universitÉ*ouverte*2012/2013*henry/coursuo2.pdf · universitÉ*ouverte*2012/2013*...
TRANSCRIPT

UNIVERSITÉ OUVERTE 2012-‐2013 Du neurone au cerveau: Une promenade dans la
complexité du système nerveux
13 Novembre 2012 : Introduc=on: le cerveau, machine électrique ou machine chimique; la cellule nerveuse 20 Novembre 2012 : Comment le neurone fabrique de l’électricité et quels signaux électriques sont échangés 27 Novembre 2012 : Comment l’informa=on passe entre les neurones; la mémoire à l’échelle cellulaire 4 Décembre 2012 : Comment le cerveau prend l’informa=on extérieure: de la vision à la lecture 11 Décembre 2012 : Comment la neurobiologie aborde les fonc=ons complexes: la mémoire, les émo=ons, l’empathie, la conscience
Jean-‐Pierre Henry

Origine des courants électriques
L’informa=on est électrique et pourtant il n’est pas nécessaire de meVre les doigts
dans la prise pour penser

UNIVERSITÉ OUVERTE 2012-‐2013 Comment le neurone fabrique de l’électricité? Quels signaux électriques sont échangés?
-‐2-‐ 20 Novembre 2012
Jean-‐Pierre Henry Laboratoire Ma=ère Et Systèmes Complexes

Résumé du premier cours
• L’unité de base est le neurone, la cellule nerveuse; c’est une cellule avec des prolongements: axone, dendrites
• Les neurones sont organisés en réseaux dans lesquels circule l’informa=on
• Dans les neurones, l’informa=on est « électrique »; elle circule dans un seul sens: des dendrites vers l’extrémité de l’axone
• Entre les neurones (synapse), l’informa=on est chimique: neurotransmeVeurs libérés par l’axone

Rappel d’électricité
• Les effets électriques sont dus à des espèces possédant une charge; dans le système nerveux: des ions, ca=ons (K+, Na+, Ca2+), anions (Cl-‐, glutamate)
• Une différence de poten=el (Volt,V) fait bouger les ions: Si V>0, les ca=ons sont repoussés, les anions acrés
• Le courant électrique i est le nombre de charges transportées par unité de temps (Ampère, A)
• La différence de poten=el V= V1-‐V2 est liée à l’intensité du courant par la Loi d’Ohm: V= R x i où R est la résistance (Ohm,Ω); l’inverse de la résistance est la conductance γ
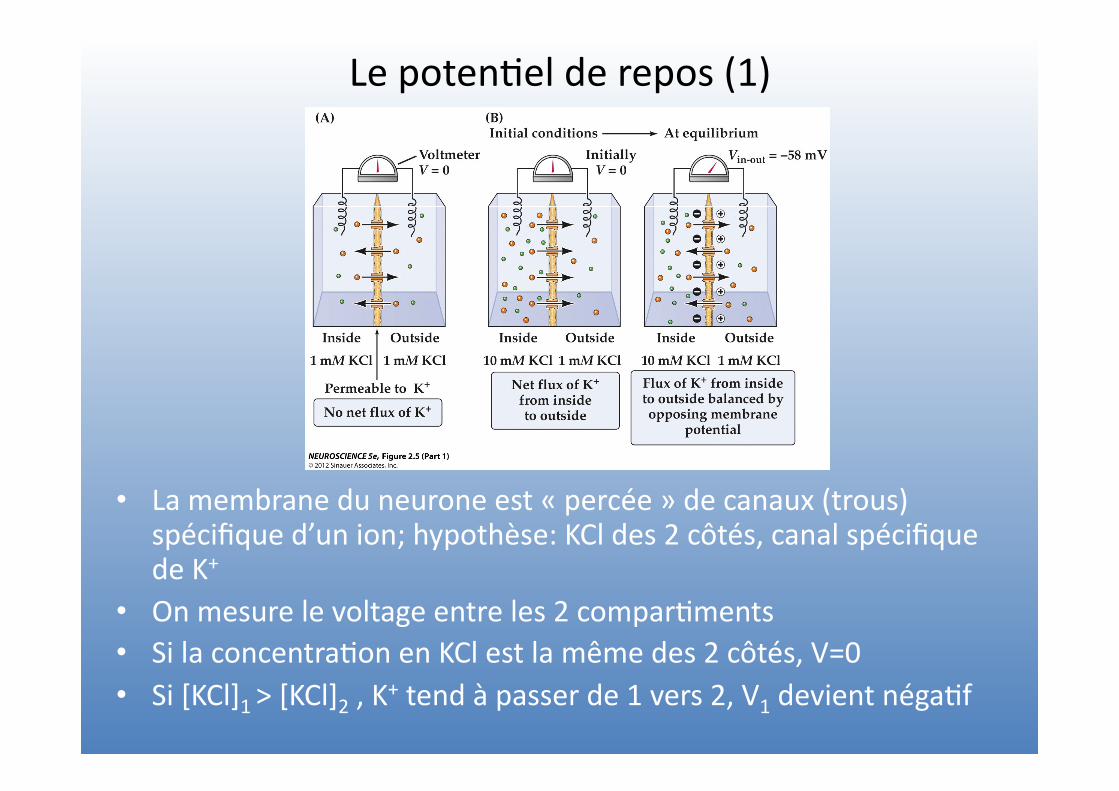
Le poten=el de repos (1)
• La membrane du neurone est « percée » de canaux (trous) spécifique d’un ion; hypothèse: KCl des 2 côtés, canal spécifique de K+
• On mesure le voltage entre les 2 compar=ments • Si la concentra=on en KCl est la même des 2 côtés, V=0 • Si [KCl]1 > [KCl]2 , K+ tend à passer de 1 vers 2, V1 devient néga=f
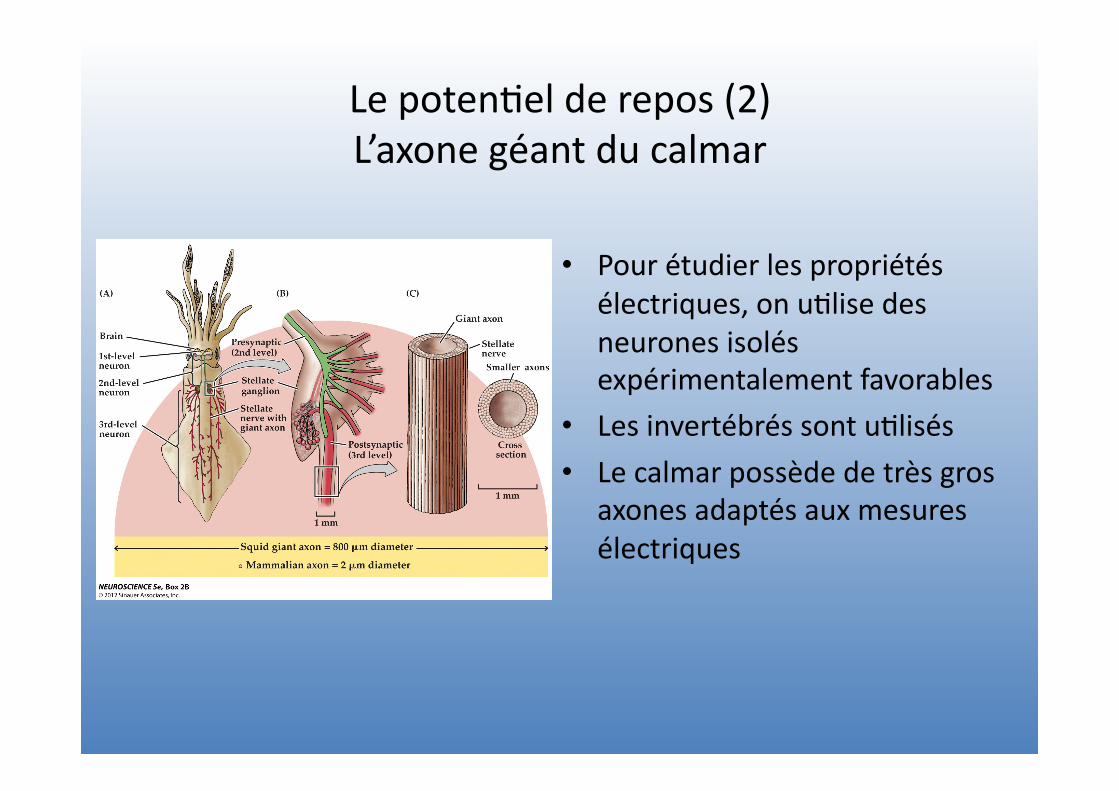
Le poten=el de repos (2) L’axone géant du calmar
• Pour étudier les propriétés électriques, on u=lise des neurones isolés expérimentalement favorables
• Les invertébrés sont u=lisés • Le calmar possède de très gros
axones adaptés aux mesures électriques

Le poten=el de repos (3)
• Chez le calmar (et de manière générale), la concentra=on de K+ est plus forte à l’intérieur et celles de Na+ et Cl-‐ plus fortes à l’extérieur
• CeVe dissymétrie a pour résultat un poten=el néga=f de – 60 mV à l’intérieur, par rapport au milieu
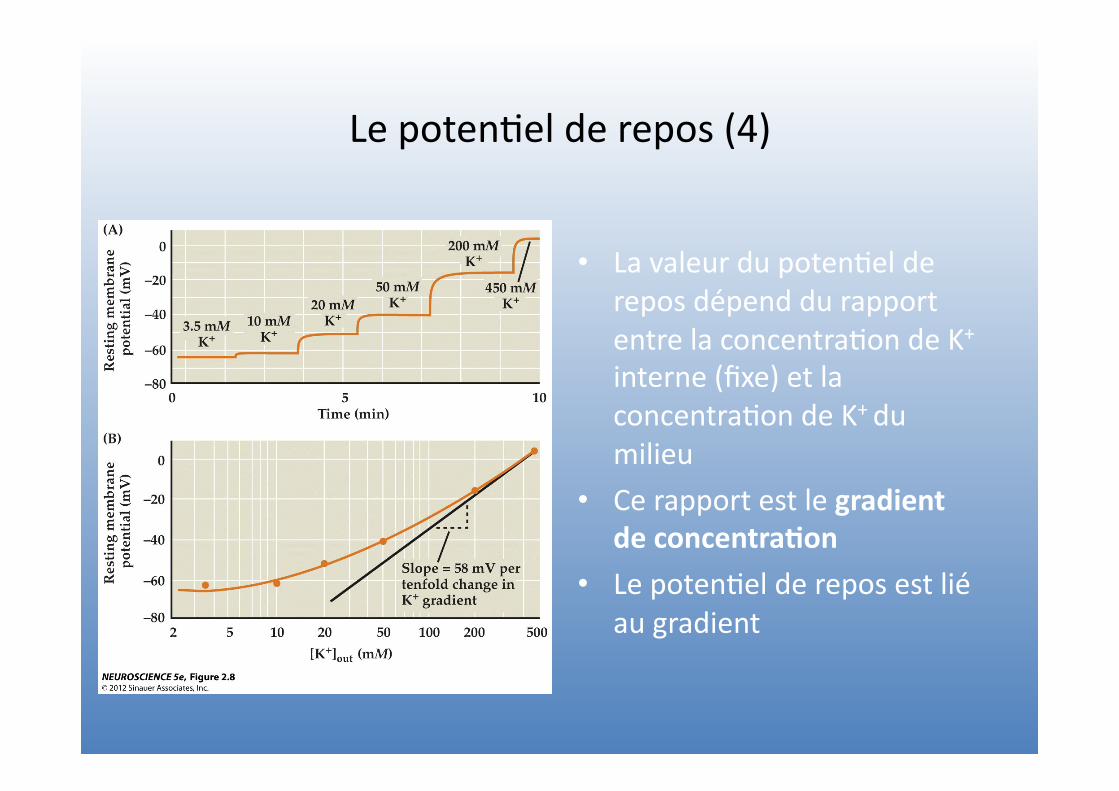
Le poten=el de repos (4)
• La valeur du poten=el de repos dépend du rapport entre la concentra=on de K+ interne (fixe) et la concentra=on de K+ du milieu
• Ce rapport est le gradient de concentra4on
• Le poten=el de repos est lié au gradient


Résumé: poten=el de repos
• L’existence d’un poten=el de repos est dû à une asymétrie dans la distribu=on des ions
• Phénomène réclamant de l’énergie (à expliquer) • Il faut aussi des perméabilités sélec=ves aux ions: la membrane est perméable (faiblement) aux ions K+
• Existence de canaux ioniques ouverts (à expliquer) • Toutes les cellules sont électriquement polarisées: ce n’est pas une propriété des neurones
• Quelle est la spécificité des neurones: le poten=el d’ac=on

