a partir de l’exploitation des documents et de vos...
TRANSCRIPT

Devoir N°409/01/08
BIOLOGIE
Épreuve B
Durée : 3 heures 30 minutes
_________
L’usage de la calculatrice, d’abaques et de tables est interdit pour cette épreuve.
Si, au cours de l’épreuve, un candidat repère ce qui lui semble être une erreur d’énoncé, il le signale sur sa copie et poursuit sa composition en expliquant les raisons des initiatives qu’il a été amené à prendre.
A partir de l’exploitation des documents et de vos connaissances, montrez que des gènes sont impliqués dans le contrôle du fonctionnement du méristème apical caulinaire et l'organogenèse végétale. Quelques aspects de leurs interventions seront précisés.
* L’exposé sera encadré par une introduction et une conclusion et sera structuré par un plan faisant apparaître explicitement les thèmes abordés et la progression suivie.
* L’exposé doit se limiter aux trois thèmes abordés. Les trois parties sont indépendantes.
* Le candidat ne doit pas rédiger de longs développements de ses connaissances sur le sujet indépendamment de l’exploitation des documents.
* Les documents peuvent être découpés et intégrés à la copie à condition d’être exploités.
Les documents proposés portent - pour la partie I sur l'intervention d'un gène (le gène PHAN) sur la morphogenèse foliaire - pour la partie II sur les interventions de gènes dans le fonctionnement du méristème apical caulinaire et les interactions entre ces gènes- pour la partie III sur l'intervention d'ARN dans les interactions entre des gènes au cours du développement floral.
Remarque : Parmi les documents proposés certains font référence à des méristèmes floraux non étudiés pour le moment en cours. Leur exploitation ne nécessite aucune connaissance au delà des connaissances les plus simples acquises en TP.
1/14
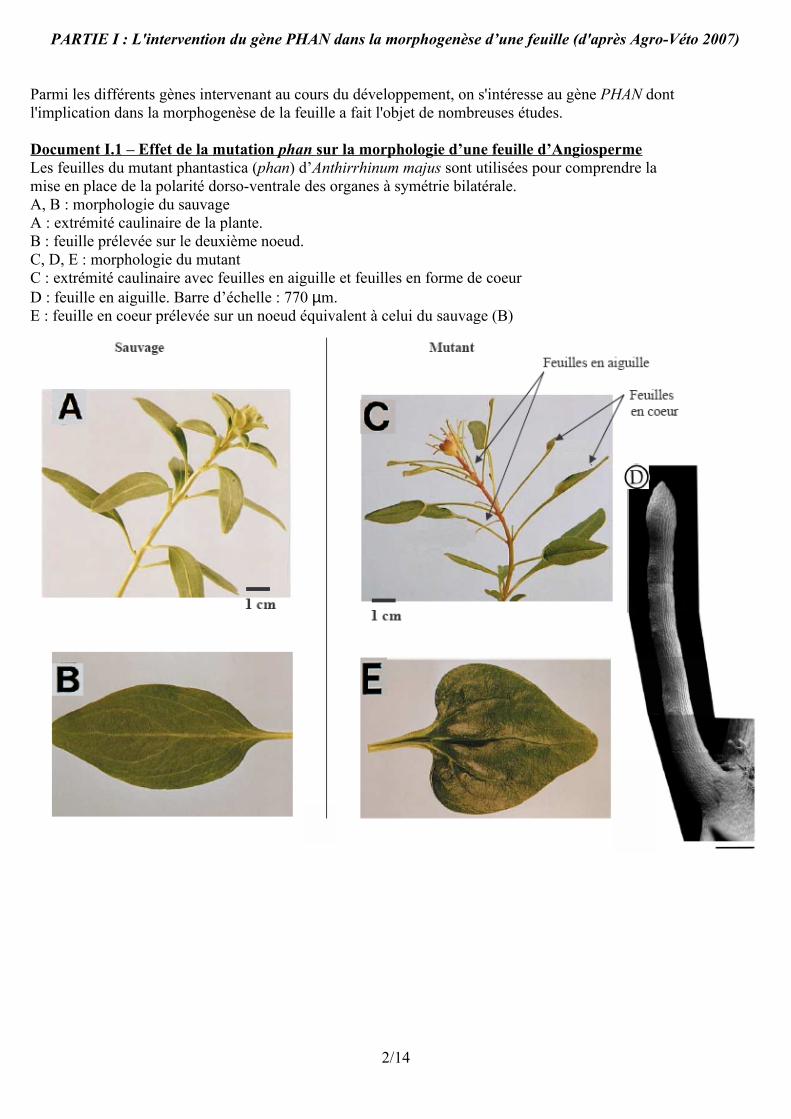
PARTIE I : L'intervention du gène PHAN dans la morphogenèse d’une feuille (d'après Agro-Véto 2007)
Parmi les différents gènes intervenant au cours du développement, on s'intéresse au gène PHAN dontl'implication dans la morphogenèse de la feuille a fait l'objet de nombreuses études.
