dynamique de la population de cerfs du domaine national de ... · photos de couverture : château...
TRANSCRIPT

Dynamique de la population de cerfs
du domaine national de Chambord :
Analyse temporelle des indicateurs de changement écologique (ICE)

2
TITRE : Dynamique de la population de cerfs du domaine national
de Chambord : analyse temporelle des indicateurs de
changement écologique (ICE)
AUTEURS : Maryline Pellerin, Christophe Bonenfant, Mathieu Garel,
Thierry Chevrier, Guillaume Queney, François Klein &
Jacques Michallet
PUBLICATION : CNERAs Cervidés-Sanglier et Faune de Montagne de
l’ONCFS & LBBE-CNRS de l’université de Lyon 1, Gières,
juillet 2014
CITATION : Pellerin M., Bonenfant C., Garel M., Chevrier T., Queney G.,
Klein F. & Michallet J. 2014. « Dynamique de la population
de cerfs du domaine national de Chambord : analyse
temporelle des ICE », Rapport d’expertise ONCFS
MOTS-CLÉS : Chambord – cerf élaphe – indicateur de changement
écologique – capture-marquage-recapture fèces – modèle
démographique – prélèvements cynégétiques
Photos de couverture : Château de Chambord (©Domaine national de Chambord) et groupe de
cerfs élaphe (©Erich Marek).

3
SOMMAIRE
I. INTRODUCTION ............................................................................................................. 4
1. Dynamique des populations d’ongulés : généralités et problématique ....................... 5
2. Utilisation des ICE : application au domaine national de Chambord ........................ 7
II. DONNÉES DISPONIBLES ET UTILISÉES ................................................................. 10
1. Description du domaine national de Chambord ........................................................ 10
2. Les ICE potentiels pour la population de cerfs du domaine de Chambord .............. 11
2.1. ICE « Abondance » ............................................................................................. 12
2.2. ICE « Performance » .......................................................................................... 14
2.2.1. La masse corporelle des jeunes .................................................................. 14
2.2.2. La gestation des femelles .......................................................................... 15
2.2.3. Intégration des conditions environnementales ............................................ 16
2.3. ICE « Impact » sur le milieu ............................................................................... 17
3. Capture-Marquage-Recapture non-invasif ................................................................ 18
III. ANALYSES ET DISCUSSION ....................................................................................... 20
1. Analyse des trois familles d’ICE ................................................................................. 20
1.1. Variations des ICE « Abondance » ..................................................................... 20
1.2. Variations des ICE « Performance » ................................................................... 27
1.2.1. La masse corporelle des jeunes .................................................................. 27
1.2.2. La gestation des femelles .......................................................................... 33
1.3. Variations des ICE « Impact » sur le milieu ....................................................... 40
1.4. Interprétation combinée des ICE ........................................................................ 43
2. Effectifs estimés de la population de cerfs par CMR non-invasif ............................. 44
3. Modélisation et plans de prélèvements ....................................................................... 49
IV. CONCLUSIONS .............................................................................................................. 55
BIBLIOGRAPHIE ................................................................................................................. 58

4
I. INTRODUCTION
L’histoire récente du cerf élaphe (Cervus elaphus) en France est étroitement liée au
domaine national de Chambord (DNC). Plusieurs événements et particularités ont contribué à
faire de ce domaine un site unique en Europe, auquel l’image du cerf élaphe est souvent associée.
D’une part, une majorité des animaux relâchés dans le cadre du renforcement des populations
françaises de cerfs provenait de ce site (Leduc & Klein 2004). D’autre part, la population de cerfs
du domaine a été isolée démographiquement depuis plusieurs siècles par un mur d’enceinte, tout
en faisant l’objet de chasses de renommée internationale. Enfin, le DNC contribue depuis
longtemps à sensibiliser le grand public à la conservation de la biodiversité, en favorisant
l’observation de la faune en général et du cerf en particulier, et en organisant des sessions de
captures de cerfs. L’intérêt patrimonial et cynégétique de cette espèce a conduit les gestionnaires
du domaine à construire, depuis plus de 30 ans, une base de données unique en France dont la
valeur scientifique et l’intérêt pour la gestion de la population de cerfs est indiscutable. C’est
cette base de données que nous nous proposons de valoriser ici afin de statuer sur l’état actuel de
la population de cerfs de Chambord. Ce travail sera la première réalisation concrète de la volonté
conjointe du domaine national de Chambord et de l’Office National de la Chasse et de la Faune
Sauvage (ONCFS) de mettre en place un programme scientifique à long terme sur la
démographie et la chasse des ongulés sauvages à Chambord.
Dans ce travail d’expertise, les informations analysées et exploitées portent sur :
les six modalités de comptages mises en place depuis les années 1980,
le plan de chasse,
la masse corporelle des faons et le taux de gestation des femelles prélevées à la
chasse,
le suivi de la pression des animaux sur la forêt de 1999 à 2008,
et les fèces échantillonnées en 2009 et 2010.
Après un bref rappel théorique sur le fonctionnement des populations de grands mammifères
herbivores, nous détaillerons la notion d’indicateur de changement écologique (ICE), développée
par le consortium ONCFS - CNRS - INRA - IRSTEA -ONF - FNC pour le suivi et la gestion
raisonnée de la faune sauvage (Morellet et al. 2007, 2008). Ensuite, après une synthèse des
données disponibles, les données pertinentes seront analysées dans le cadre d’une gestion par ICE

5
afin de ré-interpréter l’histoire de la dynamique de la population de cerfs du DNC. Nous
proposerons également une estimation des effectifs de cette population en 2009 et 2010 à l’aide
des techniques de capture-marquage-recapture et d’échantillonnage non-invasif (ADN ; voir
Valière et al. 2007 pour une étude pilote sur le domaine). Puis, à l’aide d’un modèle
démographique intégrant un indice d’abondance et les données cynégétiques, nous déterminerons
dans quelle mesure la chasse a été un facteur de variation d’abondance de la population de cerfs.
Enfin, nous conclurons le rapport d’expertise sur les rôles relatifs des facteurs écologiques et
cynégétiques à l’origine des variations d’abondance observées depuis près de 30 ans, et ouvrirons
la réflexion sur de nouvelles pistes de gestion du cerf sur le domaine pour les années à venir. Les
objectifs de gestion cynégétique du DNC ayant été un maintien de la population de cerfs à un
niveau relativement élevé depuis plusieurs décennies, on s’attend à ce que les ICE montrent que
les ressources disponibles sont en quantité limitée et que l’espèce a un impact marqué sur son
habitat.
1. Dynamique des populations d’ongulés : généralités et problématique
Les populations de grands mammifères herbivores sont caractérisées par une très forte
structuration en âge des paramètres démographiques que sont les taux de survie, de reproduction
et de dispersion. Par exemple chez le chevreuil (Capreolus capreolus), il a été montré que la
survie des faons lors des 6 premiers mois de vie (60% en moyenne) est nettement plus faible que
la survie des adultes entre en 2 et 8 ans (92% en moyenne), tout comme la survie des animaux
âgés qui diminue progressivement à partir de 9 ans chez cette espèce (Gaillard et al. 1996).
L’importance de la structure en âge des populations de grands mammifères pour la dynamique de
populations tient à ce que certains paramètres démographiques vont avoir plus ou moins d’impact
sur le taux d’accroissement des populations (noté r), selon que l’on s’intéresse aux individus
jeunes ou adultes. En conséquence, un résultat majeur des recherches récentes conduites sur les
grands herbivores a été de mettre en évidence que les variations, dans l’espace et dans le temps,
des paramètres de survie et de reproduction sont beaucoup plus importantes pour les juvéniles
que pour les adultes (Gaillard et al. 2000). On parle alors d’une dynamique des populations à
recrutement variable et survie constante (Gaillard et al. 1998). De fait, les variations d’abondance
observées dans les populations naturelles de grands mammifères herbivores sont essentiellement

6
dues à des variations de la survie juvénile et de la reproduction des femelles. De par ses
variations moindres, la survie des adultes a un rôle plus limité, sauf cas d’épizootie, de crash de
population ou de prélèvements par la chasse dans cette classe d’âge.
Les causes de variation des paramètres démographiques des populations de grands
mammifères herbivores sont fortement liées à la quantité et à la qualité des ressources
alimentaires (mais voir Garel et al. 2005). De fait, en l’absence de grands prédateurs carnivores,
le type d’habitat (Pettorelli et al. 2002), les conditions météorologiques (Gaillard et al. 1993) et la
densité d’individus (Bonenfant et al. 2002) sont les principales sources de variation des
paramètres démographiques rapportées dans la littérature (voir Bonenfant et al. 2009 pour une
synthèse). Ces variations environnementales génèrent des effets cohortes, c’est-à-dire des effets à
long terme des conditions de vie sur les paramètres démographiques et les caractéristiques
phénotypiques qui sont partagés par les individus nés la même année (Albon et al. 1992, Rose et
al. 1998, Gaillard et al. 1997, Toïgo et al. 1999). Ces effets cohortes ont pour origine des
conditions environnementales différentes, d’une année sur l’autre, durant la phase tardive de
gestation et durant la lactation. Au sein d’une même population, jusqu’à 15 cohortes peuvent
coexister (Gaillard et al. 2003a) dont les contributions à la dynamique des populations est très
hétérogène - les bonnes cohortes ayant un rôle plus important car elles possèdent les taux de
survie et de reproduction les plus élevés. De plus, l’habitat, le climat, et la densité interagissent
sur les paramètres démographiques. Un exemple classique est l’interaction climat – densité, mise
en évidence chez le cerf élaphe et le mouton de Soay (Ovis aries), et où l’effet des précipitations
hivernales sur la survie des faons et des agneaux est plus important à forte qu’à faible densité
(Coulson et al. 2001). Une fois encore, la structure en âge des populations est cruciale puisque les
effets de l’habitat, du climat et de la densité sur les paramètres démographiques diffèrent selon
les classes d’âges considérées. Par exemple, si les faons de chevreuils vivant dans un habitat de
mauvaise qualité survivent moins bien que ceux vivant dans un habitat de bonne qualité, les
adultes ont des taux de survie comparables quel que soit l’habitat (Pettorelli et al. 2005). Les
effets de la densité sur les paramètres démographiques n’échappent pas à cette règle et sont aussi
fortement dépendants de l’âge des individus et du paramètre démographique considéré (survie ou
reproduction). La séquence des paramètres démographiques qui diminuent avec l’augmentation
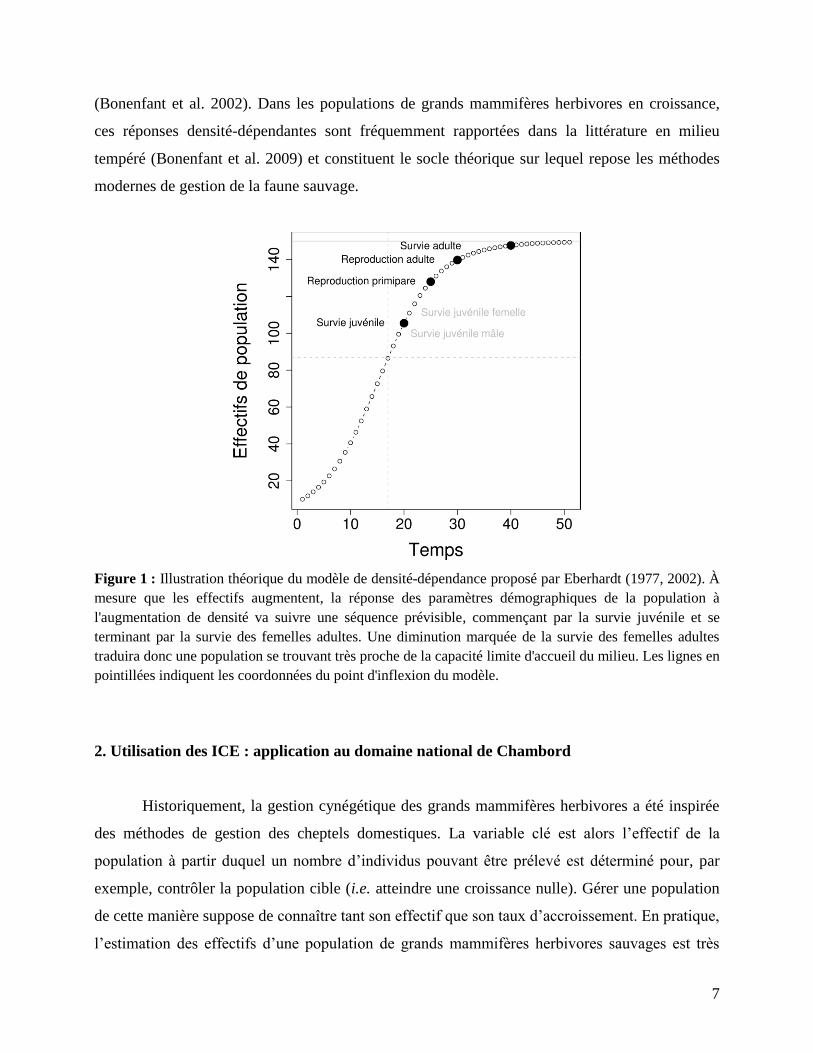
de la densité de population est même prévisible (Fig. 1) et connue sous le nom de modèle
d’Eberhardt (1977, 2002). Ce modèle, très général, s’applique également au cas du cerf élaphe
7
(Bonenfant et al. 2002). Dans les populations de grands mammifères herbivores en croissance,
ces réponses densité-dépendantes sont fréquemment rapportées dans la littérature en milieu
tempéré (Bonenfant et al. 2009) et constituent le socle théorique sur lequel repose les méthodes
modernes de gestion de la faune sauvage.
Figure 1 : Illustration théorique du modèle de densité-dépendance proposé par Eberhardt (1977, 2002). À
mesure que les effectifs augmentent, la réponse des paramètres démographiques de la population à
l'augmentation de densité va suivre une séquence prévisible, commençant par la survie juvénile et se
terminant par la survie des femelles adultes. Une diminution marquée de la survie des femelles adultes
traduira donc une population se trouvant très proche de la capacité limite d'accueil du milieu. Les lignes en
pointillées indiquent les coordonnées du point d'inflexion du modèle.
2. Utilisation des ICE : application au domaine national de Chambord
Historiquement, la gestion cynégétique des grands mammifères herbivores a été inspirée
des méthodes de gestion des cheptels domestiques. La variable clé est alors l’effectif de la
population à partir duquel un nombre d’individus pouvant être prélevé est déterminé pour, par
exemple, contrôler la population cible (i.e. atteindre une croissance nulle). Gérer une population
de cette manière suppose de connaître tant son effectif que son taux d’accroissement. En pratique,
l’estimation des effectifs d’une population de grands mammifères herbivores sauvages est très

8
difficile à obtenir, nécessite des protocoles spécifiques et coûteux (transect linéaire, capture-
marquage-recapture ; voir Schwarz & Seber 1999 pour une synthèse) et, dans le meilleur des cas,
ne permet pas d’avoir une précision inférieure à 30% (Caughley 1977). Une telle incertitude
limite fortement les conclusions quant à l’augmentation, la diminution ou la stabilisation d’une
population d’une année à l’autre. De plus, un effectif n’est pas un indicateur pertinent de la
qualité d’un habitat (vanHorne 1983) et ne renseigne en rien sur la quantité de ressources
disponibles pour les herbivores (et donc sur la relation entre les animaux et leur milieu).
Afin de contourner ces problèmes, un ensemble d’outils de suivi appelés indicateurs de
changement écologique (ICE ; Morellet et al. 2007, 2008) ont été développés. Ces ICE découlent
des recherches réalisées sur des sites de référence (par exemple : Chizé et Trois-Fontaines, pour
le chevreuil ; La Petite Pierre, pour le cerf) propices à la mise au point de méthodes de suivi des
populations de grands herbivores (Klein et al. 2007) et à partir desquelles les différents ICE ont
été pensés et validés. L’étape de validation, indispensable, consiste à démontrer que les outils
proposés reflètent effectivement l’effet d’une variation de l’effectif réel de la population et, de
fait, constituent un moyen fiable de suivre les populations. Les ICE traduisent alors les variations
relatives, au cours du temps, de l’abondance des populations de grands herbivores, de leur impact
sur leur milieu de vie, et de la quantité de ressources alimentaires qui leur est disponible. In fine,
les ICE permettent une gestion raisonnée, objective et adaptative du système « population-
environnement » (Eberhardt 2002), tout en s’affranchissant des difficultés inhérentes à
l’estimation absolue des effectifs. En pratique, le suivi d’un seul ICE ne permet pas de trancher
entre différents scénarii démographiques caractérisant le système étudié (Morellet et al. 2007).
C’est bien l’analyse conjointe des trois familles d’ICE (indicateurs d’abondance, indicateurs de
performance et les indicateurs de pression sur le milieu) qui permettra aux gestionnaires de
statuer sur l’état du système « population-environnement » (Fig. 1). Différents ICE ont donc été
proposés selon les espèces ciblées (Michallet & Chevrier 2013). Pour le cerf, l’indice nocturne
comme indicateur d’abondance (Garel et al. 2010), la masse corporelle des faons et la gestation
des bichettes comme indicateurs de performance (Bonenfant et al. 2002) ont été validés. L’indice
de consommation qui a été validé comme indicateur de pression sur le milieu pour le chevreuil
(Morellet et al. 2003), est en cours de validation pour le cerf et montre d'ores et déjà des résultats
biologiquement pertinents sur les sites où il est utilisé (Michallet & Chevrier 2013,
www.oncfs.gouv.fr/OGFH). Les ICE supposent également de pouvoir délimiter des unités de

9
gestion biologiquement cohérentes, correspondant à des unités de population fonctionnelles
(Zannèse et al. 2006). Dans le cas du DNC, l’unité de gestion fonctionnelle de la population de
cerfs correspond à l’unité de gestion administrative que représente le DNC.
L’utilisation des ICE est en pleine phase de démocratisation en France (Morellet et al.
2008) et à l’étranger (Strickland et al. 2008). Le principe d’une gestion cynégétique par ICE
consiste à ajuster les prélèvements par un processus d’essais et d’erreurs qui, à travers l’examen
des variations temporelles des ICE, doit permettre d’atteindre les objectifs de gestion initialement
définis. Les variations du niveau de prélèvements doivent donc être suffisantes et durables pour
engendrer une réponse détectable des ICE suivis, ce qui peut nécessiter des variations
importantes des plans de chasse sur plusieurs années. La mise en œuvre des ICE ne se justifie que
sur le moyen terme et pour laquelle des suivis de 5 à 10 années constituent un minimum pour une
espèce comme le cerf. Grâce aux données collectées sur le long terme sur le DNC, les ICE sont
tout à fait applicables à sa population de cerfs de manière rétrospective, mais ils seront également
proposés pour la gestion cynégétique à venir du site.

10
II. DONNÉES DISPONIBLES ET UTILISÉES
1. Description du domaine national de Chambord
Le DNC, situé à la porte nord-ouest de la Sologne dans le Loir-et-Cher (41), créé par
l’article 230 de la loi du 23 février 2005 relative au développement des territoires ruraux, est un
établissement public national à caractère industriel et commercial. Propriété de l’État, le domaine
s’étend sur 5 439,88 ha d’un territoire boisé à hauteur de 4 330 ha environ (soit 79%),
entièrement ceint par un mur de 32 km de long. Classé « Réserve Nationale de Chasse et de
Faune Sauvage » (RNCFS) depuis 1947 dans sa partie intra-muros, il est également inscrit au
réseau écologique européen Natura 2000 portant désignation d’une Zone de Protection Spéciale
(ZPS) via l’arrêté du 7 mars 2006 et d’une Zone Spéciale de Conservation (ZSC) via l’arrêté du
13 avril 2007. La gestion sylvo-cynégétique est actuellement assurée par un service spécialisé de
Chambord. Aujourd’hui, à l’exception de 200 ha de terres agricoles, de 160 ha de prairies à gibier
et des 200 ha d’emprise du village, l’ensemble est couvert principalement en peuplements de
chênes (Quercus sp.) et de pins sylvestres (Pinus sylvestris). L’ensemble de la forêt compte
désormais 499 parcelles, d’une superficie moyenne de 10 ha (Fig. 2).
Caractérisé par une topographie peu mouvementée (128 m pour le point culminant), le
domaine de Chambord présente une légère dépression d’orientation est-ouest empruntée par la
rivière « Cosson », qui le traverse sur une longueur de 8 km environ. En raison de sa situation
géographique, Chambord connaît un climat de transition sous double influence entre l’ouest
océanique et l’est plus continental. Ce climat est caractérisé par des précipitations moyennement
abondantes et des amplitudes thermiques limitées. L’importance de la surface couverte par les
feuillus à Chambord (chêne pédonculé Quercus robur et chêne sessile Quercus petraea
principalement), et l’abondance de leurs fruits sont très favorables aux Cervidés ainsi qu’aux
sangliers (Sus scrofa). En période d’absence de fructification forestière, une distribution
quotidienne de céréales en grain est réalisée en 16 points du parc, appelés « centres
d’agrainage ». Les années de glandées, l’agrainage peut être arrêté pendant une durée qui sera
fonction de l’importance de la fructification, et qui pourra, selon le cas, se prolonger sur plus d’un
trimestre.

