synthèse translésionnelle translesion synthesis...
TRANSCRIPT

A. Cordonnier Equipe: Modifications post-traductionnelles et Cancérogenèse UMR7242-IREBS Ecole supérieure de biotechnologie de Strasbourg-Illkirch E-mail [email protected]
Synthèse translésionnelle Translesion Synthesis (TLS)

Synthèse translésionnelle Translesion Synthesis (TLS)
Plan du cours:
1. Réplication de l’ADN en présence de lésions: Recombinaison homologue et synthèse translésionnelle
1. La famille Y des ADN Polymérases translésionnelles chez les procaryotes et les eucaryotes
3. Régulation de la synthèse translésionnelle (TLS) 4. Rôle des ADN polymérases translésionnelles dans l’hypermutation somatique et l’induction du checkpoint de la réplication

1
Réplication de l’ADN en présence de lésions
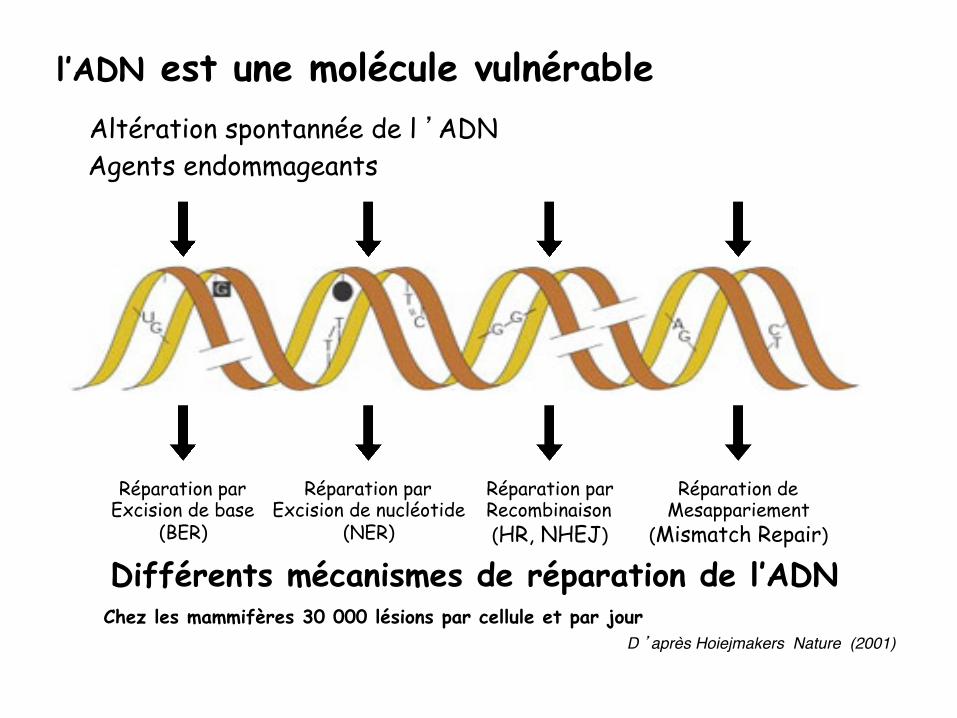
Différents mécanismes de réparation de l’ADN Chez les mammifères 30 000 lésions par cellule et par jour
D ’après Hoiejmakers Nature (2001)"
Réparation par Excision de base
(BER)
Réparation par Excision de nucléotide
(NER)
Réparation par Recombinaison (HR, NHEJ)
Réparation de Mesappariement
(Mismatch Repair)
Agents endommageants Altération spontannée de l ’ADN
l’ADN est une molécule vulnérable

Chez E.coli, Une cellule sauvage (WT) peut tolérer 3700 lésions UV (CPD) par cellule
Mécanismes de tolérance chez E.coli
Une cellule défective pour la réparation (NER -) ne peut tolérer que 60 lésions => Importance de la réparation par excision de nucléotides Une cellule défective pour la réparation (NER -) et mutée dans le gène RecA: une seule lésion est létale !!!!!
En plus de la réparation, il existe un mécanisme (dépendant de RecA chez E.coli) qui permet de « tolérer des lésions » c.a.d. de répliquer le matériel génétique malgré la présence de lésions Les mécanismes de tolérance des lésions sont aussi appelés mécanismes de réparation post-réplicative
Conclusion
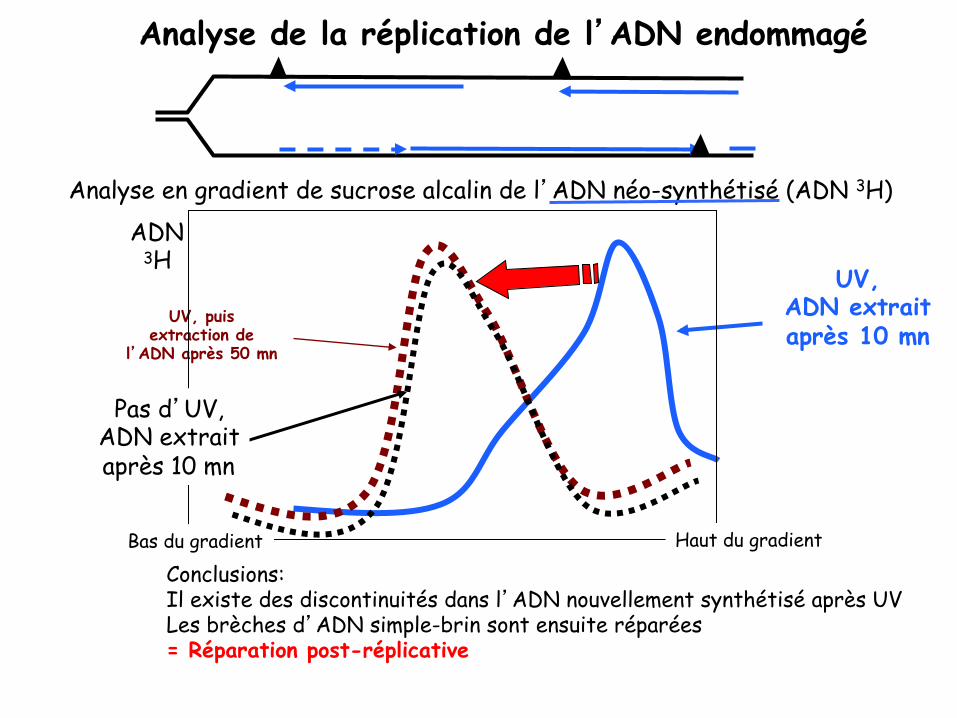
Analyse de la réplication de l’ADN endommagé
UV, ADN extrait après 10 mn
Conclusions: Il existe des discontinuités dans l’ADN nouvellement synthétisé après UV Les brèches d’ADN simple-brin sont ensuite réparées = Réparation post-réplicative
UV, puis extraction de
l’ADN après 50 mn
Analyse en gradient de sucrose alcalin de l’ADN néo-synthétisé (ADN 3H)
Haut du gradient Bas du gradient
ADN 3H
Pas d’UV, ADN extrait après 10 mn

Peignage moléculaire de l’ADN
IdU=iododeoxyuridine CldU=chlorodeoxyuridine
Analyse statistique de la dynamique de la réplication au niveau de fibres d’ADN
.Incorporation de IdU et CldU
.Lyse des cellules
.Etirement de l’ADN déprotéinisé sur des lamelles .Détection des nucléotides incorporés par immunofluorescence
WT XPV lignées cellulaires
Les cellules XPV ont un défaut de réplication de l’ADN après traitement aux rayons UV
UV