Document I.1 – Effet de la mutation phan sur la morphologie d’une feuille d’Angiosperme Les feuilles du mutant phantastica (phan) d’Anthirrhinum majus sont utilisées pour comprendre lamise en place de la polarité dorso-ventrale des organes à symétrie bilatérale.A, B : morphologie du sauvageA : extrémité caulinaire de la plante.B : feuille prélevée sur le deuxième noeud.C, D, E : morphologie du mutantC : extrémité caulinaire avec feuilles en aiguille et feuilles en forme de coeurD : feuille en aiguille. Barre d’échelle : 770 µm.E : feuille en coeur prélevée sur un noeud équivalent à celui du sauvage (B)
2/14
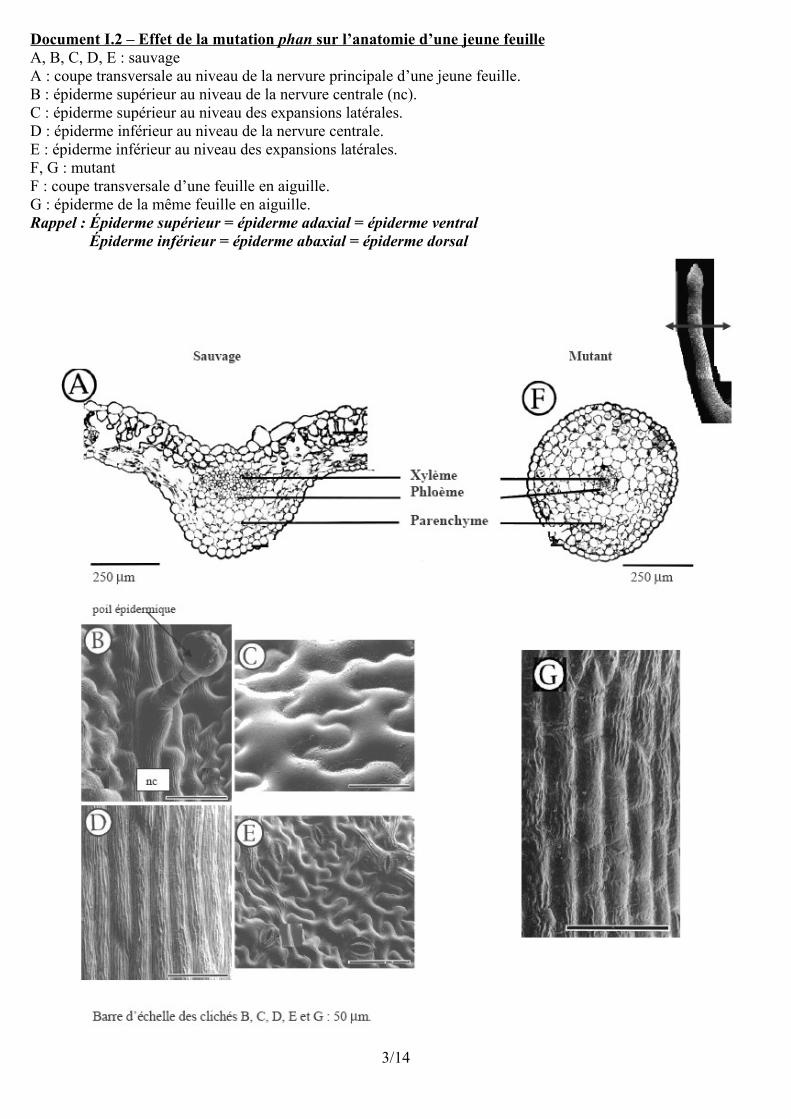
Document I.2 – Effet de la mutation phan sur l’anatomie d’une jeune feuille A, B, C, D, E : sauvageA : coupe transversale au niveau de la nervure principale d’une jeune feuille.B : épiderme supérieur au niveau de la nervure centrale (nc).C : épiderme supérieur au niveau des expansions latérales.D : épiderme inférieur au niveau de la nervure centrale.E : épiderme inférieur au niveau des expansions latérales.F, G : mutantF : coupe transversale d’une feuille en aiguille.G : épiderme de la même feuille en aiguille.Rappel : Épiderme supérieur = épiderme adaxial = épiderme ventral
Épiderme inférieur = épiderme abaxial = épiderme dorsal
3/14
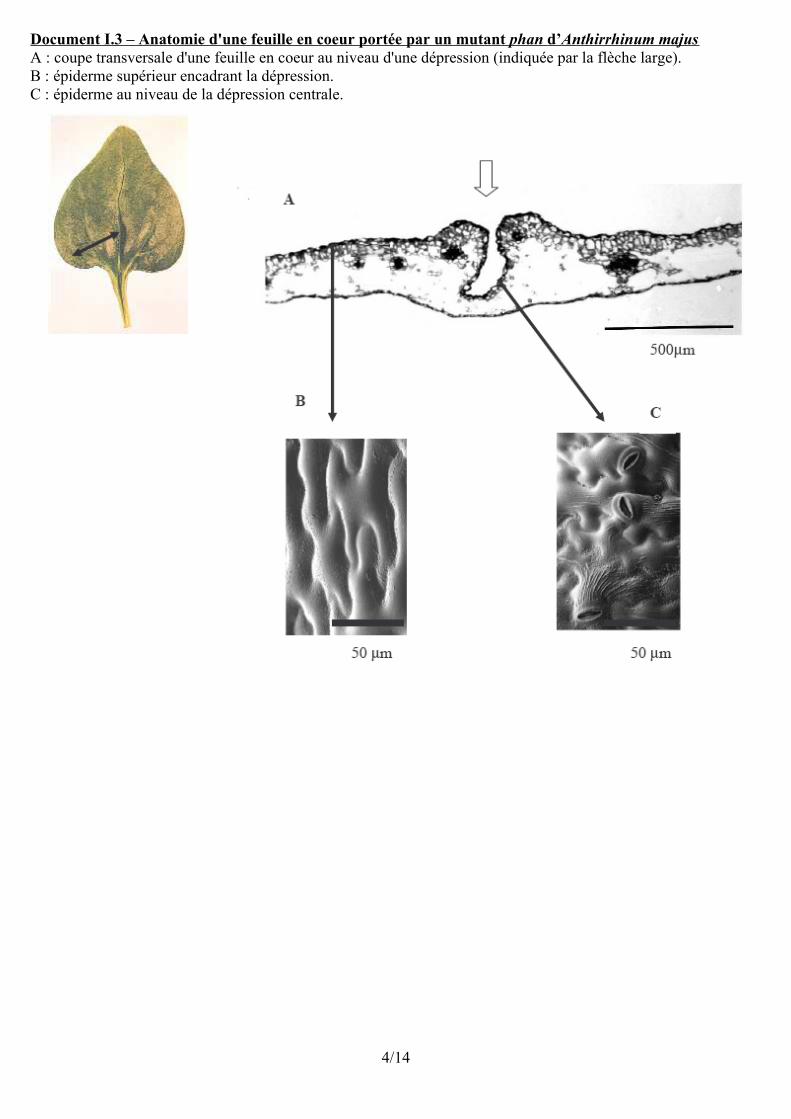
Document I.3 – Anatomie d'une feuille en coeur portée par un mutant phan d’ Anthirrhinum majus A : coupe transversale d'une feuille en coeur au niveau d'une dépression (indiquée par la flèche large).B : épiderme supérieur encadrant la dépression.C : épiderme au niveau de la dépression centrale.