11
Figure 2 : Cartographie des différents types d’habitat du domaine national de Chambord.
2. Les ICE potentiels pour la population de cerfs du domaine de Chambord
La démarche ICE, qui est un suivi du système « plante – herbivores », demande de
mesurer au moins un indicateur d’abondance, un indicateur de performance et un indicateur de
pression sur le milieu (Morellet et al. 2007). Les données recueillies sur le domaine depuis près
de 30 ans sont nombreuses, mais surtout diverses et hétérogènes en termes de quantité et de

12
qualité. Or, un nombre restreint d’indicateurs judicieusement choisis est toujours préférable à un
nombre plus important mais dont la réponse aux variations de densité est inconnue ou
imparfaitement connue. Un choix s’impose donc et, parmi tous les ICE potentiels que l’on peut
dériver d’une telle quantité de données, certains sont plus ou moins pertinents eu égard aux
travaux antérieurs sur le cerf ou le chevreuil. Le choix d’utiliser une mesure en tant qu’ICE ou
non sera motivé ici par (i) la validation de la mesure sur un site de référence garantissant une
interprétation juste des résultats, (ii) la longueur de la chronique afin de pouvoir remonter le plus
loin possible dans le temps et (iii) la précision de la mesure conditionnée par la taille
d’échantillon.
2.1. ICE « Abondance »
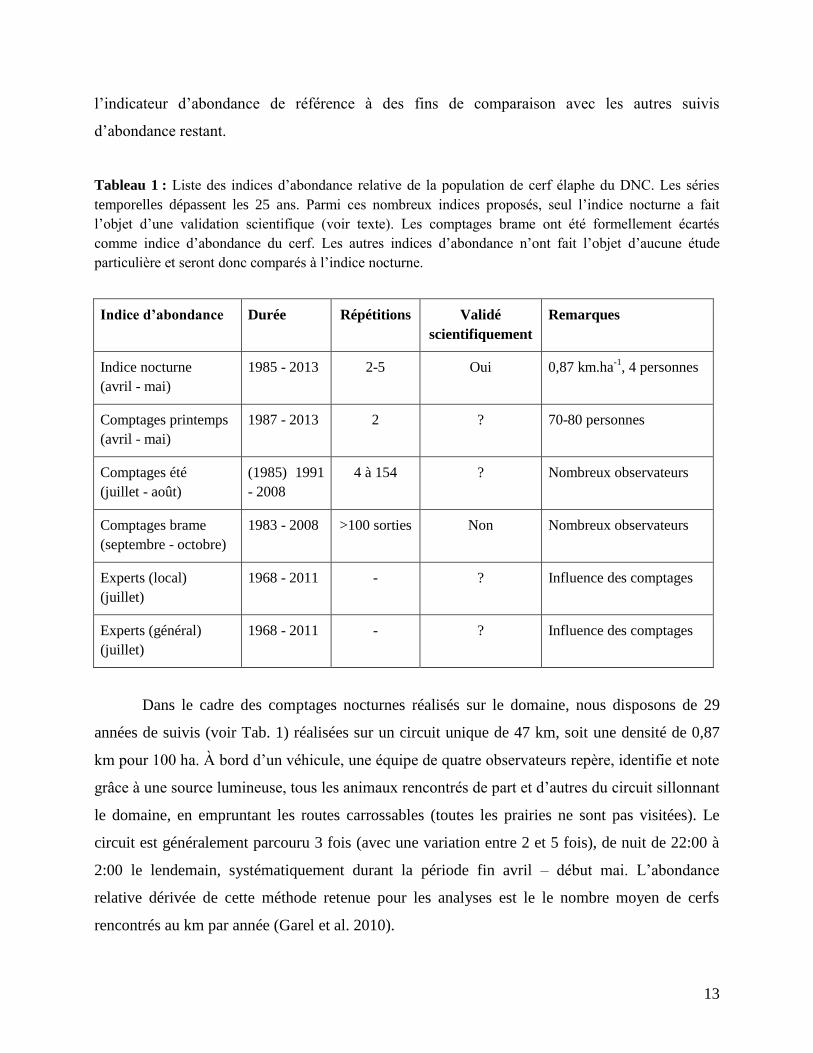
Pas moins de six mesures d’abondance relative de la population de cerfs ont été collectées
annuellement sur le domaine. Cet investissement humain et financier atteste indubitablement de
l’importance portée à l’abondance par les gestionnaires, souvent utilisée a tort comme une
estimation d’effectif. On retrouve :
(1) l’indice nocturne,
(2) les comptages de printemps,
(3) les comptages estivaux,
(4) les comptages brame,
(5-6) ainsi que deux estimations d’abondance à dires d’experts (voir Tab. 1).
Parmi cet ensemble d’indices d’abondance relative de la population de cerfs mis en place sur le
domaine, l’indice nocturne est l’ICE abondance le plus pertinent et le plus à même de donner les
meilleurs résultats. Ce suivi d’abondance, mis en place depuis 1985 dans la réserve de chasse de
Chambord, est très largement utilisé dans le monde de la gestion cynégétique (Hamann et al.
2011). Toutefois, le véritable atout de l’indice nocturne réside dans sa validation en tant qu’ICE.
En effet, une relation entre les estimations d’effectifs par capture-marquage-recaptures et l’indice
nocturne a clairement été mise en évidence sur le site de référence de La Petite Pierre (Garel et al.
2010). C’est à l’heure actuelle l’unique ICE abondance validé pour cette espèce qui est disponible
sur le DNC. Sur la base de ces études précédentes, les comptages brame ne seront donc pas
considérés dans ce travail (Douhard et al. 2013), et l’indice nocturne sera utilisé comme
13
l’indicateur d’abondance de référence à des fins de comparaison avec les autres suivis
d’abondance restant.
Tableau 1 : Liste des indices d’abondance relative de la population de cerf élaphe du DNC. Les séries
temporelles dépassent les 25 ans. Parmi ces nombreux indices proposés, seul l’indice nocturne a fait
l’objet d’une validation scientifique (voir texte). Les comptages brame ont été formellement écartés
comme indice d’abondance du cerf. Les autres indices d’abondance n’ont fait l’objet d’aucune étude
particulière et seront donc comparés à l’indice nocturne.
Indice d’abondance Durée Répétitions Validé
scientifiquement
Remarques
Indice nocturne
(avril - mai)
1985 - 2013 2-5 Oui 0,87 km.ha-1
, 4 personnes
Comptages printemps
(avril - mai)
1987 - 2013 2 ? 70-80 personnes
Comptages été
(juillet - août)
(1985) 1991
- 2008
4 à 154 ? Nombreux observateurs
Comptages brame
(septembre - octobre)
1983 - 2008 >100 sorties Non Nombreux observateurs
Experts (local)
(juillet)
1968 - 2011 - ? Influence des comptages
Experts (général)
(juillet)
1968 - 2011 - ? Influence des comptages
Dans le cadre des comptages nocturnes réalisés sur le domaine, nous disposons de 29
années de suivis (voir Tab. 1) réalisées sur un circuit unique de 47 km, soit une densité de 0,87
km pour 100 ha. À bord d’un véhicule, une équipe de quatre observateurs repère, identifie et note
grâce à une source lumineuse, tous les animaux rencontrés de part et d’autres du circuit sillonnant
le domaine, en empruntant les routes carrossables (toutes les prairies ne sont pas visitées). Le
circuit est généralement parcouru 3 fois (avec une variation entre 2 et 5 fois), de nuit de 22:00 à
2:00 le lendemain, systématiquement durant la période fin avril – début mai. L’abondance
relative dérivée de cette méthode retenue pour les analyses est le le nombre moyen de cerfs
rencontrés au km par année (Garel et al. 2010).

14
2.2. ICE « Performance »
Le terme de performance est pris ici au sens de performance démographique. Un individu
avec une performance démographique forte est un animal qui, relativement à ses congénères,
élèvera la majorité des jeunes qu’il aura produit et bénéficiera d’une forte longévité. La
performance démographique est très souvent une fonction des conditions environnementales
(notamment celles rencontrées l'année de naissance des animaux, appelées effets cohortes). Une
performance démographique faible peut faire suite à une dégradation des conditions
environnementales, comme cela se produit lorsque la population rentre en densité-dépendance.
Les ICE performance doivent donc permettre d'apporter une information robuste et simple de la
performance démographique des individus.
2.2.1. La masse corporelle des jeunes
La masse corporelle est, chez les grands mammifères, une variable biométrique de
première importance puisqu’elle exerce un rôle clé dans la dynamique des populations (Saether
1997). À titre d’exemple, il a été démontré que la masse corporelle conditionne la probabilité de
gestation des jeunes femelles (pour le cerf : Albon et al. 1983, Bonenfant et al. 2002 ; pour le
chevreuil : Gaillard et al. 1992) ou encore la survie hivernale des nouveaux nés dans les
populations de cervidés (pour le cerf : Loison et al. 1999a ; pour le chevreuil : Gaillard et al.
1993). De manière plus générale, la masse corporelle est un indicateur fiable de la qualité
phénotypique d’un individu et représente une bonne approximation de sa performance
individuelle. Réagissant promptement aux changements des conditions environnementales, des
études ont permis de mettre en évidence les réponses de la masse corporelle à l’augmentation de
densité (Mysterud et al. 2001a, Bonenfant et al. 2002, Bonenfant 2004), aux conditions
météorologiques (Mysterud et al. 2001b, Toïgo et al. 2006) ou encore aux modifications de
l’habitat (Pettorelli et al. 2003). Néanmoins, il est important de noter que ces effets agissent sur la
masse corporelle à travers une réduction (directe ou indirecte) de la quantité, de la qualité et de
l’accès aux ressources alimentaires.
À l’échelle de la population, la masse corporelle peut tout à fait être utilisée avec profit
comme un ICE car sa mesure est aisément accessible avec un coût relativement faible. À

15
Chambord, cette information est recueillie chaque année depuis 1982 lors de l’analyse du tableau
de chasse. Actuellement, plusieurs études ont montré que la masse corporelle des jeunes
individus s’avère être plus sensible aux restrictions alimentaires par rapport aux autres classes
d’âge (Gaillard et al. 1998, 2000). En effet, les priorités de croissance ne sont pas les mêmes
entre un animal qui a atteint sa taille adulte qui peut donc investir dans des réserves énergétiques
lui permettant de faire face à des épisodes de restrictions alimentaires, et un jeune animal dont la
stratégie est avant tout d'investir dans sa croissance. Dans le cas qui nous intéresse, la relation
entre la masse corporelle et la densité est la plus pertinente : la relation attendue est une relation
négative, c’est-à-dire une diminution de la masse corporelle avec l’augmentation de la densité.
Dans le cas précis du cerf, il est possible de détecter une augmentation de densité de manière plus
précoce en focalisant les suivis sur l’évolution de la masse corporelle des jeunes faons mâles
(Bonenfant 2004). Cette particularité, que l’on peut attendre pour l’ensemble des espèces
sexuellement dimorphiques en taille est, chez les mâles, la conséquence directe d’une allocation
préférentielle des ressources énergétiques à la croissance plutôt qu’à leur mise en réserve, et donc
à leur propre survie.
2.2.2. La gestation des femelles
Plus spécifique au cerf, le taux de gestation des jeunes femelles (pourcentage de bichettes
gestantes) est un autre ICE performance pertinent (voir Bonenfant et al. 2007 pour une
application au cerf élaphe sur le domaine du Donon). Il a en effet été largement démontré que la
performance reproductive chez les femelles d'ongulés était une fonction de leur masse corporelle.
Ainsi, tout facteur susceptible d'entraîner un retard de croissance aura pour conséquence de
limiter la proportion de femelles se reproduisant pour la première fois à un âge donné. Dans
certains cas, il arrive même que l'âge de reproduction soit décalé d'un an ou plus pour permettre
aux femelles d'atteindre la masse corporelle nécessaire pour assumer les coûts de la reproduction.
Ce retard est l’illustration des compromis entre croissance et reproduction (Green & Rothstein
1991, Jorgenson et al. 1993) qui sont attendus en biologie évolutive (Stearns 1992). En fonction
de la densité et du climat, les délais de première reproduction peuvent atteindre chez le cerf
élaphe jusqu’à 2 ou 3 ans. Ainsi, sur l’île de Rum, les femelles se reproduisent pour la première

16
fois à l’âge de 3,5 ans alors que dans les populations étudiées en France et en Belgique, de 30 à
80% des femelles se reproduisent à 1,5 an (Bonenfant et al. 2002, Bertouille & de Crombrugghe
2002). D’un point de vue pratique, puisque cet ICE est lié à la masse corporelle des jeunes classes
d'âge, il répondra tout aussi rapidement aux variations de densité. Dans la Réserve Nationale de
Chasse et de Faune Sauvage de La Petite Pierre, le poids des jeunes et la proportion de bichettes
gestantes ont en effet réagi à la forte augmentation des plans de chasse lancée en 1983
(Bonenfant et al. 2002). D’autre part, il a été suggéré que chez les espèces de grande taille
comme le cerf, l’âge de primiparité est le trait d’histoire de vie le plus sensible à la densité
(Gaillard et al. 2000). Théoriquement, cet ICE devrait réagir pour des niveaux de densité
relativement faibles pour le cerf et est donc de ce fait particulièrement bien adapté dans le cadre
de la gestion. À Chambord, les informations relatives au statut reproducteur des femelles
(gestante ou non, par classe d’âge) sont également recueillies dans les tableaux de chasse depuis
1982.
2.2.3. Intégration des conditions environnementales
D’un point de vue biologique, l’intégration des données météorologiques constitue un
point crucial puisque nous avons vu que le climat agissait sur la masse corporelle des herbivores
(Weladji et al. 2002, Loison et al. 1999b, Mysterud et al. 2001b). Pour cette étude, nous avons
obtenu les données météorologiques, depuis 1980, d’une station Météo France située sur la
commune de Cheverny, adjacente au Domaine. À partir de ces informations, deux indices de
sécheresse ou indices de Gaussen (GI ; Dajoz 1973) ont été calculés, l’un au printemps (mars –
avril – mai) et l’autre en été (juin – juillet – août), en soustrayant deux fois la température
moyenne à la somme des précipitations des mois considérés. D’une manière générale, l’indice de
Gaussen représente une mesure de la disponibilité en eau pour la végétation (Dajoz 1973), et
plusieurs études ont mis en évidence l’influence de cette variable climatique sur la performance
des populations d’ongulés sauvages (Gaillard et al. 1997, Garel et al. 2004). Au printemps,
l’indice de Gaussen permet d’appréhender la croissance du couvert végétal, il est donc un bon
indicateur de la quantité de ressources disponibles à la fin de l’hiver et au cours du printemps (fin
de la gestation et début de la lactation). En été, l’indice de Gaussen renseigne également sur la

17
quantité mais aussi sur la qualité des ressources (Becker et al. 1994), et constitue donc un bon
indicateur de la disponibilité alimentaire pendant la période de lactation et jusqu’au sevrage.
D’un point de vue technique, la prise en compte de ces variables climatiques permet
éventuellement de détecter les phénomènes de densité-dépendance de manière plus efficace. En
plus des données météorologiques, nous disposons du niveau des fructifications dans le DNC
pour chaque année : 1 = absente, 2 = faible, 3 = moyenne, 4 = importante et 5 = très importante.
2.3. ICE « Impact » sur le milieu
Deux indicateurs ont été utilisés pour suivre l'évolution de la pression de consommation
exercée par les cervidés sur les espèces forestières du domaine : l'IPF (Indice de Pression
Floristique ; Guibert 1997) et l'IC (Indice de consommation ; Morellet et al. 2001). Aucun de ces
indicateurs n'a été validé chez le cerf, ils doivent donc être interprétés avec prudence surtout que
son régime alimentaire est majoritairement constitué d'herbacées (Gebert & Verheyden-Tixier
2001). De plus, sur un massif où plusieurs espèces cohabitent (comme c’est le cas sur le DNC
avec cerf, mouflon, chevreuil et sanglier), il est très difficile de quantifier la part de chaque
espèce dans les variations du niveau d'abroutissement. Il sera donc important de considérer aussi
cet élément au moment de l'interprétation des résultats.
Sans rentrer dans le détail des mises en œuvre de ces approches, il faut retenir que l'IC a
été proposé comme une amélioration de l'IPF qui souffrait notamment d'un manque de robustesse
face à l'effet « observateurs » et présentait plus de difficultés quant à sa mise en œuvre et son
analyse. Dans les deux cas, la mesure de l'abroutissement est réalisée en fin d'hiver/début de
printemps (période de repos végétatif), ce qui signifie que les traces de consommation mesurées
l'année t correspondent donc à l'abroutissement de l'année de végétation t-1. Contrairement aux
autres indicateurs, les indicateurs d'impact ne sont pas spécifiques mais concerne les herbivores
au sens large.
Arrêtée depuis près de cinq ans, la mesure de l’impact des cervidés sur la flore du DNC a
été réalisée de 1999 à 2006 pour l’IPF, et de 2005 à 2008 pour l’IC. L’objectif de la prise en
compte de ces données est essentiellement de pouvoir comparer les résultats obtenus avec les
tendances observées pour les autres catégories d’ICE. Par ailleurs, il est important de noter que la

18
méthode d'engrillagement (parcelles de végétation de 50 ha) progressivement mise au point ces
30 dernières années par le personnel de Chambord permet à la fois la régénération et le maintien
de la diversité en espèces ligneuses et semis ligneuses du domaine. Toutefois, cette protection qui
entraine une réduction temporelle des zones de gagnage peut déplacer et concentrer l’impact des
animaux vers les zones accessibles.
2.4. Capture-Marquage-Recapture non-invasif
En opposition aux indices d’abondance, les estimateurs d’effectifs1 utilisant le principe de
Lincoln - Petersen, reposent sur l’identification d’une certaine proportion d’individus dans la
population (Seber 1982, Schwarz & Seber 1999). La mise en œuvre de ces méthodes passe le
plus souvent par des sessions de captures d’animaux au cours desquelles les animaux non-connus
sont marqués, et les animaux déjà marqués sont identifiés. Au moins deux sessions de captures
sont nécessaires pour obtenir une estimation d’effectif. Récemment, l’avènement de l’outil
génétique a ouvert la possibilité de s’affranchir des captures physiques et du marquage,
remplacées alors par l’identification des individus à partir de leur empreinte génétique. Cette
identification n’impliquant pas le marquage des animaux fait partie des méthodes dites non-
invasives (Kohn & Wayne 1997). Pour cela, des échantillons contenant de l’ADN (tissus, poils,
sang, fèces) collectés sur le terrain lors d’une première campagne fournissent un lot d’individus
connus dont la présence est recherchée dans les échantillons de la deuxième campagne. Ainsi,
nous disposons des données permettant l’estimation d’un effectif de population (Lincoln 1930,
Petersen 1896). De telles estimations d’effectifs à partir des méthodes non-invasives ont déjà été
faites avec succès pour plusieurs espèces de mammifères, notamment le coyote (Canis latrans,
Kohn et al. 1999), le loup (Canis lupus, Creel et al. 2003) ou encore l’éléphant de forêt d’Afrique
(Loxodonta cyclotis, Eggert et al. 2003).
1 Un estimateur d’effectif permet d’estimer la probabilité de détection des animaux, ce que ne font pas les
indicateurs d’abondance comme l’indice nocturne (qui supposent cette probabilité constante durant la
période de suivi). Un estimateur d’effectif repose sur une théorie statistique et fournit, outre l’effectif
attendu de la population, un intervalle de confiance de cette estimation. On reconnaît deux grandes
familles d’estimateurs d’abondance : le transect linéaire est ses dérivés qui estiment une densité, et les
méthodes de capture-marquage-recaptures qui estiment un effectif.

19
À Chambord, un protocole d’échantillonnage de fèces de cerfs a été mis en place en 2009
et 2010 pour permettre une estimation non-invasive de l’effectif de la population (par capture-
marquage-recapture (CMR) génétique). Notons qu’une étude pilote ayant pour but d’évaluer
l’effectif de cerfs à Chambord par CMR non-invasive avait déjà été conduite en 2003 (Valière et
al. 2007) mais n’avait pas abouti. Les échantillons de fèces ont été récoltés chaque année sur 8
zones différentes, en deux séries (2009 : 24 mars et 8 avril ; 2010 : 8 avril et 21 avril) espacées
dans le temps d’environ 15 jours afin de s’assurer que l'effectif de la population était stable
durant la période de collecte (aucune naissance et mortalité des animaux supposée négligeable).
Les analyses génétiques (extraction, amplification et séquençage) ont été réalisées par le
laboratoire Antagène (http://www.antagene.com/fr). En fonction de la quantité et de la qualité de
l’ADN extrait des échantillons, entre 0 et 8 marqueurs microsatellites ont pu être amplifiés. Parmi
les 1406 échantillons génotypés, 789 présentaient une empreinte génétique exploitable avec au
moins 5 marqueurs amplifiés sur les 8. En effet, les conditions d’échantillonnage rencontrées sur
le terrain et le délai de conservation des fèces avant l’extraction d’ADN peuvent affecter la
qualité de l’ADN extrait et sont susceptibles de biaiser les estimations d’abondance qui en
résultent. Pour les mêmes années, des échantillons de tissus prélevés sur les animaux tués à la
chasse ont également été analysés au laboratoire afin de produire une empreinte génétique
similaire à celle obtenue à partir des fèces. Un total de 307 échantillons de tissus portant sur
autant d’individus différents a pu être génotypé.

