rapport final de l’aci mocemy mod elisation de la …ljk.imag.fr/mocemy/mocemy-rapport.pdf ·...
TRANSCRIPT

Rapport Final de l’ACI MoCeMy
Modelisation de la Cellule et du Myocarde
(ACI NIM 04 5 290)
Participants au projet
Parmi les participants au projet, il est a noter que le financement obtenu a permis d’embaucherV. MILISIC en Post-doc. Celui-ci a integre le laboratoire en tant que CR CNRS l’annee suivante,et naturellement la liste des membres de l’ACI. Par ailleurs le laboratoire LMC est devenu LJKdurant le deroulement du projet.
Laboratoire Jean Kuntzmann ex Laboratoire LMCEric BONNETIER, PR UJF, Georges-Henri COTTET, PR UJF, Emmanuel MAITRE, MCF UJF(Coordinateur), Vuk MILISIC, Post-Doctorant finance par l’ACI (depuis nov. 2004), puis CR CNRS(depuis sept. 2005), Annie RAOULT, PR UJF (PR Paris Descartes depuis oct. 2005), ThomasMILCENT, Doctorant (depuis 2004), Ayman MOURAD Enseignant-chercheur, Universite Liba-naise de Beyrouth et Libanese International University, Jawad ZAAROUR, Doctorant (depuis oct.2004).
Laboratoire de Spectrometrie Physique (LSP)Thierry BIBEN, CR CNRS, Chaouqi MISBAH, DR CNRS, Philippe PEYLA, MCF UJF, ThomasPODGORSKI, CR CNRS, Julien BEAUCOURT, Doctorant (depart juin 2005), Maude MADER,Doctorante, Victoria VITKOVA, Contrat CNES.
Laboratoire Techniques de l’Ingenierie Medicale et de la Complexite (TIMC)Pierre-Simon JOUK, PU-PH TIMC-CHU Grenoble, Gabrielle MICHALOWICZ, Attachee Scienti-fique TIMC-CHU, Jacques OHAYON, PR U. Savoie, Angelique STEPHANOU, CR CNRS, PhilippeTRACQUI, DR CNRS, Yves USSON, CR CNRS.
Laboratoire Sols, Solides, Structures (3S)Denis CAILLERIE, PR INPG
Laboratoire de Mathematiques U. Paris-SudBertrand MAURY, PR Orsay
1

1 Introduction generale
L’approche que nous proposons distingue trois niveaux de modelisation - la membrane cellulaire,la cellule, le tissu cardiaque - qui sont successivement imbriques en vue de la realisation d’unmodele global explicitant les modalites des contractions rythmiques du myocarde. Cette demarcheintegrative s’appuie sur des outils mathematiques modernes (techniques variationnelles pour desenergies non convexes, methodes ”level set”, homogeneisation discrete, flot optique ...) dont ledeveloppement dans le cadre de ce projet ont permis de surmonter les difficultes inherentes a lamodelisation multi-echelles. Nous passons en revue les divers points abordes, en renvoyant auxarticles publies dans le cadre du projet, dont une liste figure page 25.
Nous avons d’abord developpe et valide un modele biomecanique d’une cellule a interieur fluide,constitue d’un systeme d’equations aux derivees partielles decrivant les proprietes rythmiquescontractiles du cytosquelette. Cette thematique est developpee dans l’equipe DynaCell, sous undouble aspect theorique (modelisation des proprietes rheologiques et contractiles des cellules) etexperimental (imagerie video-microscopique des deformations cellulaires). Les travaux de l’equipedu LSP se situent en amont et concernent la modelisation physique des membranes phospholipi-diques (Biben et al 2005, Mader et al 2006, Mader et al 2007 , Misbah 2006, Danker et Misbah 2006,Danker et al 2007). Le LJK a propose pour une cellule a paroi membranaire un modele d’interactionfluide-structure base sur une extension de la methode level-set, plus facilement implementable queles methodes classiques (Cottet et Maitre 2004, Cottet et Maitre 2006).
La cellule myocardique a une organisation propre. Elle est cylindrique, constituee de sarcomeres,qui en sont les elements contractiles : le glissement des filaments d’actine et de myosine les uns parrapport aux autres permet la contraction. Nous disposons de premieres mesures experimentales surle comportement en etirement des cardiomyocytes isoles. En particulier, nous sommes en relationdirecte avec une equipe de l’Inserm de Montpellier qui peut effectuer certaines mesures a notre de-mande et surtout nous preciser dans quel environnement d’activation electrique (ou chimique) lesmesures sont effectuees. Ce dernier point est souvent peu explicite dans les articles experimentauxque nous connaissons. Par ailleurs, la partie fluide du cardiomyocyte lui confere un caractere in-compressible. Nous avons mis en oeuvre deux approches pour l’etude du cardiomyocyte.
La premiere est au niveau de la cellule et etudie la dynamique contractile de celle-ci. Une contribu-tion de l’equipe du laboratoire TIMC porte sur la modelisation de la contraction du cardiomyocyteinduite par la dynamique auto-oscillante du calcium. L’accent est mis sur l’utilisation d’une methoded’elements finis pour resoudre les equations de l’elasticite incompressible et anisotrope du cardio-myocyte (Pustoc’h et al 2005, Tracqui et Stephanou 2007, Tracqui et Ohayon 2006 et 2007). Unecontribution de l’equipe du laboratoire LJK s’est portee sur une formulation eulerienne generalisantl’approche operee pour l’elasticite d’une membrane 2D immergee au cas d’un corps elastique 3Dimmerge. Cette formulation qui utilise une approche de type level-set permet d’utiliser des ou-tils relativement simples comme la methode des differences finies tout en prenant en compte desgeometries de cardiomyocyte issues de l’experimentation (Cottet et Maitre 2007, Cottet et al 2006).
Dans une deuxieme approche nous avons utilise une description mecanique tres simplifiee du car-diomyocyte assimile a un segment elastique, interagissant avec les autres cardiomyocytes par desmoments. Bien que primitif sur les aspects d’incompressibilite et d’activation electrique, ce modele
2

de cellule isolee transforme en un modele de tissu grace a une technique d’homogeneisation nous apermis d’obtenir un modele de tissu cardiaque qui est en accord avec les resultats experimentaux dela litterature bio-mecanique (Caillerie, Mourad et Raoult 2005, Caillerie, Mourad et Raoult 2006,Jouk et al 2007, Milisic 2006, Milisic 2007, Mourad 2003, Raoult 2005, 2007, 2008).
A long terme, nous esperons que la realisation de ces differentes etapes aura des implicationsdirectes en physio-pathologie cardiaque : comprehension des pathologies cardiaques associees ades modifications structurelles (epaississement, fibroses locales, ...) affectant la biomecanique dumyocarde, simulation d’interventions chirurgicales, remodelage chirurgical effectif apres insuffisancecoronarienne ou encore therapie par implantation cellulaire a differenciation myocardique. D’autrepart la meilleure comprehension des deformations cellulaires devrait avoir des retombees sur lestechnologies prometteuses du tri cellulaire et de la conception de cellules artificielles.
Dans les pages qui suivent figure une description plus detaillee des contributions decrites ci-dessus.Nous avons fait le choix editorial de developper particulierement les contributions concernant lecardiomyocyte comme cellule isolee, cette partie etant par certains aspects plus graphique que lesautres. Ainsi nous esperons donner un apercu du travail accompli sans perdre notre lecteur dansdes equations qui se trouvent par ailleurs dans les publications du projet, page 25.
Pour conclure signalons deux faits marquants de notre projet : tout d’abord les credits allouesont permis de recruter V. Milisic en post-doc. Recrutement heureux puisqu’il est entre au CNRSdans notre laboratoire l’annee suivante. D’autre part nous avons organise en mai 2005, des journeesde l’ACI ou nous avons pu prendre le temps entre biologistes, physiciens, et mathematiciens ap-pliques d’echanger sur nos approches parfois tres differentes de la modelisation. Le programme deces journees se trouve sur le site http ://www-lmc.imag.fr/MOCEMY/journee aci.html.
2 Modelisation des deformations de membranes phospholipidiques
Ce sujet a ete amene par les membres de l’equipe GREPHE du laboratoire LSP. Puis une interactiona eu lieu avec les membres du laboratoire LJK sur une formulation level-set/champ de phase deces objets. Un groupe de travail a eu lieu regulierement. Nous presentons ci-dessous les avanceesoperees.
2.1 Methode de champ de phase (LSP)
L’equipe a porte un interet particulier aux problemes de la dynamique des vesicules en milieuetendu et en microfluidique. En outre, des travaux ont vu le jour sur la rheologie des suspensionsde vesicules.
3

2.1.1 Introduction et contexte
Les vesicules sont des membranes fermees composee d’une bicouche phospholipidique. Elles consti-tuent un systeme modele pour les membranes cytoplasmiques de cellules reelles, et plus parti-culierement les globules rouges. Pour cette raison, une suspension de vesicules peut etre vue commeune representation simplifiee pour l’etude de la rheologie du sang. Une vesicule a l’equilibre a uneforme qui minimise son energie de courbure, en respectant les contraintes de surface constante (lamembrane est un fluide bidimensionnel incompressible) et a volume interne constant (le fluide en-clos par la membrane est incompressible). Helfrich a developpe un modele ou l’energie s’ecrit sousla forme
E =∫
(H −H0)2dA+ µ1A+ µ2V
L’integrale porte sur la surface de la vesicule, H est la courbure moyenne, H0 est la courburespontanee (en quelque sorte rendant compte du fait que les deux cotes de la membrane peuventetre differents, de sorte qu’une membrane ouverte aurait comme curbure naturelle H0 et non zero),et µ1 et µ2 sont des multiplicateurs de Lagrange fixant l’aire et le volume.
Les vesicules peuvent etre plus au moins gonflees. Leur taux de degonflement depend des additifsrajoutes dans la solution ; le degonflement est d’origine osmotique. Il peut egalement etre d’originethermique : l’expansion thermique des phospholipides est plus facile que celle de l’eau enclose sousla membrane. Ainsi on peut augmenter la surface de la membrane plus rapidement que celle duvolume interieur. La quantite qui mesure le taux de degonflement est appele le volume reduit. C’estle rapport entre le volume reel sur celui de du volume d’une sphere ayant la meme surface :
ν =V/(4π/3)
(A/4π)32
ou V est le volume, et A la surface. ν est un nombre sans dimension. Il vaut 1 pour une sphere etest toujours < 1 pour tout autre forme. Pour le globule rouge humain, par exemple, il vaut environ0.64. En faisant varier ν la forme qui minimise l’energie de courbure peut varier d’une ellipsoıdeallongee, a une forme biconcave (representant le globule rouge, cf figure 1), vers des formes varieescomme celle donnant lieu a la forme de type « anemie falciforme ». Les travaux portant sur lesformes d’equilibre se sont developpes entre les annees 1970 et 1995, environ.
Depuis les milieux des annees 1990 les phenomenes hors equilibre ont connu un veritable engoue-ment. Mis a part la force de Helfrich, il existe un couplage non trivial entre la forme et l’ecoulement,qui genere des proprietes nonlocales et nonlineaires de la dynamique. Un des premiers travaux(etude theorique) de notre equipe a porte sur l’autopropulsion d’une vesicule induit par un gradientd’adhesion (mouvement dit de haptotaxie). L’equipe a ensuite developpe des theories sur le com-portement des vesicules sous cisaillement hydrodynamique pres d’une paroi. Plus particulierement,on a analyse la force de portance d’origine visqueuse. Ces travaux ont donne lieu a des etudesexperimentales au laboratoire. Les travaux qui ont suivi ont porte sur la transition entre le mou-vement dit chenille de char vers le mouvement de bascule (ou tumbling) tant sur le plan theoriquequ’experimental.
4

