universite paris est creteil faculte de …doxa.u-pec.fr/theses/th634245.pdf · figure 4: image en...
TRANSCRIPT

UNIVERSITE PARIS EST CRETEIL
FACULTE DE MEDECINE DE CRETEIL
******************
ANNEE 2012 N° 1056
THESE
POUR LE DIPLOME D'ETAT
DE
DOCTEUR EN MEDECINE
Discipline :
OPHTALMOLOGIE
Présentée et soutenue publiquement le à
PARIS
Par M. Sylvain Michée
Né le 23/04/1982 à Corbeil-Essonnes
TITRE : ANALYSE IN VITRO DES INTERACTIONS ENTRE LES MACROPHAGES ET LES CELLULES EPITHELIALES DE LA CONJONCTIVE SOUS L’INFLUENCE DU CHLORURE DE BENZALKONIUM
PRESIDENT DE THESE : LE CONSERVATEUR DE LA
M. le Pr. Bahram Bodaghi BIBLIOTHEQUE UNIVERSITAIRE
DIRECTEUR DE THESE : M. le Dr. Antoine Labbé
Signature du Cachet de la bibliothèque Président de thèse universitaire

2
Remerciements
Au président du jury,
Monsieur le Professeur Bahram Bodaghi,
Pour m’avoir fait l’honneur d’accepter de présider la soutenance de cette thèse, vos qualités
médico-chirurgicales ainsi que votre implication dans la recherche sont un atout en
ophtalmologie.
Vos connaissances, mais aussi vos qualités pédagogiques et votre disponibilité suscitent
l’admiration de vos élèves.
Soyez assuré de ma très sincère gratitude et de mon profond respect.

3
A mon directeur de thèse,
Monsieur le Docteur Antoine Labbé,
Tu as su me soutenir, m’aiguiller dans le chemin de la recherche, me donner de précieux
conseils par ton expérience passée, ta gentillesse et ta sympathie. Tu as pu prendre le temps
nécessaire pour examiner en détail tout ce travail avec une minutie qui te fait honneur.
Ta rigueur et ton sérieux sont un exemple pour moi, ton aide a été incalculable dans la
réalisation de cette thèse.
Sois assuré de toute ma reconnaissance en acceptant ce travail.

4
A mon maître, Monsieur le Professeur Christophe Baudouin,
Vous m’avez accueilli dans votre laboratoire au sein de votre équipe devenue une des plus
performantes du monde dans l’analyse de la surface oculaire pendant une année de recherche
qui restera une expérience forte. C’est aussi au cours de mon cursus d’interne que j’ai pu
apprécier vos qualités médicales et chirurgicales. L’étendue de vos connaissances et du travail
que vous avez accompli me laissent admiratif.
Vous m’avez fait confiance et soutenu dans les décisions que j’ai pu prendre et c’est avec un
réel plaisir et enthousiasme que je vais aborder les futures années à travailler dans votre
service.
Veuillez accepter ce travail comme la preuve de ma gratitude et de mon plus profond respect.

5
Monsieur le Professeur Marc Labetoulle
Vous m’avez fait l’honneur d’accepter d’être l’un des membres du jury.
J’ai été très touché que vous accordiez une part de votre précieux temps à la lecture de mon
travail
Veuillez trouver ici le témoignage de ma profonde reconnaissance et de mon respect.

6
Au Docteur Françoise Baudouin,
Vous avez toujours su trouver les mots et les encouragements nécessaires au quotidien. Par
votre gentillesse et bienveillance, vous m’avez guidé tout au long de ce travail.
Vous m’avez fait l’honneur d’accepter de siéger dans ce jury et d’avoir consacré du temps
pour évaluer ce travail.
Recevez ce travail en témoignage de mon immense reconnaissance et de mon profond respect.

7
Au Professeur Phuc Le Hoang,
Au Professeur Alain José Sahel,
Au Professeur Laurent Laroche,
Au Professeur Vincent Borderie,
Au Professeur Alain Gaudric,
Au Professeur Pascale Massin,
Au Professeur Gilles Chaine,
Au Docteur Catherine Bourreau,
Au Docteur Sylvia Buffet-Nghiem
Au Docteur Franck Fajnkuchen
Au Docteur Ramin Tadayoni,
Au Docteur Yves Lachkar,
Au Docteur Pascal Dureau,
Au Docteur Georges Caputo,
Au Docteur Stanislas Roman,
Au Docteur Djawed Bensmail,
Au Docteur François Auclin,
Au Docteur Alexandre Denoyer,
Au Docteur Bénédicte Dupas,
Au Docteur Cécile Allouch,
Au Docteur Pascale Hamard,
Au Docteur William Rostène,
Votre exemple a été un modèle,
Recevez avec ce travail un témoignage de mon admiration et de mon respect.

8
A Luisa Riancho,
Qui a été ma maman canard au cours de mon stage en laboratoire, ce qui n’est pas rien.
A mes collègues passés et présents et amis
Michelle Nguyen, Nacim Bouheraoua, Pierre Chouraqui, Julien Bullet, Adil Darugar, Julien
Pérol, Chrysanthi Basdekidou, Eric Pérez-Campagne, Feng Liang, Hong Liang, Mohammed
El-Hammdaoui, Rachid Tahiri, Jean-Rémi Fenolland, Sylvia Cayette, Julie Frugier, Aurélie
Brolly, Jonathan Benesty, Déborah Benisty, Charles Van Went, Raphaël De Nicola et ceux
que j’oublie.
Recevez avec ce travail un témoignage de mon amitié et de ma reconnaissance.
Vous côtoyer au quotidien a été un grand plaisir.

9
A ma famille,
Mes grands parents,
Mon père et ma mère,
Mes deux frères,
Mes cousines, oncles et tantes,
Merci pour votre soutien depuis le premier jour.

10
TABLE DES MATIERES
TABLE DES ILLUSTRATIONS .......................................................................................... 13
ABREVIATIONS ................................................................................................................... 15
INTRODUCTION .................................................................................................................. 16
CHAPITRE 1 : PREAMBULE ............................................................................................. 18
1 PHYSIOPATHOLOGIE DE LA CONJONCTIVE .......................................................... 18
1.1 Généralités ................................................................................................................. 18
1.2 Interactions cellulaires épithéliales ............................................................................ 19
1.3 Interactions au sein du stroma ................................................................................... 23
2 GLAUCOME ET CONSERVATEURS ........................................................................... 24
2.1 Généralités sur le glaucome ....................................................................................... 24
2.2 Traitements du glaucome ........................................................................................... 25
2.2.1 Principes ............................................................................................................. 25
2.2.2 Les collyres antiglaucomateux ........................................................................... 26
2.2.3 La chirurgie filtrante et la fibrose conjonctivale ................................................ 27
2.3 Les conservateurs ...................................................................................................... 27
2.3.1 Historique ........................................................................................................... 27
2.3.2 Principaux conservateurs .................................................................................... 28
2.3.3 Le chlorure de benzalkonium (BAK) ................................................................. 29
3 LIGNEES CELLULAIRES .............................................................................................. 31
3.1 Généralités ................................................................................................................. 31
3.2 La lignée THP-1 ........................................................................................................ 32
3.2.1 Historique ........................................................................................................... 32
3.2.2 THP-1 en tant que lignée macrophagique .......................................................... 33
3.3 Les cellules épithéliales Wong Kilbourne dérivées de Chang (WKD) ..................... 34
CHAPITRE 2 : ETUDE IN VITRO DES EFFETS DU CHLORURE DE
BENZALKONIUM SUR LES MACROPHAGES .............................................................. 36
1 INTRODUCTION ............................................................................................................ 36
2 MATERIELS ET METHODES ....................................................................................... 37
2.1 Lignées cellulaires ..................................................................................................... 37
2.1.1 Lignée monocytaire ............................................................................................ 37
2.1.2 Lignée épithéliale conjonctivale ......................................................................... 38
2.2 Stimulations ............................................................................................................... 38
2.3 Expression des marqueurs de surface ........................................................................ 39
2.4 Immunohistochimie ................................................................................................... 39
2.5 Etude de la mort cellulaire en CMF à l’aide du test Annexine V / 7-AAD ............... 40
2.6 Etude de la phagocytose ............................................................................................ 40
2.7 Production de cytokines ............................................................................................. 41
2.8 Etude de la migration ................................................................................................. 41

11
2.9 Analyses statistiques .................................................................................................. 42
3 RESULTATS .................................................................................................................... 42
3.1 Les cellules THP-1 ont été différenciées en macrophages ........................................ 42
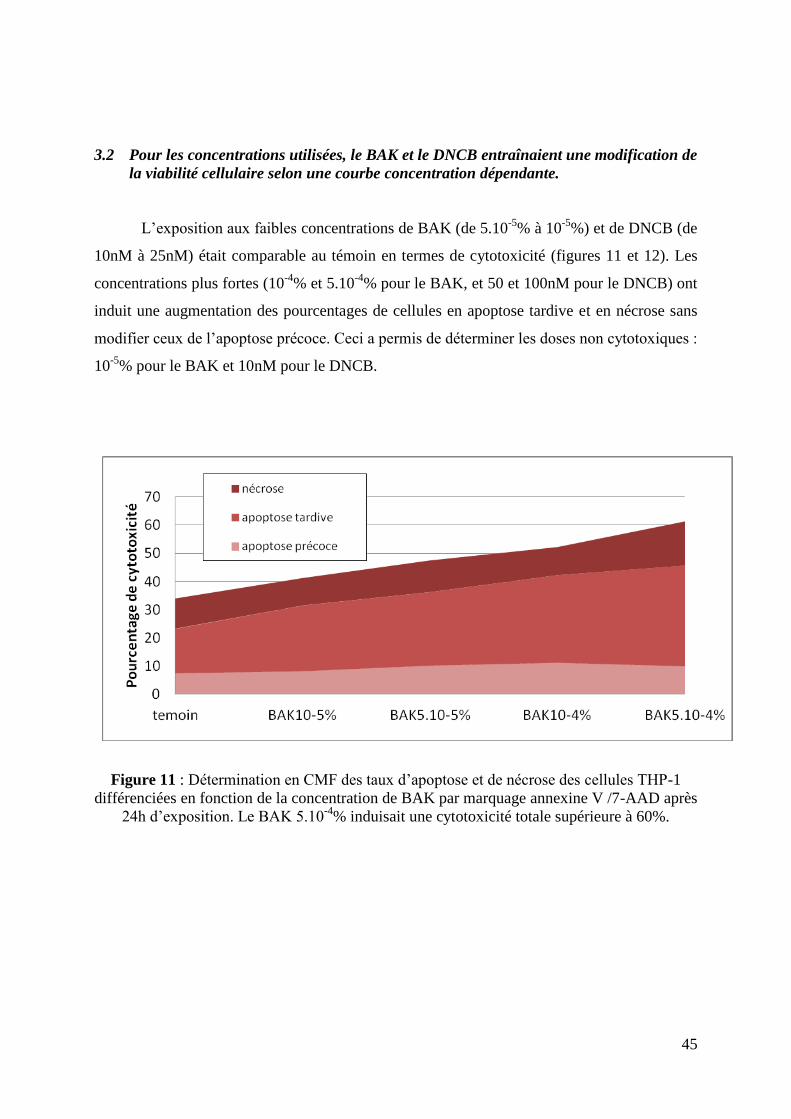
3.2 Pour les concentrations utilisées, le BAK et le DNCB entraînaient une modification
de la viabilité cellulaire selon une courbe concentration dépendante.. .................................... 45
3.3 L’exposition au LPS et au TNF-α n’a pas entraîné d’effet cytotoxique aux
concentrations étudiées ............................................................................................................ 46
3.4 L’expression des marqueurs de différenciation des cellules THP-1 différenciées a été
modifiée après exposition au BAK, DNCB, LPS et TNF-α. ................................................... 48
3.5 Les cellules THP-1 exposées au BAK, LPS et TNF-α ont augmenté leur capacité de
phagocytose .............................................................................................................................. 51
3.6 Les cellules THP-1 différenciées (macrophages) ont présenté un accroissement de
leurs capacités de migration sous l’effet du BAK et du TNF-α.. ............................................. 52
3.7 L’expression des cytokines par les cellules THP-1 a été modifiée en fonction de la
différenciation et de l’exposition aux différents facteurs de stimulation ................................. 53
4 DISCUSSION ................................................................................................................... 56
5 CONCLUSION ................................................................................................................. 59
CHAPITRE 3 : ANALYSE IN VITRO DES RELATIONS ENTRE LES
MACROPHAGES ET LES CELLULES EPITHELIALES CONJONCTIVALES ....... 61
1 INTRODUCTION ............................................................................................................ 61
2 MATERIELS ET METHODES ....................................................................................... 61
2.1 Lignées cellulaires ..................................................................................................... 61
2.1.1 Lignée monocytaire ............................................................................................ 61
2.1.2 Lignée épithéliale conjonctivale ......................................................................... 62
2.2 Stimulations ............................................................................................................... 62
2.3 Analyse de l’effet des facteurs stimulants sur les cellules épithéliales conjonctivales
en cytométrie en flux ............................................................................................................... 62
2.4 Production de cytokines ............................................................................................ 63
2.5 Interactions entre les cellules WKD et les cytokines produites par les macrophages 63
2.6 Interactions entre les macrophages et les cytokines produites par les cellules WKD 64
2.7 Analyses statistiques .................................................................................................. 64
3 RESULTATS .................................................................................................................... 65
3.1 Parmi l’expression des marqueurs membranaires des cellules WKD, seul EMMPRIN
a été modifiée après exposition au BAK et au DNCB ............................................................. 65
3.2 L’expression des chimiokines par les cellules WKD a été modifiée par l’exposition
aux facteurs stimulants. ............................................................................................................ 66
3.3 Augmentation de l’expression du CD54 par les cellules conjonctivales WKD
exposées aux surnageants des macrophages exposés au LPS .................................................. 69
3.4 Augmentation de l’expression du CD54 par les macrophages exposés aux
surnageants des cellules conjonctivales exposées au BAK. ..................................................... 70
4 DISCUSSION ................................................................................................................... 71
4.1 Stimulation directe sur les cellules conjonctivales .................................................... 71
4.2 Influence des macrophages sur les conjonctivales. ................................................... 72

12
4.3 Les effets du BAK dans les interactions entre les cellules conjonctivales et les
macrophages ............................................................................................................................. 74
5 CONCLUSION ................................................................................................................. 75
CONCLUSION ET PERSPECTIVES ................................................................................. 76
REFERENCES ....................................................................................................................... 77
RESUME EN ANGLAIS ....................................................................................................... 85
RESUME EN FRANCAIS ..................................................................................................... 86

13
TABLE DES ILLUSTRATIONS
Liste des Figures
Figure 1: Coupe histologique de la conjonctive (page 18).
Figure 2: Image en microscopie confocale de cellules à mucus après immunomarquage (page
21).
Figure 3: Image en microscopie confocale de cellules de Langerhans après immunomarquage
(page 22).
Figure 4: Image en microscopie confocale de cellules à mucus et de Langerhans après double
immunomarquage (page 23).
Figure 5: Formule chimique du chlorure de benzalkonium (page 30).
Figure 6: Image en microscopie optique des cellules THP-1 (page 33).
Figure 7: Image en microscopie optique des cellules WKD (page 35).
Figure 8: Image en microscopie optique des cellules THP-1 non différenciées et différenciées
(page 43).
Figure 9: Expression membranaire par les cellules THP-1 indifférenciées et différenciées du
CD86, CD54, CD11b, CD11c et CD33 (page 44).
Figure 10: Expression membranaire du CD11c à la surface des cellules THP-1
indifférenciées et différenciées en immunofluorescence (page 44).
Figure 11: Analyse de la cytotoxicité du BAK sur les macrophages (page 45).
Figure 12: Analyse de la cytotoxicité du DNCB sur les macrophages (page 46).
Figure 13: Analyse de la cytotoxicité du LPS sur les macrophages (page 47).
Figure 14: Analyse de la cytotoxicité du TNF-α sur les macrophages (page 47).
Figure 15: Expression du CD11b par les cellules THP-1 et les macrophages (page 49).
Figure 16: Expression du CD11c par les cellules THP-1 et les macrophages (page 50).
Figure 17: Expression du CD54 par les cellules THP-1 et les macrophages (page 50).
Figure 18: Expression du CD33 par les cellules THP-1 et les macrophages (page 51).
Figure 19: Image en microscopie à fluorescence des microsphères de carboxylate
phagocytées par les cellules THP-1 et les macrophages (page 52).
Figure 20: Analyse des capacités de phagocytose des cellules THP-1 et des macrophages
(page 52).
Figure 21: Analyse de la migration des cellules THP-1 et des macrophages (page 53).
Figure 22: Dosage des surnageants de culture des cellules THP-1 sur membranes de
nitrocellulose (page 56).
Figure 23: Expression des marqueurs membranaires CD 40, CD 54, CD 63, CD 95, HLA-DR
et EMMPRIN par les cellules épithéliales conjonctivales (page 65).
Figure 24: Dosage des surnageants de culture des cellules WKD sur membranes de
nitrocellulose (page 67).
Figure 25: Expression des marqueurs membranaires CD 40, CD 54, CD 63, CD 95, HLA-DR
et EMMPRIN par les cellules épithéliales conjonctivales après exposition aux surnageants de
culture des macrophages (page 69).
Figure 26: Expression des marqueurs membranaires CD54, CD11b et CD11c par les
macrophages sous l’influence des surnageants de culture des cellules WKD (page 70).

14
Liste des tableaux
Tableau I: Intensité des spots correspondant aux cytokines présentes dans les surnageants de
culture des cellules THP-1 (page 55).
Tableau II: Intensité des spots correspondant aux cytokines présentes dans les surnageants de
culture des cellules WKD (page 68).

15
Principales abréviations utilisées
7-AAD : 7-aminoactinomycin D
ADN : Acide désoxyribonucléique
ARNm : Acide ribonucléique messager
BAK : Chlorure de benzalkonium
C3b : Complément 3 type b
C5a : Complément 5 type a
CCL : Cystéine cystéine ligand
CD : Cluster de différenciation
CFU : Colony-forming unit
CMF : Cytométrie en flux
CXCL : Cystéine X cystéine ligand
DAPI : Di-amidino-phénylindole
DMEM : Milieu Eagle modifié de Dulbecco
DNCB : Di-nitro-chloro-benzène
EDTA : Acide éthylène diamine tétraacétique
EGF : Epidermal growth factor
EMA : European Medicine Agency
EMMPRIN : Extracellular Matrix Metalloproteinase Inducer
Fas : Fragment inducteur d’apoptose
FBS : Sérum de veau foetal
Fc : Fragment constant des immunoglobulines
FITC : Fluorescéine-isothiocyanate
G-CSF : Granulocyte colony-stimulating factor
GM-CSF : Granulocyte macrophage colony-stimulating factor
HA: Humeur aqueuse
HLA: Human leukocyte antigen
HTO: Hypertonie intraoculaire
ICAM : Intercellular adhesion molecule
IF : Immunofluorescence
IFN : Interferon
IL : Interleukine
LPS : Lipopolysaccharide
MCP : Methyl-accepting Chemotaxis Protein
MEC : Matrice extra cellulaire
MIF : Macrophage migration inhibitory factor
NF-κB : Nuclear factor-kappa B
PAI : Plasminogen activator inhibitor
PBS : Phosphate buffer saline
PE : Phycoérythrine
PFA : Paraformaldéhyde
PIO : Pression intra oculaire
PMA : Phorbol myristate acetate
RPMI : Roswell Park Memorial Institute
SDF-1 : Stromal cell derived factor 1
sTREM : Soluble triggering receptor expressed on myeloid cells
TLR : Toll like receptor
TNF-α : Tumor necrosis factor alpha
WKD : Wong-Kilbourne derivated

16
INTRODUCTION
La surface oculaire constitue la partie la plus externe de l’œil, comprenant les
paupières, le film lacrymal et les glandes lacrymales, la conjonctive et la cornée. Elle est
maintenant identifiée comme une entité fonctionnelle intégrée. La surface oculaire est en
contact permanent avec le milieu extérieur, source d’agressions et d’agents pathogènes, tels
que les virus, les bactéries, champignons, amibes, parasites, agents allergiques, polluants ou
encore les collyres. Les maladies de la surface oculaire représentent un ensemble de
pathologies extrêmement fréquentes allant de la simple irritation oculaire à de véritables
maladies cécitantes. Ainsi, la prévalence de la sécheresse oculaire serait de 5 à 35% en
fonction de l’âge dans les pays développés (34), l’allergie oculaire toucherait 7 à 10% de la
population en Europe (34), chaque personne ferait au moins un épisode de conjonctivite
infectieuse durant sa vie et enfin on estime qu’environ 65 millions de patients sont traités par
collyres antiglaucomateux dans le monde (56).
L’inflammation et les cellules inflammatoires sont au cœur des mécanismes
physiopathologiques impliqués au niveau de la surface oculaire (12). Parmi les cellules de
l’immunité, les macrophages et les cellules dendritiques ont un rôle de pivot notamment par
leurs fonctions de présentation des antigènes et la production de cytokines (41). L’utilisation
des collyres antiglaucomateux au long cours entraîne, au-delà des manifestations cliniques
classiques de l’allergie et de la toxicité, de nombreuses modifications des tissus qui
composent la surface oculaire. Ces changements tissulaires ont été démontrés et analysés par
de nombreuses études sur des empreintes conjonctivales, sur des biopsies conjonctivales ou
encore sur des modèles expérimentaux, confirmant la toxicité et l’expression de marqueurs
inflammatoires au niveau de la surface oculaire induites par ces collyres (11, 17, 29). La
réponse cicatricielle excessive de l’œil après chirurgie du glaucome est caractérisée au niveau
de la conjonctive par une inflammation et une prolifération fibroblastique qui conduit à une
production excessive de matrice extracellulaire (MEC) venant bloquer le site de filtration
(18). L’utilisation au long cours de collyres antiglaucomateux et de leurs conservateurs a une
influence directe sur les résultats chirurgicaux (46). Bien que les macrophages aient été
impliqués dans de nombreuses pathologies de la surface oculaire, les mécanismes précis du
recrutement de ces cellules inflammatoires, les interactions avec les cellules épithéliales de la
surface oculaire mais aussi les moyens de réguler l’inflammation pathologique ne sont pas

17
encore totalement expliqués. Néanmoins, les conservateurs associés au principe actif, en
particulier le chlorure de benzalkonium (BAK), semblent jouer un rôle majeur dans ces
changements tissulaires et leurs complications (9).
Largement étudiés sur les cellules épithéliales conjonctivales, les effets du BAK n’ont
été que peu analysés sur les cellules inflammatoires et notamment sur les macrophages.
L’objectif de la première partie de cette thèse a donc été de définir les effets directs in vitro du
BAK sur les macrophages. Les interactions entre les cellules épithéliales conjonctivales et les
macrophages sous l’effet du BAK ont été ensuite analysées dans une deuxième partie.
Ce travail a ainsi permis de préciser les interactions entre deux acteurs cellulaires
majeurs de la surface oculaire, les cellules épithéliales et les macrophages, sous l’effet du
principal conservateur utilisés dans les collyres, le BAK.

18
CHAPITRE 1 : PREAMBULE
1. PHYSIOPATHOLOGIE DE LA CONJONCTIVE
1.1 Généralités
La cornée et la conjonctive sont exposées de manière permanente aux agressions
mécaniques toxiques et microbiennes de l’environnement extérieur. La protection palpébrale
et le lavage lacrymal éliminent la majeure partie des corps étrangers irritants. La conjonctive
est une membrane muqueuse vascularisée qui couvre la surface antérieure du globe et la face
postérieure des paupières supérieures et inférieures. Elle est habituellement séparée en 3
parties, la conjonctive palpébrale (marginale, tarsale et orbitaire), les culs de sacs
conjonctivaux et la conjonctive bulbaire (50). Histologiquement, la conjonctive est constituée
d’un épithélium et d’un chorion sous-jacent ou substantia propria (figure1).
Figure 1: Coupe histologique de la conjonctive en coloration hématéine-éosine. On distingue
l’épithélium et le chorion sous-jacent. Au sein de l’épithélium apparaissent des cellules claires
de plus grande taille, les cellules à mucus (Image du Professeur Christophe Baudouin).

