sciences de la vie et de la terreb.fichera.free.fr/ancien/mperez/livreprofsvt.pdf · 3 la...
TRANSCRIPT

1re SSciences de la Vie et de la Terre
Liv
re
du
pro
fesse
ur
Dangerle photocopillagetue le livre
Le photocopillage, c'est l'usage abusif et collectifde la photocopie sans l'autorisation des auteurs et des éditeurs.Largement répandu dans les établissements d'enseignement, le photocopillage menace l'avenir du livre, car il met en dangerson équilibre économique. Il prive les auteurs d'une juste rémunération.En dehors de l'usage privé du copiste, toute reproduction totale ou partielle de cet ouvrage est interdite.
48 7621 5
:HSMCLI=\X[ZYY:
www.editions-hatier.fr

Sciences de la Vie et de la Terre
1sre
LIVRE DU PROFESSEUR
Sous la direction deJacques Bergeron
AuteursPierre BeaujardProfesseur à Ancenis
Jacques BergeronProfesseur à Versailles
Bernard DavidProfesseur à Châteaubriant
Annick HyonProfesseur à Meudon
Isabelle Bednarek-Maitrepierre
Professeur à Saint-Nazaire
Dominique MargerieProfesseur à Saint-Germain-en-Laye
Martine MargerieProfesseur à Saint-Germain-en-Laye
Odile MoutauxProfesseur à Saint-Germain-en-Laye

© HATIER, PARIS, SEPTEMBRE 2001 ISBN 2-218-73654-3Toute représentation, traduction, adaptation ou reproduction même partielle, par tous procédés, en tous pays, faite sans autorisation préalable est illicite et exposerait lecontrevenant à des poursuites judiciaires. Réf. : loi du 11 mars 1957, alinéas 2 et 3 de l’article 41.Une représentation ou reproduction sans autorisation de l’éditeur ou du Centre Français d’exploitation du droit de Copie (20, rue des Grands-Augustins 75006 Paris) consti-tuerait une contrefaçon sanctionnée par les articles 425 et suivants du Code pénal.
Maquette et Mise en pages : Graphismes
Illustrations : Coredoc – Laurent Blondel

SommaireDes phénotypes à différents niveaux d’organisation
du vivantunité 1 Du génotype au phénotype, relations avec l’environnement ................................ 7
1 Diversité des phénotypes...................................................................................................... 8
2 Les enzymes........................................................................................................................ 12
3 La biosynthèse des protéines .......................................................................................................... 18
4 Complexité du phénotype .................................................................................................. 24
unité 2 Génotype, phénotype et régulation............................................................................ 29
5 Régulation de la glycémie .................................................................................................. 30
6 Génotype, environnement et phénotype diabétique .......................................................... 35
unité 3 Génotype, phénotype et système nerveux ................................................................ 39
7 Un phénotype comportemental : le réflexe myotatique .................................................... 40
8 Fonctionnement des centres nerveux et mémoire génétique ............................................ 45
9 Cerveau, génotype et environnement ................................................................................ 53
unité 4 Génotype, phénotype et morphogenèse végétale.................................................. 59
10 La diversité phénotypique des végétaux ........................................................................ 60
11 Les méristèmes et la croissance des végétaux ................................................................ 66
12 Le contrôle de la croissance chez les végétaux .............................................................. 74
Structure, composition et dynamique de la Terreunité 5 Structure et composition chimique de la Terre interne ........................................ 81
13 Structure et composition de la croûte terrestre .............................................................. 82
14 Modèles de la structure de la Terre.................................................................................. 89
15 Un modèle de la dynamique lithosphérique .................................................................... 94
unité 6 La Terre : machinerie thermique .................................................................................. 99
16 Marges passives et océanisation .................................................................................. 100
17 Magmatisme et convection mantellique ...................................................................... 105
Partie 2
Part ie 1

•Un exercice corrigé permet aux élèves de s’entraîner auxactes mentaux essentiels et de saisir peu à peu ce que l’onattend d’eux : analyser un document, en tirer des informa-tions signifiantes, les mettre en rapport avec les connais-sances acquises et, à partir de là, construire un raisonnement.
Les quest ions d’aujourd’huiElles constituent une ouverture, permettant de dépasser lesstrictes connaissances de base, et offrent des pistes derecherche et d’information pour les élèves en apportantdes compléments à des ensembles de chapitres (unités).
Les f iches techniquesElles apportent des compléments d’informations sur despoints techniques ou des protocoles expérimentaux. Ellespeuvent être utilisées au cours de séances de travaux pratiques.
Lexique et indexLe manuel est complété par un lexique, précisant le sensdes mots clés rencontrés tout au long des chapitres, et unindex conçu comme le lexique à partir des termes essen-tiels du programme officiel.
Object i fs recherchés•Contribuer à donner du sens aux notions essentielles duprogramme par l’intermédiaire des activités ; à travers lesdémarches explicatives que celles-ci proposent, lesnotions apparaissent comme les solutions des problèmes àrésoudre.•Contribuer à la formation au raisonnement scientifique àtravers la sélection des problèmes envisagés et la perti-nence des documents retenus.•Aider les élèves à repérer et mémoriser les connaissancesexigibles.•Aider les élèves à surmonter leurs difficultés éventuellesde compréhension grâce à un texte explicatif ciblé sur l’es-sentiel tout en insistant sur les points cruciaux du raison-nement.•Aider les élèves à s’auto-évaluer et à préparer leurscontrôles sommatifs en leur proposant de nombreux exer-cices dont certains, corrigés, leur précisent les actes men-taux à pratiquer.
Le manuel de l’élève : les choix pédagogiques
Organisation généraleLe manuel a été conçu afin que, dans chaque chapitre,l’élève, seul ou guidé par le professeur, soit mis en situa-tion de recherche (d’où les activités) mais puisse égale-ment trouver les notions essentielles à maîtriser présentéesde façon argumentée (d’où les connaissances) en fin dechapitre. Des exercices lui permettent de tester ses acquis.Les chapitres sont regroupés en unités (4 pour les sciencesde la Vie, 2 pour les sciences de la Terre) ; chacune d’ellesse termine par une ouverture sur des problèmes voisins oudes données complémentaires proposées dans les ques-tions d’aujourd’hui.À la fin de l’ouvrage, des fiches techniques, un lexique etun index fournissent les éléments complémentaires à unebonne maîtrise de l’ensemble du programme.
Les act ivi tés•La signification de chaque activité, et donc la notionqu’elle aide à construire, est indiquée dans le problème initial et précisée, en bas de page, par l’objectif nettement exprimé, c’est-à-dire la compétenceque l’élève doit avoir acquise en fin d’activité.•Elles sont conçues de manière à permettre au professeurde faire des choix ou de les intégrer dans d’autresdémarches, leur enchaînement n’étant que suggéré par lesquestions d’exploitation.•En ce qui concerne les documents présentés, les seulsrenseignements fournis sont des clés indispensables à leurlecture, à leur compréhension mais ils ne donnent enaucun cas de conclusions. Les documents permettent unvéritable travail personnel des élèves.
Les connaissances• Développées à la fin de chaque chapitre, elles occupent3 ou 4 pages.•C’est un texte qui explique, argumente et complète lesnotions prioritaires du chapitre. Il en facilite ainsi la com-préhension. Il a été conçu pour être lu, travaillé et comprismême si certaines activités n’ont pas été réalisées.• Les documents qui l’illustrent sont accompagnés,contrairement à leur présentation dans les activités, delégendes explicatives pour permettre une meilleure appro-priation de la notion.•L’essentiel des notions à mémoriser apparaît dans desencadrés terminant chacun des paragraphes et se retrouvedans un schéma bilan.
Les exercicesIls terminent chacun des chapitres.•De types variés, les uns sont axés sur une évaluation desconnaissances, les autres sur l’aptitude à utiliser lesconnaissances au cours de raisonnements. Les premierspermettent une évaluation formative ciblée et rapide.

SCIENCES DE LA VIE DES PHÉNOTYPES À DIFFÉRENTS NIVEAUX D’ORGANISATION DU VIVANT
mètre de l’organisme étant un aspect de son phénotype aumême titre que ses aspects morphologiques ou anato-miques.Les notions de base sur les caractéristiques d’un systèmede régulation sont précisées et l’accent est mis sur l’im-portance de la communication hormonale dans la régula-tion de la glycémie. Au terme de l’étude, la perception quede multiples gènes interviennent au sein des diversgroupes cellulaires impliqués dans la régulation de la gly-cémie est soulignée : ces phénotypes sont multigéniques.Le chapitre 6 est centré sur les phénotypes diabétiques etsur le siège des déficiences de la régulation de la glycémiechez les diabétiques de type 1 et de type 2. L’objectif n’estpas d’approfondir l’étude des gènes impliqués dans la réa-lisation des génotypes diabétiques (lesquels, pour la plu-part, ne sont pas encore isolés) mais de faire réfléchir surdes données statistiques pour faire saisir le sens de l’ex-pression : «gènes de susceptibilité» et la complexité desinteractions génotype-milieu. À ce propos, les données surl’évolution du diabète de type 2 chez les Indiens Pimassont particulièrement pertinentes.
Unité 3 Génotype, phénotype etsystème nerveux
Chapi tres 7 à 9Ces chapitres envisagent les phénotypes liés à l’organi-sation du système nerveux.Les chapitres 7 et 8 ont trait au réflexe myotatique. Placéedans le cadre de la réalisation du phénotype postural, cetteétude vise d’abord à la connaissance des structures neuro-niques et de leurs propriétés fonctionnelles, supports duréflexe. Elle sensibilise aussi à l’idée que cette organisa-tion médullaire résulte de la mise en jeu d’un grandnombre de gènes tant au cours de la construction desréseaux neuroniques que durant leur activité. Ce phéno-type comportemental que constitue la contraction d’unmuscle en réponse à son étirement résulte ainsi essentiel-lement d’une mémoire génétique propre à l’espèce. Pource phénotype, il existe sans doute une variabilité entreindividus liée au polymorphisme des gènes impliqués.Probablement aussi existe-t-il une certaine plasticité duréflexe en fonction des apprentissages moteurs dans les-quels il est impliqué.Mais cette plasticité, cette action de l’environnement sur lastructure du tissu nerveux est particulièrement développéeau niveau du cortex ; elle est l’objet du chapitre 9, quis’appuie sur des données récentes relatives aux modifica-tions du cortex somatosensoriel, y compris chez l’adulte,en réponse à des changements dans les messages afférents.Il montre comment l’influence de l’histoire personnellesur la construction cérébrale conduit à une individuali-sation marquée des comportements.
Unité 1Des phénotypes à différentsniveaux d’organisation du vivant
Chapi tres 1 à 4Le chapitre 1, à partir de l’analyse des relations entre lesdivers niveaux de définition du phénotype, a pour objectifd’aboutir à l’idée qu’un gène s’exprime au sein d’une cel-lule en dirigeant la synthèse d’un polypeptide. Un autreobjectif est de sensibiliser à l’idée que les propriétés d’unpolypeptide dépendent de sa structure spatiale laquelle estliée à sa séquence d’acides aminés. Cette idée est renfor-cée dans le chapitre 2 avec la notion de site actif envi-sagée avec les propriétés des enzymes. L’importance de laséquence des acides aminés étant ainsi acquise, l’étude dela synthèse des protéines (chapitre 3) peut être centrée surla façon dont une séquence de nucléotides du gène déter-mine la séquence des acides aminés du polypeptide.Le chapitre 4 revient sur l’analyse des relations entregénotype et phénotype en mobilisant les connaissancesacquises sur la synthèse des protéines ; l’objectif étant desaisir comment des différences dans le génotype peuventse traduire ou non dans le phénotype. Il remet en cause laliaison un gène-un caractère en envisageant des cas –comme celui, classique, des groupes sanguins – où plu-sieurs gènes interviennent dans la réalisation d’un ca-ractère. L’accent est mis, avant tout, sur l’idée que mêmedans les cas où le phénotype est considéré comme mono-génique (maladies héréditaires par exemple), sa réa-lisation dépend de plusieurs gènes. Des différences géno-typiques au niveau de gènes modulateurs (modulaires)font qu’un même génotype pour un gène majeur peut setraduire par des différences phénotypiques (gravité plus oumoins accusée de la phénylcétonurie, de la drépanocy-tose…).L’accent est enfin mis sur l’idée que très généralement lephénotype ne dépend pas que de l’expression du génotyped’un individu mais résulte d’une interaction complexeentre le gène et le milieu. Différents sites d’action dumilieu sur la réalisation du phénotype sont envisagés.Au terme de cette étude, la réalisation d’un phénotype doitapparaître comme multifactorielle, dépendante de plu-sieurs gènes et de divers facteurs du milieu. Cette idée vaêtre développée à propos des phénotypes diabétiques (cha-pitre 6) et de la plasticité cérébrale (chapitre 9).
Unité 2 Génotype, phénotype etrégulat ion
Chapi tres 5 et 6Le chapitre 5, qui traite de la régulation de la glycémie estd’abord situé dans le cadre général de la relation géno-type-phénotype ; les caractéristiques d’un individu liées aufonctionnement d’un système de régulation d’un para-
Programme officiel et organisation du manuel

Unité 4 Génotype, phénotype et morphogenèse végétale
Chapi tres 10 à 12En conformité avec l’esprit du programme, le chapitre 10qui introduit cette unité sensibilise d’abord à l’idée de fac-teurs génétiques et de facteurs du milieu dans la réali-sation du phénotype d’un végétal. Il met l’accent sur l’im-portance des méristèmes dans la construction de ce phé-notype.L’analyse du fonctionnement de ces méristèmes au cha-pitre 11 amène à découvrir les processus : multiplication,élongation, différenciation cellulaires qui aboutissent à lagenèse de nouveaux organes et donc à la réalisation duphénotype.
Les mécanismes de conservation de l’information géné-tique au cours des cycles cellulaires méristématiques sontensuite envisagés : ils permettent de comprendre la per-manence du phénotype engendré par les méristèmes.Le chapitre 12 d’abord axé sur les mécanismes en œuvreau cours de l’élongation des cellules réinvestit la notiond’hormone. Par l’étude de l’action du milieu et notammentde l’éclairement sur la répartition (auxine) ou la synthèse(gibbérelline) des hormones végétales, on explique com-ment le phénotype du végétal est influencé par l’environ-nement.Les implications biotechnologiques de ces connaissancessont alors abordées lorsqu’on envisage commentl’Homme peut orienter la réalisation du phénotype enmanipulant les concentrations en hormones du milieu deculture.
Unité 5 Structure et composit ionchimique de la Terre interne
Chapi tres 13 à15Dans le programme, le chapeau introductif aux sciencesde la Terre insiste, d’une part, sur « la démarche scienti-fique qui vise en permanence à enrichir les modèles de laTerre, à les critiquer et donc à les faire évoluer» et, d’autrepart, sur la classe de terrain «moyen privilégié d’aborderla géologie à une échelle aisément accessible aux élèves».Pour respecter cette double orientation, le chapitre 13, atrait au modèle de la Terre accessible, c’est-à-dire au mo-dèle de la croûte terrestre qu’on peut construire à partirdes observations faites en milieu continental et en milieuocéanique. Ce chapitre débouche sur une notion géolo-gique majeure, à savoir l’opposition entre la croûte conti-nentale et la croûte océanique. Il a l’objectif de permettrel’acquisition des notions de base sur l’origine des rochesconstitutives de ces croûtes ainsi que sur la nature et lacomposition chimiques des minéraux qui les constituent.Ces connaissances seront constamment réinvesties.Le chapitre 14 conduit au modèle de la Terre à symétriesphérique caractérisé par l’hétérogénéité verticale duglobe (croûte, manteau, noyau) et l’homogénéité horizon-tale en dehors des croûtes. La distinction entre les donnéeselles-mêmes et le modèle interprétatif est clairement réa-lisée. Ainsi le profil des vitesses des ondes sismiques P etS en fonction de la profondeur est finalement présentécomme un modèle rendant compte des faits observables àsavoir les temps d’arrivée des ondes P et S aux diversesstations sismiques à la suite d’un séisme. De même, l’ac-cent est mis sur la façon dont on peut construire un mo-dèle minéralogique de la Terre à partir de faits d’observa-tion et de données expérimentales.Le chapitre 15, à travers les données sur la dynamique duglobe, conduit à remettre en cause le modèle à symétriesphérique préalablement établi. Ce dernier, en effet, nepermet pas d’expliquer la répartition de la séismicité et duvolcanisme. L’exploitation des informations sur ces zones
actives conduit au modèle de la tectonique des plaques etdonc à mettre l’accent sur une géographie des profondeursdu globe. Les calculs des vitesses d’expansion des fondsocéaniques à partir de données diverses focalisent sur ladynamique de surface du globe et invitent à rechercher laliaison avec ce qui se passe à l’intérieur. Le chapitre 13 aciblé sur l’opposition croûte océanique-croûte continen-tale et le chapitre 15 sur les frontières des plaques.
Unité 6 La Terre : machineriethermique
Chapi tres 16 et 17Le chapitre 16 conduit à relier les approches des chapitres13 et 15 par une réflexion sur la signification des fron-tières océan-continent : marges actives (frontières deconvergence) ou marges passives ayant enregistré la signa-ture de la déchirure continentale. Le chapitre développealors les caractéristiques de ces marges passives en s’ap-puyant sur les données sismiques, sur les modèles analo-giques et sur l’analyse de régions actuelles à différentsstades du processus d’océanisation. Au terme de ce cha-pitre, l’idée que la déchirure continentale s’achève par lamise en place d’une dorsale dont l’activité va engendrer lalithosphère océanique est bien dégagée.Le chapitre 17 vise à préciser les caractéristiques d’unedorsale, notamment la remontée de l’asthénosphère parconvection, afin de comprendre comment ce processuspeut engendrer le magma de composition basaltique à l’origine de la croûte océanique ; là encore, il y a une mo-dification du modèle préalablement établi. L’évolution dela lithosphère océanique depuis sa genèse aux dorsalesjusqu’aux plaines abyssales met l’accent sur l’augmenta-tion de son épaisseur et de sa densité. Il reste alors à ins-crire toute cette dynamique, à savoir l’ascension del’asthénosphère aux dorsales et la subduction de la litho-sphère océanique aux frontières de convergence, dans uneperspective plus large : les mouvements de convectionglobaux du manteau qui font de la Terre une machine thermique.
SCIENCES DE LA TERRE STRUCTURE, COMPOSITION ET DYNAMIQUE DE LA TERRE

Pa
rt
ie
1 Des phénotypes à différents niveaux d’organisation du vivant
Du génotype au phénotype, relationsavec l’environnement
1 Diversité des phénotypes ...................................... 8
2 Les enzymes ...................................................................... 12
3 La biosynthèse des protéines ........................ 18
4 Complexité du phénotype .................................. 24
Unité 1

8
◆ Problèmes scientifiques à résoudreL’observation d’une biche blanche, différente des bichesnon albinos permet de poser le problème des relationsentre le phénotype d’un organisme et son génotype.
P o s e r l e s p r o b l è m e s
Le DOCUMENT 1, en s’appuyant sur les connaissances de laclasse de seconde, rappelle que l’œuf contient l’informa-tion génétique nécessaire à la mise en place de l’orga-nisme et de ses caractères, que cette information estcontenue dans les chromosomes et se retrouve dans toutesles cellules, y compris celles qui sont impliquées dans laconstitution et la taille des ailes et dans la constitution etla couleur des yeux.Quelles correspondances peut-on établir entre des diffé-rences génotypiques et des différences phénotypiques?
A c t i v i t é s 1
Différents niveaux de définition du phénotypeIl s’agit de définir les divers niveaux d’analyse des phé-notypes : phénotype au niveau de l’organisme (macro-scopique ou clinique), au niveau cellulaire et au niveau
moléculaire. Une relation causale entre ces différentsniveaux est établie. Cette analyse n’est possible que sion s’appuie sur deux phénotypes alternatifs : ici nondrépanocytaire et drépanocytaire.
ExploitationL’élève doit faire une analyse des différences d’abord auniveau clinique (DOCUMENT 2) puis au niveau du phénotypecellulaire, en repérant (DOCUMENT 3) les différences entreles hématies.Il doit réinvestir ses connaissances sur le rôle des globulesrouges dans le transport du dioxygène pour établir com-ment les différences notées au niveau du phénotype cellu-laire entraînent des différences au niveau du phénotypeclinique.L’analyse du DOCUMENT 4 permet d’établir que les diffé-rences constatées au niveau cellulaire sont dues à des dif-férences dans le comportement de l’hémoglobine desdrépanocytaires et des non drépanocytaires donc à des dif-férences au niveau des phénotypes moléculaires.En conclusion, l’élève doit, dès ce moment là, saisir que legénotype détermine le phénotype moléculaire, lequeldétermine le phénotype cellulaire. Ce dernier étant res-ponsable du phénotype au niveau (à l’échelle) de l’orga-nisme.
Programme: notions et contenus• Le phénotype peut se définir à différentes échelles : de l’organisme à la molécule• Les phénotypes alternatifs sont dus à des différences dans les protéines concernées.
Objectifs à atteindreUn premier objectif consiste, à partir de l’analyse des relations entre les divers niveaux de définition du phénotype, àdégager l’idée qu’un gène s’exprime au sein d’une cellule en dirigeant la synthèse d’un polypeptide.
Un autre objectif est de dégager les connaissances de base relatives à la structure des polypeptides et de sensibiliserà l’idée que les propriétés d’un polypeptide dépendent de la séquence des acides aminés qui le constituent.
Un troisième objectif consiste à montrer que les différences dans le phénotype moléculaire sont dues à des différencesdans la séquence des nucléotides des allèles d’un gène.
Diversité des phénotypes
C H A P I T R E 1

Chapitre 1 : Diversité des phénotypes
9
entre la fonction acide de la valine et la fonction amine del’histidine avec départ d’une molécule d’eau.Les DOCUMENTS 8 et 9 traduisent que la seule différenceentre l’hémoglobine S drépanocytaire et l’hémoglobine Anon drépanocytaire résulte de la nature du 6e acide aminédes chaînes β et que la structure spatiale n’est pas modifiéepar la substitution (DOCUMENT 8) de l’acide glutamique6 parde la valine 6. Cependant, la valine est un acide aminéhydrophobe alors que l’acide glutamique est hydrophile.Le DOCUMENT 9 montre que les deux molécules d’hémoglo-bine S se lient au niveau d’une valine 6. L’élève doit com-prendre que le système étant répétitif, les dimères ainsiformés peuvent s’associer les uns aux autres pour formerdes fibres rigides qui allongent et déforment les hématies.Remarque : les molécules de HbS polymérisent car desliaisons faibles s’établissent entre la valine 6 des chaînesβ des unes et un site accepteur (phénylalanine 85 et leu-cine 88) des chaînes β des autres.
A c t i v i t é s 3
Du phénotype phénylcétonurique à la protéine responsableLe but poursuivi est triple :
1. à partir d’un nouvel exemple (phénotype phénylcéto-nurique et phénotype non phénylcétonurique), retrou-ver la relation causale entre le phénotype moléculaireet le phénotype macroscopique avec l’introduction d’unnouveau niveau phénotypique, le niveau biochimique ;
2. retrouver également que le phénotype moléculairedéterminé par le gène est toujours de nature protéique ;
3. retrouver enfin que les différences au niveau de laséquence des acides aminés sont à l’origine des diffé-rents phénotypes, à tous les niveaux.
ExploitationLes DOCUMENTS 10 et 11 permettent d’établir deux niveauxphénotypiques : le phénotype au niveau de l’organisme,clinique et le phénotype biochimique (concentration plas-matique de phénylalanine) et d’établir la relation causaleentre les deux niveaux : c’est la phénylalanine qui à forteconcentration est toxique pour le cerveau et entraîne leretard mental, principal signe clinique de la maladie.Le DOCUMENT 12 permet d’introduire dans l’exemple choisiles notions de phénotype cellulaire (aptitude des celluleshépatiques à transformer ou non la phénylalanine en tyro-sine) et de phénotype moléculaire (enzyme PAH fonction-nelle ou non). Bien que l’étude des enzymes soit abordéeseulement dans le chapitre 2, nous avons conservé cetexemple car la notion d’enzyme a été, sous une formesimple, abordée au collège.L’élève doit, à ce stade, comprendre que des différences auniveau de l’activité enzymatique, entraînent des diffé-rences dans la transformation de la phénylalanine en tyro-sine par les cellules du foie et de là des différences dans laconcentration plasmatique de la phénylalanine qui sont à
A c t i v i t é s 2
Protéine et phénotypeIl s’agit, en poursuivant l’étude de l’exemple de la dré-panocytose, d’élucider la nature des différences entrephénotype drépanocytaire et phénotype non drépanocy-taire. Pour cela, il est nécessaire d’acquérir desconnaissances sur l’hémoglobine (association de poly-peptides), sur la nature d’un polypeptide (chaîned’acides aminés) et sur la façon dont les acides aminéssont enchaînés.
Cela aboutit à établir que la différence entre les phéno-types moléculaires étudiés provient d’une différenceentre les deux chaînes � des deux types d’hémoglobineau niveau d’un seul acide aminé. C’est une première sen-sibilisation à l’idée qu’un gène s’exprime à travers uneprotéine et que les caractéristiques fonctionnelles d’unpolypeptide sont liées à sa séquence d’acides aminés.
Les données présentées ont été établies à l’aide du logi-ciel RasMol dont l’adaptation française a été réaliséepar l’INRP, logiciel disponible gratuitement auprès decet organisme.
ExploitationLe DOCUMENT 5 permet une vision globale de la moléculed’hémoglobine. Plutôt qu’une représentation en «boules»,on a choisi une représentation limitée au squelette poly-peptidique (…CH-CONH-CH-CONH-CH-CONH…) quine tient pas compte des chaînes latérales des acides ami-nés constitutifs mais respecte le repliement de cette molé-cule.Le DOCUMENT 6 prolonge cette vision en montrant que lachaîne polypeptidique β est constituée par un enchaîne-ment d’acides aminés ; la numérotation permet de souli-gner le caractère unidirectionnel de la molécule (les acidesaminés sont disposés dans un ordre précis de 1 à 146).En analysant le DOCUMENT 7a, NH2
l’élève doit retrouver que le groupement CHCOOH
est commun aux deux acides aminés valine et histidine etque, par contre, le reste de la molécule (R) est différent.On peut dès lors schématiser les molécules d’acides ami-nés suivant les modèles ci-dessous :
R1 R2
NH2 – CH – COOH ; NH2 – CH – COOH ;
R3
NH2 – CH – COOH etc.
Le DOCUMENT 7b permet une investigation de la liaison pep-tidique entre les deux acides aminés valine et histidine.L’élève doit d’abord repérer qu’à droite se trouve l’histi-dine et à gauche la valine, puis le groupement NH2 libre dela valine et le groupement COOH libre de l’histidine. Leproblème est alors de repérer la liaison peptidique CONHet de comprendre que le dipeptide résulte de la liaison

10
l’origine des différences entre les phénotypes macrosco-piques.Le DOCUMENT 13 présente le squelette polypeptidique del’enzyme PAH constituée de 451 acides aminés.Le DOCUMENT 14 doit permettre à l’élève d’établir que ladifférence d’activité entre l’enzyme d’une personne phé-nylcétonurique et celle d’une personne non phénylcétonu-rique est liée à une seule différence : l’acide glutamique estremplacé par la lysine en position 280. En regroupant lesinformations relatives à la drépanocytose et à la phénylcé-tonurie, l’élève peut conclure que le génotype détermine lephénotype moléculaire, c’est-à-dire la séquence d’acidesaminés d’un polypeptide.Remarque : alors que le phénotype drépanocytaire corres-pond à un seul allèle du gène codant pour la chaîne �(�S) ; il existe plus de 300 allèles à l’origine du phénotypephénylcétonurique. Dans ce chapitre introductif nousn’avons envisagé qu’une variante mutée et non fonction-nelle de l’enzyme PAH (E280K).
A c t i v i t é s 4
ADN et phénotypeOn admet ici, contrairement à ce qui sera développédans le chapitre 4, que les phénotypes drépanocytaireou non, phénylcétonurique ou non, dépendent unique-ment d’une protéine, produit de l’activité d’un seulgène.
Ces activités permettent de réinvestir les connaissancessur l’ADN acquises en seconde : un gène est uneséquence de nucléotides d’une molécule d’ADN et cesont des différences dans la séquence des nucléotidesdu gène qui sont à l’origine des différences dans laséquence des acides aminés des protéines concernées.Le problème de la correspondance gène-protéine peutêtre posé et sera résolu dans le chapitre 3.
ExploitationLe DOCUMENT 15 reprend les connaissances acquises dansles activités 1 à 3 ; les phénotypes au niveau de l’orga-nisme sont déterminés par des différences phénotypiquesau niveau de polypeptides. Il met en relation dans les deuxcas, drépanocytose et phénylcétonurie, les différences auniveau des phénotypes moléculaires et les différences auniveau des allèles des gènes impliqués.Cela doit conduire à l’idée qu’un gène ne s’exprime qu’ausein d’une cellule et détermine uniquement, au sein descellules où il s’exprime, la synthèse d’un polypeptide. Lesdifférences entre les allèles portent sur des différencesentre les séquences des nucléotides constitutifs et, aufinal, elles se traduisent par des différences dans laséquence des acides aminés des polypeptides synthétisés.La compréhension de cette notion est testée par les DOCU-MENTS 16 à 19 où des différences génotypiques conduisentà des différences au niveau du phénotype macroscopique :une différence entre les allèles d’un gène qui code pour lepigment « rouge » des cônes entraîne une différence dans
la séquence des acides aminés de la protéine pigmentaire(phénotype moléculaire) et, par là, une différence dansl’absorption des radiations par les cônes rouges (phéno-type cellulaire) laquelle entraîne la différence dans la per-ception des couleurs (phénotype au niveau del’organisme).
E x e r c i c e s
Évaluer ses connaissances
Vrai ou faux ?1. a., b., d. et f. sont des affirmations exactes.c. Les acides aminés ne diffèrent que par leurchaîne latérale.e. Il existe 20 acides aminés essentiels constitutifsde toutes les protéines.
2. a., b. et e. sont des affirmations exactes.c. La liaison a lieu entre le groupement aminé de l’unet le groupement carboxyle (acide) de l’autre.d. Les protéines ont des formes spatiales diffé-rentes.
3. a., c. et f. sont des affirmations exactes.b. Non (voir les exemples choisis dans les activités).d. Un allèle ne détermine au sein d’une cellule qu’unphénotype moléculaire (polypeptide).e. Les allèles d’un même gène sont situés au mêmelocus sur des chromosomes homologues (voir laproposition f).
Phrases à corriger« Le gène de la drépanocytose et celui de la phénylala-nine… »Connaissances à mobiliser par l’élève : il n’existe pasde gène d’une maladie mais un gène qui code pourun polypeptide et dont certains allèles peuvent êtreà l’origine de symptômes morbides.
«… sont constitués par une séquence d’acides aminés. »Connaissances à mobiliser par l’élève : ne pasconfondre les acides aminés constitutifs des poly-peptides et les séquences de nucléotides constitu-tifs de l’ADN.
« Ces derniers diffèrent par leurs fonctions amine etacide… »Connaissances à mobiliser par l’élève : les fonctionsamine et acide sont communes à tous les acidesaminés.
«… sont spécifiques d’une protéine donnée. »Connaissances à mobiliser par l’élève : les mêmesacides aminés (20) se retrouvent dans toutes lesprotéines ; c’est leur ordre de succession (séquence)qui est différent d’une protéine à l’autre.
2
1

Chapitre 1 : Diversité des phénotypes
11
ce cas, « incapacité à utiliser les LDL ». Le phéno-type cellulaire alternatif (protéine non mutée) estcaractérisé par la capacité à internaliser et à utiliserles LDL.Conséquences au niveau du phénotype à l’échellede l’organisme (macroscopique, clinique) : l’incapa-cité des cellules possédant des récepteurs mutés àinternaliser les LDL se traduit par le phénotypehypercholestérolémique. En effet, le cholestérolplasmatique des LDL n’étant pas utilisé, sa concen-tration dans le sang est importante (2 à 4 fois supé-rieure à la normale), il s’accumule alors au niveaudes tendons et des artères et peut-être à l’origined’accidents cardio-vasculaires. Chez les individusde phénotype non cholestérolémique, le cholestéroldes LDL est utilisé par les cellules, sa concentrationdans le sang reste modérée.
La mucoviscidose1. L’allèle muté se caractérise par la perte de troispaires de nucléotides complémentaires situés enposition 1519, 1520 et 1521 ou 1520, 1521 et1522.Remarques : pour les mêmes raisons que cellesexposées à propos de l’exercice 4, l’élève ne peutétablir les conséquences de la mutation du gène surle polypeptide : les deux triplets du brin transcrit ATCTTT sont, par délétion de TCT (ou de CTT), réduits àun seul, ATT ; le couple d’acides aminés isoleucine(AUC) – phénylalanine (UUU) de la protéine nonmutée est remplacé par un seul acide aminé, l’iso-leucine (AUU) dans le polypeptide muté.2. Phénotype moléculaire : le phénotype molécu-laire mucoviscidose est constitué par la protéinemutée caractérisée par l’absence d’un acide aminésitué en position 508 dans le polypeptide non muté.Phénotype cellulaire : la protéine mutée s’intègretrès mal à la membrane des cellules bronchiques etpancréatiques ; l’absence ou la rareté de ce récep-teur membranaire conduit les cellules à produire unmucus visqueux, épais au contraire de celui fabriquépar les cellules normales qui est beaucoup plusfluide.Phénotype macroscopique : le mucus sécrété parles cellules privées de récepteurs CFTR obstrue lesbronches et provoque peu à peu la destruction despoumons par des infections associées. La mucovis-cidose est également à l’origine d’atteintes auniveau du pancréas.
5
«Le gène responsable… toutes les cellules de l’orga-nisme... »Connaissances à mobiliser par l’élève : il n’existe pasde gène d’une maladie. Un gène ne s’exprime pasobligatoirement dans toutes les cellules de l’orga-nisme ; ainsi le gène responsable de la synthèse dela globine � ne s’exprime que dans les cellulessouches des hématies.
«… est responsable du phénotype clinique. »Connaissances à mobiliser par l’élève : un gène nedétermine pas un phénotype clinique (à l’échelle del’organisme) mais uniquement un phénotype molé-culaire qui, lui, déterminera le phénotype clinique àtravers les différents niveaux phénotypiques.
Exercice corrigé
AlbinismeVoir correction dans le manuel pages 24 et 25.
Évaluer ses compétences
L’hypercholestérolémie famil ialeLa différence entre les allèles du gène concernéporte sur le huitième nucléotide du fragment pré-senté. Un nucléotide à guanine présent chez l’indi-vidu non atteint est remplacé par un nucléotide àadénine chez un individu atteint.Remarques : à ce stade, l’élève, qui ne possède pasde notions sur le code génétique, ne peut recon-naître que cette substitution sur le brin non transcritconduira à l’apparition d’un codon stop (UAG) à laplace du codon UGG codant pour le tryptophane. Ils’agit de la mutation non sens FH 683.Conséquence au niveau du phénotype molécu-laire : le phénotype moléculaire est constitué par lerécepteur aux LDL c’est-à-dire la protéine membra-naire résultant de l’expression du gène. Chez lesindividus hypercholestérolémiques présentant lamutation, la partie cytoplasmique du polypeptide estpratiquement absente : elle est formée de 2 acidesaminés seulement contre 50 pour la protéine résul-tant de l’expression de l’allèle non muté.Conséquences au niveau du phénotype cellulaire :les cellules pourvues de récepteurs dont le domainecytoplasmique est absent sont incapables d’interna-liser les LDL. Le phénotype cellulaire est donc, dans
4
3

12
font intervenir des enzymes. Le DOCUMENT 1b. montre, enprenant appui sur les réactions chimiques de la digestion,que celles-ci ne peuvent pas se dérouler sans enzyme bienque les réactifs soient placés dans les mêmes conditions.La comparaison des deux documents permet de s’interro-ger sur le mode d’action des enzymes et sur la nature decelles-ci.
◆ Problèmes scientifiques à résoudreQuelles sont les caractéristiques de l’action enzymatique?
P o s e r l e s p r o b l è m e s
Le DOCUMENT 1a. permet de faire le point sur les acquis de3e au niveau de la digestion et de l’assimilation. Il intègrela réaction chimique vue dans le chapitre 1, à savoir laréaction d’hydroxylation de la phénylalanine en tyrosine.Cela permet de mettre en évidence la multiplicité des réac-tions au sein de l’organisme et que toutes ces réactions
Programme: notions et contenus• Des protéines actives dans la catalyse : les enzymes.• Les protéines enzymatiques sont des catalyseurs biologiques. Elles présentent une double spécificité : spécificité d’action et de substrat. Les moda-lités de leur action reposent sur la formation du complexe enzyme-substrat. Les propriétés des enzymes dépendent de leur structure spatiale. Desmodifications de structure spatiale, déterminées soit par des changements de la séquence des acides aminés, soit par des conditions du milieu (pH,température, ions…), modifient leur activité.• L’activité des enzymes contribue à la réalisation du phénotype.Limites : L’étude des coenzymes, l’étude de l’allostérie, les lois de la cinétique enzymatique, ne sont pas au programme.
Objectifs à atteindreLes élèves ont découvert la notion d’enzyme en classe de 3e à travers l’étude de la digestion. « Au cours de la diges-tion, la fragmentation de molécules de grosse taille et de nature variée (glucides, protides, lipides) contenues dans lesaliments, aboutit à un nombre réduit de types de molécules petites et non spécifiques. Cette simplification s’effectuesous l’action d’enzymes digestives. »
Le terme d’enzyme a également été employé dans le chapitre 1. Ce chapitre établit le lien entre l’ADN, support de l’in-formation génétique et le phénotype. On montre qu’une relation causale plus ou moins complexe relie le phénotypemoléculaire au phénotype macroscopique via le phénotype cellulaire et parfois le phénotype biochimique. Un gènedétermine uniquement le phénotype moléculaire, à savoir une protéine déterminée.
Certaines protéines, les protéines enzymatiques, mettent en place le phénotype biochimique.
À partir de ces acquis, ce chapitre se propose, dans un premier temps, de définir l’enzyme comme un catalyseur, pourconstruire ensuite la notion de double spécificité de l’enzyme.
Il est nécessaire, dans un deuxième temps, d’expliquer leur mode d’action, pour ensuite montrer comment la connais-sance des structures moléculaires permet de comprendre les propriétés des enzymes.
Puis, on explique les liens entre l’activité biologique de l’enzyme et son environnement de façon à bien comprendrele passage du génotype au phénotype et les relations avec l’environnement. « L’importance des facteurs de l’environ-nement comme modulateurs de l’activité des protéines enzymatiques est rapprochée de la participation des protéinesà la réalisation du phénotype. »
Les enzymes
C H A P I T R E 2

Chapitre 2 : Les enzymes
13
pris l’exemple du glucose dans deux réactions chi-miques différentes : une réaction de dégradation et uneréaction de synthèse. Le choix de la réaction de syn-thèse est très important car souvent les élèves, au tra-vers de l’exemple prégnant des enzymes digestives,associent les enzymes à des réactions de dégradation.Le protocole est décrit dans le document 5, mais il estnécessaire d’ajouter un point important : il faut utiliserdes tubercules de pommes de terre sortant du réfrigéra-teur, de l’eau distillée fraîche et tout le matériel (cou-teau, entonnoir, papier filtre, mixeur, bêcher) doit êtrerefroidi préalablement. Tout cela est nécessaire pouréviter la dégradation de l’enzyme amidon-synthétase.(Il est préférable d’utiliser des pommes de terre à chairblanche.)
Un même type de réaction, l’hydrolyse par exemple,peut affecter des substances différentes. Pour illustrercette affirmation, nous avons pris comme exemple lessubstrats des enzymes digestives : trois diosides et uneprotéine.
Exploitation� La glucose-oxydase ne réalise qu’un seul type de réac-tion chimique (DOCUMENT 4), à savoir l’oxydation aérobiedu glucose. Cette oxydation produit de l’acide gluconiqueet du peroxyde d’hydrogène. Sur le même substrat, glu-cose, l’amidon-synthétase ne réalise qu’un seul type deréaction chimique (DOCUMENT 5), à savoir une polymérisa-tion du glucose en amidon. Faire remarquer aux élèves quel’enzyme (amidon-synthétase) est obtenue par broyage decellules de pomme de terre, que l’amidon est arrêté par lefiltre et que le substrat est le glucose-1-phosphate. Deuxréactions chimiques différentes sont catalysées par deuxenzymes différentes : il y a spécificité d’action de l’en-zyme ; elle ne catalyse qu’un seul type de réaction chi-mique.
Dans le DOCUMENT 7, on remarque que la maltase n’hydro-lyse que le maltose, la lactase que le lactose, et la saccha-rase que le saccharose. Pourtant, les trois molécules sontassez proches : ce sont des diosides. À la spécificité d’ac-tion, s’ajoute celle de substrat. L’enzyme exerce son actionsur un substrat déterminé. Cette notion est renforcée par leDOCUMENT 6. Le fructose et le galactose sont des oses struc-turellement très proches du glucose. En présence de glu-cose-oxydase, il n’y a pas de produit formé avec lefructose, ni le galactose, contrairement au glucose. La glu-cose-oxydase est spécifique du substrat glucose.
Le DOCUMENT 8 permet de préciser que la spécificité d’ac-tion peut être étroite. La chymotrypsine ou la trypsinepeuvent hydrolyser des substrats appartenant à une mêmefamille de substances (chaîne polypeptidique), mais l’hy-drolyse ne s’effectue pas sur les mêmes liaisons pepti-diques. On dit que la spécificité est étroite.
A c t i v i t é s 1
L’enzyme : un catalyseur biologiqueL’objectif de ces activités est de construire la notion decatalyseur et de définir l’enzyme comme catalyseurbiologique. Les élèves rencontrent souvent ici une diffi-culté, leur conception sur les enzymes est que celles-ciagissent comme des ciseaux qui vont couper les molé-cules de grosse taille. Il y a alors obstacle pour com-prendre que les enzymes permettent la réalisation deréactions chimiques. La notion de catalyseur estconstruite dans la question 1 du guide d’exploitation, àpartir de la comparaison de deux réactions chimiquesdans lesquelles intervient HCl, dans l’une comme cata-lyseur, dans l’autre comme réactif. La question 2 per-met de définir l’enzyme comme biocatalyseur.
Exploitation� Comparaison des rôles de HCl dans les deux réactionschimiques.Dans l’hydrolyse acide de l’amidon (DOCUMENT 2), onconstate qu’il y a une réaction chimique qui est une hydro-lyse : les réactifs sont l’amidon et l’eau ; le produit forméest le glucose.La constance du pH indique que la concentration d’ionsH3O + est la même au début et à la fin de la réaction. Del’empois d’amidon à 5 ‰ porté à ébullition à 100 °C, sansHCl, ne se transforme pas en glucose. L’expérience témoinn’a pas été représentée sur cette page.Dans la réaction où HCl agit sur du magnésium, onconstate, là aussi, qu’il y a une réaction chimique : lesréactifs sont le magnésium et HCl, les produits forméssont le dihydrogène qui se dégage et le chlorure de magné-sium. La mesure du pH au début et à la fin de la réactionmet bien en évidence la disparition de HCl.Dans la 1re réaction, HCl n’intervient pas comme réactif,mais il est absolument nécessaire pour que celle-ci sedéroule. On dit que HCl est un catalyseur. Un catalyseurest une molécule chimique qui, par sa présence dans lemilieu réactionnel, permet la formation des produits d’uneréaction, sans apparaître dans l’équation-bilan.� Dans cette réaction d’hydrolyse de l’amidon (DOCUMENT 3),avec l’amylase salivaire, les réactifs sont l’amidon et l’eau,le produit est le maltose.L’expérience témoin avec l’eau distillée nous montre quel’amylase salivaire est indispensable pour que l’hydrolysede l’amidon ait lieu. Pourtant sa concentration reste lamême entre le début et la fin de la réaction. L’amylasen’est pas un réactif, mais elle est nécessaire au déroule-ment de cette réaction : c’est un catalyseur.
A c t i v i t é s 2
La double spécificité d’une enzymeUne même substance peut entrer dans de multiplesréactions. Pour illustrer cette affirmation, nous avons

A c t i v i t é s 3
La réaction enzymatiqueL’objectif de ces activités est d’expliquer le mode d’ac-tion de l’enzyme. L’étude de l’évolution de la vitesse dela réaction enzymatique en fonction de la concentrationen substrat pour une concentration en enzymeconstante et celle de l’action de la concentration enenzyme sur la vitesse de réaction enzymatique avec uneconcentration en substrat constante a permis de propo-ser une modélisation de l’activité enzymatique. L’élèvepeut alors vérifier la conformité du modèle aux résul-tats expérimentaux obtenus.
Exploitation� Le graphe (DOCUMENT 9) permet de suivre la quantité deproduit formé en fonction du temps. Chaque courbe pré-sente une pente puis un palier. La pente de la courbe(dP/dt) correspond à la vitesse d’apparition du produit,c’est-à-dire à la vitesse de réaction. Le palier correspond àl’arrêt de la production du produit, c’est-à-dire à unevitesse nulle. La vitesse de la réaction n’est donc pasconstante. Elle est maximale au départ ; elle décroîtensuite au fur et à mesure de la disparition du substrat.Plus on augmente la concentration en substrat, plus laquantité de produit formé à un temps t (par exemple : 1minute) est importante.� Dans le DOCUMENT 10, on suit la vitesse initiale de laréaction en fonction de la quantité de substrat. La vitesseinitiale augmente d’abord avec la concentration du sub-strat, puis garde une valeur constante. Même en augmen-tant la quantité de substrat, la vitesse de réactionn’augmente plus. C’est la vitesse maximale. La concen-tration en glucose cesse d’être le facteur limitant. On peut émettre l’hypothèse que le dioxygène et/ou l’en-zyme sont les facteurs limitants lorsque le glucose est àforte concentration. Le professeur peut indiquer que lacourbe Vi = f [S] obtenue avec des enzymes n’ayant qu’unseul substrat est de même type. Cela permet de conclureque l’enzyme est le facteur limitant. On peut envisager sonmode d’action, notamment avec l’idée d’une saturation del’enzyme.Le graphe du DOCUMENT 11 permet de suivre l’action de laconcentration en enzyme sur la vitesse de la réactionenzymatique. La pente de chaque courbe correspond à lavitesse d’apparition du produit, c’est-à-dire à la vitesse deréaction. Plus on augmente la concentration en enzyme,plus la pente de la courbe est importante. La vitesse de laréaction augmente avec la concentration en enzyme, donc,la concentration en enzyme est un facteur limitant de lavitesse de réaction.La mise en relation des informations livrées par les gra-phiques des DOCUMENTS 10 et 11 permet de concevoir que lapremière étape de la catalyse enzymatique consiste en laformation d’un complexe enzyme-substrat et que dans unedeuxième étape a lieu la réaction chimique.On peut alors vérifier la conformité des modèles et desrésultats obtenus. Le modèle analogique du DOCUMENT12 a. permet de se représenter la variation de la vitesse de
14
la réaction enzymatique en fonction de la concentration ensubstrat. Le grain de blé, le concasseur et la farine repré-sentent respectivement le substrat, l’enzyme et le produit.S’il arrive moins de 20 kg. min-1 de grains de blé, cetapport constitue un facteur limitant de la production pos-sible de farine, car le concasseur pourrait effectuer un tra-vail plus important. S’il arrive 20 kg. min-1, la vitesse deproduction est maximale. S’il arrive plus de 20 kg. min-1,la capacité du concasseur est insuffisante et constitue alorsun facteur limitant de la production. Nous retrouvons lesconclusions des DOCUMENTS 10 et 11.Le modèle interprétatif de l’activité enzymatique (DOCU-MENT 12 b.) repose sur la formation d’un complexe entrel’enzyme et le substrat, puis sur la réalisation de la réac-tion chimique proprement dite avec conservation de l’en-zyme en fin de réaction. Le professeur peut proposer icil’exercice 7 du manuel, qui montre également la conser-vation de l’enzyme au cours d’une réaction chimique,ainsi qu’une activité sur logiciel de simulation. Ce modèleinterprétatif rend bien compte que le substrat est un fac-teur limitant, tant que l’enzyme n’est pas saturée.Lorsqu’elle est saturée, la quantité d’enzyme devient unfacteur limitant. La vitesse maximale de la réaction enzy-matique dépend du temps de liaison entre l’enzyme et sonsubstrat.
A c t i v i t é s 4
Le complexe enzyme-substratPour expliquer les variations de la vitesse de la réac-tion enzymatique en fonction du temps, nous avons étéconduits à concevoir la formation d’un complexe tem-poraire enzyme-substrat. L’objectif de ces activités estde préciser la nature de la relation entre l’enzyme etson substrat, lors de la formation du complexe.
Exploitation� Les enzymes sont des protéines. Elles adoptent sponta-nément dans l’espace une conformation tridimensionnellequi dépend de la structure primaire. On observe sur lesDOCUMENTS 13 et 15 que le substrat s’associe à une régionlimitée de l’enzyme en forme de creux : le site actif. Onconstate une étroite complémentarité entre le site actif etune zone précise du substrat. Le site actif a deux fonc-tions : fixer le substrat spécifique et permettre la réactionchimique sur ce substrat.� L’enzyme B diffère de l’enzyme A par quatre acidesaminés seulement. On observe que cette différence modi-fie la structure spatiale de la forme B au niveau du siteactif. On peut supposer que, suite à cette modification, iln’y a plus complémentarité entre le site actif et le substratN-acétylgalactosamine, mais complémentarité avec unnouveau substrat, le galactose.� La trypsine et la chymotrypsine ont en commun, auniveau de leur site actif, un groupe de trois acides aminés :l’histidine, l’acide aspartique et la sérine. Ces acides ami-nés sont responsables de la spécificité de la réaction : unehydrolyse.