4/14
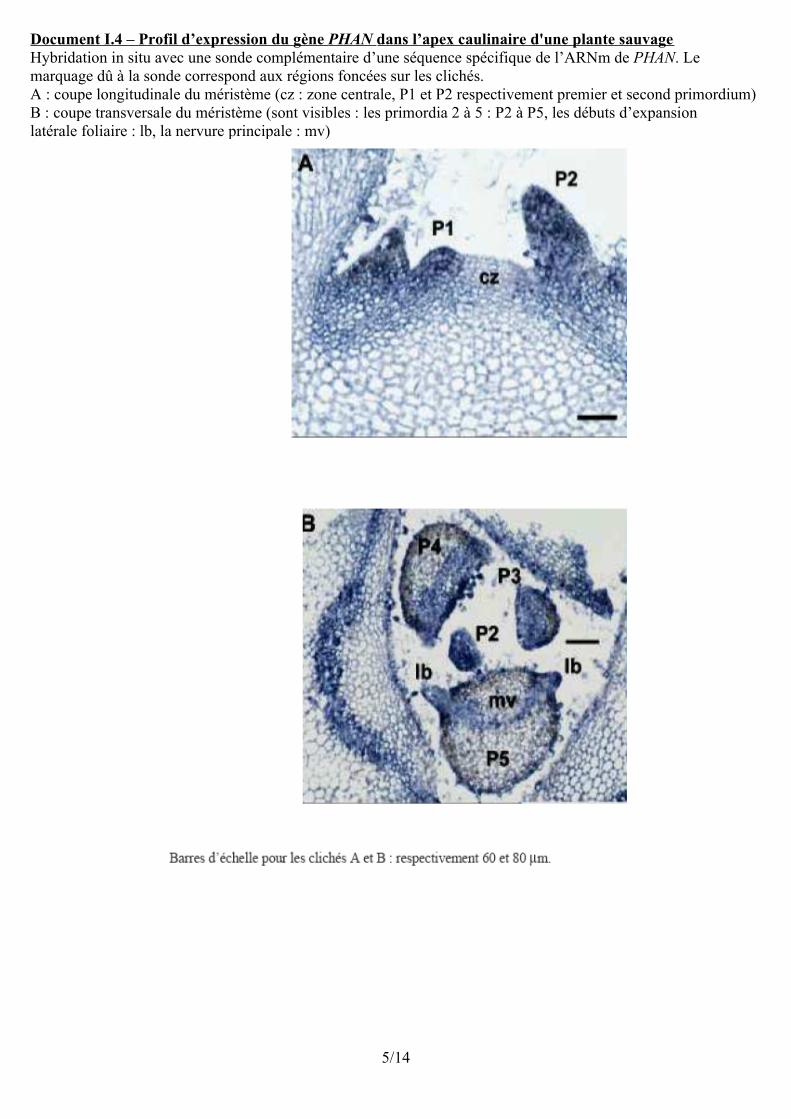
Document I.4 – Profil d’expression du gène PHAN dans l’apex caulinaire d'une plante sauvage Hybridation in situ avec une sonde complémentaire d’une séquence spécifique de l’ARNm de PHAN. Lemarquage dû à la sonde correspond aux régions foncées sur les clichés.A : coupe longitudinale du méristème (cz : zone centrale, P1 et P2 respectivement premier et second primordium)B : coupe transversale du méristème (sont visibles : les primordia 2 à 5 : P2 à P5, les débuts d’expansionlatérale foliaire : lb, la nervure principale : mv)
5/14
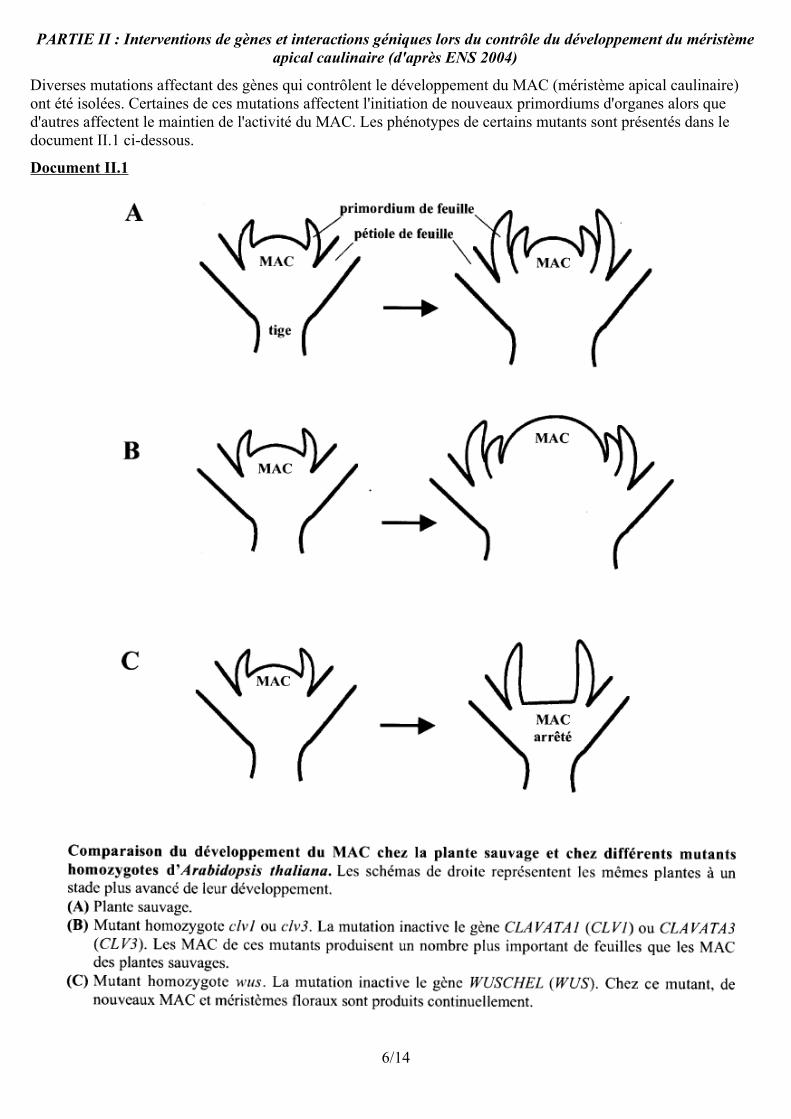
PARTIE II : Interventions de gènes et interactions géniques lors du contrôle du développement du méristème apical caulinaire (d'après ENS 2004)
Diverses mutations affectant des gènes qui contrôlent le développement du MAC (méristème apical caulinaire) ont été isolées. Certaines de ces mutations affectent l'initiation de nouveaux primordiums d'organes alors que d'autres affectent le maintien de l'activité du MAC. Les phénotypes de certains mutants sont présentés dans le document II.1 ci-dessous.
Document II.1
6/14

Afin de mieux comprendre le rôle de CLV3 au sein du MAC, des mesures de l'index mitotique ont été réalisées dans différents domaines de la couche L1 sur des MAC provenant d'une population sauvage d'Arabidopsis thaliana (tableau 1) et sur des méristèmes provenant de plantes mutantes clv3 (tableau 2).Les domaines sont définis sur le document II-2.Document II-2
7/14

Afin de mettre en évidence d'éventuelles interactions entre les gènes WUS et CLV dans le méristème sauvage, les phénotypes de doubles mutants ont été analysés et comparés aux phénotypes des simples mutants. De telles analyses permettent de dégager des relations d'épistasie entre gènes. Un gène est dit épistatique par rapport à un autre lorsque le produit d'expression du premier exerce un effet activateur ou inhibiteur sur l'expression du second. Pour cela, différentes lignées de mutants homozygotes wus, clv1 et clv3 présentant tous des allèles non fonctionnels, ont été croisées et les phénotypes des fleurs des doubles et simples mutants analysés. Les résultats sont présentés dans le tableau 3
Afin de préciser la fonction du gènes WUS, des plantes transgéniques (PANT-WUS) surexprimant le gène WUS ont été réalisées. Au bout de deux semaines de développement, le phénotype du MAC des plantes transgéniques a été analysé et les ARNm du gène CLV3 ont été détectés par hybridation in situ. Les résultats sont présentés sur le document II-3.