Lopes M. et al., 2006, Mol Cell, 21, 15-27.
Analyse en microscopie électronique de la réplication de l’ADN chez S. cerevisiae après traitement aux UV
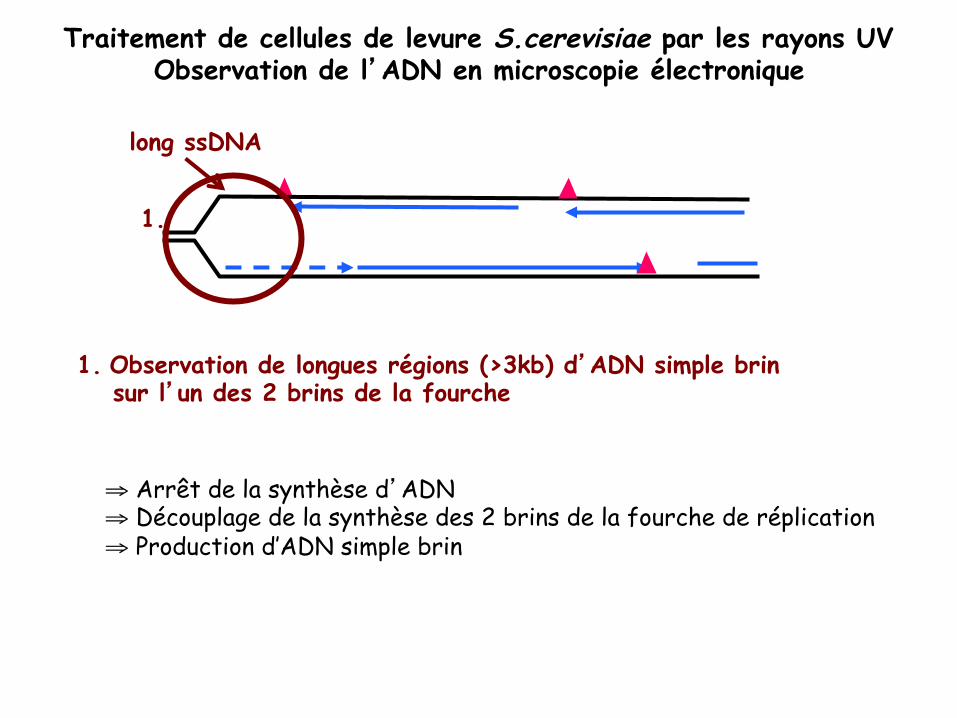
Traitement de cellules de levure S.cerevisiae par les rayons UV Observation de l’ADN en microscopie électronique
1. Observation de longues régions (>3kb) d’ADN simple brin sur l’un des 2 brins de la fourche
⇒ Arrêt de la synthèse d’ADN ⇒ Découplage de la synthèse des 2 brins de la fourche de réplication ⇒ Production d’ADN simple brin
long ssDNA
1.
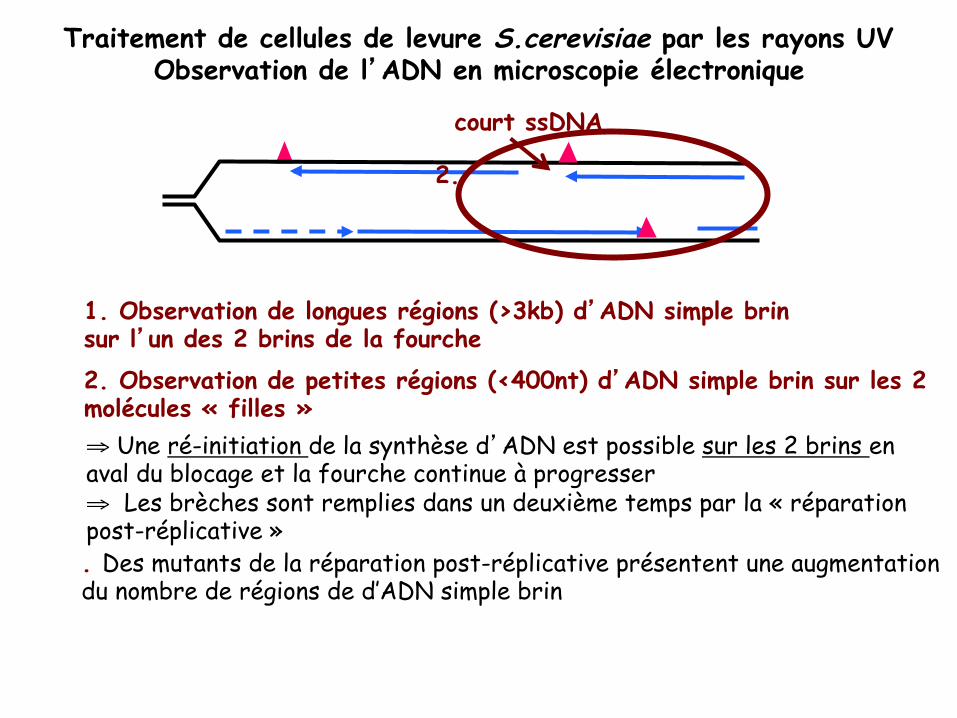
Traitement de cellules de levure S.cerevisiae par les rayons UV Observation de l’ADN en microscopie électronique
1. Observation de longues régions (>3kb) d’ADN simple brin sur l’un des 2 brins de la fourche 2. Observation de petites régions (<400nt) d’ADN simple brin sur les 2 molécules « filles » ⇒ Une ré-initiation de la synthèse d’ADN est possible sur les 2 brins en aval du blocage et la fourche continue à progresser ⇒ Les brèches sont remplies dans un deuxième temps par la « réparation post-réplicative »
2.
court ssDNA
. Des mutants de la réparation post-réplicative présentent une augmentation du nombre de régions de d’ADN simple brin
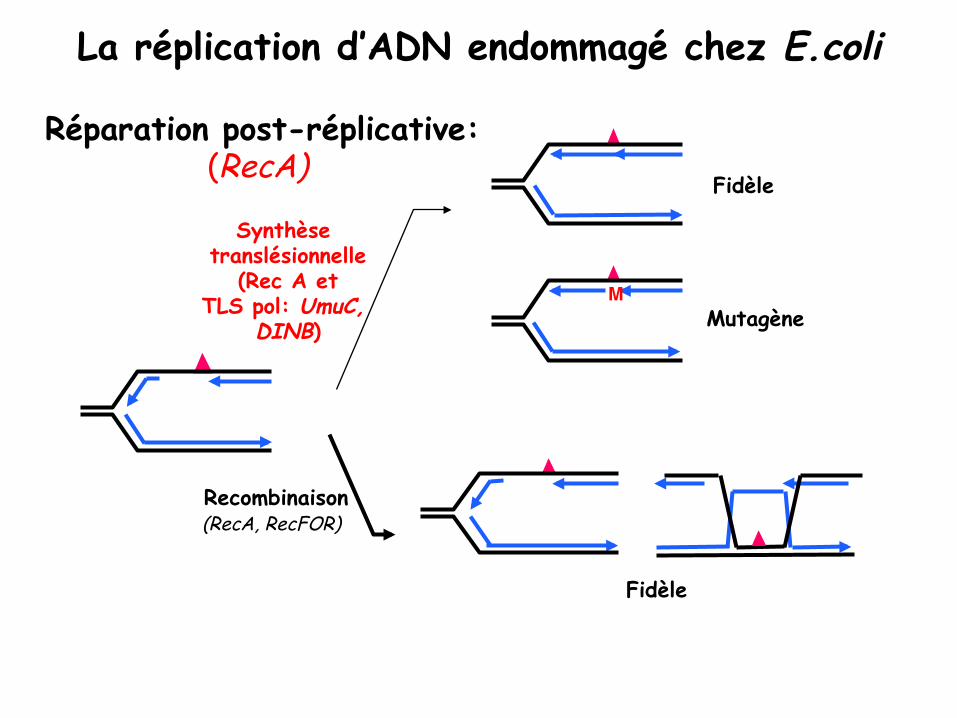
La réplication d’ADN endommagé chez E.coli"
Recombinaison (RecA, RecFOR) "
Fidèle
Réparation post-réplicative: (RecA)
Mutagène"
Fidèle
Synthèse translésionnelle
(Rec A et TLS pol: UmuC,
DINB)"M"
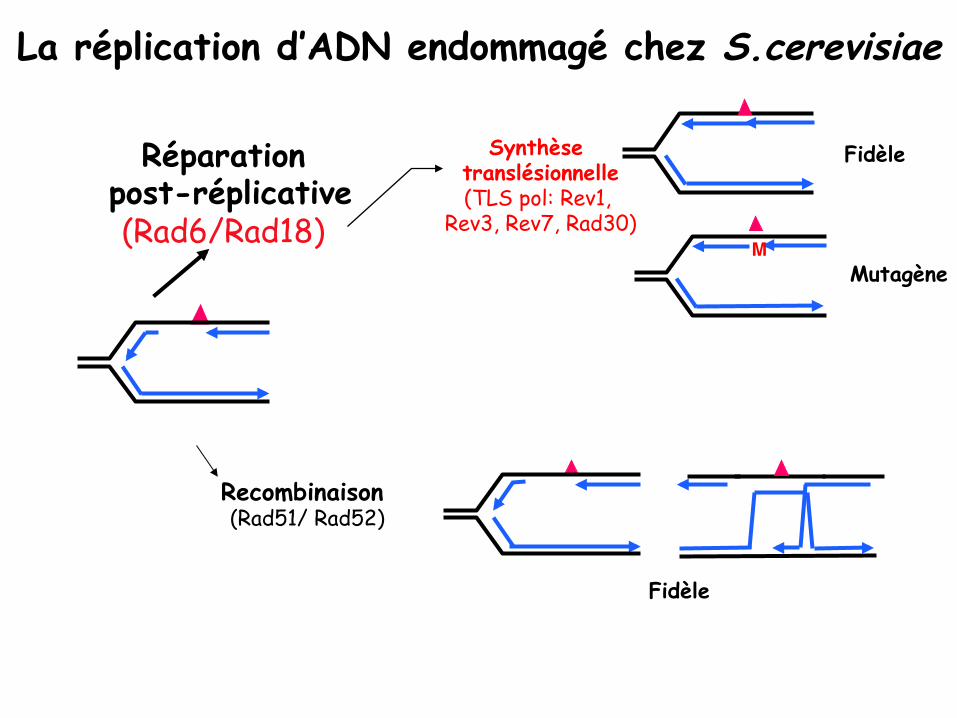
La réplication d’ADN endommagé chez S.cerevisiae"
Synthèse translésionnelle (TLS pol: Rev1,
Rev3, Rev7, Rad30)"
Mutagène"
Fidèle
M"
Recombinaison (Rad51/ Rad52)"
Fidèle
Réparation post-réplicative
(Rad6/Rad18)

Xeroderma Pigmentosum: Maladie autosomale récessive (1/106) . probabilité de développer un cancer de la peau 4000 fois plus importante que la population normale . Hypermutabilité des cellules après irradiation UV
XP classiques: 7 groupes de complémentation Déficient dans les gènes de réparation par excision de nucléotides
L’analyse en gradient de sucrose alcalin de la synthèse d’ADN après irradiation montre que les cellules des malades XPV ont un défaut dans la réparation post-réplicative
Mécanismes de tolérance chez l’homme:
XP variant:XPV 8ème groupe de complémentation Pas de déficience dans les gènes de la réparation Défaut dans la réplication de l ’ADN endommagé

