mémoire de stage de master deuxième année année ... · maxime ragué master 2 biologie,...
TRANSCRIPT

Mémoire de stage de Master deuxième année
Année universitaire 2017-2018
Le contexte paysager influence l’efficacité de la pollinisation entomophile
du colza en fonction des conditions locales.
Par :
Maxime Ragué
Master 2 Biologie, Écologie, Évolution
Parcours Dynamique et Conservation de la Biodiversité
Sous la direction de :
Sabrina Gaba, Directrice de recherche
Centre d'Études Biologiques de Chizé
Février – Juin 2018

Remerciements :
Ce travail de Master a été conduit dans le cadre du projet BIOSERV, financé par le
méta programme EcoServ (INRA).
Sabrina Gaba, merci de m’avoir permis de travailler sur cette thématique que j’ai
particulièrement apprécié. Merci pour votre confiance, j’ai rapidement pu m’approprier le
sujet et conduire ces travaux de recherche de façon autonome. Je tiens également à vous
remercier pour votre disponibilité durant ce stage, vous avez pris le temps de suivre l’avancée
de mes travaux, de m’aider dans mes réflexions autour des analyses statistiques et dans la
compréhension des dynamiques communautaires et de la fonction de pollinisation dans les
paysages agricoles. Enfin, merci pour tous vos conseils concernant la rédaction de ce rapport
de stage.
Vincent Bretagnolle, merci d’avoir rendu possible mon travail sur les données de la
Zone Atelier Plaine & Val de Sèvre et ainsi de m’inscrire dans le projet global de recherche
de cette zone. Merci également pour vos commentaires sur mon rapport.
Thomas Perrot, merci pour tes remarques et conseils concernant le protocole
expérimental auquel tu as participé, les analyses statistiques et mon rapport.
Olivier Martin, merci d’avoir pris le temps de m’expliquer et de m’aider à appliquer
sur mes données la méthode d’analyse spatiale Siland que tu as développé avec Florence
Carpentier.
Je tiens également à remercier l’ensemble des personnes qui ont participé à la récolte
des données. Je suis conscient de la quantité de travail qu’il y a derrière les tableaux de
données que j’ai eu à analyser. Je remercie particulièrement Jean-Luc Gautier pour la récolte
mais aussi pour la saisie des données.
Merci à mes collègues de bureau Béranger, Coline et Lucie de m’avoir permis de
travailler dans une ambiance sympathique et agréable.

Sommaire
A. INTRODUCTION ....................................................................................................................................... 1
B. MATÉRIEL ET MÉTHODES ................................................................................................................... 5
1. Zone d’étude ............................................................................................................................................ 5
2. Échantillonnage ....................................................................................................................................... 6
a. Choix des paysages .............................................................................................................................................. 6
b. Mesure de la fonction de pollinisation ............................................................................................................... 6
3. Mesure des traits impliqués dans le rendement du colza .................................................................... 8
4. Calcul des contributions absolues et relatives des processus de pollinisation ................................... 8
5. Variables locales et paysagères .............................................................................................................. 8
6. Analyses statistiques ............................................................................................................................... 9
a. Comment le trait impliqué dans le rendement est-t-il sélectionné ?................................................................ 9
b. Étude de l’influence des biais expérimentaux ................................................................................................. 10
c. Le succès de fructification et le rendement dépendent-ils des conditions locales ? ...................................... 10
d. Étude de l’influence du contexte paysager sur la production de grains........................................................ 11
C. RÉSULTATS ............................................................................................................................................. 12
1. Recherche du trait impliqué dans le rendement ................................................................................ 12
a. Contribution des processus de pollinisation aux traits liés au rendement .................................................... 12
b. Compromis entre traits impliqués dans le rendement ................................................................................... 12
2. Influence du protocole expérimental sur les résultats ....................................................................... 13
3. Le succès de fructification et le rendement dépendent-ils des conditions locales ? ......................... 14
4. Le succès de fructification et le rendement dépendent-ils du contexte paysager ? ......................... 15
D. DISCUSSION ............................................................................................................................................ 20
1. Les insectes pollinisateurs contribuent au rendement du colza ........................................................ 20
2. La culture testée n’influence pas la pollinisation entomophile du colza .......................................... 21
3. Le contexte paysager influence la pollinisation entomophile du colza ............................................. 22
E. CONCLUSION & PERSPECTIVES ..................................................................................................... 24
F. RÉFÉRENCES ......................................................................................................................................... 26
G. ANNEXES ................................................................................................................................................. 32
1. Différences interannuelles des protocoles ........................................................................................... 32
2. Sélection des covariables ...................................................................................................................... 33
3. Compromis entre traits (fleurs ouvertes en serres vs sur le terrain) ................................................ 33
4. Contribution des processus de pollinisation (fleurs ouvertes en serres vs sur le terrain) ............... 34
5. Etudes résiduelles .................................................................................................................................. 35
6. Corrélations entre variables paysagères ............................................................................................. 36

1
A. INTRODUCTION
Le concept de services écosystémiques, les bénéfices directs et indirects que les
humains tirent des écosystèmes, introduit l’interdépendance entre la conservation de la
biodiversité et le bien être humain (Millennium Ecosystem Assessment 2005). L’un de ces
services, central dans les agroécosystèmes, est la pollinisation, indispensable à la reproduction
sexuée des angiospermes et donc, entre autres, à la production agricole. Les rendements
mondiaux sont ainsi dépendants du transfert de pollen, soit au sein d’une même fleur
(autogamie), au sein d’une même plante (géitonomogamie) ou entre plantes par des vecteurs,
le vent (anémogamie) ou les animaux (zoogamie ; Gallai et al. 2009). Plus de 70% des
principales cultures mondiales destinées à l’alimentation humaine (Klein et al. 2007), soit
30% de la surface totale cultivée des pays développés (Aizen et al. 2008), sont dépendantes
dans différentes mesures de la zoogamie, et plus particulièrement de l’entomogamie
(pollinisation par les insectes). Une culture est dite dépendante de la pollinisation entomophile
si elle bénéficie de l’entomogamie en termes de rendement et/ou de valeur marchande (qualité
de la production ; Klein et al. 2007). Le service de pollinisation a ainsi été évalué à 153
milliards de dollars, soit 9,5% de la valeur de l’ensemble des cultures produites à des fins de
consommations humaines (Gallai et al. 2009). Malgré son importance économique, le service
de pollinisation des cultures est menacé. Ainsi, en Europe, 9% des hyménoptères sont
catégorisés comme menacés d’extinction selon les critères UICN (IUCN 2012 ; Potts et al.
2016) et la biomasse d’insectes volants a diminué de 82% en 27 ans en Allemagne (Hallmann
et al. 2017). Le déclin des pollinisateurs sauvages dans les paysages agricoles est en partie
attribuable à l’agriculture conventionnelle, via l’utilisation d’intrants chimiques, la diminution
de la surface d’habitats dits semi-naturels (HSN), constitués de communautés végétales
permanentes contrairement aux cultures (Wezel et al. 2014), et l’homogénéisation des
paysages agricoles (Tscharntke et al. 2005 ; Potts et al. 2010b). En parallèle, les pollinisateurs
domestiques (abeilles et certains bourdons) sont incapables d’assurer le service de
pollinisation seuls et durablement. En effet, il a été reporté des déclins d’abeilles domestiques
en Europe (baisse de 16% du nombre de colonies entre 1985 et 2005 ; Potts et al. 2010a) et
aux États-Unis (deux fois moins de colonies en 2008 qu’en 1935 ; Ellis et al. 2010). De plus,
l’augmentation du « stock » d’abeilles domestiques au niveau mondial (+40% entre 1961 et
2007 ; Aizen & Harder 2009) est insuffisante relativement à l’augmentation des surfaces
mondiales occupées par les cultures dépendantes de la pollinisation entomophile (+ 70 %
entre 1961 et 2006 ; Aizen et al. 2008 ; Aizen & Harder 2009 ; Breeze et al. 2014).
Le colza (Brassica napus L) participe à la croissance générale des surfaces cultivées

2
dépendantes de la pollinisation entomophile (de 22,7 à 33,7 millions d’hectares en colza entre
1994 et 2016 ; FAO 2018). Cette culture, majoritairement autogame (Becker et al. 1992), est
également dépendante de la pollinisation par le vent et les insectes (diptères, hyménoptères
sauvages et domestiques). Les insectes pollinisateurs contribuent à la valeur marchande des
grains de colza (d’environ 20% relativement à quand les insectes ne pollinisent pas le colza ;
Bommarco et al. 2012 ; Bartomeus et al. 2014 ; Marini et al. 2015) mais également au
nombre de siliques (+24% ; Kamler & Jas 2003), au succès de fructification (+ 30% environ ;
Perrot et al. en révision), au nombre de grains par silique (+ 20% et + 10% ; Kamler & Jas
2003 ; Stanley et al. 2013) et à la masse des grains (+ 18% et 8% ; Bommarco et al. 2012 ;
Stanley et al. 2013). La contribution de la pollinisation entomophile au rendement du colza
est finalement comprise entre 12% (Zou et al. 2017) et 51% (Kamler & Jas 2003). Il y a une
variabilité importante dans cette contribution et dans les composantes du rendement
influencées. Cela peut être expliqué par la variété de colza (Steffan-Dewenter 2003 ;
Hudewenz et al. 2014 ; Marini et al. 2015 ; Lindström et al. 2016), les pratiques agricoles
(Marini et al. 2015 ; Lindström et al. 2018) et le profil des communautés de pollinisateurs
(Rader et al. 2012). Le rendement du colza est positivement lié à l’abondance en diptères
(Jauker & Wolters 2008 ; Jauker et al. 2012 ; Perrot et al. en révision), en hyménoptères
sauvages (Lerin & Rivault 1982 ; Steffan-Dewenter 2003 ; Jauker et al. 2012 ; Zou et al.
2017 ; Perrot et al. 2018), en hyménoptères domestiques (Lindström et al. 2016) et à la
richesse spécifique en hyménoptères sauvages (Zou et al. 2017 ; Perrot et al. en révision). Des
absences de relations entre le rendement et la richesse en diptères (Perrot et al. en révision),
l’abondance en hyménoptères domestiques (Zou et al. 2017) ou la diversité en hyménoptères
(Bommarco et al. 2012) sont également mises en évidence. Cela pourrait traduire une forme
de redondance fonctionnelle des pollinisateurs. Par exemple dans des paysages abritant une
diversité de pollinisateurs sauvages importante, l’augmentation de l’abondance en
pollinisateurs domestiques n’influence pas significativement les rendements du colza (Zou et
al. 2017).
L’assemblage des communautés de pollinisateurs dépend de facteurs locaux, c’est-à-
dire de la qualité de l’habitat, caractérisable et quantifiable à l’échelle de la parcelle agricole.
La probabilité de présence des pollinisateurs dans un habitat est positivement liée à la richesse
spécifique en adventices (ou espèces florales) dans cet habitat (Sydenham et al. 2017). Donc,
la diversité en ressources florales gouverne la diversité en insectes pollinisateurs (Potts et al.
2003 ; Bretagnolle & Gaba 2015 ; Rollin et al. 2015 ; Cole et al. 2017). La diversité en
ressources florales est dépendante du type de culture et de l’emplacement dans la parcelle. Les

3
prairies abritent une diversité en espèces florales importante (Cole et al. 2017) et le colza
fournit une abondance en fleurs importante (Westcott & Nelson 2001). La diversité en
adventices est plus importante dans les bordures que dans les pleins champs (Henckel et al.
2015). En relation avec leurs capacités de dispersion, la diversité de pollinisateurs dépend du
contexte paysager en termes de composition et de configuration. La composition rend compte
des différents types d’habitats présents dans un paysage et la configuration considère la
complexité de l’agencement spatial des habitats (Fahrig et al. 2011 ; Pufal et al. 2017).
Différents habitats dans les paysages agricoles influencent la diversité locale de pollinisateurs
(Encadré 1). Les mécanismes proximaux et l’échelle d’influence, c’est-à-dire l’échelle
spatiale à laquelle les pollinisateurs dépendent le plus d’une variables paysagère considérée
(Holland et al. 2004), diffèrent entre variables paysagères. De plus l’influence d’une variable
dépend du contexte local (influence du colza dans le paysage ; Encadré 1), de la guilde de
pollinisateurs étudiée (influence des HSN ; Encadré 1), et des traits des pollinisateurs. Par
exemple les gros pollinisateurs et les pollinisateurs généralistes sont affectés de manière plus
importante et à plus large échelle spatiale que les petits pollinisateurs ou les pollinisateurs
spécialistes d’un habitat (Benjamin et al. 2014 ; Hopfenmuller et al. 2014).
Encadré 1 : Impact du contexte paysager sur la diversité de pollinisateurs :
Des études ont quantifié l’effet des HSN, de l’agriculture biologique (AB), du colza,
et de la configuration dans le paysage en quantifiant l’abondance ou la richesse spécifique
des pollinisateurs le long de gradients de ces variables paysagères. Les tendances globales de
leur influence sur la diversité de pollinisateurs sont présentées en Figure 1.
Figure 1 : Influence des densités A. d’HSN (Steffan-Dewenter et al. 2002 ; Westphal et al.2003 ; Holzschuh
et al. 2010 ; Bommarco et al. 2012 ; Bartomeus et al. 2014 ; Diekötter et al. 2014 ; Hopfenmuller et al. 2014 ;
Nicholson et al. 2017) B. d’AB (Holzschuh et al. 2008 ; Gabriel et al. 2010) C. de colza (Westphal et al. 2003 ;
Holzschuh et al. 2011 ; Diekötter et al. 2014 ; Holzschuh et al. 2016) et D. de la longueur des bordures entre
les habitats (Holzschuh et al. 2010 ; Hass et al. 2018) sur la diversité de pollinisateurs. Les traits pleins
représentent les tendances prédites dans la littérature (aucune taille de l’effet n’est indiquée, ici seul le sens
global de la relation est présenté sans considération de la guilde de pollinisateurs ou de l’habitat local) tandis
que les traits pointillés sont des inférences. L’échelle d’influence (EI) présentée est la moyenne des échelles
d’influence des études mettant en évidence une relation significative entre la diversité de pollinisateurs et une
variable paysagère.

4
A. HSN Il y a une relation positive entre la proportion d’HSN et la diversité de
pollinisateurs (Figure 1A). Cela a été mis en évidence par la majorité des études (2/3 des
études environ) et le long du gradient de proportion d’HSN. Ces habitats fournissent des
ressources florales diverses et continues ainsi que des sites de nidification (Bukovinszky et
al. 2017). Les bénéfices associés débordent dans les habitats voisins (Tscharntke et al. 2012).
Certains pollinisateurs moins dépendants des fleurs à toutes les étapes de leur cycle de vie
sont néanmoins moins dépendants des HSN (les diptères pollinisateurs par exemple ;
Steffan-Dewenter et al. 2002).
B. AB Holzschuh et al. (2008) mettent en évidence une relation positive entre la
diversité de pollinisateurs et le pourcentage d’AB (0-35%) dans un rayon de 500m. Ce
système de production agricole induit un plus faible taux de mortalité des pollinisateurs
(Hole et al. 2004) et une augmentation de la diversité d’adventices relativement à
l’agriculture conventionnelle (Henckel et al. 2015). Néanmoins, Gabriel et al. (2010) ne
mettent pas en évidence ce lien dans un rayon de 1750m pour une plus faible gamme d’AB
(2-17% ; Figure 1B).
C. Colza L’augmentation de la proportion de colza (entre 4 et 15% dans un rayon de
1750m) dans le paysage influence positivement la diversité de pollinisateurs dans les HSN
(Diekötter et al. 2014). Cela traduit l’effet de facilitation par effet de débordement, la
diversité de pollinisateurs présente dans les champs de colza déborde sur les habitats voisins
(Blitzer et al. 2012). La relation inverse (entre 0 et 30% de colza dans un rayon de 1000m)
est présentée dans les parcelles en colza (Holzschuh et al. 2011). Cela rend compte de la
compétition à travers l’effet de dilution, où le colza attire les pollinisateurs présents dans les
habitats adjacents (Holzschuh et al. 2011).
D. Longueur des bordures La configuration peut être quantifiée par la somme de la
longueur des bordures entre les habitats des paysages. Il y a une relation de type monod entre
cette variable et la diversité de pollinisateurs (Figure 1D), la configuration étant liée à une
interspersion plus importante, c’est-à-dire à une diminution de la distance entre habitats
complémentaires (Fahrig et al. 2011), et à une augmentation de la connectivité permettant les
mouvements des individus entre habitats (Tscharntke et al. 2012).
La compréhension des liens entre l’organisation des paysages agricoles et la diversité
de pollinisateurs associée à la fonction de pollinisation est une étape clé vers une optimisation
durable de la fonction de pollinisation. Dans ce contexte, notre étude a pour objectif de mettre
en évidence l’influence des facteurs locaux, paysagers, et leur interaction sur l’efficacité de la
pollinisation entomophile du colza. Une approche semi-expérimentale a été mise en place via
la pose de pieds de colza dans différentes localités le long de gradients paysagers dans un
territoire agricole. Nous avons deux objectifs : 1/ Quantifier l’influence de la pollinisation
entomophile sur le rendement du colza ; 2/ Étudier l’influence de facteurs locaux et paysagers
ainsi que leur interaction sur le succès de pollinisation entomophile du colza.
Il est attendu que la contribution des insectes au rendement du colza soit inférieure à
celle de l’autogamie mais supérieure à celle de l’anémogamie et que le succès de
fructification soit le trait impliqué dans le rendement le plus dépendant de ce processus
comme cela a été montré sur des plantes de colza en plein champ (Perrot et al. en révision). Il
est également attendu que le succès de pollinisation entomophile dépende des facteurs locaux,

