réponse immunitaire de l'hôte dans la symbiose...
TRANSCRIPT

N° d’ordre 2006-ISAL-00107 Année 2006
Thèse
Réponse immunitaire de l'hôte dans la symbiose bactérienne intracellulaire du
charançon Sitophilus zeamais
présentée devant L’Institut National des Sciences Appliquées de Lyon
pour obtenir
le grade de docteur
Ecole doctorale : Evolution, Ecosystèmes, Microbiologie et Modélisation Spécialité : Analyse et Modélisation des Systèmes biologiques
Par
Caroline ANSELME
Soutenue le 6 Décembre 2006 devant la Commission d’examen
Jury
LEMAITRE Bruno Rapporteur BOUCHON Didier Rapporteur FLEURY Frédéric Examinateur MOYA Andrés Examinateur VOLKOFF Anne-Nathalie Examinateur HEDDI Abdelaziz Directeur
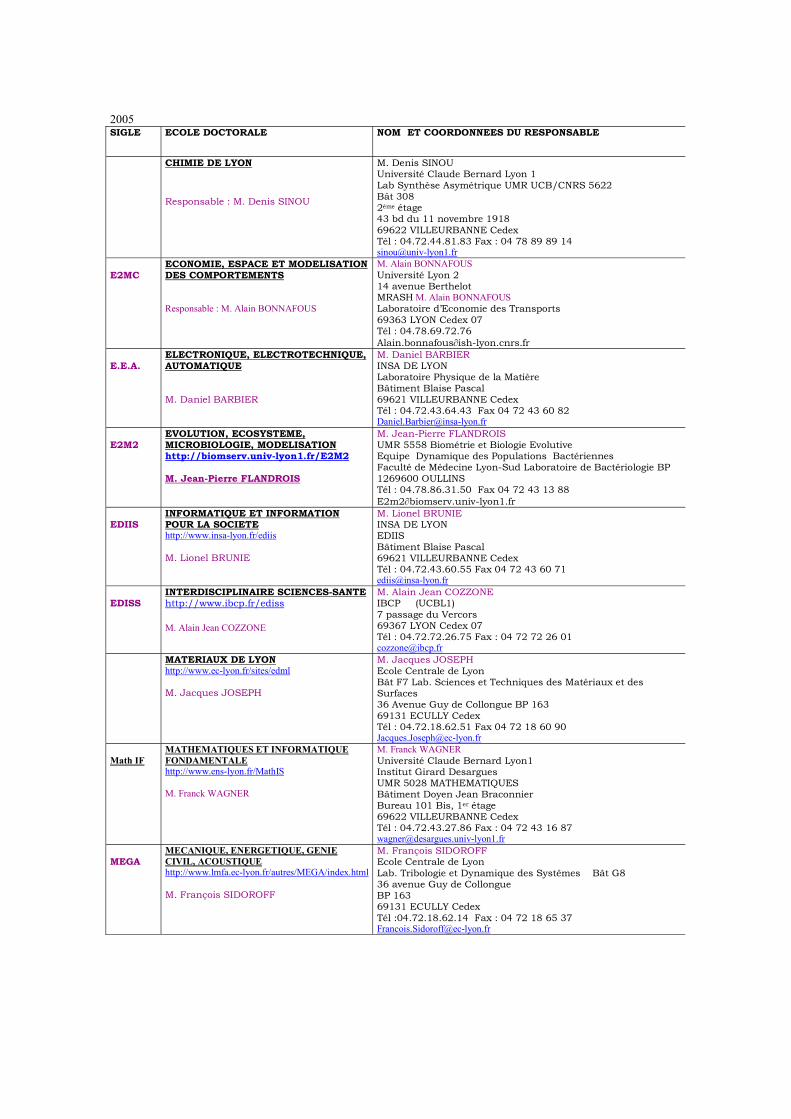
2005 SIGLE ECOLE DOCTORALE NOM ET COORDONNEES DU RESPONSABLE
CHIMIE DE LYON Responsable : M. Denis SINOU
M. Denis SINOU Université Claude Bernard Lyon 1 Lab Synthèse Asymétrique UMR UCB/CNRS 5622 Bât 308 2ème étage 43 bd du 11 novembre 1918 69622 VILLEURBANNE Cedex Tél : 04.72.44.81.83 Fax : 04 78 89 89 14 [email protected]
E2MC
ECONOMIE, ESPACE ET MODELISATION DES COMPORTEMENTS Responsable : M. Alain BONNAFOUS
M. Alain BONNAFOUS Université Lyon 2 14 avenue Berthelot MRASH M. Alain BONNAFOUS Laboratoire d’Economie des Transports 69363 LYON Cedex 07 Tél : 04.78.69.72.76 Alain.bonnafous∂ish-lyon.cnrs.fr
E.E.A.
ELECTRONIQUE, ELECTROTECHNIQUE, AUTOMATIQUE M. Daniel BARBIER
M. Daniel BARBIER INSA DE LYON Laboratoire Physique de la Matière Bâtiment Blaise Pascal 69621 VILLEURBANNE Cedex Tél : 04.72.43.64.43 Fax 04 72 43 60 82 [email protected]
E2M2
EVOLUTION, ECOSYSTEME, MICROBIOLOGIE, MODELISATION http://biomserv.univ-lyon1.fr/E2M2 M. Jean-Pierre FLANDROIS
M. Jean-Pierre FLANDROIS UMR 5558 Biométrie et Biologie Evolutive Equipe Dynamique des Populations Bactériennes Faculté de Médecine Lyon-Sud Laboratoire de Bactériologie BP 1269600 OULLINS Tél : 04.78.86.31.50 Fax 04 72 43 13 88 E2m2∂biomserv.univ-lyon1.fr
EDIIS
INFORMATIQUE ET INFORMATION POUR LA SOCIETE http://www.insa-lyon.fr/ediis M. Lionel BRUNIE
M. Lionel BRUNIE INSA DE LYON EDIIS Bâtiment Blaise Pascal 69621 VILLEURBANNE Cedex Tél : 04.72.43.60.55 Fax 04 72 43 60 71 [email protected]
EDISS
INTERDISCIPLINAIRE SCIENCES-SANTE http://www.ibcp.fr/ediss M. Alain Jean COZZONE
M. Alain Jean COZZONE IBCP (UCBL1) 7 passage du Vercors 69367 LYON Cedex 07 Tél : 04.72.72.26.75 Fax : 04 72 72 26 01 [email protected]
MATERIAUX DE LYON http://www.ec-lyon.fr/sites/edml M. Jacques JOSEPH
M. Jacques JOSEPH Ecole Centrale de Lyon Bât F7 Lab. Sciences et Techniques des Matériaux et des Surfaces 36 Avenue Guy de Collongue BP 163 69131 ECULLY Cedex Tél : 04.72.18.62.51 Fax 04 72 18 60 90 [email protected]
Math IF
MATHEMATIQUES ET INFORMATIQUE FONDAMENTALE http://www.ens-lyon.fr/MathIS M. Franck WAGNER
M. Franck WAGNER Université Claude Bernard Lyon1 Institut Girard Desargues UMR 5028 MATHEMATIQUES Bâtiment Doyen Jean Braconnier Bureau 101 Bis, 1er étage 69622 VILLEURBANNE Cedex Tél : 04.72.43.27.86 Fax : 04 72 43 16 87 [email protected]
MEGA
MECANIQUE, ENERGETIQUE, GENIE CIVIL, ACOUSTIQUE http://www.lmfa.ec-lyon.fr/autres/MEGA/index.html M. François SIDOROFF
M. François SIDOROFF Ecole Centrale de Lyon Lab. Tribologie et Dynamique des Systêmes Bât G8 36 avenue Guy de Collongue BP 163 69131 ECULLY Cedex Tél :04.72.18.62.14 Fax : 04 72 18 65 37 [email protected]

À ma famille, mes parents et ma sœur qui m’ont soutenue toutes ces années et qui m’ont permis d’en arriver là… Merci...
Remerciements
Mes remerciements vont aux rapporteurs : Didier Bouchon et Bruno Lemaitre ainsi qu’aux membres du
jury : Frédéric Fleury, Andrés Moya et Anne-Nathalie Volkoff pour avoir accepté de juger ce travail.
Merci également à Pierre Couble, Bernard Duvic, Gérard Febvay et Phillipe Normand pour les discus-
sions constructives menées lors des réunions du comité de pilotage.
Je tiens à remercier tout particulièrement Aziz Heddi, mon directeur de thèse, pour ces quatre années
de recherche passionnantes, pour son optimisme et surtout pour tout ce que j’ai appris…
Un grand merci à Agnès Vallier, sans qui ce travail ne serait pas ce qu’il est. Merci pour son encadre-
ment technique, son aide, son soutien et toutes les heures qu’elle a passées sous la loupe…
Merci aussi à Carole Vincent-Monégat pour son aide, ses précieux conseils -surtout sa technique de
“digestion micro-onde”- et pour toutes nos discussions enrichissantes...
Merci également à Séverine Balmand pour son aide en histologie et Marjolaine Rey pour son aide lors
de l’identification des peptides antibactériens du charançon... en espérant qu’elles ne m’en voudront
pas trop de les avoir embarquées avec moi dans cette galère…
Mes remerciements vont aussi à Gabrielle Duport pour les pucerons aposymbiotiques, Yvan Rahbé pour
son aide et ses conseils à de nombreuses reprises, Paul Nardon pour ses conseils en histologie et
pour avoir partagé avec moi ses connaissances sur le modèle charançon, Anne-Marie Grenier pour la
correction de ce manuscrit et Hubert Charles pour son aide précieuse en statistiques, la lecture criti-
que de cette thèse et son soutien en période de crise…
Merci également à Marie-Odile Fauvarque et toute l’équipe pour nous avoir accueillis à Grenoble, moi et
mes charançons, pour le travail avec Pseudomonas.
Merci à Vicente Pérez-Brocal, Amparo Lattore et Andrés Moya pour le séquençage des EST de charan-
çon et de puceron et à Delphine Charif pour le travail qu’elle a effectué sur ces séquences.
Enfin, je tiens à remercier Gérard Febvay et tous les membres du laboratoire pour leur accueil et leur
soutien au cours de ces quatre années… avec une mention spéciale pour toutes les petites mains qui
m’ont aidée à dissequer les grains de blé, pour Gaby, Isa et Heidi pour les séances “anti-stress” à la
salle de sport et pour Alain qui a régulièrement fleuri mon bureau.
Merci aussi à tous mes amis pour leur soutien,
Merci à Federica pour avoir été là quand j’en avais besoin,
Merci à Jeremy, mon complice,
Et enfin merci à Frédéric, tout simplement.

Sommaire
ANSELME Caroline 4 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Sommaire
ABREVIATIONS ................................................................................................................................................................8
INTRODUCTION ...............................................................................................................................................................9
SYMBIOSE ET IMMUNITE INNEE ...............................................................................................................................12 1 CONTEXTE BIOLOGIQUE : LA SYMBIOSE................................................................................ 13
1.1 Les différentes définitions et classifications de la symbiose .......................................... 13 1.2 La symbiose intracellulaire chez les insectes................................................................ 14
1.2.1 Biologie de la symbiose intracellulaire chez les insectes .....................................................14 1.2.1.1 Niveau d’intégration des endocytobiotes .............................................................................14 1.2.1.2 Rôle des endocytobiotes primaires, impact physiologique ...................................................16 1.2.1.3 Aspects moléculaires des interactions hôte-symbiote...........................................................17
1.2.2 Evolution des génomes des bactéries symbiotiques intracellulaires......................................19 1.2.2.1 Augmentation de la vitesse d’évolution...............................................................................19 1.2.2.2 Réduction de la taille des génomes des endocytobiotes........................................................20
1.3 Caractéristiques du modèle Sitophilus spp. .................................................................. 22 1.3.1 Développement de l’insecte et description des structures symbiotiques ...............................23
1.3.1.1 Transmission des symbiotes et embryogenèse .....................................................................24 1.3.1.2 Le développement du bactériome larvaire ...........................................................................24 1.3.1.3 Développement des bactériomes des cæcums mésentériques................................................25
1.3.2 Quelques aspects de l’interaction hôte-symbiote .................................................................26 1.3.2.1 Devenir des structures symbiotiques chez les insectes aposymbiotiques...............................26 1.3.2.2 Contrôle de la densité bactérienne par l’hôte .......................................................................26
1.3.3 Aspects évolutifs de la symbiose chez Sitophilus ................................................................27 1.3.3.1 Caractéristiques du génome de SOPE..................................................................................27 1.3.3.2 Histoire évolutive de la symbiose chez les Dryophthoridae..................................................27
2 L’IMMUNITE INNEE .............................................................................................................. 30 2.1 La réponse cellulaire ................................................................................................... 33
2.1.1 La phagocytose par les plasmatocytes.................................................................................33 2.1.2 L’encapsulation par les lamellocytes ..................................................................................34 2.1.3 Les cellules à cristaux et la mélanisation ............................................................................34
2.2 La réponse humorale : synthèse des peptides antimicrobiens........................................ 34 2.2.1 Le corps gras : la réponse systémique .................................................................................35
2.2.1.1 La voie Toll .......................................................................................................................35 2.2.1.2 La voie Imd .......................................................................................................................38
2.2.2 Les épithéliums : la réponse locale .....................................................................................39 2.2.3 Reconnaissance des microorganismes .................................................................................40
2.2.3.1 Les GNBP..........................................................................................................................40 2.2.3.2 Les PGRP ..........................................................................................................................41
2.2.4 Elimination des microorganismes par les peptides antimicrobiens .......................................48 2.3 La réaction de mélanisation ......................................................................................... 50 2.4 Conclusion .................................................................................................................. 52
MATERIEL ET METHODES ..........................................................................................................................................53 1 INSECTES : ELEVAGE ET MATERIEL BIOLOGIQUE.................................................................... 54
1.1 Le charançon Sitophilus zeamais ................................................................................. 54 1.1.1 Elevage..............................................................................................................................54 1.1.2 Matériel biologique ............................................................................................................54
1.1.2.1 Larves du quatrième stade et nymphes ................................................................................54 1.1.2.2 Embryons...........................................................................................................................54 1.1.2.3 Ovocytes et bactériomes larvaires .......................................................................................54
1.2 Le puceron Acyrthosiphon pisum ................................................................................. 55 1.2.1 Élevage..............................................................................................................................55 1.2.2 Matériel biologique ............................................................................................................55
2 TECHNIQUES HISTOLOGIQUES............................................................................................... 55 2.1 Fixation des échantillons ............................................................................................. 55 2.2 Double inclusion et coupe ............................................................................................ 56
ANSELME Caroline 5 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Sommaire
2.3 “Fluorescence In Situ Hybridization” (FISH) sur coupes histologiques........................ 56 3 INFECTION BACTERIENNE ..................................................................................................... 57
3.1 Culture des souches bactériennes utilisées ................................................................... 57 3.2 Infection des larves de charançon ................................................................................ 58 3.3 Mesure de la croissance bactérienne in larva............................................................... 58 3.4 Infection des pucerons ................................................................................................. 58
4 TECHNIQUES DE BIOLOGIE MOLECULAIRE ............................................................................. 59 4.1 Extraction des acides nucléiques et électrophorèse sur gel d’agarose........................... 59
4.1.1 Extraction d’ADN génomique ............................................................................................59 4.1.2 Extraction d’ARN totaux....................................................................................................59
4.1.2.1 Extraction au Trizol®..........................................................................................................60 4.1.2.2 Extraction "RNeasy Mini kit" (QIAGEN) ...........................................................................60 4.1.2.3 Traitement des échantillons à la DNase...............................................................................60
4.1.3 Electrophorèse sur gel d’agarose ........................................................................................60 4.1.4 Dosage des échantillons .....................................................................................................61
4.2 Northern Blot .............................................................................................................. 61 4.2.1 Préparation des membranes ................................................................................................62 4.2.2 Préparation des sondes radioactives ....................................................................................62
4.2.2.1 Amplification par PCR des fragments d’ADN .....................................................................62 4.2.2.2 Purification des fragments d’ADN amplifiés .......................................................................63 4.2.2.3 Marquage radioactif de la sonde .........................................................................................63
4.2.3 Hybridation et révélation....................................................................................................64 4.2.3.1 Préhybridation-hybridation .................................................................................................64 4.2.3.2 Lavages, exposition, révélation et normalisation .................................................................64
4.3 Soustraction d’ADNc (“Suppressive Subtractive Hybridization”) ................................. 65 4.3.1 Synthèse, amplification et purification des ADNc ...............................................................65 4.3.2 Digestion des ADNc, ligation aux adaptateurs et hybridations .............................................67 4.3.3 Amplification des ADNc soustraits.....................................................................................67
4.4 Obtention des séquences des ARNm à partir des EST (RACE)....................................... 68 4.4.1 Obtention des ADNc (RT-PCR)..........................................................................................68 4.4.2 Obtention des fragments d’ADNc (RACE PCR)..................................................................69
4.5 Amplification de fragment d’ADN génomique par longue PCR ..................................... 70 4.6 Marche sur le chromosome (“Genome Walking”) ........................................................ 70 4.7 Clonage des produits de PCR, préparation de l’ADN plasmidique et séquençage. ........ 71
4.7.1 Clonage par “TOPO cloning” (Invitrogen) ..........................................................................71 4.7.2 Transformation des cellules par électroporation ..................................................................72 4.7.3 Préparation d’ADN plasmidique .........................................................................................73 4.7.4 Séquençage des fragments d’ADN......................................................................................73
4.8 RT-PCR quantitative en temps réel .............................................................................. 73 4.8.1 Rétrotranscription des ARNm.............................................................................................74 4.8.2 PCR en temps réel..............................................................................................................75
4.9 Macroarrays................................................................................................................ 76 4.9.1 Synthèse des sondes radioactives par rétrotranscription.......................................................76 4.9.2 Préparation des membranes ................................................................................................76
5 TECHNIQUES D’ANALYSE DE SEQUENCES .............................................................................. 77 5.1 Analyse des séquences à l’aide du programme Mac Molly Tetra (GeneSoft) ................. 77 5.2 Analyse des séquences par comparaison aux banques de données ............................... 77 5.3 Analyse des EST des banques soustraites ..................................................................... 78
6 TECHNIQUES DE BIOCHIMIE .................................................................................................. 78 6.1 Collecte d’hémolymphe et extraction des peptides ........................................................ 78 6.2 Séparation des peptides de l’hémolymphe par chromatographie liquide à haute performance (HPLC) .............................................................................................................. 79 6.3 Test d’activité antibactérienne en milieu liquide .......................................................... 80
ANSELME Caroline 6 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Sommaire
RESULTATS.....................................................................................................................................................................81 1 IDENTIFICATION DES GENES DE LA REPONSE IMMUNITAIRE DE L’HOTE ................................... 82
1.1 Gènes de la réponse immunitaire de Sitophilus zeamais ............................................... 82 1.2 Gènes de la réponse immunitaire d’Acyrthosiphon pisum ............................................ 85 1.3 Analyse et caractérisation des GRI .............................................................................. 86
1.3.1 Analyse globale des banques soustraites .............................................................................86 1.3.1.1 Expression différentielle des EST de Sitophilus zeamais .....................................................87 1.3.1.2 Analyse de la banque d’Acyrthosiphon pisum ......................................................................90
1.3.2 Caractérisation des GRI de Sitophilus zeamais ....................................................................90 1.3.2.1 Obtention et analyse des séquences complètes des transcrits des GRI ..................................90 1.3.2.2 Etude du profil d’expression des GRI .................................................................................97 1.3.2.3 Mise en place d’une procédure adaptée à l’analyse des séquences des banques soustraites. ..99 1.3.2.4 Tentative d’identification des peptides antibactériens par une approche biochimique .........100
2. SYMBIOSE ET IMMUNITE CHEZ SITOPHILUS ZEAMAIS ..........................................................105 2.1 Expression des GRI de Sitophilus zeamais dans le bactériome larvaire .......................105 2.2 Caractérisation moléculaire du gène PGRP1 ..............................................................106
2.2.1 Expression du gène PGRP1 en relation avec la densité bactérienne. ..................................106 2.2.1.1 Mesure de la croissance bactérienne dans les larves ..........................................................107 2.2.1.2 Etude de l’expression du gène PGRP1 ..............................................................................108
2.2.2 Expression du gène PGRP1 au cours de développement de l’insecte..................................110 2.2.2.1 Expression du gène PGRP1 à différents stades de développement de l’hôte .......................110 2.2.2.2 Externalisation des endocytobiotes au stade nymphal ........................................................111
2.2.3 Epissage alternatif du gène PGRP1..........................................................................................113 2.2.3.1 Séquençage du gène PGRP1 .............................................................................................115 2.2.3.2 Quantification des deux transcrits alternatifs du gène PGRP1 ............................................116
DISCUSSION .................................................................................................................................................................. 118 1 LA REPONSE IMMUNITAIRE DE L’HOTE AUX BACTERIES GRAM (-) .........................................119 2 LA REPONSE IMMUNITAIRE DE S. ZEAMAIS AUX BACTERIES SYMBIOTIQUES ............................122
2.1 Réponse immunitaire et maintien de la symbiose dans le bactériome ...........................122 2.2 Externalisation des endocytobiotes et activation de la réponse immunitaire de l’hôte au
stade nymphal ...............................................................................................................128 2.3 Une réponse immunitaire spécifique............................................................................129
CONCLUSION................................................................................................................................................................ 131
REFERENCES BIBLIOGRAPHIQUES......................................................................................................................... 135
ANNEXES ....................................................................................................................................................................... 149
ANSELME Caroline 7 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Abréviations
aa : acide aminé ADN : Acide DésoxyriboNucléique ADNc : Acide DésoxyriboNucléique
complémentaire ADNg : Acide DésoxyriboNucléique génomique ADNr : Acide DésoxyriboNucléique ribosomal ANOVA : ANalysis Of Variance ARN : Acide RiboNucléique ARNm : Acide RiboNucléique messager BET : Bromure d’EThidium BF2I : Biologie Fonctionnelle, Insectes et
Interactions BLAST : Basic Local Alignment Search Tool BSA : Bovine Serum Albumine DAP : acide DiAminoPimélique DEPC : DiEthylPyroCarbonate DMSO : DiMéthylSulfOxyde DNase : DésoxyriboNucléase DTT : DiThioThréitol EDTA : acide EthylèneDiamineTétraAcétique EST : Expressed Sequence Tag gapdh : glycéraldéhyde-3-phosphate
déshydrogénase gb : GenBank GNBP: Gram-Negative bacteria Binding Protein GRI : Gène de la Réponse Immunitaire GSP: Gene Specific Primer HPLC : High Performance Liquid Chromatography hr : humidité relative IAGC : International Aphid Genomic Consortium IL-1R : InterLeukin-1 Receptor imd : immune deficiency IRAK : IL-1 Receptor Associated Kinase kb : kilobase LPS : LipoPolySaccharide MA : Million d’Années Mb : Mégabase NF-κB : Nuclear Factor-Kappa B
ORF : Open Reading Frame p : poids PAMP : Pathogen Associated Molecular Pattern PASS : Pea Aphid Secondary Symbiont PBS: Phosphate Buffer Saline PCR : Polymerase Chain Reaction PGN : PeptidoGlycaNPGRP : PeptidoGlycan Recognition Protein PO : PhénolOxydase PPAE (F) : ProPhenoloxydase Activating
Enzyme (Factor) PRR : Pattern-Recognition Receptor qsp: quantité suffisante pour RACE: Rapid Amplification of cDNA Ends RNAi : RiboNucleic Acid interference RNase : RiboNucléase RT : Reverse-Transcriptase SDS : Sodium Dodécyl Sulfate SOPE : Sitophilus oryzae Primary Endosymbiont sp : Swiss-Prot SPE : Sitophilus Primary Endosymbiont spp. : abréviation utilisée pour désigner plusieurs
espèces d’un même genre SSC : Saline Sodium Citrate SSPE : Saline Sodium Phosphate EDTA SSTT : Système de Sécrétion de Type Trois SZPE: Sitophilus zeamais Primary
Endosymbiont TAE : Tris Acétate EDTA TLR : Toll-Like Receptor TNF-α : Tumor Necrosis Factor alpha UPM : Universal Primers Mix v: volume
8

Introduction
ANSELME Caroline 9 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

INTRODUCTION
“On peut concevoir la symbiose, à l’origine, comme une infection bactérienne au sens pathologique du mot. La lente adaptation de ces bactéries à un même organisme a progressivement diminué leur virulence jusqu’à les rendre complètement inoffensives pour l’hôte.”
A. PAILLOT, “L’infection chez les insectes :
immunité et symbiose” (1933).
L’intérêt de cet entomologiste lyonnais pour l’étude des “maladies des in-sectes” est né en 1912, dans le cadre d’un projet visant à utiliser des parasi-tes microbiens dans la lutte contre les insectes nuisibles. Pensant qu’aucun résultat ne serait obtenu avec les microorganismes sans la connaissance des phénomènes pathologiques qu'ils entraînent, A. Paillot entreprend alors l’étude de l'immunité des insectes. Au cours de ses recherches, il constate “l’existence de processus différents de ceux qui ont été mis en évidence chez les vertébrés supérieurs”, processus qu’il décrit dans son livre “L'in-fection chez les Insectes” (Paillot, 1933).
A la même époque, P. Buchner publie un ouvrage recouvrant l’ensemble des connaissances acquises sur la symbiose chez les insectes (Buchner, 1930). Fasciné par l’origine bactérienne des symbiotes chez le puceron, A. Paillot consacre une partie de ses recherches à l’étude de la symbiose dans ce groupe d’insectes ravageurs des cultures. Non convaincu par le rôle des symbiotes dans le métabolisme de leur hôte, A. Paillot rap-proche les processus symbiotiques des processus infectieux. Il pense que la symbiose “ne devrait plus être considérée, chez les pucerons tout au moins, comme une association étroite de deux êtres vivants retirant chacun un égal bénéfice de la vie en commun, mais comme une infection bactérienne normale, équilibrée par des réactions d’immunité semblables à celles qui se produisent dans l’organisme des insectes inoculés avec des bacilles peu pathogènes”.
Bien qu’A. Paillot ait très largement sous-estimé l’impact des symbiotes sur les caractères de fitness de leur hôte, il a eu le mérite d’introduire dans la symbiose les notions d’infection et d’équilibre assuré par les réactions immunitaires. Ces notions, qui ont ouvert des perspectives nouvelles sur la conception générale de la symbiose, ont néanmoins été très peu abordées jusqu’à nos jours.
Chez les insectes, de nombreuses espèces nécessitent pour leur survie et leur reproduction, la présence de bactéries symbiotiques intracel-lulaires (ou endocytobiotes). Les bactéries sont localisées dans des cellules spécialisées, appelées bactériocytes, qui sont parfois groupées entre elles pour former un organe, le bactériome (Buchner, 1965). Ces vingt dernières
ANSELME Caroline 10 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

INTRODUCTION
années, la recherche sur les symbioses d’insectes s’est principalement foca-lisée sur la physiologie des interactions, la phylogénie des symbiotes et l’évolution moléculaire des génomes bactériens, en relation avec les contraintes du milieu intracellulaire. L’étude des endocytobiotes d’insectes a notamment permis d’étayer l’hypothèse de leur origine “pathogène”1 en raison de leur proximité phylogénétique avec des bactéries parasites ou pa-thogènes (Charles et al., 2001 ; Dale et al., 2002 ; Lefèvre et al., 2004).
Par ailleurs, deux études sur les interactions moléculaires hôte-symbiote, menées récemment chez le charançon Sitophilus zeamais et le puceron Acyrthosiphon pisum, ont mis en évidence, pour la première fois dans les symbioses d’insectes, la surexpression de gènes de la réponse im-munitaire dans les bactériocytes (Heddi et al., 2005 ; Nakabachi et al., 2005). La surexpression de ces gènes dans l’organe symbiotique suscite de nombreuses interrogations concernant le mode de perception des endocyto-biotes, leur persistance dans les bactériocytes, et le rôle éventuel de l’immunité innée dans le contrôle des endocytobiotes.
L’objectif de ma thèse était de caractériser les aspects immunitai-res de l’interaction hôte-symbiote, et leur évolution en relation avec l’évolution des capacités infectieuses des endocytobiotes. Dans ce travail, nous nous sommes d’abord attachés à identifier et caractériser, chez le cha-rançon S. zeamais et le puceron A. pisum, les gènes de la réponse immuni-taire par une approche de soustraction d’ADNc. L’analyse du profil d’expression des gènes de la réponse immunitaire dans le bactériome du charançon a ensuite permis d’aborder les mécanismes immunitaires qui permettraient le maintien de la symbiose. Cette analyse nous a également permis d’émettre l’hypothèse d’un contrôle de la localisation des endocyto-biotes par l’immunité de l’hôte. Enfin, j’ai complété cette étude par l’analyse d’un gène de la famille des “PeptidoGlycan Recognition Pro-teins” (PGRP), qui pourrait représenter un gène essentiel dans l’adaptation de la réponse immunitaire de l’hôte au maintien de la symbiose.
Pour introduire ce travail sur la symbiose et l’immunité de l’insecte, il me paraît nécessaire de présenter d’abord certains aspects de la symbiose intracellulaire et de décrire les mécanismes généraux de la ré-ponse immunitaire des insectes.
1 On désigne ici par “pathogène”, un microorganisme possédant les gènes nécessaires à l’invasion de
l’hôte et éventuellement, à la pathogénicité.
ANSELME Caroline 11 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
ANSELME Caroline 12 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
1 Contexte biologique : La symbiose
Le terme de symbiose a été introduit pour la première fois en biologie par Frank (1877) et De Bary (1879) pour désigner une association entre deux organismes spécifiquement distincts. Cette définition, très générale, inclut donc aussi bien les associations mutualistes, où les deux partenaires bénéfi-cient de l’association, que les associations parasitaires, où l’un des parte-naires vit aux dépens de l’autre. Depuis cette définition, le terme de sym-biose a engendré de nombreuses confusions, probablement du fait de l’utilisation de ce terme dans des expressions socioculturelles pour évoquer des relations plutôt harmonieuses. Il en résulte qu’au sein de la communau-té scientifique, la notion de symbiose est souvent rapprochée de celle de mutualisme et, de ce fait, opposée à celle de parasitisme (Nardon et Gre-nier, 1993). Dans cette thèse, je désignerai donc par le terme de symbiose, les associations qui se placent dans un continuum phénotypique allant du parasitisme au mutualisme.
1.1 Les différentes définitions et classifications de la symbiose
La symbiose est un phénomène largement répandu dans la nature et qui se manifeste par une grande diversité aussi bien dans les partenaires impli-qués, que dans les formes d’associations. Elle implique généralement des organismes phylogénétiquement distants, et l’association peut se limiter à deux partenaires (monosymbiose) ou impliquer plusieurs partenaires (pluri-symbiose). Il existe aussi des situations d’hypersymbiose, comme chez les aleurodes qui hébergent une β-protéobactérie à l’intérieur de laquelle vit une γ-protéobactérie (von Dohlen et al., 2001). Par ailleurs, on peut distin-guer les ectosymbioses, où le symbiote est à l’extérieur de son hôte, et les endosymbioses où le symbiote vit à l’intérieur de l’hôte. La localisation du symbiote dans l’organisme hôte donne lieu à une seconde distinction entre les endosymbioses extracellulaires comme la symbiose digestive, et les en-docytobioses, où le symbiote vit à l’intérieur des cellules de l’hôte (Schwemmler, 1980). Toutefois, le terme “endosymbiont” regroupe, dans la littérature anglophone, à la fois les endosymbiotes et les endocytobiotes, alors que les symbiotes du tube digestif qui sont extracellulaires sont quali-fiés de “gut symbionts”.
ANSELME Caroline 13 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
1.2 La symbiose intracellulaire chez les insectes
Il est difficile de dire avec précision quand les premiers insectes sont appa-rus, mais les éléments fossiles apportent la preuve de leur existence dès le dévonien (de -408 MA à -360 MA environ). Les insectes ont traversé plu-sieurs âges géologiques et constituent aujourd’hui le groupe animal le plus représenté sur terre. Il existe actuellement plus de 1,4 millions d’espèces d’insectes décrites, ce qui représente environ 80% de la biodiversité totale. Ce nombre qui est certainement sous-estimé, pourrait atteindre entre 30 et 50 millions d’espèces selon Erwin (1997).
Cette forte représentation des insectes résulte de leur capacité de diversification et d’adaptation à différentes niches écologiques, y compris celles qui sont nutritionnellement déficientes. La conquête de ces niches déficientes, comme la sève phloémienne des végétaux, les grains de céréa-les, ou encore le sang des mammifères, aurait été facilitée par la présence de bactéries symbiotiques intracellulaires (Buchner, 1965). Grâce aux nou-velles capacités physiologiques apportées par la bactérie, l’endocytobiose constitue un élément essentiel dans le pouvoir adaptatif des insectes. Au-jourd’hui, on estime à plus de 10% le nombre d’espèces qui dépendent d’endocytobiotes pour leur viabilité et leur reproduction (Buchner, 1965 ; Moran et Telang, 1998 ; Moran et Baumann, 2000).
Dans cette partie, il ne s’agit pas de développer de manière ex-haustive tous les aspects de la symbiose intracellulaire d’insectes, mais de présenter les aspects en relation avec le travail réalisé dans cette thèse. Ces aspects seront abordés essentiellement à travers le modèle Sitophilus-“Sitophilus Primary Endosymbiont” (SPE), mais les caractéristiques d’autres modèles de symbioses seront également évoquées.
1.2.1 Biologie de la symbiose intracellulaire chez les insectes
1.2.1.1 Niveau d’intégration des endocytobiotes
Chez les insectes, on distingue différents types d’endocytobioses bactériennes en fonction de la représentation des symbiotes dans les popu-lations d’insectes et de leur impact sur la physiologie et la reproduction de l’hôte.
ANSELME Caroline 14 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
1.2.1.1.1 Les symbioses primaires
La symbiose est qualifiée de primaire lorsque l’hôte dépend du symbiote pour sa survie et sa reproduction, et lorsque le symbiote est représenté dans toutes les populations de l’espèce d’insecte.
Parmi les caractéristiques de ces symbioses, il existe une localisa-tion des bactéries dans des cellules spécialisées appelées bactériocytes. La localisation des bactériocytes et leur organisation présentent une grande va-riabilité entre les différents groupes d’insectes (Buchner, 1965). Ainsi, chez les glossines (Aksoy, 2000) et les charançons (Nardon et Wicker, 1981), les bactériocytes sont regroupés pour former un organe, le bactériome. On ob-serve également un regroupement des bactériocytes chez les pucerons, mais la structure reste plus lâche (Buchner, 1965). Chez les fourmis et les blat-tes, en revanche, les bactériocytes ne sont pas regroupés en un organe mais disséminés entre les cellules intestinales et les cellules adipeuses, respecti-vement (Buchner, 1965). A l’intérieur des bactériocytes, les bactéries sont soit libres dans le cytoplasme comme chez les charançons, soit entourées d’une membrane de type vacuolaire appelée M3 comme chez les pucerons ; M1 et M2 étant les deux membranes des bactéries Gram (-).
L’autre caractéristique des endocytobiotes primaires est leur transmission maternelle. Le mode le plus fréquent est la transmission par voie ovocytaire, mais il existe d’autres modes comme la transmission par contamination intra-utérine chez les larves de glossines (Aksoy, 2000) ou encore la réinfection des jeunes embryons parthénogénétiques à un stade très précoce chez les pucerons (Buchner, 1965 ; Braendle et al., 2003 ; Miura et al., 2003). Les mécanismes de ces réinfections embryonnaires res-tent pour l’instant mal caractérisés. Il pourrait s’agir d’un phénomène pas-sif, par endocytose, ou actif, par l’utilisation d’un système bactérien impli-qué dans l’invasion cellulaire. Chez le puceron, la présence de la membrane M3 suggère que ce processus d’infection implique l’endocytose de Buchne-ra. Cette hypothèse semble confirmée par les résultats de microscopie élec-tronique obtenus récemment par l’équipe de T. Fukatsu (Koga et al., 2006).
1.2.1.1.2 Les symbioses secondaires
En plus des endocytobiotes primaires obligatoires, les insectes peuvent pos-séder des symbiotes dont la fonction et l’impact sur la physiologie de l’hôte restent mal élucidés. Ces symbiotes, dits secondaires ou facultatifs, ne sont pas présents dans toutes les populations d’insectes, sont transmis aussi bien verticalement qu’horizontalement et ne possèdent pas toujours de spécifici-té tissulaire. Chez les pucerons, les symbiotes secondaires occupent des
ANSELME Caroline 15 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
bactériocytes dits secondaires, différents des bactériocytes renfermant le symbiote primaire Buchnera (Fukatsu et al., 2000 ; Koga et al., 2003). Chez la glossine, le symbiote secondaire, Sodalis, infecte les cellules de di-vers tissus, principalement l’intestin, et peut aussi se retrouver libre dans l’hémolymphe de l’insecte.
1.2.1.1.3 Les symbioses à Wolbachia
De très nombreuses espèces d’insectes (15-20%), ainsi que d’autres inver-tébrés, vivent également en symbiose facultative avec Wolbachia. Cette bactérie, qui appartient au groupe des α-protéobactéries et qui est très pro-che des Rickettsia, est connue pour son impact sur la reproduction des in-vertébrés (Werren, 1997 ; Jeyaprakash et Hoy, 2000). Elle manipule la re-production de son hôte en faveur de sa transmission à la descendance, via quatre mécanismes connus : l’incompatibilité cytoplasmique, la parthéno-genèse, la féminisation des mâles génétiques et le “male-killing” (Breeuwers et Werren, 1990 ; Rigaud et al., 1991 ; Stouthamer et al., 1999 ; Fialho et Stevens, 2000).
Contrairement aux endocytobiotes primaires, Wolbachia infecte l’ensemble des tissus de son hôte. Elle se trouve néanmoins souvent à forte densité dans la lignée germinale, où elle exerce son rôle sur la reproduction (Dobson et al., 1999). Elle est transmise verticalement aux descendants, comme les symbiotes primaires, mais elle peut utiliser également des trans-ferts horizontaux pour infecter de nouvelles populations (Vavre et al., 1999). Bien que Wolbachia soit généralement sans effet sur la fitness de l’hôte, il existe toutefois quelques exemples de souches indispensables à la reproduction de leur hôte comme chez Asobara tabida (Hymenoptera, Braconidae) où Wolbachia est nécessaire à l’ovogenèse (Dedeine et al., 2001). Il existe également des souches pathogènes qui entraînent la mort de l’hôte (McGraw et O'Neill, 2004).
1.2.1.2 Rôle des endocytobiotes primaires, impact physiologique
Les premières études consacrées à la symbiose chez les insectes étaient es-sentiellement centrées sur des travaux histologiques. La symbiose était une curiosité biologique et les chercheurs, fascinés par les structures symbioti-ques, tentaient de décrire ces structures et de comprendre le processus de transmission des symbiotes à la descendance, ainsi que leur interaction avec l’embryogenèse.
Les études du rôle des endocytobiotes sur la physiologie et la reproduction de leur hôte, se sont développées plus récemment grâce à
ANSELME Caroline 16 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
l’obtention, en laboratoire, de souches dépourvues artificiellement de sym-biote (souches aposymbiotiques). La comparaison des souches apo- et sym-biotique a ainsi permis de mettre en évidence l’impact du symbiote sur les caractères de fitness de l’hôte. Chez le charançon Sitophilus oryzae par exemple, l’élimination de l’endocytobiote entraîne une augmentation de la durée de développement, une baisse de la fertilité et la perte de l’aptitude au vol chez les adultes (Nardon et Wicker, 1981 ; Grenier et al., 1994). Des perturbations physiologiques similaires ont aussi été observées chez d’autres insectes symbiotiques comme les blattes, les punaises ou les glos-sines. Chez le puceron, l’élimination des endocytobiotes entraîne, en plus, la stérilité de l’hôte (Sasaki et al., 1991 ; Rahbé et al., 1993 ; Iimura et al., 1995).
Puisque la plupart des insectes symbiotiques se développent dans des milieux pauvres ou déséquilibrés nutritionnellement, les causes de ces perturbations physiologiques ont été recherchées au niveau d’éventuelles complémentations nutritionnelles par le symbiote. Ainsi, grâce à des appro-ches biochimiques, il a été montré que les endocytobiotes fournissaient à leur hôte des nutriments comme les vitamines ou les acides aminés. Buchnera, le symbiote du puceron, fournit à son hôte les acides aminés es-sentiels comme la thréonine, la lysine et l’isoleucine dont est dépourvue la sève phloémienne (Sasaki et Ishikawa, 1994 ; Febvay et al., 1995). Wigglesworthia, le symbiote de la glossine, fournit également à son hôte les vitamines sous représentées dans le sang des mammifères (Aksoy, 2000). Chez Sitophilus, les endocytobiotes fournissent à leur hôte des vita-mines indispensables telles que la riboflavine, l’acide pantothénique et la biotine (Wicker, 1983). Par le biais de ces vitamines, il a été montré que le symbiote influence le métabolisme mitochondrial en augmentant les activi-tés enzymatiques spécifiques du cycle de Krebs et de la chaîne respiratoire (Heddi et al., 1993). Ces améliorations du métabolisme énergétique ont un impact direct sur l’aptitude au vol des souches symbiotiques (Grenier et al., 1994).
1.2.1.3 Aspects moléculaires des interactions hôte-symbiote
La première étude de l’interaction moléculaire hôte-symbiote au niveau des bactériocytes a été réalisée, chez le charançon, par une approche de sous-traction d’ADNc (Heddi et al., 2005). Grâce à cette approche, qui consiste à identifier les gènes de l’hôte surexprimés dans le bactériome, il a été montré qu’une proportion importante de gènes surexprimés correspond à des gènes impliqués dans le transport et le métabolisme des sucres. Ceci est probablement lié à une absorption importante de sucres par le bactériome qui est accolé au tube digestif. Une analyse par HPLC a permis d’étayer
ANSELME Caroline 17 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
cette hypothèse grâce à la mise en évidence d’une accumulation de fructose et de polyols (sorbitol et mannitol) dans le bactériome (Heddi et al., 2005). L’assimilation de ces sucres serait réalisée en partie par l’endocytobiote SPE qui a conservé les gènes impliqués dans le catabolisme des sucres dé-rivés des plantes et les gènes impliqués dans le transport des polyols (Rio et al., 2003 ; séquençage en cours du génome). L’approche soustractive a éga-lement mis en évidence la surexpression de gènes de stress et de gènes im-pliqués dans le trafic vésiculaire. La surexpression des premiers a été inter-prétée comme un moyen de limiter les effets nocifs du métabolisme intense des sucres et de la synthèse de réactifs oxygénés. Pour les seconds, ils se-raient impliqués dans l’exportation, vers l’insecte, des métabolites synthéti-sés dans les bactériocytes. Enfin, contrairement à ce qui a été décrit jusqu’à présent dans les autres modèles de symbiose, le bactériome des charançons du genre Sitophilus surexprime au moins un gène de la réponse immuni-taire. Il s’agit d’un gène de la famille des “PeptidoGlycan Recognition Pro-teins” (PGRP) homologue du PGRP-LB de Drosophila melanogaster (Heddi et al., 2005).
Une autre étude du profil transcriptionnel des bactériocytes a également été réalisée chez le puceron, par l’équipe de Fukatsu (Nakabachi et al., 2005), à travers une analyse des EST bactériocytaires par PCR quantitative en temps réel. En accord avec le rôle nutritionnel de Buchnera, plusieurs gènes impliqués dans l’utilisation des acides aminés essentiels synthétisés par le symbiote sont surexprimés dans les bactériocytes. En revanche, puisque le puceron fournit à Buchnera des acides aminés non es-sentiels que le symbiote ne peut pas synthétiser, ce sont les gènes impliqués dans la synthèse et non dans l’utilisation de ces derniers qui sont surexpri-més dans ces cellules. Les bactériocytes surexpriment également des gènes impliqués dans le transport mitochondrial et le transport vésiculaire, ce qui souligne le rôle des bactériocytes dans l’échange de métabolites et de subs-trats entre l’hôte et le symbiote. Enfin, le gène le plus exprimé dans les bactériocytes code un lysozyme de type i, une enzyme bactériolytique qui pourrait jouer un rôle dans le contrôle et le maintien des endocytobiotes (Nakabachi et al., 2005).
Les études d’Heddi et al. (2005) et de Nakabachi et al. (2005) rapportent des résultats similaires qui soulignent la surexpression de gènes impliqués dans le transport et dans le métabolisme des sucres et des acides aminés, respectivement, en relation avec le rôle nutritif des symbiotes. Ces deux études ont également mis en évidence la surexpression de gènes im-pliqués dans la réponse immunitaire de l’hôte, ce qui suggère l’implication des voies immunitaires dans le contrôle et le maintien des symbiotes dans le bactériome.
ANSELME Caroline 18 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
1.2.2 Evolution des génomes des bactéries symbiotiques intracellulaires
Ces dix dernières années, les données issues de la génomique et de la post-génomique ont permis de mettre en évidence que le génome des bactéries strictement intracellulaires est soumis à une évolution différente de celle du génome des bactéries libres.
1.2.2.1 Augmentation de la vitesse d’évolution
Le mode de vie strictement intracellulaire conduit à un isolement des endo-cytobiotes qui empêche toute recombinaison avec des bactéries libres. Par ailleurs, le mode de transmission maternel des endocytobiotes est responsa-ble d’un fort goulot d’étranglement : à chaque génération, seul un petit nombre de bactéries est transmis à la génération suivante pour engendrer la population bactérienne de la descendance. Associés au “cliquet de Muller” (c’est-à-dire l’accumulation de mutations de manière irréversible du fait de l’absence de recombinaison), ces goulots d’étranglement successifs vont accélérer les effets de la dérive génétique et favoriser la fixation de muta-tions délétères (Moran, 1996).
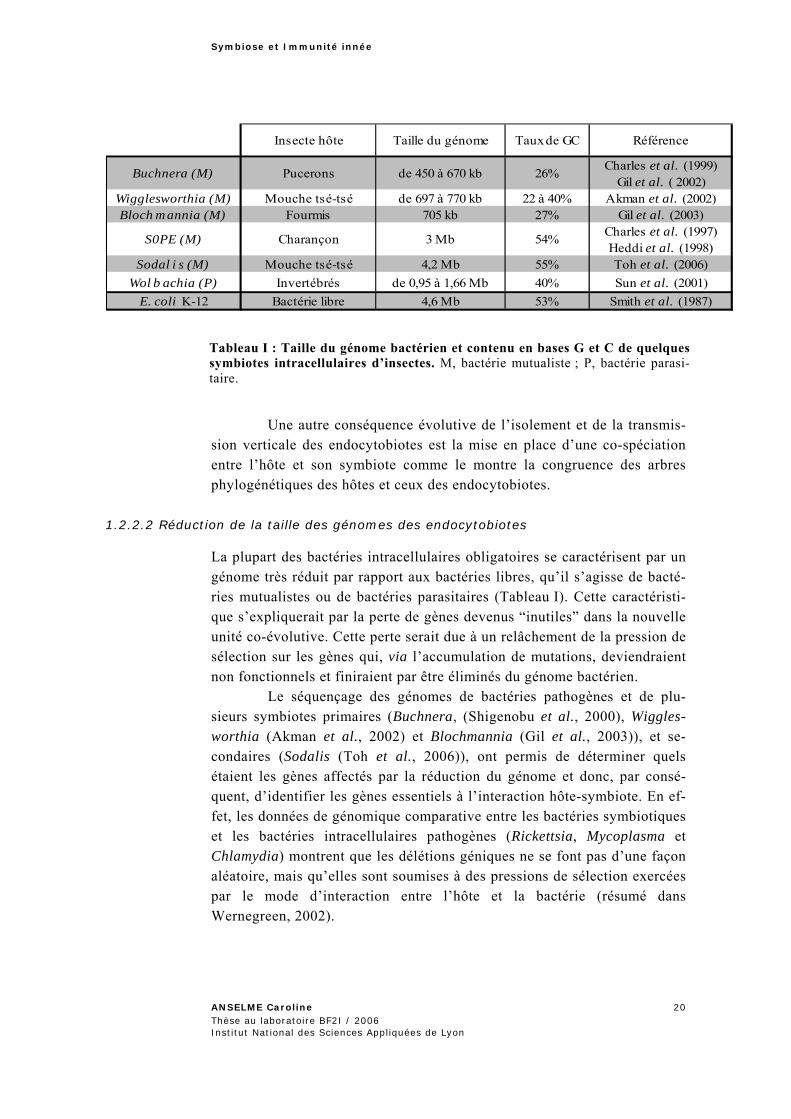
L’isolement des endocytobiotes dans les bactériocytes et leur transmission strictement maternelle augmentent donc de façon drastique la vitesse d’évolution des génomes, ce qui augmente l’influence des biais mu-tationnels. On observe ainsi chez toutes les bactéries intracellulaires, des compositions en bases A et T très fortes (Tableau I). Ce biais de composi-tion vers les bases A et T s’expliquerait par le moindre coût métabolique pour la synthèse de ces deux bases (Rocha et Danchin, 2002). Par ailleurs, on observe également une réduction de la taille des génomes chez ces bac-téries (Tableau I).
ANSELME Caroline 19 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
Symbiose et Immunité innée
(Charles et Ishikawa, 1999) (Gil et al., 2002) (Akman et al., 2002) (Gil et al., 2003) (Charles et al., 1997) (Heddi et al., 1998) (Toh et al., 2006) (Sun et al., 2001) (Smith et al., 1987)
I T T Rnsecte hôte aille du génome aux de GC éférence
arles et al. (1999)l et al. ( 2002)
Wigglesworthia (M) Mouche tsé-tsé de 697 à 770 kb 22 à 40% Akman et al. (2002)Bloch mannia (M) Fourmis 705 kb 27% Gil et al. (2003)
Charles et al. (1997) Heddi et al. (1998)
Sodal i s (M) Mouche tsé-tsé 4,2 Mb 55% Toh et al. (2006)Wol b achia (P) Invertébrés de 0,95 à 1,66 Mb 40% Sun et al. (2001)
E. coli K-12 ctérie libre 4,6 Mb 53% Smith et al. (1987)
Buchnera (M)
S0PE (M)
ucerons e 450 à 670 kb
Charançon 3 Mb 54%
ChP d 26%Gi
Ba
Tableau I : Taille du génome bactérien et contenu en bases G et C de quelques symbiotes intracellulaires d’insectes. M, bactérie mutualiste ; P, bactérie parasi-taire.
Une autre conséquence évolutive de l’isolement et de la transmis-
sion verticale des endocytobiotes est la mise en place d’une co-spéciation entre l’hôte et son symbiote comme le montre la congruence des arbres phylogénétiques des hôtes et ceux des endocytobiotes.
1.2.2.2 Réduction de la taille des génomes des endocytobiotes
La plupart des bactéries intracellulaires obligatoires se caractérisent par un génome très réduit par rapport aux bactéries libres, qu’il s’agisse de bacté-ries mutualistes ou de bactéries parasitaires (Tableau I). Cette caractéristi-que s’expliquerait par la perte de gènes devenus “inutiles” dans la nouvelle unité co-évolutive. Cette perte serait due à un relâchement de la pression de sélection sur les gènes qui, via l’accumulation de mutations, deviendraient non fonctionnels et finiraient par être éliminés du génome bactérien.
Le séquençage des génomes de bactéries pathogènes et de plu-sieurs symbiotes primaires (Buchnera, (Shigenobu et al., 2000), Wiggles-worthia (Akman et al., 2002) et Blochmannia (Gil et al., 2003)), et se-condaires (Sodalis (Toh et al., 2006)), ont permis de déterminer quels étaient les gènes affectés par la réduction du génome et donc, par consé-quent, d’identifier les gènes essentiels à l’interaction hôte-symbiote. En ef-fet, les données de génomique comparative entre les bactéries symbiotiques et les bactéries intracellulaires pathogènes (Rickettsia, Mycoplasma et Chlamydia) montrent que les délétions géniques ne se font pas d’une façon aléatoire, mais qu’elles sont soumises à des pressions de sélection exercées par le mode d’interaction entre l’hôte et la bactérie (résumé dans Wernegreen, 2002).
ANSELME Caroline 20 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
- gènes des voies métaboliques
Contrairement aux bactéries pathogènes dont les capacités métaboliques ont été largement altérées puisqu’elles vivent aux dépens de l’hôte, les endocytobiotes mutualistes ont conservé les fonctions de biosyn-thèse des constituants nécessaires à la survie et à la reproduction de leur hôte. Ils auraient, en revanche, perdu les gènes des voies métaboliques re-dondantes avec le métabolisme de l’hôte, ou inutiles par rapport au milieu nutritionnel dans lequel l’insecte se développe (Shigenobu et al., 2000 ; Rio et al., 2003 ; Toh et al., 2006). A titre d’exemple, Buchnera a perdu les gè-nes codant les voies de biosynthèse des acides aminés non essentiels pour le puceron, mais elle a conservé les voies de biosynthèse des acides aminés essentiels que le puceron ne peut pas synthétiser (Shigenobu et al., 2000). Un autre exemple concerne le symbiote secondaire de la glossine qui a per-du les gènes codant des enzymes glycolytiques, comme la galactosidase et la glucosidase, qui sont inutiles compte tenu du faible taux des substrats de ces enzymes dans le sang des mammifères (Toh et al., 2006).
La perte de gènes métaboliques contribue grandement à la dépen-dance de l’endocytobiote vis-à-vis de son hôte, ce qui explique, en partie, que la plupart des endocytobiotes ne soient pas cultivables in vitro.
- gènes impliqués dans les processus d’invasion cellulaire
Dans toutes les symbioses d’origine ancienne, les génomes bacté-riens semblent être dépourvus des systèmes de sécrétion et des gènes de virulence qui leur sont associés (Shigenobu et al., 2000 ; Akman et al., 2002 ; Gil et al., 2003). La délétion de ces gènes serait le résultat d’un relâchement de la pression de sélection sur le pouvoir infectieux de la bactérie. Toutefois, la transmission de certains symbiotes d’insecte d’une génération à l’autre, implique des phases d’infection (ou de réinfection) de certaines cellules comme chez les glossines et les pucerons. Il a de ce fait été suggéré que chez ces symbiotes, le système flagellaire pourrait assurer le rôle de système de sécrétion (Young et al., 1999 ; Akman et al., 2002 ; Majander et al., 2005). En effet, malgré son génome très réduit, Wigglesworthia aurait conservé l’ensemble des gènes constituant le système flagellaire, alors que ni flagelle, ni motilité n’ont été observés chez ce symbiote. Chez Buchnera, le système flagellaire est également conservé mais il est dépourvu du filament responsable de la motilité, et il a été suggéré que ce système pourrait servir de transporteur (Shigenobu et al., 2000). Chez les endocytobiotes, le système flagellaire pourrait donc sécréter des protéines impliquées dans les processus d’infection des bactériocytes.
ANSELME Caroline 21 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Dans le cas d’associations plus récentes, où les symbiotes peuvent présenter des phases extracellulaires, le génome bactérien possède des sys-tèmes d’invasion similaires aux systèmes employés par les bactéries patho-gènes. A titre d’exemple, le génome du symbiote secondaire de la glossine, Sodalis, code trois systèmes de sécrétion de type III (SSTT), similaires des SSTT de Yersinia enterocolitica et Salmonella spp., comprenant les protéi-nes de structure, les régulateurs, ainsi que certains effecteurs. Chez Sodalis, une étude préliminaire de deux SSTT a mis en évidence le rôle du système codé par la “Sodalis symbiosis Region-1” (SSR-1) dans l’invasion cellu-laire, et celui du système codé par la SSR-2 dans la survie intracellulaire du symbiote, deux processus nécessaires au maintien de cette symbiose se-condaire (Dale et al., 2005 ; Toh et al., 2006).
- gènes de la paroi bactérienne
Les composants de la paroi bactérienne constituent les principaux éléments qui sont reconnus par le système immunitaire de l’hôte. Ces élé-ments varient d’une bactérie à l’autre et, de ce fait, peuvent engendrer dif-férentes réponses immunitaires (voir chapitre “Immunité”). Ainsi, pour échapper au système immunitaire de l’hôte, les pathogènes intracellulaires investissent une proportion considérable de leur génome réduit pour synthé-tiser une paroi élaborée et limiter leur reconnaissance par l’hôte.
Dans le cas des endocytobiotes, le séquençage du génome de Wigglesworthia révèle que cet endocytobiote primaire de la glossine a conservé les gènes de la synthèse de la paroi bactérienne (Akman et al., 2002) alors que chez Buchnera, les gènes codant certains éléments de la pa-roi comme les lipopolysaccharides et les phospholipides, sont absents du génome (Shigenobu et al., 2000). Shigenobu et al. (2000) suggèrent que la délétion de ces gènes pourrait être liée à la séquestration de Buchnera dans une membrane vacuolaire d’origine eucaryotique (M3) qui isole la bactérie de l’environnement extérieur.
1.3 Caractéristiques du modèle Sitophilus spp.
Les charançons (Curculionidea) constituent la plus grande superfamille de coléoptères. Le genre Sitophilus appartient à la famille des Dryophthoridae qui compte plus de 140 genres et 500 espèces. Les insectes de cette famille se développent sur diverses parties des plantes comme les tiges, les stipes ou les graines. Bien que la symbiose soit très répandue dans ce groupe d’insectes (Lefèvre et al., 2004), les charançons des céréales du genre Sito-philus sont les mieux caractérisés de par leur caractère cosmopolite et leur importance économique. En effet, les trois espèces céréalières, Sitophilus oryzae, Sitophilus zeamais et Sitophilus granarius, sont responsables d’une
ANSELME Caroline 22 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
perte de denrées stockées qui peut atteindre 40% dans les pays tropicaux (Grenier et al., 1986).
La symbiose dans le genre Sitophilus a été découverte par Pieran-toni en 1927. Depuis, de nombreux auteurs ont participé à la caractérisation de ce modèle de symbiose (Mansour, Scheinert, Murray et Tiggs, Mus-grave, Nardon…) (Nardon et Grenier, 1988). Les charançons du genre Sito-philus, à l’exception d’une espèce, Sitophilus linearis, vivent tous en sym-biose intracellulaire avec une γ-protéobactérie nommée SPE pour “Sitophilus Primary Endosymbiont” (Heddi et al., 1998). Les bactéries sont localisées, chez les larves, dans le bactériome situé à la jonction de l’intestin antérieur et de l’intestin moyen, et chez les adultes, dans les bac-tériomes des cæcums mésentériques. On les trouve également chez les fe-melles, dans des bactériomes situés à l’apex des ovarioles (Figure 1).
Les différentes études moléculaires n’ont pas identifié de sym-biote secondaire chez Sitophilus, mais plus de la moitié des souches étu-diées hébergent Wolbachia (Heddi et al., 1999). Contrairement au symbiote SPE, Wolbachia est présente dans tous les tissus de l’insecte, y compris les bactériocytes. Cette bactérie, comme chez la plupart des insectes, provoque des incompatibilités cytoplasmiques partielles (Heddi et al., 1999).
1.3.1 Développement de l’insecte et description des structures symbiotiques
Les trois espèces céréalières de Sitophilus présentent des cycles de déve-loppement similaires. Grâce à leur rostre, les femelles perforent le grain de céréale et y déposent leurs œufs. Le trou est ensuite rebouché par une sé-crétion mucilagineuse de l’oviducte qui durcit rapidement à l’air. Le déve-loppement des quatre stades larvaires et du stade nymphal se déroule à l’intérieur du grain et l’adulte émerge quelques jours après la nymphose. Pendant les vies larvaire et nymphale, le grain apporte à l’insecte à la fois nourriture et protection.
ANSELME Caroline 23 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Figure 1 : Localisation des endocytobiotes chez Sitophilus. Chez le charançon, les symbiotes sont localisés dans un organe spécialisé appelé bactériome. ia, intes-tin antérieur ; im, intestin moyen ; ip, intestin postérieur ; cm, cæcums mésentéri-ques ; o, ovarioles ; r, rostre.
1.3.1.1 Transmission des symbiotes et embryogenèse
L’endocytobiote SPE est présent d’une façon permanente dans les ovaires du charançon (Pierantoni, 1927 ; Mansour, 1930 ; Murray et Tiegs, 1935 ; Nardon et Wicker, 1981). Au cours de l’embryogenèse, la lignée germinale qui est située au pôle postérieur de l’embryon est contaminée très tôt par des symbiotes (Nardon, 1971). Ainsi, au moment de leur formation, les cel-lules entraînent des symbiotes mêlés au déterminant germinal. Si les sym-biotes persistent dans la lignée germinale femelle, ils sont en revanche éli-minés de la lignée germinale mâle par un processus encore méconnu (Musgrave et Miller, 1953 ; Nardon et al., 1998). En plus de la lignée ger-minale, les symbiotes contaminent également une lignée somatique qui se différencie, au troisième jour de développement (à 27,5°C et 70% hr), en bactériocytes localisés au niveau du tube digestif antérieur de la future larve.
1.3.1.2 Le développement du bactériome larvaire
Chez la larve, le bactériome apparaît sous forme d’un collier bilobé à la jonction de l’intestin antérieur (stomodeum) et de l’intestin moyen (mésen-téron) (Figure 1 et Figure 2-A). Il est constitué de bactériocytes et de cellu-les interstitielles, dont la cohésion est assurée par une enveloppe constituée de cellules aplaties. Bien que le bactériome soit accolé à l’intestin, il ne semble pas posséder de connexion avec le tube digestif (Nardon et Wicker, 1981).
Au cours des quatre stades larvaires, la taille du bactériome aug-mente par mitose (Murray et Tiegs, 1935) et polyploïdisation, d’environ
ANSELME Caroline 24 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
quatre à cinq fois. Dans le même temps, le nombre de bactéries symbioti-ques passe de 2×104 dans l’œuf à 3×106 dans la larve de quatrième stade (Nardon, 1978). Ceci correspond à sept cycles de divisions bactériennes dans une période de vingt jours, si on considère que toutes les bactéries persistent durant la vie larvaire. La multiplication des symbiotes semble donc très limitée, en comparaison avec une bactérie libre comme E. coli. Toutefois, ce faible nombre de divisions est à relativiser du fait de l’augmentation de la taille des bactéries. En effet, les endocytobiotes pri-maires chez S. oryzae (SOPE) et S. granarius (SGPE) sont très pleïomor-phes, avec une taille variant de 5 à 30 µm de longueur. Ils sont plus ou moins flexueux et peuvent aussi se présenter sous forme de chaînettes (Figure 2-B). Chez S. zeamais (SZPE), le caractère flexueux s’accentue et les symbiotes se présentent sous forme hélicoïdale, la spirale pouvant dé-passer 10 µm de long (Figure 2-B).
Le pleïomorphisme de taille et la forme des endocytobiotes pour-raient être liés à une inhibition de la septation des endocytobiotes au cours de leur développement dans le bactériome larvaire. Cette hypothèse n’a ce-pendant pas encore été vérifiée.
trachée
100 µm
10 µm
10 µm
A) B)trachée
100 µm
10 µm
10 µm
A) B)
Figure 2 : Bactériome larvaire et pleïmorphisme des endocytobiotes chez Sito-philus. A) Bactériome larvaire observé sous la loupe binoculaire. B) Endocytobio-tes de S. zeamais (en haut) et S. oryzae (en bas) après coloration de Gram.
1.3.1.3 Développement des bactériomes des cæcums mésentériques
Pendant la nymphose, le bactériome se désintègre et libère les bactériocytes dont une partie va dégénérer, et l’autre va s’insinuer entre l’intestin imagi-nal en cours de formation et sa tunique musculaire (Nardon et Grenier, 1989). Les bactériocytes vont ainsi migrer vers l’arrière du corps de l’insecte et s’incorporer dans les cæcums mésentériques au cours de leur formation (Pierantoni, 1927 ; Tarsia In Curia, 1933 ; Murray et Tiegs, 1935 ; Nardon et Nardon, 1998).
ANSELME Caroline 25 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Très abondants chez les jeunes adultes, les endocytobiotes dispa-raissent chez les adultes âgés de plus de trois semaines par un mécanisme inconnu. Néanmoins, ils demeurent chez les femelles dans des bactériomes situés à l’apex des ovarioles et dans les ovocytes, ce qui assure leur trans-mission à la génération suivante.
1.3.2 Quelques aspects de l’interaction hôte-symbiote
1.3.2.1 Devenir des structures symbiotiques chez les insectes aposymbiotiques
Comme cela a déjà été évoqué, il existe certaines souches de charançons naturellement non symbiotiques, mais il est aussi possible d’obtenir en la-boratoire des souches aposymbiotiques par traitement à la chaleur ou à l’aide d’antibiotique.
Dans une souche non symbiotique de S. granarius découverte en Egypte, Mansour (1927, 1935) décrit la présence d’un bactériome parfaite-ment formé, mais moins volumineux que celui des souches symbiotiques. Nous avons également constaté la présence d’un bactériome larvaire rési-duel dans une souche aposymbiotique de S. oryzae obtenu récemment au laboratoire. Toutefois, il semblerait que ce bactériome se perde au cours des générations puisque les souches aposymbiotiques plus anciennes sont totalement dépourvues de bactériome larvaire. La présence des symbiotes ne semble donc pas être nécessaire à la différenciation du bactériome lar-vaire mais plutôt au maintien de cet organe au cours des générations suc-cessives. Néanmoins, les endocytobiotes sont indispensables à la différen-ciation de l’organe symbiotique dans les phases précoces de l’association. Aussi, la différenciation des bactériocytes observées en leur absence pour-rait être due à un “signal moléculaire”, qui serait transmis maternellement au cours des premières générations puis qui finirait par disparaître.
Des observations similaires ont été faites chez les pucerons non symbiotiques où les bactériocytes se différencient et persistent à l’échelle de l’individu, mais disparaissent au cours de l’évolution (Braendle et al., 2003).
1.3.2.2 Contrôle de la densité bactérienne par l’hôte
Le dénombrement des bactéries symbiotiques dans les ovaires de S. oryzae révèle que le nombre d’endocytobiotes varie d’une souche à l’autre mais reste relativement stable dans une souche donnée. A partir de cette consta-tation, Nardon et al. (1998) ont émis l’hypothèse selon laquelle le nombre de symbiotes serait contrôlé génétiquement par l’hôte. Des expériences de
ANSELME Caroline 26 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
croisements entre une souche à forte densité bactérienne (110 000 ± 4 000) et une souche à faible densité bactérienne (41 200 ± 3 000) ont montré que les individus de la génération F1 présentaient une densité bactérienne avoi-sinant la moyenne des souches parentales (75 000 ± 5 000). Ce résultat, ap-puyé par des expériences de rétrocroisements et d’irradiation, suggère que l’hôte contrôle la densité de la population bactérienne par un facteur chro-mosomique (Nardon et al., 1998).
1.3.3 Aspects évolutifs de la symbiose chez Sitophilus
1.3.3.1 Caractéristiques du génome de SOPE
L’étude des endocytobiotes de charançon du genre Sitophilus a principale-ment été réalisée chez l’espèce S. oryzae. Contrairement aux autres symbio-tes primaires d’insectes, le génome de SOPE semble présenter des caracté-ristiques proches de celles des bactéries libres. En effet, le génome de SOPE est peu réduit, avec une taille estimée à 3 Mb (Charles et al., 1997), et un taux en bases GC de 54% (Heddi et al., 1998). Par ailleurs, le génome de SOPE code un système de sécrétion de type III fonctionnel, similaire à celui de la bactérie pathogène Salmonella (Dale et al., 2002).
Compte tenu de ces particularités, un projet de séquençage com-plet du génome de SOPE a été entrepris en collaboration avec A. Latorre et A. Moya à l’université de Valence (Espagne). Plus de la moitié du génome est actuellement séquencée. Une analyse préliminaire montre que SOPE au-rait conservé plusieurs gènes impliqués dans les voies métaboliques, la vi-rulence, la motilité, le stress… Par ailleurs, plus de 20% des séquences correspondent à des séquences d’insertion ce qui témoigne d’une forte plas-ticité génomique dans les phases précoces de la symbiose intracellulaire.
1.3.3.2 Histoire évolutive de la symbiose chez les Dryophthoridae
Les caractéristiques du génome de SOPE, proches de celles des bactéries libres, suggèrent une association récente entre SPE et Sitophilus. En revan-che, sa transmission maternelle, sa localisation dans les bactériomes et son rôle dans la physiologie de l’hôte évoquent plutôt une longue co-évolution entre les deux partenaires. L’apparente contradiction entre les caractéristi-ques du génome de SOPE et celles de l’association S. oryzae-SOPE a été clarifiée par l’étude des aspects évolutifs de la symbiose chez les Dryoph-thoridae (Lefèvre et al., 2004).
