responsable de stage : jean-pascal torreton · stage d’élève –ingénieur sciences et...
TRANSCRIPT

Institut des Sciences de l’Ingénieur de Montpellier
Université Montpellier II
Stage d’élève –ingénieur Sciences et Technologies de l’Eau
Rapport de stage 2ème année
Présenté par Bérengère HERSANT
Septembre 2003
Responsable de stage : Jean-Pascal TORRETON
Stage effectué dans le cadre de la thèse de Séverine Jacquet
Institue de Recherche pour
le Développement
Institut des Sciences de l’Ingénieur
de Montpellier

Remerciements
Je tiens à remercier Mr COLIN pour m’avoir accueillie au sein de l’Unité de Recherche CAMELIA
de l’IRD de Nouvelle-Calédonie.
Je remercie également Jean-Pascal TORRETON, mon maître de stage, et Séverine JACQUET pour
tous les enseignements qu’elle m’a apportés.
J’associe à ces remerciements Alain LAPETITE, Sandrine CHIFFLET et Philippe GERARD pour
leur aide, ainsi que Miguel et Sam, les « pilotes » du bateau.
Je tiens aussi à exprimer toute ma gratitude à Pascal DOUILLET, Sylvain OUILLON et Olivier
PRINGAULT pour leur bonne humeur.
Enfin, je remercie tous les lofteurs et les autres: Héléna, Vincent, Ali, Aymeric, Romain (3),
Aurore, Barbara, Xavier (2), Kirti, K’ro, Audrey, Anaïs, Eric, Charles, Alexandre, Charlotte, Pierre
et Nicolas.

Sommaire
Sommaire 1. Introduction 1
1.1. IRD Nouvelle-Calédonie 1 1.2. Unité de Recherche CAMELIA 2 1.3. Contexte scientifique de l’étude 3 1.4. Description du site d’étude 4 1.5. Objectif du stage 6
2. Matériel et Méthodes 7
2.1. Stratégie d’échantillonnage 7 2.1.1. Choix des sites d‘échantillonnage 7 2.1.2. Echantillonnage 7
2.2. Bioessais 9 2.2.1. Enrichissements en nutriments 9 2.2.2. Cycles de mesures sur 24 heures 9 2.2.3. Validation du protocole 11
2.3. Variables biologiques mesurées 12 2.3.1. Chlorophylle a 12 2.3.2. Production primaire 12 2.3.3. Production bactérienne 15 2.3.4. Abondance bactérienne 17
2.4. Eléments nutritifs 18 2.4.1. Ammonium 18 2.4.2. Eléments nutritifs 18
2.5. Test-t sur populations hétéroscédastiques 18
3. Résultats – Discussion 19
3.1. Cycles de 24 heures 19 3.1.1. Caractéristiques physico-chimiques des stations échantillonnées 19 3.1.2. Station N12 19 3.1.3. Station M33 21 3.1.4. Mise en place du protocole 23
3.2. Test de validation 24 3.2.1. Caractéristiques physico-chimiques des stations échantillonnées 24 3.2.2. Validation sur la station N12 25 3.2.3. Validation sur la station M33 27 3.2.4. Validation sur la station N04 29 3.2.5. Choix des stations de mesures 30 3.2.6. Vérification des temps d’incubation 30 3.2.7. Nutriments limitants 31
4. Conclusion 33 Bibliographie 35 Annexes 37
- 1 -

Introduction
1 INTRODUCTION 1.1 IRD Nouvelle-Calédonie
L'Institut de recherche pour le développement (IRD) est un établissement à caractère scientifique et
technologique et dépend des ministères chargés de la Recherche et de la Coopération. Il satisfait
trois missions fondamentales : la recherche, l’expertise et la formation. Des programmes de
recherche scientifique, centrés sur l'étude des relations entre l'homme et son environnement, sont
développés à travers le monde (Afrique, Océan Indien, Amérique Latine, Asie et Pacifique), avec
comme cible le développement durable de ces régions. Ces recherches sont menées en coopération
avec des institutions d'enseignement supérieur et de recherche française ainsi qu'avec des
partenaires étrangers.
Le centre IRD de Nouméa en Nouvelle-Calédonie, créé en 1946 sous l'appellation IFO (Institut
Français d'Océanie), est le premier du Pacifique. En 1964, l'IFO prend le nom d'ORSTOM (Institut
Français de Recherche Scientifique pour le développement en Coopération), intitulé qu'il
conservera jusqu'en 1998. Par décret ministériel du 5 novembre 1998, l'ORSTOM devient IRD
(Institut de Recherche pour le Développement), et le centre de Nouméa devient centre IRD de
Nouvelle-Calédonie. Plusieurs disciplines scientifiques y sont représentées : océanographie,
écologie marine, géologie, géophysique, pharmacologie, agronomie, botanique, entomologie,
archéologie. Les recherches sont conduites en partenariat avec des institutions locales (Université
de la Nouvelle-Calédonie, Institut Agronomique Calédonien, IFREMER, Institut Pasteur, CNRS
…) ou régionales (Secrétariat général de la Communauté du Pacifique, University of the South
Pacific, Agence Universitaire de la Francophonie…). Actuellement, le centre est constitué de 13
Unités de Recherche (UR) et de 5 Unités de Service (US). Les premières ont pour mission
d'améliorer les connaissances scientifiques de la zone intertropicale, les secondes ont pour but
d’appliquer les résultats de la Recherche en répondant aux demandes d'expertise des partenaires
locaux. L’IRD remplit également un rôle de formation, le centre accueille des étudiants dans le
cadre de la formation continue et de la formation à la recherche. Intégrés dans les équipes, les
étudiants peuvent avoir le statut de stagiaires (du DEUG au DEA, IUT, école d’ingénieurs), de
thésards ou de post-doctorants.
Pour ma part, j’effectue un stage de deux mois d’élève-ingénieur S.T.E-I.S.I.M (Sciences et
Technologies de l’Eau à l’Institut des Sciences de l’Ingénieur de Montpellier) de 2e année au
sein de l’Unité de Recherche CAMELIA (Caractérisation et Modélisation des Echanges dans
les Lagons soumis aux Influences terrigènes et Anthropiques)
- 2 -

Introduction
1.2 Unité de Recherche CAMELIA Les écosystèmes tropicaux littoraux du pacifique sont soumis à des pressions anthropiques
croissantes. Les conséquences écologiques occasionnées ne sont pas sans effet sur l’économie des
états concernés. En effet, outre l’intérêt écologique, ces écosystèmes ont une importance
économique, sociale et culturelle majeure pour les pays qu’ils bordent. La pêche, et notamment la
pêche de subsistance, est vitale pour de nombreuses populations du pacifique. Le tourisme, lié
notamment aux récifs, est une source très importante de devises et constitue l'un des enjeux majeurs
de développement économique pour la plupart des états de cette zone. La bonne gestion de ces
environnements côtiers est donc une obligation écologique nécessaire à un maintien, voire un
développement économique.
L'Unité de Recherche CAMELIA s'inscrit dans le prolongement de la dynamique de recherche mise
en œuvre depuis 1996 avec le programme ECOTROPE (Ecosystèmes Côtiers et Tropicaux sous
influences terrigènes et anthropiques, 1996-2000) sur le thème général de l'influence des activités
humaines sur les écosystèmes côtiers tropicaux. Les recherches menées par CAMELIA portent sur
les mécanismes de transport et de transformation des apports en particules responsables de
l'hypersédimentation, des apports organiques et inorganiques responsables de l'eutrophisation et des
apports en métaux potentiellement inhibiteurs. Prévu sur une période de quatre ans, entre 2001 et
2004, l’UR étudie principalement deux sites lagonaires présentant des degrés d’eutrophisation
différents : le Lagon Sud Ouest de Nouvelle-Calédonie et le Lagon de Suva à Fidji.
Le Lagon Sud-Ouest de Nouvelle-Calédonie est sous influence anthropiques et terrigène, liée à la
présence de la ville de Nouméa, aux exploitations minières (nickel essentiellement) à ciel ouvert et
aux rivières (La coulée, la rivière des pirogues, la Dumbéa). On peut distinguer différentes sources
de pollution : les rejets d’eaux usées et les rejets liés aux activités industrielles et agricoles.
Différentes thématiques sont abordées au sein de l’Unité de Recherche CAMELIA :
1. Organisation des caractéristiques physico-chimiques des eaux et mécanismes de transport
2. Dynamique des échanges à l'interface eau-sédiment et influence des apports
3. Réponses des biocénoses pélagiques aux apports
4. Intégration des métaux dans la chaîne trophique et recherche de bio-indicateurs
5. Modélisation de la circulation, du transport et du fonctionnement biogéochimique du lagon
- 3 -

Introduction
1.3 Contexte scientifique de l’étude A ce jour, rares sont les études sur les impacts de l’eutrophisation en milieu côtier oligotrophe. La
structure et le fonctionnement des réseaux trophiques diffèrent fondamentalement entre milieux
oligotrophes, dominés par le réseau trophique microbien, et les milieux eutrophes, dominés par un
réseau trophique « herbivore » (Legendre et Rassoulzadegan, 1995 ; Joint et al, 2002). Les systèmes
oligotrophes fonctionnent au moyen d’un recyclage important de sels nutritifs, ce qui explique en
partie la prépondérance de la biomasse hétérotrophe. Dans ces conditions, les bactéries
hétérotrophes représentent des compétiteurs efficaces du phytoplancton vis-à-vis des nutriments
(Cotner et al, 2000 ; Joint et al, 2002). A l’opposé, les milieux eutrophes sont dominés par le
phytoplancton. Dans ces milieux, l’activité conjuguée des bactéries et de leurs consommateurs peut
amener un relargage des éléments nutritifs contenus dans la matière organique allochtone qui, en
s’ajoutant aux apports directs, augmenteront l’eutrophisation. Les expériences menées sur ce thème
montrent des modifications de la composition et du spectre de taille des communautés
planctoniques en fonction des apports en nutriments (Hein et Riemann, 1995 ; Eilers et al, 2000 ;
Henriksen et al, 2002 ; Carlson et al, 2002). On peut donc s’attendre à ce que l’eutrophisation d’un
système oligotrophe occasionne des modifications structurales des populations planctoniques.
Situés dans un environnement océanique oligotrophe tropical, les milieux récifo-lagonaires sont
généralement oligotrophes hors de l’influence des apports d’origine anthropique en raison du fort
taux de renouvellement de leurs eaux. Les peuplements planctoniques des milieux lagonaires non
perturbés sont limités par les ressources nutritives, ce qui les rend particulièrement sensibles aux
apports eutrophisants. Il existe peu de données sur la nature de cette limitation nutritive : celle-ci
varie selon les sites, les saisons et les communautés étudiées (Fisher et al, 1992 ; Holmboe et al,
1999 ; Lebaron et al, 2001). Il est donc impossible d’établir a priori la nature de cette limitation en
milieu lagonaire.
Les effets des apports eutrophisants sur le réseau trophique planctonique du Lagon Sud Ouest de
Nouvelle-Calédonie sont étudiés selon trois approches complémentaires au sein l’UR CAMELIA :
1. Etablir expérimentalement et comparer les réponses à des additions en éléments nutritifs des
différentes communautés planctoniques dans différentes situations trophiques allant de
l’oligotrophie à l’eutrophie.
2. Déterminer expérimentalement et comparer les caractéristiques cinétiques d’incorporation d’
NH4+ et de PO4
2- pour les bactéries hétérotrophes et le phytoplancton dans ces mêmes situations.
3. Etablir les relations in situ entre nutriments, abondance et activité des communautés
planctoniques au moyen de prélèvements périodiques.
- 4 -

Introduction
1.4 Description du site d’étude La Nouvelle-Calédonie est située au nord du tropique du Capricorne (22°S et 166°E). D'une
superficie totale de 18 575,5 km², l'archipel est constitué de "la Grande Terre", des îles Loyauté
(Ouvéa, Lifou, Maré, Tiga), de l'archipel des Belep, de l'Ile des Pins et de quelques îlots lointains
(figure 1). La Grande Terre se présente comme une bande d'environ 400 km de long sur 40 à 70 km
de large, traversée du nord au sud par une chaîne montagneuse. La chaîne centrale coupe l'île en
deux régions distinctes : la côte Est exposée aux alizés est la région la plus humide et la plus
chaude ; une végétation luxuriante y couvre les pentes abruptes ; la côte Ouest est plus découpée,
avec des plaines propres à la culture et à l'élevage, surplombées par des massifs riches en minerais.
La Nouvelle-Calédonie est caractérisée par un climat tropical humide où l’on distingue deux
saisons : la saison chaude (mi-novembre, mi-avril) de 25 à 27°C, marquée par une forte
pluviométrie avec des dépressions tropicales qui se produisent de manière discontinue ; la saison
fraîche (mi-mai, mi-septembre) de 20 à 23 °C. Deux régimes de vent prédominent : les vents d’Est
ou Alizés de direction ESE et SE prépondérants toute l’année et des coup d’Ouest plus occasionnels
de direction NW et SW généralement associés au passage des dépressions et des cyclones. Les
vents conditionnent largement le renouvellement et la circulation des eaux lagonaires (Douillet,
1998)
Figure 1 - localisation géographique de la Nouvelle-Calédonie
- 5 -