5
paysagers et de leur interaction, en relation avec la taille des pollinisateurs (Benjamin et al.
2014). Ainsi, le succès de pollinisation entomophile du colza devrait être plus important dans
les bordures de parcelles, les prairies et les cultures de colza car ces habitats fournissent
respectivement une diversité en plantes adventices plus importante que le plein champ
(Henckel et al. 2015), des ressources florales diverses, et un pulse de ressources florales
(Westcott & Nelson 2001) qui attire massivement les insectes pollinisateurs (Riedinger et al.
2015). Les proportions d’HSN, d’AB, de colza et la longueur des bordures dans les paysages
devraient influencer l’efficacité de la pollinisation entomophile en relation avec leur effet sur
la diversité de pollinisateurs suivant les relations décrites dans la Figure 1. Étant donné que
l’assemblage des communautés de pollinisateurs dépend de l’interaction entre les facteurs
locaux et paysagers (Kennedy et al. 2013), on s’attend également à ce que les facteurs
paysagers, dans leur échelle d’influence et dans leur capacité à diluer ou concentrer la
fonction de pollinisation entomophile du colza, dépendent de l’attractivité des localités
(Riedinger et al. 2015).
B. MATÉRIEL ET MÉTHODES
1. Zone d’étude
La Zone Atelier
Plaine & Val de Sèvre (ZA
PVS ; Figure 2) couvre une
surface de 435 kms² dans le
département des Deux-
Sèvres. Ce territoire peut
être divisé en trois
catégories couvrant des
densités différentes : 1/ Le
système productif
regroupe les parcelles agricoles destinées à la production de denrées et couvre 76,9% du
territoire. Les cultures céréalières (45,6%) couvrent la majorité de ce système, les cultures de
colza (7,5%) et l’AB (13,7%) représentent une surface non négligeable de ce système. 2/
10,4% de la ZA PVS correspondent au système urbain (bâtiments, routes) ; 3/ Les HSN
(bois, haies, prairies permanentes et temporaires) couvrent 12,7% du territoire. Un suivi
annuel des variables paysagères et des pratiques agricoles depuis 1994 et 2006 respectivement
y est effectué. Les recherches de la ZA PVS visent à trouver des solutions afin de concilier
Figure 2 : Assolement de la Zone Atelier Plaine & Val de Sèvre en 2017.

6
une production agricole suffisante ainsi que la conservation de la biodiversité et des services
écosystémiques associés dans les paysages agricoles. Les approches mobilisées pour étudier
l’influence des pratiques agricoles et des caractéristiques des paysages sur la biodiversité,
certaines fonctions écologiques et certains services écosystémiques sont à la fois des analyses
empiriques et des études expérimentales in situ dans des parcelles des agriculteurs
(Bretagnolle et al. 2018). Le suivi de la fonction de pollinisation du colza a débuté en 2013
en parcelle et en 2014 avec des plantes phytomètres (voir section B.2.a), dans le but de mettre
en évidence l’importance des insectes pollinisateurs sur la production agricole et de
comprendre comment les pratiques et les paysages agricoles affectent à la fois les
pollinisateurs et la pollinisation des cultures.
2. Échantillonnage
a. Choix des paysages
L’étude des effets du contexte paysager sur la pollinisation du colza est réalisée à
l’aide de phytomètres de colza : 832 plantes ont été posées depuis 2014. Les phytomètres sont
placés dans des parcelles agricoles de 6,09 ha de moyenne se situant dans des micro-paysages
d’un km² répartis le long de gradients de proportions d’AB et d’HSN connues pour influencer
les communautés de pollinisateurs (Figure 1A et 1B). La sélection des fenêtres paysagères
d’un km² est réalisée de manière à minimiser la corrélation entre les deux gradients et ainsi
pouvoir étudier leurs effets sur la fonction de pollinisation indépendamment l’un de l’autre.
Pour cela, de nombreux micro-paysages sont représentés graphiquement le long des gradients
de densités d’AB et d’HSN. Puis les paysages sont sélectionnés de manière à minimiser la
corrélation entre les deux variables paysagères (Fahrig et al. 2011).
b. Mesure de la fonction de pollinisation
Les phytomètres sont des plants de colza (variété DK EKSTROM) récupérés dans des
parcelles de la ZA PVS à la fin de l’hiver et rempotés dans des pots de trois litres de terre.
Ceux-ci sont placés dans des abris « insect-proof » empêchant l’accès aux insectes. Ensuite,
entre avril et juin (période de floraison du colza), les phytomètres sont placés pendant quatre
jours dans les paysages sélectionnés de la ZA PVS et dans différentes cultures (prairie, colza
ou céréale) à deux emplacements (en bordure ou en plein champ), avant d’être replacés sous
abris jusqu’à leur maturité (moment où le colza aurait été récolté par un agriculteur). Ce
protocole a connu des modifications entre années, celles-ci sont présentées en Annexe 1A.
L’expérimentation consiste en la pose sur trois hampes choisies aléatoirement de
sachets permettant la pollinisation autogame (traitement « osmolux »), la pollinisation
autogame et anémogame (traitement « petite maille »), la pollinisation autogame,
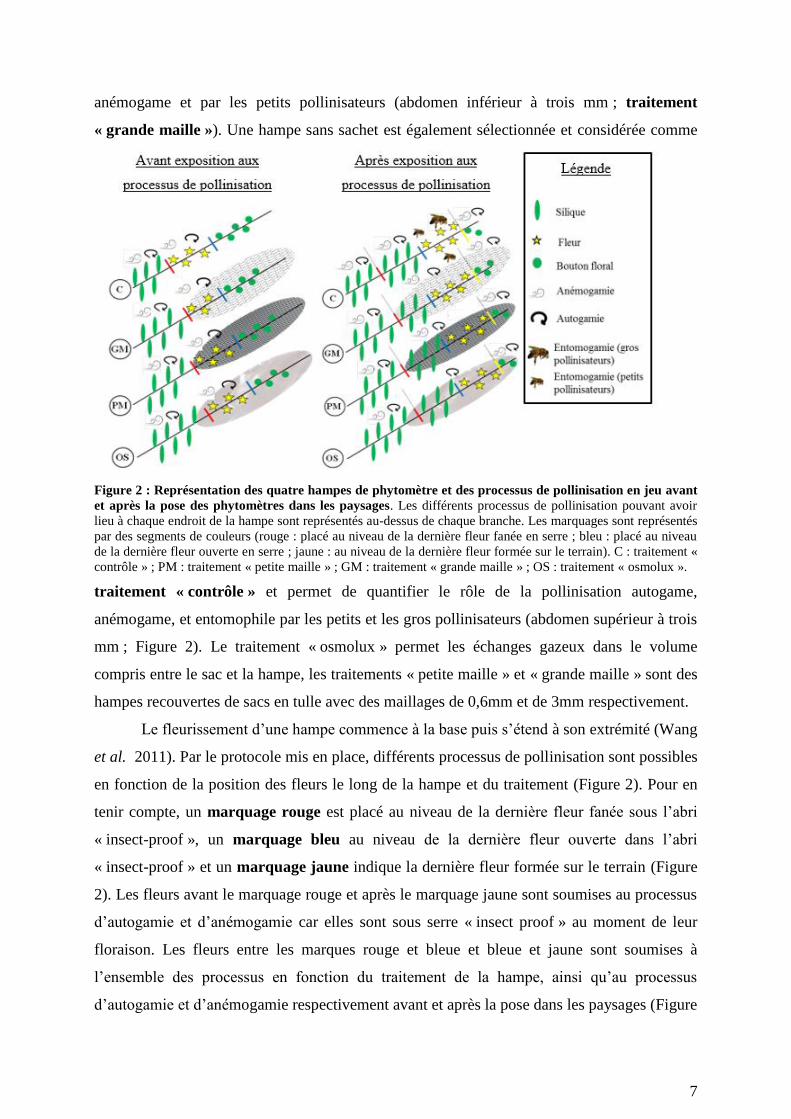
7
anémogame et par les petits pollinisateurs (abdomen inférieur à trois mm ; traitement
« grande maille »). Une hampe sans sachet est également sélectionnée et considérée comme
Figure 2 : Représentation des quatre hampes de phytomètre et des processus de pollinisation en jeu avant
et après la pose des phytomètres dans les paysages. Les différents processus de pollinisation pouvant avoir
lieu à chaque endroit de la hampe sont représentés au-dessus de chaque branche. Les marquages sont représentés
par des segments de couleurs (rouge : placé au niveau de la dernière fleur fanée en serre ; bleu : placé au niveau
de la dernière fleur ouverte en serre ; jaune : au niveau de la dernière fleur formée sur le terrain). C : traitement «
contrôle » ; PM : traitement « petite maille » ; GM : traitement « grande maille » ; OS : traitement « osmolux ».
traitement « contrôle » et permet de quantifier le rôle de la pollinisation autogame,
anémogame, et entomophile par les petits et les gros pollinisateurs (abdomen supérieur à trois
mm ; Figure 2). Le traitement « osmolux » permet les échanges gazeux dans le volume
compris entre le sac et la hampe, les traitements « petite maille » et « grande maille » sont des
hampes recouvertes de sacs en tulle avec des maillages de 0,6mm et de 3mm respectivement.
Le fleurissement d’une hampe commence à la base puis s’étend à son extrémité (Wang
et al. 2011). Par le protocole mis en place, différents processus de pollinisation sont possibles
en fonction de la position des fleurs le long de la hampe et du traitement (Figure 2). Pour en
tenir compte, un marquage rouge est placé au niveau de la dernière fleur fanée sous l’abri
« insect-proof », un marquage bleu au niveau de la dernière fleur ouverte dans l’abri
« insect-proof » et un marquage jaune indique la dernière fleur formée sur le terrain (Figure
2). Les fleurs avant le marquage rouge et après le marquage jaune sont soumises au processus
d’autogamie et d’anémogamie car elles sont sous serre « insect proof » au moment de leur
floraison. Les fleurs entre les marques rouge et bleue et bleue et jaune sont soumises à
l’ensemble des processus en fonction du traitement de la hampe, ainsi qu’au processus
d’autogamie et d’anémogamie respectivement avant et après la pose dans les paysages (Figure

8
2).
3. Mesure des traits impliqués dans le rendement du colza
Les phytomètres arrivés à maturité, les hampes sous traitement sont coupées, placées
individuellement dans des sachets puis dans une étuve à 70°C pendant 48h. Différents traits
impliqués dans le rendement sont concernés à l’échelle de la branche et quantifiés entre
chaque marque (Diepenbrock 2000) : le nombre de fleurs, le nombre de siliques, le succès
de fructification qui est le ratio entre le nombre de siliques et le nombre de fleurs formées, le
nombre moyen de grains par silique qui est le rapport entre le nombre total de grains et le
nombre de siliques et la masse moyenne des grains estimée par la masse totale des grains
divisée par le nombre total de grains. Le rendement est la masse moyenne de grains produite
sur une branche, c’est le produit du nombre de siliques, du nombre moyen de grains par
silique et de la masse moyenne des grains. Les traits liés au rendement ont été quantifiés
différemment entre années et entre positions le long des hampes (Annexe 1B).
4. Calcul des contributions absolues et relatives des processus de pollinisation
A partir des traitements sur un même phytomètre, les contributions absolues et
relatives des processus de pollinisation aux traits impliqués dans le rendement sont calculées.
Si la valeur d’un trait associée à un processus de pollinisation est calculée indépendamment
des autres processus, la contribution du processus de pollinisation au trait est dite absolue
sinon elle est relative (Tableau 1).
Tableau 1 : Calcul des contributions absolue et relative de l’autogamie (AU), de l’anémogamie (AN), des
petits pollinisateurs (ENp) et des gros pollinisateurs (ENg) à un trait impliqué dans le rendement.
Processus Contribution absolue Contribution relative
Autogamie Osmolux
AU AU% =
AU
𝐴𝑈+𝐴𝑁+ 𝐸𝑁𝑃+𝐸𝑁𝑔
Anémogamie Petite maille – Osmolux
AN = (AU + AN) – AU AN% =
AN
𝐴𝑈+𝐴𝑁+ 𝐸𝑁𝑃+𝐸𝑁𝑔
Petits pollinisateurs Grande maille – Petite maille
ENp = (AU + AN + ENp) – (AU + AN) ENp% =
𝐸𝑁𝑝
𝐴𝑈+𝐴𝑁+ 𝐸𝑁𝑃+𝐸𝑁𝑔
Gros pollinisateurs Contrôle – Grande maille
ENg = AU + AN + ENp + ENg – (AU + AN + ENp) ENg% =
𝐸𝑁𝑔
𝐴𝑈+𝐴𝑁+ 𝐸𝑁𝑃+𝐸𝑁𝑔
5. Variables locales et paysagères
Les variables locales sont déterminées à l’échelle de la parcelle agricole.
L’emplacement dans la parcelle (en bordure ou en plein champ) et la culture testée (colza,
céréale, prairie) sont considérés car ces variables locales sont suspectées d’influencer les
communautés de pollinisateurs locales.
Les variables paysagères sont les proportions d’AB, de colza, d’HSN ainsi que la
longueur des bordures entre les éléments du paysage, connues pour influencer la structure

9
des communautés de pollinisateurs dans les paysages agricoles (Figure 1). La proportion
d’HSN est décomposée en proportions de haie, de prairie et de forêt afin de prendre en
compte la variabilité dans la diversité de ressources florales entre les HSN. Ces variables sont
quantifiées dans des rayons d’étude de 500m, 750m, 1000m, 1250m, 1500m, 1750m et
2000m autour des centroïdes des parcelles d’étude car les insectes pollinisateurs couvrent des
distances entre quelques centaines de mètres et quelques kilomètres (Pufal et al. 2017) et qu’à
partir d’un rayon de 500m, les variables paysagères en dehors des parcelles dans lesquelles les
phytomètres sont placés sont caractérisables.
6. Analyses statistiques
L’ensemble des analyses a été effectué avec le logiciel R (version 3.3.2 ; R Core Team
2016). Les packages « car » (Fox & Weisberg 2011), « ggplot2 » (Wickham 2009), « Hmisc »
(Harrell 2017), « lme4 » (Bates et al. 2015), « lmerTest » (Kuznetsova et al. 2017),
« maptools » (Bivand & Lewin-Koh 2017), « nlme » (Pinheiro et al. 2016), « raster »
(Hijmans 2017), « rgeos » (Bivand & Rundel 2017) et « rgdal » (Bivand et al. 2018) ont été
utilisés pour les analyses. L’ensemble des données de l’année 2014 a été retiré car seuls deux
traitements ont été appliqués aux phytomètres (contre quatre les autres années ; annexe 1A).
a. Comment le trait impliqué dans le rendement est-t-il sélectionné ?
Les intervalles de confiance à 95% des contributions relatives des gros, des petits
pollinisateurs, de l’anémogamie et de l’autogamie à l’ensemble des traits impliqués dans le
rendement sont calculés par une méthode de bootstrapping (1000 tirages avec remises ; Efron
1979). Cette méthode donne lieu à des valeurs de contributions relatives négatives (par
exemple quand le nombre de siliques est plus important sur la hampe soumise au traitement
« grande maille » que sur la hampe « contrôle »). Des valeurs nulles sont assignées pour les
contributions relatives dont la valeur calculée est négative, et cela, pour l’ensemble des traits
liés au rendement du colza. Cela concerne respectivement de 29% à 44%, de 31% à 54%, et
de 45% à 54% des contributions relatives des gros, des petits pollinisateurs et du vent en
fonction des traits impliqués dans le rendement étudié. Pour étudier la similarité des
contributions relatives des gros (n = 337) et des petits pollinisateurs (n = 337) entre les traits
impliqués dans le rendement, des χ² de contingence sont appliqués. Le trait lié au rendement
du colza qui dépend le plus des insectes pollinisateurs est sélect000ionné par la comparaison
des moyennes et des intervalles de confiance à 95% à l’œil (Cumming & Fich 2005) des
contributions relatives des petits et des gros pollinisateurs. Les relations entre traits impliqués
dans le rendement du colza sont étudiées par des corrélations de Spearman (normalité des