19
La conjonctive est également responsable de la sécrétion du mucus essentiel pour le
film lacrymal par l’intermédiaire de glandes muqueuses, constituées par des ensembles de
cellules caliciformes ou à cellules à mucus, regroupées pour la plupart dans des cryptes de
Henlé.
Très richement vascularisée par les artères palpébrales et ciliaires antérieures, la
conjonctive joue un rôle primordial dans la défense de la surface oculaire grâce à de
nombreuses cellules immunocompétentes capables d’initier une réaction inflammatoire, d’y
participer et de synthétiser des immunoglobulines. Le drainage lymphatique est assuré par un
réseau profond et un réseau superficiel sous-épithélial. Les vaisseaux lymphatiques
conjonctivaux se drainent vers les ganglions pré-tragiens, parotidiens et sous-maxillaires. La
conjonctive est particulièrement impliquée dans les réactions de la surface oculaire aux
traitements médicaux du glaucome, mais aussi dans son traitement chirurgical.
1.2 Interactions cellulaires épithéliales
La conjonctive est constituée de cellules épithéliales pluristratifiées composée de 2 à
8-10 couches de cellules selon la localisation. Associées aux lymphocytes intra-épithéliaux
(essentiellement des lymphocytes T), les cellules épithéliales conjonctivales ne sont
maintenant plus considérées comme une simple barrière inerte, mais elles sont directement
impliquées dans de nombreuses réactions biologiques, particulièrement dans les processus
inflammatoires et apoptotiques. En cas d’inflammation, les cellules épithéliales surexpriment
CD54 ou ICAM-1 (Intercellular adhesion molecule-1), molécule d’adhésion qui en se liant au
LFA-1 des lymphocytes et des cellules phagocytaires permet leur migration à travers
l’épithélium (55). L’interféron γ (IFN-γ) semble induire cette expression d’ICAM-1 en cas
d’inflammation locale (15). Les cellules épithéliales, également sous l’effet de l’interféron
gamma (IFN-γ), expriment les antigènes de classe II, HLA-DR (Human leukocyte antigen-
DR), qui leur confèrent des propriétés d’immunocompétence propre. Les cellules épithéliales
synthétisent également des cytokines, l’IL-1 (Interleukine-1), l’IL-6 et l’IL-8, qui jouent
probablement un rôle d’amplification des réactions inflammatoires locales. Leur synthèse est

20
également augmentée chez les patients glaucomateux traités au long cours (8).
Présentes dans l’épithélium, les cellules caliciformes ou cellules à mucus sont
réparties de façon inégale sur la conjonctive (figure 1 et 2). Ces cellules jouent un rôle
particulièrement important dans les pathologies de la surface oculaire et la perte de ces
cellules est considérée comme un indicateur sensible, quoique non spécifique, d’atteinte de la
surface oculaire (51). Des cellules immunocompétentes, en particulier les cellules
dendritiques et de Langerhans, caractérisées par des expansions cytoplasmiques, sont
également présentes au sein de l’épithélium de toutes les régions conjonctivales (Figure 3). La
densité de celles-ci est estimée à 250 à 300 par mm2 chez l’homme au niveau de la
conjonctive limbique (3). Ces cellules se déplacent entre les cellules épithéliales en fonction
de l’état inflammatoire local. En cas d’inflammation de surface, leur densité augmente
considérablement et une impressionnante migration vers la superficie conjonctivale peut être
observée. Du fait de leurs prolongements les cellules dendritiques sont en contact avec de
nombreuses cellules. Un double immunomarquage des cellules à mucus et des cellules
dendritiques permet d’observer les rapports étroits qu’entretiennent ces cellules, chaque
cellule à mucus (de forme arrondie) étant bordée par des prolongements étroits de cellules
dendritiques (figure 4).
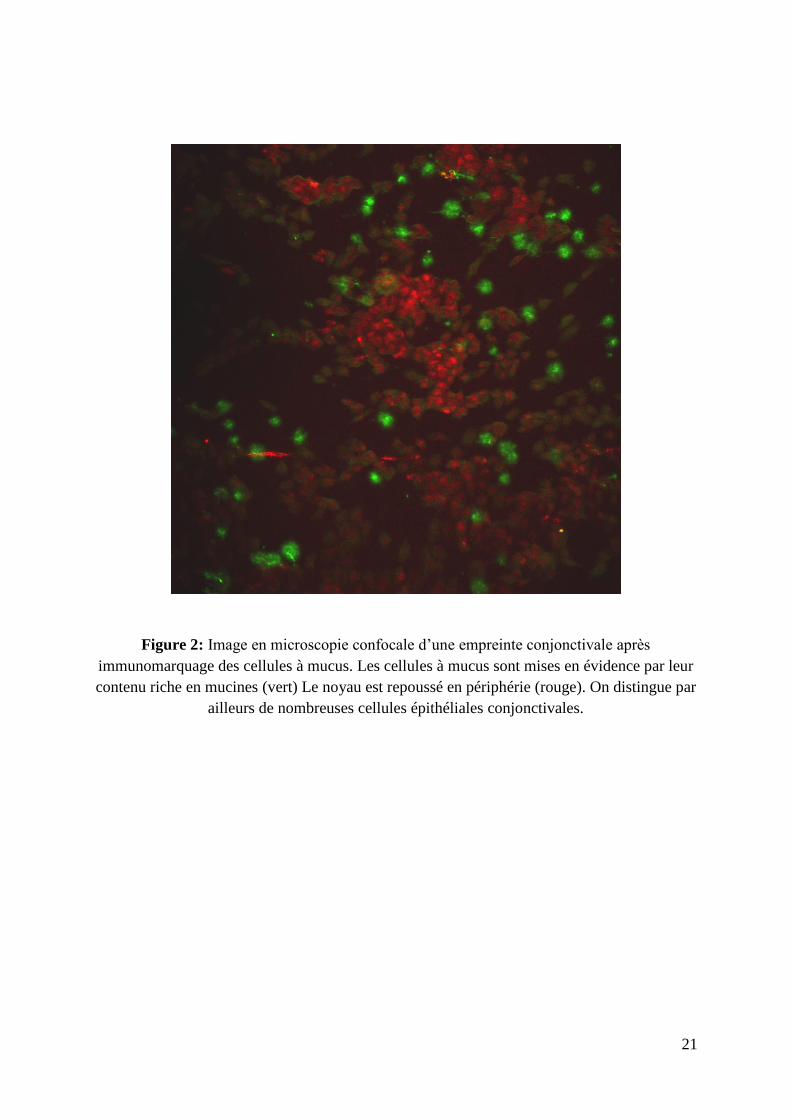
21
Figure 2: Image en microscopie confocale d’une empreinte conjonctivale après
immunomarquage des cellules à mucus. Les cellules à mucus sont mises en évidence par leur
contenu riche en mucines (vert) Le noyau est repoussé en périphérie (rouge). On distingue par
ailleurs de nombreuses cellules épithéliales conjonctivales.
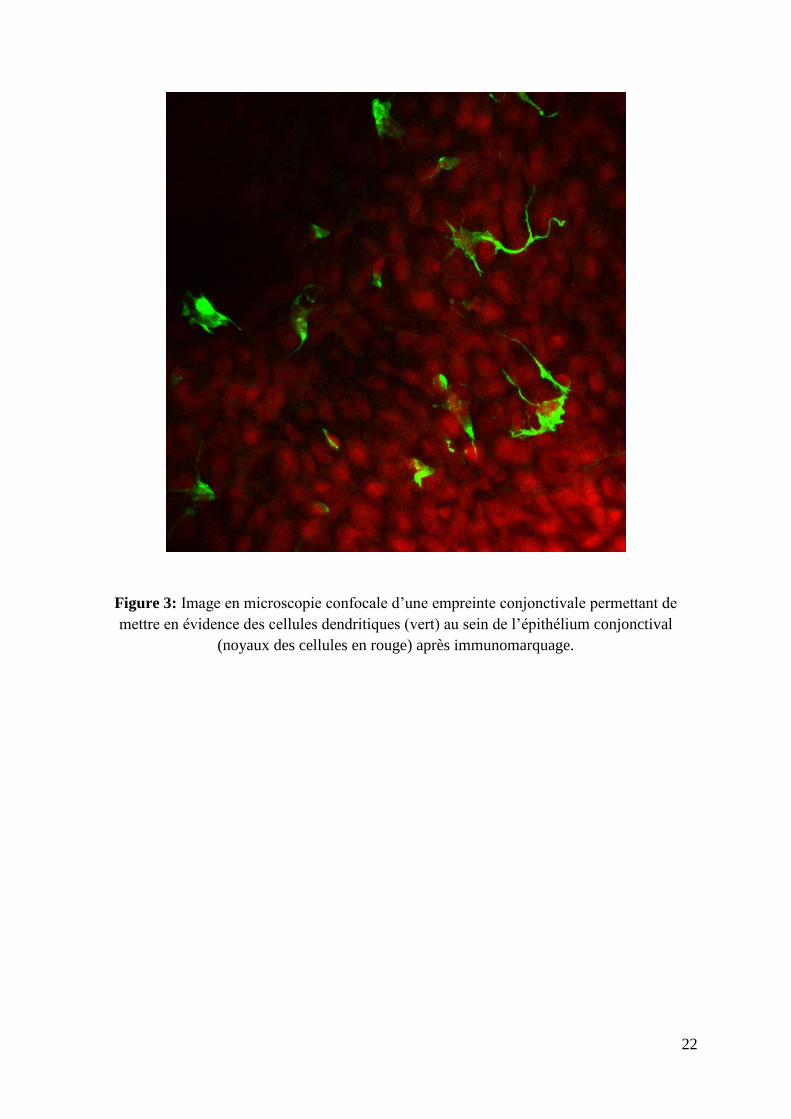
22
Figure 3: Image en microscopie confocale d’une empreinte conjonctivale permettant de
mettre en évidence des cellules dendritiques (vert) au sein de l’épithélium conjonctival
(noyaux des cellules en rouge) après immunomarquage.
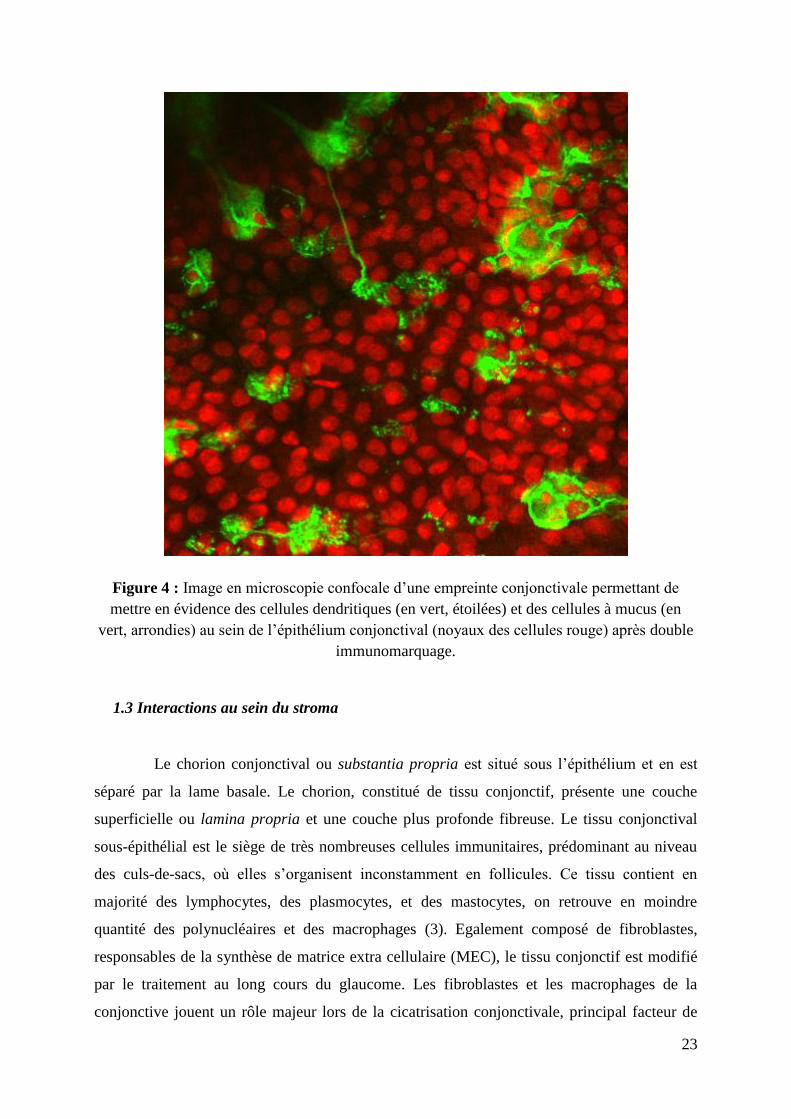
23
Figure 4 : Image en microscopie confocale d’une empreinte conjonctivale permettant de
mettre en évidence des cellules dendritiques (en vert, étoilées) et des cellules à mucus (en
vert, arrondies) au sein de l’épithélium conjonctival (noyaux des cellules rouge) après double
immunomarquage.
1.3 Interactions au sein du stroma
Le chorion conjonctival ou substantia propria est situé sous l’épithélium et en est
séparé par la lame basale. Le chorion, constitué de tissu conjonctif, présente une couche
superficielle ou lamina propria et une couche plus profonde fibreuse. Le tissu conjonctival
sous-épithélial est le siège de très nombreuses cellules immunitaires, prédominant au niveau
des culs-de-sacs, où elles s’organisent inconstamment en follicules. Ce tissu contient en
majorité des lymphocytes, des plasmocytes, et des mastocytes, on retrouve en moindre
quantité des polynucléaires et des macrophages (3). Egalement composé de fibroblastes,
responsables de la synthèse de matrice extra cellulaire (MEC), le tissu conjonctif est modifié
par le traitement au long cours du glaucome. Les fibroblastes et les macrophages de la
conjonctive jouent un rôle majeur lors de la cicatrisation conjonctivale, principal facteur de

24
risque des échecs de la chirurgie filtrante du glaucome (18). Le drainage lymphatique
s’effectue vers les ganglions pré-tragiens qui mettent en communication les cellules
immunitaires oculaires avec les autres structures immunologiques de l’organisme. La
conjonctive profonde contient, comme l’épithélium, des cellules dendritiques présentatrices
d’antigènes.
Pourtant peu nombreux dans le stroma dans les conditions physiologiques, les
macrophages se manifestent surtout en cas d’agression de la surface oculaire. Ils agissent ainsi
en tant que cellules présentatrices d’antigène pendant le développement des conjonctivites
allergiques (49). Ils sont également retrouvés en grande quantité dans la conjonctive des
patients qui présentent une kératoconjonctivite vernale, une forme grave d’allergie oculaire
(70). Dans les atteintes infectieuses de la surface oculaire, les macrophages jouent un rôle de
défense en surexprimant le toll like receptor (TLR)-4 qui se lie au lipopolysaccharide (LPS)
constituant des membranes bactériennes (48). Ils ont aussi un rôle dans la cicatrisation
conjonctivale, augmentant la production locale de facteurs de croissance endothéliaux,
fibroblastiques et matriciels dans le trachome (1). Dans la pemphigoïde oculaire cicatricielle,
une accumulation de macrophages est également retrouvée associée à une concentration
locale élevée de leurs facteurs de croissance (72). De même, lors de l’utilisation de collyres
antiglaucomateux et de leurs conservateurs au long cours, une infiltration de cellules
inflammatoires (macrophages, cellules dendritiques et lymphocytes) est observée dans la
conjonctive (17, 63, 80).
2. GLAUCOME ET CONSERVATEURS
2.1 Généralités sur le glaucome
Le glaucome est une des causes majeures de cécité et un problème de santé publique
au niveau mondial. Le glaucome représente en réalité un groupe de neuropathies optiques
caractérisées par une dégénérescence progressive des cellules ganglionnaires par apoptose
(92). L’apoptose des cellules ganglionnaires de la rétine entraîne une excavation
caractéristique de la tête du nerf optique et une atteinte du champ visuel. L’élévation de la
pression intraoculaire (PIO) est le principal facteur de risque de cette neuropathie optique, liée
à une augmentation des résistances à l’élimination de l’humeur aqueuse (HA) de la chambre

25
antérieure de l’œil (60).
En fonction de l’examen clinique ophtalmologique, de l’anatomie de l’angle
iridocornéen, de l’aspect du nerf optique et de l’existence d’une atteinte du champ visuel de
type glaucomateuse, les glaucomes sont classés en (39) :
Glaucomes congénitaux
Glaucomes à angle ouvert :
o Glaucomes primitifs à angle ouvert et l’hypertonie oculaire (HTO),
o Glaucomes secondaires à angle ouvert.
Glaucome par fermeture de l’angle iridocornéen :
o Glaucomes primitifs à angle fermé,
o Glaucomes secondaires à angle fermé.
2.2 Traitements du glaucome
2.2.1 Principes
L’objectif du traitement du glaucome est de ralentir la progression de la neuropathie
optique afin de maintenir la fonction visuelle et la qualité de vie des patients à un coût
raisonnable. Le seul traitement aujourd’hui disponible et cliniquement efficace pour prévenir
le développement d’un glaucome chez un patient présentant une HTO, et pour ralentir
l’évolution du glaucome, consiste à faire baisser la PIO afin d’obtenir une PIO dite « cible »
(39). Cette PIO cible correspond à la PIO moyenne obtenue sous traitement permettant de
ralentir la progression ou de prévenir l’apparition des déficits glaucomateux. Le choix du
traitement du glaucome ou de l’HTO est fait en fonction du patient, de son glaucome et des
bénéfices et risques attendus de la thérapeutique choisie. L’arsenal thérapeutique pour
diminuer la PIO comporte actuellement des traitements médicaux avec des collyres
antiglaucomateux, des traitements laser (trabéculoplastie sélective au laser) et des traitements
chirurgicaux : chirurgie filtrante (sclérectomie profonde non perforante et trabéculectomie) et
la pose de valve. Parmi cet arsenal thérapeutique, les collyres antiglaucomateux et la chirurgie
filtrante ont des conséquences sur la conjonctive et seront donc développés.

26
2.2.2 Les collyres antiglaucomateux
Selon la pharmacopée française, un collyre est une solution ou une suspension stérile,
aqueuse ou huileuse, contenant une ou plusieurs substances médicamenteuses destinées à
l’instillation oculaire. Il existe de très nombreux collyres antiglaucomateux et le choix du
traitement doit tenir compte, au-delà de la baisse de la PIO, de la tolérance, de l’observance et
du coût du traitement. Les molécules utilisées dans le traitement du glaucome ont toutes pour
effet d’abaisser la PIO. Le traitement médical du glaucome ou de l’HTO commence le plus
souvent avec une monothérapie. Il y a actuellement 5 différentes classes de médicaments
hypotonisants oculaires : les antagonistes adrénergiques ou β-bloquants, les agonistes
adrénergiques α-2 sélectifs et non sélectifs, les dérivés de la prostaglandine et les prostamides,
les inhibiteurs de l’anhydrase carbonique et les parasympathomimétiques (cholinergiques)
(39). Ces traitements diminuent la PIO par diminution de la production d’HA (les β-
bloquants, les inhibiteurs de l’anhydrase carbonique, les agonistes adrénergiques non sélectifs
et les agonistes α-2 adrénergiques), ou par augmentation de l’évacuation de l’HA par la voie
trabéculaire (parasympathomimétiques) ou par la voie uvéosclérale (les agonistes
adrénergiques non sélectifs, les agonistes adrénergiques α-2 sélectifs, les analogues des
prostaglandines et les prostamides).
Les β-bloquants sont utilisés depuis de nombreuses années en première intention dans le
traitement du glaucome ou de l’HTO car ils sont bien tolérés, efficaces et peu onéreux (76).
Ensuite, les analogues des prostaglandines et les prostamides ont été approuvés comme
traitement de première intention, et sont de plus en plus utilisés comme premier traitement de
l’HTO ou du glaucome. En cas d’efficacité insuffisante, différentes classes thérapeutiques
peuvent être associées et il existe maintenant de nombreuses combinaisons fixes de
traitements. Si plus de deux ou trois molécules sont nécessaires pour contrôler la maladie,
d’autres thérapeutiques comme le laser ou la chirurgie filtrante sont alors discutées.

27
2.2.3 La chirurgie filtrante et la fibrose conjonctivale
La trabéculectomie et la sclérectomie profonde non perforante sont les interventions les
plus fréquentes pratiquées en France. La chirurgie filtrante augmente la conductance de
l’écoulement de l’humeur aqueuse entre le site de production et les voies de drainages
extraoculaires, afin de diminuer durablement la PIO. La principale problématique de la
chirurgie filtrante est la cicatrisation conjonctivale. L’excès de cicatrisation est la principale
cause d’échec du bon contrôle de la PIO (73). Il est ainsi indispensable que la cicatrisation
conjonctivale et sclérale au cours de la période postopératoire soient parfaitement contrôlées.
Des agents antimitotiques, le 5-fluorouracile et la mitomycine C, sont utilisés pour contrôler
l’inflammation, la prolifération fibroblastique et la production excessive de MEC afin
d’améliorer le taux de succès de la chirurgie filtrante (52). Les modifications tissulaires
provoquées par l’utilisation au long cours de collyres antiglaucomateux contenant des
conservateurs et entrainant une stimulation de la fibrose conjonctivale du site de filtration sont
une des principales raisons des échecs de la chirurgie filtrante (18).
2.3 Les conservateurs
2.3.1 Historique
Le rôle des conservateurs est d’éviter et de limiter la contamination bactérienne des
flacons contenant les collyres. Ils peuvent être aussi employés pour stabiliser le principe actif
des flacons multidoses. La contamination peut être multiple, par contact de l’embout du
flacon sur les paupières, les cils, la conjonctive ou les larmes. Sans conservateur, le contenu
d’un flacon multidose utilisé deux fois par jour est habituellement contaminé en 1 à 2
semaines (77). Le risque est multiplié et la transmission bactérienne croisée lors de
l’utilisation d’un même flacon par plusieurs personnes d’une même famille ou en milieu
hospitalier. Dans les années soixante, devant la multiplication de transmissions bactériennes
croisées, l’industrie pharmaceutique a été poussée à développer des préparations en unidose et
de limiter la durée d’utilisation d’un flacon après son ouverture (22). La prépondérance des

28
flacons multidoses engendra le développement des conservateurs dès les années soixantes,
même si les recommandations n’ont exigé l’addition d’un agent antimicrobien que vers le
milieu des années soixante-dix. L’efficacité des conservateurs est basée sur un test
d’efficacité, qui consiste en l’inoculation de 1x106
CFU (colon- forming unit) par ml d’un
mélange de bactéries et de champignons. Il doit être observé une réduction de la concentration
bactérienne de 1 Log en une semaine et de 3 Log en deux semaines, sans prolifération des
survivants entre deux et quatre semaines, ainsi que l’absence de prolifération des
champignons entre zéro et quatre semaines (75). De cette règlementation découle deux
conséquences, la durée d’utilisation des flacons est garantie jusqu’à 1 mois et de par leur
action non spécifique, les conservateurs sont toxiques sur les tissus employés. C’est ainsi que
l’EMA (European Medicines Agency) a publié un rapport afin d’éviter l’utilisation des
conservateurs chez les patients intolérants et ceux traités au long cours, et d’utiliser la
concentration minimale efficace des conservateurs (35).
2.3.2 Principaux conservateurs
Il existe un grand nombre de conservateurs utilisés en association avec un ou plusieurs
principes actifs dans les collyres. On les distingue habituellement par leur mode d’action:
chimique, antioxydant ou complexant afin de renforcer l’action d’autres conservateurs.
Les conservateurs chimiques. Les plus utilisés, ils altèrent les membranes cellulaires et
ainsi provoquent une lyse cellulaire :
Les ammoniums quaternaires: Ce sont des agents surfactants dont les
principaux représentants sont le chlorure de benzalkonium (BAK) et le
polyquaternium. Ce sont des composés bipolaires, très hydrosolubles qui présentent
des propriétés tensioactives. Ils agissent principalement par le biais de leurs propriétés
détergentes en dissolvant les parois bactériennes et les membranes cellulaires,
détruisant la couche cytoplasmique semi-perméable. Leur activité est maximale à
37°C en milieu alcalin. Excellents fongicides, leur spectre d’activité comprend aussi
les bactéries Gram + mais aussi sur les bactéries Gram -, en particulier lorsqu’ils sont
couplés à l’acide éthylène diamine tétraacétique (EDTA). On retrouve le BAK dans
une majorité de collyres à des concentrations comprises entre 0,004% et 0,02%, et le

29
plus couramment à 0,01%. Ils sont aussi employés en tant que spermicide et dans de
nombreuses préparations à visée extraoculaire (savon, cosmétiques, produits
nettoyants et désinfectants). D’autres ammoniums quaternaires sont également utilisés
comme: le bromure de benzododécinium, le bromure de cétrimonium ou encore le
chlorure de cétylpyridinium.
Les dérivés mercuriels doivent leur efficacité aux propriétés thioloprives de
l’ion mercuriel. Le thiomersal est le principal représentant de ces dérivés, mais ce type
de conservateur est de moins en moins utilisé en raison d'une fréquence élevée
d'allergies et surtout des problèmes écologiques liés à l'industrie du mercure.
Les alcools comme le chlorobutanol, le phényléthanol, le phénoxyéthanol.
Les parabens ou esters de l’acide parahydroxybenzoïque comme le
parahydroxybenzoate de méthyle, le méthylparaben et le propylparaben.
Les amidines comme le digluconate de chlorhexidine ou le
polyhexaméthylène biguanide peuvent également être utilisés comme conservateurs
dans certains collyres.
Les conservateurs oxydants. Ils pénètrent dans la cellule et interférent avec les
fonctions cellulaires vitales des microorganismes en modifiant les lipides, les protéines ou
l’ADN. On distingue le perborate de sodium (NaBO3), le chlorite de sodium (NaClO2), le
complexe oxychloropurite stabilisé ou encore le stabilized oxyborate complex.
Les conservateurs antioxydants et autres conservateurs. La présence de conservateurs
antioxydants est utile lorsque la formulation contient des corps gras saturés ou des actifs
éminemment oxydables. L’EDTA est généralement utilisé pour renforcer le pouvoir
conservateur des ammoniums quaternaires ou du thiomersal. On peut également citer le
sorbate (acide sorbique) et le Sodium silver chloride complex (complexe à base de chlorure de
sodium d'argent et de sulfate d'argent).
2.3.3 Le chlorure de benzalkonium (BAK)
Le BAK est le conservateur le plus fréquemment retrouvé dans les collyres utilisés
pour le traitement du glaucome. Il possède trois grandes fonctions, comme biocide, comme
surfactant cationique et comme agent de transfert de phase dans l’industrie chimique.
Mélange de chlorure d’alkylbenzyldimethylammonium, sa formule chimique est la

30
suivante : C6H5CH2N(CH3)2RCl, R représentant les radicaux alkyles allant de C8 à C18. Le
BAK a déjà démontré son importante toxicité sur une variété d’organes et de systèmes, selon
un mode temps et dose-dépendant (31).
Figure 5 : formule chimique du chlorure de benzalkonium
Activité bactéricide et fongicide
In vitro, en comparaison avec les autres conservateurs chimiques (paraben,
chlorobutanol) et antioxydants (chlorure d’argent et oxychloropurite stabilisé) le BAK associé
à l’EDTA a été le seul conservateur répondant aux critères d’efficacité antimicrobienne. Il a
de plus une action d’inhibition de la prolifération du Staphylococcus aureus (20). Par contre,
certaines souches de Pseudomonas aeruginosa ont développé une résistance acquise au BAK,
éliminée par l’adjonction d’EDTA (66).
In vivo, l’efficacité du BAK pourrait être moindre, la contamination des flacons
conservés par le BAK après 15 jours d’utilisation répétée a atteint 34,8% (83). Le taux de
contamination est sans doute lié à la stabilité de la molécule qui décroit avec le temps.
BAK et pénétration oculaire
Les effets du BAK sur les membranes cellulaires ont été un argument expliquant
l’augmentation de la pénétration de la molécule active des collyres contenant ce conservateur.
La pénétration transcornéenne de l’aciclovir et de différents antibiotiques s’est révélée
augmentée en présence de BAK (58). La pénétration de la cyclosporine A a également été
améliorée lors de l’utilisation d’un véhicule contenant du BAK (94). Néanmoins, les études
sont controversées, Pellinen et Lokkila ont ainsi montré l’absence de différence de
concentration du principe actif dans l’humeur aqueuse lors de l’utilisation d’une forme
conservée et non conservée du tafluprost (66).

