rapport bibliographique : biologie et ecologie du requin ... · ... (l’âge à première...
TRANSCRIPT

Rapport bibliographique :
Biologie et Ecologie du requin tigre (Galeocerdo cuvier) et
du requin bouledogue (Carcharhinus leucas)
Antonin BLAISON, mai 2012
Programme CHARC (Connaissances de l’écologie et de l’HAbitat de deux espèces de
Requins Côtiers sur la côte Ouest de La Réunion), Phase I
Requin tigre, © http://listnation.blogspot.com/2012/03/10‐most‐badass‐sharks‐in‐world.html Requin bouledogue, © Zeppy, 1969

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
1
SOMMAIRE
Résumé ................................................................................................................. 2
Synthèse de l’étude bibliographique ................................................................. 5
Le requin tigre ..................................................................................................... 6
Introduction ................................................................................................................................6
1. Distribution et habitat............................................................................................................7
2. Alimentation..........................................................................................................................8
3. Reproduction .......................................................................................................................10
4. Age et développement.........................................................................................................11
Fécondation........................................................................................................................11
Taille à la naissance ............................................................................................................12
Taux de croissance..............................................................................................................12
Maturité sexuelle ...............................................................................................................13
Taille maximale ..................................................................................................................13
5. Patterns de mouvements et utilisation de l’habitat..............................................................14
6. Bibliographie........................................................................................................................17
Le requin bouledogue........................................................................................ 22
Introduction ..............................................................................................................................22
1. Distribution..........................................................................................................................23
2. Alimentation........................................................................................................................24
3. Reproduction .......................................................................................................................26
Gestation............................................................................................................................26
Cycle Reproducteur ............................................................................................................26
4. Age et développement.........................................................................................................27
Fécondation et taille à la naissance.....................................................................................27
Taux de croissance..............................................................................................................28
Maturité sexuelle ...............................................................................................................29
Taille maximale ..................................................................................................................29
5. Patterns de mouvements et utilisation de l’habitat..............................................................30
Saisonnalité et migration....................................................................................................30
Utilisation de l’habitat et mouvements à court terme.........................................................32
6. Bibliographie........................................................................................................................36
Annexe ................................................................................................................ 42

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
2
Résumé
Ce rapport bibliographique fait la synthèse et la critique de 125 publications et rapports
scientifiques publiés sur la biologie et l’écologie des requins tigres et des requins bouledogues entre 1943 et 2012. Requin tigre (Galeocerdo cuvier)
Le requin tigre (Galeocerdo cuvier) est le deuxième plus grand poisson prédateur au monde après le requin blanc (Carcharodon carcharias). Le requin tigre mesure généralement entre 3 et 4 mètres pour un poids compris entre 350 et 700 kilos. Le plus grand requin tigre pêché est une femelle de 7 mètres 40 pour un poids de plus de 3 tonnes. L’âge maximum estimé est de 27 ans.
Le requin tigre est une espèce côtière et pélagique. Son aire de répartition comprend les eaux côtières et hauturières des mers tropicales et subtropicales. Le caractère hautement migratoire de cette espèce suggère que son aire de répartition est encore sous-estimée.
Le régime alimentaire des requins tigres est le plus diversifié parmi les élasmobranches. Il s’alimente principalement de poissons, de dauphins, de dugongs, de tortues de mer, de serpents de mer, d’oiseaux de mer, de crustacés, d’élasmobranches voire même d’autres mammifères marins et terrestres. Il a des proies préférentielles qui varient d’une région géographique à l’autre. Ses habitudes alimentaires peuvent être influencées par plusieurs facteurs : la disponibilité en proie, l’expérience de l’individu et son stade de maturité (en rapport avec la taille).
Il est l’unique ovovivipare de la famille des Carcharhinidés. Les œufs incubent et éclosent à l’intérieur de la mère. Avec une période de gestation relativement longue (entre 15 et 16 mois), le cycle reproducteur serait d’au moins deux ans. Certains auteurs émettent l’hypothèse d’un cycle trisannuel.
Les requins tigres ont des portées importantes (une trentaine de nouveaux nés en moyenne), une taille à la naissance qui les protège de la majorité des prédateurs, un taux de croissance élevé et une maturité relativement précoce par rapport aux autres requins de la même famille (l’âge à première maturité varie entre 7 et 10 ans selon les zones du globe). Ces informations suggèrent un potentiel de reproduction plus grand chez cette espèce par rapport aux autres Carcharhinidés. Néanmoins, les études sur les jeunes suggèrent un taux de mortalité important des nouveaux nés (entre 27% et 62% selon les observations). Le requin tigre est l’un des élasmobranches avec les plus forts taux de croissance. Ce taux diminue avec l’age et varie de 50 à 10 cm.an-1 entre 1 an et 10 ans.
L’utilisation de l’habitat est très variable entre les individus qu’ils soient originaires ou non d’une même région. Dans les eaux côtières, les requins semblent limiter leurs mouvements à des zones peu étendues, ont des déplacements circulaires et nagent principalement près du fond. Les mouvements des requins tigres à l’échelle d’une baie pourraient être corrélés avec ceux de ses proies favorites. Au contraire, dans les eaux hauturières, les individus suivis ont des nages orientées à des profondeurs moyennes comprises entre 60 et 80 mètres (profondeur maximum enregistré : 335 mètres). Plusieurs individus peuvent rester dans une même zone insulaire toute leur vie, d’autres vont étendre leur domaine vital à l’ensemble d’archipels, et d’autres encore sont capables de faire des

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
3
migrations transocéaniques. La plus grande migration observée chez les requins tigres est celle d’un requin entre l’Australie et l’Afrique du Sud. La disponibilité en proies et la température de l’eau semble jouer un rôle important dans ces déplacements et leurs variations individuelles. Requin bouledogue (Carcharhinus carcharias)
Le requin bouledogue a une aire de répartition très étendue comprenant les eaux chaudes tropicales et sub-tropicales. C’est une espèce côtière et semi-pélagique. Il a longtemps été décrit comme vivant dans les eaux peu profondes mais des profondeurs jusqu’à 140 mètres ont pu être mesurées récemment ce qui a remis en cause son affinité exclusive à des zones côtières peu profondes. Il est souvent observé dans les lacs, les zones d’estuaires et remonte régulièrement les fleuves. C’est l’une des rares espèces de requin euryhalines. Comme les autres espèces de requins « évoluées », le requin bouledogue possède une mâchoire hyostylique. La mâchoire supérieure n’est pas soudée à la mâchoire inférieure. Les estimations de taille maximale indiquent une longueur de 340 cm (LT) mais un spécimen a été capturé mesurant 400 cm.
Les requins bouledogues ont un régime alimentaire diversifié mais bien défini. Il comprend la majorité des poissons (y compris les autres requins), les céphalopodes, les cétacés, les tortues, les oiseaux et certains mammifères terrestres. Les requins bouledogues ne s’attaquent pas aux baleines vivantes mais ils sont régulièrement signalés autour de carcasses dérivantes en pleine mer. Le type de proie ne change pas en fonction de l’âge, mais la taille de celle-ci augmente proportionnellement à la taille du requin. A la Réunion, la présence de tortues, de dauphins et saisonnièrement de baleine en plus des proies généralement décrites pourraient jouer un rôle sur la composition du régime alimentaire du requin bouledogue dans cette région de l’Océan Indien.
Le cycle de reproduction des requins bouledogues est encore peu connu. Les requins bouledogues ont une maturité tardive. Les femelles arrivent à maturité à 10 ans pour une taille supérieure à 200 cm et les mâles à 9-10 ans pour une taille comprise entre 190 et 200 cm. Le temps de gestation de 10 à 11 mois pourrait inclure un cycle annuel, bisannuel ou triannuel. Le requin bouledogue est une espèce vivipare. L’embryon est nourri par la mère par l’intermédiaire d’une liaison placentaire. La femelle donne naissance à des portées de 1 à 9 nouveaux nés. La taille à la naissance varie de 50 à 80 cm (LT).
Le taux de croissance des requins bouledogues est lent, variable entre individus et dépend du stade de maturité (entre 4 et 20 cm.an-1). Il varie d’une région à l’autre. Pour expliquer cette différence, les auteurs émettent l’hypothèse du rôle majeur de l’habitat et de ses caractéristiques dans le développement biologique et comportementale.
Dans les zones côtières, la répartition des requins adultes semble dépendre majoritairement de la structure bathymétrique des fonds qu’ils occupent, mais trois autres facteurs ont également été mis en évidence, la température de l’eau, la salinité et la turbidité. Le requin peut vivre dans des eaux peu salées mais lorsque l’apport d’eau douce est trop important, les requins quitteraient les rivières. Cette observation suggère que les mouvements à court terme des nouveaux nés et des jeunes de l’année sont davantage influencés par la salinité que par le risque de prédation. Bien que les requins bouledogues semblent évoluer

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
4
préférentiellement dans des eaux à forte turbidité, très peu d’études ont été réalisées sur l’influence de ce facteur.
A l’heure actuelle, il est bien admis que cette espèce montre une grande fidélité au site, particulièrement les nouveaux nés, les jeunes de l’année et les juvéniles. Les comportements des sub-adultes et des adultes apparaissent plus complexes à définir. Les différentes études ont montré que si ces derniers passent la majorité de l’année au même endroit, durant l’hiver beaucoup disparaissent pour réapparaitre quelques mois plus tard. L’hypothèse acceptée est que les requins bouledogues en âge de se reproduire migrent pendant l’hiver dans d’autres régions du monde pour l’accouplement ou la mise bas.
Les requins bouledogues auraient un changement ontogénique dans leur pattern de mouvement avec une augmentation de la taille de leur territoire et de leur déplacement migratoire au cours de leur croissance. Ainsi, les nouveaux nés et les jeunes de l’année naissent et restent pendant au moins un an dans les zones côtières à proximité d’un estuaire (nurserie). En devenant sub-adultes, ils migrent progressivement vers des habitats moins côtiers où ils rejoignent les individus adultes. A ce stade, ils restent la plupart de leur vie au même endroit migrant saisonnièrement dans d’autres régions du monde pour s’accoupler et reviennent pour mettre bas.
Les distances maximales parcourues par les requins bouledogues marqués sont faibles à 2 exceptions près, n’excédant pas 650 kilomètres. Les mouvements migratoires sont donc relativement restreints en comparaison des autres espèces. Si ce scénario est bien accepté, il manque néanmoins de validation scientifique.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
5
Synthèse de l’étude bibliographique
Une étude sur l’écologie et l’habitat des requins tigres (Galeocerdo cuvier) et des requins
bouledogues (Carcharhinus leucas) près des côtes réunionnaises a débuté suite aux attaques
de requins à La Réunion en 2011. Une des premières étapes importantes de ce projet est de
réaliser un bilan des connaissances sur ces deux espèces à travers le monde. Les résultats de
ce rapport permettront de fournir une base de connaissances pour améliorer notre capacité à
analyser les résultats et des éléments pour comparer l’écologie comportementale de ces
espèces avec d’autres régions du monde.
Ce rapport bibliographique fait la synthèse et la critique de 125 publications et rapports
scientifiques sur les deux espèces, soit 54 études sur le requin tigre entre 1948 et 2012 et 67
sur le requin bouledogue entre 1943 et 2012 (annexe 1). Ces dernières sont synthétisées selon
5 grands thèmes : distribution géographique, alimentation, reproduction, âge &
développement et utilisation de l’habitat. Ce rapport ne fait pas état de toutes les publications
compulsées existantes à l’heure actuelle sur le sujet mais présente l’ensemble des
connaissances majeures disponibles.
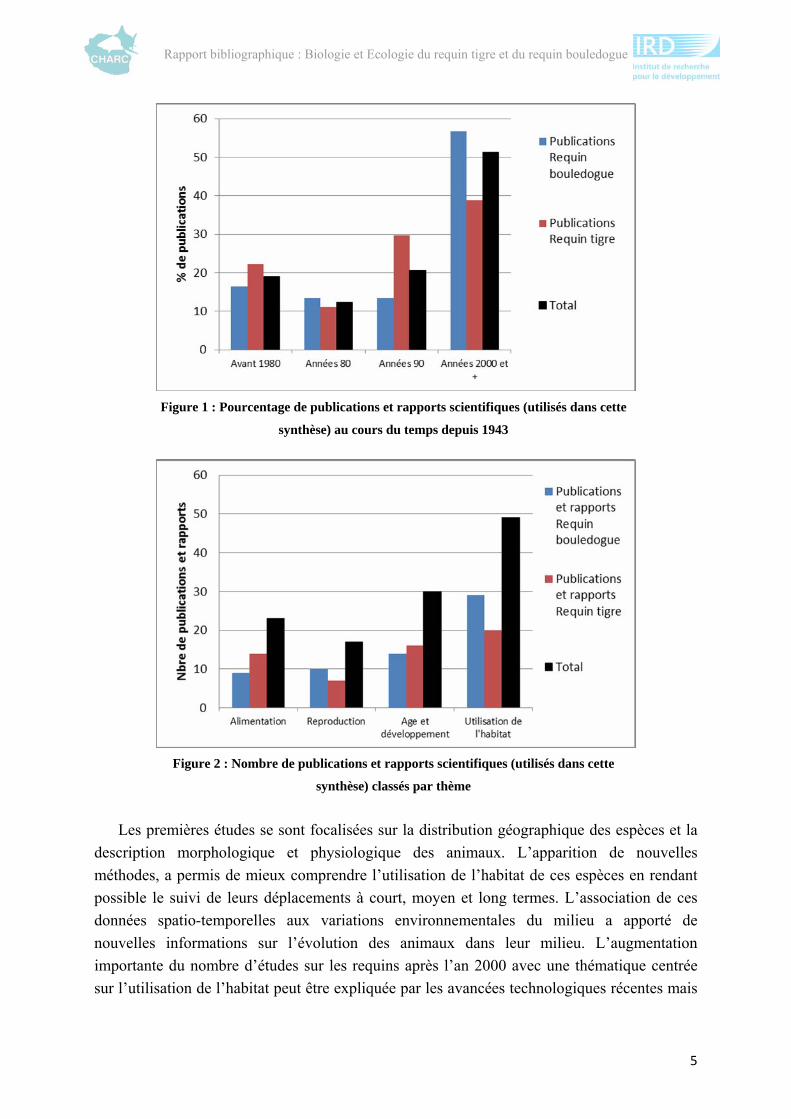
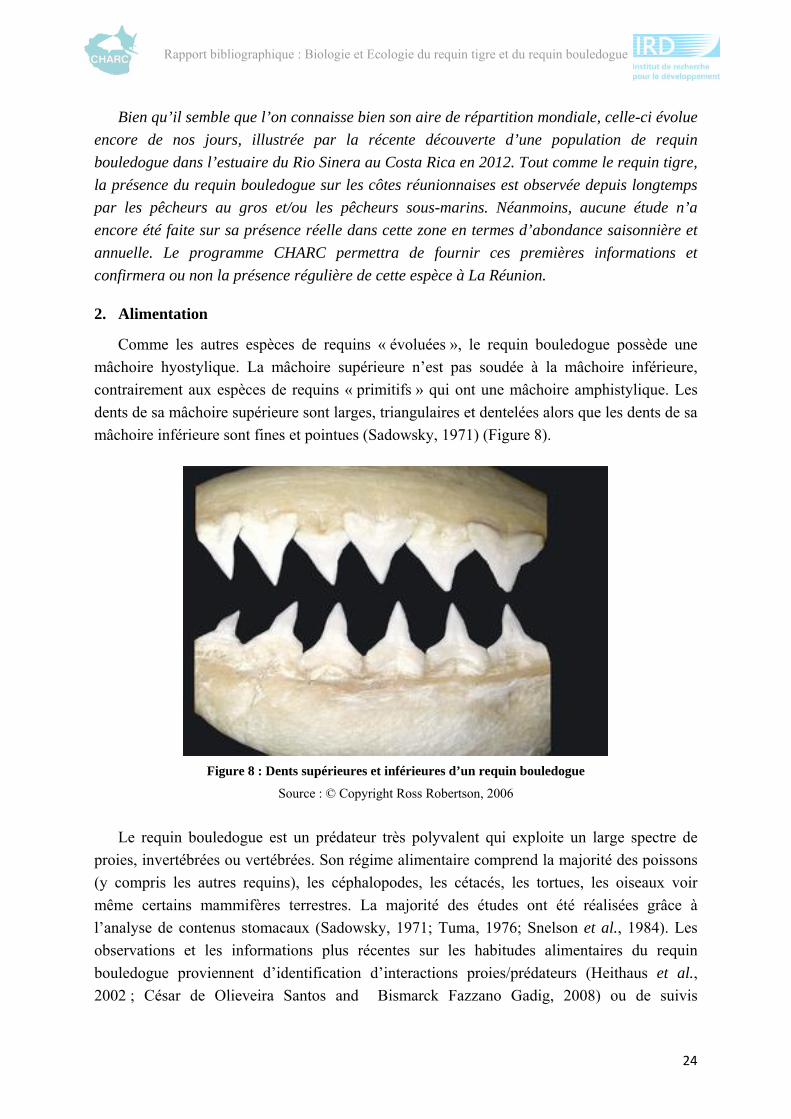
Une rapide analyse de l’évolution des études sur ces deux espèces met en évidence trois
phénomènes majeurs : une nette augmentation du nombre d’études après les années 2000
(figure 1) en particulier sur l’utilisation de l’habitat (figure 2) et un manque de connaissance
sur la reproduction des deux espèces (figure 2). En comparaison aux espèces terrestres, les
animaux marins sont difficilement observables, en particulier ceux qui viennent rarement en
surface. Pour pallier à cette contrainte, les auteurs utilisaient majoritairement la pêche, la
méthode de capture/recapture et l’observation visuelle en plongée. Cependant, ces techniques
ne donnent qu’une idée ponctuelle du comportement des requins en limitant le temps
d’observation par individu. La télémétrie a permis de faire avancer nos connaissances dans ce
domaine mais elle est encore peu développée sur ces espèces.
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
5
Figure 1 : Pourcentage de publications et rapports scientifiques (utilisés dans cette
synthèse) au cours du temps depuis 1943
Figure 2 : Nombre de publications et rapports scientifiques (utilisés dans cette
synthèse) classés par thème
Les premières études se sont focalisées sur la distribution géographique des espèces et la
description morphologique et physiologique des animaux. L’apparition de nouvelles
méthodes, a permis de mieux comprendre l’utilisation de l’habitat de ces espèces en rendant
possible le suivi de leurs déplacements à court, moyen et long termes. L’association de ces
données spatio-temporelles aux variations environnementales du milieu a apporté de
nouvelles informations sur l’évolution des animaux dans leur milieu. L’augmentation
importante du nombre d’études sur les requins après l’an 2000 avec une thématique centrée
sur l’utilisation de l’habitat peut être expliquée par les avancées technologiques récentes mais

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
6
également par une prise de conscience collective sur le rôle majeur des requins dans
l’équilibre des écosystèmes (Stevens, 2000 ; Baum et al., 2003 ; Myers et al., 2007).
Le requin tigre
Introduction
Le requin tigre (Galeocerdo cuvier) est le deuxième plus grand poisson prédateur au
monde après le requin blanc (Carcharodon carcharias) (Campagno, 1984). Il a été décrit, la
première fois, en 1822 par Péron et Lesueur sont le nom de Squalus cuvier. Ce n’est qu’en
1837 qu’il est reclassé parmi les carcharhinidés dans le genre Galeocerdo dont il est le seul
représentant actuel (Müller and Henle, 1837). Le requin tigre est l’unique ovovivipare de la
famille des carcharhinidés. Les œufs incubent et éclosent à l’intérieur de la mère.
Contrairement aux vivipares, les embryons n’ont pas de lien placentaire avec l’utérus et se
nourrissent grâce à une poche vitelline qui leur fournit tous les éléments essentiels à leur
croissance.
Le requin tigre a un corps massif de couleurs variables, du bleu au gris-marron en passant
par le gris vert. Il possède des rayures de couleur bleu-gris sur tout le corps qui sont
particulièrement visibles chez les juvéniles. Ces marques distinctives de l’espèce tendent à
disparaitre chez les adultes. Sa tête est large et son nez carré, ce qui le différencie facilement
des autres espèces de requin. Il a une mâchoire symétrique avec des dents plates, crénelées et
pointues, en forme de scie ou « crête de coq ».
Le requin tigre mesure généralement entre 300 et 400 cm pour un poids compris entre 350
et 700 kilogrammes. Cependant, il n’est pas rare d’observer des individus dépassant 500 cm.
Le plus grand requin tigre pêché est une femelle de 740 cm pour un poids de plus de 3 tonnes.
Bien qu’il soit possible que ces mesures aient été surestimées, ce record suggère que cette
espèce pourrait atteindre des tailles et des poids supérieurs à ceux du requin blanc (le plus
grand requin blanc pêché mesurait 640 cm pour un poids de 3,2 tonnes). Ils atteignent leur
maturité entre 7 et 10 ans pour des tailles comprises entre 250 et 350 cm (Whitney and Crow,
2006).
Son régime alimentaire est très diversifié ce qui lui a valu son surnom de « poubelle des
mers ». Il s’alimente principalement de poissons, de dauphins, de dugongs, de tortues de
mer, de serpents de mer, d’oiseaux de mer. Néanmoins, les dernières études sur ses habitudes
alimentaires décrivent un régime alimentaire beaucoup moins opportuniste que ce que l’on
pensait (Lowe et al., 1996).

