chapitre v: le reploiement des protéines · l’énergie libre energie totale = energie...
TRANSCRIPT
Chapitre V: Le reploiement des protéines
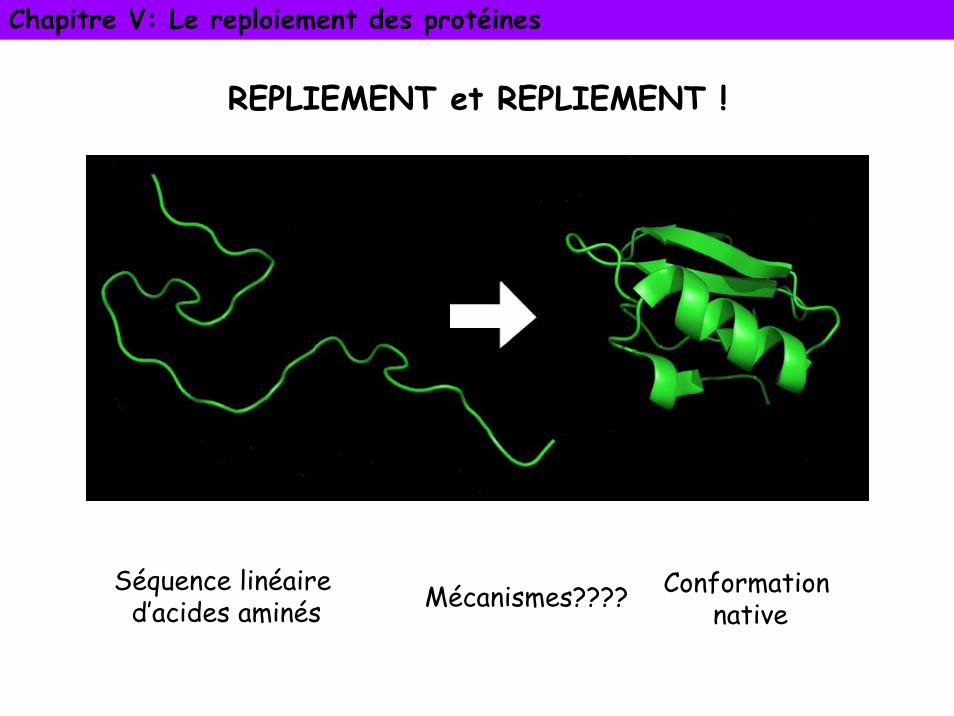
Séquence linéaire d’acides aminés
Conformation nativeMécanismes????
REPLIEMENT et REPLIEMENT !

Pourquoi s’intéresser au repliement des protéines?
Si on pouvait comprendre le chemin de repliement des protéines on pourrait:
Prédire la structure 3D d’une protéine à partir de sa séquence primaire
Créer des protéines plus stables qui se conforment plus facilement
Identifier les résidus essentiels pour le repliement (qui sont à exclure des expériences de mutagénèse dirigée)
Comprendre les maladies liées à l’altération de la conformation des protéines (Kuru et Creutzfeldt-Jakob chez l’homme, tremblante du mouton, encéphalopathie bovine et prion)
Chapitre V: Le reploiement des protéines
Chapitre V: Le reploiement des protéines
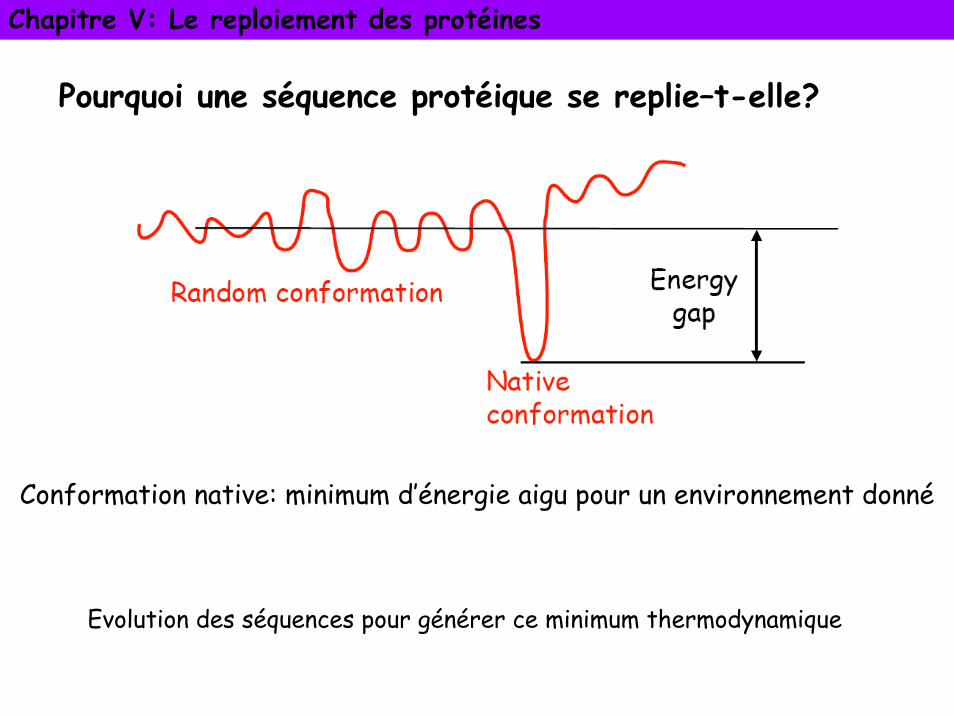
Pourquoi une séquence protéique se replie–t-elle?
Conformation native: minimum d’énergie aigu pour un environnement donné
Evolution des séquences pour générer ce minimum thermodynamique
Chapitre V: Le reploiement des protéines
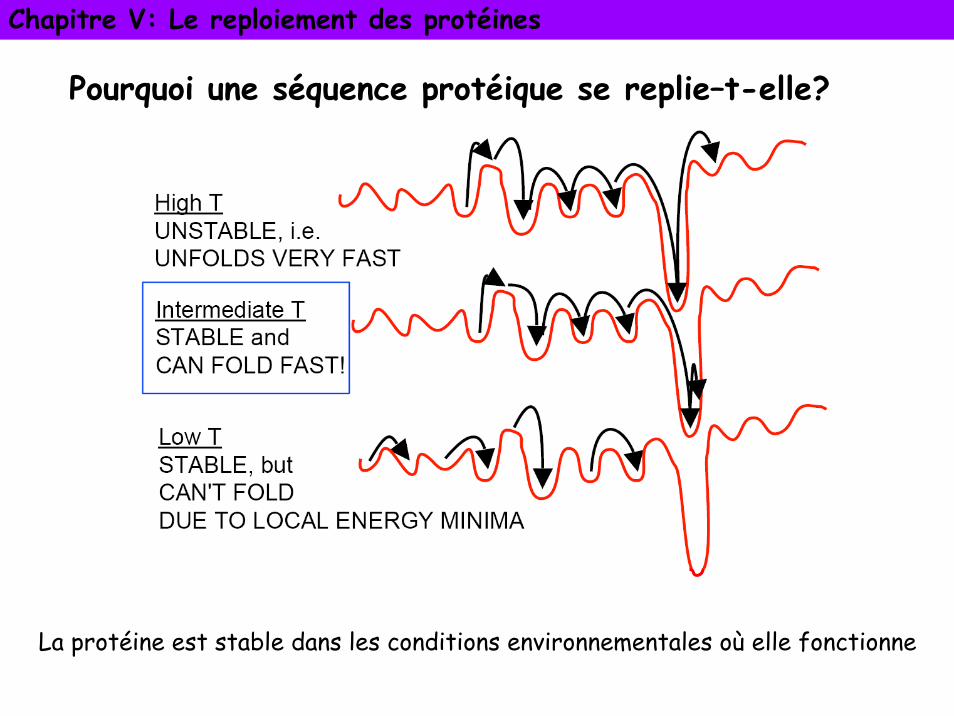
Pourquoi une séquence protéique se replie–t-elle?
La protéine est stable dans les conditions environnementales où elle fonctionne
Chapitre V: Le reploiement des protéines
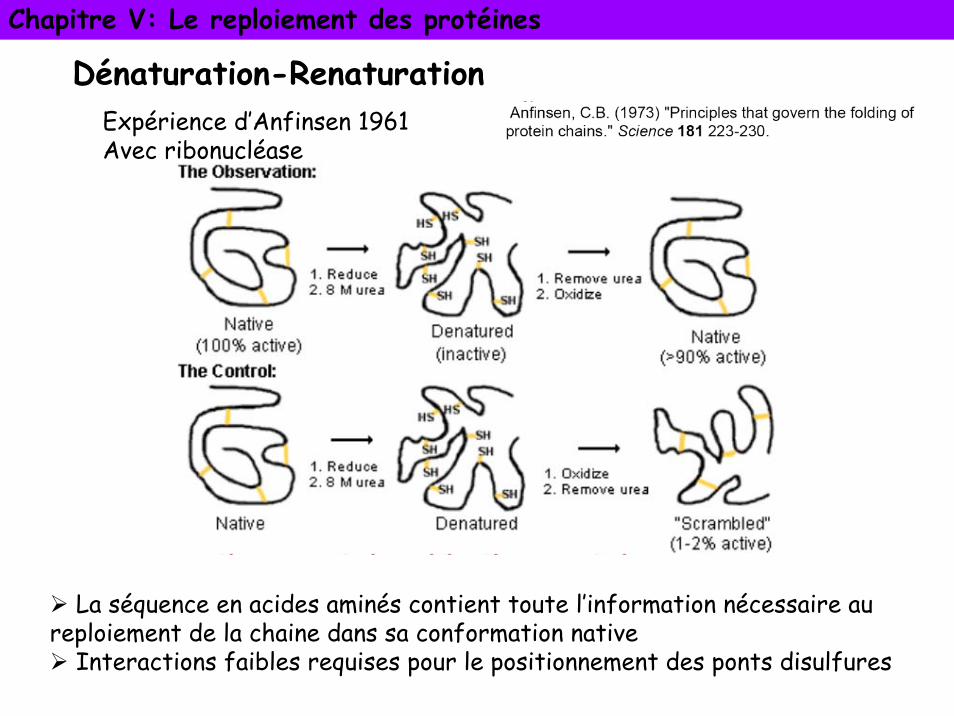
Dénaturation-RenaturationExpérience d’Anfinsen 1961Avec ribonucléase
La séquence en acides aminés contient toute l’information nécessaire au reploiement de la chaine dans sa conformation native
Interactions faibles requises pour le positionnement des ponts disulfures

