le facteur de croissance vasculaire endothéliale induit les protéines de la matrice...
TRANSCRIPT

Sciences fondamentales
Le facteur de croissance vasculaireendoth�eliale induit les prot�eines de la matriceextracellulaire et l’ost�eopontine dans l’artereombilicale
Manfred Infanger,1 Jirka Grosse,2 Kriss Westphal,2 Annekatrin Leder,2 Claudia Ulbrich,2
Martin Paul,2 Daniela Grimm,2 Berlin, Allemagne
Le facteur de croissance vasculaire endoth�eliale (VEGF) est un m�ediateur mitogene, an-giog�enique, et puissant de la perm�eabilit�e vasculaire. Il joue un role dans les l�esions, contribuea l’oedeme pendant la phase aigue des l�esions tissulaires, et favorise le remodelage pendant lar�ecup�eration. Nous avons r�ecemment montr�e que les taux s�eriques de VEGF des brul�es pr�esen-tant un nombre important de l�esions vasculaires �etaient significativement augment�es. Ici, nous�etudions les effets de VEGF sur des vaisseaux sains trait�es avec une concentration de VEGFcomparable a celle des brul�es graves. Du VEGF 165 (0,2 mL d’une solution a 10 ng/mL) ou unesolution de transport (s�erum physiologique a 0,9%) �etaient mis en contact avec des arteres om-bilicales pendant 90 min a 37 �C. Le cordon �etait ensuite perfus�e pendant 4 hr. Pendant la per-fusion, les parametres fonctionnels et biochimiques �etaient maintenus dans les margesphysiologiques. Les vaisseaux �etaient ensuite analys�es en morphom�etrie, apres coloration aurouge sirius, en microscopie a polarisation, par analyse Western blot, et en immunohistochimie.Par ailleurs, des cellules endoth�eliales humaines cultiv�ees de veine ombilicale (CEVOH) �etaienttrait�ees avec du VEGF ou su s�erum physiologique pendant 90 min et 5,5 hr pour �etudier lesprot�eines de la matrice extracellulaire (MEC) et les r�ecepteurs de la tyrosine kinase. Les arteresombilicales trait�ees au VEGF avaient un oedeme tissulaire significatif et une facilitation de la la-minine et du collagene de type I, III, et IV par comparaison avec les arteres controle. Nous avonsd�etect�e une augmentation de Flt-1, de Flk-1, de l’ost�eopontine, et de la ß1-int�egrine. En cytom�etriede flux, VEGF induisait pr�ecocement la laminine dans les CEVOH. De meme, VEGF induisait unequantit�e plus �elev�ee d’ost�eopontine, de ß1-int�egrine, et des deux kinases du r�ecepteur de latyrosine par les cellules endoth�eliales en 90 min. L’application endoluminale de VEGF augmenteles prot�eines de MEC, l’ost�eopontine, et la production de ß1-int�egrine par l’endoth�elium, tout enentraınant un oedeme tissulaire. VEGF initie le remodelage vasculaire des qu’il produit del’oedeme, meme si le vaisseau cible n’est pas l�es�e. L’ost�eopontine et la ß1-int�egrine, tous deuxinduits par VEGF, peuvent jouer un role important dans le processus de remodelage vasculaire.
DOI of original article: 10.1016/j.avsg.2007.11.002.
M. I. and J. G. ont contribu�e �egalement a ce travail.1Department of Trauma and Reconstructive Surgery, Charit�e-
Universitatsmedizin, Berlin, Allemagne.2Institute of Clinical Pharmacology and Toxicology, Charit�e-
Universitatsmedizin, Berlin, Allemagne.
Correspondance : Daniela Grimm, MD, Institute of ClinicalPharmacology and Toxicology, Charit�e-University Medicine Berlin,CBF, Garystrasse 5, D-14195 Berlin, Allemagne, E-mail: [email protected]
Ann Vasc Surg 2008; 22: 273-284DOI: 10.1016/j.acvfr.2008.05.001� Annals of Vascular Surgery Inc.�Edit�e par ELSEVIER MASSON SAS
296

Vol. 22, No. 2, 2008 VEGF et ost�eopontine 297
INTRODUCTION
Le facteur de croissance vasculaire endoth�elial
(VEGF) est un mitogene sp�ecifique de la cellule
endoth�eliale in vitro et un m�ediateur principal de
l’angiog�enese et de la perm�eabilit�e vasculaire in
vivo.1-6 La prot�eine de 45 kDa se compose de deux
sous-unit�es, qui induisent presque exclusivement la
prolif�eration des cellules endoth�eliales. Cinq iso-
formes humains avec diff�erents nombres d’acides
amin�es (VEGF 121, 145, 165, 189, 206) ont �et�ed�ecrits.7 VEGF 121 et VEGF 165 sont des isoformes
solubles et sont les plus abondants. VEGF 165 est un
stimulateur important de la prolif�eration des cellules
endoth�eliales,8 et VEGF 121 est responsable de
l’augmentation de la perm�eabilit�e vasculaire.9
L’expression de VEGF est control�ee par la
diff�erentiation, la transformation, et la fourniture
d’oxygene. L’effet de VEGF est m�edi�e par deux
kinases du r�ecepteur de la tyrosine, VEGFR-1/flt-1
(fms-like tyrosine kinase-1) et VEGFR-2/flk-1
(kinase de foie foetal)/KDR, qui sont presque
exclusivement sur les cellules endoth�eliales.
Les cellules endoth�eliales de l’intima sont haute-
ment sp�ecialis�ees, avec des fonctions endocrines, exo-
crines, d’adh�esion des cellules, de coagulation, et des
fonctions de transport, et jouent un role important
dans les m�ecanismes du flux sanguin, de la r�egulation
de la coagulation, de l’adh�erence des leucocytes, et de
la croissance des cellules musculaires lissesvasculaires
; elles servent �egalement de barriere a la diffusion
trans-vasculaire des liquides et des corps dissous. Elles
peuvent s’adapter rapidement aux changements
d’environnement.7,10 Quand, par exemple, les tissus
avoisinants sont l�es�es par des brulures ou par l�esions
c�er�ebrales traumatiques, la perm�eabilit�e vasculaire
est augment�ee, avec pour r�esultat un oedeme tissu-
laire local et g�en�eral. VEGF semble impliqu�e dans ces
processus, et Harrigan et coll.11 ont montr�e que la
perfusion intraventriculaire de VEGF favorise l’an-
giog�enese avec un oedeme c�er�ebral minime. Notre
groupe a montr�e que le niveau de VEGF est si-
gnificativement augment�e chez les patients pr�esen-
tant une brulure grave.12
En plus du VEGF, des int�egrines et des consti-
tuants de la matrice extracellulaire (MEC) sont
impliqu�es dans les fuites vasculaires et le remode-
lage. Nous avons montr�e que l’application endolu-
minale de VEGF est salutaire au processus de
cicatrisation en microchirurgie vasculaire et qu’une
augmentation des prot�eines de la MEC accompagne
la cicatrisation d’anastomoses microvasculaires.13
Beaucoup de prot�eines de la MEC, y compris divers
types de collagene, la fibronectine, la laminine, et la
vitronectine, sont d�etectables dans les vaisseaux
sanguins.14-16 Certaines sont des ligands pour les
int�egrines, qui sont des mol�ecules h�et�erodim�eriques
transmembranaires qui se composent de sous-unit�es
a et b. Les r�ecepteurs d’adh�esion de la ß1-int�egrine
controlent la fixation des cellules endoth�eliales a la
MEC et facilitent leur croissance, leur migration, et
leur capacit�e a d�eposer des prot�eines de matrice. Ils
sont des facteurs importants de la rest�enose et de
l’ath�eroscl�erose.17 Le mode d’action de VEGF sur
l’expression des int�egrines et les diff�erents compo-
sants de la MEC reste inconnu.
On sait cependant que les l�esions d’un nombre
consid�erable de vaisseaux, comme cela est observ�echez les patients ayant une brulure d’au moins
30% de leur surface corporelle, d�eclenchent une
expression majeure de VEGF.12 Entre les premiere
et troisieme semaines apres l’accident, les taux
s�eriques de VEGF sont au moins octuples du niveau
normal. Ceci peut etre une raison importante du
d�eces de beaucoup de patients. Par cons�equent,
nous �etions int�eress�es a �etudier les effets des
concentrations �elev�ees de VEGF sur un vaisseau
normal.
