caractérisation précoce de la tolérance à la salinité et à la sécheresse de deux hybrides...
TRANSCRIPT

This article was downloaded by: [Van Pelt and Opie Library]On: 22 October 2014, At: 02:21Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Acta Botanica Gallica: BotanyLettersPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tabg20
Caractérisation précoce de latolérance à la salinité et à lasécheresse de deux hybridessomatiques de Pomme deterre (Solanum tuberosum L.,Solanaceae) et de leurs parentsMarie-Luce Akossiwoa Quashie a , Aimé Nato b &Koffi Akpagana aa Département de Botanique et Biologie végétale,Faculté des Sciences , Université de Lomé , B.P.1515, Lomé , Togob Laboratoire de Morphogenèse végétaleexpérimentale , Université Paris-Sud , Bat. 360,F-91405 , Orsay CedexPublished online: 26 Apr 2013.
To cite this article: Marie-Luce Akossiwoa Quashie , Aimé Nato & Koffi Akpagana(2004) Caractérisation précoce de la tolérance à la salinité et à la sécheresse dedeux hybrides somatiques de Pomme de terre (Solanum tuberosum L., Solanaceae)et de leurs parents, Acta Botanica Gallica: Botany Letters, 151:2, 127-138, DOI:10.1080/12538078.2004.10516028
To link to this article: http://dx.doi.org/10.1080/12538078.2004.10516028
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and views

expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

Acta Bot. Gallica, 2004, 151 (2), 127-138.
Caracterisation precoce de Ia tolerance a Ia salinite et a Ia secheresse de deux hybrides somatiques de Pomme de terre (Solanum tuberosum L., Solanaceae) et de leurs parents
par Marie-Luce Akossiwoa Quashie(1), Aime Natoe) et Koffi Akpagana( 1)
(1) Departement de Botanique et Biologie vegetale, Faculte des Sciences, Universite de Lome, B.P.
1515, Lome, Togo
(2) Laboratoire de Morphogenese vegetale experimentale, Bat. 360, Universite Paris-Sud, F-91405
Orsay Cedex
arrive le 28 juillet 2003, accepte le 23 decembre 2003
Resume.- La culture de Ia Pomme de terre au Togo est affectee par deux stress abiotiques : Ia salinite et Ia secheresse. La recherche de variates adaptees aux conditions ecologiques et pedo-climatiques passe par une creation varietale. Des hybrides obtenus par fusion somatique ainsi que leurs parents peuvent eire caracterises precocement au slade de vitroplants grace a des proteines marqueurs telles que les aquaporines. Des ce slade, Jes reponses des aquaporines different selon Je type de stress applique et selon Ia variate consideree.
Mots cles : Togo - Solanum tuberosum - stress abiotiques - proteines marqueurs - aquaporines.
Abstract.- The potato growing in Togo suffers from two abiotical stress: salinity and drought. The research of adapted varieties to ecological and pedo-climatic conditions goes through a variety creation. Hybrids issued from somatic fusions as well as parents can be early characterized at the vitro-plants stage through marking proteins such as aquaporins. At this stage, aquaporins responses vary according to the kind of applied stress and variety considered.
Key words : Togo - Solanum tuberosum - abiotic stress - proteins markers -aquaporins.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

128
I. INTRODUCTION
L'extension de la culture de la Pomme de terre au Togo passe par Ia production et !'utilisation de varietes adaptees aux conditions pedo-climatiques locales et done a un certain nombre de contraintes biotiques et abiotiques. Les limites rencontrees pour la production de Pomme de terre au Togo sont celles des pays en developpement de Ia sous-region ouestafricaine ; outre des problemes d'approvisionnement en semences, il existe aussi un impact du climat et des eaux d'arrosage et d'irrigation qui entrainent respectivement stress hydrique et stress salin (Sidikou, 2002). La relation est directe et inseparable entre stress hydrique et stress salin, mais il est important de souligner que le stress salin a en plus une composante ionique introduisant des effets qu'on ne retrouve pas chez les plantes en deficit hydrique (Pasternak, 1987 ; Bray, 1997). Au Togo, la creation de genotypes reellement adaptees necessite des programmes d'amelioration a long terme. II est done interessant et meme indispensable aujourd'hui de disposer de techniques permettant de caracteriser precocement mais egalement in vitro les genotypes existants, d'evaluer les premieres etapes de Ia selection par des voies conventionnelles ou biotechnologiques.
Les stress environnementaux abiotiques affectant une culture peuvent occasionner des pertes de rendements considerables (Buchanan et a/., 2000), I' elucidation des mecanismes de reponse est primordiale pour ameliorer la tolerance aux stress aux niveaux genetique et environnemental. Pour permettre a Ia plante de survivre et de s'adapter, une des strategies mises en place au niveau cellulaire semble etre aussi la synthese de proteines specifiques. Parmi elles on cite les proteines de stress et les canaux a eau ou aquaporines.
Le but de notre etude a ete de rechercher des tests physiologiques et biochimiques, en particulier les expressions de ces proteines que sont enzymes du metabolisme du carbone, de !'azote et aquaporines, susceptibles d'etre des marqueurs potentiels du caractere sensible ou resistant au stress salin et au stress hydrique en vue de pouvoir caracteriser precocement et in vitro des especes sauvages, des varietes et des hybrides de Pomme de terre issus de fusion somatique.