L’étude phylogénétique basée sur l’ADN 16S des bactéries sym-biotiques révèle que, dans la plupart des familles d’insectes, la symbiose
ANSELME Caroline 27 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
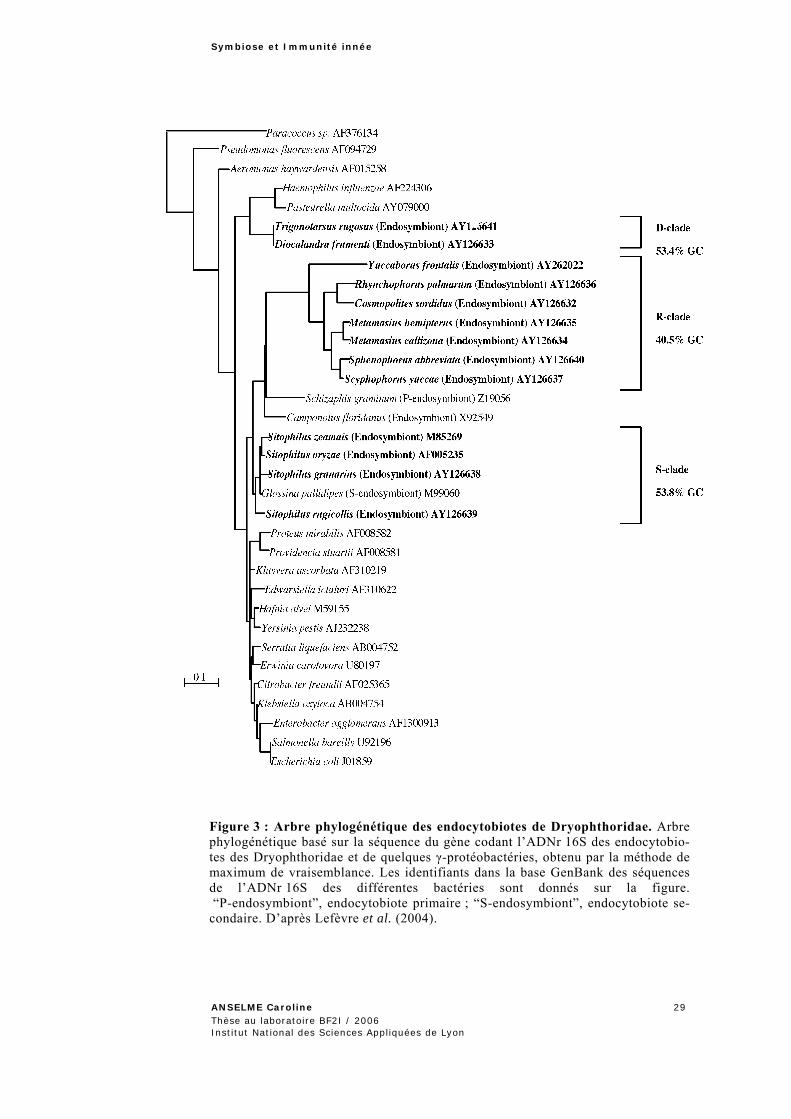
s’est établie lors d’une infection bactérienne ancestrale unique qui a ensuite conduit à une divergence parallèle des hôtes et des symbiotes (co-spéciation). Dans la famille des Dryophthoridae, en revanche, l’étude phylogénétique des bactéries symbiotiques a mis en évidence l’existence de trois clades d’endocytobiotes nommés D (Diocalandra), R (Rhynchophorus) et S (Sitophilus) (Lefèvre et al., 2004). Les endocyto-biotes des Dryophthoridae dériveraient donc de trois espèces bactériennes distinctes ayant établi une symbiose avec trois espèces ancêtres distinctes (Figure 3).
Les endocytobiotes du clade R montrent les caractéristiques évolu-tives associées aux symbioses anciennes. Ils présentent un taux de substitu-tion relatif élevé et leur génome est riche en bases AT (59,6% au niveau de l’ADN 16S). Par ailleurs, la congruence entre l’arbre des hôtes et celui des endocytobiotes de ce clade traduit une évolution parallèle entre les insectes et leur symbiote. Les endocytobiotes du clade R seraient de ce fait les des-cendants actuels directs de l’endocytobiote ancestral. D’après les données fossiles d’insectes, la symbiose entre l’ancêtre des endocytobiotes du clade R et l’ancêtre des Dryophthoridae se serait établie il y a près de 100 MA (O’Meara et Farell, communication personnelle). Le nom de Candidatus Nardonella a été proposé pour cet endocytobiote ancestral, en l’honneur du Professeur Nardon (Lefèvre et al., 2004).
Les endocytobiotes des clades D et S présentent, quant à eux, des taux de GC proches des bactéries libres (Figure 3). Les endocytobiotes de ces clades se sont donc associés plus récemment, probablement suite au remplacement d’endocytobiotes plus anciens. Dans le cas du clade S, l’âge de la symbiose a été estimé autour de 25 MA.
Le remplacement des endocytobiotes pourrait être une consé-quence de la réduction drastique des génomes bactériens intracellulaires, et donc de la perte de leurs capacités évolutives qui pourrait favoriser l’acquisition de nouveaux potentiels évolutifs à travers le recrutement d’autres symbiotes. La présence de symbiotes secondaires plus compétitifs pourrait alors aboutir à l’élimination du symbiote primaire.
ANSELME Caroline 28 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
Symbiose et Immunité innée
Figure 3 : Arbre phylogénétique des endocytobiotes de Dryophthoridae. Arbre phylogénétique basé sur la séquence du gène codant l’ADNr 16S des endocytobio-tes des Dryophthoridae et de quelques γ-protéobactéries, obtenu par la méthode de maximum de vraisemblance. Les identifiants dans la base GenBank des séquences de l’ADNr 16S des différentes bactéries sont donnés sur la figure. “P-endosymbiont”, endocytobiote primaire ; “S-endosymbiont”, endocytobiote se-condaire. D’après Lefèvre et al. (2004).
ANSELME Caroline 29 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Il existe de plus en plus d’arguments en faveur de l’existence de remplacement des symbiotes primaires par les symbiotes secondaires dans d’autres modèles. Par exemple, Koga et al. (2003) ont clairement montré que chez les pucerons, l’endocytobiote secondaire PASS (“Pea Aphid Secondary Symbiont”) pouvait, en conditions de laboratoire, remplacer Buchnera. En effet, des injections de PASS dans des pucerons mono-symbiotiques ont mis en évidence une compétition entre les symbiotes qui aboutit à l’élimination de Buchnera. De plus, lors d’une injection dans une souche dépourvue de Buchnera, PASS infecte le cytoplasme des bactériocytes et compense partiellement l’absence du symbiote primaire, en termes de survie et de reproduction de l’insecte, pendant quelques générations. L’étude phylogénétique menée par Lefèvre et al. (2004) révèle également la proximité phylogénétique des endocytobiotes du clade D et des bactéries parasitaires des genres Haemophilus et Pasteurella, ce qui étaye l’hypothèse de l’origine “pathogénique” de la symbiose. En effet, l’acquisition de symbiotes sous-entend que le futur endocytobiote ait des capacités “pathogéniques” pour pouvoir envahir le milieu cellulaire sans être éliminé par le système immunitaire de l’hôte (Charles et al., 2001). Cette hypothèse est également soutenue par la présence d’un système de sécrétion de type III fonctionnel chez Sodalis et SOPE qui pourraient donc avoir conservé des capacités infectieuses (Dale et al., 2002).
L’étude de la symbiose chez les Dryophthoridae révèle donc que
la symbiose est très ancienne dans cette famille d’insectes, mais que l’association Sitophilus-SPE est néanmoins plus récente, probablement suite à un remplacement de symbiotes. Les structures symbiotiques obser-vées chez Sitophilus spp. sont donc le résultat d’une longue co-évolution avec une bactérie symbiotique ancêtre, aujourd’hui remplacée par SPE. Ce remplacement ayant eu lieu il y a moins de 25 MA, le génome de SPE ne présente pas une réduction de taille et un biais vers les bases A+T aussi drastiques que ceux des génomes des endocytobiotes établis depuis plus de 100 MA.
2 L’immunité innée
Les êtres vivants évoluent dans des milieux environnementaux “hostiles” comportant de nombreux microorganismes, véritables menaces pour leur survie. Aussi, les mécanismes visant à les protéger des invasions par les microorganismes sont d’une importance primordiale pour leur développe-ment et leur évolution. Ces mécanismes sont regroupés au sein d’une fonc-
ANSELME Caroline 30 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
tion assurée par le système immunitaire. L’immunité comporte d’une part la reconnaissance des éléments étrangers, ce qui implique la capacité à re-connaître le Soi du Non-Soi, et d’autre part, la mise en place de voies cellu-laires ou humorales, visant à éliminer de l’organisme ces éléments étran-gers. Deux types d’immunité sont à distinguer : l’immunité innée et l’immunité acquise (ou adaptative). La première fait intervenir, pour la re-connaissance et l’élimination des corps étrangers, des facteurs codés dans la lignée germinale, alors que la seconde produit, par des réarrangements géniques somatiques, des récepteurs qui reconnaissent des antigènes spéci-fiques et qui permettent à l’organisme de développer une mémoire immuni-taire (Fearon, 1997). Ces deux types d’immunité ne sont pas indépendants puisque l’immunité innée est essentielle pour l’activation des premières étapes de l’immunité acquise (Fearon, 1999).
L’immunité acquise a suscité beaucoup d’intérêt dans le passé du fait de sa spécificité et de son efficacité. Seulement, étant apparue chez les ancêtres des poissons cartilagineux (Hoffmann et Reichhart, 2002), l’immunité acquise est absente chez de nombreux organismes (au moins 80% des espèces) qui ne possèdent pas les enzymes nécessaires aux réar-rangements géniques somatiques (Ferrandon et Royet, 2002). Bien que ces organismes ne disposent que de l’immunité innée, ils présentent une effica-cité remarquable dans la lutte contre les agents pathogènes, ce qui laisse supposer que l’immunité innée joue un rôle plus important que celui qui lui a été attribué par le passé. Récemment, les études des mécanismes de la ré-ponse immunitaire de la drosophile ont permis d’identifier de nombreuses familles de protéines qui s’avèrent être bien conservées au cours de l’évolution, puisque de nombreuses homologies ont été observées notam-ment avec les mammifères (Kang et al., 1998 ; Horng et Medzhitov, 2001 ; Vasselon et Detmers, 2002) et les végétaux (Kistner et Parniske, 2002).
Chez les invertébrés, les premières défenses immunitaires sont assurées par des barrières physiques telles que la cuticule et les épithéliums de l’intestin, de la trachée ou du tractus génital (Figure 4). Certaines cellu-les épithéliales ont également la capacité de sécréter des peptides antimi-crobiens qui permettent d’éliminer les microorganismes pathogènes (Ferrandon et al., 1998 ; Tzou et al., 2000). Lorsque ces derniers réussis-sent à échapper à ces peptides et qu’ils pénètrent dans l’hémocoele, deux voies de défense se mettent en place : une voie cellulaire et une voie humo-rale systémique qui coopèrent pour combattre l’infection (Braun et al., 1998 ; Basset et al., 2000 ; Elrod-Erickson et al., 2000 ; Lavine et Strand, 2002) (Figure 4).
La voie cellulaire implique les hémocytes, parmi lesquels se dis-tinguent les plasmatocytes qui phagocytent les bactéries, les lamellocytes
ANSELME Caroline 31 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
qui forment une capsule autour des particules trop volumineuses pour être phagocytées, et les cellules à cristaux qui interviennent dans la synthèse de mélanine lors de l’encapsulation et de la cicatrisation (Meister, 2004). La voie systémique humorale, quant à elle, comprend d’une part des cascades protéolytiques non spécifiques (coagulation et mélanisation locales), et d’autre part, la synthèse et la sécrétion dans l’hémolymphe de peptides an-timicrobiens par le corps gras (analogue fonctionnel du foie des vertébrés) (Hoffmann, 2003).
Compte tenu de la diversité et de la complexité des mécanismes de l’immunité innée, j’ai choisi de n’exposer dans cette partie bibliographique que les voies immunitaires en relation avec les travaux réalisés dans cette thèse. Je décrirai donc principalement la réponse humorale, systémique ou locale, et plus particulièrement la synthèse des peptides antibactériens régu-lée par les voies Toll et Imd (“Immune deficiency”) décrites chez la droso-phile. Je rappellerai néanmoins succinctement quelques notions sur la ré-ponse cellulaire et j’évoquerai autant que possible les éléments de la réponse immunitaire décrits chez d’autres insectes.
Figure 4 : La réponse immunitaire chez la drosophile. Lorsqu’un corps étranger comme un microorganisme franchit les premières barrières physiques de l’insecte et pénètre dans l’hémocoele, il induit un ensemble de réactions de la réponse immuni-taire. Une coagulation et une mélanisation locale au niveau des tissus endommagés vont permettre de limiter la perte d’hémolymphe. Des peptides antimicrobiens syn-thétisés par les épithéliums, le corps gras et les hémocytes vont assurer l’élimination du microorganisme. Ces microorganismes peuvent également être phagocytés par les plasmatocytes. Si le corps étranger est trop volumineux pour être phagocyté, comme dans le cas d’un œuf de parasitoïde, celui-ci est encapsulé par les lamellocytes et la capsule est ensuite mélanisée. Ce schéma général de la ré-ponse immunitaire est probablement similaire pour l’ensemble des insectes holométaboles.
ANSELME Caroline 32 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
2.1 La réponse cellulaire
Chez la drosophile, la réponse cellulaire repose sur trois types d’hémocytes : les plasmatocytes, équivalents des macrophages des mammi-fères, les lamellocytes et les cellules à cristaux qui n’auraient, quant à eux, pas d’équivalents (Meister, 2004).
Une première population d’hémocytes se différencie au cours de l’embryogenèse puis, une deuxième population se différencie, chez la larve, à partir de l’organe hématopoïétique. Cet organe disparaît ensuite pendant la métamorphose et seuls les plasmatocytes d’origines embryonnaire et lar-vaire persistent chez l’adulte (Lanot et al., 2001 ; Holz et al., 2003). Chez les larves, la population hémocytaire comprend majoritairement des plas-matocytes et, en plus faible quantité, des cellules à cristaux (5 %). En re-vanche, les lamellocytes sont rarement représentés dans les larves saines. Ces cellules plates et adhérentes se différencient massivement à la suite d’une infection par des parasites (Lanot et al., 2001). Chez les adultes, les hémocytes sont peu nombreux du fait de l’absence d’organe hématopoïéti-que.
2.1.1 La phagocytose par les plasmatocytes
Les plasmatocytes seraient responsables de la phagocytose, non seulement des cellules étrangères (Figure 4), mais également des cellules apoptotiques et des débris cellulaires. Ce processus emprunterait des mécanismes simi-laires aux macrophages des mammifères (Ramet et al., 2002b). Les plasma-tocytes joueraient donc un rôle important dans la réponse immunitaire, le développement et l’homéostasie des tissus.
La phagocytose est initiée par la reconnaissance de la cellule (ou de la particule) cible et sa liaison à la cellule phagocytaire. Il s’ensuit une réorganisation du cytosquelette permettant l’internalisation de la cible et son transport vésiculaire jusqu’au phagosome, où elle est éliminée. L’étape de reconnaissance implique des récepteurs qui peuvent se lier directement aux éléments de la surface du microorganisme ou se lier à des opsonines, qui sont des protéines qui entourent et, de ce fait, “marquent” la cible (Stuart et Ezekowitz, 2005).
ANSELME Caroline 33 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
2.1.2 L’encapsulation par les lamellocytes
L’encapsulation semble être une réponse spécifique des invertébrés. Elle se met en place lorsque des corps étrangers, trop volumineux pour être phago-cytés, sont présents dans l’organisme (Meister, 2004). C’est notamment le cas de l’encapsulation des œufs d’hyménoptères parasitoïdes par les diptè-res et les lépidoptères (Schmidt et al., 2001). Quand les plasmatocytes cir-culants détectent la présence d’un œuf d’hyménoptère dans la larve, il s’ensuit une différenciation massive des lamellocytes (Lanot et al., 2001). Les lamellocytes libérés dans l’hémolymphe forment une capsule autour de l’œuf qui va ensuite noircir, probablement suite à l’accumulation de méla-nine (Figure 4). L’élimination du parasite serait alors due soit à une asphyxie, soit aux intermédiaires toxiques synthétisés lors de la production de mélanine (§ 2.3).
2.1.3 Les cellules à cristaux et la mélanisation
Les cellules à cristaux jouent un rôle de stockage à l’interface des réponses cellulaire et humorale (Meister, 2004). En effet, les inclusions cristallines de ces cellules sont en fait une forme cristallisée de la prophénoloxydase. Cette enzyme intervient dans la synthèse de mélanine dans l’hémolymphe (§ 2.3), au niveau d’une blessure ou de la capsule formée par les lamellocy-tes. Bien que la phagocytose, l’encapsulation et la synthèse de la prophéno-loxydase constituent les fonctions majeures des hémocytes, ces cellules in-terviennent également dans d’autres fonctions, souvent en interaction avec la réponse humorale. Les hémocytes interviennent notamment dans la réac-tion de coagulation qui permet de limiter la perte d’hémolymphe et la dis-sémination de microorganismes dans l’hémocoele.
2.2 La réponse humorale : synthèse des peptides antimicrobiens
La synthèse des peptides antimicrobiens constitue actuellement la réponse immunitaire la mieux caractérisée, du fait de la spécificité et de l’importance de ces peptides dans la résistance des insectes aux infections microbiennes (Lemaitre et al., 1997 ; Tzou et al., 2002). Les peptides anti-microbiens sont sécrétés dans l’hémolymphe par les cellules du corps gras, des épithéliums et par les hémocytes, principalement en réponse à une in-
ANSELME Caroline 34 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
fection. Toutefois, comme cela sera discuté plus loin, ces peptides peuvent également être sécrétés constitutivement par les épithéliums.
Les gènes codant les peptides antimicrobiens peuvent être induits lors d’une réponse systémique quand le microorganisme est présent dans l’hémocoele, comme lors de piqûres septiques utilisées pour l’étude de la réponse immunitaire. Ils peuvent également être induits lors d’une réponse locale, au niveau des épithéliums, généralement lors des infections naturel-les de l’intestin, de la trachée ou du tractus génital.
2.2.1 Le corps gras : la réponse systémique
La synthèse des peptides antimicrobiens en réponse à une infection systé-mique se fait principalement dans le corps gras, suite à l’activation des voies appelées Toll et Imd. Ces deux voies comportent de nombreuses pro-téines homologues aux protéines des voies “Nuclear Factor-Kappa B” (NF-κB) des mammifères impliquées dans la réponse inflammatoire. La voie Toll présente des homologies avec les voies des “Toll-Like Receptors” (TLR)/“InterLeukine-1 Receptor” (IL-1R) alors que la voie Imd est plus proche de la cascade du “Tumor Necrosis Factor Receptor” (TNFR1).
L’activation de l’une ou l’autre des voies Toll et Imd est fonction du microorganisme infectant. La voie Toll est principalement activée en ré-ponse à la présence de champignons ou de bactéries Gram (+), alors que la voie Imd est activée lors d’une infection par des bactéries Gram (-) (Hoffmann, 2003).
2.2.1.1 La voie Toll
D’abord connue pour son rôle dans la mise en place de l’axe dorso-ventral lors du développement embryonnaire de la drosophile (St Johnston et Nus-slein-Volhard, 1992), elle a ensuite été étudiée pour son rôle dans la ré-ponse immunitaire de l’insecte (Lemaitre et al., 1996). La protéine Toll est un récepteur membranaire présentant un domaine extracellulaire riche en leucine (LRR, “Leucine-Rich-Repeats”), et un domaine intracellulaire simi-laire au récepteur de l’interleukine-1 des mammifères (TIR, “Toll-IL-1 Receptor domain”).
Toll est le récepteur de la protéine Spätzle, issue du clivage de la protéine pro-Spätzle par une cascade de protéases à sérine activées en ré-ponse à une infection (Figure 5). Si le ligand du récepteur Toll et certaines protéines de la cascade sont activés quel que soit le microorganisme (bacté-ries Gram (+) ou champignons), d’autres protéines de cette cascade sont ac-tivées soit par les bactéries Gram (+), soit par les champignons, assurant
ANSELME Caroline 35 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
ainsi la spécificité de la réponse (Lemaitre et al., 1996 ; Levashina et al., 1999 ; Ligoxygakis et al., 2002a ; Robertson et al., 2003 ; Kambris et al., 2006). La liaison Spätzle-Toll induit la dimérisation du récepteur et la for-mation d’un complexe récepteur-adaptateur composé des protéines dMyD88, Tube et de la protéine kinase Pelle (Horng et Medzhitov, 2001 ; Sun et al., 2002). La transduction du signal se poursuit par la phosphoryla-tion de l’inhibiteur Cactus, un analogue de la protéine I-κB (“Inhibitor of NF-κB”) des mammifères, par des intermédiaires encore non caractérisés. Cette phosphorylation est suivie par la dégradation de l’inhibiteur par le protéasome (Figure 5), la libération et la translocation nucléaire du facteur de transcription de la famille NF-κB : DIF (“Dorsal related Immune Factor”). Une fois dans le noyau, DIF assure la transcription des gènes co-dant certains peptides antimicrobiens, ainsi que d’autres gènes de la ré-ponse immunitaire (De Gregorio et al., 2002a).
ANSELME Caroline 36 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
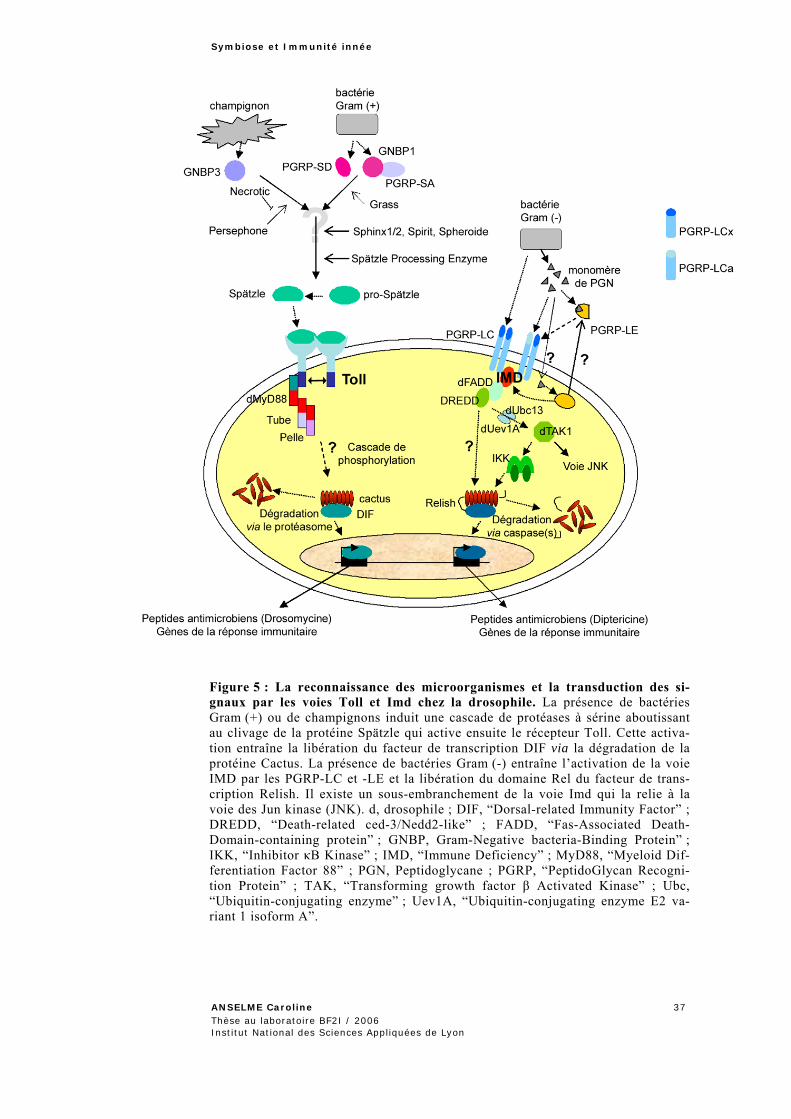
Symbiose et Immunité innée
Figure 5 : La reconnaissance des microorganismes et la transduction des si-gnaux par les voies Toll et Imd chez la drosophile. La présence de bactéries Gram (+) ou de champignons induit une cascade de protéases à sérine aboutissant au clivage de la protéine Spätzle qui active ensuite le récepteur Toll. Cette activa-tion entraîne la libération du facteur de transcription DIF via la dégradation de la protéine Cactus. La présence de bactéries Gram (-) entraîne l’activation de la voie IMD par les PGRP-LC et -LE et la libération du domaine Rel du facteur de trans-cription Relish. Il existe un sous-embranchement de la voie Imd qui la relie à la voie des Jun kinase (JNK). d, drosophile ; DIF, “Dorsal-related Immunity Factor” ; DREDD, “Death-related ced-3/Nedd2-like” ; FADD, “Fas-Associated Death-Domain-containing protein” ; GNBP, Gram-Negative bacteria-Binding Protein” ; IKK, “Inhibitor κB Kinase” ; IMD, “Immune Deficiency” ; MyD88, “Myeloid Dif-ferentiation Factor 88” ; PGN, Peptidoglycane ; PGRP, “PeptidoGlycan Recogni-tion Protein” ; TAK, “Transforming growth factor β Activated Kinase” ; Ubc, “Ubiquitin-conjugating enzyme” ; Uev1A, “Ubiquitin-conjugating enzyme E2 va-riant 1 isoform A”.
ANSELME Caroline 37 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
2.2.1.2 La voie Imd
La seconde voie de signalisation impliquée dans la synthèse des peptides antimicrobiens a été identifiée lors de l’étude du phénotype imd caractérisé par une diminution de l’inductibilité des peptides après une infection par des bactéries Gram (-) (Lemaitre et al., 1995).
L’activation de la voie Imd implique la reconnaissance directe (contrairement à la voie Toll) du peptidoglycane (PGN) bactérien par le ré-cepteur membranaire de cette voie, le PGRP-LC (Figure 5). La voie de si-gnalisation aboutissant à la libération du facteur de transcription n’est pas encore parfaitement caractérisée, mais elle impliquerait un complexe com-prenant les protéines Imd, dFADD et la caspase DREDD (Leulier et al., 2000 ; Leulier et al., 2002). Ce complexe serait responsable de l’activation ubiquitine-dépendante (dUbc13/dUev1A) d’une cascade de kinases incluant la protéine dTAK1 et le complexe IKK (Wang et al., 2001 ; Zhou et al., 2005). Cette cascade aboutit à la phosphorylation du facteur de transcrip-tion Relish. Une fois phosphorylé, Relish serait clivé, soit directement par la caspase DREDD, soit par une autre caspase activée par cette protéine (Stoven et al., 2000). La dissociation des domaines Iκ-B-like et Rel (do-maine de liaison à l’ADN) permet la translocation nucléaire du facteur de transcription et l’induction de peptides antibactériens et de gènes de la ré-ponse immunitaire (De Gregorio et al., 2002a). Il existe également d’autres sous-embranchements de la voie Imd permettant notamment d’activer les mécanismes de l’apoptose (Georgel et al., 2001) et la voie des JNK (“Jun N-terminal kinase”) impliquée dans le réarrangement du cytosquelette et la réparation des tissus (Boutros et al., 2002 ; Ramet et al., 2002a).
Les voies décrites ci-dessus récapitulent les étapes générales des
voies Toll et Imd. Toutefois, il est intéressant de signaler qu’il existe éga-lement des interactions et des recouvrements entre ces dernières, puisque certains gènes impliqués dans une voie sont régulés par l’autre, et qu’il existe des gènes co-régulés par les deux voies. La distinction entre les bac-téries Gram (+)/champignons et les bactéries Gram (-) au niveau de l’activation des voies Toll et Imd est donc parfois trop restrictive, et la ré-ponse à certains pathogènes peut impliquer une activation simultanée des deux voies (De Gregorio et al., 2002a). Par ailleurs, d’autres facteurs se-raient susceptibles d’activer les voies Toll et Imd. Par exemple, tous les peptides antimicrobiens sont faiblement induits lors d’une blessure stérile, probablement pour prévenir une infection par un microorganisme opportu-niste qui pourrait profiter de la rupture des barrières physiques. Cette in-duction préventive suggère l’existence de signaux indépendants des micro-
ANSELME Caroline 38 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
organismes, probablement liés à l’altération des tissus, qui pourraient acti-ver les voies Toll et/ou Imd ou activer d’autres voies de signalisation en-core non identifiées (Boutros et al., 2002 ; De Gregorio et al., 2002a).
2.2.2 Les épithéliums : la réponse locale
Bien que la synthèse des peptides antimicrobiens par le corps gras ait lar-gement été décrite, très peu d’études se sont consacrées à la synthèse de ces peptides par les épithéliums. En effet, les études de la réponse immunitaire ont principalement été réalisées par l’introduction des microorganismes di-rectement dans l’hémocoele (piqûre septique) et ne font donc pas intervenir les premières défenses assurées par les épithéliums. Pourtant, hormis quel-ques souches bactériennes dont certaines espèces d’Erwinia, la plupart des bactéries n’induisent pas de réponse systémique lors d’infections naturelles par ingestion (Basset et al., 2000). Il semble donc que la réponse locale constitue un mécanisme de défense efficace pour lutter contre la majorité des infections naturelles chez les insectes.
Chez la drosophile, les épithéliums en contact avec l’environnement externe (trachée, intestin, tractus génital) sont capables d’exprimer des peptides antimicrobiens en réponse à une infection naturelle (Tzou et al., 2000). Toutefois, contrairement à l’induction observée dans le corps gras, l’induction des peptides dans ces épithéliums dépendrait uni-quement de la voie Imd, et ce, même dans le cas de la drosomycine, pour-tant sous le contrôle de la voie Toll dans la réponse systémique (Ferrandon et al., 1998). D’autre part, puisque la réponse épithéliale n’est pas induite par l’injection de bactéries dans la cavité abdominale, elle pourrait impli-quer des mécanismes d’activation tissu-spécifiques, probablement via des récepteurs spécifiques des cellules épithéliales (Tzou et al., 2000). Toute-fois, l’équipe de B. Lemaitre a récemment mis en évidence le rôle du PGRP-LC dans l’activation de la réponse locale de l’intestin (Zaidman-Remy et al., 2006). Par ailleurs, un autre membre de la famille des PGRP impliqué dans l’activation de la voie Imd, le PGRP-LE, pourrait être impli-qué dans la réponse de la trachée, en synergie avec le PGRP-LC (Takehana et al., 2004). Il semblerait donc que les mêmes récepteurs soient impliqués dans les réponses systémique et locale.
Tzou et al. (2000) ont également mis en évidence l’expression constitutive de peptides dans certains épithéliums, indépendamment des gènes Toll ou imd. Cette expression ne serait donc pas sous le contrôle des facteurs de transcription NF-κB, mais d’autres facteurs. Ryu et al. (2004) ont ainsi mis en évidence le rôle du gène homéotique Caudal dans l’expression constitutive de deux peptides antimicrobiens : la cécropine
ANSELME Caroline 39 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
dans le canal éjaculatoire et la drosomycine dans les glandes salivaires. Toutefois, dans le tractus génital femelle, l’expression de la drosomycine est indépendante de Caudal, suggérant l’existence d’autres voies de signali-sation.
2.2.3 Reconnaissance des microorganismes
Bien que la notion de “non spécificité” de la réponse immunitaire soit lar-gement répandue, la majorité des mécanismes immunitaires des insectes sont non constitutifs, et le spectre de peptides antimicrobiens synthétisés diffère selon l’agent pathogène. Il existe donc des éléments fonctionnels qui permettent non seulement de détecter la présence des microorganismes, mais également de les différencier pour activer les réponses adaptées à la nature des microorganismes détectés. Cette fonction serait assurée par un ensemble de récepteurs (PRR, “Pattern-Recognition Receptors”) capables de reconnaître certaines composantes spécifiques des microorganismes (PAMP, “Pathogen Associated Molecular Patterns”) (Janeway, 1989).
De nombreux facteurs pouvant jouer le rôle de PRR ont été identi-fiés chez les insectes : l’hémoline, les “Thiolester containing proteins” (TEP), les lectines, les “β-1,3-Glucan Recognition Proteins” (βGRP), les “Gram-Negative bacteria-Binding Proteins” (GNBP) et les PGRP. Les PRR peuvent jouer le rôle d’opsonines dans l’activation de la réponse cellulaire comme l’hémoline chez les lépidoptères (Kanost et al., 2004) ou les TEP chez l’anophèle (Levashina, 2004), et probablement aussi la drosophile (De Gregorio et al., 2001). Les PRR jouent également un rôle important dans l’activation de la réponse humorale. Chez les lépidoptères, les lectines et les βGRP interviennent dans l’activation de la réaction de mélanisation en réponse aux LipoPolySaccharides (LPS) des bactéries Gram (-), et aux β-1,3-glucanes des champignons, respectivement (Kanost et al., 2004). Les rôles des deux derniers types de récepteurs, les GNBP et surtout les PGRP, ont été relativement bien décrits chez la drosophile.
2.2.3.1 Les GNBP
La famille des GNBP, spécifique des invertébrés et proche de la famille des βGRP, a été initialement identifiée chez Bombyx mori pour sa capacité de liaison aux bactéries Gram (-) (Lee et al., 1996). Toutefois, il semblerait que les protéines de cette famille soient capables de se lier à différents composants.
Chez la drosophile, trois gènes GNBP et deux gènes GNBP-like ont été identifiés. Un de ces gènes, le GNBP1 (osiris), avait été décrit dans
ANSELME Caroline 40 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
des cellules immunocompétentes comme un élément de reconnaissance des LPS des bactéries Gram (-) et des β-1,3-glucanes des champignons (Kim et al., 2000). Pourtant, Pili-Floury et al. (2004) ont ensuite montré in vivo, en utilisant le RNAi (ARN interférence), que la protéine GNBP1 était en fait impliquée dans la reconnaissance des bactéries Gram (+) et l’activation de la voie Toll (Figure 5). En ce qui concerne les autres GNBP(-like), seul le GNBP3 a été étudié pour son rôle dans l’activation de la voie Toll par les champignons (Ferrandon, 2006).
Les GNBP constituent donc une famille encore relativement mé-connue, mais les résultats apparemment contradictoires des études évo-quées précédemment soulignent la complexité des interactions PRR/PAMP et l’implication probable des GNBP dans la reconnaissance de différentes classes de microorganismes.
2.2.3.2 Les PGRP
Les PGRP, comme leur nom l’indique, sont des protéines de reconnaissance des éléments composant le peptidoglycane (PGN) bactérien. Largement étudiée depuis la caractérisation d’un PGRP chez B. mori en 1996 par Yoshida et al. (1996), la famille des PGRP est représentée aussi bien chez les invertébrés que chez les vertébrés, et elle comporte une grande variété d’isoformes et de fonctions (Dziarski, 2004). Certains PGRP sont impliqués dans l’activation de la cascade de mélanisation et/ou des voies de synthèse des peptides antimicrobiens (Michel et al., 2001 ; Takehana et al., 2002 ; Takehana et al., 2004). D’autres assurent la régulation de ces voies de syn-thèse (Bischoff et al., 2006 ; Zaidman-Remy et al., 2006). Enfin, certains PGRP auraient un rôle dans la phagocytose (Ramet et al., 2002b ; Garver et al., 2006). Chez les mammifères, les PGRP peuvent posséder des activités bactéricides ou bactériostatiques (Liu et al., 2000 ; Tydell et al., 2006).
Les protéines de la famille PGRP se caractérisent par la présence d’un domaine très conservé de 160-165 acides aminés qui présente une ho-mologie avec le lysozyme du bactériophage T7, une N-acétyl-muramyl-L-alanine amidase (Kang et al., 1998). Toutefois, si tous les PGRP ont apparemment conservé leur capacité de liaison au PGN, ce n’est pas le cas pour leur activité amidasique. De plus, lorsqu’elle est conservée, l’activité amidasique des PGRP est moins forte que celle du lysozyme phagique (Yoshida et al., 1996 ; Kang et al., 1998 ; Michel et al., 2001 ; Kim et al., 2003 ; Mellroth et al., 2003 ; Wang et al., 2003).
ANSELME Caroline 41 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Bien que la question de la reconnaissance des bactéries Gram (-) par les PGRP via les LPS soit restée longtemps en suspens du fait de la contamination des préparations commerciales de LPS par du PGN, il est au-jourd’hui admis que la reconnaissance des bactéries se fait plutôt au niveau de leur PGN (Leulier et al., 2003). En effet, le PGN des bactéries Gram (+) diffère de celui des bactéries Gram (-) et de celui de quelques espèces de Bacillus au niveau du troisième acide aminé du pont peptidique. Il s’agit d’une lysine pour les premières, et d’un acide meso-diaminopimélique (DAP) pour les autres (Leulier et al., 2003). On distingue de ce fait deux types de PGN, le PGN de type Lysine et le PGN de type DAP (Figure 6). Ainsi, la réponse immunitaire dirigée contre les bactéries Gram (+) est ac-tivée par les PGRP présentant une affinité pour le PGN de type Lysine, alors que ceux ayant une affinité pour le PGN de type DAP activeront la réponse anti-Gram (-).
Figure 6 : Structure du peptidoglycane (PGN) bactérien. Le PGN est un poly-mère constitué de chaînes polysaccharidiques composées d’une alternance de N-acétylglucosamine et d’acide N-acétylmuramique, reliées par des ponts peptidi-ques dont l’unité de base est un tétrapeptide (cadre pointillé). Dans le PGN des bac-téries Gram (-) et des bactéries Gram (+) du genre Bacillus, le tétrapeptide com-porte en troisième position un acide meso-diaminopimélique (PGN de type DAP, à gauche). Les tétrapeptides sont reliés entre eux par une liaison directe entre le rési-du DAP d’une unité et le résidu terminal d’une autre. Dans le cas des bactérie Gram (+), le résidu DAP est remplacé par un résidu lysine (PGN de type Lysine, à droite) et les liaisons interpeptidiques sont assurées par des résidus variables en fonction de l’espèce bactérienne. Par exemple, le peptide de liaison du PGN de Staphylococcus aureus (représenté à droite) comporte un chaînon de cinq résidus glycine.
ANSELME Caroline 42 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Chez la drosophile, treize gènes, dont certains possèdent des transcrits alternatifs, codent des protéines de la famille des PGRP (Werner et al., 2000 ; Werner et al., 2003). Bien que cette famille soit très diversi-fiée tant par les profils d’expression des gènes, que par la distribution tissu-laire des protéines, elle peut être divisée en deux sous-familles : les formes courtes (PGRP-S), sécrétées dans l’hémolymphe, et les formes longues (PGRP-L), dont certaines possèdent des domaines transmembranaires (PGRP-LA,-LC, -LD, -LF). Cette première classification basée sur la struc-ture des transcrits reste encore très utilisée. Toutefois, avec les progrès ré-alisés ces trois dernières années, il est également possible de classer les protéines de cette famille en fonction de leur activité, PGRP récepteurs ou PGRP enzymatiques.
2.2.3.2.1 Les PGRP récepteurs, activateurs de la réponse immunitaire
C’est à partir de l’année 2001 que les résultats des cribles génétiques de mutants de drosophile ont permis d’attribuer le rôle de récepteurs à certains membres de la famille des PGRP.
Le PGRP-SA (semmelweis) est le premier PGRP caractérisé chez la drosophile pour son implication dans la résistance aux bactéries Gram (+) (Michel et al., 2001). La mutation de ce gène, qui n’affectait ni la réponse aux champignons ni la réponse aux bactéries Gram (-), avait mis en évidence, non seulement le rôle de ce PGRP, mais également la spécificité de reconnaissance des microorganismes par les PGRP. Il a ensuite été mon-tré que la protéine PGRP-SA active la voie Toll en interaction avec la pro-téine GNBP1 qui hydrolyserait le PGN et faciliterait ainsi sa reconnais-sance par le PGRP-SA (Pili-Floury et al., 2004 ; Filipe et al., 2005). Toutefois, Bischoff et al. (2004) ont montré que la voie Toll était activée par certaines bactéries Gram (+) indépendamment du complexe PGRP-SA/GNBP1. L’implication du PGRP-SD dans la réponse à ces bac-téries a alors été mise en évidence, attribuant à un autre PGRP le rôle de ré-cepteur.
Si les PGRP-SA et -SD sont des protéines sécrétées dans l’hémolymphe et induites par une infection bactérienne Gram (+), le PGRP-LC, en revanche, est une protéine transmembranaire constitutive. Ce dernier est en fait le récepteur de la voie Imd impliquée dans la réponse aux infections bactériennes Gram (-) (Choe et al., 2002 ; Gottar et al., 2002 ; Choe et al., 2005) (Figure 4). Le gène PGRP-LC présente également la particularité de posséder trois transcrits alternatifs dont deux, PGRP-LCx et -LCa, ont été impliqués dans la réponse aux bactéries Gram (-). Ces
ANSELME Caroline 43 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
transcrits présentent des domaines intracellulaire et transmembranaire communs mais un domaine PGRP extracellulaire variable. Ceci permet d’assurer une variabilité supplémentaire dans les mécanismes de reconnais-sance du PGN (Werner et al., 2003 ; Chang et al., 2005). Parallèlement à son rôle dans l’activation de la voie Imd, le PGRP-LC pourrait également intervenir dans la phagocytose des bactéries Gram (-) (Ramet et al., 2002b). Toutefois, des résultats obtenus in vivo suggèrent que la phagocytose des bactéries Gram (-) impliquerait plutôt d’autres récepteurs, notamment le PGRP-SA (Garver et al., 2006), ce qui pourrait remettre en cause les résul-tats obtenus en culture cellulaire par Ramet et al. (2002).
La reconnaissance du PGN du type DAP des bactéries Gram (-) fait également intervenir une autre protéine exprimée constitutivement : le PGRP-LE (Takehana et al., 2002 ; Takehana et al., 2004 ; Kaneko et al., 2006). Bien que la protéine ait été initialement prédite intracellulaire sur la base de la séquence nucléotidique (Werner et al., 2000), celle-ci peut être détectée dans l’hémolymphe et serait donc sécrétée par un processus encore non élucidé (Takehana et al., 2004). De plus, Takehana et al. (2004) ont montré que ce PGRP agit à la fois en amont et en parallèle du PGRP-LC, dans l’activation de la voie Imd. Le mode d’action du PGRP-LE, en rela-tion avec sa localisation cellulaire, a alors été caractérisé par l’étude de Kaneko et al. (2006). La protéine PGRP-LE peut être soit intracellulaire, soit extracellulaire, sous une forme tronquée comportant uniquement le domaine PGRP. Ainsi, le PGRP-LE pourrait fonctionner, parallèlement au PGRP-LC, comme un récepteur intracellulaire du PGN monomérique, et en amont du PGRP-LC, en se liant au PGN de l’hémolymphe et en facilitant sa liaison au PGRP-LC (Kaneko et al., 2006).
Le PGRP-LE pourrait également jouer un rôle dans l’activation de la cascade de mélanisation en réponse à une infection par E. coli (Takehana et al., 2004).
2.2.3.2.2 Les PGRP enzymatiques, régulateurs de la réponse immunitaire
Compte tenu de l’homologie de séquence entre les PGRP et les N-acétyl-muramyl-L-alanine amidases, l’activité enzymatique des premiers PGRP identifiés chez les insectes a bien entendu été testée. Ces tests se sont révé-lés négatifs jusqu’aux travaux de Mellroth et al. (2003) qui ont montré que le PGRP-SC1 clive les peptidoglycanes bactériens (Figure 7) avec une ef-ficacité variable selon l’origine bactérienne. Toutefois, le PGRP-SC1 ne présente pas d’activité bactéricide in vitro et ne pourrait donc pas cliver le PGN de la paroi bactérienne, à moins d’une synergie in vivo avec une autre
ANSELME Caroline 44 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
protéine immunitaire. Sachant que le clivage du PGN diminue son pouvoir immunogène, les auteurs ont suggéré que le PGRP-SC1 serait impliqué dans le contrôle de l’infection bactérienne, plutôt que dans l’élimination des bactéries.
Figure 7 : Sites de coupure du peptidoglycane (PGN) bactérien par quelques enzymes. Les protéines de la famille PGRP présentent une homologie avec le lyso-zyme du bactériophage T7, une N-acétyl-muramyl-L-alanine amidase. Contraire-ment au lysozyme du blanc d’œuf qui clive la chaîne polysaccharidique (liaison osidique entre le NAM et NAG), le lysozyme phagique et certains membres de la famille des PGRP clivent le PGN au niveau de la liaison entre le pont peptidique et le NAM. L’activité des PGRP varie en fonction du type de PGN (lysine ou DAP) et de l’espèce bactérienne. A titre d’exemple, le PGRP-LB serait 100 à 1000 fois plus actif sur le PGN de type DAP que sur le PGN de type Lysine (Zaidman-Remy et al., 2006), alors que les différences d’activité observées sur ces deux types de PGN sont relativement faibles dans le cas du PGRP-SC1 (Mellroth et al., 2003).
L’analyse de la séquence du PGRP-SC1 montre qu’il a conservé,
contrairement aux PGRP-SA et -LCx, quatre des cinq résidus du site actif du lysozyme du bactériophage T7. Au niveau du cinquième résidu, il pré-sente une mutation d’une lysine en thréonine qui permet de conserver une activité, toutefois réduite par rapport à l’activité du lysozyme phagique (Cheng et al., 1994). C’est donc une mutation de l’un de ces résidus qui en-traîne la perte d’activité amidasique observée chez les PGRP récepteurs. Grâce à cette étude, Mellroth et al. (2003) ont mis en évidence une nou-velle classe de PGRP ayant conservé les résidus du site actif, et ont suscité ainsi un nouvel intérêt pour l’activité enzymatique potentielle des PGRP.
ANSELME Caroline 45 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
L’importance de la conservation des cinq résidus dans l’activité amidasique a ensuite été confirmée au niveau structural, grâce à la cristallisation du PGRP-LB (Kim et al., 2003). L’obtention de la structure de ce PGRP a confirmé l’homologie des PGRP avec le lysozyme phagique au niveau du site de liaison au PGN, et a révélé, sur la face opposée à ce site de liaison, la présence d’un segment N-terminal spécifique des PGRP. Compte tenu de la conformation des PGRP au niveau de ce segment, cette zone pourrait constituer un site de liaison protéique qui conférerait aux PGRP leur(s) fonction(s) supplémentaire(s), comme l’activation de la synthèse des peptides antimicrobiens (Kim et al., 2003).
tiques d’insectes.
Concernant le rôle in vivo de ces PGRP enzymatiques, deux études ont permis de confirmer la fonction immuno-modulatrice des PGRP amidasiques, PGRP-SC1/2 et -LB (Bischoff et al., 2006 ; Zaidman-Remy et al., 2006). Ces deux études ont mis en évidence une suractivation spécifique de la voie Imd après une infection bactérienne, locale ou systémique, en l’absence des PGRP-SC1/2 et -LB par rapport aux témoins. Elles ont également montré que la régulation n’était pas due à l’élimination des bactéries, confirmant ainsi l’absence d’activité bactéricide observée in vitro, mais à la diminution de l’immunogénicité du PGN dégradé. Les PGRP-SC1/2 et -LB sont principalement exprimés dans l’intestin, où l’épithélium est en contact permanent avec la flore bactérienne. Ils pourraient donc limiter l’activation de la réponse immunitaire de l’hôte. Sur la base de leurs derniers résultats, Zaidman-Remy et al. (2006) ont proposé un modèle pour expliquer le rôle du PGRP-LB dans la régulation de la réponse locale de l’intestin. L’activation de la voie Imd serait réalisée en fonction de la croissance de la population bactérienne de l’intestin via la détection du PGN libéré par les bactéries en division (Figure 8). Il est intéressant de mentionner ici que ce modèle, qui est décrit pour expliquer le maintien de la flore intestinale, serait potentiellement adapté à la régulation et au maintien des bactéries symbio
ANSELME Caroline 46 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
Figure 8 : Modèle de l’activation de la voie Imd par les bactéries Gram (-). Lorsque les bactéries se divisent, le PGN libéré est dégradé par le PGRP-LB pour bloquer l’activation de la voie Imd par le PGRP-LC (1). En cas d’infection, la quantité de PGN libérée lors de la multiplication bactérienne dépasse les capacités du PGRP-LB et le PGN se lie au PGRP-LC (2). L’activation de la voie Imd entraîne l’induction de la synthèse des peptides antibactériens pour éliminer les bactéries (3), et l’induction du PGRP-LB pour éliminer le PGN libre (1) et ainsi terminer la réaction immunitaire. Dans l’intestin, le PGRP-LB jouerait un rôle important dans la prévention de l’activation des réponses immunitaires locale et systémique par des bactéries de la flore commensale. D’après Zaidman-Remy et al. (2006).
Une autre étude par mutagenèse a mis en évidence le rôle de
l’activité amidasique du gène PGRP-SC1a dans la phagocytose de Staphy-lococcus aureus (Garver et al., 2006). Cette étude montre également la sen-sibilité des mutants à une infection par S. aureus (>80 % de mortalité en 24 heures) et leur incapacité à activer la voie Toll, ce qui est en contradic-tion avec les résultats de Bischoff et al. (2006). L’implication du PGRP-SC1 dans l’activation de la voie Toll reste donc controversée et il se-rait intéressant de déterminer si les différences observées sont liées à l’approche technique (mutation vs RNAi), à la présence de la protéine PGRP-SC2 dans l’étude de Garver et al. (2006) ou à d’autres différences dans les souches utilisées ou dans les techniques d’infection et de détection.
Pour conclure ce paragraphe sur les PGRP enzymatiques, il faut signaler l’existence d’une autre activité enzymatique, l’activité L,D-carboxypeptidase, mise en évidence pour la protéine PGRP-SA
ANSELME Caroline 47 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
(Figure 7) par Chang et al. (2004). La protéine PGRP-SA peut se lier aux deux types de PGN, mais la liaison au PGN de type lysine entraîne l’activation de la voie Toll alors que la liaison au PGN de type DAP en-traîne la dégradation de ce PGN. La protéine PGRP-SA assure donc des fonctions différentes en fonction du type de PGN auquel elle se lie, ajoutant ainsi un autre degré dans la diversité et la complexité des fonctions assu-rées par les PGRP.
Les protéines PGRP, qu’il s’agisse des PGRP récepteurs ou des PGRP enzymatiques, permettent à la drosophile de différencier les micro-organismes et d’induire une réponse immunitaire adaptée, permettant ainsi l’élimination des pathogènes tout en limitant les effets létaux occasionnés par une suractivation de la réponse immunitaire (Bischoff et al., 2006). Les nombreuses données apportées par les études structurales devraient permet-tre de caractériser les interactions PGN-PGRP et d’éclairer les processus de reconnaissance et de régulation des voies immunitaires.
2.2.4 Elimination des microorganismes par les peptides antimicrobiens
La production de peptides antimicrobiens est un moyen de défense très conservé depuis les microorganismes jusqu’aux plantes et aux mammifères (Bulet et al., 2004). Chez les insectes, la synthèse de ces peptides au niveau des réponses locale et systémique constitue un moyen rapide et efficace pour l’élimination des microorganismes. Cette efficacité est liée au large spectre de peptides exprimés par l’hôte, à la forte concentration des pepti-des sécrétés dans l’hémolymphe (jusqu’à 300 µM), mais également à la spécificité de la réponse engendrée par les voies Toll et Imd (Hoffmann et Reichhart, 2002).
Les peptides antimicrobiens sont de petites protéines (moins de 100 acides aminés) dont l’activité requiert une charge globale cationique à pH physiologique et une proportion suffisante de résidus hydrophobes (>50%), pour permettre une interaction avec la membrane anionique des microorganismes. Une fois l’interaction peptide-membrane établie, deux mécanismes d’action ont été décrits jusqu’à présent : (i) la création de po-res qui déstabilisent la membrane et entraînent la mort du microorganisme, (ii) la translocation cytoplasmique du peptide et son interaction avec une cible intracellulaire (Powers et Hancock, 2003 ; Ganz, 2004).
Les peptides antimicrobiens peuvent présenter des propriétés bac-téricides et/ou fongicides avec un spectre d’action plus ou moins large. Par ailleurs, il est souvent difficile d’attribuer une activité globale à un peptide. Son activité peut en effet varier en fonction de sa concentration et de la physiologie du microorganisme. A titre d’exemples, les défensines sont
ANSELME Caroline 48 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
moins efficaces vis-à-vis des bactéries métaboliquement inactives (manque de nutriments, inhibiteurs métaboliques) que vis-à-vis des bactéries actives (Ganz, 2004) ; la coléoptéricine A d’Allomyrina dichotoma présente une activité bactéricide contre S. aureus et B. subtilis mais inhibe la division d’E. coli (Sagisaka et al., 2001) ; la cécropine A peut induire chez la bacté-rie, à des concentrations sub-létales, des changements transcriptionnels (Hong et al., 2003). L’activité d’un peptide antimicrobien nécessite donc souvent d’être évaluée au cas par cas.
Les peptides antimicrobiens actifs sont généralement obtenus suite au clivage in vivo d’un précurseur comportant un peptide signal et parfois un propeptide (motifs R-x-(K/R)-R) en amont ou en aval du peptide mature (Hosaka et al., 1991 ; Lazzaro et Clark, 2003). Au niveau structural, ces peptides sont regroupés en familles : les peptides à hélice-α comme la cé-cropine ou la sarcotoxine, les peptides de la famille des défensines stabili-sés par des ponts disulfures, et les peptides linéaires sans structure se-condaire caractéristique du fait de leur richesse en résidus proline et/ou glycine comme la drosocine, la metchnikowine ou les coléoptéricines (Powers et Hancock, 2003 ; Bulet et Stocklin, 2005).
Chez la drosophile, on distingue actuellement sept types différents de peptides inductibles, d’une taille allant de 20 à 50 acides aminés et clas-sés selon leur spectre d’activité : antibactérien Gram (+), antibactérien Gram (-) et antifongique (Tableau II). Le rôle et l’importance des différents peptides antimicrobiens dans la réponse immunitaire ont été mis en évi-dence en exprimant constitutivement, par transgenèse, ces différents pepti-des dans des double-mutants Imd-Spätzle, où les deux voies de biosynthèse des peptides antimicrobiens sont non fonctionnelles. La défensine et la dro-somycine sont les principaux peptides assurant la défense, respectivement, contre les bactéries Gram (+) et les champignons, alors que dans le cas des bactéries Gram (-), aucun peptide n’est capable, à lui seul, de restaurer une résistance similaire à celle observée chez les insectes sauvages (Tzou et al., 2002).
Chez les autres insectes, de nombreux peptides ont également été caractérisés avec parfois des spécificités au niveau de l’ordre, comme les lébocines décrites chez les lépidoptères, ou encore les coléoptéricines dé-crites chez les coléoptères. Actuellement, plus de 300 peptides différents ont été caractérisés essentiellement chez les diptères et les lépidoptères. En revanche, très peu de données sont disponibles sur d’autres ordres, notam-ment sur les insectes hétérométaboles comme les homoptères ou les orthoptères.
ANSELME Caroline 49 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
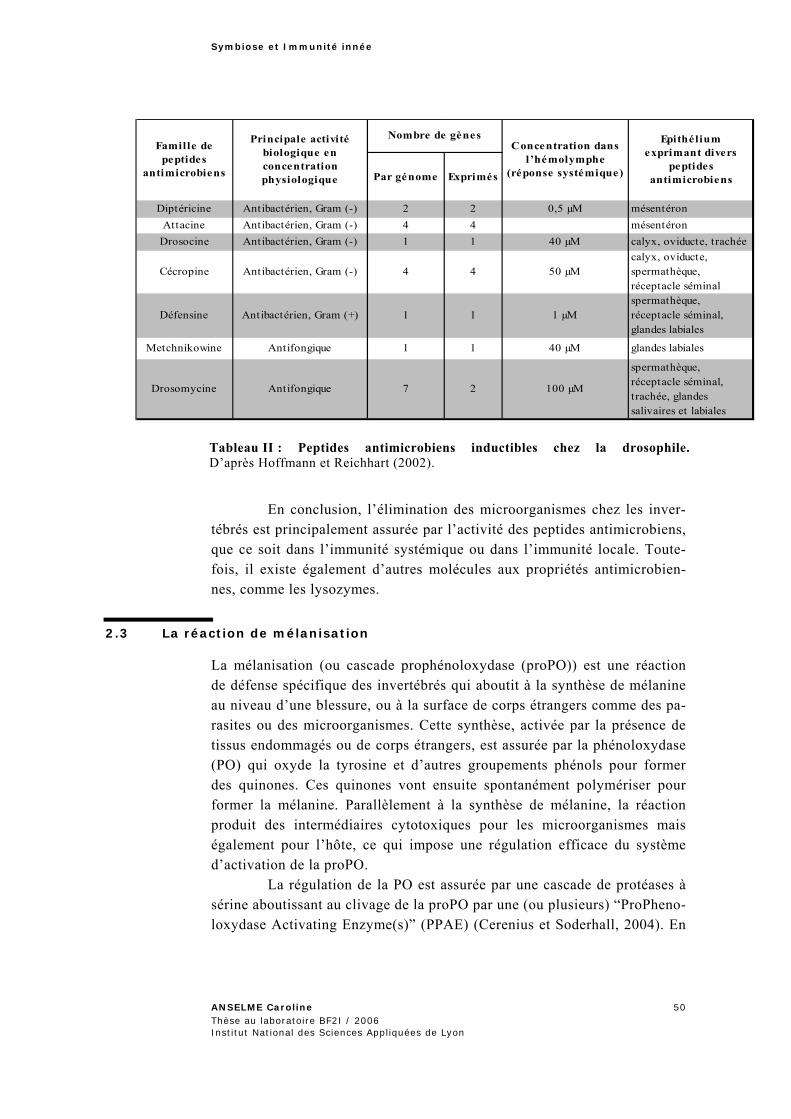
Symbiose et Immunité innée
Par génome Exprimés
Diptéricine Antibactérien, Gram (-) 2 2 0,5 µM mésentéronAttacine Antibactérien, Gram (-) 4 4 mésentéron
Drosocine Antibactérien, Gram (-) 1 1 40 µM calyx, o
s la
viducte, trachée
Cécropine Antibactérien, Gram (-) 4 4 50 µMcalyx, oviducte, spermathèque, réceptacle séminal
Défensine Antibactérien, Gram (+) 1 1 1 µMspermathèque, réceptacle séminal, glande biales
Metchnikowine Antifongique 1 1 40 µM glandes labiales
Drosomycine Antifongique 7 2 100 µM
spermathèque, réceptacle séminal, trachée, glandes salivaires et labiales
Famille de peptides
antimicrobiens
Principale activité biologique en concentration physiologique
Nombre de gènes Epithélium exprimant divers
peptides antimicrobiens
Concentration dans l’hémolymphe
(réponse systémique)
Tableau II : Peptides antimicrobiens inductibles chez la drosophile. D’après Hoffmann et Reichhart (2002).
En conclusion, l’élimination des microorganismes chez les inver-
tébrés est principalement assurée par l’activité des peptides antimicrobiens, que ce soit dans l’immunité systémique ou dans l’immunité locale. Toute-fois, il existe également d’autres molécules aux propriétés antimicrobien-nes, comme les lysozymes.
2.3 La réaction de mélanisation
La mélanisation (ou cascade prophénoloxydase (proPO)) est une réaction de défense spécifique des invertébrés qui aboutit à la synthèse de mélanine au niveau d’une blessure, ou à la surface de corps étrangers comme des pa-rasites ou des microorganismes. Cette synthèse, activée par la présence de tissus endommagés ou de corps étrangers, est assurée par la phénoloxydase (PO) qui oxyde la tyrosine et d’autres groupements phénols pour former des quinones. Ces quinones vont ensuite spontanément polymériser pour former la mélanine. Parallèlement à la synthèse de mélanine, la réaction produit des intermédiaires cytotoxiques pour les microorganismes mais également pour l’hôte, ce qui impose une régulation efficace du système d’activation de la proPO.
La régulation de la PO est assurée par une cascade de protéases à sérine aboutissant au clivage de la proPO par une (ou plusieurs) “ProPheno-loxydase Activating Enzyme(s)” (PPAE) (Cerenius et Soderhall, 2004). En
ANSELME Caroline 50 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
l’absence d’infection, ces enzymes, comme la PO, sont présentes sous forme inactive, ce qui permet de limiter la synthèse de mélanine. Toutefois, ces enzymes sont également régulées par des inhibiteurs de protéases de la famille des serpines (inhibiteurs de protéases à sérine) qui jouent un rôle important dans le contrôle et la terminaison de la réaction pour éviter une mélanisation excessive (Zhu et al., 2003 ; Cerenius et Soderhall, 2004 ; Tong et Kanost, 2005).
Différents modèles d’activation de la proPO ont été décrits chez les insectes. Chez Manduca sexta, les études menées par l’équipe de M. Kanost (Kanost et al., 2004), ont montré que l’hydrolyse de la proPO implique un complexe protéique comprenant trois “Prophenol Activating Proteinases” (PAP) et des cofacteurs de la famille des Homologues des Sérines Protéases (SPH), dépourvus d’activité enzymatique. Un complexe similaire serait également impliqué chez le coléoptère Holotrichia diom-phalia (Lee et al., 1998). Chez M. sexta, ces cofacteurs pourraient se lier aux immulectines, des protéines de reconnaissance, et aux PPAE pour for-mer un complexe à la surface des microorganismes. En ce qui concerne les éventuelles protéases à sérine et serpines impliquées dans l’activation et la régulation de la cascade, 22 ADNc codant des protéases nommées “hemolymph proteinases” (HP) et cinq gènes codant des serpines ont été identifiés. Si l’implication et le rôle des diverses HP nécessitent encore d’être caractérisés, l’inhibition de la voie proPO par les serpines, au niveau des PPAE ou d’autres protéases encore non identifiées, a clairement été démontrée (Zhu et al., 2003 ; Tong et Kanost, 2005). Chez H. diomphalia, il existe deux proPO dont l’activation est assurée en deux étapes de protéo-lyse impliquant trois “ProPhenoloxydase Activating Factors” (PPAF) (Kim et al., 2002).
Chez la drosophile, trois gènes, dont deux sont exprimés dans les cellules à cristaux, codent la proPO. Ces cellules libèrent facilement la proPO inactive dans l’hémolymphe (Meister, 2004). Le contrôle de la cas-cade proPO est assuré par la serpine Spn27A, dont la mutation se traduit par une mélanisation excessive des insectes (De Gregorio et al., 2002b ; Ligoxygakis et al., 2002b ). Compte tenu de l’interaction de la Spn27A avec les PPAE d’autres insectes, cette serpine assurerait l’inhibition d’une PPAE encore non identifiée chez la drosophile (De Gregorio et al., 2002b).
Lors d’une infection, la serpine Spn27A est éliminée de l’hémolymphe, suite à l’activation de la voie Toll, par une protéine synthé-tisée de novo (Ligoxygakis et al., 2002b). Cette élimination serait due à la formation d’un complexe PPAE-Spn27A instable, rapidement éliminé de l’hémolymphe (Ligoxygakis et al., 2002b). Il est donc probable que l’activation de la voie Toll augmente le taux de PPAE active dans
ANSELME Caroline 51 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Symbiose et Immunité innée
l’hémolymphe, ce qui entraîne l’élimination de la Spn27A. L’excès de PPAE active alors la prophénoloxydase. La fin de la réaction serait assurée par la protéine Spn27A qui réapparaît dans l’hémolymphe 17 heures après l’infection (Ligoxygakis et al., 2002b), suite à l’induction du gène par la voie Toll (De Gregorio et al., 2002b). La Spn27A interviendrait également dans la limitation de la réaction à une zone proche du microorganisme (De Gregorio et al., 2002b).
2.4 Conclusion
La réponse immunitaire des insectes implique de nombreuses réactions cel-lulaires et humorales, qui, par leurs interactions et leur complémentarité, assurent une réponse efficace contre les agents infectieux. Bien que ces réactions soient du domaine de l’immunité innée, les insectes sont capables de contrôler et d’adapter les réactions immunitaires en fonction du micro-organisme grâce à la mise en place de systèmes de régulations complexes. La caractérisation des mécanismes de reconnaissance a notamment souligné l’importance des familles multigéniques et l’intérêt de l’épissage alternatif pour assurer la reconnaissance des agents infectieux et activer/réguler une réponse adaptée.
ANSELME Caroline 52 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
ANSELME Caroline 53 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
1 Insectes : élevage et matériel biologique
1.1 Le charançon Sitophilus zeamais
1.1.1 Elevage
Les charançons Sitophilus zeamais de la souche Lagoa (Açores) sont élevés au laboratoire sur grains de blé à 27,5°C et à 70% d’humidité relative (hr) (Laviolette et Nardon, 1963). La souche aposymbiotique a été obtenue par traitement de la souche symbiotique un mois à 35°C et à 90% hr (Nardon, 1973). Elle est viable, fertile et élevée dans les mêmes conditions que la souche symbiotique. Afin d’obtenir des cohortes d’individus relativement synchrones, les charançons sont mis à pondre sur de nouveaux grains de blé tous les deux jours.
1.1.2 Matériel biologique
1.1.2.1 Larves du quatrième stade et nymphes
Les charançons se développant à l’intérieur des grains de blé, les larves de quatrième stade et les nymphes sont récoltées par dissection des grains de blé, respectivement 18 et 22 jours après la ponte pour les insectes symbio-tiques et 21 et 25 jours pour les insectes aposymbiotiques.
1.1.2.2 Embryons
Les embryons ne pouvant être récoltés à partir des grains de blé, de jeunes adultes (deux à quatre semaines) sont conservés deux à quatre heures sur des grains d’amidon pour que les femelles y déposent leurs œufs. Les em-bryons sont récoltés après trois jours par dissolution à l’eau des grains d’amidon sur un tamis fin.
1.1.2.3 Ovocytes et bactériomes larvaires
Les ovocytes et les bactériomes larvaires sont obtenus par dissection res-pectivement des femelles adultes et des larves du quatrième stade dans un tampon isoosmotique A (25 mM KCl, 10 mM MgCl2, 250 mM saccharose, 35 mM Tris-HCl, pH7,5) sous la loupe binoculaire. Les échantillons sont
ANSELME Caroline 54 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
centrifugés (10 000 g, 1 min) et les organes du culot sont utilisés immédia-tement ou, au besoin, conservés à -80°C jusqu’à utilisation.
1.2 Le puceron Acyrthosiphon pisum
1.2.1 Élevage
Les pucerons symbiotiques sont issus du clone parthénogénétique LL01 ob-tenu lors d’une infestation de luzerne à Lusignan en 1986. Les pucerons sont élevés sur des plants de fèves dans des cages en plexiglas. Ces cages sont maintenues en conditions contrôlées (21°C, 70% hr, photopériode de 16 h) pour obtenir des pucerons se reproduisant par parthénogenèse. Afin d’obtenir des cohortes relativement synchrones d’individus, les pucerons adultes sont mis à pondre 24 h sur plante hôte avant d’être retirés des plan-tes.
1.2.2 Matériel biologique
Les pucerons aposymbiotiques stériles sont obtenus, pour chaque expéri-mentation, à partir des pucerons symbiotiques par un traitement à la rifam-picine selon la procédure décrite par Rahbé et al. (1994). Les bactériocytes sont disséqués à partir de larves symbiotiques du quatrième stade dans le tampon A et sont traités comme décrit précédemment pour les bactériomes du charançon.
2 Techniques histologiques
2.1 Fixation des échantillons
Les larves de quatrième stade et les nymphes sont fixées immédiatement après récolte dans une solution alcoolique de Bouin dont la formule a été modifiée au laboratoire (20 ml d’éthanol 80° contenant 0,7% d’acide picri-que, 5 ml d’une solution de formol neutre à 40% (addition de CaCO3 au formol commercial jusqu’à saturation et filtration), 2 ml d’acide acétique glacial et H2O qsp 50 ml). Afin de faciliter la pénétration du fixateur dans les tissus de l’insecte, les larves et les nymphes sont piquées respective-ment du coté postérieur et derrière la capsule céphalique après une pre-mière étape de fixation de 4 h pour éviter toute réaction consécutive à la piqûre. Les individus sont ensuite conservés dans le fixateur à 4°C pour une durée minimale de 36 h.
ANSELME Caroline 55 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
2.2 Double inclusion et coupe
Après fixation, les individus sont lavés dans trois bains successifs de 15 min dans l’éthanol 70° puis réhydratés dans trois bains d’eau. L’excédent d’eau est éliminé à l’aide d’un papier absorbant et les individus sont inclus une première fois dans de la gélose (1,3% agar). Les blocs de gélose sont ensuite découpés et déshydratés par des bains successifs de 30 min dans l’éthanol 70° puis 95° et enfin 100°. Les blocs sont ensuite la-vés par 2 bains de butan-1-ol et conservés dans le butan-1-ol à 4°C en at-tendant l’inclusion en paraffine. Cette étape permet ainsi de ramollir les tis-sus pour faciliter les coupes ultérieures.
Avant la seconde inclusion, les blocs de gélose subissent d’abord un bain de 30 min à 60°C (50% butan-1-ol, 50% paraffine) puis deux bains successifs d’une heure dans la paraffine pure. A la sortie du deuxième bain, les blocs sont emparaffinés et, après refroidissement, les blocs de paraffine sont conservés à 4°C.
