cours 4 - accueill2bichat2012-2013.weebly.com/uploads/1/3/9/0/13905422/rono_4__ue7... · ce sont...
TRANSCRIPT

UE 7 GYNECO-ENDOCRINOLOGIE Professeur DE ROUX Le 05/10/2012 8h30-10h30Ronéotypeur : Yousra Kherabi Ronéolecteur :Thomas Brion
COURS 4 : Introduction à l'hormonologie
Rythmes biologiques
Ronéo n°4 d'UE 7 1/22

PLAN
• I) Classification biochimique des hormones 1) Définition d'une hormone 2) Règles
• II) Synthèse et catabolisme 1) Sources et nature des hormones : a) Généralités b) Les hormones peptidiques et l'exemple de la GnRH I c) Les hormones stéroïdes 2) La synthèse hormonale : un mécanisme répondant à une action très précise 3) Transport des hormones dans le sang 4) Métabolisme des hormones a) Transformation des hormones b) Élimination des hormones
• III) Mécanismes d’action 1) Généralités 2) Les hormones des récepteurs nucléaires a) Mécanismes d'action des hormones stéroïdes b) Mécanismes d'action des hormones thyroïdiennes 3) Les récepteurs hormonaux membranaires a) Présentation générale b) Les récepteurs couplés à des protéines G (RCPG) c) Le récepteur à l'hormone de croissance
• IV) Rétro-contrôle
• V) Exploration hormonale 1) Généralités 2) Méthodes
• VI) La pathologie hormonale
• VII) Take Home : petit résumé du cours à bien connaître
Ronéo n°4 d'UE 7 2/22

I) Classification biochimique
1) Définition d'une hormone
Les hormones sont des molécules informatives agissant via des récepteurs spécifiques.
• Il y a une triade historique qui a permis de décrire l'hormone. L'hormone est donc généralement caractérisée par :
- Une sécrétion à partir d’une glande endocrine - Un transport par voie sanguine - Une action à distance de sa sécrétion
Il s'agit de la définition même d'un système endocrine. Il existe également les systèmes paracrines qui agissent « à côté de » et les systèmes autocrines (action de l'hormone sur la cellule responsable de sa synthèse).
• Toutes les hormones ne répondent donc pas à cette triade (puisqu'il existe des hormones à action paracrine ou autocrine).
2) Règles
• Toutes les hormones interagissent avec les cellules cibles en se fixant sur un récepteur qui peut être membranaire ou intra-cellulaire. C'est une notion fondamentale. En effet les récepteurs sont des transducteurs d'une information extra-cellulaire vers une information intra-cellulaire ; leur fonction est de transformer un signal biochimique extra-cellulaire en un signal biochimique intra-cellulaire.
• Les récepteurs des hormones sont liés à des partenaires protéiques capables d’activer des voies de transduction.
• Une substance qui mime l’action d’une hormone sur un récepteur est un agoniste.
• Une substance qui prévient l’activation d’un récepteur par une hormone est un antagoniste.
Ce sont des notions utilisées en exploration fonctionnelle : on ajoute des agonistes qui vont activer des récepteurs et on va étudier la fonction de ces agonistes sur les axes endocriniens.
II) Synthèse et catabolisme
1) Sources et nature des hormones a) Généralités
• Les sources des hormones sont :
- Les glandes endocrines : hypophyse, thyroïde, surrénales, pancréas (qui a la particularité d'être une glande endocrine et exocrine. La sécrétion de l'insuline est endocrine).
Ronéo n°4 d'UE 7 3/22

- Les cellules endocrines au sein d’un tissu non endocrinien : l'estomac possède un certain nombre de cellules capables de synthétiser la grheline (qui joue un rôle sur la prise alimentaire et la synthèse de l'hormone de croissance).
- Les adipocytes qui synthétisent de la leptine.
• La nature biochimique de l’hormone détermine son mode de synthèse. Il y a des hormones peptidiques, des hormones stéroïdes et des dérivés des acides aminés (comme les catécholamines). Voici des exemples de classification de différentes hormones :
– Les hormones polypeptidiques : elles peuvent être des neuropeptides comme ACTH et la vasopressine, ou encore des protéines comme l'insuline et le glucagon. D'autres exemples sont sur la diapo (le prof n'en a pas parlé) : somatotropine, FSH,LH Oxytocine, Ghréline et TSH. Les hormones polypetidiques peuvent également être des glycoprotéines (de haut poids moléculaire, environ 30 kDa, et qui ont la particularité d'être glycosylées).
– Les hormones stéroïdes : il y a des hormones sexuelles comme les oestrogènes et la testostérone, des hormones de l'axe corticotrope (le cortisol synthétisé par la surrénale), il y a aussi l’aldostérone, la corticostérone et la progestérone.
– Les dérivés d'acides aminés : il y a les hormones thyroïdiennes : la Thyroxine, T3 et T4 dont la synthèse est de nature assez complexe, il y a aussi l'épinephrine, la norépinephrine, la dopamine, la mélatonine, la sérotonine…
• Les événements cellulaires conduisant à la biosynthèse d'une hormone peptidique sont très complexes et surtout, à chaque étape, la régulation de la synthèse hormonale peut survenir.
b) Les hormones peptidiques et l' exemple de la GnRH I
• Par exemple pour la synthèse d'une hormone peptidique, ces événements sont beaucoup plus longs que la simple régulation de l'expression d'un gène et la production d'une protéine (comme on serait tenté de le penser de prime abord). Il va y avoir un ARNm (messager) qui va être produit dans le noyau, il va ensuite être traduit puis il va y avoir un certain nombre d'étapes biochimiques extrêmement importantes.
Ronéo n°4 d'UE 7 4/22

