mémoire chedzer-clarc clément maîtrise en sols et
TRANSCRIPT

Nutrition minérale de la canneberge au Québec
Mémoire
Chedzer-Clarc Clément
Maîtrise en sols et environnement Maître ès sciences (M. Sc.)
Québec, Canada
© Chedzer-Clarc Clément, 2017

Nutrition minérale de la canneberge au Québec
Mémoire
Chedzer-Clarc Clément
Sous la direction de :
Léon‐Étienne Parent, directeur de recherche

iii
Résumé
Les recommandations d’engrais dans la production de canneberge sont guidées par les
analyses foliaires et de sols. Cependant, les intervalles critiques de concentrations
utilisés à des fins de diagnostic sont biaisés par les interactions entre les éléments
nutritifs et l’âge physiologique de la plante. L’utilisation des ratios logarithmiques
isométriques (ilr) pour l’analyse des données compositionnelles peut éviter les biais
relatifs aux interactions. L’objectif de ce mémoire est de développer des normes
nutritives foliaires non biaisées pour la canneberge à haut rendement au Québec en
utilisant la méthode des balances nutritives (ilr).
Des essais de fertilisation ont été conduits pendant deux années consécutives avec des
traitements variés de N, P, K, Mg, Cu, B sur quatre sites de production de canneberge
dans la région Centre-du-Québec. Les tissus foliaires ont été analysés pour leur
concentration en N, P, K, Mg, Ca, B, Cu, Mn, Fe et Zn. Les données ont été jumelées
aux données des analyses foliaires de six essais de P réalisés par Marchand et al. (2013)
entre 2001 et 2003 dans la même région et sur le même cultivar « Stevens ». Les
résultats des analyses foliaires ont été transformés en balances nutritives entre
composantes ou groupes de composantes, puis partitionnés selon la procédure Cate-
Nelson et des arbres de classification.
Des intervalles de balances nutritives de référence, ainsi que des intervalles de
concentration en soutien, ont été calculés pour la canneberge à haut rendement (> 35
Mg ha-1). Les spécimens à faible rendement ont montré des concentrations en K et en
Mg plus faibles, ce qui peut expliquer le déséquilibre des balances [Ca |Mg] et [K |Ca,
Mg] par rapport aux spécimens à haut rendement. Comparés aux intervalles de
concentration retenus dans la littérature, ceux de cette étude sont plus restreints
partiellement en raison de l’exclusion des cas de consommation de luxe et de
contamination nutritive par cette méthode de calcul. Les intervalles de concentration de
N et K sont les plus fiables, étant donné que les fertilisations N et K sont les seules
auxquelles la culture a répondu de façon significative (P < 0.05). Pour tous les autres
éléments, les intervalles caractérisent de bons rendements sans forcément être des
valeurs critiques, étant le résultat des conditions de sol et de la gestion générale de la
fertilisation.

iv
Tables des Matières RESUME .................................................................................................................................................... III
TABLES DES MATIERES ......................................................................................................................... IV
LISTE DES TABLEAUX ............................................................................................................................ V
LISTE DES FIGURES ................................................................................................................................ VI
REMERCIEMENTS ................................................................................................................................. VII
INTRODUCTION ....................................................................................................................................... 1
CHAPITRE 1 : REVUE DE LITTERATURE ............................................................................................. 4
1.1.-LA CANNEBERGE ........................................................................................................................... 4 1.2.-FERTILISATION ET ANALYSES FOLIAIRES DE LA CANNEBERGE .................................................... 5
1.2.1.-Période d’échantillonnage foliaire ........................................................................................ 5 1.2.2.-Parties du plant visées par l’analyse foliaire ......................................................................... 6 1.2.3.-Concentration foliaire critique en éléments nutritifs ............................................................ 7 1.2.4.-Recommandations en éléments fertilisants ........................................................................... 7
1.3.-APPROCHE COMPOSITIONNELLE OU CONCEPT DES BALANCES .................................................... 9 1.3.2.-Principes de l’analyse compositionnelle .............................................................................. 10 1.3.3.-Transformations des données compositionnelles : les ratios logarithmiques (logs ratios) .. 11
CHAPITRE 2 : HYPOTHESES ET OBJECTIFS ..................................................................................... 13
2.1.-HYPOTHESE ................................................................................................................................ 13 2.2.-OBJECTIFS .................................................................................................................................. 13
CHAPITRE 3 : MÉTHODOLOGIE .......................................................................................................... 14
3.1.-ENSEMBLE DE DONNÉES .............................................................................................................. 14 3.2.-SITES EXPÉRIMENTAUX .............................................................................................................. 14 3.3.-STRUCTURE DES TRAITEMENTS ET DISPOSITIF EXPÉRIMENTAL ................................................. 14 3.4.-ANALYSES DE SOLS ET FOLIAIRES ............................................................................................... 17 3.5.- RÉCOLTE .................................................................................................................................... 17 3.6.-EXPÉRIENCE ANTÉRIEURE : MARCHAND ET AL. (2013) ............................................................. 17 3.7.-ANALYSE DES DONNÉES .............................................................................................................. 18
3.7.1.-Définition des balances nutritives ....................................................................................... 18 3.7.2.-Transformation des données foliaires en log ratio isométriques (ilr).................................. 18 3.7.3.-Partition de Cate-Nelson ..................................................................................................... 19 3.7.4.-Interprétation du test de diagnostic ..................................................................................... 20 3.7.5.-Apprentissage automatique ................................................................................................. 21
CHAPITRE 4 : RÉSULTATS ET DISCUSSIONS .................................................................................... 22
4.1.-PROPRIÉTÉS DES SOLS ET PRODUCTIVITÉ ................................................................................... 22 4.2.-DIAGNOSTIC FOLIAIRE................................................................................................................ 23
4.2.1.-Partition de Cate-Nelson ..................................................................................................... 23 4.2.2.-Intervalles de suffisance nutritive ....................................................................................... 25 4.2.3.-Comparaison entre TN et TP ............................................................................................... 30
CONCLUSION .......................................................................................................................................... 35
RÉFÉRENCES BIBLIOGRAPHIQUES ................................................................................................... 36

v
Liste des tableaux
Tableau 1: Intervalles de suffisance nutritive des tissus de canneberge ....................................... 7 Tableau 2: Recommandations en azote, phosphore et potassium au Québec (Parent et Gagné, 2010) ............................................................................................................................................. 8 Tableau 3: Structure des traitements de l'expérience .................................................................. 15 Tableau 4: Calendrier d'application des engrais .......................................................................... 16 Tableau 5: Contrastes logarithmiques et partition séquentielle binaire des analyses foliaires de la canneberge ................................................................................................................................... 18 Tableau 6. Propriétés des sols des sites de canneberge en Mai 2014 (Mai 2001 pour les sites de Marchand et al., 2013)................................................................................................................. 22 Tableau 7. Intervalles de confiance (P < 0,01) des balances (ilr) pour les vrais négatifs (TN) .. 25 Tableau 8. Intervalles de suffisance nutritive des tissus de canneberge ...................................... 28

vi
Liste des figures
Figure 1.- Contenus saisonniers en azote et en potassium dans les tissus de canneberge au Massachussetts (DeMoranville, 1992) .......................................................................................... 6 Figure 2. Disposition des parcelles expérimentales : cas d'un site. ............................................. 16 Figure 3. Partition de Cate-Nelson entre la distance de Mahalanobis et le rendement en fruits de la canneberge. .............................................................................................................................. 24 Figure 4. Arbre de Classification des rendements élevés (HY) et bas (LY) au niveau de 35 Mg ha-1. .............................................................................................................................................. 31

vii
Remerciements
Je tiens à remercier mon directeur de recherche, Mr Léon-Étienne Parent, professeur au
département des sols et génie agroalimentaire, pour son temps et sa patience, et de
m’avoir fourni toute l’aide nécessaire pour mener à bien ce projet.
Mes remerciements vont aussi à Serge-Étienne Parent qui m’a beaucoup aidé dans les
analyses statistiques, et à Reza Jamaly qui a été mon collègue de travail tout au long du
projet.
Je témoigne aussi une particulière gratitude au Dr Lotfi Khiari et au Dr. Antoine Karam.
Qu'ils trouvent ici l'expression de mes remerciements les plus sincères.
Ce projet a reçu un appui financier du Conseil de Recherche en Sciences Naturelles et
en Génie du Canada (CRDPJ 385199 09) et des partenaires industriels suivants : Atocas
de l’érable Inc., Atocas Blandford et la Cannebergière.
À tous ceux qui, d’une manière ou d’une autre, m’ont apporté une aide dans la
réalisation de ce travail : Je vous dis un grand merci.

1
Introduction
La canneberge (Vaccinium macrocarpon Ait.) est une éricacée sauvage domestiquée en
Amérique du Nord à partir du XIXe siècle. Elle est depuis cultivée pour la production et
la qualité de ses fruits (Roper et Vorsa, 1997). Le petit fuit rouge, sec ou transformé, est
très apprécié pour sa teneur élevée en antioxydant. Le Québec est le second producteur
mondial de canneberge après l’État du Wisconsin aux États-Unis. La canneberge est
cultivée sur des sols acides sableux ou organiques dans des bassins de 1 à 2 ha endigués
pour faciliter la gestion de l’eau. Un ponçage régulier de la surface du support et
l’élagage des cultures permettent de maintenir le rendement de la canneberge en limitant
une fructification biennale (Roper, 2006).
Malgré sa récente domestication, la nutrition de la canneberge est plus proche de celle
des espèces fruitières domestiques que celle des espèces fruitières sauvages (Parent et
al., 2013). La canneberge est sensible à l’ajout d’éléments nutritifs. Elle l’est beaucoup
plus à l’ajout d’azote que de phosphore (Parent et Marchand, 2006). L’ajout d’éléments
nutritifs est guidé par les analyses de sols et foliaires (Yarborough et al., 1993 ;
Marchand et al., 2013). Un diagnostic nutritif des tissus des plantes doit être conduit de
façon à intégrer tous les facteurs d’interaction, et par conséquent, doit refléter à la fois
les facteurs génétiques et environnementaux (Munson et Nelson, 1990). Selon la loi de
l’optimum de Liebscher (De Wit, 1992), l’effet d’un élément nutritif sur le rendement
d’une culture dépend de la proximité des autres éléments de leur optimum. L’hypothèse
derrière l’interprétation des analyses de plantes est qu’une courbe de réponse nette peut
être dérivée de la relation entre le rendement et la concentration des tissus en un élément
nutritif où tous les autres éléments sont maintenus dans leurs plages optimales de
concentration pour éviter de modifier l’effet de l’élément nutritif cible sur le rendement
par un effet nuisible d’un autre élément nutritif (Ulrich et Hills, 1967). Une
concentration critique est généralement interpolée là où la croissance des plantes
diminue de 5-10%.
Comme toutes les autres données compositionnelles (Aitchison, 1986), les analyses
foliaires sont des données strictement positives et intrinsèquement multivariées. Elles
font partie d’un simplex compositionnel dont les composantes expriment une
information relative à l’unité ou à l’échelle de mesure telle que le g kg-1 ou le