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
7
1. Distribution et habitat
Le requin tigre est une espèce côtière et pélagique. Son aire de répartition comprend les
eaux côtières et hauturières des mers tropicales et subtropicales (figure 3).
Figure 3 : Aire de répartition mondiale du requin tigre (d’après les données publiées)
Source : Walter inc. original work by Chris
On le retrouve notamment sur le continent américain, du Canada à l’Uruguay, dans le
Golfe du Mexique, aux Bermudes et dans la mer des Caraïbes (Kohler et al., 1996 ;
Branstetter, 1987 ; Menni and Lucifora, 2007) ainsi que du Sud de la Californie au Pérou
incluant Hawaii, les îles Galápagos et Revillagigedo (Briggs, 1961 ; Holland et al., 1999 ;
Sandoval-Castillo et al., 2006 ; Lundberg et al., 2010 ; Meyer et al., 2010) ; sur le continent
européen, en Islande, au Royaume-Uni et en France (Campagno, 1984). Ces informations sont
issues de données de pêches non publiées, et ne sont pas intégrées à l’aire de répartition
mondiale illustrée sur la figure 3) ; sur le continent africain, du Maroc à l’Angola, ainsi que de
l’Afrique du Sud jusqu’en Mer Rouge (Gohar and Mazhar, 1964 ; Smale and Cliff, 1998) ;
dans l’Océan Indien, du canal du Mozambique jusqu’en Inde en passant par La Réunion
(Wintner and Dudley, 2000) ; et en Océanie, notamment en Australie, en Nouvelle Zélande,
en Nouvelle Calédonie et au Japon (Rancurel and Intes, 1982 ; Francis, 1998 ; Simpfendorfer
et al., 2001 ; Haraguchi et al., 2009).
Les requins tigres sont présents dans les eaux côtières de beaucoup de pays à travers le
monde. Cependant, le caractère hautement migratoire de cette espèce suggère que son aire de
répartition est encore sous-estimée notamment dans les eaux hauturières. L’utilisation de
nouvelles technologies comme les balises satellites permettront de mieux définir l’aire de
répartition globale du requin tigre (Meyer et al., 2010). A La Réunion, le requin tigre est
observé depuis de nombreuses années, notamment par les pêcheurs au gros qui en capturent

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
8
ou en observe de temps en temps sur le sec de Saint Gilles. Cependant, aucune donnée ne
permet de définir si cette région de l’Océan Indien constitue une partie de son aire de
répartition ou juste un lieu de passage d’une migration à plus grande échelle. Au cours du
programme CHARC, 5 à 6 requins tigres seront équipés de balises satellites (MiniPat,
Wildlife Computers).Une meilleure connaissance des déplacements à grande échelle des
requins tigres fréquentant les eaux réunionnaises permettra de définir si cette zone fait partie
ou non l’aire de répartition mondiale des requins tigres.
2. Alimentation
Le requin tigre est un prédateur opportuniste avec un régime alimentaire très diversifié
comprenant: les téléostéens, les tortues marines, les serpents de mers, les oiseaux de mer, les
crustacés, les élasmobranches voir même d’autres mammifères marins et terrestres (Gudger,
1949 ; Bass et al., 1975 ; Rancurel and Intes, 1982 ; Stevens 1984 ; Stevens and Mc Loughlin,
1991). Cette capacité à s’alimenter sur un grand nombre d’espèces différentes est expliquée
par sa grande taille (Randall, 1992), sa dentition adaptée pour attraper de grosses proies
(Witzell, 1987) (figure 4), la puissance de sa mâchoire (> 30 kg.mm-2, Evans and Gilbert,
1971) et sa large répartition géographique.
Figure 4 : Dents supérieures et inférieures d’un requin tigre
Source : © Copyright Ross Robertson, 2006
Le requin tigre a des proies préférentielles qui varient d’une région géographique à l’autre,
en fonction de l’abondance des différentes espèces présentes. Au Queensland et à Hawaii, les
requins s’alimentent principalement de téléostéens (Simpfendorfer, 1992 ; Lowe et al., 1996)
alors qu’en Nouvelle Calédonie, les proies les plus fréquentes dans l’estomac des requins
étudiés sont des serpents de mer et des oiseaux marins (Rancurel and Intes, 1982). Cette
différence de régime alimentaire semble se retrouver à des échelles très réduite comme celle
d’une baie. Ainsi, à l’Ouest de Shark Bay (Australie occidentale), Simpfendorfer et al. (2001)
démontrent que les requins tigres s’alimentent principalement de serpents de mer, et en

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
9
moindre quantité de tortues marines et de dugongs. A l’Est de cette baie, Heithaus (2001a)
trouve dans l’estomac des requins essentiellement des dugongs puis en plus petite quantité des
serpents de mers et des tortues marines. Ce dernier suggère que cette différence dans les
proportions de proies trouvées est liée à la différence de substrat entre l’Ouest, où les roches
et les coraux prédominent, et l’Est caractérisé par l’abondance d’herbiers. Le régime
alimentaire des requins tigres dépendrait de l’abondance des proies disponibles, elle-même
influencée par la nature du substrat.
La disponibilité des proies ne semble pas être le seul facteur jouant sur l’alimentation de
ces requins. A proximité d’Hawaii, au banc de la Frégate Française, certains requins tigres ont
été observés tout au long de l’année alors que d’autres ne sont présents que pendant la saison
des premiers envols des jeunes albatros de Laysan (Phoebastria immutabilis) sur lesquels ils
s’alimentent (Meyer et al., 2010). Les auteurs en déduisent que chez cette espèce de requin,
chaque individu aurait un comportement alimentaire propre, sur plusieurs sites d’alimentation,
définis par son expérience migratoire passée.
Dès les premières études sur le régime alimentaire, il a été observé un changement
ontogénique dans la taille et les espèces consommées. En Nouvelle Calédonie, plus les
requins disséqués sont de grande taille plus la proportion de serpents de mer et de téléostéens
diminue au profit de plus grosses proies comme les tortues marines, les mammifères marins et
les élasmobranches (Rancurel and Intes, 1982). De même, en Australie, l’étude du régime
alimentaire démontre une corrélation positive entre la taille des proies et celles des requins
examinés (Simpfendorfer, 1992). A Hawaii, aucune tortue verte et aucun élasmobranche n’ont
été trouvés dans l’estomac des jeunes individus alors que ces espèces sont fréquemment
observées dans ceux d’individus sub-adultes et adultes (Lowe et al., 1996). Heithaus (2001b)
arrive au même constat concernant la prédation sur les dauphins à Shark Bay. La majorité des
cicatrices laissées sur les dauphins provenait de requins tigres adultes. Lowe et al. (1996)
suggèrent trois raisons possibles pour expliquer ce changement ontogénique d’alimentation : -
les plus grands individus peuvent s’attaquer à de plus grandes proies, - les individus de
différentes tailles occupent des habitats différents, - les plus grands individus sont devenus de
meilleurs chasseurs capables de capturer des proies plus rapides.
Le régime alimentaire des requins tigres est le plus diversifié parmi les élasmobranches.
Ses habitudes alimentaires ne semblent pas pour autant aléatoires et sont influencées par
plusieurs facteurs : la disponibilité en proie, la localisation géographique, l’expérience de
l’individu et le stade de maturité (la taille) de celui-ci. A La Réunion, aucune étude sur le
régime alimentaire des requins tigres n’a été réalisée. L’analyse isotopique devrait permettre
d’avoir une première idée sur l’écologie alimentaire de ces requins et pourrait confirmer la
présence d’une ou deux espèces préférentielles avec un changement ontogénique. Ces
nouvelles données seront essentielles pour comprendre la dynamique de répartition spatio-
temporelle des requins tigres près des côtes réunionnaises et leur rôle dans les écosystèmes
marin de cette région.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
10
3. Reproduction
Le requin tigre est une espèce ovovivipare. Les embryons sont indépendants de la mère.
Ils se nourrissent et se développent grâce à une poche vitelline. La période de gestation est
encore mal définie mais elle est estimée à plus d’un an. En Floride, lors d’une étude sur la
reproduction des requins tigres, les dissections ont révélé la présence d’embryons en début de
développement chez certaines femelles et des embryons presque à terme chez d’autres.
L’ensemble des observations se déroulant sur une même période, les auteurs en déduisent que
la période de gestation doit être d’un peu plus d’un an. Ils l’estiment entre 13 et 16 mois
(Clark et Von Schmidt, 1965). Au Brésil, Alves (1977) retrouve les mêmes résultats suite aux
dissections de requins tigres pêchés au large de Ceará. Une étude plus récente, réalisée à Oahu
(Hawaii) suggère une période de gestation similaire, entre 15 et 16 mois (Withney and Crow,
2006). Cette étude est considérée comme la plus complète et la plus précise à l’heure actuelle,
avec un grand nombre d’individus étudiés et un échantillonnage étalé sur 10 mois. La
difficulté à déterminer une période de gestation précise s’explique par les caractéristiques
physiologiques des embryons. En effet, ces derniers absorbent leur poche vitelline
relativement tôt au cours de leur développement (Longueur Totale > 60cm, Withney and
Crow, 2006). Il est donc difficile de différencier des embryons arrivés à terme, d’embryons en
cours de développement (Figure 5).
Figure 5 : Requin tigre âgé de moins d’un an
Source : George Burgess
Il n’existe que peu d’informations sur le cycle reproducteur des requins tigres. Branstetter
et al. (1987) propose un cycle de reproduction d’au moins 2 ans. Ils se basent sur la période
de gestation définie par Clark et Von Schmidt (1965) et l’observation d’indices
d’accouplement sur les requins (morsures ou blessures au niveau du cloaque ou des
ptérygopodes) qui apparaissent avant la saison des mises bas. Withney and Crow (2006)

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
11
proposent un cycle trisannuel, se basant sur les études précédentes et leurs propres travaux et
observations sur les requins tigres à Hawaii. L’accouplement aurait lieu en janvier et février.
Les femelles, n’ayant pas d’ovocytes matures à cette époque de l’année, stockeraient le
sperme dans une glande oviducale (Pratt, 1993). En juin-juillet, la fécondation et le
développement de l’embryon commenceraient. Il y aurait une période de gestation de 15-16
mois suivie d’une mise bas en septembre-octobre de l’année suivante (Vatter, 2003). La
femelle passerait ensuite un an avant de s’accoupler à nouveau.
Malgré la précision dans la description de ce cycle reproducteur, il reste encore beaucoup
d’incertitude. Si les périodes d’accouplement et de parturition commencent à être mieux
connues, la question de l’annualité du cycle est encore non résolue : Est-il bisannuel ?
Trisannuel ? Est-il constant dans le temps et l’espace ? Dans le cas d’un cycle trisannuel,
pourquoi les femelles attendent un an après la parturition pour s’accoupler à nouveau ? Les
mâles ont-ils le même cycle ou se reproduisent-ils chaque année ? Où s’accouplent-ils ?
A l’heure actuelle, la reproduction des requins tigres nécessite d’avantage de
connaissances et d’études pour être mieux définie et comprise. A La Réunion, une meilleure
compréhension du cycle reproducteur des requins tigres et des comportements associés
permettrait de mieux appréhender les questions d’abondances saisonnières et/ou annuelles,
d’utilisation de l’habitat et de changement de comportements. Ces questions peuvent être
cruciales en termes de variation du « risque requin ». En effet, si les femelles requins tigres se
rapprochent de la côte pour mettre bas une cinquantaine de nouveaux nés (par femelle), le
nombre d’interactions humains/requins sera potentiellement plus élevé l’année de la
reproduction. La même logique peut s’appliquer si l’accouplement se déroule prêt des côtes à
une période donnée selon une fréquence bis ou trisannuelle. Au cours du programme
CHARC, l’observation de femelles gestantes, qui ont mis bas, qui ont des marques
d’accouplement et/ou l’observation de nouveaux nés, devraient permettre de définir la ou les
zones de reproduction (lieu de mise bas et/ou d’accouplement). Cependant, la définition d’un
cycle reproducteur sera difficile à définir étant donné son caractère pluriannuel. Il faudra
pour cela réaliser une étude spécifique sur au moins trois ans.
4. Age et développement
Fécondation
Les femelles donnent naissance à des portées d’une trentaine de nouveaux nés en
moyenne (Whitney and Crow, 2006). Cependant, ce nombre varie beaucoup d’un individu à
l’autre. A Hawaii, sur 23 femelles gestantes étudiées, le nombre d’embryons par femelle serait
compris entre 3 et 57 avec une moyenne de 33 (Whitney and Crow, 2006). En Australie,
Simpfendorfer (1992) retrouve cette grande variabilité associée au même nombre moyen
d’embryons par femelle. En Floride, ce nombre serait compris entre 37 et 56 (Kohler et al.,
1996).

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
12
Au vu du grand nombre de nouveaux nés par portée, Branstetter et al. (1987) suggère un
taux de mortalité important pour cette classe d’âge. Une étude sur la mortalité des jeunes
requins tigres au Nord du Golfe du Mexique et sur les côtes Sud-Est des Etats-Unis a
confirmé cette hypothèse (Driggers et al., 2008). Le taux de mortalité pour les jeunes de
l’année (YOY : Young of the Year) serait de 51% dans le Golfe et 39% sur les côtes Est
américaines. Ce taux est légèrement différent pour les requins juvéniles (individus de plus
d’un an) avec 62% dans le Golfe et 27% sur les côtes Est américaines. Au-delà de la
confirmation d’un taux de mortalité très élevé, les auteurs mettent en évidence la grande
variation de ce taux en fonction de la localisation géographique. Ce phénomène n’est toujours
pas bien compris à l’heure actuelle.
Le taux de mortalité élevé mais variable géographiquement peut avoir des conséquences
sur l’estimation de la population de cette espèce à la Réunion. Si le taux de mortalité se
rapproche de celui du Golfe du Mexique, la population de nouveaux nés et de jeunes de
l’année sera potentiellement plus faible que si ce taux se rapproche de celui estimé sur les
côtes Est américaines.
Taille à la naissance
La taille à la naissance varie entre 80 et 90 centimètres (Simpfendorfer, 1992 ; Schwartz,
1994 ; Whitney and Crow, 2006). Les premières études sur le sujet estiment une taille à la
naissance beaucoup plus petite. Bigelow et Schroeder (1948) et Baughman et Springer (1950)
ont suggéré une taille à la naissance comprise entre 45 et 50 centimètres. Ces résultats ont été
déduits d’observations de nouveaux nés nageant dans les eaux du Golfe du Mexique. En
1965, Clark et Von Schmidt ont observé des embryons beaucoup plus grands et concluent que
les individus observés dans les précédentes études devaient être prématurés. L’observation
d’embryons de tailles supérieures à 80 centimètres dans les études plus récentes semble
confirmer cette hypothèse.
Taux de croissance
Le requin tigre est l’un des élasmobranches avec les plus forts taux de croissance. Les
nouveaux nés et les juvéniles ont un taux de croissance très élevé jusqu’à ce qu’ils deviennent
matures. Les individus matures ont leur taux de croissance qui diminue progressivement avec
l’augmentation de leur taille. Branstetter et al. (1987) estiment que les requins tigres du Golfe
du Mexique et de l’Atlantique Nord-Ouest ont un taux de croissance de 50 cm.an-1 la première
année, de 40 cm.an-1 jusqu’à 3 ans, de 30 cm.an-1 entre 4 et 7 ans, puis de 20 à 10 cm.an-1 au-
delà de 7 ans. Natanson et al. (1999) ont retrouvé des taux de croissance assez proches chez
les populations de requins tigres en Floride. Ils estiment que les requins grandissent de 40 à 45
cm.an-1 la première année et de 35 à 45 cm.an-1 la deuxième. Les études plus récentes
montrent un taux de croissance plus faible même dans les premiers stades de maturité.
Wintner et Dudley (2000) en Afrique du Sud, puis Wirsing et al. (2006) en Australie, trouvent

