repartition et facteurs de croissance des...
TRANSCRIPT

1
REPARTITION ET FACTEURS DE CROISSANCE DES LENTILLES D’EAU
DANS LE MARAIS POITEVIN
RAPPORT D’ETUDES 2011
INSTITUTION INTERDEPARTEMENTALE DU BASSIN DE LA SEVRE NIORTAISE (IIBSN) Maison du Département - BP 531 - 79 021 NIORT

2
Sommaire
INTRODUCTION ...................................................................................................................... 3 I. Répartition des lentilles d’eau en fonction des sites de prélèvement ...................................... 4
1.1 Protocole de suivi des lentilles d’eau ............................................................................... 4
1.1.1 Sites de prélèvement étudiés ..................................................................................... 4 1.1.2 Méthodologie des prélèvements ................................................................................ 4 1.1.3 Présentation morphologique des espèces de lentilles d’eau étudiées ........................ 5
1.2 Répartition des espèces de lentilles d’eau dans le Marais poitevin en mai 2011 ............. 7 1.2.1 Comparaison des biomasses moyennes de lentilles par site...................................... 7
1.2.2 Comparaison des pourcentages de recouvrement par espèce et par site ................... 8 1.2.2 1 Recouvrements moyens des espèces de lentilles d’eau (Site Moyen).................... 8 1.2.2 2 Résultats pour le site d’Arçais .............................................................................. 10
1.2.2 3 Résultats pour le site de Damvix .......................................................................... 10 1.2.2 4 Résultats pour le site de Magné ............................................................................ 11 1.2.2 5 Résultats pour le site de Maillezais ...................................................................... 11 1.2.2 6 Résultats pour le site du Mazeau .......................................................................... 12
1.2.2 7 Résultats pour le site de St Hilaire La Palud ........................................................ 12 1.2.2 8 Résultats pour le site du Vanneau ........................................................................ 13
1.2.2 9 Conclusion ............................................................................................................ 13 1.3.3 Répartition des espèces de lentilles d’eau par site depuis 2008 .............................. 13
II. Répartition des espèces de lentille d’eau en fonction des paramètres physico-chimiques de
l’eau .......................................................................................................................................... 16 2.1 Méthodes d’analyse des paramètres physico-chimiques ................................................ 16
2.2 Analyse des paramètres physico-chimiques au cours du temps ..................................... 17 2.3 Analyses multi-variées des paramètres physico-chimiques de l’eau et de la répartition
des lentilles ........................................................................................................................... 18
2.3.1 Projection des variables selon le plan F1-F2 pour les différentes dates .................. 18
2.3.2 Projection des points de prélèvements sur l’axe F1-F2 pour chaque date .............. 20 III. Etude de la croissance des lentilles d'eau à partir de sédiments ......................................... 21
3.1 Mise en place de l'expérience ......................................................................................... 21 3.1.1 Récolte des sédiments ............................................................................................. 21 3.1.2 Mises en culture ...................................................................................................... 22
3.1.3 Principe du suivi de la croissance des lentilles ....................................................... 23 3.2 Résultats ......................................................................................................................... 23
3.2.1 Cinétique de croissance des lentilles ....................................................................... 23 Figure 34 : ........................................................................................................................ 23 culture de sédiments ......................................................................................................... 23 en chambre chaude (mésocosmes) ................................................................................... 23 3.2.2 Observation des propagules de lentilles dans le sédiment ...................................... 25
3.2.3 Analyse des différentes expériences réalisées sur 7 sites de prélèvements ............. 27 3.2.4 Comparaison des résultats de croissance avec les recouvrements observés sur le
terrain ............................................................................................................................... 30 CONCLUSION ........................................................................................................................ 34 ANNEXES ............................................................................................................................... 36

3
INTRODUCTION
Au printemps, les lentilles d’eau se multiplient dans les canaux et les fossés situés en
partie amont du marais mouillé du Marais poitevin. Leur développement et leur prolifération
dans ces zones de production dépendent principalement des contextes météorologiques et des
conditions de milieu (qualité et quantité d’eau, ombrage, substrat, interventions humaines,…).
En été, ces végétaux flottants peuvent se déplacer au gré des vents et des courants pour se
concentrer vers l’aval dans les grandes voies d’eau du réseau principal (contours de Sèvre
niortaise, ports,…) perturbant ainsi les usages et le milieu.
Une action de gestion de ces végétaux flottants visant la préservation du milieu et des
activités est engagée depuis 1992 par l’Institution Interdépartementale du Bassin de la Sèvre
Niortaise (IIBSN). Elle a pour objectif d’éviter les recouvrements totaux des voies d’eau du
réseau principal par ces végétaux et leur accumulation.
Parallèlement, une collaboration est mise en place depuis 2006 entre le Département
Génie Biologique de l'IUT de La Roche sur Yon et l'Institution Interdépartementale sur le
thème des lentilles d'eau dans le Marais poitevin. Ce partenariat a pour but d’améliorer la
connaissance des différentes espèces de lentilles, de connaître leur répartition géographique,
leur relation avec la qualité de l’eau et leur développement à partir des sédiments.
Depuis 2006, les étudiants de l'IUT suivent les peuplements de lentilles dans
différentes secteurs du Marais poitevin (annexe 1), analysent la qualité de l’eau de chaque site
et mesurent la croissance de ces végétaux à partir de prélèvements de vase.
Plusieurs axes d’études ont été définis : suivis deux fois par an de la répartition des
lentilles d’eau sur 35 sites situés dans le marais mouillé (zones de production endogène),
corrélation de la répartition des différentes espèces de lentilles avec les principaux facteurs
physico-chimiques et essais de reprise de croissance en mésocosmes à partir de sédiments.
Ces différents suivis visent à connaître annuellement les peuplements de lemnacées
présents sur le territoire (espèces et répartition), à mieux comprendre les conditions de
développement de ces végétaux flottants et à analyser si ces végétaux peuvent servir
d’indicateurs de la qualité de l’eau.
Ce rapport présente les résultats des répartitions des lentilles d’eau pour le printemps
2011 et une comparaison avec ceux du printemps 2010. Les données des analyses physico
chimiques réalisées sur les sites sont également présentées dans l’objectif d’essayer d’établir
un lien entre la répartition des espèces de lentille et les paramètres étudiés. Enfin, un bilan des
cultures en mésocosmes (écosystèmes miniatures pour le laboratoire) est dressé ainsi que des
essais de comparaison avec les résultats des recouvrements sur les mêmes sites de
prélèvement.
Ce rapport d’études a été rédigé par Myriam BOU (enseignante en écologie à l’IUT) et
Nicolas PIPET (technicien à l’Institution). Pour les prélèvements, les analyses des
échantillons et les cultures en laboratoire plusieurs étudiants de l’IUT ont collaboré à ce
travail : Jérôme MICHENAUD, Loïse SAVARY, Manon JEAN DROSSON, Mellie
GRATREAU, Jodie THENARD, Mélusine DUHAMEL, Lolita ANTIER, Clémence
BARBIER, Rébecca BAUCHET, Juliette CHASSAGNAUD et Olivia MERCIER.

4
I. Répartition des lentilles d’eau en fonction des sites de
prélèvement
1.1 Protocole de suivi des lentilles d’eau
1.1.1 Sites de prélèvement étudiés
Sept sites de prélèvement ont été suivis lors d’une campagne de prélèvements le 19
mai 2011. Ils correspondent à 7 zones de production endogène de lentilles situées en milieux
tourbeux : Arçais, Damvix, Magné, Maillezais, Mazeau, St Hilaire la Palud, Vanneau (annexe
1). Les résultats de ces prélèvements seront comparés aux résultats obtenus l’année
précédente lors de la campagne du 18 mai 2010 effectuée sur les mêmes sites.
1.1.2 Méthodologie des prélèvements
Echantillonnage
Cinq échantillons par site de prélèvement sont réalisés à partir de la berge avec une
épuisette (quadrat 20 cm x 20 cm) puis déposés dans un récipient. Le nombre total
d’échantillons prélevés est de 35 (7 sites x 5 échantillons). L’annexe 2 présente les sites pour
la campagne de prélèvement de mai 2011.
Figure 1 : Épuisette servant aux prélèvements
Estimation du recouvrement par espèce pour chaque site
Les cinq échantillons sont pesés au laboratoire une fois égouttés (poids frais total). Les
déchets (feuilles, branches ou autres) sont retirés de l’échantillon et pesés. Il reste ainsi la
biomasse fraîche de lentilles échantillonnées.
Le contenu d’un échantillon est ensuite déversé dans une bassine (34 cm x 49 cm) avec 5 cm
d’eau pour obtenir un recouvrement total de l’eau et une seule épaisseur de lentilles. Pour un
échantillon important (plus de 100g), plusieurs observations peuvent être réalisées avec un
étalement d’au maximum 100g de végétaux à chaque fois.
Les recouvrements sont évalués en pourcentage et par espèce de lentilles, pour chaque
étalement et repris dans un tableau général (annexe 3). Les observations sont effectuées dans
les 15 jours qui suivent le prélèvement afin d’éviter tout phénomène de dessèchement.

5
Figure 2 : Exemple d’étalement de lentilles dans une bassine pour l’estimation du
recouvrement des espèces de lentille
1.1.3 Présentation morphologique des espèces de lentilles d’eau étudiées
Spirodela polyrhiza
Figure 3 : Photographie de
Spirodela polyrhiza
- Les frondes sont rondes, flottantes, de couleur
vert foncé et brillantes sur la face supérieure,
souvent pourprée sur le revers.
- Chaque fronde possède au moins deux racines et
trois à quinze nervures plates ou assez renflées.
- Elle vit libre ou rattachée par deux ou cinq
frondes.
- La taille varie de un à cinq mm. C’est l’espèce la
plus grande.
Lemna gibba
Figure 4 : Photographie de
Lemna gibba
- Les frondes sont flottantes, de couleur vert gris,
prenant parfois une teinte brun rougeâtre.
- Les frondes sont de forme ovale à arrondie, assez
renflées sur la face inférieure.
- Leur taille varie de 1 à 5 mm de long.
- Elles sont liées entre elles par un pétiole
insignifiant.
- Chaque fronde possède une seule longue racine.

6
Wolffia arrhiza
Figure 5 : Photographie de
Wolffia arrhiza
- Les frondes dépassent rarement la longueur du
millimètre.
- Elles ont une forme bombée.
- Elles ne possèdent pas de racines.
Lemna minuta
Figure 6 : Photographie de
Lemna minuta
- Les frondes ont une face supérieure un peu
convexe et carénée : « toit » en lumière rasante.
- Elles ont une longueur de 1,5 à 3 mm et
possèdent une seule nervure.
- Leur bord est très mince (observé par
transparence).
Lemna minor
- Elles ont une fronde flottante vert pâle, de forme
arrondie à ovale, opaque et aplatie, de petite taille :
1,5 à 4 mm de diamètre.
- Elles possèdent 3 à 5 nervures de 1,5 à 5mm de
long, contrairement à L. minuta qui ne possède
qu’une seule nervure.
- Les frondes sont soit reliées entre elles par des
pétioles insignifiants, soit libres Figure 7 : Photographie de
Lemna minor
Les espèces Lemna minor et Lemna minuta sont très proches l’une de l’autre et sont très
difficiles à distinguer visuellement lors des tris. Elles seront par la suite traitées comme un
seul complexe. Une autre espèce de lentille (Lemna trisulca) a été signalée par le passé dans
le Marais poitevin mais n’a jamais été observée dans nos prélèvements.
Quelques autres espèces végétales ont été observées ponctuellement durant les suivis des
échantillons, elles représentent habituellement de très faibles pourcentages par rapport aux
lentilles d’eau observées. L’espèce principale trouvée est une fougère aquatique, Azolla
filiculoïdes (observations depuis octobre 2009).
7
1.2 Répartition des espèces de lentilles d’eau dans le Marais poitevin
en mai 2011
Sept sites de prélèvement ont été suivis en 2011 lors d’une campagne de prélèvement
réalisée mi-mai (annexe 3). Les résultats seront comparés avec ceux de mai 2010 (annexe 4)
issus d’une campagne dans les mêmes conditions et sur les mêmes sites. Les résultats obtenus
concernant les répartitions des lentilles à chaque date sont détaillés ci-après.
Pour avoir une tendance globale sur le territoire et pour comparer chaque site avec un résultat
moyen, un « site moyen » est présenté également dans les graphiques suivants. Il s’agit de la
moyenne des résultats des 7 sites étudiés. Il donne une idée de la répartition des lentilles à
l’échelle du territoire du Marais poitevin (marais mouillé).
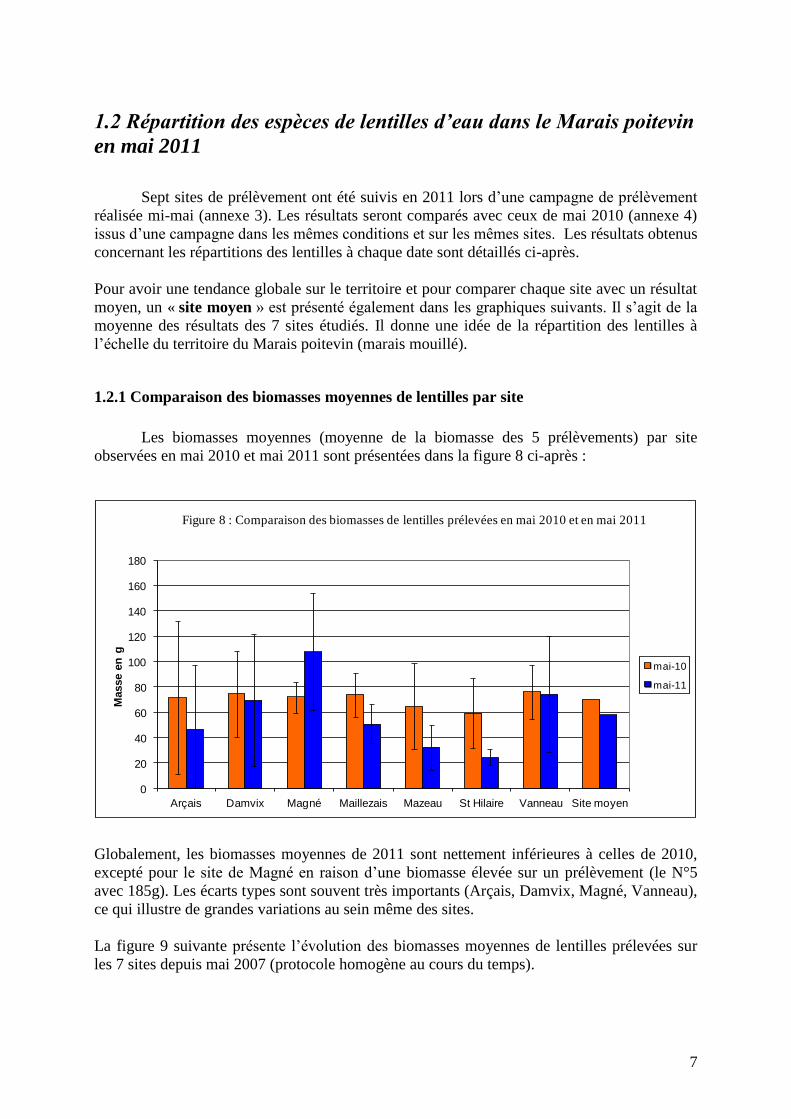
1.2.1 Comparaison des biomasses moyennes de lentilles par site
Les biomasses moyennes (moyenne de la biomasse des 5 prélèvements) par site
observées en mai 2010 et mai 2011 sont présentées dans la figure 8 ci-après :
0
20
40
60
80
100
120
140
160
180
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Masse e
n g
Figure 8 : Comparaison des biomasses de lentilles prélevées en mai 2010 et en mai 2011
mai-10
mai-11
Globalement, les biomasses moyennes de 2011 sont nettement inférieures à celles de 2010,
excepté pour le site de Magné en raison d’une biomasse élevée sur un prélèvement (le N°5
avec 185g). Les écarts types sont souvent très importants (Arçais, Damvix, Magné, Vanneau),
ce qui illustre de grandes variations au sein même des sites.
La figure 9 suivante présente l’évolution des biomasses moyennes de lentilles prélevées sur
les 7 sites depuis mai 2007 (protocole homogène au cours du temps).
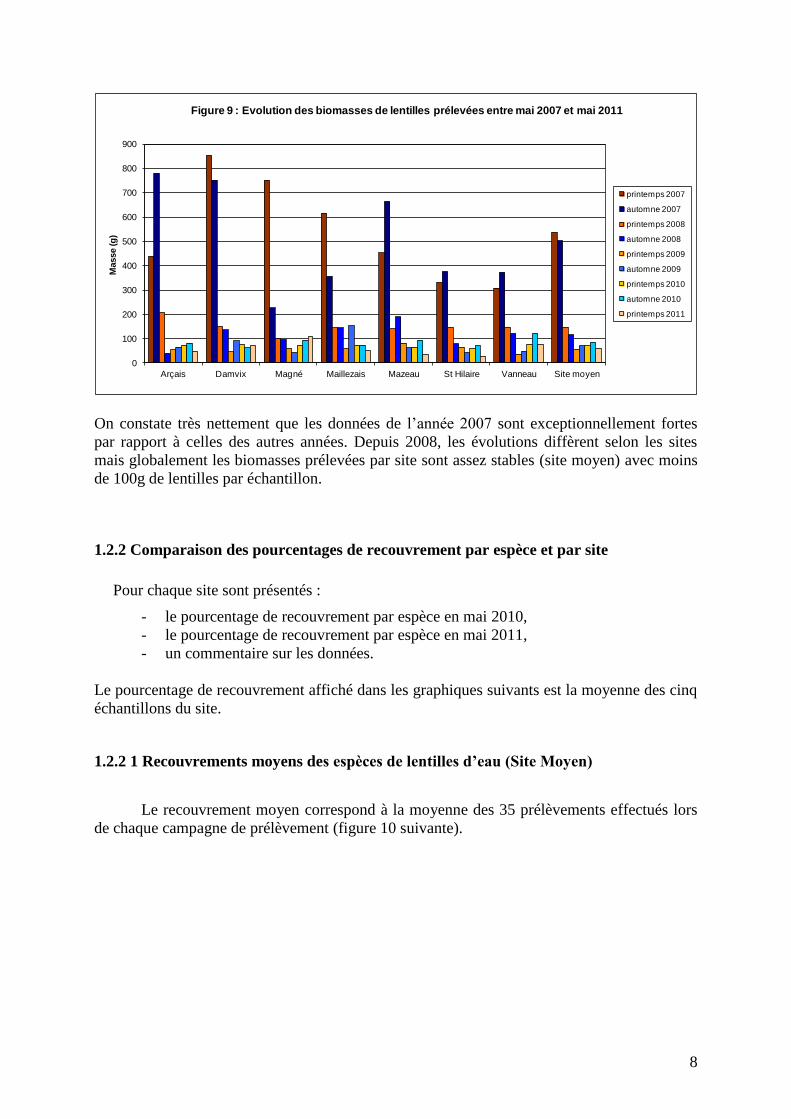
8
0
100
200
300
400
500
600
700
800
900
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Masse (g
)Figure 9 : Evolution des biomasses de lentilles prélevées entre mai 2007 et mai 2011
printemps 2007
automne 2007
printemps 2008
automne 2008
printemps 2009
automne 2009
printemps 2010
automne 2010
printemps 2011
On constate très nettement que les données de l’année 2007 sont exceptionnellement fortes
par rapport à celles des autres années. Depuis 2008, les évolutions diffèrent selon les sites
mais globalement les biomasses prélevées par site sont assez stables (site moyen) avec moins
de 100g de lentilles par échantillon.
1.2.2 Comparaison des pourcentages de recouvrement par espèce et par site
Pour chaque site sont présentés :
- le pourcentage de recouvrement par espèce en mai 2010,
- le pourcentage de recouvrement par espèce en mai 2011,
- un commentaire sur les données.
Le pourcentage de recouvrement affiché dans les graphiques suivants est la moyenne des cinq
échantillons du site.
1.2.2 1 Recouvrements moyens des espèces de lentilles d’eau (Site Moyen)
Le recouvrement moyen correspond à la moyenne des 35 prélèvements effectués lors
de chaque campagne de prélèvement (figure 10 suivante).