En revanche, au niveau du site actif, la trypsine possède unacide aminé qui fixe la chaîne latérale de la lysine ou del’arginine. Cet acide aminé ne se retrouve pas dans la chy-motrypsine. À la place de ce dernier, on trouve un autreacide aminé qui fixe les chaînes latérales d’acides aminés,comme la tyrosine, la phénylalanine et le tryptophane.Cela explique la différence dans la spécificité de substratentre la chymotrypsine et la trypsine mise en évidencedans le DOCUMENT 8 page 31.
A c t i v i t é s 5
Actions des conditions du milieu sur l’activité enzymatiqueL’objectif de ces activités est d’expliquer les liens entrel’activité biologique d’une enzyme et son environne-ment. Elles vont permettre de bien recentrer ce chapitresur la thématique générale de cette partie : du génotypeau phénotype, relation avec l’environnement. Les pro-téines participent à la réalisation du phénotype. Nousallons dans cette double page, montrer l’importancedes facteurs de l’environnement comme modulateurs del’activité des protéines enzymatiques.
Exploitation� La vitesse de la réaction enzymatique (DOCUMENT 17) estquasi nulle pour une température de 0 °C ; elle augmenteensuite progressivement avec l’élévation de la tempéra-ture, pour atteindre une vitesse optimale vers 45 °C (ce quicorrespond à la température maximale) ; elle décroîtensuite très rapidement, si la température augmente tou-jours. Elle est nulle lorsque la température avoisine les60 °C.� Le DOCUMENT 16 montre que la courbe représentative dela vitesse initiale en fonction de la température présentedeux parties distinctes de part et d’autre d’une températureoptimale pour laquelle la vitesse est maximale.Dans la première partie, la vitesse augmente avec la tem-pérature. L’énergie fournie au système augmente l’agita-tion moléculaire et augmente ainsi la probabilité derencontre entre l’enzyme et son substrat, et par voie deconséquence facilite la formation du complexe enzyme-substrat. Dans la deuxième partie, il y a dénaturation pro-gressive de l’enzyme qui perd son activité. Cettedénaturation correspond à un changement irréversible dela conformation des molécules d’enzyme qui ne peuventplus fixer les molécules de substrat. Le DOCUMENT 17 per-met de confirmer ces conclusions.� La truite arc-en-ciel, lorsqu’elle vit naturellement dansles eaux relativement chaudes, synthétise une acétylcho-line estérase de type 1. Si elle synthétisait toujours cetteenzyme à basse température, celle-ci serait inactivée, cequi aurait pour conséquence un mauvais fonctionnementmusculaire. Par contre, en synthétisant une acétylcholineestérase de type 2, ayant une température d’action opti-male plus basse, cette enzyme a une activité suffisantepour permettre un bon fonctionnement musculaire.
� Sur le DOCUMENT 19, chaque courbe représentative de lavitesse de multiplication des bactéries en fonction de latempérature présente deux parties distinctes de part etd’autre d’une température optimale, pour laquelle lavitesse de multiplication est maximale. On peut supposerque cette température optimale corresponde à la tempéra-ture optimale de l’activité des enzymes de la bactérie.On peut expliquer le développement des bactéries dansune eau très chaude par le fait que leurs enzymes ont unetempérature optimale élevée. Les enzymes de ces bacté-ries sont suffisamment rigides pour résister à des tempéra-tures élevées (proches de 100 °C) pour ne pas se dénaturer.� On constate que suivant les enzymes, la vitesse de réac-tion est maximale pour une valeur de pH dite pH optimal.Elle décroît rapidement dès que l’on s’éloigne de cettevaleur. Les variations de pH modifient l’ionisation de cer-tains acides aminés, correspondant au site actif de la molé-cule d’enzyme et modifient ainsi sa conformation spatiale.Ces modifications altèrent la liaison enzyme-substrat.Elles sont irréversibles : si l’on s’éloigne trop du pH opti-mal, l’enzyme est alors dénaturée.
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Catalyseur : c’est une molécule chimique qui, parsa présence dans le milieu réactionnel, augmente lavitesse de formation des produits d’une réaction,sans apparaître dans l’équation-bilan.b. Enzyme : protéine synthétisée par une celluleconformément à son information génétique, quiintervient dans une réaction chimique du vivant enaugmentant la vitesse de réaction, tout en se retrou-vant intacte à la fin de celle-ci. C’est un biocataly-seur.c. Substrat : molécule sur laquelle s’exerce l’actiond’une enzyme.d. Complexe enzyme-substrat : association tempo-raire entre une zone spécifique de la molécule desubstrat et le site actif de l’enzyme, étape néces-saire à la réalisation de la réaction chimique.e. Site actif d’une enzyme : région particulière d’uneenzyme en forme de creux qui a deux fonctions :fixer le(s) substrat(s) spécifique(s) et permettre laréaction chimique sur ce(s) substrat(s).
Questions à réponses cour tesa. Dans une réaction chimique où une enzyme inter-vient, elle en augmente la vitesse de réaction, touten se retrouvant intacte à la fin de celle-ci. Cetteaction est possible grâce à la formation temporaired’un complexe entre une zone spécifique de lamolécule de substrat et le site actif de l’enzyme. Cesite permet de fixer le substrat spécifique et permetla réaction chimique sur ce substrat. L’enzyme estun biocatalyseur.
2
1
Chapitre 2 : Les enzymes
15

Phrases à corrigera. Une enzyme catalyse une réaction lorsqu’elle estcombinée aux réactifs de la réaction. Ces réactifssont les substrats spécifiques de l’enzyme.b. Le site actif de l’enzyme est la partie de la molé-cule d’enzyme qui se lie au substrat.c. Une enzyme participe à la réaction qu’elle cata-lyse sans entrer dans la composition des produitsissus de cette réaction.
Choisir les bonnes réponses1. Une enzyme…a.… est un catalyseur biologique.c.… favorise la réalisation d’une réaction chimiquesans participer à celle-ci.d.… est une molécule qui favorise la réalisation deréactions chimiques spécifiques.2. Le site actif d’une enzyme…b.… dépend du programme génétique.c.… est la partie de la molécule sur laquelle se fixele substrat.d.… peut être modifié par différents facteurs : tem-pérature, pH.
Exercice corrigé
Les enzymes de la levure de boulangerVoir correction dans le manuel pages 44 et 45.
Action d’un filtrat de grainsd’orge germés sur l’empois d’amidon1. En A, on constate un changement de couleur aubout de 15 minutes. Le réactif iodo-ioduré ne donneplus une coloration bleu violacé, caractéristique del’amidon. Ce changement de couleur met donc enévidence la disparition de l’amidon dans le flacon Aau bout de 15 minutes, alors qu’il est toujours pré-sent dans le flacon B.2. La disparition de l’amidon dans le flacon A peuts’expliquer par la présence dans le filtrat d’uneenzyme qui hydrolyse l’amidon. La non disparitionde l’amidon dans le flacon B peut s’expliquer par ladénaturation de cette enzyme suite à l’ébullition dufiltrat.
Rechercher une explicat ionL’équation bilan est :C6H12O6 + O2 + H2O Glucose-oxydase C6H12O7 + H2O2
Glucose dioxygène eau acide gluconique péroxyde d’hydrogène
1. Lorsque l’on regarde l’équation-bilan, on penseimmédiatement au glucose comme substrat, mais ledioxygène est également substrat. Cela se vérifie enconstatant qu’avec une concentration constante etélevée de glucose, la vitesse de la réaction enzyma-tique augmente avec la concentration de dioxygène.Il est donc nécessaire d’être à la fois précis sur la
7
6
5
4
3b. Une enzyme a pour caractéristiques spécifiques :- une spécificité de substrat,- une spécificité d’action,- une température optimale d’action,- un pH optimal d’action.La spécificité de substrat et la spécificité d’actiondépendent de l’étroite complémentarité entre le siteactif de l’enzyme et une zone précise du substrat.Elles dépendent plus particulièrement des acidesaminés présents au niveau du site actif.Chaque enzyme a une activité maximale pour unevaleur donnée de la température et pour une valeurdonnée du pH. Lorsque l’on s’écarte de ces valeurs,la température ou le pH entraîne une modification dela conformation spatiale de l’enzyme qui aboutit àune perte de sa spécificité.c. Les enzymes sont des protéines. Elles adoptentspontanément dans l’espace une conformation tridi-mensionnelle qui dépend de leur structure primaire,déterminée par la séquence des acides aminés. Lesubstrat s’associe à une région limitée de l’enzymeen forme de creux : le site actif.L’activité de l’enzyme dépend dans un premiertemps de la formation du complexe enzyme-sub-strat, ce qui nécessite, d’une part, une complémen-tarité de forme entre ce site actif et une zonespécifique de la molécule de substrat et, d’autrepart, la présence au niveau du site actif des acidesaminés qui se lient au substrat. Après cette étape, ily a la réalisation de la réaction chimique. Elledépend de la présence au niveau du site actif decertains acides aminés qui participent à la catalyse.Ces acides aminés (de liaison ou catalytiques) sontsouvent éloignés dans la séquence de la protéineenzymatique, mais rapprochés dans la structurespatiale.d. Les facteurs qui déterminent la vitesse d’uneréaction chimique catalysée par une enzyme sont :- la concentration en substrat,- la concentration en enzyme,- la température.La vitesse croît proportionnellement à la concentra-tion en substrat jusqu’à une certaine concentration,puis elle est constante, indépendante de la concen-tration en substrat. La vitesse maximale est atteinte,toutes les molécules d’enzyme sont engagées à unmoment donné, dans le complexe enzyme-substrat.L’enzyme est saturée et devient un facteur limitantde la réaction. Pour augmenter la vitesse, il fautrajouter des molécules d’enzyme.L’élévation de température augmente l’agitationmoléculaire ; elle augmente ainsi la probabilité derencontre entre les molécules d’enzymes et lesmolécules de substrat : la vitesse de la réactionenzymatique augmente. À basse température, l’en-zyme est inactivée ; à haute température, l’enzymeest dénaturée.
16

concentration de glucose et sur la concentration endioxygène. En agitant le milieu, on le sature endioxygène.2. Les deux courbes sont superposées. On peutdonc en conclure que les deux solutions présententla même activité enzymatique. L’activité enzyma-tique de la solution « usagée » n’a pas été modifiée,malgré le fait qu’elle ait déjà servi. C’est donc qu’il ya conservation de l’enzyme au cours d’une réactionenzymatique.Remarque : Lors de la mesure de l’activité de cesdeux solutions enzymatiques, on a pris une solutionen glucose dix fois plus concentrée que celle utiliséepour préparer la solution de glucose-oxydase « usa-gée ». Cela permet de diminuer l’erreur dans le casoù il resterait du glucose issu du premier temps del’expérience, lors de la préparation de la solutiond’enzyme « usagée ».
Action de la glucose-oxydase1. En comparant les résultats, on constate que suiteà l’injection de l’enzyme, il n’y a consommation dedioxygène qu’avec l’amidon hydrolysé par del’acide. L’activité de l’enzyme n’a donc lieu qu’aveccette solution.2. Dans les activités 1 p. 29, nous avons constatéque l’hydrolyse de l’amidon par la salive donne dumaltose (C12H22O11), que l’hydrolyse de l’amidon parl’acide donne du glucose (C6H12O6).L’enzyme se trouve donc en présence de trois sub-strats : le maltose, l’amidon et le glucose. L’activitéde l’enzyme n’est présente qu’avec le glucose ; onvérifie ainsi la spécificité de substrat.
8
Activi té enzymatique et structurede l ’enzyme1.Le repliement de la chaîne polypeptidique dépendde la structure primaire de cette chaîne. Des liaisons(ponts disulfures) s’établissent entre deux cystéinesqui sont plus ou moins éloignées le long de lachaîne. Ces liaisons imposent le repliement de lachaîne.2. L’action de l’urée puis du �-mercaptoéthanolsupprime les ponts disulfures. Le �-mercaptoétha-nol est un agent réducteur qui peut rompre les pontsS-S et les réduire en groupes SH. Cela modifie doncla configuration spatiale de l’enzyme et par voie deconséquence son site actif : il n’y a donc plus d’ac-tivité enzymatique.L’élimination du �– mercaptoéthanol permet (formeC) la reformation des ponts disulfures 65-72 et 58-110. Mais deux ponts disulfures sont modifiés : 84-95 au lieu de 40-95 (forme A) et 26-40 au lieu de26-84 (forme A). L’enzyme retrouve donc uneconformation tridimensionnelle, mais différente de laforme A. Cette différence doit affecter le site actif, cequi explique que cette enzyme soit très peu active.3. La disparition de l’acide aminé soufré 84 modifie-rait la configuration spatiale de l’enzyme. Si cettemodification affecte la région limitée de l’enzyme quicorrespond au site actif, elle aura pour conséquenceune inactivation de cette enzyme.Si cette modification affecte une région qui ne cor-respond pas au site actif, elle n’aura pas de consé-quence sur l’activité de l’enzyme.
9
Chapitre 2 : Les enzymes
17

18
phénotypes individuels, cellulaires et moléculaires d’unepart, et le génotype d’autre part, avant d’étudier les méca-nismes qui permettent l’expression de ce dernier.L’enzyme dont la structure tridimensionnelle est présentéeici, est une enzyme du suc pancréatique, la carboxypepti-dase, telle qu’elle est proposée par le logiciel Rasmol. Legène du précurseur inactif de la carboxypeptidase, possède1310 paires de nucléotides. Les 27 premières paires debases de la partie codante du gène sont indiquées en regarddes acides aminés correspondants, à partir du codon d’ini-tiation (AUG) et de la méthionine. Le constat de l’exis-tence de deux molécules séquencées, ADN et protéine,pose le problème d’un système de correspondance.Remontant les documents de (d) à (a), il est possible derelier une digestion efficace des aliments à une séquencede nucléotides, et de s’interroger sur les modalités de l’ex-pression du gène codant cette enzyme pancréatique.
◆ Problèmes scientifiques à résoudreSachant que l’information génétique est contenue dans lenoyau, la question de la localisation de la synthèse desprotéines doit être posée. L’observation de celle-ci dans lecompartiment cytoplasmique légitime une nouvelle pro-blématique : comment l’information codée au niveau desgènes peut-elle s’exprimer au sein du cytoplasme par l’as-semblage d’acides aminés prélevés dans le milieu extra-cellulaire?
P o s e r l e s p r o b l è m e s
Le DOCUMENT 1 permet de relier entre eux différentsniveaux de réalisation du phénotype. Le phénotype macro-scopique correspond ici à l’aptitude à bien digérer ses ali-ments. On rappelle ainsi l’existence de relations entre les
Programme: notions et contenus• L’expression de l’information génétique se fait en deux étapes : transcription et traduction.• Au cours de la transcription, un ARN messager complémentaire du brin transcrit de l’ADN est synthétisé.• La traduction permet la synthèse cytoplasmique de chaînes polypeptidiques. La séquence des acides aminés est gouvernée par celle des nucléo-tides de l’ARN messager suivant un système de correspondance, le code génétique.• Ce code génétique est universel et dégénéré.• La traduction débute au codon d’initiation et s’arrête au codon stop.
Objectifs à atteindreAprès avoir mis en évidence la diversité des phénotypes, l’importance toute particulière des protéines enzymatiquesdans la réalisation du phénotype aura été établie, ce qui justifie l’étude des mécanismes responsables de la synthèsedes protéines. Les objectifs cognitifs de ce chapitre, la connaissance de l’existence d’un ARN messager comme celledu code génétique, visent à faire acquérir aux élèves des éléments d’une culture scientifique. La maîtrise de cesconnaissances permet d’appréhender les conséquences des mutations, tant dans le domaine de la santé que dans celuide l’évolution des espèces. Dans cet esprit, l’universalité du code génétique doit être soulignée. Bien que ne se prê-tant guère à la réalisation d’expériences dans le cadre de la classe, ce chapitre offre la possibilité d’un apprentissageà la démarche expérimentale. Avec l’utilisation de logiciels, les documents du manuel permettent cet apprentissage.La mise en relation des informations tirées de l’analyse des résultats d’expériences historiques, de l’observation declichés de microscopie électronique, de la comparaison de séquences d’ADN et d’ARN messager, vise à faire élabo-rer progressivement aux élèves, et non à leur imposer, un modèle de l’expression de l’information génétique.
La biosynthèse des protéines
C H A P I T R E 3

A c t i v i t é s 1
Localisation de la synthèse des protéinesL’observation de cellules du pancréas exocrine a mon-tré la présence de protéines enzymatiques au niveau deleur cytoplasme. Les clichés de microscopie électro-nique analysés ici, d’une part, prolongent ces observa-tions avec deux moments de la synthèse d’une protéinedu suc pancréatique, d’autre part, élargissent lesconclusions auxquelles on peut aboutir, au cas de pro-téines non sécrétées, comme l’hémoglobine, dont lecodage sera envisagé au cours des activités suivantes.
Exploitation� Le DOCUMENT 2 présente deux autoradiographies obte-nues lors d’une expérience de traçage d’un acide aminérendu radioactif (J. D. Jamieson et G. E. Palade, 1967).Des tranches de pancréas de cobaye ont été mises à incu-ber pendant 3 minutes dans un milieu contenant de la leu-cine tritiée, puis placées dans un milieu non radioactifpendant des temps de chasse de durées variables. En find’expérience, les échantillons ont été fixés au tétraoxyded’osmium, puis inclus dans une résine dans laquelle on aréalisé des coupes de 0,02 µm d’épaisseur, et sur les-quelles a été coulée une émulsion photographique. Lespréparations ont été observées au microscope électroniqueaprès 3 semaines d’exposition, la présence de grains d’ar-gent révélant la présence de radioactivité. Les clichés met-tent en évidence un transport intracellulaire des acidesaminés, puis des protéines sécrétées par les cellules aci-neuses du pancréas. Bien que l’assemblage des acidesaminés soit observé au niveau du réticulum (DOCUMENT 2a),ainsi que les vésicules golgiennes à proximité de lalumière de l’acinus (DOCUMENT 2b), les rôles du réticulumet de l’appareil de Golgi ne sont pas l’objet de cette étude.Les grains de zymogène (zy) sont des vésicules de sécré-tion renfermant les enzymes du suc pancréatique. La pré-sence de grains d’argent dans la lumière de l’acinus(DOCUMENT 2b) montre que des polypeptides contenant dela leucine tritiée ont été rejetés par les cellules pancréa-tiques. La localisation cytoplasmique de la synthèse desprotéines est confirmée par l’observation, sur le DOCUMENT3, des ribosomes associés à la synthèse de l’hémoglobine.La formation des polysomes ne pouvant être encore expli-quée, on se limite ici à montrer la présence de ribosomesà l’intérieur du cytoplasme.� Dans tous les cas, un acide aminé prélevé dans lemilieu extracellulaire est incorporé au sein d’un polypep-tide au niveau des ribosomes. Les polypeptides constituentensuite des protéines qui sont, soit exportées (DOCUMENT 2),soit rendues fonctionnelles dans le compartiment intracel-lulaire (DOCUMENT 3).
A c t i v i t é s 2
Un messager de l’information génétiqueLa localisation cytoplasmique de la synthèse des pro-téines conduit à s’interroger sur la transmission de l’in-formation génétique jusqu’au cytoplasme et sur lafonction des ribosomes. Ces activités ont pour objectifde mettre en évidence le rôle de l’ARN messager. Lesdocuments proposés permettent, dans un premiertemps, de montrer que les protéines peuvent être syn-thétisées en l’absence d’ADN, et dans un second tempsque l’ARN messager, synthétisé au niveau du noyau,migre ensuite dans le cytoplasme. Sa fonction estdécouverte à partir de l’exploitation de résultats expé-rimentaux. C’est donc de la mise en relation de donnéesobtenues au cours d’expériences différentes qu’estdéduite l’existence des deux étapes, transcription et tra-duction, de la synthèse des protéines.
Exploitation� Le DOCUMENT 4a met en évidence l’incorporation de leu-cine tritiée, même en l’absence du noyau. On remarqueque l’énucléation n’a précédé que de quelques minutes lamise en culture des cellules sur un milieu contenant unacide aminé rendu radioactif car la synthèse de protéinesne peut se poursuivre durablement en l’absence d’ADN.Le DOCUMENT 4b et son interprétation, au DOCUMENT 4c, per-mettent de visualiser l’ARN messager, les ribosomes et unpolypeptide en formation, ici une protéine fibreuse de lasoie. Bien que le grossissement permette leur observation(le diamètre d’un ribosome ne dépasse guère 0,02 µm), lessous-unités ribosomiales ne doivent pas être distinguées àce niveau.Les résultats expérimentaux apportés par le DOCUMENT 5montrent que la présence d’ARN messager suffit à provo-quer la synthèse d’hémoglobine de Lapin par des œufs deXénope privés de leur ADN. Les données du DOCUMENT 6permettent une comparaison entre les structures de l’ADNet de l’ARN, au cours de laquelle la différence de tailleentre ces molécules doit être soulignée.Le DOCUMENT 7 représente deux autoradiographies d’unprotozoaire, Tétrahyména, après son exposition à unnucléotide de l’ARN rendu radioactif. La totalité de laradioactivité incorporée dans l’ARN au cours des 15 pre-mières minutes a quitté le noyau à la fin de l’expérience,pour se retrouver dans le cytoplasme. On peut doncconclure que l’ARN est synthétisé à l’intérieur du noyau(DOCUMENT 7a), puis migre dans le cytoplasme (DOCUMENT7b), où il représente une information nécessaire à la syn-thèse d’une protéine (DOCUMENT 5). L’assemblage desacides aminés est réalisé par les ribosomes fixés sur unemolécule d’ARN messager (DOCUMENT 4).
Chapitre 3 : La biosynthèse des protéines
19

La transcription, dont la vitesse est de l’ordre de 1 µm parminute est généralement unidirectionnelle : un seul desdeux brins d’ADN est transcrit. Cependant, ce n’est pastoujours le même brin qui est transcrit, la direction detranscription variant d’une région du chromosome à uneautre, mais toujours dans la direction 5’➞ 3’. Bien querare, la transcription bilatérale existe pour plusieurs gènesde virus ou de bactéries, ainsi que pour l’ADN mitochon-drial.
� Les DOCUMENTS 11 et 12 permettent d’envisager un trans-port des molécules d’ARN du noyau vers le cytoplasme. Ilest alors possible de relier les informations extraites dedocuments différents pour une courte synthèse, de latranscription d’une molécule d’ARN à partir d’unematrice, le brin d’ADN transcrit, jusqu’à son transfertdans le noyau et la fixation des ribosomes. À ce stade,l’ARN messager doit être compris comme une copie, maisen négatif, de la partie codante d’un gène.La structure de l’enveloppe nucléaire, une double mem-brane de 7 nm d’épaisseur, oblige les échanges à se faire àtravers les pores nucléaires. Ceux-ci contrôlent activementle transport des matériaux entre les compartimentsnucléaire et cytoplasmique. Il s’agit de structures pro-téiques à symétrie radiale d’ordre 8, dont le diamètreexterne est d’environ 120 nm, et l’orifice lui-même de 60à 90 nm. Au fur et à mesure de leur synthèse, les filamentsd’ARN sont continuellement extrudés par chaque pore, àune vitesse d’environ 1 µm par minute. Le DOCUMENT 12montre une molécule d’ARN messager d’une cellule deglande salivaire d’insecte, saisie au cours de son déplace-ment vers le cytoplasme.On peut indiquer que les matériaux nécessaires à la syn-thèse de l’ARN, protéines et nucléosides triphosphates,traversent les pores nucléaires en sens inverse ; même lesprotéines qui sont fonctionnelles à l’intérieur du noyausont élaborées dans le cytoplasme.
A c t i v i t é s 4
Traduction de l’information génétiqueLe principe du codage d’une séquence de nucléotidesen une séquence d’acides aminés est acquis au cours deces activités par une approche historique, propre à sus-citer une démarche expérimentale. On détermine lenombre minimal de nucléotides susceptibles de coderensemble un acide aminé, puis on étudie le résultatd’une expérience fondamentale réalisée par Nirenberget Matthéi. Alors que l’expérimentation directe n’estguère envisageable en classe, une démarche scienti-fique rigoureuse demeure possible en s’appuyant surles expériences réalisées à l’aide de plusieurs polynu-cléotides de synthèse. Les caractéristiques du codegénétique sont ensuite étudiées par les élèves, endehors de tout contexte historique. À cette fin, les docu-ments du manuel peuvent être utilisés seuls, ou associésà l’exploitation d’une banque de données sur différentsgènes dont les séquences sont connues. Les séquences
A c t i v i t é s 3
Transcription au sein du noyauLes mécanismes qui permettent le transfert de l’infor-mation d’un gène à une molécule d’ARN messager aucours de sa synthèse constituent le support de ces acti-vités. La fonction de l’ARN messager étant connue,c’est au niveau moléculaire qu’une explication doit êtrerecherchée. La connaissance de la structure de l’ARNmessager est également acquise. Il reste à expliquer icien quoi les modalités de la synthèse d’une moléculed’ARN messager lui permettent d’être une copie dugène transcrit. On envisage ensuite la possibilité d’unemigration de l’ARN messager et, avec lui, de l’informa-tion qu’il représente désormais, vers le cytoplasme. Lacomplémentarité des bases entre l’ARN et la portioncorrespondante du brin non transcrit de l’ADN, la simi-litude entre l’ARN transcrit et l’ADN non transcrit, laspécificité de l’uracile, peuvent être découvertes àl’aide de logiciels. Les documents du manuel, utilisésseuls ou en complément d’un logiciel, permettent desituer les phénomènes étudiés au sein de la cellule, etde s’assurer de leur compréhension.
Exploitation� Les connaissances acquises précédemment permettentd’identifier les structures visibles sur le document 8 : lesfilaments A sont des molécules d’ADN, les filaments B etC des molécules d’ARN.Il n’est sans doute pas judicieux de proposer la figure,sans doute plus classique, de complexes de transcriptionsde l’ARN ribosomial contenu dans le nucléole d’un ovo-cyte d’Amphibien, car le cas de nombreuses transcriptionssimultanées donnant l’apparence d’une fronde de fougère,ne correspond pas à celui de la plupart des gènes. En effet,la majeure partie de la chromatine d’une cellule eucaryotene contient pas de gènes actifs, et se trouve donc dépour-vue de transcrits. De plus, un gène n’est, le plus souvent,transcrit simultanément qu’en quelques molécules d’ARNmessager seulement.� et � Le DOCUMENT 9 a été obtenu à l’aide du logicielAnagène, qui permet d’aligner des séquences d’ADN etd’ARN messager d’un même gène. L’ARN messagerapparaît identique au brin 1 de l’ADN, à l’exception del’uracile qui se substitue à la thymine. On peut ainsi insis-ter sur les caractères du brin non transcrit, représentatif dugène dans la plupart des données. Le DOCUMENT 10 montrecomment une molécule d’ARN peut être synthétisée, l’undes brins d’ADN lui servant de matrice : les nucléotidesmanquants sont, de gauche à droite, un nucléotide à gua-nine, un nucléotide à uracile, un nucléotide à adénine. Ondoit en conclure que le brin d’ADN transcrit est le brin 2du DOCUMENT 9. C’est pour ne pas détourner l’attention desélèves de l’objectif des activités que les ARN polymérasesn’ont pas été représentées. Elles trouvent naturellementleur place dans le texte et les schémas des«Connaissances », page 57.
20

proposées dans les documents du manuel permettent deconstruire partiellement un tableau du code génétique.Le caractère dégénéré du code peut être établi. Lesconséquences d’une mutation devraient alors être aisé-ment comprises dans le cas d’une substitution.L’exemple proposé, plus complexe, permet d’appréhen-der les effets d’une délétion. Il ne s’agit donc pas del’utilisation du code génétique pour déterminer unenchaînement d’acides aminés, mais d’une appropria-tion progressive par les élèves, mis en situation derecherche, de ses principaux caractères. Les exercices 4(corrigé), 6 et 7 permettent de s’assurer, en utilisant letableau du code génétique donné page 59, de la maî-trise des connaissances et des compétences acquises aucours des activités.
Exploitation� Trois nucléotides au moins sont nécessaires pour coderun acide aminé. Le DOCUMENT 13 ne suffit pourtant pas,seul, à apporter une réponse certaine à cette question. Cesont les informations fournies par les DOCUMENTS 14 et 15qui permettent de valider cette hypothèse.L’approche historique choisie ne retrace que quelques unsdes travaux qui ont abouti à la découverte du code géné-tique. Le DOCUMENT 14 rend compte des résultats de l’expé-rience de Nirenberg et Matthéi, qui recherchèrent lepolypeptide synthétisé in vitro par un polynucléotide desynthèse, le poly-U. Celui-ci sélectionne exclusivementdes molécules de phénylalanyl-ARNt, formant ainsi unechaîne polypeptidique ne contenant que de la phénylala-nine.Des expériences analogues effectuées avec le poly-C et lepoly-A ont pu désigner CCC comme codant la proline etAAA, la lysine (les résidus guanine des poly-G établissententre eux des liaisons qui les empêchent de se fixer sur lesribosomes). Au cours de ces premières expériences, des
concentrations élevées en magnésium ont permis à lachaîne de biosynthèse de démarrer en l’absence du facteurd’initiation.Les autres codons ont été déterminés par une méthode dif-férente, utilisant le fait que seules des molécules d’ami-noacyl-ARNt spécifiques peuvent se fixer aux complexesribosomes-ARNm. Ainsi, en présence de poly-U, seul lephénylalanyl-ARNt peut s’attacher aux ribosomes. Demême, le lysyl-ARNt se fixe quand on utilise le trinucléo-tide AAA. Cette méthode a permis de déterminer directe-ment l’ordre des nucléotides au sein de codons tels queACA, CAC, CAA.Les données du DOCUMENT 15 confirment ici que le code estlinéaire, non chevauchant et univoque, c’est-à-dire qu’à uncodon ne correspond qu’un seul acide aminé. Une analyseplus approfondie montre le caractère dégénéré du codegénétique : la proline, sélectionnée par un poly-C (DOCU-MENT 14), correspond au codon CCU des portions repré-sentées des ARNm des chaînes de l’hémoglobine(DOCUMENT 15).� Les DOCUMENTS 15 et 16 montrent que toutes lesséquences d’ARN messager commencent par le mêmecodon, AUG. La méthionine est ensuite excisée au coursde la maturation de la protéine, quelle qu’elle soit. En fait,seule la partie strictement codante de l’ARN messager estreprésentée, en amont et en aval de cette partie codante ilexiste d’autres nucléotides. La traduction doit donc com-mencer et s’arrêter en des positions internes, correspon-dant à des signaux présents dans l’ADN lui-même, et parsuite dans l’ARN messager : ce sont le codon d’initiation(AUG) et les codons stop (UAA, UAG et UGA).Le DOCUMENT 16 représente les premiers codons, et lesacides aminés correspondants, de l’ARNm de la chaîne �de l’hémoglobine dans le cas d’une forme grave d’une tha-lassémie. On constate que la délétion d’un nucléotide àcytosine en position 51 entraîne un décalage du cadre delecture, modifiant le codon concerné et le codon suivant,
Chapitre 3 : La biosynthèse des protéines
21
UUU phénylalanineUUC phénylalanineUUAUUG leucine
UCU sérineUCC sérineUCAUCG
UAU tyrosineUAC tyrosineUAA stopUAG
UGUUGCUGA stopUGG tryptophane
CUUCUCCUACUG leucine
CCU prolineCCC prolineCCACCG
CAU histidineCAC histidineCAA glutamineCAG glutamine
CGU arginineCGCCGACGG
AUUAUC isoleucineAUAAUG méthionine
ACU thréonineACC thréonineACA thréonineACG
AAU asparagineAAC asparagineAAA lysineAAG lysine
AGU sérineAGC sérineAGA arginineAGG arginine
GUU valineGUC valineGUAGUG valine
GCU alanineGCC alanineGCAGCG
GAU acide aspartiqueGAC acide aspartiqueGAA acide glutamiqueGAG acide glutamique
GGU glycineGGC glycineGGA glycineGGG
U C A G
U
C
A
G
� Les données extraites des DOCUMENTS 14, 15 et 16 permettent de construire un tableau partiel du code génétique.

Exercice corrigé
L’élaboration des protéines dulaitCet exercice peut être proposé en classe, en com-plément des activités 4, dans la mesure où il permetde vérifier la maîtrise de l’utilisation du code géné-tique. Il peut aussi constituer, dans une période detravail personnel des élèves, un entraînement effi-cace à une analyse rigoureuse de documents. Lescompétences acquises au cours des activités sontdirectement réinvesties. Il s’agit de saisir les don-nées pertinentes par rapport aux questions posées,puis de les mettre en relation avec les connais-sances. Celles-ci devraient s’en trouver consoli-dées. Une lecture de graphiques, sans difficultémajeure à ce niveau, conduit à retracer le devenird’un acide aminé incorporé dans une cellule, puisdans une protéine exportée hors de celle-ci. Lesrésultats exprimés par le document c. peuvent êtreinterprétés à l’aide des connaissances acquises surla localisation de la synthèse des protéines.Conformément au programme, on ne demande pasici de distinguer les rôles des différents comparti-ments cytoplasmiques (réticulum, appareil de Golgi)dans le transit des acides aminés, puis des pro-téines synthétisées.
Évaluer ses compétences
Synthèse protéique in vi tro1. La seconde injection d’ARN entraîne immédiate-ment la synthèse de protéines. On peut penser qu’ilen est de même à la suite de la première injection,ou après la synthèse d’ARN endogène. Les ribo-somes et les enzymes présents dans le milieu (aux-quels ont été ajoutés de l’ATP et du GTP) nesuffisent donc pas à réaliser l’assemblage desacides aminés radioactifs présents dans le milieu.2. Les molécules d’ARN injectées sont rapidementdétruites, et ne permettent donc la synthèse qued’une quantité limitée de protéines. L’ARN messagerne constitue qu’une copie éphémère d’un gène.3. Une cellule privée de noyau ne possède plusd’ADN. Elle ne peut donc plus transcrire celui-ci enARN messager. Les ARN messagers étant rapide-ment dégradés, la synthèse de protéines ne semblepouvoir se poursuivre au-delà d’une quinzaine deminutes. Les informations apportées par le docu-ment b de cet exercice peuvent être rapprochées dudocument 4 a., page 50.
La synthèse d’une protéine de lacoagulat ion du sang1. L’enzyme TaqI reconnaît la séquence TCGA d’unemolécule d’ADN, et non la séquence TTGA. Il s’agitdonc d’une mutation par substitution. Un nucléotideà thymine est substitué à un nucléotide à cytosine.2. Au triplet CGA du brin non transcrit correspond lecodon CGA, qui code l’arginine, alors qu’au triplet
6
5
4
provoquant l’apparition prématurée d’un codon stop,UGA. Pour l’ensemble de ces documents, l’attention desélèves est attirée par la coloration rouge des lettres repré-sentant les nucléotides placés en position significative.
E x e r c i c e s
Évaluer ses connaissances
Une usine imaginairePlans de fabrication : ADN; coffre : chromosomes ;bureau directorial : noyau ; photocopies : ARN mes-sager ; atelier de fabrication : ribosomes.
Vrai ou faux ?Les affirmations exactes sont a. et d.b. Le code génétique est un système de correspon-dance entre une séquence de nucléotides de lamolécule d’ARN et une séquence d’acides aminésd’un polypeptide.c. Au cours de la transcription, un ARN messagercomplémentaire de l’un des brins d’ADN est synthé-tisé.e. La séquence de nucléotides d’une moléculed’ARN messager est identique à celle du brin nontranscrit de la molécule d’ADN, à l’exception del’uracile qui remplace la thymine.
Des explicat ions brèvesa. Le même acide aminé peut être codé par plu-sieurs codons. La substitution d’un nucléotide peutdonc entraîner, pour le gène muté, un codon qui,tout en étant différent de celui du gène non muté,code pourtant le même acide aminé. La mutation estalors sans effet sur la séquence polypeptidique.b. Tous les phénotypes dépendent des phénotypesmoléculaires, donc soit des protéines de structure,soit de substances synthétisées grâce à la présenced’enzymes, qui sont des protéines.c. Dire que le code génétique est dégénéré signifieque plusieurs codons peuvent coder le même acideaminé. Tous les êtres vivants possèdent le mêmecode génétique, qui est donc universel.d. La copie d’une molécule d’ADN en une moléculed’ARN est une transcription. La copie d’une molé-cule d’ARN en une molécule d’ADN constitue l’opé-ration inverse (transcription inverse).e. Une mutation par insertion ou par délétion d’unnucléotide entraîne un décalage du cadre de lecture,qui modifie tous les codons situés en aval de lamutation. Elle peut en outre entraîner l’apparitionprécoce d’un codon stop, et donc la synthèse d’unpolypeptide tronqué.
3
2
1
22

TGA correspond le codon UGA. Or, celui-ci est uncodon stop. La protéine synthétisée est donc tron-quée et non fonctionnelle : le sang de la personneporteuse de cette mutation ne peut donc jamaiscoaguler, ce qui caractérise une forme grave d’hé-mophilie.
Les phénotypes du syndrome deDown1. Plusieurs séquences de nucléotides peuvent êtreproposées. Un ARN messager possible est :GAA UUU CGA CAU GAU UCU GGUou encore, par exemple :GAG UUC CGU CAC GAC UCC GGCEn terme de nucléotides d’ADN, on obtient :GAA TTT CGA CAT GAT TCT GGT brin non transcrit
CTT AAA GCT GTA CTA AGA CCA brin transcrit
ou, dans le second cas :GAG TTC CGT CAC GAC TCC GGC brin non transcrit
CTC AAG GCA GTG CTG AGG CCG brin non transcrit
La pluralité des réponses possibles provient du faitque le code génétique est dégénéré, c’est-à-direque plusieurs codons, en terme d’ARN messager,ou plusieurs triplets différents, en terme d’ADN, peu-vent coder le même acide aminé.2. Au niveau du phénotype moléculaire, chaqueallèle d’un gène s’exprime. Étant situé sur le chro-mosome 21, le gène codant la protéine P se trouveprésent en trois exemplaires, aboutissant à une syn-thèse accrue de cette protéine.3. Si on admet que la superoxyde dismutase n’estcodée que par un seul gène, chacun de ses allèlesserait responsable de la synthèse d’environ 500 uni-tés de cette enzyme. Les individus atteints du syn-drome de Down auraient donc 3 allèles du gène,situé probablement sur le chromosome 21.
7
E x e r c i c e c o m p l é m e n t a i r e
Un phénotype de la mucovisci-doseLa mucoviscidose est la maladie génétique la plusfréquente en Europe. Les sécrétions bronchiques etpancréatiques des individus atteints sont anormale-ment épaisses, entraînant des difficultés respira-toires et une insuffisance pancréatique. La viscositéde ces sécrétions est liée à une modification de laforme d’une protéine membranaire, nommée CFTR.La séquence de nucléotides suivante représenteune portion du brin non transcrit de l’allèle normal :
… AAA GAA AAT ATC ATC TTT GGT GTT TCC TAT…
Dans 70 % des cas de mucoviscidose, cet allèleprésente, pour la même portion de gène, laséquence suivante :
… AAA GAA AAT ATC ATT GGT GTT TCC TAT…
Comparer les séquences polypeptidiques corres-pondant à cette portion de gène, pour les deuxallèles codant la protéine CFTR. Utiliser le tableaudu code génétique, page 59.Sachant que le début de la séquence proposée cor-respond à l’acide aminé de position 503, identifier lamutation, dite F 508, à l’origine de la mucoviscidose.
Réponses attenduesSéquence partielle d’acides aminés de la protéineCFTR fonctionnelle :lysine-acide glutamique-asparagine-isoleucine-isoleucine-phénylalanine-glycine-valine-sérine-tyrosineSéquence partielle de la protéine CFTR non fonc-tionnelle, codée par le gène muté :lysine-acide glutamique-asparagine-isoleucine-isoleucine-gly-cine-valine-sérine-tyrosine
Les deux triplets ATC et TTT du gène non muté sontpar délétion de TCT (ou CTT) réduit à un seul tripletATT.
Délétion
… AAA GAA AAT ATC ATC TTT GGT GTT TCC TAT…
Le couple d’acides aminés (507) isoleucine (AUC) et(508) phénylalanine (UUU) du polypeptide non mutéest remplacé par un seul acide aminé, l’isoleucine(AUU), dans le polypeptide muté. C’est la phénylala-nine en position 508 qui a disparu.
8
Chapitre 3 : La biosynthèse des protéines
23

24
la relation génotype-phénotype doit être plus complexeque celle envisagée jusqu’ici.
A c t i v i t é s 1
Dominance et récessivitéIl s’agit d’aboutir à la notion de dominance et de réces-sivité et de montrer qu’il peut exister un même phéno-type pour deux génotypes différents.
ExploitationLe caractère héréditaire de la drépanocytose est connu(chapitre 1).
◆ Problèmes scientifiques à résoudreToutes les cellules de la peau du Lapin nain ont le mêmegénotype et pourtant la couleur du pelage n’est pas uni-forme. Comment le même génotype peut-il se traduire pardes caractéristiques différentes? Il faut, pour répondre,envisager des facteurs génétiques complexes et/ou l’actiondu milieu, de l’environnement.
P o s e r l e s p r o b l è m e s
Il s’agit, en confrontant les DOCUMENTS 1 et 2, de sensibiliserà l’idée que ce qui a été vu jusqu’à présent (DOCUMENT 1)n’est pas suffisant pour expliquer la diversité de la couleurdes yeux dans l’espèce humaine (DOCUMENT 2) et donc que
Programme: notions et contenus• Un phénotype macroscopique donné résulte de processus biologiques gouvernés par l’expression de plusieurs gènes. La mutation de l’un seule-ment de ces gènes peut altérer ce phénotype.• Un même phénotype macroscopique peut donc correspondre à plusieurs génotypes.• Chez un individu donné, l’effet des allèles d’un gène va dépendre également de l’environnement.
Objectifs à atteindreUn premier objectif est de montrer que la relation génotype-phénotype est plus complexe que celle envisagée dans lechapitre 1, du fait de l’hétérozygotie possible et du fait que plusieurs gènes peuvent être directement impliqués dansla réalisation d’un phénotype (chaîne de biosynthèse par exemple).
Un deuxième objectif est d’envisager le polymorphisme d’un gène qui peut se traduire par un polymorphisme du phé-notype moléculaire et, dans une certaine mesure, ce polymorphisme moléculaire peut entraîner une variabilité phé-notypique au niveau macroscopique.
Un troisième objectif est de dégager l’idée que l’effet phénotypique d’un gène peut varier en fonction de son contextegénétique. L’élève doit savoir qu’un gène ne s’exprime qu’au sein d’une cellule et sous la forme du polypeptide dontil dirige la synthèse. Au sein de la cellule, ce polypeptide est confronté aux produits de l’expression d’autres gènes etla même chose est vraie à d’autres niveaux de l’organisation du vivant. Entre l’effet cellulaire d’un gène et sa tra-duction au niveau de l’organisme interviennent de nombreux phénomènes qui sont, eux-mêmes sous le contrôle demultiples gènes pour lesquels le polymorphisme existe également.
On doit donc aboutir ainsi à la notion de gènes majeurs et de gènes modulateurs.
Dernier objectif, enfin, montrer que la réalisation d’un phénotype dépend souvent de l’environnement qui peut agirsoit directement sur l’expression des gènes, soit en interaction avec les différents niveaux phénotypiques.
À la fin de ce chapitre, un élève doit avoir saisi que, en dehors de cas particuliers où le phénotype est à déterminismeuniquement génétique (groupes sanguins par exemple), tout phénotype est multifactoriel, résultant d’une interactionavec les différents niveaux phénotypiques.
Complexité du phénotype
C H A P I T R E 4

L’arbre généalogique (DOCUMENT 3) soulève le problème decomprendre comment des parents non drépanocytairespeuvent avoir un enfant atteint de drépanocytose.Le DOCUMENT 4 permet l’analyse des phénotypes molécu-laires des trois individus concernés et conduit à l’idée queles deux parents fabriquent les deux types d’hémoglobineHbA et HbS alors que l’enfant ne fabrique que l’hémoglo-bine HbS. À partir de cette conclusion, il est possible dedéduire les génotypes HbA/HbS pour les parents etHbS/HbS pour l’enfant. Cela permet donc, en réponse auproblème posé, d’introduire la notion de dominance auniveau de l’organisme et au niveau cellulaire. Cette domi-nance est nettement apparente au niveau cellulaire, parcontre, elle n’existe pas au niveau moléculaire puisque lesdeux allèles s’expriment.Remarque : en toute rigueur, on devrait dire que c’est lephénotype moléculaire HbA qui est dominant sur le phé-notype HbS dans la mesure où il empêche celui-ci de setraduire dans le phénotype cellulaire. La présence d’hé-moglobine HbA empêche l’hémoglobine HbS de polyméri-ser.Pour l’élève, la notion de dominance, pour être claire, doitêtre reliée au phénotype de l’organisme, macroscopique.Le DOCUMENT 5 montre les limites, au niveau du phénotypecellulaire, de la notion de dominance qui peut varier enfonction des conditions de l’environnement. Les hématiesd’un individu hétérozygote pouvant, dans un milieu trèspauvre en oxygène, se déformer. Dans ces conditions, eneffet, l’hémoglobine se trouve essentiellement sous laforme de désoxyhémoglobine et tend à polymériser. Celapermet d’expliquer que des individus hétérozygotes peu-vent éprouver des troubles lorsqu’ils se trouvent placerdans des conditions où la pression en dioxygène est plusbasse (en altitude par exemple).
A c t i v i t é s 2
Phénotype, polymorphisme, interac-tion entre gènesDeux objectifs sont poursuivis :
1. introduire l’idée qu’un gène peut exister au sein despopulations non pas uniquement sous la forme de deuxallèles mais sous la forme de plusieurs allèles et queces différences entre allèles peuvent se traduire non paspar deux phénotypes seulement mais par plusieurs phé-notypes différents ; autrement dit, un gène sous ses dif-férentes formes peut être à l’origine de plusieursphénotypes (allèles du gène ABO);
2. montrer qu’un même phénotype peut dépendre deplusieurs gènes (en considérant ici le cas simple de lachaîne de biosynthèse impliquée dans l’établissementdes groupes sanguins).
Exploitation� Le DOCUMENT 8 permet de comparer deux allèles B dugène ABO à un allèle A de référence. Les allèles B101 et B102
diffèrent de l’allèle A101 par 6 nucléotides ce qui se traduit
par 6 triplets différents. L’utilisation du code conduit à éta-blir que cela se traduit par 4 différences au niveau desacides aminés ; cela permet de rappeler aux élèves qu’unemutation génique ne se traduit pas obligatoirement auniveau du polypeptide. Par référence au DOCUMENT 6, onvoit que deux des acides aminés différents (266 et 268)font partie du site actif de l’enzyme ; l’utilisation desconnaissances sur le site actif des enzymes (chapitre 2)permet à l’élève d’émettre l’hypothèse que ces différencessont à l’origine de l’utilisation de substrats différents parles enzymes de type A (N-acétyl-galactosamine) et B(galactose).Les allèles de type O sont caractérisés par la délétion dunucléotide G en position 261 ; l’élève doit exploiter laséquence de nucléotides de l’allèle A101 (le nucléotide Gétant repéré) pour en établir les conséquences : la délétion,en décalant le cadre de lecture, transforme le triplet 118 enun triplet (TAA) à l’origine d’un codon stop (UAA). Lesenzymes codées par l’allèle O sont donc constituées par117 acides aminés (y compris la méthionine initiatrice) aulieu de 354 pour les enzymes de type A ou B et donc deuxacides aminés du site actif (266 et 268) n’existent pas : uneenzyme de type O est inactive et ne peut catalyser la fixa-tion d’une substance sur la substance H.L’exploitation des informations relatives aux allèles A(A101, A102 et A103) permet de montrer qu’un changement denucléotide entraînant un changement d’acide aminé (pro-line de A101, en position 156, remplacée par la leucine chezA102 et A103) est sans conséquence sur le caractère fonction-nel de l’enzyme et que, en raison du caractère dégénéré ducode génétique, le remplacement en position 188 d’un tri-plet CGC de A101 par un triplet CGT chez A103 est sansconséquence sur la séquence des acides aminés (mise enplace de l’arginine dans les deux cas).
� La deuxième question demande le réinvestissement desnotions de dominance et de récessivité pour établir lesgénotypes des individus testés dans le DOCUMENT 7 :- l’individu 1 est de phénotype A et de génotype A/A ouA/O;- l’individu 2 est de phénotype B et de génotype B/B ouB/O;- l’individu 3 a pour phénotype AB et pour génotype A/B ;- l’individu 4 a pour phénotype O et pour génotype O/O.
� Le DOCUMENT 9 peut être traduit sous la forme d’un arbregénéalogique. L’élève doit trouver une contradiction entrele génotype de groupe O (O/O) établi en utilisant le docu-ment 7 et les données proposées. En utilisant ses connais-sances de la classe de troisième, il doit dire qu’un individune peut transmettre qu’un allèle de chaque gène ; le père degroupe O transmet donc à son enfant l’allèle A101 et l’allèleh et la mère également de groupe O, l’allèle O101 et l’allèleH ; l’enfant aura donc pour génotype H/h, A101/O101.Par suite de la dominance de H sur h et de A sur O, l’en-fant fabriquera une enzyme fonctionnelle H et fabriquerala substance H en fixant le fucose sur le précurseur et,grâce à son enzyme A fonctionnelle, fixera la N-acétyl-galactosamine sur la substance H : il sera de groupe A.
Chapitre 4 : Complexité du phénotype
25

ont des phénotypes différents au niveau de l’organisme,l’un (A) présentant un retard mental accusé par rapport àson frère (B).Les élèves doivent alors utiliser leurs connaissances (cha-pitre 1) sur le fait que le phénotype macroscopique estessentiellement dû à la toxicité de la phénylalanine vis-à-vis des cellules nerveuses et donc être sensibilisés à l’im-portance de la concentration de la phénylalanine dans lecerveau. Cela permet de faire l’hypothèse que les deuxfrères diffèrent par la valeur de cette concentration céré-brale.Le DOCUMENT 13 montre, en effet, que des individus ayantla même concentration plasmatique de phénylalanine peu-vent avoir des concentrations cérébrales très différentes ;pour certains, la valeur de cette concentration est voisinede celle d’individus de génotype non muté pour PAH.La conséquence, au niveau de l’organisme, d’une concen-tration plasmatique élevée de phénylalanine, semble doncdépendre du passage de cet acide aminé du plasma dans lecerveau, à travers la paroi des vaisseaux de la barrièrehémato-encéphalique.Ce passage dépend de protéines membranaires ; on peutsupposer que les gènes codant pour ces protéines mem-branaires possèdent un polymorphisme tel que le passagede la phénylalanine dans le cerveau est variable. Ces gènesne sont pas actuellement connus.Là encore, au niveau de l’organisme, les conséquences dela présence d’allèles morbides du gène PAH (majeur)dépendent de la présence d’allèles d’autres gènes (modu-lateurs) sans rapport fonctionnel apparent avec le gènePAH.Ainsi la notion de phénotype non monofactoriel est à nou-veau précisée.
A c t i v i t é s 4
Génotype, environnement et phéno-typeL’objectif est de montrer que le phénotype d’un orga-nisme ne dépend pas que de son génotype mais, le plussouvent, de l’interaction entre son génotype et l’envi-ronnement.
La question peut être envisagée de deux façons :
- un même génotype entraîne un phénotype variable enfonction de facteurs de l’environnement (document 14,15 et 16) ;
- un phénotype causé par l’environnement (par exempleune maladie d’origine virale) peut être différent selon legénotype que l’on possède (document 17).
ExploitationLe DOCUMENT 14 permet de montrer qu’un même génotypePAH normalement à l’origine de la phénylcétonurie peutse traduire par des phénotypes différents selon l’apport ali-mentaire. Cela sensibilise nettement au fait que, dans unenvironnement différent, le même génotype peut se tra-
A c t i v i t é s 3
Phénotype, gène majeur et gènes modulateursLe but est de montrer que les conséquences au niveauphénotypique de l’expression des allèles d’un gènepeuvent être différentes en fonction de l’expression desallèles d’autres gènes dont les produits ne sont pourtantpas impliqués dans une chaîne de biosynthèse com-mune (ces gènes dits modulateurs n’interviennent pasdirectement dans la réalisation d’un phénotype qui estdéterminé fondamentalement par le premier, le gènemajeur).
ExploitationLe premier élément à repérer grâce au document 11a etqu’il n’existe qu’un seul allèle à l’origine de l’hémoglo-bine S ; en conséquence, tous les individus drépanocy-taires ont le même génotype HbS/HbS. Pourtant,phénotypiquement, ils peuvent être différents puisque ran-gés en deux groupes, les uns à phénotype drépanocytairegrave, les autres à phénotype drépanocytaire bénin (DOCU-MENT 11).Les DOCUMENTS 11a et 11b permettent de rechercher uneexplication à cette différence phénotypique. Le document11 a permet de déterminer les hémoglobines des individusdrépanocytaires des deux groupes : le groupe 1 à drépano-cytose grave ne fabrique que de l’hémoglobine S, alorsque le groupe 2 à drépanocytose moins grave fabriquedeux types d’hémoglobine : HbS et HbF.Le DOCUMENT 11b montre qu’il y a une corrélation entre lenombre d’hématies qui contiennent un mélange d’hémo-globine F et d’hémoglobine S et la gravité de la maladie.Les individus du groupe 2 possèdent un nombre de glo-bules rouges contenant de l’hémoglobine F beaucoup plusimportant que celui des individus du groupe 1. C’est doncla présence fréquente de l’hémoglobine F dans les hématiesqui est responsable de la forme bénigne de la maladie; leshématies prenant le faciès falciforme étant beaucoup moinsnombreuses chez les individus du groupe 2.Remarque : il est possible d’aller un peu plus loin dansl’explication ; en effet, les molécules d’hémoglobine F,comme les molécules d’hémoglobine A empêchent lesmolécules d’hémoglobine S de polymériser.En conclusion, on voit donc que les conséquences, de laprésence d’hémoglobine S due à l’expression des allèlesd’un gène sont variables en fonction de la présence ou nond’hémoglobine fœtale, résultat de l’expression des allèlesd’un autre gène, totalement indépendant du premier. Dansle cas de la drépanocytose, le gène �, impliqué dans lasynthèse de l’hémoglobine de l’adulte (DOCUMENT 10), estun gène majeur dont les variants peuvent être à l’originedu phénotype drépanocytaire mais les effets de ce gènemajeur peuvent être modulés par l’expression des allèlesdu gène γ, gène modulateur, impliqués dans la synthèse del’hémoglobine fœtale (DOCUMENT 10).La lecture du texte (DOCUMENT 12) permet d’établir que lesfrères A et B ont le même phénotype biochimique, ce quilaisse à penser qu’ils ont le même génotype, et pourtant ils
26

duire par un phénotype différent. L’environnement agis-sant ici sur le phénotype biochimique.
Le DOCUMENT 15 permet de retrouver le problème scienti-fique posé par la photographie d’introduction à ce chapitre4 et donc de rechercher une explication aux différences decoloration du pelage du Lapin hymalayen et à leur locali-sation.
Il est nécessaire de faire un rappel de connaissances (exer-cice corrigé pages 25-26 et DOCUMENT 15b.) concernant ledéterminisme génétique de la pigmentation de la peaudonc la genèse de la mélanine qui dépend d’une chaîne debiosynthèse dont l’enzyme fondamentale est codée par ungène, celui de la tyrosinase, dont certains allèles sont nonfonctionnels.
Le DOCUMENT 15 a. indique que la production de mélaninechez le Lapin dépend de la température.
Le DOCUMENT 15 b. indique que les individus concernés ontun génotype présentant un gène muté codant pour la tyro-sinase ; dans la tyrosinase mutée, l’arginine en position442 est remplacée par la glutamine.
On a établi que chez le Lapin la fabrication de la mélaninedépendait de la température ; pour interpréter cela, on uti-lise des données chez l’Homme où existent des individusqui présentent un phénotype comparable à celui de ceslapins car ne produisant de la mélanine que dans lesrégions froides du corps. Ces individus possèdent tous unmême génotype caractérisé par un allèle qui code pour unetyrosinase dont l’activité dépend de la température (DOCU-MENT 15 c.). On voit donc ici qu’un facteur de l’environne-ment (la température) agit sur le phénotype moléculaire (latyrosinase) produit de l’expression directe d’un gène.
Le DOCUMENT 16 permet de présenter un exemple où l’envi-ronnement agit directement sur l’activité de gènes. Eneffet, l’entraînement se traduit par une augmentation desmyofibrilles c’est-à-dire par le nombre de molécules d’ac-tine et de myosine, protéines résultant de l’expression degènes : l’entraînement stimule bien, directement, l’activitédes gènes impliqués dans la synthèse de l’actine et de lamyosine.
Le DOCUMENT 17 montre un dernier aspect de l’importancede l’environnement sur l’établissement d’un phénotype.Les renseignements apportés permettent d’établir que levirus du SIDA, pour pénétrer, doit utiliser des récepteursmembranaires CCR5 et les individus dont le génotype neleur permet pas de synthétiser cette protéine membranairene seront jamais atteints. Toute personne possédant desrécepteurs CCR5 peut être atteinte du SIDA si elle estinfectée par le V.I.H. et il n’a jamais été observé d’indivi-dus atteints de la maladie et ne possédant pas de récepteursCCR5.
C’est donc bien du génotype (présence d’allèles permet-tant ou non la synthèse de certaines protéines membra-naires) que dépendent les effets de l’environnement(présence du virus) sur l’établissement du phénotypemacroscopique (maladie ou non).
E x e r c i c e s
Évaluer ses connaissances
Vrai ou fauxb., d., e. et g. sont des affirmations exactes.a. Un gène code pour un polypeptide est n’est doncresponsable que du phénotype moléculaire.c. Le polyallélisme est un fait général.f. Le polymorphisme ne concerne pas tous lesgènes (environ 30 % chez l’Homme).
Phrases à corrigera., b., c., d. et f. sont des phrases exactes.e. Si A domine B, alors l’hétérozygote AB aura unphénotype macroscopique identique à celui d’unindividu de génotype AA.
Phrases à corrigerb., c., e. et g. sont des phrases exactes.a. Il n’existe pas de gène d’une maladie mais ungène qui code pour un polypeptide donné et dontcertains allèles peuvent être à l’origine de symp-tômes morbides.d. Le plus souvent, deux ou plusieurs gènes inter-viennent dans la réalisation d’un phénotype cellu-laire, biochimique ou macroscopique.e. L’exemple du SIDA (page 73) montre que le phé-notype résultant d’un facteur de l’environnementpeut dépendre du génotype.
Exercice corrigé
La phénylcétonurieVoir correction dans le manuel pages 80 et 81.
Évaluer ses compétences
La mucoviscidoseIl s’agit de repérer la nature de la mutation au niveaude l’ADN puis d’utiliser le code génétique.Mutation DF508 : il s’agit d’une délétion de troisnucléotides affectant les triplets 507 et 508 (ATCTTT) qui conduit à un triplet ATT en position 507. Laconséquence pour le polypeptide CFTR est la dis-parition de la phénylalanine en position 508 ; lecouple isoleucine (AUC) et phénylalanine (UUU)étant remplacé par l’isoleucine seule (AUU). La délé-tion peut être localisée de deux façons : soitATCTTT, soit ATCTTT.Mutation R334W: il s’agit d’une substitution, le pre-mier nucléotide à guanine du triplet 334 (CGG) étantremplacé par un nucléotide à thymine (TGG) ; enconséquence, le 334e acide aminé de la CFTR nonmutée, l’arginine, est remplacé par le tryptophanedans la protéine mutée.
5
4
3
2
1
Chapitre 4 : Complexité du phénotype
27

ponsable de la maladie ; l’injection de BH4 rétablis-sant une concentration normale de phénylalanine,on peut conclure que le gène PAH est représentéchez lui par des allèles codant pour une enzymefonctionnelle.On met ainsi en évidence le fait qu’un même phéno-type au niveau de l’organisme peut être l’expressionfinale de deux génotypes totalement différents :PAH –/PAH–, DHPR+/ DHPR+ pour le premier (courbebleue) et PAH+/ PAH+, DHPR–/DHPR– pour le second (courberouge).
L’enzyme G6PDEn Afrique, trois allèles du gène G6PD (G6PDA-,G6PDA et G6PDB) étant représentés avec une fré-quence supérieure à 1 % (de 15 à 65 %) le gène estpolymorphe ; il en est de même dans le bassin médi-terranéen où deux allèles (G6PDB et G6PDM) ontune fréquence supérieure ou égale à 1 %.Par contre, en Europe, seul l’allèle G6PDB étantreprésenté à plus de 1 % (99,7 %) le gène n’est paspolymorphe.Le phénotype macroscopique est normal quel quesoit le génotype si on fait abstraction de l’influencede l’environnement. L’intervention de facteursexternes comme la prise de médicaments antipalu-déens ou l’ingestion de certains aliments fait appa-raître le phénotype malade. Le phénotype est doncbien, dans ce cas, la résultante de l’interaction dugénotype et de l’environnement.
Drépanocytose et paludismeOn constate que l’allèle HbS a une fréquence élevéedans les régions où sévit le paludisme (documentsa. et b.). Le paludisme est dû à un protozoaire qui sedéveloppe dans les hématies. Le document c.montre que les hétérozygotes, HbA/HbS, vivantdans les régions paludéennes sont très peu touchéspar le paludisme (1 décès pour 20,6 attendus) aucontraire des autres individus (HbA/HbA) dont lamortalité est environ 100 fois plus élevée. La pré-sence d’hémoglobine HbS dans les hématiesempêche l’apparition du phénotype paludéen. C’estdonc, ici, du génotype (présence ou non de l’allèleHbS) que dépendent les effets de l’environnement(pénétration de l’Hématozoaire) sur l’établissementdu phénotype (non paludéen ou paludéen).On retrouve ici des conclusions comparables àcelles tirées de l’exemple du SIDA, dans les activités4 page 73.
8
7
Les six enfants suivis ont tous le même génotypeR334W/DF508 et possèdent donc le même phéno-type moléculaire.Au niveau de l’organisme :- le phénotype pulmonaire est le même dans lesdeux familles ;- le phénotype pancréatique est différent ; dans lafamille A, aucun enfant n’est atteint alors que dansla famille B, ils sont tous les trois touchés ;- les premiers symptômes de la maladie se manifes-tent à des âges très divers, entre la naissance et11ans.Le phénotype moléculaire semble donc se traduireinévitablement au niveau des poumons ; on peutpenser qu’il n’existe aucune interaction avecd’autres gènes ou l’environnement. Cependant,l’âge variable d’apparition des premiers symptômeslaisse à penser que certains facteurs génétiquesindividuels peuvent retarder l’apparition de la mala-die ; la variabilité au sein de la même famille (donc àconditions de vie identiques) amenant à privilégierl’hypothèse génique au détriment de l’hypothèseenvironnementale.Le phénotype clinique au niveau pancréatique diffé-rent dans les deux familles peut s’expliquer en ima-ginant la présence d’allèles de certains gènes(inconnus), empêchant l’établissement du phéno-type mucoviscidose dans la famille A (hérédité fami-liale) en interagissant sur les produits de l’expressiondu gène CFTR ou sur le phénotype cellulaire lié à laprésence de CFTR mutée. Les trois enfants de lafamille B ne possédant pas ces allèles sont atteintsau niveau pancréatique.
PAH et DHPRChez les individus phénylcétonuriques, l’injection dephénylalanine se traduit par une forte hausse de laconcentration plasmatique de cet acide aminé qui,déjà élevée avant l’expérience (x20 par rapport à unindividu non atteint), est multipliée par 4.L’injection de BH4 :- est sans effet chez l’individu courbe bleue ; sonphénotype phénylcétonurique n’est pas lié à unmanque de cette substance et son phénotype cli-nique est donc dû à une enzyme PAH déficiente.Rien ne permet cependant de conclure sur sa capa-cité à fabriquer BH4 ;- entraîne une baisse immédiate et rapide de laconcentration en phénylalanine chez l’individucourbe rouge ; sa phénylcétonurie est donc due àl’absence de BH4 ; dans son cas, c’est une défi-cience de l’activité de l’enzyme DHPR qui est res-
6
28