Le poten=el d’ac=on
Une propriété des neurones Huxley et Hodgkin,
Prix Nobel, 1963
Andrew Huxley 1917-‐2012
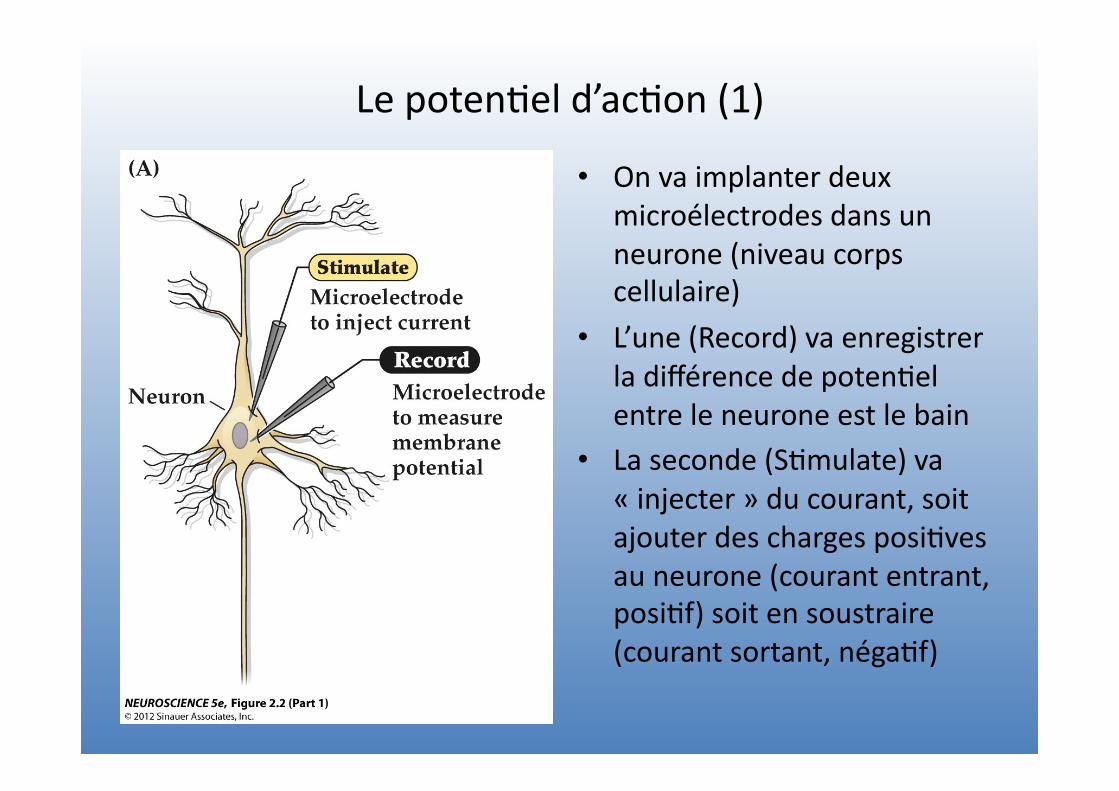
Le poten=el d’ac=on (1)
• On va implanter deux microélectrodes dans un neurone (niveau corps cellulaire)
• L’une (Record) va enregistrer la différence de poten=el entre le neurone est le bain
• La seconde (S=mulate) va « injecter » du courant, soit ajouter des charges posi=ves au neurone (courant entrant, posi=f) soit en soustraire (courant sortant, néga=f)
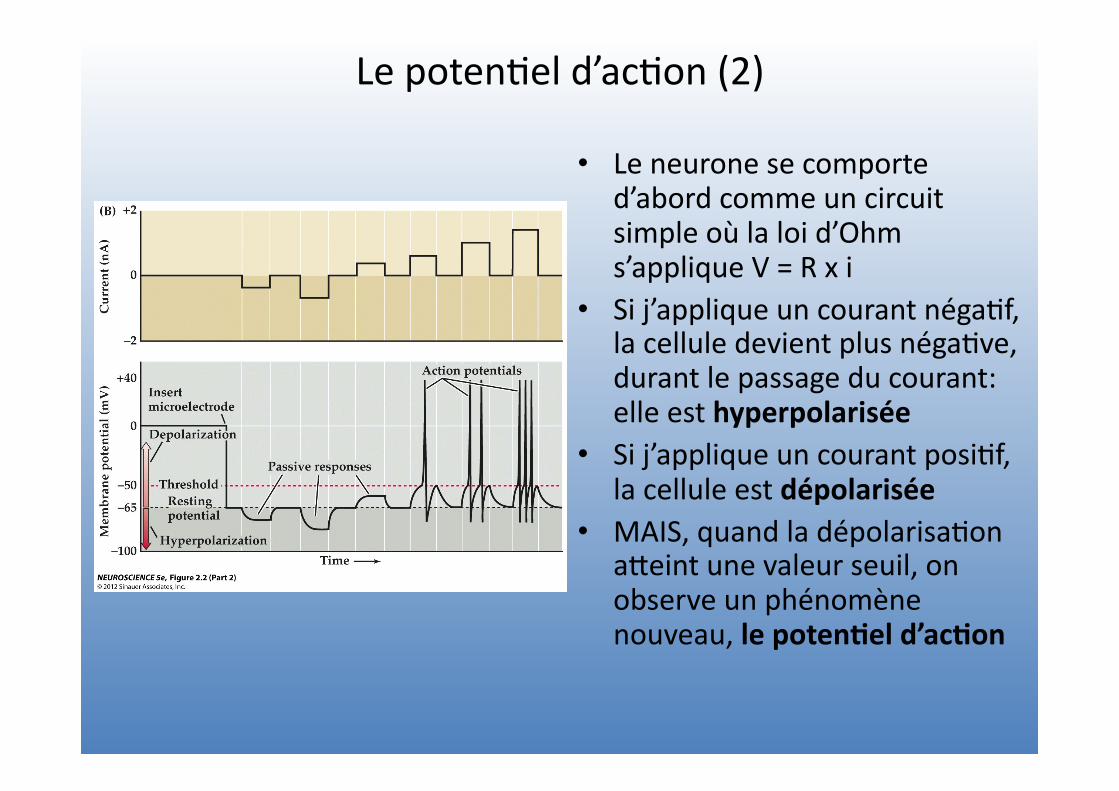
Le poten=el d’ac=on (2)
• Le neurone se comporte d’abord comme un circuit simple où la loi d’Ohm s’applique V = R x i
• Si j’applique un courant néga=f, la cellule devient plus néga=ve, durant le passage du courant: elle est hyperpolarisée
• Si j’applique un courant posi=f, la cellule est dépolarisée
• MAIS, quand la dépolarisa=on aVeint une valeur seuil, on observe un phénomène nouveau, le poten4el d’ac4on
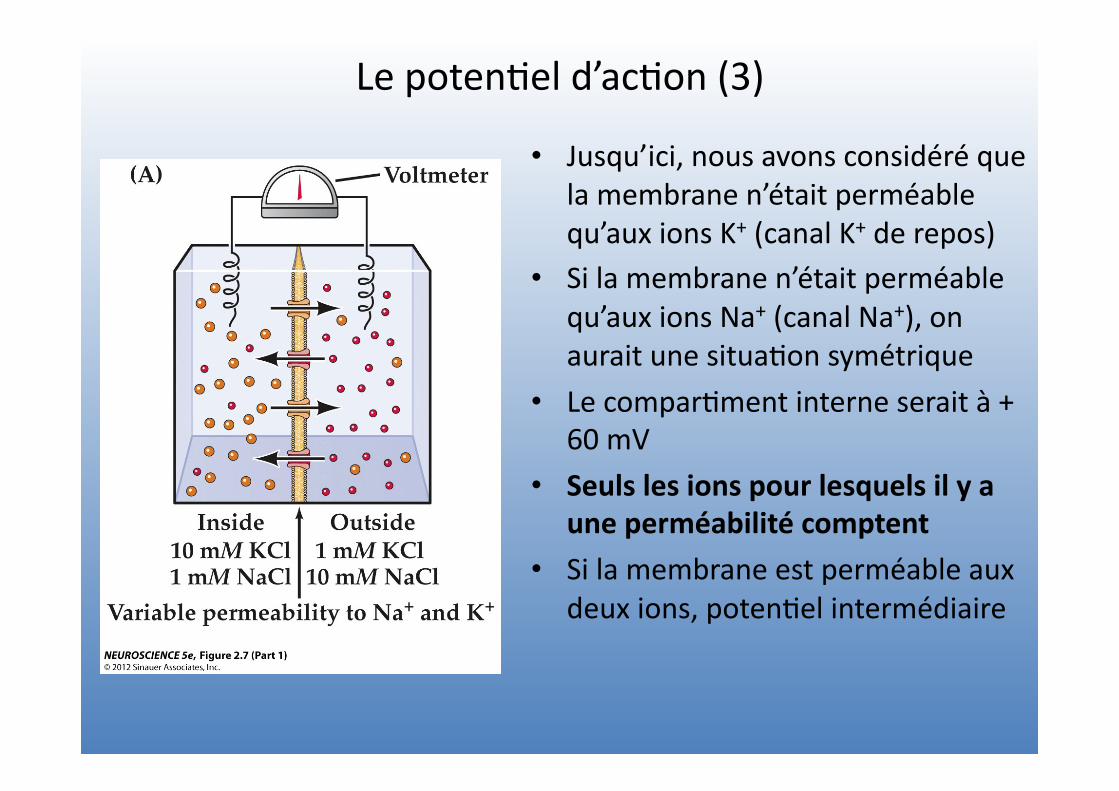
Le poten=el d’ac=on (3)
• Jusqu’ici, nous avons considéré que la membrane n’était perméable qu’aux ions K+ (canal K+ de repos)
• Si la membrane n’était perméable qu’aux ions Na+ (canal Na+), on aurait une situa=on symétrique
• Le compar=ment interne serait à + 60 mV
• Seuls les ions pour lesquels il y a une perméabilité comptent
• Si la membrane est perméable aux deux ions, poten=el intermédiaire
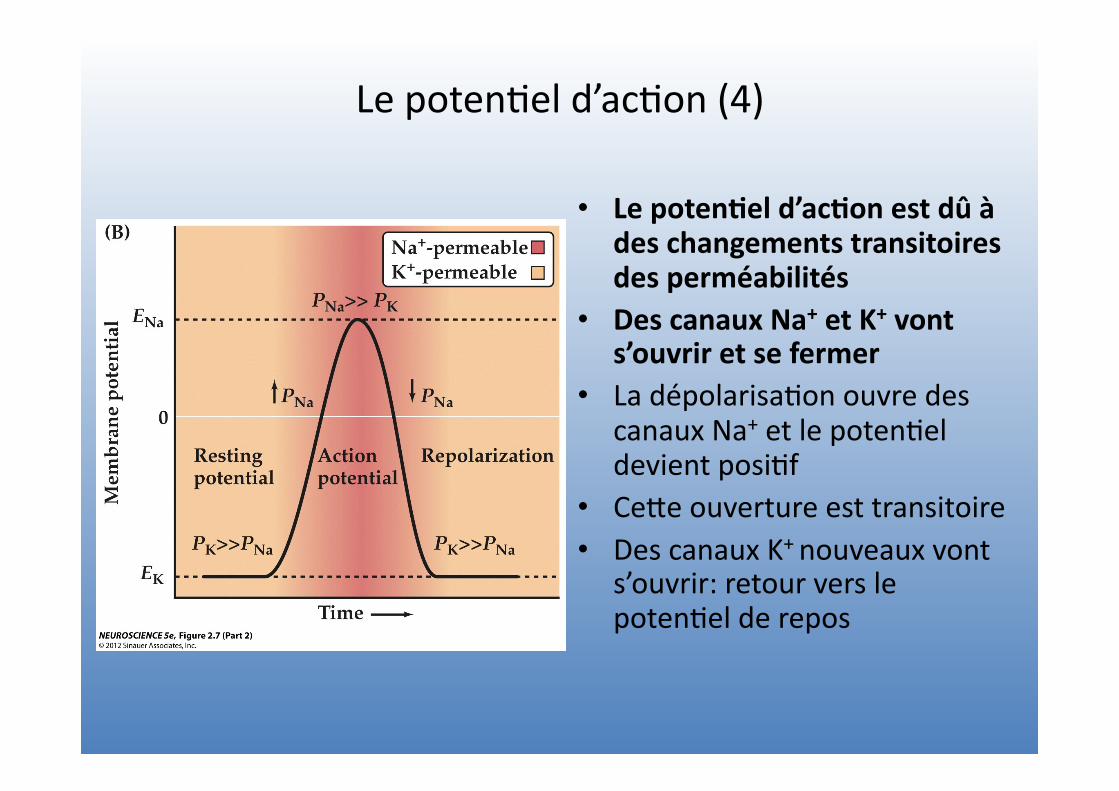
Le poten=el d’ac=on (4)
• Le poten4el d’ac4on est dû à des changements transitoires des perméabilités
• Des canaux Na+ et K+ vont s’ouvrir et se fermer
• La dépolarisa=on ouvre des canaux Na+ et le poten=el devient posi=f
• CeVe ouverture est transitoire • Des canaux K+ nouveaux vont
s’ouvrir: retour vers le poten=el de repos


Le poten=el d’ac=on (5)
• Le poten=el d’ac=on requiert une dépolarisa=on locale (V devient moins néga=f)
• Origine de ceVe dépolarisa=on ? (à expliquer) • Sa forma=on est associée avec l’ouverture de canaux Na+, puis K+
• L’ouverture (et la fermeture) de certains canaux est régulée par la différence de poten=el (canaux dépendants du voltage) (à expliquer)
• Le poten=el d’ac=on est un phénomène de tout ou rien, se formant au delà d’une dépolarisa=on seuil
• Sa forme est constante, quand les concentra=ons de Na+ et K+ sont fixées

Propaga=on du poten=el d’ac=on (1)
• L’axone est un milieu conducteur
• Si on injecte du courant en un point, la différence de poten=el avec le milieu se propage
• Mais c’est un conducteur mal isolé et le courant « fuit » vers l’extérieur : la différence de poten=el s’aVénue rapidement
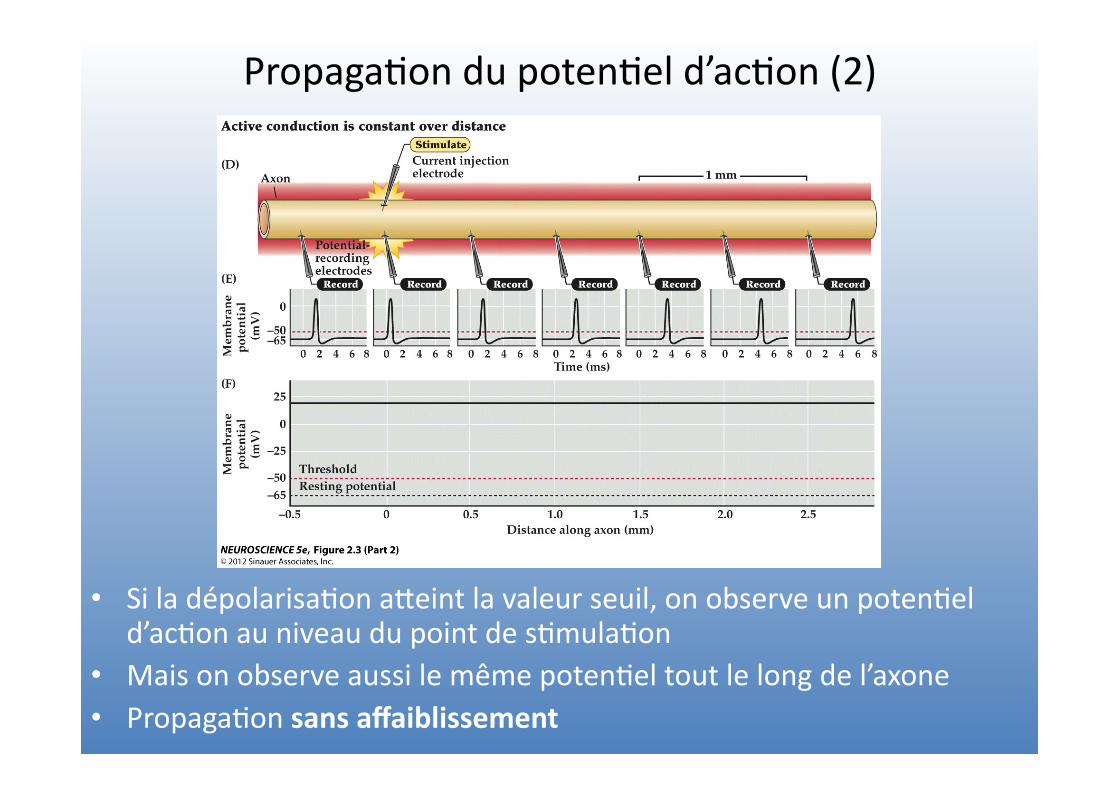
Propaga=on du poten=el d’ac=on (2)
• Si la dépolarisa=on aVeint la valeur seuil, on observe un poten=el d’ac=on au niveau du point de s=mula=on
• Mais on observe aussi le même poten=el tout le long de l’axone • Propaga=on sans affaiblissement


Propaga=on du poten=el d’ac=on (3)
• L’influx nerveux (ou l’ac=va=on des neurones) correspond à la propaga=on d’un poten=el d’ac=on
• Le poten=el d’ac=on ne s’aVénue pas et se propage sans modifica=on de forme
• L’informa=on portée par l’influx est stéréotypée, elle n’est pas contenue dans la forme des signaux
• Seulement deux types de signaux: le poten=el de repos (modulable) et le poten=el d’ac=on (fixe)
• Comment est modulé le poten=el de repos ? (à voir)

Propaga=on du poten=el d’ac=on (3)
• Si la forme du poten=el d’ac=on est invariable, sa vitesse de propaga=on varie
• Un axone moteur peut mesurer 1 mètre de long: ces axones sont myélinisés
• Une vitesse de 1 m/s correspond à 3,6 km/h et 100 m/s à 360 km/h
• La repolarisa=on (ouverture des canaux K+) introduit une période réfractaire, pendant laquelle un poten=el d’ac=on ne peut pas circuler (environ 5 ms)
• Cela signifie qu’un train de s=mula=on ne peut pas dépasser 0,2 kHz, ce qui est ridicule par rapport à un ordinateur (3 gigaHz, soit plus d’un million de fois plus rapide!)