• Le premier événement dans la synthèse d'une hormone peptidique, c'est qu'il y a un signal peptide qui permet d'orienter la protéine dans la voie dite « de sécrétion ».Ce signal peptique est clivé quand la protéine entre dans le réticulum, ce qui permet à la protéine (ou au peptide) de continuer sa synthèse. Au sein du réticulum, il va y avoir la formation de ponts disulfures, la protéine va commencer à obtenir une conformation qui va être fondamentale pour la suite de la synthèse puis, après, pour son activité biologique. Dans le réticulum, il commence à apparaître des glycosylations qui sont également importantes pour la fonction biologique du peptide (ou de la protéine). Ça fait peu de temps que l'ARNm a été traduit (ces étapes ont donc été assez rapides).
Puis la protéine ou le peptide va passer dans l'appareil de Golgi où les modifications biochimiques de la protéine ou du peptide vont être très importantes. Il va y avoir changement des glycosylations, remplacement de certains sucres par d'autres, des modifications post-traductionnelles …
A la sortie de l'appareil de Golgi, la protéine pourra avoir subi un clivage, ce clivage est fondamental : souvent les peptides sont synthétisés sous la forme d'une pro-hormone qui subit un clivage de façon à libérer uniquement la zone active du peptide (l'autre partie du peptide soit peut maturer, soit c'est une protéine dont on ne connaît pas bien la fonction). Après cette coupure, il va y avoir de nouveau des modifications biochimiques et la synthèse du peptide est parfois très éloignée de ce qu'on pouvait imaginer si on avait juste lu la séquence de l'ARNm…
Par exemple la séquence de l'ARN m peut donner une hormone de 130 acides aminés (aa) et in fine le peptide va faire seulement 15 aa et c'est lui qui sera actif (phrase pas très claire du prof mais en gros, quand tu vois la séquence de ton ARNm relativement longue tu te dis « ça va faire une grosse protéine, ça » et en fait avec toutes les modifications post-traductionnelles, ça te donne un tout p'tit peptide au final).
• Prenons l'exemple de la GnRH ( Gonadotropin Releasing Hormon OH YEAH! ) qui est l'hormone qui régule la synthèse des gonadotropines. C'est un décapeptide (il fait 10 aa).
-pGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2
Comme on peut le voir, il a subi des modifications : une en N-Ter et l'autre en C-Ter. Il ne se termine pas par une fonction carboxylique mais par une fonction amine. Comment expliquer cette modification C-terminale ?La GnRH est exprimée essentiellement dans l'hypothalamus mais aussi l'utérus, les ovaires et le placenta et est synthétisée à partir d'un gène qui comprend 4 exons (le gène est localisé sur le chromosome 8 en position 8p11.2. Il comprend quatre exons et trois introns).
L'exon 1 n'est pas codant, les exons 2 et 3 sont codants, l'exon 4 a un tout petit bout codant. Cette organisation génique en 4 exons ne peut pas expliquer un peptide en 10 aa, inévitablement l'ARN m qui est synthétisé est « plus grand que ces 10 aa ».
Ronéo n°4 d'UE 7 5/22
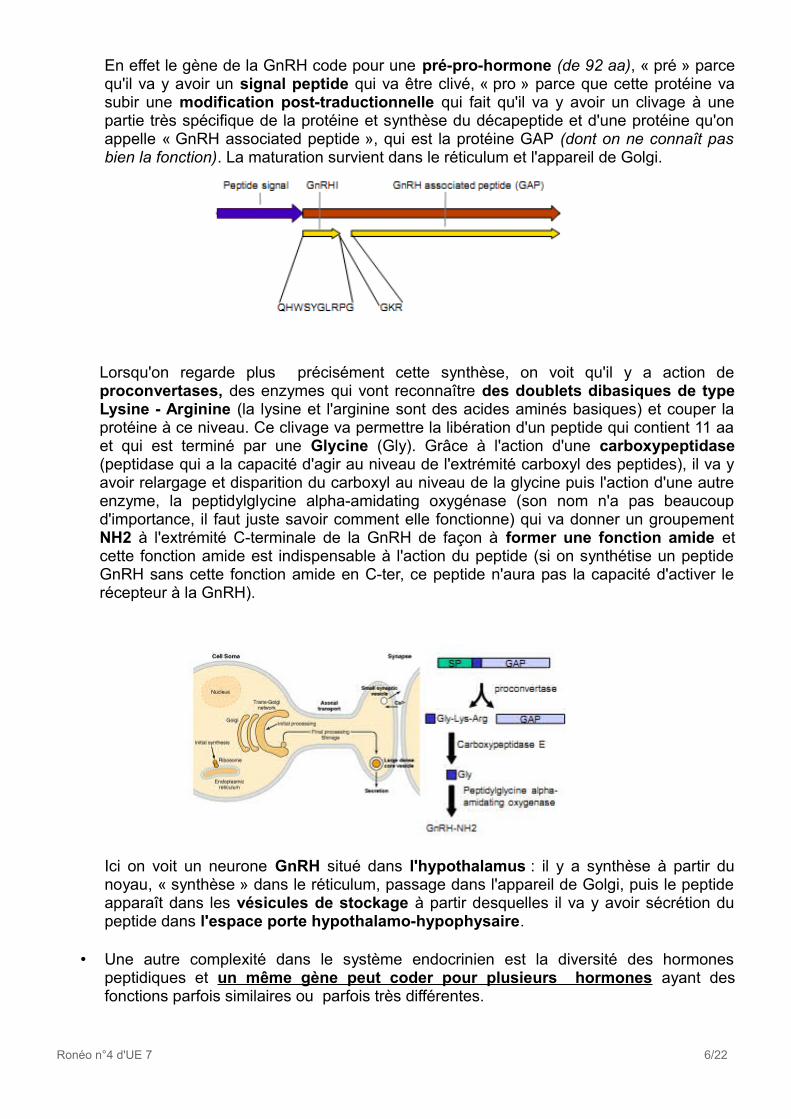
En effet le gène de la GnRH code pour une pré-pro-hormone (de 92 aa), « pré » parce qu'il va y avoir un signal peptide qui va être clivé, « pro » parce que cette protéine va subir une modification post-traductionnelle qui fait qu'il va y avoir un clivage à une partie très spécifique de la protéine et synthèse du décapeptide et d'une protéine qu'on appelle « GnRH associated peptide », qui est la protéine GAP (dont on ne connaît pas bien la fonction). La maturation survient dans le réticulum et l'appareil de Golgi.
Lorsqu'on regarde plus précisément cette synthèse, on voit qu'il y a action de proconvertases, des enzymes qui vont reconnaître des doublets dibasiques de type Lysine - Arginine (la lysine et l'arginine sont des acides aminés basiques) et couper la protéine à ce niveau. Ce clivage va permettre la libération d'un peptide qui contient 11 aa et qui est terminé par une Glycine (Gly). Grâce à l'action d'une carboxypeptidase (peptidase qui a la capacité d'agir au niveau de l'extrémité carboxyl des peptides), il va y avoir relargage et disparition du carboxyl au niveau de la glycine puis l'action d'une autre enzyme, la peptidylglycine alpha-amidating oxygénase (son nom n'a pas beaucoup d'importance, il faut juste savoir comment elle fonctionne) qui va donner un groupement NH2 à l'extrémité C-terminale de la GnRH de façon à former une fonction amide et cette fonction amide est indispensable à l'action du peptide (si on synthétise un peptide GnRH sans cette fonction amide en C-ter, ce peptide n'aura pas la capacité d'activer le récepteur à la GnRH).
Ici on voit un neurone GnRH situé dans l'hypothalamus : il y a synthèse à partir du noyau, « synthèse » dans le réticulum, passage dans l'appareil de Golgi, puis le peptide apparaît dans les vésicules de stockage à partir desquelles il va y avoir sécrétion du peptide dans l'espace porte hypothalamo-hypophysaire.
• Une autre complexité dans le système endocrinien est la diversité des hormones peptidiques et un même gène peut coder pour plusieurs hormones ayant des fonctions parfois similaires ou parfois très différentes.
Ronéo n°4 d'UE 7 6/22