20
III. ANALYSES ET DISCUSSION
Le même type d’approche sera employé dans cette étude, avec la recherche d’une
convergence et d’une cohérence globale des différents ICE analysés : comptages nocturnes, plans
de chasse réalisés, masse corporelle des faons et taux de gestation des femelles, et enfin impact
sur le milieu (de 1999 à 2008 uniquement).
1. Analyse des trois familles d’ICE
1.1. Variations des ICE « Abondance »
Pour rappel, en ce qui concerne les ICE d’abondance, nous nous intéressons tout
particulièrement à l’indice nocturne puisqu’il s’agit du seul indicateur d’abondance validé pour
l’espèce cerf (Garel et al. 2010). Dans un premier temps, nous décrivons la variabilité temporelle
de l’indice nocturne et recherchons l’existence de facteurs confondants potentiels. Ensuite, nous
étudions la relation de l’indice nocturne avec les autres mesures d’abondance disponibles ainsi
que les prélèvements cynégétiques.
Les comptages nocturnes réalisés depuis 1985 montrent une forte variabilité inter-
annuelle (Tab. 2; Fig. 3). Un minimum est observé en 2002, et trois pics d’abondance sont
enregistrés en 1996, en 2008 puis en 2013 (Fig. 3). La décroissance récente entre 2008 et 2012
est surtout due à une année 2012 particulièrement basse et fortement influencée par 2 sorties avec
très peu d’animaux observés (Fig. 3). L’indice nocturne a été multiplié par 3,4 entre 2002 et
2008, puis a été divisé par 2,0 entre 2008 et 2012. Notons tout de suite que les accroissements de
l’indice nocturne observés entre les années 1987-1988 (r = 0,281), 1994-1995 (r = 0,336), 2003-
2004 (r = 0,391), 2007-2008 (r = 0,343) et 2012-2013 (r = 0,505) ne sont pas compatibles avec la
biologie du cerf pour lequel un taux d’accroissement r > 0,30 reste exceptionnel et ne concerne
que des populations en phase de colonisation occupant des milieux riches. Cette remarque illustre
les limites de l’interprétation qu’imposent les indicateurs d’abondance, au contraire de ce que
l’on peut obtenir d’un estimateur d’abondance. Sur la période 1985 - 2013, nous détectons un
accroissement significatif de l’indice nocturne au cours du temps de l’ordre de 1,6% par an (β =
0,016 +/- 0,005, z = 3,153, P = 0,002). Bien que cette tendance exprime 15,4% seulement de la

21
variabilité annuelle moyenne de l’indice nocturne, cela suggère que l’abondance relative de la
population de cerfs est aujourd’hui plus élevée que lors de la mise en place de l’indice en 1985.
L’indice nocturne établit sans ambiguïté que l’abondance des cerfs s’est fortement accrue depuis
2002, toutefois, il n’est pas possible de quantifier l’augmentation réelle de l’effectif de population
puisque l’on se base sur un indicateur d’abondance uniquement. On peut cependant distinguer
nettement deux phases dans l’histoire récente du cerf sur le domaine : (i) une première phase de
1985 à 2002 avec une abondance relative faible mais plutôt stable dans le temps (indice nocturne
moyen : 136,9) ; (ii) et une deuxième phase de 2003 à aujourd’hui qui a vu l’abondance du cerf
augmenter fortement pour atteindre un plateau depuis 2008 (indice nocturne moyen : 180,4) et
qui s’apparente à une croissance de type logistique.
Figure 3 : Variation temporelle de l’indice nocturne mis en place sur le domaine national de Chambord.
Les points blancs représentent les valeurs de comptage de chaque sortie, et les points noirs les valeurs
moyennes annuelles. L’augmentation progressive de l’indice nocturne de 1985 à 2013, de l’ordre de 1,6%
par an, est statistiquement significative, mais traduit surtout une augmentation importante de l’abondance
depuis 2002.
22
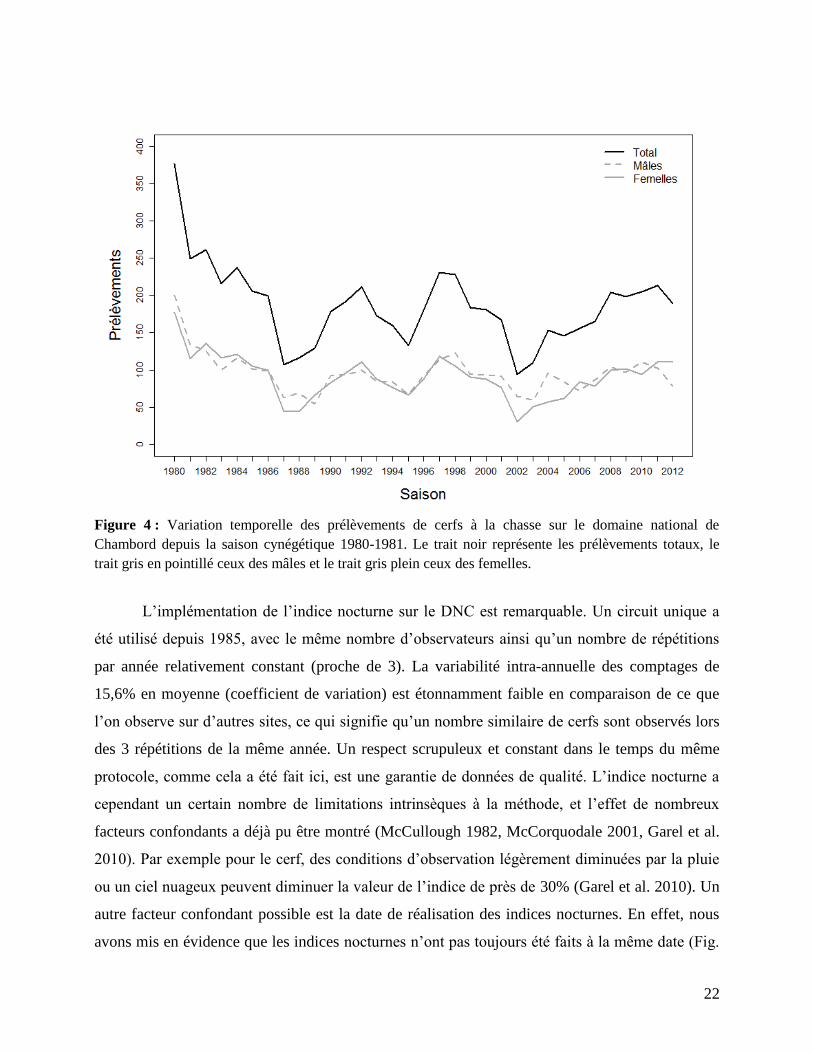
Figure 4 : Variation temporelle des prélèvements de cerfs à la chasse sur le domaine national de
Chambord depuis la saison cynégétique 1980-1981. Le trait noir représente les prélèvements totaux, le
trait gris en pointillé ceux des mâles et le trait gris plein ceux des femelles.
L’implémentation de l’indice nocturne sur le DNC est remarquable. Un circuit unique a
été utilisé depuis 1985, avec le même nombre d’observateurs ainsi qu’un nombre de répétitions
par année relativement constant (proche de 3). La variabilité intra-annuelle des comptages de
15,6% en moyenne (coefficient de variation) est étonnamment faible en comparaison de ce que
l’on observe sur d’autres sites, ce qui signifie qu’un nombre similaire de cerfs sont observés lors
des 3 répétitions de la même année. Un respect scrupuleux et constant dans le temps du même
protocole, comme cela a été fait ici, est une garantie de données de qualité. L’indice nocturne a
cependant un certain nombre de limitations intrinsèques à la méthode, et l’effet de nombreux
facteurs confondants a déjà pu être montré (McCullough 1982, McCorquodale 2001, Garel et al.
2010). Par exemple pour le cerf, des conditions d’observation légèrement diminuées par la pluie
ou un ciel nuageux peuvent diminuer la valeur de l’indice de près de 30% (Garel et al. 2010). Un
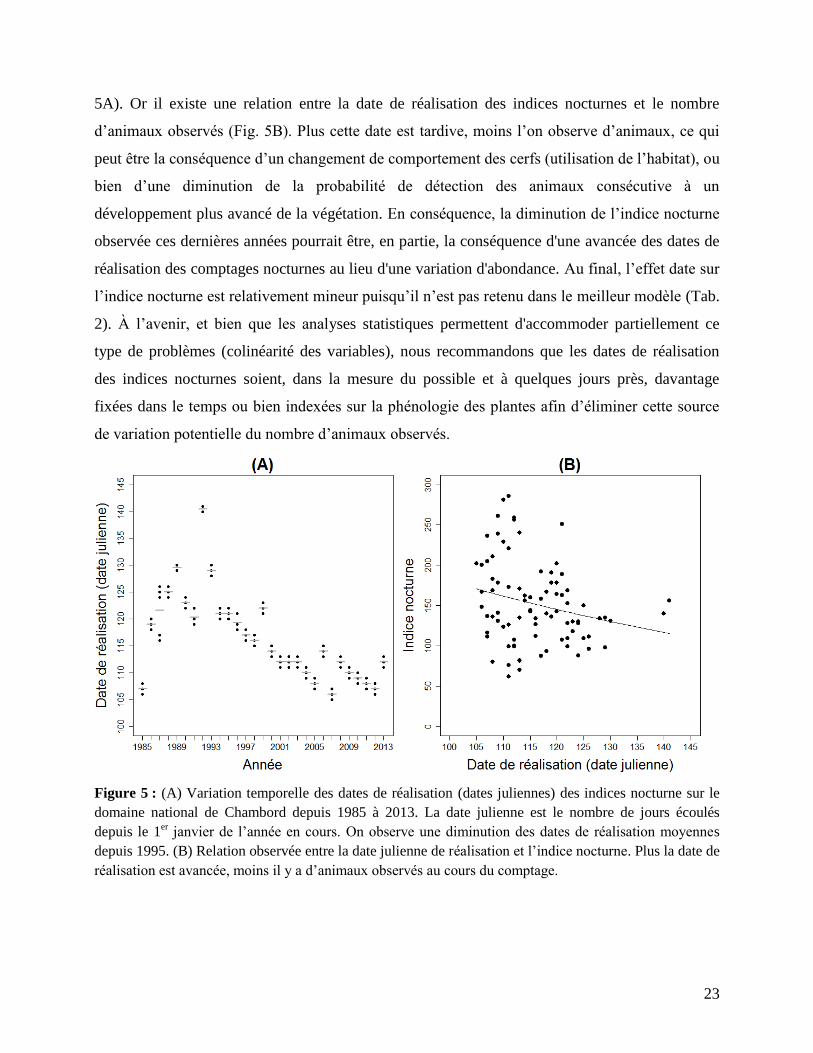
autre facteur confondant possible est la date de réalisation des indices nocturnes. En effet, nous
avons mis en évidence que les indices nocturnes n’ont pas toujours été faits à la même date (Fig.
23
5A). Or il existe une relation entre la date de réalisation des indices nocturnes et le nombre
d’animaux observés (Fig. 5B). Plus cette date est tardive, moins l’on observe d’animaux, ce qui
peut être la conséquence d’un changement de comportement des cerfs (utilisation de l’habitat), ou
bien d’une diminution de la probabilité de détection des animaux consécutive à un
développement plus avancé de la végétation. En conséquence, la diminution de l’indice nocturne
observée ces dernières années pourrait être, en partie, la conséquence d'une avancée des dates de
réalisation des comptages nocturnes au lieu d'une variation d'abondance. Au final, l’effet date sur
l’indice nocturne est relativement mineur puisqu’il n’est pas retenu dans le meilleur modèle (Tab.
2). À l’avenir, et bien que les analyses statistiques permettent d'accommoder partiellement ce
type de problèmes (colinéarité des variables), nous recommandons que les dates de réalisation
des indices nocturnes soient, dans la mesure du possible et à quelques jours près, davantage
fixées dans le temps ou bien indexées sur la phénologie des plantes afin d’éliminer cette source
de variation potentielle du nombre d’animaux observés.
Figure 5 : (A) Variation temporelle des dates de réalisation (dates juliennes) des indices nocturne sur le
domaine national de Chambord depuis 1985 à 2013. La date julienne est le nombre de jours écoulés
depuis le 1er janvier de l’année en cours. On observe une diminution des dates de réalisation moyennes
depuis 1995. (B) Relation observée entre la date julienne de réalisation et l’indice nocturne. Plus la date de
réalisation est avancée, moins il y a d’animaux observés au cours du comptage.
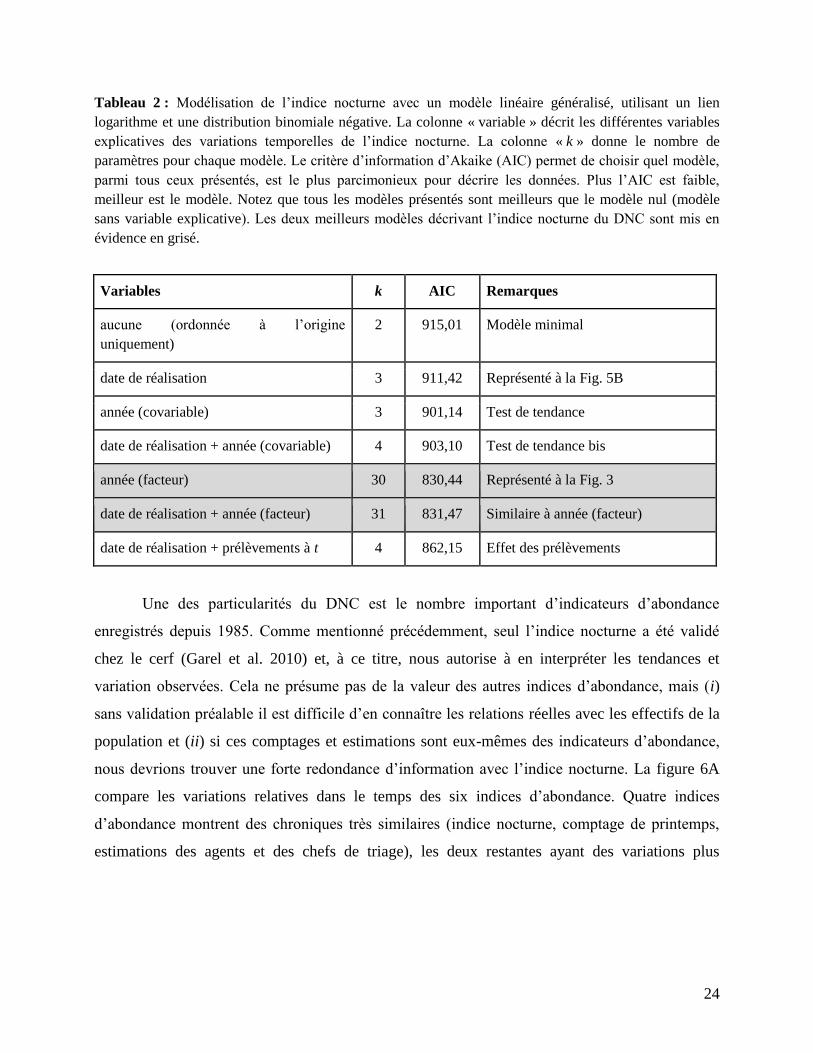
24
Tableau 2 : Modélisation de l’indice nocturne avec un modèle linéaire généralisé, utilisant un lien
logarithme et une distribution binomiale négative. La colonne « variable » décrit les différentes variables
explicatives des variations temporelles de l’indice nocturne. La colonne « k » donne le nombre de
paramètres pour chaque modèle. Le critère d’information d’Akaike (AIC) permet de choisir quel modèle,
parmi tous ceux présentés, est le plus parcimonieux pour décrire les données. Plus l’AIC est faible,
meilleur est le modèle. Notez que tous les modèles présentés sont meilleurs que le modèle nul (modèle
sans variable explicative). Les deux meilleurs modèles décrivant l’indice nocturne du DNC sont mis en
évidence en grisé.
Variables k AIC Remarques
aucune (ordonnée à l’origine
uniquement)
2 915,01 Modèle minimal
date de réalisation 3 911,42 Représenté à la Fig. 5B
année (covariable) 3 901,14 Test de tendance
date de réalisation + année (covariable) 4 903,10 Test de tendance bis
année (facteur) 30 830,44 Représenté à la Fig. 3
date de réalisation + année (facteur) 31 831,47 Similaire à année (facteur)
date de réalisation + prélèvements à t 4 862,15 Effet des prélèvements
Une des particularités du DNC est le nombre important d’indicateurs d’abondance
enregistrés depuis 1985. Comme mentionné précédemment, seul l’indice nocturne a été validé
chez le cerf (Garel et al. 2010) et, à ce titre, nous autorise à en interpréter les tendances et
variation observées. Cela ne présume pas de la valeur des autres indices d’abondance, mais (i)
sans validation préalable il est difficile d’en connaître les relations réelles avec les effectifs de la
population et (ii) si ces comptages et estimations sont eux-mêmes des indicateurs d’abondance,
nous devrions trouver une forte redondance d’information avec l’indice nocturne. La figure 6A
compare les variations relatives dans le temps des six indices d’abondance. Quatre indices
d’abondance montrent des chroniques très similaires (indice nocturne, comptage de printemps,
estimations des agents et des chefs de triage), les deux restantes ayant des variations plus

25
aléatoires. Les coefficients de corrélation croisée2 entre l’indice nocturne et les comptages de
printemps, les estimations des agents et les estimations des chefs de triage sont respectivement de
τ = 0,61, τ = 0,53, et τ = 0,47 (tous sont statistiquement significatifs). Il existe donc une relation
très forte entre tous ces indices et donc, une redondance importante dans les informations
collectées sur le terrain. D’autre part, il est difficile d’imaginer des estimations très différentes
entre les agents et les chefs de triage, ce que confirme le très fort coefficient de corrélation
croisée entre ces 2 indices (τ = 0,86 ; très proche du maximum de 1). Par comparaison, le
coefficient de corrélation croisée entre l’indice nocturne et les comptages estivaux est de τ = 0,22
et de τ = 0,27 avec l’indice brame. Cette dernière valeur est en parfait accord avec les analyses
similaires à La Petite Pierre qui n’ont pas pu mettre en évidence de relation entre l’indice brame
et l’indice nocturne (Douhard et al. 2013). Quant aux comptages estivaux, ils n’offrent pas de
réalité biologique évidente et, surtout, montrent des variations temporelles très importantes par
rapport aux autres indices d’abondance qui eux, sont plutôt en phase les uns avec les autres. À la
lumière de ces analyses, nous concluons que le maintien de l’ensemble des six indicateurs
d’abondance n’est absolument pas nécessaire pour la gestion, et que le suivi par indice nocturne
sur le long terme est amplement suffisant et justifié pour appréhender les variations d’abondance
de la population de cerfs du DNC.
Une dernière analyse pertinente concerne la relation entre l’indice nocturne et les
prélèvements par la chasse (voir Fig. 4 pour la variation temporelle des prélèvements), pour
laquelle nous pouvons poser deux hypothèses alternatives : (1) Sous l’hypothèse que les
prélèvements conduisent à une diminution de l’indice nocturne, alors le coefficient de corrélation
croisée entre les prélèvements et l’indice nocturne devrait être négatif ; (2) Au contraire, si le
niveau de prélèvements est déterminé en fonction de l’indice nocturne, alors on devrait observer
un coefficient de corrélation croisé positif. Afin de tester ces deux alternatives, nous avons
calculé le coefficient de corrélation croisée entre l’indice nocturne et les prélèvements avec des
effets retard allant de -2 à 2 ans (voir Fig. 6B). L’effet des prélèvements au cours de l’automne /
2 Comme les variables analysées sont des séries temporelles (chroniques), un coefficient de corrélation
simple entre deux indices peut être surestimé. Pour tester une relation entre 2 chroniques, on utilise plutôt
l’indice de corrélation croisée, qui met en regard les variations entre t et t+1 de la première variable avec
les variations de la deuxième variable sur le même pas de temps. Ce coefficient varie entre -1 et 1 et est
noté τ.
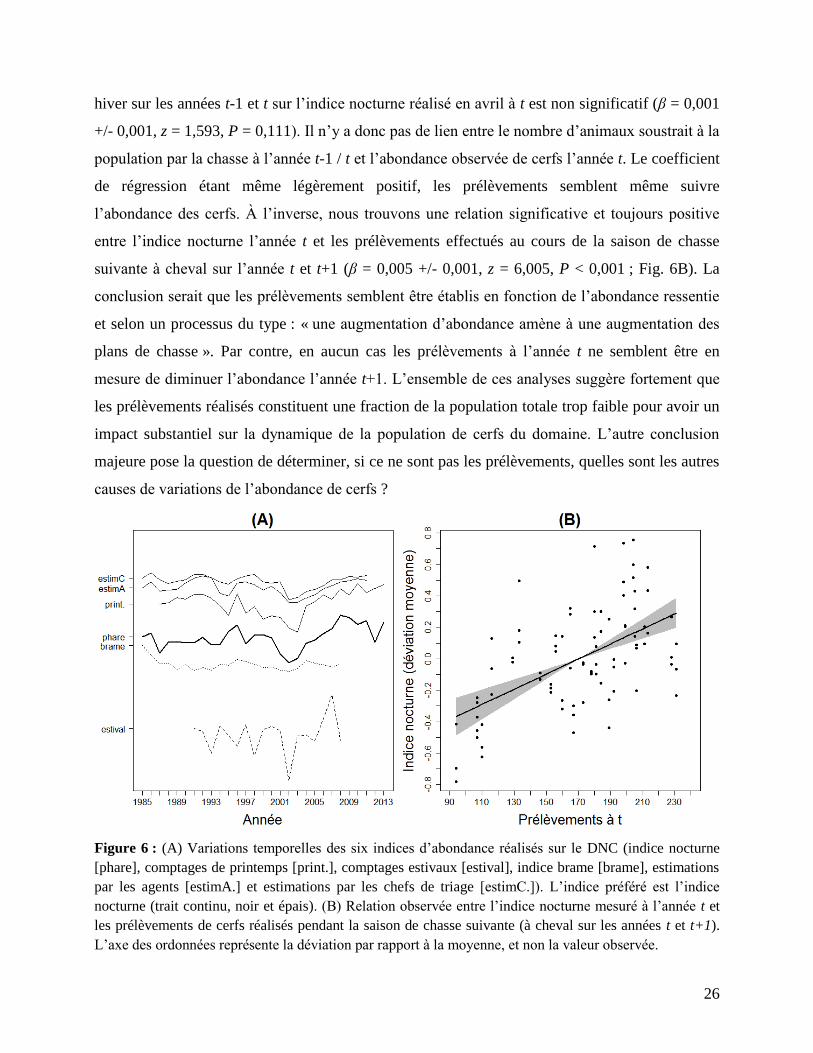
26
hiver sur les années t-1 et t sur l’indice nocturne réalisé en avril à t est non significatif (β = 0,001
+/- 0,001, z = 1,593, P = 0,111). Il n’y a donc pas de lien entre le nombre d’animaux soustrait à la
population par la chasse à l’année t-1 / t et l’abondance observée de cerfs l’année t. Le coefficient
de régression étant même légèrement positif, les prélèvements semblent même suivre
l’abondance des cerfs. À l’inverse, nous trouvons une relation significative et toujours positive
entre l’indice nocturne l’année t et les prélèvements effectués au cours de la saison de chasse
suivante à cheval sur l’année t et t+1 (β = 0,005 +/- 0,001, z = 6,005, P < 0,001 ; Fig. 6B). La
conclusion serait que les prélèvements semblent être établis en fonction de l’abondance ressentie
et selon un processus du type : « une augmentation d’abondance amène à une augmentation des
plans de chasse ». Par contre, en aucun cas les prélèvements à l’année t ne semblent être en
mesure de diminuer l’abondance l’année t+1. L’ensemble de ces analyses suggère fortement que
les prélèvements réalisés constituent une fraction de la population totale trop faible pour avoir un
impact substantiel sur la dynamique de la population de cerfs du domaine. L’autre conclusion
majeure pose la question de déterminer, si ce ne sont pas les prélèvements, quelles sont les autres
causes de variations de l’abondance de cerfs ?
Figure 6 : (A) Variations temporelles des six indices d’abondance réalisés sur le DNC (indice nocturne
[phare], comptages de printemps [print.], comptages estivaux [estival], indice brame [brame], estimations
par les agents [estimA.] et estimations par les chefs de triage [estimC.]). L’indice préféré est l’indice
nocturne (trait continu, noir et épais). (B) Relation observée entre l’indice nocturne mesuré à l’année t et
les prélèvements de cerfs réalisés pendant la saison de chasse suivante (à cheval sur les années t et t+1).
L’axe des ordonnées représente la déviation par rapport à la moyenne, et non la valeur observée.