9
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
50,00
Lemna gibba Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza Azolla
% d
e r
ec
ou
vre
me
nt
Figure 10 : Pourcentage moyen de recouvrement par espèce pour l'ensemble des sites
étudiés en mai 2010 et en mai 2011
mai-10
mai-11
Les pourcentages de recouvrement par espèce ne sont pas identiques sur les deux années. Les
espèces majoritaires sont Spirodela polyrhiza et Lemna minor/minuta. A elles deux, elles
représentent 73% du recouvrement en 2010 et 94% en 2011. La proportion de Lemna
minor/minuta a augmenté en 2011 et cette espèce est en quantité équivalente à Spirodela
polyrhiza. Au contraire Lemna gibba et Wolffia arrhiza sont en nette diminution entre 2010 et
2011.
La figure 11 suivante montre l’évolution du recouvrement moyen de chaque espèce de
lentilles depuis 2008.
0
10
20
30
40
50
60
70
80
90
Lemna gibba Spirodela polyrhiza Lemna minor/minuta Wolf f ia arrhiza Azolla f iliculoïdes
% d
e r
eco
uvre
men
t
Figure 11 : Evolution des pourcentages moyens de recouvrement par espèce de lentille entre 2008 et 2011
mai 2008
octobre 2008
mai 2009
octobre 2009
mai 2010
octobre 2010
mai 2011
10
Figure 12 : Répartition par espèces en % de recouvrement pour le site d'Arçais entre
mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
Figure 13 : Répartition par espèces en % de recouvrement pour le site de Damvix
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
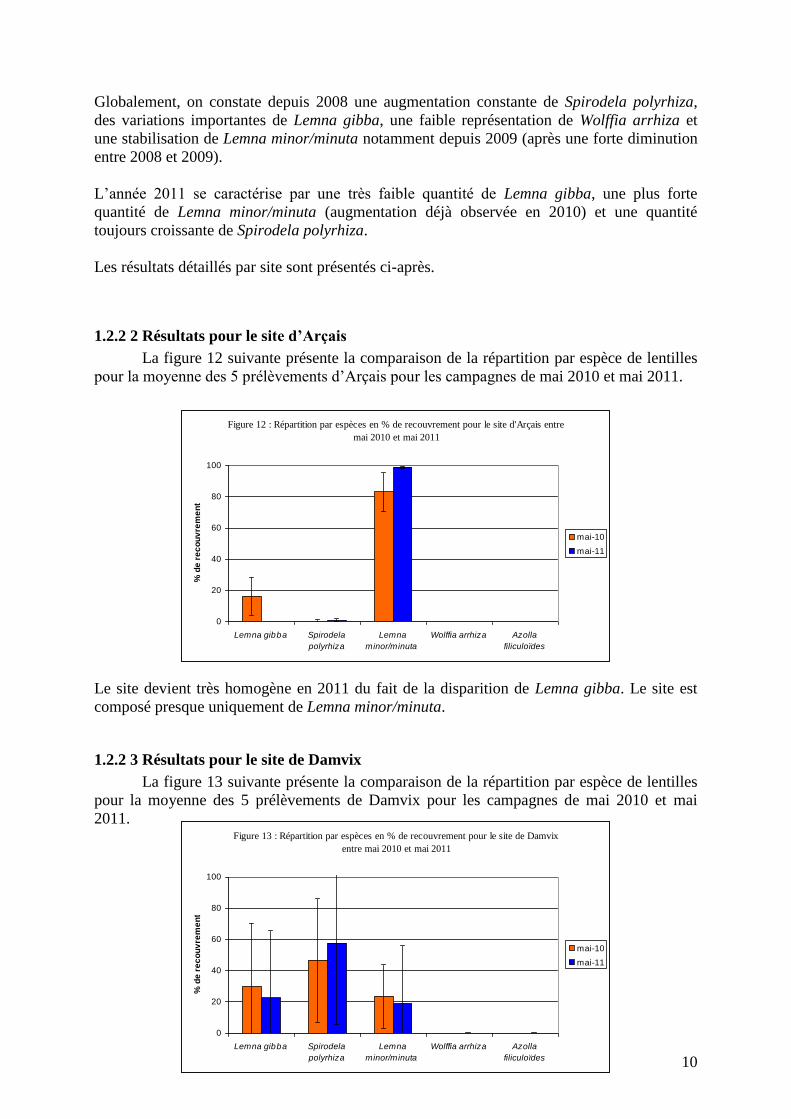
Globalement, on constate depuis 2008 une augmentation constante de Spirodela polyrhiza,
des variations importantes de Lemna gibba, une faible représentation de Wolffia arrhiza et
une stabilisation de Lemna minor/minuta notamment depuis 2009 (après une forte diminution
entre 2008 et 2009).
L’année 2011 se caractérise par une très faible quantité de Lemna gibba, une plus forte
quantité de Lemna minor/minuta (augmentation déjà observée en 2010) et une quantité
toujours croissante de Spirodela polyrhiza.
Les résultats détaillés par site sont présentés ci-après.
1.2.2 2 Résultats pour le site d’Arçais
La figure 12 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements d’Arçais pour les campagnes de mai 2010 et mai 2011.
Le site devient très homogène en 2011 du fait de la disparition de Lemna gibba. Le site est
composé presque uniquement de Lemna minor/minuta.
1.2.2 3 Résultats pour le site de Damvix
La figure 13 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements de Damvix pour les campagnes de mai 2010 et mai
2011.
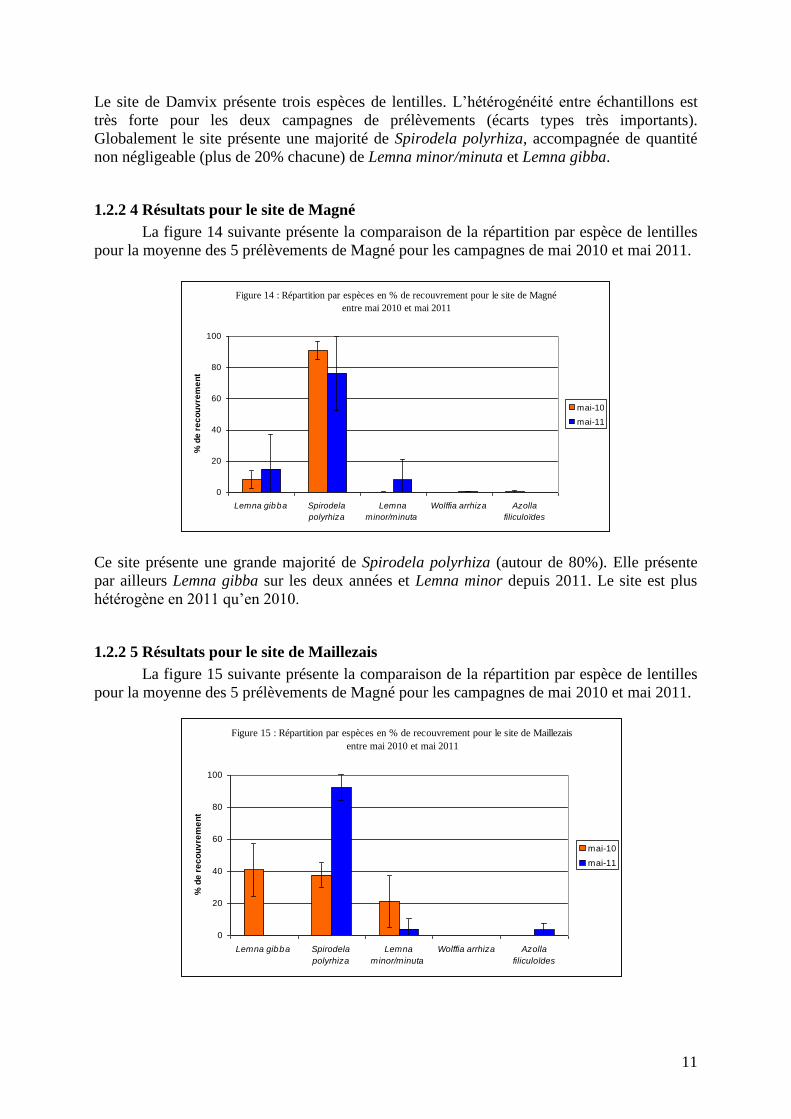
11
Figure 14 : Répartition par espèces en % de recouvrement pour le site de Magné
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
Figure 15 : Répartition par espèces en % de recouvrement pour le site de Maillezais
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
Le site de Damvix présente trois espèces de lentilles. L’hétérogénéité entre échantillons est
très forte pour les deux campagnes de prélèvements (écarts types très importants).
Globalement le site présente une majorité de Spirodela polyrhiza, accompagnée de quantité
non négligeable (plus de 20% chacune) de Lemna minor/minuta et Lemna gibba.
1.2.2 4 Résultats pour le site de Magné
La figure 14 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements de Magné pour les campagnes de mai 2010 et mai 2011.
Ce site présente une grande majorité de Spirodela polyrhiza (autour de 80%). Elle présente
par ailleurs Lemna gibba sur les deux années et Lemna minor depuis 2011. Le site est plus
hétérogène en 2011 qu’en 2010.
1.2.2 5 Résultats pour le site de Maillezais
La figure 15 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements de Magné pour les campagnes de mai 2010 et mai 2011.
12
Figure 16 : Répartition par espèces en % de recouvrement pour le site du Mazeau
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
Figure 17 : Répartition par espèces en % de recouvrement pour le site de St Hilaire
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
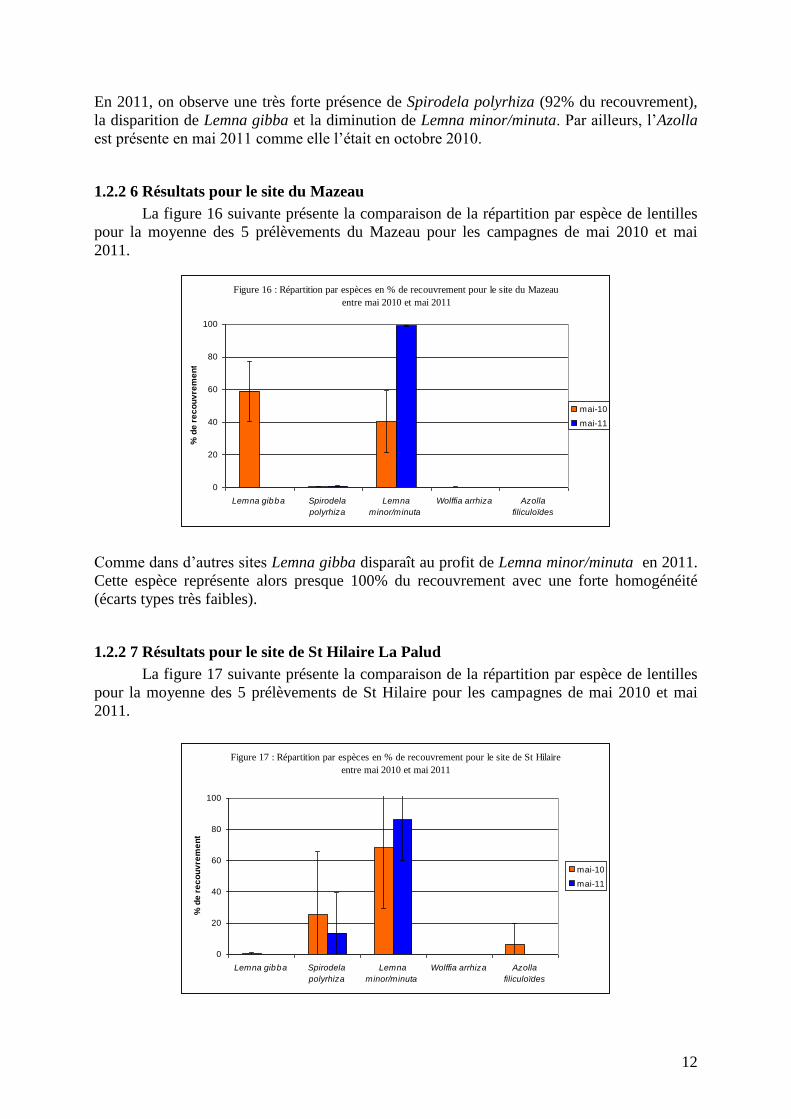
En 2011, on observe une très forte présence de Spirodela polyrhiza (92% du recouvrement),
la disparition de Lemna gibba et la diminution de Lemna minor/minuta. Par ailleurs, l’Azolla
est présente en mai 2011 comme elle l’était en octobre 2010.
1.2.2 6 Résultats pour le site du Mazeau
La figure 16 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements du Mazeau pour les campagnes de mai 2010 et mai
2011.
Comme dans d’autres sites Lemna gibba disparaît au profit de Lemna minor/minuta en 2011.
Cette espèce représente alors presque 100% du recouvrement avec une forte homogénéité
(écarts types très faibles).
1.2.2 7 Résultats pour le site de St Hilaire La Palud
La figure 17 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements de St Hilaire pour les campagnes de mai 2010 et mai
2011.

13
Figure 18 : Répartition par espèces en % de recouvrement pour le site du Vanneau
entre mai 2010 et mai 2011
0
20
40
60
80
100
Lemna gibba Spirodela
polyrhiza
Lemna
minor/minuta
Wolffia arrhiza Azolla
filiculoïdes
% d
e r
ec
ou
vre
me
nt
mai-10
mai-11
Le site est marqué par l’augmentation de Lemna minor/minuta au détriment de Spirodela
polyrhiza. Par ailleurs, l’Azolla a disparu en 2011. L’hétérogénéité de ce site reste très forte.
1.2.2 8 Résultats pour le site du Vanneau
La figure 18 suivante présente la comparaison de la répartition par espèce de lentilles
pour la moyenne des 5 prélèvements du Vanneau pour les deux campagnes de mai 2010 et
mai 2011.
Le nombre d’espèce présent sur le site à fortement diminué en raison de la disparition de
Lemna gibba et Wolffia arrhiza entre les deux dates. L’espèce majoritaire est Spirodela
polyrhiza. L’hétérogénéité reste modérée.
1.2.2 9 Conclusion
Globalement, les sites en mai 2011 présentent une quantité moindre de Lemna gibba
sauf pour Magné. Il n’y a pas de schéma général de répartition des lentilles dans les différents
sites entre les deux dates (mai 2010 et mai 211) : certains sites présentent des augmentations
de Spirodela polyrhiza (Maillezais, Vanneau), d’autres des augmentations de Lemna
minor/minuta (Vanneau, Mazeau), d’autres au contraire présentent des diminutions de
recouvrement pour ces deux espèces. Chaque site et même chaque prélèvement se comporte
de façon bien spécifique.
1.3.3 Répartition des espèces de lentilles d’eau par site depuis 2008
Le recouvrement de chaque espèce depuis mai 2008 sera représenté dans les
graphiques suivants (figure 19 à 23) afin d’étudier les modifications au cours du temps.