• Prenons l'exemple de la POMC (pro-opiomelanocortine), qui permet la synthèse de l'ACTH (axe corticotrope). Il va y avoir des modifications traductionnelles, des clivages au sein de cette grosse pro hormone donc synthèse d'autres peptides ; c'est d'autant plus complexe que la synthèse de ces peptides dépend de la cellule qui exprime la POMC. La POMC est exprimée dans l'hypophyse mais aussi dans l'hypothalamus : les peptides exprimés dans l'hypothalamus à partir de la POMC ne sont pas les mêmes que ceux exprimés dans l'hypophyse. La maturation d'une pro hormone en plusieurs peptides est donc source de diversité. Il y a d'autres exemples comme l' épissage alternatif qui pourra être cellule et tissu spécifique ; dans certaines situations il va y avoir des promoteurs alternatifs : un promoteur alternatif est un promoteur situé dans un intron du gène et qui va pouvoir transcrire un ARNm qui sera inévitablement tronqué dans sa partie « N-ter » (en fait dans sa partie 5'). Donc la protéine résultant de ce promoteur alternatif sera une protéine de petit poids moléculaire par rapport à la protéine principale. Il y a aussi la glycosylation différente, c'est-à-dire qu'il y a des niveaux de glycosylation différents en fonction du tissu ou du type cellulaire qui exprime l'hormone.
En définitive, la synthèse des hormones peptidiques est extrêmement complexe et source de régulations aussi complexes et qui participent à l'homéostasie du système endocrinien.
c) Les hormones stéroïdes
NB : une hormone stéroïde n'est PAS un peptide ! Elle n'est pas synthétisée à partir d'un ARN messager qui a été traduit mais à partir de modifications biochimiques du cholestérol.
• Les hormones stéroïdes se fixent sur des récepteurs nucléaires
• En fonction de la structure biochimique des hormones stéroïdes, on définit :
- des glucocorticoïdes comme le cortisol (glucides)
- des minéralocorticoïdes comme l'aldostérone (Na+)
- des oestrogènes comme l'oestradiol
- des androgènes comme la testostérone (il existe aussi des androgènes surrénaliens) - des progestatifs comme la progestérone (grossesse)
• La synthèse des hormones stéroïdes se fait à partir du cholestérol (les étapes seront vues dans un cours ultérieur). On voit que la structure biochimique de l'aldostérone diffère de la structure biochimique du cortisol qui elle même diffère de celle de la testostérone qui diffère de celle de l'oestradiol.
2) La synthèse hormonale : un mécanisme répondant à une action très précise
• La notion de rythme est importante dans la synthèse hormonale. Le rythme circadien est un rythme sur 24 heures, c'est le rythme nuit-jour. Le système endocrinien dépend de ce rythme et participe à l'élaboration de ce rythme, il y a un échange permanent.
Ronéo n°4 d'UE 7 7/22
Les concentrations plasmatiques des hormones peuvent donc varier au cours de la journée ( vous l'aurez compris , c'est le rythme circadien et cette variation est fortement conservée entre espèces, au cours de l'évolution) ou encore sur plusieurs semaines, comme pour le cycle ovarien .
• Les variations peuvent être sur quelques heures ou quelques dizaines de minutes : c'est ce qu'on appelle le rythme ultradien et c'est le cas de la GnRH par exemple qui a une synthèse/sécrétion toutes les 90 minutes à peu près à partir des neurones hypothalamiques. Il existe des pathologies associées à une perte de rythme de la synthèse hormonale.
• La synthèse hormonale peut dépendre du rythme circadien. Le cycle circadien rythme la journée. Il dépend (sur le plan moléculaire) d’un réseau de neurones hypothalamiques situés dans le noyau suprachiasmatique participant à la régulation de la synthèse de la mélatonine par exemple. Il est synchronisé par l’alternance nuit-jour. Pour la mélatonine, la lumière agit par l'intermédiaire de la rétine et envoie une information au noyau suprachiasmatique situé dans l'hypothalamus qui va envoyer une information jusqu’à la glande pinéale qui va synthétiser la mélatonine. C'est un réseau complexe qui passe par la moelle épinière. La mélatonine participe au rythme biologique, notamment chez l'enfant, en favorisant le sommeil. C'est donc un système qui dépend de l'alternance nuit-jour, qui entraîne la synthèse d'une hormone, cette hormone participant directement à l'endormissement et donc au rythme biologique.
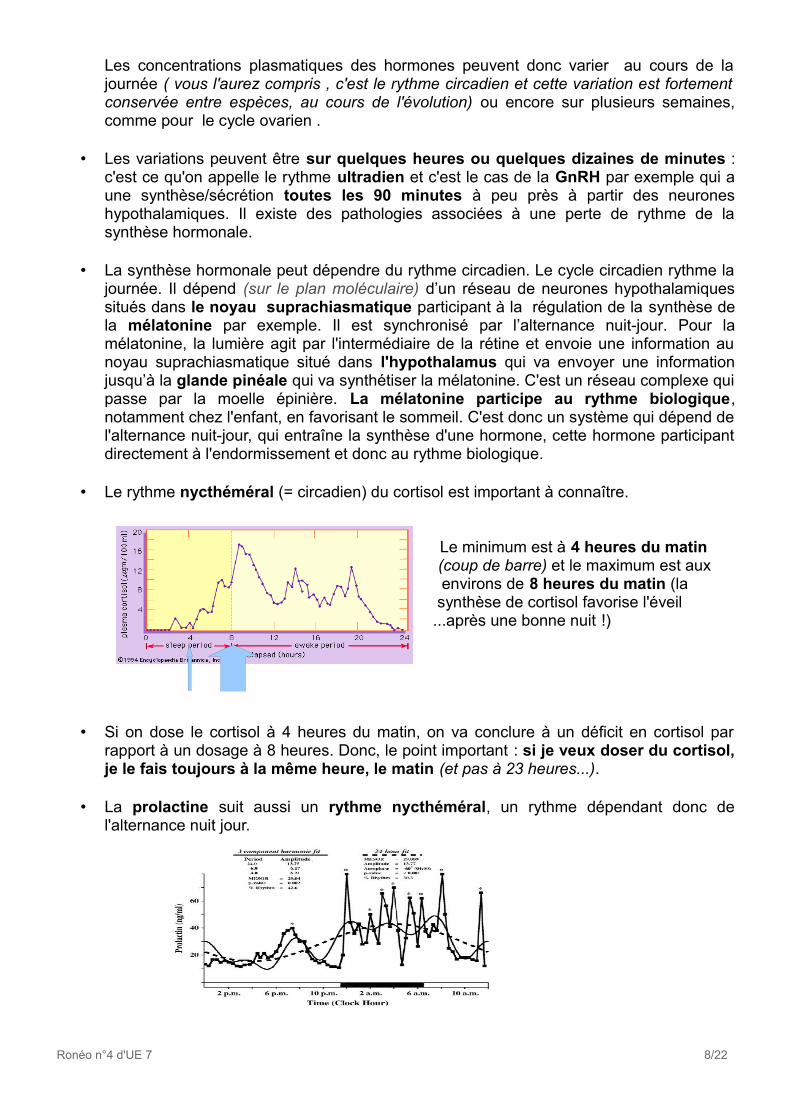
• Le rythme nycthéméral (= circadien) du cortisol est important à connaître.
MiiM Le minimum est à 4 heures du matin (coup de barre) et le maximum est aux environs de 8 heures du matin (la synthèse de cortisol favorise l'éveil ...après une bonne nuit !)
• Si on dose le cortisol à 4 heures du matin, on va conclure à un déficit en cortisol par rapport à un dosage à 8 heures. Donc, le point important : si je veux doser du cortisol, je le fais toujours à la même heure, le matin (et pas à 23 heures...).
• La prolactine suit aussi un rythme nycthéméral, un rythme dépendant donc de l'alternance nuit jour.
Ronéo n°4 d'UE 7 8/22

