ue8 - de l’agent infectieux à l’hôte j.j ... -...
TRANSCRIPT

UE8 - De l’agent infectieux à l’hôte J.J. Hoarau
Date : 10/02/2016 Plage horaire : 8h30-10h30Promo : P2 2015/2016 Enseignant : JJHRonéoiste : CELESTIN Kevin
La cellule bactérienne - Physiologie bactérienne
I. Métabolisme (généralités) 1. Besoins nutritifs non carbonés 2. Absorption des nutriments
A. Diffusion passive B. Diffusion facilitée C. Transport actif
3. L’assimilation des métaux (exemple du fer) 4. Les facteurs environnementaux
A. La pression osmotique (les solutés et la disponibilité en eau) B. Effet du pH C. Effet de la température D. Métabolisme respiratoire E. La pression (atmosphérique et hydrostatique) F. Les radiations (UV, X, γ)
II. Conditions d’analyse et conditions environnementales 1. Croissance in vitro
A. Les milieux de culture B. La croissance bactérienne C. Courbe de croissance D. Mathématiques de la croissance
2. Croissance in vivo : « la vraie vie » A. Les biofilms B. Coopération et conflits chez P. Aeruginosa (qui fait du quorum sensing)
III. Identification 1. Utilisation du métabolisme pour l’identification bactérienne 2. Identification moléculaire
A. Par PCR B. Par sondes fluorescentes C. Par électrophorèse D. Par séquençage
3. Spectrométrie de masse

I. Métabolisme (généralités)
Savoir de quoi a besoin une bactérie revient à savoir de quoi elle est constituée. Composition des bactéries (en termes de % de poids sec) :
Macroéléments (95% du poids sec des bactéries = constituants présents en grande quantité dans les bactéries)
- C, O, H, N, S, P (les 6 principaux) : Constituants de base des glucides, lipides, protéines et des acides nucléiques
- K, Ca, Mg, Fe : Aussi requis en grande quantité car indispensables au métabolisme (exemple : cofacteurs enzymatiques)
Ces macroéléments sont indispensables aux bactéries et vont énormément impacter leur développement. Ils seront un facteur limitant de ce dernier s’ils ne sont pas présents dans le milieu.
Oligo-éléments ou microéléments (5% du poids sec des bactéries) : Mn, Zn, Co, Mb, Ni, Cu.
Ils sont tout aussi importants pour le développement bactérien et indispensables pour leur métabolisme, mais en très petite quantité. La seule grande différence c’est que ces oligo-éléments sont présents uniquement à l’état de trace dans le milieu. Nécessaires qu’en très petite quantité, les bactéries n'auront donc aucun problème à en trouver dans leur milieu. De ce fait, ce ne sont pas des facteurs limitants de leur développement.
Les bactéries ont besoin de ces éléments pour construire les nouveaux constituants cellulaires et pour en tirer des sources d’énergie. Ce qui a amené à classifier les bactéries en fonction de leur besoins trophiques selon 3 critères :
- La nature de la source de carbone utilisée pour leur développement Bactéries autotrophes Source minérale (exemple : CO2) Bactéries hétérotrophes Source organique (exemple : glucose)
- Les sources d’énergie qu’elles vont utiliser Bactéries chimiotrophes Source chimique (d’origine organique généralement)
Le CO2 ne pouvant pas constituer une source d’énergie, très souvent, l’autre possibilité de source d’énergie est lumineuse.
Bactéries phototrophes Source lumineuse (cyanobactérie notamment)
- La source du pouvoir réducteur Bactéries lithotrophes Source minérale (souvent inorganique, réduite, provenant notamment de
roches) Bactéries organotrophes Source de molécules organiques

Toutes les combinaisons existent. Néanmoins, la majeure partie des bactéries font parties du groupe deschimio-organo-hétéro-trophes et où retrouve notamment, les bactéries pathogènes humaines et animales.
1. Besoins nutritifs non carbonés
Azote (indispensable pour la synthèse des acides aminés, les glucides, les lipides, les cofacteurs enzymatiques…), puisé dans le milieu sous forme :
- Acide aminé, ammoniaque (souvent la forme minérale), nitrate, N2 (azote atmosphérique dans certains cas).
Phosphore (essentiel pour les acides nucléiques, phospholipides, ATP) :- Puisé dans le milieu extérieur, sous forme organique qui va être hydrolysé dans le périplasme et être
importé dans la cellule sous forme de phosphate inorganique Toujours du Pi qui est transféré dans la cellule.
Soufre (certains acides aminés et glucides), puisé dans le milieu extérieur sous forme de :- Sulfate réduit (principalement)- Cystéine (notamment quand il provient d'autres organismes vivants).
Sels : Sodium, Potassium, Magnésium, puisés dans le milieu extérieur.
Métaux : Fer, Zinc, Manganèse, puisés sous la forme très souvent d'ions dans le milieu extérieur.
Facteurs de croissance (constituants nécessaires, non synthétisés par certaines bactéries par incapacité enzymatique) doivent être puisés dans le milieu extérieur sous forme de :
- Certains acides aminés (Exemple : la leucine)- Certaines bases azotées : purines et pyrimidines,- Les vitamines (acide folique, biotine, B1, B2, B6, B12…).
2. Absorption des nutriments
Ces mécanismes d’absorption se font toujours de façon sélective au niveau de la membrane plasmique de la cellule. Ils seront énormément influencés par les gradients de concentration. Les 3 principaux connus sont la diffusion passive, la diffusion facilitée et le transport actif.

A. Diffusion passive
Elle dépend essentiellement du gradient de concentration entre le milieu extracellulaire et le cytoplasme. Il ne nécessite pas d’énergie et ne pourra concerner que de très petites molécules (eau, O2, CO2).
B. Diffusion facilitée
Elle accroit la diffusion sélective de solutés à de faibles concentrations en faisant appel à des transporteurs membranaires (exemple : perméases). La source d’énergie est uniquement liée aux gradients de concentration.
C. Transport actif
Il se justifie lors du transport de grosses molécules. Nous verrons 3 systèmes (les mieux décrits mais pas les seuls).
Transporteurs ABC (ATP binding cassette transporteurs)
Ces transporteurs transmembranaires formant un pore, possèdent 2 domaines de liaisons à l’ATP au niveau cytoplasmique. Il nécessite l’utilisation de protéines fixatrices de soluté (il en existe des différentes en fonction du soluté qu’elles sont capables de fixer) qu’on va retrouver dans l’espace périplasmique (GRAM-) ou attachées à la face externe de la membrane cytoplasmique (GRAM+). Une fois le soluté fixé, elles vont venir se mettre en contact avec la face externe du transporteur membranaire et utiliser l’énergie de l’ATP pour permettre la translocation du soluté dans le cytoplasme. C’est un mécanisme qui nécessite un coup important.
Système symport/antiport
L’énergie utilisée n’est pas sous forme d’ATP. Elle est puisée grâce à des pompes H+ au niveau de la membrane cytoplasmique, initiant le mécanisme en transférant les H+ du cytoplasme vers le milieu extérieur (par couplage avec des électrons) créant ainsi un gradient de H+ (extracellulaire) permettant ensuite d’exporter du Na+ du cytoplasme vers le milieu extérieur grâce à un antiport, créant un gradient de Na+ (extracellulaire).
Le Na+ exporté va se fixer sur des symports (transporteurs membranaires) induisant un changement conformationel leur permettant de fixer un soluté (spécifique d’un symport).
Une fois le Na+ et le soluté fixés sur le symport, celui- ci va subir un changement conformationel transférant aussi bien le Na+ que le soluté sera dans la cellule.
L’énergie est essentiellement fournie par un mécanisme de pompe à proton.