Chapitre V: Le reploiement des protéines
Point de vue thermodynamique
Les protéines peuvent se dénaturer et se renaturer
L’information nécessaire est contenue dans la séquence
Le repliement naturel correspond au minimum d’énergie libre
Mais
Plusieurs protéines ne peuvent adopter une conformation native in vitro
Nécessité pour certaines protéines d’un reploiement assisté
Protéines avec différents états stables (Alzheimer, Prion…)

Chapitre V: Le reploiement des protéines
L’énergie libreEnergie totale = Energie d’interaction + désordre
à température constante T
Enthalpie: ∆H caractérise la stabilité. Si ∆H<0 les produits sont plus stables que les réactants
Entropie: ∆S caractérise le désordre. Si ∆S>0 les produits sont plus désordonnés que les réactants
Dénaturé (D) Natif(N)A l’équilibre ∆G = 0 Balance entre les plus fortes interactions cohésives de N (plus basse énergie) et la plus grande liberté de conformation de D (plus haute entropie).
Une augmentation de T augmente l’influence de ∆S
Une diminution de T augmente l’influence de ∆H

Chapitre V: Le reploiement des protéines
Dénaturé (D) Natif(N)
Etat natif stable: ΔG<0
Dans conditions physiologiques, équilibre déplacé vers N
Grandes valeurs négatives de ∆S : perte de liberté conformationnelle de D avec plusieurs conformations vers N avec une seule conformation
La protéine dans l’état natif doit trouver de fortes interaction inter-résidus pour atteindre grande valeur négative de ∆H
L’énergie libre
ΔG++
ΔG
G
Réactants
produits
États de transition
Passage état initial à état final avec intermédiaires instables
ΔG ≈ 20-50kJ mol-1

Chapitre V: Le reploiement des protéines
Rôle de la séquence protéique
Rnase bovine

Chapitre V: Le reploiement des protéines
Dans le vide le repliement permet la formation des liaisons H et des ponts salinsLa chaine repliée est beaucoup plus organisée que la chaine dépliée
Interactions polarisantes entre chaines latérales et solvant plus fortes dans l’état dépliéPlus d’interactions solvant/solvant que solvant/chaines latérales en état repliéLes groupes non polaires entrainent un accroissement de l’ordre des molécules d’eau dans l’état déployé
Plus d’interactions solvant/solvants en absence de la solvatation des groupes polaires (partiellement) enfouiLes molécules d’eau sont relativement ordonnées autour des groupes polaires
ΔGtotal= ΔHchaine+ΔHsolvant-T(ΔSchaine+ΔSsolvant)
Protéine déployée: chaines latérales sont en interaction avec les molécules d’eauProtéine reployée est hautement ordonnée/à structure déployée:
Biochimie, R.H. Garrett, C.M. Grisham, B. Lubochinsk
Contribution des résidus polaires et hydrophobes au repliement
Chapitre V: Le reploiement des protéines
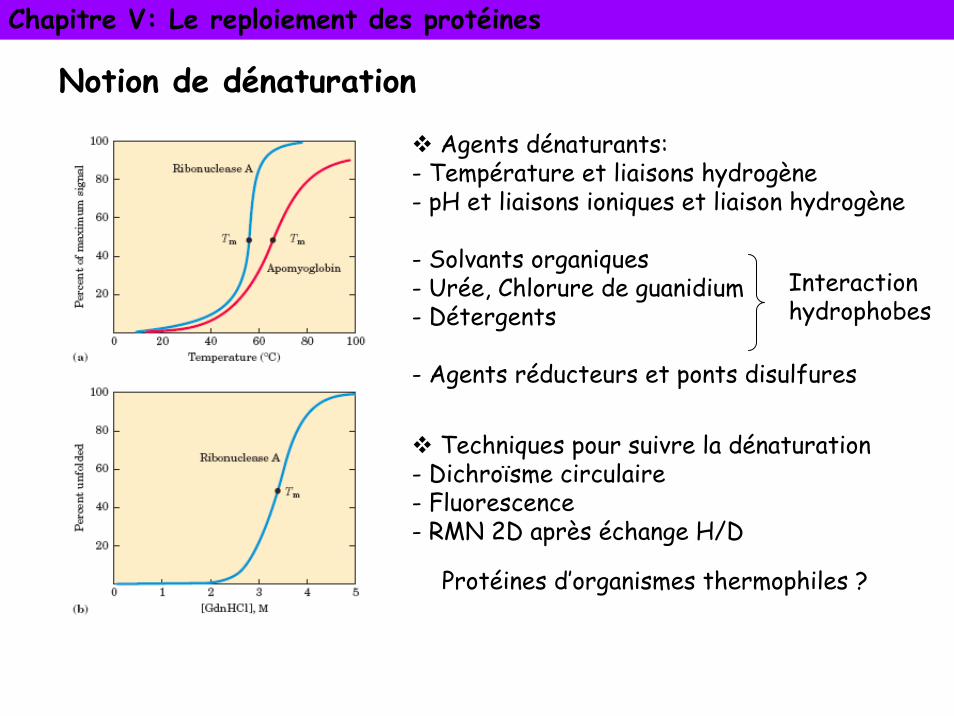
Notion de dénaturation
Agents dénaturants:- Température et liaisons hydrogène- pH et liaisons ioniques et liaison hydrogène
- Solvants organiques- Urée, Chlorure de guanidium- Détergents
- Agents réducteurs et ponts disulfures
Interactionhydrophobes
Techniques pour suivre la dénaturation- Dichroïsme circulaire- Fluorescence- RMN 2D après échange H/D
Protéines d’organismes thermophiles ?

Chapitre V: Le reploiement des protéines
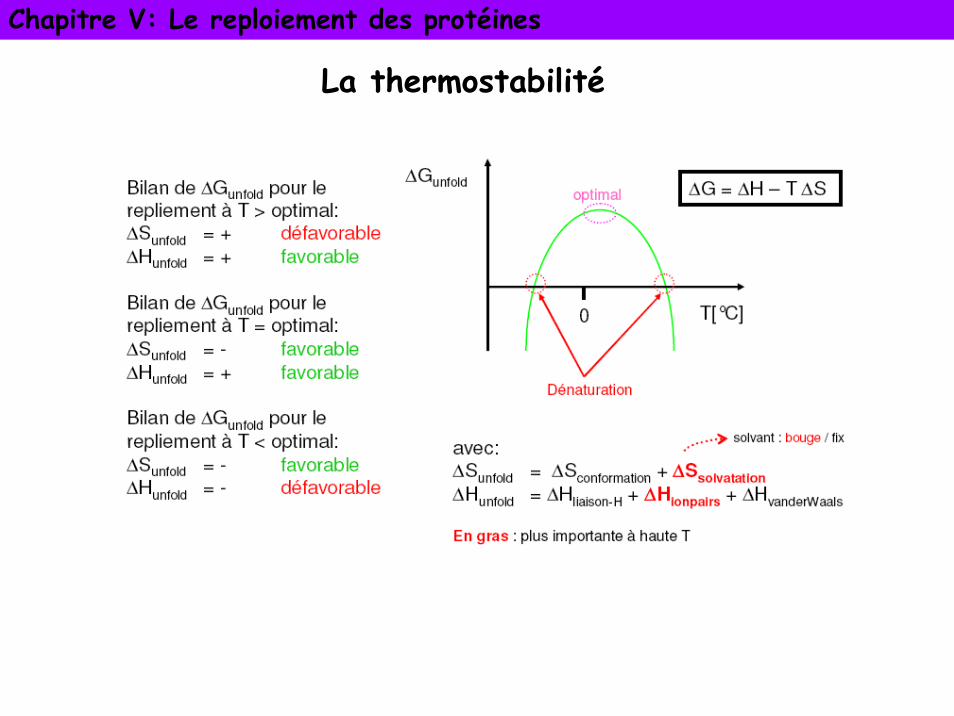
L’énergie de dépliement (ΔG unfold)
La réaction se déplace dans le sens ou ΔG est négatif
Chapitre V: Le reploiement des protéines
La thermostabilité