• La GnRH est une hormone qui est synthétisée toutes les 90 minutes chez l'adulte. Toutes les 90 minutes il y a donc un relargage de GnRH par l'hypothalamus.
3) Transport des hormones dans le sang
• Lorsque l'hormone est synthétisée, elle arrive dans des vésicules de sécrétion et est relarguée dans le sang en fonction des besoins. Les hormones peptidiques sont (plus ou moins) hydrophiles, elles sont donc transportées dans le sang sans problème. Le propre d'une hormone stéroïde, c'est d'être hydrophobe ; dans le sang elle va donc devoir se lier à des protéines de liaison pour pouvoir être transportée. Certaines hormones sont donc libres, et d'autres sont liées à des protéines de transport (par des liaisons de faible énergie). C’est important car on peut doser ou bien l'hormone libre ou bien l'hormone totale. Les protéines de transport ont des actions contre les dégradations dans le sang et la capacité de réservoir tampon (c'est notamment vrai pour les hormones thyroïdiennes) et elles peuvent faciliter le transfert membranaire.
• Il y a différentes protéines de transport et elles sont spécialisées pour les hormones. Par exemple, les glucocorticoides sont portés par la CBG (Corticosteroid binding protein).
• Elles ne sont pas toutes à apprendre par-coeur-même-les-couleurs mais il y en a « quelques unes plus importantes que d'autres » comme la TBG (Thyroxine Binding Globulin) qui joue vraiment le rôle de réservoir pour la T3 et la T4. Les IGF BPs sont importantes aussi, on a aujourd'hui la possibilité de doser ces protéines : en fonction des situations pathologiques le taux d'IGF BP va varier. La Neurophysine a la capacité de transporter de la vasopressine et de l'ocytocine, quand il y a des mutations sur les neurophysines, on va avoir des déficits en vasopressine et en ocytocine.
4) Métabolisme des hormones
a) Transformation des hormones
Les hormones sont synthétisées sous la forme d'un peptide ou alors elles sont transformées comme les hormones stéroïdes en différents métabolites et ces métabolites ont plus ou moins une activité sur le récepteur.
Ronéo n°4 d'UE 7 9/22

• On peut avoir des transformations des hormones sous forme active :
- On a la possibilité de transformer les hormones en une forme active atténuée par exemple le cortisol en cortisone. La cortisone a toujours la capacité d'activer le récepteur des glucocorticoïdes mais avec une affinité moins importante que le cortisol, de même pour l'estrone par rapport à l'oestradiol.
- La modification biochimique de certaines de ces hormones entraîne une augmentation locale de l'activité. Par exemple, la transformation de la testostérone en 5β-dihydroandrogènes est une transformation qui survient localement notamment au niveau des testicules et cette transformation augmente l’affinité pour le récepteur. Mais ce que l'endocrinologue est capable de doser et qu'il dose facilement, c'est la testostérone qui est alors le reflet de l'imprégnation androgènique (on ne dose pas ou exceptionnellement 5β-dihydroandrogènes).
• Les hormones peuvent être transformées en formes inactives :
- Les hormones peptidiques peuvent subir des protéolyses dans le sang. Par exemple, la GnRH va activer un récepteur situé au niveau des cellules hypophysaires et la GnRH a une durée de vie de 10 minutes dans le sang, c'est trop court donc on ne peut pas doser la GnRH (et pourtant elle a été active). Il y a une enzyme qui coupe ce peptide de 10 aa qu'est la GnRH en 2 peptides de 5 aa qui ne peuvent pas activer le récepteur. Donc la protéolyse va entraîner l'inactivation du peptide.
- Les hormones stéroïdiennes vont aussi subir des transformations sous la forme d'oxydoréductases ou d'hydrolases et qui vont entraîner une perte d'activité pour ces hormones stéroïdes. - L’action des désiodases est très importante en clinique car elles ont la capacité d'enlever l'iode à partir des hormones thyroïdiennes (T3 et T4). En enlevant un iode, on va former une hormone qui va dans un cas être active (lorsqu'on va passer de T4 à T3 comme on le verra dans un prochain cours) mais dans d'autres situations, par l'action d'autres désiodases, qui va être inactive (la diapo mentionne l'action de décarboxylases pour les hormones thyroïdiennes mais le prof n'en a pas parlé).
- Pour les catécholamines, on va avoir des modifications biochimiques (avec notamment des monoamines oxydases mentionnées sur la diapo et dont le prof ne parle pas) qui vont faire que l'hormone ne sera plus active mais par contre c'est elle qui sera dosée. Et celle qui sera dosée sera le reflet de l'hormone synthétisée, de l'hormone active donc.
b) Élimination des hormones
Après avoir été transformées, les hormones vont être éliminées (hormones stéroïdes ou thyroïdiennes) mais il y a des hormones qui vont être éliminées sans modification (LH, FSH, hCG, aldostérone). Cette élimination peut être rénale ou hépatique. On a donc la possibilité de doser les hormones dans les urines… (par exemple certaines équipes estiment que pour la LH et la FSH, le dosage dans les urines est un très bon reflet de l'activité de l'axe gonadotrope alors que d'autres équipes, beaucoup plus nombreuses, pensent que le dosage dans le sang est un meilleur reflet et plus facile d'accès).
Ronéo n°4 d'UE 7 10/22

III) Mécanismes d’action
1) Généralités
• Les hormones de la même classe biochimique ont des mécanismes d ’action similaires.
NB :Il y a des exceptions à cette grande idée. Aujourd'hui, on sait qu'il existe des hormones stéroïdes qui ont la capacité d'activer des récepteurs membranaires comme l'Oestradiol ou la Progestérone (on avait vu que les hormones stéroïdes se fixent sur des récepteurs nucléaires classiquement). C’est intéressant parce que ça permet de voir comment une hormone stéroïde peut avoir une action rapide sur une cellule, et non via des récepteurs nucléaires qui sont des facteurs de transcription hormono-inductibles (il y a plusieurs heures entre la synthèse de l'ARNm et la sécrétion de la protéine donc l'effet de l'oestradiol n'a pas un effet rapide a priori, mais par l’intermédiaire d'un récepteur membranaire on commence à comprendre comment l'oestradiol peut avoir un effet rapide sur les cellules).
• Les cellules doivent posséder le récepteur spécifique de l'hormone.Pour agir, une hormone doit se fixer sur un récepteur donc les cellules cibles de cette hormone doivent posséder ce récepteur. C'est important car parfois en pharmacologie, le but du traitement est justement de bloquer le récepteur, c'est de le faire disparaître de la cellule. C'est un principe utilisé en endocrinologie notamment dans le traitement de la puberté précoce.
• Les hormones se lient sur les récepteurs avec une très forte affinité.
• Les récepteurs hormonaux sont facilement saturables. C'est-à dire qu'on peut se retrouver dans une situation de désensibilisation du système parce qu'il n'y a plus de récepteur à la surface de la cellule ou dans la cellule.
• Le signal hormonal peut subir une désensibilisation.Dans certaines situations, cette désensibilisation n'arrive pas et cela peut entraîner une pathologie.
La transduction du signal est la suite des événements qui participent à la transmission du signal extra-cellulaire vers l'intérieur de la cellule.
Ronéo n°4 d'UE 7 11/22
Il s'agit alors de quelque chose de complètement coordonné. Le récepteur peut être cytosolique ou nucléaire, il va y avoir initiation du signal puis traduction du signal et enfin activation de la molécule cible. Si le récepteur est absent tout cela n'a pas lieu,il y a des situations où la traduction du signal (c'est-à-dire sa transduction) n'est pas normale, et dans ce cas là on se retrouve notamment dans des cas de pathologies tumorales où il y a une activation anormale du signal et donc augmentation du nombre de cellules.