8/14

Afin de poursuivre la détermination des interactions entre les gènes WUS et CLV, l'expression des ARNm du gène WUS a été analysé dans les méristèmes floraux chez les mutants clv1 et clv3, par une technique d'hybridation in situ.Ces profils d'expression sont présentés dans ce document II-4 ci-dessous.
Par ailleurs des plantes transgéniques (P35S-CLV3) exprimant de façon continue et anormalement importante le gène CLV3 ont été obtenues. Ces plantes présentent un MAC qui initie la formation de premières feuilles, mais qui s'arrête précocement de fonctionner. Enfin, l'expression du gène WUS a été analysée dans les MAC de ces plantes. Les résultats sont présentés dans le document II-5
Le transgène P35S-CLV3 a également été introduit dans le mutant clv1 d'Arabidopsis thaliana. Les plantes transgéniques obtenues expriment une quantité élevée d'ARNm CLV3 et présentent le même phénotype que les mutants clv1.
9/14

PARTIE III : L'intervention d'ARN dans les interactions géniques au cours du développement floral (d'après ENS 2007)
Le ‘RNA silencing’ correspond à l’extinction de l’expression génique via des interactions séquence-spécifiques impliquant de petites molécules d’ARN simple-brin de 21 à 26 nucléotides. Découvert récemment chez les plantes, ce processus affecte différentes étapes de l’expression du génome selon des mécanismes conservés. Il peut être induit notamment par des ARN non codants exprimés par le génome, appelés microARN.Au cours du développement floral d’Arabidopsis thaliana, l’identité des pièces florales est déterminée par des gènes homéotiques dont la mutation conduit au remplacement de certains organes par d’autres.Parmi ces gènes, les gènes de classe A sont impliqués dans la spécification de l’identité des pièces du périanthe (sépales et pétales) et les gènes de classe C dans la spécification de l’identité des pièces fertiles (étamines et carpelles). Les mutations apetala2 et agamous, inactivant les gènes APETALA2 (AP2) et AGAMOUS (AG) qui codent respectivement les protéines AP2 et AG, ont été isolées. Les phénotypes des fleurs mutantes sont donnés à la Figure III-1.
Chez les plantes apetala2, la protéine AG s’exprime dans les ébauches de toutes les pièces florales alors qu’elle n’est présente que dans celles des pièces fertiles des plantes sauvages. Chez les plantes agamous, la protéine AP2 s’exprime dans les ébauches de toutes les pièces florales, mais elle n’est présente que dans celles des pièces du périanthe des plantes sauvages.