II. MATERIELS ET METHODES
A. Materiel vegetal Les genotypes choisis sont :
- un clone dihaploi:de deS. tuberosum (2n = 2x = 24 chromosomes) cv. BF15 (Belle de Fontenay 15) sensible a Ia secheresse eta Ia salinite obtenu de I'Institut national de la recherche agronomique (INRA) de Landerneau; - un clone (PI234013) diploi"de sauvage deS. stenotomum (Sst) (2n = 2x = 24 chromosomes), resistante au fletrissement bacterien (Martin, 1979) et fourni par le Centre international de Ia pomme de terre (CIP) de Lima (Perou) ; - un clone diploi"de Sauvage s. vernei 3 (V3) resistant a Ia salinite ; - un clone tetraploi"de de S. tuberosum cv Sahel considere tolerant a Ia secheresse.
Les deux hybrides etudies issus de Ia fusion somatique des deux parents BFI5 et Sst baptises BS31 et BS42 tetraploides (2n = 4x = 48 chromosomes) obtenus par electrofusion au laboratoire de Morphogenese experimentale vegetale (Orsay).
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

B. Methodes Conditions de culture
129
Les vitroplants utilises sont cultives sur milieu liquide de Murashige & Skoog (1962) additionne de 30 g.I-1 de saccharose et des vitamines de Morel & Wetmore (1951) durant 4 a 6 semaines. Les conditions environnementales sont une photoperiode joumaliere de 16 heures de lumiere d'intensite 60 JlE. m-2 .s-1, une temperature de 21 °C et une humidite relative de 60%.
Application du stress sa/in 1 ml de milieu liquide de Murashige & Skoog (1962) enrichi en NaCl est ajoute de
fa<;:on a avoir dans le tube 6 ml de milieu liquide a 200 mM de NaCl ; 1 ml de milieu liquide de base est ajoute pour le lot temoin. Les plants sont ensuite remis en chambre de culture sous une photoperiode de seize heures d'intensite 80 JlE.m-2 .s-1, a 21 oc. 48 vitroplants repartis en quatre lots ont ete utilises pour chaque variete et hybride. Le premier lot a servi de temoin, les trois autres soumis a un stress et places au hasard dans la chambre de culture. Les analyses sont effectuees 2 heures, 24 heures, et 48 heures apres !'application du stress.
Application du stress hydrique 1 ml de milieu liquide de Murashige & Skoog (1962) enrichi en poly-ethylene glycol
(PEG 8000 Sigma) est ajoute de fa<;:on a avoir dans le tube 6 ml de milieu liquide a 10% de PEG ; 1 ml de milieu liquide de base est ajoute pour le lot temoin. Les plants sont ensuite remis en chambre de culture dans les memes conditions que celles precedemment decrites. Les analyses sont effectuees 2 heures, 24 heures, et 48 heures apres !'application du stress.
Extraction des proteines solubles totales Les extractions sont realisees a partir de vitroplants ages de quatre a six semaines. 500
mg par echantillon sont broyes dans 0,5 ml de tampon d'extraction ( 100 mM Tris pH 8, 10 mM MgC12, 5 mM DTT). Les homogenats sont centrifuges a 4 oc (15 000 g en centrifugeuse Sigma 1Kl5 pendant dix minutes). Les sumageants sont mesures et stockes a 4 °C. Les culots conserves sont repris, laves deux fois et remis en suspension dans 0,5 ml de tampon d'extraction des proteines insolubles ( 100 mM Tris pH 8, 10 mM MgC12, 5 mM DTT, 1 g SDS, 0,25 g CHAPS). Apres 30 minutes d'incubation et une centrifugation a 15 000 g pendant 10 minutes, les proteines de chaque echantillon sont recuperees et dosees.
Dosage des proteines Le principe du dosage repose sur la methode de Sedmak & Grossberg ( 1977). Le com
plexe proteine-colorant presente une absorption caracteristique a 610 nm. La quantite de proteines dans un echantillon est determinee par reference a une gamme etalon preetablie avec une solution de serum albumine SAB (0 a 50 Jlg). La quantite de proteines totales est exprimee en mg.g-1 MF.
Analyse de l'activite enzymatique de Ia phosphoenolpyruvate carboxylase (E.C.4.1.1.31) par electrophorese en conditions non denaturantes
L'etude de l'activite de la PEPC (E.C.4.1.1.31) est effectuee par detection et caracterisation apres separation des proteines solubles par electrophorese en conditions non denaturantes. 30 11g d'extrait sont deposes par puits, la migration s'effectue en chambre froide.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

130
Les gels sont ensuite incubes a 37 oc pendant 20 a 30 minutes (0,1 M, Tris-HCI, pH 8 ; NaHC03, 0,3% ; MgC12, 0,6% ; PEP, 0,2%). Un gel temoin est incube dans les memes conditions en absence du PEP. Apres !'incubation, l'ajout dans les boites de 10 a 20 mg de Fast Violet Blue (4 benzoylamino-2-methoxy-5-methylbenzenediazonium chloride hemi zinc chloride C15H14N302 Sigma) en poudre permet de visualiser l'acide oxaloacetique. L'oxaloacetate colore en rouge orange, produit de la reaction enzymatique, materialise les endroits ou se trouve la PEPC ; Ia presence des isoformes de Ia PEPC se traduit par Ia materialisation de plusieurs bandes.
Electrophorese SDS-PAGE Pour les electrophoreses denaturantes (SDS-PAGE a 15% d'acrylamide), 300 jlg de pro
teines par echantillon sont preleves et ajoutes au melange denaturant (0,5 M Iris HCI pH 6,8 ; glycerol ; SDS 1 0%; ~-mercaptoethanol et quelques grains de bleu de bromophenol) de far;:on a obtenir une solution contenant 300 jlg de proteines dans un volume final de 300 jll. Avant d'etre deposes, les echantillons additionnes de melange denaturant sont denatures par chauffage au bain-marie a 90 oc pendant trois minutes. Pour effectuer des comparaisons valables entre les echantillons, 15 a 30 jlg de proteines denaturees sont deposes par puits.