Pour �etudier les effets de concentrations patholo-
giques de VEGF sur un vaisseau sain, nous avons
appliqu�e une perfusion ex vivo, qui s’est d�eja av�er�ee
etre un modele utile de perfusion pour un coeur
h�emoperfus�e isol�e fonctionnant ex vivo.18,19
L’appareil permet des �etudes dans des conditions
physiologiques d’�ecoulement et permet l’enregis-
trement en ligne du d�ebit et des pressions. Nous
avons ensuite recherch�e l’oedeme et �evalu�e le
remodelage potentiel. Nous avons d�etermin�e l’ex-
pression des int�egrines, des prot�eines de la MEC, et
des kinases du r�ecepteur de la tyrosine dans l’artere
ombilicale sous l’influence de VEGF. Par ailleurs,
nous avons �etudi�e les effets du VEGF sur les cellules
endoth�eliales humaines de veine ombilicale (CE-
VOH) pour �etudier les changements des prot�eines de
la MEC et prouver que l’agent agit sur l’endoth�elium
ombilical ex vivo et in vitro.
METHODES
Obtention et transport des sp�ecimens
Vingt cordons ombilicaux ont �et�e obtenus de l’Unit�ede N�eonatologie de l’Hopital Universitaire de la
Charit�e (Berlin, Allemagne). Ces cordons ombilicaux
ont �et�e obtenus de femmes en bonne sant�e et normo-
tendues (tension art�erielle normale, absence d’infec-
tion ou de diabete) apres accouchement a terme par
c�esarienne et employ�es pour les exp�eriences de perfu-
sion. Les criteres d’exclusion �etaient les l�esions vascu-
laires et les anomalies du cordon ombilical. Seuls les

298 Infanger et al. Annales de chirurgie vasculaire
segments d’une longueur de 12 cm �etaient choisis.
Juste apres la naissance, le cordon ombilical �etait
soigneusement pr�epar�e et plac�e dans une solution
isotonique. Les cordons ombilicaux �etaient trans-
port�es au laboratoire dans un d�elai de 1 hr (pr�eserv�es
du s�erum physiologique froid [4 �C]).
Pr�eparation des cordons
Apres examen (exclusion d’anomalies, par exemple
une artere ombilicale simple, une plicature ou un
noeud, partiels ou complets), 20 cordons ombilicaux
ont �et�e pr�epar�es. Une partie de ces sp�ecimens a �et�epr�elev�ee en tant que controles non perfus�es (Cnp,
n¼ 20). A l’aide de la microscopie, une artere om-
bilicale et la veine �etaient pr�epar�ees et canul�ees
(cath�eter d’alimentation p�ediatrique). Les cath�eters�etaient fix�es aux vaisseaux avec des sutures (Vicryl
3-0 ; Ethicon, St-Stevens-Woluwe, Belgique). Puis
0,2 mL de VEGF 165 (groupe VEGF n¼ 10, 10 ng/
mL ; Chemicon, Hofheim, Allemagne) ou d’une
solution v�ehicule (groupe Cp n¼ 10, s�erum a 0,9%)
et 0,2 ml de bleu d’Evans (Merck, Darmstadt, Alle-
magne ; 2% dilu�es dans du s�erum physiologique)�etaient inject�es dans le cath�eter art�eriel (groupes
perfus�e). Ensuite, les arteres de cordon ombilical�etaient incub�ees a 37 �C pendant 90 min dans une
chambre de perfusion (temps d’adaptation).
Modele de perfusion du cordon ombilical
Apres adaptation (90 min), le groupe Cp et les cor-
dons ombilicaux du groupe VEGF �etaient reli�es a
l’appareil de perfusion r�ecemment d�ecrit, 18,19 tan-
dis que les 20 sp�ecimens de cordon ombilical non
perfus�es (controles z�ero) servaient de controles
(Cnp). Ensuite, une temp�erature de 37 �C �etait
r�etablie et la perfusion commencait avec une solu-
tion de dialysat humain (1872,3 mL d’eau bidistill�ee
st�erile, 57,1 mL de solut�e d’h�emodialyse SW 239 A9
; Braun, Melsungen, Allemagne) et 70 mL d’une
solution de bicarbonate pour h�emodialyse (Bic
8,4%, Braun). La solution (pH 7,4) contenait
140 mmol/L Na+, 2 mmol/L K+, 1,75 mmol/L Ca2+,
0,75 mmol/L Mg2+, 112 mmol/L Cl-, 32 mmol/L
HCO3-, 84 g/L NaHCO3, 3 mmol/g/L CH3COO-, et
2 g/L de glucose. Selon le protocole, la circulation de
perfusion contenait 750 ml de dialysat et la circu-
lation de dialysat 1250 ml de solution de dialysat.
Tandis que la solution de perfusion �etait perfus�ee
dans le cordon ombilical pendant 4 hr, elle �etait
maintenue a 37 �C par un �echangeur de chaleur
reli�e a un bain d’eau et oxyg�en�e par un d�ebitmetre
(DK800R ; Krohne, Duisburg, Allemagne) avec un
m�elange d’oxygene et d’anhydride carbonique
(95% d’O2 et 5% de CO2). Les donn�ees
h�emodynamiques �etaient surveill�ees avec un mi-
crocontroleur reli�e a un PC avec un logiciel adapt�e(Mediport Biotechnik, Berlin, Allemagne), �evaluant
sans interruption ces parametres : d�ebit, pressions de
perfusion art�erielles et veineuses. Nous avons men�eune analyse continue de la solution en ce qui con-
cerne son pH et sa temp�erature (37 �C), ainsi que des
analyses des gaz en solution pendant l’exp�erience
(ABL 555 ; Radiometer, Copenhague, Danemark).
Par ailleurs, les taux de sodium, de potassium, de
calcium, et de bicarbonate �etaient d�etermin�es dans
le milieu de perfusion.18,19 A 1, 2, et 3 hr, 1 ml de
bleu d’Evans �etait inject�e dans l’artere ombilicale
par l’interm�ediaire d’un robinet a trois voies.
Exp�eriences de culture de cellules
Les CEVOH et le milieu de culture complet pour cel-
lules endoth�eliales (pret a employer) �etait achet�echez Promocell (Heidelberg, Allemagne). Trente
cultures de cellules ont �et�e trait�ees avec le v�ehicule
(s�erum a 0,9%) ou du VEGF 165 (10 ng/mL)
pendant 90 min ou 5,5 hr. Cinq �echantillons de
chaque groupe ont �et�e fix�es avec de l’�ethanol
(70%), et cinq autres �etaient congel�es rapidement
pour l’analyse Western blot.
Etude histologique et microscopie a
polarisation
Pour l’histologie, des coupes transversales du cor-
don ombilical (n¼ 10, chaque groupe) �etaient fai-
tes et color�ees a l’h�ematoxyline �eosine et au rouge
sirius. Une coloration sp�ecifique du tissu conjonctif�etait �egalement faite.13,20 Les coupes �etaient
immerg�es dans du fixatif methacarn, incluses en
paraffine, et sectionn�es en coupe de 3 mm
d’�epaisseur. Une coloration normalis�ee �etait r�ealis�ee
grace a un systeme de Robot-Stainer (Robot-Stainer
HMS 760 ; Microm International, Walldorf, Alle-
magne). Les vaisseaux �etaient examin�es dans un
microscope Zeiss Axiophot (Oberkochen, Allema-
gne). L’endoth�elium, la structure vasculaire, la gel�ee
de Wharton, et l’�epith�elium amniotique �etaient
document�es. La microscopie a polarisation a �et�eex�ecut�ee selon Junqueira et coll.20 Le collagene de
type I apparaıt jaune, orange, ou rouge, alors que le
collagene de type III semble vert.