Introduction
Le Lagon Sud-Ouest de Nouvelle-Calédonie a une superficie de 2000 km². Les rivières des
Pirogues, de La Coulée, et de Dumbéa représentent les principaux apports en eau douce dans le
lagon. La présence de trois passes (Boulari, Dumbéa, Uitoé) au niveau du récif barrière permet des
échanges entre le lagon et l’océan. Le Lagon comprend le site urbain de Nouméa, et les zones
estuariennes environnantes, soit une superficie de 600 km² (figure 2). Nouméa regroupe 60 % de la
population sur 1 % du territoire. Au cours des trois dernières décennies, la ville a connu un essor
extrêmement rapide et compte 100 000 habitants. Ainsi les baies peu profondes (environ 10m),
autour de Nouméa sont soumises à une forte pression anthropique et terrigène.
30
5
8
34
166°16'E 166°26'E 166°36'E 166°46'E
22°10'S
22°20'S
22°30'S
22°40'S
TerreRécifs
N
Noum
éa
Piro
gues
Coul
éeDumbéa
1020
20
10
30
20
20
30
20
10
20
2030
40
20
20
2030
20
20
Nouméa
20° S
21° S
22° S
23° S
164° E 165° E 166° E 167° E 168° E
OCEANPACIFIQUE
NOUVELLE - CALEDONIE
ILES LOYAUTESOuvéa
Lifou
Maré
Figure 2 – Lagon Sud-Ouest de Nouvelle-Calédonie
- 6 -

Introduction
1.5 Objectifs du stage Le stage que j’effectue au sein de l’UR CAMELIA s’inscrit dans le travail de thèse de Séverine
Jacquet (dirigée par J-P Torréton). Cette thèse traite de « l’influence des apports en nutriments sur
le fonctionnement du réseau trophique microbien dans le Lagon Sud-Ouest de Nouvelle-
Calédonie » et s’intègre essentiellement dans le thème d’action de CAMELIA : réponses des
biocénoses pélagiques.
Un premier travail réalisé dans le cadre d’un stage de DEA (Jacquet, 2001) a permis d’évaluer
l’importance comparée des productions bactérienne et primaire planctoniques dans un gradient
d’eutrophisation dans le Lagon Sud-Ouest de Nouvelle-Calédonie et leurs relations avec les
variables physico-chimiques. Cette étude a mis en évidence le contrôle potentiel par l’azote minéral
dissous de la production et de la biomasse phytoplanctonique, et un contrôle potentiel par l’azote
minéral dissous de la production et du taux de croissance bactérien. Elle a également montré
l’augmentation de la taille des producteurs primaires en réponse à un apport en azote minéral
dissous. Les objectifs de la thèse faisant suite à ce DEA sont : (1) d’étendre cette étude sur une plus
large échelle spatiale et temporelle, (2) d’approfondir l’étude des changements des communautés
planctoniques et de leurs activités avec l’eutrophisation.
L’aspect limitant de l’azote minéral dissous a été mis en évidence au travers de l’étude des relations
in situ entre nutriments d’une part, et biomasses et productions phytoplanctonique et bactérienne,
d’autre part. Il s’agit maintenant de compléter cette étude par une approche in vitro des réponses des
communautés planctoniques à des enrichissements. Une première expérience sur l’addition de
nutriments a été réalisée sur le lagon par J-P Torréton en 2000 (com. pers.). Dans les conditions
d’expérimentations (grazing, longues durées d’incubation), l’azote apparaît comme le premier
facteur limitant des activités planctoniques. Toutefois, des biais dans les résultats (exemple : forte
augmentation de la production bactérienne dans les témoins) montrent la nécessité de mettre en
place un protocole permettant d’avoir un temps d’incubation plus court.
L’objectif de mon stage est de mettre en place ce protocole. Il s’agit plus précisément de déterminer
un temps d’incubation minimal permettant d’avoir une réponse suffisante et limitant les biais liés au
confinement (augmentation de la production bactérienne et modification des communautés
planctoniques dans les contrôles non enrichis) et de tester ensuite ce protocole sur des situations
trophiques contrastées pour en vérifier la validité.
- 7 -

Matériel et méthodes
2 MATERIEL ET METHODES 2.1. Stratégie d’échantillonnage
2.1.1. Choix des sites d’échantillonnage
Pour la mise en place du protocole, deux stations d’échantillonnage ont été choisies : N12 et M33
(figure 3). Ces stations, étudiées au cours d’un suivi saisonnier dans le cadre de la thèse de S.
Jacquet, sont représentatives de situations trophiques contrastées et pérennes.
M33 est située dans le chenal lagonaire hors influence anthropique.
N12 est située dans la baie de Sainte-Marie où sont déversées les eaux usées non traitées de
l’agglomération de Nouméa.
Pour les tests de routine, les stations choisies sont N04, N12, M33 (figure 3) afin d’avoir un
gradient trophique.
N04 est située en fond de baie de Sainte-Marie à proximité des zones de rejets.
Figure 3 – Répartition des stations d’échantillonnage dans le lagon Sud-Ouest de Nouvelle-Calédonie
- 8 -

Matériel et méthodes
2.1.2 Echantillonnage
Les opérations en mer sont effectuées à bord du « Coris », navire de l’IRD. Les prélèvements sont
réalisés aux mêmes heures, le matin (7h00 - 8h00) pour la phase de mise en place du protocole et en
début d’après-midi (~13 heures) pour les tests de routine. L’eau est prélevée à l’aide de bouteilles
NISKIN de 5 litres (figure 4 a) à une profondeur de 3 m pour éviter l’éventuelle couche de
dessalure après de fortes pluies.
Chaque prélèvement est accompagné de mesures des paramètres physico-chimiques (température,
salinité, conductivité, pression) à l’aide de la sonde CTD (Conductivity Temperature Depth)
Seabird SBE 19 (figure 4 b). Des capteurs supplémentaires sont utilisés :
un fluorimètre permettant la détermination in situ de la concentration en chlorophylle
un capteur PAR (Photosynthetically Active Radiation) permettant l’estimation de
l’atténuation lumineuse en fonction de la profondeur
un néphélomètre mesurant la turbidité des eaux traversées par la sonde.
La sonde, fixée à un boute, est descendue à vitesse régulière pour acquérir ces paramètres sur
l’ensemble de la colonne d’eau.
Figure 4 : (a) bouteille Niskin de cinq litres, (b) Sonde CTD Seabird SBE 19
- 9 -

Matériel et méthodes
2.2. Bioessais Les bioessais ont pour objet d’établir et comparer les réponses à des additions en éléments nutritifs
des différentes communautés planctoniques : les bactéries hétérotrophes et le phytoplancton.
2.2.1 Enrichissements en nutriments
On choisit d’enrichir les milieux en azote (N), phosphore (P) et carbone (C) fournis respectivement
sous la forme chlorure d’ammonium (H4ClN), phosphate di-sodium hydrogéné (HNa2O4P) et
glucose (C6H12O6). Ces nutriments sont ceux utilisés dans la plupart des expériences
d’enrichissements (Cotner et al., 2000 ; Fisher et al., 2002 ; Henriksen et al., 2002 ; Joint et al.,
2002). L’azote sous forme ammonium présente l’avantage d’être directement assimilable par les
compartiments bactériens et phytoplanctoniques, contrairement au nitrate qui nécessite un
catabolisme supplémentaire chez les bactéries hétérotrophes entraînant un coût énergétique élevé.
Le glucose est aussi facilement et rapidement dégradable par les bactéries (Joint et al., 2002). Les
flacons destinés aux mesures d’activités phytoplanctoniques ne sont pas enrichis en glucose car la
production primaire est déterminée par incorporation de 14C-bicarbonate.
La quantité de nutriments ajoutée est calculée sur la base des concentrations moyennes en NH4+
mesurées au cours du suivi saisonnier (avril 2002-avril 2003) sur les stations N12 (0,163 µM) et
M33 (0,045 µM). On choisit d’ajouter une concentration en NH4+ environ cinq fois supérieure à
celle mesurée in situ en N12. La concentration en NH4+ ajoutée est 1 µM. Les concentrations des
autres nutriments sont calculées en se basant sur le rapport de Redfield, 1963 (C:N:P = 106:16:1).
Ce rapport correspond à une première approximation de la composition de la matière organique
océanique nécessaire à une croissance équilibrée du phytoplancton. Chez les bactéries hétérotrophes
des communautés côtières, le ratio est très proche de celui du phytoplancton (Kirchman, 2001). La
quantité de phosphore ajoutée est donc de 0,625 µM (10 fois supérieure à la moyenne annuelle en
N12) et celle de carbone 6,625 µM. Il n’existe pas de règle en matière de quantité ajoutée dans les
expériences d’enrichissement, en effet le choix des concentrations dans la littérature est assez
variable et pas toujours justifié (Fisher et al., 1992 ; Cotner et al, 2000). Se reporter à l’annexe 1
pour les préparations des solutions mère de nutriments ajoutés.
2.2.2 Cycles de mesures sur 24 heures
Cette étape constitue la phase de mise en place du protocole de mesure des réponses des
communautés planctoniques à des enrichissements. L’objectif étant de déterminer un temps optimal
d’incubation et donc un signal réponse conséquent, les milieux seront enrichis de manière à ce
qu’ils ne soient pas limités (C, N, P pour le compartiment bactérien et N, P pour le compartiment
- 10 -

Matériel et méthodes
phytoplanctonique). Les activités bactériennes et phytoplanctoniques sont mesurées selon un pas de
temps de 5 heures jusqu’à 24 heures d’incubation totale.
Les cycles de mesures sont réalisés le 22 juillet 2003 pour la station N12 et le 31 juillet 2003 pour la
station M33.
Variables mesurées avant ajout de nutriments (eau brute) : production bactérienne, abondance
bactérienne, concentrations en chlorophylle a, ammonium, nitrate, phosphate, azote et phosphore
organique dissous.
Etude du compartiment bactérien : 250 µl d’une solution mère CNP (C : glucose ; N : NH4Cl ;
P : HNa2O2P ) sont injectés dans un flacon de 500 ml (réplicat). Deux flacons témoins non enrichis
de même volume sont soumis aux mêmes conditions expérimentales. Les flacons sont mis à incuber
à température in situ, à l’obscurité pour empêcher un développement du compartiment
phytoplanctonique. On prélève de l’eau dans chacun des flacons à t = 5, 10, 15, 20 et 24 heures pour
la mesure de la production bactérienne (protocole cf. § 2.1.3). Les variables mesurées à t = 24
heures sont la production bactérienne, l’abondance bactérienne et la concentration en NH4+.
Etude du compartiment phytoplanctonique : 250 µl d’une solution mère NP (N : NH4Cl ; P :
Hna2O2P) sont injectés dans 5 flacons de 76 ml. 5 témoins non enrichis seront soumis aux mêmes
conditions expérimentales. Les flacons sont placés à incuber à température in situ et à lumière
constante 200 µE.m-2.s-1 (tubes fluorescents) dans un bac en plexiglas crée pour les expériences in
vitro (figure 5). Une circulation d’eau dans le bac permet de maintenir une température constante
palliant à l’échauffement engendré par les tubes fluorescents. Un flacon enrichi et un non enrichi
sont retirés du bac à t = 5, 10, 15, 20 et 24 heures pour des mesures de production primaire
(protocole cf. § 2.1.2).
Tubes fluorescents
200 µE.m-2.s-1
Bac en plexiglas contenant les flacons de 76 ml : 5 enrichis avec 250 µl de la solution mère NP et 5 témoins
: sens de circulation de l’eau
Figure 5 : dispositif expérimental pour l’étude du compartiment phytoplanctonique
- 11 -

Matériel et méthodes
2.2.3 Validation du protocole
Une fois le protocole mis en place, il sera appliqué aux stations N04, N12 et M33 pour validation.
Les expériences sont réalisées le 7 août 2003 pour N12, le 12 août 2003 pour M33 et le 14 août
2003 pour N04.
Variables mesurées avant ajout de nutriments (eau brute) : production bactérienne, abondance
bactérienne, concentrations en chlorophylle a, ammonium, nitrate, phosphate, azote et phosphore
organique dissous.
Etude du compartiment bactérien : afin de tester les réponses des communautés bactériennes à
des enrichissements, différents milieux seront élaborés (figure 6). Pour chaque milieu, 250 µl de la
solution mère sont injectés (réplicats). Deux flacons témoins non enrichis de même volume sont
soumis aux mêmes conditions expérimentales. L’incubation se fera dans les conditions déterminées
par les cycles de mesures. Les variables mesurées au terme de l’incubation sont la production
bactérienne et la concentration en ammonium pour chacun des milieux et l’abondance bactérienne
pour le milieu qui montre la réponse la plus importante à l’ajout de nutriments.
s
ph
m
de
da
m
Le
ét
co
Témoin
Figure 6 : enrichis
250 µ 250 µl 250 µl m
250 µ
Etude du com
ytoplanctoniques
ère P), et NP (sol
ux témoins non e
ns flacons de 76
esures.
s cycles de 24
endus à d’autres
mmandes en reta
+ CNP l de la solution
mère CNP
sements effectués pour l’étu
partiment phytoplancto
à des enrichissements,
ution mère NP) seront e
nrichis. Pour chaque m
ml. L’incubation se f
heures et les validation
stations pour des rais
rd).
+ N de la solution
ère N
de des réponses du comparl
nique : afin de tester
des milieux enrichis e
ffectués (2 flacons par
ilieu, 250 µl de la solu
era dans les conditio
s n’ont pu être répété
ons de temps et de
- 12 -
+ P de la solutionmère P
timent bactérien. Flacon
les réponses des
n N (solution mère N
enrichissement), av
tion mère considéré
ns déterminées par
s sur les stations c
moyens matériels (
+ NP l de la solutionmère NP
Flacons V = 500 m
s de V = 500 mlcommunautés
), P (solution
ec en parallèle
e sont injectés
les cycles de
onsidérées ou
problèmes de