2
pourcentage, et sont généralement exprimées sur la base de matière fraîche ou de
matière sèche (Jones et al., 1990) ou sur une autre base comme l’azote (Ingestad, 1987).
Bien avant qu’Aitchison (1986) ait développé, pour des raisons mathématiques et
géométriques, le concept de « l’espace clos » derrière l’analyse des données
compositionnelles, Lagatu et Maume (1934), les fondateurs des analyses et diagnostics
foliaires, intégrèrent dans une relation le N-P-K dans les feuilles de pomme de terre en
utilisant un diagramme ternaire. Des concepts d’interprétation ultérieurs tels que les
seuils ou les plages de concentration critique (Macy, 1936 ; Ulrich, 1952) et les ratios
binaires conçus pour refléter des interactions importantes entre les éléments (Redfield,
1936 ; Beaufils, 1973) ont ignoré le problème de « l’espace clos » conduisant ainsi à des
distorsions à l’étape du diagnostic (Parent et Dafir, 1992 ; Parent et al., 2012). En plus
des biais méthodologiques liés aux analyses des données compositionnelles tels que la
redondance, l’incohérence de sous-composition et la distribution non normale, la
délimitation des intervalles d’éléments nutritifs par des moyennes graphiques (Ulrich et
Hills, 1967) ou des intervalles de confiance (Beaufils, 1973) est assujettie à des cas de
concentrations sous-optimales, de consommation de luxe et de contamination (Parent et
al., 2012).
Les diagrammes de performance des cultures en fonction des indices nutritifs peuvent
être partitionnés en quadrants suivant la procédure de Cate-Nelson afin d’isoler les vrais
négatifs (true negative) et d’en déduire les normes nutritives associées (Parent et al.,
2012). Les ratios logarithmiques, notamment le ratio logarithmique centré (centered log
ratio) (Aitchison, 1986 ; Parent et Dafir, 1992) et le ratio logarithmique isométrique
(isometric log ratio) (Egozcue et al., 2003 ; Parent, 2011), sont des techniques d’analyse
compositionnelle très utiles ayant déjà été utilisées dans le diagnostic nutritif des tissus
des plantes. La transformation logarithmique ordinaire et le ratio logarithmique centré
donnent exactement les mêmes résultats de l'analyse statistique si et seulement si les
moyennes géométriques des compositions testées sont les mêmes (Lovell et al., 2011).
Cependant, les moyennes géométriques diffèrent à cause de l’hypothèse ceteris paribus
(tous les autres facteurs autres que ceux étudiés sont égaux) qui est rarement applicable
en raison de la redondance des données à l’intérieur de l’espace clos compositionnel
(Aitchison, 1986) ou à la suite d’interactions entre les éléments nutritifs (Bates, 1971)
qui contribuent aussi à la redondance des données (toute variation dans la concentration
d’un élément modifie celle d’un ou de plusieurs autres éléments).

3
Il est donc devenu impératif d’utiliser des méthodes appropriées pour analyser les
données des analyses foliaires en vue d’un diagnostic nutritif non biaisé. L’approche des
ilr est la plus appropriée pour décrire des modèles naturels et les balances nutritives
dans les tissus des plantes (Parent, 2011). C’est dans ce contexte que s’inscrit ce travail
de recherche orienté vers un diagnostic nutritif non biaisé des tissus de canneberge en
utilisant la technique du ratio logarithmique isométrique afin d’établir des normes
nutritives foliaires caractéristiques de haut rendement.

4
Chapitre 1 : Revue de littérature
1.1.-La canneberge
La canneberge appartient à la famille des Ericacées, qui colonisent en général les sols
très acides et mal pourvus en éléments nutritifs. En plus d’être peu exigeante en
éléments nutritifs, la canneberge vit en association avec un champignon mycorhizien
endophyte (Bajwa et Read, 1986 ; Stribley et Read, 1974). Cette symbiose améliore son
approvisionnement en éléments minéraux.
Le pH d’un sol sous culture de canneberge doit se situer entre 4,0 et 5,5 (Eck, 1990).
Cette fourchette de pH assure à la canneberge un approvisionnement en azote
ammoniacal, la forme d’azote que la canneberge préfère (Barak et al., 1996 ; Greidanus
et al., 1972 ; Rosen et al., 1990).
L’azote ammoniacal provenant de la matière organique ou des engrais peut se
transformer sous la forme nitrique par le processus de nitrification (Alexander, 1980).
La canneberge utilise très mal le nitrate. Vu que dans les conditions d’aérobie et
d’acidité dans lesquelles elle évolue, l’azote ammoniacal est la forme d’azote minéral
qui prédomine. En plus, le nitrate peut être lessivé dans le sol ou converti sous des
formes gazeuses comme N2 et N2O. C’est une réaction d’oxydation qui se fait par
catalyse enzymatique reliée à des bactéries nitrifiantes (nitrosomonas et nitrobacter)
(Schmidt, 1982). La nitrification peut être évitée par la gestion du pH afin de limiter
l’activité des bactéries nitrifiantes (Foy, 1984). Ces bactéries sont sensibles aux
conditions du milieu dans lesquelles elles vivent. Leur croissance ainsi que les enzymes
qu’elles produisent à chaque séquence du processus de nitrification subissent l’influence
des facteurs du milieu. Parmi les facteurs d’influence, le pH est un facteur capital qui
affecte la nitrification (Alexander, 1980 ; Paul et Clark, 1989).
Le pH optimal favorisant la nitrification varie entre 6.6 et 8 (Paul et Clark, 1989). Dans
les sols sableux ou les sols de canneberge (M. O<5%), il est recommandé de maintenir
un pHeau entre 4 et 5. Si le sol contient plus de 5 % de matière organique, un pHeau
inférieur à 4,5 est recommandé (DeMoranville, 2010).

5
1.2.-Fertilisation et analyses foliaires de la canneberge
Les plants de canneberge nécessitent l’apport d’éléments nutritifs pour assurer une
bonne croissance végétative et une production élevée de fruits. Quand ces éléments se
trouvent en quantité insuffisante, la croissance ainsi que le rendement peuvent être
affectés. Des éléments fertilisants comme l’azote, le phosphore et le potassium sont
apportés aux plants de canneberge pour suppléer les quantités déjà disponibles dans le
sol. Les analyses foliaires permettent d’évaluer l’adéquation entre les éléments
minéraux prélevés par la plante et ses besoins nutritifs. Les résultats de ces analyses
aident à diagnostiquer d’éventuels problèmes de nutrition de la plante, à prévoir les
besoins en éléments fertilisants pour l’année suivante et à ajuster un programme de
fertilisation d’une plante pérenne comme la canneberge (Davenport et al., 1995).
Des recherches ont montré des relations entre la fertilisation et la concentration des
éléments dans les tissus de canneberge. Ainsi, la concentration en azote dans les tissus
augmente avec l’augmentation de la fertilisation azotée dans la culture de la canneberge
(Davenport et Provost, 1994 ; Eaton, 1971 ; Eaton et Meehan, 1973; Eck, 1971). Il n’y a
pas que la concentration en azote qui augmente avec l’application de fertilisant azoté, la
concentration du P et du K augmente aussi pendant que celle du Fe diminue (Davenport
et Prevost, 1994). Parent et Marchand (2006) ont trouvé une augmentation linéaire de la
concentration de P dans les tissus suite à l’ajout de P dans les sols ayant un ratio molaire
P/(Al+Fe) MIII compris entre 0,024 à 0,044.
1.2.1.-Période d’échantillonnage foliaire
Lors du prélèvement des échantillons foliaires, les feuilles et la tige ne sont pas
séparées. Les analyses foliaires incluent donc la tige et les feuilles. Des changements
importants dans la concentration en certains éléments sont observés durant la période de
croissance active (mai à août) comme illustré à la figure 1. Les concentrations en N et K
dans les tissus varient rapidement durant la saison de croissance, soit de mai à août,
pour atteindre un niveau constant entre mi-août et mi-septembre (fig.1). Il est préférable
de prélever les échantillons lorsque les éléments nutritifs contenus dans les tissus sont
stabilisés (Davenport et al., 1995). L’échantillonnage doit être effectué quand la
concentration des éléments nutritifs dans les tissus est stable (Davenport et al., 1995).
La figure 1 illustre le risque émanant de la collecte des tissus tard en septembre. La
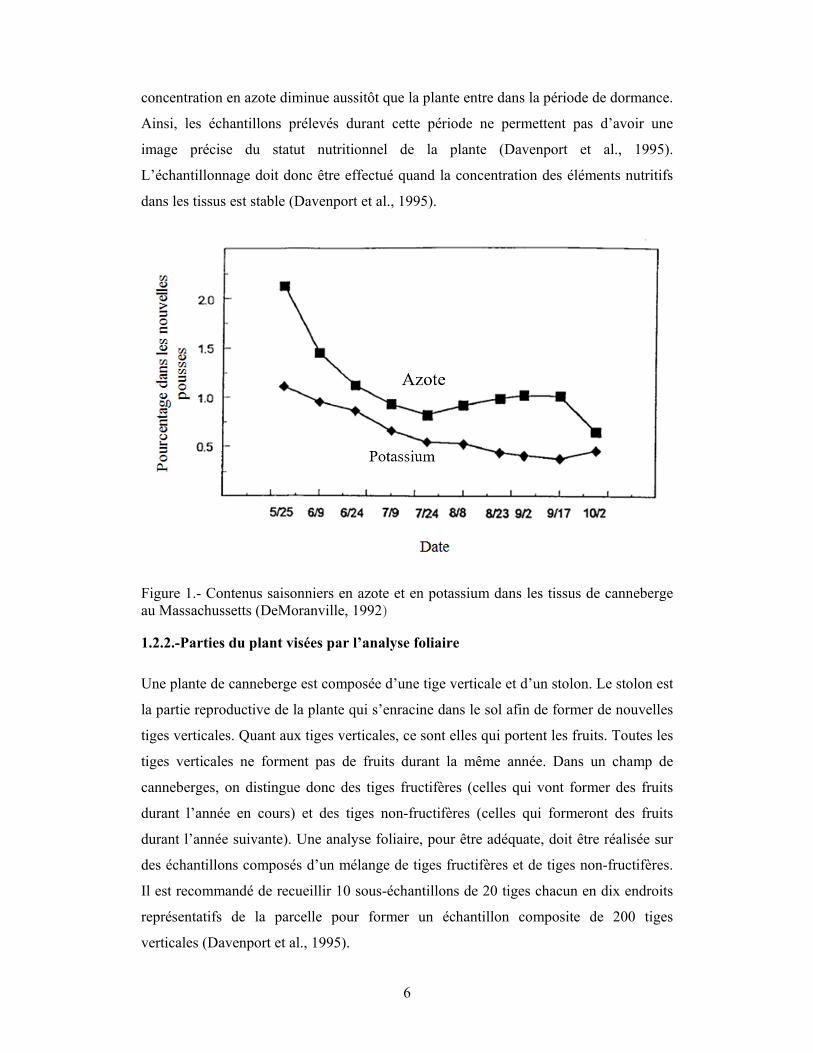
6
concentration en azote diminue aussitôt que la plante entre dans la période de dormance.
Ainsi, les échantillons prélevés durant cette période ne permettent pas d’avoir une
image précise du statut nutritionnel de la plante (Davenport et al., 1995).
L’échantillonnage doit donc être effectué quand la concentration des éléments nutritifs
dans les tissus est stable (Davenport et al., 1995).
Figure 1.- Contenus saisonniers en azote et en potassium dans les tissus de canneberge au Massachussetts (DeMoranville, 1992)
1.2.2.-Parties du plant visées par l’analyse foliaire
Une plante de canneberge est composée d’une tige verticale et d’un stolon. Le stolon est
la partie reproductive de la plante qui s’enracine dans le sol afin de former de nouvelles
tiges verticales. Quant aux tiges verticales, ce sont elles qui portent les fruits. Toutes les
tiges verticales ne forment pas de fruits durant la même année. Dans un champ de
canneberges, on distingue donc des tiges fructifères (celles qui vont former des fruits
durant l’année en cours) et des tiges non-fructifères (celles qui formeront des fruits
durant l’année suivante). Une analyse foliaire, pour être adéquate, doit être réalisée sur
des échantillons composés d’un mélange de tiges fructifères et de tiges non-fructifères.
Il est recommandé de recueillir 10 sous-échantillons de 20 tiges chacun en dix endroits
représentatifs de la parcelle pour former un échantillon composite de 200 tiges
verticales (Davenport et al., 1995).

