e, 1 zoo, -...
TRANSCRIPT

[.
l~ 1.
UNIVERSITEDE OUAGADOUGOU
N° d'ordre:
Faculté des Scienceset Techniques
Laboratoire de Botaniqueet Biologie Végétale
1!-~SEfl-;;~~~~" ~-;- ----lPOUR l'ENSelGNEt. rA;~~~H!:1
, C A \-\f:NT .JUj~i·V ..IJ" 1/ - _,_~, ' . . M. E, S. _ .' - ""... c, .A .' . OUAGADOUGn i i
. rr'vee 1f.\ nr ~,
i Enregistré' ~~~tn~C.·ZOO, '.,THE5E 1.. -. .. .......-. .-~~-~~-'~9
Présentée pour obtenir le titre de :
DOCTEUR DE TROISIEME CYCLE
Spécialité : Sciences Biologiques AppliquéesOption : Biologie et Ecologie Végétales
Par : SouJeymane GANABA
i .1î
ROLE DES STRUCTURES RACINAIRES DANS LA DYNAMIQUEDU PEUPLEMENT LIGNEUX DE LA REGION DE LA MARE
D'OURSI (BURKINA FASO) EI/TRE 1980 ET 1992
soutenue le 6 Décembre 1994 devant la Oommlsslon d'Examen :
PréSident : Mahamane SAADOU, Maitre de Conférences,Université ABDOU Moumouni, Niamey
Examinateurs: Koffi AKPAGANA, MaItre de Conférences, Université du Bénin-Togo",(1
Sita GUINKO, Professeur, Université de OuagadougouJeanne MILLOGO-RASOLODIMBY, Maitre-Assistant, Université de OuagadougouJean Louis DEVINEAU, Docteur d'Etat Es Sciences, ORSTOM Bobo·Dioulasso
•

Nous dédions ce travail :
Aux vaillantes populations du Sahel, pourles souffrances endurées par les difficilesconditions de vie ;
A notre nouveau-né,Kiswendsida Sidi Mohamed

Avant-propos
Nous voudrions tout d'abord témoigner nos sentimentsde sincère reconnaissance à tous ceux quiont contribué de quelque manière que ce soit à l'aboutissement de ce travail.Nous voudrions adresser tout particulièrement nos remerciements:
Aux responsables du projet RUG/FAST/Ecologie et du projet RC.S Sahel pour leurs aidesmultiformes dans l'exécution de ce travail.
Au Professeur Sita GUINKO, pour avoir su avec grande lucidité entrevoir la nécessité de menerdes travaux de recherche sur la dynamique du peuplement ligneux de la région de la mared'Oursi. Nous avons beaucoup appris de sa parfaite connaissance de la région qu'il parcourtdepuis 1978. Il a dirigé avec rigueur ses travaux malgré les multiples difficultés qui ont jalonnéleur exécution. Qu'il nous soit permis de lui témoigner ici notre reconnaissance.
A Monsieur Daniel Yves ALEXANDRE, Directeur de Recherche, Conseiller Technique àl'IRBET (C.N.RS.T), pour sa disponibilité fréquente et sa courtoisie. Il a mis à notre dispositionune documentation scientifique importante, nous aidé matériellement et financièrement, a apportéune grande contribution scientifique.Nous lui exprimons le témoignage d'une reconnaissance.
Aux Professeurs Antoine NONGONIERMA de l'IFAN (Dakar) et Mahamane SAADOU del'Université de Niamey, qui, au cours de leurs brèves missions ont bien voulu accepter, aveccourtoisie, échanger leurs expériences et émettre des suggestions. Le Professeur SAADOU acorrigé notre manuscrit et accepté de présider notre jury.Nous leur exprimons notre gratitude.
A Monsieur Jean Louis DEVINEAU, Directeur de Recherche à l'ORSTOM, Antenne de BoboDioulasso, qui a fait une correction détaillée de notre manuscrit et suggéré de nombreusescorrections. Qu'il trouve ici l'expression de notre respectueuse gratitude.
A Monsieur Koffi AKPAGANA, Maître de Conférences à l'Université du Bénin (TOGO), quia accepté faire partie de notre jury pour nous juger. Qu'il soit vivement remercié.
Notre gratitude va également :
Au Professeur Issaka ZOUNGRANA ex Directeur de l'I.D.R pour sa contribution dans laconduite des travaux et dans l'acquisition de documents.
A Madame Jeanne MILLOGO-RASOLODIMBY, Maître-Assistant à la FAST qui nous a aidédans l'étude anatomique des coupes microscopiques et dans la recherche bibliographique. Ellea aussi corrigé notre manuscrit.

A Monsieur Jean Marie OUADBA, Directeur de l'IRBET qui à travers le projet R.C.S. Sahela financé les sorties de la première année. Il a par ailleurs apporté une aide fmancière etmatérielle dans l'exécution de nos travaux de recherche.
Monsieur Amadou BA du laboratoire de Microbiologie de l'IRBET nous a apporté une aidematérielle appréciable. De même Monsieur Arzouma OUATTARA de l'IRSSH (C.N.R.S.T) aaccepté de corriger notre manuscrit. Nous leur adressons nos remerciements sincères.
Nous sommes redevables aux enseignants de la rive droite de la FA.S.T. et de l'I.D.R. pour leurcollaboration et leurs contributions multiformes. Les remerciements vont en particulier:A Monsieur Dorosso A. SAMATE, Maître Assistant au Laboratoire de Chimie OrganiqueAppliquée à la FAST, qui a effectué le screening phytochimique ;A Messieurs Urbain WENNMINGA et François BOURGES Maîtres Assistants au départementde Géologie pour leur apport matériel notamment la mise à notre disposition d'un microscopebiologique photographique;A Messieurs Jean Didier ZONGO, Maître de Conférences, Abdoulaye OUEDRAOGO, PhilippeSANKARA, Henri Y. YE et Prosper ZOMBRE, Maîtres Assistants à l'I.D.R. qui nous ont aussiapporté leur aide matérielle.
Nos remerciements vont également à Messieurs Ibrahim TOE et Isidore ZONGO respectivementancien et actuel Directeurs du projet FAO Mare d'Oursi, qui durant les 3 années de terrain ontmis à notre disposition un logement décent et ont apporté une contribution logistique. Nous leuradressons ainsi qu'au personnel du projet et services associés toute notre reconnaissance.
Monsieur Nouhoun SOW du c.R. T.O. s'est dévoué pour la cartographie, mais malheureusementla non acquisition de documents de télédétection n'a pas permis la production de carte de lavégétation.
Nos pensées vont également à l'endroit de nos collègues de 3e cycle.
Nous gardons une amitié sincère des manoeuvres qui ont tour à tour travaillé avec nous dansl'excavation du sol: Yahia ABDOULAYE, Ruddouane AG AGHALI, Hama Amadou BAHDIO,Moussa BOURAIMA, Boukary BONDIERE, Yoro GANAME et Mounkaylou HAMADOU..
Nous ne saurions oublier notre épouse Ramata, pour sa participation active et son soutien moral.Nous lui renouvellons notre attachement affectif.
Nous remercions enfin nos parents, amis et connaissances pour leur soutien multiforme.

TABLE DE MATIERES
LISTE DES ABREVIATIONS UTILISEES . . . . . . . . . . . . . . . . . . . . . . .....7
ABSTRACT . . . . . . . . . . . . . . . . . . . . . . . . . . r . . . . . . . . . . . 8
INTRODUCTION GENERALE " . 9
Première partie :
GENERALITES SUR LA REGION DE LA MARE D'OURSI
PRESENTATION DE LA REGION D'ETUDE 14
1 - LOCALISATION GEOGRAPHIQUE 14
2 - CLIMAT 14
3 - GEOMORPHOLOPHIE 15
3. 1 - Les massifs rocheux et les buttes cuirassées . . . . . . . . . . . . . . . . . . . . . . 17
3.2 - Les elacis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 17
3.3 - Les dépressions et les talwees . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
3.4 - Les systèmes dunaires . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
4 - PEDOLOGIE .19
5 - HYDROGRAPHIE , 21
6 - VEGETATION 21
7 - LES HOMMES ET LEURS ACTIVITES 23
7.1 - Les hommes 23
7.2 - Les activités , 24
Deuxième partie :
DESCRIPTION ET ANALYSE DES STRUCTURES RACINAIRES
DE ACACIA RADDIANA SAVI ET PTEROCARPUS LUCENS LEPR.
INTRUDUCTION 26
1 - BIOLOGIE ET ECOLOGIE DE ACACIA RADDIANA ET PTEROCARPUS
LUCENS 26
1.1 - Distribution eéoKI'aphique. . . . . . . . . . . . . . . . .. . . . . . . . . 26
1.2 - Ecolo&ie et description de leurs peuplements 28

1.3 - Fruits et mode de dispersion des auaines . . . . . . . . . . . . . . . . . . . . . . . . 29
1.4 - Essai de eennination des waines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
2 - METHODOLOGIE 31
2. 1 - Choix des esPtces . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
2.2 - Choix des sites et des pieds 32
2.3 - Matériel et méthodes de déterrage des racines 33
2.4 - Analyses pédolo&iques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 34
2.5 - Analyses morpholo&iques et structurales des racines 34
2.5.1 - Matériel végétal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 34
2.5.2 - Méthodes d'observations 36
2.5.2.1 - Observations macroscopiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
2.5.2.2 - Observations à la loupe binoculaire . . . . . . . . . . . . . . . . . . . . . . . . . . 37
2.5.2.3 - Observations microscopiques 38
2.6 - Quelques difficultés méthodologiques 38
3 - RESULTATS ET DISCUSSIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.1 - Caractéristiques des racines de Acacia raddiana et de Pterocarpus lucens ... 39
3.2 - Oreanisation morpholo&igue des systèmes racinaires . . . . . . . . . . . . . . . . .41
3.3 - Oreanisation structurale des systèmes racinaires . . . . . . . . . . . . . . . . . . . 44
3.4 - Analyses du développement et du fonctionnement des systèmes racinaires ... 46
3.4.1 - Relations entre l'enracinement et la hauteur du pied
chez Acacia raddiana 46
3.4.2 - Relations entre l'enracinement et le diamètre à la base du pied
chez Acacia raddiana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 50
3.4.3 - Relations entre l'enracinement et l'âge estimé du pied
chez Acacia raddiana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 52
3.4.4 - Relations entre la profondeur de la zone d'absorption et la hauteur du pied
chez Acacia raddiana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 58
3.4.5 - Relations entre d'une part l'enracinement et d'autre part la hauteur et le
diamètre à la base du pied chez Pterocarpus lucens . . . . . . . . . . . . . . . . 58
3.4.6 - Formes d'absorption de l'eau et des substances dissoutes 61
3.5 - Relations entre les systèmes racinaires 65
3.5.1 - Compétition spatiale 65
3.5.2 - Compétition trophique 67

3.5.2.1 - Morphologie des anastomoses 67
3.5.2.2 - Structures et mécanismes des anastomoses 69.
3.5.2.3 - Caractérisation phytochimique des anastomoses . . . . . . . . . . . . . . . . . . . 70
3.5.2.4 - Nature des anastomoses 71i
3.6 - Associations symbiotiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .72
3.6.1 - Plantes-bactéries _ . . . . . . . . . . . . . . . . . . . . . . . . .. 72
3.6.1.1 - Nodules d'Acacia raddiana 73
3.6.1.2 - Nodules de Pterocarpus lucens 73
3.6.1.3 - Quelques réflexions sur l'association plante-bactéries 74
3.6.2 - Plantes-termites 74
3.6.2.1 - Observations in situ 74
3.6.2.2 - Rôle des termites dans les écosystèmes 75
CONCLUSION 76
Troisième partie :
ESSAI D'ETUDE DESCRIPTIVE ET DYNAMIQUE DU PEUPLEMENT LIGNEUX
1 - RAPPELS BmLIOGRAPHIQUES . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 80
1.1 - Subdivisions phyto&éo2faphiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
1.2 - Formations et groupements véeétaux . . . . . . . . . . . . . . . . . . . . . . . . . . .80
2 - METHODOLOGIE &2
2.1 - Carto2faphie 82
2.2 - Inventaire floristique 83
2.2.1 - Mesures qualitatives 84
2.2.2 - Mesures quantitatives 85
2.3 - Dynamique du peuplement lieneux 87
3 - RESULTATS ET DISCUSSIONS 88
3.1 - Etude descriptive du peuplement liameux . . . . . . . . . . . . . . . . . . . . . . . . 88
3.1.1 - Composition floristique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
3.1.2 - Structure " 90
3.1.2.1 - Peuplement ligneux des dépressions et talwegs 91
3.1.2.2 - 'Peuplement ligneux des glacis 93
3.1.2.3 - Peuplement ligneux des buttes cuirassées et reliefs rocheux 9.3

3. 1.2.4 - Peuplement ligneux des dunes et ensablements . . . . . . . . . . . . . . . . . . . 94
3.1.2.5 - Peuplement ligneux des champs et jachères . . . . . . . . . . . . . . . . . . . . . 94
3.1.2.6 - Interprétation des résultats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .97
3. 1.2.7 - Discussions sur la structure du peuplement ligneux . . . . . . . . . . . . . . . . .9.8
3.1.3 - Analyse de la répartition en fonction du diamètre . . . . . . . . . . . . . . . . 101
3.1.4 - Degré d'appétabilité 101
3.2 - Dynamique du peuplement lilmeux . . . . . . . . . . . . . . . . . . . . . . . . . . . 102
3.2.1 - Analyse qualitative et quantitative entre 1980 et 1992 102
3.2.2 - La régénération. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 106
3.2.3 - La mortalité 107
3.2.4 - Les coupes 108
3.2.5 - Discussions sur la vitalité. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .108
3.2.6 - Les jachères en région sahélienne 110
3.2.6.1 - Nature et mode de défrichement . . . . . . . . . . . . . . . . . . . . . . . . . . . 110
3.2.6.2 - Dynamique spatiale et mode d'installation des espèces ligneuses . . . . . . . . 111
3.2.6.3 - Importance et gestion des jachères 112
3.2.7 - Discussions sur les causes et les mécanismes de la dégradation
du peuplement ligneux . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
3.2.7.1 - Les facteurs climatiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
3.2.7.2 - Les facteurs mésologiques induits par le climat. . . . . . . . . . . . . . . . . . . 114
3.2.7.3 - Les facteurs anthropiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116
3.2.7.4 - Cas particulier des deux espèces . . . . . . . . . . . . . . . . . . . . . . . . . . . 118
3.2.8 - Conséquences de la dynamique 122
3.2.9 - Localisation de la dégradation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
CONCLUSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .... 126
CONCLUSION GENERALE ET PERSPECTIVES 129
BIBLIOGRAPHIE D2
LISTE DES ANNEXES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 141
LISTE DES CARTES " 143
LISTE DES FIGURES . . . .. 1M

A.C.C.- LAT =
BU.NA.SOLS =
C.N.RS.T. =
C.N.S.F. =
C.R.T.O. =
C.T.F.T. =
CILSS =
FA.S.T. =
FAO =
GERDAT =
GRIZA =
I.D.R. =
I.S.N.-I.D.R. =
IRBET =
ORSTOM =
PEPS Oursi =
RC.S.- Sahel =
SPOT =
7
LISTE DES ABREVIATIONS UTILISEES
Action Complémentaire Coordonnée - Lutte contre l'Aridité en milieu
Tropical
Bureau National des Sols (Burkina Faso)
Centre National de Recherche Scientifique et Technologique
(Burkina Faso)
Centre National des Semences Forestières (Burkina Faso)
Centre Régional de Télédétection de Ouagadougou
Centre Technique Forestier Tropical
Comité Inter-Etats de Lutte contre la Sécheresse dans le Sahel
Faculté des Sciences et Techniques de l'Université de Ouagadougou
Organisation des Nations Unies pour l'Alimentation et l'Agriculture
Groupement d'Etudes et de Recherches pour le Développement de
l'Agronomie Tropicale
Groupe de Recherches Interdisciplinaires en Zones Arides
Institut de Développement Rural (Université de Ouagadougou)
Institut de Développement Rural - Institut des Sciences de la Nature
(ancien I.S.P.)
Institut de Recherche en Biologie et Ecologie Tropicale du C.N .RS.T.
Institut français de recherche scientifique pour le développement en
coopération
Programme d'Evaluation Préliminaire SPOT à Oursi
Renforcement des Capacités Scientifiques dans le Sahel
Satellite Probatoire d'Observation de la Terre
Langues nationales:
(f) = foulfouldé ; (t) = tamachek ; (s) = sonraï ; (m) = mooré

8
ABSTRACT
A selective mortality of woody plants is observed in the region of the Swamp of
Oursi, sinee the high drought in 1972-1973.
The tentative to explain such selectivity of mortality led to the study of the root
system of two woody plants speeies eharacteristics of the region, Acacia raddiana Savi, an
invading plant and Pterocarpus lucens Lepr., a perishing plant.
Their root system is both tap-rooted and up-running. The difference is the ability of Acacia
raddiana to have deep and extensive root system and also large root vessels which resulted
in inereased water absorption. But the other species, Pterocarpus lucens, have a less
developped root system because of soil texture, aerial euttings by humans and frequent roots
eut by termites. The low water table explained the high mortality of this species, so that it
is in danger to disappear.
In addition, root anastomosis were identified between Balanites aegyptiaca and Acacia
raddiana or Bauhinia rufescens. This phenomenon quite new and related to sorne kind of
parasitism.
Furthennore, analysis of the dynamie of woody vegetation between 1980 and 1992,
indicated floristie graduaI withdrawal of soudanian speeies (Anogeissus leiocarpus, Entada
africana and SC!erocarya birrea) in favor of sahelian speeies (Acacia raddiana, Balanites
aegyptiaca, Calotropis procera, Leptadenia pyrotechnica and Maerua crassifoliay. and
disappearance or relocation of high-water table species (Anogeissus leiocarpus, Diospyros
mespiliformis, Ficus gnaphalocarpa, Mitragyna inermis and Tamarindus indicaï,
The pseudoclimax seems to be evolving toward Acacia raddiana savannah bushland.
Keys words : Burkina Faso - Oursi Swamp - drought - Acacia raddiana - Pterocarpus lucens
- roots system - root anastomosis - woody vegetation - dynamic - mortality - savannah
bushland.

9
INTRODUCTION GENERALE
Depuis plusieurs décennies, le Sahel est confronté à de graves déficits
pluviométriques, sources de sécheresses (1911, 1929, 1944, 1968, 1973 et 1984). Celles-ci
se traduisent par une mortalité de certaines plantes ligneuses. Le phénomène touche de vastes
espaces (des milliers d'ha) sur lesquels les arbres se dessèchent et meurent. Si certaines
espèces présentent en effet une certaine résistance comme Acacia raddiana Savi en pleine
expansion, d'autres en revanche sont très touchées comme Pterocarpus lucens Lepr. dont les
peuplements régressent fortement.
La mortalité des plantes s'est manifestée dans la partie méridionale de la zone
subsahélienne (région de Tougouri à Yalgo) et jusque dans la zone soudanienne du Plateau
Central (DIALLO, 1990).
Pour parvenir à une utilisation rationnelle des ressources renouvelables du Sahel, il
est nécessaire de bien connaître la situation actuelle, de cerner les tendances évolutives et
aussi de rechercher les causes de la forte mortalité des plantes.
Diverses causes notamment climatiques et anthropiques, ont déjà été largement
évoquées par divers auteurs (DEPIERRE et GILLET, 1971; TOUTAIN et PlOT, 1980;
PlOT et al., 1980; BARRY et al., 1983; BORTOU, 1983; GUINKO, 1984) pour expliquer
la dégradation du peuplement ligneux. Toutefois, les causes biologiques (adaptation, système
racinaire, âge... ) et hydrogéologiques (niveau des nappes d'eau souterraine) sont moins bien
connues. Peu est par ailleurs connu sur les divers comportements des ligneux vis-à-vis des
facteurs du milieu.
Nous avons choisi d'approcher le phénomène de la dynamique du peuplement ligneux
par l'étude comparative des systèmes racinaires de 2 espèces: Acacia raddiana Savi dont les
populations s'accroissent et Pterocarpus lucens Lepr. subissant une forte mortalité. Ces 2
espèces résument la dynamique du peuplement ligneux de la région.

10
La région de la Mare d'Oursi qui est le cadre de la présente étude est considérée
comme représentative des espaces sahéliens. En effet, elle concentre en un espace restreint
toutes les caractéristiques de la région sahélienne comme le montrent les communications au
Colloque International sur la Mare d'Oursi (C.N.R.S.T., 1992). Elle est par ailleurs bien
connue grâce aux nombreux travaux scientifiques qui y ont déjà été menés dans divers
programmes de recherche.
Ces programmes correspondaient à un élan de solidarité internationale qui s'est
manifesté à la suite des effets catastrophiques des épisodes de sécheresse climatique qui ont
affecté le Sahel. Celui des années 1972-73 a eu des conséquences écologiques, économiques
et sociales graves (BARRAL et BENOIT, 1976). Ces programmes avaient en outre pour
objectif une meilleure connaissance des problèmes de la région par l'étude du milieu
physique, des écosystèmes et des systèmes de production, utiles pour des actions
d'intervention futures. Ils devaient permettre de juguler les effets des crises climatiques dans
cette frange sahélienne. Ce sont : le programme A.C.C. Lutte contre l'Aridité en milieu
Tropical (LAT) de 1976 à 1981 ; Simulation SPOT, 1981-1983 ; Programme d'Evaluation
Préliminaire SPOT (PEPS) Oursi, 1985-1987 ; Groupe de Recherches Interdisciplinaires en
Zones Arides (GRIZA), 1980.
Le colloque tenu au Burkina Faso, du 17 au 21 Février 1992, a été l'occasion surtout
de mettre en relief les acquis des études réalisées. Il a aussi permis de définir des orientations
pour les futurs programmes de recherche dans la région sahélienne en général. Les axes
d'orientation sont le suivi des systèmes écologiques et la recherche expérimentale pour le
développement.
Notre thème intitulé :
"Rôle des structures racinaires dans la dynamique du peuplement ligneux
de la région de la Mare d'Oursi (Burkina Faso) entre 1980 et 1992"
s'inscrit dans le cadre de ces programmes. Il sera présenté selon le plan suivant:
- Pour mieux comprendre et situer la région d'étude, la première partie du travail présentera
les généralités sur les facteurs physiques et socio-économiques,

11
- La deuxième partie exposera la description et l'analyse des structures racinaires des deux
espèces ligneuses choisies.
- Elle nous conduira à la troisième partie, qui est un essai de description de la physionomie
et d'analyse de la dynamique du peuplement ligneux de la région.

Première partie :
GENERALITES SUR LA REGION DE
LA MARE D'OURSI
Carte nO 1
SITUATION DE LA MARE D'OURSI, BURKINA FASO( D'après CHEVALIER et 01. 1 ~985)
12" N--....,.=-----+l-----i---..-+~---~---+_'~L-------~,__
BENIN
GHANA
10° N ---k--f---b......;;;;;;~~T---+--------t_--~::------
a 50 100 150 200 250km1 1 1 1 1 1
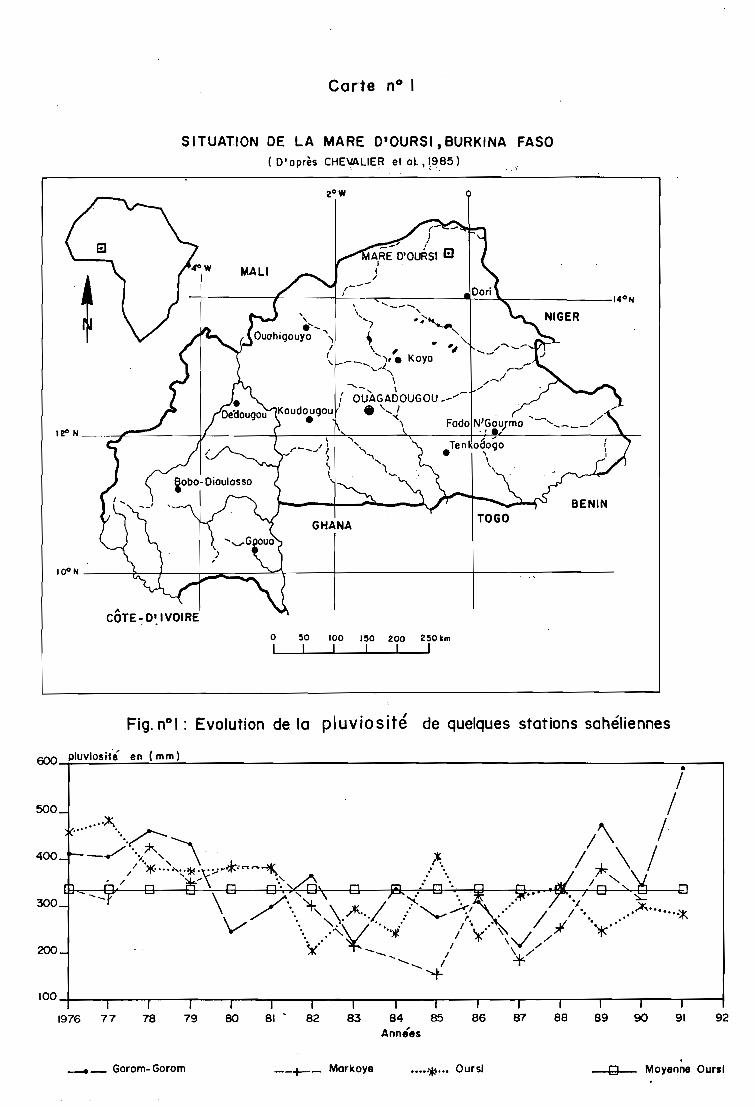
Fig. nOI : Evolution de la pluviosité de quelques stations sahéliennes
600 pluviosité en (mm)•/
//
/\ ..,/' -, ~. ./ ~
\ J'/ ~'''\'~'''/ / '-....-'(/>~V..<\·. ///./1 "'" )I:. •••••••~V .,<.\ / ....'i .,.. 'y 7f' l'
~., ~ --- / \"-- / .:..1,./
-.... 1 1-....--+
........,):.-.~:;'" ~ -..... ..- ....... ~" \
/ "'f{,'::-"~~'\~.*""~~. / ' .~"
200
300
400
500
100 -f---r-----r---r-----r--T"""'---r---r--......,,......--.---r----r--,...----r--,...---,.--1
__ Gorom- Gorom .....*... Oursl__+-_ Markoye.
Moyenne Oursl
929190
g
8988878683 84 85Années
81' 828079781976 77

14
PRESENTATION DE LA ZONE D'ETUDE
1 - LOCALISATION GEOGRAPHIQUE
Oursi est une localité située au nord du Burkina Faso, dans la province sahélienne de
l'Oudalan, à environ 360 km de Ouagadougou la capitale. La mare du même nom est un lac
éolien endoréique reposant sur un socle précambrien. Elle est pérenne en années successives
de pluviosité normale.
La région d'étude qui inclut le bassin versant de la mare couvre une superficie de 1
200 km'. Elle est comprise entre les latitudes 140 32' et 140 50' nord et les longitudes 0 0
16' et 0 0 40' ouest (carte n° 1).
La région est dominée par une pénéplaine granitique dont le stade d'évolution est
avancé. Les reliefs correspondant à des affleurements de roches endogènes cristallines et
éruptives émergent çà et là.
L'agencement de ces différents éléments du paysage conduit à distinguer 2 parties:
- une plaine méridionale constituée d'une mosaïque de buttes rocheuses, de glacis et d'un
réseau hydrographique très ramifié convergeant vers les dépressions ;
- au nord, le socle est recouvert de sable formant des successions de cordons dunaires
parallèles hauts de 20 à 50m.
2 - CLIl\fAT
Le régime climatique de l'Oudalan est caractérisé par une longue saison sèche qui
dure 8 à 10 mois suivie d'une courte saison de pluies marquée par un changement brusque
du paysage. Ces pluies qui redonnent vivacité à la végétation ont pour la plupart un caractère
orageux très prononcé et une répartition spatiale très hétérogène.

15
Les analyses de température, d'humidité, de vitesse et direction des vents, de
l'évapotranspiration faites par ALBERGEL et al., (1984); CHEVALIER et al., (1985),
confirment le caractère aride du climat.
Les moyennes interannuelles des précipitations mensuelles et annuelles observées aux
postes de référence de Gorom-Gorom et Markoye, situés respectivement à 40 km au sud-est
et à 70 km au nord-est d'Oursi, montrent une variation interannuelle des quantités de pluies.
Des périodes fortement déficitaires ont conduit à des années de crise climatique et écologique
comme celles de 1983 et 1987. D'autres ont une pluviosité supérieure à la moyenne d'Oursi
1976-78, 1986, 1989 et 1991.
Cette analyse montre aussi une diminution générale de la pluviosité annuelle depuis
1982 (figure n° 1).
Tous les auteurs ayant travaillé sur le climat sahélien (PUECH, 1983; ALBERGEL
et al., 1984; CHEVALIER et al., 1985; HUBERT et CARBONNEL, 1987; CLAUDE et
al., 1991) soulignent le caractère aléatoire des flux atmosphériques et océaniques à l'origine
des pluies. Ils concluent par ailleurs à l'impossibilité d'établir leur caractère cyclique et
régulier.
Les auteurs admettent cependant, l'évolution vers une aridification. Cette évolution
se traduit par un allongement de la saison sèche, une diminution de la disponibilité en eau
pour les végétaux et une répercussion sur leur répartition et leur phénologie.
3 - GEOMORPHOLOGIE
Les auteurs ayant travaillé dans la région (TOUTAIN, 1976; JOLY, DEWOLF et
RIOU, 1980; PlOT et al., 1980; GROUZIS, 1988 et C.R.T.O, 1989) distinguent
globalement 4 ensembles géomorphologiques :
- les talwegs et les dépressions,
- le système dunaire et les placages sableux,
- les glacis,
- les massifs rocheux et les buttes cuirassées.

Corte nO 2
NN
Echelle: 1/200 000
21
1-376\ Point géodésique
LEGENDE
Site d'Acacia
Villages
B Route secondaire--
~ Piste
14° 0 Zone inondabler ......:
BJ Cours d'eau
~ More
15
~~1
Source: CR.T.O., 1989
D' OURSI
~J'----' -_T--_l-
MARE
Z ormoko
LA
,
"\
--,-~L .-.:-----343' 1 ... '-',-.--'- " -,-<.._.«("-- Timataltal
........ ), '-- .>-- \
,',' \--._---)
• .-.--- ).--<,
........, -.-/./. ........ ..... _- ...' •• __l._ Tirnbolo ".
\111L.. .__--J .....---_
~\ '-.,. \ <,~.>, " _,•.
' .._ ...

17
3. 1 - Les massifs rocheux et les buttes cuirassées
Les reliefs rocheux et les buttes cuirassées sont constitués de :
- massifs de gabbros formant les reliefs les plus élevés du bassin: Tin Edjar (518 m) et Kolel
(372 m) riches en magnétite et Warga (365 m) ;
- reliefs granitiques élevés sur 5 à 20 fi, dispersés dans la région et dont l'altération forme
des chaos et des boules (Tondaré, Tounté ... );
- bunes cuirassées au sud du bassin: Gouba (350 à 380 m).
3.2 - Les Klacis
Les glacis désignent des surfaces relativement planes à pente faible 1 englobant une
grande variété de matériaux, d'évolution et de dynamique de modelés. C'est pourquoi leur
classification est différente chez les géomorphologues et les pédologues (CLAUDE et al.,
1991).
Ce sont:
- des glacis arénacés liés aux formations granitiques à aspect de surface en mosaïque de
plages glacées et de sable d'arène en transit;
- des glacis gravillonnaires occupant les versants nord-ouest et sud-ouest du bassin ainsi que
le centre et dont l'aspect unitaire de surface est lié à la présence remarquable d'un pavage
de cailloux et de graviers ;
- des glacis cuirassés peu nombreux (cas du piémont sud de Tin Edjar) qui sont souvent
recouverts d'une fine couche de sable éolien.
Ces différents types de glacis ont une caractéristique commune : la présence de
formations sous-jacentes compactes qui empêchent l'infiltration de l'eau dans le sol.

19
3.3 - Les talwee;s et les dépressions
Les cours d'eau exploitent quelques failles et fractures du socle cristallin. Ces cours
d'eau prenant naissance dans les buttes rocheuses et les glacis se concentrent dans le sud. Ils
se gonflent seulement après les orages et coulent pour alimenter les bassins fermés dont les
cordons dunaires barrent l'écoulement.
Des mares d'étendues inégales dont certaines sont pérennes jalonnent le cordon
dunaire sur toute sa longueur; la plus importante, celle d'Oursi, est longue de 7 km et large
de 3 km. A l'est, se trouve la mare de Yomboli et à l'ouest celles de Tin Edjar et de Dibissi.
Toutes ces mares se caractérisent par leur faible profondeur. Zones privilégiées de
pâturage et d'abreuvement pour le bétail en saison sèche, les mares sont progressivement
comblées par le sable dunaire qui y est entraîné quotidiennement par le piétinement des
animaux et le passage des véhicules de transport (carte n° 2).
3.4 - Les systèmes dunaires
Les systèmes dunaires se composent de 4 grands ensembles:
- l'erg récent en cordon rectiligne à pente raide vers le nord et douce vers le sud, se
remobilisant au niveau du village d' Oursi au passage quotidien du bétail pour la mare ;
- l'erg ancien est occupé surtout par des champs de petit mil (cas de la dune de Ganadawri);
- les dunes de placages sableux forment les piémonts nord des massifs gabbroïques de Kolel
et de Warga et sont exploitées pour la culture de petit mil et de sorgho ;
- les placages amorphes peu développés et à faible profondeur (2 m environ) présents sur le
bassin voisin du Gountouré entre Kolel et Pétoy sont aussi exploités pour l'agriculture.
4 - PEDOLOGIE
On distingue une grande diversité de sols liés au climat soudano-sahélien et à la
diversité des roches mères (AUBREVILLE, 1940). Pour la région de la Mare d'Oursi, 7
classes de sol sont définies (carte n° 3 et annexe II).

87 mm. IK =15
mm ~ massifs rocheuxmm E3 bas- fonds
mm Ij--d-- mores en eau0 1 2km1 1 1
< LNI !C:: 60"""
<LNI tE;; 70
<LNI !C:: 75"""
50
60
70
1\
ruissellement et
fréquence décennale pour P =
mm
Corte des aptitudesd' Oursi
4~
5~
6~
mm 7 E:3 75 <LNI
--
LN! ~ 30 mm
30 <LN! ~40
._- ",.
LNI = lame non- infiltrée deSource: DEVINEAU et 01. 1 1992
1 m:i!!${.12 r:~';;:'i::::J
3~ 40 <LN! ~50
Carte n04

21
Les sols de la région se distinguent par une carence à des degrés divers en potassium
échangeable, en phosphore total et assimilable et en azote total. Ils ont également une teneur
en matière .organique faible à très faible, à l'exception des sols hydromorphes. En outre,
presque tous les sols ont une capacité de rétention en eau très faible et des propriétés
physiques défavorables: structures massives, porosité intergranulaire élevée. La teneur en
fer présent sous forme de taches d'amas de concrétions est très élevée (BUNASOLS, 1981).
Les termites y sont abondants avec des formations en termitières cathédrales. Il existe
aussi des termitières hypogées qui ne sont pas visibles.
5 - HYDROGRAPHIE
Le réseau hydrographique de la région appartient au bassin du Niger à l'exception des
zones d'endoréisme.
Le Gountouré est le seul cours d'eau capable de franchir les cordons dunaires pour
remonter à l'extrême nord et alimenter le Béli. Il forme sur son parcours la mare de
Ganadawri.
L'écoulement des eaux, actif seulement après les orages. est discontinu et intermittent.
Les sols des axes de drainage constituent en saison des pluies des prairies à fonio sauvage
(Panicum anabaptistum, Panicum laetum et Panicum subalbidumï,
Trois traits caractérisent le réseau hydrographique: la faiblesse générale des pentes
d'écoulement, la brièveté de la période d'écoulement et l'importance de l'évaporation
(BARRAL, 1977; carte n° 4).
6 - VEGETATION
Les principales formations végétales naturelles de la région sont les steppes et les
"brousses tigrées" dominées par les espèces épineuses. Acacia raddiana est l'espèce la plus
répandue.

Carte nO 5
+++
L1PTAKO
EXTENSION DE LA BROUSSE TIGREE PAR
RAPPORT AUX FORMATIONS GEOLOGIQUES
(D'après schéma géologique de J. GALLAISmodifié 1 BARRY el 01, 1983 )
o1
~ Socle ~ Série d' Yaoubo ~ Serie d'Isma
[]]]] Grès de Firgoun ~ Serie d' Hombori- - Brousse ti9réeDouenlza
m Orthogneiss (Bourré) ~ Primaire. .. .
• Série d'Ansango ~ C. terminal-'\ Limite du ",U
~de Hambori-
Série de Labezanga 0 Quorlernaire ,.... Goundam

23
Les steppes constituent les formations des dunes, des cordons sableux, des piémonts de
buttes et des glacis. Dans les dépressions et les talwegs, les steppes forment des fourrés ou
galeries ripicoles plus ou moins ouverts.
Les "brousses tigrées", formées d'une alternance de bandes sinueuses de végétation
dense et de plages dénudées de glacis encroûtés, se rencontrent dans la partie septentrionale
(Férérillo, Zarmakoy et Tin Atan ; carte n° 5). Ces formations végétales se caractérisent par
une dégradation consécutive à la forte mortalité de Pterocarpus lucens.
La végétation de la région se singularise par une abondance d'espèces herbacées parmi
lesquelles Schoenefeldia gracilis tient une place importante. Ces espèces constituent des
pâturages riches et variés. Par contre, les ligneux sont faiblement représentés tant en nombre
de pieds qu'en espèces. Parmi les ligneux, l'espèce dominante est Acacia raddiana alors que
Pterocarpus lucens est en voie de disparition.
Les feux de brousse sont absents de cette région.
7 - LES HOMMES ET LEURS ACTIVITES
7.1 - Les hommes
L'Oudalan a été envahi vers 1760 par les Kel Tamachek puis par les Peul Gaobé, et
les Peul du Djelgodji. Les premiers allaient instaurer une hégémonie dans la région qui
prendra désormais leur nom. Des relations de dépendance et une organisation de la société
en classes de nobles et d'esclaves se sont établies. Cela allait reléguer au second plan, la
recherche de la "codification" dans l'utilisation et l'exploitation des ressources du milieu.
Cela expliquerait l'absence de droit sur le sol et l'eau (BARRAL, 1977; GROUZIS, 1988).
Quatre principaux groupes ethniques ayant abandonné ce type de rapports de
dépendance entre hommes libres (nobles) et captifs tributaires (esclaves) coexistent. Ce sont
les Bella, les Peul, les Sonraï et les Touareg (Al kasseibaytan principalement) (BARRAL,
1977, GROUZIS, 1988).

24
Les facteurs de concentration humaine sont la présence d'eau et l'existence de terres
arables.
6 2 - Les activités
Les activités socio-économiques des sociétés sahéliennes sont basées sur l'exploitation
des ressources naturelles locales. La nature des droits sur le milieu, la pression
démographique et les événements socio-politiques antérieurs à l'indépendance du pays
(esclavage), ont marqué de manière significative l'exploitation et l'évolution des ressources
du milieu par la cueillette, l'élevage et l'agriculture (GROUZIS, 1988; MILLEVILLE,
1990).
L'élevage constitue l'activité principale de la population de cette région. II assure la
plus grande partie de l'alimentation et constitue la principale source de revenus.
Dans l'Oudalan, les hommes se déplacent avec leurs troupeaux pendant la saison des
pluies, le regroupement s'imposant en saison sèche à cause du manque d'eau.
Il n'y a pas de doute que le troupeau constitue un capital pour l'éleveur sahélien. Mais
ce capital est précaire du fait de la menace permanente : le bétail croît lentement, peut
décroître rapidement et dans le pire des cas disparaître totalement. Les causes en sont les
variations du niveau des ressources fourragères et hydrauliques d'une année à l'autre
(THEBAUD, 1990).
La gestion de ce capital est peu productive. Ainsi non seulement elle ne permet pas de
couvrir les besoins en biens de consommation des éleveurs, mais encore elle fragilise
beaucoup le milieu qui est sensé le faire fructifier.
En définitive, la région de la Mare d'Oursi est caractérisée par un climat aride, des
sols pauvres et une végétation riche en herbacées annuelles mais pauvre en plantes ligneuses.
Ces ligneux subissent une forte et sélective mortalité et un taux élevé d'exploitation. Nous
avons tenté une description et une analyse des structures racinaires afm d'expliquer leurs
propriétés et leur rôle dans la vie des plantes étudiées.

25
Deuxième partie :
DESCRIPTION ET ANALYSE DES
STRUCTURES RACINAIRES DE ACACIA
RADDIANA SAVI ET PTEROCARPUS
LUCENS LEPR.