Matériel et méthodes
2.3. Variables biologiques mesurées 2.3.1 Chlorophylle a
La concentration en chlorophylle a a été mesurée par le laboratoire de Chimie Marine de l’IRD. Les
mesures ont été effectuées après filtration de 200 ml d’échantillon sur filtres Whatman GF/F
(porosité 0.7 µm). Les concentrations en chlorophylle a ont ensuite été déterminées par fluorimétrie
(fluorimètre TURNER TD 700) selon la méthode de Yentsch et Menzel (1963). Les concentrations
des pigments sont données par les formules de Lorenzen (1966), modifiées par Jeffrey et Humphrey
(1975) (formule 1) :
[ ]( ) ( ) DKKV
FFlµgachla
a ×⋅××
×⋅=−
1000)(. 0
1 ν et [ ]( ) DKKKV
FKFKlgPhéoaaa
aa ×−×××
×−××=−
)(1000. 01 νµ
Formule 1 – Formules de Lorenzen (1966) modifiées
Avec :
ν : volume de l’extrait (7 ml) V : volume filtré (ml) D : facteur de dilution
Ka : facteur d’étalonnage du fluorimètre pour la phéophytine a pure
K : facteur d’étalonnage du fluorimètre pour la Chlorophylle a pure
F0, Fa : fluorescence de l’échantillon avant et après acidification
La teneur en Chlorophylle a peut être considérée comme un estimateur de la biomasse
phytoplanctonique. Néanmoins, la relation entre la concentration en chlorophylle et le carbone
phytoplanctonique n’est pas constante (Hein & Riemann, 1995). Cette relation dépend de l’espèce
et de l’état physiologique des organismes. La concentration en Chlorophylle a est donc un
indicateur de la biomasse chlorophyllienne plutôt qu’une valeur de référence.
2.3.2 Production primaire
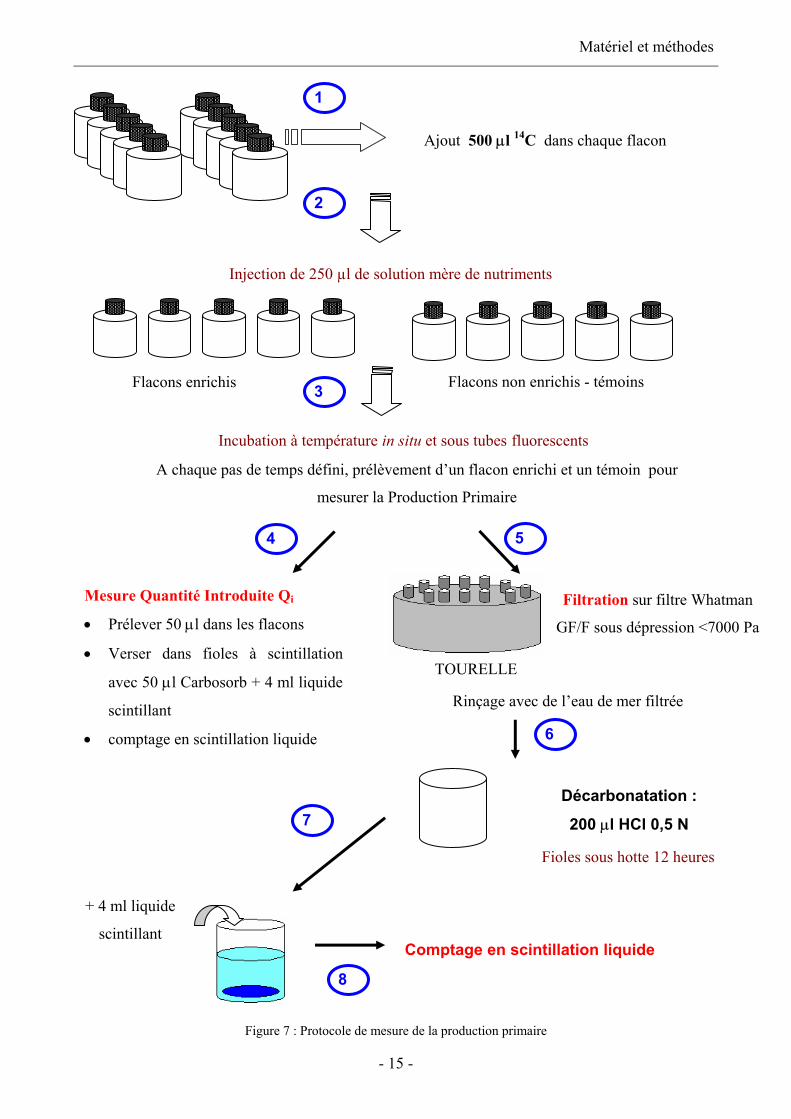
Protocole (figure 7)
La production primaire est déterminée par incorporation de 14C-bicarbonate (Steeman-Nielsen,
1992). On mesure ainsi la quantité de carbone incorporée dans le compartiment autotrophe au cours
de la photosynthèse. Dans chaque flacon de V = 76 ml, on injecte 500 µl de 14C-bicarbonate
correspondant à 13,33 µCi final. Les flacons sont ensuite placés dans le bac d’incubation en
plexiglas (cf. § 2.1.2). La quantité totale de radioactivité introduite est vérifiée dans chaque flacon.
Pour cela et avant toute autre manipulation, 50 µl sont prélevés dans chaque flacon et placés dans
des fioles à scintillation contenant 4 ml de liquide scintillant (Packard Ultima Gold) et 50 µl de
Carbosorb. Après incubation, le phytoplancton est collecté par filtration (<7000 Pa) sur filtre
Whatman GF/F préalablement mouillé avec de l’eau de mer filtrée. Les filtres sont ensuite rincés
- 13 -

Matériel et méthodes
avec 5 ml d’eau de mer filtrée. Les filtres sont ensuite placés dans des fioles à scintillation et
immédiatement décarbonatés par ajout de 250 µl d’acide chlorhydrique HCl 0,5 N. Les fioles sont
placées sous la hotte pendant la nuit et la quantité de radioactivité incorporée est mesurée après
ajout de 4 ml de liquide scintillant. Se reporter à l’annexe 2 pour le principe du marquage par un
isotope radioactif et du comptage par scintillation.
Calcul de la production primaire (formule 2)
WV
Vdpm
dpmdpmhmmgCPPf
P
t
be ×××
×−
=−− 05,11000)..( 13
Formule 2 - calcul de la production primaire
Avec :
dpme : désintégration par min dans l’échantillon, et dpmb à l’obscurité
Vp : volume prélevé pour la mesure de radioactivité introduite (µl)
Vf : volume d’échantillon filtré (ml)
1,05 : fractionnement isotopique entre 12C et 14C
W : concentration en carbonates (mgC.m-3)
Remarque : La concentration en carbonates a été obtenue par des mesures d’alcalinité totale par le
laboratoire de chimie de l’IRD sur des situations contrastées du lagon Sud Ouest. La valeur
moyenne est de 28215 mgC.m-3 (σ = 215 mgC.m-3).
- 14 -
Matériel et méthodes
1
Rinçage avec de l’eau de mer filtrée
2
Injection de 250 µl de solution mère de nutriments
3 Flacons non enrichis - témoins Flacons enrichis
4 5
Filtration sur filtre Whatman
GF/F sous dépression <7000 Pa
TOURELLE
6
A chaque pas de temps défini, prélèvement d’un flacon enrichi et un témoin pour
mesurer la Production Primaire
Incubation à température in situ et sous tubes fluorescents
Ajout 500 µl 14C dans chaque flacon
Mesure Quantité Introduite Qi
• Prélever 50 µl dans les flacons
• Verser dans fioles à scintillation
avec 50 µl Carbosorb + 4 ml liquide
scintillant
• comptage en scintillation liquide
7
+ 4 ml liquide
scintillant
Fioles sous hotte 12 heures
Décarbonatation :
200 µl HCl 0,5 N
8
Comptage en scintillation liquide
Figure 7 : Protocole de mesure de la production primaire
- 15 -

Matériel et méthodes
2.3.3. Production bactérienne
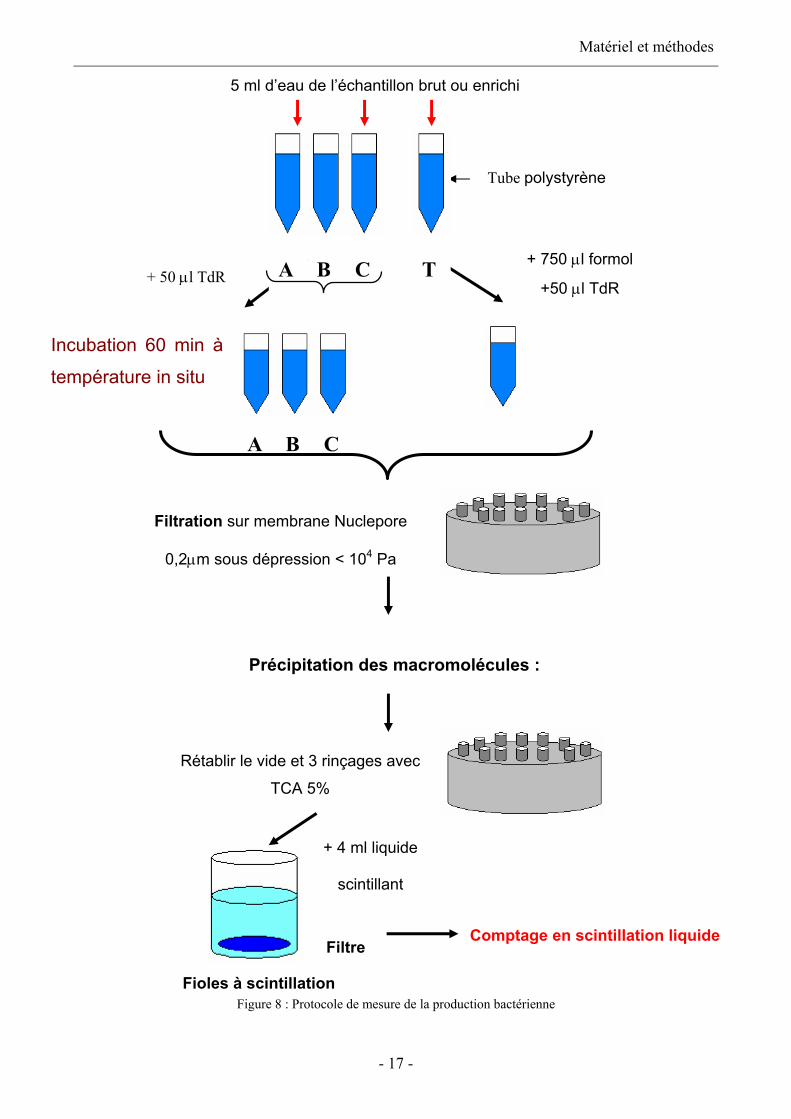
Protocole (figure 8)
La production bactérienne est mesurée par incorporation de [menthyl-3H] thymidine (TdR)
(Amersham, 1.92 TBq.mmol-1) dans le TCA précipité suivant le protocole de Torréton et Dufour
(1996). La thymidine, précurseur de la synthèse d’ADN, permet d’obtenir une estimation de la
synthèse d’ADN bactérien.
Des réplicats (parfois triplicatas) de 5 ml sont réalisés dans lesquels 50 µl de TdR sont injectés, soit
une concentration finale de 7.5 nM (un essai réalisé dans le cadre du DEA S. Jacquet, 2001 montre
que l’incorporation saturait dès 2 nM). Les tubes sont mis à incuber pendant 60 minutes à
température in situ ( ± 1°C). L’incorporation est stoppée en injectant 750 µl de formol (37%
tamponné au borate de sodium). Un témoin est réalisé pour chaque réplicat. Ce témoin est formolé
avant addition de TdR, puis est placé au froid. Après incubation, les bactéries sont collectées par
filtration sous faible dépression (104 Pa) sur filtres en polycarbonate (Nuclepore) de porosité 0,2 µm
reposant sur des membranes en microfibres de verre Whatman GF/D, préalablement imbibés avec
250 µl de thymidine non radioactive. Une fois la filtration faite, le vide est rompu et les
macromolécules sont précipitées avec de l’acide TriChloroAcétique (TCA 5% w/v) froid dans les
tourelles de filtration. Après 15 minutes à 4°C, le vide est rétabli et on rince les tourelles de
filtration trois fois avec du TCA 5% froid. Les filtres sont mis dans des fioles à scintillation, dans
lesquelles on ajoute 4 ml de liquide scintillant. Les fioles sont alors placées dans le compteur à
scintillation afin de déterminer la quantité de radioactivité incorporée par les bactéries.
L’autoabsorption dans le précipité TCA des radiations faiblement énergétiques du tritium a été
déterminée au cours d’une précédente étude par incubation d’une série d’échantillons en double
(Jacquet, 2001). L’un des lots a subi une hydrolyse acide à chaud (HCl 0 ;5 N, 100°C, 30 min) et
comparé au traitement standard. Le rapport entre les échantillons hydrolysés et non hydrolysés est
de 1,3 en moyenne (σ= 0,1). Un facteur correctif a donc été appliqué aux mesures. Se reporter à
l’annexe 2 pour le principe du marquage par un isotope radioactif et du comptage par scintillation.
Calcul de la production bactérienne
ASKTVdpmdpmhpMPB
fblancréplicats ×
×××−=−12
1 1060100)().(
Formule 4 – calcul la production bactérienne
dpmréplicats : radioactivité moyenne des réplicats T : temps d’incubation (minutes)
dpmblanc : radioactivité du témoin Vf : volume d’échantillon filtré (ml)
AS : activité spécifique de la solution mère de TdR (1920 TBq.mol-1)
K : 60.1012 TBq.mol-1
- 16 -
Matériel et méthodes
i
R
Incubation 60 min à
température in situ
e
A B C
A B C T
F
Précipitation des macromolécules :
Rétablir le vide et 3 rinçages avec
TCA 5%
ee
igure 8 : Proto
Filtr
n
Fioles à scintillatio+ 4 ml liquide
scintillant
cole de mesure de la produ
- 17 -
Comptage en scintillation liquid
+ 50 µl Td
ction bacté
+ 750 µl formol
+50 µl TdR
Tube polystyrèn
Filtration sur membrane Nuclepore
0,2µm sous dépression < 104 Pa
5 ml d’eau de l’échantillon brut ou enrich
rienne