14
Figure 19 : Evolution par site du recouvrement de Spirodela polyrhiza depuis 2008
0
10
20
30
40
50
60
70
80
90
100
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Reco
uvre
men
t (%
)
mai-08
sept-08
mai-09
sept-09
mai-10
sept-10
mai-11
Figure 20 : Evolution par site du recouvrement de Lemna minor/minuta depuis 2008
0
10
20
30
40
50
60
70
80
90
100
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Reco
uvre
men
t (%
)
mai-08
sept-08
mai-09
sept-09
mai-10
sept-10
mai-11
Figure 21 : Evolution par site du recouvrement de Lemna gibba depuis 2008
0
10
20
30
40
50
60
70
80
90
100
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Reco
uvre
men
t (%
)
mai-08
sept-08
mai-09
sept-09
mai-10
sept-10
mai-11

15
Figure 22 : Evolution par site du recouvrement de Wolffia arrhiza depuis 2008
0
5
10
15
20
25
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Reco
uvre
men
t (%
)
mai-08
sept-08
mai-09
sept-09
mai-10
sept-10
mai-11
Figure 23 : Evolution par site du recouvrement de Azolla filiculoïdes depuis 2008
0
5
10
15
20
25
Arçais Damvix Magné Maillezais Mazeau St Hilaire Vanneau Site moyen
Reco
uvre
men
t (%
)
mai-08
sept-08
mai-09
sept-09
mai-10
sept-10
mai-11
La figure 20 montre qu’après plusieurs années de diminution du recouvrement, le complexe
Lemna minor/minuta réaugmente au printemps 2011. Ce complexe reste dominant sur le
marais avec une autre espèce, Spirodela polyrhiza (figure 19). Cette dernière est en
augmentation tout au long de la période. Elle n’est que très faiblement représentée
uniquement sur deux sites : Arçais et Mazeau. Sur ces deux sites, Lemna minor/minuta est
dominante.
Les quantités de Lemna gibba (figure 21) sont très variables en fonction des sites et des
années. Elle est relativement abondante sur le site du Mazeau en permanence. Sa quantité
maximale a été observée en mai 2010.
La dernière espèce de lentille, Wolffia arrhiza, est toujours faiblement représentée (figure 22).
Sa présence n’est pas stable dans les sites.
Enfin, une autre plante aquatique flottante a été observée Azolla filiculoïdes (figure 23) en
raison de son abondance depuis septembre 2009. Elle est depuis en déclin. Elle est surtout
présente en automne et beaucoup moins au printemps.
Afin de comprendre les observations sur les répartitions des lentilles, un lien avec les
paramètres physico-chimiques va être étudié.

16
II. Répartition des espèces de lentille d’eau en fonction des
paramètres physico-chimiques de l’eau
Les données précédentes montrent que les différentes espèces de lentilles d’eau ne
sont pas réparties de la même manière sur l’ensemble du Marais poitevin. Les raisons de leur
répartition pourraient être liées aux paramètres physico-chimiques du milieu dans lequel elles
évoluent.
2.1 Méthodes d’analyse des paramètres physico-chimiques
Plusieurs paramètres physico-chimiques influencent la croissance des végétaux
aquatiques : température, pH, dioxygène, conductimétrie, turbidité, profondeur, concentration
en nitrates, concentration en phosphates.
Les mesures de conductimétrie, dioxygène, pH, température et turbidité ont été réalisées à
l’aide de sondes électroniques. Elles ont une précision au centième sauf pour la turbidité où la
précision est au dixième.
La profondeur est évaluée à l’aide d’une règle graduée.
Ces différents paramètres ont été mesurés le jour de la réalisation des prélèvements et pour
chaque échantillon prélevé (soit 5 mesures par site de prélèvement).
Pour les nitrates (NO3-) et les phosphates (PO4
3-) les mesures sont faites en laboratoire grâce à
des méthodes Hach depuis 2009 (auparavant, on utilisait des méthodes colorimétriques
classiques). Pour les phosphates, on utilise la gamme 0 à 5 mg/L de phosphates et la précision
est au centième de mg. Pour les nitrates, on utilise une gamme 0-30 mg/L et la précision est
au dixième de mg/L. Les analyses ont été réalisées dans les 48h qui ont suivi le prélèvement
(temps maximal préconisé afin d’avoir des données conformes au terrain). Certaines données
sont manquantes dans les années précédentes en raison de pannes matérielles.
Figure 24 : Mallettes de mesures pour le terrain
(turbidimètre et sondes multi-paramètres)

17
Figure 25 : Concentration moyenne en dioxygène dans le marais Poitevin au cours du temps (mg/L)
0
1
2
3
4
5
6
Mai 2008 Oct 2008 Mai 2009 Sep 2009 Mai 2010 Sep 2010 mai-11
Figure 26 : Conductimétrie moyenne dans le marais Poitevin au cours du temps (mS/cm)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
Mai 2008 Oct 2008 Mai 2009 Sep 2009 Mai 2010 Sep 2010 mai-11
Figure 27 : Concentration moyenne en nitrates dans le marais Poitevin au cours du temps (mg/L)
0
2
4
6
8
10
12
Mai 2008 Oct 2008 Mai 2009 Sep 2009 Mai 2010 Sep 2010 mai-11
Figure 28 : Concentration moyenne en phosphates dans le marais Poitevin au cours du temps
(mg/L)
0
0,5
1
1,5
2
2,5
3
Mai 2008 Oct 2008 Mai 2009 Sep 2009 Mai 2010 Sep 2010 mai-11
2.2 Analyse des paramètres physico-chimiques au cours du temps De grandes tendances ayant été observées au niveau des répartitions des lentilles, les
paramètres physico-chimiques moyens pour chaque campagne de prélèvements seront
représentés pour essayer de trouver des corrélations. Les résultats bruts sont dans les annexes
3 et 4.

18
Les figures 25 à 28 montrent des évolutions globales des paramètres dans le marais poitevin
depuis mai 2008. L’année 2011 est marquée par un faible taux de dioxygène, certainement lié
au printemps très chaud qu’il y a eu (température moyenne de l’eau en mai 2010 est de 15.2
contre 18.5 en mai 2011). Les autres valeurs sont conformes à ce qui avait été observé lors des
printemps précédents.
Les modifications observées sur les répartitions de lentilles en mai 2011 sont donc peut être
liées au printemps très chaud qu’il y a eu associé à un hiver froid.
2.3 Analyses multi-variées des paramètres physico-chimiques de
l’eau et de la répartition des lentilles
L’analyse en composante principale (ACP) permet d’analyser de nombreux
paramètres présentant des caractères quantitatifs. Pour notre étude, elle permet de projeter sur
une carte en deux dimensions, les relations entre toutes les données mesurées par point de
prélèvement (données physico-chimiques et répartition des espèces). Plus les points sont
proches sur la carte, plus ils possèdent des caractéristiques communes.
L’analyse est réalisée grâce au logiciel XLstat, macro d’Excel avec un coefficient de
corrélation de Pearson (n). Un code est attribué à chaque site, dont les prélèvements sont
déclinés de 1 à 5. Les variables sont en rouge sur les cartes.
2.3.1 Projection des variables selon le plan F1-F2 pour les différentes dates
L’ACP permet de projeter les données suivant 35 axes, autant que le nombre de site.
Ces axes de projection sont classés en fonction de la quantité d’information qu’ils permettent
de représenter. L’axe F1 est le plus significatif. Pour obtenir une projection contenant le
maximum d’informations, le plan de projection F1-F2 est utilisé.
La figure 29 suivante montre les projections selon l’axe F1-F2 des variables étudiées pour
chaque date de prélèvement et pour 2 années (2010 et 2011).
On constate que sur l’ensemble des figures observées, les axes F1-F2 représentent des
pourcentages faibles (de 39,5% à 51%), ce qui montre qu’au maximum la moitié de
l’information contenue dans les données est représentée.
Les pourcentages obtenus sont pour :
- mai 2009 : 51%
- septembre 2009 : 41%
- mai 2010 : 39,6%
- septembre 2010 : 40,7%
- mai 2011 : 44.5%
L’année 2011 se situe dans la moyenne au niveau du pourcentage représenté par les axes F1-
F2.

19
Variables mai 2010 (ax es F1 et F2 : 39,56 %)
Cond
pH
Turb
Nit
Phos
M tot
L. gibba
Spirodela
L. minor
Wolffia
Autres
-1
-0,75
-0,5
-0,25
0
0,25
0,5
0,75
1
-1 -0,75 -0,5 -0,25 0 0,25 0,5 0,75 1
F1 (20,36 %)
F2
(1
9,2
0 %
)Variables s ept 2010 (ax es F1 et F2 : 40,70 %)
Cond
pH
Turb
Nit
Phos
M tot
L. gibba
SpirodelaL. minor
Wolffia
Autres
-1
-0,75
-0,5
-0,25
0
0,25
0,5
0,75
1
-1 -0,75 -0,5 -0,25 0 0,25 0,5 0,75 1
F1 (20,84 %)
F2
(1
9,8
6 %
)Variables mai 2011 (axes F1 et F2 : 44,52 %)
Azolla
Wolff ia
L. minor
Spirodela
L. gibba
M asse
Phos
Nit
Turb
pH
Cond
O2
-1
-0,75
-0,5
-0,25
0
0,25
0,5
0,75
1
-1 -0,75 -0,5 -0,25 0 0,25 0,5 0,75 1
F1 (25,09 %)
F2 (
19,4
3 %
)
Figure 29 : Projections selon l’axe F1-F2 des
différentes dates de prélèvements en 2010 et 2011
La projection des variables de mai 2011, montre une fois de plus que la variable Spirodela est
opposée à Lemna minor/minuta. De même qu’en septembre 2010, Lemna gibba se situe à la
perpendiculaire des deux espèces précédentes assez proche de Wolffia.
Par ailleurs, ni Lemna minor/minuta, ni Spirodela, ne sont corrélées à aucune variable
physicochimique. Ceci est confirmé par la matrice de corrélation située en annexe 5. Lemna
gibba est assez bien corrélée à la turbidité et à la concentration en phosphates mais ce n’était
pas le cas les autres années. Wolffia est corrélée à la masse de lentilles : l’espèce étant rare, on
peut supposer que lorsque les échantillons sont importants, les probabilités de l’observer sont
meilleures.
En comparant les résultats des deux années consécutives et des années précédentes, nous ne
pouvons pas mettre en évidence de corrélations stables entre les variables. La structuration
des données est donc très faible entre les recouvrements des espèces de lentilles et les
paramètres physicochimiques mesurés.
20
Figure 30 : Projection des différents prélèvements selon l'axe
F1-F2 de l'ACP
VA5
VA4VA3
VA2VA1
HI5HI4
HI3
HI2HI1 MZ5
MZ4MZ3
MZ2
MZ1
MI5
MI4
MI3
MI2
MI1MG5
MG4
MG3
MG2MG1
DA5
DA4
DA3
DA3
DA1
AR5
AR4
AR3AR2AR1
Azolla
Wolffia
L. minor
Spirodela
L. gibba
Masse
Phos
Nit
Turb
pH
Cond
O2
-3
-2
-1
0
1
2
3
4
-5 -4 -3 -2 -1 0 1 2 3 4 5 6
F1 (25,09 %)
F2
(1
9,4
3 %
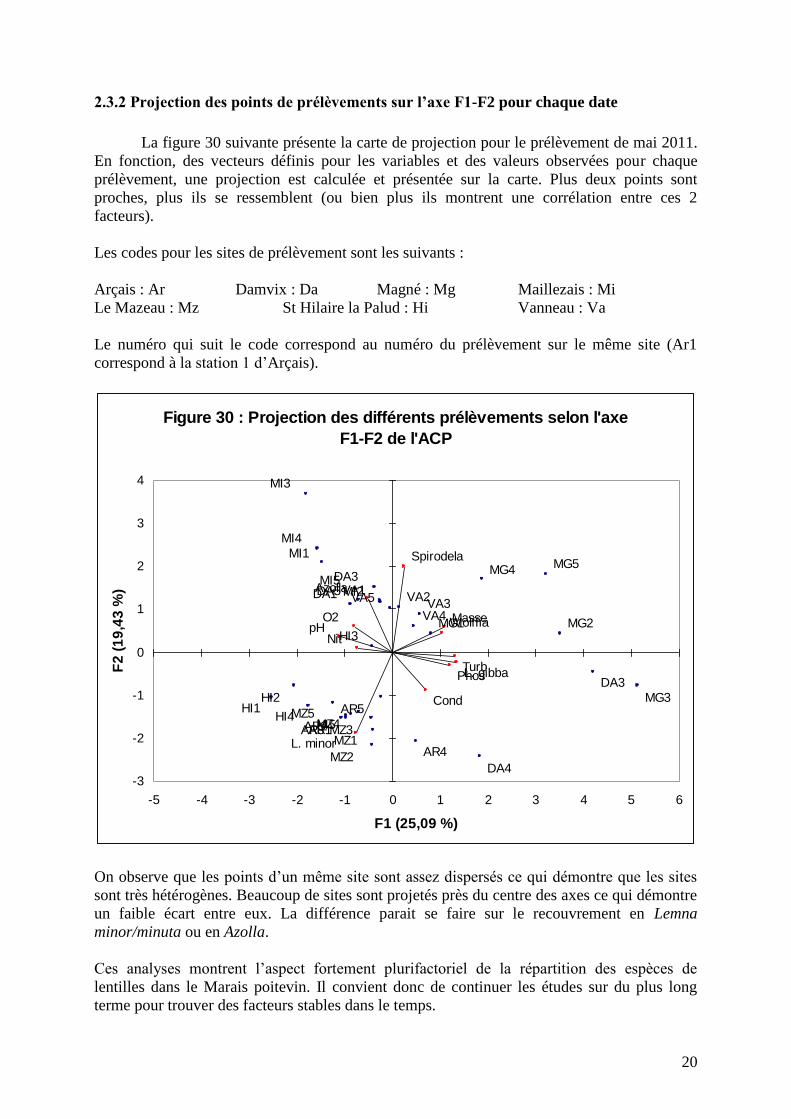
)2.3.2 Projection des points de prélèvements sur l’axe F1-F2 pour chaque date
La figure 30 suivante présente la carte de projection pour le prélèvement de mai 2011.
En fonction, des vecteurs définis pour les variables et des valeurs observées pour chaque
prélèvement, une projection est calculée et présentée sur la carte. Plus deux points sont
proches, plus ils se ressemblent (ou bien plus ils montrent une corrélation entre ces 2
facteurs).
Les codes pour les sites de prélèvement sont les suivants :
Arçais : Ar Damvix : Da Magné : Mg Maillezais : Mi
Le Mazeau : Mz St Hilaire la Palud : Hi Vanneau : Va
Le numéro qui suit le code correspond au numéro du prélèvement sur le même site (Ar1
correspond à la station 1 d’Arçais).
On observe que les points d’un même site sont assez dispersés ce qui démontre que les sites
sont très hétérogènes. Beaucoup de sites sont projetés près du centre des axes ce qui démontre
un faible écart entre eux. La différence parait se faire sur le recouvrement en Lemna
minor/minuta ou en Azolla.
Ces analyses montrent l’aspect fortement plurifactoriel de la répartition des espèces de
lentilles dans le Marais poitevin. Il convient donc de continuer les études sur du plus long
terme pour trouver des facteurs stables dans le temps.

