regulation of sterol content in membranes by … · lnstitut de biologie moléculaire des plantes,...
TRANSCRIPT

Plant Physiol. (1994) 105: 509-518
Regulation of Sterol Content in Membranes by Subcellular Compartmentation of Steryl-Esters Accumulating in a
Sterol-Overprod ucing Tobacco Mutant
Laurence Condet*, Roberte Bronner, and Pierre Benveniste
lnstitut de Biologie Moléculaire des Plantes, Centre National de Ia Recherche Scientifique, Département d’Enzymologie Cellulaire et Moléculaire, lnstitut de Botanique, 28 rue Goethe, 67083 Strasbourg Cédex,
France (L.G., P.B.); and Département de Biologie Cellulaire, 12 rue du Général Zimmer, 67084 Strasbourg Cédex, France (R.B.)
lhe study of sterol overproduction in tissues of LAB 1-4 mutant tobacco (Nicotiana tabacum L. cv Xanthi) (P. Maillot-Vernier, H. Schaller, P. Benveniste, C. Belliard [1989] Biochem Biophys Res Commun 165: 125-130) over several generations showed that the overproduction phenotype i s stable in calli, with a 10-fold stimu- lation of sterol content when compared with wild-type calli. How- ever, leaves of LAB 1-4 plants obtained after two steps of self- fertilization were characterized by a mere 3-fold stimulation, whereas calli obtained from these plants retained a typical sterol- overproducing mutant phenotype (i.e. a 10-fold increase of sterol content). lhese results suggest that the expression of the LAB 1-4 phenotype i s dependent on the differentiation state of cells. Most of the sterols accumulating in the mutant tissues were present as steryl-esters, which were minor species in wild-type tissues. Sub- cellular fractionation showed that in both mutant and wild-type tissues, free sterols were associated mainly with microsomal mem- branes. In contrast, the bulk of steryl-esters present in mutant tissues was found in the soluble fraction of cells. Numerous lipid droplets were detected in the hyaloplasm of LAB 1-4 cells by cytochemical and cytological techniques. After isolation, these lipid granules were shown to contain steryl-esters. These results show that the overproduced sterols of mutant tissues accumulate as steryl-esters in hyaloplasmic bodies. The esterification process thus allows regulation of the amount of free sterols in membranes by subcellular compartmentation.
~~ ~
Recently, three classes of mutant tobacco (Nicotiana taba- cum L. cv Xanthi) calli exhibiting resistance to LAB 170250F have been isolated in our laboratory (Maillot-Vemier et al., 1990). LAB 170250F is a phytotoxic sterol biosynthesis inhib- itor of the triazole type that has been shown to inhibit strongly the obtusifoliol-14a-methyl-demethylase, a Cyt P450-dependent enzyme of the phytosterol biosynthesis pathway (Fig. 1) (Taton et al., 1988).
Among these resistant calli, the LAB 1-4 genotype has been extensively studied. Its main characteristic is a 10-fold sterol overproduction in comparison with wild-type calli, with a concomitant accumulation of several biosynthetic intermedi- ates (Maillot-Vemier et al., 1989). Sterol esterification is also
* Corresponding author; fax 33-88-35-84-84.
a typical feature of this mutant: 80% of the sterols are found as steryl-esters (Fig. 2) (Maillot-Vemier et al., 1991). More- over, genetic studies showed that this phenotype is due to a single nuclear and semidominant mutation (Maillot-Vemier et al., 1991). Finally, it was shown that sterol overproduction was the consequence of the increased activity of HMGR, a key enzyme in isoprenoid biosynthesis. Indeed, the activity of this enzyme was about 3-fold higher in mutant tissues than in wild-type tissues (Gondet et al., 1992). The latter result emphasized the limiting role of the HMGR-catalyzed step in phytosterol biosynthesis, which supports a potential regulatory role of this enzyme in sterol metabolism in higher plants.
Taken together, these characteristics raised a question of prime importance: what is the location of this spectacular m a s of sterols in mutant cells? Free sterols (with a free /3- hydroxyl in the C3 position) (Fig. 2) are membrane constitu- ents and are found mainly in the plasma membrane (for review, see Hartmann and Benveniste, 1987). On the other hand, little is known about the localization of steryl-esters, which are described either in membranes (for review, see Dyas and Goad, 1993) or in the soluble cellular fraction (Dupéron and Dupéron, 1973). The LAB 1-4 tissues, charac- terized by a very high steryl-ester content, were an ideal material with which to increase our knowledge in this field. Furthermore, the determination of the subcellular localization of sterols in mutant tissues was a prerequisite for a complete understanding of the LAB 1-4 mutant phenotype.
The present paper reports on the subcellular localization of sterols (both free and esterified) in LAB 1-4 and wild-type tissues by two complementary approaches: first, a biochem- ical determination of the sterol content in microsomal and soluble subcellular fractions; and second, cytochemical and cytological analyses, with the aim of determining the in situ localization of sterols. As a prerequisite, we determined the sterolic composition of the wild-type and LAB 1-4 tissues over several successive generations to evaluate the stability of the LAB 1-4 phenotype.
Abbreviation: HMGR, 3-hydroxy-3-methylglutaryl-coenzyme A reductase.
509
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

510 Gondet et al. Plant Physiol. Vol. 105, 1994
ACETYL-COA
i CYCLOARTENOL
I 24-METHYLENE CYCLOARTANOL
1 i
LAB 170250F OBTUSIFOLIOL
I CYTOCHROME P450 OBTUSIFOLIOL
14-DEMETHYLASE
1 1 1
24-ETHY LIDENE 24-METHYLENE LOPHENOL LOPHENOL
1 1 I A’- AVENASTEROL
4 I 1 ZCMETHYLENE
CHOLESTEROL ISOFUCOSTEROL I I
I CAMPESTEROL SITOSTEROL 1
I finalphyrosrerols STIGMASTEROL 1
Figure 1. Simplified phytosterol biosynthesis from cycloartenol, the first sterol precursor specific to higher plants. The biosynthetic intermediates accumulating in LAB 1-4 tissues are the only ones represented.