27
Les points clés
1. L’indice nocturne est l’ICE d’abondance préféré sur le DNC.
2. L’indice nocturne met en évidence deux périodes contrastées en termes de variabilité et
d’abondance, et se situant avant et après 2002.
3. L’indice nocturne gagnerait à être réalisé à la même date tous les ans.
4. Les prélèvements semblent être basés sur l’indice nocturne or l’idéal serait, à l’inverse, de
contrôler les variations de l’indice nocturne par les prélèvements pour avoir un levier d’action
sur la population de cerfs.
1.2. Variations des ICE « Performance »
1.2.1. La masse corporelle des jeunes
Pour le cerf comme pour de nombreuses autres espèces de grands herbivores, la masse
corporelle des faons est un indicateur de performance particulièrement sensible aux effets de la
densité et du climat sur la qualité et la quantité de ressource disponibles (Bonenfant et al. 2002).
La première étape pour rendre les mesures de masse corporelle comparables au cours de la saison
de chasse, et d’une année sur l’autre, est de modéliser la croissance pondérale des faons en
fonction de la date de tir de l’animal (date julienne). Nous avons ensuite testé spécifiquement les
effets suivants sur le poids des faons : différence entre mâles et femelles, présence d’une
tendance linéaire au cours du temps, et enfin rôle de l’abondance de la population (indice
nocturne), des prélèvements annuels et des conditions environnementales (fructification et indices
de Gaussen au printemps et en été).
Au cours de la saison de chasse, les faons mâles et femelles prennent en moyenne 6 kg
entre le mois d’octobre et le mois de mars, et cette croissance est la même pour les deux sexes (β
= 0,015 +/- 0,005, t = 2,487, P = 0,004). Cette croissance n’est pas linéaire entre le début et la fin
de la saison de chasse puisque vers le milieu de l’hiver les faons arrêtent quasiment de prendre du
poids. Par contre, le dimorphisme sexuel de taille est évident puisque les faons femelles pèsent en
moyenne 3,6 ± 0,47 kg de moins que les faons mâles. Une fois la croissance pondérale prise en
compte, nous observons que la masse corporelle moyenne des faons, corrigée pour la date de tir

28
et le sexe, varie d’une année sur l’autre de manière significative (Tab. 3; Fig. 7), avec des masses
corporelles variant de 27,8 kg à 44,5 kg pour les femelles (soit 62% de variation relative) et de
31,0 kg à 47,7 kg pour les mâles (soit 65% de variation relative). Cette forte variation de la masse
corporelle moyenne des faons de cerfs d’une année à l’autre est une illustration très parlante des
effets cohortes discutés en introduction (cf page 5). La masse corporelle des faons réagit donc
très rapidement aux changements de qualité et de quantité des ressources alimentaires, induisant
des variations du taux de croissance pondérale. La question importante est donc de savoir quelles
ont été les causes biologiques ou environnementales à l’origine de ces différences très marquées
de la masse corporelle des faons de cerfs du DNC.
Figure 7 : Variation temporelle de la masse corporelle des faons prélevés sur le domaine national de
Chambord (cohorte et année sont confondues pour les faons). Les points représentent les prédictions par le
meilleur modèle des moyennes annuelles par sexe et corrigée par la date de tir, et les barres verticales les
intervalles de confiance à 95% des prédictions. Les nombres à l’intérieur du graphique donnent les tailles
d’échantillons pour les faons mâles (en haut) et les faons femelles (en bas).
29
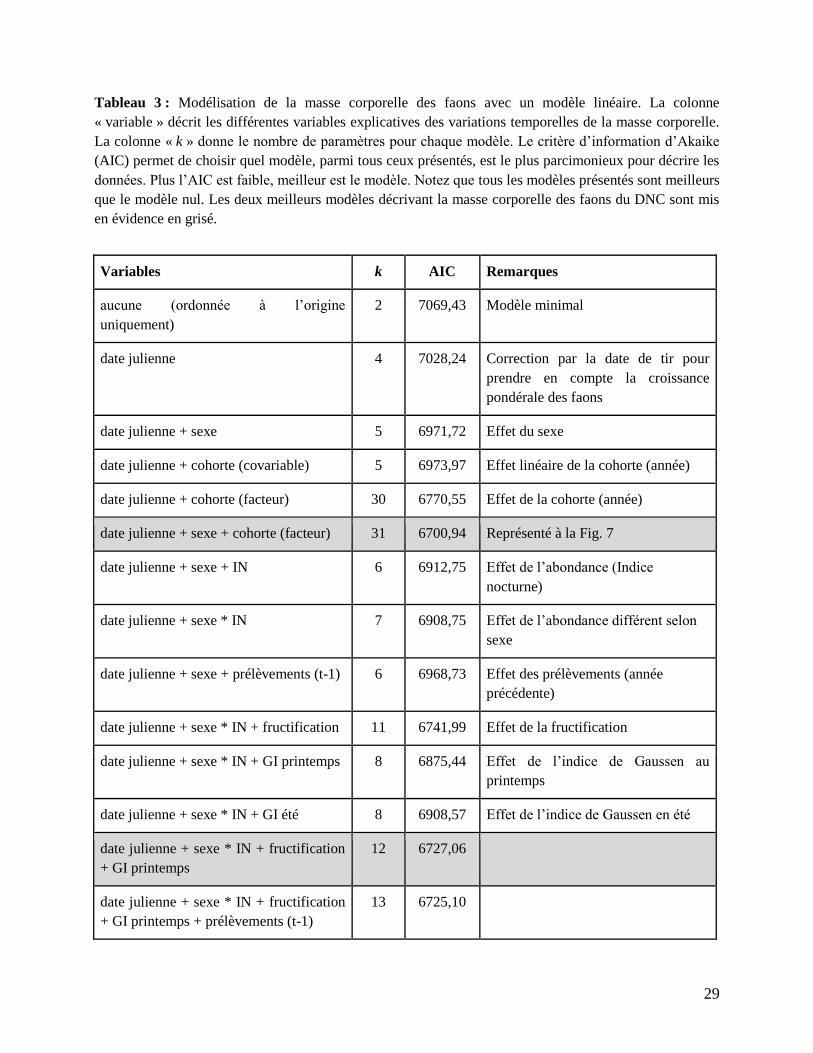
Tableau 3 : Modélisation de la masse corporelle des faons avec un modèle linéaire. La colonne
« variable » décrit les différentes variables explicatives des variations temporelles de la masse corporelle.
La colonne « k » donne le nombre de paramètres pour chaque modèle. Le critère d’information d’Akaike
(AIC) permet de choisir quel modèle, parmi tous ceux présentés, est le plus parcimonieux pour décrire les
données. Plus l’AIC est faible, meilleur est le modèle. Notez que tous les modèles présentés sont meilleurs
que le modèle nul. Les deux meilleurs modèles décrivant la masse corporelle des faons du DNC sont mis
en évidence en grisé.
Variables k AIC Remarques
aucune (ordonnée à l’origine
uniquement)
2 7069,43 Modèle minimal
date julienne 4 7028,24 Correction par la date de tir pour
prendre en compte la croissance
pondérale des faons
date julienne + sexe 5 6971,72 Effet du sexe
date julienne + cohorte (covariable) 5 6973,97 Effet linéaire de la cohorte (année)
date julienne + cohorte (facteur) 30 6770,55 Effet de la cohorte (année)
date julienne + sexe + cohorte (facteur) 31 6700,94 Représenté à la Fig. 7
date julienne + sexe + IN 6 6912,75 Effet de l’abondance (Indice
nocturne)
date julienne + sexe * IN 7 6908,75 Effet de l’abondance différent selon
sexe
date julienne + sexe + prélèvements (t-1) 6 6968,73 Effet des prélèvements (année
précédente)
date julienne + sexe * IN + fructification 11 6741,99 Effet de la fructification
date julienne + sexe * IN + GI printemps 8 6875,44 Effet de l’indice de Gaussen au
printemps
date julienne + sexe * IN + GI été 8 6908,57 Effet de l’indice de Gaussen en été
date julienne + sexe * IN + fructification
+ GI printemps
12 6727,06
date julienne + sexe * IN + fructification
+ GI printemps + prélèvements (t-1)
13 6725,10

30
Une première observation est que la masse corporelle des faons montre une tendance à la
diminution au cours du temps (-0,21 kg par an en moyenne), bien qu’elle semble ré-augmenter
ces cinq dernières années après une forte diminution en 2008 (Fig. 7). Cette tendance rappelle
fortement celle observée pour l’indice nocturne (Fig. 3) suggérant que l’abondance de la
population pourrait être un facteur de variation de la masse corporelle des faons. De fait, le
nombre moyen de cerfs comptés au kilomètre par an sur le domaine (i.e., la valeur annuelle de
l’indice nocturne), explique 17% de la variation interannuelle de la masse corporelle des faons
sur le DNC. L’effet de l’indice nocturne sur la masse corporelle des faons étant négatif (β = -
0,040 +/- 0,005), ce résultat décrit un phénomène de densité dépendance très classique (Fig. 8A).
Les faons perdent en moyenne 1 kg pour une augmentation de 27 cerfs observés lors des
comptages nocturnes (Fig. 8A). La diminution de masse corporelle est par contre plus prononcée
chez les faons mâles que chez les faons femelles (βmâle = -0,053 +/- 0,010 vs. βfemelle = -0,028 +/-
0,007 ; Tab. 3), ce qui est bien attendu pour une espèce présentant un dimorphisme sexuel en
taille aussi marqué que le cerf (Bonenfant et al. 2002, 2009). L’explication biologique à cela est
que les faons mâles allouent davantage d’énergie à leur croissance que les femelles mais, en
contrepartie, souffrent plus de la compétition pour les ressources alimentaires (Clutton-Brock et
al. 1982, Bonenfant et al. 2009). La plus forte diminution de la masse corporelle des faons mâles
avec l’indice nocturne confirme également que cette dernière est un ICE particulièrement
pertinent pour le cerf. Dans la séquence de réponse des indicateurs aux changements de densité,
on s’attend donc à voir réagir en premier le poids des faons mâles, puis celui des femelles,
permettant ainsi de préciser la position de la population sur le gradient de densité-dépendance (le
poids des animaux étant étroitement lié à leur survie ; modèle d’Eberhardt, Fig. 1). Au-delà des
différences de réponses entre les sexes, la diminution de masse corporelle avec l’indice nocturne
indique surtout que les ressources alimentaires ont été limitantes pour le cerf depuis 1982 sur le
DNC et donc, que la population s’est approchée une ou plusieurs fois de la capacité limite
d’accueil du milieu (modèle d’Eberhardt). Les périodes de probable densité-dépendance les plus
fortes sont les années 1982-1983 et 2008-2012, la masse corporelle étant restée relativement
stable entre temps (42,7 kg en moyenne pour les mâles et 39,5 kg pour les femelles). Depuis
2008, la masse corporelle moyenne des faons mâles est de 37,1 kg seulement et celle des faons
femelles 33,9 kg, soit des valeurs très proches de celles enregistrées à La Petite Pierre (30,6-34,8
kg pour les mâles et 29,3-31.2 kg pour les femelles ; Bonenfant et al. 2002) ou sur les zones
31
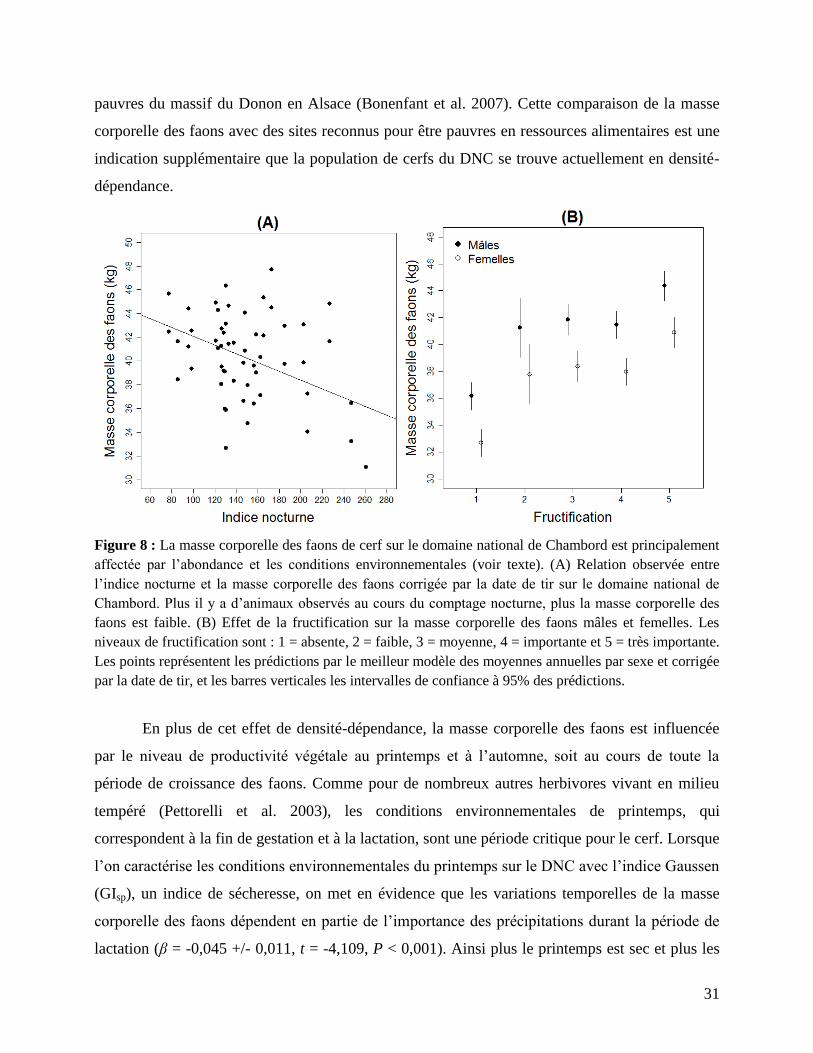
pauvres du massif du Donon en Alsace (Bonenfant et al. 2007). Cette comparaison de la masse
corporelle des faons avec des sites reconnus pour être pauvres en ressources alimentaires est une
indication supplémentaire que la population de cerfs du DNC se trouve actuellement en densité-
dépendance.
Figure 8 : La masse corporelle des faons de cerf sur le domaine national de Chambord est principalement
affectée par l’abondance et les conditions environnementales (voir texte). (A) Relation observée entre
l’indice nocturne et la masse corporelle des faons corrigée par la date de tir sur le domaine national de
Chambord. Plus il y a d’animaux observés au cours du comptage nocturne, plus la masse corporelle des
faons est faible. (B) Effet de la fructification sur la masse corporelle des faons mâles et femelles. Les
niveaux de fructification sont : 1 = absente, 2 = faible, 3 = moyenne, 4 = importante et 5 = très importante.
Les points représentent les prédictions par le meilleur modèle des moyennes annuelles par sexe et corrigée
par la date de tir, et les barres verticales les intervalles de confiance à 95% des prédictions.
En plus de cet effet de densité-dépendance, la masse corporelle des faons est influencée
par le niveau de productivité végétale au printemps et à l’automne, soit au cours de toute la
période de croissance des faons. Comme pour de nombreux autres herbivores vivant en milieu
tempéré (Pettorelli et al. 2003), les conditions environnementales de printemps, qui
correspondent à la fin de gestation et à la lactation, sont une période critique pour le cerf. Lorsque
l’on caractérise les conditions environnementales du printemps sur le DNC avec l’indice Gaussen
(GIsp), un indice de sécheresse, on met en évidence que les variations temporelles de la masse
corporelle des faons dépendent en partie de l’importance des précipitations durant la période de
lactation (β = -0,045 +/- 0,011, t = -4,109, P < 0,001). Ainsi plus le printemps est sec et plus les

32
faons sont maigres, traduisant la relation bien établie en milieu tempéré entre précipitations et
productivité végétale (Langvatn et al. 1996). L’indice de Gaussen estival (GIsu) n’apporte quant à
lui aucune explication concernant la variabilité annuelle de la masse corporelle des faons pour
cette étude (Tab. 3), laissant penser que cette période de l’année n’est pas limitante pour la
croissance des faons. Un autre résultat très intéressant est que la masse corporelle des faons
augmente avec le niveau de fructification des chênes (catégories allant de 1 à 5, 5 étant le niveau
de fructification forestière maximal) passant de 36,1 kg/32,7 kg (mâles/femelles) en l’absence de
fructification à 44,3 kg/40,9 kg (mâles/femelles) pour les années avec très forte fructification
(Fig. 8B). De manière générale, les glands de chêne sont une ressource alimentaire très
recherchée par les grands mammifères herbivores au sens large. Par exemple la fructification agît
sur la reproduction du sanglier (Gamelon et al. 2013), avec des tailles de portées plus importantes
les années à forte glandée. Pour le cerf, il s’agit à notre connaissance de l’une des premières
mises en évidence claire de ce phénomène et du rôle important que peuvent jouer les conditions
environnementales d’automne. Ces dernières ont souvent été omises à la préférence des
conditions printanières, supposées plus critiques. D’un point de vue pratique, le fait que la masse
corporelle des faons augmente avec le niveau de fructification en automne malgré l’agrainage
intensifié lors des faibles fructifications et inversement lors des fortes fructifications, laisse
penser que l’agrainage ne permet pas de compenser complètement l’absence de fructification à
l’automne. La question de la pertinence de cette pratique mérite donc d’être posée. Pratiquement,
ces effets des conditions environnementales printanières et automnales sur la masse corporelle
des faons corroborent notre interprétation d’une limitation de la performance démographique du
cerf par les ressources alimentaires sur le DNC.
La dernière source de variation de la masse corporelle des faons pour laquelle on pouvait
attendre un effet est le prélèvement par la chasse à t-1. Le raisonnement est le suivant : si les
prélèvements conduisent à une diminution de l’abondance, alors la quantité de ressources par
individu devrait aussi être plus importante et donc être favorable pour la croissance des faons. La
résultante serait un effet positif des prélèvements à t-1 sur la masse corporelle des faons au temps
t. Contrairement à cette attente, l’effet des prélèvements de l’année précédente sur la masse
corporelle des faons est négatif sur le DNC (β = -0,016 +/- 0,007, t = -2,230, P = 0,026 ; Tab. 3).
Donc plus les prélèvements sont importants, plus la masse corporelle des faons diminue. Ce
résultat, plutôt inattendu, s’explique au moins en partie à la lumière des analyses des comptages