Mise au point d’un test in vitro de synthèse d’ADN utilisant une matrice ADN endommagé : Les extraits de cellules de Xeroderma Pigmentosum Variant sont déficients en synthèse translésionnelle
Purification du facteur XPV: A partir de cellules normales, purification du facteur qui permet de complémenter la déficience des cellules XPV
Séquençage de la protéine et du cDNA: Le facteur XPV est une ADN polymérase translésionnelle L’ADN polymérase η"
Découverte des ADN polymérases translésionnelles dans les cellules humaines

TLS et XPV
Propriété de Pol η : Pol η est capable de répliquer un dimère de TT induit par les UV (CPD) en incorporant AA en face.
En absence de Pol η (dans un contexte XPV): Une autre polymérase translésionnelle incorpore une mauvaise base en face de lésion induite par les UV mutations
cancer de la peau dans les zones exposées
Le facteur XPV est une ADN polymérase translésionnelle L’ADN polymérase η"
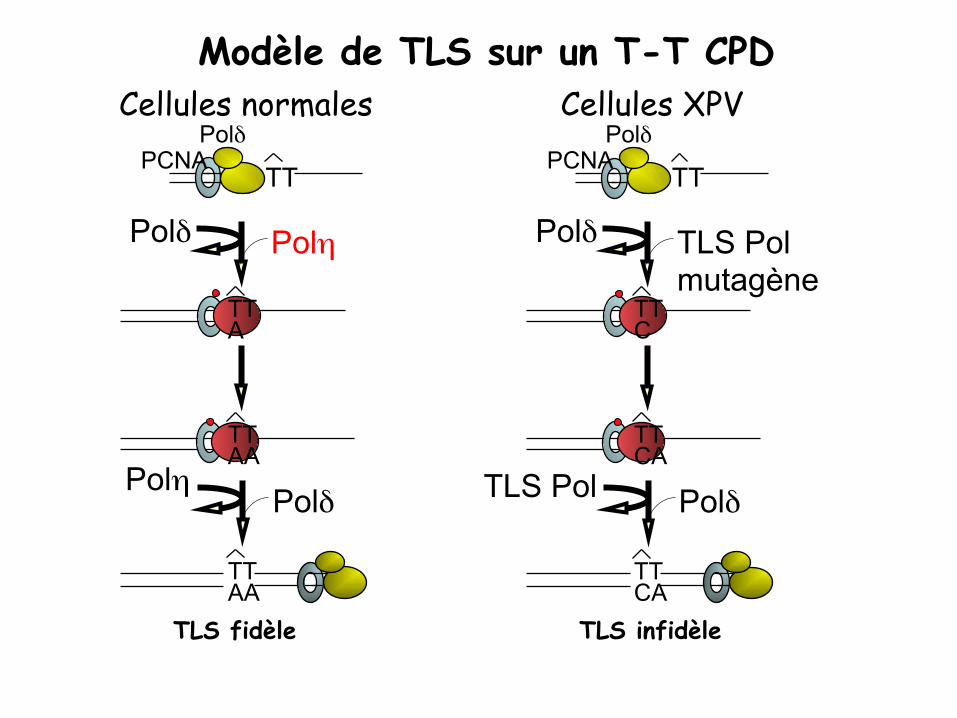
Modèle de TLS sur un T-T CPD
Polδ PCNA
TT
TLS fidèle
Polδ Polη
Polη Polδ
TT AA
TT AA
TT A
Cellules normales Polδ
PCNA TT
TLS infidèle
Polδ TLS Pol mutagène
TLS Pol Polδ
TT CA
TT CA
TT C
Cellules XPV

1
La famille Y des ADN polymérases
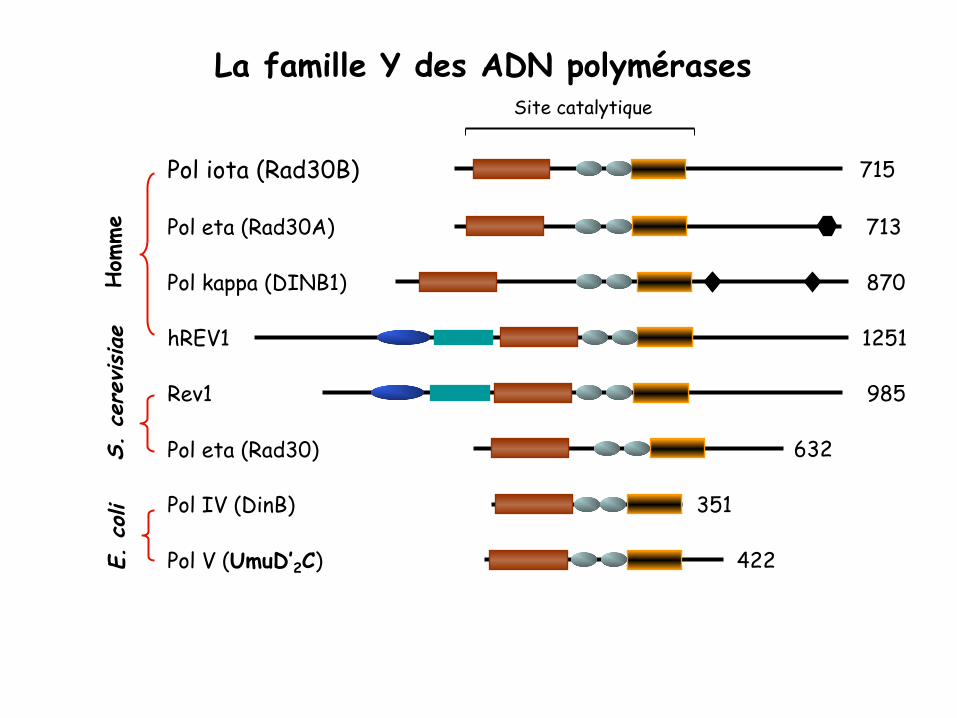
La famille Y des ADN polymérases
Pol V (UmuD’2C) 422
Pol IV (DinB) 351
Pol eta (Rad30) 632
Rev1 985
hREV1 1251
Pol kappa (DINB1) 870
Pol eta (Rad30A) 713
Pol iota (Rad30B) 715
Hom
me
S. c
erev
isiae
E. c
oli
Site catalytique

Translesion synthesis DNA
polymerases Origines de la mutagenèse dans une cellule normale: . Erreurs de la réplication corrigées par le mécanisme de réparation des mésappariements . Lésions de l’ADN dues au métabolisme cellulaire ou à des agents environnementaux
Sélectivité du nucleotide entrant élevée Activité de correction de lecture
Taux de mutation
(substitution de base)
Sélectivité du nucleotide entrant faible Pas d’activité de correction de lecture
Replicative DNA polymerases
Caractéristiques des ADN polymérases réplicatives et ADN polymérases translésionnelles
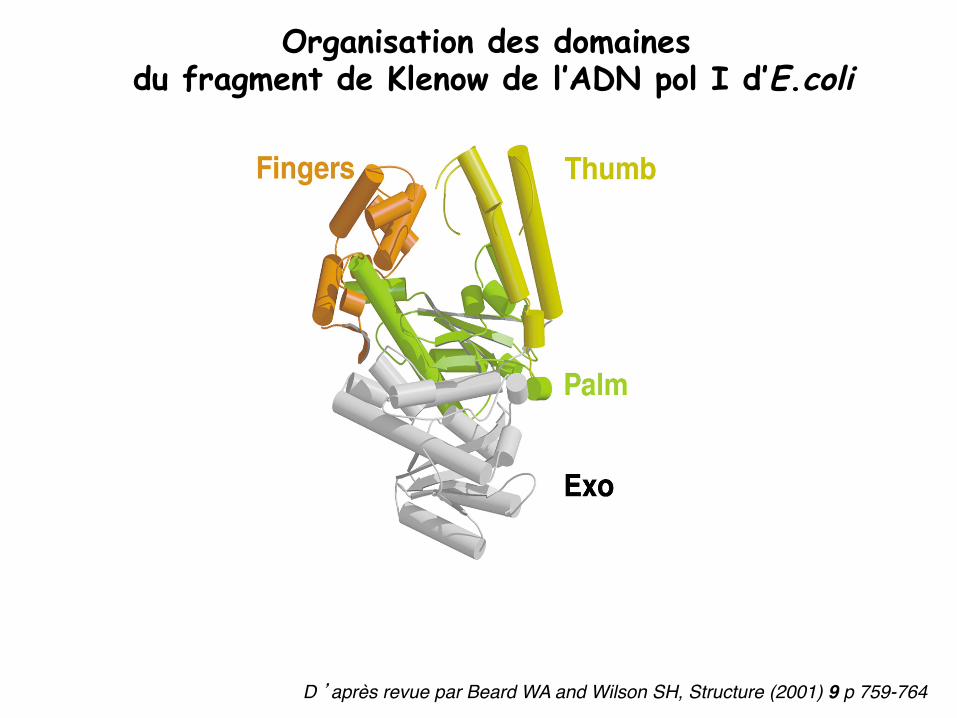
Organisation des domaines du fragment de Klenow de l’ADN pol I d’E.coli
D ’après revue par Beard WA and Wilson SH, Structure (2001) 9 p 759-764"
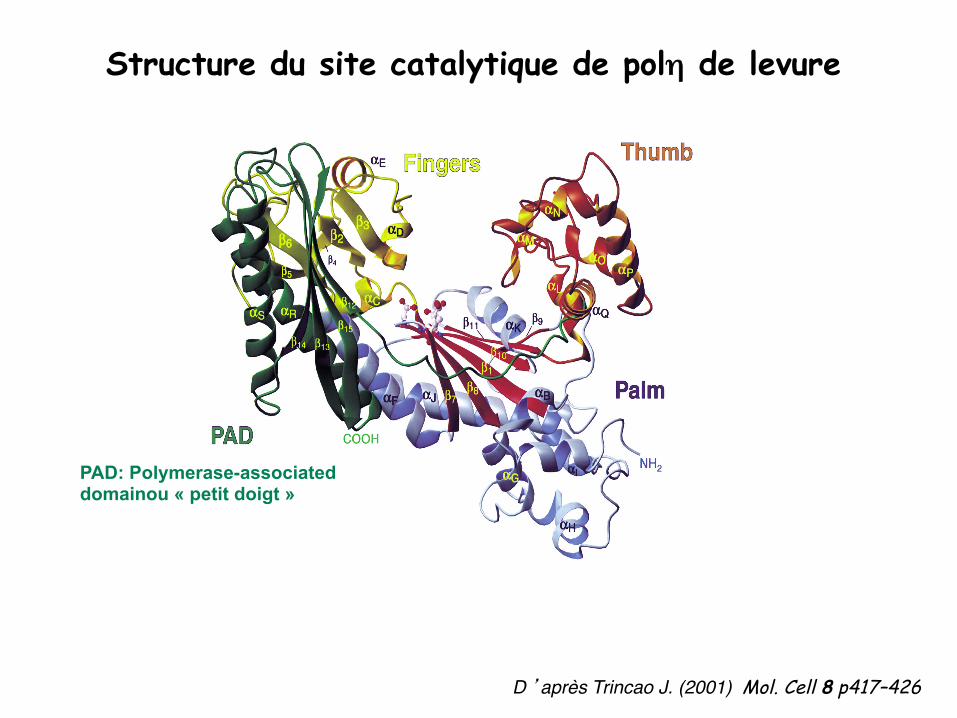
Structure du site catalytique de polη de levure
D ’après Trincao J. (2001) Mol. Cell 8 p417–426
PAD: Polymerase-associated domainou « petit doigt »