10
données non respectée). L’intervalle de confiance à 95% du coefficient de corrélation ρ est
obtenu par une méthode de bootstrapping (1000 tirages avec remises ; Efron 1979). Les traits
associés à ces tests sont calculés entre les marques rouge et jaune étant donné que les liens
entre traits (Annexes 3) et les contributions relatives des différents processus de pollinisation
(Annexes 4) sont similaires qu’ils soient étudiés entre les marques rouge et bleue ou bleue et
jaune. Étant donné le nombre important de valeurs de contributions relatives transformées, la
suite des analyses s’intéresse aux traitements et non pas aux contributions relatives des
processus de pollinisation.
b. Étude de l’influence des biais expérimentaux
Afin de vérifier que les hampes sont similaires morphologiquement entre les
différentes localités dans lesquelles les phytomètres sont placés et les traitements, le nombre
de fleurs entre les marques rouge et bleue est étudié en fonction du traitement, de la culture
testée et de l’emplacement dans la parcelle par un modèle linéaire généralisé (n = 1909 ;
distribution de type « poisson » ; étude résiduelle en annexes 5Aa et 5Ba). Les hampes ne sont
pas morphologiquement similaires selon les cultures qui ont été expérimentées (voir section
C.2). L’allocation de l’énergie à la production de fleurs dépend de la hampe (Dienpenbrock
2000 ; Wang et al. 2011) mais également de la position le long de la hampe (Wang et al.
2011). L’influence de la position verticale de la hampe (numéro de hampe) sur le nombre de
fleurs entre les marques rouge et bleue est étudiée par un modèle linéaire généralisé (n =
1838 ; distribution de type « poisson » ; étude résiduelle en annexes 5Ab et 5Bb). Un χ² de
contingence permet d’étudier la dissimilarité des numéros de hampes entre les trois cultures
locales (n = 1830). L’effet de la position de la première marque (nombre de fleurs avant la
marque rouge) sur le nombre de fleurs entre les marques rouge et jaune est étudié. Pour cela,
un modèle mixte généralisé (n = 1919 ; distribution de type « poisson » ; analyse résiduelle en
annexes 5Ac et 5Bc) avec le traitement, la culture testée et l’année comme effets fixes et le
phytomètre comme effet aléatoire expliquant le nombre de fleurs avant la marque rouge a été
construit. Puis l’influence de la position de la première marque sur le nombre de fleurs entre
les marques rouge et jaune a été étudiée par une corrélation de Spearman entre ces deux
variables (l’intervalle de confiance à 95% du coefficient de corrélation ρ est donnée suite à
1000 tirages avec remise ; Efron 1979). L’ensemble des modèles a été analysé avec des
Anova de type II.
c. Le succès de fructification et le rendement dépendent-ils des conditions locales ?
L’analyse des facteurs locaux et paysagers s’applique sur le succès de fructification
(section C. 1) et sur le rendement. Le rendement a subi une transformation logarithmique

11
afin de respecter les conditions de normalité et d’homoscédasticité des modèles linéaires.
L’influence des facteurs locaux (n = 1722) sur le succès de fructification et sur le rendement
(n = 1010) est étudiée par des modèles linéaires mixtes avec le phytomètre comme effet
aléatoire (analyse résiduelle en annexes 5Ad et 5Bd pour le succès de fructification et en
annexes 5Ae et 5Be pour le rendement) puis des Anova de type II sont appliquées sur les
modèles sélectionnés. Les variables explicatives sont la culture testée, l’emplacement dans la
parcelle, le traitement et les interactions entre ces variables. L’année d’étude et la date de
pose des phytomètres (nombre de jours depuis la floraison du colza niché dans la variable
culture car la date de pose des phytomètres est plus précoce dans les cultures de colza ;
données sur la commune de Saint-Pierre-D’Amilly proche de la ZA PVS extraites de
http://apibotanica.inra.fr/) sont incluses dans le modèle afin de prendre en compte les
dynamiques temporelles intra et interannuelles des communautés de pollinisateurs (Petanidou
et al. 2008 ; Rollin et al. 2014) et des paysages (Annexe 2). La position de la première
marque sur la hampe est incluse comme covariable afin de prendre en compte la variabilité du
nombre de fleurs entre les marques rouge et jaune. Une procédure de sélection de modèle
basée sur le critère d'information d'Akaike (AIC ; Aho et al. 2014) est effectuée.
d. Étude de l’influence du contexte paysager sur la production de grains
L’effet du contexte paysager sur le succès de fructification et sur le rendement est
étudié indépendamment pour les trois cultures locales. Dans les cultures de colza (n = 697 et n
= 439 respectivement pour le succès de fructification et le rendement), les cultures de céréales
(n = 740 et n = 405) et les prairies (n = 285 et n = 160), un modèle linéaire mixte avec le
phytomètre en effet aléatoire est construit pour chaque variable dépendante (analyses
résiduelles en annexes 5A et B f à k). Les variables explicatives sont le traitement, les
variables paysagères et les interactions deux à deux entre ces variables. Les variables
paysagères, correspondant à des proportions, subissent une transformation de type arcsin car
de nombreux paysages ont une proportion de variable paysagère proche de zéro. L’année
d’étude, la date de pose du phytomètre et la position de la première marque sont incluses dans
les modèles car elles influencent le succès de fructification et le rendement (voir analyse ci-
dessous). La surface de la parcelle est également ajoutée comme covariable car plusieurs
phytomètres sont posés dans une même parcelle d’étude. Pour chaque culture testée et chaque
variable dépendante, 4096 combinaisons de modèles sont testées. Celles-ci correspondent aux
variations des rayons d’études des six variables paysagères. Les échelles d’influence des
variables paysagères sont déterminées par l’AIC. La part de variance expliquée par chacune
des variables explicatives est analysée par des Anova de type II. L’indice de sensibilité de

12
chaque variable explicative est calculé en fonction de la culture testée et de la variable
dépendante en divisant la somme des carrés d’une variable par la somme totale des carrés de
l’ensemble des variables (Ginot et al. 2006).
C. RÉSULTATS
1. Recherche du trait impliqué dans le rendement
a. Contribution des processus de pollinisation aux traits liés au rendement
L’autogamie contribue significativement plus aux traits impliqués dans le rendement
que les autres processus de pollinisation (Tableau 2). La contribution de l’entomogamie n’est
pas similaire suivant le trait lié au rendement étudié, que ce soit pour les gros (χ² = 1523,53 ;
ddl = 1245 ; p = 8,67.10-8) ou les petits pollinisateurs (χ² = 1354,73 ; ddl = 1119 ; p = 1,40.10-
6). D’après le Tableau 2, les contributions relatives des gros et des petits pollinisateurs sont
plus importantes pour le succès de fructification et le nombre de siliques.
Tableau 2 : Contributions relatives des processus de pollinisation aux traits impliqués dans le rendement
du colza. La moyenne ainsi que l’intervalle de confiance à 95% de ces contributions sont calculés à partir d’une
méthode de bootstrapping (1000 tirages avec remise).
Succès de
fructification Nombre de
siliques Nombre moyen de
grains par silique Masse moyenne
des grains
Entomogamie
(> 3mm) 0,246
[0,210 ; 0,280] 0,250
[0,220 ; 0,278] 0,187
[0,154 ; 0,216] 0,080
[0,063 ; 0,098]
Entomogamie
(< 3mm) 0,170
[0,147 ; 0,194] 0,183
[0,156 ; 0,208] 0,137
[0,107 ; 0,159] 0,092
[0,072 ; 0,116]
Anémogamie 0,123
[0,090 ; 0,149] 0,130
[0,105 ; 0,161] 0,121
[0,088; 0,154] 0,102
[0,075 ; 0,123]
Autogamie 0,456
[0,423 ; 0,488] 0,434
[0,402 ; 0,464] 0,557
[0,520 ; 0,587] 0,720
[0,688 ; 0,752]
b. Compromis entre traits impliqués dans le rendement
Le succès de fructification, le nombre de siliques et le nombre moyen de grains par
silique sont corrélés positivement (Figure 3). L’augmentation d’un de ces traits associés au
Figure 3 : Représentation des relations entre les traits impliqués dans le rendement. ρ correspond à
l’intervalle de confiance à 95% du coefficient de corrélation de Spearman. * : p-value < 0,05 ; ** : p-value <
0,01 ; *** : p-value < 0,001.
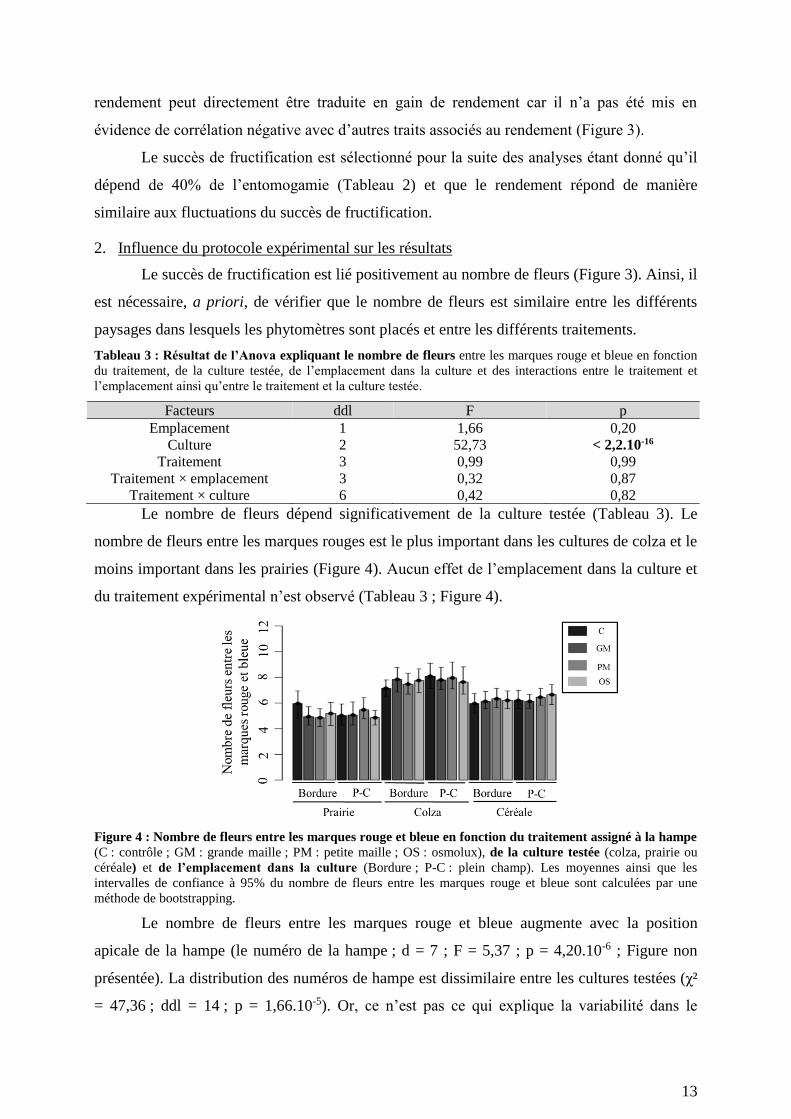
13
rendement peut directement être traduite en gain de rendement car il n’a pas été mis en
évidence de corrélation négative avec d’autres traits associés au rendement (Figure 3).
Le succès de fructification est sélectionné pour la suite des analyses étant donné qu’il
dépend de 40% de l’entomogamie (Tableau 2) et que le rendement répond de manière
similaire aux fluctuations du succès de fructification.
2. Influence du protocole expérimental sur les résultats
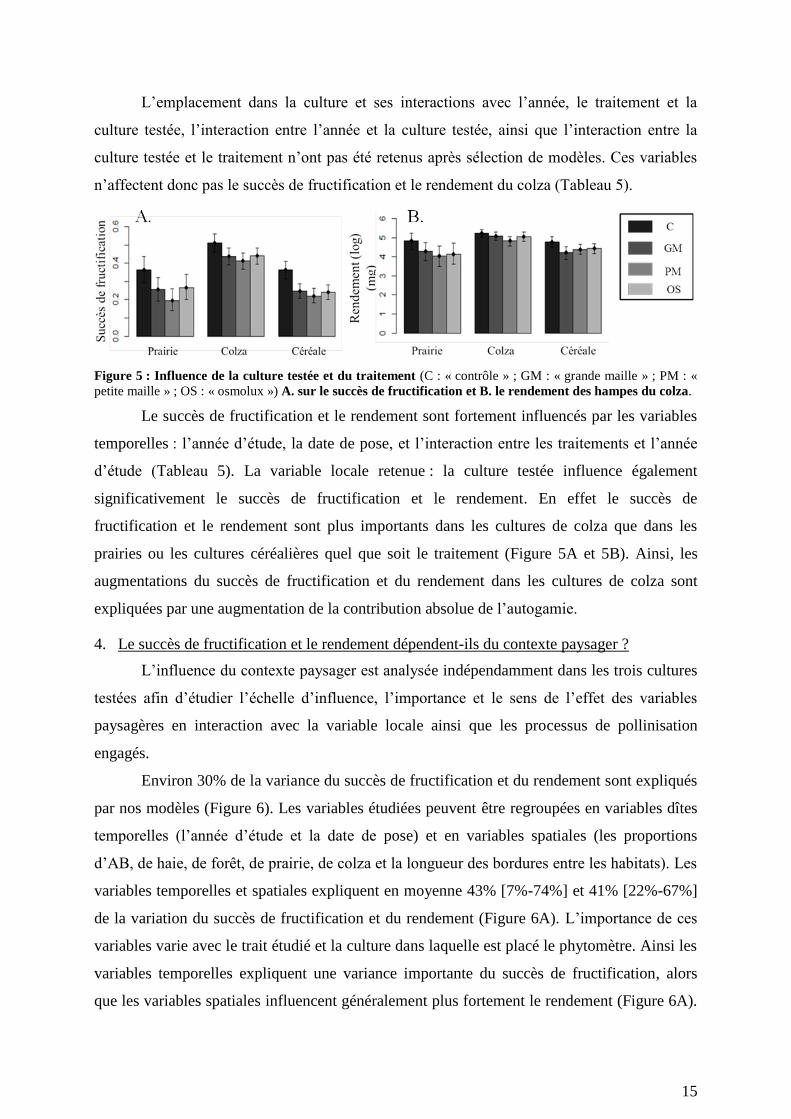
Le succès de fructification est lié positivement au nombre de fleurs (Figure 3). Ainsi, il
est nécessaire, a priori, de vérifier que le nombre de fleurs est similaire entre les différents
paysages dans lesquels les phytomètres sont placés et entre les différents traitements.
Tableau 3 : Résultat de l’Anova expliquant le nombre de fleurs entre les marques rouge et bleue en fonction
du traitement, de la culture testée, de l’emplacement dans la culture et des interactions entre le traitement et
l’emplacement ainsi qu’entre le traitement et la culture testée.
Facteurs ddl F p
Emplacement 1 1,66 0,20
Culture 2 52,73 < 2,2.10-16
Traitement 3 0,99 0,99
Traitement × emplacement 3 0,32 0,87
Traitement × culture 6 0,42 0,82
Le nombre de fleurs dépend significativement de la culture testée (Tableau 3). Le
nombre de fleurs entre les marques rouges est le plus important dans les cultures de colza et le
moins important dans les prairies (Figure 4). Aucun effet de l’emplacement dans la culture et
du traitement expérimental n’est observé (Tableau 3 ; Figure 4).
Figure 4 : Nombre de fleurs entre les marques rouge et bleue en fonction du traitement assigné à la hampe
(C : contrôle ; GM : grande maille ; PM : petite maille ; OS : osmolux), de la culture testée (colza, prairie ou
céréale) et de l’emplacement dans la culture (Bordure ; P-C : plein champ). Les moyennes ainsi que les
intervalles de confiance à 95% du nombre de fleurs entre les marques rouge et bleue sont calculées par une
méthode de bootstrapping.
Le nombre de fleurs entre les marques rouge et bleue augmente avec la position
apicale de la hampe (le numéro de la hampe ; d = 7 ; F = 5,37 ; p = 4,20.10-6 ; Figure non
présentée). La distribution des numéros de hampe est dissimilaire entre les cultures testées (χ²
= 47,36 ; ddl = 14 ; p = 1,66.10-5). Or, ce n’est pas ce qui explique la variabilité dans le