26
INTRODUCTION
De nombreux travaux ont déjà été consacrés aux deux espèces qui font l'objet de la
présente étude. Toutefois, ces travaux n'ont pas intéressé la partie souterraine de ces plantes.
Or, le rôle du système racinaire reste, pour tout végétal, primordial dans la fixation
et dans la recherche des éléments nutritifs. Dans les conditions naturelles, le développement
des végétaux dépend à la fois de leurs possibilités d'absorption par les racines et de la
disponibilité d'éléments minéraux et d'eau pour satisfaire les besoins de la plante. La
disponibilité de ces éléments est d'abord fonction de l'importance des réserves, de leur
dynamique dans le sol et de la distribution des racines dans ce dernier.
En milieu sahélien, caractérisé par une aridité climatique, l'importance de la
colonisation du sol par les racines apparaît donc comme un facteur très important dans la
dynamique des peuplements ligneux. Nous nous sommes intéressés aux 2 espèces Acacia
raddiana et Pterocarpus lucensdont l'opportunité du choix est mentionnée dans le paragraphe
suivant.
1 - BIOLOGIE ET ECOLOGIE DE ACACIA RADDIANA ET PTEROCARPUS LUCENS
Acacia raddiana Savi ou Acacia de Raddi (BERHAUT, 1975) ou Acacia tortilis
(Forsk) Hayne ssp. raddiana (Savi) Brenan (AUBREVILLE, 1950 ; FAHN, 1959 ;
VASSAL, 1972 ; BERHAUT, 1976; NONGONIERMA, 1978 ; VON MAYDELL, 1983)
et Pterocarpus lucens Lepr. ex Guill et Perr. ou Ptérocarpe luisant, sont deux légumineuses
appartenant respectivement aux familles des Mimosaceae et des Papilionaceae.
1.1 - Distribution .:éoarraphique
Acacia raddiana, espèce ligneuse des régions arides et semi-arides (isohyètes 50 à
500 mm) au sud et au nord du Sahara, a une aire de distribution qui va du Sénégal à
l'Afrique Orientale et l'Arabie du Sud. Elle se rencontre au Proche et Moyen Orient et en
Afrique du Sud.
BURKINA FASO
AIRE DE DISTRIBUTION DE PTEROCARPUS LUCENS
ET ACACIA RADDIANA
850
,/
Car1è nO 6
Dm Groupement sahélien
~ Groupement soudono-sohelien
~ Groupement soudonien vrai
~ Groupement sud-soudonien
CARTOGRAPHIE DU CNRST ~.o. c. KI1° 20
NA
_. _ Limite sud Pterocorpus lucens
.. sud~ roddiono•• ••• Llm,te nord Butyrospermum parkii
Pterocorpus lucens hors de son aire
H
Source: 80RTOLI, 198320 10
G
VOl R E4"
1001
SC)1
o1
LI MITE DES GROUPEMENTS.~
C 0 T E
'\.
-"--e.,J;\1
/(

28
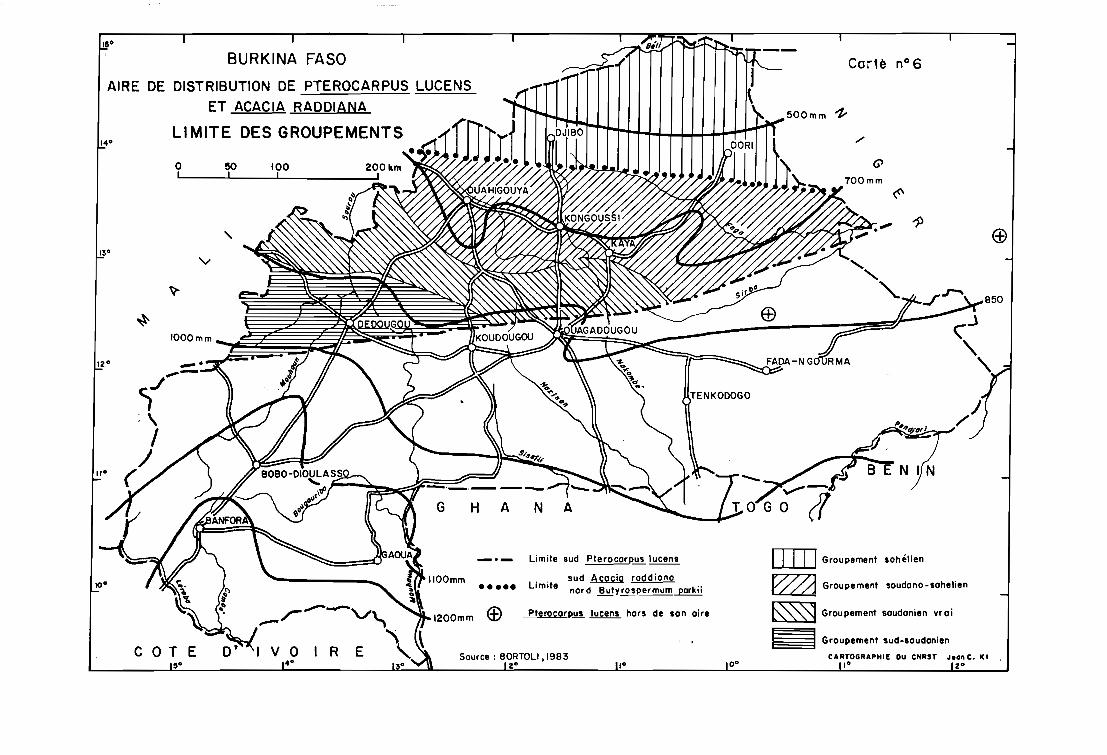
Au Burkina Faso, son aire de répartition est incluse dans le domaine sahélien. Sa
limite sud correspond à la barrière naturelle constituée par la chaîne de buttes cuirassées du
Birrimien (carte n° 6).
Pterocarpus lucens est une espèce de l'Afrique sèche sahélo-soudanienne dont l'aire
de répartition s'étend du Sénégal et de la Mauritanie à la Come de l'Afrique. Mais elle serait
absente du Niger (BORTOLI, 1983) où par contre Acacia raddiana donnerait des formes
arborescentes dans les vallées sèches de l'Aïr, vers 700 m d'altitude,dans des conditions
particulières de pédologie sableuse et de drainage abondant (communication personnelle du
Professeur SAADOU). Cette aire déborde largement le Sahel burkinabè vers le sud, en deçà
de la chaîne birrimienne, jusqu'aux rives du Nakambé et dans la province de la Kossi (carte
n° 6).
1.2 - EcoloKÏe et description des peuplements
Dans le Sahel burkinabè, Acacia raddiana ou affagak (t), gonponsgo (m), forme des
peuplements arbustifs bas (2 à 3 m) souvent monospécifiques sur les placages sableux (photo
n° 1). On retrouve des pieds plus grands (7 à 8 m) sur les dunes. Sur les glacis, les pieds
sont arbustifs et dispersés parmi d'autres espèces.
Sa régénération est grande en bordure des mares temporaires (photo n° 2).
Se présentant sous la forme d'arbuste de 3 à 5 m de haut, atteignant rarement la taille
de 7 à 10 m, mais pratiquement jamais les 15 m, selon la description de AUBREVILLE
(1950), le Ptérocarpe luisant (alibunis (t), penperga (m)) pousse sur des terrains secs. Sa
taille dépend à la fois de la nature et de la profondeur du sol, des conditions pluviométriques
et d'exploitation. Cette espèce constitue encore de nos jours des fourrés arbustifs (bushs)
entre Dori et Kaya (DIALLO, 1990) mais dans lesquels elle est en forte régression.
Dans l'Oudalan, cette espèce est dominante surtout dans les brousses tigrées au nord
des cordons dunaires en compagnie de Combretum micranthum, Commiphora africana et
Grewia bicolor. Au sud, on rencontre des reliques d'anciens groupements à proximité des
massifs et reliefs parsemés de restes d'anciens hauts fourneaux et souvent de bois morts.

29
Cette espèce est très recherchée comme combustible mais aussi comme arbre
fourrager. C'est pourquoi elle a une croissance lente liée à la coupe fréquente des parties
aériennes (photo n° 3).
De plus de nombreux jeunes plants de l'espèce poussent sous les pieds touffus de ligneux de
ces groupements.
Le ptérocarpe est fortement soumis à de multiples contraintes de développement dans
son milieu naturel (photo n° 4). Il est en déséquilibre avec son milieu. Cette situation laisse
penser, selon BORTOLI (op. cit.), que certains glacis à Acacia laeta, Balanites aegyptiaca
et Commiphora africana seraient d'anciens peuplements de Pterocarpus lucens.
1.3 - Fruits et mode de dispersion des graines
Les fruits de Pterocarpus lucens sont des gousses ovées, allongées et aplaties
apparaissant en fin de saison pluvieuse. Ces fruits se maintiennent sur le pied pendant la
défoliation en saison sèche. Par contre, les gousses typiquement spiralées d'Acacia raddiana
sont abondantes de février à mai (AUBREVILLE, 1950 ; VON MAYDELL, 1983).
Les fruits d'Acacia raddiana peuvent tomber sous l'action du vent mais peuvent
également être arrachés par les bergers, les chèvres ou les dromadaires.
Tandis que les sucs digestifs des animaux altèrent les graines du ptérocarpe, ils
augmentent en revanche le pouvoir germinatif de celles de l'acacia en levant la dormance due
au tégument (MAYDELL, 1983; TYBIRK, 1991). Ceci explique l'abondance des jeunes
pousses dans les parcs àchèvres et à moutons. La levéede cette dormance physique, exogène
ou inhibition tégumentaire (BELLEFONTAINE, 1992) est aussi réalisée par plusieurs
techniques. Chez les acacias elle est réalisée par ébouillantage (ou cuisson de 5 à 10 rnn)
suivi d'un refroidissement lent, par traitement à l'acide sulfurique concentré suivi d'un
rinçage à l'eau, et par abrasion ou scarification manuelle ou mécanique du tégument
(KARSCHüN, 1975; LE GRAND, 1979; PATHAK et al., 1980; VON MAYDELL, 1983;
TYBIRK, 1991).

30
Dans la nature, elle s'effectuerait par l'action des insectes notamment les termites et
par l'alternance chaleur/fraîcheur. En effet, les graines non traitées ont un pouvoir de
germination lent et faible, comme l'a indiqué PATHAK et al., (1980). Acacia raddiana est
une espèce endozoochore à l'opposé de Pterocarpus lucens, espèce anémochore.
Les fruits verts d'Acacia et du ptérocarpe sont l'objet d'attaque de la part des insectes:
des Coléoptères (Coleoptera) dont la famille des Bruchidés (Bruchidae) constitue la plus
importante des prédateurs, des punaises (Hemiptera), des papillons et mites (Lepidoptera),
des guêpes et fourmis (Hymenoptera) (TYBIRK, 1991). Cette attaque peut engendrer une
perte du pouvoir germinatif des graines. Ainsi KARSCHüN (1961, 1965, 1975),
DONAHAYE et al., (1966), HEDQVIST (1967), ARAP et al., (1985), SINGH et
BHANDARI (1987), ont montré que l'infestation des graines d'Acacia raddiana en Israël et
en Algérie, par des larves de bruchidés (Bruchidius albonatatus, Bruchidius albosparsus,
Caryedon serratus, Pseudopachymerus laillemantîï, réduisait de 40 % leur viabilité.
Par ailleurs les gousses d'Acacia raddiana et les samares de Pterocarpus lucens
n'arrivent à maturité que hors de portée du bétail qui les broute.
1.5 - Essai de Kermination des I:raines
La germination des graines de ces 2 plantes est de type épigée et phanérocotylaire
caractéristique des espèces d'acacias africains. Les cotylédons s'échappent de l'enveloppe
séminale durant la germination comme l'ont indiqué VASSAL (1972), NONGUIERMA
(1978) et SOME (1991). Nous avons par ailleurs observé, qu'au stade cotylédonnaire , le
collet redressé se distinguait grâce à la nette différence de diamètre entre les organes
racinaires et hypocotylaires (future tige). Cette différence est marquée par une crête annulaire
nette chez Acacia raddiana (SOME, 1991) et un hypocotyle à diamètre réduit à partir du
collet chez Pterocarpus lucens.
La radicule apparait au bout de 1 à 2 jours chez l'acacia, après prétraitement des
graines, et au bout de 5 à 7 jours chez le ptérocarpe, après trempage.

31
2 - METHODOLOGIE
2. 1 - Choix des espèces
L'importance des 2 espèces, Acacia raddiana et Pterocarpus lucens dans les
écosystèmes sahéliens du Burkina Faso, est grande.
L'inventaire tloristique montre qu'elles marquent de manière singulière la dynamique
de la végétation ligneuse de la région sahélienne : la première par la forte expansion du
nombre de ses pieds et la seconde par leur grande régression.
Ces 2 espèces ont des aires de distribution qui se chevauchent et couvrent près de la
moitié nord du pays. Elles sont très utilisées par les populations sahéliennes : feuilles. fruits,
branches, bois, écorces et racines.
Acacia raddiana est bien appétée par les caprins (feuilles fraîches, fleurs, fruits) et
les camelins (feuilles fraîches et fruits).
Pterocarpus lucens est un excellent fourrage ligneux bien apprécié des caprins, des ovins,
des bovins et des camelins (feuilles fraîches et sèches, fruits). Les feuilles sont aussi
consommées par les populations samo de la province de la Kossi et mossi du Yatenga.
Ces 2 espèces répondent bien aux objectifs de recherche de l'IRBET et du C.N.S.F.
qui les classent parmi les 23 espèces prioritaires à protéger et à améliorer pour leur
transformation en vue d'une meilleure satisfaction des besoins des populations.
En effet, Acacia raddiana est la seule Mimosaceae sahélienne qui colonise presque
tous les milieux exondés: bordures de mare, dépressions interdunaires, glacis, talwegs et
même les dunes. Elle est la plus abondante et forme parfois des formations monospécifiques.
C'est elle qui résiste le mieux aux conditions de dégradation du milieu avec une faible
mortalité et une forte régénération. Elle s'adapte parfaitement à l'aridité, elle a une
croissance rapide et de multiples utilisations comme en Inde (ROY et al., 1973 cité par
CORNET, 1981). Le bois donne un excellent charbon (4360 kcal/kg) et le système racinaire
à la fois traçant et pivotant stabilise très bien les sols dunaires.

32
Pterocarpus lucens est par contre la seule Papilionaceae arborescente du Sahel qui
forme des boisements encore denses dans la région. Mais c'est aussi l'espèce qui a le plus
souffert ces dernières années de la sécheresse au Sahel, au point qu'il est difficile de la
trouver en bon état. Les pieds du Ptérocarpe sont encombrés de bois mort et sont entre la vie
et la mort. Ils rejetent plus ou moins vigoureusement de la souche ou des branches étalées
au sol.
Cette espèce occupe des milieux les plus ingrats sur le plan de l'économie de l'eau
(BORTOLI, 1983).
Tout cela justifie le choix de l'étude de ces 2 espèces.
2.2 - Choix des sites et des pieds
Dans les conditions du milieu, les sites d'investigation du système racinaire sont
choisis en fonction de différents types de sol par calage du calque d'interprétation sur les
cartes pédologiques du BUNASOLS (1981 et 1989) sur la région à la même échelle que celle
de la végétation.
Les critères de choix des sites sont: la proximité par rapport au village, la praticabilité des
pistes en saison pluvieuse et la nature du sol (carte n° 2).
Les sites retenus pour Acacia raddiana sont :
- les sols sableux bruns subarides de Tondaré, entre Gountouré et Oursi (1);
- les sols isohumiques modaux de Pétel Borti, à la sortie sud-ouest du village (2);
- les sols ferrugineux tropicaux lessivés à concrétions des glacis arénacés de Kolangal, au
nord-est de la Mare d'Oursi (3);
- les sols peu évolués d'apport alluvial vertiques des axes de drainage de Bossia, à l'est de
la Mare d'Oursi (4);
- les sols argileux hydromorphes peu humifères à gley peu profonds de la bordure (30 m)
nord-ouest de la Mare d'Oursi (5).

33
Pour Pterocarpus lucens, ce sont :
- les sols sableux peu épais de Timataltal, à 10 km au nord d'Oursi ;
- les sols ferrugineux tropicaux lessivés sur cuirasse des axes de drainage de Pété Yaï au sud- 1
ouest de Tin Edjar ,
- les sites des brousses tigrées de Férérillo, de Zarmakoy et de Tin Atan hors de la région
de la Mare d' Oursi.
Le choix des pieds de chaque espèce est opéré selon des hauteurs croissantes
inférieures à 150 cm. Cela permet d'apprécier l'influence de la hauteur du pied sur
l'organisation du système racinaire en rapport avec l'âge. L'âge du pied est estimé par
comparaison avec les jeunes pousses de l'année facilement identifiables et par comparaison
des semis dans le jardin botanique de la FA.S.T. La méthode de détermination de l'âge par
les cernes évoquée dans le paragraphe 2.5.2.2 n'est pas fiable.
Nous veillons dans la mesure du possible à retenir des pieds d'individus non
émondés. Ils sont choisis à l'intérieur ou à la périphérie de leurs formations afin d'analyser
les interférences éventuelles de leurs racines.
Nous cherchons enfin à déterminer le rapport des hauteurs des appareils aérien et
souterrain chez les pieds étudiés in situ. Il permet d'apprécier le niveau des nappes d'eau
souterraine. D'autre part il renseigne sur l'influence des coupes des tiges aériennes sur
l'enracinement des ligneux.
2.3 - Matériel et méthodes de déterra2e des racines
Nous avons utilisé la méthode d'excavation décrite par BOHM (1979) et utilisée dans
la forêt guyanaise par ALEXANDRE (1990) (annexe III). Elle permet de mesurer la
profondeur de l'enracinement et le degré d'étalement, en un mot l'intensité de la colonisation
du sol par le système racinaire du ligneux. Elle a aussi le mérite de permettre une
observation directe dans les conditions naturelles du système souterrain. Elle est cependant
coûteuse.

34
Des échantillons de racines, de nodules et d'organismes associés sont en outre
prélevés et conservés dans l'alcool à 75° pour observations ultérieures au laboratoire.
2.4 - Analyses pédoloKiques
Les analyses des échantillons des horizons de sol ont porté sur la couleur avec le
"Munsell Soil Color Chart" et les teneurs en N, P, K par analyse chimique au laboratoire
d'écologie. La texture est appréciée selon la méthode rapide de terrain dont la technique est
décrite en annexe IV.
Nous nous sommes référés à quelques données piézométriques de la Direction de
l'Inventaire des Ressources Hydrauliques du Ministère de l'eau, aux études de l'ORSTOM
(CLAUDE et al., 1978, 1979 et 1980 en annexe V) et aux données de 1980 (RICOLVI,
1989) et actuelles du B.R.G.M pour apprécier les variations de niveaux des nappes
souterraines: données de Gorom-Gorom et d'Aribinda (département situé à 66 km au sud
ouest de la région d'étude).
2.5 - Analyses morpholoiPques et structurales des racines
Les observations de terrain (annexes VI et VII) et de préparations microscopiques de
racines, permettront de mettre en évidence:
- l'organisation morphologique des systèmes racinaires,
- la profondeur et l'organisation des structures racinaires d'absorption minérale,
- la structure anatomique des racines,
- le mode et la structure des anastomoses racinaires,
- l'organisation des associations racinaires.
2.5.1 - Matériel végétal
Nous avons réalisé une culture au laboratoire par germination de graines des 2 espèces
dans des germoirs contenant de la vermiculite et dans des bacs contenant du sable.

35
Les plantules sont utilisées pour l'observation du type de germination de chaque espèce et
les coupes microscopiques.
Cela permet de disposer d'échantillons d'organes frais compte tenu de l'éloignement de la
région d'étude.
Des cultures de plants sont en outre réalisées en pépinière dans des pots de
polyéthylène. Dans ce cas, nous avons utilisé:
- du sol brun rouge subaride de Gountouré et du sol isohumique modal de Pétel Borti pour
Acacia raddiana ;
- du sol ferrugineux tropical lessivé sur cuirasse de Tin Edjar pour Pterocarpus /ucens.
Les cultures sont faites simultanément à la pépinière du Ministère de l'Environnement
et du Tourisme à Oursi et dans la serre de la Faculté. Les racines sont mises à nu
successivement 3 et 8 semaines après la germination (CORNET, 1981).
Elles servent à compléter les observations de terrain sur les capacités de leur
association symbiotique et à remplacer les pieds déracinés in situ pendant les travaux
d'excavations du sol.
Auparavant, les graines d'Acacia raddiana sont prétraitées par scarification à l'acide
sulfurique concentré par agitation manuelle pendant 30 mn suivi d'un trempage à l'eau de
robinet en 24 h.
Celles du Pterocarpus /ucens sont seulement trempées à l'eau pendant 24 h. Ces
graines sont obtenues auprès du C.N.S.F ..
Les nodules prélevés sur les différents pieds sont déshydratés avec du chlorure de
calcium anhydre et livrés au laboratoire de microbiologie de l'IRBET.

36
2.5.2 - Méthodes d'observations
2.5.2.1 - Observations macroscopiques
Les observations macroscopiques faites pendant l'excavation permettent d'une part
d'étudier la morphologie et l'organisation spatiale du système racinaire : disposition des
racines les unes par rapport aux autres, leur mode d'étalement et leur comportement dans les
différents horizons du sol (annexes VI et VII).
Les paramètres d'enracinement sont:
-l'enracinement en profondeur mesuré du collet à l'extrémité de la racine pivotante;
- l'enracinement latéral ou extension racinaire de la plus longue racine latérale située dans
les cinquante premiers centimètres du sol.
L'horizon superficiel du sol est généralement à dominance sableuse. De ce fait il est
perméable et facilement débitable.
La mesure de ces 2 paramètres de l'enracinement généralement utilisés (POUPON,
1980 et ALEXANDRE, 1990) suffit à expliquer la dynamique des espèces ligneuses de ce
milieu. La mesure de la profondeur d'enracinement permet de définir le niveau des nappes
d'eau souterraine en fonction des sols. L'extension racinaire latérale permet d'apprécier la
superficie de sol colonisée dans la collecte des eaux superficielles.
De ce point de vue, il est judicieux de corréler l'enracinement avec des paramètres
dendrométriques, facilement mesurables, tels que la hauteur du fût et le diamètre à la base
du tronc du pied. La prise en compte des propriétés pédologiques et de l'âge estimé permet
de défmir le temps de protection nécessaire au peuplement naturel ou planté dans les unités
géomorphologiques de la région.
L'identification des horizons d'absorption hydrique est faite. Pour cela, nous nous
sommes intéressés particulièrement aux poils absorbants car ils constituent la structure de
nutrition minérale par excellence de la plante. La profondeur et les caractères pédologiques
de cet horizon sont mesurés et décrits.

37
Un traitement global des données est réalisé avec le logiciel STAT ITeF pour la
définitiondes équations de régression. Le choix de chaque équation est basé sur la recherche
de la fonction, linéaire ou non, à ajuster en fonction des données observées. Nous
déterminons les valeur,s des paramètres qui doivent figurer dans l'équation préalablement
choisie dans une série de fonctions proposées. Il s'agit d'identifier celle qui est la meilleure.
L'identification est faite par appréciation de la valeur du coefficient de détermination r,significative si sa valeur est supérieure à 0,65 ou inférieure à -0,65. Une comparaison est
aussi faite entre les valeurs calculées, estimées et celles observées, mesurées.
2.5.2.2 - Observations à la loupe binoculaire
Elles visaient à déterminer l'âge des pieds par l'observation des cernes des tiges et
des racines prélevées pendant les excavations. Les fragments secs d'organes sont taillés en
rondelles de 0.1 cm d'épaisseur à la scie à disque.
Les fragments sont éclaircis dans l'hypochlorite de sodium "pur" par trempage à
froid. Le rinçage abondant à l'eau distillée suivi de séchage donne une meilleure visibilité
de la structure.
Mais comme d'autres (FAHN, 195,9 et MARIAUX, 1975) l'ont montré bien avant,
la détermination de l'âge des ligneux par la séparation des cernes de bois (ou
dendrochronologie) au sahel est aléatoire. Les raisons de cet échec sont :
- d'une part la variabilité de la largeur des cernes due à plusieurs facteurs dont les variations
de fertilité du sol, l'enracinement de l'arbre, le traumatismes subis etc.,
- d'autre part, I'activité du cambium responsable de la formation des cernes n'est pas
forcément liée à la hauteur annuelle des pluies mais à des conditions telles la durée et hl
continuitéde la saison sèche, les variations d'humidité de l'air, l'effet cumulatif de plusieurs
années sèches etc.
Des photographies d'anastomoses entre racines de Balanites aegyptiaca et d'Acacia
raddiana sont réalisées au stéréomicroscope WI;LD 3 B.

38
2.5.2.3 - Observations microscopiques
Pour les plants cultivés, les coupes sont réalisées manuellement dans la zone
subéreuse où les tissus sont bien différenciés (2 cm du collet). Les coupes intéressent des
plants de 4 et 14 jours. Ceci permet de mettre en évidence les tissus des racines des 2
espèces.
Les racines anastomosées d'Acacia raddiana et de Balanites aegyptiaca sont coupées
en séries au microtome selon la méthode d'inclusion à la paraffine (annexe VIII).
Les coupes sont montées dans de la glycérine sur microscope photonique pour
observations et microphotographies au microscope biologique NIKON.
2.6 - Quelques difficultés méthodolo2iques
Ces difficultés sont liées :
- au transport du matériel d'excavation du sol à moto et à l'absence fréquente d'eau pour
mouiller les horizons secs
- aux difficultés d'analyse texturale des horizons du fait qu'il n'a pas été possible de
collaborer avec un pédologue ;
- aux contraintes financières qui n'ont pas permis de respecter le programme de travail défini
tel le suivi phénologique ;
- la nécessité d'effectuer en 1991 un certain nombre d'excavations afin de mettre en place
une méthodologie fiable d'investigation.

39
3 - RESULTATS ET DISCUSSIONS
3.1 - Caractéristiques des racines de Acacia raddiana et de Pterocarpus lucens
Les racines d'Acacia raddiana sont de couleur rougeâtre sur sol sableux à jaune pâle
sur sol argileux. Elles ont une odeur caractéristique et piquante d'ail à l'état frais. Cette
odeur permet de les distinguer des autres racines, notamment de celles des herbacées. Elles
sont en outre fibreuses et tendres, d'où leur utilisation comme lien dans l'artisanat et dans
la construction des habitations. On rencontre quelquefois des racines asséchées et cassantes
dans les sols argileux des glacis se débitant en blocs pendant les excavations. Certaines
pourrissent dans les sols à immersion prolongée des zones d'écoulement. D'autres enfin sont
rongées et coupées par des larves d'insectes qu'elles hébergent.
Aux extrémités coupées apparaissent des bourgeons en période humide pour
reconstituer la racine en formant plusieurs brins en "queue de cheval". VARTANIAN (1972)
citée par PICARD (1984) montre en effet que le stress hydrique entraîne l'apparition chez
les Dicotylédones de structures racinaires particulières en massue à partir desquelles se fait
la reprise après réhumectation.
Les racines dans la colonisation du sol évitent les obstacles qu'elles contournent
jusqu'à rencontrer une voie de passage. Parfois encore, elles forment un réseau dense de
radicelles s'incrustant dans les blocs de latérite.
Elles possèdent enfin des vaisseaux conducteurs avec des canaux larges et visibles à
l'oeil nu dans la section transversale. Un fragment de racine long de 50 cm permet d'aspirer
par la bouche l'eau d'un récipient dans lequel plonge l'autre bout. Or, selon PASSOURA
(1982) cité par CHOPART (1984), le nombre et le diamètre des vaisseaux de xylème jouent
un rôle important dans les capacités de transfert de l'eau des sites d'absorption vers les
parties aériennes. Les racines d'Acacia raddiana seraient alors plus compétitives dans la
nutrition minérale que celles de Pterocarpus lucens, par l'amélioration de la conduction
hydraulique.

Figure n° 2
H= 127cm
0= 2,4 cm
Prof cm Couleur Cohesion Texture Observ.
a 7,5YR4/4 sons cohesion sables
12 10YR 4/4 bonne sabie-aroliie cavités
85 10YR 4/4 forte oroiie
125 5YR bonne oroiie
gravillons
215 10YR 8/1 tres forte sable dalleQrossier rocheuse
Ocm
20
50
100
150
200
,Partie souterraine dt Acacia raddiana a Bassia
(Axe de drainage)

41
En ce qui concerne le ptérocarpe, il possède des racines de couleur blanc laiteux. Son
odeur est douce. Les racines ont une consistance plus ferme que celles de l'acacia et sont
moins fragiles que celles de Balanites aegyptiaca. Le dessèchement des racines suite aux
assèchements périodiques des sols peu profonds est plus fréquent chez cette espèce. Il
apparait alors de multiples radicelles de remplacement des racines coupées, touffues en
"queue de cheval".
En outre, la nature des sols est à l'origine des multiples déviations des racines au
contact des horizons compacts et durs chez toutes les 2 espèces.
3.2 - Ora:anisation morpholoWque des ~stèmes racinaires
Acacia raddiana et Pterocarpus lucens présentent un appareil souterrain comprenant
une racine principale pivotante et des racines secondaires. Ces dernières sont plus ou moins
étendues latéralement, grosses, ramifiées et disposées autour de la racine principale.
Chez Acacia raddiana comme chez Pterocarpus lucens , les premières racines latérales
apparaissent dans les trente premiers centimètres. Elles sont alors superficielles.
Chez la première espèce, les racines latérales sont nombreuses et s'étalent dans toutes
les directions perpendiculairement au pivot (figure n° 2 et photo n° 5). En outre, elles sont
très ramifiées. Mais on rencontre parfois des ramifications, de gros diamètre dans les
horizons profonds des différents types de sol.
Chez la deuxième surtout, l'extension racinaire latérale est moins étalée et s'enfonce
parallèlement au pivot (figure n° 3 et photo n° 6). Ces racines présentent en outre de
nombreux replis formant parfois des noeuds, car beaucoup de forces d'inertie s'opposent à
leur développement normal : la texture du sol et la dessiccation.
Les ramifications racinaires latérales sont assimilables à des racines pivotantes. Elles
laissent penser aux racines latérales primitives ayant évolué dans ce sens à la suite de
l'inhibition de la croissance de la racine pivotante initiale. Cette ramification de
remplacement du pivot semble toutefois s'effectuer du même côté.

Figure nO 3
H=60cm
0= 1,7cm
Ocm
10
20
50
enroulement de poilsfin dans unecavité termitique
100
racine pivotantede substitutionsectionnée parles termites
150
o 20 cm Prof cm Couleur Cohesion Texture Observ.L-..-J
0 7,5YR 6/4 sons cohesion sable
17 7,5YR 6/6 faible ar9Ïle-soblè cavites
70 7,5YR6/6 bonne arqiJe-sable
125 IQYRs/e forte arQile
135 7,5 YR 6/6 très forte cuirasse dalle
Partie souterraine de Pterocarpus lucens à Timataltal
( Mi crodune de brousse tigrée dégradée)

43
En effet, le pivot est toujours présent mais il est fréquemment sectionné chez
Pterocarpus lucens. Il est alors court et souvent prolongé par des ramifications multiples de
longueurs inégales ou simplement substitué par une racine latérale.
L'extension racinaire latérale des deux espèces forme aussi des filaments fins et
tendres, non ramifiés qui s'enroulent et tapissent les parois des cavités abandonnées de
termites hypogés.
Les associations racinaires avec les termites sont plus fréquentes avec Pterocarpus
lucens. Les sols sous les pieds de cette espèce abrite aussi des scorpions se nourrissant de
termites.
Le tronc racinaire généralement renflé se prolonge par un pivot racinaire qui pénètre
plus ou moins en profondeur chez les 2 espèces. Le pivot est souvent unique, parfois divisé,
plus ou moins en profondeur, en 2 ou 3 fourches portant chacune des racines latérales.
L'allure et la morphologie du pivot des deux espèces sont aussi fortement influencées
par la nature et la structure des horizons pédologiques traversés. Ainsi, les racines sont
droites et lisses dans les horizons sableux, moins ramifiées dans les horizons argileux,
tortillantes et étranglées en chapelets dans les horizons concrétionnaires à gravillonnaires.
Au contact des obstacles (horizon compact, boules, dalle inférieure rocheuse ou
cuirassée), la racine se courbe à la recherche d'un passage. Parfois, elle peut s'enrouler en
plusieurs spires avant de trouver un passage vers les horizons inférieurs. C'est le cas de la
racine d'Acacia raddiana rencontrée au piémont des buttes rocheuses à Tounté (photo n° 5).
Au delà des horizons compacts, la racine reprend son aspect normal lisse et vertical.
Le système racinaire des 2 espèces ligneuses est à extension mixte (GORENFLüT,
1986). Cette morphologie permet à la fois d'atteindre des profondeurs où le sol reste
longtemps humide et de récupérer l'eau superficielle.
Ce type morphologique caractérise les arbustes des régions arides.

44
Chez Acacia albida, le système racinaire est fortement pivotant et profond, parfois
horizontal et superficiel sur sol compact (ALEXANDRE et OUEDRAOGO, 1990).
La structure profonde du pivot et la dense ramification des racines latérales caractérisent la
plupart des acacias sahéliens comme Acacia ni/otica var. adansonii, Acacia raddiana et
Acacia senegal, décrits par CAZET (1989).
Balanites aegyptiaca possède une système pivotant et profond avec la présence
d'anastomoses entre les racines du même pied (POUPON, 1980).
Par contre en région soudanienne plus humide, la morphologie racinaire est différente.
BAMBA (1985) a décrit dans la région de Saponé (Burkina Faso) :
- un système racinaire superficiel extensif dans lequel le pivot est absent chez Combretum
ghasalense, Dichrostachys cinerea, Desmodium gangeticum, Erythrina senegalensis,
Securinega virosa et Stereospermum kunthianum ;
- un système racinaire à pivot très court avec un réseau latéral plus ou moins étendu chez
Azadirachta indica, Annona senegalensis, Cassia siamea, Feretia apodanthera; Mitragyna
inermis, Piliostigma reticulatum, Piliostigma thonningii, Pseudocedrela kotschyi et
Terminalia avicennioides ;
- un système racinaire avec un pivot profond et des racines latérales étendues et plus ou
moins profondes chez Acacia dudgeonii, Acacia gounnaensis, Butyrospermum paradoxum
et Gardenia temifolia. L'extension racinaire augmente avec l'âge contrairement au pivot qui
disparaît parfois. C'est le cas de Parkia biglobosa dont le pivot est remplacé par une cavité
dans le tronc.
La morphologie du système racinaire chez les ligneux varie avec la pluviométrie.
3.3 - Ora:anisation structurale des systèmes racinaires
Les structures racinaires primaires de la zone pilifère chez Acacia raddiana et chez
Pterocarpus lucens sont presque identiques. Elles sont constituées de l'extérieur vers
l'intérieur d'un épiderme formé d'une couche continue de cellules parenchymateuses non
rangées. Elles assurent la protection contre la dessiccation et les agressions externes
(parasites... ) .