2.1.2 Les principaux resultats de l’equipe et les publications correspondantes sur lesujet pour la periode 2005-2007
1. Developpement d’un modele de champ de phase pour l’etude de la dynamique des vesicules a3D. On a egalement realise une etude asymptotique du modele pour montrer que les equationsse reduisent a celles du modele d’interface mince (Biben et al, 2005)
2. Etude experimentale detaillee de la dynamique des vesicules en fonction du taux de cisaille-ment. Decouverte d’effets importants lies a la deformabilite de la vesicule (Mader et al, 2006)
3. Elaboration d’un modele simple pour expliquer l’effet de la deformabilite de la vesicule sur lemouvement de bascule. En particulier nous avons pu expliquer tous les resultats qualitatifsdu regime de bascule (Mader et al, 2007)
4. Etude analytique d’une vesicule dans un champ de cisaillement, et decouverte d’un no-veau mode baptise vacillation-respiration (Misbah, 2006). Ce mode a depuis ete observeexperimentalement.
5. Etude theorique de la rheologie d’une suspension diluee de vesicules. On a derive une loiconstitutive pour une suspension diluee et decrit le lien entre la dynamique microscopique etla rheologie (Danger et Misbah, 2007)
6. Nous avons etendu le calcul develope en 4) aux ordres superieurs (i.e. en incluant des deformationsplus fortes de la vesicule). Nous avons etabli un diagramme de phase general pour fixer ledomaine (dans l’espace des parametres) associe a chacun des trois modes dynamique desvesicules (mouvement chenille de char, mouvement de bascule et mouvement de vacillation-respiration) (Danger et al, a paraıtre).
7. Nous avons etabli un parallele entre la physique d’une vesicule et celle d’une goutte (Dankeret al, 2007).
8. Nous avons etendu le modele de champ de phase 1) pour lui donner une forme compatibleavec le second principe de la thermodynamique (Jamet et Misbah, a paraıtre).
9. Nous avons realise une etude experimentale en microgravite et au sol, ainsi qu’une etudeanalytique pour expliquer l’influence de la gravite sur la dynamique des vesicules (Mader etal, 2006).
2.2 Methode level-set (LJK)
Dans le cadre de la modelisation des vesicules phospholipidiques en dimension 3, nous avonsdeveloppe un code de calcul permettant d’obtenir la forme de ceux-ci en fonction de leur tauxde remplissage (Cottet et Maitre 2006, Maitre et al, a paraıtre) Mathematiquement, ils s’agit deresoudre un probleme de couplage fluide-structure entre une structure elastique exercant a la foisune force due a son etirement mais aussi a sa courbure, ce qui complique notablement son ex-pression. L’objectif a plus long terme et qui interesse les physiciens du laboratoire LSP (et quifigure dans les objectifs d’un projet finance par l’ESA) est de simuler en 3D cette vesicule dansun ecoulement en cisaillement qui reproduit la situation d’un ecoulement dans un vaisseau san-guin. Cette etude complete donc celle menee en dimension deux par le LSP avec une technique dechamp de phase et developpee au LJK avec la methode level-set (cf http ://www-lmc.imag.fr/lmc-edp/Emmanuel.Maitre/simulation3.html). L’objectif est de retrouver des comportements observes parles experimentalistes : lorsque le rapport entre la viscosite du fluide interieur a la vesicule et celle
5

Fig. 1 – Formes optimales de vesicules phospholipidiques. En haut : ν = 0.8. En bas : ν = 0.586. Del’initialisation a l’etat stationnaire, de gauche a droite.
du fluide exterieur est inferieur a un certain seuil, la (pseudo-)cellule trouve un regime d’equilibreou sa membrane glisse sur elle-meme avec un mouvement de chenille de char (tank-treading). Audela de ce seuil, la cellule dans son ensemble connaıt un mouvement de rotation (tumbling). Latransition entre ces deux etats se produit pour un ration de viscosites qui correspond grosso modoa celui que l’on rencontre pour les globules rouges in vivo. Ceci expliquerait donc l’observationexperimentale qui est faite des deux mouvements pour ces objets biologiques.
Les resultats numeriques presentes ci-dessous montrent deux situations typiques de formes opti-males de vesicules 3D. La forme finale depend du volume reduit ν defini ci-dessus. Le premier cascorrespond a ν = 0.8, et donne une surface optimale en forme de cacahouete (figure 1, en haut).La seconde correspond quant a elle a ν = 0.586 ce qui produit une forme optimale en forme deglobule rouge (figure 1, en bas). Pour chacune des formes nous avons fait figurer la forme initialea gauche et une suite de formes avant convergence vers la forme optimale, image de droite. Cesformes correspondent qualitativement a celle observees par l’equipe du LSP.
Un article comparant les approches de level-set et de champ de phase d’un point de vue theoriqueest en cours de redaction (Maitre et al, en preparation).
3 Modelisation de la dynamique cellulaire et tissulaire, applica-tion au myocarde.
Deux approches ont ete explorees pour cette contribution sur l’etude de la contraction de la cellulemyocardique. La premiere considere le cardiomyocyte comme un corps elastique isole, dans lequelune onde calcique provoque une contraction active. La geometrie est issue de l’experience en 2Dou axisymetrique en 3D. Le calcul numerique utilise la methode des elements finis. Une deuxieme
6

approche utilise une methode de type frontiere immergee ou le cardiomyocyte isole est modeliseainsi que le fluide qui l’entoure. La geometrie complexe est prise en compte grace a l’utilisationd’une fonction level-set pour localiser l’interface. L’utilisation de cette fonction permet d’utiliser lamethode des differences finies malgre la complexite geometrique.
3.1 Contribution TIMC
La cellule myocardique a une organisation specifique. De forme quasi-cylindrique, elle est constitueede sarcomeres, qui en sont les elements contractiles. Cette contraction est controlee par la concentra-tion cytosolique locale de calcium qui, par fixation sur des proteines specifiques, induit le glissementdes filaments d’actine et de myosine les uns par rapport pour provoquer le raccourcissement rapidede la cellule. La regulation des interactions entre les filaments d’actine et de myosine est etroitementdependante des concentrations en calcium a l’exterieur et a l’interieur de la cellule, ainsi que dansles differents compartiments intracellulaires. Des nombreux modeles mathematiques de la celluleindividuelle myocardique ont ete developpes, mais la grande majorite de ceux-ci sont dedies soita la representation des phenomenes electriques qui modulent les flux calciques membranaires, soita l’etude de la diffusion du calcium intracellulaire, sous forme d’une vague calcique parcourant lacellule. L’originalite de l’approche que nous avons propose ici est de coupler la dynamique spatio-temporelle complexe du calcium intracellulaire aux contraintes mecaniques locales engendrees parle raccourcissement des sarcomeres. L’objectif est de mettre en evidence les couplages nonlineairesqui existent au sein de la cellule cardiaque entre les processus de reaction-diffusion d’une part, lesprocessus biomecaniques actifs et passifs dans un regime de grandes deformations d’autre part.Soulignons que les proprietes mecaniques de la cellule myocardique sont tout a fait determinantesvis a vis des performances du tissu cardiaque. Des modifications des proprietes mecaniques passives(diminution de son elasticite, augmentation de la viscosite du cytoplasme, ...) ou actives (tonuscontractile) de la cellule cardiaque ont en effet une influence directe sur la frequence des batte-ments cardiaques et sur l’eventuelle apparition d’un comportement asynchrone des cellules quine se contracteraient plus en phase. Ce couplage entre activite mecanique et electrique du tissucardiaque est implique dans l’initiation de differentes pathologies comme les arythmies ou les fibril-lations. Il est donc essentiel de pouvoir disposer d’un modele cytomecanique de la cellule cardiaquequi permette d’analyser l’influence de contraintes mecaniques sur la dynamique et l’efficacite decontraction cellulaire. Le travail realise dans le cadre de ce projet associe approches theoriques etexperimentales, avec pour l’analyse des donnees le couplage necessaire de methodes specifiques dequantification de processus dynamiques cellulaires a partir de sequences d’images. Ces sequences,acquises par video-microscopie, permettent d’analyser le comportement contractile rythmique spon-tane de cardiomyocytes isoles. A partir de l’analyse de ces observations, nous avons developpe unmodele realiste et original de la contraction spontanee de la cellule cardiaque qui prend en compte :– La possibilite d’oscillations calciques non lineaires auto-entretenues generees par le mecanisme
d’auto-stimulation de la liberation du calcium (mecanisme CICR, Calcium-Induced Calcium Re-lease). Il s’agit d’un processus non lineaire dans lequel le calcium, entrant par les canaux voltage– dependants de la membrane du sarcolemne, active les canaux calciques recepteurs a la ryano-dine du reticulum sarcoplasmique (RS), ce qui induit alors un relargage massif de calcium dansle cytosol.
– Le couplage de ces flux calciques locaux avec une diffusion anisotrope du calcium dans le cardio-myocyte, du fait de l’architecture interne specifique de cette cellule
– Les proprietes mecaniques passives de la cellule cardiaque dans le domaine des grandes deformations
7
– Le developpement de contraintes actives anisotropes, selon des directions donnees par l’orien-tation des sarcomeres, et dependant de facon non-lineaire des concentrations cytosoliques decalcium.
L’ensemble de ces elements est integre au sein d’un modele mecano-biochimique de la cellulecardiaque, decrit par un systeme d’equations aux derivees partielles non-lineaires dont les ca-racteristiques et les proprietes sont detaillees dans les paragraphes suivants. La validation de cesresultats theoriques a ete realisee sur la base des differents resultats experimentaux obtenus dansle cadre de ce projet ou a travers des collaborations externes.
3.1.1 Caracterisation experimentale des proprietes macroscopiques emergentes ducardiomyocyte isole.
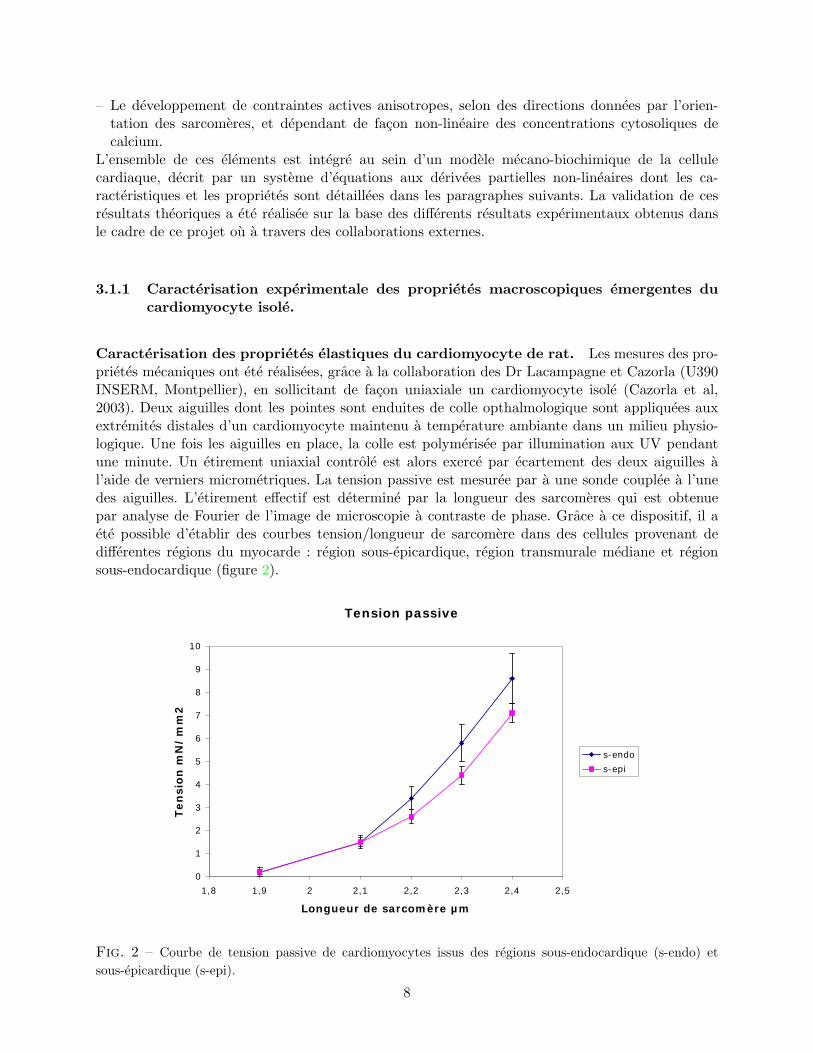
Caracterisation des proprietes elastiques du cardiomyocyte de rat. Les mesures des pro-prietes mecaniques ont ete realisees, grace a la collaboration des Dr Lacampagne et Cazorla (U390INSERM, Montpellier), en sollicitant de facon uniaxiale un cardiomyocyte isole (Cazorla et al,2003). Deux aiguilles dont les pointes sont enduites de colle opthalmologique sont appliquees auxextremites distales d’un cardiomyocyte maintenu a temperature ambiante dans un milieu physio-logique. Une fois les aiguilles en place, la colle est polymerisee par illumination aux UV pendantune minute. Un etirement uniaxial controle est alors exerce par ecartement des deux aiguilles al’aide de verniers micrometriques. La tension passive est mesuree par a une sonde couplee a l’unedes aiguilles. L’etirement effectif est determine par la longueur des sarcomeres qui est obtenuepar analyse de Fourier de l’image de microscopie a contraste de phase. Grace a ce dispositif, il aete possible d’etablir des courbes tension/longueur de sarcomere dans des cellules provenant dedifferentes regions du myocarde : region sous-epicardique, region transmurale mediane et regionsous-endocardique (figure 2).
Tension passive
0
1
2
3
4
5
6
7
8
9
10
1,8 1,9 2 2,1 2,2 2,3 2,4 2,5
Longueur de sarcomère µm
Ten
sio
n m
N/
mm
2
s-endo
s-epi
Fig. 2 – Courbe de tension passive de cardiomyocytes issus des regions sous-endocardique (s-endo) etsous-epicardique (s-epi).
8