14
nombre de fleurs entre les cultures locales car plus de hampes en position apicale sont
sélectionnées dans les prairies. Respectivement 7,3%, 11,5% et 13,3% des numéros de
hampes sélectionnés dans les cultures de colza, les cultures céréalières et les prairies sont
supérieurs à cinq.
La position de la première marque varie significativement avec la culture testée et le
traitement assigné à la hampe (Tableau 4). Ainsi, le nombre de fleurs avant la marque rouge
est plus faible quand le traitement « osmolux » est assigné à la hampe et quand les
phytomètres sont placés dans les cultures de colza (Figure non présentée).
Tableau 4 : Résultat de l’Anova expliquant le nombre de fleurs avant la première marque en fonction de
la culture testée et du traitement, de leur interaction et du phytomètre (en effet aléatoire).
Facteurs ddl F p
Culture 2 13,03 3,10.10-6
Traitement 3 6,86 1,37.10-4
Traitement × culture 6 0,45 0,84
La position de la première marque est plus proche de la position basale de la hampe
dans les cultures de colza car les phytomètres sont placés plus tôt dans celles-ci. Ainsi, les
phytomètres sont respectivement posés 30, 40, et 44 jours après la floraison du colza dans les
cultures de colza, dans les parcelles en céréales et dans les prairies. Il y a une corrélation
négative entre le nombre de fleurs avant la marque rouge et le nombre de fleurs entre les
marques rouge et jaune (ρ = [-0,59 ; - 0,25] ; ddl = 1933 ; p <2,2.10-16).
Afin de prendre en compte la variabilité du nombre de fleurs entre cultures locales et
la corrélation positive entre le nombre de fleurs et le succès de fructification, la position de la
première marque est incluse comme covariable pour la suite des analyses.
3. Le succès de fructification et le rendement dépendent-ils des conditions locales ?
Par des procédures de sélection de modèles basées sur l’AIC, des modèles similaires
ont été sélectionnés afin d’expliquer l’influence des facteurs locaux sur le succès de
fructification et le rendement (Tableau 5).
Tableau 5 : Résultat de l’Anova expliquant le succès de fructification et le rendement en fonction du
traitement, de la culture, de l’année, de la date de pose, de la position de la première marque, de
l’interaction entre l’année et le traitement et du phytomètre en effet aléatoire. Les deux modèles ont été
sélectionnés suite à des procédures de sélection de modèle basées sur l’AIC.
Traits Succès de fructification Rendement
Facteurs ddl F p ddl F P
Culture 2 14,71 6,40.10-7 2 11,16 2,00.10-5
Traitement 3 30,95 < 2,2.10-16 3 11,52 2,18.10-7
Année 2 108,25 < 2,2.10-16 2 33,01 6,56.10-14
Date de pose 3 43,21 < 2,2.10-16 3 12,27 1,12.10-7
Position de la première marque 1 26,33 3,33.10-7 1 28,39 1,29.10-7
Traitement × année 6 46,21 7,65.10-11 6 3,47 2,20.10-3
15
L’emplacement dans la culture et ses interactions avec l’année, le traitement et la
culture testée, l’interaction entre l’année et la culture testée, ainsi que l’interaction entre la
culture testée et le traitement n’ont pas été retenus après sélection de modèles. Ces variables
n’affectent donc pas le succès de fructification et le rendement du colza (Tableau 5).
Figure 5 : Influence de la culture testée et du traitement (C : « contrôle » ; GM : « grande maille » ; PM : «
petite maille » ; OS : « osmolux ») A. sur le succès de fructification et B. le rendement des hampes du colza.
Le succès de fructification et le rendement sont fortement influencés par les variables
temporelles : l’année d’étude, la date de pose, et l’interaction entre les traitements et l’année
d’étude (Tableau 5). La variable locale retenue : la culture testée influence également
significativement le succès de fructification et le rendement. En effet le succès de
fructification et le rendement sont plus importants dans les cultures de colza que dans les
prairies ou les cultures céréalières quel que soit le traitement (Figure 5A et 5B). Ainsi, les
augmentations du succès de fructification et du rendement dans les cultures de colza sont
expliquées par une augmentation de la contribution absolue de l’autogamie.
4. Le succès de fructification et le rendement dépendent-ils du contexte paysager ?
L’influence du contexte paysager est analysée indépendamment dans les trois cultures
testées afin d’étudier l’échelle d’influence, l’importance et le sens de l’effet des variables
paysagères en interaction avec la variable locale ainsi que les processus de pollinisation
engagés.
Environ 30% de la variance du succès de fructification et du rendement sont expliqués
par nos modèles (Figure 6). Les variables étudiées peuvent être regroupées en variables dîtes
temporelles (l’année d’étude et la date de pose) et en variables spatiales (les proportions
d’AB, de haie, de forêt, de prairie, de colza et la longueur des bordures entre les habitats). Les
variables temporelles et spatiales expliquent en moyenne 43% [7%-74%] et 41% [22%-67%]
de la variation du succès de fructification et du rendement (Figure 6A). L’importance de ces
variables varie avec le trait étudié et la culture dans laquelle est placé le phytomètre. Ainsi les
variables temporelles expliquent une variance importante du succès de fructification, alors
que les variables spatiales influencent généralement plus fortement le rendement (Figure 6A).
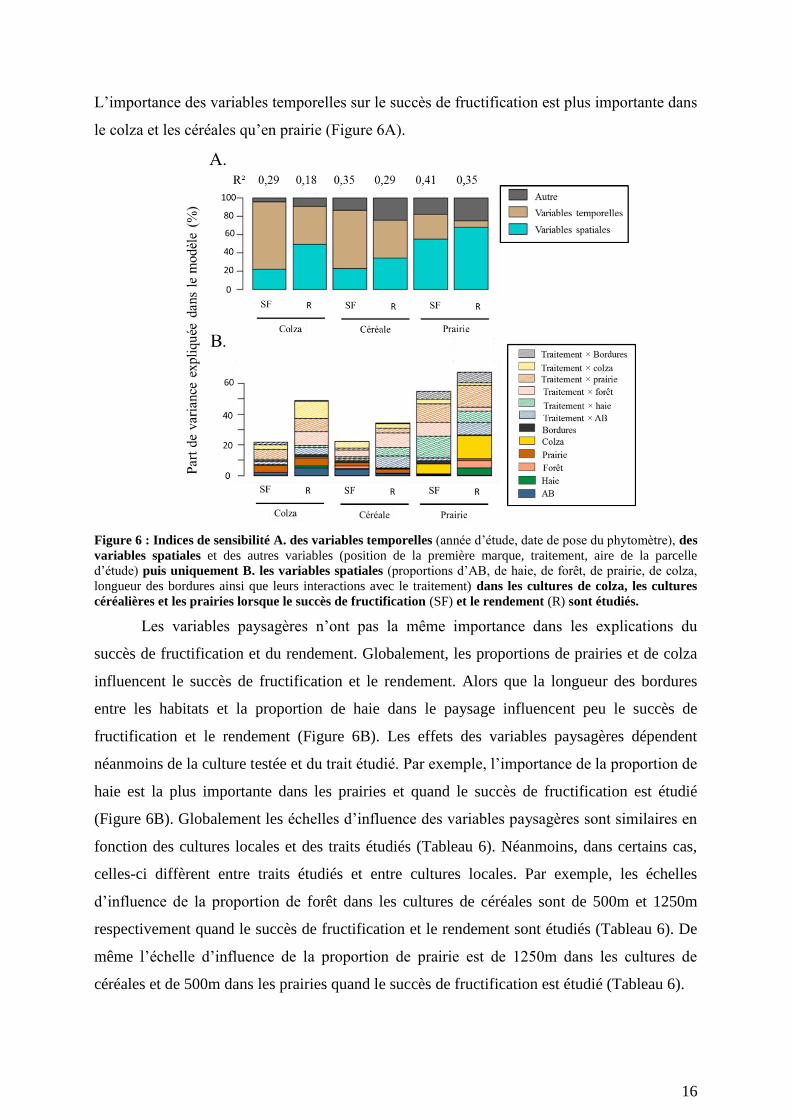
16
L’importance des variables temporelles sur le succès de fructification est plus importante dans
le colza et les céréales qu’en prairie (Figure 6A).
Figure 6 : Indices de sensibilité A. des variables temporelles (année d’étude, date de pose du phytomètre), des
variables spatiales et des autres variables (position de la première marque, traitement, aire de la parcelle
d’étude) puis uniquement B. les variables spatiales (proportions d’AB, de haie, de forêt, de prairie, de colza,
longueur des bordures ainsi que leurs interactions avec le traitement) dans les cultures de colza, les cultures
céréalières et les prairies lorsque le succès de fructification (SF) et le rendement (R) sont étudiés.
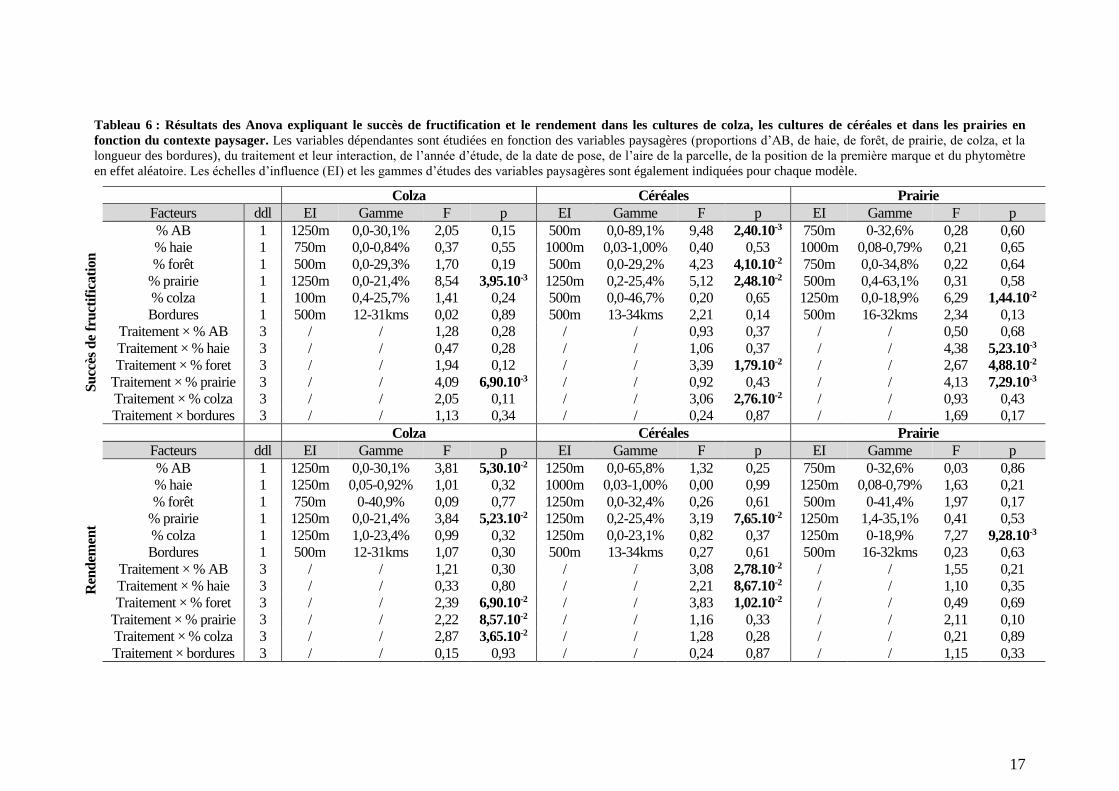
Les variables paysagères n’ont pas la même importance dans les explications du
succès de fructification et du rendement. Globalement, les proportions de prairies et de colza
influencent le succès de fructification et le rendement. Alors que la longueur des bordures
entre les habitats et la proportion de haie dans le paysage influencent peu le succès de
fructification et le rendement (Figure 6B). Les effets des variables paysagères dépendent
néanmoins de la culture testée et du trait étudié. Par exemple, l’importance de la proportion de
haie est la plus importante dans les prairies et quand le succès de fructification est étudié
(Figure 6B). Globalement les échelles d’influence des variables paysagères sont similaires en
fonction des cultures locales et des traits étudiés (Tableau 6). Néanmoins, dans certains cas,
celles-ci diffèrent entre traits étudiés et entre cultures locales. Par exemple, les échelles
d’influence de la proportion de forêt dans les cultures de céréales sont de 500m et 1250m
respectivement quand le succès de fructification et le rendement sont étudiés (Tableau 6). De
même l’échelle d’influence de la proportion de prairie est de 1250m dans les cultures de
céréales et de 500m dans les prairies quand le succès de fructification est étudié (Tableau 6).
17
Tableau 6 : Résultats des Anova expliquant le succès de fructification et le rendement dans les cultures de colza, les cultures de céréales et dans les prairies en
fonction du contexte paysager. Les variables dépendantes sont étudiées en fonction des variables paysagères (proportions d’AB, de haie, de forêt, de prairie, de colza, et la
longueur des bordures), du traitement et leur interaction, de l’année d’étude, de la date de pose, de l’aire de la parcelle, de la position de la première marque et du phytomètre
en effet aléatoire. Les échelles d’influence (EI) et les gammes d’études des variables paysagères sont également indiquées pour chaque modèle.
Colza Céréales Prairie
Facteurs ddl EI Gamme F p EI Gamme F p EI Gamme F p
Su
ccès
de
fru
ctif
icati
on
% AB 1 1250m 0,0-30,1% 2,05 0,15 500m 0,0-89,1% 9,48 2,40.10-3 750m 0-32,6% 0,28 0,60
% haie 1 750m 0,0-0,84% 0,37 0,55 1000m 0,03-1,00% 0,40 0,53 1000m 0,08-0,79% 0,21 0,65
% forêt 1 500m 0,0-29,3% 1,70 0,19 500m 0,0-29,2% 4,23 4,10.10-2 750m 0,0-34,8% 0,22 0,64
% prairie 1 1250m 0,0-21,4% 8,54 3,95.10-3 1250m 0,2-25,4% 5,12 2,48.10-2 500m 0,4-63,1% 0,31 0,58
% colza 1 100m 0,4-25,7% 1,41 0,24 500m 0,0-46,7% 0,20 0,65 1250m 0,0-18,9% 6,29 1,44.10-2
Bordures 1 500m 12-31kms 0,02 0,89 500m 13-34kms 2,21 0,14 500m 16-32kms 2,34 0,13
Traitement × % AB 3 / / 1,28 0,28 / / 0,93 0,37 / / 0,50 0,68
Traitement × % haie 3 / / 0,47 0,28 / / 1,06 0,37 / / 4,38 5,23.10-3
Traitement × % foret 3 / / 1,94 0,12 / / 3,39 1,79.10-2 / / 2,67 4,88.10-2
Traitement × % prairie 3 / / 4,09 6,90.10-3 / / 0,92 0,43 / / 4,13 7,29.10-3
Traitement × % colza 3 / / 2,05 0,11 / / 3,06 2,76.10-2 / / 0,93 0,43
Traitement × bordures 3 / / 1,13 0,34 / / 0,24 0,87 / / 1,69 0,17
Colza Céréales Prairie
Facteurs ddl EI Gamme F p EI Gamme F p EI Gamme F p
Ren
dem
ent
% AB 1 1250m 0,0-30,1% 3,81 5,30.10-2 1250m 0,0-65,8% 1,32 0,25 750m 0-32,6% 0,03 0,86
% haie 1 1250m 0,05-0,92% 1,01 0,32 1000m 0,03-1,00% 0,00 0,99 1250m 0,08-0,79% 1,63 0,21
% forêt 1 750m 0-40,9% 0,09 0,77 1250m 0,0-32,4% 0,26 0,61 500m 0-41,4% 1,97 0,17
% prairie 1 1250m 0,0-21,4% 3,84 5,23.10-2 1250m 0,2-25,4% 3,19 7,65.10-2 1250m 1,4-35,1% 0,41 0,53
% colza 1 1250m 1,0-23,4% 0,99 0,32 1250m 0,0-23,1% 0,82 0,37 1250m 0-18,9% 7,27 9,28.10-3
Bordures 1 500m 12-31kms 1,07 0,30 500m 13-34kms 0,27 0,61 500m 16-32kms 0,23 0,63
Traitement × % AB 3 / / 1,21 0,30 / / 3,08 2,78.10-2 / / 1,55 0,21
Traitement × % haie 3 / / 0,33 0,80 / / 2,21 8,67.10-2 / / 1,10 0,35
Traitement × % foret 3 / / 2,39 6,90.10-2 / / 3,83 1,02.10-2 / / 0,49 0,69
Traitement × % prairie 3 / / 2,22 8,57.10-2 / / 1,16 0,33 / / 2,11 0,10
Traitement × % colza 3 / / 2,87 3,65.10-2 / / 1,28 0,28 / / 0,21 0,89
Traitement × bordures 3 / / 0,15 0,93 / / 0,24 0,87 / / 1,15 0,33

18
Dans les cultures de colza,
le succès de fructification et le
rendement augmentent
significativement avec la
proportion de prairie dans un
rayon de 1250m (Tableau 6). Il y
a une interaction entre la
proportion de prairie dans le
paysage et le traitement (Tableau
6) : le succès de fructification et
le rendement augmentent
seulement pour les traitements
« contrôle » et « grande maille »
avec l’augmentation de la
proportion de prairie dans un
rayon de 1250m (Figure 7A). Ce
résultat suggère que la
contribution des petits
pollinisateurs au succès de
fructification et au rendement
augmente avec la proportion de
prairie dans le paysage quand les
phytomètres sont posés dans les
cultures de colza.
Dans les cultures de
céréales, il y a une interaction
entre la proportion d’AB dans un
rayon de 1250m et le traitement
quand le rendement est considéré (Tableau 6). Cette interaction est liée à l’augmentation de la
contribution absolue de l’anémogamie couplée à une diminution de la contribution absolue
des petits pollinisateurs avec la proportion d’AB dans le paysage (Figure 8A).
L’augmentation de la proportion de colza dans un rayon de 1250m influence le
rendement des phytomètres placés dans le colza en interagissant avec le traitement (Tableau
Figure 7 : A. Succès de fructification en fonction du pourcentage
de prairie dans un rayon de 1250m. Rendement en fonction des
pourcentages B. de colza dans un rayon de 1250m et C. de forêt
dans un rayon de 750, et cela dans les cultures de colza. Les
proportions ont subi une transformation arcsin. Les points représentent
les données brutes tandis que les intervalles de prédiction du succès de
fructification et du rendement sont représentés. Les traitements sont
également indiqués : C (contrôle), GM (Grande maille), PM (Petite
maille), OS (osmolux).