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
13
des taux de croissance de respectivement 32 cm.an-1 et 35 cm.an-1 jusqu’à ce que les animaux
atteignent leur taille adulte. Ils estiment le taux de croissance des adultes à respectivement 4
cm.an-1 et 12 à 15 cm.an-1. Cette différence peut être due à plusieurs facteurs : - la localisation
géographique : Branstetter et al. (1987) met en évidence une différence de vitesse de
croissance entre les populations du Golfe du Mexique et de l’Atlantique Nord-Ouest ; - la
taille considérée pour les nouveaux nés : Natanson et al. (1994) a considéré une taille à la
naissance de 65 cm alors que l’on sait aujourd’hui que cette taille est sous-estimée et serait
comprise entre 80 et 90 cm ; - et la méthodologie : bien que dès les premières études,
l’analyse des anneaux de croissance sur les vertèbres était couplée avec des campagnes de
capture/recapture, cette technique a beaucoup évoluée et est désormais beaucoup plus précise.
Maturité sexuelle
Les requins tigres ont une maturité relativement précoce en comparaison avec les autres
requins de la même famille (Carcharhinus plumbeus : 15–30 ans ; Casey and Natanson, 1992
; Carcharhinus falciformis : 6,5–12 ans ; Bonfil et al., 1993 ; Carcharhinus obscurus : 19–20
ans ; Natanson, 1994). Dans l’Atlantique Nord-Ouest, l’âge à première maturité était de 10
ans pour les mâles et les femelles (Branstetter et al., 1987). Dans le Golfe du Mexique,
(Branstetter et al., 1987) et en Floride (Natanson et al., 1999), l’âge à première maturité était
estimé à 7 ans pour les mâles et 8 ans pour les femelles. La taille à maturité est de 310
centimètres chez les mâles et 315-320 cm chez les femelles (Branstetter et al., 1987 ; Kohler
et al., 1996).
Taille maximale
La taille maximale enregistrée pour un requin tigre est 740 cm (LT). Cependant, la
majorité des plus grands requins tigres observés ne dépasse pas 500 cm (LT). L’étude de
Branstetter et al. (1987), indique que la plus grande femelle et le plus grand mâle mesuraient
tous les deux 381 cm (LT) pour des âges respectifs de 16 et 15 ans. Kohler et al. (1996)
observaient une taille maximum de 339 cm (LT) dans l’Atlantique Nord-Ouest. Wirsing et al.
(2006) ont recensé un requin tigre de 445 cm (LT). Les différents auteurs ont calculé un âge
maximum compris entre 20 et 37 ans. Les résultats de Natanson et al. (1999) en Floride
étaient plus précis en estimant un âge maximum de 27 ans. Branstetter et al. (1987) et
Natanson et al. (1999) suggèrent que les femelles deviennent matures à 25% de leur âge
maximum et seraient capables de se reproduire une dizaine de fois, sur la base d’un cycle de
reproduction de plus de 2 ans.
Les requins tigres ont des portées importantes, une taille à la naissance qui les protège de
la majorité des prédateurs, un taux de croissance élevé et une maturité relativement précoce.
Ces informations suggèrent un potentiel de reproduction plus grand chez cette espèce par
rapport aux autres Carcharhinidés. Cela implique une population de requins tigres

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
14
importante, composée d’individus qui atteignent rapidement de grandes tailles par rapport
aux requins bouledogues.
5. Patterns de mouvements et utilisation de l’habitat
L’utilisation de l’habitat est l’un des traits d’histoire de vie le plus étudié ces dernières
années. La majorité des premières études sur le sujet se sont basées sur des observations
visuelles opportunistes ou sur le résultat de campagnes de pêche (Springer, 1943 ; Bigelow
and Schroeder, 1948 ; Springer, 1963 ; Randall, 1967 ; Tester, 1969). Les informations étaient
fragmentaires et souvent limitées par la méthodologie. C’est en 1981 que la première étude
utilisant la technologie acoustique fut lancée (Tricas et al., 1981). Bien qu’elle s’est déroulée
sur un seul requin, elle fut le prélude à de nombreuses autres études sur les déplacements à
court et long termes des requins tigres (Holland et al., 1999 ; Heithaus et al., 2001 ; Holland
et al., 2001 ; Heithaus et al., 2002 ; Heithaus et al., 2006 ; Lowe et al., 2006 ; Heithaus et al.,
2007) (Figure 6).
Les mouvements et les habitudes migratoires des requins tigres sont complexes à définir.
En effet dès les premières études, il est apparu que cette espèce est séparée en groupes
d’individus occupant des espaces vitaux plus au moins grands et composés de zones
spécifiques qu’ils fréquentent annuellement. Alors que la majorité des requins tigres
fréquentant les côtes de la Floride sont des résidents permanents (Springer, 1963), certains
migrent plus au Nord pendant l’été longeant la côte Est des Etats-Unis (Bigelow and
Schroeder, 1948). De même, à Hawaii, certains requins tigres limitent leurs mouvements à
certaines zones alors que d’autres ne sont plus ré-observés dans la zone après le marquage
(Tester, 1969).
Figure 6 : Marquage acoustique en interne et relâche d’un requin tigre à La Réunion
Source : © Geoffrey Bertrand et Estelle Crochelet, 2011

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
15
Ces informations mettent en évidence toute la complexité de l’utilisation de l’habitat par
ces requins. La technique de capture/recapture n’étant pas adaptée à l’étude d’espèces qui font
de grands déplacements, il a fallu attendre le développement des suivis acoustiques pour
mieux comprendre les mouvements de ces requins. En 1981, dans les eaux du banc de la
Frégate Française, Tricas et al. (1981), ont réalisé le premier marquage acoustique d’un
requin tigre. Pendant les 48 heures de suivi, l’individu a limité ses mouvements aux eaux
côtières et a nagé préférentiellement au fond. Bien que cette étude a apporté des nouvelles
informations sur l’utilisation de l’habitat des requins tigres, ce n’est qu’à la fin des années
1990 que les suivis acoustiques ont été utilisés de manière systématique pour ce type d’étude.
En 1999, à Hawaii, Holland et al. ont marqué et suivi 9 requins tigres sur des durées
comprises entre 7 et 50 heures. Les résultats de cette étude ont été sensiblement différents de
ceux de Tricas et al. (1981). Les neuf requins marqués n’ont pas cantonné leurs mouvements
aux eaux côtières mais ont effectué des déplacements au large notamment entre Oahu et le
banc des Pingouins. Les auteurs ont ainsi pu définir l’utilisation de l’habitat côtier et hauturier
des requins tigres à Hawaii et ses alentours. Dans les eaux côtières, les requins semblent
limiter leurs mouvements à des zones peu étendues, ont des déplacements circulaires et
nagent principalement près du fond. Au contraire, dans les eaux hauturières, les individus
suivis nagent en ligne droite à des profondeurs moyennes comprises entre 60 et 80 mètres
avec un record à 335 mètres. Cette capacité à utiliser les milieux pélagiques pour se déplacer a
été déjà suggérée par les prises accessoires des thoniers au large d’Hawaii mais n’avait pas
encore été observée (Polovina and Lau, 1993). Pour conclure, les auteurs déduisent que la
taille de l’espace vital des requins tigres pourrait être très étendue, ne se limitant pas à une île,
un banc ou une baie mais inclurait des zones pouvant être séparées par de grandes distances.
Une étude menée en Atlantique Nord-Ouest, a montré qu’un requin tigre marqué dans cette
zone a traversé l’océan Atlantique jusqu’en Afrique du Sud (Kohler et al., 1998). Cette
observation met en évidence la capacité de certains requins tigres à effectuer des grands
déplacements à l’échelle d’un océan. En 2001, une étude utilisant 5 types de marquages
différents (Roto-tag, balises acoustiques internes et externes, balises archives acoustiques
CHAT et balises satellites) dans la même zone d’étude a confirmé et précisé ces nouvelles
informations sur les requins tigres (Holland et al., 2001). L’ensemble des résultats issus des
différentes méthodes suggère une fidélité au site variable dans l’année et confirme l’existence
d’un espace vital étendu. Les requins tigres seraient constamment en déplacement dans ces
espaces vitaux et ne resteraient pas dans un site particulier plus de quelques semaines. Plus
récemment, il a été mis en évidence que les requins tigres d’Hawaii effectuaient des
déplacements à grande échelle entre plusieurs zones, et non entre deux sites comme l’avait
suggéré Holland et al. (Oahu-banc de la Frégate Française, 1999). Plusieurs requins marqués
ont effectué des allers-retours entre le banc de la Frégate Française et l’île de Midway ou
Kona (Hawaii) (ce qui représente respectivement 1280 kilomètres et 1190 kilomètres; Lowe
et al., 2006). De même, les requins marqués à East Island ont été détectés sur l’ensemble des
îles environnantes. Ainsi, la taille de l’espace vital des requins tigres à Hawaii s’étendrait au

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
16
moins à l’ensemble des archipels (Hawaii, Penguins Island, le banc de la Frégate Française et
ses îles alentours).
Cette fidélité partielle aux sites ainsi que la capacité à effectuer de grandes distances
semblent induire une saisonnalité en termes de présence et d’abondance sur chaque site. Si au
banc de la Frégate Française les requins tigres sont observés tout au long de l’année, une
tendance saisonnière se dessine concernant les îles et îlots environnants. Des individus
marqués acoustiquement ont passé plus de temps autour d’East Island durant l’été alors qu’ils
ont été majoritairement détectés autour des îles du Nord (Tern, Trig, and Shark Islands) en
hiver (Lowe et al., 2006). Les études menées à Shark Bay en Australie, montrent des
tendances similaires. Les études de Heithaus et al. (2001) et de Wirsing et al. (2006),
montrent que le nombre de requins tigres capturés est significativement plus élevé en été
(septembre à mai) qu’en hiver (juin à août), avec un pic en février.
L’influence des paramètres environnementaux sur cette variation saisonnière reste encore
mal connue et difficile à définir. La température de l’eau semble un facteur important mais
pas déterminant. A Shark Bay, le taux de capture des requins a diminué fortement lorsque la
température a dépassé 20°C (Heithaus et al., 2001). Cependant, une autre étude sur le même
site l’année suivante n’a pas trouvé de lien évident entre la température de l’eau et la variation
d’abondance saisonnière des requins tigres (Heithaus et al., 2002).
Un autre facteur étudié est la disponibilité en proie (Heithaus et al., 2001, Heithaus et al.,
2002). Deux hypothèses ont été émises pour expliquer l’influence du comportement des
proies chez le requin tigre. Les changements d’abondance saisonnière des requins tigres
observés à Shark Bay correspondent au départ et à l’arrivée des dugongs et des serpents de
mer dans la baie (Heithaus et al., 2001). Ainsi, les mouvements des requins tigres à l’échelle
d’une baie pourraient être corrélés avec ceux de ses proies favorites. Heithaus et al. (2002) a
associé des suivis acoustiques à des crittercams (caméra embarquée) pour étudier l’utilisation
de l’habitat des requins tigres à Shark Bay. Les requins suivis ont utilisé majoritairement les
zones côtières où les herbiers dominaient (habitat préférentiel de ses proies). Il appuie ainsi
l’hypothèse d’une distribution des requins tigres influencée par la disponibilité en proies. La
variabilité saisonnière pourrait aussi être influencée par la variation de la disponibilité de
proies dans d’autres régions du monde. Par exemple, certains requins observés à Oahu
(Hawaii) quittent la zone pour se rendre aux bancs de la Frégate Française où ils s’alimentent
de juvéniles d’albatros de Laysan (Meyer et al., 2010).
Une étude récente sur les mouvements à long terme des requins bouledogues à Shark Bay
met également en évidence un nouveau comportement migratoire (Heithaus et al., 2007). Si la
majorité des requins marqués avec des balises satellites sont restés dans les eaux
australiennes, une femelle adulte est allée jusqu’en Afrique du Sud parcourant plus de 8000
kilomètres en un peu plus de 50 jours. C’est la plus grande migration observée chez les
requins tigres. C’est la deuxième observation d’une migration Est-Ouest entre l’Australie et
l’Afrique du Sud réalisée par un requin. La première était un requin blanc adulte qui, marqué

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
17
en Afrique du Sud, a fait l’aller-retour en Australie en 9 mois soit plus de 20000 kilomètres
(Bonfil et al., 2005). Les raisons de cette migration Est-Ouest reste encore inconnues.
Pour résumer, l’utilisation de l’habitat est très variable entre les individus qu’ils soient
originaires ou non d’une même région. Ainsi, plusieurs individus peuvent rester dans une
même zone insulaire toute leur vie, d’autres vont étendre leur domaine vital à l’ensemble
d’archipels, et d’autres encore sont capables de faire des migrations transocéaniques. Les
facteurs influençant ces différents comportements sont encore peu connus. Cependant, la
disponibilité en proies semble jouer un rôle important. Plusieurs questions se posent alors :
Pourquoi existe-il dans une même zone des requins « sédentaires » et d’autres plus
migrateurs ? Quels sont les facteurs qui déterminent si un individu sera résident permanent
ou temporaire ? Existe-il un déterminisme génétique ? Existe-il une ségrégation
comportementale en fonction des habitudes migratoires ? Autant de questions sans réponse
qui nécessitent de continuer les études dans ce domaine en s’aidant de nouvelles techniques
comme l’analyse génétique. A l’échelle de l’océan indien, la question de connectivité prend
tout son sens : est-ce que la population de requin de la Réunion est interconnectée avec celles
des Seychelles, de Madagascar ou de l’Afrique du Sud. L’existence d’un tel lien entre
plusieurs populations peut avoir un impact important sur la variation saisonnière et annuel
de l’abondance des requins tigres dans les eaux côtières de l’île. Elle sera étudiée à travers
les suivis satellites mais surtout grâce à une étude génétique.
6. Bibliographie
Alves, M. I. M. (1977). Algumas consideracoes sobre a reproducao do cacao jaguara,
Galeocerdo cuvieri (Le Sueur, 1822) (Selachii: Carcharhinidae). Arq Cienc Mar, 17 121–
125
Bass, A. J., D'Aubrey, J. D. and Kistnasamy, N. (1975). Sharks of the east coast of
southern Africa. III. The families Carcharhinidae (excluding Carcharhinus and Mustelus)
and Sphyrnidae. Oceanogr. Res. Int. (Durban) Invest. Rep. 38, 100 p.
Baughman, J.L. and Springer, S. (1950). Biological and economic notes on the sharks of
the Gulf of Mexico, with special reference to those of Texas, and with a key for their
identification. Amer. Mid. Natur. 44, 96-152
Baum, J. K., Myers, R. a, Kehler, D. G., Worm, B., Harley, S. J., and Doherty, P. a.
(2003). Collapse and conservation of shark populations in the Northwest Atlantic. Science,
299, 389-92. doi:10.1126/science.1079777
Bigelow, H. B., and W. C. Schroeder. (1948). Sharks. In J. Tee-Van, C. M. Breder, S. F.
Hilde- brand, A. E. Parr, and W. C. Schroeder (eds.). Fishes of the Western North Atlantic,
part one, vol. 1, p. 59–546. Mem. Sears Found. Mar. Res., Yale Univ.
Bonfil, R., Mena, R. and de Anda, D. (1993). Biological parameters of commercially
exploited silky sharks, Carcharhinus falciformis, from the Campeche Bank, Mexico. In S.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
18
Branstetter (ed.). Conservation biology of elasmobranchs, p. 73–86. U.S. Dep. Commer.,
NOAA Tech Rep. NMFS 115.
Bonfil, R., Meÿer, M., Scholl, M. C., Johnson, R., O’Brien, S., Oosthuizen, H., Swanson,
S., Deon, K. and Paterson, M. (2005). Transoceanic migration, spatial dynamics, and
population linkages of white sharks. Science, 310, 100-103. doi:10.1126/science.1114898
Branstetter, S. , Muscik, J. A. and Colvocoresses, J. A. (1987). A comparison of the age
and growth of the tiger shark, Galeocerdo cuvier, from off Virginia and from the
northwestern Gulf of Mexico. Fishery Bulletin, 85(2), 269-279.
Briggs, J. C. (1961). The East Pacific Barrier and the Distribution of Marine Shore Fishes.
Society for the study of Evolution, 15(4), 545-554
Clark, E., and Von Schmidt, K. (1965). Sharks of the central Gulf coast of Florida. Bull
Mar Sci, 15, 13–83
Casey, J. G. and Natanson, L. J. (1992). Revised estimates of age and growth of the sandbar
shark (Carcharhinus plumbeus) from the western North Atlantic. Can J Fish Aquat Sci 49,
1474–1477
Compagno, L.J.V., (1984). Sharks of the world. Part 1, Hexanchiformes to Lamniformes.
FAO Species Catalogue 4, FAO Fish. Synop. 125, Food and Agriculture Organisation of the
United Nations, Rome, 249 p.
Driggers, W., Ingram, G., Grace, M., Gledhill, C., Henwood, T., Horton, C., & Jones, C.
(2008). Pupping areas and mortality rates of young tiger sharks Galeocerdo cuvier in the
western North Atlantic Ocean. Aquatic Biology, 2, 161-170. doi:10.3354/ab00045
Francis, M. P. (1998). New Zealand shark fisheries: development, size and management.
Marine and Freshwater Research, 49(7) 579 - 591
Gohar, H. A. F. and Mazhar, F. M. (1964). The elasmobranchs of the northwestern
Red Sea. Pub. Mar. BioI. Stn. Ghardaqa, (13): 1-144.
Gudger, E.W. (1949). Natural history notes on tiger sharks, Galeocerdo tigrinis, caught at
KeyWest, Florida, with emphasis on food and feeding habits. Copeia, 39–47.
Haraguchi, K., Hisamichi, Y. Kotaki, Y., Kato, Y. and Endo, T. (2009). Halogenated
ipyrroles and Methoxylated Tetrabromodiphenyl Ethers in Tiger Shark (Galeocerdo cuvier)
from the Southern Coast of Japan. Environ. Sci. Technol., 43 (7) 2288–2294
Heithaus, M. R. (2001a). The biology of tiger sharks (Galeocerdo cuvier) in Shark Bay,
Western Australia: sex ratio, size distribution, diet, and seasonal changes in catch rates.
Environ Biol Fish, 61, 25–36
Heithaus, R. (2001b). Shark attacks on bottlenose dolphins (Tursiops aduncus) in Shark Bay,
Western Australia : Attack rate, bites scar frequencies and attack seasonality. Marine
Mammal Science, 17(3), 526-539.
Heithaus, M., Marshall, G., Buhleier, B., and Dill, L. (2001c). Employing Crittercam to
study habitat use and behavior of large sharks. Marine Ecology Progress Series, 209, 307-
310. doi:10.3354/meps209307