MATERIALS AND METHODS
Plant Material
Wild-type seeds of Nicotiana tabacum L. cv Xanthi were harvested from plants originating after severa1 selfings of a plant obtained by diploidization of the SH6 dihaploid line that was used in the previous selection experiments (Maillot- Vemier et al., 1990). Wild-type calli were derived from mesophyll protoplasts of plants descending from the original wild-type plants by successive selfings.
The original LAB 1-4 callus was selected for resistance to LAB 170250F (Maillot-Vemier et al., 1990), and a first mutant plant was regenerated from a part of this callus (Maillot- Vemier et al., 1989). Mutant plants used in the present work were obtained by successive selfings of this first plant. Mutant calli were derived from mesophyll protoplasts of these plants. The media and conditions for mesophyll protoplast prepa- ration, plant regeneration, and culture of calli and plants have been described (Maillot-Vemier et al., 1990).
Preparation of Subcellular Fractions from Tobacco Tissues
Calli or plant leaf fragments (15 g) were washed in 0.1 M Tris-HC1, pH 7.4, and homogenized in a blender (6 X 5 s) with 30 mL of MTEB buffer (0.1 M Tris-HC1, pH 7.4, 0.5 M mannitol, 10 mM EDTA, 0.4% [w/v] BSA). The homogenized
crude extract was filtered through gauze and cell debris, nuclei, and most of organelles were removed by (centrifuga- tion (6,0OOg, 10 min). The post-6,OOOg supematartt was cen- trifuged i3t 100,OOOg for 90 min. The pellet corresponded to the mimosomal membranes and the supematant corre- sponded to the soluble fraction of cells. To restrict the possible contamiriation of microsomes with supematant and vice versa, the pellet and the soluble fraction were centrifuged again (lOO,OOOg, 60 min). Microsomes were finedly diluted with 5 mL of MTEB buffer.
Determiination of the Global Sterol Content of Plant Tissues
About 100 mg of dried material were ground in dichloro- methane:methanol (2:1, v/v) with an Ultra-turrax homoge- nizer (Janke and Kunkel Ika-Work, Stanfen, Germany) and lipids were extracted at 7OoC (3 X 2 h) in this solvent mixture. The total lipid extract was dried in a Biichi rotary evaporator and saponified with 5 mL of 6% (w/v) methanolic KOH for 1 h at 90OC. This alkaline hydrolysis step allows the release of the sterol moiety of steryl-esters.
The tcital sterols were then extracted with 2 volumes of n-hexane, filtered through sodium sulfate, and analyzed by TLC in a pure dichloromethane solvent system, 0x1 0.25-mm- thick precoated silica plates (Merck 60F 254, Darmstadt, Germany). The TLC step led to the separation of sterols in three bands (RF = 0.2, 0.25, and 0.3), depending on their degree of methylation in the C4 position. Sterols were visu- alized by fluorescence (A = 366 nm) after spraying with an ethanolic Berberine sulfate solution (O. 1 %, w/v).
The sterol-containing bands were scraped off and eluted together in dichloromethane, filtered to remove the residual silica, dried in a rotary evaporator, and finally siibmitted to acetylation in a to1uene:acetic anhydride:pyridine mixture (l:l:l, v/v/v) for 2 h at 7OOC. After evaporalion of the reagents, the steryl-acetates were analyzed again by TLC in dichloroimethane. Under these conditions they a11 migrated as a single band (RF = 0.5) that was scraped off and eluted in dichloromethane to form the final extract, which was then analyzecl by GC in a Carlo Erba GC 4160 apparatus (Erba
130 ,#P A : FREE STEROL
B : STERYL ESTER
Figure 2. Structure of t h e two major forms of sterols. A, Free sterol (free 8-hydroxyl in C3 is indicated by arrow). B, Steryl-ester (the 0- hydroxyl in C, is esterified by a fatty acid); the arrow shows t h e ester bond.
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

Subcellular Localization of Steryl-Esters in Tobacco Mutant Tissues 51 1
Science, Paris, France) equipped with a glass capillary column (WCOT, 30 m long, 0.25 mm intemal diameter, coated with OV1 [J&W Scientific, Folsom, CAI). The camer gaz (H2) was delivered at 2 mL/min.
The temperature program for GC included two phases: a fast heating phase from 6OoC to 24OOC (3OoC/min) and a slow heating phase from 24OOC to 28OOC (2OC/min). Free cholesterol was used as intemal standard for qualitative and quantitative determination of the sterol content of the plant sample. The amount and the relative retention time of each sterol was determined by a Spectra Physics SP4270 integrator (Spectra Physics, Les Ullis, France) connected with a detector (constant temperature: 28OOC). When required, sterol struc- tures were identified by GC/MS (Rahier and Benveniste, 1989). The SD for the quantification of sterols was never above 15%.
Determination of the Sterol Content of Subcellular Fractions
Dichloromethane-methanol extracts of the lyophilized microsomal and soluble cellular fractions were made and processed essentially as described in the previous paragraph. Before the alkaline hydrolysis step, the extract was analyzed by TLC in dichloromethane. Free sterols were found as three distinct bands (RF = 0.2, 0.25, and 0.3) as noted previously, whereas steryl-esters migrated near the solvent front (RF
= 0.9). Free and esterified sterols were thereafter treated separately .
The free sterols were pooled and eluted in dichlorometh- ane, the solvent was evaporated, and the residue was sub- mitted to acetylation. The steryl-esters were eluted in dichlo- romethane, dried, and saponified. The resulting free sterols were extracted with n-hexane and the extract was filtered through sodium sulfate and evaporated. After TLC analysis (to check that each sterol from the esterified fraction was indeed under the free @-hydroxyl form), the sterolic residue was acetylated. The steryl-acetates from both the free and the esterified fraction were analyzed separately as described in the previous paragraph. Results presented in this work are representative of three replicates.
Cytochemical Studies
Various tobacco samples were used for these studies: wild- type and LAB 1-4 callus fragments, plant leaf epiderma, and soluble fraction of callus cells. The material was stained without any previous fixation according to Martoja and Mar- toja (1967) to detect cellular lipid compounds. Observations were made by microscopy (X650 magnification). The follow- ing staining methods were used.
Sudan 111 (Dye No. 26 100 According to the Color Index)
The material was squashed directly on the glass plate and covered with a small volume of a saturated (about 0.3%, w/v) Sudan 111 solution in 70% (v/v) ethanol. After 15 min, the sample was washed with distilled water. A11 lipids were stained red-orange.
