taxonomie, morphologie et anatomie du polysphaeria...
TRANSCRIPT

Cah. ORSTOM, sér. Biol., no 8 - juin 1969.
TAXONOMIE, MORPHOLOGIE ET ANATOMIE
DU Polysphaeria congesta (H.Bn.) A.Chev.,
FAUX CAFEIER SAUVAGE DE MADAGASCAR
PAR
H. RABl%.!HAULT*
RÉsuMF:
L’espèce étudiée éiail autrefois confondue avec celles des genres Coffea et Cremaspora. Ce faux-Caéfier s’en distingue en particulier par les poils qui ferment la gorge de la corolle et par ses graines ruminées: l’albumen est constitué par des feuillets longitudinaux.
La structure des organes végétatifs est proche le plus souvent de celle des Caféiers mais celle des graines est très différente. Ces dernières ne renferment pas de caféine.
Les essais de greffage avec les Caféiers cultivés n’ont pas donné de résultats.
SLJM~IARY
The species studied was previously mistaken for thaf of the genus Coffea and Cremaspora. The false coffee-shrub cari be distinguished from ihem by the hairs which close the throat of the corolla and by the ruminafe seeds: the albumen is formed of longitudinal layers.
The structure of the plant organs in most cases resemble those of the coffee-shrub but thaf of fhe seed is very differenf. The laffer does nof contain caffein.
Affempfs af graffing with cultivated coffee-shrubs did nof produce good results.
’ Mattre de Recherches, Services Scientifiques Centraux de I’ORSTOM, 70, 74 route d’Aulnay, 93-Bondy.

30 H. RABBCHAULT
L’un des objectifs de l’Institut français du Café et du Cacao est l’amélioration des Caféiers cultivés par la sélection voire par l’hybridation et le greffage. 11 est intéressant pour cela de disposer de nombreuses variétés et d’espèces sauvages qui présentent des gènes nouveaux, des caractères particuliers que ne possèdent pas les Caféiers cultivés. Or, depuis le début du siècle, les botanistes français, parmi lesquels le Professeur A. CHE- VALIER, ont, au cours de missions successives, découvert de nombreuses espèces sauvages à Madagascar et dans les îles voisines. J.-F. LEROY (19, 21, 22) a récemment recensé 35 espèces malgaches, et il pense qu’il en reste encore à découvrir. Certaines sont à pré- sent disparues et nous ne les connaissons que sous la forme d’échantillons d’herbier.
C’est la raison pour laquelle A. CHEVALIER (9, 12, 13), pour sauver ces richesses, avait préconisé à plusieurs reprises la création de collections vivantes ou vergers d’études. Ces collections vivantes auraient permis de cataloguer les espèces sauvages et d’étudier leur biologie.
En 1901, le savant chimiste G. BERTRAND (4, 5, 6) a montré que certaines espèces malgaches et des Ples Comores présentaient une particularité très importante : elles ne renferment pas de caféine dans leurs graines et A. CHEVALIER (9, 12) pensait que leur sélection aurait permis l’obtention naturelle d’un café sans caféine.
Ces dernières années, les idées et l’œuvre de ce savant ont été reprises par plusieurs botanistes du Muséum National d’Histoire Naturelle, notamment par le Professeur R. PORTÈRES (25) et par J.-F. LEROY (19 à 22) qui ont, en relation avec l’Institut français du Café et du Cacao, continué l’inventaire et commencé la création des premières collec- tions vivantes.
La flore malgache recèle aussi d’autres plantes très intéressantes parce qu’elles ressemblent à s’y méprendre aux Caféiers* ; ce sont en général également des Rubiacées qui appartiennent à des genres divers : Feretia, Cremaspora, Tricalysia, Polysphaeria, Belonophora, Craterispermum, Canthium, Pavetta, Psychotria, etc. La ressemblance est telle que leurs graines sont parfois utilisées localement pour la préparation d’une infusion identique au café*.
l Cette ressemblance est souvent telle que les systématiciens ont fait de nombreuses erreurs d’identitlcation. Pour indiquer cette affmite, ils ont utilise le nom d’espèce Q coffeoides n (Feretia coffeoides A. CHEV., Belo-
nophora coffeoides HOOK. f., Saldinia cofeoides AD. BR., etc.). Certains faux cafeiers ont ete cultives au même titre que les Cafeiers, c’est le cas du Café marron et du Cafe bâtard des Antilles, qui est le Rudgea caribea BENTH., du Cafeier sauvage de Sierra Leone qui serait selon HIERN le Randia grandiFora DC.. A. CHEVALIER (~OC. cif.) cite l’anecdote suivante concernant le Ferefia coffeoides appel8 Songala par les indigènes du Fouta-Djallon : les cerises jaunes de cet arbuste donnent des graines comme le Cafeier. Un echantillon ayant ete adresse a un expert d’Amsterdam, M. GRESHOFF, celui-ci a indique qu’il s’agissait d’un Cafe sans cafeine qui, apres torréfaction, donnait une boisson absolument analogue au Café.
PLANCHE 1. - Polysphaeria congesta (H. BN.) A. CHEV. : fig. A, Rameau plagiotrope avec fleurs et fruits ; B., deux boutons floraux ; C., fleur epanouie ; D, fleur en coupe longitudinale grossie, montrant l’insertion d’une étamine et la pilosité de la face interne de la corolle ; E, ovaire et calice, coupe grossie montrant, en particulier, la disposition des ovules ; F, imbrication des bractees a la base du calice des fleurs ; Fi et Fa, formes de calices ; Gr et G*, etamines vues de profil ; G8 et G”, anthères, les sacs polliniques sont parfois portes sur un receptacle charnu (connectif) plus large et plus long ; H1 et H*, details de la surface et de l’extrémité des poils qui recouvrent l’inte- rieur du tube de la corolle : 1’ et Ia, grains de pollen ; J, un fruit ; K, fruit en coupe transversale montrant la rumi- nation des graines ; L, graine vue de face montrant les sillons longitudinaux et sinueux qui penètrent l’albumen ; M, graine vue par dessus ; N, ecorché d’une graine montrant la disposition de l’embryon ; 0 et P, coupes longitu- dinales d’une graine selon deux plans : l’un passe par l’embryon qu’il sectionne transversalement, l’autre est dans
le grand axe de la graine ; Q, un embryon grossi. (Grossissements fig. A ys; B x 3; C, F, FI, F*, J, K x 4; D x 8; E x 20; G x 30; H x 200; 1 x 250;
L, M, N, 0, P x 4; Q x 16).

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 31
PLANCHE 1

32 H. RABhCHAULT
A. CHEVALIER (8, 9, 10) les a appelés (( Faux Caféiers )).
Leur étude s’avère nécessaire, ne serait-ce que pour définir les caractères taxo- nomique, morphologique, anatomique et biologique permettant de les distinguer des véritables Caféiers.
C’est une confusion entre un Caféier sauvage et un faux Caféier (le Polysphaeria congesfa) qui nous a amené à l’étude taxonomique, morphologique et anatomique de ce dernier.
1. TAXONOMIE, ORIGINE DU MATERIEL
En 1952, j’ai reçu de la part de M. CARRÉ, directeur de la Station de 1’Ivoloina (Madagascar), des échantillons (rameaux en fleurs, fruits et graines) d’un (( Caféier sauvage o sans caféine, baptisé Coffea humblofiana H. BN. et cultivé sur la station rive gauche de la rivière Ivoloina. Cette plante avait été introduite par une mission envoyée aux Comores à la recherche de Caféiers sauvages sans caféine, à la suite des travaux précités de CI. BERTRAND et de A. CHEVALIER.
Le C. humblofiana a été en effet signalé dans ces îles mais grâce à un examen som- maire, je pus me rendre à l’évidence que les échantillons dont je disposais ne correspon- daient pas à la diagnose de cette espèce et qu’il ne s’agissait pas non plus d’un Coffea.
Les graines semées dans les serres de la Section technique d’Agriculture Tropicale de Nogent-sur-Marne ont donné plusieurs jeunes plantes qui m’ont été très utiles pour avoir le matériel frais nécessaire à l’étude anatomique.
Cette plante se distinguait du genre Coffea par deux caractères essentiels : la gorge de la corolle est obturée par des poils nombreux et d’autre part la graine petite ne com- porte pas de sillon ventral comme le grain de café mais est profondément ruminée.
H. BAILLON (2, p. 283) avait décrit cette plante sous le nom de Cremaspora congesfa H. BN., en 1879.
Frufex, ramis opposifis virgafis, ufi planta fofa glabris. Folia opposifa oblongo- lanceolafa (ad 10 cent. longa, 4 cent. lafa), apice longiuscule acuminafa, basi angusfafa, v. rarius obfusiuscula, infegerrima membranacea, supra lucida laevia, subfus paulo pallidiora : nervis secundaris 12-14, inaequali-sinuafis; venis reficulafis vix prominulis. Pefioli breves (1/2-l cent.). Sfipulae inferpefiolares breves, deciduae. Flores axillares glomerafi crebri,
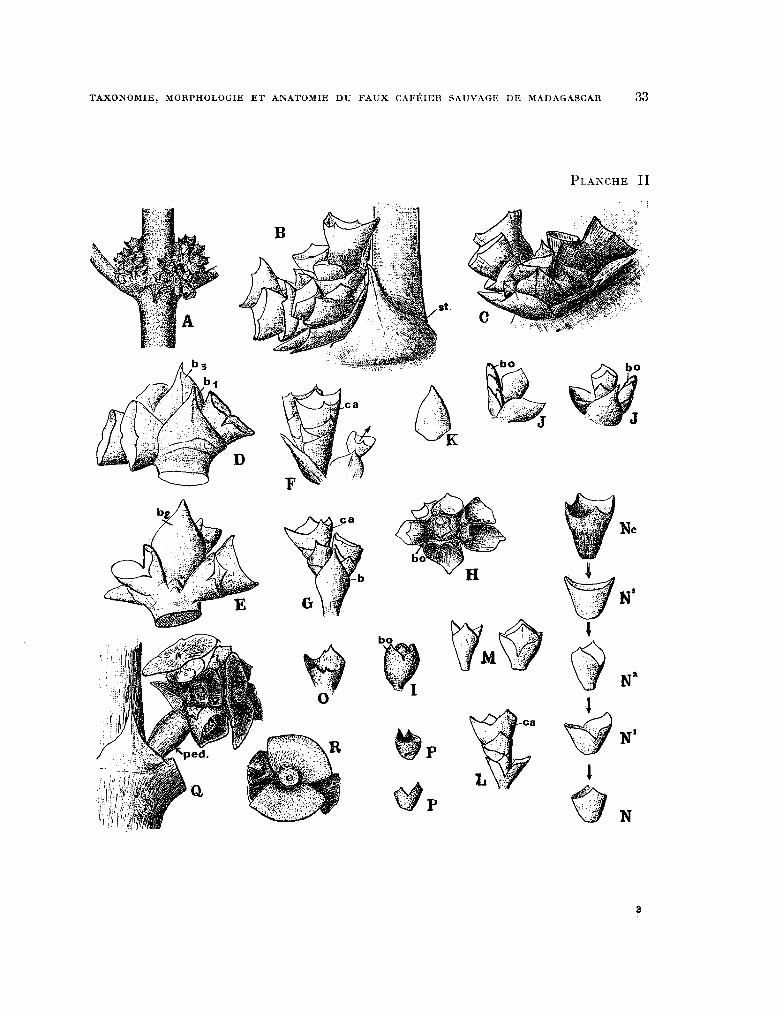
PLANCHE II. - Morphologie de l’inflorescence : fig. A, groupes de fleurs après la chute des corolles ; B, C, ensemble de calicule et calices cal. composant une inflorescence vue de profil, st. stipule b.,, Premiere bractée ; D, base de l’inflorescence vue de la face extérieure (face vers le petiole). On remarque le pedoncule aplati de l’inflores- cence, bl la premiere bractée, ba la troisieme bractée et les premiers calicules ; E, base de l’inflorescence vue de la face interne (face contre la tige), b, deuxiéme bractee , . F, calices et calicules des deux fleurs d’une subdivision de
l’inflorescence ; G, le même groupe vu de face avec sa bractée b; H, extremite de l’inflorescence montrant le bour-
geon terminal bo vu de dessus ; 1, bourgeon terminal de l’inflorescence vu de profil ; J, bractee et bourgeon terminal
de la derniere subdivision de l’inflorescence ; K, une bractee ; L, ensemble des calicules d’une fleur, ca. calice ; M, calicules 2 et 3 d’une fleur ; N, N,, N,, N,, N,, calicules grossis d’une fleur et calice disposes dans l’espace dans les positions avec lesquelles ils s’emboitent les uns dans les autres ; 0, calice a dents irregulieres ; P, calicule Q dents
irregulieres ; Q, reste des axes et des calicules de l’inflorescence après la chute des fruits. Remarquer le pedoncule ped. qui s’est allonge ; R, calicule dechire persistant apres la chute du fruit.
TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 33
PLANCHE II
NC
P
P
N”
N’
N