Des sections de 5 à 10 µm sont réalisées sur les pièces de paraf-fine à l’aide d’un microtome. Les rubans de paraffine sont étalés sur des lames recouvertes de poly-L-lysine (POLYSINETM, Menzel-Glaser®) à l’aide d’une goutte d’eau puis l’eau est éliminée et les pièces sont fixées sur la lame sur une plaque chauffante à 60°C. Les lames peuvent ainsi être conservées plusieurs mois.
2.3 “Fluorescence In Situ Hybridization” (FISH) sur coupes histologiques
La FISH est une technique d’hybridation in situ ADN/ARN (sonde/cible) très spécifique. Elle permet de détecter et de localiser sur des coupes histo-logiques ou des tissus en suspension la présence d’une ou plusieurs espèces bactériennes cibles à l’aide d’une sonde ADN marquée par un fluoro-chrome. Toutes les étapes de cette expérience sont réalisées en l’absence de RNase pour éviter toute dégradation de l’ARN cible.
La première étape consiste à déparaffiner les lames dans le mé-thylcyclohexane (2×15 min). Les lames sont ensuite déshydratées par deux bains de 10 min dans l’éthanol 100° avant d’être séchées sous hotte. Les membranes des tissus sont perméabilisées à l’aide d’une goutte d’acide acé-tique 70% (v/v) déposée sur les coupes. Après séchage complet à l’étuve (45°C), les coupes sont rincées au PBS (140 mM NaCl, 2 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH7,4) puis déshydratées à nouveau par des bains d’éthanol 100° de 2 min. Les coupes sont ensuite déprotéinisées 30 min à 37°C dans une solution d’HCl 0,01 M contenant 100 µg.ml-1 de pepsine avant de subir une nouvelle étape de rinçage-déshydratation. Les
ANSELME Caroline 56 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
lames sont enfin préhybridées 30 min à 45°C sous agitation dans le tam-pon H [0,7 M NaCl, 15,8 mM Tris-HCl, 4 mM EDTA, 0,1% SDS, Denhardt 10X (Ficoll 0,2% (p/v), polyvinylpyrrolidone 0,2% (p/v) ; BSA, 0,2% (p/v))] avant d’être hybridées entre lame et lamelle dans le tampon H addi-tionné de la sonde marquée (12,5 ng.µl-1) pendant trois heures à 45°C dans une chambre noire humide. Les lames sont finalement lavées dans deux bains de 15 min à 45°C dans un tampon de lavage (0,9 M NaCl, 29,8 mM Tris-HCl, 5 mM EDTA, 0,1% SDS) avant d’être rincées au PBS et montées entre lame et lamelle dans le VectashieldTM (Vector Laboratories) contenant 3 µg.ml-1de DAPI (4’-6-diamidino-2-phenylindole). Les lames peuvent être conservées quelques jours à 4°C à l’obscurité. Observées sous un micros-cope à épifluorescence avec les filtres adaptés, les sondes marquées à la rhodamine restituent une fluorescence rouge alors que les structures nucléi-ques marquées au DAPI restituent une fluorescence bleue.
La sonde utilisée dans cette étude est une sonde spécifique de l’ARNr 16S bactérien, dirigée contre des régions variables de l’ARNr 16S de SPE (5’-ACC-CCC-CTC-TAC-GAG-AC-3’). Cette sonde a été synthéti-sée et marquée en 5’ à la rhodamine par la société Eurogentec (Angers, France).
3 Infection bactérienne
3.1 Culture des souches bactériennes utilisées
Trois souches bactériennes ont été utilisées :
Escherichia coli (TOP 10, Invitrogen), bactérie Gram-négative, souche non pathogène.
Bacillus megaterium (souche 526.86, UMR 5557 CNRS UCBL), bactérie Gram-positive, souche non pathogène.
Pseudomonas aeruginosa (PAO1, CEA Grenoble)2, bactérie Gram-négative, souche pathogène pour D. melanogaster.
Toutes les souches bactériennes sont cultivées à 37°C sous agitation (250 rotations par minute) dans le milieu de culture de Luria Bertoni (LB).
2 Toutes les expérimentations avec P. aeruginosa ont été réalisées au CEA de Grenoble en collabora-
tion avec M.O. Fauvarque.
ANSELME Caroline 57 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Afin de tester également la réponse de l’hôte vis-à-vis de son en-docytobiote, un culot de bactéries a été préparé extemporanément à partir d’une centaine de bacteriomes de larves de S. zeamais disséquées dans le tampon A comme décrit précédemment. Après centrifugation, le culot de bactériomes est repris dans 400 µl de milieu LB, les bactériomes sont broyés au potter et centrifugés (7000 g, 10 min) afin d’obtenir le culot bac-térien.
3.2 Infection des larves de charançon
Des larves aposymbiotiques du quatrième stade extraites des grains de blé sont maintenues sur le flanc à l’aide d’une pince souple. Elles sont ensuite piquées, au niveau de l’extrémité abdominale postérieure, à l’aide d’une épingle d’entomologie stérilisée à la flamme ou d’une épingle stérilisée puis trempée dans un culot bactérien obtenu par centrifugation d’une culture d’une nuit (E. coli ou B. megaterium) ou obtenu à partir de bacté-riomes disséqués (endocytobiote). Dans le cas de l’infection par P. aeruginosa, la bactérie doit être en phase de croissance exponentielle pour que l’infection soit efficace. Les larves ont donc été infectées par les bactéries d’une culture de DO600nm=0,8. Des larves témoins, non piquées, sont également traitées en parallèle.
Les larves sont ensuite placées dans les puits d’une microplaque de titration sous atmosphère humide, dans l’étuve à 27,5°C et à 70% hr. Après une période d’incubation de 1 à 12 h, les larves sont récoltées puis congelées à -80°C dans des microtubes Eppendorfs.
3.3 Mesure de la croissance bactérienne in larva
Des lots de cinq larves ont été broyés dans 200 µl de LB et après centrifugation (300 g, 10 min) pour séparer les bactéries des débris cellulai-res, le surnageant est prélevé et dilué en série (jusqu’à 10 000 fois). 100 µl de chaque dilution sont ensuite étalés sur boîtes LB-agar et, après une nuit à 37°C, le nombre de colonies est estimé à partir de la (des) dilution(s) la (les) plus appropriée(s) pour des temps d’incubation de six et vingt-quatre heures.
3.4 Infection des pucerons
Des pucerons aposymbiotiques du quatrième stade larvaire sont piqués au niveau de l’abdomen à l’aide d’une épingle stérilisée puis trempée dans un culot bactérien d’E. coli obtenu par centrifugation d’une culture d’une nuit.
ANSELME Caroline 58 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Les pucerons sont ensuite placés sur les plants de fève, dans une cage de plexiglas à 21°C et à 70% d’hr, sous éclairage artificiel. Après une période d’incubation de 1 à 12 h, les pucerons sont récoltés puis congelés à -80°C dans des Eppendorfs.
4 Techniques de biologie moléculaire
4.1 Extraction des acides nucléiques et électrophorèse sur gel d’agarose
4.1.1 Extraction d’ADN génomique
Les extractions d’ADN génomique (ADNg) ont été réalisées à partir de lar-ves de quatrième stade pouvant être broyées en Eppendorf à l’aide de sim-ples pistons. Les tissus sont homogénéisés dans 500 µl d’une solution STE (100 mM NaCl, 1 mM Na2EDTA, 10 mM Tris HCl pH8). La lyse cellulaire est assurée par un traitement au SDS 0,5% (m/v) et à la protéinase K (120 ng.µl-1) deux heures à 55°C. Les ARN sont ensuite dégradés par trai-tement à la RNase A (50 ng.µl-1) une heure à 37°C. Les protéines sont éli-minées par précipitation au cours d’une extraction phénol/chloroforme (500 µl). Après centrifugation (10 000 g, 2 min) la phase supérieure aqueuse est transférée dans un nouvel Eppendorf et l’ADN est précipité une heure à -20°C par addition de deux volumes d’éthanol 100° et de 1/10 de volume d’acétate de sodium 3 M. Le culot d’ADN obtenu après centrifugation (10 000 g, 15 min) est ensuite lavé à l’éthanol 70° (10 000 g, 5 min) et sé-ché sous hotte. L’ADN est resuspendu dans 50 µl d’eau ultra pure (QBio-gen).
4.1.2 Extraction d’ARN totaux
Toutes les expérimentations menées sur les ARN seront réalisées avec des gants, des consommables exempts de RNase et avec de la verrerie et du ma-tériel autoclavés (115°C, 20 min) ou traités 10 minutes à la soude 0,1 M et rincés à l’eau traitée au Diéthylpyrocarbonate 0,1% (v/v) (H2O DEPC). Toutes les solutions, si elles ne sont pas fournies avec les kits utilisés, sont également traitées au DEPC 0,1% (v/v) et autoclavées (120°C, 15 min) après une incubation d’au moins six heures.
ANSELME Caroline 59 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
4.1.2.1 Extraction au Trizol®
Les échantillons congelés sont broyés dans 1 ml de Trizol® Reagent (Invi-trogen) à l’aide d’un homogénéiseur Dounce, puis le broyat est transféré dans des microtubes Eppendorf. Après 5 min, 200 µl de chloroforme sont ajoutés et après 3 min d’incubation, les échantillons sont centrifugés (10 000 g, 15 min). La phase aqueuse contenant les ARN est transférée dans un nouveau tube, puis les ARN sont précipités pendant 10 min en pré-sence de 500 µl d’isopropanol. Le culot d’ARN obtenu après centrifugation (10 000 g, 10 min) est lavé à l’éthanol 70°, séché à température ambiante, puis repris dans 50 µl d’H2O DEPC 0,1% (v/v). Les ARN sont quantifiés puis stockés à -80°C jusqu’à utilisation.
4.1.2.2 Extraction "RNeasy Mini kit" (QIAGEN)
Cette technique de purification sur colonne est préférée à l’extraction Tri-zol lorsque l’extraction est réalisée sur de faibles quantités de tissus (ovo-cytes, embryons, bactériomes).
Les ARN sont extraits à partir des tissus selon les recommanda-tions du fournisseur. Brièvement, les échantillons sont lysés et homogénéi-sés dans un volume de tampon contenant du β-mercaptoéthanol et du thiocyanate de guanidine. Après centrifugation (10 000 g, 3 min), le surna-geant, additionné d’un volume d’éthanol 70°, est déposé sur une colonne RNeasy retenant spécifiquement les ARN lors de la centrifugation (10 000 g, 15 s). Les contaminants sont éliminés par lavages, et les ARN élués par 40 µl d’H2O dépourvues de RNase (10 000 g, 1 min).
4.1.2.3 Traitement des échantillons à la DNase
A l’exception des échantillons utilisés pour les Northern blot, les échantil-lons d’ARN sont soumis à un traitement à la DNase. La digestion de 40 µg d’acides nucléiques est réalisée pendant 10 min à 37°C en présence de 40 unités de DNAse (Promega) dans le tampon de digestion de l’enzyme (H2O DEPC qsp 100 µl). Après digestion, l’enzyme et les sels du tampon sont éliminés par purification de l’ARN sur colonne à l’aide du kit “RNeasy mini kit” (Qiagen).
4.1.3 Electrophorèse sur gel d’agarose
L’électrophorèse des produits de l’extraction est effectuée sur un gel d’agarose-TAE (Tris-HCl 1,6 mM, acétate de sodium 1,6 mM, EDTA 0,04 mM pH8, agarose 1% (p/v), bromure d’éthydium (BET) 0,5 µg.ml-1).
ANSELME Caroline 60 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Une fraction des échantillons (généralement 10%) est additionnée de solu-tion de dépôt orangé G 6X (orange G 0,25% (p/v), glycérol 50% (v/v)) pour l’ADN ou de bleu de dépôt 5X (bleu de bromophénol 0,25% (p/v), xylène cyanol 0,25% (p/v), EDTA 50 mM, glycérol 50% (v/v), H20 DEPC) pour l’ARN, et déposée sur le gel. La migration se déroule dans le tampon TAE sous une tension de 100 volts et peut être suivie grâce à la présence des co-lorants.
Les fragments d’ADN (ou d’ARN) sont ensuite visualisés par ex-position du gel sur une table UV équipée d’une caméra. Dans le cas de l’ADN, la taille est estimée par comparaison à un marqueur de taille déposé sur le gel. Dans le cas de l’ARN, ce type d’électrophorèse en conditions non dénaturantes ne permet pas l’estimation de la taille des transcrits. Elle permet néanmoins d’estimer la qualité de l’ARN à partir des transcrits ribo-somaux.
4.1.4 Dosage des échantillons
Les quantités d’acides nucléiques obtenues peuvent être estimées à l’aide du gel d’électrophorèse ou dosées précisément par spectrophotométrie.
Pour une estimation des quantités relatives, l’image du gel obte-nue avec le logiciel Bio-CAPT peut être traitée à l’aide du logiciel Bio-1D (Bio-profil, Vilber Lourmat). Ce logiciel permet de quantifier l’ADN (ou l’ARN) à partir de l’intensité des bandes par comparaison à l’intensité des bandes du marqueur de taille ou d’un échantillon de référence de quantité connue.
Pour une mesure fiable de la concentration des échantillons, les solutions sont dosées à l’aide du NanoDrop® ND-1000 UV-Vis Spectro-photometer (NanoDrop Technologies, Wilmington, Delaware USA.).
4.2 Northern Blot
Le Northern blot est une technique permettant de quantifier le taux de transcrits d’un gène à un instant et dans des conditions donnés. Cette tech-nique est basée sur la dénaturation et la séparation des ARN sur gel d’agarose, le transfert et la fixation de ces ARN sur une membrane de ny-lon, et enfin sur l’hybridation d’une sonde ADN radioactive avec les trans-crits du gène étudié.
ANSELME Caroline 61 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
4.2.1 Préparation des membranes
La première étape du Northern blot consiste à séparer les ARN par électro-phorèse sur un gel d’agarose 1% (p/v) dans du tampon phosphate 0,01 M (pH7) en conditions dénaturantes. Pour cela, 15 µg d’ARN (qsp H2O DEPC 7,4 µl) sont dénaturés pendant 30 min à 55°C en présence de 24,6 µl d’une solution de dénaturation (glyoxal désionisé 1M, DMSO 50% (v/v), tampon phosphate 0,01 M pH7, H20 DEPC) puis refroidis dans la glace. Les échantillons additionnés de solution de dépôt sont déposés sur gel. Chaque gel comporte 3 µg d’ARN dénaturés du marqueur de taille (0.24-9.5 Kb RNA ladder, Invitrogen), ainsi qu’un échantillon choisi comme contrôle interne afin de normaliser les données obtenues sur diffé-rentes membranes. La migration s’effectue à 80 volts dans le tampon phos-phate 0,01M pH7 avec une recirculation constante du tampon de la cathode à l’anode pour éviter l’acidification du tampon de migration.
Après migration, la portion de gel contenant le marqueur de tailles d’ARN est colorée au BET (0,5 µg.ml-1) afin de déterminer la corres-pondance entre la taille des transcrits (en kb) et leur distance de migration (en cm) par photographie sous UV du gel en présence d’une règle fluores-cente.
Les ARN des échantillons sont transférés par capillarité sur une membrane de nylon (HybondTM-N, Amersham) en présence de SSC 20X (Sigma). Après une nuit de transfert, les ARN sont fixés sur la membrane par traitement à 80°C pendant deux heures. La membrane peut ensuite être conservée plusieurs semaines.
4.2.2 Préparation des sondes radioactives
Les sondes nucléotidiques utilisées sont obtenues par une étape de PCR et une étape de marquage, à partir des clones bactériens comportant les ADN complémentaires des transcrits étudiés.
4.2.2.1 Amplification par PCR des fragments d’ADN
Les réactions de PCR ont été effectuées dans le thermocycleur TGradient (Biometra, Goettingen, Germany) avec les amorces du plasmide pCR®2.1-TOPO® : le T7 forward (5’-TAA-TAC-GAC-TCA-CTA-TAG-GG-3’) et le M13 reverse (5’-CAG-GAA-ACA-GCT-ATG-ACC-3’) dans les conditions décrites ci-après.
ANSELME Caroline 62 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Conditions de PCR : Amorce T7 forward 300 nM Amorce M13 reverse 300 nM dNTPs 200 µM (chacun) Tampon PCR 1 X MgCl2 2 mM Taq uptitherm (Upthima) 1,25 U H20 qsp 50 µl ADN matrice : une pointe de clone bactérien
25 cycles
7’ 30’’
30’’
30’’ 5’
72°C 92°C
54°C
4.2.2.2 Purification des fragments d’ADN amplifiés
La purification des produits de PCR est réalisée sur colonne à l’aide du kit “MinElute Gel Extraction” (QIAGEN) selon les recommandations du four-nisseur. La totalité de la réaction de PCR préalablement contrôlée est dépo-sée sur gel d’agarose pour une électrophorèse (§ 4.1.3). La bande de gel contenant l’ADN d’intérêt est découpée sous UV. L’agarose contenant les fragments d’ADN est dissous, additionné d’isopropanol, et déposé sur une colonne fixant l’ADN. Après centrifugation, l’agarose est éliminé puis, après une étape de lavage, l’ADN est élué dans 10 µl d’eau ultra pure.
4.2.2.3 Marquage radioactif de la sonde
Le marquage radioactif est effectué une heure à température ambiante sur 50 ng d’ADN (H20 qsp 5 µl), dénaturés 5 min à 100°C puis refroidis immé-diatement dans la glace, selon la procédure du kit Prime-a-Gene® Labeling System (Promega).
Mélange réactionnel pour une réaction :
ADN dénaturé (10 ng.µl-1) 5 µl Tampon de marquage 5X 10 µl dATP, dGTP, dTTP (500 µM chacun) 2 µl α32P-dCTP (10µCi.µl-1) 5 µl ADN polymérase I, Large (Klenow) fragment (5U.µl-1) 1 µl BSA dépourvue de nucléase (10 mg.ml-1) 2 µl H20 dépourvue de nucléase qsp 50 µl
La sonde est ensuite purifiée par filtration sur gel sur une colonne
de Sephadex G50 (Quick SpinTM columns, Boehringer Mannheim) pour éliminer les nucléotides non incorporés.
ANSELME Caroline 63 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
4.2.3 Hybridation et révélation
4.2.3.1 Préhybridation-hybridation
La membrane de nylon est d’abord réhydratée dans du SSC 2X, enroulée avec une feuille de nylon puis introduite dans une bouteille d’hybridation. Pour saturer les sites aspécifiques de la membrane, une étape de préhybri-dation est réalisée pendant 24 h à 50°C en présence d’une solution d’hybridation (formamide désionisé 50% (v/v), SDS 0,2% (p/v), Denhardt 5X (Ficoll 0,1% (p/v), polyvinylpyrrolidone 0,1% (p/v) ; BSA, 0,1% (p/v)), SSPE 5X, H20 DEPC) additionnée d’un mélange d’ADN de sperme de ha-reng et de saumon (0,5% (p/v) chacun) dénaturé pendant 15 min à 100°C et refroidi immédiatement dans la glace.
L’hybridation est ensuite réalisée pendant 24 h à 42°C dans un nouveau tampon d’hybridation additionné d’ADN de sperme de ha-reng/saumon dénaturé extemporanément et de la totalité de la sonde ra-dioactive dénaturée extemporanément à 100°C pendant 5 min et refroidie immédiatement dans la glace.
4.2.3.2 Lavages, exposition, révélation et normalisation
Pour éliminer les hybridations non spécifiques de la sonde, la membrane subit plusieurs lavages de stringence croissante jusqu’à ce que le signal soit clairement différencié du bruit de fond. Généralement, deux lavages de 15 min à 52°C (SSC 2X, SDS 0,1%, H20 DEPC) suivis de deux lavages de 15 min à 52°C (SSC 0,2X, SDS 0,1%, H20 DEPC) et éventuellement de deux lavages de 15 min à 60°C (SSC 0,1X, SDS 0,1%, H20 DEPC) sont suffisants pour éliminer le bruit de fond et les hybridations non spécifiques. Le suivi des lavages étant réalisé grâce à un compteur Geiger. La mem-brane est ensuite enveloppée dans un film de SARAN (Dow), mise en contact 4 à 12 h avec un écran qui est ensuite scanné dans un appareil STORMTM (Amersham) pour obtenir une image numérique des signaux ra-dioactifs présents sur la membrane.
Afin de normaliser les données obtenues, la membrane est stockée pour décroissance de la radioactivité pendant environ deux mois, puis ré-hybridée comme précédemment avec un gène de normalisation. Les don-nées sont ensuite traitées à l’aide du logiciel ImageQuant (Molecular Dy-namics). Les intensités des bandes obtenues pour les transcrits d’intérêt sont d’abord corrigées par rapport au bruit de fond de chaque membrane. La normalisation entre les échantillons s’effectue ensuite en divisant l’intensité corrigée de l’échantillon par celle obtenue avec le gène de nor-
ANSELME Caroline 64 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
malisation. Enfin, la normalisation entre membranes s’effectue en divisant l’intensité normalisée de l’échantillon par celle du témoin interne.
4.3 Soustraction d’ADNc (“Suppressive Subtractive Hybridization”)
La soustraction d’ADNc permet d’isoler des séquences présentes dans une population d’ARNm, mais absentes ou faiblement représentées dans une autre. La technique consiste à mélanger une préparation d’ADNc appelée “tester” obtenue à partir d’une population d’ARNm d’intérêt, avec un excès d’ADNc obtenus à partir d’une seconde population d’ARNm appelée “dri-ver”. Dans ce type de méthode, les séquences communes entre les deux po-pulations sont éliminées (d’où le terme de soustraction) par hybridation des ADNc communs aux deux populations. En utilisant ensuite la technique de PCR associée à des jeux d'adaptateurs spécifiques, seuls les ADNc spécifi-ques de l'une des deux conditions (“tester”) sont amplifiés par PCR. La soustraction d’ADNc a été réalisée à l’aide du kit “PCR-Select™ cDNA Subtraction Kit” (BD Biosciences Clontech).
4.3.1 Synthèse, amplification et purification des ADNc
La synthèse des ADNc correspondants aux deux populations d’ARN (“tes-ter” et “driver”) est réalisée à l’aide du kit “BD SMARTTM PCR cDNA syn-thesis kit” (BD Biosciences Clontech) selon les recommandations du four-nisseur. Ce kit permet grâce à un oligo(dT) de synthétiser les ADN complémentaires des ARNm à partir d’ARN totaux et, grâce à l’activité terminale transférase de la rétrotranscriptase, d’incorporer un adaptateur synthétique spécifiquement en 5’ pendant la synthèse de l’ADNc (Figure 9). Un jeu d’amorces s’hybridant au niveau de cet adaptateur et de l’oligo(dT) permet ensuite d’amplifier les ADNc complets. Ce jeu d’adaptateurs permet ainsi de diminuer considérablement la proportion de fragments d’ADNc obtenue généralement lors de la rétrotranscription et donc, d’enrichir la population d’ADNc en ADNc complet.
ANSELME Caroline 65 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Figure 9 : Principe de la rétrotranscription des ARNm à l’aide du kit “BD SMARTTM PCR cDNA synthesis kit” (BD Biosciences Clontech). 1-L’oligo(dT) spécialisé s’amorce sélectivement aux ARNm de l’échantillon d’ARN total. L’oligo contient une séquence pour la PCR et un site de restriction RsaI. 2-La rétrotrans-criptase (RT) atteint l’extrémité 5’ de l’ARNm matrice et ajoute une série de cyto-sines grâce à son activité terminale transférase. 3-Un oligo SMARTTM spécialisé s’hybride au niveau des cytosines, créant une nouvelle matrice pour la RT. Cette dernière poursuit la synthèse d’ADNc jusqu’à l’extrémité de l’oligo SMART incor-porant la séquence de celui-ci. Ainsi, les deux extrémités de l’ADNc comportent un adaptateur. 4-Les ADNc double brins sont ensuite amplifiés par PCR avec les amorces correspondant aux séquences de l’adaptateur. L’oligo SMART et l’oligo(dT) contiennent également un site de restriction RsaI.
Les ADNc sont synthétisés à partir de 1 µg d’ARN (H20 qsp 4 µl)
incubé en présence de 1 µl d’amorce oligo(dT) (12 µM) et de 1 µl de l’adaptateur SMART (12 µM) 2 min à 70°C. Le mélange est ensuite refroi-di rapidement sur glace. Sont alors ajoutés au mélange : 2 µl du tampon 5X, 1 µl de DTT (20 mM), 1 µl de dNTP (10 mM chacun) et 1 µl de rétrotrans-criptase. La réaction est réalisée une heure à 42°C puis le mélange est re-froidi rapidement sur glace avant d’être dilué par addition de 40 µl de tam-pon TE (1 mM EDTA, 10 mM Tris pH7,6).
Les PCR sont réalisées à partir de 1 µl du produit de la rétrotrans-cription, dans un volume final de 100 µl, à l’aide du kit “BD AdvantageTM 2 PCR” (BD Biosciences Clontech). Après optimisation du nombre de cy-cles de PCR (conservation des proportions initiales d’ADNc), les PCR sont réalisées comme décrit ci-après.
ANSELME Caroline 66 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
Conditions de PCR : Amorces PCR primers 240 nM dNTPs (chacun) 200 µM Tampon PCR 1 X BD Advantage 2 polymerase mix 1 X ADNc monobrin dilué 1 µl H20 PCR qsp 99 µl
17 cycles
6’
30’’
1’ 15’’
72°C 95°C
65°C Les produits de PCR sont ensuite purifiés par addition d’un vo-
lume de phénol:chloroforme:alcool isoamylique (25:24:1). Après centrifu-gation (10 000 g, 10 min), la phase supérieure est prélevée et ajoutée à 700 µl de butanol. Après une nouvelle centrifugation (10 000 g, 1 min), la phase supérieure est purifiée par filtration sur gel sur une colonne “BD chromaspin 1000” (BD Biosciences Clontech). Après purification, les pro-duits de PCR sont collectés dans 380 µl de TNE (10 mM Tris-HCl pH8, 10 mM NaCl, 0,1 mM EDTA,).
4.3.2 Digestion des ADNc, ligation aux adaptateurs et hybridations
Après purification, les ADNc sont digérés trois heures à 37°C par 10 unités de l’enzyme RsaI puis purifiés sur colonne à l’aide du kit “GenElute PCR clean-up” (Sigma) selon les recommandations du fournisseur.
Deux échantillons de 300 ng d’ADN du “tester” digérés par l’enzyme de restriction RsaI sont ensuite liés aux adaptateurs 1 et 2R res-pectivement, une nuit à 16°C. Un contrôle de soustraction réalisé à partir d’un mélange des deux milieux réactionnels (comportant donc les deux adaptateurs) est également soumis à une ligation durant la nuit.
La soustraction proprement dite est réalisée par deux hybridations successives. La première hybridation est réalisée pour chaque échantillon (1 et 2R), entre l’ADNc “tester” et l’ADNc “driver” dans un rapport 1:30 (p/p) pendant huit heures à 68°C après 1,5 min de dénaturation à 98°C. La seconde hybridation est réalisée une nuit à 68°C entre les deux échantillons issus de la première hybridation additionnés d’ADNc “driver” dénaturé ex-temporanément (rapport final 1:25). A l’issue de la seconde hybridation, l’échantillon est dilué dans 200 µl de tampon (20 mM HEPES pH6,6, 20 mM NaCl, 0,2 mM EDTA pH8) et incubé 7 min à 68°C.
4.3.3 Amplification des ADNc soustraits
Les ADNc soustraits, ainsi que le contrôle de soustraction, sont amplifiés par deux réactions de PCR successives précédées de la synthèse de la sé-
ANSELME Caroline 67 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
quence complémentaire des adaptateurs à l’extrémité des brins hybridés, par une incubation de 5 min à 75°C. Au cours de la première PCR, seuls les ADNc comportant les deux adaptateurs sont amplifiés. La seconde PCR est une PCR nichée qui permet de réduire le bruit de fond et d’enrichir l’échantillon en ADNc soustraits.
A l’issue de la seconde PCR, les produits de PCR sont analysés sur gel d’électrophorèse pour comparer le profil des ADNc soustraits et ce-lui du contrôle non soustrait. Si les profils sont bien différents, les ADNc soustraits sont purifiés sur colonne à l’aide du kit “GenElute PCR clean-up” (Sigma) selon les recommandations du fournisseur. La banque sous-traite est ensuite obtenue par clonage des produits de PCR purifiés dans le vecteur pCR®2.1-TOPO® (§ 4.7.1).
Conditions de PCR 1: Amorces PCR primer1 400 nM dNTPs 200 µM (chacun) Tampon PCR 1 X 50X Advantage polymerase Mix 1 X ADNc dilué 1 µl H20 PCR qsp 25 µl
25’’
27 cycles
1’30’’
30’’
10’’
72°C75°C
5’ 94 °C
66°C
Conditions de PCR 2: Amorce Nested PCR primer1 400 nM Amorce Nested PCR primer 2R 400 nM dNTPs 200 µM (chacun) Tampon PCR 1 X 50X Advantage polymerase Mix 1 X PCR 1 diluée 10 fois 1 µl H20 PCR qsp 25 µl
12 cycles
1’30’’
30’’
10’’
94 °C72°C
68°C
4.4 Obtention des séquences des ARNm à partir des EST (RACE)
Les séquences complètes des ADNc sont obtenues à partir des EST à l’aide du kit “BD smart Rapid Amplification of cDNA Ends (RACE) cDNA am-plification” (BD Biosciences Clontech), grâce à la technologie SMART dé-crite précédemment (Figure 9).
4.4.1 Obtention des ADNc (RT-PCR)
La première étape de la réaction consiste à rétrotranscrire les ARNm conte-nus dans l’échantillon d’ARN utilisé comme “tester” dans la technique de PCR soustractive. Deux rétrotranscriptions sont réalisées en parallèle, l’une pour l’incorporation de l’adaptateur contenant la séquence SMART à
ANSELME Caroline 68 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
l’extrémité 5’ des ARNm (5’ADNc) comme décrit précédemment, et l’autre, pour introduire la séquence SMART à l’extrémité 3’ de l’ARNm (3’ADNc) grâce à une amorce oligo(dT)-SMART (Figure 9).
Brièvement, pour chaque réaction, 1 µg d’ARN est incubé en pré-sence des adaptateurs associés (1,2 µM chacun) puis la réaction s’effectue 1,5 h à 42°C dans le milieu réactionnel (Reverse transcriptase, tampon 1X, 2 mM DTT, 1 mM dNTP mix). Les ADNc sont ensuite dilués par addition de 100 µl de tampon Tricine-EDTA (10 mM Tricine-KOH pH8,5, 1mM EDTA) et incubés 7 min à 72°C. Ces deux échantillons, 5’ADNc et 3’ ADNc sont alors prêts pour l’amplification respective des extrémités 5’ et 3’ des ADNc d’intérêt.
4.4.2 Obtention des fragments d’ADNc (RACE PCR)
Les fragments d’ADN situés de part et d’autre de l’EST sont obtenus par PCR grâce à la combinaison d’une amorce spécifique (GSP, “Gene Specific Primer”) et d’une amorce universelle (UMP, “Universal Primer Mix”). Les PCR sont réalisées comme décrit ci-dessous.
Condition de 5’ RACE PCR (ou 3’ RACE PCR) : Amorce GSP1 (ou GSP2) 200 nM Amorce UPM 1 X Tampon PCR 1 X 50X Advantage polymerase Mix 1 X dNTPs 200 µM (chacun) 5’ ADNc (ou 3’ADNc) 2,5 µl H20 PCR qsp 50 µl
30’’
70°C
30’’
5 cycles 5 cycles
72°C 94°C
72°C
3’ 94°C
3’ 30’’
72°C
3’ 30’’
94°C
25 cycles
30’’
68°C
Les premiers cycles de PCR sont réalisés à haute température
d’hybridation (72 et 70°C) pour favoriser la spécificité de la PCR au niveau de l’hybridation de l’amorce GSP (l’amorce UMP s’hybridant à l’extrémité de tous les ADNc). Après l’amplification préférentielle des fragments d’intérêt dans le mélange d’ADNc initial, l’amplification est réalisée avec une température d’hybridation plus basse (68°C) pour favoriser l’efficacité de la PCR. Après vérification de la présence d’une bande majoritaire par électrophorèse sur gel d’agarose, celle-ci est purifiée à partir du gel et le fragment d’ADN est cloné et séquencé comme décrit plus loin (§ 4.7).
Si la première PCR ne permet pas d’obtenir une bande majoritaire facilement isolable, une seconde PCR peut être réalisée, dans les mêmes conditions, avec une amorce spécifique située à l’intérieur du fragment am-plifié lors de la première PCR et de l’amorce NUP (“Nested Universal Pri-
ANSELME Caroline 69 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
mer”). Les séquences des amorces utilisées dans ce travail sont données dans l’Annexe I.
4.5 Amplification de fragment d’ADN génomique par longue PCR
L’amplification des fragments d’ADN à partir d’ADN génomique a été ré-alisée à l’aide du kit “Expand long template PCR system” (Roche applied science). Ce système est composé d’un mélange de deux polymérases ther-mostables, une Taq polymérase classiquement utilisée en PCR et une Tgo polymérase avec une activité 3’-5’ exonucléase permettant d’augmenter la fidélité de l’amplification. De plus, la combinaison de ces deux enzymes permet d’augmenter le rendement de la PCR et d’amplifier des fragments allant jusqu’à 22 kb. Les PCR ont été réalisées selon les re-commandations du fournisseur en testant pour chaque PCR les trois milieux réactionnels disponibles.
4.6 Marche sur le chromosome (“Genome Walking”)
La marche sur le chromosome est une approche qui permet d’obtenir la sé-quence complète d’un gène, à partir d’une partie de la séquence, grâce à l’amplification, de proche en proche, des fragments d’ADN. Cette approche est basée sur la digestion de l’ADNg par une enzyme de restriction et la li-gation d’adaptateurs aux extrémités des fragments obtenus. L’utilisation d’ADNg digéré permet ainsi de réduire la taille des fragments à amplifier. Un premier fragment est amplifié à l’aide d’une amorce spécifique et d’une amorce s’hybridant au niveau de l’adaptateur. L’obtention de la séquence de ce fragment permet alors de définir une nouvelle amorce spécifique et de réaliser une nouvelle PCR sur une autre digestion d’ADNg. L’utilisation de différentes enzymes de restriction à bouts francs (“blunt ends”) permet ain-si d’amplifier et de séquencer progressivement le gène d’intérêt. Les enzy-mes utilisées sont : AluI, DraI, EcoRV, PvuII, ScaI et StuI.
2,5 µg d’ADNg sont digérés une nuit à 37°C par 80 unités d’enzyme de restriction dans 100 µl du tampon de digestion. Après vérifi-cation de l’efficacité de la digestion par électrophorèse, l’enzyme et les sels sont éliminés par une extraction phénol-chloroforme-alcool-isoamylique de l’ADNg redissous dans un volume de 20 µl (§ 4.1.1). L’adaptateur est ob-tenu par une étape d’hybridation de 30 min à 28°C des deux oligonucléoti-des (1 mM chacun) synthétisés par la société Eurogentec : adaptator1 (5’-GTA-ATA-CGA-CTC-ACT-ATA-GGG-CAC-GCG-TGG-TCG-ACG-GCC-CGG-GCT-GGT-3’) et adaptator2 (5’-ACC-AGC-CC-3’, amine C7 en 3’). La ligation à l’adaptateur (5,9 µM) est réalisée par la T4 DNA Li-gase (Promega) une nuit à 16°C dans un volume de 8 µl contenant 4 µl
ANSELME Caroline 70 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
d’ADNg. La réaction est achevée par l’inactivation de l’enzyme 5 min à 72°C et la dilution par dix du produit de ligation dans du TE (10 mM Tris, 1 mM EDTA pH7,5).
Les fragments d’ADNg sont ensuite amplifiés par PCR nichées dans les conditions décrites ci-dessous.
Conditions de PCR 1: Amorce GSP1 200 nM Amorce AP1 200 nM dNTPs 200 µM (chacun) Tampon PCR 1 X 50X Advantage polymerase Mix 1 X ADNg-adaptateur 1 µl H20 PCR qsp 50 µl
72°C
2’’ 3’
7 cycles
94 °C 67 °C
2’’
3’
32 cycles
94 °C 4’
Conditions de PCR 2:
Amorce GSP2 200 nM Amorce AP2 200 nM dNTPs 200 µM (chacun) Tampon PCR 1 X 50X Advantage polymerase Mix 1 X PCR 1 diluée 50 fois 1 µl H20 PCR qsp 50 µl
72°C
2’’ 3’
5 cycles
94 °C 67 °C
2’’
3’
20 cycles
94 °C 4’
4.7 Clonage des produits de PCR, préparation de l’ADN plasmidique et
séquençage.
4.7.1 Clonage par “TOPO cloning” (Invitrogen)
La technique de “TOPO cloning” est une technique permettant de cloner des fragments de PCR sans avoir recours à des étapes de digestion par des enzymes de restriction. Le plasmide classique pCR®2.1-TOPO® et le plas-mide pCR®-XL-TOPO® approprié au clonage des fragments obtenus par longue PCR, sont disponibles sous forme linéarisée avec, à leurs extrémités 3’, une désoxythymidine et une topoisomérase I associée de manière cova-lente. Lors de l’amplification par PCR, la polymérase utilisée ajoute une désoxyadénosine aux extrémités 3’ des produits de PCR permettant l’insertion de ces produits dans le vecteur grâce à la désoxythymidine aux extrémités 3’ de ce dernier. Lors de l’insertion, la topoisomérase est libérée et exerce alors son activité de ligase.
La ligation dans le vecteur pCR®2.1-TOPO® est réalisée 30 min à température ambiante à partir de 2 à 4 µl d’ADN purifié (H2O qsp 4 µl) et de 1 µl du tampon contenant le vecteur pCR®2.1 (10 ng.µl-1 plasmide, 50% glycérol, 50 mM Tris-HCl pH7,4, 1 mM EDTA, 1 mM DTT, 0,1% Triton X-100, 100 µg.ml-1 BSA) additionnés de 1 µl de solution saline (300 mM
ANSELME Caroline 71 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
NaCl, 15 mM MgCl2). La ligation dans le vecteur pCR®-XL-TOPO® est ré-alisée 5 min à température ambiante à partir de 4 µl d’ADN purifié et de 1 µl du tampon contenant le vecteur pCR®XL (10 ng.µl-1 plasmide, 50% gly-cérol, 50 mM Tris-HCl pH7,4, 1 mM EDTA, 1 mM DTT, 0,1% Triton X-100, 100 µg.ml-1 BSA). Après les 5 min d’incubation, la réaction est ar-rêtée par l’ajout d’1 µl de “stop solution” (300 mM NaCl, 60 mM MgCl2).
4.7.2 Transformation des cellules par électroporation
L’électroporation est une technique de transformation, utilisant un courant électrique pour perméabiliser la membrane des bactéries compétentes (E. coli, TOP10, Invitrogen) afin d’y faire pénétrer de l’ADN. Cette étape d’électroporation est réalisée grâce au système Gene Pulser II (Biorad). 4.108 bactéries compétentes sont soumises à une décharge électrique en présence de 2 µl du produit de ligation. Après l’impulsion électrique, les bactéries sont incubées 1 h à 37°C dans 250 µl de milieu SOC (2% tryp-tone, 0,5% extrait de levure, 10 mM NaCl, 2,5 mM KCl, 10 mM MgCl2, 10 mM MgSO4, 20 mM glucose) afin de permettre aux cellules transfor-mées d’exprimer le gène de résistance à un antibiotique présent sur le plasmide. Les cultures sont ensuite étalées sur milieu sélectif, LB-agar (Sigma) additionné d’antibiotique, où seules les bactéries transformées peuvent se développer. Après une nuit à 37°C, les boites sont conservées à 4°C.
Lorsque les fragments sont clonés dans le pCR®2.1 porteur d’un gène de résistance à l’ampicilline, les cultures sont étalées sur un milieu sé-lectif contenant de l’ampicilline (50 µg.ml-1) sur lequel ont préalablement été étalés 40 µl d’une solution à 40 mg.ml-1de Xgalactose (Xgal) dans du diméthylformamide. La distinction entre les cellules possédant un plasmide vide et celle possédant un plasmide porteur d’un insert est ensuite réalisée par α-complémentation. Le vecteur pCR2.1 possède un fragment du gène LacZ codant pour la protéine de la β-galactosidase, une enzyme qui dégrade le chromogène X-gal présent dans le milieu en un produit bleu. Durant l’étape de ligation, le produit de PCR cloné s’insère dans le vecteur au sein de ce gène altérant le codage du gène LacZ (clones LacZ-) : le chromogène n’est donc pas hydrolysé et les colonies apparaissent blanches. Pour obtenir les plasmides recombinants, seules les colonies blanches sont sélectionnées.
Dans le cas d’un clonage dans le vecteur pCR-XL-TOPO, la sélec-tion des bactéries possédant un plasmide recombinant est réalisée grâce à la présence d’un gène de résistance à la kanamycine et d’un gène létal, le gène ccdB d’E. coli. Lors de l’étape de ligation, le produit de PCR s’insère dans le vecteur au sein du gène létal ccdB ce qui altère le codage du gène et ainsi
ANSELME Caroline 72 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
permet à la bactérie de se développer. Ainsi, seules les bactéries recombi-nantes pourront se développer sur un milieu LB gélosé additionné de kanamicyne (50 µg.ml-1)
Les colonies bactériennes possédant les plasmides recombinants peuvent être conservées à -80° à partir d’un volume d’une culture d’une nuit additionnée d’un volume de solution de glycérol (glycérol 65%, 0,1 mM MgSO4, 25 mM Tris-HCl pH8).
4.7.3 Préparation d’ADN plasmidique
L’extraction d’ADN plasmidique est réalisée par lyse alcaline à l’aide du kit “Nucleospin plasmid” (Macherey-Nagel) sur 1,5 ml d’une culture d’une nuit. Cette technique est basée sur une dénaturation différentielle entre l’ADN plasmidique superenroulé et l’ADN chromosomique de structure re-lâchée, suivie d’une purification de l’ADN plasmidique sur colonne. Briè-vement, le culot de bactéries est suspendu dans un premier tampon addi-tionné de RNases puis les bactéries sont lysées, et l’ADN dénaturé, par l’ajout d’un tampon contenant du SDS et de la soude. Enfin, l’ajout d’une solution de neutralisation entraîne la formation d’un précipité de protéines et d’ADN chromosomique alors que l’ADN plasmidique reprend sa forme soluble native. L’ADNg et le complexe SDS-protéine sont éliminés par centrifugation (10 000 g, 10 min) et le surnageant contenant l’ADN plasmi-dique est déposé sur colonne pour purification. Après lavage, l’ADN plas-midique est élué dans 50 µl de tampon Tris-HCl 5 mM, pH8,5.
4.7.4 Séquençage des fragments d’ADN
La réaction de séquençage est généralement effectuée sur séquenceur auto-matique par la société Génome Express (Meylan, France) à l’exception des EST des banques soustraites qui ont été séquencées par Vicente Pérèz-Brocal, en collaboration avec A. Moya et A. Latorre, à “l’Institut Cavanil-les de Biodiversitat i Biologia Evolutiva”, Université de Valence (Espa-gne).
4.8 RT-PCR quantitative en temps réel
La technique de RT-PCR quantitative en temps réel est une technique per-mettant de quantifier précisément les ARN d’un échantillon. Elle comprend une étape de rétrotranscription des ARNm et une étape de PCR en temps réel qui permet, comme son nom l’indique, de suivre l’accumulation des
ANSELME Caroline 73 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
produits d’amplification. Parmi les différents systèmes de PCR en temps réel disponibles actuellement, nous avons utilisé le système LightCycler® (Roche applied science) disponible dans la plateforme du DTAMB (Déve-loppement de Techniques et Analyse Moléculaire de la Biodiversité) de l’IFR 41 (Instituts Fédératifs de Recherche, université Claude Bernard Lyon 1). Les PCR sont réalisées à l’aide du kit “LightCycler® FastStart DNA Master SYBR Green I” (Roche applied science).
Le principe de ce système est basé sur une molécule, le SYBR Green, qui n’émet de la fluorescence que lorsqu’elle est liée à de l’ADN double brin. Au cours de l’élongation, le SYBR green se lie progressive-ment à l’ADN double brin nouvellement formé. La PCR peut donc être sui-vie en temps réel par l’accumulation de la fluorescence qui est directement proportionnelle à la quantité d’ADN double brin. Pour quantifier les pro-duits de PCR, la fluorescence est mesurée à la fin de l’étape d’élongation de chaque cycle. La dénaturation de l’ADN entraîne ensuite la libération des molécules de SYBR green et la chute de la fluorescence.
L’utilisation du SYBR green permet également, à la fin de la PCR, de vérifier la spécificité de la réaction et l’absence de dimères des amorces par la réalisation d’une courbe de fusion. Pour cela, la température du mé-lange réactionnel est élevée progressivement entraînant la dénaturation des produits de PCR et donc la chute de la fluorescence à la température d’hybridation de l’ADN (Tm). Chaque Tm étant spécifique du produit de PCR (ou du dimère des amorces), une chute de fluorescence correspond à un produit de PCR donné. La mesure de la fluorescence permet ainsi de dé-terminer le nombre de produits de PCR obtenus en fin de réaction.
4.8.1 Rétrotranscription des ARNm
La rétrotranscription des ARNm est réalisée à l’aide du kit “Super ScriptTM First strand synthesis system for RT-PCR” (Invitrogen) à partir de 1 µg d’ARN. L’ARN est dénaturé 5 min à 65°C en présence de 0,5 µg d’amorce oligo(dT) et de 10 nmol de dNTP puis le mélange (10 µl) est refroidi sur glace. La rétrotranscription est ensuite réalisée 1 h à 42°C dans un volume final de 20 µl de milieu réactionnel (tampon 1X, MgCl2 5 mM, DTT 10 mM, RNaseOUTTM 1/20 (v/v), SuperScriptTM II RT 50 U). La réaction est achevée par 15 min d’incubation à 70°C. Les ARN de la matrice sont ensuite dégradés par une incubation de 20 min à 37°C en présence de deux unités de RNase H. Les ADNc sont enfin dilués dans 4 volumes d’H2O puis aliquotés pour limiter leur dégradation par congélations-décongélations.
ANSELME Caroline 74 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
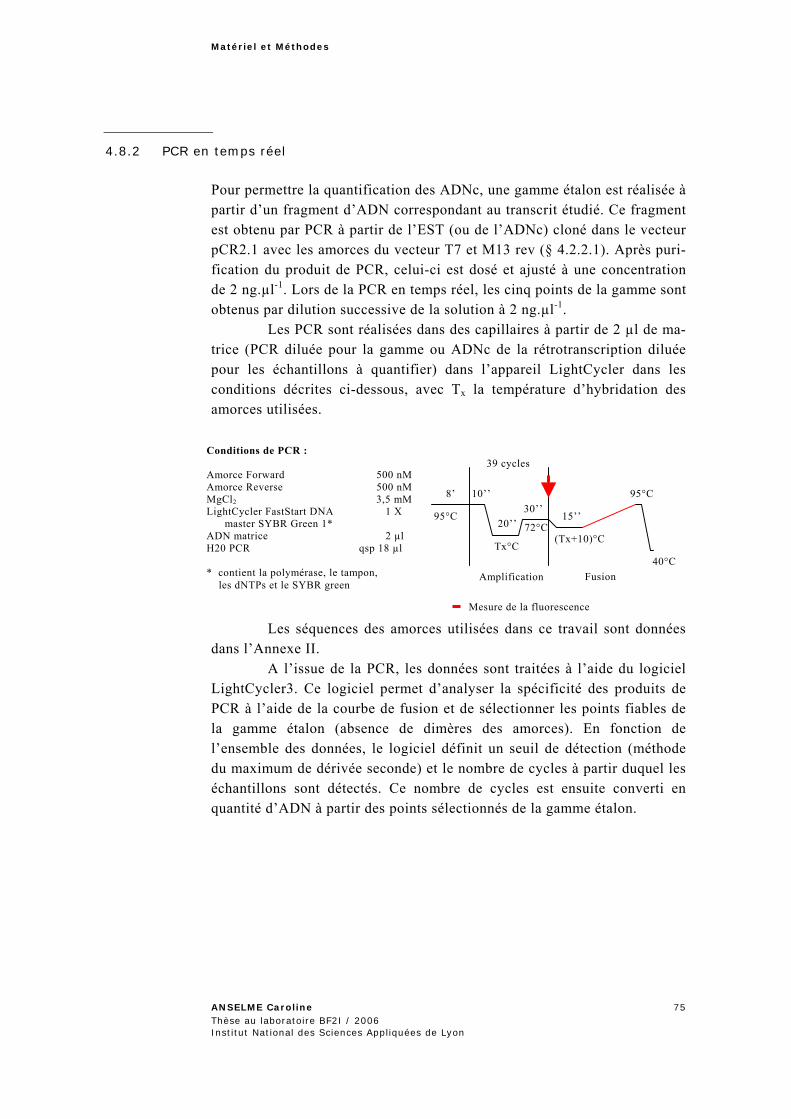
Matériel et Méthodes
4.8.2 PCR en temps réel
Pour permettre la quantification des ADNc, une gamme étalon est réalisée à partir d’un fragment d’ADN correspondant au transcrit étudié. Ce fragment est obtenu par PCR à partir de l’EST (ou de l’ADNc) cloné dans le vecteur pCR2.1 avec les amorces du vecteur T7 et M13 rev (§ 4.2.2.1). Après puri-fication du produit de PCR, celui-ci est dosé et ajusté à une concentration de 2 ng.µl-1. Lors de la PCR en temps réel, les cinq points de la gamme sont obtenus par dilution successive de la solution à 2 ng.µl-1.
Les PCR sont réalisées dans des capillaires à partir de 2 µl de ma-trice (PCR diluée pour la gamme ou ADNc de la rétrotranscription diluée pour les échantillons à quantifier) dans l’appareil LightCycler dans les conditions décrites ci-dessous, avec Tx la température d’hybridation des amorces utilisées.
ar r
Conditions de PCR : Amorce Forw d 500 nM Amorce Reve se 500 nM MgCl2 3,5 mM LightCycler FastStart DNA 1 X master SYBR Green 1* ADN matrice 2 µl H20 PCR qsp 18 µl * contient la polymérase, le tampon, les dNTPs et le SYBR green
40°C
95°C 8’ 10’’
20’’ 30’’
15’’ 95°C
Tx°C
72°C(Tx+10)°C
39 cycles
Amplification Fusion
Mesure de la fluorescence Les séquences des amorces utilisées dans ce travail sont données
dans l’Annexe II. A l’issue de la PCR, les données sont traitées à l’aide du logiciel
LightCycler3. Ce logiciel permet d’analyser la spécificité des produits de PCR à l’aide de la courbe de fusion et de sélectionner les points fiables de la gamme étalon (absence de dimères des amorces). En fonction de l’ensemble des données, le logiciel définit un seuil de détection (méthode du maximum de dérivée seconde) et le nombre de cycles à partir duquel les échantillons sont détectés. Ce nombre de cycles est ensuite converti en quantité d’ADN à partir des points sélectionnés de la gamme étalon.
ANSELME Caroline 75 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
4.9 Macroarrays
La technique de macroarray est une méthode permettant d’étudier le profil d’expression d’un ensemble de gènes dans différentes conditions physiolo-giques. Cette technique est basée sur la fixation sur une membrane de nylon des produits de PCR correspondant aux gènes étudiés puis, à l’hybridation d’une sonde radioactive constituée d’un mélange d’ADNc correspondant à une condition physiologique donnée.
4.9.1 Synthèse des sondes radioactives par rétrotranscription
La synthèse des sondes radioactives est réalisée à l’aide du kit “Super ScriptTM First strand synthesis system for RT-PCR” (Invitrogen) à partir des échantillons d’ARN utilisés dans la technique de PCR soustractive et d’ARN de bactériomes.
10 µg d’ARN sont dénaturés 5 à 10 min à 65°C en présence de 1 µg d’amorce oligo(dT) puis refroidis sur glace. La rétrotranscription est ensuite réalisée 1,5 h à 42°C en présence de dCTP radioactif dans 40 µl de milieu réactionnel (tampon 1X, MgCl2 5mM, DTT 10 mM, RNaseOUTTM 1/20 (v/v), dATP 1 mM, dTTP 1 mM, dGTP 1 mM, α32P-dCTP 100 µCi, SuperScriptTM II RT 200U). La réaction est arrêtée par 15 min d’incubation à 70°C. La sonde est ensuite purifiée par filtration sur gel (“Quick spin”, Roche Molecular Biochemicals) et la radioactivité est comptée pour vérifier l’équivalence des sondes synthétisées.
4.9.2 Préparation des membranes
Les EST des clones de la banque soustraite sont amplifiées à partir des stocks bactériens conservés à -80°C (§ 4.7.2). Les PCR sont réalisées avec les amorces de la soustraction, Nested PCR primer 1 et 2R dans les condi-tions décrites dans le paragraphe 4.2.2.1 à une température d’hybridation de 68°C. Les produits de PCR sont ensuite contrôlés par électrophorèse et un volume fixe de 10 µl de chaque PCR non purifiée a été utilisé pour chaque dépôt.
Les membranes sont réalisées à l’aide du système de microfiltra-tion Bio-Dot® (Biorad) qui permet la réalisation de membranes comportant 96 dépôts. Les membranes de nylon (HybondTM-N, Amersham) préalable-ment humidifiées dans un tampon SSC 6X sont montées dans le système Bio-Dot selon les recommandations du fournisseur. Après l’application de
ANSELME Caroline 76 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
100 µl/puits de SSC 6X et élimination sous vide, 10 µl/puits de produit de PCR sont déposés sur la membrane. La solution est ensuite éliminée sous vide alors que l’ADN est retenu sur la membrane. Après un lavage par 100 µl/puits de SSC 6X, la membrane est transférée 10 min dans de la soude 0,6 M pour dénaturer l’ADN, puis dans un tampon Tris-HCl 0,5 M pH7,5 avant d’être rincée dans des bains de SSC 2X. L’ADN est fixé sur la membrane par traitement à 80°C pendant deux heures. Les membra-nes sont ensuite préhybridées (24 heures), hybridées (48 heures), puis la-vées et révélées comme décrit dans le paragraphe 4.2.3.
L’analyse quantitative des membranes est réalisée à l’aide du logi-ciel ImageQuant (Molecular Dynamics). La normalisation des données s’effectue en divisant l’intensité du signal de l’EST par celle obtenue avec le gène de normalisation.
5 Techniques d’analyse de séquences
5.1 Analyse des séquences à l’aide du programme Mac Molly Tetra (GeneSoft)
La première étape de l’analyse des résultats du séquençage consiste à iden-tifier les différents éléments qui constituent la séquence : le vecteur de clo-nage, les amorces spécifiques ou universelles, les adaptateurs, les séquen-ces connues et les nouvelles séquences. Cette première analyse est réalisée par alignement des séquences à l’aide des applications du programme Mac Molly Tetra (GeneSoft).
Les séquences peuvent alors être nettoyées des éléments issus de la construction (vecteur, amorces universelles, adaptateurs) et intégrées aux séquences déjà identifiées dans le cas de la reconstitution d’une séquence complète d’un ADNc (RACE) ou d’un gène. Les séquences peuvent ensuite être analysées par comparaison aux banques de données.
5.2 Analyse des séquences par comparaison aux banques de données
La recherche de séquences homologues dans les banques de données est ré-alisée grâce à l’application BLAST (“Basic Local Alignment Search Tool”) disponible sur le site NCBI (“National Center of Biotechnology Informa-tion”, http://www.ncbi.nlm.nih.gov).
L’analyse au niveau des protéines théoriques (traduction, composition en acides aminés, localisation cellulaire putative…) a été réalisée à l’aide d’applications disponibles sur le site ExPASy (“Expert Protein Analysis System”, http://us.expasy.org). La recherche des motifs
ANSELME Caroline 77 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
lysis System”, http://us.expasy.org). La recherche des motifs protéiques a ensuite été réalisée à l’aide d’un programme qui combine différentes mé-thodes d’identification des domaines protéiques décrits dans les bases de données : l’InterProScan (http://www.ebi.ac.uk/InterProScan).
5.3 Analyse des EST des banques soustraites
Les séquences des EST sont nettoyées manuellement et les sites de restric-tion RsaI sont identifiés pour décomposer les séquences lorsque celles-ci sont le résultat de la ligation de plusieurs EST (chimères). Les protéines putatives définies par les six phases de lecture possibles des EST sont en-suite comparées aux banques protéiques (BLASTx) à l’aide d’une applica-tion disponible sur la plateforme du Pôle BioInformatique Lyonnais (PBIL). Contrairement aux applications du site NCBI qui ne permettent de traiter qu’une séquence à la fois, l’application utilisée, dans cette analyse, permet de traiter l’ensemble des EST en une seule requête. A partir des identifiants des protéines issues du BLASTx, les informations associées à ces protéines sont obtenues grâce au “Sequence Retrieval System” (SRS) disponible sur le site ExPaSy (http://us.expasy.org/srs5). Les résultats ob-tenus, comme l’identifiant, la E-value, ainsi que la description des protéi-nes, les mots-clés qui leur sont associés et l’organisme dont elles provien-nent, sont organisés dans un tableau de données pour visualiser l’ensemble des données et accéder à la banque Swiss-Prot-trEMBL à l’aide de liens hypertextes.
6 Techniques de biochimie
6.1 Collecte d’hémolymphe et extraction des peptides
Le système de collecte d’hémolymphe est constitué d’un cône de 1 ml pla-cé, grâce à un adaptateur (microtube 0,5 ml Eppendorf coupé), sur un tube Eppendorf contenant 30 µl d’acide trifluoroacétique (TFA) 0,1 % et 10 µl d’une solution saturée de phénylthiourée (PTU) (5 mg.ml-1). Le PTU est un inhibiteur de la phénoloxydase qui limitera la mélanisation de l’hémolymphe et qui servira d’étalon interne pour la Chromatographie Li-quide à Haute Performance (HPLC).
Des lots de 25 larves sont déposés dans les cônes et les larves sont ensuite blessées à l’aide d’une aiguille d’entomologie pour faciliter l’extraction. La collecte de l’hémolymphe est réalisée par une centrifuga-
ANSELME Caroline 78 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
tion des systèmes Eppendorf-cône (10 000 g, 1 min) permettant de broyer/presser les larves lors du passage par l’extrémité du cône et d’éliminer ensuite les débris larvaires. Le surnageant contenant l’hémolymphe est additionné de 70 µl de TFA 0,1% et centrifugé de nou-veau (15 000 g, 10 min). Le deuxième surnageant est collecté à la seringue Hamilton et conservé trois heures à 4°C pour favoriser la précipitation des protéines. L’extraction est achevée par une dernière centrifugation (15 000 g, 10 min) pour éliminer le culot protéique. Le dernier surnageant contenant les peptides de l’hémolymphe est collecté à la seringue Hamilton et évaporé sous vide d’air au Speed-Vac.
6.2 Séparation des peptides de l’hémolymphe par chromatographie
liquide à haute performance (HPLC)
La technique d’HPLC est une technique de séparation basée sur la diffé-rence de polarité des composants d’un échantillon. La séparation des pepti-des de l’hémolymphe est réalisée en phase inverse grâce à une phase sta-tionnaire apolaire (C18) et une phase mobile polaire (H20 90%, CH3CN 10%, TFA 0,1%) dont on diminue progressivement la polarité en augmen-tant la proportion d’acétonitrile (CH3CN). Les peptides sont ainsi élués pro-gressivement (et donc séparés) en fonction de leur polarité.
Les extraits d’hémolymphe sont repris dans 130 µl de phase mo-bile initiale et la solution est ensuite filtrée (0,45 µm). 100 µl sont injectés sur une colonne Nucléosil C18 (250×4,6 mm, 5 µm, 300Å). Les paramètres chromatographiques (gradient d’élution, débit et détection) sont donnés ci-dessous.
Le solvant des fractions collectées à la sortie de la chaîne est éva-
poré sous vide d’air et les échantillons sont conservés à -20°C.
ANSELME Caroline 79 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Matériel et Méthodes
6.3 Test d’activité antibactérienne en milieu liquide
Les fractions testées sont resuspendues dans 50 µl de PBS et additionnées à 50 µl d’une solution à 6×107 bactéries.ml-1 obtenue par dilution d’une culture d’E. coli (TOP 10, Invitrogen) en phase exponentielle. Le test est réalisé en plaque de titration 96 puits à 37°C sous agitation. L’absorbance à 595 nm est mesurée dans un lecteur de plaques PowerWave x (Bio-Tek) toutes les 20 min sur une durée de cinq heures.
ANSELME Caroline 80 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats
ANSELME Caroline 81 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1 Identification des gènes de la réponse
immunitaire de l’hôte
L’étude moléculaire de la perception des endocytobiotes par l’hôte et de sa réponse immunitaire exige la connaissance des gènes impliqués dans la re-connaissance et l’élimination des bactéries. Chez le charançon S. zeamais, aucune séquence de gènes de la réponse immunitaire n’était connue, à l’exception d’un gène de la famille des PGRP (Heddi et al., 2005). L’enjeu principal de cette thèse était donc d’identifier des gènes de la réponse hu-morale activée par les bactéries Gram (-). Dans le but d’intégrer notre étude de la réponse immunitaire de l’hôte dans un cadre évolutif, et de compren-dre comment évolue l’interaction hôte-symbiote parallèlement à la réduc-tion de la taille du génome bactérien, une étude comparative a été initiée entre le modèle S. zeamais-SZPE et le modèle A. pisum–Buchnera dont la symbiose remonte à 150-200 MA, et où le génome bactérien ne code ni SSTT, ni certains éléments de la paroi bactérienne comme les LPS et les phospholipides.
1.1 Gènes de la réponse immunitaire de Sitophilus zeamais
Pour identifier les Gènes de la Réponse Immunitaire (GRI) du charançon S. zeamais, j’ai réalisé une soustraction d’ADNc entre des larves aposymbiotiques infectées par E. coli et des larves aposymbiotiques témoins (non traitées). Afin d’obtenir un large spectre de gènes intervenant dans plusieurs phases de la réponse immunitaire (i.e. reconnaissance du microorganisme, transmission du signal, élimination du microorganisme et régulation de la réponse), la soustraction a été réalisée avec un ADNc “tester” synthétisé à partir d’un mélange en quantités égales de trois échantillons d’ARN extraits 3, 6 et 12 h après infection par E. coli.
Après les étapes de digestion par l’enzyme de restriction RsaI et de purification des ADNc (voir Matériel et Méthodes), la quantité d’ADNc “driver” obtenue à partir des larves aposymbiotiques témoins était infé-rieure à la quantité recommandée par le fournisseur pour poursuivre la soustraction. Les quantités d’ADNc du “tester” et du “driver” utilisées dans la suite de la soustraction ont donc été adaptées en fonction des quantités disponibles, pour pouvoir conserver la proportion entre le “tester” et le “driver”.
ANSELME Caroline 82 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
A l’issue de cette soustraction, 527 clones ont été isolés, et, après vérification de leur contenu plasmidique, 513 plasmides ont été envoyés à Valence (Espagne), où ils ont été séquencés dans le cadre d’une collabora-tion. Après élimination des séquences du vecteur et des amorces, j’ai obte-nu 507 séquences d’EST. Ces séquences ont été analysées par comparaison aux banques de données protéiques, sans limitation du degré de similitude (Evalue<10), et les EST présentant une similitude de séquence avec d’autres organismes ont été répertoriées dans le Tableau A-III présenté en annexe.
Le classement des ces EST par catégories fonctionnelles a mis en évidence l’efficacité de la soustraction puisque 12,7 % des EST de la ban-que soustraite correspondent à des peptides antibactériens et 5,7 % des EST à d’autres protéines de la réponse immunitaire (Figure 10). Il est important de signaler que dans le cas des EST présentant des similitudes avec des sé-quences de bactéries, de virus ou de champignons, ces similitudes sont le plus souvent non significatives (Evalue>1) (Tableau A-III) et sont généra-lement obtenues sur des EST correspondant à des séquences courtes ou non codantes.
Figure 10 : Distribution en catégories fonctionnelles des EST de la banque soustraite de S. zeamais.
Les EST présentant une similitude de séquence avec des peptides ou des protéines de l’immunité sont répertoriées dans le Tableau III. Ces peptides et protéines incluent des peptides antibactériens, des lysozymes, une protéine homologue de la thaumatine de plantes, des protéines de la famille PGRP, une protéine homologue d’un régulateur de la réponse im-munitaire des mammifères (Tollip) et enfin, des protéases et des inhibiteurs de protéases dont des éléments de la cascade prophénoloxydase (PPAF, Serpines).
ANSELME Caroline 83 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
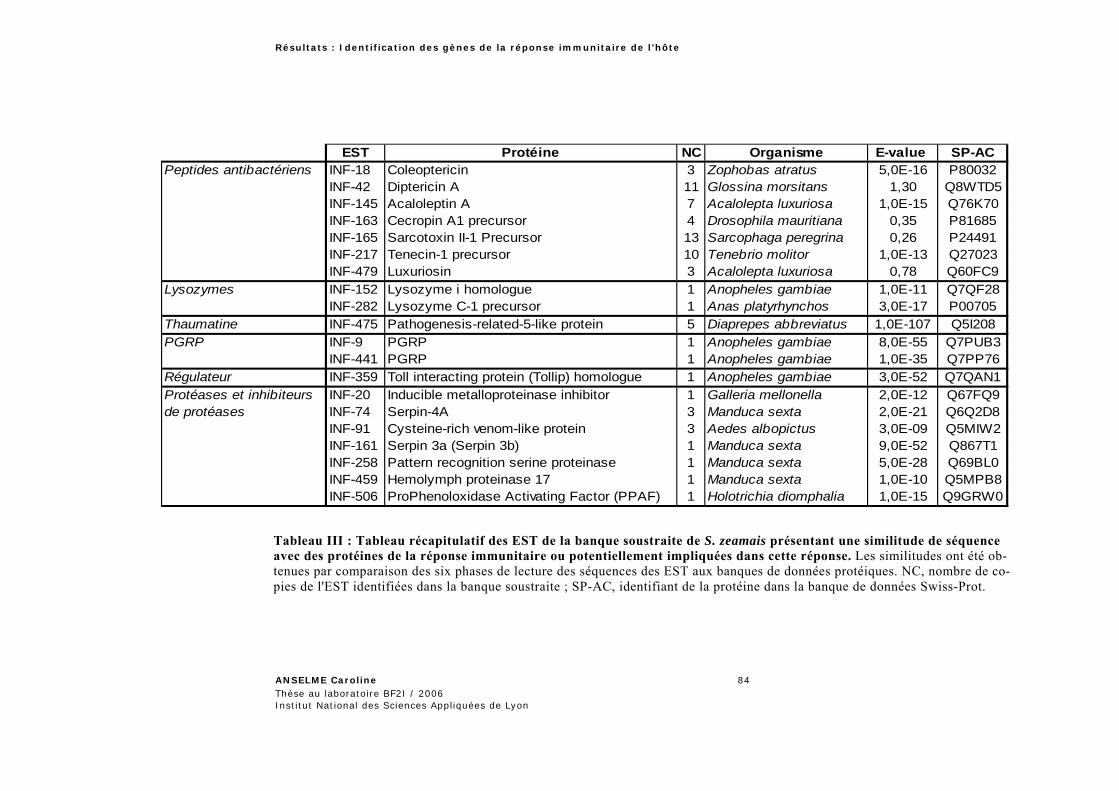
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
ANSELME Caroline 84
F-18 Coleoptericin 3 Zophobas atratus 5,0E-16 P80032F-42 Diptericin A 11 Glossina morsitans 1,30 Q8WTD5F-145 Acaloleptin A 7 Acalolepta luxuriosa 1,0E-15 Q76K70F-163 Cecropin A1 precursor 4 Drosophila mauritiana 0,35 P81685F-165 Sarcotoxin II-1 Precursor 13 Sarcophaga peregrina 0,26 P24491F-217 Tenecin-1 precursor 10 Tenebrio molitor 1,0E-13 Q27023F-479 Luxuriosin 3 Acalolepta luxuriosa 0,78 Q60FC9
Lysozymes F-152 Lysozyme i homologue 1 Anopheles gambiae 1,0E-11 Q7QF28F-282 Lysozyme C-1 precursor 1 Anas platyrhynchos 3,0E-17 P00705
Thaumatine F-475 Pathogenesis-related-5-like protein 5 Diaprepes abbreviatus 1,0E-107 Q5I208PGRP F-9 PGRP 1 Anopheles gambiae 8,0E-55 Q7PUB3
F-441 PGRP 1 Anopheles gambiae 1,0E-35 Q7PP76Régulateur F-359 Toll interacting protein (Tollip) homologue 1 Anopheles gambiae 3,0E-52 Q7QAN1Protéases et inhibiteurs F-20 Inducible metalloproteinase inhibitor 1 Galleria mellonella 2,0E-12 Q67FQ9de protéases F-74 Serpin-4A 3 Manduca sexta 2,0E-21 Q6Q2D8
F-91 Cysteine-rich venom-like protein 3 Aedes albopictus 3,0E-09 Q5MIW2F-161 Serpin 3a (Serpin 3b) 1 Manduca sexta 9,0E-52 Q867T1F-258 Pattern recognition serine proteinase 1 Manduca sexta 5,0E-28 Q69BL0F-459 Hemolymph proteinase 17 1 Manduca sexta 1,0E-10 Q5MPB8F-506 ProPhenoloxidase Activating Factor (PPAF) 1 Holotrichia diomphalia 1,0E-15 Q9GRW0
IN IN IN IN IN ININ IN IN IN IN IN IN ININ IN IN IN IN IN
EST Protéine NC Organisme E-value SP-ACPeptides antibactériens
Tableau III : Tableau récapitulatif des EST de la banque soustraite de S. zeamais présentant une similitude de séquence avec des protéines de la réponse immunitaire ou potentiellement impliquées dans cette réponse. Les similitudes ont été ob-tenues par comparaison des six phases de lecture des séquences des EST aux banques de données protéiques. NC, nombre de co-pies de l'EST identifiées dans la banque soustraite ; SP-AC, identifiant de la protéine dans la banque de données Swiss-Prot.
Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
On constate que la plupart des similitudes de séquence sont signi-ficatives (Evalue<1×10-9). En revanche, certaines similitudes avec des pep-tides antibactériens, tels que la cécropine, la diptéricine et la sarcotoxine, sont peu significatives (Evalue>0). Compte tenu de la redondance de ces EST dans la banque soustraite (4, 11 et 13 clones respectivement), nous avons choisi de poursuivre l’étude des gènes correspondants.
1.2 Gènes de la réponse immunitaire d’Acyrthosiphon pisum
La seconde soustraction d’ADNc a été réalisée entre des larves de pucerons aposymbiotiques témoins et des larves aposymbiotiques infectées par E. coli, selon le même plan expérimental que celui établi pour le charançon. A l’issue de cette soustraction, 521 clones ont été isolés et après vérifica-tion de leur contenu plasmidique, 509 plasmides ont été séquencés à Va-lence (Espagne).
Après l’analyse des 192 premières séquences, aucune EST ne pré-sentait une homologie avec un gène immunitaire connu. Aussi, afin de véri-fier l’efficacité de la soustraction avant de séquencer le reste des clones, nous avons choisi d’utiliser une méthode indirecte en comparant les 192 premières EST obtenues à la banque d’EST du Consortium Internatio-nal de la Génomique des Aphides (IAGC). Cette banque comptait près de 40 000 EST au moment de cette analyse (Sabater-Munoz et al., 2006). La comparaison de ces deux banques a révélé que seulement 50% des EST de la banque soustraite présentaient une similitude de plus de 90% avec des EST de l’IAGC. Ceci nous a laissés supposer que l’absence de similitude avec des gènes de la réponse immunitaire ne serait pas le résultat d’un pro-blème technique lié à la soustraction et nous a encouragés à poursuivre le séquençage des EST de la banque d’A. pisum. A l’issue du séquençage complet, la moitié des séquences seulement trouve leur homologue dans la banque de l’IAGC.
La comparaison des 494 séquences du puceron aux banques de données protéiques, n’a permis d’identifier aucun peptide antibactérien, ni lysozyme, ni PGRP. En revanche, certaines EST présentent des similitudes de séquence avec des éléments des voies de signalisation immunitaire des mammifères telles que les protéines “TNF Receptor Associated Factor 3 in-teracting protein” (Evalue=3×10-9) et “Immune signaling kinase MEK3” (Evalue=2×10-52) (Tableau A-IV en annexe). De plus, il existe une faible redondance des EST de la banque du puceron par rapport au nombre d’EST redondantes obtenues dans la banque soustraite du charançon. Par ailleurs, 210 séquences (soit 42% des EST) ne présentent aucune similitude signifi-cative avec des séquences des banques de données protéiques (Evalue>1).
ANSELME Caroline 85 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3 Analyse et caractérisation des GRI
1.3.1 Analyse globale des banques soustraites
Suite à l’analyse in silico des EST de la banque soustraite d’A. pisum qui n’a pas révélé de gènes de la réponse immunitaire, nous avons mesuré, par macroarray, l’expression des EST obtenues chez les insectes témoins et les insectes infectés par E. coli afin de vérifier le caractère différentiel des EST. Cette expérience a aussi été réalisée sur les EST du charançon S. zeamais pour tenter d’identifier d’éventuels gènes sans homologie avec des GRI connus mais qui seraient induits par l’infection bactérienne.
Ainsi, 268 EST non redondantes du charançon et 344 EST non re-dondantes du puceron ont été analysées. Les membranes de Dot blot ont été réalisées en double exemplaire, l’une a été hybridée avec une sonde ra-dioactive obtenue par rétrotranscription à partir des ARN extraits des insec-tes témoins (sonde “témoin”) et l’autre, avec une sonde radioactive obtenue à partir des ARN extraits des insectes infectés (sonde “infectée”).
Les résultats obtenus sur quatre des quatorze membranes sont pré-sentés dans la Figure 11. Les autres membranes sont présentées dans l’Annexe III.
ANSELME Caroline 86 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
S. zeamais A. pisum
Larves infectées par E. coli Larves infectées par E. coli
Larves témoins Larves témoins
Figure 11 : Membranes de Dot-blot comportant les EST des banques soustrai-tes de S. zeamais (à gauche) et d’A. pisum (à droite). Chaque membrane, réalisée en double exemplaire, comporte un témoin négatif correspondant à une amplifica-tion réalisée en absence d’ADN (en rouge) et un témoin de normalisation (gapdh, en bleu). Les membranes du haut ont été hybridées avec une sonde obtenue à partir des larves témoins et celles du bas, avec une sonde obtenue à partir des larves pi-quées par une aiguille infectée par E. coli. Chaque dépôt correspond à un volume fixe de 10 µl et seules les comparaisons intermembranes sont possibles.
1.3.1.1 Expression différentielle des EST de Sitophilus zeamais
Une première analyse qualitative des membranes révèle l’existence de plu-sieurs signaux d’intensité différente entre les membranes hybridées par la sonde “témoin” et celles hybridées par la sonde “infectée”. Nous avons en-suite commencé une analyse quantitative des résultats, mais nous avons été confrontés à des difficultés liées à la variabilité de l’intensité entre les si-gnaux et à l’encombrement des signaux sur la membrane. En effet, l’analyse des signaux de faible intensité nécessite l’augmentation du contraste de l’image de la membrane. Cependant, cette augmentation ampli-fie également l’intensité de signaux déjà très forts, et, du fait de l’encombrement stérique, ces signaux vont alors interférer avec les signaux environnants. Par ailleurs, sur certaines membranes, l’intensité du signal obtenu pour le gène de normalisation (glycéraldéhyde-3-phosphate déshy-drogénase) est trop faible pour permettre une quantification satisfaisante du signal et donc, une normalisation des données de ces membranes.
ANSELME Caroline 87 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Toutes les EST de la banque soustraite ne pouvant être analysées quantitativement, aucune estimation objective du pourcentage d’EST diffé-rentielles ne peut être donnée à partir de cette analyse. Nous avons donc simplement identifié qualitativement les EST correspondant aux signaux qui présentent un différentiel important entre les deux membranes. Nous avons ainsi identifié 28 signaux présentant un différentiel important entre les membranes hybridées par la sonde “témoin” et celles hybridées par la sonde “infectée”. Les EST correspondantes ainsi que les résultats de leurs comparaisons aux banques de données sont répertoriés dans le Tableau IV. Parmi les EST correspondant aux signaux différentiels, on peut distinguer des similitudes avec des peptides antibactériens et d’autres protéines asso-ciées à la réponse immunitaire, ainsi que des similitudes avec des protéines associées au stress. On peut distinguer également des EST présentant une faible similitude avec des protéines de microorganismes et enfin, des EST qui ne présentent aucune similitude de séquence dans les banques de don-nées.
L’analyse globale des EST de la banque soustraite du charançon S. zeamais nous a donc permis d’une part, de confirmer le caractère diffé-rentiel des EST qui présentaient une faible similitude de séquence avec la diptéricine et la cécropine et, d’autre part, de mettre en évidence d’autres gènes qui présentent une similitude faible (Evalue>0), ou qui ne présentent pas de similitude avec des séquences des banques de données. Pour ces derniers, des études biochimiques et moléculaires sont nécessaires pour dé-terminer s’il s’agit de nouvelles familles de GRI.
ANSELME Caroline 88 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
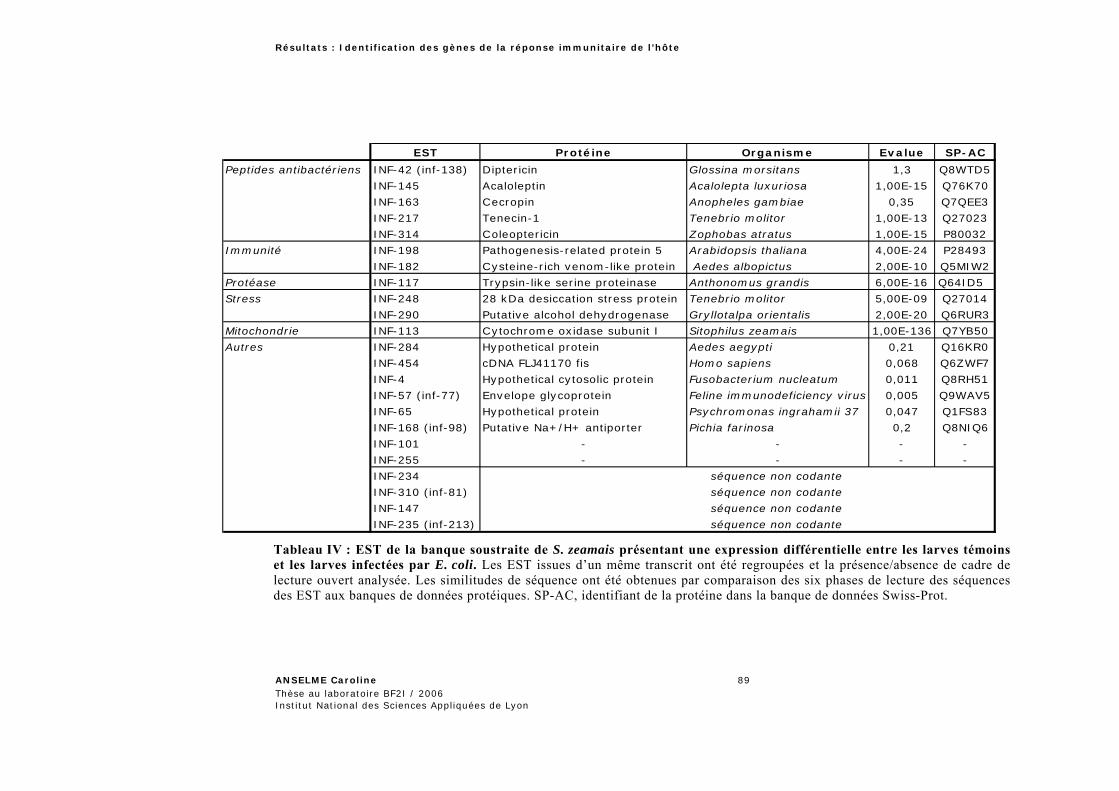
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
EST Protéine Organisme Evalue SP-AC
Peptides antibactériens INF-42 (inf-138) Diptericin Glossina morsitans 1,3 Q8WTD5
INF-145 Acaloleptin Acalolepta luxuriosa 1,00E-15 Q76K70
INF-163 Cecropin Anopheles gambiae 0,35 Q7QEE3
INF-217 Tenecin-1 Tenebrio molitor 1,00E-13 Q27023
INF-314 Coleoptericin Zophobas atratus 1,00E-15 P80032
Immunité INF-198 Pathogenesis-related protein 5 Arabidopsis thaliana 4,00E-24 P28493
INF-182 Cysteine-rich venom-like protein Aedes albopictus 2,00E-10 Q5MIW2
Protéase INF-117 Trypsin-like serine proteinase Anthonomus grandis 6,00E-16 Q64ID5
Stress INF-248 28 kDa desiccation stress protein Tenebrio molitor 5,00E-09 Q27014
INF-290 Putative alcohol dehydrogenase Gryllotalpa orientalis 2,00E-20 Q6RUR3
Mitochondrie INF-113 Cytochrome oxidase subunit I Sitophilus zeamais 1,00E-136 Q7YB50
Autres INF-284 Hypothetical protein Aedes aegypti 0,21 Q16KR0
INF-454 cDNA FLJ41170 fis Homo sapiens 0,068 Q6ZWF7
INF-4 Hypothetical cytosolic protein Fusobacterium nucleatum 0,011 Q8RH51
INF-57 (inf-77) Envelope glycoprotein Feline immunodeficiency virus 0,005 Q9WAV5
INF-65 Hypothetical protein Psychromonas ingrahamii 37 0,047 Q1FS83
INF-168 (inf-98) Putative Na+/H+ antiporter Pichia farinosa 0,2 Q8NIQ6
INF-101 - - - -
INF-255 - - - -
INF-234 séquence non codante
INF-310 (inf-81) séquence non codante
INF-147 séquence non codante
INF-235 (inf-213) séquence non codante
Tableau IV : EST de la banque soustraite de S. zeamais présentant une expression différentielle entre les larves témoins et les larves infectées par E. coli. Les EST issues d’un même transcrit ont été regroupées et la présence/absence de cadre de lecture ouvert analysée. Les similitudes de séquence ont été obtenues par comparaison des six phases de lecture des séquences des EST aux banques de données protéiques. SP-AC, identifiant de la protéine dans la banque de données Swiss-Prot.
ANSELME Caroline 89 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3.1.2 Analyse de la banque d’Acyrthosiphon pisum
Contrairement aux résultats obtenus sur le charançon, les membranes obte-nues à partir des EST du puceron paraissent très similaires et aucun signal n’apparaît nettement différentiel (Figure 11 et Annexe III). Ceci suggère que la banque soustraite comporte peu de différentiel et donc que la sous-traction réalisée chez A. pisum n’a pas permis d’obtenir de gènes surexpri-més après l’infection par E. coli. L’analyse in silico des EST de la banque soustraite d’A. pisum d’une part, et l’analyse de leur profil d’expression d’autre part, n’ont pas permis d’identifier des gènes de la réponse immunitaire chez le puceron. Nous avons donc choisi de continuer le travail de thèse sur notre premier modèle d’étude, le charançon S. zeamais.
1.3.2 Caractérisation des GRI de Sitophilus zeamais
Pour la suite de l’étude de la réponse immunitaire du charançon, nous avons choisi d’obtenir les séquences complètes des transcrits des GRI par la technique de RACE afin de confirmer les similitudes de séquence et d’obtenir la séquence des protéines. Par ailleurs, puisque les GRI des insec-tes peuvent être induits différemment à la suite d’une blessure ou d’infec-tions par divers microorganismes, nous avons également décidé de caractériser le profil d’expression des gènes de Sitophilus, par la technique de RT-PCR quantitative. Ces deux approches ne permettant pas d’analyser un grand nombre de gènes, nous avons privilégié les EST présentant une similitude de séquence avec des protéines de fonction(s) connue(s).
1.3.2.1 Obtention et analyse des séquences complètes des transcrits des GRI
Pour cette étude, notre choix s’est porté sur les EST codant des protéines dont la fonction putative présente un intérêt direct dans l’étude de la ré-ponse immunitaire de l’hôte dans le contexte de la symbiose. Nous avons sélectionné les homologues des PGRP puisqu’un PGRP est surexprimé dans le bactériome (Heddi et al., 2005), et l’homologue de Tollip (Burns et al., 2000 ; Zhang et Ghosh, 2002), qui pourrait assurer une inhibition, partielle ou totale, de la réponse immunitaire dans l’organe symbiotique. Nous avons également considéré les EST correspondant aux effecteurs de la réponse immunitaire connus pour leur activité antibactérienne : les lysozymes et les peptides antibactériens, particulièrement les EST inf-42, inf-163 et inf-165 dont la similitude de séquence est très faible.
ANSELME Caroline 90 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Les résultats de la comparaison des séquences complètes des ADNc aux bases de données, et de l’analyse in silico des séquences protéi-ques sont présentés dans le Tableau V et la Figure 12.
Figure 12 : Représentation schématique des protéines théoriques de la réponse immunitaire identifiées chez S. zeamais. Les séquences protéiques ont été déduites des séquences des ADNc obtenues par RACE. Les domaines identifiés par la compa-raison aux banques de domaines protéiques (InterProScan) ainsi que la E-value asso-ciée sont donnés pour chaque protéine (régions hachurées). Toutes les E-values pré-sentées ici sont inférieures au seuil de significativité défini pour chaque famille de domaine, dans l’application InterProScan. Les peptides signaux (en noir) ont été identifiés par le programme TargetP (http://www.cbs.dtu.dk/services/TargetP) et les domaines propeptides (en gris) ont été prédits à partir des motifs de coupure R-x-(K/R)-R. La taille des protéines et des différents domaines peut être estimée à partir de l’échelle donnée en haut de la figure. Coleoptericin, IPR009382 ; Defensin, IPR001542 ; Kunitz, IPR002223 ; Lysozyme, IPR000974 ; Destabilase, IPR008597 ; PGRP, IPR002502 ; C2, IPR000008 ; CUE, IPR003892.
ANSELME Caroline 91 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
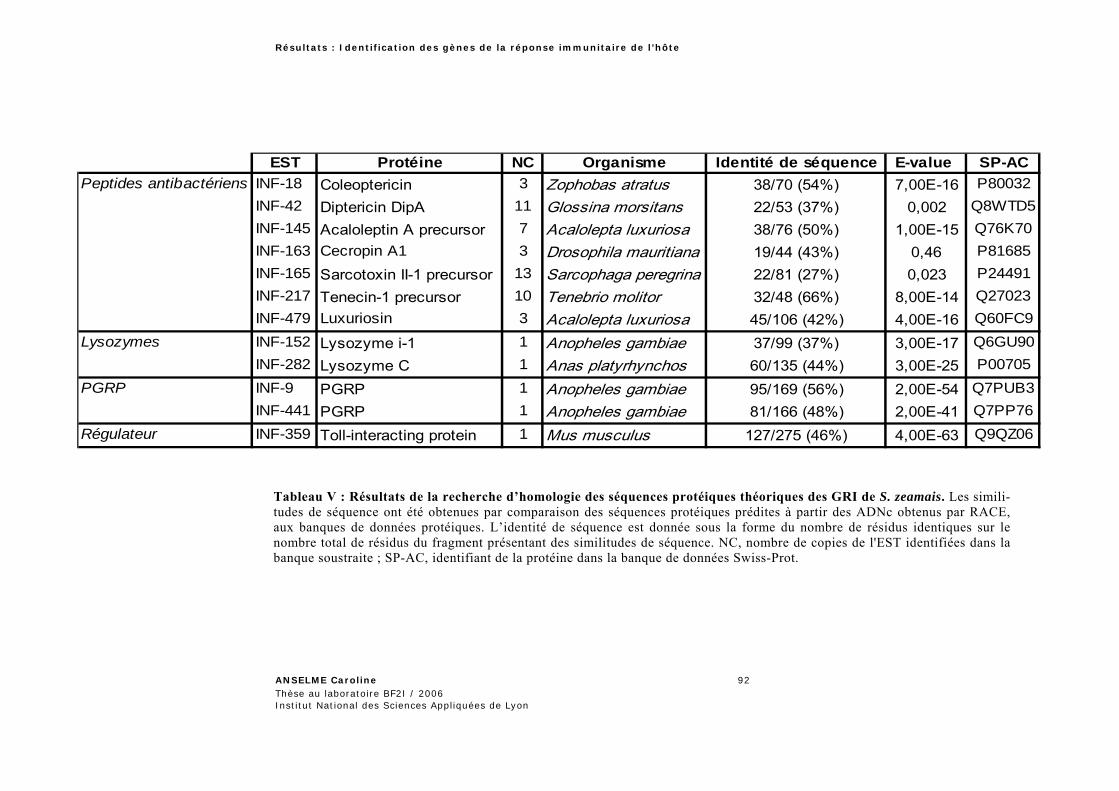
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
E
ANSELME Caroline 92
-145 Acaloleptin A precursor 7 Acalolepta luxuriosa 38/76 (50%) 1,00E-15 Q76K70IN -163 Cecropin A1 3 Drosophila mauritiana 19/44 (43%) 0,46 P81685INF-165 Sarcotoxin II-1 precursor 13 Sarcophaga peregrina 22/81 (27%) 0,023 P24491IN -217 Tenecin-1 precursor 10 Tenebrio molitor 32/48 (66%) 8,00E-14 Q27023IN -479 Luxuriosin 3 Acalolepta luxuriosa 45/106 (42%) 4,00E-16 Q60FC9
Lysozymes IN -152 Lysozyme i-1 1 Anopheles gambiae 37/99 (37%) 3,00E-17 Q6GU90INF-282 Lysozyme C 1 Anas platyrhynchos 60/135 (44%) 3,00E-25 P00705
PGRP IN -9 PGRP 1 Anopheles gambiae 95/169 (56%) 2,00E-54 Q7PUB3IN -441 PGRP 1 Anopheles gambiae 81/166 (48%) 2,00E-41 Q7PP76
Régulateur INF-359 Toll-interacting protein 1 Mus musculus 127/275 (46%) 4,00E-63 Q9QZ06
F F
F F F
F F
ST Protéine NC Organisme Identité de séquence E-value SP-ACPeptides antibactériens INF-18 Coleoptericin 3 Zophobas atratus 38/70 (54%) 7,00E-16 P80032
INF-42 Diptericin DipA 11 Glossina morsitans 22/53 (37%) 0,002 Q8WTD5IN
Tableau V : Résultats de la recherche d’homologie des séquences protéiques théoriques des GRI de S. zeamais. Les simili-tudes de séquence ont été obtenues par comparaison des séquences protéiques prédites à partir des ADNc obtenus par RACE, aux banques de données protéiques. L’identité de séquence est donnée sous la forme du nombre de résidus identiques sur le nombre total de résidus du fragment présentant des similitudes de séquence. NC, nombre de copies de l'EST identifiées dans la banque soustraite ; SP-AC, identifiant de la protéine dans la banque de données Swiss-Prot.
Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3.2.1.1 Reconnaissance des microorganismes et régulation de la
réponse immunitaire: les PGRP et Tollip
Les PGRP
Parmi les EST de la banque soustraite, deux EST présentent une similitude de séquence avec les PGRP : inf-9 dont la séquence correspond à celle de l’EST obtenue dans la banque soustraite du bactériome (Heddi et al., 2005), et inf-441 qui correspond à un deuxième gène PGRP.
L’analyse des séquences obtenues par RACE, montre que le pre-mier PGRP code pour une protéine de 262-aa sans séquence signal, et que le second code pour une protéine de 187-aa avec un peptide signal. Les deux protéines présentent 30% d’identité et possèdent toutes les deux le domaine caractéristique de la famille PGRP (IPR006619) et les cinq résidus nécessaires à l’activité amidasique (Kim et al., 2003) (Figure 13). Les pro-téines codées par les deux gènes PGRP identifiés dans la banque soustraite auraient donc une activité amidasique. En revanche, un seul des deux gènes (inf-441, PGRP2) possède une séquence de peptide signal et coderait donc une protéine sécrétée dans l’hémolymphe. L’autre protéine PGRP (inf-9, PGRP1) serait intracellulaire.
La comparaison de la séquence protéique prédite des deux PGRP par BLAST a confirmé que le gène PGRP1, identifié dans la banque sous-traite du bactériome, serait l’homologue du PGRP-LB de la drosophile puisqu’il présente un pourcentage de similitude par longueur de 51% (92/179 résidus, Evalue=1×10-52). En ce qui concerne le gène PGRP2, l’analyse de la séquence protéique par BLAST n’a pas permis d’identifier un homologue précis chez la drosophile. En effet, l’homologie la plus éle-vée est obtenue avec le PGRP-LCx (72/161 résidus, Evalue=2×10-37), mais l’alignement entre ce dernier et la protéine PGRP2 du charançon se limite au domaine PGRP (161/500 résidus). Ainsi, si la séquence complète de la protéine est considérée, le PGRP2 du charançon serait plus proche du PGRP-SC2 (71/161 résidus pour un total de 184 résidus, Evalue=2×10-35).
ANSELME Caroline 93 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
10 20 30 40 50 60 | | | | | | INF-441 MSS-LSFVIV--- LNIG- TCPAI RAEWGA P
MSSKQSRSIVPTC LNIVA TDPEI RDEWGA P
--------------IV -IVIA LK R ALSTSL INF-9 ECKFEKMEEIGEKKVE NSFVC VT K PTGVEN +++R +WGA+ P VE+ Dm PGRP-LB (32) LLSRSDWGARLPKSVEH 70 80 90 100 110 120 | | | | | | INF-441 A F LV NEVA TRLHGI DQKG I GT FE LRQEP P V HHSDTP-QCI CK QNYHM WDDIGYNFM GGD I
INF-9 V Y VI TKEE NDMQWM QNNS V NR YV P PYV+IHHSY+PA C + +C+ M+ MQ++HQ W DIGY+F +GGD IY
LT-LP P V HHSYIPAACS CI QNYHQ WCDIGYNFA GGD I
Dm PGRP-LB FQGPAPYVIIHHSYMPAVCYSTPDCMKSMRDMQDFHQLERGWNDIGYSFGIGGDGMIYT 130 140 150 160 170 180 | | | | | | INF-441 T K SL L NFQ SAA LT ES EC VKEEK Q R M INF-9 V R AR I DWT PES ML KQ NM IRDGY N K I GRG+ +GAHAP+YN +S+GIVLIGDW LPP+ + AAK LI G+ GYI YKL+ Dm PGRP-LB GRGFNVIGAHAPKYNDKSVGIVLIGDWRTELPPKQMLDAAKNLIAFGVFKGYIDPAYKLL 190 200 210 220 230 240 | | | | | |
GRGWGL GAHAV YN SIGICL G ETNP Q AL LI G IHT Y LGRGWTA GAHAP YN SIGIVL G EILP Q AA LI G ISE Y L
INF-441 DK FRVLSHM NFVRT----------------------------------- GHRQVSATACPG L P
INF-9 EA YKEIQTW HWI---DNPSLEDEVIPVVPLDKEDLKPDPREDSDASLAS GHRQVR+TECPG L+ EI +WPH+ D + PVVP
GHRQVRETECPG L P
Dm PGRP-LB GHRQVRDTECPGGRLFAEISSWPHFTHINDTEGVSSTTAPVVP (210) 250 260 | | INF-441 ----------------------- INF-9 EDNKIKRHSSTDKEESTSRSIKQ
Figure 13 : Alignement des séquences protéiques des deux PGRP de S. zeamais et du PGRP-LB de D. melanogaster. Les cinq résidus nécessaires à l’activité ami-dasique qui ont été identifiés chez le lysozyme du phage T7, et qui sont conservés chez les PGRP amidasiques, sont surlignés en bleu.
Tollip
Une des EST de la banque soustraite, inf-359, présente une homologie très significative avec un régulateur négatif de la voie des “Toll-Like Recep-tors” identifié chez les mammifères : la protéine Tollip (Burns et al., 2000 ; Zhang et Ghosh, 2002).
L’analyse du cadre de lecture ouvert (“Open Reading Frame”, ORF) obtenu par RACE prédit une protéine de 274-aa. La protéine Tollip du charançon présente les deux domaines caractéristiques du Tollip des mammifères, un domaine C2 impliqué dans les interactions avec les phos-pholipides membranaires (IPR000008) et un domaine C-terminal de liaison à l’ubiquitine (CUE, IPR003892). Aucune séquence signal n’ayant été mise en évidence, la protéine serait cytoplasmique.
ANSELME Caroline 94 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3.2.1.2 Les effecteurs : lysozymes et peptides antibactériens
Les lysozymes
Deux EST (inf-152 et inf-282) présentant des similitudes de séquence avec des lysozymes ont été identifiées dans la banque soustraite. La première présente une homologie avec les lysozymes de type i caractérisés chez les invertébrés et la seconde, une homologie avec les lysozymes de type c, dont l’un des membres a été caractérisé chez le poulet. L’analyse des ADNc ob-tenus par RACE montre que le premier code une protéine putative de 178-aa avec un peptide signal et un domaine destabilase (IPR008597) correspondant à une activité peptidase identifiée chez un lysozyme de type i (Zavalova et al., 2000). Ce même domaine est présent chez le lysozyme i-1 de A. gambiae. Le second lysozyme de S. zeamais code une protéine de 146-aa, caractérisée par la présence d’un peptide signal et d’un domaine ly-sozyme caractéristique des lysozymes de type c (IPR000974).
Les peptides antibactériens
Les coléoptéricines
Deux EST (inf-18 et inf-145) de la banque soustraite présentent une homo-logie avec des membres d’une famille de peptides antibactériens spécifique des coléoptères, les coléoptéricines. L’ADNc obtenu par 5’RACE à partir de l’EST inf-18 possède une ORF de 126-aa. L’EST inf-145 correspond à un ADNc complet avec une ORF de 115-aa. Les deux protéines présentent 40% d’identité et codent un peptide signal et un domaine propeptide (motif RxRR). Les peptides matures de 75-aa et de 67-aa comportent tous les deux un domaine coléoptéricine (IPR009382).
La luxuriosine
Parmi les EST présentant une homologie avec les peptides antibactériens, inf-479 semble être un homologue de la luxuriosine, un peptide original ca-ractérisé par la présence d’un domaine inhibiteur de protéases (Kunitz, IPR002223). La protéase cible et le rôle de ce peptide, récemment identifié chez le longicorne Acalolepta luxuriosa, n’ont pas encore été caractérisés (Ueda et al., 2005). Chez Sitophilus, le peptide inf-479 correspondrait à un peptide sécrété de 90-aa comportant, comme la luxuriosine, un domaine Kunitz.
ANSELME Caroline 95 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Les défensines
L’ADNc obtenu par 5’RACE à partir de l’EST inf-217 est un homologue de la ténécine, un peptide de la famille des défensines. L’ADNc comporte une ORF de 83-aa avec un peptide signal, un propeptide (motif RQKR) et un peptide mature de 44-aa. Les six cystéines impliquées dans les trois ponts disulfures caractéristiques des défensines (IPR001542) sont conservées chez le peptide du charançon.
Les autres peptides potentiellement antimicrobiens
Dans le cas des EST inf-42, -163 et -165, la séquence complète des ADNc n’apporte pas d’amélioration quant à leur similitude de séquence avec des peptides antibactériens (Tableau V). De plus, aucun domaine de peptide an-tibactérien connu n’a été identifié lors de l’analyse des séquences par In-terProScan.
L’ADNc obtenu à partir de l’EST inf-42 présente une ORF de 133-aa constituée d’un peptide signal et d’un propeptide (domaine RQKR). Le peptide mature prédit de 91-aa serait sécrété dans l’hémolymphe. Il est riche en résidus glycine (27,5 %) et pourrait être cationique, puisqu’il pos-sède huit résidus positifs et six résidus négatifs (pI=9,23).
En ce qui concerne l’EST inf-163, l’ADNc code un peptide de 69-aa avec une séquence signal. Un motif minimal de clivage (RxxR) a été mis en évidence en combinaison avec une arginine en position P6 (RxxRxR), ce qui pourrait favoriser la coupure du peptide. Dans ce cas, le peptide inf-163 serait sécrété dans l’hémolymphe sous forme d’un peptide mature de 26-aa, riche en résidus glycine (15,4 %).
Enfin, l’analyse in silico de l’ADNc obtenu à partir de l’EST inf-165 prédit l’existence d’une protéine de 181-aa avec une séquence si-gnal et un motif RTKR. La protéine mature de 125-aa est, là encore, riche en résidus glycine (20,8 %) et possède onze résidus positifs et seulement huit résidus négatifs (pI=9,35).
L’analyse des séquences de ces trois peptides révèle plusieurs ca-ractéristiques des peptides antibactériens en relation avec la structure pep-tide signal-propeptide-peptide et la richesse en résidus glycine. Toutefois, il n’est pas possible, sur la base de l’analyse in silico, de les classer dans la catégorie des peptides antibactériens. L’étude de leur activité devrait per-mettre de mieux caractériser ces nouveaux peptides potentiellement antimi-crobiens.
ANSELME Caroline 96 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3.2.2 Etude du profil d’expression des GRI
Pour confirmer l’induction des GRI suite à une infection bactérienne, j’ai mesuré leur taux de transcrits par RT-PCR quantitative dans les larves apo-symbiotiques témoins (non traitées) et dans les larves infectées par E. coli ou par B. megaterium, six heures après l’infection. Le taux de transcrits des GRI a également été mesuré sur des larves piquées par une aiguille stérile pour tester l’effet lié à la blessure provoquée par la piqûre.
Au total, 22 EST ont été sélectionnées pour cette analyse (Tableau VI). La spécificité de l’amplification a été testée pour chaque couple d’amorces grâce à la courbe de fusion. Lorsque plusieurs produits de PCR ont été détectés, de nouvelles amorces ont été définies sur d’autres régions des séquences.
Ainsi, sur les 22 EST sélectionnées, seuls les transcrits corres-pondant aux EST inf-475 et inf-258 n’ont pu être quantifiés. L’amplification des ADNc correspondant à l’EST inf-475 ne présentait pas de spécificité malgré plusieurs changements d'amorces, alors que le gène correspondant à l’EST inf-258 ne serait pas suffisamment exprimé pour que le taux de transcrits soit quantifiable. Les résultats obtenus sur les diffé-rents échantillons ont été normalisés par l’expression du gène gapdh. Pour l’ensemble des gènes étudiés, les résultats obtenus après l’infection par E. coli et ceux obtenus après l’infection par B. megaterium sont très simi-laires. Aussi, pour la suite de l’analyse, nous ne considérerons que l’infection par E. coli.
Les valeurs des taux de transcrits obtenues dans les différentes conditions expérimentales ont été analysées à l’aide du test non paramétri-que de Wilcoxon. Les résultats de cette analyse ainsi que les rapports entre les taux des transcrits mesurés après traitement et le taux de transcrits me-suré dans les larves témoins (taux de base) sont présentés dans le Tableau VI.
ANSELME Caroline 97 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
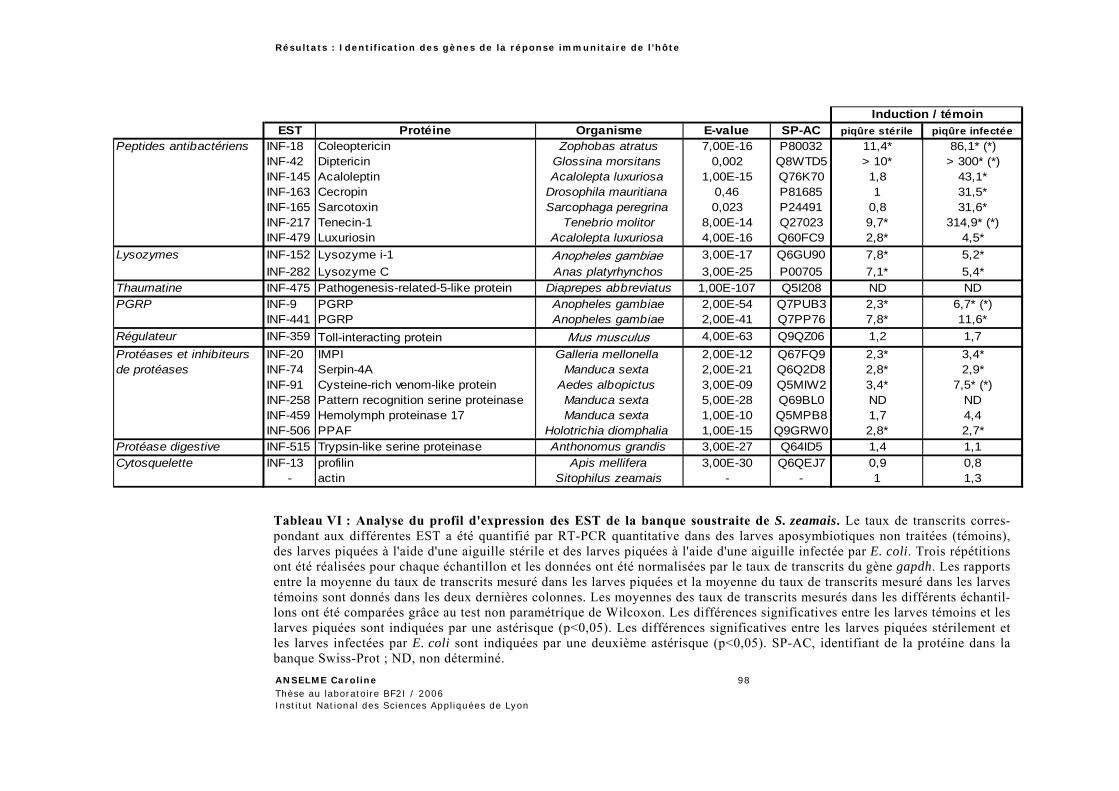
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
ANSELME Caroline 98
eoptericin Zophobas atratus 7,00E-16 P80032 11,4* 86,1* (*)INF-42 Diptericin Glossina morsitans 0,002 Q8WTD5 > 10* > 300* (*)INF-145 Acaloleptin Acalolepta luxuriosa 1,00E-15 Q76K70 1,8 43,1*INF-163 Cecropin Drosophila mauritiana 0,46 P81685 1 31,5*INF-165 Sarcotoxin Sarcophaga peregrina 0,023 P24491 0,8 31,6*INF-217 Tenecin-1 Tenebrio molitor 8,00E-14 Q27023 9,7* 314,9* (*)INF-479 Luxuriosin Acalolepta luxuriosa 4,00E-16 Q60FC9 2,8* 4,5*
Lysozymes INF-152 Lysozyme i-1 Anopheles gambiae 3,00E-17 Q6GU90 7,8* 5,2*INF-282 Lysozyme C Anas platyrhynchos 3,00E-25 P00705 7,1* 5,4*
Thaumatine INF-475 Pathogenesis-related-5-like protein Diaprepes abbreviatus 1,00E-107 Q5I208 ND NDPGRP INF-9 PGRP Anopheles gambiae 2,00E-54 Q7PUB3 2,3* 6,7* (*)
INF-441 PGRP Anopheles gambiae 2,00E-41 Q7PP76 7,8* 11,6*Régulateur INF-359 To l-interacting protein Mus musculus 4,00E-63 Q9QZ06 1,2 1,7Protéases et inhibiteurs INF-20 IMPI Galleria mellonella 2,00E-12 Q67FQ9 2,3* 3,4*de protéases INF-74 Serpin-4A Manduca sexta 2,00E-21 Q6Q2D8 2,8* 2,9*
INF-91 Cysteine-rich venom-like protein Aedes albopictus 3,00E-09 Q5MIW2 3,4* 7,5* (*)INF-258 Pattern recognition serine proteinase Manduca sexta 5,00E-28 Q69BL0 ND NDINF-459 Hemolymph proteinase 17 Manduca sexta 1,00E-10 Q5MPB8 1,7 4,4INF-506 PPAF Holotrichia diomphalia 1,00E-15 Q9GRW0 2,8* 2,7*
Protéase digestive INF-515 Trypsin-like serine proteinase Anthonomus grandis 3,00E-27 Q64ID5 1,4 1,1Cytosquelette INF-13 profilin Apis mellifera 3,00E-30 Q6QEJ7 0,9 0,8
- actin Sitophilus zeamais - - 1 1,3
Induction / témoin l l
EST Protéine Organisme E-value SP-AC piqûre stérile piqûre infectéePeptides antibactériens INF-18 Co
Tableau VI : Analyse du profil d'expression des EST de la banque soustraite de S. zeamais. Le taux de transcrits corres-pondant aux différentes EST a été quantifié par RT-PCR quantitative dans des larves aposymbiotiques non traitées (témoins), des larves piquées à l'aide d'une aiguille stérile et des larves piquées à l'aide d'une aiguille infectée par E. coli. Trois répétitions ont été réalisées pour chaque échantillon et les données ont été normalisées par le taux de transcrits du gène gapdh. Les rapports entre la moyenne du taux de transcrits mesuré dans les larves piquées et la moyenne du taux de transcrits mesuré dans les larves témoins sont donnés dans les deux dernières colonnes. Les moyennes des taux de transcrits mesurés dans les différents échantil-lons ont été comparées grâce au test non paramétrique de Wilcoxon. Les différences significatives entre les larves témoins et les larves piquées sont indiquées par une astérisque (p<0,05). Les différences significatives entre les larves piquées stérilement et les larves infectées par E. coli sont indiquées par une deuxième astérisque (p<0,05). SP-AC, identifiant de la protéine dans la banque Swiss-Prot ; ND, non déterminé.
Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Tous les peptides antibactériens putatifs, à l’exception de la luxuriosine, sont induits après l’infection par E. coli avec des variations très importantes des taux de transcrits allant de 30 à plus de 300 fois le taux de base. Dans le cas des EST inf-18, inf-42, et inf-217, on observe également une induction à la suite de la blessure occasionnée par la piqûre stérile avec des variations néanmoins plus faibles (10 à 30 fois le taux de base). Les gènes codant la luxuriosine, les lysozymes, les homologues des protéines de la voie prophénoloxydase (inf-506 et inf-74) et de la métalloprotéase (inf-20) sont induits par la blessure, indépendamment de l’infection (2 à 8 fois le taux de base). En ce qui concerne les deux gènes PGRP, ils sont induits par la blessure mais seul le gène correspondant au PGRP1 (inf-9) est induit à la suite de l’infection par E. coli. Enfin, aucune variation significative n’a été observée dans le cas des homologues de Tollip, de deux protéases (inf-459 et inf-515) et des éléments du cytosquelette.
1.3.2.3 Mise en place d’une procédure adaptée à l’analyse des séquences des banques
soustraites.
A la suite du travail réalisé au cours de la thèse, nous avons été contactés par l’équipe de F. Fleury (Biologie et Biométrie Evolutive, UCBL) qui souhaitait utiliser la technique de PCR soustractive dans leur projet de re-cherche. Nous avons profité de ces échanges pour développer avec D. Charif (IE, CNRS), une procédure d’analyse de séquences adaptée aux EST des banques soustraites. Cette procédure permet d’éliminer les sé-quences contaminantes, de séparer les séquences chimères (voir Matériel et Méthodes) et de regrouper en clusters les EST issues d’un même transcrit. Après cette étape, la banque soustraite est constituée par les singletons et par les contigs représentés par les séquences consensus associées aux grou-pes d’EST. Les séquences des singletons et les séquences consensus sont ensuite comparées aux banques de données pour rechercher des homologies de séquence. Une interface Web, accessible aux deux équipes, a également été développée pour faciliter l’accès à la description de la procédure et des outils informatiques utilisés, ainsi qu’aux résultats de l’analyse des séquen-ces.
Une deuxième analyse des banques immunitaires soustraites a donc été réalisée par la procédure mise en place par D. Charif. Dans le cas de la banque soustraite de Sitophilus, le regroupement des EST a permis de réduire le nombre de séquences à 283 avec 226 singletons et 57 contigs correspondant à des regroupements de 2 à 22 séquences. Dans le cas d’A. pisum, les séquences obtenues ont été rassemblées en 425 singletons et 21 contigs, ce qui confirme le manque de redondance de cette banque par rapport à la banque de S. zeamais. Globalement, les deux analyses, indivi-
ANSELME Caroline 99 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
duelle ou après regroupement, ont permis d’obtenir des résultats similaires avec toutefois quelques différences au niveau des EST qui composent les contigs. Ces différences se sont avérées sans conséquence sur le résultat des analyses des banques soustraites.
1.3.2.4 Tentative d’identification des peptides antibactériens par une approche biochimique
La plupart des peptides antibactériens d’insectes ont été isolés à partir de l’hémolymphe, par purification par HPLC et mesure in vitro de l’activité antibactérienne. Pour utiliser une telle approche sur les modèles charançon et puceron, où les quantités d’hémolymphe sont faibles, une méthode rapide de collecte en masse d’hémolymphe et d’extraction des peptides à l’acide trifluoroacétique (TFA) a été mise au point au laboratoire BF2I par Y. Rahbé et G. Duport.
Parmi les GRI de Sitophilus, trois gènes correspondant aux EST inf-42, inf-163 et inf-165 qui sont surexprimés après l’infection bactérienne pourraient, d’après l’analyse in silico de leur séquence, coder de nouveaux peptides antimicrobiens. Pour confirmer cette hypothèse, nous avons donc utilisé une approche biochimique pour tenter d’identifier les peptides anti-microbiens du charançon.
1.3.2.4.1 Identification des peptides induits dans l’hémolymphe des
larves infectées par E. coli.
Afin d’identifier les peptides induits dans l’hémolymphe lors de la réponse immunitaire de l’insecte, les peptides de larves non traitées (témoins) et de larves infectées par E. coli ont été extraits de l’hémolymphe 18 heures après l’infection. Ces peptides ont ensuite été analysés par HPLC et les chromatogrammes obtenus ont été superposés afin d’identifier d’éventuels pics différentiels (Figure 14). Quatre pics, un réprimé et trois induits par l’infection ont ainsi été identifiés de façon reproductible sur les trois répéti-tions réalisées pour cette étude.
ANSELME Caroline 100 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
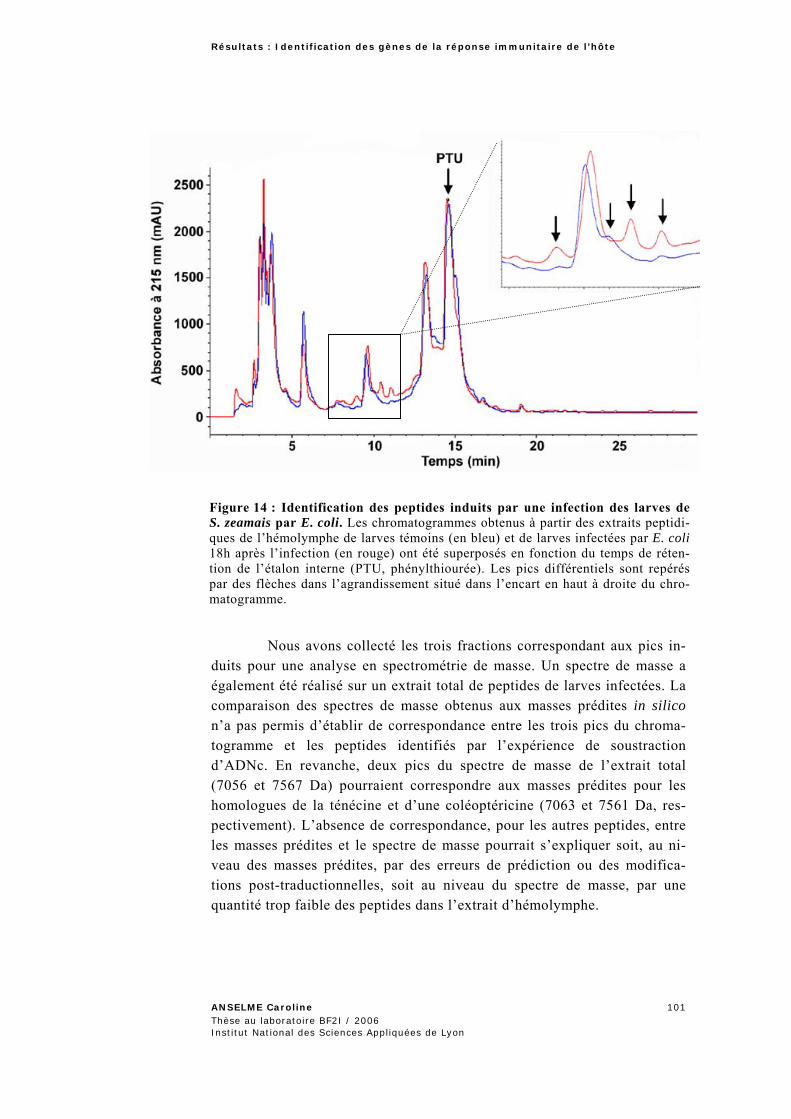
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Figure 14 : Identification des peptides induits par une infection des larves de S. zeamais par E. coli. Les chromatogrammes obtenus à partir des extraits peptidi-ques de l’hémolymphe de larves témoins (en bleu) et de larves infectées par E. coli 18h après l’infection (en rouge) ont été superposés en fonction du temps de réten-tion de l’étalon interne (PTU, phénylthiourée). Les pics différentiels sont repérés par des flèches dans l’agrandissement situé dans l’encart en haut à droite du chro-matogramme.
Nous avons collecté les trois fractions correspondant aux pics in-
duits pour une analyse en spectrométrie de masse. Un spectre de masse a également été réalisé sur un extrait total de peptides de larves infectées. La comparaison des spectres de masse obtenus aux masses prédites in silico n’a pas permis d’établir de correspondance entre les trois pics du chroma-togramme et les peptides identifiés par l’expérience de soustraction d’ADNc. En revanche, deux pics du spectre de masse de l’extrait total (7056 et 7567 Da) pourraient correspondre aux masses prédites pour les homologues de la ténécine et d’une coléoptéricine (7063 et 7561 Da, res-pectivement). L’absence de correspondance, pour les autres peptides, entre les masses prédites et le spectre de masse pourrait s’expliquer soit, au ni-veau des masses prédites, par des erreurs de prédiction ou des modifica-tions post-traductionnelles, soit au niveau du spectre de masse, par une quantité trop faible des peptides dans l’extrait d’hémolymphe.
ANSELME Caroline 101 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
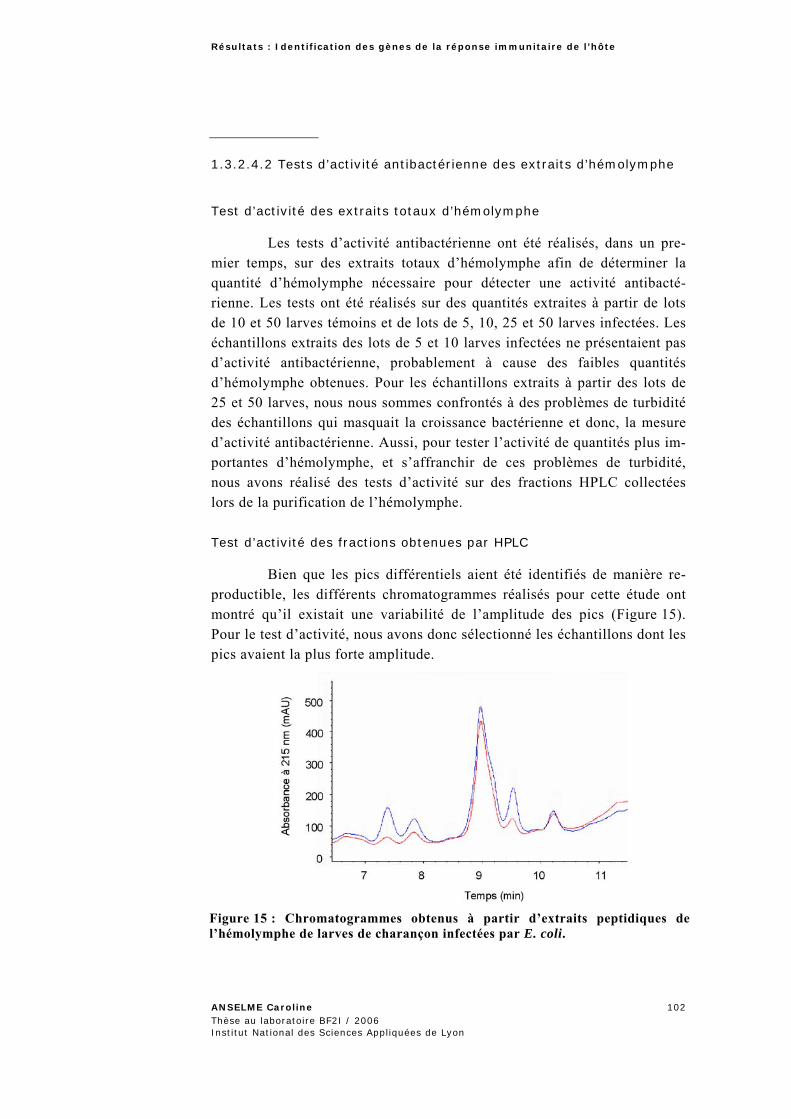
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
1.3.2.4.2 Tests d’activité antibactérienne des extraits d’hémolymphe
Test d’activité des extraits totaux d’hémolymphe
Les tests d’activité antibactérienne ont été réalisés, dans un pre-mier temps, sur des extraits totaux d’hémolymphe afin de déterminer la quantité d’hémolymphe nécessaire pour détecter une activité antibacté-rienne. Les tests ont été réalisés sur des quantités extraites à partir de lots de 10 et 50 larves témoins et de lots de 5, 10, 25 et 50 larves infectées. Les échantillons extraits des lots de 5 et 10 larves infectées ne présentaient pas d’activité antibactérienne, probablement à cause des faibles quantités d’hémolymphe obtenues. Pour les échantillons extraits à partir des lots de 25 et 50 larves, nous nous sommes confrontés à des problèmes de turbidité des échantillons qui masquait la croissance bactérienne et donc, la mesure d’activité antibactérienne. Aussi, pour tester l’activité de quantités plus im-portantes d’hémolymphe, et s’affranchir de ces problèmes de turbidité, nous avons réalisé des tests d’activité sur des fractions HPLC collectées lors de la purification de l’hémolymphe.
Test d’activité des fractions obtenues par HPLC
Bien que les pics différentiels aient été identifiés de manière re-productible, les différents chromatogrammes réalisés pour cette étude ont montré qu’il existait une variabilité de l’amplitude des pics (Figure 15). Pour le test d’activité, nous avons donc sélectionné les échantillons dont les pics avaient la plus forte amplitude.
Figure 15 : Chromatogrammes obtenus à partir d’extraits peptidiques de l’hémolymphe de larves de charançon infectées par E. coli.
ANSELME Caroline 102 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Cinq échantillons d’hémolymphe ont été extraits et purifiés à par-tir de lots de 25 larves infectées par E. coli, pour avoir, après regroupement des fractions, l’équivalent de 125 larves par fraction. Pour tester spécifi-quement les pics différentiels, mais également tous les autres constituants de l’hémolymphe, les fractions ont été collectées toutes les minutes pendant les sept premières minutes, puis, en fonction des pics du chromatogramme entre la septième et la onzième minute, et à nouveau toutes les minutes, jusqu’à la trentième minute (Figure 16). Les résultats obtenus pour le test d’activité de ces fractions sont présentés sur le chromatogramme de la Figure 16. Cinq échantillons d’hémolymphe extraits et purifiés à partir de lots de 25 larves témoins ont été traités en parallèle.
L’inhibition de la croissance bactérienne a été observée au niveau des premières (1 à 5 min) et des dernières (12 à 16 min) fractions collectées dans les larves infectées, mais également dans les larves témoins. Cette in-hibition serait donc due soit à des artefacts liés à l’expérimentation, soit à des peptides synthétisés constitutivement. Aucune activité antibactérienne n’a, en revanche, été observée sur les fractions correspondant aux trois pics induits par l’infection bactérienne. Ceci est peut-être dû à des quantités trop faibles de peptides. Toutefois, il est également possible que les pics diffé-rentiels ne correspondent pas à des peptides antibactériens mais à d’autres peptides présents dans l’hémolymphe en réponse à l’infection bactérienne. Il pourrait notamment s’agir de produits libérés à la suite du clivage d’un précurseur de la réponse immunitaire.
Bien que nous n’ayons pas pu confirmer l’activité antibactérienne
des trois peptides correspondant aux EST inf-42, inf-163 et inf-165, cette étude nous a permis de détecter une activité antibactérienne dans des frac-tions de l’hémolymphe. Nos conditions chromatographiques ne permettant pas de séparer les composés de ces fractions, une nouvelle mise au point devrait donc être envisagée pour séparer ces composés. Nous pourrons ainsi identifier les composés qui sont responsables de l’activité antibactérienne observée et déterminer s’il s’agit, ou non, d’un artefact.
ANSELME Caroline 103 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
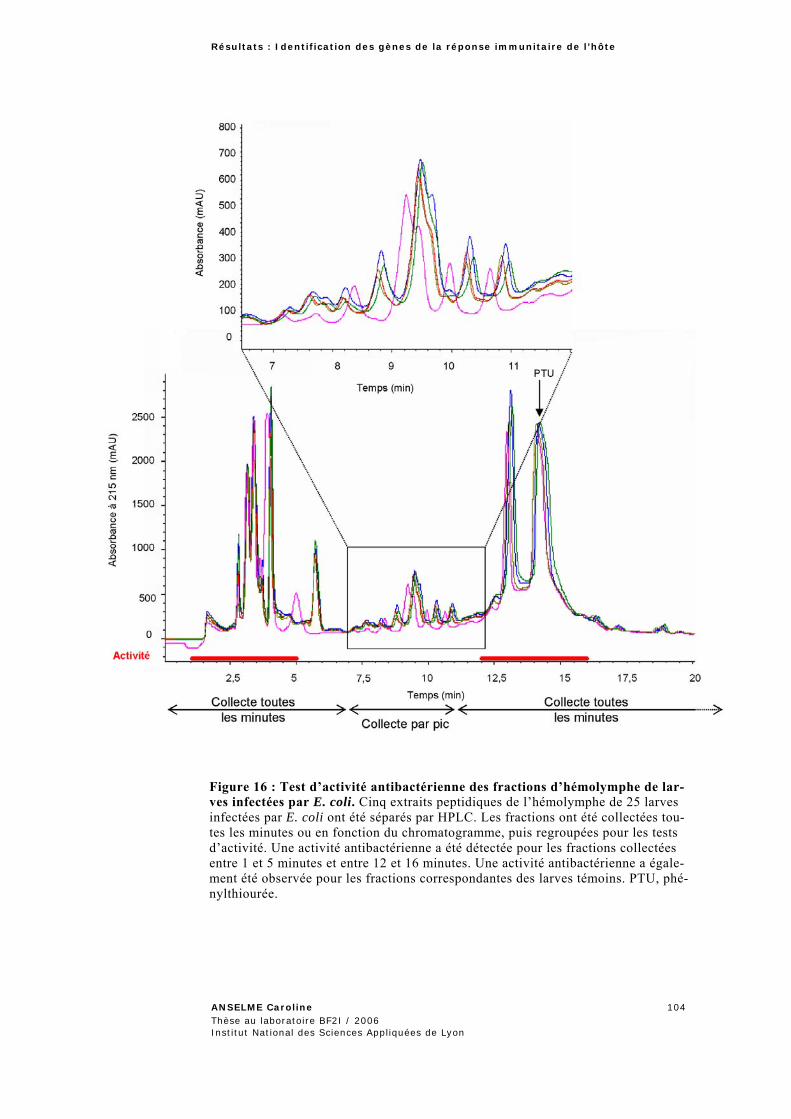
Résultats : Identification des gènes de la réponse immunitaire de l’hôte
Figure 16 : Test d’activité antibactérienne des fractions d’hémolymphe de lar-ves infectées par E. coli. Cinq extraits peptidiques de l’hémolymphe de 25 larves infectées par E. coli ont été séparés par HPLC. Les fractions ont été collectées tou-tes les minutes ou en fonction du chromatogramme, puis regroupées pour les tests d’activité. Une activité antibactérienne a été détectée pour les fractions collectées entre 1 et 5 minutes et entre 12 et 16 minutes. Une activité antibactérienne a égale-ment été observée pour les fractions correspondantes des larves témoins. PTU, phé-nylthiourée.
ANSELME Caroline 104 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
2. Symbiose et immunité chez Sitophilus
zeamais
Après avoir identifié les gènes de la réponse humorale activée par les bac-téries Gram (-) chez S. zeamais, je me suis intéressée à leur régulation dans le bactériome larvaire, en présence des bactéries symbiotiques. Cette étude avait notamment pour but de déterminer si des gènes impliqués dans l’élimination des bactéries, comme les peptides antibactériens et les lyso-zymes, étaient exprimés dans le bactériome du charançon. Parmi les gènes identifiés, un intérêt particulier a été accordé au gène PGRP1 qui est surex-primé dans le bactériome larvaire (Heddi et al., 2005), et qui appartient à une famille de gènes très diversifiés de par leur(s) fonction(s) dans la ré-ponse immunitaire innée (Dziarski, 2004 ; Steiner, 2004 ; Bischoff et al., 2006 ; Zaidman-Remy et al., 2006).
2.1 Expression des GRI de Sitophilus zeamais dans le bactériome larvaire
Dans cette partie, le taux de transcrits de chacun des 20 gènes étudiés pré-cédemment (Tableau VI) a été mesuré dans le bactériome larvaire et a en-suite été comparé au taux de transcrits des larves aposymbiotiques à l’aide du test non paramétrique de Wilcoxon. Les valeurs moyennes obtenues après normalisation des données ainsi que les différences significatives en-tre les taux de transcrits sont présentées dans la Figure 17.
Cette analyse révèle que de nombreux homologues putatifs des peptides antibactériens (inf-145, -163, -165 et inf-217), ainsi que celui de l’inhibiteur de métalloprotéase (inf-20), ne présentent pas de différence d’expression significative entre les larves aposymbiotiques et les bactério-mes. En revanche, les lysozymes (inf-152 et inf-282), le PGRP2 (inf-441), l’homologue du gène luxuriosine (inf-479) et les homologues putatifs des protéases et des inhibiteurs de protéases (inf-506, -74, -91, -459 et inf-515) sont sous-exprimés, voire non-exprimés, dans le bactériome. Une sous-expression inattendue des éléments du cytosquelette (inf-13 et β-actine) a également été mise en évidence. Enfin, parmi tous les gènes étudiés, trois seulement sont surexprimés dans le bactériome : le gène coléoptéricine (inf-18 ; 10 fois), le gène PGRP1 (inf-9 ; 39 fois) déjà identifié dans la banque soustraite du bactériome et l’homologue de Tollip (inf-359 ; 9,5 fois).
ANSELME Caroline 105 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
Figure 17 : Quantification du taux de transcrits dans les larves aposymbioti-ques et dans les bactériomes larvaires chez S. zeamais. La mesure du taux de transcrits correspond à la moyenne de trois répétitions. Les erreurs-types sont re-présentées par les barres d’erreur. Les différences significatives par rapport au té-moin (p<0,05) sont signalées par une astérisque. Les gènes surexprimés dans le bactériome larvaire sont indiqués par une flèche (voir Tableau VI pour l’identification des EST).
2.2 Caractérisation moléculaire du gène PGRP1
Parmi les trois gènes surexprimés dans le bactériome, le gène PGRP1 est le seul gène qui est induit par les bactéries Gram (-), et qui coderait une pro-téine cytoplasmique (§ 1.3.2.). La protéine PGRP1 serait donc potentielle-ment en contact avec les endocytobiotes et pourrait jouer un rôle direct dans la perception de ces derniers.
2.2.1 Expression du gène PGRP1 en relation avec la densité bactérienne.
Pour approcher l’hypothèse selon laquelle le gène PGRP1 serait impliqué dans la perception des endocytobiotes et la régulation de la réponse immu-nitaire dans le bactériome, j’ai dans un premier temps étudié la régulation de son expression en relation avec la densité bactérienne (Anselme et al., 2006). Pour cela, j’ai infecté des larves de charançons aposymbiotiques avec deux souches bactériennes sélectionnées en fonction des résultats ob-tenus chez D. melanogaster : une souche non pathogène d’E. coli qui est rapidement éliminée par l’insecte, et une souche pathogène de Pseudomo-nas aeruginosa (PAO1) qui se multiplie rapidement dans l’insecte et qui tue la drosophile en moins de 18 h (Fauvarque et al., 2002). Pour chacune des deux souches bactériennes, nous avons mesuré la croissance bacté-
ANSELME Caroline 106 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
rienne in larva et le taux de transcrits PGRP1 après différents temps d’infection.
2.2.1.1 Mesure de la croissance bactérienne dans les larves
La densité in larva de chacune des deux souches bactériennes, a été estimée à partir du nombre de colonies (cfu) formées sur boîte après l’étalement d’un broyat de larves infectées. Dans le cas de larves aposymbiotiques té-moins non infectées, aucune colonie n’a été observée sur boîte. Ceci est en accord avec des travaux réalisés au laboratoire qui ont conclu à l’absence de bactéries dans le tube digestif des larves (Nardon, communication personnelle).
Les larves ont été infectées par des bactéries en phase de crois-sance exponentielle pour permettre une activation plus rapide des systèmes de virulence bactériens. Cependant, devant l’incapacité de la souche d’E. coli à infecter l’hôte dans ces conditions, les infections par E. coli ont été réalisées avec un culot de bactéries obtenu à partir d’une culture d’une nuit, pour augmenter le nombre de bactéries. Les résultats obtenus dans ces conditions sont présentés dans la Figure 18.
Deux heures après l’infection, le nombre de colonies par larve est supérieur pour E. coli par rapport à P. aeruginosa, ce qui est cohérent avec la différence de concentration des solutions bactériennes utilisées pour l’infection des larves. Le nombre d’E. coli reste ensuite constant pendant toute la durée de l’expérimentation avec, toutefois, une légère diminution entre 12 h et 24 h. En revanche, la bactérie P. aeruginosa prolifère rapide-ment dans la larve avec un nombre de colonies multiplié par près de 104 en-tre 2 h et 24 h.
Figure 18 : Croissance bactérienne mesurée dans les larves aposymbiotiques infectées par E. coli et P. aeruginosa. La densité bactérienne a été estimée par le nombre de colonies (cfu) formées sur boîte après l’étalement d’un broyat de larves infectées. Les mesures ont été effectuées 2, 6, 12 et 24 h après l’infection. Les va-leurs correspondent à la moyenne de deux répétitions.
ANSELME Caroline 107 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
L’ensemble de ces résultats suggère que la capacité d’infection de la souche PAO1 de P. aeruginosa est bien supérieure à celle de la souche non pathogène d’E. coli. Toutefois, on notera que le taux de survie des lar-ves infectées par PAO1 est semblable au taux de survie obtenu après une piqûre stérile, pour la durée de l’expérimentation. Cette bactérie serait donc moins pathogène pour le charançon que pour la drosophile.
2.2.1.2 Etude de l’expression du gène PGRP1
Le taux de transcrits du gène PGRP1 a été mesuré par la technique de Nor-thern blot dans des larves aposymbiotiques non traitées (contrôle) et des larves piquées à l’aide d’une aiguille stérile ou d’une aiguille infectée par E. coli ou P. aeruginosa. Les variations dans le temps du taux de transcrits ont été analysées en effectuant des mesures 1, 2, 6 ou 12 h après la piqûre. Les membranes réalisées dans cette expérimentation sont présentées dans la Figure 19-A. Pour chaque échantillon, l’intensité du signal radioactif obte-nu a été corrigée par rapport au bruit de fond et normalisée par l’intensité du signal obtenu avec la β-actine. Une normalisation inter-membranes a en-suite été réalisée à l’aide d’un échantillon calibrateur.
Les résultats obtenus par cette démarche (Figure 19-B) indiquent clairement une variation du taux de transcrits PGRP1 différente pour cha-que traitement, ce qui traduit une interaction entre le facteur temps et le facteur traitement. Afin de prendre en compte cette interaction, les résultats ont été analysés par des tests paramétriques. La confirmation de la signifi-cativité de l’interaction par une ANOVA (p<3,5×10-8) a conduit à une ana-lyse des effets simples (analyse de l’effet d’un facteur pour chaque modali-té de l’autre facteur).
Les premiers résultats ne montrent aucune différence significative entre les échantillons une heure après traitement (p=0,80). L’effet du trai-tement n’est donc visible que pour des temps d’incubation de plus de deux heures. Concernant la variation du taux de transcrits dans le temps, celui-ci reste constant dans les larves non traitées (p=0,13) et les larves piquées sté-rilement (p=0,15). En revanche, il augmente jusqu’à 5,5 fois après l’infection par E. coli (p=1,7×10-6) et jusqu’à 8,5 fois après l’infection par P. aeruginosa (p=3,7×10-11). D’après un test de comparaison de Dunnett utilisant le taux de transcrits estimé une heure après l’infection comme ré-férence, le taux de transcrits estimé douze heures après l’infection par E. coli n’est plus significativement différent du taux de base. En revanche, ce même test reste significatif douze heures après l’infection par P. aeruginosa.