Figure nO 4
65 7,5YR 4/6 assez bonne orQïle- sable
200105 7,5YR 5Ai assez forte orQile
Qrovillons
135 10YR 6/4 forte cuirasse
235 lOYR 8/1 tree forte soble dolleQrOlsier rocheUH
H= 50 cm
0= 0,6 cm
H: 30 cm
0= 0,5 cm
0150
Prof. cm Couleur Cohesion Texture Observ.
0 10YR sons sablecohésion
7 IQYR 4/4 cavités
35 7YR 4/6 froQile OI'Qile-soble
100
20
Ocm
5
50
H=13cm
0=0,3 cm
Parties souterraines dt Acacia raddlana à Tondaré( Ploc cçe amorphe)

46
Sous l'épiderme se trouve le parenchyme cortical constitué de cellules volumineuses
dans la partie médiane avec des méats contrairement au parenchyme médullaire qui est jointif
et peu abondant. L'endoderme apparait plus différencié et de forme anguleuse chez le
ptérocarpe.
3.4 - Analyse du développement et du fonctionnement des systèmes racinaires
Les résultats de mesure d'enracinement des 2 espèces sont consignés en annexe IX.
3.4.1 - Les relations entre l'enracinement et la hauteur du pied chez Acacia raddiana
La profondeur de l'enracinement augmente avec la hauteur des individus dans chaque
site. Le ratio dimensionnel marque, cependant un développement relativement de moins en
moins important de l'enracinement en profondeur au cours de la croissance de la tige des
jeunes plants. Ainsi à Tondaré (figure n° 4), le ratio hauteur du pied/profondeur de la racine
est de 0,12 ; 0,19 et 0,22 respectivement pour les pieds de 13 cm, 30 cm et 50 cm.
Mais l'augmentation de l'enracinement en profondeur diffère selon les sites (figure
n° 5). Elle est plus importante dans le site de bordure de mare. Cela peut s'expliquer par la
constitution prédominante en sable et la profondeur plus importante des sols de ce site. Dans
d'autres sites la présence de dalle rocheuse plus ou moins profonde limite l'excavation.
Les équations de courbes de régression de l'enracinement en profondeur (Yen cm)
en fonction de la hauteur (H en cm) sont :
Bossia :
y = -494,17 H / (114,57 + H)
Bordure de mare :
y = -798,17 H / (93,38 + H)
Kolangal
y = -484,17 H / (77,02 + H)
(r' = 0,90)
(r' = 0,84)
(r' = 0,89)

Fig.no5 : Relation entre profondeurd'enracinement et hauteur du pied
chez Acacia roda/ana
Profondeur d enracinei'n.ent (cm) .. \ ï
0...------------------
-100
-200
-300
-400
- ..... - __dalle". ~ ----- ..
......+ :..~.."'_= ~ dalle+ p
x . .~
x
50 .100'150 200···· 250
Hauteur du pied (cm)
-500 '--__.l...-__....L..__---L .L-_---J
o
Figure n 6: Relation entre extensionracinaire latérale et hauteur du pied
chez Acacia raddiana
Extension racinaire latérale (cm)400 c----------~-----~---,
+
300
200
100
/
50 100 150 200 250
Hauteur du pied (cm)
~ Tondaré
-e- Kolangal
+ Pétel Bortl .-*_. Bosala
-- Bordure de mare

Pétel Borti :
y = -397,38 H 1 (218,91 + H)
Tondaré
y = -445,97 H 1 (50,12 + H)
avec r2 = coefficient de détermination
48
(r' = 0,94)
(r' = 0,94)
L'enracinement en profondeur atteint 4,30 m en bordure de mare chez un pied
d'acacia de 1,14 m de hauteur et 2 cm de diamètre. L'effet limitatif de la dalle à Bossia
semble réduit par 1'hydromorphie fréquente dans cette station. Dans tous les sites la racine
traverse la carapace vers la nappe souterraine.
L'enracinement latéral augmente aussi avec la hauteur du pied. L'élongation racinaire
latérale est aussi différente dans les sites (figure n06). Elle est plus importante dans les sols
à horizon supérieur sableux de bordure de mare, de Pétel Borti et de Tondaré que dans les
sols à horizon supérieur argilo-limono-sableux de Bassia et de Kolangal géographiquement
proches.
Les équations de régression de l'enracinement latéral (L en cm) en fonction de la
hauteur (H en cm) sont:
Bordure de mare:
L = -2,91 H + 9, 83
Bossia :
L = -1,17 H + 21, 06
Kolangal
L= -1,24 H + 14,97
Pétel Borti
L = -5,45 H + 156
Tondaré
L = -82281,60 H 1(85218 + H)
(r' = 0,84)
(r' = 0,98)
(r' = 0,83)
(r = 0,81)
(r' = 0,96)
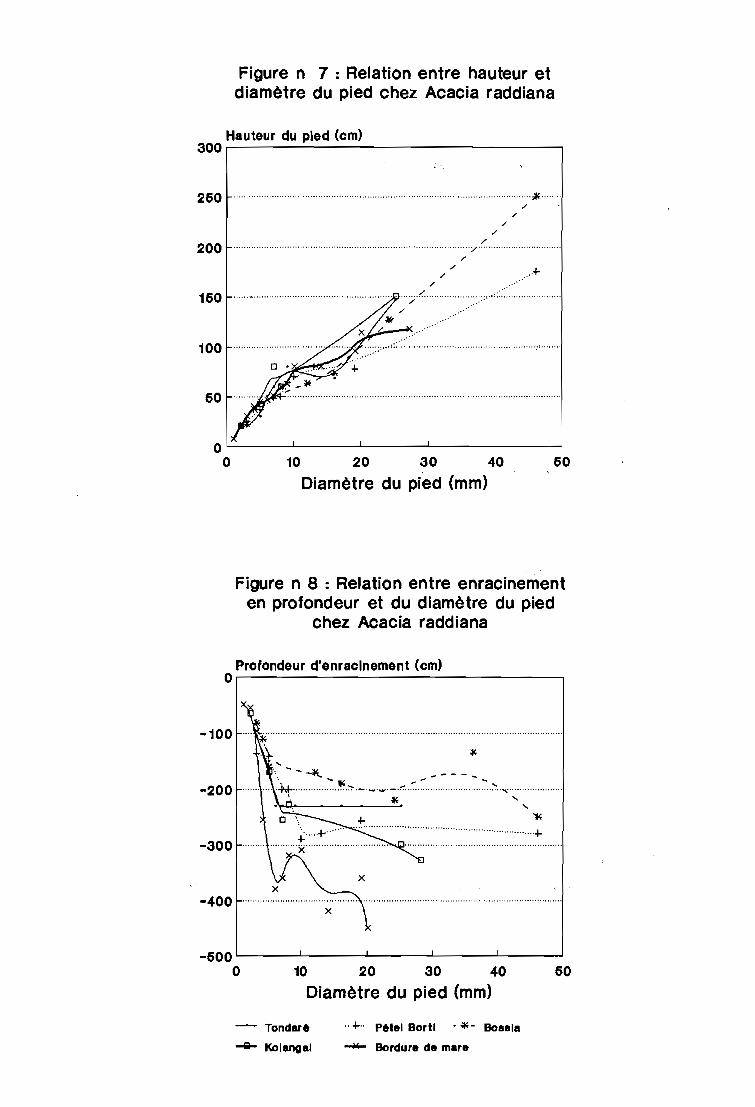
Figure n 7: Relation entre hauteur etdiamètre du pied chez Acacia raddiana
Hauteur du pied (cm)300,--------------------,
260 ...................................................... · · · · · · · · · · · · · · · · · · · · · · ·· · M · · · · · · ·
//
//
/200 ",7 ..
/
/ ...1-/
160 ..
100 ..........
60
................. ::;,.. ::::: .
....
6010 20 30 40
Diamètre du pied (mm)
O'----.......!----'----.......!----L.---......Jo
Figure n 8 : Relation entre enracinementen profondeur et du diamètre du pied
chez Acacia raddiana
Profondeur d'enracinement (cm)0.--------------------,
-100
-200
-300
.............................,......
"~
....................... +
-400 ..x
6010 20 30 40
Diamètre du pied (mm)
-600 '--__....I....-__---'- ...L.-__---L__---l
o
- Tondar6
..... Kolangal
...+.. P61el Borll . ~... Boaala
-- Bordure de mare

50
3.4.2 - Les relations entre l'enracinement et le diamètre à la base du pied chez Acacia
raddiana
La croissance de la tige en fonction du diamètre à la base du pied (figure n° 7)
montre une relation linéaire dans le premier centimètre de diamètre. La croissance des pieds
ralentit ensuite dans tous les sites pour les pieds de diamètre compris entre 1 et 2 cm
environ. Ce ralentissement serait certainement dû aux effets de la saison sèche suivant la
germination. Il y a assèchement de la région apicale des jeunes plants et destruction des pieds
par pâture. La croissance se poursuit assez régulièrement chez les pieds de diamètre
supérieur à 2 cm ; c'est comme si l'alimentation en eau était assurée chez ces pieds qui
atteignent 1 m de hauteur à ce diamètre sur presque tous les sites.
La relation entre l'enracinement en profondeur et le diamètre (figure n° 8) est
identique à celle de l'enracinement en profondeur avec la hauteur. En effet les 2 courbes sont
superposables. Cela est sans doute dû à la corrélation très significative (-0,95) qui lie la
hauteur au diamètre à la base du pied de l'espèce.
La courbe montre que la dalle sous-jacente est atteinte par les pieds de diamètre au
moins égal à 0,5 cm. Les horizons humides de la nappe souterraine sont atteints en bordure
de mare avant que le diamètre du pied n'atteigne 1 cm (figure n° 9).
L'enracinement en profondeur s'effectue ici sans contrainte majeure jusqu'à 1'horizon humide
situé à environ 3,5 m de la surface. Il est réalisé au détriment de la croissance en épaisseur.
Le développement de la racine principale doit assurer la recherche des eaux souterraines.
Les équations de régression (hyperboles) des courbes de l'enracinement en profondeur
(Yen cm) en fonction du diamètre (D en cm) sont :
Bordure de mare :
y = -584,26 D /(0.69 + D)
Bossia :
y = -243,22 D / (0, 44 + D)
(r' = 0,86)
(r' = 0,75)

Figure n° 9
150
H= 46cm
0= 0,6cm
manchon arQileux
Bouhiniorufescens
H= 80cm
0= 1,4 cm
250
200
300
350
400200
50
30
100
150
7
H: 47cm
0= 0,8 cm
Ocm __
Partie s souterraines d'Acacia raddiana à Petel Sorti (a) et
en bordure de mare ( b,c )

Kolangal :
y = -395,67 D / (0,61 + D)
Pétel Borti :
y = -363,45 D / (0,55 + D)
avec r = coefficient de détermination
52
(r' = 0,80)
(r' = 0,69)
En considérant l'extension racinaire latérale (figure n° 10), nous remarquons que son
élongation est réduite dans le site de Bossia souvent engorgé en saison pluvieuse et dans celui
de Kolangal. Ces 2 sites ont des horizons de sol superficiels à dominante argileuse. Ils sont
asphyxiants en saison pluvieuse et compacts en saison sèche.
A Kolangal, l'écoulement des eaux sur ces sols glacés avec pente empêche leur "',l;1 :' v {
infiltration (ALBERGEL et al., 1986 ; CASENAVE et VALENTIN"; DEVINEAU et al., /
1992). Les racines des ligneux dans ce site profitent peu de ces eaux et se développent peu.
Si l'enracinement en profondeur est manifeste dès la germination de la graine
d'Acacia raddiana, l'extension latérale accuse un léger retard. La croissance de la racine
pivotante constitue une priorité pour le jeune plant. Elle doit atteindre la frange capillaire du
sol avant la fin de la saison des pluies pour pouvoir traverser la saison sèche. C'est pourquoi
sa croissance s'effectue au détriment de celle des racines latérales pendant les premières
semaines après la germination, comme l'ont aussi souligné MUTHANA et al., (1980), et
SOME (1991) sur Acacia raddiana. L'enracinement latéral servira en début de saison
pluvieuse à alimenter la plante par les couches superficielles humectées par les premières
pluies.
Toute cette organisation constitue une forme d'adaptation de la plante à l'aridité
climatique.
3.4.3 - Les relations entre l'enracinement et l'âge estimé du pied chez Acacia raddiana
L'extension racinaire latérale est plus rapide chez les pieds de bordure de mare, lente
chez les pieds des sols sableux de Pétel Borti et de Tondaré et plus lente encore chez les
pieds des sols argileux en surface de Bossia et de Kolangal (figure n° 11).

Figure n 10: Relation entre extensionracinaire latérale et diamètre du pied
chez Acacia raddiana
Extension racinaire latérale (cm)600 ,-----~----------___y_.____r_-__c_o
+500
400+
x
10 20 30 40 50
Diamètre du pied (mm)
300 f- .
200
100-~----- *
* -----
Figure n 11: Relation entre extensionracinaire latérale et âge estimé du pied
chez Acacia raddiana
Extension racinaire latérale (cm)600 ~---------------,r-----,,------,
500
400
x
300
200
100
2 4 6 8 10 12
Age estimé du pied (années)
Tondaré
-e- Kolangal
+ Pétel Bortl --*- Bossla
-*""" Bordure de mare

Figure n 12: Relation entre hauteur etage es timé du pied chez Acacia raddiana
Hauteur du pied (cm)250,---------------------,
~ ",
~o ..............................................................•................................./
.//
/150 /. ..
100 .
50
124 6 8 10
Age estimé '(année)2
oL....-__.L...-__.J......__.J......__l.-__l.-__
o
Figure n 13: Relation entre profondeurd'enracinement et âge estimé du pied
chez Acacia raddiana
Profondeur d'enracinement (cm)0.-------------------------,
+. "',,,,.
. *+
~x .~ ....
-,
-~
\....
+ ..-.. *
-400
-100
-300
-200
122 4 6 8 10
Age estimé du pied (années)
- 500 L....-~__L....___L....__ _____.l______l_______"_ _
o
Tondaré
~ Kolangal
+ Pètel Bortl -*- Bossla
-- Bordure de mare

55
Les équations de régression de l'enracinement en profondeur en fonction de l'âge
estimé du pied sont :
Bordure de mare
y = 12265,33 A / (l + A)
Bossia
y = -24,50 A - 78,69
Kolangal
y = -442,85 A / (3,87 + A)
Pétel Borti
y = -225,53 A / (5,28 + A)
Tondaré
y = -997,50 A / (A + 8,5)
(r' = 0,65)
(r' = 0,91)
(r' = 0,91)
(r' = 0,73)
(r' = 0,60)
Les horizons sableux sont favorables à l'extension latérale des racines vu que leur
porosité exerce peu de contrainte et favorise l'infiltration des eaux de pluies qui sont ensuite
collectées par ces racines.
La croissance en hauteur des pieds en fonction de l'âge estimé, en régénération
naturelle (figure n° 12), est sensiblement identique dans tous les sites au cours des 4
premières années de vie de la plante, et d'environ 18 cm/an.
Au delà de cet âge, la croissance devient plus importante: 25 cm/an. Elle est plus
élevée chez les pieds du site de l'axe de drainage (Bossia). Au bout de 6 ans tous les pieds
atteignent 1 m de hauteur.
L'enracinement en profondeur (figure n° 13) est plus rapide que la croissance en
hauteur de la tige aérienne. Au bout de 2 ans les racines entrent en contact avec les horizons
humides (présence de nodules d'hydromorphie à -3,5 m) au niveau de la bordure de mare
(figure n° 9b et 9c). Il devrait en être de même au niveau des autres sites à partir de 4 ans
d'enracinement. En effet les racines des pieds de cet âge pénètrent dans la dalle (figure n°
14). Or cette carapace pourrait constituer la toiture de la nappe d'eau qui limite l'évaporation
hydrique en atmosphère sèche.
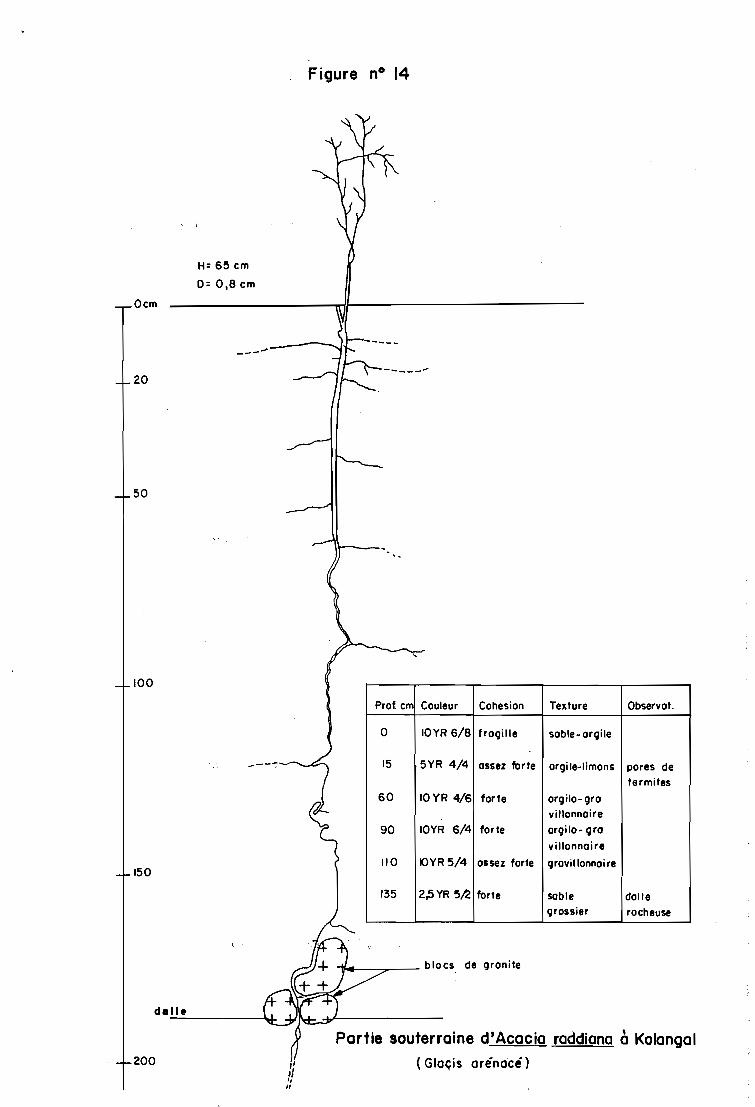
Figure nO 14
Ocm
20
H= 65 cm
0= 0,8 cm
--...,-
50
Partie souterraine d'Acacia raddiana à Kolangal
(Glaçis arénacé)
"..__~__ blocs de granite
"Il"Il
dalle
100
Prof. cm Couleur Cohesion Texture Observat.
0 10YR 6/8 fragille sable- arc~i1e
---.-. 15 5YR 4/4 assez forte argi le-limons pores determites
60 10YR 4/6 forte argilo- gravillonnoire
90 10YR 6/4 forte argilo- gro
villonnoire
110 IOYR5/4 assez forte gravillonnaire150
135 2tJYR 5~ forte sable dallegrossier rocheuse
200

Figure nO 15
H= 65 cm
0= 2,8 cm
nodule,,--
250
50
10
200
100
150
H=25cm
0= 0,5 cm
Ocm
20
50
· H =20cm
0= 0,3 cm
Ocm
100
Prof. cm Couleur cohésion Texture
0 10YR 5/8 sons cohesion sableuse300
7 7,5YR 6/6 bonne oroiie-sobie CV50 7,5 YR 5/4 for le oro Ho-concretion
aire90 5YR 5/6 forte oroi1o Orovillon-
-noire
150 5YR 5/6 bonne oroiio-concre- 350
tionnoire
0Parties souterraines de pterocarpJlJ lucens 0 Timataltal
( Axe de drainaQe de brousse tigrée dégradée)

58
3.4.4 - Les relations entre l'enracinement et la hauteur d'une part et le diamètre à la
base de la tige d'autre part chez Pterocarpus lucens
L'enracinement augmente en condition normale avec la hauteur et le diamètre à la
base du pied chez Pterocarpus lucens. Dans l'axe de drainage de Timataltal cette espèce a
des ratios dimensionnels de 0,23, 0,08 et 0,15 respectivement pour des pieds de 20 cm, 25
cm et 65 cm (figure n° 15). Il apparait que l'enracinement n'est pas proportionnel à la
hauteur de la tige qui est fréquemment appétée. En sol profond et relativement humide de
Timataltal, l'enracinement est pivotant et profond comme chez Acacia raddiana.
L'enracinement latéral et l'enracinement en profondeur chez Pterocarpus lucens donne
une représentation en "patate". De ce fait aucune relation ne peut être définie entre
l'enracinement et les paramètres dendrométriques (hauteur et diamètre) chez cette espèce.
Plusieurs raisons expliquent cela :
- la résistance du sol à la pénétration racinaire,
- les coupures fréquentes des racines par les termites hypogés,
- les influences du ralentissement de la croissance des tiges aériennes par les coupes
multiformes et très fréquentes des pieds.
Les pieds de cette espèce sont souvent rabougris et leurs racines peu développées.
Ainsi dans la zone d'épandage de Pété Yaï, le pivot est fréquemment coupé au dessous de
120 cm (figure n° 16). Les racines latérales sont elles localisées entre 25 et 60 cm et forment
des réseaux denses de chevelu racinaire s'infiltrant dans les pores des plaquettes cuirassées.
3.4.5 - Les relations entre la profondeur de la zone d'absorption et la hauteur du pied
chez les 2 espèces
La profondeur de la zone d'absorption de l'eau et des substances dissoutes du sol en
hivernage varie avec la hauteur du pied d'Acacia raddiana (figure n° 17). Elle reste
cependant superficielle (-1 m) dans l'axe de drainage de Bossia, moyenne (-1 à -1,5 m) pour
le glacis de Kolangal et les sols subarides de Pétel Borti et de Tondaré. Elle se situe par
exemple à 20,92 m en début de saison pluvieuse (18 juin 1993) à Tondaré.

H= 30cm
D= O,Gcm
Ocm
20
50
racine pivotantecoupée par les termites
100
Figure n° 16
H =35cm
D= 2,2 cm
enroulement racina ire
dans cavité termitique
o 20 cmL--..J
dalle
"\..\
Parties souterraines de Pterocarpus lucens a Pete Aï(Axe de drainage)
Prof. cm Couleur cohésion Texture Observ.
0 7,5 YR 5/6 argi le-limons faible nodules
rClci naires
GO 7,5YR 4/6 argile bonnegravillons ,
120 7,5YR 6/6 cuirosse tret farte dalle
-

60
La zone d'absorption est plus profonde (-3 à -2 m) dans les sols argileux humifères
et profonds de Bordure de mare. La nappe y est superficielle. Sa profondeur, consignée en
annexe V, était de 2,5 fi en 1977-80 selon CLAUDE et al., (1977, 1978, 1979, 1980). En
août 1992 et 1993, elle se situe actuellement à 4 m.
Le retrait progressif de la mare conduit à une colonisation de ses berges par Acacia
raddiana.
La zone d' absorption évolue avec la croissance de la tige. Elle se stabilise, dans tous
les sites, lorsque le pied a environ 40 cm de hauteur, ce qui correspond à un âge estimé à 2
ou 3 ans. A cette hauteur, la plante serait en contact avec les horizons humides du sol. La
mise en place des structures d'absorption de l'eau (poils fins) est alors effective.
Figure n 17: Relation entre profondeurde zone d'absorption et hauteur du pied
chez Acacia raddiana
Profondeur d'absorption (cm)Or---------------------,
-100
-200
-300
-400
250so 100 150 200
Hauteur du pied (cm)
-eoo L-__......I- L-__......I-__----JL-__--J
o
~ Tond."
~ Kolangal
.+ P61el Bortl . ~ - Boula
~ Bordure de mare

61
Des travaux antérieurs confirment que l'enracinement en profondeur chez les deux
espèces s'effectue en un temps plus ou moins court, fonction du niveau de la nappe. Par
contre l'extension racinaire latérale se poursuit longtemps.
Ainsi un pied d'Acacia raddiana, vieux d'environ 300 ans, dans le désert sableux du
Niger, avait un pivot atteignant la nappe d'eau profonde de 35 m (ANONYME, 1974).
MUTHANA et al., (1980) ont mesuré en Inde chez des pieds de 5 à 10 ans, des racines
latérales de 20 m de longueur.
Dans les horizons durs (carapace ou roche mère), l'enracinement s'effectue à la
faveur des fissures, des pores ou des gaines d'anciennes racines pourries.
Chez Pterocarpus lucens, la zone d'absorption est souvent diffuse et superficielle
comme dans le site de Pété Yaï. Ses racines pénètrent dans les gaines d'anciennes racines de
pieds de la même espèce, de Combretum micranthum ou d'autres espèces ligneuses mortes.
L'axe des gaines est rempli de terre, des ramifications en sortent par des fissures pour
coloniser le sol.
En empruntant ces voies, la plante s'adapte en évitant les contraintes pédologiques.
3.4.6 - Les formes d'absorption de l'eau et des substances dissoutes chez les 2 espèces
Chez les plants de 1 et 2 ans, l'alimentation en eau semble se faire sur toute la
longueur du pivot qui est hérissé de poils fins d'absorption. La racine du plant qui vient de
germer rencontre peu de résistance; ce qui fait que son allure générale est rectiligne (figure
n° 18).
Sur les sols sableux comme sur les sols argileux de bordure de mare, les pieds
d'Acacia raddiana âgés de quelques semaines ont déjà un enracinement supérieur à 1 m et
des ramifications nombreuses dirigées vers le bas.
A la profondeur de 1 m, la racine est à l'évidence très éloignée de la nappe
souterraine et de son horizon humide en saison sèche. Seuls des phénomènes d'adaptation
(défoliation par exemple) peuvent expliquer la survie des jeunes plants pendant cette période.

Figure n° 18
Prof. cm Couleur Cohesion Texture Observ.
0 10YR 6~ faible sable
37 10YR 4/6 fragile sable-argile
85 7,5YR 4Ai bonne argille-sable cavites determites
160 10YR 5/5 fragile sable
200 10YR fragile sable
272 . 10YR 5/8 bonne argile cenere--tians
315 10YR 5/6 bonne argile nodu- hydromorphelaire
360 10YR 5/4 forte argile gra-
villonnaire
Hl: 38 cm
D=O,3cm
H = 33t:m
D=O,3cm
H= 21 cm
D= 0,3cm
Bordure de more
20
50
70
H= 20cm
D= 0,2 cm
Ocm
100
Parties souterraines d'Acacia raddiana en bordure de la mare d'Oursi (a et c), ,
et a Petel Sorti (b et d )

63
Chez Acacia raddiana, la structure d'absorption de l'eau et des substances dissoutes,
en saison pluvieuse, est constituée des racines latérales ramifiées et très étendues dans les
horizons sableux superficiels; par exemple 3,8 m de longueur pour un pied de 0,8 m de
hauteur à Pétel Borti.
Dans les horizons intermédiaires, de dominante argileuse à limoneuse, les racines
latérales se terminent par des poils fins qui s'enchevêtrent et tapissent les parois des cavités
des nids hypogés abandonnés par les termites souterrains.
Ces nids sont des poches argileuses cimentées constituant des réservoirs d'eau
(BOYER, 1969; 1973; 1975a et 1975b et LEPRUN, 1976). Des mouvements hydriques sont
signalés d'une part avec les eaux des nappes souterraines et d'autre part avec l'atmosphère
par évaporation capillaire ou par infiltration des eaux de pluie.
Dans les horizons inférieurs généralement argileux plus ou moins gravillonnaires, le
système racinaire forme des poils absorbants qui restent appliqués contre le pivot le tout étant
entouré d'une couche de mucilage ou mucigel sur toute la longueur.
C'est un manchon d'une épaisseur variant entre quelques micromètres à 100
micromètres (CALLOT et al., 1981). Il est constitué d'une agrégation de particules
argileuses soudées par le mucigel secrété par les racines et constitué en majeure partie de
polysaccharides, de lipides et de peptides, autour de la racine. 11 est signalé en 1937 par
PARFITT et GREENLAND et repris par CALLOT et al., (1981). Ces auteurs montrent en
outre que ce manchon constitue en quelque sorte une protection de la racine contre le
dessèchement excessif du sol. Ce qui permet aussi d'assurer sa nutrition en conservant une
partie de la phase liquide du sol dans ses agrégats.
Nous avons observé dans ces horizons une traînée humide le long du pivot de certains
pieds d'Acacia raddiana. Les poils d'absorption forment des réseaux de mailles épousant les
contours des graviers plus ou moins grossiers de l'horizon d'absorption.
Ces observations confirment celles de POUPON (1980) qui estimait que l'eau s'écoule
préférentiellement le long du pivot qui est ainsi entouré d'une gaine humide.

64
Chez Pterocarpus lucens, l'absorption semble se faire préférentiellement dans les nids
hypogés de termites et au niveau des arborisations du chevelu racina ire. Il n'a pas été
observé de mucilage chez cene espèce et les poils d'absorption se situent généralement dans
les horizons argileux concrétionnaires à gravillonnaires de faible profondeur (60 cm à Pété
Yaï).
Malgré les coupes fréquentes de ces racines, des rejets apparaissent souvent sur la
souche en saison pluvieuse. Mais ce ne sont pas les possibilités d'absorption qui limitent la
croissance des végétaux, c'est plutôt le processus de métabolisme qui pennet l'utilisation par
la plante des éléments absorbés. En effet, selon STODDART el al., (1975), tout ce qui
affecte la photosynthèse affecte les racines car les quantités de substances carbonées produites
sont influencées par les coupes et autres prélèvements aériens.
Ainsi PARKER et SAMPSON (1931), qu'ils citent, confirment qu'une défoliation ou
une fauche provoque une réaction immédiate marquée par une cessation temporaire de la
croissance racinaire. Par exemple chez les graminées, une fauche entraîne une réduction du
cinquième de la longueur racinaire normale selon ROBERTSON (1933) cité par STODDART
el al., (op. cit.). Les diverses formes de coupe limitent alors la croissance racinaire.
CALLOT el al. (1981) par ailleurs affirment que le cinquième seulement du système
racinaire du maïs isolé et plongé dans un milieu riche suffit à satisfaire entièrement les
besoins de la plante. Par extrapolation, nous pouvons déduire que dans ce milieu sahélien,
l'éloignement du niveau de la nappe souterraine constitue la cause majeure de la mortalité
des pieds de Pterocarpus lucens. Elle situe par exemple à 35,8 m en septembre et à plus de
50 m en mars de 1993 à Timataltal (B.R.G.M non publié). Malgré le faible enracinement de
cette espèce, une pluviométrie abondante peut permettre à ces pieds de survivre.
En saison sèche, ce serait le pivot racina ire qui assurerait l'alimentation minérale des
ligneux à partir des nappes d'eau souterraines.

65
3.5 - Relations entre les systèmes racinaires
3.5.1 - Compétition spatiale
Il existe une concurrence entre les appareils souterrains des ligneux et des herbacées
d'une part et les appareils souterrains des ligneux entre eux mêmes d'autre part.
La compétition avec les herbacées est très importante chez les pieds de régénération.
Les racines de très jeunes pieds d'Acacia raddiana et celles des herbacées colonisent
ensemble le premier mètre du sol. Souvent elles se côtoient sur près de 2 m de profondeur,
sans lien apparent entre elles.
Il est aussi évident qu'une compétition existe entre les pieds de régénération de
Pterocarpus lucens et ceux des espèces ligneuses qui les protègent du broutage : Guiera
senegalensis et Combretum micranthum. Des enchevêtrements racinaires ont été observés
entre ces espèces. Dans ces cas, il se forme le plus souvent une couche horizontale et
superficielle de racines latérales entrecroisées sans liaisonsapparentes entre elles. Toutefois,
elles s'accrochent souvent par des noeuds.
Dans l'enracinement en profondeur, une fissure d'un horizon compact, peur être
exploitée par plusieurs racines d'espèces différentes. Elles présentent ou non des liaisons
entre elles.
Cette compétition spatiale dans l'exploitation des ressources du sol est observée dans
le système racinaire d'Acacia raddiana avec plusieurs autres espèces en particulier Balanites
aegyptiaca.
Les racines de Balanites aegyptiaca colonisent intensément le sol sous les pieds des
ligneux voisins jusqu'à une profondeur de 2 m. Cette compétition dans l'exploitation des
ressources du sol est surtout remarquable au niveau des restes de cendres d'un ancien foyer
rencontré à 50 cm de profondeur, sous un pied d'Acaciaraddiana. Les radicelles du "dattier"
développent un réseau dense emprisonnant les cendres.

réseou de rodicellesde Bolonites oegyptico
-,-ll---- rocine d' Acocio roddlono
étronglement de rodicellede Bolonites oegyptioco
Fig.noI9: Anastomoses racinaires de Balanites aegïptiaca
et Acacia raddiana
lmm- j- rocine d' Acocio roddiono
L- rhylidome
"- popille de pénélroliOIl de rodicellede Boloniles Oegyplioco
Fig.no20 :Mode de pénétration de radicelle de Balanites aegyp-tiacadans la racine d' Acacia raddiana

67
3.5.2 - Compétition trophique
Nous avons observé à partir d'un pied d'Acacia raddiana en cours de dessèchement,
des racines provenant d'une souche vivante de Balanites aegyptiaca. Cette souche était
constanunent piétinée par les passages réguliers des véhicules. Ces racines s'appliquent et
s'enroulent autour du tronc racinaire et des racines latérales d'Acacia raddiana, qu'elles
semblent étrangler à environ 50 cm de profondeur (figure n" 19 et photo n° 7).
Nous nous sommes intéressés à d'autres pieds morts de cette espèce dans les
formations végétales dominées par Balanites aegyptiaca, en bordure de la mare. Nous avons
encore découvert la présence de contacts entre les racines de ces 2 espèces. Il en est de
même avec les pieds d'acacia vivants mais présentant un retard de feuillaison. Par contre,
avec les pieds à phénologie normale, aucune relation racinaire n'est décelée.
3.5.2.1 - Morphologie des anastomoses
Les radicelles de Balanites aegyptiaca s'appliquent intimement sur celles d'Acacia
raddiana.
A la loupe binoculaire, les radicelles du dattier d'Egypte forment une sorte de maille
autour de la racine de l'hôte dans laquelle elles pénètrent. Au point de pénétration la radicelle
forme une papille sous forme de plaque ovale de 1 mm de diamètre (figure n° 20 et planches
n° 8). Parfois, la racine hôte forme aussi une tumeur. Les papilles présentent en outre des
cavités en forme de réservoir en relation avec les tissus de l'hôte. Des poils glandulaires sont
parfois présents en bordure du point de pénétration.
L'arrachage par ailleurs d'une racine pénétrante du tronc racinaire de l'hôte laisse une
plaie béante dans les tissus corticaux légèrement enflés. Pourtant l' écorçage des racines
latérales réceptrices ne montre aucun lien visible à l'oeil nu avec le bois interne. D'autre part
la séparation de l'écorce du liber ne laisse aucune trace visible à l'oeil nu sur celui-ci.

a
'--------- :~::';: }L- phellodeme
L- suber
b
1enlice Ile
rocine d' Acacio roddino4
Tinus de
Balanites aeQyptiaca
S3~--- rhytidome
tissus
d' AcoClo roddlono
racine de Bolanites aeQyptiaca ..Fig.no21 : Contacts tissulaires entre Acacia raddlana
Balanites aegyptiaca dans l'anastomose racinaire

69
3.5.2.2 - Structure et mécanismes des anastomoses
Les observations microscopiques de coupes transversales de racines de la plante hôte
indiquent que les radicelles de Balanites aegyptiaca pénètrent soit tangentiellement soit
perpendiculairement au niveau des lenticelles.
Ses tissus racinaires sont en contact avec le phelloderme. Le suber ouvert au niveau
de la lenticelle se replie sous sa pression.
Balanites aegyptiaca pénètre par un canal en forme de suçoir peu développé qui
aménage une cavité. C'est probablement par ce canal que les substances absorbées transitent
dans la cavité (figure n° 21a).
D'autres coupes présentent de grandes lacunes avec l'interruption du phelloderme
communiquant la cavité de la tumeur au phloème de l'hôte (figure n° 21b). Dans ce cas le
dattier sauvage absorberait alors la sève élaborée produite par les organes foliaires de son
hôte. Le transfert des substances carbonées s'effectuerait soit par aspiration soit par simple
gradient de pression osmotique.
La reconnaissance racinaire s'effectue par les lectines (glycoprotéines) émises par les
racines de l'espèce hôte (communication personnelle du Pr ANNO). En effet les racines ne
stockent pas de composés azotés, ces derniers étant exportés dès leur formation. Par contre
on observe une accumulation de protéine chez les plantes soumises à la sécheresse. Cette
accumulation représente un moyen pour la plante de maintenir une pression osmotique
supérieure à celle du sol sans faire appel à une trop grande quantité d'ions minéraux qui se
révèleraient toxiques (GUIGNARD et al., 1985).
L'arrachage des radicelles pénétrantes sur 3 pieds d'acacia en début de dessèchement
a permis la reprise de la feuillaison au bout de 2 semaines. Le dattier d'Egypte retirerait
alors des substances nutritives de la plante à laquelle il est lié, contribuant ainsi à son
dépérissement.

70
II ressort de ces observations que Balanites aegyptiaca joue peut-être un rôle
important dans la mortalité des pieds d'Acacia raddiana. Le taux de mortalité est
respectivement de 4,8 % et Il, Il % du peuplement ligneux et des effectifs de cet Acacia.
3.5.2.3 - Caractérisation phytochimique des anastomoses
Balanites aegyptiaca contient dans tous ses organes végétaux des poisons organiques
non azotés. Des tests chimiques préliminaires de screening phytochimique que nous avons
effectués sur différents organes (écorces de tronc et de racines) montrent la présence de
saponosides, de stéroïdes-terpoïdes, de quinones et l'absence de tanins, d'alcaloïdes et de
flavonoïdes. Ces substances ont déjà été signalées par KERHARO et ADAM (1974) au
Sénégal.
Ce sont des hétérosides stéroïdiques, classés dans le groupe des saponosides. Ils ont
des propriétés moussantes, d'où l'utilisation des écorces du tronc et des racines comme savon
par les populations touaregs et peuhls de la région. Ils ont en outre une action hémolytique
et sont très toxiques pour les animaux à sang froid.
Ces auteurs citent HARDMAN et al., qui ont extrait, en 1970, en Tanzanie, des
racines, 1% de la substance, et ARCHIBAL (1933-34) qui a montré l'action mortelle des-----;:--- ;,:
différents organes (fruits, amandes, écorces, racines et branches) pour les mollusques, les
miracidies, les cercaires, les poissons et les têtards. Aussi avait-il recommandé la plantation
de Balanites aegyptiaca le long des rivières pour éviter la propagation de la bilharziose.
Toutefois il est signalé l'utilisation de branches de cette espèce dans la pêche au Niger
(communication personnelle du Pr. SAADOU). En revanche on ne signale aucune action de
ces substances sur les végétaux de leur voisinage.
Comme l'ont montré JEAN-BLAIN et GRISVARD (1973), la toxicité d'une espèce
peut subir des variations considérables sous l'action du milieu et de facteurs intrinsèques.
Cela pourrait expliquer des manifestations de toxicité responsable de la mortalité des ligneux,
particulièrement en bordure de la Mare d'Oursi.

71
Cette station comporte un peuplement ligneux dense à dominante Balanites
aegyptiaca. La station abrite par ailleurs les campements de saison sèche des éleveurs. Il y
a ainsi chaque année une concentration de la fumure azotée des déjections animales dans un
sol pauvre en phosphate. Les variations entre les fumures azotée et phosphatée pourrait
favoriser l'accumulation de poison comme il a été signalé pour les alcaloïdes du sorgho et
les stéroïdes des légumineuses par KERHARO et ADAM (1974).
Il est connu d'autre part que Acacia raddiana est capable d'exsudation naturelle de
gomme arabique à la suite de traumatismes, par lyse des membranes cellulaires. La gomme
se gonfle fortement au contact de l'eau en masse gélatineuse ou solution colloïdale au niveau
des racines. KERHARO et ADAM (1973) pensent qu'en réaction à ces exsudations, Balanitesli-
aegyptiaca excréterait le poison qui formerait une mousse stable empêchant la circulation de
l'eau dans la plante hôte et/ou inhiberait les enzymes secrétées par elle. L'acacia dont les
fonctions physiologiques se trouvent perturbées finit par mourir.
Des anastomoses racinaires analogues sont aussi observées entre Balanites aegyptiaca
et Bauhinia rufescens d'une part et entre les racines d'un même pied de Balanites aegyptiaca
d'autre part. Des cas de mortalité n'ont toutefois pas été observés chez Bauhinia rufescens.
5.3.2.4 - Nature des anastomoses
,;J
De telles anastomoses ont été signalées au Sénégal entre les racines latérales de
Balanites aegyptiaca (POUPON, 1980). On les rencontre aussi chez les figuiers épiphytes
(Ficus glabra par exemple), dans les soudures racinaires du flamboyant (Delonix regia). dans
les racines de l'okoumé (Aucoumea klaineana) (DEVAL, 1974) et chez le jujubier (Ziziphus
mauritiana) (communication personnelle du Dr ALEXANDRE). Dans les plantations de pins
en Nouvelle Zélande et de pins tailladés de Floride (WILL, 1986}t SCHULTZ, 1972 cités fpar KRAMER, 1983), il existe des liaisons entre les racines d'un même pied et entre les X'
'l .
racines de pieds différents de même espèce (Pinus radiata). Le système racinaire et le pied
des arbres coupés survivent souvent à partir des sucres fournis par les arbres intacts auxquels
ils sont liés.

72
Le rapport tissulaire dattier-acacia est de nature trophique. Toutefois il n'est pas aussi
évident que chez des espèces hémiparasites du genre Cuscuta (Convolvulaceae) ou
Tapinanthus (Loranthaceae).
Par ailleurs chez les hémiparasites comme le gui (Viscum album) et les Orobanches, le suçoir
forme des cellules terminales dont l'agencement rappelle des hyphes fungiques établissant des
contacts avec le liber de l'hôte. Ces hyphes s'appliquent contre les tubes criblés sans jamais
y pénétrer. C'est l'hôte qui met en place des trachéides qui rejoignent celles du parasite.
Chez les Balanophoraceae l'association hôte-parasite est plus intime. La racine parasitaire
pénètre de place en place au niveau des tumeurs volumineuses. MANGENOT cité par
GORENFLOT (1986) montre la formation de chimères dans lesquelles vivent et se
développent en synergie et suivant un mode défini et uniforme d'association, des tissus
d'apparence extrêmement différente.
L'hypothèse peut être faite à partir de la mortalité par anastomose racinaire, que les
pieds plus anciens du dattier d'Egypte s'opposent à la pénétration des régénérations d'acacia
dans le peuplement. La logique voudrait par ailleurs que le dattier profite des performances
des pieds d'acacia déjà existants. Par comparaison à l'enracinement de Balanites aegyptiaca
défini dans les travaux de POUPON (1980), nous pouvons affirmer que le système racinaire
de l'acacia est plus profond malgré le jeune âge des pieds.
Balanites aegyptiaca et Acacia raddiana sont toutes des espèces sahéliennes qui
occupent beaucoup de stations. La première pénètre plus profondément en région soudanienne
tandis que la deuxième remonte au nord saharien. Alors Balanites aegyptiaca recherche chez
Acacia raddiana, mieux adaptée à la sécheresse les conditions permettant sa survie.
3.6 - Associations symbiotiques
3.6.1 - Plante-bactéries
La formation de nodules bactériens a été observée sur les racines des 2 espèces
ligneuses étudiées.

73
3.6.1.1 - Nodules d'Acacia radâiana
Les nodules d'Acacia raddiana n'ont été retrouvés que sur les racines des pieds en
pépinière à Oursi et à Ouagadougou dans un sol de la région d'étude. BALASUNDARAM
(1987) avait aussi constaté en Inde, l'absence de nodules in situ sur les racines d'Acacia
raddiana contrairement à Acacia tortilis. DREYFUS et DOMMERGUES (1981) et FAGG
(1981) indiquent que Acacia raddiana nodulait seulement par inoculation de lignées de
Rhizobiums à croissance rapide. DUCOUSSO (1991) a récolté au Sénégal des nodules sur
un pied de 4 ans de cette espèce, en condition expérimentale par essai d'inoculation. Par
contre DUPUY et al., (1991) en ont récolté sur des racines secondaires de cette espèce. in
situ et à 2m de profondeur, au Sénégal.
La morphologie des nodules varie selon l'âge: d'abord globuleuse chez les jeunes
pieds d'environ 3 semaines avec une taille d'environ 1 mm ils deviennent allongés en
bâtonnet, dichotomique ou coralloïde au bout de 8 semaines pour une largeur atteignant 1
cm. La couleur est brun foncé et éclaircie aux extrémités.
La forme et la couleur rappellent celles des nodules produits par rhizobium sur des
racines d'Acacia colei (Acacia holosericea) au Sénégal (CORNET et DIEM, 1982) et sur
celles d'Acacia auriculiformis au Zaïre (KHUSA et al., 1990). En effet les souches isolées
par DUPUY et al., (1991) chez Acacia raddiana appartiennent au genre Rhizobium,
caractérisé par une croissance rapide en culture pure.
3.6.1.2 - Nodules de Pterocarpus lucens
Les nodules du ptérocarpe sont appliqués contre les racines latérales ou directement
sur le pivot racinaire des pieds de pépinière et de ceux en conditions naturelles. Leur
répartition n'est pas uniforme. Ils sont isolés ou associés en grappe le long de la racine
infestée, l'englobant plus ou moins totalement. Toutes les racines d'un même pied ne portent
pas nécessairement des nodules.