Immunohistochimie
Les anticorps utilis�es pour cette �etude �etaient dirig�es
contre le collagene de type IV (1:100 ; Dako,
Hambourg, Allemagne), la laminine (kit pr�edilu�ede Sigma, Taufkirchen, Allemagne), le VEGF-A
(1:50, Chemicon), Flk-1, Flt-1 (1:50 pour les deux

Vol. 22, No. 2, 2008 VEGF et ost�eopontine 299
; Santa Cruz Biotechnology, Santa Cruz, CA, Etats-
Unis), et l’ost�eopontine (1:100 ; Developmental
Studies Hybridoma Bank, Department of Biological
Sciences, Universit�e de l’Iowa, Iowa City, IA,
Etats-Unis). Les complexes antigene/anticorps�etaient visualis�es par la technique d’immunopero-
xydase indirecte.21,22
Analyse d’image automatique
L’analyse d’image automatique a �et�e appliqu�ee pour�evaluer quantitativement les changements intersti-
tiels de l’oedeme des tissus de cordon ombilical per-
fus�e et la distribution des prot�eines de MEC avec un
analyseur d’image assist�e par ordinateur (Olympus
Optical, Hambourg, Allemagne).18,19,21,22
Analyse de la laminine en cytom�etrie de
flux
Dans chaque test, des anticorps monoclonaux�etaient ajout�es a 105 cellules qui avaient �et�epr�epar�ees et fix�ees a l’�ethanol (70%) comme
pr�ec�edemment d�ecrit.23 Les cellules �etaient incub�ees
avec les anticorps non-conjugu�es de la laminine
(laminin, Sigma) pendant 60 min a temp�erature
ambiante dans l’obscurit�e et lav�ees trois fois avec une
solution de tampon phosphate contenant du s�erum
foetal de veau a 2%. Les cellules �etaient ensuite
trait�ees pendant 60 min a temp�erature ambiante
avec un antis�erum conjugu�e avec de l’isothiocyanate
de fluoresc�eine anti- immunoglobulines G de lapin
(Dako) et lav�ees. Les suspensions de cellules �etaient
analys�ees avec un cytometre de flux Facscan (Becton
Dickinson, Heidelberg, Allemagne) �equip�e d’un laser
a argon comme pr�ec�edemment d�ecrits.23 Les cellules
exercant des intensit�es de fluorescence au-dessus de
la limite sup�erieure de la distribution controle
n�egative �etaient consid�er�ees positives.
Teneur en eau
Les coupes de cordon ombilical �etaient pes�ees et
s�ech�ees pendant 24 hr a 95 �C. Apres pesage des
coupes seches, le pourcentage de teneur en eau du
cordon ombilical �etait calcul�e comme suit :
%H2O¼ ½ðpoids humide� poids secÞ� 100�=poids humide
Analyse Western blot
A la fin de la perfusion, les arteres �etaient soigneuse-
ment diss�equ�ees de la gel�ee de Wharton avec des
instruments de microchirurgie. L’extraction de
prot�eine, l’�electrophorese sur gel de dod�ecyl
sulfate-polyacrylamide de sodium, et l’immuno-
blotting �etaient effectu�es selon des protocoles
courants.18,19,22,24 Le test �etait fait six fois. Des
anticorps contre les antigenes suivants ont �et�eemploy�es pour cette �etude : collagene de type IV
(1:1.000, Dako), laminine (1:1000, Chemicon), ß1-
int�egrine (1:1000 ; BD Transduction Laboratories,
San Jose, CA, Etats-Unis), ost�eopontine (1:1000,
Developmental Studies Hybridoma Bank), glyce-
rald�ehydes-3-phosphate d�eshydrog�enase (GAPDH,
1:1000 ; Affinity BioReagents, Golden, CO), VEGF
(1:1000, Chemicon), Flk-1, et Flt-1 (1:1000, tous les
deux de Santa Cruz Biotechnology).
Analyse statistique
L’analyse statistique a �et�e faite avec SPSS 11.5
(SPSS, Inc., Chicago, IL, Etats-Unis). Toutes les
donn�ees ont �et�e exprim�ees en tant que moyenne ±�ecart type. Nous avons examin�e tous les parametres
pour des d�eviations de distribution gaussienne par le
test de Kolmogorov-Smirnov et avons compar�e les
cas en employant le test t ou le test de Mann Whit-
ney (en fonction des r�esultats du test de normalit�e).
Les diff�erences �etaient consid�er�ees significatives au
seuil de p< 0,05.
RESULTATS
Apres 90 min d’incubation avec VEGF ou le
v�ehicule, suivie d’une p�eriode de perfusion de 4 hr,
le cordon ombilical �etait imm�ediatement employ�epour d�eterminer la morphologie, la teneur en eau,
les prot�eines de MEC, la ß1-int�egrine, l’ost�eopontine,
et le VEGF et ses kinases du r�ecepteur de tyrosine.
Morphologie, oedeme tissulaire etteneur en eau des cordons
Les �echantillons trait�es avec VEGF montraient un
oedeme tissulaire significativement augment�e avec
extravasation du bleu d’Evans ( p< 0,018, Fig. 1A-
C). Il y avait une diff�erence nette dans l’espace
interstitiel des arteres de cordon ombilical trait�ees
avec le v�ehicule ou avec VEGF et perfus�ees pendant
4 hr. Afin de montrer que l’agrandissement de l’in-
terstitium est du au d�epot d’eau, la teneur en eau des
cordons ombilicaux �etait d�etermin�ee. Les r�esultats
montraient que les cordons ombilicaux trait�es au
VEGF avaient une teneur en eau accrue (groupe
VEGF 94 ± 1%) par comparaison avec les controles
(groupe Cnp 91 ± 1%, groupe Cp 93 ± 1%).
Apres 4 hr de perfusion, la coloration a
l’h�ematoxyline �eosine montrait une morphologie

300 Infanger et al. Annales de chirurgie vasculaire
VEGFCp2 mm2 mm Cp VEGFS
urface d
'extravasatio
n d
u
bleu
d
'Evan
s (cm
2)
0.0
0.1
0.2
0.3
0.4
0.5 P<0,018A B
C
D E
Cnp
50 µm
400 µmCp
50 µm
400 µm VEGF
50 µm
400 µm
I I
IF
Fig. 1. Extravasation du bleu d’Evans de l’artere ombi-
licale trait�ee par solution v�ehicule (A) et de l’artere
trait�ee par VEGF (B). L’oedeme tissulaire a �et�e mesur�epar analyse d’image quantitative et est sensiblement
augment�e par VEGF (C). La coloration a l’h�ematoxyline
�eosine montre une histologie normale de l’artere ombi-
licale au temps z�ero (D) et apres 4 hr de perfusion (E). Le
traitement de F VEGF induit un oedeme tissulaire
mod�er�e dans l’espace interstitiel ( fleches). Grossissement
original �100, encart �400. I, intima.
normale de l’artere ombilicale (Fig. 1D, E). L’en-
doth�elium �etait inchang�e, et les structures de la
gel�ee de Wharton et de l’�epith�elium amniotique
n’�etaient pas chang�ees par la perfusion ou le trai-
tement au VEGF. Cependant, l’oedeme tissulaire�etait �evident dans l’espace interstitiel (Fig. 1F) des
parois art�erielles ombilicales humaines perfus�ees
avec le milieu contenant VEGF.
VEGF change les prot�eines de la MEC
Collagene de types I et III. Pour d�eterminer le
contenu en collagene de type I et III, nous avons
utilis�e la coloration au rouge sirius selon Junqueira
et coll.20 La microscopie optique montrait une artere
ombilicale non perfus�ee (Fig. 2A), une artere per-
fus�ee avec un oedeme interstitiel mod�er�e (Fig. 2B),
et une artere trait�ee au VEGF avec un oedeme in-
terstitiel marqu�e (Fig. 2C). La microscopie en pola-
risation montrait du collagene de types I et III sur des
cordons ombilicaux fraıchement pr�epar�es (Fig. 2D).
La quantit�e et le rapport du collagene de types I et III
n’�etaient pas affect�es par la perfusion (Fig. 2E). Du
collagene de type III �etait visible dans toute la paroi
de l’artere ombilicale dans les groupes Cnp et Cp
(Fig. 2D, E), la gel�ee de Wharton contenant une
quantit�e importante de collagene de types I et III.
Par comparaison avec Cnp et Cp, VEGF avait clai-
rement augment�e le contenu de la paroi art�erielle en
collagene de type III (Fig. 2F), alors que le collagene
de type I (jaune, orange) �etait augment�e dans le
secteur p�erivasculaire (Fig. 2F) et dans la gel�ee de
Wharton apres l’application du VEGF (Fig. 2F).
Collagene de type IV. Du collagene de type IV, autre
prot�eine de la MEC et constituant des membranes
basales, �etait �egalement �elev�e apres traitement par
VEGF ( p< 0,05 vs les controles Cnp et Cp), en
analyse Western blot (Fig. 3A, B) et en analyse
d’image quantitative (Fig. 4A-D). Du collagene de
type IV �etait �egalement d�etect�e dans l’�epith�elium
amniotique (Fig. 4C).