31
La majeure partie du BAK est absorbée par les couches superficielles mais un
certain pourcentage parvient à pénétrer les couches plus profondes qui ne bénéficient pas d’un
lavage par le film lacrymal. Le BAK pourrait ainsi s’infiltrer dans une moindre mesure dans
des tissus encore plus profond et passer dans la circulation générale par drainage veino-
lymphatique.
BAK et contrôle de la PIO
La possibilité d’un meilleur contrôle de la PIO avec un collyre conservé a été
écartée par de nombreuses études et pour la plupart des collyres. Les études sur les
bêtabloquants, puis avec les analogues des prostaglandines (45) et enfin avec l’association
bêtabloquant-inhibiteur de l’anhydrase carbonique (53) ont toutes montré l’absence de
supériorité entre la forme conservée et non conservée sur la baisse de la PIO. Seule
l’efficacité des prostamides serait influencée par la présence du BAK (40).
3. LIGNEES CELLULAIRES
3.1 Généralités
Les lignées cellulaires sont utilisées depuis plus d’un siècle. En 1907, Ross Harrison a
pour la première fois observé la croissance de neuroblastes en culture à partir d’embryons de
grenouille (62). Développée ensuite avec les cellules cancéreuses, les premières cellules mises
en culture ont été isolées par George Gey. Les cellules provenaient d’un cancer du col utérin
de Henrietta Lacks, qui a donné son nom à la lignée cellulaire, les cellules HeLa, encore
utilisées aujourd’hui (61).
Par définition, une lignée cellulaire est une population homogène de cellules qui ont
une capacité illimitée de division en culture tout en gardant les caractéristiques de leur cellule
d’origine. C’est la capacité théorique de division illimitée des cellules cancéreuses qui a
permis leur culture initiale. Actuellement, les lignées cellulaires peuvent être créées en
modifiant les gènes impliqués dans la régulation du cycle cellulaire ou en intégrant un
oncogène. Elles ont de nombreuses applications en recherche biomédicale pour évaluer les
effets de l’environnement ou d’une molécule sur la lignée employée. Elles peuvent aussi être

32
utilisées dans l’industrie pharmaceutique pour la production de molécules à visée
thérapeutique.
Grâce à la possibilité de stockage quasi-infinie dans l’azote liquide, les lignées
cellulaires sont une source de cellules illimitée. Seulement, l’utilisation de lignées cellulaires
n’est pas dénuée d’inconvénients. Les cellules peuvent posséder des caractéristiques de
cellules cancéreuses, limitant ainsi l’interprétation des résultats. La contamination des lignées
cellulaires, par des agents pathogènes ou d’autres lignées cellulaires n’est pas rare, les
techniques de culture doivent donc être parfaitement maîtrisées. En opposition aux cultures
primaires, nécessitant des moyens lourds et onéreux, leur utilisation est largement étendue et
validée dans le domaine de la recherche. De plus, techniquement, les lignées cellulaires ont
l’avantage d’avoir une origine génétique homogène, limitant les variations de phénotypes
liées aux sources des cellules primaires.
3.2 La lignée THP-1
3.2.1 Historique
Mise au point en 1980 (89), la lignée THP-1 provient de cellules d’une leucémie aigue
monocytaire d’un jeune garçon. Les premières études ont décrit cette lignée ayant des
caractères morphologiques de monocytes. L’aspect microscopique montre des cellules
arrondies indifférenciées (figure 6). La lignée exprime en surface les récepteurs au fragment
constant (Fc) des immunoglobulines et au complément (C3b), son haplotype est composé de
HLA-A2, -A9, -B5, -DRW1 et -DRW2. Le caractère monocytaire de la lignée a été confirmé
par l’activité d’estérase d’alpha-naphthyl butyrate, la production de lysozyme, la phagocytose
de particules de latex et la capacité à restaurer la réponse des lymphocytes à la concanavaline
A.

33
Figure 6 : Image en microscopie optique des cellules THP-1, arrondies, non confluentes et
non adhérentes.
3.2.2 THP-1 en tant que lignée macrophagique
Plus tard, Auwerx a décrit la différenciation de la lignée THP-1 en macrophages sous
l’influence d’esters de phorbol (6). Les cellules différenciées adhèrent à la plaque de culture et
acquièrent un phénotype de macrophages avec des changements morphologiques, une forme
aplatie et possédant des pseudopodes, un appareil de Golgi et un réticulum endoplasmique
bien développés, avec de nombreux ribosomes libres dans le cytoplasme. Une fois
différenciées, les cellules surexpriment le CD11b et le CD14, diminuent leur prolifération et
augmentent leur taux de phagocytose. En comparaison avec les principales lignées myélo-
monocytaires (71), les cellules dérivées de la lignée THP-1 sont les plus proches des
macrophages dérivés de monocytes sanguins. La lignée THP-1 a ainsi été largement utilisée
depuis comme modèle d’étude in vitro des mécanismes impliquant les macrophages. Plus
spécifiquement, les cellules THP-1 adhérentes au fond des plaques de culture ont montré des
caractéristiques de macrophages activés (87). Elles possèdent des propriétés morphologiques

34
et fonctionnelles différentes. Avec une activité de phagocytose augmentée, la surexpression
d’HLA-DR et la stimulation d’activité lymphocytaire, la lignée THP-1 confirme son potentiel
d’étude de la lignée macrophagique.
La lignée THP-1 a fait l’objet d’études afin de valider ses propriétés monocytaires et
macrophagiques avec les techniques actuelles. Les interactions cellulaires entre les cellules
THP-1 et les cellules vasculaires et immunitaires se sont révélées concordantes avec les
interactions des monocytes/macrophages dérivés du sang. De plus, l’étude des modifications
du transcriptome, du profil ARNm et des histones a permis de définir les mécanismes de
régulation des cellules THP-1 en réponse à différents médiateurs inflammatoires (71). Les
mécanismes mis en évidence ont confirmé le parallèle entre les THP-1 et les
monocytes/macrophages dérivés du sang.
3.3 Les cellules épithéliales Wong Kilbourne dérivées de Chang (WKD)
La lignée WKD a fait l’objet de nombreuses études dans le domaine de la surface
oculaire et en particulier en toxicologie. La lignée WKD a montré de nombreuses
caractéristiques communes avec les cellules épithéliales conjonctivales, par exemple la
formation de desmosomes, le développement de microvillosités, la présence de récepteur à
l’EGF (epidermal growth factor), l’expression du fragment inducteur d’apoptose (Fas) ou
CD95 et l’absence d’expression basale d’HLA-DR (28). En microscopie optique, les cellules
sont adhérentes au fond de la plaque, étalées avec une tendance à la confluence (figure 7). Les
cellules WKD sont aussi un modèle validé pour l’étude de la production de cytokines, de
facteurs de croissance et de leurs récepteurs, ainsi que dans l’étude des phénomènes
inflammatoires et allergiques.

35
Figure 7 : Image en microscopie optique des cellules WKD, cellules aplaties,
confluentes adhérentes au fond de la plaque de culture.
L’utilisation des cellules WKD a été mise en cause par la découverte d’une
contamination par les cellules de la lignée HeLa (54). La contamination s’est uniquement
manifestée au niveau du caryotype et aucune démonstration de la ressemblance entre les deux
lignées n’a été rapportée. En comparaison avec une autre lignée conjonctivale immortalisée,
non connue pour être contaminée, la lignée IOBA-NHC, les effets d’ordre toxicologique et en
particulier ceux du BAK se sont révélés similaires (16), validant de ce fait l’utilisation de la
lignée WKD.
A travers l’étude de la morphologie, du cycle cellulaire, de l’expression des mucines,
de la réponse à la stimulation au TNF-α et à l’IFN-γ, de leur potentiel présentateur d’antigène
et de la susceptibilité à l’apoptose, les cellules WKD ont été comparées aux cellules
conjonctivales primaires (28). Les résultats entre les deux types cellulaires ont été variables
sur certains points, les cellules WKD ayant une morphologie plus proche des cellules
mésenchymateuses avec un phénotype plus dédifférencié que les cellules conjonctivales
primaires. Les réactions inflammatoires ont été plus importantes dans la lignée primaire, ce
qui laisserait penser que les réactions de la lignée WKD sont amoindries. Ainsi, il faut tenir
compte du fait que les cellules WKD ne possèdent pas toutes les caractéristiques des cellules
épithéliales conjonctivales primaires dans l’interprétation des résultats. Cependant, cette
lignée reste un bon modèle d’étude en particulier dans les interactions cellulaires avec les
lymphocytes (21).

36
CHAPITRE 2 : ETUDE IN VITRO DES EFFETS DU CHLORURE DE
BENZALKONIUM SUR LES MACROPHAGES.
1. INTRODUCTION
L’inflammation et les cellules inflammatoires sont au centre des mécanismes
physiopathologiques impliqués dans les pathologies de la surface oculaire (33). Les réactions
des cellules inflammatoires sont un paramètre clé dans les nombreuses voies de signalisation
et les mécanismes immunitaires de l’inflammation. Les macrophages sont des cellules
phagocytaires mononuclées, jouant un rôle de pivot dans le maintien de l’homéostasie des
tissus dans les conditions physiologiques. Ils servent de sentinelles en éliminant sans réaction
inflammatoire les cellules mortes ou en apoptose et en facilitant la réparation des tissus
endommagés (41). Les macrophages partagent avec les cellules dendritiques le rôle de
cellules présentatrices d’antigène (41). Les macrophages tissulaires produisent des médiateurs
inflammatoires et présentent une hétérogénéité phénotypique selon leur environnement. Bien
que les macrophages conjonctivaux soient localisés dans la substantia propria, sous-jacente à
l’épithélium conjonctival, ils interagissent étroitement avec les cellules épithéliales
conjonctivales et jouent un rôle important dans les pathologies de la surface oculaire. En cas
de conjonctivite allergique, la conjonctive est infiltrée de façon excessive par les macrophages
(49). Les macrophages jouent aussi un rôle dans le trachome en augmentant la sécrétion locale
de facteurs de croissance endothéliaux, fibroblastiques et matriciels (1, 70). De façon
similaire, dans la pemphigoïde cicatricielle oculaire la substantia propria conjonctivale est
infiltrée de façon significative par les macrophages (85).
A cause de l’inflammation infraclinique provoquée par l’utilisation au long cours des
collyres antiglaucomateux, les patients traités pour un glaucome ou HTO souffrent
fréquemment d’une atteinte de la surface oculaire (9). Les conservateurs associés au
composant actif des collyres ont déjà montré leurs effets toxiques dans des études cliniques et
expérimentales, et pourraient être en partie responsables de ces changements tissulaires
inflammatoires (10). Le chlorure de benzalkonium, un ammonium quaternaire, est le
conservateur le plus communément utilisé dans les traitements antiglaucomateux (10). Le
BAK est directement responsable de la perte des cellules à mucus ainsi que de la métaplasie et
l’apoptose des cellules épithéliales conjonctivales, et d’une rupture de la barrière épithéliale
cornéenne (5). Il possède une cytotoxicité dose-dépendante aboutissant à l’apoptose et la
nécrose des cellules épithéliales conjonctivales
(29). Chez les patients traités pour le

37
glaucome, une association entre la sévérité des pathologies de la surface oculaire et le nombre
de collyres contenant du BAK a été établie (10). De même, Malvitte et al. ont montré que
l’expression de marqueurs fibroblastiques et inflammatoires, et l’intensité des réactions
inflammatoires étaient liées à la durée d’exposition et au nombre de collyres contenant des
conservateurs utilisés (59). De nombreuses cellules inflammatoires et en particulier de
macrophages ont été observées dans la substantia propria de la conjonctive après utilisation
au long cours de collyres antiglaucomateux contenant des conservateurs (17, 63, 80). Ces
modifications tissulaires responsables d’une stimulation de la fibrose conjonctivale du site de
filtration en post-opératoire pourraient être la raison principale d’échec de la chirurgie
filtrante du glaucome (18). En comparaison avec les patients traités avec des collyres sans
conservateur, les biopsies conjonctivales de patients traités avec des collyres conservés sont
infiltrées de façon anormale par des macrophages (11). De même, dans un modèle animal
exposé au timolol contenant du BAK, les biopsies conjonctivales présentaient une densité
augmentée de macrophages en comparaison avec les animaux ayant reçu du timolol sans
conservateur (11).
En considérant le rôle essentiel des macrophages dans les pathologies de la surface
oculaire et en particulier chez les patients traités pour glaucome ou une HTO, l’objectif de ce
premier travail a été de caractériser les effets du BAK sur les macrophages in vitro.
2. MATERIELS ET METHODES
2.1 Lignées cellulaires
2.1.1 Lignée monocytaire
Les cellules THP-1 ont été fournies par l’American Type Culture Collection (ATCC,
Rockville, MD). Cette lignée est issue des cellules d’un patient présentant une leucémie aiguë
monocytaire. Ces cellules sont connues pour se différencier en macrophages sous l’effet du
phorbol myristate acétate (PMA) (6). Les cellules ont été cultivées dans un milieu RPMI 1640

38
(Invitrogen life technologies Grand Island, NE) enrichi avec 10% de sérum de veau fœtal
(FBS; HyClone, Logan, USA), 1% de glutamine (Invitrogen), 50 UI/ml de pénicilline et 50
UI/ml de streptomycine (BioSource International, Camarillo, CA). Les conditions de culture
étaient standard en incubateur à 37°C avec 5% de CO2. Afin d’obtenir des cellules
différenciées, les cellules THP-1 ont été ensemencées en plaque 6 puits à 106 cellules/ml et
stimulées avec du PMA (Sigma-Aldrich St Louis, MO) pendant 72 heures à la concentration
de 100nM (6).
2.1.2 Lignée épithéliale conjonctivale
Les cellules conjonctivales Wong-Kilbourne dérivées d’une conjonctive humaine de
Chang (WKD) (clone 1 à 5c-4l, ATCC) ont été cultivées en conditions standard (atmosphère
humidifiée, 5% de CO2 et 37°C) dans un milieu DMEM (Invitrogen) enrichi avec 10% de
sérum de veau fœtal (HyClone), 1% de glutamine (Invitrogen), 50 UI/ml de pénicilline et 50
UI/ml de streptomycine (BioSource International).
2.2 Stimulations
Les cellules THP-1 ont été exposées pendant 72 heures au PMA (différenciation), puis
après renouvellement du milieu de culture, les cellules ont été exposées pendant 24 heures à
du chlorure de benzalkonium (BAK) (Sigma-Aldrich) pour les phénomènes toxiques et
iatrogéniques, du dinitrochlorobenzène (DNCB) (Sigma-Aldrich) en temps qu’allergène de
contact, du lipopolysaccharide (LPS) (Sigma-Aldrich) pour mimer une stimulation
infectieuse, du TNF-α (tumor necrosis factor) (PeproTech, Rocky Hill, NJ) pour mimer une
réaction inflammatoire ou du tampon phosphate isotonique (PBS) comme témoin. Les
surnageants ont été recueillis et stockés à -20°C. Ils ont été recueillis à l’issue de 24 heures
d’exposition aux différents facteurs de stimulation.
Le BAK a été dilué dans du PBS à quatre concentrations : 5.10-4
%, 10-4
%, 5.10-5
% et
10-5
% (3). Après 24 heures d’exposition, les concentrations plus fortes (10-3
% et au-delà) se
sont révélées extrêmement toxiques, empêchant une analyse convenable (données non

39
présentées). Le DNCB a été dilué dans du PBS à quatre concentrations : 10nM, 25nM, 50nM
et 100nM. Le LPS d’Escherichia Coli de sérotype 0111 a été dilué dans du PBS selon une
gamme de quatre concentrations 10ng/ml, 50ng/ml, 100ng/ml et 250ng/ml. Enfin, le TNF-α a
également été dilué en PBS à quatre concentrations : 1ng/ml, 2ng/ml, 5ng/ml et 10ng/ml.
2.3 Expression des marqueurs de surface
Pour déterminer le phénotype des cellules, l’expression de marqueurs membranaires a
été quantifiée en cytométrie en flux (CMF) (Beckman Coulter, Miami, FL). Les cellules
étaient recueillies au grattoir pour les cellules THP-1 et après une incubation de 10 minutes en
EDTA 0,5mM (Sigma-Aldrich) pour les cellules WKD. Les cellules ont été ensuite lavées
puis fixées dans une solution de paraformaldéhyde (PFA) (Alfa Aesar, Ward Hill, MA) à
0,5% pendant 24 heures à 4°C avant d’être marquées. Les anticorps monoclonaux utilisés ont
été : CD11b couplé au fluorescéine isothyocyanate (FITC), CD11c couplé à la phycoérythrine
(PE), CD86 couplé FITC, CD54 couplé FITC, CD33 couplé PE, (BD Biosciences, San Jose,
CA) ainsi qu’un anticorps isotypique de contrôle (IgG1 de souris) (Pharmingen). Chaque
suspension cellulaire correspondant à une condition a été incubée en présence des anticorps
primaires fluorescents pendant 35 minutes ou à l’aide d’un anticorps secondaire de chèvre
anti-souris conjugué au FITC pendant une autre incubation de 35 minutes. Après chaque
incubation les cellules ont été lavées en PBS puis analysées en CMF. Pour les cellules THP-1,
l’expression des marqueurs en CMF a été quantifiée avant et après la différenciation et après
exposition pendant 24 heures aux différents facteurs stimulants.
2.4 Immunohistochimie
Afin de visualiser l’expression des marqueurs de différenciation, les cellules THP-1
ont été mises en culture sur des lames compartimentées (Lab-TekTM
Chamber Slide, NUNC
A/S, Roskilde, Danemark) à raison de 500ml de suspension cellulaire à 106cellules/ml par
compartiment. Après exposition aux différents facteurs, les lames ont été fixées pendant 15
minutes en PFA à 4%. Les lames ont ensuite été lavées trois fois pendant 5 minutes avec du

40
PBS. Puis pendant 1 heure et à température ambiante, une moitié des compartiments a été
incubée avec le CD11c couplé à la PE (Pharmingen) dilué au 1/100ème
en PBS, l’autre moitié
a été incubée avec une solution contenant le contrôle isotypique IgG1 couplé à la PE. Après
trois lavages en PBS, les compartiments ont été démontés et les lames ont été ensuite
recouvertes de lamelles avec un milieu de montage contenant du di-amidino-phénylindole
(DAPI) (Vectashield-DAPI ; Vector Laboratories, Burlingame, USA). Les lames ont été
photographiées sous microscope (DM 6000, Leica Microsystems GmbH, Wetzlar,
Allemagne).
2.5 Etude de la mort cellulaire en CMF à l’aide du test Annexine V / 7-AAD
Ce test comprend deux marquages : le marquage annexine V et l’analyse du contenu
ADN en 7-AAD (7-aminoactinomycin D). L’appareil est équipé d’un laser Argon émettant à
488nm. L’annexine V se fixe spécifiquement aux phosphatidylsérines qui sont exprimées à la
paroi externe de la membrane plasmique dès les phases précoces de l’apoptose. Le 7-AAD est
une sonde fluorescente qui s’intercale aux acides nucléiques des cellules en nécrose ou en
apoptose tardive (91).
Après différenciation pendant 72 heures et exposition aux facteurs de stimulation
pendant les 24 heures suivantes, les cellules ont été recueillies avec un grattoir. L’analyse a
été effectuée à l’aide du kit ANNEXIN V-FITC / 7-AAD (Beckman Coulter) sans fixation des
cellules et selon les recommandations du fabricant. Un histogramme biparamétrique a été
utilisé pour déterminer les marquages en annexine V et 7-AAD. Quatre populations ont été
identifiées, négatives aux deux marqueurs (cellules viables), positives uniquement à
l’annexine V (apoptose précoce), positives à l’annexine V et au 7-AAD (apoptose tardive) et
positives uniquement au 7-AAD (nécrose).
2.6 Etude de la phagocytose
Pour étudier la fonction de phagocytose, les cellules THP-1 ont été ensemencées sur
lames compartimentées (Lab-Tek) et sur des plaques 6 puits. Elles ont été ensuite incubées
pendant 1 heure avec 100µl d’une solution contenant des billes de latex fluorescentes

41
(Invitrogen) à 50µg/ml préalablement incubées pendant une heure dans une solution de PBS
contenant 10% de sérum de veau fœtal pour entraîner une reconnaissance antigénique. Les
cellules cultivées sur lames compartimentées ont été fixées et montées avec lamelle sans
utilisation d’anticorps. Ces lames ont été analysées au microscope avec détection de la
longueur d’onde des microsphères fluorescentes. Les cellules cultivées sur plaque 6 puits ont
été recueillies et fixées (PFA à 0,5% pendant 24 heures) pour analyse en CMF. Le cytomètre
a été calibré pour la détection de la longueur d’onde des microsphères fluorescentes selon un
histogramme monoparamétrique.
2.7 Production de cytokines
L’analyse des cytokines produites a été effectuée sur les surnageants des cellules THP-
1 dans les six conditions différentes précédemment décrites : non différenciées, différenciées
et exposées aux différents facteurs de stimulation : BAK (10-5
%), DNCB (10nM), LPS
(50ng/ml), TNF-α (5ng/ml) ou PBS (1X). Les surnageants correspondaient à la production
cellulaire sur les dernières 24 heures, le milieu étant changé après les 72 heures d’exposition
au PMA. Les surnageants ont été analysés à l’aide d’un kit permettant d’identifier 38
cytokines différentes (Proteome profiler array, ARY005, R&D Systems, Minneapolis, USA).
Les expérimentations ont été effectuées selon les consignes du fournisseur. Les six conditions
d’évaluation représentaient chacune une membrane de nitrocellulose révélée sur film
radiographique (Amersham : GE Healthcare Limited, Buckinghamshire, UK) avec utilisation
d’un agent électroluminescent (Amersham : GE Healthcare Limited) selon les
recommandations du fournisseur. La densité en pixel des spots a été analysée par logiciel
informatique (Macbiophotonics ImageJ®).
2.8 Etude de la migration
Pour évaluer la migration des macrophages, les cellules THP-1 ont été cultivées sur
des membranes perméables avec des pores de 8,0 µm (inserts) (FALCON, BD Biosciences)
en suspension dans les puits. Les cellules conjonctivales WKD ont été ensemencées dans une