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
19
Heithaus, M.R., Dill, L. M., Marshall, G. J. and Buhleier B. M. (2002). Habitat use and
foraging behavior of tiger sharks (Galeocerdo cuvier) in a seagrass ecosystem. Mar Biol
140: 237–248
Heithaus, M. R., Hamilton, I. M., Wirsing, A. J., and Dill, L. M. (2006). Validation of a
randomization procedure to assess animal habitat preferences: microhabitat use of tiger
sharks in a seagrass ecosystem. The Journal of animal ecology, 75, 666-76.
doi:10.1111/j.1365-2656.2006.01087
Heithaus, M. R., Frid, A., Wirsing, A. J., Dill, L. M., Fourqurean, J. W., Burkholder, D.,
Thomson, J., et al. (2007). State-dependent risk-taking by green sea turtles mediates top-
down effects of tiger shark intimidation in a marine ecosystem. The Journal of animal
ecology, 76, 837-44. doi:10.1111/j.1365-2656.2007.01260
Holland, K. N., Wetherbee, B. M., Lowe, C. G., and Meyer, C. G. (1999). Movements of
tiger sharks (Galeocerdo cuvier) in coastal Hawaiian waters. Marine Biology, 134(4), 665-
673. doi:10.1007/s002270050582
Holland, K. N., Bush, A., Meyer, C. G., Kajiura, S., Wetherbee, B. M. and Lowe, C. G.
(2001). Five tags Applied to a single species in a single location: The tiger shark experience.
Sibert JR, Nielsen J (eds) Electronic tagging and tracking in marine fisheries (pp. 237-247).
Kohler, N. E., Casey, J. G., and Turner, P. A. (1996). Length-Length and Length-Weight
Relationships for 13 Shark Species from the Western North Atlantic. Fisheries Science (p.
29).
Kohler, N. E., Casey, J. G., Turner, P. A. (1998). NMFS cooperative shark tagging
program, 1962–93: an atlas of shark tag and recapture data. Mar Fish Rev 60:1–87
Lowe, C. G., Wetherbee, B. M., Crow, G. L. and Tester, A. L. (1996). Ontogenic dietary
shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, in Hawaiian waters.
Environmental Biology of Fishes, (47), 203-211.
Lowe, C. G., Wetherbee, B. C., Meyer, C. G. (2006). Using acoustic telemetry monitoring
techniques to quantify movement patterns and site fidelity of sharks and giant trevally
around French Frigate Shoals and Midway atoll. Atoll Research Bulletin, 281-303
Lundberg1 J. G, M. Pérez, H. S., Dahdul, W. M., Aguilera, O. A., Hoorn, C. and
Wesselingh, F. P. (2010). The Amazonian Neogene Fish Fauna. Blackwell Publishing
Menni, R. C.and Lucifora, L. O. (2007). Condrictios de la Argentina y Uruguay (p. 18).
Myers, R. a, Baum, J. K., Shepherd, T. D., Powers, S. P., and Peterson, C. H. (2007).
Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315,
1846-1850. doi:10.1126/science.1138657
Meyer, C. G., Papastamatiou, Y. P., & Holland, K. N. (2010). A multiple instrument
approach to quantifying the movement patterns and habitat use of tiger (Galeocerdo cuvier)
and Galapagos sharks (Carcharhinus galapagensis) at French Frigate Shoals, Hawaii.
Marine Biology, 157, 1857-1868. doi:10.1007/s00227-010-1457

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
20
Natanson, L. J., Casey, J. G., and Kohler, N. E. (1999). Growth of the tiger shark,
Galeocerdo cuvier, in the western North Atlantic based on tag returns and length
frequencies�; and a note on the effects of tagging. Fishery Bulletin, 97, 944-953.
Natanson, L. J. (1994). Age and growth estimates for the dusky shark, Carcharhinus
obscurus, in the western North Atlantic. Fish. Bull., 93, 116–126.
Polovina, J. J., and Lau, B. B. (1993). Temporal and Spatial Distribution of Catches of Tiger
Sharks, Galeocerdo cuvier, in the Pelagic Longline Fishery Around the Hawaiian Islands.
Marine Fisheries Review, 55(3), 1-3.
Pratt, H. L. Jr. (1993). The storage of spermatazoa in the oviducal glands of western North
Atlantic sharks. Environ Biol Fish, 38, 139–149
Rancurel, P. and Intes, A. (1982). Le requin tigre, Galeocerdo cuvier, Lacépède des eaux
Néocalédoniennes examen des contenus stomacaux. Tethys, 10, 195-199
Randall, J.E. (1967). Feed habit of the reef fishes of the West Indies. Institut of Marine
Science, Univ. Miami Stud. Trop. Oceanogr, 5: 654-847
Randall, J.E. (1992). Review of the biology of the tiger shark (Galeocerdo cuvier). Aust J
Mar Freshw Res 43, 21–31
Sandoval-castillo, J., Mariano-melendez, E and Villaciencio-Garayzar, C. (2006). New
records of albinism in two elasmobranchs: the tiger shark Galeocerdo cuvier and the giant
electric ray Narcine entemedor. Cybium, 30(2), 191-192.
Simpfendorfer, C. (1992). Biology of tiger sharks (Galeocerdo cuvier) caught by the
Queensland shark meshing program off Townsville, Australia. Aust. J. Mar. Freshw. Res.
43, 33–43
Simpfendorfer, C. A, Goodreid, A.B., and McAuley, R. B.(2001). Size, Sex And
Geographic Variation in the Diet of the Tiger Shark, Galeocerdo Cuvier, From Western
Australian Waters. Env. Bio. Of Fish., 61(1)37-46
Smale, M. J. and Cliff, G. (1998). Cephalopods in the diets of four shark species
(Galeocerdo cuvier, Sphyrna lewini, S. zygaena and S. mokarran) from KwaZulu-Natal,
South Africa. South African Journal of Marine Science, 20(1), 241-253
Springer, S., (1943). Sharks and Their Behavior, with Particular Reference to Eight Genera
Implicated in Reports of Attacks on Man, 52 pp., illus. Unpubl. rept. in files of coordinator
of Research & Development, USN, Emergency Rescue Equipment Section, Washington, D.
C.
Springer, S., (1963). Field observations on large sharks of the Florida- Caribbean region. In
P. W. Gilbert (editor), Sharks and survival, Sect. II. Ch. 3, p. 95-113. Heath and Co.,
Boston.
Stevens, J. D. (1984). Biological observation on sharks caught by sport fishermen off New
South Wales. Aust J Mar Freshw Res 35:573–590
Stevens, J.D. and Mc Loughlin, K.J. (1991). Distribution, size and sex composition,
reproductive biology and diets of sharks from northern Australia. Aust. J. Mar. Freshw. Res
42: 151–199.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
21
Stevens, J. (2000). The effects of fishing on sharks, rays, and chimaeras (chondrichthyans),
and the implications for marine ecosystems. ICES Journal of Marine Science, 57, 476-494.
doi:10.1006/jmsc.2000.0724
Schwartz, F. J. (1994). Body–organ weight relationships of near- term and newborn tiger
sharks, Galeocerdo cuvier, captured off North Carolina. J Elisha Mitchell Sci Soc, 110,
104–107
Tester, A. L. (1969). Cooperative shark research and control program, final report, 1967-69.
Univ. Hawaii Press, Honolulu, 47 p.
Tricas, T. C., Taylor, L. R. and Naftel. G. (1981). Diel behavior of the tiger shark,
Galeocerdo cuvier, at French Frigate Shoals, Hawaiian Islands. Copeia : 904-908.
Vatter, A. (2003). Bottom longline fishing for sharks in the Northwest Hawaiian Islands. US
Department of Commerce, AR-PIR-03–01, Honolulu, HI
Whitney, N. M., and Crow, G. L. (2006). Reproductive biology of the tiger shark
(Galeocerdo cuvier) in Hawaii. Marine Biology, 151(1), 63-70. doi:10.1007/s00227-006-
0476-0
Wintner, S. P. and Cliff, G. (1999). Age and growth determination of the white shark,
Carcharodon carcharias, from the east coast of South Africa. Fishery Bulletin, 153-169.
Wintner, S. P. and Dudley, S. F. J. (2000). Age and growth estimates for the tiger shark,
Galeocerdo cuvier, from the east coast of South Africa. Mar. Freshwat. Res., 51, 43-53.
Wirsing, A. J., Heithaus, M. R., & Dill, L. M. (2006). Tiger shark (Galeocerdo cuvier)
abundance and growth in a subtropical embayment: evidence from 7 years of standardized
fishing effort. Marine Biology, 149, 961-968. doi:10.1007/s00227-006-0278-4
Witzell, W. N. (1987). Selective predation on large cheloniid sea turtles by tiger sharks
(Galeocerdo cuvier). Jpn. J. Herpetol. 12, 22–29.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
22
Le requin bouledogue
Introduction
Le requin bouledogue est l’un des plus grands poissons prédateurs au monde (Campagno,
1984). Comme le requin tigre, il appartient à la famille des Carcharhinidés ou « requins
requiem », mais fait partie du genre Carcharhinus (Blainville, 1816). Ce genre regroupe le
plus grand nombre d’espèces de requin actuel avec 32 espèces distinctes (ITIS, 2012) dont des
espèces de récif, comme le requin pointes noires (Carcharhinus melanopterus, Quoy and
Gaimard, 1824), le requin pointe blanche de récif (Carcharhinus albimarginatus, Rüppell,
1837) ou le requin gris de récif (Carcharhinus amblyrhynchos, Bleeker, 1856) mais aussi des
espèces océaniques, comme le requin pointe blanche océanique (Carcharhinus longimanus,
Poey, 1861). Ces espèces sont le plus souvent vivipares (les nouveaux nés naissent déjà
complètement formés), et vivent principalement dans les eaux chaudes des tropiques.
Massif, le requin bouledogue possède un nez court et arrondi et de petits yeux ronds. Son
dos est gris, gris-bleu, ou brun avec l’extrémité des nageoires un peu plus foncé, notamment
au niveau de la nageoire caudale des jeunes individus. Sa face ventrale est beaucoup plus
claire, pouvant aller de gris clair à blanchâtre. Il a une mâchoire asymétrique avec des dents
larges, triangulaires et dentelées au niveau de la mâchoire supérieure et des dents fines, lisses
et pointues au niveau de la mâchoire inférieure.
Le requin bouledogue peut atteindre une taille maximale de 400 cm (individu pêché dans
le fleuve Zambèze, Afrique du Sud) pour un poids maximum (connu) de 320 kg. Néanmoins,
de récentes découvertes montrent l’existence d’individus dépassant cette taille (population de
requins bouledogues de l’embouchure du Rio Sinera, Costa Rica, com. pers. 2012). Les
femelles arrivent à maturité à 10 ans pour une taille supérieure à 200 cm et les mâles à 9-10
ans pour une taille comprise entre 190 et 200 cm (Martinez et al., 2004).
Son régime alimentaire est diversifié, des téléostéens aux élasmobranches et/ou aux
mammifères marins (Schmid et al., 1990).
Le requin bouledogue est l’une des seules espèces euryhalines (Montoya and Thorson,
1982 ; Last and Stevens 1994 ; Pillans et al., 2005). Il est présent dans toutes les eaux
tropicales et sub-tropicales, aussi bien à la côte qu’en milieu semi-pélagique. Il est également
observé dans les fleuves et les estuaires, où il est connu sous d’autres noms. En Afrique du
Sud, les requins bouledogues fréquentant les eaux du Zambèze prennent le nom de requin du
Zambèze ; au lac Nicaragua, il a longtemps été considéré comme une espèce endémique du
lac sous le nom de requin du Nicaragua (Carcharhinus nicaraguensis, Bigelow and
Schroeder, 1961). En Inde, il fut très souvent confondu avec le requin du Gange (Glyphis
gangeticus, Müller and Henle, 1839) qu’il côtoie, mais qui est aujourd’hui considéré comme
disparu. Dans l’Océan Indien, il est aussi souvent confondu avec le requin Java, Carcharhinus
amboinensis (Müller and Henle, 1839). Ce dernier est physiquement très proche du requin
bouledogue qui ne se différencie que par un ratio de hauteur entre la première et deuxième

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
23
nageoire dorsale plus petit et un nombre de dents plus faible (13-1-13/12-1-12 chez le requin
bouledogue et 12-1-12/11-1-11 chez le requin Java) (Smith, 1952 ; Sadowsky, 1971).
1. Distribution
Le requin bouledogue a une aire de répartition très étendue comprenant les eaux chaudes
tropicales et sub-tropicales, les rivières et les lacs (Figure 7). On le retrouve notamment sur le
continent américain, de la Caroline du Nord à l’Argentine sur la côte Est (Bigelow and
Schroeder, 1948, Menni and Lucifora,2007) et du Sud de la Californie au Nord du Pérou sur
la côte Ouest (Castro and Aguirre, 1978); sur le continent africain, du Nord du Sénégal à
l’Afrique du Sud jusqu’en Egypte (Valencienne, 1841; Coad and Papahr, 1988; Cliff and
Dudley, 1992; Trape, 2008); dans l’Océan Indien, des côtes du canal du Mozambique
jusqu’en Inde en passant par La Réunion (com. pers.); en Océanie, des côtes australiennes aux
Fidji (Heithaus and Dill, 2002; Brunnschweiler and Earle, 2006).
Il est également souvent observé dans les zones d’estuaires et remonte régulièrement les
fleuves, comme l’estuaire du Santa-Lucia et du Breede (Afrique du sud, Bass et al., 1973;
McCord and Lamberth, 2009), l’estuaire du Paraná (Brésil, César de Olieveira Santos and
Bismarck Fazzano Gadig, 2008), le Brisbane (Australie, Pillans and Franklin, 2004), l’Indien
et le Caloosahatchee (Floride, Snelson et al., 1984; Simpfendorfer et al. 2005), le Rio Bayano
(Panama, Montoya and Thorson, 1982), l’Amazone (Pérou, Myers, 1952), le Mississippi
(Illinois, Thomerson and Thorson, 1977) ou encore le Gange (Inde, com. pers.). Il est même
signalé dans certains lacs, le plus connu étant le lac Nicaragua (Nicaragua, Bigelow and
Schroeder, 1976) mais on le recense aussi dans le lac Miraflores (Colombie, Bigelow and
Schroeder, 1948), le lac Bayano (Panama, Montoya et al. 1982) et plus récemment, le lac
Pontchartrain (Louisiane, O'Connell et al., 2007).
Figure 7 : Aire de répartition mondiale du requin bouledogue (d’après les données publiées).
Source : Compagno, Dando & Fowler (2005). Sharks of the World. Collins Field Guides.
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
24
Bien qu’il semble que l’on connaisse bien son aire de répartition mondiale, celle-ci évolue
encore de nos jours, illustrée par la récente découverte d’une population de requin
bouledogue dans l’estuaire du Rio Sinera au Costa Rica en 2012. Tout comme le requin tigre,
la présence du requin bouledogue sur les côtes réunionnaises est observée depuis longtemps
par les pêcheurs au gros et/ou les pêcheurs sous-marins. Néanmoins, aucune étude n’a
encore été faite sur sa présence réelle dans cette zone en termes d’abondance saisonnière et
annuelle. Le programme CHARC permettra de fournir ces premières informations et
confirmera ou non la présence régulière de cette espèce à La Réunion.
2. Alimentation
Comme les autres espèces de requins « évoluées », le requin bouledogue possède une
mâchoire hyostylique. La mâchoire supérieure n’est pas soudée à la mâchoire inférieure,
contrairement aux espèces de requins « primitifs » qui ont une mâchoire amphistylique. Les
dents de sa mâchoire supérieure sont larges, triangulaires et dentelées alors que les dents de sa
mâchoire inférieure sont fines et pointues (Sadowsky, 1971) (Figure 8).
Figure 8 : Dents supérieures et inférieures d’un requin bouledogue
Source : © Copyright Ross Robertson, 2006
Le requin bouledogue est un prédateur très polyvalent qui exploite un large spectre de
proies, invertébrées ou vertébrées. Son régime alimentaire comprend la majorité des poissons
(y compris les autres requins), les céphalopodes, les cétacés, les tortues, les oiseaux voir
même certains mammifères terrestres. La majorité des études ont été réalisées grâce à
l’analyse de contenus stomacaux (Sadowsky, 1971; Tuma, 1976; Snelson et al., 1984). Les
observations et les informations plus récentes sur les habitudes alimentaires du requin
bouledogue proviennent d’identification d’interactions proies/prédateurs (Heithaus et al.,
2002 ; César de Olieveira Santos and Bismarck Fazzano Gadig, 2008) ou de suivis

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
25
acoustiques associés à la biodiversité présente (Heithaus et al., 2009; McCord and Lamberth,
2009).
Bien que son régime alimentaire soit très varié, les études des contenus stomacaux ont
permis de différencier les proies préférentielles des proies « accessoires ». Au Brésil, dans le
lagon de Cananéia, les requins bouledogues s’alimentent principalement de mâchoirons (Arius
spixii, Arius grandicassus…), de carangues médailles (Chloroscombrus chrysurus), de
requins (Carcharhinus porosus, Sphyrna tiburo) et de raies pastenagues (Dasyatis americana)
(Sadowsky, 1971). Au Nicaragua et au Costa Rica, les contenus stomacaux d’individus
juvéniles et adultes contiennent 85% d’Osteichtyes, 10% de crustacés et 5% de reptiles
(Cheloniidae) (Tuma, 1976). Occasionnellement, des restes d’oiseaux de la famille des
Tyrannidés et de poissons scies (famille des Pristidae) ont également été trouvés dans les
estomacs. Tout comme dans l’étude de Sadowsky (1971), aucune variation alimentaire
ontogénique n’a été observée. La taille des poissons varie proportionnellement à la taille du
requin et le pourcentage de tortues est plus élevé chez les adultes, dû certainement à
l’incapacité des juvéniles à s’attaquer à des proies plus grandes qu’eux. En Floride, dans
l’Indian River, Snelson et al. (1984) ont retrouvé ces mêmes proportions de proies dans le
régime alimentaire des requins bouledogues. Les estomacs contenaient essentiellement des
mâchoirons (Ariis), des carangues, des raies pastenagues et des requins. L’estomac d’un
requin sub-adulte (162 cm LT) comportait la tête d’un requin bouledogue juvénile, ce qui
constitue la première évidence de prédation intra-spécifique.
Bien que très peu de restes de mammifères marins soient retrouvés dans les estomacs des
requins bouledogues étudiés (Last and Stevens, 1994), plusieurs grosses cicatrices ont été
observées sur le corps de dauphins et de baleines. Au Brésil, dans l’estuaire du Paraná,
plusieurs dauphins de Guyane (Sotalia guianensis) présentent des marques de morsures sur
l’aileron (César de Oliveira Santos and Bismarck Fazzano Gadig, 2008). Certaines sont assez
marquées pour que l’on puisse identifier la mâchoire d’un requin bouledogue. Néanmoins,
aucun évènement de prédation de dauphin de Guyane par un requin bouledogue n’a encore été
observé. A La Réunion, dans la baie de Saint Paul, plusieurs grands dauphins de
l’indopacifique (Tursiops aduncus) présentent des marques identiques au niveau de la queue
et/ou de l’aileron dorsal (Rabeariosa, com. pers.). Bien que là encore aucune prédation n’ait
pu être mise en évidence, la forme de la morsure et la présence de requin bouledogue dans la
zone permet d’émettre l’hypothèse que ces blessures proviennent aussi de ces derniers.
Par ailleurs, les requins bouledogues ne s’attaquent pas à des baleines vivantes mais ils
sont régulièrement signalés autour de carcasses dérivantes en pleine mer, notamment en
Afrique du Sud et dans le canal du Mozambique (com. pers.).
Ainsi, les requins bouledogues ont un régime alimentaire diversifié mais bien défini sans
changement ontogénique. Si le type de proie de change pas en fonction de l’âge, la taille de
celle-ci augmente proportionnellement à la taille du requin. Plus le requin atteint des tailles
importantes plus le pourcentage de grosses proies, comme les tortues, les dauphins et les