Transfert par ~stern blot Le transfert des proteines par Western blot est realise sur des membranes Immobilon-P
(millipore) constituees de polyvinylidene difluoride. Les anticorps primaires utilises pour les experiences d'immunoempreintes sont, pour la PEPC, un anticorps polyclonal de lapin dirige contre la PEPC de tabac, proteine purifiee par Lavergne eta/. (1992). Pour Ia GS, un anticorps polyclonal de lapin dirige contre la GS1 et la GS2 de tabac, proteine purifiee par Hirel eta/. (1992). Pour les aquaporines, l'aquaporine isochem neosystem peptide synthetique (AQP) dont la portion de sequence correspondant ala region C-terminale determinee est la sequence
Tyr-Val-Glu-Leu-His-Ser-Pro-Gln-Ser-Leu-Pro-Arg-Gly-Thr-Lys-Ala.
III. RESULTATS ET DISCUSSION
A. Analyses des reponses de Ia PEPC Chez l'hybride BS42 et le parent cultive BF15, le stress salin entraine un renforcement
d'activite de Ia PEPC des 24 heures. Ce renforcement d'activite est encore plus marquee apres 48 heures de stress (Fig. 1 ). La salinite n'entraine aucune modification d'activite chez les clones sauvages deS. stenotomum et S. vernei 3. La reaction du metabolisme des vitroplants au stress hydrique se traduit par une augmentation transitoire d'activite de la PEPC pour le parent sauvage S. stenotomum et les hybrides BS31 et BS42 (Fig. 1 ). C'est aussi le cas du clone sauvage de S. vernei 3 et de Ia variete cultivee Sahel. On note au contraire, pour Ia variete cultivee BF15, une importante baisse d'activite de Ia PEPC.
La complexite de Ia reponse de Ia plante face au stress salin pourrait s'expliquer par le fait que Ia salinite implique a Ia fois un stress ionique et un stress osmotique (Bray, 1993). Hdider et Desjardins (1994) ont montre que !'assimilation du carbone chez les plants cultives in vitro se faisait preferentiellement par la voie de Ia PEPC. La presence de saccharose en forte concentration dans le milieu inhibe la photosynthese des vitroplants et provoque une augmentation d'activite de Ia PEPC. Cette inhibition est due a Ia repression
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4
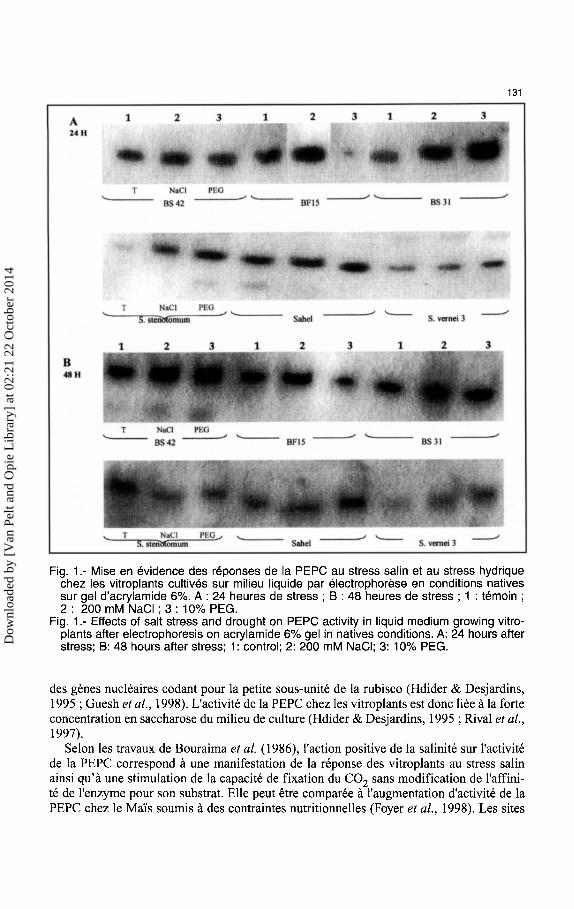
131
Fig. 1.- Mise en evidence des reponses de Ia PEPC au stress salin et au stress hydrique chez les vitroplants cultives sur milieu liquide par electrophorese en conditions natives sur gel d'acrylamide 6%. A : 24 heures de stress ; B : 48 heures de stress ; 1 : temoin ; 2: 200 mM NaCI; 3: 10% PEG.
Fig. 1.- Effects of salt stress and drought on PEPC activity in liquid medium growing vitroplants after electrophoresis on acrylamide 6% gel in natives conditions. A: 24 hours after stress; B: 48 hours after stress; 1: control; 2: 200 mM NaCI; 3: 10% PEG.
des genes nucleaires codant pour Ia petite sous-unite de Ia rubisco (Hdider & Desjardins, 1995 ; Guesh eta!., 1998). L'activite de Ia PEPC chez les vitroplants est done liee a Ia forte concentration en saccharose du milieu de culture (Hdider & Desjardins, 1995 ; Rival eta!., 1997).