Translocation de groupe (système PTS)
C’est un mécanisme qui utilise une énergie chimique qui va être libérée en métabolisant la molécule au cours de son transfert. Les sucres à importer (mannitol, glucose…), vont être modifiés et l’énergie de ces modifications va permettre la translocation de ces molécules dans la cellule.Il en existe plusieurs mais le plus connu est le système PTS (Phospho-enol-pyruvate-dependant Transport System).
C’est un système phospho-relais (à phospho-transférase) qui utilise comme point de départ, l’énergie d’une liaison phosphate puisée à partir du PEP. Au départ on part du PEP intra-cytoplasmique, qui va transférer l’énergie d’une liaison phosphate vers une 1ère enzyme, libérant du pyruvate ainsi qu’une enzyme phosphorylée.
Cette énergie va être transférée successivement de protéines en protéines. On peut avoir des phosphorylations multiples jusqu’à arriver au transporteur membranaire qui va alors phosphorylé le composé qui doit être importé.
Ce système PTS n’est pas systématiquement retrouvé chez toutes les bactéries. On le retrouve chez E. Coli, Salmonella, Staphylococcus, chez de nombreuses bactéries anaérobies facultatives et quelques anaérobies obligatoires (clostridium).
La majorité des bactéries aérobies ne possèdent pas de système PTS et ont des systèmes alternatifs à la place. Une exception : Les bactéries de type Bacillus.
4. L’assimilation des métaux (exemple du fer)
Le fer qui est retrouvé par les bactéries dans le milieu environnant est du fer ferrique (Fe3+) et elles ont besoin de fer ferreux (Fe2+).Le Fe3+, insoluble dans le milieu extérieur, a besoin d’être solubilisé pour être importé dans la cellule. Les cellules vont le solubiliser en utilisant des sidérophores (molécules sécrétées) qui vont se complexer avec les Fe3+ et des récepteurs de sidérophores vont reconnaître le complexe (sidérophore + fer ferreux). Chez les bactéries GRAM- ces récepteurs auxsidérophores se trouvent au niveau de la membrane externe permettant le transfert de tout le complexe dans le périplasme.
Il existe des canaux qui permettent ensuite soit de libérer directement le Fe2+ et permettre son transport direct dans le cytoplasme de la bactérie (souvent GRAM +) ou alors il existe des canaux au niveau de la membrane cytoplasmique permettant le transfert (consommant de l’énergie sous forme d’ATP) de l’ensemble du complexe (Exemple : transporteur de type ABC). Une fois dans la cellule, le complexe va se dissocier et le Fe3+ se transformera en Fe2+.

Un sidérophore solubilise les ions fer en formant des complexes qui peuvent être utilisés par des mécanismes de transport actif.
5. Les facteurs environnementaux
Ils vont influencer la croissance bactérienne, qu’ils soient chimiques ou physiques.
A. La pression osmotique (les solutés et la disponibilité en eau)
La nature du soluté dans lequel évoluent les bactéries et notamment la pression osmotique va conditionner indirectement la disponibilité en eau du milieu (principal constituant), en raison de la membrane plasmique à perméabilité sélective.
Le cytoplasme des bactéries est toujours maintenu à une pression osmotique légèrement supérieure à celle du milieu extérieur pour permettre à la membrane cytoplasmique de rester fermement collée à la paroi de la cellule, évitant un phénomène d’implosion. Les bactéries ont besoin et peuvent contrôler en permanence la pression osmotique de leur cytoplasme mais aussi potentiellement du milieu environnant en faisait intervenir :
- Corps d’inclusions, présents dans le cytoplasme, stockent des sources d’énergie mais aussi différents constituants tels que des acides aminés ou des ions qui vont pouvoir être relâchés dans le milieu extérieur pour contrôler cette pression osmotique.
- Vacuoles pulsatives
- Solutés compatibles : composés qui peuvent être stockés dans les corps d’inclusions, qui lorsqu’ils sont stockés dans la cellule en forte concentration n’ont pas d’impact sur le métabolisme cellulaire. Une libération dans le milieu extérieur de ces solutés compatibles sera possible pour agir sur la pression osmotique du milieu environnant :
o Choline, bétaïne, acide aminé,o K (limité),o Sucres : saccharose et polyols, qu'elle va soit absorber, soit libérer dans le milieu,o …
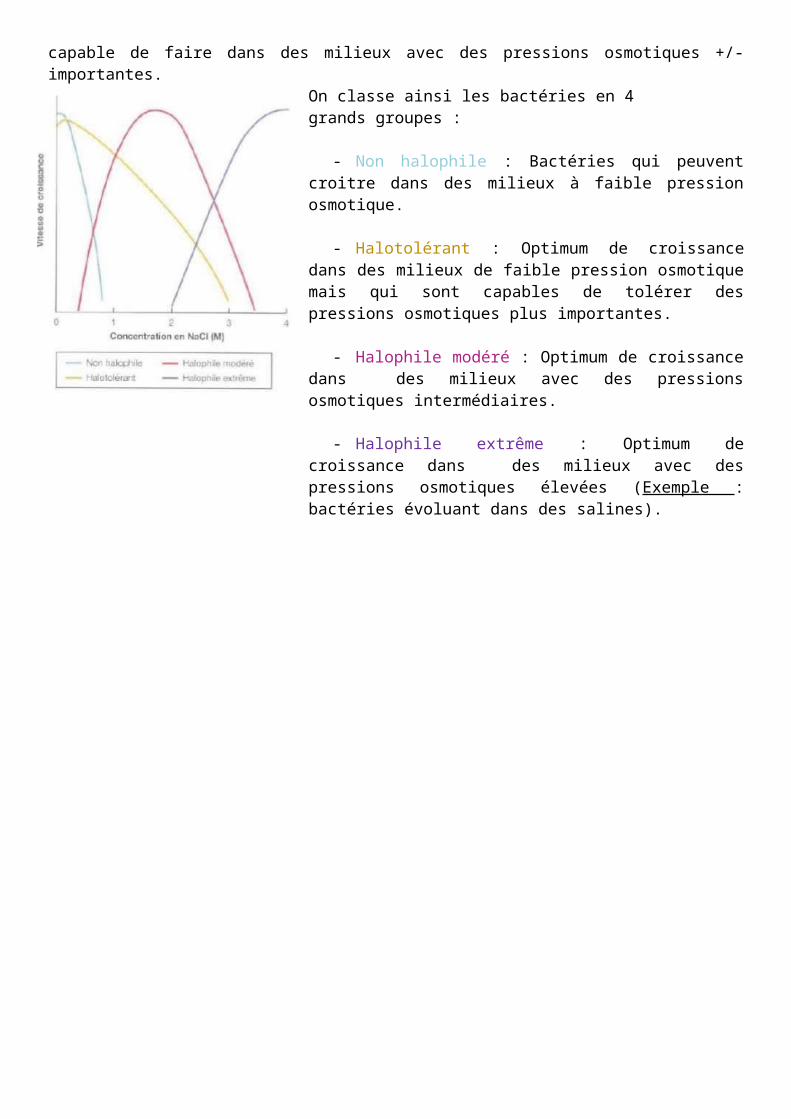
Toutes les bactéries ne sont pas capables de vivre dans des milieux avec des pressions osmotiques identiques et on peut les classer en fonction de leur capacité à résister à la pression du milieu extérieur. Chaque bactérie possède une capacité à croitre en fonction de ce qu’elle est capable de faire dans des milieux avec des pressions osmotiques +/- importantes.
On classe ainsi les bactéries en 4 grands groupes :
- Non halophile : Bactéries qui peuvent croitre dans des milieux à faible pression osmotique.
- Halotolérant : Optimum de croissance dans des milieux de faible pression osmotique mais qui sont capables de tolérer des pressions osmotiques plus importantes.
- Halophile modéré : Optimum de croissance dans des milieux avec des pressions osmotiques intermédiaires.
- Halophile extrême : Optimum de croissance dans des milieux avec des pressions osmotiques élevées (Exemple : bactéries évoluant dans des salines).