7
1.2.3.-Concentration foliaire critique en éléments nutritifs
La majorité des résultats de recherche sur la canneberge proposent des normes foliaires
établies pour chaque élément minéral individuellement. Pour chaque élément, un
intervalle de suffisance est donné en pourcentage ou en ppm par rapport au poids sec du
tissu végétal. En-dessous de la limite minimale fixée par cet intervalle, l’élément est
considéré en carence et au-dessus de la limite maximale, il est considéré en excès. Le
tableau 1 présente les intervalles de suffisance nutritive des plants de canneberge au
Maine (Yarborough et al., 1993), au Wisconsin (Roper, 2001) et au Québec (Marchand
et al., 2013).
Tableau 1: Intervalles de suffisance nutritive des tissus de canneberge
Éléments Intervalle de suffisance
Roper, 2001 Yarborough et al., 1993 Marchand et al., 2013
% massique
Azote (N) 0,90-1,10 0,95-1,05 1,09-1,24
Phosphore (P) 0,10-0,20 0,11-0,14 0,11-0,13
Potassium (K) 0,40-0,75 0,40-0,65 0,50-0,56
Calcium (Ca) 0,30-0,80 0,60-0,80 0,84-1,00
Magnésium (Mg) 0,15-0,25 0,20-0,25 0,17-0,20
Soufre (S) 0,08-0,25 ND ND
mg.kg-1
Bore (B) 15-60 30-50 30-39
Fer (Fe) >20 40-80 60-69
Manganèse (Mn) >10 150-250 493-711
Zinc (Zn) 15-30 15-30 22-26
Cuivre (Cu) 4-10 4-7 4-5
1.2.4.-Recommandations en éléments fertilisants
Les recommandations actuelles en N-P-K dans la culture de la canneberge au Québec
sont basées sur des résultats expérimentaux obtenus dans le Centre-du-Québec avec la

8
variété « Stevens » sur des sols du groupe de texture sableuse. Les doses à appliquer
dépendent de la quantité de l’élément présent dans le sol et des concentrations foliaires
visées (Parent et Gagné, 2010). Les concentrations foliaires visées au Québec entre le
15 août et le 15 septembre sont de 0,90 à 1,10 % en azote, 0,10 % en phosphore (bien
que des États américains recommandent 0,11 %) et de 0,40 à 0,75 % en potassium
(Parent et Gagné, 2010). Les doses d’application proposées au Québec sont présentées
au tableau 2.
Tableau 2: Recommandations en azote, phosphore et potassium au Québec (Parent et
Gagné, 2010)
Azote
Temps et mode d’apport Recommandation (kg N ha-1)
Dose répartie en 4 applications (au début de la floraison, à 50 % floraison, à 50 %
nouaison, Au grossissement du fruit)
20-65
Phosphore
Indice de saturation en phosphore :
(%)
Recommandation selon le P foliaire visé (kg P2O5 ha-1)
0,10 % 0,11 %
0-3,5 40 80
3,6-7,0 20 65
7,1-14,0 0 0
14,1 et plus 0 0
Potassium
Analyse (kg KM-3 ha-1)
Recommandation (kg K2O ha-1)
0-115 65-110
116-230 0-65
231 et plus 0
Ces recommandations s’appliquent à des sols de pH 4 à 5 et pour des champs en
production depuis au moins 3 ans.
Cette approche de diagnostic foliaire selon laquelle chaque élément est considéré
isolément est biaisée vu qu’elle ne prend pas en compte les interactions entre les

9
éléments ainsi que l’âge physiologique de la plante (Bates, 1971 ; Parent, 2011 ;
Marchand et al., 2013). Ainsi, une nouvelle méthode de diagnostic nutritif a été
développée au fil des années : l’approche compositionnelle qui permet de faire un
diagnostic nutritif non biaisé (Parent et Dafir, 1992 ; Parent, 2011 ; Parent et al., 2013).
1.3.-Approche compositionnelle ou concept des balances
1.3.1.-Définition et concepts de base
Les données compositionnelles sont définies comme un vecteur où toutes les
composantes sont des nombres réels strictement positifs et fournissent uniquement des
informations relatives. En effet, l’information relative réside dans l’unité de mesure de
la composition exprimée en général en pourcentage de poids ou de volume, en ppm, en
ppb ou en proportion molaire. La somme de toutes les composantes d’une composition
est égale à une constante k. À noter que les valeurs prises par k dépendent de l’unité de
mesure. Les valeurs usuelles sont 1 si l’unité de mesure est en proportion, 100 si c’est
en pourcentage, 106 si elle est en ppm et 109 si c’est en ppb (Pawlowsky-Glahn et
Buccianti, 2011).
Mathématiquement un vecteur de composition est défini par le simplex :
SD={x=[x1, x2, … xD]; avec xi>0; i=1, 2,…, D;∑ } ;
x1, x2,…, xD sont les D composantes du vecteur de composition.
À la lumière de ces définitions, on reconnaît que les données des analyses foliaires sont
des données compositionnelles :
Ce sont des nombres réels strictement positifs
Elles fournissent uniquement des informations relatives
La somme des résultats d’analyse de toutes les composantes est contrainte à k.
Si toutes les composantes du vecteur de composition ne sont pas disponibles, on doit,
soit définir une valeur de remplissage ou valeur résiduelle, ou simplement définir une
sous-composition avec les composantes disponibles (Egozcue et Pawlowsky-Glahn,
2006).
La valeur de remplissage est la différence entre la constante k et la somme de toutes les
composantes analysées. Elle représente les composantes non prises en compte dans le
vecteur de composition.

10
Pour définir une sous-composition à partir d’une composition dont toutes les
composantes ne sont pas disponibles, on procède comme suit (Pawlowsky-Glahn et
Buccianti, 2011) :
Soit z une composition avec s composantes analysées et connues et C(z) une sous-
composition de z:
z = [z1, z2, … zj]; avec zj>0; j=1, 2,…, D; ∑ ;
C(z)= [ . 1∑ 1
, . 2∑ 1
,… , .∑ 1
] avec zi>0 et i= 1,2,…,s.
1.3.2.-Principes de l’analyse compositionnelle
Selon Aitchison (1986), trois (3) conditions doivent être remplies pour une analyse non
biaisée des données compositionnelles :
a. Invariance d’échelle
La plus importante caractéristique des données compositionnelles est le fait
qu’elles apportent seulement une information relative due à l’unité de mesure
(proportion, %, ppm, ppb, etc.) (Pawlowsky-Glahn et Buccianti, 2011). Ainsi,
changer l’unité de mesure des données (par exemple de ppm à %) revient à
multiplier les données par une constante de conversion ʎ (équivalence
compositionnelle). Les analyses doivent donner les mêmes résultats peu importe
la valeur de ʎ ou l’unité de mesure des données. C’est ce qu’Aitchison (1986)
appelle invariance d’échelle.
b. Invariance de permutation
Il y a invariance de permutation si on obtient des résultats équivalents même en
changeant l’ordre des composantes dans une composition.
Pour résoudre le problème de singularité de la matrice de covariance des
données compositionnelles, la méthode classique est de réduire le nombre de
dimensions D à D-1 sans perte d’information (i.e. en évitant d’enlever une
composante redondante utile).
c. Cohérence sous-compositionnelle
Ce principe peut être résumé comme suit : projeter les données
compositionnelles dans l’espace réel sans dépasser les limites de l’espace clos.

11
Autrement dit, les sous-compositions devraient se comporter de la même façon
que les projections orthogonales dans les analyses réelles conventionnelles. La
dimension d’un segment projeté est inférieure ou égale à celle du segment lui-
même. En d’autres termes, la distance entre deux compositions complètes doit
être supérieure ou égale à leur distance quand on considère des sous-
compositions.
Pour projeter les données compositionnelles dans l’espace réel, les logs ratios ont été
conçus par des mathématiciens. Il s’agit du log ratio additif (alr), du log ratio centré
(clr) et du log ratio isométrique (ilr). En plus de libérer les données compositionnelles
de leur espace clos et de les projeter dans l’espace réel, le log ratio respecte le principe
de l’invariance d’échelle. Quel que soit l’échelle de mesure, le log ratio doit être le
même pour que l’interprétation soit cohérente.
1.3.3.-Transformations des données compositionnelles : les ratios logarithmiques
(logs ratios)
a. alr (additive log ratio)
Cette transformation a été élaborée par Aitchison (1986) et utilisée comme une
variable stoïchiométrique par Ingestad (1987) pour le suivi de la nutrition des plants
forestiers. Le nombre de degrés de liberté est réduit à D-1 en utilisant une
composante comme dénominateur commun. L’analyse multivariée est invariante au
choix du dénominateur commun (Aitchison, 1986). Dans le modèle d’Ingestad
(1987), le dénominateur commun est la concentration en N. Le jème alr est calculé
comme suit :
est le jème élément nutritif hormis le dénominateur commun, N. Par exemple, si un
tissu contient 1.8 % N et 0.12 % P, le est égal à0.121.8
2.71.
Toutefois, les alr sont obliques (60 o) l’un à l’autre dans l’espace euclidien et donc
difficiles à ajuster géométriquement à un espace orthogonal (Egozcue et
Pawlowsky-Glahn, 2005).

12
b. clr (centered log ratio)
La transformation clr a été utilisée pour l’analyse des tissus végétaux (Parent et
Dafir, 1992 ; Parent et al., 2009), des sols (Parent et al., 2012) et des solutions
nutritives (Lopez et al., 2002). Le jième clr pour la ième observation est calculé comme
suit :
ln
Où est la jième composante et est la moyenne géométrique des proportions
pour la ième observation. Comme il y a D clr pour D concentration et que la somme
des clr est égale à 0, la matrice est singulière et un clr (généralement celui de la
valeur de remplissage) doit être sacrifié pour effectuer une analyse multivariée. Par
conséquent, le clr n’est pas adapté aux mesures de corrélation (Filzmoser et Hron,
2009).
c. ilr (isometric log ratio)
L’ilr, aussi appelé « balance orthonormale » décrite dans une partition binaire
séquentielle, est défini comme le ratio logarithmique entre les moyennes
géométriques de deux composantes ou groupes de composantes. L’ilr permet de
résoudre le problème de la matrice de singularité. Contrairement à l’alr, il a
l’avantage d’être géométriquement adapté pour conduire des analyses multivariées
en raison de l’orthogonalité (Filzmoser et al., 2009). Il est calculé comme
suit (Egozcue et al., 2003) :
Où ilri est la ième balance entre deux sous-compositions, i est compris entre 1 et D-1,
r et s sont respectivement le nombre de composantes au numérateur et au
dénominateur, et et sont les moyennes géométriques des sous-
ensembles au numérateur et au dénominateur, respectivement.

13
Chapitre 2 : Hypothèses et Objectifs
2.1.-Hypothèse
Les essais de fertilisation font varier un ou plusieurs éléments nutritifs sous l’hypothèse
du « ceteris paribus » (tous les autres facteurs ont un effet égal par ailleurs), ce qui
permet de relier le rendement de la culture à l’analyse foliaire (Ulrich et Hills, 1967).
Notre hypothèse est que la fertilisation N, P, K, Mg, Cu, et B de la canneberge permet
d’élaborer des normes nutritives foliaires caractéristiques de rendements élevés compte
tenu de la variation normale de l’ensemble de la composition minérale d’une feuille.
2.2.-Objectifs
L’objectif général de ce travail est de développer des normes foliaires nutritives pour la
canneberge (cv. ‘Stevens’) à haut rendement, établie sur des sols sableux acides au
Québec en utilisant de nouvelles méthodes d’analyses de données comme l’analyse
compositionnelle et les arbres de classification et de régression. Plus spécifiquement,
nos objectifs sont de :
1. Définir des balances nutritives idéales des plants de canneberge à haut rendement ;
2. Réévaluer les normes de concentrations foliaires de la canneberge ;
3. Comparer les nouveaux intervalles de concentration aux normes de concentrations
foliaires publiées dans la littérature.