Caracterisation de la contraction rythmique spontanee du cardiomyocyte de rat isole.Nous avons enregistre par videomicroscopie les contractions de cardiomyocytes de rat en culture,maintenus a temperature ambiante dans du liquide physiologique (Fig. 3). Ces cellules nous ontete gracieusement fournies par le Dr Olivares (LBFA UJF Grenoble). Pour l’observation, la boıtede culture a ete placee sur un microscope confocal a balayage laser. Les images ont ete acquisesen microscopie de transmission a contraste interferentiel differentiel (DIC) de Nomarski. Cettetechnique ne necessite aucune coloration des cellules et est tres resolutive. Apres avoir repere unecellule viable montrant une activite contractile spontanee, un enregistrement video est realise, defacon a obtenir des images numeriques (8 images /s) sur plusieurs cycles de contraction de la cellule(Fig. 3), de facon a s’assurer du caractere periodique ou non des contractions
Fig. 3 – Enregistrement videomicroscopique de la contraction d’un cardiomyocyte isole de rat. La taille dela cellule est d’environ 117 µm x 26 µm.
Quantification de la dynamique de contraction par analyse d’images. Pour pouvoirquantifier avec suffisamment de precision les differents aspects de la dynamique contractile ducardiomyocyte, nous avons ete amenes a developper de facon originale deux methodes d’analysedans le cadre de ce projet. La premiere repose sur une analyse de Fourier de l’image qui permetd’isoler les frequences spatiales correspondant aux unites sarcomeriques de la cellule. La seconderepose sur une approche par flot optique qui caracterise le champ de deplacement dans une zoned’image a partir d’un modele parametrique affine.
a. Quantification de la contraction par analyse de Fourier. Cette premiere approche permet deconnaıtre en tout point de la cellule la deformation longitudinale. L’algorithme developpe se baseen premier lieu sur une analyse de Fourier de l’image qui permet de selectionner les frequencesspatiales correspondant aux unites sarcomeriques de la cellule. Puis les lignes de cretes delimitantces unites sont detectees. A partir de ces lignes la deformation longitudinale est donnee enchaque point par le rapport entre la distance separant deux lignes de cretes les plus proches etla longueur de sarcomere au ”repos”. Il est ainsi possible de suivre la propagation de l’onde deraccourcissement longitudinal qui traverse la cellule pendant sa contraction (figure 4).
9
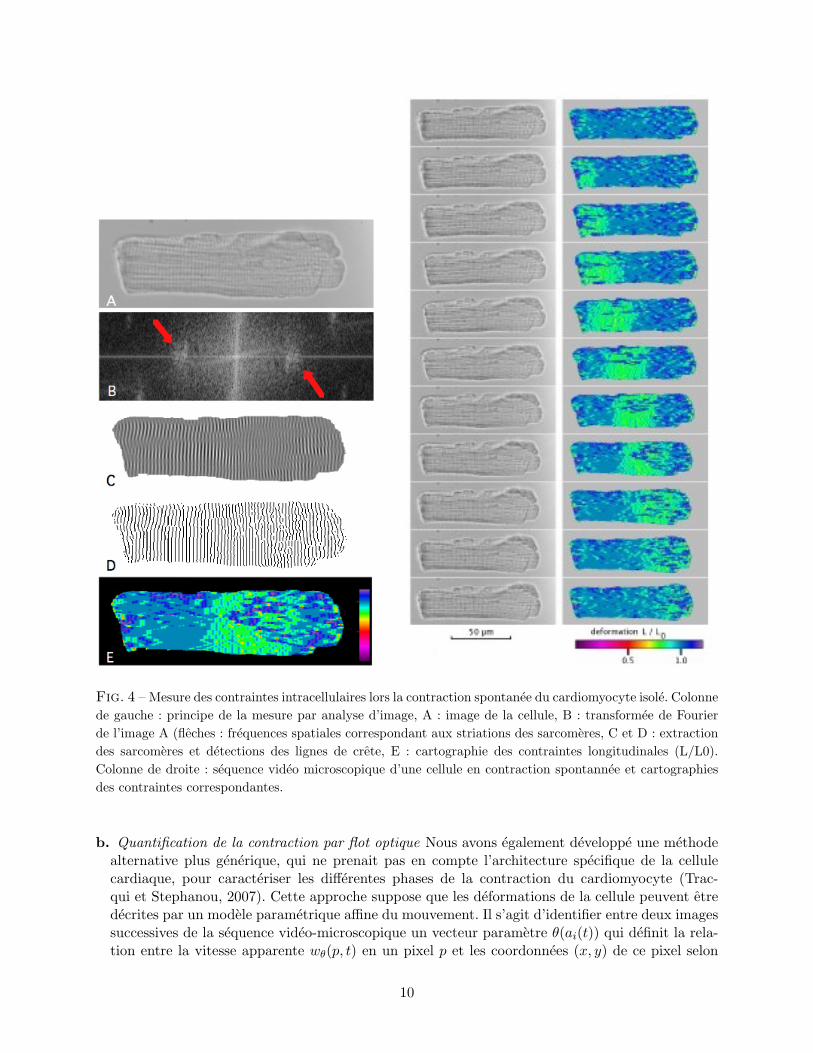
Fig. 4 – Mesure des contraintes intracellulaires lors la contraction spontanee du cardiomyocyte isole. Colonnede gauche : principe de la mesure par analyse d’image, A : image de la cellule, B : transformee de Fourierde l’image A (fleches : frequences spatiales correspondant aux striations des sarcomeres, C et D : extractiondes sarcomeres et detections des lignes de crete, E : cartographie des contraintes longitudinales (L/L0).Colonne de droite : sequence video microscopique d’une cellule en contraction spontannee et cartographiesdes contraintes correspondantes.
b. Quantification de la contraction par flot optique Nous avons egalement developpe une methodealternative plus generique, qui ne prenait pas en compte l’architecture specifique de la cellulecardiaque, pour caracteriser les differentes phases de la contraction du cardiomyocyte (Trac-qui et Stephanou, 2007). Cette approche suppose que les deformations de la cellule peuvent etredecrites par un modele parametrique affine du mouvement. Il s’agit d’identifier entre deux imagessuccessives de la sequence video-microscopique un vecteur parametre θ(ai(t)) qui definit la rela-tion entre la vitesse apparente wθ(p, t) en un pixel p et les coordonnees (x, y) de ce pixel selon
10
l’expression :
wθ(p, t) =(a0(t)a3(t)
)+(a1(t) a2(t)a4(t) a5(t)
)(xy
)sachant que le vecteur inconnu wθ(p, t) verifie l’equation de contrainte du mouvement apparent :
∂I
∂t+ wθ(p, t) · ∇I(p, t) = 0
ou I(p, t) est l’intensite lumineuse (ou luminance) projetee sur le pixel p a l’instant t. Nous avonsalors quantifie localement la contraction des cardiomyocytes observes en utilisant une approchepar fenetre glissante. En effet, a l’echelle de la cellule entiere, certaines zones restent au repos,certaines se contractent et d’autres se relaxent juste apres le passage de la vague calcique. Lamethode par fenetre glissante consiste a deplacer une fenetre etroite le long du grand axe Ox dela cellule, c’est-a-dire dans la direction parallele a l’orientation des sarcomeres, en supposant quele mouvement est homogene dans chaque fenetre (Fig. 5).
Fig. 5 – Evolution au cours du temps des descripteurs significatifs de la contraction spatialement heterogenede la cellule cardiaque E(t) et S1(t), calcules dans trois fenetres distinctes. On voit clairement que la vaguecontractile caracterisee par le descripteur S1(t) se propage, pratiquement sans deformation, de gauche adroite le long du grand axe de la cellule.
11