Matériel et méthodes
2.3.4 Abondance bactérienne
La technique de comptage bactérien utilisée est la microscopie à épifluorescence. Le marqueur
utilisé est le DAPI (4’6-diamidino-2-phenylindole), il se fixe sur l’ADN. Quand le complexe ADN-
DAPI est excité avec une lumière d’une longueur d’onde comprise entre 330 et 380 nm il fluoresce
dans le bleu. Le DAPI fixé sur du matériel non ADN peut fluorescer en jaune pâle. Cette méthode
améliore la visualisation des bactéries et leur distinction des cyanobactéries de taille inférieure à 1
µm (Porter & Feig, 1980). Se reporter à l’annexe 3 pour le principe de la microscopie à
épifluorescence.
Protocole
On filtre (< 10 mm Hg) 10 ml d’échantillon, fixé au formol (2 % final) immédiatement après le
prélèvement, sur une membrane en polycarbonate noire (Nuclepore) de porosité 0,2 µm. Lorsqu’il
reste environ 1 ml de l’échantillon dans la tourelle de filtration, le vide est rompu et on ajoute 4
gouttes de DAPI dilué. La coloration au DAPI dure 10 minutes à l’obscurité puis on reprends la
filtration jusqu’à ce que le filtre soit sec. Le filtre et le sous-filtre sont ensuite placés dans une boîte
de Pétri que l’on conserve à –20°C jusqu’au comptage.
Comptage
Les bactéries sont comptées au microscope à épifluorescence. Plus de 400 bactéries ont été
dénombrées sur au moins 20 champs pour obtenir un coefficient de variation de la moyenne par
champs d’environ 10%.
Calcul de la production bactérienne
66
101183191100)/10( ××××=
fT VcC
BmlBBactéries
Formule 5 – Calcul de l’abondance bactérienne
Avec :
BT : nombre total de bactéries comptées
C : nombre de carreaux par champs sur lesquels sont dénombrées les bactéries
c : nombre de champs comptés
Vf : volume filtré (ml)
18319 : nombre de champs de 100 carreaux sur un filtre de 25 mm
- 18 -

Matériel et méthodes
2.4. Eléments nutritifs 2.4.1 Ammonium
Principe
Le dosage des ions ammonium est fait par fluorimétrie. Pour cela, de l’OrthoPhtaldiAldehyde
(OPA) est ajouté dans chaque échantillon et forme avec l’ammonium un complexe mesurable par
fluorimètrie. Pour chaque série d’analyse, une gamme d’étalonnage complète est effectuée en début
et fin d’analyse. Un étalon interne est inséré tous les vingt échantillons pour vérifier qu’il n’y a pas
de dérive du signal.
Protocole
Quatre flacons de 40 ml sont remplis d’eau provenant des stations étudiées, 2ml de réactif OPA sont
ajoutés. Ils sont ensuite mis à incuber 6 heures à l’obscurité le temps que la réaction de
complexation se fasse entre les ions ammoniums et l’OPA. La fluorescence dans le flacon est
ensuite mesurée et convertie en concentration avec la relation établie grâce à la gamme
d’étalonnage.
2.4.2. Eléments nutritifs
Les mesures en nitrates, phosphates, et silicates ainsi que l’estimation de l’azote organique dissous
et du phosphore organique dissous ont été réalisées par le Laboratoire de Chimie Marine de l’IRD à
l’aide d’un Technicon selon une méthode colorimétrique.
2.5. Test-t sur populations hétéroscédastiques Le test de comparaison t considère deux populations normales de moyenne µ1 et µ2 et de variance
commune σ 2. Lorsque les variances s’avèrent inégales, il est préférable d’utiliser un test t modifié.
Ce test a été appliqué sur les données pour déterminer si les réponses aux enrichissements sont
significatives.
- 19 -
Résultats
3 RESULTATS - DISCUSSION 3.1 Cycles de 24 heures
3.1.1 Caractéristiques physico-chimiques des stations échantillonnées
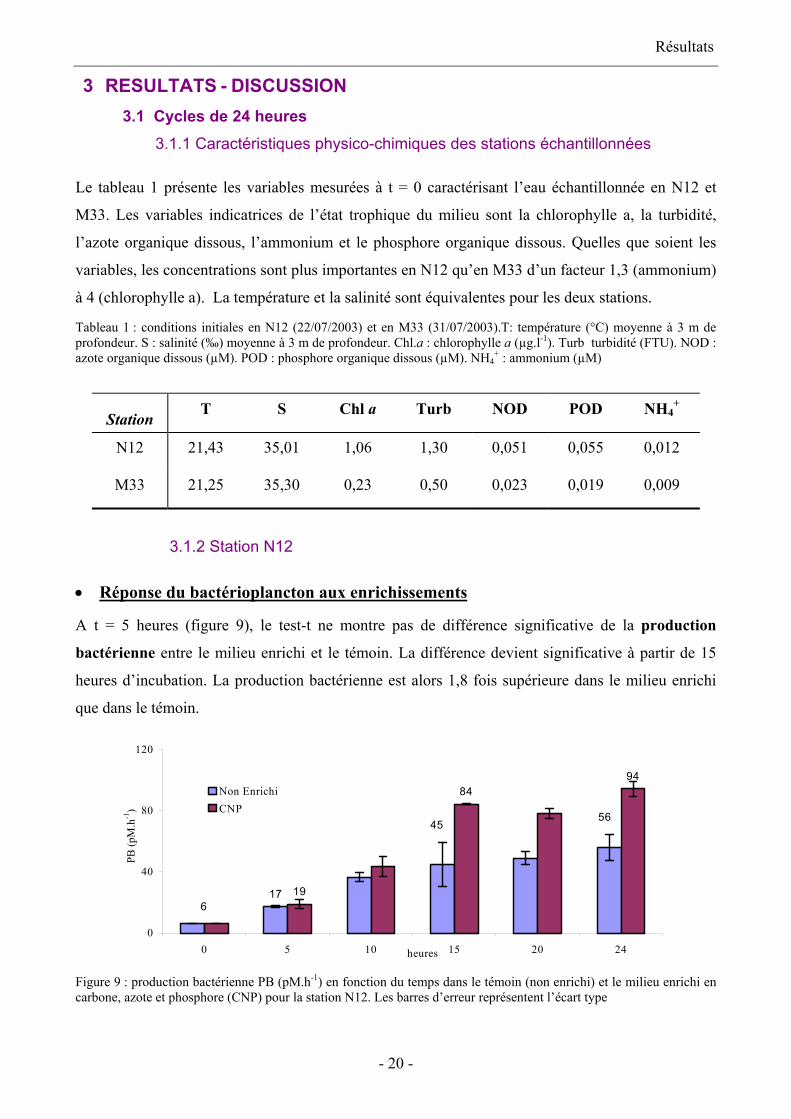
Le tableau 1 présente les variables mesurées à t = 0 caractérisant l’eau échantillonnée en N12 et
M33. Les variables indicatrices de l’état trophique du milieu sont la chlorophylle a, la turbidité,
l’azote organique dissous, l’ammonium et le phosphore organique dissous. Quelles que soient les
variables, les concentrations sont plus importantes en N12 qu’en M33 d’un facteur 1,3 (ammonium)
à 4 (chlorophylle a). La température et la salinité sont équivalentes pour les deux stations.
Tableau 1 : conditions initiales en N12 (22/07/2003) et en M33 (31/07/2003).T: température (°C) moyenne à 3 m de profondeur. S : salinité (‰) moyenne à 3 m de profondeur. Chl.a : chlorophylle a (µg.l-1). Turb turbidité (FTU). NOD : azote organique dissous (µM). POD : phosphore organique dissous (µM). NH4
+ : ammonium (µM)
Station T S Chl a Turb NOD POD NH4+
N12 21,43 35,01 1,06 1,30 0,051 0,055 0,012
M33 21,25 35,30 0,23 0,50 0,023 0,019 0,009
3.1.2 Station N12
Réponse du bactérioplancton aux enrichissements •
A t = 5 heures (figure 9), le test-t ne montre pas de différence significative de la production
bactérienne entre le milieu enrichi et le témoin. La différence devient significative à partir de 15
heures d’incubation. La production bactérienne est alors 1,8 fois supérieure dans le milieu enrichi
que dans le témoin.
8494
4556
617 19
0
40
80
120
0 5 10 15 20 24heures
PB (p
M.h
-1)
Non EnrichiCNP
Figure 9 : production bactérienne PB (pM.h-1) en fonction du temps dans le témoin (non enrichi) et le milieu enrichi en carbone, azote et phosphore (CNP) pour la station N12. Les barres d’erreur représentent l’écart type
- 20 -

Résultats
L’abondance bactérienne dans le témoin après 24 heures d’incubation est peu différente de celle
mesurée à t = 0 (tableau 2). Dans le milieu enrichi, on observe une augmentation de l’abondance
bactérienne de 28 %. Tableau 2 : Abondance bactérienne AB (106 cell.ml-1) pour la station N12 à t =0 et t= 24 heures dans le milieu témoin et le milieu enrichi en CNP (carbone, azote et phosphore)
AB
t = 0
AB
t= 24 heures
Milieu Témoin 0.99 0.96
Milieu CNP
0.99
1.27
La concentration en NH4+ ajoutée dans le flacon enrichi est de 1 µM : la concentration initiale en
NH4+ est donc de 1,012 µM dans ce milieu. La concentration en NH4
+ au bout de 24 heures
d’incubation, est de 0.037 µM dans le milieu enrichi et 0,057 µM dans le témoin.
Réponse du phytoplancton aux enrichissements •
Dès 5 heures d’incubation, la production primaire instantanée (figure 10a) dans le
milieu enrichi en NP est deux fois plus importante que dans le témoin. A partir de 20
heures d’incubation, la production primaire instantanée dans le milieu enrichi est trois
fois plus importante que dans le témoin. La production primaire cumulée (figure 10b)
est aussi trois fois plus forte dans le milieu enrichi que dans le témoin, celui-ci ayant
peu varié depuis le début de l’incubation.
- 21 -