10/14
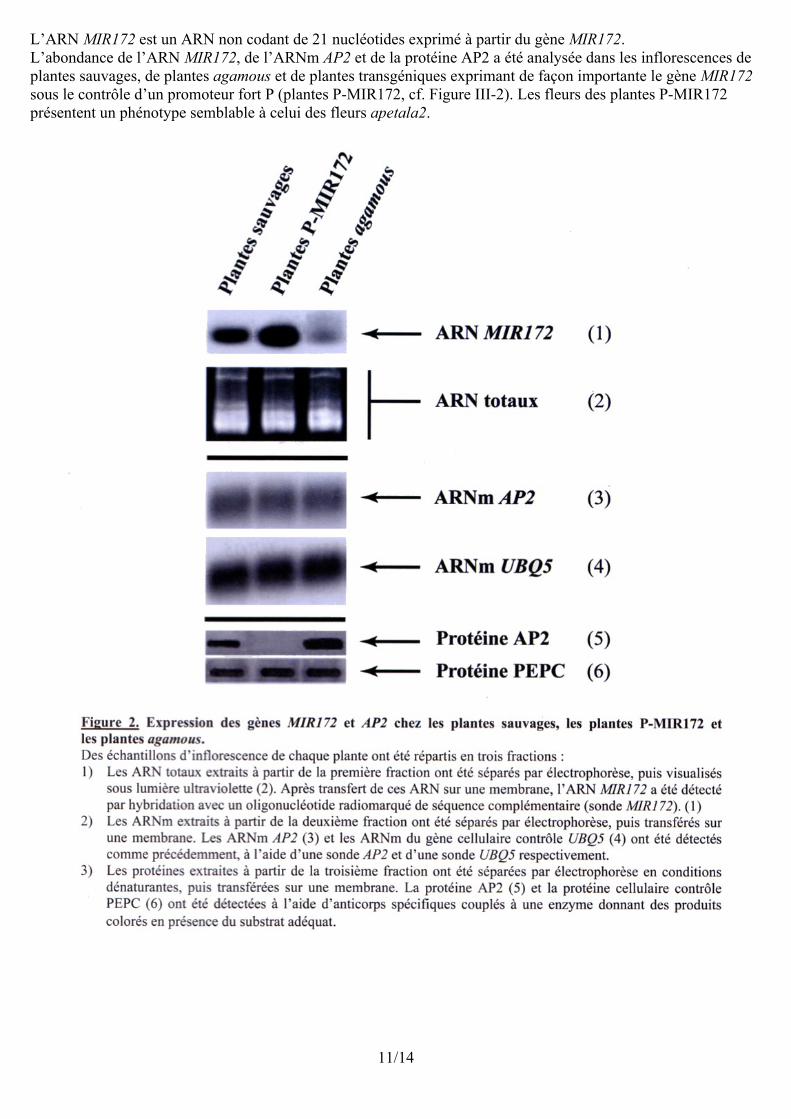
L’ARN MIR172 est un ARN non codant de 21 nucléotides exprimé à partir du gène MIR172.L’abondance de l’ARN MIR172, de l’ARNm AP2 et de la protéine AP2 a été analysée dans les inflorescences de plantes sauvages, de plantes agamous et de plantes transgéniques exprimant de façon importante le gène MIR172 sous le contrôle d’un promoteur fort P (plantes P-MIR172, cf. Figure III-2). Les fleurs des plantes P-MIR172 présentent un phénotype semblable à celui des fleurs apetala2.
11/14
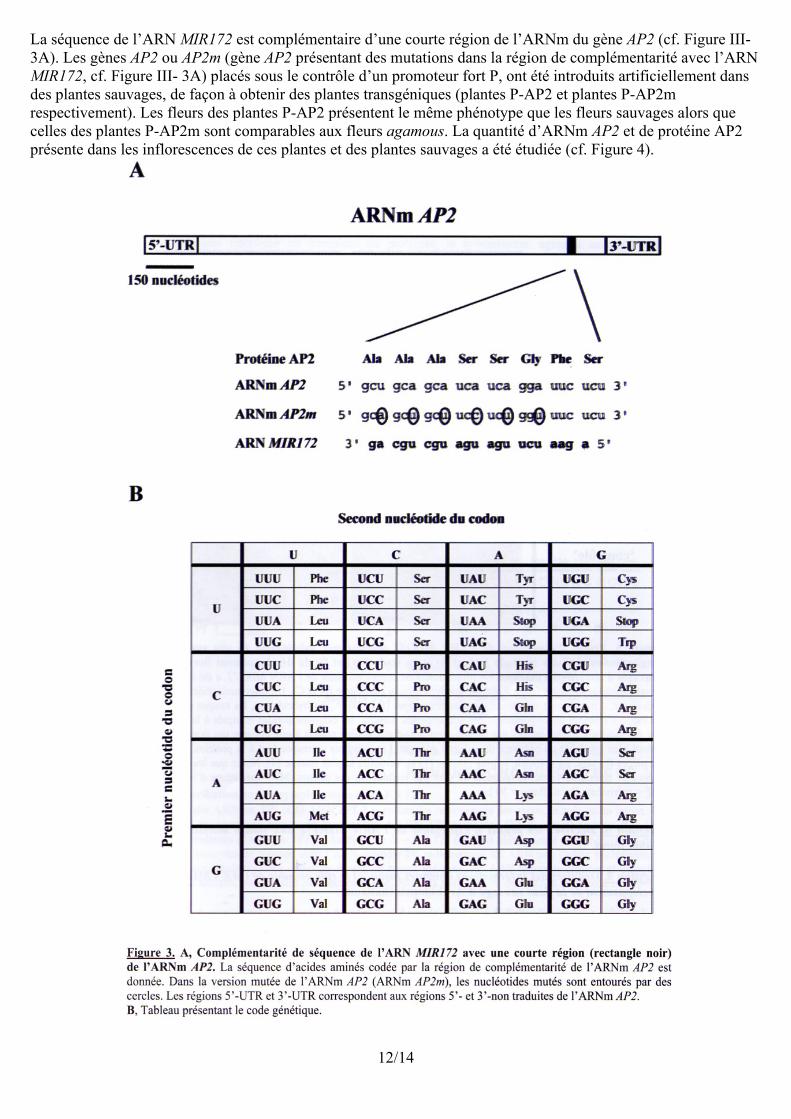
La séquence de l’ARN MIR172 est complémentaire d’une courte région de l’ARNm du gène AP2 (cf. Figure III-3A). Les gènes AP2 ou AP2m (gène AP2 présentant des mutations dans la région de complémentarité avec l’ARN MIR172, cf. Figure III- 3A) placés sous le contrôle d’un promoteur fort P, ont été introduits artificiellement dans des plantes sauvages, de façon à obtenir des plantes transgéniques (plantes P-AP2 et plantes P-AP2m respectivement). Les fleurs des plantes P-AP2 présentent le même phénotype que les fleurs sauvages alors que celles des plantes P-AP2m sont comparables aux fleurs agamous. La quantité d’ARNm AP2 et de protéine AP2 présente dans les inflorescences de ces plantes et des plantes sauvages a été étudiée (cf. Figure 4).