Chapitre V: Le reploiement des protéines
Forces contribuant à la stabilité des protéines
Contribution à l’enthalpie
Ponts salins: interactions ioniques fortes mais ne stabilisent pas fortement les protéines
Forces de van der Waals: (interactions entres dipôles permanents et/ou induits) faibles mais stabilisent significativement les structures des protéines
Liaisons hydrogène internes d’une protéine stabilisent faiblement les protéines (essentiellement locales, structures compactes locales)
Forces hydrophobes facteur déterminant pour provoquer le repliement qui leur donne conformation native
La structure des protéines est le résultat d’un équilibre fragile entre des forces compensatoires puissantes
Contribution à l’entropie
Ponts disulfures: dans la protéine repliée et non repliée. Grand effet sur ΔS
Chapitre V: Le reploiement des protéines
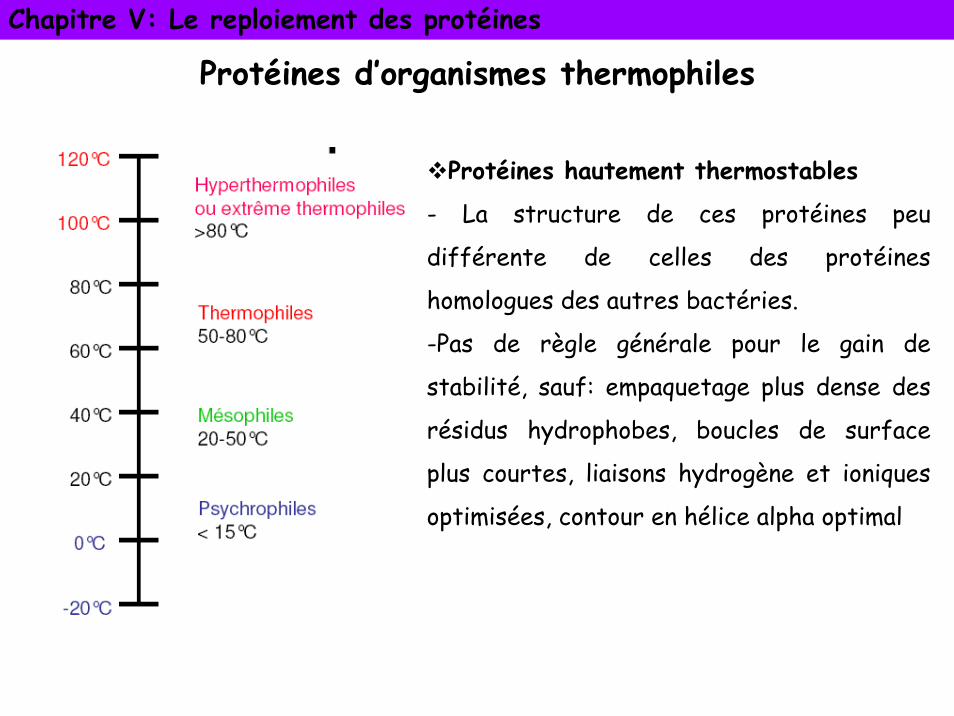
Protéines d’organismes thermophiles
Protéines hautement thermostables
- La structure de ces protéines peu
différente de celles des protéines
homologues des autres bactéries.
-Pas de règle générale pour le gain de
stabilité, sauf: empaquetage plus dense des
résidus hydrophobes, boucles de surface
plus courtes, liaisons hydrogène et ioniques
optimisées, contour en hélice alpha optimal

Chapitre V: Le reploiement des protéines
Le repliement cinétique des protéinesPlusieurs modèles construits à partir de considérations théoriques ouexpérimentales:
• Modèle framework: les structures secondaires se forment d’abord puis arrangement des structures secondaires
• Modèle diffusion-collision: Quelques résidus interagissent entre eux puis se forment les structures secondaires et le noyau hydrophobe
• Modèle d’effondrement (collapse) hydrophobe: Les résidus hydrophobes se rassemblent pour former le cœur de la protéine (état « effondré »=molten globule ou globule fondu) puis la chaine s’organise et les structures secondaires se forment
Ou mélange de toutes ces théories ou pas de règle fixe, toutes les approches sont possibles selon la protéine

Chapitre V: Le reploiement des protéines
Point de vue cinétique: Le paradoxe de Levinthal
Soit une chaine de 100 résidus
- chaque résidu n’adopte que 2 conformations (sous-estimé)
Soit 2100 ou 1030 conformations possibles
- 10-11 sec pour passer d’une conformation à une autre (sous-estimé)
1019 sec soit 1011 années!!!!!!
Hypothèse: exploration aléatoire des conformations possibles avant de trouver la conformation native
MAIS:
Temps de génération d’une bactérie: 30 minutes
Synthèse d’une protéine = quelques secondes
Passage en « molten globule » quelques micro-milliseconde
Atteinte de l’état natif en quelques millisecondes