Laminine. La laminine est normalement localis�ee
dans les membranes basales des cellules en-
doth�eliales et dans la m�edia art�erielle. Sa distribu-
tion est rest�ee semblable dans tous les cordons,
ind�ependamment du traitement. Cependant, sa
quantit�e �etait significativement �elev�ee apres traite-
ment par VEGF par comparaison avec les controles
( p< 0,05, Fig. 3A, C). Elle �etait d�etectable dans la
gel�ee de Wharton et l’�epith�elium amniotique du
cordon ombilical (Fig. 4E-H).
Impact de VEGF sur l’ost�eopontine et la
ß1-int�egrine
L’ost�eopontine �etait d�etect�ee sur tous les cordons om-
bilicaux examin�es, ind�ependamment de la perfusion
(Fig. 4I-L). Cependant, sa quantit�e �etait augment�ee
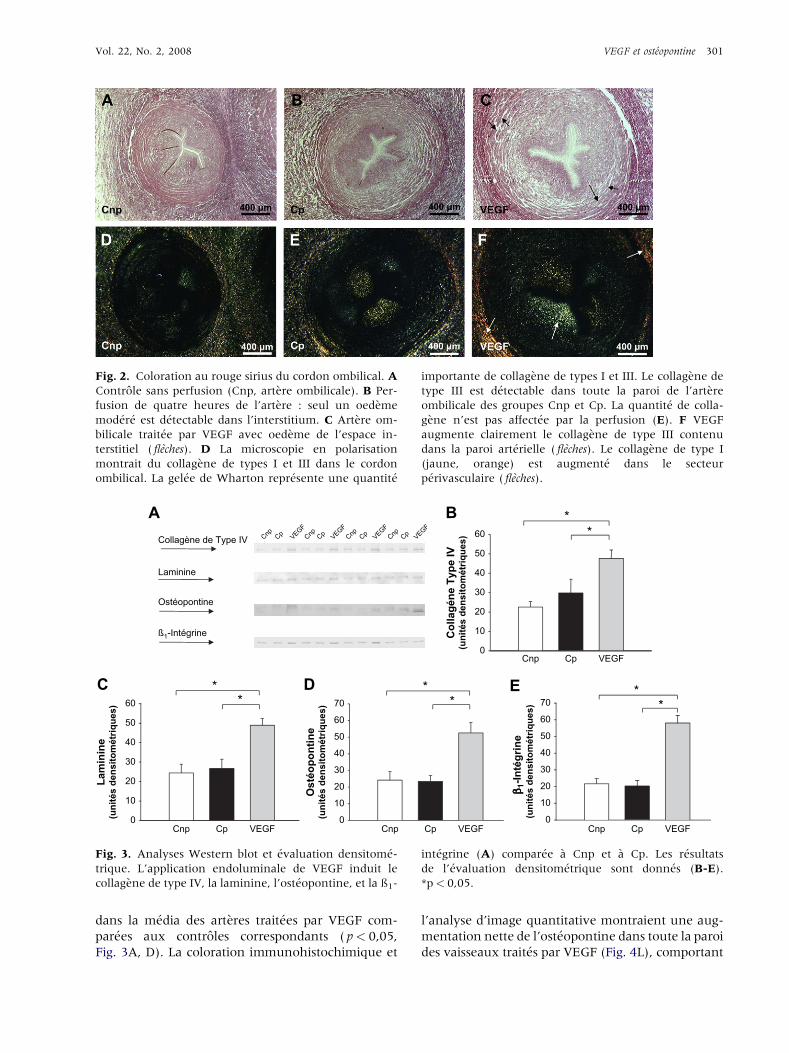
Vol. 22, No. 2, 2008 VEGF et ost�eopontine 301
Fig. 2. Coloration au rouge sirius du cordon ombilical. A
Controle sans perfusion (Cnp, artere ombilicale). B Per-
fusion de quatre heures de l’artere : seul un oedeme
mod�er�e est d�etectable dans l’interstitium. C Artere om-
bilicale trait�ee par VEGF avec oedeme de l’espace in-
terstitiel ( fleches). D La microscopie en polarisation
montrait du collagene de types I et III dans le cordon
ombilical. La gel�ee de Wharton repr�esente une quantit�e
importante de collagene de types I et III. Le collagene de
type III est d�etectable dans toute la paroi de l’artere
ombilicale des groupes Cnp et Cp. La quantit�e de colla-
gene n’est pas affect�ee par la perfusion (E). F VEGF
augmente clairement le collagene de type III contenu
dans la paroi art�erielle ( fleches). Le collagene de type I
(jaune, orange) est augment�e dans le secteur
p�erivasculaire ( fleches).
Cnp Cp VEGF
Co
llag
én
e T
yp
e IV
(u
nités d
en
sito
métriq
ues)
**
10
20
30
40
50
60
0
A B
Collagène de Type IV Cnp Cp Cnp CpVEGFVEGF
Cnp Cp VEGFCnp Cp VEGF
Laminine
Ostéopontine
ß1-Intégrine
0
10
20
30
40
50
60
70
Ostéo
po
ntin
e
(u
nités d
en
sito
métriq
ues)
**
Cnp Cp VEGF
βββ 1-In
tég
rin
e
(u
nités d
en
sito
métriq
ues)
0
10
20
30
40
50
60
70*
*
Cnp Cp VEGF
Lam
in
in
e
(u
nités d
en
sito
métriq
ues)
**
Cnp Cp VEGF
10
20
30
40
50
60
0
C ED
Fig. 3. Analyses Western blot et �evaluation densitom�e-
trique. L’application endoluminale de VEGF induit le
collagene de type IV, la laminine, l’ost�eopontine, et la ß1-
int�egrine (A) compar�ee a Cnp et a Cp. Les r�esultats
de l’�evaluation densitom�etrique sont donn�es (B-E).
*p< 0,05.
dans la m�edia des arteres trait�ees par VEGF com-
par�ees aux controles correspondants ( p< 0,05,
Fig. 3A, D). La coloration immunohistochimique et
l’analyse d’image quantitative montraient une aug-
mentation nette de l’ost�eopontine dans toute la paroi
des vaisseaux trait�es par VEGF (Fig. 4L), comportant
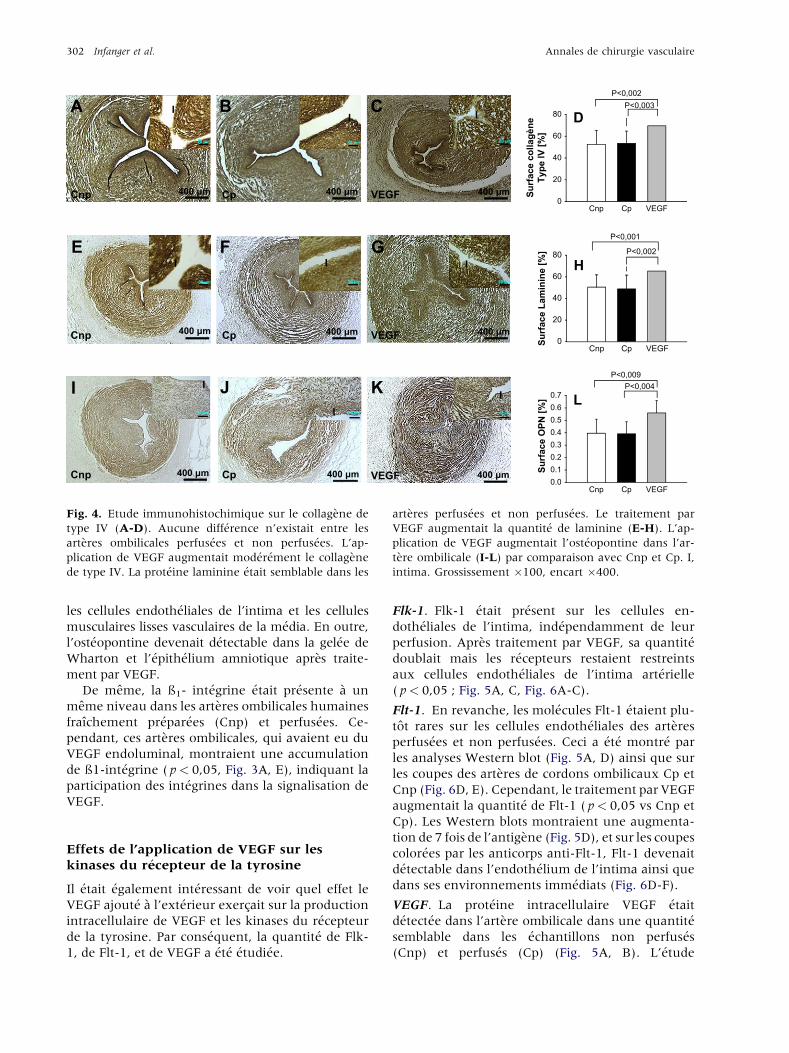
302 Infanger et al. Annales de chirurgie vasculaire
A B C
E F
JI K
G
Cnp Cp VEGF0
20
40
60
80
Su
rface co
llag
èn
e
Typ
e IV
[%
]
P<0,002P<0,003
D
Cnp Cp VEGF0
20
40
60
80
Su
rface L
am
in
in
e [%
]
P<0,001
P<0,002
H
S
urface O
PN
[%
]
0.00.10.20.30.40.50.60.7
P<0,009P<0,004
Cnp Cp VEGF
L
VEGF 400 µmCnp 400 µm
VEGF
VEGF400 µm
Cp400 µm
Cnp400 µm
400 µmCp
400 µm
Cp 400 µm
Cnp
I
II
III
I
I
I
400 µm
Fig. 4. Etude immunohistochimique sur le collagene de
type IV (A-D). Aucune diff�erence n’existait entre les
arteres ombilicales perfus�ees et non perfus�ees. L’ap-
plication de VEGF augmentait mod�er�ement le collagene
de type IV. La prot�eine laminine �etait semblable dans les
arteres perfus�ees et non perfus�ees. Le traitement par
VEGF augmentait la quantit�e de laminine (E-H). L’ap-
plication de VEGF augmentait l’ost�eopontine dans l’ar-
tere ombilicale (I-L) par comparaison avec Cnp et Cp. I,
intima. Grossissement �100, encart �400.