Il faut savoir qu’à tous les types bactériens, correspond une pression osmotique optimale où la croissance bactérienne est maximale.
Ne pas prendre le schéma au sens strict (exemple : il existe d’autre sel que le NaCl).
B. Effet du pH
L’acidité du milieu va également impacter énormément la croissance bactérienne. Ici on parle du pH du milieu extérieur. Le pH des bactéries (quel que soit le type bactérien) au niveau du cytoplasme est toujours neutre (≈ 7)
pH = -log [H+] = log (1/ [H+])
En fonction de la capacité des bactéries à croître dans un milieu +/- acide, ou +/- basique, on va les classer en :
- Acidophiles : 0 < pH < 5,5- Neutrophiles (la majorité des cas) : 5,5 < pH < 8 ; Exemple : E. Coli- Alcalophiles : 8 < pH < 11,5 ; Exemple : Pseudomonas Vibrio- Alcalophiles extrêmes : pH > 11,5
La bactérie va devoir maintenir un pH neutre vis-à-vis du milieu extérieur, notamment via des mécanismes permettant l’importation ou l’exportation de H+ :
- Système symport/antiport (par l'échange K+/Na+ il y a équilibration du milieu en H+ ; dans les 2 sens)
- ATPases (transporteurs de protons permettant de les rejeter à l’extérieur de la cellule)
- Exportation des déchets métaboliques nécessaire : Le métabolisme bactérien en interne ayant tendance à acidifier son cytoplasme.
- « Protéines chaperonnes », qui dans les conditions extrêmes où le pH cytoplasmique devient limite, empêchent la dénaturation des protéines en les protégeant momentanément.
Remarque et exemple : Une bactérie neutrophile a une croissance optimale lorsque le pH est autour de 7 ; cependant, elle pourra continuer à croitre dans un pH un peu plus acide ou basique mais moins rapidement.
C. Effet de la température
Les bactéries n'ont pas le moyen de réguler leur température, elles vont donc évoluer à la température du milieu extérieur. Elles ont la capacité à persister dans certains milieux en fonction de la température. Le gros effet de la température c’est qu’elle impacte les réactions enzymatiques.
- Si les températures sont inférieures à la température optimale de fonctionnement des enzymes, il va y avoir une diminution du métabolisme.
- Plus on va monter en température, plus on risque d’inactiver ces enzymes et notamment les dénaturés et oriente alors les bactéries vers une mort cellulaire.