42
autre plaque et les deux lignées ont été exposées aux mêmes stimulants. Les inserts contenant
les macrophages ont été ensuite cultivés pendant 4 heures dans les puits contenant les cellules
WKD. Les membranes ont été recueillies, fixées au PFA 4% et montées entre lames et
lamelles avec du DAPI. Les lames ont été ensuite photographiées en focalisant sur la face
supérieure et la face inférieure de la membrane. La migration des cellules a été mesurée en
comparant le ratio du nombre de noyaux sur la face inférieure par rapport au nombre de
noyaux sur la face supérieure de la membrane pour chaque condition. Le décompte des
noyaux a été effectué à l’aide du logiciel ImageJ® .
2.9 Analyses statistiques
Dans cette étude, toutes les expérimentations ont été réalisées trois fois. Pour les
comparaisons multiples entre les différentes conditions, le test non paramétrique de Kruskall-
Wallis avec comparaisons multiples par paires et correction de Bonferroni a été utilisé. Pour
les comparaisons simples entre deux conditions, un test t de Student non apparié a été réalisé.
Une valeur de p<0,05 a été considérée comme significative.
3. RESULTATS
3.1 Les cellules THP-1 ont été différenciées en macrophages.
Après 72h en PMA, la morphologie des cellules THP-1 était modifiée en microscopie
optique: Au départ rondes et non adhérentes, elles prenaient un aspect dendritiforme et
adhéraient au fond de la plaque (figure 8).
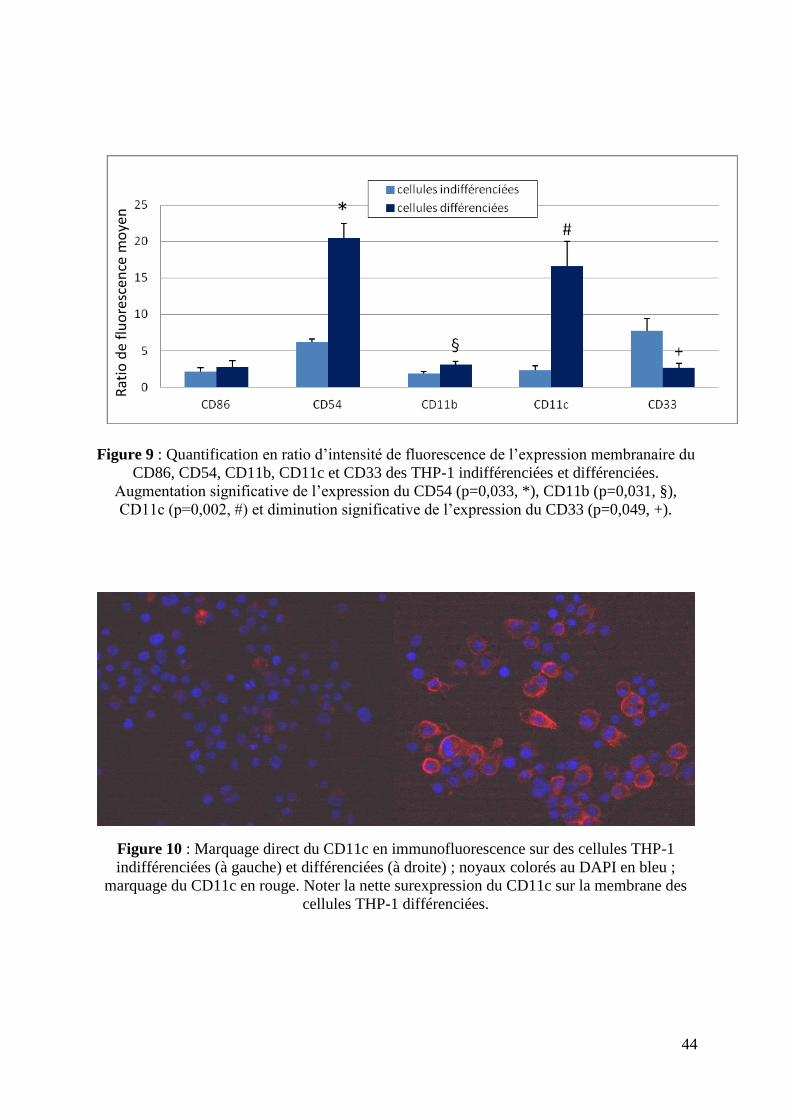
Sur le plan phénotypique, l’expression des marqueurs de différenciation analysée en
CMF a été modifiée après exposition au PMA avec une surexpression du CD11b (p=0,031),
CD11c (p=0,033), CD54 (p=0,002) ainsi qu’une diminution de l’expression du CD33

43
(p=0,049), tant en pourcentage d’expression (données non présentées) qu’en intensité de
fluorescence (figure 9).
En immunohistochimie sur lames, une forte expression membranaire du CD11c a été
observée sur les cellules différenciées en comparaison avec les cellules indifférenciées (figure
10).
Figure 8 : Aspect en microscopie optique (x400) des cellules THP-1 non différenciées (à
gauche) ayant un aspect ovalaire sans adhérence au fond de la plaque) et différenciées (à
droite) avec un aspect dendritique et adhérente au fond de la plaque.
44
Figure 9 : Quantification en ratio d’intensité de fluorescence de l’expression membranaire du
CD86, CD54, CD11b, CD11c et CD33 des THP-1 indifférenciées et différenciées.
Augmentation significative de l’expression du CD54 (p=0,033, *), CD11b (p=0,031, §),
CD11c (p=0,002, #) et diminution significative de l’expression du CD33 (p=0,049, +).
Figure 10 : Marquage direct du CD11c en immunofluorescence sur des cellules THP-1
indifférenciées (à gauche) et différenciées (à droite) ; noyaux colorés au DAPI en bleu ;
marquage du CD11c en rouge. Noter la nette surexpression du CD11c sur la membrane des
cellules THP-1 différenciées.
Rat
io d
e fl
uo
resc
ence
mo
yen
45
3.2 Pour les concentrations utilisées, le BAK et le DNCB entraînaient une modification de
la viabilité cellulaire selon une courbe concentration dépendante.
L’exposition aux faibles concentrations de BAK (de 5.10-5
% à 10-5
%) et de DNCB (de
10nM à 25nM) était comparable au témoin en termes de cytotoxicité (figures 11 et 12). Les
concentrations plus fortes (10-4
% et 5.10-4
% pour le BAK, et 50 et 100nM pour le DNCB) ont
induit une augmentation des pourcentages de cellules en apoptose tardive et en nécrose sans
modifier ceux de l’apoptose précoce. Ceci a permis de déterminer les doses non cytotoxiques :
10-5
% pour le BAK et 10nM pour le DNCB.
Figure 11 : Détermination en CMF des taux d’apoptose et de nécrose des cellules THP-1
différenciées en fonction de la concentration de BAK par marquage annexine V /7-AAD après
24h d’exposition. Le BAK 5.10-4
% induisait une cytotoxicité totale supérieure à 60%.
46
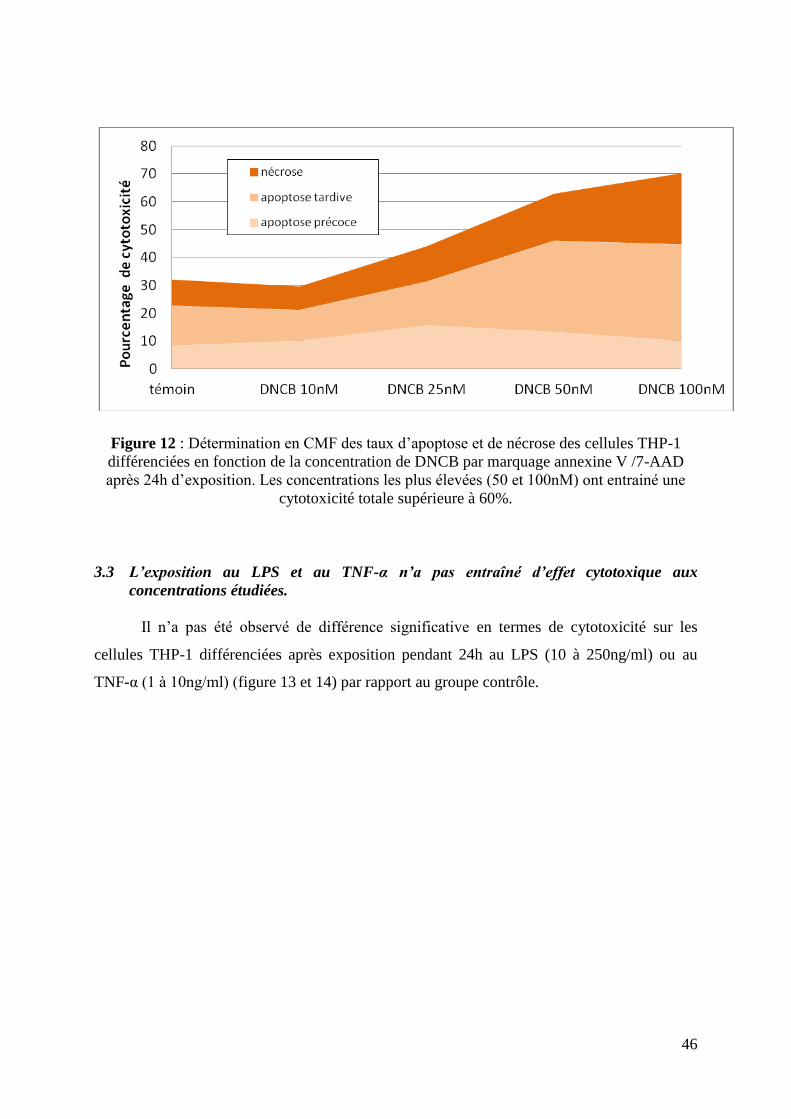
Figure 12 : Détermination en CMF des taux d’apoptose et de nécrose des cellules THP-1
différenciées en fonction de la concentration de DNCB par marquage annexine V /7-AAD
après 24h d’exposition. Les concentrations les plus élevées (50 et 100nM) ont entrainé une
cytotoxicité totale supérieure à 60%.
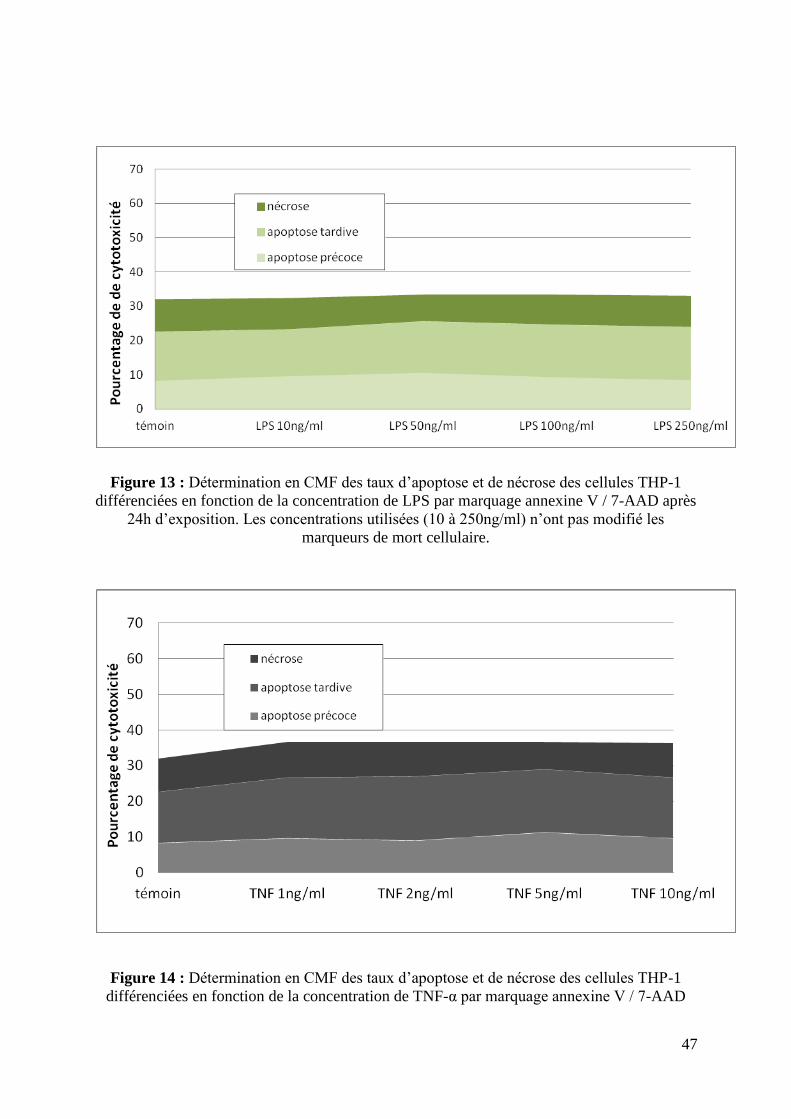
3.3 L’exposition au LPS et au TNF-α n’a pas entraîné d’effet cytotoxique aux
concentrations étudiées.
Il n’a pas été observé de différence significative en termes de cytotoxicité sur les
cellules THP-1 différenciées après exposition pendant 24h au LPS (10 à 250ng/ml) ou au
TNF-α (1 à 10ng/ml) (figure 13 et 14) par rapport au groupe contrôle.
47
Figure 13 : Détermination en CMF des taux d’apoptose et de nécrose des cellules THP-1
différenciées en fonction de la concentration de LPS par marquage annexine V / 7-AAD après
24h d’exposition. Les concentrations utilisées (10 à 250ng/ml) n’ont pas modifié les
marqueurs de mort cellulaire.
Figure 14 : Détermination en CMF des taux d’apoptose et de nécrose des cellules THP-1
différenciées en fonction de la concentration de TNF-α par marquage annexine V / 7-AAD