Organisation des domaines des ADN polymérases de type main droite pouce - paume - doigt
Structure des ADN polymérases de la famille Y pouce - paume - doigt - petit doigt
Le site actif des ADN polymérases de la famille Y tolère une base endommagée ou un mésappariement
Conclusion

Caractéristiques des ADN polymérases de la famille Y
.Distributives"
.Infidèles, pas d’activité de correction de lecture"
.Capables de répliquer l’ADN contenant des lésions"
.Interagissent avec le facteur de processivité des ADN polymérases réplicatives (PCNA chez les eucaryotes et β clamp chez les procaryotes)
.Structure du site catalytique similaire à une main droite: “palme/pouce/doigt/petit doigt”"
"
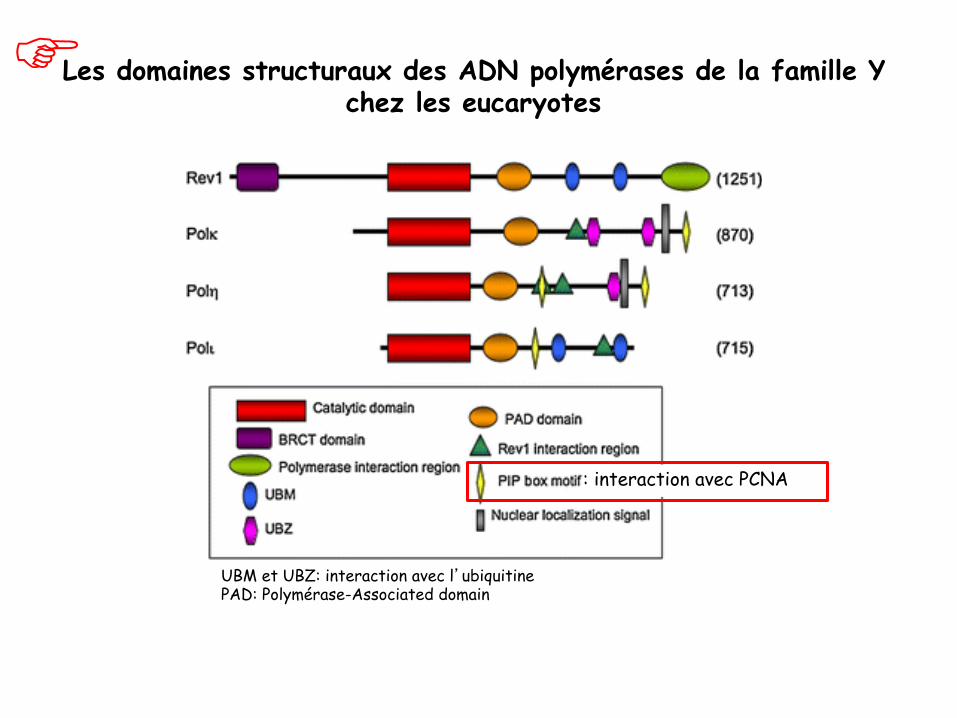
Les domaines structuraux des ADN polymérases de la famille Y chez les eucaryotes
UBM et UBZ: interaction avec l’ubiquitine PAD: Polymérase-Associated domain
: interaction avec PCNA
"
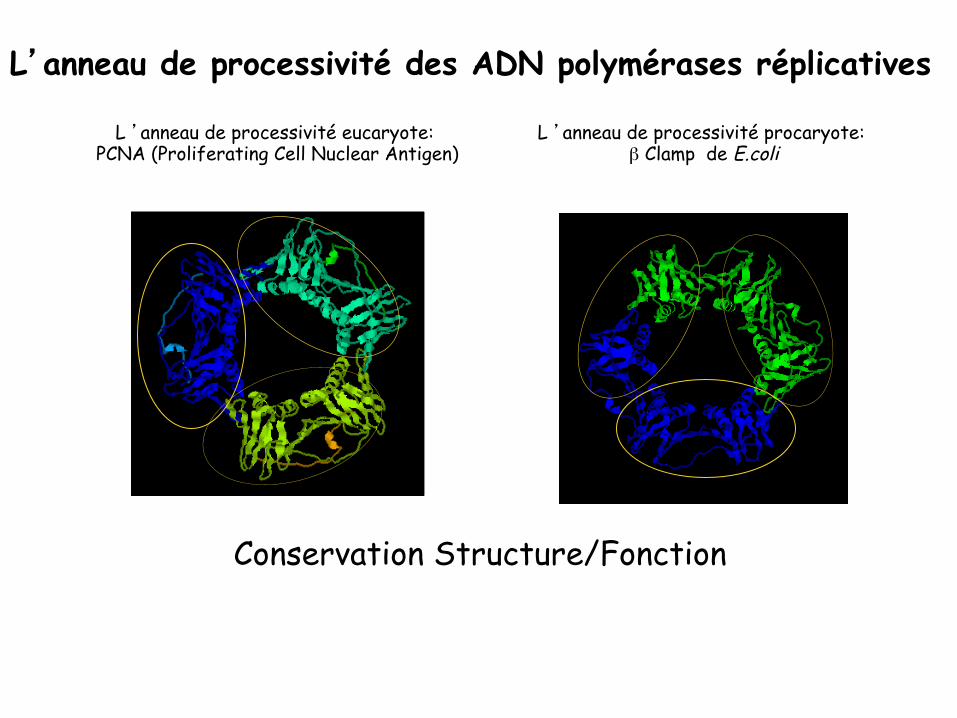
L’anneau de processivité des ADN polymérases réplicatives
L ’anneau de processivité eucaryote: PCNA (Proliferating Cell Nuclear Antigen)
L ’anneau de processivité procaryote: β Clamp de E.coli
Conservation Structure/Fonction
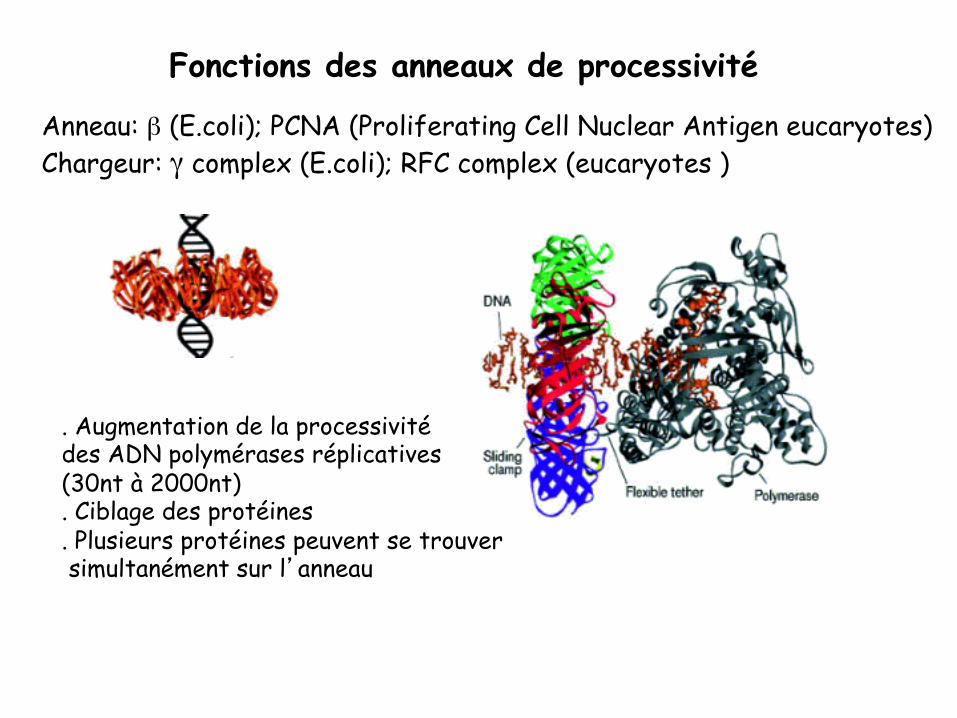
Fonctions des anneaux de processivité
Anneau: β (E.coli); PCNA (Proliferating Cell Nuclear Antigen eucaryotes) Chargeur: γ complex (E.coli); RFC complex (eucaryotes )
. Augmentation de la processivité des ADN polymérases réplicatives (30nt à 2000nt) . Ciblage des protéines . Plusieurs protéines peuvent se trouver simultanément sur l’anneau
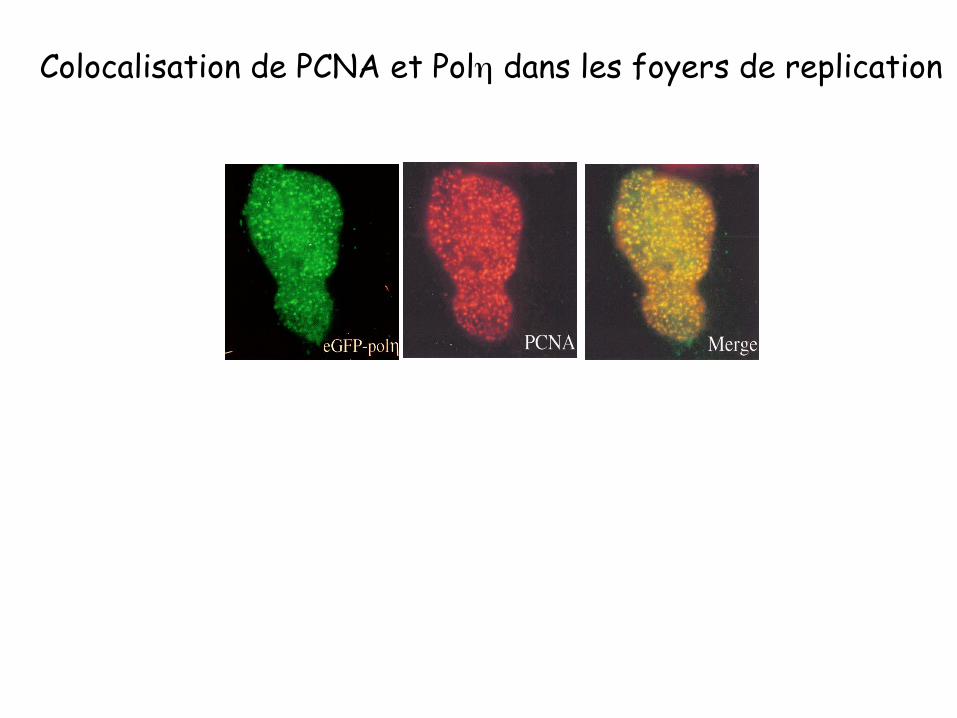
Colocalisation de PCNA et Polη dans les foyers de replication
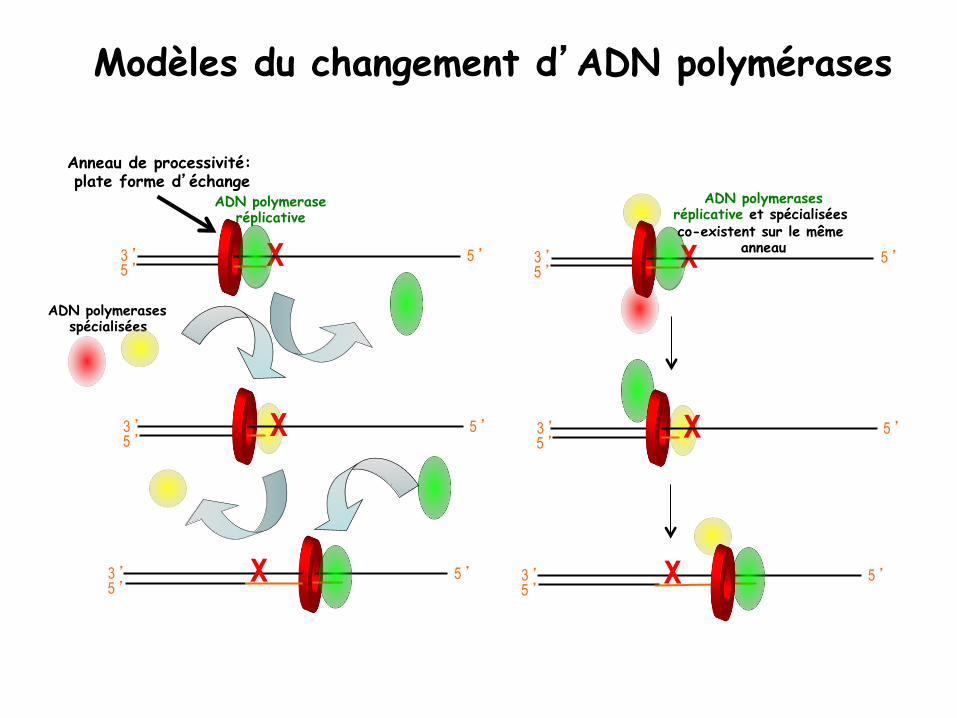
ADN polymerases spécialisées
ADN polymerase réplicative
Modèles du changement d’ADN polymérases
3 ’ 5 ’ 5 ’
X
3 ’ 5 ’ 5 ’ X
3 ’ 5 ’ 5 ’ X
Anneau de processivité: plate forme d’échange
3 ’ 5 ’ 5 ’
X
3 ’ 5 ’ 5 ’ X
3 ’ 5 ’ 5 ’ X
ADN polymerases réplicative et spécialisées co-existent sur le même
anneau