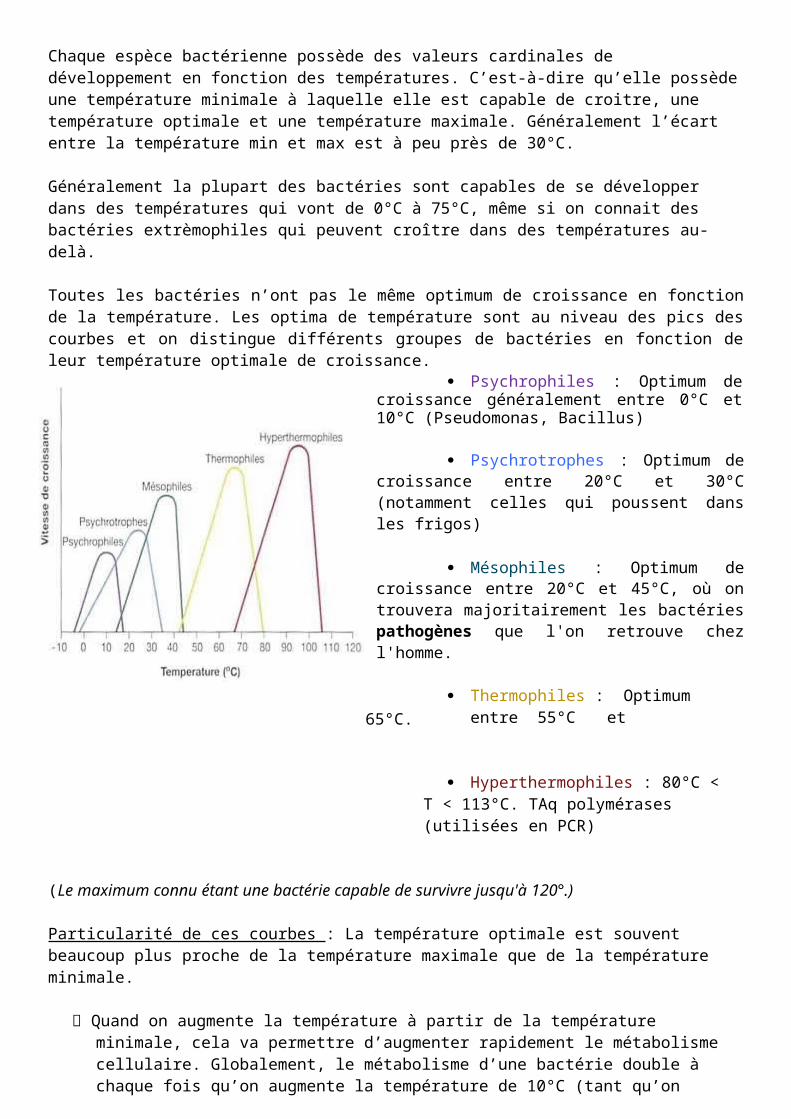
Chaque espèce bactérienne possède des valeurs cardinales de développement en fonction des températures. C’est-à-dire qu’elle possède une température minimale à laquelle elle est capable de croitre, une température optimale et une température maximale. Généralement l’écart entre la température min et max est à peu près de 30°C.
Généralement la plupart des bactéries sont capables de se développer dans des températures qui vont de 0°C à 75°C, même si on connait des bactéries extrèmophiles qui peuvent croître dans des températures au-delà.
Toutes les bactéries n’ont pas le même optimum de croissance en fonction de la température. Les optima de température sont au niveau des pics des courbes et on distingue différents groupes de bactéries en fonction de leur température optimale de croissance.
Psychrophiles : Optimum de croissance généralement entre 0°C et 10°C (Pseudomonas, Bacillus)
Psychrotrophes : Optimum de croissance entre 20°C et 30°C (notamment celles qui poussent dans les frigos)
Mésophiles : Optimum de croissance entre 20°C et 45°C, où on trouvera majoritairement les bactéries pathogènes que l'on retrouve chez l'homme.
65°C. Thermophiles : Optimum entre 55°C et
Hyperthermophiles : 80°C < T < 113°C. TAq polymérases (utilisées en PCR)
(Le maximum connu étant une bactérie capable de survivre jusqu'à 120°.)
Particularité de ces courbes : La température optimale est souvent beaucoup plus proche de la température maximale que de la température minimale.
Quand on augmente la température à partir de la température minimale, cela va permettre d’augmenter rapidement le métabolisme cellulaire. Globalement, le métabolisme d’une bactérie double à chaque fois qu’on augmente la température de 10°C (tant qu’on reste dans les gammes de température qu’une bactérie est capable d’accepter).
Par contre une fois qu’on dépasse cette température, très rapidement le métabolisme bactérien va diminuer et les bactéries vont mourir.
Cela serait une des explications à la fièvre, quand on fait une fièvre post infection bactérienne, il n’y a pas besoin de monter très haut en température, quelques degrés d’augmentation suffisent à abolir le métabolisme bactérien et donc à impacter la croissance bactérienne.
Rq : Les bactéries thermophiles et hyperthermophiles ont généralement un métabolisme qui met en place des mécanismes de stabilisations au niveau des protéines. Elles vont généralement présenter au niveau moléculaire, d’avantages de liaisons covalentes, de liaisons hydrogènes…
Rq : Les courbes donnent une idée générale. Si on fouille, on peut trouver une bactérie avec un optimum de croissance à 50°C par exemple.

D. Métabolisme respiratoire
Un autre paramètre qui influence beaucoup les bactéries c’est le métabolisme respiratoire. C’est particulièrement important car on utilise l’identification du métabolisme respiratoire pour faire de l’identification bactérienne.
Dans la chaine respiratoire des bactéries, l’O2 peut être ou non un accepteur d’électron. Tout dépend si les bactéries l’utilisent pour conserver l’énergie fournie par les différentes sources d’énergie. Comme l’O2 est un accepteur final d’électron, il va être réduit et produire des espèces oxygénées réactives qui sont extrêmement agressives pour les différents constituants cellulaires (O2-, H2O2). Les bactéries l’utilisant doivent donc posséder tout un arsenal enzymatique pour détoxifier. Parmi ces enzymes on va retrouver la SOD (super-oxyde dismutase) et la catalase.
Des bactéries capables de croitre et d’utiliser l’oxygène vont principalement disposer de ces enzymes alors que les bactériens n’en disposant pas y seront extrêmement sensibles et celui-ci sera toxique. Généralement les bactéries anaérobies ne possèdent pas ces enzymes.
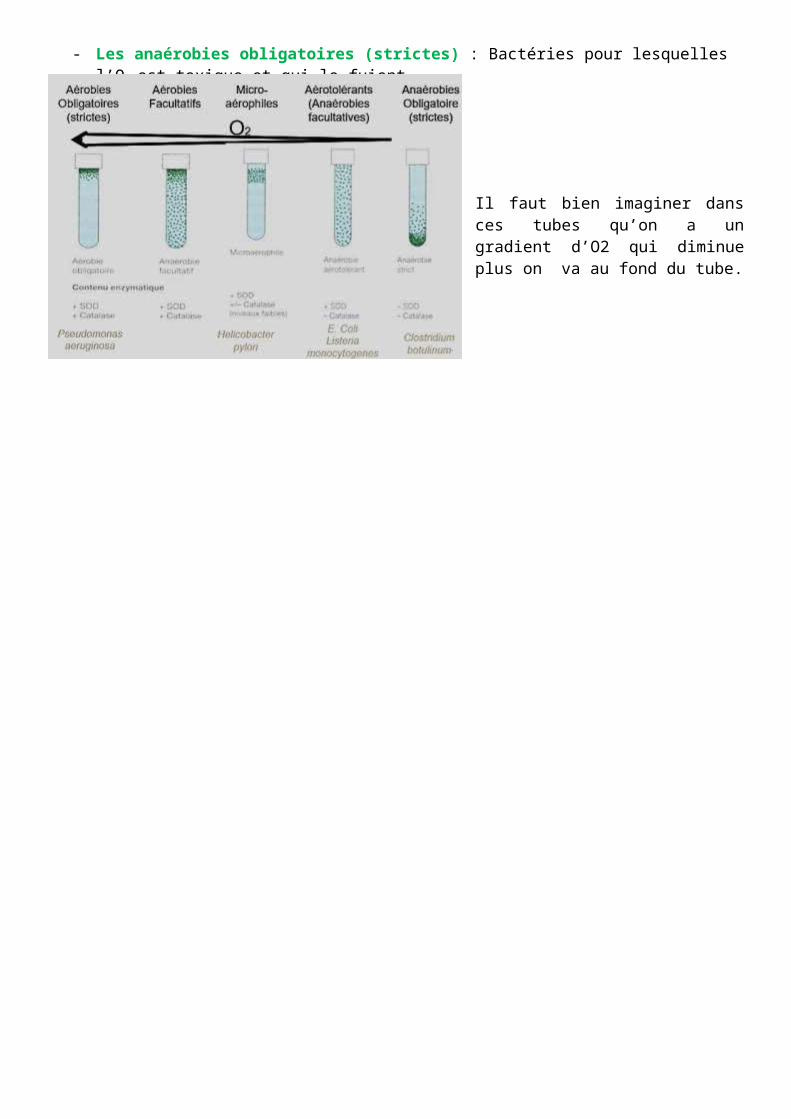
On va donc trouver des gammes de bactérie en fonction des enzymes dont elles disposent et effectuer un classement :
- Aérobie obligatoire (strictes) : Bactéries ayant un besoin absolue d’O2 pour leur métabolisme et possédant ces 2 enzymes en grande quantité (Exemple : Pseudomonas)
- Aérobies facultatives : Bactéries qui ont besoin d’O2 mais qui ont la capacité à croitre même en son absence. Néanmoins quand elles sont en présence d’O2 elles vont l’utiliser principalement comme accepteur final d’électron. Lors de test en laboratoire, on va cultiver (ensemencer) des bactéries dans des milieux liquides ou semi-liquides et constater qu’elles poussent en surface mais également dans le milieu.
- Les micro-aérophiles (cas particulier) : Bactéries qui possèdent généralement au moins la SOD mais qui parfois ne disposent pas de la catalase. Généralement elles sont capables de croître en présence d’O2 mais là où on a une concentration plus réduite (elles poussent légèrement en retrait).
Les bactéries qui n’ont pas besoin d’O2 comme accepteur final d’électron (utilisant le souffre ou autre à la place) :
- Les aérotolérants (anaérobies facultatives) : Bactéries qui disposent néanmoins des enzymes de détoxification leur permettant de croitre en présence d’O2 même si elles n’en ont pas besoin.
- Les anaérobies obligatoires (strictes) : Bactéries pour lesquelles l’O2 est toxique et qui le fuient.
Il faut bien imaginer dans ces tubes qu’on a un gradient d’O2 qui diminue plus on va au fond du tube.