48
après 24h d’exposition. Les concentrations utilisées n’ont pas modifié les marqueurs de mort
cellulaire.
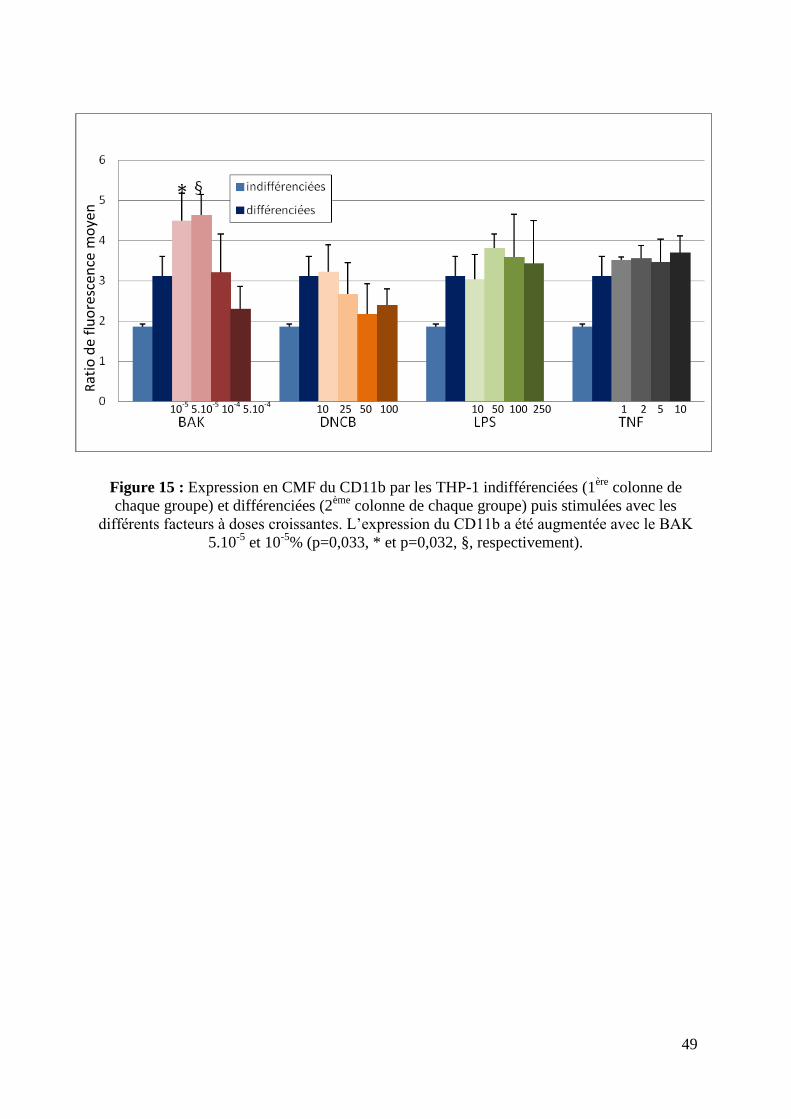
3.4 L’expression des marqueurs de différenciation des cellules THP-1 différenciées a été
modifiée après exposition au BAK, DNCB, LPS et TNF-α.
L’expression membranaire du CD11b par les cellules THP-1 différenciées n’a été
augmentée qu’après exposition au BAK à 10-5
et 5.10-5
% (p=0,033 et p=0,032,
respectivement) par rapport au PBS. Le DNCB, le LPS et le TNF-α n’ont pas modifié
l’expression du CD11b (figure 15).
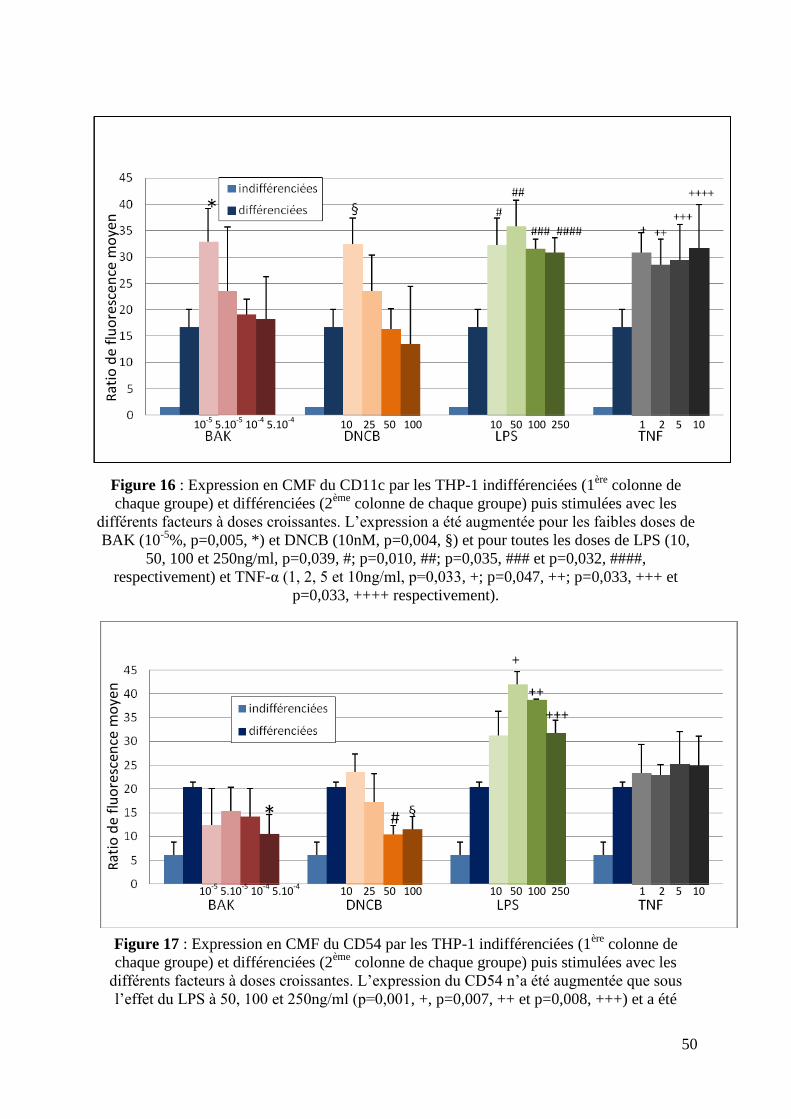
L’expression membranaire du CD11c était augmentée après exposition au LPS (10,
50, 100, et 250ng/ml ; p=0,039, p=0,010, p=0,035 et p=0,032, respectivement) et au TNF-α
(1, 2, 5 et 10ng/ml ; p=0,033, p=0,047, p=0,033 et p= 0,033, respectivement) par rapport au
PBS. L’expression du CD11c n’était augmentée après exposition au BAK ou au DNCB
qu’aux concentrations les plus faibles : 10-5
% pour le BAK (p=0,005) et 10nM pour le DNCB
(p=0,004) (figure 16).
L’expression membranaire du CD54 était augmentée avec le LPS à 50, 100 et
250ng/ml (p=0,001, p=0,007 et p=0,008, respectivement). L’exposition au TNF-α n’a pas
entraîné de modification de l’expression du CD54. Seules les fortes concentrations de BAK
(5.10-4
%) (p=0,009) et de DNCB (50 et 100nM, p=0,012 et p=0,03, respectivement) ont
entraîné une diminution de l’expression du CD54 (figure 17).
L’expression du CD33 n’était pas diminuée après exposition au LPS et au TNF-α aux
concentrations utilisées. Dans le groupe exposé au BAK, l’expression du CD33 était corrélée
à la concentration de BAK, l’expression de CD33 était diminuée aux concentrations faibles de
BAK (10-5
% et 5.10-5
%, p=0,003 et p=0,007, respectivement). Dans le groupe exposé au
DNCB, l’expression était diminuée aux fortes concentrations de DNCB (50 et 100nM)
(p=0,031) (figure 18).
49
Figure 15 : Expression en CMF du CD11b par les THP-1 indifférenciées (1ère
colonne de
chaque groupe) et différenciées (2ème
colonne de chaque groupe) puis stimulées avec les
différents facteurs à doses croissantes. L’expression du CD11b a été augmentée avec le BAK
5.10-5
et 10-5
% (p=0,033, * et p=0,032, §, respectivement).
Rat
io d
e fl
uo
resc
ence
mo
yen
10-5
5.10-5
10-4
5.10-4
10 25 50 100 10 50 100 250 1 2 5 10
50
Figure 16 : Expression en CMF du CD11c par les THP-1 indifférenciées (1ère
colonne de
chaque groupe) et différenciées (2ème
colonne de chaque groupe) puis stimulées avec les
différents facteurs à doses croissantes. L’expression a été augmentée pour les faibles doses de
BAK (10-5
%, p=0,005, *) et DNCB (10nM, p=0,004, §) et pour toutes les doses de LPS (10,
50, 100 et 250ng/ml, p=0,039, #; p=0,010, ##; p=0,035, ### et p=0,032, ####,
respectivement) et TNF-α (1, 2, 5 et 10ng/ml, p=0,033, +; p=0,047, ++; p=0,033, +++ et
p=0,033, ++++ respectivement).
Figure 17 : Expression en CMF du CD54 par les THP-1 indifférenciées (1ère
colonne de
chaque groupe) et différenciées (2ème
colonne de chaque groupe) puis stimulées avec les
différents facteurs à doses croissantes. L’expression du CD54 n’a été augmentée que sous
l’effet du LPS à 50, 100 et 250ng/ml (p=0,001, +, p=0,007, ++ et p=0,008, +++) et a été
Rat
io d
e fl
uo
resc
en
ce m
oye
n
10-5
5.10-5
10-4
5.10-4
10 25 50 100 10 50 100 250 1 2 5 10
Rat
io d
e fl
uo
resc
ence
mo
yen
10-5
5.10-5
10-4
5.10-4
10 25 50 100 10 50 100 250 1 2 5 10
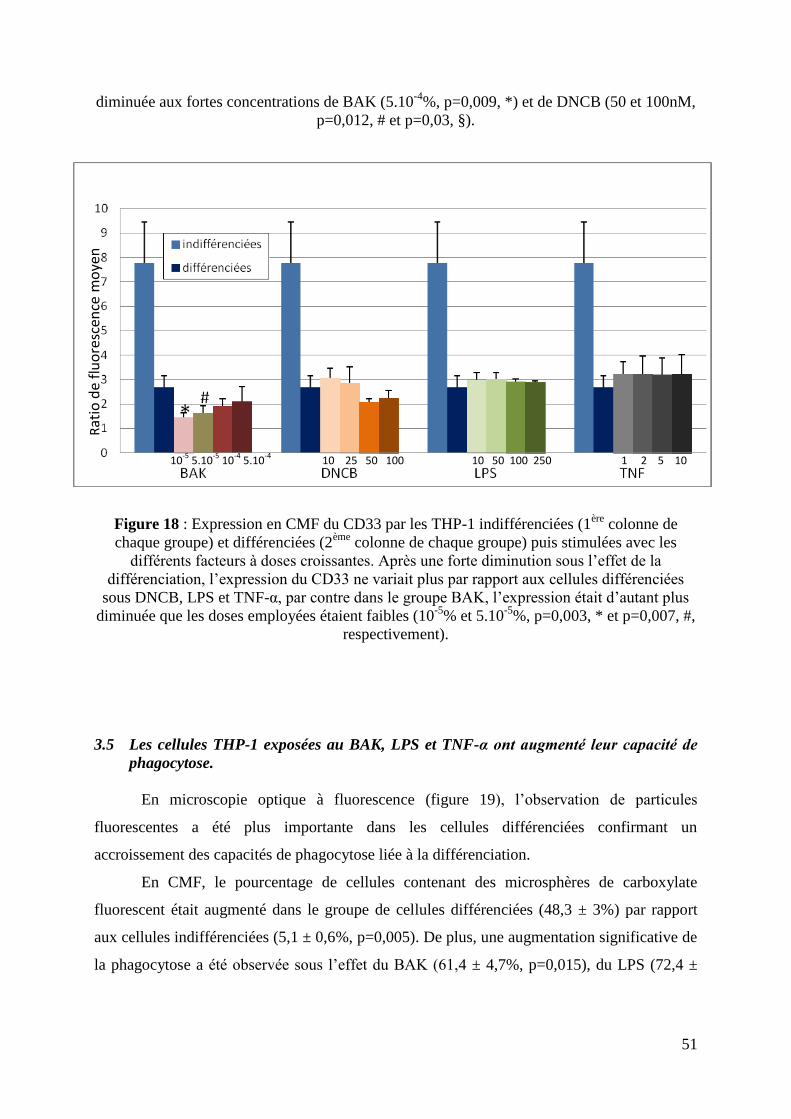
51
diminuée aux fortes concentrations de BAK (5.10-4
%, p=0,009, *) et de DNCB (50 et 100nM,
p=0,012, # et p=0,03, §).
Figure 18 : Expression en CMF du CD33 par les THP-1 indifférenciées (1ère
colonne de
chaque groupe) et différenciées (2ème
colonne de chaque groupe) puis stimulées avec les
différents facteurs à doses croissantes. Après une forte diminution sous l’effet de la
différenciation, l’expression du CD33 ne variait plus par rapport aux cellules différenciées
sous DNCB, LPS et TNF-α, par contre dans le groupe BAK, l’expression était d’autant plus
diminuée que les doses employées étaient faibles (10-5
% et 5.10-5
%, p=0,003, * et p=0,007, #,
respectivement).
3.5 Les cellules THP-1 exposées au BAK, LPS et TNF-α ont augmenté leur capacité de
phagocytose.
En microscopie optique à fluorescence (figure 19), l’observation de particules
fluorescentes a été plus importante dans les cellules différenciées confirmant un
accroissement des capacités de phagocytose liée à la différenciation.
En CMF, le pourcentage de cellules contenant des microsphères de carboxylate
fluorescent était augmenté dans le groupe de cellules différenciées (48,3 ± 3%) par rapport
aux cellules indifférenciées (5,1 ± 0,6%, p=0,005). De plus, une augmentation significative de
la phagocytose a été observée sous l’effet du BAK (61,4 ± 4,7%, p=0,015), du LPS (72,4 ±
Rat
io d
e fl
uo
resc
ence
mo
yen
10-5
5.10-5
10-4
5.10-4
10 25 50 100 10 50 100 250 1 2 5 10
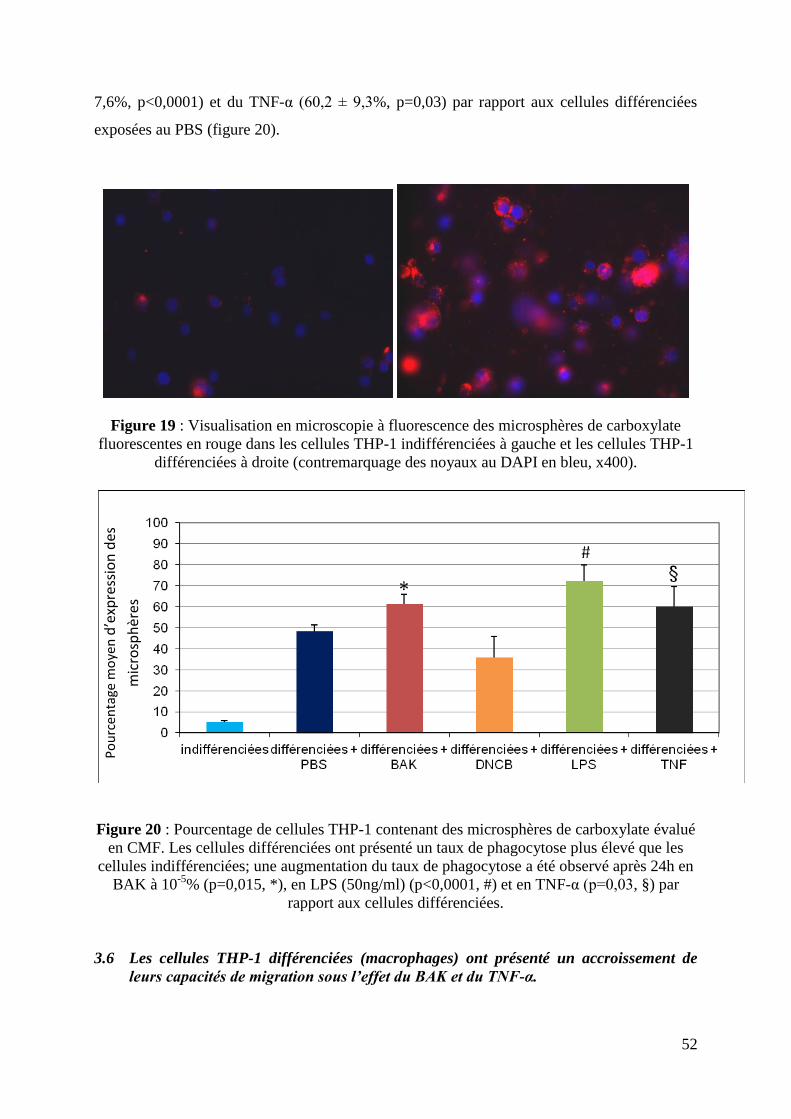
52
7,6%, p<0,0001) et du TNF-α (60,2 ± 9,3%, p=0,03) par rapport aux cellules différenciées
exposées au PBS (figure 20).
Figure 19 : Visualisation en microscopie à fluorescence des microsphères de carboxylate
fluorescentes en rouge dans les cellules THP-1 indifférenciées à gauche et les cellules THP-1
différenciées à droite (contremarquage des noyaux au DAPI en bleu, x400).
Figure 20 : Pourcentage de cellules THP-1 contenant des microsphères de carboxylate évalué
en CMF. Les cellules différenciées ont présenté un taux de phagocytose plus élevé que les
cellules indifférenciées; une augmentation du taux de phagocytose a été observé après 24h en
BAK à 10-5
% (p=0,015, *), en LPS (50ng/ml) (p<0,0001, #) et en TNF-α (p=0,03, §) par
rapport aux cellules différenciées.
3.6 Les cellules THP-1 différenciées (macrophages) ont présenté un accroissement de
leurs capacités de migration sous l’effet du BAK et du TNF-α.
Po
urc
enta
ge m
oye
n d’exp
ression
des
mic
rosp
hèr
es
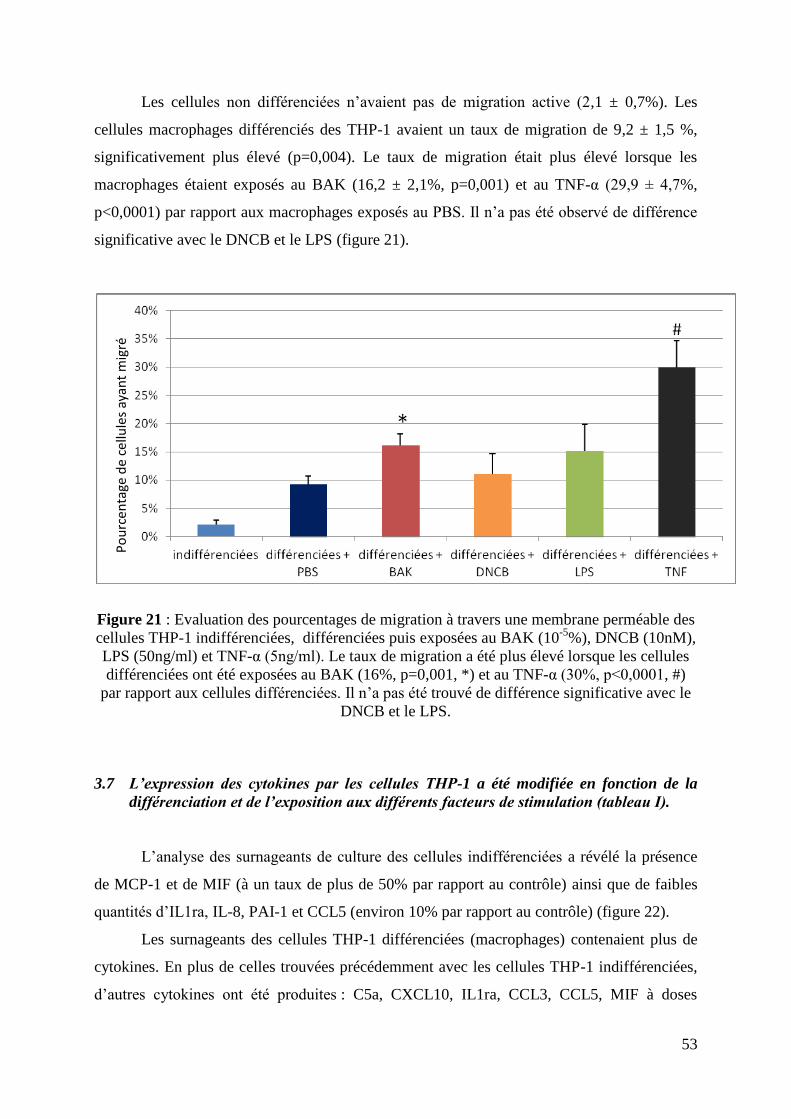
53
Po
urc
enta
ge d
e ce
llule
s ay
ant
mig
ré
Les cellules non différenciées n’avaient pas de migration active (2,1 ± 0,7%). Les
cellules macrophages différenciés des THP-1 avaient un taux de migration de 9,2 ± 1,5 %,
significativement plus élevé (p=0,004). Le taux de migration était plus élevé lorsque les
macrophages étaient exposés au BAK (16,2 ± 2,1%, p=0,001) et au TNF-α (29,9 ± 4,7%,
p<0,0001) par rapport aux macrophages exposés au PBS. Il n’a pas été observé de différence
significative avec le DNCB et le LPS (figure 21).
Figure 21 : Evaluation des pourcentages de migration à travers une membrane perméable des
cellules THP-1 indifférenciées, différenciées puis exposées au BAK (10-5
%), DNCB (10nM),
LPS (50ng/ml) et TNF-α (5ng/ml). Le taux de migration a été plus élevé lorsque les cellules
différenciées ont été exposées au BAK (16%, p=0,001, *) et au TNF-α (30%, p<0,0001, #)
par rapport aux cellules différenciées. Il n’a pas été trouvé de différence significative avec le
DNCB et le LPS.
3.7 L’expression des cytokines par les cellules THP-1 a été modifiée en fonction de la
différenciation et de l’exposition aux différents facteurs de stimulation (tableau I).
L’analyse des surnageants de culture des cellules indifférenciées a révélé la présence
de MCP-1 et de MIF (à un taux de plus de 50% par rapport au contrôle) ainsi que de faibles
quantités d’IL1ra, IL-8, PAI-1 et CCL5 (environ 10% par rapport au contrôle) (figure 22).
Les surnageants des cellules THP-1 différenciées (macrophages) contenaient plus de
cytokines. En plus de celles trouvées précédemment avec les cellules THP-1 indifférenciées,
d’autres cytokines ont été produites : C5a, CXCL10, IL1ra, CCL3, CCL5, MIF à doses

54
élevées (plus de 65% par rapport au contrôle) et CCL1, GM-CSF et TNF-α à doses plus
faibles (entre 15 et 35% par rapport au contrôle).
Après exposition aux facteurs de stimulation, les surnageants des macrophages
(cellules THP-1 différenciées) contenaient les cytokines précédemment observées sur les
cellules non stimulées mais aussi d’autres cytokines : CCL1, CD54 soluble, IL-1β, CCL4,
TNF-α à doses élevées (plus de 65% par rapport au contrôle) et CXCL11 à dose faible (moins
de 20% par rapport au contrôle). A noter la présence de CXCL1 et d’IL-16 (plus de 60% par
rapport au contrôle) dans les groupes DNCB et LPS. De plus, le surnageant des cellules
exposées au LPS (50ng/ml) contenait d’autres cytokines : G-CSF, IL-1α, IL-6 et IL-10 à
doses élevées (entre 40 et 90% par rapport au contrôle) et IFN-γ, IL-23, CXCL12 et sTREM-1
à doses faibles (environ 20% par rapport au contrôle). Il n’y avait pas de différence
d’expression des cytokines testées dans les surnageants des cellules exposées au TNF-α et au
BAK.
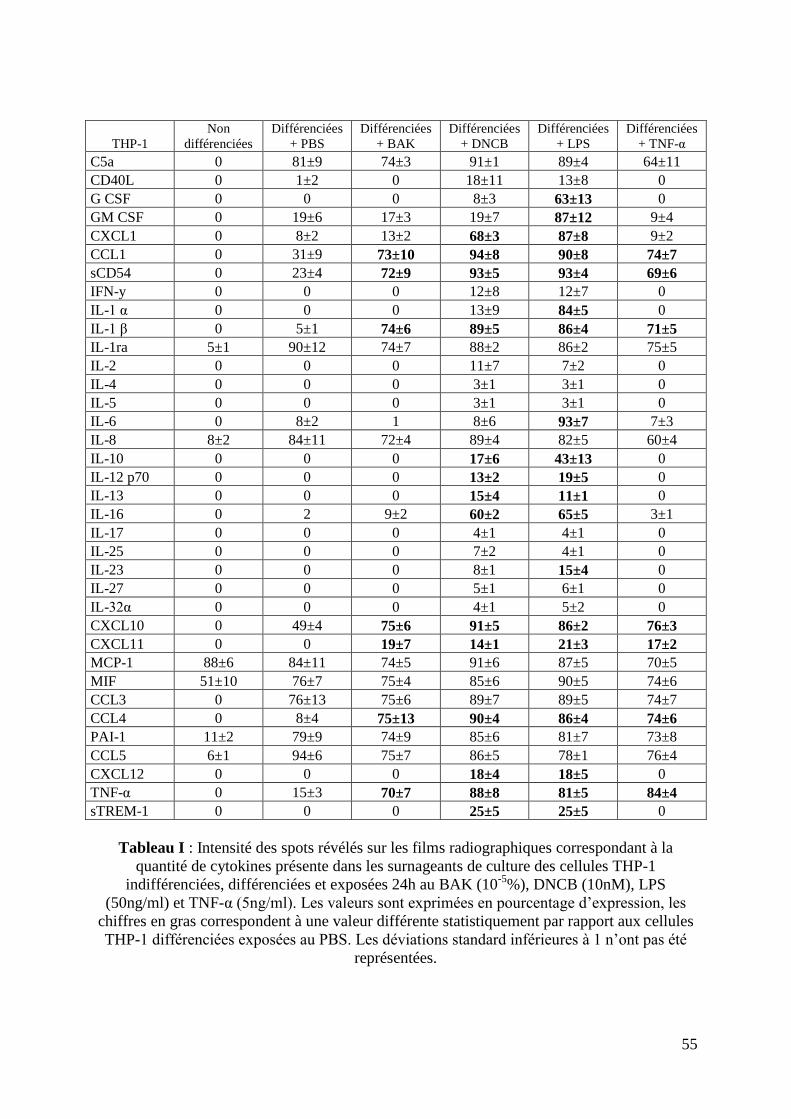
55
THP-1
Non
différenciées
Différenciées
+ PBS
Différenciées
+ BAK
Différenciées
+ DNCB
Différenciées
+ LPS
Différenciées
+ TNF-α
C5a 0 81±9 74±3 91±1 89±4 64±11
CD40L 0 1±2 0 18±11 13±8 0
G CSF 0 0 0 8±3 63±13 0
GM CSF 0 19±6 17±3 19±7 87±12 9±4
CXCL1 0 8±2 13±2 68±3 87±8 9±2
CCL1 0 31±9 73±10 94±8 90±8 74±7
sCD54 0 23±4 72±9 93±5 93±4 69±6
IFN-y 0 0 0 12±8 12±7 0
IL-1 α 0 0 0 13±9 84±5 0
IL-1 β 0 5±1 74±6 89±5 86±4 71±5
IL-1ra 5±1 90±12 74±7 88±2 86±2 75±5
IL-2 0 0 0 11±7 7±2 0
IL-4 0 0 0 3±1 3±1 0
IL-5 0 0 0 3±1 3±1 0
IL-6 0 8±2 1 8±6 93±7 7±3
IL-8 8±2 84±11 72±4 89±4 82±5 60±4
IL-10 0 0 0 17±6 43±13 0
IL-12 p70 0 0 0 13±2 19±5 0
IL-13 0 0 0 15±4 11±1 0
IL-16 0 2 9±2 60±2 65±5 3±1
IL-17 0 0 0 4±1 4±1 0
IL-25 0 0 0 7±2 4±1 0
IL-23 0 0 0 8±1 15±4 0
IL-27 0 0 0 5±1 6±1 0
IL-32α 0 0 0 4±1 5±2 0
CXCL10 0 49±4 75±6 91±5 86±2 76±3
CXCL11 0 0 19±7 14±1 21±3 17±2
MCP-1 88±6 84±11 74±5 91±6 87±5 70±5
MIF 51±10 76±7 75±4 85±6 90±5 74±6
CCL3 0 76±13 75±6 89±7 89±5 74±7
CCL4 0 8±4 75±13 90±4 86±4 74±6
PAI-1 11±2 79±9 74±9 85±6 81±7 73±8
CCL5 6±1 94±6 75±7 86±5 78±1 76±4
CXCL12 0 0 0 18±4 18±5 0
TNF-α 0 15±3 70±7 88±8 81±5 84±4
sTREM-1 0 0 0 25±5 25±5 0
Tableau I : Intensité des spots révélés sur les films radiographiques correspondant à la
quantité de cytokines présente dans les surnageants de culture des cellules THP-1
indifférenciées, différenciées et exposées 24h au BAK (10-5
%), DNCB (10nM), LPS
(50ng/ml) et TNF-α (5ng/ml). Les valeurs sont exprimées en pourcentage d’expression, les
chiffres en gras correspondent à une valeur différente statistiquement par rapport aux cellules
THP-1 différenciées exposées au PBS. Les déviations standard inférieures à 1 n’ont pas été
représentées.

56
Expression des cytokines par les cellules THP1
Indifférenciée
s
Différenciées BAK DNCB LPS TNF-α
Figure 22 : Films radiographiques correspondant aux membranes de nitrocellulose sur
lesquelles ont été dosés les surnageants des cellules THP-1 indifférenciées et différenciées
puis exposées au BAK (10-5
%), DNCB (10nM), LPS (50ng/ml) ou au TNF-α (5ng/ml).
Chaque couple de point représente la mesure d’une molécule, les résultats ont été regroupés
dans le tableau I.
4. DISCUSSION
Dans cette étude in vitro nous avons procédé à la différenciation des cellules THP-1 en
macrophages, selon une procédure largement validée (6). Les cellules THP-1 sont devenues
une des lignées cellulaires les plus employées pour étudier la fonction et la régulation des
macrophages in vitro (71). La lignée cellulaire est issue de cellules d’une leucémie aigue
monocytaire qui ne possède pas toutes les caractéristiques des cellules dérivées du sang (27).
Néanmoins, au vu de l’association de la morphologie et des marqueurs fonctionnels présentés
par les cellules THP-1 différenciées, ces cellules sont communément utilisées en tant que
cellules présentatrices d’antigène in vitro (38). Dans cette étude, nous avons sélectionné les
cellules THP-1 différenciées adhérentes au fond des puits de culture comme précédemment
décrit (89). Ces cellules avaient un aspect dendritiforme et des capacités d’adhésion, capacités
partagées par les cellules dendritiques et les macrophages (41). Ces changements
morphologiques étaient aussi associés à l’expression de CD11b, un marqueur de macrophages
conjonctivaux (41), et de CD11c, un marqueur de cellules dendritiques conjonctivales (41).

57
De plus, une diminution de l’expression de CD33, un marqueur des cellules myéloïdes qui
diminue avec la différenciation des cellules (43), a confirmé le phénotype différencié de ces
cellules. Les cellules THP-1 différenciées ont aussi démontré des capacités de migration et de
phagocytose communes aux cellules présentatrices d’antigène in vitro.
Le BAK est largement utilisé dans les collyres comme conservateur. Sa toxicité et ses
effets pro inflammatoires sur les cellules épithéliales de la surface oculaire sont connus (9, 29,
63). Exposées à une concentration de BAK à 0.0001%, la croissance des cellules épithéliales
conjonctivales et cornéennes est réduite, et les cellules deviennent apoptotiques à des
concentrations plus élevées
(29). Le BAK est aussi responsable de la production de
médiateurs de l’inflammation et de cytokines proinflammatoires par les cellules
inflammatoires cutanées selon une relation directe entre l’intensité des signes cliniques
d’irritation et la libération in vitro de médiateurs proinflammatoires (90). Dans notre étude,
une faible concentration de BAK (10-5
%) a été utilisée dans le but d’être en dessous des effets
cytotoxiques directs sur les macrophages et de mieux reproduire les effets du BAK observés
in vivo sur les tissus de la surface oculaire. Ainsi, nous avons montré que de faibles
concentrations de BAK stimulaient les macrophages sur le plan de la migration, de la
production de chimiokines, de la phagocytose et de l’expression de marqueurs cellulaires.
En utilisant la même lignée cellulaire THP-1, Hirota et al. ont observé que le BAK à 10-
4%
n’induisait pas de production de CCL4 ou MIP-1β et de CD86 in vitro (47). De façon plus
intéressante, pour Lim et al., la concentration d’inhibition de la prolifération cellulaire des
THP-1 à 50% était de 3.2x10-4
% pour le BAK. Cette concentration de BAK entrainait une
diminution de la production de CCL4 par les cellules THP-1, tandis que des concentrations
plus faibles (3.2x10-5
% and 3.2x10-6
%) n’avaient aucun effet sur la production de CCL4 (57).
Nous avons observé de façon similaire qu’une faible concentration de BAK n’induisait pas de
production de CCL4 par les cellules THP-1, toutefois, cette concentration (10-5
%) augmentait
la production de CCL4 par les macrophages (cellules THP-1 différenciées), ce qui n’avait pas
été évalué dans les deux précédentes études. Les cellules THP-1 doivent ainsi être
différenciées en macrophages pour être stimulées par de faibles concentrations de BAK. Il a
été démontré auparavant que le BAK induisait la libération de cytokines inflammatoires (IL-
1β, IL-8, CXCL1, CCL3, CCL5 and MCP-1) par le parenchyme vaginal (4, 24). Sur la peau
de l’oreille, l’application de BAK libère des chimiokines inflammatoires comme TNF-α, IL-
1β et MIP-1α et MIP-1β (74).
Sur les cellules de la surface oculaire, le BAK induit la
production d’IL-1 et de TNF-α par les cellules conjonctivales et cornéennes (37). Dans l’étude
présentée ici, les surnageants de macrophages stimulés par le BAK ont montré l’expression

58
d’ICAM-1, IL-8, CCL4, IL-1β et TNF-α. De façon intéressante, ICAM-1 et IL-8 ont aussi été
retrouvés sur les empreintes conjonctivales de patients traités avec des collyres
antiglaucomateux contenant du BAK (10). Le BAK a induit in vitro la production par les
macrophages de chimiokines similaires à celles retrouvées in vivo dans les tissus exposés au
BAK (4, 24, 74). Ces résultats soulignent les effets stimulant des macrophages in vivo.
Afin de mieux évaluer les effets du BAK sur les macrophages, d’autres facteurs
reproduisant des pathologies de la surface oculaire ont été utilisés : le DNCB, le TNF-α et le
LPS. Le DNCB est un allergène dérivé du benzène qui est responsable de dermite de contact
(97). Il possède un rôle stimulant pour les cellules présentatrices d’antigène, en augmentant
l’expression des marqueurs cellulaires (HLA-DR, CD86, CD40 et CD54), la prolifération des
lymphocytes T et la production de TNF-α, ainsi que l’expression du récepteur aux
chimiokines Cys-Cys 7 (CCR7) (13). Le DNCB est considéré comme un allergène in vivo, au
regard de son action proinflammatoire sur les macrophages (13, 97). Sur les macrophages
issus des cellules THP-1, nous avons observé que le DNCB induisait une surexpression de
CD11c et la libération de chimiokines proinflammatoires (CXCL1, IL-16, CCL1, CD54
soluble, IL-1β, CCL4 et TNF-α) mais le DNCB n’avait pas d’effet sur la migration et la
phagocytose. Le LPS et le TNF-α sont tous deux des facteurs stimulants connus pour les
fonctions de migration, de phagocytose et la production de chimiokines par les macrophages,
le LPS est exprimé sur la membrane externe des bactéries Gram -, ces effets pro
inflammatoires sur les cellules conjonctivales et les cellules présentatrices d’antigène ont été
largement démontrés (23, 78). Le TNF-α est un médiateur de l’inflammation produit par
différents types cellulaires qui agit en induisant la production de chimiokines, de molécules
d’adhésion et de co-stimulation par les cellules inflammatoires (32). Dans l’étude présentée
ici, l’expression du CD11b et du CD11c par les macrophages induite par le BAK, le LPS et le
TNF-α étaient comparables. De même, sur le plan de la phagocytose, aucune différence n’a
été observée entre les effets stimulants du BAK, du LPS et du TNF-α. Le LPS était le facteur
stimulant le plus actif sur le plan de la production de chimiokines par les macrophages. Le
TNF augmentait l’expression de CD54 par les macrophages, expression qui n’était pas
modifiée par les autres facteurs (BAK, TNF-α et DNCB). Pour la migration des macrophages,
le TNF-α a eu l’effet le plus important, en comparaison avec les effets observés par le LPS et
le BAK. Malgré quelques variations en fonction des propriétés stimulées, une faible
concentration de BAK pourrait agir comme un facteur stimulant comparable au TNF-α et au
LPS au niveau de la surface oculaire.