Nile Blue (Color Index: No. 5 1 180)
The material was immersed in a 1% (w/v) Nile blue solu- tion in water, treated with acetic acid (I%, v/v), and washed with distilled water. Steryl-esters and glycerides were stained pink.
Liebermann-Schultz Technique
Tissues were pretreated for 3 d by an aqueous iron alum solution (2.5%, w/v), then washed with distilled water and laid on a glass plate. The preparation was covered with drops of acetic acid:sulfuric acid (l:l, v/v) prepared just before use. Sterols (both free and esterified forms) were stained brown- green.
Digitonin Treatment (Lison Technique)
Tissues were immersed for several hours in a 0.5% (w/v) digitonin solution in ethanol (50%, v/v) and were observed under polarizing light. Free sterol-digitonin complexes ap- peared as birefringent crystals.
Smith-Dietrich Method
Tissues were immersed for 2 d in an aqueous potassium bichromate solution (5%, w/v), washed with distilled water, and treated for 5 h with the Kultschitzky hematoxyline solution, freshly prepared by mixing 10 mL of solution A (10% [w/v] hematoxyline [Color Index: No. 752901 in pure ethanol; this solution has to be stored for 6 months before use) and 92 mL of solution B (2 mL of acetic acid and 90 mL of distilled water). After several washes with distilled water, tissues were treated for 12 h with the Weigert mixture (4 g of sodium borate and 5 g of potassium femcyanide in 600 mL of distilled water). Finally, they were washed again with distilled water before observation. Phospholipids were stained dark blue.
Cytological Studies
LAB 1-4 callus pieces were fixed in a 2% (v/v) glutaralde- hyde solution in a 0.1 M potassium phosphate buffer, pH 7.2, for 2 h at 4OC. After an ovemight washing step in the same buffer at 4OC, the samples were treated by 1% (w/v) osmium tetroxide in 0.1 M phosphate buffer, pH 7.2, for 2 h at 4OC. The fixed material was washed in distilled water and dehy- drated in a gradual ethanol series (70-95-100% [v/v] in water). After a final treatment with propylene oxide, the sample was embedded in Araldite M and the resin was polymerized for 3 d at 6OOC.
For light microscopy, semithin sections (about 0.7 wm thick) were made with a Sorva11 microtome (Porter Blum MT2). The sections were stained with a 1% (w/v) solution of toluidine blue in 5% (w/v) sodium tetraborate (Lynn, 1965). They were also stained by Sudan black (Bronner, 1975) for lipid local- ization. For EM, thin sections (0.1 pm thick) were collected on copper grids and contrasted with uranyl acetate and lead citrate according to Reynolds (1963). Observations were made with a Phillips transmission electron microscope.
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

512 Gondet e t al. Plant Physiol. Vol. 105, 1994
RESULTS
Determination of Sterol Overproduction Leve1 in LAB 1-4 Tissues
The sterolic composition of wild-type and LAB 1-4 tissues from different generations was compared. The results show that after only two generations, sterol overproduction level in leaves of mutant plants decreased by a factor of 3 (Table I) when compared with that of the initial plant regenerated from the selected LAB 1-4 callus (Maillot-Vernier et al., 1989). Indeed, plant leaves obtained after two selfings contained only 2- to 3-fold as much sterol as did wild-type leaves (Table I). Mutant calli derived from these plants were nevertheless still characterized by a typical sterol overproduction level, i.e. 10-fold as compared with wild-type calli, with a strong accumulation (over 50%) of biosynthetic intermediates (Table 11). The differentiation state of cells might therefore have a strong influence over the expression level of the sterol-over- producing phenotype.
Biochemical Determination of Sterolic Composition in Subcellular Fractions Obtained from LAB 1-4 and Wild-Type Tissues
Wild-type and mutant calli showed notable differences in their subcellular sterolic distribution (Table 111). In wild-type calli, 80% (11 pg g-' fresh weight) of total sterols were free sterols and were localized in the microsomal fraction, whereas in LAB 1-4 calli, 75% of the sterol content (111 pg g-' fresh weight) was present as steryl-esters in the soluble fraction (post-100,000g supernatant), which is formed by cell hyalo- plasm and vacuoles. On the other hand, the quantity of free sterols in microsomes was similar for wild-type and mutant calli (1 1 and 9 pg g-' fresh weight, respectively).
These observations also apply to mutant plant leaves (Table IV). However, subcellular localization of sterols was less
marked than it was in calli, especially that of steryl-esters. Indeed, the steryl-esters accumulating in the soluble fraction (22 pg g,-' fresh weight) amounted to only one-fourth of the total sterols. This observation is certainly connecied with the lower overproducing capacity of mutant leaves as compared with calli, which was noted in the previous paragraph. Con- sequently, the quantity of steryl-esters accumuhting in the soluble fraction of mutant cells is a good indicator of sterol overproduction intensity and, therefore, of the level of the LAB 1-4L phenotype expression.