ANSELME Caroline 108 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
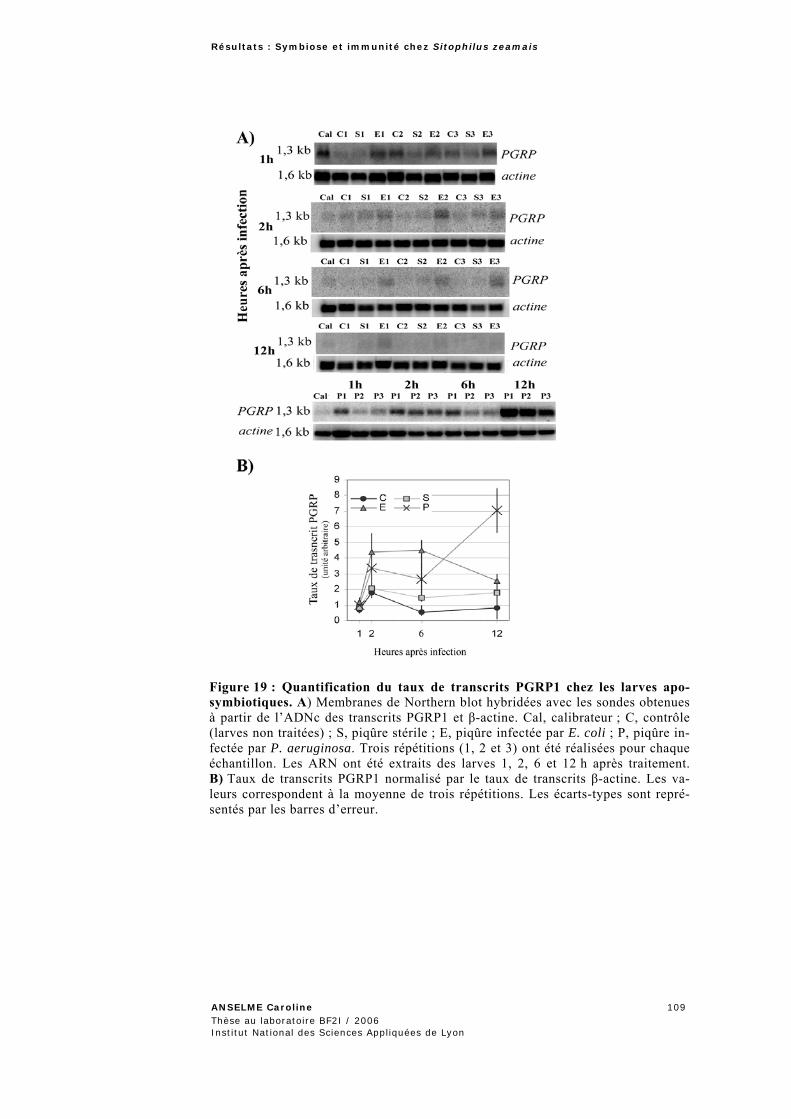
Résultats : Symbiose et immunité chez Sitophilus zeamais
Figure 19 : Quantification du taux de transcrits PGRP1 chez les larves apo-symbiotiques. A) Membranes de Northern blot hybridées avec les sondes obtenues à partir de l’ADNc des transcrits PGRP1 et β-actine. Cal, calibrateur ; C, contrôle (larves non traitées) ; S, piqûre stérile ; E, piqûre infectée par E. coli ; P, piqûre in-fectée par P. aeruginosa. Trois répétitions (1, 2 et 3) ont été réalisées pour chaque échantillon. Les ARN ont été extraits des larves 1, 2, 6 et 12 h après traitement. B) Taux de transcrits PGRP1 normalisé par le taux de transcrits β-actine. Les va-leurs correspondent à la moyenne de trois répétitions. Les écarts-types sont repré-sentés par les barres d’erreur.
ANSELME Caroline 109 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
Quelle que soit la souche bactérienne utilisée, le gène PGRP1 est donc induit par l’infection. Toutefois, une différence entre les souches est visible au niveau de la variation du taux de transcrits dans le temps. Dans le cas de l’infection par E. coli, le taux de transcrits décroît après six heures d’infection et retrouve, après douze heures, le taux de transcrits mesuré en l’absence d’infection. Suite à l’infection par P. aeruginosa, en revanche, le taux de transcrits augmente encore après six heures d’infection pour attein-dre, à douze heures, la valeur maximale obtenue dans cette expérimenta-tion.
L’analyse conjointe des résultats de la croissance bactérienne in larva (Figure 18) et du taux de transcrits PGRP1 (Figure 19) après un temps d’infection donné, suggère donc que l’expression du gène PGRP1 est régu-lée en fonction de la densité bactérienne (Anselme et al., 2006).
2.2.2 Expression du gène PGRP1 au cours de développement de l’insecte
En mesurant l’expression des gènes inv/spa du SSTT de SZPE au cours du développement du charançon, Dale et al. (2002) ont mis en évidence une forte induction des gènes du SSTT au stade nymphal. Ce résultat suggère un changement dans la physiologie bactérienne, et donc dans l’interaction hôte-symbiote au cours de la métamorphose. Aussi, pour déterminer si ce changement a des conséquences sur la réponse immunitaire de l’hôte, nous avons étudié l’expression du gène PGRP1 à différentes étapes du dévelop-pement de l’insecte.
2.2.2.1 Expression du gène PGRP1 à différents stades de développement de l’hôte
Le taux de transcrits PGRP1 a été quantifié par PCR en temps réel chez les individus apo- et symbiotiques dans les ovocytes, les embryons de trois jours (lors de la formation du bactériome), les larves du quatrième et der-nier stade, les nymphes au moment de la transition du bactériome larvaire aux bactériomes des cæcums mésentériques, et enfin, dans les adultes âgés de plus de trois semaines qui sont dépourvus de bactériomes des cæcums mésentériques. Les résultats obtenus après normalisation des données sont présentés dans la Figure 20.
ANSELME Caroline 110 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
Figure 20 : Quantification du taux de transcrits dans les insectes apo- et sym-biotiques. La mesure du taux de transcrits correspond à la moyenne de trois répéti-tions. Les écarts-types sont représentés par les barres d’erreur. Les différences si-gnificatives entre les insectes apo- et symbiotiques (p<0,05) sont signalées par les astérisques.
On constate tout d’abord un taux important de transcrits dans les
ovocytes, indépendamment du statut symbiotique de l’insecte. Ces trans-crits seraient d’origine maternelle et seraient exprimés dans les ovocytes pour assurer une fonction dans le développement et/ou l’immunité de l’embryon avant la mise en place de son propre système immunitaire.
Par une analyse des effets simples, la comparaison entre les indi-vidus apo- et symbiotiques montre que le stade nymphal est le seul stade, parmi ceux considérés, qui présente une différence significative des taux de transcrits (p=2,12×10-5). A titre de remarque, il faut signaler que malgré la surexpression du gène PGRP1 dans le bactériome des larves symbiotiques, aucune différence significative n’est observée au stade larvaire entre les in-dividus apo- et symbiotiques. Ceci est probablement dû à une faible repré-sentation des transcrits du bactériome par rapport aux transcrits de la larve, et donc à une dilution de ces transcrits.
2.2.2.2 Externalisation des endocytobiotes au stade nymphal
La surexpression du gène PGRP1 au stade nymphal chez l’insecte coïncide avec la surexpression, au même stade, des gènes du SSTT inv/spa de la bactérie symbiotique (Dale et al., 2002). Une des hypothèses qui peut ex-pliquer l’induction simultanée d’un gène immunitaire de l’hôte et de gènes bactériens impliqués dans l’invasion cellulaire est la présence de bactéries libres dans la nymphe. Afin de vérifier cette hypothèse, des expériences de
ANSELME Caroline 111 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
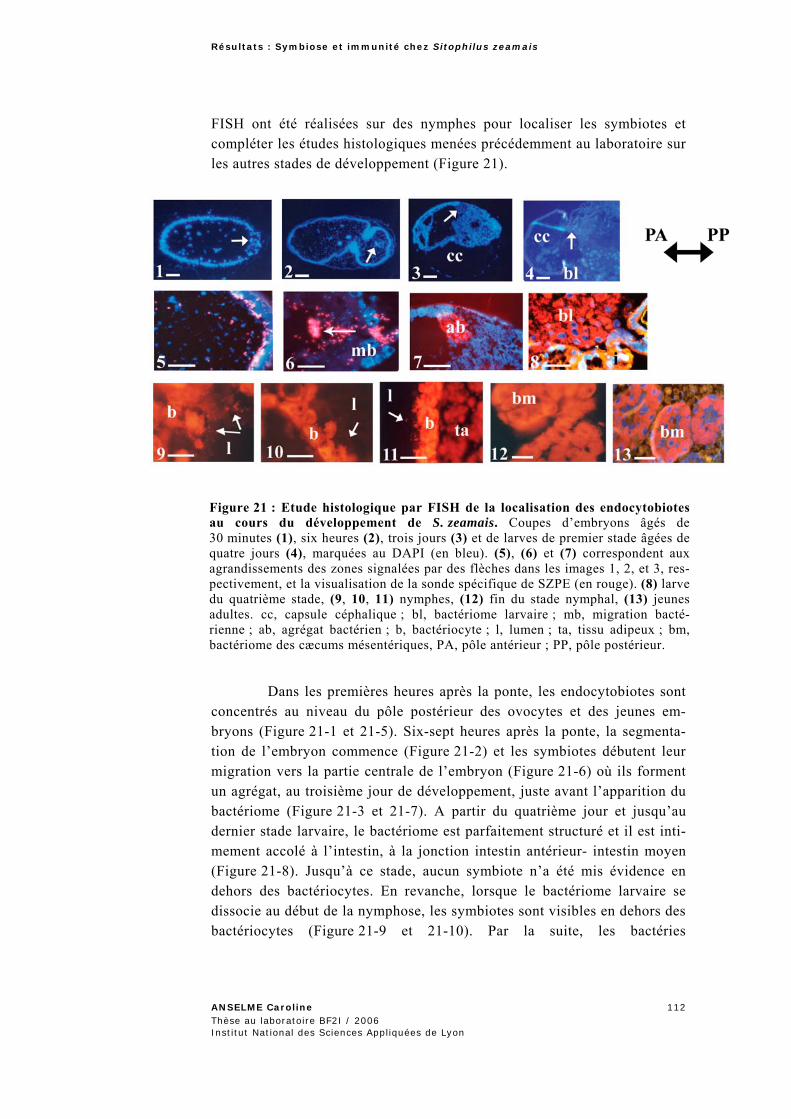
Résultats : Symbiose et immunité chez Sitophilus zeamais
FISH ont été réalisées sur des nymphes pour localiser les symbiotes et compléter les études histologiques menées précédemment au laboratoire sur les autres stades de développement (Figure 21).
Figure 21 : Etude histologique par FISH de la localisation des endocytobiotes au cours du développement de S. zeamais. Coupes d’embryons âgés de 30 minutes (1), six heures (2), trois jours (3) et de larves de premier stade âgées de quatre jours (4), marquées au DAPI (en bleu). (5), (6) et (7) correspondent aux agrandissements des zones signalées par des flèches dans les images 1, 2, et 3, res-pectivement, et la visualisation de la sonde spécifique de SZPE (en rouge). (8) larve du quatrième stade, (9, 10, 11) nymphes, (12) fin du stade nymphal, (13) jeunes adultes. cc, capsule céphalique ; bl, bactériome larvaire ; mb, migration bacté-rienne ; ab, agrégat bactérien ; b, bactériocyte ; l, lumen ; ta, tissu adipeux ; bm, bactériome des cæcums mésentériques, PA, pôle antérieur ; PP, pôle postérieur.
Dans les premières heures après la ponte, les endocytobiotes sont
concentrés au niveau du pôle postérieur des ovocytes et des jeunes em-bryons (Figure 21-1 et 21-5). Six-sept heures après la ponte, la segmenta-tion de l’embryon commence (Figure 21-2) et les symbiotes débutent leur migration vers la partie centrale de l’embryon (Figure 21-6) où ils forment un agrégat, au troisième jour de développement, juste avant l’apparition du bactériome (Figure 21-3 et 21-7). A partir du quatrième jour et jusqu’au dernier stade larvaire, le bactériome est parfaitement structuré et il est inti-mement accolé à l’intestin, à la jonction intestin antérieur- intestin moyen (Figure 21-8). Jusqu’à ce stade, aucun symbiote n’a été mis évidence en dehors des bactériocytes. En revanche, lorsque le bactériome larvaire se dissocie au début de la nymphose, les symbiotes sont visibles en dehors des bactériocytes (Figure 21-9 et 21-10). Par la suite, les bactéries
ANSELME Caroline 112 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
libres sont moins nombreuses et les bactériocytes sont alignés le long de l’intestin évoquant la migration bactériocytaire de l’ancien bactériome lar-vaire vers les cæcums mésentériques (Figure 21-11). A la fin de la méta-morphose, les bactériocytes se regroupent pour former les bactériomes des cæcums mésentériques chez les jeunes adultes, et tous les symbiotes sont à nouveau intracellulaires.
2.2.3 Epissage alternatif du gène PGRP1
Pour poursuivre la caractérisation du gène PGRP1 et aborder sa fonction, nous avons exprimé la protéine dans un système hétérologue pour obtenir des anticorps, et étudier l’activité amidasique prédite sur la base de la sé-quence protéique. Au cours du clonage du transcrit PGRP1, deux fragments d’ADN ont été amplifiés par PCR (Figure 22). Ces derniers ont été clonés et séquencés. Les deux produits de PCR correspondent à deux transcrits PGRP1 de 789 et 903 pb, respectivement, qui ne diffèrent qu’au niveau d’une séquence de 114 pb située en amont du domaine PGRP dans le trans-crit de 903 pb (Figure 23). Ceci suggère l’existence d’un épissage alternatif du gène PGRP1 et pour tester cette hypothèse, le séquençage du gène et l’analyse de ces deux transcrits ont été réalisés.
Figure 22 : Gel d’électrophorèse des produits de l’amplification de l’ADNc du gène PGRP1. (M) Marqueur de tailles, (1) Produit de PCR, (2) et (3) Produits de purification sur gel des deux fragments de PCR. La PCR a été réalisée sur les ADNc du bactériome, avec des amorces situées de part et d’autre de la séquence codante de l’ADNc obtenu à partir de l’EST inf-9.
ANSELME Caroline 113 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
5’-ATGTCCAGTA AGCAATCACG GTCGATAGTA CCCACGTGCG AGTGTAAGTT CGAAAAAATG 60 ^||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
5’-ATGTCCAGTA AGCAATCACG GTCGATAGTA CCCACGTGCG AGTGTAAGTT CGAAAAAATG 60 61 GAAGAAA
ATG AAG G AGTT? ||||
T G A
CCG |||
127 GAAATAGTTA CGAGGGACGA ATGGGGTGCC AAACCACCCA CGGGCGTGGA AAACTTAACT 186 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
187 CTACCAGTCC CTTACGTCGT CATACACCAC AGTTATATAC CTGCAGCTTG TTCAACAAAA 246 |||||||||| |||||||||| |||||||||| |||||||||| |?|||||||| ||||||||||
C
247 GAGGAATGTA TAAATGACAT GCAATGGATG CAAAACTATC ATCAACAAAA TAATTCATGG 306 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
307 TGCGATATTG GATATAATTT CGCTGTTGGA GGGGACAACA GAATTTACGT TGGACGAGGT 366 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
367 TGGACAGCTG TAGGAGCCCA CGCTCCCCGT TACAATGCCA GAAGCATTGG AATCGTCCTG 426 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
427 ATAGGAGATT GGACAGAAAT ATTACCTCCA GAGAGTCAAA TGTTAGCAGC AAAACAATTG 486 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| |||||||?||
C
487 ATTAATATGG GAATCAGAGA CGGCTATATA AGTGAAAACT ACAAATTAAT TGGTCATAGG 546 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| |||?||||||
G
GGAGA|||||||||| |||
A
CCCCA GGAC
T T C
ATGCCA
G T
AGAAAA GAATTA AATATAG 97 |||||||?|| ?|||||||| |||||| |||||||
61 GAAGAAA TG AGAGAAAA AGTTGAATTA AATATAGTAC CCCAGGTTGC TCCACATTAC 120
121 CCCCAGAATT CCTGGAGGAG TTTATTAACA AAATCAAGAA TAATTCTAAT TGTGACAGTG 180
98 TTGCGAACA GCTTCGTATG TACAGAC 126 ||||||||| |||||||||| |||||||
181 TTAGTCATCG CAGTCATCAT ATGTGCAACA TTTGCGAACA GCTTCGTATG TACAGACCCG 240
241 GAAATAGTTA CGAGGGACGA ATGGGGTGCC AAACCACCCA CGGGCGTGGA AAACTTAACT 300
301 CTACCAGTCC CTTACGTCGT CATACACCAC AGTTATATAC C GCAGCTTG TTCAACAAAA 360
361 GAGGAATGTA TAAATGACAT GCAATGGATG CAAAACTATC ATCAACAAAA TAATTCATGG 420
421 TGCGATATTG GATATAATTT CGCTGTTGGA GGGGACAACA GAATTTACGT TGGACGAGGT 480
481 TGGACAGCTG TAGGAGCCCA CGCTCCCCGT TACAATGCCA GAAGCATTGG AATCGTCCTG 540
541 ATAGGAGATT GGACAGAAAT ATTACCTCCA GAGAGTCAAA TGTTAGCAGC AAAACAA TG 600
601 ATTAATATGG GAATCAGAGA CGGCTATATA AGTGAAAACT ACAAATTAAT TGG CATAGG 660
547 CAAGTCCGAG AAACAGAATG TCCAGGAGAA GCATTATATA A TCCA AACGTGGCCT 606 ||||||| |||||||||| |||||||||| |?|||||||| ||||||||||
661 CAAGTCCGAG AAACAGAATG TCCAGGAGAA GCATTATATA A GAGATCCA AACGTGGCCT 720
607 CACTGGATAG ACAA G CCTCGA GAGGTCATTC CTGTAGTACC TCTAGACAAA 666 |||||||||| ||||?||?|| ||||||?||| |||||||||| |||||||||| ||||||||||
721 CACTGGATAG ACAA CC AG CCTCGA GAC GAGGTCATTC CTGTAGTACC TCTAGACAAA 780
667 GAAGACCTTA AACCAGATCC CAGAGAGGAT TCTG GCTTGGCATC TGAAGATAAT 726 |||||||||| |||||||||| |||||||||| ||||?|||?| |||||||||| ||||||||||
781 GAAGACCTTA AACCAGATCC CAGAGAGGAT TCTG TGC A GCTTGGCATC TGAAGATAAT 840
727 AAAATTAAAC GACATAGTTC TACAGACAAA GAGGAAAGTA CGTCGAGGAG TATAAAACAG 786 |||||||||| |||||||||| |||||||||| |||||||||| |||||||||| ||||||||||
841 AAAATTAAAC GACATAGTTC TACAGACAAA GAGGAAAGTA CGTCGAGGAG TATAAAACAG 900
787 TAA-3’ 789 |||
901 TAA-3’ 903
Figure 23 : Alignement des séquences des fragments de 789 pb (en noir) et de 903 pb (en bleu) obtenus lors de l’amplification de la séquence codante du gène PGRP1. L’alignement au niveau de la séquence codante du domaine PGRP est in-diqué en rouge.
ANSELME Caroline 114 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
2.2.3.1 Séquençage du gène PGRP1
Le séquençage du gène PGRP1 de S. zeamais a été réalisé par deux appro-ches différentes (Figure 24). Une approche par longues PCR, à partir d’amorces définies sur la séquence des transcrits, et une approche de mar-che sur le chromosome qui permet de réduire la taille des fragments de PCR par l’utilisation d’enzymes de restriction et d’adaptateurs.
Figure 24 : Epissage alternatif du gène PGRP1 de S. zeamais. Le gène (représen-té en haut) a été séquencé à partir de fragments d’ADNg obtenus par longue PCR et “Genome Walking” (GW). La taille des introns (traits) et des exons (rectangles) est donnée en nombre de paires de bases. Le séquençage de l’intron 2 n’a pas été ache-vé. Les deux transcrits obtenus par l’épissage alternatif du gène sont représentés au-dessous du gène.
Les longues PCR ont été réalisées en testant différentes combinai-
sons d’amorces, différentes températures d’hybridation, et enfin différentes polymérases. La plupart des fragments d’ADN ont été amplifiés grâce aux polymérases et aux différents milieux réactionnels du kit “Expand long template PCR system” (Roche applied science). Les premières longues PCR ont d’abord permis d’obtenir les séquences des introns 1, 3 et 4 de 1224, 392 et 3020 pb, respectivement, et de confirmer l’épissage alternatif du gène en obtenant le début de la séquence de l’intron 2 (Figure 24). Tou-tefois, le produit de PCR de 5518 pb correspondant à la fin de l’exon 2 et au début de l’intron 2 a été amplifié suite à une hybridation aspécifique de l’amorce anti-sens dans l’intron 2. D’autres amorces ont ensuite été défi-nies à partir de cette séquence mais l’approche par longues PCR s’est avé-rée inefficace pour obtenir la fin de l’intron 2.
Grâce à l’utilisation d’une seule amorce spécifique, sens ou anti-sens, définie aux extrémités des exons 1, 2 et 3, la marche sur le chromo-
ANSELME Caroline 115 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Résultats : Symbiose et immunité chez Sitophilus zeamais
some a permis d’obtenir une séquence de 1153 pb en amont du codon initiateur des transcrits et de progresser dans le séquençage de l’intron 2. La séquence obtenue à partir de l’extrémité 5’ de l’exon 3 a permis d’obtenir les 1497 dernières paires de bases de l’intron 2 (Figure 24). En ce qui concerne la séquence de 2223 pb obtenue à partir de l’extrémité 3’ de l’exon 2, une comparaison avec la séquence obtenue par longue PCR a ré-vélé de nombreuses différences entre ces deux séquences. Ces différences correspondent non seulement à des substitutions ou des insertions/délétions ponctuelles mais également à des insertions/délétions de fragments plus importants. Si la taille de ces fragments est en général de 20 à 30 pb, il existe des insertions/délétions de plusieurs centaines de nucléotides pou-vant aller jusqu’à 700 pb. Le cumul des insertions/délétions aboutit à une différence de près de 2 kb entre les deux séquences. Le séquençage de l’intron 2 a ensuite nécessité la définition des amorces au niveau de sé-quences non codantes du gène, séquences riches en bases AT. Les PCR ré-alisées avec ces amorces se sont révélées vaines, quelle que soit l’approche utilisée.
Le séquençage du gène n’a donc pas été achevé, mais le but de ce travail a toutefois été atteint puisque l’épissage alternatif du gène est confirmé. Par ailleurs, l’analyse de l’exon 2 révèle que la séquence de 114 pb code une région aux propriétés transmembranaires (Méthode SOPMA ; Geourjon et Deleage, 1995). Sur la base des séquences nucléoti-diques, l’épissage alternatif du gène conduit donc à deux protéines qui au-raient des localisations cellulaires différentes, une protéine PGRP intracel-lulaire et une protéine PGRP transmembranaire.
2.2.3.2 Quantification des deux transcrits alternatifs du gène PGRP1
Dans les études précédentes de l’expression du gène PGRP1, les amorces utilisées pour les expériences de PCR quantitative ont été définies, en aval de l’exon 2, sur une région commune aux deux transcrits PGRP1. Dans ces conditions, aucune distinction entre les deux transcrits alternatifs ne pou-vait être faite lors de la quantification du taux de transcrits du gène PGRP1. La surexpression de ce gène dans les larves infectées, le bactériome et les nymphes symbiotiques a donc été reconsidérée pour différencier les deux transcrits, et déterminer s’il existe une régulation au niveau de l’épissage du gène. Pour cela, une nouvelle amorce a été définie au niveau de l’exon 2 pour permettre la quantification spécifique du transcrit de 903 pb. Pour chaque échantillon, nous avons quantifié, comme précédemment, la totalité des transcrits PGRP1, et nous avons également quantifié spécifiquement le transcrit de 903 pb en utilisant l’amorce située dans l’exon 2.
ANSELME Caroline 116 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
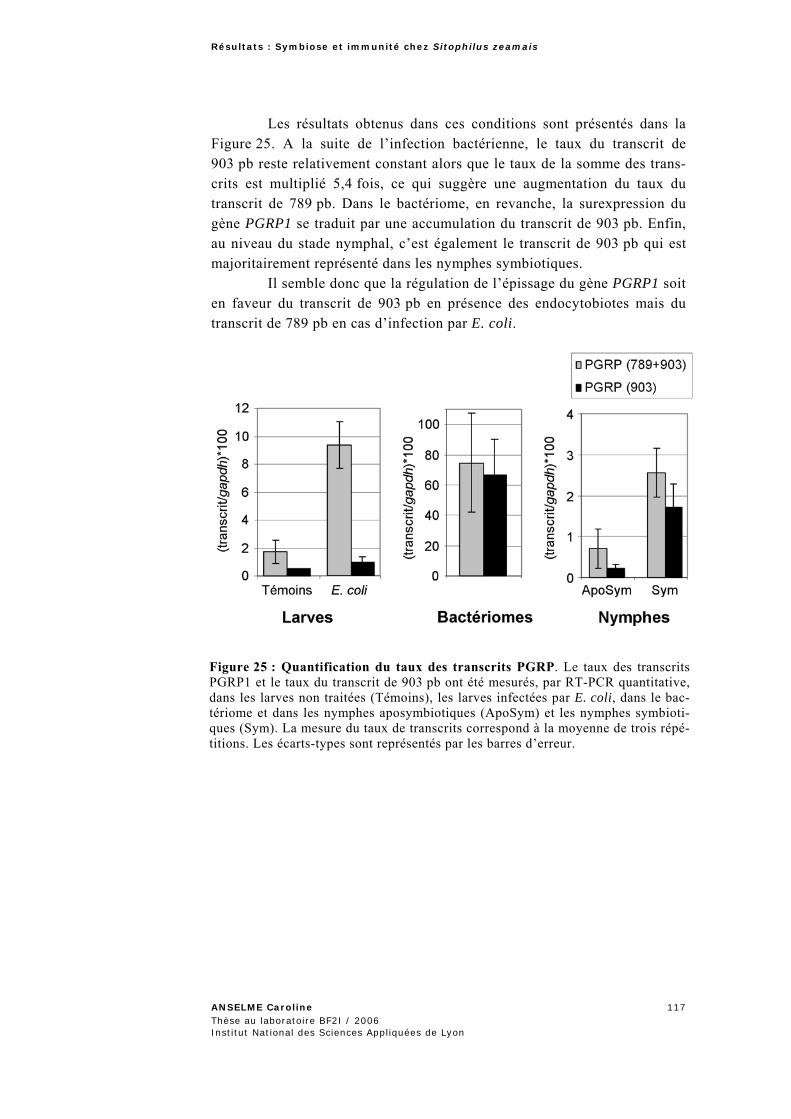
Résultats : Symbiose et immunité chez Sitophilus zeamais
Les résultats obtenus dans ces conditions sont présentés dans la Figure 25. A la suite de l’infection bactérienne, le taux du transcrit de 903 pb reste relativement constant alors que le taux de la somme des trans-crits est multiplié 5,4 fois, ce qui suggère une augmentation du taux du transcrit de 789 pb. Dans le bactériome, en revanche, la surexpression du gène PGRP1 se traduit par une accumulation du transcrit de 903 pb. Enfin, au niveau du stade nymphal, c’est également le transcrit de 903 pb qui est majoritairement représenté dans les nymphes symbiotiques.
Il semble donc que la régulation de l’épissage du gène PGRP1 soit en faveur du transcrit de 903 pb en présence des endocytobiotes mais du transcrit de 789 pb en cas d’infection par E. coli.
Figure 25 : Quantification du taux des transcrits PGRP. Le taux des transcrits PGRP1 et le taux du transcrit de 903 pb ont été mesurés, par RT-PCR quantitative, dans les larves non traitées (Témoins), les larves infectées par E. coli, dans le bac-tériome et dans les nymphes aposymbiotiques (ApoSym) et les nymphes symbioti-ques (Sym). La mesure du taux de transcrits correspond à la moyenne de trois répé-titions. Les écarts-types sont représentés par les barres d’erreur.
ANSELME Caroline 117 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
ANSELME Caroline 118 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
L’étude de l’interaction moléculaire hôte-symbiote menée au laboratoire BF2I sur le modèle Sitophilus-SPE a mis en évidence la surexpression d’un gène immunitaire de la famille des PGRP dans le bactériome (Heddi et al., 2005). Par ailleurs, les études du transcriptome des bactériocytes du puce-ron menées par l’équipe de T. Fukatsu ont révélé la surexpression d’un ly-sozyme, un autre gène de la réponse immunitaire (Nakabachi et al., 2005). Etant donné les fonctions des homologues de ces gènes chez d’autres insec-tes, leur surexpression dans l’organe symbiotique du charançon et du puce-ron suscite de nombreuses interrogations sur la persistance des bactéries, mais également sur le rôle de cette réponse immunitaire dans les bactério-cytes.
Au cours de ma thèse, j’ai entrepris l’étude des aspects immunitai-res de l’interaction hôte-symbiote chez les insectes, pour aborder les ques-tions de la régulation de la population bactérienne dans les bactériocytes, et du rôle de la réponse immunitaire dans le maintien de la symbiose intracel-lulaire. Pour étudier ces aspects et les intégrer dans le processus évolutif des bactéries intracellulaires, une étude comparative a été initiée entre les modèles S. zeamais-SZPE et A. pisum-Buchnera.
1 La réponse immunitaire de l’hôte aux
bactéries Gram (-)
La première étape de cette étude a consisté à identifier les gènes de la ré-ponse immunitaire (GRI) du charançon et du puceron. En raison de l’absence de séquences génomiques, j’ai utilisé une approche de soustrac-tion d’ADNc pour identifier les gènes surexprimés après une infection des larves par E. coli. L’objectif de ce travail n’était pas d’étudier la réponse immunitaire de ces insectes, mais d’identifier des gènes de la réponse hu-morale induits par les bactéries Gram (-) afin d’étudier ensuite leur profil d’expression dans le bactériome, et leur rôle éventuel dans la symbiose in-tracellulaire.
Grâce à la comparaison des séquences des banques soustraites aux bases de données protéiques, j’ai identifié chez S. zeamais une vingtaine d’EST présentant des similitudes de séquence avec des GRI décrits chez d’autres insectes. Pour confirmer l’expression différentielle des EST et identifier les GRI, au-delà de la simple similitude de séquence, j’ai analysé la banque soustraite par la technique de macroarray. J’ai ainsi confirmé l’expression différentielle de nombreuses EST et identifié des EST qui pourraient correspondre à des peptides ou à des protéines spécifiques de la
ANSELME Caroline 119 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
réponse immunitaire du charançon (§ 1.3.1.1 et 1.3.2.1.2). Toutefois, le ca-ractère différentiel de toutes les EST n’a pas pu être analysé car cette tech-nique ne semble pas être bien adaptée aux transcrits faiblement exprimés et/ou lorsque le différentiel est faible, du fait notamment de l’encombrement stérique des signaux sur la membrane. Aussi, pour exploi-ter pleinement la banque soustraite, une seconde analyse globale devrait être réalisée avec la technique de Reverse Northern (Heddi et al., 2005). Cette technique permet de séparer les produits de PCR par électrophorèse avant leur transfert sur membranes. Cette séparation permettrait ainsi de diminuer l’encombrement des signaux sur la membrane et d’analyser l’ensemble des EST.
Si la soustraction a permis l’identification des GRI de S. zeamais, les résultats de l’analyse de la banque soustraite d’A. pisum soulèvent, en revanche, quelques questions. Aucune EST de cette banque ne présente de similitude avec des gènes codant des récepteurs, des peptides antibacté-riens, des lysozymes ou des éléments de la cascade prophénoloxidase. De plus, la faible redondance des EST dans la banque d’A. pisum, et l’absence d’EST nettement différentielle mise en évidence par l’analyse par macroar-ray, suggèrent un manque d’efficacité de la soustraction. Ceci pourrait s’expliquer simplement par un problème technique au cours d’une des éta-pes de la soustraction, mais il est aussi possible que l’approche soustractive ne soit pas adaptée au modèle biologique. En effet, lorsque la différence entre les transcriptomes des deux conditions étudiées est faible, les gènes cibles sont peu représentés dans la banque soustraite. Ainsi, si la réponse immunitaire du puceron à l’infection par E. coli implique des gènes expri-més constitutivement, ou faiblement induits, les GRI ne peuvent être obte-nus par une approche soustractive. Par ailleurs, il faut noter que la réponse immunitaire du puceron est actuellement méconnue car il semble qu’il existe peu d’homologie entre les protéines immunitaires du puceron (ho-moptère) et celles de diptères ou de lépidoptères ; les pucerons étant phylo-génétiquement très éloignés de ces ordres d’insectes (300 MA environ). En effet, seuls un lysozyme de type i (Nakabachi et al., 2005) et un peptide an-tifongique (Megourine, swiss-prot P83417-9), qui ne présente aucune simi-litude avec une autre protéine d’insecte, ont jusqu’à présent été identifiés chez les Aphididae. De plus, chez A. pisum, les EST de l’IAGC compren-nent 59% de séquences orphelines et aucun homologue de gènes de la ré-ponse immunitaire n’a été identifié, ni parmi ces EST, ni parmi les EST d’une banque de tube digestif infecté par Erwinia chrysanthemi (Y. Rahbé, communication personnelle). Les défenses immunitaires du puceron pour-raient donc être assurées par des voies différentes de celles décrites, jus-
ANSELME Caroline 120 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
qu’à présent, dans d’autres modèles. Le séquençage complet du génome du puceron devrait permettre de confirmer ou infirmer cette hypothèse.
Quelle qu’en soit la cause, technique ou biologique, la soustrac-tion chez A. pisum n’a pas permis d’identifier les GRI du puceron. Les as-pects comparatifs de la réponse immunitaire de l’hôte en relation avec la l’évolution du génomes des endocytobiotes n’ont donc pas pu être abordés, mais cette banque d’EST nous aura tout de même permis d’obtenir de nou-velles séquences d’A. pisum, absentes des EST séquencées par le consor-tium international. Dans le cadre de cette thèse, l’étude des aspects immu-nitaires de l’interaction hôte-symbiote a donc été recentrée sur le modèle S. zeamais-SZPE. A l’issue de la soustraction menée chez le charançon, nous disposions d’un ensemble d’EST correspondant à des gènes induits suite à la piqûre des lar-ves par une aiguille infectée par E. coli. Les GRI pouvant être induits par la blessure provoquée par la piqûre et/ou par l’infection bactérienne, j’ai étu-dié plus précisément le profil d’expression des GRI du charançon. En effet, pour aborder l’hypothèse d’une éventuelle induction des GRI par l’endocytobiote SZPE dans le bactériome, il était nécessaire de distinguer dans un premier temps, parmi les GRI identifiés, les gènes induits par les bactéries Gram (-). Pour cette analyse, notre choix s’est porté sur la techni-que de PCR quantitative, et nous avons donc dû recentrer notre étude sur une vingtaine de gènes candidats. La sélection de ces gènes pouvait être guidée par l’homologie des EST et/ou par les résultats de l’analyse globale par macroarray. Nous avons cependant choisi de limiter cette première étude aux gènes présentant une homologie, même faible, à des GRI d’autres insectes.
Nous avons ainsi confirmé que tous les GRI sélectionnés étaient bien surexprimés dans les larves six heures après la blessure et/ou l’infection par E. coli, à l’exception des EST inf-515 et inf-459. Toutefois, les EST de la banque soustraite ayant été obtenues en considérant des temps post-infection de trois, six et douze heures, l’absence d’induction de ces deux gènes doit encore être vérifiée par une quantification des taux de transcrits trois et douze heures après l’infection.
L’analyse des taux de transcrits des larves piquées stérilement et des larves infectées nous a ensuite permis de définir deux catégories de gè-nes : ceux qui étaient induits par l’altération des tissus indépendamment de l’infection, comme les lysozymes (inf-152, -282), le PGRP2 (inf-441) et les homologues putatifs des protéases et des inhibiteurs de protéases (inf-20, -74, -91, -479 et inf-506), et ceux qui étaient induits par E. coli
ANSELME Caroline 121 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
comme les peptides antibactériens putatifs (inf- 18, -42, -145, -163, -165 et inf-217) et le PGRP1 (inf-9).
Pour compléter l’étude du profil d’expression des GRI de S. zeamais, nous avions également choisi de mesurer leur taux de transcrits après une infection par une bactérie Gram (+) puisque la réponse immuni-taire varie en fonction des microorganismes infectants (Hoffmann, 2003). Notre choix s’était alors porté sur une souche de Bacillus megaterium, une bactérie non pathogène, qui pouvait être manipulée dans notre laboratoire. Cependant, lorsque nous avons choisi cette souche, il n’avait pas encore été montré que l’induction différentielle des voies Toll et Imd était liée au type de peptidoglycane bactérien (Leulier et al., 2003). En accord avec les résul-tats décrits chez la drosophile, nous n’avons obtenu aucune différence si-gnificative entre l’infection par E. coli et l’infection par B. megaterium qui possède un PGN de type DAP comme les bactéries Gram (-). Ceci suggère que, si le charançon met effectivement en place une réponse adaptée au type de bactérie, la reconnaissance de ces bactéries se ferait également au niveau du type de PGN. La confirmation de cette hypothèse nécessiterait une étude du profil d’expression des GRI en réponse à une infection par une bactérie possédant un PGN de type Lysine. Cette première partie de la thèse avait pour objectif principal d’identifier les GRI de S. zeamais. Un ensemble de gènes homologues de GRI décrits dans d’autres modèles a été obtenu et la caractérisation du profil d’expression de ces gènes a permis de déterminer ceux qui étaient induits par les bactéries Gram (-). Grâce à ce travail, j’ai donc pu aborder la se-conde partie de la thèse sur les aspects immunitaires de l’interaction hôte-symbiote, dans le modèle S. zeamais, par l’étude de la réponse immunitaire de l’hôte en présence des endocytobiotes.
2 La réponse immunitaire de S. zeamais
aux bactéries symbiotiques
2.1 Réponse immunitaire et maintien de la symbiose dans le bactériome
L’analyse de l’expression des gènes de la réponse immunitaire de S. zeamais dans le bactériome a confirmé la surexpression du gène PGRP1 et a révélé la surexpression de deux autres gènes : un peptide antibactérien de la famille des coléoptéricines et un homologue d’un inhibiteur de la voie des TLR des mammifères nommé Tollip. Cette analyse met donc en évi-dence l’existence d’une réponse immunitaire dans l’organe symbiotique du
ANSELME Caroline 122 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
charançon, avec la surexpression notamment d’un peptide antibactérien. Toutefois, il s’agit d’une réponse modérée en comparaison de la réponse observée à la suite d’une infection par E. coli. En effet, en dehors de la co-léoptéricine, les peptides antibactériens induits chez la larve par E. coli ne sont pas induits dans le bactériome. A titre de remarque, en accord avec leur profil d’expression, les GRI induits par l’altération des tissus provo-quée par la piqûre, et non par les bactéries Gram (-), sont sous-exprimés voire non-exprimés dans le bactériome.
Une des questions suscitées par la mise en évidence de la surex-pression de trois gènes de la réponse immunitaire dans le bactériome, concerne la régulation de ces gènes. En effet, dans les épithéliums qui, comme le bactériome, sont en contact permanent avec des microorganis-mes, il a été montré que les gènes impliqués dans la réponse immunitaire locale pouvaient être exprimés constitutivement ou être induits par ces microorganismes (Ferrandon et al., 1998 ; Tzou et al., 2000 ; Werner et al., 2000 ; Ryu et al., 2004 ; Bischoff et al., 2006 ; Zaidman-Remy et al., 2006). Aussi, la question se pose de savoir si les gènes surexprimés dans le bactériome sont induits par les endocytobiotes ou s’il s’agit d’une expres-sion constitutive.
Dans le cas de l’homologue du gène Tollip, nous avons montré que, contrairement aux gènes coléoptéricine et PGRP1, ce gène n’est pas induit par une infection des larves aposymbiotiques par E. coli ou par B. megaterium. Par ailleurs, Tollip est exprimé constitutivement chez la souris (Burns et al., 2000 ; Li et al., 2004 ; Didierlaurent et al., 2006). Il est donc probable que l’homologue du gène Tollip soit exprimé constitutive-ment dans le bactériome du charançon. Dans le cas de la coléoptéricine et du PGRP1, en revanche, il est plus difficile de privilégier l’une ou l’autre de ces hypothèses au vu des données dont nous disposons.
Pour déterminer si l’expression des GRI dans le bactériome est in-duite ou constitutive, l’approche la plus appropriée serait de comparer leur taux de transcrits entre les bactériomes de larves symbiotiques et les bacté-riomes vides de larves aposymbiotiques. Cette approche reste toutefois dé-licate car les souches aposymbiotiques sont généralement dépourvues de bactériome (§ 1.3.2.1). De plus, les bactériomes vides présents dans une souche obtenue récemment au laboratoire sont fragiles et de taille très ré-duite. Ils sont, de ce fait, difficiles à isoler en quantité suffisante pour une étude par RT-PCR quantitative. Une alternative à la PCR quantitative et à la dissection des bactériomes vides, serait d’utiliser des approches d’hybridation in situ sur des coupes de larves aposymbiotiques pour déter-miner si ces gènes s’expriment dans le bactériome vide.
ANSELME Caroline 123 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
Une question sous-jacente à l’hypothèse d’une induction des gènes coléoptéricine et PGRP1 dans le bactériome, consiste à savoir si les endo-cytobiotes sont perçus ou non par l’hôte. Pour tenter de répondre à cette question, nous avons quantifié le taux de transcrits des GRI dans les larves aposymbiotiques après une “infection” par SZPE, pour déterminer si ces gènes sont induits par le symbiote. Aucune différence significative n’a été observée entre le taux de transcrits obtenu après une piqûre stérile et celui obtenu après une piqûre infectée par SZPE. Toutefois, nous ne pouvons pas conclure à la non induction des GRI sans vérifier l’efficacité de “l’infection” par une mesure de la densité bactérienne dans les larves. En effet, l’absence d’induction des GRI pourrait s’expliquer par une quantité trop faible de bactéries qui n’arriveraient pas à se multiplier à l’intérieur de la larve. Cette dernière hypothèse semble être confirmée par les résultats obtenus lors de l’étude du stade nymphal, qui seront discutés plus loin et qui suggèrent l’induction d’une réponse immunitaire de l’hôte par les sym-biotes.
La réponse immunitaire observée dans le bactériome pourrait donc être constitutive et/ou induite par les endocytobiotes, notamment en cas de risque d’invasion des autres tissus de l’insecte par l'endocytobiote. Dans ce cas, l’induction dans le bactériome impliquerait alors l’existence d’un ré-cepteur intracellulaire, comme le PGRP-LE de la drosophile (Kaneko et al., 2006) ou encore les récepteurs “Nucleotide-binding Oligomerization Do-main proteins” (NOD) décrits chez les mammifères (Strober et al., 2006).
En ce qui concerne le rôle de la réponse immunitaire dans les bac-
tériomes, nous ne pouvons que spéculer, dans l’état actuel de nos connais-sances, sur la fonction des trois gènes surexprimés dans cet organe en fonc-tion des données obtenues sur leurs homologues. Il est toutefois important de garder à l’esprit que, dans le contexte symbiotique, la réponse immuni-taire de Sitophilus pourrait impliquer des processus spécifiques en relation avec la localisation intracellulaire des symbiotes.
Le gène Tollip
Tollip est une protéine des voies de signalisation immunitaire si-tuées en aval des récepteurs membranaires IL-1RI (“Interleukin-1 Receptor type I”), TLR2 et TLR4 (“Toll-Like Receptors”). Elle est décrite comme un inhibiteur de la réponse immunitaire innée des mammifères, car sa surex-pression conduit à une inhibition de l’activation de ces voies (Burns et al., 2000 ; Zhang et Ghosh, 2002). En l’absence de stimulation, Tollip inhibe-rait l’activité de la protéine “Interleukin-1 Receptor Associated Kinase-1” (IRAK-1), une kinase nécessaire à la transduction du signal aboutissant à
ANSELME Caroline 124 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
l’activation de la voie NF-κB (Croston et al., 1995). Il a été suggéré que Tollip pourrait permettre d’une part, de maintenir les cellules immunitaires en état de quiescence en absence d’infection, et d’autre part, de faciliter la terminaison de la réaction immunitaire (Zhang et Ghosh, 2002). Toutefois, le mécanisme d’action de cette protéine n’a pas encore été caractérisé, et les résultats obtenus chez des souris déficientes pour le gène Tollip suggè-rent une fonction beaucoup plus complexe de ce gène (Didierlaurent et al., 2006). L’absence de Tollip résulte en une atténuation de la production de l’IL-6 et du TNF-α, ce qui suggère que cette protéine régulerait la magni-tude de la réponse inflammatoire. La concentration cellulaire de Tollip pourrait donc jouer un rôle crucial dans la régulation de la réponse immuni-taire (Didierlaurent et al., 2006).
Chez les insectes, la recherche de similitude de séquences révèle l’existence d’homologues de Tollip chez Anopheles gambiae (sp Q7QAN1, identité : 83/166), Tribolium castaneum (gb XM_970075.1, identi-té : 138/273), Apis mellifera (gb XM_624414, identité : 117/222) et donc chez S. zeamais (inf-359), mais pas chez D. melanogaster. Par ailleurs, Burns et al. (2000) ont constaté l’absence d’interaction entre la protéine Tollip de la souris et un homologue de la kinase IRAK chez la drosophile : la protéine Pelle (Hoffmann, 2003). Les auteurs suggèrent que la protéine Tollip pourrait assurer une fonction équivalente au domaine C-terminal in-hibiteur du récepteur Toll de la drosophile, qui est absent des TLR (Shen et Manley, 1998). Ceci pourrait alors expliquer l’absence d’homologue de Tollip chez cet insecte.
Compte tenu de l’ensemble de ces données, la surexpression d’un homologue de Tollip dans le bactériome de S. zeamais pourrait assurer une inhibition de l’activation de la réponse immunitaire, voire une régulation fine de cette réponse.
Le gène coléoptéricine
Les coléoptéricines sont des peptides antibactériens d’environ 70 résidus, qui sont riches en résidus glycine, et qui sont spécifiques des coléoptères (Bulet et al., 1991 ; Lee et al., 1994 ; Yang et al., 1998 ; Ima-mura et al., 1999 ; Sagisaka et al., 2001). L’activité antibactérienne anti-Gram (-) ou anti-Gram (+) de ces peptides est variable. La plupart sont actifs contre les bactéries Gram (-), mais la rhinocérosine d’Oryctes rhino-ceros présente également une activité antibactérienne anti-Gram (+) (Yang et al., 1998). Dans le cas de la coléoptéricine d’ Allomyrina dichotoma, il a même été montré que ce peptide présente une forte activité antibactérienne anti-Gram (+), mais qu’il inhibe la division bactérienne d’E. coli (Sagisaka et al., 2001). Les bactéries se présentent alors sous forme de chaînettes qui
ANSELME Caroline 125 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
rappelle les observations faites sur les endocytobiotes du charançon (Figure 26).
Si la coléoptéricine du charançon possède une activité semblable à celle de la coléoptéricine d’A. dichotoma, sa surexpression dans le bacté-riome pourrait alors expliquer en partie le pleïomorphisme des endocyto-biotes observé chez Sitophilus spp. Ce peptide pourrait inhiber la division des endocytobiotes et assurer ainsi une forme de contrôle de la population bactérienne. Bien sûr, il ne s’agit là que de spéculations dans l’attente d’une étude fonctionnelle de la coléoptéricine du charançon. D’après l’analyse in silico de sa séquence, la coléoptéricine serait sécrétée en de-hors des bactériocytes, et ne serait donc pas en contact avec les endocyto-biotes. Néanmoins, comme cela a été mis en évidence au stade nymphal, il est possible que des endocytobiotes se retrouvent à l’extérieur des bactério-cytes. La coléoptéricine pourrait alors prévenir l’invasion des tissus envi-ronnants en éliminant ces symbiotes, ce qui expliquerait qu’aucun symbiote ne soit détecté en dehors des bactériocytes chez la larve. La coléoptéricine pourrait alors permettre à l’hôte de contrôler la localisation des symbiotes.
Figure 26 : Morphologie d’E. coli avant et après traitement par la coléoptéri-cine d’A. dichotoma (en haut) et morphologie des endocytobiotes de Sitophilus spp. (en bas) . Observation en microscopie à contraste de phase d’une culture d’une nuit d’E. coli en l’absence (a) et en présence de la coléoptéricine d’A. dichotoma (1µg/50µl) (b), d’après Sagisaka et al. (2001). Coloration de Gram des endocytobiotes de S. zeamais (c) et S. oryzae (d).
Le gène PGRP
La famille des PGRP comporte une grande variété d’isoformes et de fonctions. Certains PGRP sont des activateurs de la réponse immunitaire alors que d’autres sont, au contraire, des régulateurs négatifs de cette ré-ponse, par le biais d’une activité amidasique. L’homologue du PGRP1 du charançon chez la drosophile, le PGRP-LB, est l’un de ces régulateurs né-gatifs (Zaidman-Remy et al., 2006). Les études menées sur ce PGRP et le
ANSELME Caroline 126 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
PGRP-SC1/2, un autre PGRP amidasique de la drosophile, ont montré que l’activité enzymatique de ces PGRP diminue l’immunogénicité du PGN bactérien, régulant ainsi négativement la réponse immunitaire (Bischoff et al., 2006 ; Zaidman-Remy et al., 2006). Principalement exprimés dans l’intestin, où l’épithélium est en contact permanent avec la flore bacté-rienne, ces PGRP pourraient limiter l’activation de la réponse immunitaire de l’hôte.
Le PGRP1 du charançon, comme le PGRP-LB de la drosophile, a conservé les résidus nécessaires à l’activité amidasique et pourrait donc cliver les PGN bactériens. Ces prédictions ont récemment été confirmées par des mesures d’activité enzymatique3 sur la protéine PGRP1 produite dans un système hétérologue bactérien. Aussi, le modèle proposé par Zaid-man-Remy et al. (2006) pour expliquer le rôle du PGRP-LB dans la régula-tion de la réponse locale de l’intestin chez la drosophile (Figure 8) pourrait s’appliquer au PGRP1 du charançon. Le PGRP1 pourrait, via la dégradation du PGN de SZPE, moduler la réponse immunitaire de l’hôte dans les bacté-riocytes et assurer le maintien de la symbiose chez le charançon. Cette ré-gulation serait d’autant plus efficace que le gène PGRP1 est régulé en fonc-tion de la densité bactérienne (Figure 18 et Figure 19). Toutefois, l’épissage alternatif du gène PGRP1, la localisation prédite in silico des deux protéines PGRP et la localisation intracellulaire des symbiotes susci-tent un grand nombre d’interrogations.
L’étude de l’expression du gène PGRP1 a montré que le transcrit de 789 pb, codant une amidase intracellulaire, est induit par l’infection bac-térienne des larves aposymbiotiques. Ceci suggère que la protéine codée par ce transcrit ait un rôle dans la réponse immunitaire du charançon, indé-pendamment de la symbiose. Toutefois, pour assurer ce rôle, la protéine devrait être présente dans l’hémolymphe. Il est donc probable que la pro-téine soit sécrétée à la suite d’une infection bactérienne, par un processus qu’il reste à déterminer.
Dans le bactériome, en revanche, la protéine PGRP1 intracellu-laire serait en contact direct avec SZPE, et pourrait donc dégrader le PGN libéré par les endocytobiotes. Toutefois, la prévention de l’activation de la réponse immunitaire par le PGN d’une bactérie intracellulaire, implique l’existence d’un récepteur intracellulaire ou encore que le PGN puisse tra-verser la membrane cellulaire. Ces deux hypothèses n’ont pas encore été vérifiées. Par ailleurs, la forme intracellulaire ne serait pas majoritaire dans
3 L’étude de l’activité amidasique de la protéine PGRP du charançon est réalisée par Mireille Hervé à
l’Institut de Biochimie et Biophysique Moléculaire Cellulaire, à Orsay, dans le cadre d’une collabora-tion avec l’équipe de Dominique Mengin-Lecreulx.
ANSELME Caroline 127 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
le bactériome si l’on se réfère aux résultats obtenus par RT-PCR quantita-tive (Figure 25). En effet, dans cet organe, l’épissage du gène PGRP1 gé-nère le transcrit de 903 pb, qui code une amidase dont la localisation transmembranaire suscite un certain nombre de questions. Il est peu proba-ble que ce PGRP soit un récepteur car aucun élément n’a été obtenu en ce sens lors de l’analyse in silico. Dans ce cas, quel rôle joue la localisation membranaire du PGRP en relation avec l’activité amidasique ? Le domaine PGRP est-il intra- ou extracellulaire ? D’après les prédictions in silico, ce domaine serait extracellulaire. Il ne serait donc pas impliqué dans la per-ception des symbiotes à l’intérieur des bactériocytes. Des données supplé-mentaires sont donc nécessaires pour émettre une hypothèse sur la fonction du PGRP transmembranaire, et sur son rôle dans la symbiose du charançon.
2.2 Externalisation des endocytobiotes et activation de la réponse
immunitaire de l’hôte au stade nymphal
Parallèlement aux travaux de Dale et al. (2002) qui ont mis en évidence au stade nymphal une induction des gènes du SSTT inv/spa, nous avons obser-vé la surexpression, à ce stade, du gène PGRP1. Des résultats préliminai-res, non présentés dans cette thèse, suggèrent également une expression dif-férentielle entre les nymphes symbiotiques et aposymbiotiques, de la coléoptéricine (jusqu’à 15 fois) et des peptides potentiellement antibacté-riens, inf-163 et inf-165 (jusqu’à 5 fois). Puisque la taille du bactériome n’augmente pas au-delà du dernier stade larvaire et que l’expression des gènes du bactériome n’est pas perceptible au niveau de la comparaison des larves apo- et symbiotiques (Figure 20), la surexpression des GRI dans les nymphes symbiotiques serait due à l’activation de la réponse immunitaire de l’hôte. Des expériences d’hybridation in situ permettraient de déterminer s’il s’agit d’une réponse locale, dans le bactériome, et/ou d’une réponse systémique.
Parallèlement à l’induction des GRI de S. zeamais et des gènes inv/spa de SZPE au stade nymphal, nous avons également mis en évidence, par les expériences de FISH, la présence de symbiotes en dehors des bacté-riocytes, ce qui n’avait jamais été décrit chez Sitophilus. Bien que nous ne puissions exclure un rôle actif de SZPE, il est probable que la libération des endocytobiotes soit liée à la dissociation du bactériome larvaire, et à la lyse de quelques bactériocytes, au cours des réarrangements cellulaires pendant la métamorphose. L’expression des GRI de S. zeamais et des gènes du SSTT de SZPE serait alors la conséquence de la libération des endocytobio-tes. Il semble donc que l’hôte perçoive les symbiotes en dehors des bacté-riocytes, ce que nous n’avions pas pu mettre en évidence par des expérien-ces d’infection des larves aposymbiotiques par SZPE. Les GRI de l’insecte
ANSELME Caroline 128 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
seraient alors exprimés pour éliminer les symbiotes libres. La surexpression des gènes du SSTT des symbiotes, quant à elle, pourrait être interprétée comme un changement physiologique des bactéries qui leur permettrait de réinfecter de nouvelles cellules de l’hôte. Dans ce cas, la formation des bactériomes des cæcums mésentériques chez l’adulte ne serait pas seule-ment due à la migration des bactériocytes larvaires (Nardon et Nardon, 1998), mais pourrait impliquer des phénomènes de réinfection cellulaire.
Par ailleurs, l’induction des gènes du SSTT de SZPE, au stade nymphal uniquement, suggère que l’expression des gènes impliqués dans l’invasion cellulaire est réprimée dans les bactériocytes larvaires. Cette ré-pression pourrait être due à un système de régulation bactérien à deux com-posantes impliqué dans la perception du milieu environnant (comme le sys-tème PhoPQ) qui permettrait à la bactérie de déterminer sa localisation cellulaire et de moduler, en fonction, l’expression des gènes du SSTT. Ain-si, lorsque SZPE est libéré dans l’hémolymphe au stade nymphal, les gènes du SSTT s’expriment pour permettre à SZPE de réinfecter une nouvelle cel-lule. Bien que cette hypothèse paraisse vraisemblable, il n’est pas exclu que l’hôte puisse inhiber les gènes de virulence bactériens. Leur induction au stade nymphal serait alors due à une levée de l’inhibition à la suite de la li-bération des symbiotes.
2.3 Une réponse immunitaire spécifique
Avant la mise en évidence d’un épissage alternatif du gène PGRP1, la su-rexpression de ce gène dans le bactériome larvaire avait été considérée comme une réponse immunitaire de l’hôte, semblable à celle observée après une infection bactérienne (Heddi et al., 2005). La quantification du taux des deux transcrits alternatifs par RT-PCR quantitative montre que le gène est surexprimé sous la forme d’un transcrit de 903 pb dans le bactériome alors que c’est le transcrit de 789 pb qui est épissé à la suite d’une infec-tions des larves par E. coli ou B. megaterium (Figure 25). La réponse im-munitaire de l’hôte dans l’organe symbiotique serait donc différente de celle observée après une infection bactérienne.
Bien que le rôle de ces deux transcrits ne soit pas encore élucidé, deux hypothèses pourraient expliquer cette différence. La première serait qu’il existe un mécanisme de régulation au niveau de l’épissage du gène PGRP1 qui produit le transcrit de 903 pb dans les bactériocytes, et le trans-crit de 789 pb dans les autres cellules immunocompétentes. La réponse immunitaire observée dans les bactériocytes serait alors tissu-spécifique. La seconde hypothèse repose sur l’existence d’une réponse immunitaire spéci-fique aux symbiotes, à travers une régulation de l’épissage du gène PGRP1 qui privilégierait le transcrit de 903 pb en présence de SZPE.
ANSELME Caroline 129 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Discussion
Pour déterminer quel type de régulation est à l’origine de la ré-ponse observée dans le bactériome, il faudrait déterminer quel transcrit est épissé par les cellules immunocompétentes lors d’une “infection” par SZPE. Les expériences de RT-PCR quantitative et de FISH ont mis en évi-dence l’induction du transcrit PGRP1 de 903 pb au stade nymphal, où des symbiotes sont présents en dehors des bactériocytes. L’étude de ce stade par des approches d’hybridation in situ permettrait de déterminer si le transcrit de 903 pb est épissé uniquement dans les bactériocytes (réponse locale), ou s’il est épissé dans d’autres cellules immunocompétentes à la suite de la libération des symbiotes.
L’existence de deux transcrits PGRP1, l’un épissé dans la réponse aux infections bactériennes, et l’autre épissé spécifiquement dans la ré-ponse aux symbiotes (bactériome larvaire et stade nymphal), suggère que l’épissage alternatif du gène PGRP1 constitue une forme d’adaptation de la réponse immunitaire du charançon au maintien de la symbiose. Les études histologique (immunolocalisation) et fonctionnelle (test d’activité, RNAi) menées actuellement au laboratoire devraient permettre de caractériser le rôle des deux protéines codées par le gène PGRP1 dans la réponse immuni-taire aux bactéries symbiotiques intracellulaires.
ANSELME Caroline 130 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Conclusion
ANSELME Caroline 131 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Conclusion
L’immunité est un domaine qui est resté longtemps négligé chez les insec-tes en comparaison de l’intérêt porté aux modèles de vertébrés. Cependant, depuis une dizaine d’années, ce domaine a connu un développement consi-dérable chez les insectes, notamment grâce aux données apportées par les travaux de génétique menés chez la drosophile et par le séquençage du gé-nome complet de cet insecte. Ces données ont révélé l’existence de nom-breuses familles de gènes de la réponse immunitaire souvent conservées chez les mammifères, ce qui a engendré un intérêt grandissant pour la ca-ractérisation de la fonction de ces gènes.
Chez les insectes symbiotiques, les premières évidences de l’implication de l’immunité dans la perception et le contrôle des endocyto-biotes n’ont été publiées que très récemment (Heddi et al., 2005 ; Nakaba-chi et al., 2005). En effet, très peu de travaux ont été consacrés aux aspects immunitaires de la symbiose bactérienne intracellulaire des insectes. Ceci est probablement dû à une conception qui consiste à séparer les bactéries mutualistes des bactéries pathogènes. Pourtant, la proximité phylogénétique des endocytobiotes avec des bactéries parasites ou pathogènes étaye l’hypothèse d’une origine “pathogène” de ces bactéries mutualistes (Charles et al., 2001 ; Dale et al., 2002 ; Lefèvre et al., 2004), et les don-nées cellulaires et moléculaires récentes montrent que les interactions hôte-bactérie, qu’elles soient pathogènes ou mutualistes, impliquent des gènes et des voies cellulaires très similaires.
L’étude menée au cours de cette thèse a permis d’identifier les gè-nes de la réponse immunitaire du charançon, et a mis en évidence l’existence d’une réponse immunitaire de l’hôte dans l’organe symbiotique, le bactériome. Toutefois, cette réponse ne correspond pas à la réponse im-munitaire observée chez les larves, à la suite d’une infection bactérienne. Les premiers résultats de ce travail suggèrent donc l’existence d’une ré-ponse immunitaire spécifique dans le bactériome, avec la surexpression d’un seul peptide antibactérien, une coléoptéricine, d’un homologue de Tollip, et surtout la surexpression et l’épissage alternatif du gène PGRP1. La coléoptéricine pourrait prévenir l’invasion de l’hôte par les symbiotes, et les gènes PGRP1 et Tollip auraient quand à eux un rôle immunomodula-teur qui permettrait de modérer la réponse immunitaire dans l’organe sym-biotique. Enfin, l’épissage spécifique d’un des deux transcrits du gène PGRP1 dans la réponse aux symbiotes suggère que l’épissage de ce gène constitue une forme d’adaptation de la réponse immunitaire dans le contexte symbiotique.
La fonction des trois gènes surexprimés dans le bactériome reste cependant spéculative et de nombreux aspects de la réponse immunitaire doivent encore être caractérisés (réponse induite ou constitutive, spécificité
ANSELME Caroline 132 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Conclusion
liée à l’organe ou à l’endocytobiote...). Les travaux menés au laboratoire devraient permettre, par des approches fonctionnelles (RNAi, test d’activité) et histologiques (hybridation in situ, immunolocalisation), de tester les différentes hypothèses formulées dans cette thèse.
Cette première étude des aspects immunitaires de l’interaction hôte-symbiote chez le charançon apporte des éléments en faveur d’une adaptation de la réponse immunitaire de l’hôte au maintien de la symbiose intracellulaire. Ces éléments suggèrent notamment que la différenciation du bactériome ne permet pas simplement d’isoler les endocytobiotes du sys-tème immunitaire de l’insecte mais que cet organe possède une réponse immunitaire adaptée à ces bactéries symbiotiques. L’existence de cette ré-ponse ouvre de nouvelles perspectives quant aux mécanismes qui pour-raient assurer le contrôle des endocytobiotes dans le bactériome, mais éga-lement l’élimination de ces derniers dans les bactériomes des cæcums mésentériques des adultes âgés de plus de trois semaines et dans la lignée germinale mâle, un phénomène qui demeure encore méconnu.
L’intérêt du modèle charançon, par rapport à d’autres modèles comme celui du puceron, tient au fait que SPE n’a intégré la symbiose in-tracellulaire que depuis 25 MA, probablement suite à un remplacement de symbiote (Lefèvre et al., 2004). SPE possède des structures similaires aux bactéries libres, notamment en ce qui concerne la paroi bactérienne (recon-naissance par les PRR) et le système de sécrétion de type III (invasion cel-lulaire). Les résultats obtenus dans ce modèle devraient permettre de com-prendre les aspects immunitaires impliqués dans les phases précoces de l’interaction hôte-symbiote. Une des questions qui reste posée concerne l’évolution de la réponse immunitaire de l’hôte parallèlement à la dégéné-rescence du génome des endocytobiotes qui engendre des modifications profondes dans la structure de la paroi bactérienne et dans les modes de transmission et d’invasion cellulaire. Il serait alors intéressant de mener une étude comparative entre le modèle S. zeamais-SZPE et un autre modèle appartenant au clade ancestral des Dryophthoridae, où l’endocytobiote (Candidatus Nardonella) aurait intégré la symbiose il y a près de 100 MA (Lefèvre et al., 2004).
Chez le puceron, dont la symbiose remonte à 150-200 MA, une réponse immunitaire semble subsister dans les bactériocytes avec la surex-pression d’un lysozyme dont la fonction, en relation avec la symbiose, reste encore à caractériser (Nakabachi et al., 2005). Pour déterminer si la diffé-rence observée entre le modèle puceron et le modèle charançon est due à l’absence, chez Buchnera, de système de sécrétion (excepté peut être le système flagellaire) et de certains constituants de la paroi bactérienne (LPS et phospholipides), nous avions abordé une étude comparative entre ces
ANSELME Caroline 133 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Conclusion
deux modèles. Cependant, en l’absence d’homologues de gènes de la ré-ponse immunitaire décrits chez les insectes holométaboles, comme les pep-tides antibactériens ou les PGRP, une étude comparative qui inclurait le modèle puceron semble difficilement envisageable. En effet, la différence observée entre le puceron et le charançon pourrait être due à une différence dans les gènes de la réponse immunitaire de l’insecte. Dès lors, il serait plus judicieux de considérer pour une étude comparative des modèles de symbiose primaire chez des insectes holométaboles, comme les diptères ou les coléoptères, où des gènes homologues interviennent dans la réponse immunitaire de l’hôte. Par ailleurs, il serait intéressant d’envisager une étude de la réponse immunitaire des insectes holométaboles en présence de Buchnera afin de déterminer si la dégénérescence du génome du symbiote altère sa perception par le système immunitaire de l’hôte. Cette étude nous permettrait ainsi de savoir si la réponse observée dans les bactériocytes du puceron est liée au système immunitaire de l’insecte ou aux caractéristiques de l’endocytobiote.
Les éléments apportés par ce travail de thèse suggèrent que la ré-ponse immunitaire ne se limite pas à un ensemble de processus permettant à l’hôte d’éliminer les microorganismes. Cette définition trop réductrice qui semblait incompatible avec le maintien de la symbiose, à moins que le symbiote n’échappe au système immunitaire de l’hôte, est probablement responsable du faible nombre d’études consacrées aux aspects immunitaires de la symbiose chez les insectes. Comme le suggérait A. Paillot, déjà en 1933, la symbiose devrait être considérée comme une infection bactérienne non pathogène qui résulte d’un “équilibre” entre la réponse immunitaire de l’hôte et la virulence du symbiote. Les notions d’immunité et de virulence ne devraient alors pas être restreintes à une conception où la fonction des gènes impliqués dans ces processus se limiterait à l’élimination des bacté-ries et à l’envahissement de l’hôte, respectivement.
ANSELME Caroline 134 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
ANSELME Caroline 135 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Akman, L., Yamashita, A., Watanabe, H., Oshima, K., Shiba, T., Hattori, M. and Aksoy, S. (2002)
Genome sequence of the endocellular obligate symbiont of tsetse flies, Wigglesworthia glossinidia.
Nat Genet. 32: 402-407.
Aksoy, S. (2000) Tsetse - A haven for microorganisms. Parasitology today. 16: 114-118.
Anselme, C., Vallier, A., Balmand, S., Fauvarque, M.O. and Heddi, A. (2006) Host PGRP gene
expression and bacterial release in endosymbiosis of the weevil Sitophilus zeamais. Appl Environ
Microbiol. 72: 6766-6772.
Basset, A., Khush, R.S., Braun, A., Gardan, L., Boccard, F., Hoffmann, J.A. and Lemaitre, B.
(2000) The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an
immune response. Proc Natl Acad Sci U S A. 97: 3376-3381.
Bischoff, V., Vignal, C., Boneca, I.G., Michel, T., Hoffmann, J.A. and Royet, J. (2004) Function of
the drosophila pattern-recognition receptor PGRP-SD in the detection of Gram-positive bacteria.
Nat Immunol. 5: 1175-1180.
Bischoff, V., Vignal, C., Duvic, B., Boneca, I.G., Hoffmann, J.A. and Royet, J. (2006) Downregula-
tion of the Drosophila Immune Response by Peptidoglycan-Recognition Proteins SC1 and SC2.
PLoS Pathog. 2: 139-147.
Boutros, M., Agaisse, H. and Perrimon, N. (2002) Sequential activation of signaling pathways during
innate immune responses in Drosophila. Dev Cell. 3: 711-722.
Braendle, C., Miura, T., Bickel, R., Shingleton, A.W., Kambhampati, S. and Stern, D.L. (2003)
Developmental origin and evolution of bacteriocytes in the aphid-Buchnera symbiosis. PLoS Biol.
1: 70-76.
Braun, A., Hoffmann, J.A. and Meister, M. (1998) Analysis of the Drosophila host defense in domino
mutant larvae, which are devoid of hemocytes. Proc Natl Acad Sci U S A. 95: 14337-14342.
Breeuwers, J.A.J. and Werren, J.H. (1990) Microorganisms associated with chromosome destruction
and reproductive isolation between two insect species. Nature. 346: 558-560.
Buchner, P. (1930) Tier und Pflanze in Symbiose, 2nd ed. Berlin: Gebrüder Borntraeger, 900 p.
Buchner, P. (1965) Endosymbiosis of animals with plant microorganisms. New York, London, Sydney:
Interscience Publishers, a division of Wiley & sons, Inc., 909 p.
Bulet, P. and Stocklin, R. (2005) Insect antimicrobial peptides: structures, properties and gene regula-
tion. Protein Pept Lett. 12: 3-11.
Bulet, P., Stocklin, R. and Menin, L. (2004) Anti-microbial peptides: from invertebrates to vertebrates.
Immunol Rev. 198: 169-184.
Bulet, P., Cociancich, S., Dimarcq, J.L., Lambert, J., Reichhart, J.M., Hoffmann, D., Hetru, C.
and Hoffmann, J. A. (1991) Insect immunity. Isolation from a coleopteran insect of a novel induc-
ible antibacterial peptide and of new members of the insect defensin family. J Biol Chem. 266:
24520-24525.