2) Les hormones des récepteurs nucléaires
Les récepteurs des hormones stéroïdes sont dits nucléaires parce qu'ils agissent dans le noyau, en se fixant sur l'ADN. Ce sont donc des récepteurs qui sont des facteurs de transcription hormono-inductibles.
• Les hormones stéroïdes ou thyroïdes (qui ne sont pas des hormones stéroïdes) se détachent des protéines de transport (par exemple les hormones thyroïdiennes se détachent de la TBG).• Elles passent à travers la membrane cytoplasmique et pénètrent dans la cellule
grâce à leurs propriétés hydrophobes ou lipophiles (c'est donc une diffusion passive. Dans certains cas, cela se fait par des transporteurs actifs – mais le prof n'en dit pas plus donc bon...-).
• Les récepteurs des hormones lipophiliques sont des récepteurs nucléaires (déjà localisés dans l'ADN) ou cytoplasmiques.
a) Mécanismes d'action des hormones stéroïdes
• Une fois qu'elle se trouve dans le cytoplasme et qu'elle a fixé le récepteur, l'hormone stéroïde va permettre une dimérisation du récepteur puis ce récepteur dimérisé va pénétrer dans le noyau (navette vers le noyau) où il va se fixer sur l'ADN au niveau de ce que l'on appelle des éléments de réponse.
• Il va y avoir deux effets principaux : soit il va y avoir activation de la transcription, soit il va y avoir inhibition de la transcription.Tout cela dépend de l'environnement du promoteur, des co-facteurs qui vont se fixer sur le récepteur (soit ce sont des co-facteurs activateurs soit ce sont des co-facteurs inhibiteurs de la transcription).
b) Mécanismes d'action des hormones thyroïdiennes
• Pour les hormones thyroïdiennes, c'est légèrement différent car les récepteurs sont fixés sur l'ADN en absence d’hormone et la fixation des hormones sur le récepteur (qui est donc déjà sur l'ADN) va avoir pour effet de recruter ou des co-activateurs ou des co-inhibiteurs.
3) Les récepteurs hormonaux membranaires
a) Présentation générale• Les hormones hydrophiles ne passent pas la membrane cytoplasmique.
• Les catécholamines, les polypeptides et les glycoprotéines activent donc des récepteurs situés à la surface des cellules.
• Le signal extra-cellulaire est traduit en un signal intra-cellulaire par le récepteur.
• Le second messager active de nouveaux signaux cellulaires dans le cytoplasme ou dans le noyau.
Ronéo n°4 d'UE 7 12/22

Si on prend l'exemple des RCPG (Récepteurs couplés à des protéines G), les récepteurs vont activer des protéines G (AMAZING!). En fonction de la protéine G qui est activée (et ça dépend de la nature du récepteur et de la cellule dans laquelle est exprimé ce récepteur : un même récepteur peut activer une protéine Gs et une protéine Gq, il activera Gs dans une cellule A et Gq dans une cellule B. Il y a donc une spécificité qui dépend du récepteur mais aussi de la cellule qui exprime le couple récepteur-protéine G).
En fonction de la protéine G activée, il va y avoir activation d'un effecteur qui peut être l'adénylate cyclase par exemple (et ça va dans ce cas entraîner la synthèse d'AMPc), ou de la guanylate cyclase (synthèse de GMPc). Ça peut aussi être une protéine Gq qui va activer des phospholipases qui vont permettre la synthèse d'Inositol tri-phosphate (IP3). L'IP3 a la capacité d'activer un récepteur au niveau du réticulum qui va libérer du calcium en intracellulaire ; ce calcium va avoir la capacité de se fixer sur des protéines effectrices. Gq a également la particularité de produire, en activant les phospholipases, du Diacylglycérol (DAG) qui va pouvoir aller activer des protéines kinases C (PKC) qui vont avoir pour fonction de phosphoryler d'autres kinases. Il va y avoir aussi l'activation de canaux membranaires qui vont permettre la rentrée du calcium dans la cellule.
Il y a aussi des récepteurs à activité tyrosine kinase (comme le récepteur à l'insuline, le récepteur de la GH) ou qui ont la particularité de pouvoir activer des protéines tyrosine kinases.
Le but in fine est d'activer des voies de phosphorylation, ces protéines phosphorylées vont avoir 2 effets :
– Elles vont modifier les interactions protéines-protéines et elles vont pénétrer (ou entraîner la pénétration d'autres protéines) dans le noyau, ce qui va entraîner l'activation de l'activité transcriptionnelle de certains gènes.
– Elles peuvent aussi modifier au sein du cytoplasme l'équilibre de phosphorylation et l'équilibre d'interaction entre les protéines et favoriser par exemple le relargage d'un certain nombre de peptides ou l'activation globale de la cellule.
Le signal final de l'activation des récepteurs membranaires est toujours la phosphorylation des protéines.
Ronéo n°4 d'UE 7 13/22

b) Les récepteurs couplés à des protéines G (RCPG)
• Ce qui est intéressant, c'est qu'une même structure biochimique dépend de ligands extrêmement variés qui vont se fixer plus ou moins bien sur le récepteur et activer des protéines G.
Le prof a décidé de ne pas trop s'attarder sur cette diapo parce qu'on sait normalement pas mal de choses, je vous la mets avec en gras les éléments qu'il a mentionné :
• C'est une super famille comprenant plusieurs centaines de membres (> 1000).
• Il y a une architecture moléculaire commune :- Sept domaines transmembranaires reliés par des boucles extra-cellulaires et intra-cellulaires- Un domaine extra-cellulaire et un domaine intra-cellulaire- Des modifications post-traductionnelles non spécifiques : glycosylation, sulfatation, phosphorylation, clivage.
• Des mécanismes d’activation communs : la protéine G.
• Des mécanismes de désensibilisation.
Il y a 3 types de pathologies qui ont été décrites en rapport avec les RCPG :
– La maladie de Basedow qui est une hyperthyroïdie due à des anticorps dirigés contre un RCPG.
– Des pathologies rares associées à des mutations au RCPG qui font que le récepteur est : ou bien pas activé ou bien activé en l'absence de ligand (activité constitutionnelle).Ces récepteurs sont désensibilisables, c'est donc une stratégie thérapeutique qui est utilisée pour bloquer un axe endocrinien.
• Les caractéristiques des protéines G sont importantes :
- ( /!\ QCM possible : quelle est la définition d'une protéine G ?) Les protéines G sont une famille de protéines capables de fixer un GTP et d'hydrolyser ce GTP en GDP.
- Les protéines G peuvent être hétérotrimériques (composées des 3 sous-unités α, β et γ, et qui jouent un rôle dans la transduction du signal) ou monomériques (qui sont homologues d'une certaine façon à la sous-unité α des protéines G hétérotrimériques, qui porte l'activité GTPasique).
- Elles sont activées par de très nombreux récepteurs et elles peuvent stimuler ou inhiber un signal intracellulaire. Il y a une forme active et une forme inactive, la forme active est la forme ayant fixé le GTP. Après hydrolyse du GTP en GDP, le signal est arrêté.
Ronéo n°4 d'UE 7 14/22