33
nocturnes (voir section 1.1). Il semble en effet que sur le DNC les prélèvements sont calqués sur
la valeur de l’indice nocturne conduit au printemps précédent (Fig. 6B), en conséquence de quoi,
il existe une relation positive entre l’indice nocturne et les prélèvements. Nous avons donc mis en
évidence une variable confondante, l’abondance du cerf, qui lie la masse corporelle des faons et
les prélèvements. Le mécanisme de relation indirecte entre ces deux dernières variables est donc
le suivant : (i) l’abondance augmente et induit un accroissement immédiat des prélèvements ; (ii)
l’augmentation d’abondance diminue les ressources alimentaires, de même que la masse
corporelle des faons ; (iii) il apparaît à l’analyse que la masse corporelle des faons est
négativement associée aux prélèvements. L’ensemble des résultats sur la masse corporelle sont
donc cohérents même si les prélèvements n’apportent pas plus d’information que l’abondance sur
la compréhension de la dynamique de la population de cerfs du DNC.
1.2.2. La gestation des femelles
Chez le cerf, l’entrée en œstrus au moment du brame est fortement liée à la condition
physique des femelles, dont la masse corporelle est une bonne mesure chez les mammifères
herbivores. En conséquence, la probabilité pour une femelle d’être gestante pendant la période de
la chasse dépend de sa masse corporelle. Cette relation est d’autant plus marquée pour les
bichettes que pour les biches (Bonenfant et al. 2002, Bertouille & de Crombrugghe 2002), ce qui
se vérifie très clairement pour les femelles du DNC (Fig. 9). Or, la condition physique des
animaux étant fortement dépendante de la quantité des ressources alimentaires disponibles, le
taux de gestation des femelles est, à même titre que la masse corporelle des faons, une fonction
des variables environnementales affectant la disponibilité en ressources et en premier lieu, une
fonction de l’abondance des populations (Clutton-Brock & Albon 1989, Bonenfant et al. 2002).
Comme prédit par le modèle d’Eberhardt (1977, 2002), le taux de gestation des bichettes est
rapporté être beaucoup plus variable et sensible aux variations de l’abondance des populations
que le taux de gestation des biches (Bonenfant 2004). Le taux de gestation des bichettes est donc
un ICE performance qui complète utilement l’ICE masse corporelle des faons (voir Bonenfant &
Klein 2004, Bonenfant et al. 2007 pour des applications pratiques). Un atout de cet ICE
performance est qu’il varie entre 0 (aucune bichette gestante) et 1 (toutes les bichettes sont
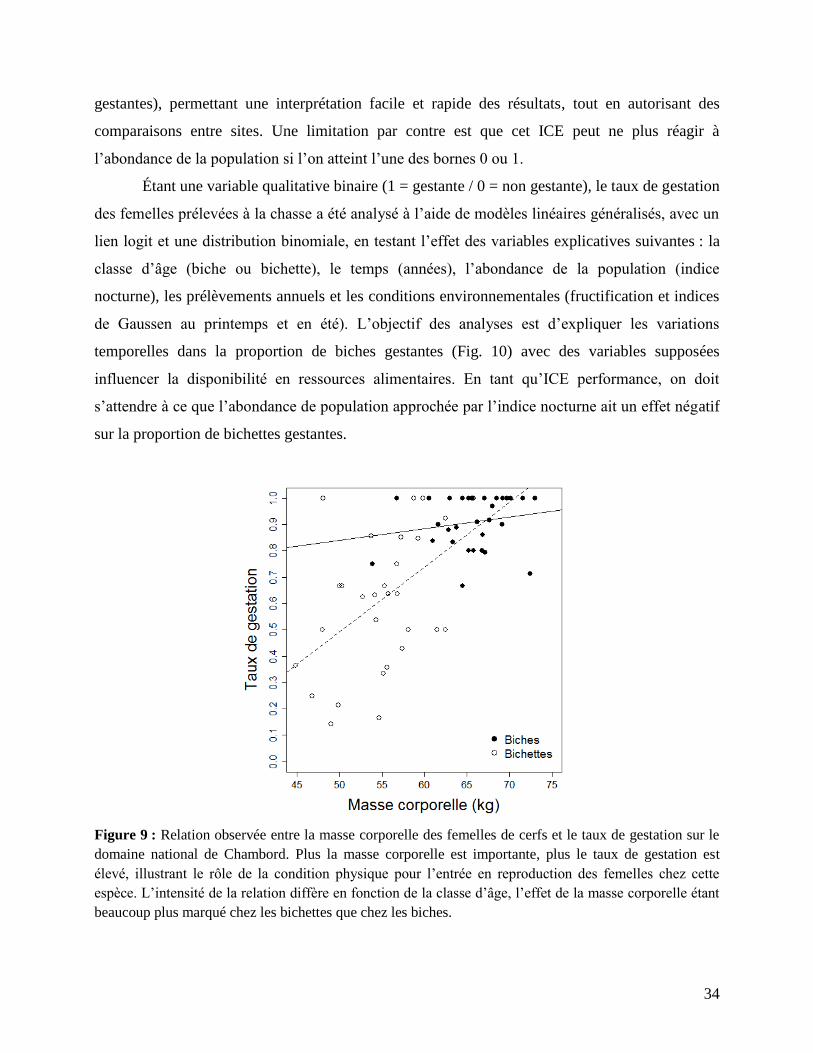
34
gestantes), permettant une interprétation facile et rapide des résultats, tout en autorisant des
comparaisons entre sites. Une limitation par contre est que cet ICE peut ne plus réagir à
l’abondance de la population si l’on atteint l’une des bornes 0 ou 1.
Étant une variable qualitative binaire (1 = gestante / 0 = non gestante), le taux de gestation
des femelles prélevées à la chasse a été analysé à l’aide de modèles linéaires généralisés, avec un
lien logit et une distribution binomiale, en testant l’effet des variables explicatives suivantes : la
classe d’âge (biche ou bichette), le temps (années), l’abondance de la population (indice
nocturne), les prélèvements annuels et les conditions environnementales (fructification et indices
de Gaussen au printemps et en été). L’objectif des analyses est d’expliquer les variations
temporelles dans la proportion de biches gestantes (Fig. 10) avec des variables supposées
influencer la disponibilité en ressources alimentaires. En tant qu’ICE performance, on doit
s’attendre à ce que l’abondance de population approchée par l’indice nocturne ait un effet négatif
sur la proportion de bichettes gestantes.
Figure 9 : Relation observée entre la masse corporelle des femelles de cerfs et le taux de gestation sur le
domaine national de Chambord. Plus la masse corporelle est importante, plus le taux de gestation est
élevé, illustrant le rôle de la condition physique pour l’entrée en reproduction des femelles chez cette
espèce. L’intensité de la relation diffère en fonction de la classe d’âge, l’effet de la masse corporelle étant
beaucoup plus marqué chez les bichettes que chez les biches.
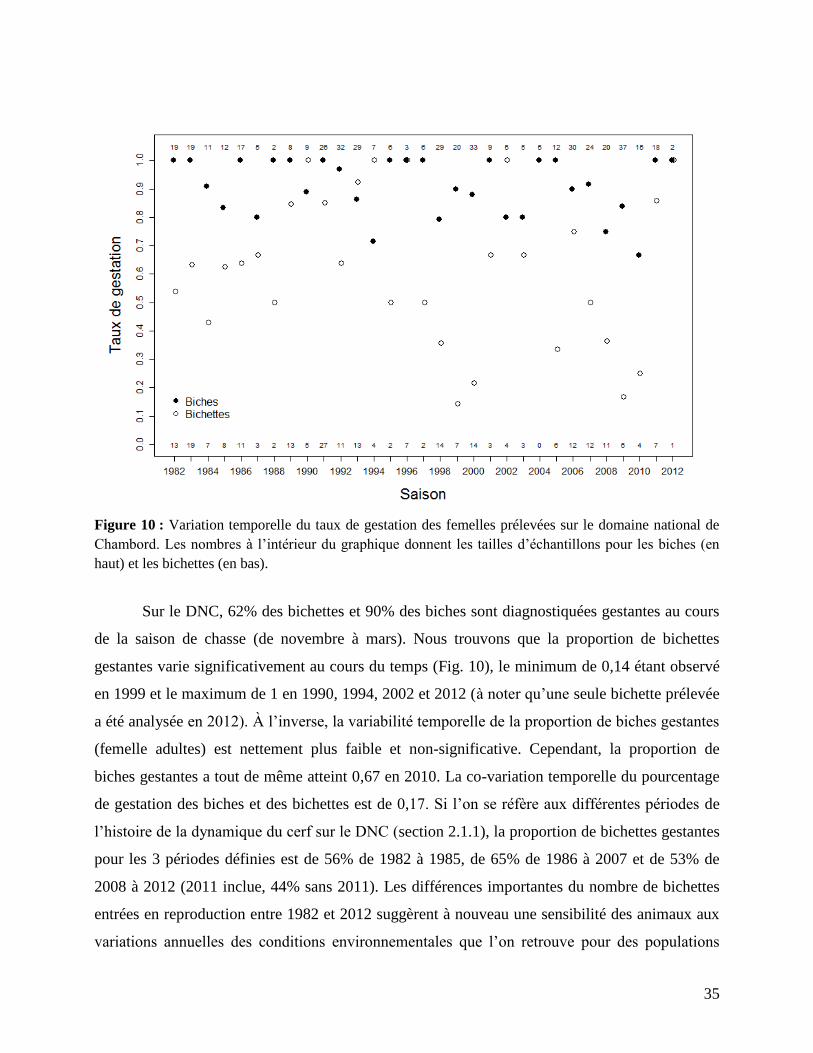
35
Figure 10 : Variation temporelle du taux de gestation des femelles prélevées sur le domaine national de
Chambord. Les nombres à l’intérieur du graphique donnent les tailles d’échantillons pour les biches (en
haut) et les bichettes (en bas).
Sur le DNC, 62% des bichettes et 90% des biches sont diagnostiquées gestantes au cours
de la saison de chasse (de novembre à mars). Nous trouvons que la proportion de bichettes
gestantes varie significativement au cours du temps (Fig. 10), le minimum de 0,14 étant observé
en 1999 et le maximum de 1 en 1990, 1994, 2002 et 2012 (à noter qu’une seule bichette prélevée
a été analysée en 2012). À l’inverse, la variabilité temporelle de la proportion de biches gestantes
(femelle adultes) est nettement plus faible et non-significative. Cependant, la proportion de
biches gestantes a tout de même atteint 0,67 en 2010. La co-variation temporelle du pourcentage
de gestation des biches et des bichettes est de 0,17. Si l’on se réfère aux différentes périodes de
l’histoire de la dynamique du cerf sur le DNC (section 2.1.1), la proportion de bichettes gestantes
pour les 3 périodes définies est de 56% de 1982 à 1985, de 65% de 1986 à 2007 et de 53% de
2008 à 2012 (2011 inclue, 44% sans 2011). Les différences importantes du nombre de bichettes
entrées en reproduction entre 1982 et 2012 suggèrent à nouveau une sensibilité des animaux aux
variations annuelles des conditions environnementales que l’on retrouve pour des populations

36
limitées par les ressources. En comparaison avec les autres sites pour lesquels ces données sont
disponibles, la proportion moyenne de bichettes gestantes de 62% est relativement élevée. Ce
taux est similaire à celui observé sur le site de la Petite Pierre (55% en moyenne, variant de 0 à
80% entre 1978 et 2012). À titre de comparaison, la proportion de bichettes gestantes est de 9,6%
(n = 18) dans le massif du Donon (Bonenfant et al. 2007, p. 63). Cependant, la période de 2008 à
2012 est marquée par une proportion de bichettes gestantes plutôt faible, hormis l’année 2011 qui
a été exceptionnelle en termes de glandée et qui se caractérise par une anomalie positive (comme
pour la masse corporelle des faons) forte de la proportion de bichettes gestantes de presque 90%.
Nous pouvons donc affirmer que ces dernières années, la population de cerfs du DNC se trouve
limitée par la nourriture. Ceci dit, le seuil de 0% de bichette gestante n’étant pas encore atteint, il
est tout à fait possible que la situation puisse se dégrader davantage à l’avenir (du point de vue du
cerf) de sorte que l’âge de première reproduction passerait à 2,5 ans pour toutes les femelles.
Lorsque l’on recherche quelles variables environnementales expliquent les variations
inter-annuelles dans la proportion de bichettes gestantes (Tab. 4), l’indice nocturne, les
prélèvements et, dans une moindre mesure, la fructification des chênes semblent les plus
pertinentes. Ensemble, ces trois variables expliquent 55% de la variation annuelle de la gestation
des bichettes. En accord avec nos prédictions sur la densité-dépendance dans les populations de
cerfs, un effet négatif de l’abondance de la population est mis en évidence sur le taux de gestation
des bichettes principalement (β = -0,002 +/- 0,001, t = -2,224, P = 0,035). Sur le domaine, il se
produit une diminution de 10% du taux de gestation quand le nombre de cerfs observés augmente
de 40 individus lors des comptages nocturnes (Fig. 11). Une réponse densité dépendante existe
également pour la proportion de biches adultes gestantes, mais l’effet est non-significatif et
beaucoup plus faible que pour les bichettes (Fig. 11), avec une diminution de l’ordre de 1%
seulement du taux de gestation quand l’indice nocturne augmente de 40 individus. Là encore, que
seules les bichettes soient affectées par l’abondance de population indique qu’en l’absence de
contrôle, le DNC pourrait supporter davantage d’animaux. En Écosse par exemple, les femelles
adultes se reproduisent une année sur deux uniquement dans certaines populations, du fait d’une
combinaison entre un milieu très pauvre et peu favorable pour le cerf et des densités relatives
élevées (Clutton-Brock & Albon 1989). Cette reproduction densité-dépendante des bichettes est
vraisemblablement à l’origine de la diminution de la proportion de bichettes gestantes que l’on
rapporte depuis 2008 (Fig. 11 ; 15% de la déviance est expliquée par l’indice nocturne).

37
Tableau 4 : Modélisation du taux de gestation des femelles avec un modèle linéaire généralisé, utilisant
un lien logit et une distribution binomiale. La colonne « variable » décrit les différentes variables
explicatives des variations temporelles du taux de gestation. La colonne « k » donne le nombre de
paramètres pour chaque modèle. Le critère d’information d’Akaike (AIC) permet de choisir quel modèle,
parmi tous ceux présentés, est le plus parcimonieux pour décrire les données. Plus l’AIC est faible,
meilleur est le modèle. Notez que tous les modèles présentés sont meilleurs que le modèle nul. Les deux
meilleurs modèles décrivant le taux de gestation du DNC sont mis en évidence en grisé.
Variables k AIC Remarques
aucune (ordonnée à l’origine
uniquement)
1 628,84 Modèle minimal
classe âge 2 568,25 Différence entre bichettes et biches
année (covariable) 2 625,08 Effet linéaire du temps (année)
année (facteur) 27 631,45 Effet du temps (année)
classe âge + année (facteur) 28 551,90
classe âge + IN 3 555,01 Effet de l’abondance (Indice
nocturne)
classe âge + masse corporelle 3 488,58 Effet de la masse corporelle
classe âge * masse corporelle 4 483,72 Effet de la masse corporelle différent
selon classe d’âge
classe âge * masse corporelle + IN 5 483,88
classe âge * masse corporelle +
fructification
8 487,51
classe âge * masse corporelle + classe
âge * fructification
12 482,13 Effet de la fructification différent
selon classe d’âge
classe âge * masse corporelle +
prélèvements (t-1)
5 479,21 Effet des prélèvements (année
précédente)
classe âge * masse corporelle +
prélèvements femelles (t-1)
5 477,78 Effet des prélèvements des femelles
(année précédente)
classe âge * masse corporelle +
prélèvements femelles (t-1) + classe âge
* fructification
13 476,34
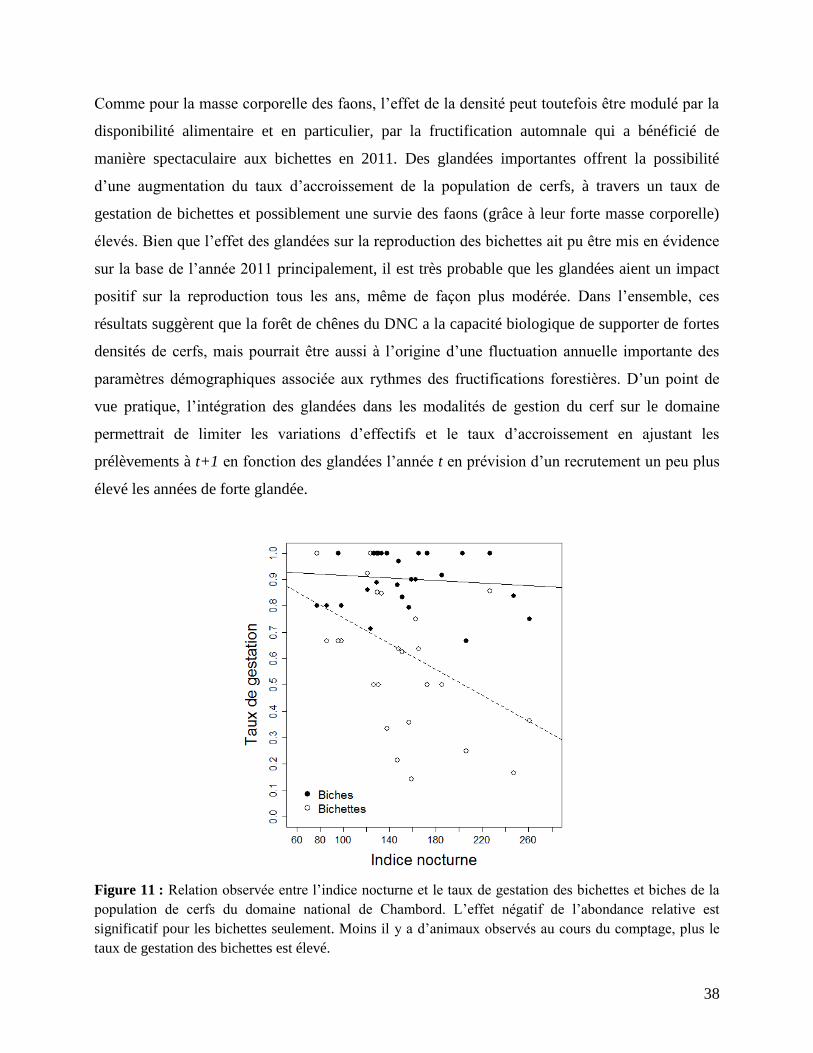
38
Comme pour la masse corporelle des faons, l’effet de la densité peut toutefois être modulé par la
disponibilité alimentaire et en particulier, par la fructification automnale qui a bénéficié de
manière spectaculaire aux bichettes en 2011. Des glandées importantes offrent la possibilité
d’une augmentation du taux d’accroissement de la population de cerfs, à travers un taux de
gestation de bichettes et possiblement une survie des faons (grâce à leur forte masse corporelle)
élevés. Bien que l’effet des glandées sur la reproduction des bichettes ait pu être mis en évidence
sur la base de l’année 2011 principalement, il est très probable que les glandées aient un impact
positif sur la reproduction tous les ans, même de façon plus modérée. Dans l’ensemble, ces
résultats suggèrent que la forêt de chênes du DNC a la capacité biologique de supporter de fortes
densités de cerfs, mais pourrait être aussi à l’origine d’une fluctuation annuelle importante des
paramètres démographiques associée aux rythmes des fructifications forestières. D’un point de
vue pratique, l’intégration des glandées dans les modalités de gestion du cerf sur le domaine
permettrait de limiter les variations d’effectifs et le taux d’accroissement en ajustant les
prélèvements à t+1 en fonction des glandées l’année t en prévision d’un recrutement un peu plus
élevé les années de forte glandée.
Figure 11 : Relation observée entre l’indice nocturne et le taux de gestation des bichettes et biches de la
population de cerfs du domaine national de Chambord. L’effet négatif de l’abondance relative est
significatif pour les bichettes seulement. Moins il y a d’animaux observés au cours du comptage, plus le
taux de gestation des bichettes est élevé.