14
Chapitre 3 : Méthodologie
3.1.-Ensemble de données
Les données utilisées dans le cadre de ce travail sont issues d’essais de fertilisation
réalisés pendant deux (2) ans dans la région Centre-du-Québec et des travaux de
Marchand et al. (2013). Au total, le jeu de données est constitué de 488 observations
dont 202 sont issues des travaux antérieurs.
3.2.-Sites expérimentaux
L’expérience a été réalisée pendant deux ans (2014 et 2015) sur quatre (4) sites (A9, 10,
45 et 9) sur les fermes Atocas Blandford (Latitude: 46o14ˈ48,63 ̎ N; Longitude: 72o
02ˈ01,20 ̎ O), la Cannebergière (Latitude: 46o19ˈ37,15 ̎ N; Longitude: 71o 44ˈ04,27 ̎ O)
et Atocas de l’érable (Latitude: 46o21ˈ33,57 ̎ N; Longitude: 71o 52ˈ34,16 ̎ O) localisées
dans la région Centre-du-Québec. Le sol est un brunisol de la série Blandford (site A9)
(Rompré et al., 1984), un podzol de la série Villeroy (sites 9 et 45) et un gleysol de la
série Lyster (site 10) (Laflamme et al.,1989). Durant la première année de l’expérience,
les quatre sites étaient en production conventionnelle. En 2015, un bassin chez Atocas
Blandford (A9) est passé en transition biologique. La deuxième année de l’expérience a
été donc réalisée en utilisant des engrais alors certifiés biologiques par Écocert sur le
site A9.
L’étude a été menée sur le cultivar ‘Stevens’. C’est un cultivar hybride de mi- saison qui
produit de gros fruits ronds de coloration moyenne (Eck, 1990). C’est le cultivar le plus
populaire au Québec étant donné son haut potentiel de rendement.
3.3.-Structure des traitements et dispositif expérimental
Au total 18 traitements ont été testés, répartis en cinq (5) doses d’azote, trois (3) doses
de phosphore, quatre (4) doses de potassium, deux (2) doses de magnésium, deux (2)
doses de cuivre et deux (2) doses de bore. Sur les sites conventionnels, les éléments
fertilisants ont été apportés sous forme de sulfate d’ammonium (21-0-0), de triple
superphosphate (0-46-0), de sulfate de potasse (0-0-50), de sulfate de potasse magnésien
(sulpomag) (0-0-22-11% Mg), du sel d’Epsom (9% Mg), de sulfate de cuivre (25% Cu)
et de solubor (21% B). Sur le site en transition biologique (A9), l’azote, le phosphore et
le potassium ont été apportés sous forme d’engrais alors certifiés biologiques par

15
Écocert, soit des acides aminés liquides (8-0-0) pour l’azote, de l’os fossile moulu (0-
13-0) pour le phosphore, du sulfate de potasse et du sulpomag biologiques pour le
potassium. Le sel d’Epsom, le sulfate de cuivre et le solubor sont aussi certifiés
biologiques. Dans le cas des traitements sans magnésium (Mg0) et sans potassium (K0),
du sel d’Epsom et du sulfate de potassium ont respectivement été utilisés.
Les traitements sont structurés de façon telle qu’une parcelle reçoit le traitement qui lui
est attribué aléatoirement et la dose des autres éléments recommandée par Parent et
Gagné (2010). Par exemple, les traitements azotés sont composés de doses différentes
d’azote mais d’une même dose de phosphore, de potassium, de magnésium, de bore et
de cuivre. Le tableau 3 présente avec plus de détails la structure des traitements.
Tableau 3: Structure des traitements de l'expérience
Composantes (kg/ha)
Traitement N P K Mg B Cu
N0 0 30 80 12 2 1
N15 15 30 80 12 2 1
N30 30 30 80 12 2 1
N45 45 30 80 12 2 1
N60 60 30 80 12 2 1
P0 45 0 80 12 2 1
P15 45 15 80 12 2 1
P30 45 30 80 12 2 1
K0 45 30 0 12 2 1
K40 45 30 40 12 2 1
K80 45 30 80 12 2 1
K120 45 30 120 12 2 1
Mg0 45 30 80 0 2 1
Mg12 45 30 80 12 2 1
B0 45 30 80 12 0 1
B1 45 30 80 12 2 1
Cu0 45 30 80 12 2 0
Cu2 45 30 80 12 2 1
Les traitements sont randomisés dans des unités expérimentales disposées en blocs
complets. Le dispositif expérimental de chacun des sites est constitué de deux (2) blocs
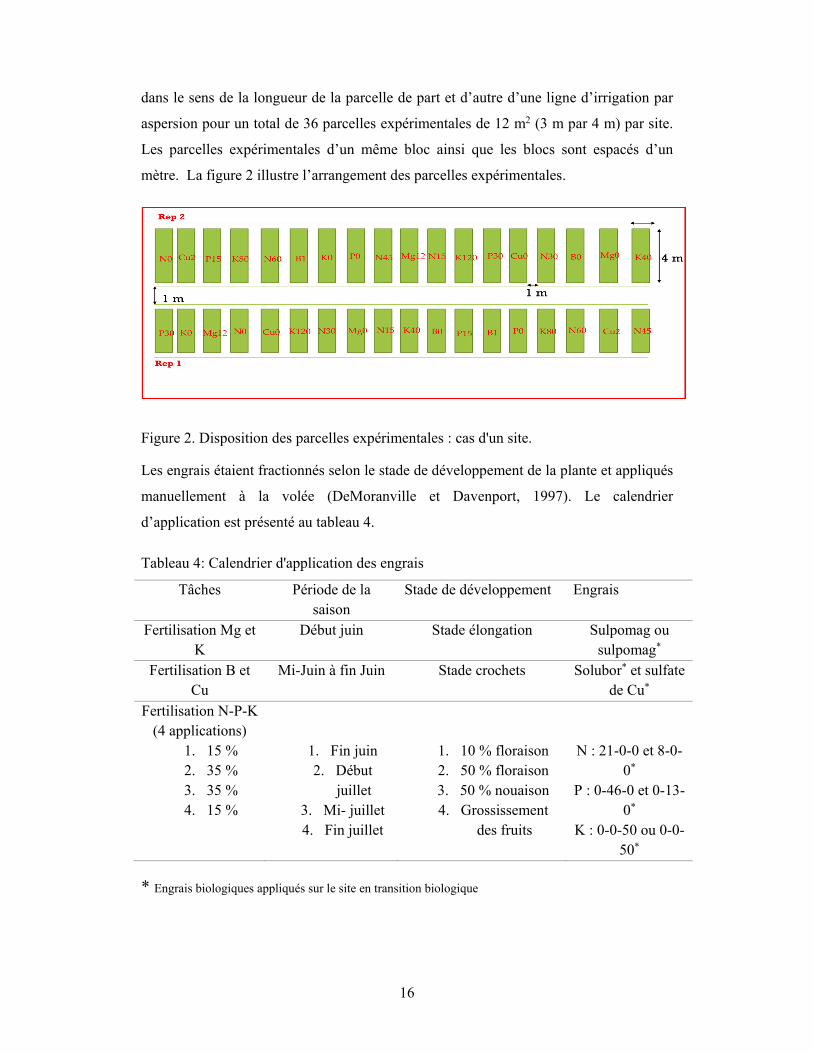
16
dans le sens de la longueur de la parcelle de part et d’autre d’une ligne d’irrigation par
aspersion pour un total de 36 parcelles expérimentales de 12 m2 (3 m par 4 m) par site.
Les parcelles expérimentales d’un même bloc ainsi que les blocs sont espacés d’un
mètre. La figure 2 illustre l’arrangement des parcelles expérimentales.
Figure 2. Disposition des parcelles expérimentales : cas d'un site.
Les engrais étaient fractionnés selon le stade de développement de la plante et appliqués
manuellement à la volée (DeMoranville et Davenport, 1997). Le calendrier
d’application est présenté au tableau 4.
Tableau 4: Calendrier d'application des engrais
Tâches Période de la saison
Stade de développement Engrais
Fertilisation Mg et K
Début juin Stade élongation Sulpomag ou sulpomag*
Fertilisation B et Cu
Mi-Juin à fin Juin Stade crochets Solubor* et sulfate de Cu*
Fertilisation N-P-K (4 applications)
1. 15 % 2. 35 % 3. 35 % 4. 15 %
1. Fin juin 2. Début
juillet 3. Mi- juillet 4. Fin juillet
1. 10 % floraison 2. 50 % floraison 3. 50 % nouaison 4. Grossissement
des fruits
N : 21-0-0 et 8-0-0*
P : 0-46-0 et 0-13-0*
K : 0-0-50 ou 0-0-50*
* Engrais biologiques appliqués sur le site en transition biologique

17
3.4.-Analyses de sols et foliaires
Des échantillons de sols composés d’un mélange de cinq (5) sous-échantillons ont été
prélevés dans la couche 0-15 cm de chaque parcelle avant la fertilisation afin de
caractériser les sites. Les échantillons ont été mis à sécher à l’air libre (20oC) et tamisés
à < 2 mm avant les analyses au laboratoire.
Le pH a été déterminé dans une solution de CaCl2 (0,01 M) en utilisant un rapport de
sol-solution de 1:2 (Hendershot et al., 1993). Les éléments assimilables (P, K, Ca, Mg,
Cu, Fe, Zn, Mn) ont été déterminés par extraction à la solution Mehlich –III (Ziadi et
Tran, 2006). Les formes amorphes d’Al et de Fe ont été extraites par l’oxalate
d'ammonium 0,2 M et l’acide oxalique 0,2 M (Schwertmann, 1973).
Des échantillons foliaires ont été prélevés à raison de 200 tiges par parcelle (Davenport
et al., 1995). Les échantillons ont été séchés à 55 oC, broyés à 2 mm et analysés à
l’acide perchlorique (P-K-Ca-Mg-Al-Cu-Mn-Zn-Fe) (Barnhisel et Bertsch, 1982).
Quant au bore, il a été extrait par calcination puisque le bore peut se volatiliser durant la
digestion humide (Wickner, 1986). Le dosage des éléments a été réalisé par spectrométrie
d’induction au plasma à haute fréquence (ICP-OES).
3.5.- Récolte
Les fruits ont été récoltés manuellement dans quatre placettes d’un pied carré par
parcelle entre les deuxième et troisième semaines de septembre. Pour chaque parcelle,
les fruits récoltés ont été pesés afin de déterminer le rendement par unité de surface.
3.6.-Expérience antérieure : Marchand et al. (2013)
Marchand et al. (2013) ont mené un essai de fertilisation de P sur six sites pendant trois
ans dans la région Centre-du-Québec. L’essai a été réalisé sur le cultivar ‘Stevens’ avec
trois (3) niveaux de P (0 à 39 ou 99 selon l’année). Au total 202 observations sur les
concentrations foliaires en N, P, K, Mg, Ca, B, Zn, Cu, Mn, Fe ainsi que les rendements
correspondants ont été rapportées par Marchand et al. (2013).