La figure 5 montre l’evolution simultanee des descripteurs de deformations E(t) (E(t) = (a1(t) +a5(t))/2) et S1(t) (S1(t) = (a1(t) − a5(t))/2) de la cellule pour les trois fenetres d’observationrepresentees sur la figure. Il apparaıt clairement que le descripteur dominant est la fonction S1(t),qui represente les contractions/expansions longitudinales. Ce resultat est une consequence directeet attendue de l’orientation anisotrope des sarcomeres selon cette direction. Plus precisement, onobserve que S1(t) est d’abord negatif, ce qui caracterise la phase de contraction de la cellule, puispositif, ce qui correspond a la relaxation des sarcomeres vers leur longueur de repos. La vaguede contraction se propage sous forme de soliton, avec une largeur (duree de contraction) et uneamplitude (intensite de contraction) qui se conserve d’une fenetre a l’autre et que cette methoded’analyse d’image permet de quantifier directement.
3.1.2 Modelisation de la contraction du cardiomyocyte induite par la dynamiqueautooscillante du calcium
Dynamique spatio-temporelle du calcium intracellulaire Les techniques d’imagerie de lacellule vivante ont permis de visualiser les oscillations rapides et spontanees du calcium intracellu-laire, ainsi que sa diffusion anisotrope (Subramanian et al., 2001). En nous basant sur le modelecinetique a un compartiment calcique de Goldbeter et al. (Goldbeter et al., 1990) decrivant lemecanisme CICR, nous avons integre cette anisotropie de la diffusion calcique au sein d’un modelecomportant deux variables qui sont respectivement les concentrations en calcium sarcoplasmiqueY (t) et en calcium cytosolique Z(t).
L’evolution temporelle de ces variables est regie par le systeme d’equations differentielles non-lineaires suivant :
∂Y
∂t= ν2(Z)− ν3(Y,Z)− kfY (1)
∂Z
∂t= ν0 + ν1β − ν2(Z) + ν3(Y,Z) + kfY − kZ +∇ · (D∇Z) (2)
Z(r, t0) = Z0; Y (r, t0) = Y0 (3)
ou D est le tenseur diagonal de diffusion de composantes Dii (Dij = 0 pour i 6= j). En accordavec les donnees experimentales (Subramanian et al., 2001), nous avons considere un rapport D22
D11
de 0.5 entre les diffusions transverse et longitudinale. L’interpretation des termes reactionnels desequations (1) et (2) est la suivante : le reticulum sarcoplasmique (RS) controle un flux constant decalcium vers le cytosol note ν1β. La concentration en calcium cytosolique Z(t) varie dans le tempset oscille entre deux phases : l’une pendant laquelle la concentration est tres faible en raison dupompage actif du calcium par le RS (ν2) et l’autre pendant laquelle la concentration Z(t) est treselevee caracterisant le relargage massif de Ca2+ (ν3) du RS dans le cytosol. Les parametres ν0 etk designent respectivement les flux d’entree et de sortie du calcium au sein de la cellule. Le termekfY represente une fuite de calcium depuis le RS dans le cytoplasme. Les flux de calcium ν2 et ν3
sont exprimes a l’aide des fonctions de Michaelis-Menten suivantes :
ν2 = VM2
Zn
Kn2 + Zn
ν3 = VM3
Y m
KmR + Y m
Zp
KpA + Zp
12
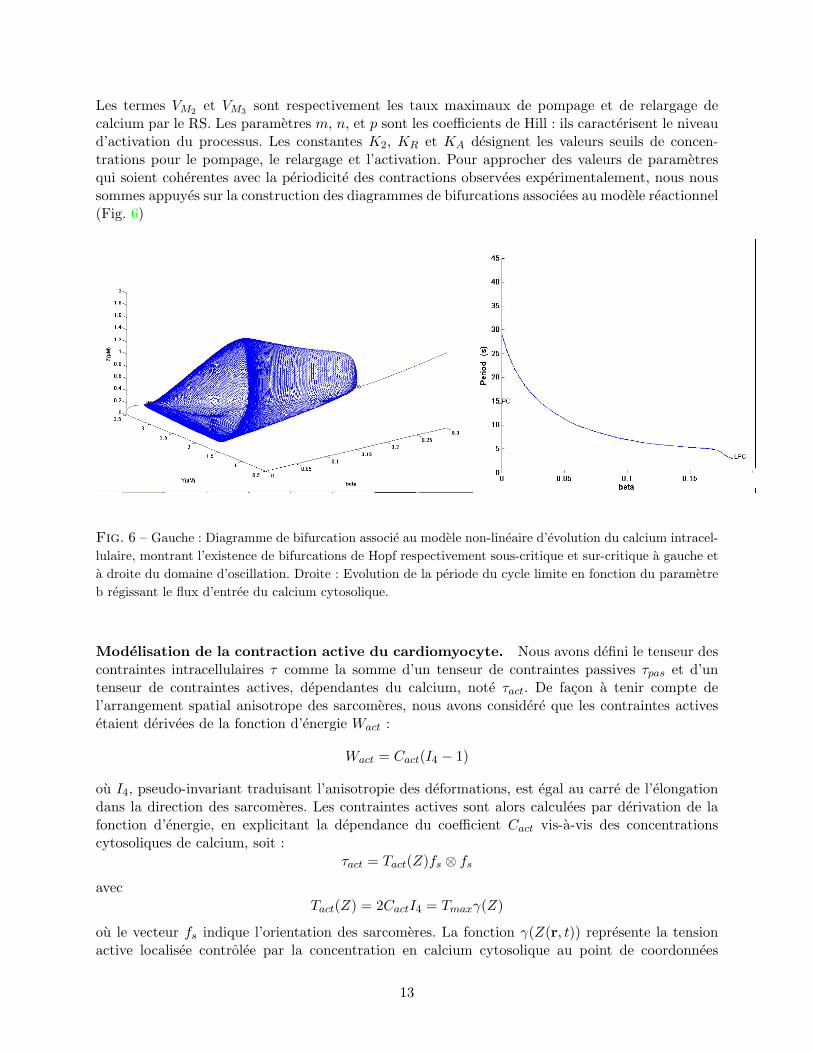
Les termes VM2 et VM3 sont respectivement les taux maximaux de pompage et de relargage decalcium par le RS. Les parametres m, n, et p sont les coefficients de Hill : ils caracterisent le niveaud’activation du processus. Les constantes K2, KR et KA designent les valeurs seuils de concen-trations pour le pompage, le relargage et l’activation. Pour approcher des valeurs de parametresqui soient coherentes avec la periodicite des contractions observees experimentalement, nous noussommes appuyes sur la construction des diagrammes de bifurcations associees au modele reactionnel(Fig. 6)
Fig. 6 – Gauche : Diagramme de bifurcation associe au modele non-lineaire d’evolution du calcium intracel-lulaire, montrant l’existence de bifurcations de Hopf respectivement sous-critique et sur-critique a gauche eta droite du domaine d’oscillation. Droite : Evolution de la periode du cycle limite en fonction du parametreb regissant le flux d’entree du calcium cytosolique.
Modelisation de la contraction active du cardiomyocyte. Nous avons defini le tenseur descontraintes intracellulaires τ comme la somme d’un tenseur de contraintes passives τpas et d’untenseur de contraintes actives, dependantes du calcium, note τact. De facon a tenir compte del’arrangement spatial anisotrope des sarcomeres, nous avons considere que les contraintes activesetaient derivees de la fonction d’energie Wact :
Wact = Cact(I4 − 1)
ou I4, pseudo-invariant traduisant l’anisotropie des deformations, est egal au carre de l’elongationdans la direction des sarcomeres. Les contraintes actives sont alors calculees par derivation de lafonction d’energie, en explicitant la dependance du coefficient Cact vis-a-vis des concentrationscytosoliques de calcium, soit :
τact = Tact(Z)fs ⊗ fsavec
Tact(Z) = 2CactI4 = Tmaxγ(Z)
ou le vecteur fs indique l’orientation des sarcomeres. La fonction γ(Z(r, t)) represente la tensionactive localisee controlee par la concentration en calcium cytosolique au point de coordonnees
13

spatiales r. En accord avec les donnees de la litterature, le terme γ(Z(r, t)) est exprime par lafonction de Hill suivante :
γ(Z(r, t)) =ZnH
ZnH50 + ZnH
ou Z50 correspond a 50% de la valeur maximale atteinte par la concentration en calcium cytosoliqueZ(r, t) et nH , coefficient de Hill, est egal a 4.
En considerant de plus que le comportement mecanique passif est decrit par la fonction d’energie hy-perelastique Wp (Lin et Yin, 1998), et en supposant que la cardiomyocyte est quasi-incompressible,le tenseur des contraintes dans le cardiomyocyte s’exprime par la relation :
τ = −pI + F∂Wp
∂EF T + τact
ou p est un multiplicateur de Lagrange introduit pour satisfaire l’hypothese d’incompressibilite.
La resolution par elements finis du probleme, implementee dans le logiciel Comsol, est realiseesur une geometrie maillee du cardiomyocyte issue de l’importation du contour reel d’une celluleobservee par video-microscopie (Fig. 7). Les conditions aux limites considerees sont :(i) des flux de calcium passifs nuls aux frontieres (conditions de Neumann)(ii) la cellule semble adherente dans sa position centrale(iii) des contraintes nulles sur les frontieres externes de la cellule
Fig. 7 – Haut : Image issue d’une sequence de video-microscopie de cardiomyocyte isole de rat. Bas :Contour cellulaire importe et maille utilise pour nos simulations.
Contraction simulee du cardiomyocyte Les valeurs des differents parametres representant ladynamique calcique ont ete choisies (Tableau 2) en accord avec la physiologie cellulaire et de manierea obtenir une periode des oscillations calciques correspondant aux mesures experimentales. A partirdes donnees de la figure 2, nous avons estime un module d’Young E egal a 30kPa. L’amplitude descontractions est controlee par le coefficient de tension active Tmax. Une amplitude de contraction dela cellule correspondant aux mesures effectuees sur les cardiomyocytes de rat permet de determinerla valeur de ce coefficient, soit 5, 5kPa dans nos conditions experimentales.
14