Selon les travaux de Bouraima eta!. ( 1986), !'action positive de Ia salinite sur 1'activite de Ia PEPC correspond a une manifestation de Ia reponse des vitrop1ants au stress salin ainsi qu'a une stimulation de Ia capacite de fixation du C02 sans modification de l'affinite de !'enzyme pour son substrat. Elle peut etre comparee a !'augmentation d'activite de Ia PEPC chez le Mai's soumis a des contraintes nutritionnelles (Foyer et a!., 1998). Les sites
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4
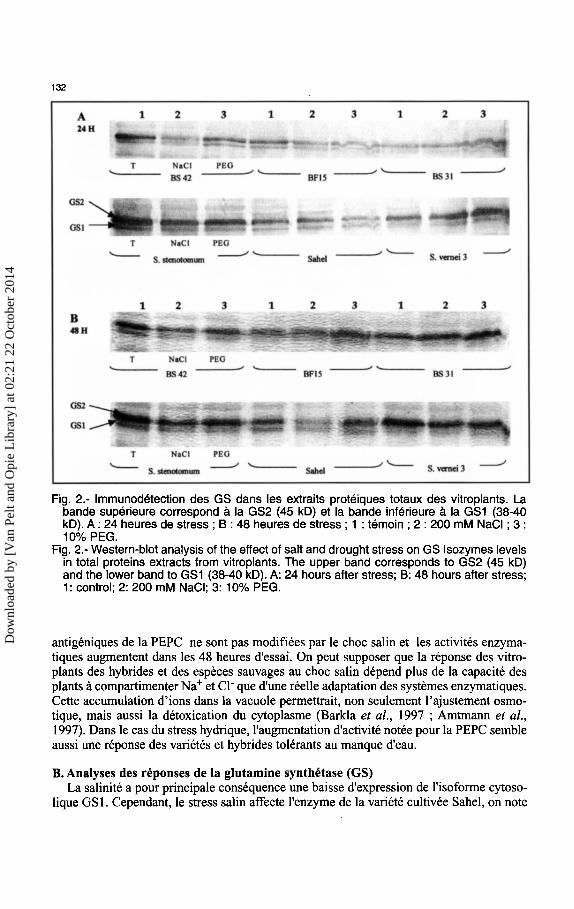
132
Fig. 2.- lmmunodetection des GS dans les extraits proteiques totaux des vitroplants. La bande superieure correspond a Ia GS2 (45 kD) et Ia bande inferieure a Ia GS1 (38-40 kD). A: 24 heures de stress ; B : 48 heures de stress ; 1 : temoin ; 2 : 200 mM NaCI ; 3 : 10% PEG.
Fig. 2.- Western-blot analysis of the effect of salt and drought stress on GS lsozymes levels in total proteins extracts from vitroplants. The upper band corresponds to GS2 (45 kD) and the lower band to GS1 (38-40 kD). A: 24 hours after stress; B: 48 hours after stress; 1: control; 2: 200 mM NaCI; 3: 10% PEG.
antigeniques de la PEPC ne sont pas modifiees par le choc salin et les activites enzymatiques augmentent dans les 48 heures d'essai. On peut supposer que la reponse des vitroplants des hybrides et des especes sauvages au choc salin depend plus de la capacite des plants a compartimenter Na+ et Cl- que d'une reelle adaptation des systemes enzymatiques. Cette accumulation d'ions dans la vacuole permettrait, non seulement l'ajustement osmotique, mais aussi la detoxication du cytoplasme (Barkla et a/., 1997 ; Amtmann et a/., 1997). Dans le cas du stress hydrique, I' augmentation d'activite notee pour la PEPC semble aussi une reponse des varietes et hybrides tolerants au manque d'eau.
B. Analyses des reponses de Ia glutamine synthetase (GS) La salinite a pour principale consequence une baisse d'expression de l'isoforme cytoso
lique GS I. Cependant, le stress sal in affecte I' enzyme de la variete cultivee Sahel, on note
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4
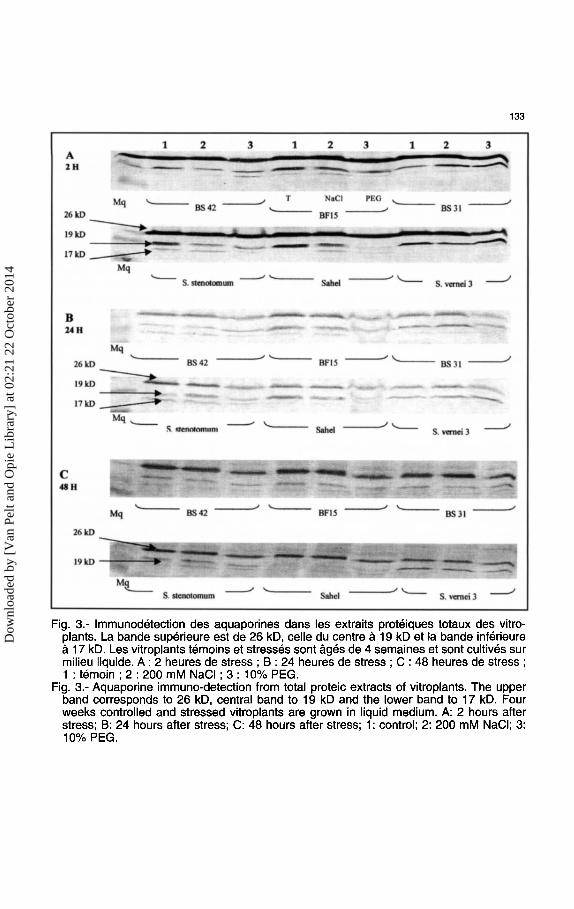
133
Fig. 3.- lmmunodetection des aquaporines dans les extraits proteiques totaux des vitroplants. La bande superieure est de 26 kD, celle du centre a 19 kD et Ia bande inferieure a 17 kD. Les vitroplants temoins et stresses sont ages de 4 semaines et sont cultives sur milieu liquide. A : 2 heures de stress ; 8 : 24 heures de stress ; C : 48 heures de stress ; 1 : temoin; 2: 200 mM NaCI; 3: 10% PEG.