ANSELME Caroline 136 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Burns, K., Clatworthy, J., Martin, L., Martinon, F., Plumpton, C., Maschera, B., Lewis, A., Ray,
K., Tschopp, J. and Volpe, F. (2000) Tollip, a new component of the IL-1RI pathway, links IRAK
to the IL-1 receptor. Nat Cell Biol. 2: 346-351.
Cerenius, L. and Soderhall, K. (2004) The prophenoloxidase-activating system in invertebrates.
Immunol Rev. 198: 116-126.
Chang, C.I., Ihara, K., Chelliah, Y., Mengin-Lecreulx, D., Wakatsuki, S. and Deisenhofer, J.
(2005) Structure of the ectodomain of Drosophila peptidoglycan-recognition protein LCa suggests a
molecular mechanism for pattern recognition. Proc Natl Acad Sci U S A. 102: 10279-10284.
Chang, C.I., Pili-Floury, S.S., Herve, M., Parquet, C., Chelliah, Y., Lemaitre, B., Mengin-Lecreulx,
D. and Deisenhofer, J. (2004) A Drosophila pattern recognition receptor contains a peptidoglycan
docking groove and unusual L,D-carboxypeptidase activity. PLoS Biol. 2: 1293-1302.
Charles, H. and Ishikawa, H. (1999) Physical and genetic map of the genome of Buchnera, the primary
endosymbiont of the pea aphid Acyrthosiphon pisum. J Mol Evol. 48: 142-150.
Charles, H., Heddi, A. and Rahbé, Y. (2001) A putative insect intracellular endosymbiont stem clade,
within the Enterobacteriaceae, infered from phylogenetic analysis based on a heterogeneous model of
DNA evolution. C R Acad Sci Paris. 324: 489-494.
Charles, H., Condemine, G., Nardon, C. and Nardon, P. (1997) Genome size characterization of the
principal endocellular symbiotic bacteria of the weevil Sitophilus oryzae, using pulsed field gel elec-
trophoresis. Insect Biochem Mol Biol. 27: 345-350.
Cheng, X., Zhang, X., Pflugrath, J.W. and Studier, F.W. (1994) The structure of bacteriophage T7
lysozyme, a zinc amidase and an inhibitor of T7 RNA polymerase. Proc Natl Acad Sci U S A.
91: 4034-4038.
Choe, K.M., Lee, H. and Anderson, K.V. (2005) Drosophila peptidoglycan recognition protein LC
(PGRP-LC) acts as a signal-transducing innate immune receptor. Proc Natl Acad Sci U S A.
102: 1122-1126.
Choe, K.M., Werner, T., Stoven, S., Hultmark, D. and Anderson, K.V. (2002) Requirement for a
peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in
Drosophila. Science. 296: 359-362.
Croston, G.E., Cao, Z. and Goeddel, D.V. (1995) NF-kappa B activation by interleukin-1 (IL-1)
requires an IL-1 receptor-associated protein kinase activity. J Biol Chem. 270: 16514-16517.
Dale, C., Jones, T. and Pontes, M. (2005) Degenerative evolution and functional diversification of
type-III secretion systems in the insect endosymbiont Sodalis glossinidius. Mol Biol Evol.
22: 758-766.
Dale, C., Plague, G.R., Wang, B., Ochman, H. and Moran, N.A. (2002) Type III secretion systems
and the evolution of mutualistic endosymbiosis. Proc Natl Acad Sci U S A. 99: 12397-12402.
De Bary, A. (1879) De la symbiose. Revue Inter Sci. 3: 301-309.
ANSELME Caroline 137 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
De Gregorio, E., Spellman, P.T., Rubin, G.M. and Lemaitre, B. (2001) Genome-wide analysis of the
Drosophila immune response by using oligonucleotide microarrays. Proc Natl Acad Sci U S A.
98: 12590-12595.
De Gregorio, E., Spellman, P.T., Tzou, P., Rubin, G.M. and Lemaitre, B. (2002a) The Toll and Imd
pathways are the major regulators of the immune response in Drosophila. EMBO J. 21: 2568-2579.
De Gregorio, E., Han, S.J., Lee, W.J., Baek, M.J., Osaki, T., Kawabata, S., Lee, B. L., Iwanaga, S.,
Lemaitre, B. and Brey, P. T. (2002b) An immune-responsive Serpin regulates the melanization cas-
cade in Drosophila. Dev Cell. 3: 581-592.
Dedeine, F., Vavre, F., Fleury, F., Loppin, B., Hochberg, E. and Boulétreau, M. (2001) Removing
symbiotic Wolbachia bacteria specifically inhibits oogenesis in a parasitic wasp. Proc Natl Acad Sci
U S A. 98: 6247-6252.
Didierlaurent, A., Brissoni, B., Velin, D., Aebi, N., Tardivel, A., Kaslin, E., Sirard, J. C., Angelov,
G., Tschopp, J. and Burns, K. (2006) Tollip regulates proinflammatory responses to interleukin-1
and lipopolysaccharide. Mol Cell Biol. 26: 735-742.
Dobson, S.L., Bourtzis, K., Braig, H.R., Jones, B.F., Zhou, W., Rousset, F. and O'Neill, S.L. (1999)
Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem
Mol Biol. 29: 153-160.
Dziarski, R. (2004) Peptidoglycan recognition proteins (PGRPs). Mol Immunol. 40: 877-886.
Elrod-Erickson, M., Mishra, S. and Schneider, D. (2000) Interactions between the cellular and hu-
moral immune responses in Drosophila. Curr Biol. 10: 781-784.
Erwin, T.L. (1997) Biodiversity at its utmost: Tropical forest beetles. In Biodiversity II: Understanding
and Protecting Our Biological Ressources. Reaka-Kudla, M.L., Wilson, D.E., and Wilson, E.O.
(eds.) Washington, D. C.: Joseph Henry Press, pp. 27-40.
Fauvarque, M.O., Bergeret, E., Chabert, J., Dacheux, D., Satre, M. and Attree, I. (2002) Role and
activation of type III secretion system genes in Pseudomonas aeruginosa-induced Drosophila killing.
Microb Pathog. 32: 287-295.
Fearon, D.T. (1997) Seeking wisdom in innate immunity. Nature. 388: 323-324.
Fearon, D.T. (1999) Innate immunity and the biological relevance of the acquired immune response.
QJM. 92: 235-237.
Febvay, G., Liadouze, I., Guillaud, J. and Bonnot, G. (1995) Analysis of energetic amino acid me-
tabolism in Acyrthosiphon pisum: a multidimentional approach to amino acid metabolism in aphids.
Arch Insect Biochem Physiol. 29: 45-69.
Ferrandon, D. (2006) Sensing infections in Drosophila. Communication orale. Conférence Jacques-
Monod, Roscoff, France. In Bangham, J., Jiggins, F. and Lemaitre, B. (2006) Insect immunity: the
post-genomic era. Immunity. 25: 1-5.
ANSELME Caroline 138 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Ferrandon, D. and Royet, J. (2002) La drosophile : un modèle pour l'étude de la réponse immunitaire
des invertébrés. Regard sur la biochimie. 1: 17-23.
Ferrandon, D., Jung, A.C., Criqui, M., Lemaitre, B., Uttenweiler-Joseph, S., Michaut, L., Reich-
hart, J. and Hoffmann, J. A. (1998) A drosomycin-GFP reporter transgene reveals a local immune
response in Drosophila that is not dependent on the Toll pathway. EMBO J. 17: 1217-1227.
Fialho, R.F. and Stevens, L. (2000) Male-killing Wolbachia in a flour beetle. Proc R Soc Lond B Biol
Sci. 267: 1469-1473.
Filipe, S.R., Tomasz, A. and Ligoxygakis, P. (2005) Requirements of peptidoglycan structure that
allow detection by the Drosophila Toll pathway. EMBO Rep. 6: 327-333.
Frank, A.B. (1877) Uber die biologischen Verhältnisse des Thallus eineger Krustenflechnten. Beitr Biol
Pflanz. 2: 123-200.
Fukatsu, T., Nikoh, N., Kawai, R. and Koga, R. (2000) The secondary endosymbiotic bacterium of the
pea aphid Acyrthosiphon pisum (Insecta : Homoptera). Appl Environ Microbiol. 66: 2748-2758.
Ganz, T. (2004) Defensins: antimicrobial peptides of vertebrates. C R Biol. 327: 539-549.
Garver, L.S., Wu, J. and Wu, L.P. (2006) The peptidoglycan recognition protein PGRP-SC1a is essen-
tial for Toll signaling and phagocytosis of Staphylococcus aureus in Drosophila. Proc Natl Acad Sci
U S A. 103: 660-665.
Georgel, P., Naitza, S., Kappler, C., Ferrandon, D., Zachary, D., Swimmer, C., Kopczynski, C.,
Duyk, G., Reichhart, J. M. and Hoffmann, J. A. (2001) Drosophila immune deficiency (IMD) is a
death domain protein that activates antibacterial defense and can promote apoptosis. Dev Cell. 1:
503-514.
Geourjon, C. and Deleage, G. (1995) SOPMA: significant improvements in protein secondary structure
prediction by consensus prediction from multiple alignments. Comput Appl Biosci. 11: 681-684.
Gil, R., Sabater-Munoz, B., Latorre, A., Silva, F.J. and Moya, A. (2002) Extreme genome reduction
in Buchnera spp.: Toward the minimal genome needed for symbiotic life. Proc Natl Acad Sci U S A.
99: 4454-4458.
Gil, R., Silva, F.J., Zientz, E., Delmotte, F., Gonzalez-Candelas, F., Latorre, A., Rausell, C., Ka-
merbeek, J., Gadau, J., Hölldobler, B., van Ham, R.C.H.J., Gross, R. and Moya, A. (2003) The
genome sequence of Blochmannia floridanus: Comparative analysis of reduced genomes. Proc Natl
Acad Sci U S A. 100: 9388-9393.
Gottar, M., Gobert, V., Michel, T., Belvin, M., Duyk, G., Hoffmann, J.A., Ferrandon, D. and
Royet, J. (2002) The Drosophila immune response against Gram-negative bacteria is mediated by a
peptidoglycan recognition protein. Nature. 416: 640-644.
Grenier, A.M., Nardon, P. and Bonnot, G. (1986) Importance de la symbiose dans la croissance des
populations de Sitophilus oryzae L. (Coleoptère Curculionidae). Etude théorique et expérimentale.
Acta Oecologica. 7: 93-110.
ANSELME Caroline 139 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Grenier, A.M., Nardon, C. and Nardon, P. (1994) The role of symbionts in flight activity of Sitophi-
lus weevils. Entomol exp appl. 70: 201-208.
Heddi, A., Lefebvre, F. and Nardon, P. (1993) Effect of endocytobiotic bacteria on mitochondrial en-
zymatic activities in the weevil Sitophilus oryzae (Coleoptera, Curculionidae). Insect Biochem Mol
Biol. 23: 403-411.
Heddi, A., Charles, H., Khatchadourian, C., Bonnot, G. and Nardon, P. (1998) Molecular charac-
terization of the principal symbiotic bacteria of the weevil Sitophilus oryzae: A peculiar G - C con-
tent of an endocytobiotic DNA. J Mol Evol. 47: 52-61.
Heddi, A., Grenier, A.M., Khatchadourian, C., Charles, H. and Nardon, P. (1999) Four intracellular
genomes direct weevil biology: Nuclear, mitochondrial, principal endosymbionts, and Wolbachia.
Proc Natl Acad Sci U S A. 96: 6814-6819.
Heddi, A., Vallier, A., Anselme, C., Xin, H., Rahbé, Y. and Wäckers, F. (2005) Molecular and cellu-
lar profiles of insect bacteriocytes: mutualism and harm at the initial evolutionary step of symbio-
genesis. Cell Microbiol. 7: 293-305.
Hoffmann, J.A. (2003) The immune response of Drosophila. Nature. 426: 33-38.
Hoffmann, J.A. and Reichhart, J.M. (2002) Drosophila innate immunity: an evolutionary perspective.
Nat Immunol. 3: 121-126.
Holz, A., Bossinger, B., Strasser, T., Janning, W. and Klapper, R. (2003) The two origins of hemo-
cytes in Drosophila. Development. 130: 4955-4962.
Hong, R.W., Shchepetov, M., Weiser, J.N. and Axelsen, P.H. (2003) Transcriptional profile of the
Escherichia coli response to the antimicrobial insect peptide cecropin A. Antimicrob Agents Chemo-
ther. 47: 1-6.
Horng, T. and Medzhitov, R. (2001) Drosophila MyD88 is an adapter in the Toll signaling pathway.
Proc Natl Acad Sci U S A. 98: 12654-12658.
Hosaka, M., Nagahama, M., Kim, W.S., Watanabe, T., Hatsuzawa, K., Ikemizu, J., Murakami, K.
and Nakayama, K. (1991) Arg-X-Lys/Arg-Arg motif as a signal for precursor cleavage catalyzed by
furin within the constitutive secretory pathway. J Biol Chem. 266: 12127-12130.
Iimura, Y., Sasaki, T., Fukatsu, T. and Ishikawa, H. (1995) Diminution of intracellular symbiont of
aphid maintained on artificial diet: a morphological study. Zool Sci. 12: 795-799.
Imamura, M., Wada, S., Koizumi, N., Kadotani, T., Yaoi, K., Sato, R. and Iwahana, H. (1999)
Acaloleptins A: inducible antibacterial peptides from larvae of the beetle, Acalolepta luxuriosa.
Arch Insect Biochem Physiol. 40: 88-98.
Janeway, C.A., Jr. (1989) Approaching the asymptote? Evolution and revolution in immunology. Cold
Spring Harb Symp Quant Biol. 54: 1-13.
Jeyaprakash, A. and Hoy, M.A. (2000) Long PCR improves Wolbachia DNA amplification: wsp
sequences found in 76% of sixty-three arthropod species. Insect Mol Biol. 9: 393-405.
ANSELME Caroline 140 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Kambris, Z., Brun, S., Jang, I.H., Nam, H.J., Romeo, Y., Takahashi, K., Lee, W. J., Ueda, R. and
Lemaitre, B. (2006) Drosophila immunity: a large-scale in vivo RNAi screen identifies five serine
proteases required for Toll activation. Curr Biol. 16: 808-813.
Kaneko, T., Yano, T., Aggarwal, K., Lim, J.H., Ueda, K., Oshima, Y., Peach, C., Erturk-Hasdemir,
D., Goldman, W. E., Oh, B. H., Kurata, S. and Silverman, N. (2006) PGRP-LC and PGRP-LE
have essential yet distinct functions in the Drosophila immune response to monomeric DAP-type
peptidoglycan. Nat Immunol. 7: 715-723.
Kang, D., Liu, G., Lundstrom, A., Gelius, E. and Steiner, H. (1998) A peptidoglycan recognition pro-
tein in innate immunity conserved from insects to humans. Proc Natl Acad Sci U S A. 95: 10078-
10082.
Kanost, M.R., Jiang, H. and Yu, X.Q. (2004) Innate immune responses of a lepidopteran insect,
Manduca sexta. Immunol Rev. 198: 97-105.
Kim, M.S., Baek, M.J., Lee, M.H., Park, J.W., Lee, S.Y., Soderhall, K. and Lee, B.L. (2002) A new
easter-type serine protease cleaves a masquerade-like protein during prophenoloxidase activation in
Holotrichia diomphalia larvae. J Biol Chem. 277: 39999-40004.
Kim, M.S., Byun, M. and Oh, B.H. (2003) Crystal structure of peptidoglycan recognition protein LB
from Drosophila melanogaster. Nat Immunol. 4: 787-793.
Kim, Y.S., Ryu, J.H., Han, S.J., Choi, K.H., Nam, K.B., Jang, I.H., Lemaitre, B., Brey, P. T. and
Lee, W. J. (2000) Gram-negative bacteria-binding protein, a pattern recognition receptor for
lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate im-
mune genes in Drosophila melanogaster cells. J Biol Chem. 275: 32721-32727.
Kistner, C. and Parniske, M. (2002) Evolution of signal transduction in intracellular symbiosis. Trends
Plant Sci. 7: 511-518.
Koga, R., Meng, X.Y., Kamagata, Y, Sakurai, M., Tsuchida, T. and Fukatsu, T. (2006) Infection
dynamics of the secondary endosymbiotic bacterium Serratia symbiotica associated with the pea
aphid Acyrthosiphon pisum. Communication orale. International Symbiosis Society congress,
Vienne, Autriche,
Koga, R., Tsuchida, T. and Fukatsu, T. (2003) Changing partners in an obligate symbiosis: a faculta-
tive endosymbiont can compensate for loss of the essential endosymbiont Buchnera in an aphid. Proc
R Soc Lond B Biol Sci. 270: 2543-2550.
Lanot, R., Zachary, D., Holder, F. and Meister, M. (2001) Postembryonic hematopoiesis in Droso-
phila. Dev Biol. 230: 243-257.
Lavine, M.D. and Strand, M.R. (2002) Insect hemocytes and their role in immunity. Insect Biochem
Mol Biol. 32: 1295-1309.
Laviolette, P. and Nardon, P. (1963) Action des rayons gamma du Cobalt 60 sur la mortalité et la ferti-
lité des adultes d'un charançon du riz. Bull Biol Fr Belg. 97: 305-333.
ANSELME Caroline 141 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Lazzaro, B.P. and Clark, A.G. (2003) Molecular population genetics of inducible antibacterial peptide
genes in Drosophila melanogaster. Mol Biol Evol. 20: 914-923.
Lee, S.Y., Moon, H.J., Kurata, S., Kurama, T., Natori, S. and Lee, B.L. (1994) Purification and mo-
lecular cloning of cDNA for an inducible antibacterial protein of larvae of a coleopteran insect,
Holotrichia diomphalia. J Biochem (Tokyo). 115: 82-86.
Lee, S.Y., Kwon, T.H., Hyun, J.H., Choi, J.S., Kawabata, S.I., Iwanaga, S. and Lee, B.L. (1998)
In vitro activation of pro-phenol-oxidase by two kinds of pro-phenol-oxidase-activating factors iso-
lated from hemolymph of coleopteran, Holotrichia diomphalia larvae. Eur J Biochem. 254: 50-57.
Lee, W.J., Lee, J.D., Kravchenko, V.V., Ulevitch, R.J. and Brey, P.T. (1996) Purification and mo-
lecular cloning of an inducible Gram-negative bacteria-binding protein from the silkworm, Bombyx
mori. Proc Natl Acad Sci U S A. 93: 7888-7893.
Lefèvre, C., Charles, H., Vallier, A., Delobel, B., Farrell, B. and Heddi, A. (2004) Endosymbiont
phylogenesis in the Dryophthoridae weevils: Evidence for bacterial replacement. Mol Biol Evol.
21: 965-973.
Lemaitre, B., Reichhart, J.M. and Hoffmann, J.A. (1997) Drosophila host defense: differential induc-
tion of antimicrobial peptide genes after infection by various classes of microorganisms. Proc Natl
Acad Sci U S A. 94: 14614-14619.
Lemaitre, B., Nicolas, E., Michaut, L., Reichhart, J.M. and Hoffmann, J.A. (1996) The dorsoventral
regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila
adults. Cell. 86: 973-983.
Lemaitre, B., Kromer-Metzger, E., Michaut, L., Nicolas, E., Meister, M., Georgel, P., Reichhart, J.
M. and Hoffmann, J. A. (1995) A recessive mutation, immune deficiency (imd), defines two distinct
control pathways in the Drosophila host defense. Proc Natl Acad Sci U S A. 92: 9465-9469.
Leulier, F., Rodriguez, A., Khush, R.S., Abrams, J.M. and Lemaitre, B. (2000) The Drosophila cas-
pase Dredd is required to resist Gram-negative bacterial infection. EMBO Rep. 1: 353-358.
Leulier, F., Vidal, S., Saigo, K., Ueda, R. and Lemaitre, B. (2002) Inducible expression of double-
stranded RNA reveals a role for dFADD in the regulation of the antibacterial response in Drosophila
adults. Curr Biol. 12: 996-1000.
Leulier, F., Parquet, C., Pili-Floury, S., Ryu, J.H., Caroff, M., Lee, W.J., Mengin-Lecreulx, D. and
Lemaitre, B. (2003) The Drosophila immune system detects bacteria through specific peptidoglycan
recognition. Nat Immunol. 4: 478-484.
Levashina, E.A. (2004) Immune responses in Anopheles gambiae. Insect Biochem Mol Biol. 34: 673-
678.
Levashina, E.A., Langley, E., Green, C., Gubb, D., Ashburner, M., Hoffmann, J.A. and Reichhart,
J.M. (1999) Constitutive activation of Toll-mediated antifungal defense in serpin-deficient Droso-
phila. Science. 285: 1917-1919.
ANSELME Caroline 142 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Li, T., Hu, J. and Li, L. (2004) Characterization of Tollip protein upon Lipopolysaccharide challenge.
Mol Immunol. 41: 85-92.
Ligoxygakis, P., Pelte, N., Hoffmann, J.A. and Reichhart, J.M. (2002a) Activation of Drosophila
Toll during fungal infection by a blood serine protease. Science. 297: 114-116.
Ligoxygakis, P., Pelte, N., Ji, C., Leclerc, V., Duvic, B., Belvin, M., Jiang, H., Hoffmann, J. A. and
Reichhart, J. M. (2002b) A serpin mutant links Toll activation to melanization in the host defence
of Drosophila. EMBO J. 21: 6330-6337.
Liu, C., Gelius, E., Liu, G., Streiner, H. and Dziarski, R. (2000) Mammalian peptidoglycan recogni-
tion protein binds peptidoglycan with high affinity, is expressed in Neutrophils, and inhibits bacterial
growth. J Biol Chem. 275: 24490-24499.
Majander, K., Anton, L., Antikainen, J., Lang, H., Brummer, M., Korhonen, T.K. and Wester-
lund-Wikstrom, B. (2005) Extracellular secretion of polypeptides using a modified Escherichia coli
flagellar secretion apparatus. Nat Biotechnol. 23: 475-481.
Mansour, K. (1930) Preliminary studies on the bacterial cell mass (accessory cell mass) of Calandra
oryzae: the rice weevil. Q J Microsc Sci. 73: 421-436.
McGraw, E.A. and O'Neill, S.L. (2004) Wolbachia pipientis: intracellular infection and pathogenesis
in Drosophila. Curr Opin Microbiol. 7: 67-70.
Meister, M. (2004) Blood cells of Drosophila: cell lineages and role in host defence. Curr Opin Immu-
nol. 16: 10-15.
Mellroth, P., Karlsson, J. and Steiner, H. (2003) A scavenger function for a Drosophila peptidoglycan
recognition protein. J Biol Chem. 278: 7059-7064.
Michel, T., Reichhart, J.M., Hoffmann, J.A. and Royet, J. (2001) Drosophila Toll is activated by
Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature. 414: 756-
759.
Miura, T., Braendle, C., Shingleton, A., Sisk, G., Kambhampati, S. and Stern, D.L. (2003) A com-
parison of parthenogenetic and sexual embryogenesis of the pea aphid Acyrthosiphon pisum (Hemip-
tera: Aphidoidea). J Exp Zoolo B Mol Dev Evol. 295: 59-81.
Moran, A.N. and Telang, A. (1998) Bacteriocyte-associated symbionts of insects. Bioscience.
48: 295-304.
Moran, N.A. (1996) Accelerated evolution and Muller's rachet in endosymbiotic bacteria. Proc Natl
Acad Sci U S A. 93: 2873-2878.
Moran, N.A. and Baumann, P. (2000) Bacterial endosymbionts in animals. Curr Opin Microbiol.
3: 270-275.
Murray, F.V. and Tiegs, O.W. (1935) The metamorphosis of Calandra oryzae. Q J Microsc Sci.
77: 404-495.
ANSELME Caroline 143 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Musgrave, A.J. and Miller, J.J. (1953) Some microoganisms associated with the weevils, Sitophilus
granarius and Sitophilus oryzae. I - Distribution and description of the organisms. Can Entomol.
85: 387-390.
Nakabachi, A., Shigenobu, S., Sakazume, N., Shiraki, T., Hayashizaki, Y., Carninci, P., Ishikawa,
H., Kudo, T. and Fukatsu, T. (2005) Transcriptome analysis of the aphid bacteriocyte, the symbi-
otic host cell that harbors an endocellular mutualistic bacterium, Buchnera. Proc Natl Acad Sci
U S A. 102: 5477-5482.
Nardon, P. (1971) Contribution à l'étude des symbiotes ovariens de Sitophilus sasakii : localisation, his-
tochimie et ultrastructure chez la femelle adulte. C R Acad Sci Paris. 272D: 2975-2978.
Nardon, P. (1973) Obtention d'une souche asymbiotique chez le charançon Sitophilus sasakii Tak : dif-
férentes méthodes d'obtention et comparaison avec la souche symbiotique d'origine. C R Acad Sci
Paris. 277D: 981-984.
Nardon, P. (1978) Etude des interactions physiologiques et génétiques entre l'hôte et les symbiotes
chez le coléoptère Curculionidae Sitophilus sasakii (= S. oryzae). Thèse de doctorat.
Lyon : INSA-Université Lyon 1. 285 p.
Nardon, P. and Wicker, C. (1981) La symbiose chez le genre Sitophilus (Coleoptère Curculionidae).
Principaux aspects morphologiques, physiologiques et génétiques. Ann Biol. 4: 329-373.
Nardon, P. and Grenier, A.M. (1988) Genetical and biochemical interactions between the host and its
endosymbiotes in the weevil Sitophilus (Coleoptera Curculionidae) and other related species.
In Cell to cell signals in plant, animal and microbial symbiosis. Scannerini S., Smith, D., Bonfante-
Fasolo, P., and Gianinazzi-Pearson, V. (eds) Berlin: Springer-Verlag, pp. 255-270.
Nardon, P. and Grenier, A.M. (1989) Endocytobiosis in Coleoptera: Biological, Biochemical and Ge-
netic Aspects. In Insect Endocytobiosis: Morphology, Physiology, Genetics, Evolution. Schwemmler,
W. and Gassner, G. (eds) Boca Raton: CRC Press, pp. 175-215.
Nardon, P. and Grenier, A.M. (1993) Symbiose et évolution. Ann Soc Entomol Fr. 29: 113-140.
Nardon, P. and Nardon, C. (1998) Morphology and cytology of symbiosis in insects. Ann Soc Entomol
Fr. 34: 105-134.
Nardon, P., Grenier, A.M. and Heddi, A. (1998) Endocytobiote control by the host in the weevil Sito-
philus oryzae, Coleoptera, Curculionidae. Symbiosis. 25: 237-250.
Paillot, A. (1933) L'infection chez les insectes : Immunité et symbiose. Trévoux: Imprimerie de G. Patis-
sier, 535 p.
Pierantoni, U. (1927) L'organo simbiotico nello sviluppo di Calandra oryzae. Rendiconti della Reale
Academia delle Scienza di Napoli. 35: 244-250.
Pili-Floury, S., Leulier, F., Takahashi, K., Saigo, K., Samain, E., Ueda, R. and Lemaitre, B. (2004)
In vivo RNAi analysis reveals an unexpected role for GNBP1 in the defense against Gram-positive
bacterial infection in Drosophila adults. J Biol Chem. 279: 12848-128453.
ANSELME Caroline 144 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Powers, J.P. and Hancock, R.E. (2003) The relationship between peptide structure and antibacterial
activity. Peptides. 24: 1681-1691.
Rahbé, Y., Delobel, B., Febvay, G. and Chantegrel, B. (1994) Aphid-specific triglycerides in symbi-
otic and aposymbiotic Acyrthosiphon pisum. Insect Biochem Mol Biol. 24: 95-101.
Rahbé, Y., Delobel, B., Febvay, G., Nardon, C. and Nardon, P. (1993) Are aphid bacterial endosym-
bionts involved in the biosynthesis of aphid peculiar triglycerides ? In Endocytobiology V. Proc. 5th
Int. Coll. on Endocytobiology & Symbiosis. Philips Univ., Uji-Kyoto (JAP), June 23-27 1992. Sato,
S., Ishida, M., Ishikawa, H. (eds.) Tubingen (GER): Tubingen University Press, pp. 139-144.
Ramet, M., Lanot, R., Zachary, D. and Manfruelli, P. (2002a) JNK signaling pathway is required for
efficient wound healing in Drosophila. Dev Biol. 241: 145-156.
Ramet, M., Manfruelli, P., Pearson, A., Mathey-Prevot, B. and Ezekowitz, R.A. (2002b) Functional
genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature.
416: 644-648.
Rigaud, T., Souty-Grosset, C., Raimond, R., Mocquard, J.P. and Juchault, P. (1991) Feminizing
endocytobiosis in the terrestrial crustacean Armadillidium vulgare Latr: recent acquisistion. Endocy-
tobiosis and Cell Res. 7: 259-273.
Rio, R.V.M., Lefèvre, C., Heddi, A. and Aksoy, S. (2003) Comparative genomics of insect-symbiotic
bacteria: Influence of host environmental on microbial genome composition. Appl Environ Micro-
biol. 69: 6825-6832.
Robertson, A.S., Belorgey, D., Lilley, K.S., Lomas, D.A., Gubb, D. and Dafforn, T.R. (2003) Char-
acterization of the necrotic protein that regulates the Toll-mediated immune response in Drosophila.
J Biol Chem. 278: 6175-6180.
Rocha, E.P. and Danchin, A. (2002) Base composition bias might result from competition for meta-
bolic resources. Trends Genet. 18: 291-294.
Ryu, J.H., Nam, K.B., Oh, C.T., Nam, H.J., Kim, S.H., Yoon, J.H., Jang, I. H., Brey, P. T. and Lee,
W. J. (2004) The homeobox gene Caudal regulates constitutive local expression of antimicrobial
peptide genes in Drosophila epithelia. Mol Cell Biol. 24: 172-185.
Sabater-Munoz, B., Legeai, F., Rispe, C., Bonhomme, J., Dearden, P., Dossat, C., et al (2006)
Large-scale gene discovery in the pea aphid Acyrthosiphon pisum (Hemiptera). Genome Biol. 7: R21.
Sagisaka, A., Miyanoshita, A., Ishibashi, J. and Yamakawa, M. (2001) Purification, characterization
and gene expression of a glycine and proline-rich antibacterial protein family from larvae of a beetle,
Allomyrina dichotoma. Insect Mol Biol. 10: 293-302.
Sasaki, T. and Ishikawa, H. (1994) Production of essential amino acids from glutamate by mycetocyte
symbionts of the pea aphid, Acyrthosiphon pisum. J Insect Physiol. 41: 41-46.
Sasaki, T., Hayasaki, H. and Ishikawa, H. (1991) Growth and reproduction of symbiotic and aposym-
biotic pea aphids, Acyrthosiphum pisum, maintained on artificial diets. J Insect Physiol. 37: 749-756.
ANSELME Caroline 145 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Schmidt, O., Theopold, U. and Strand, M. (2001) Innate immunity and its evasion and suppression by
hymenopteran endoparasitoids. Bioessays. 23: 344-351.
Schwemmler, W. (1980) Endocytobiosis: general principles. Biosystems. 12: 111-122.
Shen, B. and Manley, J.L. (1998) Phosphorylation modulates direct interactions between the Toll
receptor, Pelle kinase and Tube. Development. 125: 4719-4728.
Shigenobu, S., Watanabe, H., Hattori, M., Sakaki, Y. and Ishikawa, H. (2000) Genome sequence of
the endocellular bacterial symbiont of aphids Buchnera sp. APS. Nature. 407: 81-86.
Smith, C.L., Econome, J.G., Schutt, A., Klco, S. and Cantor, C.R. (1987) A physical map of the
Escherichia coli K12 genome. Science. 236: 1448-1453.
St Johnston, D. and Nusslein-Volhard, C. (1992) The origin of pattern and polarity in the Drosophila
embryo. Cell. 68: 201-219.
Steiner, H. (2004) Peptidoglycan recognition proteins: on and off switches for innate immunity.
Immunol Rev. 198: 83-96.
Stouthamer, R., Breeuwers, J.A.J. and Hurst, G.D.D. (1999) Wolbachia pipientis: Microbial manipu-
lator of arthropod reproduction. Annu Rev Microbiol. 53: 71-102.
Stoven, S., Ando, I., Kadalayil, L., Engstrom, Y. and Hultmark, D. (2000) Activation of the Droso-
phila NF-kappaB factor Relish by rapid endoproteolytic cleavage. EMBO Rep. 1: 347-352.
Strober, W., Murray, P.J., Kitani, A. and Watanabe, T. (2006) Signalling pathways and molecular
interactions of NOD1 and NOD2. Nat Rev Immunol. 6: 9-20.
Stuart, L.M. and Ezekowitz, R.A. (2005) Phagocytosis: elegant complexity. Immunity. 22: 539-550.
Sun, H., Bristow, B.N., Qu, G. and Wasserman, S.A. (2002) A heterotrimeric death domain complex
in Toll signaling. Proc Natl Acad Sci U S A. 99: 12871-12876.
Sun, L.V., Foster, J.M., Tzertzinis, G., Ono, M., Bandi, C., Slatko, B.E. and SL, O.N. (2001)
Determination of Wolbachia genome size by pulsed-field gel electrophoresis. J Bacteriol. 183: 2219-
2225.
Takehana, A., Yano, T., Mita, S., Kotani, A., Oshima, Y. and Kurata, S. (2004) Peptidoglycan rec-
ognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. EMBO J.
23: 4690-4700.
Takehana, A., Katsuyama, T., Yano, T., Oshima, Y., Takada, H., Aigaki, T. and Kurata, S. (2002)
Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates
imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae.
Proc Natl Acad Sci U S A. 99: 13705-13710.
Tarsia In Curia, I. (1933) Nuove osservazioni sull'organo simbiotico di Calandra oryzae. Q J Microsc
Sci. 80: 159-284.
ANSELME Caroline 146 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Toh, H., Weiss, B.L., Perkin, S.A., Yamashita, A., Oshima, K., Hattori, M. and Aksoy, S. (2006)
Massive genome erosion and functional adaptations provide insights into the symbiotic lifestyle of
Sodalis glossinidius in the tsetse host. Genome Res. 16: 149-156.
Tong, Y. and Kanost, M.R. (2005) Manduca sexta serpin-4 and serpin-5 inhibit the prophenol oxidase
activation pathway: cDNA cloning, protein expression, and characterization. J Biol Chem.
280: 14923-14931.
Tydell, C.C., Yuan, J., Tran, P. and Selsted, M.E. (2006) Bovine peptidoglycan recognition protein-S:
antimicrobial activity, localization, secretion, and binding properties. J Immunol. 176: 1154-1162.
Tzou, P., Reichhart, J.M. and Lemaitre, B. (2002) Constitutive expression of a single antimicrobial
peptide can restore wild-type resistance to infection in immunodeficient Drosophila mutants. Proc
Natl Acad Sci U S A. 99: 2152-2157.
Tzou, P., Ohresser, S., Ferrandon, D., Capovilla, M., Reichhart, J.M., Lemaitre, B., Hoffmann,
J.A. and Imler, J.L. (2000) Tissue-specific inducible expression of antimicrobial peptide genes in
Drosophila surface epithelia. Immunity. 13: 737-748.
Ueda, K., Saito, A., Imamura, M., Miura, N., Atsumi, S., Tabunoki, H., Watanabe, A., Kitami, M.
and Sato, R. (2005) Purification and cDNA cloning of luxuriosin, a novel antibacterial peptide with
Kunitz domain from the longicorn beetle, Acalolepta luxuriosa. Biochim Biophys Acta. 1722: 36-42.
Vasselon, T. and Detmers, P.A. (2002) Toll receptors: a central element in innate immune responses.
Infect Immun. 70: 1033-1041.
Vavre, F., Fleury, F., Lepetit, D., Fouillet, P. and Bouletreau, M. (1999) Phylogenetic evidence for
horizontal transmission of Wolbachia in host-parasitoid associations. Mol Biol Evol. 16: 1711-1723.
von Dohlen, C.D., Kohler, S., Alsop, S.T. and McManus, W.R. (2001) Mealybug beta-proteobacterial
endosymbionts contain gamma-proteobacterial symbionts. Nature. 412: 433-436.
Wang, C., Deng, L., Hong, M., Akkaraju, G.R., Inoue, J. and Chen, Z.J. (2001) TAK1 is a ubiqui-
tin-dependent kinase of MKK and IKK. Nature. 412: 346-351.
Wang, Z.M., Li, X., Cocklin, R.R., Wang, M., Fukase, K., Inamura, S., Kusumoto, S., Gupta, D.
and Dziarski, R. (2003) Human peptidoglycan recognition protein-L is an N-acetylmuramoyl-L-
alanine amidase. J Biol Chem. 278: 49044-49052.
Wernegreen, J. (2002) Genome evolution in bacterial endosymbionts of insects. Nat Rev Genet. 3: 850-
861.
Werner, T., Borge-Renberg, K., Mellroth, P., Steiner, H. and Hultmark, D. (2003) Functional diver-
sity of the Drosophila PGRP-LC gene cluster in the response to lipopolysaccharide and peptidogly-
can. J Biol Chem. 278: 26319-26322.
Werner, T., Liu, G., Kang, D., Ekengren, S., Steiner, H. and Hultmark, D. (2000) A family of pepti-
doglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc Nat Acad Sci U S A.
97: 13772-13777.
ANSELME Caroline 147 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Références bibliographiques
Werren, J.H. (1997) Biology of Wolbachia. Annu Rev Entomol. 42: 587-609.
Wicker, C. (1983) Differential vitamin and choline requirements of symbiotic and aposymbiotic Sito-
philus oryzae (Coleoptera : Curculionidae). Comp Biochem physiol. 76A: 177-182.
Yang, J., Yamamoto, M., Ishibashi, J., Taniai, K. and Yamakawa, M. (1998) Isolation, cDNA clon-
ing and gene expression of an antibacterial protein from larvae of the coconut rhinoceros beetle,
Oryctes rhinoceros. Eur J Biochem. 255: 734-738.
Yoshida, H., Kinoshita, K. and Ashida, M. (1996) Purification of a peptidoglycan recognition protein
from hemolymph of the silkworm, Bombyx mori. J Biol Chem. 271: 13854-13860.
Young, G.M., Schmiel, D.H. and Miller, V.L. (1999) A new pathway for the secretion of virulence
factors by bacteria: the flagellar export apparatus functions as a protein-secretion system. Proc Natl
Acad Sci U S A. 96: 6456-6461.
Zaidman-Remy, A., Herve, M., Poidevin, M., Pili-Floury, S., Kim, M.S., Blanot, D., Oh, B.H.,
Ueda, R., Mengin-Lecreulx, D. and Lemaitre, B. (2006) The Drosophila Amidase PGRP-LB
Modulates the Immune Response to Bacterial Infection. Immunity. 24: 463-473.
Zavalova, L.L., Baskova, I.P., Lukyanov, S.A., Sass, A.V., Snezhkov, E.V., Akopov, S.B., et al
(2000) Destabilase from the medicinal leech is a representative of a novel family of lysozymes.
Biochim Biophys Acta. 1478: 69-77.
Zhang, G. and Ghosh, S. (2002) Negative regulation of Toll-like receptor-mediated signaling by
Tollip. J Biol Chem. 277: 7059-7065.
Zhou, R., Silverman, N., Hong, M., Liao, D.S., Chung, Y., Chen, Z.J. and Maniatis, T. (2005)
The role of ubiquitination in Drosophila innate immunity. J Biol Chem. 280: 34048-34055.
Zhu, Y., Wang, Y., Gorman, M.J., Jiang, H. and Kanost, M.R. (2003) Manduca sexta serpin-3 regu-
lates prophenoloxidase activation in response to infection by inhibiting prophenoloxidase-activating
proteinases. J Biol Chem. 278: 46556-46564.
ANSELME Caroline 148 Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon

Annexes
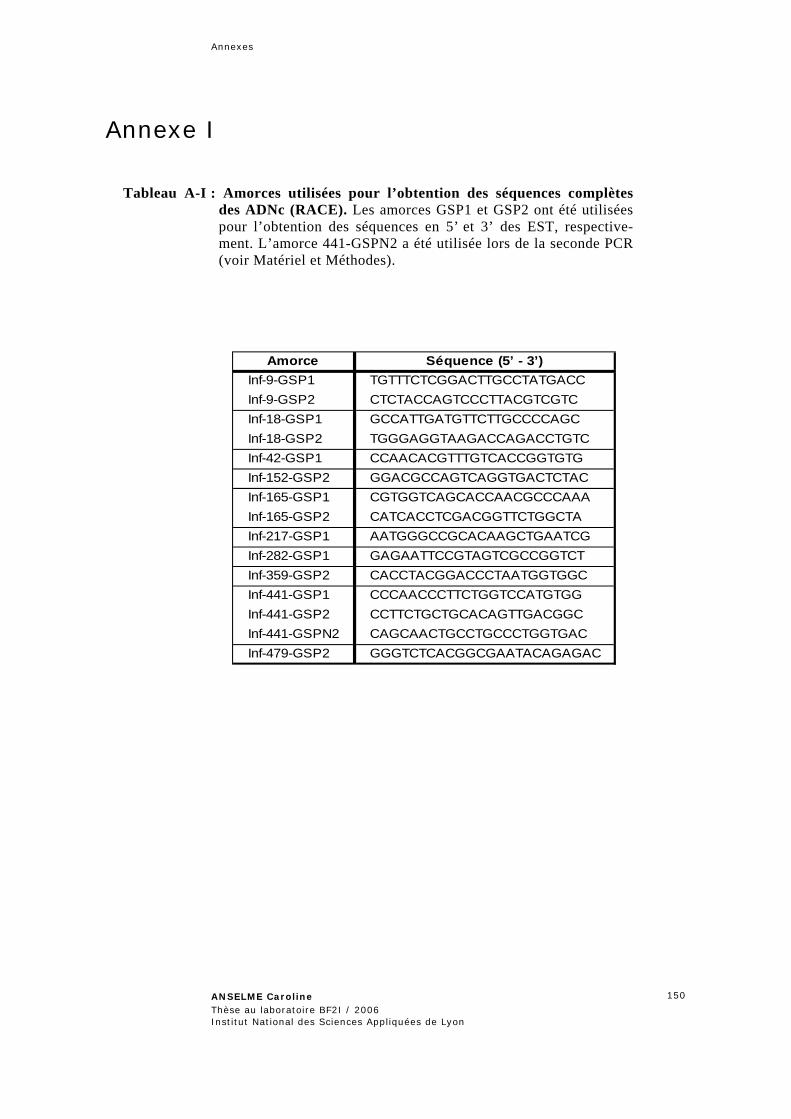
Annexes
Annexe I
Tableau A-I : Amorces utilisées pour l’obtention des séquences complètes des ADNc (RACE). Les amorces GSP1 et GSP2 ont été utilisées pour l’obtention des séquences en 5’ et 3’ des EST, respective-ment. L’amorce 441-GSPN2 a été utilisée lors de la seconde PCR (voir Matériel et Méthodes).
Amorce Séquence (5’ - 3’)
Inf-9-GSP1 TGTTTCTCGGACTTGCCTATGACCInf-9-GSP2 CTCTACCAGTCCCTTACGTCGTCInf-18-GSP1 GCCATTGATGTTCTTGCCCCAGCInf-18-GSP2 TGGGAGGTAAGACCAGACCTGTCInf-42-GSP1 CCAACACGTTTGTCACCGGTGTGInf-152-GSP2 GGACGCCAGTCAGGTGACTCTACInf-165-GSP1 CGTGGTCAGCACCAACGCCCAAAInf-165-GSP2 CATCACCTCGACGGTTCTGGCTAInf-217-GSP1 AATGGGCCGCACAAGCTGAATCGInf-282-GSP1 GAGAATTCCGTAGTCGCCGGTCTInf-359-GSP2 CACCTACGGACCCTAATGGTGGCInf-441-GSP1 CCCAACCCTTCTGGTCCATGTGGInf-441-GSP2 CCTTCTGCTGCACAGTTGACGGCInf-441-GSPN2 CAGCAACTGCCTGCCCTGGTGACInf-479-GSP2 GGGTCTCACGGCGAATACAGAGAC
ANSELME Caroline Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
150

Annexes
Annexe II
Tableau A-II : Amorces utilisées dans les expériences de PCR quantitative.
Amorce Séquence (5’- 3’) Amorce Séquence (5’- 3’)5’inf-9 ATAATTTCGCTGTTGGAGGG 5’inf-165 AGTCACAAAGGAACAATTTTGG3’inf-9 TCTCGGACTTGCCTATGACC 3’inf-165 AGAAGCCGTCACGTCCGTTC5’inf-13 GAAGAGGTGAGGATGTCCT 5’inf-217 ACGCTGTAGTGATAAGTGCA3’inf-13 ACATGAGTTGGCAGGATTAC 3’inf-217 CACACGTACAGGTCCTTCT5’inf-18 GAATAGATACAACGGGGGTCA 5’inf-282 GCGGGGAAAAAAAGTGGCGTA3’inf-18 CTACCATCTGACACTTCCTC 3’inf-282 CCATCGGGATTGTGGTTTTTAG5’inf-20 ACCCTGGACCTAACGATCC 5’inf-359 AAGAGCGACGCAATAGGGT3’inf-20 TACTGCTACAAACGAACATTG 3’inf-359 CATTAGGGTCCGTAGGTGT5’inf-42 TCAGGATGAAGATGGCCAAG 5’inf-441 TGCAAGGCCTGCCCTTAGT3’inf-42 ACACTGGTACTCTGGCATCT 3’inf-441 GTGCCATCTCCTCCAATCAT5’inf-74 TCTGTTGTATGGCATTCCGA 5’inf-459 CAACAAATCCAGACGAGCC3’inf-74 AGCGCTTGAAGTATCGAACT 3’inf-459 ACCACCGCATAGCCATCG5’inf-91 GATCGGTATACTTCAGCGAA 5’inf-479 ACCTTCACGACCCACACAT3’inf-91 AACGCATCTACCAGTTCTGT 3’inf-479 GTCATACGATACGTTTTCAGT5’inf-145 TCGGCATCCCCTAATCCAGAC 5’inf-506 CAATCGTTTATGGTCAGGAC3’inf-145 CTACCAGCGGATGGCGCCACC 3’inf-506 GGAATTCCACAACAAACGTC5’inf-152 CTCTCCGTCACCTCGTGTTAT 5’inf-515 GGTTCGAACGCCGTCAGA3’inf-152 TCCTGCTCGTTCTTTGTCAT 3’inf-515 TCACGGAGATGTCGTTAGC5’inf-163 GTCCTGTTAGTTGTGGCAGTA Actine-for CAACTTCCCTAGAAAAGAGC3’inf-163 TTATCTTCCAGGCACCAATCC Actine-rev TTCCTTCTGCATCCTGTCGG