In Situ Localization of Steryl-Esters in LAB 1-4 Cells by a Cytochemical Approach
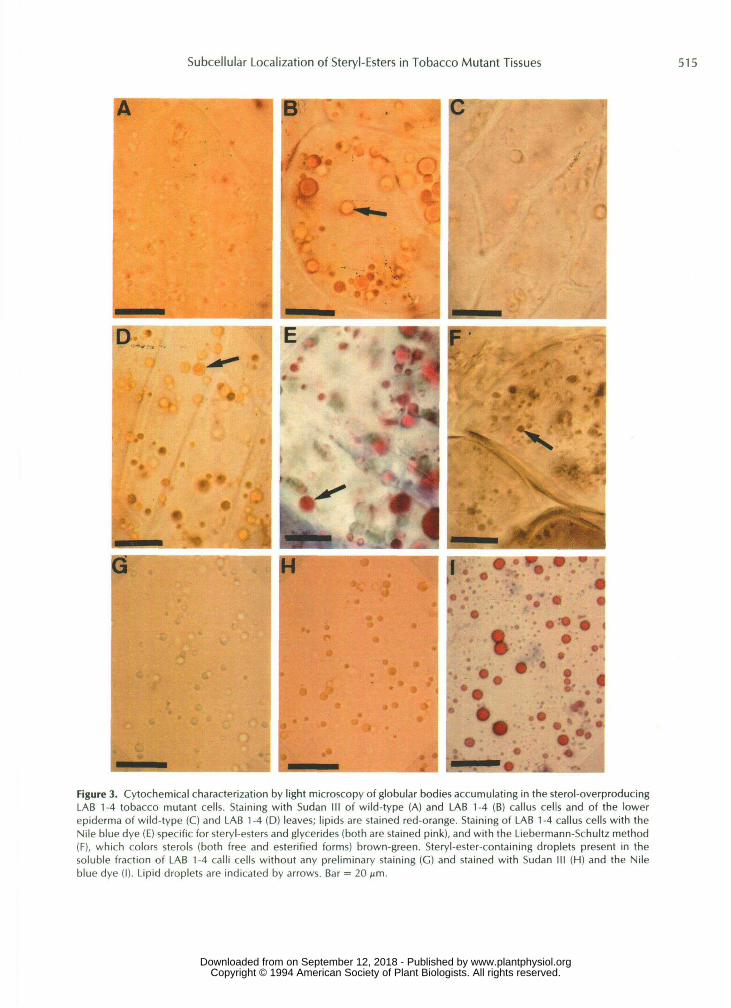
After staining with Sudan 111, a general dye iised for the detection of a11 classes of lipids, LAB 1-4 calli were shown to contain lipid droplets (Fig. 3B), which were not present in wild-type calli (Fig. 3A). Such bodies were also identified in the epidlermis of mutant leaves (Fig. 3D) but not in wild-type leaves (Fig. 3C).
These lipid granules were also detected in LAB 1-4 tissues by the Nile blue dye (specific for glycerides and steryl-esters, which are stained pink) (Fig. 3E) and the Liebermann-Schultz staining technique, which allows identification of free and esterified sterols (stained brown-green) (Fig. 3F). These drop- lets were not stained by a digitonin treatment (detection of free sterols only) or by the Smith-Dietrich method (specific for pho!jpholipids). These results show that the lipid droplets present in mutant tissues contain steryl-esters.
Characiterization of Purified Lipid Droplets frorn LAB 1-4 Callus Cells
To establish without ambiguity the link between the steryl- esters contained in the droplets and those identified in the soluble cellular fraction by previous biochemical character-
Table 1. Sterol analysis of leaves from wild-type and LAB 1-4 miitant plants obtained after severa/ selfings of the initial plants
Total sterols of 100 ma of dried material were extracted as dlescribed in "Materials and Methods." Number of Selfings from t h e Original Plant 2 3 4
Sterol composition (wt '/O of total sterols) w.t.a LAB 1-4 w.t. LAB 1-4 w.t. LAB 1-4
Cycloartenol 8 21 2 14 3 18 24-Methylene cycloartanol O 2 O 2 O 1 Obtusifoliol O 4 O 2 O 3 24-Ethylidene lophenol O 5 O 4 O 3 A7-Avenasterol O O O 4 3 2 24-Methylene cholesterol O O O O O O lsofucosterol 8 13 7 10 9 14 Percent of biosynthetic intermediates 16 45 9 36 15 41
Cholesterol 12 4 11 4 11 5
Sitosterol 16 16 15 20 16 18 Stigmasterol 35 17 43 24 37 17
Total sterol content (kg g-' dry weight) 1152 3278 1092 2374 887 2472
Campesterol 21 18 22 16 21 19
Percent of final sterols 84 55 91 64 85 59
Sterol overproduction factorb 3 2 3
a w.t., Wild-type. Ratio of t h e total sterol content of LAB 1-4 tissues to that of wild-type tissues.
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
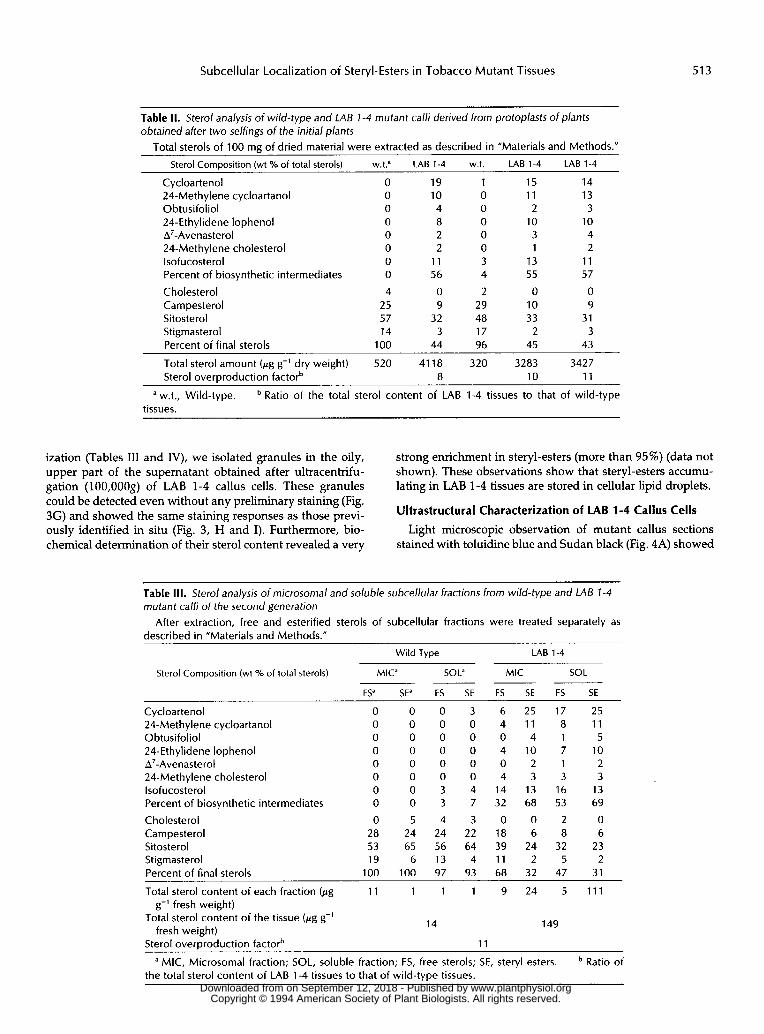
Subcellular Localization of Steryl-Esters in Tobacco Mutant Tissues 513
Table II. Sterol analysis of wild-type and LAB I-4 mutant calli derived from protoplasts of plants obtained after two selfings of the initial plants
Total sterols of 100 mg of dried material were extracted as described in ”Materials and Methods.”