Fig. 3.- Aquaporine immune-detection from total proteic extracts of vitroplants. The upper band corresponds to 26 kD, central band to 19 kD and the lower band to 17 kD. Four weeks controlled and stressed vitroplants are grown in liquid medium. A: 2 hours after stress; 8: 24 hours after stress; C: 48 hours after stress; 1: control; 2: 200 mM NaCI; 3: 10% PEG.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

134
une baisse nette d'expression des deux isoformes (Fig. 2). Brugiere et al. ( 1999) concluent, apres une etude realisee sur des tabacs transgeniques, que Ia GS1 joue un role majeur dans Ia regulation de Ia production de Ia proline lors d'un stress salin ou hydrique. Sa presence dans le cytosol des cellules compagnes du phloeme a ete mise en evidence (Carvalho et al., 1992). Faiblement concentree dans 1es feuilles, elle est abondante dans les racines (Hire! et al., 1993; Oliveira et al., 1997), suggerant ainsi son action dans ]'assimilation primaire de ]'ammonium derivant de Ia reduction du nitrate au niveau de ces organes. On remarque egalement une expression accrue de Ia forme chloroplastique GS2 de !'enzyme chez S. vernei 3 consideree comme resistante a la salinite (Ross, 1986). La GS2 interviendrait dans Ia photorespiration ; Hoshida et al. (2000), utilisant des plants transgeniques de Riz soumis pendant deux semaines a un stress salin (NaCI, 12 g.I- 1 ), suggerent qu'elle serait a l'origine de Ia protection de l'activite du photosysteme II contre Ia salinite, expliquant ainsi Ia tolerance de cette espece a la salinite.
La coordination entre le metabolisme du carbone et de ]'azote n'est pas rompue par le stress hydrique (Foyer et al., 1998) puisque l'activite de Ia glutamine synthetase ne semble pas affectee. Comme l'ont montre Wingler et al. ( 1999), Ia GS2 ne semble pas affectee par le stress hydrique. On peut toutefois remarquer qu'on retrouve en cas de stress hydrique une surexpression de Ia GS2 chez S. vernei 3. Ce resultat peut etre lie toujours au role de Ia GS2 dans Ia photorespiration et Ia protection des systemes photosynthetiques lors des stress abiotiques.
C. Analyse des reponses des aquaporines Stress sa/in
La salinite affecte les aquaporines des les deux premieres heures. Seule la forme de 19 kDa a une expression modulee en reponse au stress, alors que ]'expression de la forme de 26 kDa ne change pas (Fig. 3). Les variations d'expression des aquaporines en reponse au stress salin s'expriment pour toutes les especes par des variations dans !'expression de Ia forme a 19 kDa principalement. L'expression de Ia forme majoritaire a 26 kDa n'est manifestement pas touchee par le stress salin. Ce resultat est valable pour toutes les especes et les hybrides analyses. 11 semblerait que, dans ce cas, Ia forme d'aquaporines a 26 kDa ne soit pas impliquee dans la reponse au stress salin mais que ce soit plutot la forme de 19 kDa.
Les reponses des aquaporines en 48 heures de stress salin en milieu liquide permettent deja dans une premiere approche de caracteriser les especes et les hybrides etudies. Chez l'espece cultivee, parent de fusion, la reponse au stress salina pour consequence une baisse d'expression des aquaporines. Cette baisse d'expression se retrouve egalement chez l'un des hybrides somatiques pour Ia forme a 19 kDa. La baisse d'expression des aquaporines est une des reponses connues des canaux a eau au stress salin (Kirch et al., 2000). La baisse nette d'expression observee, tant chez les aquaporines de la membrane vacuolaire (TIP) que chez les aquaporines de la membrane plasmique (PIP), pourrait expliquer la tolerance des plantes au stress osmotique. La reduction d'expression des genes de certaines aquaporines soumises a un stress osmotique pourrait etre un mecanisme pour diminuer les pertes en eau ou pour limiter !'influx du sodium principalement dans les vacuoles. Des genes d'aquaporines du plasmalemme et surtout du tonoplaste ont ete recenses comme etant actives en reponse au stress salin chez les glycophytes (Fray et a/., 1994 ; Guererro et al., 1990 ; Yamaguchi-Shinozaki et al., 1992, 1993 ; Yamada et al., 1997). Ces memes chercheurs ont emis une hypothese selon laquelle, face au stress salin, Ia regulation de ]'expression des genes d'aquaporines se fait en deux phases, en reponse au statut hydrique de
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

135
Ia plante. Une baisse moden!e du potentiel hydrique des feuilles entraine une baisse d'expression des aquaporines, tandis qu'une baisse plus drastique au bout de 48 heures de stress salin entraine par contre une surexpression des aquaporines. Chez !'halophyte facultative, Mesembryanthemum crista/linum, !'expression des aquaporines est reprimee durant les trente premieres heures suivant un stress salin avec 400 mM de NaCl (12 g.I-1). Apres cette periode, les aquaporines retrouvent Ia meme expression que celle qu'elles avaient avant !'application du stress (Yamada et a/., 1995). On retrouve cette particularite dans le cas de notre etude, chez le parent sauvage et un hybride, soulignant ainsi une tolerance plus importante a la salinite que le parent cultive. La surexpression des aquaporines du tonoplaste a UTI stress osmotique a egalement ete signalee chez le Chou-fleur (Barrieu et al., 1999)
La tolerance au stress salin ne s'exprime pas de la meme fa<;:on chez les deux especes sauvages au regard de !'expression des aquaporines. Chez S. stenotomum resistante a R. solanacearum, agent du fletrissement bacterien, Ia resistance au stress osmotique passe en milieu liquide par une surexpression des formes d'aquaporines caracteristiques de cette espece : les formes a 19 et 17 kDa. Cette surexpression face au stress salin se retrouve chez les hybrides somatiques qui expriment plus fortement les formes a 19 kDa et a 17 kDa sans que l'on puisse noter d'alterations d'expression de Ia forme a 26 kDa. S. vernei 3 tolerante au sel exprime, queUes que soient les conditions de stress salin, les deux formes d'aquaporines presentes aussi chez le temoin (26 et 19 kDa). Apres 24 heures de stress salin, !'expression de Ia forme a 19 kDa est plus forte. La reponse des aquaporines est aussi une illustration de Ia tolerance de S. vernei 3 au sel puisque !'expression des deux formes d'aquaporines toujours retrouvee chez cette espece ne change pratiquement pas avec !'addition de sel.