les cellules endoth�eliales de l’intima et les cellules
musculaires lisses vasculaires de la m�edia. En outre,
l’ost�eopontine devenait d�etectable dans la gel�ee de
Wharton et l’�epith�elium amniotique apres traite-
ment par VEGF.
De meme, la ß1- int�egrine �etait pr�esente a un
meme niveau dans les arteres ombilicales humaines
fraıchement pr�epar�ees (Cnp) et perfus�ees. Ce-
pendant, ces arteres ombilicales, qui avaient eu du
VEGF endoluminal, montraient une accumulation
de ß1-int�egrine ( p< 0,05, Fig. 3A, E), indiquant la
participation des int�egrines dans la signalisation de
VEGF.
Effets de l’application de VEGF sur les
kinases du r�ecepteur de la tyrosine
Il �etait �egalement int�eressant de voir quel effet le
VEGF ajout�e a l’ext�erieur exercait sur la production
intracellulaire de VEGF et les kinases du r�ecepteur
de la tyrosine. Par cons�equent, la quantit�e de Flk-
1, de Flt-1, et de VEGF a �et�e �etudi�ee.
Flk-1. Flk-1 �etait pr�esent sur les cellules en-
doth�eliales de l’intima, ind�ependamment de leur
perfusion. Apres traitement par VEGF, sa quantit�edoublait mais les r�ecepteurs restaient restreints
aux cellules endoth�eliales de l’intima art�erielle
( p< 0,05 ; Fig. 5A, C, Fig. 6A-C).
Flt-1. En revanche, les mol�ecules Flt-1 �etaient plu-
tot rares sur les cellules endoth�eliales des arteres
perfus�ees et non perfus�ees. Ceci a �et�e montr�e par
les analyses Western blot (Fig. 5A, D) ainsi que sur
les coupes des arteres de cordons ombilicaux Cp et
Cnp (Fig. 6D, E). Cependant, le traitement par VEGF
augmentait la quantit�e de Flt-1 ( p< 0,05 vs Cnp et
Cp). Les Western blots montraient une augmenta-
tion de 7 fois de l’antigene (Fig. 5D), et sur les coupes
color�ees par les anticorps anti-Flt-1, Flt-1 devenait
d�etectable dans l’endoth�elium de l’intima ainsi que
dans ses environnements imm�ediats (Fig. 6D-F).
VEGF. La prot�eine intracellulaire VEGF �etait
d�etect�ee dans l’artere ombilicale dans une quantit�esemblable dans les �echantillons non perfus�es
(Cnp) et perfus�es (Cp) (Fig. 5A, B). L’�etude
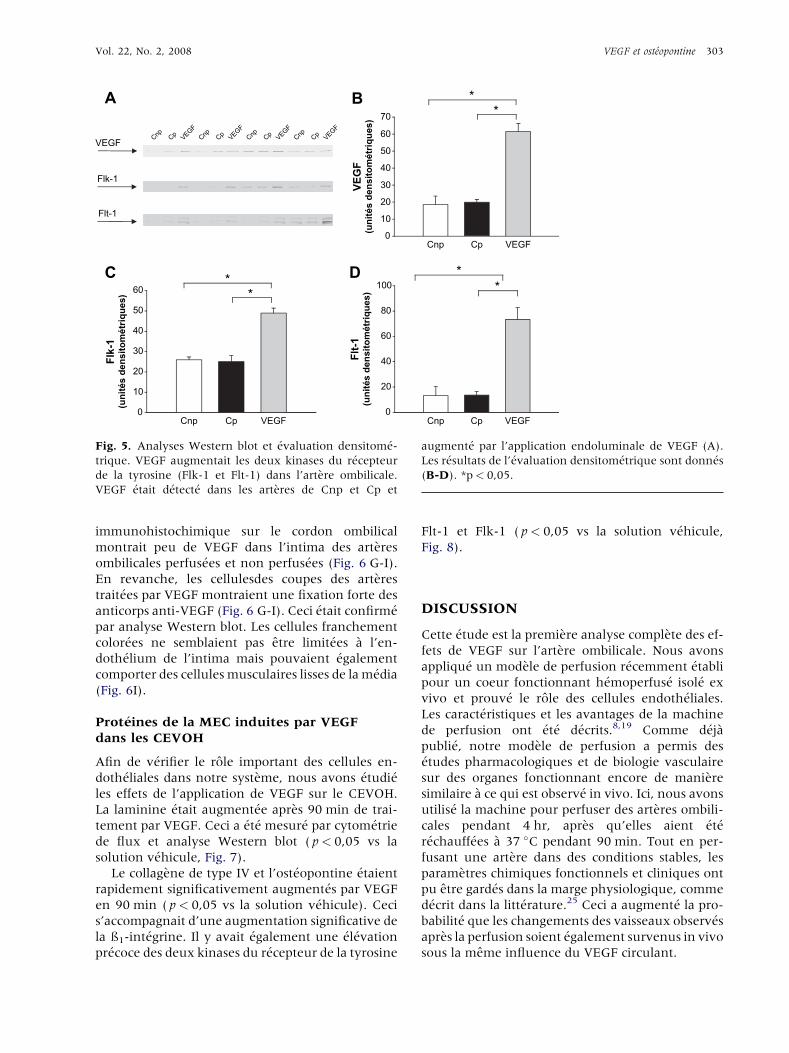
Vol. 22, No. 2, 2008 VEGF et ost�eopontine 303
Cnp Cp VEGF0
10
20
30
40
50
60*
Flk-1
(u
nité
s d
en
sito
mé
triq
ue
s) *
C
Cnp Cp VEGF0
20
40
60
80
100
Flt-1
(u
nité
s d
en
sito
mé
triq
ue
s)
**D
*
Cnp Cp VEGF0
10
20
30
40
50
60
70
VE
GF
(u
nité
s d
en
sito
mé
triq
ue
s)
*
VEGF
Flk-1
Flt-1
A B
Cp VEGFCnp Cp VEGF
Cnp Cp VEGFCp VEGF
CnpCnp
Fig. 5. Analyses Western blot et �evaluation densitom�e-
trique. VEGF augmentait les deux kinases du r�ecepteur
de la tyrosine (Flk-1 et Flt-1) dans l’artere ombilicale.
VEGF �etait d�etect�e dans les arteres de Cnp et Cp et
augment�e par l’application endoluminale de VEGF (A).
Les r�esultats de l’�evaluation densitom�etrique sont donn�es
(B-D). *p< 0,05.
immunohistochimique sur le cordon ombilical
montrait peu de VEGF dans l’intima des arteres
ombilicales perfus�ees et non perfus�ees (Fig. 6 G-I).