59
Le BAK est considéré comme un agent cytotoxique au regard de ces effets sur les cellules
épithéliales conjonctivales (29, 31), mais peu considéré comme un facteur stimulant direct sur
les cellules inflammatoires (9). De faibles concentrations de BAK, telles qu’observées chez
les patients traités au long cours avec des collyres hypotonisants contenant des conservateurs
pourraient être directement responsables du recrutement des macrophages et de
l’augmentation de leurs capacités migratoires. De façon intéressante, dans notre étude, les
macrophages exposés au BAK ont davantage migré en présence de cellules épithéliales
conjonctivales. Il a été précédemment démontré que le TNF-α induisait un effet pro-
migratoire sur les cellules dendritiques de la cornée (30). Cet effet similaire sur la migration
provoqué par le BAK pourrait être lié à une production de TNF-α induite par le BAK et
démontrée dans notre étude. Peu d’études ont analysé les effets du BAK sur les fonctions de
phagocytose des macrophages. Les cellules épithéliales conjonctivales exposées au BAK
contiennent des phagosomes et des autolysosomes (19), conduisant à leur phagocytose par les
autres cellules. In vitro, nous avons observé que la fonction de phagocytose des macrophages
était augmentée par l’exposition au BAK. La surexpression du CD11b et du CD11c facilitant
la phagocytose de particules recouvertes de complément (81) pourrait expliquer cet effet du
BAK sur les macrophages. Ainsi, dans les tissus de la surface oculaire, le BAK pourrait
activer la phagocytose en induisant l’apoptose et l’autophagie des cellules épithéliales
conjonctivales et en stimulant directement les macrophages.
5. CONCLUSION
Ces résultats suggèrent que les macrophages sont en partie responsables des réactions
inflammatoires observées lorsque les tissus de la surface oculaire sont exposés au BAK. Des
concentrations infra toxiques de BAK se sont révélées stimulantes pour certaines fonctions
macrophagiques. Il a été montré dans la littérature que le BAK était responsable d’une
infiltration de macrophages dans la conjonctive et de l’expression de marqueurs
inflammatoires et de fibrose chez les patients traités pour glaucome ou HTO (11, 17, 63, 80).
Dans cette étude in vitro, nous avons utilisé une concentration plus faible de BAK (10-5
%)
que celles utilisées dans les collyres antiglaucomateux (0,025 à 0,004%). Cependant, chez les
patients recevant des collyres hypotonisants contenant des conservateurs, les macrophages et
les cellules dendritiques de la surface oculaire devraient être exposés à des concentrations plus

60
faibles de BAK à cause de la dilution dans le film lacrymal et la localisation profonde des
macrophages en comparaison avec la localisation superficielle des cellules épithéliales
conjonctivales. Les effets proinflammatoires induits par les collyres conservés et le BAK en
particulier, sont responsables de symptômes d’irritation de la surface oculaire constituant un
véritable obstacle à l’observance des traitements du glaucome. De plus, l’infiltration de
macrophages dans la conjonctive a été mise en cause, au moins en partie, dans les processus
de cicatrisation pathologiques observés après chirurgie filtrante (18, 46). Nos résultats mettent
une fois de plus en avant le rôle du BAK dans ces changements tissulaires conjonctivaux. En
considérant le pouvoir stimulant du BAK sur les macrophages, le BAK devrait être évité
avant chirurgie filtrante au moins pendant la période pré-opératoire immédiate.

61
CHAPITRE 3 : ANALYSE IN VITRO DES RELATIONS ENTRE LES
MACROPHAGES ET LES CELLULES EPITHELIALES
CONJONCTIVALES
1. INTRODUCTION
Les cellules épithéliales conjonctivales, par l’intermédiaire des marqueurs
proinflammatoires qu’elles expriment lorsqu’elles sont soumises à des substances irritantes,
posséderaient un rôle immunomodulateur sur la surface oculaire (96). Dans la première partie
de ce travail, nous avons démontré que de faibles concentrations de BAK avaient un effet
stimulant sur les macrophages in vitro. Il existe probablement des mécanismes de recrutement
des macrophages induits par le BAK, directement liés à l’effet du BAK sur les macrophages
comme démontré précédemment mais aussi par l’intermédiaire de l’effet du BAK sur les
cellules épithéliales conjonctivales. L’objectif de cette deuxième partie a donc été d’analyser
les interactions entre les cellules épithéliales conjonctivales et les macrophages lors d’une
stimulation par le BAK.
2. MATERIELS ET METHODES
2.1 Lignées cellulaires
2.1.1 Lignée monocytaire
Les cellules de la lignée THP-1 ont été utilisées après différenciation en macrophages
selon la méthode employée en première partie.

62
2.1.2 Lignée épithéliale conjonctivale
Les cellules conjonctivales Wong-Kilbourne dérivées d’une conjonctive humaine de
Chang (WKD) (clone 1 à 5c-4l, ATCC) ont été cultivées en conditions standard (atmosphère
humidifiée, 5% de CO2 et 37°C) dans un milieu DMEM (Invitrogen) enrichi avec 10% de
sérum de veau fœtal (HyClone), 1% de glutamine (Invitrogen), 50 UI/ml de pénicilline et 50
UI/ml de streptomycine (BioSource International).
2.2 Stimulations
Les cellules WKD ont été ensemencées dans des plaques 6 puits à la concentration de
80 000 cellules/ml à raison de 2 ml par puits. Après 48 heures de culture et renouvellement du
milieu, les cellules ont été exposées pendant 24 heures au BAK à 10-5
%, au DNCB à 10nM,
au LPS à 50ng/ml ou au TNF-α à 5ng/ml. Comme précédemment décrit, les cellules THP-1
ont été exposées pendant 72 heures au PMA (différenciation), puis après renouvellement du
milieu de culture, les cellules ont été exposées pendant 24 heures à du BAK, du DNCB, du
LPS, du TNF-α ou PBS. Seules les concentrations infra toxiques déterminées dans la première
partie de ce travail ont été utilisées, soit le BAK 10-5
%, le DNCB 10nM, le LPS 50ng/ml et le
TNF-α 5ng/ml. Les surnageants ont été recueillis et stockés à -20°C. Ils ont été recueillis suite
à 24 heures d’exposition aux différents facteurs de stimulation.
2.3 Analyse de l’effet des facteurs stimulants sur les cellules épithéliales conjonctivales en
cytométrie en flux.
L’analyse en CMF a été réalisée après 24 heures d’exposition aux différents facteurs
de stimulation. Les cellules WKD ont été recueillies après une incubation de 10 minutes en
EDTA 0,5mM (Sigma-Aldrich) pour favoriser le décollement des cellules de la plaque. Les
cellules ont été ensuite lavées puis fixées dans une solution de PFA (Alfa Aesar, Ward Hill,
USA) à 0,5% pendant 24 heures à 4°C avant d’être marquées. Les anticorps monoclonaux

63
utilisés étaient : CD54 (FITC), CD40 (FITC), CD63 (FITC), CD95 (FITC) (Pharmingen, San
Diego, USA), CD33 (PE), (BD Biosciences, San Jose, USA), EMMPRIN (Extracellular
Matrix Metalloproteinase Inducer) (Santa Cruz Biotechnology, Santa Cruz, USA) ainsi qu’un
anticorps isotypique de contrôle (IgG1 de souris) (Pharmingen). Chaque suspension cellulaire
correspondant à une condition a été incubée en présence des anticorps primaires directement
fluorescents pendant 35 minutes ou contremarquée à l’aide d’un anticorps secondaire de
chèvre anti-souris conjugué au FITC pendant une autre incubation de 35 minutes. Après
chaque incubation les cellules ont été lavées en PBS puis finalement analysées en CMF.
2.4 Production de cytokines
Les surnageants des cellules conjonctivales ont été analysés à l’aide d’un kit
permettant d’identifier 38 cytokines différentes (R&D Systems) selon une méthode décrite
précédemment dans ce travail. Les six conditions d’évaluation représentaient chacune une
membrane de nitrocellulose révélée sur film radiographique (GE Healthcare Limited) avec
utilisation d’un agent électroluminescent (GE Healthcare Limited) selon les recommandations
du fournisseur. La densité en pixel des spots a été analysée par logiciel informatique (ImageJ).
L’intensité de production de cytokine est exprimé en pourcentage, ce pourcentage est calculé
en rapportant la mesure à l’intensité du contrôle négatif, 100% correspondant à l’intensité du
contrôle positif.
2.5 Interactions entre les cellules WKD et les cytokines produites par les macrophages
Les cellules WKD ont été cultivées avec les surnageants des cellules THP-1 selon les
conditions présentées précédemment. Les milieux de culture des THP-1 et des cellules WKD
étant différents, les surnageants ont été filtrés avec une centrifugeuse (AvantiR J-E, Beckman
Coulter) à haute vélocité à 4000G, à 4°C pendant 10 minutes dans des tubes de filtration
(Merck Milipore). L’ultrafiltrat de surnageant contenant les cytokines (200µL à partir d’une
base de 2 mL) a été ensuite complété par du milieu DMEM et utilisé comme milieu de culture
pour les cellules WKD. Les six conditions précédemment décrites ont été évaluées. Après 24

64
heures d’exposition, les cellules WKD ont été recueillies après décollement de la plaque par
de l’EDTA (Sigma-Aldrich) et fixées en PFA 0,5% pendant 24 heures à 4°C avant analyse en
CMF. Les marqueurs utilisés étaient le CD40 (FITC), CD63 (FITC), CD95 (FITC) HLA-DR
(FITC) (Pharmingen) et EMMPRIN (Santa Cruz Biotechnology).
2.6 Interactions entre les macrophages et les cytokines produites par les cellules WKD
Les macrophages issus de la lignée THP-1 ont été cultivés avec les surnageants des
cellules WKD soumises aux différentes conditions de stimulation précédemment décrites. Les
surnageants des WKD ont été filtrés selon les mêmes conditions. L’ultrafiltrat de surnageant
contenant les cytokines (200µL à partir d’une base de 2 mL) a été ensuite complété par du
milieu RPMI et utilisé comme milieu de culture pour les macrophages. Les six conditions
précédemment décrites ont été évaluées. Après 24 heures d’exposition, les cellules THP-1 ont
été recueillies et fixées en PFA 0,5% pendant 24 heures à 4°C avant analyse en CMF. Les
marqueurs utilisés étaient le CD11b couplé au FITC, CD11c couplé à la PE, et le CD54
couplé FITC (Pharmingen).
2.7 Analyses statistiques
Dans cette étude, toutes les expérimentations ont été réalisées trois fois sauf pour le
dosage des cytokines sur membranes de nitrocellulose qui n’a été réalisé qu’une fois. Pour les
comparaisons multiples entre les différentes conditions, le test non paramétrique de Kruskall-
Wallis avec comparaisons multiples par paires et correction de Bonferroni a été utilisé. Pour
les comparaisons simples entre deux conditions, un test t de Student non apparié a été réalisé.
Une valeur de p<0,05 a été considérée comme significative.
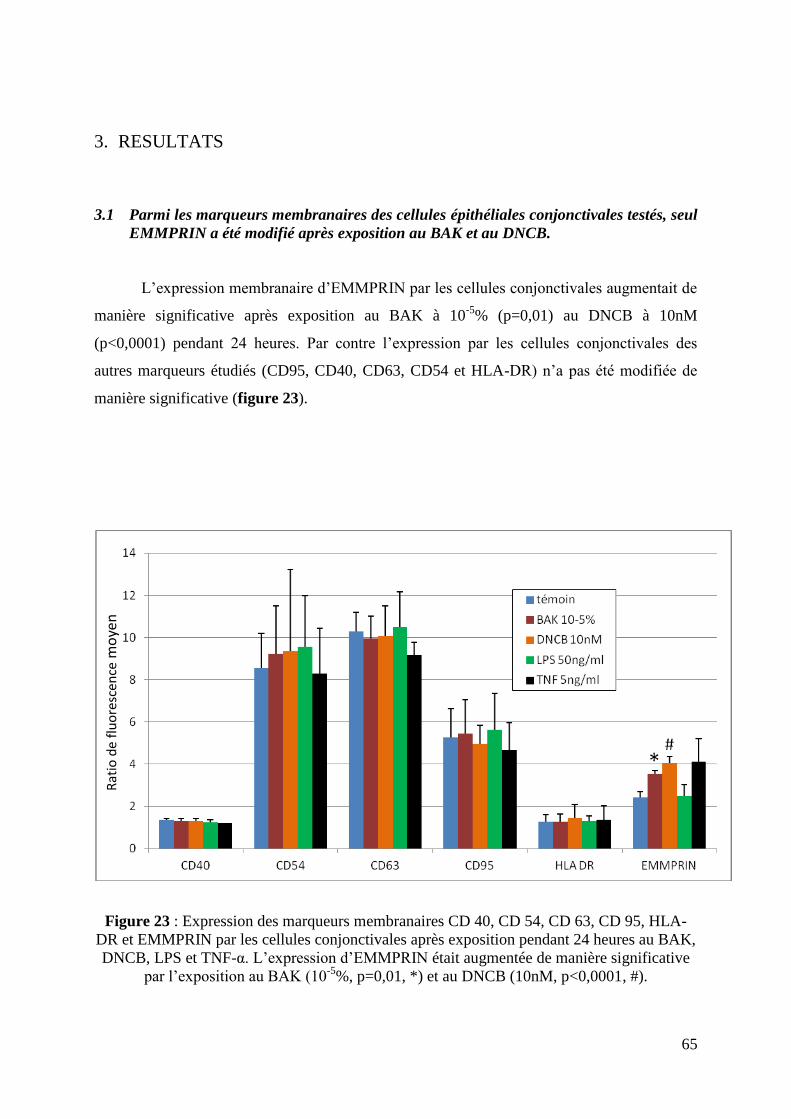
65
3. RESULTATS
3.1 Parmi les marqueurs membranaires des cellules épithéliales conjonctivales testés, seul
EMMPRIN a été modifié après exposition au BAK et au DNCB.
L’expression membranaire d’EMMPRIN par les cellules conjonctivales augmentait de
manière significative après exposition au BAK à 10-5
% (p=0,01) au DNCB à 10nM
(p<0,0001) pendant 24 heures. Par contre l’expression par les cellules conjonctivales des
autres marqueurs étudiés (CD95, CD40, CD63, CD54 et HLA-DR) n’a pas été modifiée de
manière significative (figure 23).
Figure 23 : Expression des marqueurs membranaires CD 40, CD 54, CD 63, CD 95, HLA-
DR et EMMPRIN par les cellules conjonctivales après exposition pendant 24 heures au BAK,
DNCB, LPS et TNF-α. L’expression d’EMMPRIN était augmentée de manière significative
par l’exposition au BAK (10-5
%, p=0,01, *) et au DNCB (10nM, p<0,0001, #).
Rat
io d
e fl
uo
resc
ence
mo
yen

66
3.2 L’expression des chimiokines par les cellules épithéliales conjonctivales a été modifiée
par l’exposition aux facteurs stimulants. (figure 24 et tableau II).
Sans stimulation, les surnageants des cellules WKD contenaient une concentration
élevée de MIF et de PAI-1 (77 et 72% par rapport au contrôle) et une concentration faible
d’IL-6 (12% par rapport au contrôle). Les autres chimiokines testées étaient à l’état de traces.
Après stimulation par le BAK, l’expression de PAI-1 n’était pas modifiée mais la
concentration de MIF était augmentée (91% par rapport au groupe contrôle). La présence
d’IL-6 et d’IL-8 a également été retrouvée à des concentrations faibles après stimulation par
le BAK (21% et 11% par rapport au contrôle).
L’action du DNCB sur la présence de chimiokines dans les surnageants de culture des
cellules WKD était modérée, des concentrations modérées de MIF et PAI-1 (57 et 33% par
rapport au contrôle) ont été mises en évidence.
L’exposition des cellules WKD au LPS a mis en évidence une concentration élevée de
MIF (83% par rapport au contrôle) restant inférieure aux surnageants des cellules exposées au
BAK, une concentration équivalente de PAI-1 (74% par rapport au contrôle) et des
concentrations modérées d’IL-6 et d’IL-8 (20 et 10% par rapport au contrôle) dans leurs
surnageants.
Les surnageants des cellules WKD exposées au TNF-α ont révélé la présence de plus
de chimiokines qu’avec les autres stimulants. Des concentrations élevées de MIF, PAI-1 IL-6,
IL-8 et de TNF-α ont été retrouvées (respectivement 65, 55, 75, 77 et 93% par rapport au
contrôle). Le CCL-1 a été mis en évidence en quantité modérée (15% par rapport au contrôle).
Le MIF et le PAI-1 étaient retrouvés également, mais plus faiblement qu’après exposition au
BAK.