• Dans certaines pathologies, la sous-unité α est mutée et est incapable de fixer le GTP et dans ces cas là le système n'est pas activable. Beaucoup plus intéressant, d'autres mutations entraînent une perte de l'activité GTPasique : si la sous-unité α ne peut plus hydrolyser le GTP, le système est activé en permanence. La conséquence de ça : c'est une tumeur (hypophysaire, thyroïdienne…). Une mutation somatique de la sous-unité α avec perte de l'activité GTPasique entraîne une activation permanente du système ; si ce système participe à la prolifération cellulaire, il y a tumeur. Si on voit un patient arriver avec un tableau d'acromégalie - une synthèse anormale de GH - ou un tableau d’hyperthyroïdie - une synthèse anormale de TSH -, une des hypothèses à évoquer est qu'il ait une mutation perte de fonction de l'activité GTPasique donc activation permanente du système.
• Les protéines G sont situés sous la membrane plasmique à laquelle elles sont accrochées par l'intermédiaire de lipides : ce ne sont donc pas des protéines membranaires mais qui sont fixées à la membranes plasmique et qui peuvent entrer en interaction avec les RCPG assez facilement sous l'effet de l'activation et du changement de conformation de ces récepteurs.
c) Le récepteur à l'hormone de croissance
• Il s'agit d'un récepteur avec un seul domaine transmembranaire, qui a un grand domaine extra cellulaire et qui a un domaine intracellulaire.
• Le domaine extra cellulaire fixe l'hormone de croissance, ce qui entraîne la dimérisation de 2 molécules du récepteur. Le fait qu'il y ait cette dimérisation va entraîner la phosphorylation de tyrosines situées dans le domaine intra-cellulaire, cette phosphorylation de tyrosines se fait par un mécanisme croisé (une molécule de récepteur phosphoryle l'autre et vice-versa). Cette phosphorylation du récepteur va entraîner le recrutement de protéines intra-cellulaires qui ont la particularité d'aller se fixer spécifiquement sur les tyrosines phosphorylées : cette fixation va entraîner une cascade d'activation, de phosphorylations. In fine, une protéine phosphorylée va aller se fixer sur un élément de réponse au sein du noyau et entraîner une synthèse d'une protéine donnée.
Ronéo n°4 d'UE 7 15/22
βγ
α
GDP
βγ
βγ
α +RGS
GTP
GTP
Effecteur Effecteur
Pi
α
EffecteurAdé nylate cyclasePLC β 1-3
Echangeur Na+/H+
Adé nylate cyclase (+) Adé nylate cyclase (-)Ras-Map KinasePLC β 1-3
GDP
Une perturbation de ce cycle peut entraîner des pathologies (notamment en lien avec le calcium, les pseudoaldostéronismes par exemple). Les 3 sous-unités α, β, γ forment la protéine G hétérotrimérique. L'α a la capacité de fixer du GDP. L'activation du récepteur par le ligand entraîne un relargage du GDP et la fixation du GTP sur la sous-unité α : ceci va être responsable d'une dissociation de la sous-unité α-GTP et des 2 sous-unités β et γ. C'est α-GTP qui va avoir la capacité d'activer un effecteur (adénylate cyclase, phospholipase). β et γ ont également des fonctions au sein de la cellule d'activation ou d'inhibition des effecteurs. La sous-unité α a une activité GTPasique intrinsèque, elle a donc la capacité d'hydrolyser le GTP en GDP (et relargage d'un phosphate inorganique).

• Donc le récepteur à l'hormone de croissance est un mécanisme complètement différent de celui qui est décrit pour les RCPG, même si in fine le but est la phosphorylation de protéines intra-cellulaires.
IV) Rétro-contrôle
• Le rétro contrôle dans le système endocrinien a une fonction principale qui est d'établir un équilibre entre la synthèse de l'hormone et l'effet de l'hormone. Toujours et en permanence, il faut que cette synthèse soit adaptée aux besoins du système. Il y a des rétro-contrôles négatifs (dans la très grande majorité) mais également des rétro-contrôles positifs (notamment dans l'axe gonadotrope).
• Un axe endocrinien est composé de 3 étages :- Un étage cérébral dans le cerveau, au sein de l'hypothalamus (région situé à la base du cerveau).- L'hypothalamus envoie une information à l'hypophyse (qui elle est située en dehors du cerveau – elle est en dehors de la barrière hémato-encéphalique).- L'hypophyse synthétise des hormones qui envoient des informations aux « glandes endocrines » proprement dites, dites « glandes périphériques ».Elles vont elles-mêmes synthétiser une hormone qui va agir sur les cellules cibles (c'est l'étage final).Il y a donc trois étages dans un axe endocrinien :Hypothalamus → hypophyse → glande périphérique et un étage final constitué par les cellules cibles (par exemple testicule ovaire foie etc...).
• Il existe un système porte entre l'hypothalamus et l'hypophyse : le système porte hypothalamo-hypophysaire.
Ronéo n°4 d'UE 7 16/22
Il est composé d'un réseau de capillaires qui vont recevoir des peptides de l'hypothalamus et les transporter au niveau de l'hypophyse antérieure et il va y avoir une réponse spécifique pour chaque neuropeptide. Ce système est en lien avec la circulation générale mais un peptide synthétisé par l'hypothalamus va agir tout de suite sur l'hypophyse puis va être dégradé très rapidement, ce qui fait que dans le sang périphérique (sang veineux au niveau du bras), on ne pourra pas doser ce peptide. Pour doser ce peptide, il faut faire un cathétérisme au niveau du système porte donc prélever du sang directement à la sortie de l'hypothalamus.

• On définit finalement 5 grands axes endocriniens.
- L'axe gonadotrope qui synthétise la LH et la FSH, qui va permettre la synthèse d'Oestradiol et de Testostérone.- L'axe lactotrope dont la fonction est la synthèse de la prolactine par l'hypophyse qui elle va avoir une action directe sur le tissu périphérique sans passer par une glande endocrine.
• Il y a dans les axes endocriniens une partie qui dépend de la post hypophyse (et pas de l'anté hypophyse comme les axes précédents). Ce sont des neurones qui ont pour origine l'hypothalamus mais dont l'extrémité est localisée dans l'hypophyse. Ces neurones-là sont donc différents des axes qui dépendent de l'hypophyse antérieure dans le sens où il n'y a pas de relargage de peptides dans le système porte hypothalamo-hypophysaire. Le peptide est relargué à partir de la post hypophyse qui est constituée de tissu nerveux (ce sont des extrémités axonales dans la post hypophyse contrairement à l'anté hypophyse qui est constituée de cellules endocriniennes). La post hypophyse synthétise la vasopressine et l'ocytocine qui vont avoir des fonctions par exemple sur le rein ou sur l'utérus.
• Les pathologies des hormones peuvent être d'origine hypothalamique, hypophysaire ou périphérique. En fonction du niveau de l'anomalie le traitement sera différent.
• Un peu par analogie avec le système enzymatique, le système endocrinien subit un rétrocontrôle par le produit final. L'hormone finale à la capacité de réguler au niveau de l'hypothalamus ou de l'hypophyse la synthèse des hormones hypothalamiques ou hypophysaires. Prenons l'exemple de l'axe corticotrope. Il va y avoir synthèse du CRF (Corticotropin releasing hormon) qui va aller réguler la synthèse, par l'anté hypophyse, de l'ACTH (ou béta-corticotropin). Cette ACTH va agir au niveau du cortex surrénalien et va réguler au niveau des cellules surrénaliennes la synthèse du cortisol. Donc pour l'instant, on a un axe uniquement activateur.
Ronéo n°4 d'UE 7 17/22
- L'axe thyréotrope a pour fonction de synthétiser les hormones thyroïdiennes. Il est composé d'un étage hypothalamique qui synthétise la TRH qui va agir sur l'hypophyse (antérieure) qui va synthétiser la thyrotropine ou TSH qui elle même va agir sur la thyroïde.
- L'axe corticotrope : il y a synthèse de CRF, puis d 'ACTH qui va agir sur les surrénales.
- L'axe somatotrope qui synthétise la GH (le prof précise qu'il y a une erreur dans le tableau ci-contre pour l'axe somatotrope – ça ne devait pas être somatotropine mais GH – mais on reverra tout ça dans un autre cours donc bon...)
NB : Pituitary c'est l' hypophyse , les enfants .