Pa
rt
ie
1 Des phénotypes à différents niveaux d’organisation du vivant
Génotype, phénotype et régulation
5 Régulation de la glycémie ................................ 30
6 Génotype, environnement et phénotypediabétique ............................................................................ 35
Unité 2

30
◆ Problèmes scientifiques à résoudreLe document de la page d’entrée évoque des organes inter-venant dans la régulation de la glycémie. Quel est leur rôleau sein du système de régulation ?
P o s e r l e s p r o b l è m e s
Il s’agit de mettre en relation des phénotypes au niveau del’organisme (DOCUMENT 1) : hyperglycémique, hypoglycé-mique, euglycémique (= ni hyper ni hypoglycémique),avec des phénotypes biochimiques : concentrations plas-matiques de glucose. On peut alors envisager comment lephénotype biochimique détermine le phénotype macro-scopique ; par exemple, si le taux de glucose sanguintombe au-dessous de 0,7 g.L-1 les cellules nerveuses nesont pas convenablement approvisionnées en glucose d’oùles symptômes visibles de l’hypoglycémie (phénotypes’établissant à court terme) ; inversement, une concentra-tion supérieure à 1,26 g.L-1 entraîne à plus ou moins longterme des anomalies au niveau des vaisseaux sanguins(DOCUMENTS 2 et 3), le glucose apparaissant ainsi commetoxique à forte concentration.
Quels sont les mécanismes qui sous-tendent ce phénotypebiochimique (concentration de glucose sanguin) dontdépend tout le reste ?
A c t i v i t é s 1
Réalité d’une régulation de la glycé-mieL’objectif est de mettre en évidence l’existence d’un sys-tème de régulation ce qui permet de déboucher surl’hypothèse de l’existence d’organes stockeurs de glu-cose et d’organes producteurs de glucose. Cela revientà démontrer que le phénotype biochimique dépend d’unsystème de régulation.
ExploitationDans le DOCUMENT 4 l’élève doit repérer qu’à la suite d’unrepas, la glycémie augmente et qu’elle baisse après unexercice physique mais que ses fluctuations restent relati-vement faibles (de 0,8 à 1,4 pour le sujet A par exemple).Intuitivement, la faiblesse de l’amplitude des fluctuations
Programme : notions et contenus• Malgré des variations importantes (prise alimentaire discontinue, consommation énergétique variable), la glycémie (grandeur réglée de l’homéo-stat) oscille en permanence autour d’une valeur physiologique voisine de 1 g.L-1 (grandeur consigne).• Cette homéostasie glycémique nécessite une gestion des réserves de l’organisme.• Les cellules � et � du pancréas endocrine sont des capteurs de la glycémie. En fonction des variations de la glycémie, elles émettent des mes-sagers chimiques, les hormones glucagon et insuline.• Le message hormonal est codé par la concentration plasmatique de l’hormone.• Les cellules cibles expriment les récepteurs spécifiques à ces hormones.• Sous l’action de l’insuline, le glucose est stocké sous forme de glycogène dans le foie et les cellules musculaires squelettiques, ainsi que sousforme de triglycérides dans le foie et les adipocytes.• Sous l’action du glucagon, le glucose est libéré par le foie dans le plasma.• Cellules pancréatiques (� et �), hormones (glucagon et insuline) et cellules cibles constituent le système réglant de l’homéostat glycémique.
Objectifs à atteindreArriver à établir que le phénotype biochimique «concentration plasmatique de glucose» résulte du fonctionnementd’un système de régulation qui fait intervenir des phénotypes cellulaires multiples : capteurs pancréatiques de la gly-cémie, sécréteurs d’insuline et de glucagon et cellules effectrices répondant aux messages hormonaux pancréatiques :cellules hépatiques, musculaires et adipeuses.
Régulation de la glycémie
C H A P I T R E 5

Chapitre 5: Régulation de la glycémie
31
L’hépatotectomie (DOCUMENT 14) s’accompagnant d’unehypoglycémie rapide, semble indiquer que l’utilisation duglycogène hépatique est publique : le foie livre du glucoseau milieu intérieur. Le glycogène musculaire, quantitative-ment le plus important (DOCUMENT 8) ne semble pas pouvoirêtre utilisé pour enrichir le milieu intérieur en glucose.Le DOCUMENT 15, expérience du foie lavé montrant que cedernier est capable de fournir du glucose à un liquide deperfusion alors qu’il n’en contenait plus confirme le rôledu foie dans la production de glucose.Le DOCUMENT 16 permet de démontrer que les cellules adi-peuses sont capables de synthétiser des triglycérides à par-tir du glucose.
A c t i v i t é s 3Cellules hépatiques et glycémieL’objectif principal est de démontrer que le foie est leseul producteur de glucose et cela par deux processus :glycogénolyse et néoglucogenèse.
Un autre objectif est de montrer qu’à jeun, le foie livreau milieu intérieur, donc à l’ensemble de l’organisme,une dizaine de grammes de glucose par heure ; ce glu-cose est à 75 % utilisé par les cellules glucodépen-dantes et le reste par les autres cellules notammentmusculaires.
Lors d’un apport alimentaire, tous les organes prélè-vent du glucose (la consommation des cellules gluco-dépendantes restant la même qu’à jeun), foie, muscleset tissu adipeux stockant le glucose.
Ces systèmes effecteurs sont à l’origine du maintien dela glycémie aux alentours de 1 g.L-1. Les phénotypescellulaires impliqués dans la régulation dépendent dephénotypes moléculaires et, par là même, des gènes quicodent pour les enzymes qui interviennent dans les dif-férentes réactions de stockage et de production.
ExploitationLe DOCUMENT 17 est relatif à des expériences effectuéeschez le Chien ; le protocole expérimental complexe n’estpas rappelé ici pour ne pas compliquer et notamment iln’est pas fait allusion au montage permettant, en particu-lier, de relier la veine porte directement à la veine caveinférieure donc de court-circuiter le foie. Le sang arriveainsi uniquement par l’artère hépatique et en sort par lesveines sus-hépatiques. Le glucose étant injecté dans lesystème circulatoire, il est non ingéré.Ce document montre que, à jeun, la glycémie du sang veineux quittant le foie est nettement supérieure à celle du sang artériel qui y arrive : le foie livre du glucose (DOCUMENT 17a.).En revanche (DOCUMENT 17b.), la glycémie du sang veineuxdu muscle est légèrement inférieure à celle du sang arté-riel : le muscle, à jeun, consomme un peu de glucose maissurtout il n’en produit pas.
laisse à penser qu’il existe des mécanismes qui empêchentdes fluctuations importantes qui, en particulier, n’attei-gnent jamais les valeurs caractéristiques des phénotypeshyper ou hypoglycémique (DOCUMENT 1).Constance glycémique ne signifie donc pas valeur fixemais fluctuations relativement faibles autour d’une valeurde l’ordre de 1 g.L-1.Ce document permet en outre de mettre en évidence l’ex-trême variabilité entre des individus de même phénotype(euglycémique) qui présentent des glycémies à jeun diffé-rentes et des fluctuations d’amplitude plus ou moinsimportante (de 22,1 à 59,7)Le DOCUMENT 5 permet d’approcher quantitativementl’existence d’un système de régulation et de postuler celled’organes stockeurs de glucose. En effet, pendant les troisheures, la consommation de glucose (DOCUMENT 7) est de45 g (3 � 15) ; donc 30 g environ, sur les 75 g absorbés,sont restés dans les 15 litres de milieu intérieur ; dans ces conditions, la glycémie aurait dû augmenter de30/15 = 2 g.L-1, c’est-à-dire atteindre des valeurs de l’ordrede 3. Or elle atteint au maximum 1,4 g.L-1. On est doncconduit à supposer que le surplus a été soutiré, sans êtremétabolisé, et a été stocké.Le DOCUMENT 6 montre que la glycémie reste constantealors que la consommation de glucose est de 10 g.h-1 ; or,la quantité totale de glucose disponible, à un momentdonné, dans le milieu intérieur est d’environ 15 g ; la gly-cémie devrait donc être nulle au bout de 1 h 30 environ.Cela permet d’envisager l’existence probable d’organescapables de produire du glucose et de le livrer au milieuintérieur.
A c t i v i t é s 2
Les organes effecteurs, stockeurset/ou producteurs de glucoseDémontrer que les muscles et le foie stockent le glucosesous forme de glycogène et que le tissu adipeux stockele glucose sous forme de triglycérides.
Approcher le rôle du foie en tant que producteur de glu-cose (sans le foie, il y a hypoglycémie).
ExploitationLes cellules musculaires (DOCUMENT 10 a.), hépatiques(DOCUMENT 11) renferment du glycogène et la molécule deglycogène est formée d’un grand nombre de molécules deglucose (DOCUMENT 12).Le DOCUMENT 9 permet de montrer que la synthèse de glycogène dépend de l’apport glucidique alimentaire (le 0correspond au début des observations).Au cours d’un exercice, la teneur en glycogène diminue.Pour la cellule musculaire, le glycogène est une forme de réserve puisque stocké et utilisé ce que confirme leDOCUMENT 10 b.Le DOCUMENT 13 montre que, durant le jeune, la teneur englycogène du foie diminue ce qui confirme son rôle deréserve et de producteur.

L’élève doit repérer la méthode utilisée pour la mise enévidence du rôle hormonal d’un organe : ablation (doc.22a.), greffe (doc. 22b.) et injection d’extraits (doc. 26) etconfirmer ainsi l’idée que le pancréas agit sur la glycémiepar l’intermédiaire d’un messager chimique. Seuls lesextraits pancréatiques étant capables d’abaisser la glycé-mie, les autres extraits étant sans effet.Le DOCUMENT 24, associé au DOCUMENT 25, permet d’établirque les îlots de Langerhans suffisent à eux seuls pourmaintenir une glycémie normale et que leur destructionentraîne une hyperglycémie. Les cellules sécrétant lesmessagers hormonaux sont donc celles des îlots deLangerhans.Remarque : l’originalité de Banting et Best est d’avoirobtenu les extraits actifs à partir de pancréas dont la par-tie acineuse avait été détruite après ligature du canal pan-créatique. Contrairement aux autres expérimentateurs quitravaillaient avec des extraits de pancréas complet, leurméthode leur permettait d’obtenir des extraits dont leshormones, de nature protéique, n’étaient pas hydrolyséespar les enzymes du pancréas exocrine.
A c t i v i t é s 5
Hormones pancréatiques : action sur les cellules ciblesLes objectifs visés par ces activités sont relatifs à la foisau rôle joué par l’insuline et le glucagon dans la régu-lation de la glycémie et à une généralisation de lanotion d’hormone. Il s’agit :
– d’arriver à la conclusion que l’insuline a une actionhypoglycémiante en inhibant la production de glucosepar le foie et en stimulant son utilisation (stockage) parles muscles, le foie et le tissu adipeux ;
– de montrer que l’insuline agit au niveau de cellulescibles qui possèdent des récepteurs spécifiques ;
– de sensibiliser l’élève à l’idée que la liaison récep-teur-insuline au niveau des cellules musculaires ethépatiques entraîne une modification du profil enzyma-tique de ces cellules qui se traduit par l’activation del’enzyme impliquée dans la synthèse du glycogène etl’inactivation de celle qui est impliquée dans sa dégra-dation. Les critères caractérisant un messager hormo-nal peuvent donc être établis ;
– de montrer que le glucagon a un effet hyperglycé-miant en activant les enzymes hépatiques impliquésdans la glycogénolyse et la néoglucogenèse.
ExploitationLe DOCUMENT 27 montre que l’injection d’insuline provoqueune hypoglycémie (DOCUMENT 27a.) et les DOCUMENTS 27b. et27 c. permettent d’expliquer cette baisse de la glycémie ;on voit, en effet, sur ce DOCUMENT 27c., que l’insuline,même à faible concentration, bloque la production de glu-cose qui devient nulle pour une insulinémie égale à 50mU.mL–1 environ, valeur rapidement atteinte lors de l’in-
Remarque : il peut être intéressant de compléter cette acti-vité en exploitant les graphes relatifs à d’autres organesde l’exercice 2 page 108 permettant de montrer que, mis àpart le foie, aucun autre organe n’est producteur de glu-cose. En réalité, il existe un autre organe producteur deglucose, le rein dont le rôle ne devient important que lorsde jeunes prolongés.Après injection de glucose, la glycémie s’élève et onconstate, au bout de quelque temps, que foie et muscle ontle même profil métabolique qui correspond à un stockagedu glucose (DOCUMENTS 17a. et 17b.).Le DOCUMENT 18 permet de mettre en évidence la néoglu-cogenèse : 8 heures après un repas, il n’y a plus d’entréede glucose exogène, l’organisme est à jeun. L’utilisationdu glucose (2e colonne) permet de satisfaire les besoinsénergétiques de l’organisme (il n’est évidemment pasquestion de stockage). La glycémie à jeun reste constantecar la production de glucose par le foie (4e colonne) et sonutilisation sont égales.Au bout de 24 heures (et même avant), le foie continue àproduire 10 grammes de glucose à l’heure alors que sesréserves sont quasiment épuisées. Cela implique l’exis-tence d’un autre processus que la glycogénolyse pour pro-duire du glucose : la néoglucogenèse.En réalité, dès le début de la période de jeune (environ4 heures après un repas), cette néoglucogenèse assure25 % de la production contre 75 % pour la glycogénolyse.Les DOCUMENTS 19 et 20 permettent à l’élève de réfléchiraux phénotypes cellulaires hépatique et musculaire liésaux phénotypes moléculaires constitués par les enzymesimpliqués dans la glycogénolyse, la néoglucogenèse et lasynthèse du glycogène. Ils indiquent, qu’au sein des cel-lules musculaires et hépatiques, les enzymes peuvent pas-ser d’une forme active à une forme inactive et vice-versa.Le DOCUMENT 21 insiste sur l’originalité du phénotype cel-lulaire hépatique liée à un phénotype moléculaire particu-lier, la présence d’une enzyme active qui n’existe pas dansles autres cellules de l’organisme, la glucose-6-phospha-tase.
A c t i v i t é s 4
Pancréas et glycémieL’objectif est de sensibiliser l’élève à l’idée que la miseen jeux des organes effecteurs est sous la dépendancede sécrétions internes du pancréas. Il s’agit donc éga-lement de dégager les notions de glande endocrine etd’hormone.
ExploitationL’exploitation du DOCUMENT 22 peut laisser à penser que lepancréas est un organe stockeur de glucose : son ablationentraîne en effet une hyperglycémie. Or, le DOCUMENT 26montre que de simples extraits de pancréas font baisser laglycémie alors qu’il n’y a plus de pancréas (animal pan-créatectomisé totalement). En conséquence, le pancréasn’agit pas directement sur la glycémie, il n’est pas unorgane effecteur de la glycémie.
32

jection d’insuline (doc. 27b.). En même temps (doc. 27c.),cette augmentation de l’insulinémie entraîne celle de l’uti-lisation du glucose. La baisse de la glycémie est donc dueau blocage de sa production par le foie et à l’augmentationde son prélèvement et donc de son stockage par le foie, lesmuscles et le tissu adipeux.Le retour à une glycémie normale (DOCUMENT 27a.) au boutd’une heure et demi environ peut-être expliqué en partiepar la disparition de l’insuline injectée (DOCUMENT 27b.) ;cela indique que l’insuline est rapidement dégradée parl’organisme ce qui peut permettre d’introduire la notion dedemi-vie d’une hormone (de l’ordre de 5 minutes pourl’insuline et le glucagon).Le DOCUMENT 28 est relatif à un cas très rare de diabètecaractérisé par une glycémie très élevée à jeun (de l’ordrede 200 mg.dL-1). Dans ce cas, l’injection d’insulinen’abaisse pas la glycémie, alors que la même injection àun individu non atteint provoque une hypoglycémie.L’injection d’insuline est donc sans effet sur les cellulescibles effectrices ; une explication possible est que ces cel-lules cibles ne possèdent pas de récepteurs à l’insuline.En réunissant les conclusions tirées de l’exploitation desDOCUMENTS 27, 28 et 29, on peut établir les différentesétapes de l’action de l’insuline : sécrétion par les cellules� des îlots de langerhans → fixation aux récepteurs descellules cibles hépatiques, musculaires et adipeuses →modification du profil enzymatique de ces cellules cibles→ synthèse de glycogène (le glucose est soutiré du milieuintérieur).Le DOCUMENT 30 permet le même travail à propos du gluca-gon : sécrétion par les cellules � → fixation aux récepteursdes cellules hépatiques → modification du profil enzyma-tique de ces cellules (activation de la glycogène phospho-rylase) → dégradation du glycogène (du glucose est libérédans le milieu intérieur).
A c t i v i t é s 6
Le système de régulation de la glycé-mieIl s’agit :
– d’établir que les cellules � et � du pancréas sont descapteurs de la glycémie ;– d’établir la notion de message hormonal ;– de rassembler toutes les informations vues jusqu’icipour arriver à réunir les différentes composantes dusystème de régulation et ainsi de faire le bilan de tousles phénotypes cellulaires impliqués dans l’établisse-ment du phénotype biochimique (glycémie) et, par làmême, de réfléchir à la pluralité des gènes à l’originedes divers phénotypes moléculaires (hormones, récep-teurs, enzymes), tous de nature protéique.
ExploitationL’exploitation du DOCUMENT 31, doit permettre à l’élèved’établir que les cellules � et � sont des capteurs de la gly-cémie. Le pancréas étant isolé, la seule chose qui varie est
la teneur en glucose du liquide de perfusion. Les résultatsindiquent que la sécrétion d’insuline et celle de glucagonvarient en sens inverse suivant la concentration en glucosedu liquide de perfusion. Les cellules pancréatiques réagis-sent bien aux variations de la glycémie.L’analyse du DOCUMENT 32 permet d’établir que :– même à jeun, il y a une sécrétion insulinique de base del’ordre de 10 mU environ ;– à la suite d’un repas, l’insulinémie augmente durant 40minutes pour atteindre une valeur de 140 mU ;– ce n’est donc pas seulement la présence d’insuline quiintervient mais sa concentration : le message hormonal estcodé en amplitude, en quantité d’hormone. La mêmechose peut être établie pour le glucagon ;– les cellules effectrices, à jeun, sont soumises à un rap-port insuline/glucagon faible et à la suite d’un repas, cerapport augmente. Le véritable message hormonal agis-sant sur les cellules effectrices est le rapport concentrationd’insuline/concentration de glucagon.Le DOCUMENT 33 permet à l’élève de faire fonctionner lesystème de régulation lors d’un apport exogène importantde glucose : 1 : augmentation ; 2 : diminution ; 3 et 4 : aug-mentation ; 5 : diminution ; 7, 8, 9, et 10 : augmentation ;11 : diminution (retour à une glycémie normale) ; 12 : sanschangement (entrée constante de glucose).Le DOCUMENT 34 permet d’étudier le fonctionnement dusystème de régulation dans des conditions particulières :un injection intraveineuse d’insuline entraîne une hypo-glycémie. Cette hypoglycémie, captée par les cellules �,entraîne une augmentation de la sécrétion de glucagon quilimite la baisse de la glycémie et la fait revenir plus rapi-dement à sa valeur initiale ; en effet, si on supprime lasécrétion de glucagon, l’hypoglycémie est accentuée et leretour à la normale beaucoup plus tardif.
E x e r c i c e s
Évaluer ses connaissanses
Choisir la bonne réponse1. a., b., e. sont des affirmations exactes.c. C’est l’inverse.d. Les cellules glucodépendantes (donc les cellulesnerveuses) ont une consommation constante deglucose.
2. Aucune réponse exacte.a. Les récepteurs doivent être spécifiques de l’hor-mone considérée.b. En nanogrammes ou picogrammes.c. Endocrine.d. Voir a.e. Non ; se fixe à un récepteur spécifique et le com-plexe hormone-récepteiur modifie le comportementde la cellule.
1
Chapitre 5: Régulation de la glycémie
33

Données médicalesChez les malades privés de pancréas, on constateque l’insulinémie est nulle mais que la glucagonémieest élevée ce qui explique l’hyperglycémie. L’élèvedevrait s’étonner de trouver du glucagon chez unindividu dépourvu de pancréas. Dans ce cas-là, ilfaut conclure à une autre origine du glucagon. Ilexiste des cellules intestinales capables de sécréter,dans certaines circonstances, du glucagon (entéro-glucagon).
5Comportement vis-à-vis du glucoseOrgane A : prélève du glucose de manière abon-dante mais n’en libère pas à jeun ; il s’agit d’unmuscle.Organe B : utilise autant de glucose à jeun et aprèsl’injection ; il s’agit de l’encéphale.Organe C : cède du glucose à jeun et en stockeaprès l’injection ; il s’agit du foie.Organe D : consomme du glucose à jeun un peuplus (très peu) après l’injection ; il s’agit d’un organenon glucodépendant et non effecteur de la glycémie,la rate par exemple.
Exercice corrigé
Système de régulat ion de la calcémieVoir correction dans le manuel pages 108 et 109.
Évaluer ses compétences
Séquence d’événements1. L’injection d’insuline au temps 0 entraîne :– de 0 à 20 minutes : un arrêt quasi total de la pro-duction de glucose et une augmentation rapide deson utilisation (stockage du glucose) ; en consé-quence, on constate une baisse importante de laglycémie (de 95 à 40 mg.dL–1) qui entraîne une aug-mentation de la sécrétion de glucagon ;– au bout de 20 minutes : la destruction de l’insulineet la production accrue de glucagon font que la pro-duction de glucose par le foie augmente considéra-blement ; en même temps, il y a une baisse del’utilisation liée à la diminution de l’insulinémie : enconséquence, la glycémie qui était descendue jus-qu’à 35 mg.dL–1 commence à remonter pour retrou-ver sa valeur initiale au bout de 75 minutes environ.
2. La perfusion du glucagon entraîne :– une augmentation immédiate de la production deglucose alors que l’utilisation de celui-ci resteconstante dans un premier temps ; en conséquence,la glycémie augmente (de 100 à 120 mg.dL–1) ;– l’augmentation plus tardive (au bout de 15minutes) de l’utilisation du glucose ; en effet, la gly-cémie élevée a entraîné une sécrétion accrue d’in-suline responsable de cette utilisation accrue et quis’oppose à la production de glucose ; en consé-quence, malgré une glucagonémie élevée, la glycé-mie retrouve sa valeur initiale.
4
3
2
34

35
Il s’agit donc, dans le cas de diabète, d’émettre des hypo-thèses sur les phénotypes cellulaires possibles à l’originedu phénotype diabétique.
A c t i v i t é s 1
Les diabètesIl s’agit de définir les deux grands types de phénotypediabétique, d’établir les caractéristiques communes àces deux types, d’en voir les différences et en particu-lier montrer que, dans le type 2, le phénotype diabé-tique est très variable au contraire du phénotypediabétique de type 1.
◆ Problèmes scientifiques à résoudreLe document de la page d’entrée évoque une populationoù la fréquence du diabète de type 2 est particulièrementimportante et amène à s’interroger sur les causes de cenombre élevé de cas de diabètes et d’une façon plus géné-rale sur les causes du diabète.
P o s e r l e s p r o b l è m e s
On veut rappeler ici les caractéristiques du métabolisme àjeun et celles du métabolisme qui suit un repas et l’actiondes messagers hormonaux pancréatiques dans ce métabo-lisme. Ce qui revient à dire qu’il faut maintenir uneconcentration de glucose permettant de satisfaire lesbesoins des cellules glucodépendantes tout en évitant quecette concentration soit trop élevée ce qui à long termeserait nocif.
Programme : notions et contenus• Au niveau métabolique, le phénotype diabétique est défini par une hyperglycémie (glycémie à jeun supérieure à 1,26 g.L-1). Sur le plan clinique, ondistingue deux phénotypes : le diabète de type 1 et le diabète de type 2.• Au niveau cellulaire, le diabète de type 1 est caractérisé par la destruction totale des cellules � sécrétrices d’insuline. Le diabète de type 2 est dûà une insulinorésistance des cellules cibles de l’insuline ainsi qu’à un déficit de l’insulinosécrétion.• De nombreux gènes sont impliqués dans le développement des diabètes. On peut avoir une prédisposition génétique à un phénotype diabétique.Les diabètes résultent de l’interaction entre ces gènes et des facteurs de l’environnement, en particulier l’alimentation. Dans la majorité des cas, lediabète de type 2 se développe à la suite d’une obésité.• La connaissance précise des gènes de susceptibilité aux diabètes et de leur polymorphisme entre dans le cadre de la médecine prédictive.L’utilisation de cette connaissance soulève des problèmes éthiques importants.
Objectifs à atteindreDéfinir les deux grands types de phénotype diabétique au niveau de l’organisme et au niveau biochimique.
Rechercher les anomalies au niveau des phénotypes cellulaires à l’origine des phénotypes biochimiques diabétiques.L’élève doit aboutir à établir que le phénotype cellulaire diabétique de type 1 est caractérisé au niveau pancréatique,par la destruction des cellules � alors que les cellules � sécrétrices de glucagon restent fonctionnelles. Pour le dia-bète de type 2, il doit établir que le phénotype cellulaire pancréatique est caractérisé par une insuffisance de l’insu-linosécrétion et que le phénotype cellulaire des effecteurs est caractérisé par une insulinorésistance.
Rechercher les facteurs à l’origine de ces phénotypes cellulaires, ce qui conduit à établir que plusieurs gènes sont,dans les deux cas, obligatoirement concernés et que l’environnement joue un rôle important ; autrement dit, qu’ils’agit de phénotypes multifactoriels.
Génotype, environnement et phénotype diabétique
C H A P I T R E 6

On peut alors émettre des hypothèses à partir des diversesobservations.L’insulinémie à jeun, d’un diabétique au stade 3 est prati-quement égale à celle d’un individu non diabétique (envi-ron 10 mU. mL–1) alors que sa glycémie, très élevée, estsupérieure à 300 mg. dL–1 ; il est possible d’émettre l’hy-pothèse que l’insuline est inefficace sur ses cellulescibles : il y a insulinorésistance.Mais avec cette glycémie élevée, un individu non diabé-tique sécréterait beaucoup plus d’insuline ; d’où unedeuxième hypothèse : il y a déficit de l’insulinosécrétion.Chez l’individu au premier stade de la maladie, la sécré-tion insulinique de base est un peu plus élevée et surtout,la réponse est plus forte que la normale à la suite d’unrepas ; dans ce cas, l’insulinosécrétion ne paraît pas affec-tée mais il faut une concentration plus élevée d’insulinepour maintenir la glycémie à une valeur proche de la nor-male ; cela traduit une insulinorésistance qui est la carac-téristique précoce d’un diabète de type 2 (et permetd’ailleurs de le dépister).Le DOCUMENT 10 permet de tester ces diverses hypothèses ;utilisation du glucose signifiant ici prise de glucose dansle sang. C’est la technique du clamp euglycémique hyper-insulinique qui est utilisée ; elle consiste en une perfusiond’insuline à une vitesse constante avec simultanément uneperfusion de glucose afin de maintenir la glycémieconstante (= euglycémie) ; on « clampe » la glycémie entre80 et 90 mg. dL– 1. Le calcul de la consommation globalede glucose permet d’évaluer la réponse de l’organismedans son ensemble à un taux spécifique d’insuline.Pour une insulinémie de 50 mU. mL– 1 l’utilisation du glu-cose (DOCUMENT 10a.) est nettement plus faible chez l’indi-vidu diabétique que chez le non diabétique ; cela revient àdire que les organes effecteurs stockent moins le glucosechez le diabétique que chez le non diabétique, ils sontmoins sensibles à l’insuline, ils sont insulinorésistants.Inversement, pour cette même concentration d’insuline, laproduction de glucose par le foie est totalement stoppéechez le non diabétique alors qu’elle est encore élevée chezle diabétique. L’insuline est donc moins efficace pour inhi-ber la glycogénolyse et la néoglucogenèse chez le diabé-tique, cela confirme l’insulinorésistance des celluleshépatiques dans ce diabète de type 2.
A c t i v i t é s 3
Les causes du diabète de type 2L’objectif est de mettre en évidence l’intervention defacteurs génétiques et environnementaux cause d’obé-sité dans l’établissement du phénotype diabétique detype 2.
ExploitationLe DOCUMENT 11 permet de montrer que la fréquence dudiabète varie beaucoup suivant les populations et que desindividus de même origine éthnique sont plus ou moinstouchés suivant leur mode de vie (urbain ou rural) ; que,d’autre part, à milieu de vie identique, des individus
ExploitationLes DOCUMENTS 1 et 2 permettent d’établir les caractèrescommuns aux deux formes de diabètes :– une hyperglycémie à jeun supérieure à 1,26 g. L–1,– une hyperglycémie exagérée à la suite d’une ingestion deglucose.Il est alors possible de conduire l’élève à émettre deshypothèses pour localiser ces anomalies qui affectent :– soit les cellules effectrices,– soit les cellules émettrices des messages hormonaux.À noter que le DOCUMENT 2 peut être interprété soit commetrois phénotypes biochimiques diabétiques de personnesdifférentes, soit comme trois phénotypes successifs chezla même personne.
A c t i v i t é s 2
Diabètes et système de régulationIl s’agit d’aboutir aux caractéristiques des phénotypescellulaires responsables des phénotypes diabétiques detype 1 et 2.
ExploitationLa comparaison des DOCUMENTS 5a. et 5b. montre que chezl’individu diabétique de type 1 par rapport à un individunon diabétique, il y a absence d’insuline à jeun et absencede sécrétion d’insuline en réponse à une ingestion de glu-cose. Cette absence d’insuline pouvant être compensée parune injection de cette hormone (DOCUMENT 4b.). L’absencede sécrétion d’insuline est donc un facteur important del’hyperglycémie observée dans un cas de diabète de type 1.Le DOCUMENT 6 révèle une autre caractéristique de ce dia-bète de type 1 : une sécrétion exagérée de glucagon à jeun ;malgré l’hyperglycémie, il y a une glucagonémie élevée.À jeun, ce diabète s’explique donc non seulement par l’ab-sence d’insuline mais également par une sécrétion accruede glucagon.L’utilisation du système de régulation (document 2a.) doitpermettre à l’élève de saisir que, par rapport à un individunon diabétique, la production de glucose est augmentée etson utilisation plus faible.Le DOCUMENT 7 confirmé par le DOCUMENT 8 permet d’établirl’origine de l’anomalie principale : l’absence totale de cel-lules �. On ne constate pas, d’autre part, une augmentationdu nombre de cellules � qui, au contraire, diminue.Cependant, les cellules � présentes ont une activité plusgrande (hyperglucagonémie, DOCUMENT 6) ce qui est dû à lalevée de l’inhibition que l’insuline exerce sur les cel-lules �.Le DOCUMENT 9 permet une étude du même ordre pour lediabète de type 2. Il permet de constater :– une sécrétion insulinique de base quelle que soit la gra-vité du diabète ;– une certaine réponse insulinique à la suite d’un repas.Par ailleurs, le DOCUMENT 7 montre que les cellules � ne sontpas détruites. Les causes du diabète de type 2 sont donc dif-férentes de celles responsables du diabète de type 1.
36

appartenant à la même population (hispanique parexemple) sont plus ou moins touchés suivant leur niveaude vie, c’est-à-dire leur type d’alimentation. Cela laissesoupçonner des facteurs génétiques et environnementaux.Le DOCUMENT 12 montre des populations ayant une simili-tude génétique importante et chez lesquelles la fréquencedu diabète est différente ; la comparaison des deux popu-lations de Pimas permet d’établir qu’il y a une différencede fréquence entre ces deux groupes et que l’évolution dudiabète chez les Pimas d’Arizona depuis leur changementde mode de vie semble indiquer que c’est ce dernier,source d’obésité, qui est responsable.Le DOCUMENT 13, étant admis que les deux populationscomparées ont le même mode de vie, permet à nouveau demettre en évidence l’influence de facteurs génétiques dansune population, les Pimas étant beaucoup plus touchés parle diabète que les habitants de Rochester.Le DOCUMENT 14 permet d’établir l’influence de l’obésité.Quelle que soit la tranche d’âge, au sein de la populationdes Pimas d’Arizona, les individus obèses (indice demasse corporelle supérieur à 35), sont beaucoup plus fré-quemment touchés par le diabète de type 2 que les autres.L’environnement, cause de cette obésité, est donc un fac-teur très influent dans l’établissement du phénotype diabé-tique.Le DOCUMENT 15 conduit à démontrer cette fois, le rôle dugénotype dans la mise en place de ce phénotype. En effet,à partir de la tranche d’âge 15-24 ans, les individus ayantdes parents diabétiques (précoces ou tardifs) sont plus fré-quemment atteints que les individus n’ayant pas lesmêmes antécédents.Le DOCUMENT 16, enfin, fait la synthèse des deux précé-dents. Il montre en effet que les individus de masse cor-porelle élevée et dont un des parents ou les deux sontdiabétiques sont atteints de diabète beaucoup plus fré-quemment que les individus ne présentant pas ces caracté-ristiques.Il confirme bien que le phénotype diabétique de type 2 estmultifactoriel.
A c t i v i t é s 4
Les facteurs de risqueL’objectif est de dégager la notion de gènes de suscep-tibilité c’est-à-dire de gènes dont certains allèles pré-sents chez un individu sont susceptibles de favoriserl’apparition d’une maladie mais ne sont ni nécessairesni suffisants ; leurs effets étant le plus souvent liés à desfacteurs environnementaux.
ExploitationLe DOCUMENT 17, relatif au diabète de type 1, montre quelorsque l’un des parents est diabétique, 3 à 6 % des enfantsle sont également. Cela est très supérieur à la fréquenced’apparition du diabète chez des enfants sans antécédentsqui ne sont touchés que dans 0,2 % des cas. Cela indiqueque des facteurs génétiques transmissibles favorisent l’ap-parition du diabète. Cela est confirmé par les cas de dia-
bète chez les enfants ayant un frère ou une sœur diabétiqueet très nettement par l’étude des jumeaux vrais : si l’un estdiabétique, l’autre l’est dans 30 à 40 % des cas.Toutefois, bien que les jumeaux vrais aient le même géno-type, dans 60 à 70 % des cas, la concordance n’existe pas.Cela permet d’établir que les facteurs génétiques à euxseuls ne peuvent expliquer l’apparition du diabète. Il y aseulement prédisposition génétique au diabète.Le DOCUMENT 18 sensibilise aux résultats des recherchesmenées pour déterminer les gènes impliqués dans lagenèse du diabète de type 1. Celui-ci étant une maladieauto-immune, les recherches se sont orientées vers lesgènes du système HLA dont le polymorphisme permet de penser que certains allèles pourraient prédisposer audiabète.Le document teste cette idée : la probabilité pour que deuxenfants d’un même couple possède le même HLA est de25 %; or lorsqu’ils sont diabétiques, dans 57 % des cas, ilsont le même système HLA et, dans 38 % des cas, un HLAà demi-identique (contre 25 % attendus). Seulement 5 %des enfants diabétiques d’un couple ont un système HLAdifférent contre 50 % attendus. Autrement dit, les propor-tions 57, 38 et 5 sont extrêmement différentes des propor-tions 25, 25 et 50 et confirment l’implication des gènes duCMH dans le diabète de type 1.Le DOCUMENT 19 permet de préciser quels sont parmi lesallèles des gènes du système HLA ceux qui seraient plusprécisément impliqués ; cela signifie que ces allèles diri-gent la synthèse de protéines membranaires qui, pour desraisons encore inconnues, favoriseraient l’apparition dudiabète. Ici encore, le document montre que chez les dia-bétiques, les allèles DR3 et DR4 sont présents avec unefréquence beaucoup plus grande que chez les non diabé-tiques.On peut ajouter que des études récentes ont montré que90 % des diabétiques possèdent l’un des allèles DR3 ouDR4 contre 40 à 50 % chez les non diabétiques et que lamoitié des diabétiques ont le génotype DR3/DR4 contre5 % chez les non diabétiques.Tout cela confirme que si les gènes du système HLA nesont pas « les gènes du diabète », certains de leurs allèlesconfèrent une prédisposition à cette maladie.Les DOCUMENTS 20 et 21 confirment le caractère multifacto-riel du diabète de type 2.Les données familiales présentées dans le document 20confirment l’importance des facteurs génétiques déjà éta-blie pour les indiens Pimas et montrent même que les don-nées génétiques semblent plus conséquentes dans lediabète de type 2 que dans celui de type 1 puisque chez lesjumeaux monozygotes, la concordance atteint 90 %.Le DOCUMENT 21 confirme, dans une autre population quecelle des Pimas, l’importance de l’obésité dans l’appari-tion du diabète de type 2.Le DOCUMENT 22 précise que la répartition même de cetteobésité représente un facteur de risque.
Chapitre 6 : Génotype, environnement et phénotype diabétique
37

Évaluer ses compétences
Exploitation du document a. :– de 0 à 2 ans (stade 1), la glycémie à jeun nechange que très peu et reste normale alors que l’in-sulinémie augmente de plus en plus. Cela traduitune résistance des effecteurs à l’action de l’insulinecompensés par l’augmentation de l’insulinémie quipermet de maintenir une glycémie à jeun à peu prèsnormale ;– de 2 à 6 ans (stade 2), la glycémie à jeun augmentenettement alors que l’insulinémie, qui reste stable etélevée, ne peut s’opposer à la résistance des effec-teurs à l’action de l’insuline;– de 6 à 12 ans (stade 3), la glycémie à jeun devienttrès élevée car l’insulinémie s’effondre, ce qui traduitun déficit de l’insulinosécrétion.Exploitation du document b. :Normalement, l’état diabétique se manifeste par uneglycémie supérieure à 200 mg.dL–1 deux heuresaprès l’ingestion de 75 g de glucose (hyperglycémieprovoquée). En réponse à une décharge glucidique,la glycémie de A à B ne peut être maintenueconstante que grâce à une augmentation de l’insuli-némie ; cette sécrétion d’insuline atteint un maxi-mum puis s’effondre en C entraînant l’apparition del’état diabétique.
E x e r c i c e s
Évaluer ses connaissances
Choisir la bonne réponse1. Affirmations exactes : a., b., c.d. Pas de sécrétion d’insulinee. Cellules � détruites.
2. Affirmations exactes : a., d., e.b. La sécrétion d’insuline n’est jamais nulle.c. Les cellules b sont intactes.
Choisir la bonne réponse1. Affirmation exacte : a.b. Vrai pour le type 1, mais toujours supérieure ouégale à la normale dans le diabète de type 2.c. Non.d. Vrai seulement pour le type 1 (insulinodépen-dant) ; faux pour le type 2 (non insulinodépendant).
2. Affirmation exacte : b.a. Non.c. Non, il existe des allèles de prédisposition (pas degène du diabète).d. Non.e. Non.
Exercice corrigé
Recherche d’un diabèteVoir correction dans le manuel pages 126 et 127.
3
2
1
38

Pa
rt
ie
1 Des phénotypes à différents niveaux d’organisation du vivant
Génotype, phénotype et système nerveux
7 Un phénotype comportemental :le réflexe myotatique .............................................. 40
8 Fonctionnement des centres nerveuxet mémoire génétique ............................................ 45
9 Cerveau, génotype et environnement .. 53
Unité 3

40
◆ Les acquisCette partie du programme permet l’acquisition desnotions de base sur la communication nerveuse chez lesMammifères et plus particulièrement chez l’Homme.Dans les classes antérieures, les élèves ont acquis :– en cinquième, dans la partie de programme : « Le mou-vement et sa commande » : contractions et relâchementscoordonnés des muscles assurent les mouvements. Lefonctionnement des muscles est commandé par les centresnerveux. Les messages nerveux sont transmis par lescentres nerveux (cerveau et moelle épinière) et les nerfs ;– en troisième, dans la partie du programme : « Relationsà l’environnement et activité nerveuse » : l’activité desrécepteurs sensoriels est déclenchée par un stimulus spé-cifique, provoquant la naissance de messages nerveux. Lapropagation des messages nerveux vers le cerveau se fait
le long de fibres nerveuses en relation avec les récepteurssensoriels. Les messages nerveux sont élaborés et transmispar des cellules spécialisées : les neurones. Les neuronescommuniquent entre eux au niveau des dispositifs spécia-lisés appelés synapses, par l’intermédiaire de messageschimiques. L’étude du neurone est limitée à son identifi-cation dans le tissu nerveux, mais ne vise pas à localiserles corps cellulaires et les synapses sur le schéma fonc-tionnel demandé. On se contente d’indiquer que les neu-rones sont juxtaposés et de constater qu’ils communiquententre eux au niveau des synapses. Ce constat permet d’ex-pliquer les disfonctionnements du système nerveux parl’usage de certaines substances chimiques ;– en seconde, dans la partie du programme : « L’organismeen fonctionnement » : le fonctionnement automatique ducœur est modulé par le système nerveux. L’activité ryth-mique des muscles respiratoires est commandée par le
Programme : notions et contenus• La part du génotype et la part de l’expérience individuelle dans le fonctionnement du système nerveux (durée indicative : 6 semaines).• Cette partie du programme présente un double objectif :– d’une part, permettre l’acquisition de notions de base sur la communication nerveuse chez les Mammifères et plus particulièrement chez l’Homme ;– d’autre part, élargir la compréhension des relations entre le phénotype et le génotype d’un organisme.• Les réactions comportementales, les représentations du monde que se construit un organisme grâce à son système nerveux, sont des aspects deson phénotype au même titre que ses caractéristiques physiques.• Le réflexe myotatique fournit un exemple du déterminisme génétique impliqué dans l’organisation du système nerveux et les propriétés des neu-rones.• Les approches suggérées de la plasticité du cortex cérébral attirent l’attention sur le fait que, depuis le tout début de sa mise en place jusqu’à lamort, l’organisation cérébrale inscrit, dans sa structure, l’histoire individuelle de l’organisme. Cette épigenèse, permise par les gènes, ouvre l’archi-tecture corticale sur l’environnement physique et social. Elle fait de chaque individu – même les vrais jumeaux – un être cérébralement unique, caren constante reconstruction.• L’outil informatique est particulièrement utile pour aborder l’étude de ces sujets de neurophysiologie. Outre l’expérimentation assistée par ordina-teur, il existe plusieurs logiciels de simulation, complémentaires les uns des autres, permettant de mettre les élèves en situation d’investigation.• Les propriétés intégratrices des centres nerveux et le fonctionnement des neurones.• Les circuits neuroniques médullaires mobilisés au cours du réflexe myotatique.• Le réflexe myotatique assure le tonus musculaire nécessaire au maintien de la posture.• Le réflexe myotatique repose sur des populations neuronales :– les neurones afférents qui ont leurs corps cellulaires dans les ganglions des racines dorsales ; les extrémités de ces neurones afférents sont enliaison avec des récepteurs sensoriels : les fuseaux neuromusculaires ;– les motoneurones des muscles étirés et les motoneurones des muscles antagonistes dont les axones aboutissent aux fibres musculaires effec-trices ;– les interneurones inhibiteurs assurant les connexions entre les neurones afférents et les motoneurones des muscles antagonistes.• Limites : l’étude détaillée du récepteur sensoriel et de la plaque motrice, l’étude détaillée de la structure et du fonctionnement des fibres musculairesne sont pas au programme.
Un phénotype comportemental : le réflexe myotatique
C H A P I T R E 7

Chapitre 7 : Un phénotype comportemental : le réflexe myotatique
41
A c t i v i t é s 1
Posture et tonus musculaireL’objectif de ces activités est d’expliquer la notion detonus et sa relation avec la posture.
Exploitation� Pendant l’état de veille et l’état de sommeil lent, nousobservons une activité électrique enregistrable qui traduitun état de contraction des muscles de la nuque des indivi-dus allongés. Ces individus n’effectuent pas de mouve-ment, les muscles de la nuque sont donc au repos, maissont légèrement contractés. Le tonus musculaire est donccet état de contraction des muscles, permanent et de faibleintensité.� Le personnage du tableau a une position du corps com-plètement relâchée ; sa tête et ses bras « tombent » passi-vement sous l’effet de la pesanteur. On peut supposer queses muscles squelettiques sont au repos, c’est-à-dire noncontractés.� Sur le DOCUMENT 4, on constate que, pendant le sommeilparadoxal, l’amplitude de l’électromyogramme est trèsfaible, ce qui traduit un très faible état de contraction desmuscles, donc une forte diminution du tonus musculaire.Cette atonie musculaire explique la position du corps relâ-chée observée précédemment. Le tonus musculaire, étatpermanent de contraction des muscles, ne produit aucunmouvement, mais il est nécessaire au maintien de la pos-ture.
A c t i v i t é s 2
Tonus musculaire et réflexe myota-tiqueDans la rubrique « Poser des problèmes », nous avonsdéfini le réflexe myotatique en observant que l’étire-ment du biceps entraîne une contraction de ce muscle.Cette contraction permet de maintenir la posture pré-sente avant la pose de l’haltère. Dans les activités 1,nous avons établi la relation entre tonus et posture.L’objectif est donc maintenant d’expliquer en quoi leréflexe myotatique constitue une composante du tonusmusculaire. Pour atteindre cet objectif, deux supportsd’activités sont proposés : une expérience historique etune expérience assistée par ordinateur.
Exploitation� L’étirement du muscle lors de l’expérience deSherrington (DOCUMENT 5) a pour conséquence la contrac-tion de ce muscle. L’étirement du muscle extenseur dupied suite au choc du marteau réflexe (DOCUMENT 7)entraîne la contraction de celui-ci. Dans l’expérience deSherrington, l’animal utilisé a sa moelle épinière isolée del’encéphale, ce qui élimine toute possibilité de réponsevolontaire. Dans le cas du réflexe achilléen, nous obser-vons également une réaction involontaire et qui se pré-sente toujours de la même façon. Le réflexe myotatique est
système nerveux. Cette étude du fonctionnement du sys-tème nerveux s’effectue au niveau organe et non au niveaucellulaire. Les variations des messages nerveux sont tra-duites par des flèches ascendantes ou descendantes. Lesnotions de potentiel d’action et de codage de l’informationsont hors programme.
Objectifs à atteindrePour permettre l’acquisition de notions de base sur lacommunication nerveuse chez les Mammifères et plusparticulièrement chez l’Homme, l’élève va découvrirdans ce chapitre :
– les structures impliquées au niveau des organes et descellules dans un exemple de communication nerveusequ’est le réflexe myotatique ;
– le sens de communication des messages nerveux dansces structures.
Dans le chapitre suivant, l’élève approfondira lescaractéristiques du message nerveux, la transmissionde ces messages entre les neurones, et mettra en évi-dence la complexité du réseau de neurones, complexitésous la dépendance du génome.
P o s e r l e s p r o b l è m e s
Nous avons voulu, dans cette rubrique, éviter le côté arti-ficiel lié à l’expérimentation comme dans les réflexesachilléen ou rotulien. Pour cela, nous proposons une situa-tion naturelle comme celle qui consiste à tendre légère-ment l’avant-bras afin de recevoir quelque chose dans lamain (exemple : lorsque, lors d’un cocktail, on tend sonverre pour le faire remplir).Pour éviter l’activité volontaire, l’élève a les yeux bandéset est surpris par la pose de l’haltère dans la main. Sonavant-bras s’abaisse et retrouve presque instantanément saposition initiale.Nous observons une réaction involontaire, stéréotypée,déclenchée par une stimulation de l’environnement : c’estun réflexe. Lors de la pose de l’haltère, on observe un éti-rement du biceps. Si l’avant-bras retrouve sa position ini-tiale, c’est que le biceps s’est contracté. Cette contractions’observe sur l’électromyogramme. On peut donc définirle réflexe myotatique. Sur l’électromyogramme, onconstate qu’avant la pose de l’haltère, le biceps présentaitun certain degré de contraction : cela correspond au tonus.On peut donc s’interroger sur la relation qui existe entre letonus musculaire observé et la position initiale, c’est-à-dire la posture de l’élève (en a.), puis sur la relation entrele tonus musculaire et le réflexe myotatique. Alors le pro-blème des organes et des cellules impliquées dans ceréflexe apparaît facilement.

rentes et efférentes. Ces fibres se retrouvent dans le nerfrachidien dont le nerf musculaire n’est qu’une ramifica-tion. Il reste donc à préciser comment le nerf rachidien seraccorde à la moelle épinière et comment s’effectue lacommunication entre les capteurs et les effecteurs.
A c t i v i t é s 4
Des fuseaux neuromusculaires auxfibres musculaires contractilesLes activités 4 sont le prolongement des activités 3 etvont permettre de préciser les relations entre les nerfsrachidiens et la moelle épinière, et d’expliquer com-ment circule l’information dans les structures impli-quées dans le réflexe myotatique.
Exploitation� Le nerf rachidien se raccorde à la moelle épinière pardeux racines, une racine postérieure ou dorsale et uneracine antérieure ou ventrale.– Conséquences de la section de la racine antérieure : sen-sibilité conservée et motricité impossible.– Conséquences de la section de la racine postérieure :sensibilité abolie et motricité possible.Le message nerveux afférent emprunte la racine dorsale,alors que le message nerveux efférent emprunte la racineventrale.� Une stimulation du nerf musculaire provoque, sur lesfibres nerveuses de ce nerf, la naissance d’un messagenerveux aussi bien sur les fibres nerveuses afférentes quesur les fibres nerveuses efférentes. Les messages nerveuxconduits par ces fibres sont à l’origine de phénomènesélectriques que l’on peut enregistrer sur la surface du nerf :c’est le potentiel global, potentiel qui sera étudié dans lesactivités 2 du chapitre 8. La section évite que la réceptionsur la racine ventrale soit perturbée par des messages véhi-culés par des fibres motrices vers la moelle épinière. LeDOCUMENT 15a. met en évidence qu’un message empruntela racine dorsale et est à l’origine du message que l’onenregistre sur la racine ventrale. Par contre, le DOCUMENT15b. montre que le message créé sur la racine ventrale,suite à la stimulation, ne se transmet pas à la racine dor-sale. Il y a unidirectionalité dans la conduction du messagenerveux lors du réflexe myotatique.En résumé de toutes ces activités, l’élève peut construireun schéma fonctionnel. Celui-ci sera tel qu’il est repré-senté sur le schéma-bilan pages 146-147, avec deux diffé-rences. Le neurone afférent est remplacé par un trait bleuqui part du capteur et qui va jusqu’à la moelle : ce traitbleu symbolise la fibre nerveuse afférente. Le neuroneefférent est remplacé par un trait rouge qui part de lamoelle épinière vers les fibres musculaires : ce trait rougesymbolise la fibre nerveuse efférente. À la jonction entreces deux types de fibres, on place un point d’interrogation(le problème de la jonction est posé par le DOCUMENT 15b.).On s’interroge alors sur la nécessité de préciser la naturedes fibres nerveuses et la nature de la jonction entre cesfibres.
donc une réaction involontaire et stéréotypée qui entraînela contraction du muscle en réponse à son propre étire-ment. Nous observons (DOCUMENT 6) qu’après la section dunerf innervant le muscle quadriceps du chat, ce musclen’exerce plus, pour un étirement identique, qu’une forcebeaucoup plus faible due aux propriétés de sa composanteélastique parallèle. La différence entre les courbes P et Mreprésente la contribution du réflexe à la tension mesurée.Le système nerveux est donc nécessaire dans la réalisationde ce réflexe.� L’observation des enregistrements de la tension dumuscle dans l’expérience de Sherrington, montre que latension développée croît rapidement et atteint son maxi-mum dès que l’étirement est maximal. À ce sommet suc-cède un plateau qui persiste autant que dure l’étirement.Dans le cas du réflexe achilléen, l’étirement bref provoqueun réflexe de courte durée (DOCUMENT 8b.). Lorsque l’étire-ment est bref, la réponse musculaire est de courte durée :c’est la réponse phasique du réflexe myotatique. Lorsquel’étirement est permanent, la contraction se maintient pen-dant toute la durée de cet étirement : c’est la réponsetonique ou statique du réflexe myotatique. Le réflexe myo-tatique présente donc deux aspects : un aspect phasique(réflexes rotulien, achilléen) et un aspect tonique (élémentimportant du tonus musculaire). Recherchons maintenantles supports du réflexe d’étirement.
A c t i v i t é s 3
Les relations nerf-muscleDans la continuité de l’activité précédente, cette acti-vité va permettre à l’élève de préciser les relationsentre le nerf et les structures réceptrices et effectricesdu muscle.
Exploitation� Chez des malades présentant des lésions des fuseauxneuromusculaires d’un muscle (DOCUMENT 11), on constatela disparition du réflexe myotatique et la persistance de lamotricité volontaire : on peut donc supposer que lesfuseaux neuromusculaires sont les structures réceptricesou capteurs, sensibles à l’étirement. Par contre, la capacitéde contracter ses muscles disparaît chez une personnemyopathe : les fibres musculaires engendrent donc lacontraction du muscle, ce sont les structures effectrices.Sur le DOCUMENT 9b., on observe que le nerf musculaire(nerf qui innerve un muscle) est constitué de fibres ner-veuses. Sur le DOCUMENT 10b., on observe qu’une fibre ner-veuse pénètre dans chaque fuseau neuromusculaire et s’yramifie. L’élève peut émettre l’hypothèse que ces fibresvont conduire le message nerveux issu du capteur jusqu’àla moelle épinière : ce sont des fibres afférentes. Le DOCU-MENT 10a. permet de préciser la relation entre les fibresnerveuses et les fibres musculaires, et de proposer l’hypo-thèse que ces fibres vont conduire un message nerveuxvers les fibres musculaires contractiles : ce sont les fibresefférentes.Le muscle contient à la fois, les capteurs et les effecteursdu réflexe. Le nerf musculaire est constitué de fibres affé-
42

A c t i v i t é s 5
Cellules nerveuses et réflexe myota-tiqueAprès avoir construit la notion de cellule nerveuse ouneurone, cette double page va permettre de connaîtrel’organisation neuronique, support du réflexe myota-tique.
Exploitation� Le DOCUMENT 16 présente une culture de neurones. Onobserve des cellules avec des prolongements. Cetensemble, corps cellulaire (noyau + cytoplasme) et sesprolongements (fibres) constitue un neurone.Les fibres nerveuses observées dans les activités précé-dentes sont donc des prolongements de corps cellulaires.Les DOCUMENTS 17 et 18b. vont permettre de localiser lecorps cellulaire du neurone afférent dans le ganglion spi-nal. La fibre afférente issue du fuseau neuromusculaire estun prolongement (prolongement périphérique), qui, à l’in-térieur du ganglion spinal, bifurque vers le corps cellulaireet se prolonge vers la moelle (prolongement central). Auniveau du neurone afférent, classiquement, on appelaitdendrite, le prolongement périphérique qui conduit lessignaux nerveux du fuseau vers le corps cellulaire. Cettedistinction est aujourd’hui abandonnée : on utilise le termede fibre afférente pour les deux prolongements. Les DOCUMENTS 17 et 18a. vont permettre de localiser lecorps cellulaire du neurone efférent (motoneurone) dans lasubstance grise des cornes antérieures ou ventrales de lamoelle épinière.La fibre efférente est le prolongement du corps cellulairesitué dans la corne antérieure de la moelle épinière. Lecorps cellulaire présente d’autres petits prolongements. Àson arrivée dans le muscle, la fibre efférente se divise enun certain nombre de ramifications, chacune d’elles inner-vant une seule fibre musculaire (DOCUMENT 19 b.).L’ensemble formé par le neurone efférent et l’ensembledes fibres musculaires qu’il innerve constitue une unitéfonctionnelle appelée unité motrice.La cellule nerveuse est donc une cellule très longue quipermet d’établir des liaisons entre des points éloignés del’organisme et elle conduit des messages nerveux.� À partir de ces nouvelles données, le schéma fonction-nel élaboré dans les activités 4 peut être complété enconstruisant la chaîne de neurones, support du réflexemyotatique. Le DOCUMENT 19 permet de lever l’interroga-tion sur la jonction entre les neurones afférents et effé-rents, ainsi que celle entre le neurone efférent et la fibremusculaire. Le schéma bilan de la rubrique « Essentiel »est ainsi construit.
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Tonus musculaire : C’est l’état permanent decontraction des muscles qui ne produit aucun mou-vement. Il est nécessaire au maintien de la posture.b. Réflexe myotatique : C’est une réaction involon-taire et stéréotypée qui entraîne la contraction dumuscle en réponse à son propre étirement.c. Fuseau neuromusculaire : Capteur du stimulusétirement, enfoui dans la masse musculaire. C’estune capsule de tissu conjonctif de quelques milli-mètres de longueur contenant de 3 à 10 fibres mus-culaires modifiées et dans laquelle pénètre une fibrenerveuse.d. Fibres musculaires contractiles : Effecteurs dumouvement. Cellules de grande taille, de formeallongée, dont la longueur atteint plusieurs centi-mètres. Elles constituent l’essentiel de la massemusculaire.e. Nerf musculaire : Nerf qui innerve un muscle etqui correspond à une ramification d’un nerf rachi-dien. Il contient à la fois les fibres afférentes et effé-rentes.f. Neurone afférent : Neurone dont le corps cellulairese trouve dans le ganglion spinal de la racine posté-rieure de la moelle épinière. À partir du corps cellu-laire, il possède un seul prolongement qui bifurque àl’intérieur même du ganglion spinal. Le prolonge-ment périphérique constitue une fibre afférente d’unfuseau neuromusculaire, tandis que le prolongementcentral pénètre dans la moelle épinière. Le terme defibre afférente est utilisé pour les deux prolonge-ments.g. Neurone efférent : Neurone dont le corps cellu-laire se trouve dans la substance grise des cornesantérieures ou ventrales de la moelle épinière, géné-ralement appelé motoneurone. Son long prolonge-ment quitte la moelle épinière, passe par la racineventrale d’un nerf rachidien et constitue une fibreefférente qui va jusqu’au muscle innervé.
Questions à réponses cour tesa. Le tonus musculaire correspond à l’état perma-nent de contraction des muscles. Cet état decontraction ne produit aucun mouvement mais per-met de stabiliser les différents segments osseux depart et d’autre des articulations. Sans cette forceantagoniste à la pesanteur exercée par le tonusmusculaire, on observerait l’effondrement du sujet.Le tonus musculaire est nécessaire au maintien de laposture.b. L’action de la pesanteur étire nos muscles exten-seurs. Le réflexe myotatique, c’est-à-dire la contrac-tion du muscle en réponse à son propre étirement,se produit donc en permanence. Il est la compo-sante essentielle du tonus musculaire, qui estnécessaire au maintien de la posture.
2
1
Chapitre 7 : Un phénotype comportemental : le réflexe myotatique
43

c. Le réflexe myotatique est un réflexe qui entraînela contraction d’un muscle en réponse à son propreétirement. Cette réaction réflexe implique, pour saréalisation, la mise en jeu :– de capteurs, ce sont les fuseaux neuromuscu-laires, qui réagissent spécifiquement au stimulus éti-rement ;– de fibres afférentes issues des capteurs quiconduisent le message jusqu’au centre nerveux,– d’un centre nerveux, la moelle épinière, quiexploite le message qui lui parvient et émet un mes-sage nerveux efférent ;– de fibres efférentes qui conduisent ce messageefférent jusqu’aux organes effecteurs ;– d’effecteurs, les fibres musculaires, qui réagissentau message efférent en se contractant.Le nerf musculaire contient à la fois les fibres affé-rentes et les fibres efférentes. Les fibres afférentessont le prolongement du corps cellulaire d’un neu-rone afférent. Les fibres efférentes sont le prolonge-ment du corps cellulaire d’un neurone efférent.d. Voir schéma « Essentiel » du réflexe myotatiquepages 146-147.
Choisir les bonnes réponses1. a., b., d.2. a., b.3. a., d.4. a.
Exercice corrigé
Le ref lexe d’évitementVoir correction dans le manuel pages 148 et 149.
Évaluer ses compétences
Deux réf lexes différentsLors du réflexe d’évitement, le stimulus « piqûre »provoque, au niveau des capteurs cutanés à la dou-leur, la naissance d’un message nerveux afférentconduit par des fibres nerveuses afférentes jusqu’àla moelle épinière. Celle-ci émet un message ner-veux efférent conduit par des fibres nerveuses effé-rentes jusqu’aux cellules musculaires du biceps, quiréagissent au message nerveux en se contractant.La contraction de ces fibres entraîne le raccourcis-sement du muscle biceps qui tire, grâce aux ten-dons, sur les os de l’avant-bras : le mouvement deflexion se réalise.
5
4
3
44
Tension Longueur du biceps du biceps
Position 1 de référence de référence
Position 2 diminuée augmentée
Position 3 augmentée de référence
L’augmentation de la tension observée dans la posi-tion 3 est le résultat du réflexe myotatique.Ces deux types de réflexes présentent trois diffé-rences :– la nature du stimulus (douleur ou étirement) ;– les récepteurs spécifiques associés (capteurcutané à la douleur ou fuseau neuromusculaire) ;– dans le cas du réflexe myotatique, le capteur etl’effecteur sont dans le même organe : le muscle.
« Tomber de sommeil »Lors du sommeil profond ou sommeil paradoxal, onobserve une annulation du tonus musculaire. Lesmuscles extenseurs de la tête ne sont plus contrac-tés et n’exercent plus une force antagoniste à lapesanteur, la tête bascule alors en avant.
Les conséquences d’un accidentLa section du nerf musculo-cutané entraîne la sec-tion des fibres afférentes et efférentes de ce muscle.Cette section a pour conséquence la disparition duréflexe myotatique et celle de la motricité volontaire.Ce muscle ne se contracte plus jamais, il n’y a plusde tonus. L’absence d’activité musculaire expliquel’atrophie importante de ce muscle.
7
6
Lors du réflexe myotatique, le stimulus « étirement »provoque au niveau des fuseaux neuromusculaires,la naissance d’un message nerveux afférent conduitpar des fibres nerveuses afférentes jusqu’à la moelleépinière. Celle-ci émet un message nerveux efférentconduit par des fibres nerveuses efférentes jus-qu’aux cellules musculaires du biceps qui réagissentau message nerveux en se contractant. La contrac-tion de ces fibres entraîne le raccourcissement dumuscle biceps qui tire sur les os de l’avant-bras : lemouvement de flexion se réalise.