74
Leur forme est globuleuse, généralement aplatie. Leur aspect de surface est froissé
par la présence de sortes de petites pointes. La taille est de 1 mm environ chez les sujets de
3 semaines et de 5 mm chez ceux de 8 semaines. La couleur varie du blanc laiteux au bleu
foncé en passant par le violet.
Or, il est connu que seuls les nodules de couleur foncée sont fixateurs d'azote
atmosphérique. Leur nombre dépasse généralement celui des nodules clairs.
3.6.1.3 - Quelques réflexions sur l'association plante-bactérie
Nous pensons que la formation de nodules chez Acacia raddiana est favorisée en
pépinière par l'humidité assurée par un arrosage régulier (tous les jours) et/ou par l'aération
du sol des pots de germination ou la levée d'autres facteurs inhibiteurs de la nodulation.
Comme le ptérocarpe porte des nodules fixateurs d'azote in situ, on devrait
s'intéresser à ses possibilités d'apport de cet élément au sol. Dans les pays de la sous- région
ouest africaine, il existe des phosphates en minerais dont le traitement peut donner l'élément
phosphore complémentaire. L'engrais azoté est importé et coûte relativement cher. Nous
avons cependant constaté peu de différences entre la végétation sous ces arbres et celle des
alentours. L'azote rejeté serait dans ces conditions peu important.
Mais même si l'azote apporté au sol est faible, il migre dans les feuilles et enrichi les
écosystèmes par l'alimentation des animaux et par la décomposition des débris végétaux.
3.6.2 - PJante-termites
3.6.2.1 - Observations in situ
Nos observations in situ montrent la présence dans les sols de tous les sites
d'excavation du sol de nids tennitiques. Ces cavités souterraines sont situées entre 0,3 et 1
m, rarement jusqu'à 2 m dans le sol sans édifice externe et contiennent fréquemment des
meules (annexe X et photo n° 9).

75
Ils sont généralement localisés dans les horizons à dominante argileuse peu
gravillonnaire, au dessus de la carapace ou cuirasse ferrugineuse ou de la dalle rocheuse. En
effet, JOSENS (1982) cité par SIEBER (1983) confirme que la texture sableuse du sol est peu
favorable au creusement des galeries et des loges souterraines. D'autre part son pF est
difficilement maintenu à une valeur proche de celle souhaitée, égale à 3,5.
Les insectes adultes eux sont présents dans de minuscules galeries en dessous des
nids abandonnés ou remontent dans les chambres à meules.
Les meules ou gâteaux de termites sont de couleur jaune à bleu et de diamètre
variable de 2 à 7 cm. Leur forme est variable: circulaire et concave, ou en forme de
croissant, ou bombée et épais de 1,5 à 5 cm plus ou moins perforés d'alvéoles en ruches
d'abeilles. Cette architecture délicate donne des formes plus ouvertes, moins épaisse et plus
plates avec des excroissances. Un emboîtement de deux meules a été observé dans une loge
à Timataltal (photo n° 9).
Les racines sont dépourvues de poils absorbants indiquant la présence probable
d'endomycorhizes. Ce phénomène se rencontre chez les deux espèces végétales.
Deux genres (Microtermes et Odontotermesy de termites ont été déterminés à partir
des échantillons récoltés.
Toutefois, l'affinité des termites est plus grande pour Pterocarpus lucens dont ils
sectionnent les racines des pieds même vivants. Ces insectes contribuent activement à la
dégradation du peuplement de cette espèce par la consommation de substances ligneuses des
racines. Les capacités de colonisation du sol par les racines attaquées se trouvent limitées,
d'où une faible performance dans la recherche de l'eau des nappes profondes en saison sèche.
3.6.2.2 - Rôle des termites dans les écosystèmes
Les termites se nourrissent de la végétation vivante, morte récemment ou en
décomposition, de l'humus et des champignons. Ils se comportent au niveau trophique
comme des consommateurs de premier ordre (herbivores) et des décomposeurs (détritivores).

76
Ils libèrent par cette dégradation des débris végétaux, des substances organiques et minérales,
notamment de l'azote (ABBADIE et LEPAGE, 1989), au sol.
D'autre part, les termites creusent des galeries dans le sol jusqu'à 40 à 55 fi de
profondeur pour atteindre la nappe, dans le Ferlo sénégalais, comme l'ont montré LEPAGE
et al., cités par CASENAVE et VALENTIN, 1989. Ils jouent un rôle important dans
l'évolution des écosystèmes sahéliens aération, humidification, réserves
alimentaires, ... (HEIM, 1977; BRIAN, 1978; ABBADIE et LEPAGE, 1989). Un des rôles
importants est la désintégration des croûtes cuirassées de surface (MAIGNIEN, 1966;
GRASSE, 1950; TRICART, 1957 et BOYER, 1959 cités par BRIAND, 1978). MANDa et
al., (1993) confirment que l'apport de matière végétale sèche (paillis composite de bois et
d'herbacées) favorisait le mieux la réinstallation des termites, la destruction des croûtes et
l'aération des sols en région sahélienne de Bourzanga, Burkina Faso. Un autre rôle est la
stimulation de la croissance végétale rarement observée et décrite par KAISER (1953). II
affirme que les nids de Anoplotennes pacijicus de la forêt humide du Brésil sont envahis par
un réseau dense de radicelles de Mapairia cephalantha dont les apex racinaires sont
préférentiellement consommés par les termites. Dans les termitières abandonnées les
radicelles sont mortes.
Certaines espèces de termites joueraient aussi un rôle positif, contrairement à l'idée
répandue selon laquelle elles détruisent les écosystèmes forestiers.
CONCLUSION
Le système racinaire des arbustes sahéliens montre un réseau superficiel et extensif
colonisant une superficie importante du sol, ce qui permet une utilisation maximale des
pluies.
Les racines latérales apparaissent dans les trente premiers centimètres du sol. Elles s'étalent
dans toutes les directions mais pénètrent sous forme de racine pivotante de substitution chez
Pterocarpus lucens. La racine principale pivotante, plus ou moins longue et ramifiée en
fonction des propriétés pédologiques du sol, existe toujours.

77
L'enracinement du ptérocarpe est cependant limité par le sectionnement par les
termites hypogés, les contraintes pédologiques et les influences négatives des coupes
fréquentes et sévères de l'appareil aérien.
Dans l'anastomose racinaire, Balanites aegyptiaca utilise des substances prélevées
dans le milieu ou élaborées par son hôte. En pénétrant dans les tissus de son hôte. cette
espèce provoque des proliférations cellulaires localisées. Elle pourrait l'intoxiquer en libérant
des substances. Elle possède les aspects fondamentaux du phénomène parasitaire défini par
GORENFLOT (1986). Elle se comporte en hémiparasite facultatif.
Or selon cet auteur, l'hémiparasitisme est caractéristique uniquement des 3 familles de
Scrophulariaceae, de Santalaceae et de Loranthaceae.
Les moyens d'investigation ne nous permettent pas de définir de manière précise la
nature de relation trophique observée.
Plusieurs hypothèses d'association tissulaire dans ces anastomoses racinaires peuvent
être émises :
- l'existence de lacunes dans le phelloderme mettant en commun le canal parasitaire avec les
vaisseaux conducteurs et les tubes criblés ;
- l'existence de prolongements fins ressemblant aux hyphes fongiques, peu visibles reliant
le parasite aux vaisseaux ;
- l'échange de substances entre le liber de l'hôte et les tissus parasitaires par migration à
travers les cellules à paroi pectocellulosique du phelloderme.
Les nodules symbiotiques racinaires améliorants de la fertilité des sols sont présents
en conditions naturelles chez Pterocarpus lucens et en pépinière seulement pour Acacia
raddiana. Leurs racines sont par ailleurs en relation avec les cavités tennitiques dont les poils
fins tapissent les parois. Elles profitent de l' humidité crée par les remontées de ces insectes
des nappes profondes. Les termites joueraient aussi un rôle positif dans l'évolution des
écosystèmes sahéliens.

78
Pterocarpus lucens présente un système racina ire organisé suivant le même mode que
celui de Acacia raddiana mais dont le développement est réduit par des facteurs extrinsèques:
termites hypogés, contraintes pédologiques et coupes de la tige. La plante n'arrivant pas à
profiter des apports en eau des premières pluies, il s'en suit une saison sèche prolongée pour
cette espèce.
L'acacia a un pouvoir de colonisation du sol plus élevé que le ptérocarpe. II pourra
profiter davantage des rares pluies de début et de fin de saison pluvieuse. Par ailleurs il
possède un mucilage protecteur contre le dessèchement racinaire et des capacités de transfert
plus grandes des substances des sites d'absorption vers les parties aériennes. Toutefois, en
présence de quantité suffisante d'eau, Pterocarpus lucens pourrait se développer
normalement.
Les systèmes racinaires des ligneux assurent une fonction de fixation mécanique et
jouent un rôle dans la nutrition minérale. Ils jouent à travers ces fonctions et l'intervention
de leurs structures spécialisées (poils absorbants, nodules et anastomoses) un rôle important
dans la croissance, la productivité et les mécanismes d'adaptation à la sécheresse. Comme
les conditions mésologiques évoluent rapidement, le peuplement ligneux change de facto dans
sa composition floristique et sa structure.

Troisième partie:
ESSAI D'ETUDE DESCRIPTIVE ET
DYNAMIQUE
DU PEUPLEMENT LIGNEUX

80
1 - RAPPELS BffiLIOGRAPHIQUES
1.1 - Subdivisions phytogéographiques
D'après les subdivisions phytogéographiques d'AUBREVILLE (1957), le Burkina
septentrional appartient à la zone sahélienne du domaine atlantico-nilotique. Quant à la région
de la Mare d'Oursi en particulier, GUINKO (1984) la classe dans le secteur sahélien strict
du domaine sahélien. Cette classification sera utilisée dans le présent rapport.
Les limites de ce secteur coïncident avec:
- la limite septentrionale de Combretum glutinosum et Adansonia digitata (TROCHAIN,
1940) ;
- la limite méridionale des aires de répartition d'Acacia laeta, de Leptadenia pyrotechnica
et de Maerua crassifolia ;
- l'isohyète 400 mm séparant les 2 secteurs sahélien strict et subsahélien;
- enfin la ligne rejoignant les stations où le nombre annuel de jours pluvieux est voisin de 30
jours.
Le Sahel, comme l'indique RIOU (1990), couvre une bande riveraine du grand désert.
Elle s'étend de la côte atlantique sénégalo-mauritanienne (Ferlo sénégalais) aux littoraux
coralliens de la Mer Rouge et de l'Océan Indien, entre l'Ethiopie et le Kenya (steppes
Masaï).
1.2 - Formations et groupements végétaux
TOUTAIN a réalisé en 1976 un inventaire cartographique de la végétation de la
région dans laquelle il distingue 23 unités sur des bases tloristiques. GROUZIS (1988) les
a regroupées en 4 grands ensembles correspondant à des conditions géomorphologiques et
édaphiques différentes:
- les unités de végétation liées aux dunes et aux ensablements,
- les unités de végétation liées aux cuirasses ferrugineuses,
- les unités de végétation liées aux affleurements rocheux,

81
- les formations inondables.
Puis PlOT et al., (1980), dans une étude quantitative des ligneux dans la zone sud de
la région de la Mare d 'Oursi, ont regroupé les 23 formations végétales en 3 grands ensembles
structuraux: les formations de bas-fonds, celles de glacis et celles des systèmes dunaires.
Dans une cartographie des unités écologiques de la région, le C.R.T.O. (1989) a
réparti la végétation en 9 groupements, en fonction de leur physionomie et de leur écologie
(caractères pédologiques et géomorphologiques des unités). Une synthèse est proposée par
le tableau n° 1.
Tableau n° 1 : Principales unités écologiques de la région de la Mare d'Oursi (C.R.T.O.,1989)
Ensembles Unités Formations Espèces ligneuses dominantesgéomorphologi cartographiques végétalesques
- Oueds des - Fourré - Bauhinia rufescensensembles Balanites aegyptiacasableux
- Steppe - Bauhinia rufescensherbeuse Pterocarpus lucens
- "
Formations dedépressions et de - Steppe - Bauhinia rufescenstalwegs - Bordure de herbeuse à Combretum aculeatum
mare arbustive
- mare - Prairieaquatique àgraminées

82
- dune vive - Leptadenia pyrotechnica
- dune (erg -Steppe - Acacia senegalrécent) arbustive Leptadenia pyrotechnica
Formations dedunes etensablements -dune (erg - Steppe - Acacia raddiana
ancien) arbustive Combretum glutinosumdégradée
- .. -Steppe - Acacia raddianaarbustive plus Combretum glutinosumou moinsdégradée
- Glacis - Steppe - Acacia laetapierreux arbustive à Acacia seyal
herbeuse
-Dépression - Brousse - Pterocarpus lucensFormations de et/ou plaine tigrée plus ou Acacia senegalglacis latéritique moins
dégradée
-glacis d'oued -Fourré- Balanites aegyptiaca
Acacia senegal
Formations de -buttes et - Steppe - Boscia angustifoliabuttes cuirassées et massifs arbustive Combretum glutinosumreliefs rocheux
2 - METHODOLOGIE
2.1 - Carto2l'aphie
Nous avons utilisé des cartes et des photographies aériennes de 1981 pour réaliser
l'inventaire floristique de la région. En effet, les recherches de documents de télédétection
ont permis d'acquérir seulement des photographies de décembre 1981 auprès du Projet FAü
Mare d'Oursi et une image SPOT d'août 1986 au 1/200 000 au C.R.T.O.

83
Dans ces conditions la production d'une carte floristique était difficile. Il n'est alors
pas possible de réaliser une étude diachronique de la végétation comme celles de
DEWISPALAERE et TOUTAIN (1976 et 1981) de la même région pour les périodes 1955
et 1975 puis celles de 1975 et 1981. Une telle approche aurait renseigné sur la localisation,
l'ampleur et la vitesse de l'évolution globale de la végétation ligneuse de la région.
Les cartes utilisées sont :
- la carte topographique de l'I.G.N. (1960) feuille de Dori au 11200 000;
- la carte pédologique de reconnaissance de la région de la mare d'Oursi du BUNASOLS
(1981) au 1/50000 ;
- la carte des sols et des aptitudes de la région Nord de la mare d'Oursi du BUNASOLS
(1989) au 1150000 ;
- la carte toponymique de l'ORSTOM (1979) au 1/50000;
- la carte de situation du bassin de la mare d'Oursi, du C.N.R.S. (1980) au 1150000 ;
- la carte des ressources fourragères A. C .C. LAT de l'I. E.M.V. T. (1976) au 1/50 000 ;
- enfin celle des unités écologiques de la mare d'Oursi du C.R.T.O. (1989) au 1150000.
2.2 - Inventaire floristigue
Nous avons effectué un sondage stratifié aléatoire au taux de 10-3 . Il présente
l'avantage de réduire le coût et le temps du sondage (GANABA, 1990).
Le taux d'échantillonnage est déterminé par unité géomorphologique en référence aux
cartographies de TOUTAIN (1976) et du C.R.T.O (1989), tel que défini sur le tableau n02.
Ensuite les placeaux sont positionnés de manière orienté en choisisant dans l'unité définie au
préalable sur notre calque d'interprétation de photos aériennes, une station qui nous semble
représentative.

84
Tableau n° 2 : Définition du nombre de placeaux d'échantillonnage du peuplement ligneuxpar unité géomorphologique
Formations végétales TOUTAIN C.R.T.O. Nombre de(1976) (1989) placeaux
retenus
Dépressions et talwegs 12 14 15
Glacis 29 38 321
Buttes et reliefs - 5 4
Dunes et ensablements 13 8 8
Champs et jachères 12 - 9
Total 66 65 68
Sur chaque unité de l'échantillonnage, les relevés de terrain vont porter sur des
placeaux de 0,1 ha. Les placeaux correspondent à des rectangles de 50 m de longueur sur
20 m de largeur (FONTES et GUINKO, 1993). La forme allongée et la taille des placeaux
sont conformes aux recommandations du Séminaire, tenu en 1988 à Koudougou, sur
l'Aménagement des Forêts Naturelles, qui préconise une taille optimum de 0,125 ha retenue
ailleurs comme au Cameroun (GANABA, 1990).
Chaque placeau est numéroté et décrit par une fiche de reconnaissance où l'accent est
mis sur le peuplement ligneux et les conditions stationnelles (fiches d'inventaires en annexes
XI et XII). Les informations sur le peuplement herbacé concernent la composition floristique
dominante, la hauteur et le taux de recouvrement.
A titre de comparaison, nous avons sondé les anciennes parcelles de végétation de
GROUZIS (1979 et 1988) dont les caractéristiques sont définies en annexe XIII.
2.2.1 - Mesures qualitatives
Ces mesures permettent de caractériser la végétation dans sa composition, sa
physionomie et ses tendances évolutives.

85
Tous les ligneux du placeau sont comptés et leurs espèces identifiées. Tout ligneux
de hauteur supérieure ou égale à 7 m est considéré comme arbre. Si la hauteur est comprise
entre 7 m et 1 m c'est un arbuste ou un arbrisseau. Les régénérations représentent les pieds
de moins de 1 m de hauteur avec un diamètre à la base du tronc inférieure ou égale à 2,5 cm
(FONTES et GUINKO, 1993).
Par ailleurs, la mesure de la vitalité (régénération, coupes et mortalité) est prise en
compte. La coupe regroupe les individus ayant subi des prélèvements des tiges aériennes ou
des racines. La mortalité concerne les individus (pieds isolés) en dessèchement total en début
de saison sèche et les bois morts sur pieds ou couchés.
Des difficultés d'évaluation de la mortalité sont apparues à cause du ramassage des
pieds morts sur certaines stations.
2.2.2 - Mesures quantitatives
Les paramètres de la végétationà mesurer sont: le diamètre à la base (environ 30 cm
du sol), la hauteur totale du fût, le nombre de pieds et de tiges. La mesure de ces paramètres
permet d'apprécier les potentialités en ressources ligneuses du peuplement à l'état actuel et
leur évolution.
Selon KALA (1977) cité par POUPON (1980), l'utilisation des 2 paramètres (hauteur
et circonférence) permet d'envisager la dynamique des peuplements ligneux.
Certes, la circonférence plus précise que le diamètre permet de mieux caractériser la
végétation. Mais la structure buissonnante et la nature épineuse des ligneux de cette région,
nous a fait abandonné la méthode de mesure de la circonférence du tronc au ruban mètre au
profit de celle du diamètre au compas forestier.
Le diamètre du fût se mesure à la base du tronc (30 cm du sol) pour la plus grosse
tige, à cause de la présence de ramifications chez les espèces sahéliennes en dessous de la
hauteur de poitrine (fixée conventionnellement à 1,30 m) (POUPON, 1980; NEBOUT, 1978
et PIOT et al., 1980).

86
La hauteur totale du fût est appréciée par mesure à la perche dendrométrique.
Nous avons caractérisé chaque espèce par son indice de présence, son degré
d'appétabilité et son taux d'exploitation dans le milieu.
Pour définir les indices de présence des ligneux, nous nous sommes référé à la
fréquence centésimale utilisée par GROUZIS (1988) et GUINKO et al. (1989).
La fréquence spécifique (F.S.) d'une espèce représente le nombre de points où cette
espèce a été rencontrée. La fréquence centésimale (FoC.] est égale au rapport de la F.S. au
nombre de points échantillonnés (N) :
F.S.
F.C. = ------ X 100 (%)
N
Pour enfin être conforme au document de référence pour la comparaison, nous avons adopté
la classification suivante pour le degré d'abondance:
- espèces très répandues: 100 à 50% de F.C.,
- espèces moyennement répandues: 50 à 10% de F.C.,
- espèces pas ou rarement rencontrées: 10 à 0% de F.C.
Les taux d'exploitation et de mortalité sont définis par le rapport entre les pieds
coupés ou morts et le nombre de pieds vivants intacts de l'espêce considérée.
Les ligneux constituent un fourrage dont il est nécessaire d'apprécier le degré
d'appétabilité. Aussi avons-nous regroupé, à partir de nos propres observations et des études
agrostologiques de la région et d'autres régions similaires, les données sur l'appétabilité des
principales espèces ligneuses (TOUTAIN et DEWISPALAERE, 1978; POUPON, 1980;
CISSE, 1987; GUINKO et al., 1989).

87
2.3 - Dynamique du peuplement lieneux
Pour savoir comment se manifeste la dynamique et son importance, il est nécessaire
de suivre les changements de la végétation sur des sites-échantillons pendant une période
assez longue. Il est aussi possible de comparer les résultats d'inventaire d'une situation de
départ relativement éloignée dans le temps (1980) avec ceux de la situation présente. Cela
nécessite une similitude de méthodes de collecte et de traitement des données. Or, les
informations précises sur cette période ne concernent que des zones limitées et sont en outre
orientées vers des approches pastoralistes.
Face à ces limites, l'approche méthodologique de la dynamique est indirecte: une
comparaison est faite entre les résultats de notre inventaire et ceux de PlOT et al. (1980) et
de GROUZIS (1983). Puis, des indices de toute nature permettant de reconstituer des
situations antérieures ou d'apprécier le mouvement évolutif actuel sont recherchés: ce sont
le degré de vitalité des différentes espèces ligneuses et leur importance écologique (utilisation
en pâturage ou par l'homme).
Nous pourrons ainsi définir sur le plan floristique et physionomique, les zones de
relative stabilité celles de dégradation plus ou moins avancée, enfin celles de repeuplement.
Pour permettre une comparaison de nos résultats à ceux de PlOT et al. (op. cit) et
pour caractériser le peuplement ligneux, nous avons subdivisé le paysage végétal en 5 types
de peuplement:
- le peuplement ligneux des dépressions et talwegs,
- le peuplement ligneux des glacis,
- le peuplement ligneux des buttes cuirassées et reliefs rocheux,
- le peuplement ligneux des dunes et ensablements,
- le peuplement ligneux des champs et jachères.
La distinction d'un peuplement des champs et jachères permettra d'appréhender le
sens de l'artificialisation du milieu par les activités agricoles et la sélection opérée au niveau
des espèces ligneuses.

88
Les résultats d'inventaire sont traités au logiciel REFLEX.
3 - RESULTATS ET DISCUSSIONS
3.1 - Etude descriptive du peuplement ligneux
La description floristique et structurale du peuplement ligneux représente son état en
décembre 1992.
3.1.1 - Composition floristlque
L'inventaire floristique des surfaces exondées de la région de la Mare d'Oursi
comporte 59 espèces végétales ligneuses, consignées en annexes XIV, XV et XVI.
D'après les inventaires floristiques établis par GASTON et BOTTE (1971),
TOUTAIN (1976) puis GROUZIS et NGARSARI (1981), la flore de cette région comprend
près de 400 espèces végétales. Ils distinguent en outre 61% de thérophytes, 17% de
phanérophytes et 22% de chaméphytes, géophytes et hémicryptophytes.
L'examen de la liste floristique ligneuse de la région conduit aux remarques suivantes:
1- l'importance spécifique de 4 familles: les Asclepiadaceae (9%), les Capparaceae
(Capparidaceae) (13%), les Combretaceae (9%) et les Mimosaceae (23%). Comme l'indique
MICHEL et a/. (1969) cité par POUPON (1984), les familles des Capparaceae et
Mimosaceae sont caractéristiques du domaine sahélien et plus particulièrement de sa partie
septentrionale, ce qui correspond au secteur sahélien strict de GUINKO (1984). Il poursuit
en montrant que la famille des Combretaceae marque généralement la transition entre les
zones sahélienne et soudanienne.
2- l'abondance des espèces épineuses d'où le nom de "steppe armée" ou "brousse à
épineux" donné par certains auteurs à ce type de formation. La plus abondante est Acacia
raddiana qui, par son extrême plasticité écologique, colonise pratiquement tous les milieux
non inondés.

89
3- la présence d'espèces soudaniennes représentant certainement des vestiges
d'anciennes formations d'ailleurs relatées dans les récits des populations.
En résumé, nous distinguons dans la flore des groupes d'espèces liées aux 3 étages
bioclimatiques: sahélien, subsahélien et soudanien (tableau n° 3).
Tableau n° 3 : Tableau de répartition bioclimatique des principales espèces ligneuses de larégion de la Mare d' Oursi
Etage bioclimatique Espèces ligneuses caractéristiques Total d'espèces
Acacia erhenbergianaAcacia laetaAcacia raddiana
Sahélien Acacia senegal 6Commiphora africanaLeptadenia pyrotechnica
Acacia nilotica var. adansoniiBalanites aegyptiacaBauhinia rufescensBoscia senegalensis
Subsahélien Combretum .micranthumDalbergia melanoxylon 9Maerua crassifoliaPterocarpus lucensZiziphus mauritiana
Acacia albidaAdansonia digitataAnogeissus leiocarpus
Soudanien Celtis integrifolia 7Entada africanaMitragyna inermisSclerocarya birrea
L'analyse de la structure des peuplements des unités géomorphologiques permet de
mieux caractériser le peuplement ligneux.

90
3.1.2 - Structure
Les résultats de l'inventaire concernent les pieds vivants de toutes les espèces
ligneuses, tous diamètres compris. Ils sont exprimés en fonction des unités
géomorphologiques définies auparavant. Les mesures de densité et des paramètres de vitalité
du peuplement ligneux par unité géomorphologique sont donnés en annexe XVII
3.1.2.1 - Peuplement ligneux des dépressions et talwegs
Cette unité géomorphologique renferme les formations végétales les plus denses de
la région, ayant parfois la physionomie des fourrés ou "bushs" plus ou moins impénétrables
des galeries forestières. Autour des mares d'eau, plus ou moins pérennes selon la
pluviométrie des années successives, les ligneux s'organisent en zones concentriques formées
de groupes d'espèces de moins en moins exigeantes en eau au fur et à mesure qu'on s'en
éloigne (tableau n° 4).
Tableau n" 4 . Répartition zonale du peuplemeœ ligneux autour des mares naturelles de la région de la Mare d'Oursi
Zone Zone à longue inondation Zone à inondation temporaire Zone rarement illllOl1œ
Mare
Acacia nilonca var. nilouca Acacia nilonca var. adansoniiAcacia seyal Balanites aegyptiaca
Piiiostigma reticularumAcacia seyal
Ours! Bauhinia rufescensAcacia raddiana
Ficus gnafalocarpa
Acacia nilotica var. nilotica Acacia nilonca var. adansonii Balanites aegyptiac.aAcacia nilotica var. IOmentosa Acacia seyal Acacia raddiana
Tin Edjar Miuag)na rœrmts Bauhinia rufescens Combrecum glutinosum
Acacia nilcoca var. nilccca Bauhinia rufesœns Acacia nilotica var. adansoniiAcacia seyal Acacia raddiana
Yomboli Ceins integrifolia Acacia seyalBalanites aegyptiacaOxystehna bomouense
Mais cette organisation végétale est diffuse le long des talwegs.
Au regard de la composition floristique de ce peuplement et par rapport à l'inventaire de
PlOT et al. (1980), nous remarquons qu'il est le plus riche en espèces.

91
Les plus nombreuses sont: Balanites aegyptiaca (24% des individus), Combretum aculeatum
(15%), Acacia raddiana (12%), Acacia nilotica (10%), Acacia seyal (9%), Boscia
senegalensis (6%), Combretum micranthum (4%), Ziziphus mauritiana (3%) et Pterocarpus
lucens (3%).
Certaines espèces ne se rencontrent que dans cette unité. Ce sont: Acacia nilotica
var. nilotica, Acacia nilotica var. tomentosa, Combretum micranthum, Feretia apodanthera,
Piliostigma reticulatum, Anogeissus leiocarpus, Grewia bicolor, Grewia vil/osa et Grewia
tenax .
D'autres sont absentes des bordures de mares: Anogeissus leiocarpus, Combretum
micranthum, Feretia apodanthera, Pterocarpus lucens, Grewia bicolor, Grewia flavescens,
Grewia tenax et Grewia vil/osa.
Ces formations de dépressions constituent en outre les réserves en potentiel ligneux
de la région malgré des cas de mortalité qu'elles présentent.
D'autre part, beaucoup d'espèces sont buissonnantes ou ramifiées dans cette région.
Ce sont: Acacia ataxacantha, Combretum micranthum, Feretia apodanthera, Bauhinia
rufescens et Dalbergia melanoxylon respectivement 15,6,5,4 et 3 tiges/pied en moyenne.
Guiera senegalensis possède aussi en moyenne 4 rejets/pied, Mitragyna inermis et
Pterocarpus lucens 3 tiges/pied. Cette allure des pieds s'explique par les coupes fréquentes
qui les affectent.
En outre, la densité élevée et le caractère ramifié de la grande majorité des espèces
contribuent à expliquer la nature plus ou moins fermée de ces steppes arborées.
Les pieds de régénération de hauteur inférieure à 0,5 m et ceux dont la hauteur se
situe entre 2 et 5 m représentent respectivement 51 % et 29 % des effectifs de l'unité.

92
Les petits arbres de hauteur comprise entre 7 et 10 m en représentent 4 %. Les arbres
moyens, de hauteur égale ou supérieure à 10 m dans ce cas, forment 1 % de cene
population. Les plus grands arbres recensés sont par ordre décroissant : Anogeissus
leiocarpus, Acacia seyal, Acacia nilotica, Pterocarpus lucens et Combretum micranthum.
Les ligneux les plus vigoureux (diamètre supérieur à 22,5 cm) sont: Acacia nilotica,
Acacia raddiana, Acaciaseyal, Balanites aegyptiaca, Piliostigma reticulatum et Pterocarpus
lucens.
3.1.2.2 - Peuplement ligneux des glacis
L'analyse de la composition tloristique du peuplement de cene unité (tableau n° 5)
indique une pauvreté spécifique avec Acacia raddiana (49% des individus) comme espèce
dominante dans cene unité. Balanites aegyptiaca représente 15% des pieds.
Les glacis présentent entre eux une différence importante dans la densité et dans la
composition floristique des peuplements ligneux qu'ils portent. En effet, le glacis d'épandage
de Windé a une densité de 290 pieds/ha contre 36 pour le glacis gravillonnaire de Djalafanka.
Les espèces sont peu ramifiées à l'exception d'Acacia laeta (2 tiges/pied en moyenne) et
Dalbergia melanoxylon environ 7 tiges/pied en moyenne.
Là encore, les ligneux se concentrent dans les classes de hauteur de 0 à 0,5 m et de
2 à 5 m avec respectivement 57 % et 28 % des effectifs. Les pieds ont une hauteur maximale
de 6 m. Les plus grands. Acacia raddiana, Balanites aegyptiaca et Acacia laeta sont aussi
les plus fréquents.
Quatre vingt six pour cent des ligneux ont un diamètre inférieur à 10 cm. Les plus
gros pieds sont encore représentés par Acacia raddiana et Balanites aegyptiaca, qui sont
aussi représentées dans toutes les classes.

93
3.1.2.3 - Peuplement ligneux des buttes cuirassées et des reliefs rocheux
Le peuplement ligneux de cette unité est dominé par Acacia raddiana (23 % des
individus), Pterocarpus lucens (18%) et Leptadenia hastata (15%).
La hauteur maximale des pieds est de 8 m rencontrée essentiellement chez
Pterocarpus lucens. Cinquante sept pour cent des pieds ont une hauteur inférieure à 2 m.
Par ailleurs 73 % des ligneux ont un diamètre inférieur à 7,5 cm et le plus gros diamètre est
de 24 cm.
3.1.2.4 - Peuplement ligneux des dunes et ensablements
Les espèces les plus abondantes sont: Acacia raddiana (40 % des pieds) Balanites
aegyptiaca (21%) Acacia senegal (9%), Maerua crassifolia (8%) et Guiera senegalensis
(7%).
Certaines de ces espèces sont ramifiées : Acacia laeta et Guiera senegalensis avec en
moyenne 3 tiges/pied, Acacia raddiana et Maerua crassifolia 2 tiges/pied.
La hauteur maximale du peuplement dans cette unité est de 9 m. Près de la moitié
(49%) des ligneux ont une hauteur comprise entre 2 et 5 m avec 34% des pieds dans celle
de 0 à 0,5 m. Seulement 1% des pieds ont une hauteur comprise entre 7 et 10 m. Les plus
hauts ligneux de cette unité sont: Combretum glutinosum, Balanites aegyptiaca et Acacia
raddiana. Cette formation constitue une steppe arbustive basse.
La répartition des ligneux par classe de diamètre, représentée par la figure n° 8,
révèle une forte proportion d'individus de diamètre compris entre 0 et 7,5 cm soit 70 % de
la population, le plus gros diamètre étant de 32,5 cm.
3.1.2.5 - Peuplement ligneux des champs et jachères
Les principales espèces sont: Balanites aegyptiaca (39% des pieds, Acacia raddiana
(17%), Leptadenia hastata (15%).

94
Les ramifications y sont rares à cause des élagages effectués par les paysans. Les
espèces parfois épargnées sont Ca/otropis procera, espèce secondaire des jachères, et
Bauhinia rufescens. Chacune possède en moyenne 5 tiges/pied.
Tableau n° 5 : Densité moyenne par ha et par unité géomorphologique du peuplementligneux de la région de la Mare d'Oursi
Densité moyenne des ligneux par unitéEspèce ligneuse géomorphologique
Dépressions Glacis Buttes Dunes Champs
Acacia nilotica var. adansonii 37 1Acacia laeta 5 8 1 8Acacia raddiana 54 50 75 118 21Acacia senegal 3 27Acacia seyal 37Balanites aegyptiaca 103 15 17 62 48Boscia senegalensis 25 25Calotropis procera 5Combretum aculeatum 66Combretum glutinosum 11 4Combreturn micranthum 19 13Grewia bicolor 8Grewia villosa 10Guiera senegalensis 6 30 20Leptadenia hastata 11 50 17 18Maerua crassifolia 10 25Piliostigma reticulatum 5Pterocarpus lucens 11 60Ziziphus mauritiana 15
Autres 53 19 39 7 14
Densité moyenne/ha 434 103 327 ~99 123
Ecart-type densité 61,40 18,85 114,99 58,56 28,33
Total espèces 32 28 19 15 18
Ca/otropis procera est abondante dans les jachères de piémont des buttes de Kolel et
de Warga et sur quelques jachères dunaires de Ganadawri.
Les pieds épargnés dans les champs sont utilisés comme bois de service.

24,16"10
Acacia raddiana
4,88% 8,74%
41,39%
Balanites aegyptiaca
16,61%
Toutes espèces
7,59% 8,95%
44,57%
Leptadenia hastata
• Buttes etreliefs
22,56% ŒJ Glacis
~ Bas-fonds
mDunes elensablements
D Champs etjachères
Pterocarpus lucens
12,61"10
6,31%
40,00%
60,00%
Répartition par ha et par unité géomorphologique du peuplementligneux de la région de la mare d' Oursi en 1992

96
La majorité des ligneux sont des arbustes. Les arbres sont représentés essentiellement
par Acacia albida assez répandue dans les terroirs de la région. Ils sont protégés par la
population qui récolte les fruits pour son bétail.
3.1.2.6 - Interprétation des résultats
La comparaison des moyennes arithmétiques des densités par unité géomorphologique
permet de faire la classification décroissante suivante: dépressions, buttes - dune, champs
et glacis.
La densité des ligneux augmente progressivement des glacis vers les champs et
jachères, identique sur les dunes et les piémonts de buttes et reliefs, plus élevée dans les
dépressions. La composition floristique et la physionomie varient aussi avec la topographie
(figure n° 22).
Acacia raddiana comme Balanites aegyptiaca et Leptadenia hastata s'installent dans
toutes les unités géomorphologiques à l'exception des stations à immersion prolongée (annexe
XVII).
Ces espèces tolèrent les variations de conditions pédologiques. Elles ont une amplitude
écologique large et sont ubiquistes dans cette zone.
Au contraire Mitragyna inermis, Acacia nilotica var. nilotica et Acacia nilotica var.
tomentosa affectionnent uniquement les sols hydromorphes des axes de drainage.
D'autres espèces enfin sont présentes de façon préférentielle dans des unités
géomorphologiques déterminées. Ainsi Boscia salicifolia, Euphorbia balsamifera et
Leptadenia pyrotechnica sont présentes uniquement sur les sols dunaires et les ensablements.
Leptadenia pyrotechnica est répandue sur la dune vive d'Oursi et Boscia salicifolia sur la
dune fixée de Zarmakoy. Pterocarpus lucens aussi ne se rencontre actuellement que sur les
plateaux cuirassés et les zones d'écoulement d'eau qui les séparent. Cette "rigidité" aux
conditions mésologiques restreint son amplitude écologique et fait d'elle une espèce
relativement exclusive.

97
Acacia raddiana et Balanites aegyptiaca sont les espèces les plus abondantes dans
la région
3.1. 2.7 - Discussions sur la structure du peuplement ligneux
Une différence significative de 31 à 78 % existe entre les résultats des densités du
peuplement de J'inventaire général et ceux des parcelles de végétation (anciennes parcelles
de GROUZIS) situées sur ces unités (tableau n° 6).
Tableau n° 6 : Comparaison des densités et des nombres d'espèces du peuplement ligneuxentre l'inventaire général et celui des parcelles de végétation
Inventaire Différence engénéral/parcelles de pourcentage (%)
Unités végétationgéomorphologiques
Nombre Densité Nombre Densitéd'espèces pieds/ha d'espèces pieds/ha
Dépressions et talwegs 32/8 434/95 75 78
Glacis 28/6 103171 79 31
Bunes et reliefs 19 327 - -
Dunes et ensablements 15/9 299/180 40 39
Champs et jachères 18/13 123/47 28 61
NB : La parcelle de végétation au niveau des buttes cuirassées de Kolel piémont esttransformée en champ et celle de Windé incluse dans les dépressions et talwegs.
La différence de densité à la même époque (1992) entre les deux entités peut
s'expliquer par la différence de taille entre les placeaux de sondage (placeaux de 0,1 ha
contre des parcelles de 1 ha). Des constats révèlent d'autre part des différences
structurales entre les parcelles de végétation et les formations végétales environnantes. En
effet, des dégradations importantes ont affecté ces parcelles.
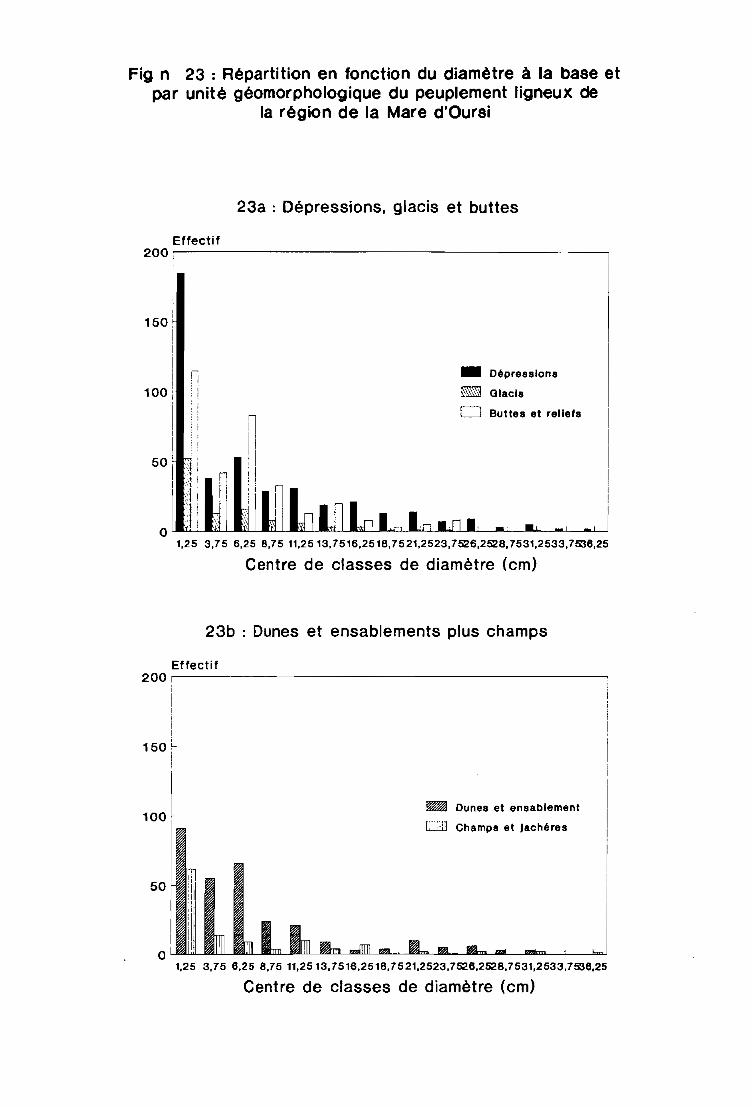
Fig n 23: Répartition en fonction du diamètre à la baae etpar unité géomorphologique du peuplement ligneux de
la région de la Mare d'Ourai
23a : Dépressions. glacis et buttes
Effectif200
1
!
1
i150
100
50
nIl! 1
il: 1
_ Dépressions
_ Glacis
o Bulles et rellels
1,25 3,75 6,25 8,75 11,2513,7516,2518,7521,2523,7526,2528,7531,2533,7536,25
Centre de classes de diamètre (cm)
23b : Dunes et ensablements plus champs
Ellecti 1200 ri-------
150
100_ Dunes et ensablement
o Chsmps et Jachéres
50
o1,25 3,75 6,25 8,75 11,2513,7516,2518,7521,2523,7526,2528,7531,2533,7536,25
Centre de classes de diamètre (cm)

Fig ri 23c: Toutes unités
Effectif700,----------------------------,
600
500
400
300
200
100
o
o Autres espèces
_ Balanites aegyptlaca
_ Acacia raddlana
1,25 3,75 6,25 8,75 11,2513,7516,2518,7521,2523,7526,2528,7531,2533,7536,25
Centre de classes de; diamètre (cm).