34 H. RABtiCHAULT
bracteis 2 in calyculum brevem sub flore connatis. Calyx brevis, obtuse 4-dentatus; corolla in alabastro conico acutato torta, 4-loba, Stamina 4, inclusa. Germen obconicum; loculis 2; ovulo in singulis descendente. Fructus pisiformi glaber, obtuse ad dissepimentum sulcatus, indehiscens; seminibus 2, subglobosis descendentibus; hilo lato (fuscato); albumine carti- lagineo dense sulcato ruminatoque; integumento sericeo; embryonis obliqui v. subverticalis cotyledonibus inferioribus ellipticis membranaceis; radicula tereti supera v. laterali.
Arbrisseau, rameaux opposés fins, plante entièrement glabre. Feuilles opposées oblongues-lancéolées (de 10 c,rn de longueur sur 4 cm de large), apex longuement acuminé, base décurrente ou rarement obtuse, limbe membraneux, le dessus plus brillant, le dessous un peu plus pâle nervures secondaires 12-14, inégalement sinuées ; veines réti- culées à peine proéminentes. Pétiole court (1/2-l cm). Stipules interpétiolaires courtes,
* caduques. Fleurs axillaires en glomérules serrées, 2 bractées en calicule court opposées sous la fleur. Calice court à 4 dents obtuses. Corolle en bouton floral, conique aiguë, tordue et à 4 lobes. Etamines 4, incluses. Ovaire obconique ; deux loges ; ovule unique descendant. Fruit pisiforme glabre, obtus à cloisons cannelées longitudinalement, indéhiscent ; hile large ; albumen cartilagineux très strié ou ruminé. Téguments sémi- naux soyeux ; embryon oblique ou sub-vertical, cotylédons inférieurs, elliptiques mem- braneux ; radicule arrondie super ou latérale.
J’ai retrouvé au Muséum National d’Histoire Naturelle de Paris les échantillons ayant servi à cette description : a Types )) : échantillons collectés par BOYER, no 281, et par RICHARD, n 0 658 à Mohilla, appartenant à l’herbier E. DRAKE ; o Cotypes 1) (exsicata no 2419 et no 3175) ; échantillons collectés par BO~VIN à Mayotte en 1847-1852, lequel a noté sa fréquence sur les côteaux boisés au-dessus de Moussapéré, et par PERVILLE. Une note de ce dernier collecteur signale que la plante aurait été cultivée au Muséum où elle fleurit en 1843.
Les échantillons en ma possession correspondaient bien aux types et co-types ci- dessus, mais également aux n 08 238 (ou 298) et 375 de l’herbier E. DRAKE collectés par le jardinier RICHAR» à l’île de Moheli (Comores) et au no 33 récolté par M. LAVRANCHE à Anjouan (Comores) qui a noté Q arbuste très commun sous toutes les latitudes, fleurit de février à mars 1).
En réalité, nos échantillons comme l’espèce décrite ci-dessus par H. BAILLON ne sont pas des Cremaspora, car l’un des caractères des espèces de ce genre est l’absence de rumi- nation de l’albumen. Les Cremaspora ont un albumen corné (cartilagineux) plus ou moins strié mais non pénétré profondément par les téguments séminaux. L’espèce décrite par BAILLON est un Polysphaeria, genre que nous devons à HOOKER fils (3, p. 108). D’ailleurs, A. CHEVALIER, dans ses travaux sur les Rubiacées, l’a désigné sous le nom de Poly- sphaeria congesta ou congestum. Il faut donc l’appeler Polysphaeria congesta (H. BN.) A. CHEV. = Cremaspora congesta H. BN. = Saldinia coffeoides AD. BR., Schizospermum congestum BVN.
Je pense qu’il y a trois causes à la confusion de cette plante, cultivée à la Station agronomique de l’Ivoloina, avec le Coffea Humblotiana. On peut évoquer sa ressemblance avec un Caféier, mais aussi le fait qu’elle serait cultivée par les Malgaches dans certaines régions pour la fabrication d’une infusion dont le goût serait analogue au café ; ce fait a été rapporté par plusieurs collecteurs, notamment par BOYER* et A. CHEVALIER (8, 11). Il faut ajouter enfin que les échantillons de 1’Ivoloina sont absolument identiques à ceux qui ont été collectés par HUMBLOT aux Comores, et que l’on trouve au Muséum dans l’herbier E. DRAKE, sous le no 1506.
l Voir note infrapaginale, p. 30.

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 35
Il existe malgré leurs liens de parenté très nette chez les spécimens des herbiers du Muséum, comme chez les Caféiers cultivés, une certaine variabilité. Ainsi le no 375 (herbier DRAKE), RIC~ARD Hortul. collect., a des feuilles subcoriaces et sur le no 658 id. M. RICHAR» recolté en 1840 à l’île Mohilla, on peut remarquer que des groupes de fleurs sont identiques à ceux des autres échantillons, mais qu’ils sont subdivisés en deux glomé- rules portés à l’aisselle de chaque feuille par deux pédoncules. Ce caractère a servi aux systématiciens pour subdiviser le genre en deux groupes ; nous en reparlerons au chapitre de la morphologie des inflorescences (page 46).
D’autres échantillons récoltés par M. FRANÇOIS, en mars 1937, sont cités par A. CHEVALIER qui a noté à leur sujet (( Une autre espèce de l’archipel des Comores, P. congestum H. BN. a une telle ressemblance avec un Caféier, qu’on a tenté de la cultiver dans les jardins d’essais de Madagascar ef de La Réunion O. Les échantillons de FRANC~IS, accompagnés d’un lot de graines, proviennent de plantes cultivées dans la région de Coconi (Madagascar) et l’on sait qu’elles sont originaires des Comores. Or, ces plantes ont des feuilles moitié plus petites (long. 54 mm x larg. 15 mm) qlie celles des plantes de l’Ivoloina, et le dessin joint à l’herbier par BEILLE fait ressortir d’autres différences avec nos échantillons, notamment dans la forme de certaines pièces florales.
II. MORPHOLOGIE ET ANATOMIE
A. Les Rameaux et les Racines (Photographie 2 et Planches 1 et III).
a. MORPHOLOGIE.
Parmi les plantes voisines des Caféiers, cet arbuste est certainement celui qui ressemble le plus à ces derniers. Par son port et sa charpente, la forme et la couleur du feuillage, on peut le rapprocher du Coffea arabica L. ou du Coffea congensis FROEHNER. Les pieds qui sont maintenus en culture en serre comprennent deux sortes de rameaux. La flèche comporte un axe principal sur lequel sont disposées des feuilles et rameaux opposés décussés (Phot. 2). Tandis que les rameaux latéraux sont plagiotropes et ne peuvent pas donner de rameaux érigés. Si l’on supprime l’axe principal, un axe axillaire se développe au-dessus de l’aisselle du rameau latéral plagiotrope comme chez le Coffea arabica L.
Le tronc est grisâtre, et les jeunes branches vertes deviennent marron rougeâtre clair tout en restant lisses. L’exfoliation de l’écorce se fait par lambeaux. Les rameaux plagiotropes sont divisés en palmettes légèrement graciles.
b. ANATOMIE (Planche III).
Je n’ai pas eu, hélas, à ma disposition de rameaux, de troncs ou de racines âgés. Il ne m’a donc pas été possible d’en étudier la structure. Malgré tout, l’examen de coupes de rameaux aoûtés, montre peu de différence de structure avec les Caféiers. Je suppose donc qu’il ne m’aurait pas été permis d’en trouver davantage à l’examen de sections d’organes âgés, si je m’en rapporte aux observations que j’ai faites sur les Caféiers (26).
Le rameau orthotrope sectionné (Pl. III, fig. A et B) était aoûté, de couleur brune, et avait un grand diamètre de cinq millimètres.