45
Programme, notions et contenus• Les potentiels d’action et les messages nerveux.• Les signaux émis par les neurones sont des potentiels d’action.• La genèse de potentiels d’action repose sur l’existence d’un potentiel dit de repos, propriété commune à toutes les cellules.• Un potentiel d’action est une inversion transitoire de la polarisation membranaire. Au cours de sa propagation le long d’une fibre, le potentiel d’ac-tion conserve toutes ses caractéristiques.• Limites : les mécanismes ioniques sous-tendant la genèse du potentiel d’action, la propagation des potentiels d’action par les courants locaux, nesont pas au programme.• Les messages nerveux : les messages nerveux (afférents et efférents) se traduisent au niveau d’une fibre par des trains de potentiels d’action,d’amplitude constante. Les messages nerveux sont codés par la fréquence des potentiels d’action et le nombre de fibres nerveuses mises en jeu.• Limites : les mécanismes de la transduction (potentiels de récepteurs, potentiels générateurs) ne sont pas au programme.• Caractéristiques du fonctionnement des synapses : un message nerveux est transmis d’un neurone à d’autres neurones ou à des cellules effec-trices par des synapses.• Au niveau d’une synapse, le message nerveux présynaptique, codé en fréquence de potentiels d’action, est traduit en message chimique codé enconcentration de neurotransmetteur. Les molécules de neurotransmetteur se fixent sur des récepteurs de la membrane postsynaptique : cette modi-fication induit une modification de l’activité du neurone postsynaptique. Ce changement d’activité est à l’origine d’un nouveau message.• Limites : les mécanismes ioniques liés à l’activité des synapses ne sont pas au programme.• Activité du centre nerveux : le traitement des messages afférents, en réponse au stimulus d’étirement à l’origine du réflexe myotatique, modifie lafréquence des potentiels d’action des motoneurones. Celle des motoneurones du muscle étiré est augmentée alors que celle des motoneurones desmuscles antagonistes est diminuée, voire annulée.• Les motoneurones et les interneurones du réflexe myotatique sont en connexion avec d’autres neurones que les neurones afférents issus desfuseaux neuromusculaires.• Dans certaines limites, la stimulation d’autres récepteurs sensoriels (par exemple, les récepteurs nociceptifs) ou une commande volontaire peu-vent inhiber le réflexe myotatique.• Limites : les notions de potentiel postsynaptique excitateur et de potentiel postsynaptique inhibiteur ne sont pas au programme.• La part du génotype dans le fonctionnement du système nerveux : le phénotype comportemental des réflexes (par exemple le réflexe myotatiqueet le réflexe nociceptif d’évitement) est la conséquence de la mise en place, au cours du développement, des chaînes de neurones, sous le contrôlede l’information génétique.
Objectifs à atteindreDans le chapitre précédent, les organes et les cellules impliqués dans le réflexe myotatique ont été mis en place. Lesens de circulation des messages nerveux dans ces structures a été découvert par les élèves.
Dans ce chapitre, les notions à construire sont celles qui concernent :
– les caractéristiques du message nerveux au niveau de la fibre (message afférent, message efférent) ;
– les caractéristiques du message nerveux au niveau du nerf ;
– la transmission des messages entre les neurones ;
– le rôle intégrateur des neurones ;
– la notion d’inhibition ;
– la mise en place de chaînes de neurones au cours du développement et par là comprendre que le phénotype com-portemental d’un individu est sous le contrôle de l’information génétique.
Fonctionnement des centres nerveux et mémoire génétique
C H A P I T R E 8

tion, il faut que celle-ci soit d’intensité suffisante pourdéclencher un potentiel d’action. Pour une stimulationd’intensité I1, il n’y a pas de déclenchement du potentield’action (PA), pour les intensités I2 et I3 croissantes, il y adéclenchement d’un potentiel d’action dont l’amplitudeest constante.Cette caractéristique est confirmée par le DOCUMENT 2c. quimontre que l’amplitude et la durée du potentiel d’action nevarient pas quand celui-ci apparaît à la suite d’une stimu-lation (I2, I3, I4). On peut ainsi constater que le potentield’action ne code pas l’intensité du stimulus qui lui a donnénaissance.L’intérêt de ce document est aussi de montrer que lorsquel’on diminue la vitesse de balayage du faisceau d’électronsde l’oscilloscope, les potentiels d’action vont se présentersous forme de traits verticaux, aspect que l’on retrouveradans le DOCUMENT 3 page 154.
� Calcul de la vitesse de propagation du signal nerveux :v (m.s-1) = d (m)/t (s), v = 3.10-3/0,6.10-3 = 5 m.s-1.Cette valeur de vitesse de propagation du signal n’est pasdu même ordre de grandeur que la vitesse du courant élec-trique. Le potentiel d’action n’est pas un courant élec-trique, mais s’accompagne de phénomènes de natureélectrique qui sont enregistrables.
A c t i v i t é s 2
Le message nerveux afférentLe potentiel d’action (PA) est le signal nerveux ou mes-sager nerveux. C’est l’élément de base du message ner-veux. Le PA ne codant pas l’intensité du stimulus qui luia donné naissance, il est nécessaire de rechercher com-ment le message nerveux peut traduire l’intensité du sti-mulus.
On se remettra dans le cadre du réflexe myotatique etl’on recherchera dans un premier temps comment lemessage nerveux issu d’un fuseau neuromusculaire tra-duit l’intensité de l’étirement lorsque celle-ci aug-mente. Dans un deuxième temps, sachant que, dans lemuscle, il y a de nombreux fuseaux neuromusculaires,et que l’ensemble des fibres issues de ces fuseauxconstitue le nerf musculaire, on cherchera comment, auniveau du nerf, est codé le message nerveux.
Exploitation� Dans le DOCUMENT 3a, on constate que l’enregistrement dumessage nerveux sur une fibre issue d’un fuseau neuro-musculaire se traduit par une suite de PA, un train de PAd’amplitude constante, mais dont la fréquence varie enfonction de l’intensité de l’étirement : pour un étirement de2 g, la fréquence des PA est de 35 PA.s-1 ; pour un étirementde 15 g, la fréquence des PA est de 55 PA.s-1 ; pour un éti-rement de 35 g, la fréquence des PA est de 65 PA.s-1.L’augmentation de l’intensité du stimulus est traduite auniveau de la fibre par une augmentation de la fréquence desPA : le codage du message nerveux au niveau d’une fibreissue du fuseau neuromusculaire se fait en fréquence de PA.
◆ Problèmes scientifiques à résoudreComment l’organisation fonctionnelle du système ner-veux permet-elle la réalisation du réflexe myotatique ?
P o s e r l e s p r o b l è m e s
Les DOCUMENTS 1a. et 1b. permettent de constater que plusl’intensité du coup, c’est-à-dire du stimulus porté sur letendon, est grande et plus l’amplitude de l’électromyo-gramme est importante, montrant que la contraction dumuscle permettant l’extension de la jambe est plus forte.Se pose alors le problème de savoir comment le messagenerveux afférent issu des fuseaux neuromusculaires peuttraduire l’intensité de la stimulation (étirement).
A c t i v i t é s 1
Le signal nerveuxL’objectif de ces activités est de construire les notionsde potentiel de repos, de potentiel d’action et de déga-ger leurs caractéristiques.
Pour faire l’étude de ces notions, il est nécessaire quel’élève s’approprie les conditions d’enregistrement deces différents potentiels : pour cela, il sera aidé par leschéma du dispositif expérimental du document 2a.
Exploitation� Le DOCUMENT 2a. montre le dispositif expérimental per-mettant les enregistrements des différents potentiels. Undispositif d’excitation est en place (stimulateur) avec,parallèlement, un dispositif d’enregistrement (une micro-électrode disposée à la surface ou enfoncée dans la fibre etune électrode de référence). Ces deux électrodes récep-trices sont reliées aux plaques horizontales d’un oscillo-scope cathodique. Il est à préciser que la fibre nerveusegéante doit se trouver dans des conditions physiologiquespermettant sa survie (la fibre doit baigner dans du sérumphysiologique).Le potentiel de repos de la fibre correspond à la différencede potentiel enregistrée, ici – 72 mV lorsque l’on enfoncela microélectrode dans la fibre. En effet, le DOCUMENT 2b.indique que, lorsque la microélectrode est en position 1 eten l’absence de stimulation, aucune différence de potentieln’est enregistrée entre la microélectrode et l’électrode deréférence. Lorsque la microélectrode est enfoncée dans lafibre (position 2) et en l’absence de stimulation, unedéviation brusque vers le bas du spot de l’oscilloscope estobservée ; cette déviation s’installe à une valeur de – 72 mV, valeur du potentiel de repos montrant que la fibrenerveuse est polarisée positivement à l’extérieur et négati-vement à l’intérieur.La deuxième partie du DOCUMENT 2b. montre que le poten-tiel d’action de la fibre correspond, lorsque l’on stimulecette fibre, à une inversion brutale et brève du potentiel derepos. L’amplitude du potentiel d’action de cette fibre estd’environ 115 mV; elle a une durée d’environ 2 ms. Lepotentiel d’action ne se déclenche pas à chaque stimula-
46

Cette observation est confirmée par le DOCUMENT 3b. quimontre graphiquement qu’il y a une relation de propor-tionnalité entre l’intensité de l’étirement et le nombre dePA par seconde émis par le fuseau neuromusculaire.
� Le DOCUMENT 4 montre que pour 3 fuseaux neuromus-culaires localisés de façon différente dans le muscle, lecodage se fait toujours en fréquence de PA. Mais, pourchaque fuseau, et pour une intensité d’étirement donnée,la fréquence est différente. Cela indique que la sensibilitéd’un fuseau neuromusculaire est variable et que cette sen-sibilité semble être fonction de sa place dans le muscle.Cette observation semble indiquer que dans le nerf mus-culaire constitué d’un ensemble de fibres nerveuses issuesdes fuseaux neuromusculaires circule un message pluscomplexe sans doute que le message qui circule au niveaud’une fibre.Les DOCUMENTS 5 et 6 vont permettre d’expliquer commentse fait le codage du message au niveau du nerf musculaire.Si on étire de façon très brève le muscle (étirement équi-valent à une stimulation) et que l’on enregistre les phéno-mènes électriques sur la racine dorsale du nerf, on constateque, pour une intensité d’étirement A, on n’enregistreaucun phénomène électrique ; pour une intensité d’étire-ment B, on observe un phénomène électrique dont l’am-plitude est de l’ordre de 2 mV; pour des intensitésd’étirement plus grandes (de C à G) les phénomènes élec-triques enregistrés ont une amplitude qui augmente pouratteindre environ 8 mV. Ces phénomènes électriques quel’on enregistre quand les électrodes réceptrices sontposées sur la racine d’un nerf constituent le potentiel glo-bal de nerf. On peut noter que les conditions d’enregistre-ment ne sont pas les mêmes que celles de l’enregistrementd’un potentiel d’action de fibre. L’amplitude du potentielglobal de nerf augmente avec l’intensité de la stimulation(ici, l’étirement). Pour l’intensité de stimulation H, plusgrande que G, l’amplitude du potentiel global n’augmenteplus. L’enregistrement du potentiel global d’un nerf deCrabe, d’Étrille ou d’Araignée montre le même phéno-mène. Le dispositif est de même type, le nerf est déposésur les deux électrodes réceptrices reliées à un dispositifEx.A.O. On constate que pour une faible intensité(courbes bleu foncé) on n’enregistre pas de potentiel glo-bal ; ensuite lorsque l’intensité de la stimulation augmente(courbes rouge, bleue, violette, verte) on enregistre unpotentiel global dont l’amplitude augmente et se stabilisevers 8 mV. Cette amplitude augmente en fonction dunombre de fibres nerveuses stimulées et donc mises en jeusuite à leur recrutement lors de la stimulation. Le potentielglobal correspond à la somme des PA de chacune desfibres mises en jeu lors de la stimulation.Dans le cas du réflexe myotatique, le codage du messagenerveux afférent au niveau du nerf musculaire se fait enamplitude en fonction du nombre de fuseaux neuromuscu-laires mis en jeu lors de l’étirement (leur nombre dépen-dant de leur place dans le muscle) et de l’intensité del’étirement (utilisation du logiciel SYNAPSES, CRDP,Versailles). Lorsque tous les fuseaux neuromusculairessont mis en jeu, l’amplitude du potentiel global est maxi-male.Le message nerveux véhiculé par un nerf est donc codépar un potentiel global dont l’amplitude est liée au nombre
de fibres nerveuses recrutées et à la fréquence des poten-tiels d’action qui parcourent chacune de ces fibres.
A c t i v i t é s 3
Émission des messages par le motoneuroneLes messages nerveux afférents qui circulent dans le nerfmusculaire sont codés par la fréquence des potentielsd’action qui parcourent chacune des fibres et par le nombre de fibres nerveuses mises en jeu. Dans le cha-pitre 7, nous avons vu que chaque fibre nerveuse affé-rente issue d’un fuseau neuromusculaire est connectéeau niveau de la moelle épinière avec au moins un moto-neurone. Le problème qui se pose est de savoir ce qui sepasse au niveau de la moelle épinière, si la moelle épi-nière n’est qu’un simple lieu de passage du message affé-rent ou s’il se produit des modifications du message auniveau de ce centre. On recherche dans ces activitéscomment se fait le traitement des messages qui y arriventet dans quelles conditions les motoneurones émettent unmessage à l’origine de la contraction musculaire.
L’ensemble des activités permettra progressivement demontrer :– qu’il y a traitement des messages afférents au niveaudu centre nerveux ;– que plusieurs fibres afférentes issues des fuseaux neu-romusculaires entrent en contact avec le même moto-neurone ;– que le motoneurone, par les afférences qu’il reçoit,fait la somme des signaux qui lui arrivent (sommationsspatiale et temporelle) et émet un nouveau message.
Exploitation� Le DOCUMENT 7a. précise les conditions d’enregistre-ment des phénomènes électriques présentés dans le DOCU-MENT 7b. mais aussi dans le DOCUMENT 8.Comme dans le DOCUMENT 5 des activités 2, les électrodesréceptrices R1 et R2 sont déposées sur les racines du nerf.Ce sont donc, comme il est précisé dans le texte, despotentiels globaux qui sont enregistrés. En E1 et E2, la sti-mulation est telle (un seul stimulus électrique) que chaquefibre n’émet qu’un seul potentiel d’action.Si on stimule un groupe de fibres G1 en E1 avec une inten-sité I1, on observe en O1 un potentiel global dont l’ampli-tude est de l’ordre de 10 mV. On n’enregistre aucunpotentiel global en O2 montrant qu’aucun motoneuronen’a émis de message. Ceci montre que les synapses situéesentre les neurones afférents et les neurones efférents nefonctionnent pas au coup par coup et que par là, la moelleépinière n’est pas un simple lieu de passage du messagemais un centre où l’information est traitée.Si on stimule un groupe de fibres G2 en E2 avec une inten-sité I2 > I1, on observe en O1 un potentiel global dont l’am-plitude est de l’ordre de 25 mV suggérant qu’un plusgrand nombre de fibres a été recruté que dans la stimula-
Chapitre 8 : Fonctionnement des centres nerveux et mémoire génétique
47

tion convergente du centre nerveux. Le nombre de moto-neurones n’est pas inférieur au nombre de fibres affé-rentes, ce qui implique une organisation divergente.Le DOCUMENT 10 montre que si l’on stimule un nerf muscu-laire avec toujours la même intensité, mettant en jeu tou-jours le même nombre de fibres afférentes et que l’onaugmente la fréquence des stimulations, on constate quepour une fréquence donnée F1, le motoneurone n’émet pasde potentiel d’action ; par contre, pour la fréquence F2, lemotoneurone émet un potentiel d’action ; pour F3, il émet2 potentiels d’action. Dans ce cas, les fibres afférentesrecrutées ont véhiculé un train de potentiels d’action.Lorsque la fréquence est trop faible, chaque potentield’action est inefficace ; par contre, à plus grande fré-quence, le motoneurone fait la somme des potentiels d’ac-tion qui lui arrivent : ce phénomène est nommé sommationtemporelle. Il est à remarquer dans ce DOCUMENT 10, qu’il ya à la fois sommation temporelle et spatiale car on consi-dère plusieurs fibres nerveuses qui sont connectées avec lemotoneurone.
A c t i v i t é s 4
Synapses et transmission du messageIl a été montré précédemment que la synapse neuro-neuronique ne fonctionnait pas au coup par coup. Sepose alors le problème de savoir quelle est la structurede la synapse, quel est son fonctionnement et surtoutcomment ce fonctionnement permet d’expliquer les pro-priétés de sommation du neurone.
Dans ces activités, seront comparées les structures de la synapse neuroneuronique et de la synapse neuro-musculaire. Puis, en s’appuyant sur le fonctionnementde la synapse neuromusculaire, le fonctionnement de lasynapse neuroneuronique sera abordé.
ExploitationLes DOCUMENTS 11 et 12 montrent l’ultrastructure d’unesynapse neuroneuronique et d’une synapse neuromuscu-laire. Le tableau ci-dessous donne la comparaison desdeux types de synapses.
tion I1. On enregistre en O2 un potentiel global dont l’am-plitude est de l’ordre de 10 mV. Un ou plusieurs motoneu-rones ont émis un message qui circule dans la racineventrale. Si on stimule en E1 et E2 en même temps, onenregistre en O1 un potentiel global dont l’amplitude estd’environ 30 mV montrant qu’un plus grand nombre defibres afférentes du nerf musculaire est recruté ; on enre-gistre en O2 un potentiel global dont l’amplitude est d’en-viron 20 mV montrant qu’un plus grand nombre demotoneurones a été sollicité.Ces résultats peuvent être expliqués grâce au DOCUMENT 9 quischématise l’organisation convergente mais aussi diver-gente des neurones. Un neurone afférent est connecté à plu-sieurs motoneurones (divergence) et chaque motoneuronereçoit des afférences de plusieurs neurones afférents(convergence). Stimulé par un petit nombre de fibres affé-rentes, le motoneurone n’émet pas de message. Enrevanche, activé par un plus grand nombre de fibres affé-rentes, il émet un message. Ceci étant valable pour plusieursmotoneurones, on peut enregistrer, au niveau de la racineventrale, un potentiel global dont l’amplitude varie en fonc-tion du nombre de motoneurones mis en jeu. Il y a bien auniveau du centre un traitement des informations issues desfuseaux neuromusculaires lors du réflexe myotatique.
� Le DOCUMENT 8, dont le dispositif expérimental se trouvedans le DOCUMENT 7a., montre que lorsque l’on stimule enE1 (par exemple), avec des stimulations croissantes I1, I2, I3
et I4, on enregistre en O1 tout d’abord aucun potentiel glo-bal puis un potentiel global dont l’amplitude augmenteavec l’intensité de la stimulation montrant que, pour I1,aucune fibre n’est recrutée mais que pour les autres inten-sités, un nombre de plus en plus grand de fibres issues desfuseaux neuromusculaires est mis en jeu.Au niveau de O3 où l’enregistrement ne se fait que sur unefibre issue d’un seul motoneurone, d’où l’enregistrementd’un éventuel potentiel d’action, on constate que pour lesstimulations I1, I2 et I3, le motoneurone n’émet pas depotentiel d’action. Par contre, pour l’intensité I4, le moto-neurone émet un potentiel d’action. Le motoneurone, enI4, est activé par un plus grand nombre de fibres affé-rentes : il fait la somme des potentiels d’action qui lui par-viennent provenant de chacune des fibres stimulées etrépond par un potentiel d’action : ce phénomène est appelésommation spatiale. L’explication repose sur l’organisa-
48
Synapse neuroneuronique Synapse neuromusculaire
Élément présynaptique (cellule) Neurone présynaptique Arborisation terminale de la fibre nerveuse
Élément postsynaptique (cellule) Neurone postsynaptique Fibre musculaire
Dans l’élément présynaptique Présence de vésicules synaptiques Présence de vésicules synaptiques
Limite de l’élément présynaptique Présence d’une membrane Présence d’une membrane présynaptique présynaptique
Limite de l’élément postsynaptique Membrane postsynaptique Une membrane postsynaptique existe mais elle est difficilementvisible sur cette électronographie
Discontinuité entre les deux Présence d’une fente synaptique Présence d’une fente synaptiquemembranes plasmiques

La synapse est polarisée structurellement et ce caractère seretrouve sur le plan fonctionnel.On fait l’hypothèse que les molécules d’acétylcholinecontenues dans les vésicules synaptiques du neurone pré-synaptique interviennent dans le fonctionnement de lasynapse neuromusculaire.Le DOCUMENT 13 montre que lorsque l’on dépose de l’acé-tylcholine sur une fibre musculaire, au niveau d’unesynapse, on constate que le potentiel membranaire de cettecellule, au départ de – 80 mV passe à + 50 mV. Cetteinversion brutale et brève du potentiel membranaire derepos de la cellule musculaire correspond au potentield’action musculaire. Ce potentiel d’action est à l’originede la contraction de la fibre musculaire. Ce potentiel d’ac-tion n’apparaît pas si on injecte l’acétylcholine à l’inté-rieur de la fibre musculaire montrant ainsi quel’acétylcholine agit au niveau de la fente synaptique etplus particulièrement au niveau de récepteurs membra-naires spécifiques situés sur la membrane postsynaptique(DOCUMENT 14). Cette substance chimique contenue dansles vésicules synaptiques, déversée dans la fente synap-tique et agissant sur des récepteurs situés dans la mem-brane synaptique modifiant l’activité de cette cellule estun neurotransmetteur.On peut penser que l’arrivée d’un potentiel d’action ner-veux au niveau du neurone présynaptique provoque lalibération de l’acétylcholine dans la fente synaptique,déclenchant un potentiel d’action musculaire à l’originede la contraction musculaire.Le DOCUMENT 15 confirme que, dans le cas de la synapseneuromusculaire, l’arrivée d’un potentiel d’action nerveuxà l’extrémité de la fibre du motoneurone est suffisantepour stimuler la fibre musculaire : la synapse neuromus-culaire fonctionne au coup par coup.Dans le cas des synapses neuroneuroniques, un autre neu-rotransmetteur peut être contenu dans les vésicules synap-tiques. La libération du neuromédiateur dans la fentesynaptique à la suite de l’arrivée d’un potentiel d’actionnerveux n’est pas suffisante pour déclencher un potentield’action dans le neurone postsynaptique. Cela confirmel’observation faite dans les activités 3 et qui montre que lasynapse neuroneuronique ne fonctionne pas au coup parcoup.On peut expliquer les propriétés de sommation des moto-neurones en sachant que l’effet du neuromédiateur sur lemotoneurone persiste pendant 10 à 20 millisecondes. Eneffet, un train de PA dont la fréquence est élevée et quiarrive au niveau d’une synapse va entraîner les libérationssuccessives et rapprochées de plusieurs quantités de neu-romédiateur dont les effets vont s’additionner permettantle déclenchement d’un potentiel d’action dans le neuronepostsynaptique. Le message nerveux présynaptique, codéen fréquence de potentiels d’action, est traduit en un mes-sage chimique codé en concentration de neurotransmet-teur. Ceci permet d’expliquer la sommation temporelle.De la même façon, on peut expliquer la sommation spa-tiale : il peut y avoir au même moment addition des effetsd’une libération simultanée de neurotransmetteur par plu-sieurs fibres sur le corps cellulaire du neurone et déclen-chement d’un potentiel d’action postsynaptique.
Les sommations temporelle et spatiale résultent bien descaractéristiques du fonctionnement synaptique.
A c t i v i t é s 5
Inhibition et réponse coordonnée des musclesLors du réflexe myotatique, l’étirement d’un muscleprovoque, par voie réflexe, sa contraction. Le muscleantagoniste, parallèlement, se relâche. Que se produit-il au niveau de ce muscle et plus particulièrement quelmessage les motoneurones véhiculent-ils juqu’à cemuscle permettant le relâchement?
Dans ces activités seront mis en évidence :– le phénomène d’inhibition ;– le lieu où s’exerce cette inhibition ;– les neurones inhibiteurs et le fonctionnement synap-tique de ces neurones.
ExploitationD’après le DOCUMENT 16, on constate que lorsque l’on étirevivement le muscle fléchisseur antagoniste d’un muscleextenseur (quadriceps) déjà contracté, la contraction duquadriceps cesse brusquement. Le réflexe myotatique duquadriceps est donc inhibé par les messages afférents issusdes fuseaux neuromusculaires du muscle antagoniste.À quel niveau se situe l’inhibition ? Comment circulent lesmessages dans les différentes fibres issues des fuseauxneuromusculaires de ces deux muscles antagonistes etquel est le type de messages partant des motoneurones etarrivant aux deux muscles?Le DOCUMENT 17 montre que le message afférent qui circuledans une fibre issue d’un fuseau neuromusculaire dumuscle extenseur est le même avant et au cours de l’étire-ment : l’inhibition ne se fait pas sur la voie afférente. Lemessage nerveux afférent qui circule dans une fibre issued’un fuseau neuromusculaire du muscle fléchisseur cor-respondant à une fréquence de potentiels d’action qui estaugmentée, s’explique par l’augmentation de l’intensité dela stimulation lors de l’étirement.Pour le message efférent : la fréquence des potentiels d’ac-tion du message qui circule dans une fibre issue d’unmotoneurone allant au muscle extenseur est importanteavant l’étirement, c’est ce message qui permet la contrac-tion musculaire du muscle extenseur. La fréquence despotentiels d’action de ce message est diminuée au cours del’étirement montrant que l’inhibition se fait au niveau desmotoneurones qui conduisent le message efférent aumuscle extenseur. La fréquence des potentiels d’action dumessage nerveux efférent qui circule dans une fibre issued’un motoneurone allant au muscle fléchisseur est nulleavant l’étirement du fléchisseur, elle est augmentée aucours de l’étirement à l’origine de la contraction de cemuscle.L’inhibition de la contraction du muscle extenseur se faitdonc au niveau des motoneurones innervant le muscleextenseur, et cette inhibition s’effectue par l’intermédiairedu message nerveux afférent issu du muscle fléchisseur.
Chapitre 8 : Fonctionnement des centres nerveux et mémoire génétique
49

Le GRAPHIQUE 19b. indique que lorsque le fléchisseur estrelâché, le réflexe myotatique se produit normalement ;lorsque ce fléchisseur est contracté, le réflexe myotatiquene se produit pas.Le DOCUMENT 20 montre que, quel que soit l’état de contrac-tion (volontaire) ou de décontraction du muscle fléchis-seur, quand on étire le tendon d’Achille, la fréquence despotentiels d’action du message qui est véhiculé par lesvoies nerveuses afférentes vers la moelle épinière est tou-jours la même : ce n’est pas sur la voie afférente que se faitl’inhibition. La fréquence des potentiels d’action qui cir-culent dans les fibres efférentes est diminuée indiquantque l’inhibition se fait au niveau des motoneurones desmuscles extenseurs.Le DOCUMENT 21 permet de comprendre les mécanismesmis en jeu. On constate que les motoneurones du muscleextenseur du pied sont en relation par l’intermédiaire d’in-terneurones avec des neurones corticaux moteurs d’oùpartent des messages commandant ce muscle extenseur.Lors d’une flexion volontaire du pied, les motoneuronesdu muscle extenseur sont inhibés par les messages desinterneurones inhibiteurs mis en jeu volontairement.Lorsque le réflexe myotatique achilléen est déclenché dansla situation où le pied est fléchi, les motoneurones condui-sant le message nerveux aux muscles extenseurs reçoiventà la fois des messages excitateurs venant des fuseaux neu-romusculaires et des messages inhibiteurs venant des neu-rones corticaux via les interneurones. Les motoneuronesréalisent alors la somme de ces deux types d’informationset si l’inhibition l’emporte, partiellement ou totalement,l’extension du pied peut être partielle ou inexistante (celadépendra de la force de contraction volontaire de départ).
� Le DOCUMENT 22 montre un autre type de réflexe : leréflexe nociceptif d’évitement dans lequel les neuronesafférents sont connectés aux motoneurones de façon com-plexe par l’intermédiaire d’interneurones excitateurs ouinhibiteurs.Dans le cas du réflexe myotatique et du réflexe nociceptif,chez tous les individus humains, on trouve le même typed’organisation, le même type de fonctionnement suggé-rant que la mise en place des neurones est sous la dépen-dance du génome. Le DOCUMENT 23 complète cetteinformation. Dans le cas d’une maladie congénitale d’ori-gine génétique d’insensibilité à la douleur, les fibres ner-veuses, et par là les neurones impliqués dans le réflexenociceptif, présentent une dégénérescence. Cette maladie,sous contrôle génétique, indique que la mise en place desneurones est sous la dépendance du génome.
L’explication est à rechercher au niveau du fonctionne-ment des synapses : certaines synapses sont inhibitrices,d’autres excitatrices.Le DOCUMENT 18a. précise le circuit neuronique du réflexemyotatique, c’est la mise en jeu d’un interneurone qui per-met cette inhibition. La fibre du neurone afférent en rela-tion avec un fuseau neuromusculaire se ramifie. Une deces ramifications établit un contact synaptique excitateuren libérant le glutamate comme neurotransmetteur avec lemotoneurone du même muscle, alors qu’une autre de cesramifications forme une synapse excitatrice (c’est tou-jours du glutamate qui est sécrété par la fibre au niveau dela synapse) avec l’interneurone inhibiteur. La fibre issuede cet interneurone forme une synapse inhibitrice (dont leneurotransmetteur est de la glycine) avec le motoneuronedu muscle antagoniste.Ainsi l’étirement d’un muscle active les motoneurones dumême muscle (par une voie ne comportant qu’une seulesynapse neuroneuronique : voie monosynaptique) etinhibe ceux du muscle antagoniste (par une voie où inter-vient un interneurone donc une synapse neuroneuroniquesupplémentaire : voie disynaptique).Le DOCUMENT 18b. montrera que le message nerveux issu dumuscle extenseur du pied inhibe les motoneurones dumuscle fléchisseur antagoniste et que, inversement, lemessage afférent de ce muscle fléchisseur inhibe les moto-neurones du muscle extenseur : ceci correspond à uneinnervation réciproque. Cette organisation assure la miseen jeu coordonnée des muscles.Il est à remarquer que ce sont des populations de neuronesqui interviennent et non un seul neurone à chaque fois.
A c t i v i t é s 6
Réseaux nerveux et déterminisme génétiquePar l’ensemble des cinq activités, il a été répondu auproblème suivant : comment l’organisation fonction-nelle du système nerveux permet-elle la réalisation duréflexe myotatique ?
Le réflexe myotatique peut être partiellement ou totale-ment inhibé par la commande volontaire ou par la sti-mulation d’autres récepteurs.
Dans cette dernière partie, on verra comment la com-mande volontaire va intervenir sur le réflexe myota-tique.
Le dernier point abordé permettra de montrer que lamise en place du système nerveux, et plus particulière-ment les chaînes de neurones, se trouvent sous lecontrôle de l’information génétique.
Exploitation� Le GRAPHIQUE 19a. montre que lorsque l’on fléchitvolontairement le pied, l’amplitude de l’électromyo-gramme enregistré à la suite de la percussion du tendond’Achille est diminuée.
50

E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Potentiel de repos : différence de potentiel quiexiste entre l’extérieur et l’intérieur de toute cellulevivante. Cette différence est de l’ordre de – 70 mV.Le potentiel de repos indique que la membraneplasmique est polarisée positivement à l’extérieur,négativement à l’intérieur.b. Potentiel d’action : inversion brutale et brève dupotentiel de repos à la suite d’une stimulation de lafibre nerveuse. Il est propageable à la surface de lafibre. À partir du moment où il se déclenche, l’ampli-tude est de l’ordre de 100 mV et est constantecomme sa durée (de l’ordre de 1 à 2 ms). Le PA estle messager nerveux ou signal de base du messageneveux.c. Potentiel global : phénomène électrique enregis-tré lorsqu’un nerf disposé sur 2 électrodes récep-tives est stimulé. L’amplitude du potentiel global estvariable et dépend du nombre de fibres du nerfrecrutées lors de la stimulation.d. Message nerveux afférent : message nerveux quicircule dans les fibres nerveuses afférentes (dans lecas du réflexe myotatique, il circule des fuseauxneuromusculaires au centre nerveux), ce messageest codé dans chaque fibre en fréquence de poten-tiels d’action.e. Message nerveux efférent : message nerveux quicircule dans les fibres nerveuses efférentes (dans lecas du réflexe myotatique, il circule du centre ner-veux aux fibres musculaires contractiles), ce mes-sage est codé dans chaque fibre en fréquence depotentiels d’action.f. Synapse : zone de contact entre l’extrémité d’unneurone et une autre cellule qui peut être soit unneurone (on a alors une synapse neuroneuronique),soit une cellule musculaire (on a alors une synapseneuromusculaire). La synapse est polarisée structu-rellement et fonctionnellement. La synapse neuro-neuronique ne fonctionne pas au coup par coupcontrairement à la synapse neuromusculaire. Dansle cas de la synapse neuroneuronique, il existe dessynapses excitatrices et des synapses inhibitrices.g. Sommation spatiale : propriété d’un neuronepostsynaptique de réagir à l’ensemble des informa-tions qui lui parvient. Dans le cas de la sommationspatiale, c’est la capacité pour le neurone de faire lasommation de tous les potentiels d’action qui luiarrivent au même moment.h. Sommation temporelle : propriété d’un neuronepostsynaptique de réagir à l’ensemble des informa-tions qui lui parvient. C’est la capacité pour le neu-rone de faire la sommation, au niveau d’une synapsedonnée, des potentiels d’action arrivant à quelquesmillisecondes d’intervalle.i. Neurotransmetteur : molécule libérée dans la fentesynaptique par le neurone présynaptique enréponse à l’arrivée d’un message nerveux et res-
1
ponsable de la transmission d’un message de ceneurone à une autre cellule excitable. Certains neu-rotransmetteurs sont excitateurs, d’autres sont inhi-biteurs.
Questions à réponses cour tesa. Le codage du message nerveux au niveau d’unefibre se fait en fréquence de potentiels d’action. Lecodage du message nerveux au niveau d’un nerf sefait par le nombre de fibres recrutées (ce qui se tra-duit à la surface du nerf par un potentiel global gra-duable en amplitude).Au cours de l’étirement d’un muscle, les motoneu-rones innervant ce muscle font une sommation spa-tiale et temporelle des messages afférents nés auniveau des fuseaux neuromusculaires. Le messagenerveux qui parcourt chaque fibre en relation avecun motoneurone est donc codé en fréquence depotentiels d’action. Au cours de l’étirement dumuscle, chaque motoneurone augmente l’émissiondes potentiels d’action vers les fibres musculairescontractiles.b. À la suite de l’arrivée d’un potentiel d’action, l’élé-ment présynaptique libère dans la fente synaptiquedes molécules de neurotransmetteur. En se fixantsur les récepteurs membranaires postsynaptiques,le neurotransmetteur est le stimulus de l’élémentpostsynaptique. Dans le cas de la synapse neuro-musculaire, un potentiel d’action nerveux déclencheà chaque fois un potentiel d’action musculaire àl’origine de la contraction. Dans le cas de la synapseneuroneuronique, la quantité de neurotransmetteurlibéré par l’arrivée d’un seul potentiel d’action estinsuffisante pour que le neurone postsynaptique yréponde par un potentiel d’action. Cela induit unemodification de l’activité du neurone postsynaptiquemais ce changement d’activité est insuffisant pourêtre à l’origine d’un potentiel d’action. En revanche,si en divers endroits de la surface du neurone post-synaptique, il y a libération simultanée de neuro-transmetteur, alors il y aura émission d’un potentield’action ; de même, si au niveau d’une seulesynapse, les potentiels d’action se suivent à une fré-quence élevée, alors les quantités de neurotrans-metteur s’ajoutent. De plus, suivant la nature duneurotransmetteur, le neurone postsynaptique eststimulé ou inhibé.c. L’étirement d’un muscle est à l’origine d’un mes-sage nerveux afférent qui active les motoneuronesdu même muscle et qui inhibe ceux du muscle anta-goniste. Cette double voie, simultanément mise nejeu, rend compte de la réponse coordonnée deseffecteurs musculaires.
Phrases à corrigera. Le message nerveux issu d’un fuseau neuromus-culaire se traduit, au niveau de la fibre afférentelorsque l’intensité de la stimulation augmente, parun train de potentiels d’action dont la fréquenceaugmente et dont l’amplitude reste constante.b. La synapse neuroneuronique ne fonctionne pasau coup par coup.
3
2
Chapitre 8 : Fonctionnement des centres nerveux et mémoire génétique
51

motoneurone à la suite de l’arrivée simultanée d’unplus grand nombre de potentiels d’action. Le moto-neurone a fait sommation des informations qui sontarrivées à sa surface.
4. Le phénomène mis en évidence dans cette expé-rience est le phénomène de sommation spatiale.
E x e r c i c e c o m p l é m e n t a i r e
Deux bactéries pathogènes,deux modes d’act ion différentsLe tétanos est une maladie extrêmement grave quidébute par une contracture douloureuse, invincible,permanente des muscles masticateurs, qui s’op-pose à toute ouverture de la bouche et gêne l’ali-mentation et l’élocution. Les contractures segénéralisent en quelques jours, atteignant l’en-semble de la musculature. Les troubles respira-toires, par contracture des muscles respiratoires, quibloquent la cage thoracique, peuvent entraîner l’as-phyxie. Cette maladie est due à une bactérie, lebacille tétanique, qui végète dans le sol et quipénètre dans l’organisme à la faveur d’une plaie nonsoignée.La toxine produite par ce bacille perturbe le fonc-tionnement des synapses inhibitrices sur les moto-neurones qui innervent les muscles squelettiques.
1. Expliquer comment le bacille tétanique peutentraîner une augmentation de la contracture mus-culaire dans l’organisme.La toxine de la bactérie botulique, bactérie que l’onpeut trouver dans des nourritures avariées, inhibe lalibération de l’acétylcholine au niveau de la synapseneuromusculaire. Elle provoque une paralysie desmuscles, souvent fatale, lorsque les muscles respi-ratoires sont atteints.
2. Expliquer comment la bactérie botulique peutentraîner la mort par asphyxie.
Réponses attendues1. Les axones d’un grand nombre de neurones éta-blissent des synapses sur un motoneurone pourréguler son activité. En permanence, les motoneu-rones reçoivent des influences excitatrice et inhibi-trice. La toxine tétanique élimine l’action inhibitriceexercée sur les motoneurones et laisse la seuleinfluence excitatrice. Il en résulte une activité exces-sive des motoneurones, qui provoque une contrac-tion involontaire des muscles squelettiques.2. La toxine botulique inhibe la libération d’acétyl-choline, elle empêche donc la transmission dusignal chimique entre le motoneurone et la fibremusculaire. L’arrivée du message nerveux à l’extré-mité de la fibre nerveuse n’entraîne donc pas la for-mation de potentiel d’action au niveau de la fibremusculaire. En l’absence de PA, celle-ci ne secontracte pas, les muscles sont paralysés, notam-ment les muscles intercostaux et du diaphragme.L’inspiration ne peut plus s’effectuer, la personneintoxiquée meurt par asphyxie.
7
c. Les messages nerveux nés de l’étirement d’unmuscle ont une action inhibitrice par l’intermédiaired’interneurones sur les motoneurones du muscleantagoniste.
Choisir les bonnes réponses1. Affirmations exactes : b. et c.2. Affirmations exactes : a., d. et e.
Exercice corrigé
Action du curare sur la synapseVoir correction dans le manuel pages 170 et 171.
Évaluer ses compétences
Sommation spatiale et temporel le1. Les enregistrements a correspondent aux mes-sages nerveux qui circulent dans la fibre afférente àla suite de la stimulation. Le stimulus correspond audéplacement de la soie dont l’angle de déplacementaugmente traduisant ainsi l’intensité de la stimula-tion. Les enregistrements correspondent à despotentiels d’action. Plus l’intensité de la stimulationaugmente et plus la fréquence des potentiels d’ac-tion est grande (pendant 120 ms, 2 pour a1, 4 poura2, 7 pour a3). Le codage du message au niveau dela fibre afférente se fait en fréquence de potentielsd’action.Les enregistrements b correspondent aux messagesnerveux qui circulent dans la fibre afférente à la suitede la stimulation. Les enregistrements correspon-dent à des potentiels d’action. Dans le cas de lapremière stimulation, on ne constate la présenced’aucun potentiel d’action. Lorsque la fréquencedes potentiels d’action augmente en b3, la fréquenceélevée des potentiels d’action déclenche l’émissiond’un potentiel d’action par le neurone géant. Le neu-rone géant a fait la somme des informations qui luiarrivent : c’est le phénomène de sommation tempo-relle.
2. On considère dans cette expérience un nerfdéposé sur des électrodes réceptrices (enregistre-ment c) que l’on stimule grâce à des électrodesexcitatrices. Au niveau des enregistrements, onconstate que, après un artefact de stimulation, onenregistre un potentiel dont l’amplitude varie de 3 à15 mV. Ce potentiel est un potentiel global de nerfdont l’amplitude varie en fonction de l’intensité de lastimulation. Cette augmentation de l’amplitude estdue au nombre de plus en plus grand de fibres ner-veuses mises en jeu.
3. Lors des stimulations I1 et I2, aucun potentiel d’ac-tion n’est enregistré au niveau du neurone géant :cela signifie que le neurone géant n’est pas suffi-samment stimulé par l’ensemble des potentielsd’action qui lui arrivent simultanément par les diffé-rentes fibres recrutées. L’enregistrement b3 corres-pond à un potentiel d’action émis par le
6
5
4
52

53
Programme : notions et contenus• Thème général : la part du génotype et la part de l’expérience individuelle dans le fonctionnement du système nerveux.• Des phénotypes à différents niveaux d’organisation du vivant.• Le cortex sensoriel et la plasticité du système nerveux central.• L’information sensorielle générée à la périphérie est transmise au cortex sensoriel.• Dans le cortex somatosensoriel, chaque territoire de l’organisme est représenté. Cette représentation est déformée par rapport à la surface desterritoires corporels.• Les zones corticales concernées sont constituées de neurones interconnectés et organisés en colonnes.• Des modifications de l’activité neuronale à la périphérie régulent l’organisation dynamique du cortex. Elles se traduisent par un remodelage desconnexions synaptiques, témoin de la plasticité neuronale.• La neuroplasticité est une propriété générale du système nerveux central.• Limites : les détails de l’organisation anatomique du cortex cérébral ne sont pas au programme.
Objectifs à atteindreLe phénotype comportemental dépend du génome et de l’environnement. Il évolue toute la vie par suite des interac-tions permanentes de l’organisme avec le milieu dans lequel il vit. Cette interaction permanente est à l’origine del’identité biologique de l’individu.
Les notions auxquelles ce chapitre fait référence ont été peu abordées dans les classes de collège et de lycée.Cependant, l’enseignant peut s’appuyer sur certaines d’entre elles vues en classe de troisième : « Relations à l’envi-ronnement et activité nerveuse ».
Les notions à construire sont les suivantes :
– le phénotype comportemental perceptif dépend de l’activité de zones particulières de notre cerveau : la perceptiondu monde est une création de notre cerveau ;
– les représentations du cortex somatosensoriel dépendent du génome de l’espèce. Elles sont modulées par les fac-teurs de l’environnement ;
– le développement du système nerveux résulte d’une collaboration entre le génome et l’environnement : c’est la plas-ticité cérébrale ;
– chez l’adulte, cette plasticité est maintenue, elle est liée à la plasticité synaptique et est à l’origine de notre indivi-dualité.
Cerveau, génotype et environnement
C H A P I T R E 9

Exploitation� Le DOCUMENT 2a. montre que lorsque deux doigts de lamain droite sont stimulés, une zone précise du cortexpariétal gauche est activée. Un message nerveux né auniveau des doigts de la main droite à la suite de leur sti-mulation est à l’origine d’une information sensoriellevéhiculée par des voies nerveuses afférentes jusqu’au cor-tex pariétal gauche. On peut noter que les voies afférentesdoivent se croiser.Le DOCUMENT 2b. montre que l’information sensorielle néeau niveau des yeux est transmise à une zone précise ducerveau : le cortex occipital.Il existe des aires corticales spécialisées dans le traitementdes informations venant des récepteurs périphériques.
� Dans le cas du DOCUMENT 3, des lésions dans des zonesprécises du cerveau entraînent des modifications de per-ception de l’environnement.La perte de la sensibilité de la moitié gauche du corps lors-qu’une zone du cortex pariétal de l’hémisphère cérébraldroit est lésée indique qu’il existe, au niveau du cortex,une aire spécialisée dans la sensibilité générale. Les récep-teurs périphériques sont intacts et pourtant la perceptionne se fait pas, montrant ainsi que cette perception se faitau niveau du cerveau.Pour Madame M. P., la perception des objets en mouve-ment est abolie. Cette modification de la perception est àmettre en relation avec une lésion bilatérale du cortex. Ilexiste, au niveau du cortex, dans chaque hémisphère céré-bral, une zone spécialisée dans la vision des mouvements.L’absence de cette perception est due à cette lésion : la per-ception naît bien au niveau du cerveau.Le DOCUMENT 1c. montre qu’à la suite d’une amputation,alors même que le membre ne peut plus être stimulé, dessensations semblent naître au niveau de ce membre : lesperceptions ne peuvent être qu’une création du cerveaupuisque des informations ne peuvent provenir de cemembre.La perception est une création de notre cerveau.
A c t i v i t é s 2
Les cartes corticalesSachant que les messages nerveux issus des organessensoriels arrivent dans des aires spécialisées du cer-veau, on s’intéresse à certaines de ces aires : celles quiconcernent la sensibilité générale : ce sont les airessomatosensorielles ou somesthésiques.
Deux objectifs sont poursuivis dans cette série d’activités :– la notion de carte ou représentation somatosenso-rielle qui correspond à la projection sur le cortex de lasensibilité générale des différents territoires de l’orga-nisme. Cette carte porte le nom d’homoncule. Cethomoncule n’est pas homothétique, il est déformé parrapport à la surface des territoires corporels et cettedéformation est à mettre en relation avec la densité desrécepteurs sensoriels présents à la surface de l’orga-nisme ;
◆ Problèmes scientifiques à résoudreLes structures du système nerveux central sont-elles inva-riantes au cours de la vie?Quelles sont les parts respectives du génome et de l’envi-ronnement dans la construction de notre cerveau?
P o s e r l e s p r o b l è m e s
Les vrais jumeaux ont le même génome ; du point de vuemorphologique et physiologique, ils sont très proches l’unde l’autre. Leurs cerveaux sont-ils les mêmes? Sont-ilsconstruits de la même façon?Les cerveaux 1 et 2 des jumeaux ont le même volume, cequi n’est pas le cas du cerveau 3 non apparenté. On peutproposer à l’élève de faire le contour du cerveau 1 avec unfeutre sur un transparent, puis de superposer celui-ci sur le2 et le 3. On constate un volume cérébral identique pourles cerveaux 1 et 2 et différent du cerveau 3. Pour les cer-veaux 1 et 2, on observe que les circonvolutions cérébralessont différentes. Un apport du professeur permet unegénéralisation : l’examen de dix paires de jumeaux mono-zygotes effectué par Bartley et ses collègues duNeuroscience Center de Washington montre que le volumedu cerveau est presque entièrement déterminé par des fac-teurs génétiques tandis que le dessin des circonvolutionsdépendrait surtout de facteurs environnementaux (LaRecherche, p. 77, juillet-août 1998).On peut donc s’interroger sur la part respective de la géné-tique et de l’environnement sur la construction du cerveau.L’illusion que le membre amputé soit toujours présent per-met de s’interroger sur l’intervention du cerveau dans laperception du monde.
A c t i v i t é s 1
Perception du monde et cerveauL’objectif de ces activités est de montrer que la percep-tion de notre environnement dépend de l’activité denotre cerveau.
Pour atteindre cet objectif, on utilise :– des données de l’imagerie cérébrale révélant leszones du cortex activées par les stimuli externes ;– d’autres données relatives à des troubles liés à deslésions de zones corticales bien définies.Les documents permettent de montrer que :– le cerveau, et plus particulièrement le cortex senso-riel, reçoit de nombreux messages issus de nos récep-teurs sensoriels périphériques ;– ces messages nerveux sont traités dans des airescérébrales spécialisées ;– la perception du monde qui nous entoure dépend del’interprétation qu’en fait notre cerveau : la perceptiondu monde est donc une création de notre cerveau.
54