Adapted from Bunting et al. (2003)
Position off
Interaction de Pol IV avec l’anneau de processivité de E.coli
Position on
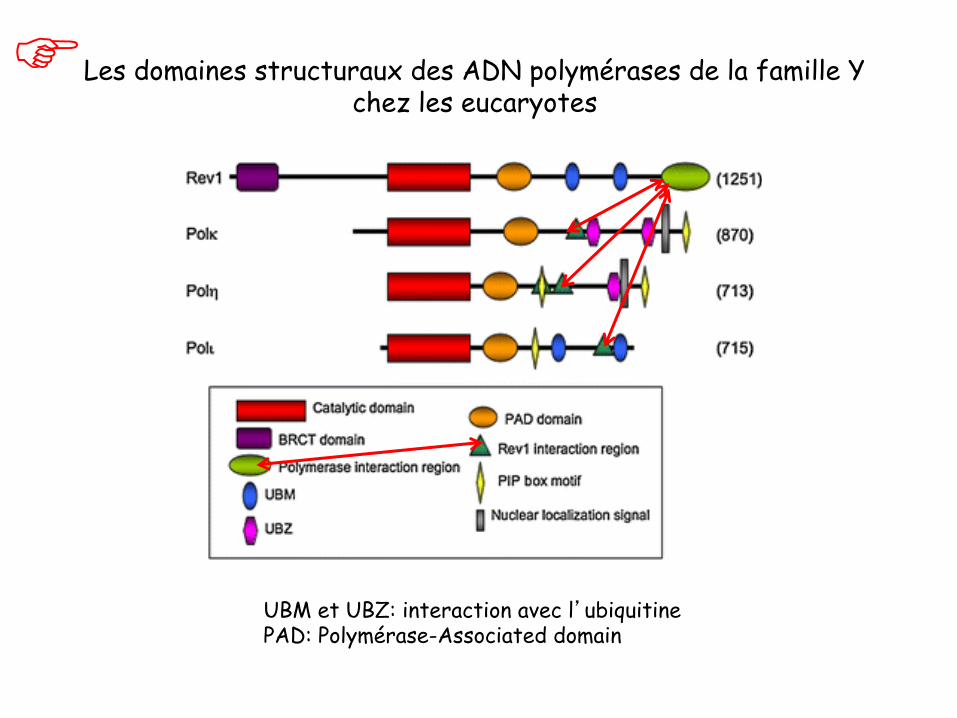
Les domaines structuraux des ADN polymérases de la famille Y chez les eucaryotes
UBM et UBZ: interaction avec l’ubiquitine PAD: Polymérase-Associated domain
"
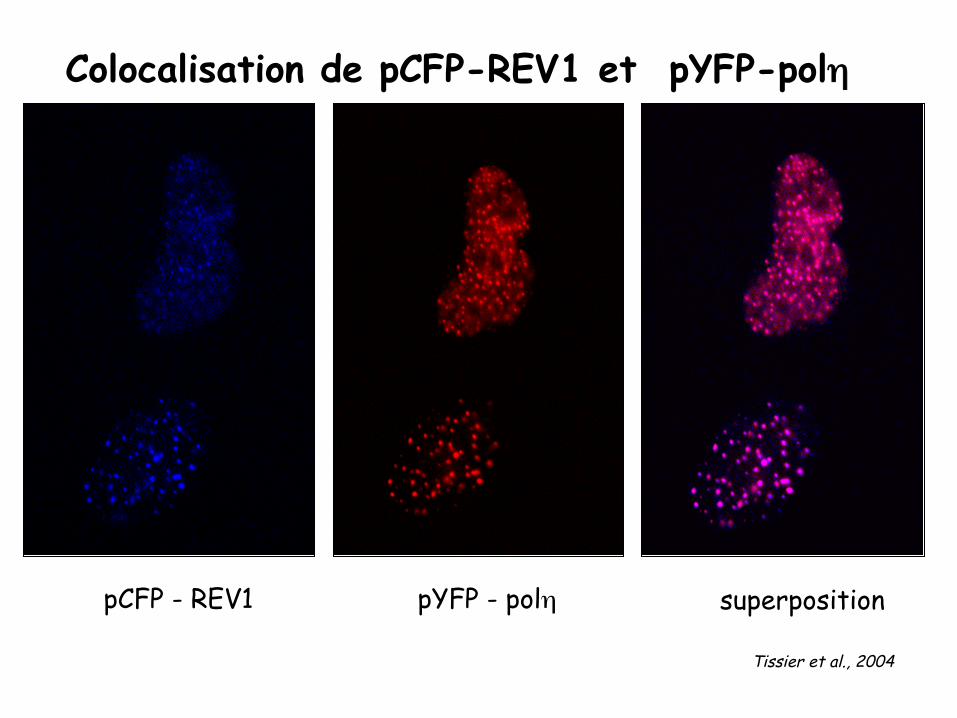
Colocalisation de pCFP-REV1 et pYFP-polη
pYFP - polη pCFP - REV1 superposition
Tissier et al., 2004
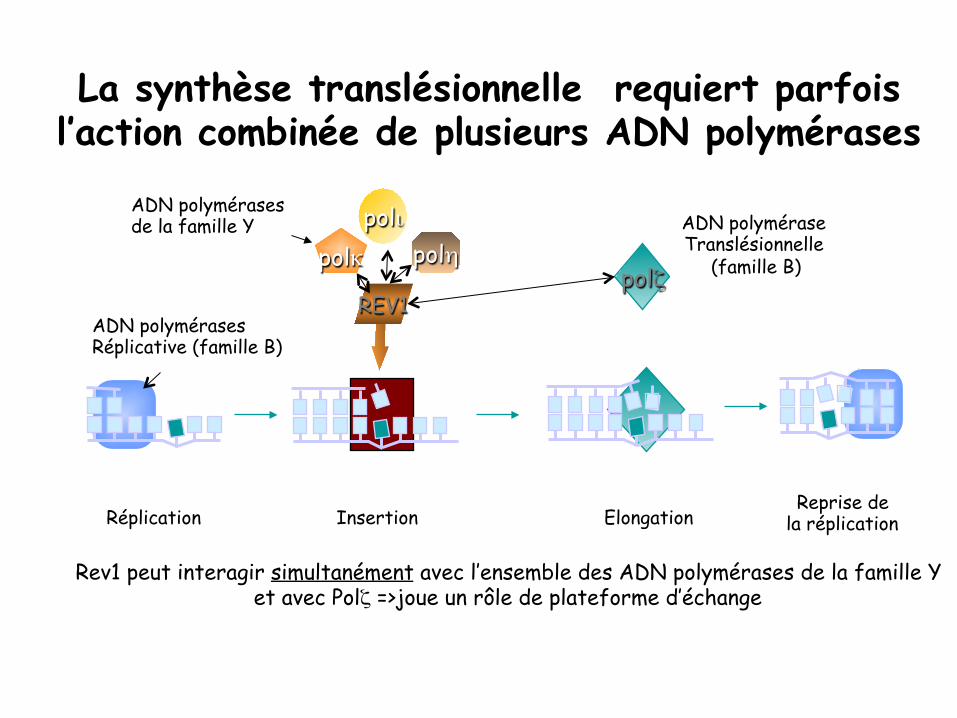
La synthèse translésionnelle requiert parfois l’action combinée de plusieurs ADN polymérases
ADN polymérases Réplicative (famille B)
polζ
Elongation
polι polη polκ
REV1
Insertion Réplication Reprise de
la réplication
ADN polymérases de la famille Y
Rev1 peut interagir simultanément avec l’ensemble des ADN polymérases de la famille Y et avec Polζ =>joue un rôle de plateforme d’échange
ADN polymérase Translésionnelle
(famille B)