100
3.1.3 - Analyse de la répartition en fonction du diamètre
Les histogrammes relatifs à la répartition par classe de diamètre mesuré à la base et
exprimée par unité géomorphologique du peuplement ligneux sont représentés par la figure
n° 23.
Ils se caractérisent par une structure plurimodale (3 à 4 pics) marquée par un grand
pic pour les individus de petit diamètre [0-1,25 cm] et une série de pics irréguliers et moins
élevés pour les ligneux plus gros. Cette structure est similaire pour les peuplements des 5
unités géomorphologiques. Mais elle diffère très sensiblement des courbes théoriques de type
exponentielle décroissante caractérisant la structure des peuplements naturels en équilibre
comme celle de Balanites aegyptiaca étudiée par POUPON (1980).
Tout se passe comme si la population avait subi des périodes de faible régénération
ou de forte mortalité de ses jeunes plants responsables des "classes creuses" observées.
En dépit des difficultés d'estimation de l'âge de ces ligneux sahéliens, on est tenté de
faire le lien entre les diamètres des classes creuses avec la série des années à déficit
pluviométrique. Ainsi les creux des classes [2,5-5 cm] et [15-17-20 cm] seraient sans doute
provoqués respectivement par les déficits pluviométriques des années 1984, 1987 et 1990.
Les creux des classes de [20-25 cm] et [27-30 cm] seraient liées aux épisodes antérieurs,
probablement celui de 1973.
3.1.4 - Degré d'appétabilité
Le taux d'espèces ligneuses appétées est consigné sur le tableau n° 7 suivant.
Nous pouvons déduire des données de ce tableau, le rôle important assuré
effectivement par les ligneux, en complément des herbacées, dans l'alimentation du bétail.
En effet, 59% des espèces sont bien appétées. Seule l'espèce CaraLluma retrospiciens
est reconnue toxique. Le latex de cette espèce est utilisé pour empoisonner les hyènes.

101
Tableau n° 7 : Taux d'appétabilité des principales espèces ligneuses de la région de la mared'Oursi
Degré d'appétabilité Nombre d'espèces Pourcentage (%)
Très appété (TA) 17 30Appété (A) 16 29Peu appété (PA) 18 32Non appété (NA) 4 7Toxique (T) 1 2
Total 56 100
Comme l'ont indiqué RIPPSTEN et PEYRE De FABREGUES (1972, cités par
BERNUS, 1983), les ligneux constituent non seulement un fourrage de relais des pâturages
herbacés mais aussi une ressource fourragère stable pendant tout le cycle annuel. Ils sont
moins tributaires de la répartition des pluies de la saison précédente.
La plupart des ligneux sahéliens possède en effet une valeur fourragère très bonne à
assez bonne pendant toute I' année. Les feuilles toujours vertes de certaines espèces (Balanites
aegyptiaca, Boscia senegalensis, Salvadora persica et Ziziphus mauritiana) et les fruits des
espèces à feuilles caduques (Acacia spp. Maerua crassifolia, Pterocarpus lucens , Celtis
integrifoliaï sont très recherchés en saison sèche.
A cette période de l'année ils constituent les seules sources de matières azotées
indispensables à une ration équilibrée.
3.2 - Dynamique du peuplement ligneux
3.2.1 - Analyses qualitative et quantitative entre 1980 et 1992
Un accroissement sensible se remarque entre les densités des ligneux de notre
inventaire et de celui de PlOT et al. (1980,) dont la comparaison est consignée sur la figure
n° 24.

Fig n 24: Comparaison par unité géomorphologique desdensités du peuplement ligneux de la région de la Mare
d'Oursi entre 1980 et 1992
Légende 24a : Dépressions et talwegs
Densité/ha120,-~~~~------------------
Espèces ligneuses
Acacia nilot/caAcacle laetaAcacia raddienaAcacia lenegalAcacia aeyalBalanltea aegyptiacaCalotropia prcceraCombretum aculeatumCombretum glutinoaumCombretum micranthumQuiers aenegalenlilLeptadenia hastataPllioaligma reticulatumZlziphua maurilians
code
AniAisArsAagAsyBaeCprCacCglCmiOaeLhaPrelma
100
80
60
40
20
J---=-Ala Ara Asy Bae Csc Cml
Espèces ligneuses
.1980 ~1992
Cal lms
24 b : Glacis
Densité/ha
::~-------------------------.,
24 C : Dunes et ensablements
Denaité/ha140,-----'--------------------------,
60
40
20
Ala Ani Ara
Espèces ligneuses
.1980 ~1992
Bse Ala Ara Bae Cac Cgl Cal Ose Pre lma
Espèces ligneuses
.1980 ~1992

103
Il apparait qu'en 1980 certaines espèces étaient faiblement représentées et d'autres ne
figurent pas dans la liste des nombres d'arbres mesurés par ha (annexe XVIII). C'est le cas
pour Acacia senegal et Leptadenia hastata dans les formations des dunes et ensablements et
dans les glacis. Il en est de même pour Maerua crassifolia dans les dunes et ensablements,
Acacia nilotica, Acacia ataxacantha et Grewia spp. dans les bas-fonds.
Par ailleurs, la taille inférieure à 1 haet le sondage pourraient aussi expliquer les écarts. De
plus l'inventaire du C.T. F. T est relativement plus proche de la grande sécheresse des années
1972-73 qui avait entamé sérieusement le potentiel ligneux.
Nos valeurs de densité des ligneux sont en deçà de celles de POUPON (1978) dans
le sahel sénégalais (Fêté-Olé), qui est toutefois influencé par la proximité de la mer. La
répartition par éléments de reliefs des densités par hectare des ligneux est la suivante :
sommets et versants de dune, 296 et 492 ; replats 1004 ; bas de versants, 1497 ; et
dépressions, 3499. En région subsahélienne de Banh (Burkina Faso), COUTERON,
AQUINO et OUEDRAOGO (1992) ont mesuré en fourré de bas-fonds, en steppe arbustive
claire, en steppe arbustive dense et brousse tigrée respectivement environ 100, 125, 150 et
175 pieds/ha.
Ces constats rendent toute analyse quantitative indicative. Aussi insisterons-nous sur
l'analyse qualitative de la dynamique du peuplement ligneux entre 1980 et 1992.
L'analyse qualitative de la flore montre l'absence de certaines espèces rencontrées en
1980 par PlOT et al.. Ce sont: Acacia polyacantha, Craieva religiosa, Cordia myxa (sic!
Cordia myxa probablement), Diospyros mespilifonnis et Ziziphus spina-christi.
La comparaison des degrés d'abondance permet de classer les espèces ligneuses en:
- espèces abondantes en 1980 devenues moyennement fréquentes: Acacia laeta, Acacia
nilotica var. adansonii, Acacia seyal, Anogeissus leiocarpus, Combretum glutinosum,
Combretum aculeatum, Guiera senegalensis, Pterocarpus lucens et Ziziphus mauritiana;
- espèces abondantes rarement rencontrées maintenant : Commiphora africana, Dalbergia
melanoxylon, Euphorbia balsamifera, Grewia bicolor et Piliostigma reticulatum ;

104
- espèces moyennement fréquentes devenues peu fréquentes: Acacia ataxacantha, Adansonia
digitata, Boscia senegalensis, Dichrostachys cinerea, Feretia apodanthera, Grewia tenax et
Grewia villosa.
D'autres espèces conservent leur niveau d'abondance initial:
- espèces toujours abondantes: Acacia raddiana et Balanites aegyptiaca ;
- espèces toujours moyennement fréquentes: Acacia senegal et Maerua crassifolia.
D'autres enfin ont vu leur distribution augmenter sensiblement. Rarement rencontrées
auparavant, elles sont devenues moyennement fréquentes. Ce sont: Calotropis procera,
Leptadenia hastata et Leptadenia pyrotechnica.
La comparaison des valeurs des densités des principales espèces végétales entre 1980
et 1992 confirme la prédominance de Acacia raddiana et Balanites aegyptiaca. Leur
abondance est en hausse sensible dans les' peuplements de toutes les unités
géomorphologiques.
Dans le cas de Combretum aculeatum, Commiphora africana et Piliostigma
reticulatum, nous observons une diminution de leurs effectifs.
D'autres espèces deviennent plus nombreuses dans certaines unités. C'est le cas de Guiera
senegalensis, Leptadenia hastata et Maerua crassifolia sur les dunes et ensablements.
Par contre Pterocarpus lucens ne ressort pas dans les listes des principales espèces des unités
géomorphologiques de PlOT et al. (1980).
Par ailleurs la comparaison des densités des parcelles de végétation, ayant un état de
dégradation élevée, avec celles de l'inventaire floristique du C. T. F. T., indique aussi des
augmentations de densité de 134 et 35 respectivement pour celle de dunes et de glacis. Il y
a une diminution avec 68 pieds/ha pour celle de Windé. Cela confirme soit un repeuplement
en ligneux soit une sous-estimation des potentialités ligneuses dans l'inventaire du C.T.F.T.
Les variations de la distribution des espèces peuvent être illustrées par l'analyse des
paramètres d'évolution à savoir la régénération, les coupes et la mortalité.

Figure n 25: Taux de régénératlon/hades espèces ligneuses
Taux de régénération200 ~:::C-_--""'-- _
Unité géomorphologique
Hauteur 1 1m et dIamètre ~. 2,15 cm
Figure n 27: Taux de coupe en % Ipiedvivants des espèces ligneuses
T.a.:::u::.x::.d:::e=--=c::.o.::.u"-p.::.e -t40 -
Figure n 26: Taux de mortalité/hades espéces ligneuses
Taux de mortalité140 r=--'--.::..::..:.:..:::.:..:..:...-------------,
Unité géomorphologiquePied. I.CI en tin de ••laon pluvleu••
Figure n 28: Croissance des ligneuxde la plantation du glacis de Djalafanka
Hauleur du pied (cm) Diamétre (cm)1BO r=-==--=--=.::..::.='------'--===.:;3
_ Hauteur _ DiamétreUnités gèomorphologiques
A. seyal o A. seyal A. raddiana A. senegal
Espéce ligneuse
2,5
2
1,5
1 .
-. ~Acacia. aeyal~ • pieds d'une année
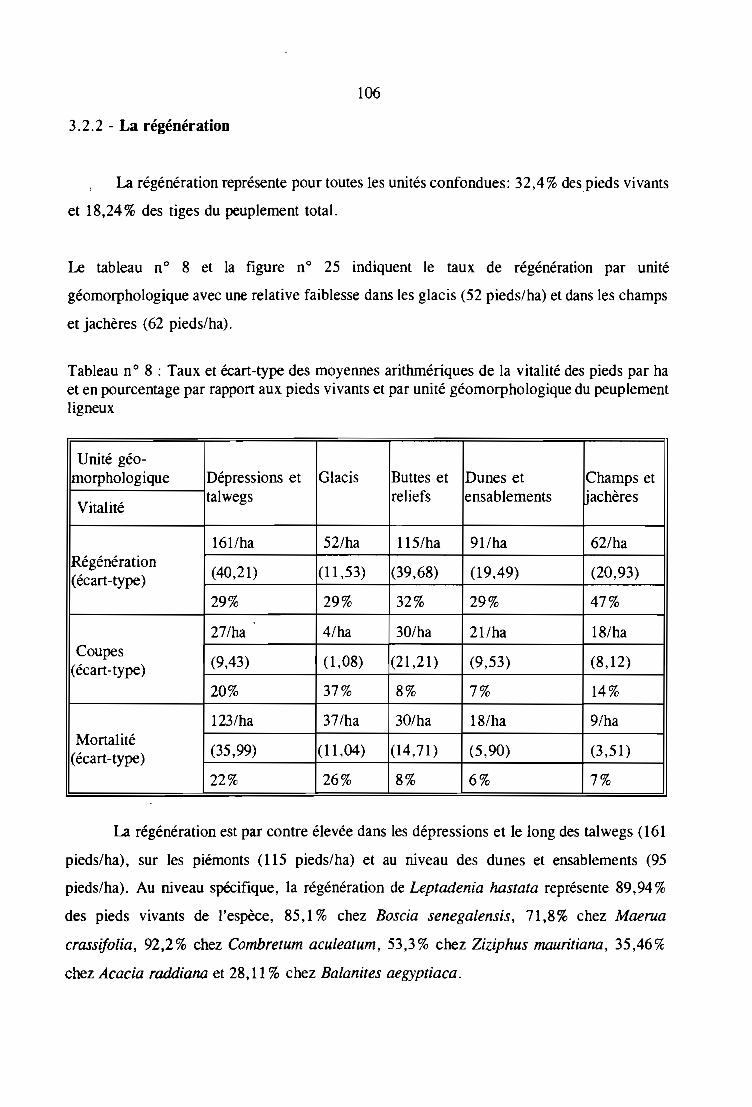
106
3.2.2 - La régénération
La régénération représente pour toutes les unités confondues: 32,4% despieds vivants
et 18,24% des tiges du peuplement total.
Le tableau n° 8 et la figure n° 25 indiquent le taux de régénération par unité
géomorphologique avec une relative faiblesse dans les glacis (52 pieds/ha) et dans les champs
et jachères (62 pieds/ha).
Tableau n° 8 : Taux et écart-type des moyennes arithmériques de la vitalité des pieds par haet en pourcentage par rapport aux pieds vivants et par unité géomorphologique du peuplementligneux
Unité géo-morphologique Dépressions et Glacis Buttes et Dunes et Champs et
Vitalitétalwegs reliefs ensablements jachères
16l/ha 52/ha 115/ha 911ha 62/haRégénération
(40,21) (11 ,53) (39,68) (19,49) (20,93)(écart-type )
29% 29% 32% 29% 47%
27/ha 4/ha 30/ha 21/ha 18/haCoupes
(9,43) (1,08) (21,21) (9,53) (8,12)(écart-type)
20% 37% 8% 7% 14%
123/ha 37/ha 30/ha 18/ha 9/haMortalité
(35,99) (11,04) (14,71) (5,90)(écart-type) (3,51)
22% 26% 8% 6% 7%
La régénération est par contre élevée dans les dépressions et le long des talwegs (161
pieds/ha), sur les piémonts (115 pieds/ha) et au niveau des dunes et ensablements (95
pieds/ha). Au niveau spécifique, la régénération de Leptadenia hastata représente 89,94%
des pieds vivants de l'espèce, 85,1 % chez Boscia senegalensis, 71,8 % chez Maerua
crassifolia, 92,2% chez Combretum aculeatum, 53,3% chez Ziziphus mauritiana, 35,46%
chez Acacia raddiana et 28, Il % chez Balanites aegyptiaca.

107
D'une part, Acacia raddiana est l'espèce la plus représentée en effectif, d'autre part
sa régénération équivaut au quart de la régénération totale de la région. Elle tend de ce fait
à dominer davantage les autres espèces.
La régénération de cette espèce est surtout localisée en bordure exondée de la Mare
d'Oursi (photo n° 7) et le long des couloirs d'écoulement des eaux des versants de dune
(Pétel Borti par exemple) et des piémonts de buttes rocheuses (Kolel et Bombourou par
exemple).
De nombreux jeunes plants de Pterocarpus lucens poussent sous les pieds touffus de
ligneux de ces groupements: jusqu'à 85 semis sous un pied de Guiera senegalensis. Très peu
cependant survivront pendant la saison sèche.
3.2.3 - La mortalité
Parmi les individus morts sur pied ou couchés, il est parfois difficile de reconnaître
la plante. Cela explique le nombre élevé des pieds indéterminés qui représentent 34,2 % des
pieds morts.
Cette mortalité est paradoxalement importante dans les dépressions et talwegs, les
dernières zones de retranchement des espèces ligneuses après la mortalité des années 1972-73
(photo n° 8). La poursuite des épisodes de sécheresse concentre la mortalité dans cette unité
qui contient les espèces soudaniennes, C'est pourquoi le taux de mortalité est actuellement
élevé dans cette unité géomorphologique. Le tableau n° 8 et la figure n° 26 nous donnent les
taux par unité géomorphologique.
Nous avons mesuré un pourcentage de mortalité de 65,62% des pieds de Grewia
bicolor, 60% chez Pterocarpus lucens, 57,28 % chez Acacia nilotica et 28,57% chez Acacia
senegal. Elle concerne seulement 4,18% des pieds de Acacia raddiana et 3,83% de ceux de
Balanites aegyptiaca (tableau n° 8).

108
3.2.4 - Les coupes
Le tableau n° 8 et la figure n° 27 montrent que ce phénomène est faible dans les
glacis à cause du nombre réduit de ligneux de cette unité. Il est uniformément réparti dans
les autres unités géomorphologiques.
Les espèces concernées sont essentiellement les arbustes épineux.
3.2.5 - Discussions sur la vitalité
La comparaison des moyennes arithmétiques des paramètres d'évolution (régénération,
mortalité et coupes) par unité géomorphologique permet de réaliser les classifications
décroissantes suivantes (annexe XIX) :
La régénération est plus élevée dans les dépressions, moins élevée et presqu'identique sur
les buttes et les dunes, faible et identique dans tes champs et les glacis.
L'émondage est similaire dans les buttes, les dépressions, les dunes et les champs et moins
importante dans les glacis.
La mortalité est plus élevée dans les dépressions, indentique dans les buttes et les glacis,
moins élevée sur les dunes et plus réduite dans tes champs.
Il ressort de ses observations que les dunes et en moindre mesure les piémonts de buttes et
reliefs constituent les unités de repeuplement ligneux.
Les espèces les plus exploitées par les coupes sont: Bauhinia rufescens avec 31,25%
des pieds, 6,79% de Acacia nilotica var. adansonii, 6,89% de Acacia raddiana, 6,38% de
Balanites aegyptiaca, Il % de Pterocarpus lucens et 10% de Ziziphus mauritiana (tableau
n° 9).
Avec une régénération faible, les espèces à fort taux de mortalité Grewia bicolor,
Pterocarpus lucens, Acacia nilotica et Acacia senegal sont menacées de disparition si le
mouvement de régression se poursuit. Le taux de régression du nombre de pieds est
respectivement de 62,77%,59%,54,37% et 16,07% (tableau n° 9).

109
Tableau n° 9 : Caractéristiques dynamiques par rapport aux pieds vivants de quelquesespèces ligneuses de la région de la mare d'Oursi
Taux de Taux de Taux de Taux deEspèce ligneuse régénération mortalité coupes dynamique
(R) (M) (R-M)
Acacia raddiana 35,46% 4,18% 6,89% + 31,28%Pterocarpus lucens 1% 60% 11% - 59%Acacia nilotica * 2,91% 57,28% 6,79% - 54,37%Acacia senegal 12,5% 28,57% 7,69% - 16,07%Balanites aegyptiaca 28,11 % 3,83% 6,38% + 24,28%Bauhinia rufescens 8,33% 8,33% 31,25% 0%Grewia bicolor 2,85% 65,62% 5,71 % + 62,77%Leptadenia hastata 89,94%
Acacia nilotica * = Acacia nilotica var. adansonii
Les espèces appétées sont aussi les plus exploitées ce qui prouve bien la vocation
pastorale de la région.
Les conséquences des modes d'exploitation des ligneux sur la dégradation des peuplements
seront analysées dans les paragraphes suivants.
3.2.6 - Les jachères en région sahélienne
3.2.6.1 - Nature et mode défrichement
Avec l'aridification croissante et l'augmentation des surfaces cultivées consécutives
à la croissance démographique, les défrichements de nouveaux champs se localisent dans les
creux interdunaires et dans les dépressions argileuses. Ces zones étaient autrefois réservées
au pâturage de saison des pluies et à la récolte du fonio. En effet MILLEVILLE (CLAUDE
et al., 1991) montre que les champs étaient absents des dépressions dans les photographies
aériennes de 1955.
La préparation d'un champ cultivé défriché un an plus tôt consiste à détruire au
coupe-coupe ou à la hache certaines repousses arbustives.

110
Les pieds de Balanites aegyptiaca sont par ailleurs élagués et parfois brûlés purement et
simplement vu qu'ils abritent les oiseaux granivores.
Dans le cas d'un nouveau champ, la préparation s'effectue par coupe à ras de certains
pieds et l'élagage d'autres. Les branches détachées sont utilisées pour la confection de haie
morte.
Par exemple la mise en culture de l'ancienne parcelle du piémont de Kolel en 1992
a provoqué les modifications suivantes (tableau n° 10) : la coupe à ras du quart des ligneux
soit 83% de Mimosaceae et 16,5% de Balanites aegyptiaca et l'élagage d'un autre quart.
Tableau n° la : Taux de coupe à ras des ligneux dans l'ancienne parcelle de piémont deKolel
Taux absolu de Taux relatif deEspèce ligneuse coupe (%) coupe (%)
Acacia nilotica var. adansonii 100 16,5Acacia raddiana 19 25Acacia seyal 67 16,5Balanites aegyptiaca 20 16,5Dichrostachys cinerea 75 25
Total 100
En définitive, le choix des espèces ligneuses épargnées pendant le défrichement est
fonction de leur utilité, de leur répartition spatiale par la jonction des couronnes et de leur
fréquentation par les oiseaux granivores répandus dans la région.
Ainsi les pieds de Calotropis procera sont souvent maintenus dans les champs du fait
de leur utilisation comme cordage des toiles de tentes des Bella.
3.2.6.2 - Dynamique spatiale et mode d'installation des espèces ligneuses
Le rôle des "arbres pionniers" (FLORET et PONTANIER, 1991) ou arbres laissés
par le paysan durant le cycle cultural, est important dans la reprise de la végétation des
jachères.

111
L'action des sucs digestifs des arumaux pennet la germination des graines et la
dissémination des espèces endozoochores comme les acacias, le dattier d'Egypte et le jujubier
. Les zones privilégiées sont les bordures de .mares humides et ombragées en saison sèche.
Les pieds de Calotropis procera produisent de nombreuses graines à aigrette
éparpillées par les vents et piégées par divers obstacles. Les sols relativement découverts et
meubles des champs et des jachères favorisent leur germination et l'expansion de l'espèce.
3.2.6.3 - Importance et gestion des jachères
La mise en jachère représente un phénomène presque accidentel chez les agriculteurs
de la région comme l'a souligné MILLEVILLE (CLAUDE et al., 1991). Elle est souvent
liée au déplacement de la famille ou à une réduction de la main d'oeuvre. En effet, le
caractère réduit des surfaces cultivées et le respect des zones de parcours fourrager
expliquent que des champs soient cultivés pendant 40 ans sans jachère.
La pauvreté chimique des sols tout comme le non emploi des engrais chimiques dans
cette région sont bien connus. Ainsi dès la fin des récoltes, les champs sont ouverts au bétail
qui broute les tiges et y laissent ses déchets.
3.2.7 - Discussions sur les causes et les mécanismes de la dynamique du peuplement
ligneux
La discussion porte sur la dynamique du peuplement ligneux en général et sur celle
des espèces choisies Acacia raddiana et Pterocarpus lucens en particulier.
Au niveau du peuplement ligneux, les causes de la dégradation sont de 3 ordres: d'abord
climatiques, puis mésologiques induits par le climat et enfin anthropiques.
3.2.7.1 - Les facteurs climatiques
Les principaux facteurs climatiques sont la baisse de la pluviosité, l'action du•
ruissellement des eaux et des vents.

112
La mort accidentelle des ligneux en région sahélienne est considérée par habitude
comme le résultat de la sécheresse. La baisse de la pluviosité et le recul des isohyètes
annuelles est un phénomène déjà connudans les pays sahéliens. Ce phénomène s'est accentué
ces dernières années et se traduit en fait par des mouvements de va et vient de la pluviosité.
Comme l'ont montré TOUTAIN et PlOT (1980), la succession des années déficitaires
en pluviométrie engendre des conditions naturelles difficiles pour la survie et la régénération
des végétaux. Ce fut le cas pendant les sécheresses de 1972-73 et 1984 qui ont provoqué la
mort de nombreux ligneux et l'absence de régénération. TOUTAlN (1980) a montré que la
pluviométrie excédentaire de 1977 a conduit à une forte activité ligneuse avec une faible
régénération qui s'est plutôt manifestée l'année suivante malgré une pluviométrie moyenne.
Mais la pluviométrie médiocre de 1979 a entraîné une réduction de la feuillaison et la mort
d'une partie de cette régénération.
La régénération nécessite alors 2 années consécutives de bonne pluviométrie. II en est
de même pour sa pérennité. Cela fait en tout 4 années successives de pluviométrie non
déficitaire, ce qui est rarement enregistré dans la région.
Les espèces peu exigeantes en eau font preuve d'une plasticité écologique particulière,
ce qui justifie leur abondance dans presque toutes les stations.
L'aridité fait que l'eau disponible pour les besoins de la plante est souvent si rare que
les fonctions indispensables à la survie se trouvent perturbées. Le phénomène se manifeste
malgré les possibilités élevées de limitation de la perte d'eau par réduction des surfaces
foliaires, par défoliation, malgré la vitesse de croissance, de ramification et d'extension en
profondeur et en surface des racines. Le niveau bas des nappes souterraines n'arrive pas à
combler ces pertes par remontées capillaires. Les jeunes plants à pivot peu profond doivent
entrer en quiescence pour survivre.
II arrive que toutes les feuilles des ligneux s'étiolent. La plante finit par mourir, si
cet état de stress hydrique perdure, d'autant plus qu'il favorise l'action des insectes
phytophages.

113
En saison des pluies, l'érosion en nappe des eaux de ruissellement entraîne les
horizons de surface et emporte les semences végétales. Il s'en suit des phénomènes de
glaçage des sols en pente (glacis). Les bandes dénudées à ruissellement fort existant des
brousses tigrées septentrionales, s'agrandissent davantage. L'eau qui ruisselle sur ces bandes
s'infiltre au niveau de la végétation (CORNET et al., 1983 ; GROTEN, 1990) mais ne
semble pas permettre une compensation totale des pertes en eau. Le déplacement de fertilité
et de l'humidité serait responsable de leur dégradation.
La violence de l'écoulement des eaux dans les talwegs provoque souvent le
déracinement des ligneux plus ou moins grands.
Dans la dynamique éolienne, le vent déchausse et déracine les ligneux (photo n° 10),
balaie les surfaces nues et évacue les graines qui ne peuvent s'y maintenir. Elles sont piégées
préférentiellement dans les zones sableuses meubles. Ces zones ont une capacité d'infiltration
en eau relativement élevée et une évaporation réduite du fait du manque de capillaires fins
capables de remonter l'eau jusqu'en surface. En conservant l'humidité, ces sables permettent
leur germination.
Le vent augmente l'évapotranspiration et provoque une dessiccation des végétaux.
3.2.7.2 - Les facteurs mésologiques induits par le climat
La baisse du niveau des nappes, les modifications des propriétés pédologiques et
l'action des termites sont prépondérantes.
La diminution de la pluviosité provoque la baisse du niveau des nappes d'eau qui ne sont pas
rechargées au fil des années. Des variations de niveau de 9 à 14 m en une décennie sont
observées dans cette région: de 19 à 29 m entre 1977 et 1988 à Kolel, de 3,6 à 12,74 m
entre 1980 et 1993 à Tin Edjar, de 15,9 à 30,46 m entre 1980 et 1993 à Férérillo.
Comme la présence de nappes est liée à l'existence des fissures dans les roches
sous-jacentes et en outre comme ces dernières présentent des structures variables, il n'est pas
prudent de faire des extrapolations de ces mesures à d'autres sites même voisines.

•
114
Dans ces conditions de baisse de niveau, la remontée capillaire ne suffit pas à
alimenter les ligneux à racines peu profondes (DUPRIEZ et DE LEENER, 1990).
Les espèces ligneuses à système racinaire à la fois pivotant, étendu et intact pourront
s'alimenter. Les jeunes plants doivent avoir une croissance racinaire élevée ou développer
des formes d'adaptation à l'aridité de la saison sèche pour survivre.
La diminution de la pluviosité provoque aussi des modifications sur la nature
pédologique des sols. Par ailleurs les termitières cathédrales écroulées sont de plus en plus
nombreuses en région sahélienne. Or les termites sont actifs dans l'aération, l'humification
et l'augmentation de la profondeur, de la porosité et de la fertilité des sols (HEIM, 1977;
BACHELIER, 1978; BRIAND, 1978; ABBADIE et LEPAGE, 1989).
Selon GUINKO (1984), en zone sahélienne, les sols des termitières durcissent et
asphyxient toute racine qui s'y trouve. Non seulement les ligneux des termitières meurent
mais aussi aucune germination n'est possible sur ces sols nus. Les sols des termitières
abandonnées, à surface dure, s'arasent et s'assimilent par la suite à une croûte d'érosion
(CASENAVE et VALENTIN, 1989 ; photo n° Il).
Plusieurs espèces de termites ayant des actions contradictoires sur la vie des plantes
ligneuses sont en cause.
L'écorçage des pieds des acacias par les rats (Arvicanthis niloticusï est présent dans
les formations végétales à strate herbacée abondante. Les pieds écorcés à la base de la tige
finissent par mourir.
Ainsi la protection par grillage (GROUZIS, 1988) ou par gardiennage (glacis restauré
par la FAO) a favorisé la strate herbacée et, par contre coup, a engendré une pullulation de
rats qui ont tué de nombreux pieds d'Acacia seyal et d'Acacia raddiana essentiellement.
Des anastomoses racinaires sont identifiées entre Balanites aegyptiaca et Bauhinia
rufescens. Ce phénomène qui s'apparente à un parasitisme racinaire est localisé .

115
Toutefois il contribue de manière naturelle à la dynamique des espèces ligneuses dans les
conditions sahéliennes. Il n'est pas possible actuellement de prévoir l'évolution de ce
processus de sélection naturelle.
La mort naturelle est le terme ultime du cycle végétatif. La plante meurt après avoir
atteint un âge plus ou moins avancé : près de 5 000 ans pour Adansonia digitata
(GUILLAUMIN, MOREAU et MOREAU; 1955). Ce phénomène de déchéance progressive
est perceptible dans la région, chez les 2 espèces précédemment citées.
3.2.7.3 - Les facteurs anthropiques
La recherche du fourrage aérien est très répandue dans la région et constitue le
principal facteur. Son exploitation s'effectue par émondage, élagage, ébranchage ou étêtage
(annexes XX et XXI).
Les ovins et les caprins constituent les agents de pâture des ligneux les plus nocifs et
les moins sélectifs. Heureusement que les ligneux sont peu sensibles à cette action au dessus
de 1 m de hauteur selon DEPIERRE et GILLET (1977).
Cette hauteur est atteinte dans la région au bout de 5 ans en condition de régénération
naturelle et en 4 ans en reboisement protégé (figure n° 28). PlOT et al., (1980) et TYBIRK
(1991) recommandent une protection des espèces ligneuses pendant une période de 5 ans au
moins dans cette région.
Les formes de prélèvement aérien sont nombreuses et loin d'exercer les mêmes effets
sur les espèces.
Selon BERNUS (op.cit), l'émondage "en parapluie" par les bergers, plus répandu,
pour nourrir le petit bétail est préjudiciable aux sujets.
En effet il ne s'agit pas d'une coupe franche mais d'une entaille qui provoque l'affaissement
des branches et la déchirure des tissus jusqu'au tronc (photo n° 2).

116
Des modes d'ébranchage sont effectués à titre d'essai sur Acacia seyal et Pterocarpus
lucens en zone sahélienne et en zone nord soudanienne au Mali de 1978 à 1983 par CISSE
(1987). Ils révèlent que suivant les espèces, l'ébranchage répété a un effet dépressif de 90%
chez les 2 espèces précédentes et stimulateur sur la production foliaire cumulée chez Acacia
albida.
L'ébranchage pratiqué après la saison des pluies tant chez Acacia albida que chez
Pterocarpus lucens fournit des biomasses foliaires cumulées les plus élevées. Par contre
aucune de ces espèces ne supporte un rythme d'exploitation de 2 fois par an qui entraîne, à
plus ou moins longue échéance, la mort des sujets.
TOUTAIN et PlOT (1980) ont montré, dans notre région d'étude, qu'un ébranchage
trop important ou répété plusieurs années de suite sur un même pied d'Acacia seyal entraîne
la mort du sujet. L'ébranchage total ou étêtage cause une mortalité de 60% des pieds. 18
mois après le traitement. La coupe au ras du sol ou à 1 m de hauteur entraîne la mort des
souches au cours de la saison sèche suivant l'intervention.
Le traitement le moins préjudiciable au développement du pied est l'ébranchage au
tiers du houppier, les branches étant totalement détachées du ligneux.
La coupe des ligneux verts constitue une cause importante de la mortalité qui s'ajoute
à la mortalité climatique.
D'autres types d'exploitation courantes dans la région concourent à la mortalité des
ligneux. C'est le cas des déterrages des racines de Acacia laeta, Acacia raddiana, Acacia
senegal et Combretum glutinosum sur les dunes et ensablements (photos n° 12 et 13).
Ces racines sont utilisées dans la confection des corbeilles de récolte de fonio sauvage et
comme charpente dans la construction des tentes (photos n° 14 et 15). Il en est de même de
la récolte des fibres par écorçage des pieds de Acacia nilotica, Acacia raddiana et surtout
Adansonia digitata qui réduit la résistance des troncs dénudés des pieds concernés, à l'aridité
du milieu.

117
Les épineux sont en outre utilisés pour la confection d'enclos d'animaux et de haie
des champs. D'autres sont coupés et utilisés comme bois d'oeuvre ou de service.
3.2.7.4 - Cas particulier des deux espèces
* Cas de Acacia raddiana
La dynamique de cette espèce se caractérise par une forte expansion due
essentiellement à une résistance et une croissance rapide des pieds. Son système racinaire
pivotant et traçant à la fois permet à la plante d'atteindre au cours de la saison pluvieuse de
la germination de la graine, les horizons humides du sol . Au cours de cette saison
l'enracinement est plus rapide que la croissance de la tige. En outre rallongement en
profondeur du pivot s'effectue au détriment de l'extension racinaire latérale. La jeune pousse
pourra alors résister à l'aridité de la saison sèche.
L'enracinement de la plante se poursuit afin d'atteindre la nappe souterraine et de collecter
les eaux des premières pluies. Ainsi le pivot de cette espèce peut atteindre 35 m et
l'extension latérale 20 m (ANONYME, 1974 et MUTHANA et al., 1980).
La mortalité, quoique faible (4,18 % des pieds vivants) est cependant présente chez cette
espèce. Elle peut être liée aux facteurs climatiques, mésologiques et anthropiques.
Une pluviométrie déficitaire peut en effet provoquer la mort des régénérations de
l'année précédente tout comme les vents violents de débuts de saison pluvieuse déracine de
nombreux pieds d'Acacia raddiana des glacis d'épandage et des versants de dune.
Les pieds des zones à herbacées denses sont par ailleurs écorcés par les rats.
La mort naturelle est assez tardive chez cette espèce: 300 ans pour Acacia raddiana
du Ténéré (ANONYME, 1974). Cet Acacia aurait été en réalité bousculé par un touriste
imprudent selon le Pr SAADOU (communication personnelle).

118
Une autre forme de mortalité est causée par les anastomoses racinaires de cette espèce
avec Balanites aegyptiaca. Ce phénomène est localisé et peu important (5 %du peuplement
ligneux).
A l'instar des autres épineux de la région, Acacia raddiana est beaucoup exploitée par
coupes aériennes, par écorçage et par déterrage des racines. Elle est par ailleurs très appétée.
* Cas de Pterocarpus lucens
La forte mortalité de l'espèce (60% des pieds vivants) provoque une dégradation de
ses formations. Les mécanismes de ces transformations sont:
Le système racinaire de cette plante est aussi pivotant et traçant à la fois. Cependant
le niveau des nappes souterraines a beaucoup baissé au cours des dernières décennies : par
exemple de 15,9 m à 30,46 m entre 1980 et 1993 à Férérillo.
L'enracinement est limité par les coupes aériennes fréquentes et sévères influencent
négativement la croissance racinaire comme l'a montré STODDART et al., (1975).
En outre, les contraintes pédologiques réduisent ses capacités d'enracinement de sorte que
la colonisation du sol se fait en suivant les anciennes racines ou en pénétrant dans leur gaine.
Les racines de cette espèce même fraîches sont aussi sectionnées par les termites hypogés.
Les termites, à la recherche de débris organiques mais aussi d 'humidité rare en saison sèche,
s'attaquent aux racines. Il y a réduction des capacités de colonisation du sol engendrée par
l'action des termites.
Les formations à Pterocarpus lucens sont caractérisées par l'abondance de termitières
épigées dégradées dont le colmatage des débris argileux glacent les sols, asphyxie leurs
racines et empêchent les germinations de leurs graines.
Les jeunes plants sont aussi particulièrement broutés par le bétail de sorte que seuls les pieds
dissimulés sous les branchages sont épargnés : 93 %des pieds du ptérocarpe de moins de 50
cm de haut en steppe arbustive de Banh (CüUTERüN, AQUINO et OUEDRAOGO, 1992).

119
Ces auteurs montrent toutefois que le broutage n'est pas à l'origine de leur mortalité. Il
provoquerait tout simplement une perte de la dominance apicale et un rabougrissement de ces
pieds.
* Comparaison entre Acacia raddiana et Pterocarpus lucens
Acacia raddiana et Pterocarpus lucens sont 2 légumineuses fourragères bien appétées
dans la région. Elles possèdent toutes les un système racinaire pivotant et traçant à la fois.
Il existe aussi au voisinage de leurs pieds, des nids termitiques souterrains dans
lesquelles s'enchevêtrent des poils racinaires qui tapissent les parois des cavités. Ces poches
argileuses cimentées constitueraient des réservoirs d'absorption d'eau remontée par les
termites en saison sèche notamment (LEPRUN, 1976; BOYER 1973, 1975a et 1975b cité
par ABBADIE et LEPAGE, 1989; CALLOT et al., 1981).
Leur pivot s'entoure d'une couche de mucilage qui constitue une protection de la racine
contre le dessèchement excessif du sol (POUPON, 1980).
Des différences importantes existent cependant entre les 2 espèces et expliquent
l'expansion de l'une et la régression de l'autre.
Pterocarpus lucens est présente sur les terrains à bilan hydrique médiocre : massifs
rocheux, buttes cuirassées, sols pierreux gravillonnaire ou latéritique (MAYDELL, 1983).
Le niveau des nappes d'eau souterraine est plus bas dans les sols des formations à
Pterocarpus lucens (30,8 m le 10/9/93,66,27 met 50,37 m le 31/3/94 à Timataltal et 30,46
m le 28/6/93 à Férérillo) que celles à Acacia raddiana dominant ( 20,92 m le 18/6/93 à
Tondaré, 21,92 m le 27/6/93 à Kolel et 24,97 le 31/3/94 à Tayaret (piémont de Tin Edjar)
selon les données du B.R.G.M. (RICOLVI, 1989 et données récentes non publiées).
L'enracinement du ptérocarpe est aussi limité par les contraintes pédologiques, les
coupes aériennes fréquentes et sévères et les attaques des termites hypogés qui les
sectionnent.