18
3.7.-Analyse des données
3.7.1.-Définition des balances nutritives
Les prélèvements des tissus foliaires ont été analysés en adoptant l’approche
compositionnelle vu qu’il s’agit de données compositionnelles, c’est-à-dire des données
strictement positives faisant partie d’un système et que chaque composante peut être
calculée par la différence entre le système et la somme des autres composantes
(Egozcue et Pawlowsky-Glahn, 2005). Les ratios logarithmiques isométriques (ilr) est
l’approche la plus appropriée pour décrire les balances nutritives des plantes (Parent,
2011). Ainsi, dix (10) contrastes logarithmiques ont été définis à partir des 10 éléments
(C-N-P-K-Ca-Mg-Mn-Cu-Zn-Fe-B) analysés dans les tissus foliaires (Tableau 5).
Tableau 5: Contrastes logarithmiques et partition séquentielle binaire des analyses
foliaires de la canneberge
Ilr N P K Ca Mg B Cu Zn Mn Fe Fv
1 -1 -1 -1 -1 -1 -1 -1 -1 -1 -1 1 1 10
2 1 1 1 1 1 1 -1 -1 -1 -1 0 6 4
3 1 1 1 1 1 -1 0 0 0 0 0 5 1
4 1 1 -1 -1 -1 0 0 0 0 0 0 2 3
5 0 0 1 -1 -1 0 0 0 0 0 0 1 2
6 0 0 0 1 -1 0 0 0 0 0 0 1 1
7 1 -1 0 0 0 0 0 0 0 0 0 1 1
8 0 0 0 0 0 0 1 1 -1 -1 0 2 2
9 0 0 0 0 0 0 1 -1 0 0 0 1 1
10 0 0 0 0 0 0 0 0 1 -1 0 1 1
3.7.2.-Transformation des données foliaires en log ratio isométriques (ilr)
Le calcul des ilr a été réalisé en utilisant le package « compositions » de R (R
Development Core Team, 2013). Pour éviter toute redondance, le nombre de balances

19
entre les éléments nutritifs (ilr) doit être égal au nombre d’éléments nutritifs D analysés
dans le tissu foliaire moins un (D-1) pour ainsi assurer l’orthogonalité des contrastes
logarithmiques. Le calcul des ilr est effectué comme suit (Egozcue et Pawlowsky-
Glahn, 2005) :
Où est le nombre de composantes au numérateur, est le nombre de composantes
au dénominateur, est la moyenne géométrique des composantes au numérateur et
est la moyenne géométrique des composantes au dénominateur. Le coefficient
permet de normaliser les balances orthogonales en balances orthonormales.
Comme les balances sont orthogonales dans l’espace euclidien, la distance de
Mahalanobis peut être calculée comme indice de balance nutritive comme suit (Parent et
al., 2013) :
∗ ∗
Où ∗ est l’ilr de la composition de la population de référence et COV est la matrice
de covariance. Cette population est isolée à l’aide de la procédure itérative de Cate-
Nelson (Nelson et Anderson, 1977) où les paramètres ∗ et COV sont recalculés à
chaque itération jusqu’à ce que le nombre de spécimens vrais négatifs ne change plus.
3.7.3.-Partition de Cate-Nelson
La procédure de partition de Cate-Nelson se fait en plusieurs étapes (Nelson et
Anderson, 1977). La première consiste à ordonner les observations en ordre croissant
selon leur distance de Mahalanobis calculée avec des normes nutritives provisoires
(moyennes et écart-types) en utilisant une sous-population à haut rendement
(initialement, le quartile supérieur basé sur les rendements) comme composition de
référence. La seconde étape est de maximiser la somme des carrés des valeurs de
rendement comme suit :

20
∑ ∑
Où Y est le rendement, k est un compteur d’incrémentation allant de 1 à n, le nombre
total d’observations, et CF un facteur de correction calculé comme suit : ∑ ⁄ .
La partition donne lieu à quatre (4) groupes d’observations. Les TN (True Negative en
anglais appelés Vrais Négatifs en français) sont les observations à haut rendement qui
sont diagnostiquées balancées (c’est-à-dire en dessous de la distance multivariée
critique) ; les FN (False Negative en anglais) appelés « Faux Négatifs » en français sont
les observations à faible rendement incorrectement diagnostiquées comme balancées en
raison de facteurs limitants autres que ceux pris en compte. Ce groupe représente
l’erreur de type II. Les FP (False Positive, « Vrais Positifs » en français) représentent
l’erreur de type I. Ce sont les observations à haut rendement incorrectement classées
dans le groupe des spécimens non balancés (c’est-à-dire au-dessus de la distance
multivariée critique) à cause de cas de consommation de luxe ou de contamination. Les
TP (True Positive, « Faux Positifs » en français) sont les observations à faible
rendement et correctement diagnostiquées non balancées parce qu’au moins un élément
nutritif est limitant pour la croissance.
3.7.4.-Interprétation du test de diagnostic
Le test de diagnostic est interprété à l’aide des critères de performance suivant (Parent et
al., 2013) :
NPV (Negative Predictive Value) est la probabilité pour qu’un diagnostic équilibré
retourne un haut rendement. Elle est calculée comme suit : TN/(TN+FN).
PPV (Positive Predictive Value) est la probabilité pour qu’un diagnostic
déséquilibré retourne un faible rendement. Elle est égale au rapport suivant :
TP/(TP+FP).
Précision (Accuracy) est la probabilité pour qu’une observation soit correctement
diagnostiquée comme équilibrée ou déséquilibrée. Elle est calculée comme suit :
(TN+TP)/(TN+FN+TP+FP).
Spécificité (Specificity) est la probabilité pour qu’une observation à haut rendement
soit équilibrée et est calculée comme suit : TN/(TN+FP).

21
Sensibilité (Sensitivity) est la probabilité pour qu’une observation à faible
rendement soit diagnostiquée en déséquilibre nutritif. Ce critère est calculé comme
suit : TP/(TP+FN).
Le NPV, la précision et la sensibilité sont des critères qui permettent d’identifier des
carences nutritives potentielles et d’indiquer si d’autres facteurs autres que ceux pris en
compte dans le diagnostic limitent la croissance de la plante. Par conséquent, le modèle
avec le NPV, la précision et la sensibilité les plus élevés sera considéré comme le plus
approprié à des fins diagnostiques. Le PPV et la spécificité permettent de détecter de
problèmes potentiels de consommation de luxe, de concentration sous-optimale et de
contamination.
La distribution des ilr des TN a été examinée graphiquement avec le logiciel Orange
3.2. Les intervalles de confiance des ilrs des TN (P < 0,01) ont ensuite été soumis à
100 000 combinaisons de Monte Carlo dans R (Parent, 2016) puis transformés en
valeurs de concentration (fonction ilrInv du module compositions de R). Les valeurs
maximales et minimales des concentrations ont été retenues comme intervalles de
concentrations. Ceci évite de calculer des statistiques sur des valeurs de concentration
biaisées par des interactions.
3.7.5.-Apprentissage automatique
Cette méthode appelée « apprentissage par arbre de décision », utilisée notamment en
fouilles de données (data mining) et en apprentissage automatique (machine learning),
consiste à utiliser un arbre de décision comme modèle prédictif permettant d’évaluer la
valeur d’une caractéristique d’un système depuis l’observation d’autres caractéristiques
du système en question (Rokach et Maimon, 2008). Un arbre de décision de type «
classification » est utilisé afin de différencier les observations équilibrées à haut
rendement et celles non équilibrées à faible rendement. Pour ce faire, le module
orngtree du logiciel Orange Canvas a été utilisé. Le modèle de régression k nearest
Neighbors du logiciel Orange Canvas a permis de prédire la performance du système
(rendement) à partir des valeurs de ses différentes composantes (ilr).

22
Chapitre 4 : Résultats et Discussions
4.1.-Propriétés des sols et productivité
Les sols sont des sols acides (pH CaCl2 < 4,5), contenant plus de 85 % de sable, tous de
la même classe de fertilité potassique (classe 1 : 0-115 kg KM-3 ha-1) et leurs indices de
saturation en P (ratios molaires [P/(Al+Fe)]M-3 sont inférieurs à la valeur limite
environnementale de 11 % (Parent et Gagné, 2010) (Tableau 6).
Tableau 6. Propriétés des sols des sites de canneberge en Mai 2014 (Mai 2001 pour les
sites de Marchand et al., 2013)
Propriété de sol
Site
A9
10
45
9
Marchand et al.
(2013)†
Sable, % 91,5 92,7 93,8 92,8 89,3
Limon, % 4,9 5 3,7 4,4 7,1
Argile, % 3,6 2,3 2,6 2,8 3,6
Al, mg kg-1 1488 594 1240 1510 1847
Ca, mg kg-1 15 32 70 106 ND
Cu, mg kg-1 0,9 1,8 1,1 1,7 ND
Fe, mg kg-1 238,7 177,4 304,3 281,5 248,6
K, mg kg-1 10,7 8,5 26,1 16,6 ND
Mg, mg kg-1 3,9 10,7 6,6 9,8 ND
Mn, mg kg-1 0,4 1,5 0,7 1,2 ND
P, mg kg-1 90,8 62,9 100,3 164,3 119
Zn, mg kg-1 0,4 0,6 1 1,9 ND
pH (CaCl2) 4,1 4,2 4,2 4,3 4,3
P/ (Al+Fe) M-3, % 4,9 8,2 6,4 8,8 5,3
ND Valeur non disponible
† Moyenne de cinq sites
Les rendements des nouveaux essais variaient de 6 à 74 Mg ha-1. La culture n’a répondu
de façon significative qu’aux fertilisations azotée et potassique (Jamaly, 2017). Pour les
deux (2) années de l’expérience, le rendement a augmenté suivant un effet linéaire

23
significatif avec l’augmentation de la dose d’azote (P < 0,0001 en 2014 et 2015). Dans
le cas du potassium, une augmentation linéaire significative a été obtenue en 2014 (P <
0,05). En 2015, aucune différence significative n’a été décelée entre les doses de
potassium appliquées. Les traitements se sont comportés de la même façon sur tous les
sites (interaction site*traitement non significative). Les variations de rendement sont
attribuables en partie au statut nutritif différentiel des plantes de canneberge sous les
divers régimes nutritifs.
4.2.-Diagnostic foliaire
4.2.1.-Partition de Cate-Nelson
La partition de Cate-Nelson a divisé les observations en deux (2) sous-populations à un
seuil de rendement de 35 Mg ha-1. Le compromis optimal entre spécificité et sensibilité
a été trouvé à 80 % pour les deux (spécificité et sensibilité), ce qui correspond à un seuil
de prédiction (distance de Mahalanobis) de 4,65. En somme, quatre groupes ont découlé
de la partition (Fig. 3). Les observations ont été réparties comme suit : 180 vrais
négatifs (TN), 207 vrais positifs (TP), 55 faux négatifs (FN) et 47 faux positifs (FP).
Les TN regroupent les observations à haut rendement (>35 Mg ha-1) qui sont
correctement diagnostiquées en équilibre nutritif (distance de Mahalanobis inférieure à
4,65). Les FP sont les observations à haut rendement incorrectement diagnostiquées en
déséquilibre nutritif (distance de Mahalanobis supérieure à 4,65). Ces observations
indiquent une consommation de luxe de nutriments ou des cas de nutrition sous-
optimale ou de contamination. Les cas de consommation de luxe par la canneberge
concernent surtout le potassium (Roper, 2010). D’autre part, les contaminations (Cu, Zn
et Mn) proviennent des applications de fongicides. De plus, la canneberge est cultivée
sur une large variété de sols, des sols organiques aux podzols sableux, avec de larges
gammes de Mn et de Fe disponibles (Marchand et al., 2013).