36 H. RABBCHAULT
A un faible grossissement on remarque (fig. A), comme chez les caféiers, au centre la moelle (m.) ou parenchyme interne entouré d’un anneau de xylème (z. bois), lui-même entouré d’un anneau de phloème (ph. liber) recouvert par du parenchyme cortical (par.), limité enfin à l’extérieur par un épiderme (ep.). La moelle ou parenchyme interne, au centre, est constituée de cellules arrondies et ne présente pas de caractères anato- miques particuliers. Le xylème, de même, est assez semblable à celui du Coffea arabica, mais présente cependant une proportion moins grande de vaisseaux ligneux, ceci au profit des fibres ligneuses de petit diamètre et à lumen presque ponctuel. Les rayons ligneux sont constitués de grandes cellules parallélépipédiques à grand lumen, plus allongées que chez le Caféier et disposées par 1 à 2 files radiales, de façon irrégulière, toutes les 1 à 6 files fibre-vasculaires.
Le cambium (c) est très irrégulier. Le liber secondaire (phloème) ne présente pas de caractère particulier. Le protophloème est important. Il est constitué de nombreuses cellules polygonales de forme et de diamètre irréguliers et dont les angles sont brusque- ment épaissis.
Dans le liber, les cellules des rayons libériens, peu nombreuses, ont des parois fines. Leur diamètre est comme chez les Caféiers, supérieur à celui des cellules libériennes. Elles renferment peu ou pas de sable d’oxalate de calcium. La fréquence des rayons libériens correspond à celle des rayons ligneux. Le phloème et le protophloème repré- sentent environ la moitié de l’épaisseur de l’écorce.
A la limite du liber, on remarque des groupes irréguliers (1 à 10 fibres) de fibres péri- cycliques (f.) arrondies, à lumen ponctuel. Le péricycle (pe.) ne présente aucun caractère particulier mais l’endoderme (en.) est toujours constitué de cellules plus grandes que chez le Caféier et dont les parois sont souvent épaissies.
Une assise génératrice subérophellodermique se forme au niveau de l’endoderme comme chez les Caféiers.
Le parenchyme cortical et l’épiderme sont semblables à ceux des Caféiers. Le collen- chyme est en faible proportion.
La racine (Pl. III, fig. D et E) examinée était jeune (diamètre 2,5 mm) aussi je n’ai hélas pu faire une comparaison de grande portée avec celle des caféiers. D’une façon générale, on remarquera que le diarnètre des cellules est beaucoup plus grand que celui des cellules du jeune rameau dont l’étude précède. On remarque l’allongement tangentiel général des cellules.
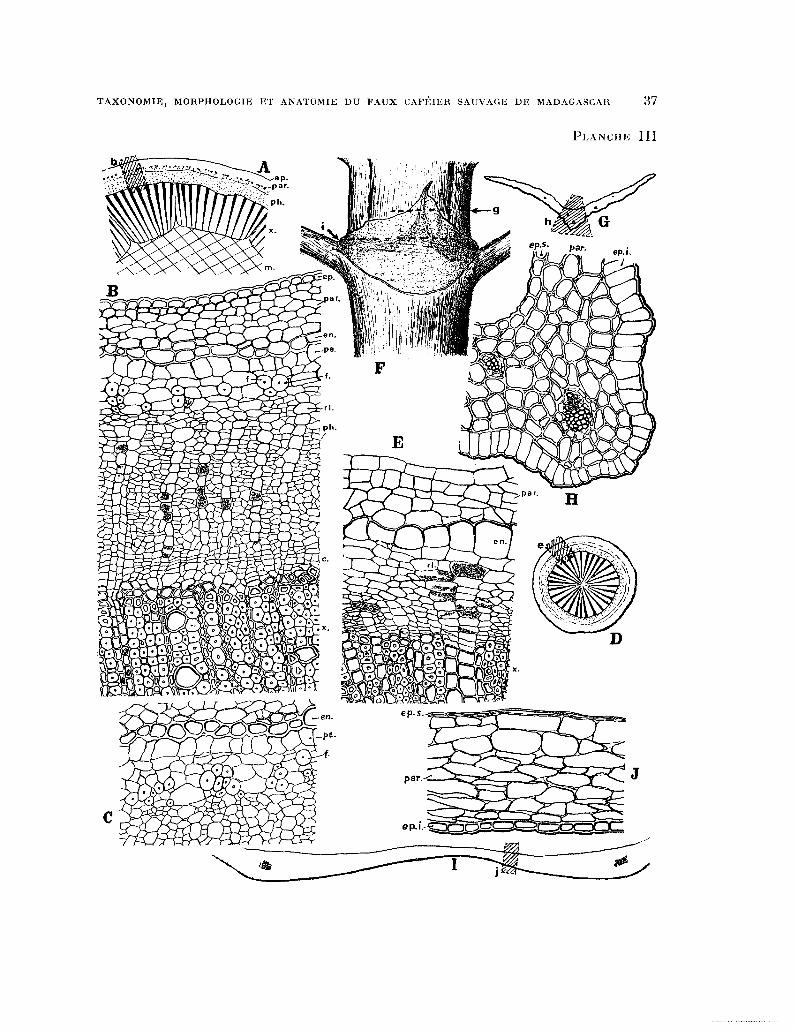
PLANCHE III. - Anatomie d’un rameau, d’une racine ct d’une stipule : fig. A, schema d’une partie de la section transversale d’une tige ; B, partie grossie de la section transversale de la tige representee par la zone hachu- rée b de la figure precédente, en.. epiderme, par. parenchyme cortical, en. endoderme, f. fibres libériennes, ph. phloème, rl. rayons libériens avec cellules renfermant du sable d’oxalate de calcium, e. cambium, z. xyleme (quel- ques cellules d’un rayon ligneux en bas a gauche renferment des cristaux d’oxalate de calciumj ; C, une autre partie de la section transversale de la tige dans la région du pericycle et de l’endoderme ; les Bbres sont plus nombreuses que dans la section précédente ; D, schema de la section transversale d’une jeune racine ; E, partie de la section transversale d’une racine. On remarque la faible epaisseur du parenchyme cortical par., les grandes cellules de l’endoderme en. et les rayons libériens rl. plus larges que chez la tige avec leurs cellules aplaties tangentiellement ; F, nœud d’un rameau orthotrope montrant les niveaux g. et i. auxquels ont et& effectuees les sections transversales dans la stipule ; G, schema de la section transversale d’une stipule au niveau g de la figure precedente ; H, section transversale grossie representee par la zone hachuree h de la figure précedente, en. s. Bpiderme superieur, ep. i. epiderme inferieur, par. parenchyme avec cellules a parois epaisses ; 1, schema de la section transversale de la
stipule au niveau i de la figure F ; J, section transversale grossie représentée par la zone hachuree j de la figure precedente.
(Grossissements fig. A, D x 15 ; F x 6 ; B, C, H, E, J x 150).
t
a
1
TAXONOMIE, MORPHOLOGIR ET ANATOMIE DU FAUX ChFbIliH Y.4UVAGE DE MADhGASChH 37
PLANCHE III
ep.s.

38 H. RABÉCHAULT
L’endoderme (en.) est constitué de cellules encore plus grandes que dans la tige. Le sable d’oxalate de calcium est beaucoup moins abondant (voire rare) dans les rayons libériens (rd.) que chez les caféiers.
En résumé, les rameaux et racines s’identifient donc morphologiquement comme anatomiquement à ceux des caféiers, à quelques détails près.
B. La Feuille (Planche IV).
Je donnerai les dimensions des feuilles prises sur matériel d’herbier en provenance directe du lieu de culture car j’ai observé qu’en serre, les feuilles étaient plus longues par suite d’un léger étiolement de la plante. Sur 20 feuilles choisies au hasard, j’ai relevé les dimensions en mm suivantes : longueur totale, variation 82 à 116 mm, moyenne 95,4 ;
longueur du limbe (73-106) X86,5 ; longueur de l’acumen (7-16) X 10,7 mm ; largeur du limbe (22-31) Y? 25,6 mm ; longueur du pétiole (7-10) ? 8,l mm ; diamètre du pétiole (0,8-1,9) X 0,98 mm.
1. Le Pétiole (Planche IV, fig. A, B, C).
a. MORPHOLOGIE.
Le pétiole est court (5 à 20 mm de long, sur 0,8 à 2 mm environ de diamètre), arrondi, glabre, et possède une rainure longitudinale peu accentuée à la face supérieure. Sa partie distale est ailée par les bords décurrents du limbe et analogue par conséquent à celle des Caféiers. C’est à ce point dit G caractéristique )) par PETIT (cf. 23), que j’ai effectué une coupe transversale.
b. ANATOMIE (Pl. IV, fig. A, B, C).
L’anneau principal libéro-ligneux (fig. B) est presque fermé à sa partie supérieure. Il entoure un parenchyme interne à cellules arrondies dont la paroi est épaisse. L’anneau ligneux est peu épais. Les vaisseaux ligneux sont nombreux. Les fibres ligneuses plus épaisses que chez le Coffea arabica L. ont malgré tout un grand lumen et emplissent les espaces entre les files de vaisseaux. De même, le liber est constitué d’amas de vaisseaux libériens de faible diamètre et de cellules polygonales irrégulières à angles vifs. Elles renferment peu ou pas de sable d’oxalate de calcium. On n’observe pas ou très peu de fibres péricycliques.
Le parenchyme cortical (par.) est épais, à cellules arrondies, laissant entre elles de nombreux et grands méats. L’épaisseur de leur paroi augmente progressivement du liber vers l’extérieur de l’organe où l’on observe un important collenchyme (col.). L’épiderme (ep.) est constitué de petites cellules arrondies sans caractère particulier. Une deuxième coupe transversale effectuée vers le milieu du pétiole, montre un arc libéro-ligneux plus épais que l’anneau ci-dessus. Les extrémités de cet arc sont repliées en dehors sur elles- mêmes.
Une troisième coupe transversale (fig. A) effectuée dans la partie proximale du pétiole ne montre pas de différence sensible de forme du système libero-ligneux. Cepen- dant, nous remarquons la présence de fibres (f.) due sans doute à la proximité de la tige.
I
I
I
!
\,
I

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 39
2. Les Nervures (Pl. 1, fig. A et Pl. IV, fig. D).
a. MORPHOLOGIE.
Les nervures glabres font saillie à la face inférieure ou dorsale du limbe.
On a tenté d’introduire dans les diagnoses des Caféiers les caractères de la nervation. Mais ces caractères sont extrêmement variables notamment le nombre de (( paires o de nervures secondaires ou (t paries H*. Pour ne rien omettre cependant, on peut indiquer que les feuilles de mes échantillons comportent 6 à 12 Q paires )) de nervures secondaires, ce qui est légèrement inférieur au nombre donné par BAILLON dans sa diagnose.
A l’aisselle des nervures secondaires, je n’ai pas constaté la présence de domaties ni sur mes échantillons, ni sur ceux de BO~VIN, de RICHARD, de LAGRANGE et de HUMBLOT. Ces nervures sont saillantes à la face dorsale comme la nervure principale et font un angle de 700 environ avec cette dernière. Des nervures tertiaires les relient entre elles vers la marge par leur extrémité et en faisant des arcs très ouverts et réguliers dont la courbure augmente cependant progressivement en allant vers la nervure située au-dessus, c’est-à- dire vers l’apex.
La nervation tertiaire et quaternaire peu procminente et réticulée, est souvent plus visible à la face supérieure qu’à la face inférieure lorsque la feuille sèche. Ce qui provient de ce que ces fines nervures ou veines sont peu ou pas saillantes sur la feuille verte car elles sont exactement dans le plan du limbe. Le réseau formé est constitué de grandes mailles polygonales irrégulières, souvent allongées dans le sens des nervures secondaires.
b. ANATOMIE (Pl. IV, fig. D).
Ainsi que l’a montré MARIANI (23) pour le caféier, le point (( caractéristique 1) de la nervure principale est sa région moyenne ; c’est à cet endroit que j’ai effectué une coupe transversale.
Le système libéro-ligneux y est fermé, c’est-à-dire annulaire. On peut remarquer, en comparant les figures A, B, et D (Pl. IV) que la formation de cet anneau se poursuit de la partie proximale du pétiole à la nervure principale. Le mot (c anneau )) laisse supposer que l’ensemble des éléments du xylème et du phloème épouse la forme de deux couronnes arrondies concentriques ; or, il n’en est rien. En effet, l’anneau est aplati vers la partie supérieure. Il est constitué par l’arc que nous avons observé dans le pétiole, dont les deux extrémités sont reliées par une partie droite qui apparaît par la suite vers la région distale de cet organe.
Autre caractère : on remarque la présence de nombreux sclérites et fibres péri- cycliques ; phénomène que j’avais observé déjà chez les Caféiers et qui traduit une structure (( plus âgée )) de la nervure principale par rapport au pétiole.
Les autres caractères anatomiques de la nervure sont semblables à ceux du pétiole ; c’est la raison pour laquelle je n’ai pas jugé nécessaire d’en donner un dessin de détail.
Les très jeunes nervures (n.) (Pl. IV, fig. E), appelées encore nervules ou veines, qui innervent les tissus du limbe, sont constituées de quelques vaisseaux ligneux, et de quelques éléments libériens. Le tout est entouré par une couche de cellules péricycliques. Les fibres péricycliques apparaissent très tôt, alors que le volume de la nervure ne se traduit pas encore par une excroissance de la face inférieure du limbe comme chez le Caféier.
* Terme erronk puisque les nervures secondaires sont alternes sur la principale. On a voulu par 1~4 simplifier
le comptage des nervures en se les reprbsentant par groupes de 2.