Stress hydrique Les deux premieres heures apres !'addition de PEG apportent deja une reponse metabo
lique des aquaporines. Pour toutes les varietes, c'est Ia forme a 26 kDa qui est affectee (Fig. 3). La baisse d'expression de Ia forme a 26 kDa notee des les deux premieres heures ne s'accentue pas et, au bout de 48 heures, on retrouve environ le meme taux d'expression pour les hybrides BS31 et BS42. Les deux autres formes sont presentes chez les hybrides avec 1a meme intensite chez les vitroplants temoins et les vitroplants stresses. Chez le parent Sst, seule Ia forme a 26 kDa est affectee mais !'expression de cette forme chute de moins de 30% 48 heures apres !'application du stress. La baisse d'expression de la forme a 26 kDa est bien nette chez BF15 apres 48 heures de stress hydrique. Les aquaporines sont impliquees dans les mecanismes de Ia plante pour eviter UTI deficit en eau (Clarkson et al., 2000). La reponse au stress hydrique chez les vitroplants se traduit par une baisse d'expression de Ia forme majoritaire presente chez Ia Pomme de terre d'un poids moleculaire voisin de 26 kDa. Cette baisse d'expression est aussi notee chez S. stenotomum et S. vernei 3. Au contraire du stress salin, le stress hydrique affecte la forme principale d'aquaporines retrouvee chez Ia Pomme de terre. On enregistre une baisse de cette forme principale tout au long du stress. Cette baisse est cependant transitoire chez les especes sauvages et les hybrides.
Un nombre important d'aquaporines est present chez les plantes. Certaines sont localisees dans le tonoplaste, d'autres au niveau de Ia membrane plasmique et enfin certaines au niveau des endomembranes (NOD26 aquaporine de Ia membrane peribacteroi'de du nodule du Soja) (Weaver et al., 1994 ; Weig et al., 1997; Kjelbom et al., 1999). II est clairement etabli aujourd'hui que les aquaporines sont impliquees dans la reponse des plantes au stress
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

136
hydrique. L'activite des aquaporines peut etre regulee par phosphorylation (Johansson et al., 1998). Dans le cas de l'aquaporine PM28 de l'Epinard, la phosphorylation n'est active que dans les conditions de potentiel hydrique eleve (Maurel et al., 1995 ; Johansson et al., 1998). La phosphorylation est assuree par une proteine kinase Ca2+ dependante liee a la membrane. Cette aquaporine peut etre dephosphorylee et inactivee dans les conditions de stress hydrique, ceci pour conserver l'eau dans la plante (Johansson et al., 1996, 1998). La meme observation a ete faite chez Mesembryanthemum crystal/inurn (Yamada et al., 1995). Chez le Toumesol, les mRNA du gene Sun TIP 18 declinent apres 24 heures de stress hydrique (Sarda et al., 1999) et, chez Nicotiana glauca, trois aquaporines du plasmalemme voient leur expression chuter en conditions de stress hydrique (Smart et al., 2001). 11 semblerait que ce soit le cas de l'aquaporine de 26 kDa trouvee ici chez la Pomme de terre et dont le poids moleculaire semble la rapprocher des aquaporines du plasmalemme.
IV CONCLUSION
La maitrise de la fusion de protoplastes et de la regeneration de vitroplants de Pomme de terre (Solanum tuberosum L.) est l'outil indispensable a !'obtention plus rapide d'hybrides associant les caracteres avantageux des deux parents. La creation ou !'amelioration varietale fait appel a un pool genique issu d'especes dites "sauvages" souvent diploides. La fusion somatique est maintenant un moyen efficace et incontoumable d'arriver plus rapidement aux varietes "cultivees" possedant les caracteres recherchees. La fusion somatique a pour avantage un gain de temps significatif par rapport aux techniques traditionnelles de croisement. Cependant l'introgression des caracteres avantageux apres retrocroisements necessite deux a trois generations. Ces retrocroisements restent utiles dans le cas des hybrides pour arriver a une nouvelle variete.
I.; utilisation pour la creation de nouveau materiel vegetal des biotechnologies, et notamment de la culture in vitro associee a la fusion de protoplastes, requiert des tests precoces et adaptes. Dans I' evaluation de la resistance des hybrides, il est souvent plus aise d'operer sur des vitroplants. 11 est en effet indispensable de pouvoir evaluer la resistance des hybrides issus de fusion directement sur le vitroplant afin de pouvoir operer rapidement un tri sur les produits de fusion obtenus. La caracterisation precoce au bout de 48 heures des hybrides somatiques et des parents de fusion, par !'addition d'une solution saline ou d'une solution de PEG dans le milieu de culture liquide, a apporte des elements de reponse interessants. L'augmentation de l'activite de la PEPC est mise en evidence chez les vitroplants stresses des differentes especes et hybrides analysees. Elle semble etre une reaction generale d'activation de !'assimilation du carbone face aux deux contraintes abiotiques majeures. Le stress salin affecte !'expression de la GS chez les especes sensibles a la salinite (BF 15 et Sahel). Chez les especes tolerantes, la reponse a la secheresse et a la salinite passe par une meilleure compartimentation des sels. Elle fait egalement suite a une protection des photosystemes de la photosynthese, permettant ainsi de preserver la coordination entre les metabolismes d'assimilation du carbone et de l'azote.