Les canaux ioniques
Bases moléculaires des phénomènes électriques

Les canaux sensibles au poten=el
• Les travaux de Hodgkin et Huxley (1945-‐ 1955) ont établi la nature du poten=el d’ac=on en postulant des changements de conductance, sans connaître l’existence des canaux ioniques
• Leurs travaux consistent en équa=ons qui rendent parfaitement compte des phénomènes observés
• La base des changements de conductance est due à l’ouverture de canaux, spécifiques des neurones
• La principale caractéris=que des canaux du poten=el d’ac=on est d’être dépendants du poten=el
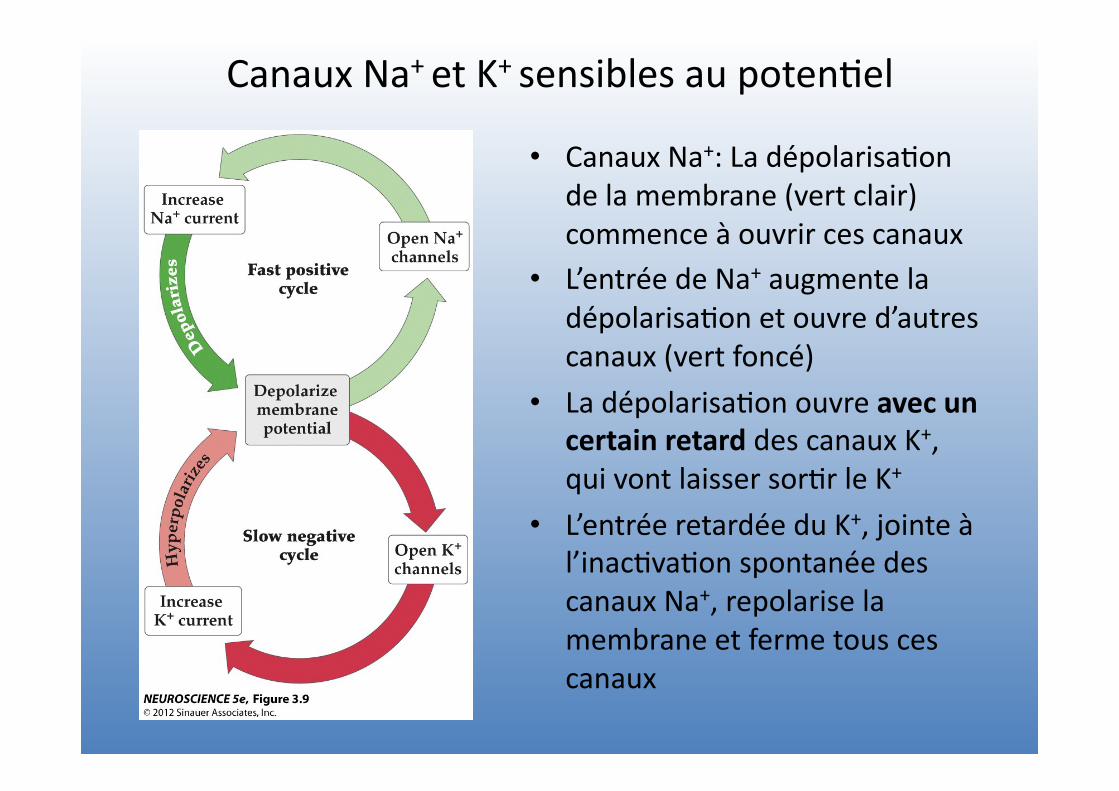
Canaux Na+ et K+ sensibles au poten=el
• Canaux Na+: La dépolarisa=on de la membrane (vert clair) commence à ouvrir ces canaux
• L’entrée de Na+ augmente la dépolarisa=on et ouvre d’autres canaux (vert foncé)
• La dépolarisa=on ouvre avec un certain retard des canaux K+, qui vont laisser sor=r le K+
• L’entrée retardée du K+, jointe à l’inac=va=on spontanée des canaux Na+, repolarise la membrane et ferme tous ces canaux
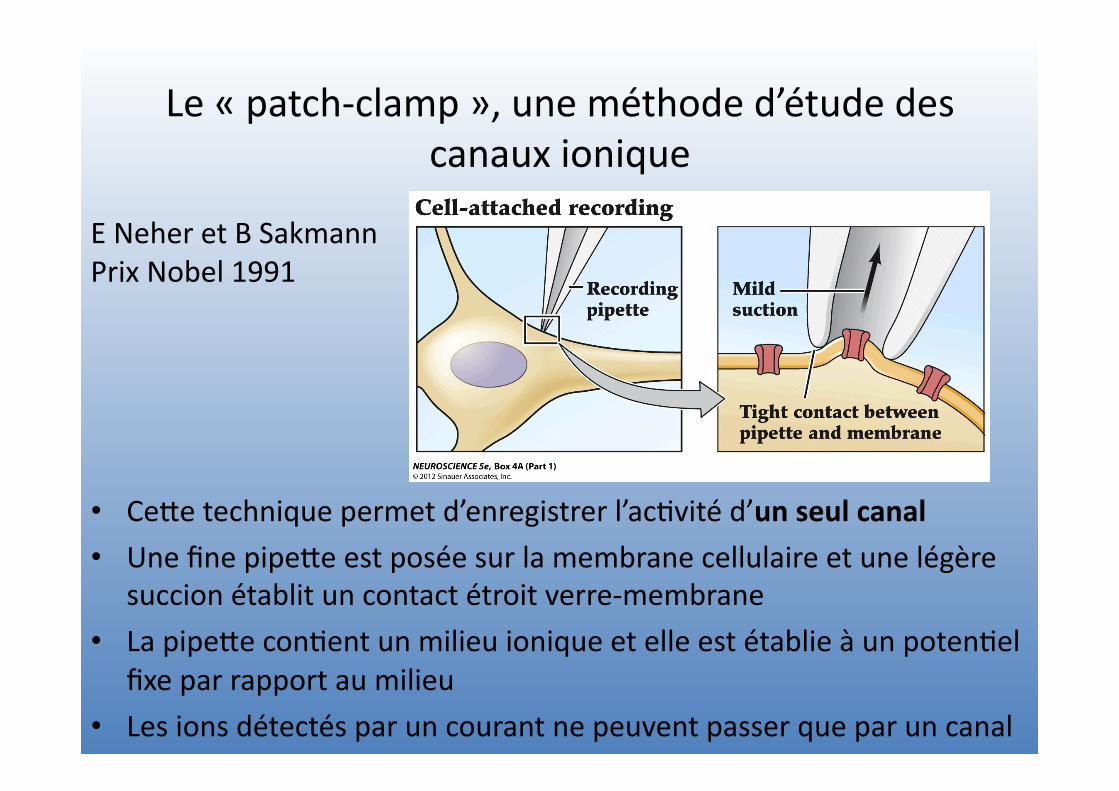
Le « patch-‐clamp », une méthode d’étude des canaux ionique
• CeVe technique permet d’enregistrer l’ac=vité d’un seul canal
• Une fine pipeVe est posée sur la membrane cellulaire et une légère succion établit un contact étroit verre-‐membrane
• La pipeVe con=ent un milieu ionique et elle est établie à un poten=el fixe par rapport au milieu
• Les ions détectés par un courant ne peuvent passer que par un canal
E Neher et B Sakmann Prix Nobel 1991
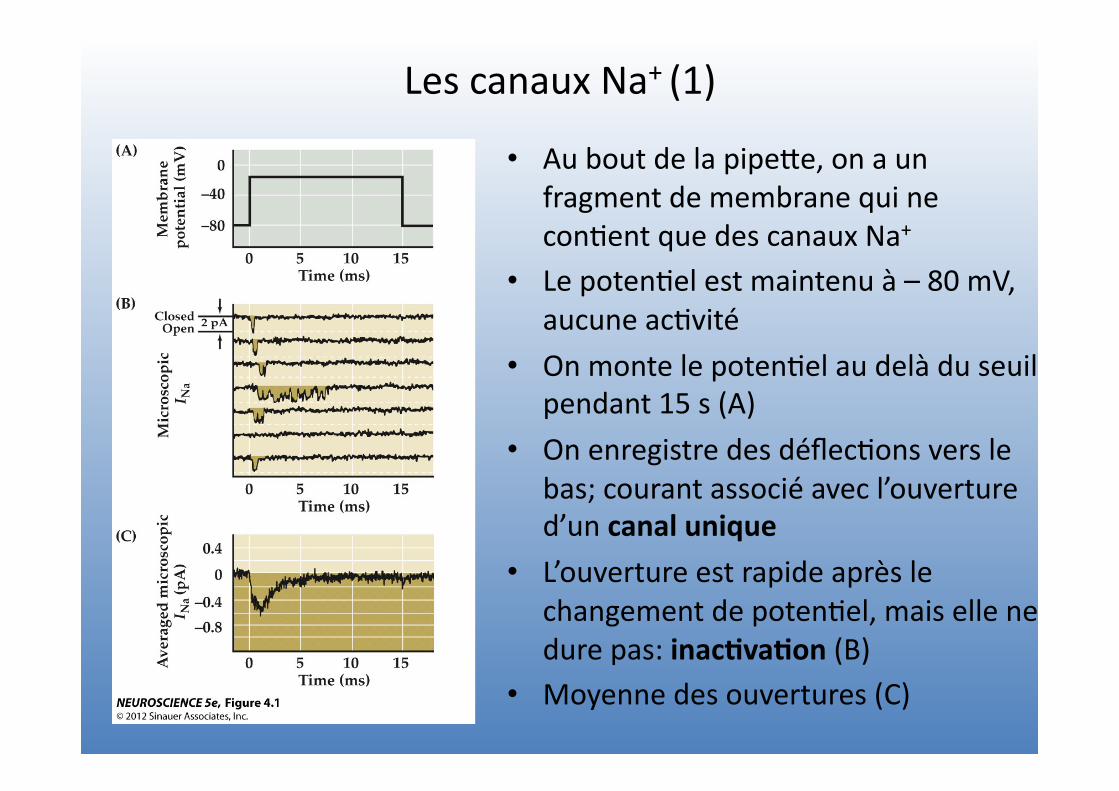
Les canaux Na+ (1)
• Au bout de la pipeVe, on a un fragment de membrane qui ne con=ent que des canaux Na+
• Le poten=el est maintenu à – 80 mV, aucune ac=vité
• On monte le poten=el au delà du seuil pendant 15 s (A)
• On enregistre des déflec=ons vers le bas; courant associé avec l’ouverture d’un canal unique
• L’ouverture est rapide après le changement de poten=el, mais elle ne dure pas: inac4va4on (B)
• Moyenne des ouvertures (C)
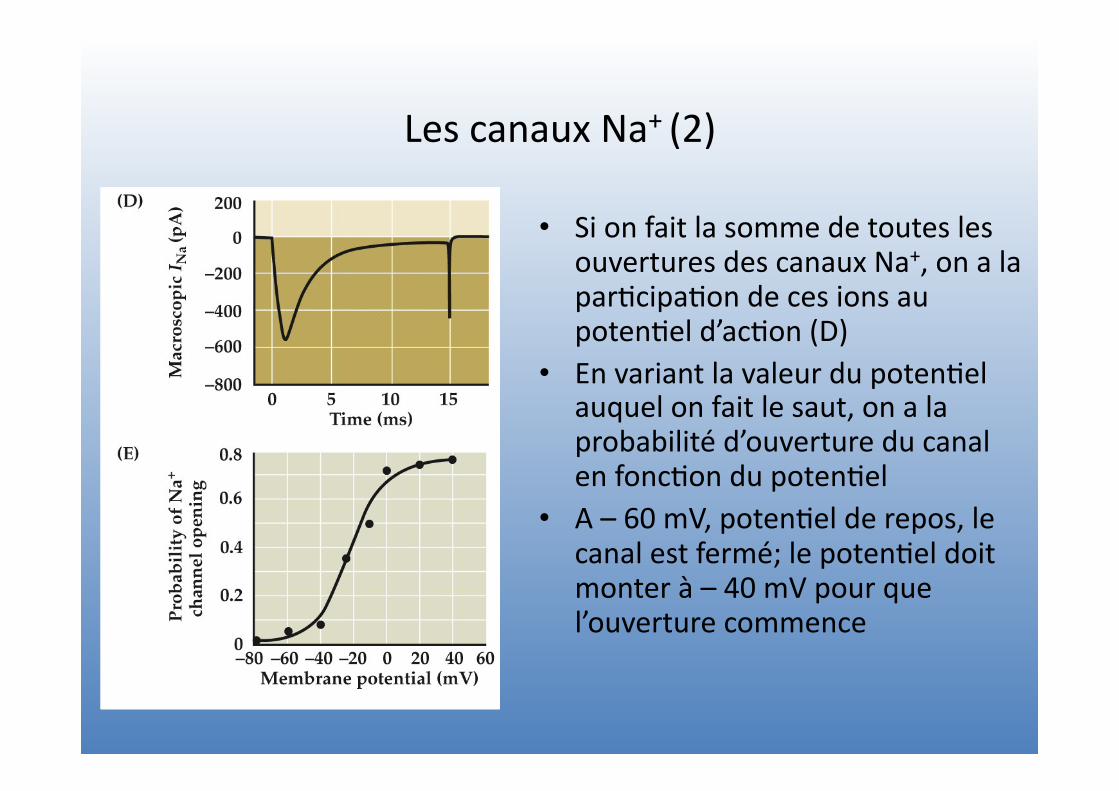
Les canaux Na+ (2)
• Si on fait la somme de toutes les ouvertures des canaux Na+, on a la par=cipa=on de ces ions au poten=el d’ac=on (D)
• En variant la valeur du poten=el auquel on fait le saut, on a la probabilité d’ouverture du canal en fonc=on du poten=el
• A – 60 mV, poten=el de repos, le canal est fermé; le poten=el doit monter à – 40 mV pour que l’ouverture commence
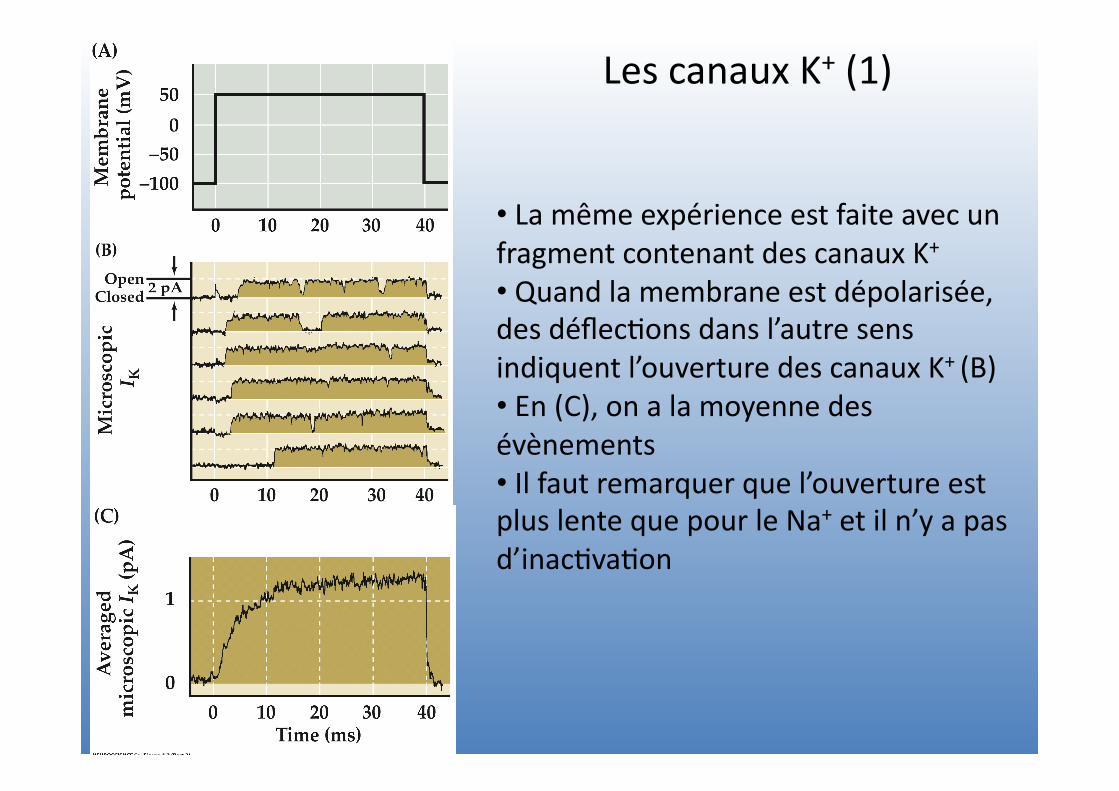
Les canaux K+ (1)
• La même expérience est faite avec un fragment contenant des canaux K+
• Quand la membrane est dépolarisée, des déflec=ons dans l’autre sens indiquent l’ouverture des canaux K+ (B) • En (C), on a la moyenne des évènements • Il faut remarquer que l’ouverture est plus lente que pour le Na+ et il n’y a pas d’inac=va=on
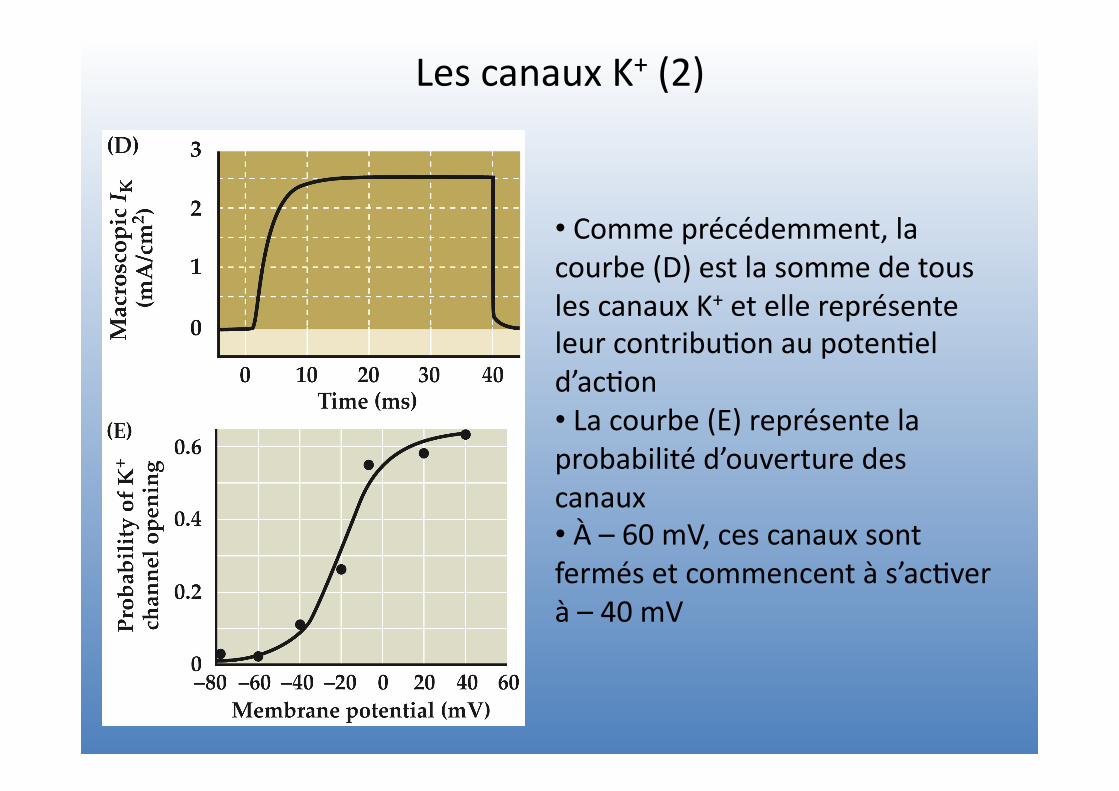
Les canaux K+ (2)
• Comme précédemment, la courbe (D) est la somme de tous les canaux K+ et elle représente leur contribu=on au poten=el d’ac=on • La courbe (E) représente la probabilité d’ouverture des canaux • À – 60 mV, ces canaux sont fermés et commencent à s’ac=ver à – 40 mV