Rq : Dans le cadre d’E. Coli qu’on étudiera en TP, il faut retenir que c’est une bactérie aérotolérante (anaérobies facultatives). Lors des manipulations, on assurera une anaérobie en recouvrant l’échantillon avec de l’huile de paraffine qui va chasser l’air et empêcher le contact avec l’O2.
E. La pression (atmosphérique et hydrostatique)
Généralement pour les bactéries qui poussent à la surface du sol, la pression atmosphérique est de 1 atm et la majeure partie des bactéries sont calibrées pour fonctionner au niveau de cette pression atmosphérique, Toujours est-il qu’il existe des bactéries capables de vivre dans des milieux extrêmes avec des pressions bien supérieures :
- Les barotolérants qui généralement vont se développer avec un optimum à une pression de 1 atm mais qui sont capables d’accepter des pressions supérieures
- Les barophiles, au fond des océans et qui ne poussent qu’à des pressions élevées (600-1000 atm).
F. Les radiations (UV, X, γ)
Faibles doses = mutation ADN / Fortes doses = létales
Classiquement, les méthodes de stérilisation les plus simple sont les UV (exposition de matériel à des lampes UV), mais généralement les bactéries disposent de mécanismes leur permettant de résister à des doses d’UV limitées. Par contre, rayonnement X et gamma à forte dose, pas de bactérie capable de survivre très longtemps.
Utilité pour la stérilisation (inactivation des spores…)
II. Conditions d’analyse et conditions environnementales
1. Croissance in vitro
Pour faire du diagnostic, on a besoin d’isoler les bactéries, de les ensemencer et de les cultiver. On ne sait pas le faire pour toutes les bactéries, mais on dispose d’un certain nombre de milieu de culture qui permettent d’assurer le développement d’une large gamme de bactéries.
A. Les milieux de culture
Lorsque l’on veut cultiver des bactéries, il faut leur apporter les conditions idéales de croissance en termes notamment de nutriments. Ainsi pour faire de la croissance in vitro on va utiliser des « milieux de cultures ». Lors de l’élaboration d’un milieu de culture, on tient compte de trois paramètres :
1) La composition chimique (où on va leur apporter tous les éléments nécessaires) Milieux définis purement synthétique dont on connait parfaitement la composition : Pour des
espèces bactériennes qui sont bien caractérisées.
Milieux complexes dont la composition n’est pas exactement connue : Pour les bactéries dont on ne connait pas exactement les besoins.
2) Le support de cultures Milieux liquides, généralement ces cultures se font en flacons (tube à essai qu’on appelle
flasque).

Milieux solides ou semi solides (boite de pétri)
3) Le type fonctionnel (en fonction des exigences de bactéries et de ce que l’on veut faire) : Milieux de base (peu d’exigences)
Milieux enrichis : besoins particuliers (exemple : bactéries incapables de synthétiser certains acides aminés, qu’on va donc leur apporter si on veut les sélectionner)
Milieux sélectifs ou différentiels : Pour sélectionner ou favoriser une bactérie par rapport à une autre.
1) Composition chimique des milieux
Les milieux synthétiques ou définis sont de composition connue qualitativement et quantitativement correspondant à l’optimum de croissance de la bactérie que l’on veut cultiver.
- Liquide (bouillon) ou solide
- Source de carbone :CO2 (sous forme de Na2CO3, …),Glucose (élément nécessaire à la
croissance)
- Source d’azote (Nitrate, ammonium)
- Sulfate, phosphate
- Minéraux
- Milieu tamponné (pH ajusté)
Adapté aux microorganismes font les besoins sont parfaitement définis
Les milieux complexes (composants indéfinis), riches et complets, adaptés aux microorganismes dont les besoins nutritifs sont inconnus :
- Peptones (hydrolysats de protéines obtenu par digestion partielle de viande, caséine, soja, gélatine et autre protéines)
- Extraits de viande
- Extraits de levure
Ces extraits et ces hydrolysats ont toujours la même fonction : source de carbone, d’énergie, d’azote, d’acides aminés, de facteurs de croissance, de vitamines, de minéraux.
Généralement, qu’on fait une mise en culture d’une bactérie, on le teste toujours sur un milieu synthétique. Si les résultats ne sont pas satisfaisants, on bascule sur des milieux complexes.
Qu’on utilise des milieux définis ou complexes, il existe des bactéries qui ne poussent pas car elles ont vraiment des besoins très particuliers.
2) Nature physique des milieux
Lorsqu’on travaille avec des bactéries de type anaérobie, on préfère travailler en milieu liquide. Lorsqu’on travaille avec des bactéries de type aérobie on travaille sur un milieu solide.
Le passage de l’état liquide à solide est obtenu en ajoutant de l’agar (ou du gel de silice dans le cas des bactéries autotrophes). Ce sont des composés qui restent neutre (les bactéries ne vont pas les utilisées lors de leur croissance). Le composé le plus couramment utilisé reste l’agar, à une concentration de 1% à 2%.
3) Les types fonctionnels
Pour certain type bactérien particulièrement exigeant, on peut utiliser des milieux fonctionnels (en fonction de ce que l’on veut faire) :
- Milieu de base = utilité générale (exemple : bouillon ou gélose de soja)
- Milieu enrichi (pour les organismes exigeants : ajout d’aliments spéciaux aux milieux de base)
- Milieu sélectif (Exemple : ajout de fuscine basique ou cristal violet Favoriser les GRAM- au GRAM +)
- Milieu différentiel (Pour distinguer différents groupes bactériens) Exemple : La gélose au sangOn va rajouter au milieu des hématies pour visualiser les bactéries hémolytiques. Les plages de lyses indiquent qu’il y a des bactéries qui poussent.