Sterol Composition (wt % of total sterols) w.t? LAB 1-4 w.t. LAB 1-4 LAB 1-4
Cycloartenol O 19 1 15 14 24-Methylene cycloartanol O 10 O 11 13 Obtusifoliol O 4 O 2 3 24-Ethylidene lophenol O 8 O 10 10 A’-Avenasterol O 2 O 3 4 24-Methylene cholesterol O 2 O 1 2 lsofucosterol O 11 3 13 11 Percent of biosynthetic intermediates O 56 4 55 57
Cholesterol 4 O 2 O O Campesterol 25 9 29 10 9 Sitosterol 57 32 48 33 31 Stigmasterol 14 3 17 2 3 Percent of final sterols 1 O0 44 96 45 43
Total sterol amount (pg g-’ dry weight) 520 41 18 320 3283 3427 Sterol overproduction factorb 8 10 11
a w.t., Wild-type. Ratio of the total sterol content of LAB 1-4 tissues to that of wild-type tissues.
ization (Tables I11 and IV), w e isolated granules in the oily, upper part o f the supernatant obtained after ultracentrifu- gation (100,OOOg) of LAB 1-4 callus cells. These granules could be detected even without any preliminary staining (Fig. 3G) and showed the Same staining responses as those previ- ously identi f ied in situ (Fig. 3, H and I). Furthermore, bio- chemical determination o f their sterol content revealed a very
strong enrichment in steryl-esters (more than 95%) (data not shown). These observations show that steryl-esters accumu- lat ing in LAB 1-4 tissues are stored in cellular lipid droplets.
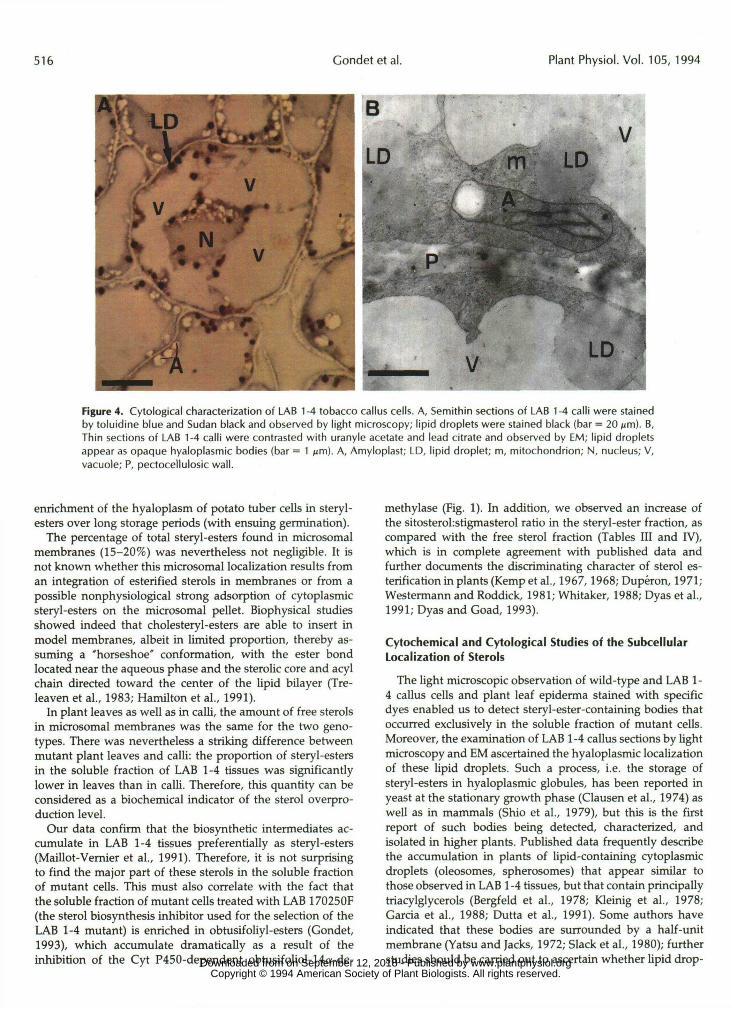
Ultrastructural Characterization of LAB 1-4 Callus Cells Light microscopic observation o f mutant callus sections
stained with toluidine blue and Sudan black (Fig. 4A) showed
Table 111. Sterol analysis of microsomal and soluble subcellular fractions from wild-type and LAB 1-4 mutant calli of the second generation
described in ”Materials and Methods.” After extraction, free and esterified sterols of subcellular fractions were treated separately as
Wild Type LAB 1-4
Sterol Composition (wt O/O of total sterols) MIC” SOL“ MIC SOL _ _ _ ~
FS” SE” FS SE FS SE FS SE ~~
Cycloartenol 24-Methylene cycloartanol Obtusifoliol 24-Ethylidene lophenol A’-Avenasterol 24-Methylene cholesterol lsofucosterol Percent of biosynthetic intermediates
C holesterol Campesterol Sitosterol Stigmasterol Percent of final sterols
O O O O O O O O
O 28 53 19
1 O0
O 0 O 0 O 0 O 0 O 0 O 0 o 3 o 3
5 4 24 24 65 56
6 13 100 97
3 O O O O O 4 7
3 22 64 4
93
6 25 17 25 4 11 8 11 o 4 1 5 4 10 7 10 o 2 1 2 4 3 3 3
14 13 16 13 32 68 53 69
O 0 2 O 1 8 6 8 6 39 24 32 23
2 11 2 5 68 32 47 31
Total sterol content of each fraction (pg 11 1 1 1 9 24 5 111
Total sterol content of the tissue (pg g-’ g-’ fresh weight)
fresh weight)
a MIC, Microsomal fraction; SOL, soluble fraction; FS, free sterols; SE, steryl esters.
14 149
Sterol overproduction factorb 11
Ratio of the total sterol content of LAB 1-4 tissues to that of wild-type tissues.