67
Expression des cytokines par les cellules WKD
PBS BAK DNCB LPS TNF-α
Figure 24 : Films radiographiques correspondant aux membranes de nitrocellulose sur
lesquelles ont été dosés les surnageants des cellules WKD exposées et au BAK (10-5
%),
DNCB (10nM), LPS (50ng/ml) ou au TNF-α (5ng/ml). Chaque couple de point représente la
mesure d’une molécule, les résultats ont été regroupés dans le tableau II.
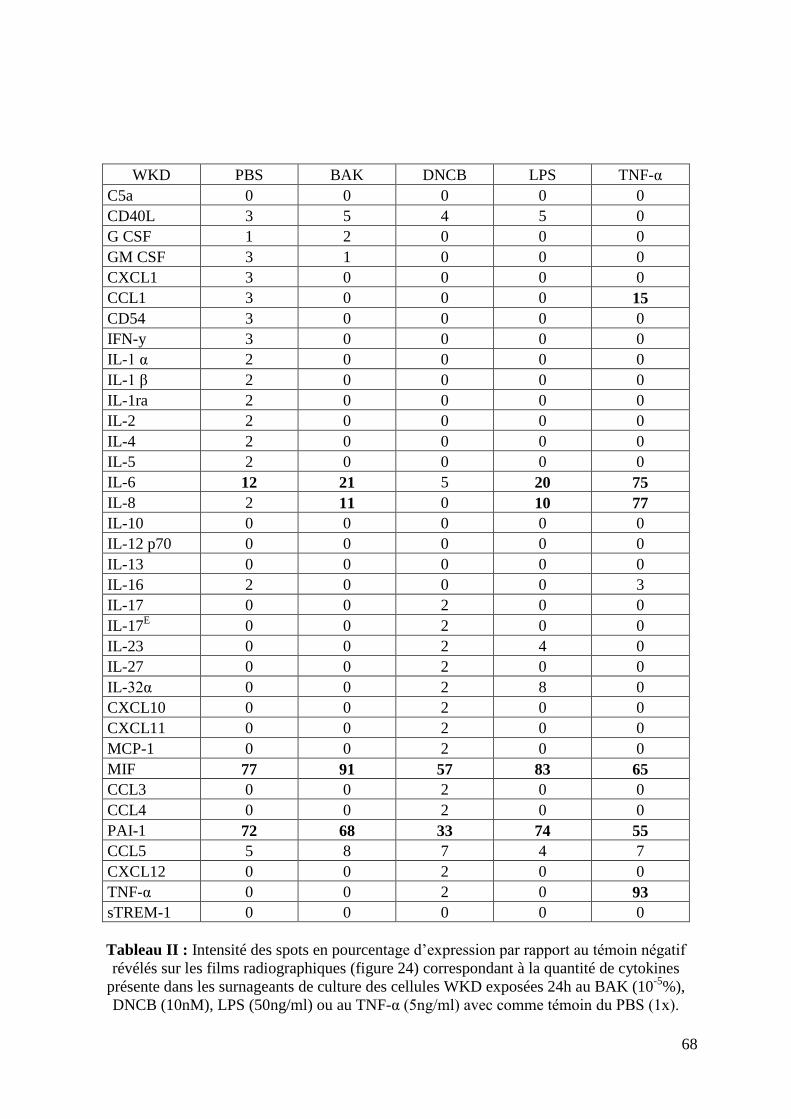
68
WKD PBS BAK DNCB LPS TNF-α
C5a 0 0 0 0 0
CD40L 3 5 4 5 0
G CSF 1 2 0 0 0
GM CSF 3 1 0 0 0
CXCL1 3 0 0 0 0
CCL1 3 0 0 0 15
CD54 3 0 0 0 0
IFN-y 3 0 0 0 0
IL-1 α 2 0 0 0 0
IL-1 β 2 0 0 0 0
IL-1ra 2 0 0 0 0
IL-2 2 0 0 0 0
IL-4 2 0 0 0 0
IL-5 2 0 0 0 0
IL-6 12 21 5 20 75
IL-8 2 11 0 10 77
IL-10 0 0 0 0 0
IL-12 p70 0 0 0 0 0
IL-13 0 0 0 0 0
IL-16 2 0 0 0 3
IL-17 0 0 2 0 0
IL-17E 0 0 2 0 0
IL-23 0 0 2 4 0
IL-27 0 0 2 0 0
IL-32α 0 0 2 8 0
CXCL10 0 0 2 0 0
CXCL11 0 0 2 0 0
MCP-1 0 0 2 0 0
MIF 77 91 57 83 65
CCL3 0 0 2 0 0
CCL4 0 0 2 0 0
PAI-1 72 68 33 74 55
CCL5 5 8 7 4 7
CXCL12 0 0 2 0 0
TNF-α 0 0 2 0 93
sTREM-1 0 0 0 0 0
Tableau II : Intensité des spots en pourcentage d’expression par rapport au témoin négatif
révélés sur les films radiographiques (figure 24) correspondant à la quantité de cytokines
présente dans les surnageants de culture des cellules WKD exposées 24h au BAK (10-5
%),
DNCB (10nM), LPS (50ng/ml) ou au TNF-α (5ng/ml) avec comme témoin du PBS (1x).
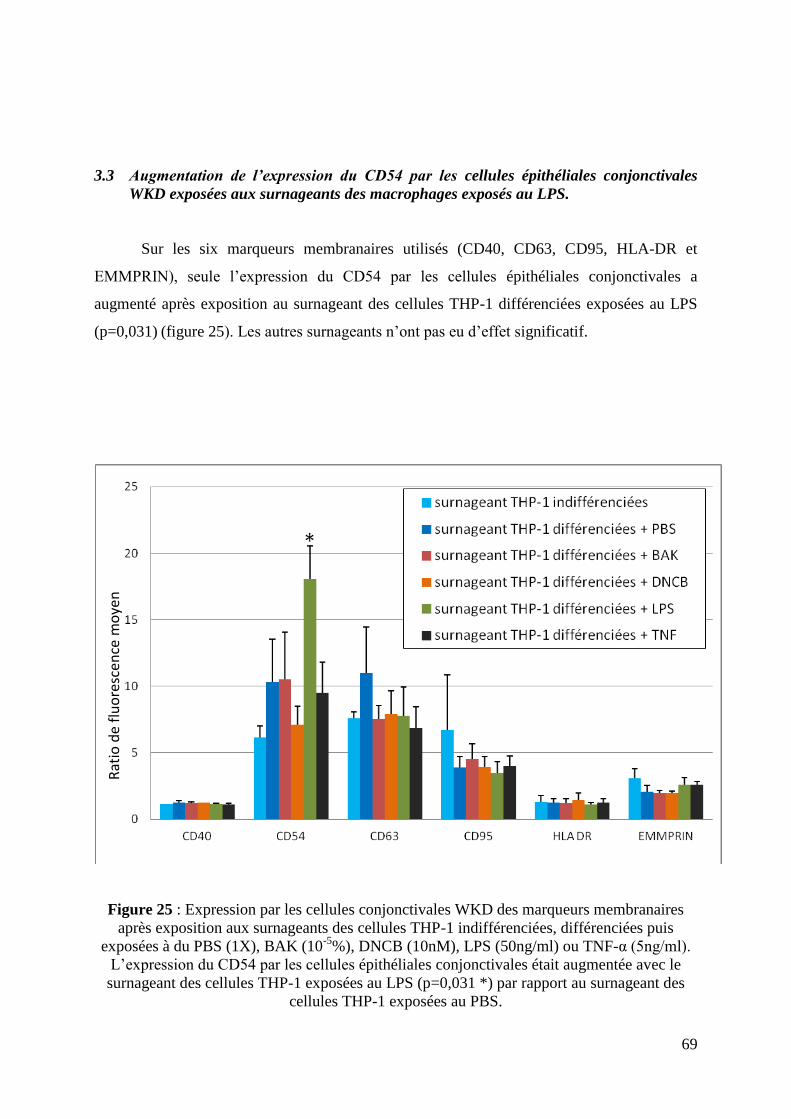
69
3.3 Augmentation de l’expression du CD54 par les cellules épithéliales conjonctivales
WKD exposées aux surnageants des macrophages exposés au LPS.
Sur les six marqueurs membranaires utilisés (CD40, CD63, CD95, HLA-DR et
EMMPRIN), seule l’expression du CD54 par les cellules épithéliales conjonctivales a
augmenté après exposition au surnageant des cellules THP-1 différenciées exposées au LPS
(p=0,031) (figure 25). Les autres surnageants n’ont pas eu d’effet significatif.
Figure 25 : Expression par les cellules conjonctivales WKD des marqueurs membranaires
après exposition aux surnageants des cellules THP-1 indifférenciées, différenciées puis
exposées à du PBS (1X), BAK (10-5
%), DNCB (10nM), LPS (50ng/ml) ou TNF-α (5ng/ml).
L’expression du CD54 par les cellules épithéliales conjonctivales était augmentée avec le
surnageant des cellules THP-1 exposées au LPS (p=0,031 *) par rapport au surnageant des
cellules THP-1 exposées au PBS.
Rat
io d
e fl
uo
resc
ence
mo
yen
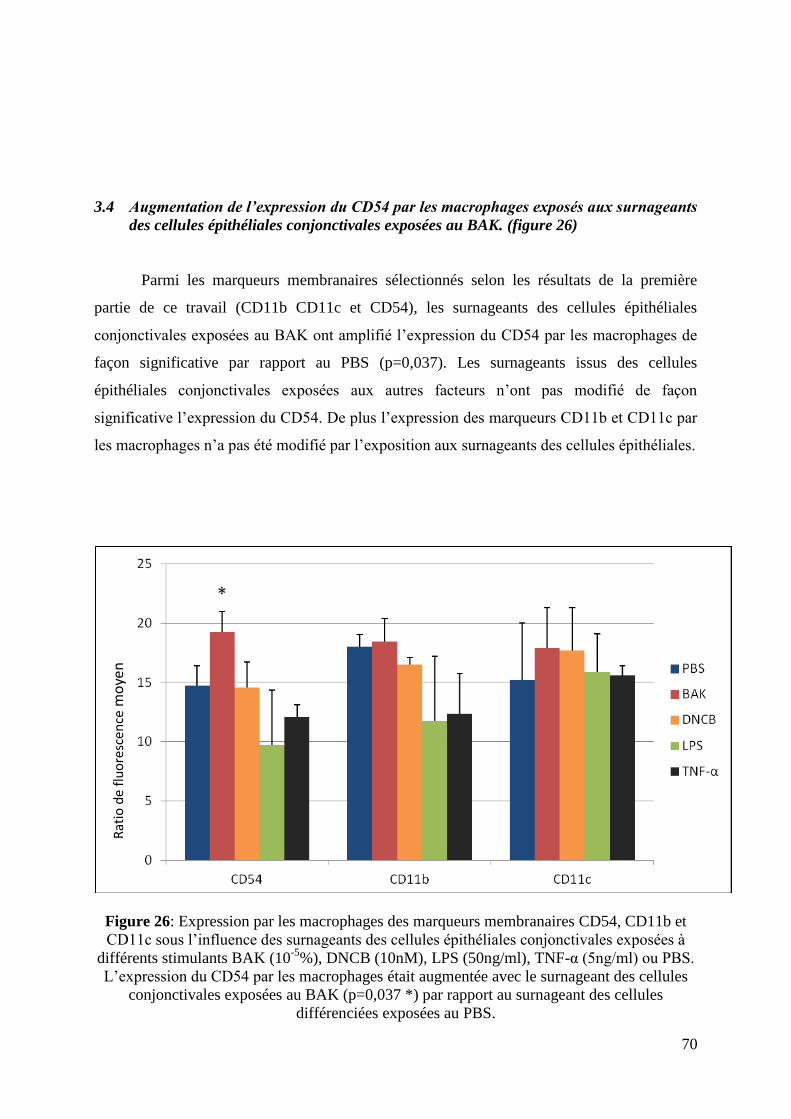
70
3.4 Augmentation de l’expression du CD54 par les macrophages exposés aux surnageants
des cellules épithéliales conjonctivales exposées au BAK. (figure 26)
Parmi les marqueurs membranaires sélectionnés selon les résultats de la première
partie de ce travail (CD11b CD11c et CD54), les surnageants des cellules épithéliales
conjonctivales exposées au BAK ont amplifié l’expression du CD54 par les macrophages de
façon significative par rapport au PBS (p=0,037). Les surnageants issus des cellules
épithéliales conjonctivales exposées aux autres facteurs n’ont pas modifié de façon
significative l’expression du CD54. De plus l’expression des marqueurs CD11b et CD11c par
les macrophages n’a pas été modifié par l’exposition aux surnageants des cellules épithéliales.
Figure 26: Expression par les macrophages des marqueurs membranaires CD54, CD11b et
CD11c sous l’influence des surnageants des cellules épithéliales conjonctivales exposées à
différents stimulants BAK (10-5
%), DNCB (10nM), LPS (50ng/ml), TNF-α (5ng/ml) ou PBS.
L’expression du CD54 par les macrophages était augmentée avec le surnageant des cellules
conjonctivales exposées au BAK (p=0,037 *) par rapport au surnageant des cellules
différenciées exposées au PBS.
Rat
io d
e fl
uo
resc
ence
mo
yen

71
4. DISCUSSION
4.1 Stimulation directe des cellules épithéliales conjonctivales
Pour valider le modèle et afin de croiser les effets directs et indirects du BAK sur les
cellules épithéliales conjonctivales et les macrophages, les cellules épithéliales ont été
exposées de façon isolée aux différents stimulants utilisés en première partie (BAK, DNCB,
LPS et TNF-α) à des concentrations infra toxiques. Par l’intermédiaire de l’expression
d’EMMPRIN, le BAK et le DNCB avaient une action directe sur les cellules épithéliales
conjonctivales. EMMPRIN (CD147), en plus d’être un inducteur des métallo-protéinases
(MMPs), possède aussi des capacités de régulation de la réponse lymphocytaire (95), ce qui
en fait un modulateur et témoin d’une activité inflammatoire. Ces fonctions sont expliquées
en partie par le rôle d’adressage du transporteur monocarboxylate (MCT 1 et 3) à la
membrane plasmique (68). L’activation d’EMMPRIN est impliquée dans les interactions
entre les cellules épithéliales et le stroma sous jacent notamment dans le remodelage
pathologique mais aussi physiologique de la matrice extra cellulaire (42). Les cellules
épithéliales conjonctivales exposées au BAK et au DNCB pourraient ainsi stimuler la
dégradation de la MEC même à des concentrations infra toxiques. Sur le plan des cytokines
produites par les cellules épithéliales, le DNCB n’a pas modifié le profil étudié, par contre le
BAK induisait la production d’IL-6 et d’IL-8. Epstein et al. ont montré que sous l’influence
du BAK, les cellules épithéliales conjonctivales produisaient du TNF-α et de l’IL-1 (37). En
comparaison avec notre étude, les concentrations de BAK utilisées par ces auteurs étaient bien
plus importantes. De plus, Baudouin et al. ont observé une quantité d’IL-8 significativement
plus importante dans les empreintes conjonctivales de patients traités par collyres
hypotonisants avec conservateur par rapport aux patients traités avec des collyres sans
conservateur (8). Ces résultats confirmerait in vitro le rôle pro-inflammatoire du BAK à des
concentrations non toxiques sur les cellules épithéliales conjonctivales à travers la production
de cytokines et l’expression d’EMMPRIN.
Le LPS est exprimé dans la paroi des bactéries Gram -, c’est une des principales voies de
reconnaissance de ces bactéries par les membranes cellulaires, par l’intermédiaire du TLR-4.
Ces effets proinflammatoires sur les cellules épithéliales conjonctivales et les cellules
présentatrices d’antigènes sont connus (23). Par ces effets stimulants sur les deux lignées

72
cellulaires il a ainsi joué le rôle de témoin positif. Le LPS est un moyen simple de mimer une
partie de l’action de certaines bactéries. In vivo, il produit une inflammation conjonctivale liée
à une infiltration de monocytes avec production d’IL1-α (79). Dans notre étude, malgré
l’absence de modification d’expression des marqueurs membranaires testés (CD40, 54, 63,
95, HLA-DR et EMMPRIN), les cellules conjonctivales ont produit de l’IL-6 et de l’IL-8 sous
l’influence directe du LPS. Même si cela n’a pas été mis en évidence dans notre étude,
l’exposition des cellules conjonctivales au LPS pourrait aussi induire une production d’IL-1
(37, 79), confirmant son rôle activateur des cellules épithéliales conjonctivales.
Le TNF-α est un médiateur de l’inflammation produit par de nombreux types cellulaires
différents. Son rôle dans les maladies inflammatoires auto-immunes est reconnu (32). Il agit
en induisant l’expression de cytokines, de molécules d’adhésion et de co-stimulation, en
activant la voie neutrophile, par l’intermédiaire de la voie NF-κB (nuclear factor-kappa B).
Plus spécifiquement, il a aussi une action favorisant la migration des cellules dendritiques
cornéennes (30). Dans la première partie de ce travail, il s’est révélé comme stimulant de
façon directe les macrophages. Bien qu’il n’ait pas modifié l’expression membranaire du
CD40, 54, 63, 95, HLA-DR et EMMPRIN, l’exposition des cellules conjonctivales au TNF-α
a induisait une forte production d’IL-6 et d’IL-8, et une production plus modérée de CCL-1.
Enriquez-de-Salamanca et al. ,ont montré que la stimulation par du TNF-α des cellules
épithéliales conjonctivales in vitro induisait la production de IL-12p40, GM-CSF, MCP-1, IL-
7, RANTES, IFN-γ, IL-1α, IL-1β, IL-3 et CXCL-10 (36). Bien que le panel de cytokines
testées, tout comme le kit de détection ou encore la lignée cellulaire étaient différentes entre
nos deux études, la production d’IL-6 et d’IL-8 a été retrouvée stimulée dans les deux cas.
4.2 Influence des macrophages sur les cellules conjonctivales
Les surnageants provenant de la culture des macrophages exposés au BAK et au
DNCB n’ont pas modifié l’expression des marqueurs étudiés sur les cellules épithéliales
conjonctivales. Ceci est surprenant car l’expression d’EMMPRIN par les cellules épithéliales
était augmentée sous l’influence directe du BAK et du DNCB. Nous pouvons en déduire que
le BAK et le DNCB n’avaient un effet sur l’expression d’EMMPRIN par les cellules
épithéliales conjonctivales qu’en cas d’exposition directe. Ceci pourrait confirmer ce que l’on

73
observe in vivo, où les cellules les plus exposées sont les cellules épithéliales, avec des
conséquences connues (29, 31, 37).
Bien que stimulant actif direct des cellules conjonctivales des macrophages, la
stimulation des macrophages par le TNF-α n’a pas produit d’effet sur les cellules épithéliales.
Les surnageants des macrophages exposés au LPS ont augmenté l’expression du CD54
par les cellules conjonctivales, alors que l’exposition directe au LPS n’avait pas eu d’effet sur
les cellules conjonctivales. Bien que le CD54 soit exprimé en majorité par les leucocytes et
les cellules endothéliales, il est aussi exprimé par les cellules épithéliales conjonctivales. Son
expression est augmentée en cas d’inflammation et d’allergie. Il a été montré que son
expression est endogène et ne provient pas d’un captage de CD54 soluble (7). Dans la
première partie de ce travail, nous avions montré que les macrophages exposés au LPS
produisaient du CD54 soluble, ce n’est donc pas du CD54 soluble qui s’est fixé sur les
cellules conjonctivales mais bien l’influence des cytokines produites par les macrophages
exposés au LPS qui a entrainé une production endogène de CD54 membranaire par les
cellules conjonctivales. Le CD54 joue un rôle important dans l’immunité innée et adaptative.
Son expression est augmentée par les cytokines pro inflammatoires (55). Il a été montré que
l’interaction entre les cellules cornéennes et les CPA liée au LPS entrainait une production de
TNF-α et d’IL-6 liée aux TLR 2 et 4 (23). La surexpression du CD54 par les cellules
conjonctivales lorsqu’elles étaient cultivées en présence des surnageants de macrophages
exposés au LPS suggérait que les macrophages aient un effet d’activation sur les cellules
conjonctivales en cas d’infection par les bactéries produisant du LPS. Il a été montré que les
macrophages activés par le LPS permettent le déclenchement de la voie des MAP kinases en
l’absence de corécepteur au LPS (86). Il a été aussi montré que l’IFN-γ était nécessaire afin
d’obtenir une réaction conjonctivale en cas d’exposition au LPS (82). De l’IFN-γ a été
retrouvé en faible quantité dans les surnageants des macrophages exposés au LPS. Dans notre
modèle in vitro, les macrophages posséderaient donc un rôle indispensable dans la réponse
aux infections liées au LPS. Les cellules épithéliales conjonctivales ne réagiraient au LPS
qu’en présence d’un cofacteur, elles n’activeraient donc pas les macrophages en cas
d’exposition non agressive au LPS. Par contre, en cas d’exposition invasive au LPS,
atteignant les couches plus profondes contenant des macrophages, le LPS aurait un effet
stimulant sur toute la surface oculaire, par l’intermédiaire de l’expression du CD54, entrainant
une cascade inflammatoire.

74
4.3 Les effets du BAK dans les interactions entre les cellules conjonctivales et les
macrophages
Précédemment nous avions observé que le BAK avait un effet stimulant direct sur les
macrophages à des concentrations infra toxiques.
Par l’intermédiaire des cellules conjonctivales (les plus exposées au BAK), les
macrophages subissent l’influence indirecte du BAK. Présents de façon assez rare dans la
conjonctive en conditions physiologiques, la conjonctive des patients exposés au BAK
contient un plus grand nombre de macrophages. Cette infiltration de macrophages présente
chez les patients exposés au BAK peut être expliquée de deux manières. La première est un
recrutement direct des cellules inflammatoires par le BAK comme cela a été démontré dans la
première partie de ce travail. La deuxième est un recrutement des cellules inflammatoires en
réponse à une stimulation par la première ligne de cellules exposées au BAK, les cellules
épithéliales. Si les surnageants des macrophages exposés au BAK n’ont pas eu d’effet sur
l’expression des marqueurs de surface des cellules épithéliales, les surnageants des cellules
conjonctivales exposées au BAK ont amplifié l’expression du CD54 par les macrophages.
Ceci est d’autant plus intéressant que le BAK n’a pas eu d’effet direct sur l’expression du
CD54 par les macrophages aussi bien dans notre étude (première partie) que dans la littérature
(47, 57). C’est donc bien par l’intermédiaire d’un facteur produit par les cellules épithéliales
que l’expression du CD54 a été augmentée sur les macrophages. Le CD54 joue un rôle
important dans la diapédèse des leucocytes et leur transmigration (2). D’autres fonctions lui
ont été attribuées, en particulier l’activation de cascades inflammatoires qui propagent la
réaction inflammatoire (55). Dans la première partie de ce travail, l’exposition au BAK des
macrophages avait entraîné une sécrétion de CD54 soluble dans les surnageants de ces
cellules. Il semblerait donc que l’exposition au BAK induise une production de CD54 soluble
par les macrophages et parallèlement que les cellules conjonctivales exposées au BAK
induisent l’expression membranaire de CD54 par les macrophages. Les macrophages de la
conjonctive seraient ainsi doublement activés.

75
5. CONCLUSION
Les cellules épithéliales conjonctivales et les macrophages ont montré des interactions par
l’intermédiaire de leurs surnageants en cas d’exposition croisée à des facteurs toxiques,
allergiques, inflammatoires et infectieux. Avec des capacités de réaction directe face aux
stimulations pro inflammatoires, en produisant des cytokines, en augmentant l’expression de
marqueurs membranaires, mais aussi par des capacités d’activation des macrophages, le rôle
immunomodulateur des cellules épithéliales conjonctivales est confirmé. Les macrophages
exposés au LPS ont augmenté l’expression du CD54 par les cellules épithéliales
conjonctivales tout comme les cellules épithéliales conjonctivales exposées au BAK ont
augmenté l’expression du CD54 par les macrophages. Le BAK pourrai avoir ainsi un
potentiel de recrutement de cellules inflammatoires au même titre que le LPS.

76
CONCLUSION ET PERSPECTIVES
Il a été intéressant d’observer que les facteurs utilisés, BAK, DNCB, LPS et TNF-α, ont
eu un effet sur les macrophages mais aussi sur les cellules épithéliales conjonctivales in vitro.
Ceci est d’autant plus remarquable que la concentration des stimulants utilisés était en dessous
des seuils de cytoxicité. En particulier pour le BAK, qui s’est révélé un stimulant pro-
inflammatoire direct et indirect des macrophages comparable aux facteurs pro-inflammatoires
classiques comme le LPS. Les macrophages sont moins exposés que les cellules épithéliales
conjonctivales aux agressions environnementales, leur recrutement s’explique donc aussi par
un recrutement indirect. C’est ce recrutement indirect par l’intermédiaire des cellules
épithéliales conjonctivales qui pourrait expliquer en partie l’infiltration conjonctivale
retrouvée chez les patients traités au long cours par des collyres contenant des conservateurs.
Le BAK dans les préparations ophtalmologiques ne devrait pas être considéré seulement
comme un élément toxique mais aussi comme un facteur pro-inflammatoire direct.
Il existe néanmoins de très nombreuses limites à ces deux travaux réalisés in vitro.
L’ensemble des acteurs cellulaires et leurs interactions au sein de la surface oculaire n’ont pas
été pris en compte notamment les autres cellules immunitaires (lymphocytes et cellules
dendritiques), les cellules à mucus et les fibroblastes mais aussi les constituants extra-
cellulaires ou encore le film lacrymal. Actuellement, des modèles animaux des pathologies
iatrogènes de la surface oculaire (rat, souris et lapin essentiellement) permettent de prendre en
compte l’ensemble des acteurs de la surface oculaire, mais la multiplication des éléments qui
composent la surface oculaire multiplie aussi les différences phénotypiques entre les modèle
animaux et l’homme. Parallèlement, des modèles in vitro humains d’étude de la surface
oculaire ont été développés (modèle 3D-HCE : Laboratoires SkinEthic®, Nice, France),
offrant une large possibilité d’expérimentations toxicologiques (65). Cependant, ces modèles
in vitro ne considèrent actuellement qu’un seul type cellulaire ou des interactions simples
entre deux types cellulaires (21). La caractérisation et l’obtention progressive des différents
constituants cellulaires de la surface oculaire comme les macrophages permettront sans doute
dans le futur de voir se développer un modèle d’étude in vitro mimant les réactions
impliquées chez l’homme.

77
Références bibliographiques
1. ABU EL-ASRAR AM, AL-KHARASHI SA, MISSOTTEN L, GEBOES K. Expression of growth
factors in the conjunctiva from patients with active trachoma. Eye (Lond). 2006 ; 20 : 362-9.
2. ADAMSON P, ETIENNE S, COURAUD PO, CALDER V, GREENWOOD J: Lymphocyte
migration through brain endothelial cell mono layers involves signaling through endothelial
ICAM-1 via a Rho-dependent pathway. J Immunol. 1999 ; 162 : 2964–2973.
3. ALLANSMITH MR, GREINER JV, BAIRD RS. Number of inflammatory cells in the normal
conjunctiva. Am J Ophthalmol. 1978 ; 86 : 250-9.
4. ALT C, HARRISON T, DOUSMAN L, FUJITA N, SHEW K, TRAN TT, et al. Increased CCL2
expression and macrophage/monocyte migration during microbicide-induced vaginal irritation.
Curr HIV Res. 2009 ; 7 : 639-49.
5. AMMAR DA, NOECKER RJ, KAHOOK MY. Effects of benzalkonium chloride- and polyquad-
preserved combination glaucoma medications on cultured human ocular surface cells. Adv Ther.
2011 ; 28 : 501-10.
6. AUWERX J. The human leukemia cell line, THP-1: a multifacetted model for the study of
monocyte-macrophage differentiation. Experientia. 1991 ; 47 : 22-31.
7. BAGNASCO M, PESCE G, FIORINO N, RICCIO AM, CIPRANDI G, BUSCAGLIA S, et al. In
situ hybridization analysis of ICAM-1 (CD54) mRNA on conjunctival epithelium during allergic
inflammation. Clin Exp Allergy. 1997 ; 27 : 737-43.
8. BAUDOUIN C, HAMARD P, LIANG H, CREUZOT-GARCHER C, BENSOUSSAN L,
BRIGNOLE F. Conjunctival epithelial cell expression of interleukins and inflammatory markers
in glaucoma patients treated over the long term. Ophthalmology. 2004 ; 111 : 2186-92.
9. BAUDOUIN C, LABBÉ A, LIANG H, PAULY A, BRIGNOLE-BAUDOUIN F. Preservatives in
eyedrops: the good, the bad and the ugly. Prog Retin Eye Res. 2010 ; 29 : 312-34.
10. BAUDOUIN C, LIANG H, HAMARD P, RIANCHO L, CREUZOT-GARCHER C, WARNET
JM, et al. The ocular surface of glaucoma patients treated over the long term expresses
inflammatory markers related to both T-helper 1 and T-helper 2 pathways. Ophthalmology. 2008 ;
115 : 109-15
11. BAUDOUIN C, PISELLA PJ, FILLACIER K, GOLDSCHILD M, BECQUET F, DE SAINT
JEAN M, et al. Ocular surface inflammatory changes induced by topical antiglaucoma drugs:
human and animal studies. Ophthalmology. 1999 ; 106 : 556-63.
12. BAUDOUIN C. A new approach for better comprehension of diseases of the ocular surface. J Fr
Ophtalmol. 2007 ; 30 : 239-46.