Exemple : la gélose en chocolatRajout de sang mais cuit qui donne cet aspect chocolat. La cuisson des hématies entraine la libération de facteurs de croissance indispensable à la croissance des bactéries.
B. La croissance bactérienne
Elle se fait par scissiparité c’est-à-dire qu’à partir d’une bactérie (à matériel génétique cyclique), on en obtient deux.
En condition optimale de croissance, les bactéries sont quasiment constamment en situation de division cellulaire.
Celle-ci commence toujours par un allongement de la cellule qui s’étire. Puis, la réplication du génome démarre au niveau de l’ORI qui est unique et qui se termine à l’autre extrémité (origine de terminaison).
Généralement, le chromosome bactérien se positionne de façon à ce que chacune des extrémités (origine de réplication et origine de terminaison) se retrouvent à des extrémités opposées de la cellule.
Une fois la réplication terminée, un septum va se mettre en place qui va progresser et permettre de scinder la cellule en deux conduisant à la division cellulaire.
Le mécanisme de croissance bactérienne peut être divisé en 3 étapes : la réplication, la répartition et la cytocinèse.

La réplication thêta : 1 fourche de réplication avance dans les 2 sens de sorte que les ORI puissent évoluer et aller à chaque extrémité de la cellule.- 1 chromosome circulaire- 1 origine de réplication- 1 site de terminaison
La répartition : mécanisme actifFait intervenir les protéines MreB du cytosquelette pour pouvoir répartir les chromosomes bactériens.
La cytocinèse : partition de la cellule en 2- Sélection la région de formation du septum. (position centrale séparation équitable de la cellule
en 2 mais pas dans le cas de la sporulation ou la séparation est asymétrique)
- Assemblage d’un anneau Z (protéines FtsZ) lié à la membrane plasmique et venant pincer le cytoplasme, permettant ensuite l’assemblage d’une machinerie permettant de synthétiser la paroi.
o Constriction de l’anneau Z entrainant une invagination de la membrane cellulaire puis synthèse de la paroi du septum
- Scission de la cellule en 2 cellules filles.
Ces 3 étapes sont +/- longues en fonction des bactéries auquelles on a à faire. Généralement, en milieu naturel, E. Coli a une croissance relativement lente (≈ 1h avec réplication ≈ 40 min, mise en place du septum+ cytocinèse ≈ 20 min). Cultivée en milieu synthétique dans des conditions optimales fait état d’une croissance beaucoup plus rapide (1 cycle ≈ 20min). E. Coli n’attend pas que son chromosome soit entièrement répliqué pour entamer une nouvelle réplication.
Souvent on peut avoir 2, 3 voire même 4 complexes réplicatifs qui se mettent en pace simultanément, Ce qui fait que lorsqu’une cellule se divise et qu’on a 2 cellules filles, ces cellules filles sont déjà en train de répliquer leur génome. C’est la raison pour laquelle on peut trouver plus d’un chromosome bactérien dans la cellule même si ce chromosome n’est pas forcément complet.
Le doublement de la population cellulaire peut se faire toutes les 20 minutes, ce qui est extrêmemnt rapide.
C. Courbe de croissance
Quel que soit néanmoins la bactérie, lorsqu’on regarde sa courbe de croissance, dans un milieu qui ne contient qu’une seule source nutritive (lien avec les cas de dioxie : quand il y a plusieurs sources nutritives on peut avoir des courbes bi-phasiques) on a toujours une courbe de croissance comme celle-ci-dessous et qui est divisée en 6 phases.
Phase 1 : phase de latence : la croissance est nulle, adaptation des bactéries aux ressources (production et activation des enzymes).
Phase 2 : phase d’accélération avec augmentation de la vitesse de croissance.
Phase 3 : phase de croissance exponentielle durant laquelle le taux de croissance est maximal (doublement du nombre de bactéries à chaque génération).
Phase 4 : phase de ralentissement où la vitesse de croissance diminue. Epuisement des nutriments du milieu de culture et accumulation des déchets. En milieu artificiel comme en milieu naturel, devient facteur limitant. Sporulation, développement du pouvoir pathogène ou de la virulence.
Phase 5 : phase stationnaire : arrêt de la multiplication bactérienne due à un facteur limitant dans l’environnement, le taux de croissance est nul taux de division = taux d’autolyse.Rq : généralement, en milieu in vitro, on rentre dans la phase stationnaire lorsqu’on arrive à saturation (densité cellulaire ≈ 109 cellules/ml, saturation entrainant un épuisement du milieu).
Phase 6 : phase de déclin (sénescence et mort) : les ressources sont épuisées et le nombre de bactéries diminue.
Rq : il ne faut pas s’imaginer qu’elle est toujours rapide. Elle peut durer de quelques jours, à quelque mois, à quelques années car au sein de la colonie bactérienne, les cellules qui meurent vont être utilisées par les autres. Même s’il n’y a plus de croissance, certaines bactéries vont pouvoir survivre.
D. Mathématiques de la croissance
Quand on fait de la culture, en fonction de la quantité de bactéries qu’on ensemence, si on veut suivre la croissance bactérienne, il faut bien savoir à quel moment elles vont être en phase exponentielle de croissance (seul moment où on peut faire des mathématiques) pour pouvoir obtenir les quantités de bactéries dont on a besoin.
En phase exponentielle de croissance : Chaque bactérie se divise à intervalle de temps constant
La population double à intervalle de temps régulier Temps de génération ou temps de doublement
Au bout de n génération, l’augmentation de la population est de 2n
Nt = N0 x 2n
Le développement de la population est exponentiel.Si on passe en échelle logarithmique, on va obtenir une linéarité qui permet de déterminer le temps de génération.
2. Croissance in vivo : « la vraie vie »
In-vitro, des conditions optimales de croissance sont artificiellement fournies.
Dans la vraie vie, le milieu naturel est limitant :- Pauvre en nutriment (milieu oligotrophe) ou qui vont s’épuiser rapidement- Complexe (plusieurs sources de facteurs de croissance et de nutriments différents avec des
gradients chevauchants)- En changement perpétuel (variation de pH, disponibilité en eau…)
Globalement, plusieurs études ont été mise en place pour essayer de comprendre, de façon synthétique et simple, les facteurs qui influencent de façon naturelle ou même artificiel le développement bactérien et finalement il y a 2 grands éléments qui vont limiter la croissance bactérienne qui sont :
1) La disponibilité du milieu en nutriment2) La tolérance aux conditions environnementales