3
Régulation de la synthèse translésionnelle

L’activité des ADN polymérases translésionnelles doit être régulée pour éviter l’introduction de mutations dans le génome
Chez les eucaryotes: Modification post traductionnelle de PCNA "
Chez les procaryotes: Régulation par le système SOS
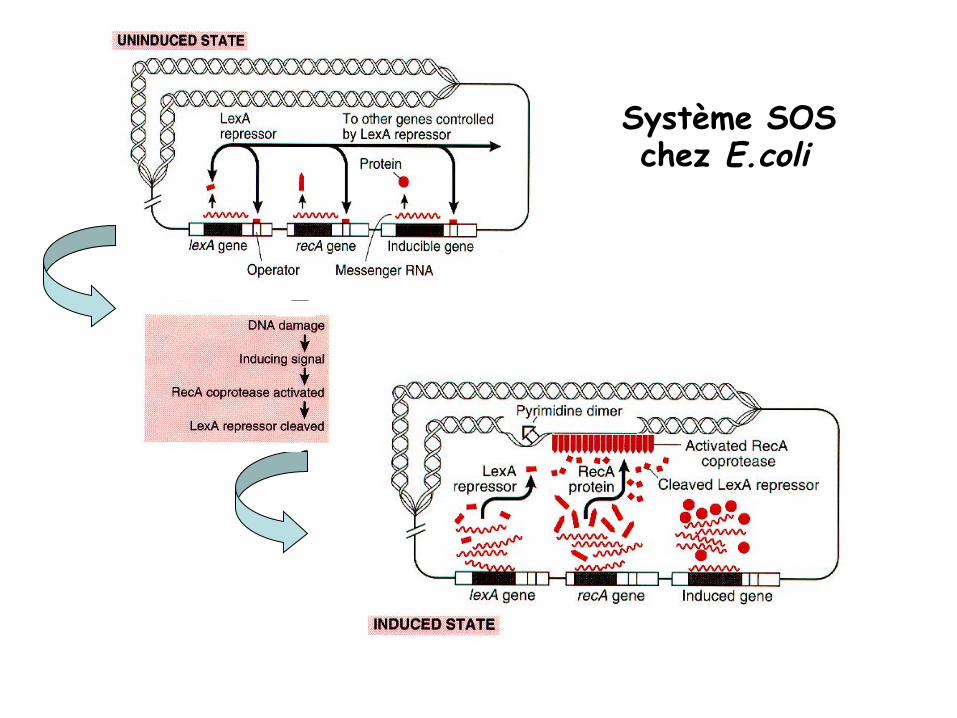
Système SOS chez E.coli
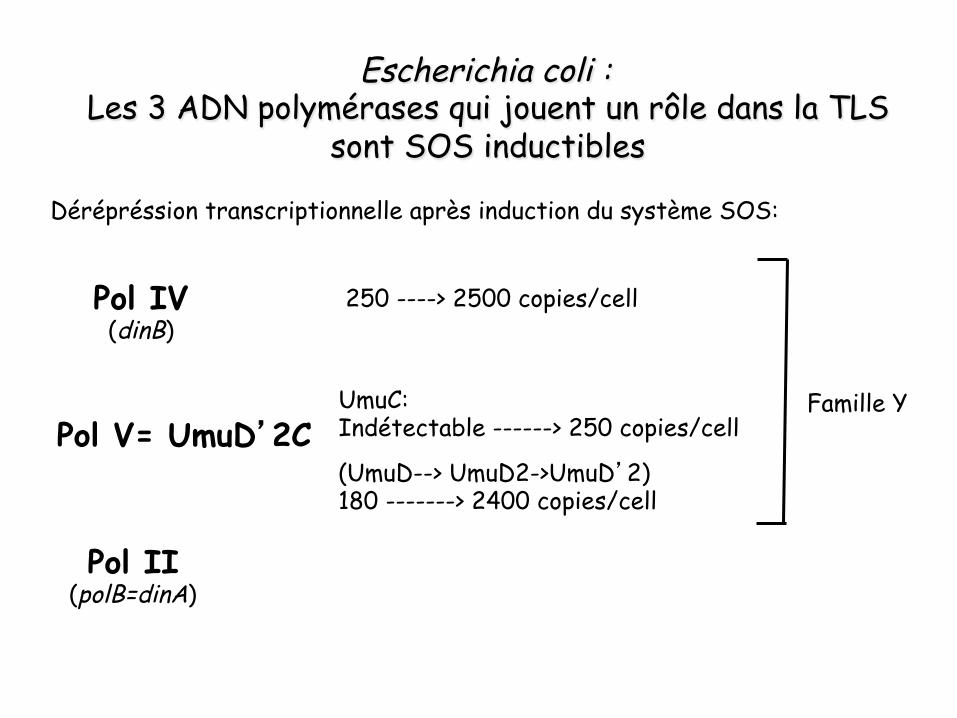
Pol II (polB=dinA)
Pol IV (dinB)
Pol V= UmuD’2C
Escherichia coli : Les 3 ADN polymérases qui jouent un rôle dans la TLS
sont SOS inductibles
Famille Y UmuC: Indétectable ------> 250 copies/cell
(UmuD--> UmuD2->UmuD’2) 180 -------> 2400 copies/cell
250 ----> 2500 copies/cell
Dérépréssion transcriptionnelle après induction du système SOS:
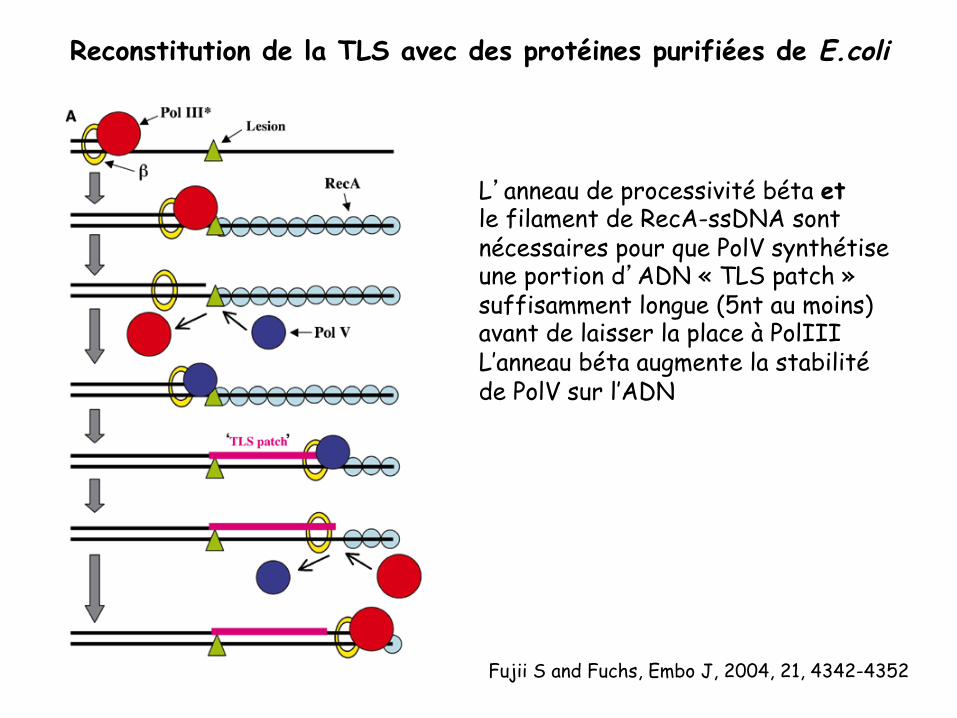
Fujii S and Fuchs, Embo J, 2004, 21, 4342-4352
Reconstitution de la TLS avec des protéines purifiées de E.coli
L’anneau de processivité béta et le filament de RecA-ssDNA sont nécessaires pour que PolV synthétise une portion d’ADN « TLS patch » suffisamment longue (5nt au moins) avant de laisser la place à PolIII L’anneau béta augmente la stabilité de PolV sur l’ADN