19
6). Le rendement des branches
en traitement « petite maille »
augmente avec la proportion de
colza dans le paysage ; aucun
effet n’est observé sur le
rendement des branches soumis
aux trois autres traitements
(Figure 7B). Ceci suggère que la
contribution absolue de
l’anémogamie augmente tandis
que celle des petits pollinisateurs
diminue lorsque la proportion de
colza autour des parcelles de
colza augmente. Dans les
cultures de céréales, le succès de
fructification dépend également
de l’interaction entre le
traitement et la proportion de
colza à une petite échelle
spatiale (Tableau 6). La
contribution absolue des gros
pollinisateurs augmente avec la
proportion de colza car seul le
succès de fructification du
traitement « contrôle » augmente
avec la proportion de colza
(Figure 8B).
La proportion de forêt
autour des parcelles influence le rendement dans les cultures de colza en interagissant avec le
traitement (Tableau 6). La contribution absolue de l’anémogamie au rendement augmente
tandis que celles de l’autogamie et des petits insectes diminuent avec la proportion de forêt
dans le paysage (Figure 7C). Dans les cultures de céréales, le succès de fructification dépend
significativement de la proportion de forêt dans un rayon de 500m et de l’interaction avec le
traitement tandis que seule l’interaction entre le traitement et la proportion de foret dans un
Figure 8 : Dans les cultures de céréales, A. Rendement en fonction
du pourcentage d’AB dans un rayon de 1250m, succès de
fructification en fonction des proportions B. de colza dans un
rayon de 500m et C. de forêt dans un rayon de 500m. Une
transformation arcsin est appliquée aux proportions. Les points
représentés sont issus des données brutes alors que les intervalles de
prédiction du succès de fructification et du rendement sont représentés.
Les traitements sont également indiqués : C (contrôle), GM (Grande
maille), PM (Petite maille), OS (osmolux).

20
rayon de 1250m influence le rendement (Tableau 6). Que le succès de fructification ou le
rendement soient étudié, la contribution absolue des petits insectes diminue avec la proportion
de forêt autour des cultures de céréales (Figure 8C).
D. DISCUSSION
1. Les insectes pollinisateurs contribuent au rendement du colza
L’autogamie est le principal processus de pollinisation du colza, suivi de
l’entomogamie et de l’anémogamie (Tableau 2). Le colza est une plante cultivée
majoritairement autogame (Becker et al. 1992) et plus adaptée à la dispersion du pollen par
les insectes que par le vent (Cresswell et al. 2014). La contribution des insectes pollinisateurs
est dépendante de leur taille et du trait impliqué dans le rendement du colza (Tableau 2). Les
pollinisateurs dont l’abdomen mesure plus de trois mm (par exemple les bourdons, les abeilles
domestiques) contribuent plus au rendement que les pollinisateurs dont l’abdomen mesure
moins de trois mm (les diptères pollinisateurs par exemple ; Tableau 2) car ce sont des
pollinisateurs du colza globalement plus efficaces (notamment l’abeille domestique Apis
mellifera) en termes de charge pollinique déposée sur le stigmate, de temps passé au contact
du stigmate et du nombre de fleurs visitées par unité de temps (Rader et al. 2009). C’est sur
le succès de fructification (fortement corrélé au nombre de siliques) que la contribution des
insectes est la plus importante (Tableau 2). Cela est cohérent avec une étude faite en plein
champ dans la ZA PVS (Perrot et al. en révision). Dans certaines études, le succès de
fructification est corrélé négativement avec le nombre de grains par silique (Perrot et al. en
révision) ou la masse des grains (Marini et al. 2015 ; Zou et al. 2017) par des compromis dans
l’allocation des ressources (Diepenbrock 2000). Ce n’est pas le cas dans notre étude (Figure
3). Ainsi l’augmentation du succès de fructification est directement liée à un gain de
rendement. In fine, les rendements du colza mesurés à l’échelle de la branche dépendent de
l’importance de l’entomogamie entre 28 et 35%. La transformation des valeurs de
contributions relatives (les valeurs négatives remises à zéro) n’induit pas une surestimation de
la contribution des insectes pollinisateurs au rendement du colza. Par exemple, sans
transformation, le succès de fructification dépend de [23 ; 36%] des gros pollinisateurs et de
[12-36%] des petits. Ce n’est pas significativement différent de ce qui est obtenu à la suite de
la transformation des données (Tableau 2). Mais, la contribution des insectes pollinisateurs au
rendement du colza est sous-estimée car les phytomètres sont placés dans les paysages
seulement quatre jours. Dans les études utilisant un protocole expérimental similaire
(Bommarco et al. 2012 ; Stanley et al. 2013 ; Bartomeus et al. 2014), les phytomètres sont
laissés dans les paysages de la floraison de la première fleur à la fane de la dernière fleur. La

21
durée de floraison d’une fleur de colza est comprise entre trois et cinq jours (Mesquida &
Renard 1981) mais, dans notre étude, certaines fleurs sont présentes un temps dans les
paysages et un autre temps dans les abris où seuls les processus d’autogamie et d’anémogamie
peuvent contribuer au rendement. Malgré cela, notre quantification est cohérente avec les
résultats de nombreuses autres études (par exemple Bommarco et al. 2012 ; Bartomeus et al.
2014 ; Perrot et al. en révision) où la contribution des insectes pollinisateurs au rendement du
colza est estimée entre 12% (Zou et al. 2017) et 51% (Kamler & Jas 2003).
2. La culture testée n’influence pas la pollinisation entomophile du colza
Le succès de fructification et le rendement sont plus importants dans les cultures de
colza que dans les cultures de céréales ou les prairies (Tableau 5 ; Figure 5) car la contribution
absolue de l’autogamie y est plus importante. Mais, contrairement à nos hypothèses, la
pollinisation entomophile est indépendante des variables locales, soit la culture testée et
l’emplacement dans les cultures (Tableau 5 ; Figure 5). La diversité de pollinisateurs dépend
de la qualité de l’habitat, et plus particulièrement de la densité et de la richesse en ressources
florales (Albrecht et al. 2007 ; Cole et al. 2017 ; Sydenham et al. 2017). Malgré les
différences d’attractivités entre les prairies, les cultures de colza et les cultures céréalières, la
pollinisation du colza par les insectes est similaire entre ces trois cultures locales (Tableau 5).
A notre connaissance aucune étude n’a quantifié le lien entre la culture testée et le succès de
pollinisation entomophile du colza. Seules certaines mettent en évidence que la culture testée
influence la diversité de pollinisateurs. Ainsi, la richesse spécifique (Gabriel et al. 2010 ;
Kennedy et al. 2013) et l’abondance (Concepción et al. 2012 ; Kennedy et al. 2013) en
hyménoptères sont plus importantes dans les parcelles en AB qui abritent une diversité en
adventices plus importante (Henckel et al. 2015) que dans celles en agriculture
conventionnelle. De plus, en dépit du gain d’adventices dans les bordures par rapport au plein
champ (Henckel et al. 2015) et d’une abondance en pollinisateurs décroissante en allant vers
le centre des cultures (Ricketts et al. 2008), la pollinisation du colza est similaire entre la
bordure et le plein champ (Tableau 5). C’est contraire à ce qui a été mis en évidence par
Stanley et al. (2013), la contribution des insectes pollinisateurs au rendement du colza à
l’échelle de la branche étant plus importante dans les bordures que dans les pleins champs.
Néanmoins dans leur étude, une population importante de phytomètres (85 en moyenne contre
quatre maximum dans notre étude) est placée dans les parcelles d’études. Dans notre cas, une
relation positive entre la qualité de l’habitat et l’importance de la compétition entre les
phytomètres et les autres ressources florales pour la pollinisation expliquerait l’absence

22
d’effet des facteurs locaux sur la pollinisation du colza.
3. Le contexte paysager influence la pollinisation entomophile du colza
Les variables paysagères étudiées n’expliquent pas de façon similaire les variations du
succès de fructification et du rendement. Par exemple, le colza explique en moyenne 10,1%
de la variance expliquée par le modèle alors que la longueur des bordures en explique en
moyenne 3,5% (Figure 6B). La longueur des bordures entre les habitats n’influence pas la
pollinisation des cultures de colza (Tableau 6), cela est cohérent avec d’autres études. La
configuration du paysage n’influence ni la diversité d’abeilles nicheuses (Holzschuh et al.
2010), ni la diversité de pollinisateurs, ni le taux de visites des myrtilliers (Nicholson et al.
2017). Récemment, Hass et al. (2018) ont montré que malgré l’influence positive de
l’augmentation de la longueur des bordures dans le paysage sur l’abondance en abeilles
sauvages, la pollinisation des radis noirs en est indépendante. Par l’augmentation de
l’interspersion (Fahrig et al. 2011) et de la connectivité des paysages (Lindenmayer et al.
2008 ; Tscharntke et al. 2012), la configuration influence positivement la diversité de
pollinisateurs. Ce n’est pas le cas de la fonction de pollinisation associée dans notre étude
parce que cette variable configurationnelle est fortement corrélée à d’autres variables
paysagères (Annexe 6 A, B, C).
Les HSN dans le paysage influencent globalement le succès de fructification et le
rendement du colza (Figure 6B). Peu d’études ont quantifié l’importance des HSN dans le
paysage sur la pollinisation des cultures. Ricketts et al. (2008) mettent en évidence dans une
méta-analyse que la pollinisation des cultures est indépendante de la distance aux HSN. Plus
récemment, deux études montrent que la proportion d’HSN dans les paysages favorise la
pollinisation des cultures. L’efficacité de la pollinisation des fraisiers suit une relation de type
monod entre 0 et 16% d’HSN dans un rayon de 5000m (Bukovinszky et al. 2017) et
l’efficacité de la pollinisation du radis noir augmente linéairement entre 1,5 et 10% d’HSN
dans un rayon de 1000m (Hass et al. 2018). Les HSN fournissent des ressources florales
diversifiées et des sites de nidification nécessaires à la survie et à la reproduction des
pollinisateurs (Bukovinszky et al. 2017). Les HSN sont constitués de différents habitats qui
diffèrent dans le type et la quantité de ressources florales disponible (Cole et al. 2017). Notre
étude a permis de mettre en évidence que dans les cultures de colza, augmenter la proportion
de prairie de 10% dans le paysage augmente le succès de fructification des traitements
contrôle de colza de 10% grâce aux petits pollinisateurs (Figure 7A). Au contraire, à une
échelle d’étude similaire (rayon de 1000m) mais en considérant une gamme de proportion de

23
prairie moins importante (0,6 – 12,9%), Lindström et al. (2016) montrent que les rendements
du colza sont indépendants de la proportion de prairie dans le paysage. Les prairies sont des
habitats avec une ressource en fleurs importante en termes de richesse ou d’abondance (Cole
et al. 2017). Il en découle une diversité en pollinisateurs importante dans ces milieux qui
déborde (Tscharntke et al. 2012) dans les cultures de colza. A l’inverse, les forêts sont des
milieux ombragés avec de faibles diversités de ressources florales et de pollinisateurs (Cole et
al. 2017). Nous avons ainsi montré un effet négatif de la proportion de forêt environnant les
parcelles en colza et en céréales dans un rayon de 750m environ sur la pollinisation des
phytomètres par les petits insectes (Tableau 6 ; Figure 7C ; Figure 8C). Les forêts n’agissent
donc pas comme source de pollinisateurs pour les cultures de colza et de céréale (Figure 9A).
Les haies abritent des diversités de ressources florales et de pollinisateurs importantes
relativement aux autres habitats de la matrice agricole (Cole et al. 2017). Dans notre étude, la
proportion de haie dans le paysage n’influence pas la pollinisation entomophile du colza
(Tableau 6). Étant donné la corrélation entre les proportions de prairie et de haie dans les
paysages (Annexes 7A) et la similarité des échelles d’influences, ces deux habitats peuvent
expliquer de façon synergique l’influence positive de la proportion de prairie dans le paysage
sur la pollinisation du colza dans les cultures de colza.
L’importance des variables paysagères dépend aussi de la culture testée. Le
pourcentage de variance expliquée par la proportion de prairie dans les modèles varie entre
3,5% et 12,5% quand le succès de fructification est considéré (Figure 6B). L’impact positif
des prairies sur la pollinisation entomophile n’est vrai que dans les cultures de colza. Cela
signifie qu’il y a une interaction entre la capacité de débordement (Tscharntke et al. 2012) de
la fonction de pollinisation des prairies vers les habitats voisins et la culture testée (Figure
9B). La culture de colza par le pulse de ressources florales que cet habitat fournit attire
massivement les insectes des prairies, contrairement aux cultures de céréales ou les prairies
qui attirent moins les insectes (Holzschuh et al. 2011 ; Riedinger et al. 2015).
Globalement, l’AB influence peu la pollinisation du colza (Figure 6B). La proportion
d’AB dans le paysage autour des cultures de céréales influence négativement la contribution
absolue des petits pollinisateurs au rendement du colza (Figure 8A). Ce n’est pas le cas dans
les autres cultures locales (Tableau 6). Ce résultat est surprenant étant donné que l’AB est
connue pour influencer positivement la diversité locale en hyménoptères (Gabriel et al. 2010 ;
Concepción et al. 2012 ; Kennedy et al. 2013 ; Tuck et al. 2014 ; Lichtenberg et al. 2017). Ce
système de production agricole s’affranchit de l’utilisation de pesticides (Norton et al. 2009)
et abrite une diversité de ressources florales relativement plus importante que l’agriculture

24
conventionnelle (Henckel et al. 2015) par la non utilisation d’herbicides. Les effets positifs de
l’AB au niveau local en termes d’abondance et de richesse débordent dans les habitats voisins
(Gabriel et al. 2010 ; Tuck et al. 2014 ; Lichtenberg et al. 2017).
Le sens de l’effet de la proportion de colza sur la pollinisation des phytomètres diffère
en fonction de la culture testée. Avec l’augmentation de la proportion de colza dans le
paysage, la contribution des petits pollinisateurs au rendement dans les cultures de colza
diminue (Figure 7B), tandis que la contribution des gros pollinisateurs au succès de
fructification des phytomètres augmente dans les cultures céréalières (Figure 8B). Les
cultures de colza sont en compétition pour les pollinisateurs et la fonction de pollinisation
associée. Cela rend compte d’un effet de dilution de la diversité de pollinisateurs associée à la
fonction de pollinisation dans les cultures de colza (Holzschuh et al. 2011). Un effet de
facilitation par effet de débordement (Blitzer et al. 2012) est également mis en évidence car la
fonction de pollinisation dans les cultures de colza déborde dans les cultures de céréales
(Figure 9C).
Différents points liés à l’approche utilisée consistant à quantifier chaque variable dans
des buffers de rayons différents (Holland et al. 2004) sont critiquables. 1/ La quantification de
nombreuses variables paysagères conduit à des corrélations entre certaines de ces variables
(Annexe 6) ; 2/ Les échelles d’influence des variables paysagères n’étant pas testées
statistiquement, elles ne peuvent pas être comparées entre cultures locales ; 3/ L’effet d’une
variable paysagère sur la pollinisation du colza est supposé constant dans l’espace, alors qu’il
est attendu que cet effet décroisse avec la distance (Chandler & Hepinstall-Cymerman 2016).
Afin de soustraire des corrélations entre variables paysagères, la méthode de sélection des
paysages décrite par Fahrig et al. (2011 ; section B.2.a) doit être appliquée en prenant en
compte l’ensemble des variables paysagères. Pour pallier aux deux autres contraintes, une
méthode récemment développée par Carpentier et Martin (2017) permet de quantifier la force
et la distance d’influence des variables paysagères sur une variable dépendante en s’abstenant
de l’approche buffer.
E. Conclusion & Perspectives
Notre étude est la première à mettre en évidence le lien direct entre contexte paysager
et pollinisation entomophile du colza par une approche expérimentale. Différents habitats
dans le paysage agricole gouvernent la dynamique spatiale de la fonction de pollinisation
entomophile dépendamment de l’attractivité des cultures testées : les prairies, les forêts, les
cultures de colza et l’AB (Figure 9). Nos travaux s’ancrent dans l’intensification écologique

25
des systèmes agricoles, c’est-à-
dire la valorisation et
l’optimisation des services
écosystémiques dans les systèmes
de production agricole, afin de
réduire la dépendance aux intrants
chimiques par le rétablissement de
la biodiversité et de ses fonctions
(Bommarco et al. 2013 ; Tittonell
2014). En effet, nos travaux sont
nécessaires à l’élaboration de
schémas d’aménagements des
territoires agricoles dans lesquels
les habitats constituants la matrice
agricole sont agencés de manière à
abriter une diversité de
pollinisateurs importante, résiliente, qui disperse vers les cultures dépendantes de la
pollinisation. Ces schémas d’aménagement doivent tenir compte du fait que la fonction de
pollinisation est majoritairement gouvernée par la dispersion aléatoire des pollinisateurs dans
la matrice agricole, avec un rôle important des processus stochastiques (Figure 6A ;
Sydenham et al. 2017). La dynamique temporelle des pollinisateurs dans les paysages
agricoles est également à considérer, néanmoins, rares sont les études à la prendre à compte
(Senapathi et al. 2017). Différents habitats interviennent dans la dynamique temporelle des
pollinisateurs associée à la fonction de pollinisation. Les HSN favorisent les mouvements des
pollinisateurs et réduisent la dissimilarité temporelle des communautés de pollinisateurs dans
la matrice agricole (Beduschi et al. 2018). La diversité de pollinisateurs présente dans les
cultures de colza déborde dans les habitats voisins après la période de floraison de cette
culture (Diekötter et al. 2014). Ces effets positifs sont également visibles d’une année à
l’autre : la diversité de pollinisateurs l’année n étant positivement liée à la proportion de colza
de l’année n-1 (Riedinger et al. 2015). La compréhension de la dynamique temporelle
associée à la dynamique spatiale des pollinisateurs permettrait d’assurer les mesures de
conservation des pollinisateurs tout en optimisant les rendements des cultures dépendantes de
la pollinisation dans les paysages agricoles.
Figure 9 : Dynamique spatiale de la fonction de pollinisation
dans les territoires agricoles. Différentes paires de paysages sont
schématisées en fonction de la proportion de A. forêt B. prairie et
C. colza dans les paysages. Plus le nombre de points est important
dans un habitat, plus la fonction de pollinisation y est importante.