•
120
Par contre l'accessibilité plus grande de l'eau pour Acacia raddiana, la facilité de
circulation de l'eau et des substances dissoutes dans ses tissus racinaires, la moindre
importance des coupes aériennes et le croissance rapide des pieds favorisent l'expansion de
cette espèce.
Il en est de même pour sa forme de dissémination des graines essentiellement par les
animaux qui donne une large répartition et une bonne germination par rapport à la
dissémination par les vents des samares de Pterocarpus lucens sont les graines sont digérées
par les animaux (TYBIRK, 1979).
Dans l'étude de la dynamique, il ressort que Acacia raddiana est en expansion de
31,28% et Pterocarpus lucens est en régression de 59% de leurs pieds vivants actuels. Cette
différence d'évolution entre les espèces ligneuses a pour conséquences des changements
physionomiques, structurales et floristiques du peuplement
3.2.8 - Conséquences de la dynamique
Les conséquences de la dynamique du peuplement sont :
l'apparition de vastes "cimetières de bois morts" et de vastes clairières soumises à
l'érosion, à proximité des zones d'habitation, après ramassage du bois;
- la dispersion des ligneux vivants et une tendance à leur concentration dans les bas niveaux
des toposéquences (mares, talwegs et creux interdunaires), lieux privilégiés de la régénération
qui comble à peine la dégradation du peuplement par mortalité;
- l'appauvrissement floristique par la disparition de certaines espèces tandis que celles qui se
maintiennent et qui sont bien appétées par le bétail se rabougrissent avec une perturbation du
cycle végétatif et une baisse du pouvoir reproducteur des pieds, comme chez Balanites
aegyptiaca, Boscia angustifolia, Boscia senegalensis et Maerua crassifolia ;
- la tendance à la dominance de certaines espèces, dont Acacia raddiana ; l'expansion
progressive de cette espèce dans le milieu est la preuve de l'évolution vers un nouvel
équilibre biologique différent de celui qui existait auparavant et qui fut rompu suite aux
épisodes de sécheresse et aux conditions d'exploitation qui ont affecté les écosystèmes ;

121
- l'avivement des dunes fixées auparavant dont les sables comblent les dépressions,
notamment les mares naturelles ;
- la réduction des ressources végétales de la région engendrant parfois des situations de
désolation chez les populations par des pertes importantes de bétail.
Par ailleurs, certaines espèces envahissantes et caractéristiques du lessivage se sont
propagées ces 4 dernières années de manière extraordinaire dans la région. Ce sont:
- Calotropis procera seulement signalée en 1979 dans un champ du piémont de Kolel
(TOUTAIN et PlOT, 1980) et qui occupe les champs et jachères dunaires et de piémont
souvent en formation monospécifique et dense,
- Leptadenia hastata et Leptadenia pyrotechnica dans une moindre mesure sont présentes
dans toutes les unités géomorphologiques.
La mortalité intéresse principalement des espèces utilitaires :
- Acacia nilotica var. adansonii, utilisée dans le traitement des peaux d'animaux pour
l'artisanat ;
- Acacia senegal qui produit de la gomme arabique ;
- Grewia bicolor utilisée comme bois de service ;
- Adansonia digitata écorcée pour ses fibres et élaguée pour ses feuilles (sauces de repas) ;
- et Pterocarpus lucens très appétée et utilisée comme bois de feu (présence de restes
d'anciens hauts fourneaux de fonte de la magnétite extraite des buttes) et de service à l'état
frais. De nos jours, des chargements de perches fraîches de ptérocarpe sortent toujours des
formations restantes, pour la construction de tente et de grenier. Il est par conséquent difficile
de trouver encore un pied intact de cette espèce.
Les tiges des pieds desséchés servent d'abris de reproduction pour de nombreux
insectes: Cerambicidae, Curculionidae et Scolytidae.
Les espèces soudaniennes ou à affinités soudaniennes, à l'instar de Anogeissus
leiocarpus, Entada africana et Scferocarya birrea, deviennent rares. Il en est de même des
espèces sahéliennes dont la répartition dépasse la limite sud : Cadaba farinosa, Boscia
senegalensis, Grewia bicolor et Commiphora africana.

122
D'autres espèces ligneuses indicatrices de la proximité du ruveau de la nappe
phréatique sont localisées au bord des mares pérennes qui deviennent semi-pérennes
iAnogeissus leiocarpus, Ficus gnaphalocarpa, Mitragyna inermis, Tamarindus indicaï ou ont
disparues (Diospyros mespiliformis et Acacia sieberianay. Cela confmne l'aridisation
croissante de la région.
Al'opposé les espèces dont la répartition remonte au nord du domaine sahélien se
répandent : Acacia raddiana, Combretum aculeatum, Balanites aegyptiaca, Calotropis
procera, Leptadenia pyrotechnica. En effet, le repeuplement de la dernière décennie est
marquée par l'expansion d'espèces caractéristiques des zones sèches comme Acacia raddiana,
et envahissantes telles que Calotropis procera, Leptadenia hastata et Leptadenia
pyrotechnica. La dernière espèce constitue selon la population le véritable messager du
désert. Les espèces à tendance hygrophile font place à une végétation de région sèche.
l..e pseudoclimax évolue vers un paratype de substitution dans lequel Acacia raddiana
et Calotropis procera sont dominants car leur régénération est abondante et leur mortalité
faible par rapport à toutes les autres espèces ligneuses de la région.
Par contre Pterocarpus lucens est menacée de disparition dans toute sa zone de
répartition (GUINKO, ZOUNGRANA et ZOUNGRANA, 1989 ; GROTEN, 1990), malgré
ses capacités d'adaptation. Rappelons qu'elle rejette abondamment de la souche et qu'elle
développe des "racines aériennes". L'alimentation des branches survivantes qui restent
accolées au tronc desséché reste possible par une mince couche de rhytidome (DIALLO,
1990).
Si la tendance se maintient, ce sont les espèces moins exigeantes en eau et moins
utilisées par l'homme et son bétail qui domineront le peuplement futur de la région. Il se
distinguera par une diminution des potentialités fourragères arborées.
La dynamique c'est-à-dire le taux de régénération (R) moins celui de la mortalité (M),
(R-M), est seulement très positive sur les buttes et reliefs et sur les dunes et ensablements.

123
Elle est davantage réduite dans les dépressions, les champs et jachères et encore plus réduire
sur les glacis.
3.2.9 - Localisation de la dégradation
Dans la région de la Mare d'Oursi en particulier, la dégradation du peuplement
ligneux est, selon les témoignages, manifeste depuis la sécheresse de 1972-73.
La dégradation est essentiellement localisée dans les axes de drainage, sur les sols
squelettiques des glacis d'épandage. Ce sont la partie nord-ouest de la Mare de Tin Edjar
(Pété Yaï, photo n° 4), les glacis de transit de Djalafanka et ceux d'épandage de Windé. les
brousses tigrées septentrionales de Férérillo, Zarmakoy et Tin Atan.
Les formations dégradées proches des campements d 'habitation sont débarrassées de
leurs bois morts. Par contre les anciennes formations de fourrés plus éloignées, se
caractérisent par la présence de "cimetières de bois morts".
La proximité des villages et des campements par ailleurs favorise la dégradation des
peuplements ligneux par les défrichements de champs, les coupes diverses et la pâture.
La dégradation du peuplement ligneux s'est maintenant déplacée en région
subsahélienne.
Elle y progresse en un front de mortalité perceptible dans les localités de Yalgo et Tougouri
avec 87,6% et 20% de mortalité respectivement de Pterocarpus lucens et Balanites
aegyptiaca (DIALLO, 1990). A l'ouest de cette zone à Banh, COUTERON, AQUINO et
OUEDRAOGO (1992) relèvent des taux de mortalité de 8%, 35%, 20% et 28 % respectifs
pour la steppe arbustive claire, la steppe arbustive dense, le fourré de bas-fonds et la brousse
tigrée.

124
CONCLUSION
Après les phénomènes de mortalité des ligneux liés aux déficits pluviométriques des
années 1972-73 et 1984, nous assistons à une repeuplement en ligneux caractérisée par une
composition et une structure différentes de celles qui existaient auparavant.
Les brousses tigrées sont constituées d'espèces subsahéliennes (Boscia senegalensis,
Combretum micranthum, Grewia bieolor, Pteroearpus lucens) et d'espèces soudaniennes
(Adansonia digitata, Diehrostaehys cinerea, Entada afrieana.) qui se maintiennent ainsi en
région sahélienne. Ces espèces à écologie humide trouvaient dans le biotope privilégié que
constituaient les bandes boisées en position basse, des conditions qui ressemblaient à celles
dont elles disposaient plus au sud (CORNET, 1992). Les brousses tigrées équivalaient alors
à un écosystème humide en milieu subaride.
Avec la diminution de la pluviosité annuelle, la baisse du niveau des nappes
phréatiques qui s'ensuit et les coupes trop fréquentes, les conditions de survie de ces espèces
sont remises en cause. Une mortalité massive mais sélective affecte ces brousses tigrées qui
se rétractent par élargissement des bandes dénudées des microdunes.
La mortalité a rendu disponible le bois de chauffe dont le ramassage et la commercialisation
laisse des surfaces dénudées soumises à l'érosion éolienne et hydrique au voisinage des zones
d'habitation.
En zone sahélienne, Pteroearpus lucens est réduit à quelques pieds isolés dans les zones
d'écoulement des eaux. Les espèces relictuelles des brousses tigrées sont Combretum
micranihum et Guiera senegalensis.
La mortalité des arbres accroît provisoirement la possibilité d'approvisionnement de
la population en bois de chauffe. Mais le ramassage du bois favorise l'érosion hydrique et
éolienne des sols et a un effet néfaste sur la régénération (DEWISPALAERE, 1980 et
BAUMER, 1987). Qu'en sera-t-il lorsque ces réserves en bois seront épuisées?

125
Dans le milieu sahélien, les ligneux jouent un rôle essentiel dans la vie de la
population. Ils constituent non seulement un fourrage de relais pour les animaux pendant la
période de soudure, mais aussi une ressource fourragère stable, pendant tout le cycle annuel
(PlOT et al., 1980). De plus ils apportent un complément alimentaire pour l'homme et sont
utilisés comme bois de feu, bois de service et bois d'oeuvre. En un mot la population retire
les ressources nécessaires à sa vie du milieu naturel avec des pratiques ancestrales
d'exploitation.
Même si les conditions hydrogéologiques, climatiques et édaphiques jouent un rôle
évident dans cette dynamique, l'homme et les animaux domestiques accélèrent de manière
très significative le processus de dégradation du peuplement ligneux. L'homme y prélève des
perches et du bois d'artisanat. Les épineux sont utilisés pour la construction de parcs à bétail
et des haies mortes des champs. Les fibres de cordage, les médicaments et le bois de chauffe
y sont prélevés. Même les racines sont déterrées pour la confection des corbeilles et des
armatures de tente. Le surpâturage nuit aux ligneux en interrompant leur croissance et en
modifiant leur pouvoir reproducteur.
Le peuplement ligneux en reconstitution est localisé essentiellement en bordure des
mares, sur les dunes et ensablements, sur les piémonts de relief rocheux ou cuirassé et le
long des axes de drainage des eaux. Ces axes correspondent aux zones de fracture.
L'analyse descriptive et dynamique du peuplement ligneux montre une expansion de
Acacia raddiana et une régression de Pterocarpus lucens. Il y a remplacement de la
végétation.

126
CONCLUSION GENERALE ET PERSPECTIVES
L'aridification du milieu se traduit sur le plan hydrogéologique par la localisation, en
bordure des mares endoréiques, des espèces indicatrices de la présence de nappe phréatique
ou phréatophytes : Anogeissus leiocarpus, Ficus gnaphalocarpa, Mitragyna inermis,
Tamarindus indica; et la disparition d'autres: Diospyros mespiliformis et Acacia sieberiana.
La raréfaction de ces phréatophytes, la transformation des mares pérennes en mares semi
pérennes et l'abondance des régénérations ligneuses dans les zones de fractures traduisent une
désertification croissante. Elle se traduit par une baisse du niveau des nappes d'eau
souterraine de la région.
Les relevés météorologiques confirment par ailleurs une diminution générale de la
pluviométrie annuelle. Or le cumul de ses déficits successifs joue un rôle déterminant dans
les variations de niveau des nappes et dans la stagnation ou la disparition des espèces
végétales.
Les termites résistent assez bien à la sécheresse, en creusant des galeries profondes
et en aménageant des réservoirs d'eau. C'est pourquoi au Burkina Faso, certains
hydrogéologues s'appuient sur la répartition spatiale des termitières épigées et sur la
reconnaissance des phréatophytes pour orienter leur prospection des eaux souterraines
(SAVADOGO, 1987). Alors l'érosion des termitières et la raréfaction des phréatophytes
peuvent corroborer l'affirmation de l'abaissement du niveau des eaux souterraines.
La meilleure résistance d'Acacia raddiana à la sécheresse s'explique par la capacité
à éviter les pertes en eau par une réduction foliaire et à augmenter l'absorption en eau par
un système racinaire plus extensif et intacte et une amélioration de la conduction hydraulique
dans la racine (VARTANIAN et LEMEE, 1984) ..
L'enracinement de Pterocarpus lucens pourtant aussi pivotant et traçant est toutefois
très limité par les coupes aériennes fréquentes et l'action des termites souterrains.

127
Il Ya alors réduction du pouvoir colonisateur du sol par la plante, ce qui ne lui permet plus
d'exploiter les rares eaux des premières pluies précédant la saison sèche longue et
desséchante pour les racines.
L'abaissement du niveau de la nappe d'eau souterraine constitue la cause majeure de
la mortalité des pieds de cette espèce.
Les nombreuses contraintes pédologiques des sols (cuirasse ou roche mère
superficielle) du peuplement de cette espèce, l'abondance et la fréquence des coupes des tiges
aériennes influencent négativement la croissance des racines des pieds généralement
encombrés de bois mort.
Pterocarpus lucens qui subit une forte mortalité dans toute son aire de répartition, en
zone sahélienne et subsahélienne malgré le développement de diverses formes de résistance:
formation de racines aériennes et alimentation par une mince couche de rhytidome des
branches survivantes. Elle est appelée à disparaître si le processus se poursuit.
Nous avons mis en évidence l'existence d'une compétition trophique poussée
jusqu'aux anastomoses racinaires chez Balanites aegyptiaca avec des pieds voisins. Ces
fusions tissulaires sont sources de mortalité chez Acacia raddiana.
Contrairement à l'idée répandue d'une action destructrice des termites, certaines
espèces créeraient des conditions favorables à la nutrition minérale des racines chez les 2
espèces ligneuses.
Pour compléter ces données et permettre un meilleur suivi de la dynamique du
peuplement ligneux dans une région aussi sensible que le Sahel, nous envisageons un suivi
par inventaire des parcelles de végétation pendant 10 ans.
Finalement, à défaut de pouvoir maîtriser les facteurs climatiques, on peut atténuer
leurs effets dégradants sur le peuplement ligneux par les quelques mesures suivantes :

128
- Protéger la régénération naturelle, de manière limitée dans le temps des zones déterminées
avec la participation de la population pour permettre une restauration des surfaces dénudées
et/ou un maintien du potentiel végétal. Cette méthode est plus efficace, moins, coûteuse,
écologiquement adaptée et permet une gestion participative de la population de l'espace agro
sylvo-pastoral tour à tour utilisé pour la production de la nourriture, du bois et pour
entretenir le bétail.
- Codifier l'exploitation des ressources du milieu est nécessaire car la conception répandue
de n J'appartenance à tous et à personne" contribue énormément à la dégradation des
ressources ligneuses par la coupes du bois et les défrichements anarchiques des champs. Cette
exploitation se localise dans les bordures de mare, les dépressions d'écoulement des eaux et
les creux interdunaires qui sont les zones de retranchement des espèces menacées de
disparition.
- Associer la pâture contrôlée ou la fauche des herbacées pendant la période de protection
maximale de 4 ans des zones de régénération, pour éviter la pullulation des rongeurs
responsables de la mortalité de nombreux ligneux par écorçage dans la région.
- Répandre des matières végétales (arbres morts, pailles et herbes sèches) pour favoriser la
réimplantation des termites sur les sols et leur destruction des encroûtements.
En effet, TOUTAIN (1977) affirmait que cette méthode de récupération des surfaces
dégradées et nues aurait les mêmes effets qu'une scarification ou un sous-solage. MANDa
et al., (1993) confirment qu'elle est une alternative pour le Sahel, car peu coûteuse et basée
sur les ressources naturelles. Toutefois des investigations sur les effets durables et l'efficacité
en milieu sahélien des termites sont à poursuivre.
- Dévier la voie routière de Gorom-Gorom à Oursi vers Djalafanka contribuerait énormément
à freiner la remobilisation continuelle du sable dunaire et l'invasion de la mare par celui-ci.

128
- Protéger la régénération naturelle, de manière limitée dans le temps des zones déterminées
avec la participation de la population pour permettre une restauration des surfaces dénudées
et/ou un maintien du potentiel végétal. Cette méthode est plus efficace, moins coûteuse,
écologiquement adaptée et permet une gestion participative de la population de l'espace agro
sylvo-pastoral tour à tour utilisé pour la production de la nourriture, du bois et pour
entretenir le bétail.
- Codifier l'exploitation des ressources du milieu est nécessaire car la conception répandue
de "l'appartenance à tous et à personne" contribue énormément à la dégradation des
ressources ligneuses par la coupes du bois et les défrichements anarchiques des champs. Cette
exploitation se localise dans les bordures de mare, les dépressions d'écoulement des eaux et
les creux interdunaires qui sont les zones de retranchement des espèces menacées de
disparition.
- Associer la pâture contrôlée ou la fauche des herbacées pendant la période de protection
maximale de 4 ans des zones de régénération, pour éviter la pullulation des rongeurs
responsables de la mortalité de nombreux ligneux par écorçage dans la région.
- Répandre des matières végétales (arbres morts, pailles et herbes sèches) pour favoriser la
réimplantation des termites sur les sols et leur destruction des encroûtements.
En effet, TOUTAIN (1977) affirmait que cette méthode de récupération des surfaces
dégradées et nues aurait les mêmes effets qu'une scarification ou un sous-solage. MANDü
et al., (1993) confirment qu'elle est une alternative pour le Sahel, car peu coûteuse et basée
sur les ressources naturelles. Toutefois des investigations sur les effets durables et l'efficacité
en milieu sahélien des termites sont à poursuivre.
- Dévier la voie routière de Gorom-Gorom à Oursi vers Djalafanka contribuerait énormément
à freiner la remobilisation continuelle du sable dunaire et l'invasion de la mare par celui-ci.

129
La mise en évidence d'anastomoses racinaires entre Balanites aegyptiaca et Acacia
raddiana ou Bauhinia rufescens mérite des investigations plus poussées afin de préciser les
liens tissulaires entre les racines fusionnées. Des analyses chimiques par screening
phytochimique permettraient d'identifier la nature des substances de transfert. Des recherches
sur la spécificité de ce phénomène aboutirait à indiquer son importance et son rôle dans la
dynamique du peuplement ligneux.
Cette approche de la dynamique par l'étude des structures racinaires a le mérite de
rechercher dans les propriétés biologiques des plantes ligneuses, les causes de leur expansion
ou de leur régression. Elle n'a intéressé cependant que 2 espèces : Acacia raddiana et
Pterocarpus lucens. Or chaque espèce possède des particularités biologiques auxquelles
s'ajoutent les influences édaphiques et anthropiques sur l'enracinement.
PERSPECTIVES
Il s'agira dans des études ultérieures de prendre en compte un plus grand nombre
d'espèces ou mieux celles-là.
Une approche pluridisciplinaire, associant botaniste, écologue, hydrogéologue,
sociologue, peut aussi permettre de mieux appréhender le phénomène de la dynamique du
peuplement ligneux dans cette région fortement artificialisée.
Cette approche peut se concrétiser par l'exécution du projet de recherche sur la mortalité des
ligneux dans le sahel, initié par le Laboratoire d'Hydrogéologie de la Faculté des Sciences.
L'utilisation de la télédétection (photographies aériennes et images satellitaires) est
indispensable pour une étude complète de la dynamique du peuplement ligneux (étude
diachronique sur une longue période).
Par ailleurs des travaux de recherche sur la production des ligneux, leur régénération
in situ et les relations entre leur âge, leurs paramètres dendrométriques, les conditions
pédologiques et les conditions pluviométriques pourraient fournir des informations nécessaires
à la connaissance de la biologie des espèces ligneuses et de leur potentiel de régénération.

•
BIBLIOGRAPHIE

131
ABBADIE L. et LEPAGE M., 1989 - The role of subterranean fungus comb chambers(Isoptera, Macrotermitinaeï in soil nitrogen cycling in a preforest savana (CôteD'Ivoire). Soil biol. biochem., 21, (8) : 1067-1071.
ALBERGEL J. CARBONNEL J.P. et GROUZIS M., 1984 - Pluies, eaux de surface,productions végétales en Haute-Volta 1920-1983. ORSTOM-DGRST, Ouagadougou, 64p.
ALBERGEL J., RIBSTEN P. et VALENTIN C., 1986 - L'infiltration: quels facteursexplicatifs? Analyse des résultats acquis sur 48 parcelles soumises à des simulations de pluiesau Burkina Faso. Journées hydrologiques de l'ORSTOM à Montpellier, Coll. ORSTOM,Colloques et Séminaires : 25 - 28.
ALEXANDRE D.Y., 1990 - Morphologie racina ire en relation avec l'organisation du sol.Cas de deux espèces pionnières de Guyane française: Goupia glabra et Yismia guianensis.Cahier ORSTOM, sér, pédol., XXV, (4), 1989-1990 : 417-422.
ALEXANDRE D.Y. et OUEDRAOGO S.J., 1991 - Variations de la morphologieracinaire de Faidherbia albida en relation avec le sol et effets agronomiques de l'arbre:quelques observations au Burkina Faso. Note présentée à l'atelier Faidherbia albida,Niamey 22-26 avril 1991, 8p.
AMIN (El) H. M., 1973 - Sudan acacias. Forest Research, Institute Bulletin n 01,
Khartoum, 1973, 57p.
ANONYME, 1974 - L'arbre du Ténéré est mort. Bois et Forêts des Tropiques, n0153,Janv.-Fév. 1974 : 61-65.
ARAP SANG F.K., HOEKSTRA D.A. et MWENDANDU R., 1985 - Rehabilitation ofBalanites aegyptiaca, Acacia tortilis and Commiphora ajricana. In the grazing land :perlimary results. In FAGG et GREA VES, 1990 : 16.
BACHELIER G., 1978 - La faune des sols. Son écologie et son action.Initiations-Documentations Techniques n° 38, ORSTOM, 391p.
BALASUNDARAM V. R., 1987 - Studies on the native nodulation and biomass productionof sorne tree legumes. In FAGG et GREAVES, 1990 : 25.
BAMBA K., 1985 - Systèmes aériens et racinaires de quelques essences spontanées etexotiques dans la région de Saponé. Mémoire IDR. Eaux et Forêts, Université deOuagadougou, 135p.
BARRAL H., 1977 - Les populations nomades de 1'00dalan et leur espace pastoral.Travaux et Documents de l'ORSTOM n077, 119p.
BARRAL H. et BENOIT M., 1976 - Nature et genre de vie au Sahel. L'année 1973 dansle nord de la Haute-Volta, ORSTOM, Ouagadougou, 2Op.

132
BARRY J. P., BOUDET G., BOURGEOT A., CELLES J.-C., COULffiALY A.M.,LEPRUN J.C. et MANIERE R., 1983 - Etudes des potentialités pastorales et de leurévolution en milieu sahélien au Mali. ACC-GRIZA-LAT, 114p.
BAUMER M., 1987 - Le rôle possible de l'Agroforesterie dans la lutte contre ladésertification et la dégradation de l'environnement. ICRAF, UNIVERSA, WEITEREN,Belgique, 26Op.
BELLEFONTAINE R., 1992 - Prétraîtrements des Semences Forestières. SymposiumIUFRO sur les problèmes de semences forestières. Ouagadougou, 23-27 Novembre1992, 12p.
BERNUS E., 1992 - Hydraulique pastorale et gestion des parcours. in L'aridité, unecontrainte au développement. Editions ORSTOM, 1992 : 555-563.
BOHM W., 1979 - Methods of Studying Root Systems. Springer-Verlag Heidelberg, NewYork, 188p.
BORTOLIH., 1983 - Pterocarpus lucens en Haute-Volta. In TOUTAIN et al., 1983 : 5766.
BOUILLON A. et MATHOT G., 1965 - Quel est ce termite africain? Edition del'Université de Léopoldville "ZOOLEO" n°l. Recueil.
BOUSSIM I.J., 1991 - Contribution à l'étude des tapinanthus parasites du karité au BurkinaFaso. Thèse de Doctorat 3e cycle, FAST, Université de Ouagadougou n01, 152p.
BRIAND M. V., 1978 - Production ecology of ants and termites. International BiologicalProgramme 13, Cambridge University Press, 407p.
BRUNCK F., COLONA J.-P., DOMMERGUES Y., DUCOUSSO M., GALIANA A.,PRIN Y., ROEDERER Y. et SOUGOUFARA B., 1990 - La maîtrise de l'inoculation desarbres avec leurs symbiotes racinaires. Synthèse d'une sélection d'essais au champ en zonetropicale. Bois et Forêts des Tropiques, n° 223, 1990 : 24-42.
BUNASOLS, 1981 - Etude pédologique de reconnaissance de la région de la Mare d' Oursi.Echelle 1150000. Rapport technique n025. Projet BKF/74/007, 69p.
BUNASOLS, 1989 - Etude pédologique de la région nord de la mare d'Oursi. Echelle 1150000. Rapport technique n° 63 + carte. Projet BKF/87/020, 136p.
CALLOT G., CHAMAYOU H., MAERTENS C. et SALSAC L., 1981 - Mieuxcomprendre les interactions sol-racine: incidence sur la nutrition minérale, INRA, 325p.
CASENAVE A. et VALENTIN C., 1989 - Les états de surface de la zone sahélienne.Influence sur l'infiltration. ORSTOM, Paris, Didactiques, 23Op., 196 photos.

133
CAZET M., 1989 - Les plantations linéaires denses sur les sols sableux dégradés de la zonecentre-nord du Sénégal. Comportement et effets sur les cultures adjacentes de quelquesespèces locales et introduites. Bois et Forêts des Tropiques, n° 222, (4) 1989: 27-37.
CHEVALIER P., CLAUDE J., POUYAUD B. et BERNARD A., 1985 - Pluies et Cruesau Sahel. Hydrologie de la Mare d'Oursi, Burkina Faso (1976-1981). Collection Travauxet Documents de l'ORSTOM n 019O, Paris, 251p.
CHOPART J.- L., 1984 - Développement racinaire de quelques espèces annuelles cultivéesen Afrique de l'Ouest et résistantes à la sécheresse. Actes du colloque "Résistance à lasécheresse en milieu intertropical: quelles recherches pour le moyen terme?" Dakar,24-27 Septembre 1984, Paris, 591p : 145-154.
CISSE M. 1., 1987 - Aménagements sylopastoraux au Sahel : rôle des ligneux pourl'élevage. Rapport final du Séminaire Régional sur l'Aménagement des ForêtsNaturelles, Bamako (Mali), 21-28 mai 1987, 189 P : 148-166.
CLAUDE J., GROVZIS M. et MILLEVILLE P., 1991 - Un espace sahélien. La Mared'oursi. Burkina Faso. Editions ORSTOM, 241p. + cartes.
CLAUDE J., SAADOUN N. et BERNARD A., 1978 - Etude hydrologique de septbassins versants alimentant la Mare d'Oursi. Rapport de campagne 1976 et 1977,oRSTOl\f, 76p.
CLAUDE J., SAADOUN N. et BERNARD A., 1979 - Etude hydrologique de septbassins versants alimentant la Mare d'Oursi. Rapport de campagne 1978, ORSTOM,67p.
CLAUDE J., BERNARD A. et BARDIN E., 1980 - Etude hydrologique de sept bassinsversants alimentant la mare d'Oursi. Rapport de campagne 1979, ORSTOM, 23p.
CNRST, 1992 - Les actes du Colloque Scientifique International sur la mare d'Oursi.Ouagadougou (Burkina Faso), 17-21 février 1992. Edition CNRST, 137p.
CORNET A., 1992 - Relations entre la structure spontanée des peuplements végétaux et lebilan hydrique des sols de quelques phytocénoses en zone aride. L'aridité, une contrainteau développement : 245-265.
CORNET A., DELHOUME J. P. et MONTANA C., 1983 - Vegetation patterns and soilsin the Mapimi Bolson (Chihuahuan Desert Mexico)1. Vegetation arcs. Second ChihuahuanDesert Symposium, Alpin Texas, V.S.A, 19p.
CORNET F. et DJEM H. G., 1982 - Etudes comparatives de l'efficacité des souches deRhizobium d'acacia isolées de sols de Sénégal et effet de la double symbiose Rhizobium Glomus mosseae sur la croissance Acacia holosericee et Acacia raddiana. Bois et forêts desTropiques, n0198 (4), 1982 : 3 - 15.

134
COUTERON P., AQUINO (de) P. et OUEDRAOGO M. I. O., 1992 - Pterocarpuslucens Lepr. dans la région de Banh (nord-ouest du Burkina Faso, Afrique de l'Ouest).Importance pastorale et état actuel des peuplements. Revue Elev. Méd. Vét. Pays Trop.,1992, 45 (2) : 179-190.
C.R.T.O, 1989 - Cartographie. Unités écologiques. Projet PNUD/FAO/BKF/OO3 "Maredl Oursi", 58p.
DAGNELIE P., 1992 - Statistique Théorique et Appliquée Tome 1. Les bases théoriques.Les presses agronomiques de Gembloux, A.S.B.L, 492p.
DEPIERRE D. et GILLET H., 1971 - Désertification de la zone sahélienne au Tchad.Bilan de dix années de mise en défens. Bois et Forêts des Tropiques n0139, 1971 : 3-25.
DEVAL J.L., 1974 - Structure dynamique de rhizosphère de l'okoumé dans les rapportsavec la sylviculture. C.T.F.T, 113p.
DEVINEAU J. L., FOURNIER A., LAMACHERE J.M et MALEK C., 1992 - Leprogramme d'évaluation préliminaire SPOT n° 149 - SPOT Oursi. Observation au sol,photointerprétation et traitements numériques d' images satellitaires sur le bassin versant dela mare d' Oursi. Colloque Scientifique International sur la Mare d' Oursi.Ouagadougou, 17-21 février 1992, 20p.
DIALLO A., 1990 - Contribution à l'étude de la mortalité des plantes dans le secteursubsahélien du Burkina Faso : cas de la zone située entre Tougouri et Yalgo. DEA,Université de Ouagadougou, 68p.
OONAHAYE E., NAVARRO S., et CALDERON M., 1966 - Observations on the lifecycle of Caryedon gonagra (F) on its natural hosts in Israel, Acacia spirocarpa and Acaciatortilis. In FAGG et GREAVES A. : 29.
DUCOUSSO M., 1991 - Importance et rôle des symbioses racinaires pour l'utilisation desacacias des zones sèches en Afrique de l'Ouest. Thèse de Doctorat, Université ClaudeBernard de Lyon, 205p.
DUPRIEZ H. et DE LEENER P., 1990 - Les chemins de l'eau: ruissellement, irrigation,drainage (manuel tropical). Terres et Vies, Nivelles, Belgique, 380p.
DUPUY N., DETREZ C., NEYRA M., DE LAJUDIE P. et DREYFUS B., 1991 - Lesacacis fixateurs d'azote du Sahel. La Recherche 233, 1991, vol., 22 : 803 - 804.
ESAU K., 1977 - Anatomy of Seed Plants, 2e Edition, California, U.S.A., 55Op.
FAGG C.W., 1991 - Acacia tortilis : Fodder Tree for Desert Sands. NFf Highlights,NFTA 91-01, Waimanalo, USA, 2p.

135
FAGG C. W; et GREAVES A., 1990 - Acacia tortilis 1925-1988. CABI/OFI annotedbibliography N° F41. CAB International, Wallingford, Oxon, UK., 36p.
FAHN A., 1959 - Xylem structure and annual rhythm of development in trees and shrubsof the desert. II Acacia tortilis and Acacia radâiana. In FAGG et GREAVES : 31.
FLOC'H (LE) E., GROUZIS M., CORNET A. et BILLE J.-C., 1992 Caractérisation, réponses biologiques, stratégies des sociétés. L'aridité, une contrainte audéveloppement. Editions de l'ORSTOM, Paris.
FLORET C. et PONTANIER R., 1991 - Recherches sur la jachère en Afrique Tropicale.Atelier" La jachère en Afrique de l'Ouest" ORSTOM, Montpellier, 3-5 décembre1991, 28p.
FONTES J. et GUINKO S., 1993 - Inventaire cartographique des ressources renouvelablesdu Burkina Faso par télédétection. Actes des journées scientifiques de Montréal 1991.Edition AUPELF-UREF: 227 - 238.
GANABA S., 1990 - Approche des méthodes d'inventaire des ressources ligneuses à petiteset moyennes échelles. Application d'une méthode par télédétection à une région-test duBurkina Faso. Mémoire DEA, Université de Ouagadougou, l00p.
GASTON A. et BOTTE F., 1971 - Etude agrostologique de la réserve de Tin Arkachem(République de Haute-Volta). I.E.M.V.T, Rapp. Multigr. 146p + 1 carte.
GIFFARD P. L., 1971 - Evolution des peuplements forestiers au Sénégal, C.T.F.T, Dakar,59p.
GORENFLOT R., 1986 - Biologie végétale. Plantes supérieures. 1 Appareil végétatif. 2eédition, Masson, Paris, 238p.
GOURLAY 1. D. et KANOWSKI P. J., 1991 - Marginal parenchyma bands andcristalloferous chains as indicators of age in african Acacia species. lAWA Bulletin n. s.,Vol. 12 (2), 1991 : 187-194.
GRAND (LE) E., 1979 - Etude expérimentale des propriétés germinatives de quelquessemences sahéliennes. A.C.C LAT, ORSTOM, 39p.
GROTEN S. M. E., 1990 - Analyse et lutte contre la désertification à l'aide de latélédétection. Etude comprenant l'analyse de cinq ans de données de l'index de végétation duBurkina Faso. Symposium International sur le suivi de la dynamique de ladésertification, Niamey, 20-22 Mars 1990, CILSS, 23p.
GROUZIS M., 1979 - Structure, Composition tloristique et Dynamique de la Productivitéde matière sèche des formations végétales sahéliennes (Mare d'Oursi, Haute-Volta).ORSTOM, Ouagadougou, 55p.

"136
GROUZIS M., 1988 - Structure, Productivité et Dynamique des systèmes écologiquessahéliens (mare d'Oursi, Burkina Faso). Etudes et Thèses de l'ORSTOM, Paris, 336p.
GROUZIS M. et NGARSARI M., 1988 - Herbier du centre ORSTOM : liste des espèces.Rapport multigr., ORSTOM Ouagadougou, 25p.
GIDGMA P., 1992 - Impact du semis direct dans la régénération des formations naturellesdu Nazinon. Communication au Symposium IUFRO sur les problèmes de Semencesforestières, Ouagadougou, 23-27 novembre 1992, 5p.
GIDGNARD J.-L., COSSON L. et HENRY M., 1985 - Abrégé de phytochimie. Masson,224p.
GUILLAUMIN A., MOREAU F. et MOREAU C., 1955 - La vie des plantes. LibrairieLarousse, Paris, 472p.
GUINKO S., 1984 - Végétation de la Haute-Volta. Thèse de Doctorat ès-SciencesNaturelles, Université de Bordeaux III, 2 tomes.
GUINKO S., 1992 - Rôle des acacias dans le développement rural au Burkina Faso et auNiger, Afrique de l'Ouest. Etudes flor, véga Burkina Faso 1, 3-16,Francfort/Ouagadougou, Nov. 1992.
GUINKO S., ZOUNGRANA 1. et ZOUNGRANA C. Y., 1989 - Etude des pâturages dela région de la mare d'Oursi, Burkina Faso. Projet BKF/87/003, 51p.
GUNN B. V. et VERCOE T. K., 1992 - Acacias australiens et leurs applications enAfrique Occidentale Subsahélienne. Symposium IUFRO sur les problèmes de SemencesForestières, Ouagadougou 23-27 novembre 1992, 12p.
HEDQVIST K.J., 1967 - Notes on sorne chalcid t1ies reared from Buprestidae andBostrychidae, in furious to Acacia raddiana Savi in Algeria (Sahara) and North Tchad. InFAGG et GREAVES : 29.
HEIM R., 1977 - Termites et Champignons. Les champignons termitophiles d' Afrique Noireet d'Asie Méridionale. Société Nouvelle des Editions BOUBEE, Paris, 190p.
HUBERT P. et CARBONNEL J.P., 1987 - Approche statistique de l'aridification del'Afrique de l'Ouest. Journal of Hydrology n° 95 : 165-183.
HUMPHREYS L. R., 1987 - Tropical Pastures and Fodder Crops. Second Edition.Intennediate Tropical Agriculture Séries, Longman Scientific et Technical, New York,155p.

137
HUTCHINSON J. et DALZIEL J. M., 1954, 1958, 1968, 1972 - Flora of West TropicalAfrica'. Second Edition.Vol 1 Part 1 (1954), Vol 1 Part 2 (1958), Vol III Part 1 (1968), Vol III Part 2 (1972).Crown Agents For Oversea Governements And Administrations, Millbank, London,S.Wl.
IRBET, 1987 - Recueil des communications présentées au Séminaire national sur lesessences forestières locales, Ouagadougou 6-16 juillet 1987, 222p.
JEAN-BLAIN C. et GRISVARD M., 1973 - Les plantes vénéneuses. Leur toxicologie.La Maison Rustique, Paris, 139p.
JOLY F., DEWOLF Y. et RIOU G., 1980 - Le bassin de la Mare d'Oursi (Haute-Volta).Etude géomorphologique et géodynamique. Contraintes naturelles. Université Paris VII DGRST, n07, 65p.
JOSENS G., 1982 - Le bilan énergétique de Trinervitermes geminatus (Wasmann)(Terminitidae, Nasutitermitinae). 2. Mesures de consommation en laboratoire. InsectesSociaux, Paris, 1982, vol. 29, n04 : 511-523.
KARSCHON R., 1961 - Contribution to the arboreal fIora of Israel: Acacia raddiana Saviand Acacia tortilis Hayne. In FAGG et GREAVES : 7.
KARSCHON R., 1975a - On the concurence of Acacia raddiana Savi and Acacia albidaDehle in Jericho. In FAGG et GREAVES : 7.
KARSCHON R., 1975b - Seed germination of Acacia raddiana Savi and Acacia tortilisHayne as related to infestation by bruchids. In FAGG et GREAVES : 14.
KAUL R.N. et NAMBIAR K.T.N., 1966 - Exotic trees and shrubs for arid tracts. InFAGG et GREAVES : 19.
KERHARO J. et ADAM J. G., 1974 - La pharmacopée sénégalaise traditionnelle. Plantesmédecinales et toxiques. Edition VIGOT FRERES, Paris, 1011p.
KHAN M.I.Ur-R., 1960 - A note on exotic Acacia spp. introduced into West Pakistan. InFAGG et GREAVES : 19.
KHUSA P., FURLAN V. et LUMANDE K., 1990 - Symbioses racinaires chez quelquesessences forestières importantes au Zaïre. Bois et Forêts des Tropiques n° 224 : 27-33.
KRAMER P. J. et KOZWWSKIT.T., 1979 - Physiology of Woody Plants. AcademiePress Ine, USA, 811p.
KUMMEROW J., KRAUSE D. et JOW W., 1977 - Roots systems of Chaparaal Shrubs.Oecologia (BerI.) 29, 1977: 163-177.

138
LANGLOIS M., 1983 - Les sociétés agro-pastorales de la région de la mare d'Oursi. Etudesocio-économique. ORSTOM, Ouagadougou, 101p.
LEPRUN J. C., 1992 - Etudes des brousses tigrées sahéliennes: structure, dynamique,écologie. L'aridité, une contrainte au développement, 1992 : 221-244.
LEPRUN J. C. et SILVEIRA (Da) O., 1992 - Sahel et Nordeste : analogies etparticularités. L'aridité, une contrainte au développement, 1992 : 555-563.
LEVANG P. et GROUZIS M., 1980 - Méthodes d'étude de la biomasse herbacée deformations sahéliennes: application à la Mare d'Oursi, Haute-Volta. Acta OEcologica,OEcol. Plant., vol. 1, 15, n03 : 231 - 244.
MAIGNIEN R., 1969 - Manuel de prospection pédologique. Iniations-DocumentationsTechniques n011, ORSTOM, Paris, 132p.
MALEK C., 1989 - Diagnostic du paysage à partir des données satellitaires. Application auSahel- Oudalan (Burkina Faso). Thèse de Doctorat nouveau régime, Paris, 20Sp.
MANDO A., VAN DRIEL W. et ZOMBRE P., 1993 - Les termites dans la restaurationdes sols ferrugineux tropicaux encroûtés au Sahel. Communication au Colloque AOCASS" Gestion durable des sols et environnement en Afrique intertropicale", 6 - 10 déc.1993, Ouagadougou, 18p.
MARIAUX A., 1975 - Essaide dendroclimatologie en climat sahélien sur Acacia raddiana.Bois et Forêts des Tropiques, 1975, n° 163: 27-35.
MAYDELL (Von) H. J., 1983 - Arbres et arbustes du Sahel. Leurs caractéristiques et leursutilisations. Eschborn, 531p.
METCALFE C. R. et CHALK. L., 1979 - Anatomy of the Dicotyledons. Second Edition,Vol. 1. Systematic Anatomy of the Leaf and Stem. Oxford Science Publications, 276p.
MUTHANA K.D. et ARORA G.D., 1980 - Acacia tortilis (Forsk.) - a promising fastgrowing tree for India arid zones. In FAGG et GREAVES : 20.
NEBOUT J. P. et TOUTAIN B., 1978 - Etude sur les arbres fourragers dans la zonesahélienne (Oudalan voltaïque). C.T.F.T- I.E.M.V.T, 119p.
NONGONIERMA A., 1978 - Contribution à l'étude biosystématique du genre AcaciaMiller (Mimosaceae) en Afrique Occidentale. Thèse de Doctorat d'Etat ès-SciencesNaturelles, Dakar, 3 tomes.
OUEDRAOGO J. S., 1993 - La multiplication végétative de Faidherbia albida. Evolutioncomparée des parties souterraines et aériennes de plants issus de semis et bouturage. Bois etForêts des Tropiques, 1993, n0237. (3), 31-43.