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
26
élasmobranches. Peu de données sont disponibles sur la part des baleines dans le régime
alimentaire de cette espèce. Il semble que ces évènements de prédation soient rares et très
opportunistes. Les données prises dans le cadre du programme CHARC chercheront à
confirmer ou infirmer ces conclusions. Comme pour le requin tigre, une étude isotopique sera
réalisée pour connaître l’évolution du régime alimentaire des requins bouledogues en
fonction des saisons et l’âge. Il sera donc possible de déterminer si les requins bouledogues
s’alimentent sur les mêmes familles d’espèces que précédemment observées. A la Réunion, la
présence de tortues, de dauphins et saisonnièrement de baleine en plus des proies
généralement décrite pour l’espèce pourraient jouer un rôle sur la composition du régime
alimentaire du requin bouledogue dans cette région de l’Océan Indien.
3. Reproduction
Gestation
Le requin bouledogue est une espèce vivipare. L’embryon est nourri par la mère par
l’intermédiaire d’une liaison placentaire. Le temps de gestation est de 10 à 11 mois
(Rodriguez de La Cruz et al., 1996). Les nouveau-nés naissent complètement formés et
indépendants.
Si beaucoup d’auteurs décrivent le requin bouledogue comme vivipare, cette espèce est
encore aujourd’hui décrite par certains comme ovovivipare. Une espèce est ovovivipare
lorsque les œufs incubent et éclosent dans le ventre de la femelle, sans relation nutritive avec
celle-ci. Chez les requins vivipares, le lien placentaire avec l’utérus maternel ne se crée pas
immédiatement. Au premier stade de développement, l’embryon est indépendant et se nourrit
grâce à une poche vitelline. Après quelques semaines, cette poche va s’attacher à la paroi
utérine créant ainsi un lien placentaire entre l’embryon et la mère. Le placenta apportera à
l’embryon l’eau, les nutriments et le dioxygène nécessaire à son bon développement (Price
and Daiber, 1967).
Il y a pu avoir une confusion entre ces deux modes de reproduction. Si les embryons sont
observés en tout début de développement, ils possèdent des poches vitellines généralement
typiques d’un mode de reproduction ovovivipare. C’est l’observation d’embryons plus âgés
qui a permis de déterminer que chez cette espèce la liaison placentaire avec la mère se faisait
plus tardivement. Bien que cela ne soit pas corrigé dans la littérature, on est actuellement sûr
que le requin bouledogue est bien vivipare et non ovovivipare.
Cycle Reproducteur
De nos jours, il existe peu d’information sur le cycle reproducteur de cette espèce. On ne
connaît pas avec exactitude leurs périodes et leurs zones de reproduction et aucun
accouplement n’a été observé. Les principales informations sont issues d’études sur leurs
déplacements et d’observations visuelles de femelles gestantes, de cicatrices laissées par des
accouplements récents ou de marques au niveau du cloaque signalant une mise bas dans la
zone. Au Brésil, dans le lagon de Cananéia, les femelles adultes en fin de gestation ne sont

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
27
présentes qu’entre novembre et février (Sadowsky, 1971). Cette période a ainsi été définie
comme la période de parturition. L’absence de mâle mature dans le lagon quel que soit la
période de l’année suggère que l’accouplement a lieu hors de cette zone. En Floride, dans
l’Indian River, les femelles viennet mettre bas dans l’estuaire dès la fin du printemps (avril)
jusqu’en été (juillet) (Snelson et al., 1984) avant de quitter la zone. D’après Dodrill (1977), la
saison d’accouplement aurait lieu juste après durant les mois de juin et juillet. Le nombre de
mâle mature présent sur les côtes de Floride étant limité, les auteurs pensent que
l’accouplement doit se passer soit dans une autre zone côtière soit plus au large. Au Nicaragua
et au Costa Rica, Jenson (1976) trouve sensiblement les mêmes résultats. La parturition a lieu
dans l’estuaire du Rio Colorado entre avril et juin. L’accouplement, bien que n’ayant jamais
été observé, se déroulerait juste après la mise bas dans une zone proche de l’estuaire.
Ce cycle de reproduction annuel est différent du cycle bisannuel décrit pour les requins
bouledogues au Fidji (Brunnschweiler and Baensch, 2011). Les auteurs proposent le cycle
suivant : l’accouplement se passe en début d’année, la mise bas en fin d’année et les femelles
s’accouplent de nouveau un an après leur mise bas. Ce cycle bisannuel est assez fréquent chez
les requins, notamment pour les espèces qui ont une durée de gestation longue. Il a
notamment était décrit chez des espèces de la même famille que le requin bouledogue comme,
le requin à Nez Rude (Carcharhinus macloti), le requin taupe (Lamna nasus) ou le requin
peau bleu (Prionace glauca) (Stevens and Mcloughlin 1991 ; Pratt and Carrier, 2001).
Ainsi, le cycle de reproduction des requins bouledogues est encore peu connu et beaucoup
de questions restent encore sans réponse. Ont-ils un cycle annuel, bisannuel voir triannuel ?
Est-ce que l’ensemble de la population mondiale a le même cycle ou diffère-t-il en fonction de
la localisation géographique ? Où se déroule l’accouplement ? A quelle période de l’année ?
Comme chez beaucoup d’autres espèces de requins, la reproduction du requin bouledogue est
l’un des traits d’histoire de vie les moins bien connus, et ce malgré son importance pour la
gestion, la conservation et la protection des espèces. Comme pour le requin tigre, aucune
étude spécifique à la reproduction ne sera réalisée dans le cadre de CHARC. Néanmoins,
l’observation de femelles gestantes, ayant mis bas ou portant des marques d’accouplement
récent et de nouveaux nés devraient apporter les premières informations sur le lieu et la
période de mise bas et/ou d’accouplement. A la suite de CHARC, ces données pourront servir
à une étude plus spécifique sur le cycle de reproduction de cette espèce.
4. Age et développement
Fécondation et taille à la naissance
La femelle requin bouledogue donne naissance à des portées de 1 à 9 nouveaux nés
(Sadowsky, 1971 ; Jenson, 1976) (Figure 9). La variation du nombre de nouveaux nés par
portée peut s’expliquer par la présence d’embryons qui ne se développent pas ou
anormalement. On observe alors du cannibalisme intra-utérin par les embryons les mieux
développés (Jenson, 1976). La taille à la naissance varie de 50 à 80 cm (LT). Il existe des

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
28
différences de taille entre les individus d’une même portée et entre populations de zones
géographiques différentes. En Afrique du Sud, Cliff et Dudley (1991) ont observé des
nouveaux nés dont la taille ne dépasse pas 62 cm (LT). Au Nicaragua et au Costa Rica, dans
le lac Nicaragua et dans le réseau fluvial du Rio San Juan, les nouveaux nés mesurent
majoritairement 65 à 70 cm (Jenson, 1976). Au Brésil, dans le lagon de Cananéia, ils sont
sensiblement plus grands, avec des tailles comprises entre 76 et 81 cm (LT) (Sadowsky,
1971). Ces différences de tailles, relativement importantes, peuvent s’expliquer par la
variabilité de vitesse de croissance propre à chaque individu. Dodrill (1977) a observé la
présence associée de trois embryons de petites tailles à un de grande taille dans le même
utérus. Il conclut à un phénomène de compétition alimentaire intra-utérine qui favoriserait le
développement d’un ou plusieurs embryons par rapport aux autres. La méthode
d’échantillonnage peut, elle aussi, influencer cette différence. Sadowsky (1971) ou Jenson
(1976) ont mesuré la taille d’embryons qu’ils considéraient sur le point de naître alors que
Cliff et Dudley (1991) ont observé des nouveaux nés nageant en pleine eau. Ainsi, il se peut
que ces individus n’aient pas le même âge. Par ailleurs, le taux de croissance des premières
semaines ou des premiers mois semble être important.
Figure 9 : Nouveaux nés requins bouledogues
Source : ©SeaPics
A La Réunion, deux requins juvéniles de 50 à 60 centimètres ont été observés dans la
ravine sud de Saint Leu. Un mois plus tard, leurs tailles ont été estimées à 70-80 centimètres.
Cette observation qui demande à être vérifiée sur un plus grand nombre d’individus
soutiendrait l’hypothèse d’un taux de croissance élevé dans les premières semaines après la
naissance.
Taux de croissance
Plusieurs études ont été réalisées sur l’âge et la croissance de ces requins (Brandstetter et
Stiles, 1987; Jenson, 1976; Cruz-Martínez et al., 2004; Neer et al. 2005). Bien qu’effectuées
sur des populations différentes, les résultats obtenus sont relativement similaires. Le taux de
croissance des requins bouledogues est lent, variable entre individu et dépendant du stade de
maturité de l’individu. Au Nord du Golfe du Mexique, la croissance et l’âge des requins
bouledogue ont été étudiés en comptant les anneaux de croissance qui se forment sur les
vertèbres (Brandstetter et Stiles, 1987). Indifféremment du sexe, le taux de croissance est de

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
29
15-20 cm.an-1 entre 0 et 5 ans, de 10 cm.an-1 de 6 à 10 ans, de 5-7 cm.an-1 de 11 à 16 ans et
moins de 4-5 cm.an-1 pour les années suivantes. Cinq requins bouledogues élevés en aquarium
et mesurés plusieurs fois par an pendant 7 ans ont présenté des taux de croissances proches de
ceux observés par Brandstetter et Stiles (1987) (Schimd and Muru, 1994). Les trois premières
années de leur vie, ils ont eu un taux de croissance de 23 cm.an-1 puis de 7 cm.an-1 sur les 4
d’après.
Par contre, la différence de croissance en fonction du sexe reste confuse. Brandstetter et
Stiles (1987) n’ont pas trouvé de différence significative en fonction des sexes. A l’inverse,
Cruz-Martínez et al. (2004) ont mis en évidence une différence de croissance entre les mâles
et les femelles en utilisant le même protocole. Les mâles avaient une croissance plus rapide
que les femelles et ces dernières atteignaient des tailles plus importantes que les mâles. Ces
résultats se rapprochent de ceux observés chez d’autres espèces de requins, comme le requin
blanc (Carcharodon carcharias, Dudley et Simpfendorfer, 2006) ou le requin taupe (Lamna
nasus, Natanson et al., 2002) dont les mâles ont un taux de croissance plus élevé dans les
premiers stades de leur vie mais ont des tailles adultes généralement plus faibles que les
femelles.
Maturité sexuelle
Les requins bouledogues ont une maturité tardive. Dans le sud du Golfe du Mexique, à
Veracruz et Campeche, l’âge à première maturité est estimé à 10 ans avec une taille moyenne
de 204 cm (LT) pour les femelles et entre 9 et 10 ans avec une taille moyenne de 190 cm (LT)
pour les mâles (Cruz-Martínez et al., 2004). L’étude menée dans le Nord du Golfe du
Mexique par Brandstetter et Stiles (1987) décrit une maturité encore plus tardive, 18 ans voir
plus avec une taille moyenne supérieure à 225 cm (LT) pour les femelles et entre 14 et 15 ans
pour une taille moyenne comprise entre 210 et 220 cm (LT) pour les mâles. A l’inverse, les
requins bouledogues présents dans le lac Nicaragua atteignent leur maturité plus rapidement.
Jenson (1976) décrit des individus matures, femelles comme mâles, ne dépassant pas 170 cm
(LT).
Taille maximale
Campagno (1984) a estimé la taille maximale des requins bouledogues à 340 cm (LT).
Lors de l’étude de Brandstetter et Stiles (1987), la plus grande femelle mesurait 268 cm à 24
ans et le plus grand mâle mesurait 245 cm à 21 ans. Cruz-Martínez et al. (2004) ont enregistré
des tailles maximales encore plus faibles mais correspondant à des âges plus avancés. La plus
grande femelle mesurait 256 cm (LT) à 28 ans et le plus grand mâle mesurait 243 cm (LT) à
23 ans. Les plus vieux individus ont été capturés en Afrique du Sud, une femelle de 32 ans
mesurant 221 cm (LT) et un mâle de 29 ans mesurant 216 cm (LT) (Wintner et al., 2002).
Cette limite d’âge et de taille n’a plus été contestée jusqu’en 2009. Cette année-là, une équipe
Sud-Africaine, cherchant à confirmer la présence de requin bouledogue dans l’estuaire du
Breede, a pêché une femelle de 400 cm (McCord and Lamberth, 2009). Cette première
mondiale a été confirmée par la découverte d’autres spécimens de plus de 4 mètres au Costa

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
30
Rica (com. pers.). Cette découverte laisse à penser que la longévité du requin bouledogue est
encore aujourd’hui sous-estimée.
Pour conclure, il semble que les caractéristiques biologiques des requins bouledogues
varient en fonction de leur localisation géographique. En effet, le taux de croissance entre
individu semble similaire au sein d’une même région mais varie d’une région à l’autre. Pour
expliquer cette différence, les auteurs émettent l’hypothèse du rôle majeur de l’habitat et de
ses caractéristiques dans le développement biologique et le comportement du requin
bouledogue. Si aucune étude sur la croissance et le développement n’est prévue dans
CHARC, il sera possible de définir une taille moyenne et maximale des requins bouledogues à
La Réunion. En comparant, en associant cette moyenne aux caractéristiques de l’habitat puis
en le comparant aux résultats des études précédentes, il sera possible d’apporter des
nouveaux éléments de compréhension de cette différence géographique et intra-spécifique des
caractéristiques biologiques de cette espèce.
5. Patterns de mouvements et utilisation de l’habitat
L’utilisation de l’habitat par les requins bouledogues a été beaucoup étudiée dans le
monde. Si les comportements des nouveaux nés, des jeunes de l’année et des juvéniles
commencent à être bien connus (Snelson et al., 1984; Neer et al., 2005; Pillans et al., 2005;
Simpfendorfer et al., 2005; Curtis, 2008; Heupel and Simpfendorfer, 2008; Heithaus et al.,
2009 ; Ortega et al., 2009), il existe très peu d’études sur le comportement des sub-adultes et
des adultes (McCord and Lamberth, 2009; Brunnschweiler et al., 2010; Carlson et al., 2010).
Saisonnalité et migration
Les requins bouledogues semblent avoir une fidélité au site très prononcée dans des
milieux côtiers ou semi-pélagiques, ponctuée par des migrations saisonnières.
La majorité des études menées sur le sujet a mis en œuvre la méthode de
marquage/recapture (roto-tag ou marque spaghettis), généralement associée à des
observations visuelles en plongée (Thorson, 1971; Kohler et al., 1998; Kohler and Turner,
2001; Curtis 2008). Une des premières études utilisant cette méthode a été réalisée par
Thorson (1971) entre le lac Nicaragua et le réseau de rivières de San Juan. Prêt de 2200
requins ont été marqués et 220 recapturés. C’est à l’heure actuelle la plus grande campagne de
marquage de requin bouledogue jamais réalisée. L’auteur a montré que la population de
requin du lac Nicaragua n’était pas isolée et qu’un requin pouvait traverser les 175 kilomètres
du réseau de rivières de San Juan pour rejoindre la mer des Caraïbes en un peu moins d’une
journée. Ainsi, les requins passeraient la majorité de l’année dans le lac mais effectueraient
ponctuellement des allers-retours dans la mer des Caraïbes. Thorson (1971) émet alors
l’hypothèse d’une grande fidélité de cette espèce au site malgré leur capacité à faire des
déplacements sur de grande distance. A Tampa Bay, à l’Ouest de la Floride, des jeunes
requins bouledogue de l’année (YOY) ont été recapturés à moins de 5 km du site de marquage

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
31
après plus de 113 jours (Hueter and Manire, 1984). En Afrique du sud, des requins
bouledogues juvéniles ont été observés sur le site de marquage plus d’un an après (Bullen and
Mann, 2000). Ces deux dernières études confirment la forte fidélité des requins bouledogues
au site, comme suggéré par Thorson (1971).
Les distances maximales parcourues par les requins bouledogues capturés sur les côtes
Est/Sud-Est des Etats-Unis sont faibles, n’excédant pas 235 kilomètres selon Kolher et al.
(1998) et 643 kilomètres selon Kolher et Turner (2001). En comparaison, les résultats
d’études similaires sur des requins babosses (Carcharhinus altimus), des requins gris
(Carcharhinus plombeus) ou des requins tigres (Galeocerdo cuvier) ont montré des distances
maximum parcourues respectivement de 3343 kilomètres, 3776 kilomètres et 6747 kilomètres
(Kolher and Turner, 2001). Il apparait donc que les mouvements migratoires des requins
bouledogues sont relativement restreints en comparaison avec d’autres espèces. Néanmoins,
la technique de marquage-recapture ne donne qu’une image instantanée de la localisation
temporelle et spatiale des requins marqués (Heupel et al., 2007).
Il faudra attendre 2008 pour que la première étude utilisant la télémétrie acoustique voit le
jour. En Floride, à Pine Island Sound, Yeiser et al. (2008) ont déployé un réseau passif
d’écoute acoustique pour suivre des requins bouledogues sub-adultes. Les individus marqués
sont restés dans une zone restreinte pendant la majorité du suivi (89 jours) avec des courtes
périodes d’absence. Les requins auraient quitté cette zone côtière préférentielle pour aller dans
des zones pélagiques. Bien que ces nouveaux résultats confirment une haute fidélité au site,
les déplacements des requins n’ont pu être étudiés qu’à l’échelle d’une île et ne donnent pas
encore d’idée précise sur le comportement migratoire de cette espèce.
En 2010, J.K. Carlson et son équipe ont posé les premières balises satellites sur des
requins bouledogues dans la partie Nord du Golfe du Mexique (Carlson et al., 2010). Dix-huit
individus sub-adultes ont été marqués en 2 ans avec des balises programmées pour enregistrer
leurs mouvements pendant 90 à 120 jours. Bien que la majorité des requins marqués a limité
ses déplacements aux zones proches de celles du lieu de marquage, 2 individus marqués en
septembre à Tampa sur la côte Ouest de la Floride, ont effectué des déplacements beaucoup
plus importants. Un des deux requins a migré de la Floride au Texas en 85 jours en passant
par des eaux plus profondes au large du Golfe. Bien qu’aucun pattern journalier n’ait pu être
mis en évidence, l’individu a effectué de nombreuses plongées en profondeur avec un
maximum enregistré de 128 mètres. Le deuxième requin est parti de l’Ouest de La Floride, a
longé la côte jusqu’à la pointe Sud puis est remonté le long de la côte Est des Etats-Unis
jusqu’en Caroline du Sud où il est arrivé 30 jours plus tard. Il n’a quitté les eaux côtières
qu’après avoir dépassé le Nord de la Floride où il a commencé à migrer vers le large. A la
différence du premier requin, il a nagé à des profondeurs ne dépassant pas 30 mètres. Les
résultats de cette première étude utilisant le suivi satellite confirme la fidélité du requin
bouledogue au site mais met en évidence des comportements migratoires nouveaux. Les
auteurs émettent l’hypothèse d’une migration hivernale et ainsi une utilisation de l’habitat qui
diffère entre les saisons. Ils suggèrent également que les requins bouledogues ont un