– les cartes somatosensorielles sont spécifiques de l’es-pèce ce qui indique qu’elles sont sous la dépendance defacteurs génétiques.
Exploitation� DOCUMENT 4a. : la représentation en coupe du cortexsomatosensoriel montre une projection des différents ter-ritoires corporels sur ce cortex. Toutes les parties du corpssont projetées sur le cortex, mais on constate que les dif-férentes régions du corps sont déformées, que l’étenduedu cortex somatosensoriel représentant les mains et la faceest beaucoup plus importante que leur surface relative parrapport au reste du corps. La face et les mains sont surdi-mensionnées par rapport au torse et aux segments proxi-maux des membres.De plus, on constate que les différentes régions du corpsne sont pas contiguës : la face se trouve près du pouce parexemple.DOCUMENT 4b. : l’importance des différents territoires auniveau du cortex est confirmée par l’homoncule (homon-culus = petit homme). Les régions correspondant à la faceet à la main sont très étendues.Ces observations sont à mettre en relation avec le DOCUMENT 5 qui montre que la sensibilité tactile est impor-tante (discrimination tactile fine) au niveau de la face :nez, joue, lèvre supérieure et au niveau des différentsdoigts de la main.La surface de l’homoncule est directement proportionnelleà la densité des récepteurs sensoriels présents à la surfacedu corps.
� L’homoncule ou homonculus, représentation corticalesomatosensorielle de l’Homme et le ratonculus, représen-tation somatosensorielle du Rat sont différents. Dans lesdeux cas, on reconnaît « l’image » corporelle de l’espèce.Dans le cas du ratonculus, la représentation de la tête etplus particulièrement des vibrisses du museau occupe unesurface du cortex importante : les vibrisses qui se trouventà la surface du museau sont essentielles pour la détectionde l’environnement du rat.Les afférences sensorielles qui sont particulièrementimportantes pour une espèce donnée ont une représenta-tion corticale proportionnellement plus étendue.Les différences constatées sont sans doute dues au fait quel’on considère deux espèces différentes chez lesquellesl’importance relative des capteurs sensoriels à la surfacede l’organisme est différente. L’hypothèse que l’on peutformuler est que ces différences constatées sont dues à desdifférences de génomes, le cortex somatosensoriel dépenddonc du génome caractéristique de l’espèce.
� Le DOCUMENT 7a. correspond à une reproduction duschéma du tissu nerveux du cortex visuel du rat dessinépar Cajal en 1888. Une observation au microscope optiquemontrerait le même type d’architecture. (voir doc. 3p. 184). Le cortex cérébral est une fine couche de sub-stance grise d’environ 5 mm d’épaisseur qui recouvre lesdeux hémisphères cérébraux du cerveau. Lorsqu’onobserve ce cortex, on constate l’existence de six couchessuperposées dans lesquelles se trouvent des neuronesconnectés grâce à leurs prolongements. La mise en placede ces six couches au cours du développement s’effectue
toujours de la même façon suggérant que cette construc-tion est sous la dépendance du génome de l’espèce. Cetteorigine génétique est confirmée par le texte du DOCUMENT7b. relatant les conséquences d’une mutation sur l’organi-sation du cortex dans le cas des souris reeler : l’allèle dugène responsable a été identifié, l’expression de ce gèneaffecte le développement des neurones du cortex cérébral.
A c t i v i t é s 3
La plasticité cérébrale au cours du développementL’objectif de ces activités est de montrer que les repré-sentations corticales sont bien sous la dépendance defacteurs génétiques mais que, au cours du développe-ment, des facteurs environnementaux vont intervenirsur la construction des représentations corticales.Cette construction du cerveau est malléable en fonctiondes conditions extérieures ; il y a au sein de l’espèceune variabilité individuelle indépendante du génome etfonction de l’expérience individuelle : c’est la plasticitécérébrale. Pour mettre en évidence cette plasticité céré-brale, deux types d’observation peuvent être faites :
– celles correspondant à la suppression des entréespériphériques ;– celles correspondant à des augmentations de cesentrées périphériques : stimulations de certains organesdes sens.
Exploitation� L’observation des DOCUMENTS 8b. et 8c. permet deconstater que le motif précis des vibrisses observé sur lemuseau est projeté sur le cortex somatosensoriel et quechaque tonneau correspond à une vibrisse. Le fait de véri-fier qu’à chaque vibrisse du museau correspond un ton-neau du cortex permettra ensuite à l’élève de constaterque, s’il y a absence de vibrisse, il y a absence de tonneau.
� DOCUMENT 9 : Si on effectue l’ablation d’une rangéehorizontale de vibrisses chez un raton, on constate qu’ilmanque une rangée horizontale de tonneaux correspon-dant à la projection de la rangée de vibrisses chez l’animaltémoin (DOCUMENT 8). De même, si on effectue l’ablationd’une rangée verticale de vibrisses, on constate l’absenced’une rangée verticale de tonneaux, tonneaux correspon-dant à la projection de la rangée de vibrisses chez l’animaltémoin, et l’occupation de l’espace laissé vaccant par lestonnaux non développés.Cette observation montre que la présence des récepteurssensoriels à la naissance et leurs stimulations jouent unrôle important dans la construction de la structure céré-brale. L’absence de la circulation de messages des récep-teurs sensoriels à certaines zones du cortex ne permet pasla construction de représentations corticales normales : ily a une réorganisation corticale.Il semble exister une période critique (le 3e jour) en deçàde laquelle les facteurs de l’environnement (ici l’ablationdes vibrisses) entraînent des modifications irréversibles.
Chapitre 9 : Cerveau, génotype et environnement
55

On montrera, dans un dernier temps, que cette plasti-cité cérébrale est liée à la plasticité synaptique.
Exploitation� DOCUMENT 14 : Chez les singes dont les racines dorsalesen relation avec les membres antérieurs ont été section-nées, c’est-à-dire chez des singes désafférentés, onconstate une modification des représentations corticales,l’aire du visage proche des aires du bras et de la main s’estétendue. Il y a donc une réorganisation corticale, à la suitede l’absence d’arrivée de messages nerveux afférents pro-venant de la main et du bras.DOCUMENT 15 : Cette précédente observation peut être miseen relation avec l’observation faite par Ramachandran en1993. L’observation réalisée par Ramachandran montreque chez le sujet observé, l’absence de main et d’avant-bras a sans doute conduit à une réorganisation corticalecomme chez le singe. Si on regarde l’homoncule du cor-tex somatosensoriel, on constate que les aires du visage,de la main et de l’avant-bras sont proches les unes desautres montrant qu’il y a un degré important de plasticitéfonctionnelle dans le cortex cérébral humain.
� DOCUMENT 12 : Chez le singe, on constate qu’après uneutilisation préférentielle d’un doigt (le singe est soumis àla même tâche qu’il répète des milliers de fois) la repré-sentation corticale de la zone du doigt stimulée augmentede surface, s’étend au dépend des autres doigts montrantune réorganisation corticale qui confirme la plasticitécérébrale.L’exercice 7 page 189 pourra compléter cette étude. Desvariations des surfaces des aires somatosensorielles peu-vent être constatées à la suite d’amputation d’un doigtchez le singe hibou. Chez l’Homme, on peut observer lemême type de réorganisation corticale dans le cortex desviolonistes : voir l’exercice 6 page 188 dont le documentpeut être utilisé comme texte d’activité.DOCUMENT 13 : Chez l’Homme, les personnes qui présen-tent un syndrome de syndactylie à la naissance et chez les-quelles une opération chirurgicale permet la séparationdes doigts montrent des modifications sensibles et rapidesdes cartes corticales liées à ces doigts.Il y a bien dans ce cas, là encore, une malléabilité du cor-tex cérébral : une plasticité cérébrale.
� Lorsque l’on stimule le siphon de l’Aplysie, un mes-sage nerveux circule dans le neurone sensoriel puis dans leneurone moteur provoquant la contraction des muscles dusiphon, du manteau et des branchies. Lors du conditionne-ment, un autre message circule parallèlement entre le neu-rone sensoriel de la queue, l’interneurone et lemotoneurone. La mise en jeu des deux circuits en mêmetemps entraîne une contraction plus forte des muscles. Siau bout d’un certain nombre de fois, seul le siphon est sti-mulé, on constate que la contraction est aussi importanteque si la stimulation du siphon avait été suivie de la sti-mulation de la queue. Cette observation montre que lasynapse présente entre le neurone sensitif du siphon et lemotoneurone a été modifiée. Il y a une augmentation del’efficacité de la synapse entre le neurone sensoriel prove-nant du siphon et le neurone moteur entraînant un change-ment dans son fonctionnement. Cette augmentation de
DOCUMENT 10 : L’apparition chez un rat nouveau-né de ton-neaux après la transplantation d’un fragment de cortexvisuel dans lequel ne se forment jamais de tonneaux, dansla région du cortex somatosensoriel montre que le cortexvisuel placé dans la région du cortex somatosensoriel,évolue différemment. Dans cette région, il reçoit des affé-rences différentes. Le tissu nerveux est plastique puisquecapable d’acquérir de nouvelles structures dans un envi-ronnement particulier.DOCUMENT 11 : Les expérimentations effectuées chez lesanimaux sont confirmées par les observations faites chezl’Homme. Chez les aveugles de naissance, le cortex visuelqui n’est pas sollicité par la fonction visuelle est mis enactivité lors d’exercices de lecture du braille mettant en jeula sensibilité des doigts. Le cortex visuel est mis à profitpour la reconnaissance des lettres en braille par le toucher.Chez l’individu témoin, seul le cortex somatosensoriel estactivé.Ces trois documents montrent que la structure cérébrale aucours du début de la vie de l’individu est malléable, on ditque cette structure cérébrale est plastique. La structurecérébrale dépend du génome, mais elle se modifie aucours du développement de l’individu sous l’influence del’environnement.
A c t i v i t é s 4
La plasticité cérébrale chez l’adulteDans cette double page, on montrera que la plasticitécérébrale observée au cours du développement de l’in-dividu est maintenue chez l’adulte. L’individualité dechacun d’entre nous est à relier à cette plasticité.L’architecture du cerveau de chaque individu est modi-fiée de façon unique. L’environnement est différent pourchaque individu et celui-ci n’intervient pas de la mêmefaçon en fonction des allèles des gènes que l’individupossède.
On pourra à la fin de ce chapitre revenir sur la page« Poser les problèmes » et répondre aux questionsposées au départ. Les vrais jumeaux qui ont le mêmegénome n’ont pas le même cerveau car la part de l’ex-périence individuelle dans la construction du cortex(somatosensoriel en particulier) n’est pas la mêmepour chacun des jumeaux.
Chez l’adulte, la plasticité est maintenue, des modifica-tions des représentations corticales peuvent résulter del’entraînement ou au contraire résulter de la perte oude la dénervation des membres.
Pour construire ces notions, là encore on fera deuxtypes d’observations :– des stimulations préférentielles dans le cas d’expé-riences chez l’animal ou de constats chez l’Homme;– les conséquences de désafférentations lors d’expéri-mentations chez l’animal ou d’accidents chezl’Homme.
56

l’efficacité est due à l’action préalable de la synapse pro-venant de l’interneurone et qui se connecte au niveau de laterminaison présynaptique du neurone sensoriel. L’activitéaccrue de la synapse entraîne des changements dans sonfonctionnement : libération plus importante de neurotrans-metteur. Il y a donc acquisition d’une nouvelle caractéris-tique de cette synapse : c’est la plasticité synaptique.
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Aires cérébrales sensorielles : ce sont les surfacesdu cortex cérébral recevant des messages prove-nant des récepteurs sensoriels périphériques. Ilexiste plusieurs aires cérébrales sensorielles corres-pondant aux différents sens.b.Représentations corticales : dans le cas du cortexsomatosensoriel (correspondant à l’aire de la sensi-bilité générale) ce sont les projections de chaquerégion du corps donnant un homoncule déformédont la surface est proportionnelle à la densité desrécepteurs sensoriels présents à la surface de l’or-ganisme.c.Plasticité cérébrale : capacité du système ner-veux, dont la construction est sous la dépendancedu génome, à se modifier en fonction des facteursde l’environnement (expérience individuelle) aucours du développement ou chez l’adulte ; les modi-fications pouvant être fonctionnelles et/ou structu-rales.d. Plasticité synaptique : modifications desconnexions entre les neurones par renforcement,formation, disparition de synapse. La plasticité céré-brale est une conséquence de la plasticité synap-tique.
Questions à réponses cour tesa. La perception de notre environnement dépenddes récepteurs sensoriels périphériques que nouspossédons, du traitement des messages nerveuxissus de ces récepteurs sensoriels et du traitementdes messages nerveux issus de ces récepteurs parles aires cérébrales sensorielles spécialisées.b. Le cortex somatosensoriel correspond à cer-taines aires corticales sensorielles qui traitent desinformations concernant le toucher et les sensationsde pression en provenance de la surface du corps,des muscles et des articulations. Il représentechaque territoire de l’organisme. Le cortex somato-sensoriel reste sous la dépendance du génomecaractéristique de l’espèce, il est modulé par desfacteurs de l’environnement.c. La construction des représentations corticales(somatosensorielles) dépend du génome de l’es-pèce : l’homonculus est différent du ratonculus.Cette construction est modulée par des facteurs del’environnement : dans le cas du rat par exemple, lasuppression d’une rangée de vibrisses à la nais-
2
1
sance entraîne une modification de la représentationsomatosensorielle dans la mesure où certains motifscorrespondant à des tonneaux situés normalementà un endroit bien précis du cortex et à la projectionde cette rangée de vibrisses sont dans ce cas,absents.d. Le développement du système nerveux d’un indi-vidu est sous la dépendance du génome et estmodulé par les facteurs de l’environnement. Cettecapacité du système nerveux de se modifier sousles facteurs de l’environnement (l’expérience indivi-duelle) constitue la plasticité cérébrale. Cette plasti-cité cérébrale est due à la plasticité synaptique.Cette plasticité est à la fois fonctionnelle et structu-rale : augmentation du nombre des connexions,activité augmentée des synapses.
Phrases à corrigera. Au cours du développement chez le jeune, lastructure du cerveau dépend de facteurs génétiqueset de facteurs de l’environnement.b. La plasticité chez l’adulte est importante car ilpeut y avoir de nouvelles connexions synaptiquesmais de nouveaux neurones ne peuvent se mettreen place.c. Chez l’adulte, après un accident cérébral vascu-laire, une récupération est possible car de nouvellesconnexions synaptiques peuvent se faire.
Choisir la bonne réponse1. Affirmations exactes : c. et d.2. Affirmations exactes : d.
Évaluer ses compétences
Des représentat ions cor t icalessomatosensoriel les différentesOn constate que les représentations corticales chezles différentes espèces n’ont pas le même aspect.Cette variabilité est due au génome des différentesespèces. Chaque représentation rappelle bien lamorphologie de chacune des espèces mais cesreprésentations présentent des déformations impor-tantes en fonction de l’importance de la sensibilitésomatosensorielle des différentes régions du corps.Dans chaque cas, les régions ayant les innervationssensorielles les plus denses sont les plus représen-tées : c’est le cas du singe araignée dont la surfacede la représentation corticale des membres et de laqueue préhensile est grande car ces organes jouentun rôle important dans le déplacement de cet ani-mal ; c’est le cas du raton laveur dont la représenta-tion corticale des membres antérieurs estimportante car ceux-ci jouent un rôle important dansla préhension des objets ; c’est le cas du rat dont lemuseau est très représenté au niveau cortical car lesvibrisses ont un rôle dans la détection de l’environ-nement. Dans le cas du mouton, aucune région enparticulier ne semble être privilégiée ; de plus, cettereprésentation corticale présente des discontinuitéscomme dans le cas de l’homonculus.
5
4
3
Chapitre 9 : Cerveau, génotype et environnement
57

Apprentissage chez les s inges et chez les rats1. On constate que plus l’ablation du cortex frontala été réalisée tôt dans la vie du singe et plus lenombre d’erreurs au test d’apprentissage par rap-port au témoin non opéré est faible. Les lésionscérébrales précoces entraînent un déficit comporte-mental moins important que lorsque la lésion se faitplus tard.On peut expliquer cette observation par le faitqu’une réorganisation neuronique laissant peu deséquelles peut se faire plus facilement quand lalésion a été réalisée tôt, avant même la naissance dusinge dans ce cas précis. La neuroplasticité est plusimportante chez les très jeunes singes que chez lessinges plus âgés.
2. Les rats qui ont subi la pseudo-opération peuventêtre considérés comme des rats témoins : ils ontsubi le traumatisme de l’opération sans subir lalésion du cortex occipital. Or, ce sont les consé-quences de cette lésion qui intéressent les cher-cheurs et le non traumatisme.
3. Comparaisons des lots A et C : pas de lésionchez ces deux lots, différences au niveau de l’envi-ronnement : on constate que les rats qui ont subiune lésion corticale ont un nombre moyen d’erreursélevé au test d’apprentissage par rapport aux ratsn’ayant pas subi de lésion. Lorsque les rats sont pla-cés dans un milieu pauvre en stimulations, lenombre d’erreurs augmente par rapport aux rats quisont dans les mêmes conditions (lésion ou absencede lésion). Cette étude a permis de montrer que l’en-vironnement joue un rôle important dans la récupé-ration après une atteinte cérébrale.
8Le cerveau des violonistesChez les violonistes, la représentation corticale del’auriculaire gauche est augmentée par rapport à unsujet témoin. L’auriculaire gauche est un doigt utilisépréférentiellement par les violonistes et son utilisa-tion répétée entraîne une modification par augmen-tation de la surface au niveau de la représentationcorticale du cortex somatosensoriel montrant laplasticité cérébrale. Plasticité cérébrale plus impor-tante avant l’âge de 13 ans ce qui correspond à lafin de la mise en place du système nerveux chezl’homme : le cortex n’acquiert son organisation défi-nitive que vers l’âge de 10 ans. Cette plasticité estmaintenue chez l’adulte puisque, même après cetâge, l’agrandissement de la représentation corticalede l’auriculaire est encore substantiel.
Amputation chez un singe hibou1. Surface des aires corticales des cinq doigts avantamputation : 4,53 mm2.Surfaces des aires corticales des cinq doigts aprèsamputation : 4,67 mm2.
2. Pourcentage d’augmentation des différentsdoigts après amputation du doigt 3.Doigt 1 : + 4 %.Doigt 2 : + 80 %.Doigt 4 : + 64 %.Doigt 5 : – 16 %.On constate que, pour les doigts, 1, 2 et 4, il y a uneaugmentation de la surface de la représentation cor-ticale. En revanche, il y a une diminution de cettesurface pour le doigt 5. La représentation fonction-nelle corticale s’étend le plus pour les doigts 2 et 4qui, dans les tâches courantes doivent compenserl’absence du doigt 3 amputé. La réduction de lareprésentation corticale du doigt 5 peut peut-êtres’expliquer par une moindre utilisation de ce doigt àla suite de l’amputation.
7
6
58

Pa
rt
ie
1 Des phénotypes à différents niveaux d’organisation du vivant
Génotype, phénotype et morphogenèsevégétale
10 La diversité phénotypique des végétaux ...................................................................... 60
11 Les méristèmes et la croissancedes végétaux ...................................................................... 66
12 Le contrôle de la croissancechez les végétaux ........................................................ 74
Unité 4

60
◆ Problèmes scientifiques à résoudreLes variations phénotypiques observées au sein d’unemême espèce posent le problème de leur déterminisme.Quels sont les rôles du génome et de l’environnement dansla réalisation du phénotype d’un végétal ? Comment peut-on expliquer qu’un végétal puisse modifier sa forme aucours de sa croissance et continuer celle-ci, même lors-qu’il est devenu adulte ?
P o s e r l e s p r o b l è m e s
L’observation d’arbres d’espèces différentes, vivant dansdes environnements variés, mais tous soumis à un fort ventdominant, montre qu’ils possèdent tous une morphologiesemblable, leur port étant communément dit « endrapeau ». La présence de la mer, visible sur une des pho-tographies proposées, laisse penser que leurs ramifications
ne peuvent se développer contre le vent dominant. L’idéed’une influence des facteurs du milieu dans l’établisse-ment du phénotype se trouve renforcée par l’observationde deux hêtres soumis à des conditions environnementalesdifférentes. D’autres observations peuvent être effectuées au cours dela sortie de terrain, telle celle d’écotypes d’une mêmeespèce, présentant un port en rosette en altitude, et un portdressé en plaine. Les problèmes scientifiques à résoudreseront alors les mêmes que dans les exemples précédents.Dans tous les cas, cette approche doit faire surgir des inter-rogations scientifiques, en se gardant de conclusionshâtives, souvent erronées à partir des seules observations.En cela, les documents proposés, associés aux observa-tions de terrain, ou les rappelant, permettent de susciterune attitude scientifique face au monde végétal, dont lefonctionnement est encore méconnu au début de ce cha-pitre.
Programme : notions et contenus• La morphologie d’un végétal dépend en partie des caractéristiques génétiques de l’espèce.• En fonction de leur environnement, des individus d’une même espèce peuvent avoir des morphologies différentes.• Des réponses morphologiques semblables peuvent être obtenues avec des végétaux d’espèces différentes placées dans un même environnement.
Objectifs à atteindreL’étude de la morphogenèse chez un végétal permet, probablement plus aisément que chez les animaux, de montrerque la réalisation du phénotype ne saurait dépendre de la seule expression de son génotype. Des observations réali-sées au cours de la sortie sur le terrain, dans le cadre du programme de géologie, auront mis en évidence la grandeplasticité phénotypique de végétaux de la même espèce placés dans des conditions différentes de milieu, ou celle devégétaux d’espèces différentes ayant grandi dans le même environnement. Il s’agit donc ici d’établir la réalisation duphénotype comme le résultat d’interactions constantes entre l’expression d’un génotype et son contrôle par l’envi-ronnement. Ce premier chapitre consacré à la morphogenèse végétale permet de dégager des problèmes scientifiquesà partir de la mise en relation de données issues d’observations, macroscopiques et microscopiques, de pratiquesagricoles ou de résultats expérimentaux. Les problèmes ainsi dégagés seront résolus au cours des deux chapitres sui-vants, avec l’étude des mécanismes de la croissance et du rôle joué par les hormones végétales, en interaction avecles facteurs de l’environnement. Il convient donc, avec ce chapitre, de situer cette étude dans le cadre plus vaste desrelations entre l’environnement et le génotype dans l’établissement du phénotype individuel. Les activités proposéespermettent à la fois d’établir les connaissances indispensables à la compréhension des activités ultérieures et, à par-tir d’observations simples, de montrer que la formulation des problèmes à résoudre constitue une étape essentielle dela démarche scientifique.
La diversité phénotypique des végétaux
C H A P I T R E 1 0

Chapitre 10 : La diversité phénotypique des végétaux
61
dépende de l’environnement, permet de formuler, à unniveau simple, une hypothèse quant à l’action d’un fac-teur du milieu dans l’établissement du phénotype. Lesobservations initiales de variétés de Pois nains et nor-maux mettent en évidence un déterminisme génétiquede l’élongation des entre-nœuds. Le problème peutalors être posé du rôle de l’environnement chez desespèces où l’élongation des tiges dépend de l’allonge-ment de la durée journalière d’éclairement. Les résul-tats expérimentaux proposés à propos de l’élongationde la tige, indispensable à la floraison des épinards,entraînent à une démarche scientifique rigoureuse. S’iln’est pas utile, à ce stade, de définir la gibbérellinecomme une hormone végétale, les conditions de sa syn-thèse doivent être envisagées en mobilisant les connais-sances acquises sur l’expression des gènes. À la fin deces activités, l’action d’un facteur de l’environnementdoit être située, non au niveau du génome lui-même, entant que facteur mutagène, mais à celui de son expres-sion.
Exploitation� Le DOCUMENT 5 présente deux variétés de Pois, dontl’une est génétiquement naine. Leur appartenance à lamême espèce doit être reliée à l’existence de nombreuxcaractères morphologiques communs, comme la forme etla coloration des feuilles, bien visibles sur les documents5b. et 5c. Il en est de même pour les feuilles d’épinardsobservées sur les DOCUMENTS 6 et 8. Ces caractères dépen-dent d’un programme génétique commun à tous les indi-vidus d’une même espèce. Chez le Pois, seule l’élongation des entre-nœuds dépendde la nature des allèles possédés par la plante. En fait, plu-sieurs gènes sont impliqués dans la synthèse de gibbérel-line, principale hormone responsable de l’élongation desentre-nœuds. Parmi ceux-ci, on trouve les gènes «Le»,dont la transmission de certains allèles a permis à Mendeld’établir les bases des lois de l’hérédité, et «Na». Lamutation en double exemplaire d’un seul de ces gènesconduit à l’absence de gibbérelline active, et donc à unevariété naine.
� Le DOCUMENT 6 montre que, chez l’Épinard, l’élongationde la tige ne se produit qu’au printemps dans les condi-tions naturelles, différentes de celles de la culture maraîchère lorsqu’on ne désire pas obtenir de graines.L’hypothèse du rôle joué par l’allongement de la duréequotidienne d’éclairement peut être formulée, puis validéepar les données du DOCUMENT 7. On doit cependant se garder de conclusions qui auraient une portée plus générale : si, chez l’Épinard, la photopériode semble responsable de l’allongement des entre-nœuds, chez leChou (Brassica sp.), c’est une combinaison de plusieursfacteurs, température et photopériode, qui déclenche l’allongement de la hampe florale.
� Le DOCUMENT 8 permet un premier niveau d’explicationdu mode d’action de la photopériode. On ne peut conclureici qu’à l’induction de la synthèse de gibbérelline par lesjours longs, au-delà d’une certaine durée, et à l’action dela gibbérelline sur l’élongation des pétioles et des tiges.
A c t i v i t é s 1
Environnement et phénotypeL’observation d’un Hêtre en lisière de forêt prolonge,mais en faisant appel à un facteur environnemental dif-férent, les observations faites dans «Poser les pro-blèmes». Elle permet de prendre conscience de ladiversité des facteurs du milieu susceptibles de modifierle phénotype d’un végétal. L’exemple de la Sagittairemontre l’influence d’un autre facteur du milieu sur ledéveloppement des feuilles. Cet exemple permet d’envi-sager les influences respectives du milieu et de l’envi-ronnement : si la morphologie des feuilles de Sagittairen’est pas génétiquement déterminée... elle n’en n’estpas moins celle de feuilles de Sagittaire. L’objectif estdonc de dégager un problème scientifique majeur, laplace tenue par le milieu et celle tenue par l’hérédité,dans la réalisation du phénotype individuel. Uneapproche véritablement scientifique de ce problèmesera proposée avec les activités suivantes.
Exploitation
� Le DOCUMENT 3 montre un développement plus impor-tant des ramifications du hêtre du côté le mieux éclairé.Même si l’hypothèse d’une photosynthèse plus importantepour les feuilles les plus exposées à la lumière peut êtreproposée, cette observation, assez commune, ne permetpas de conclure quant au mécanisme responsable de cettedissymétrie, mais seulement d’envisager une influence del’éclairement sur les mécanismes participant à la crois-sance du Hêtre. Ce document doit être rapproché du DO-CUMENT 1, et peut-être de l’observation d’une futaie ren-contrée au cours de la sortie, pour une comparaison desdifférents phénotypes du Hêtre. Le DOCUMENT 4 permet dedistinguer l’importance d’un facteur du milieu, la hauteurde l’eau, par rapport à celle des facteurs génétiques. Eneffet, les différents types de feuilles apparaissant sur lemême individu, les feuilles possèdent toutes le mêmegénome. Cependant, les observations ne rendent pascompte de la manière, au niveau moléculaire, dont peuts’exercer l’influence des facteurs de l’environnement.
A c t i v i t é s 2
Le rôle du génome et celui de l’environnementAvant d’envisager l’influence d’un facteur de l’environ-nement, il apparaît nécessaire de s’assurer que la mor-phologie d’un végétal ne dépende pas uniquement deson génome. Le choix de plantes naines, et de plantesde la même espèce dont les entre-nœuds sont allongés,se justifie par le fait que le caractère considéré neconcerne qu’une partie limitée du génome, dont l’ex-pression est aisément observable. Par ailleurs, le faitque, chez d’autres espèces, l’élongation des tiges

différents moments de l’éclosion d’un bourgeon. Alorsque les activités précédentes avaient permis d’appré-hender le rôle de l’environnement sur la morphogenèsevégétale, on met ici en évidence le fait que cette mor-phogenèse se réalise tout au long de la vie d’une plante.Ces deux aspects de la croissance des végétaux ayantété abordés au cours de ce chapitre, l’étude de l’activitédes méristèmes, puis celle du contrôle de la croissancecellulaire, pourront être entreprises au cours des cha-pitres suivants.
Exploitation� Les différentes opérations d’une greffe en écusson sontreprésentées sur le DOCUMENT 9 dans leur continuité. Onprélève un rameau (a.), puis la base d’une feuille compor-tant un seul bourgeon axillaire, accompagné d’un lambeaud’écorce et de vaisseaux conducteurs de sève (b.). Onincise l’écorce du porte-greffe (d.) et on installe le greffonen réalisant une ligature afin de permettre l’établissementd’une continuité entre les vaisseaux conducteurs du gref-fon et ceux du porte-greffe (e.). L’observation du greffonisolé présenté sur la paume d’une main (c.), doit être com-parée à celle d’un poirier adulte, ici greffé sur un cognas-sier (g.). On en déduit qu’un seul bourgeon axillairerenferme potentiellement toutes les ramifications qui sedévelopperont ultérieurement. On peut alors penser queles ramifications naturelles d’un arbre dépendent du déve-loppement, ou non, de ses bourgeons axillaires.
� L’étude du DOCUMENT 10 suit logiquement celle du DOCU-MENT 9. Le choix de ce document a été dicté par le souci deproposer une image d’un bourgeon axillaire d’une espèceligneuse : le bourgeon axillaire d’Érable est fort semblableau bourgeon de Poirier utilisé pour la greffe en écusson.On remarque la taille réduite du bourgeon axillaire et sasituation, à l’aisselle d’une feuille de l’année. De nou-velles feuilles sont déjà présentes, à l’état d’ébauche, àl’intérieur du bourgeon, de part et d’autre du méristèmeaxillaire. Le DOCUMENT 11 présente un bourgeon terminal, ou apical,d’une plante herbacée, le Coléus. Si ce bourgeon ne pos-sède pas les organes nécessaires à sa pérennité, onretrouve cependant des structures communes à tous lesbourgeons, un méristème et l’ébauche des feuilles.L’existence de bourgeons axillaires doit également êtresoulignée, car elle élargit la présence des méristèmes à dif-férents endroits, bien localisés, de l’anatomie d’un végé-tal. À ce stade, il ne paraît pas possible de conclure quantau rôle des méristèmes dans la croissance des feuilles, dela tige ou du rameau. Tout au plus, la coloration rougesombre, identique à celle des noyaux des cellules différen-ciées, suggère que les cellules qui les constituent sontnombreuses, de petite taille par rapport à celle de leurnoyau, et possèdent des parois minces, alors que celles desautres cellules, plus larges, apparaissent colorées en vertsombre. Une analyse du DOCUMENT 12 permet de penser que lesméristèmes sont bien à l’origine du développement desfeuilles et, ici, de la tige. On remarque la persistance duméristème en position apicale, alors que les feuilles sedéveloppent, posant ainsi à la fois le problème du renou-
L’hypothèse d’une action de la lumière sur la productionde gibbérelline doit donc être proposée.En fait, il existe plusieurs gibbérellines, nommées GA1 àGA53 selon l’ordre de leurs découvertes. Ces moléculessont ainsi nommées, car elles furent mises d’abord en évi-dence chez un champignon parasite du Riz, Fusariumheterosporum, nommé au Japon Gibberella fujikori, et quiprovoque le gigantisme chez son hôte. Étant rapidementisolée à partir d’une culture fungique, GA3 est la forme laplus fréquemment commercialisée, et dont les effets ontété les plus étudiés. En particulier, on a pu montrer chez leRiz que GA3 permet la transcription de gènes codant desprotéines responsables du processus de la mitose, histoneset cyclines, et ainsi l’activité des méristèmes intercalaires(Van der Knapp et al. , 2000). Chez l’Orge, GA3 initie lasynthèse de l’ARNm dirigeant celle d’une enzyme, l’�-amylase, utilisée lors de la germination (Jacobsen etBeach, 1985). Mais le problème étudié ici est celui de la synthèse d’unegibérelline active, en relation avec l’éclairement.L’Épinard possède 6 gibbérellines. Celle utilisée au coursde l’expérimentation décrite par le DOCUMENT 8 est GA3,naturellement synthétisée à partir d’autres gibbérellines,dont GA19, abondante en situation de jours courts, et GA20,abondante en situation de jours longs (Metzger etZeevaart, 1980). Le photocontrôle a pu être situé, à l’inté-rieur de la chaîne de biosynthèse des gibbérellines, auniveau de la conversion de GA19 en GA20, en marquant leurprécurseur commun, GA53, à l’aide de deutérium(Gianfragna et al. , 1983).
A c t i v i t é s 3
Des cellules responsables de la croissance des plantesLes objectifs de ces activités sont, au niveau cognitif,d’établir la persistance de méristèmes au sein du végé-tal, et au niveau méthodologique, la mise en relation dedonnées afin de proposer une explication à la crois-sance indéfinie des végétaux. L’acquisition de ces don-nées peut donner lieu à des observations depréparations microscopiques, coupes de bourgeons ter-minaux et axillaires. Parmi les techniques utilisées enarboriculture fruitière, la greffe en écusson est celle quipermet le mieux d’appréhender le rôle des bourgeonsaxillaires dans le développement ultérieur desrameaux. En effet, on ne prélève alors qu’un seul bour-geon appartenant à une espèce productive, pour le gref-fer sur un arbre moins productif, ou de moins bonnequalité, mais mieux adapté aux conditions locales deculture. L’obtention de nombreux rameaux donnant desfruits de la variété, voire de l’espèce greffée, pose leproblème des propriétés des tissus contenus dans lebourgeon transplanté. Il reste alors à rechercher, à l’in-térieur du bourgeon, les structures responsables de sacroissance. Leur dynamique ne peut être abordée auniveau cellulaire que par l’étude de coupes réalisées à
62

vellement cellulaire en son sein, et celui du devenir descellules qui ont quitté le méristème.Il n’est pas envisagé d’étudier ici, ni l’organisation duméristème apical, ni le contrôle de son fonctionnement.Toutefois, le développement latéral des feuilles, l’aligne-ment des cellules issues du méristème médullaire, pour-ront être remarqués. En effet, les méristèmes caulinairesconstituent des territoires bien structurés : à la partieexterne du dôme méristématique, les cellules de la« tunica » formeront les feuilles, alors que les cellules plusinternes du « corpus » donneront naissance aux différentstissus différenciés de la tige (document). Aux extrémitéslatérales du méristème, les cellules de la tunica produisentdes unités comprenant chacune une feuille, un bourgeonaxillaire et un entre-noeud, les phytomères. La zone cen-trale du méristème n’est pas organogène, mais elle produitdes cellules-souches, à l’origine des cellules qui se diffé-rencieront au fur et à mesure que les mitoses successivesles auront éloignées de cette zone. Le méristème est ainsiune structure en équilibre dynamique : les cellules pro-duites par mitose au niveau de la zone centrale sont aussinombreuses que les cellules repoussées vers la périphérieet recrutées pour l’organogenèse. De ce fait, les dimen-sions du méristème apical du Coléus ne paraissent pasmodifiées sur le DOCUMENT 12.
E x e r c i c e s
Évaluer ses connaissances
Choisir les bonnes réponsesb. et c. sont les affirmations exactes.a. Une variation des conditions environnementalesne peut avoir de conséquence immédiate sur laforme d’un végétal, celle-ci dépendant du fonction-nement de ses méristèmes.d. Les facteurs du milieu ne peuvent modifier le phé-notype d’un végétal que dans les limites imposéespar ses potentialités génétiques.e. Un végétal peut modifier sa forme en fonction desvariations de son environnement, sans que songénome ne soit modifié. Remarque : il convient de bien distinguer un accom-modat, variation phénotypique due à une croissanceorientée des organes végétaux, des effets d’unemutation somatique.
Exercice corrigé
La culture des nénupharsCet exercice permet aux élèves de s’assurer de lacompréhension de notions abordées au cours del’ensemble des activités proposées. La croissancedes nénuphars dépend en effet, à la fois de la hau-teur de l’eau recouvrant leur rhizome, et de leurscaractéristiques génétiques. Il s’agit donc de distin-guer les rôles respectifs du génome et d’un facteurde l’environnement dans la réalisation du phénotypedes nénuphars. Pour cela, les connaissancesacquises sur les méristèmes doivent être mobili-sées, afin d’expliquer à un niveau ne dépassant pascelui des activités, comment des cellules au génomeidentique peuvent donner naissance à des feuillesde morphologies différentes. Différentes étapes du raisonnement scientifiquesont mises en œuvre. La première questiondemande de saisir les données pertinentes danschacun des documents proposés. La difficulté tientà la diversité des informations fournies, les donnéesne devant être retenues qu’en fonction du problèmeposé. La seconde question conduit à envisager desmodèles différents de nénuphar selon que le phéno-type est totalement déterminé, ou non, par le pro-gramme génétique, puis à les confronter auxdonnées concernant les conditions nécessaires àleur survie. Enfin, la dernière question associe la sai-sie de données à leur mise en relation entre elles,puis aux données acquises concernant l’accommo-dation au milieu. Elle peut donc constituer un entraî-nement à la réalisation d’une courte synthèse.
3
1
Chapitre 10 : La diversité phénotypique des végétaux
63
Méristème apical caulinaire
Méristèmemédullaire
Bourgeon axillaire
Feuille
Nœud
Entre-nœud
Zonecentrale
Zonepériphérique Primordia
Tunica
Corpus
Le méristème apical caulinaire est composé de différentes zones,dont chacune possède une fonction.
On peut consulter à ce sujet : Marianne Delarue et JeanTraas , L’Architecture des plantes, dans le Dossier Horssérie Pour la Science «De la graine à la plante», janvier2000.

3. Les feuilles exposées à l’ombre ont une capacitéphotosynthétique inférieure à celles qui sont expo-sées au Soleil. Cependant, une plus grande surfacefoliaire, associée à une teneur plus élevée en chloro-phylle, compense partiellement le fait qu’elles reçoi-vent une plus faible quantité d’énergie par unité desurface. Le document (c.) montre que l’intensité deleur photosynthèse est environ deux fois moindreque celle des feuilles de lumière, alors que le rayon-nement solaire reçu par unité de surface est environ500 fois moins élevé. Le rendement photosynthé-tique, exprimé en quantité de matière produite pourune même quantité d’énergie reçue, est donc plusélevé pour les feuilles d’ombre que pour les feuillesexposées à la lumière.
Environnement et photosynthèse1. Toutes les cellules du même individu possédant lemême génome, les différences observées entre lesfeuilles de Hêtre les plus exposées au rayonnementsolaire, en périphérie de la frondaison, et celles lesmoins éclairées, à l’intérieur de la frondaison, nepeuvent provenir que des conditions du milieu. Cesdifférences portent essentiellement sur l’épaisseurdes feuilles, mais aussi sur leur surface moyenne etleur teneur en chlorophylle.
2. L’intensité de la photosynthèse, variable selon laplace tenue par les feuilles au sein de la frondaison,est donnée par unité de surface foliaire. Il ne s’agitdonc pas de la photosynthèse totale réalisée parl’ensemble des feuilles, soit d’ombre, soit delumière, mais de la capacité photosynthétique dechaque type de feuille. On constate que celle desfeuilles qui se sont développées à la périphérie de lafrondaison est plus importante que celle des feuillesqui se sont développées à l’intérieur de celle-ci.
5Évaluer ses compétences
Le por t des genêts sur le l i t toral1. Deux explications paraissent également pos-sibles : soit le caractère prostré est inscrit dans legénome des pieds de genêt qui expriment ce carac-tère, soit la forme prostrée résulte de l’action du ventsur l’expression de leur génome. Dans ce derniercas, le génome des genêts à port rampant ne seraitpas différent, pour les gènes mis en cause, de celuides genêts à port dressé.2. Le fait que des genêts à port rampant puissentavoir des descendants ayant le même port qu’eux,bien que cultivés en milieu abrité, laisse penser quela forme prostrée peut être due à une mutation. Lecaractère prostré serait donc inscrit dans leurgénome. 3. La forme prostrée des genêts du littoral peut avoirdeux origines différentes. Il peut s’agir, soit d’unemutation, qui peut donc être transmise à leurs des-cendants, soit d’un accommodat, c’est-à-dire d’unevariation phénotypique sans modification dugénome, mais liée à l’action d’un facteur de l’envi-ronnement. Dans ce dernier cas, les graines issuesdes genêts présentant sur le littoral un port rampantdonnent des genêts à port dressé.
La mise en forme d’un arbre fruitier1. Un pommier en cordon est obtenu en arquant lajeune tige, puis en supprimant les rameaux qui sedévelopperaient du côté opposé à celle-ci. En réalité,ce sont les méristèmes des jeunes rameaux qui setrouvent ainsi régulièrement supprimés. Les méris-tèmes non supprimés, celui du bourgeon terminal etceux des bourgeons axillaires situés à la face supé-rieure de la tige, continuent à se développer.2. La similitude entre la forme d’un arbre taillé pourobtenir un cordon et celle d’un arbre ne développantses ramifications que du seul côté abrité du vent,s’explique par une cause commune. Du côté exposéau vent dominant, les méristèmes sont régulière-ment détruits, par effet mécanique, ou en bord demer, en raison de la salinité trop élevée des embrunsqu’ils reçoivent.
4
2
64
Feuilles exposées Feuilles exposéesà la lumière à l’ombre
Capacité 3,5 1,3photosynthétique,en mg de CO2
absorbé.dm-2.h-1

E x e r c i c e c o m p l é m e n t a i r e
Le chancre du Hêtre
La maladie du chancre du Hêtre est provoquée parun champignon (Nectria ditissima) dont les spores,transportées par le vent, pénètrent dans les tissusde l’arbre par des orifices naturels, comme les cica-trices foliaires, ou par des blessures accidentelles,causées par des piqûres d’insectes, par exemple.Le mycélium issu de la germination des spores colo-nise les tissus vivants superficiels de l’hôte, provo-quant la formation de chancres à la périphériedesquels de nouvelles spores apparaissent au boutde deux ans. Le ruissellement de la rosée ou de lapluie le long du tronc entraîne leur dissémination,contribuant à la multiplication des chancres sur lemême arbre.
1. Quels sont les effets de la maladie sur la crois-sance du Hêtre ?2. Identifier les facteurs de l’environnement qui sontintervenus dans la morphogenèse d’un hêtre atteintde la maladie du chancre. Distinguer les facteursbiotiques et abiotiques.
6
Réponses attendues1. Le mycélium du champignon parasite provoquedes déformations des tissus de son hôte.Cependant, ces déformations ne peuvent être direc-tement causées par le parasite. Elles représententune réponse de l’arbre à la présence du mycélium.C’est donc bien la croissance du Hêtre qui se trouvemodifiée sous l’influence d’un facteur de son envi-ronnement.
2. Les facteurs biotiques sont représentés par lechampignon lui-même (Nectria ditissima), mais aussipar les insectes qui, par leurs piqûres, favorisent sadissémination. La chute des feuilles du Hêtre, enautomne, constitue un facteur biotique favorisant,par les cicatrices laissées, la propagation du para-site d’un arbre à un autre. Le ruissellement de l’eau,de rosée ou de pluie, est un facteur abiotique favo-risant la dissémination des spores sur le mêmearbre. Le vent, lorsqu’il transporte les spores d’unarbre à un autre, est un facteur abiotique intervenantdans la morphogenèse d’un arbre atteint.
Chapitre 10 : La diversité phénotypique des végétaux
65
Un hêtre atteint de la maladie du chancre.
ph
© S
outr
enon
A.

66
P o s e r l e s p r o b l è m e s
Dans des conditions strictes de culture, des massifs cellu-laires issus d’embryons de graines de pin peuvent donnerplusieurs embryons qui, séparés, deviendront des plantsgénétiquement identiques. Le fait qu’un massif cellulaireembryonnaire donne, au départ, des cellules toutes iden-tiques pose le problème de la conservation du matérielgénétique au cours des divisions cellulaires. On peutétendre ce problème aux cellules différenciées puisque,
◆ Problèmes scientifiques à résoudreLa morphogenèse d’un végétal dépend en grande partie dufonctionnement des méristèmes du végétal. Ce fonction-nement permanent permet la croissance continue desorganes du végétal (voir chapitre précédent). Comment unméristème assure-t-il à la fois sa pérennité (dans l’espaceet dans le temps) et la mise en place, dans son voisinage,de nouveaux organes ? Comment, d’autre part, le maintiendu patrimoine génétique, qui s’exprime partiellement outotalement en fonction du devenir des cellules, peut-il êtreassuré au sein de chacune des cellules méristématiques ?
Programme : notions et contenus• La morphogenèse associe la division et la croissance cellulaire au niveau de territoires spécialisés.• La mitose est localisée dans les méristèmes. Elle permet de produire :– des cellules qui vont ensuite se différencier et participer à la croissance et à la structuration de l’organisme (feuilles, tiges, racines) ;– des cellules qui restent indifférenciées et qui vont à leur tour constituer des méristèmes (apical ou axillaire).• La mitose est un processus commun aux cellules eucaryotes.• Au cours de l’interphase du cycle cellulaire, la réplication de l’ADN s’effectue selon un mécanisme semi-conservatif, fondé sur la complémentaritédes bases.• Les structures cellulaires se modifient lors de la mitose.• Chaque cellule fille issue de la mitose contient le même patrimoine génétique que la cellule initiale.
Objectifs à atteindreLes activités de ce chapitre permettent de localiser les zones de croissance du végétal et de déterminer les mécanismesqui assurent cette croissance. Cette étape est essentielle pour bien montrer que les multiplications cellulaires ont lieudans des territoires spécialisés et que, étant toujours identiques, leurs mécanismes pourront être généralisés. Pourcela, on partira d’observations faites sur le végétal à l’échelle de l’organe, puis après avoir localisé expérimentale-ment les zones de croissance, on s’attachera à déterminer les mécanismes cellulaires qui assurent la croissance del’organe :
– la multiplication cellulaire (ou mitose) de cellules indifférenciées ;
– l’élongation cellulaire (début de la différenciation cellulaire).
Cette dernière étape, sous la dépendance d’hormones, sera étudiée avec plus de détails dans le chapitre 12. Ce cha-pitre, consacré essentiellement à la multiplication cellulaire, permet de dégager des problèmes scientifiques à partirde la mise en relation de données issues d’observations macroscopiques et microscopiques, puis de les résoudre àl’aide de résultats expérimentaux.
Étudiée avec le fonctionnement des méristèmes, où elle est le plus aisément mise en évidence, la mitose doit êtreensuite généralisée à l’ensemble des cellules eucaryotes. Sa signification, le maintien du patrimoine génétique au fildes générations cellulaires, constitue un objectif notionnel important de ce chapitre.
Les méristèmes et la croissance des végétaux
C H A P I T R E 1 1

chez les végétaux, on peut reproduire des clones d’indivi-dus à partir de boutures de tiges ou de feuilles d’un mêmeplant. Ce qui signifie que ces cellules conservent le maté-riel génétique de départ, et sont capables de la même toti-potence que les cellules embryonnaires. La secondeinterrogation concerne le devenir de chacune des micro-boutures. En effet, chaque îlot méristématique donnera descellules différenciées structurées en organe, dont le déve-loppement commencera par la croissance. La comparaisondes DOCUMENTS 1a. et 1b. doit faire prendre conscience,qu’après une phase de multiplication cellulaire, des amasuniformes vont donner des plants dont la morphologiedépend à la fois de la mise en place des organes et de lacroissance de ces derniers.
A c t i v i t é s 1
La croissance d’un organe végétalL’observation de la croissance de plantules de petitsPois, matériel facile à se procurer, va permettre deconclure que celle-ci commence surtout par une impor-tante croissance en longueur de la racine et une crois-sance plus tardive de la tigelle. Il sera donc plus aiséd’étudier la croissance de la racine que celle de latigelle et c’est le choix qui a été opéré ici. On seraensuite amené à se demander si la croissance affectetoute la longueur de la racine ou au contraire une ouplusieurs zones spécialisées. S’interroger sur lesmoyens de parvenir à une réponse peut être un préludeà l’activité 1.
ExploitationLe DOCUMENT 2 présente le développement in situ de plan-tules de petits Pois. La coupe de sol montre que le déve-loppement commence par la croissance de la racine, suiviede celle de la tigelle. On pourra noter que la position de laradicule au moment de la germination importe peu, car lacroissance de la racine est toujours orientée vers le sol.Cette observation pourra éventuellement trouver des élé-ments d’explication après l’étude du chapitre 12 sur le rôlede facteurs externes. La réponse (gravitropisme) ne peutêtre demandée à ce stade. Ce document permet aussi d’in-troduire une démarche expérimentale sur la recherche dela (ou des) zones de croissance. Avec le DOCUMENT 3, on serestreint à la réalisation d’une des démarches, à savoir, lemarquage équidistant fait à l’aide d’un fil trempé dansl’encre de Chine sur la jeune racine. On remarque ainsique l’extrémité (apex racinaire) n’entre jamais en crois-sance ; la zone du 2e millimètre initial est la zone qui s’al-longe le plus ; le 3e millimètre a une croissance pluslimitée ; enfin les 4e et 5e millimètres initiaux ne s’allon-gent pas. La zone de croissance est donc localisée à lazone située juste en arrière de l’apex (donc après le 1er mil-limètre). En classe, d’autres germinations, blé, soja, per-mettent d’obtenir des résultats identiques. Lagénéralisation des résultats pourra être affirmée.
A c t i v i t é s 2
Les mécanismes cellulaires de la croissanceL’étude de la croissance, ici croissance en longueur, nepermet pas, à l’issue des activités 1 d’en connaître lesmécanismes. Il faut donc passer à l’échelle cellulaire(préparation microscopique) afin d’établir si crois-sance est synonyme de multiplication cellulaire, d’élon-gation cellulaire ou des deux à la fois. D’autre part, unezone peut évoluer au cours du temps et présenter desmécanismes successifs différents. Le suivi dans l’es-pace, grâce au marquage, doit être complété par unsuivi dans le temps grâce à la comparaison de la mêmezone à quelques jours d’intervalle (deux jours parexemple). Les résultats d’observation directe menéeavec rigueur (évaluation de la taille des cellules, pour-centage de cellules en mitose) peuvent être confirméspar des résultats expérimentaux obtenus en laboratoire.Cela permettra aux élèves d’être véritablementconfrontés à la recherche d’une réponse au problèmeposé et d’être confortés dans leur démarche.
Exploitation� Le DOCUMENT 4 est un rappel des zones marquées. Il per-met de repérer la zone de croissance (zone B) de la racinede Blé. Le choix du Blé est préférable à celui du petit Pois.En effet, les mécanismes cellulaires sont beaucoup plusvisibles chez le Blé ; en particulier, les mitoses y sont plusnombreuses dans la zone A. La taille des cellules apparte-nant à chacune des zones peut être mesurée directementsur les DOCUMENTS 5a. à 5e. et avec plus d’incertitude surles DOCUMENTS 6a. et 6b. Dans la zone A (5a et 5b.), les cel-lules sont petites, de l’ordre de 10 à 15 µm et leur contenucytoplasmique est indifférencié. Dans la zone B (5c. et5d.), les cellules sont plus allongées (20 à 30 µm pour c.et 90 à 100 µm pour d.). L’élongation cellulaire a donc lieudans la zone B. Les cellules de la zone C (5e.) peuventatteindre 120 à 130 µm, elles sont donc plus grandes quedans la zone B ; en revanche, dans le temps, elles ne gran-dissent plus. Les cellules présentes dans la zone C sontdonc des cellules ayant atteint leur taille définitive. Toutela croissance cellulaire a eu lieu dans la zone B.
� Le DOCUMENT 6a. permet de situer la zone X à l’apex dela racine et de constater qu’il existe dans cette zone méris-tématique de nombreuses cellules en division (DOCUMENT6b.) : chromosomes visibles, rapport nucléocytoplasmiquefaible, taille réduite des cellules. La zone méristématiquecomporte environ 430 cellules dont environ 70 présententdes figures de division. Le pourcentage est donc d’environ15 % sur le cliché.Remarque : Les alignements de cellules en périphérie nesont pas à prendre en compte dans la mesure où ils subis-sent très tôt une élongation cellulaire.
� À l’aide des constats précédents, et du DOCUMENT 7, onpeut préciser le fonctionnement du méristème et expliquersa régénération. Il y a des zones de division et des zonesd’élongation. Les divisions ont lieu à l’extrémité de la
Chapitre 11 : Les méristèmes et la croissance des végétaux
67

et non pas à son renouvellement. En effet, à l’aide du DOCU-MENT 9, on constate que pour les 10 % de cellules qui sonten mitose, 92 % d’entre elles ont un contenu en ADNsupérieur à la quantité 2C correspondant à la quantitéd’ADN de n paires de chromosomes. Cette quantité pas-sant du simple (2C) au double (4C), on peut affirmer quela synthèse d’ADN se fait progressivement et aboutit audoublement de la quantité initiale. Par contre, on peutnoter que, dans la zone d’élongation, toutes les cellules ontune quantité égale à 2C et qu’aucune cellule n’est entréeen mitose. Augmentation de la quantité d’ADN et mitosessont donc liées et, dans la zone d’élongation, il n’y a nisynthèse d’ADN, ni division cellulaire, ce qui avait étédéjà noté dans l’activité 2. Le DOCUMENT 10 permet de com-pléter l’étude. On y voit par endroits (DOCUMENT 10b.) laduplication d’un chromosome. On peut ainsi réaliser quel’augmentation progressive de la quantité d’ADN d’unecellule méristématique correspond en fait à la duplicationprogressive du chromosome, donc à la synthèse d’ADN.La structure de la molécule d’ADN (DOCUMENT 11) corres-pond à une séquence donnée de nucléotides.L’incorporation de nucléotides à thymine, mais bienentendu l’incorporation possible d’autres nucléotides nonradioactifs, doit aussi se faire selon un ordre donné. Ledédoublement du chromosome vu en DOCUMENT 10b. sug-gère que celui-ci ait lieu au moins en parallèle à la molé-cule d’ADN initiale. En trouvant une quantité finale égaleà deux fois la quantité initiale (quantité 4C du DOCUMENT 9),on peut penser que la duplication est totale et s’effectuesur toute la longueur du chromosome. Cette hypothèsesera validée au cours des activités suivantes.
A c t i v i t é s 4
Les modalités de la réplication del’ADNL’observation au microscope électronique de la dupli-cation partielle des chromosomes des cellules méristé-matiques ne permet pas de connaître les modalités de laréplication de l’ADN. Malgré tout, les réponses élabo-rées précédemment amènent à penser que la moléculed’ADN conserve à la fois la succession de ses nucléo-tides, mais aussi le nombre de nucléotides puisque l’in-formation génétique est conservée. Il convient doncdésormais de faire des hypothèses sur les modalités decette réplication et de les confronter aux résultats d’ex-périences menées historiquement par Meselson etStahl. Leur démarche, présentée ici dans son intégra-lité, permet un raisonnement scientifique rigoureux. Enl’absence d’activités pratiques réalisables en classe,elle est l’occasion d’un apprentissage au mode de pen-sée hypothético-déductif.
ExploitationBien que les clichés du DOCUMENT 12 représentent un plas-mide bactérien et non pas un chromosome bactérien, ils’agit avant tout du matériel génétique de Bactéries et desa réplication. Le chromosome bactérien, circulaire et fili-
racine (au niveau du premier millimètre) sans qu’il y aitcroissance. Puis, à partir de 1 mm, le nombre de mitosesdiminue progressivement, et la croissance en longueurcommence. Cette croissance s’effectue tant que les cel-lules sont situées dans la zone comprise entre 1 et 3 mm.Ensuite elle cesse. Le fonctionnement de la zone méristé-matique permet donc la multiplication des cellules dans lazone du premier millimètre. Les cellules repoussées dufait des divisions des cellules de ce premier millimètrecommencent à croître. Elles repoussent ainsi vers l’avantla zone méristématique (zone X). En cours d’allongement,les cellules qui sont en fait alignées parallèlement à l’axede la racine, conservent une place qui détermine leur dif-férenciation en fin de croissance : cellules des vaisseauxconducteurs, de l’épiderme, du parenchyme, de la zonepilifère… Les cellules méristématiques sont donc des cel-lules totipotentes dont une partie est capable de régénéreren permanence le méristème lui-même, et l’autre partied’assurer la croissance et la mise en place des tissus et desorganes.
A c t i v i t é s 3
L’ADN dans les méristèmesÀ la faveur de l’étude précédente, on peut affirmer quele méristème est la zone de multiplications cellulaires.Sachant d’autre part que toute cellule méristématiquepossède la totalité de l’information génétique spéci-fique du végétal étudié (voir la rubrique « Poser lesproblèmes »), il convient de s’intéresser, dans les acti-vités 3 à 5, au devenir du matériel nucléaire au coursde ces divisions cellulaires. La première étape (activi-tés 3) permet de noter qu’au sein du noyau existent desmécanismes qui ne s’apparentent pas à la synthèsed’ARNm (voir la première partie du programme) maisqui concernent par contre l’augmentation de la quan-tité d’ADN. Ainsi, avant même d’étudier les modalitésde la transmission du matériel génétique, l’élève pren-dra conscience que ce matériel subit des transforma-tions.
ExploitationEn suivant l’incorporation ou non de nucléotides à thy-mine, nucléotide spécifique de l’ADN, on peut discernerles lieux de synthèse d’ADN. Ainsi observe-t-on au niveaudes méristèmes de racine (DOCUMENT 8b.), quelques cel-lules centrales qui ont incorporé de la thymine. Les cel-lules de la coiffe et les cellules situées au-dessus de lazone méristématique ne présentent pas de radioactivité.L’ADN est donc synthétisé seulement au sein du méris-tème racinaire. Afin de généraliser ce phénomène auxméristèmes caulinaires, un méristème de bourgeon est pré-senté au DOCUMENT 8a.. Là aussi la thymine radioactive estvisible au sein des cellules du méristème, mais aussi à labase des primordium foliaires. Par conséquent, tous lesméristèmes racinaire et caulinaire et quelques cellules deprimordium présentent une synthèse d’ADN. Celle-ci cor-respond en fait à une augmentation de la quantité d’ADN
68