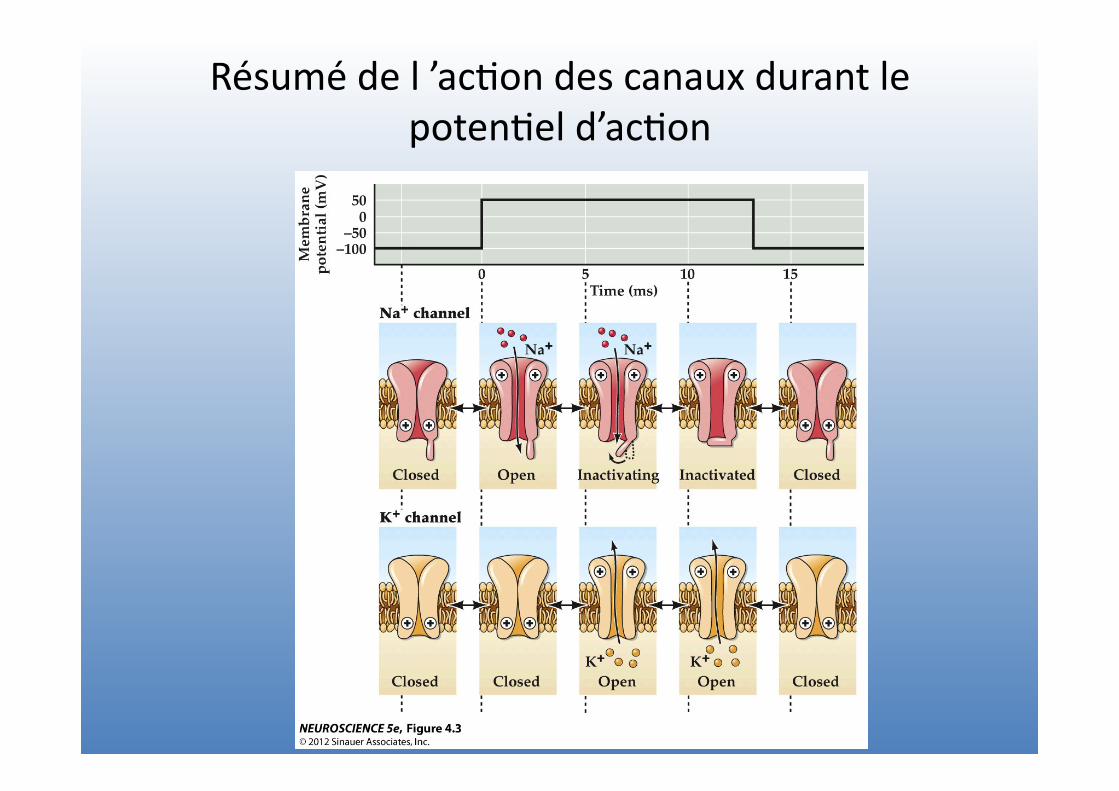
Résumé de l ’ac=on des canaux durant le poten=el d’ac=on

Structure des canaux ioniques
Des protéines membranaires enfin bien connues

Comment passer de la fonc=on à la structure ? (1)
• Les canaux sont des protéines enchâssées dans la membrane; leur purifica=on requiert leur solubilisa=on. Ce sont des procédures difficiles
• Pour suivre leur ac=vité après solubilisa=on, on peut u=liser des toxines de forte affinité (Ex.: Tétrodotoxine)
• On passe de la protéine purifiée au gène par les techniques de la géné=que moléculaire
• Le gène donne de nombreuses informa=ons: séquence de la protéine, repliement de la chaine pep=dique, diversité protéique, localisa=on,…
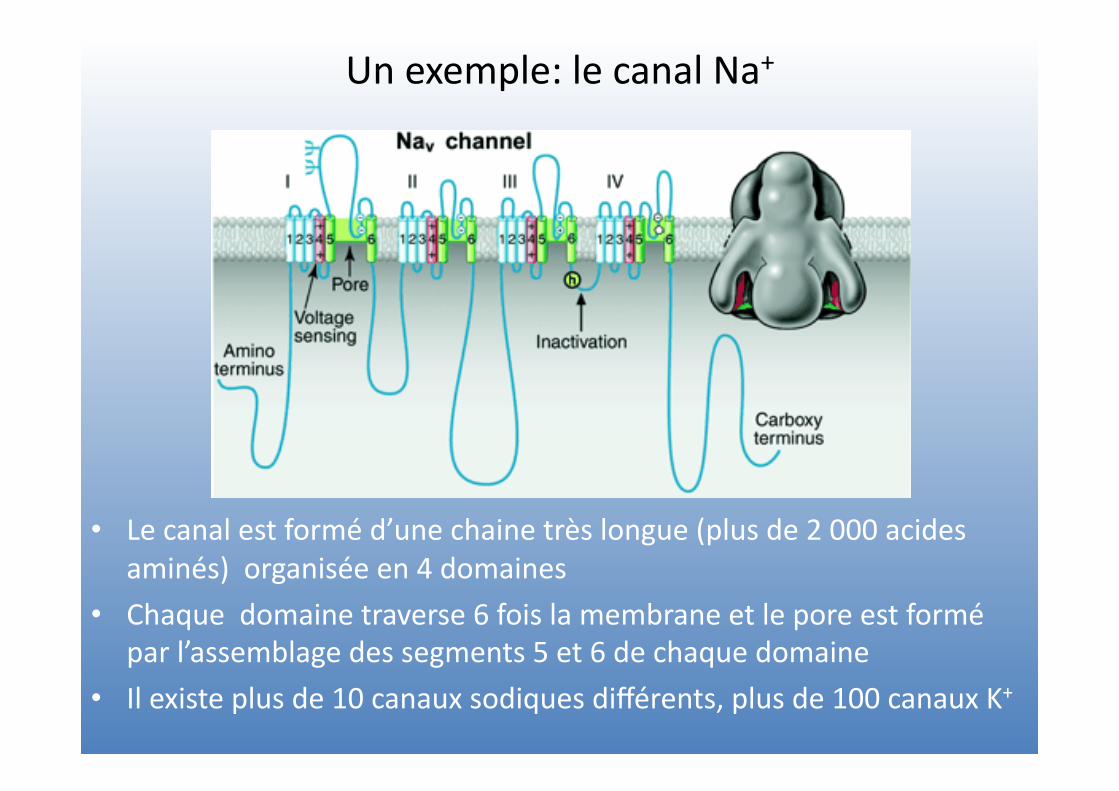
Un exemple: le canal Na+
• Le canal est formé d’une chaine très longue (plus de 2 000 acides aminés) organisée en 4 domaines
• Chaque domaine traverse 6 fois la membrane et le pore est formé par l’assemblage des segments 5 et 6 de chaque domaine
• Il existe plus de 10 canaux sodiques différents, plus de 100 canaux K+