En revanche, les cellulesdes coupes des arteres
trait�ees par VEGF montraient une fixation forte des
anticorps anti-VEGF (Fig. 6 G-I). Ceci �etait confirm�epar analyse Western blot. Les cellules franchement
color�ees ne semblaient pas etre limit�ees a l’en-
doth�elium de l’intima mais pouvaient �egalement
comporter des cellules musculaires lisses de la m�edia
(Fig. 6I).
Prot�eines de la MEC induites par VEGF
dans les CEVOH
Afin de v�erifier le role important des cellules en-
doth�eliales dans notre systeme, nous avons �etudi�eles effets de l’application de VEGF sur le CEVOH.
La laminine �etait augment�ee apres 90 min de trai-
tement par VEGF. Ceci a �et�e mesur�e par cytom�etrie
de flux et analyse Western blot ( p< 0,05 vs la
solution v�ehicule, Fig. 7).
Le collagene de type IV et l’ost�eopontine �etaient
rapidement significativement augment�es par VEGF
en 90 min ( p< 0,05 vs la solution v�ehicule). Ceci
s’accompagnait d’une augmentation significative de
la ß1-int�egrine. Il y avait �egalement une �el�evation
pr�ecoce des deux kinases du r�ecepteur de la tyrosine
Flt-1 et Flk-1 ( p< 0,05 vs la solution v�ehicule,
Fig. 8).
DISCUSSION
Cette �etude est la premiere analyse complete des ef-
fets de VEGF sur l’artere ombilicale. Nous avons
appliqu�e un modele de perfusion r�ecemment �etabli
pour un coeur fonctionnant h�emoperfus�e isol�e ex
vivo et prouv�e le role des cellules endoth�eliales.
Les caract�eristiques et les avantages de la machine
de perfusion ont �et�e d�ecrits.8,19 Comme d�eja
publi�e, notre modele de perfusion a permis des�etudes pharmacologiques et de biologie vasculaire
sur des organes fonctionnant encore de maniere
similaire a ce qui est observ�e in vivo. Ici, nous avons
utilis�e la machine pour perfuser des arteres ombili-
cales pendant 4 hr, apres qu’elles aient �et�er�echauff�ees a 37 �C pendant 90 min. Tout en per-
fusant une artere dans des conditions stables, les
parametres chimiques fonctionnels et cliniques ont
pu etre gard�es dans la marge physiologique, comme
d�ecrit dans la litt�erature.25 Ceci a augment�e la pro-
babilit�e que les changements des vaisseaux observ�es
apres la perfusion soient �egalement survenus in vivo
sous la meme influence du VEGF circulant.

304 Infanger et al. Annales de chirurgie vasculaire
Fig. 6. Des cellules endoth�eliales positives pour Flk-1
�etaient d�etect�ees dans l’intima de toutes les arteres dans
tous les groupes (A-C). En parallele, nous avons
�egalement trouv�e des cellules endoth�eliales positives
pour Flt-1 dans l’intima du groupe VEGF mais pas dans
les arteres Cnp et Cp (D-F). La prot�eine VEGF �etait
d�etectable dans les cellules endoth�eliales de toutes les
arteres mais clairement augment�ee apres application de
VEGF (G-I). I, intima. Grossissement �400.
Impact de VEGF sur la perm�eabilit�evasculaire
Les diverses �etudes ont montr�e que l’ARNm et la
prot�eine VEGF sont largement distribu�es dans les
tissus, les organes, ou les s�erums normaux. Elles
soutiennent le concept que VEGF peut jouer un
role important dans l’entretien et le controle de la
fonction vasculaire.12,26 Dans cette �etude, nous
avons appliqu�e une solution physiologique de VEGF
a l’artere ombilicale. Le temps d’incubation �etait de
90 min. L’application endoluminale de VEGF a in-
duit un oedeme tissulaire du cordon ombilical sans
dommages �evidents. L’extravasation �etait d�etectable
apres 4 hr de perfusion et d�ependait fortement de la
pr�ecession du traitement par VEGF, mais elle n’�etait
pas induite par la machine de perfusion. Ainsi, il
devient de plus en plus �evident qu’un oedeme
g�en�eral peut etre provoqu�e par VEGF seul chez les
patients pr�esentant un taux s�erique �elev�e de VEGF
du a une l�esion importante mais localis�ee.
Effets de VEGF sur les kinases du
r�ecepteur de la tyrosine
Nous avons �egalement d�emontr�e que le milieu de
perfusion contenant du VEGF d�eclenche l’expres-
sion de la prot�eine VEGF par les cellules
endoth�eliales de l’intima de l’artere ombilicale.
L’observation s’accorde avec un premier r�esultat
obtenu a partir de l’incubation des cellules en-
doth�eliales en pr�esence de VEGF externe sous mi-
crogravit�e.24 VEGF se lie aux kinases du r�ecepteur
de surface des cellules pour la tyrosine principale-
ment exprim�ees sur les cellules endoth�eliales, dont
les plus importantes sont VEGFR-1 (Flt-1) et
VEGFR-2 (Flk-1). La coloration immunohisto-
chimique de Flk-1 et de Flt-1 a clairement montr�e

Vol. 22, No. 2, 2008 VEGF et ost�eopontine 305
VEH 90 min VEGF 5.5 h VEGF0
10
20
30
40
50
60 **
CEVOHVEH39%
CEVOH90 min VEGF
45%
CEVOH5,5 h VEGF
44%
0
5
10
15
20
25
Lam
in
in
e
(u
nités d
en
sito
métriq
ues)
Laminine
VEH 90 min VEGF 5.5 h VEGF
**
*
Lam
in
in
e
(%
d
e cellu
les p
ositives)
A
B
veh veh 90 min 90 min 5.5h 5.5h
C
Num
ber
Num
ber
R1 R2 R1 R2 R1 R2
Fig. 7. Recherche en cytom�etrie de flux de la laminine
dans les CEVOH (A, B). VEGF augmentait pr�ecocement
la quantit�e de laminine. Cette observation �etait
confirm�ee par l’analyse Western blot (C). VEH, v�ehicule.
*p< 0,05.
VEGF 165 sur les cellules endoth�eliales de l’intima
des arteres ombilicales. En outre, l’application de
VEGF a augment�e l’expression de ses r�ecepteurs.
Flk-1 �etait d�etectable dans les arteres ombilicales
dans une certaine mesure au d�ebut de l’exp�erience,
alors que Flt-1 n’�etait pas trouv�e a ce moment.
Apres 4 hr de perfusion, Flk-1 �etait clairement
stimul�e dans l’intima et Flt-1 �etait devenu
d�etectable.
Pendant longtemps, on a largement suppos�e que
VEGF agit en tant que facteur de croissance sp�eci-
fique des cellules endoth�eliales et att�enue la mala-
die vasculaire en induisant la prolif�eration
endoth�eliale principalement par Flk-1. Des preuves
r�ecentes suggerent que Flt-1 et Flk-1 fonctionnels
sont aussi exprim�es sur les cellules musculaires
lisses de la m�edia apres que les vaisseaux sanguins
aient �et�e l�es�es. Dans ce travail, l’application endolu-
minale de VEGF augmente les deux kinases du
r�ecepteur de la tyrosine au niveau de l’en-
doth�elium de l’artere ombilicale, indiquant que
notre appareil de perfusion fonctionne sans cr�eer
de l�esions de perfusion. VEGF et Flt-1 jouent un
role essentiel dans la formation de nouvelles
intimas apres des l�esions.27
VEGF, Flk-1, et Flt-1 peuvent stimuler la pro-
lif�eration, la migration, et l’activit�e prot�eolytique
des cellules endoth�eliales. VEGF induit �egalement
la perm�eabilit�e, la vasodilatation, et l’angiog�enese
vasculaires.28 Par ailleurs, la quantit�e de VEGF
exprim�e est control�ee par la diff�erentiation, la
transformation, et la fourniture d’oxygene. Flt-1
et Flk-1 sont presque exclusivement situ�es sur les
cellules endoth�eliales.26,29,30 La densit�e des deux
kinases du r�ecepteur de la tyrosine est basse dans le
tissu normal mais augment�ee dans les tissus tumo-
raux ou a proximit�e en cas de n�eoangiog�enese et/ou
de perm�eabilit�e vasculaire augment�ee comme
observ�e dans l’angiog�enese tumorale.31 Comme
d’autres32 nous avons d�emontr�e que VEGF lui-
vmeme est un facteur important menant a l’ac-
croissement des r�ecepteurs de VEGF.