forme sur le DOCUMENT 12a., présente un dédoublementpartiel sur le DOCUMENT 12b.. Le dédoublement devienttotal sur le DOCUMENT 12c.. La conservation de la longueurdu chromosome, donc du nombre de nucléotides, doit êtreprise en compte pour la représentation schématique de laréplication. D’autre part, l’épaisseur entre A et B des deuxsegments de chromosomes étant identiques, on doit consi-dérer que la molécule d’ADN a la même structure. On peut ainsi émettre deux hypothèses :a. le doublement progressif de la molécule d’ADN,b. le doublement progressif de chaque brin d’ADN.L’hypothèse (a) correspond à l’hypothèse de la réplicationconservative de la molécule d’ADN, et l’hypothèse (b) àcelle d’une réplication semi-conservative de la moléculed’ADN (DOCUMENT 13). L’emploi par Meselson et Stahl del’azote lourd 15N et/ou de l’azote léger 14N (DOCUMENT 14a.et 14 b.) au cours de deux divisions successives de bacté-ries permet de distinguer, après isolement et centrifugationde l’ADN, l’ADN ne contenant que du 14N, de l’ADN necontenant que de l’azote 15N, et même de l’ADN mixte à14N et 15N. Les résultats donnés dans le DOCUMENT 14 c. cor-respondent à l’hypothèse d’une réplication semi-conserva-tive, alors que dans le cas d’une réplication uniquementconservative, on obtiendrait déjà de l’ADN de densité 1.7et de l’ADN de densité 1.65 au temps t1. Au temps t2, lesrésultats seraient maintenus identiques à ceux de t1. Cettedernière explication peut être demandée aux élèves dans lecadre d’un exercice de vérification de la compréhensionde la méthode utilisée par Meselson et Stahl.
A c t i v i t é s 5
Conservation de l’information aucours des cycles cellulairesAprès avoir établi qu’il existe une duplication du maté-riel génétique, il reste à étudier comment celui-ci estéquitablement réparti entre les deux cellules filles. Eneffet, la première doit recevoir un matériel génétiqueidentique à celui de la seconde. Les modalités phy-siques (condensation des chromosomes et transmissiondes chromatides de ces derniers) et la succession desétapes (la réplication précédant toujours la mitose)permettent de construire la notion de cycle cellulaire.Et après avoir étudié la réplication de la moléculed’ADN, se rendre compte que chaque nouvelle molé-cule d’ADN formera, après condensation, une des chro-matides d’un chromosome permet d’établir sansambiguïté qu’il y a bien transmission équitable desmolécules d’ADN entre les cellules filles. Dans cetteactivité, il est encore fait appel à un raisonnementscientifique rigoureux : observations et hypothèses surla répartition des molécules d’ADN, confrontées auxrésultats d’expériences, permettent de proposer uneexplication à la conservation de l’information géné-tique.
Exploitation� Grâce au DOCUMENT 15, on constate que la transmissiondu matériel génétique passe par sa condensation puis lepositionnement de tous les chromosomes au centre de lacellule avant toute séparation. On obtient donc, aprèscondensation, en prophase de mitose, des chromosomes àdeux chromatides et on peut noter que chacune des chro-matides partira vers un pôle de la cellule à l’anaphase : lepartage équitable en quantité doit donc avoir lieu à cemoment-là. On doit alors se demander si chaque chroma-tide est constituée d’une des deux molécules d’ADN for-mées au cours de la réplication, celle-ci précèdant lamitose (DOCUMENT 16).
� Au cours du cycle cellulaire (DOCUMENT 16), en phase G1,les chromosomes sont à une chromatide, donc à une seulemolécule d’ADN (1 exemplaire de chaque gène). En phasede réplication (phase S), les chromosomes sont partielle-ment à une ou deux chromatides : les gènes sont donc soità un exemplaire (segment d’ADN non répliqué), soit àdeux exemplaires (segment d’ADN répliqué). De la phaseG2 à la métaphase de mitose incluse, les chromosomessont à deux chromatides (deux molécules d’ADN), doncchaque gène est en deux exemplaires. De l’anaphase demitose à la télophase, les chromosomes sont de nouveau àune chromatide : les gènes sont donc à ce moment, à unseul exemplaire dans chaque chromosome. Par contre, iln’y a pas encore eu de cytodiérèse et la cellule conserveles deux exemplaires de chaque gène. En fin de télophase,la cytodiérèse a eu lieu, chaque nouvelle cellule en phaseG1 possède un exemplaire de chaque gène. Conclusion : àl’issue de la mitose, chaque cellule fille possède le mêmenombre de gènes, identiques à ceux de la cellule mère.
� Les chromosomes visibles sur les clichés du DOCUMENT17 sont à deux chromatides : ce sont des chromosomes enfin de prophase ou en métaphase. En fait, la légende sou-ligne qu’il s’agit de chromosomes bloqués en métaphase.La radioactivité que certains chromosomes présententtémoigne de l’incorporation de thymine radioactive entout début de culture, en phase S du cycle cellulaire (DOCU-MENT 17a.). Les résultats (chromatides toutes radioactives)permettent de réactualiser les connaissances sur la réplica-tion semi-conservative et d’affirmer que chaque chroma-tide est formée de ces nouvelles molécules d’ADN. Àl’étape suivante, c’est-à-dire en métaphase de deuxièmedivision (DOCUMENT 17b.), une seule chromatide sur deuxest radioactive : on doit donc considérer que la chromatidenon radioactive est constituée d’une molécule d’ADNdont les deux brins sont non radioactifs. La réplicationétant semi-conservative, il existe ainsi une associationentre un brin non radioactif (étape S de l’interphase 2) etun brin initial non radioactif (celui de la cellule mère dudébut de l’expérience). D’où la correspondance : « unechromatide = une molécule d’ADN ». Avec ce schéma entête, il est aisé de considérer que la deuxième chromatideest formée d’une molécule d’ADN dont un brin est nonradioactif (phase S de la mitose 2) et d’un brin radioactif(phase S de la mitose 1). À la mitose suivante, du fait dela réplication semi-conservative et de la correspondance«chromatide = molécule d’ADN», on observera : la moi-tié des chromosomes à deux chromatides non radioactives(ils proviennent des chromatides neutres précédentes), et
Chapitre 11 : Les méristèmes et la croissance des végétaux
69

b. Les chromosomes se condensent en prophasede mitose.d. Les cellules différenciées ne subissent plus decycle cellulaire. Seules celles qui se dédifférencie-ront pourront à nouveau amorcer un nouveau cyclecellulaire : ce seront alors des cellules totipotentesnon différenciées.
Proposer une explicat iona. Dans un même méristème, de nombreusesmitoses ont lieu. Toutes les cellules filles issuesd’une cellule mère de départ ont toutes le mêmeprogramme génétique (aux mutations près dont lafréquence est 10–6) car la duplication de l’ADN et lamitose conservent l’information génétique au seinde chaque cellule. Ces cellules constituent un clone.En prélevant des microboutures d’un même méris-tème, on peut régénérer des plantes entières quitoutes posséderont donc la même information géné-tique.b. La tonte d’une pelouse raccourcit essentiellementdes feuilles. Les bourgeons terminaux des plantesacaules, situés au ras du sol ne subissent pas decoupe. Par conséquent, malgré les tontes, la forma-tion de nouvelles feuilles peut avoir lieu à partir de ceméristème terminal.c. Les cellules végétales sont totipotentes. Dansdes conditions favorables, elles peuvent se dédiffé-rencier et subir des mitoses. Les cellules issues deces mitoses peuvent à leur tour se différencier.
Exercice corrigé
Évolut ion de la quanti té d’ADNdans une culture cel lulaireCet exercice permet d’insister sur les variations dela quantité d’ADN cellulaire mais aussi tissulaire aucours du temps, donc sur la notion de cycle cellu-laire. La construction du graphique « Évolution de laquantité d’ADN en fonction du temps » permetensuite d’évaluer la durée d’un cycle cellulaire,durée qui servira à déterminer le nombre de mitosesqui ont lieu au terme de 37 heures après la mise enculture.La difficulté de l’exercice résulte du fait que les pre-mières données sur les quantités d’ADN débutenten phase G2. L’élève doit donc bien avoir enmémoire les différentes phases du cycle cellulaire.L’autre difficulté réside dans l’appréhension desquantités d’ADN d’une culture de cellules. Aprèsmitose de toutes les cellules de la culture, la quan-tité d’ADN de la culture est le double de la quantitéde départ, le nombre de cellules aussi. Par contre,au terme de 37 heures, la quantité d’ADN peut avoirdoublé alors que les cellules ne sont pas encoreentrées en mitose. Il faut donc dissocier l’étape de laduplication de l’ADN pendant l’interphase de lamitose proprement dite.
4
3E x e r c i c e s
Évaluer ses connaissances
Vrai ou fauxa. Faux.Remarque : On peut toutefois admettre que desmutations peuvent intervenir au cours de la duplica-tion de l’ADN. Toute cellule issue d’une cellulemutée sera porteuse de la mutation et ne posséderapas le programme génétique de la plante mère.b. Vrai.c. Faux. Certaines cellules méristématiques se diffé-rencient et d’autres cellules méristématiques ne sedifférencient pas. Ces dernières assurent la perma-nence du méristème.d. Faux. La croissance en longueur d’une racinedépend, d’une part, des multiplications cellulairesqui ont lieu au sein du méristème racinaire, d’autrepart, de l’élongation des parois de certaines de sescellules.e. Vrai. Toutefois, les lignées cellulaires se différen-ciant à des vitesses différentes, on peut encoreobserver quelques mitoses dans les ébauches d’or-ganes, feuilles par exemple.
Choisir la bonne réponse1. b. est la bonne réponse.a. La duplication de l’information génétique a lieupendant l’interphase, avant la mitose proprementdite.c. En début d’anaphase, les chromatides de chaquechromosome se séparent. Chaque chromosome està une chromatide en fin d’anaphase.d. Chaque cellule fille hérite de l’ensemble des chro-mosomes. Chaque chromosome est à une chroma-tide.2. b. est la bonne réponse.a. La totalité de l’information génétique est trans-mise aux cellules filles.c. Les chromosomes sont tous conservés et vontêtre formés de deux chromatides au lieu d’uneseule.d. Chaque chromatide est formée d’un brin d’ADNconservé et d’un brin d’ADN néoformé. La réplica-tion ne peut conserver une chromatide entière.
3. c. est la bonne réponse.a. Les chromatides se séparent en début d’ana-phase de mitose.
2
1
70
la moitié de chromosomes à une chromatide neutre etl’autre radioactive (ils proviennent des chromatides radio-actives précédentes).

Évaluer ses compétences
Méristème et cycle cel lulaire1. La présence de nombreuses figures de mitose, lafaible taille des cellules et l’absence de développe-ment du système vacuolaire font que ce tissu végé-tal est un méristème.2. Pour chaque stade, le matériel chromosomiqueprésente les caractéristiques suivantes :D : les chromosomes ne sont pas condensés (noyauquiescent) ;B : condensation de la chromatine ; les chromo-somes deviennent visibles ;C : chromosomes placés au centre de la cellule(plaque équatoriale) ;A : on observe deux lots de chromosomes. Unechromatide de chaque chromosome migrant à l’undes deux pôles de la cellule.Les stades identifiés sont donc : A (fin d’anaphase),B (prophase), C (métaphase) et D (interphase). Aucours du cycle cellulaire, ils se succèdent dansl’ordre suivant : D, B, C et A… puis D,…3. Les chromosomes sont à deux chromatides pourles stades B et C. Ils sont à une seule chromatidepour les stades D et A. On notera toutefois qu’austade D, les chromosomes sont formés de chroma-tine non condensée. L’ordre chronologique est lesuivant : D, B, C et A.
ADN et cycle cel lulaire1. Les chromosomes présentent tous une colorationbicolore jaune et orange. La coloration orange tra-duit la présence de chromatide dont les deux brinsd’ADN possèdent de la BrdU. La coloration jaunecorrespond au contraire à une chromatide dont unseul brin possède de la BrdU. Ces chromosomessont des chromosomes à deux chromatides doncprésents en prophase ou métaphase de division cel-lulaire.En suivant le devenir d’un chromosome au coursdes deux cycles cellulaires, on peut dire qu’à la finde la première interphase et du fait que la réplicationest semi-conservative, chaque chromatide a intégréde la BrdU sur le brin d’ADN néoformé. L’autre brin,conservé, ne contient pas de BrdU. Chaque chro-matide est donc jaune. À l’anaphase, chaque chro-mosome sera formé d’une seule chromatide jaune.À la fin de l’interphase suivante, chaque brin d’ADNde la chromatide aura un brin complémentaire néo-formé, toujours avec de la BrdU puisque le milieu deculture en contient. Le brin d’ADN sans BrdU auraun brin complémentaire avec de la BrdU : la chro-matide apparaîtra jaune. Le brin d’ADN initial avecde la BrdU aura un brin complémentaire avec de laBrdU : la chromatide apparaîtra orange.L’observation de chromosomes bicolores ne peutdonc avoir lieu qu’à la deuxième division cellulaire,en prophase ou métaphase, lorsque les chromo-somes sont condensés et à deux chromatides.
6
5
2. Après l’anaphase de la deuxième mitose, leschromosomes seront à une chromatide : soit jaune,soit orange après transfert sur un milieu normal. Leschromosomes de couleur orange dont les deuxbrins d’ADN possèdent de la BrdU auront, aprèsduplication de l’ADN, deux chromatides. Chacunedes chromatides possédera un brin initial à BrdU etun brin néoformé sans BrdU : les chromosomesauront deux chromatides jaunes.Les chromosomes jaunes dont un seul brin possèdede la BrdU auront, après duplication de l’ADN, deuxchromatides. Une chromatide possédera un brin ini-tial sans BrdU et un brin néoformé lui aussi sansBrdU. La chromatide ne sera pas colorée. L’autrechromatide possédera un brin avec BrdU et un brinnéoformé sans BrdU : elle sera colorée en jaune. Leschromosomes présenteront donc une chromatidejaune et l’autre non colorée.
ADN et différenciat ion cel lulaireEn s’éloignant de l’apex de la racine, les cellules ont,d’une part, un volume cellulaire qui augmente(+ 50 % à une distance de 2 cm de la zone méristé-matique), d’autre part, un volume nucléaire qui, luiaussi, augmente (dans ce cas, une augmentationd’environ 30 % du volume nucléaire a lieu à une dis-tance de 2 cm).En fait, les cellules de l’apex quittent progressive-ment la zone méristématique (zone de multiplication)pour accéder à la zone d’élongation cellulaire (voiractivité 2) : leur volume, réduit lorsque les cellules necessent de se diviser, augmente très rapidementlorsque les cellules passent dans la zone de crois-sance. On observe parallèlement une augmentationdu volume nucléaire, mais dans une moindre pro-portion que le reste de la cellule. Le rapport nucléo-cytoplasmique diminue.Remarque : L’augmentation du volume nucléaireprovient en fait d’un phénomène fréquent dans lerègne végétal, l’existence de la polyploïdie. Laconservation du nombre de chromosomes n’en-traîne pas de variation du volume nucléaire. Parcontre, l’augmentation du nombre des chromo-somes provoque une augmentation de la taille dunoyau.
Le déroulement de la mitose1. L’ordre chronologique des différentes phases dela mitose est le suivant :c. Métaphase : les chromosomes bien visibles sontpositionnés en cercle (formation de la plaque équa-toriale au centre de la cellule).d. Métaphase : les chromosomes visibles au centrede la cellule forment la plaque équatoriale.b. Début d’anaphase : les chromosomes situés aucentre de la cellule viennent de se séparer en deux.Chaque lot de chromatides commence sa migrationvers un des pôles de la cellule.e. Fin d’anaphase : il existe deux lots de chromo-somes présents chacun à un des pôles de la cellule ;début d’étranglement de la cellule.
8
7
Chapitre 11 : Les méristèmes et la croissance des végétaux
71

mer ou non dans les cellules dans lesquelles ils ontété introduits. Mais ces gènes n’ont pas pour rôled’acquérir le rôle des gènes des cellules animales :les gènes permettant la régénération des cellules dela peau humaine peuvent ne pas être les mêmesque ceux de la multiplication cellulaire chez lesvégétaux.2. Les molécules d’ADN et d’ARN ne doivent pasêtre déstructurées pour être fonctionnelles. Or,extraites des cellules vivantes, ces molécules nepourront pas conserver leur structure car la durée devie de nombreux ARN par exemple n’est que dequelques dizaines de minutes. D’autre part, pourêtre fonctionnelles, les molécules d’ADN doiventêtre intégrées au noyau cellulaire, et les moléculesd’ARN au cytoplasme.
Structure du matériel chromosomique avant la mitoseOn peut rendre visible la structure des chromo-somes de cellules parvenues aux stades précédantla mitose en les fusionnant avec des cellules encours de mitose. Celles-ci induisent une condensa-tion prématurée du matériel chromosomique despremières cellules. Les documents a. et b. montrentle résultat de telles fusions entre cellules deCervidés dont la formule chromosomique est 2n = 6.D’autre part, on réalise un étalement de matérielchromosomique afin de visualiser l’ADN cellulaire aumicroscope électronique (doc c.).
10
a. Début de télophase : les deux lots de chromo-somes sont encore visibles, et la cytodiérèse estpresque achevée. Les deux cellules filles sont bien-tôt formées.Le classement chronologique est donc le suivant : c. ou d., b., e. et a.2. Les photos correspondent aux divisions cellu-laires de cellules humaines en culture. En comparantavec les photos de l’activité 5 qui correspondent auxdifférentes phases des divisions cellulaires de cel-lules végétales, on retrouve les mêmes étapes, àsavoir : métaphase, anaphase et télophase. Étapesqui correspondent au partage du matériel nucléaireen deux lots identiques et à la formation de deuxcellules filles à partir d’une cellule mère.3. Les chromosomes du cliché f. sont à deux chro-matides ; ce sont des chromosomes de prophase oude métaphase. Le prélèvement a donc pu être effec-tué aux stades c. et d.
E x e r c i c e s c o m p l é m e n t a i r e sProduits cosmétiques et ADN
– Publicité 1Produits de beauté à base d’ADN. Très fortementdosé en extrait de cellules végétales dont l’ADN(14 %) active les fonctions de la peau et stimule leprocessus naturel d’oxygénation cellulaire. Ce fluideréparateur a une action dynamisante sur toutes lesfonctions de la peau.– Publicité 2Produits de beauté à base d’ADN. Convient auxfemmes de tout âge tant pour son rôle de préventiondes rides que pour celui de soin. De formulation trèsélaborée, les produits de la ligne contiennent unpourcentage important d’ADN ou d’ARN (moléculesdu noyau de la cellule vivante, jouant un rôle essen-tiel dans la régénération cellulaire).1. Quelles affirmations sont en accord ou désaccordavec vos connaissances sur l’ADN et l’ARN ?2. Quelles conditions, non citées dans ces textes,doivent être remplies pour que ces molécules puis-sent agir ?
Réponses attendues1. Affirmations en accord avec les connaissancesacquises : les molécules d’ADN et d’ARN sont desmolécules d’origine nucléaire. Ces molécules jouentun rôle essentiel dans le renouvellement cellulaire etmoléculaire (régénération cellulaire). Elles permet-tent aux cellules d’être fonctionnelles, en particulierelles permettent le fonctionnement de certains orga-nites (les mitochondries : oxygénation cellulaire).Affirmations en désaccord avec les connaissancesacquises : les molécules d’ADN et d’ARN permet-tent le fonctionnement des cellules et de leurs mito-chondries, mais elles n’activent pas et ne stimulentpas les mécanismes naturels. Les gènes portés parl’ADN, spécifiques d’une espèce, peuvent s’expri-
9
72
a. Structures des chromosomes à deux moments du cycle cellulaire.
b. Structures des chromosomes à deux moments du cycle cellulaire.

À quelles phases du cycle cellulaire se trouvaient lescellules dont les chromosomes n’étaient pas enmétaphase?
Réponses attenduesSur les documents a. et b., les chromosomes méta-phasiques sont facilement reconnaissables : ils sont formés de deux chromatides condenséesreliées par un centromère. Les autres chromosomespartiellement condensés ne peuvent être que deschromosomes d’interphase. Sur le document a., ces chromosomes sont à unechromatide : ils se trouvent en phase G1 du cyclecellulaire. Les chromosomes du document b. possè-dent deux chromatides : ils se trouvent en phase G2
du cycle cellulaire, la duplication est totale.La phase reconnaissable sur le document c. est laphase S d’interphase : on distingue une moléculed’ADN partiellement dupliquée (la réplication estamorcée mais non terminée).
Chapitre 11 : Les méristèmes et la croissance des végétaux
73
c. ADN cellulaire observé au microscope électronique à une certaine phase du cycle cellulaire.
Ph © Extrait de A. Berkaloff, Biologie et physiologie cellulaires-tome 4,Ed.Hermann, 1989.
0,2 µm

74
Programme : notions et contenus• Dans la tige, la croissance cellulaire est contrôlée par une hormone, l’auxine.• La paroi des cellules végétales en extension est essentiellement composée de polysaccharides, dont la cellulose et les hémicelluloses.• La pression de turgescence cellulaire et la plasticité pariétale permettent la croissance cellulaire.• L’auxine, facteur de croissance ou une hormone végétale, contrôle la croissance cellulaire.• L’auxine est synthétisée à l’apex des tiges. Elle possède une double action :– une action à court terme sur la plasticité pariétale ;– une action à plus long terme sur l’expression des gènes qui participent aux divers événements du métabolisme nécessaires à la croissance.• Le développement du végétal est influencé par la répartition des hormones en interaction avec les facteurs de l’environnement.• La répartition inégale de l’auxine dans les tissus, conséquence d’un éclairement anisotrope, permet une croissance orientée.• Les ramifications naturelles ou provoquées sont sous la dépendance d’un changement de répartition des hormones dans le végétal qui conduit àun changement de morphologie.• La totipotence des cellules végétales permet le clonage. Les proportions des différentes hormones (rapport des concentrations d’auxine et de cyto-kinine) contrôlent l’organogenèse (tige, racines).
Objectifs à atteindreCe chapitre termine l’étude de l’établissement du phénotype chez les végétaux. Il vise à apporter des explicationsscientifiques aux faits mis en évidence au cours des chapitres précédents. La notion d’hormone, établie à l’occasionde la réalisation des phénotypes diabétiques, est ici réinvestie, pour une généralisation de la communication inter-cellulaire à l’ensemble du monde vivant. L’activité des méristèmes, dont l’existence est désormais connue, est en effetsous le contrôle d’hormones végétales, dont la répartition est elle-même soumise à l’influence de facteurs externes.Les notions à construire concernent donc les mécanismes de la croissance cellulaire, les modes d’action de l’auxineet le rôle des facteurs environnementaux dans la morphogenèse. Le choix a été fait de traiter l’ensemble de ces notionsdans un même chapitre afin de lui conserver sa cohérence, l’objectif étant d’expliquer, dans un cadre intégré, la plas-ticité phénotypique des végétaux. L’existence de différentes hormones végétales doit être signalée afin d’envisager leurutilisation dans la réalisation de cultures in vitro. Si ce dernier point se prête particulièrement à des activités pra-tiques, c’est l’ensemble des activités proposées qui peuvent être l’occasion de travaux pratiques, d’observations etd’expérimentations. Associées aux documents du manuel, elles permettent aux élèves de pratiquer une démarche expé-rimentale. Bien que la plupart des activités soient réalisables en classe, certaines expériences nécessitent une duréeou une technicité qui les rendent délicates à réaliser avec les élèves. Dans le cas où des choix doivent être faits, lesdonnées du manuel, parfois tirées de travaux récents, permettent d’accomplir la totalité des étapes du raisonnementscientifique.
Le contrôle de la croissance chez les végétaux
C H A P I T R E 1 2

◆ Problèmes scientifiques à résoudreComment les cellules issues du fonctionnement d’unméristème peuvent-elles grandir ? Comment les facteursde l’environnement peuvent-ils exercer leur action sur lamorphologie d’un végétal ?
P o s e r l e s p r o b l è m e s
Les documents rendent compte d’observations qui peu-vent avoir été déjà faites par les élèves : les activités duchapitre, à dominante expérimentale, ont pour but d’ap-porter une explication scientifique à des faits rencontrésdans la vie courante. Ainsi, le DOCUMENT 1 rappelle l’exis-tence du phototropisme, souvent observé chez les plantesd’intérieur. Le DOCUMENT 2, montrant différents phénotypesréalisés par des individus d’une même espèce, conduit às’interroger sur la manière dont s’exercent les facteursenvironnementaux dans la morphogenèse d’un arbre frui-tier. Le développement d’un méristème axillaire après lamort d’un rameau peut être remarqué sur le DOCUMENT 2 c.,introduisant l’idée d’une inhibition possible, exercée aupa-ravant sur ce méristème. Les constats d’une relation entreun facteur externe, comme l’éclairement unidirectionnelou la taille sélective de certains rameaux, et la croissanced’un végétal, ligneux ou non, permettent la formulationd’hypothèses quant aux mécanismes responsables d’unecroissance orientée.
A c t i v i t é s 1
La paroi des cellules végétalesDans le chapitre précédent, l’étude du fonctionnementdes méristèmes a permis d’identifier deux mécanismesresponsables de la croissance, la division cellulaire etl’élongation cellulaire, alors que la présence d’uneparoi rigide semble interdire cette dernière. L’étude descaractéristiques de la paroi primaire constitue donc unpréalable à celle de l’action de l’auxine. Elle pose clai-rement le problème à résoudre au niveau moléculaire,celui-ci permettant la compréhension de phénomènesenvisagés au niveau macroscopique précédemment. Lacroissance d’un végétal est ainsi continuellementappréhendée à différentes échelles d’observation, decelle de l’individu à celle de la cellule et de ses consti-tuants biochimiques.
Exploitation� La forme sphérique des protoplastes observés sur leDOCUMENT 3 contraste avec la forme parallélépipédique descellules foliaires en place. Bien que ces protoplastes neproviennent pas de cellules de feuilles d’élodée, le choixde ces dernières a été dicté, d’une part, par le souci demontrer à la fois la présence de chloroplastes et celle de laparoi cellulosique, d’autre part, par le fait que ce type decellules est aisément observable en classe. L’observationdirecte peut alors être complétée par l’étude des autresdocuments du manuel. On constate que les protoplastes
sont des cellules chlorophylliennes qui ne diffèrent descellules foliaires que par leur forme. Celle-ci n’est doncque celle de la paroi cellulosique.� Le DOCUMENT 4 montre que les fibres de cellulose peu-vent être synthétisées par des cellules végétales, dès lorsqu’elles sont privées de leur paroi et cultivées sur unmilieu nutritif favorable. Cependant, le DOCUMENT 5 montreque ces fibres sont strictement agencées au sein de la paroiprimaire. On doit souligner le maintien en place des fibresde cellulose par des fibres plus petites d’hémicellulose,disposées transversalement, et faire comprendre que si lesliaisons unissant les fibres de cellulose et d’hémicellulosene sont pas rompues, la synthèse de nouvelles fibres decellulose ne pourrait qu’épaissir la paroi sans l’allonger.La paroi primaire est un réseau hydraté composé essen-tiellement de microfibrilles de cellulose, étroitemententourées par les molécules de la matrice, hémicelluloseset pectines, auxquelles elles sont unies par des liaisonshydrogène. La distinction entre ces deux derniers typesmoléculaires porte sur des différences de solubilité, lespectines seules pouvant être extraites dans l’eau à 80 °C.Ce sont des hétéropolysaccharides dont l’hydrolyse libèredes pentoses (arabinose, xylose), des hexoses (glucose,galactose, mannose) et des oses méthylés (rhamnose,fucose). Ces résidus sont unis entre eux par des liaisonscovalentes, les chaînes ainsi constituées étant reliées auxmicrofibrilles de cellulose par des liaisons hydrogène àl’axe de xyloglucanes. C’est la matrice, réseau macromo-léculaire déformable, qui confère à la paroi sa plasticité.Pour une étude plus détaillée de la structure de la paroi, onpeut consulter :– P. Mazliak, Physiologie végétale, tome 2,«Développement et croissance», Hermann.– D. Robert et J.-C. Roland, Biologie végétale, tome 1,«Organisation cellulaire», Doin.– C. Brett et K. Waldron, Physiology and Biochemistry ofPlant Cell Walls, Unwin Hyman, Londres.
A c t i v i t é s 2
Les mécanismes de l’élongation cellulaireLa mise en évidence d’une pression de turgescence peutdonner lieu à la réalisation d’activités pratiques.Toutefois, s’il peut être commode d’opposer la formed’une cellule adulte plasmolysée à la même cellule enturgescence, il s’agit surtout ici d’envisager ces phéno-mènes chez une cellule jeune, en phase d’élongation.L’observation d’un accroissement des vacuoles au furet à mesure que les cellules s’allongent justifie l’étudeultérieure des échanges hydriques et de leur consé-quence sur l’élongation cellulaire. L’analyse ou la réa-lisation d’expériences sur le coléoptile montre ensuitel’influence exercée par une sécrétion hormonale. Lecoléoptile de blé représente ici un modèle expérimental,historique, mais également utilisable au cours d’uneséance de travaux pratiques, et dont la croissance ne
Chapitre 12 : Le contrôle de la croissance chez les végétaux
75

A c t i v i t é s 3
Actions de l’auxineLa notion d’hormone végétale est explicitée au coursd’activités mettant en évidence le rôle de l’auxine dansl’élongation des parois cellulaires et la rhizogenèse.Des mesures réalisées sur des pieds de Tabac et deFévier montrent une diffusion de l’auxine à partir del’apex des tiges, sa forte concentration à ce niveau lais-sant penser qu’elle y est produite comme à l’apex descoléoptiles. Le constat de son inégale répartition peutdéjà être fait, en l’absence de variation des facteursexternes. Les effets de l’auxine sur l’allongement cellu-laire et sur la rhizogenèse s’exercent par deux modesd’action différents, en fonction des cellules ciblesatteintes et de sa concentration instantanée. Modifiantl’activité de cellules éloignées de son lieu de produc-tion, l’auxine apparaît bien comme une hormone végé-tale, dont les modalités d’action peuvent êtrerapprochées de celles de l’insuline, par exemple.
Exploitation� Le DOCUMENT 11 montre une forte concentrationd’auxine à l’apex des tiges, ainsi que son inégale réparti-tion dans les conditions naturelles. Dans les conditionsexpérimentales du DOCUMENT 12, on observe une diffusionde l’auxine à partir de l’apex et son accumulation auniveau des nœuds, et donc des méristèmes intercalaires, enparticulier au niveau du premier bourgeon. Les informa-tions tirées de ces deux documents fournissent des argu-ments en faveur d’une diffusion de l’auxine à partir duméristème du bourgeon terminal. L’étude du DOCUMENT 13permet d’affirmer une implication de l’auxine dans lesmécanismes de l’élongation cellulaire, et le DOCUMENT 14de proposer un mode d’action de l’auxine, à savoir sonintervention dans la synthèse de protéines. L’auxine agitdonc sur la croissance des plantes en modifiant l’activitédes cellules en élongation.� Une action directe sur l’extensibilité de la paroi neserait pas inhibée par la présence de la cycloheximide(CHI). On peut remarquer que la vitesse d’élongationdiminue de moitié, 16 minutes après son application, cequi correspond à la courte demi-vie d’une « protéine limi-tante de croissance ». C’est une action de l’auxine sur l’ex-pression de gènes codant des protéines enzymatiquesnécessaires à la plasticité de la paroi qui est ici mise enévidence.Une action à long terme peut également être observéelorsqu’on lave le segment de coléoptile afin de supprimertoute trace d’auxine (document ci-après, d’après les tra-vaux d’Edelmann et Schopfer, 1989), la vitesse de crois-sance étant encore égale à la moitié de sa valeur maximale,40 minutes après le retrait de l’auxine. Au contraire, l’ad-dition de KCN, inhibiteur de la respiration cellulaire, à 1 mmol.L–1, réduit rapidement les effets de l’auxine, maisn’a pas d’effet sur la pression de turgescence.En fait, l’auxine n’agit pas directement sur les parois iso-lées. Ses récepteurs ont été mis en évidence au niveau desmembranes du plasmalemme et du réticulum. Par ailleurs,
constitue pas, en elle-même, l’objet de cette étude. À cestade, on se limite à une appréhension au niveau cellu-laire, et non moléculaire, des mécanismes de l’élonga-tion.
Exploitation� Des cellules en élongation peuvent être observées àl’extrémité de jeunes racines de blé. Le DOCUMENT 6 pré-sente de jeunes cellules racinaires colorées au rougeneutre, sur lesquelles des mesures relativement précisesmontreront, non seulement la fusion des vacuoles et l’aug-mentation de leur volume, mais aussi leur élongation. Afinde visualiser l’effet de la pression de turgescence, il estpossible de provoquer la plasmolyse de cellules adultesdont la vacuole est naturellement pigmentée. Il est alorssouhaitable d’observer les modifications de diamètred’une même cellule, montée successivement dans de l’eaudistillée et dans une solution molaire de saccharose, soit342 g.L–1 (voir la fiche technique 2, page 365). Le DOCU-MENT 7 montre une cellule d’épiderme d’oignon rouge tur-gescente (a.), puis plasmolysée (b.). La variation observée,environ 1 mm en largeur et 2 mm en longueur, entre cesdeux stades, fait percevoir l’existence d’une pression exer-cée sur les parois lorsque la vacuole est emplie d’eau. LeDOCUMENT 8 permet de relier l’entrée d’eau dans la vacuoleà l’élongation de la paroi. L’enchaînement des phéno-mènes qui peut être proposé est donc :– une augmentation du volume vacuolaire, liée à uneentrée d’eau croissante au cours de la phase d’élongation ;– l’étirement de la paroi sous l’action de la pression de tur-gescence exercée sur elle par la vacuole emplie d’eau ;– la synthèse de nouvelles microfibrilles de cellulose per-mettant à la paroi amincie de retrouver son épaisseur ini-tiale.� Le DOCUMENT 9 permet de situer le coléoptile, organevégétal encore inconnu des élèves. Les mitoses y sont trèséphémères, et sa croissance est essentiellement assurée parl’élongation de ses cellules. Le résultat obtenu après lasection de l’apex, et présenté sur le DOCUMENT 10 ne peutdonc s’interpréter par l’arrêt des mitoses, la partie décapi-tée ne possédant plus de méristème. Les expériences his-toriques (Boysen-Jensen, 1913) dessinées sur lesDOCUMENTS 10 c. et d. permettent de conclure à la diffusiond’une substance à partir de l’apex du coléoptile, et res-ponsable de l’élongation des cellules sous-jacentes.L’auxine est ici introduite comme étant une hormone végé-tale, en dehors du contexte de sa découverte, qui impli-querait une étude prématurée du phototropisme.Des protocoles expérimentaux peuvent être consultésdans :– D. Pol, Travaux pratiques de biologie, Bordas, 1994.– R. Prat, L’Expérimentation en physiologie végétale,Hermann, 1993.
76
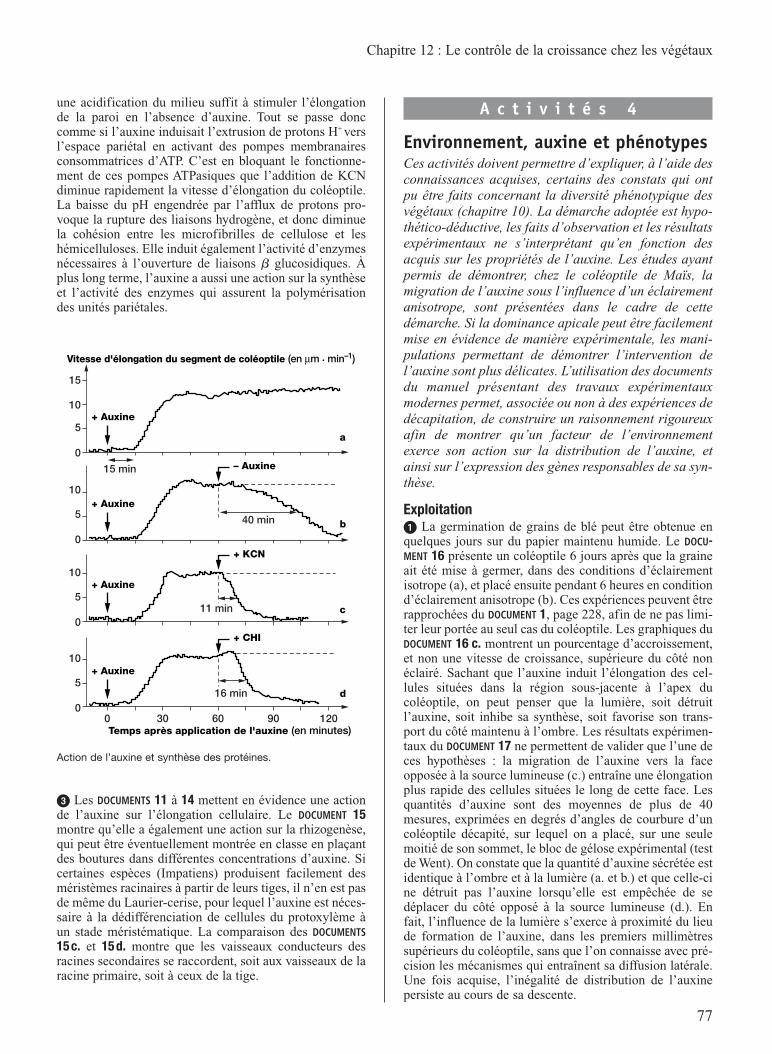
une acidification du milieu suffit à stimuler l’élongationde la paroi en l’absence d’auxine. Tout se passe donccomme si l’auxine induisait l’extrusion de protons H+ versl’espace pariétal en activant des pompes membranairesconsommatrices d’ATP. C’est en bloquant le fonctionne-ment de ces pompes ATPasiques que l’addition de KCNdiminue rapidement la vitesse d’élongation du coléoptile.La baisse du pH engendrée par l’afflux de protons pro-voque la rupture des liaisons hydrogène, et donc diminuela cohésion entre les microfibrilles de cellulose et leshémicelluloses. Elle induit également l’activité d’enzymesnécessaires à l’ouverture de liaisons � glucosidiques. Àplus long terme, l’auxine a aussi une action sur la synthèseet l’activité des enzymes qui assurent la polymérisationdes unités pariétales.
A c t i v i t é s 4
Environnement, auxine et phénotypesCes activités doivent permettre d’expliquer, à l’aide desconnaissances acquises, certains des constats qui ontpu être faits concernant la diversité phénotypique desvégétaux (chapitre 10). La démarche adoptée est hypo-thético-déductive, les faits d’observation et les résultatsexpérimentaux ne s’interprétant qu’en fonction desacquis sur les propriétés de l’auxine. Les études ayantpermis de démontrer, chez le coléoptile de Maïs, lamigration de l’auxine sous l’influence d’un éclairementanisotrope, sont présentées dans le cadre de cettedémarche. Si la dominance apicale peut être facilementmise en évidence de manière expérimentale, les mani-pulations permettant de démontrer l’intervention del’auxine sont plus délicates. L’utilisation des documentsdu manuel présentant des travaux expérimentauxmodernes permet, associée ou non à des expériences dedécapitation, de construire un raisonnement rigoureuxafin de montrer qu’un facteur de l’environnementexerce son action sur la distribution de l’auxine, etainsi sur l’expression des gènes responsables de sa syn-thèse.
Exploitation� La germination de grains de blé peut être obtenue enquelques jours sur du papier maintenu humide. Le DOCU-MENT 16 présente un coléoptile 6 jours après que la graineait été mise à germer, dans des conditions d’éclairementisotrope (a), et placé ensuite pendant 6 heures en conditiond’éclairement anisotrope (b). Ces expériences peuvent êtrerapprochées du DOCUMENT 1, page 228, afin de ne pas limi-ter leur portée au seul cas du coléoptile. Les graphiques duDOCUMENT 16 c. montrent un pourcentage d’accroissement,et non une vitesse de croissance, supérieure du côté nonéclairé. Sachant que l’auxine induit l’élongation des cel-lules situées dans la région sous-jacente à l’apex ducoléoptile, on peut penser que la lumière, soit détruitl’auxine, soit inhibe sa synthèse, soit favorise son trans-port du côté maintenu à l’ombre. Les résultats expérimen-taux du DOCUMENT 17 ne permettent de valider que l’une deces hypothèses : la migration de l’auxine vers la faceopposée à la source lumineuse (c.) entraîne une élongationplus rapide des cellules situées le long de cette face. Lesquantités d’auxine sont des moyennes de plus de 40mesures, exprimées en degrés d’angles de courbure d’uncoléoptile décapité, sur lequel on a placé, sur une seulemoitié de son sommet, le bloc de gélose expérimental (testde Went). On constate que la quantité d’auxine sécrétée estidentique à l’ombre et à la lumière (a. et b.) et que celle-cine détruit pas l’auxine lorsqu’elle est empêchée de sedéplacer du côté opposé à la source lumineuse (d.). Enfait, l’influence de la lumière s’exerce à proximité du lieude formation de l’auxine, dans les premiers millimètressupérieurs du coléoptile, sans que l’on connaisse avec pré-cision les mécanismes qui entraînent sa diffusion latérale.Une fois acquise, l’inégalité de distribution de l’auxinepersiste au cours de sa descente.
Chapitre 12 : Le contrôle de la croissance chez les végétaux
77
Action de l’auxine et synthèse des protéines.
� Les DOCUMENTS 11 à 14 mettent en évidence une actionde l’auxine sur l’élongation cellulaire. Le DOCUMENT 15montre qu’elle a également une action sur la rhizogenèse,qui peut être éventuellement montrée en classe en plaçantdes boutures dans différentes concentrations d’auxine. Sicertaines espèces (Impatiens) produisent facilement desméristèmes racinaires à partir de leurs tiges, il n’en est pasde même du Laurier-cerise, pour lequel l’auxine est néces-saire à la dédifférenciation de cellules du protoxylème àun stade méristématique. La comparaison des DOCUMENTS15c. et 15d. montre que les vaisseaux conducteurs desracines secondaires se raccordent, soit aux vaisseaux de laracine primaire, soit à ceux de la tige.
Temps après application de l'auxine (en minutes)
+ Auxine
Vitesse d'élongation du segment de coléoptile (en µm . min–1)
00
5
10
0
5
10
0
5
10
0
5
10
15
30 60 90 120
15 min
40 min
16 min
11 min
+ Auxine
+ Auxine
+ Auxine
+ CHI
+ KCN
– Auxine
a
b
c
d

végétaux in vitro (voir les « Questions d’aujourd’hui »,page 247). L’étape du raisonnement scientifique à pri-vilégier est alors l’élaboration d’un protocole expéri-mental, que celui-ci soit ou non mis en œuvre entravaux pratiques.
Exploitation� Le DOCUMENT 21 indique que les effets de l’auxine surl’organogenèse dépendent de sa concentration. On remarque qu’une forte concentration d’auxine (> 10–5 g.mL–1) favorise l’émission de racines secondaires.Ces données sont confirmées par les informations tiréesdu DOCUMENT 22, puisqu’une concentration d’auxine de3 mg.L–1 (ou 3.10–3 g.mL–1) est utilisée pour induire la for-mation des racines à partir d’un cal issu d’un fragment detige. Dans ces conditions, une faible concentration decytokinine (0,02 mg.L–1) est également nécessaire. Enfin,le DOCUMENT 23 montre que le développement des racinesest obtenu à partir d’un explant foliaire sur un milieucontenant 1 mg.L–1 d’auxine, en l’absence de cytokinines.On peut conclure qu’un rapport élevé des concentrationsauxine/cytokinine favorise la formation des racines.Cependant, leur croissance nécessite ensuite une concen-tration beaucoup plus faible en auxine, de l’ordre de 10–8 g.mL–1 (ou 10–5 mg.L–1).
� Les résultats expérimentaux du DOCUMENT 20 sont ex-traits d’une publication de D. Reinhardt, Th. Mandel et C. H. Kuhlemeier, de l’Université de Berne (« AuxinRegulates the Initiation and Radial Position of PlantLateral Organs », The Plant Cell, volume 12, n° 4, avril2000). Ils démontrent la nécessité de la présence d’auxinepour induire des ébauches foliaires in vivo. Le DOCUMENT21 permet de penser que la concentration d’auxine (0,1mM.L–1) est alors très faible. En effet, le DOCUMENT 22indique que l’obtention de feuilles demande, dans lesconditions de culture in vitro, une concentration faible enauxine (0,003 mg.L–1), mais forte en cytokinine (1 mg.L–1).Des données du même ordre sont fournies par le DOCUMENT23. Au contraire de celle des racines, la formation desfeuilles est donc favorisée par un rapport auxine/cytoki-nine faible. Leur croissance est ensuite stimulée par uneconcentration plus élevée en auxine (DOCUMENTS 21 et 23).
� Les réponses aux questions précédentes permettent deproposer un protocole pouvant être éventuellement mis enœuvre en classe. Sa réalisation exige de travailler dans desconditions parfaites de stérilité. Un explant doit être pré-levé sur un organe végétal, tige, feuille ou tubercule, puismis en culture sur un milieu contenant plus d’auxine quede cytokinine, ou même de l’auxine seule, pour l’obten-tion d’un cal. Le cal doit être transplanté sur un milieucontenant moins de cytokinine pour le développement desracines, ou moins d’auxine et plus de cytokinine pourobtenir de nombreux bourgeons axillaires. Les modifica-tions successives de milieu permettent d’obtenir uneplante entière possédant de nombreux bourgeons axil-laires, à l’origine d’une multiplication des plants obtenuspar culture de méristèmes.
� Chez le Haricot, l’ablation du bourgeon terminalentraîne, comme le montre le DOCUMENT 18, le développe-ment rapide d’un bourgeon axillaire. L’hypothèse d’uneinhibition due à l’auxine sécrétée par le méristème apicaldoit alors être proposée. Elle se trouve validée par lesrésultats des expériences présentées par le DOCUMENT 19 etréalisées chez le Févier : le fait d’inhiber la diffusion del’auxine à l’aide d’un manchon réfrigéré entraîne le déve-loppement du premier bourgeon axillaire (et dans unemoindre mesure, du second). Au cours de la croissance dela plante, l’auxine sécrétée par le bourgeon terminal dif-fuse dans la plante, et, s’accumulant temporairement auniveau des méristèmes axillaires (DOCUMENT 12), s’oppose àleur activité. L’absence du bourgeon terminal prive lesbourgeons axillaires d’auxine, levant ainsi l’inhibitionqu’il exerçait auparavant. On peut remarquer que lerameau obtenu possède à son tour un méristème terminalen activité sécréteur d’auxine... Une explication peut alorsêtre apportée aux morphologies des arbres fruitiers ayantsubi une taille de mise en forme (DOCUMENT 2, page 228).Les données expérimentales des DOCUMENTS 12 et 19 pro-viennent d’une publication de A. Everat-Bourbouloux etG. Roblin (Some Questions about the Release fromInhibition of Axillary Buds in Vicia faba, INRA, 1990).Le rôle de l’auxine dans la dominance apicale avait déjàété mis en évidence par Thimann et Skoog en 1934, enrétablissant l’inhibition des bourgeons axillaires par l’ap-plication, au sommet d’une tige décapitée de Févier, d’unbloc de lanoline imprégné d’auxine. Cependant, l’inhibi-tion exercée par l’auxine pourrait être indirecte, en indui-sant par exemple la sécrétion d’éthylène ou d’acideabscissique, substances inhibitrices du développement desbourgeons.
A c t i v i t é s 5
La différenciation des organes végé-tauxUn objectif notionnel de ces activités concerne l’uni-versalité des modalités de la communication intercellu-laire, l’organogenèse dépendant des rapports deconcentrations instantanées entre les différentes hor-mones végétales présentes dans le milieu extracellu-laire. Leur connaissance a permis de mettre au pointdes techniques de culture in vitro qui font appel à desassociations de plusieurs hormones, présentes à desconcentrations variées à différents moments de la cul-ture. Il s’agit moins, ici, d’enrichir les connaissancesdes élèves des rôles de chacune de ces hormones, quede sensibiliser à des applications biotechnologiquesactuelles. On s’appuie donc d’abord sur une expéri-mentation établissant le rôle de l’auxine, seule hor-mone végétale connue des élèves, dans l’induction desébauches foliaires. L’existence des cytokinines estensuite signalée, sans que leurs modes d’action nesoient étudiés au cours de ces activités, afin de com-prendre les principes qui régissent la multiplication de
78

E x e r c i c e s
Évaluer ses connaissances
Des relat ions logiquesLes affirmations a, d, et e sont exactes. Les affirma-tions b, c et f sont fausses.a. L’auxine est une hormone végétale, car l’auxinesécrétée par des cellules est transportée vers descellules cibles qui répondent en s’allongeant.d. Les bourgeons axillaires ne se développent pasen présence du bourgeon terminal, car l’auxine libé-rée par le bourgeon terminal inhibe l’activité desméristèmes sous-jacents.e. Les ramifications naturelles d’un arbre dépendentde son exposition à la lumière, car un éclairementanisotrope provoque une migration de l’auxine ducôté opposé à la source lumineuse.
Choisir la bonne réponse
1. Le mécanisme cellulaire responsable du photo-tropisme est...b. ... une élongation plus importante des cellules ducôté opposé à la lumière.2. L’influence de la lumière sur la morphologie d’unvégétal s’exerce par une modification...a. ... de la répartition de l’auxine au sein de la plante.
Exercice corrigé
Environnement, phénotype et hormone végétaleL’exercice proposé consiste, à propos des gibbérel-lines, à réinvestir les connaissances acquises dansle domaine des hormones végétales et la manièredont s’exerce l’influence d’un facteur du milieu.Cependant, si la lumière modifie la répartition del’auxine, elle induit la synthèse d’une gibbérellineactive. Le transfert des connaissances et des com-pétences acquises ne saurait donc être immédiatdans la résolution de cet exercice. Celle-ci supposeégalement la compréhension du fonctionnementd’une chaîne de biosynthèse. En cela, cet exercicepeut aussi être considéré comme un exercice desynthèse. À partir d’une saisie complexe de don-nées, il propose de construire un modèle possiblede chaîne de biosynthèses comportant l’interventiond’un éclairement de jours longs, puis de confronterce modèle aux données fournies. Il peut être inté-ressant de faire résoudre les problèmes posés enprenant appui sur les activités 2 du chapitre 10,pages 196 et 197.
3
2
1
Évaluer ses compétences
Auxine et cytokinines1. Les travaux de Miller et Skoog montrent que,pour une même concentration de cytokinine, l’ajoutd’auxine inhibe d’autant plus la formation des bour-geons que sa concentration est élevée. Les travauxde Vanderhoef montrent que l’auxine favorise l’élon-gation cellulaire, et que celle-ci est, à tous les stadesde l’élongation, inhibée par l’ajout de cytokinine.L’auxine favorise donc l’élongation cellulaire etinhibe la formation des bourgeons ; la cytokininefavorise la formation des bourgeons et inhibe l’élon-gation cellulaire.
2. Pour obtenir de nombreux bourgeons, il faut aug-menter le taux de cytokinines, et réduire celuid’auxine (< 5.10–6 g.mL–1). Pour obtenir la croissancedes tiges, il faut au contraire réduire le taux de cyto-kinine et augmenter celui d’auxine.
E x e r c i c e c o m p l é m e n t a i r e
Une technique tradit ionnel le, le bouturageAfin de multiplier les plantes d’une même variété, onprélève des fragments de végétaux, que l’on traitede manière à provoquer l’émission de racines à labase de la tige.Les boutures sont le plus souvent issues derameaux prélevés sur le pied de la plante mère justeau-dessous d’un nœud, c’est-à-dire du point d’in-sertion d’une feuille.Il est recommandé de recouvrir la base du rameauavec une poudre contenant des hormones végétalesde synthèse, voisines de l’auxine naturelle (ABA, ouacide � indolbutyrique à 0,056 %), mais dont leseffets sont plus durables (voir « Questions d’aujour-d’hui », page 248).En outre, l’ablation de la partie terminale du rameaupeut s’avérer nécessaire.Justifier chacune de ces pratiques à l’aide desconnaissances acquises.
Réponses attendues– La section d’un rameau au-dessous d’un nœudgarantit la présence d’au moins un bourgeon axil-laire sur le fragment de tige prélevé. Les traitementsproposés ont pour but de lever l’inhibition du méris-tème de ce bourgeon, resté inactif sur le pied de laplante mère.– La dédifférenciation de cellules de la tige en cel-lules méristématiques, à l’origine de méristèmesracinaires secondaires, ne peut être induite que pardes concentrations élevées en auxine. La teneur enauxine étant, le plus souvent, naturellement faible auniveau des entre-noeuds, une application localiséed’auxine de synthèse peut y favoriser l’émission deracines.
5
4
Chapitre 12 : Le contrôle de la croissance chez les végétaux
79

– L’auxine endogène sécrétée par le bourgeon ter-minal peut inhiber l’activité du méristème du bour-geon axillaire. Sa disparition peut lever cetteinhibition.Remarque : La circulation de l’auxine étant polarisée,les auxines de synthèse appliquées à la base durameau ne peuvent provoquer à leur tour une inhibi-tion.
80

Pa
rt
ie
2 Structure, composition et dynamique de la Terre
Structure et composition chimiquede la Terre interne
13 Structure et composition de la croûte terrestre .............................................. 82
14 Modèles de la structure de la Terre.......... 89
15 Un modèle de la dynamiquelithosphérique .................................................................. 94
Unité 5

82
Programme : notions et contenus• La classe sur le terrain, une démarche scientifique.La sortie de terrain a pour objectif d’observer des affleurements, de s’approprier logiquement l’information géologique et les questions qu’elle sou-lève, de sensibiliser l’élève à l’importance de la collecte de données de terrain. Le contexte et les problèmes géologiques choisis doivent se ratta-cher au contenu du programme.Cette sortie géologique a également pour objectif une approche concrète de la diversité morphologique des végétaux en liaison avec leur environ-nement.• Composition chimique de la Terre : des échantillons naturels aux matériaux inaccessibles.• Seuls les matériaux de la croûte et du manteau supérieur sont observables à la surface de la Terre. Les enveloppes de la Terre, accessibles paréchantillonnage, ont des compositions chimiques différentes que l’on détermine à partir de l’étude de roches représentatives. Ces roches sont for-mées de minéraux et/ou de verre.• La composition chimique des enveloppes de la Terre est dominée par un nombre limité d’éléments dits “ majeurs ” (Si, O, Mg, Fe, Ca, Na, K, Al).• Les principaux minéraux qui hébergent ces éléments sont : olivines, pyroxènes, feldspaths, quartz, amphiboles et micas.– La composition chimique des roches est présentée en % massique d’éléments chimiques.– Les matériaux du manteau profond et du noyau sont inaccessibles.
Objectifs à atteindreL’objectif visé par ce chapitre introductif est de mettre en place des connaissances concernant la structure et la com-position des croûtes continentale et océanique, connaissances qui serviront de base aux chapitres suivants.
Il est également destiné à montrer que la géologie est, à la base, une science de terrain qui privilégie les faits direc-tement accessibles ; l’observation d’affleurements et d’échantillons de roches prélevées sur les continents ou au fonddes océans :
– permet une saisie d’informations concernant la composition et la structure des croûtes ;
– aboutit à la formulation de problèmes relatifs à la composition et à la genèse de ces roches et par là même à la dyna-mique du globe.
Ce chapitre à caractère descriptif motive ainsi l’étude dynamique du globe : ainsi la présence de basalte et de gab-bro d’origine magmatique au niveau de la croûte océanique induit la recherche de l’origine du magma ainsi que celledes conditions de mise en place des roches.
La classe sur le terrain permet également de saisir les limites d’exploration directe de la structure du globe et motivel’étude des méthodes indirectes, en particulier sismiques, à l’échelle de la croûte dans ce chapitre puis à l’échelle duglobe dans le chapitre suivant.
Cette première approche permet également de sensibiliser à la notion d’échelle en passant de l’affleurement (la sor-tie), à l’échantillon (la roche) puis au minéral (la molécule) et enfin à l’élément chimique. Ces différentes informa-tions devant être à nouveau intégrées dans un cadre de plus en plus général passant de l’échelle locale (la sortie), àcelle d’un pays (la France) puis à celle du globe.
Structure et composition de la croûte terrestre
C H A P I T R E 1 3