40 H. RABkHAULT
3. Le Limbe (Pl. 1, fig. A et Pl. IV, fig. E).
a. MORPHOLOGIE.
Le limbe est simple, uni, glabre, lisse, brillant cn dessus, un peu plus pâle en dessous, subcoriace à membraneux, lancéolé à oblong, parfois assez longuement acuminé, à base cunée décurrente. Il est plus ou moins ondulé vers la marge et rappelle ainsi celui de la feuille du Coffea arabica L.
En l’observant par transparence avec une loupe, on remarque de nombreux petits points clairs que l’on identifiera plus loin à autant de poches à mucilage.
b. ANATOI\IIE (Pl. IV, fig. E).
L’épiderme supérieur sans stomates, est constitué de cellules aplaties à contour très sinueux (Pl. IV, fig. F).
L’épiderme inférieur (ep. i.) (Pl. IV, fig. E), au contraire, comporte de nombreux stomates caractéristiques de la famille des Rubiacées (Pl. IV, fig. G, 1, H). Les cellules épidermiques aplaties ont un contour également sinueux, mais leur paroi est fine et présente souvent de nombreux étranglements.
En coupe transversale (Pl. IV, fig. E), la structure du limbe se montre assez diffé- rente de la structure du limbe de la feuille des Caféiers.
L’épiderme supérieur (ep. s.) est constitué de cellules de section quadrangulaire. Le parenchyme palissadique (par. pal.) comprend une seule couche de cellules allongées courtes, à parois fines et parfois légèrement sineuses. Ces cellules, à l’inverse de celles des Caféiers, ne renferment pas de sable d’oxalate de calcium, lequel est d’ailleurs en très faible quantité dans les autres tissus.
Au-dessous, le parenchyme lacuneux (par. lac.), contrairement à celui des feuilles de Caféier, est constitué de cellules arrondies serrées les unes contre les autres, laissant entre elles de très petits méats. Ce tissu est très lâche chez les Caféiers, ce qui donne à la feuille l’aspect anatomique d’une feuille de plante aquatique (RABÉCHAULT, 26, p. 173).
Les couches de cellules du parenchyme lacuneux, situées immédiatement au-dessous du parenchyme palissadique, comprennent de grandes poches dc 21 à 83 p de diamètre (moyenne 44 p) pleines d’une matière mucilagineuse brune (r.). En face des poches parfois très vastes, les cellules palissadiques sont jusqu’à moitié plus courtes que leurs voisines ou bien sont écartées de part et d’autre.
Chez certains Caféiers, la dernière couche de parenchyme lacuneux est formée de cellules serrées les unes contre les autres. Cette couche rappelle, comme je l’ai fait remar- quer dans l’anatomie des Caféiers (26, p. 172), le parenchyme palissadique placé à la face
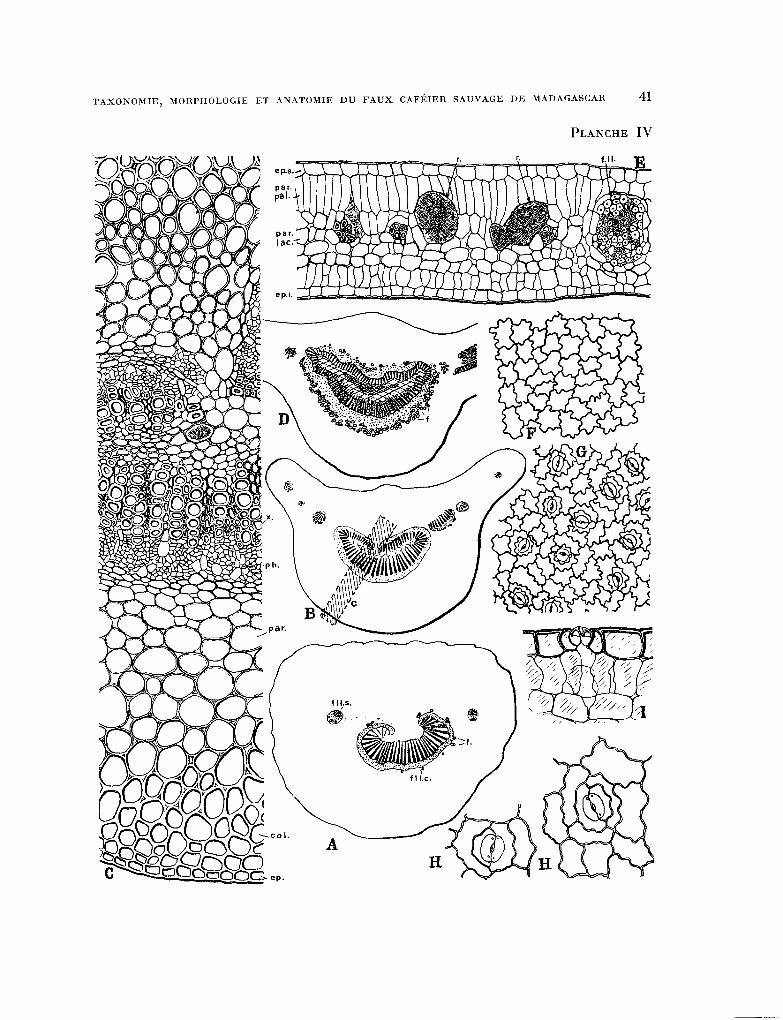
PLANCHE IV. - Anatomie de la feuille : fig. A, schkma de la section transversale du pktiole avec son fais-
ceau libéro-ligneux central (f.1.Z.c.) en forme de croissant et deux faisceaux libkro-ligneux secondaires (f.Z.l.s.),f. fibres libkriennes ; B, schkma de la section transversale du pbtiole à l’extrémité distale (point * critique * de PETIT) prés du limbe ; D, schéma de la section transversale effectuée a mi-longueur de la nervure principale. On remarquera l’abondance des fibres libkriennes f. ; C, partie grossie de la section transversale du pétiole au point * critique * repr6senUe par la région hachurbe c de la figure B : ep., epiderme, col. collenchyme, par. parenchyme cortical, ph. phelloderme, z. xylème ; E, section transversale du limbe : ep. s. Epiderme supkrieur, par. pal. parenchyme palissa- dique, f. 1. 1. faisceau libéro-ligneux (nervule), P. poche à mucilages et tanins, ep. i. bpiderme infbrieur ; F, cellules de 1’6piderme vues perpendiculairement & la surface du limbe ; G, cellules de l’bpiderme infbrieur vues perpendi- culairement à la surface du limbe ; 1, cellules d’un stomate dans l’épiderme inférieur en coupe t,ransversale ; H, deux stomates grossis vus perpendiculairement & la surface de l’bpiderme infkieur.
(Grossissements fig. A, B, D x 30 ; C, E x 150 ; F, G x 200 ; H, 1 x 400).
I
I
!
i
c. i
TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 41
PLANCHE IV

42 H. RABÉCHAULT
supérieure de la feuille. Chez le Pofysphaeria, cette ressemblance est encore plus parfaite, car ces cellules sous-épidermiques (épiderme inférieur) ont une forme allongée (une à deux fois plus courtes que les cellules palissadiques), leur paroi est fine et très légèrement sinueuse. Elles forment comme un deuxième tissu palissadique inférieur.
En ce qui concerne la forme générale, le limbe des feuilles de Polysphaeria tend donc vers une symétrie bifaciale.
4. Les Stipules (Pl. III, fig. F, G, H, 1, J).
a. MORPHOLOGIE.
On compte deux stipules glabres embrassant la tige dc part et d’autre des points d’insertion de chaque paire de feuilles (interpétiolaires). Il y a donc une paire de stipules par paire de feuilles.
Chaque stipule est acuminée et surbaissée, c’est-à-dire souvent plus large que longue, acumen compris. J’ai relevé sur matériel d’herbier les dimensions suivantes :
hauteur 3,5 à 4,0 mm, largeur 5 à 8 mm.
Les stipules comprennent : une base large, arrondie, grisâtre et striée transversale- ment jusqu’à la moitié environ de la hauteur totale, surmontée d’une partie supérieure plus épaisse acuminée (Pl. III, fig. F).
b. ANATOMIE (Pl. III, fg. G, H, 1, J).
Une section transversale (fig. G et H, Pl. III), effectuée vers l’acumen, présente une forme générale en V très ouvert. Les deux branches de ce V sont dirigées vers la tige et la pointe occupe la face dorsale de la stipule ; dans cette arête limitée à la partie acumi- née, passe la nervure principale.
La nervation de la stipule est mal définie. La nervure principale, en allant vers la base, se divise bientiX en un nombre variable de nervures secondaires dont les deux plus importantes rejoignent le système vasculaire de la tige à la base de chaque pétiole. La plupart du temps, la nervure principale s’annule ainsi par division et on ne retrouve plus vers la base de la stipule que des faisceaux libéro-ligneux divergents.
Les tissus de la stipule (Pl. III, fig. H) peuvent être divisés en trois parties : les épidermes supérieur (ep. s.) (face ventrale) et inférieur (ep. i.) (face dorsale) et un parenchyme à cellules allongées (par.) dans l’épaisseur duquel passe le système vascu- laire.
Le parenchyme (par.) a une allure collenchymateuse car les méats intercellulaires sont réduits et ses cellules sont arrondies à ovales et avec une paroi épaisse. L’épiderme ventral ou supérieur est sans intérêt. Il est constitué de cellules à parois épaisses de section carrée à rectangulaire. L’épiderme dorsal ou inférieur présente lui, plus de caractères. Ses cellules, de section quadrangulaire, ont une membrane épaisse vers l’exté- rieur qui devient fine sur les parois radiales et intérieure.
Une section transversale (Pl. III, fig. Fi et I), ef’fectuée dans la base d’une stipule a la forme d’une bande très étroite plus épaisse aux deux extrémités et par conséquent déprimée dans la partie médiane. Dans chaque partie épaisse passe l’une des deux nervures secondaires précitées issues de la nervure principale unique rencontrée dans la section précédente (fig. G, H). Tant et si bien que la stipule semble formée en cet endroit par
I
I
I
t
! I

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAH 43
deux stipules axillaires accolées dont les bords les plus proches se seraient soudés (concres- cence marginale) ; les deux nervures sont représentées par les deux nervures secondaires issues de la nervure principale de la région acuminée décrite précédemment qui divergent vers la base de l’organe.
La section transversale (Pl. III, fig. J), portion grossie d’une zone située entre les deux nervures secondaires permet de reconnaître : un épiderme inférieur (ep. i) (face dorsale) avec des cellules à parois épaisses, un parenchyme (pur.) à cellules allongées et un épiderme supérieur (ep. s.) (face ventrale) constitué de cellules pavimenteuses très aplaties. L’épiderme supérieur se détache très facilement des autres tissus sous la forme d’une très fine pellicule.
C. Floraison. .
A Madagascar (Ivoloina), le Polysphaeria congesla (H. BN) A. CHEV. fleurit en avril- mai et de février à mars selon LAVRANCHE, dans l’fle d’Anjouan (Comores).
Pendant cette floraison saisonnière, les fruits se forment aussitôt après la féconda- tion puis mûrissent et il faut attendre une autre période favorable pour assister à une nouvelle initiation florale. Au moment de la floraison, les vingt à trente premiers centi- mètres de l’extrémité des rameaux présentent des nœuds pourvus de fleurs. Celles-ci sont plus jeunes et non encore épanouies vers le bourgeon terminal (Pl. 1, fig. A). Le développement se poursuit au fur et à mesure que l’on s’éloigne de l’extrémité du rameau.
Les fleurs sont groupées en glomérules ou cymes contractées, situées à l’aisselle de chaque feuille (axillaires) vers l’extrémité des rameaux.
Le rameau comprend ensuite une zone où les nœuds ne présentent ni fleurs ni fruits (Pl. 1, fig. A). Cette portion est celle qui s’est développée pendant la saison non favorable à la floraison (saison sèche). Puis nous trouvons une zone à fruits, résultat de la floraison précédente. Là, les petits fruits sont groupés par 2-3, le plus souvent 4 (plus rarement 5 ou 6) à l’aisselle des deux feuilles de chaque paire.
Ainsi, le long des jeunes rameaux on peut, observer tous les stades de la floraison à la fructification.
D. L’inflorescence (Pl. 1 et II, fig. A).
On distingue deux glomérules par nœud. Chacun d’eux comprend 10 à 20 fleurs (moyenne 14-18) serrées les unes contre les autres. Lorsque les glomérules ont un grand nombre de fleurs ou vers l’extrémité des branches lorsque le rameau diminue de diamètre, les deux glomérules se rejoignent et semblent n’en former qu’un seul entourant chaque noeud.
Les glomérules sont sessiles, mais cependant les plus âgés vers la base des rameaux florifères sont parfois pédonculés (Pl. II, fig. Q). J ‘ai observé également un glomérule pédonculé sur le dernier entre-nœud au bas de l’échantillon no 658, Herbier E. DRAKE (M. RICHARD, collect.) du Muséum National d’Histoire Naturelle de Paris.