Ces dépendances ont donné lieu à 2 lois :
Loi du minimum de Liebig : « La biomasse totale d’un organisme sera déterminée par l’élément nutritif présent à la concentration la plus faible par rapport aux exigences de l’organisme. »
Dans un milieu, pour savoir ce qui va limiter la croissance bactérienne, il faut regarder le réactif limitant c’est-à-dire celui qui est en plus petite quantité et ceci même si les autres réactifs sont en très grande concentration.
Loi de tolérance de Shelford : « Il existe des limites dans les facteurs environnementaux au-dessus et au- dessous desquelles un organisme ne peut survivre et se développer quel que soit l’apport en nutriment »
On a vu que certaines bactéries ont besoin d’un pH précis ou encore la température. Si on les écarte de cette gamme de développement optimum, on peut leur donner des nutriments ces dernières ne pousseront pas.
En milieu naturel, le développement bactérien consommera très vite les nutriments et libérera des déchets toxiques qui limiteront sa croissance.
Les bactéries devront s’adapter à ce milieu, optimiser leur croissance et établir des stratégies de survie. L’une de ces stratégies de survie est la formation de biofilm.
A. Les biofilms
Dès 1940, observation de microorganismes se développant au sein de communautés complexes enveloppées dans un mucus appelées biofilms (ressemble à des couches muqueuses, MEC), omniprésent dans la nature (surface des pierres, eau…).
Voici un biofilm de Staphylocoque doré à la surface d’un pacemaker. Il s’est formé une couche homogène
Ici, des colonies bactériennes (chaque boule est un ensemble de bactérie) qui peuvent regrouper des bactéries d’origines différentes, à l’intérieur desquelles elles vont coopérer, établir des communications pour permettre de s’échanger des nutriments, des gènes.

Différentes stratégies sont étudiées pour lutter contre ces biofilms car ces matrices filtrent les échanges entre ces communautés bactériennes et le milieu extérieur (antibiotiques, blocage du passage des cellules phagocytaires (macrophage, PNN…)). C’est un moyen de protection qu’utilisent les bactéries.
Inconvénient :- Ils rendent les bactéries résistantes aux UV, peu perméables aux biocides et protègent donc les bactéries.- Ils permettent aussi d’assurer l’alimentation correcte de l’ensemble des bactéries qui s’y trouvent (transport de O2, de nutriments…) ce qui fait que même les bactéries du centre ont une nutrition correcte. Non seulement il protège, mais il participe au développement de la communauté bactérienne.
Le biofilm se développe et certain morceau peuvent même se détacher (érosion/écaillement) et se disséminer. Ces biofilms peuvent être à l’origine d’échecs d’implantation, de maladies graves… Malheureusement le système immunitaire a très peu de moyen d’enlever ces biofilms. Le seul moyen est donc d’enlever le dispositif médical.
Comment se forme un biofilm ?
Le biofilm nécessite toujours que le substrat au niveau duquel il va se développer soit conditionné par le dépôt préalable de molécules (polysaccharides, protéines, ADN…) pour que les bactéries puissent s’y fixer de façon ferme. A partir de ce moment, les bactéries commencent alors à se développer.
A un moment donné, les bactéries vont s’activer afin de développer une coopération entres elles. Elles vont commencer à établir des communications intercellulaires et à produire des molécules (exo-polymères) venant former ce mucus au sein duquel les bactéries vont se développer.
Comment cette communication s’établie entre les bactéries ?
Quorum sensing et développement de biofilms : Dans un biofilm, la communication intercellulaire des différents types bactériens permet de maintenir les canaux de solutés ouverts.
Toutes les cellules du biofilm (même celles situées au centre) peuvent poursuivre leur croissance sans épuisement du milieu.Le Quorum sensing est la capacité de certaines bactéries à sentir leur densité au sein d’une colonie bactérienne. Lorsque ces bactéries vont atteindre une certaine densité, elles vont déclencher l’activation de certains gènes permettant d’établir la coopération entre bactéries (biofilm, mécanismes de virulence…).
Le Quorum sensing est lié à la capacité de certaines bactéries à produire de façon constitutive, à un niveau de base et de façon constante des molécules activatrices.Toutes les bactéries ne sont pas capables de faire du quorum sensing.
Quorum sensing : senseur de densité bactérienne Expression constitutive d’un activateur (HSL : homoserine
lactone, AHL….)
Augmentation de concentration (par augmentation du nombre de bactéries)
Au-delà d’un seuil activation de promoteurs transcriptionnels Luminescence, virulence, biofilms, sporulation…
Rq: Les bactéries possédant les récepteurs de ces molécules activatrices peuvent donc connaitre la densité bactérienne.
Rq : Ces molécules activatrices tel que l’HSL, peuvent traverser la membrane cellulaire par diffusion passive ou facilitée et agir au niveau de certain opéron et notamment déclencher l’activation de ces cellules et de différents gènes.
Exemple : le gène LasR qui va activer d’autres gènes déclenchant l’expression de gènes intervenant dans la virulence, la production de biofilms…
Virulence par quorum sensing obtenue par : auto- induction ou par action sur d’autres types cellulaires qui seront responsable de la virulence.
Dans une communauté bactérienne, on a des bactéries qui produisent de façon constitutive ces inducteurs et qui possèdent eux-mêmes les récepteurs de ces inducteurs (autocrine), et on a d’autres bactéries qui ne participent pas à la production des activateurs mais qui possèdent les récepteurs et qui vont pouvoir participer à la communauté bactérienne.
Tout cela n’est pas toujours équitable car produire de l’activateur a un coût énergétique pour la cellule alors que celles qui n’en produit pas économise ce coût, mais cela peut être compensé par le fait qu’elle va participer à la production de différentes enzymes. Le cout de cette coopération n’est pas forcément le même entre toutes les bactéries.
Communication intercellulaire et coordination: QUORUM SENSING
Développement de la virulence et de biofilms chez Staphylococcus aureus et Pseudomonas aeruginosa
Développement de la compétence naturelle (capacité à absorber du matériel génétique du milieu extérieur pour aboutir à de la recombinaison) et de la sporulation chez Bacillus subtilis
Déclenchement de la conjugaison chez Enterococcus faecalis
Absorption d’ADN chez Staphylococcus aureus et Bacillus subtilis