Chapitre 13 : Structure et composition de la croûte terrestre
83
(Bassins parisien et aquitain, Limagne, Alsace et Bresseen particulier), la présence des trois catégories de rochesdans les massifs et chaînes récentes avec une prédomi-nance des affleurements de roches métamorphiques et degranites dans les massifs (anciennes chaînes érodées) ;– l’importance relative des affleurements ; ainsi les rochessédimentaires semblent être les roches continentales pré-dominantes si on considère l’étendue des bassins et leurprésence au sein des massifs et chaînes (à ce niveau, il fautinsister sur le fait qu’il manque la dimension verticale dela croûte qu’il faudra étudier par des méthodes directes,les forages ou indirectes, sismiques réfraction ouréflexion).
A c t i v i t é s 2
La classe sur le terrainCette activité peut être située au tout début du chapitreafin de motiver l’intérêt de la collecte de données sur leterrain et de rechercher les différentes activités qui peu-vent être réalisées au niveau d’un affleurement ainsique le matériel nécessaire à leur réalisation.
Exploitation� La lecture du DOCUMENT 4 (photographie et texte) cou-plée au DOCUMENT 5 permet de mettre en correspondanceune activité et l’utilisation du matériel ; ainsi on peut suc-cessivement :– localiser le site et se repérer (orientation) à l’aide decartes topographiques et géologiques et d’une boussole(DOCUMENT 5b.) ;– saisir des informations en prenant des photographies(aspect général ou détails de l’affleurement) et en réalisantdes croquis légendés de l’affleurement (DOCUMENT 5c. et5d.) en veillant à ce que l’échelle soit repérée c’est-à-direla dimension des objets ;– prélever des échantillons représentatifs de roches ou defossiles, en place et non transportés, les moins altérés pos-sible en veillant à bien repérer le lieu de la collecte et à nepas piller le site (DOCUMENTS 5e. et 5 f.) ;– classer les échantillons dans des sachets ou des boîtes etles situer sur le croquis de l’affleurement à l’aide de numé-ros repères.� Les activités sur le terrain amènent le géologue àobserver et à décrire les objets géologiques (affleurements,roches ainsi que leurs caractéristiques : disposition,aspect, contenus à différentes échelles). Cette saisie d’in-formation peut permettre de répondre à un problème déjàformulé ou au contraire permettre la formulation de pro-blèmes qui nécessiteront la recherche d’éléments nou-veaux pour les résoudre ; ainsi l’observation de rochesdifférentes, certaines en strates plus résistantes à l’érosion(DOCUMENT 5a.) ou la présence de fossiles d’huîtres dansune roche (DOCUMENT 5 f.) peuvent amener l’élève à formu-ler la problématique suivante : « Comment expliquer laprésence de roches disposées en strates, de nature diffé-rente et la présence de fossiles d’huîtres dans certaines ? »Une recherche d’explication peut alors être engagée avec
◆ Problèmes scientifiques à résoudreDans ce chapitre, il s’agit plutôt d’une interrogation sur lastructure et la composition de la partie superficielle duglobe et la correspondance entre la géomorphologie et lagéologie sous-jacente.
P o s e r l e s p r o b l è m e s
L’existence d’affleurements naturels de roches sur lescontinents ou au fond des océans permet de saisir l’im-portance de la récolte de données de terrain et de formulerun certain nombre de questions relatives à la nature et à lacomposition chimique des roches de la croûte continentaleet de la croûte océanique.
A c t i v i t é s 1
Matériaux accessibles de la croûtecontinentaleL’objectif à partir du réel (vue aérienne, affleurements)ou de sa représentation (cartes géologiques) est derepérer la diversité, la localisation et l’importancerelative des matériaux affleurant au niveau d’un payscomme la France.
Exploitation� La carte géologique de Clermont-Ferrand (DOCUMENT3b.) associée à une observation aérienne (DOCUMENT 3a.) età celle de deux affleurements (DOCUMENTS 3c. et 3d.) per-met de retrouver les trois catégories de roches envisagéesen classe de 4e ainsi que certaines de leurs caractéristiquessur le terrain :– les roches magmatiques (plutonique comme le granite :en massif sur la carte et en blocs séparés par des diaclasessur le terrain ; volcanique avec des coulées qui recouvrent,le plus souvent, les autres terrains) ;– les roches métamorphiques (DOCUMENT 3b.) ;– les roches sédimentaires disposées en strates (DOCUMENT3d.).Le choix de cette carte a été motivé par le fait qu’elle estune des seules à représenter de façon simple les trois caté-gories de roches présentes sur les continents.La comparaison des DOCUMENTS 3a. et 3b. permet égale-ment un travail d’orientation avec la mise en correspon-dance des édifices volcaniques et donc d’une orientationde la photographie (est à gauche avec la Limagne, ouest àdroite avec les autres puys).
� Le passage à la carte géologique de la France (rabat decouverture du manuel) permet de passer à une échelle plusgrande et de généraliser l’existence des trois catégories deroches. Deux autres objectifs peuvent également êtreatteints :– la localisation des catégories de roches dans les grandsdomaines morphologiques français : ainsi l’importancedes roches sédimentaires dans les bassins et fossés

Les roches sédimentaires représentent donc une très faiblepart des roches de la croûte continentale malgré leursaffleurements très importants.
A c t i v i t é s 4
Les roches volcaniques : des roches mises en place en surfaceL’objectif de cette activité est de réinvestir des acquisde la classe de 4e mais également d’exprimer l’abon-dance relative des roches volcaniques continentales parrapport aux autres roches de la croûte continentale.
Exploitation� Le DOCUMENT 11 rappelle le mode de mise en place desroches volcaniques continentales (écoulement de laves leplus souvent fluides à haute température suivi d’un refroi-dissement qui aboutit à une roche de couleur foncée, lebasalte dans ce cas).Le DOCUMENT 10 représente les affleurements volcaniquessur les continents, soit ponctuels (volcans isolés ou for-mant des guirlandes), soit constituant des nappesimmenses appelées encore trapps (ainsi les trapps duDeccan représentent la superficie de la France avec uneépaisseur moyenne de 2 km !). Cependant, si on comparela surface et l’épaisseur des trapps à celle des continents,on constate que les roches volcaniques continentales nereprésentent qu’une très faible part de la croûte continen-tale.� Les roches volcaniques sont des roches issues durefroidissement de laves d’origine magmatique de hautetempérature (plus de 1 000 °C) (DOCUMENT 11).Le basalte (DOCUMENT 12) est une roche cristallisée quicomporte des cristaux de taille variable (olivine etpyroxène de grande taille et feldspath plagioclase de pluspetite taille appelé microlites ) noyés dans un verre noncristallisé : la structure de la roche est qualifiée de struc-ture microlitique.Le DOCUMENT 14 confronté au DOCUMENT 12 permet deconclure que les cristaux se forment à partir du refroidis-sement d’un liquide magmatique et que la taille est liée àla vitesse de refroidissement (plus la vitesse est rapide,plus les cristaux sont petits). On peut ainsi déduire que lesgros cristaux d’olivine et de pyroxène du basalte se sontformés au cours d’un refroidissement lent précoce, suivid’un refroidissement plus rapide donnant les microlites defeldspath puis d’un refroidissement très rapide qui a figéle magma en un verre non cristallisé emprisonnant lescristaux préexistants.Le DOCUMENT 13 sera exploité plus à fond avec les DOCU-MENTS 18, 22, 23, 28 qui permettront de définir la constitu-tion et la composition des croûtes continentales etocéaniques.
la formulation d’hypothèses pouvant être étayées par desobservations de terrains (le DOCUMENT 5g. pouvant suggérerune solution à l’existence de coquilles dans une roche).Les éléments collectés sur le terrain pourront faire l’objetd’une étude plus précise au laboratoire (à la loupe ou aumicroscope) permettant ainsi, avec d’autres documents, derésoudre les problèmes formulés précédemment.
A c t i v i t é s 3
Les roches sédimentaires : des roches de surfaceL’objectif de cette activité est de rappeler les caracté-ristiques des roches sédimentaires déjà étudiées aucycle central du collège (5e ou 4e) et surtout de montrerque les roches sédimentaires ne représentent qu’unetrès faible part de la croûte continentale malgré leursgrands affleurements à cause de leur faible épaisseurpar rapport aux autres roches tels que granite et rochesmétamorphiques.
Exploitation� Le DOCUMENT 6 permet de dégager les critères caractéri-sant les roches sédimentaires, leur disposition en strates auniveau de l’affleurement et la présence de fossiles danscertaines d’entre elles.Le DOCUMENT 7 permet de classer les roches de la carrièrede Cormeilles-en-Parisis en roches :– d’origine détritique : les sables et les marnes contenantrespectivement des grains de quartz et des particules argi-leuses issus de l’érosion du continent (les marnes renfer-mant également du calcaire) ;– d’origine biochimique : les calcaires qui sont souvent lerésultat de l’accumulation de squelettes calcaires élaboréspar des organismes planctoniques marins.Le gypse quant à lui a une origine chimique, sa précipita-tion étant provoquée par l’évaporation de l’eau de mer quiconcentre les ions ; cette roche est ainsi qualifiée d’éva-porite.
� Le DOCUMENT 8 montre l’existence, dans la partie sud duBassin parisien, d’une épaisseur voisine de 1 000 m deroches sédimentaires (strates de roches de nature diffé-rente et comportant des fossiles caractéristiques d’unmilieu marin de profondeur variable) surmontant desroches métamorphiques (gneiss) de plus de 2 500 m, leforage s’arrêtant à cette profondeur.Le DOCUMENT 9, interprétation d’un profil de sismiqueréfraction, montre :– que la croûte continentale a une épaisseur qui oscilleentre 25 km sous le Massif central et plus de 40 km sousles Alpes (soit une épaisseur moyenne de 30 km) ;– que les roches sédimentaires n’excèdent pas 5 kmd’épaisseur et qu’elles représentent un plaquage au-dessusd’une croûte essentiellement constituée de roches grani-tiques et métamorphiques.
84

A c t i v i t é s 5
Le granite, une roche fondamentale de la croûte continentaleCette activité est destinée à montrer l’importance dugranite dans la constitution de la croûte continentaleainsi qu’à rechercher son origine par comparaison àcelle du basalte.
Exploitation� Le DOCUMENT 15 montre un affleurement à grandeéchelle (plusieurs centaines de km) qui présente des mas-sifs granitiques anciens érodés inclus dans des rochesmétamorphiques. Le DOCUMENT 16 rappelle que la croûtecontinentale est épaisse de 30 km en moyenne et qu’elleest constituée essentiellement de granite et de rochesmétamorphiques. Les granites peuvent être inclus dans lesroches métamorphiques ou remonter à travers les rochessus-jacentes constituant des batholites intrusifs que l’éro-sion pourra mettre à jour.
� Le DOCUMENT 17 montre que le granite est constitué dela juxtaposition de cristaux tous visibles à l’œil nu donnantun aspect granuleux à la roche, d’où son nom. Ces cristauxsont de nature différente ; on rencontre du quartz, du micaet du feldspath qui peuvent être identifiés à l’œil nu ou enmicroscopie optique en lumière polarisée analysée (parleur forme et leur couleur).La structure grenue de la roche et la présence de gros cris-taux sous-tendent que le granite est une roche d’originemagmatique et que le magma s’est refroidi en remontantlentement vers la surface mais sans l’atteindre aboutissantà la formation de gros cristaux : la roche est dite pluto-nique.La comparaison des DOCUMENTS 12-13 et 17-18 montre quebasalte et granite sont toutes deux des roches magma-tiques mais que leur composition et leur structure étantdifférentes, leur origine (composition des magmas) et leurmise en place sont différentes (volcanique dans un cas,plutonique dans l’autre).
A c t i v i t é s 6
La croûte continentaleCe chapitre, en incluant l’étude des roches métamor-phiques, est destiné à faire le point sur la compositionde la croûte continentale et en particulier à montrerque la composition globale de la croûte continentale estvoisine de celle du granite.
Exploitation� Les DOCUMENTS 19 et 20 montrent des roches métamor-phiques, à deux échelles différentes, qui présentent desdéformations (plis d’épaisseurs variables suggérant uneorigine dans un contexte compressif avec un comporte-ment souple et non cassant des roches et donc à une pro-fondeur et à une température relativement élevées) ; le
DOCUMENT 21 montre que la roche est entièrement cristalli-sée et que les cristaux sont disposés en lits ou feuillets(micacés et quartzo-feldspathiques). Le gneiss n’est doncpas une roche magmatique ayant pour origine un magmaen fusion.� Le gneiss et le granite comportent les mêmes minérauxet montrent une composition chimique voisine (DOCUMENTS17-18 et 21-22). La seule différence concerne la disposi-tion des minéraux qui est orientée dans le gneiss.� La composition chimique globale de la croûte conti-nentale est très voisine de celle du granite. En effet, leDOCUMENT 23 montre que le granite représente 40 % duvolume de la croûte continentale et que le gneiss de mêmecomposition chimique que le granite représente 55 % duvolume de la croûte continentale. La croûte continentale adonc, pour 95 % de son volume, une composition grani-tique.
A c t i v i t é s 7
Les matériaux de la croûte océaniqueL’objectif de cette activité est de comparer l’organisa-tion de la croûte océanique à celle de la croûte conti-nentale et d’opposer la nature et la composition desroches qui les constituent.
Exploitation� Le basalte et le gabbro présentent la même compositionminéralogique mais une structure différente (le basalteprésente une structure microlitique avec des cristaux noyésdans un verre non cristallisé alors que le gabbro présenteune structure grenue, roche entièrement cristallisée for-mée de cristaux de grande taille).Ces deux roches ont pour origine un même magma, l’uneétant volcanique (le basalte) subissant une phase finale derefroidissement rapide par effusion de lave en surface (aufond de l’océan prenant la forme de coussin au contact del’eau), l’autre plutonique subissant un refroidissementplus lent en profondeur (sous les basaltes en coussins et enfilons).
� Les croûtes continentales et océaniques s’opposent surplusieurs points :– Leurs épaisseurs sont différentes : la croûte continentaleest plus épaisse (30 km en moyenne contre 7 km pour lacroûte océanique) et son épaisseur est variable (elle estplus épaisse à l’aplomb des reliefs pouvant atteindre 70-80 km).– La croûte continentale est hétérogène horizontalement etverticalement (épaisseur et nature des roches variablesselon les régions) alors que la croûte océanique est homo-gène sur tous les fonds océaniques (épaisseur constante etsuperposition de basaltes et de gabbros).– La composition minéralogique et donc la compositionchimique des roches sont différentes entre les deuxcroûtes. La croûte océanique est ainsi plus riche en cer-tains éléments chimiques tels que le fer, le magnésium etle calcium et plus pauvre en silicium, sodium et potas-sium.
Chapitre 13 : Structure et composition de la croûte terrestre
85

E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Granite : roche magmatique plutonique consti-tuée de trois catégories de minéraux, quartz, mica,feldspath. Elle est présente uniquement au niveaudes continents.b. Basalte : roche magmatique volcanique consti-tuée de minéraux, olivine, pyroxène et feldspath pla-gioclase noyés dans un verre. Elle constitue le fondde tous les océans et se trouve également au niveaudu volcanisme de points chauds en milieu continen-tal ou océanique.c. Roche sédimentaire : roche caractérisée par unaspect stratifié à l’affleurement résultant d’un dépôtde particules et par la présence fréquente de fos-siles.
1
86
d. Roche métamorphique : roche souvent plissée àl’affleurement et présentant une disposition enfeuillets des minéraux. Roche issue de transforma-tions (métamorphose), à l’état solide, de roches pré-existantes sous l’effet de contraintes subies enprofondeur.e. Roche volcanique : roche issue du refroidisse-ment rapide de laves issues d’un magma et émisesà la surface des continents ou au fond des océans.f. Roche plutonique : roche issue du refroidissementlent d’un magma qui ne parvient pas à la surface.g. Roche magmatique : roche issue du refroidisse-ment d’un magma (soit volcanique, soit plutonique).h. Bassin sédimentaire : dépression plus ou moinsvaste remplie d’une succession de couches sédi-mentaires.i. Massif : zone continentale érodée qui laisse appa-raître les roches formées en profondeur à la suite del’érosion des roches sus-jacentes.j. Minéral : substance chimique naturelle, solide, decomposition chimique définie et qui présente unestructure cristalline.k. Cristal : édifice comportant une structure géomé-trique précise qui dépend de la nature et du moded’assemblage d’éléments chimiques présents dansles minéraux des roches.
Vrai ou faux ?1. La croûte océanique : vrai : b., e., f.2. la croûte continentale : vrai : b., c., e.
2
Catégories de roches Disposition sur le terrain Structure de la roche Contenu de la roche
A – MAGMATIQUES
1 – volcaniques coulées ou projections microlitique cristaux noyés dans aériennes un verrecoulées en coussinsen milieu aquatique
2 – plutoniques aspect massif grenue gros cristaux jointifs
B – MÉTAMORPHIQUES plissées (plis d’épaisseur feuilletée cristaux disposés variable et souples) en feuillets
C – SÉDIMENTAIRES strates superposées variable selon la nature présence fréquente de nature souvent différente des roches de fossiles
– La croûte continentale se raccorde à la croûte océaniquenon pas à la limite océan-continent mais au niveau du taluscontinental qui peut se trouver à plusieurs centaines dekilomètres du littoral. Il faut donc opposer le couple terresémergées (30 %) et surface océanique (70 %) au couplecontinent au sens géologique (40 %) et océan au sens géo-logique (60 %).
Expliquer brièvementa.
3

Exercice corrigé
Les ophiol i tes alpinesVoir correction dans le manuel, pages 274 et 275.
Évaluer ses compétences
Enclaves rocheuses dans unbasalte1. Le basalte est une roche cristalline à structuremicrolitique qui atteste d’une origine magmatiquevolcanique (magma d’origine profonde qui remonteet donne naissance à des laves qui s’écoulent à lasurface et qui aboutissent à des roches en se refroi-dissant rapidement).2. La présence d’une enclave de granite à l’intérieurdu basalte prouve que le granite est antérieur aubasalte. En remontant à travers les autres roches dela croûte continentale, le magma basaltique a dûarracher un fragment de granite ; ce dernier s’estretrouvé emprisonné dans la lave puis dans la rochebasaltique après refroidissement de cette lave (Voirschéma page 88).3. Le basalte n’est pas d’origine océanique car lesfonds océaniques étant constitués uniquement debasalte et de gabbro, les enclaves ne pourraient pascontenir de granite caractéristique des continents.4. La péridotite est une roche qui ne se trouve nidans la croûte continentale ni dans la croûte océa-nique. Le fait de la retrouver en enclave dans dubasalte peut s’expliquer par l’existence de cetteroche au-dessous de la croûte (ce serait la rocheconstitutive du manteau). Dans ce cas, le magmabasaltique proviendrait de la fusion partielle de cettemême péridotite ; le magma devant se former dansou au-dessous de la région contenant les péridotitespour que certains fragments de celles-ci se retrou-vent en enclaves (Voir schéma page 88).
5
4
Chapitre 13 : Structure et composition de la croûte terrestre
87
b. Si on prend l’exemple du quartz de formule SiO2,ce minéral est constitué de l’association detétraèdres tous constitués des mêmes éléments chi-miques, un cation Si4+ étant entouré de 4 anions O2–.En revanche, dans le cas des feldspaths, le siliciumpeut être remplacé par l’aluminium (Al3+) au cœur decertains tétraèdres ; dans ce cas, un certain nombrede charges négatives persistent au niveau destétraèdres, des cations tels que K+, Na+ ou Ca2+ s’in-tercalent entre les tétraèdres. Selon la nature et lenombre de ces cations, on considère deux famillesde feldspaths soit potassiques, soit plagioclasesdont la composition oscille entre un pôle sodique etun pôle calcique.c. Les roches métamorphiques ont une origine liée àla formation des chaînes de montagnes et donc auxcollisions continentales car elles présentent des plisà l’échelle de l’affleurement et à l’échelle de l’échan-tillon attestant ainsi de l’action de forces compres-sives. Leur formation se déroule en profondeur carles plis les affectant ne sont pas cassants maissouples avec une épaisseur variable attestant d’unplissement en conditions thermiques élevées maissans atteindre la fusion. Un autre argument est queles roches métamorphiques affleurent largement aucœur des massifs anciens très érodés au même titreque les granites.d. La structure cristalline grenue du granite ne peuts’expliquer que par une origine magmatique, donc àune certaine profondeur nécessaire à la fusion decertaines roches à l’origine du magma de composi-tion granitique. Le refroidissement du magma s’estfait très lentement expliquant la grande taille descristaux sans arrivée à la surface (absence de verre).C’est l’érosion des terrains sus-jacents qui a dégagéle granite ce qui est confirmé par l’abondance desaffleurements granitiques dans les massifs très éro-dés tels que le Massif armoricain ou le Massif cen-tral par rapport aux chaînes de montagnes récentes.e. Le fond de la Manche présente, au niveau litholo-gique, toutes les caractéristiques d’un continent ;c’est donc une partie déprimée envahie par la mermais qui est en continuité avec le continent euro-péen dont fait également partie la Grande-Bretagne.Si la Manche était un océan au sens géologique, ondevrait trouver un fond constitué d’une superposi-tion de basaltes et de gabbros.
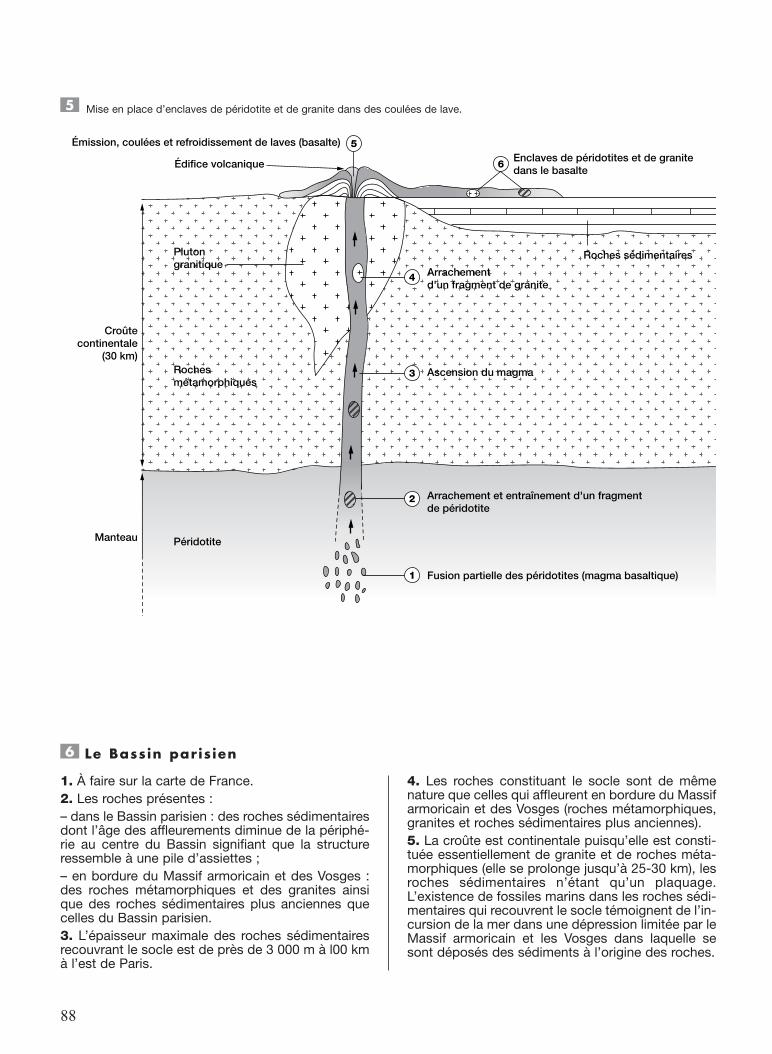
4. Les roches constituant le socle sont de mêmenature que celles qui affleurent en bordure du Massifarmoricain et des Vosges (roches métamorphiques,granites et roches sédimentaires plus anciennes).5. La croûte est continentale puisqu’elle est consti-tuée essentiellement de granite et de roches méta-morphiques (elle se prolonge jusqu’à 25-30 km), lesroches sédimentaires n’étant qu’un plaquage.L’existence de fossiles marins dans les roches sédi-mentaires qui recouvrent le socle témoignent de l’in-cursion de la mer dans une dépression limitée par leMassif armoricain et les Vosges dans laquelle sesont déposés des sédiments à l’origine des roches.
1. À faire sur la carte de France.2. Les roches présentes :– dans le Bassin parisien : des roches sédimentairesdont l’âge des affleurements diminue de la périphé-rie au centre du Bassin signifiant que la structureressemble à une pile d’assiettes ;– en bordure du Massif armoricain et des Vosges :des roches métamorphiques et des granites ainsique des roches sédimentaires plus anciennes quecelles du Bassin parisien.3. L’épaisseur maximale des roches sédimentairesrecouvrant le socle est de près de 3 000 m à l00 kmà l’est de Paris.
88
Mise en place d’enclaves de péridotite et de granite dans des coulées de lave.
3
4
5
2
1
6
Fusion partielle des péridotites (magma basaltique)
Arrachement et entraînement d'un fragmentde péridotite
Arrachementd'un fragment de granite
Plutongranitique
Édifice volcanique
Rochesmétamorphiques
Croûtecontinentale
(30 km)
Manteau Péridotite
Roches sédimentaires
Enclaves de péridotites et de granitedans le basalte
Émission, coulées et refroidissement de laves (basalte)
Ascension du magma
Le Bassin paris ien6
5

89
engendre des ondes qui se déplacent dans toutes lesdirections, et qui pourront atteindre différents endroitsde la planète. On cherche à montrer ici que les troistypes d’ondes enregistrées par les sismographes n’ontpas les mêmes caractéristiques.
Cette activité permet de motiver les activités suivantes,car c’est grâce à l’étude de la propagation des ondessismiques que l’on peut connaître la structure internedu globe.
ExploitationLe DOCUMENT 3 permet de présenter les différents typesd’ondes sismiques. Les schémas représentent la déforma-tion des roches près de la surface lors du passage desondes P, S et L. Les deux derniers schémas correspondentaux ondes de Love (oscillations dans un plan horizontalperpendiculaire à la direction de propagation) et deRayleigh (oscillations dans un plan vertical).Le DOCUMENT 4 montre qu’un séisme ayant eu lieu à Kobé(île de Honshu, au Japon) a pu être enregistré à Hawaii et
◆ Problème scientifique à résoudreComment peut-on connaître la structure et la compositioninterne de la Terre ? À quels modèles du globe aboutit-on ?
P o s e r l e s p r o b l è m e s
L’observation de météorites, qui ont le même âge que lesystème solaire, et d’enregistrements d’ondes sismiques àl’aide de sismographes peut amener l’élève à s’interrogersur l’étude (forcément indirecte) des structures profondesdu globe.
A c t i v i t é s 1
Les ondes sismiquesUn séisme provoqué au foyer par rupture et déplace-ment des roches de part et d’autre du plan de faille,
Programme : notions et contenus• Structure et composition chimique de la Terre interne.• Origine, différenciation et structure interne de la Terre.• L’étude de la propagation des ondes sismiques montre que la Terre est structurée en enveloppes concentriques de tailles, masses et masses volu-miques différentes : la croûte (continentale ou océanique), le manteau et le noyau. Les enveloppes sont séparées par des discontinuités physiqueset/ou chimiques. La lithosphère se distingue de l’asthénosphère sous-jacente par un comportement rigide.• La température, la pression et la masse volumique varient avec la profondeur.• Cette structure de la Terre résulte, d’une part, de sa formation par accrétion de petits corps dont les météorites de type chondrite sont les vestigeset, d’autre part, de sa différenciation.• Les matériaux du manteau profond et du noyau sont inaccessibles. On peut néanmoins, par des modèles et des raisonnements qui tiennent comptede la formation de la Terre, à partir des chondrites, préciser leurs compositions.
Objectifs à atteindreDans le chapitre 13, nous avons vu que les enveloppes accessibles de la Terre, croûte et manteau supérieur, ont descompositions chimiques différentes, déterminées à partir de l’étude des roches qui les constituent.
Pour connaître la structure et la composition de l’intérieur de la Terre, on doit en revanche utiliser des méthodes indi-rectes, telle que la sismologie. C’est effectivement grâce à l’étude de la propagation des ondes sismiques que l’on peutaboutir à un modèle sismologique du globe : les enveloppes concentriques de la Terre (croûte, manteau, noyau) sontséparées par des discontinuités.
Ce modèle sismologique sera enrichi en indiquant la nature et la composition chimique des roches (dont une partie aété étudiée précédemment) et les limites entre les différentes couches seront précisées.
Modèles de la structure de la Terre
C H A P I T R E 1 4

sés, plus ces matériaux (roches et minéraux constitutifs)sont denses et plus la vitesse des ondes P est élevée. Onpeut donc en déduire que la densité des matériaux de laTerre augmente avec la profondeur.
A c t i v i t é s 3
Zone d’ombre et structure interne du globeL’analyse des temps d’arrivée des ondes sismiques amontré une augmentation de la densité avec la profon-deur. Nous allons voir que cette densité n’augmente pasrégulièrement mais qu’il existe des discontinuités àl’intérieur du globe.
Donc, après avoir montré dans l’activité précédente,que la structure du globe est hétérogène, on cherchemaintenant à construire un modèle de la structureinterne du globe.
Exploitation� Le DOCUMENT 9 nous montre que pour un séisme donné,il existe une région du globe où les enregistrements sontnuls, c’est la zone d’ombre. Elle se situe entre 103° et 142°de distance angulaire de l’épicentre pour les ondes P, quine sont donc pas reçues entre 11 500 km et 14 500 km del’épicentre, et pour les ondes S, la zone d’ombre est bienplus importante puisque ces ondes ne sont pas reçues au-delà de 103° (11 500 km). Donc on ne peut expliquer ceszones d’ombre avec le modèle d’une sphère dans lequel ladensité augmente régulièrement avec la profondeur,comme on l’a vu dans l’activité précédente.� On a vu que l’on pouvait appliquer aux ondes sis-miques les lois de l’optique (activités 2) : les rais sis-miques de même que les rayons lumineux, sont destrajectoires qui divergent à partir d’une source (foyer sis-mique ou source lumineuse). On réalise donc ici unmodèle analogique en utilisant une source lumineuse pourvisualiser le trajet des ondes P dans un milieu homogènepuis dans un milieu hétérogène.Grâce au modèle, on peut expliquer la zone d’ombre car ilpermet de visualiser celle-ci et de comprendre qu’elle estliée à l’existence d’une surface de discontinuité, ici maté-rialisée par le petit cristallisoir.Dans le modèle de la Terre du DOCUMENT 11, pour la zoned’ombre des ondes P, on voit que le rai sismique qui arriveà 103° est tangent à la discontinuité (il effleure le noyau)alors que le rai plus profond se réfracte deux fois, à l’en-trée et à la sortie du noyau. L’existence de cette zoned’ombre repose donc sur la présence d’une discontinuitésituée à 2 900 km de profondeur, qui sépare le noyau, cen-tral, du manteau, périphérique.
� On a vu que la zone d’ombre des ondes S est bien plusimportante que celle des ondes P, puisque les ondes S nesont plus du tout enregistrées à partir de 103°. La zoned’ombre des ondes S indique que celles-ci ne sont pastransmises dans le noyau ; sachant que les ondes S,contrairement aux ondes P, ne se propagent pas en milieu
en Australie, dans des lieux situés respectivement à6630 km et 7870 km de la ville de Kobé.Les ondes sismiques partent toutes du même endroit, orelles présentent des temps d’arrivée différents. Les sismo-grammes montrent que pour les deux stations, ce sont lesondes P qui arrivent les premières et qu’elles sont suiviesdes ondes S puis des ondes L : les ondes sismiques n’arri-vent donc pas en même temps.On note que les ondes sismiques arrivent plus tard pour lastation située plus loin du lieu du séisme, ici en Australie.On peut donc penser que les ondes sismiques ont unevitesse de propagation différente ou que le trajet qu’ellesempruntent n’est pas le même (se déplaçant dans lescouches superficielles du globe pour les unes, dans lescouches profondes pour les autres).On peut faire calculer par l’élève la vitesse de propagationdes ondes P, S et L d’après le sismogramme enregistré àHawaii par exemple : on obtient une vitesse d’environ10 km/s pour les ondes P, 5,6 km/s pour les ondes S et de3,9 km/s pour les ondes L.On retrouve ensuite les mêmes valeurs pour le sismo-gramme enregistré en Australie.
A c t i v i t é s 2
Informations apportées par les ondessismiquesL’activité précédente a permis de présenter les caracté-ristiques des ondes sismiques. On cherche maintenant àmontrer que l’étude de ces ondes apporte des rensei-gnements sur la structure de la Terre. L’objectif est demontrer que la vitesse des ondes sismiques P et Sindique que l’intérieur du globe n’est pas homogène.
Exploitation� Le DOCUMENT 5 présente les stations du réseau sismolo-gique mondial géoscope. Chaque station est équipée de 3sismographes permettant l’enregistrement des mouve-ments du sol dans les 3 directions E-W, N-S et verticale.D’après le DOCUMENT 6, on voit que la vitesse de propaga-tion des ondes L est représentée par une droite, donc estconstante, contrairement aux vitesses des ondes P et S, quiaugmentent avec la distance à l’épicentre. C’est-à-dire queles ondes P et S arrivent plus rapidement aux stations éloi-gnées que les ondes L ; or les ondes P et S, contrairementaux ondes L, se propagent en profondeur.Donc les ondes P et S traversent un milieu qui n’est pashomogène, ce qui correspond au modèle b du document 7.De ce fait, les rais sismiques subissent une succession deréfractions, leur donnant une trajectoire courbe commel’indique le DOCUMENT 7c. Ceux qui arrivent aux stations lesplus éloignées de l’épicentre ont parcouru de plus grandesprofondeurs que ceux arrivant aux stations proches del’épicentre, donc la vitesse de propagation des ondes P etS augmente avec la profondeur.
� Le DOCUMENT 8 nous montre que la vitesse des ondes Pdépend essentiellement de la densité des matériaux traver-
90

liquide, on peut en déduire que le noyau présente (dans sapartie externe tout au moins) les propriétés d’un liquide.La discontinuité située à 2 900 km est donc une interfaceentre un milieu solide (le manteau) et un milieu liquide (lenoyau).Les trajectoires des ondes P sont déviées à la frontière2 900 km, car elles subissent une double réfraction, ce quiimplique une diminution de la vitesse de propagation deces ondes.
A c t i v i t é s 4
Un modèle sismologique du globeUn modèle simple de la structure du globe, lié à l’exis-tence de zones d’ombre, a été élaboré dans l’activitéprécédente. La zone d’ombre des ondes P et S étant lapreuve de l’existence d’une surface de discontinuité quisépare le manteau, solide, du noyau, liquide (au moinsdans sa partie externe).
L’objectif de cette activité sera d’affiner le modèle destructure du globe élaboré dans l’activité précédente enutilisant des données sismiques.
Exploitation� Les vitesses des ondes P et S ne sont pas constantes, lemilieu parcouru change donc de caractéristiques, de den-sité notamment. Les variations brutales de la vitesse depropagation des ondes sismiques permettent de repérerdes discontinuités.On remarque qu’à 2 900 km de profondeur, les ondes Pralentissent brutalement et les ondes S s’arrêtent, révélantla discontinuité mise en évidence dans l’activité précé-dente, appelée discontinuité du Gutenberg, séparant lemanteau du noyau externe. Celui-ci est à l’état liquide.À 5 100 km de profondeur, on observe un autre ralentisse-ment de la vitesse des ondes P correspondant à une dis-continuité séparant le noyau externe du noyau interne ougraine, la discontinuité de Lehman. Cette graine est solide(on n’a pas d’information ici permettant de le prouver).L’évolution de la vitesse des ondes sismiques dans unezone plus superficielle révèle la présence d’une disconti-nuité située aux alentours de 30 km sous les continents etde 10 km sous les océans, elle sépare la croûte du man-teau, on l’appelle la discontinuité de Mohorovicic (ouMoho). Elle est repérée par une augmentation de la vitessedes ondes.On met en évidence un ralentissement des ondes sis-miques entre 100/120 et 700 km de profondeur, corres-pondant à l’asthénosphère. Ce ralentissement estinterprété comme la conséquence d’une moindre rigiditédu manteau à ces profondeurs. Entre 0 et 100/120 km deprofondeur, la croûte et la partie supérieure du manteausupérieur forment la lithosphère, plus rigide.Au-delà de 670 km de profondeur, la légère augmentationde vitesse des ondes sismiques révèle un milieu, le man-teau inférieur, rigide lui aussi.
On note une diminution de la vitesse des ondes sismiquesaux alentours de 120 km jusqu’à environ 300 km de pro-fondeur, révélant une zone à moindre vitesse (ou LVZ),c’est-à-dire que le milieu est moins rigide que les zonesadjacentes.Les variations de vitesse observées sont graduelles et cor-respondent à des changements très progressifs de naturephysique. Ainsi, tous les niveaux situés au-dessus de120 km de profondeur sont rigides et forment la litho-sphère, puis jusqu’à 700 km de profondeur, le milieu,quoique solide est un peu moins rigide et appelé asthéno-sphère. À partir de 700 km de profondeur, le manteau infé-rieur redevient rigide. Entre 2 900 km de profondeur et lecentre de la Terre, le noyau est formé de deux couches : unnoyau externe, liquide, jusqu’à 5 100 km de profondeur etun noyau interne, ou graine, solide.Les données sismiques ont donc permis de proposer unmodèle d’organisation interne du globe, à symétrie sphé-rique.
A c t i v i t é s 5
Un modèle minéralogique et chimiquedu globeOn a pu aboutir dans l’activité précédente à un modèlesismologique du globe, caractérisé par l’existenced’enveloppes concentriques séparées les unes desautres par des discontinuités.
On cherche maintenant à connaître la nature des rochesconstituant le manteau et le noyau, la composition descroûtes continentale et océanique ayant été étudiée auchapitre 13. C’est à l’aide de méthodes indirectes quel’on pourra connaître la composition du manteau et dunoyau.
Exploitation� Quand la lave basaltique remonte des profondeurs de laTerre, elle ramone les parois du manteau, entraînant avecelle des fragments des matériaux du manteau. Ceux-ci for-ment donc des enclaves dans le basalte et on les retrouveen surface. Ces enclaves sont de la péridotite, rocheconstituée d’olivine, de pyroxène et de grenat ; le manteaupourrait donc être constitué de péridotites.Le DOCUMENT 16 indique une augmentation importante detempérature entre 150 et 300 km de profondeur, montrantque la péridotite est presque fondue mais toujours solide.Or cette zone correspond à la zone de moindre vitesse quiappartient au manteau, c’est donc que le manteau est denature péridotitique.Le changement de rigidité indique une discontinuité denature physique qui sépare la lithosphère de l’asthéno-sphère.Le DOCUMENT 17 indique en parallèle l’évolution de lavitesse des ondes P et l’interprétation minéralogique deces variations de vitesse. Jusqu’à 400 km de profondeur,les principaux minéraux du manteau péridotitique sontl’olivine et le pyroxène, puis à partir de 400 km, ces miné-
Chapitre 14 : Modèles de la structure de la Terre
91

aurait été de même pour la Terre, puisque ces deux objetsdu système solaire, très proches, se sont formés en mêmetemps.Donc les planètes, dont la Terre, tout comme la Lune sesont formées par accrétion c’est-à-dire par agglomérationde matériaux en orbite autour du Soleil ; les planètes ontvu leur taille augmenter en attirant par gravité la matièreavoisinante, en l’occurrence les météorites qui les bom-bardaient.Ainsi les planètes résultent de l’agglomération de météo-rites, donc d’une accrétion.
� On voit que les chondrites ont même composition quela Terre dans son ensemble : elles sont formées de sili-cates, de fer et de nickel.Les achondrites quant à elles, sont des météorites diffé-renciées qui pourraient être l’équivalent des différentesenveloppes de la Terre : les achondrites, constituées de feret de nickel, correspondraient au noyau alors que lesachondrites silicatées pourraient correspondre aux enve-loppes externes de la Terre.Ces données permettent de proposer comme scénario quela Terre aurait été une gigantesque chondrite qui auraitfondu et se serait complètement différenciée.
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Asthénosphère : Zone du manteau supérieursituée sous la lithosphère et moins rigide qu’elle. Elles’étend à environ 120 à 700 km de profondeur.b. Noyau : Enveloppe terrestre comprise entre 2 900et 5 100 km de profondeur pour le noyau externe,liquide, et entre 5 100 km et le centre de la Terrepour le noyau interne, solide.c. Ondes S : Ondes transversales, dites de cisaille-ment, elles ne se propagent pas dans les milieuxliquides.d. Surface de discontinuité : Frontière séparant deuxenveloppes du globe dont les propriétés, physiquesou chimiques, sont différentes.
Questions à réponse cour tea. La Terre n’étant pas un milieu homogène, les raissismiques subissent une succession de réfractions,qui conduisent à une trajectoire incurvée.b. Les ondes sismiques, lorsqu’elles atteignent unesurface de discontinuité, peuvent se réfléchir ou seréfracter.c. Parmi les météorites, on distingue les chondriteset les achondrites qui peuvent être pierreuses,métalliques ou mixtes.d. Une zone d’ombre sismique est une zone carac-térisée par l’absence totale de réception d’ondessismiques.
2
1
raux changent de nature : on passe à des spinelles et desgrenats. Vers 700 km de profondeur, une nouvelle struc-ture, la pérovskite, remplace les structures précédentes etconstitue tout le manteau inférieur.La densité de ces structures augmente en fonction de laprofondeur, alors que la composition chimique de la péri-dotite ne change pas.On voit bien le lien entre les modifications de nature desminéraux de la péridotite et les variations de vitesse desondes sismiques, donc des changements de nature phy-sique des différentes enveloppes, la péridotite étant moinsrigide pour l’asthénosphère.Le manteau est donc bien constitué de péridotites, vis-queuses parce que froides pour la lithosphère, moins vis-queuses parce que plus chaudes pour l’asthénosphère.
� Les météorites se sont formées en même temps que lesystème solaire, elles ont donc le même âge que la Terre etpossèdent les mêmes constituants, ce qui permet de les uti-liser pour connaître la composition de l’intérieur de laTerre. Comme elles possèdent du fer dans leur composi-tion, on peut donc penser que le noyau est lui aussi consti-tué de fer.On voit que le manteau a une masse volumique inférieureà celle de la Terre dans son ensemble, et on sait que lamasse volumique de la croûte continentale est de 2,7 etcelle de la croûte océanique est de 2,8. Il faut doncadmettre que le noyau possède une masse volumiquebeaucoup plus importante. Celle-ci pourrait correspondreà celle du fer, dont la densité est indiquée (12).– La masse volumique de la Terre est de 5,5 g/cm3 soit 5,5 gigatonnes/km3, la masse de la Terre est donc de 5,5 x 1,1.1012 = 6,05.1012 Gt.– La masse du manteau est : 4,5 x 0,92.1012 = 4,14.1012 Gt.– La masse du noyau est donc : 6,05.1012 – 4,14.1012 = 1,91.1012 Gt.– La masse volumique du noyau est donc :1,91.1012/1,8.1011 = 10,6 g/cm3, ce qui correspond approxi-mativement à celle du fer.
A c t i v i t é s 6
Formation et différenciation du globe terrestreOn a pu obtenir la composition du manteau et du noyaugrâce à l’activité précédente. Le manteau est de naturepéridotitique, alors que le noyau est essentiellementconstitué de fer.
Dans cette activité, on cherche à montrer comment laTerre s’est formée et si elle était déjà différenciée dès saformation.
Exploitation� La majorité des cratères se sont formés sur la Luneentre – 4 et – 3 milliards d’années, c’est-à-dire lors du pre-mier milliard d’années de son histoire. Ils sont dus à desbombardements météoritiques. On peut penser qu’il en
92

Expliquer brièvementa. La distinction entre lithosphère et asthénosphèreest de nature physique car la lithosphère, qui com-prend la croûte et la partie supérieure du manteauest très rigide, alors que l’asthénosphère, qui estsituée en dessous, est moins rigide.b. Le manteau et la croûte sont constitués de rochessurtout formées de minéraux silicatés alors que lenoyau est constitué d’un alliage de fer et de nickel.c. Le manteau est constitué de péridotite, roche quicomprend surtout des minéraux ferro-magnésienstels que l’olivine et les pyroxènes ainsi que le grenat,alors que les croûtes océanique et continentale sontmoins riches en minéraux ferro-magnésiens.
Phrases logiquesa. Les ondes sismiques sont reçues par des sismo-graphes qui enregistrent les vibrations du sol. Ellessont traduites sous forme de sismogrammes.b. La vitesse de propagation des ondes sismiquesdiminue si la rigidité des matériaux traversés estfaible. C’est le cas entre 120 et 300 km de profondeur.c. Les météorites ont permis d’établir l’idée que laplanète Terre est une planète qui a subi une diffé-renciation en enveloppes concentriques ; on ditqu’elle s’est différenciée au cours de sa formation.d. Le noyau comprend le noyau externe, liquide, etle noyau interne, solide, encore appelé graine.
Phrases à corrigera. Les ondes L, superficielles, se déplacent à vitesseconstante alors que les ondes P et S ont une vitessed’autant plus élevée que la distance entre l’épicentreet la station est importante.b. Le Moho n’est pas toujours situé à la même pro-fondeur : il est situé à 30 km environ sous les conti-nents et à 6 km environ sous les océans.c. L’asthénosphère est située sous la lithosphère etest moins rigide que cette dernière.d. Les chondrites sont des météorites formées enmême temps que le système solaire, et donc que laplanète Terre et permettent ainsi de connaître lacomposition interne du globe.
Exercice corrigé
La discontinuité de Mohorovic icVoir correction dans le manuel, pages 294 et 295.
Évaluer ses compétences
Une caractérist ique de la propagation des ondes L1. Sachant que 2 cm correspondent à 3 000 km, etcomme la distance entre Tokyo et l’épicentre est de6 cm sur le papier, la distance à l’épicentre pourTokyo est de 6 x 3 000 / 2 = 9 000 km en réalité.
7
6
5
4
3 Pour les autres stations, on suit le même raisonne-ment et on obtient 6 300 km pour Papeete, 4 350 kmpour Montréal et 3 150 km pour Atlanta.2. Les ondes L arriveront en premier à la stationd’Atlanta car c’est la plus proche de l’épicentre.3. On convertit les temps d’arrivée des ondes ensecondes, soit pour Tokyo 1 h 1 min et 32 secondessoit 3 692 secondes ; la vitesse de propagation desondes L est donc de 9 000/3 692 = 2,43 km.s-1.Pour Papeete : 2,52 km/s, pour Montréal : 2,06 km/set pour Atlanta : 1,96 km/s.On constate que la vitesse des ondes est d’environ2,4 km/s pour les stations de Tokyo et Papeete etd’environ 2 km/s pour les stations d’Atlanta etMontréal. Ceci peut s’expliquer par le fait que lesondes L se propagent au niveau du continent pourles stations de Montréal et Atlanta (vitesses proches)alors qu’elles se propagent au niveau de l’océanpour les stations de Tokyo et Papeete (vitessesproches l’une de l’autre mais éloignées de celles desdeux autres stations). Les ondes L ont donc traversédeux milieux de nature différente.
Nature de la croûte terrestre1. On remarque que la vitesse des ondes P aug-mente en fonction de la profondeur que ce soit endomaine continental ou océanique. C’est donc quela nature des roches est différente.L’épaisseur de la croûte est de 40 km sous l’Écosse,de 55 km sous les Andes et de presque 70 km sousles Alpes. Comme le Moho marque la limite infé-rieure de la croûte, il est donc localisé à 40 km sousl’Écosse, 55 km sous les Andes et environ 70 kmsous les Alpes. On constate que la profondeur duMoho est plus importante sous les chaînes de mon-tagnes jeunes comme les Alpes.En domaine océanique, la croûte est bien moinsépaisse, la limite qui sépare la croûte du manteaun’est localisée qu’à environ 7 km de profondeur. LeMoho est donc situé à environ 7 km sous les océansalors qu’il est plus profond sous les continents.2. Dans les deux cas, les Vp d’une valeur d’environ8 correspondent au manteau supérieur, formé depéridotites. On remarque qu’il y a trois couches dematériaux différents sous les océans et deuxcouches sous les continents.En ce qui concerne la croûte océanique, Vp = 2 km/s, ce qui correspond à des sédiments nonconsolidés. Puis on trouve Vp = 5,1 ce qui corres-pond à des basaltes (avec sédiments consolidés) etVp = 6,7 indique des gabbros (ou roches métamor-phiques, mais c’est impossible en domaine océa-nique) : donc la croûte océanique comprend, de hauten bas, des sédiments non consolidés, desbasaltes, puis des gabbros.Pour la croûte continentale, les Vp de 5 à 6,1 cor-respondent à des granites, puis les Vp de 6,5 à 7,4correspondent à des gabbros ou à des roches méta-morphiques. Les gabbros sont caractéristiques de lacroûte océanique ; donc la croûte continentale estconstituée de granites et de roches métamor-phiques.
8
Chapitre 14 : Modèles de la structure de la Terre
93

94
caractéristiques du relief. Cette activité motive les sui-vantes en ce sens que l’on recherchera ensuite ce qui sepasse au niveau de ces zones actives du globe, et lasignification du relief qui leur est associé.
Exploitation� En utilisant dans un premier temps le DOCUMENT 1, quireprésente la carte de répartition mondiale des séismes etdu volcanisme, on localise les zones géologiquementactives du globe, situées en frontière de vastes surfacesgéologiquement peu ou pas actives. Dans un deuxièmetemps, on utilise en plus la carte du fond des océans(située sur le rabat), et le DOCUMENT 3 pour associer lesreliefs aux zones actives du globe.Ainsi, les dorsales sont les reliefs des fonds océaniques(exemple : dorsale Nord Atlantique), les chaînes de mon-tagnes sont des reliefs continentaux (exemple : chaînehimalayenne). Il existe un troisième type de relief(exemple : relief sous-marin du Pacifique) représenté parles fosses océaniques, bordant ici un continent.
◆ Problèmes scientifiques à résoudreComment expliquer la dynamique du globe?À quel modèle de la dynamique lithosphérique, appelémodèle de la tectonique des plaques, arrive-t-on?
P o s e r l e s p r o b l è m e s
La superposition de la répartition mondiale des séismes etcelle du volcanisme, sur le DOCUMENT 1, montre que devastes surfaces sont limitées par des frontières géologi-quement très actives.La photo du DOCUMENT 2 évoque une déchirure continentale.On peut donc s’interroger sur la signification des zonesgéologiquement actives, et leur relation avec la dynamiquede la lithosphère.
A c t i v i t é s 1
Zones actives et formes associéesIl s’agit ici de localiser les manifestations en surface del’activité interne du globe, donc les zones géologique-ment actives du globe, et de les associer à des formes
Programme : notions et contenus• La lithosphère et la tectonique des plaques.• Découpage de la lithosphère en plaques d’épaisseur variable, peu déformables à l’exception de leurs limites.• Mouvements relatifs des plaques : divergence au niveau des dorsales océaniques où elles se forment, convergence dans les zones de subductionet de collision où elles disparaissent, coulissage le long des failles transformantes.
Objectifs à atteindreLa structure verticale de la Terre a été étudiée : le chapitre 14 a permis d’aboutir à un modèle du globe correspon-dant à la synthèse de plusieurs modèles (sismologique, chimique, minéralogique et thermique). On va maintenantmettre en évidence une régionalisation dynamique de la lithosphère, permettant d’élaborer un modèle de la cinéma-tique globale des plaques.
Le modèle va révéler des hétérogénéités latérales du manteau, à mettre en relation avec les phénomènes de surfacequi affectent la croûte. Ce modèle sera ensuite enrichi grâce aux deux chapitres suivants.
Un modèle de la dynamique lithosphérique
C H A P I T R E 1 5