21
III. Etude de la croissance des lentilles d'eau à partir de
sédiments
En hiver, les lentilles se déposent dans les sédiments ce qui leur permet de survivre
durant la période de froid. La reprise de la croissance a lieu au printemps dès que les
conditions météorologiques redeviennent favorables. Le but de nos manipulations est de
reproduire ce phénomène en laboratoire.
3.1 Mise en place de l'expérience
3.1.1 Récolte des sédiments
Trois campagnes de prélèvements ont été réalisées dans le Marais poitevin durant
l’hiver 2010-2011 : le 30 septembre 2010, le 22 novembre 2010 et le 10 février 2011. Les
prélèvements de sédiments ont été effectués dans sept stations : Vanneau 4, Vanneau 5,
Arçais 5, Damvix 1, Damvix 5, Mazeau 1 et Magné 2.
Les résultats détaillés des deux premières campagnes ont été présentés dans le rapport de
2010.
Dans ce rapport, nous analyserons la totalité des résultats obtenus depuis janvier 2009 (date de
la première expérience réalisée sur mésocosmes).
Les différents prélèvements réalisés au cours du temps sont résumés dans la figure 31 ci-après
(cf. cartes de localisation, annexe 6).
Figure 31 : Dates et localisation des prélèvements de sédiments
Commune
N°site
N°
Prélèv
15 Janv
2009
30 Avril
2009
6 Nov
2009
20 Janv
2010
30 Sept
2010
22 Nov
2010
10 Fev
2011
Arçais 5 P1 X X X X X X X
Damvix 3 P2 X
Damvix 5 P3 X X X X X X
Damvix 1 P4 X X X X X
Le Mazeau 5 P5 X
Le Mazeau 3 P6 X X X X X X
Le Vanneau 1 P7 X
Le Vanneau 4 P8 X X X X X
Le Vanneau 5 P9 X X X X X
Magné 5 P10 X
Magné 2 P11 X X X X X X X
Maillezais 5 P12 X
Maillezais 3 P13 X
Maillezais 2 P14 X
Saint Hilaire la
Palud
5
P15
X

22
3.1.2 Mises en culture
Afin d'observer le développement de lentilles d'eau à partir de ces sédiments, le
protocole de mise en culture est le suivant :
- mettre dans chaque bassine environ 4cm de vermiculite (minéral naturel formé
d’argile expansée, qui est un support neutre de culture),
- ajouter 1,5 L d’eau prélevée dans le marais,
- laisser gonfler la vermiculite une nuit,
- le lendemain, déposer une moustiquaire qui empêchera le mélange entre sédiments et
vermiculite
- déposer environ 2 cm de sédiment sur la moustiquaire,
- ajouter 4 cm d’eau de rivière afin de recouvrir les sédiments,
- mettre les bassines en chambre chaude sous un éclairement de 14 à 16h par jour,
- compléter avec de l'eau distillée au fur et à mesure l'expérience pour maintenir un
niveau d'eau de 4 cm et afin de ne pas concentrer le milieu en ions.
Figure 33 : Schéma de la mise en culture des sédiments
Les sédiments ont été prélevés à l’aide d’une pelle
à sédiment. Ils sont stockés en chambre froide
entre le moment où ils sont prélevés et le moment
où ils sont mis en culture. Ce temps est
généralement assez court (<10 jours). Le seul
prélèvement à être resté longtemps en chambre
froide est celui du 30 septembre 2010, qui a été
mis en culture le 10 novembre 2010. En effet, les
sédiments ayant été prélevés tôt, une levée
d’éventuelles dormances a été effectuée par le
passage en chambre froide.
Figure 32 : Récupération des
sédiments en vue d'une mise en
culture en laboratoire

23
3.1.3 Principe du suivi de la croissance des lentilles
La croissance des lentilles est suivie pendant au moins 1 mois et jusqu’à l’arrêt de
l’apparition de nouvelles lentilles (figure 34 suivante).
Les différentes espèces de lentilles d'eau sont donc dénombrées pour chaque site puis
enlevées du récipient de culture. Elles sont ensuite déposées dans des récipients contenant de
l’eau du marais afin de les laisser se développer et de vérifier leur détermination.
3.2 Résultats
Les résultats de toutes les expériences de culture de sédiments sont présentés dans ce
rapport et ils sont détaillés dans les tableaux récapitulatifs en annexe 7. Globalement, la
présence de lentilles a pu être observée très rapidement après la mise en culture des
sédiments, excepté pour le prélèvement d’avril 2009 où aucune lentille ne s’est développée.
Plusieurs axes de travail vont être présentés :
- la cinétique de croissance des lentilles sur mésocosmes (à l’aide de la première
série d’expériences en 2009),
- la recherche des éléments issus de lentilles contenus dans le sédiment,
- les résultats des différentes expériences et comparés par site (à partir du
prélèvement de novembre 2009),
- les corrélations possibles entre les croissances observées en laboratoire et les
recouvrements observés dans le milieu naturel.
3.2.1 Cinétique de croissance des lentilles
Afin d’étudier la cinétique de croissance des lentilles à partir des sédiments, seules les
données du 15 janvier 2009 seront utilisées. Les sédiments ont été mis en culture le 23 janvier
2009. Cette expérience a permis d’étudier la faisabilité du protocole et la durée de suivi.
Figure 34 :
culture de sédiments
en chambre chaude (mésocosmes)

24
Figure 35 : Cinétique de la croissance des lentilles sur mésocosmes
pour le site de Damvix 3
0
100
200
300
400
500
600
0 10 20 30 40 50 60No
mb
re c
um
un
é d
e f
ron
des o
bserv
ées
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolff ia arrhiza
Figure 36 : Cinétique de la croissance des lentilles sur mésocosmes
pour le site de Magné 2
0
100
200
300
400
500
600
0 10 20 30 40 50 60No
mb
re c
um
un
é d
e f
ron
des o
bserv
ées
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolff ia arrhiza
Figure 37 : Cinétique de la croissance des lentilles sur mésocosmes
pour le site de Arçais 5
0
100
200
300
400
500
600
0 10 20 30 40 50 60No
mb
re c
um
un
é d
e f
ron
des o
bserv
ées
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolff ia arrhiza

25
Figure 38 : Cinétique de la croissance des lentilles sur mésocosmes
pour le site de Maillezais 3
0
100
200
300
400
500
600
0 10 20 30 40 50 60No
mb
re c
um
un
é d
e f
ron
des o
bserv
ées
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolff ia arrhiza
Commentaires Figures 35 à 38
Pour les sites de Magné 2 et Arçais 5, les lentilles apparaissent au bout de 13 jours de culture.
Pour les deux autres sites, il faut attendre 40 jours pour observer une croissance de lentilles.
D’ailleurs, pour les sites de Maillezais 3 et Damvix 3, la quantité de lentilles observée est
beaucoup plus faible (moins de 100 frondes) et seule l’espèce Lemna minor est représentée.
Pour le site d’Arçais 5, seule Lemna minor est représentée mais un grand nombre de frondes a
poussé (594 au total). La croissance s’étale sur deux mois puis s’arrête. L’expérience a été
poursuivie après 52 jours mais aucune nouvelle fronde n’est apparue.
Le site de Magné 2 est très satisfaisant puisque 3 espèces se développent en grande quantité :
Lemna minor (390 frondes), Lemna gibba (64 frondes) et Spirodela polyrhiza (66 frondes).
Ce site démontre que l’on arrive à faire pousser depuis les réserves de propagules contenues
dans les sédiments 3 des 4 espèces observées dans le marais.
Cette expérience préliminaire montre l’intérêt d’étudier la reprise de la croissance des lentilles
à partir de sédiments. Pour ce faire, le sédiment doit être choisi correctement afin de contenir
potentiellement des propagules de lentille et être mis en culture sur un temps assez long (entre
1 et 2 mois).
3.2.2 Observation des propagules de lentilles dans le sédiment
Les résultats des expériences précédentes semblent indiquer la présence et la
conservation des lentilles dans les sédiments lorsque les conditions du milieu leurs deviennent
défavorables durant l’hiver. Pour connaître sous quelle forme celles-ci se conservent durant
cette période de froid, des sédiments ont été observés à la loupe binoculaire, préalablement
filtré avec un tamis de maille 200µm, permettant de retenir les éléments susceptibles d’être
intéressants.
Tout d’abord, nous avons observé dans nos bassines de cultures des lentilles décolorées, que
nous avons supposées mortes. Ces débris végétaux ont été prélevés et observés à la
loupe (figure 39 suivante).

26
Figure 39 : Photographie d’une lentille morte observée à la loupe binoculaire (x20)
Nous pouvons voir sur cette photographie qu’il semble manquer une partie à la fronde de
lentille. Cette partie correspondrait sur les lentilles vivantes à une zone de couleur plus
foncée, d’où est issue la racine : le turion. On observe la disparition de celui-ci sur plusieurs
lentilles mortes étudiées à la loupe binoculaire.
Figure 40 : Photographie d’une lentille Spirodela polyrhiza à turion
Les observations à la loupe laissent supposer que c’est donc le turion des lentilles qui se
conserve dans les sédiments. Nous avons donc ciblé les éléments susceptibles d’être un turion
lors de nos observations à la loupe, c’est-à-dire de petite taille et de forme plutôt circulaire.
A l’issue de cette recherche, quelques éléments ont été isolés, et placés en chambre chaude
dans des boîtes remplies d’eau et hermétiquement fermées (afin d’éviter l’évaporation), dans
le but d’observer une éventuelle croissance à partir des éléments sélectionnés.
Turion
27
Figure 41 : Nombre de frondes observées pour les
différentes expériences pour le site d'Arçais 4
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 42 : Nombre de frondes observées pour les
différentes expériences pour le site de Damvix 1
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 41 : Photographie de quelques éléments sélectionnés après observation
à la loupe binoculaire (x20)
Cependant, aucune croissance à partir de ces éléments n’a pu être mise en évidence. Aucune
conclusion certaine ne peut donc être tirée de ces expériences. Néanmoins, il serait intéressant
de renouveler le protocole sur un échantillonnage plus important.
3.2.3 Analyse des différentes expériences réalisées sur 7 sites de prélèvements
Sept sites ont été choisis pour les prélèvements de sédiments dans des zones qui
présentent des pourcentages de recouvrements variés in situ : Arçais 5, Damvix 1 et 5, Magné
2, Mazeau 3, Vanneau 4 et 5 (figures 41 à 48). Entre 2009 et 2011 (hiver 2009-2010 et hiver
2010-2011), 5 expériences de croissance de lentilles à partir de sédiments ont été réalisées.
Elles s’arrêtent lorsque l’on n’observe plus aucune nouvelle fronde. Les durées sont très
variables (de 26 à 55 jours).
Légende des figures 41 à 47 * Passage des échantillons au froid pendant 40 jours
Lemna gibba Spirodela polyrhiza
Lemna minor/minuta Wolffia arrhiza

28
Figure 43 : Nombre de frondes observées pour les
différentes expériences pour le site de Damvix 5
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 44 : Nombre de frondes observées pour les
différentes expériences pour le site de Magné 2
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 45 : Nombre de frondes observées pour les
différentes expériences pour le site du Vanneau 4
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 46 : Nombre de frondes observées pour les
différentes expériences pour le site du Vanneau 5
0
50
100
150
200
250
300
350
400
450
500
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées
Figure 47 : Nombre de frondes observées pour les
différentes expériences pour le site du Mazeau 3
Attention l'échelle est différente
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Prélèvement
06-11-2009
Prélèvement
20-1-2010
Prélèvement
30-9-2010*
Prélèvement
22-11-2010
Prélèvement
10-2-2011
No
mb
re c
um
ulé
de f
ron
des o
bserv
ées

29
Commentaires Figures 41 à 47
L’expérience sur le site de Arçais 4 (figure 41) ne montre que le développement de Lemna
minor/minuta. Plus l’hiver avance, plus le nombre de fronde qui redémarre est élevé et ce
pour les deux années. Le passage au froid sur l’expérience de septembre 2010 n’a pas
d’influence notable ou au contraire une inhibition des lentilles.
Pour le site de Damvix 1 (figure 42), les résultats sont très différents entre les deux années.
Les 4 espèces ont été observées. Il semblerait que le passage au froid est favorisé l’apparition
de Wolffia arrhiza qui n’est pas présente sur les autres dates à part en très faible quantité en
novembre 2010. L’année 2010 a permis d’observer des croissances de Lemna gibba. L’année
2011 est marquée par des croissances importantes de Spirodela polyrhiza. Lemna
minor/minuta est toujours en faible quantité.
Le site de Damvix 5 (figure 43) présente les mêmes évolutions mais avec une quantité de
lentilles bien moindre. Pourtant les masses de lentilles observées sur les deux sites sont
comparables au mois d’octobre voire plus importantes à Damvix 5. Il semblerait que la
quantité de lentilles observées sur le site ne soit pas corrélée avec le nombre de propagules
dans le sédiment.
Pour le site de Magné 2 (figure 44), on observe une évolution semblable : Lemna gibba en
2009-2010, Spirodela polyrhiza en 2010-2011. Là encore le nombre de frondes observé est
relativement faible (moins de 150 frondes pour une espèce par prélèvement).
Le site du Vanneau 4 (figure 45) présente toujours de très faible quantité de frondes qui
croissent depuis les sédiments (maximum 100 frondes pour une date). Là encore, si on
compare par rapport à Vanneau 5 (figure 46) qui présente un nombre de frondes beaucoup
plus important, il n’y a pas de masse de lentilles observées in situ plus faible au contraire.
Pour le Vanneau 5 (figure 46), on observe sur le prélèvement de février 2011, trois espèces
simultanément. C’est le seul prélèvement pour lequel on observe des Lemna gibba en 2010-
2011. Ce site présente d’assez grandes quantités de Lemna minor/minuta.
Enfin, le site du Mazeau 3 (figure 47) présente une très grande quantité de Lemna
minor/minuta pour l’expérience de novembre 2010 (1830 frondes) alors que pour celle de
septembre 2010 avec passage au froid, il n’y avait quasiment pas de croissance (36 frondes).
Il semblerait que le froid artificiel ait diminué la reprise de croissance des Lemna
minor/minuta. Même si les quantités sont beaucoup plus faibles, l’effet semble inversé pour
Spirodela polyrhiza.
Globalement, les croissances sont très différentes entre l’hiver 2009-2010 et l’hiver 2010-
2011. Durant l’hiver 2009-2010, on observe des Lemna gibba alors qu’elles sont ensuite
quasiment absentes. Au contraire, la quantité de Spirodela polyrhiza est plus importante
durant l’hiver 2010-2011. Le froid artificiel (40 jours à 4°C en chambre froide) semble
inhiber la reprise de croissance des Lemna minor/minuta. Par contre, ce froid a permis
l’observation de Wolffia arrhiza jusque là non observée. Enfin, la quantité de lentilles
observées ne semble pas du tout corrélée à la masse de lentilles observée sur le terrain. Ce
sont donc d’autres facteurs qui interviennent dans la mise en sommeil des lentilles pendant
l’hiver. La comparaison des résultats obtenus avec les recouvrements observés sur le terrain
va permettre d’affiner cette conclusion.
30
Figure 48 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Arçais 4
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
Figure 49 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Damvix 1
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
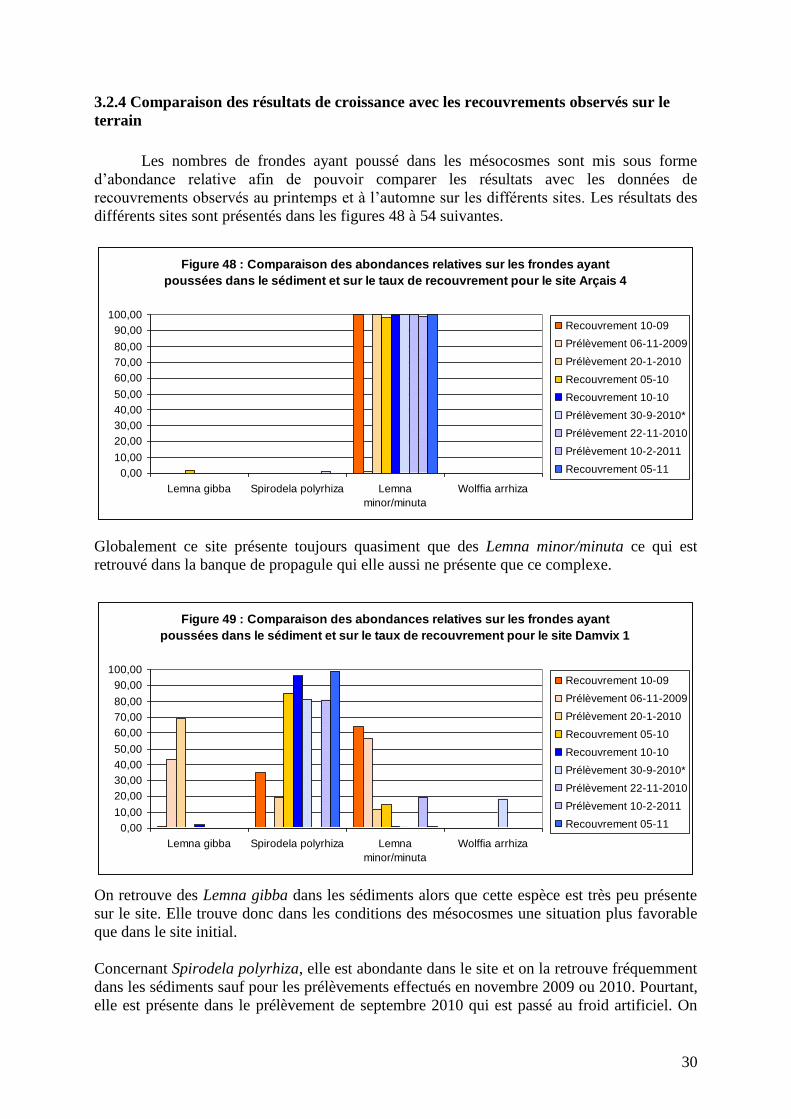
3.2.4 Comparaison des résultats de croissance avec les recouvrements observés sur le
terrain
Les nombres de frondes ayant poussé dans les mésocosmes sont mis sous forme
d’abondance relative afin de pouvoir comparer les résultats avec les données de
recouvrements observés au printemps et à l’automne sur les différents sites. Les résultats des
différents sites sont présentés dans les figures 48 à 54 suivantes.
Globalement ce site présente toujours quasiment que des Lemna minor/minuta ce qui est
retrouvé dans la banque de propagule qui elle aussi ne présente que ce complexe.
On retrouve des Lemna gibba dans les sédiments alors que cette espèce est très peu présente
sur le site. Elle trouve donc dans les conditions des mésocosmes une situation plus favorable
que dans le site initial.
Concernant Spirodela polyrhiza, elle est abondante dans le site et on la retrouve fréquemment
dans les sédiments sauf pour les prélèvements effectués en novembre 2009 ou 2010. Pourtant,
elle est présente dans le prélèvement de septembre 2010 qui est passé au froid artificiel. On