12/14

13/14
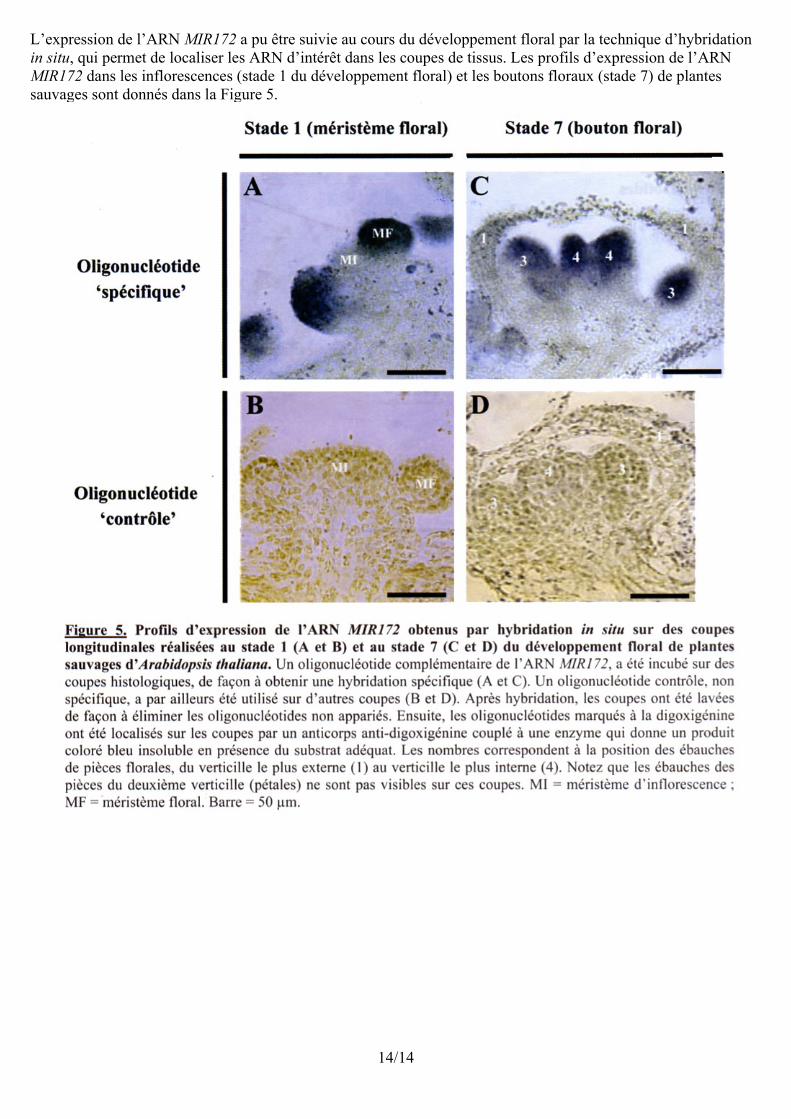
L’expression de l’ARN MIR172 a pu être suivie au cours du développement floral par la technique d’hybridation in situ, qui permet de localiser les ARN d’intérêt dans les coupes de tissus. Les profils d’expression de l’ARN MIR172 dans les inflorescences (stade 1 du développement floral) et les boutons floraux (stade 7) de plantes sauvages sont donnés dans la Figure 5.
14/14

Corrigé du devoir n°4 du mercredi 9 janvier 2008Épreuve de type B : Implication des gènes dans le contrôle du fonctionnement du MAC et la morphogenèse Sur 80
Introduction : (on n'écrit pas ce qui entre parenthèse mais on construit trois petits paragraphes)(Approche générale du sujet :) La formation d'organes ou organogenèse est un phénomène continu au cours de la vie de la plante. Elle s'initie au niveau des méristèmes apicaux, zones de mérèse situées aux extrémités des tiges et des racines, et notamment au niveau du méristème apical caulinaire (MAC). S'y mettent place deux catégories d'organes végétatifs, tige et feuille, et les différentes pièces florales lors de sa transformation en méristème floral.(Problématique :) Grâce à l'étude de la morphologie et de l'anatomie de mutants, à des profils d'expression géniques, et des analyses quantitatives de différentes molécules, sera mise en évidence et précisée l'implication de gènes, appartenant à la catégorie des gènes homéotiques, dans le contrôle du fonctionnement du méristème apical caulinaire et la morphogenèse végétale.(Présentation des thèmes :) Dans le premier thème, on montrera comment un gène intervient dans l'un des aspects de la morphogénèse de la feuille, puis dans un deuxième thème, comment le fonctionnement du MAC est contrôlé par des gènes qui interagissent entre eux puis, dans une troisième thème, un type particulier d'interaction entre gènes homéotiques faisant intervenir un ARN sera mis en évidence.