PGRP-903-For TTACCCCAGGTTGCTCCACPGRP-903-Rev ATTGCATGTCATTTATACATTCCTC
ANSELME Caroline Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
151

Annexes
Annexe III Figure A-1 : Membranes de Dot-blot comportant les EST des banques soustraites de
S. zeamais (A) et d’A. pisum (B). Chaque membrane, réalisée en double exemplaire, comporte un témoin négatif correspondant à une amplification réalisée en absence d’ADN (en rouge) et un témoin de normalisation (gapdh, en bleu). Les membranes du haut ont été hybridées avec une sonde obtenue à partir des larves témoins (T) et celles du bas, avec une sonde obtenue à par-tir des larves piquées par une aiguille infectée par E. coli (i). Chaque dépôt correspond à un volume fixe de 10 µl et seules les comparaisons intermem-branes sont possibles.
ANSELME Caroline Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
152

Annexes
Annexe IV Tableau A-III : Tableau récapitulatif des EST de la banque soustraite de S. zeamais. Les similitudes de séquence ont été obtenues par comparaison des six phases de lecture des séquences des EST aux banques de données protéiques. SP-AC, identifiant de la protéine dans la banque de données Swiss-Prot.
ANSELME Caroline Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
153
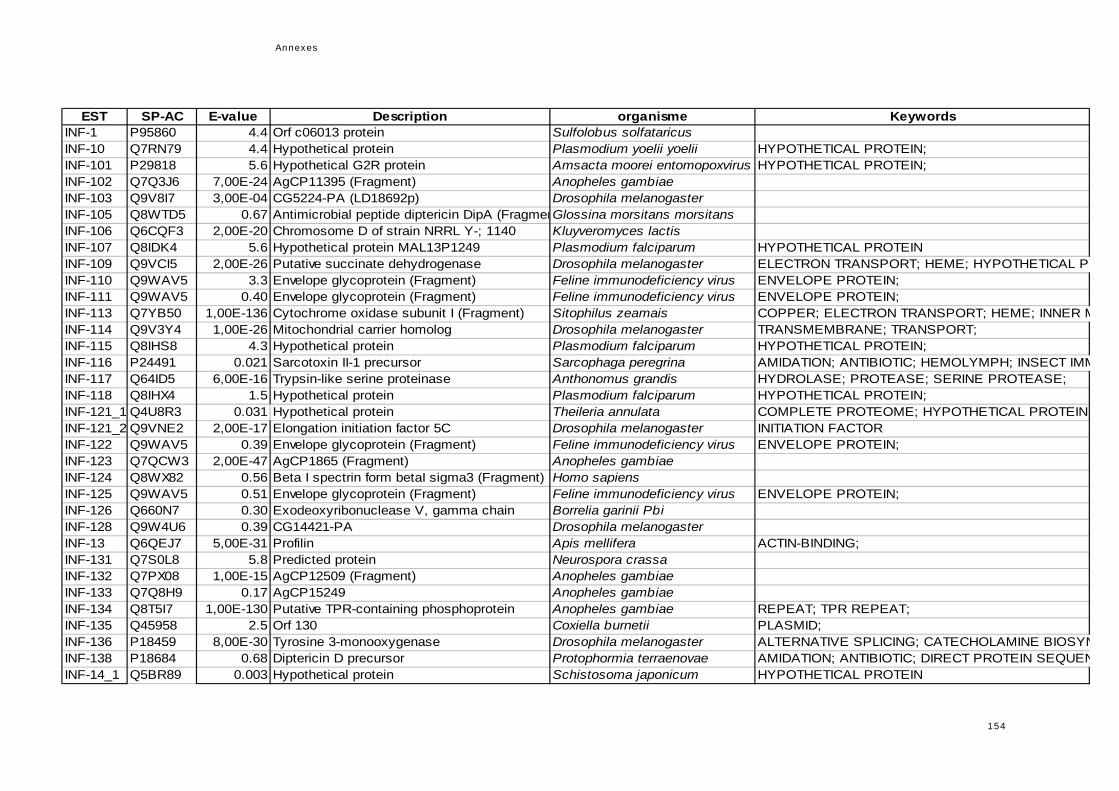
Annexes
154
C E-value Description organisme KeywordsINF-1 P95860 4.4 Orf c06013 protein Sulfolobus solfataricus INF-10 Q7RN79 4.4 Hypothetical protein Plasmodium yoelii yoelii HYPOTHETICAL PROTEIN; INF-101 P29818 5.6 Hypothetical G2R protein Amsacta moorei entomopoxvirus HYPOTHETICAL PROTEIN; INF-102 Q7Q3J6 7,00E-24 AgCP11395 (Fragment) Anopheles gambiae INF-103 Q9V8I7 3,00E-04 CG5224-PA (LD18692p) Drosophila melanogasterINF-105 Q8WTD5 0.67
A
EST SP-
Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-106 Q6CQF3 2,00E-20 Chromosome D of strain NRRL Y-; 1140 Kluyveromyces lactis INF-107 Q8IDK4 5.6 Hypothetical protein MAL13P1249 Plasmodium falciparum HYPOTHETICAL PROTEININF-109 Q9VCI5 2,00E-26 Putative succinate dehydrogenase Drosophila melanogaster ELECTRON TRANSPORT; HEME; HYPOTHETICAL PINF-110 Q9WAV5 3.3 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-111 Q9WAV5 0.40 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-113 Q7YB50 1,00E-136 Cytochrome oxidase subunit I (Fragment) Sitophilus zeamais COPPER; ELECTRON TRANSPORT; HEME; INNER MINF-114 Q9V3Y4 1,00E-26 Mitochondrial carrier homolog Drosophila melanogaster TRANSMEMBRANE; TRANSPORT; INF-115 Q8IHS8 4.3 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-116 P24491 0.021 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-117 Q64ID5 6,00E-16 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASE; INF-118 Q8IHX4 1.5 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-121_1 Q4U8R3 0.031 Hypothetical protein Theileria annulata COMPLETE PROTEOME; HYPOTHETICAL PROTEININF-121_2 Q9VNE2 2,00E-17 Elongation initiation factor 5C Drosophila melanogaster INITIATION FACTORINF-122 Q9WAV5 0.39 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-123 Q7QCW3 2,00E-47 AgCP1865 (Fragment) Anopheles gambiaeINF-124 Q8WX82 0.56 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-125 Q9WAV5 0.51 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-126 Q660N7 0.30 Exodeoxyribonuclease V, gamma chain Borrelia garinii Pbi INF-128 Q9W4U6 0.39 CG14421-PA Drosophila melanogaster INF-13 Q6QEJ7 5,00E-31 Profilin Apis mellifera ACTIN-BINDING; INF-131 Q7S0L8 5.8 Predicted protein Neurospora crassa INF-132 Q7PX08 1,00E-15 AgCP12509 (Fragment) Anopheles gambiae INF-133 Q7Q8H9 0.17 AgCP15249 Anopheles gambiae INF-134 Q8T5I7 1,00E-130 Putative TPR-containing phosphoprotein Anopheles gambiae REPEAT; TPR REPEAT; INF-135 Q45958 2.5 Orf 130 Coxiella burnetii PLASMID; INF-136 P18459 8,00E-30 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-138 P18684 0.68 Diptericin D precursor Protophormia terraenovae AMIDATION; ANTIBIOTIC; DIRECT PROTEIN SEQUENINF-14_1 Q5BR89 0.003 Hypothetical protein Schistosoma japonicum HYPOTHETICAL PROTEIN
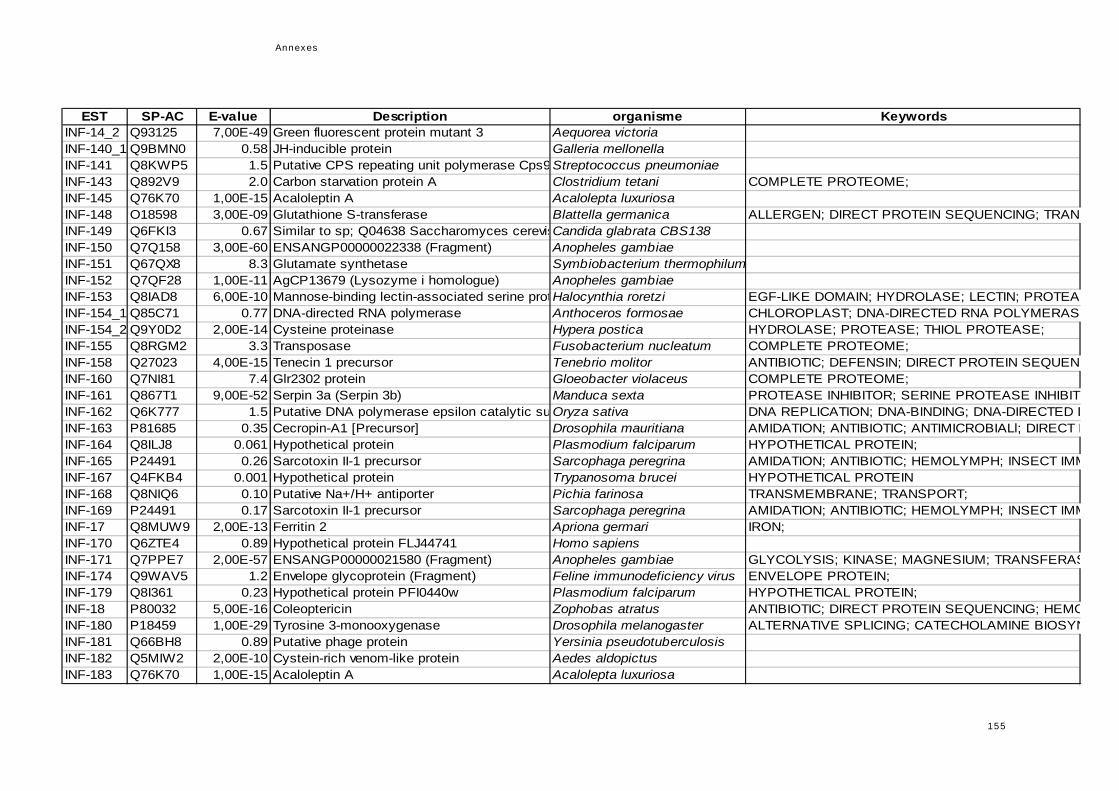
Annexes
155
EST SP- C E-value Description organisme KeywordsINF-14_2 Q93125 7,00E-49 Green fluorescent protein mutant 3 Aequorea victoriaINF-140_1 Q9BMN0 0.58 JH-inducible protein Galleria mellonella INF-141 Q8KWP5 1.5 Putative CPS repeating unit polymerase Cps
A
9Streptococcus pneumoniae INF-143 Q892V9 2.0 Carbon starvation protein A Clostridium tetani COMPLETE PROTEOME; INF-145 Q76K70 1,00E-15 Acaloleptin A Acalolepta luxuriosa INF-148 O18598 3,00E-09 Glutathione S-transferase Blattella germanica ALLERGEN; DIRECT PROTEIN SEQUENCING; TRANSINF-149 Q6FKI3 0.67 Similar to sp; Q04638 Saccharomyces cerevisCandida glabrata CBS138 INF-150 Q7Q158 3,00E-60 ENSANGP00000022338 (Fragment) Anopheles gambiaeINF-151 Q67QX8 8.3 Glutamate synthetase Symbiobacterium thermophilum INF-152 Q7QF28 1,00E-11 AgCP13679 (Lysozyme i homologue) Anopheles gambiaeINF-153 Q8IAD8 6,00E-10 Mannose-binding lectin-associated serine protHalocynthia roretzi EGF-LIKE DOMAIN; HYDROLASE; LECTIN; PROTEASINF-154_1 Q85C71 0.77 DNA-directed RNA polymerase Anthoceros formosae CHLOROPLAST; DNA-DIRECTED RNA POLYMERASINF-154_2 Q9Y0D2 2,00E-14 Cysteine proteinase Hypera postica HYDROLASE; PROTEASE; THIOL PROTEASE; INF-155 Q8RGM2 3.3 Transposase Fusobacterium nucleatum COMPLETE PROTEOME; INF-158 Q27023 4,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-160 Q7NI81 7.4 Glr2302 protein Gloeobacter violaceus COMPLETE PROTEOME; INF-161 Q867T1 9,00E-52 Serpin 3a (Serpin 3b) Manduca sexta PROTEASE INHIBITOR; SERINE PROTEASE INHIBITOINF-162 Q6K777 1.5 Putative DNA polymerase epsilon catalytic suOryza sativa DNA REPLICATION; DNA-BINDING; DNA-DIRECTED DINF-163 P81685 0.35 Cecropin-A1 [Precursor] Drosophila mauritiana AMIDATION; ANTIBIOTIC; ANTIMICROBIALl; DIRECT PINF-164 Q8ILJ8 0.061 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-165 P24491 0.26 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-167 Q4FKB4 0.001 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEININF-168 Q8NIQ6 0.10 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORT; INF-169 P24491 0.17 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-17 Q8MUW9 2,00E-13 Ferritin 2 Apriona germari IRON; INF-170 Q6ZTE4 0.89 Hypothetical protein FLJ44741 Homo sapiens INF-171 Q7PPE7 2,00E-57 ENSANGP00000021580 (Fragment) Anopheles gambiae GLYCOLYSIS; KINASE; MAGNESIUM; TRANSFERASINF-174 Q9WAV5 1.2 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-179 Q8I361 0.23 Hypothetical protein PFI0440w Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-18 P80032 5,00E-16 Coleoptericin Zophobas atratus ANTIBIOTIC; DIRECT PROTEIN SEQUENCING; HEMOINF-180 P18459 1,00E-29 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-181 Q66BH8 0.89 Putative phage protein Yersinia pseudotuberculosis INF-182 Q5MIW2 2,00E-10 Cystein-rich venom-like protein Aedes aldopictusINF-183 Q76K70 1,00E-15 Acaloleptin A Acalolepta luxuriosa
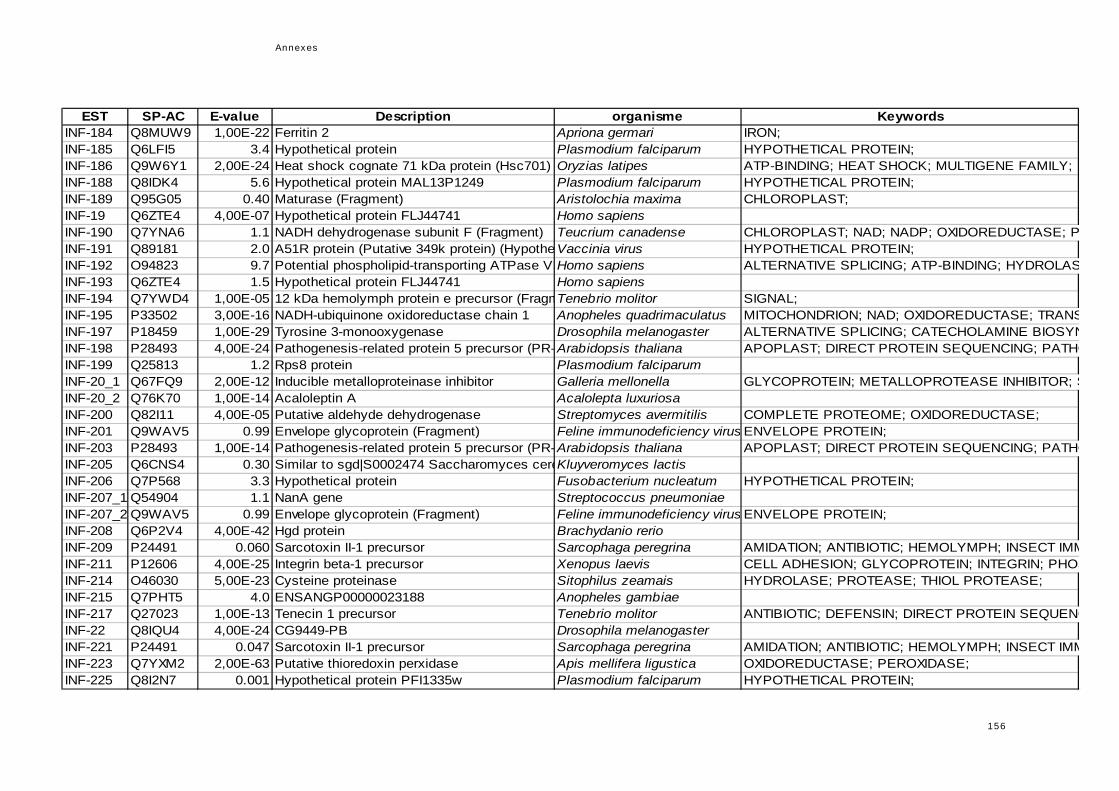
Annexes
156
C E-value Description organisme KeywordsINF-184 Q8MUW9 1,00E-22 Ferritin 2 Apriona germari IRON; INF-185 Q6LFI5 3.4 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-186 Q9W6Y1 2,00E-24 Heat shock cognate 71 kDa protein (Hsc701) Oryzias latipes ATP-BINDING; HEAT SHOCK; MULTIGENE FAMILY; INF-188 Q8IDK4 5.6 Hypothetical protein MAL13P1249 Plasmodium falciparum HYPOTHETICAL PROTEIN; INF-189 Q95G05 0.40 Maturase (Fragment) Aristolochia maxima CHLOROPLAST; INF-19 Q6ZTE4 4,00E-07 Hypothetical protein FLJ44741 Homo sapiens INF-190 Q7YNA6 1.1 NADH dehydrogenase subunit F (Fragment) Teucrium canadense CHLOROPLAST; NAD; NADP; OXIDOREDUCTASE;
A
EST SP-
PINF-191 Q89181 2.0 A51R protein (Putative 349k protein) (HypotheVaccinia virus HYPOTHETICAL PROTEIN; INF-192 O94823 9.7 Potential phospholipid-transporting ATPase VBHomo sapiens ALTERNATIVE SPLICING; ATP-BINDING; HYDROLASINF-193 Q6ZTE4 1.5 Hypothetical protein FLJ44741 Homo sapiens INF-194 Q7YWD4 1,00E-05 12 kDa hemolymph protein e precursor (FragmTenebrio molitor SIGNAL; INF-195 P33502 3,00E-16 NADH-ubiquinone oxidoreductase chain 1 Anopheles quadrimaculatus MITOCHONDRION; NAD; OXIDOREDUCTASE; TRANSINF-197 P18459 1,00E-29 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-198 P28493 4,00E-24 Pathogenesis-related protein 5 precursor (PR-Arabidopsis thaliana APOPLAST; DIRECT PROTEIN SEQUENCING; PATHOINF-199 Q25813 1.2 Rps8 protein Plasmodium falciparumINF-20_1 Q67FQ9 2,00E-12 Inducible metalloproteinase inhibitor Galleria mellonella GLYCOPROTEIN; METALLOPROTEASE INHIBITOR; SINF-20_2 Q76K70 1,00E-14 Acaloleptin A Acalolepta luxuriosaINF-200 Q82I11 4,00E-05 Putative aldehyde dehydrogenase Streptomyces avermitilis COMPLETE PROTEOME; OXIDOREDUCTASE; INF-201 Q9WAV5 0.99 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-203 P28493 1,00E-14 Pathogenesis-related protein 5 precursor (PR-Arabidopsis thaliana APOPLAST; DIRECT PROTEIN SEQUENCING; PATHOINF-205 Q6CNS4 0.30 Similar to sgd|S0002474 Saccharomyces cereKluyveromyces lactis INF-206 Q7P568 3.3 Hypothetical protein Fusobacterium nucleatum HYPOTHETICAL PROTEIN; INF-207_1 Q54904 1.1 NanA gene Streptococcus pneumoniae INF-207_2 Q9WAV5 0.99 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-208 Q6P2V4 4,00E-42 Hgd protein Brachydanio rerio INF-209 P24491 0.060 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-211 P12606 4,00E-25 Integrin beta-1 precursor Xenopus laevis CELL ADHESION; GLYCOPROTEIN; INTEGRIN; PHOSINF-214 O46030 5,00E-23 Cysteine proteinase Sitophilus zeamais HYDROLASE; PROTEASE; THIOL PROTEASE; INF-215 Q7PHT5 4.0 ENSANGP00000023188 Anopheles gambiaeINF-217 Q27023 1,00E-13 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-22 Q8IQU4 4,00E-24 CG9449-PB Drosophila melanogaster INF-221 P24491 0.047 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-223 Q7YXM2 2,00E-63 Putative thioredoxin perxidase Apis mellifera ligustica OXIDOREDUCTASE; PEROXIDASE; INF-225 Q8I2N7 0.001 Hypothetical protein PFI1335w Plasmodium falciparum HYPOTHETICAL PROTEIN;
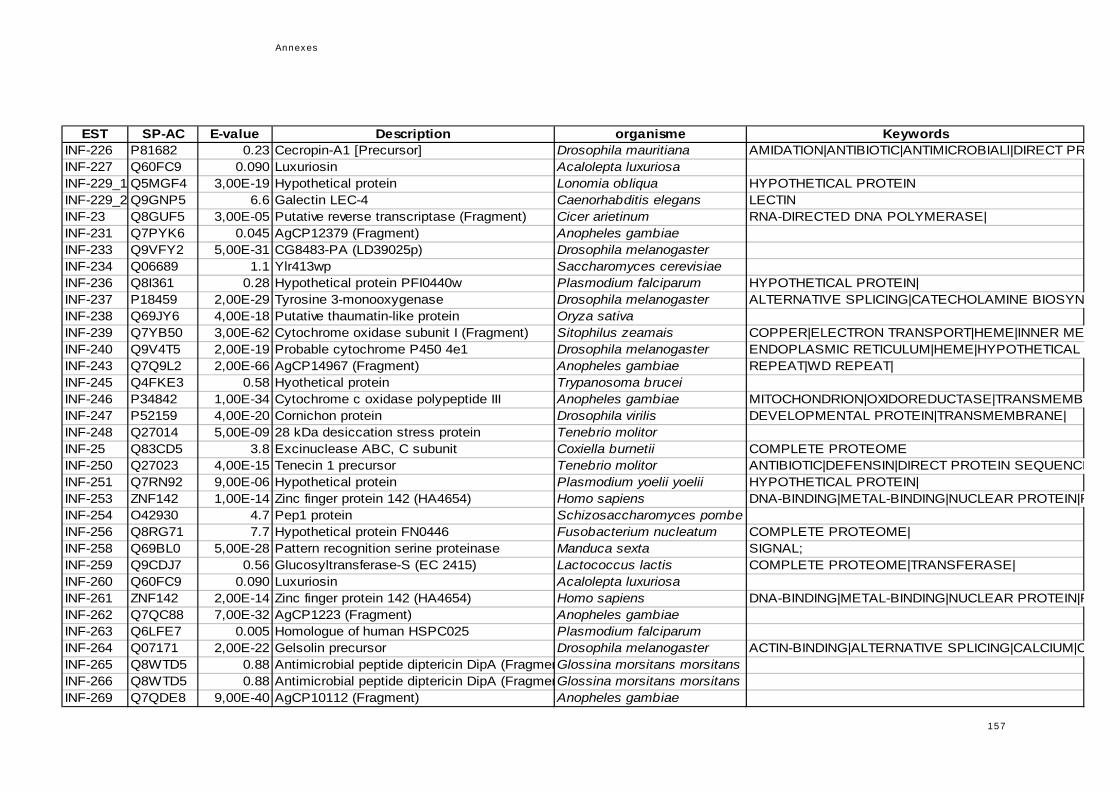
Annexes
A
EST SP- C E-value Description organisme KeywordsINF-226 P81682 0.23 Cecropin-A1 [Precursor] Drosophila mauritiana AMIDATION|ANTIBIOTIC|ANTIMICROBIALl|DIRECT PRINF-227 Q60FC9 0.090 Luxuriosin Acalolepta luxuriosaINF-229_1 Q5MGF4 3,00E-19 Hypothetical protein Lonomia obliqua HYPOTHETICAL PROTEININF-229_2 Q9GNP5 6.6 Galectin LEC-4 Caenorhabditis elegans LECTININF-23 Q8GUF5 3,00E-05 Putative reverse transcriptase (Fragment) Cicer arietinum RNA-DIRECTED DNA POLYMERASE|INF-231 Q7PYK6 0.045 AgCP12379 (Fragment) Anopheles gambiae INF-233 Q9VFY2 5,00E-31 CG8483-PA (LD39025p) Drosophila melanogaster INF-234 Q06689 1.1 Ylr413wp Saccharomyces cerevisiae INF-236 Q8I361 0.28 Hypothetical protein PFI0440w Plasmodium falciparum HYPOTHETICAL PROTEIN|INF-237 P18459 2,00E-29 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING|CATECHOLAMINE BIOSYNINF-238 Q69JY6 4,00E-18 Putative thaumatin-like protein Oryza sativa INF-239 Q7YB50 3,00E-62 Cytochrome oxidase subunit I (Fragment) Sitophilus zeamais COPPER|ELECTRON TRANSPORT|HEME|INNER MEINF-240 Q9V4T5 2,00E-19 Probable cytochrome P450 4e1 Drosophila melanogaster ENDOPLASMIC RETICULUM|HEME|HYPOTHETICAL INF-243 Q7Q9L2 2,00E-66 AgCP14967 (Fragment) Anopheles gambiae REPEAT|WD REPEAT|INF-245 Q4FKE3 0.58 Hyothetical protein Trypanosoma brucei INF-246 P34842 1,00E-34 Cytochrome c oxidase polypeptide III Anopheles gambiae MITOCHONDRION|OXIDOREDUCTASE|TRANSMEMBINF-247 P52159 4,00E-20 Cornichon protein Drosophila virilis DEVELOPMENTAL PROTEIN|TRANSMEMBRANE|INF-248 Q27014 5,00E-09 28 kDa desiccation stress protein Tenebrio molitor INF-25 Q83CD5 3.8 Excinuclease ABC, C subunit Coxiella burnetii COMPLETE PROTEOMEINF-250 Q27023 4,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC|DEFENSIN|DIRECT PROTEIN SEQUENCIINF-251 Q7RN92 9,00E-06 Hypothetical protein Plasmodium yoelii yoelii HYPOTHETICAL PROTEIN|INF-253 ZNF142 1,00E-14 Zinc finger protein 142 (HA4654) Homo sapiens DNA-BINDING|METAL-BINDING|NUCLEAR PROTEIN|RINF-254 O42930 4.7 Pep1 protein Schizosaccharomyces pombe INF-256 Q8RG71 7.7 Hypothetical protein FN0446 Fusobacterium nucleatum COMPLETE PROTEOME|INF-258 Q69BL0 5,00E-28 Pattern recognition serine proteinase Manduca sexta SIGNAL;INF-259 Q9CDJ7 0.56 Glucosyltransferase-S (EC 2415) Lactococcus lactis COMPLETE PROTEOME|TRANSFERASE|INF-260 Q60FC9 0.090 Luxuriosin Acalolepta luxuriosaINF-261 ZNF142 2,00E-14 Zinc finger protein 142 (HA4654) Homo sapiens DNA-BINDING|METAL-BINDING|NUCLEAR PROTEIN|RINF-262 Q7QC88 7,00E-32 AgCP1223 (Fragment) Anopheles gambiaeINF-263 Q6LFE7 0.005 Homologue of human HSPC025 Plasmodium falciparumINF-264 Q07171 2,00E-22 Gelsolin precursor Drosophila melanogaster ACTIN-BINDING|ALTERNATIVE SPLICING|CALCIUM|CINF-265 Q8WTD5 0.88 Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-266 Q8WTD5 0.88 Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-269 Q7QDE8 9,00E-40 AgCP10112 (Fragment) Anopheles gambiae
157
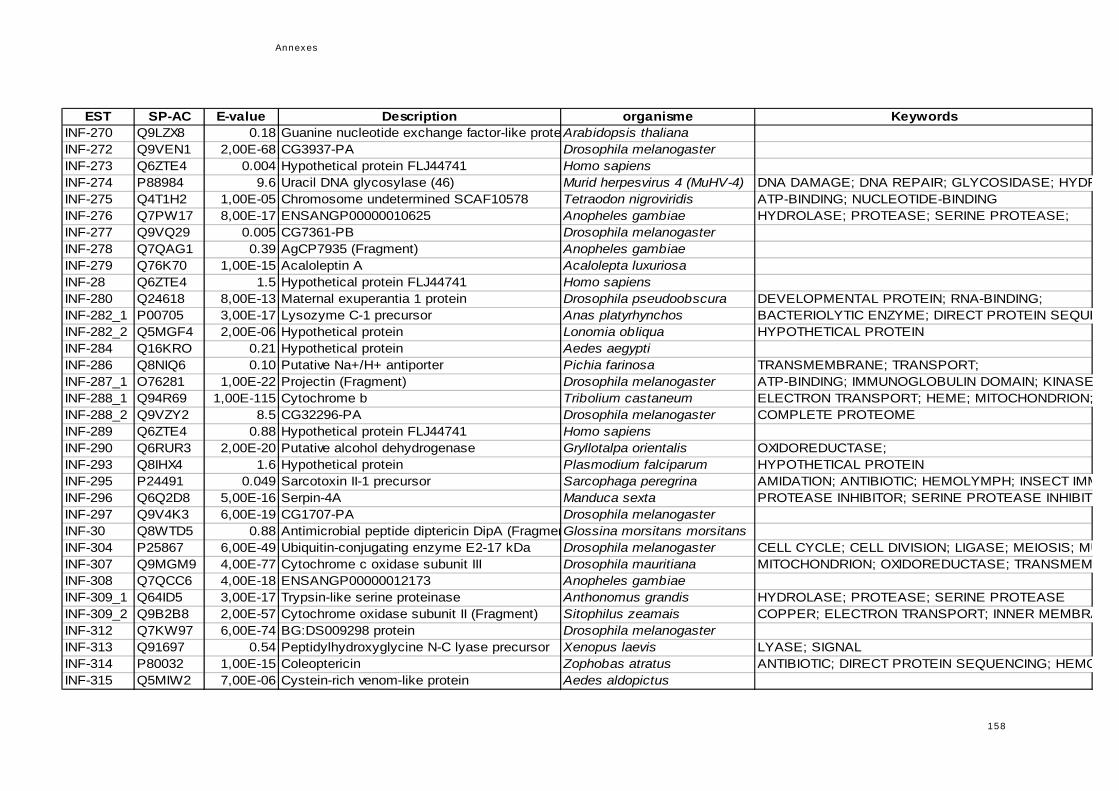
Annexes
158
AC E-value Description organisme KeywordsINF-270 Q9LZX8 0.18 Guanine nucleotide exchange factor-like prot
-
EST SPeArabidopsis thaliana
INF-272 Q9VEN1 2,00E-68 CG3937-PA Drosophila melanogasterINF-273 Q6ZTE4 0.004 Hypothetical protein FLJ44741 Homo sapiensINF-274 P88984 9.6 Uracil DNA glycosylase (46) Murid herpesvirus 4 (MuHV-4) DNA DAMAGE; DNA REPAIR; GLYCOSIDASE; HYDRINF-275 Q4T1H2 1,00E-05 Chromosome undetermined SCAF10578 Tetraodon nigroviridis ATP-BINDING; NUCLEOTIDE-BINDINGINF-276 Q7PW17 8,00E-17 ENSANGP00000010625 Anopheles gambiae HYDROLASE; PROTEASE; SERINE PROTEASE; INF-277 Q9VQ29 0.005 CG7361-PB Drosophila melanogasterINF-278 Q7QAG1 0.39 AgCP7935 (Fragment) Anopheles gambiaeINF-279 Q76K70 1,00E-15 Acaloleptin A Acalolepta luxuriosaINF-28 Q6ZTE4 1.5 Hypothetical protein FLJ44741 Homo sapiensINF-280 Q24618 8,00E-13 Maternal exuperantia 1 protein Drosophila pseudoobscura DEVELOPMENTAL PROTEIN; RNA-BINDING; INF-282_1 P00705 3,00E-17 Lysozyme C-1 precursor Anas platyrhynchos BACTERIOLYTIC ENZYME; DIRECT PROTEIN SEQUEINF-282_2 Q5MGF4 2,00E-06 Hypothetical protein Lonomia obliqua HYPOTHETICAL PROTEININF-284 Q16KRO 0.21 Hypothetical protein Aedes aegyptiINF-286 Q8NIQ6 0.10 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORT; INF-287_1 O76281 1,00E-22 Projectin (Fragment) Drosophila melanogaster ATP-BINDING; IMMUNOGLOBULIN DOMAIN; KINASEINF-288_1 Q94R69 1,00E-115 Cytochrome b Tribolium castaneum ELECTRON TRANSPORT; HEME; MITOCHONDRION;INF-288_2 Q9VZY2 8.5 CG32296-PA Drosophila melanogaster COMPLETE PROTEOMEINF-289 Q6ZTE4 0.88 Hypothetical protein FLJ44741 Homo sapiensINF-290 Q6RUR3 2,00E-20 Putative alcohol dehydrogenase Gryllotalpa orientalis OXIDOREDUCTASE; INF-293 Q8IHX4 1.6 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEININF-295 P24491 0.049 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-296 Q6Q2D8 5,00E-16 Serpin-4A Manduca sexta PROTEASE INHIBITOR; SERINE PROTEASE INHIBITOINF-297 Q9V4K3 6,00E-19 CG1707-PA Drosophila melanogaster INF-30 Q8WTD5 0.88 Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-304 P25867 6,00E-49 Ubiquitin-conjugating enzyme E2-17 kDa Drosophila melanogaster CELL CYCLE; CELL DIVISION; LIGASE; MEIOSIS; MUINF-307 Q9MGM9 4,00E-77 Cytochrome c oxidase subunit III Drosophila mauritiana MITOCHONDRION; OXIDOREDUCTASE; TRANSMEMINF-308 Q7QCC6 4,00E-18 ENSANGP00000012173 Anopheles gambiae INF-309_1 Q64ID5 3,00E-17 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-309_2 Q9B2B8 2,00E-57 Cytochrome oxidase subunit II (Fragment) Sitophilus zeamais COPPER; ELECTRON TRANSPORT; INNER MEMBRAINF-312 Q7KW97 6,00E-74 BG:DS009298 protein Drosophila melanogaster INF-313 Q91697 0.54 Peptidylhydroxyglycine N-C lyase precursor Xenopus laevis LYASE; SIGNALINF-314 P80032 1,00E-15 Coleoptericin Zophobas atratus ANTIBIOTIC; DIRECT PROTEIN SEQUENCING; HEMOINF-315 Q5MIW2 7,00E-06 Cystein-rich venom-like protein Aedes aldopictus
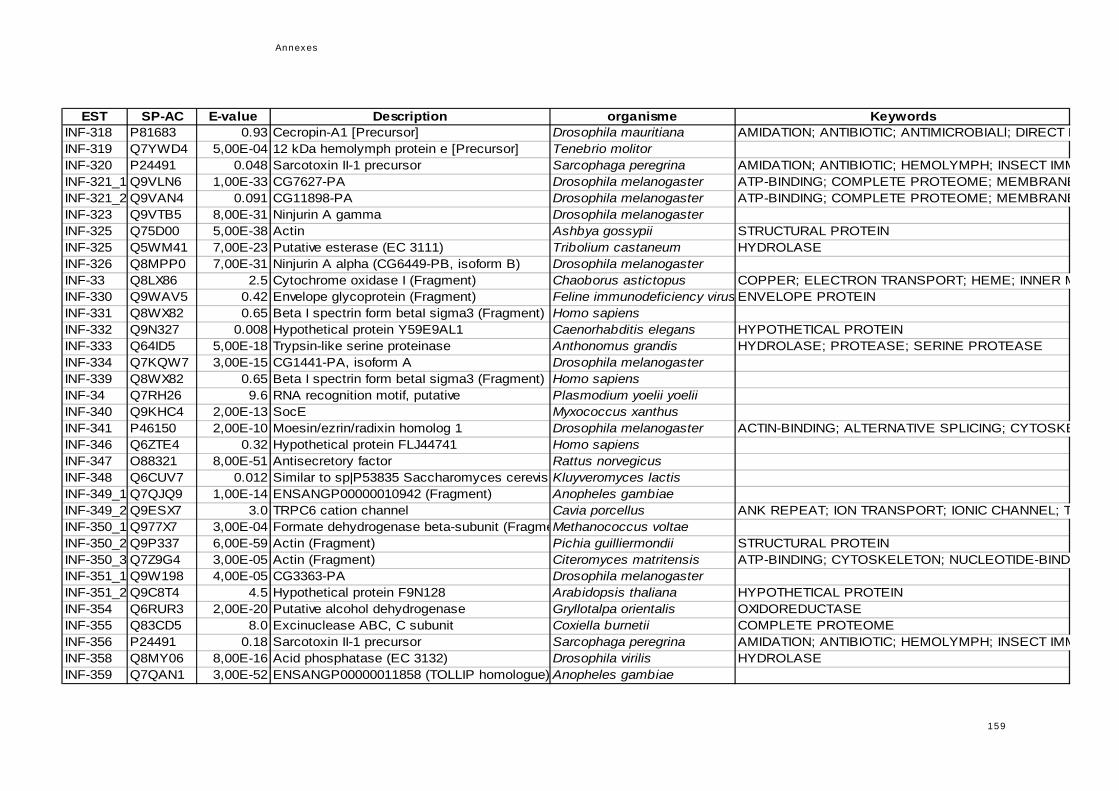
Annexes
159
C E-value Description organisme KeywordsINF-318 P81683 0.93 Cecropin-A1 [Precursor] Drosophila mauritiana
A
EST SP-AMIDATION; ANTIBIOTIC; ANTIMICROBIALl; DIRECT P
INF-319 Q7YWD4 5,00E-04 12 kDa hemolymph protein e [Precursor] Tenebrio molitorINF-320 P24491 0.048 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-321_1 Q9VLN6 1,00E-33 CG7627-PA Drosophila melanogaster ATP-BINDING; COMPLETE PROTEOME; MEMBRANEINF-321_2 Q9VAN4 0.091 CG11898-PA Drosophila melanogaster ATP-BINDING; COMPLETE PROTEOME; MEMBRANEINF-323 Q9VTB5 8,00E-31 Ninjurin A gamma Drosophila melanogasterINF-325 Q75D00 5,00E-38 Actin Ashbya gossypii STRUCTURAL PROTEININF-325 Q5WM41 7,00E-23 Putative esterase (EC 3111) Tribolium castaneum HYDROLASEINF-326 Q8MPP0 7,00E-31 Ninjurin A alpha (CG6449-PB, isoform B) Drosophila melanogasterINF-33 Q8LX86 2.5 Cytochrome oxidase I (Fragment) Chaoborus astictopus COPPER; ELECTRON TRANSPORT; HEME; INNER MINF-330 Q9WAV5 0.42 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEININF-331 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-332 Q9N327 0.008 Hypothetical protein Y59E9AL1 Caenorhabditis elegans HYPOTHETICAL PROTEIN INF-333 Q64ID5 5,00E-18 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-334 Q7KQW7 3,00E-15 CG1441-PA, isoform A Drosophila melanogasterINF-339 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-34 Q7RH26 9.6 RNA recognition motif, putative Plasmodium yoelii yoeliiINF-340 Q9KHC4 2,00E-13 SocE Myxococcus xanthusINF-341 P46150 2,00E-10 Moesin/ezrin/radixin homolog 1 Drosophila melanogaster ACTIN-BINDING; ALTERNATIVE SPLICING; CYTOSKEINF-346 Q6ZTE4 0.32 Hypothetical protein FLJ44741 Homo sapiens INF-347 O88321 8,00E-51 Antisecretory factor Rattus norvegicus INF-348 Q6CUV7 0.012 Similar to sp|P53835 Saccharomyces cerevisiKluyveromyces lactis INF-349_1 Q7QJQ9 1,00E-14 ENSANGP00000010942 (Fragment) Anopheles gambiaeINF-349_2 Q9ESX7 3.0 TRPC6 cation channel Cavia porcellus ANK REPEAT; ION TRANSPORT; IONIC CHANNEL; TINF-350_1 Q977X7 3,00E-04 Formate dehydrogenase beta-subunit (FragmeMethanococcus voltae INF-350_2 Q9P337 6,00E-59 Actin (Fragment) Pichia guilliermondii STRUCTURAL PROTEININF-350_3 Q7Z9G4 3,00E-05 Actin (Fragment) Citeromyces matritensis ATP-BINDING; CYTOSKELETON; NUCLEOTIDE-BINDINF-351_1 Q9W198 4,00E-05 CG3363-PA Drosophila melanogaster INF-351_2 Q9C8T4 4.5 Hypothetical protein F9N128 Arabidopsis thaliana HYPOTHETICAL PROTEININF-354 Q6RUR3 2,00E-20 Putative alcohol dehydrogenase Gryllotalpa orientalis OXIDOREDUCTASEINF-355 Q83CD5 8.0 Excinuclease ABC, C subunit Coxiella burnetii COMPLETE PROTEOMEINF-356 P24491 0.18 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-358 Q8MY06 8,00E-16 Acid phosphatase (EC 3132) Drosophila virilis HYDROLASEINF-359 Q7QAN1 3,00E-52 ENSANGP00000011858 (TOLLIP homologue) Anopheles gambiae
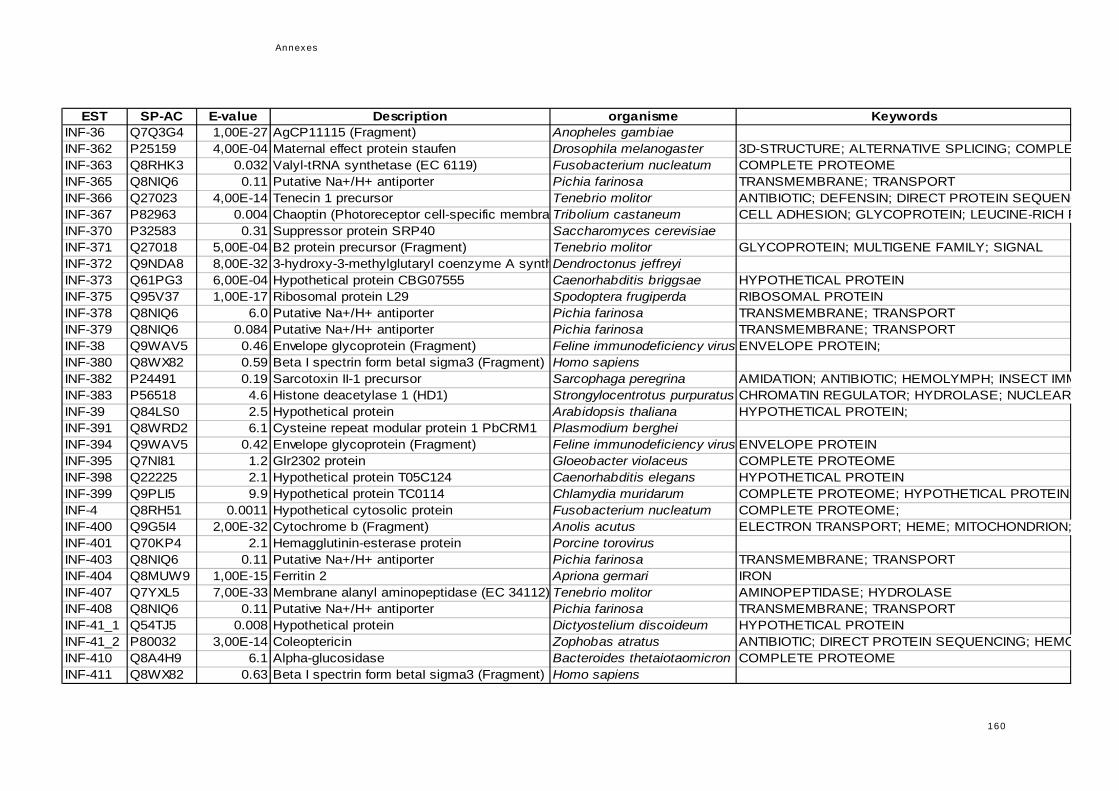
Annexes
160
EST SP-AC E-value Description organisme KeywordsINF-36 Q7Q3G4 1,00E-27 AgCP11115 (Fragment) Anopheles gambiaeINF-362 P25159 4,00E-04 Maternal effect protein staufen Drosophila melanogaster 3D-STRUCTURE; ALTERNATIVE SPLICING; COMPLEINF-363 Q8RHK3 0.032 Valyl-tRNA synthetase (EC 6119) Fusobacterium nucleatum COMPLETE PROTEOMEINF-365 Q8NIQ6 0.11 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-366 Q27023 4,00E-14 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-367 P82963 0.004 Chaoptin (Photoreceptor cell-specific membra Tribolium castaneum CELL ADHESION; GLYCOPROTEIN; LEUCINE-RICH RINF-370 P32583 0.31 Suppressor protein SRP40 Saccharomyces cerevisiae INF-371 Q27018 5,00E-04 B2 protein precursor (Fragment) Tenebrio molitor GLYCOPROTEIN; MULTIGENE FAMILY; SIGNALINF-372 Q9NDA8 8,00E-32 3-hydroxy-3-methylglutaryl coenzyme A synthDendroctonus jeffreyi INF-373 Q61PG3 6,00E-04 Hypothetical protein CBG07555 Caenorhabditis briggsae HYPOTHETICAL PROTEININF-375 Q95V37 1,00E-17 Ribosomal protein L29 Spodoptera frugiperda RIBOSOMAL PROTEININF-378 Q8NIQ6 6.0 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-379 Q8NIQ6 0.084 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-38 Q9WAV5 0.46 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-380 Q8WX82 0.59 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-382 P24491 0.19 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-383 P56518 4.6 Histone deacetylase 1 (HD1) Strongylocentrotus purpuratus CHROMATIN REGULATOR; HYDROLASE; NUCLEAR INF-39 Q84LS0 2.5 Hypothetical protein Arabidopsis thaliana HYPOTHETICAL PROTEIN; INF-391 Q8WRD2 6.1 Cysteine repeat modular protein 1 PbCRM1 Plasmodium bergheiINF-394 Q9WAV5 0.42 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEININF-395 Q7NI81 1.2 Glr2302 protein Gloeobacter violaceus COMPLETE PROTEOMEINF-398 Q22225 2.1 Hypothetical protein T05C124 Caenorhabditis elegans HYPOTHETICAL PROTEININF-399 Q9PLI5 9.9 Hypothetical protein TC0114 Chlamydia muridarum COMPLETE PROTEOME; HYPOTHETICAL PROTEININF-4 Q8RH51 0.0011 Hypothetical cytosolic protein Fusobacterium nucleatum COMPLETE PROTEOME; INF-400 Q9G5I4 2,00E-32 Cytochrome b (Fragment) Anolis acutus ELECTRON TRANSPORT; HEME; MITOCHONDRION;INF-401 Q70KP4 2.1 Hemagglutinin-esterase protein Porcine torovirus INF-403 Q8NIQ6 0.11 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-404 Q8MUW9 1,00E-15 Ferritin 2 Apriona germari IRONINF-407 Q7YXL5 7,00E-33 Membrane alanyl aminopeptidase (EC 34112) Tenebrio molitor AMINOPEPTIDASE; HYDROLASEINF-408 Q8NIQ6 0.11 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-41_1 Q54TJ5 0.008 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEININF-41_2 P80032 3,00E-14 Coleoptericin Zophobas atratus ANTIBIOTIC; DIRECT PROTEIN SEQUENCING; HEMOINF-410 Q8A4H9 6.1 Alpha-glucosidase Bacteroides thetaiotaomicron COMPLETE PROTEOMEINF-411 Q8WX82 0.63 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiens
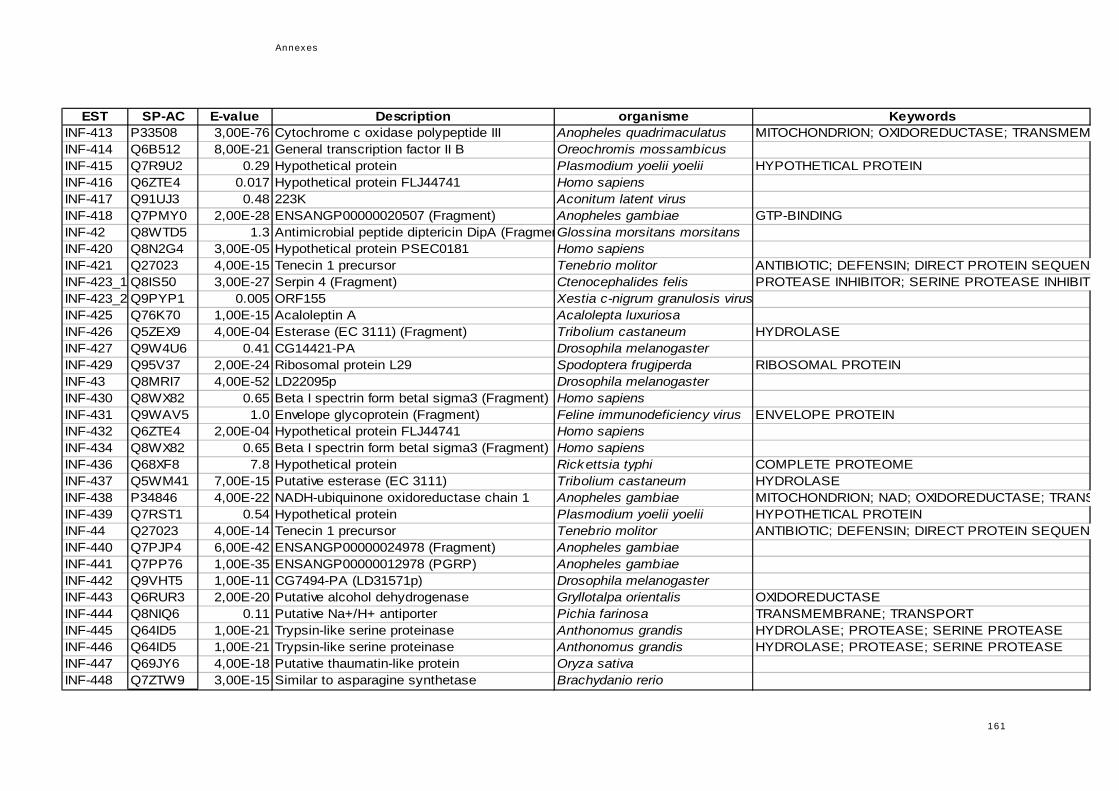
Annexes
161
C E-value Description organisme KeywordsINF-413 P33508 3,00E-76 Cytochrome c oxidase polypeptide III Anopheles quadrimaculatus MITOCHONDRION; OXIDOREDUCTASE; TRANSMEMINF-414 Q6B512 8,00E-21 General transcription factor II B Oreochromis mossambicus INF-415 Q7R9U2 0.29 Hypothetical protein Plasmodium yoelii yoelii HYPOTHETICAL PROTEININF-416 Q6ZTE4 0.017 Hypothetical protein FLJ44741 Homo sapiens INF-417 Q91UJ3 0.48 223K Aconitum latent virus INF-418 Q7PMY0 2,00E-28 ENSANGP00000020507 (Fragment) Anopheles gambiae GTP-BINDINGINF-42 Q8WTD5 1.3
A
EST SP-
Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-420 Q8N2G4 3,00E-05 Hypothetical protein PSEC0181 Homo sapiens INF-421 Q27023 4,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-423_1 Q8IS50 3,00E-27 Serpin 4 (Fragment) Ctenocephalides felis PROTEASE INHIBITOR; SERINE PROTEASE INHIBITOINF-423_2 Q9PYP1 0.005 ORF155 Xestia c-nigrum granulosis virus INF-425 Q76K70 1,00E-15 Acaloleptin A Acalolepta luxuriosa INF-426 Q5ZEX9 4,00E-04 Esterase (EC 3111) (Fragment) Tribolium castaneum HYDROLASEINF-427 Q9W4U6 0.41 CG14421-PA Drosophila melanogaster INF-429 Q95V37 2,00E-24 Ribosomal protein L29 Spodoptera frugiperda RIBOSOMAL PROTEININF-43 Q8MRI7 4,00E-52 LD22095p Drosophila melanogasterINF-430 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-431 Q9WAV5 1.0 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEININF-432 Q6ZTE4 2,00E-04 Hypothetical protein FLJ44741 Homo sapiens INF-434 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-436 Q68XF8 7.8 Hypothetical protein Rickettsia typhi COMPLETE PROTEOMEINF-437 Q5WM41 7,00E-15 Putative esterase (EC 3111) Tribolium castaneum HYDROLASEINF-438 P34846 4,00E-22 NADH-ubiquinone oxidoreductase chain 1 Anopheles gambiae MITOCHONDRION; NAD; OXIDOREDUCTASE; TRANSINF-439 Q7RST1 0.54 Hypothetical protein Plasmodium yoelii yoelii HYPOTHETICAL PROTEININF-44 Q27023 4,00E-14 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-440 Q7PJP4 6,00E-42 ENSANGP00000024978 (Fragment) Anopheles gambiaeINF-441 Q7PP76 1,00E-35 ENSANGP00000012978 (PGRP) Anopheles gambiaeINF-442 Q9VHT5 1,00E-11 CG7494-PA (LD31571p) Drosophila melanogasterINF-443 Q6RUR3 2,00E-20 Putative alcohol dehydrogenase Gryllotalpa orientalis OXIDOREDUCTASEINF-444 Q8NIQ6 0.11 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-445 Q64ID5 1,00E-21 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-446 Q64ID5 1,00E-21 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-447 Q69JY6 4,00E-18 Putative thaumatin-like protein Oryza sativa INF-448 Q7ZTW9 3,00E-15 Similar to asparagine synthetase Brachydanio rerio
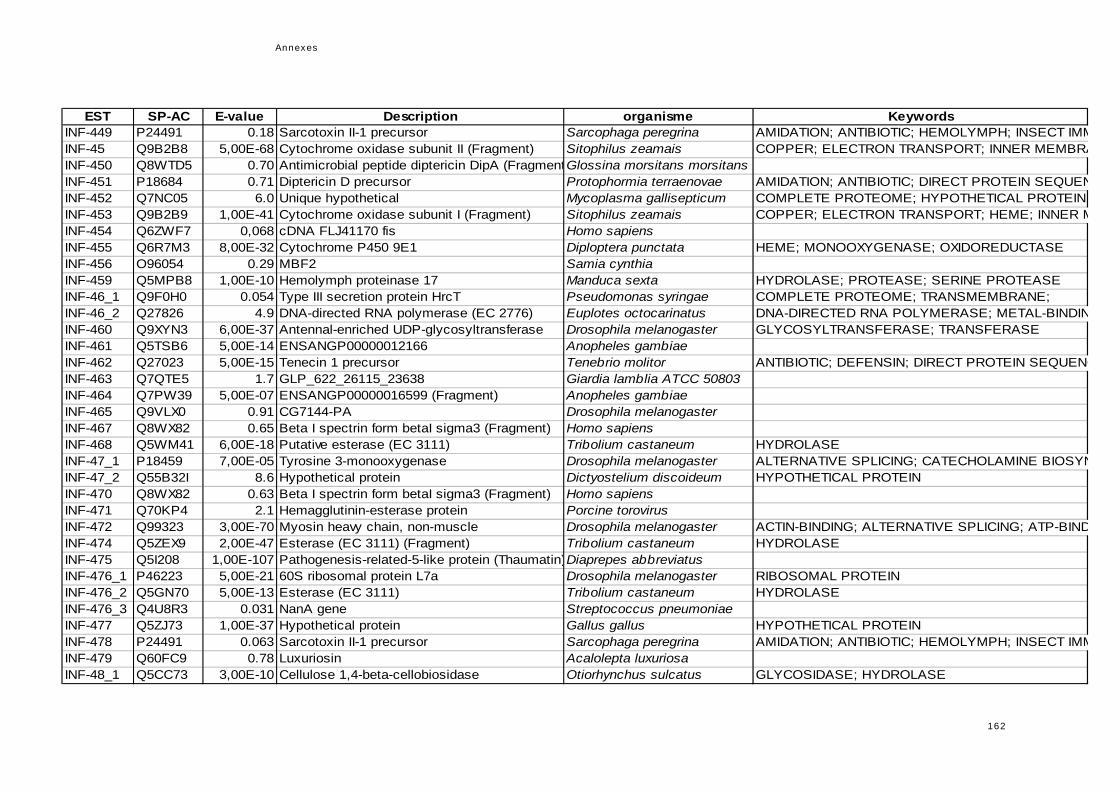
Annexes
162
AC E-value Description organisme KeywordsINF-449 P24491 0.18 Sarcotoxin II-1 precursor Sarcophaga peregrina
-
EST SPAMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMM
INF-45 Q9B2B8 5,00E-68 Cytochrome oxidase subunit II (Fragment) Sitophilus zeamais COPPER; ELECTRON TRANSPORT; INNER MEMBRAINF-450 Q8WTD5 0.70 Antimicrobial peptide diptericin DipA (FragmentGlossina morsitans morsitansINF-451 P18684 0.71 Diptericin D precursor Protophormia terraenovae AMIDATION; ANTIBIOTIC; DIRECT PROTEIN SEQUENINF-452 Q7NC05 6.0 Unique hypothetical Mycoplasma gallisepticum COMPLETE PROTEOME; HYPOTHETICAL PROTEININF-453 Q9B2B9 1,00E-41 Cytochrome oxidase subunit I (Fragment) Sitophilus zeamais COPPER; ELECTRON TRANSPORT; HEME; INNER MINF-454 Q6ZWF7 0,068 cDNA FLJ41170 fis Homo sapiensINF-455 Q6R7M3 8,00E-32 Cytochrome P450 9E1 Diploptera punctata HEME; MONOOXYGENASE; OXIDOREDUCTASEINF-456 O96054 0.29 MBF2 Samia cynthia INF-459 Q5MPB8 1,00E-10 Hemolymph proteinase 17 Manduca sexta HYDROLASE; PROTEASE; SERINE PROTEASEINF-46_1 Q9F0H0 0.054 Type III secretion protein HrcT Pseudomonas syringae COMPLETE PROTEOME; TRANSMEMBRANE; INF-46_2 Q27826 4.9 DNA-directed RNA polymerase (EC 2776) Euplotes octocarinatus DNA-DIRECTED RNA POLYMERASE; METAL-BINDININF-460 Q9XYN3 6,00E-37 Antennal-enriched UDP-glycosyltransferase Drosophila melanogaster GLYCOSYLTRANSFERASE; TRANSFERASEINF-461 Q5TSB6 5,00E-14 ENSANGP00000012166 Anopheles gambiaeINF-462 Q27023 5,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-463 Q7QTE5 1.7 GLP_622_26115_23638 Giardia lamblia ATCC 50803 INF-464 Q7PW39 5,00E-07 ENSANGP00000016599 (Fragment) Anopheles gambiae INF-465 Q9VLX0 0.91 CG7144-PA Drosophila melanogaster INF-467 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-468 Q5WM41 6,00E-18 Putative esterase (EC 3111) Tribolium castaneum HYDROLASEINF-47_1 P18459 7,00E-05 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-47_2 Q55B32I 8.6 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEININF-470 Q8WX82 0.63 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-471 Q70KP4 2.1 Hemagglutinin-esterase protein Porcine torovirus INF-472 Q99323 3,00E-70 Myosin heavy chain, non-muscle Drosophila melanogaster ACTIN-BINDING; ALTERNATIVE SPLICING; ATP-BINDINF-474 Q5ZEX9 2,00E-47 Esterase (EC 3111) (Fragment) Tribolium castaneum HYDROLASEINF-475 Q5I208 1,00E-107 Pathogenesis-related-5-like protein (Thaumatin)Diaprepes abbreviatus INF-476_1 P46223 5,00E-21 60S ribosomal protein L7a Drosophila melanogaster RIBOSOMAL PROTEININF-476_2 Q5GN70 5,00E-13 Esterase (EC 3111) Tribolium castaneum HYDROLASEINF-476_3 Q4U8R3 0.031 NanA gene Streptococcus pneumoniae INF-477 Q5ZJ73 1,00E-37 Hypothetical protein Gallus gallus HYPOTHETICAL PROTEININF-478 P24491 0.063 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-479 Q60FC9 0.78 Luxuriosin Acalolepta luxuriosaINF-48_1 Q5CC73 3,00E-10 Cellulose 1,4-beta-cellobiosidase Otiorhynchus sulcatus GLYCOSIDASE; HYDROLASE
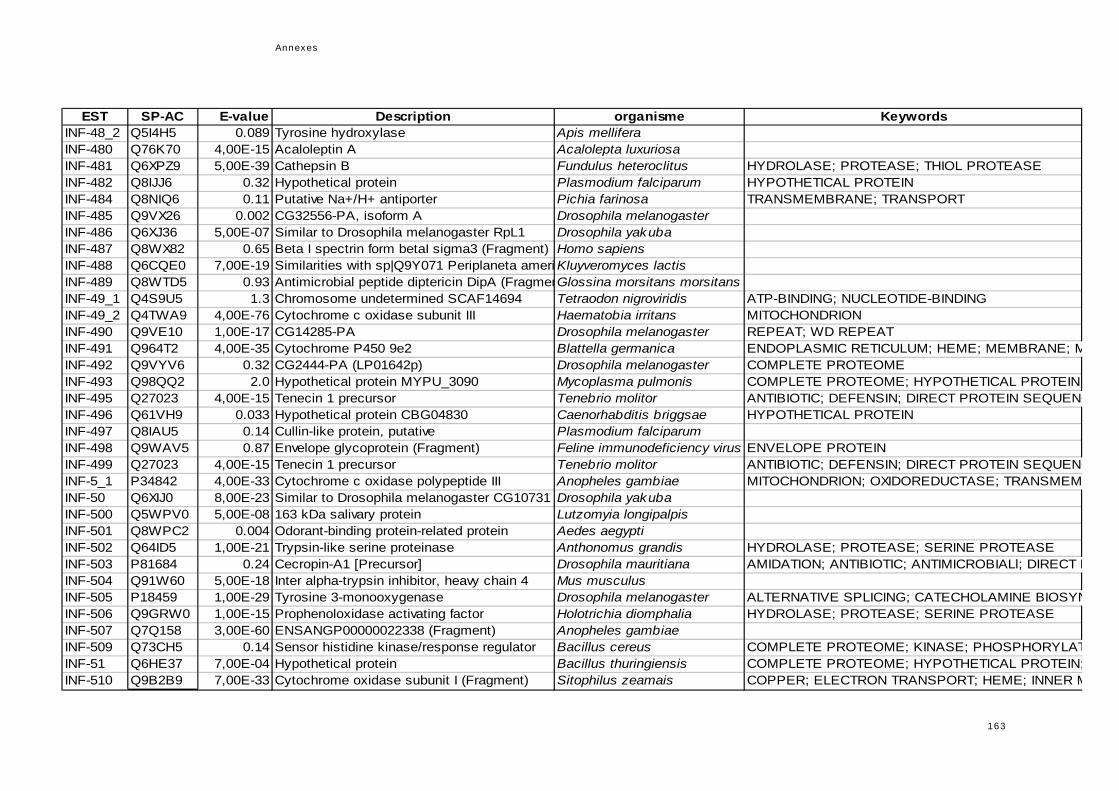
Annexes
163
EST SP- C E-value Description organisme KeywordsINF-48_2 Q5I4H5 0.089 Tyrosine hydroxylase Apis mellifera INF-480 Q76K70 4,00E-15 Acaloleptin A Acalolepta luxuriosa INF-481 Q6XPZ9 5,00E-39 Cathepsin B Fundulus heteroclitus HYDROLASE; PROTEASE; THIOL PROTEASEINF-482 Q8IJJ6 0.32 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEININF-484 Q8NIQ6 0.11 Putative Na+/H+ antiporter Pichia farinosa TRANSMEMBRANE; TRANSPORTINF-485 Q9VX26 0.002 CG32556-PA, isoform A Drosophila melanogaster INF-486 Q6XJ36 5,00E-07 Similar to Drosophila melanogaster RpL1 Drosophila yakubaINF-487 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-488 Q6CQE0 7,00E-19 Similarities with sp|Q9Y071 Periplaneta ameri Kluyveromyces lactisINF-489 Q8WTD5 0.93
A
Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-49_1 Q4S9U5 1.3 Chromosome undetermined SCAF14694 Tetraodon nigroviridis ATP-BINDING; NUCLEOTIDE-BINDINGINF-49_2 Q4TWA9 4,00E-76 Cytochrome c oxidase subunit III Haematobia irritans MITOCHONDRIONINF-490 Q9VE10 1,00E-17 CG14285-PA Drosophila melanogaster REPEAT; WD REPEATINF-491 Q964T2 4,00E-35 Cytochrome P450 9e2 Blattella germanica ENDOPLASMIC RETICULUM; HEME; MEMBRANE; MINF-492 Q9VYV6 0.32 CG2444-PA (LP01642p) Drosophila melanogaster COMPLETE PROTEOMEINF-493 Q98QQ2 2.0 Hypothetical protein MYPU_3090 Mycoplasma pulmonis COMPLETE PROTEOME; HYPOTHETICAL PROTEININF-495 Q27023 4,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-496 Q61VH9 0.033 Hypothetical protein CBG04830 Caenorhabditis briggsae HYPOTHETICAL PROTEININF-497 Q8IAU5 0.14 Cullin-like protein, putative Plasmodium falciparum INF-498 Q9WAV5 0.87 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEININF-499 Q27023 4,00E-15 Tenecin 1 precursor Tenebrio molitor ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-5_1 P34842 4,00E-33 Cytochrome c oxidase polypeptide III Anopheles gambiae MITOCHONDRION; OXIDOREDUCTASE; TRANSMEMINF-50 Q6XIJ0 8,00E-23 Similar to Drosophila melanogaster CG10731 Drosophila yakubaINF-500 Q5WPV0 5,00E-08 163 kDa salivary protein Lutzomyia longipalpis INF-501 Q8WPC2 0.004 Odorant-binding protein-related protein Aedes aegypti INF-502 Q64ID5 1,00E-21 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-503 P81684 0.24 Cecropin-A1 [Precursor] Drosophila mauritiana AMIDATION; ANTIBIOTIC; ANTIMICROBIALl; DIRECT PINF-504 Q91W60 5,00E-18 Inter alpha-trypsin inhibitor, heavy chain 4 Mus musculus INF-505 P18459 1,00E-29 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-506 Q9GRW0 1,00E-15 Prophenoloxidase activating factor Holotrichia diomphalia HYDROLASE; PROTEASE; SERINE PROTEASEINF-507 Q7Q158 3,00E-60 ENSANGP00000022338 (Fragment) Anopheles gambiaeINF-509 Q73CH5 0.14 Sensor histidine kinase/response regulator Bacillus cereus COMPLETE PROTEOME; KINASE; PHOSPHORYLATINF-51 Q6HE37 7,00E-04 Hypothetical protein Bacillus thuringiensis COMPLETE PROTEOME; HYPOTHETICAL PROTEIN;INF-510 Q9B2B9 7,00E-33 Cytochrome oxidase subunit I (Fragment) Sitophilus zeamais COPPER; ELECTRON TRANSPORT; HEME; INNER M
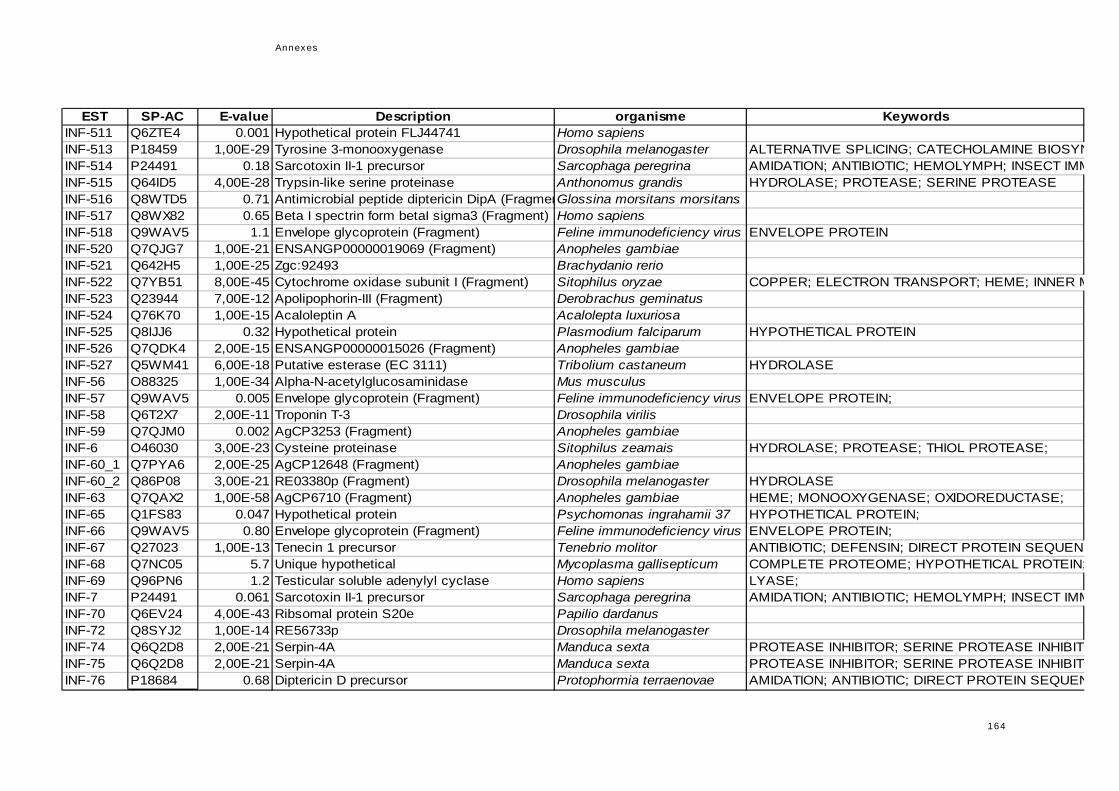
Annexes
164
EST SP- C E-value Description organisme KeywordsINF-511 Q6ZTE4 0.001 Hypothetical protein FLJ44741 Homo sapiens INF-513 P18459 1,00E-29 Tyrosine 3-monooxygenase Drosophila melanogaster
A
ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-514 P24491 0.18 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-515 Q64ID5 4,00E-28 Trypsin-like serine proteinase Anthonomus grandis HYDROLASE; PROTEASE; SERINE PROTEASEINF-516 Q8WTD5 0.71 Antimicrobial peptide diptericin DipA (FragmenGlossina morsitans morsitansINF-517 Q8WX82 0.65 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-518 Q9WAV5 1.1 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEININF-520 Q7QJG7 1,00E-21 ENSANGP00000019069 (Fragment) Anopheles gambiaeINF-521 Q642H5 1,00E-25 Zgc:92493 Brachydanio rerio INF-522 Q7YB51 8,00E-45 Cytochrome oxidase subunit I (Fragment) Sitophilus oryzae COPPER; ELECTRON TRANSPORT; HEME; INNER MINF-523 Q23944 7,00E-12 Apolipophorin-III (Fragment) Derobrachus geminatusINF-524 Q76K70 1,00E-15 Acaloleptin A Acalolepta luxuriosa INF-525 Q8IJJ6 0.32 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEININF-526 Q7QD 4 2,00E-15 ENSANGP00000015026 (Fragment) Anopheles gambiaeINF-527 Q5WM41 6,00E-18 Putative esterase (EC 3111) Tribolium castaneum HYDROLASEINF-56 O88325 1,00E-34 Alpha-N-acetylglucosaminidase Mus musculus INF-57 Q9WAV5 0.005 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-58 Q6T2X7 2,00E-11 Troponin T-3 Drosophila virilis INF-59 Q7QJM0 0.002 AgCP3253 (Fragment) Anopheles gambiae INF-6 O46030 3,00E-23 Cysteine proteinase Sitophilus zeamais HYDROLASE; PROTEASE; THIOL PROTEASE; INF-60_1 Q7PYA6 2,00E-25 AgCP12648 (Fragment) Anopheles gambiaeINF-60_2 Q86P08 3,00E-21 RE03380p (Fragment) Drosophila melanogaster HYDROLASEINF-63 Q7QAX2 1,00E-58 AgCP6710 (Fragment) Anopheles gambiae HEME; MONOOXYGENASE; OXIDOREDUCTASE; INF-65 Q1FS83 0.047 Hypothetical protein Psychomonas ingrahamii 37 HYPOTHETICAL PROTEIN; INF-66 Q9WAV5 0.80 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-67 Q27023 1,00E-13 Tenecin 1 precursor Tenebrio molitor
K
ANTIBIOTIC; DEFENSIN; DIRECT PROTEIN SEQUENCINF-68 Q7NC05 5.7 Unique hypothetical Mycoplasma gallisepticum COMPLETE PROTEOME; HYPOTHETICAL PROTEIN;INF-69 Q96PN6 1.2 Testicular soluble adenylyl cyclase Homo sapiens LYASE; INF-7 P24491 0.061 Sarcotoxin II-1 precursor Sarcophaga peregrina AMIDATION; ANTIBIOTIC; HEMOLYMPH; INSECT IMMINF-70 Q6EV24 4,00E-43 Ribsomal protein S20e Papilio dardanus INF-72 Q8SYJ2 1,00E-14 RE56733p Drosophila melanogaster INF-74 Q6Q2D8 2,00E-21 Serpin-4A Manduca sexta PROTEASE INHIBITOR; SERINE PROTEASE INHIBITOINF-75 Q6Q2D8 2,00E-21 Serpin-4A Manduca sexta PROTEASE INHIBITOR; SERINE PROTEASE INHIBITOINF-76 P18684 0.68 Diptericin D precursor Protophormia terraenovae AMIDATION; ANTIBIOTIC; DIRECT PROTEIN SEQUEN
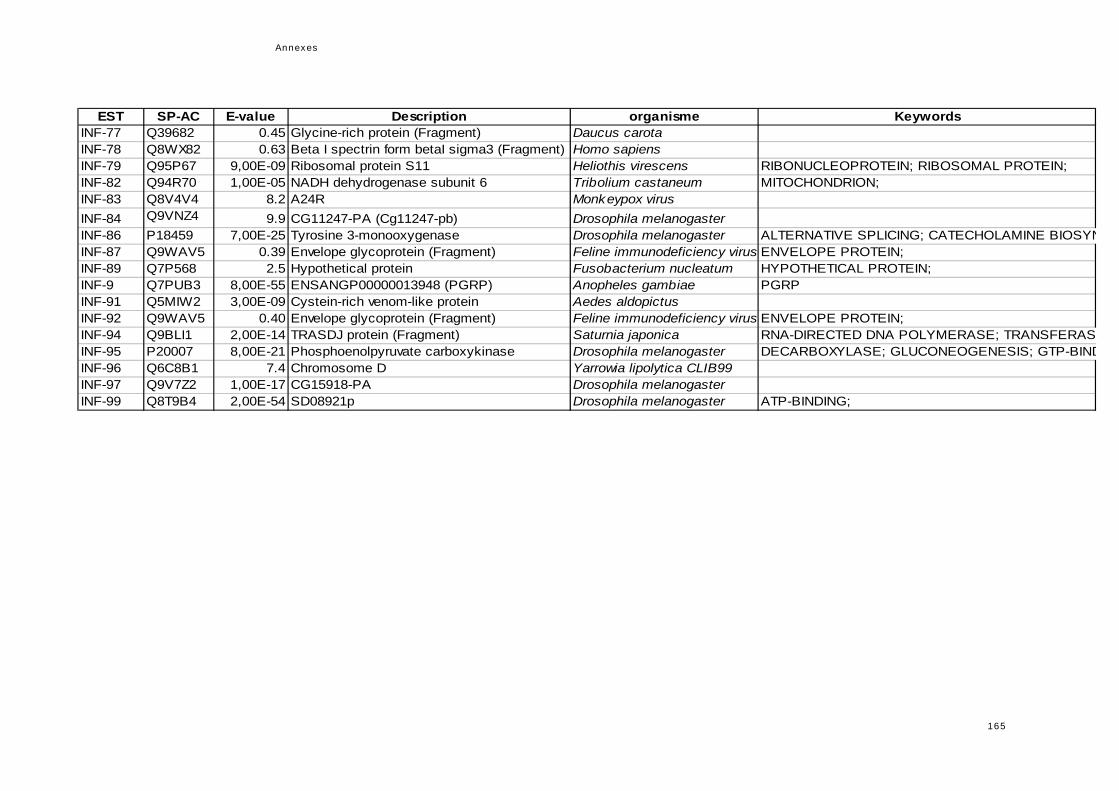
Annexes
P
EST S -AC E-value Description organisme KeywordsINF-77 Q39682 0.45 Glycine-rich protein (Fragment) Daucus carotaINF-78 Q8WX82 0.63 Beta I spectrin form betaI sigma3 (Fragment) Homo sapiensINF-79 Q95P67 9,00E-09 Ribosomal protein S11 Heliothis virescens RIBONUCLEOPROTEIN; RIBOSOMAL PROTEIN; INF-82 Q94R70 1,00E-05 NADH dehydrogenase subunit 6 Tribolium castaneum MITOCHONDRION; INF-83 Q8V4V4 8.2 A24R Monkeypox virus INF-84 Q9VNZ4 9.9 CG11247-PA (Cg11247-pb) Drosophila melanogaster INF-86 P18459 7,00E-25 Tyrosine 3-monooxygenase Drosophila melanogaster ALTERNATIVE SPLICING; CATECHOLAMINE BIOSYNINF-87 Q9WAV5 0.39 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-89 Q7P568 2.5 Hypothetical protein Fusobacterium nucleatum HYPOTHETICAL PROTEIN; INF-9 Q7PUB3 8,00E-55 ENSANGP00000013948 (PGRP) Anopheles gambiae PGRPINF-91 Q5MIW2 3,00E-09 Cystein-rich venom-like protein Aedes aldopictusINF-92 Q9WAV5 0.40 Envelope glycoprotein (Fragment) Feline immunodeficiency virus ENVELOPE PROTEIN; INF-94 Q9BLI1 2,00E-14 TRASDJ protein (Fragment) Saturnia japonica RNA-DIRECTED DNA POLYMERASE; TRANSFERASINF-95 P20007 8,00E-21 Phosphoenolpyruvate carboxykinase Drosophila melanogaster DECARBOXYLASE; GLUCONEOGENESIS; GTP-BINDINF-96 Q6C8B1 7.4 Chromosome D Yarrowia lipolytica CLIB99 INF-97 Q9V7Z2 1,00E-17 CG15918-PA Drosophila melanogasterINF-99 Q8T9B4 2,00E-54 SD08921p Drosophila melanogaster ATP-BINDING;
165

Annexes
Annexe V Tableau A-IV : Tableau récapitulatif des EST de la banque soustraite d’A. pisum. Les similitudes de séquence ont été obtenues par comparaison des six phases de lecture des sé-quences des EST aux banques de données protéiques. SP-AC, identifiant de la protéine dans la banque de données Swiss-Prot.