24
Figure 3. Partition de Cate-Nelson entre la distance de Mahalanobis et le rendement en fruits de la canneberge.
Les TP sont les observations à faible rendement (< 35 Mg ha-1) et qui sont correctement
diagnostiquées en déséquilibre nutritif. Les FN représentent les bas rendements qui sont
incorrectement diagnostiquées en équilibre nutritif. Au moins un autre facteur que ceux
pris en compte dans le diagnostic nutritif limite le rendement de ces observations
(Marchand et al., 2013). Un grand nombre de FN sont souvent dus à de petites
variations climatiques et à des changements naturels ou pathologiques au niveau de la
plante. Ils peuvent aussi avoir pour cause le caractère biennal de la canneberge qui
alterne des années à haut rendement (on-year) et à faible rendement (off-year)
(Monselise and Goldschmidt, 1982 ; Parent et al., 2013). La taille et le sablage servent à
réduire l’effet biennal sur le rendement (Roper, 2006).
La majorité des spécimens sont correctement diagnostiqués (précision ou accuracy de
80 %) par la distance de Mahalanobis comme indice de diagnostic. Plus de 80 % des
spécimens diagnostiquées non balancés avaient un rendement inférieur au seuil de
rendement critique de 35 Mg ha-1 (PPV=80 %). D’autre part, plus de trois quarts des

25
spécimens balancés ont eu un rendement supérieur au seuil de rendement critique
(NPV=76%).
4.2.2.-Intervalles de suffisance nutritive
Le groupe des TN étant le groupe équilibré à haut rendement, on peut établir à partir de
ces observations des intervalles de suffisance nutritive pour avoir de bon rendement.
Des intervalles de référence pour les balances nutritives sont proposés dans le tableau 7.
Tableau 7. Intervalles de confiance (P < 0,01) des balances (ilr) pour les vrais négatifs (TN)
Balance Minimum Moyenne Maximum
[Fv | B,Ca,Cu,Fe,K,Mg,Mn,P,Zn,N] 7,47 7,50 7,53
[Cu,Fe,Mn,Zn | B,Ca,K,Mg,P,N] -6,32 -6,23 -6,15
[Fe,Mn | Cu Zn] 3,07 3,13 3,20
[Fe | Mn] -0,78 -0,68 -0,57
[Cu | Zn] -1,33 -1,24 -1,16
[B | Ca,K,Mg,P,N] -3,99 -3,93 -3,87
[P,N | Ca,K,Mg] -0,33 -0,31 -0,29
[P | N] -1,73 -1,71 -1,69
[K | Ca,Mg] 0,17 0,19 0,22
[Ca | Mg] 1,03 1,05 1,08
Les balances nutritives décrivent les interactions qui peuvent survenir entre les éléments
nutritifs au niveau du sol ou des tissus des plantes. Ces interactions interviennent quand
l’ajout ou l’absorption d’un élément affecte l’absorption ou l’utilisation d’autres
éléments (Fageria, 2001). La balance [P | N] décrit l’interaction positive entre le N et P.
Wilkinson et al. (2000) ont rapporté que le N peut augmenter le prélèvement du P par la
plante par une augmentation de la croissance racinaire et de la capacité des racines à
absorber et à acheminer le P vers les autres parties et par une baisse du pH du sol
résultant de l’absorption de NH4+ augmentant ainsi la solubilité du P. Le N et le P
interagissent aussi avec le Ca, le Mg et le K (balance [P, N | Ca,K,Mg]) (Wilkinson et
al., 2000).

26
L’augmentation du niveau de N dans la plante peut augmenter (si le niveau de K est
élevé dans le milieu) ou diminuer (si K est faible dans le milieu) la concentration de K
dans les tissus des plantes (Dibb et Thompson, 1985). Le Mg est un activateur des
enzymes kinases et active la plupart des réactions impliquant le transfert des
phosphates. De ce fait, on s’attend à une interaction positive entre le P et le Mg (Fageria
et al., 2011) dans les tissus de la plante.
Quant au K, à concentration élevée, il a des effets antagonistes potentiels sur
l’absorption du Ca et du Mg (Fageria, 2001). La diminution du prélèvement du Ca avec
l’augmentation de la concentration du K est liée à la compétition entre les deux
éléments due aux propriétés physiologiques des ions Ca2+ et K+ (Johansen et al., 1968).
L’effet antagoniste du K à concentration élevée sur le prélèvement du Mg peut être
interprété comme le résultat de la compétition entre le K et le Mg pour se lier à des
composés métaboliques (Omar et Kobbia, 1966). Ces effets sont décrits par la balance
[K | Ca,Mg].
Les interactions entre les éléments majeurs et les éléments mineurs ([Cu,Fe,Mn,Zn |
B,Ca,K,Mg,P,N]; ainsi que celles entre les éléments mineurs ([Cu | Zn]; [Fe | Mn];
[Fe,Mn | Cu Zn]) sont aussi décrites par les balances. Dans le cas où de grandes
quantités de phosphore sont appliquées, une consommation de luxe de P (Tagliavini et
al., 1991) peut faire augmenter les ratios P/Fe (De Kock et al., 1965) et P/Zn
(Loneragan et al., 1979 ; Loneragan et al., 1982) dans les tissus de plante et a souvent
été associée à des symptômes de déficience en ces deux éléments mineurs (Murphy et
al., 1981).
Les interactions de N avec les micronutriments sont dues aux changements de pH dans
la rhizosphère et de la forme de N utilisée (Fageria, 2001). Dans le cas de la canneberge,
l’azote ammoniacal NH4+ est la forme utilisée. L’absorption cationique du NH4
+ fait
baisser le pH du sol et du coup augmente le prélèvement de certains micronutriments
notamment le fer (Wilkinson et al., 2000). D’autre part, quand l’azote stimule la
croissance, cela peut causer une augmentation de la demande pour certains
micronutriments (bore, manganèse, molybdène) et une possible déficience (Fageria,
2001). Il peut aussi avoir une compétition cationique entre le NH4+, Cu2+, Zn2+ et Mn2+
pour les sites d’absorption, de transport ou dans les tissus de la plante (Robson et

27
Pitman,1983) vu que la somme des cations dans les tissus de la plante est à peu près
constante (Dibb et Thompson, 1985).
La balance [Fe | Mn] décrit l’importance de maintenir un ratio Mn/Fe équilibré. Un
excès de Mn augmente ce ratio et peut induire une déficience en fer (Lee, 1972) et dans
les sols où des problèmes de toxicité due au fer existe, le prélèvement du Mn peut être
fortement réduit (Fageria, 2001).
Les « ilr » sont utilisées à des fins de diagnostic nutritif. Pour avoir une appréciation en
termes de carence nutritive, d’adéquation, de consommation de luxe ou d’excès, les
concentrations nutritives sont plus familières. En effet, l’utilisation des ratios
logarithmiques isométriques ne permet pas de savoir si le niveau d’un élément nutritif
est élevé, adéquat ou insuffisant. Ceci est une faiblesse apparente de l’interprétation des
ratios nutritifs (Walworth and Summer, 1987 ; Wilkinson et al., 2000 : Marschner,
2011).
L’approche des balances intègre les balances et les concentrations nutritives dans un
modèle physiologiquement cohérent et statistiquement non biaisé, où les concentrations
nutritives peuvent aider à apprécier les résultats des analyses statistiques effectuées sous
forme de ratios logarithmiques isométriques (Parent et al., 2013).
L’utilisation des ilr permet un diagnostic nutritif non biaisé avec des valeurs de
concentrations nutritives en soutien pour fournir une appréciation des résultats (carence
relative, adéquation ou excès par rapport aux TN). Les intervalles de concentrations
dérivées des simulations de Monte Carlo sont présentés au tableau 8 à titre de
comparaison aux normes nutritives foliaires déjà établies qui ont été présentées au
tableau 1.
28
Tableau 8. Intervalles de suffisance nutritive des tissus de canneberge
Éléments
Minimum Moyenne Maximum
%
Azote (N) 1,00 1,08 1,17
Phosphore(P) 0,090 0,097 0,100
Potassium (K) 0,46 0,50 0,54
Calcium (Ca) 0,76 0,83 0,90
Magnésium (Mg) 0,17 0,19 0,20
mg kg-1
Bore (B) 46,27 51,4 57,04
Fer (Fe) 84,58 98,83 116,53
Manganèse (Mn) 218,07 256,76 301,82
Zinc (Zn) 14,47 16,73 19,33
Cuivre (Cu) 2,5 2,9 3,3
Contrairement aux méthodes traditionnelles de diagnostic qui incluent toutes les
observations à haut rendement (équilibrées et non équilibrées) pour établir des normes
nutritives foliaires, la méthode utilisée dans ce travail inclut uniquement les
observations à haut rendement et équilibrées (TN). Les cas de consommation de luxe,
de concentration sous-optimales et de contamination (FP) ne sont pas pris en compte
dans les intervalles proposés. Ainsi, les intervalles de concentrations présentés au
tableau 8 sont plus restreints que ceux retrouvés dans la littérature et présentés au
tableau 1. Toutefois, seuls les intervalles de N et K sont fiables vu que les fertilisations
azotée et potassique sont les seules auxquelles la culture a répondu de façon
significative. Les intervalles nutritifs pour les autres éléments caractérisent de bons
rendements sans nécessairement montrer des valeurs critiques.
Un élément important qui ne figure pas dans le diagnostic est le soufre. C’est l’un des
trois (3) éléments secondaires requis par la plante pour s’assurer d’une croissance
normale et saine ; les deux autres étant le calcium et le magnésium. Il existe un équilibre
significatif entre l'azote et le soufre : sans une quantité suffisante de soufre, les plantes
ne peuvent pas utiliser l'azote et les autres nutriments de façon efficace afin d'atteindre
leur plein potentiel. De ce fait, les engrais apportés à la canneberge (sulfate de K et de

29
Mg, sulfate d’ammonium, sulfate de cuivre, sulfate de K, sel d’Epsom) ont des teneurs
non négligeables en soufre.
Vu que le soufre n’était pas pris en compte dans les travaux de Marchand et al. (2013),
il a été enlevé du modèle de diagnostic initial. Toutefois, dans le diagnostic initial
réalisé seulement avec les données foliaires des expériences de 2014 et 2015 (excluant
les données de Marchand et al. (2013)), on a obtenu des intervalles de suffisance
nutritive de 0,08 % à 0,09 % pour cet élément. Cet intervalle de suffisance se trouve à
l’intérieur de la norme établie par Roper (2011) qui est de 0,08 % à 0,25 %.
Les écarts observés entre les intervalles de concentration de la littérature (tableau 1) et
ceux de ce travail (tableau 8) peuvent résider dans divers facteurs qui prévalaient lors
des expériences. L’un des premiers facteurs qui peut affecter les intervalles de
concentrations est le cultivar sur lequel l’expérience a été réalisée. Il est connu que les
concentrations critiques varient d’une espèce à une autre (Chapman, 1966) même si les
mêmes organes sont comparés au même stade physiologique (Marschner, 2011). De
même, les cultivars d’une même espèce fruitière donnée varient considérablement en
leur capacité à extraire et à utiliser les éléments nutritifs du sol ou des milieux nutritifs
(Vose, 1963). Ces variations sont principalement basées sur des différences dans le
métabolisme et la composition des plantes (Marschner, 2011). Par exemple, Kessler
(1961) a montré des différences marquées dans les concentrations critiques du zinc pour
deux variétés de pommes, et Holford (1968) dans les concentrations critiques de l’azote
dans des cultivars de canne à sucre. Bien que plus d’informations puissent sembler
nécessaires avant de valider ces conclusions pour la canneberge, il y a lieu de penser
que les différences variétales pourraient causer des écarts entre les intervalles de
concentration quand on sait que la grande majorité des recherches ayant conduit à
l’élaboration des normes nutritives pour la canneberge ont été réalisées aux États-Unis
où un grand nombre de cultivars sont utilisés (Early Blacks, Howes, Stevens, Searles,
Ben Lear, McFarlin). Au Québec, le cultivar Stevens, sur lequel cette expérience a été
réalisée, est de loin, le plus populaire.
L’utilisation d’engrais liquides (concentrations élevées en bore et zinc), un drainage
inadéquat du sol (teneur en Mn dépasse 300 mg Mn kg-1), des sols saturés ou secs, une
température élevée, des dommages liés à l’utilisation d’herbicides sont parmi les autres

30
conditions expérimentales pouvant affecter les intervalles de concentrations des
éléments nutritifs (Davenport et al., 1995).
Les fluctuations dans les facteurs environnementaux tels que la température et la teneur
en eau du sol peuvent aussi affecter considérablement les concentrations en éléments
nutritifs des tissus foliaires (Marschner, 2011 ; Bates, 1971). Ces facteurs influencent à
la fois la disponibilité et le prélèvement des éléments nutritifs par les racines ainsi que le
taux de croissance des racines (Marschner, 2011). Divers auteurs ont relié l’effet des
facteurs environnementaux aux concentrations foliaires de certaines cultures. Burr
(1961) a montré que les températures du sol et de l’air ainsi que l’intensité de la lumière
affectent la concentration de l’azote dans les tissus foliaires de la canne à sucre. Les plus
faibles concentrations en azote ont été enregistrées pour une forte intensité lumineuse,
des sols froids et un air chaud et les concentrations les plus élevées dans le cas de sols
chauds, un air froid et une faible intensité lumineuse. Des tendances similaires ont été
retrouvées pour le phosphore foliaire de la canne à sucre. Les variations des facteurs
environnementaux sont beaucoup plus importantes d’une région à une autre (Bates,
1971). Ces variations sont susceptibles d’être à la base des écarts qui existent entre les
intervalles de concentrations élaborés ici au Québec et les standards de concentrations
nutritives qui ont été élaborés aux États-Unis.
4.2.3.-Comparaison entre TN et TP
L’arbre de classification conçu à l’aide du module Orngtree du logiciel Orange 3.2 a
permis de détecter les balances qui diffèrent entre les TN et les TP (Figure 4). La
précision de la classification est très élevée à 92 %. Cet arbre est un outil permettant
d’identifier les balances nutritives critiques pour l’obtention d’un rendement > 35 Mg
ha-1.