V) Exploration hormonale
1) Généralités
• On considère aujourd'hui que la concentration plasmatique ou urinaire des hormones est un reflet de leur activité. La concentration plasmatique des hormones est le reflet de la sécrétion de ces hormones (c'est également un reflet de leur catabolisme, pour certaines hormones c'est le reflet de la quantité de protéines porteuses). Globalement, s'il y a trop d'hormone dans le sang, c'est qu'il y a eu une synthèse anormale, trop de synthèse. Quand il n'y en a pas assez, on considère qu'il y a eu une synthèse insuffisante.
• La demi-vie d'une hormone est le temps requis pour diminuer de moitié la concentration hormonale. En général, la demi vie de l'hormone est suffisamment longue pour permettre son dosage dans le sang sans difficulté (sauf pour la GnRH par exemple comme on l'a vu précédemment) . Dans certaines situations où la demi-vie est particulièrement diminuée, il va y avoir diminution de la concentration plasmatique des hormones.
• Les concentrations physiologiques sont responsables des effets physiologiques.
On considère que ce qu'on est capable de doser dans le sang est le reflet des effets physiologiques. Par exemple devant un tableau d'hyperthyroïdie, il est logique de voir une augmentation de la concentration d'hormones thyroïdiennes dans le plasma. Si on se retrouve face à un tableau d’hypothyroïdie avec des concentrations plasmatiques élevées, il faut évoquer un problème au niveau des récepteurs de ces hormones.
• Les variations physiologiques de la concentration hormonale modifient la réponse des effecteurs.
Cette propriété physiologique est logique et est utilisée en exploration fonctionnelle. Si on injecte un agoniste qui mime les effets de l'hormone et si l'axe est fonctionnel, on doit observer une augmentation de l'effet de cette hormone qui est généralement vue par l'augmentation de la synthèse et la sécrétion à partir de la glande cible de cette hormone.
Ronéo n°4 d'UE 7 18/22
= CRF
= ACTH
Le cortisol a pour capacité de réguler négativement la synthèse du CRF et a également une action sur l'hypophyse antérieure.
- Quand il y a trop de cortisol, il y a moins de CRF et moins d'ACTH. - Quand il n'y a pas suffisamment de cortisol, il y a plus de CRF et plus d'ACTH.
En clinique on dose l'ACTH, le cortisol et exceptionnellement le CRF.

• La concentration des hormones peut avoir plusieurs effets :
- L'effet de « priming » :
On va donner un petit peu d'hormone à un patient, la réponse va être modérée et quand on va en redonner une dose à peu près équivalente quelques heures ou quelques jours plus tard, on va avoir un effet très important. On a donc dans un premier temps stimulé l'axe de façon infra-clinique (ou infra-biologique) et dans un second temps l'axe est prêt à recevoir une deuxième stimulation.
Il a été montré que cet effet de « priming » est dû à une augmentation du nombre de récepteurs dans les cellules cibles (mais aussi une capacité de la cellule à mieux répondre à une stimulation hormonale) d'où une augmentation de la réponse cellulaire.
Dans l'axe gonadotrope,en physiologie, durant la fin de la phase folliculaire avant le pic ovulatoire de LH, il y une préparation du système comme si tout était organisé pour qu'au moment où il y a un relargage de LH l'ovulation puisse survenir. Le système subit une maturation, une activation, un « priming » qui fait que le relargage brutal de LH permet l'ovulation au 14ème jour du cycle.
- La désensibilisation :
Si on administre trop d'hormone à un patient, il y aura à l'inverse un effet de désensibilisation, de perte de la réponse hormonale (c'est une stratégie utilisée dans certaines situations pathologiques).
Il y a diminution du nombre de récepteurs à la surface de la cellule et donc diminution de la réponse cellulaire.
La sécrétion pulsatile de certaines hormones prévient la désensibilisation et est absolument indispensable à la synthèse hormonale.
Si on administre de la GnRH à un patient en continu (et non par pics toutes les 90 minutes), on bloque son axe gonadotrope complètement. C'est utilisé pour les cancers hormono-dépendants, par exemple le cancer de la prostate où il faut tout faire pour que la testostérone soit au plus bas, on administre des agonistes de la GnRH en continu. Le patient va avoir une désensibilisation de l'axe gonadotrope et un arrêt de synthèse de testostérone. C'est ce qu'on appelle une castration chimique. C'est aussi utilisé dans le cas du traitement de la puberté précoce chez le garçon et chez la fille.
• Il y a deux moyens d'évaluer l'activité d'une hormone : soit on regarde la quantité qui est présente dans le sang, soit on regarde sa bio activité.
- On a la possibilité de doser une hormone de façon statique donc d'évaluer le niveau sécrétoire du tissu endocrine.
Le patient vient à 8 heures, on lui fait une prise de sang, on regarde son taux de cortisol, de T3, de T4… et on est capable de dire si le patient est dans une situation d'hyper ou d'hypo activité endocrinienne de façon chronique.
Ronéo n°4 d'UE 7 19/22