Parametre Valeur Unite Parametre Valeur Unite Parametre Valeur Uniteν0 0.45 µM.s−1 KA 0.92 µM D11 300 µm2s−1
k 2.2 s−1 KR 3.5 µM D22 150 µm2s−1
kf 0.1 s−1 Y0 0.1 µM D12 0 µm2s−1
ν1 4 µM.s−1 Z0 10 µM D21 0 µm2s−1
VM2 65 µM.s−1 Z50 2.5 µM E 30 kPaVM3 500 µM.s−1 β 0.05 – ν 0.49 –K2 1.2 µM T0 5.5 kPa
Tab. 2 – Valeurs des parametres rheologiques et mecaniques utilisees dans le modele.
Une sequence typique de contraction du cardiomyocyte virtuel est representee sur la figure 8,lorsque l’on suppose que le pic (spark) de calcium declencheur est situe dans le coin superieurgauche de la cellule. L’onde calcique se propage de la gauche vers la droite de la cellule et induitune augmentation progressive de la contraction dans la zone centrale, avant la relaxation de lacellule vers sa morphologie initiale non contrainte. Cette dynamique simulee est en tres bon accordavec les donnees experimentales que nous avons enregistrees en marquant le calcium par une sondefluorescente (Fluo-3, colonne de gauche sur la figure 8).
Fig. 8 – Propagation d’une vague de calcium dans un cardiomyocyte isole (l’intervalle de temps entre deuximages successives est de 268 ms) et dynamique correspondante simulee par le modele.
15

En tenant compte de la morphologie quasi-cylindrique de la cellule cardiaque, nous avons purepresenter ce comportement contractile dans une cellule 3D reconstruite en considerant pour li-miter les temps de calculs une geometrie et des conditions axisymetriques. La figure 9 montre lavague calcique simulee et la contraction associee lorsque que le spark declencheur est cette fois cilocalise au centre de la cellule. Dans ce cas, deux vagues calciques emergent, qui se propagent ensens contraire dans la cellule, ce qui provoque la contraction simultanee de deux regions differentesdu cardiomyocyte.
Fig. 9 – Simulation de la contraction d’un cardiomyocyte avec une geometrie idealisee presentantune symetrie de revolution. Le declenchement de la vague calcique par un « spark » de calcium sefait a partir d’un region centrale de la cellule.
Cette hypothese simplificatrice d’axisymetrie n’est pas contraignante pour etudier la dynamiquecontractile de la cellule cardiaque. Neanmoins, elle ne permet pas de prendre en compte la geometrie3D reelle de la cellule, contrairement a l’approche level-set developpee dans ce projet (cf. § suivants).
Validation du modele mecano-biochimique du cardiomyocyte. Nos resultats de simu-lations ont ete compares aux donnees issues des sequences de video-microscopie que nous avonsacquises sur des cardiomyocytes de rat, avec ou sans perfusion avec un indicateur fluorescent ducalcium (Fluo-3). Les cellules observees presentent a temperature ambiante une periode de contrac-tion de 17 ± 5, 8 s, avec une duree de contraction tres breve de 1, 5 ± 0, 4 s. Les simulations de lacontraction du cardiomyocyte virtuel donnent une duree de contraction, une periode des oscillationstout a fait en accord avec ces valeurs experimentales. La contraction/relaxation simulee se repro-duit toutes les 18, 8 s, avec une amplitude maximale de la contraction de la cellule cardiaque reelled’environ 7, 3%, soit approximativement 8µm, ce qui est tout a fait en accord avec les observations.
3.1.3 Conclusions et perspectives
A travers ce travail, mene aux interfaces biologie cellulaire–mathematiques–biomecanique, nousavons pu proposer de facon originale une cellule cardiaque virtuelle modelisee par un oscilla-teur cytomecanique autonome qui reproduit de maniere satisfaisante un ensemble de donneesexperimentales obtenues a l’echelle cellulaire. En permettant de quantifier les contraintes et les
16

deformations locales subies par la cellule cardiaque, cette approche ouvre la voie a des analysesplus fines, menees au niveau moleculaire, et qui devraient permettre de mieux comprendre le rolejoue par certaines proteines, comme la titine, dans la modulation des proprietes mecaniques du car-diomyocyte. Plus globalement, c’est une meilleure connaissance des voies de mecanotransductionau sein de la cellule cardiaque qui peut etre envisagee dans la mesure ou ce modele de cellule vir-tuelle un pont entre la description de processus subcellulaires et l’emergence de proprietes contrac-tiles macroscopiques. L’etude du comportement mecanique genere par un ensemble de cellules vir-tuelles contractiles organisees en couche tissulaire reste un objectif prometteur, mais les resultatspreliminaires que nous avons obtenus ont montre les limites de l’approche numerique de resolutionpar elements finis, ou un nombre limite de cellules peut etre considere pour des temps de calculsraisonnables. Une approche generalisee basee sur une description des cellules par level-set devraitpermettre de contourner ce probleme.
3.2 Contribution LJK
Dans un article recent, Okada et al (2005) etudient le mecanisme de la propagation d’une vaguecalcique dans un cardiomyocyte et la contraction qui en resulte. Ils presentent des simulations 3Dutilisant le modele de propagation de vagues calciques introduit par Subramanian et al (2002), etle modele de contraction de Negroni et Lascano (1996). Le cardiomyocyte est represente comme unmilieu hyperelastique obeissant a la loi de comportement hyperelastique de St-Venant-Kirchoff. Sastructure microscopique est decrite par un choix d’elements finis correspondant a la geometrie desobjets biologiques : myofibrilles, lignes Z, ... Le resultat est une simulation certes en adequationavec l’experience, mais au prix d’un calcul de 37 heures pour une seconde de simulation. Nousproposons ici une modelisation alternative, basee sur une formulation eulerienne de l’elasticite ducardiomyocyte, qui fait suite a une formulation particulaire de l’elasticite incompressible proposeepar G.-H. Cottet (Cottet, 2004).
Les mesures effectuees a Montpellier sur des cardiomyocytes de rat par les medecins collaborant avecTIMC nous permettent de nous baser sur une geometrie reelle. Nous considerons donc le systemeconstitue d’un cardiomyocyte immerge dans un fluide et decrivons le probleme d’interaction fluidestructure en variables euleriennes. D’autre part nous utilisons la dynamique calcique introduite auparagraphe precedent qui intervient dans la partie active du tenseur des contraintes (cf ci-dessous).
3.2.1 Description du modele
Formulation eulerienne du mileu continu passif Le cardiomyocyte est immerge dans unfluide qui occupe un domaine borne Ω ⊂ R3. On note u le champ de vitesse, a divergence nulle, dumilieu continu incompressible dans son ensemble. On le suppose regulier et nul au bord :
(H) u ∈ C1(Ω× [0, T ]) and u = 0 on ∂Ω× [0, T ]
La frontiere du cardiomyocyte est decrite de la maniere suivante : initialement le cardiomycocyteoccupe une domaine Ω0
c = x ∈ Ω, ψ0(x) < 0, o˘ ψ0 est donne (il n’est pas necessaire que cesoit une fonction distance). Cette fonction peut-etre donnee par une formule mathematique dans le
17

cas d’un cardiomoycyte cylindrique, ou tiree directement des donnees experimentale obtenues par(Usson et al, 2005), figure 10.
Fig. 10 – Image d’un cardiomyocyte obtenue par microscopie confocale (TIMC) et utilisee commefonction level-set ψ.
Nous remarquons ensuite dans (Cottet et Maitre, 2006) que les caracteristiques retrogrades de usont des quantites euleriennes X(x, t) solutions de
Xt + u(x, t) · ∇X = X(0) = x (4)
Une fois ces caracteristiques calculees, toute quantite transportee peut facilement se calculer. Parexemple la frontiere du cardiomyocyte est la ligne de niveau 0 de la fonction ψ(x, t) = ψ0(X(0;x, t)).Le tenseur de Cauchy-Green a gauche est donne par (Ciarlet, 1985, p. 15, Ciarlet, 1988, p. 43)
B = FF t ou F (x, t) = (∇X)−1(x, t)
Pour un materiau dont la reponse verifie le principe d’indifference materielle et est isotropique aupoint ξ = X(x, t), le tenseur des contraintes de Cauchy en x est donne par (Ciarlet, 1985, p. 50,Ciarlet, 1988, p. 115)
T (x) = TD(x,B(x, t))
Si ce materiau est incompressible, alors cette loi constitutive devient
ΣS(x) = TD(x,B(x, t))− p(x)I
ou I est le tenseur identite. Si nous considerons le fluide environnant le cardiomyocyte commenewtonnien, le tenseur de contrainte dans la partie fluide du milieu continu (soit x, ψ(x, t) > 0)est
ΣF (x) = −p(x)I + µD(u)
18

ou µ est la viscosite et D(u) = 12(∇u +∇ut). La conservation de la quantite de mouvement dans
tout le milieu continu s’ecrit alors
ρ(ut + u · ∇u)− div σ +∇p = f
ou σ = σSχφ<0 + σFχφ>0 et σS = TD(x,B(x, t)), σF = µD(u). Dans ce cas l’equation ci-dessus s’entend au sens des distributions et des conditions de saut de contraintes sur l’interfacefluide-solide existent et est compensee par un saut de pression. Une autre maniere de poser leprobleme est d’introduire une regularisation de χ utilisant la fonction niveau ψ. Notons r → H( rε)une approximation de la fonction de Heaviside lorsque ε→ 0. Alors la contrainte regularisee
σε = σS(1−H(ψ
ε)) + σFH(
ψ
ε)
varie regulierement au passage de l’interface ψ = 0 et l’equation de conservation de mouvementpeut etre prise au sens classique, la pression devenant alors continue. Un schema differences finiespeut donc etre utilise pour resoudre de telles equations.
3.2.2 Isotropie transverse
Dans le cardiomyocyte, les myofibrilles sont alignes selon une direction privilegiee, qui correspondd’ailleurs a celle des fibres cardiaque, a l’echelle superieure. On peut donc s’attendre a un compor-tement elastique anisotropique. Plus precisement, comme indique dans (Mourad, 2003), p. 80, lecardiomyocyte peut etre considere comme isotropique transverse (Ogden, 2003). Soit τ la directiondu grand axe du cardiomyocyte au repos. Un materiau de ce type peut etre caracterise par uneenergie qui depend de F et de τ ⊗ τ . Le tenseur des contraintes a alors l’expression suivante
ΣS = −pI + 2α1B + 2α2(tr(B)B −B2) + 2α4Fτ ⊗ Fτ + 2α5(Fτ ⊗BFτ +BFτ ⊗ Fτ) (5)
ou αi est la derivee de l’energie par rapport a l’invariant i, avec
I1 = tr(B), I2 =12
[tr(B)2 − tr(B2)], I4 = |Fτ |2, I5 = (BFτ) · (Fτ). (6)
Une interpretation de I124 en termes d’elongation dans le sens privilegie est aisee. L’interpretation
de I5 est moins intuitive : il s’agit de l’elongation dans la direction orthogonale aux directions nonprivilegiee dans la configuration de depart. Ces deux notions sont evidemment liees et coincidents’il n’y a pas de cisaillement tranverse. Nous avons donc suppose ci-dessous que α5 = 0.
3.2.3 Contraction active
Pour la partie active du comportement du myocarde, nous nous sommes bases sur le modele de(Stephanou et Tracqui, 2002), ou la contrainte active est ajoutee a σS dans la direction privilegiee,ce qui dans nos notations correspond a ajouter dans (5) T0γ(Z(x, t)) ‡ α4, ou Z est la concentrationintracellulaire en Ca2+. Z est donc la solution du systeme de reaction diffusion (1)-(2).
19