**
**
**I. L'intervention du gène PHAN dans la morphogenèse d’une feuille
A. Gène PHAN et morphogenèse de la feuille Les feuilles d'Anthirrhinum majus sont des feuilles à limbe ovale, à nervation pennée.Celles des mutants phan sont, soit en forme de coeur à nervation pédalée avec un pétiole beaucoup plus long, soit en forme d'aiguille. Dans ce dernier cas les feuilles sont beaucoup plus courtes (de l'ordre de 1cm alors que les autres feuilles mesurent de l'ordre de 5cm).Conclusion : la mutation phan se traduit donc par l'absence de mise en place du limbe de la feuille ou par la mise en place d'un limbe réduit. Le gène PHAN joue donc un rôle indispensable dans la morphogenèse foliaire : mise en place du limbe caractéristique de l'espèce. Cette mise en place du limbe passe notamment par le maintien d'une activité méristématique en bordure de l'ébauche foliaire. On peut supposer que le gène PHAN contribue à ce maintien.
B. Gène PHAN et histogenèse au sein de la feuilleComparaison des anatomies (tableau envisageable)
**
**
+
Feuille de l'individu sauvage Feuille de l'individu muté phan- Symétrie bilatérale- xylème et phloème superposé- parenchyme palissadique vers la face supérieureparenchyme lacuneux en dessous- épiderme supérieur et épiderme inférieur- épiderme à cellules allongées au niveau de la nervurepoils épidermiques au niveau de la face sup de la nervureépid. à cel. en forme de pièces de puzzle au niveau du limbestomates nombreux au niveau de l'épiderme inférieur
Symétrie axialexylème central et phloème périphériqueparenchyme homogène (+ ou moins lacuneux)
un seul type d'épidermeépiderme à cellules allongées tout autour(sans poils épidermiques ?)
**
*
**
Conclusion : La feuille des mutants a une organisation tissulaire voisine de celle de la nervure de la feuille des individus sauvages. Il n'y a pas de polarité dorso-ventrale dans la disposition des tissus. Le gène PHAN joue donc un rôle indispensable dans la détermination des types de différenciation cellulaire à l'origine de la polarité dorso-ventrale de la feuille. **
C. Gène PHAN et polarité de l'épidermeDans le cas d'une feuille en coeur une polarité dorso-ventrale est perceptible au niveau anatomique : parenchyme palissadique du côté supérieur et lacuneux en dessous.Au niveau du limbe l'épiderme supérieur est conforme à celui d'une feuille normale (cellules en forme de pièces de puzzle) sans stomates.Au niveau de dépressions présentes sur la face sup., l'épid. est de type « limbe inférieur » avec des stomates.Conclusion : Au niveau des feuilles en coeur, la polarisation dorso-ventrale des tissus est imparfaite.Ceci confirme le rôle du gène PHAN dans la déterm. des types de diff. cell à l'origine de la polarité dorso-ventrale.
**
**D. Localisation de l'expression du gène PHAN
L'hybridation in situ permet grâce au repérage des ARNm de PHAN la déterm. des zones où ce gène s'exprime.La coupe A révèle une expression limitée aux cellules des primordiums foliaires.La coupe transversale montre une expression dans l'ensemble des primordiums 2 et 3 puis réduite aux zones périphériques (début d'expansions latérales et futures zones épidermiques) pour les primordiums 4 et 5. Conclusion : le gène PHAN s'exprime très précocement dans le primordium foliaire. Il s'exprime alors dans l'ensemble du primordium puis son expression se restreint aux zones méristématiques marginales à l'origine du limbe, et aux épidermes. On peut supposer que, sous son action, ces zones acquièrent leur polarité dorso-ventrale.
**
**
15/14

II. Interventions de gènes et interactions géniques lors du contrôle du développement du méristème apical caulinaire
A. Gènes CLV et WUS et développement du MACLes mutants clv ayant un MAC hypertrophié, on peut en déduire que ces gènes exercent un rôle inhibiteur sur le développement du MAC.Le mutant wus ayant un MAC dont l'activité s'arrête, on peut en déduire que ce gène exerce un rôle indispensable dans le maintien de l'activité du MAC.D'autres MAC et des méristème floraux se mettant en place, le gène WUS peut être mis en cause dans l'inhibition de la mise en place d'autres méristèmes.
B. Gène CLV3 et activité mitotique au sein du MACChez les mutants clv3 la zone médiane caractérisées par un index mitotique de l'ordre de 1,8% assimilable à la zone centrale occupe une proportion plus importante du méristème car son rayon est égal à 0,5R au lieu de 0,25R.Les index mitotiques mesurés par ailleurs ne présentent pas de différences significatives.Conclusion : On en déduit que le gène CLV3 exerce son action inhibitrice sur le développement du MAC en stimulant la transformation des « cellules de la zone centrale » en « cellules de la zone périphérique ».
C. Interactions entre gènes CLV et gènes WUS1. Action de WUS sur CLV
- Chez les mutants wus le nombre d'organes floraux initiés ne varie pas en ce qui concerne les sépales et les pétales et est réduit en ce qui concerne les pièces fertiles. On peut en déduire que WUS exerce une action augmentant le nombre de pièces fertiles.- Chez les mutants clv, toutes les pièces florales augmentent en nombre. On peut donc supposer que les gènes CLV exercent une action réduisant le nombre de pièces florales.- Les doubles mutants wus.clv présentent les mêmes résultats que les mutants simples wus chez lesquels on pourrait s'attendre à observer moins de pièces florales.Conclusion : Si lorsque le gène WUS est muté, on observe une absence d'effet des mutations clv, c'est que les gènes CLV sont déjà neutralisésLe gène WUS parait donc nécessaire à l'expression des gènes CLV. Le gène WUS exerce donc un effet épistatique activateur sur les gènes CLV.(ou sur l'un avec répercussion sur l'autre)- Chez les plantes surexprimant WUS, on constate un domaine d'expression de CLV3 bien plus important que chez les plantes sauvages.Conclusion : Ceci confirme l'effet activateur de WUS sur l'expression du gènes CLV3.