139
PATHAK P.S., GUPTA S.K et ROY R.D., 1980 - Studies on seed polymorphism,germination and seedling growth of Acacia tortilis Hayne. In FAGG et GREAVES : 14.'
PERRIN R., 1990 - Mycorhizes et protection phytosanitaire. in Les mycorhyzes desarbres et plantes cultivées. Editions Techniques et Documentation, 250p : 93-130.
PICARD D., 1984 - Etudes racinaires et résistances à la sécheresse. Synthèse introductive.Actes du colloque "Résistance à la sécheresse en milieu intertropical : quellesrecherches pour le moyen terme?" Dakar, 24-27 septembre 1984, 591p.
PlOT J., NEBOUT J.P., NANOT R. et TOUTAIN B., 1980 - Utilisation des ligneuxsahéliens par les herbivores domestiques. Etude quantitative dans la zone sud de la mared'Oursi (Haute-Volta), C.T.F.T - I.E.M.V.T.
POUPON H., 1980 - Structure et dynamique de la strate ligneuse d'une steppe sahélienneau nord du Sénégal. Travaux et Documents de l'ORSTOM, Paris, 351p.
PUECH C., 1983 - Persistance de la sécheresse au Sahel : Conséquences sur les normeshydrologiques et pluviométriques. C.LE.H, série hydrologie, 24p.
RAPHAEL L. et MWALYOSE B. B., 1990 - The dynamic ecology of Acacia tortiliswoodland in Lake Manyara National Park, Tanzania. Afr. J. Ecol. 1990, Vol. 28 :189-199.
RICOLVI M., 1989 - Actualisation de factibilité d'un programme d'hydraulique villageoisedans les provinces du Sourn, du Séno et de l'Oudalan. B.R.G.M - BURGEAP, 2ü9p.
RIOU G., 1990 - L'eau et les sols dans les géosystèmes tropicaux. Masson, CollectionGéographie, 221p.
SAVADOGO N. A., 1987 - Localisation des eaux souterraines par l'étude des indicesbiologiques dans les régions cristallines du Burkina Faso. Atelier International sur lagestion de l'eau et des ressources naturelles pour intensifier la production agricole enAfrique. FIS/Univ. 9 - 14 mars 1987, Niamey, 22p.
SIEBER R., 1983 - Establishment of fungus comb in laboratory colonies of Macrotermesmichailseni and Ondontotermes montanus (Isoptera, Macrotermitinae). Insectes Sociaux,1983, Vol. 30, n02, : 204-209.
SINGH P., et BHANDARI R.S., 1987 - Insect pests of Acacia tortilis in India. In FAGGet GREAVES : 28.
SOME A.N., 1991 - Etude des phénomènes germinatifs et des plantules de quelquesessences locales de Mimosaceae. Mémoire I.D.R, Université de Ouagadougou, l06p.
STODDART L. A., SMITH A. D. et BOX T. W., 1975 - Range Management. Thirdedition, Mc Graw-Hill Book Company, U.S.A; 532p.

140
STRULLU D. G., 1990 - Les relations entre les plantes et les champignons. In Lesmycorhizes des arbres et plantes cultivées. Editions Techniques et Documentation,250p : 9-49.
THEBAUD B., 1990 - Sociétés pastorales et développement. Cahiers des SciencesHumaines, Vol. 26, nOl-2, ORSTOM, Paris.
TOUTAIN B. et DEWISPALAERE G., 1978 - Paturages de l'O.R.D. du Sahel et de lazone de délestage au nord-est de Fada N'gounna (Haute-Volta). 3 tomes. I.E.M.V.T.
TOUTAIN B. et PlOT J., 1980 - Mise en défens et possibilités de régénération desressources fourragères sahéliennes. Etudes expérimentales dans le bassin de la mare d'Oursi(Haute-Volta). I.E.M.V.T - C.T.F.T, 15Op.
TOUTAIN B., BORTOLI L., DULIEU D., FORGIANI G., MENAUT J.C. et PlOTJ., 1983 - Espèces ligneuses et herbacées dans les écosystèmes pâturés sahéliens deHaute-Volta. Synthèse des résultats du programme ACC-GRIZA (LAT), 124p.
TROCHAIN J., 1940 - Contribution à l'étude de la végétation du Sénégal. Mémoires del'Institut Français dlAfrique Noire, n02, SWETS et ZEITLINGER N. V., Amsterdam,433p.
TYBIRK K., 1991 - Régénération des légumineuses ligneuses du Sahel. AAU REPORTS28, Aarhus University Press, Danemark, 86p.
VARTANIAN N. et LEMEE G., 1984 - La notion d'adaptation à la sécheresse. Bull.Soc. Bot. Fr., 131. Actual. bot., (1) : 7-13.
VASSAL J., 1972 - Apport des recherches ontogéniques et séminologiques à l'étudemorphologique, taxonomique et phylogénique du genre Acacia. Thèse de Doctorat ès-SciencesNaturelles, U.P.S Toulouse, n0485, 128p.

141
LISTE DES ANNEXES
Annexe X:
Annexe XI :
Annexe xn .Annexe XTII :
Annexe VI :
Annexe VII:
Annexe vmAnnexe IX :
Annexe 1 :
Annexe TI:
Annexe m :Annexe IV :
Annexe V:
Planches photographiques.
Propriétés pédologiques des sols de la région de la Mare d'Oursi
Techniques d'excavation des sols
Techniques d'analyse des propriétés pédologiques des sols
Variations des hauteurs d'eau (en cm) de la mare et de la nappe d'eau
souterraine autour de la Mare d'Oursi au cours de la période 1977-1979
Fiche de description des sites d'excavation des sols
Fiche d'analyse du système racinaire des ligneux
Techniques de préparation des coupes microscopiques
Résultats de mesure des systèmes racinaires d'Acacia raddiana et
de Pterocarpus lucens et de leur traitement statistique
Nomenclature des termes concernant les diverses parties des Agarics
termitophiles associés aux meules des Macrotermites
Fiche de reconnaissance des unités d'échantillonnage
Fiche d'inventaire floristique
Principales caractéristiques des parcelles de végétation de la région
de la Mare d'Oursi
Annexe XIV : Liste des espèces ligneuses naturelles rencontrées
Annexe XV : Liste des espèces herbacées naturelles rencontrées
Annexe XVI : Appelation en langues nationales des espèces ligneuses naturelles dans la
région de la Mare d'Oursi
Annexe XVII : Variation de la densité et de la vitalité du peuplement ligneux de la région
de la Mare d'Oursi en 1992 et par unité géomorphologique
Annexe xvm : Résultats d'inventaire floristique du peuplement ligneux de la région de la
Mare d'Oursi de PlOT et al. (1980)
Annexe XIX : Comparaison des moyennes arythmétiques des densités et des paramètres
d'évolution du peuplement ligneux de la région de la Mare d'Oursi

142
Annexe XX : Distribution, indice de présence et paramètres d'évolution des espèces
ligneuses de la région de la Mare d'Oursi
Annexe XXI : Utilisations humaine et animale des principales espèces ligneuses de la
région de la 'Mare d'Oursi

143
LISTE DES CARTES
Carte n° 1 : Situation de la Mare d'Oursi, Burkina Faso 13!
Carte n° 2 : La Mare d'Oursi 16
Carte n° 3 : Carte de reconnaissance des unités pédologiques de la région
de la Mare d'Oursi 18
Carte n° 4 : Carte des aptitudes au ruissellement et à l' infiltration des sols
de la région d'Oursi 20
Carte n° 5 : Aire de distribution de Pterocarpus lucens et d'Acacia raddiana
au Burkina Faso 22
Carte n° 6 : Carte pédologique de reconnaissance de la région de la Mare d'Oursi 27

Figure n° 1 :
Figure n° 2 :
Figure n° 3 :
Figure n° 4 :
Figure n° 5 :
Figure n° 6 :
Figure n° 7 :
Figure n° 8 :
Figure n° 9 :
Figure n° 10 :
Figure n° 11 :
Figure n° 12 :
Figure n° 13 :
Figure n° 14 :
Figure n° 15 :
Figure n° 16 :
Figure n° 17 :
Figure n° 18 :
Figure n° 19 :
144
LISTE DES FIGURES
Evolution de la pluviométrie annuelle dans quelques stations
sahéliennes 13
Partie souterraine d'Acacia raddiana à Bossia 40
Partie souterraine de Pterocarpus lucens à Timataltal 42
Partie souterraine d'Acacia raddiana à Tondaré 45
Relations entre profondeur d'enracinement et hauteur du pied
chez Acacia raddiana 47
Relations entre extension racinaire latérale et hauteur du pied
chez Acacia raddiana 47
Relations entre hauteur et diamètre du pied chez Acacia raddiana 49
Relations entre enracinement en profondeur d'enracinement et diamètre
du pied chez Acacia raddiana 49
Parties souterraines d'Acacia raddiana à Pétel Borti et en bordure
de la Mare d'Qursi 51
Relations entre extension racina ire latérale et diamètre du pied
chez Acacia raddiana 53
Relations entre extension racinaire latérale et âge estimé du pied
chez Acacia raddiana 53
Relations entre hauteur et âge estimé du pied chez Acacia raddianâa
Relations entre profondeur d'enracinement et âge estimé du pied
chez Acacia raddiana 54
Partie souterraine d'Acacia raddiana à Kolangal (glacis arénacé) 56
Parties souterraines de Pteracarpus lucens à Timataltal (axe de drainage
de brousse tigrée dégradée) 57
Parties souterraines de Pterocarpus lucens à Pété Yaï 59
Relations entre profondeur de zone d'absorption et hauteur du pied
chez Acacia raddiana 60
Parties souterraines d'Acacia raddiana en bordure de la Mare d'Qursi
et à Pétel Borti 62
Anastomoses racinaires entre Acacia raddiana et Balanites aegyptiaca

Figure 00 18 :
Figure 00 19 :
Figure 00 20 :
Figure 00 21 :
Figure 00 22 :
Figure 00 23 :
Figure 00 24 :
Figure 00 25 :
Figure 00 26 :
Figure 00 27 :
Figure 00 28 :
145
Parties souterraines d'Acacia raddiana en bordure de la Mare d'Oursi
et à Pétel Borti 62
Anastomoses racinaires entre Acacia raddiana et Balanites aegyptiaca66
Mode de pénétration de radicelle de Balanites aegyptiaca dans la racine
d'Acacia raddiana 66
Contacts tissulaires entre Acacia raddiana et Balanites aegyptiaca
dans l'anastomose racinaire 68
Répartition par unité géomorphologique du peuplement ligneux
de la région de la Mare d'Oursi . 95
Répartition en fonction du diamètre à la base et par unité
géomorphologique du peuplement ligneux de la région de la Mare
d'Oursi 98
Comparaison par unité géomorphologique des densités du peuplement
ligneux de la région de la Mare d'Oursi entre 1980 et 1992 102
Taux de régénération/ha et par unité géomorphologique
des espèces ligneuses 105
Taux de mortalité/ha et par unité géomorphologique
des espèces ligneuses 105
Taux de coupes en % par rapport aux pieds vivants
des espèces ligneuses 105
Croissance des ligneux de la plantation du glacis de Djalafanka
(projet FAO/Mare d'Oursi) 105

Tableau 00 1 :
Tableau 00 2 :
Tableau 00 3 :
Tableau 00 4 :
Tableau 00 5 :
Tableau 00 6 :
Tableau 00 7 :
Tableau 00 8 :
Tableau 00 9 :
Tableau 00 10 :
146
LISTE DES TABLEAUX
Principales unités écologiques de la région de la Mare d'Oursi 81
Définition du nombre de placeaux d'échantillonnage du peuplement
ligneux par unité géomorphologique de la région de la Mare d'Oursi 84
Tableau de répartition bioclimatique des principales espèces
ligneuses de la région de la Mare d'Oursi 89
Répartition zonale du peuplement ligneux autour des mares naturelles
de la région de la Mare d'Oursi 90
Densité moyenne par ha et par unité géomorphologique du peuplement
ligneux de la région de la Mare d'Oursi 94
Comparaison des densités et des nombres d'espèces du peuplement
ligneux entre l'inventaire général et celui des parcelles de végétation 97
Taux d'appétabilité des principales espèces ligneuses de la région
de la Mare d'Oursi 101
Taux de vitalité par ha et en pourcentage par rapport aux pieds vivants,
et par unité géomorphologique du peuplement ligneux de la région
de la Mare d'Oursi 106
Caractéristiques dynamiques de quelques espèces ligneuses de la région
de la Mare d'Oursi 109
Taux de coupe à ras des ligneux dans l'ancienne parcelle de piémont
de Kolel 110

ANNEXES

Annexe 1
PLANCHES PHOTOGRAPHIQUES
\ 1 _ . : :.

· ;
PLANCHE N° 1
Physionomie et dynamique des peuplements ligneux à Acacia raddiana et à Pterocarpuslucens dominantes "
1 : Formation monospécifique à Acaciaraddiana sur placages sableux à Gountouré (décembre1992)
, I~'
2 : Formation arbustive de bordure nord-ouest de la mare d' Oursi où est découvert lephénomène des fusions racinaires entre Acacia raddiana et Balanites aegyptiaca avoisinantes(août 1993)Ar = Acacia raddiana en retard de feuillaison en juilletBa = Balanites aegyptiaca avoisinants
3 : Emondage en "parapluie" des rares pieds vigoureux de Pterocarpus lucens dans l'axe dedrainage à Pété Yaï (juillet 1992)
4 : Mortalité en cours de Pterocarpus lucensdans l'axe de drainage de Pété Yaï (juillet 1992)
, .'

8

, "
Morphologie du système racinaire et anastomoses racinaires
5 : Système racinaire pivotant et traçant d'Acacia raddiana montrant quelques racineslatérales en extension et une racine pivotante présentant un enroulement autour d'une boulede sable solidifié et pénètre en profondeur en fm filament (août 1991)rl = racine latérale .er = enroulement racinaire
6 : Système racinaire avec racines pivotantes de susbtitution et association avec les termiteschez Pterocarpus lucens à Tirnataltal (microdune ; août 1992) ,,'rpi = racine pivotante principale initialerps = racine pivotante de substitutionec = enroulement racinaire dans une cavité termitique
7 : Morphologie des anastomoses racina ires entre Acacia raddiana et Balanites aegyptiacapar application intime et par enroulement de racines du dattier d'Égypte sur celles de l'acacia(août 1993)tr = tronc racinaire d'Acacia raddianarb = racines de Balanites aegyptiaca
8 : Mode de pénétration des racines de Balanites aegyptiaca (dattier d'Égypte) par formationde papille ovale appliquée contre la racine d'Acacia raddiana dans laquelle elles pénètrent
(x 64) , 1
po = papille ovale de pénétration .e
rb = radicelle de Balanites aegyptiacara = racine hôte ct 1Acacia raddiana

PLANCHE II

; ". , ~
Rôle du climat et des termites dans la dynamique du peuplement ligneux
9 : Meules (= association symbiotique champignons-termites)' emboîtées dans une cavitétermitique souterraine (- 60 cm) à Timataltal (août 1993)ID = meule
la : Formation de brousse tigrée dégradée avec des termitières cathédrales mortes induisantdes plages stériles autour d' elles 1
(août 1993) ,
Il : Racines de Guiera senegalensis déchaussées par les vents et les eaux de ruissellement surune vielle dune à Tirnataltal (août 1993)

PLANCHE DI
®

PLANCHE N° IV
Déterrage et utilisations des racines des ligneux par-la population de la région de la Mared'Oursi
12 : Déterrage des racines de Acacia laeta et Acacia senegal sur la dune fixée d' Oursi àTondaré (août 1993)rd = racine déterrée
13 : Racines de Acacia laeta et Acacia senegal déterrées et saisies pa:.- le servicedépartemental de l'Environnement et du Tourisme d' Oursi en juin 1992 (août 1992)
14 : Utilisation des racines déterrées de Acacia sp. et Combretum glutinosum dans laconstruction de tentes d'habiartion à Timbolo (octobre 1992)
15 : Utilisation de racines déterrées de Acacia sp. dans la confection de corbeille de récoltede fonio sauvage (Panicum laetum, Panicum subalbidum et Panicum dnabaptistum desprairies; juillet 1992)

• 1
. l
@

Annexe n° fi
SYNTHESE DES PROPRIETES DES UNITES PEDOLOGIQUES DE LA REGIONDE lA MARE D'OURSI (BUNASOLS, 1981)
Valeur de Définition Propriétés Contraintes Utilité IVégétation LocalisatioUnité pédologique 'unité agronomique ~ominante n
sols inaptes à toute massifsaffleurement Aucune valeur culture rocheux et
lithosols rocheux ou de agricole buttescuirasse latéritique cuiras-sées
Sols texture sableuse sols maptes à toute ...eptadenia hastata dune viveminéraux régosols 6.6% sable vif dunaire ~rande culturebruts Œ>erméabilité
!COuleur ururorme, sols assez arbustes épineux axes desols pauvres en convenanles draînage
d'apport 2.96% sols lirnono- m.o. et K capacitéalluvial vertique argileux d'échange élevée Hydromorphie peu
accentuée
sols moyennement sols peu Acacias spp. plateau!profonds, convenables Balanites aegyptiaca
Sols peu d'érosion Œ>erméabilité kJravillonne-mentévolués régosolique sur 2.3% sols limona- ~Ievée, capacité et cuirasse
cuirasse sableux d'échange faible
réserves minérales Acacia nilotica plainetopomorphes 1.12% bonnes, réserves Acacia seyal alluviale
hydriquesAcacia seyal axes de
sols argileux à insuffisantes Sans contrainte sols convenablesAcacia nilotica draînage
Vertisolstopomorphessur texture lourde majeure aux culturesmatériaux 0.08% Cornbretum
aculeatum
sols très tragiles sols assez Balanites aegyptiaca bas delet pauvres en m. onvenables: mises dune,o. et en réserves [Texture très en valeur limitée glacis
sols bruns sub- 32.74% sols sableux minérales sableuse par réservesarides modaux hydriques
insuffisantes
sols profonds orêts galeries terrasseSols sols bruns sols limono-argilo- pauvres en m. o. sols assez alluvialeisohurni-ques rouges subarides 28.34% sableux et en réserves Texture sableuse convenables
minérales ethydriques
7.9% sols pauvres en sols convenables ~cacla raddiana ~Iacls
sols bruns m.o. en P, K Sans contrainte lo\cacia laeta
subarides sols argilo-sableu~ échangeables majeurevertiques mauvaise structure
taux élevé en sols peu plateauconcrétions convenables
sols ferrugineux ferrugineuses, Taux detropicaux carapace à moins gravillonne-ment
lessivés à 13.7% de l m, teneur élevéconcrétions faible en m.o. et
Sols à en réservessexquioxy- minéralesdes sols ferrugineux présence de !plaine
tropicaux sols sablo-argileux cuirasse savane arbustive alluviale
indurés sur 0.92% ferrugineuse, Cuirasseuirasse pauvreté en
m.o.,N,P
hydromorphie très conviennent à la bordure deprononcée, sol riziculture et au mare,
lsols hydromorphespeu profond et maraîchage prairie talweg
humifères à gley peu profond 0.86% sols argileux himiquement Hydromorphieriche
. pauvrete en Phénomènes sols peu I&lacls
~ols sodiques à structure en 2.48% éléments d'alcalinisa-tion convenables rares épineux
colonette minéraux et m.o. possible
Légende m.o. = matière organique K = potassium N = azote P = phosphore

Annexe ID
TECHNIQUES D'EXCAVATION DU SOL
Le travail est effectué par une équipe de 2 manoeuvres avec pour matériel essentiel
un pic, une barre à mine, deux grandes pèles (bêche et ronde) et une petite pelle ronde, un
seau et une corde. IL s'est déroulé de juillet à septembre, de 1991 à 1993, période humide
favorable à l'excavation du sol et à l'observation de structures symbiotiques associées. En
effet, l'intensité de nodulation dépend des conditions écologiques (KHUSA et al., 1990) et
les plantes perdent leurs nodules en réponse au froid, à la sécheresse, à l'inondation et à
l'ombrage (HUMPHREYS, 1987).
D'abord les caractéristiques dendrométriques (hauteur du fût et diamètre du tronc au
pied) sont mesurées. Puis la phase d'excavation intervient.
Elle débute par un dégagement superficiel du sol jusqu'à 50 cm de profondeur autour
du pied afin de mettre en évidence le système racinaire latéral. Le creusement manuel se
poursuit de manière horizontale et centrifuge à partir du tronc racinaire afm de dégager les
terminaisons de toutes les racines latérales.
Ensuite, une tranchée est creusée dans le sol, le long du pivot racinaire, à une distance
de 30-40 cm de celui-ci pour éviter toute rupture éventuelle pendant l'excavation en cas de
déviation de ce pivot vers la tranchée. Cette coupe transversale de 3 m de longueur et 1 m
de largeur permettra le dégagement à la pelle puis au seau du sol creusé, jusqu'à l'extrémité
de la racine pivotante si possible.
La progression s'effectue par horizons de sol successifs.
Les racines sont dégagées à l'aide d'instruments mécaniques et de jets d'eau à la
pissette. Les mesures concernent d'une part l'épaisseur, la couleur et la texture des horizons
de sol traversés, la longueur et le diamètre moyen de système racinaire. Ce dernier est illustré
par un dessin à l'échelle de 1/10 et par des prises de vue photographiques.

Annexe IV
TECHNIQUES D'ANALYSE DES HORIZONS DE SOL
La texture des horizons de sol est déterminée en écrasant et en roulant une fraction
entre les doigts à l'état sec et/ou à l'état humide. D'après les réactions au toucher on apprécie
la texture en appliquant les critères suivants :
- l'argile mouillée forme une pâte collante aux doigts et s'enroule en un filament pliable ;
- le limon à l'état sec donne une impression douce et à l'état humide forme une pâte qui ne
colle pas aux doigts et un filament qui se casse au pliage;
- les sables fins crissent sous les doigts et ne peuvent pas s'agglomérer en boule;
- les sables grossiers s'observent "de visu".
Pour les mélanges argile-sable, la fraction est dite argilo-sableuse si la boule peut être
obtenue et sablo-argileuse dans le cas contraire.

Annexe V
VARIATION EN CM DE LA HAUTEUR D'EAU DE LA'MARE ET DU NIVEAU DELA NAPPE D'EAU SOUTERRAINE AUTOUR DE LA MARE D'OURSI AU COURSDE LA PERIODE ~977-1979 SELON CLAUDE ET AL., (1977, 1978 ET 1979)
Date Pl (450) P2(444) P3(426) P4(598) Puits(578) Mare
03/05/77 125 94 59 105 15611/05/77 118 84 55 74 106 14630/05/77 85 69 40 58 108 11718/06/77 94 60 40 18126/06/77 84 60 38 113 17202/07/77 88 54 34 95 16706/07/77 71 52 32 105 16314/07/77 86 47 31 103 18922/07/77 124 60 33 98 23306/08/77 184 84 54 116 24217/08/77 204 99 54 118 24824/08/77 212 114 50 128 24629/08/77 218 114 52 127 25601/09/77 289 161 57 140 30006/09/77 297 204 64 164 30017/09/77 320 263 86 76 189 32327/09/77 342 324 114 78 34304/10/77 339 320 136 80 232 33809/10/77 336 309 152 96 277 33506/10/77 329 303 211 102 256 32923/10/77 323 297 194 104 248 32530/10/77 318 292 189 111 254 32006/11/77 313 278 178 143 254 31613/11/77 309 272 174 139 253 31220/11/77 306 266 171 135 245 30904/12/77 303 254 164 138 250 30018/12/77 287 238 156 128 232 29511/01/78 270 213 152 120 226 28223/01/78 265 205 146 123 220 27312/02/78 255 216 147 128 220 26723/02/78 250 154 133 113 215 26005/03/78 231 182 130 119 218 255,312/03/78 279 179 130 116 206 25219/03/78 274 176 127 115 201 248,326/03/78 266 166 125 114 193 243,302/04/78 260 162 122 113 188 238,416/04/78 233 151 112 98 168 228,230/04/78 230 147 108 96 154 221,818/05/78 220 132 99 88 138 209,728/05/78 127 101 86 116 210,512/06/78 138 120 93 82 114 19326/06/78 132 111 82 74 118 179,812/07/78 126 101 79 67 134 160,827/07/78 125 42 77 66 123 146,311/08/78 242 157 86 23 160 274

26/08/78 255 185 109 92 183 27810/09/78 249 185 119 95 192 273,525/09/78 248 187 118 95 190 27010/10/78 237 177 87 101 198 26025/10/78 226 165 125 103 199 25009/11/78 229 165 116 100 189 24224/11/78 217 149 109 93 188 23309/12/78 140 97 90 177 22624/12/78 134 94 84 163 21808/01/79 130 93 82 162 21223/01/79 124 85 81 149 20507/02/79 173 111 75 71 138 19422/02/79 171 100 71 68 136 18304/03/79 175 94 86 61 96 17424/03/79 80 80 57 116 16618/04/79 66 73 106 15824/04/79 54 62 98 13718/05/79 85 10007/06/79 7623/06/79
Légende : Pn (x) n nO de puitsx côte du niveau de la nappe en cm par rapport
à l'échelle
" ",

Annexe VI
Etude des structures racinaires du peuplementligneux de la région de la Mare d'Oursi
Fiche n03
Pied n° .Date .
FICHE DE DESCRIPTION DU SITE D'EXCAVATION
LOCALISATIONLOCAL ITE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . PHOTO N° .UNITE CARTOGRAPHIQUE DE VEGETATION .
Nature du rel ief . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. Pente................... .Affl eurement rocheux: Oui Non Eros ion .
PEDOLOGIETYPE DE SOL .UNITE CARTOGRAPHIQUE ET ETUDE .
Horizon Profondeur Couleur Texture Stucture Nbe de racineslatérales
sèche humide
ZONE D'ABSORPTION:Organisation des poils .Profondeur et nature de 1'horizon .
VEGETATIONType de formation: 1 Steppe arborée 2 Steppe arbustive 3 Steppe herbeuse
l l 4 Brousse tigrée 5 Fourré 6 Autres
Ligneux domi nants .
Autres .
Herbacées .
REGENERATION MORTALITE
Annexe VII
Etude des structures racinaires du peuplementligneux de la région de la Mare d'Oursi
Espèce: .Pi ed n": .Date: , .. '"
1
FICHE D'ANALYSE DU SYSTEME RACINAIRE
PARAMETRES DENDROMETRIQUESHauteur du fOt .Diamètre du tronc .
OBSERVATIONS DE L'EXCAVATION
Couronne N-S .E-W .
Horizon Aspect de la racine Longueur racinaire Diamètre moyen Observ.
pivotante latérale pivotante latérale pivot. latét.

Fiche n04 ( suite )Espèce: .Pi ed n": .Date: , .
Horizon Aspect de la racine Longueur raci na ire Il Di amètre moyen Il Observ. Il
pivotante latérale pivotante latérale pivot. latér.

Annexe VIII
TECHNIQUES DE PREPARATION DES COUPES MICROSCOPIQUES
1 - La fixation et la déshydratation
La fixation consiste à tuer le tissu en solidifiant les protéines cellulaires soit par
coagulation grâce aux agents physiques ou par précipitation par des agents chimiques par
phénomène d'osmose.
Nous avons utilisé de l'éthanol 70 0 qui conserve le matériel sans dommage.
La déshydratation des racines longueur d'environ 0,5 cm est faite dans des bains alcooliques
de 30 mu dans de l'éthanol à 95 0 et 3 fois dans de l'alcool absolu en agitant fréquemment
les récipients.
2 - L'éclaircissement
Les objets sont plongés dans un mélange toluène-éthanol absolu puis successivement dans 3
bains de 25 mu de solvant pur de toluène pour éliminer totalement l'alcool : c'est
l'éclaircissement.
3 - L'inclusion
Les objets sont plongés dans un mélange toluène-paraffine à chaud pendant 25 mn à l'étuve
maintenue à la température légèrement supérieure au point de fusion de la paraffine (56°).
Le toluène soluble dans l'alcool et dans la paraffine permet l'imprégnation des objets par la
paraffine.
Ils subissent en outre 3 bains de 20 mn dans la paraffine pure fondue pour éliminer
complètement le solvant et éviter leur durcissement.
L'inclusion définitive des échantillons est enfin réalisée dans la parrafine pure avec
des barres de Leuckart.

4 - Confection des coupes
Les coupes sont réalisées au microtome à rasoir rotatif de type Leitz Wetzlar. La
microtomisation libère des coupes recueillies sur des lames. Puis les coupes sont étalées à
l'eau gélatinée et mises à sécher à l'étuve à 45 0 pendant 24h.
Après cela on procède au déparaffinage par immersion dans le toluène puis passage
2 fois dans de l'alcool 96 0 pour éliminer l'excès de toluène et enfin une réhydratation dans
une solution alcoolique 50 0 et dans de l'eau distillée quelques mn.
Les lames sont prêtes pour la coloration.
Les coupes à la main sont immergées 30 mn dans de l'eau de javel contenant de
l'hypochlorite de sodium et rincées à l'eau puis immergées pendant 5 mn dans de l'acide
acétique à 50 % et rincées de nouveau.
5 - La coloration
Toutes les coupes réalisée sont colorées pendant 5 mn au cannino-vert acétique de
MIRANDE.
Le vert d'iode colore en vert la lignine donc le xylème et le sclérenchyme, en bleu
vert la subérine (subéroïde des Monocotylédones et suber des Dicotylédones) tandis que le
carmin colore en rose la cellulose et l'hémicellulose ce qui correspond aux parenchymes non
sc1érifiés et non lignifiés et au collenchyme.
L'étape finale est le montage des coupes déjà soigneusement rincées.

Annexe IX
MESURES D'ENRACINEMENT DE ACACIA RADDIANA ET PTBROCARPUS LUCENS
Espèce Site Sol Age Haut. Diam. Prof. Extens. Absorp.ans cm cm obs. cm cm
1 Pétel Borti 5a 1 21 0.3 -110 2 -451 Pétel Borti 5a 1 30 0.3 -125 6 -451 Pétel Borti 5a 2 30 0.5 -140 101 Pétel Borti 5a 3 37 0.7 -170 341 Pétel Borti 5a 1 38 0.3 -175 40 -1051 Pétel Borti 5a 2 47 0.8 -170 100 -1051 Pétel Borti 5a 2 48 0.8 -200 1251 Pétel Borti 5a 3 60 0.7 -230 1301 Pétel Borti 5a 3 60 0.8 -230 150 -1101 Pétel Borti 5a 4 70 1 -290 115 -1901 Pétel Borti 5a 4 78 1.9 -257 3001 Pétel Borti 5a 4 80 1.3 -280 3801 Pétel Borti 5a 10 175 4.6 -280 5201 Bossia 2b 1 22 0.3 -82 4 -521 Bossia 2b 1 32 0.4 -85 8 -521 Bossia 2b 2 40 0.4 -134 15 -521 Bossia 2b 3 45 0.5 -160 30 -401 Bossia 2b 3 63 1.2 -170 701 Bossia 2b 4 73 1.6 -190 761 Bossia 2b 5 127 2.4 -220 125 -1251 Bossia 2b 8 200 4.6 -250 210 -1101 Tonda ré 5b 1 13 0.3 -105 15 -551 Tondaré 5b 2 30 0.5 -150 25 -701 Tondaré 5b 2 50 0.6 -230 50 -1451 Tondaré 5b 3 60 0.7 <-230 2001 Tondaré 5b 4 68 1.6 <-230 170 -1301 Tonda ré 5b 4 70 1.3 <-230 240 -1101 Tondaré 5b 3 80 0.9 <-230 70 -1551 Tondaré 5b 5 95 1.9 <-230 240 -1551 Tondaré 5b 8 147 2.5 <-230 350 -1151 Kolangal 5c 1 20 0.2 -65 10 -401 Kolangal 5c 2 35 0.5 -108 20 -521 Kolangal 5c 3 50 0.5 -230 90 -1011 Kolangal 5c 4 55 0.8 -230 100 -1001 Kolangal 5c 4 65 0.8 -225 125 -1101 Kolangal 5c 4 80 0.7 -255 155 -1241 Kolangal 5c 10 150 2.5 -300 185 -1101 Kolangal 5c 11 160 2.8 -328 2001 Bordure mare lIa 1 1 0.5 -13.5 01 Bordure mare lIa 1 6 0.1 -36 01 Bordure mare IIa 1 9 0.1 -65 11 Bordure mare IIa 1 20 0.2 -55 19 -371 Bordure mare IIa 1 27 0.3 101 Bordure mare lIa 1 33 0.3 -97 50 -371 Bordure mare IIa 2 40 0.4 -255 841 Bordure mare lIa 2 46 0.6 -380 200 -3151 Bordure mare lIa 2 50 0.7 -360 176 -3151 Bordure mare lIa 3 60 0.8 -320 230 -711 Bordure mare lIa 4 80 1 -310 1631 Bordure mare lIa 3 80 1.4 -420 255 -2501 Bordure mare lIa 4 95 1.9 -360 200 -250

1 Bordure mare IIa 5 114 2 -430 3402 Féréri 110 2 22 D,57 -142 442 Féréri 11 0 4 33 0,99 -56 282 Féréri 11 0 4 33 1.2 -78 412 Féréri 11 0 4 36 0,88 -189 682 Féréri 11 0 5 37 . 1.7 -87 902 Féréri 11 0 6 44 2,8 -220 153:2 Férér t l lo 3 48 0.78 -195 842 Féréri 11 0 3 54 0,78 -80 1202 Tin Atan 2 25 0.76 -130 58 -402 Tin Atan 3 44 0,86 -109 48 -392 Pété Yaï 8b 5 10 2.2 -85 1002 Pété Yaï 8b 4 15 1.9 -240 190 -502 Pété Yaï 8b 2 24 0,6 -146 80 -502 Pété Yaï 8b 2 30 0,8 -190 210 -602 Pété Yaï 8b 2 32 0.6 -75 1102 Pété Yaï 8b 5 35 2,2 -230 2002 Pété Yaï 8b 4 56 1.9 -108 802 Pété Yaï 8b 5 67 2,2 -95 1302 Timataltal 5b 1 6 0.1 -35 02 Timataltal 5b 1 7 0,1 -37 12 Ti mata Hal 5b 2 10 0,25 -85 202 Ti mata Hal 5b 2 20 0,3 -85 302 Ti mata Hal 5b 3 25 0.5 -270 100 -1402 Timataltal 5b 2 27 0.7 -155 80? Ti mata Hal 5b 2 45 0.7 -160 1202 Timataltal 5b 5 50 1.4 -180 170 -1352 Ti mata Ha l 5b 4 60 1 -300 100 -1502 Ti mata Hal 5b 6 60 1.7 -200 190 -1252 Ti mata Hal 5b 8 65 2.8 -380 270 -135
Légende : 1 = Acacia raddiana2 = Pterocarpus lucens
Haut. = Hauteur du piedDiam. = Diamètre à la baseProf. = Profondeur d'enracinementExtens. = Extension racinaire latéraleAbsorp. = Profondeur de la zone d'absorption
Sol : unités cartographi ques du BUNASOLS (1981)

ESTIMATIONS STATISTIQUES (STAT ITCF) DES VALEURS OBSERVEES OU MESUREES DES RACINESD'ACACIA RADDI~ PAR LES COURBES DE REGRESSION PROPOSEES
Pétel Borti
Hauteur Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
21 -110 -102 -7,99 2 41.58 41.5830 -140 -137 -2.41 6 7.46 -1.4630 -125 -137 12.58 10 7,46 2,5337 -170 -162,6 -7,37 34 45.60 -11.6038 -175 -166 8.96 40 51.06 -11,0547 -170 -194,98 24.98 100 100,10 -0,1048 -200 -198.01 -1.98 125 105,55 19.4460 -230 -231,98 1. 98 130 170.95 -40.9560 -230 -231 1.98 150 170.95 -20,9570 -290 -257,19 -32,8 115 225,95 -110.4478 -257 -275 18,62 300 269.04 30,9580 -280 -280.01 0 380 279,94 100,05
Pétel Borti
Diamètre Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
0.3 -110 -127.61 17.61 2 8.78 -6.770.3 -125 -127,61 2,61 6 8.78 -2.770.3 -175 -127.61 -47.38 10 53.14 -43,140.5 -140 -172,34 32.34 34 97,51 -63.510.7 -170 81 32.81 40 8.77 31.220.7 -230 -202.81 -27.18 100 19,6 -19,690,8 -230 -214.67 -15.32 125 19.69 5.300.8 -200 -214,67 14,67 130 97.51 32,480.8 -170 -214.67 44,67 150 119,69 50,30
1 -290 -233 -56.18 115 164.06 -49.061.3 -280 -254.79 -25.2 300 363.72 -63.721.9 -257 -281 24.35 380 250.61 149,38
Pétel Borti
Age Pro obs. Pro est. Résidus
1 -110 -129,53 19,532 -125 -129.53 4.633 -140 -175.23 35,234 -170 -220.93 50,935 -175 -129,53 -45.466 -170 -175.23 5,237 -200 -175,23 -24,769 -230 -220,93 -9.06
10 -290 -266,63 -23,3611 -257 -266,63 9,6312 -280 -266,63 -13,36

Bossia
Age Pro obs. Pro est. Résidus
1 -82 -103,19 21.191 -85 -103.19 18.192 -134 -127.58 6.423 -160 -152.18 -7.823 -170 152.18 -17,824 -190 -176.68 13.325 -220 -201.18 -18.828 -250 -274.67 24.67
Kolangal
Hauteur Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
20 -65 -99.81 34,81 10 -39.85 29.8535 -108 -151. 28 43.28 20 58.51 38.5150 -230 -190.59 -39.4 90 77 .18 12.8155 -230 -201,71 -28,28 100 83.41 16,5965 -225 -221.6 -3.39 125 95.84 29.1580 -255 -246.68 -8.31 155 114.51 40,48
150 -300 -319.91 19.91 185 201. 61 16.61160 -328 -326.84 -1.15 200 214.06 14.06
Kolangal
Diamètre Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
0.2 -65 -97 32.36 10 22.97 12.970.5 -230 -177.78 -52.21 20 39.27 19.270.5 -108 -177,78 69.78 90 88.22 -1. 770.7 -255 -210,98 -44.01 100 71.89 -28.110.8 -225 -224.05 -0.94 125 104.52 -20.470.8 -230 -224,05 -5.94 155 164.35 9.352.5 -300 -317.78 17.78 185 196.89 11.892.8 -328 -324.62 -3.37 200 196.87 -3.12
Kolangal
Age est. Pro obs. Pro est. Résidus
1 -55 -90.93 25.932 -108 -150.89 42.893 -230 -193.39 36.604 -255 -225.10 -29,894 -225 -225.10 04 -230 -225.10 -4.89
10 -300 -319.33 19.3311 -328 -327,64 -0.35
Tondaré
Hauteur Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
13 -105 -91 -13.83 15 12.55 2.4530 -150 -166.97 -16.97 25 28.95 3.9550 -230 -222.7 -7.29 50 48.24 1. 7560 <-230 -242.99 12.99 200 57.97 142.0368 <-230 -256.74 26.74 170 65.70 104.370 <-230 -259.89 29.89 240 67.64 172.3680 <-230 -274.19 44.19 70 77 .31 -7.3195 <-230 -291. 95 61.95 240 91.82 148.18
147 <-230 -332.58 102.58 350 142.17 207.83
Tondaré
Age est. Pro obs. Pro est. Résidus
1 -105 -104 -02 -150 -189 392 -250 -189 -40
Bossia
Hauteur Pro obs. Pro est. Rés idus Ext. obs. Ext. est. Résidus
22 -82 -79 -2.39 -4 -4.76 0.7632 -85 -107.88 22.88 -8 -16.5 8.4940 -134 -127 -6.12 -15 -25.88 10.8845 -160 -139.35 -20.64 -30 -31.75 1. 7563 -170 -175.32 5.32 -70 -52.87 -17 .1373 -190 -192.32 2.32 -76 -64.61 -11.39
127 -220 -259.80 39.8 -125 -127 2,98200 -250 -314.19 64.19 -210 -213.64 3.64
Bossia
Diamètre Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
0.3 -82 -97.67 15.67 4 13.67 9.670.4 -85 -114.85 -19.14 8 18.43 10.430.4 -134 -114.85 29.85 15 18.43 3.430.5 -160 -128.41 -31.59 30 23.19 6.81.2 -170 -177.2 7.2 70 56.53 13.461.6 -190 -190.1 0.1 76 75.58 0.412.4 -220 -205.45 14.55 125 113.68 11.314.6 -250 -221. 98 28.02 210 218.46 8.46