A c t i v i t é s 2
Les dorsales et l’expansion des fondsocéaniquesAprès avoir étudié ce qui caractérise morphologique-ment les zones actives du globe, on va, dans les activi-tés 2 et 3, comprendre la signification dynamique deces zones.
On cherche ici à montrer que tous les fonds océaniquesont été fabriqués au niveau des dorsales, frontières dedivergence.
Exploitation� L’axe de la dorsale est caractérisé par des basaltes encoussins, traduisant un volcanisme sous-marin, et l’exis-tence de failles dans le plancher océanique basaltiqueindique un écartement des terrains (DOCUMENT 4). Le DOCU-MENT 5 confirme le précédent, et on observe en plus la pré-sence de failles normales, qui attestent de mouvements enextension : tout se passe comme si la croûte océaniqueétait tirée de part et d’autre de l’axe de la dorsale.D’après le DOCUMENT 6, la zone d’activité tectonique et sis-mique n’est située qu’à une distance de 20 km autour del’axe de la dorsale, et l’activité volcanique est concentréeexclusivement aux abords de cet axe. Elle est à l’originedes coulées volcaniques basaltiques qui forment les fondsocéaniques. Les foyers des séismes sont superficiels, cesséismes témoignent de contraintes à l’origine de rupturesdans la croûte océanique (plus généralement la litho-sphère), engendrée par l’activité magmatique.On note l’absence de sédiments sur 30 km environ de partet d’autre de l’axe de la dorsale, ce qui confirme la nais-sance de la lithosphère océanique à l’axe de la dorsale.Les sédiments reposent sur le basalte des fonds océa-niques au-delà de 30 km de l’axe, et on peut les dater grâceaux fossiles qu’ils contiennent (DOCUMENT 7).On remarque ainsi que l’âge des fonds océaniques augmenteen s’éloignant de l’axe de la dorsale (médio-atlantique ici).On peut donc conforter l’idée de l’expansion océaniqueémise par Hess : la lithosphère océanique se forme au niveaude l’axe de la dorsale, et s’en éloigne progressivement depart et d’autre, engendrant les fonds océaniques.Les dorsales apparaissent donc bien comme des frontièresde divergence, permettant la création de tous les fondsocéaniques.
A c t i v i t é s 3
Fosses, montagnes et frontières de convergenceOn a vu dans l’activité précédente que la lithosphèreocéanique résulte de l’activité des dorsales, zones dedivergence.
On cherche maintenant à comprendre la significationdynamique des fosses océaniques et des chaînes demontagnes, et à montrer que ce sont des zones deconvergence.
Exploitation
� D’après le DOCUMENT 8, on observe que les foyers sis-miques peuvent atteindre jusque 800 km de profondeursous l’archipel du Japon ; or les séismes ne peuvent appa-raître que dans des matériaux rigides, et l’asthénosphèreest moins rigide dès 100 km de profondeur.Le DOCUMENT 9 montre que les foyers des séismes situéssous l’archipel du Japon sont d’autant plus profonds qu’ilssont éloignés de la fosse océanique qui borde l’Est duJapon (voir la carte du fond des océans sur le rabat).Ces observations nous permettent de supposer un enfon-cement de la lithosphère océanique sous la lithosphèrecontinentale (= subduction). Ainsi les foyers des séismesauraient pour origine des libérations brutales d’énergie,liées aux cassures affectant la lithosphère océanique rigidequi plonge.Les données tomographiques du DOCUMENT 9 révèlent lasubduction de la plaque Pacifique dans l’asthénosphère,sous la plaque Eurasienne. L’ensemble des séismes maté-rialise le plan incliné suivant lequel la plaque Pacifiqueplonge sous la plaque Eurasienne (plan de Benioff-Wadati).� Les régions situées entre l’Inde et l’Asie sont le sièged’une sismicité intense. Ces régions correspondent à unegigantesque chaîne de montagnes : l’Himalaya.On a retrouvé des basaltes en coussins faisant partie desophiolites, dans une région de l’Himalaya à 5000 mètresd’altitude. Or, les coulées basaltiques sous-marines pré-sentent ce même épanchement en coussins : c’est donc lapreuve qu’un océan a jadis existé dans cette région.Le DOCUMENT 12 nous montre le plongement vers le Nordde la plaque indienne, correspondant à une subduction dela plaque indienne sous la plaque asiatique.Lorsque l’Inde a commencé à s’enfoncer sous le Tibet, unfragment de lithosphère océanique est resté coincé entreles deux blocs continentaux, comme en témoignent lesophiolites, prouvant que l’Inde était alors séparée del’Asie par un océan. Les ophiolites indiquent donc qu’il ya eu une suture entre l’Inde et l’Asie.Le fait que la plaque indienne devienne verticale et se ren-verse, traduit un blocage de la subduction : il y a alors col-lision entre les deux blocs continentaux, ce qui a permisd’ériger l’Himalaya.Cette gigantesque chaîne de montagnes correspond donc àune frontière de convergence entre deux plaques.
Chapitre 15 : Un modèle de la dynamique lithosphérique
95

teau : on voit la lithosphère qui plonge dans l’asthéno-sphère au niveau des zones de subduction.
A c t i v i t é s 5
Vitesses et mouvements des plaquesLe modèle de la cinématique du globe a pu être élaborédans l’activité précédente, à l’aide des informationsobtenues au cours des activités 1 à 4. Les plaques litho-sphériques sont en mouvement, elles se déplacent lesunes par rapport aux autres.
L’objectif de cette activité est de calculer la vitesse dedéplacement des plaques lithosphériques et d’évaluerle sens de leur déplacement.
Exploitation� D’après le DOCUMENT 18, un point chaud fixe émet dumagma qui perfore régulièrement la lithosphère qui sedéplace au-dessus, formant des alignements de volcans,comme c’est le cas dans l’océan Pacifique.L’activité volcanique la plus récente (actuelle), se situe ausud-est de la chaîne, alors que les volcans éteints sontd’autant plus anciens qu’on s’éloigne d’Hawaii en allantvers le nord-ouest (DOCUMENT 19), ce qui confirmed’ailleurs bien le sens du déplacement de la plaquePacifique indiqué sur le document.On remarque, d’après le DOCUMENT 20, que sur la plaquePacifique, les trois alignements volcaniques présentent lemême coude, indiquant un changement d’orientation il y aquarante millions d’années, ce qui prouve que le déplace-ment de la plaque Pacifique a changé de direction au coursdu temps.– Calcul de la vitesse moyenne du déplacement de laplaque Pacifique : V = d/t ; sachant que l’on mesure 14 cmentre le volcan sur Niihau et Kilauea et que 2,5 cm cor-respondent à 100 km.V = 56 x 106/4,9 x 106 = 11,4 cm/an.– Pour l’océan Atlantique, en utilisant les données dudocument 7, page 301 (activités 2), on obtient la vitesse dudéplacement de la plaque Sud-américaine :1) En utilisant le forage 15 : d = 400 km = 40 x 106 cm;t = 22 millions d’années = 22 x 106 ans ;V = 40/22 = 1,8 cm/an.2) En utilisant le forage 19 : d = 1000 km = 100 x 106 cm;t = 47 millions d’années = 47 x 106 ans ;V = 100/47 = 2 cm/an.
A c t i v i t é s 4
Modèle de la tectonique des plaquesNous avons vu dans les activités 2 et 3 que les plaquesdivergent au niveau des dorsales océaniques, etqu’elles convergent dans les zones de subduction et decollision où elles disparaissent. Cette activité va per-mettre d’envisager un autre type de frontières deplaques, où les plaques coulissent le long de faillestransformantes.
Tous les phénomènes géologiques ayant lieu au niveaudes zones actives du globe ont été envisagés, on peutmaintenant établir le modèle de la tectonique desplaques.
Exploitation� On voit que les zones de fractures sont ici parallèles àleur sens de déplacement, il existe ici un coulissement quis’effectue bord à bord. Ces zones de fracture, ou faillestransformantes peuvent être localisées en milieu océa-nique ou en milieu continental. Elles peuvent joindre dessegments de dorsales, par exemple dans l’Atlantique cen-tral (DOCUMENT 13), ou peuvent relier, comme c’est le caspour la faille de San Andreas (DOCUMENT 14) une frontièreconvergente (la zone de subduction au Nord) à une fron-tière divergente (au Sud).Le DOCUMENT 14 nous permet de voir que seule la zone cor-respondant à la faille transformante est sismiquementactive : de nombreux séismes, dont les foyers sont super-ficiels (essentiellement entre 0 et 10 km de profondeur)sont enregistrés au niveau de la faille de San Andreas,créant des décalages horizontaux permettant d’étendreencore la longueur de cette faille, qui est un réseau de cas-sures. Ce réseau sépare l’Ouest de la Californie, qui glissevers le Nord, du reste des États-Unis. Les failles transfor-mantes sont ainsi nommées parce qu’elles transforment lemouvement de divergence ou de convergence en un mou-vement de coulissement ; il n’y a ni création ni disparitionde matière à leur niveau.� En utilisant toutes les données précédentes, un modèlede la cinématique du globe peut être construit. La super-position de la répartition mondiale des séismes et du vol-canisme a permis de repérer les plaques, limitées par desfrontières géologiquement très actives.Le DOCUMENT 15 place les différentes frontières de plaquesétudiées dans les activités précédentes : frontières diver-gentes (dorsales), frontières convergentes (zones de sub-duction et de collision) et frontières coulissantes (faillestransformantes).Les mouvements d’une plaque par rapport à une autre ontété indiqués, et on remarque que certaines plaques sontentièrement océaniques (comme la plaque Pacifique),alors que d’autres présentent à la fois une partie continen-tale et une partie océanique (plaque Eurasienne). De plus,leurs limites ne correspondent pas aux bordures établiesentre océans et continents.Le modèle de la figure 16 est enrichi par rapport à celuiqui a été élaboré dans le chapitre 14, car il révèle cette foisdes hétérogénéités latérales, qui sont situées dans le man-
96

A c t i v i t é s 6
Vitesses passées et vitesses actuellesVitesse et sens de déplacement des plaques lithosphé-riques ont pu être évalués dans l’activité précédentegrâce aux alignements volcaniques dus aux pointschauds, ainsi qu’en utilisant les données fournies parl’âge des sédiments surmontant les basaltes de lacroûte océanique. Nous verrons dans cette dernièreactivité que la vitesse des mouvements des plaques a puaussi être calculée grâce aux anomalies magnétiques,et que les techniques actuelles permettent d’en retrou-ver les valeurs.
Exploitation� Les anomalies magnétiques permettent de dater lesfonds océaniques, donc de dresser une carte géologique del’âge des fonds océaniques (DOCUMENT 22).On remarque d’après la carte que la vitesse d’expansion del’océan Atlantique est moins rapide que celle de l’océanPacifique, ce qui confirme les vitesses calculées à l’acti-vité précédente.� Le calcul de la vitesse à l’aide des anomalies magné-tiques donne le résultat suivant : V = 10 x 106/9 x 106 = 1,1 cm/an.On peut retrouver les valeurs de vitesses établies sur desmillions d’années en calculant la vitesse d’expansionactuelle grâce aux mesures instantanées fournies par lessatellites. Si on prend le coefficient directeur sur la courbedu DOCUMENT 23b. entre 1989 et 1991, on trouve 50 mmpour une période de 2 ans donc 25 mm pour 1 an,l’Amérique s’éloigne donc de l’Europe à une vitesse d’en-viron 2 cm/an.
E x e r c i c e s
Évaluer ses connaissances
Questions à réponses cour tesa. La lithosphère océanique formée au niveau desdorsales s’en éloigne de part et d’autre à la manièred’un tapis roulant : c’est l’expansion des fondsocéaniques.b. La complémentarité des formes entre certainscontinents suggère qu’ils étaient autrefois soudés,et qu’ils se sont éloignés l’un de l’autre au cours destemps géologiques : c’est la dérive des continents.c. La subduction correspond à l’enfoncement d’uneplaque océanique sous la lithosphère continentale(cas le plus fréquent, exemple : Amérique du Sud)ou sous la lithosphère océanique (exemple : îlesMariannes).d. L’expansion des fonds océaniques a pu être miseen évidence grâce à la datation des roches sédi-mentaires au contact direct du basalte, à l’enregis-
1
trement des anomalies magnétiques, à l’étude desalignements de volcans de points chauds et plusrécemment, à l’aide des techniques géodésiques.
Expliquer brièvementa. La lithosphère océanique, froide et dense, dispa-raît par plongement dans l’asthénosphère, selon unplan incliné au niveau d’une fosse océanique : c’estla subduction.b. Les trois types de frontières qui animent lesplaques sont des frontières de divergence, repré-sentées par les dorsales, de convergence, représen-tées par les zones de subduction et de collision, decoulissement, représentées par les failles transfor-mantes.c. La lithosphère océanique dérive de part et d’autrede l’axe de la dorsale, puis disparaît par subductiondans les zones de convergence des plaques ; et lasubduction peut aboutir à la collision de deuxmasses continentales et à la formation des chaînesde montagnes.
Phrases logiquesa. La présence d’ophiolites dans les chaînes demontagnes témoigne de la fermeture d’un océan.b. Les dorsales sont le siège d’une activité volca-nique et de nombreux séismes, dont le foyer est peuprofond.c. La mesure des anomalies magnétiques montreune symétrie parallèle à l’axe des dorsales ettémoigne de l’expansion des fonds océaniques.
Phrases à corrigera. Les anomalies magnétiques sont disposées enbandes alignées parallèlement à l’axe de la dorsaleocéanique.b. Au niveau des fosses océaniques, la lithosphèrefroide et dense s’enfonce dans l’asthénosphèreselon un plan incliné.c. Les dorsales ne possèdent pas toujours de pro-fonde vallée axiale, ou rift.d. Les points chauds permettent de mettre en évi-dence le mouvement absolu des plaques, car ilssont fixes et situés en dessous de la plaque litho-sphérique qui se déplace au-dessus d’eux.
Exercice corrigé
Anomalies magnétiques etexpansion des fonds océaniquesVoir correction dans le manuel pages 314 et 315.
5
4
3
2
Chapitre 15 : Un modèle de la dynamique lithosphérique
97

Évaluer ses compétences
Les î les de la polynésie française1. L’âge des îles volcaniques est de plus en plusancien quand on s’éloigne de Méhétia, on peut doncpenser qu’elles résulteraient de l’activité d’un pointchaud fixe, localisé sous l’île de Méhétia.
2. Les volcans de l’archipel de la société sont ali-gnés selon la direction NW-SE ; leur origine peuts’expliquer par le déplacement de la plaquePacifique au-dessus d’un point chaud (Méhétia), cedéplacement s’effectuant vers l’Ouest.Le mouvement de la plaque Pacifique traduit bienl’expansion des fonds océaniques.
3. V = d/t.t = 4,80 + 4,05/2 = 4,425 x 106 ans.d = 480 km, soit 48 x 106 cm.V = (48 x 106)/(4,425 x 106) = 10,84 cm/an.
Foyers s ismiques en Amérique du Sud1. Les foyers sismiques sont d’autant plus profondsque l’on s’éloigne de l’océan Pacifique et donc de lafosse du Chili.
2. La représentation graphique correspond au docu-ment ci-dessous.
7
6
98
Foyer sismique
100
300
200
100
0
200 300 400 500 600 700
B
Profondeur (km)
Axe de la fosse du Chili
Distance (km)
Mobil i té de la l i thosphère1. Au fur et à mesure que l’on s’éloigne de la dor-sale, on remarque que la nature et l’épaisseur dessédiments changent : il y a d’abord dépôt de sédi-ments métallifères, puis de boues carbonatées,d’argiles rouges des grands fonds et enfin de bouesà radiolaires. Ces dernières se trouvent entre deuxcouches d’argiles rouges pour le dernier forage.
2. Les boues à radiolaires indiquent la présence devie, on voit que le dépôt de cette couche correspondà une zone de haute productivité équatoriale ; laplaque passe dans cette zone et quand la plaquequitte cette zone, il se produit de nouveau un dépôtd’argiles rouges. Cela indique que la plaque sedéplace, donc la sédimentation est ici un indiceprouvant la dynamique lithosphérique.
8
3. Sachant que les séismes n’apparaissent quedans des matériaux rigides, ceux-ci ne peuventconcerner l’asthénosphère. On peut donc supposerqu’il y a ici une subduction, la lithosphère océaniques’enfoncerait sous la lithosphère continentale.L’ensemble des foyers sismiques matérialise le plansuivant lequel la plaque Nazca glisse sous la plaqueSud-américaine (voir le modèle page 313).
Répartition des foyers sismiques en fonction de la profondeurselon la coupe AB.

Pa
rt
ie
2 Structure, composition et dynamique de la Terre
La Terre : machinerie thermique
16 Marges passives et océanisation .............. 100
17 Magmatisme et convection mantellique105
Unité 6

100
Programme : notions et contenusMarges passives des continents : elles sont structurées par des failles normales et sont le siège d’une sédimentation importante. Elles ont enregis-tré l’histoire précoce de la rupture continentale et de l’océanisation. L’activité des failles normales, héritage de rifts continentaux, témoigne de l’amin-cissement de la lithosphère et de sa subsidence.
Objectifs à atteindreLe chapitre 15 partant des manifestations de surface de la dynamique de la Terre aboutit à un modèle où la lithosphèreest découpée en plaques séparées par des frontières (de divergence, de convergence ou coulissantes).
On peut alors s’interroger sur la signification des marges passives comme celles de l’Atlantique sans activité sismiqueet volcanique et ne correspondant pas à des frontières de plaques.
Sachant que les fonds océaniques résultent de l’activité des dorsales, on peut remonter dans le temps et rapprocherles marges passives d’un océan comme l’Atlantique ; ainsi ces marges apparaissent comme le résultat de la fractura-tion d’un continent initial qui s’est poursuivie par la naissance et la croissance d’un océan. L’étude des marges pas-sives doit alors fournir des renseignements sur les modalités de la déchirure continentale et le début de l’océanisation.
Marges passives et océanisation
C H A P I T R E 1 6
◆ Problèmes scientifiques à résoudreQuelles sont les caractéristiques des marges passives etquels enseignements apportent-elles sur la formation etl’évolution des océans?
P o s e r l e s p r o b l è m e s
Les supports choisis permettent d’opposer les deux typesde marges continentales, les unes actives associées à laprésence de séismes, volcanisme et fosses (cas du pour-tour du Pacifique), d’autres dépourvues d’activité tecto-nique (marge Atlantique à l’exception de la zone Caraïbe).Cette opposition aboutit à la formulation de problèmesconcernant le lien existant entre la structure des margespassives et la genèse d’un océan à partir de la fracturationinitiale d’un continent.
A c t i v i t é s 1
La morphologie des marges passivesL’objectif de cette activité est de préciser les caracté-ristiques morphologiques d’une marge et ses relationsavec le domaine océanique.
Exploitation� Le DOCUMENT 3a. permet de repérer les différentsdomaines morphologiques continentaux et océaniquesainsi que la structure d’une marge passive bien visible surla côte Est des États-Unis et du Canada ; le DOCUMENT 3b.précise les différents domaines de la marge qui constituela partie immergée du continent :– le plateau ou plate-forme en pente très douce de faibleprofondeur (200 m) et large de 70 à 80 km en moyenne,– le talus s’étageant de 200 m à 3000 m avec une pente de7 % souvent entamé par des canyons sous-marins,– le glacis entre 3000 et 5000 m qui se raccorde à la plaineabyssale pratiquement horizontale ; le glacis reçoit trèssouvent des avalanches de sédiments appelés courants deturbidités qui se détachent du talus et qui s’étalent en for-mant des deltas sous-marins.� Une marge passive au sens géologique corresponddonc au prolongement immergé du continent. Le domainecontinental se raccorde ainsi au domaine océanique loindu littoral (parfois à près de 100 km).

Chapitre 16 : Marges passives et océanisation
101
leur disposition en éventail à la surface de chaque bloc nepeut s’expliquer que par un basculement progressif desblocs dans le sens horaire, celui-ci ayant commencé avecle dépôt des premiers sédiments à leur surface et se termi-nant avec le dépôt des derniers sédiments déformés sur-montés par les sédiments non déformés plus ou moinshorizontaux (3).Pour reconstituer l’état initial, il faut donc remettre lesblocs en place en les redressant et en les faisant pivoter sureux-mêmes dans le sens anti-horaire ; on aboutit ainsi àune croûte continentale plus ou moins plane qui va se frac-turer lors de la formation de la marge délimitant des blocsqui vont basculer le long de failles normales. Des sédi-ments sont ainsi piégés et suivent le basculement des blocsjusqu’à ce que le phénomène s’arrête ; la sédimentationdevient alors horizontale (3) et recoupe la précédentedéformée par le basculement (2).On peut ainsi dater la période de formation de la marge quicorrespond à la période de dépôt des sédiments contem-porains du basculement des blocs (sédimentation en éven-tail) : elle correspond ici pour la marge de Galice auxdépôts aptien et albien du crétacé inférieur datés entre – 114 et – 96 millions d’années soit une durée de 18 mil-lions d’années. Le début de l’ouverture de l’AtlantiqueNord s’est réalisé durant cette période (qualifiée de rif-ting), la marge de Galice correspondant à celle de TerreNeuve.
A c t i v i t é s 3
Du rifting à l’océanisationL’activité 2 a montré que l’amincissement de la croûtecontinentale peut conduire à sa rupture aboutissant à lacréation d’un océan, les marges de part et d’autre d’unocéan constituant les témoins de la déchirure initiale. Ils’agit de rechercher des témoignages actuels qui per-mettent de reconstituer l’histoire d’une marge passiveavant d’envisager les mécanismes responsables (objetdu chapitre 4)
Exploitation� Le DOCUMENT 6a. montre que le rift rhénan correspond àun vaste fossé d’effondrement de 30 km de largeur et deplusieurs centaines de kilomètres de longueur (seule unepartie est représentée sur le document).Le DOCUMENT 6b. issu de la sismique réflexion montre :– un amincissement de la croûte continentale à l’aplombdu fossé d’environ 8 km pour deux raisons : un effondre-ment de 3 km en surface rempli de sédiments et uneremontée du manteau supérieur (le moho se trouve à25 km de profondeur au lieu de 30 km environ en dehorsdu rift) ;– un amincissement cassant lié à l’existence de failles nor-males qui accompagnent l’effondrement dans la partiesupérieure de la croûte ;
A c t i v i t é s 2
Les caractéristiques d’une marge pas-siveL’objectif de cette activité est de préciser la structureprofonde d’une marge passive (nature des roches, dis-position et épaisseur) et de dater la période de forma-tion d’une marge à l’aide des sédiments qui surmontentle socle.
Exploitation� Le DOCUMENT 4c. issu d’un profil de sismique réfraction(voir la fiche technique à la fin du manuel), montre :– une stratification des vitesses sismiques allant desvitesses les plus basses en surface voisines de 2 km. s– 1
aux vitesses les plus rapides en profondeur voisines de 8 km. s – 1; l’utilisation du tableau 4b. couplé aux symbolesde couleurs du DOCUMENT 4c. permet de retrouver la naturedes roches au niveau de la marge (sédiments en surfaceconsolidés ou non, socle granitique puis péridotite dumanteau).Remarque : On constate en réalité deux catégories devitesses au niveau du socle : les géologues subdivisent lacroûte continentale en deux parties, la croûte supérieureet la croûte inférieure.– un amincissement important de la croûte continentalequi passe de 35 km à l’Est à 10 km environ au niveau deson raccordement avec la croûte océanique soit un facteurd’amincissement de 3,5.Le DOCUMENT 4d. issu des techniques de sismique réflexionà un niveau voisin du profil précédent confirme l’amin-cissement de la croûte continentale et précise de quellefaçon cet amincissement s’est produit :– dans la partie supérieure de la croûte, des failles nor-males limitent des blocs d’une dizaine de kilomètres delargeur et qui semblent de moins en moins épais lorsqu’ons’éloigne du continent ; ces blocs semblent avoir glissé etbasculé les uns contre les autres du continent vers l’océan.Certaines des failles normales ne sont pas rectilignes maisincurvées en forme de cuiller d’où leur nom de faille listrique ;– dans la partie inférieure, la croûte s’amincit sans pré-sence de failles c’est-à-dire sans rupture des roches.La croûte continentale présente donc un comportementcassant en surface et souple ou ductile en profondeur.
� Les DOCUMENTS 5a. (profil sismique brut) et 5b. (profilsismique avec un repérage des limites lithologiques et desfailles) au niveau de la marge de Galice permettent d’iden-tifier les mêmes structures que celles rencontrées auniveau de la marge de Bretagne : un enfoncement du fondqui passe de – 2900 m à – 4500 m, la présence de blocsbasculés limités par des failles normales au niveau dusocle (1) ainsi que celle de sédiments (2 et 3) surmontantle socle. On peut remarquer que les sédiments 2 sont dis-posés en éventail alors que les sédiments 3 sont plus oumoins parallèles.La disposition des sédiments notés 2 pose problème étantdonné que la sédimentation s’effectue horizontalement ;

DOCUMENT 7 c.) suivie d’une océanisation (le rift continentalétant relayé par un rift océanique de dorsale : DOCUMENT 7d.).
A c t i v i t é s 4
Les mécanismes de la rupture continentaleLe chapitre précédent permet d’émettre l’hypothèse demouvements extensifs à l’origine de la déchirure conti-nentale et de la création d’un océan dont les margespassives représentent les premiers témoins.
Ce chapitre est destiné à confronter des modélisationsaux données de terrains afin de conforter les hypo-thèses explicatives.
Exploitation� Le DOCUMENT 8 confirme que des mouvements extensifs(écartement) exercés sur un matériau cassant simulant lacroûte supérieure entraînent :– la rupture du matériau selon des failles normales (deuxprincipales au départ suivies de nombreuse autres lorsquel’extension se poursuit) ;– le déplacement des blocs le long des failles ;– l’amincissement de l’ensemble par formation d’un fosséd’effondrement.
� Ce résultat peut alors être comparé à la formation d’unrift continental. Cependant, ceci n’explique pas l’amincis-sement de façon ductile de la partie inférieure de la croûteni la remontée du manteau inférieur.
� Le DOCUMENT 9 montre une prise en compte plus précisedes propriétés physiques des différentes couches superfi-cielles du globe. En particulier, la résistance des matériauxà la déformation (la rhéologie) varie : on considère que lacroûte et la partie supérieure du manteau présentent unepartie superficielle cassante et une partie plus profondeductile expliquant l’alternance de sable et silicone super-posés dans la modélisation. Enfin l’asthénosphère plusductile est simulée par l’utilisation du miel à la base dudispositif.Lors de l’extension du dispositif, on peut remarquer :– un amincissement de la croûte selon deux modalités :cassant pour la partie supérieure avec formation de faillesnormales limitant des blocs basculés entraînant la forma-tion d’un fossé d’effondrement comparable à un rift, duc-tile pour la partie inférieure selon l’effet « carambar » ;– un amincissement de la partie supérieure du manteauselon des modalités comparables à celles des deux partiesde la croûte ;– une remontée passive de l’asthénosphère.Ainsi l’étirement de la croûte continentale par des mouve-ments extensifs sous-jacents (en réalité la lithosphère) suf-fit à provoquer un amincissement qui s’accompagne del’effondrement de la partie centrale superficielle et de laremontée passive de l’asthénosphère : on parle dans ce casde rifting passif, la remontée de l’asthénosphère étantdavantage une conséquence qu’une cause de l’extension.
– un amincissement non cassant mais souple dans la par-tie inférieure de la croûte.Indications complémentaires : des profils sismiques réali-sés en Alsace (à la hauteur de Colmar) ou dans le fossé dela Bresse plus au Sud montrent des structures comparablesà celle du DOCUMENT 6b.� La comparaison des DOCUMENTS 4, 5 et 6 permet demettre en évidence des similitudes entre la structure d’unrift et celle d’une marge passive (amincissement de lacroûte, failles normales dans sa partie supérieure, amin-cissement ductile dans sa partie inférieure). On peut à ceniveau, compte tenu des informations, imaginer que lesdeux marges symétriques d’un océan sont les bords sépa-rés d’un ancien rift continental qui a poursuivi son ouver-ture. L’amincissement de la croûte continentale serait alorsle résultat d’un étirement de la croûte c’est-à-dire d’unmouvement d’extension.
� Le DOCUMENT 7c. montre que la rive ouest de la merRouge a une structure comparable à celle d’une margepassive (blocs basculés limités par des failles normalesrecouvertes par des sédiments contemporains ou posté-rieurs au rifting). L’effondrement du rift continental de lamer Rouge avec basculement des blocs le long des faillescommence à l’éocène et se termine à la fin du miocènemoyen (daté par les sédiments qui accompagnent le bas-culement des blocs et ceux qui reposent sur ces dernierssans être déformés).Le DOCUMENT 7d. montre la présence d’une jeune croûteocéanique à 1,5 km de profondeur et large de 50 km envi-ron avec des basaltes jeunes (< 0,5 mA) dans la partieaxiale et de plus en plus vieux lorsqu’on s’éloigne de cettezone (de 0,5 à 3 MA). À ce niveau, on peut parler de riftocéanique avec présence de failles normales qui résultentdu fonctionnement d’une jeune dorsale. Celle-ci élaboreune croûte océanique qui va repousser et séparer, au fur età mesure de sa formation, les bords devenus passifs desdeux marges continentales.Le DOCUMENT 7e. confirme que dans la partie Nord de lamer Rouge, en tout début d’océanisation (le document 7b.montrant quelques rares émissions basaltiques), le man-teau supérieur remonte et accompagne ainsi un amincisse-ment de la croûte continentale confirmé en surface par leDOCUMENT 7c. (effondrement avec failles normales).Le DOCUMENT 7a. (complémentarité des rivages) laisse pen-ser que les deux bords de la mer Rouge résultent d’unedéchirure initiale d’un continent unique liée à des mouve-ments d’extension (avec création d’un rift continental :
102

� La genèse d’un océan nécessite plusieurs étapes liées àdes phénomènes extensifs profonds : formation d’un riftcontinental par amincissement de la lithosphère (effondre-ment central et remontée de l’asthénosphère), rupture de lalithosphère à la fin du rifting coïncidant avec le début del’océanisation : la dorsale commençant à fonctionnergénère un fond océanique qui repousse alors les deuxbords de l’ancien rift qui deviennent des marges passives.Au début de son existence, la marge passive est peu pro-fonde puis celle-ci va s’enfoncer et se recouvrir plus oumoins de sédiments.
A c t i v i t é s 5
Histoire des marges passives après la rupture continentaleL’objectif de cette activité est de montrer que les margesévoluent et de façon différente selon leur situation géo-graphique.
Exploitation� La marge de Saint-Pierre-et-Miquelon au large deTerre-Neuve est recouverte d’une épaisseur considérablede sédiments (DOCUMENT 11c.) pouvant atteindre près de15 km d’épaisseur alors que les marges européennes(Bretagne et Galice) sont recouvertes d’une faible épais-seur de sédiments (3-4 km) ; la marge américaine est ditegrasse ou nourrie alors que les marges européennes sontqualifiées de marges maigres. L’épaisseur des sédimentsest liée en particulier aux apports continentaux, ainsi lamarge de Saint-Pierre-et-Miquelon au débouché du fleuveSaint-Laurent a reçu de nombreux sédiments (au mêmetitre que la marge Ouest Africaine équatoriale qui reçoitdes quantités considérables de sédiments charriés par lesgrands fleuves).� Le DOCUMENT 11b. couplé au DOCUMENT 12 atteste que lesroches sédimentaires qui recouvrent la marge de Saint-Pierre-et-Miquelon se sont formées dans un océan peuprofond sous une épaisseur d’eau n’excédant pas quelquescentaines de mètres voire moins pour les évaporites. Il fautdonc admettre que le socle de la marge jeune peu profondau départ s’est enfoncé régulièrement et que, au fur et àmesure, sous une faible épaisseur d’eau, des sédiments sesont déposés aboutissant à une épaisseur considérable. Cetenfoncement progressif est qualifié de subsidence (onconsidère actuellement qu’elle est due à un refroidisse-
ment progressif de la lithosphère à l’aplomb des marges ;elle est ainsi qualifiée de subsidence thermique).
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Plateau continental : Encore appelé plate-forme ;correspond à la partie du continent qui se prolongeen pente douce sous l’océan jusqu’au talus conti-nental.b. Glacis : Zone immergée à pente très faible, situéeentre 4000 et 5000 m de profondeur qui raccorde letalus continental à la plaine abyssale.c. Marge passive : Partie immergée du continent denature granitique qui fait la transition avec le fondocéanique de nature basaltique et qui ne montreaucune activité volcanique ou sismique car située ausein d’une même plaque lithosphérique.d. Rift : Fossé d’effondrement continental ou océa-nique (au niveau de certaines dorsales dans ce der-nier cas) marqué par l’existence de failles normalesrésultant d’un étirement de la croûte qui s’amincit.e. Faille normale : Cassure affectant un terrain avecun mouvement relatif des deux parties séparéescorrespondant à un allongement de l’ensemble etrésultant de mouvement de distension (rift).f. Ductile : Se dit d’un matériau qui peut se déformerpar étirement sans se casser.g. Subsidence : Enfoncement progressif de cer-taines parties de l’écorce terrestre sous l’action dupoids des matériaux (sédiments ou glaces) ou parsuite d’un refroidissement entraînant une augmenta-tion de densité.h. Marge grasse : Marge passive comportant unegrande épaisseur de sédiments.
Questions à réponses cour tesa. Une marge passive est caractérisée par une mor-phologie comportant un plateau continental large àpente douce suivi d’un talus à pente raide qui abou-tit à un glacis se raccordant à la plaine abyssale ; sastructure se caractérise par une épaisseur de lacroûte continentale qui diminue progressivement etfinit par atteindre celle de la croûte océanique et parla présence de failles normales qui séparent desblocs basculés, le tout étant recouvert de sédi-ments.b. Un rift est le résultat d’une extension de la croûtequi entraîne la formation de failles normales auniveau desquelles des blocs coulissent et basculent.On date la période de formation du rift en datant ledébut et la fin de dépôt des sédiments disposés enéventail sur les blocs c’est-à-dire contemporains dubasculement des blocs.c. La formation d’un rift continental par suite d’uneextension de la croûte a pour conséquence l’amin-cissement et la rupture du continent qui aboutit à la
2
1
Chapitre 16 : Marges passives et océanisation
103

– un retour de la mer au pliocène avec reprise de lasédimentation (2 km d’épaisseur).La Méditerranée présente la structure d’un jeuneocéan qui n’a pas poursuivi sa croissance.
Une marge continentale grasse1. 22 mm correspondent à 100 km au niveau del’échelle horizontale (soit 4,5 km pour 1 mm) alorsque pour l’échelle verticale, 10 mm correspondent à5 km soit 0,5 km pour 1 mm. L’exagération del’échelle verticale par rapport à l’échelle horizontaleest donc de : 4,5/0,5 = 9. Ainsi le récif corallien quia une épaisseur de 5 km devrait avoir une largeur 9fois plus importante à la même échelle.2. Il s’agit d’une marge passive avec un amincisse-ment de la croûte qui passe de 35 km au Nord-Ouest à 7 km d’épaisseur en se raccordant à lacroûte océanique. La phase d’extension de la croûtecontinentale coïncide avec le dépôt des sédimentscontemporains du basculement des blocs, elle datedonc du trias.3. L’épaisseur maximale des sédiments est voisinede 10 km au niveau de la formation récifale ; il s’agitdonc d’une marge grasse. La formation récifale estépaisse de 5 km et large de 30 à 40 km.4. L’ensemble des dépôts sédimentaires correspon-dent à des dépôts de milieu peu profonds doncimmergés sous une faible épaisseur d’eau. Les éva-porites se formant par évaporation de l’eau peu pro-fonde et les récifs ne pouvant exister que sous unefaible tranche d’eau, on peut conclure que la marges’est enfoncée progressivement (subsidence) et queles sédiments se sont déposés sous une faibleépaisseur d’eau mais se sont accumulés sur desépaisseurs considérables.
6
remontée de l’asthénosphère à l’origine de magmabasaltique.d. Les marges maigres comportent une faible épais-seur de sédiments (au large de la Bretagne ou de laGalice ) ; les marges grasses comportent une trèsgrande épaisseur de sédiments (marge de Terre-Neuve).e. Les mouvements d’extension provoquent deseffondrements (formation d’un rift bordé d’épaules)et une remontée de l’asthénosphère.
Une marge passivea. Les arguments qui permettent de reconnaître unestructure de marge passive sont :– la présence de blocs séparés par des failles ;– la présence de sédiments disposés en éventailattestant du basculement des blocs ;– la présence de sédiments horizontaux postérieursau basculement des blocs.b. Les sédiments disposés en éventail correspon-dent à la période de basculement des blocs et ledébut de la sédimentation horizontale coïncide avecla fin du rifting.
Exercice corrigé
Marge de l ’éperon de GobanVoir correction dans le manuel, pages 338-339.
Évaluer ses compétences
La marge du golfe du Lion1. La marge du golfe du Lion est une marge passivecar elle présente :– un amincissement progressif de la croûte conti-nentale depuis sa partie émergée jusqu’à son rac-cordement avec le fond basaltique ;– la présence de failles normales dans la partiesupérieure de la croûte.2. La période d’extension continentale (rifting) estd’âge oligocène-début miocène car elle correspondau remplissage sédimentaire de la partie supérieuredes blocs lors de leur basculement. On peut remar-quer que le début du volcanisme basaltique date decette même période (basaltes superposés au socleet non inclus dans le miocène).3. La suite de l’histoire géologique correspond à :– une période de sédimentation au miocène inférieuret moyen ayant déposé plus de 5 km de sédimentsau Sud-Est accompagnée de la poursuite du volca-nisme basaltique (basaltes inclus dans le miocèneinférieur et moyen) ;– une période d’émersion sur les bords du bassinresponsable de la surface d’érosion et d’évaporationintense au centre du bassin (présence d’évaporitesd’1 km d’épaisseur environ) au miocène supérieur ;
5
4
3
104

105
Programme : notions et contenus• Divergence et phénomènes liés– Formation et divergence des plaques lithosphériques au niveau des dorsales océaniques.– Activités tectoniques et magmatiques associées.– Tectonique : la morphologie, la présence de séismes et les failles normales qui structurent les dorsales océaniques attestent de mouvements enextension.– Magmatique : les dorsales océaniques sont le siège d’une production importante de magma : de l’ordre de 20 km3 par an. Ces magmas sont issusde la fusion partielle des péridotites du manteau induite par décompression. Ils sont de nature basaltique. La fusion partielle leur donne une compo-sition chimique différente de celle de la roche source. Le refroidissement plus ou moins rapide des magmas conduit à des roches de texture diffé-rente (basalte/gabbros).– En s’éloignant de la dorsale, la lithosphère océanique se refroidit, s’hydrate et s’épaissit.• La machinerie thermique de la Terre– Dissipation de l’énergie interne.– Le flux de chaleur en surface en est la manifestation principale. La chaleur interne a pour origine essentielle la désintégration de certains isotopesradioactifs.– La convection du manteau terrestre.– La fabrication de la lithosphère océanique, la subduction et les mouvements des plaques lithosphériques sont les manifestations d’une convectionthermique à l’état solide du manteau (transport de chaleur par mouvement de matière). Les dorsales océaniques traduisent des courants montantschauds de matériel du manteau. Les plaques en subduction traduisent des courants descendants froids.– Points chauds.Le magmatisme lié aux points chauds marque la remontée ponctuelle de matériel du manteau profond. Il s‘exprime par des éruptions massives delaves basaltiques (plateaux océaniques, trapps, alignements insulaires).
Objectifs à atteindreLe chapitre 15 a montré que les fonds océaniques résultent de l’activité des dorsales au niveau desquelles se formentdes roches magmatiques déjà identifiées dans le chapitre 13 (basalte et gabbro de textures différentes mais de mêmecomposition).
L’objectif de ce chapitre est de comprendre la genèse de la croûte et de la lithosphère océaniques au niveau d’une dor-sale à partir d’une activité magmatique. Il s’agit, dans un premier temps, d’identifier l’origine du magma (rochesource) et de préciser les conditions de genèse de ce magma basaltique. La compréhension de la genèse du magmanécessite la prise en compte préalable :
– du flux de chaleur terrestre en particulier à l’aplomb des dorsales ;
– de la mobilité du manteau à l’état solide en particulier des remontées d’asthénosphère chaude à l’aplomb des dorsales.
Dans un second temps, le devenir de la lithosphère océanique doit être envisagé au fur et à mesure qu’elle s’éloignede la dorsale (hydrothermalisme, refroidissement, épaississement, subsidence) en liaison avec la morphologie desfonds océaniques.
Magmatisme et convection mantellique
C H A P I T R E 1 7

A c t i v i t é s 2
Dynamique du globe au niveau des dorsalesL’objectif de cette activité est de montrer qu’il existeune hétérogénéité thermique du manteau en profondeuret que celle-ci entraîne des mouvements lents dematière à l’état solide en particulier des ascendancesde péridotites chaudes à l’aplomb des dorsales.
Exploitation� Le DOCUMENT 4 montre qu’à l’aplomb des dorsales, à100 km de profondeur, les vitesses des ondes S sont infé-rieures aux vitesses attendues (anomalies négatives) ; celasignifie que les péridotites du manteau sont moins rigides àce niveau et que leur température est plus élevée qu’ailleurs(sous les plaines abyssales ou sous les continents).Le DOCUMENT 5 confirme l’existence d’un matériel chaud àtrès faible profondeur à l’aplomb de la dorsale alors que lematériel froid en surface s’épaissit lorsqu’on s’éloigne dela dorsale.Le DOCUMENT 6 confirme la structure de la croûte océanique(superposition de basalte et gabbro) et montre la remontéed’un matériel moins dense à l’aplomb de la dorsale direc-tement sous la croûte : il s’agit de l’asthénosphère. Lalithosphère à ce niveau est donc réduite à la croûte océa-nique.Le DOCUMENT 7 montre que les lignes d’égale température(isogéothermes) remontent et convergent vers la dorsale cequi signifie une remontée de matériel chaud à ce niveau.L’isogéotherme 1 200 °C correspond à la limite entre lalithosphère (péridotites rigides) et l’asthénosphère sous-jacente (péridotites ductiles).L’ensemble des documents suggère qu’un matériel chaudremonte jusque sous la croûte à l’aplomb de la dorsale ; ils’agit de l’asthénosphère dont la température atteint1 200 °C.� Le DOCUMENT 8 montre que l’huile réchauffée locale-ment monte à travers l’huile plus froide qui la surmonte(on parle de diapir thermique). On peut ainsi supposerqu’une hétérogénéité thermique du manteau avec desrégions chaudes et des régions froides est à l’origine demouvements très lents, à l’état solide ; les matériaux pluschauds et donc moins denses auraient ainsi tendance àmonter ce qui expliquerait l’existence des dorsales (ouplus ponctuellement des points chauds).
A c t i v i t é s 3
Genèse des magmas au niveau des dorsalesL’objectif de cette activité est de montrer que la tempé-rature élevée à l’aplomb des dorsales n’est pas la seuleen cause pour expliquer l’origine des magmas et que labaisse de pression consécutive à l’ascension de l’asthé-nosphère intervient également.
◆ Problèmes scientifiques à résoudreComment expliquer la genèse des magmas à l’origine dela croûte océanique au niveau des dorsales et des édificesvolcaniques énormes des points chauds ?
P o s e r l e s p r o b l è m e s
Les supports choisis permettent de mobiliser les acquisconcernant la structure et la dynamique superficielle duglobe, de situer certaines roches et de rappeler l’originemagmatique des roches de la croûte océanique et despoints chauds.Un certain nombre de problèmes peuvent alors être formulés concernant la genèse des magmas à partir de la péridotite solide du manteau ainsi que le devenir de lalithosphère océanique qui s’éloigne lentement de la dor-sale.
A c t i v i t é s 1
Le flux de chaleur terrestreL’origine magmatique des roches de la croûte océa-nique permet de supposer que la température du globeest plus élevée à l’aplomb des dorsales. L’objectif decette activité est de repérer les régions du globe quidégagent le plus de chaleur par unité de surface et detemps.
Exploitation� Le DOCUMENT 2 permet de constater que les régions quiperdent le plus de chaleur sont effectivement les dorsales.Cette perte de chaleur est maximale à l’aplomb des dor-sales et diminue de part et d’autre (dégradé de couleursallant du rouge ou de l’orangé au bleu foncé). La perte dechaleur est plus importante au niveau de la dorsalePacifique (jusqu’à 350 mW.m–2) et celle-ci est inégaleselon les régions d’une dorsale (ainsi la partie Sud de ladorsale Atlantique perd plus de chaleur que sa partieNord). Les plaines abyssales émettent environ 4 à 5 foismoins de chaleur par m2 et par seconde que les dorsales(70 mW.m–2 contre 350 mW.m–2 au niveau de la dorsale).Le DOCUMENT 3 confirme l’existence d’un flux thermiqueélevé à l’aplomb de la dorsale, ce dernier s’abaisse lors-qu’on se dirige vers les plaines abyssales (le document 3représente une « coupe » à travers la dorsale Pacifiquefigurant sur le DOCUMENT 2)Ces valeurs sont bien en accord avec la tectonique desplaques et avec l’origine des magmas ; le flux de chaleurimportant au niveau des dorsales sous-entend qu’une pro-duction de chaleur importante existe en profondeur et quecelle-ci permet la fusion des péridotites à l’origine dumagma basaltique.
� Le flux de chaleur élevé à l’aplomb des dorsales signi-fie qu’un matériel chaud existe à faible profondeur.
106

Exploitation� Le DOCUMENT 9 montre qu’à l’aplomb d’une dorsale, latempérature augmente très vite avec la profondeur jusqu’à30-40 km environ (elle atteint 1 400 °C contre 750 °C à lamême profondeur au niveau d’une plaine abyssale).Ensuite la température reste stable et voisine de 1 450 °Clorsqu’on passe à des profondeurs supérieures. Le géo-therme de dorsale traduit en fait le comportement ther-mique du diapir asthénosphérique qui remonte en gardantà peu près la même température jusqu’à 20 à 30 km deprofondeur.Le DOCUMENT 10 montre l’état d’une péridotite en fonctionde la température et de la pression. On constate que pourla faire fondre partiellement, il faut franchir une limiteappelée solidus ; pour franchir le solidus, on peut :– augmenter la température pour une pression donnée (onconstate qu’il faut augmenter la température lorsque lapression augmente) ;– diminuer la pression à condition qu’on se trouve, audépart, à une température supérieure à 1 250 °C (et en sup-posant que la diminution de pression ne s’accompagne pasd’une chute de la température).Le DOCUMENT 11 permet de montrer que pour une tempéra-ture égale, la fusion d’un corps solide est d’autant plusrapide que la pression exercée sur ce corps est faible.Le DOCUMENT 12 correspond à la superposition du géo-therme de dorsale et des résultats expérimentaux de fusiondes péridotites (solidus). Pour exploiter ce document, ilfaut situer les péridotites asthénosphériques dans uncontexte dynamique c’est-à-dire comprendre que le géo-therme correspond au chemin suivi par ces roches. Ainsiune péridotite solide à 160 km de profondeur constituée detrois minéraux a, b, c va remonter ; au cours de son ascen-sion, elle garde presque la même température alors que lapression à laquelle elle est soumise diminue. Elle va ainsifranchir son solidus entraînant une fusion partielle de laroche. Le magma liquide et chaud issu de la fusion de cer-tains minéraux va migrer vers la surface et donner, parrefroidissement rapide, des basaltes en surface ou, parrefroidissement plus lent, des gabbros à la base desbasaltes. La partie résiduelle non fondue de la péridotitereste en dessous de la croûte nouvellement formée etforme la partie mantellique de la lithosphère océanique.
� Le DOCUMENT 13a. montre que la composition minéralo-gique des basaltes et gabbros est la même que celle de lapéridotite source mais que la proportion des minérauxn’est pas la même, les basaltes et gabbros étant plus richesen pyroxènes calciques et en feldspaths plagioclases. LeDOCUMENT 13b. confirme l’enrichissement en aluminiumfer et calcium des basaltes et gabbros.La fusion partielle de la péridotite source affecte donc pré-férentiellement les minéraux riches en aluminium et cal-cium. Ceci explique que la péridotite résiduelle soitenrichie de façon relative en magnésium et appauvrie enaluminium, fer et surtout calcium.
A c t i v i t é s 4
Formation et devenir de la litho-sphère océaniqueCette activité a pour objectif de situer la mise en placedes roches de la croûte océanique et de suivre le deve-nir de la croûte et de la lithosphère océanique quis’éloignent lentement de la dorsale.
Exploitation� Le DOCUMENT 14 résulte de l’interprétation de profil desismique réflexion. On constate l’existence d’une chambremagmatique de petite taille (1 km de largeur) et située àfaible profondeur (1,5 km) à la base des basaltes en filonset au sommet des gabbros. Cependant, une vaste zone de10 km de large à la base s’étale sous la chambre magma-tique ; elle correspond à la présence de magma à l’étatplus pâteux en cours de cristallisation.On peut ainsi imaginer que le magma qui s’extrait de lapéridotite source après fusion partielle remonte et ali-mente régulièrement la chambre magmatique ainsi que lapartie sous-jacente. Le magma présent dans la chambremagmatique peut s’injecter dans le toit de la chambre,engendrant des filons ou des coulées de basaltes en cous-sin alors que le magma sous-jacent va refroidir plus lente-ment engendrant les gabbros.
� Le DOCUMENT 17 montre que l’eau de mer peut s’infiltrerà travers la croûte et se réchauffer fortement au voisinagede la chambre magmatique avant de ressortir à une tempé-rature pouvant atteindre 350 °C. La comparaison de lacomposition de l’eau de mer avec celle du fluide hydro-thermal montre une soustraction de certains éléments(magnésium en particulier) et un enrichissement trèsimportant en d’autres éléments en particulier métalliques(fer, manganèse, zinc et cuivre) dont une partie se déposesous forme de sulfures métalliques au voisinage desfumeurs noirs. Ceci reflète l’existence de réactions chi-miques entre l’eau de mer et les minéraux des roches faci-litées par les hautes températures ; les roches subissent unvéritable métamorphisme ; ainsi les minéraux ferroma-gnésiens des basaltes et gabbros (olivine et pyroxène) sonttransformés en serpentine, silicate hydraté de fer et demagnésium de couleur verdâtre.
� Le DOCUMENT 15a. montre :– que le fond océanique s’enfonce de part et d’autre d’unedorsale, la profondeur passant de 2 km à l’aplomb de ladorsale à 5 km au niveau des plaines abyssales ;– une différence entre les profils des dorsales Atlantique etPacifique ( la pente est plus abrupte dans le cas de la dor-sale Atlantique et la profondeur de 5 km est atteinte plusrapidement que dans le cas de la dorsale Pacifique).Le DOCUMENT 15b. montre que la lithosphère est très minceau niveau de la dorsale, l’asthénosphère chaude de faibledensité (3,25) remontant pratiquement jusque sous lacroûte. Au fur et à mesure de l’expansion océanique, lacroûte et l’asthénosphère s’éloignent de la dorsale et serefroidissent par conduction thermique. Le sommet del’asthénosphère se refroidissant se transforme en litho-sphère de densité plus élevée (3,3 contre 3,25).
Chapitre 17 : Magmatisme et convection mantellique
107

d. La plus grande partie de l’énergie dissipée par laTerre provient de la désintégration de certains iso-topes radioactifs tels que thorium, uranium et potas-sium qui libèrent de la chaleur en se désintégrant.Une autre partie de la chaleur provient de la chaleurinitiale accumulée lors de l’accrétion à l’origine de laTerre ainsi que de la cristallisation du fer liquide à lalimite noyau externe-noyau interne qui est très exo-thermique.e. La fusion partielle des péridotites solides du man-teau peut être provoquée par une élévation de latempérature (c’est le cas des panaches à l’originedes points chauds) ou par abaissement de pressionsans variation de température (c’est ce qui se passeà l’aplomb des dorsales lors de la remontée de l’as-thénosphère entraînée dans les courants de convec-tion).f. La présence de basaltes et de gabbros surmontésde sédiments renfermant des gisements de sulfuresde cuivre est une indication de l’existence d’unancien fond océanique proche d’une dorsale. Eneffet, actuellement, les dorsales sont les lieux de for-mation des basaltes et gabbros ; la circulation del’eau de mer entraîne un hydrothermalisme quiaboutit à la formation de fumeurs noirs au niveaudesquels sont rejetées de grandes quantités de sul-fures de fer, de zinc et de cuivre.
Choisir la bonne réponse1. Réponse exacte : c.2. Réponse exacte : a.3. Réponse exacte : b.
Exercice corrigé
Fusion part iel le et magma basalt iqueVoir correction dans le manuel pages 356 et 357.
Évaluer ses compétences
Circulat ion d’eau dans la croûteocéanique1. La durée pour que toute l’eau de mer traverse lacroûte océanique est de : 1,4 x 1021 kg/2,6x1014 kg.an–1 = 5 384 615 années soitplus de 5 millions d’années ce qui est très court àl’échelle des temps géologiques.2. Le volume d’eau de mer incorporé dans lesbasaltes et les gabbros chaque année :– volume de basalte émis chaque année : 20 km3 soit 20 x 1 0003 m3 ;– masse de basalte émis chaque année : (20 x 1 0003 m3) x (2 900 kg.m–3) = 58 x 1012 kg ;– masse d’eau incorporée dans les basaltes et gab-bros : (58 x 1012) x (3,5 x 10–2) = 203 x 1010 kg, soitl’équivalent de 2 milliards de tonnes d’eau.
5
4
3
L’épaississement progressif de la lithosphère au détrimentde l’asthénosphère et l’augmentation de sa densité entraî-nent un enfoncement progressif du fond océanique. La dif-férence de profils des fonds de l’océan Atlantique et del’océan Pacifique est liée au fonctionnement des dorsales,la dorsale Pacifique responsable d’une expansion rapidede la plaque Pacifique entraîne loin la jeune lithosphèreencore chaude, donc peu subsidente.
E x e r c i c e s
Évaluer ses connaissances
Définit ionsa. Flux thermique : Quantité de chaleur émise parunité de surface d’un matériau exprimée le plus sou-vent en W.m–2 qui représente en réalité un débitd’énergie exprimé en J.m–2.s–1.b. Solidus : Courbe obtenue expérimentalementdéfinissant la limite de température et de pressionséparant l’état solide d’une roche de l’état de débutde fusion.c. Convection : Déplacement lent de matière sousl’effet de différences de densité liées à des diffé-rences de température.d. Gradient géothermique : Augmentation de la tem-pérature avec la profondeur au sein du globe ter-restre.e. Panache mantellique : Remontée magmatiqued’origine profonde qui traverse le manteau et vientperforer la croûte terrestre.f. Fusion partielle : Processus qui aboutit à l’appari-tion d’une faible quantité de liquide magmatique àpartir d’une roche à l’état solide lors d’une augmen-tation de température ou/et d’une diminution depression.
Questions à réponses cour tesa. Le géotherme océanique montre que la tempéra-ture s’accroît très rapidement lorsque la profondeuraugmente et ce jusqu’à 40 km environ puis aug-mente très peu au-delà de cette profondeur dumoins dans le manteau supérieur.b. Les mouvements du manteau qui permettent lamobilité des plaques lithosphériques sont appelésmouvements de convection. Ces mouvements sontdes déplacements lents de matière à l’état solidesous l’effet des différences de densité liées à desdifférences de température. Les parties pluschaudes moins denses ont tendance à remontervers la surface où elles se refroidissent, deviennentplus denses et redescendent avant de se réchaufferà nouveau ; le cycle recommence ensuite.c. Les zones chaudes du manteau correspondent àdes remontées mantelliques chaudes situées àl’aplomb des dorsales alors que les zones froidescorrespondent à des plongées (subductions) delithosphère océanique refroidie.
2
1
108

L’ Is lande : une dorsale émergéeet un point chaud1. L’Islande est une dorsale émergée car elle estsituée dans le prolongement de la dorsale médio-Atlantique et présente les caractéristiques d’un riftde dorsale lente avec émission discontinue debasaltes accompagnée de phénomènes d’exten-sion.2. La tomographie sismique montre une zone relati-vement étroite d’anomalies négatives qui atteste dela présence, à l’aplomb de l’Islande, d’un matérielchaud qui part de la limite noyau externe-manteau
inférieur soit à 2 900 km de profondeur. Cette zoneplus chaude présente toutes les caractéristiquesd’un panache mantellique qui alimente un volca-nisme de point chaud.3. La topographie actuelle de l’Islande s’explique àla fois par son caractère de dorsale (rift et extensionaccompagnée de failles normales et d’émissionsbasaltiques) et par son caractère de point chaud (laremontée de matériel chaud, donc moins denses’accompagne d’une émersion de la dorsale.
6
E x e r c i c e c o m p l é m e n t a i r e
Topographie des fonds océaniques et dynamique du globeLe document ci-dessous présente le profil topographique des fonds océaniques de l’Atlantique entre Saint-Christophe aux Antilles (point A) et Dakar au Sénégal (point B).
7
1. Repérer la situation de ce profil sur la carte desfonds océaniques et calculer l’exagération del’échelle verticale par rapport à l’échelle horizontale.2. Repérer les principaux éléments topographiquesde ce profil en précisant leurs caractéristiques.3. Rechercher une explication à ce profil en vousaidant de vos connaissances sur la structure et ladynamique du globe.
Réponses attendues1. Voir la carte : 14 mm sur l’échelle verticale repré-sentent 6 km soit 1 mm pour 0,43 km alors que29 mm représentent 1 000 km sur l’échelle horizon-tale soit 1 mm pour 34,5 km. L’exagération del’échelle verticale est donc de 34,5/0,43 = 80 foisenviron (les reliefs devraient être réduits 80 fois pourêtre en correspondance avec l’échelle horizontale).
Chapitre 17 : Magmatisme et convection mantellique
109
0
1 000 2 000 3 000 4 000 5 0006 000
Profondeur (m)
Saint-Christophe DakarA B
Distance (km)
Profil topographique.
2. Les principaux éléments topographiques repé-rables sont :– la dorsale Atlantique profonde de 1,5 km en sonaxe qui se raccorde aux plaines abyssales à environ700 km de son axe ;– les plaines abyssales profondes de 5 km ;– une fosse avant les Antilles, îles volcaniques àl’ouest ;– un relief important correspondant aux îles du Capvert ;– une marge passive à la hauteur du Sénégal à l’est.3. Ce profil s’explique par la géodynamique interne :– la dorsale est due à la remontée de matériel asthé-nosphérique chaud qui explique le relief de la dor-sale. Cette remontée aboutit à la genèse de lalithosphère océanique par fusion partielle des péri-dotites à l’origine d’un magma basaltique ;

110
1
1
2
D
PAPA
ZS
F
A PC B
MP C
P
DPAFMPPCZSCP
: Dorsale: Plaine abyssale: Fosse: Marge continentale passive: Point chaud: Zone de subduction: Continent: Panache mantellique
Lithosphère jeune, mince, encore chaude, moins dense, en relief
Lithosphère agée, épaisse, refroidie, plus dense qui s'est enfoncée
Croûte continentale(granite-roches métamorphiques)
Croûte océanique (basalte et gabbro)
Manteau lithosphériquePéridotite
Manteau asthénosphérique
Volcanisme de point chaud (basalte)
Fusion partielle
Ascension dela péridotite
2
Structure et dynamique du globe au niveau de la dorsale Atlantique.
– le profil situé de part et d’autre de la dorsale s’ex-plique par la subsidence thermique liée au refroidis-sement et à l’épaississement de la lithosphère quidevient plus dense en s’éloignant de la dorsale ;– la fosse profonde de 6 000 m avant les Antillescorrespond à une subduction de la lithosphèreocéanique de l’Atlantique ;
– les îles du Cap Vert sont liées à un volcanisme depoint chaud car isolé et sans relation avec une zonede subduction ;– la marge passive du Sénégal est un témoin de ladéchirure continentale à l’origine de l’Atlantique.
73654_01_p099a112 01/04/2005 14:19 Page 110