Comment passer de la fonc=on à la structure ? (2)
• Pour avoir la structure atomique (place des atomes), la technique est l’analyse aux rayons X de la structure atomique
• Elle suppose d’obtenir des cristaux de la protéine (empilements réguliers de la protéine); la posi=on des atome est alors fixe
• Pour les protéines membranaires, ce stade est par=culièrement difficile
• Il a été franchi par Roderick McKinnon, Prix Nobel 2003
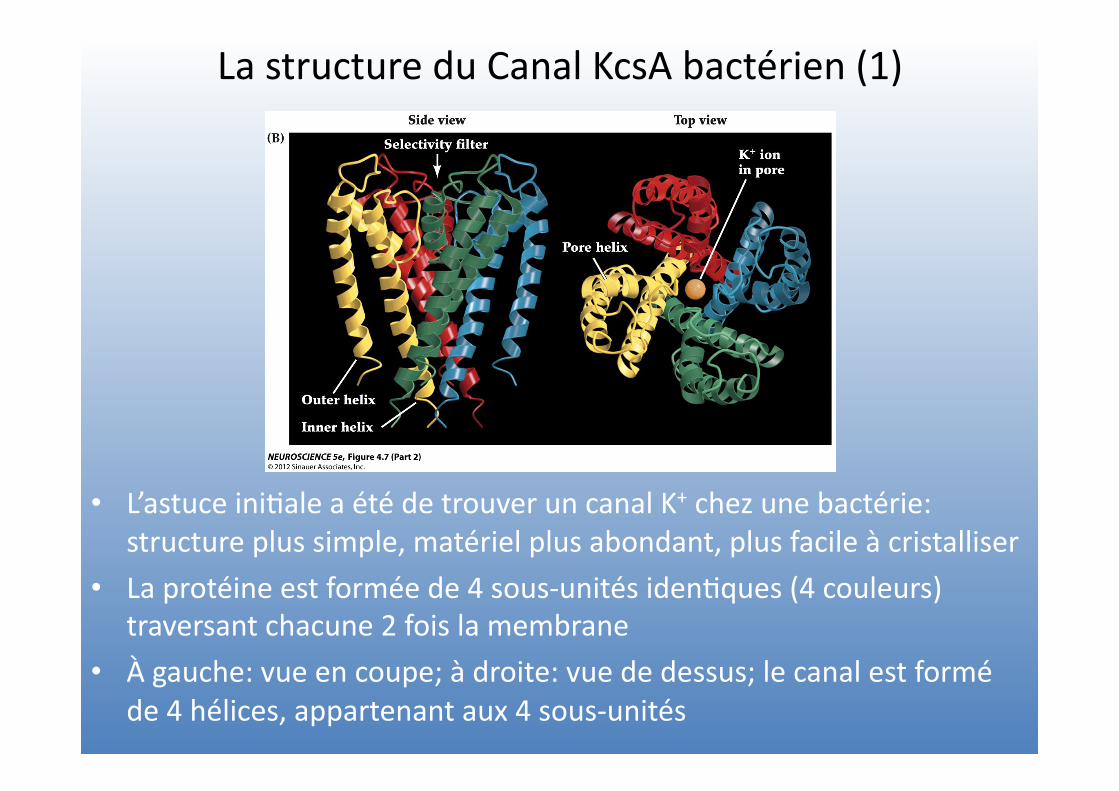
La structure du Canal KcsA bactérien (1)
• L’astuce ini=ale a été de trouver un canal K+ chez une bactérie: structure plus simple, matériel plus abondant, plus facile à cristalliser
• La protéine est formée de 4 sous-‐unités iden=ques (4 couleurs) traversant chacune 2 fois la membrane
• À gauche: vue en coupe; à droite: vue de dessus; le canal est formé de 4 hélices, appartenant aux 4 sous-‐unités
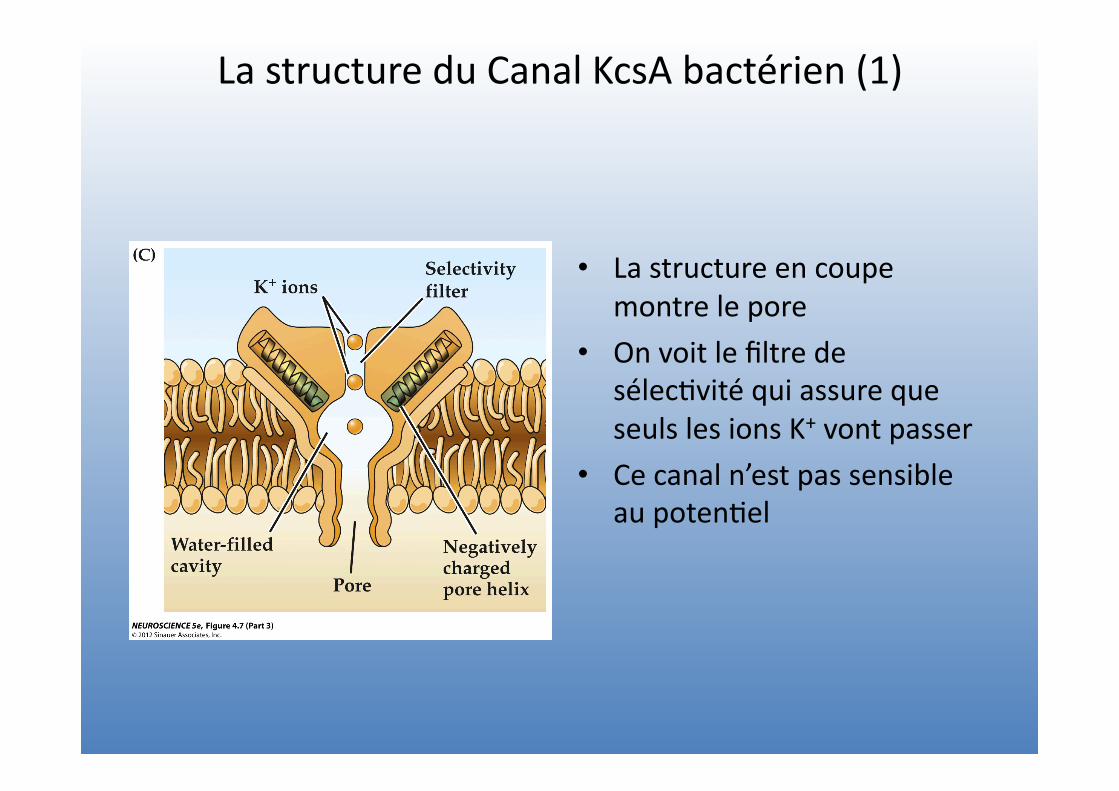
La structure du Canal KcsA bactérien (1)
• La structure en coupe montre le pore
• On voit le filtre de sélec=vité qui assure que seuls les ions K+ vont passer
• Ce canal n’est pas sensible au poten=el
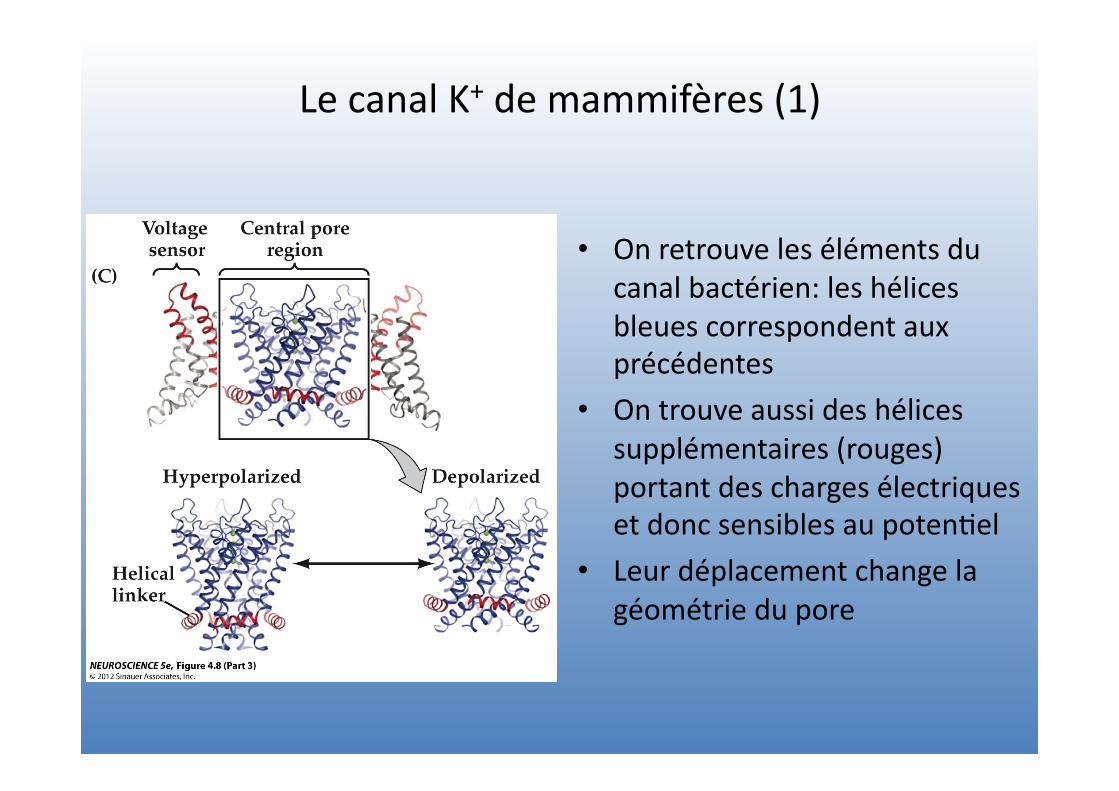
Le canal K+ de mammifères (1)
• On retrouve les éléments du canal bactérien: les hélices bleues correspondent aux précédentes
• On trouve aussi des hélices supplémentaires (rouges) portant des charges électriques et donc sensibles au poten=el
• Leur déplacement change la géométrie du pore
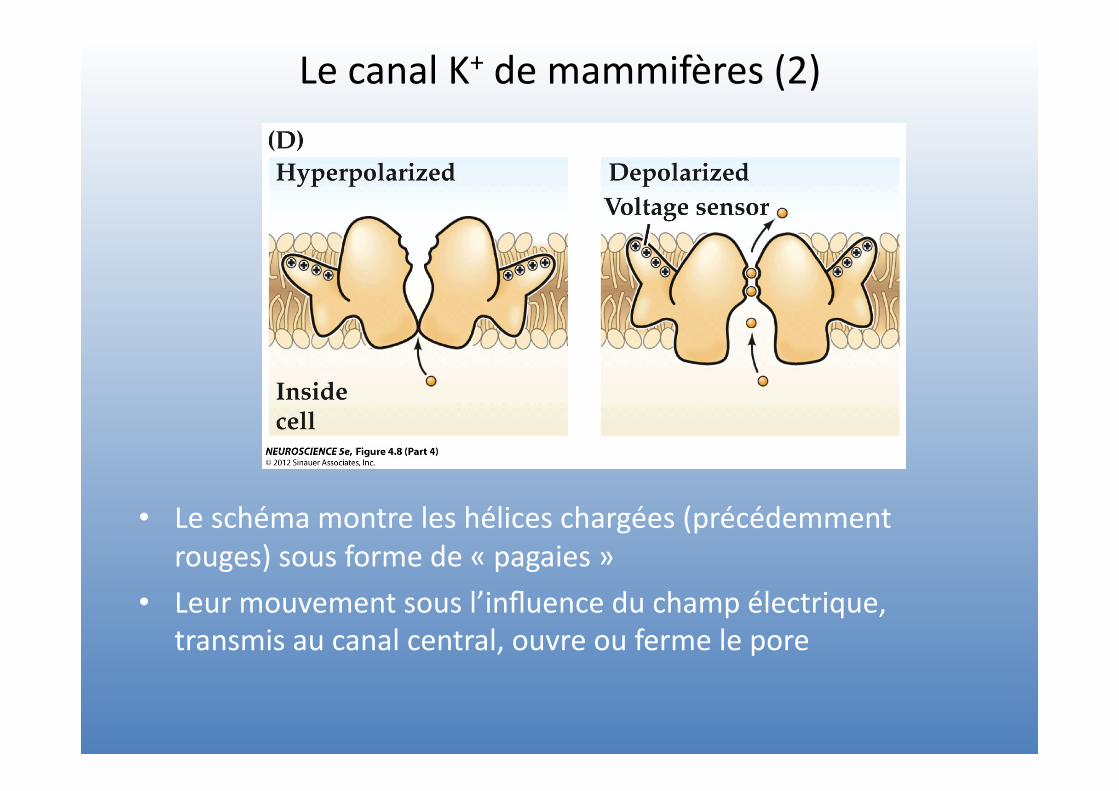
Le canal K+ de mammifères (2)
• Le schéma montre les hélices chargées (précédemment rouges) sous forme de « pagaies »
• Leur mouvement sous l’influence du champ électrique, transmis au canal central, ouvre ou ferme le pore

Comment naissent les poten=els d’ac=on?
Les canaux ioniques ac=vés par des ligands