Bordure de mare
Hauteur Pro obs. Pro est. Résidus Ext. obs. Ext. est. Résidus
1 -13 -8.46 -5.04 0 6,91 -6,91 .6 -36 -48.18 12.18 0 7,64 7.649 -65 -70,16 5.16 1 16,37 15,37
20 -55 -140.79 85,79 19 48.39 29,3933 -97 -208,41 111.41 50 86,24 36,2440 -255 -239,36 -15,63 84 106.62 22,6246 -380 -263,42 -116.57 200 124,09 75.950 -360 -278.34 -81. 65 176 135.73 -40,2660 -320 -312,23 -7,76 230 164,85 -65.1580 -420 -368.28 -61.71 163 223.07 60,0780 -310 -368,28 58,28 255 223,07 31.9295 -360 -402.51 42.51 200 266.74 66.74
114 -430 -438.76 8.76 340 322.05 -17,94
Bordure de ma re
Diamètre Pro obs. Pro est. Résidus Ext obs.
0.1 -13 -73,14 8,14 00.1 -36 -73,14 37,14 00.1 -65 -73.14 59.64 10,2 -55 -130 75 190.3 -97 -175,48 78,48 500,4 -255 -212.68 -42.3 1840.6 -380 -269.9 -110,09 2000.7 -360 -292,37 -67,62 1760,8 -320 -311,85 -8.14 230
1 -420 -343.92 33.92 1631,4 -310 -389.72 -30.27 2551.9 -360 -427.15 67,15 2002 -430 -432.97 2,97 340
Bordure de mare
Age est. Pro obs. Pro est. Résidus
1 -13,5 -99.33 85.831 -55 -99,33 44.331 -65 -99.33 34,331 -36 -99.33 63.331 -97 -99.33 2.332 -360 -199,47 -166.522 -380 -199,47 -180.522 -255 -199,47 -55.523 -420 -300.42 -119,583 -320 -300,42 -19.584 -360 -402.18 42.184 -310 -402,18 92.185 -430 -504.78 74.78
Pro obs. = profondeur observée ou mesurée (en cm)Pro est. = profondeur estimée ou calculée (en cm)Ext. obs. = Extension racinaire latérale observée ou mesurée (cm)Ext. est. = Extension racinaire latérale estimée ou calculée (cm)

Annexe X
~------- perfora1orium
7-----anneou supère descendant
~----débris cil bléma1o~ne
!\ collier perioileiqoe~ille
) ou anneau oscendont
~-pseudorhize
loge
~-- coussinet basal
n----- primordium,meule
/ .~~~~~~~u--;. mycotete
Nomenclature des termes concernant les diversesparties des Agarics termitophiles associés auxmeules des Macrotermites. ( HEIM, 1977)

Annexe XI
Etude de la dynamique du peuplementligneux de la région de la Mare d'Oursi
N° .Date .
FICHE DE RECONNAISSANCE
STATION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . PLACEAU .W PVA.............................. N° DE PHOTO .
TYPE DE VEGETATION: 1 Steppe arborée 2 Steppe arbustive 3 Steppe herbeuse.1 1 4 Brousse tigrée 5 Fourré 6 Autres
TAUX DE RECOUVREMENT: 1 = Nul à très flaible: 0 à 10 % 2 = Flaible: la à 25 %des ligneux 1 1 3 = Moyen: 25 à 50 % 4 = Elevé : 50 à 75 %des herbacées-r- 1 5 = Très élevé: > 75 %
HAUTEUR MOYENNE DU PEUPLEMENT .. ARBORE ARBUSTIF , HERBACE .
ESPECES DOMINANTES Strate arborée 1 .2 .
Strate arbustive 1 .2 .
Strate herbacée 1 : .2 .
ARTIFICIALISATION 1 11 Surpâturage~ Ebranchange et coupe 3 Cultures et/ou jachères
SITUATION TOPOGRAPHIQUE 1 Butte4 Talweg
2 Glacis 3 Bordure de mare5 Dune 6 Dépression interdunaire
NATURE DE LA ROCHE. . . . . . . . . . . . . . . . . .. NATURE DU SOL .CUIRASSE... OUI NON Texture ... sableuse limoneuse argileuseEROSION Gravillonnaire Oui Non
ANALYSE DU PEUPLEMENT LIGNEUX ET HERBACE
PEUPLEMENT LIGNEUX1 .2 .3 .4 .5 .6 .7 , .8 .9 .10 .Il .12 .13 .14 .15
PEUPLEMENT HERBACE

Annexe XII
Etude de la dynamique du peuplementligneux de la région de la Mare d'Oursi
Fiche N°2FICHE D'INVENTAIRE
N° .Date .
Genre Espèce Nb de Hauteur de Di amètre Di amètre de Vi ta -ti ges 1 1 i nd ividu du fût 1a couronne 1i té
12345678910111213141516171819202122232425262728293031323334353637383940
Régénération = diamètre < 2,5 cm ou hauteur < lm
Vitalité: néant= individu sain 0= malade 1= coupes2= déssèchement partiel 3= déssèchement total 4= bois mort
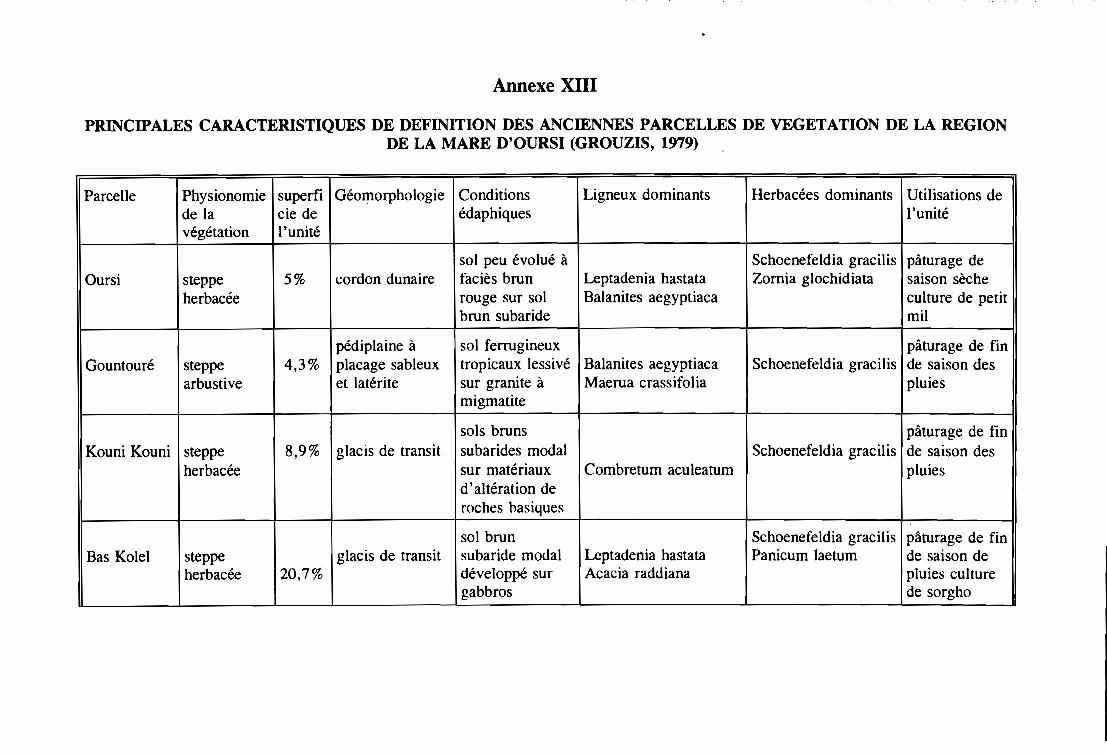
Annexe XIII
PRINCIPALES CARACTERISTIQUES DE DEFINITION DES ANCIENNES PARCELLES DE VEGETATION DE LA REGIONDE LA MARE D'OURSI (GROUZIS, 1979)
Parcelle Physionomie superfi Géomorphologie Conditions Ligneux dominants Herbacées dominants Utilisations dede la cie de édaphiques l'unitévégétation l'unité
sol peu évolué à Schoenefeldia gracilis pâturage deOursi steppe 5% cordon dunaire faciès brun Leptadenia hastata Zomia glochidiata saison sèche
herbacée rouge sur sol Balanites aegyptiaca culture de petitbrun subaride mil
pédiplaine à sol ferrugineux pâturage de finGountouré steppe 4,3% placage sableux tropicaux lessivé Balanites aegyptiaca Schoenefeldia gracilis de saison des
arbustive et latérite sur granite à Maerua crassifolia pluiesmigmatite
sols bruns pâturage de finKouni Kouni steppe 8,9% glacis de transit subarides modal Schoenefeldia gracilis de saison des
herbacée sur matériaux Combretum aculeatum pluiesd'al tération deroches basiques
sol brun Schoenefeldia gracilis pâturage de finBas Kolel steppe glacis de transit subaride modal Leptadenia hastata Panicum laetum de saison de
herbacée 20,7% développé sur Acacia raddiana pluies culturegabbros de sorgho

sols ferrugineux jachères etKolel steppe piémont de tropicaux peu Acacia raddiana Schoenefeldia gracilis champs depiémont arbustive 2,4% gabbros léssivé Acacia nilotica Cassia obtusifolia petit mil
pâturage desaison sèche
sol brun Acacia seyal Panicum laetum pâturage deWindé galerie 10% glacis subaride vertique Acacia nilotica Cassia obtusifolia début de
forestière d'épandage peu épais sur saison desmigmatites pluies

Annexe n° XIV
LISTE DES ESPECES LIGNEUSES NATURELLES RENCONTREES DANS LAREGION DE LA MARE D'OURSI
N° D'ORDRE GENRE ET ESPECE - N° D'ECHANTILLON
1 Acacia albida Del. syn. Faidherbia albida (Del.)A. Chev.
2 Acacia ataxacantha DC. - 283 Acacia ehrenbergiana Hayne - 1114 Acacia laeta R. Br. ex Benth. -305 Acacia nilotica var. adansonii (Guill. et Perr.)
O. Ktze - 1076 Acacia nilotica var. nilotica (Linn.) Willd ex Del. - 1147 Acacia nilotica var. tomentosa (Benth.) A. F. Hill.- 1138 Acacia pennata (Linn.) Willd. - 29, 1029 Acacia raddiana Savi -103
10 Acacia senegal (Linn.) Willd. - 5511 Acacia seyal Del.12 Adansonia digitata Linn.13 Anogeissus leiocarpus (DC.) Guill. et Perr.14 Balanites aegyptiaca (Linn.) Del.15 Bauhinia rufescens Lam.16 Boscia angustifolia A. Rich. - 23, 6617 Boscia salicifolia Olîv. - 7018 Boscia senegalensis (Pers.) Lam. ex. Poir. - 4519 Cadaba farinosa Forsk. - 9220 Cadaba glandulosa Forsk. -9321 Calotropis procera (Ait.) Ait. F.22 Capparis corymbosa Lam. - 47, 6423 Caralluma retropiscens (Ehrenb.) N. E. Br.24 Celtis integrifolia Lam.25 Combretum aculeatum Vent. - 8026 Combretum glutinosum Perr. ex. DC. - 4427 Combretum micranthum G. Don.28 Commiphora africana (A. Rich.) Engl.29 Dalbergia melanoxylon Guill.et Perr. - 1830 Dichrostachys cinerea (Linn.) Wight et Am.31 Entada africana Guill. et Perr. - 2232 Euphorbia balsamifera Ait. Hort. Kew33 Feretia apodanthera Del. - 5834 Ficus gnaphalocarpa (Miq.) Stend. ex A. Rich. -7935 Grewia bicolor Juss. -27, 5736 Grewia flavescens Juss. - 11537 Grewia tenax (Forsk.) - 19, 3138 Grewia villosa Willd. - 45
FAMILLE
MIMOSACEAEMIMOSACEAEMIMOSACEAEMIMOSACEAE
MIMOSACEAEMIMOSACEAEMIMOSACEAEMIMOSACEAEMIMOSACEAEMIMOSACEAEMIMOSACEAE
BOMBACACEAECOMBRETACEAE
ZYGOPHYLLACEAECAESALPINIACEAE
CAPPARACEAECAPPARACEAECAPPARACEAECAPPARACEAECAPPARACEAE
ASCLEPIADACEAECAPPARACEAE
ASCLEPIADACEAEULMACEAE
COMBRETACEAECOMBRETACEAECOMBRETACEAE
BURSERACEAEPAPILIONACEAE
MIMOSACEAEMIMOSACEAE
EUPHORBIACEAERUBIACEAEMORACEAETILIACEAETILIACEAETILIACEAETILIACEAE

39 Guiera senegalensis J. F. Gmel - 2440 Hyphaene thebaica (Linn.) Mart.41 Leptadenia hastata (Pers.) Decne. - 69, 7142 Leptadenia pyrotechnica (Forsk.) Decne. - 6843 Loeseneriella africana (Willd.) R. WiLczek - 4444 Maerua angolensis DC. - 6345 Maerua crassifolia Forsk. - 25, 11046 Mitragyna inermis (Willd.) O. Ktze. - 10647 Momordica balsamina L. - 6748 Oxystelma bomouense R. Br. - 9449 Piliostigma reticulatum (DC.) Hoch.50 Pterocarpus lucens Lepr. ex Guil 1. et Perr. - 32, 7651 Salvadora persica Linn. - 21, 4952 Sclerocarya birrea (A. Rich.) Hochst53 Stereospermum kunthianum Cham.54 Tamarindus indica Linn.55 Tapinanthus globiferus (A. Rich.) Van Tiegh - 11256 Tinospora bakis (A. Rich.) Miers - 26, 10557 Ziziphus mauritiana Lam.
COMBRETACEAEPALMACEAE
ASCLEPIADACEAEASCLEPIADACEAE
HYPPOCRATEACEAECAPPARACEAECAPPARACEAE
RUBIACEAECUCURBITACEAE
ASCLEPIADACEAECAESALPINIACEAE
PAPILIONACEAESALVADORACEAEANACARDIACEAE
BIGNONIACEAETAMARICACEAELORANTHACEAE
MENISPERMACEAERHAMNACEAE

Annexe XV
LISTE DES ESPECES HERBACEES RENCONTREES DANS LA REGION DE LAMARE D'OURSI
N° GENRE ET ESPECE - N° D'ECHANTILLON
1 Abutilon pannosum (Forst. f.) Sehl.2 Aehyrantes aspera Linn. - 473 Adenia gracilis Harms4 Aerva persiea (Burm. f.) Merrill - 465 Althernanthera nodiflora R. Br.6 Alysiearpus ovalifolius (Sehum. et Thonn.) J. Léonard7 Amaranthus graeeizans Linn.8 Andropogon gayanus Kunth - 719 Aristida adseensionis Linn.10 Aristida sieberana Trin. - 8811 Aristida mutabilis Trin et Rupr. - 612 Aristida hordeaeea Kunth - 4012 Aristida funieulata Trin. et Rupr. - 9813 Asparagus afrieanus Lam.14 Bergia suffrutieosa (Del.) Fenzl. - 3415 Blepharis linariifolia Pers.16 Borreria filifolia (Sehum. et Thonn.) K. Sehum. - 117 Braehiara ramosa (L.) Stapf.18 Braehiaria distiehopylla (Trin.) Stapf.19 Bulbostylis barbata (Rottb.) C. B. Cl.20 Bulbostylis hispidula subsp. sengalensis (Cherm.)
Napper syn. Fimbristymis exilis (Kunth) Rormeret Sehultes var. senegalensis Cherm. - 72
21 Caralluma dalzielii N. E; Br.22 Cassia nigricans Vahl23 Cassia obtusifolia Linn. syn. Cassia tora L. - 4324 Cassia oceidentalis Linn. - 1725 Cassia mimosoides Linn.26 Cenehrus biflorus Roxb. - 10127 Cenehrus prieuri (Kunth) Maire - 9628 Chrozophora broehiana Vis. - 5029 Cleome viseosa Linn.30 Colocynthis citrullus (Linn.) O. Ktze - 84, 9131 Commelina forskalaei Vahl32 Corehorus tridens Linn. - 3733 Corchorus olitorus Linn.34 Crinum omatum (Ait) Bury35 Cueumus metuliferus E. Mey. ex Naud - 936 Cynodon daetylon (Linn.) Pers. - 7437 Cyperus iria Linn. n° 838 Cyperus digitatus Roxb.
FAMILLE
MALVACAEEAMARANTHACEAECAESALPINIACEAEAMARANTHACEAEAMARANTHACEAE
PAPILIONACEAEARANTHACEAE
POACEAEPOACEAEPOACEAEPOACEAEPOACEAEPOACEAE
LILIACEAEELATINACEAE
ACANTHACEAERUBIACEAE
POACEAEPOACEAE
CYPERACEAE
CYPERACEAEASCLEPIADACEAE
CEASALPINIACEAECAESALPINIACEAECAESALPINIACEAECAESALPINIACEAE
POACEAEPOACEAE
EUPHORBIACEAECAPPARACEAE
CUCURBITACEAECOMMELINACEAE
TILIACEAETILIACEAE
AMARYLLIDACEAECUCURBITACEAE
POACEAECYPERACEAE
CYPERCEAE

39 Cyperus amabilis Vahl40 Dactyloctenium aegyptium (Linn.) P. Beauv.41 Digitaria lecardii (Pilger) Stapf - 4142 Diheteropogon amplectan (Nees) W. D. Clayton - 9743 Echinoc1oa colona (Linn.) Link44 Echinocloa stagnina (Retz.) P. Beauv.45 Eleocarys dulcis (Bunn. f.) Henschel46 Eragrostis elegantissima Chiov. - 7847 Eragrostis tremula Hoschst. ex Stend. - 7348 Eragrostis pilosa (Linn.) P. Beauv. - 3949 Euphorbia aegyptiaca Boiss.50 Gynandropsis gynandra (Linn.) Briq.51 Hackelochloa granularis (Linn.) O. Ktze - 10052 Hibiscus sidiformis Baill. - 5453 Hibiscus micranthus Linn.54 Indigofera diphylla Vent. - 4955 Indigofera microcarpa Desv.56 Indigofera pilosa Poir. - 8657 Indigofera stenophylla Guill. et Perr. - 3358 Ipomoea aquatica Forsk. - 3859 Ipomoea coptica (Linn.) Roth ex Roem. et Schult.60 Ipomoea heterotricha F. Didr.61 Ipomoea eriocarpa R. Br. - 7562 Ipomoea vagans Bak.63 Ipomoea kotschyana Hoscht. ex Choisy64 Jacquemontia tamnifolia (Linn.) Griseb. - 465 Limeum viscosum (Gay) Fenzl66 Merremia tridentata (Linn.) Hallier f.67 Merremia pinnata (Hoscht. ex Choisy) Hallier f. - 8968 Mitracarpus scaber Zucc.69 Neptunia oleracea Lour. - 3670 Pancratium thrianthum Herb. - 7771 Pandiaka involucrata (Moq.) HooK72 Panicum laetum Kunth - 4273 Panicum subalbidum Kunth - 9974 Panicum anabaptistum Steud.75 Pennisetum pedicellatum Trin.76 Pergularia tomentosa Linn. (Mant.) - 8277 Peristrophe bicalyculata (Retz.) Nees - 1078 Polycarpea linearifolia (DC.) DC. - 4879 Polycarpea eriantha Hoscht. ex A. Rich.80 Polygonum lirnbatum Meisn. - 9581 Schoenefeldia gracilis Kunth - 582 Scirpus brachyceras Hoscht. ex A. Rich.83 Sesamum alatum Thonning - 8784 Sesbania leptocarpa DC.85 Sida ovata Forsk. - 1686 Sida alba Linn. - 9087 Sphenoclea zeylanica Gaertn.
CYPERACEAEPOACEAEPOACEAEPOACEAEPOACEAEPOACEAE
CYPERACEAEPOACEAEPOACEAEPOACEAE
EUPHORBIACEAECAPPARACEAE
POACEAEMALVACEAEMALVACEAE
PAPILIONACEAEPAPILIONACEAEPAPILIONACEAEPAPILlONACEAE
CONCOLVULACEAECONVOLVULACEAECONVOLVULACEAECONVOLVULACEAECONVOLVULACEAECONVOLVULACEAECONVOLVULACEAE
MOLLUGINACEAECONVOLVULACEAE
POACEAERUBIACEAE
MIMOSACEAEAMARYLLIDACEAE
AMARANTHANCEAEPOACEAEPOACEAEPOACEAEPOACEAE
ASCLEPIADACEAEACANTHACEAE
CARYOPHYLLACEAECARYOPHYLLACEAE
POACEAEPOACEAE
CYPERACEAEPEDALIACEAE
PAPILlONACEAEMALVACEAEMALVACEAE
SPHENOCLEACEAE

88 Stylochiton hypogaeus Lepr. - 1189 Tephrosia linearis (Willd.) Pers. - 3590 Thephrosia noctitlora Bojer ex Bak. - 391 Tragus racemosus (L.) AU92 Tribulus terrestris Linn.93 Urochloa trichopus (Hoscht.) Staff94 Vossia cuspidata (Roxb.) Griff.95 Waltheria indica Linn.96 Zornia glochidiata Reichb. ex De. - 52
ARACEAEPAPILIONACEAEPAPILIONACEAE
POACEAEZYGOPHYLLACEAE
POACEAEPOACEAE
STERCULIACEAEPAPILIONACEAE

Annexe XVI
APPELATIONS EN FRANCAIS ET EN LANGUES NATIONALES DES PRINCIPALESESPECES LIGNEUSES DE LA REGION DE LA MARE D'OURSI .
Espèce ligneuse Français Peul Sonraï Tamachek
Acacia albida kad kaïki kokoya assanana
Acacia ataxacantha wonraï kubo bichu
Acacia erhenbergiana kiluki tamat
Acacia laeta patuki ndingare ewarwar
Acacia nilotica var. adansonii nèb-nèb ngaudi baano taïdjart
Acacia nilotica var. nilotica gonaade " "
Acacia nilotica var. tomentosa gommier " " "rouge
Acacia pennata gurm
Acacia raddiana acacia de kiluki bissa affagagRaddi
Acacia senegal gommier patuki ndingaré ewarwarblanc
Acacia seyal mimosa bulbe saaga orafépineux
Adansonia digitata baobab boki koa tekudis
Anogeissus leiocarpus bouleau koguioli wya akarkarad'Afrique
Balanites aegyptiaca dattier tanni garba taboraksauvage oumyroboland'Egypte
Bauhinia rufescens nammadi namaro tédéini
Boscia angustifolia daranéhi hascara agardahant
Boscia salicifolia tiréhi ingalemum
Boscia senegalensis riguile hora tadahant
Cadaba farinosa balami assona agar
Cadaba glandulosa wadagohi tahahistiCalotropis procera arbre à soie bamarndi saga torchaou pomme deSodome
Capparis corymbosa gulujaabi kodara kukukowalel
Caralluma retropiscens 00110 taïbru
Celtis integrifolia micocoulier nganki siad'Afrique
Combretum aculeatum laugi burbura agrizigil
Combretum glutinosum dooki kokorba akalaf
Combretum micranthum kinkéliba ngumui tinguéla ewan
Commiphora africana bdélium à mbadaadi gomba adarasrésineodorante
Dalbergia melanoxy lon ébénier du gelhelani gobia gelganaliSénégal gelgélani
Dichrostachys cinerea mimosa mburi taglchubaïclochette
Entada africana
Euphorbia balsamifera bédréi bèèfra tiharan
Feretia apodanthera brudéhi filirgo taouhat
Ficus gnaphalocarpa figuier gaïgaï ou nbé iwi diwi ou baurigia
Grewia bicolor kéli déiga guégué
Grewia flavescens kibolé grusuma gaïsamum
Grewia tenax jingeli bendicrila tarakat
Grewia villosa gursohi grusuma agarsemum
Guiera senegalensis nger gueloki kanfula ou sabarasaabra
Hyphaene thebaica palmier doum gélègé kanga
Leptadenia hastata sabatrogé dula tatolat
Leptadenia pyrothecnica pibeteki saabra anna
Loeseneriella africana jebukal oboka
Maerua angolensis éléfitaï
Maerua crassifolia hasu agar
Mitragyna inermis kooli haso, kaba taguialalt
Momordica balsamina asaksagaral
Oxystelma bomouense

Piliostigma reticulatum barkelehi kosora tafagargat
Pterocarpus lucens ptérocarpe kiami tinta alibunisluisant
Salvadora persica bromagé bramadanga tadumunttadanunt
Sclerocarya birrea prunier heedi dina tuhilaafricain
Securinega virosa sankara
Stereospermum kunthianum ngolobi danegobo anmit
Tamarindus indica tamarinier guétèbé boosa bososo
Tinospora bakis tinderia
Ziziphus mauritiana jujubier djabé daraï
NB: "u" en langue nationale transcrit le son "ou" du français

Annexe XVII
VARIATION DE LA DENSITE ET DES PA.R.AMETRES DE VITALITE PAR HA DU
PEUPLEMENT LIGNEUX DE LA REGION DE LA MARE DOURSI EN 1992.
Unité géomorphologique Densité Régénération Coupes Mortalité
Buttes et reliefs 610 110 90 50260 200 la
60 la 60380 140 30
Glacis 470 310 la la80 20 150
380 140 la 80240 90160 60 la
90 la 7070 50 la
130 120la 3040 20 lala
150 130 la 2020
170 100 3040 la la 4070 30 7060 la 2030 8090 40 20 2030 , 2070 40 21 32030 30 60la la40 la20
110 60 lala la
180 130 la70 6030 la la 20
190 80 la 30200 100 10 80
Dépressions et 400 180 40 40talwegs 420 20 130 190
180 80 la la810 30 60 la540 280 250150:260 100 10 50790 410 la 20290 20 80450 140 la 20

410 230 10 160890 540 20 160210 100 10 70210 80 290510 230 80 500
Dunes et 170 80ensablements 680 170 40 30
210 90 10 40360 50 80 10270 130 10290 150 10210 10 20 40200 50 10 10
Champs et 100 40 10 10jachères 220 160
90 20 4020
120 90 20110 90 10
70 10 30300 150 70 10
80 10 30
-, :

Annexe XVIll
RESULTATS D'INVENTAIRE FLORISTIQUE DU PEUPLEMENT LIGNEUX DE
LA REGION DE LA MARE D'OURSI DE PlOT ET AL. (1980)
Les espèces ligneuses des formations des dunes et ensablements
Nombre d'arbres mesurés/haEspèces ligneuses
Densité %
Balanites aegyptiaca 21 59Guiera senegalensis 5 14Combretum aculeatum 2 5Combretum glutinosum 2 5Acacia laeta 1 3Acacia raddiana 2 5Commiphora africana 1 3Ziziphus mauritiana 1 3Piliostigma reticulatum 1 3
Total 36 100
Les espèces ligneuses des formations ligneuses des glacis
Nombre d'arbres mesurés/haEspèces ligneuses
Nombre %
Acacia laeta 30 83Balanites aegyptiaca 3 8,5
Divers 3 8,5Acacia raddianaAcacia nilotica
Total 36 100

Les espèces ligneuses des formations de bas-fonds
Nombre dl arbres mesurés/haEspèces ligneuses
Nombre %
Combretum aculeatum 61 37,5Balanites aegyptiaca 58 35,5Acacia seyal 16 1~Combretum micranthum 13Ziziphus mauritiana 6 3,5Acacia laeta 5 3Commiphora africana 3 2Acacia raddiana 1 0,5
Total 163 100
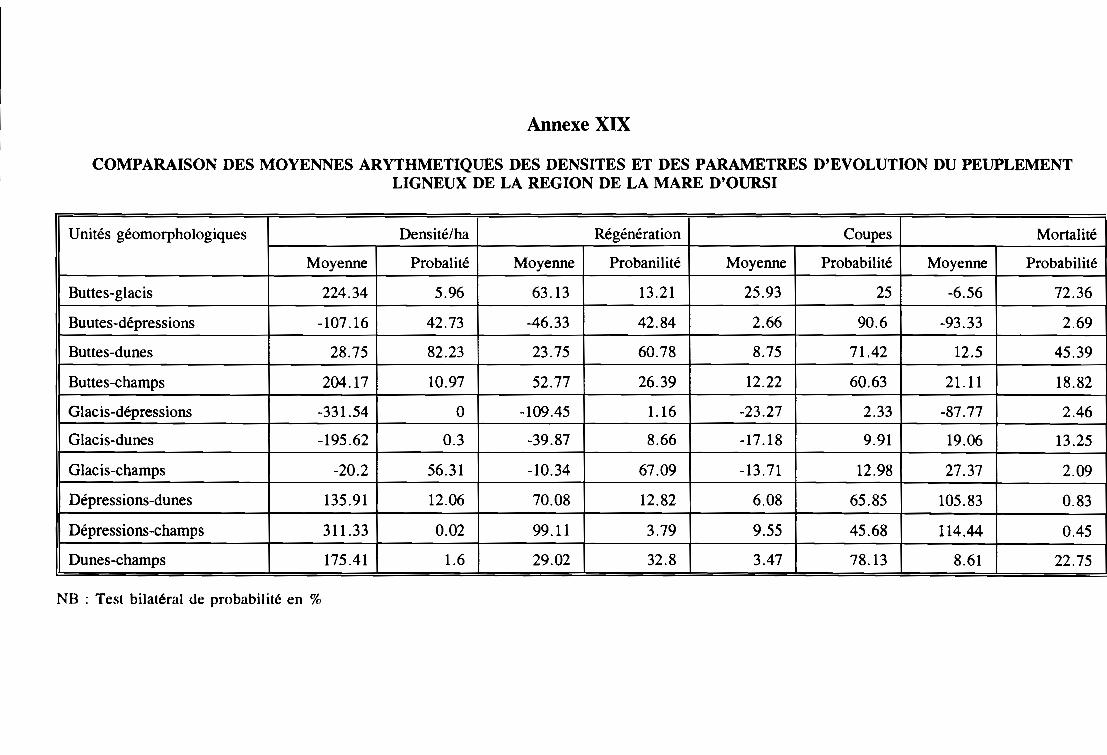
Annexe XIX
COMPARAISON DES MOYENNES ARYTHMETIQUES DES DENSITES ET DES PARAMETRES D'EVOLUTION DU PEUPLEMENTLIGNEUX DE LA REGION DE LA MARE D'OURSI
Unités géomorphologiques Densité/ha Régénération Coupes Mortalité
Moyenne Probalité Moyenne Probanilité Moyenne Probabilité Moyenne Probabilité
Buttes-glacis 224.34 5.96 63.13 13.21 25.93 25 -6.56 72.36
Buutes-dépressions -107.16 42.73 -46.33 42.84 2.66 90.6 -93.33 2.69
Buttes-dunes 28.75 82.23 23.75 60.78 8.75 71.42 12.5 45.39
Buttes-champs 204.17 10.97 52.77 26.39 12.22 60.63 21.11 18.82
Glacis-dépressions -331.54 0 -109.45 1.16 -23.27 2.33 -87.77 2.46
Glacis-dunes -195.62 0.3 -39.87 8.66 -17.18 9.91 19.06 13.25
Glacis-champs -20.2 56.31 -10.34 67.09 -13.71 12.98 27.37 2.09
Dépressions-dunes 135.91 12.06 70.08 12.82 6.08 65.85 105.83 0.83
Dépressions-champs 311.33 0.02 99.11 3.79 9.55 45.68 114.44 0.45
Dunes-champs 175.41 1.6 29.02 32.8 3.47 78.13 8.61 22.75
NB : Test bilatéral de probabilité en %

Annexe XX
DISTRIBUTION, INDICE DE PRESENCE ET PARAMETRES D'EVOLUTION DESESPECES LIGNEUSES DE LA REGION DE LA MARE D'OURSI
Espèce Station Indice de Appéta- Degré Régéné Cou Mortaliécologique présence bilité d'exploi ration -pes -té
-tation % % %
Acacia albida C,D + TA 2 1
Acacia ataxacantha T + A
Acacia erhenbergiana D + TA 2
Acacia laeta D,G,P + + A 3 0,17 2 1,18
Acacia nilotica var. D,G,T + + + PA 3 0,5 7 11adansonii
Acacia nilotica var. T + PA 2nilotica
Acacia nilotica var. T + PA 2tomentosa
Acacia penneta D + A
Acacia raddiana C,D,G,P,T + + + TA 3 24,7 528,3
Acacia senegal D,G + + A 3 1,2 2 4,8
Acacia seyal C,G,T + + TA 2 0,3 4 1,2
Adansonia digitata D,T + PA 3
Anogeissus leiocarpus T + PA 1
Balanites aegyptiaca G,P,T + + + TA 3 15 20 3,5
Bauhinia rufescens T + + TA 2 0,3 5 0,6
Boscia angustifolia D,G,P + + A 3 1 0,6
Boscia salicifolia D + A 1
Boscia senegalensis P,T + PA 2 6
Cadaba farinosa D + TA 3 0,8 1
Cadaba glandulosa D + TA 3 0,5
Calotropis procera C,D,G,P + + PA 1 0,8 1 2,1
Capparis corymbosa + A 2
Caralluma retropiscens D + NA 0

Celtis integrifolia T + TA 2
Combretum aculeatum T + + TA 3 16 1
Combretum glutinosum D + PA 2 1 1
Combretum micranthum P,T + + PA 1 1,5 1 2,4
Commiphora africana D,G,T + TA 3 0,3
Dalbergia melanoxylon D,G,P + A 2 2 1,2
Dichrostachys cinerea C,D,T + A 3
Entada africana D + PA 1
Euphorbia balsamifera D,C + NA 0
Feretia apodanthera T + TA 2 1
Ficus gnafalocarpa T + PA 0
Grewia bicolor T + A 2 0,2 2 6
Grewia tlavescens T + A 1 0,2
Grewia villosa T + PA 1 0,2 1
Grewia tenax T + PA 1 0,3
Guiera senegalensis D,G,P + + PA 1 2,7 4 0,3
Hyphaene thebaica T + NA 1
Leptadenia hastata C,D,G,P,T + + + A 1 15,7 2
Leptadenia pyrotechnica D,P + A 1
Loeseneriella africana T + PA 1
Maerua angolensis D + TA 2
Maerua crassifolia D,T + + TA 3 4
Mitragyna inermis T + PA 2 0,34
Momordica balsamina D + NA 1
Oxyxtelma bornouense T + A 1
Piliostigma reticulatum T + PA 1 1 1
F1erocarpus lucens G,P,T + + TA 3 0,2 11 18
Salvadora persica T + TA 3
Sclerocarya birrea D + A 1
Securinega virosa T + PA 2

Annexe XXI
PRINCIPALES UTILISATIONS DES ~SPECES LIGNEUSES DANS LA REGION DE LAMARE D'OURSI
Espèce Artisanat Alimentation Autres
humaine animale
Acacia albida manches feuilles fraîches fertilisation dud'outils, fruits solconstructions
Acacia laeta louches gomme feuilles fraîches clôture,pharmaAcacia senegal charpente crue fruits copée (gomme)
vannerie(racines)
Acacia erhenbergiana cordage, feuilles fraîches teintureriespaterie fleurs, fruits (écorces),(racines et clôtureécorces) pharmacopée
(feuilles)
Acacia nilotica charpente feuilles fraîches teintureriecordage fleurs, fruits (écorces,
gousses),pharmacopée(fruits)
Acacia raddiana manches feuilles fraîches clôtured'outils, fleurs, fruitsvannerie,cordage(racines)
Acacia seyal charpente feuilles fraîches et pharmacopéesèches, fleurs, fruits (écorces),
clôture
Adansonia digitata cordage feuilles, feuilles, fleurs cuisson(écorces), pulpe, (poterie)vannerie graines
Anogeissus leiocarpus feuilles, rameaux teinturerie devêtements(écorces)

Stereospennum T + PA 1lcunthianum
Tamarindus indica T + A 2
Tinospora bakis D + NA 0
Ziziphus mauritiana T + + TA 3 2,7 3 0,3
Légende du tableau:
Station écologique ou milieu de distribution de l'espèce :C = Champs et jachèresD = Dunes et ensablementsG = GlacisP = Piémonts de relief rocheux ou cuirasséT = Dépressions (bordure de mare et talweg)
Indice de présence :+ + + = espèce très répandue
+ + = espèce moyennement fréquente+ = espèce peu ou rarement rencontrée
Degré d'exploitation :3 = espèce très exploitée2 = espèce moyennement exploitée1 = espèce peu exploitéeo = espèce non exploitée
Degré d' appétabilité :TA = espèce très appétée
A = espèce appétéePA = espèce peu appétée
NA = espèce non appétée
Régénération: pourcentage des individus de diamètre ~ 2,5 cm de l'espèce/ensemble des pieds derégénération
Coupe : Pourcentage des pieds coupés/ensemble des pieds coupés
Mortalité : Pourcentage des individus morts sur pied ou couchés de cette espèce/ensemble desindividus morts
Dichrostachys cinerea gourdins feuilles fraîches,fleurs, jeunespousses
Entada africana feuilles et fruits
Feretia apodanthera feuilles fraîches etsèches
Ficus gnaphalocarpa fruits, fruitsfeuilles(disette)
Grewia bicolor cuillères, fruits feuilles fraîches etliterie, sèches, fruitsbâtons,pilons
Guiera senegalensis literie jeunes feuilles etfeuilles fraîches,jeunes pousses
Hyphaene thebaica cordage, fruitsnattes(palmes),poutres
Leptadenia hastata fruits
Leptadenia pyrotechnica cordage fleurs cure-dents(fibres) pharmacopée
(suc, graines)
Loeseneriella africana feuilles
Maerua angolensis feuilles fraîches
Maerua crassifolia fruits, feuilles fraîches, cure-dentsfeuilles fleurs, fruits,(disette) rameaux
Mitragyna inennis piquet de feuillestente literie,vannerie
Momordica balsamina fruits feuilles pharmacopée(racines)
Oxystelma bomouense feuilles
~erocarpus lucens charpente feuilles fraîches et bois de chauffegrenier sèches, fruits
Salvadora persica fruits feuilles fraîches cure-dents
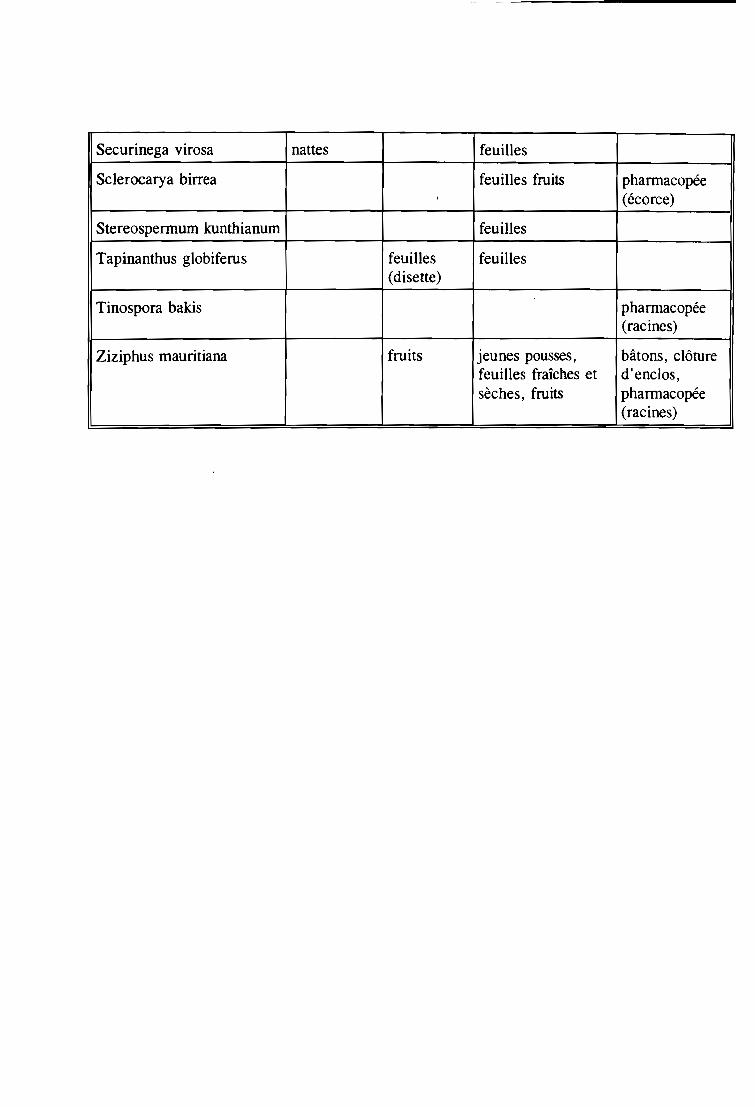
Securinega virosa nattes feuilles
Sclerocarya birrea feuilles fruits pharmacopée, (écorce)
Stereospennum kunthianum feuilles
Tapinanthus globiferus feuilles feuilles(disette)
Tinospora bakis pharmacopée(racines)
Ziziphus mauritiana fruits jeunes pousses, bâtons, clôturefeuilles fraîches et d'enclos,sèches, fruits pharmacopée
(racines)