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
32
changement ontogénique dans leur pattern de mouvement avec une augmentation de la taille
de leur territoire et de leur déplacement migratoire en relation avec leur croissance.
Une étude similaire menée aux Fidji et aux Bahamas sur des requins bouledogues adultes
n’a cependant pas pu confirmer ces hypothèses (Brunnschweiler et al., 2010). Les requins
marqués avaient des déplacements très limités dans l’espace et le temps. Seule une femelle a
effectué un déplacement plus important. Marquée aux Bahamas, elle a été suivie jusque dans
les lagons de l’Indian River en Floride. Cette zone étant considérée comme une nurserie pour
cette espèce (Curtis, 2008), il se pourrait que les femelles requins bouledogues des Bahamas
migrent en Floride pour mettre bas.
Ainsi, les comportements migratoire et saisonnier des requins bouledogues restent encore
mal connus et nécessitent d’avantage d’études. A l’heure actuelle, il est bien admis que cette
espèce montre une grande fidélité au site, particulièrement les nouveaux nés, les jeunes de
l’année et les juvéniles. Les comportements des sub-adultes et des adultes apparaissent plus
complexes à définir. Les différentes études ont montré que si ces derniers passent la majorité
de l’année au même endroit, durant l’hiver beaucoup disparaissent pour réapparaitre 3 mois
plus tard. Ces trois mois d’absence ne sont toujours pas expliqués et nous ne savons pas où et
pourquoi ces individus quittent leur zone habituelle. Les dernières études utilisant les suivis
satellites commencent à donner les premiers éléments de réponse. L’hypothèse acceptée à
l’heure actuelle serait que les requins bouledogues en âge de se reproduire migrent pendant
l’hiver dans d’autres régions du monde pour l’accouplement ou la mise bas. L’utilisation de
balises satellites devrait permettre au programme CHARC de confirmer ou d’infirmer cette
hypothèse concernant les requins bouledogue des eaux réunionnaises.
Si le comportement des requins bouledogues pendant les périodes d’absence reste mal
connu, l’utilisation de l’habitat et des mouvements à court terme est, lui, beaucoup mieux
compris.
Utilisation de l’habitat et mouvements à court terme
Le requin bouledogue est une espèce côtière et semi-pélagique (Brunnschweiler et al.,
2010). Il a longtemps été décrit comme vivant dans les eaux peu profondes à proximité des
côtes et dans les estuaires à des profondeurs dépassant rarement les 30 mètres (Campagno,
1984). La majorité des études portant sur l’habitat du requin bouledogue, s’est intéressé aux
comportements d’individus immatures souvent observés à proximité des estuaires. En Floride,
les populations de requins bouledogues juvéniles, recensés par l’intermédiaire de pêches et/ou
d’observations visuelles sous-marine, ont évoluées le plus souvent à des profondeurs
n’excédant pas 10 à 5 mètres (Simpfendorfer et al., 2005; Curtis, 2008). L’utilisation de suivi
acoustique (VEMCO V13P et VR100) mesurant la profondeur de nage des requins marqués
confirme ces résultats, avec des profondeurs moyennes de 1 mètre en dessous de la surface
(Ortega et al., 2009). Cependant, il s’agissait d’individus immatures.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
33
La première étude sur l’habitat de requin bouledogue adulte a été menée par
Bunnschweiler et Buskirk en 2006 à Walker’s Cay (Bahamas). Six individus adultes ont été
équipés de balises satellites pop-up et ont été suivis entre 4 et 24 jours. Bien que le pattern de
mouvements est proche de celui des juvéniles avec 80% du temps passés dans des eaux peu
profondes (< 20 mètres), deux individus se sont distingués. Ils ont temporairement quitté
Walker’s Cay et ont nagé à une profondeur moyenne de 25 mètres. Durant cette période, ils
ont effectué régulièrement des plongées entre 50 et 90 mètres, avec un record à 140 mètres
pour l’un d’entre eux. C’est la première fois que l’on a observé un requin bouledogue à ces
profondeurs, ce qui a remis en cause son affinité exclusive à des zones côtières peu profondes.
La même année, le suivi acoustique de requins bouledogues adultes dans la Réserve Naturelle
de Shark Reef aux îles Fidji, a mis en évidence que les individus passaient la majorité de leur
temps à des profondeurs supérieures à 30 mètres (Bunnschweiler, 2009). Cette différence
d’habitat sera confirmée par une étude comparative des populations de requins bouledogues
aux Bahamas et aux Fidji utilisant des balises satellites (Bunnschweiler et al., 2010). Bien que
les deux populations de requins adultes ont été capables de sonder à des profondeurs
supérieures à 100 mètres (avec des records à plus de 200 mètres), les requins des Bahamas
sont restés majoritairement à des profondeurs supérieures à 30 mètres alors que les requins
fidjiens étaient essentiellement entre 30 et 100 mètres.
Ainsi, l’habitat des requins bouledogues juvéniles est défini par une zone côtière peu
profonde et/ou dans un estuaire où ils restent jusqu’à l’âge adulte (âge à première maturité),
habitat que l’on nommera par la suite « nurserie ». L’habitat des requins adultes est beaucoup
moins bien défini dans la littérature. Alors qu’il est essentiellement côtier dans certaines
régions, il est semi-pélagique dans d’autres. Lors de l’analyse comparative des requins adultes
des iles Bahamas et Fidji (Bunnschweiler et al., 2010), l’étude de la structure des fonds sous-
marins a montré qu’à Walker’s Cay, les eaux environnantes restent peu profondes sur
plusieurs kilomètres alors qu’aux Iles Fidji, les fonds tombent rapidement à plus de 250
mètres. Les auteurs concluent que la répartition des requins adultes dépend majoritairement de
la structure bathymétrique des fonds de l’habitat qu’ils occupent. Enfin, il faut noter l’absence
de donnée sur l’habitat des sub-adultes même s’il est pressenti qu’ils utilisent majoritairement
les zones côtières très peu profondes et qu’ils migrent progressivement vers des zones plus
profondes voir semi-pélagique, au fur et à mesure qu’ils atteignent leur taille adulte.
Si la bathymétrie joue un rôle important dans l’utilisation de l’habitat par les requins
bouledogues, trois grands autres facteurs ont été mis en évidence, la température de l’eau, la
salinité et la turbidité.
Les requins bouledogues semblent évoluer dans des eaux entre 20°C et 32°C en moyenne
(Simpfendorfer et al. 2005; Bunnschweiler and Buskirk, 2006; Curtis, 2008; Carlson et al.,
2010; Brunnschweiler et al., 2010). La température minimale et maximale de l’eau
enregistrées sont de 12°C et 37°C. Néanmoins, les individus observés à ces deux extrêmes de
températures étaient soit maigres soit faibles (Curtis, 2008; Carlson et al., 2010). Comme la
majorité des requins, les requins bouledogues sont ectothermiques, c'est-à-dire que la

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
34
température de leurs corps est la même que celle du milieu extérieur, ce qui pourrait expliquer
cette faible tolérance à des températures extrêmes. De plus, selon les régions concernées, cette
espèce semble préférer des intervalles de température encore plus spécifiques. En Afrique du
Sud, les requins semblent évoluer principalement dans des eaux entre 20 et 24°C (McCord
and Lamberth, 2009). A Walker’s Cay au Bahamas, les requins passaient plus de 72% du
temps dans des eaux à 25-26°C (Bunnschweiler and Buskirk, 2006; Brunnschweiler et al.,
2010). Aux Fidji, ils préféraient les eaux dont la température dépassait les 26°C
(Brunnschweiler et al., 2010) et dans le Nord du Golfe du Mexique, l’intervalle de
température était compris entre 30 et 32°C (Curtis, 2008; Carlson et al., 2010). Cette
préférence à des intervalles de température restreints montre l’importance de ce facteur dans
l’utilisation de l’habitat par les requins bouledogues. Dans les zones tempérées, où la
température de l’eau varie en fonction des saisons, ce facteur jouerait même un rôle majeur
dans le cycle de vie de ces animaux. Castro (1993) émet l’hypothèse qu’en Caroline du Sud,
les eaux froides de l’hiver forcent les requins bouledogues juvéniles à quitter les eaux côtières
refroidies par l’apport d’eaux froides du continent, pour les eaux plus chaudes du large. Ces
individus ne reviendraient qu’au printemps une fois que les eaux côtières se sont réchauffées.
On retrouve ce même comportement d’évitement chez les requins bouledogues juvéniles en
Floride (Hueter and Tyminski 2002; Wiley and Simpfendorfer 2007). Pendant longtemps, on
a cru que l’absence de requins bouledogues adultes en hiver dans la plupart des régions du
monde était due à une période d’hibernation. La baisse de température de l’eau et donc de leur
température corporelle diminuerait leur métabolisme et leur activité jusqu’à un état proche de
l’hibernation. Cette hypothèse est maintenant abandonnée par la plupart des scientifiques
même si l’on ne sait toujours pas où vont les requins bouledogues adultes pendant l’hiver
(Curtis, 2008 ; Carlson et al., 2010). La même hypothèse avait été formulée concernant
l’absence d’observation de requin pèlerin (Cethorinus maximus) en hiver sur les côtes Nord-
Ouest de l’Europe. En 2009, la découverte d’une migration trans-équatorial par des requins
pèlerins équipés de balises satellites a infirmé cette théorie et a donné des premiers éléments
d’explication sur le comportement hivernal de ces requins (Skomal et al., 2009).
Le programme CHARC mettra en place une zone test de suivi acoustique durant l’hiver
2012 afin de collecter les premières données comportementales des requins bouledogues à La
Réunion durant cette période. De plus, la pause de balises satellites sur des requins
bouledogues adultes pourrait apporter des éléments de réponse sur l’éventuel comportement
migratoire de cette espèce pendant l’hiver.
L’utilisation de l’habitat par les requins bouledogues est aussi influencée par la salinité de
l’eau. Ils peuvent être observés en eau douce, en eau saumâtre et en eau salée (voir en milieu
hypersalé). Leur présence en eaux saumâtres ou en eaux douces semble être liée à leur
capacité d’osmorégulation. En effet, ce mécanisme par lesquels les organismes effectuent le
contrôle de la pression osmotique interne, a un coût énergétique important en milieu salé
(Kidder et al., 2006). Ce coût énergétique serait suffisamment important pour influencer le
comportement des animaux. Les requins bouledogues, bien que capables de faire cette

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
35
osmorégulation dans des conditions de salinité très variables (Pillans and Franklin, 2004;
Pillans et al., 2005; Pillans et al., 2006) semblent rester préférentiellement dans des eaux avec
une salinité comprise entre 7 à 29‰ et éviter les eaux où la salinité est inférieure à 7‰
(Heupel and Simpfendorfer, 2008; Curtis, 2008). Les nouveau nés et les jeunes de l’année
tendent à être dans des milieux à plus faible salinité que les juvéniles, les sub-adultes et les
adultes (Simpfendorfer et al., 2005; Curtis, 2008). Les différents auteurs expliquent cette
ségrégation en fonction de la taille et de la salinité, par le besoin des plus jeunes individus à
éviter la prédation des individus plus âgés et/ou l’avantage physiologique d’évoluer dans des
milieux saumâtre pour réduire le coût de l’osmorégulation. L’évitement de la prédation
associé à la capacité des requins bouledogues à osmoréguler dans des milieux saumâtres voir
d’eaux douces confèreraient un avantage pour leur survie en comparaison à des espèces moins
euryhaline. Il faut toutefois noter que Simpfendorfer et al. (2005) ont observé que lorsque
l’apport d’eau douce dans le Caloosahatchee River était trop important, les requins quittent la
rivière pour aller dans des eaux plus salées. Cette dernière observation suggère que les
mouvements à court terme des nouveaux nés et des jeunes de l’année sont plus influencés par
la salinité que par le risque de prédation. Néanmoins, il est nécessaire de faire d’autres études
pour comprendre plus en détail le rôle de la salinité dans la distribution et la ségrégation
spatio-temporelle des requins bouledogues.
Au cours du programme CHARC, les mouvements des requins bouledogues seront
associés aux paramètres abiotiques et biotiques pour étudier l’influence et l’importance de
ces différents paramètres sur le comportement des individus suivis. La présence de requins de
maturité différentes et la variabilité de la salinité dans la zone étudiée devrait permettre de
tester l’hypothèse avancer par Simpfendorfer et al. (2005).
Bien qu’il soit admis que les requins bouledogues évoluent préférentiellement dans des
eaux à forte turbidité, très peu d’études ont été réalisées sur l’influence de ce facteur. En
Afrique du Sud, un requin bouledogue adulte suivi acoustiquement est resté dans des eaux
légèrement trouble pendant les heures de la journée (15 < Nephelometric Turbidity Unit < 20)
alors que durant la nuit et au levée du soleil, il a évolué dans des eaux claires (NTU<5)
(McCord and Lamberth, 2009). Ortega et al. (2009) ont aussi trouvé une influence de la
turbidité sur le comportement de requins bouledogues juvéniles dans le Caloosahatchee River.
Si la turbidité semble être un facteur influençant le comportement des requins bouledogues, il
existe très peu d’informations permettant d’évaluer son importance en comparaison avec
d’autres facteurs comme la profondeur, la température de l’eau ou la salinité.
En résumé, l’utilisation de l’habitat par les requins bouledogues est le résultat de
l’influence de plusieurs facteurs qu’ils soient physique, physiologique ou comportemental. De
plus, comme chez beaucoup d’espèces de requins, cette utilisation de l’habitat connait un
changement ontogénique : les nouveaux nés et les jeunes de l’année naissent et restent
pendant au moins un an dans les zones côtières à proximité d’un estuaire, zone appelée
« nurserie ». En devenant sub-adultes, ils migrent progressivement vers des habitats moins

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
36
côtiers où ils finiront par rejoindre la population d’individus adultes. A ce stade, ils resteront
la plupart de leur vie au même endroit migrant saisonnièrement dans d’autres régions du
monde pour s’accoupler et reviendront pour mettre bas. Si ce scénario est bien accepté, il
manque cruellement de validation scientifique. Les zones de nurseries sont assez bien connues
mais le niveau de connaissances sur les habitats des sub-adultes et des adultes reste encore
faible et limité à certaines régions du monde. Ainsi, il est nécessaire de continuer les études
dans ce domaine pour mieux comprendre l’écologie comportementale de cette espèce. A La
Réunion, les résultats de la première campagne de marquage acoustique sur cette espèce
réalisée par Squal’Idée et l’IRD donne les premiers informations sur l’utilisation de l’habitat
par des individus adultes et sub-adultes (Blaison, 2012). Cependant, ces données sont limitées
dans le temps et l’espace (3 mois de suivis entre la Baie de Saint-Paul et Les Brisants) et
basées sur un nombre de requins marqués réduit. Le but du programme CHARC sera donc
d’augmenter le nombre de requins marqués, et d’étendre les suivis à l’ensemble de la côte
Ouest sur au moins un an. En associant ces suivis aux paramètres environnementaux, il sera
alors possible de définir l’utilisation de l’habitat des requins bouledogues à l’échelle d’une
île selon les périodes de l’année et leur stade de maturité. Il s’agira de mettre en évidence
l’existence ou non de micro-habitat, telle que les nurseries, et de les décrire. A La Réunion, la
présence d’individus adultes, sub-adultes et juvéniles dans une zone restreinte donne
l’opportunité d’étudier l’évolution de l’utilisation de l’habitat des requins bouledogues dès
leurs plus jeunes âges.
6. Bibliographie
Bass, A.J., D’Aubrey, J.D. and Kistnasamy, N. (1973). Sharks of the east coast of southern
Africa. I. The genus Carcharhinus (Carcharhinidae). Invest. Rep. Oceanogr. Res. Inst.
Durban, 33. 168 pp.
Bigelow, H. B., and W. C. Schroeder. (1948). Sharks. In J. Tee-Van, C. M. Breder, S. F.
Hildebrand, A. E. Parr, and W. C. Schroeder (eds.). Fishes of the Western North
Atlantic, part one, vol. 1, p. 59–546. Mem. Sears Found. Mar. Res., Yale Univ.
Bigelow, H. B., and Schroeder, W. C. (1961). Carcharhinus Nicaraguensis , a Synonym of
the Bull Shark , C . Leucas. Copeia, (3), 511.
Bigelow, H. B., and Schroeder, W. C. (1976). Investigations of the Ichthyofauna of
Nicaraguan Lakes. in Investigations of the Ichtyofauna of Nicaraguan lakes, ed. Thomas
B. Thorson (University of Nebraska-Lincoln, 1976). Copyright © 1976 School of Life
Sciences, University of Nebraska-Lincoln.
Blainville, H. M. (1816). Prodrome d'une nouvelle distribution systématique
du règne animal. Bull. Soc. Philomath. Paris, 3 (3), 113-124
Blaison, A. (2012). Bilan de la première campagne de marquage de requins bouledogues
(Carcharhinus leucas) et de requins tigres (Galeocerdo cuvier) sur la côte Ouest de La
Réunion. Rapport Squal’Idées/DEAL. 40p