39
En principe, les prélèvements à la chasse réalisés l’année t-1 devraient diminuer
l’abondance de la population à l’année t et conduire à des animaux avec une plus forte masse
corporelle par le phénomène de densité-dépendance. Des femelles ayant une masse corporelle
plus élevée devraient également montrer une fécondité accrue (voir introduction). En
conséquence nous devrions observer une relation positive entre le taux de gestation des biches et
les prélèvements réalisés à la chasse l’année précédente. Il s’agit ici du même raisonnement que
celui présenté pour l’effet des prélèvements sur la masse corporelle des faons. Contrairement au
raisonnement théorique proposé ici, l’effet des prélèvements à la chasse de l’année précédente
affecte négativement le taux de gestation des femelles (bichettes et biches) l’année suivante (β = -
0,009 +/- 0,004, z = -2,499, P = 0,012). Cet effet des prélèvements sur la reproduction des
femelles est même davantage marqué lorsque l’on considère le nombre de femelles (β = -0,017
+/- 0,006, z = -2,739, P = 0,006) au lieu du nombre total d’animaux chassés. Suite à l’analyse et à
l’interprétation de la masse corporelle des faons, il n’est pas surprenant de trouver que la
gestation des femelles diminue avec les prélèvements, soit le contraire de nos attentes. Ici encore,
les mêmes mécanismes que pour la masse corporelle sont en jeux, l’abondance de la population
jouant le rôle de variable confondante. Lorsque les prélèvements augmentent, l’abondance fait de
même, et la masse corporelle des animaux diminue. Et lorsque la masse corporelle des femelles
diminue, la probabilité qu’elles entrent en œstrus diminue également (Fig. 9). Les analyses de la
masse corporelle et de la gestation des femelles sont donc parfaitement cohérentes de ce point de
vue. Le plus fort impact des prélèvements femelles par rapport aux prélèvements totaux (incluant
mâles et femelles) peut s’expliquer par l’utilisation de l’espace qui est différente selon les sexes.
À l’exception du brame, les mâles et les femelles forment des groupes distincts la plupart du
temps pour de nombreuses espèces de mammifères herbivores, un phénomène dénommé
ségrégation sexuelle (McCullough et al. 1989, Ruckstuhl & Neuhaus 2005). La ségrégation
sexuelle s’explique par une sélection de l’habitat propre à chaque sexe, les femelles cherchant
plutôt des habitats qui offrent une certaine protection vis-à-vis du dérangement et de la prédation
alors que les mâles utilisent des milieux plus exposés mais souvent plus riches en ressources
également (Clutton-Brock et al. 1982, Bonenfant et al. 2004). La compétition pour les ressources
n’est donc pas symétrique pour les mâles et les femelles. À l’échelle de la population, du fait de
la ségrégation sexuelle marquée chez le cerf, les femelles sont plus impactées par les individus du
même sexe que par l’abondance totale ou l’abondance des mâles, car elles partagent les mêmes

40
zones géographiques et sont plus grégaires que ces derniers (Clutton-Brock et al. 1982, Ruckstuhl
& Clutton-Brock 2005). Ce résultat est donc en accord avec la biologie du cerf et devrait porter
l’attention des gestionnaires sur le fait que pour cette espèce, la gestion des mâles et des femelles
n’a pas à être obligatoirement identique, et devrait faire l’objet d’une attention particulière.
Les points clés
1. L’analyse des ICE de performance démontre l’existence d’épisodes de densité-dépendance
dans l’histoire récente du cerf sur le DNC. La période 2008-2012 est l’une d’elle avec des
paramètres biologiques pour les bichettes proches des minima observés sur des sites de
référence connus.
2. La population de cerfs du DNC semble avoir été contrôlée essentiellement entre 1984 et 2007.
3. Le taux de gestation des biches n’ayant pas beaucoup varié, la population semble être dans les
premières phases de densité-dépendance.
1.3. Variations des ICE « Impact »
L’étude de l’évolution de la pression des cervidés sur la flore forestière du domaine s’est
appuyée sur la mise en place de deux indicateurs : l’IPF de 1999 à 2006 puis l’IC de 2005 à 2008.
Le premier, développé par le CEMAGREF (Guibert 1997) a été abandonné en 2006 au profit de
l’IC (Morellet et al. 2001) afin de profiter de la meilleure robustesse du protocole et de conditions
d’application plus accessibles pour le personnel. À partir d’un réseau de placettes fixes (n = 189),
ces deux indicateurs ont permis de mesurer dans le temps la consommation des animaux sur le
cortège floristique ligneux et semi-ligneux du domaine. Compte tenu de l’impossibilité
d’attribuer la consommation à une espèce précise (cerfs, chevreuils et mouflons sont présents) les
résultats présentés seront considérés comme étant d’origine multi-spécifique.
À défaut de pouvoir travailler sur les données brutes, nous avons pris en compte dans nos
analyses les valeurs intrinsèques des indices annuels. Comme il s'agit de probabilité / proportion
(valeurs bornées entre 0 et 1 ou 0% et 100%), nous avons transformé les données pour les rendre
compatibles avec des modèles linéaires classiques. Ceci explique le terme de « variable
transformée » utilisé sur la figure 12 (A&B). Cela ne modifie pas l'interprétation : une forte

41
valeur transformée représente bien une forte valeur d'impact. Il faut noter que le faible nombre de
données (8 pour l'IPF et 4 pour l'IC) limite la puissance statistique et les résultats doivent être
considérés avec précaution. Nous avons d'ailleurs principalement porté nos efforts sur l'IPF pour
privilégier une meilleure série temporelle. Les analyses sur l'IC sont uniquement données à titre
indicatif. Par ailleurs, l'IPF et l'IC étant mesurés durant le repos végétatif (mars - avril), ils
reflètent les abroutissements réalisés au cours de la saison de végétation précédente. Par exemple
l'IPF / l'IC réalisés en 2005 mesurent l'abroutissement de la saison de végétation 2004. Si l'on
souhaite comparer ces deux indices avec des informations d'abondance ou de pression de chasse,
c'est donc l'indice nocturne de 2004 et les prélèvements de la saison 2003-2004 qui devront être
confrontés aux mesures d'impact mesurées en 2005.
Figure 12 : L’indice de pression sur la flore (IPF) sur le domaine national de Chambord est
principalement affecté par les prélèvements de femelles à la chasse la saison précédente (voir texte). (A)
Variations temporelles de l’IPF (cercles noirs) et des prélèvements de femelles à la chasse (croix) sur le
domaine national de Chambord. (B) Relation observée entre l’IPF et le nombre de femelles prélevées à la
chasse l’année précédente sur le domaine national de Chambord. Plus il y a de femelles prélevées, plus
l’IPF est faible.
Dans l'absolu, les valeurs d'IPF (entre 0,388 et 0,920) et les valeurs d'IC (entre 0,690 et
0,960) sont particulièrement fortes et proches du niveau maximum d'abroutissement (valeur de
1). Ces valeurs pourraient signifier que les indices saturent, c'est-à-dire qu'ils ont atteint des
valeurs ne pouvant plus détecter les variations du système « population-environnement ». Dans
un tel cas, ils perdent en pertinence et une analyse fine traitant séparément l'abroutissement sur

42
les différentes espèces végétales suivies plutôt qu'en regroupant toutes les espèces est nécessaire.
On pourrait notamment suivre le report d'abroutissement des végétaux les plus appétents vers les
moins appétents. Cependant, pour les raisons évoquées plus haut (défaut d'accès aux données
brutes), cette approche n'a pas été possible ici mais devra être envisagée dans le futur. Toutefois,
les données « IC » accessibles montrent que la ronce (Rubus sp.) présente sur 30% des placettes
examinées a un taux moyen de consommation compris entre 70 et 97% traduisant une pression
importante des herbivores sur cette espèce.
Les analyses montrent qu'il existe une augmentation linéaire marquée de l'IPF sur la
période 1998-2005 (valeur prédite minimum = 0,36 et valeur maximum = 0,80, soit une
augmentation par 2,2 ; β = 0,066 +/- 0,021, t = 3,148, P = 0,020 ; Fig. 12A). Cette tendance au
cours du temps explique une forte proportion de la variabilité de l'IPF (62%) indépendamment de
tout autre facteur. Si les variations d'abondance montrée par l'indice nocturne ne sont pas
corrélées à l'IPF (β = 0,001 +/- 0,002, t = 0,300, P = 0,775), une diminution du nombre de
femelles prélevées à la chasse pourrait en être à l'origine (β = -0,004 +/- 0,002, t = -1,976, P =
0,096 ; Fig. 12B). Ainsi une augmentation de la proportion de biches prélevées permettrait de
réduire l'impact de l'espèce sur le milieu. Même si la relation entre l’IPF et le nombre de mâles
prélevés à la chasse va dans le même sens, la tendance est loin d'être significative et confirme
l'effet prédominant du prélèvement des femelles adultes sur la dynamique de la population. Le
modèle intégrant l'effet des prélèvements femelles explique 40% de la variabilité de l'IPF. Cette
valeur est proche de celle du modèle considérant une simple augmentation temporelle de l'IPF et
suggère qu'une part importante de celle-ci pourrait être liée à la réduction du plan de chasse.
Aucun résultat comparable n'a été obtenu pour l'IC. À noter que si l'on associe les deux jeux de
données (IPF et IC) en faisant l'hypothèse que ces mesures sont comparables, on continue
d'observer une relation négative entre nombre de femelles prélevées et IPF (β = -0,005 +/- 0,003,
t = -1,969, P = 0,085). Ces résultats soulignent un réel intérêt à remettre en place sur le domaine
un suivi de l'impact des herbivores sur la végétation forestière. Même si les deux indicateurs
étudiés ne sont pas validés pour le cerf qui a un régime alimentaire estival plus orienté vers les
plantes herbacées que celui du chevreuil, il n'en reste pas moins vrai que cette espèce est la plus
représentée sur le DNC. De ce fait, on doit pouvoir lui attribuer une majorité de l'impact sur la
flore forestière. La présence localisée du mouflon et, dans une moindre mesure celle du chevreuil,

43
appelle à approfondir les analyses présentées ici en spatialisant les données d'impact pour tenter
de déterminer le rôle de chaque ongulé.
Les points clés
1. L'impact des herbivores sur le milieu a fortement augmenté au début des années 2000.
2. Cette augmentation pourrait être le résultat d'une diminution du plan de chasse.
3. Les indices atteignent des valeurs proches de 1 et pourraient saturer quant à leur capacité à
suivre l'état du système « population-environnement ».
4. De plus, le faible nombre de données ne permet pas forcément de relier tous les indices entre
eux avec la puissance statistique nécessaire et appelle à remettre en place un suivi de l'impact
(type IC) et à approfondir les analyses en traitant les données brutes.
1.4. Interprétation combinée des ICE
L'interprétation conjointe des 3 familles d'ICE permet de conclure clairement à l'existence
d'une régulation de la population par un processus de densité-dépendance. Régulation
« naturelle » qui est donc capable de prendre place malgré un contrôle anthropique de la
population par la chasse. Ce processus de densité-dépendance se traduit par une diminution
immédiate et linéaire des indicateurs de performance (poids des faons et taux de bichettes
gestantes) à mesure que l'abondance de la population augmente ; et ce dès les plus faibles niveaux
d'abondance observés. Dans la mesure où seuls les jeunes individus sont touchés (pas de réponse
détectée sur le taux de biches gestantes), on peut considérer que les gestionnaires ont su maintenir
le système population-environnement à un faible niveau de densité-dépendance. Ce maintien tire
probablement son origine d'un ajustement assez réactif des plans de chasse et d'un apport
supplémentaire de nourriture (apport qui n'est pas à l'origine ciblé « cervidés » uniquement). Cet
apport explique aussi que l'on puisse observer, même en période de faible abondance, des
niveaux d'abroutissement très importants (proches du maximum théorique) sans que cela soit
associé à une forte réponse démographique de la population. Cependant, si le niveau de
prélèvement a été jusqu’aujourd'hui globalement adapté à la situation écologique du DNC, il
pourrait malgré tout souffrir d'une sous-estimation croissante de l'effectif réel à mesure que la

44
population gagne en abondance. C'est ce que nous montrerons par la suite grâce la modélisation
démographique et aux estimations d'effectif par la génétique. Les réponses marquées des
indicateurs ces dernières années (forte abondance, faible poids et faible reproduction) pourraient
donc appeler à une certaine vigilance et encouragent d’autant plus à une définition précise des
objectifs de gestion (compromis qualité des animaux/opportunité de chasse).
2. Effectifs estimés de la population de cerfs par CMR non-invasif
Sauf conditions très particulières, le dénombrement direct d’animaux est impossible car
tous ne sont pas observables au moment du comptage, soit parce qu’ils ne se trouvent
physiquement pas sur la zone échantillonnée, soit par ce qu’ils sont bien présents mais non
détectés par l’observateur. L’objet de toutes les méthodes d’estimations d’effectifs est donc
d’estimer une probabilité de détection des animaux (notée p) afin de prendre en compte ces
observations imparfaites (Schwarz & Seber 1999). La méthode de Petersen-Lincoln estime ce
paramètre p sur la base d’individus marqués et recapturés lors de 2 échantillonnages successifs
(Seber 1982). Une fois la probabilité de détection estimée, il devient possible d’obtenir un effectif
de la population. Bien que proposée il y a plus de 50 ans, la méthode de Petersen-Lincoln reste à
ce jour un des estimateurs d’effectifs de référence, et pour lequel il existe de nombreuses
variantes mais reposant toutes sur ce même principe. Le frein majeur à une utilisation généralisée
aux grands herbivores tient à la nécessité de capturer et marquer les animaux. Aujourd’hui, la
génétique permet de s’affranchir de cette capture physique puisqu’il suffit « simplement » de
récupérer de l’ADN à partir de n’importe quelles traces biologiques (poils, fèces) laissées par les
animaux. Toutefois, plusieurs sources de biais à l’application de telles méthodes ont été
rapportés. Par exemple, les erreurs de génotypages sont susceptibles de biaiser les estimations
d’effectifs en faisant apparaître deux prélèvements comme appartenant à des individus différents
alors qu’il s’agit d’un seul et même individu (Waits & Leberg 2000). On surestime alors l’effectif
de la population en sous-estimant la probabilité de recapturer un même animal (Valière et al.
2007). Aujourd’hui, grâce aux développements statistiques, il est possible de s’affranchir en
partie de ce problème en intégrant explicitement dans les analyses une probabilité d’erreur
d’identification d’un individu par son génotype (notée 𝜓).
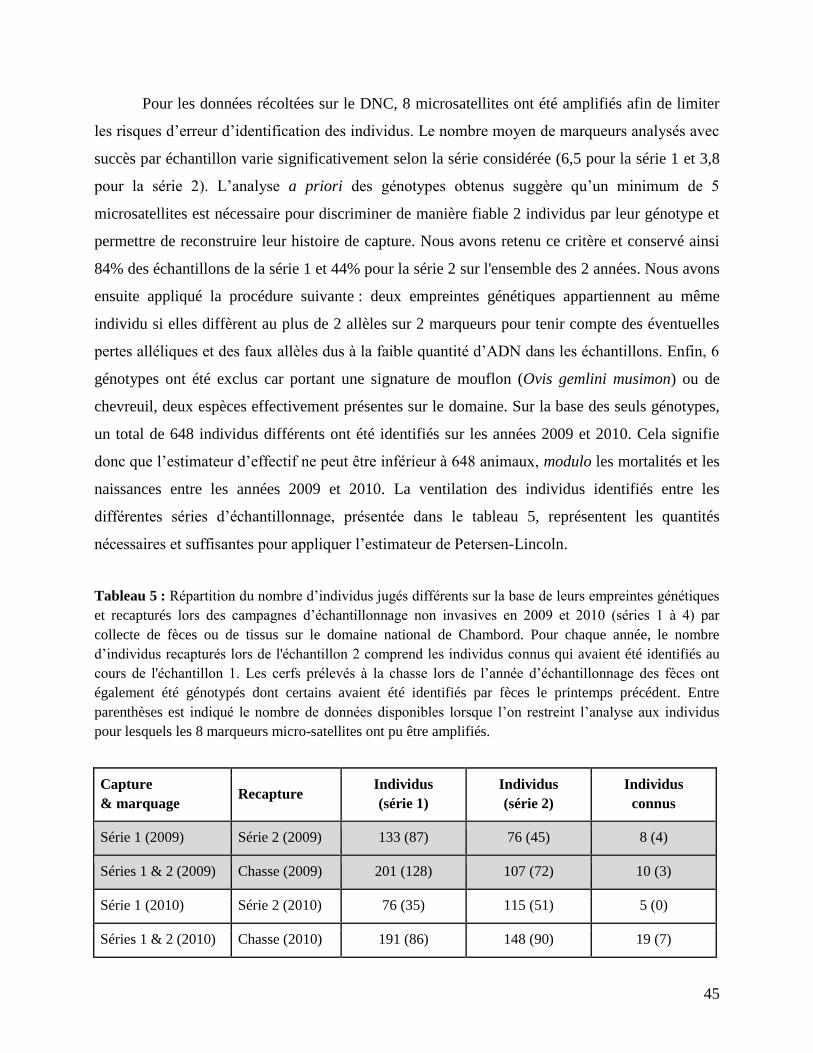
45
Pour les données récoltées sur le DNC, 8 microsatellites ont été amplifiés afin de limiter
les risques d’erreur d’identification des individus. Le nombre moyen de marqueurs analysés avec
succès par échantillon varie significativement selon la série considérée (6,5 pour la série 1 et 3,8
pour la série 2). L’analyse a priori des génotypes obtenus suggère qu’un minimum de 5
microsatellites est nécessaire pour discriminer de manière fiable 2 individus par leur génotype et
permettre de reconstruire leur histoire de capture. Nous avons retenu ce critère et conservé ainsi
84% des échantillons de la série 1 et 44% pour la série 2 sur l'ensemble des 2 années. Nous avons
ensuite appliqué la procédure suivante : deux empreintes génétiques appartiennent au même
individu si elles diffèrent au plus de 2 allèles sur 2 marqueurs pour tenir compte des éventuelles
pertes alléliques et des faux allèles dus à la faible quantité d’ADN dans les échantillons. Enfin, 6
génotypes ont été exclus car portant une signature de mouflon (Ovis gemlini musimon) ou de
chevreuil, deux espèces effectivement présentes sur le domaine. Sur la base des seuls génotypes,
un total de 648 individus différents ont été identifiés sur les années 2009 et 2010. Cela signifie
donc que l’estimateur d’effectif ne peut être inférieur à 648 animaux, modulo les mortalités et les
naissances entre les années 2009 et 2010. La ventilation des individus identifiés entre les
différentes séries d’échantillonnage, présentée dans le tableau 5, représentent les quantités
nécessaires et suffisantes pour appliquer l’estimateur de Petersen-Lincoln.
Tableau 5 : Répartition du nombre d’individus jugés différents sur la base de leurs empreintes génétiques
et recapturés lors des campagnes d’échantillonnage non invasives en 2009 et 2010 (séries 1 à 4) par
collecte de fèces ou de tissus sur le domaine national de Chambord. Pour chaque année, le nombre
d’individus recapturés lors de l'échantillon 2 comprend les individus connus qui avaient été identifiés au
cours de l'échantillon 1. Les cerfs prélevés à la chasse lors de l’année d’échantillonnage des fèces ont
également été génotypés dont certains avaient été identifiés par fèces le printemps précédent. Entre
parenthèses est indiqué le nombre de données disponibles lorsque l’on restreint l’analyse aux individus
pour lesquels les 8 marqueurs micro-satellites ont pu être amplifiés.
Capture
& marquage Recapture
Individus
(série 1)
Individus
(série 2)
Individus
connus
Série 1 (2009) Série 2 (2009) 133 (87) 76 (45) 8 (4)
Séries 1 & 2 (2009) Chasse (2009) 201 (128) 107 (72) 10 (3)
Série 1 (2010) Série 2 (2010) 76 (35) 115 (51) 5 (0)
Séries 1 & 2 (2010) Chasse (2010) 191 (86) 148 (90) 19 (7)

46
Les estimations d’effectifs de cerfs ont été réalisées pour 2009 et 2010 indépendamment à
l’aide de modèles de Capture-Marquage-Recaptures pour populations fermées (Pollock 1991).
Ces modèles supposent que la population est démographiquement close (mortalité = natalité =
immigration = émigration = 0) entre deux sessions d’échantillonnage. C’est très probablement le
cas lorsque l’on travaille sur les fèces puisque le domaine est enclos et que les données ont été
prélevées en mars-avril sur un intervalle de temps inférieur à 16 jours. Ce n’est par contre plus le
cas lorsque l’on compare les individus retrouvés par les fèces avec les individus prélevés à la
chasse puisque des naissances et de la mortalité interviennent entre les deux périodes
d’échantillonnage. Les analyses ont été effectuées avec le programme MARK version 7.1 (White
& Burnham 1999). Les modèles avec identification imparfaite ont été employés en fixant la
probabilité d’identification correcte d’un individu à 𝜓 = 0,95. La probabilité de recapture (p) a été
maintenue constante dans le temps pour les échantillonnages fèces (pserie1 = pserie2 = p), mais
différente pour les 2 pas de temps lorsque les données de chasse ont été ajoutées (pfèces ≠ pchasse).
Les modèles avec hétérogénéité de capture (ou modèle de mélange) ont également été ajustés et
comparés avec les modèles sans hétérogénéité de capture. Les mêmes analyses ont été répliquées
pour les données sans incertitude (8 microsatellites amplifiés) et pour les données avec
incertitude (génotype présentant au moins 5 microsatellites amplifiés).
Les estimations de l’effectif de la population de cerfs du DNC sont très variables selon les
séries et les années considérées (entre 885 et 3108 animaux, voir Tab. 6). La seule constante est
l’intervalle de confiance à 95% des estimations qui est toujours très grand et traduit la faible
précision des estimations d’effectifs. Le coefficient de variation minimale des différentes
estimations est de 20%. Les estimations d’effectifs les plus fortes proviennent systématiquement
des comparaisons entre les échantillons de fèces et les échantillons obtenus à la chasse (Tab. 6).
Outre que des effectifs supérieurs à 2000 animaux semblent peu réalistes biologiquement, ces
valeurs traduisent davantage un non-respect des hypothèses sous-jacentes à la méthode, et en
particulier l’hypothèse d’une population fermée. En effet, entre les récoltes des fèces au
printemps et pendant la saison de chasse en automne, se produisent des naissances et de la
mortalité qui vont diminuer la probabilité de recapture des individus et donc, amener à surestimer
l’estimation de l’effectif. En égard au non-respect de cette hypothèse, les estimations d’effectifs
faisant intervenir les échantillons prélevés sur les animaux chassés ne doivent être ni considérées,
ni interprétées. Si l’on se restreint aux données fèces récoltés spécifiquement pour appliquer la