Résultats
3,7
1 ,31,7
6 ,4
3 ,84 ,3
0
2
4
6
5 10 15 20 24 he u re s
PP (µ
gC.l-
1.h-1
)
n o n en r ich i
N P
1925
32
77
0
30
60
90
5 10 15 20 24 heures
PP (µ
gC.l-1
)
non enrichi
NP
b
a
Figure 10: (a) production primaire instantanée PP (µgC.l-1.h-1); (b) production primaire cumulée PP (µgC.l-1.h-1) en fonction du temps dans les témoins (non enrichi) et les milieux enrichis en azote et phosphore (NP) pour la station N12
3.1.3 Station M33
• Réponse du bactérioplancton aux enrichissements
La figure 11 représente la production bactérienne dans les milieux témoin et enrichis en CNP au
cours du temps pour la station M33. Le test-t ne montre pas de différence significative entre le
témoin et l’enrichi jusqu’à t=10 heures. A partir de 15 heures d’incubation la différence est
significative jusqu’à la fin de l’expérience. L’activité dans le témoin reste faible en comparaison de
celle des flacons enrichis. A t = 20 heures, la production bactérienne dans le témoin représente 9%
de celle dans le milieu CNP. La production bactérienne décroît dans le milieu enrichi après 24
heures, ce qui pourrait être dû à la consommation totale des nutriments du milieu. Ce qui est
confirmée par la concentration en ions ammonium proche de zéro dans le milieu enrichi au bout de
24 heures d’incubation.
- 22 -
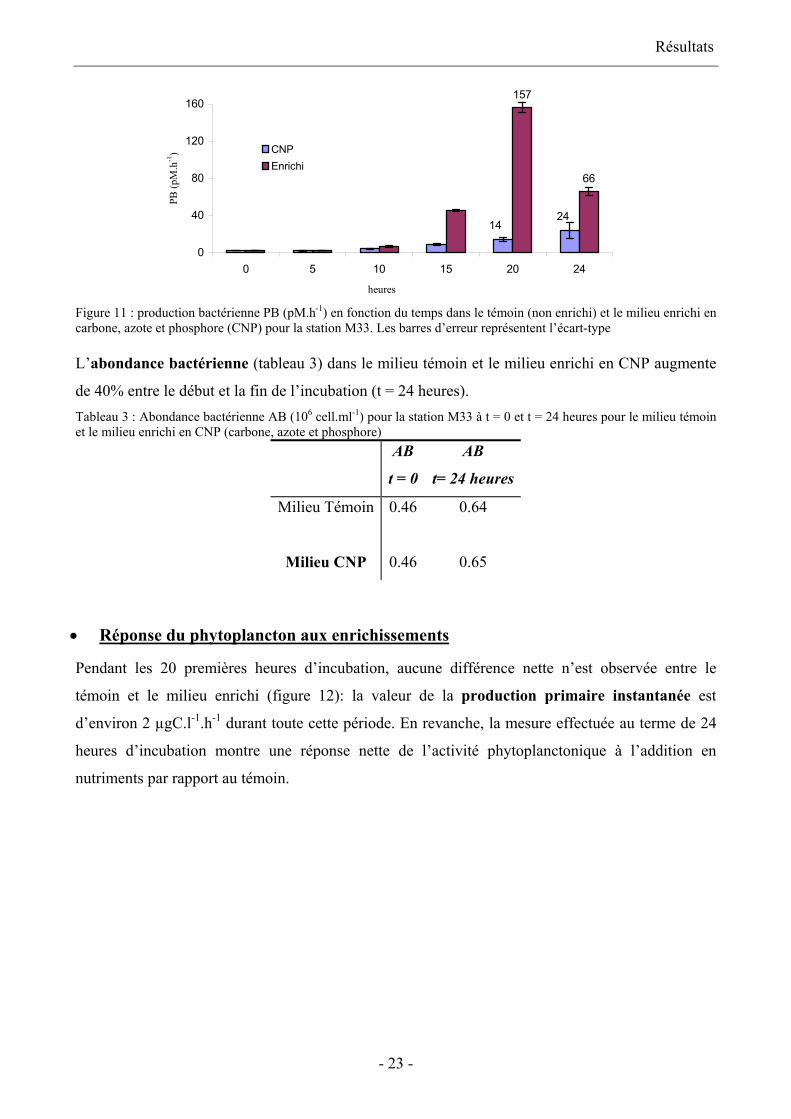
Résultats
157
66
2414
0
40
80
120
160
0 5 10 15 20 24
heures
PB (p
M.h
-1) CNP
Enrichi
Figure 11 : production bactérienne PB (pM.h-1) en fonction du temps dans le témoin (non enrichi) et le milieu enrichi en carbone, azote et phosphore (CNP) pour la station M33. Les barres d’erreur représentent l’écart-type
L’abondance bactérienne (tableau 3) dans le milieu témoin et le milieu enrichi en CNP augmente
de 40% entre le début et la fin de l’incubation (t = 24 heures). Tableau 3 : Abondance bactérienne AB (106 cell.ml-1) pour la station M33 à t = 0 et t = 24 heures pour le milieu témoin et le milieu enrichi en CNP (carbone, azote et phosphore)
AB
t = 0
AB
t= 24 heures
Milieu Témoin 0.46 0.64
Milieu CNP
0.46
0.65
Réponse du phytoplancton aux enrichissements •
Pendant les 20 premières heures d’incubation, aucune différence nette n’est observée entre le
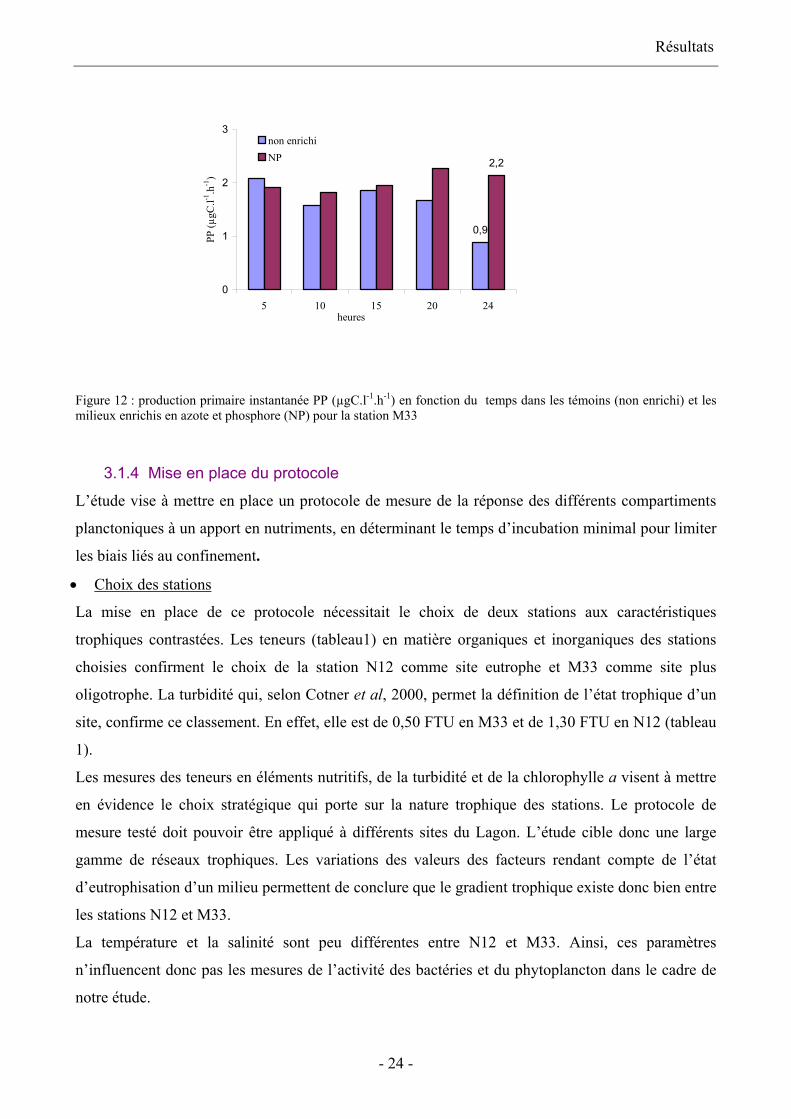
témoin et le milieu enrichi (figure 12): la valeur de la production primaire instantanée est
d’environ 2 µgC.l-1.h-1 durant toute cette période. En revanche, la mesure effectuée au terme de 24
heures d’incubation montre une réponse nette de l’activité phytoplanctonique à l’addition en
nutriments par rapport au témoin.
- 23 -
Résultats
0,9
2,2
0
1
2
3
5 10 15 20 24 heures
PP (µ
gC.l-1
.h-1
)
non enrichiNP
Figure 12 : production primaire instantanée PP (µgC.l-1.h-1) en fonction du temps dans les témoins (non enrichi) et les milieux enrichis en azote et phosphore (NP) pour la station M33
3.1.4 Mise en place du protocole
L’étude vise à mettre en place un protocole de mesure de la réponse des différents compartiments
planctoniques à un apport en nutriments, en déterminant le temps d’incubation minimal pour limiter
les biais liés au confinement.
• Choix des stations
La mise en place de ce protocole nécessitait le choix de deux stations aux caractéristiques
trophiques contrastées. Les teneurs (tableau1) en matière organiques et inorganiques des stations
choisies confirment le choix de la station N12 comme site eutrophe et M33 comme site plus
oligotrophe. La turbidité qui, selon Cotner et al, 2000, permet la définition de l’état trophique d’un
site, confirme ce classement. En effet, elle est de 0,50 FTU en M33 et de 1,30 FTU en N12 (tableau
1).
Les mesures des teneurs en éléments nutritifs, de la turbidité et de la chlorophylle a visent à mettre
en évidence le choix stratégique qui porte sur la nature trophique des stations. Le protocole de
mesure testé doit pouvoir être appliqué à différents sites du Lagon. L’étude cible donc une large
gamme de réseaux trophiques. Les variations des valeurs des facteurs rendant compte de l’état
d’eutrophisation d’un milieu permettent de conclure que le gradient trophique existe donc bien entre
les stations N12 et M33.
La température et la salinité sont peu différentes entre N12 et M33. Ainsi, ces paramètres
n’influencent donc pas les mesures de l’activité des bactéries et du phytoplancton dans le cadre de
notre étude.
- 24 -