31
Figure 50 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Damvix 5
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
Figure 51 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Magné 2
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
peut supposer que cette espèce a une période de repos minimum avant de se développer à
partir du sédiment et/ou qu’il lui faut une quantité minimale de froid.
La quantité de Lemna minor/minuta décroit dans le site ce qui s’observe aussi dans les
croissances obtenues sur sédiment.
Enfin, on observe Wolffia arrhiza dans les croissances sur sédiments passés au froid artificiel
pourtant cette espèce n’est pas présente sur le site. Néanmoins, cette espèce est observable sur
d’autres sites de Damvix situés à proximité.
Ce site présente le même type d’évolution que Damvix 1. En octobre 2009, il n’y avait pas de
Spirodela polyrhiza observée sur site et il n’y en a pas eu dans les sédiments. Par contre, cette
espèce est observée en mai et octobre 2010 et cette fois elle est aussi abondante dans les
sédiments. La présence à partir de mai 2010 sur le site ne semble pas provenir des sédiments
eux-mêmes mais plutôt de sites adjacents qui présentent cette espèce.
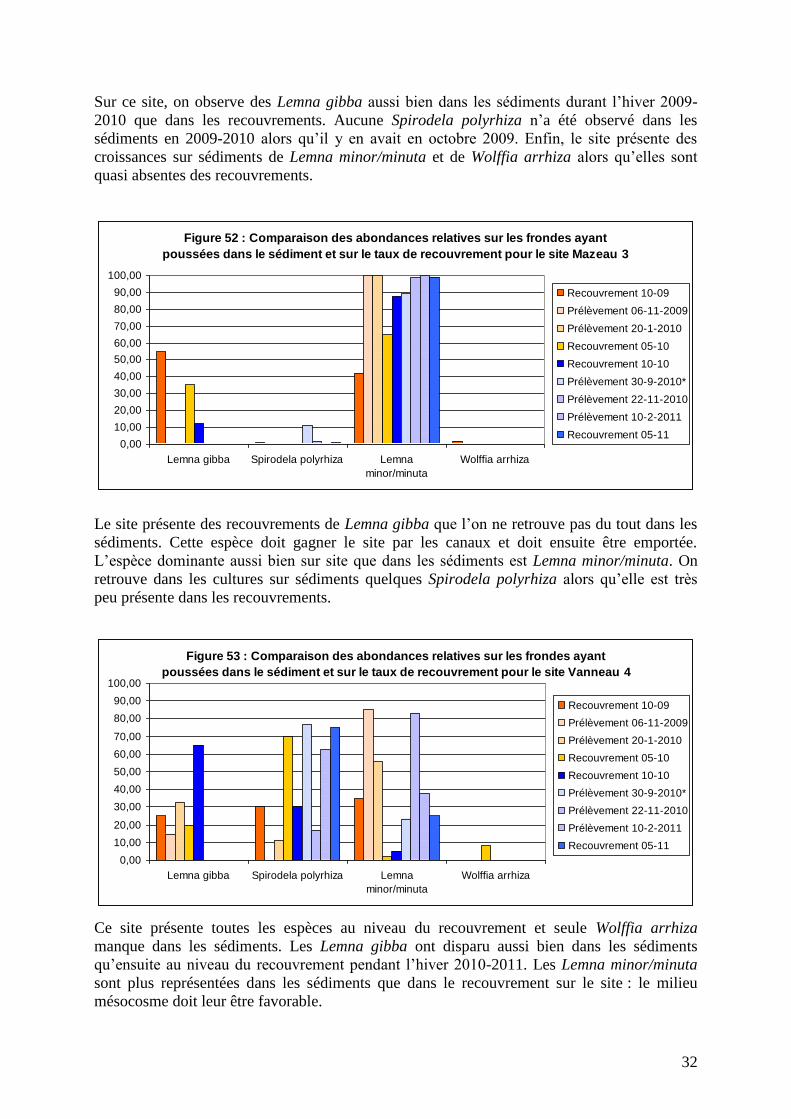
32
Figure 52 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Mazeau 3
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
Figure 53 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Vanneau 4
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
Sur ce site, on observe des Lemna gibba aussi bien dans les sédiments durant l’hiver 2009-
2010 que dans les recouvrements. Aucune Spirodela polyrhiza n’a été observé dans les
sédiments en 2009-2010 alors qu’il y en avait en octobre 2009. Enfin, le site présente des
croissances sur sédiments de Lemna minor/minuta et de Wolffia arrhiza alors qu’elles sont
quasi absentes des recouvrements.
Le site présente des recouvrements de Lemna gibba que l’on ne retrouve pas du tout dans les
sédiments. Cette espèce doit gagner le site par les canaux et doit ensuite être emportée.
L’espèce dominante aussi bien sur site que dans les sédiments est Lemna minor/minuta. On
retrouve dans les cultures sur sédiments quelques Spirodela polyrhiza alors qu’elle est très
peu présente dans les recouvrements.
Ce site présente toutes les espèces au niveau du recouvrement et seule Wolffia arrhiza
manque dans les sédiments. Les Lemna gibba ont disparu aussi bien dans les sédiments
qu’ensuite au niveau du recouvrement pendant l’hiver 2010-2011. Les Lemna minor/minuta
sont plus représentées dans les sédiments que dans le recouvrement sur le site : le milieu
mésocosme doit leur être favorable.
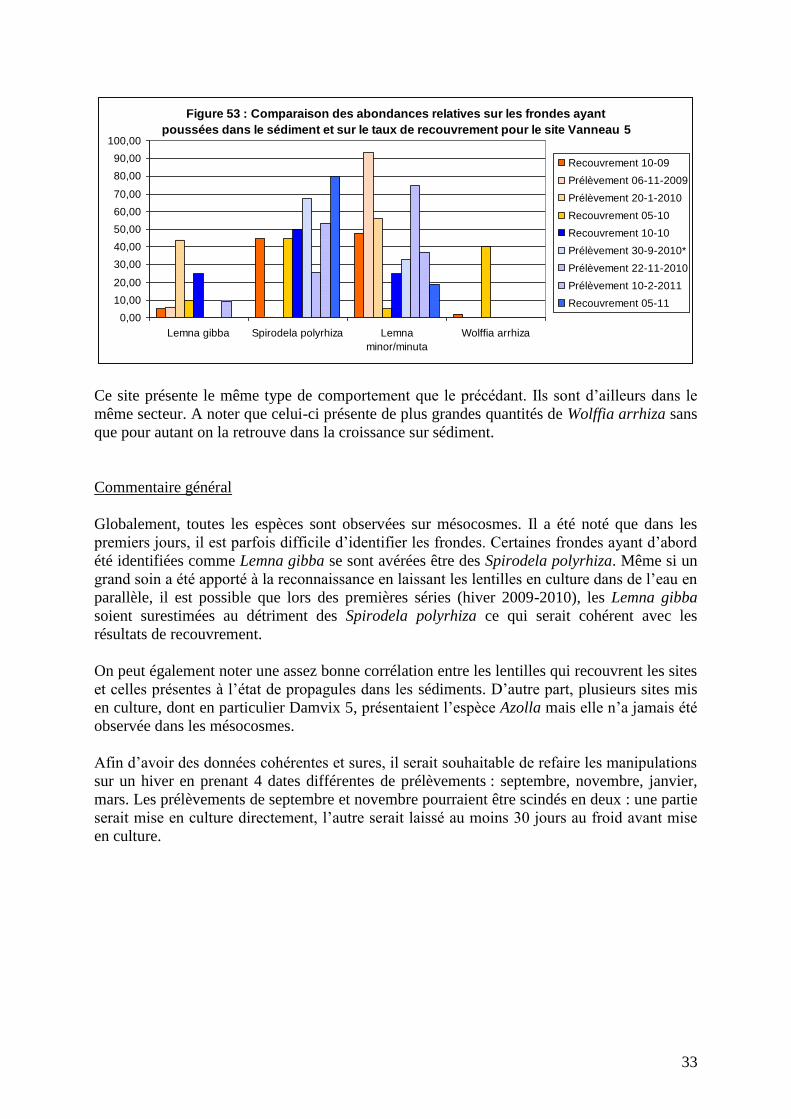
33
Figure 53 : Comparaison des abondances relatives sur les frondes ayant
poussées dans le sédiment et sur le taux de recouvrement pour le site Vanneau 5
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Lemna gibba Spirodela polyrhiza Lemna
minor/minuta
Wolffia arrhiza
Recouvrement 10-09
Prélèvement 06-11-2009
Prélèvement 20-1-2010
Recouvrement 05-10
Recouvrement 10-10
Prélèvement 30-9-2010*
Prélèvement 22-11-2010
Prélèvement 10-2-2011
Recouvrement 05-11
Ce site présente le même type de comportement que le précédant. Ils sont d’ailleurs dans le
même secteur. A noter que celui-ci présente de plus grandes quantités de Wolffia arrhiza sans
que pour autant on la retrouve dans la croissance sur sédiment.
Commentaire général
Globalement, toutes les espèces sont observées sur mésocosmes. Il a été noté que dans les
premiers jours, il est parfois difficile d’identifier les frondes. Certaines frondes ayant d’abord
été identifiées comme Lemna gibba se sont avérées être des Spirodela polyrhiza. Même si un
grand soin a été apporté à la reconnaissance en laissant les lentilles en culture dans de l’eau en
parallèle, il est possible que lors des premières séries (hiver 2009-2010), les Lemna gibba
soient surestimées au détriment des Spirodela polyrhiza ce qui serait cohérent avec les
résultats de recouvrement.
On peut également noter une assez bonne corrélation entre les lentilles qui recouvrent les sites
et celles présentes à l’état de propagules dans les sédiments. D’autre part, plusieurs sites mis
en culture, dont en particulier Damvix 5, présentaient l’espèce Azolla mais elle n’a jamais été
observée dans les mésocosmes.
Afin d’avoir des données cohérentes et sures, il serait souhaitable de refaire les manipulations
sur un hiver en prenant 4 dates différentes de prélèvements : septembre, novembre, janvier,
mars. Les prélèvements de septembre et novembre pourraient être scindés en deux : une partie
serait mise en culture directement, l’autre serait laissé au moins 30 jours au froid avant mise
en culture.

34
CONCLUSION
Les résultats des suivis de 2011 et les comparaisons avec les données des années
précédentes confirment plusieurs tendances déjà observées.
Les biomasses moyennes prélevées en 2011 sont inférieures à celles de 2010 et les
variations au sein même des sites sont souvent importantes. Excepté 2007 où les valeurs de
biomasses sont exceptionnellement fortes par rapport à celles des autres années, les évolutions
depuis 2008 diffèrent principalement selon les sites. Généralement les biomasses prélevées
par site sont assez stables et donc comparables, avec moins de 100g de lentilles par
échantillon.
Les suivis à partir des estimations des recouvrements ont permis d’observer quatre
espèces de lentilles en 2011 et l’évolution par rapport à 2010 est la suivante : augmentation
des Spirodela polyrhiza et des Lemna minor/minuta (94% des recouvrements en 2011 contre
73% en 2010) et diminution des Lemna gibba et des Wolffia arrhiza. Depuis 2008, on
constate globalement une augmentation constante de Spirodela polyrhiza, des variations
importantes de Lemna gibba, une faible représentation de Wolffia arrhiza et une stabilisation
de Lemna minor/minuta (notamment depuis 2009). Il existe toujours une hétérogénéite dans la
répartition des espèces entre les sites (secteurs de marais) et également à l’intérieur d’un site
(zone de prélèvement = fossé).
Les résultats des analyses physicochimiques réalisées pour chaque prélèvement
montrent des variations globales des facteurs mais ne permettent pas de démontrer un lien
entre espèce de lentille et qualité de l’eau. Les facteurs pouvant expliquer les répartitions
d’espèce varient pour tous les prélèvements ce qui ne permet pas de déterminer un schéma
général.
De grandes tendances se dessinent et semblent plutôt en lien avec les conditions
météorologiques de ces dernières années (hivers rigoureux entre autre) : biomasses en
diminution, augmentation de Spirodela polyrhiza au détriment de Lemna minor/minuta (dont
on a démontré par ailleurs sur les expériences sur mésocosmes qu’elles n’aimaient pas
beaucoup le froid), quantité de Lemna gibba très variable et faible quantité de Wolffia arrhiza.
Les résultats des expériences de cultures réalisées depuis 2009 nous permettent de dire
que des lentilles peuvent se développer à partir de sédiments et que seule l’observation sur site
de présence ou non de lentilles à la surface de l’eau n’est pas suffisante pour conclure sur la
notion de présence/absence de lemnacées. On remarque toutefois des résultats différents selon
les sites de prélèvements, les espèces de lentille et la date du prélèvement. Les observations
faites cette année à la loupe laissent supposer que c’est le turion des lentilles qui se conserve
dans les sédiments (pas de confirmation cette année lors des cultures de turions identifiés).

35
Concernant les banques de propagules de lentilles contenues dans les sédiments, il
semble qu’elles soient assez bien corrélées avec les recouvrements observés sur le terrain. Il
est à noter que toutes les espèces ne sont pas aptes à reprendre leur croissance en même
temps, le froid ayant apparemment un effet significatif. Ce rythme pourrait expliquer en partie
certains recouvrements observés en décalage dans le temps tout en faisant le lien avec la
compétition interspécifique existante et aux mouvements d’eau dans le marais (quantitatifs et
qualitatifs).
Les résultats et les données issus depuis 2007 des suivis de la répartition des lentilles,
des analyses de la qualité de l’eau, des éléments biologiques sur les espèces,...démontrent
qu’il s’agit bien d’un système très complexe et qu’il est difficile de mettre en évidence des
facteurs structurants stables malgré l’ampleur des prélèvements réalisés. Beaucoup
d’informations nouvelles sur ces végétaux flottants sont à ce jour connues mais des
observations similaires devront être poursuivies dans le temps ainsi que d’autres essais
méritent d’être mis en œuvre pour affiner et consolider certaines tendances.

36
ANNEXES
Annexe 1 : Répartition des sites de prélèvements de lentilles en 2011
Annexe 2 : Cartes des sites de prélèvements pour mai 2011
Annexe 3 : Résultats bruts par échantillon pour chaque site en mai 2011
Annexe 4 : Résultats bruts par échantillon pour chaque site en mai 2010
Annexe 5 : Coefficients de corrélation des ACP pour les résultats de mai 2011
Annexe 6 : Localisation des prélèvements de sédiments sur la période 2009-2011
Annexe 7 : Suivi de la croissance du nombre de frondes à partir des sédiments
37
Annexe 1 :
Répartition des sites de prélèvements de lentilles en 2011
dans le Marais poitevin
38
Annexe 2 : Cartes des sites de prélèvements de lentilles en 2011
39

40
41

42
Annexe 3 :
Résultats bruts par échantillon pour chaque site en mai 2011
Site de prélèvement ARCAIS
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 16,7 16 17 17 16,5 16,64 0,42
Concentration O2
(mg/L) 0,36 0,21 0,17 1,21 0,92
0,57 0,47
Conductimétrie
(µS/cm) 453 453 447 783 428
512,80 151,39
pH 8,23 8,32 8,24 8,26 8 8,21 0,12
Turbidité 6,78 11,4 5,48 6,68 3,28 6,72 2,97
Profondeur (cm) 10 10 20 10 25 15,00 7,07
Nitrates (mg/L) 0,6 0,7 0,6 0 0,9 0,56 0,34
Phosphates (mg/L) 0,11 0,09 0,06 1,19 0,02 0,29 0,50
Quantité de lentilles
Masse totale (g) 30,90 29,10 40,30 79,40 136,80 63,30 45,85
Masse de déchets (g) 8,00 5,50 19,80 50,80 0,10 16,84 20,30
Masse de lentilles (g) 22,90 23,60 20,50 28,60 136,70 46,46 50,53
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Spirodela polyrhiza 1,00 2,00 1,00 0,00 1,00 1,00 0,71
Lemna minor/minuta 99,00 98,00 99,00 100,00 99,00 99,00 0,71
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

43
Site de prélèvement DAMVIX
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 18,2 16,7 13,7 14,4 18 16,20 2,06
Concentration O2
(mg/L) 0,39 3,53 0,18 0,16 3,42
1,54 1,77
Conductimétrie
(µS/cm) 453 631 736 1955 461
847,20 630,65
pH 8,3 8,1 7,58 8,25 8,41 8,13 0,33
Turbidité 2,57 1,52 12 16,9 9,52 8,50 6,48
Profondeur (cm) 20 30 80 25 50 41,00 24,60
Nitrates (mg/L) 1,6 1,4 0,6 0 0,4 0,80 0,68
Phosphates (mg/L) 0,11 0,1 0,06 0,04 0,16 0,09 0,05
Quantité de lentilles
Masse totale (g) 50,90 110,10 127,10 141,40 51,00 96,10 42,68
Masse de déchets (g) 31,70 17,50 50,00 1,60 34,00 26,96 18,27
Masse de lentilles (g) 19,20 92,60 77,10 139,80 17,00 69,14 52,01
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 1,00 98,00 15,00 0,00 22,80 42,52
Spirodela polyrhiza 99,00 93,00 1,00 0,00 95,00 57,60 52,17
Lemna minor/minuta 1,00 5,00 0,00 85,00 5,00 19,20 36,85
Wolffia arrhiza 0,00 0,00 1,00 0,00 0,00 0,20 0,45
Autres espèces 0,00 1,00 0,00 0,00 0,00 0,20 0,45
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

44
Site de prélèvement MAGNE
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 17,1 23,5 18,1 18,1 16,1 18,58 2,87
Concentration O2
(mg/L) 0,11 0,15 0,23 1,95 0,63
0,61 0,77
Conductimétrie
(µS/cm) 382 500 647 605 539
534,60 102,58
pH 7,89 7,75 7,85 8,01 7,86 7,87 0,09
Turbidité 21,9 45,4 88,6 9,07 10,1 35,01 33,34
Profondeur (cm) 30 20 10 45 60 33,00 19,87
Nitrates (mg/L) 0,400 0,100 0,6 0,1 0,3 0,30 0,21
Phosphates (mg/L) 0,16 1,04 1,65 0,21 0,65 0,74 0,62
Quantité de lentilles
Masse totale (g) 90,50 115,00 68,20 96,20 185,20 111,02 44,70
Masse de déchets (g) 5,00 6,10 4,10 1,80 0,00 3,40 2,47
Masse de lentilles (g) 85,50 108,90 64,10 94,40 185,20 107,62 46,31
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 25,00 50,00 0,00 0,00 15,00 22,36
Spirodela polyrhiza 70,00 75,00 40,00 99,00 97,00 76,20 23,99
Lemna minor/minuta 30,00 0,00 10,00 0,00 2,00 8,40 12,76
Wolffia arrhiza 0,00 0,00 0,00 1,00 1,00 0,40 0,55
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