31
Figure 4. Arbre de Classification des rendements élevés (HY) et bas (LY) au niveau de
35 Mg ha-1.
La première balance qui semble limiter le rendement des TP est la balance [Ca | Mg].
La totalité des 72 observations de l’échantillon considéré (271 observations) dont la
balance [Ca | Mg] est inférieure ou égale à 0,881 (sous la borne inférieure de 1,03 de
l’intervalle optimal, tableau 7) sont des TP. Quand la balance [Ca | Mg] est supérieure à
0,881, la probabilité d’avoir des observations TN est de 59,3 %. Cette probabilité
dépend des autres balances critiques. La deuxième balance dans la hiérarchie des
balances limitantes est [K | Ca, Mg]. Quand cette balance est inférieure ou égale à 0,343
(près de la borne supérieure au tableau 7), 87,2 % des observations (dont [Ca | Mg] >
0,881 et [K | Ca, Mg] < ou = 0,343) sont équilibrées et à haut rendement. 80,5 % des
observations à [Ca | Mg] > 0,881 et [K | Ca, Mg] > 0,343 sont du groupe des TP. Une
autre balance importante dans l’obtention d’un bon rendement est la balance [Cu | Zn].
Lorsqu’elle est supérieure à -1,498 (près de la borne inférieure de l’intervalle optimal,
tableau 7), les observations respectant les critères de bon rendement des autres balances
critiques ont près de 97 % de probabilité pour être du groupe des TN. L’arbre de
classification peut être un bon outil d’aide à la décision en permettant d’identifier les

32
balances critiques à équilibrer afin de maximiser ses chances d’obtenir le plus
d’observations possibles du groupe des TN.
Un arbre de classification en fonction des concentrations nutritives peut être informatif
pour savoir quels éléments nutritifs sont à la base du déséquilibre des balances. Cet
arbre permet de différencier les concentrations nutritives du groupe des TN de celles des
TP (Figure 5).
Figure 5. Arbre de classification des vrais négatifs et des vrais positifs selon les
concentrations.
La classification des concentrations renseigne sur les éléments nutritifs à considérer
pour équilibrer les balances critiques. Par exemple, la balance [Ca | Mg] plus élevée
dans les TN que les TP peut s’expliquer par une concentration de Mg plus élevée (>
0,188 % ou 1880 mg kg-1) dans les TN. Cette valeur limite de 0,188 % est d’autant plus
fiable qu’elle est sensiblement égale à la moyenne de l’intervalle de suffisance établi
pour le magnésium (tableau 8). Ceci n’est toutefois pas le cas de Ca et K. Comme les

33
arbres de classification et de régression sont utilisés pour la première fois pour effectuer
des diagnostics nutritifs de cultures fruitières, des recherches additionnelles sont
nécessaires pour intégrer l’information pertinente provenant de la classification Cate-
Nelson et ces nouveaux outils numériques d’auto-apprentissage.
En s’inspirant des arbres de classification, on peut identifier les mesures appropriées
pour corriger les balances nutritives. Les mesures correctives incluant un élément
peuvent influencer toutes les balances qui intègrent cet élément. Par conséquent, l’effet
des mesures correctives sur un système si complexe doit être confirmé par
expérimentations (Parent et al., 2013).
Les méthodes de Cate-Nelson et des arbres de classification ne produisent donc pas
toujours les mêmes résultats. Il devient alors intéressant de vérifier si le rendement peut
être prédit par une régression (Figure 6).
Figure 6. Rendement réel vs rendement prédit se basant sur les ilr des groupes TN et
TP.
Les résultats de la figure 6 montrent que l’arbre de régression par les k= 5 voisins les
plus près peuvent bien prédire (R2 = 0,829) le rendement de la canneberge avec les
balances nutritives, confirmant ainsi que celles-ci constituent une voie prometteuse pour
les producteurs dans leur quête d’optimiser la fertilisation et d’augmenter le rendement
0
10
20
30
40
50
60
70
0 10 20 30 40 50 60 70 80
Ren
dem
ent p
rédi
t (M
g/ha
)
Rendement (Mg/ha)
R2=0,829

34
de la canneberge. Les producteurs ont tout intérêt à atteindre et à maintenir l’équilibre
au niveau des différentes balances nutritives en vue d’obtenir des rendements supérieurs
au seuil de 35 Mg ha-1. Pour cela, ils doivent connaître les doses d’engrais à appliquer
pour atteindre cet équilibre. D’où la nécessité d’effectuer des recherches additionnelles
sur la sensibilité conjointe des rendements et des balances nutritives à la fertilisation
afin de faire des recommandations d’engrais.

35
Conclusion
Cette étude a permis d’établir des normes nutritives foliaires dans la canneberge en
utilisant de nouvelles méthodes de diagnostic nutritif. Les données de concentrations
foliaires ont été transformées en indices non biaisés de balances nutritives (ilr). Des ilr
standards ont été définis à partir de la sous-population équilibrée à haut rendement (> 35
Mg ha-1).
Les ilr, valeurs non biaisées utilisées à des fins de diagnostic, ont permis de générer des
concentrations plus familières pour apprécier les résultats en termes de pénurie, d’excès,
de carence ou d’adéquation nutritive. Toutefois, seuls les intervalles de concentrations
pour le K et le N sont fiables vu que les fertilisations K et N sont les seules auxquelles
la culture a répondu. Pour les autres éléments, les intervalles caractérisent de bons
rendements sans nécessairement montrer des valeurs critiques.
Cette méthode de diagnostic exclut les cas de consommation de luxe et de
contamination nutritive des intervalles. Ainsi, ces derniers sont plus restreints que les
intervalles de concentrations de la littérature. En plus, les différences entre les
intervalles obtenus et ceux de la littérature reflètent les conditions édaphiques locales et
la gestion de la fertilisation.
Les spécimens à faible rendement (TP) ont montré des concentrations en K et Mg
faibles par rapport aux spécimens à haut rendement (TN). Des mesures correctives
devraient donc viser ces éléments afin d’équilibrer principalement les balances [Ca |
Mg] et [K | Ca, Mg] qui constituaient les principales balances limitantes à l’obtention
d’un bon rendement (> 35 Mg ha-1). Ces mesures correctives peuvent toutefois
influencer toutes les balances incluant les éléments K et Mg. Il est donc impératif de
confirmer leurs effets par expérimentation.
Bien que cette étude fournisse des intervalles de balances nutritives et de concentrations
pour obtenir de bons rendements, elle n’indique pas les doses d’engrais à appliquer pour
un équilibre nutritif. D’ailleurs, la culture n’a pas répondu à la majorité des fertilisations
(P, Mg, B, Cu). Une étude additionnelle sur la sensibilité conjointe des rendements et
des balances nutritives à la fertilisation devrait être menée afin de recommander les
doses d’engrais pour atteindre l’équilibre nutritif.

36
Références bibliographiques
Aitchison, J. (1986). The Statistical Analysis of Compositional Data. London: Chapman and Hall.
Alexander, M. (1980). “Effects of acidity on microorganisms and microbial processes in soil,” in Effects of acid precipitation on terrestrial ecosystems, eds. T.C. Hutchinson et M. Havas (Boston, Massachussetts: Springer US), 363-374.
Bajwa, R., et D. J. Read. (1986). Utilization of mineral and amino N sources by the ericoid mycorrhizal endophyte Hymenoscyphus ericae and by mycorrhizal and non-mycorrhizal seedlings of Vaccinium. Transactions of the British Mycological Society 87, 269-277.
Barak, P., J. D. Smith, A. R. Krueger et L. A. Peterson. (1996). Measurement of short‐term nutrient uptake rates in cranberry by aeroponics. Plant, Cell & Environment 19, 237-242.
Barnhisel, R. et P. Bertsch (1982). “Digestion with perchloric-nitric acids,” in Methods of soil analysis, ed. Page, A.L. (Madison, Wisconsin: American Society of Agronomy 9), 279-280.
Bates, T. E. (1971). Factors affecting critical nutrient concentrations in plants and their evaluation: a review. Soil Science 112, 116-130.
Beaufils, E. R. (1973). “Diagnosis and recommendation integrated system (DRIS),” in Soil Science, Bulletin, 1 (Pietermaritzburg, South Africa: University of Natal).
Burr, G. O. (1961). “Growth and composition of sugar cane as influenced by nitrogen,” in Plant Analysis and Fertilizer Problems, ed. W. Reuther (Washington: American Institute of Biologics Sciences), 327-337.
Chapman H.D. (1966). “Zinc”, in Diagnostic criteria for plants and soils, ed. H. Chapman (Riverside: University of California, Division of Agricultural Sciences), 484-499.
Davenport, J., C. DeMoranville, K Hart, L. Patten, T. Peterson, T. Planer, A. Poole, T. Roper, et J. Smith. (1995). Cranberry Tissue Testing for producing beds in North America. Oregon State University, Corvallis.
Davenport, J.R. et J. Provost. (1994). Cranberry tissue nutrient levels as impacted by three levels of nitrogen fertilizer and their relationship to fruit yield and quality, Journal of Plant Nutrition 17, 1625-1634.
De Kock, P.C. et A. Wallace. (1965). Excess phosphorus and iron chlorosis. California Agriculture 19, 3-4.
De Wit, C.T. (1992). Resource use in agriculture. Agricultural Systems 40, 125-151.
DeMoranville, C. (1992). Cranberry nutrients, phenology, and N-P-K fertilization. Doctoral Dissertation, Department of Plant and Soil Science, University of Massachusetts, Amherst, Massachusetts.