3.2.4 Simulation numerique
Methode La methode numerique utilisee est basee principalement sur deux ingredients : l’utili-sation d’un schema differences finies explicite d’ordre deux de type projection sur un maillage MACpour la resolution des equations de Navier-Stokes (Brown et al, 2001), couple avec un schema WENOd’ordre 5 (cf http://techreports.larc.nasa.gov/icase/1997/icase-1997-65.pdf) pour la resolution destermes de transport (dans Navier-Stokes et dans l’equation 4). Comme indique dans (Cottet etMaitre, 2004), aucune procedure de reinitialisation n’est implementee pour redresser la fonction ψa une distance. Le systeme de reaction-diffusion est resolu sur le meme maillage en utilisant unedircretisation standard des termes diffusifs.
Contraction uniforme d’un cardiomyocyte cylindrique incompressible Pour tester notrecode de calcul, nous avons commence par resoudre le probleme dans le cas d’un cardiomyocytecylindrique. La simulation presentee ici est fait sur une grille MAC de taille 1003 et une contractiondemande 3 heures de calcul sur un ordinateur de type PC AMD 64 a 3 GHz tournant sous linux64 bits.
Simulation en geometrie reelle Un interet fort de notre methode est que l’interface compliqueedu cardiomyocyte n’est pas maillee de maniere adaptative. Une fonction niveau la represente. Iln’y a donc aucun cout supplementaire a considerer une forme initiale de cardiomyocyte issue demesures experimentales par microscopie (figure 11).
Fig. 11 – Contraction uniforme d’un cardiomyocyte resultant d’un largage homogene de calcium
20

De meme, on pourrait pour le meme cout considerer plusieurs cardiomyocytes en reseau et etudierla contraction de l’ensemble. Bien entendu, la resolution de la geometrie de cardiomyocyte estconditionnee par le pas de la grille de calcul. Mais il semble peu probable que les details a petiteechelle de la forme du myocyte jouent un role determinant dans la contraction de celui-ci.
Dans le cas de la propagation d’une vague calcique, la contraction est notamment moins importante.On note aussi l’anisotropie de la propagation de l’onde due a la difference dans les coefficients dediffusion du modele CICR. Elle reflete le fait que la propagation est effectivement plus rapide dansla direction des fibres (figure 12). Les donnees biologiques sont celles du tableau 2.
Fig. 12 – Vague calcique se propageant dans un cardiomyocyte. La couleur represente la concen-tration calcique. Temps total de la simulation : 1s
4 Modelisation continue d’un reseau de cardiomyocytes.
Lois de comportement. La modelisation du mouvement cardiaque necessite la connaissancede lois de comportement du tissu myocardique. Les lois classiques reposent pour la plupart surune hypothese d’isotropie transverse, hypothese qui traduit l’existence de fibres. Dans une etudeprealable a l’ACI MoCeMy, nous avions developpe (Mourad, 2003) une approche nouvelle dansce domaine : en effet, nous etions partis de lois de comportement de la cellule myocardique pourconstruire par homogeneisation une loi globale. Des donnees experimentales sur les deformationsen elongation des myocytes sont en effet disponibles et servent de support a la construction de laloi globale. Ces donnees sont, par exemple, obtenues par une manipulation de cellules de rat aumoyen de pinces optiques.
21

Durant les annees 2004-2007, nous avons poursuivi nos travaux sur les lois de comportement ob-tenues de maniere directe comme homogeneisee. D’une part, nous avons decrit de maniere aussisimple que possible la methode d’homogeneisation discrete en l’utilisant dans un domaine autre quela modelisation biologique, celui de la modelisation de graphenes (Caillerie et al, 2006) qui sont desreseaux d’atomes de carbone. Le systeme d’equilibre mecanique que nous ecrivons pour un treillis– myocardique ou carbone – tient compte aussi bien de l’equilibre des forces que des moments,et il se revele – dans le cas carbone – etre le systeme d’Euler de l’energie classique utilisee parles physico-chimistes : l’energie contient un terme du aux changements de longueurs et un termedu aux changements d’angles. Le probleme homogeneise obtenu est un probleme membranaire engrandes deformations. Comme dans le cas du myocarde, la loi de comportement n’est pas expli-cite. Connaıtre les contraintes associees a une deformation necessite la resolution d’un systemenon lineaire, dont les inconnues sont des vecteurs de R3. Un des interets de l’exemple du reseauperiodique hexagonal de carbone est que dans le cas linearise les lois de comportement sont expli-citables et que les parametres mecaniques microscopiques et macroscopiques peuvent etre trouvesdans la litterature physico-chimiste. Nous avons constate que les valeur predites par la methoded’homogeneisation sont en accord avec les valeurs experimentales.
La validation mathematique de la methode d’homogeneisation de treillis par des resultats de conver-gence fait partie de notre programme de recherche. La difficulte technique a surmonter est la priseen compte des changements d’angles qui s’expriment par des produits scalaires de vecteurs unitairesportes par les “myocytes” ou “liens chimiques” deformes. Lorsque la deformation (non physique)tend a superposer des anastomoses ou des atomes, les denominateurs des vecteurs unitaires tendentvers 0 et generent, sans que l’on en soit surpris, des complications mathematiques. Notons que lamodelisation mathematique des reseaux est, independamment des applications citees ici, un do-maine actif (travaux de Braides, Weinan E., Friesecke, Le Bris, Le Dret, P.-L. Lions,...) parmibeaucoup d’autres.
Par ailleurs, nous avons examine les lois de comportement anisotropes utilisees par les biomecaniciens.Une premiere etude bibliographique a permis de constater que les restrictions sur les formes de ceslois bien que correctes dans le cas isotrope transverse sont obtenues – au moins dans une cer-taine ecole (un peu “ancienne”) de biomecaniciens – par des arguments ad hoc qui ne sont pas desconsequences directes de l’existence d’un groupe de symetries donne. Nous avons montre que l’onpeut obtenir de maniere rigoureuse ces resultats et surtout nous avons considere des lois pour desmateriaux a deux familles de fibres (Raoult, en preparation). La, il est interessant de constater queles lois de comportement que nous obtenons ne sont pas des lois a sept ou huit invariants, comme ilest d’usage (et maladroit, sinon absurde, puisque ce sont des fonctions de six variables reelles), maisdes fonctions de six variables assujetties a certaines conditions de symetrie. L’exploration biblio-graphique des travaux, tout juste decouverts, d’une autre mouvance de chercheurs (Boehler, Liu,Smith, Wang) est au programme, et nous decouvrirons peut-etre que nous n’avons rien invente...
Dans le domaine applicatif qui nous concerne, nous nous sommes pose la question des symetries deslois de comportement construites par homogeneisation. Nous avons demontre que la loi non lineairedu graphene restreinte au cas planaire est non isotrope, contrairement a la croyance repandue quidit qu’“un reseau hexagonal a necessairement une loi de comportement isotrope”. Et nous avonsexplique la croyance repandue en demontrant que la loi linearisee d’une loi admettant une rotationautre que 0, π/2, π ou 3π/2 (ici, π/3) comme symetrie est necessairement isotrope. Jawad Zaaroura dans le cadre de son contrat doctoral travaille sur une cellule comportant une direction principale
22

(celle de la fibre cardiaque) et 12 autres branches, qui represente un myocyte. Il a demontre quepour cette cellule les rotations d’angle π/3 et d’axe la direction principale sont dans le groupede symetrie. Et il a verifie numeriquement que des rotations d’angle inferieur ne sont pas dans legroupe de symetrie. Ainsi, la loi homogeneisee n’est pas transverse isotrope.
Resultats numeriques.
Ainsi qu’il a ete dit plus haut, la loi de comportement homogeneisee du myocarde n’est pas explicite.Pour chaque calcul des vecteurs de contraintes associes a un gradient de deformation, il faut resoudreun systeme non lineaire par une methode de Newton. Ceci avait ete realise dans (Mourad, 2003)et est appele dans la suite “code micro”. Vuk Milisic a implemente le code micro dans un codeelements finis en grandes deformations. Il a ensuite construit un des maillages d’une geometrieidealisee du ventricule gauche et y a inclus l’information sur l’orientation des fibres ainsi que leurvecteurs normaux et binormaux. Des outils de visualisation (GMSH, MEdit, Vtk, Toolkit) ont eteincorpores. Ce travail numerique a mis en evidence le caractere instable de la loi homogeneisee. Eneffet, pour certaines deformations les matrices de raideur necessaires a la resolution des problemesiteres de la methode de Newton (pour le probleme global) sont singulieres. L’analyse du spectredes matrices linearisees dans le code micro montre un phenomene de flambage.
En parallele, une simplification du modele de deformation a ete proposee fondee sur la symetrie derevolution du ventricule gauche : on ramene le probleme 3D a un probleme pose sur une sectiontransverse. Jawad Zaarour travaille a l’obtention de simulations numeriques sur ce cas simplifie enutilisant certains des programmes ecrits par Vuk Milisic.
Au chapitre visualisation, indiquons aussi que des traces de geodesiques sur des tores emboı tes ontete realises illustrant les travaux de (Mourad, 2003) sur les nombres de tours et de spires que lesfibres cardiaques peuvent realiser. Le code decrit ci-dessus a ete utilise pour differents nombres detours et/ou spires et l’on peut voir des changements quantitatifs sur la deformation engendree.
Perspectives.
Rappelons que le point de depart de notre travail a ete une sollicitation de la part de Pierre-SimonJouk, clinicien, et d’Yves Usson, biologiste, du laboratoire TIMC qui possedent des releves carto-graphiques des fibres cardiaques. Nous avons eu la satisfaction de constater que notre travail etaitbien recu dans la communaute medicale (Jouk et al, 2007). Les releves (cartes d’elevation et d’azi-mut) ont ete jusqu’a present utilises pour la validation de l’hypothese de Streeter sur l’organisationgeometrique des fibres cardiaques. Nous souhaitons les utiliser pour la generation automatique demaillages du ventricule gardant l’information sur l’orientation des fibres la ou on en dispose ef-fectivement. En effet, les interpolations des vecteurs de direction sont non fiables. Ce travail serapropose en stage de M2R et pourrait faire l’objet d’une these.
Par ailleurs, nous souhaitons ameliorer la loi de comportement homogeneisee dont nous avonsconstate le caractere instable. Nous cherchons donc un motif de base evitant le flambage. Nouscomptons, d’une part, etudier la sensibilite par rapport aux parametres de raideur en tensions et enmoments dans la cellule sur laquelle on resout le probleme microscopique et d’autre part modifierla configuration geometrique des barres de cette cellule.
23