78
13. BOISLÈVE F, KERDINE-RÖMER S, ROUGIER-LARZAT N, PALLARDY M. Nickel and
DNCB Induce CCR7 Expression on Human Dendritic Cells Through Different Signalling
Pathways:Role of TNF-a and MAPK. J Invest Dermatol. 2004 ; 123 : 494-502.
14. BORA NS, GOBLEMAN CL, ATKINSON JP, PEPOSE JS, KAPLAN HJ. Differential
expression of the complement regulatory proteins in the human eye. Invest Ophthalmol Vis Sci.
1993 ; 34 : 3579-84.
15. BOUCHARD CS, LASKY JB, CUNDIFF JE, SMITH BS. Ocular surface upregulation of
intercellular adhesive molecule-1 (ICAM-1) by local interferon-gamma (IFN-gamma) in the rat.
Curr Eye Res. 1996 ; 15 : 203-8.
16. BRASNU E, BRIGNOLE-BAUDOUIN F, RIANCHO L, WARNET JM, BAUDOUIN C.
Comparative study on the cytotoxic effects of benzalkonium chloride on the Wong-Kilbourne
derivative of Chang conjunctival and IOBA-NHC cell lines. Mol Vis. 2008 ; 14 : 394-402.
17. BROADWAY DC, GRIERSON I, O'BRIEN C, HITCHINGS RA. Adverse effects of topical
antiglaucoma medication. I. The conjunctival cell profile. Arch Ophthalmol. 1994 ; 112 : 1437-45.
18. BROADWAY DC, GRIERSON I, O'BRIEN C, HITCHINGS RA. Adverse effects of topical
antiglaucoma medication. II. The outcome of filtration surgery. Arch Ophthalmol. 1994 ; 112 :
1446-54.
19. BURON N, MICHEAU O, CATHELIN S, LAFONTAINE PO, CREUZOT-GARCHER C,
SOLARY E. Differential mechanisms of conjunctival cell death induction by ultraviolet
irradiation and benzalkonium chloride. Invest Ophthalmol Vis Sci. 2006 ; 47 : 4221-30.
20. CHARNOCK, C. Are multidose over-the-counter artificial tears adequately preserved? Cornea.
2006 ; 25 : 432-437.
21. CHASSIGNOL A, BRASNU E, BAUDOUIN C, RIANCHO L, WARNET JM, BRIGNOLE-
BAUDOUIN F. In vitro interactions between peripheral blood lymphocytes and the Wong-
Kilbourne derivative of Chang conjunctival cells. Invest Ophthalmol Vis Sci. 2012 ; 53 : 1492-8.
22. CHIBRET H. Conservateurs et préparations ophtalmiques: réalités et perspectives. Ann Pharm fr.
1997 ; 55 : 228-31
23. CHUNG SH, KWEON MN, LEE HK, CHOI SI, YANG JY, KIM EK. Toll-like receptor 4
initiates an innate immune response to lipopolysaccharide in human conjunctival epithelial cells.
Exp Eye Res. 2009 ; 88 : 49-56.
24. CONE RA, HOEN T, WONG X, ABUSUWWA R, ANDERSON DJ, MOENCH TR. Vaginal
microbicides: detecting toxicities in vivo that paradoxically increase pathogen transmission. BMC
Infect Dis. 2006 ; 6 : 90.
25. COUTANT KD, DE FRAISSINETTE AB, CORDIER A, ULRICH P. Modulation of the activity
of human monocyte-derived dendritic cells by chemical haptens, a metal allergen, and a
staphylococcal superantigen. Toxicol Sci. 1999 ; 52 : 189-98.

79
26. CRUIKSHANK W, LITTLE F. lnterleukin-16: the ins and outs of regulating T-cell activation.
Crit Rev Immunol. 2008 ; 28 : 467-83.
27. DAIGNEAULT M, PRESTON JA, MARRIOTT HM, WHYTE MK, DOCKRELL DH. The
Identification of Markers of Macrophage Differentiation in PMA-Stimulated THP-1 Cells and
Monocyte-Derived Macrophages. PLoS One. 2010 ; 5:e8668.
28. DE SAINT JEAN M, BAUDOUIN C, DI NOLFO M, ROMAN S, LOZATO P, WARNET JM,
BRIGNOLE F. Comparison of morphological and functional characteristics of primary-cultured
human conjunctival epithelium and of Wong-Kilbourne derivative of Chang conjunctival cell line.
Exp Eye Res. 2004 ; 78 : 257-74.
29. DE SAINT JEAN M, BRIGNOLE F, BRINGUIER AF, BAUCHET A, FELDMANN G,
BAUDOUIN C. Effects of Benzalkonium Chloride on Growth and Survival of Chang
Conjunctival Cells. Invest Ophthalmol Vis Sci. 1999 ; 40 : 6l9-630
30. DEKARIS I, ZHU SN, DANA MR. TNF-{alpha} Regulates Corneal Langerhans Cell Migration.
J. Immunol. 1999 ; 162 : 4235-4239
31. DEUTSCHLE T, PORKERT U, REITER R, KECK T, RIECHELMANN H. In vitro genotoxicity
and cytotoxicity of benzalkonium chloride. Toxicol In Vitro. 2006 ; 20 : 1472-1477.
32. DINARELLO CA. Inflammatory cytokines: interleukin-1 and tumor necrosis factor as effector
molecules in autoimmune diseases. Curr Opin Immunol. 1991 ; 3 : 941-8.
33. DRY EYE WORK SHOP. The definition and classification of dry eye disease: report of the
Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007). Ocul
Surf. 2007 ; 5 : 75-92.
34. DRY EYE WORK SHOP. The epidemiology of dry eye disease: report of the Epidemiology
Subcommittee of the International. Ocul Surf. 2007 ; 5 : 93-107.
35. EMEA public statement on antimicrobial preservatives in ophthalmic preparations for human use.
EMEA/622721/2009, London, 2009.
36. ENRÍQUEZ-DE-SALAMANCA A, CALDER V, GAO J, GALATOWICZ G, GARCÍA-
VÁZQUEZ C, FERNÁNDEZ I, et al. Cytokine responses by conjunctival epithelial cells: an in
vitro model of ocular inflammation. Cytokine. 2008 ; 44 : 160-7.
37. EPSTEIN SP, CHEN D, ASBELL PA. Evaluation of biomarkers of inflammation in response to
benzalkonium chloride on corneal and conjunctival epithelial cells. J Ocul Pharmacol Ther. 2009 ;
25 : 415-24.
38. ESTRELLA JL, KAN-SUTTON C, GONG X, RAJAGOPALAN M, LEWIS DE, HUNTER RL,
et al. A Novel in vitro Human Macrophage Model to Study the Persistence of Mycobacterium
tuberculosis Using Vitamin D(3) and Retinoic Acid Activated THP-1 Macrophages. Front
Microbiol. 2011 ; 2 : 67.

80
39. EUROPEAN GLAUCOMA SOCIETY. Terminology and guidelines for glaucoma (3rd edition).
Savona : Dogma, 2008 ; 183 p.
40. FOGAGNOLO P, ROSSETTI L. Medical treatment of glaucoma: present and future. Expert Opin
Investig Drugs. 2011 ; 20 : 947-959.
41. FORRESTER JV, XU H, KUFFOVÁ L, DICK AD, MCMENAMIN PG. Dendritic cell
physiology and function in the eye. Immunological Reviews. 2010 ; 234 : 282–304.
42. GABISON EE, HUET E, BAUDOUIN C, MENASHI S. Direct epithelial-stromal interaction in
corneal wound healing: Role of EMMPRIN/CD147 in MMPs induction and beyond. Prog Retin
Eye Res. 2009 ; 28 : 19-33.
43. GARNACHE-OTTOU F, CHAPEROT L, BIICHLE S, FERRAND C, REMY-MARTIN JP,
DECONINCK E, et al. Expression of the myeloid-associated marker CD33 is not an exclusive
factor for leukemic plasmacytoid dendritic cells. Blood. 2005 ; 105 : 1256-64.
44. GILL KS, YANNARIELLO-BROWN J, PATEL J, NAKAJIMA N, RAJARAMAN S, TROCMÉ
SD. ICAM-1 expression in corneal epithelium of a patient with vernal keratoconjunctivitis: case
report. Cornea. 1997 ; 16 : 107-11.
45. GROSS RL,PEACE JH, SMITH SE, WALTERS TR, DUBINER HB, WEISS MJ, et al. Duration
of IOP reduction with travoprost BAK-free solution. J Glaucoma. 2008 ; 17 : 217-222.
46. HELIN M, RÖNKKÖ S, PUUSTJÄRVI T, TERÄSVIRTA M, OLLIKAINEN M, UUSITALO H.
Conjunctival inflammatory cells and their predictive role for deep sclerectomy in primary open-
angle glaucoma and exfoliation glaucoma. J Glaucoma. 2011 ; 20 : 172-8.
47. HIROTA M, MORO O. MIP-1beta, a novel biomarker for in vitro sensitization test using human
monocytic cell line. Toxicol In Vitro. 2006 ; 20 : 736-42.
48. HU J, WANG Y, XIE L. Potential role of macrophages in experimental keratomycosis. Invest
Ophthalmol Vis Sci. 2009 ; 50 : 2087-94.
49. ISHIDA W, FUKUDA K, KAJISAKO M, TAKAHASHI A, SUMI T, VAN ROOIJEN N, et al.
Conjunctival macrophages act as antigen-presenting cells in the conjunctiva during the
development of experimental allergic conjunctivitis. Mol Vis. 2010 ; 16 : 1280-5.
50. KANTELIP B, CREUZOT-GARCHER C. Anatomie descriptive in HOANG XUAN T.
Inflammation chronique de la conjonctive. Marseille : Lamy, 1998 : p. 21-45.
51. KINOSHITA S, KIORPES TC, FRIEND J, THOFT RA. Goblet cell density in ocular surface
disease. A better indicator than tear mucin. Arch Ophthalmol. 1983 ; 101 : 1284-7.
52. LABBÉ A, KHAMMARI C, BAUDOUIN C. Wound healing modulation in glaucoma surgery. J
Fr Ophtalmol. 2007 ; 30 : 631-46.
53. LAIBOVITZ R, ZIMMERMAN, K, GETSON A, SHEDDEN A, LAURENCE J, ADAMSONS I.
Comparison of the efficacy and tolerability of preservative-free 2.0% dorzolamide / 0.5% timolol
and 2.0% dorzolamide / 0.5% timolol. Invest Ophthalmol Vis Sci. 1999 ; 40 : S665.

81
54. LAVAPPA KS. Survey of ATCC stocks of human cell lines for HeLa contamination. In Vitro.
1978 ; 14 : 469-475.
55. LAWSON C, WOLF S. ICAM-1 signaling in endothelial cells. Pharmacol Rep. 2009 ; 61 : 22-32.
56. LEUNG EW, MEDEIROS FA, WEINREB RN. Prevalence of ocular surface disease in glaucoma
patients. J Glaucoma. 2008 ; 17 : 350-5.
57. LIM YM, MOON SJ, AN SS, LEE SJ, KIM SY, CHANG IS, et al. Suitability of macrophage
inflammatory protein-1beta production by THP-1 cells in differentiating skin sensitizers from
irritant chemicals. Contact Dermatitis. 2008 ; 58 : 193-8.
58. MAJUMDAR S, HIPPALGAONKAR K, REPKA MA. Effect of chitosan, benzalkonium chloride
and ethylenediaminetetraacetic acid on permeation of acyclovir across isolated rabbit cornea. Int J
Pharm. 2008 ; 348 : 175-178.
59. MALVITTE L, MONTANGE T, VEJUX A, BAUDOUIN C, BRON AM,
CREUZOT‐GARCHER C, et al. Measurement of inflammatory cytokines by multicytokine assay
in tears of patients with glaucoma topically treated with chronic drugs. Br J Ophthalmol. 2007 ; 91
: 29–32.
60. MANSBERGER SL, CIOFFI GA. The probability of glaucoma from ocular hypertension
determined by ophthalmologists in comparison to a risk calculator. J Glaucoma. 2006 ; 15 : 426-
31.
61. MASTERS JR. Hela cells 50 years on: The good, the bad and the ugly. Nature reviews. 2002 ; 2 :
315-319.
62. MATHER JP, ROBERTS PE. Introduction to cell and tissue culture, theory and technique, 1st
edition. New York : Springer, 1998 : 241 p.
63. MIETZ H, NIESEN U, KRIEGLSTEIN GK. The effect of preservatives and antiglaucomatous
medication on the histopathology of the conjunctiva. Graefes Arch Clin Exp Ophthalmol. 1994 ;
232 : 561-5.
64. NUZZI R, VERCELLI A, FINAZZO C, CRACCO C. Conjunctiva and subconjunctival tissue in
primary open-angle glaucoma after long-term topical treatment: an immunohistochemical and
ultrastructural study. Graefes Arch Clin Exp Ophthalmol. 1995 ; 233 : 154-62.
65. PAULY A, MELONI M, BRIGNOLE-BAUDOUIN F, WARNET JM, BAUDOUIN C. Multiple
endpoint analysis of the 3D-reconstituted corneal epithelium after treatment with benzalkonium
chloride: early detection of toxic damage. Invest Ophthalmol Vis Sci. 2009 ; 50 : 1644-52.
66. PAWAR PK, MAJUMDAR DK. Effect of formulation factors on in vitro permeation of
moxifloxac in from aqueous drops through excised goat, sheep, and buffalo corneas. AAPS Pharm
Sci Tech. 2006 ; 7 : e13.
67. PELLINEN P, LOKKILA J. Corneal penetration into rabbit aqueous humor is comparable
between preserved and preservative-free tafluprost. Ophthalmic Res. 2009 ; 41 : 118-122.

82
68. PHILP NJ, OCHRIETOR JD, RUDOY C, MURAMATSU T, LINSER PJ. Loss of MCT1, MCT3,
and MCT4 expression in the retinal pigment epithelium and neural retina of the 5A11/basigin-null
mouse. Invest Ophthalmol Vis Sci. 2003 ; 44 : 1305–11.
69. PISELLA PJ, DEBBASCH C, HAMARD P, CREUZOT-GARCHER C, RAT P, BRIGNOLE F,
et al. Conjunctival proinflammatory and proapoptotic effects of latanoprost and preserved and
unpreserved timolol: an ex vivo and in vitro study. Invest Ophthalmol Vis Sci. 2004 ; 45 : 1360-8.
70. PONG JC, CHU CY, LI WY, TANG LY, LI L, LUI WT, et al. Association of hemopexin in tear
film and conjunctival macrophages with vernal keratoconjunctivitis. Arch Ophthalmol. 2011 ; 129
: 453-61.
71. QIN Z. The use of THP-1 cells as a model for mimicking the function and regulation of
monocytes and macrophages in the vasculature. Atherosclerosis. 2012 ; 221 : 2-11.
72. RAZZAQUE MS, FOSTER CS, AHMED AR. Role of enhanced expression of m-CSF in
conjunctiva affected by cicatricial pemphigoid. Invest Ophthalmol Vis Sci. 2002 ; 43 : 2977-83.
73. RICHTER CU, SHINGLETON BJ, BELLOWS AR, HUTCHINSON BT, O'CONNOR T, BRILL
I. The development of encapsulated filtering blebs. Ophthalmology. 1988 ; 95 : 1163-8.
74. ROEBROCK K, WOLF M, BOVENS S, LEHR M, SUNDERKÖTTER C. Inhibition of
benzalkonium chloride-induced skin inflammation in mice by an indol-1-ylpropan-2-one inhibitor
of cytosolic phospholipase A2 α. Br J Dermatol. 2012 ; 166 : 306-16.
75. ROSENTHAL RA, BUCK SL, HENRY CL, SCHLECH BA. Evaluation of the preserving
efficacy of lubricant eye drops with a novel preservative system. J Ocul Pharmacol Ther. 2006 ;
22 : 440-8.
76. ROULAND JF. [Initial medical treatment: beta-blockers or prostaglandins? The beta-blocker is
the Swiss Army knife of the glaucoma physician]. J Fr Ophtalmol. 2004 ; 27 : 716-7.
77. SCHEIN OD, HIBBERD PL, STARCK T, BAKER AS, KENYON KR. Microbial contamination
of in-use ocular medications. Arch Ophthalmol. 1992 ; 110 : 82-5.
78. SCHULTZ CL, MORCK DW, MCKAY SG, OLSON ME, BURET A. Lipopolysaccharide
induced acute red eye and corneal ulcers. Exp Eye Res. 1997 ; 64 : 3-9.
79. SCHUMACHER C, CLARK-LEWIS I, BAGGIOLINI M, MOSER B. High- and low-affinity
binding of GRO alpha and neutrophil-activating peptide 2 to interleukin 8 receptors on human
neutrophils. Proc Natl Acad Sci USA. 1992 ; 89 : 10542-6.
80. SHERWOOD MB, GRIERSON I, MILLAR L, HITCHINGS RA. Long-term morphologic effects
of antiglaucoma drugs on the conjunctiva and Tenon's capsule in glaucomatous patients.
Ophthalmology. 1989 ; 96 : 327-35.
81. STEWART M, THIEL M, HOGG N. Leukocyte integrins. Curr Opin Cell Biol. 1995 ; 7 : 690-6.
82. TAKAHASHI N. Quantitative cytotoxicity of preservatives evaluated in cell culture with Chang's
human conjunctival cells - effect of temperature on cytotoxicity. Jpn J Ophthalmol. 1982 ; 26 :
234-8.

83
83. TASLI H, COSAR G. Microbial contamination of eye drops. Cent Eur J Public Health. 2001 : 9 :
162-164.
84. TERAI N, SCHLÖTZER-SCHREHARDT U, LAMPEL J, BÖHM AG, RUMMELT C,
SCHMIDT E, et al. Effect of latanoprost and timolol on the histopathology of the human
conjunctiva. Br J Ophthalmol. 2009 ; 93 : 219-24.
85. TESAVIBUL N, DORFMAN D, SANGWAN VS, CHRISTEN W, PANAYOTIS Z, ROJAS B, et
al. Costimulatory molecules in ocular cicatricial pemphigoid. Invest Ophthalmol Vis Sci. 1998 ; 39
: 982–988.
86. THORLEY AJ, GRANDOLFO D, LIM E, GOLDSTRAW P, YOUNG A, TETLEY TD. Innate
Immune Responses to Bacterial Ligands in the Peripheral Human Lung - Role of Alveolar
Epithelial TLR Expression and Signaling. PLoS One. 2011 ; 6 : e21827
87. TOMINAGA T, SUZUKI M, SAEKI H, MATSUNO S, TACHIBANA T, KUDO T.
Establishment of an activated macrophage cell line, A-THP-1, and its properties. Tohoku J Exp
Med. 1998 ; 186 : 99-119.
88. TSUBOTA K, FUKAGAWA K, FUJIHARA T, SHIMMURA S, SAITO I, SAITO K, et al.
Regulation of Human Leukocyte Antigen Expression in Human Conjunctival Epithelium. Invest
Ophthalmol Vis Sci. 1999 ; 40 : 28-34.
89. TSUCHIYA S, YAMABE M, YAMAGUCHI Y, KOBAYASHI Y, KONNO T, TADA K.
Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int J
Cancer. 1980 ; 26 : 171-6.
90. VAN DE SANDT JJM, MAAS WJM, DOORNINK PC, RUTTEN AAJJL. Release of arachidonic
and linoleic acid metabolites in skin organ cultures as characteristics of in vitro skin irritancy.
Fundam Appl Toxicol. l995 ; 25 : 20-28.
91. VERMES I, HAANEN C, STEFFENS-NAKKEN H, REUTELINGSPERGER C. A novel assay
for apoptosis: flow cytometric detection of phosphatidylserine expression on early apoptotic cells
using fluorescein labelled annexin-V. J Immunol Methods. 1995 ; 184 : 39-51.
92. WEINREB RN, KHAW PT. Primary open-angle glaucoma. Lancet. 2004 ; 363 : 1711-20.
93. WILLIS CM, STEPHENS CJ, WILKINSON JD. Differential effects of structurally unrelated
chemical irritants on the density and morphology of epidermal CD 1+ cells. J Invest Dermatol.
1990 ; 95 : 711-6.
94. YENICE I, MOCAN MC, PALASKA E, BOCHOT A, BILENSOY E, VURAL I, et al.
Hyaluronic acid coated polyepsilon caprolactone nanospheres deliver high concentrations of
cyclosporine A into the cornea. Exp Eye Res. 2008 ; 87 : 162-167.
95. YURCHENKO V, CONSTANT S, EISENMESSER E, BUKRINSKY M. Cyclophilin–CD147
interactions : a new target for anti-inflammatory therapeutics. Clin Exp Immunol. 2010 ; 160 : 305-
17.

84
96. ZHAN H, TOWLER HM, CALDER VL. The immunomodulatory role of human conjunctival
epithelial cells. Invest Ophthalmol Vis Sci. 2003 ; 44 : 3906-10
97. ZHANG EY, CHEN AY, ZHU BT. Mechanism of dinitrochlorobenzene-induced dermatitis in
mice: role of specific antibodies in pathogenesis. PLoS One. 2009 ; 4 : e7703.

85
YEAR : 2012
AUTHOR’S NAME AND SURNAME : MICHEE SYLVAIN
MEMOIRE’S PRESIDENT: Pr BAHRAM BODAGHI
MEMOIRE’S DIRECTOR : Dr ANTOINE LABBE
TITLE : IN VITRO ANALYSIS OF INTERACTIONS BETWEEN MACROPHAGES AND CONJUNCTIVAL EPITHELIAL CELLS INFLUENCED BY
BENZALKONIUM CHLORIDE.
The purpose of the study was to establish an experimental model of interactions
between conjunctival epithelial cells and macrophages. Two cell lines were
studied, a monocytic line, THP-1 cells, differentiated in macrophages, and a
conjunctival cell line, Wong Kilbourne derivative of conjunctival Chang cells. Both
cell lines were exposed to stimulating factors mimicking pathophysiological
conditions implied in the ocular surface : benzalkonium chloride (BAK),
dinitrochlorobenzen (DNCB), lipopolysaccharide (LPS) and TNF-alpha (tumor
necrosis factor).
The direct effects of these factors on conjunctival cells and macrophages, also
their interactions with their surfactants, were studied through the expression of
cell markers, cell death, migration and phagocytosis behavior and cytokine
production. In this study, the stimulating factors reproducing ocular surface
pathologies activated phagocytosis, migration and cytokine production by this
cells. Compared with well known stimulating factors such as TNF-alpha or LPS,
the exposure to an infra toxic concentration of BAK had also stimulated
macrophages. In order to integrate macrophages in an ocular surface model,
conjunctival epithelial cells were associated to the study of macrophages. The
interactions between both cell lines showed that BAK activated macrophages as
well through the surfactants from conjunctival cells. This study showed that BAK
stimulated an inflammatory response and this effect was produced not only by a
direct effect on macrophages but also mediated by conjunctival epithelial cells.
KEY WORDS : INFLAMMATION, MACROPHAGES, CONJONCTIVA, PHARMACEUTICAL PRESERVATIVES, GLAUCOMA.
ADRESSE DE L’U.F.R. : 8, Rue du Général Sarrail 94010 CRETEIL

86
ANNEE : 2012
NOM ET PRENOM DE L’AUTEUR : MICHEE SYLVAIN
PRESIDENT DE THESE : Pr BAHRAM BODAGHI
DIRECTEUR DE THESE : Dr ANTOINE LABBE
TITRE DE LA THESE : ANALYSE IN VITRO DES INTERACTIONS ENTRE LES MACROPHAGES ET LES CELLULES EPITHELIALES DE LA CONJONCTIVE
SOUS L’INFLUENCE DU CHLORURE DE BENZALKONIUM
L’objectif de ce travail était de mettre au point un modèle d’étude des
interactions entre les cellules épithéliales de la conjonctive et les macrophages
sous l’effet du chlorure de benzalkonium. Deux lignées cellulaires humaines ont
été étudiées, une lignée monocytaire, les cellules THP-1, différenciée en
macrophages, et une lignée de cellules conjonctivales, les cellules de Wong
Kilbourne dérivées de la conjonctive de Chang. Les cellules ont été exposées à
différents stimulants reproduisant des conditions physiopathologiques impliquées
au niveau de la surface oculaire : le chlorure de benzalkonium (BAK), le
dinitrochlorobenzene (DNCB), le lipopolysaccharide (LPS) et le TNF-αlpha (tumor
necrosis factor).
Les effets directs des stimulants sur les cellules conjonctivales et les
macrophages ainsi que leurs interactions au travers de leurs surnageants, ont
été étudiés à travers l’expression de marqueurs membranaires, la mortalité
cellulaire, les capacités de migration et de phagocytose et la production de
cytokines. Dans cette étude, les facteurs de stimulation mimant des pathologies
de la surface oculaire ont activé la phagocytose, la migration et la production de
cytokines par ces cellules. En comparaison avec les effets stimulants de facteurs
biens connus comme le TNF-αlpha ou le LPS, une exposition à une concentration
infra toxique de BAK a également eu un effet stimulant sur les macrophages. Afin
d’intégrer les macrophages dans un modèle de surface oculaire, des cellules
épithéliales conjonctivales ont été associées à l’étude des macrophages. Les
interactions entre ces deux populations cellulaires ont montré que le BAK activait
également les macrophages par l’intermédiaire des surnageants des cellules
épithéliales. Cette étude montrait que le BAK stimule la réponse inflammatoire
non seulement par un effet direct sur les macrophages mais aussi par
l’intermédiaire des cellules épithéliales conjonctivales.
MOTS-CLES :
INFLAMMATION, MACROPHAGES, CONJONCTIVE, CONSERVATEURS PHARMACEUTIQUES,
GLAUCOME.
ADRESSE DE L’U.F.R. : 8, Rue du Général Sarrail 94010 CRETEIL