2. Action de CLV sur WUS Les mutants clv1 et clv3 expriment le gène WUS sur une zone plus grande. Conclusion : On en déduit que les gènes CLV1 et CLV3 exercent un effet épistatique inhibiteur sur l'expression du gène WUS.Chez l'individu transgénique surexprimant CLV3, on observe la disparition de la zone d'expression de WUS.Ceci confirme la conclusion précédente.
3. Action de CLV3 sur CLV1 Les effets similaires des mutations des 2 gènes suggèrent une intervention en chaîne des 2 gènes.Il n'y pas de différence entre le phénotype des simples mutants clv1 et des doubles mutants P35S-CLV3. La mutation clv1 se traduit par l'absence d'effet de la surexpression de CLV3. Conclusion : C'est donc CLV3 qui agit sur CLV1 (et non l'inverse) et CLV3 exerce un effet épistatique positif sur CLV1
Bilan : WUS exerce effet + sur CLV3 qui exerce effet + sur CLV1 qui exerce effet – sur WUS
*
*
+
***
**
**
**
*
**
**
*
****
*
**
+
16/14

III. L'intervention d'ARN dans les interactions géniques au cours du développement floralA. Des gènes impliqués dans l'identité des pièces florales
- Chez les mutants apetala2, il n'y a ni sépales, ni pétales, il manque des étamines mais il y a des carpelles surnuméraires.Chez les mutants agamous, il n'y a pas de pièces fertiles (ni étamines, ni carpelles) mais des pétales surnuméraires.Conclusion : Les gènes AP2 et AG sont des gènes impliqués dans l'identité des pièces florales. Le gène AP2 est impliqués dans l'identité des pièces du périanthe. C'est un gène de classe A.Le gène AG est impliqué dans l'identité des pièces fertiles. C'est un gène de classe C.
B. Des interactions entre ces gènesLa mutation apetala2 entraîne une expression d'AGAMOUS dans toutes les pièces florales et la mutation agamous entraîne l'expression d'APETALA dans toutes les pièces florales alors que normalement ces gènes ne s'expriment que dans la catégorie d'organes qu'ils spécifient.Conclusion : Le gène APETALA2 inhibe l'expression du gène AGAMOUS et réciproquement.
C. Une interaction entre gènes exercée par un ARNLa comparaison des quantités d'ARN et de protéines chez les plantes agamous et les plantes sauvages montre - que les quantités d'ARNm AP2 sont identiques mais que la protéine AP2 est plus abondante chez les plantes agamous- une quantité plus faible d'ARN MIR172 chez les plantes agamous que chez les plantes sauvages.- aucun changement concernant les ARN totaux, les ARNm et protéines témoins.Chez les plantes surexprimant MIR172, corrélativement à l'abondance de l'ARN MIR172, la quantité d'ARNmAP2 est identique aux plantes précédentes mais la protéine AP2 est absente.Conclusion : L'action du gène AGAMOUS sur l'expression du gène APETALA s'effectue par l'intermédiaire de l'ARN MIR172 qui exerce une action inhibitrice sur la traduction de l'ARNm AP2 en protéine AP2. Il s'agit d'un « RNA silencing »
D. Une action de l'ARN silencing sur l'ARNmLa redondance du code génétique fait que l'ARNm AP2m conduit à la même protéine que l'ARNm AP2.L'absence de complémentarité entre l'ARNm AP2 pour les plantes P-AP2m a pour conséquence une synthèse plus active de protéine AP2 (environ deux fois plus abondante).Conclusion : C'est la fixation de l'ARN silencing MIR172 sur l'ARNm AP2 qui inhibe sa traduction en protéine(division par 2 de la traduction)
E. Une expression de l'ARN silencing variable au sein du méristème florall'hybridation in situ permet de localiser l'ARN MIR 172 dans l'ensemble des méristèmes floraux à un stade précoce. Plus tard lors de la formation des verticilles floraux, on ne le localise plus que dans les pièces des verticilles à l'origine des pièces fertiles. Conclusion : Dans les ébauches de pièces fertiles, le gène AGAMOUS s'exprime et active la production d'ARN MIR 172 qui en retour inhibe la production de protéine AP2 à partir du gène APETALAUn autre processus similaire doit intervenir dans l'inhibition par le gène APETALA de la synthèse de protéine AG.
Conclusion bilan : L'ensemble de cette étude montre que chaque gène homéotique s'exprime à un moment déterminé de l'organogenèse et dans une zone déterminée du méristème ou de l'ébauche d'organe. Ces gènes déterminent, pour certains l'identité de l'organe, pour d'autres permettent l'acquisition de ses caractéristiques morphologiques et anatomiques. Des interactions positives ou négatives ont lieu entre ces gènes. Si des études récentes évoquées dans le thème III montrent que ces interactions peuvent faire intervenir des ARN, il existe aussi d'autres mécanismes connus depuis plus longtemps. En effet par exemple, dans le cas des interactions entre les gènes CLAVATA, une protéine issue de l'expression d'un gène est le ligand de celle issue de l'expression de l'autre gène. PlanRigueur du raisonnementPrésentation des documents, orthographe, rédaction, clarté et soin
**
**
**
***
*
**
*
*
**
**
**+
***********
17/14