44 H. RABÉCHAULT
J’insiste sur ce caractère car il fut pris pour base par S. MOORE pour scinder le genre Polysphaeria en deux sections : les Ephedranthae comprenant les espèces à glomérules sessiles et les Cladanthae comprenant les espèces à glomérules pédonculés.
BRENAN (7) a revu cette question et a confirmé et étendu la conception de S. MOORE à l’ensemble du genre. Cependant, ayant observé comme moi la variabilité de ce caractère et que cette subdivision n’avait par conséquent rien d’absolu, comme je l’ai noté chez l’espèce qui nous préoccupe ici, il a du modifier légèrement les subdivisions de MOORE. BRENAN a adjoint des caractères relatifs à la forme et aux dimensions du calice et de la corolle qui lui permirent de créer trois sections au lieu de deux et de subdiviser la première section en deux séries nouvelles.
Le Polysphaeria congesta (H. BN.) A. CHEV. appartiendrait à la SECTION 1 Ephe- dranthae S. MOORE, qui renferme les espèces dont le calice des boutons floraux est ouvert, court (l-3 mm de long), cupuliforme, à marge toujours tronquée à denticulée et dont la corolle petite dépasse largement le calice dans le bouton floral (la corolle dépasse le calice de 3 à 4 mm chez P. congesta), et à la série b-ephedranthae (S. MOORE) BRIXNAN caracté- risée par des inflorescences sessiles ou subsessiles.
Le pédoncule (Pl. II, fig. Q), lorsqu’il existe, est court, 2 à 4 mm. Sa section trans- versale est aplatie, elliptique, plus rarement quadrangulaire (carrée). Lorsqu’il est plus large qu’épais, la largeur est tangentielle à la section transversale de la tige.
La constitution des glomérules (cymes) est assez difficile à observer du fait de la contraction de l’inflorescence et par conséquent, du fait que les axes de cette dernière sont extrêmement courts ; les fleurs semblent attachées sur un point commun ou sur un réceptacle (Pl. II, fig. B, C, D, E). Seules, les bractées qui se développent à chaque sub- division des axes peuvent guider dans cet examen morphologique. Elles sont toutes glabrescentes sur la face extérieure ou dorsale mais sont garnies de nombreux poils courts et apprimés sur leur face ventrale ou intérieure.
A la base de l’inflorescence, les deux premières bractées alternes sont dans l’axe foliaire, c’est-à-dire que leurs axes sont dans un plan passant par les grands axes de la tige et de la feuille.
La plus grande, la première, est dirigée vers l’extérieur, c’est-à-dire vers la feuille. Elle vient s’attacher au sommet du pédoncule, sur la face extérieure. Sa base est donc à peu près égale à la largeur du pédoncule. Elle s’élargit légèrement et s’applique aux groupes de fleurs situés au-dessus d’elle. Elle marque la première subdivision de l’axe principal de l’inflorescence.
En soulevant cette bractée, on reconnaît à droite et à gauche deux groupements floraux correspondant à ces deux subdivisions opposées contractées, comme l’est l’ensem- ble de l’inflorescence.
Retournons le glomérule. Sur la face de l’inflorescence tournée vers la tige, nous retrouvons les deux groupes floraux observés sur l’autre face et au milieu, un peu plus haut, la deuxième bractée qui marque la seconde subdivision de l’axe de l’inflorescence.
Cette bractée est moins développée que la première. Elle est aussi moins large, moins ventrue, plus cochléaire (*) et subovale.
Lorsque le glomérule est petit, ces subdivisions de l’axe sont seules aisément obser- vables.
(‘) Du latin cochleorium = cuiller.
1
!
I
L
i
I

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 45
Les gros glomérules présentent quelques autres subdivisions mais leur nombre est toujours limité car la croissance de l’axe principal est inhibée en général très tôt au profit des deux ou trois premières subdivisions (axes secondaires) qui s’allongent et, se subdi- visent à leur tour avec plus de vigueur.
Or, nous avons vu d’après la position des deux premières bractées (axe tige-feuille) que les deux premières paires d’axes secondaires sont dans un plan perpendiculaire au plan tige-feuille et tangentiel à la section de la tige. La dernière subdivision observable des axes est celle constituée par deux fleurs et le bourgeon terminal de l’axe floral, à la base de laquelle on trouve une bractée comme à toutes les subdivisions de l’inflorescence.
L,e bourgeon terminal (Pl. II, bo, fig. H, 1 et bo fig. J) est de forme ovoïde à ellipsoïde. Parfois légèrement courbé, il suit la face intérieure du calicule contre laquelle il est pressé par le développement des deux dernières fleurs et pourrait être pris pour une fleur non développée. On l’observe facilement sur les vieux glomérules où les corolles des fleurs sont toutes tombées. Sa dissection montre qu’il est constitué de nombreux calicules et bractées qui se recouvrent. Une inclusion et des coupes en séries ont permis d’observer l’agencement des ébauches en cyme bipare.
Les deux dernières fleurs dont je viens de parler prennent naissance dans un calicule commun court qui embrasse la base de leurs pédoncules : on appellera calicule, comme l’a fait DUBARD, un ensemble formé de deux bractées opposées qui se sont soudées par leurs bords (concrescence marginale) de manière à former une collerette obconique ressem- blant au calice gamosépale (Pl. II, fig. N). Le calicule, par suite de son origine, ne pré- sente donc que deux dents opposées au lieu de 4 chez le calice de la fleur. DC plus, la soudure des bords des bractées est plus courte chez les calicules de la base des pédoncules, que sur ceux de l’axe floral, ce qui se traduit par deux échancrures plus profondes. Ces échancrures diminuent sur les calicules au fur et à mesure que l’on s’approche de la fleur (Pl. 1, fig. N, N,, N,, NS). Do même, les deux dents deviennent de moins en moins pro- noncées.
La fleur n’est pas portée directement au bout d’un axe floral, mais on observe que l’ovaire, surmonté de son calice à quatre dents, est logé dans un premier calicule à dents peu prononcées (Pl. 1, fig. C, F et Pl. II, fig. N, N,, N,, N,, N,), lui-même logé dans un second calicule en forme d’entonnoir dont les dents plus prononcées sont décussées par rapport aux dents du premier calicule. La base de ce second calicule est enfin logée dans un troisième et le tout dans un quatrième calicule, plus petit, dont les deux dent,s sont décussées par rapport aux deux dents du calicule supérieur.
Les calicules 2 et 3 sont parfois si bien emboîtés l’un dans l’autre, qu’ils semblent n’en former qu’un seul à 4 dents - dont deux plus petites, comme le calice.
Les deux fleurs dans leur calicule commun sont munies de 3 (parfois 4) calicules superposées que l’on peut considérer comme les vestiges de bractées opposées décussées qui marquent les subdivisions (inhibées) également opposées décussées des deux axes floraux à l’image de la décussation des rameaux sur la tige principale.
Ainsi, cette plante est caractérisée dans son ensemble par une parfaite symétrie basée sur le chiffre quatre. Les jeunes rameaux orthotropes sont quadrangulaires aux angles arrondis ; sur leurs faces opposées, on trouve quatre files de feuilles opposées décussées. Dans l’inflorescence, cyme bipare, les premières ramifications de l’axe princi- pal sont aberrantes. Elles sont opposées et superposées par suite de la compression feuille-tige. Mais les autres subdivisions sont opposées décussées et enfin, sur les axes floraux, les dents des calicules opposées décussées reproduisent la règle des feuilles opposées décussées des tiges. Dans la fleur, le calice lui-même est du type 4 avec ses

46 H. RARltCHAIJLT
4 dents dont les deux plus grandes sont décussées par rapport aux deux dents du dernier calicule. Le calice ressemble ainsi à deux calicules soudés l’un à l’autre. La corolle a 4 lobes et enfin les étamines sont au nombre de quatre.
Cette symétrie régulière est caractéristique du genre Polysphaeria, ce qui le diffé- rencie nettement du genre Cremaspora avec lequel il était autrefois confondu. Ainsi, chez les espèces du genre Cremaspora, les fleurs comportent en général cinq à six pétales et étamines. Il en est de même pour les Caféiers (Coffea) q ui ont également des fleurs avec cinq, parfois six pétales et étamines.
E. La Prefloraison.
Dans la fleur en bouton, la corolle dépasse largement le calice (3 à 4 mm). Elle se rétrécit au niveau de ce dernier puis augmente de diamètre et se termine enune forme ellipsoïde. Cependant, j’ai observé à 1’Herbier du Muséum de Paris des corolles coniques plus courtes, diminuant par conséquent de diamètre, de façon régulière jusqu’à l’extré- mité.
La préfloraison est tordue. Vue de l’extérieur, la corolle se tord en effet légèrement sur la gauche ; chaque lobe de la corolle recouvre par son bord de gauche le bord droit du lobe qui est à sa gauche. Dans cette position, on remarque que chaque lobe est presque auriculé à la base (Pl. 1, fig. B).
F. La Fleur (Pl. 1, fig. C, D, E, F, G, H, 1).
MORPHOLOGIE.
La fleur (Pl. 1, fig. C, D) est gamosépale et gamopétale comme chez toutes les Rubiacées et du type 4 (quatre dents au calice, 4 lobes au tube de la corolle). Ovaire compris, elle mesure en moyenne 6 à 9 mm de haut. Le tube de la corolle mesure à lui seul 4,7 à 5,5 mm. Le diamètre de la corolle est de 4,7-5,0 mm environ lorsque les lobes (pétales) sont dans leur position normale, repliés vers le bas (rédupliqués) et de 5,0 à 5,7 mm lorsque les lobes sont étalés dans un plan horizontal.
Nous examinerons d’abord les pièces du périanthe.
Le calice (0,7 mm de haut sur 1,5 mm de diamètre environ), identique comme texture au dernier calicule au fond duquel vient s’insérer la fleur, est vert. Il est formé d’une collerette tronconique située au sommet de l’ovaire et entoure la base du tube de la corolle. Le bord supérieur présente quatre dents, parfois très atténuées, dont deux opposées, sont plus petites que les deux autres, par suite des compressions dans le glomérule au moment de la formation des fleurs. J’ai rarement constaté une malfor- mation du calice. Planche II, figure 0, j’ai dessiné un calice anormal avec les dents peu prononcées, très aiguës, réparties inégalement tout autour. Le calice est légèrement pubescent à l’extérieur, mais présente de nombreux poils courts sur sa face interne. Le calice persiste sur le fruit.
La corolle, de couleur blanc à blanc-crême comprend deux parties : un tube conique (longueur 4,7 à 5,5 mm environ) moins évasé que le calice et quatre lobes (longueur 2 à 3 mm et largeur 2 à 2,5 mm environ), subcunés, correspondant aux quatre pétales de la fleur, qui sont autant de divisions du bord supérieur du tube.