Résultats
Choix du temps d’incubation
Dans le compartiment bactérien, une réponse significative à l’addition de nutriments apparaît à
partir de 15 heures d’incubation pour les stations N12 et M33. L’abondance bactérienne a été
contrôlée afin de vérifier que l’augmentation d’activité était bien liée à l’apport en nutriment plutôt
qu’à un développement cellulaire lié à l’expérimentation (ex : confinement). En M33, le témoin et
le milieu enrichi ne présentent pas de différences d’abondance en fin d’incubation. En N12,
l’abondance bactérienne dans le milieu enrichi augmente de 28 % après 24 heures d’incubation
alors que dans le témoin elle reste inchangée. Cette augmentation reste tout de fois insignifiante en
comparaison de celle de la production bactérienne (1500% d’augmentation par rapport à t = 0).
L’eau prélevée en n’a été ni filtrée ni diluée selon la méthode de Landry (1995) afin d’éliminer les
prédateurs (Landry & al, 1995 ; Cotner & al, 2000 ; Lebaron & al, 2000). Ainsi, l’augmentation de
l’abondance bactérienne dans les milieux enrichis pour N12 et M33 a favorisé la croissance des
prédateurs, d’où une intensification du grazing (Joint & al, 2002). L’abondance bactérienne est donc
sujet à un contrôle top-down.
Les 15 heures d’incubation ont donc été retenues pour le protocole de mesures de la réponse du
compartiment bactérien aux enrichissements. Ce temps d’incubation est testé lors de la phase de
validation du protocole.
Dans le compartiment phytoplanctonique, une réponse significative et suffisante apparaît au bout de
20 heures pour la station N12 et 24 heures pour la station M33. Ainsi, Le temps d’incubation retenu
pour l’étude de la communauté phytoplanctonique est 24 heures, il est testé lors de la phase de
validation du protocole.
3.2 Test de validation 3.2.1 Caractéristiques physico-chimiques des stations échantillonnées
La turbidité, la concentration en chlorophylle a et en ammonium sont plus importantes en N12
qu’en M33 d’un facteur compris entre 2,6 (chlorophylle a) et 4,5 (ammonium). On observe
également que ces variables sont plus fortes en N04 qu’en N12. En N04, la chlorophylle a est 1,4
fois supérieure qu’en N12 ; la turbidité, 1,7 fois ; et la concentration en NH4+, 4,5 fois. En M33, la
chlorophylle a et la turbidité sont en moyenne 4,2 fois inférieures et l’ammonium est 20,5 fois
inférieur qu’en N04. Par contre, la salinité et la température ne varient pas entre les trois stations.
- 25 -
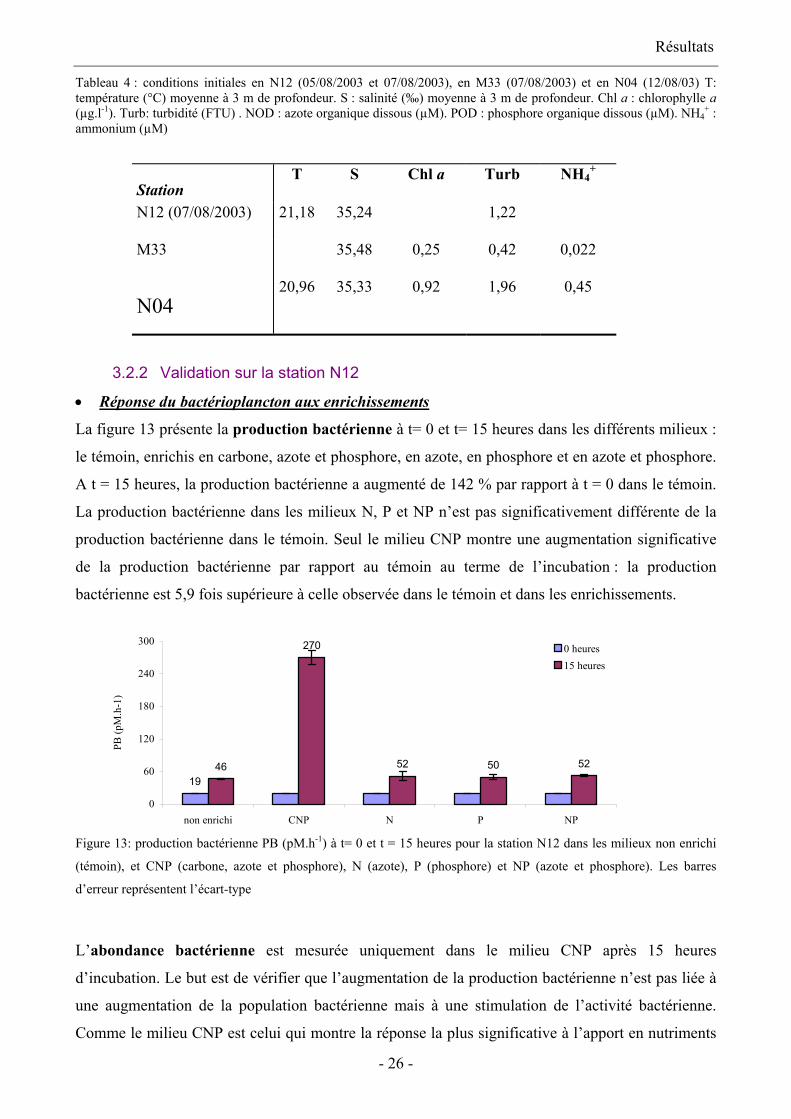
Résultats
Tableau 4 : conditions initiales en N12 (05/08/2003 et 07/08/2003), en M33 (07/08/2003) et en N04 (12/08/03) T: température (°C) moyenne à 3 m de profondeur. S : salinité (‰) moyenne à 3 m de profondeur. Chl a : chlorophylle a (µg.l-1). Turb: turbidité (FTU) . NOD : azote organique dissous (µM). POD : phosphore organique dissous (µM). NH4
+ : ammonium (µM)
Station T S Chl a Turb NH4
+
N12 (07/08/2003) 21,18 35,24 1,22
M33 35,48 0,25 0,42 0,022
N04 20,96 35,33 0,92 1,96 0,45
3.2.2 Validation sur la station N12
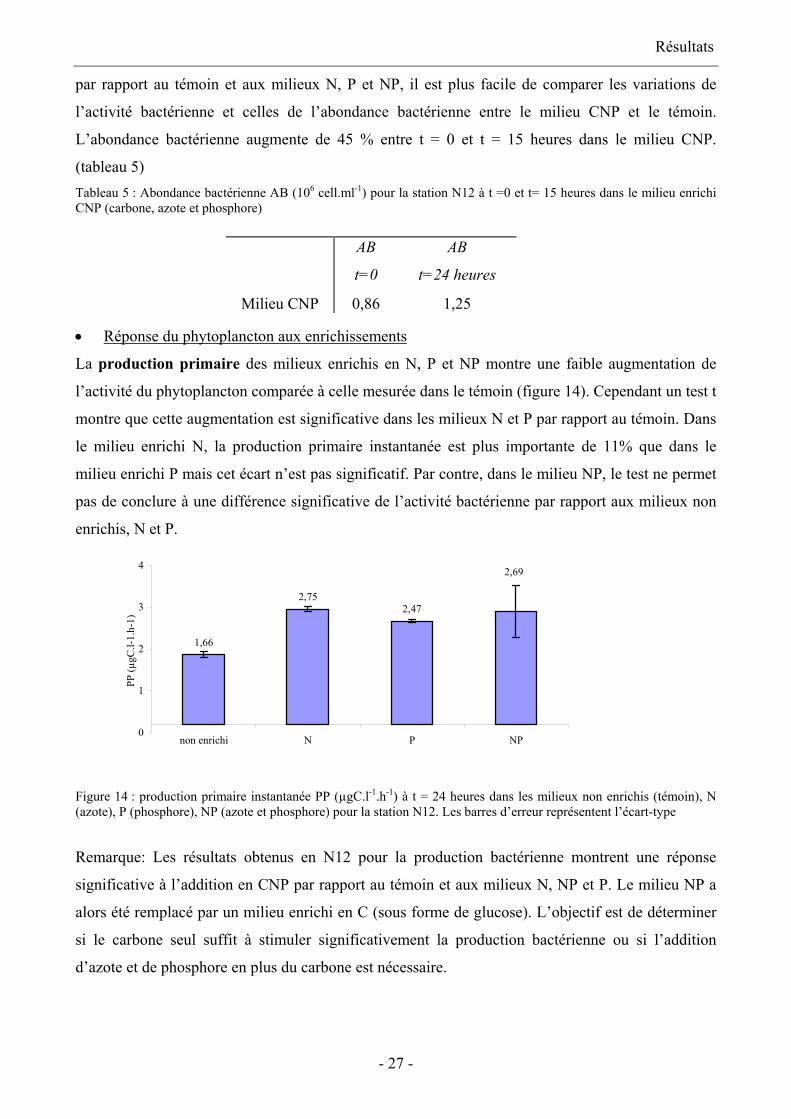
Réponse du bactérioplancton aux enrichissements •
La figure 13 présente la production bactérienne à t= 0 et t= 15 heures dans les différents milieux :
le témoin, enrichis en carbone, azote et phosphore, en azote, en phosphore et en azote et phosphore.
A t = 15 heures, la production bactérienne a augmenté de 142 % par rapport à t = 0 dans le témoin.
La production bactérienne dans les milieux N, P et NP n’est pas significativement différente de la
production bactérienne dans le témoin. Seul le milieu CNP montre une augmentation significative
de la production bactérienne par rapport au témoin au terme de l’incubation : la production
bactérienne est 5,9 fois supérieure à celle observée dans le témoin et dans les enrichissements.
1946
270
52 50 52
0
60
120
180
240
300
non enrichi CNP N P NP
PB (p
M.h
-1)
0 heures15 heures
Figure 13: production bactérienne PB (pM.h-1) à t= 0 et t = 15 heures pour la station N12 dans les milieux non enrichi
(témoin), et CNP (carbone, azote et phosphore), N (azote), P (phosphore) et NP (azote et phosphore). Les barres
d’erreur représentent l’écart-type
L’abondance bactérienne est mesurée uniquement dans le milieu CNP après 15 heures
d’incubation. Le but est de vérifier que l’augmentation de la production bactérienne n’est pas liée à
une augmentation de la population bactérienne mais à une stimulation de l’activité bactérienne.
Comme le milieu CNP est celui qui montre la réponse la plus significative à l’apport en nutriments
- 26 -
Résultats
par rapport au témoin et aux milieux N, P et NP, il est plus facile de comparer les variations de
l’activité bactérienne et celles de l’abondance bactérienne entre le milieu CNP et le témoin.
L’abondance bactérienne augmente de 45 % entre t = 0 et t = 15 heures dans le milieu CNP.
(tableau 5) Tableau 5 : Abondance bactérienne AB (106 cell.ml-1) pour la station N12 à t =0 et t= 15 heures dans le milieu enrichi CNP (carbone, azote et phosphore)
Milie
• Réponse du phytoplancton
La production primaire des
l’activité du phytoplancton com
montre que cette augmentation
le milieu enrichi N, la produc
milieu enrichi P mais cet écart
pas de conclure à une différen
enrichis, N et P.
1,66
0
1
2
3
4
non enrichi
PP (µ
gC.l-
1.h-
1)
Figure 14 : production primaire insta(azote), P (phosphore), NP (azote et p
Remarque: Les résultats obte
significative à l’addition en CN
alors été remplacé par un milie
si le carbone seul suffit à sti
d’azote et de phosphore en plus
AB
t=0
AB
t=24 heures
u CNP 0,86 1,25
aux enrichissements
milieux enrichis en N, P et NP montre une faible augmentation de
parée à celle mesurée dans le témoin (figure 14). Cependant un test t
est significative dans les milieux N et P par rapport au témoin. Dans
tion primaire instantanée est plus importante de 11% que dans le
n’est pas significatif. Par contre, dans le milieu NP, le test ne permet
ce significative de l’activité bactérienne par rapport aux milieux non
2,752,47
2,69
N P NP
ntanée PP (µgC.l-1.h-1) à t = 24 heures dans les milieux non enrichis (témoin), N hosphore) pour la station N12. Les barres d’erreur représentent l’écart-type
nus en N12 pour la production bactérienne montrent une réponse
P par rapport au témoin et aux milieux N, NP et P. Le milieu NP a
u enrichi en C (sous forme de glucose). L’objectif est de déterminer
muler significativement la production bactérienne ou si l’addition
du carbone est nécessaire.
- 27 -
Résultats
3.2.3 Validation sur la station M33
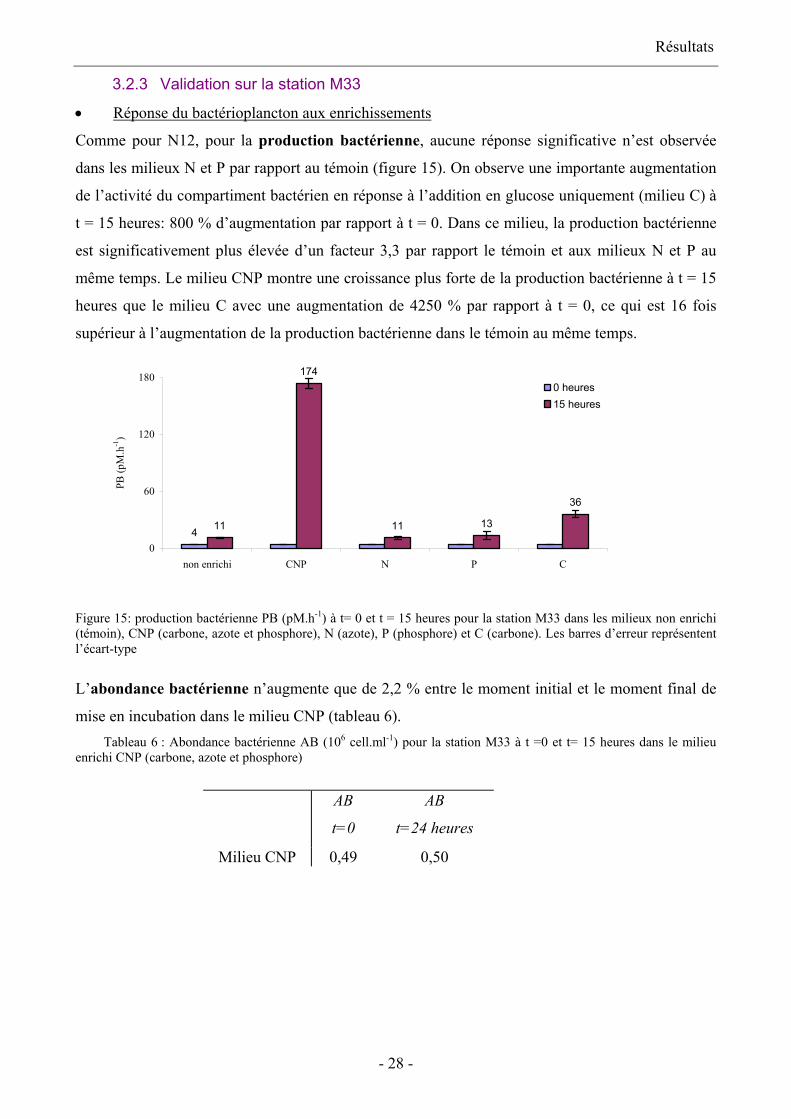
• Réponse du bactérioplancton aux enrichissements
Comme pour N12, pour la production bactérienne, aucune réponse significative n’est observée
dans les milieux N et P par rapport au témoin (figure 15). On observe une importante augmentation
de l’activité du compartiment bactérien en réponse à l’addition en glucose uniquement (milieu C) à
t = 15 heures: 800 % d’augmentation par rapport à t = 0. Dans ce milieu, la production bactérienne
est significativement plus élevée d’un facteur 3,3 par rapport le témoin et aux milieux N et P au
même temps. Le milieu CNP montre une croissance plus forte de la production bactérienne à t = 15
heures que le milieu C avec une augmentation de 4250 % par rapport à t = 0, ce qui est 16 fois
supérieur à l’augmentation de la production bactérienne dans le témoin au même temps.
4 11
174
11 13
36
0
60
120
180
non enrichi CNP N P C
PB (p
M.h
-1)
0 heures15 heures
Figure 15: production bactérienne PB (pM.h-1) à t= 0 et t = 15 heures pour la station M33 dans les milieux non enrichi (témoin), CNP (carbone, azote et phosphore), N (azote), P (phosphore) et C (carbone). Les barres d’erreur représentent l’écart-type
L’abondance bactérienne n’augmente que de 2,2 % entre le moment initial et le moment final de
mise en incubation dans le milieu CNP (tableau 6). Tableau 6 : Abondance bactérienne AB (106 cell.ml-1) pour la station M33 à t =0 et t= 15 heures dans le milieu
enrichi CNP (carbone, azote et phosphore)
AB
t=0
AB
t=24 heures
Milieu CNP 0,49 0,50
- 28 -