Chapitre V: Le reploiement des protéines
Le repliement cinétique des protéines
Les protéines ne peuvent pas explorer l’espace des configurations de façon aléatoire
Existence de un ou plusieurs chemins privilégiés pour arriver à l’état natif stable qui correspond au
minimum thermodynamique stable
Combinaison de mécanismes sous contrôle mixte: cinétique et thermodynamique
Chapitre V: Le reploiement des protéines
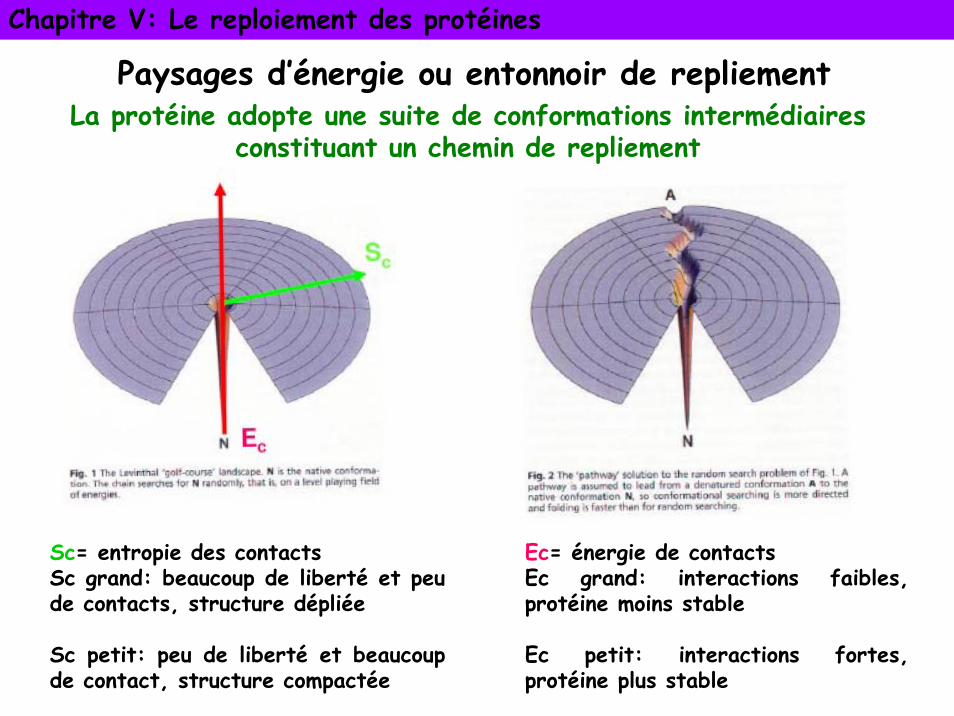
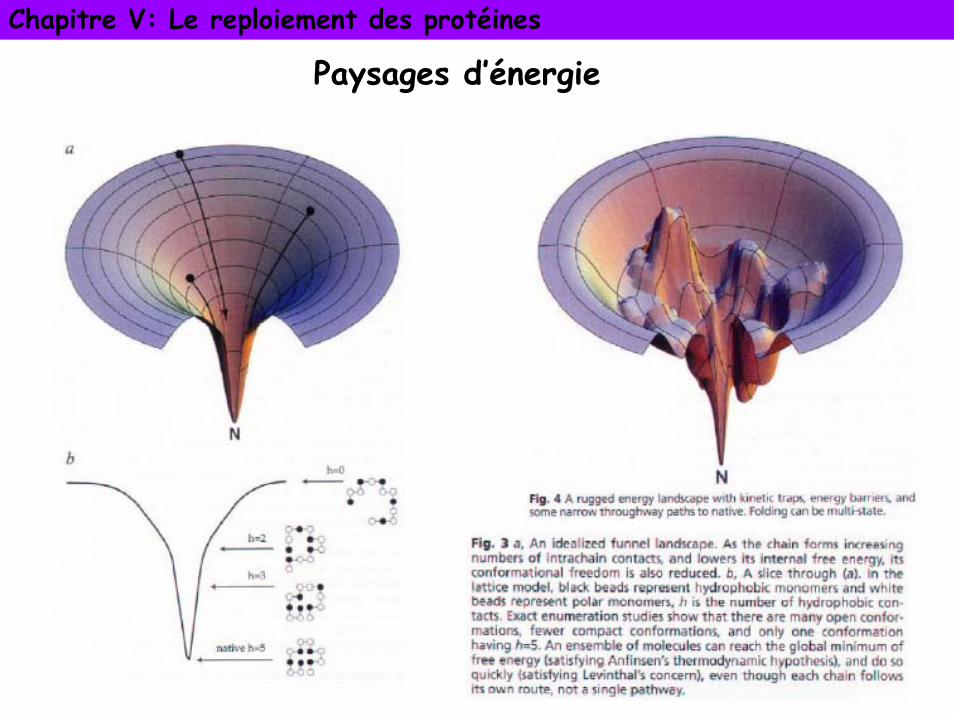
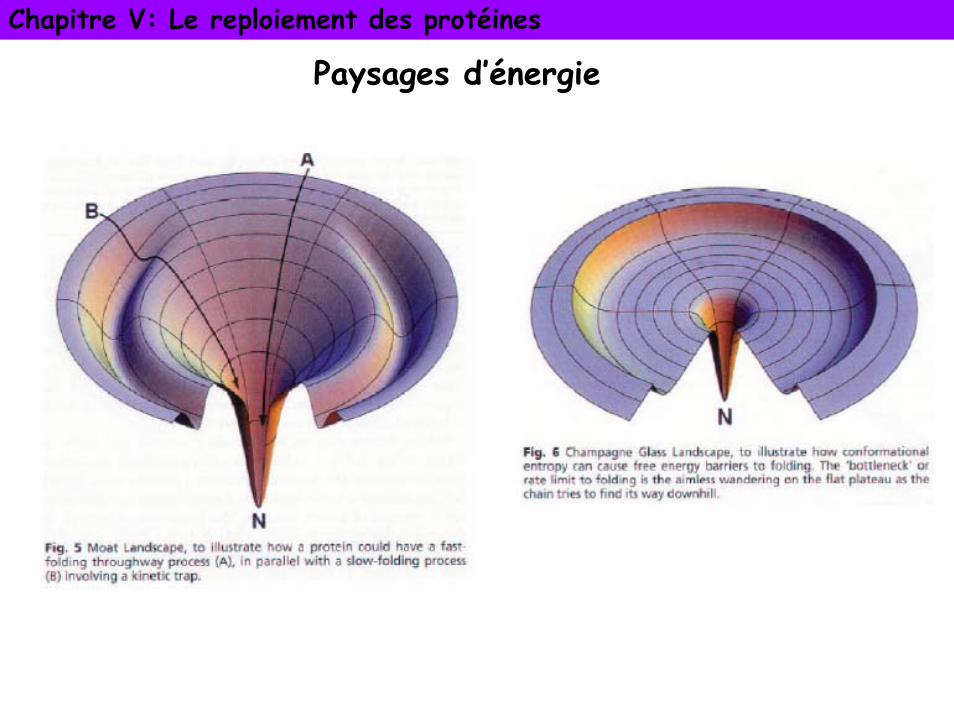
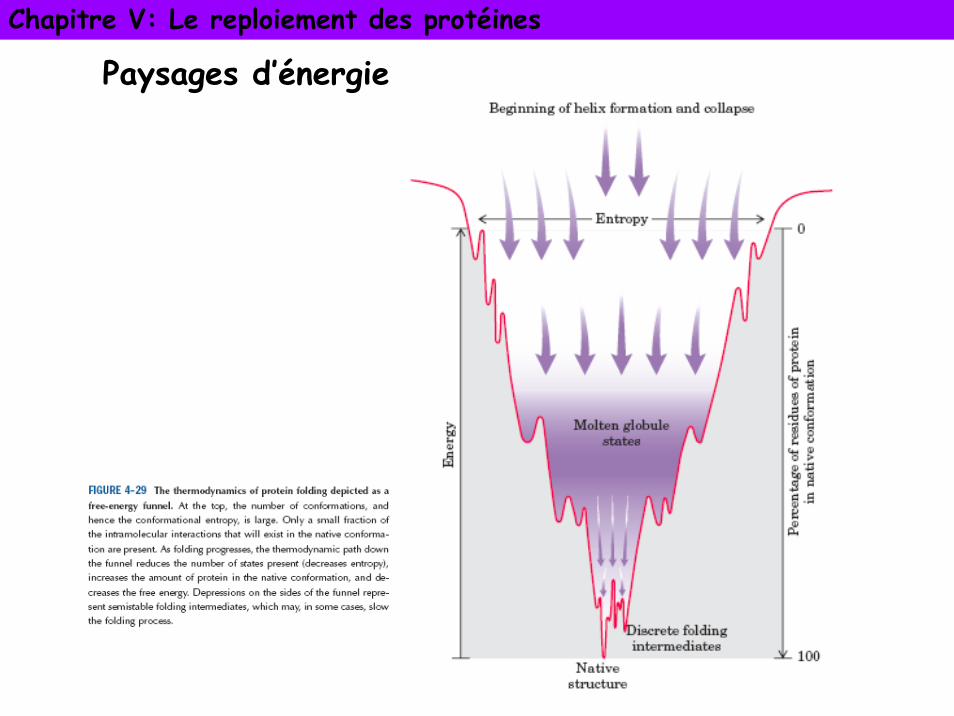
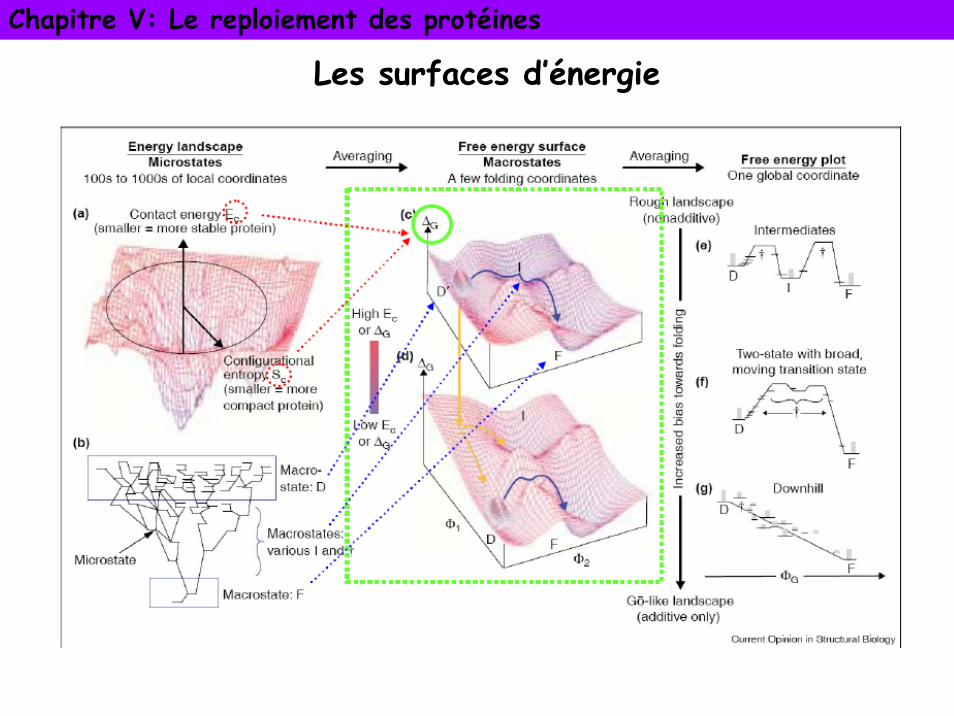
Paysages d’énergie ou entonnoir de repliement
Sc= entropie des contactsSc grand: beaucoup de liberté et peu de contacts, structure dépliée
Sc petit: peu de liberté et beaucoup de contact, structure compactée
Ec= énergie de contactsEc grand: interactions faibles, protéine moins stable
Ec petit: interactions fortes, protéine plus stable
La protéine adopte une suite de conformations intermédiaires constituant un chemin de repliement
Chapitre V: Le reploiement des protéines
Paysages d’énergie
Chapitre V: Le reploiement des protéines
Paysages d’énergie
Chapitre V: Le reploiement des protéines
Paysages d’énergie
Chapitre V: Le reploiement des protéines
Les surfaces d’énergie
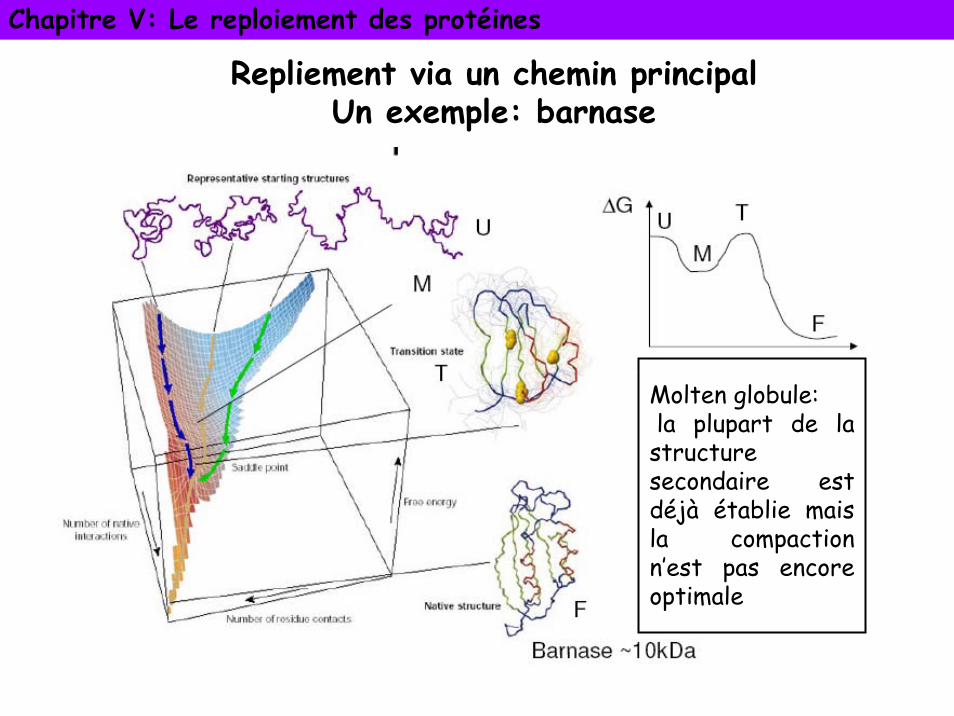
Chapitre V: Le reploiement des protéines
Repliement via un chemin principalUn exemple: barnase
Molten globule:la plupart de la
structure secondaire est déjà établie mais la compaction n’est pas encore optimale
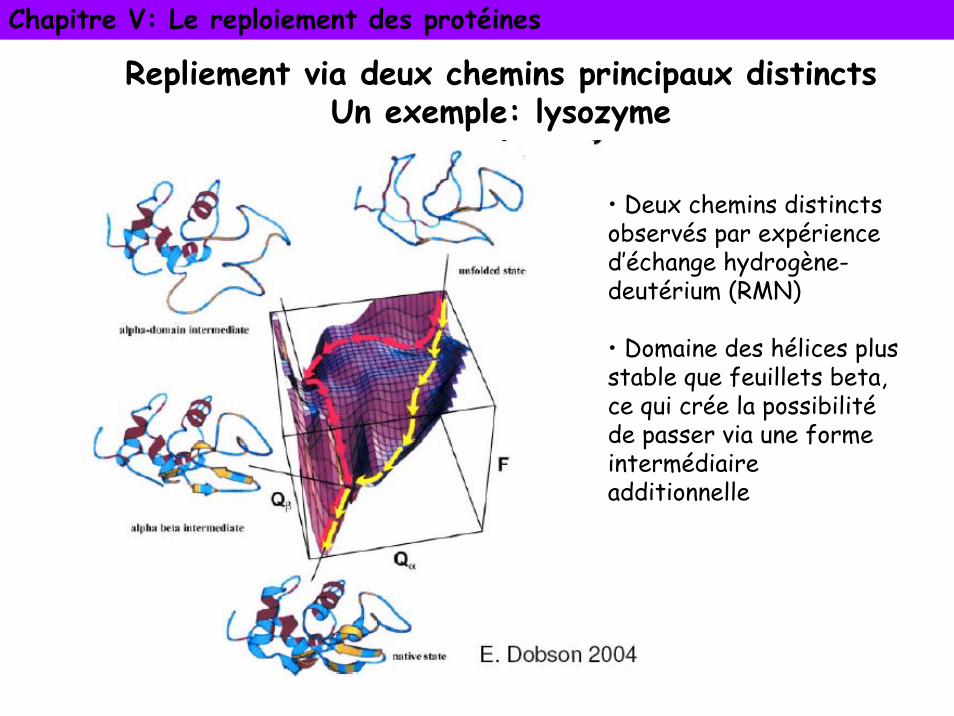
Chapitre V: Le reploiement des protéines
Repliement via deux chemins principaux distinctsUn exemple: lysozyme
• Deux chemins distincts observés par expérience d’échange hydrogène-deutérium (RMN)
• Domaine des hélices plus stable que feuillets beta, ce qui crée la possibilitéde passer via une forme intermédiaire additionnelle