26
F. RÉFÉRENCES
Aho, K., Derryberry, D. et Peterson, T. (2014). Model selection for ecologists: the
worldviews of AIC and BIC. Ecology 95, 631–636.
Aizen, M. A. et Harder, L. D. (2009). The global stock of domesticated honey bees is
growing slower than agricultural demand for pollination. Current Biology 19, 915–918. doi:
10.1016/j.cub.2009.03.071.
Aizen, M. A., Garibaldi, L. A., Cunningham, S. A. et Klein, A. M. (2008). Long-Term
Global Trends in Crop Yield and Production Reveal No Current Pollination Shortage but Increasing
Pollinator Dependency. Current Biology 18, 1572–1575. doi: 10.1016/j.cub.2008.08.066.
Albrecht, M., Duelli, P., Müller, C., Kleijn, D. et Schmid, B. (2007) The Swiss agri-
environment scheme enhances pollinator diversity and plant reproductive success in nearby
intensively managed farmland. Journal of Applied Ecology 44, 813–822. doi: 10.1111/j.1365-
2664.2007.01306.x.
Bartomeus, I., Potts, S. G., Steffan-Dewenter, I., Vaissière, B. E., Woyciechowski, M.,
Krewenka, K. M., Tscheulin, T., Roberts, S. P. M., Szentgyörgyi, H., Westphal, C. et Bommarco,
R. (2014). Contribution of insect pollinators to crop yield and quality varies with agricultural
intensification. PeerJ 2, e328. doi: 10.7717/peerj.328.
Bates, D., Maechler, M., Bolker, B., et Walker, S. (2015). Fitting Linear Mixed-Effects
Models Using lme4. Journal of Statistical Software 67, 1-48. doi:10.18637/jss.v067.i01.
Becker, H. C., Damgaard, C. et Karlsson, B. (1992). Environmental variation for
outcrossing rate in rapeseed (Brassica napus). Theoretical and Applied Genetics 84, 303–306. doi:
10.1007/BF00229487.
Beduschi, T., Kormann, U. G., Tscharntke, T. et Scherber, C. (2018). Spatial community
turnover of pollinators is relaxed by semi-natural habitats, but not by mass-flowering crops in
agricultural landscapes. Biological Conservation 221, 59–66. doi: 10.1016/j.biocon.2018.01.016.
Benjamin, F. E., Reilly, J. R. et Winfree, R. (2014). Pollinator body size mediates the scale
at which land use drives crop pollination services. Journal of Applied Ecology 51, 440–449. doi:
10.1111/1365-2664.12198.
Bivand, R. et Lewin-Koh, N. (2017). maptools: Tools for Reading and Handling Spatial
Objects. R package version 0.9-2. https://CRAN.R-project.org/package=maptools.
Bivand, R. et Rundel, C. (2017). rgeos: Interface to Geometry Engine – Open Source
('GEOS'). R package version 0.3-26. https://CRAN.R-project.org/package=rgeos.
Bivand, R., Keitt, T. et Rowlingson, B. (2018). rgdal: Bindings for the 'Geospatial' Data
Abstraction Library. R package version 1.2-18. https://CRAN.R-project.org/package=rgdal.
Blitzer, E. J., Dormann, C. F., Holzschuh, A., Klein, A.-M., Rand, T. A. et Tscharntke, T.
(2012). Spillover of functionally important organisms between managed and natural habitats.
Agriculture, Ecosystems & Environment 146, 34–43. doi: 10.1016/j.agee.2011.09.005.
Bommarco, R., Kleijn, D. and Potts, S. G. (2013). Ecological intensification: harnessing
ecosystem services for food security. Trends in Ecology & Evolution 28, 230–238. doi:
10.1016/j.tree.2012.10.012.
Bommarco, R., Marini, L. et Vaissière, B. E. (2012). Insect pollination enhances seed yield,
quality, and market value in oilseed rape. Oecologia 169, 1025–1032. doi: 10.1007/s00442-012-2271-
6.
Breeze, T. D., Vaissière, B. E., Bommarco, R., Petanidou, T., Seraphides, N., Kozák, L.,
Scheper, J., Biesmeijer, J. C., Kleijn, D., Gyldenkærne, S., Moretti, M., Holzschuh, A., Steffan-
Dewenter, I., Stout, J. C., Pärtel, M., Zobel, M. et Potts, S. G. (2014). Agricultural Policies
Exacerbate Honeybee Pollination Service Supply-Demand Mismatches Across Europe. PLOS ONE 9,
e82996. doi: 10.1371/journal.pone.0082996.
Bretagnolle, V. et Gaba, S. (2015). Weeds for bees? A review. Agronomy for Sustainable
Development 35, 891–909. doi: 10.1007/s13593-015-0302-5.
Bretagnolle, V., Berthet, E., Gross, N., Gauffre, B., Plumejeaud, C., Houte, S.,
Badenhausser, I., Monceau, K., Allier, F., Monestiez, P. et Gaba, S. (2018). Towards sustainable
and multifunctional agriculture in farmland landscapes: Lessons from the integrative approach of a

27
French LTSER platform. Science of The Total Environment 627, 822–834. doi:
10.1016/j.scitotenv.2018.01.142.
Bukovinszky, T., Verheijen, J., Zwerver, S., Klop, E., Biesmeijer, J. C., Wäckers, F. L.,
Prins, H. H. T. et Kleijn, D. (2017). Exploring the relationships between landscape complexity, wild
bee species richness and reproduction, and pollination services along a complexity gradient in the
Netherlands. Biological Conservation 214, 312–319. doi: 10.1016/j.biocon.2017.08.027.
Carpentier, F., et Martin, O. (2017). siland: Spatial Influence of Landscape. R package
version 1.0. https://CRAN.R-project.org/package=siland
Chandler, R. et Hepinstall-Cymerman, J. (2016). Estimating the spatial scales of landscape
effects on abundance. Landscape Ecology 31, 1383–1394. doi: 10.1007/s10980-016-0380-z.
Cole, L. J., Brocklehurst, S., Robertson, D., Harrison, W. et McCracken, D. I. (2017).
Exploring the interactions between resource availability and the utilisation of semi-natural habitats by
insect pollinators in an intensive agricultural landscape. Agriculture, Ecosystems & Environment 246,
157–167. doi: 10.1016/j.agee.2017.05.007.
Concepción, E. D., Díaz, M., Kleijn, D., Báldi, A., Batáry, P., Clough, Y., Gabriel, D.,
Herzog, F., Holzschuh, A., Knop, E., Marshall, E. J. P., Tscharntke, T. et Verhulst, J. (2012).
Interactive effects of landscape context constrain the effectiveness of local agri-environmental
management: Landscape constrains the effectiveness of local management. Journal of Applied
Ecology 49, 695–705. doi: 10.1111/j.1365-2664.2012.02131.x.
Cresswell, J. E., Davies, T. W., Patrick, M. A., Russell, F., Pennel, C., Vicot, M. et
Lahoubi, M. (2014). Aerodynamics of wind pollination in a zoophilous flower, Brassica napus.
Functional Ecology 18, 861–866. doi: 10.1111/j.0269-8463.2004.00917.x.
Cumming, G. et Finch, S. (2005). Inference by eye: confidence intervals and how to read
pictures of data. The American Psychologist 60, 170–180. doi: 10.1037/0003-066X.60.2.170.
Diekötter, T., Peter, F., Jauker, B., Wolters, V. et Jauker, F. (2014). Mass-flowering crops
increase richness of cavity-nesting bees and wasps in modern agro-ecosystems. GCB Bioenergy 6,
219–226. doi: 10.1111/gcbb.12080.
Diepenbrock, W. (2000). Yield analysis of winter oilseed rape (Brassica napus L.): a review.
Field Crops Research 67, 35–49. doi: 10.1016/S0378-4290(00)00082-4.
Efron, B. (1979). Bootstrap Methods: Another Look at the Jackknife. Ann Statist 7, 1-26.
1979 Ellis, J. D., Evans, J. D. et Pettis, J. (2010). Colony losses, managed colony population
decline, and Colony Collapse Disorder in the United States. Journal of Apicultural Research 49, 134–
136. doi: 10.3896/IBRA.1.49.1.30.
Fahrig, L., Baudry, J., Brotons, L., Burel, F. G., Crist, T. O., Fuller, R. J., Sirami, C.,
Siriwardena, G. M. et Martin, J.-L. (2011). Functional landscape heterogeneity and animal
biodiversity in agricultural landscapes: Heterogeneity and biodiversity. Ecology Letters 14, 101–112.
doi: 10.1111/j.1461-0248.2010.01559.x.
FAO. (2018). http://www.fao.org/faostat/fr/#data/QC/visualize. Consulté le 4 mai 2018.
Fox, J. and Weisberg, S. (2011). An {R} Companion to Applied Regression, Second Edition.
Thousand Oaks CA: Sage. URL: http://socserv.socsci.mcmaster.ca/jfox/Books/ Companion
Gabriel, D., Sait, S. M., Hodgson, J. A., Schmutz, U., Kunin, W. E. et Benton, T. G.
(2010). Scale matters: the impact of organic farming on biodiversity at different spatial scales: Scale
matters in organic farming. Ecology Letters 13, 858–869. doi: 10.1111/j.1461-0248.2010.01481.x.
Gallai, N., Salles, J.-M., Settele, J. et Vaissière, B. E. (2009). Economic valuation of the
vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68, 810–
821. doi: 10.1016/j.ecolecon.2008.06.014.
Ginot, V., Gaba, S., Beaudouin, R., Aries, F. et Monod, H. (2006). Combined use of local
and ANOVA-based global sensitivity analyses for the investigation of a stochastic dynamic model:
Application to the case study of an individual-based model of a fish population. Ecological Modelling
193, 479–491. doi: 10.1016/j.ecolmodel.2005.08.025.
Hallmann, C. A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., Stenmans,
W., Müller, A., Sumser, H., Hörren, T., Goulson, D. et de Kroon, H. (2017). More than 75 percent
decline over 27 years in total flying insect biomass in protected areas. PLOS ONE 12, e0185809. doi:
10.1371/journal.pone.0185809.

28
Harrell, E. (2017). Hmisc: Harrell Miscellaneous. R package version 4.0-3. https://CRAN.R-
project.org/package=HmiscH.
Hass, A. L., Kormann, U. G., Tscharntke, T., Clough, Y., Baillod, A. B., Sirami, C.,
Fahrig, L., Martin, J.-L., Baudry, J., Bertrand, C., Bosch, J., Brotons, L., Burel, F., Georges, R.,
Giralt, D., Marcos-García, M. Á., Ricarte, A., Siriwardena, G. et Batáry, P. (2018). Landscape
configurational heterogeneity by small-scale agriculture, not crop diversity, maintains pollinators and
plant reproduction in western Europe. Proceedings of the Royal Society B: Biological Sciences 285,
526-544. doi: 10.1098/rspb.2017.2242.
Henckel, L., Borger, L., Meiss, H., Gaba, S. et Bretagnolle, V. (2015). Organic fields
sustain weed metacommunity dynamics in farmland landscapes. Proceedings of the Royal Society B:
Biological Sciences 282. doi: 10.1098/rspb.2015.0002.
Hijmans, R. (2017). raster: Geographic Data Analysis and Modeling. R package version 2.6-
7. https://CRAN.R-project.org/package=raster.
Hole, D. G., Perkins, A. J., Wilson, J. D., Alexander, I. H., Grice, P. V. et Evans, A. D.
(2005). Does organic farming benefit biodiversity? Biological Conservation 122, 113–130. doi:
10.1016/j.biocon.2004.07.018.
Holland, J. D., Bert, D. G. et Fahrig, L. (2004). Determining the Spatial Scale of Species’
Response to Habitat. BioScience 54, 227–233.
Holzschuh, A., Dainese, M., González‐Varo, J. P., Mudri‐Stojnić, S., Riedinger, V.,
Rundlöf, M., Scheper, J., Wickens, J. B., Wickens, V. J., Bommarco, R., Kleijn, D., Potts, S. G.,
Roberts, S. P. M., Smith, H. G., Vilà, M., Vujić, A. et Steffan‐Dewenter, I. (2016). Mass-flowering
crops dilute pollinator abundance in agricultural landscapes across Europe. Ecology Letters 19, 1228–
1236. doi: 10.1111/ele.12657.
Holzschuh, A., Dormann, C. F., Tscharntke, T. et Steffan-Dewenter, I. (2011). Expansion
of mass-flowering crops leads to transient pollinator dilution and reduced wild plant pollination.
Proceedings of the Royal Society B: Biological Sciences 278, 3444–3451. doi:
10.1098/rspb.2011.0268.
Holzschuh, A., Steffan-Dewenter, I. et Tscharntke, T. (2008). Agricultural landscapes with
organic crops support higher pollinator diversity. Oikos 117, 354–361. doi: 10.1111/j.2007.0030-
1299.16303.x.
Holzschuh, A., Steffan‐Dewenter, I. et Tscharntke, T. (2010). How do landscape
composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps
and their parasitoids? Journal of Animal Ecology 79, 491–500. doi: 10.1111/j.1365-
2656.2009.01642.x.
Hudewenz, A., Pufal, G., BöGeholz, A.-L. et Klein, A.-M. (2014). Cross-pollination
benefits differ among oilseed rape varieties. The Journal of Agricultural Science 152, 770–778. doi:
10.1017/S0021859613000440.
IUCN (2012). Iucn Red List Categories And Criteria: Version 3.1. Second Edition. Zenodo.
Jauker, F. et Wolters, V. (2008). Hover flies are efficient pollinators of oilseed rape.
Oecologia 156, 819–823. doi: 10.1007/s00442-008-1034-x.
Jauker, F., Bondarenko, B., Becker, H. C. et Steffan-Dewenter, I. (2012). Pollination
efficiency of wild bees and hoverflies provided to oilseed rape. Agricultural and Forest Entomology
14, 81–87. doi: 10.1111/j.1461-9563.2011.00541.x.
Kamler, F. and Jas, S. (2003). Influence of pollination by honey bee on seed yield on
selected cultivars of winter rape. Journal of Apicultural Science 47, 119–125.
Kennedy, C. M., Lonsdorf, E., Neel, M. C., Williams, N. M., Ricketts, T. H., Winfree, R.,
Bommarco, R., Brittain, C., Burley, A. L., Cariveau, D., Carvalheiro, L. G., Chacoff, N. P.,
Cunningham, S. A., Danforth, B. N., Dudenhöffer, J.-H., Elle, E., Gaines, H. R., Garibaldi, L. A.,
Gratton, C., Holzschuh, A., Isaacs, R., Javorek, S. K., Jha, S., Klein, A. M., Krewenka, K.,
Mandelik, Y., Mayfield, M. M., Morandin, L., Neame, L. A., Otieno, M., Park, M., Potts, S. G.,
Rundlöf, M., Saez, A., Steffan-Dewenter, I., Taki, H., Viana, B. F., Westphal, C., Wilson, J. K.,
Greenleaf, S. S. et Kremen, C. (2013). A global quantitative synthesis of local and landscape effects
on wild bee pollinators in agroecosystems. Ecology Letters 16, 584–599. doi: 10.1111/ele.12082.
Klein, A.-M., Vaissière, B. E., Cane, J. H., Steffan-Dewenter, I., Cunningham, S. A.,
Kremen, C. et Tscharntke, T. (2007). Importance of pollinators in changing landscapes for world