L’activation electrique n’est pas presente dans la modelisation effectuee jusqu’a present. Nous pen-sons la modeliser par un parametre d’activation dans les lois de comportement des myocytes. Onpeut par exemple changer la longueur a vide des myocytes. Le temps effectif n’intervient pas danscette modelisation locale. Par contre, il devra intervenir dans les lois globales reliant la pression surl’endocarde au volume de la cavite.
Un dernier point est celui de l’estimation des coefficients mecaniques du myocarde a partir dedonnees images. Des methodes statistiques peuvent se reveler utiles. Mais la faisabilite de cetteapproche n’a pas encore ete exploree.
5 Conclusion generale et perspectives
Pour conclure ce rapport sur une collaboration pluridisciplinaire, nous passons en revue les differentescontributions.
L’etude d’une membrane elastique couplee avec un fluide a abouti a la mise au point d’une methodegenerale pour le couplage fluide-structure en coordonnees euleriennes, etendue ensuite au cas d’unsolide elastique. Cette methode semble tres prometteuse en sciences du vivant notamment. L’inter-action de ces methodes de type level-set avec la methode du champ de phase developpee au LSP estun sujet actuel de recherche, qui interesse aussi d’autres modelisations. Nous avons eu l’occasion departiciper en tant que formateur dans une ecole de printemps du GDR Champ de Phase et avons puconstater que les demandes d’expertise dans le domaine des methodes champ de phase/level set esttres forte de la part des physiciens notamment. D’un point de vue biomecanique, une perspectiveest l’etude des vesicules phospholipidiques 3D en cisaillement. Une these est en cours au laboratoireLJK dont une partie y est consacree (T. Milcent).
Au dela du comportement d’un cardiomyocyte isole qui a ete cerne par plusieurs approches, l’etudedu comportement mecanique genere par un ensemble de cellules virtuelles contractiles organiseesen couche tissulaire reste un objectif prometteur.
A une echelle superieure la methode d’homogenisation discrete apporte une reponse complementaireen delivrant une loi de comportement macroscopique base sur un modele simplifie de cellule. Lesrecherches actuelles portent sur l’etude de la sensibilite par rapport aux parametres de raideuren tensions et en moments dans la cellule sur laquelle on resout le probleme microscopique et laconfiguration geometrique des barres de cette cellule.
Au final, nous pensons avoir propose des avancees significatives dans plusieurs directions annonceesdans ce projet, comme l’indique entre autres les publications liees au projet (cf ci-dessous). Au delade cet aspect quantitatif, l’interaction avec des collegues d’autres disciplines est un enrichissementindeniable et une experience d’ouverture qui justifie a elle seule la poursuite d’une telle collaboration.
24

6 Publications associees au projet
6.1 Articles dans des revues
Biben T., Kassner K. et Misbah C., Phase-field approach to three-dimensional vesicle dynamics,Phys. Rev. E. 72, 041921, 2005
Caillerie D., Mourad A., Raoult A., Discrete homogenization in graphene sheet modeling, J. Elas.,84, 33-68, 2006.
Caillerie D., Mourad A., Raoult A., Confronting a mathematically derived constitutive law for themyocardium with experimental data, Third MIT Conference Computational Fluid and SolidMechanics, 2005, K.J. Bathe Ed., 127-130, 2005.
Cottet G.-H. et Maitre E., A level-set formulation of immersed boundary methods for fluid-structureinteraction problems, C. R. Acad. Sci. Paris, Ser. I 338, 581-586, 2004
Cottet G.-H. et Maitre E., A level-set method for fluid-structure interactions with immersed sur-faces, M3AS, 16 (3), 415–438, 2006
Danker G. et Misbah C., Rheology of a dilute suspension of vesicles, Phys. Rev. Lett. 98, 088104,2007
Danker G., Biben T., Podgorski T., Verdier C., et Misbah C., Dynamics and rheology of a dilutesuspension of vesicles : higher order theory, a paraıtre dans Phys. Rev. E
Danker G., Verdier C., et Misbah C., Rheology and dynamics of vesicle suspension in comparisonwith droplet emulsion, J. Non. Newontian Fluid. Mech. 2007
Jamet D. et Misbah C., Towards a thermodynamically consistent picture of the phase-field modelof vesicles, a paraıtre dans Phys. Rev. E
Jouk P.-S., Mourad A., Milisic V., Michalowicz G., Raoult A., Caillerie D., Usson Y., Analysisof the fibre architecture of the heart, European J. of Cardiac Thoracic Surgery, 31, 915-921,2007.
Mader M.-A., Vitkova V., Abkarian M., Vialix A., et Podgorski T., Dynamics of viscous vesiclesin shear flow, Eur. Phys. J. E, 19, 389-397, 2006.
Mader M.-A., Misbah C., Podgorski T., Dynamics and rheology of vesicles under a shear flow undergravity and microgravity, Microgravity sci. technol. XVIII-3/4, 2000-2003, 2006
Mader M.-A., Ez-Zahraouy H., Misbah C., Podgorski T., On coupling between the orientation andthe shape of a vesicle under shear flow, Eur. Phys. J. E 79 , 54002, 2007
Maitre E., Milcent T., Cottet G.-H., Raoult A., Usson Y., Applications of level set methods incomputational biophysics, a paraıtre dans Mathematical and Computer Modelling.
Maitre E., Misbah C., Peyla P., et Raoult A., Comparison between advected-field and level-setmethods in the study of vesicle dynamics, en preparation.
Misbah C., Vacillating-breathing and tumbling of vesicles under shear flow, Phys. Rev. Lett., 96,028104, 2006
Mourad A., “Description topologique de l’architecture fibreuse et modelisation mecanique du myo-carde”, These INPG, 2003.
Pustoc’h A., Ohayon J., Usson Y., Kamgoue A. et Tracqui P., An integrative model of the sel–sustained oscillating contractions of cardiac myocytes, Acta Biotheor. 53(4) :277–293, 2005
Raoult A., Symmetry groups in nonlinear elasticity. An exercise in vintage mathematics, en coursde redaction.
Tracqui P. et Stephanou A., Description integree de processus cellulaires spatio-temporels basee surun modele geometrique de transformation, a paraıtre dans Tech. Sci. Inf., 2007
25

6.2 Communications a des congres (liste non exhaustive)
Cottet G.-H., Maitre E. et Milcent T. An eulerian method for fluid-structure interaction withbiophysical applications, ECCOMAS CFD conference, 2006
Cottet G.-H., Maitre E. et Milcent T. New methods in fluid-structure coupling with application tobiomechanics, iCIAM, Zurich, 2007
Milisic V., Modelling and simulation of lattice structures by homogenization, poster presente lorsdu workshop Successes and failures of continuous models for discrete systems, 5-8 septembre2006, a Bristol
Milisic V., Modelling and simulation of fibrous biological tissues via dicrete homogeneization me-thods, conferencier invite pour le minisymposium “Tissue deformation and cell : a new frontierin cancer modelling” au congres iCIAM 2007, a Zurich
Milisic V., Modelisation et simulation multi-echelle de la contraction du myocarde, workshop or-ganise par le Gdr CNRS MaBeM (Modelisation mathematique en biologie et medecine) endecembre 2007, a Bordeaux.
Raoult A., Session Shells and other thin-walled structures, 9th US National Congress on Compu-tational Mechanics, San Francisco, Etats-Unis, juillet 2007
Raoult A., Mini-symposium From discrete to continuous, and back’, SIAM Conference on Mathe-matical Aspects of Materials Science, Philadelphie, Etats-Unis, mai 2008,
Raoult A., Session Multi-physics coupling in material processing, Third MIT conference on compu-tational solid and fluid mechanics, Cambridge, Etats-Unis, juin 2005
Raoult A., Ecole d’ete Poly-, quasi-, and rank-one convexity in applied mechanics, InternationalCentre for Mechanical Sciences, Udine, Italie, 24-28 septembre 2007
Raoult A., Sejour au Tata institute, Bangalore, janvier 2005,Tracqui P. and Ohayon J., Modelling and Simulation of Cardiac Tissue Deformations 2nd Workshop
on Computer Assisted Diagnosis and Surgery, 8–10 mars, 2006, Santiago (Chili) (Conferencierinvite)
Tracqui P. and Ohayon J., Coupling calcium wave propagation to anisotropic contraction in amulti–scale model of cardiac myocyte, 5th World Congress of Biomechanics, 29 juillet–4aoutMunich (Allemagne) 2006 (Conferencier invite)
Tracqui P. and Ohayon J., Approche theorique de la mecanobiologie de la cellule cardiaque : in-fluence des deformations cellulaires sur la regulation autobarique de la contraction anisotropedu cardiomyocyte, 2nde Conf. Int. Societe Francophone de Biologie Theorique, 4–6 juin 2007,Winnipeg (Canada),
Tracqui P. and Ohayon J., Multiphysic Modelling of the Self–sustained Anisotropic Contraction ofCardiac Myocytes, Comsol European Conference 23–24 octobre 2007, Grenoble.
7 Bibliographie
Biben T., Kassner K. et Misbah C., Phase-field approach to three-dimensional vesicle dynamics,Phys. Rev. E. 72, 041921, 2005
Brown D., Cortez R. et Minion L., Accurate Projection Methods for the Incompressible Navier-Stokes Equations, J. Comp. Phys., 168, 464–499 (2001)
Ciarlet P.G., Elasticite tridimensionnelle, Masson (1985)Ciarlet P.G., Mathematical elasticity, Volume 1 : Three-dimensional elasticity, North-Holland Publ.
(1988)Cai H., Loi de comportement en gandes deformations du muscle a fibres actives. Application a la
mecanique du cœur humain et a sa croissance, These de l’Universite de Savoie (1998)
26