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 47
.Je n’ai pas repéré de différence de grandeur sensible entre les lobes, comme on peut en observer parmi les dents du calice. La paroi du tube de la corolle n’est pas droite ; elle présente un léger renflement ou galbe vers son tiers supérieur (diamètre 2,l à 2,5 mm environ à la gorge contre 1 à 1,5 mm à la base). Comme le calice, la corolle est poilue sur sa face interne. Dans la partie la plus évasée ou gorge (Pl. 1, fig. D), on trouve de nombreux poils blancs effilés, caractéristiques du genre Polysphaeria. Ces poils nombreux sont dirigés radialement et leur extrémité touche le style qui passe au centre de la corolle. Ils forment comme une bourre qui obstrue le tube et déborde légèrement au dehors en un dôme blanc.
Au grossissement de deux cents fois (Pl. 1, fig. H,, I-I*), la paroi de ces longs poils (2-3 mm) présente une ornementation particulière. On distingue, orientées dans le sens du grand axe du poil, de nombreuses petites élévations en forme de tirets et des traits d’union parallèles, légèrement arqués. Ces élévations sont réguliêrement réparties mais plus on s’avance vers l’extrémité du poil, plus elles sont courtes. L’extrémité de chaque poil est ainsi comme nucamentée. Dans le sens de la longueur, on observe parfois des bandes étroites, claires, ou les élévations sont des tirets droits équidistants et transver- saux (Pl. 1, fig. H,). Les élévations longitudinales sont plus ou moins prononcées, parfois irrégulières, sinueuses et très longues. Enfin, sur les gros poils on peut observer une cer- taine irrégularité dans la distribution des tirets longitudinaux dont l’ensemble est interrompu par des élévations transversales longues, fines, cn forme de lignes brisées. Entre ces dernières, les petits tirets longitudinaux se font plus rares.
A l’extérieur, le tube de la corolle présente une fine et légère pubescence, résultat de la présence de poils très courts et apprimés.
Au bas, la partie rétrécie du tube de la corolle est glabre aussi bien à l’intérieur qu’à l’extérieur.
Les pétales ou lobes de la corolle, presque aussi longs que larges (longueur 2 à 2,5 mm, largeur 2,4 à 3 mm environ), sont plus épais et plus charnus que le tube. Ils sont courts, pointus, cunés, obtus et rarement dans un plan horizontal perpendiculaire au grand axe du tube de la corolle. Au contraire, ils s’inclinent et recourbent très t6t leur extrémité vers le bas (révoluté).
Les étamines (Pl. 1, fig. D) ne sont pas visibles de l’extérieur et ne sortent en aucun cas par la gorge de la corolle. En effet, après avoir déchiré le tube de la corolle, on trouve les anthères presque sessiles, noyées dans la masse des poils, fixées par leur filet très court et arqué vers le bas, à la paroi interne du tube entre chaque pétale. On compte ainsi autant d’étamines que de pétales, soit quatre.
Sur les fleurs fraîches prélevées directement sur l’arbre, les anthères (Pl. 1, fig. G,, G,, G,, G4) sont constituées par deux loges boudinées de 1,9 mm environ, d’un jaune pâle, dont le diamètre va en diminuant vers le haut, fixées sur un connectif charnu de forme générale triangulaire lorsqu’elles sont jeunes, puis presque linéaire par la suite et arrondi vers le bas. En haut, le connectif dépasse nettement les loges comme un mucron et se termine graduellement en pointe. Le filet trapu, arqué, prend naissance vers le milieu du connectif.
Les grains de pollen (21 à 27 p de diamètre environ), jaune pâle, ne présentent pas d’ornementation particulière sur le matériel frais (Pl. 1, fig. 1,). Toutefois, en les exami- nant avec attention, on observe de grands ports germinatifs arrondis, reliés entre eux par deux bandes aussi larges que le diamètre de ces ports et plus réfringentes. On compte

48 H. RABÉCHAULT
ainsi deux bandes par port, qui s’entrecroisent à angle droit. Le plus souvent, ces bandes s’estompent et se confondent avec la paroi avant d’avoir atteint le port germinatif voisin. Sur les échantillons d’herbiers, les grains de pollen semblent cloisonnés (Pl. 1, fig. 1,). En tout cas, ils présentent un ou plusieurs méridiens fortement marqués. Certains grains sont aplatis dans le sens d’un méridien et d’autres semblent séparés en deux par cette ligne médiane et les deux moitiés ainsi délimitées sont d’inégale grandeur.
Le gynécée comporte un ovaire infère ob-conique. Le calice qui le surmonte dépasse le dernier calicule de l’inflorescence et ressemble à ce dernier. La partie de la fleur compre- nant l’ovaire, muni de son calice à la partie supérieure, mesure 2 mm environ de hauteur.
L’ovaire (Pl. 1, fig. E) présente deux loges dans chacune desquelles pend, accroché par son funicule au sommet du placenta, un ovule orthotrope : chaque ovule donnera naissance à une graine et il y aura ainsi deux graines par fruit.
Au sommet de l’ovaire est inséré un style filiforme de 5 à 7 mm de longueur, glabre à la base puis poilu lorsqu’il sort de la corolle, situé dans le grand axe de la fleur et qui passe au milieu de la masse des poils de la gorge de la corolle. Le style est billde au sommet ; ses deux subdivisions (0,9 mm de long) s’écartent l’une de l’autre et SC, placent dans un plan horizontal. Les papilles stigmatiques sont nombreuses.
G. Le huit (Pl. 1, fig. J, K et Pl. V).
a. MORPHOLOGIE.
Le fruit sessile (Pl. 1, fig. J) (76-86 mm de hauteur sur 64-80 mm de diamètre) est une drupe ou cerise comme chez le Caféier, mais moins charnue. Il est ovoïde à sphérique, biloculaire, légèrement déprimé selon un méridien marquant la limite de séparation des deux loges, glabre, sessile et surmonté d’une petite couronne en forme de tronc de cône renversé représentant le calice acrescent qui entoure le (< disque H ou partie supérieure de l’ancien ovaire de la lleur. Le disque, légèrement bombé, est arrondi avec une petite concavité également arrondie, en son centre qui représente le point d’attache de la base du style.
En ouvrant le fruit (Pl. 1, fig. K), on trouve une graine dans chaque loge. Les deux graines de chaque fruit peuvent être facilement extraites surtout lorsque le fruit est sec. Elles restent cependant très souvent adhérentes l’une à l’autre par leurs faces planes collées par la cloison très fine de séparation des loges du fruit. Leur ensemble épouse alors une forme subsphérique.
On remarque que l’endocarpe uni, lisse, grisâtre et résistant, qui tapisse l’intérieur de chaque loge, reste adhérent à la paroi du fruit (au mésocarpe). 11 correspond à ce que les technologistes appellent la (t parche )) chez le grain de café.
b. ANATOMIE (Pl. V).
k
k
k
I
k
1
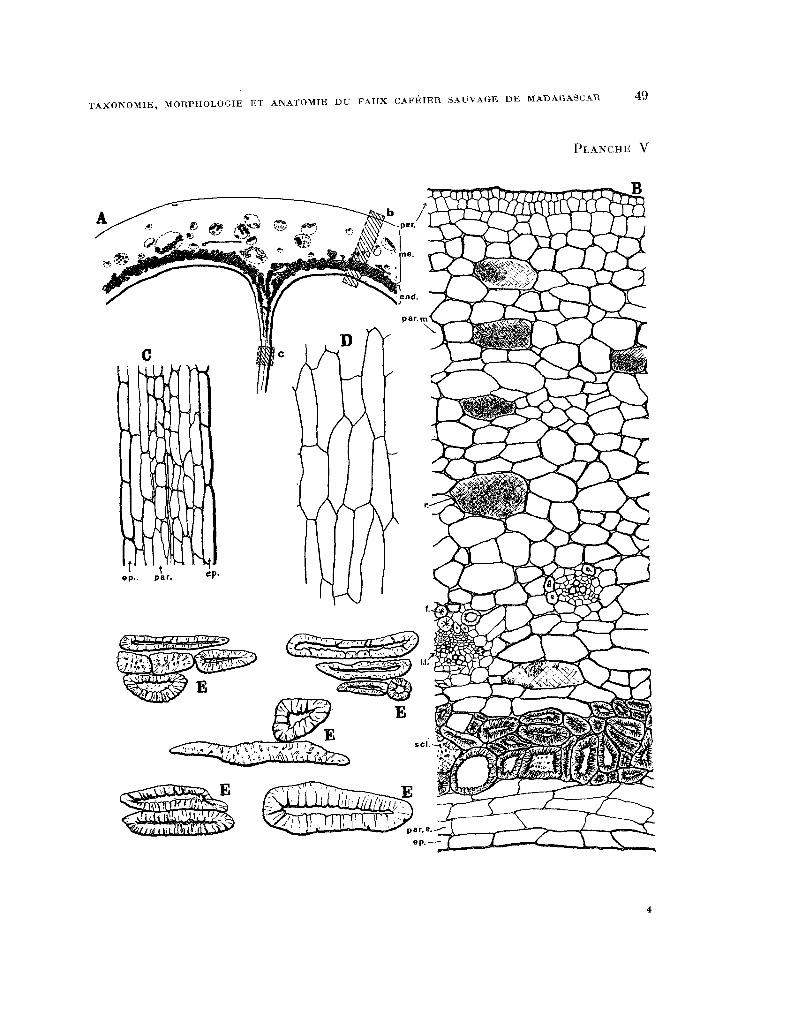
PLANCHE V. - Anatomie du fruit : fig. A, schéma d’une partie de la section transversale de la paroi du fruit ; per. péricarpe, me. mésocarpe, end. endocarpe ; B, portion grossie de la section transversale de la paroi du
fruit representée par la zone hachurée b de la figure precedente, per. péricarpe (deux couches de cellules), par. m.
parenchyme du mesocarpe, P. cellule a mucilages et tanins, 1.1. faisceau libero-ligneux, f. fibres libériennes, sel. sclérites, par.e. parenchyme endocarpique, ep. epiderme interne ; C, portion grossie de la section transversale de la paroi du fruit représentee par la zone hachuree c du schema general ; D, epiderme a l’interieur des loges en examen
perpendiculaire a la surface ; E, sclerites de l’endocarpe du fruit. (Grossissements : Bg. A x 25, les autres fig. x 200).
TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER S.4UVAG1-3 DE MADAGASCAR 49
PLANCHE V