29
crops. Proceedings of the Royal Society of London B: Biological Sciences 274, 303–313. doi:
10.1098/rspb.2006.3721.
Kuznetsova, A., Brockhoff, P. B. et Christensen, R.H.B. (2017). “lmerTest Package: Tests
in Linear Mixed Effects Models.” Journal of Statistical Software 8, 1-26. doi: 10.18637/jss.v082.i13
Lerin, J. et Rivault, E. (1982). Effets de la pollinisation entomophile sur le colza dans une
expérience en cage. Agronomie 2, 249–256.
Lichtenberg, E. M., Kennedy, C. M., Kremen, C., Batáry, P., Berendse, F., Bommarco,
R., Bosque-Pérez, N. A., Carvalheiro, L. G., Snyder, W. E., Williams, N. M., Winfree, R., Klatt,
B. K., Åström, S., Benjamin, F., Brittain, C., Chaplin-Kramer, R., Clough, Y., Danforth, B.,
Diekötter, T., Eigenbrode, S. D., Ekroos, J., Elle, E., Freitas, B. M., Fukuda, Y., Gaines-Day, H.
R., Grab, H., Gratton, C., Holzschuh, A., Isaacs, R., Isaia, M., Jha, S., Jonason, D., Jones, V. P.,
Klein, A.-M., Krauss, J., Letourneau, D. K., Macfadyen, S., Mallinger, R. E., Martin, E. A.,
Martinez, E., Memmott, J., Morandin, L., Neame, L., Otieno, M., Park, M. G., Pfiffner, L.,
Pocock, M. J. O., Ponce, C., Potts, S. G., Poveda, K., Ramos, M., Rosenheim, J. A., Rundlöf, M.,
Sardiñas, H., Saunders, M. E., Schon, N. L., Sciligo, A. R., Sidhu, C. S., Steffan-Dewenter, I.,
Tscharntke, T., Veselý, M., Weisser, W. W., Wilson, J. K. et Crowder, D. W. (2017). A global
synthesis of the effects of diversified farming systems on arthropod diversity within fields and across
agricultural landscapes. Global Change Biology 23, 4946–4957. doi: 10.1111/gcb.13714.
Lindenmayer, D., Hobbs, R. J., Montague-Drake, R., Alexandra, J., Bennett, A.,
Burgman, M., Cale, P., Calhoun, A., Cramer, V., Cullen, P., Driscoll, D., Fahrig, L., Fischer, J.,
Franklin, J., Haila, Y., Hunter, M., Gibbons, P., Lake, S., Luck, G., MacGregor, C., McIntyre,
S., Nally, R. M., Manning, A., Miller, J., Mooney, H., Noss, R., Possingham, H., Saunders, D.,
Schmiegelow, F., Scott, M., Simberloff, D., Sisk, T., Tabor, G., Walker, B., Wiens, J., Woinarski,
J. et Zavaleta, E. (2008). A checklist for ecological management of landscapes for conservation.
Ecology Letters 11, 78–91. doi: 10.1111/j.1461-0248.2007.01114.x.
Lindström, S. A. M., Herbertsson, L., Rundlöf, M., Smith, H. G. et Bommarco, R. (2016).
Large-scale pollination experiment demonstrates the importance of insect pollination in winter oilseed
rape. Oecologia 180, 759–769. doi: 10.1007/s00442-015-3517-x.
Lindström, S. A. M., Klatt, B. K., Smith, H. G. et Bommarco, R. (2018). Crop
management affects pollinator attractiveness and visitation in oilseed rape. Basic and Applied Ecology
26, 82–88. doi: 10.1016/j.baae.2017.09.005.
Marini, L., Tamburini, G., Petrucco-Toffolo, E., Lindström, S. A. M., Zanetti, F., Mosca,
G. et Bommarco, R. (2015). Crop management modifies the benefits of insect pollination in oilseed
rape. Agriculture, Ecosystems & Environment 207, 61–66. doi: 10.1016/j.agee.2015.03.027.
Mesquida, J. and Renard, M. (1981). Pollinisation du colza d’hiver male-fertile et male-
sterile (Brassica napus L. var. oleifera Metzger) par l’abeille domestique (Apis m. mellifica L.). Effets
sur la phenologie et le rendement. Apidologie 12, 345–362.
Millennium Ecosystem Assessment (Program) ed. (2005). Ecosystems and human well-
being: synthesis. Island Press, Washington, DC.
Nicholson, C. C., Koh, I., Richardson, L. L., Beauchemin, A. et Ricketts, T. H. (2017).
Farm and landscape factors interact to affect the supply of pollination services. Agriculture,
Ecosystems & Environment 250, 113–122. doi: 10.1016/j.agee.2017.08.030.
Norton, L., Johnson, P., Joys, A., Stuart, R., Chamberlain, D., Feber, R., Firbank, L.,
Manley, W., Wolfe, M., Hart, B., Mathews, F., Macdonald, D. et Fuller, R. J. (2009).
Consequences of organic and non-organic farming practices for field, farm and landscape complexity.
Agriculture, Ecosystems & Environment 129, 221–227. doi: 10.1016/j.agee.2008.09.002.
Perrot Thomas, Gaba Sabrina, Roncoroni Maryline, Gautier Jean-Luc et Vincent
Bretagnolle (en révision). Experimental evidence in farmers’ fields shows that bees can increase
oilseed rape yield up to 35%. Agriculture Ecosystem and the Environment.
Petanidou, T., Kallimanis, A. S., Tzanopoulos, J., Sgardelis, S. P. et Pantis, J. D. (2008).
Long-term observation of a pollination network: fluctuation in species and interactions, relative
invariance of network structure and implications for estimates of specialization: High plasticity in
plant-pollinator networks. Ecology Letters 11, 564–575. doi: 10.1111/j.1461-0248.2008.01170.x.
Pinheiro J., Bates D., DebRoy S., Sarkar D. et R Core Team. (2016). _nlme: Linear and
Nonlinear Mixed Effects Models_. R package version 3.1-128, <URL: http://CRAN.R-

30
project.org/package=nlme>.
Potts Simon G., Vulliamy Betsy, Dafni Amots, Ne’eman Gidi et Willmer Pat (2003).
Linking bees and flowers: how do floral communities structure pollinator communities? Ecology 84,
2628–2642. doi: 10.1890/02-0136.
Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O. et Kunin, W. E.
(2010b). Global pollinator declines: trends, impacts and drivers. Trends in Ecology & Evolution 25,
345–353. doi: 10.1016/j.tree.2010.01.007.
Potts, S. G., Imperatriz-Fonseca, V., Ngo, H. T., Aizen, M. A., Biesmeijer, J. C., Breeze,
T. D., Dicks, L. V., Garibaldi, L. A., Hill, R., Settele, J. et Vanbergen, A. J. (2016). Safeguarding
pollinators and their values to human well-being. Nature 540, 220–229. doi: 10.1038/nature20588.
Potts, S. G., Roberts, S. P. M., Dean, R., Marris, G., Brown, M. A., Jones, R., Neumann,
P. et Settele, J. (2010a). Declines of managed honey bees and beekeepers in Europe. Journal of
Apicultural Research 49, 15–22. doi: 10.3896/IBRA.1.49.1.02.
Pufal, G., Steffan-Dewenter, I. et Klein, A.-M. (2017). Crop pollination services at the
landscape scale. Current Opinion in Insect Science 21, 91–97. doi: 10.1016/j.cois.2017.05.021.
Rader, R., Howlett, B. G., Cunningham, S. A., Westcott, D. A., Newstrom-Lloyd, L. E.,
Walker, M. K., Teulon, D. A. J. et Edwards, W. (2009). Alternative pollinator taxa are equally
efficient but not as effective as the honeybee in a mass flowering crop. Journal of Applied Ecology 46,
1080–1087. doi: 10.1111/j.1365-2664.2009.01700.x.
Rader, R., Howlett, B. G., Cunningham, S. A., Westcott, D. A. et Edwards, W. (2012).
Spatial and temporal variation in pollinator effectiveness: do unmanaged insects provide consistent
pollination services to mass flowering crops? Journal of Applied Ecology 49, 126–134. doi:
10.1111/j.1365-2664.2011.02066.x.
R Core Team (2016). R: A language and environment for statistical computing. R
Ricketts, T. H., Regetz, J., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C.,
Bogdanski, A., Gemmill-Herren, B., Greenleaf, S. S., Klein, A. M., Mayfield, M. M., Morandin,
L. A., Ochieng’, A. et Viana, B. F. (2008). Landscape effects on crop pollination services: are there
general patterns? Ecology Letters 11, 499–515. doi: 10.1111/j.1461-0248.2008.01157.x.
Riedinger, V., Mitesser, O., Hovestadt, T., Steffan-Dewenter, I. et Holzschuh, A. (2015).
Annual dynamics of wild bee densities: attractiveness and productivity effects of oilseed rape. Ecology
96, 1351–1360. doi: 10.1890/14-1124.1.
Rollin, O., Bretagnolle, V., Fortel, L., Guilbaud, L. et Henry, M. (2015). Habitat, spatial
and temporal drivers of diversity patterns in a wild bee assemblage. Biodiversity and Conservation 24,
1195–1214. doi: 10.1007/s10531-014-0852-x.
Senapathi, D., Goddard, M. A., Kunin, W. E. et Baldock, K. C. R. (2017). Landscape
impacts on pollinator communities in temperate systems: evidence and knowledge gaps. Functional
Ecology 31, 26–37. doi: 10.1111/1365-2435.12809.
Stanley, D. A., Gunning, D. et Stout, J. C. (2013). Pollinators and pollination of oilseed rape
crops (Brassica napus L.) in Ireland: ecological and economic incentives for pollinator conservation.
Journal of Insect Conservation 17, 1181–1189. doi: 10.1007/s10841-013-9599-z.
Steffan-Dewenter, I., Münzenberg, U., Bürger, C., Thies, C. et Tscharntke, T. (2002).
Scale‐dependent effects of landscape context on three pollinator guilds. Ecology 83, 1421–1432. doi:
10.1890/0012-9658(2002)083[1421:SDEOLC]2.0.CO;2.
Steffan-dewenter, I. (2003). Seed set of male-sterile and male-fertile oilseed rape (Brassica
napus) in relation to pollinator density. Apidologie 34, 227–235. doi: 10.1051/apido:2003015.
Sydenham, M. A. K., Moe, S. R., Kuhlmann, M., Potts, S. G., Roberts, S. P. M., Totland,
Ø. et Eldegard, K. Disentangling the contributions of dispersal limitation, ecological drift, and
ecological filtering to wild bee community assembly. Ecosphere 8. doi: 10.1002/ecs2.1650.
Tittonell, P. (2014). Ecological intensification of agriculture—sustainable by nature. Current
Opinion in Environmental Sustainability 8, 53–61. doi: 10.1016/j.cosust.2014.08.006.
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I. et Thies, C. (2005).
Landscape perspectives on agricultural intensification and biodiversity – ecosystem service
management. Ecology Letters 8, 857–874. doi: 10.1111/j.1461-0248.2005.00782.x.
Tscharntke, T., Tylianakis, J. M., Rand, T. A., Didham, R. K., Fahrig, L., Batáry, P.,
Bengtsson, J., Clough, Y., Crist, T. O., Dormann, C. F., Ewers, R. M., Fründ, J., Holt, R. D.,

31
Holzschuh, A., Klein, A. M., Kleijn, D., Kremen, C., Landis, D. A., Laurance, W., Lindenmayer,
D., Scherber, C., Sodhi, N., Steffan-Dewenter, I., Thies, C., van der Putten, W. H. et Westphal,
C. (2012). Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biological
Reviews 87, 661–685. doi: 10.1111/j.1469-185X.2011.00216.x.
Tuck, S. L., Winqvist, C., Mota, F., Ahnström, J., Turnbull, L. A. et Bengtsson, J. (2014).
Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis.
Journal of Applied Ecology 51, 746–755. doi: 10.1111/1365-2664.12219.
Wang, X., Mathieu, A., Cournède, P.-H., Allirand, J.-M., Jullien, A., de Reffye, P. et
Zhang, B. G. (2011). Variability and regulation of the number of ovules, seeds and pods according to
assimilate availability in winter oilseed rape (Brassica napus L.). Field Crops Research 122, 60–69.
doi: 10.1016/j.fcr.2011.02.008.
Westcott, L. et Nelson, D. (2001). Canola pollination: an update. Bee World 82, 115–129.
doi: 10.1080/0005772X.2001.11099514.
Westphal, C., Steffan-Dewenter, I. et Tscharntke, T. (2003). Mass flowering crops enhance
pollinator densities at a landscape scale: Flowering crops enhance pollinator densities. Ecology Letters
6, 961–965. doi: 10.1046/j.1461-0248.2003.00523.x.
Wezel, A., Casagrande, M., Celette, F., Vian, J.-F., Ferrer, A. et Peigné, J. (2014).
Agroecological practices for sustainable agriculture. A review. Agronomy for Sustainable
Development 34, 1–20. doi: 10.1007/s13593-013-0180-7.
Wickham, H. (2009). ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New
York.
Zou, Y., Xiao, H., Bianchi, F. J. J. A., Jauker, F., Luo, S. et van der Werf, W. (2017).
Wild pollinators enhance oilseed rape yield in small-holder farming systems in China. BMC Ecology
17, 109–116. doi: 10.1186/s12898-017-0116-1.
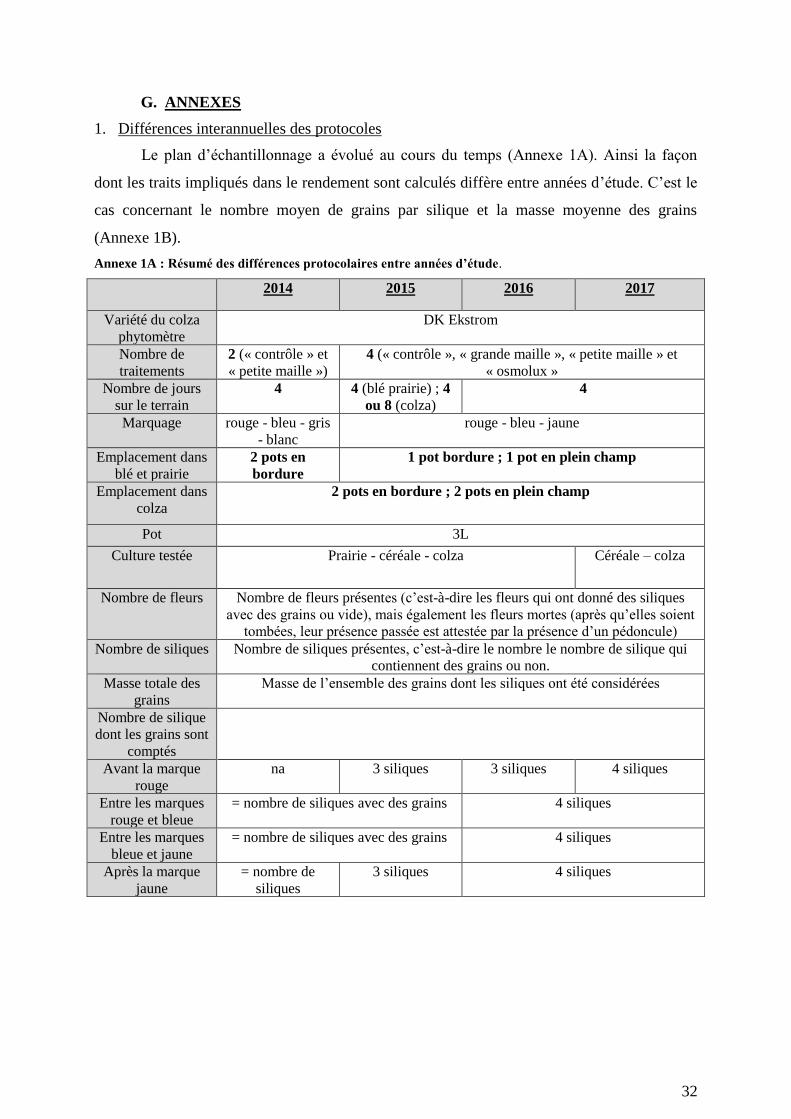
32
G. ANNEXES
1. Différences interannuelles des protocoles
Le plan d’échantillonnage a évolué au cours du temps (Annexe 1A). Ainsi la façon
dont les traits impliqués dans le rendement sont calculés diffère entre années d’étude. C’est le
cas concernant le nombre moyen de grains par silique et la masse moyenne des grains
(Annexe 1B).
Annexe 1A : Résumé des différences protocolaires entre années d’étude.
2014 2015 2016 2017
Variété du colza
phytomètre
DK Ekstrom
Nombre de
traitements
2 (« contrôle » et
« petite maille »)
4 (« contrôle », « grande maille », « petite maille » et
« osmolux »
Nombre de jours
sur le terrain
4 4 (blé prairie) ; 4
ou 8 (colza)
4
Marquage rouge - bleu - gris
- blanc
rouge - bleu - jaune
Emplacement dans
blé et prairie
2 pots en
bordure
1 pot bordure ; 1 pot en plein champ
Emplacement dans
colza
2 pots en bordure ; 2 pots en plein champ
Pot 3L
Culture testée Prairie - céréale - colza Céréale – colza
Nombre de fleurs Nombre de fleurs présentes (c’est-à-dire les fleurs qui ont donné des siliques
avec des grains ou vide), mais également les fleurs mortes (après qu’elles soient
tombées, leur présence passée est attestée par la présence d’un pédoncule)
Nombre de siliques Nombre de siliques présentes, c’est-à-dire le nombre le nombre de silique qui
contiennent des grains ou non.
Masse totale des
grains
Masse de l’ensemble des grains dont les siliques ont été considérées
Nombre de silique
dont les grains sont
comptés
Avant la marque
rouge
na 3 siliques 3 siliques 4 siliques
Entre les marques
rouge et bleue
= nombre de siliques avec des grains 4 siliques
Entre les marques
bleue et jaune
= nombre de siliques avec des grains 4 siliques
Après la marque
jaune
= nombre de
siliques
3 siliques 4 siliques