Rôle de RecA dans la TLS chez les procaryotes 1. Induction transcriptionnelle des gènes codant pour 3 ADN polymérases
translésionnelles
2. Activité coprotéase de RecA: activation de UmuD2 en UmuD’2= polV 3. Interaction de RecA avec UmuD’2C => Ciblage de PolV sur la lésion
=> Stimulation de l’activité de PolV
"
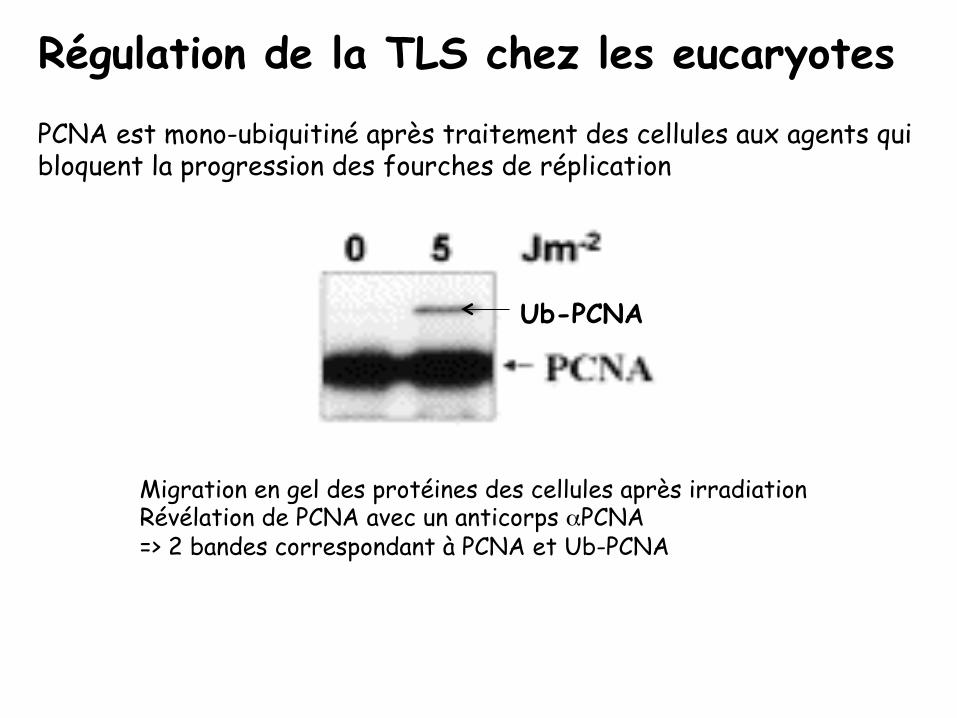
PCNA est mono-ubiquitiné après traitement des cellules aux agents qui bloquent la progression des fourches de réplication
Ub-PCNA
Migration en gel des protéines des cellules après irradiation Révélation de PCNA avec un anticorps αPCNA => 2 bandes correspondant à PCNA et Ub-PCNA
Régulation de la TLS chez les eucaryotes