Chapitre V: Le reploiement des protéines
Repliement des protéines in vivoDifférences entre repliement des protéines in vivo par rapport aux
expériences in vitro ou in silicoConcentration: dans la cellule concentration est estimée à 350 mg/mL. In vitro, on travaille à 10 mg/mL
chemin de repliement: les protéines sont synthétisées du N-ter vers C-ter. La partie N-ter naissante peut se replier dans des formes intermédiaires uniquement accessibles en raison de la taille réduite de la séquence naissante.
Repliements intermédiaires: des protéines sont déplacées dans les différents organelles et environnements de la cellule. Font-ils le passage dans leur forme native?
Repliement assisté: Des cellules mutées, soit défectueuses pour certaines protéines soit produisant des protéines hétérologues accumulent des protéines natives mal repliées. Existence de protéines auxiliaires du repliement dont la fonction est d’aider les polypeptides à se replier pour prendre leur conformation native

Chapitre V: Le reploiement des protéines
Protéines disulfure isomérases (PDI)
PDI : enzyme du RE chez les eucaryotes ou du périplasme chez les procaryotes. Appartiennent à la super famille des thioredoxines
• Homodimère. Chaque SU constituée de 4 domaines dont 2 sont homologues à la thioredoxine (protéine redox) et sont catalytiquement actifs.
• Activité oxydase/réductase qui catalyse l’appariement correct des ponts disulfures des protéines et activité foldase qui catalyse le repliement des séquences non natives grâce aux domaines thioredoxine.
• PDI fixe des peptides avec des cystéines mais aussi des séquences hydrophobes sans ponts disulfure. Spécificité de substrat très large
• En échangeant les ponts disulfures sur une protéine partiellement repliée (activité isomérase), permet de trouver rapidement la forme la plus thermostable

Chapitre V: Le reploiement des protéines
Protéines disulfure isomérases (PDI)
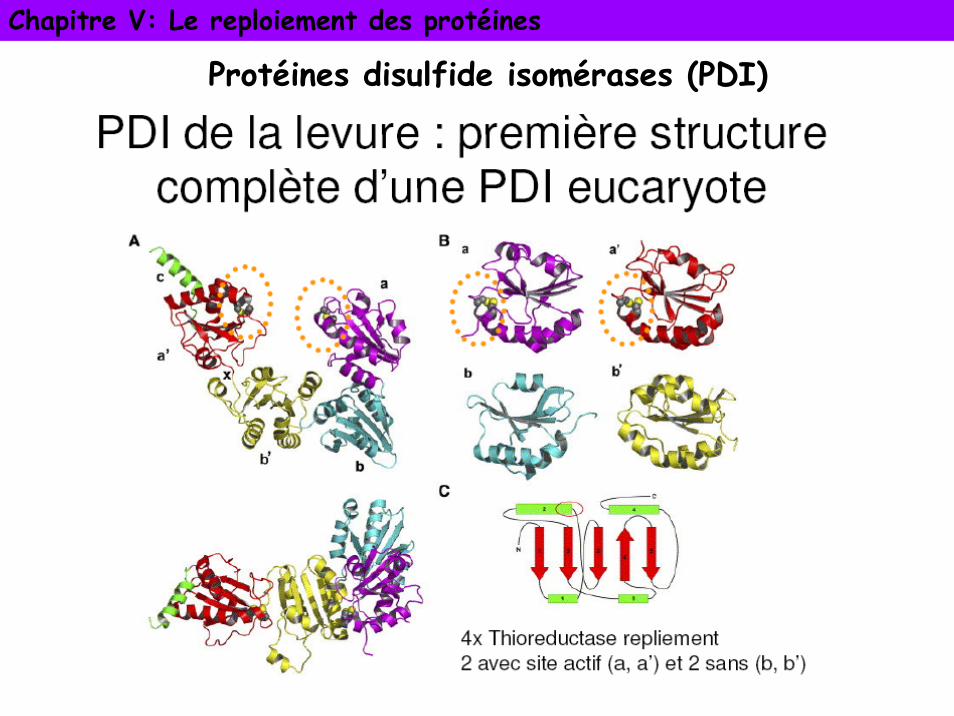
Chapitre V: Le reploiement des protéines
Protéines disulfide isomérases (PDI)