Les comportements sociaux chez les bactéries : un équilibre entre coopérateurs (produits les stimulateurs) et tricheurs (ne font que bénéficier des effets du Quorum sensing).
Ceci peut être intéressant en thérapeutique: On pourrait chercher à interférer avec les communications intercellulaires plutôt que de donner des
antibiotiques.
On pourrait aussi essayer de favoriser le développement des bactéries tricheuses au détriment des coopérateurs (pas mal d’études infructueuses, mais la recherche continue)
B. Coopération et conflits chez P. Aeruginosa (qui fait du Quorum sensing)
Pseudomonas aeruginosa (Bacille pyocyanique) responsable d’infections nosocomiales est une des bactéries les plus difficiles à traiter. Les cellules produisent un auto inducteur (HSL) qui s’accumule dans le milieu.
Au-delà d’une concentration seuil, ce signal déclenche : La formation d’un biofilm L’expression de gènes de virulence La production de protéases (élastase) qui dégradent les protéines de l’hôte (élastine,
fibrine, collagène…) au bénéfice de la communauté bactérienne (biofilm).
On constate que dans les colonies, on a des variants de pseudomonas aeruginosa. Certains ne produisent pas de HSL et bénéficient de l’apport de ces facteurs par les autres. Certains variant ne produisent pas les fameuses protéases. L’idée était de voir comment il est possible d’agir sur cet équilibre pour favoriser les pseudomonas ne produisant pas d’elastase et qui n’agressent pas les tissus des patients vis-à-vis de ceux qui en produisent.
III. Les méthodes d’identification bactérienne Comment utiliser la physiologie bactérienne pour faire de l’identification bactérienne ?
1. Utilisation du métabolisme pour l’identification bactérienne
Pour tester leur métabolisme, on utilise des galeries et notamment les galeries API. Il y a de nombreux types de galerie selon ce que l'on veut faire. Ces galeries sont une série de tests biochimiques. Chaque cupule correspond à un test.
Dans chacun de ces tubes on va tester différents caractères qui vont nous permettre d'identifier une propriété associée à un agent bactérien particulier.

Certaines cupules vont être associées au métabolisme carboné, à l'absence ou présence de certaines enzymes, au caractère aérobie ou anaérobie (on ferme la cupule et les bactéries anaérobies ne pourront pas pousser).
Ce genre de tests nécessite deux choses : de disposer d’une galerie convenable par rapport à l'agent que l'on veut mettre en évidence et un inoculum. Pour cela il nous faut la bactérie et la bactérie pure.
On suit le protocole du fabriquant. On ensemence chaque cupule avec une solution bactérienne qui aura été préalablement titrée pour avoir une densité en bactérie correcte pour le développement (trop riche on risque de négativer le test trop rapidement).
Ces tests durent entre 24h et 48h. A la fin on révèle les galeries, les puits vont être d'une couleur. Les résultats sont donnés par la couleur des puits. Une couleur représente le positif, une autre le négatif
Il y a un code d’identification : la lecture se fait par triplet. Si un puits est négatif le résultat est de 0Mais si un résultat est positif, cela dépend de sa position au sein du triplet :
- La positivité du premier puits aura pour valeur 1- La positivité du deuxième puits : valeur 2- La positivité du troisième puits : valeur 4 (et non pas 3)
Ensuite on fait la somme du triplet et on obtient alors un code d’identification qui correspond à une valeur précise pour un triplet qui permet d’identifier les bactéries avec en plus un seuil de fiabilité. Si le pourcentage de fiabilité est de 50 % soit il y a eu des erreurs dans les manipulations, soit c'est une nouvelle bactérie qui possède 50 % de ressemblance avec une bactérie existante.
Ces tests sont toutefois à relativiser à l'aide des observations déjà faites comme la morphologie de la bactérie que l'on aura auparavant déterminée au microscope. Ces techniques sont toujours en laboratoire car très exhaustives mais de moins en moins utilisées.
2. Identification moléculaire
A. Par PCR
Il s’agit d’amplifier le génome d’une bactérie par PCR. Les amorces utilisées sont spécifiques de la bactérie et nous indiquent donc sa nature, si la bactérie s'est effectivement amplifiée. Tout ceci est facilité par les banques de données dans lesquels on va faire des alignements. On aura des taux d’identité en % par rapport à tel séquence.Là encore il faut des colonies pures. Le résultat est obtenu plus rapidement que la technique précédente : 4h. La PCR en temps réel peut être interessante en clinique s’il y a un traitement à mettre en place car on peut savoir s’il faut agir à forte dose ou pas.
PCR multiplex : on peut mettre plusieurs amorces et identifier 2/3 ou 4 types bactériens différents dans un échantillon.

i.ii.
iii. iv.v.
vi. vii. viii. ix.
x.xi.
xii. xiii.
B. Par sondes fluorescentes
xiv.C. Par électrophorèse
D. Par séquençage
Aujourd’hui, le plus simple est de se baser sur le séquençage du fragment d’ADN ribosomique 16S (1500pb) qui est utilisé comme référence. On utilise des sondes fluorescentes plutôt que l'électrophorèse avec la technique de PCR en temps réel.
1) Amplification par PCR d’un gène bactérien : ADNr 16S (~1500pb)
2) Séquençage du produit d’amplification (amplicon)
3) BLAST : Basic Local Alignment Search Tool

Le séquençage permet une plus grande précision dans l’identification bactérien pour un coût modéré, notamment quand on a des variants (bactéries qui mutent).
Les tests d’identification moléculaire sont de plus en plus utilisés face aux test API. Mais la spectrométrie de masse est en train de révolutionner l’identification bactérienne.
3. Spectrométrie de masse
Elle permet d’analyser les protéines totales d’un échantillon et d’identifier directement des bactéries. Le temps nécessaire pour l’identification à partir du moment où la souche est pure est en générale de 6 à 8 minutes. Malheureusement c’est extrêmement couteux mais cette technique nécessite qu’une machine et un technicien.