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

514 Gondet et al. Plant Physiol. Vol. 105, 1994
Table IV. Sterol analysis of microsomal and soluble subcellular fractions of leaves from wild-type and LAB 1-4 plants (obtained after four selfings from the initial plants)
described in “Material5 and Methods.” After extraction, free and esterified sterols of subcellular fractions were treated separately as
Wild Type LAB 1-4
Sterol Composition (wt % of total sterols) MIC” SOL’ MIC SOL ~ - - _ _ _ _ ~ FS” SE” FS SE FS SE FS SE
Cycloartenol 24-Methylene cycloartanol Obtusifoliol 24-Ethylidene lophenol A’-Avenasterol 24-Methylene cholesterol lsofucosterol Percent of biosynthetic intermediates C holesterol Campesterol Sitosterol Stigmasterol Percent of final sterols
2 O 5 0 9 2 4 1 2 1 8 0 0 0 0 1 3 2 2 0 0 0 0 1 4 1 4 O 0 O 0 1 5 2 4 O 0 O 0 1 4 1 4 0 0 3 0 4 0 0 3 5 O 9 O 11 12 12 15 7 O 17 O 28 52 30 50
11 O 11 O 6 3 5 4 23 O 19 O 15 16 15 14 16 O 19 O 20 22 24 24 43 O 34 O 31 7 26 8 93 O 83 O 72 48 70 50
Total sterol content of each fraction (pg 42 O 1 O 47 19 6 22 g-’ fresh weight)
Total sterol amount in the tissue (pg g-’ fresh weinht) 43 94
Sterol overiroduction factorb 2
MIC, Microsomal fraction; SOL, soluble fraction; FS, free sterols; SE, steryl esters. Ratio of the total sterol content of LAB 1-4 tissues to that of wild-type tissues.
that the lipid granules were located in the thin parietal cytoplasmic layer of cells. This observation was confirmed by EM (Fig. 4B): the lipid droplets appeared as slight opaque globular todies in the hyaloplasm of cells. It is worth noting that they did not seem to be surrounded by a typical phospholipid membrane and were approximately 1 pm in diameter.
DISCUSSION
Sterol Overproduction Leve1
The quantitative analysis of sterols in wild-type and LAB 1-4 tissues from different generations showed that the sterol overproducing capacity is dependent on the differentiation state of the tissues. Indeed, calli were characterized by a stable overproduction factor, whereas overproduction level decreased in plant leaves to reach, after two selfings, a stable value that was 3- to 4-fold lower than in mutant calli. It is worth noting that this reduction of sterol overproduction was already apparent after the first selfing of the initial plant (Maillot-Vemier, 1990).
The fact that we were able to obtain calli with a typical LAB 1-4 overproduction factor (about 10-fold) from leaves of mutant plants with a low phenotype intensity proves that we were not dealing with a reversion phenomenon. These results suggest that a regulation process is involved in the expression of sterol overproduction in LAB 1-4 tissues, and that this regulation depends on the differentiation state of cells. In a previous work (Gondet et al., 1992), we established
the corrc4ation between sterol overproduction and the in- creased activity of HMGR in mutant tissues. Since sterol overprocluction level is different in LAB 1-4 calli and leaves, it is surprising that both types of tissues were characterized by the same stimulation factor of HMGR (3-fold ai: compared with wild-type tissues). Further studies will be required to shed more light on this lack of correlation between the activity of HMGR and sterol overproduction intensity in the different types of mutant tissues.
Distribution of Sterols in Subcellular Compartmcmts
Great tiifferences in the subcellular distribution of free and esterifiedl sterols were noted in wild-type and LAB 1-4 calli. In mutarit calli, the major part (75%) of sterols accumulated as steryl-esters in the soluble fraction of cells, whereas in wild-type calli, 80% of the sterols remained unesterified and were located in the microsomal fraction (which contains the majority of cellular membranes such as ER, Gol@ apparatus, plasma membrane, and tonoplast).
The arnount of free sterols in microsomes was similar for the two genotypes (about 10 pg g-’ fresh weight), implying a tight control of this amount. This is in agreement with the unesterif ied state and the membrane localization of sterols being reqpirements of their main physiological functions: the regulation of membrane permeability and fluidity (Demel and De IOUyff, 1976). In addition, our results demonstrate that steryl-esters are preferentially (about 80%) located in the soluble fraction of cells. This is in accordance with the results obtained by Dupéron and Dupéron (1973), who noticed an
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
Subcellular Localization of Steryl-Esters in Tobacco Mutant Tissues 515
A
Figure 3. Cytochemical characterization by light microscopy of globular bodies accumulating in the sterol-overproducingLAB 1-4 tobacco mutant cells. Staining with Sudan III of wild-type (A) and LAB 1-4 (B) callus cells and of the lowerepiderma of wild-type (C) and LAB 1-4 (D) leaves; lipids are stained red-orange. Staining of LAB 1-4 callus cells with theNile blue dye (E) specific for steryl-esters and glycerides (both are stained pink), and with the Liebermann-Schultz method(F), which colors sterols (both free and esterified forms) brown-green. Steryl-ester-containing droplets present in thesoluble fraction of LAB 1-4 calli cells without any preliminary staining (C) and stained with Sudan III (H) and the Nileblue dye (I). Lipid droplets are indicated by arrows. Bar = 20 urn.
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
516 Condet et al. Plant Physiol. Vol. 105, 1994
V
Figure 4. Cytological characterization of LAB 1-4 tobacco callus cells. A, Semithin sections of LAB 1-4 calli were stainedby toluidine blue and Sudan black and observed by light microscopy; lipid droplets were stained black (bar = 20 jim). B,Thin sections of LAB 1-4 calli were contrasted with uranyle acetate and lead citrate and observed by EM; lipid dropletsappear as opaque hyaloplasmic bodies (bar = 1 ̂ m). A, Amyloplast; LD, lipid droplet; m, mitochondrion; N, nucleus; V,vacuole; P, pectocellulosic wall.
enrichment of the hyaloplasm of potato tuber cells in steryl-esters over long storage periods (with ensuing germination).