Les reponses des aquaporines de la Pomme de terre sont caracteristiques du type de stress abiotique applique aux vitroplants. Le stress salin n'affecte pas !'expression de la forme majoritaire d'aquaporines a 26 kDa presentes chez la Pomme de terre. Les reponses varietales a la salinite passent principalement par des variations d'expression de la forme a 19 kDa. Le stress hydrique a pour principale consequence la baisse d'expression des aquaporines et principalement de la forme la plus abondante.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

137
BIBLIOGRAPHIE Amtmann A., S. Laurie, R.A. Leigh & D. Sanders, 1997.
Multiple inward channels provide flexibility in Na+fK+ discrimination at the plasma membrane of barley suspension culture cells. J. Exp. Bot., 48, 481-497.
Barkla B.J., R. Vera-Estrella & 0. Pantoja, 1997.Aquaporin localization. How valid are the TIP and PIP labels? Trends Plant Sci., 4, 86-88.
Barrieu F., F. Chaumont & M.J. Chrispeels, 1999.- High expression of the tonoplast aquaporin ZmTIP1 in epidermial and conducting tissues of maize. Plant Physiol., 117, 1153-1163.
Barrieu F., D. Marty-Mazars, D. Thomas, F. Chaumont, M. Charbonnier & F. Marty, 1999.- Dessication and osmotic stress increase the abundance of mRNA of the tonoplast aquaporin BobTIP26-1 in cauliflower cells. Planta, 209, 77-86.
Bouraima S., D. Lavergne & M.L. Champigny, 1986.Etude comparee de Ia tolerance au sel de differents mils : croissance, activites phosphoenol-pyruvate carboxylase et enzyme malique il NADP. Agronomie, 6 (7), 675-682.
Bray E., 1993.- Molecular responses to water deficit. Plant Physiol., 103, 1035-1040.
Bray E., 1997.- Plant responses to water deficit. Trends Plant Science, 2, 48-54
Brugiere N., F. Dubois, A.M. Limani, M. Lelandais, Y. Roux, R.S. Sangwan & B. Hire!, 1999.- Glutamine synthetase in the phloem plays a major role in controlling proline production. The Plant Cell, 11, 1995-2011.
Buchanan B., W. Gruissem & R. Jones (eds), 2000.Biochemistry and molecular biology of plants. Am. Soc. Plant Physiol., 1 341 p.
Carvalho H., S. Pereira, C. Sunkel & R. Salema, 1992.Detection of a cytosolic glutamine synthetase in leaves of Nicotiana tabacum L. by immunocytochemical methods. Plant Physiol., 100, 1591-1594.
Clarkson D.T., M. Carvajal, T. Henzler, R.N. Waterhouse, A.J. Smyth, D.T. Cooke & E. Steudle, 2000.- Root hydraulic conductance: diurnal expression and the effects of nutrient stress. J. Exp. Bot., 51 (342), 61-70.
Foyer C., M.H. Valadier, A. Migge & T. Becker, 1998.Drought-induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physio/., 117, 283-292.
Fray R.G., A. Wallac., D. Grierson & G.W. Lycett, 1994.Nucleotide sequence and expression of a ripening and water stress-related eDNA from tomato with homology to the MIP class of membrane channel protein. Plant. Mol. Bioi., 24, 539-543.
Guererro F.D., J.T. Jones & J.E. Mullet, 1990.- Turgorresponsive gene transcription and RNA levels increase rapidly when pea shoots are wilted - sequence and expression of 3 inducible genes. Plant Mol. Bioi., 15, 11-26.
Guesch R.W., K.J. Boote, J.V.C. Vu, L.H.A. Allen & G. Bowed, 1998.- Changes in growth C02 result in rapid adjustments of ribulose-1 ,5-bisphosphate carboxylase/oxygenase small subunit gene expression in expanding and mature leaves of rice. Plant Physiol., 118, 521-529.
Hdider C. & Y. Desjardins, 1994.- Effects of sucrose on photosynthesis and phosphoenolpyruvate carboxyla-
se activity of in vitro cultured strawberry plantlets. Plant Cell. Tissue and Organ Cult., 36, 27-33.
Hdider C. & Y. Desjardins, 1995.- Reaction of ribulose-1 ,5-bisphosphate carboxylase/oxygenase efficiency by the presence of sucrose during the tissue culture of strawberry plantlets. In vitro Cell Dev. Bioi. Plant, 31, 165-170.
Hire! B., M.C. Marsolier, J. Hoarau, J. Brangeon, R. Schafer & D.P.S. Verma, 1992. Forcing expression of a soybean root glutamine synthetase gene in tobacco leaves induces a native gene encoding cytosolic enzyme. Plant. Mol. Bioi., 20, 207-218.
Hirel B., G.H. Miao & D.P.S. Verma, 1993.- Metabolic and developmental control of glutamine synthetase genes in legurne and non-legurne plants. In: Plant Gene Expression. D.P.S. Verma (ed)., CRC Press Inc., 443-458.
Hoshida H., Y. Tanaka, T. Hibino, Y. Hayashi, A. Tanaka, T. Takabe & T. Takabe, 2000.- Enhanced tolerance to salt stress in transgenic rice that overexpressed chloroplast glutamine synthetase. Plant. Mol. Bioi., 43 (1), 103-111.