50 H. RABlkHAULT
Une coupe transversale effectuée dans la paroi du fruit (Pl. V, fig. A) permet, à faible grossissement, d’observer les tissus suivants : à l’extérieur un épiderme, le péricarpe (per.) ; dans la partie moyenne un parenchyme, le mésocarpe (me.) ; à l’intérieur, un tissu sclérifié, l’endocarpe (end.).
Le péricarpe (Pl. V, fig. B) est en général recouvert à l’extérieur par une cuticule et une fine couche de pruine. Il est constitué par une ou deux couches de petites cellules à section carrée assez régulières et à parois fines (per.).
Le mésocarpe, avec ses grandes cellules (par. m.), forme la partie charnue de la drupe. Certaines cellules renferment des inclusions analogues à celles rencontrées dans le limbe (P.). On observe, disséminés dans l’épaisseur du tissu des cordons libéro-ligneux (11.) plus ou moins importants, entourés chacun par leur péricycle et quelques fibres péricycliques (f.).
Les dernières couches de cellules parenchymateuses, situées immédiatement au contact des sclérites de l’endocarpe ci-après, sont remplacées chez plusieurs espèces de Caféiers, notamment les Coff ea canephora et excelsa par une couche de cellules à parois trés fines appelée couche mucilagineuse par les technologistes, ou couche palissadique par les anatomistes (RABÉCHAULT, 27). Les cellules de ce tissu sont en effet très allongées et disposées côte à cote perpendiculairement à la paroi du fruit, comme le tissu palissadique du limbe des feuilles. Ici, ce tissu particulier n’existe pas.
L’endocarpe représente ce que l’on appelle la (( parche )) chez le grain de café. Il comprend : une assise de 1 à 4 couches de gros sclérites à grand lumen tapissant la face intérieure de la paroi du fruit (sel.), plusieurs couches de cellules parenchymateuses (par. e.) à parois fines de section transversale rectangulaire et un épiderme de cellules pavimenteuses (ep.) de section rectangulaire à parois fines qui donne à l’intérieur de chaque loge du fruit un aspect lisse.
En examen longitudinal, les sclérites de l’endocarpe sont très courts, souvent isodia- métriques. Leur paroi est pénétrée par de nombreux plasmodesmes comme ceux de la parche des caféiers cultivés (Pl. V, fig. E).
Dans la paroi fine de séparation des loges (Pl. V, fig. A, C), la plus grande partie du mésocarpe a disparu. Les sclérites de l’endocarpe disparaissent complètement dans le premier millimètre voisin de la paroi du fruit. A l’intérieur de la cloison on trouve un parenchyme formé de cellules allongées à section rectangulaire aux angles arrondis, limité extérieurement de part et d’autre par un épiderme. La fig. D, Pl. V, représente les cellules de cet épiderme en examen perpendiculaire.
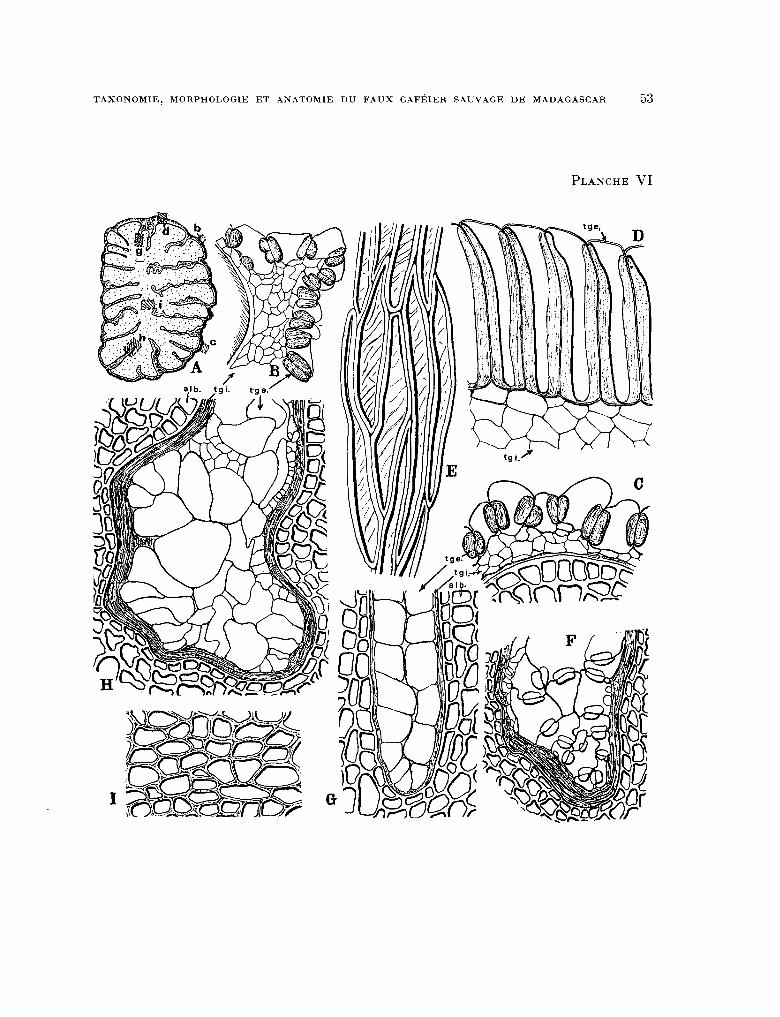
H. La Graine (Pl. 1 et Pl. VI).
a. MORPHOLOGIE (Pl. 1, fig. L, M, N, 0, P, Q).
Les dimensions d’une graine, calculées à partir de 100 graines sèches sont les sui- vantes : longueur moyenne X = 5,71 mm (4,4-6,3 mm), largeur x = 6,63 mm (5,1-7,5 mm, épaisseur Z = 4,02 (3,1-4,6 mm).
Une graine bouillie dans l’eau mesurait : L = 7,3 mm, 1 = 8,O mm, ép. = 4,9 mm. Sur son embryon, j’ai relevé les dimensions suivantes : longueur totale 3,5 mm, longueur des cotylédons 2,0 mm, largeur 1,9 mm, diamètre de la radicule 0,6 mm. Cent graines sèches pèsent 9,6 g.
La graine arrondie, de couleur brun clair (marron clair), comporte comme le grain de café, une face plane, la face ventrale, et une face bombée, la face dorsale. Vue de profil
i
I
i

TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 51
(Pl. 1, fig. 0, P) la graine a donc une forme hémi-ovoïde, plus rarement hémisphérique. La partie inférieure dirigée vers le pédoncule du fruit, est légèrement pointue, alors qu’au contraire, la partie supérieure est plus obtuse et arrondie.
Vue de face (Pl. 1, fig. L, N), on retrouve les mêmes caractères, forme arrondie à la partie supérieure et amincissement vers le bas conduisant à une forme cunée.
Vue de dessus (extrémité arrondie), la forme est hémisphérique (Pl. 1, fig. M).
La surface de la graine (Pl. 1, fig. L) est divisée par des stries ou sillons longitudi- naux que l’on reconnaît bientôt comme autant de séparations des feuillets de l’albumen aboutissant au centre de la graine (Pl. 1, fig. K et Pl. VI, fig. A). Entre ces stries longitu- dinales et sineuses se trouvent d’autres stries plus fines, peu profondes, de même sens et de même forme, qui subdivisent chaque feuillet en autant de feuillets secondaires. Les feuillets sont plus larges vers la partie équatoriale de la graine. En règle générale, l’amin- cissement des feuillets est graduel au fur et à mesure que l’on s’avance vers les extrémités de la graine où ils convergent tous.
La pointe de la graine ne présente aucune caractéristique, mais à la partie opposée arrondie, les feuillets semblent s’interrompre brusquement autour du hile, sorte de demi- pastille nucamentée, demi-circulaire Pl. 1, fig. M. Au centre de cette pastille, on trouve une partie arrondie, deux tiers plus petite et d’aspect fibreux.
En coupe transversale, la graine montre une rumination accentuée et profonde (Pl. VI, fig. A) que laissait prévoir la striation de la surface de la graine. Les feuillets qui divisent l’albumen convergent tous vers une partie centrale que nous appellerons (t âme de l’albumen >. L’âme de l’albumen (Pl. VI, fig. A) aplatie dans le sens dorso-ventral de la graine, est parfois légèrement excentrée vers la face ventrale de cette dernière.
Les téguments séminaux bordent les nombreuses invaginations de l’albumen (stries). On dit que les téguments séminaux 0 pénètrent >) l’albumen. C’est là un caractère du genre Polysphaeria. La graine des Cremaspora, genre voisin, ne serait en effet pas pénétrée du tout (non ruminée) et l’on se rappelle que le grain de café possède un seul sillon longitudinal sur la face ventrale plane.
L’embryon (Pl. 1, fig. N, 0, Q) est logé dans l’âme de l’albumen vers la partie supérieure de la graine. Il est orienté dans un plan parallèle à la face ventrale (plane). Lorsque l’on place cette face vers soi, l’embryon est légèrement à gauche du grand axe de la graine. De plus, il est incliné ; sa radicule, dirigée vers le haut, est normale (perpen- diculaire) par rapport à la limite extérieure de la graine.
L’embryon (Pl. 1, fig. Q) présente une radicule allongée cylindrique, brusquement arrondie à l’extrémité. Un peu au-dessous des cotylédons, la radicule s’amincit un peu. Les cotylédons sont ovales, presque cordiformes à la base, cunés à leur extrémité, tri-ou
pentanervés et appliqués l’un contre l’autre par leur face ventrale.
b. ANATOMIE.
Bien que divisé en feuillets, l’albumen de la graine est dur et corné, de couleur blanche translucide, avec parfois une très légère teinte verte. Ses cellules sont identiques à celles du grain de café (Pl. VI, fig. 1) mais renferment peu de substances de réserve. Leur lumen est en effet vide ou très clair. Elles ont une forme polygonale irrégulière, aux angles arrondis. Les parois sont très épaisses et régulières, avec peu de plasmodesmes.
L’albumen est limité à l’extérieur par la pellicule fibreuse et soyeuse (correspondant à la pellicule argentée chez le Café) dont j’ai parlé précédemment et qui représente les
i
1
1