Les canaux ac=vés par des ligands (1)
• Les canaux Na+ et K+ assurent la propaga=on du poten=el d’ac=on; ils sont localisés sur l’axone et le corps cellulaire
• Au niveau des synapses, principalement sur les dendrites et le corps cellulaire, on trouve d’autres canaux dont l’ouverture est contrôlée chimiquement
• Les molécules ac=ves sont les neurotransmeVeurs: ces canaux sont des récepteurs des neurotransmeVeurs (ligands = se liant)
• Remarque: il existe d’autres récepteurs des neurotransmeVeurs qui ne sont pas canaux ioniques (cours 3)
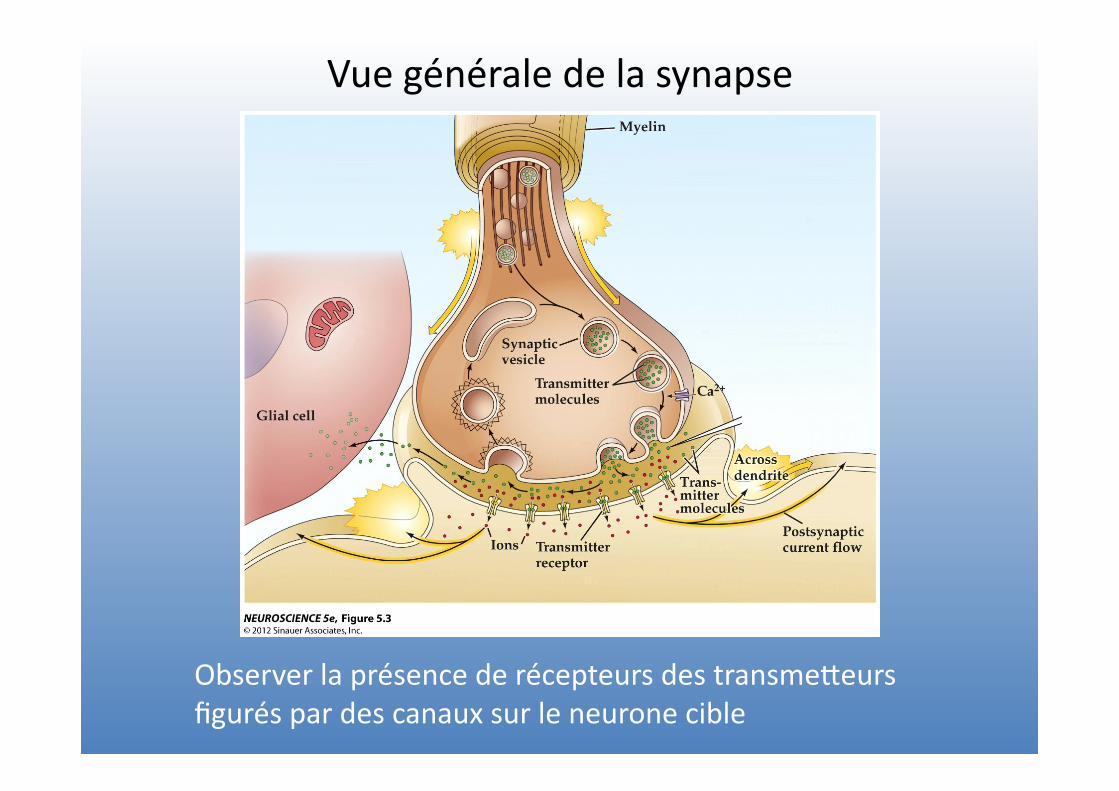
Vue générale de la synapse
Observer la présence de récepteurs des transmeVeurs figurés par des canaux sur le neurone cible
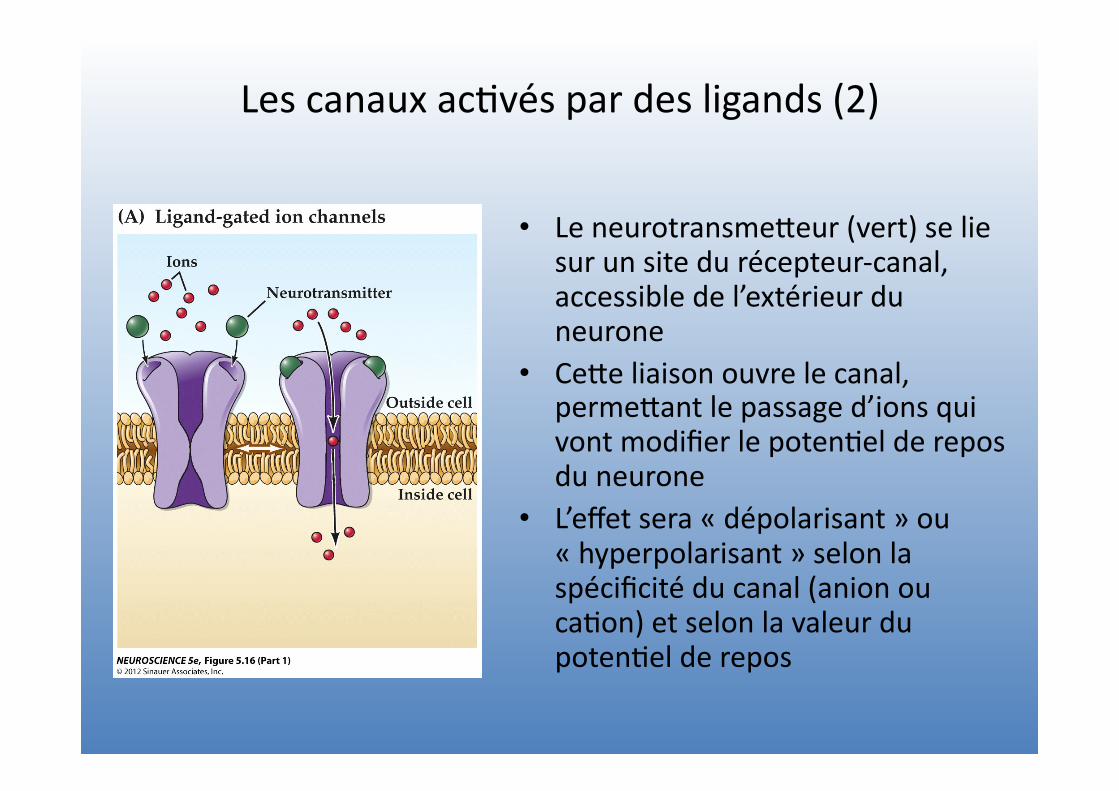
Les canaux ac=vés par des ligands (2)
• Le neurotransmeVeur (vert) se lie sur un site du récepteur-‐canal, accessible de l’extérieur du neurone
• CeVe liaison ouvre le canal, permeVant le passage d’ions qui vont modifier le poten=el de repos du neurone
• L’effet sera « dépolarisant » ou « hyperpolarisant » selon la spécificité du canal (anion ou ca=on) et selon la valeur du poten=el de repos
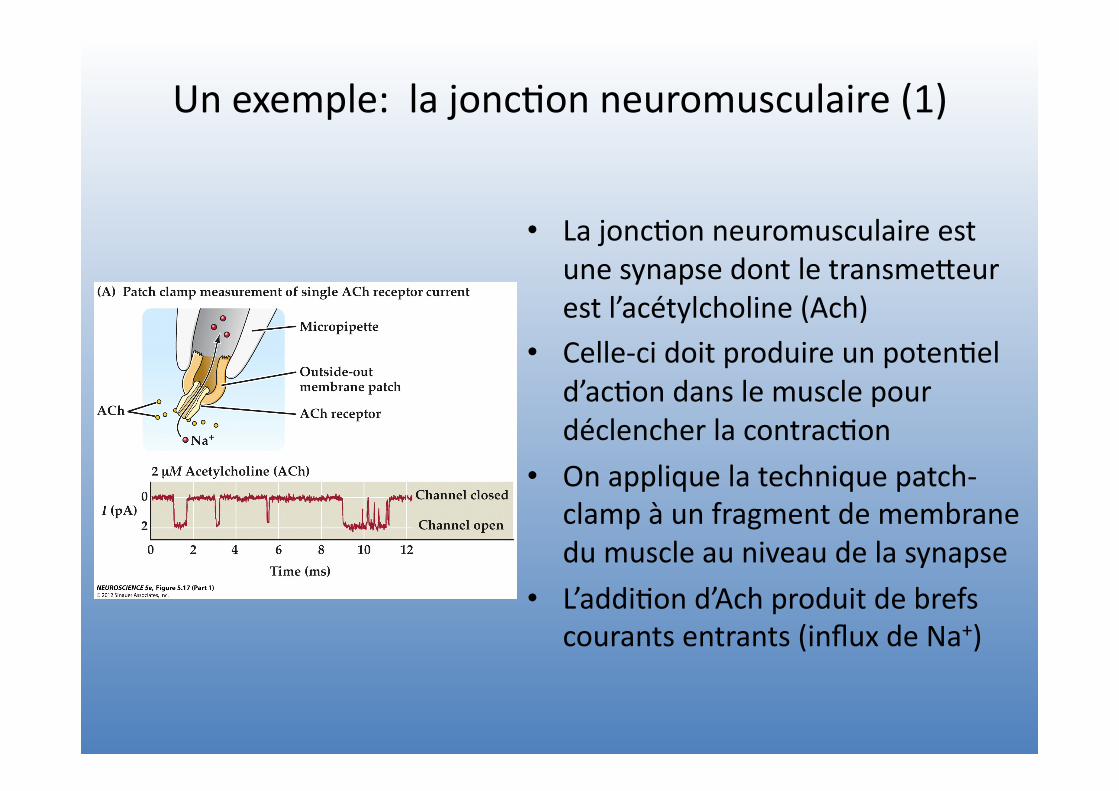
Un exemple: la jonc=on neuromusculaire (1)
• La jonc=on neuromusculaire est une synapse dont le transmeVeur est l’acétylcholine (Ach)
• Celle-‐ci doit produire un poten=el d’ac=on dans le muscle pour déclencher la contrac=on
• On applique la technique patch-‐clamp à un fragment de membrane du muscle au niveau de la synapse
• L’addi=on d’Ach produit de brefs courants entrants (influx de Na+)
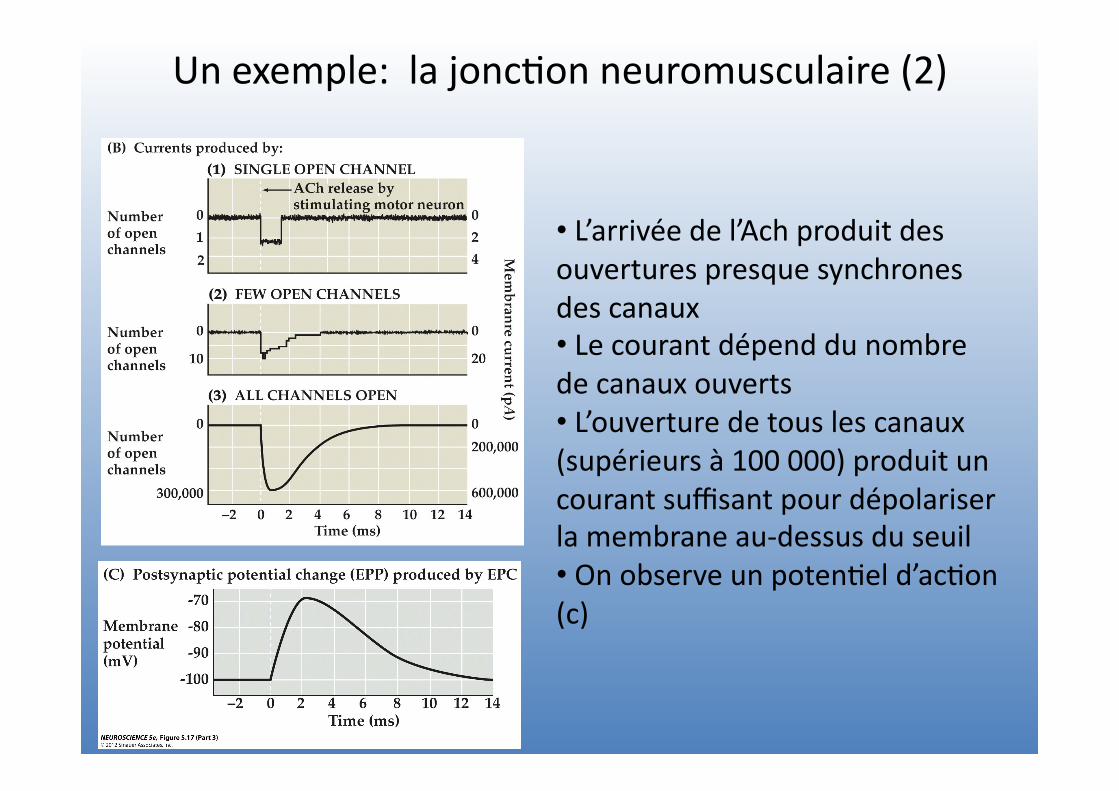
Un exemple: la jonc=on neuromusculaire (2)
• L’arrivée de l’Ach produit des ouvertures presque synchrones des canaux • Le courant dépend du nombre de canaux ouverts • L’ouverture de tous les canaux (supérieurs à 100 000) produit un courant suffisant pour dépolariser la membrane au-‐dessus du seuil • On observe un poten=el d’ac=on (c)
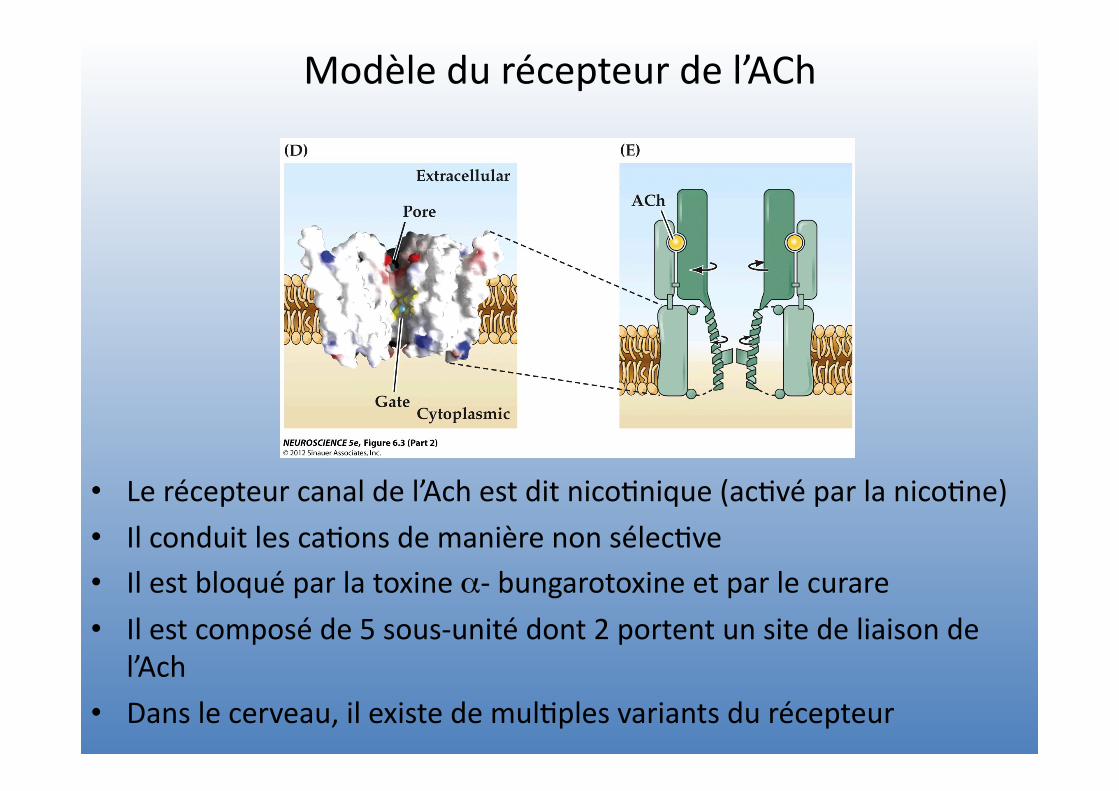
Modèle du récepteur de l’ACh
• Le récepteur canal de l’Ach est dit nico=nique (ac=vé par la nico=ne) • Il conduit les ca=ons de manière non sélec=ve • Il est bloqué par la toxine α-‐ bungarotoxine et par le curare • Il est composé de 5 sous-‐unité dont 2 portent un site de liaison de
l’Ach
• Dans le cerveau, il existe de mul=ples variants du récepteur
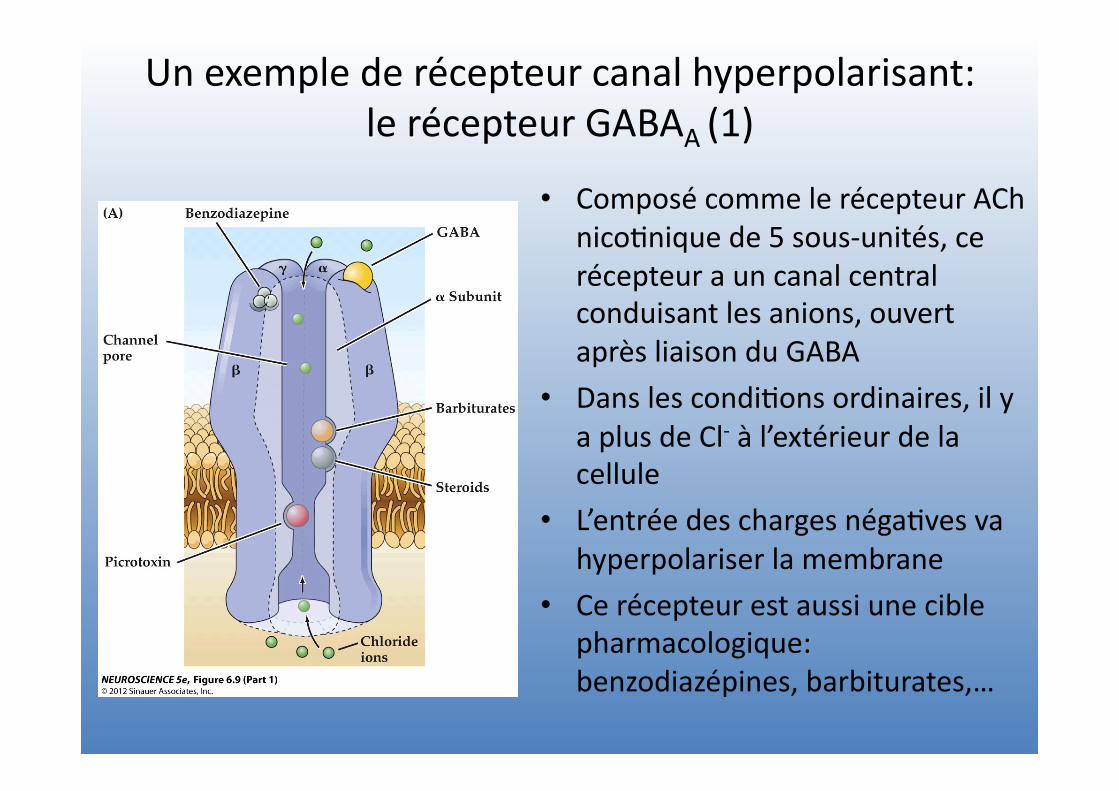
Un exemple de récepteur canal hyperpolarisant: le récepteur GABAA (1)
• Composé comme le récepteur ACh nico=nique de 5 sous-‐unités, ce récepteur a un canal central conduisant les anions, ouvert après liaison du GABA
• Dans les condi=ons ordinaires, il y a plus de Cl-‐ à l’extérieur de la cellule
• L’entrée des charges néga=ves va hyperpolariser la membrane
• Ce récepteur est aussi une cible pharmacologique: benzodiazépines, barbiturates,…
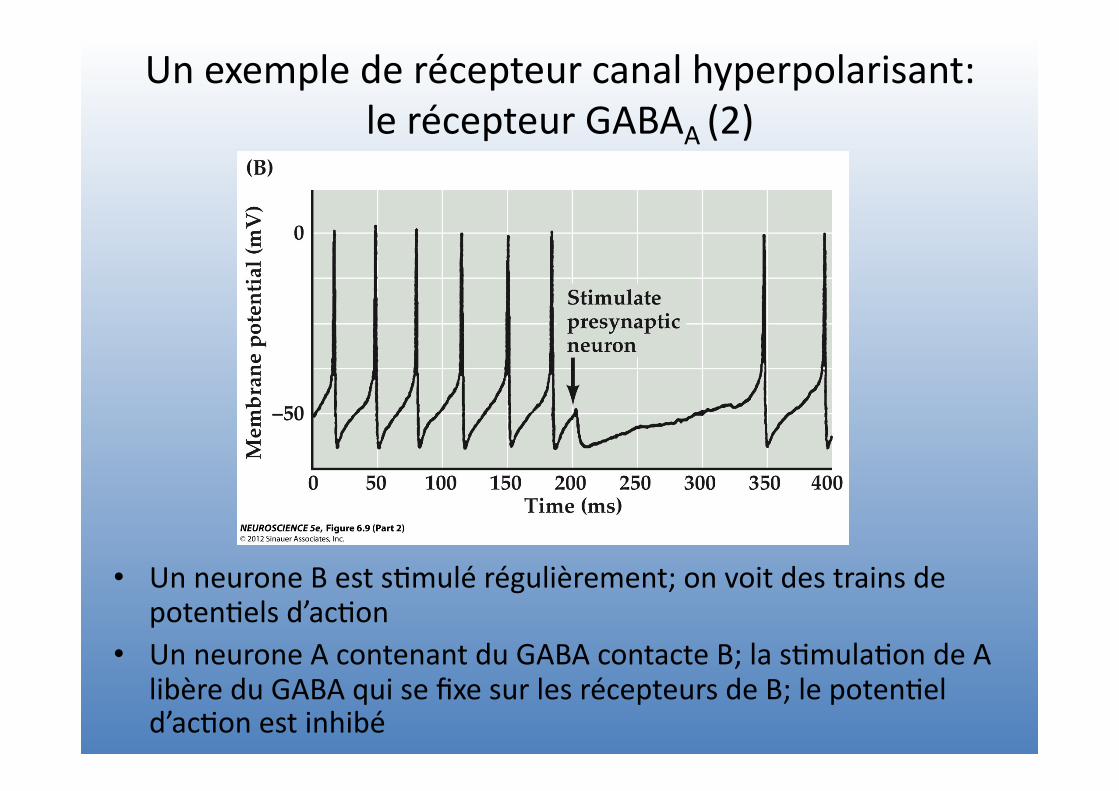
Un exemple de récepteur canal hyperpolarisant: le récepteur GABAA (2)
• Un neurone B est s=mulé régulièrement; on voit des trains de poten=els d’ac=on
• Un neurone A contenant du GABA contacte B; la s=mula=on de A libère du GABA qui se fixe sur les récepteurs de B; le poten=el d’ac=on est inhibé