The percentage of total steryl-esters found in microsomalmembranes (15-20%) was nevertheless not negligible. It isnot known whether this microsomal localization results froman integration of esterified sterols in membranes or from apossible nonphysiological strong adsorption of cytoplasmicsteryl-esters on the microsomal pellet. Biophysical studiesshowed indeed that cholesteryl-esters are able to insert inmodel membranes, albeit in limited proportion, thereby as-suming a 'horseshoe* conformation, with the ester bondlocated near the aqueous phase and the sterolic core and acylchain directed toward the center of the lipid bilayer (Tre-leaven et al., 1983; Hamilton et al., 1991).
In plant leaves as well as in calli, the amount of free sterolsin microsomal membranes was the same for the two geno-types. There was nevertheless a striking difference betweenmutant plant leaves and calli: the proportion of steryl-estersin the soluble fraction of LAB 1-4 tissues was significantlylower in leaves than in calli. Therefore, this quantity can beconsidered as a biochemical indicator of the sterol overpro-duction level.
Our data confirm that the biosynthetic intermediates ac-cumulate in LAB 1-4 tissues preferentially as steryl-esters(Maillot-Vernier et al., 1991). Therefore, it is not surprisingto find the major part of these sterols in the soluble fractionof mutant cells. This must also correlate with the fact thatthe soluble fraction of mutant cells treated with LAB 170250F(the sterol biosynthesis inhibitor used for the selection of theLAB 1-4 mutant) is enriched in obtusifoliyl-esters (Gondet,1993), which accumulate dramatically as a result of theinhibition of the Cyt P450-dependent obtusifoliol-14a-de-
methylase (Fig. 1). In addition, we observed an increase ofthe sitosterol:stigmasterol ratio in the steryl-ester fraction, ascompared with the free sterol fraction (Tables III and IV),which is in complete agreement with published data andfurther documents the discriminating character of sterol es-terification in plants (Kemp et al., 1967,1968; Duperon, 1971;Westermann and Roddick, 1981; Whitaker, 1988; Dyas et al.,1991; Dyas and Goad, 1993).
Cytochemical and Cytological Studies of the SubcellularLocalization of Sterols
The light microscopic observation of wild-type and LAB 1-4 callus cells and plant leaf epiderma stained with specificdyes enabled us to detect steryl-ester-containing bodies thatoccurred exclusively in the soluble fraction of mutant cells.Moreover, the examination of LAB 1-4 callus sections by lightmicroscopy and EM ascertained the hyaloplasmic localizationof these lipid droplets. Such a process, i.e. the storage ofsteryl-esters in hyaloplasmic globules, has been reported inyeast at the stationary growth phase (Clausen et al., 1974) aswell as in mammals (Shio et al., 1979), but this is the firstreport of such bodies being detected, characterized, andisolated in higher plants. Published data frequently describethe accumulation in plants of lipid-containing cytoplasmicdroplets (oleosomes, spherosomes) that appear similar tothose observed in LAB 1-4 tissues, but that contain principallytriacylglycerols (Bergfeld et al., 1978; Kleinig et al., 1978;Garcia et al., 1988; Dutta et al., 1991). Some authors haveindicated that these bodies are surrounded by a half-unitmembrane (Yatsu and Jacks, 1972; Slack et al., 1980); furtherstudies should be carried out to ascertain whether lipid drop- www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from
Copyright © 1994 American Society of Plant Biologists. All rights reserved.

Subcellular Localization of Steryl-Esters in Tobacco Mutant Tissues 51 7
lets contained in LAB 1-4 tissues are surrounded by such a membrane.
In contrast with the oleosomes and spherosomes, whose function is to store triacylglycerols until their future metabolic use, the steryl-ester-containing droplets of LAB 1-4 tissues might have a role in sequestering the overproduced sterols. Therefore, esterification should be considered a detoxification process taking place when the sterol amount in membranes becomes higher than a physiological threshold, resulting in the hyaloplasmic localization of nonrequired sterols.
Global Understanding of the LAB 1-4 Phenotype
Studies performed on the LAB 1-4 tobacco mutant (Maillot- Vemier et al., 1989, 1991; Gondet et al., 1992; work reported here) allow us to better understand its sterol metabolism. The LAB 1-4 mutation leads to an increase of the apparent activity of HMGR, thereby raising the leve1 of isoprenoid precursors and thus the metabolic flux to sterols. Severa1 enzymes of the sterol pathway thus become saturated by the enonnous amount of available substrates, and this results in the accu- mulation of n u m e r o u biosynthetic intermediates in mutant tissues. The major part of these sterol precursors are then esterified by a sterol-acyltransferase, whose characterization is in progress (P. Bouvier-Nave, unpublished results), to be finally confined to hyaloplasmic globules. This process allows the maintenance of a correct amount of free sterols in the membranes of mutant cells (comparable with that of wild- type cells), in spite of the massive sterol production in LAB 1-4 tissues, especially in calli. A similar chain of events may occur in another tobacco mutant genotype, which has been selected in our group for resistance to a y-keto triazole herbicide (Schaller et al., 1992).
These results confirm the great importance of biochemical mutants in achieving a better understanding of metabolic processes. Indeed, the characterization of the LAB 1-4 phe- notype allowed us to obtain valuable information on the regulation of sterol metabolism in higher plants.
ACKNOWLEDCMENTS
This work was supported by a doctoral fellowship from the French Ministry of Research and Technology (MRT) to L.G. We are grateful to B. Bastian for the skillful typing of the manuscript. Finally, we thank P. Bouvier-Navé, P. Maillot-Vemier, H. Schaller, and J. Broadhvest for valuable discussion, and T. Colbum for critica1 read- ing of the manuscript.
Received November 23, 1993; accepted February 17, 1994. Copyright Clearance Center: 0032-0889/94/105/0509/10.