Caillerie D., Mourad A. et Raoult A., Cell-to-muscle homogenization. Application to a constitutivelaw for the myocardium, Math. Mod. Numer. Anal., 37(4) :681–698 (2003)
Caillerie D., Mourad A. et Raoult A., A discrete homogenization technique for graphene sheets, InThird MIT Conference on Computational Fluid and Solid Mechanics, Boston, USA (2005)
Caillerie D., Mourad A. et Raoult A., Discrete homogenization in graphene sheet modeling, J. Elas.,84, 33-68, 2006.
Caillerie D., Mourad A., Raoult A., Confronting a mathematically derived constitutive law for themyocardium with experimental data, Third MIT Conference Computational Fluid and SolidMechanics, 2005, K.J. Bathe Ed., 127-130, 2005.
Cazorla O., Lacampagne A., Fauconnier J. et Vassort G., SR 33805 a Ca2+antagonist with length–dependent Ca2+-sensitizing properties in cardiac myocytes, Brit. J. Pharmaco, 139, 99–108(2003)
Chadwick R.S., Mechanics of the left ventricle, Biophys. J., 39, 279–288 (1982)Chang Y.C., Hou T.Y., Merriman B. et Osher S., A Level Set Formulation of Eulerian Interface
Capturing Methods for Incompressible Fluid Flows, J. Comp. Phys., 124, 449–464 (1996)Caulfield J.B. et Janicki J.S., Structure and function of myocardial fibrillar collagen, Technology
and Health Care, IOS Press 5, 95–113 (1997)Cortez R., Peskin C.S., Stockie J. et Varela D.A., Parametric resonance in immersed elastic boun-
daries, SIAM Journal on Applied Mathematics, 65(2) :494-520 (2004)Cottet G.-H. et Maitre E., A level-set formulation of immersed boundary methods for fluid-structure
interaction problems, C. R. Acad. Sci. Paris, Ser. I 338, 581-586 (2004)Cottet G.-H., Elasticite incompressible par methode particulaire, non publieCottet G.-H. et Maitre E., A level-set method for fluid-structure interactions with immersed sur-
faces, M3AS, 16 (3), 415–438, 2006Danker G. et Misbah C., Rheology of a dilute suspension of vesicles, Phys. Rev. Lett. 98, 088104,
2007Danker G., Biben T., Podgorski T., Verdier C., et Misbah C., Dynamics and rheology of a dilute
suspension of vesicles : higher order theory, a paraıtre dans Phys. Rev. EDanker G., Verdier C., et Misbah C., Rheology and dynamics of vesicle suspension in comparison
with droplet emulsion, J. Non. Newontian Fluid. Mech. 2007Dolber P.C. et Spach M.S., Conventional and confocal fluorescence microscopy of collagen fibers in
the heart, J. Histochem Cytochem.,41, 465–469 (1993)Fabiato A., Calcium–induced release of calcium from the cardiac sarcoplasmic reticulum, Am. J.
Physio., 245, C1–C14 (1983)Goldbeter A., Dupont G. et Berridge M.J., Minimal model for signal–induced Ca2+oscillations and
for their frequency encoding through protein phosphorylation, Proc .Natl. Acad. Sci. USA, 87,1461–1465 (1990)
Holzapfel G.A., Nonlinear Solid Mechanics, John Wiley & Sons, Ltd (2000)Jamet D. et Misbah C., Towards a thermodynamically consistent picture of the phase-field model
of vesicles, a paraıtre dans Phys. Rev. EJouk P.S., Usson Y., Michalowicz G. et Grossi L., Three-dimensional cartography of the pattern of
the myofibres in the second trimester fœtal human heart, Anal. Embryol, 202, 103–118 (2000)Jouk P.-S., Mourad A., Milisic V., Michalowicz G., Raoult A., Caillerie D., Usson Y., Analysis of
the fibre architecture of the heart, European J. of Cardiac Thoracic Surgery, 31, 915-921, 2007Lacampagne A., INSERM, Montpellier, communication privee.Lambrechts A., Van Troys M., Ampe Ch., The actin cytoskeleton in normal and pathological cell
motility, The International Journal of Biochemistry and Cell Biology, 36, 1890–1909 (2004)Lin D.H.S., et Yin F.C.P., A multiaxial constitutive law for mammalian left ventricular myocardium
27

in steady–state barium contracture or tetanus, ASME, Journal of Biomechanical Engineering,Vol. 120, 504–517 (1998)
Mader M.-A., Vitkova V., Abkarian M., Vialix A., et Podgorski T., Dynamics of viscous vesiclesin shear flow, Eur. Phys. J. E, 19, 389-397, 2006.
Mader M.-A., Misbah C., Podgorski T., Dynamics and rheology of vesicles under a shear flow undergravity and microgravity, Microgravity sci. technol. XVIII-3/4, 2000-2003, 2006
Mader M.-A., Ez-Zahraouy H., Misbah C., Podgorski T., On coupling between the orientation andthe shape of a vesicle under shear flow, Eur. Phys. J. E 79 , 54002, 2007
Marcy Y., Prost P., Carlier M.-F. et Sykes C., Forces generated during actin-based propulsion : Adirect measurement by micromanipulation, Proc. Natl. Acad. Sci. U. S. A. 101 : 5992-5997(2004)
Misbah C., Vacillating-breathing and tumbling of vesicles under shear flow, Phys. Rev. Lett., 96,028104, 2006
Mourad A., Biard L., Caillerie D., Jouk P.-S., Raoult A., Szafran N., et Usson Y., Geometricalmodelling of the fibre organization in the human left ventricle, In T. Katila, I.E. Magnin,P. Clarysse, J. Montagnat, et J. Nenonen, editors, Functional Imaging and Modeling of theHeart, volume 2230 in Lecture Notes in Computer Sciences, pages 32–38, Helsinki, novembre2001. Springer (2001)
Mourad A., Description topologique de l’architecture fibreuse et modelisation mecanique du myo-carde, These de doctorat, Institut National Polytechnique de Grenoble, decembre 2003.
Nash M.P. et Hunter P.J., Computational mechanics of the heart, Journal of Elasticity, Vol. 61,113–141 (2000)
Ogden R.W., Nonlinear elasticity, anisoptropy, material staility and residual stresses in soft tissue,Biomechanics of Soft Tissue in Cardiovascular Systems (edited by G.A. Holzapfel and R.W.Ogden), CISM Course and Lectures Series 441, Springer, Wien, 65–108 (2003)
Ohayon J., Cai H., Usson Y., Jouk P.S. et Azancot A., A model of the structural and functionaldevelopment of the normal human fetal left ventricle based on a global growth law, ComputerMethods in Biomechanics and Biomedical Engineering, 5, 113–126 (2002)
Ohayon J. et Chadwick R.S., Effect of collagen microstructure on the mechanics of the left ventricle,Biophys. J. 54 1077–1088 (1988)
Okada J., Sugiura S., Nishimura S., and Hisada T., Three-dimensional simulation of calcium wavesand contraction in cardiomyocytes using the finite element method, Am. J. Physiol. Cell.Physiol. 288 : C510–C522 (2005)
Osher S. et Fedkiw R. P., Level set methods, ULCA CAM Report 00-08 (2000)Peskin C.S., The immersed boundary method Acta Numerica, 11, 479–517 (2002)Pustoc’h A., J. Ohayon, Y. Usson et P. Tracqui, Modelling the oscillating calcium-induced aniso-
tropic contraction of cardiac myocytes, dans Mecanotransduction 2004, I. VinAon, C. Oddou,J.P. Renaudeaux, C. Ribreau et M.R. Moreau, eds., GAMC, ISBN2-9516368-1-4, p. 71-76(2004a)
Pustoc’h A., Ohayon J., Usson Y., Kamgoue A. and Tracqui P., An integrative model of the self–sustained oscillating contractions of cardiac myocytes, Acta Biotheor. 53(4) :277–293, 2005
Pustoc’h A., T. Boudou, J. Ohayon, Y. Usson et P. Tracqui, Finite element modelling of the calcium-induced contraction of cardiomyocytes based on time-lapse videomicroscopy, in WSEAS Tran-sactions Information Science & Applications, ISBN 1790-0832, 1(1) :376-378 (2004b)
Raoult A., Symmetry groups in nonlinear elasticity. An exercise in vintage mathematics, en coursde redaction.
Robinson T.F., Factor S.M. et Sonnenblick E.H., The heart as a suction pump, Scientific American,254, 84–91 (1986)
28

Schuster S., Marhl M., Hofer T., Modelling of simple and complex calcium oscillations, Eur. J.Biochem., 269, 1333–1355 (2002)
Stephanou A. et Tracqui Ph., Cytomechanis of cell deformation and migration : from models toexperiments, C.R. Biologie, 325, 295–308 (2002)
Stuyvers B.D, McCulloch A.D. Guo J., Duff H.J, et Keurs H.E.D.J., Effect of stimulation rate,sarcomere length and Ca2+on force generation by mouse cardiac muscle, J. Physiol., 544.3,817–830 (2002)
Subramanian S., Viatchenko–Karpinski S., Lukyanenko V., Gyork S. et Wiesner T.F, Underlyingmechanisms of symmetric calcium wave propagation in rat ventricular myocytes, Biophys. J,80, 1–11 (2001)
Taber L.A. and Perucchio R., Modeling heart development, Journal of Elasticity, Vol. 61, 165–197(2000)
Tollenaere H. et Caillerie D., Continuous modeling of lattice structures by homogenization, Advancesin Engineering Softwares. Special Issue, BHV Topping Ed. Vol 29, n 7–9, 699–705 (1998)
Tracqui P. et Stephanou A., Description integree de processus cellulaires spatio-temporels basee surun modele geometrique de transformation, a paraıtre dans Tech. Sci. Inf. (2007).
Usyk T.P., Omens J.H. et McCulloch A.D., Regional septal dysfunction in a three–dimensionalcomputational model of focal myofiber dissaray, Am. J. Physiol. Heart Circ. Physiol. , 281,506–514 (2001)
Vidrascu M., Solution of Non-Linear Elasticity Problems Using the Continu Software, Rapport derecherche de l’INRIA - Rocquencourt, RR-4128, (2001)
Zile M.R., Cowles M.K., Buckley J.M., Richardson K., Cowles B.A., Baicu C.F., Cooper G., etGharpuray V., Gel stretch method : a new method to measure constitutive properties of cardiacmuscle cells, Am. J. Physiol., 274, H2, 188–202 (1998)
29