Effets de VEGF sur la MEC
Dans une �etude r�ecente,13 nous avons montr�e que
VEGF appliqu�e par voie endoluminale acc�el�erait la
r�eendoth�elialisation et am�eliorait la cicatrisation
vasculaire apres traumatisme. Plusieurs prot�eines de
la MEC, telles que la fibronectine et le collagene de
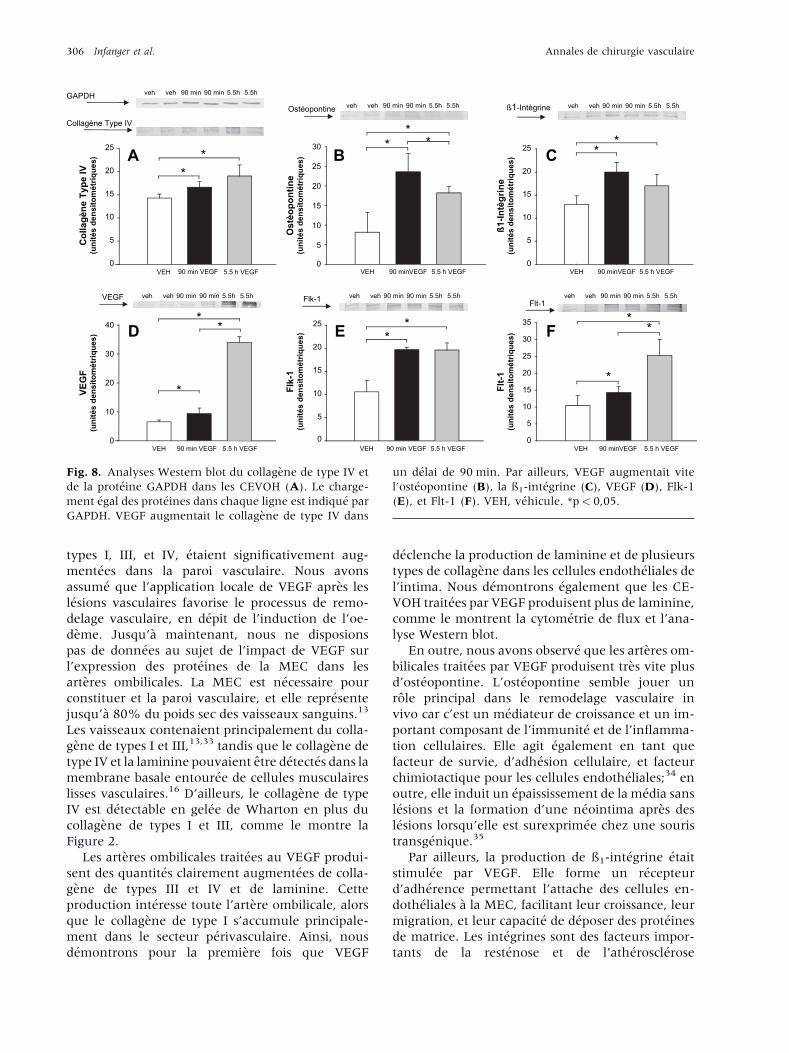
306 Infanger et al. Annales de chirurgie vasculaire
0
5
10
15
20
25
*
Co
lla
gè
ne
T
yp
e IV
(u
nités d
en
sito
métriq
ues)
VEH 90 min VEGF 5.5 h VEGF
VEH 90 min VEGF 5.5 h VEGF
Collagène Type IV
GAPDH
*A B
0
5
10
15
20
25 *
Flk-1
(u
nités d
en
sito
métriq
ues)
Flk-1
*
VEH 90 min VEGF 5.5 h VEGF
E
0
5
10
15
20
25
30
35 *
Flt-1
(u
nités d
en
sito
métriq
ues)
Flt-1
*
*
VEH 90 minVEGF 5.5 h VEGF
F
0
5
10
15
20
25
ß1
-In
tè
grin
e
(u
nités d
en
sito
métriq
ues)
ß1-Intègrine
**
0
10
20
30
40
VE
GF
(u
nités d
en
sito
métriq
ues)
VEGF
C
*
**
veh veh 90 min 90 min 5.5h 5.5h
D
0
5
10
15
20
25
30
Os
tè
op
on
tin
e
(u
nités d
en
sito
métriq
ues)
Ostéopontine
VEH 90 minVEGF 5.5 h VEGF VEH 90 minVEGF 5.5 h VEGF
* **
veh veh 90 min 90 min 5.5h 5.5h veh veh 90 min 90 min 5.5h 5.5h
veh veh 90 min 90 min 5.5h 5.5h veh veh 90 min 90 min 5.5h 5.5h veh veh 90 min 90 min 5.5h 5.5h
Fig. 8. Analyses Western blot du collagene de type IV et
de la prot�eine GAPDH dans les CEVOH (A). Le charge-
ment �egal des prot�eines dans chaque ligne est indiqu�e par
GAPDH. VEGF augmentait le collagene de type IV dans
un d�elai de 90 min. Par ailleurs, VEGF augmentait vite
l’ost�eopontine (B), la ß1-int�egrine (C), VEGF (D), Flk-1
(E), et Flt-1 (F). VEH, v�ehicule. *p< 0,05.
types I, III, et IV, �etaient significativement aug-
ment�ees dans la paroi vasculaire. Nous avons
assum�e que l’application locale de VEGF apres les
l�esions vasculaires favorise le processus de remo-
delage vasculaire, en d�epit de l’induction de l’oe-
deme. Jusqu’a maintenant, nous ne disposions
pas de donn�ees au sujet de l’impact de VEGF sur
l’expression des prot�eines de la MEC dans les
arteres ombilicales. La MEC est n�ecessaire pour
constituer et la paroi vasculaire, et elle repr�esente
jusqu’a 80% du poids sec des vaisseaux sanguins.13
Les vaisseaux contenaient principalement du colla-
gene de types I et III,13,33 tandis que le collagene de
type IV et la laminine pouvaient etre d�etect�es dans la
membrane basale entour�ee de cellules musculaires
lisses vasculaires.16 D’ailleurs, le collagene de type
IV est d�etectable en gel�ee de Wharton en plus du
collagene de types I et III, comme le montre la
Figure 2.
Les arteres ombilicales trait�ees au VEGF produi-
sent des quantit�es clairement augment�ees de colla-
gene de types III et IV et de laminine. Cette
production int�eresse toute l’artere ombilicale, alors
que le collagene de type I s’accumule principale-
ment dans le secteur p�erivasculaire. Ainsi, nous
d�emontrons pour la premiere fois que VEGF
d�eclenche la production de laminine et de plusieurs
types de collagene dans les cellules endoth�eliales de
l’intima. Nous d�emontrons �egalement que les CE-
VOH trait�ees par VEGF produisent plus de laminine,
comme le montrent la cytom�etrie de flux et l’ana-
lyse Western blot.
En outre, nous avons observ�e que les arteres om-
bilicales trait�ees par VEGF produisent tres vite plus
d’ost�eopontine. L’ost�eopontine semble jouer un
role principal dans le remodelage vasculaire in
vivo car c’est un m�ediateur de croissance et un im-
portant composant de l’immunit�e et de l’inflamma-
tion cellulaires. Elle agit �egalement en tant que
facteur de survie, d’adh�esion cellulaire, et facteur
chimiotactique pour les cellules endoth�eliales;34 en
outre, elle induit un �epaississement de la m�edia sans
l�esions et la formation d’une n�eointima apres des
l�esions lorsqu’elle est surexprim�ee chez une souris
transg�enique.35
Par ailleurs, la production de ß1-int�egrine �etait
stimul�ee par VEGF. Elle forme un r�ecepteur
d’adh�erence permettant l’attache des cellules en-
doth�eliales a la MEC, facilitant leur croissance, leur
migration, et leur capacit�e de d�eposer des prot�eines
de matrice. Les int�egrines sont des facteurs impor-
tants de la rest�enose et de l’ath�eroscl�erose

Vol. 22, No. 2, 2008 VEGF et ost�eopontine 307
art�erielles17 et peuvent etre important dans le re-
modelage vasculaire.