45
Site de prélèvement MAILLEZAIS
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 20,6 19,4 18,1 20,9 19,5 19,70 1,11
Concentration O2
(mg/L) 0,14 0,15 6,73 6,13 0,14
2,66 3,45
Conductimétrie
(µS/cm) 365 393 365 355 364
368,40 14,38
pH 9,5 8,25 8,18 8,39 8,44 8,55 0,54
Turbidité 4,82 5,31 8,51 3,84 2,21 4,94 2,32
Profondeur (cm) 80 40 100 70 80 74,00 21,91
Nitrates (mg/L) 0,5 0,4 1 0,5 0,6 0,60 0,23
Phosphates (mg/L) 0,12 0,06 0,03 0,05 0,03 0,06 0,04
Quantité de lentilles
Masse totale (g) 75,80 43,40 66,60 61,60 36,10 56,70 16,49
Masse de déchets (g) 2,20 5,90 11,80 7,70 1,80 5,88 4,14
Masse de lentilles (g) 73,60 37,50 54,80 53,90 34,30 50,82 15,77
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Spirodela polyrhiza 96,00 99,00 88,00 80,00 98,00 92,20 8,07
Lemna minor/minuta 2,00 1,00 2,00 15,00 1,00 4,20 6,06
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 2,00 0,00 10,00 5,00 1,00 3,60 4,04
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

46
Site de prélèvement MAZEAU
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 18,5 18,7 21 24,6 20,6 20,68 2,46
Concentration O2
(mg/L) 0,18 1,08 0,27 1,46 0,3
0,66 0,58
Conductimétrie
(µS/cm) 724 974 378 355 384
563,00 275,85
pH 8,18 7,85 8,1 8,15 8,86 8,23 0,38
Turbidité 7,72 3,15 16,8 10,9 5,69 8,85 5,27
Profondeur (cm) 5 15 15 2 10 9,40 5,86
Nitrates (mg/L) 0,5 0,5 0,3 0,5 0,1 0,38 0,18
Phosphates (mg/L) 0,25 0,12 0,15 0,28 0,15 0,19 0,07
Quantité de lentilles
Masse totale (g) 61,50 50,40 42,90 48,90 62,10 53,16 8,37
Masse de déchets (g) 27,00 42,00 14,20 18,00 4,60 21,16 14,15
Masse de lentilles (g) 34,50 8,40 28,70 30,90 57,50 32,00 17,50
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Spirodela polyrhiza 1,00 1,00 1,00 1,00 0,00 0,80 0,45
Lemna minor/minuta 99,00 99,00 99,00 99,00 100,00 99,20 0,45
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

47
Site de prélèvement ST HILAIRE LA PALUD
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 17,4 16,2 18,3 17,3 15,8 17,00 1,00
Concentration O2
(mg/L) 3,99 9,56 1,53 2,13 0,17
3,48 3,67
Conductimétrie
(µS/cm) 454 457 444 431 431
443,40 12,30
pH 8,16 8,25 8,14 8,34 8,28 8,23 0,08
Turbidité 1,29 2,18 10,2 5,06 6,9 5,13 3,62
Profondeur (cm) 30 10 15 30 30 23,00 9,75
Nitrates (mg/L) 3,5 0,6 0,5 1,5 0,1 1,24 1,36
Phosphates (mg/L) 0,02 0,21 0,1 0,03 0,04 0,08 0,08
Quantité de lentilles
Masse totale (g) 52,30 32,80 43,30 58,00 47,10 46,70 9,54
Masse de déchets (g) 26,50 16,00 10,00 38,50 20,40 22,28 10,89
Masse de lentilles (g) 25,80 16,80 33,30 19,50 26,70 24,42 6,49
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Spirodela polyrhiza 2,00 2,00 60,00 2,00 3,00 13,80 25,83
Lemna minor/minuta 98,00 98,00 40,00 98,00 97,00 86,20 25,83
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00
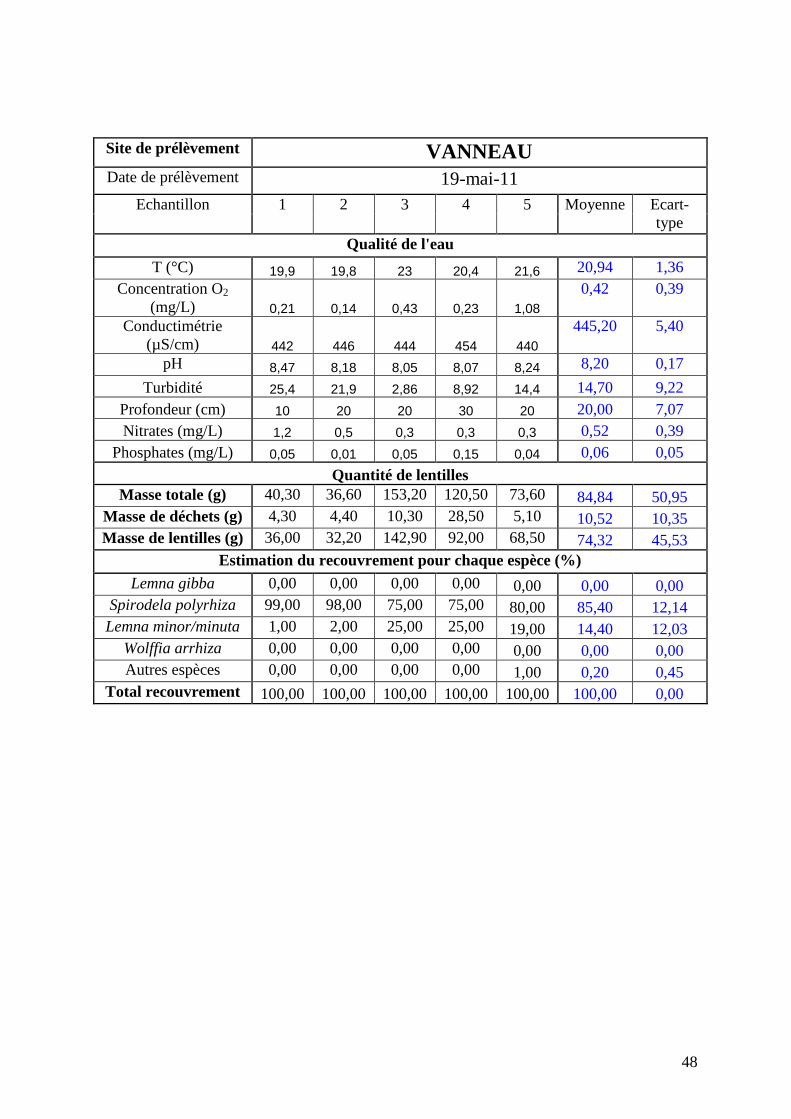
48
Site de prélèvement VANNEAU
Date de prélèvement 19-mai-11
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 19,9 19,8 23 20,4 21,6 20,94 1,36
Concentration O2
(mg/L) 0,21 0,14 0,43 0,23 1,08
0,42 0,39
Conductimétrie
(µS/cm) 442 446 444 454 440
445,20 5,40
pH 8,47 8,18 8,05 8,07 8,24 8,20 0,17
Turbidité 25,4 21,9 2,86 8,92 14,4 14,70 9,22
Profondeur (cm) 10 20 20 30 20 20,00 7,07
Nitrates (mg/L) 1,2 0,5 0,3 0,3 0,3 0,52 0,39
Phosphates (mg/L) 0,05 0,01 0,05 0,15 0,04 0,06 0,05
Quantité de lentilles
Masse totale (g) 40,30 36,60 153,20 120,50 73,60 84,84 50,95
Masse de déchets (g) 4,30 4,40 10,30 28,50 5,10 10,52 10,35
Masse de lentilles (g) 36,00 32,20 142,90 92,00 68,50 74,32 45,53
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Spirodela polyrhiza 99,00 98,00 75,00 75,00 80,00 85,40 12,14
Lemna minor/minuta 1,00 2,00 25,00 25,00 19,00 14,40 12,03
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 1,00 0,20 0,45
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00
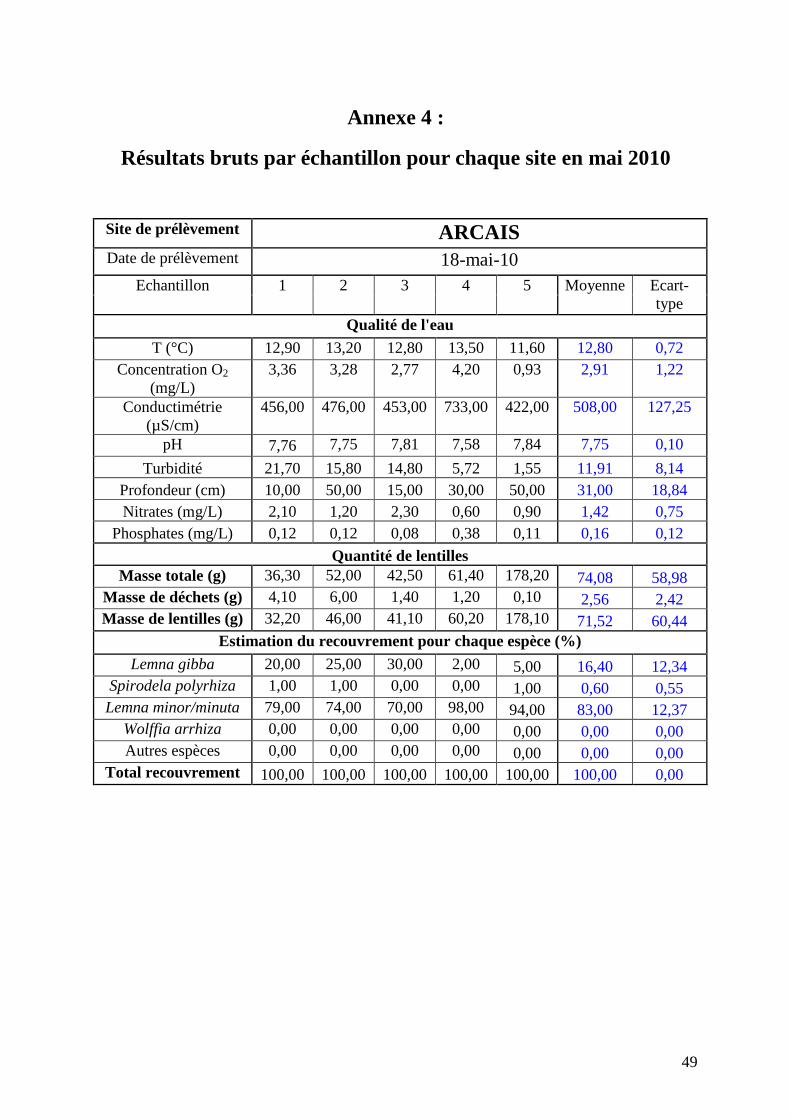
49
Annexe 4 :
Résultats bruts par échantillon pour chaque site en mai 2010
Site de prélèvement ARCAIS
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 12,90 13,20 12,80 13,50 11,60 12,80 0,72
Concentration O2
(mg/L)
3,36 3,28 2,77 4,20 0,93 2,91 1,22
Conductimétrie
(µS/cm)
456,00 476,00 453,00 733,00 422,00 508,00 127,25
pH 7,76 7,75 7,81 7,58 7,84 7,75 0,10
Turbidité 21,70 15,80 14,80 5,72 1,55 11,91 8,14
Profondeur (cm) 10,00 50,00 15,00 30,00 50,00 31,00 18,84
Nitrates (mg/L) 2,10 1,20 2,30 0,60 0,90 1,42 0,75
Phosphates (mg/L) 0,12 0,12 0,08 0,38 0,11 0,16 0,12
Quantité de lentilles
Masse totale (g) 36,30 52,00 42,50 61,40 178,20 74,08 58,98
Masse de déchets (g) 4,10 6,00 1,40 1,20 0,10 2,56 2,42
Masse de lentilles (g) 32,20 46,00 41,10 60,20 178,10 71,52 60,44
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 20,00 25,00 30,00 2,00 5,00 16,40 12,34
Spirodela polyrhiza 1,00 1,00 0,00 0,00 1,00 0,60 0,55
Lemna minor/minuta 79,00 74,00 70,00 98,00 94,00 83,00 12,37
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00
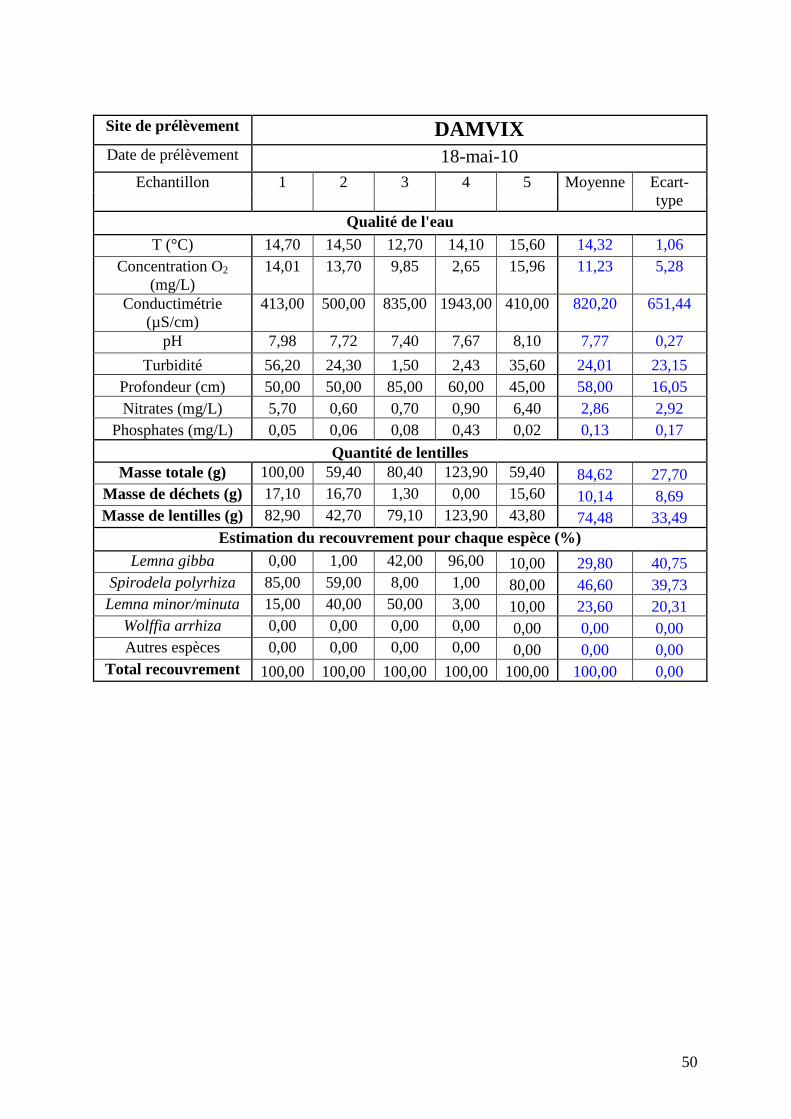
50
Site de prélèvement DAMVIX
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 14,70 14,50 12,70 14,10 15,60 14,32 1,06
Concentration O2
(mg/L)
14,01 13,70 9,85 2,65 15,96 11,23 5,28
Conductimétrie
(µS/cm)
413,00 500,00 835,00 1943,00 410,00 820,20 651,44
pH 7,98 7,72 7,40 7,67 8,10 7,77 0,27
Turbidité 56,20 24,30 1,50 2,43 35,60 24,01 23,15
Profondeur (cm) 50,00 50,00 85,00 60,00 45,00 58,00 16,05
Nitrates (mg/L) 5,70 0,60 0,70 0,90 6,40 2,86 2,92
Phosphates (mg/L) 0,05 0,06 0,08 0,43 0,02 0,13 0,17
Quantité de lentilles
Masse totale (g) 100,00 59,40 80,40 123,90 59,40 84,62 27,70
Masse de déchets (g) 17,10 16,70 1,30 0,00 15,60 10,14 8,69
Masse de lentilles (g) 82,90 42,70 79,10 123,90 43,80 74,48 33,49
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 1,00 42,00 96,00 10,00 29,80 40,75
Spirodela polyrhiza 85,00 59,00 8,00 1,00 80,00 46,60 39,73
Lemna minor/minuta 15,00 40,00 50,00 3,00 10,00 23,60 20,31
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00
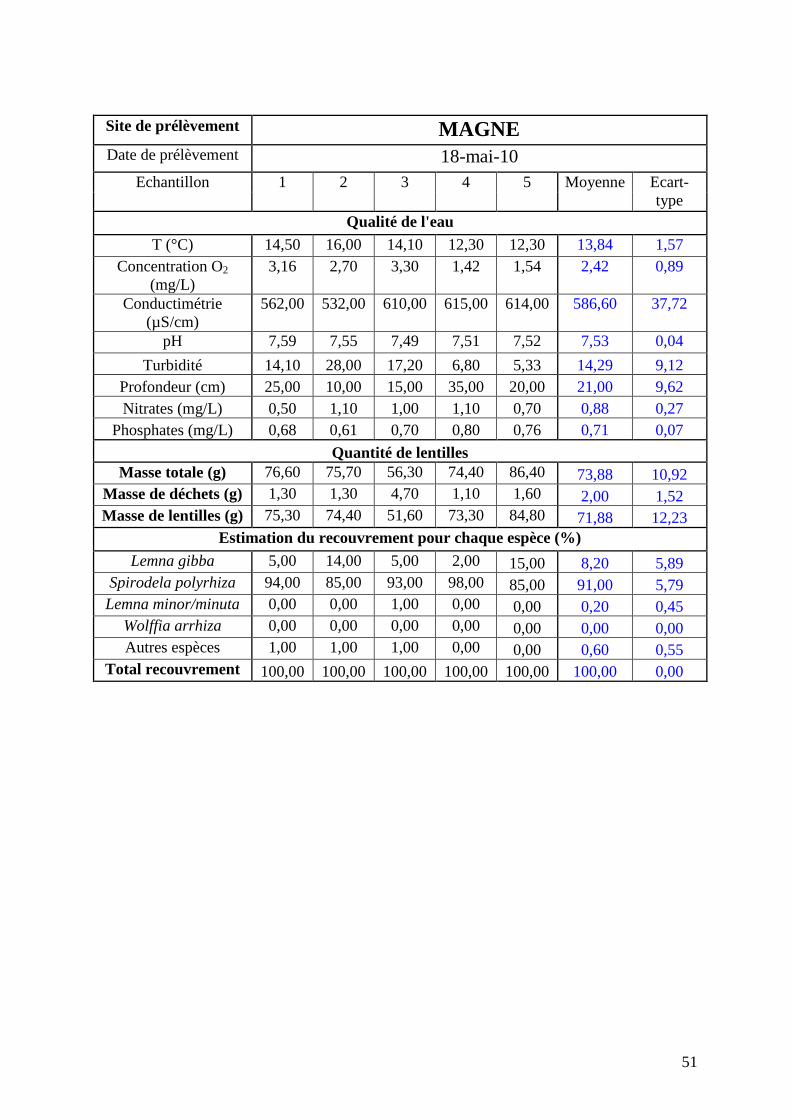
51
Site de prélèvement MAGNE
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 14,50 16,00 14,10 12,30 12,30 13,84 1,57
Concentration O2
(mg/L)
3,16 2,70 3,30 1,42 1,54 2,42 0,89
Conductimétrie
(µS/cm)
562,00 532,00 610,00 615,00 614,00 586,60 37,72
pH 7,59 7,55 7,49 7,51 7,52 7,53 0,04
Turbidité 14,10 28,00 17,20 6,80 5,33 14,29 9,12
Profondeur (cm) 25,00 10,00 15,00 35,00 20,00 21,00 9,62
Nitrates (mg/L) 0,50 1,10 1,00 1,10 0,70 0,88 0,27
Phosphates (mg/L) 0,68 0,61 0,70 0,80 0,76 0,71 0,07
Quantité de lentilles
Masse totale (g) 76,60 75,70 56,30 74,40 86,40 73,88 10,92
Masse de déchets (g) 1,30 1,30 4,70 1,10 1,60 2,00 1,52
Masse de lentilles (g) 75,30 74,40 51,60 73,30 84,80 71,88 12,23
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 5,00 14,00 5,00 2,00 15,00 8,20 5,89
Spirodela polyrhiza 94,00 85,00 93,00 98,00 85,00 91,00 5,79
Lemna minor/minuta 0,00 0,00 1,00 0,00 0,00 0,20 0,45
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 1,00 1,00 1,00 0,00 0,00 0,60 0,55
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