Johansson 1., M. Karlsson, V.K. Shukla, M.J. Chrispeels, C. Larsson & P. Kjelbom, 1998.- Water transport activity of the plasma membrane aquaporin PM28 is regulated by phosphorylation. The Plant Cell, 10,451-459.
Johansson 1., C. Larsson, B. Ek& P. Kjelbom, 1996.- The major integral proteins of spinach leaf plasma membranes are putative aquporins and are phosphorylated in response to Ca2+ and apoplastic water potential. The Plant Cell, 8, 1181-1191.
Kjelbom P., C. Larsson, I. Johansson, M. Karlsson & U. Johansson, 1999.- Aquaporin and water homeostasis in plants. Trends Plant Science, 4 (8), 308-314.
Kirch H. H., R. Veraestrella, D. Golldack, F. Quigley, C.B. Michalowski, B.J. Barkla & H.J. Bohnert, 2000.Expression of water channel proteins in Mesembryanthemum crystal/inurn. Plant Physiol., 123, 111-124.
Lavergne D., A. Nato, J.M. Dupuis, M. Pean & P. Chagvardieff, 1992.- Evidence for the expression of morphological and biochemical characteristics of C3-photosynthesis in chlorophyllous callus cultures of Zea mays. Physiologia Plantarum, 84, 292-300.
Martin C., 1979.- Sources of resistance to Pseudomonas solanacearum. In: Developments in control of potato bacterial diseases. Report of a planning conference held at CIP, Lirna, Peru, June 12-15, 49-53.
Maurel C., Kado R. T., Guern J. and M. J. Chrispeels, 1995.- Phosphorylation regulates the water channel activity of the seed-specific aquaporin a - TIP. EMBO J., 14, 3028-3035.
Morel G. and R. H. Wetmore, 1951.- Fern callus tissue culture. Am. J. Bot., 38, 141-143.
Murashige T. & F. Skoog, 1962.- A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant, 15, 473-497.
Oliveira J.C., H.M. Lam, K. Coschigano, R. Melo-Oiiveira & G. Coruzzi, 1997.- Molecular-genetic dissection of ammonium assimilation in Arabidopsis thaliana. Plant Physiol. Bioch., 35, 185-198.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4

138
Pasternak D., 1987.- Salt tolerance and crop production: a comprehensive approach. Ann. Rev. Phytopathol., 25, 271-291.
Rival A., T. Beule, D. Lavergne, A. Nato, M. Havaux & M. Puard, 1997.- Development of photosynthetic characteristics in oil palm during in vitro micropropagation. J. Plant Physiol., 150, 520-527.
Ross H., 1986.- Potato breeding. Problems and pespectives. Advances in Plant Breeding, 13, Paul Parey (eds}, Berlin and Hamburg, 1-132.
Sarda X., D. Tausch, K. Ferrare, F. Cellie., C. Alcon, J.M. Dupuis, F. Casse & T. Lamaze, 1999.Characterization of closely related ll-TIP genes encoding aquaporins which are differentially expressed in sunflower roots upon water deprivation through exposure to air. Plant Mol. Bioi., 40, 179-191.
Sedmak J.J. & S.E. Grossberg, 1977.- A rapid, sensitive and versatile assay for protein using coomassie brilliant blue G250. Analytical Biochemistry, 79, 544-552.
Sidikou R.D. S., 2002.- Contribution des biotechnologies vegetales a /'adaptation de Ia pomme de terre (Solanum tuberosum L.) au Niger. These de doctoral es Sciences, Universite de Niamey (Niger), 473 p.
Smart LB., W.A. Moskal, K.D. Cameron & A.B. Bennett, 2001.- MIP genes are down-regulated under drought stress in Nicotiana glauca. Plant Cell Physiol., 42 (7), 686-693.
Weaver C. D., N.H. Shomer, C. F. Louis & D.M. Roberts, 1994.- Nodulin 26, a nodule specific symbiosome
membrane protein from soybean, is an ion channel. J. Bioi. Chemistry, 268, 17858-17862.
Weig A., C. Deswarte & M.J. Chrispeels, 1997.- The major Intrinsic protein family of Arabidopsis has 23 members that form three distinct groups with functional aquaporines in each group. Plant Physiol., 57, 1347-1357.
Wingler A., W.P. Quick, R.A. Bungard, P.J. Bailey, P.J. Lea & R.C. Leegood, 1999.- The role of photorespiration during drought stress: an analysis utilizing barley mutants with reduced activities of photorespiratory enzymes. Plant Cell Env., 22, 361-373.
Yamada S., M. Katsuhara, W.B. Kelly, G.B. Michalowski & H. J. Bonhert, 1995.- A family of transcripts encoding water channel proteins: tissue specific expression in the common ice plant. Plant Cell, 7, 1129-1142.
Yamada S., D.E. Nelson, E. Ley, S. Marquez S. & H.J. Bonhert, 1997.- The expression of an aquaporin promoter for Mesembryanthemum crystal/inurn in tobacco. Plant Cell Physiol., 38 (12), 1326-1332.
Yamaguchi-Shinozaki K., M. Koizumi, S. Urao & K. Shinozaki, 1992.- Molecular cloning and characterization of 9 cDNAs for genes that are responsive to desiccation in Arabidopsis thaliana: sequence analysis of one eDNA clone that encodes a putative transmembrane channel protein. Plant Cell Physiol., 33, 217-224.
Yamaguchi-Shinozaki K. & K. Shinozaki, 1993.Arabidopsis-DNA encoding two desiccation-responsive rd29genes. PlantPhysiol.,101, 1119-1120.
Dow
nloa
ded
by [
Van
Pel
t and
Opi
e L
ibra
ry]
at 0
2:21
22
Oct
ober
201
4