Chapitre V: Le reploiement des protéines
Protéines disulfide isomérases (PDI)
Chapitre V: Le reploiement des protéines
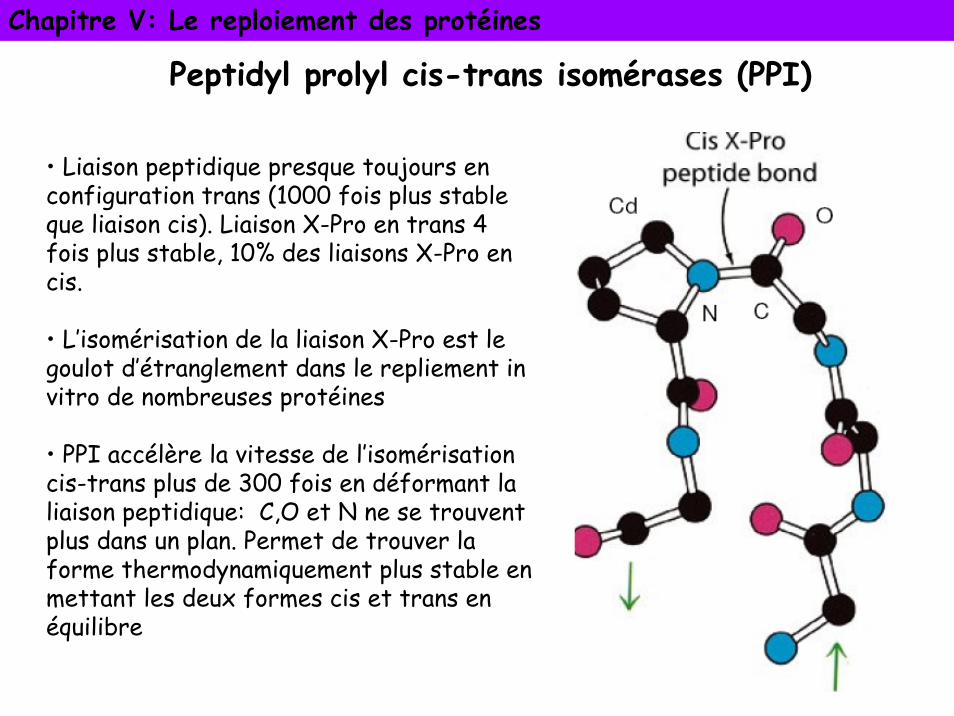
Peptidyl prolyl cis-trans isomérases (PPI)
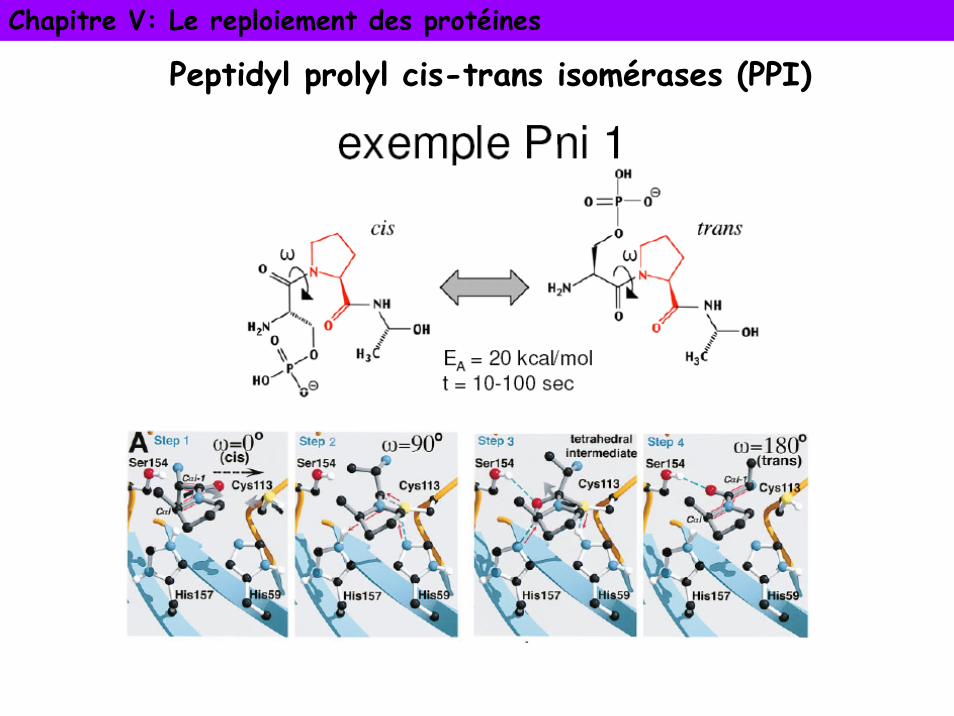
• Liaison peptidique presque toujours en configuration trans (1000 fois plus stable que liaison cis). Liaison X-Pro en trans 4 fois plus stable, 10% des liaisons X-Pro en cis.
• L’isomérisation de la liaison X-Pro est le goulot d’étranglement dans le repliement in vitro de nombreuses protéines
• PPI accélère la vitesse de l’isomérisation cis-trans plus de 300 fois en déformant la liaison peptidique: C,O et N ne se trouvent plus dans un plan. Permet de trouver la forme thermodynamiquement plus stable en mettant les deux formes cis et trans en équilibre
Chapitre V: Le reploiement des protéines
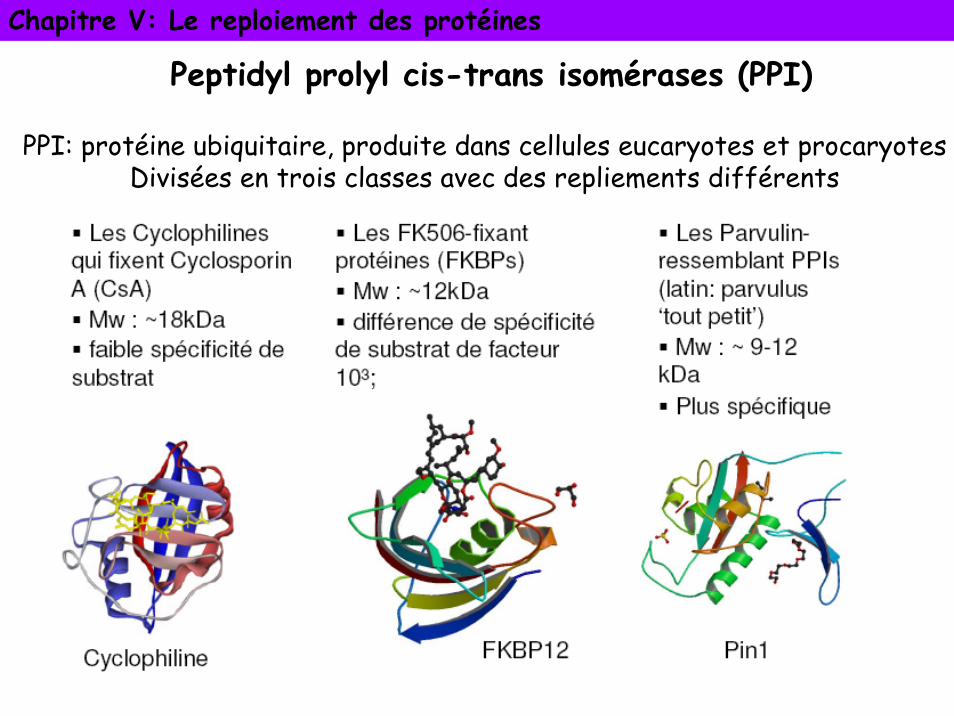
Peptidyl prolyl cis-trans isomérases (PPI)
PPI: protéine ubiquitaire, produite dans cellules eucaryotes et procaryotesDivisées en trois classes avec des repliements différents
Chapitre V: Le reploiement des protéines
Peptidyl prolyl cis-trans isomérases (PPI)

Chapitre V: Le reploiement des protéines
Les molécules chaperonnes
In vivo, toutes les protéines ne se replient pas spontanément.
Chaperons moléculairesProtéines spécialisées qui facilitent le repliement de beaucoup de protéines

Chapitre V: Le reploiement des protéines
Les molécules chaperonnes
Plusieurs classes de protéines indépendantes avec fonctions assez
différentes
-Hsp 70
-Chaperonines ou Hsp60 et Hsp10-Hsp 90: impliquées dans repliement de protéines qui jouent un rôle dans la
transduction du signal
-Nucléoplasmines: protéines nucléaires acides nécessaires pour assemblage
correct des nucléosomes (ADN et histones)
Chaperons moléculaires: protéines qui interagissent avec polypeptides
partiellement ou mal conformés, facilitant les chemins de repliement
correct ou en produisant microenvironnements favorables au repliement. La
plupart ont une synthèse fortement augmentée à température élevée: heat
shock proteins (Hsp)