Structure de l’ubiquitine (76aa)
L’ubiquitine est liée à PCNA de façon covalente par les enzymes RAD6/RAD18
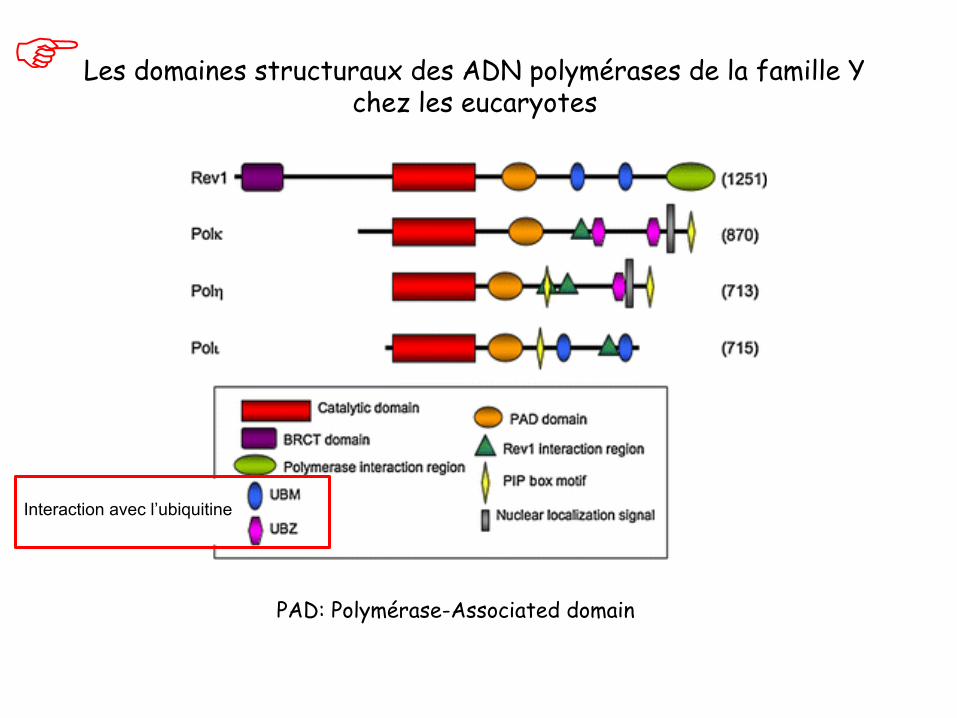
Les domaines structuraux des ADN polymérases de la famille Y chez les eucaryotes
PAD: Polymérase-Associated domain
Interaction avec l’ubiquitine
"
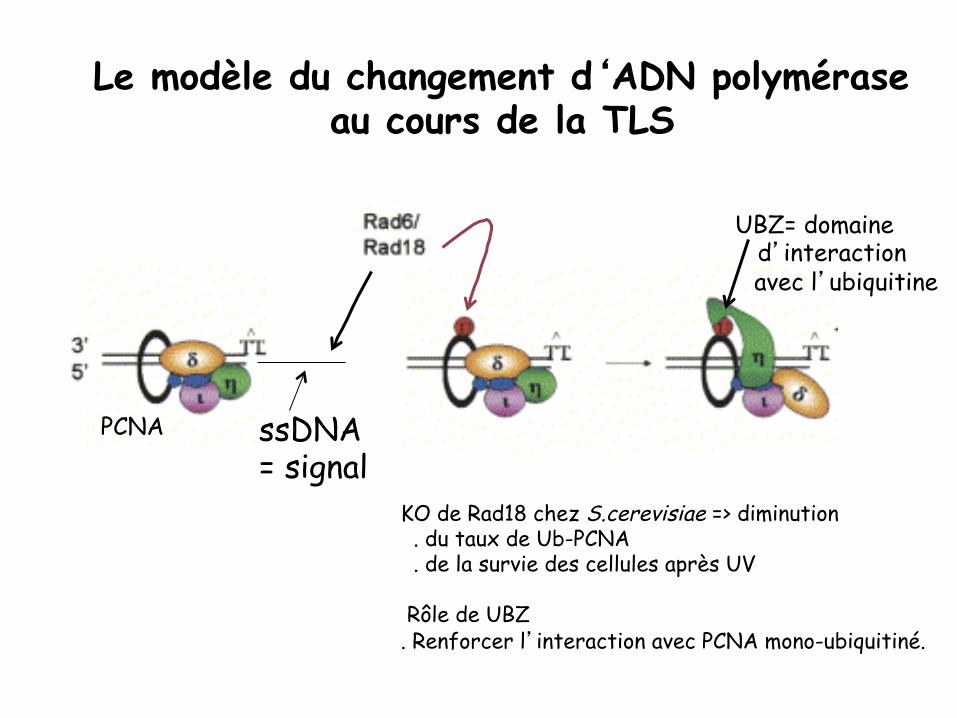
Le modèle du changement d‘ADN polymérase au cours de la TLS
KO de Rad18 chez S.cerevisiae => diminution . du taux de Ub-PCNA . de la survie des cellules après UV Rôle de UBZ . Renforcer l’interaction avec PCNA mono-ubiquitiné.
PCNA
UBZ= domaine d’interaction
avec l’ubiquitine
ssDNA = signal
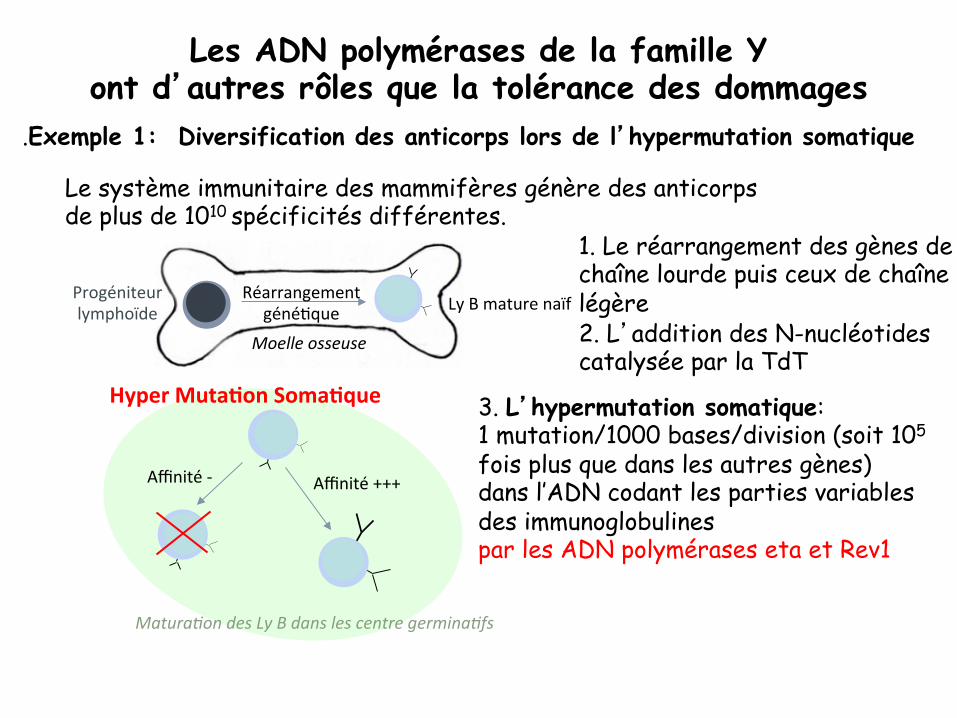
Les ADN polymérases de la famille Y ont d’autres rôles que la tolérance des dommages
.Exemple 1: Diversification des anticorps lors de l’hypermutation somatique
Le système immunitaire des mammifères génère des anticorps de plus de 1010 spécificités différentes.
1. Le réarrangement des gènes de chaîne lourde puis ceux de chaîne légère 2. L’addition des N-nucléotides catalysée par la TdT
3. L’hypermutation somatique: 1 mutation/1000 bases/division (soit 105 fois plus que dans les autres gènes) dans l’ADN codant les parties variables des immunoglobulines par les ADN polymérases eta et Rev1
Moelle osseuse
Ly B mature naïf Progéniteur lymphoïde
Réarrangement géné8que
Matura+on des Ly B dans les centre germina+fs
Hyper Muta+on Soma+que
Affinité +++ Affinité -‐
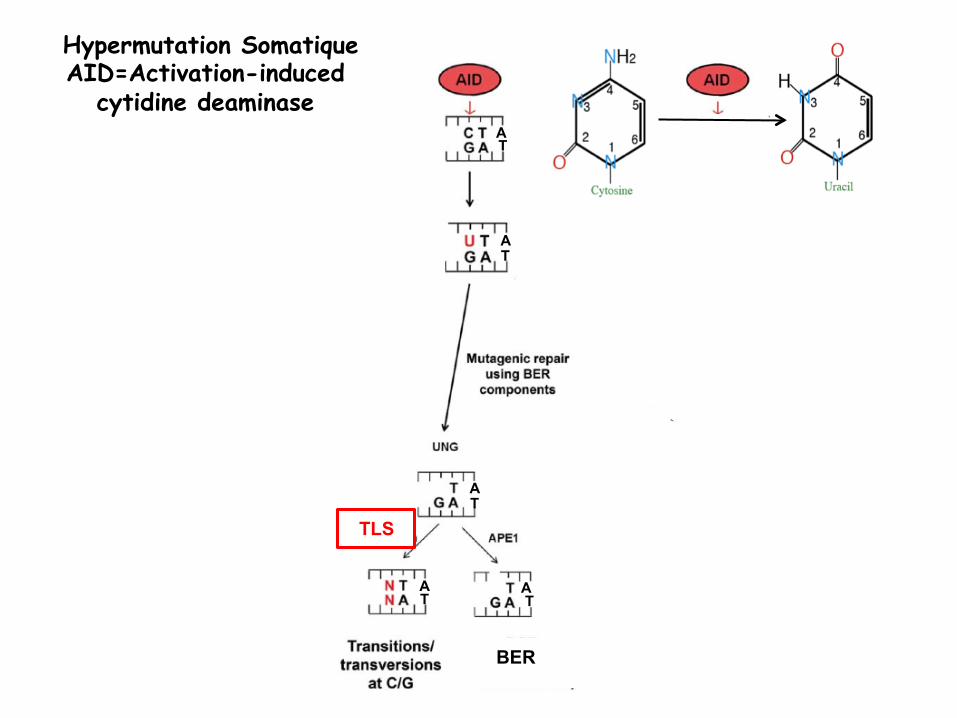
Hypermutation Somatique AID=Activation-induced
cytidine deaminase T A
BER
TLS TLS:Poleta et Ub-PCNA
T
T T
T
T
T
CT G T
Transitions at A/T
A A
A
A
A
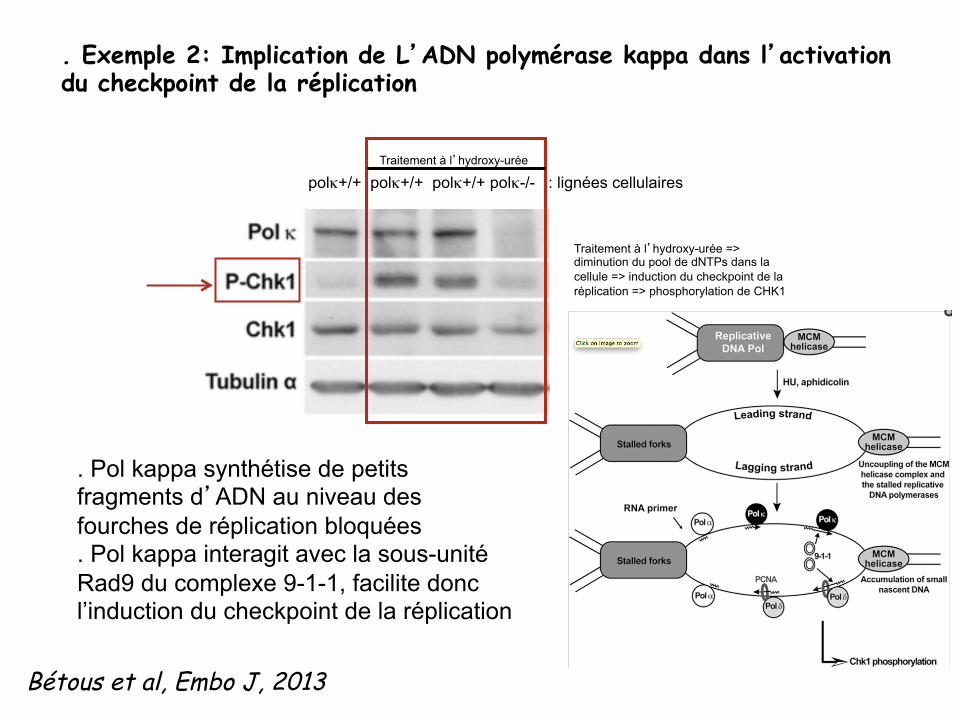
. Pol kappa synthétise de petits fragments d’ADN au niveau des fourches de réplication bloquées . Pol kappa interagit avec la sous-unité Rad9 du complexe 9-1-1, facilite donc l’induction du checkpoint de la réplication
. Exemple 2: Implication de L’ADN polymérase kappa dans l’activation du checkpoint de la réplication
Bétous et al, Embo J, 2013
polκ+/+ polκ+/+ polκ+/+ polκ-/- : lignées cellulaires Traitement à l’hydroxy-urée
Traitement à l’hydroxy-urée => diminution du pool de dNTPs dans la cellule => induction du checkpoint de la réplication => phosphorylation de CHK1

Réplication de l’ADN endommagé: résumé
Caractéristiques des ADN polymérases de la famille Y . Distributives, fidélité réduite, pas d’activité de correction de lecture, structure ouverte du site actif, capables de répliquer l’ADN contenant des lésions (seules ou de façon combinées)
. Interagissent avec le facteur de processivité (PCNA et β clamp) des ADN polymérases réplicatives
Régulation de l’activité des ADN polymérases de la famille Y . Chez les procaryotes: . Induction transcriptionnelle par le système SOS . Clivage de UmuD2 . Rôle de RecA dans le ciblage de UmuD’2C (PolV)
. Chez les eucaryotes: Interaction avec PCNA monoubiquitiné via les motifs UBM et UBZ
"La TLS est une des voies de tolérance des dommages
Rôle des ADN polymérases de la famille Y dans l’hypermutation somatique et dans l’induction du checkpoint de la réplication