Résumé sur les récepteurs canaux
• Un neurotransmeVeur se lie aux récepteurs portés par la membrane du neurone cible
• Certains de ces récepteurs sont des canaux et la liaison du neurotransmeVeur les ouvre
• Selon la valeur du poten=el de repos et la spécificité du canal, le poten=el de repos va être modifié
• Le récepteur de l’Ach nico=nique et les récepteurs du glutamate de type NMDA et AMPA sont ac=vateurs
• Les récepteurs GABA de type A et C sont inhibiteurs
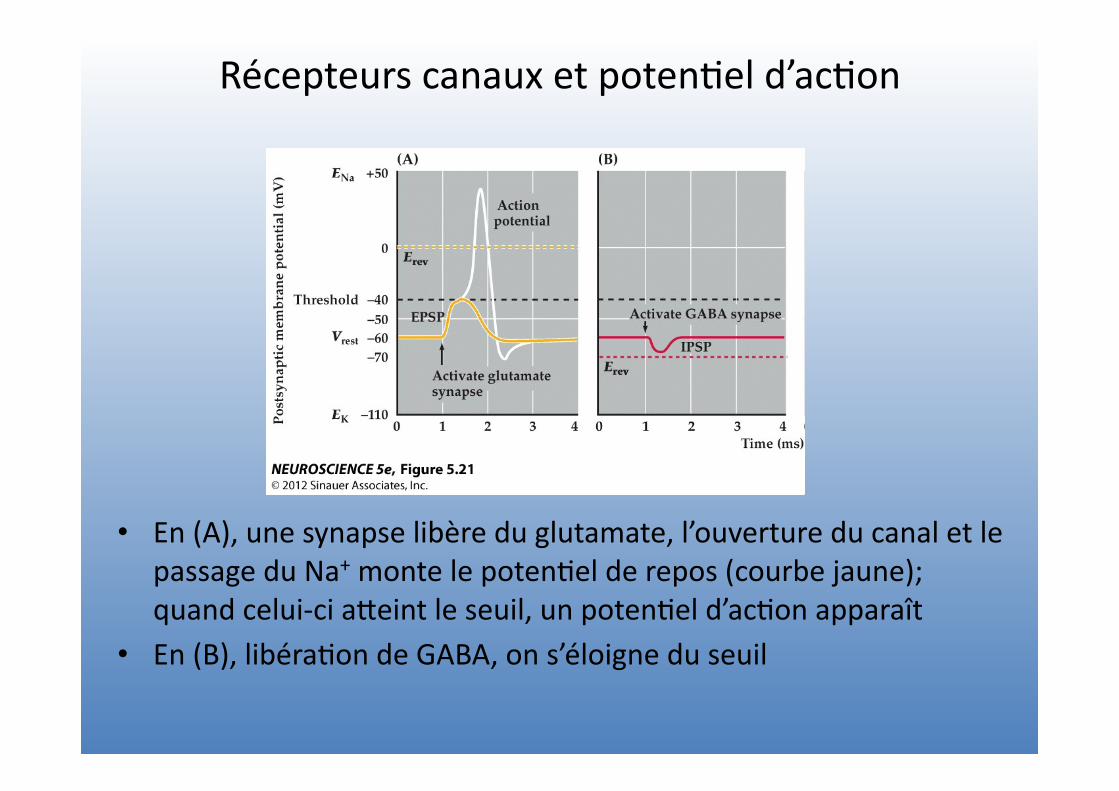
Récepteurs canaux et poten=el d’ac=on
• En (A), une synapse libère du glutamate, l’ouverture du canal et le passage du Na+ monte le poten=el de repos (courbe jaune); quand celui-‐ci aVeint le seuil, un poten=el d’ac=on apparaît
• En (B), libéra=on de GABA, on s’éloigne du seuil
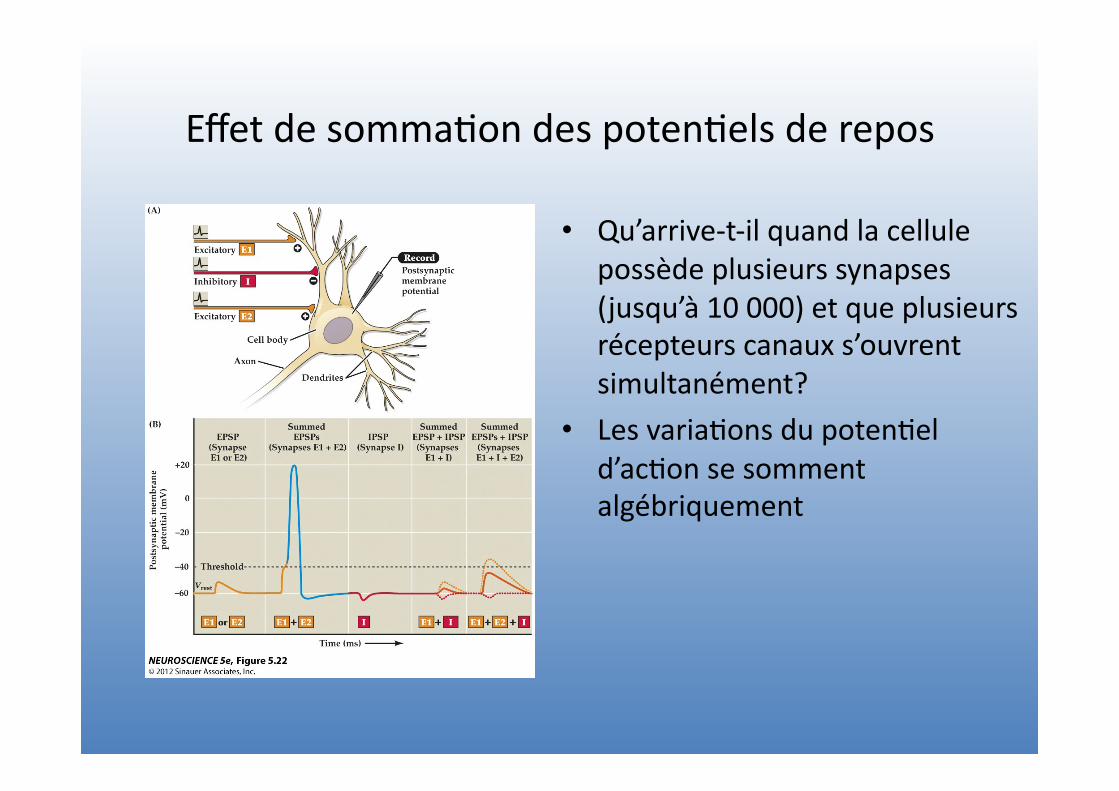
Effet de somma=on des poten=els de repos
• Qu’arrive-‐t-‐il quand la cellule possède plusieurs synapses (jusqu’à 10 000) et que plusieurs récepteurs canaux s’ouvrent simultanément?
• Les varia=ons du poten=el d’ac=on se somment algébriquement


Quelques conclusions
• L’informa=on traversant un neurone est électrique • Elle a la forme d’un poten=el d’ac=on « stéréotypé » • Le poten=el d’ac=on nait à la base de l’axone et se propage vers la terminaison
• L’induc=on du poten=el d’ac=on dépend de varia=ons du poten=el de repos
• Celles-‐ci sont induites par les synapses des neurones afférents (ou des récepteurs sensoriels, cours 4)
• La mul=plicité des synapses permet l’intégra=on des signaux électriques
• Il existe une intégra=on chimique des signaux (cours 3)

Comment sont entretenus les gradients ioniques?
Un coût énergé=que bien u=le
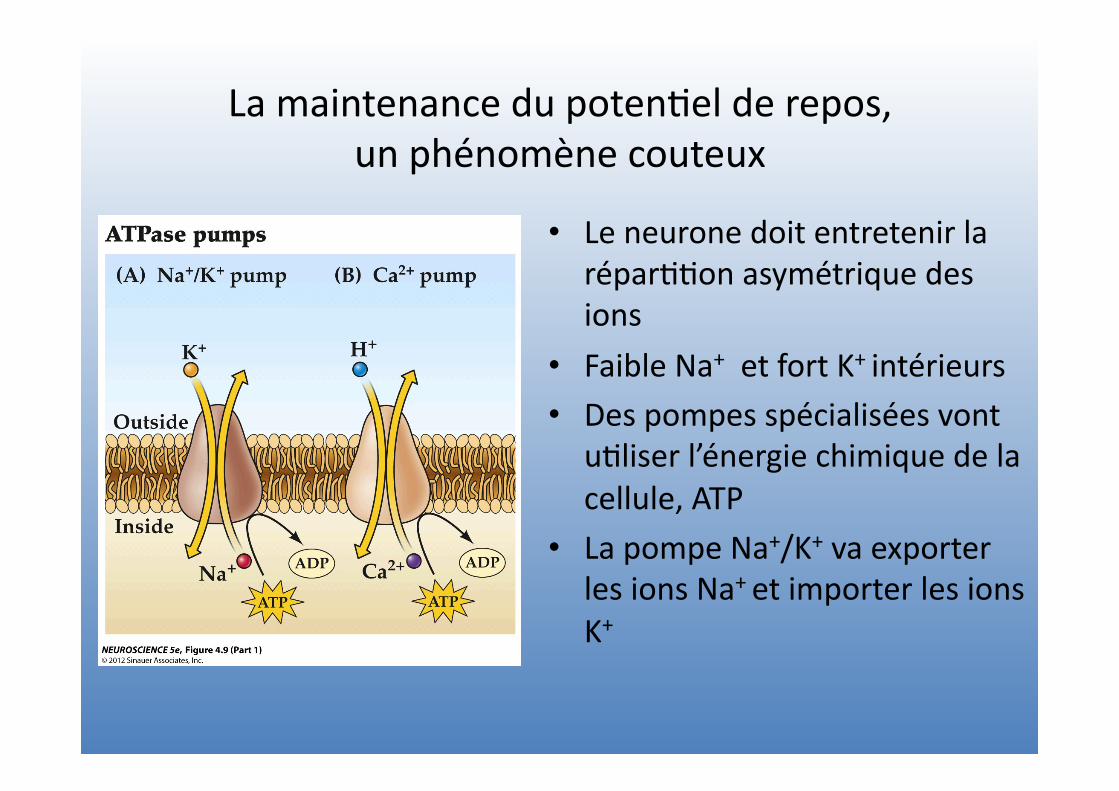
La maintenance du poten=el de repos, un phénomène couteux
• Le neurone doit entretenir la répar==on asymétrique des ions
• Faible Na+ et fort K+ intérieurs • Des pompes spécialisées vont
u=liser l’énergie chimique de la cellule, ATP
• La pompe Na+/K+ va exporter les ions Na+ et importer les ions K+