52 H. RABÉCHAULT
téguments séminaux. Le tégument séminal interne (tgi.) (Pl. VI, fig. B) est constitué de cellules polygonales irrégulières. Il est difficilement observable au fond des rumina- tions (sillons) de l’albumen. Il est souvent très comprimé et réduit à une couche aplatie brune, comme inorganisée.
Le tégument séminal externe (tge) (Pl. VI, fig. B, C, D, E, F, G, H) donne à la pelli- cule recouvrant la graine son aspect fibreux. Très grossi en examen superficiel, ce tissu se montre constitué de cellules sclériflées accolées les unes aux autres (Pl. VI, fig. E)et rappelant un peu les sclérites que l’on trouve dans la pellicule argentée du Café (RABÉ- CHAULT, 27). Cependant, contrairement aux sclérites des Caféiers, les sclérites du Poly- sphaeria ont une paroi continue régulière épaisse sans plasmodesmes. Ces cellules se colorent vivement par le bleu de méthylène ou le vert d’iode.
En coupe transversale, les cellules du tégument séminal externe montrent une grande variabilité dans la forme, les dimensions et l’épaississement de leurs parois, selon qu’elles se sont formées à l’extérieur au sommet d’un feuillet de l’albumen (Pl. VI, fig. B, C, D) ou au fond d’un sillon (rumination) (Pl. VI, fig. G, H).
Au sommet des feuillets, les cellules sont très allongées perpendiculairement à l’albumen sur le bord de la graine - zone hachurée d sur le schéma A, Pl. VI -- tandis que sur le sommet des feuillets du dos de la graine les cellules sont arrondies, irrégulières (zone hachurée b et c du même schéma).
Dans la zone d représentée Pl. VI, fig. D, les parois latérales radiales des cellules sont très épaissies, tandis que les parois tangentielles ne le sont pas du tout.
Dans les zones b et c représentées Pl. VI, fig. B et C, l’épaississement des parois latérales radiales des cellules, très limité, est globulaire. Il en est de même pour les cellules du tégument séminal externe situées au fond des sillons peu profonds (Pl. V, fig. F).
Dans les zones hachurées g et h du schéma A représentées par les figures G et H de la Pl. VI, les cellules du tégument externe qui se trouvent au fond de chaque rumination sont subarrondies, souvent très irrégulières et ne présentent aucun épaississement de leurs parois. ,
PLANCHE VI. - Anatomie de la graine : fig. A, schema de la section transversale d’une graine, les zones hachurees b, e, d, g, h, i sont les parties de l’albumen et des téguments seminaux représentees grossies dans les figures suivantes ; B, section transversale grossie des teguments seminaux dans la region b de la figure précedente. tgi. tegument seminal interne, tge. tegument seminal externe ; C, section identique a celle representee par la figure précedente mais observee dans la région c du schema general ; D, section transversale effectuee dans la region d du schema genéral ; E, tegument seminat externe de la region d du schéma géncral en examen perpendiculaire à la surface ; F, section transversale effectuee dans la région située a l’extremité d’un sillon peu profond et trés ouvert ; A comparer avec les deux B$ures suivantes qui representent les téguments au fond de sillons profonds ; G, section transversale effectuee dans la region g du schema géneral ; H, section transversale effectuee dans la region h du schema general ; 1, section transversale effectuee dans la region i du schéma général (albumen).
(Grossissements fig. A x 8 ; E x 100 ; les autres fig. x 200).
TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIER SAUVAGE DE MADAGASCAR 53
PLANCHE VI
tg i./

54 H. RABÉCHAULT
III. ESSAIS DE GREFFAGE DU POLYSPHAERIA CONGESTA (H. BN) A. CHEV. AVEC LES CAFfiIERS CULTIVfiS
MOTHES (cf. 28) avait montré en 1928 que la nicotine ne pouvait se former dans les feuilles isolées de Tabac. Puis BERNHARDINI (cf. 28) en greffant un tabac sur une tomate a observé que le tabac ne pouvait pas biosynthétiser la nicotine parce que celle-ci se formait dans les racines. En 1955, aucune recherche n’avait encore été effectuée sur la biosynthèse de la caféine. Mais j’avais pensé que si la caféine se formait dans les racines comme la nicotine, nous aurions la possibilité d’obtenir du Café sans caféine de façon naturelle en greffant un Caféier cultivé sur un Caféier sauvage de Madagascar ou un faux Caféier.
Un dosage effectué sur les graines de Polysphaeria envoyé par la Station Agronomique de 1’Ivoloina ne m’avait pas permis de détecter la présence de l’alcaloïde. Aussi j’ai commencé dès le début de l’année 1955 des greffes de Polysphaeria congesta sur Coffea arabica et C. canephora var. robusta (Phot. 1) et inversement des greffes de ces Caféiers sur un pied de Polysphaeria (Phot. 2). J’ai effectué des greffes par approche et des greffes de tête en fente simple.
Les greffes, maintenues sous un capuchon en matière plastique, ont manifesté un début de croissance et ont formé des feuilles ; un bourrelet cicatriciel est, apparu (Photo- graphies 1 et 2). Malheureusement, au bout de huit mois les sacs en polyét,hylène ayant été enlevés, les greffes sont mortes par dessèchement. L’examen anatomique du bourrelet cicatriciel a montré qu’il ne s’était pas formé de vaisseaux libéro-ligneux intermédiaires
D’autres tentatives effectuées les années suivantes n’ont pas eu plus de succès (RABÉCHAULT, 28).

’ \
’ \
’ I
TAXONOMIE, MORPHOLOGIE ET ANATOMIE DU FAUX CAFÉIEH SAUVAGE DE MADAGASCAR 55
CONCLUSIONS
La plante, introduite sous le nom de (l Coffea humblofiana H à la Station de Recherches Agronomiques de 1’Ivoloina (République Malgache) est un faux Caféier Polysphaeria congesfa (H. BN) A. CHEV.
L’espèce ne peut être confondue avec les Coffea et les Cremaspora parce que ses fleurs n’ont que 4 pétales, 4 étamines au lieu de 5 et à cause des poils nombreux de la gorge de la corolle et de la rumination profonde de l’albumen des graines.
La décussation des feuilles se retrouve dans tous les organes et jusque dans les inflorescences et les pièces florales.
Les différences de structure entre le Polysphaeria congesfa et les Coffea sont peu importantes en ce qui concerne les organes végétatifs à l’exception des feuilles, dont le limbe en particulier, comporte de nombreuses poches remplies d’une matière mucila- gineuse brune, et tend vers une symétrie bifaciale par la formation d’un second paren- chyme palissadique à la face inférieure.
L’anatomie du fruit n’a pas révélé la présence de la couche de cellules en palissade ou couche mucilagineuse rencontrée chez les Caféiers.
La graine présente des caractères anatomiques très particuliers concernant son tégument séminal externe.
Enfin, il n’a pas été trouvé de caféine dans les graines et les tentatives de greffage de cette plante avec les Caféiers cultivés Coffea arabica L. et C. canephora var. Robusfa ont échoué.
BIBLIOGRAPHIE
1. BAILLON (H.), 1880. - Histoire des Plantes : Rubiacées, VII, pp. 303-304, 385-386, 429-430 (Hachette et Cie fidit.), Paris.
2. BAILLON (H.), 1876-1879. - Stirpes exoticae novae. - Adansonia, XII, p. 283, Paris.
3. BENTHAM (G.) et HOOKER (J. D.), 1873. - Genera Plantarum, II, p. 108 (Reeve L. Edit.), London.
4. BERTRAND (G.), 1901. - Sur la composition chimique du Café de la Grande Comore. - Bull. Soc. Chim., 3e sér., XXV, pp. 379-380.
5. BERTRAND (G.), 1902. - Recherches et dosage de la caféine dans plusieurs espèces de Café. - Agric. Praf. des Pays chauds, 2, pp. 211-213.
6. BERTRAND (G.), 1905. - Sur les Cafés sans caféine. - C. R. Acad. SC., Paris, 141,
pp. 209-211. 7. BRENAN (J.-P. M.), 1949. - A contribution to the Flora of East Tropical Africa. -
Kew Bulletin, no 1, pp. 81-86.
8. CHEVALIER (A.), 1929, 1942, 1947. - Les Caféiers du Globe : 1. Généralités sur les Caféiers, 196 p., 32 fig., 1929 ; II. Iconographie des Caféiers sauvages et cultivés, 36 p., 158 pl. ; III. Systématique des Caféiers et des faux Caféiers, Maladies et Insectes nuisibles, 357 p., 17 pl. (206 fig.). (P. Lechevalier fidit.), Paris, 1947.
9. CHEVALIER (A.), 1931. - L’amélioration des Caféiers et la production des types sauvages devant l’Association Internationale des Sélectionneurs de Plantes de grande culture. - Rev. Bof. Appl. et Agric. trop., pp. 355-358, 1931. (Extr. du (( Bull. Assoc. Internation. des Sélect. de Plantes de grande culture o).

56 H. RABkCHAULT
10. CHEVALIER (A.), 1937. - Un nouveau Caféier sauvage de Madagascar à grains sans caféine. - Reu. Bot. Appf. et Agr. Trop., 17e année, no 195, pp. 821-826.
11. CHEVALIER (A.), 1938. - Sur quelques faux Caféiers d’Afrique Tropicale. - Reu. Bof. Appl. et Agr. Trop., 18e année, no 202, pp. 413-419.
12. CHEVALIER (A.), 1938. - Essai d’un groupement systématique des Caféiers sau- vages de Madagascar et des îles Mascareignes. - Rev. Bof. Appl. et Agr. Trop., pp. 825-843.
13. CHEVALIER (A.), 1946. - Écologie et distribution des Caféiers sauvages et cultivés. - Rev. Bof. Appl. et Agric. Trop., pp. 81-94.
14. COSTE (R.), 1966. - Les Caféiers sylvestres de Madagascar, Café, Cacao, Thé, X, no3 pp. 207-208.
15. FRIDMANN (M.) et VIANNEY (J.), 1966. - Contribution à l’étude des Caféiers sylvestres de Madagascar. - Café, Cacao, Thé, X, no 4, pp. 209-215.
16. GAFF~ER (M.-L.), 1933. - L’anatomie des feuilles des Caféiers sauvages. - Ann. Musée Colon. Marseille, 41e, 5e série, 1, no 4, pp. 15-27.
17. JUMELLE (H.), 1933. - Caféiers sauvages de Madagascar. - Ann. Mus. Colon. Marseille, 41e, 5e série, fasc. 4, pp. 5-14.
18. LEBRUN (J.), 1941. - Recherches morphologiques et systématiques sur les Caféiers du Congo. - Public. Insf. Naf. Etude Agron. Congo Belge (I.N.E.A.C.), vol. H. S.
19. LEROY (J.-F.), 1961. - Coffeae novae madagascarienses. - J. Agric. Trop. ef Bof. Appl., VIII, no l-2-3, pp. l-20.
20. LEROY (J.-F.), 1961. - Les faux Caféiers du genre Argocoffea (Pierre Lebrun). - J. Agric. Trop. et Bof. Appl., VIII, no l-2-3, p. 65.
21. LEROY (J.-F.), 1962. - Prospection des Caféiers sauvages : Rapport préliminaire sur une mission scientifique à Madagascar et aux Ples Mascareignes (27 avril- 15 juillet 1962). - J. Agric. Trop. et Bof. Appf., IX, nO3-6, pp. 211-244.
22. LEROY (J.-F.), 1962. - Coffea novae madagascarienses et mauritanae. - J. Agric. Trop. et Bof. Appl., IX, no 11-12, pp. 525-530.
23. MARIANI (J.), 1908. - Les Caféiers. Structure anatomique de la feuille. - Thèse Docf. Univ., 137 p., Lons-le-Saunier.
24. METCALFE (C. R.), CHALK (L.), 1950. - Anatomy of the dicotyledons, II, pp. 759- 776 . - (Clarendon Press Edit.), Oxford.
25. PORTÈRES (R.), 1962. - Sur quelques Caféiers sauvages de Madagascar. - J. Agric. Trop. et Bof. Appl., IX, no 3-6, pp. 201-210.
26. RABÉCHAULT (H.), 1954. - I%udc anatomique comparée des organes végétatifs des Caféiers de la Côte d’ivoire, in (( H. JACQUES-FÉLIX, Contributions à l’l%ude du Caféier en Côte d’ivoire 1). - L’Agron. Trop., Bull. Scient., no 5, pp. 151-177 (Sect. Techn. Agr. Trop. fidit.), Nogent-sur-Marne.
27. RABÉCHAULT (H.), 1959. - Anatomie comparée des fruits et des grains de Caféiers cultivés in (( R. COSTE, Les Caféiers et les Cafés dans le Monde )), II, pp. 97-128 (Larose adit.), Paris.
28. RABÉCHAULT (H.), CAMBRONY (H.), 1964. - Physiologie du Caféier. De la germi- nation à la morphogénèse. - Physiologie des Plantes Tropicales cultivées. Cah. ORSTOM, 1, no 1, 80 p., 291 réf. bibl.
I
\
I,
I