LITERATURE ClTED
Bergfeld R, Hong YN, Kühnl T, Schopfer P (1978) Formation of oleosomes (storage lipid bodies) during embryogenesis and their breakdown during seedling development in cotyledons of Sinapis alba L. Planta 143 297-307
Bronner R (1975) Simultaneous differential characterization of lipids and starch in plant tissues. Stain Technol 50 1-4
Clausen MK, Christiansen K, Jensen PK, Behnke O (1974) lsolation of lipid particles from baker's yeast. FEBS Lett 43: 176-179
Demel RA, De Kruyff B (1976) The function of sterols in mem- branes. Biochim Biophys Acta 457: 109-132
Duperon P (1971) Nature et comportement des stérols 'libres" et estérifiés, au cours de la germination de divers types de semences. Hypothèses sur le rÔle de ces substances chez les végétaux. Physiol Veg 9 373-399
Duperon P, Duperon R (1973) Evolution des différentes catégories de composés stéroliques au cours de la 'germination" du tubercule de pomme de terre. Localisation intracellulaire de ces substances durant cette période. Physiol Veg 11: 487-505
Dutta PC, Appelqvist LA, Gunnarsson S, Von Hofsten A (1991) Lipid bodies in tissue culture, somatic and zygotic embryo of Daucus carota L.: a qualitative and quantitative study. Plant Sci 7 8
Dyas L, Goad LJ (1993) Steryl fatty acyl esters in plants. Phytochem- istry 3 4 17-29
Dyas L, Prescott MC, Evershed RP, Goad LJ (1991) Steryl esters in a cell suspension culture of celery (Apium graveolens). Lipids 26
Garcia JM, Quintero LC, Mancha M (1988) Oil bodies and lipid synthesis in developing soybean seeds. Phytochemistry 27:
Gondet L (1993) Caractérisation biochimique et cytochimique d'un mutant de tabac (Nicotiana tabacum L.) surproducteur de stérols. PhD thesis. Université Louis Pasteur, Strasbourg, France
Gondet L, Weber T, Maillot-Vernier P, Benveniste P, Bach TJ (1992) Regulatory role of microsomal 3-hydroxy-3-methyl glu- taryl-coenzyme A reductase in a tobacco mutant that overproduces sterols. Biochem Biophys Res C o m " 186 888-893
Hamilton JA, Fujito DT, Hammer CF (1991) Solubilization and localization of weakly polar lipids in unsonicated egg phosphati- dylcholine: a "C MAS NMR study. Biochemistry 30 2894-2902
Hartmann MA, Benveniste P (1987) Plant membrane sterols: iso- lation, identification and biosynthesis. Methods Enzymol 148
Kemp RJ, Goad LJ, Mercer E1 (1967) Changes in the levels and composition of the esterified and unesterified sterols of maize seedlings during germination. Phytochemistry 6 1609-1615
Kemp RJ, Hamman ASA, Goad LJ, Goodwin TW (1968) Studies on phytosterol biosynthesis: observations on the esterified sterols of higher plants. Phytochemistry 7: 447-450
Kleinig H, Steinki C, Kopp C, Zaar K (1978) Oleosomes (sphero- somes) from Daucus carota suspension culture cells. Planta 140
Lynn JA (1965) Rapid toluidine blue staining of epon-embedded and mounted adjacent sections. Am J Clin Pathol44: 57-58
Maillot-Vernier P (1990) Sélection in vitro et caractérisation de mutants résistants i des inhibiteurs de Ia biosynthbe des phyto- stérols i partir de protoplastes de Nicotiana tabacum dihaploide. PhD thesis. Universiti Louis Pasteur, Strasbourg, France
Maillot-Vernier P, Gondet L, Schaller H, Benveniste P, Belliard G (1991) Genetic study and further biochemical characterization of a tobacco mutant that overproduces sterols. Mo1 Gen Genet
Maillot-Vernier P, Schaller H, Benveniste P, Belliard G (1989) Biochemical characterization of a sterol mutant plant regenerated from a tobacco callus resistant to a triazole cytochrome P-450- obtusifoliol-14-demethylase inhibitor. Biochem Biophys Res Com- mun 165 125-130
Maillot-Vernier P, Schaller H, Benveniste P, Belliard G (1990) In vitro selection of calli resistant to a triazole cytochrome P-450- obtusifoliol- 14-demethylase inhibitor from protoplasts of Nicotiana tabacum L. cv Xanthi. Plant Physiol93 1190-1195
Martoja R, Martoja M (1967) Initiation aux Techniques de l'Histo- logie Animale. Masson Cie, Paris
Rahier A, Benveniste P (1989) Mass spectral identification of phy- tosterols. Zn WD Nes, E Parish, eds, Analysis of Sterols and Other Significant Steroids. Academic Press, New York, pp 223-250
Reynolds M (1963) The use of lead citrate at high pH as electron- opaque stain in electron microscopy. J Cell Biol 17: 209-212
Schaller H, Maillot-Vernier P, Belliard G, Benveniste P (1992) Increased sterol biosynthesis in tobacco calli resistant to a triazole herbicide which inhibits demethylation of l4a-methyl sterols. Planta 187: 315-321
259-267
536-541
3083-3087
632-650
233-237
231: 33-40
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

518 Gondet
Shio H, Haley NJ, Fowler S (1979) Characterization of lipid-laden aortic cells from cholesterol-fed rabbits. 111. Intracellular localiza- tion of cholesterol and cholesteryl ester. Lab Invest 41: 160-167
Slack CR, Bertaud WS, Shaw BD, Holland R, Browse JA, Wright H (1980) Some studies on the composition and surface properties of oil bodies from oil seed cotyledons. Biochem J 190 551-561
Taton M, Ullmann P, Benveniste P, Rahier A (1988) Interaction of triazole fungicides and plant growth regulators with microsomal cytochrome P450-dependent obtusifoliol 14a-methyl demethyl- ase. Pestic Biochem Physiol30 178-189
et al. Plant Physiol. Vol. 105, 1994
Tre1eave:n WD, Wassall SR, Cushley RJ (1983) Carbon-13 nuclear magneiic resonance study of spin labelled cholesberyl ester in m s e l imembranes.Chem Phys Lipids 33: 223-23 1
WesJerm,an L, Roddick JG (1981) Annual variation in sterol levels in leaves of Tarasumof f i c ina le Weber. Plant Physiol 6 8 872-875
Whitakeir BD (1988) Changes in the steryl lipid content and com- position of tomato fruit during ripening. Phytochemistry 27:
Yatsu LY, Jacks TJ (1972) Spherosome membranes (half unit-mem- 341 1-3'4 16
branes)~. Plant Physiol49 937-943
www.plantphysiol.orgon September 12, 2018 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.