47
méthode de Petersen-Lincoln, les effectifs de cerfs estimés se situent entre 885 et 1503 animaux
pour le domaine, soit une estimation passant du simple au double (Tab. 6). La qualité des
estimations dépend de la taille des échantillons, mais surtout du nombre d’individus marqués
dans la série 1 qui sont ensuite recapturés dans la série 2. En deçà de 7 recaptures, les estimations
ne sont pas fiables (Robson & Regier 1964). Or, le tri des meilleures données (génotypes avec 8
microsatellites amplifiés) est bien trop restrictif et seulement 4 et 0 individus ont été recapturés
dans la série 2 en 2009 et 2010 respectivement (Tab. 5) - de fait, aucune estimation n’est possible
pour 2010 avec ces données. Avec les données optimisées (génotypes avec au moins 5
microsatellites amplifiés), le problème existe toujours mais s’avère moins marqué avec 8
animaux recapturés en 2009 et 5 en 2010 (Tab. 5), ce qui doit permettre des estimations de
meilleure qualité. Les effectifs de la population de cerfs du DNC seraient ainsi de 1352 (758 -
2579) animaux en 2009 et 1503 (1038 - 2268) animaux en 2010, soit des densités
correspondantes de 24,8 et 27,6 individus aux 100ha et un taux d’accroissement r de 0,11 entre
les deux années. A titre de comparaison, dans les Hautes Terres d’Écosse, les densités de cerfs
observées ont varié entre 3,4 et 34,3 individus aux 100ha (Clutton-Brock & Albon 1989). À La
Petite Pierre, population française vivant dans un milieu relativement pauvre, les densités de cerfs
ont pu atteindre des valeurs de l’ordre de 20 individus aux 100ha (Garel et al. 2010). Les
estimations de densité faites sur le DNC sont donc des valeurs biologiquement réalistes, quoique
relativement élevées.
L’estimation de l’effectif de la population de cerfs à dires d’experts en 2009 et 2010 est de
l’ordre de 700 individus environ, soit près de la moitié de la valeur estimée par la méthode de
Petersen-Lincoln. À ce stade de l’analyse, plusieurs arguments plaident en faveur d’une sous-
estimation de l’effectif à dires d’experts. Sur les années 2009 et 2010, le nombre total d’individus
différents retrouvés via les fèces ou la chasse est sensiblement identique (648) donc, sauf à
imaginer que les fèces de tous les individus de la population aient été collectés, le nombre de
cerfs vivants sur le DNC est nécessairement bien supérieur à 700. Bien que ce chiffre de 648
fasse fit des mortalités et des naissances, il sous-estime nettement le nombre d’individus
différents puisque seulement 46% des échantillons de fèces ont pu être analysés. Un autre
argument nous est apporté par l’indice nocturne qui suggère que les années 2009 et 2010 sont,
historiquement, des années de très forte abondance (Fig. 3), avec une détérioration marquée des
indicateurs de performance que sont la masse corporelle des faons et le taux de gestation des
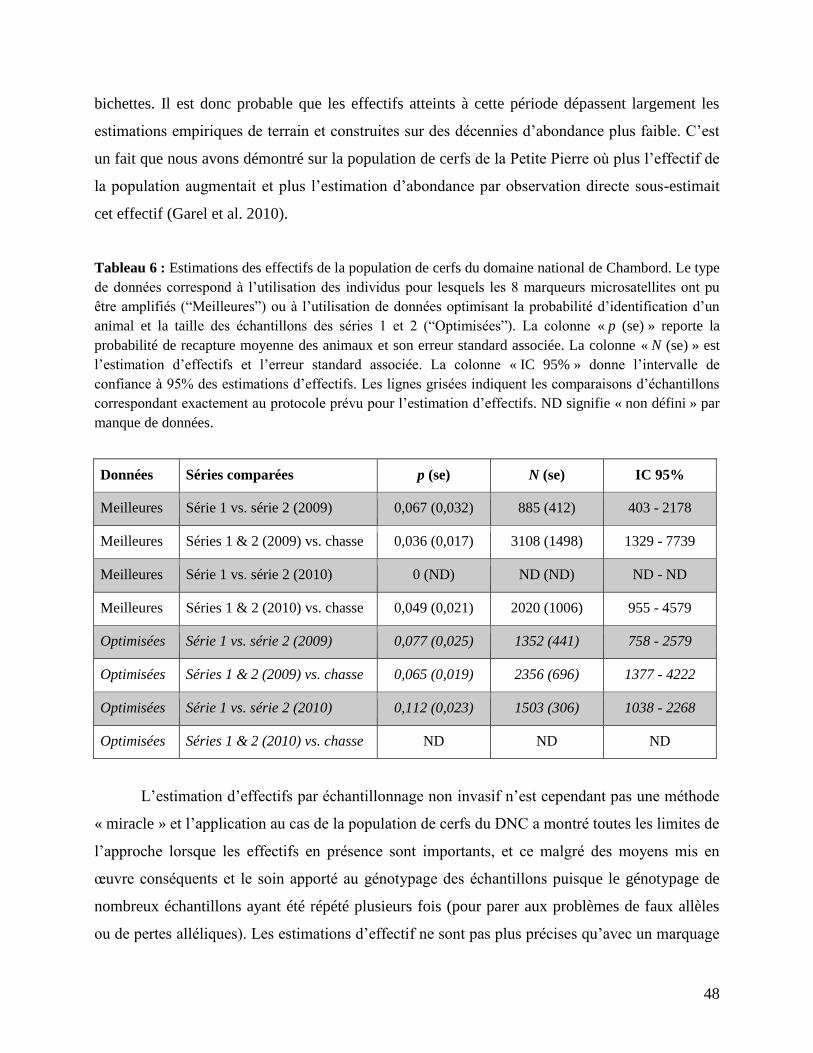
48
bichettes. Il est donc probable que les effectifs atteints à cette période dépassent largement les
estimations empiriques de terrain et construites sur des décennies d’abondance plus faible. C’est
un fait que nous avons démontré sur la population de cerfs de la Petite Pierre où plus l’effectif de
la population augmentait et plus l’estimation d’abondance par observation directe sous-estimait
cet effectif (Garel et al. 2010).
Tableau 6 : Estimations des effectifs de la population de cerfs du domaine national de Chambord. Le type
de données correspond à l’utilisation des individus pour lesquels les 8 marqueurs microsatellites ont pu
être amplifiés (“Meilleures”) ou à l’utilisation de données optimisant la probabilité d’identification d’un
animal et la taille des échantillons des séries 1 et 2 (“Optimisées”). La colonne « p (se) » reporte la
probabilité de recapture moyenne des animaux et son erreur standard associée. La colonne « N (se) » est
l’estimation d’effectifs et l’erreur standard associée. La colonne « IC 95% » donne l’intervalle de
confiance à 95% des estimations d’effectifs. Les lignes grisées indiquent les comparaisons d’échantillons
correspondant exactement au protocole prévu pour l’estimation d’effectifs. ND signifie « non défini » par
manque de données.
Données Séries comparées p (se) N (se) IC 95%
Meilleures Série 1 vs. série 2 (2009) 0,067 (0,032) 885 (412) 403 - 2178
Meilleures Séries 1 & 2 (2009) vs. chasse 0,036 (0,017) 3108 (1498) 1329 - 7739
Meilleures Série 1 vs. série 2 (2010) 0 (ND) ND (ND) ND - ND
Meilleures Séries 1 & 2 (2010) vs. chasse 0,049 (0,021) 2020 (1006) 955 - 4579
Optimisées Série 1 vs. série 2 (2009) 0,077 (0,025) 1352 (441) 758 - 2579
Optimisées Séries 1 & 2 (2009) vs. chasse 0,065 (0,019) 2356 (696) 1377 - 4222
Optimisées Série 1 vs. série 2 (2010) 0,112 (0,023) 1503 (306) 1038 - 2268
Optimisées Séries 1 & 2 (2010) vs. chasse ND ND ND
L’estimation d’effectifs par échantillonnage non invasif n’est cependant pas une méthode
« miracle » et l’application au cas de la population de cerfs du DNC a montré toutes les limites de
l’approche lorsque les effectifs en présence sont importants, et ce malgré des moyens mis en
œuvre conséquents et le soin apporté au génotypage des échantillons puisque le génotypage de
nombreux échantillons ayant été répété plusieurs fois (pour parer aux problèmes de faux allèles
ou de pertes alléliques). Les estimations d’effectif ne sont pas plus précises qu’avec un marquage

49
physique, et des problèmes techniques et statistiques apparaissent rapidement. Si la méthode
devait être reconduite, un nouveau plan d’échantillonnage, spatialisé, devra être proposé et le
nombre de marqueurs micro-satellite augmenté pour accroître sensiblement la probabilité de
recapture (p). De plus, et contrairement aux captures physiques, il n’est pas possible de connaître
en temps réel le nombre d’animaux différents que l’on capture, de sorte que l’échantillonnage
pourrait être ajusté rapidement. En conséquence, lorsque l’on ne dispose pas d'à priori sur
l’effectif de la population lors de la mise en place du protocole, le risque d’avoir un taux de
recapture très faible est important, compromettant alors l’objectif initial d’une estimation
d’effectif juste et précise.
Les points clés
1. Six cent quarante-huit (648) individus différents ont été identifiés sur la base des génotypes
récoltés en 2009 et 2010.
2. Les estimations de l’effectif de la population de cerfs du DNC par capture-marquage-recapture
non invasif varient entre 1352 et 1503 animaux en 2009 et 2010 respectivement, toujours avec
une très forte incertitude (min = 750, max = 2600).
3. Les effectifs estimés sont cohérents avec l’analyse des indicateurs de changement écologique.
3. Modélisation et plans de prélèvements
Afin de compléter la démarche de gestion adaptative des populations fondée sur les ICE,
nous avons élaboré un modèle démographique en combinant des informations sur la démographie
des cerfs avec des données de prélèvements et un indice d’abondance. Lorsque l'on dispose de
toutes les informations sur les valeurs des paramètres démographiques de la population étudiée,
ce modèle permet de reconstruire a posteriori les variations passées de l'abondance de cerfs. D'un
point de vue appliqué, cette approche permet de prévoir l’évolution des populations sur le cours
terme et, en fonction de différents scénarii de prélèvements, d’aider à établir les plans de
prélèvements.
Pour ce faire, nous nous sommes inspirés des résultats du travail de modélisation réalisé
sur la population de cerfs de la RNCFS de la Petite-Pierre pour laquelle de nombreuses données

50
sont disponibles depuis une trentaine d’années (Pellerin et al. 2013). Dans un premier temps,
nous avons développé un modèle matriciel de Leslie-Lewis structuré en âge (Leslie 1954) qui
intègre les paramètres démographique de la population : la survie naturelle et la fécondité. Afin
que ce modèle soit facilement utilisable par les gestionnaires et qu’il puisse être confronté avec
les indices d'abondance pour lesquels l'âge exact des animaux n'est pas connu, nous l’avons
structuré en gardant 3 classes d’âge : faon, bichette et biche (Fig. 13). Concernant la survie, nous
ne disposons actuellement pas des valeurs pour la population de cerfs du domaine de Chambord.
Par conséquent, nous avons utilisé des valeurs communes pour cette espèce (survie = 0,90 pour
faons, bichettes et femelles adultes ; Gaillard et al. 2000) et considéré les survies constantes au
cours du temps. La fécondité des femelles (taux de gestation) du domaine de Chambord a pu être
estimée grâce aux femelles prélevées à la chasse (voir section 1.2.2). Dans le modèle, ce taux de
gestation varie annuellement (de 1982-1983 à 2012-2013), avec des variations plus marquées
pour les bichettes (entre 14 et 100%) que pour les biches (entre 67 et 100%). On obtient alors une
matrice de Leslie annuelle. Le modèle est femelle centrée, c'est-à-dire qu'il ne considère que le
segment femelle de la population. Le cerf étant fortement polygyne, les mâles sont considérés
comme non limitant pour la croissance de la population. Ce modèle simple à croissance
exponentielle permet alors de calculer pour chaque année le taux d’accroissement annuel (rt) et
moyen (moyenne géométrique) de la population de cerfs.
Appliqué au cas du domaine de Chambord, nous trouvons un taux d’accroissement moyen
de la population de cerfs depuis 1982 de r = +17,0%, avec une variation observée entre r =
+10,7% (en 2010) et r = +20,6% (en 1996). Aucune des valeurs de r que l'on a calculées ne
tombe en dehors de valeurs biologiques possibles pour cette espèce monotoque (un seul petit par
portée).
Les données de prélèvements par classe d’âge n’étant disponibles que depuis la saison
1983-1984, l’année 1983 a été choisie comme année de départ pour le modèle. Après l’estimation
de l’effectif initial (en 1983) pour les trois classes d’âge de femelles à partir des données d’indice
nocturne et des connaissances sur la structure en âge d’une population de cerfs (Gaillard et al.
2000, Gaillard et al. 2003b), nous avons tenté de reconstruire les trajectoires passées de la
population (Fig. 13&14).

51
Figure 13 : Estimation de l’effectif de femelles (faons, bichettes, biches) à la saison t de la population de
Chambord à partir de l’effectif de la saison précédente, du modèle matriciel et du nombre de femelles
prélevées à la chasse la saison précédente.
Figure 14 : Variation temporelle de l’effectif de la population de cerfs du domaine national de Chambord
prédit (par le modèle de Leslie, avant et après prélèvement) de 1983-1984 à 2012-2013. Par comparaison,
l’indice nocturne mis en place depuis 1985 (correspondant à la saison 1984-1985) a été ajouté.
Les prédictions d’effectif de femelles est en moyenne de 455 +/- 50, avec deux valeurs
minimum en 1987 (361 femelles) et en 2000 (390 femelles), et un maximum de 546 en 2007.
L'effectif total comprenant les mâles et les femelles est obtenu en doublant ces chiffres (en
supposant un équilibre des sexes dans la population), donnant des valeurs de 722 animaux en
1987 et 1092 en 2007. Notons que l'effectif de 1092 animaux en 2007 est tout à fait compatible
avec les estimations d'effectifs obtenues par capture-marquage-recapture non-invasif les années

52
suivantes (voir section III.2), puisque tombant dans l'intervalle de confiance des estimations (min
= 750, max = 2600). Les effectifs prédits (avant et après prélèvements par la chasse) sont bien
corrélés avec l’indice nocturne (P < 0,001 et r = 0,70 et 0,62 ; respectivement avant et après
prélèvements ; Fig. 15). Ce résultat signifie que les prélèvements et la variation temporelle de la
fécondité des femelles permettent d'appréhender, dans les grandes lignes, les structures observées
depuis 1985 dans la variation temporelle d'abondance de la population de cerfs du DNC. En
particulier, on retrouve bien deux épisodes marquant que sont l'abondance minimale du début des
années 2000 et le pic atteint vers 2008-2009. Enfin, plus qu'une diminution, le modèle prédit une
stabilisation de l'abondance depuis 2010 liée à une reproduction des bichettes plutôt faible et un
fort prélèvement à la chasse depuis 2008-2009. Une autre conclusion majeure de cet exercice de
modélisation est que l'effectif de population à dires d'experts (environ 700 individus) ne permet
pas à la population de cerfs du domaine de soutenir les prélèvements annuels réalisés. En d'autres
termes, partant d'effectifs plus proches de ceux à dires d'experts, les prélèvements réalisés
amènent la population de cerfs du DNC à l'extinction avant 2012. Comme pour de nombreux cas
(voir Bonenfant & Klein 2004 pour un exemple dans les Cévennes), l'effectif de la population de
cerfs du domaine a donc probablement été sous-estimée de manière chronique par les
gestionnaires, au moins depuis la mise en place des indices de suivi au début des années 1980.
Reste que la faiblesse de l'approche proposée ici est que l'on ne connaît pas tous les paramètres
démographiques de la population du DNC et que l'on a du se reposer sur les estimations faites à
La Petite Pierre, en particulier pour les paramètres de survie et de structure en âge. Des
imprécisions sont donc possibles, et les résultats pourront être affinés avec l'utilisation des
estimations de survie réalisées sur le domaine de Chambord. Pour la structure en âge (faons,
biches, bichettes), les données de capture par panneautage seraient particulièrement pertinentes à
exploiter, sous l'hypothèse qu'il n'y a pas de biais de capture des individus selon leur âge.
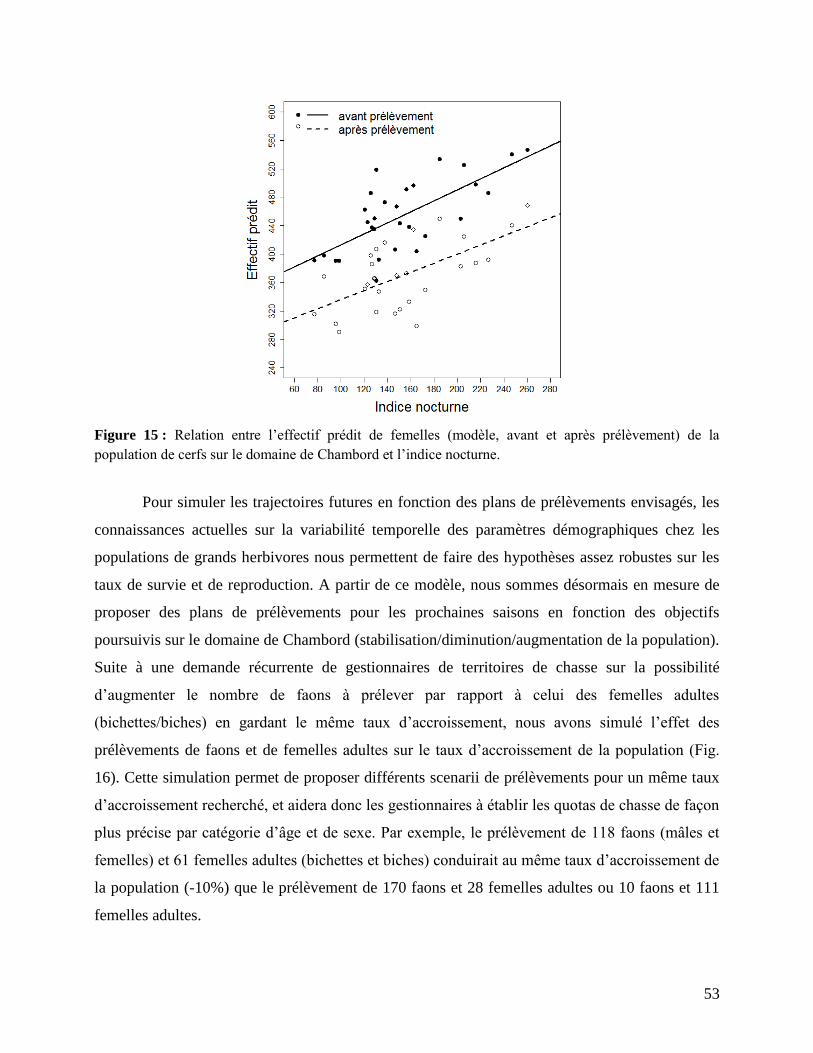
53
Figure 15 : Relation entre l’effectif prédit de femelles (modèle, avant et après prélèvement) de la
population de cerfs sur le domaine de Chambord et l’indice nocturne.
Pour simuler les trajectoires futures en fonction des plans de prélèvements envisagés, les
connaissances actuelles sur la variabilité temporelle des paramètres démographiques chez les
populations de grands herbivores nous permettent de faire des hypothèses assez robustes sur les
taux de survie et de reproduction. A partir de ce modèle, nous sommes désormais en mesure de
proposer des plans de prélèvements pour les prochaines saisons en fonction des objectifs
poursuivis sur le domaine de Chambord (stabilisation/diminution/augmentation de la population).
Suite à une demande récurrente de gestionnaires de territoires de chasse sur la possibilité
d’augmenter le nombre de faons à prélever par rapport à celui des femelles adultes
(bichettes/biches) en gardant le même taux d’accroissement, nous avons simulé l’effet des
prélèvements de faons et de femelles adultes sur le taux d’accroissement de la population (Fig.
16). Cette simulation permet de proposer différents scenarii de prélèvements pour un même taux
d’accroissement recherché, et aidera donc les gestionnaires à établir les quotas de chasse de façon
plus précise par catégorie d’âge et de sexe. Par exemple, le prélèvement de 118 faons (mâles et
femelles) et 61 femelles adultes (bichettes et biches) conduirait au même taux d’accroissement de
la population (-10%) que le prélèvement de 170 faons et 28 femelles adultes ou 10 faons et 111
femelles adultes.

54
Figure 16 : Simulation des prélèvements de femelles faons et adultes (bichettes+biches) et effet sur le
taux d’accroissement de la population (les proportions indiquées sont exprimées par rapport à la
population existante de femelles faons et adultes).
Les points clés
1. Les estimations d’effectif de femelles de la population de cerfs du DNC par modélisation
varient de 361 à 546 entre 1983-1984 et 2012-2013.
2. Les effectifs prédits par le modèle démographique sont compatibles avec les variations
d’abondance relative observées avec l’indice nocturne et avec les estimations par capture-
marquage-recapture non invasif.
3. La modélisation de la population du DNC devrait permettre d’améliorer l’adéquation entre les
plans de prélèvements et les objectifs de gestion recherchés.