Chapitre V: Le reploiement des protéines
La fonction des Hsp
Minimisent l’endommagement de la cellule provoqué par la chaleur ou le stress (renaturation/dégradation)
Aident à l’obtention de protéines correctement repliées en minimisant l’agrégation et d’autres erreurs apparaissant pendant le repliement des protéines
Se fixent sur les protéines naissantes pour empêcher leur repliement prématuré
Facilitent la translocation/l’importation à travers la membrane en empêchant la protéine de se replier avant son transport
Assistent l’assemblage/dissolution des complexes multiprotéiques

Chapitre V: Le reploiement des protéines
La famille Hsp70
PM d’environ 70000 daltons
Se fixent sur chaines polypeptidiques naissantes
Se lient à des régions riches en résidus hydrophobes empêchant l’agrégation
Bloquent le repliement de protéines qui doivent rester non conformées jusqu’à ce qu’elles traversent les membranes
Sont actives en monomères
Ont une activité ATPase
Interviennent dans un cycle qui impliquent plusieurs autres protéines comme Hsp40
Hsp70 = DnaK chez E. coli Hsp40= DnaJ chez E.coli
Chapitre V: Le reploiement des protéines
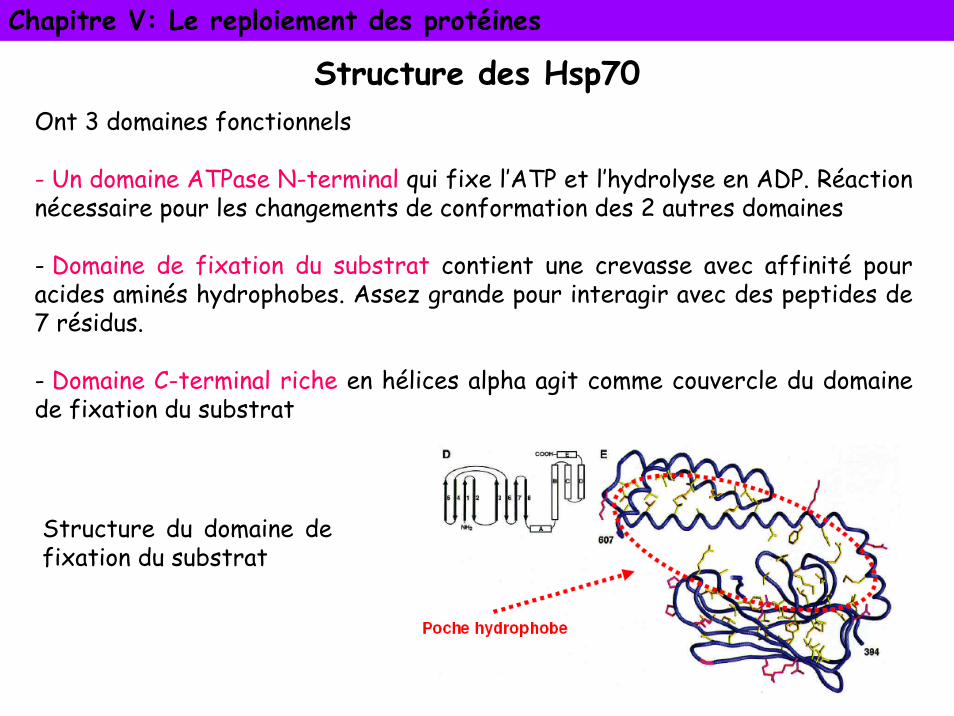
Structure des Hsp70Ont 3 domaines fonctionnels
- Un domaine ATPase N-terminal qui fixe l’ATP et l’hydrolyse en ADP. Réaction nécessaire pour les changements de conformation des 2 autres domaines
- Domaine de fixation du substrat contient une crevasse avec affinité pour acides aminés hydrophobes. Assez grande pour interagir avec des peptides de 7 résidus.
- Domaine C-terminal riche en hélices alpha agit comme couvercle du domaine de fixation du substrat
Structure du domaine de fixation du substrat
Chapitre V: Le reploiement des protéines
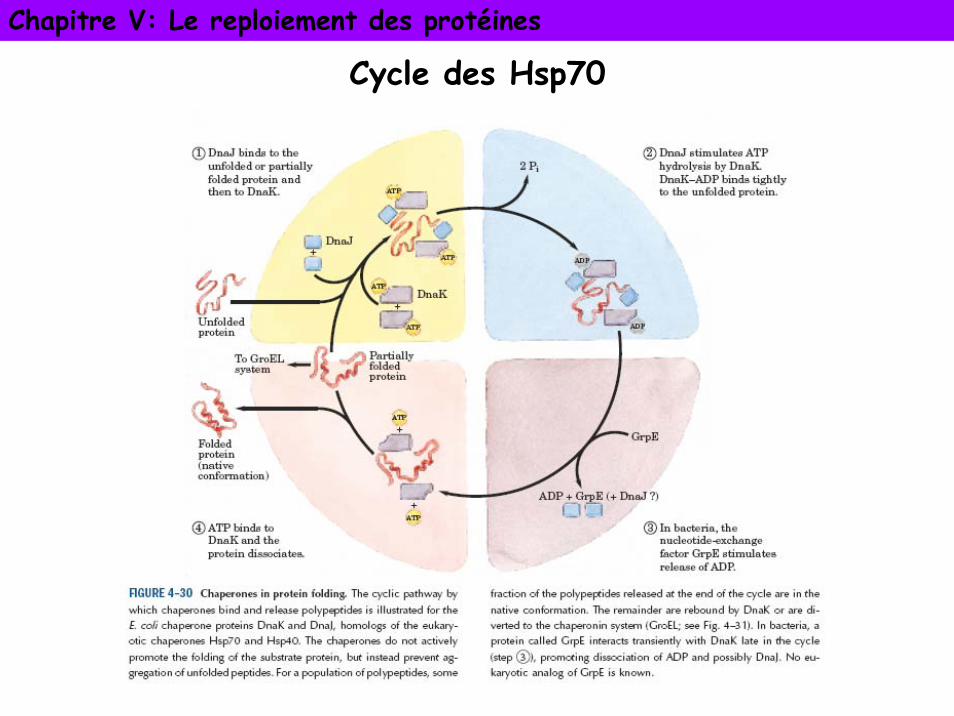
Cycle des Hsp70