33
Annexe 1B : Equations permettant le calcul du nombre moyen de grains par silique et de la masse
moyenne des grains pour les différentes années d’étude.
Année Nombre moyen de grains par silique Masse moyenne des grains
2014 = Nombre total de grains
Nombre de silique avec des grains =
Masse totale des grains
Nombre total de grains
2015 = Nombre total de grains
Nombre de silique avec des grains =
Masse totale des grains
Nombre total de grains
2016 = Nombre de grains
n avec n le nombre
de siliques considérés (1 ≤ n ≤ 4)
= Masse totale des grains
n
avec n le nombre de siliques considérés (1 ≤ n
≤ 4)
2017 = Nombre de grains
n avec n le nombre
de siliques considérés (1 ≤ n ≤ 4)
= Masse totale des grains
n
avec n le nombre de siliques considérés (1 ≤ n
≤ 4)
2. Sélection des covariables
L’année d’étude et la date de pose du phytomètre sont incluses comme covariables lors
de l’étude des effets des facteurs locaux et paysagers sur le succès de pollinisation du colza.
En effet, par la construction d’un modèle linéaire expliquant le succès de fructification par le
traitement, de l’année et du nombre de jours depuis la floraison du colza, il a été mis en
évidence que ces deux variables influencent le succès de fructification (Annexe 2).
Annexe 2 : Résultat de l’Anova du modèle expliquant le succès de fructification entre les marques rouge et
jaune en fonction du traitement, de l’année d’étude et du nombre de jours depuis la floraison du colza.
Facteurs d F p
Traitement 3 20,17 7,56.10-13
Année 2 242,81 <2,2.10-16
Date de pose 1 450,23 <2,2.10-16
Traitement × année 6 7,10 1,75.10-7
Traitement × date de pose 3 0,98 0,40
Année × date de pose 2 3,93 1,97.10-2
Ainsi, le succès de fructification est de 0,23, 0,38, 0,54 respectivement pour les années
2015, 2016 et 2017, tandis que des phytomètres placés sur le terrain 30 jours plus tard que
d’autres ont un succès de fructification 26 à 35% plus faible. Cet effet année peut être
expliqué par des dynamiques interannuelles des communautés de pollinisateurs, par des effets
du paysages ou des effets locaux (les phytomètres ont été placés dans différentes cultures ou
différents paysages en fonction des années) mais également par des effets du paysage (par
exemple moins de colza cultivé en 2015 et 2016 et donc moins de pollen disponible). L’effet
temps depuis la floraison du colza peut s’apparenter à des dynamiques intra annuelles des
pollinisateurs (pic d’émergence des pollinisateurs au début de la floraison du colza par
exemple).
3. Compromis entre traits (fleurs ouvertes en serres vs sur le terrain)
Les liens entre les traits impliqués dans le rendement du colza sont similaires en
fonction de la position le long de la hampe étudiée (les intervalles de confiance à 95% des

34
tailles de l’effet se recoupent ; Annexe 3A ; Annexe 3B). Etant donné que les relations entre
traits ne s’opposent pas en fonction de la position le long de la hampe étudiée, l’étude des
relations entre traits impliqués dans le rendement est faite entre les marques rouge et jaune.
Annexe 3A : Représentation graphique des relations entre les traits impliqués dans le rendement entre les
marques rouge et bleue. ρ correspond à l’intervalle de confiance à 95% du coefficient de corrélation de
Spearman. * : p-value < 0,05 ; ** : p-value < 0,01 ; *** : p-value < 0,001 ; sinon p > 0,05.
Annexe 3B : Corrélations entre les traits impliqués dans le rendement entre les marques bleue et jaune. ρ
correspond à l’intervalle de confiance à 95% du coefficient de corrélation de Spearman. * : p-value < 0,05 ; ** :
p-value < 0,01 ; *** : p-value < 0,001 ; sinon p > 0,05.
4. Contribution des processus de pollinisation (fleurs ouvertes en serres vs sur le terrain)
La contribution des différents processus de pollinisation aux traits impliqués dans le
rendement diffère en fonction de la position de la hampe étudiée. En effet, la contribution de
l’autogamie est moins importante et celle de l’entomogamie plus importante entre les marques
bleue et jaune qu’entre les marques rouge et bleue (Annexe 4A ; Annexe 4B). Les
pollinisateurs ont ainsi accès moins longtemps aux fleurs entre les marques rouge et bleue
qu’entre les marques bleue et jaune étant donné que ces premières se trouvent dans un premier
temps dans la serre « insect proof » où elles ne peuvent être pollinisées que par autogamie et
anémogamie. Ce patron est néanmoins similaire quel que soit le trait lié au rendement étudié,
l’étude du différentiel de contribution des insectes pollinisateurs en fonction du trait étudié est
donc faite entre les marques rouge et jaune.
35
Annexe 4A : Contribution des processus de pollinisation pour les traits impliqués dans le rendement du
colza seulement entre les marques rouge et bleue. La moyenne ainsi que l’intervalle de confiance à 95% de
ces contributions sont calculés à partir d’une méthode de bootstrapping (100 tirages avec remises).
Succès de
fructification
Nombre de
silique
Nombre moyen
de grains par
silique
Masse moyenne
des grains
Entomogamie
(>6mm)
0,224
[0,192 ; 0,257]
0,236
[0,211 ; 0,267]
0,161
[0,133 ; 0,189]
0,085
[0,064 ; 0,106]
Entomogamie
(<6mm)
0,170
[0,144 ; 0,199]
0,185
[0,155 ; 0,217]
0,144
[0,113 ; 0,173]
0,095
[0,074 ; 0,120]
Anémogamie 0,133
[0,106 ; 0,131]
0,135
[0,108 ; 0,168]
0,153
[0,115 ; 0,195]
0,110
[0,082 ; 0,138]
Autogamie 0,473
[0,436 ; 0,508]
0,446
[0,412 ; 0,482]
0,543
[0,497 ; 0,584]
0,710
[0,679 ; 0,746]
Annexe 4B : Contribution des processus de pollinisation pour les traits impliqués dans le rendement du
colza uniquement entre les marques bleue et jaune. La moyenne ainsi que l’intervalle de confiance à 95% de
ces contributions sont calculés à partir d’une méthode de bootstrapping (100 tirages avec remises)
Succès de
fructification
Nombre de
silique
Nombre moyen
de grains par
silique
Masse moyenne
des grains
Entomogamie
(>6mm)
0,290
[0,242 ; 0,331]
0,294
[0,250 ; 0,330]
0,189
[0,139 ; 0,233]
0,097
[0,068 ; 0,131]
Entomogamie
(<6mm)
0,189
[0,163 ; 0,223]
0,199
[0,167 ; 0,232]
0,136
[0,099 ; 0,176]
0,094
[0,058 ; 0,143]
Anémogamie 0,144
[0,110 ; 0,177]
0,161
[0,122 ; 0,199]
0,181
[0,121 ; 0,242]
0,089
[0,056 ; 0,128]
Autogamie 0,377
[0,335 ; 0,410]
0,353
[0,310 ; 0,386]
0,500
[0,441 ; 0,557]
0,721
[0,673 ; 0,763]
5. Etudes résiduelles
Globalement, les modèles utilisés respectent les conditions de normalité (Annexe 5A)
et d’homoscédasticité (Annexe 5B) requis pour l’application des modèles linéaires, les
modèles linéaires généralisés ainsi que les modèles linéaires à effet mixtes.
36
Annexe 5A : Résidus standardisés en fonction des quantiles théoriques de l’ensemble des modèles utilisés
dans cette étude.
Annexe 5B : Résidus en fonction des valeurs ajustées de l’ensemble des modèles linéaires appliqués dans
l’étude.
6. Corrélations entre variables paysagères
Dans le but de vérifier que les variables paysagères sont indépendantes les unes des
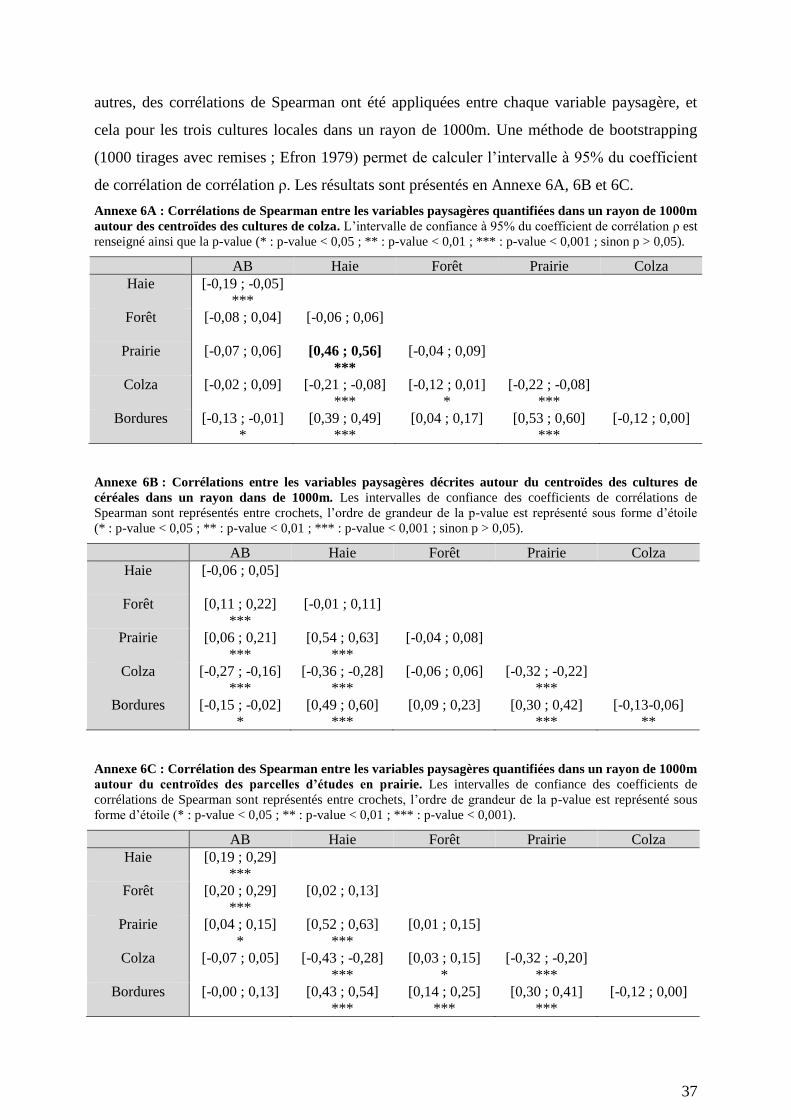
37
autres, des corrélations de Spearman ont été appliquées entre chaque variable paysagère, et
cela pour les trois cultures locales dans un rayon de 1000m. Une méthode de bootstrapping
(1000 tirages avec remises ; Efron 1979) permet de calculer l’intervalle à 95% du coefficient
de corrélation de corrélation ρ. Les résultats sont présentés en Annexe 6A, 6B et 6C.
Annexe 6A : Corrélations de Spearman entre les variables paysagères quantifiées dans un rayon de 1000m
autour des centroïdes des cultures de colza. L’intervalle de confiance à 95% du coefficient de corrélation ρ est
renseigné ainsi que la p-value (* : p-value < 0,05 ; ** : p-value < 0,01 ; *** : p-value < 0,001 ; sinon p > 0,05).
AB Haie Forêt Prairie Colza
Haie [-0,19 ; -0,05]
***
Forêt [-0,08 ; 0,04]
[-0,06 ; 0,06]
Prairie [-0,07 ; 0,06]
[0,46 ; 0,56]
***
[-0,04 ; 0,09]
Colza [-0,02 ; 0,09]
[-0,21 ; -0,08]
***
[-0,12 ; 0,01]
*
[-0,22 ; -0,08]
***
Bordures [-0,13 ; -0,01]
*
[0,39 ; 0,49]
***
[0,04 ; 0,17] [0,53 ; 0,60]
***
[-0,12 ; 0,00]
Annexe 6B : Corrélations entre les variables paysagères décrites autour du centroïdes des cultures de
céréales dans un rayon dans de 1000m. Les intervalles de confiance des coefficients de corrélations de
Spearman sont représentés entre crochets, l’ordre de grandeur de la p-value est représenté sous forme d’étoile
(* : p-value < 0,05 ; ** : p-value < 0,01 ; *** : p-value < 0,001 ; sinon p > 0,05).
AB Haie Forêt Prairie Colza
Haie [-0,06 ; 0,05]
Forêt [0,11 ; 0,22]
***
[-0,01 ; 0,11]
Prairie [0,06 ; 0,21]
***
[0,54 ; 0,63]
***
[-0,04 ; 0,08]
Colza [-0,27 ; -0,16]
***
[-0,36 ; -0,28]
***
[-0,06 ; 0,06] [-0,32 ; -0,22]
***
Bordures [-0,15 ; -0,02]
*
[0,49 ; 0,60]
***
[0,09 ; 0,23] [0,30 ; 0,42]
***
[-0,13-0,06]
**
Annexe 6C : Corrélation des Spearman entre les variables paysagères quantifiées dans un rayon de 1000m
autour du centroïdes des parcelles d’études en prairie. Les intervalles de confiance des coefficients de
corrélations de Spearman sont représentés entre crochets, l’ordre de grandeur de la p-value est représenté sous
forme d’étoile (* : p-value < 0,05 ; ** : p-value < 0,01 ; *** : p-value < 0,001).
AB Haie Forêt Prairie Colza
Haie [0,19 ; 0,29]
***
Forêt [0,20 ; 0,29]
***
[0,02 ; 0,13]
Prairie [0,04 ; 0,15]
*
[0,52 ; 0,63]
***
[0,01 ; 0,15]
Colza [-0,07 ; 0,05]
[-0,43 ; -0,28]
***
[0,03 ; 0,15]
*
[-0,32 ; -0,20]
***
Bordures [-0,00 ; 0,13]
[0,43 ; 0,54]
***
[0,14 ; 0,25]
***
[0,30 ; 0,41]
***
[-0,12 ; 0,00]

38
Résumé
Le service de pollinisation est menacé alors que plus de 70% des cultures mondiales
sont dépendantes de la pollinisation entomophile et que la surface mondiale de ces cultures
augmente. Le service de pollinisation est dépendant des communautés de pollinisateurs qui
s’assemblent en fonction des facteurs locaux (niche écologique) et du contexte paysager
(dispersion). Notre objectif est de comprendre comment les facteurs locaux et paysagers
interagissent pour influencer l’efficacité de la pollinisation entomophile du colza (Brassica
napus L).
Pour cela, des phytomètres de colza (539 depuis 2015) ont été disposés dans la “Zone
Atelier Plaine & Val de Sèvre” dans différents types de cultures abritant des ressources
florales différentes. Les parcelles sont localisées dans des paysages le long des gradients de
longueur des bordures entre les habitats et de proportions d’agriculture biologique, de prairies,
de haie, de forêt et de colza. Ces variables paysagères, connues pour influencer la diversité de
pollinisateurs dans les paysages agricoles, sont quantifiées dans des rayons allant de 500m à
2000m.
Notre expérimentation a mis en évidence que le contexte paysager influence la
pollinisation entomophile du colza en fonction de la localité. Ainsi, la fonction de
pollinisation associée à une diversité de pollinisateurs présente dans les prairies déborde vers
les cultures de colza, tandis que cette fonction est diluée entre les cultures de colza et qu’elle
déborde des cultures de colza vers les cultures de céréales. Notre étude est la première à
mettre en évidence l’importance du contexte paysager sur la pollinisation entomophile du
colza. Les schémas d’aménagement du paysage peuvent s’en inspirer afin d’optimiser la
dispersion des insectes pollinisateurs associée à la fonction de pollinisation vers les cultures
dépendantes de la pollinisation.
Mots clés :
Pollinisation | Colza | Phytomètre | Contexte paysager | Facteurs locaux