ANSELME Caroline Thèse au laboratoire BF2I / 2006 Institut National des Sciences Appliquées de Lyon
166
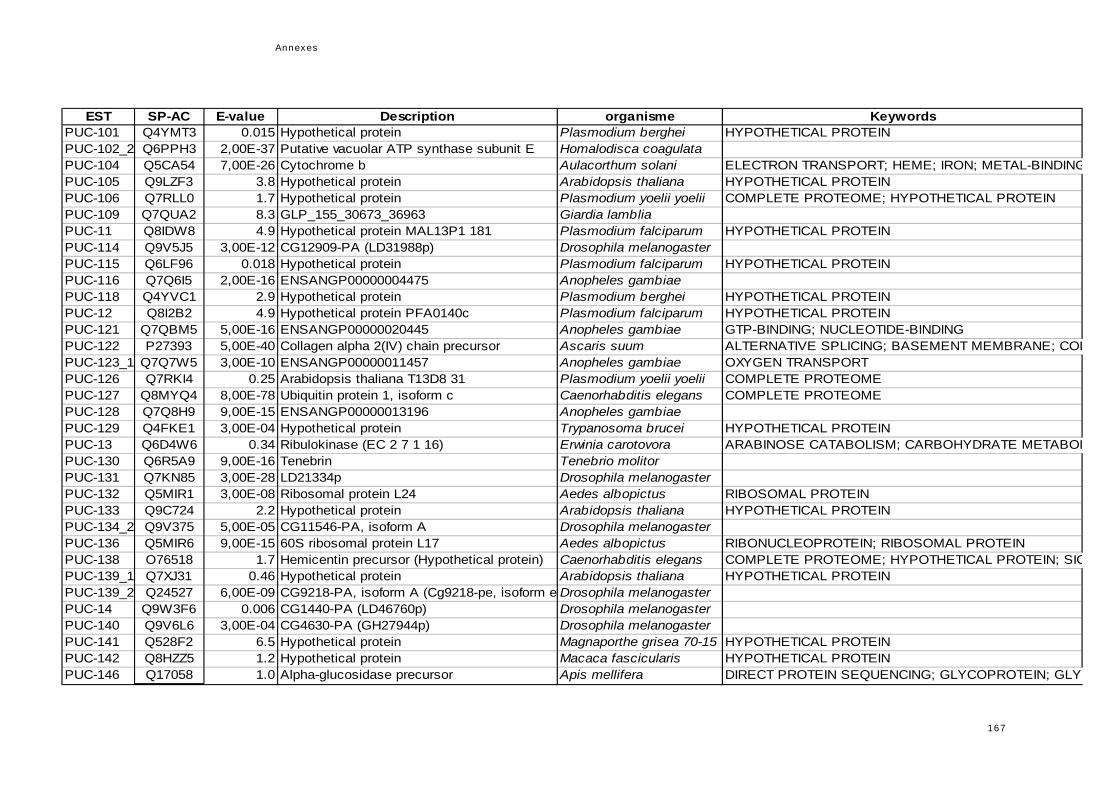
Annexes
167
AC E-value Description organisme KeywordsPUC-101 Q4YMT3 0.015 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-102_2 Q6PPH3 2,00E-37 Putative vacuolar ATP synthase subunit E Homalodisca coagulata PUC-104 Q5CA54 7,00E-26 Cytochrome b Aulacorthum solani ELECTRON TRANSPORT; HEME; IRON; METAL-BINDIN
-
EST SP
GPUC-105 Q9LZF3 3.8 Hypothetical protein Arabidopsis thaliana HYPOTHETICAL PROTEINPUC-106 Q7RLL0 1.7 Hypothetical protein Plasmodium yoelii yoelii COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-109 Q7QUA2 8.3 GLP_155_30673_36963 Giardia lamblia PUC-11 Q8IDW8 4.9 Hypothetical protein MAL13P1 181 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-114 Q9V5J5 3,00E-12 CG12909-PA (LD31988p) Drosophila melanogasterPUC-115 Q6LF96 0.018 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-116 Q7Q6I5 2,00E-16 ENSANGP00000004475 Anopheles gambiaePUC-118 Q4YVC1 2.9 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-12 Q8I2B2 4.9 Hypothetical protein PFA0140c Plasmodium falciparum HYPOTHETICAL PROTEINPUC-121 Q7QBM5 5,00E-16 ENSANGP00000020445 Anopheles gambiae GTP-BINDING; NUCLEOTIDE-BINDINGPUC-122 P27393 5,00E-40 Collagen alpha 2(IV) chain precursor Ascaris suum ALTERNATIVE SPLICING; BASEMENT MEMBRANE; COLPUC-123_1 Q7Q7W5 3,00E-10 ENSANGP00000011457 Anopheles gambiae OXYGEN TRANSPORTPUC-126 Q7RKI4 0.25 Arabidopsis thaliana T13D8 31 Plasmodium yoelii yoelii COMPLETE PROTEOMEPUC-127 Q8MYQ4 8,00E-78 Ubiquitin protein 1, isoform c Caenorhabditis elegans COMPLETE PROTEOMEPUC-128 Q7Q8H9 9,00E-15 ENSANGP00000013196 Anopheles gambiaePUC-129 Q4FKE1 3,00E-04 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-13 Q6D4W6 0.34 Ribulokinase (EC 2 7 1 16) Erwinia carotovora ARABINOSE CATABOLISM; CARBOHYDRATE METABOLPUC-130 Q6R5A9 9,00E-16 Tenebrin Tenebrio molitor PUC-131 Q7KN85 3,00E-28 LD21334p Drosophila melanogasterPUC-132 Q5MIR1 3,00E-08 Ribosomal protein L24 Aedes albopictus RIBOSOMAL PROTEINPUC-133 Q9C724 2.2 Hypothetical protein Arabidopsis thaliana HYPOTHETICAL PROTEINPUC-134_2 Q9V375 5,00E-05 CG11546-PA, isoform A Drosophila melanogasterPUC-136 Q5MIR6 9,00E-15 60S ribosomal protein L17 Aedes albopictus RIBONUCLEOPROTEIN; RIBOSOMAL PROTEINPUC-138 O76518 1.7 Hemicentin precursor (Hypothetical protein) Caenorhabditis elegans COMPLETE PROTEOME; HYPOTHETICAL PROTEIN; SIGPUC-139_1 Q7XJ31 0.46 Hypothetical protein Arabidopsis thaliana HYPOTHETICAL PROTEINPUC-139_2 Q24527 6,00E-09 CG9218-PA, isoform A (Cg9218-pe, isoform e Drosophila melanogasterPUC-14 Q9W3F6 0.006 CG1440-PA (LD46760p) Drosophila melanogasterPUC-140 Q9V6L6 3,00E-04 CG4630-PA (GH27944p) Drosophila melanogasterPUC-141 Q528F2 6.5 Hypothetical protein Magnaporthe grisea 70-15 HYPOTHETICAL PROTEINPUC-142 Q8HZZ5 1.2 Hypothetical protein Macaca fascicularis HYPOTHETICAL PROTEINPUC-146 Q17058 1.0 Alpha-glucosidase precursor Apis mellifera DIRECT PROTEIN SEQUENCING; GLYCOPROTEIN; GLYC
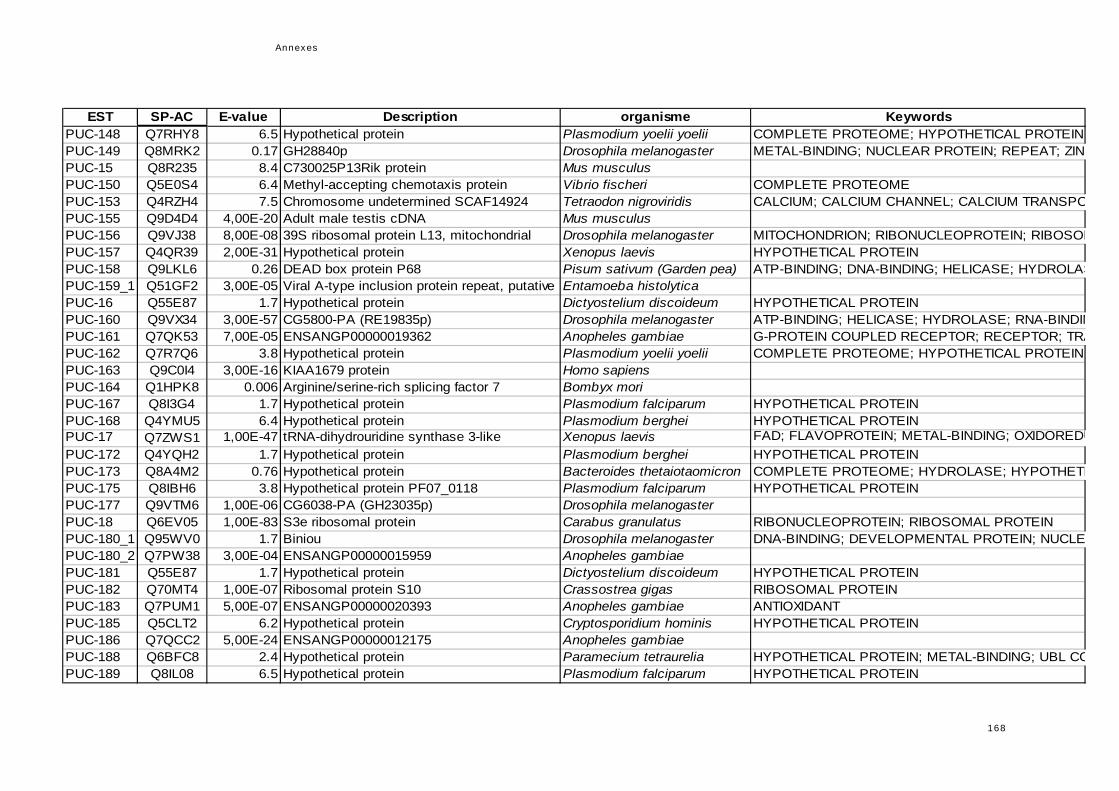
Annexes
168
AC E-value Description organisme KeywordsPUC-148 Q7RHY8 6.5 Hypothetical protein Plasmodium yoelii yoelii COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-149 Q8MRK2 0.17 GH28840p Drosophila melanogaster METAL-BINDING; NUCLEAR PROTEIN; REPEAT; ZIN
-
EST SP
CPUC-15 Q8R235 8.4 C730025P13Rik protein Mus musculusPUC-150 Q5E0S4 6.4 Methyl-accepting chemotaxis protein Vibrio fischeri COMPLETE PROTEOMEPUC-153 Q4RZH4 7.5 Chromosome undetermined SCAF14924 Tetraodon nigroviridis CALCIUM; CALCIUM CHANNEL; CALCIUM TRANSPOPUC-155 Q9D4D4 4,00E-20 Adult male testis cDNA Mus musculusPUC-156 Q9VJ38 8,00E-08 39S ribosomal protein L13, mitochondrial Drosophila melanogaster MITOCHONDRION; RIBONUCLEOPROTEIN; RIBOSOMPUC-157 Q4QR39 2,00E-31 Hypothetical protein Xenopus laevis HYPOTHETICAL PROTEINPUC-158 Q9LKL6 0.26 DEAD box protein P68 Pisum sativum (Garden pea) ATP-BINDING; DNA-BINDING; HELICASE; HYDROLASPUC-159_1 Q51GF2 3,00E-05 Viral A-type inclusion protein repeat, putative Entamoeba histolyticaPUC-16 Q55E87 1.7 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-160 Q9VX34 3,00E-57 CG5800-PA (RE19835p) Drosophila melanogaster ATP-BINDING; HELICASE; HYDROLASE; RNA-BINDINPUC-161 Q7QK53 7,00E-05 ENSANGP00000019362 Anopheles gambiae G-PROTEIN COUPLED RECEPTOR; RECEPTOR; TRAPUC-162 Q7R7Q6 3.8 Hypothetical protein Plasmodium yoelii yoelii COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-163 Q9C0I4 3,00E-16 KIAA1679 protein Homo sapiensPUC-164 Q1HPK8 0.006 Arginine/serine-rich splicing factor 7 Bombyx moriPUC-167 Q8I3G4 1.7 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-168 Q4YMU5 6.4 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-17 Q7ZWS1 1,00E-47 tRNA-dihydrouridine synthase 3-like Xenopus laevis FAD; FLAVOPROTEIN; METAL-BINDING; OXIDOREDUPUC-172 Q4YQH2 1.7 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-173 Q8A4M2 0.76 Hypothetical protein Bacteroides thetaiotaomicron COMPLETE PROTEOME; HYDROLASE; HYPOTHETIPUC-175 Q8IBH6 3.8 Hypothetical protein PF07_0118 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-177 Q9VTM6 1,00E-06 CG6038-PA (GH23035p) Drosophila melanogasterPUC-18 Q6EV05 1,00E-83 S3e ribosomal protein Carabus granulatus RIBONUCLEOPROTEIN; RIBOSOMAL PROTEINPUC-180_1 Q95WV0 1.7 Biniou Drosophila melanogaster DNA-BINDING; DEVELOPMENTAL PROTEIN; NUCLEAPUC-180_2 Q7PW38 3,00E-04 ENSANGP00000015959 Anopheles gambiaePUC-181 Q55E87 1.7 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-182 Q70MT4 1,00E-07 Ribosomal protein S10 Crassostrea gigas RIBOSOMAL PROTEINPUC-183 Q7PUM1 5,00E-07 ENSANGP00000020393 Anopheles gambiae ANTIOXIDANTPUC-185 Q5CLT2 6.2 Hypothetical protein Cryptosporidium hominis HYPOTHETICAL PROTEINPUC-186 Q7QCC2 5,00E-24 ENSANGP00000012175 Anopheles gambiaePUC-188 Q6BFC8 2.4 Hypothetical protein Paramecium tetraurelia HYPOTHETICAL PROTEIN; METAL-BINDING; UBL COPUC-189 Q8IL08 6.5 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN
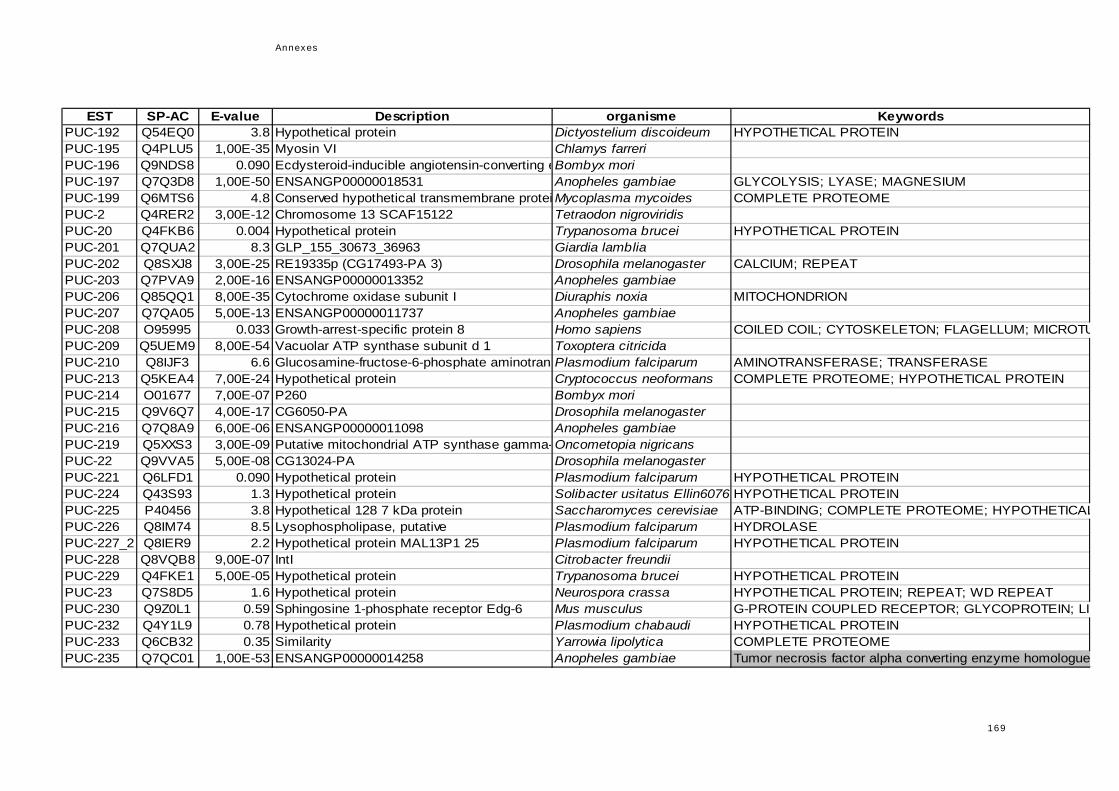
Annexes
169
EST SP-AC E-value Description organisme KeywordsPUC-192 Q54EQ0 3.8 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-195 Q4PLU5 1,00E-35 Myosin VI Chlamys farreri PUC-196 Q9NDS8 0.090 Ecdysteroid-inducible angiotensin-converting eBombyx mori PUC-197 Q7Q3D8 1,00E-50 ENSANGP00000018531 Anopheles gambiae GLYCOLYSIS; LYASE; MAGNESIUMPUC-199 Q6MTS6 4.8 Conserved hypothetical transmembrane protei Mycoplasma mycoides COMPLETE PROTEOMEPUC-2 Q4RER2 3,00E-12 Chromosome 13 SCAF15122 Tetraodon nigroviridisPUC-20 Q4FKB6 0.004 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-201 Q7QUA2 8.3 GLP_155_30673_36963 Giardia lamblia PUC-202 Q8SXJ8 3,00E-25 RE19335p (CG17493-PA 3) Drosophila melanogaster CALCIUM; REPEATPUC-203 Q7PVA9 2,00E-16 ENSANGP00000013352 Anopheles gambiaePUC-206 Q85QQ1 8,00E-35 Cytochrome oxidase subunit I Diuraphis noxia MITOCHONDRIONPUC-207 Q7QA05 5,00E-13 ENSANGP00000011737 Anopheles gambiaePUC-208 O95995 0.033 Growth-arrest-specific protein 8 Homo sapiens COILED COIL; CYTOSKELETON; FLAGELLUM; MICROTUPUC-209 Q5UEM9 8,00E-54 Vacuolar ATP synthase subunit d 1 Toxoptera citricida PUC-210 Q8IJF3 6.6 Glucosamine-fructose-6-phosphate aminotransPlasmodium falciparum AMINOTRANSFERASE; TRANSFERASEPUC-213 Q5KEA4 7,00E-24 Hypothetical protein Cryptococcus neoformans COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-214 O01677 7,00E-07 P260 Bombyx mori PUC-215 Q9V6Q7 4,00E-17 CG6050-PA Drosophila melanogasterPUC-216 Q7Q8A9 6,00E-06 ENSANGP00000011098 Anopheles gambiaePUC-219 Q5XXS3 3,00E-09 Putative mitochondrial ATP synthase gamma-Oncometopia nigricans PUC-22 Q9VVA5 5,00E-08 CG13024-PA Drosophila melanogasterPUC-221 Q6LFD1 0.090 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-224 Q43S93 1.3 Hypothetical protein Solibacter usitatus Ellin6076 HYPOTHETICAL PROTEINPUC-225 P40456 3.8 Hypothetical 128 7 kDa protein Saccharomyces cerevisiae ATP-BINDING; COMPLETE PROTEOME; HYPOTHETICALPUC-226 Q8IM74 8.5 Lysophospholipase, putative Plasmodium falciparum HYDROLASEPUC-227_2 Q8IER9 2.2 Hypothetical protein MAL13P1 25 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-228 Q8VQB8 9,00E-07 IntI Citrobacter freundiiPUC-229 Q4FKE1 5,00E-05 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-23 Q7S8D5 1.6 Hypothetical protein Neurospora crassa HYPOTHETICAL PROTEIN; REPEAT; WD REPEATPUC-230 Q9Z0L1 0.59 Sphingosine 1-phosphate receptor Edg-6 Mus musculus G-PROTEIN COUPLED RECEPTOR; GLYCOPROTEIN; LIPUC-232 Q4Y1L9 0.78 Hypothetical protein Plasmodium chabaudi HYPOTHETICAL PROTEINPUC-233 Q6CB32 0.35 Similarity Yarrowia lipolytica COMPLETE PROTEOMEPUC-235 Q7QC01 1,00E-53 ENSANGP00000014258 Anopheles gambiae Tumor necrosis factor alpha converting enzyme homologue
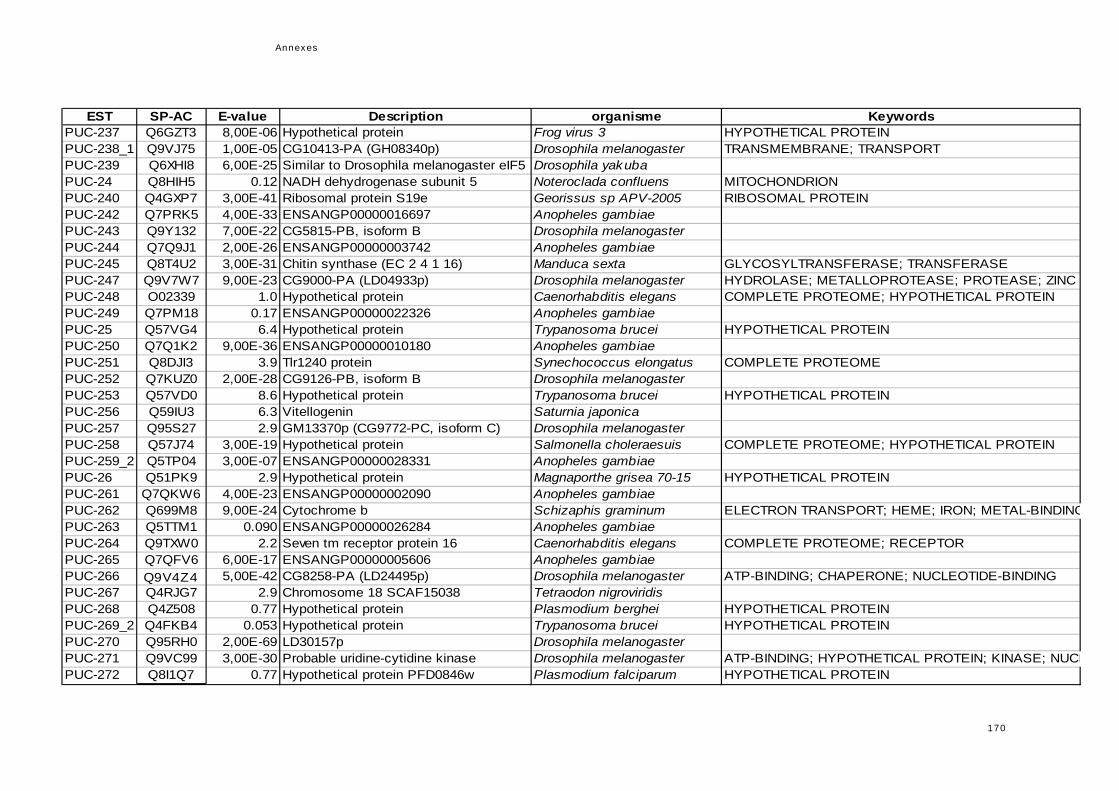
Annexes
170
AC E-value Description organisme KeywordsPUC-237 Q6GZT3 8,00E-06 Hypothetical protein Frog virus 3 HYPOTHETICAL PROTEINPUC-238_1 Q9VJ75 1,00E-05 CG10413-PA (GH08340p) Drosophila melanogaster TRANSMEMBRANE; TRANSPORTPUC-239 Q6XHI8 6,00E-25 Similar to Drosophila melanogaster eIF5 Drosophila yakubaPUC-24 Q8HIH5 0.12 NADH dehydrogenase subunit 5 Noteroclada confluens MITOCHONDRIONPUC-240 Q4GXP7 3,00E-41 Ribosomal protein S19e Georissus sp APV-2005 RIBOSOMAL PROTEINPUC-242 Q7PRK5 4,00E-33 ENSANGP00000016697 Anopheles gambiaePUC-243 Q9Y132 7,00E-22 CG5815-PB, isoform B Drosophila melanogasterPUC-244 Q7Q9J1 2,00E-26 ENSANGP00000003742 Anopheles gambiaePUC-245 Q8T4U2 3,00E-31 Chitin synthase (EC 2 4 1 16) Manduca sexta GLYCOSYLTRANSFERASE; TRANSFERASEPUC-247 Q9V7W7 9,00E-23 CG9000-PA (LD04933p) Drosophila melanogaster HYDROLASE; METALLOPROTEASE; PROTEASE; ZINCPUC-248 O02339 1.0 Hypothetical protein Caenorhabditis elegans COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-249 Q7PM18 0.17 ENSANGP00000022326 Anopheles gambiaePUC-25 Q57VG4 6.4 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-250 Q7Q1K2 9,00E-36 ENSANGP00000010180 Anopheles gambiaePUC-251 Q8DJI3 3.9 Tlr1240 protein Synechococcus elongatus COMPLETE PROTEOMEPUC-252 Q7KUZ0 2,00E-28 CG9126-PB, isoform B Drosophila melanogasterPUC-253 Q57VD0 8.6 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-256 Q59IU3 6.3 Vitellogenin Saturnia japonicaPUC-257 Q95S27 2.9 GM13370p (CG9772-PC, isoform C) Drosophila melanogasterPUC-258 Q57J74 3,00E-19 Hypothetical protein Salmonella choleraesuis COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-259_2 Q5TP04 3,00E-07 ENSANGP00000028331 Anopheles gambiaePUC-26 Q51PK9 2.9 Hypothetical protein Magnaporthe grisea 70-15 HYPOTHETICAL PROTEINPUC-261 Q7QKW6 4,00E-23 ENSANGP00000002090 Anopheles gambiaePUC-262 Q699M8 9,00E-24 Cytochrome b Schizaphis graminum ELECTRON TRANSPORT; HEME; IRON; METAL-BINDIN
-
EST SP
GPUC-263 Q5TTM1 0.090 ENSANGP00000026284 Anopheles gambiaePUC-264 Q9TXW0 2.2 Seven tm receptor protein 16 Caenorhabditis elegans COMPLETE PROTEOME; RECEPTORPUC-265 Q7QFV6 6,00E-17 ENSANGP00000005606 Anopheles gambiaePUC-266 Q9V4Z4 5,00E-42 CG8258-PA (LD24495p) Drosophila melanogaster ATP-BINDING; CHAPERONE; NUCLEOTIDE-BINDINGPUC-267 Q4RJG7 2.9 Chromosome 18 SCAF15038 Tetraodon nigroviridisPUC-268 Q4Z508 0.77 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-269_2 Q4FKB4 0.053 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-270 Q95RH0 2,00E-69 LD30157p Drosophila melanogasterPUC-271 Q9VC99 3,00E-30 Probable uridine-cytidine kinase Drosophila melanogaster ATP-BINDING; HYPOTHETICAL PROTEIN; KINASE; NUCLPUC-272 Q8I1Q7 0.77 Hypothetical protein PFD0846w Plasmodium falciparum HYPOTHETICAL PROTEIN
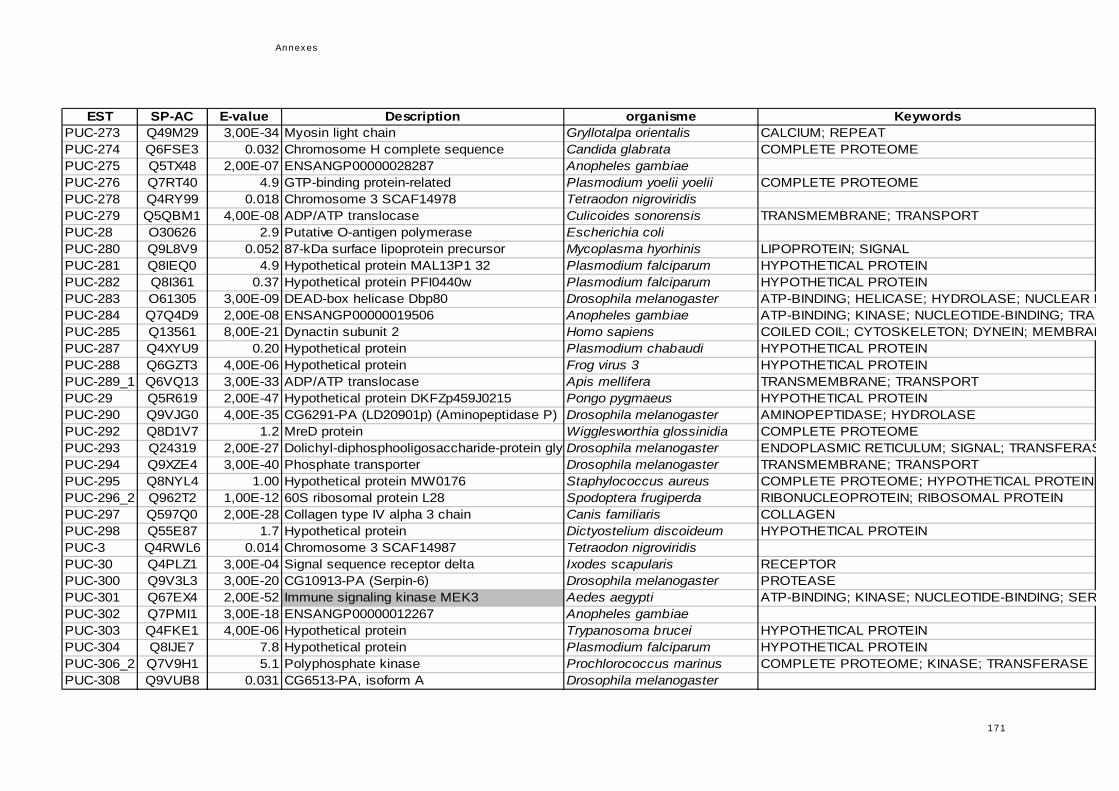
Annexes
171
EST SP-AC E-value Description organisme KeywordsPUC-273 Q49M29 3,00E-34 Myosin light chain Gryllotalpa orientalis CALCIUM; REPEATPUC-274 Q6FSE3 0.032 Chromosome H complete sequence Candida glabrata COMPLETE PROTEOMEPUC-275 Q5TX48 2,00E-07 ENSANGP00000028287 Anopheles gambiaePUC-276 Q7RT40 4.9 GTP-binding protein-related Plasmodium yoelii yoelii COMPLETE PROTEOMEPUC-278 Q4RY99 0.018 Chromosome 3 SCAF14978 Tetraodon nigroviridisPUC-279 Q5QBM1 4,00E-08 ADP/ATP translocase Culicoides sonorensis TRANSMEMBRANE; TRANSPORTPUC-28 O30626 2.9 Putative O-antigen polymerase Escherichia coliPUC-280 Q9L8V9 0.052 87-kDa surface lipoprotein precursor Mycoplasma hyorhinis LIPOPROTEIN; SIGNALPUC-281 Q8IEQ0 4.9 Hypothetical protein MAL13P1 32 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-282 Q8I361 0.37 Hypothetical protein PFI0440w Plasmodium falciparum HYPOTHETICAL PROTEINPUC-283 O61305 3,00E-09 DEAD-box helicase Dbp80 Drosophila melanogaster ATP-BINDING; HELICASE; HYDROLASE; NUCLEAR PPUC-284 Q7Q4D9 2,00E-08 ENSANGP00000019506 Anopheles gambiae ATP-BINDING; KINASE; NUCLEOTIDE-BINDING; TRAPUC-285 Q13561 8,00E-21 Dynactin subunit 2 Homo sapiens COILED COIL; CYTOSKELETON; DYNEIN; MEMBRANPUC-287 Q4XYU9 0.20 Hypothetical protein Plasmodium chabaudi HYPOTHETICAL PROTEINPUC-288 Q6GZT3 4,00E-06 Hypothetical protein Frog virus 3 HYPOTHETICAL PROTEINPUC-289_1 Q6VQ13 3,00E-33 ADP/ATP translocase Apis mellifera TRANSMEMBRANE; TRANSPORTPUC-29 Q5R619 2,00E-47 Hypothetical protein DKFZp459J0215 Pongo pygmaeus HYPOTHETICAL PROTEINPUC-290 Q9VJG0 4,00E-35 CG6291-PA (LD20901p) (Aminopeptidase P) Drosophila melanogaster AMINOPEPTIDASE; HYDROLASEPUC-292 Q8D1V7 1.2 MreD protein Wigglesworthia glossinidia COMPLETE PROTEOMEPUC-293 Q24319 2,00E-27 Dolichyl-diphosphooligosaccharide-protein gly Drosophila melanogaster ENDOPLASMIC RETICULUM; SIGNAL; TRANSFERASPUC-294 Q9XZE4 3,00E-40 Phosphate transporter Drosophila melanogaster TRANSMEMBRANE; TRANSPORTPUC-295 Q8NYL4 1.00 Hypothetical protein MW0176 Staphylococcus aureus COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-296_2 Q962T2 1,00E-12 60S ribosomal protein L28 Spodoptera frugiperda RIBONUCLEOPROTEIN; RIBOSOMAL PROTEINPUC-297 Q597Q0 2,00E-28 Collagen type IV alpha 3 chain Canis familiaris COLLAGENPUC-298 Q55E87 1.7 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-3 Q4RWL6 0.014 Chromosome 3 SCAF14987 Tetraodon nigroviridisPUC-30 Q4PLZ1 3,00E-04 Signal sequence receptor delta Ixodes scapularis RECEPTORPUC-300 Q9V3L3 3,00E-20 CG10913-PA (Serpin-6) Drosophila melanogaster PROTEASEPUC-301 Q67EX4 2,00E-52 Immune signaling kinase MEK3 Aedes aegypti ATP-BINDING; KINASE; NUCLEOTIDE-BINDING; SERPUC-302 Q7PMI1 3,00E-18 ENSANGP00000012267 Anopheles gambiaePUC-303 Q4FKE1 4,00E-06 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-304 Q8IJE7 7.8 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-306_2 Q7V9H1 5.1 Polyphosphate kinase Prochlorococcus marinus COMPLETE PROTEOME; KINASE; TRANSFERASEPUC-308 Q9VUB8 0.031 CG6513-PA, isoform A Drosophila melanogaster
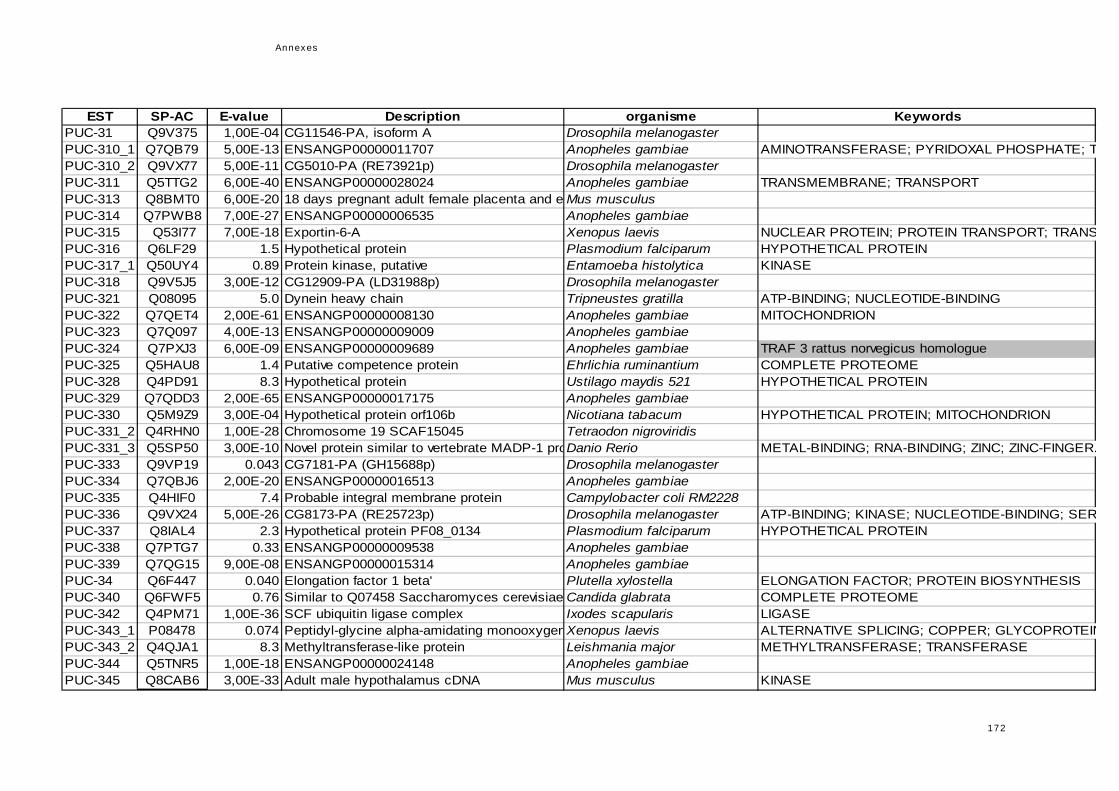
Annexes
172
EST SP-AC E-value Description organisme KeywordsPUC-31 Q9V375 1,00E-04 CG11546-PA, isoform A Drosophila melanogasterPUC-310_1 Q7QB79 5,00E-13 ENSANGP00000011707 Anopheles gambiae AMINOTRANSFERASE; PYRIDOXAL PHOSPHATE; TPUC-310_2 Q9VX77 5,00E-11 CG5010-PA (RE73921p) Drosophila melanogasterPUC-311 Q5TTG2 6,00E-40 ENSANGP00000028024 Anopheles gambiae TRANSMEMBRANE; TRANSPORTPUC-313 Q8BMT0 6,00E-20 18 days pregnant adult female placenta and e Mus musculusPUC-314 Q7PWB8 7,00E-27 ENSANGP00000006535 Anopheles gambiaePUC-315 Q53I77 7,00E-18 Exportin-6-A Xenopus laevis NUCLEAR PROTEIN; PROTEIN TRANSPORT; TRANSPUC-316 Q6LF29 1.5 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-317_1 Q50UY4 0.89 Protein kinase, putative Entamoeba histolytica KINASEPUC-318 Q9V5J5 3,00E-12 CG12909-PA (LD31988p) Drosophila melanogasterPUC-321 Q08095 5.0 Dynein heavy chain Tripneustes gratilla ATP-BINDING; NUCLEOTIDE-BINDINGPUC-322 Q7QET4 2,00E-61 ENSANGP00000008130 Anopheles gambiae MITOCHONDRIONPUC-323 Q7Q097 4,00E-13 ENSANGP00000009009 Anopheles gambiaePUC-324 Q7PXJ3 6,00E-09 ENSANGP00000009689 Anopheles gambiae TRAF 3 rattus norvegicus homologuePUC-325 Q5HAU8 1.4 Putative competence protein Ehrlichia ruminantium COMPLETE PROTEOMEPUC-328 Q4PD91 8.3 Hypothetical protein Ustilago maydis 521 HYPOTHETICAL PROTEINPUC-329 Q7QDD3 2,00E-65 ENSANGP00000017175 Anopheles gambiaePUC-330 Q5M9Z9 3,00E-04 Hypothetical protein orf106b Nicotiana tabacum HYPOTHETICAL PROTEIN; MITOCHONDRIONPUC-331_2 Q4RHN0 1,00E-28 Chromosome 19 SCAF15045 Tetraodon nigroviridisPUC-331_3 Q5SP50 3,00E-10 Novel protein similar to vertebrate MADP-1 proDanio Rerio METAL-BINDING; RNA-BINDING; ZINC; ZINC-FINGER.PUC-333 Q9VP19 0.043 CG7181-PA (GH15688p) Drosophila melanogasterPUC-334 Q7QBJ6 2,00E-20 ENSANGP00000016513 Anopheles gambiaePUC-335 Q4HIF0 7.4 Probable integral membrane protein Campylobacter coli RM2228PUC-336 Q9VX24 5,00E-26 CG8173-PA (RE25723p) Drosophila melanogaster ATP-BINDING; KINASE; NUCLEOTIDE-BINDING; SERPUC-337 Q8IAL4 2.3 Hypothetical protein PF08_0134 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-338 Q7PTG7 0.33 ENSANGP00000009538 Anopheles gambiaePUC-339 Q7QG15 9,00E-08 ENSANGP00000015314 Anopheles gambiaePUC-34 Q6F447 0.040 Elongation factor 1 beta' Plutella xylostella ELONGATION FACTOR; PROTEIN BIOSYNTHESISPUC-340 Q6FWF5 0.76 Similar to Q07458 Saccharomyces cerevisiae Candida glabrata COMPLETE PROTEOMEPUC-342 Q4PM71 1,00E-36 SCF ubiquitin ligase complex Ixodes scapularis LIGASEPUC-343_1 P08478 0.074 Peptidyl-glycine alpha-amidating monooxygenXenopus laevis ALTERNATIVE SPLICING; COPPER; GLYCOPROTEINPUC-343_2 Q4QJA1 8.3 Methyltransferase-like protein Leishmania major METHYLTRANSFERASE; TRANSFERASEPUC-344 Q5TNR5 1,00E-18 ENSANGP00000024148 Anopheles gambiaePUC-345 Q8CAB6 3,00E-33 Adult male hypothalamus cDNA Mus musculus KINASE
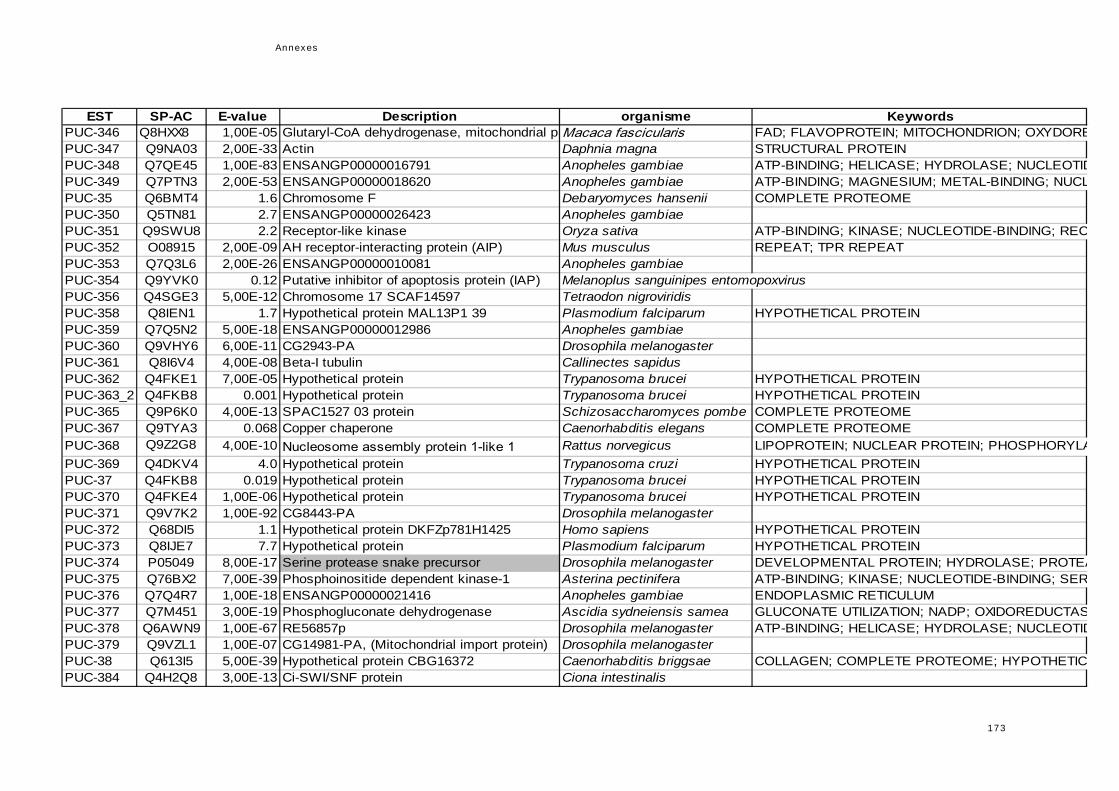
Annexes
173
EST SP AC E-value Description organisme KeywordsPUC-346 Q8HXX8 1,00E-05 Glutaryl-CoA dehydrogenase, mitochondrial p
-rMacaca fascicularis FAD; FLAVOPROTEIN; MITOCHONDRION; OXYDORE
PUC-347 Q9NA03 2,00E-33 Actin Daphnia magna STRUCTURAL PROTEINPUC-348 Q7QE45 1,00E-83 ENSANGP00000016791 Anopheles gambiae ATP-BINDING; HELICASE; HYDROLASE; NUCLEOTIDPUC-349 Q7PTN3 2,00E-53 ENSANGP00000018620 Anopheles gambiae ATP-BINDING; MAGNESIUM; METAL-BINDING; NUCLPUC-35 Q6BMT4 1.6 Chromosome F Debaryomyces hansenii COMPLETE PROTEOMEPUC-350 Q5TN81 2.7 ENSANGP00000026423 Anopheles gambiaePUC-351 Q9SWU8 2.2 Receptor-like kinase Oryza sativa ATP-BINDING; KINASE; NUCLEOTIDE-BINDING; RECPUC-352 O08915 2,00E-09 AH receptor-interacting protein (AIP) Mus musculus REPEAT; TPR REPEATPUC-353 Q7Q3L6 2,00E-26 ENSANGP00000010081 Anopheles gambiaePUC-354 Q9YVK0 0.12 Putative inhibitor of apoptosis protein (IAP) Melanoplus sanguinipes entomopoxvirusPUC-356 Q4SGE3 5,00E-12 Chromosome 17 SCAF14597 Tetraodon nigroviridisPUC-358 Q8IEN1 1.7 Hypothetical protein MAL13P1 39 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-359 Q7Q5N2 5,00E-18 ENSANGP00000012986 Anopheles gambiaePUC-360 Q9VHY6 6,00E-11 CG2943-PA Drosophila melanogasterPUC-361 Q8I6V4 4,00E-08 Beta-I tubulin Callinectes sapidus PUC-362 Q4FKE1 7,00E-05 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-363_2 Q4FKB8 0.001 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-365 Q9P6K0 4,00E-13 SPAC1527 03 protein Schizosaccharomyces pombe COMPLETE PROTEOMEPUC-367 Q9TYA3 0.068 Copper chaperone Caenorhabditis elegans COMPLETE PROTEOMEPUC-368 Q9Z2G8 4,00E-10 Nucleosome assembly protein 1-like 1 Rattus norvegicus LIPOPROTEIN; NUCLEAR PROTEIN; PHOSPHORYLAPUC-369 Q4DKV4 4.0 Hypothetical protein Trypanosoma cruzi HYPOTHETICAL PROTEINPUC-37 Q4FKB8 0.019 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-370 Q4FKE4 1,00E-06 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-371 Q9V7K2 1,00E-92 CG8443-PA Drosophila melanogasterPUC-372 Q68DI5 1.1 Hypothetical protein DKFZp781H1425 Homo sapiens HYPOTHETICAL PROTEINPUC-373 Q8IJE7 7.7 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-374 P05049 8,00E-17 Serine protease snake precursor Drosophila melanogaster DEVELOPMENTAL PROTEIN; HYDROLASE; PROTEAPUC-375 Q76BX2 7,00E-39 Phosphoinositide dependent kinase-1 Asterina pectinifera ATP-BINDING; KINASE; NUCLEOTIDE-BINDING; SERPUC-376 Q7Q4R7 1,00E-18 ENSANGP00000021416 Anopheles gambiae ENDOPLASMIC RETICULUMPUC-377 Q7M451 3,00E-19 Phosphogluconate dehydrogenase Ascidia sydneiensis samea GLUCONATE UTILIZATION; NADP; OXIDOREDUCTASPUC-378 Q6AWN9 1,00E-67 RE56857p Drosophila melanogaster ATP-BINDING; HELICASE; HYDROLASE; NUCLEOTIDPUC-379 Q9VZL1 1,00E-07 CG14981-PA, (Mitochondrial import protein) Drosophila melanogasterPUC-38 Q613I5 5,00E-39 Hypothetical protein CBG16372 Caenorhabditis briggsae COLLAGEN; COMPLETE PROTEOME; HYPOTHETICAPUC-384 Q4H2Q8 3,00E-13 Ci-SWI/SNF protein Ciona intestinalis
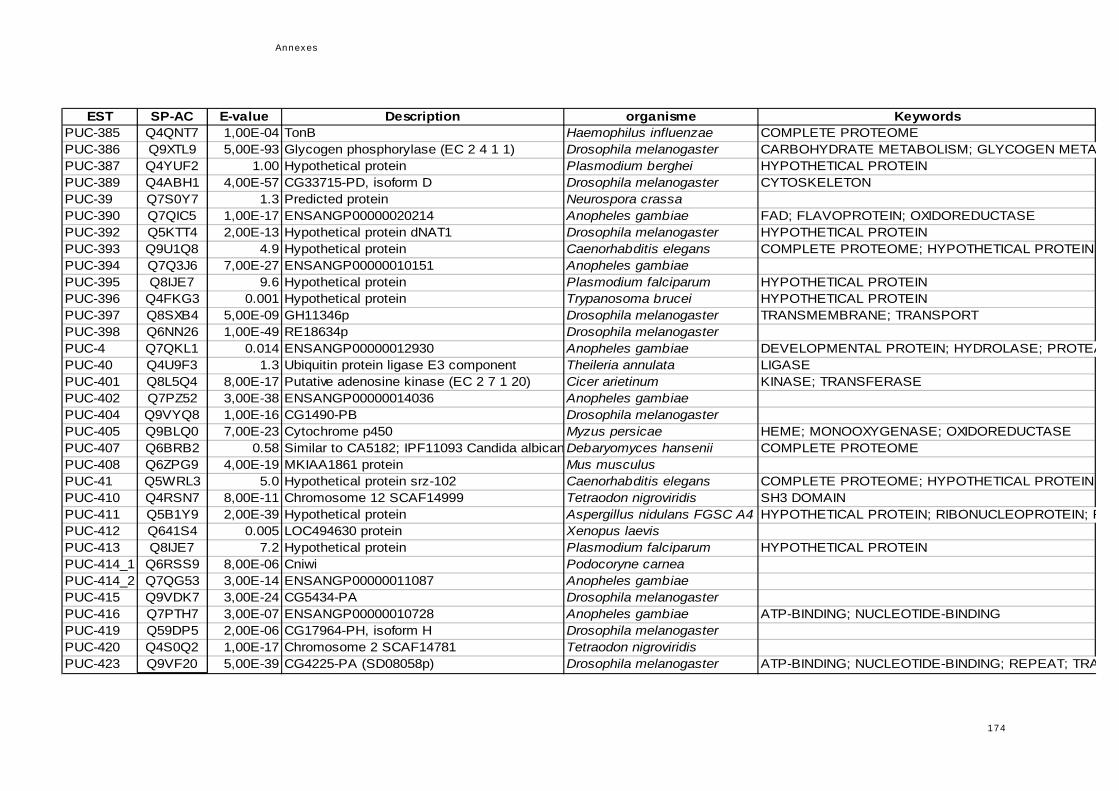
Annexes
174
AC E-value Description organisme KeywordsPUC-385 Q4QNT7 1,00E-04 TonB Haemophilus influenzae COMPLETE PROTEOMEPUC-386 Q9XTL9 5,00E-93 Glycogen phosphorylase (EC 2 4 1 1) Drosophila melanogaster CARBOHYDRATE METABOLISM; GLYCOGEN MET
-
EST SP
APUC-387 Q4YUF2 1.00 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-389 Q4ABH1 4,00E-57 CG33715-PD, isoform D Drosophila melanogaster CYTOSKELETONPUC-39 Q7S0Y7 1.3 Predicted protein Neurospora crassaPUC-390 Q7QIC5 1,00E-17 ENSANGP00000020214 Anopheles gambiae FAD; FLAVOPROTEIN; OXIDOREDUCTASEPUC-392 Q5KTT4 2,00E-13 Hypothetical protein dNAT1 Drosophila melanogaster HYPOTHETICAL PROTEINPUC-393 Q9U1Q8 4.9 Hypothetical protein Caenorhabditis elegans COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-394 Q7Q3J6 7,00E-27 ENSANGP00000010151 Anopheles gambiaePUC-395 Q8IJE7 9.6 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-396 Q4FKG3 0.001 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-397 Q8SXB4 5,00E-09 GH11346p Drosophila melanogaster TRANSMEMBRANE; TRANSPORTPUC-398 Q6NN26 1,00E-49 RE18634p Drosophila melanogasterPUC-4 Q7QKL1 0.014 ENSANGP00000012930 Anopheles gambiae DEVELOPMENTAL PROTEIN; HYDROLASE; PROTEAPUC-40 Q4U9F3 1.3 Ubiquitin protein ligase E3 component Theileria annulata LIGASEPUC-401 Q8L5Q4 8,00E-17 Putative adenosine kinase (EC 2 7 1 20) Cicer arietinum KINASE; TRANSFERASEPUC-402 Q7PZ52 3,00E-38 ENSANGP00000014036 Anopheles gambiaePUC-404 Q9VYQ8 1,00E-16 CG1490-PB Drosophila melanogasterPUC-405 Q9BLQ0 7,00E-23 Cytochrome p450 Myzus persicae HEME; MONOOXYGENASE; OXIDOREDUCTASEPUC-407 Q6BRB2 0.58 Similar to CA5182; IPF11093 Candida albicanDebaryomyces hansenii COMPLETE PROTEOMEPUC-408 Q6ZPG9 4,00E-19 MKIAA1861 protein Mus musculusPUC-41 Q5WRL3 5.0 Hypothetical protein srz-102 Caenorhabditis elegans COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-410 Q4RSN7 8,00E-11 Chromosome 12 SCAF14999 Tetraodon nigroviridis SH3 DOMAINPUC-411 Q5B1Y9 2,00E-39 Hypothetical protein Aspergillus nidulans FGSC A4 HYPOTHETICAL PROTEIN; RIBONUCLEOPROTEIN; RPUC-412 Q641S4 0.005 LOC494630 protein Xenopus laevisPUC-413 Q8IJE7 7.2 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-414_1 Q6RSS9 8,00E-06 Cniwi Podocoryne carneaPUC-414_2 Q7QG53 3,00E-14 ENSANGP00000011087 Anopheles gambiaePUC-415 Q9VDK7 3,00E-24 CG5434-PA Drosophila melanogasterPUC-416 Q7PTH7 3,00E-07 ENSANGP00000010728 Anopheles gambiae ATP-BINDING; NUCLEOTIDE-BINDINGPUC-419 Q59DP5 2,00E-06 CG17964-PH, isoform H Drosophila melanogasterPUC-420 Q4S0Q2 1,00E-17 Chromosome 2 SCAF14781 Tetraodon nigroviridisPUC-423 Q9VF20 5,00E-39 CG4225-PA (SD08058p) Drosophila melanogaster ATP-BINDING; NUCLEOTIDE-BINDING; REPEAT; TRA
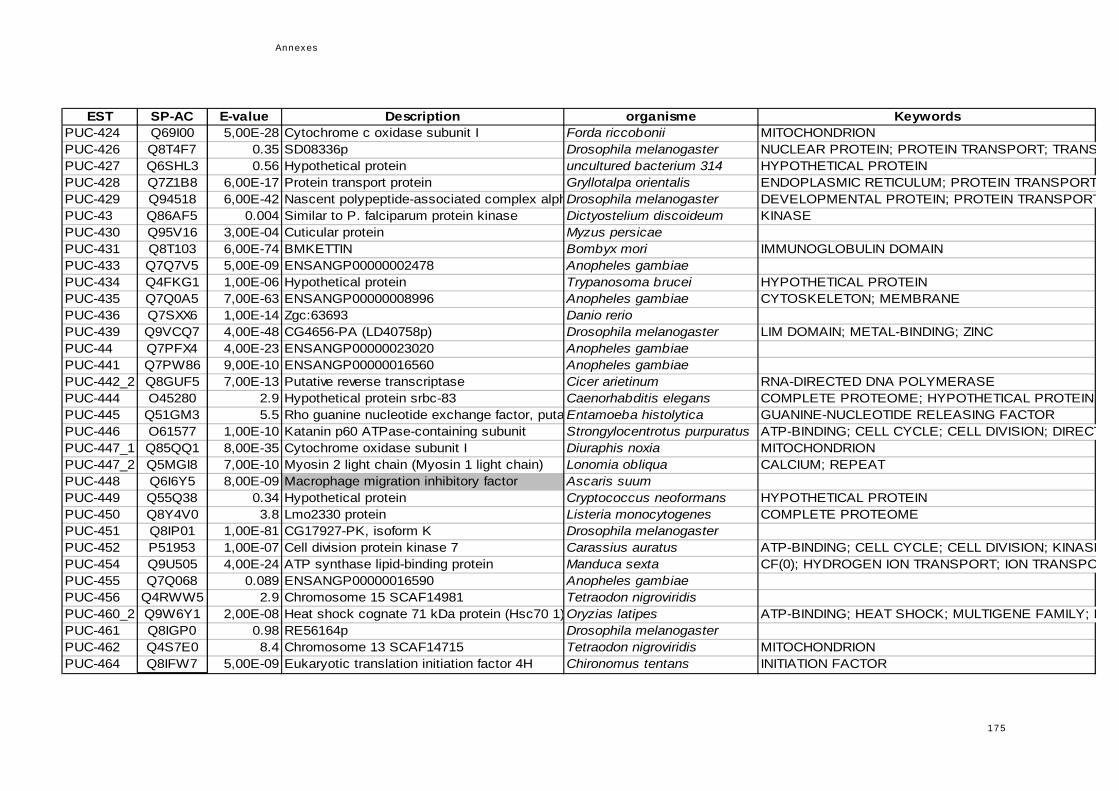
Annexes
175
EST SP-AC E-value Description organisme KeywordsPUC-424 Q69I00 5,00E-28 Cytochrome c oxidase subunit I Forda riccobonii MITOCHONDRIONPUC-426 Q8T4F7 0.35 SD08336p Drosophila melanogaster NUCLEAR PROTEIN; PROTEIN TRANSPORT; TRANSPUC-427 Q6SHL3 0.56 Hypothetical protein uncultured bacterium 314 HYPOTHETICAL PROTEINPUC-428 Q7Z1B8 6,00E-17 Protein transport protein Gryllotalpa orientalis ENDOPLASMIC RETICULUM; PROTEIN TRANSPORTPUC-429 Q94518 6,00E-42 Nascent polypeptide-associated complex alphDrosophila melanogaster DEVELOPMENTAL PROTEIN; PROTEIN TRANSPORTPUC-43 Q86AF5 0.004 Similar to P. falciparum protein kinase Dictyostelium discoideum KINASEPUC-430 Q95V16 3,00E-04 Cuticular protein Myzus persicae PUC-431 Q8T103 6,00E-74 BMKETTIN Bombyx mori IMMUNOGLOBULIN DOMAINPUC-433 Q7Q7V5 5,00E-09 ENSANGP00000002478 Anopheles gambiaePUC-434 Q4FKG1 1,00E-06 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-435 Q7Q0A5 7,00E-63 ENSANGP00000008996 Anopheles gambiae CYTOSKELETON; MEMBRANEPUC-436 Q7SXX6 1,00E-14 Zgc:63693 Danio rerio PUC-439 Q9VCQ7 4,00E-48 CG4656-PA (LD40758p) Drosophila melanogaster LIM DOMAIN; METAL-BINDING; ZINCPUC-44 Q7PFX4 4,00E-23 ENSANGP00000023020 Anopheles gambiaePUC-441 Q7PW86 9,00E-10 ENSANGP00000016560 Anopheles gambiaePUC-442_2 Q8GUF5 7,00E-13 Putative reverse transcriptase Cicer arietinum RNA-DIRECTED DNA POLYMERASEPUC-444 O45280 2.9 Hypothetical protein srbc-83 Caenorhabditis elegans COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-445 Q51GM3 5.5 Rho guanine nucleotide exchange factor, putaEntamoeba histolytica GUANINE-NUCLEOTIDE RELEASING FACTORPUC-446 O61577 1,00E-10 Katanin p60 ATPase-containing subunit Strongylocentrotus purpuratus ATP-BINDING; CELL CYCLE; CELL DIVISION; DIRECTPUC-447_1 Q85QQ1 8,00E-35 Cytochrome oxidase subunit I Diuraphis noxia MITOCHONDRIONPUC-447_2 Q5MGI8 7,00E-10 Myosin 2 light chain (Myosin 1 light chain) Lonomia obliqua CALCIUM; REPEATPUC-448 Q6I6Y5 8,00E-09 Macrophage migration inhibitory factor Ascaris suum PUC-449 Q55Q38 0.34 Hypothetical protein Cryptococcus neoformans HYPOTHETICAL PROTEINPUC-450 Q8Y4V0 3.8 Lmo2330 protein Listeria monocytogenes COMPLETE PROTEOMEPUC-451 Q8IP01 1,00E-81 CG17927-PK, isoform K Drosophila melanogasterPUC-452 P51953 1,00E-07 Cell division protein kinase 7 Carassius auratus ATP-BINDING; CELL CYCLE; CELL DIVISION; KINASEPUC-454 Q9U505 4,00E-24 ATP synthase lipid-binding protein Manduca sexta CF(0); HYDROGEN ION TRANSPORT; ION TRANSPOPUC-455 Q7Q068 0.089 ENSANGP00000016590 Anopheles gambiaePUC-456 Q4RWW5 2.9 Chromosome 15 SCAF14981 Tetraodon nigroviridisPUC-460_2 Q9W6Y1 2,00E-08 Heat shock cognate 71 kDa protein (Hsc70 1)Oryzias latipes ATP-BINDING; HEAT SHOCK; MULTIGENE FAMILY; NPUC-461 Q8IGP0 0.98 RE56164p Drosophila melanogasterPUC-462 Q4S7E0 8.4 Chromosome 13 SCAF14715 Tetraodon nigroviridis MITOCHONDRIONPUC-464 Q8IFW7 5,00E-09 Eukaryotic translation initiation factor 4H Chironomus tentans INITIATION FACTOR
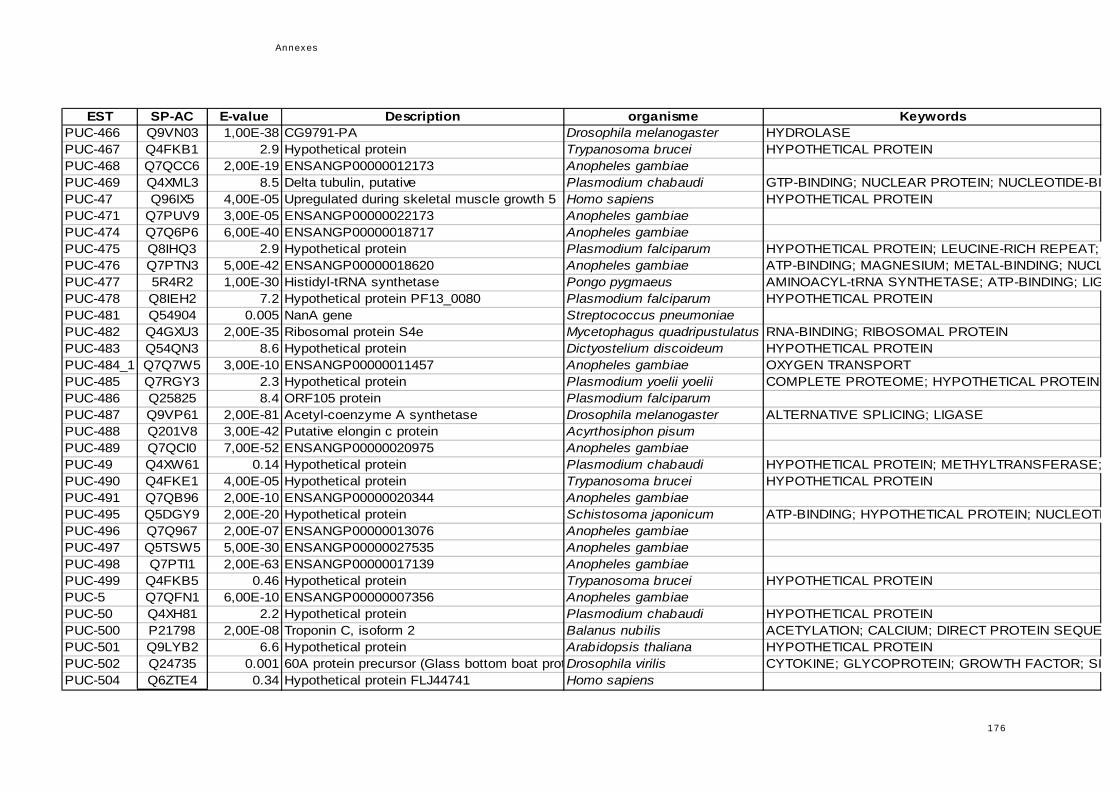
Annexes
176
AC E-value Description organisme KeywordsPUC-466 Q9VN03 1,00E-38 CG9791-PA Drosophila melanogaster HYDROLASEPUC-467 Q4FKB1 2.9 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-468 Q7QCC6 2,00E-19 ENSANGP00000012173 Anopheles gambiaePUC-469 Q4XML3 8.5 Delta tubulin, putative Plasmodium chabaudi GTP-BINDING; NUCLEAR PROTEIN; NUCLEOTIDE-BIPUC-47 Q96IX5 4,00E-05 Upregulated during skeletal muscle growth 5 Homo sapiens HYPOTHETICAL PROTEINPUC-471 Q7PUV9 3,00E-05 ENSANGP00000022173 Anopheles gambiaePUC-474 Q7Q6P6 6,00E-40 ENSANGP00000018717 Anopheles gambiaePUC-475 Q8IHQ3 2.9 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEIN; LEUCINE-RICH REPEAT; PUC-476 Q7PTN3 5,00E-42 ENSANGP00000018620 Anopheles gambiae
-
EST SP
ATP-BINDING; MAGNESIUM; METAL-BINDING; NUCLPUC-477 5R4R2 1,00E-30 Histidyl-tRNA synthetase Pongo pygmaeus AMINOACYL-tRNA SYNTHETASE; ATP-BINDING; LIGPUC-478 Q8IEH2 7.2 Hypothetical protein PF13_0080 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-481 Q54904 0.005 NanA gene Streptococcus pneumoniae PUC-482 Q4GXU3 2,00E-35 Ribosomal protein S4e Mycetophagus quadripustulatus RNA-BINDING; RIBOSOMAL PROTEINPUC-483 Q54QN3 8.6 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-484_1 Q7Q7W5 3,00E-10 ENSANGP00000011457 Anopheles gambiae OXYGEN TRANSPORTPUC-485 Q7RGY3 2.3 Hypothetical protein Plasmodium yoelii yoelii COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-486 Q25825 8.4 ORF105 protein Plasmodium falciparumPUC-487 Q9VP61 2,00E-81 Acetyl-coenzyme A synthetase Drosophila melanogaster ALTERNATIVE SPLICING; LIGASEPUC-488 Q201V8 3,00E-42 Putative elongin c protein Acyrthosiphon pisum PUC-489 Q7QCI0 7,00E-52 ENSANGP00000020975 Anopheles gambiaePUC-49 Q4XW61 0.14 Hypothetical protein Plasmodium chabaudi HYPOTHETICAL PROTEIN; METHYLTRANSFERASE;PUC-490 Q4FKE1 4,00E-05 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-491 Q7QB96 2,00E-10 ENSANGP00000020344 Anopheles gambiaePUC-495 Q5DGY9 2,00E-20 Hypothetical protein Schistosoma japonicum ATP-BINDING; HYPOTHETICAL PROTEIN; NUCLEOTIPUC-496 Q7Q967 2,00E-07 ENSANGP00000013076 Anopheles gambiaePUC-497 Q5TSW5 5,00E-30 ENSANGP00000027535 Anopheles gambiaePUC-498 Q7PTI1 2,00E-63 ENSANGP00000017139 Anopheles gambiaePUC-499 Q4FKB5 0.46 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-5 Q7QFN1 6,00E-10 ENSANGP00000007356 Anopheles gambiaePUC-50 Q4XH81 2.2 Hypothetical protein Plasmodium chabaudi HYPOTHETICAL PROTEINPUC-500 P21798 2,00E-08 Troponin C, isoform 2 Balanus nubilis ACETYLATION; CALCIUM; DIRECT PROTEIN SEQUEPUC-501 Q9LYB2 6.6 Hypothetical protein Arabidopsis thaliana HYPOTHETICAL PROTEINPUC-502 Q24735 0.001 60A protein precursor (Glass bottom boat protDrosophila virilis CYTOKINE; GLYCOPROTEIN; GROWTH FACTOR; SIPUC-504 Q6ZTE4 0.34 Hypothetical protein FLJ44741 Homo sapiens
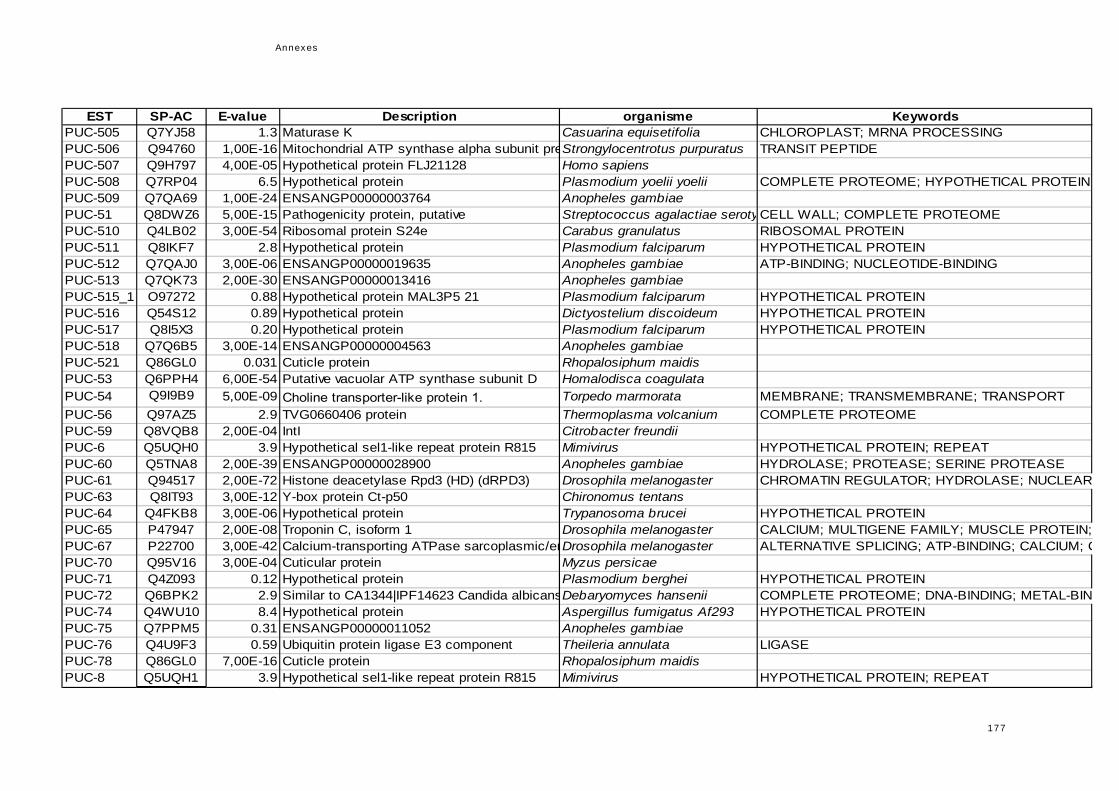
Annexes
177
EST SP AC E-value Description organisme KeywordsPUC-505 Q7YJ58 1.3 Maturase K Casuarina equisetifolia CHLOROPLAST; MRNA PROCESSINGPUC-506 Q94760 1,00E-16 Mitochondrial ATP synthase alpha subunit pr
-
eStrongylocentrotus purpuratus TRANSIT PEPTIDEPUC-507 Q9H797 4,00E-05 Hypothetical protein FLJ21128 Homo sapiens PUC-508 Q7RP04 6.5 Hypothetical protein Plasmodium yoelii yoelii COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-509 Q7QA69 1,00E-24 ENSANGP00000003764 Anopheles gambiaePUC-51 Q8DWZ6 5,00E-15 Pathogenicity protein, putative Streptococcus agalactiae serotyCELL WALL; COMPLETE PROTEOMEPUC-510 Q4LB02 3,00E-54 Ribosomal protein S24e Carabus granulatus RIBOSOMAL PROTEINPUC-511 Q8IKF7 2.8 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-512 Q7QAJ0 3,00E-06 ENSANGP00000019635 Anopheles gambiae ATP-BINDING; NUCLEOTIDE-BINDINGPUC-513 Q7QK73 2,00E-30 ENSANGP00000013416 Anopheles gambiaePUC-515_1 O97272 0.88 Hypothetical protein MAL3P5 21 Plasmodium falciparum HYPOTHETICAL PROTEINPUC-516 Q54S12 0.89 Hypothetical protein Dictyostelium discoideum HYPOTHETICAL PROTEINPUC-517 Q8I5X3 0.20 Hypothetical protein Plasmodium falciparum HYPOTHETICAL PROTEINPUC-518 Q7Q6B5 3,00E-14 ENSANGP00000004563 Anopheles gambiaePUC-521 Q86GL0 0.031 Cuticle protein Rhopalosiphum maidis PUC-53 Q6PPH4 6,00E-54 Putative vacuolar ATP synthase subunit D Homalodisca coagulata PUC-54 Q9I9B9 5,00E-09 Choline transporter-like protein 1. Torpedo marmorata MEMBRANE; TRANSMEMBRANE; TRANSPORTPUC-56 Q9 Z5 2.9 TVG0660406 protein Thermoplasma volcanium COMPLETE PROTEOMEPUC-59 Q8VQB8 2,00E-04 IntI Citrobacter freundiiPUC-6 Q5UQH0 3.9 Hypothetical sel1-like repeat protein R815 Mimivirus HYPOTHETICAL PROTEIN; REPEATPUC-60 Q5TNA8 2,00E-39 ENSANGP00000028900 Anopheles gambiae HYDROLASE; PROTEASE; SERINE PROTEASEPUC-61 Q94517 2,00E-72 Histone deacetylase Rpd3 (HD) (dRPD3) Drosophila melanogaster CHROMATIN REGULATOR; HYDROLASE; NUCLEA
7A
R PUC-63 Q8IT93 3,00E-12 Y-box protein Ct-p50 Chironomus tentans PUC-64 Q4FKB8 3,00E-06 Hypothetical protein Trypanosoma brucei HYPOTHETICAL PROTEINPUC-65 P47947 2,00E-08 Troponin C, isoform 1 Drosophila melanogaster CALCIUM; MULTIGENE FAMILY; MUSCLE PROTEIN; PUC-67 P22700 3,00E-42 Calcium-transporting ATPase sarcoplasmic/enDrosophila melanogaster ALTERNATIVE SPLICING; ATP-BINDING; CALCIUM; CPUC-70 Q95V16 3,00E-04 Cuticular protein Myzus persicae PUC-71 Q4Z093 0.12 Hypothetical protein Plasmodium berghei HYPOTHETICAL PROTEINPUC-72 Q6BPK2 2.9 Similar to CA1344|IPF14623 Candida albicansDebaryomyces hansenii COMPLETE PROTEOME; DNA-BINDING; METAL-BINPUC-74 Q4WU10 8.4 Hypothetical protein Aspergillus fumigatus Af293 HYPOTHETICAL PROTEINPUC-75 Q7PPM5 0.31 ENSANGP00000011052 Anopheles gambiaePUC-76 Q4U9F3 0.59 Ubiquitin protein ligase E3 component Theileria annulata LIGASEPUC-78 Q86GL0 7,00E-16 Cuticle protein Rhopalosiphum maidis PUC-8 Q5UQH1 3.9 Hypothetical sel1-like repeat protein R815 Mimivirus HYPOTHETICAL PROTEIN; REPEAT
Annexes
178
EST SP AC E-value Description organisme KeywordsPUC-80 Q95V16 3,00E-09 Cuticular protein Myzus persicae PUC-81 Q7P2A6 2.9 Ser/Thr and Tyr protein phosphatase Fusobacterium nucleatumPUC-82 Q9VZ57 2,00E-05 CG1637-PA, isoform A Drosophila melanogasterPUC-86 Q7QDW9 4,00E-06 ENSANGP00000013749 Anopheles gambiaePUC-87 Q6YPV6 1.1 Type I restriction-modification system methylt Onion yellows phytoplasma COMPLETE PROTEOMEPUC-88 Q9VYM3 2.5 CG15730-PA Drosophila melanogasterPUC-89 Q6BGY0 6.5 Similar to CA3283|IPF6274 Candida albicans Debaryomyces hansenii COMPLETE PROTEOMEPUC-9 Q7PHW7 1,00E-06 ENSANGP00000025159 Anopheles gambiae TRANSMEMBRANE; TRANSPORTPUC-90 Q7Q5L5 5,00E-06 ENSANGP00000013033 Anopheles gambiaePUC-91 Q7PTH7 3,00E-07 ENSANGP00000010728 Anopheles gambiae ATP-BINDING; NUCLEOTIDE-BINDINGPUC-92 Q6PPH4 5,00E-39 Putative vacuolar ATP synthase subunit D Homalodisca coagulata PUC-93 Q7PHW7 1,00E-06 ENSANGP00000025159 Anopheles gambiae TRANSMEMBRANE; TRANSPORTPUC-94 Q8IJF3 6.6 Glucosamine-fructose-6-phosphate aminotran
-
sPlasmodium falciparum AMINOTRANSFERASE; TRANSFERASEPUC-95 Q57J74 3,00E-19 Hypothetical protein Salmonella choleraesuis COMPLETE PROTEOME; HYPOTHETICAL PROTEINPUC-96_2 Q4H450 6,00E-09 Ribosomal protein S18 Crassostrea gigas RIBONUCLEOPROTEIN; RIBOSOMAL PROTEINPUC-97 Q8IPZ7 0.018 CG31950-PA Drosophila melanogasterPUC-98 Q7RHV7 0.16 TAP1 protein Plasmodium yoelii yoelii COLLAGEN; COMPLETE PROTEOME

FOLIO ADMINISTRATIF THESE SOUTENUE DEVANT L'INSTITUT NATIONAL DES SCIENCES APPLIQUEES DE LYON
NOM : ANSELME DATE de SOUTENANCE : 06/12/06 Prénoms : Caroline
TITRE : Réponse immunitaire de l’hôte dans la symbiose bactérienne intracellulaire du charançon Sitophilus zeamais. NATURE : Doctorat Numéro d’ordre : 06-ISAL-00107 Ecole doctorale : Evolution, Ecosystèmes, Microbiologie et Modélisation (E2M2) Spécialité : Analyse et Modélisation des Systèmes Biologiques
Cote B.I.U. – Lyon : T 50/210/19 / et bis CLASSE :
RESUME : De nombreuses espèces d’insectes dépendent de bactéries symbiotiques intracellulaires (ou endocytobiotes) pour leur survie et leur reproduction. Ces bactéries sont transmises maternellement et sont localisées dans le bactériome, un organe spécialisé qui se différencie chez l’embryon en présence des symbiotes. Bien que la symbiose ait largement été étudiée depuis les années trente, le mode de perception des endocytobiotes par l’hôte et les aspects immunitaires de ces associations mutualistes demeurent inconnus. Pour aborder ces aspects, nous avons utilisé chez le charançon Sitophilus zeamais une approche de soustraction d’ADNc (SSH) pour identifier les gènes de la réponse immunitaire (GRI) de l’insecte. Cette approche a permis d’obtenir un large spectre de gènes présentant des similitudes de séquence avec des peptides antibactériens, des lysozymes, des éléments de la cascade prophénoloxydase et des régulateurs de la réponse immunitaire. La surexpression de ces gènes après une infection des larves par des bactéries Gram (-) a ensuite été confirmée par RT-PCR en temps réel. La quantification du taux de transcrits des GRI a révélé l’existence d’une réponse immunitaire de l’hôte dans le bactériome larvaire. Cette réponse est toutefois modérée puisqu’à l’exception d’une coléoptéricine, les peptides antibactériens ne sont pas induits par la présence des endocytobiotes. On observe en revanche la surexpression d’un homologue de Tollip, un inhibiteur de la réponse immunitaire décrit chez les mammifères, et la surexpression d’un gène de la famille des “Peptidoglycan recognition proteins” (PGRP) dont l’homologue chez la drosophile (PGRP-LB) est un régulateur négatif de la réponse immunitaire. L’étude du gène PGRP montre qu’il est induit par les bactéries Gram (-), et que son niveau d’expression dépend de la densité bactérienne. Par ailleurs, au cours du développement de l’insecte, ce gène est induit au stade nymphal. Des expériences de FISH ont révélé que cette induction serait liée à la libération d’endocytobiotes à la suite de l’altération du bactériome au cours de la métamorphose. Enfin, nous avons montré l’existence de deux transcrits alternatifs du gène PGRP. Un des transcrit coderait une protéine intracellulaire, et l’autre une protéine transmembranaire. L’analyse du taux de ces deux transcrits révèle que le premier est épissé après une infection des larves par E. coli, alors que le second est épissé majoritairement dans le bactériome larvaire, en présence des endocytobiotes. L’ensemble des données présentées dans ce travail révèle l’existence d’une réponse immunitaire modérée dans l’organe symbiotique, et suggère que la régulation moléculaire de cette réponse serait adaptée au maintien de la symbiose intracellulaire. MOTS CLES : Symbiose intracellulaire / Immunité innée / Insectes / Sitophilus spp. / PGRP Laboratoire(s) de recherches : Laboratoire de Biologie Fonctionnelle, Insectes et Interactions UMR INRA/INSA de Lyon 0203 Directeur de thèse : A. Heddi Président du jury : Composition du jury Rapporteurs : D. Bouchon, B. Lemaitre Examinateurs : F. Fleury, A. Moya, A.N. Volkoff Directeur : A. Heddi