Résultats
• Réponse du phytoplancton aux enrichissements
La production primaire dans les milieux enrichis N et P est significativement plus importante que
la production primaire instantanée dans le témoin après 24 heures d’incubation (figure 16). La
production primaire instantanée dans le témoin représente 67 % de celle dans le milieu N, 86 % de
celle dans le milieu P. Le test t montre également une différence de la production primaire
significative de 22 % favorable au milieu N par rapport au milieu P. En revanche, il ne montre
aucun écart significatif entre l’activité phytoplanctonique dans le milieu NP par rapport aux milieux
enrichis N et P et le témoin.
0,25
0,37
0,29
0,32
0,0
0,1
0,2
0,3
0,4
non enrichi N P NP
PP (µ
gC.l-
1.h-
1)
Figure 16 : production primaire instantanée PP (µgC.l-1.h-1) à t = 24 heures dans les milieux non enrichis (témoin), N (azote), P (phosphore), NP (azote et phosphore) pour la station M33. Les barres d’erreur représentent l’écart-type
- 29 -
Résultats
3.2.4 Validation sur la station N04
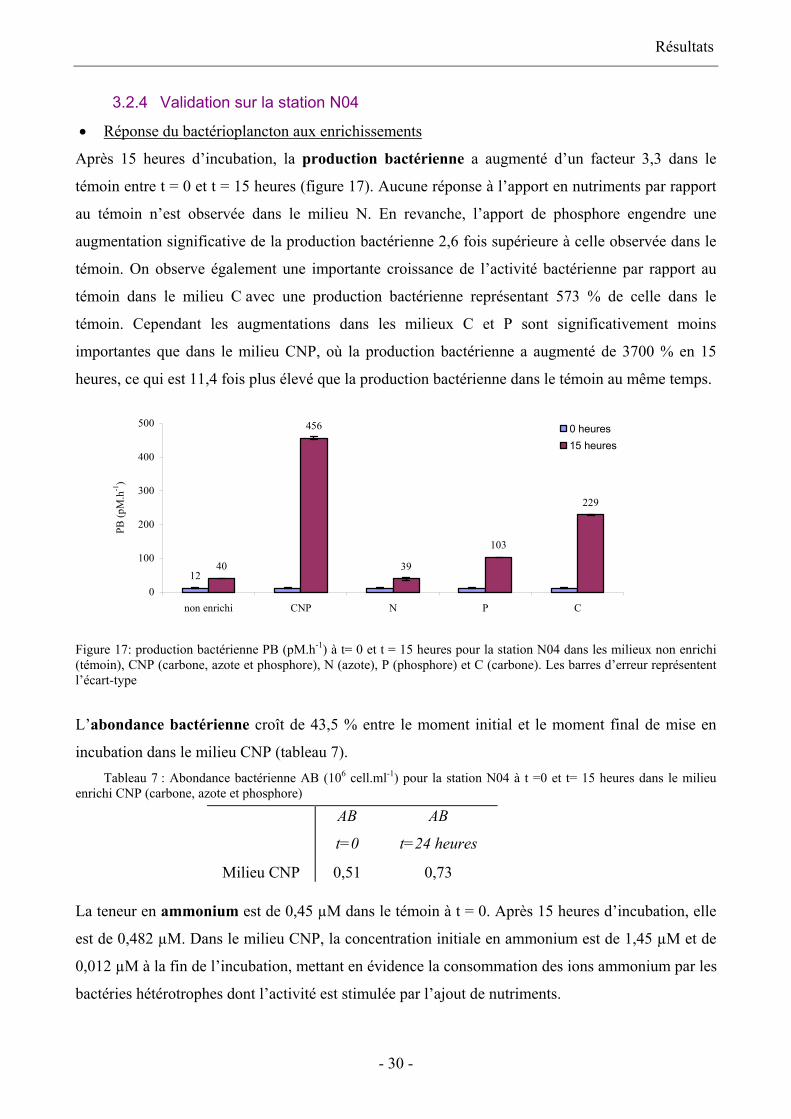
• Réponse du bactérioplancton aux enrichissements
Après 15 heures d’incubation, la production bactérienne a augmenté d’un facteur 3,3 dans le
témoin entre t = 0 et t = 15 heures (figure 17). Aucune réponse à l’apport en nutriments par rapport
au témoin n’est observée dans le milieu N. En revanche, l’apport de phosphore engendre une
augmentation significative de la production bactérienne 2,6 fois supérieure à celle observée dans le
témoin. On observe également une importante croissance de l’activité bactérienne par rapport au
témoin dans le milieu C avec une production bactérienne représentant 573 % de celle dans le
témoin. Cependant les augmentations dans les milieux C et P sont significativement moins
importantes que dans le milieu CNP, où la production bactérienne a augmenté de 3700 % en 15
heures, ce qui est 11,4 fois plus élevé que la production bactérienne dans le témoin au même temps.
1240
456
39
103
229
0
100
200
300
400
500
non enrichi CNP N P C
PB (p
M.h
-1)
0 heures15 heures
Figure 17: production bactérienne PB (pM.h-1) à t= 0 et t = 15 heures pour la station N04 dans les milieux non enrichi (témoin), CNP (carbone, azote et phosphore), N (azote), P (phosphore) et C (carbone). Les barres d’erreur représentent l’écart-type
L’abondance bactérienne croît de 43,5 % entre le moment initial et le moment final de mise en
incubation dans le milieu CNP (tableau 7). Tableau 7 : Abondance bactérienne AB (106 cell.ml-1) pour la station N04 à t =0 et t= 15 heures dans le milieu
enrichi CNP (carbone, azote et phosphore)
Milie
La teneur en ammonium es
est de 0,482 µM. Dans le m
0,012 µM à la fin de l’incub
bactéries hétérotrophes dont
AB
t=0
AB
t=24 heures
u CNP 0,51 0,73
t de 0,45 µM dans le témoin à t = 0. Après 15 heures d’incubation, elle
ilieu CNP, la concentration initiale en ammonium est de 1,45 µM et de
ation, mettant en évidence la consommation des ions ammonium par les
l’activité est stimulée par l’ajout de nutriments.
- 30 -
Résultats
• Réponse du phytoplancton aux enrichissements
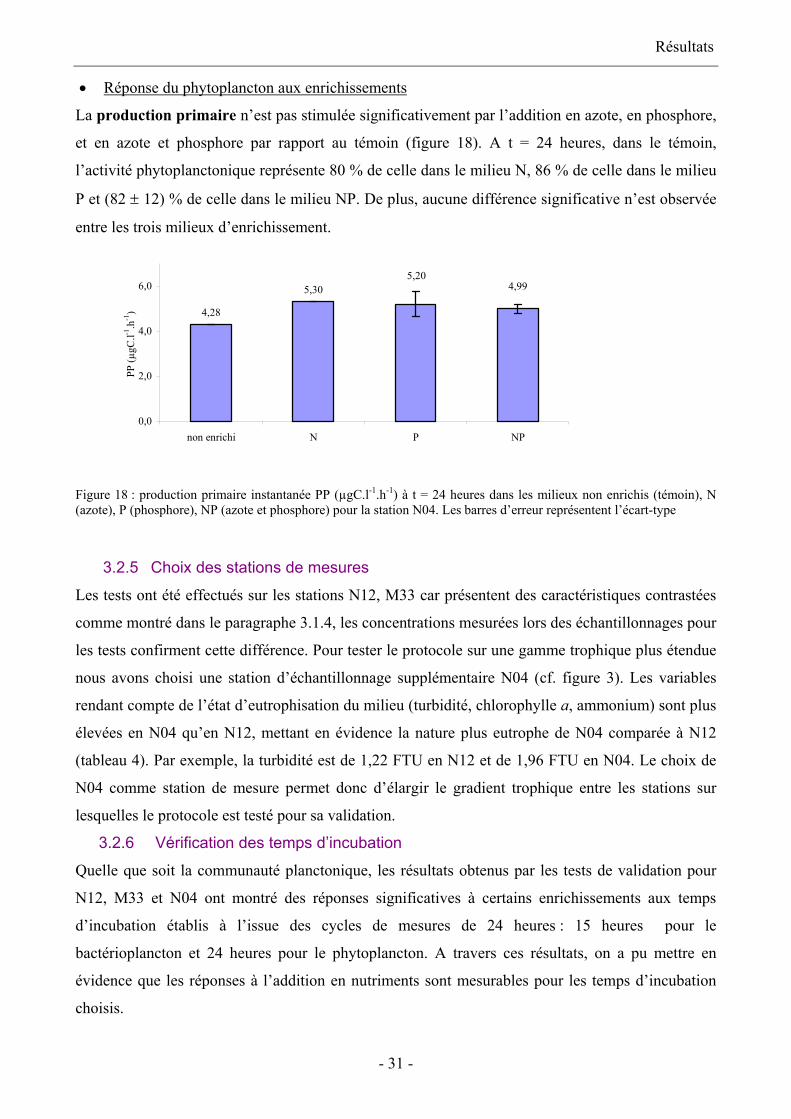
La production primaire n’est pas stimulée significativement par l’addition en azote, en phosphore,
et en azote et phosphore par rapport au témoin (figure 18). A t = 24 heures, dans le témoin,
l’activité phytoplanctonique représente 80 % de celle dans le milieu N, 86 % de celle dans le milieu
P et (82 ± 12) % de celle dans le milieu NP. De plus, aucune différence significative n’est observée
entre les trois milieux d’enrichissement.
4,28
5,305,20
4,99
0,0
2,0
4,0
6,0
non enrichi N P NP
PP (µ
gC.l-1
.h-1
)
Figure 18 : production primaire instantanée PP (µgC.l-1.h-1) à t = 24 heures dans les milieux non enrichis (témoin), N (azote), P (phosphore), NP (azote et phosphore) pour la station N04. Les barres d’erreur représentent l’écart-type
3.2.5 Choix des stations de mesures
Les tests ont été effectués sur les stations N12, M33 car présentent des caractéristiques contrastées
comme montré dans le paragraphe 3.1.4, les concentrations mesurées lors des échantillonnages pour
les tests confirment cette différence. Pour tester le protocole sur une gamme trophique plus étendue
nous avons choisi une station d’échantillonnage supplémentaire N04 (cf. figure 3). Les variables
rendant compte de l’état d’eutrophisation du milieu (turbidité, chlorophylle a, ammonium) sont plus
élevées en N04 qu’en N12, mettant en évidence la nature plus eutrophe de N04 comparée à N12
(tableau 4). Par exemple, la turbidité est de 1,22 FTU en N12 et de 1,96 FTU en N04. Le choix de
N04 comme station de mesure permet donc d’élargir le gradient trophique entre les stations sur
lesquelles le protocole est testé pour sa validation.
3.2.6 Vérification des temps d’incubation
Quelle que soit la communauté planctonique, les résultats obtenus par les tests de validation pour
N12, M33 et N04 ont montré des réponses significatives à certains enrichissements aux temps
d’incubation établis à l’issue des cycles de mesures de 24 heures : 15 heures pour le
bactérioplancton et 24 heures pour le phytoplancton. A travers ces résultats, on a pu mettre en
évidence que les réponses à l’addition en nutriments sont mesurables pour les temps d’incubation
choisis.
- 31 -

Résultats
On observe également des réponses aux apports nutritifs significatives par rapport au témoin pour
N04 au niveau de la population bactérienne. En revanche, pour cette même station, l’addition de
nutriments ne stimule pas l’activité phytoplanctonique. En raison des fortes concentrations en
éléments nutritifs en N04 et de l’exposition du milieu à la lumière pendant 24 heures, la production
primaire dans le témoin à t = 24 heures (figure 18) est considérablement favorisée. Le temps
d’incubation semble dans ce cas trop long. Cependant, N04 est une station de fond de baie (baie de
Sainte-Marie), recevant beaucoup d’apports d’origine anthropique (eaux usées non traitées de
Nouméa) et il y a peu de stations similaires en terme de richesse trophique d’après les données
existant au sein de l’UR CAMELIA : les mesures porteront donc essentiellement sur des stations
moins eutrophes. Les réponses à l’addition de nutriments observées permettent donc de valider les
temps d’incubation fixés pour l’utilisation du protocole.
3.2.7 Nutriments limitants
Les tests de validation ont permis de montrer les limitations nutritives dans le Lagon. Quelle que
soit la station étudiée, les résultats obtenus pour la production bactérienne montrent que ni l’azote ni
le phosphore ne sont limitants dans les conditions rencontrées. Par contre, l’addition de carbone a
largement favorisé l’activité bactérienne. L’augmentation de la production bactérienne stimulée par
le carbone s’accompagne d’un besoin croissant en nutriments inorganiques (Joint & al, 2002). La
production bactérienne est donc plus forte lorsque aucun nutriment est limitant (Carlson & al,
2002). Cela est confirmé par l’augmentation significative de la production bactérienne dans le
milieu CNP par rapport aux autres milieux enrichis et au témoin. Les expériences menées par Déliat
en octobre 1998 montrent que la concentration moyenne de carbone dissous dans le Lagon s’élève à
77 ± 5µM et que 16 ± 3 % de ce COD est labile, soit 13 µM de COD dégradable disponible pour les
bactéries hétérotrophes. On apporte 6,625 µM de carbone labile au milieu, soit seulement la moitié
de la quantité déjà disponible dans les eaux prélevées. On peut émettre l’hypothèse que la
proportion de COD labile serait peut-être plus faible au moment de notre étude (juillet-août) que
celle estimée par Déliat. Cela nous amène à nous poser la question de la variabilité saisonnière des
limitations nutritives dans le Lagon. Selon la saison, des limitations nutritives existeraient dans le
Lagon et seraient de nature différentes. Dans notre cas, le carbone est limitant pour la production
bactérienne. Or, les expériences menées sur le Lagon par Jean-Pascal Torréton en novembre 2000
montrent une limitation par l’azote. L’étude des relations in situ entre variables du compartiment
planctoniques d‘une part et les nutriments d’autre part montre aussi l’aspect limitant de l’azote
organique dissous dans le Lagon (Jacquet, 2001). Ces deux études désignent donc l’azote comme
facteur limitant des activités du plancton. Cela montrerait donc la possibilité d’une limitation
saisonnière…
- 32 -

Résultats
Les mesures sur la production primaire montrent une augmentation de l’activité phytoplanctonique
en réponse à l’apport en azote et en phosphore par rapport au témoin pour les stations N12 et M33.
Cela indique une limitation en azote et en phosphore. Il existe aussi peut-être une limitation en
carbone, mais la mesure de la production primaire se faisant par incorporation de 14C-bicarbonate,
cela n’a pas pu être vérifié. Ces résultats rejoignent ceux de Torréton et Jacquet en terme de limite
nutritive pour l’azote. Ils montrent également une limitation nutritive en phosphore et permettent de
soutenir l’idée d’une éventuelle limitation saisonnière.
- 33 -

Conclusion
4 Conclusion
Le stage avait pour objectif de mettre en place un protocole de mesures des réponses des
communautés planctoniques à l’addition en nutriments, en déterminant le temps d’incubation
minimal pour limiter les biais liés au confinement. A partir d’eau provenant de deux sites présentant
un gradient trophique (une station oligotrophe et une station eutrophe), les activités des bactéries
hétérotrophes et du phytoplancton de milieux enrichis en nutriments (carbone, azote, phosphore) ont
été mesurées pendant des cycles de 24 heures. Une fois le temps d’incubation minimal fixé, la suite
du stage a consisté à appliquer ce protocole sous la forme de test de validation sur les deux stations
d’échantillonnage déjà considérées et sur une troisième présentant un caractère plus eutrophe. Cette
deuxième étape a permis de vérifier que le protocole est applicable à des sites ayant des
caractéristiques trophiques contrastées.
Pour le compartiment bactérioplanctonique, 250 µl de solutions de nutriments sont ajoutés aux eaux
prélevées dans des flacons de 500ml. Pour le compartiment phytoplanctonique, 250 µl de solutions
de nutriments sont ajoutés aux eaux prélevées dans des flacons de 76 ml puis placés sous une
lumière constante 200 µE.m-2.s-1. Les flacons sont placés à température in situ.
Dans ces conditions expérimentales, les temps d’incubation utilisés sont les suivants :
• Pour l’étude du bactérioplancton : 15 heures
• Pour l’étude du phytoplancton : 24 heures
Suite à la validation du protocole, la réponse des communautés planctoniques à l’addition de
nutriments sera analysée à une échelle temporelle avec la mise en place d’un suivi saisonnier
bimensuel en M33, dans le cadre de la thèse de Séverine Jacquet, afin de voir s’il existe une
saisonnalité dans la limitation nutritive.
Le choix de la station d’échantillonnage s’est porté sur M33 en raison de sa nature oligotrophe.
M33 n’est pas sous l’influence des apports nutritifs d’origine anthropique et terrigène. Elle semble
donc bien adaptée pour l’étude du fonctionnement du lagon, contrairement à N12 qui, étant
eutrophe, ne subit pas de limitation au niveau du phytoplancton
Sur le plan personnel, ce stage m’a permis de m’initier au monde de la recherche. Cela m’a offert
une vision plus concrète de manières dont sont menées les études et des contraintes auxquelles les
chercheurs doivent faire face : temps, utilisation du matériel (éviter tout gaspillage), commandes. Le
temps a d’ailleurs été un paramètre très important au cours de ce stage : deux mois m’ont semblé un
peu courts pour pouvoir apprendre des techniques de mesures et mettre en place le protocole de
façon la plus rigoureuse possible (un seul cycle de mesure et un seul test de validation pour chaque
station ont pu être réalisés). Il serait donc préférable de prévoir des stages plus longs. En revanche,
- 34 -

Conclusion
les efforts de communication entre les différents « acteurs » de l’UR CAMELIA (réunions pour la
répartition du matériel et s’informer sur l’évolution des études de chacun) permettent une bonne
coordination des différents laboratoires et m’ont donné un bon aperçu du travail en équipe.
- 35 -