37
DeMoranville, C. (2010). Nutrient management in cranberry production, University of Massachusetts-Amherst. Available at: http://scholarworks.umass.edu/cgi/viewcontent.cgi?article=1018&context=cranberrybmp.
DeMoranville, C.J., et J.R. Davenport. (1997). Phosphorus forms, rates and timing in Massachussets cranberry production. Acta Horticulturae 446, 381-388.
Dibb, D. W. et W. R. Thompson. (1985). “Interaction of potassium with other nutrients,” in Potassium in agriculture, ed. R.D. Munson (Madison, Wisconsin: American Society of Agronomy, Crop Science Society of America and Soil Science Society of America), 515-533.
Eaton, G. W. (1978). Floral induction and biennial bearing in the cranberry. Fruit Varieties Journal 32, 58–60.
Eaton, G.W. (1971). Effect of N, P, and K fertilizer applications on cranberry leaf nutrient composition, fruit color and yield in a mature bog. American Society of. Horticultural Sciences 96, 430-433.
Eaton, G.W., et C.N. Meehan. (1973). Effects of N, P, and K fertilizer on leaf composition, yield, and fruit quality of bearing 'Ben Lear' cranberries. American Society of Horticultural Sciences 98, 89-93.
Egozcue, J. J., et V. Pawlowsky-Glahn (2006). “Simplicial geometry for compositional data,” in Compositional Data Analysis: Theory and Applications, eds V. Pawlowsky-Glahn, G. Mateu-Figueras, et A. Buccianti (London: Geological Society of London), 145–160.
Egozcue, J. J., V. Pawlowsky-Glahn, G. Mateu-Figueras, et C. Barceló-Vidal, (2003). Isometric log-ratio transformations for compositional data analysis. Mathematical Geology 35, 279-300.
Egozcue, J.J., et V. Pawlowsky-Glahn. (2005). Groups of parts and their balances in compositional data analysis. Mathematical Geology 37, 795–828.
Fageria, N.K.; V.C. Baligar et C.A. Jones. (2011). “Diagnostic techniques for nutritional disorders,” in Growth and Mineral Nutrition of Crop Plants, 3rd Edn, ed. CRC Press (Boca Raton, Florida: Taylor & Francis Group), 81-123.
Fageria, V. D. (2001). Nutrient interactions in crop plants. Journal of Plant Nutrition 24, 1269-1290.
Filzmoser, P. et K. Hron, (2009). Correlation analysis for compositional data. Mathematical Geoscience 41, 905–919.
Filzmoser, P., K. Hron, et C. Reimann. (2009). Univariate statistical analysis of environmental (compositional) data : problems and possibilities. Science of the Total Environnement 407, 6100–6108.

38
Foy, C.D. (1984). “Physiological effects of hydrogen, aluminum, and manganese toxicities in acid soil,” in Soil acidity and liming, ed. F. Adams (Madison, WI: Agronomy Monograph), 57-97.
Greidanus, T., L. A. Peterson, L. E. Schrader et M. N. Dana. (1972). Essentiality of ammonium for cranberry nutrition. Journal of the American. Society of Horticultural Science 97, 272-277.
Hendershot, W. H., H. Lalande, et M. Duquette. (1993). “Ion exchange and exchangeable cations,” in Soil sampling and methods of analysis, ed. Carter, M. R. (Boca Raton, FL: Lewis Publishers), 167-176.
Holford, I. C. R. (1968). Nutrient status of sugar cane in relation to leaf nutrient concentration. Animal Production of Science 8, 606-614.
Ingestad, T. (1987). New concepts on soil fertility and plant nutrition as illustrated by research on for-est trees and stands. Geoderma 40, 237-252.
Jamaly, R. (2017). Effet de la fertilisation N, P et K sur le rendement et les paramètres du rendement de la canneberge. Mémoire de maîtrise, Département des Sols et de Génie agroalimentaire, Université Laval, Québec, Canada.
Johansen, C., D. G. Edwards, et J. F. Loneragan. (1968). Interactions between potassium and calcium in their absorption by intact barley plants: Effects of potassium on calcium absorption. Plant Physiology 43, 1717-1721.
Jones, J. B. J., et V. W. Case. (1990). “Sampling, handling, and analyzing plant tissue samples,” in Soil Testing and Plant Analysis, ed. R. L. Westerman (Madison, WI: Soil Science Society of America), 389–427.
Kessler, B. (1961). Ribonuclease as a guide to the determination of zinc deficiency in orchard trees. Plant Analysis and Fertilizer Problems 3, 314-322.
Laflamme, G., M. Rompré, D. Carrier et L. Ouellet. (1989). Étude pédologique du comté de Mégantic. Division de pédologie, service de recherche en sols, Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec, Québec, Canada.
Lagatu, H., et L. Maume. (1935). The variations of the sum N + P205 + K20 per 100 parts of dry material of the leaf of a cultivated plant. Comptes rendus de l’Académie d’Agriculture (Paris, France) 21, 85-92.
Lee, C.R. (1972). Interrelationships of aluminum and manganese on the potato plant. Agronomy Journal 64, 546-549.
Loneragan, J.F.; D.L. Grunes; R.M. Welch; E.A. Aduayi; A. Tengah; V.A. Lazar et E.E. Cary. (1982). Phosphorus accumulation and toxicity in leaves in relation to zinc supply. Soil Science Society of America Journal 46, 45-352.
Loneragan, J.F.; T.S. Grove; A.D. Robson et K. Snowball. (1979). Phosphorus toxicity as a factor in zinc-phosphorus interactions in plants. Soil Science Society of America Journal 43, 966-972.

39
Lopez, J., L.É. Parent, N. Tremblay, et A. Gosselin, (2002). Sulfate accumulation and Ca balance in hydroponic tomato culture. Journal of Plant Nutrition 25, 1585-1597.
Lovell, D., W. Müller, J. Taylor, A. Zwart, et C. Helliwell. (2011). “Proportions, Percentages, PPM: do the molecular bioscience treat compositional data right?” in Compositional data analysis: Theory and Applications, eds. V. Pawlowsky- Glahn et A. Buccianti (New York, NY: John Wiley and Sons), 193–207.
Macy, P. (1936). The quantitative mineral nutrient requirements of plants. Plant Physiology 11, 749-764.
Marchand S., S.É. Parent., J.P. Deland., et L.É. Parent. (2013). Nutrient signature of Quebec cranberry. Revista Brasileira de Fruticultura 35, 292 – 304.
Marschner, H. (2011). Marschner's mineral nutrition of higher plants. London: Academic press.
Monselise, S.P., et E.E. Goldschmidt. (1982). ”Alternate bearing in fruit trees,” in Horticultural reviews 4, ed. J. Janick (Hoboken, New Jersey: John Wiley and sons, Inc.), 128-173.
Munson, R. D., et W. L. Nelson. (1990). "Principles and practices in plant analysis,” in Soil testing and plant analysis, ed. R.L Westerman (Stillwater, Oklahoma: Soil Science Society of America), 359-388.
Murphy, L.S. ; R. Ellis Jr et D.C. Adriano. (1981). Phosphorus-micronutrient interaction effects on crop production. Journal of Plant Nutrition 3, 593-613.
Nelson, L. A. et R. L Anderson. (1977). “Partitioning of soil test-crop response probability,” in Soil testing: correlating and interpreting the analytical results, eds. Peck T.R., J.P. Cope Jr et D.A. Whitney (Madison, WI: American Society of Agronomy), 19-38.
Omar, M.A., et T.E. Kobbia, (1966). Some observations on the interrelationships of potassium and magnesium. Soil Science 101, 437-439.
Parent, L. É. et M. Dafir. (1992). A theoretical concept of compositional nutrient diagnosis. Journal of the American Society for Horticultural Science 117, 239-242.
Parent, L.É. (2011). Diagnosis of the nutrient compositional space of fruit crops. Revista Brasileira de Fruticultura 33, 321-334.
Parent, L.-É. et S. Marchand. (2006). Response to phosphorus of cranberry on high phosphorus testing acid sandy soils. Soil Science of America Journal 70, 1914-1921.
Parent, L.-É., et G. Gagné. (2010). Guide de référence en fertilisation, 2nd ed. Québec, Canada : Centre de référence en agriculture et agroalimentaire du Québec (CRAAQ).
Parent, L.É., W. Natale, et N. Ziadi. (2009). Compositional nutrient diagnosis of corn using the Mahalanobis distance as nutrient imbalance index. Canadian Journal of Soil Science (Ottawa) 89, 383-390.

40
Parent, S.É. (2016). https://github.com/essicolo/AgFun/blob/master/compci.R.
Parent, S.É., L.É. Parent, D.E. Rozane, et W. Natale. (2013). Nutrient balance ionomics: case study with mango (Mangifera indica). Frontiers Plant Science 4, article 449.
Parent, S.-É., L. E. Parent, , D.-E. Rozane, A. Hernandes et W. Natale (2012). “Nutrient balance as paradigm of soil and plant chemometrics,” in Soil Fertility, ed. R. N. Issaka (New York, NY: InTech), 83–114.
Paul, E.A. et F.E. Clark. (1989). Soil microbiology and biochemistry. New York: Academic Press.
Pawlowsky-Glahn, V., et A. Buccianti. (2011). Compositional data analysis: Theory and Applications. New York: John Wiley and Sons.
R Development Core Team. (2013). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Redfield, A. C. (1936). An ecological aspect of the Gulf Stream. Nature 138, 1013.
Robson, A.D., et J.B. Pitman. (1983). “Interactions between nutrients in higher plants,” in Inorganic plant nutrition, eds. Pirson, A. et M.H. Zimmermann (New York: Springer Verlag), 147-180.
Rokach L. et O. Maimon. (2008). Data mining with decision trees: theory and applications. New Jersey, USA: World Scientific Publishing Co Inc.
Rompré M., G. Laflamme, D. Carrier et L. Ouellet. (1984). Étude pédologique du comté d’Arthabaska. Division de pédologie, service de recherche en sols, Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec, Québec, Canada.
Roper, T. (2001). Taking and interpreting soil and tissue samples. Cranberry School 11, 16-23.
Roper, T. (2006). The physiology of cranberry yield. Department of Horticulture, University of Wisconsin, Madison, Wisconsin.
Roper, T. (2010). How much potassium is needed? Cranberry School Proceedings 18, 23-26.
Roper, T.R., et N. Vorsa. (1997). Cranberry: Botany and Horticulture. Horticultural Reviews 21, 215-249.
Rosen, C. J., D. L. Allan, et J. J. Luby. (1990). Nitrogen form and solution pH influence growth and nutrition of two Vaccinium clones. Journal of the American Society for Horticultural Science 115, 83-89.
Schmidt, E.L. (1982). Nitrification in soil. Nitrogen in Agricultural soils. American Society of Agronomy 22, 253-288.

41
Schwertmann, U. (1973). Use of oxalate for Fe extraction from soils. Canadian Journal of Soil Science 53, 244-246.
Stribley, D. P., et D. J. Read. (1974). The biology of mycorrhiza in the ericaceae: the effect of mycorrhizal infection on uptake of 15N from labelled soil by Vaccinium Macrocarpon AIT. New Phytologist 73, 1149-1155.
Tagliavini, M., E.J. Hogue et G.H. Neilsen. (1991). Influence of phosphorus nutrition and root zone temperature on growth and mineral uptake of peach seedlings. Journal of Plant Nutrition 14, 1267-1275.
Ulrich, A. (1952). Physiological bases for assessing the nutritional requirements of plants. Annual Review of Plant Physiology 3, 207-228.
Ulrich, A., et F.J. Hills. (1967). “Principles and practices of plant analysis,” in Soil testing and plant analysis: Part II, ed. G. W. Hardy (Madison, WI: Soil Science Society of America), 11-24.
Vose, P.F. (1963). Varietal differences in plant nutrition. Herbage Abstracts Journal 33, 1-13.
Walworth, J.L., et M.E. Summer. (1987). The Diagnosis and Recommendation Integrated System (DRIS). Advances in Soil Science 6, 149-188.
Wikner, B. (1986). Pretreatment of plant and soil samples a problem in boron analysis: Part I: Plants. Communications in Soil Science and Plant Analysis 17, 1-25.
Wilkinson, S.R.; D.L. Grunes et M.E. Sumner. (2000). “Nutrients interactions in soil and plant nutrition,” in Handbook of Soil Science, ed. M.E. Sumner (Boca Raton, FL: CRC Press), 89-112.
Yarborough, D., J. Harker, et J.D Leiby (1993). Cranberry agriculture in Maine: a grower’s guide. Orono, Maine: University of Maine.
Ziadi, N. et T. S. Tran (2006). “Mehlich 3- extractable elements,” in Soil sampling and methods of analysis, eds. M.R. Carter et E.G Gregorich (Boca Raton, FL: CRC press, Taylor and Francis group), 107-120.