52
Site de prélèvement MAILLEZAIS
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 20,70 17,80 18,30 18,50 16,80 18,42 1,43
Concentration O2
(mg/L)
25,00 6,09 4,75 7,88 4,16 9,58 8,74
Conductimétrie
(µS/cm)
324,00 409,00 413,00 417,00 435,00 399,60 43,41
pH 9,19 7,80 7,97 8,04 7,65 8,13 0,61
Turbidité 51,00 17,50 11,70 11,60 3,61 19,08 18,51
Profondeur (cm) 130,00 90,00 40,00 10,00 60,00 66,00 46,15
Nitrates (mg/L) 1,30 3,60 2,90 2,70 1,20 2,34 1,05
Phosphates (mg/L) 0,18 0,07 0,09 0,03 0,10 0,09 0,06
Quantité de lentilles
Masse totale (g) 84,00 80,60 58,10 116,90 106,80 89,28 23,16
Masse de déchets (g) 19,00 10,70 6,70 27,70 13,00 15,42 8,18
Masse de lentilles (g) 65,00 69,90 51,40 89,20 93,80 73,86 17,55
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 40,00 65,00 45,00 35,00 20,00 41,00 16,36
Spirodela polyrhiza 50,00 29,00 40,00 35,00 35,00 37,80 7,85
Lemna minor/minuta 10,00 6,00 15,00 30,00 45,00 21,20 16,12
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

53
Site de prélèvement MAZEAU
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 16,20 15,90 16,10 16,50 22,20 17,38 2,70
Concentration O2
(mg/L)
5,95 7,58 6,59 2,34 3,79 5,25 2,14
Conductimétrie
(µS/cm)
505,00 502,00 415,00 419,00 417,00 451,60 47,41
pH 7,90 8,14 7,77 7,61 7,72 7,83 0,20
Turbidité 11,00 49,40 86,50 71,40 14,60 46,58 33,56
Profondeur (cm) 10,00 10,00 8,00 10,00 10,00 9,60 0,89
Nitrates (mg/L) 0,60 0,60 0,20 1,00 0,30 0,54 0,31
Phosphates (mg/L) 0,14 0,15 0,16 0,09 0,07 0,12 0,04
Quantité de lentilles
Masse totale (g) 55,30 61,10 32,50 69,10 122,30 68,06 33,24
Masse de déchets (g) 5,20 6,30 1,50 3,40 0,60 3,40 2,40
Masse de lentilles (g) 50,10 54,80 31,00 65,70 121,70 64,66 34,27
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 70,00 80,00 35,00 45,00 65,00 59,00 18,51
Spirodela polyrhiza 1,00 1,00 0,00 0,00 0,00 0,40 0,55
Lemna minor/minuta 28,00 19,00 65,00 55,00 35,00 40,40 19,10
Wolffia arrhiza 1,00 0,00 0,00 0,00 0,00 0,20 0,45
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

54
Site de prélèvement ST HILAIRE LA PALUD
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 13,40 12,90 14,90 12,50 12,40 13,22 1,02
Concentration O2
(mg/L)
4,49 0,87 3,91 2,29 1,80 2,67 1,50
Conductimétrie
(µS/cm)
442,00 480,00 449,00 534,00 563,00 493,60 53,12
pH 7,91 7,73 7,74 7,75 7,86 7,80 0,08
Turbidité 12,40 19,70 6,53 5,00 5,05 9,74 6,35
Profondeur (cm) 60,00 30,00 30,00 40,00 25,00 37,00 13,96
Nitrates (mg/L) 4,80 0,60 2,10 0,60 0,70 1,76 1,81
Phosphates (mg/L) 0,15 0,57 0,15 0,21 0,36 0,29 0,18
Quantité de lentilles
Masse totale (g) 98,80 102,30 64,20 106,30 42,60 82,84 28,06
Masse de déchets (g) 50,10 47,50 8,40 1,00 12,60 23,92 23,11
Masse de lentilles (g) 48,70 54,80 55,80 105,30 30,00 58,92 27,92
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 0,00 0,00 0,00 2,00 0,00 0,40 0,89
Spirodela polyrhiza 20,00 5,00 97,00 3,00 2,00 25,40 40,69
Lemna minor/minuta 80,00 94,00 3,00 65,00 98,00 68,00 38,58
Wolffia arrhiza 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Autres espèces 0,00 1,00 0,00 30,00 0,00 6,20 13,31
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00
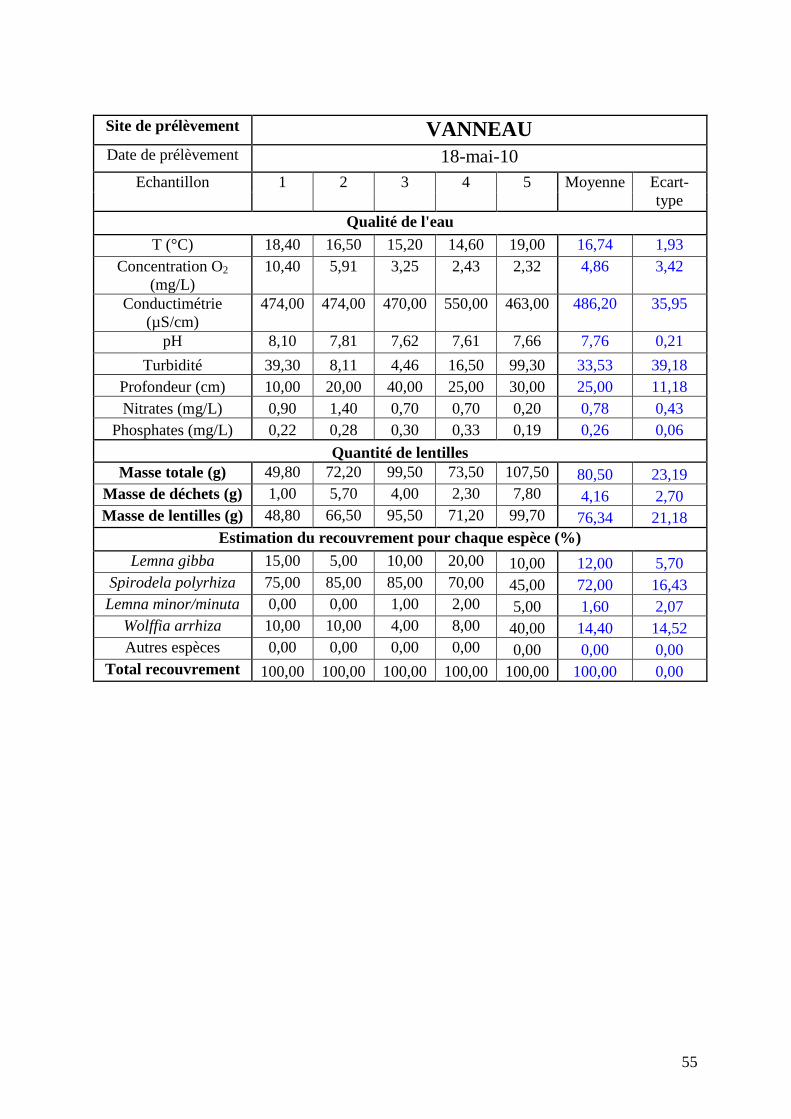
55
Site de prélèvement VANNEAU
Date de prélèvement 18-mai-10
Echantillon 1 2 3 4 5 Moyenne Ecart-
type
Qualité de l'eau
T (°C) 18,40 16,50 15,20 14,60 19,00 16,74 1,93
Concentration O2
(mg/L)
10,40 5,91 3,25 2,43 2,32 4,86 3,42
Conductimétrie
(µS/cm)
474,00 474,00 470,00 550,00 463,00 486,20 35,95
pH 8,10 7,81 7,62 7,61 7,66 7,76 0,21
Turbidité 39,30 8,11 4,46 16,50 99,30 33,53 39,18
Profondeur (cm) 10,00 20,00 40,00 25,00 30,00 25,00 11,18
Nitrates (mg/L) 0,90 1,40 0,70 0,70 0,20 0,78 0,43
Phosphates (mg/L) 0,22 0,28 0,30 0,33 0,19 0,26 0,06
Quantité de lentilles
Masse totale (g) 49,80 72,20 99,50 73,50 107,50 80,50 23,19
Masse de déchets (g) 1,00 5,70 4,00 2,30 7,80 4,16 2,70
Masse de lentilles (g) 48,80 66,50 95,50 71,20 99,70 76,34 21,18
Estimation du recouvrement pour chaque espèce (%)
Lemna gibba 15,00 5,00 10,00 20,00 10,00 12,00 5,70
Spirodela polyrhiza 75,00 85,00 85,00 70,00 45,00 72,00 16,43
Lemna minor/minuta 0,00 0,00 1,00 2,00 5,00 1,60 2,07
Wolffia arrhiza 10,00 10,00 4,00 8,00 40,00 14,40 14,52
Autres espèces 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Total recouvrement 100,00 100,00 100,00 100,00 100,00 100,00 0,00

56
Annexe 5 : Coefficients de corrélation des ACP pour les données de mai 2011
Matrice de corrélation (Pearson (n)) :
Variables O2 Cond pH Turb Nit Phos Masse L. gibba Spirodela L. minor Wolffia Azolla
O2 1 -0,141 0,033 -0,246 0,289 -0,122 -0,182 -0,165 0,024 0,023 -0,071 0,532
Cond -0,141 1 -0,194 0,108 -0,191 0,114 0,280 0,258 -0,286 0,178 0,105 -0,166
pH 0,033 -0,194 1 -0,307 0,019 -0,276 -0,262 -0,433 0,129 0,049 -0,370 0,166
Turb -0,246 0,108 -0,307 1 -0,164 0,734 0,130 0,467 0,074 -0,260 -0,031 -0,106
Nit 0,289 -0,191 0,019 -0,164 1 -0,233 -0,254 -0,064 -0,046 0,069 -0,144 0,079
Phos -0,122 0,114 -0,276 0,734 -0,233 1 0,156 0,321 -0,039 -0,089 0,077 -0,146
Masse -0,182 0,280 -0,262 0,130 -0,254 0,156 1 0,171 0,266 -0,331 0,436 0,001
L. gibba -0,165 0,258 -0,433 0,467 -0,064 0,321 0,171 1 -0,170 -0,247 0,452 -0,089
Spirodela 0,024 -0,286 0,129 0,074 -0,046 -0,039 0,266 -0,170 1 -0,912 0,134 0,287
L. minor 0,023 0,178 0,049 -0,260 0,069 -0,089 -0,331 -0,247 -0,912 1 -0,319 -0,282
Wolffia -0,071 0,105 -0,370 -0,031 -0,144 0,077 0,436 0,452 0,134 -0,319 1 -0,094
Azolla 0,532 -0,166 0,166 -0,106 0,079 -0,146 0,001 -0,089 0,287 -0,282 -0,094 1