Annexes
BIBLIOGRAPHIE
Craig A. (2002) Effect of nutrient amendments on bacterioplankton production, community structure, and DOC utilization in the North western Sargasso Sea. Aquat Microb Ecol 30: 19-36 Cotner J.B (2000) Nutrient Limitation of Heterotrophic Bacteria in Florida Bay. Estuaries 23 : 611-620 Déliat G(1998) La matière organique dissoute des zones côtières : Sources, Distribution et Biodégradabilité. Rapport de thèse de Doctorat Spécialité Océanographie, de l’Université Paris VI Douillet P (1998) Tidal dynamics of the south-west lagoon of New Caledonia: observations and 2Dnumératical modelling. Oceanol. Acta. 21 : 69-79 Eilers H (2000) Succession of pelagic marine bacteria during enrichment : a close look at cultivation-induced shifts. Applied and Environmental Microbiology 66 : 4634-4640 Fisher T. R (1992) Nutrient limitation of phytoplankton in Chesapeake Bay. Mar Ecol. Prog. Ser. 82 : 51-63 Hein M, B R (1995) Nutrient limitation of phytoplankton biomass or growth rate: an experimental approach using marine enclosures. Journal of Experimental Marine Biology and Ecology 188:167-180 Henriksen P (2002) Effects of nutrient-limitation and irradiance on marine phytoplankton pigments. J. Plankton Res. 24: 835-858 Holmboe (1999) Nutrient addition bioassays as indicators of nutrient limitation of phytoplankton in an eutrophic estuary. Mar Ecol Prog Ser. 186 : 95-104 Jacquet S (2001) Importance comparée des biomasses et des productions bactérienne et primaire planctoniques et leurs relations avec les variables physico-chimiques dans le Lagon Sud Ouest de Nouvelle Calédonie. Rapport de stage de DEA Environnement marin et biogéochimie, Université Pierre et Marie Curie – Paris VI Joint I (2002) Competition for inorganic nutrients between phytoplankton and bacterioplankton in nutrient manipulated mesocosms. Aquatic Microbial Ecology 29:145-159 Kirchman (2001) Microbial Ecology of the Oceans. Landry M.R (1995) A Refined dilution technique for measuring the community grazing impact of microzooplankton, with experimental tests in the central equatorial Pacific. Marine Ecology Progress Series. 120 : 53-63 Lebaron P (2001) Microbial community dynamics in Mediterranean nutrient-enriched seawater mesocosm: changes in abundance, activity and composition. FEMS Microbiology Ecology. 34 : 255-266 Legendre L., Rassoulzadegan F. (1995) Plankton and nutrients dynamics in marine waters. Ophelia 41 : 153-172
- 36 -

Annexes
Annexes
Annexe 1 - Solutions Mère utilisées pour les enrichissements
Solution CNP :
V = 50 ml d’EMQ
5,35 mg de NH4Cl, M = 53.49 g.mol-1
0,88 mg de HNa2O2P, M =141,96 g.mol-1
19,9 mg de glucose, M = 180.2 g.mol-1
Solution NP :
V= 50 ml d’EMQ
5,35 mg de NH4Cl, M = 53.49 g.mol-1
0,88 mg de HNa2O2P, M =141,96 g.mol-1
Solution N :
V= 50 ml d’EMQ
5,35 mg de NH4Cl, M = 53.49 g.mol-1
Solution P :
V= 50 ml d’EMQ
0,88 mg de HNa2O2P, M =141,96 g.mol-1
Solution C :
V = 50 ml d’EMQ
19,9 mg de glucose, M = 180.2 g.mol-1
- 37 -

Annexes
Annexe 2 - Techniques de mesures des productions primaire et bactérienne
On a recours au marquage par des isotopes radioactifs puis au comptage par scintillation au cours de
l’étude afin d’estimer la production bactérienne et la production primaire dans les différents milieux
étudiés.
Marquage par un isotope radioactif
Les techniques de détection des activités bactério- et phytoplanctoniques reposent sur le même
principe : il existe une entité ayant la propriété de se fixer de façon plus ou moins spécifique à ce
qu'on cherche à mettre en évidence. L’objectif reste de fixer un marqueur, c'est à dire, un élément
qui va rendre visible ces molécules biologiques au matériel (microscope, scintillateur) dont dispose
l’expérimentateur. Les marqueurs sont divers(molécule colorée ou fluorescente, atome radioactif,
métaux lourds, enzyme) et permettent de suivre l’évolution de la teneur d’une espèce chimique, une
particule, sans en perturber le comportement physique, chimique et biologique. Au cours des
expériences, pour la production bactérienne, on utilise 3H-thymidine pour pouvoir suivre la
production d’ADN. On peut également estimer la production bactérienne avec 14C-leucine (Joint &
al, 2002), incorporé pour la production de protéines, dont celles des cyanobactéries non recherchées
dans notre cas. Pour la production primaire, on utilise 14C-bicarbonate qui permet de suivre la
photosynthèse.
Il s’agit de techniques très sensibles et les productions primaire et bactériennes constituent ainsi des
variables très représentatives du degré trophique d’un milieu.
Comptage par scintillation
On peut dénombrer le nombre de molécules radioactives en mesurant la quantité de leur produit de
décomposition. En biochimie, on se sert principalement de la mesure des particules par scintillation
en milieu liquide.
Principe
Lorsque les radio-isotopes se désintègrent, ils émettent des particules d'un niveau d'énergie moyen.
Ces particules, et éventuellement leur niveau d'énergie, peuvent facilement être mesurées. Les
mesure des particules fournissent des valeurs en "coups par minute" ou CPM, plus faible que le
nombre réel de désintégrations, c’est-à-dire, « désintégrations par minute » ou DPM. Le procédé ne
décèle donc qu'une partie des décompositions, et la proportion dépend de l'efficacité du processus
de détection, d’où la relation : CPM = DPM x eff
- 38 -

Annexes
Le comptage en milieu liquide est employé pour mesurer la quantité d'émetteurs dans un
échantillon. Pour se faire, on place l'échantillon à mesurer dans une fiole avec un liquide à
scintillation. On récolte l’échantillon sur un filtre, ce qui permet de retenir les macromolécules tout
en éliminant les précurseurs radioactifs de petites tailles (acides aminés, nucléotides, etc.).
Méthodologie et appareillage
- Le liquide à scintillation
Dans la scintillation en milieu liquide, on mesure la quantité de photons émis dans un mélange
scintillant suite à une décomposition radioactive. Le liquide à scintillation contient trois
composantes de base intervenant dans le phénomène de scintillation: un solvant organique, capable
d'absorber des radiations β et d'émettre des photons, un scintillant primaire et un scintillant
secondaire. Le solvant organique joue deux rôles: il solubilise les scintillants primaires et
secondaires ; il capte l'énergie émise par les particules β.
Une décomposition d'un radio-isotope provoque l'émission de particules de différents niveaux
d'énergie. Les particules β entrent en collision avec un certain nombre de molécules du solvant du
liquide à scintillation. Ces molécules de solvant vont alors être excitées. Un électron de la couche
périphérique de la molécule passe à un niveau d'énergie supérieur. Lors du retour de cet électron à
son état fondamental, il y aura émission d'énergie, sous forme de photons généralement assez
énergétiques. Les photons peuvent alors entrer en collision avec une molécule du scintillant
primaire et l'exciter, provoquant, lors du retour à l'état fondamental de ce dernier, la production d'un
photon d'énergie moins grande. Ce photon va à son tour exciter une molécule de scintillant
secondaire qui émettra un photon faiblement énergétique.
Pour procéder aux comptages pour la production primaire et la production bactérienne, le liquide à
scintillation utilisé est : Ultima Gold tm MV (Packard)
- Les fioles
Les fioles (10 ml) sont transparentes aux photons émis lors du processus de comptage. Il est
également important que le matériau des fioles ne contienne pas des radio-isotopes susceptibles
d’interférer avec le comptage. C’est pourquoi le verre ordinaire n’est pas préconisé, car il peut
renfermer du potassium-40, un émetteur β. On utilise donc surtout des fioles en plastique,
polyéthylène ou polypropylène.
- 39 -

Annexes
- Le compteur à scintillation
Le compteur à scintillation est muni d'un dispositif photosensible : deux cellules
photomultiplicatrices déclenchent une impulsion électrique à chaque fois que des photons atteignent
sa surface, à la suite des décompositions radioactives. Le nombre et l'intensité de ces signaux sont
enregistrés. Ces signaux peuvent être classifiés selon leur intensité ("pulse height analyzer") et
comptabilisés dans des canaux correspondant à divers niveaux d'énergie. Le nombre de ces
impulsions (coups) est enregistré sur une période plus ou moins longue de temps. Connaissant le
nombre de coups et le temps d'enregistrement, les circuits électroniques de l'appareil peuvent donc,
pour chaque échantillon, donner le nombre correspondant de CPM.
Le compteur se programme pour n'enregistrer que les décompositions radioactives d'un isotope
spécifique (14C ou 3H), c’est-à-dire, d'un niveau d'énergie correspondant à l'atome qu'on veut
compter.
Le compteur utilisé pour l’étude des productions primaire et bactérienne est le Tri-CarbTM liquid
Scintillation Analyser 1600 TR (Packard).
- Le problème du quenching
La mesure obtenue en CPM n'est pas forcément égale au nombre total de désintégrations par minute
(DPM), ni à leur niveau d'énergie réel, car le processus n’apparaît pas efficace à 100%. L'efficacité
est en effet empesée par le grand nombre d'étapes du processus. Elle dépend aussi du niveau
d'énergie des particules β, et de la fiabilité de la capture des photons par les cellules
photodétectrices.
De plus cette relative inefficacité peut être accrue par le quenching qui interagit avec le mécanisme
de conversion des particules β en photons décelables par les cellules photosensibles. Le masquage
peut occasionner un amoindrissement de la vague de photons, d’où une désintégration semblant
avoir un niveau énergétique inférieur à la réalité. Ce phénomène peut aller jusqu'à la disparition
complète du signal engendré par des émissions faibles, et donc diminue le compte obtenu.
- Détermination des DPM
La quantité de radioactivité est déduite à partir de l’équation: CPM = DPM x eff
On peut construire une courbe de quenching (quenching curve) qui donne l’efficacité en
pourcentage en fonction d'une mesure du quenching. Cette mesure dépend du modèle de compteur.
En faisant cette même mesure sur l'échantillon, on peut déduire son efficacité en pourcentage
d'après la courbe de masquage, puis avec le CPM obtenu, reconvertir en DPM.
- 40 -

Annexes
Annexe 3 - Microscopie à épifluorescence
On utilise cette technique pour évaluer l’abondance bactérienne. Elle est basée sur les qualités
fluorescentes de certains composés. Cette fluorescence peut avoir plusieurs origines :
elle peut être d’origine naturelle : pigments •
•
•
elle peut être induite par l’utilisation d'une substance fluorescente se fixant spécifiquement
sur une structure. On parle alors de test de type cytochimique.
on peut également introduire dans la cellule une substance non spécifique fluorescente
naturellement.
L’outil utilisé est le microscope à épifluorescence. Il s’agit d’un microscope photonique équipé de
deux lampes : une lampe ordinaire pour une observation classique par transmission et une lampe à
arc pour la fluorescence. Des filtres d'excitation permettent de choisir la longueur d'onde incidente
et des filtres d'émission permettent de sélectionner les radiations émises par l'entité excitée.
En effet, un tel microscope possède plusieurs jeux de filtres correspondant aux fluorochromes les
plus habituellement utilisés. Chaque jeu de filtres est constitué d'un filtre d'excitation, d'un miroir
dichroïque et d'un filtre d'émission. Les fluorochromes les plus classiques sont la rhodamine et ses
dérivés, la fluorescéine et ses dérivés et le DAPI (Di Aminido Phenyl lndol). Ils émettent
respectivement dans le rouge, le vert et le bleu.
Dans un premier temps, la fluorescence des composés est excitée par la lumière. La longueur
d’onde d'excitation désirée est sélectionnée grâce aux filtres. Après excitation, les composés
émettent une fluorescence qui permet la visualisation et le comptage des cellules.
Pour déterminer l’abondance bactérienne des échantillons de notre analyse, on se sert du DAPI :
Le DAPI se caractérise par sa propriété à se fixer spécifiquement sur l'ADN : excité à une longueur
d’onde de 372nm, il émet une fluorescence bleue ( 456nm).
Le microscope est doté d'un jeu de filtres correspondant aux caractéristiques du fluorochrome: un
filtre d'excitation permettant la sélection des radiations absorbées par le fluorochrome (environ
380nm) ; un miroir dichroïque réfléchissant les radiations absorbables vers l'échantillon et ne
laissant passer par transmission que les radiations bleues et au-dessus (ici >400nm) ; un filtre
d'émission ne laissant passer par transmission que les radiations bleues et au-dessus (>400nm).
On peut également distinguer au microscope à fluorescence les cyanobactéries des autres bactéries
car elles possèdent un pigment caractéristique, la phycoérythrine qui fluoresce en orange lorsqu’elle
est excitée dans le bleu, soit à une longueur d’onde de 500 nm
- 41 -

Résumé
Les effets des apports d’origine anthropique et terrigène sur le réseau trophique planctonique
du Lagon Sud-Ouest de Nouvelle-Calédonie sont étudiés selon plusieurs approches
complémentaires au sein de l’unité de recherche CAMELIA de l’IRD de Nouvelle-Calédonie.
Une d’elle vise à établir expérimentalement et à comparer les réponses à des additions en
éléments nutritifs des différentes communautés planctoniques dans différentes situations
trophiques allant de l’oligotrophie à l’eutrophie.
Réalisé dans le cadre de la thèse de S. Jacquet qui traite de « l’influence des apports en
nutriments sur le fonctionnement du réseau trophique microbien dans le Lagon Sud-Ouest de
Nouvelle-Calédonie », mon stage avait pour objectif la mise en place d’un protocole. Il
s’agissait de déterminer un temps d’incubation permettant d’avoir une réponse suffisante à
l’addition de nutriments des communautés planctoniques et limitant les biais liés au
confinement.
Testés sur des stations d’échantillonnage présentant un gradient trophique, les temps
d’incubation sont de 15 heures pour l’étude du bactérioplancton et de 24 heures pour l’étude
du phytoplancton dans nos conditions expérimentales,.
Ce travail aura pour suite la mise en place d’un suivi saisonnier de la réponse des
communautés planctoniques aux apports nutritifs sur une station oligotrophe du Lagon.