55
IV. CONCLUSIONS
La population de cerfs du domaine national de Chambord semble se trouver à des niveaux
de densité pour lesquels une manifestation de densité dépendance n’est perceptible que sur les
classes d’âge des faons et des bichettes. A ce stade de l’analyse, deux hypothèses pourraient être
formulées :
1. La plus parcimonieuse est l’hypothèse que la population a atteint environ 60% de la
capacité limite du milieu, ce qui entraîne alors une diminution séquentielle des 3 premiers
paramètres démographiques montrant une réponse à la densité que sont : (1) la masse
corporelle des faons mâles, (2) la masse corporelle des faons femelles, (3) la proportion
de bichettes gestantes. En effet, chez les grands mammifères, cette valeur théorique
correspond au point d’inflexion de la courbe de croissance d’une population, i.e., au point
à partir duquel le taux de multiplication annuel instantané devient négatif (r ≤ 0 ; voir
Delorme & Gaillard 1999).
2. La seconde hypothèse que l’on peut formuler serait que les prélèvements annuels
permettent un contrôle efficace des effectifs prévenant ainsi l’augmentation de la densité.
Cette dernière alternative est cependant peu compatible avec les variations annuelles des
différents ICE mesurés.
Deux causes peuvent être à l’origine de la diminution de masse corporelle des faons qui
aboutissent apparemment aux mêmes résultats. Il a pu se produire une dégradation progressive de
l’environnement, soit par des modifications anthropiques ou bien par des modifications de la
qualité et de la quantité des ressources par le cerf lui-même (Caughley 1977). Une autre
explication serait que la densité de cerfs a augmenté durant toutes ces années. Dans les deux cas,
la quantité de ressources disponibles par individu est amoindrie, conduisant à une diminution de
la masse corporelle moyenne. Cependant, en l’absence de suivi de longue durée par indicateur de
changement écologique sur le compartiment végétale du domaine, il n’est pas possible de
véritablement trancher entre ces deux hypothèses.
D’autre part l’existence d’effets cohortes suggère que les jeunes cerfs sont sujets à des
contraintes environnementales diverses et de fait, ne se trouvent pas dans des conditions les plus
favorables pour leur croissance. On s’attend à ce que les effets cohortes soient plus marqués dans

56
les populations soumises à une forte variabilité environnementale et/ou des populations se
trouvant à forte densité, ce qui exacerbe les effets du climat (Albon et al. 1992, Gaillard et al.
2003b). Nous avons pu détecter l’influence du climat (Gaussen Index - GI) que sur la masse
corporelle des faons (Mysterud et al. 2001b, Post & Stenseth 1998 ; pour des exemples sur les
ongulés). Compte tenu du caractère local de cette variable, le GI est très représentatif du climat
local que l’on trouve en région Centre. Il semblerait que les différents épisodes de sécheresse
printanièrs rencontrés ponctuellement au cours de ces dernières années affectent directement les
classe d’âge les plus jeunes, et que cet effet ne se limite pas toujours à l’ICE masse corporelle
puisque nous avons également détecté une baisse de la reproduction des femelles primipares pour
certaines années. L’amélioration sensible de la qualité phénotypique des faons est probablement,
au cours des dernières années de suivi, la conséquence d’une augmentation de la qualité et de la
quantité des ressources (notamment en graminées) liée à une diminution de la pression exercée
sur le milieu, ou de l’absence d’épisode de sécheresse climatique sévère pour les jeunes classes
d’âge.
En résumé le domaine national de Chambord démontre qu'il est bien un site unique pour
le cerf, le travail accompli a permis une analyse approfondie de la dynamique de sa population de
cerfs sur des durées rarement atteintes ailleurs. Les principaux résultats mettent en évidence que :
● Les ICE montrent une fois de plus leur intérêt pratique et la possibilité de les appliquer à
des conditions écologiques variées. Avec la conclusion que ces outils de suivi ont permis
de comprendre à rebours la dynamique de la population de cerfs du DNC depuis 1982.
● Certaines mesures de terrain, surtout celles relatives aux indices d'abondance relative, ne
sont pas nécessaires car souvent redondantes ou bien démontrées comme peu fiables. Il
est donc possible d'alléger les protocoles de suivi annuels en conservant uniquement
l’indice nocturne.
● Au contraire, certains suivis comme celui de la reproduction des biches et bichettes
devraient être poursuivi et intensifié (effectifs trop faibles les dernières années) ainsi que
les mesures de pression des animaux sur la flore forestière (abandonnées depuis 2009).
● La population a traversé des épisodes de relative stabilité mais la dernière décennie se
caractérise par une forte augmentation de l'abondance et une population probablement
bien entrée en phase de densité-dépendance depuis les 5 dernières années.

57
● Tout indique que l'abondance du cerf est certainement plus forte que supposée jusqu'alors,
en particulier à partir des estimations par capture-marquage-recaptures non invasif.
● Une augmentation substantielle des prélèvements est nécessaire pour un objectif de
gestion qui envisagerait un retour à des conditions écologiques similaires à celles des
années 1990.
Enfin les données récoltées sont d'une qualité remarquable et méritent d'être valorisées
scientifiquement pour un exemple d'utilisation des ICE sur le cerf. Cette connaissance souligne
l'intérêt de poursuivre en optimisant les suivis ICE déjà en place, et en pérennisant le programme
scientifique débuté en 2013 (« Chasse et dynamique des populations d’ongulés sauvages ») pour
faire du domaine de Chambord un site d’étude de référence sur le cerf en France et en Europe.

58
BIBLIOGRAPHIE
Albon, S. D., T. H. Clutton-Brock, and R. Langvatn. 1992. Cohort variation in reproduction and
survival: implications for population demography. Pages 15-21 in The biology of deer,
2nd int. symp. Springer-Verlag, Mississippi.
Albon, S. D., B. Mitchell, and B. W. Staines. 1983. Fertility and body weight in female red deer:
a density-dependent relationship. Journal of Animal Ecology 52:969-980.
Becker, M., T. M. Nieminen, and F. Geremia. 1994. Short term variations and long term changes
in oak productivity in northeastern France. The role of climate and atmospheric CO2.
Annales des Sciences Forestières 51:477-492.
Bertouille, S. B. and S. A. de Crombrugghe. 2002. Fertility of red deer in relation to area, age,
body mass, and mandible length. Zeitschrift Fur Jagdwissenschaft 48:87-98.
Bonenfant, C. 2004. Rôle des contraintes évolutives dépendantes du sexe en biologie des
populations. L'exemple du Cerf élaphe (Cervus elaphus). PhD Thesis. Université Claude
Bernard - Lyon 1, Lyon.
Bonenfant, C., J. M. Gaillard, T. Coulson, M. Festa-Bianchet, A. Loison, M. Garel, L. E. Loe, P.
Blanchard, N. Pettorelli, N. Owen-Smith, J. Du Toit, and P. Duncan. 2009. Empirical
evidence of density-dependence in populations of large herbivores. Advances in
Ecological Research 41:313-357.
Bonenfant, C., J. M. Gaillard, F. Klein, and A. Loison. 2002. Sex- and age-dependent effects of
population density on life history traits of red deer Cervus elaphus in a temperate forest.
Ecography 25:446-458.
Bonenfant, C. and F. Klein. 2004. Évolution de la population de Cerfs (Cervus elaphus L.) du
Parc National des Cévennes. Rapport ONCFS.
Bonenfant, C., L. E. Loe, A. Mysterud, R. Langvatn, N. C. Stenseth, J.-M. Gaillard, and F. Klein.
2004. Multiple causes of sexual segregation in European red deer: enlightenments from
varying breeding phenology at high and low latitude. Proceedings of the Royal Society of
London B 271:883-892.
Bonenfant, C., M. L. Schwoerer, B. Boisaubert, and F. Klein. 2007. Observatoire du Donon -
Analyses des données. Rapport ONCFS.
Caughley, G. 1977. Analysis of vertebrate populations. John Wiley & Sons, New York.

59
Clutton-Brock, T. H. and S. D. Albon. 1989. Red deer in the highlands. Blackwell Scientific
Publication, Oxford.
Clutton-Brock, T. H., F. E. Guinness, and S. D. Albon. 1982. Red deer - Behaviour and ecology
of two sexes. Edinburgh University Press, Edinburgh, UK.
Coulson, T. N., E. A. Catchpole, S. D. Albon, B. J. T. Morgan, J. M. Pemberton, T. H. Clutton-
Brock, M. J. Crawley, and B. T. Grenfell. 2001. Age, sex, density, winter weather, and
population crashes in Soay sheep. Science 292:1528-1531.
Creel, S., G. Spong, J. L. Sands, J. Rotella, J. Zeigle, L. Joe, K. M. Murphy, and D. Smith. 2003.
Population size estimation in Yellowstone wolves with error-prone noninvasive
microsatellite genotypes. Molecular Ecology 12:2003-2009.
Dajoz, R. 1973. Précis d'écologie. Gauthier Villars.
Delorme, D. and J.-M. Gaillard. 1999. Modèles de croissance des populations de chevreuils : de
la théorie à la pratique. Bulletin Mensuel de l'ONC 244:12-16.
Douhard, M., C. Bonenfant, J. M. Gaillard, J. L. Hamann, M. Garel, J. Michallet, and F. Klein.
2013. Roaring counts are not suitable for the monitoring of red deer Cervus elaphus
population abundance. Wildlife Biology 19:94-101.
Eberhardt, L. L. 1977. Optimal policies for conservation of large mammals, with special
references to marine ecosystems. Environmental Conservation 4:205-212.
Eberhardt, L. L. 2002. A paradigm for population analysis of long-lived vertebrates. Ecology
83:2841-2854.
Eggert, L. S., J. A. Eggert, and D. S. Woodruff. 2003. Estimating population sizes for elusive
animals: the forest elephants of Kakum National Park, Ghana. Molecular Ecology
12:1389-1402.
Gaillard, J. M., J. M. Boutin, D. Delorme, G. VanLaere, P. Duncan, and J. D. Lebreton. 1997.
Early survival in roe deer: causes and consequences of cohort variation in two contrasted
populations. Oecologia 112:502-513.
Gaillard, J. M., D. Delorme, J. M. Boutin, G. VanLaere, and B. Boisaubert. 1996. Body mass of
roe deer fawns during winter in 2 contrasting populations. Journal of Wildlife
Management 60:29-36.

60
Gaillard, J. M., D. Delorme, J. M. Boutin, G. Vanlaere, B. Boisaubert, and R. Pradel. 1993. Roe
deer survival patterns: a comparative analysis of contrasting populations. Journal of
Animal Ecology 62:778-791.
Gaillard, J. M., M. Festa-Bianchet, and N. G. Yoccoz. 1998. Population dynamics of large
herbivores: variable recruitment with constant adult survival. Trends in Ecology and
Evolution 13:58-63.
Gaillard, J. M., M. Festa-Bianchet, N. G. Yoccoz, A. Loison, and C. Toigo. 2000. Temporal
variation in fitness components and population dynamics of large herbivores. Annual
Review of Ecology and Systematics 31:367-393.
Gaillard, J. M., A. Loison, C. Toigo, D. Delorme, and G. Van Laere. 2003a. Cohort effects and
deer population dynamics. Ecoscience 10:412-420.
Gaillard, J. M., A. Loison, and C. Toïgo. 2003b. Variation in life history traits and realistic
population models for wildlife management: the case of ungulates. Pages 115-132 in M.
Festa-Bianchet and M. Apollonio, editors. Animal Behavior and Wildlife Conservation.
Island Press, Washington.
Gaillard, J. M., A. J. Sempéré, J. M. Boutin, G. Vanlaere, and B. Boisaubert. 1992. Effects of age
and body weight on the proportion of females breeding in a population of roe deer
(Capreolus Capreolus). Canadian Journal of Zoology 70:1541-1545.
Gamelon, M., M. Douhard, E. Baubet, O. Gimenez, S. Brandt, and J. M. Gaillard. 2013.
Fluctuating food resources influence developmental plasticity in wild boar. Biology
Letters 9.
Garel, M., C. Bonenfant, J. L. Hamann, F. Klein, and J. M. Gaillard. 2010. Are abundance
indices derived from spotlight counts reliable to monitor red deer Cervus elaphus
populations? Wildlife Biology 16:77-84.
Garel, M., J. M. Cugnasse, J. M. Gaillard, A. Loison, P. Gibert, P. Douvre, and D. Dubray. 2005.
Reproductive output of female mouflon (Ovis gmelini musimon x Ovis sp.): a comparative
analysis. Journal of Zoology, London 266:65-71.
Garel, M., A. Loison, J.-M. Gaillard, J.-M. Cugnasse, and D. Maillard. 2004. The effects of a
severe drought on mouflon lamb survival. Proceedings of the Royal Society of London B
271 (Suppl.):S471-S473.

61
Gebert, C. and H. Verheyden-Tixier. 2001. Variations of diet composition of red deer (Cervus
elaphus L.) in Europe. Mammal Review 31:189-201.
Green, W. C. H. and A. Rothstein. 1991. Trade-offs between growth and reproduction in female
bison. Oecologia 86:521-527.
Guibert, B. 1997. Une nouvelle approche des populations de chevreuils en forêt : l'« Indice de
pression sur la flore ». Bulletin Technique de l'ONF 32:5-13.
Hamann, J. L., C. Bonenfant, J. Michallet, H. Holveck, F. Klein, and M. Garel. 2011. L’indice
nocturne : un indicateur des variations d’abondance des populations de cerfs. Faune
Sauvage 292:17-22.
Jorgenson, J. T., M. Festa-Bianchet, M. Lucherini, and W. D. Wishart. 1993. Effects of body
size, population density, and maternal characteristics on age at first reproduction in
bighorn ewes. Canadian Journal of Zoology 71:2509-2517.
Klein, F., D. Maillard, A. Loison, and J. M. Gaillard. 2007. Les réserves, des territoires de
référence pour la mise au point d'outils de gestion des populations. Faune Sauvage
278:52-56.
Kohn, M. H. and R. K. Wayne. 1997. Facts from feces revisited. Trends in Ecology and
Evolution 12:223-227.
Kohn, M. H., E. C. York, D. A. Kamradt, G. Haugt, R. M. Sauvajot, and R. K. Wayne. 1999.
Estimating population size by genotyping faeces. Proceedings of the Royal Society of
London B 266:657-663.
Langvatn, R., S. D. Albon, T. Burkey, and T. H. Clutton-Brock. 1996. Climate, plant phenology
and variation in age at first reproduction in a temperate herbivore. Journal of Animal
Ecology 65:653-670.
Leduc, D. and F. Klein. 2004. Origine des populations de cerfs présentes en France. Faune
Sauvage 264:27-28.
Lincoln, F. C. 1930. Calculating waterfowl abundance on the basis of banding returns. Circular
118, U.S. Department of Agriculture.
Loison, A., J.-M. Jullien, and P. Menaut. 1999a. Relationship between chamois and isard survival
and variation in global and local climate regimes: contrasting examples from the Alps and
Pyrenees. Ecological Bulletins 47:126-136.

62
Loison, A., R. Langvatn, and E. J. Solberg. 1999b. Body mass and winter mortality in red deer
calves: disentangling sex and climate effects. Ecography 22:20-30.
McCorquodale, S. M. 2001. Sex-specific bias in helicopter surveys of elk: Sightability and
dispersion effects. Journal of Wildlife Management 65:216-225.
McCullough, D. R. 1982. Evaluation of night spotlighting as a deer study technique. Journal of
Wildlife Management 46:963-973.
McCullough, D. R., D. H. Hirth, and S. J. Newhouse. 1989. Resource partitioning between sexes
in white-tailed deer. Journal of Wildlife Management 53:277-283.
Michallet, J. and T. Chevrier. 2013. Gestion des ongulés par les indicateurs de changement
écologique. Forêt Entreprise 210:15-17.
Morellet, N. 2008. La gestion des grands herbivores par les indicateurs de changement
écologique. Faune Sauvage 282:9-18.
Morellet, N., P. Ballon, Y. Boscardin, and S. Champely. 2003. A new index to measure roe deer
(Capreolus capreolus) browsing pressure on woody flora. Game and Wildlife Science
20:155-173.
Morellet, N., S. Champely, J. M. Gaillard, P. Ballon, and Y. Boscardin. 2001. The browsing
index: new tool uses browsing pressure to monitor deer populations. Wildlife Society
Bulletin 29:1243-1252.
Morellet, N., J. M. Gaillard, A. J. M. Hewison, P. Ballon, Y. Boscardin, P. Duncan, F. Klein, and
D. Maillard. 2007. Indicators of ecological change: new tools for managing populations
of large herbivores. Journal of Applied Ecology 44:634-643.
Mysterud, A., N. C. Stenseth, N. G. Yoccoz, R. Langvatn, and G. Steinheim. 2001a. Nonlinear
effects of large-scale climatic variability on wild and domestic herbivores. Nature
410:1096-1099.
Mysterud, A., N. G. Yoccoz, N. Stenseth, and R. Langvatn. 2001b. Effects of age, sex and
density on body weight of Norwegian red deer: evidence of density-dependent
senescence. Proceedings of the Royal Society of London B 268:911-919.
Pellerin, M., C. Bonenfant, J. L. Hamann, F. Klein, J. Michallet, and S. Saïd. 2013. Combiner
données cynégétiques et modélisation pour améliorer la gestion des populations de cerfs.
Rapport Scientifique ONCFS 2012:31.

63
Petersen, C. G. J. 1896. The yearly immigration of young plaice into the Limfjord from the
German sea. Report of the Danish Biological Station 6:1-48.
Pettorelli, N., S. Dray, J. M. Gaillard, D. Chessel, P. Duncan, A. Illius, N. Guillon, F. Klein, and
G. Van Laere. 2003. Spatial variation in springtime food resources influences the winter
body mass of roe deer fawns. Oecologia 137:363-369.
Pettorelli, N., J. M. Gaillard, G. Van Laere, P. Duncan, P. Kjellander, O. Liberg, D. Delorme, and
D. Maillard. 2002. Variations in adult body mass in roe deer: the effects of population
density at birth and of habitat quality. Proceedings of the Royal Society of London B
269:747-753.
Pettorelli, N., J. M. Gaillard, N. G. Yoccoz, P. Duncan, D. Maillard, D. Delorme, G. Van Laere,
and C. Toïgo. 2005. The response of fawn survival to changes in habitat quality varies
according to cohort quality and spatial scale. Journal of Animal Ecology 74:972-981.
Pollock, K. H. 1991. Modeling capture, recapture, and removal statistics for estimation of
demographic parameters for fish and wildlife population: past, present, and future. Journal
of the American Statistical Association 86:225-238.
Post, E. and N. C. Stenseth. 1998. Large-scale climatic fluctuation and population dynamics of
moose and white-tailed deer. Journal of Animal Ecology 67:537-543.
Robson, D. S. and H. A. Regier. 1964. Sample size in Petersen mark–recapture experiments.
Transactions of the American Fisheries Society 93:215-226.
Rose, K. E., T. H. Clutton-Brock, and F. E. Guinness. 1998. Cohort variation in male survival
and lifetime breeding success in red deer. Journal of Animal Ecology 67:979-986.
Ruckstuhl, K. E. and T. H. Clutton Brock. 2005. Sexual segregation and the ecology of the two
sexes. Pages 3-10 in K. E. N. Ruckstuhl, P., editor. Sexual segregation in vertebrates:
ecology of the two sexes. Cambridge University Press.
Ruckstuhl, K. E. and P. Neuhaus. 2005. Sexual segregation in vertebrates: ecology of the two
sexes. Cambridge University Press.
Sæther, B.-E. 1997. Environmental stochasticity and population dynamics of large herbivores: a
search for mechanisms. Trends in Ecology and Evolution 12:143-149.
Schwarz, C. J. and G. A. F. Seber. 1999. Estimating animal abundance: review III. Statistical
Science 14:427-456.

64
Seber, G. A. F. 1982. The estimation of animal abundance and related parameters. 2nd edition.
Griffin, London.
Stearns, S. C. 1992. The evolution of life histories. Oxford University Press, Oxford.
Strickland, B. K., S. Demarais, and P. D. Gerard. 2008. Variation in mass and lactation among
cohorts of white-tailed deer Odocoileus virginianus. Wildlife Biology 14:263-271.
Toïgo, C., J.-M. Gaillard, and J. Michallet. 1999. Cohort affects growth of males but not females
in alpine ibex (Capra ibex ibex). Journal of Mammalogy 80:1021-1027.
Toïgo, C., J. M. Gaillard, G. Van Laere, M. Hewison, and N. Morellet. 2006. How does
environmental variation influence body mass, body size, and body condition? Roe deer as
a case study. Ecography 29:301-308.
Valière, N., C. Bonenfant, C. Toïgo, G. Luikart, J. M. Gaillard, and F. Klein. 2007. Importance of
a pilot study for non-invasive genetic sampling: genotyping errors and population size
estimation in red deer. Conservation Genetics 8:69-78.
Van Horne, B. 1983. Density as a misleading indicator of habitat quality. Journal of Wildlife
Management 47:893-901.
Waits, J. L. and P. L. Leberg. 2000. Biases associated with population estimation using molecular
tagging. Animal Conservation 3:191-199.
Weladji, R., D. R. Klein, O. Holand, and A. Mysterud. 2002. Comparative response of Rangifer
tarandus and other northern ungulates to climatic variability. Rangifer 22:29-46.
White, G. C. and K. P. Burnham. 1999. Program MARK: survival estimation from populations of
marked animals. Bird Study 46 (Suppl.):120-139.
Zannèse, A., N. Morellet, C. Targhetta, A. Coulon, S. Fuser, A. J. M. Hewison, and M.
Ramanzin. 2006. Spatial structure of roe deer populations: towards defining management
units at a landscape scale. Journal of Applied Ecology 43:1087-1097.