57
Annexe 6 : Localisation des prélèvements (P) de sédiments sur la période 2009-2011
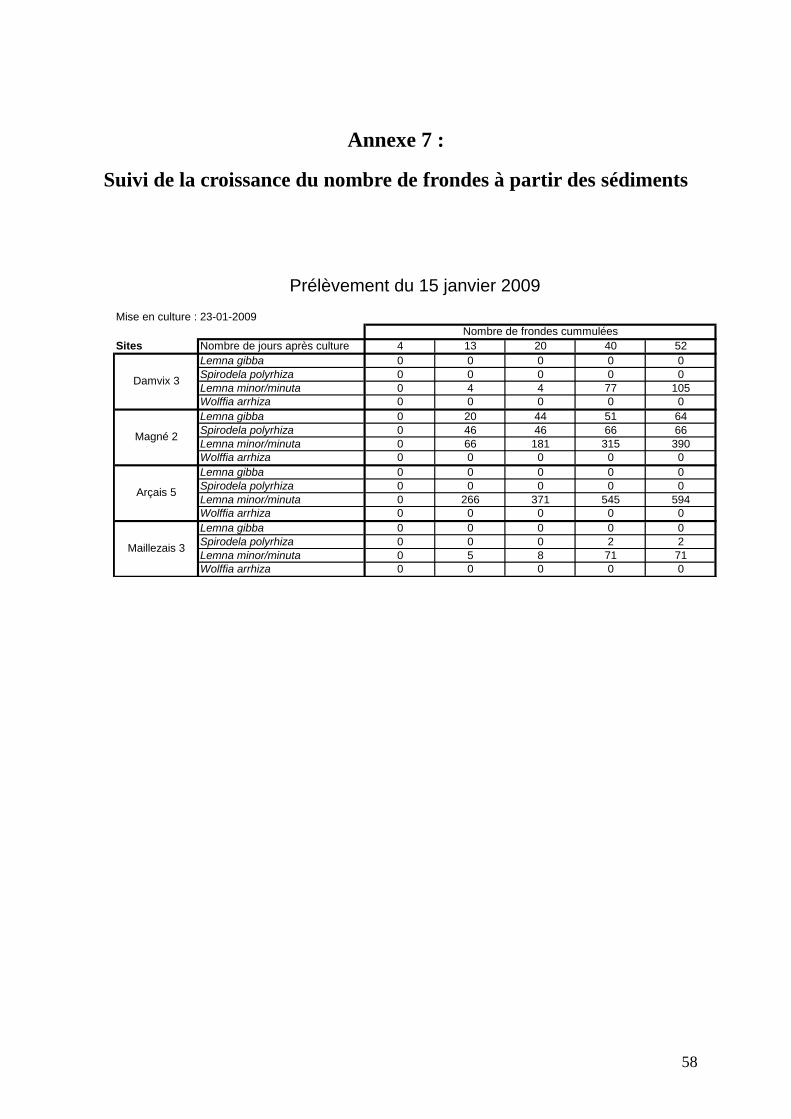
58
Annexe 7 :
Suivi de la croissance du nombre de frondes à partir des sédiments
Mise en culture : 23-01-2009
Sites Nombre de jours après culture 4 13 20 40 52
Lemna gibba 0 0 0 0 0
Spirodela polyrhiza 0 0 0 0 0
Lemna minor/minuta 0 4 4 77 105
Wolffia arrhiza 0 0 0 0 0
Lemna gibba 0 20 44 51 64
Spirodela polyrhiza 0 46 46 66 66
Lemna minor/minuta 0 66 181 315 390
Wolffia arrhiza 0 0 0 0 0
Lemna gibba 0 0 0 0 0
Spirodela polyrhiza 0 0 0 0 0
Lemna minor/minuta 0 266 371 545 594
Wolffia arrhiza 0 0 0 0 0
Lemna gibba 0 0 0 0 0
Spirodela polyrhiza 0 0 0 2 2
Lemna minor/minuta 0 5 8 71 71
Wolffia arrhiza 0 0 0 0 0
Maillezais 3
Prélèvement du 15 janvier 2009
Nombre de frondes cummulées
Damvix 3
Magné 2
Arçais 5
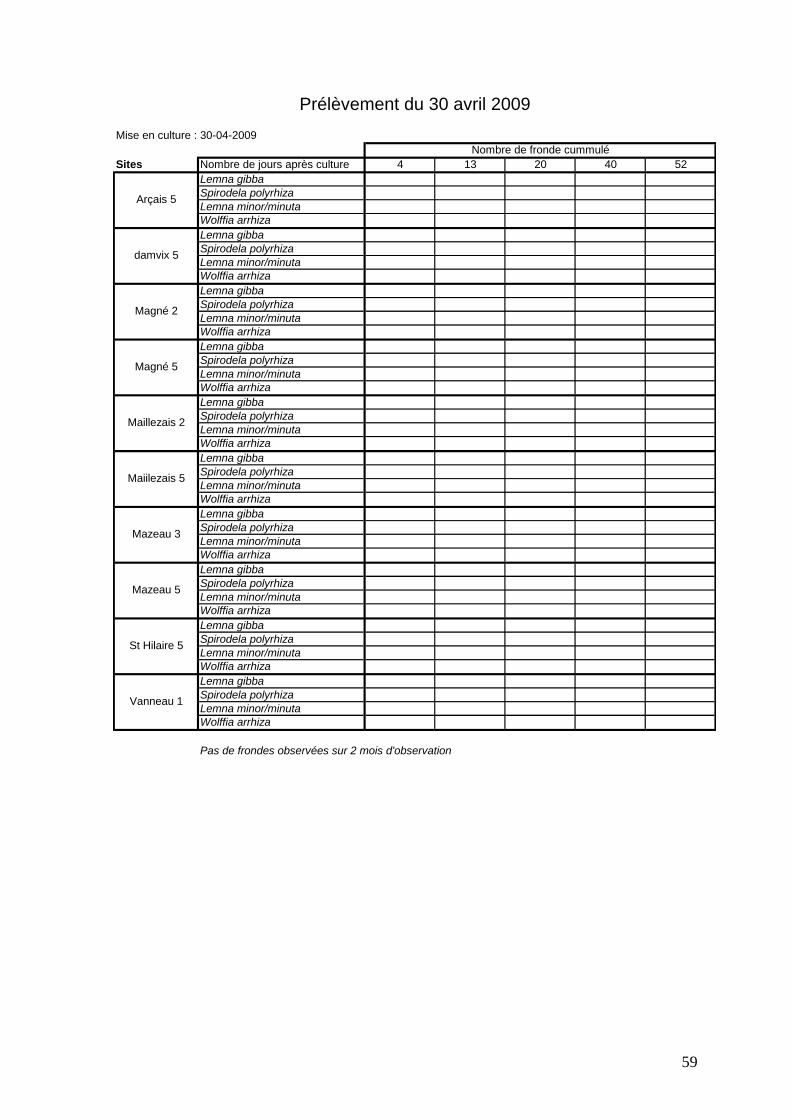
59
Mise en culture : 30-04-2009
Sites Nombre de jours après culture 4 13 20 40 52
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Lemna gibba
Spirodela polyrhiza
Lemna minor/minuta
Wolffia arrhiza
Pas de frondes observées sur 2 mois d'observation
Prélèvement du 30 avril 2009
Maillezais 2
Maiilezais 5
Nombre de fronde cummulé
Arçais 5
damvix 5
Magné 2
Magné 5
Mazeau 3
Mazeau 5
St Hilaire 5
Vanneau 1
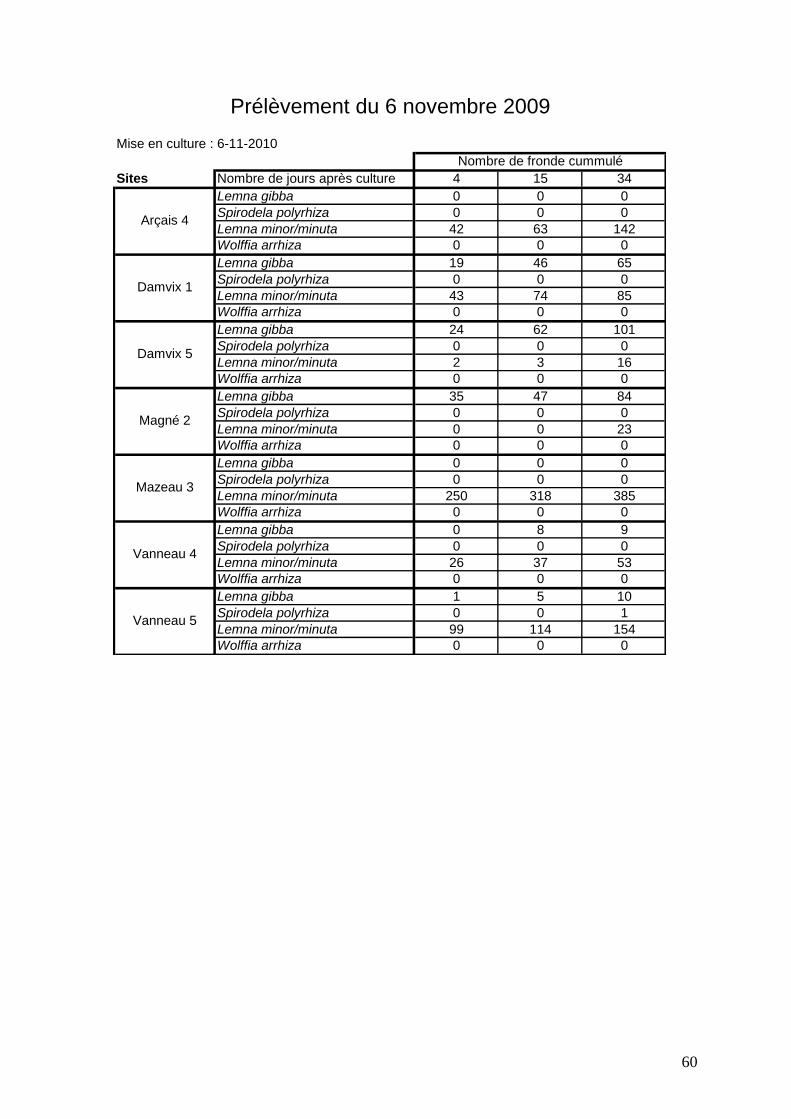
60
Mise en culture : 6-11-2010
Sites Nombre de jours après culture 4 15 34
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 42 63 142
Wolffia arrhiza 0 0 0
Lemna gibba 19 46 65
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 43 74 85
Wolffia arrhiza 0 0 0
Lemna gibba 24 62 101
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 2 3 16
Wolffia arrhiza 0 0 0
Lemna gibba 35 47 84
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 0 0 23
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 250 318 385
Wolffia arrhiza 0 0 0
Lemna gibba 0 8 9
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 26 37 53
Wolffia arrhiza 0 0 0
Lemna gibba 1 5 10
Spirodela polyrhiza 0 0 1
Lemna minor/minuta 99 114 154
Wolffia arrhiza 0 0 0
Prélèvement du 6 novembre 2009
Vanneau 5
Mazeau 3
Vanneau 4
Nombre de fronde cummulé
Arçais 4
Damvix 1
Damvix 5
Magné 2
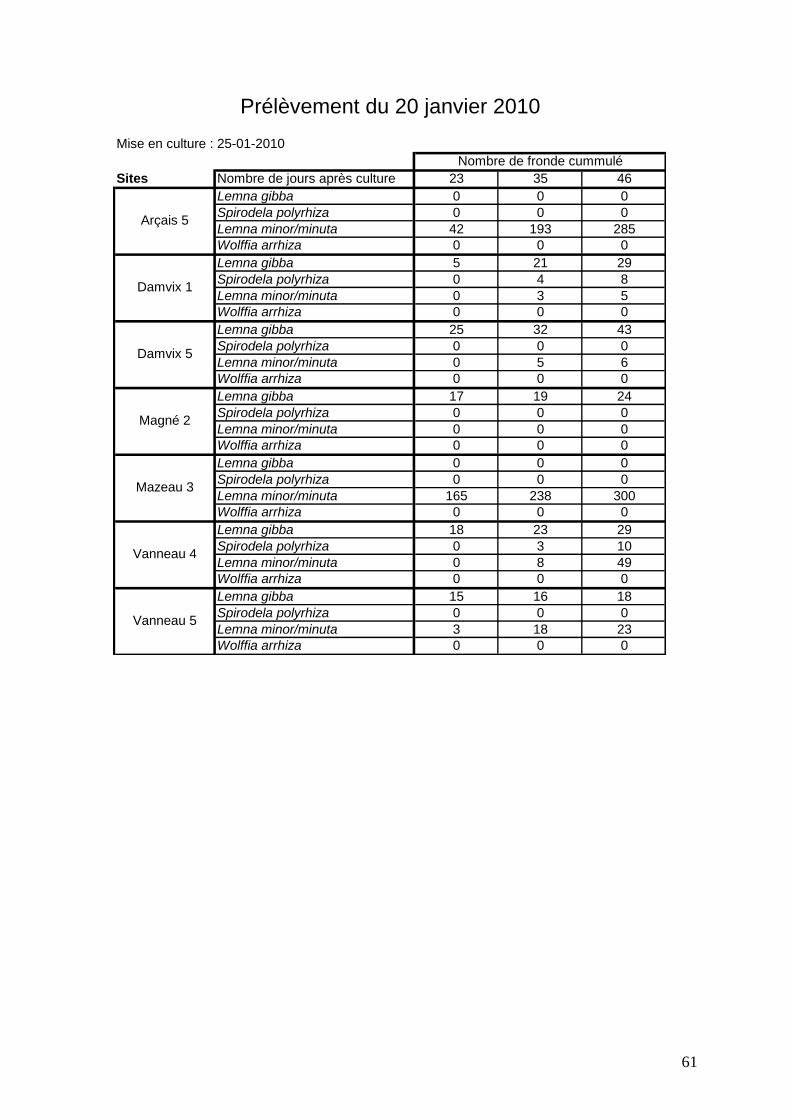
61
Mise en culture : 25-01-2010
Sites Nombre de jours après culture 23 35 46
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 42 193 285
Wolffia arrhiza 0 0 0
Lemna gibba 5 21 29
Spirodela polyrhiza 0 4 8
Lemna minor/minuta 0 3 5
Wolffia arrhiza 0 0 0
Lemna gibba 25 32 43
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 0 5 6
Wolffia arrhiza 0 0 0
Lemna gibba 17 19 24
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 0 0 0
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 165 238 300
Wolffia arrhiza 0 0 0
Lemna gibba 18 23 29
Spirodela polyrhiza 0 3 10
Lemna minor/minuta 0 8 49
Wolffia arrhiza 0 0 0
Lemna gibba 15 16 18
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 3 18 23
Wolffia arrhiza 0 0 0
Prélèvement du 20 janvier 2010
Vanneau 5
Mazeau 3
Vanneau 4
Nombre de fronde cummulé
Arçais 5
Damvix 1
Damvix 5
Magné 2

62
Mise en culture : 10-11-2010
Sites Nombre de jours après culture 8 14 20 26
Lemna gibba 0 0 0 0
Spirodela polyrhiza 0 0 0 0
Lemna minor/minuta 0 3 13 71
Wolffia arrhiza 0 0 0 0
Lemna gibba 0 0 0 0
Spirodela polyrhiza 62 94 128 175
Lemna minor/minuta 0 0 1 1
Wolffia arrhiza 0 0 0 39
Lemna gibba 0 0 0 0
Spirodela polyrhiza 148 196 219 265
Lemna minor/minuta 1 1 2 3
Wolffia arrhiza 0 0 0 7
Lemna gibba 0 0 0 0
Spirodela polyrhiza 14 38 63 96
Lemna minor/minuta 0 0 0 0
Wolffia arrhiza 0 0 0 4
Lemna gibba 0 0 0 0
Spirodela polyrhiza 2 6 0 7
Lemna minor/minuta 30 42 50 58
Wolffia arrhiza 0 0 0 0
Lemna gibba 0 0 0 0
Spirodela polyrhiza 13 26 32 40
Lemna minor/minuta 1 6 8 12
Wolffia arrhiza 0 0 0 0
Lemna gibba 0 0 0 0
Spirodela polyrhiza 7 24 37 57
Lemna minor/minuta 8 11 13 28
Wolffia arrhiza 0 0 0 0
Prélèvement du 30 septembre 2010
Vanneau 5
Mazeau 1
Vanneau 4
Nombre de fronde cummulé
Arçais 5
Damvix 1
Damvix 5
Magné 2

63
Mise en culture : 10-12-2010
Sites Nombre de jours après culture 7 21 28
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 59 99 162
Wolffia arrhiza 0 0 0
Lemna gibba 1 1 1
Spirodela polyrhiza 120 158 226
Lemna minor/minuta 9 10 16
Wolffia arrhiza 6 6 6
Lemna gibba 0 0 0
Spirodela polyrhiza 7 8 12
Lemna minor/minuta 0 0 0
Wolffia arrhiza 0 0 0
Lemna gibba 2 3 3
Spirodela polyrhiza 114 121 141
Lemna minor/minuta 16 29 36
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 20 24 24
Lemna minor/minuta 1400 1706 1831
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 0 2 8
Lemna minor/minuta 29 31 39
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 31 43 97
Lemna minor/minuta 188 254 285
Wolffia arrhiza 0 0 0
Prélèvement du 22 novembre 2010
Vanneau 5
Mazeau 1
Vanneau 4
Nombre de fronde cummulé
Arçais 5
Damvix 1
Damvix 5
Magné 2
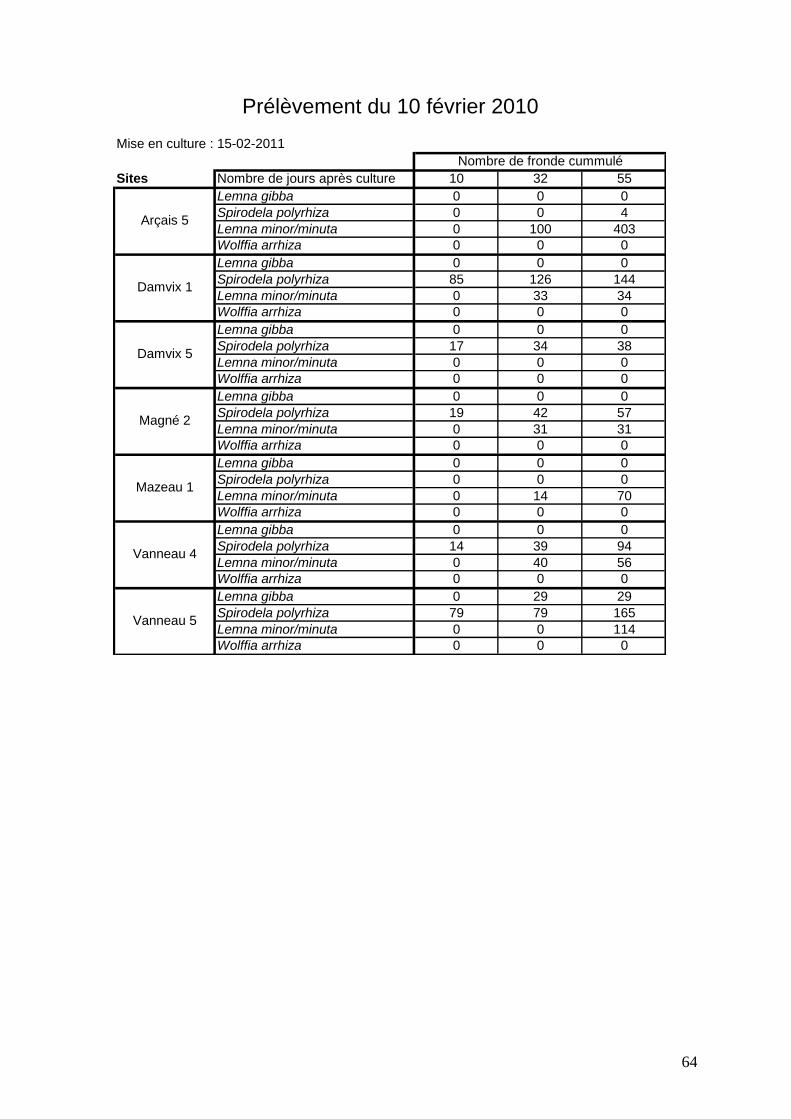
64
Mise en culture : 15-02-2011
Sites Nombre de jours après culture 10 32 55
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 4
Lemna minor/minuta 0 100 403
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 85 126 144
Lemna minor/minuta 0 33 34
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 17 34 38
Lemna minor/minuta 0 0 0
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 19 42 57
Lemna minor/minuta 0 31 31
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 0 0 0
Lemna minor/minuta 0 14 70
Wolffia arrhiza 0 0 0
Lemna gibba 0 0 0
Spirodela polyrhiza 14 39 94
Lemna minor/minuta 0 40 56
Wolffia arrhiza 0 0 0
Lemna gibba 0 29 29
Spirodela polyrhiza 79 79 165
Lemna minor/minuta 0 0 114
Wolffia arrhiza 0 0 0
Prélèvement du 10 février 2010
Vanneau 5
Mazeau 1
Vanneau 4
Nombre de fronde cummulé
Arçais 5
Damvix 1
Damvix 5
Magné 2