Chapitre V: Le reploiement des protéines
Les chaperonines Hsp60 et Hsp10 (groEL et GroES chez E.coli
Cpn60 et Cpn10 dans les chloroplastes)
Se fixent sur protéines libérées des ribosomes ou après transport dans les organelles
2 classes- classe 1: dans bactéries, chloroplastes et mitochondries. Structure
commune à GroES ou GroEL (1 ou 2 anneaux de 7 monomères identiques)- classe 2: archaebactéries et cytoplasme des cellules eucaryotes. 2
anneaux de 8 ou 9 sous-unités par forcément identiques
Cavité de GroEL couverte de surfaces hydrophobespermettent le dépliement et le repliement de la protéine.
Fixation et hydrolyse de l’ATP induit changement de conformation des boucles dans la cavité
masquent surfaces hydrophobes et relâchement de la protéine non repliée
Poursuite du processus le long de la cavité jusqu’à libération de la protéine dans conformation native
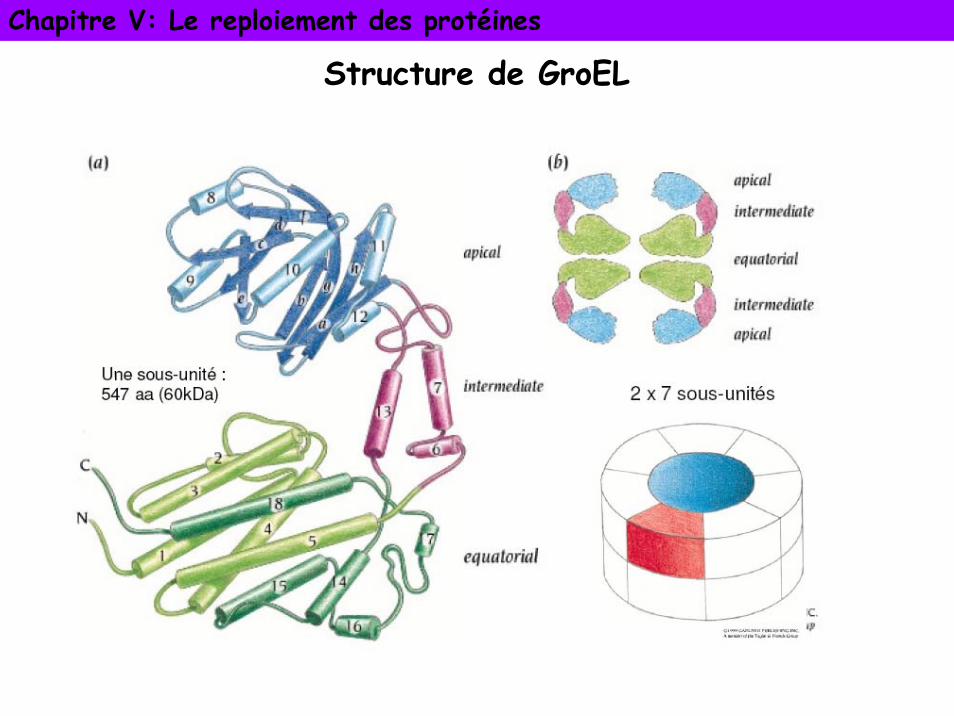
Chapitre V: Le reploiement des protéines
Structure de GroEL
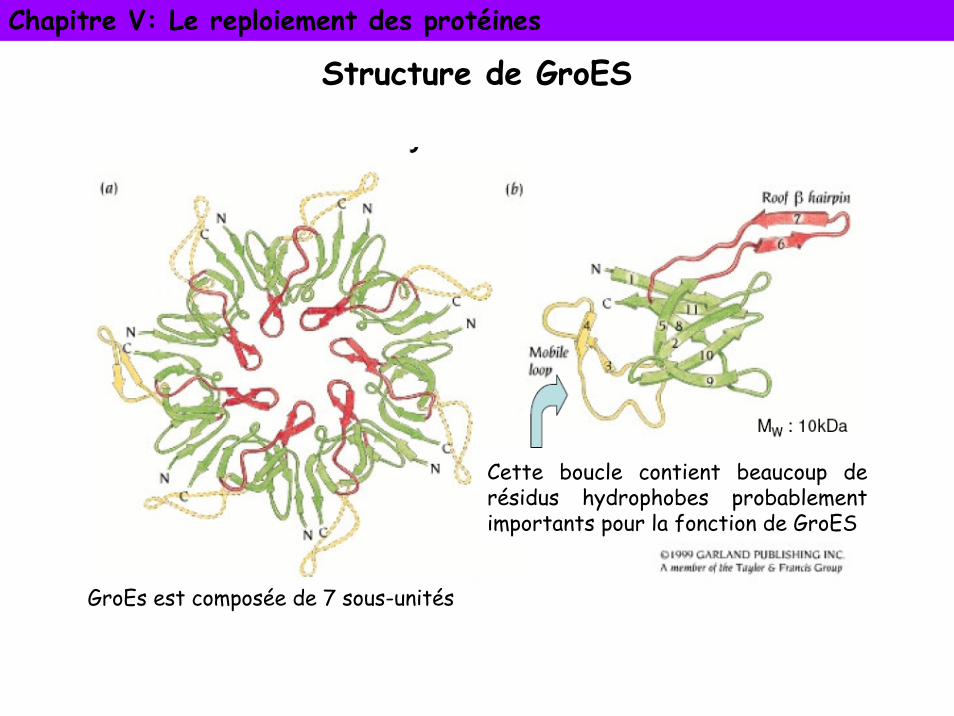
Chapitre V: Le reploiement des protéines
Structure de GroES
GroEs est composée de 7 sous-unités
Cette boucle contient beaucoup de résidus hydrophobes probablement importants pour la fonction de GroES
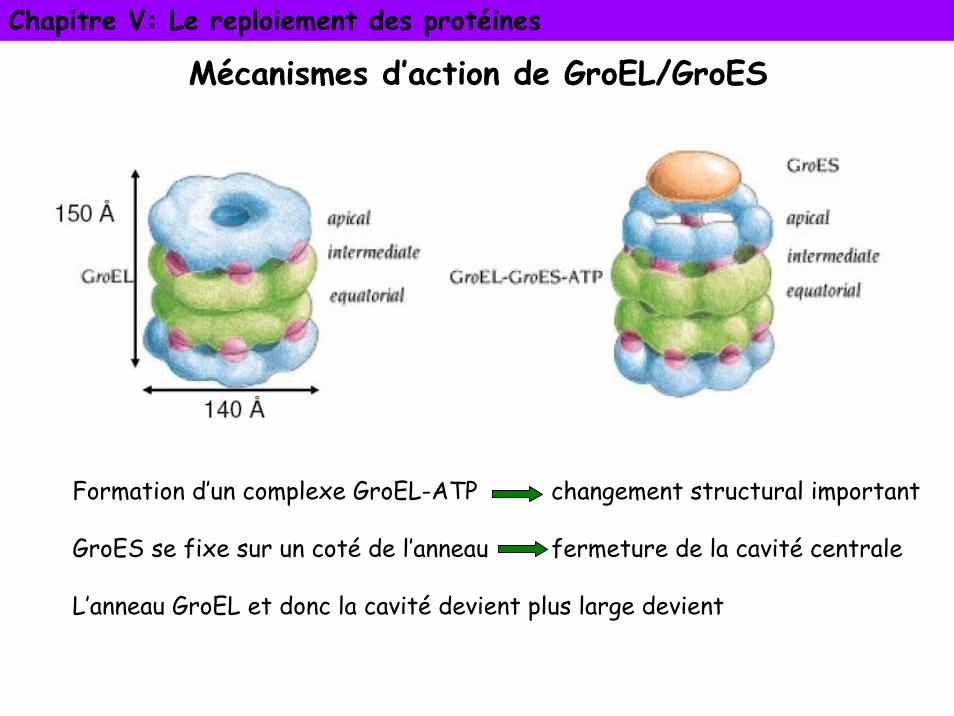
Chapitre V: Le reploiement des protéines
Mécanismes d’action de GroEL/GroES
Formation d’un complexe GroEL-ATP changement structural important
GroES se fixe sur un coté de l’anneau fermeture de la cavité centrale
L’anneau GroEL et donc la cavité devient plus large devient
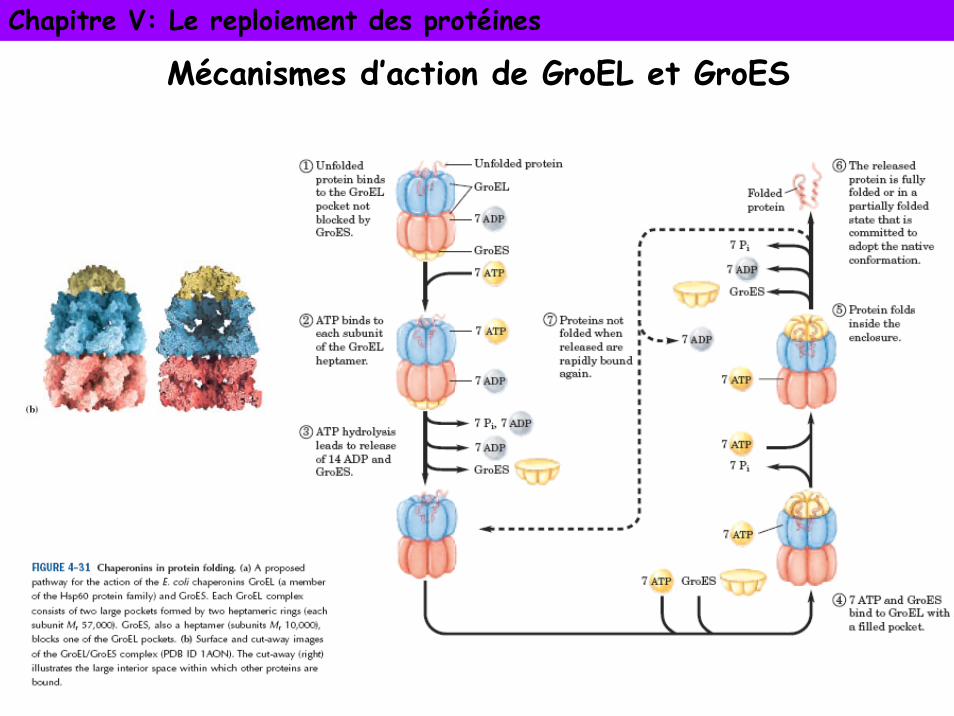
Chapitre V: Le reploiement des protéines
Mécanismes d’action de GroEL et GroES

Chapitre V: Le reploiement des protéines
Repliement des protéines et maladies18 maladies létales associées à l’altération de la structure secundo-
tertiaire des protéines: Maladie d’Alzheimer, encéphalopathies spongiformes transmissibles (TSE) ou maladies à prion, amyloïdoses.
Agrégats de protéines normalement solubles (amyloïdes):-Protéines concernées non apparentées et structures natives différentes. - Formes amyloïdes présentent des similitudes - 2 conformations stables: forme native et forme amyloïde
Prion: 2 conformations PrPc (normale) et PrPsc (altérée)
Conversion autocatalytique de PrPc en PrPsc

Chapitre V: Le reploiement des protéines
300000 séquences connues, 20000 structures 3D déterminées, 40% des ORF codent pour des protéines dont la fonction est inconnue.
Conception de protéines sur mesure
Interventions thérapeutiques
Conclusions
Livres:Introduction à la structure des protéines: carl Branden et John ToozeBiochimie: Reginald H. Garrett,Charles M. GrishamBiochimie: Donald Voet,Judith G. VoetBiologie moléculaire de la cellule: David Baltimore, Harvey Lodish,Arnold Berk, Laurence-S Zipursky, Paul Matsudaira, James DarnellBiochimie: Lubert Stryer, Jérémy M. Berg, John L. Tymoczko
Prédiction du repliement et de la structure 3D