La dépense énergé=que croît pendant l’ac=vité neuronale (1)
• La libéra=on des neurotransmeVeurs (cours 3), la régénéra=on des poten=els de repos, et d’autres phénomènes augmentent la dépense
• A poids égal, le cerveau consomme 20 fois plus d’ATP que le muscle
• La réalisa=on d’une tâche implique une dépense dans une zone déterminée du cerveau
• Le cerveau s’adapte en augmentant sa microcircula=on dans la zone ac=vée
• Observa=ons anciennes: A Mosso (19ème siècle): augmenta=on pulsa=ons; Walter K (1928) : bruit, vision
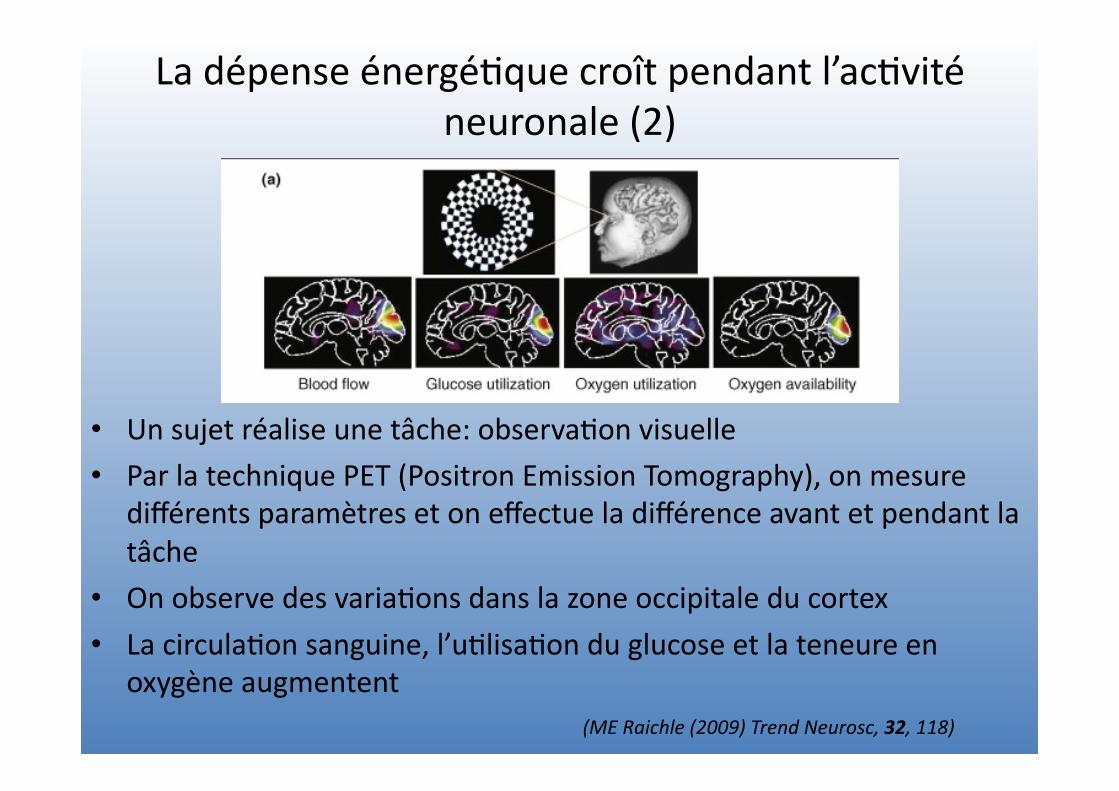
La dépense énergé=que croît pendant l’ac=vité neuronale (2)
• Un sujet réalise une tâche: observa=on visuelle • Par la technique PET (Positron Emission Tomography), on mesure
différents paramètres et on effectue la différence avant et pendant la tâche
• On observe des varia=ons dans la zone occipitale du cortex • La circula=on sanguine, l’u=lisa=on du glucose et la teneure en
oxygène augmentent (ME Raichle (2009) Trend Neurosc, 32, 118)
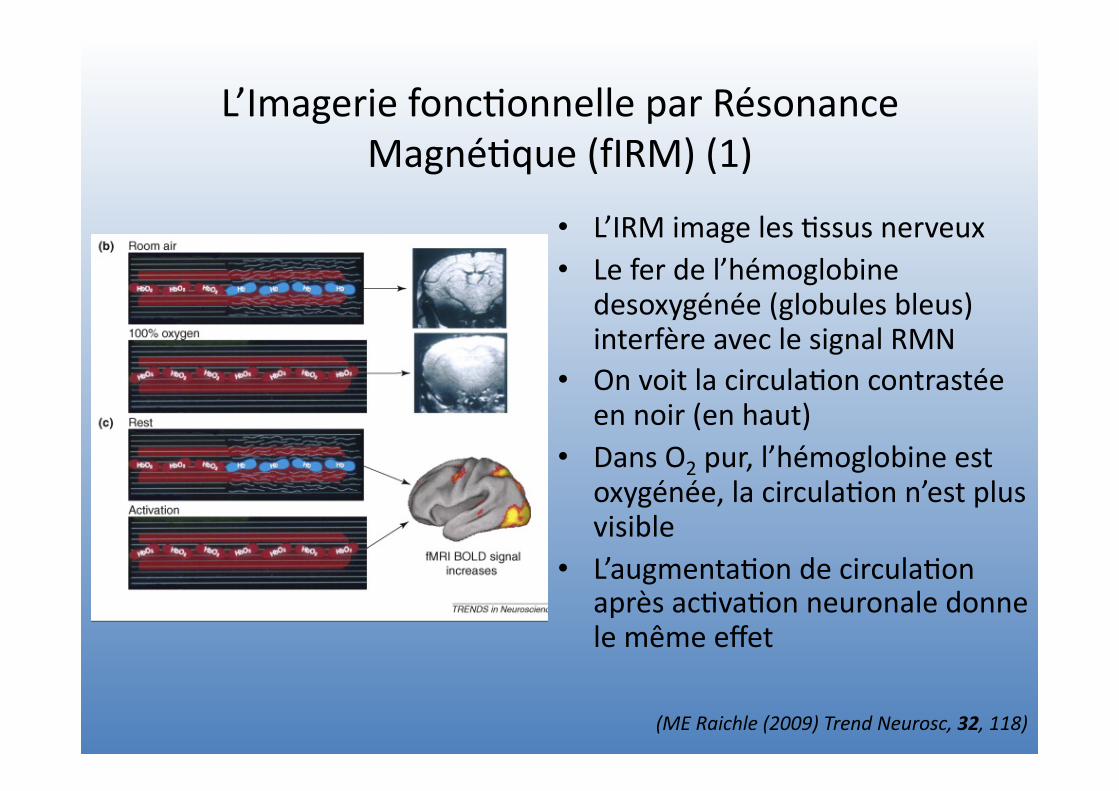
L’Imagerie fonc=onnelle par Résonance Magné=que (fIRM) (1)
• L’IRM image les =ssus nerveux • Le fer de l’hémoglobine
desoxygénée (globules bleus) interfère avec le signal RMN
• On voit la circula=on contrastée en noir (en haut)
• Dans O2 pur, l’hémoglobine est oxygénée, la circula=on n’est plus visible
• L’augmenta=on de circula=on après ac=va=on neuronale donne le même effet
(ME Raichle (2009) Trend Neurosc, 32, 118)
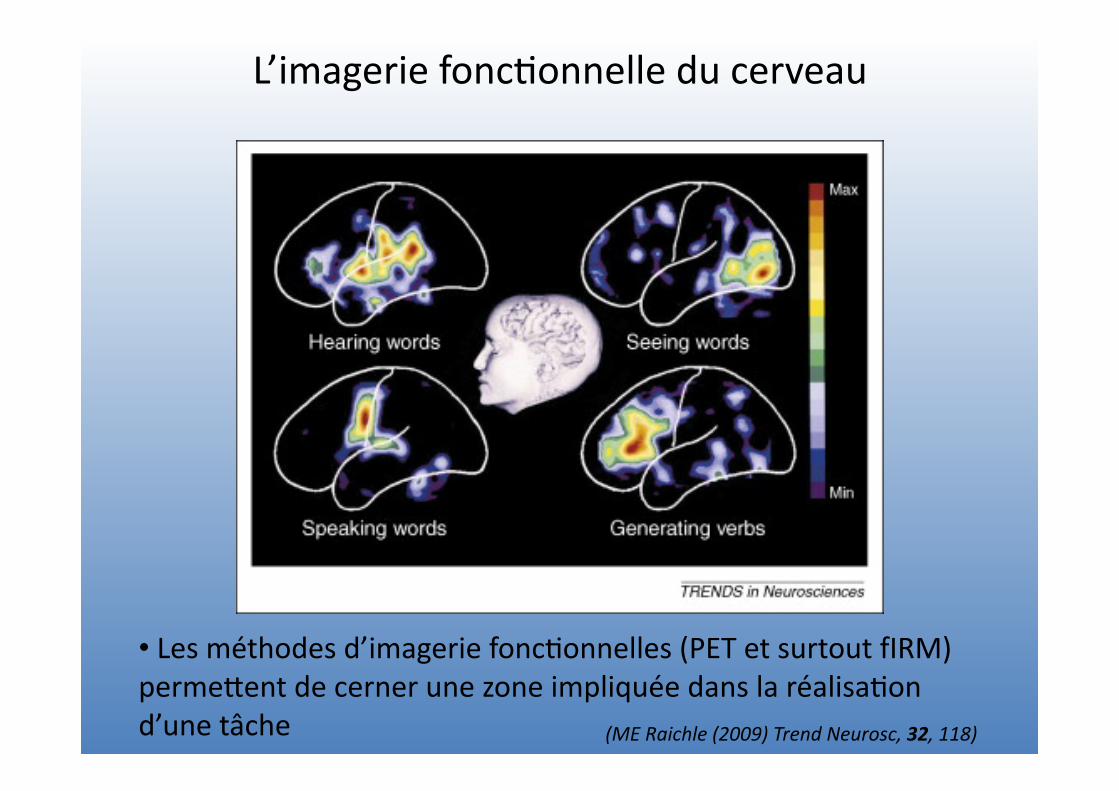
L’imagerie fonc=onnelle du cerveau
• Les méthodes d’imagerie fonc=onnelles (PET et surtout fIRM) permeVent de cerner une zone impliquée dans la réalisa=on d’une tâche (ME Raichle (2009) Trend Neurosc, 32, 118)
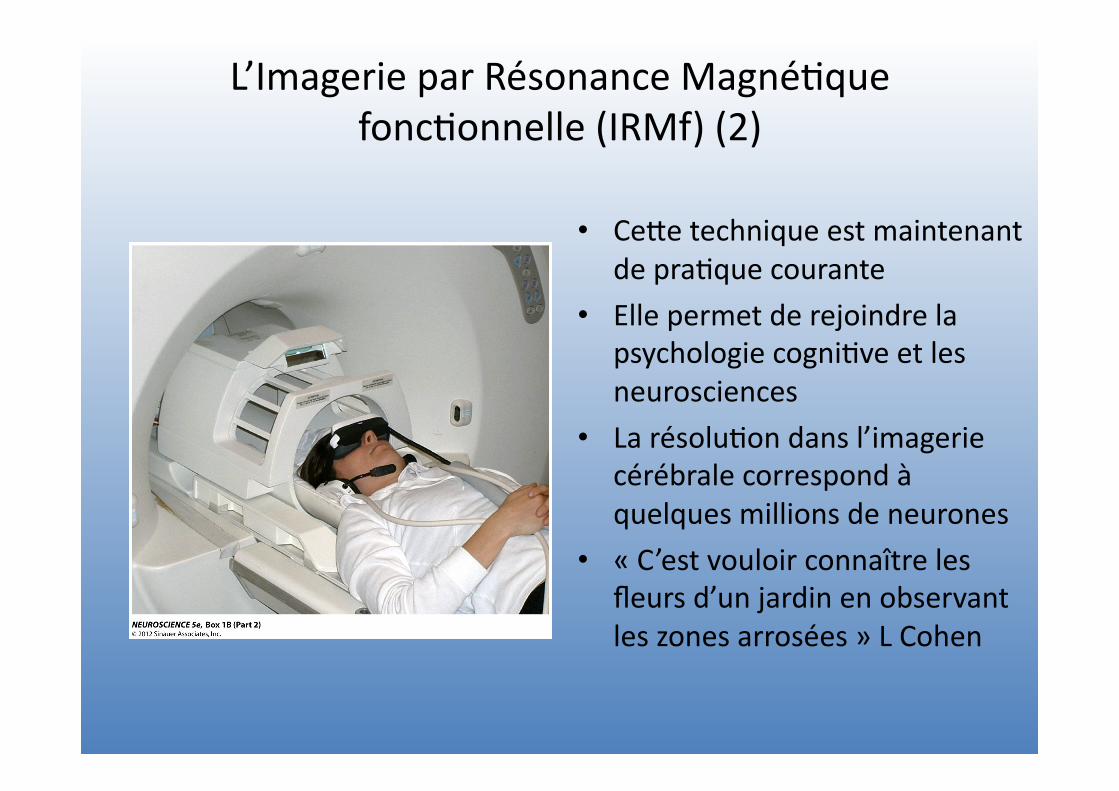
L’Imagerie par Résonance Magné=que fonc=onnelle (IRMf) (2)
• CeVe technique est maintenant de pra=que courante
• Elle permet de rejoindre la psychologie cogni=ve et les neurosciences
• La résolu=on dans l’imagerie cérébrale correspond à quelques millions de neurones
• « C’est vouloir connaître les fleurs d’un jardin en observant les zones arrosées » L Cohen

Cerveau et ordinateur
Une comparaison sauvage!

Comparaison des performances (1)
• Le cerveau possède de l’ordre de 1011 (100 milliards) de neurones; nous supposons que chacun fait en moyenne 10 synapses, soit un total de 1012 synapses
• Un neurone fait jusqu’à 100 opéra=ons/s (100 Hz) • Au total, si tout marche ensemble, le cerveau fait 1014 (1 million de milliards) d’opéra=ons/s !
• Un ordinateur traite 64 à 128 opéra=ons simultanément (64 à 128 bits) à une vitesse de 3 GigaHz (3 milliards d’opéra=ons/s)
• L’ordinateur traite 64 x 3 x 109 = 2 1011 opéra=ons/s, soit 200 milliards d’opéra=ons/s
• Avantage au cerveau, mais pas de KO!

Comparaison des performances (2)
• Pour comprendre où est vraiment la différence • Le cerveau reconnaît une image complexe en 100 ms (0,1 s)
• Sa vitesse de travail étant de 100 Hz, il a accompli seulement 10 opéra=ons dans chacun des circuits impliqués
• L’ordinateur sera obligé de faire beaucoup plus d’opéra=ons
• Le cerveau fonc4onne sur un mode massivement parallèle
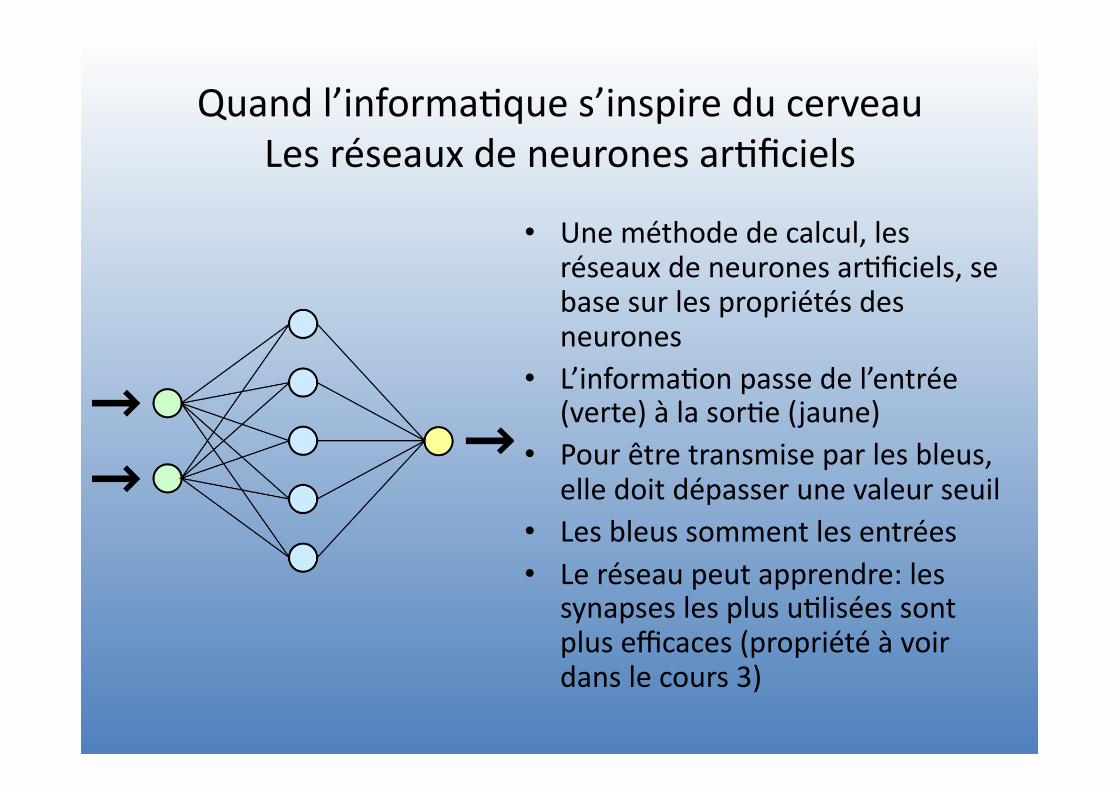
Quand l’informa=que s’inspire du cerveau Les réseaux de neurones ar=ficiels
• Une méthode de calcul, les réseaux de neurones ar=ficiels, se base sur les propriétés des neurones
• L’informa=on passe de l’entrée (verte) à la sor=e (jaune)
• Pour être transmise par les bleus, elle doit dépasser une valeur seuil
• Les bleus somment les entrées • Le réseau peut apprendre: les
synapses les plus u=lisées sont plus efficaces (propriété à voir dans le cours 3)

Conclusions du cours 2
• Les phénomènes électriques du système nerveux sont d’origine ionique
• Leur nature est bien comprise jusqu’à l’échelle moléculaire
• Leur principale manifesta=on est le poten=el d‘ac=on, une informa=on très conservée
• Le poten=el d’ac=on se propage dans le neurone et peut passer à un autre neurone après filtrage par la synapse (cours 3)
• Les phénomènes électriques consomment de l’énergie, ce qui est la base de l’IRMf