CONCLUSIONS
En r�esum�e, le modele automatis�e de perfusion
utilis�e offre des possibilit�es tres int�eressantes pour
la recherche sur les interactions complexes entre
VEGF et la paroi de l’artere ombilicale. Ainsi, nous
avons montr�e pour la premiere fois que VEGF sti-
mule les cellules endoth�eliales de l’intima de l’artere
ombilicale pour produire des concentrations aug-
ment�ees des prot�eines de la MEC. VEGF influence�egalement la m�edia et l’adventice adjacents et en-
traıne un oedeme vasculaire pr�ecoce. VEGF stimule
la production de la prot�eine ost�eopontine, ainsi que
celle du VEGF intracellulaire et ses kinases du
r�ecepteur de la tyrosine en 90 min ex vivo et in vi-
tro. Toutes les mol�ecules mentionn�ees ci-dessus
sont connues pour etre impliqu�ees dans le processus
de remodelage vasculaire. Le remodelage vasculaire
commence des la formation d’un oedeme d’ac-
compagnement. Cependant, ce sera un futur travail
que d’�etudier comment il est possible d’empecher
l’oedeme induit par VEGF dans des vaisseaux sains a
distance des l�esions. Un traitement par des inhibi-
teurs de VEGF commencant imm�ediatement apres
une brulure ou des l�esions traumatiques peut
r�eduire l’oedeme tissulaire �etendu. La cons�equence
peut etre une am�elioration des r�esultats apres des
l�esions graves.
Nous remercions Jessie Webb pour la pr�eparation du manuscrit
et Mme Ursula Schwikowski pour son aide avec les figures.
REFERENCES
1. Senger DR, Galli SJ, Dvorak AM, et coll. Tumor cells secrete
a vascular permeability factor that promotes accumulation
of ascites fluid. Science 1983;219:983-985.
2. Ferrara N, Henzel WJ. Pituitary follicular cells secrete a
novel heparin binding growth factor specific for vascular
endothelial cells. Biochem Biophys Res Commun 1989;161:
851-858.
3. Plouet J, Schilling J, Gospodarowicz D. Isolation and cha-
racterization of a newly identified endothelial cell mitogen
produced by At T20 cells. EMBO J 1989;8:3801-3808.
4. Vincenti V, Cassano C, Rocchi M, et coll. Assignement of the
vascular endothelial growth factor gene to human chro-
mosome 6p21.3. Circulation 1996;93:1493-1495.
5. Leung DW, Cachianes G, Kuang WJ, et coll. Vascular en-
dothelial growth factor is a secreted angiogenic mitogen.
Science 1989;246:1306-1309.
6. Keck PJ, Hauser SD, Krivi G, et coll. Vascular permeability
factor, an endothelial cell mitogen related to PDGF. Science
1989;246:1309-1312.
7. Aird WC. Spatial and temporal dynamics of the endo-
thelium. J Thromb Haemost 2005;3:1392-1406.
8. Houck KA, Ferrara N, Winer J, et coll. The vascular endo-
thelial growth factor family: identification of a fourth mo-
lecular species and characterization of alternative splicing of
RNA. Mol Endocrinol 1991;5:1806-1814.
9. Devries C, Escobedo JA, Ueno H, et coll. The fms-like
tyrosine kinase, a receptor for vascular endothelial growth
factor. Science 1992;255:989-991.
10. Schiffrin EL. The endothelium and control of blood-vessel
function in health and disease. Clin Invest Med 1994;17:
602-620.
11. Harrigan MR, Ennis SR, Masada T, et coll. Intraventricular
infusion of vascular endothelial growth factor promotes
cerebral angiogenesis with minimal brain edema. Neuro-
surgery 2002;50:589-598.
12. Infanger M, Schmidt O, Kossmehl P, et coll. Vascular en-
dothelial growth factor serum level is strongly enhanced
after burn injury and correlated with local and general tissue
edema. Burns 2004;30:305-311.
13. Infanger M, Shakibaei M, Kossmehl P, et coll. Intraluminal
application of vascular endothelial growth factor enhances
healing of microvascular anastomosis in a rat model. J Vasc
Res 2005;42:202-213.
14. Sixma JJ, van Zanten GH, Saelman EU, et coll. Platelet ad-
hesion to collagen. Thromb Haemost 1995;74:454-459.
15. Polanowska-Grabowska R, Simon CG, Jr, Gear AR. Platelet
adhesion to collagen type I, collagen type IV, von Wille-
brand factor, fibronectin, laminin and fibrinogen: rapid ki-
netics under shear. Thromb Haemost 1999;81:118-123.
16. Hallmann R, Horn N, Selg M, et coll. Expression and func-
tion of laminins in the embryonic and mature vasculature.
Physiol Rev 2005;85:979-1000.
17. Xu H, Zeng L, Peng H, et coll. HMG-CoA reductase inhibitor
simvastatin mitigates VEGF-induced ‘‘inside-out’’ signaling
to extracellular matrix by preventing RhoA activation. Am J
Physiol Renal Physiol 2006;291:F995-F1004.
18. Kossmehl P, Schonberger J, Shakibaei M, et coll. Increase of
fibronectin and osteopontin in porcine hearts following
ischemia and reperfusion. J Mol Med 2005;83:626-637.
19. Kossmehl P, Kurth E, Faramarzi S, et coll. Mechanisms of
apoptosis after ischemia and reperfusion: role of the renin-
angiotensin system. Apoptosis 2006;11:347-358.
20. Junqueira LCU, Cossermelli W, Brentani R. Differential
staining of collagens type I, II and III by sirius red and po-
larization microscopy. Arch Histol Jpn 1978;41:267-274.
21. Grimm D, Cameron D, Griese DP, et coll. Differential effects
of growth hormone on cardiomyocyte and extracellular
matrix protein remodeling following experimental myo-
cardial infarction. Cardiovasc Res 1998;40:297-306.
22. Grimm D, Huber M, Jabusch HC, et coll. Extracellular ma-
trix proteins in cardiac fibroblasts derived from rat hearts
with chromic pressure-overload: effects of beta-receptor
blockade. J Mol Cell Cardiol 2001;33:487-501.
23. Infanger M, Ulbrich C, Baatout S, et coll. Modeled gravita-
tional unloading induced downregulation of endothelin-1
in human endothelial cells. J Cell Biochem 2007;101:
1439-1455.
24. Infanger M, Kossmehl P, Shakibaei M, et coll. Induction of
three- dimensional assembly and increase in apoptosis of
human endothelial cells by simulated microgravity: impact
of vascular endothelial growth factor. Apoptosis 2006;11:
749-764.
25. Gan L, Sjogren LS, Doroudi R, et coll. A new computerized
biomechanical perfusion model for ex vivo study of fluid

308 Infanger et al. Annales de chirurgie vasculaire
mechanical forces in intact conduit vessels. J Vasc Res
1999;36:68-78.
26. Jakeman LB, Winer J, Bennett GL, et coll. Binding sites for
vascular endothelial growth factor are localized on endo-
thelial cells in adult rat tissues. J Clin Invest 1992;89:244-253.
27. Zhao Q, Egashira K, Hiasa K, et coll. Essential role of vas-
cular endothelial growth factor and Flt-1 signals in neoin-
timal formation after periadventitial injury. Arterioscler
Thromb Vasc Biol 2004;24:2284-2289.
28. Ferrara N, Davis-Smyth T. The biology of vascular endo-
thelial growth factor. Endocr Rev 1997;18:4-25.
29. Shibuya M, Yamguchi S, Yamane A, et coll. Nucleotide
sequence and expression of a novel human receptor-type
tyrosine kinase gene (FLT) closely related to the fms family.
Oncogene 1990;5:519-524.
30. Terman BI, Dougher VM, Carrion ME, et coll. Identification
of the KDR tyrosine kinase as a receptor for vascular en-
dothelial growth factor. Biochem Biophys Res Commun
1992;187:1579-1586.
31. Barleon B, Siemeister G, Martiny-Baron G, et coll. Vascular
endothelial growth factor up-regulates its receptor fms-like
tyrosine kinase 1 (FLT-1) and a soluble variant of FLT-1 in
human vascular endothelial cells. Cancer Res 1997;57:
5421-5425.
32. Plate KH, Breier G, Millauer B, et coll. Up-regulation of
vascular endothelial growth factor and its cognate receptors
in a rat glioma model of tumor angiogenesis. Cancer Res
1993;53:5822-5827.
33. Farquharson C, Robins SP. Immunolocalization of collagens
type I and III in the arterial wall of the rat. Histochem J
1989;21:172-178.
34. Leali D, Dell’Era P, Stabile H, et coll. Osteopontin (Eta-1)
and fibroblast growth factor-2 cross-talk in angiogenesis.
J Immunol 2003;171:1085-1093.
35. Isoda K, Nishikawa K, Kamezawa Y, et coll. Osteopontin
plays an important role in the development of medial
thickening and neointimal formation. Circ Res 2002;91:
77-82.