- On a aussi la possibilité de faire des explorations dynamiques donc d'évaluer l'activité de l'hormone au niveau des tissus cibles.
Dans ce cas, on rajoute des agonistes des axes endocriniens et on regarde l'effet de ces agonistes ou au contraire on bloque l'axe en rajoutant le produit final et on regarde si l’administration de l'hormone périphérique a bien un effet de rétro-contrôle négatif sur l'hypothalamus et sur l'hypophyse. Dans certaines situations pathologiques, comme par exemple la maladie de Cushing qui consiste en une sécrétion anormale d'ACTH par l'hypophyse, lorsqu'on rajoute un agoniste du cortisol, on ne peut pas bloquer l'axe corticotrope : l'ACTH reste élevée alors qu'on a rajouté des quantités importantes de glucocorticoide. Cela témoigne d'une autonomie de l'hypophyse par rapport à l'hormone périphérique. C'est le témoin d'un dérèglement hypophysaire qui explique cette maladie de Cushing qui est une synthèse trop importante de cortisol (et qui entraîne un tableau clinique grave et compliqué).
• Les milieux biologiques où l'on dose les hormones sont :
- Le sang : veineux périphérique, exceptionnellement on fait un cathétérisme dans le sinus pétreux pour prélever du sang dans le système porte hypothalamo-hypophysaire (notamment pour doser le CRF).
- Les urines : sur la diurèse des 24 heures, on a la capacité de doser les hormones qui ne sont pas transformées ou les métabolites de certaines hormones.
- La salive : plus rare en clinique (dosage de la testostérone par exemple), surtout pour les études de cohortes.
- Et exceptionnellement (à l’hôpital) dans le LCR, le liquide d'ascite, de ponction pleurale ou encore de kyste. On commence aussi à savoir doser les stéroïdes dans les cheveux (ça permet de voir l'imprégnation stéroïdienne au cours du temps), mais c'est anecdotique.
Donc en clinique c'est surtout le sang et les urines.
2) Méthodes • On peut faire des dosages biologiques pour analyser l’activité biologique d’une
hormone (bioactivité). C'est rare et difficile à mettre en œuvre (réservé à des laboratoires très spécialisés). Ça manque de spécificité (ça dépend de l'hormone en fait) et globalement de sensibilité. Cette méthode est plutôt utilisée dans le domaine de la recherche sauf pour les anticorps dirigés contre le récepteur de la TSH, et la majorité des dosages se fait selon une méthode de radio immunologie pour quantifier la quantité de récepteur. On a la possibilité de montrer que ces anticorps sont bien activateurs sur le récepteur (dans le sens où il y a aussi des anticorps inhibiteurs).
• La majorité des dosages en biologie sont faits par quantification de l'antigène par une méthode de radio immunologie ou par une méthode de type ELISA (dites froides).On a fabriqué un anticorps spécifique polyclonal ou monoclonal contre l'antigène à doser, l'antigène (ou l'anticorps si on est pas dans une méthode par compétition) est marqué soit de manière radioactive (radioimmunodosage, par exemple l'iode), soit avec des molécules fluorescentes, luminescentes ou encore avec des enzymes, avec la biotine...
Ronéo n°4 d'UE 7 20/22

- Soit c'est une méthode par compétition : méthode de radio immunodosage (RIA)
- Soit c'est une méthode beaucoup plus sensible, c'est la méthode dite de(par) sandwich (méthode immunométrique) :
Pour les petites molécules, il n'est pas facile de trouver 2 épitopes donc on préfère la méthode par compétition. Cependant la méthode par sandwich est automatisable alors que la méthode par RIA reste manuelle.
VI) La pathologie hormonale
Il y a deux grands groupes de pathologies. On observe un défaut de régulation de la synthèse ou de l'activité hormonale.
Ronéo n°4 d'UE 7 21/22
Dans ce cas-là l'anticorps est fixé sur le plastique du tube à essai, l'antigène marqué est mélangé avec l'antigène à doser et il y a compétition pour l'anticorps : une partie de l'antigène marqué ne pourra pas se fixer (on essaye de faire en sorte que l'antigène non marqué se fixe le mieux possible à l'anticorps). Plus il y a d'antigène non marqué dans le sérum, et moins il y aura d'antigène marqué sur l'anticorps. Donc plus le signal diminue, et plus il y a d'hormone dans le plasma (cf petit graphique ci-contre). Les anticorps sont en défaut , pour que la compétition puisse se faire.
Il a deux anticorps polyclonaux ou monoclonaux dont l'un est marqué ( par les méthodes qu'on a vu précédemment). Les anticorps sont en excès et on dose l'hormone dans l'échantillon. C'est une méthode de choix car elle est très spécifique ( lle utilise 2 anticorps) et elle peut être très sensible si les anticorps utilisés sont très sensibles. L'anticorps est fixé sur le tube à essai, l'antigène a la capacité de fixer l'anticorps, l'anticorps marqué se fixe sur un autre épitope de l'antigène. On quantifie le signal. Plus le signal est important, plus la quantité d'hormone dans l'échantillon est importante. On n'est pas dans un système de compétition. C'est la méthode de choix aujourd'hui utilisée.
C'est une méthode simple pour les petites molécules, l'échantillon comporte l'hormone à doser. La diapo mentionne une amplification mais le prof n'en parle pas.
(H)
IS NB ici c'est un anticorps qui est marqué et non pas un antigène comme dans la méthode par compétition

• Soit il y a un excès de synthèse (ou excès d'activité) :
- Soit il y a une augmentation anormale de la production de l'hormone (hyperactivation). - Soit il y a (le plus souvent) une croissance anormale de la glande endocrine (augmentation des cellules qui entraîne une augmentation de la synthèse hormonale).
• Soit il y a un défaut de synthèse (ou d'activité au niveau du récepteur) :
- Il peut y avoir une anomalie du développement qui fait que la glande endocrine n'est pas développée.- Il peut y avoir une anomalie enzymatique dans la synthèse hormonale d'où une diminution du produit final et donc il y aura une hypoactivité endocrinienne.
- Il peut aussi y avoir une atrophie de la glande (maladies auto-immunes par exemple : la glande s'est développée normalement mais il y a eu l'action d'auto anticorps notamment chez l'adulte qui entraîne une diminution de la taille de la glande ) donc il y aura une hypoactivité de cette glande endocrine et un tableau en rapport avec une baisse de la synthèse...
- Il y des situations iatrogènes (traitements qui entraînent des défauts de synthèse).
VII) Take Home : petit résumé du cours à bien connaître
• Une hormone permet la transmission de l’information entre deux tissus• Les hormones ont des structures physico-chimiques bien précises (encore une fois, une
hormone stéroïde c'est pas une hormone peptidique !)• Les hormones sont synthétisées par les glandes endocrines (qui subissent des régulations
extrêmement fines)• Elles agissent sur des tissus cibles périphériques ou endocriniens (cf. les 3 étages)• Elles subissent un catabolisme spécifique ou non (quand le catabolisme est spécifique, le
dosage du catabolite est le reflet de la molécule active ; s'il est non spécifique il n'y a pas d’intérêt à doser)• Toutes les hormones agissent par l’intermédiaire d’un récepteur• Les structures des récepteurs dépendent en partie de la structure biochimique de
l’hormone (exemple de la progestérone et de l'oestradiol qui activent un récepteur membranaire)• Les récepteurs sont membranaires pour les hormones hydrophiles et
cytoplasmiques ou nucléaires pour les hormones lipophiles (il y a des exceptions) • Les récepteurs nucléaires sont des facteurs de transcription hormono-dépendant• Les récepteurs membranaires activent des voies de transduction intra-cellulaires• Les hormones constituent des axes endocriniens régulés par des rétro-contrôles
négatifs (il existe aussi des rétro-contrôles positifs)• La concentration plasmatique des hormones est le reflet combiné de la sécrétion
et du catabolisme des hormones• Le bilan hormonal est basé sur le dosage des concentrations hormonales de base
et après stimulation des axes endocriniens (+++ source de QR +++)• La pathologie hormonale signe un défaut de production hormonale (excès ou défaut)
ou de réceptivité hormonale (hyper ou hypoactivation des récepteurs)
~ THE END ~
Une (très ) petite dédicace à tous mes lotus (mon petit surtout ♥ mais aussi les grands, et les moins grands aussi).
Ronéo n°4 d'UE 7 22/22