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
37
Branstetter, S., and Stiles, R. (1987). Age and growth estimates of the bull shark ,
Carcharhinus leucas , from the northern Gulf of Mexico. Environmental Biology of
Fishes, 20(3), 169-181.
Bunnschweiler, J and Buskirk, J. V. (2006). Satellite tagging of bull shark at Walker’s Cay.
Bahamas Naturalist & Journal of Science, 1(1), 30-34.
Brunnschweiler, J. M., and Earle, J. L. (2006). A contribution to marine life conservation
efforts in the South Pacific�: The Shark Reef Marine Reserve , Fiji by. Cybium, 30(4),
133-139.
Brunnschweiler, J. M. (2009). Tracking free-ranging sharks with hand-fed intra-gastric
acoustic transmitters. Marine and Freshwater Behaviour and Physiology, 42(3), 201-
209. doi:10.1080/10236240903033519
Brunnschweiler, J. M., Queiroz, N., and Sims, D. W. (2010). Oceans apart? Short-term
movements and behaviour of adult bull sharks Carcharhinus leucas in Atlantic and
Pacific Oceans determined from pop-off satellite archival tagging. Journal of fish
biology, 77, 1343-58. doi:10.1111/j.1095-8649.2010.02757.x
Brunnschweiler, J. M., and Baensch, H. (2011). Seasonal and Long-Term Changes in
Relative Abundance of Bull Sharks from a Tourist Shark Feeding Site in Fiji. (B.
Gratwicke, Ed.) PLoS ONE, 6(1), 5. Public Library of Science. Retrieved from
http://dx.plos.org/10.1371/journal.pone.0016597
Bullen, E. and Mann, B.Q. (2000). Sedgewick’s/ORI/WWF Tagging Programme: Zambezi
shark (Carcharhinus leucas). Data Report, Oceanographic Research Institute 2000(11). 6
pp.
Carlson, J. K., Ribera, M. M., Conrath, C. L., Heupel, M. R., and Burgess, G. H. (2010).
Habitat use and movement patterns of bull sharks Carcharhinus leucas determined using
pop-up satellite archival tags. Journal of fish biology, 77, 661-75. doi:10.1111/j.1095-
8649.2010.02707.x
Castro, J. I. (1993). The shark nursery of Bulls Bay , South Carolina , with a review of the
shark nurseries of the southeastern coast of the United States. Environmental Biology of
Fishes, 38, 37-48.
Castro and Aguirre, J.L. (1978). Catalogo sistematico de 1os peces marinos que penetran
alas aguas continentales de Mexico con aspectos zoogeograficos y ecol6gicos.
Direcci6n General del Instituto Nacional de Pesca, Serie Cientifica 19, 1-298
César de Olieveira Santos, M. and Bismarck Fazzano Gadig, O. (2008). Evidence of a
failed predation attempt of a bull shark (Carcharinhus leucas) on a Guiana dolphin
(Sotalia guianensis). Arquivos de Ciências do Mar, 43(1), 1-6.
Cliff, G., and Dudley, S. F. J. (1991). Sharks caught in the protective gill nets off Natal,
South Africa. 4. The bull shark Carcharhinus leucas (Valenciennes). South African
Journal of Marine Science, 10, 253-70.
Cliff, G., and Dudley, S. F. J. (1992). Protection against Shark Attack in South Africa ,
1952-90. Aust. J. Mar. Freshwater Res., (43), 263-272

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
38
Coad, B. W., & Papahr, F. (1988). Shark attacks in the rivers of southern Iran.
Environmental Biology of Fishes, 23(1-2), 131-134.
Compagno, L.J.V., (1984). Sharks of the world. Part 1, Hexanchiformes to Lamniformes.
FAO Species Catalogue 4, FAO Fish. Synop. 125, Food and Agriculture Organisation
of the United Nations, Rome, 249 p.
Cruz-Martínez, A., Chiappa-Carrara, X., and Arenas-Fuentes, V. (2004). Age and
Growth of the Bull Shark, Carcharhinus leucas, from Southern Gulf of Mexico. Journal
of Northwest Atlantic Fishery Science, 35, 367-374. doi:10.2960/J.v35.m481
Curtis, T. H. (2008). Distribution, Movements and Habitat use of Bull Sharks (Carcharhinus
leucas, Müller and Henle 1839) in the Indian River lagoon system, Florida. Wildlife
Conservation (p. 130).
Dodrill, J.W. (1977). A hook and line survey of the sharks of Melbourne Beach, Brevard
County, Florida. Masters Thesis, Florida Institute of Technology, Melbourne.
Dudley, S.F.J and Simpfendorfer, C.A. (2006). Population status of 14 shark species caught
in the protective gillnets off KwaZulu-Natal beaches, South Africa, 1978-2003. Marine
and Freshwater Research, 57, 225-240.
Heithaus, M., Frid, A., and Dill, L. (2002). Shark-inflicted injury frequencies, escape
ability, and habitat use of green and loggerhead turtles. Marine Biology, 140, 229-236.
doi:10.1007/s00227-001-0712-6
Heithaus, M. R., Delius, B. K., Wirsing, A. J., and Dunphydaly, M. M. (2009). Physical
factors influencing the distribution of a top predator in a subtropical oligotrophic estuary.
Limnol. Oceanogr., 54(2), 472-482.
Heupel, M. R., Carlson, J. K. and Simpfendorfer, C. A. (2007). Shark nursery areas:
concepts, definition, characterization and assumptions. Mar Ecol Prog Ser, 337, 287-297
Heupel, M. R. and Simpfendorfer, C. A. (2008). Movements and distribution of young bull
sharks (Carcharhinus leucas) in a variable estuarine environment. Aquat Biol, 1, 277–
289. doi:10.3354/ab00030
Hueter, R. E. and Manire, C. A. (1994). Bycatch and catch-release mortality of small sharks
in the Gulf coast nursery grounds of Tampa Bay and Charlotte Harbor. Mote Marine
Laboratory Tech Rep No 368. 183 pp.
Hueter, R. E. and Tyminski, J.P. (2002). Center for Shark Research (CSR) U.S. shark
nursery research overview 1991-2001. Pages 116-135 in McCandless CT, Pratt HL Jr.,
Kohler NE (eds). Shark nursery grounds of the Gulf of Mexico and the east coast waters
of the United States: an overview. NMFS Internal Report to NOAA’s Highly Migratory
Species Office.
ITIS (2012). Integrated Taxonomic Information System.
http://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=160
275
Jenson, N. H. (1976). Reproduction of the Bull Shark, Carcharhinus leucas, in the Lake
Nicaragua-Rio San Juan System. in Investigations of the Ichthyofauna of

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
39
NicarNVESTIGATIONS OF THE ICHTHYOFAUNA OF NICARAGUAN LAKES, ed.
Thomas B. Thorson (University of Nebraska-Lincoln, 1976). Copyright © 1976 School
of Life Sciences, University of Nebraska-Lincoln, 539-559.
Kidder, G. W., Petersen, C. W. and Preston R. L. (2006). Energetics of osmoregulation: II.
water flux and osmoregulatory work in the euryhaline fish, Fundulus heteroclitus. J Exp
Zool Part A Comp Exp Biol, 305A:318–327. doi:10.1002/jez.a.252
Kohler, N. E., Casey, J. G., and Turner, P. A. (1998). NMFS Cooperative SharkTagging
Program, 1962-93: An Atlas of Shark Tag and Recapture Data. Marine Fisheries Review,
60(2), 1-87.
Kohler, N. E. and Turner, P. A. (2001). Shark tagging: a review of conventional methods
and studies. Environ. Biol. Fish., 60, 191-223.
Last, P.R. and Stevens, J.D. (1994). Sharks and Rays of Australia CSIRO, Melbourne,
Australia. 513 pp.
McCord, M., and Lamberth, S. (2009). Catching and tracking the world’s largest Zambezi
(bull) shark Carcharhinus leucas in the Breede Estuary, South Africa: the first 43 hours.
African Journal of Marine Science, 31(1), 107-111. doi:10.2989/AJMS.2009.31.1.11.782
Menni, R. C.and Lucifora, L. O. (2007). CONDRICTIOS DE LA ARGENTINA Y
URUGUAY (p. 18).
Montoya, R. V., and Thorson, T. B. (1982). The bull shark (Carcharhinus leucas) and
largetooth sawfiish (Pristis perotteti) in Lake Bayano , a tropical man-made
impoundment in Panama. Environmental Biology of Fishes, 7(4), 341-347.
Myers. G. S. (1952). Sharks and sawfishes in the Amazon. Copeia, 268-269.
Natanson, L. J., Mello, J. J., and Campana, S. E. (2002). Validated age and growth of the
porbeagle shark, Lamna nasus, in the western North Atlantic Ocean. Fisheries Bulletin,
100, 266–278.
Neer, J. A., Thompson, B. A. and Carlson, J. K. (2005). Age and growth of the bull shark,
Carcharhinus leucas, in the northern Gulf of Mexico: incorporating variability in size at
birth. Journal of Fish Biology, 67, 370–383.
O’Connell, M. T, Sheperd, T. D., O'Connell, A. M. U. and Myers, R. A. (2007). Long-
term declines in two apex predators Bull Sharks and Alligator Gar, in Lake
Pontchartrain, an Oligohaline Estuary in Southern Louisana. Estuaries and Coasts,
30(4), 567-574.
Ortega, L. A., Heupel, M. R., Beynen, P. V., and Motta, P. J. (2009). Movement patterns
and water quality preferences of juvenile bull sharks (Carcharhinus leucas) in a Florida
estuary. Environmental Biology of Fishes, 84, 361-373. doi:10.1007/s10641-009-9442-2
Pillans, R. D., and Franklin, C. E. (2004). Plasma osmolyte concentrations and rectal gland
mass of bull sharks Carcharhinus leucas, captured along a salinity gradient.
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology,
138, 363-71. doi:10.1016/j.cbpb.2004.05.006

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
40
Pillans, R. D., Good, J. P., Anderson, W. G., Hazon, N., and Franklin, C. E. (2005).
Freshwater to seawater acclimation of juvenile bull sharks (Carcharhinus leucas):
plasma osmolytes and Na+/K+-ATPase activity in gill, rectal gland, kidney and intestine.
Journal of comparative physiology. B, Biochemical, systemic, and environmental
physiology, 175, 37-44. doi:10.1007/s00360-004-0460-2
Pillans, R. D., Anderson, W. G., Good, J. P., Hyodo, S., Takei, Y., Hazon, N., and
Franklin, C. E. (2006). Plasma and erythrocyte solute properties of juvenile bull sharks,
Carcharhinus leucas, acutely exposed to increasing environmental salinity. Journal of
Experimental Marine Biology and Ecology, 331, 145-157.
doi:10.1016/j.jembe.2005.10.013
Pratt, H. L., & Carrier, J. C. (2001). A review of elasmobranch reproductive behavior with
a case study on the nurse shark , Ginglymostoma cirratum. Environmental Biology of
Fishes, (60), 157-188.
Price, K. S. Jr. and Daiber, F. C. (1967). Osmotic environments during fetal development of
dogfish, Mustelus canis (Mitchill) and Squalus acanthias Linnaeus, and some
comparisons with skates and rays. Physiol. Zoot. 40, 248-260.
Rodriguez de la Cruz, C., Castillo- Géniz, J. L. and Marquez – Farias, F. (1996).
Evaluación de la Pesquería de Tiburón en el Golfo de México. Consejo Nacional de
Ciencia y Tecnología. Informe final del proyecto de Investigación 116002–5–
1314N9206, 199 p.
Sadowsky, V. (1971). Notes on the Bull Shark, Carcharhinus leucas, in the lagoon region of
Canania, Brazil. Bolm. Inst. oceanogr. S Paulo, 20(2), 71-78.
Schmid, T. H., Murru, F. L. and McDonald, F. (1990). Feeding habits and growth rates of
bull (Carcharhinus leucas (Valenciennes)), sandbar (Carcharhinus plumbeus (Nardo)),
sandtiger (Eugomphodus taurus (Rafinesque)), and nurse (Ginglymostoma cirratum
(Bonnaterre)) sharks maintained in captivity. J. Aquac. Aquat. Sci. 5, 100–105.
Schmid, T. H. and Murru, F. L. (1994). Bioenergetics of the bull shark, Carcharhinus
leucas, maintained in captivity. Zoo Biology, 2(13), 177–185
Skomal, G. B., Zeeman, S. I., Chisholm, J. H., Summers, E. L., Walsh, H. J., McMahon, K.
W. and Thorrold, S. R. (2009). Transequatorial migrations by basking sharks in the
western Atlantic Ocean. Current Biology, 19, 1019–1022.
Simpfendorfer, C. A., Freitas, G. G., Wiley, T. R. and Heupel, M. R. (2005). Distribution
and Habitat Partitioning of Immature Bull Sharks ( Carcharhinus leucas ) in a Southwest
Florida Estuary. Estuaries, 28(1), 78-85.
Snelson, F. F., Mulligan, T. J. and Williams, S. E. (1984). Food habits, occurrence and
population structure of the Bull Shark, Carcharhinus leucas, in Florida coastal lagoons.
Bulletin of Marine Science, 34(1), 71-80.
Stevens, J. D. and McLoughlin, K. J. (1991). Distribution, size, and sex composition,
reproductive biology and diet of sharks from northern Australia. Aust. J. Mar.
Freshwater Res., 40:129–146.

Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
41
Thomerson, J. E., Thorson, T. B. and Hempel, R. L. (1977). The bull shark, Carcharhinus
leucas, from the upper Mississippi River near Alton, Illinois. Copeia, 166-168.
Thorson, T. B. (1971). Movement of Bull Sharks , Carcharhinus leucas , Between Carribean
Sea and Lake Nicaragua Demonstrated by Tagging. Copeia, 2, 336-338.
Trape, S. (2008). Shark Attacks in Dakar and the Cap Vert Peninsula, Senegal: Low
Incidence despite High Occurrence of Potentially Dangerous Species. (W. Cresswell,
Ed.) PLoS ONE, 3(1), 1-6.
Tuma, R. E. (1976). An Investigation of the Feeding Habits of the Bull Shark , Carcharhinus
leucas , in the Lake Nicaragua- Rio San Juan System. in Investigation of the
Ichthyofauna of Nicaraguan lakes, ed. Thomas B. Thorson (University of Nebraska-
Lincoln, 1976). Copyright © 1976 School of Life Sciences, University of Nebraska-
Lincoln., 533-538.
Wintner, S. P., Dudley, S. F. J., Kistnasamy, N. and Everett, B. (2002). Age and growth
estimates for the Zambezi shark, Cacharhinus leucas, from the east coast of South
Africa. Mar Freshwater Res, 53, 557–566
Wiley, T. R. and Simpfendorfer, C. A. (2007). The ecology of elasmobranchs occurring in
the Everglades National Park, Florida: Implications for conservation and management.
Bull. Mar. Sci. 80: 171–189.
Yeiser, B. G., Heupel, M. R. and Simpfendorfer, C. A. (2008). Occurrence, home range
and movement patterns of juvenile bull (Carcharhinus leucas) and lemon (Negaprion
brevi- rostris) sharks within a Florida estuary. Marine and Freshwater Research, 59,
489–501.
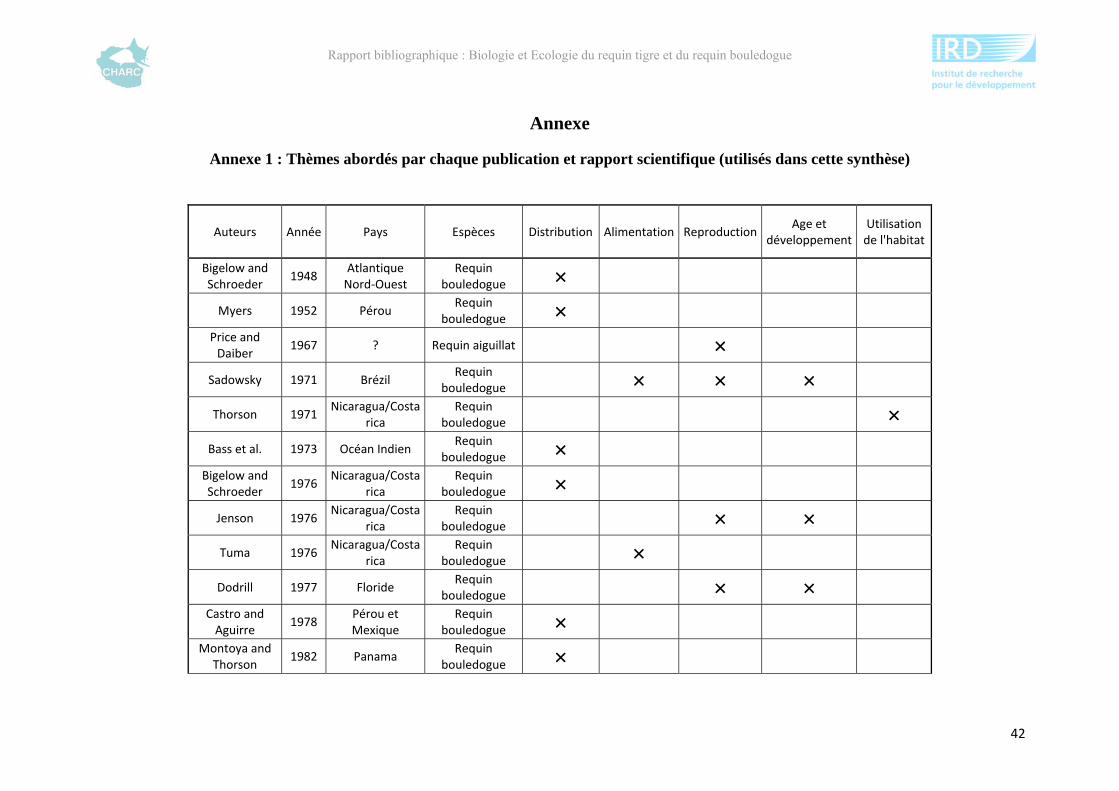
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
42
Annexe
Annexe 1 : Thèmes abordés par chaque publication et rapport scientifique (utilisés dans cette synthèse)
Auteurs Année Pays Espèces Distribution Alimentation ReproductionAge et
développementUtilisation de l'habitat
Bigelow and Schroeder
1948 Atlantique Nord‐Ouest
Requin bouledogue ×
Myers 1952 Pérou Requin
bouledogue ×
Price and Daiber
1967 ? Requin aiguillat ×
Sadowsky 1971 Brézil Requin
bouledogue × × ×
Thorson 1971 Nicaragua/Costa
rica Requin
bouledogue ×
Bass et al. 1973 Océan Indien Requin
bouledogue ×
Bigelow and Schroeder
1976 Nicaragua/Costa
rica Requin
bouledogue ×
Jenson 1976 Nicaragua/Costa
rica Requin
bouledogue × ×
Tuma 1976 Nicaragua/Costa
rica Requin
bouledogue ×
Dodrill 1977 Floride Requin
bouledogue × ×
Castro and Aguirre
1978 Pérou et Mexique
Requin bouledogue ×
Montoya and Thorson
1982 Panama Requin
bouledogue ×
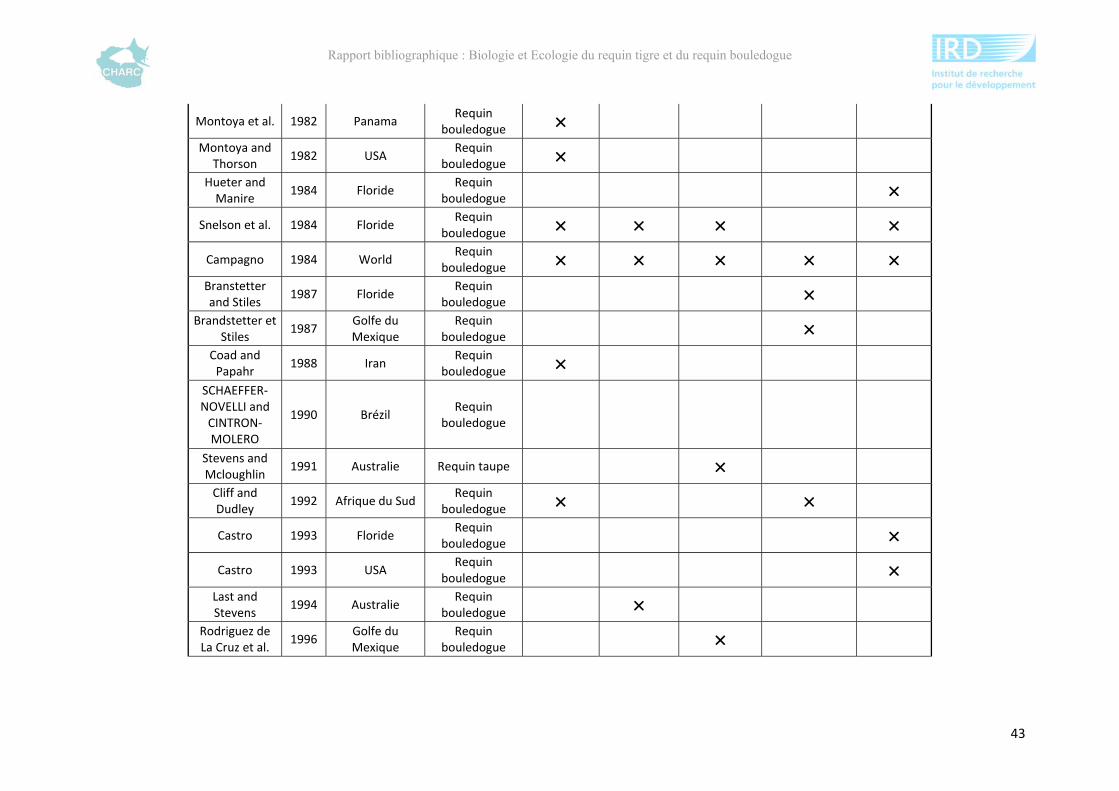
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
43
Montoya et al. 1982 Panama Requin
bouledogue ×
Montoya and Thorson
1982 USA Requin
bouledogue ×
Hueter and Manire
1984 Floride Requin
bouledogue ×
Snelson et al. 1984 Floride Requin
bouledogue × × × ×
Campagno 1984 World Requin
bouledogue × × × × × Branstetter and Stiles
1987 Floride Requin
bouledogue ×
Brandstetter et Stiles
1987 Golfe du Mexique
Requin bouledogue
×
Coad and Papahr
1988 Iran Requin
bouledogue ×
SCHAEFFER‐NOVELLI and CINTRON‐MOLERO
1990 Brézil Requin
bouledogue
Stevens and Mcloughlin
1991 Australie Requin taupe ×
Cliff and Dudley
1992 Afrique du Sud Requin
bouledogue × ×
Castro 1993 Floride Requin
bouledogue ×
Castro 1993 USA Requin
bouledogue ×
Last and Stevens
1994 Australie Requin
bouledogue ×
Rodriguez de La Cruz et al.
1996 Golfe du Mexique
Requin bouledogue
×
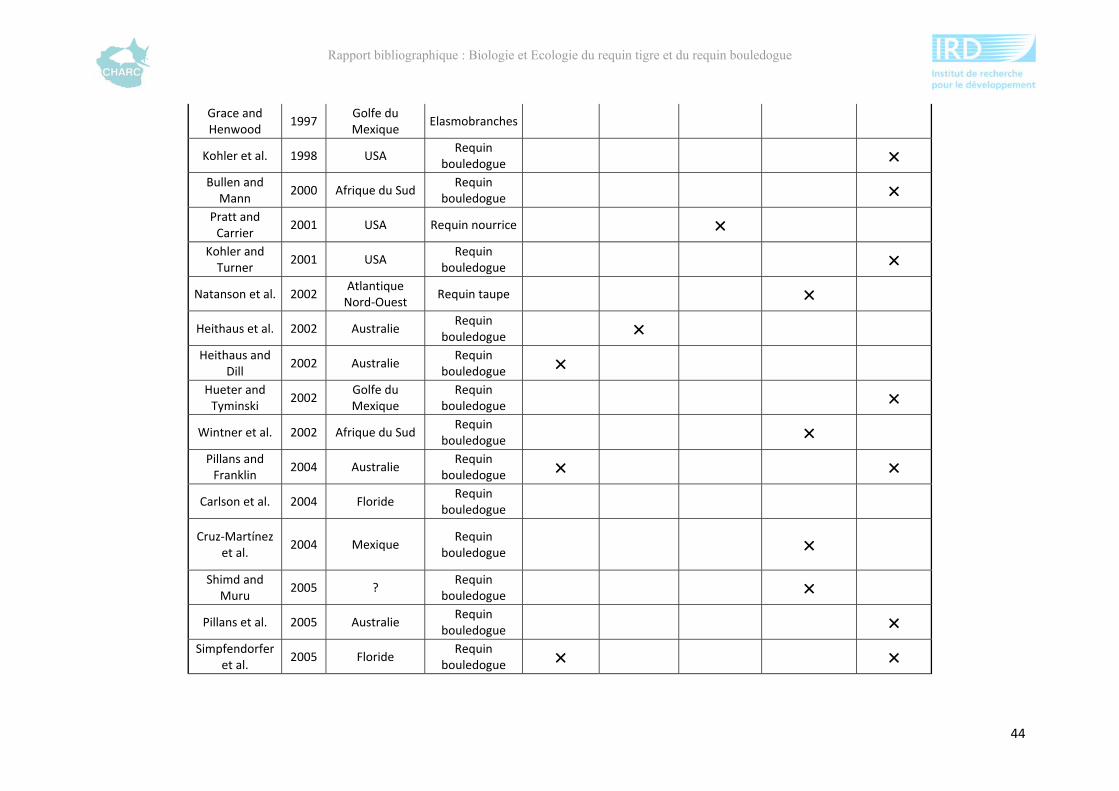
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
44
Grace and Henwood
1997 Golfe du Mexique
Elasmobranches
Kohler et al. 1998 USA Requin
bouledogue ×
Bullen and Mann
2000 Afrique du Sud Requin
bouledogue ×
Pratt and Carrier
2001 USA Requin nourrice ×
Kohler and Turner
2001 USA Requin
bouledogue ×
Natanson et al. 2002 Atlantique Nord‐Ouest
Requin taupe ×
Heithaus et al. 2002 Australie Requin
bouledogue ×
Heithaus and Dill
2002 Australie Requin
bouledogue ×
Hueter and Tyminski
2002 Golfe du Mexique
Requin bouledogue
×
Wintner et al. 2002 Afrique du Sud Requin
bouledogue ×
Pillans and Franklin
2004 Australie Requin
bouledogue × ×
Carlson et al. 2004 Floride Requin
bouledogue
Cruz‐Martínez et al.
2004 Mexique Requin
bouledogue ×
Shimd and Muru
2005 ? Requin
bouledogue ×
Pillans et al. 2005 Australie Requin
bouledogue ×
Simpfendorfer et al.
2005 Floride Requin
bouledogue × ×
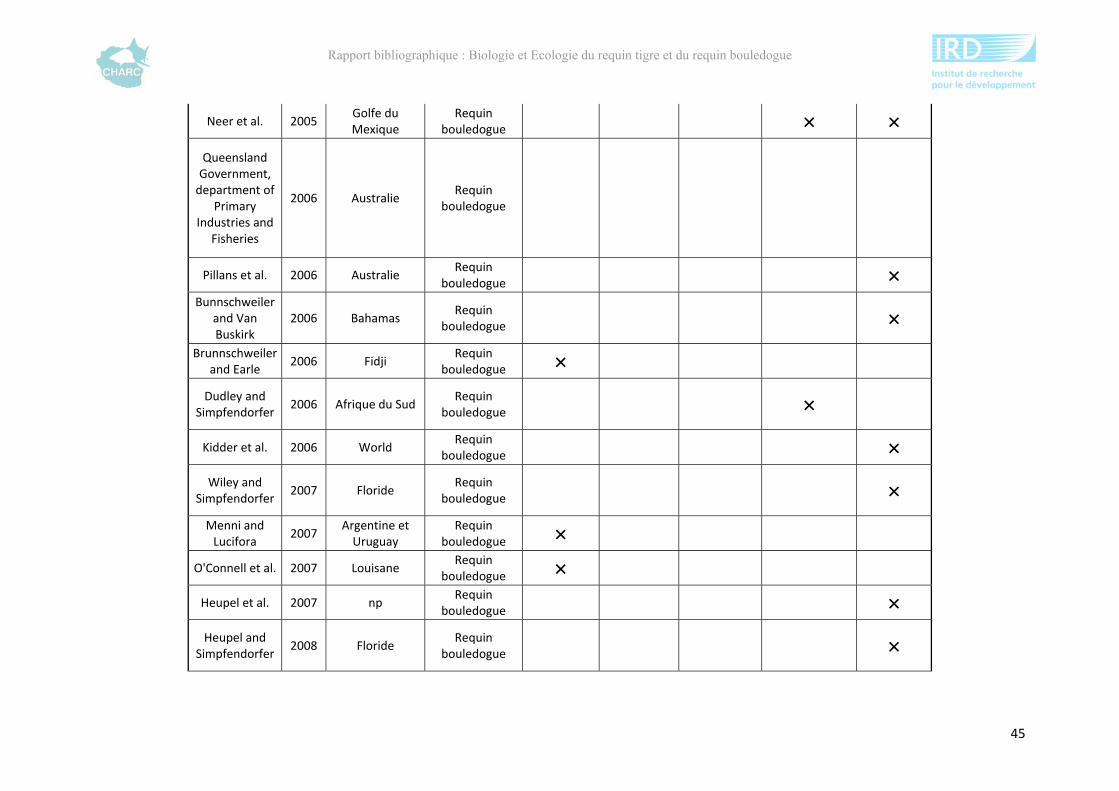
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
45
Neer et al. 2005 Golfe du Mexique
Requin bouledogue
× ×
Queensland Government, department of
Primary Industries and
Fisheries
2006 Australie Requin
bouledogue
Pillans et al. 2006 Australie Requin
bouledogue ×
Bunnschweiler and Van Buskirk
2006 Bahamas Requin
bouledogue ×
Brunnschweiler and Earle
2006 Fidji Requin
bouledogue ×
Dudley and Simpfendorfer
2006 Afrique du Sud Requin
bouledogue ×
Kidder et al. 2006 World Requin
bouledogue ×
Wiley and Simpfendorfer
2007 Floride Requin
bouledogue ×
Menni and Lucifora
2007 Argentine et Uruguay
Requin bouledogue ×
O'Connell et al. 2007 Louisane Requin
bouledogue ×
Heupel et al. 2007 np Requin
bouledogue ×
Heupel and Simpfendorfer
2008 Floride Requin
bouledogue ×
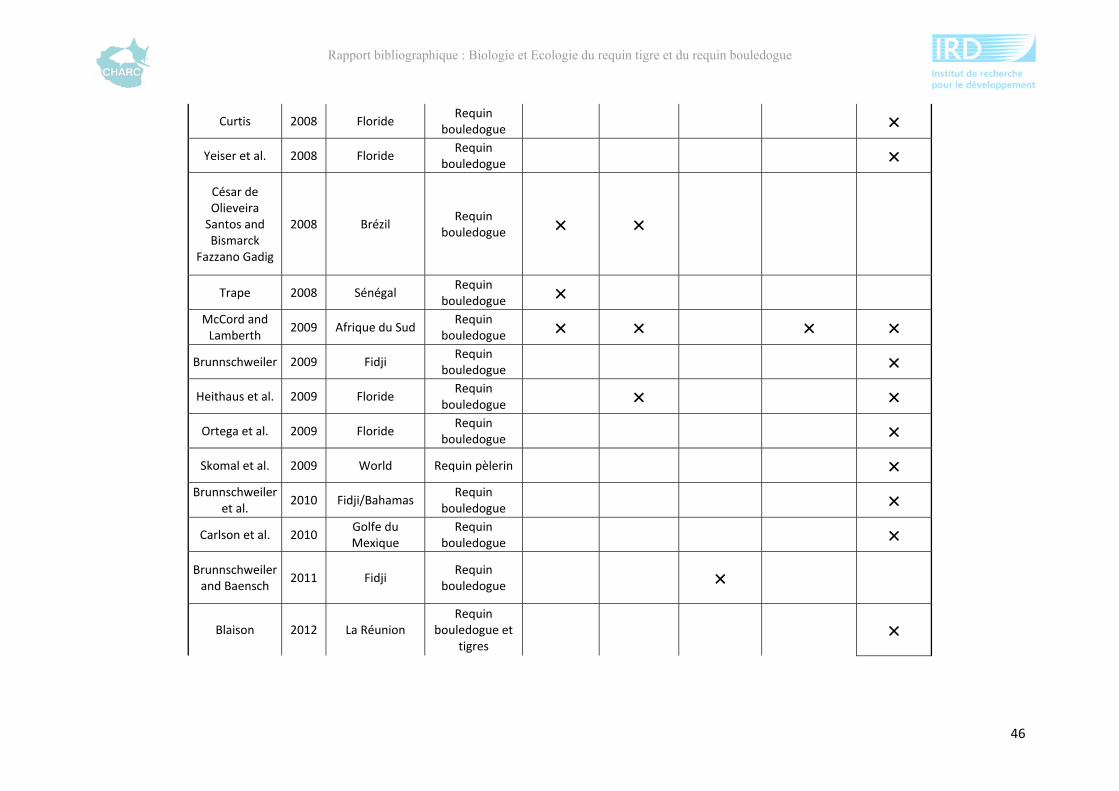
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
46
Curtis 2008 Floride Requin
bouledogue ×
Yeiser et al. 2008 Floride Requin
bouledogue ×
César de Olieveira Santos and Bismarck
Fazzano Gadig
2008 Brézil Requin
bouledogue × ×
Trape 2008 Sénégal Requin
bouledogue ×
McCord and Lamberth
2009 Afrique du Sud Requin
bouledogue × × × ×
Brunnschweiler 2009 Fidji Requin
bouledogue ×
Heithaus et al. 2009 Floride Requin
bouledogue × ×
Ortega et al. 2009 Floride Requin
bouledogue ×
Skomal et al. 2009 World Requin pèlerin × Brunnschweiler
et al. 2010 Fidji/Bahamas
Requin bouledogue
×
Carlson et al. 2010 Golfe du Mexique
Requin bouledogue
×
Brunnschweiler and Baensch
2011 Fidji Requin
bouledogue ×
Blaison 2012 La Réunion Requin
bouledogue et tigres
×
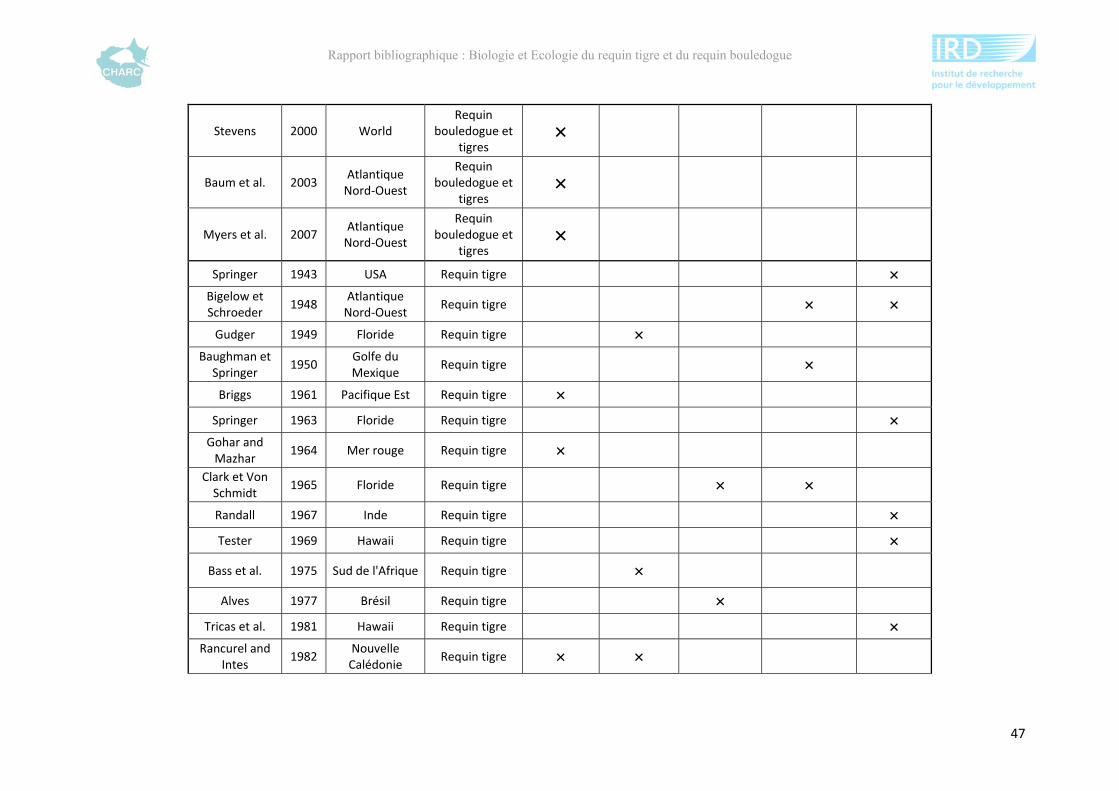
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
47
Stevens 2000 World Requin
bouledogue et tigres
×
Baum et al. 2003 Atlantique Nord‐Ouest
Requin bouledogue et
tigres ×
Myers et al. 2007 Atlantique Nord‐Ouest
Requin bouledogue et
tigres ×
Springer 1943 USA Requin tigre × Bigelow et Schroeder
1948 Atlantique Nord‐Ouest
Requin tigre × ×
Gudger 1949 Floride Requin tigre × Baughman et Springer
1950 Golfe du Mexique
Requin tigre ×
Briggs 1961 Pacifique Est Requin tigre ×
Springer 1963 Floride Requin tigre × Gohar and Mazhar
1964 Mer rouge Requin tigre ×
Clark et Von Schmidt
1965 Floride Requin tigre × ×
Randall 1967 Inde Requin tigre ×
Tester 1969 Hawaii Requin tigre ×
Bass et al. 1975 Sud de l'Afrique Requin tigre ×
Alves 1977 Brésil Requin tigre ×
Tricas et al. 1981 Hawaii Requin tigre × Rancurel and
Intes 1982
Nouvelle Calédonie
Requin tigre × ×
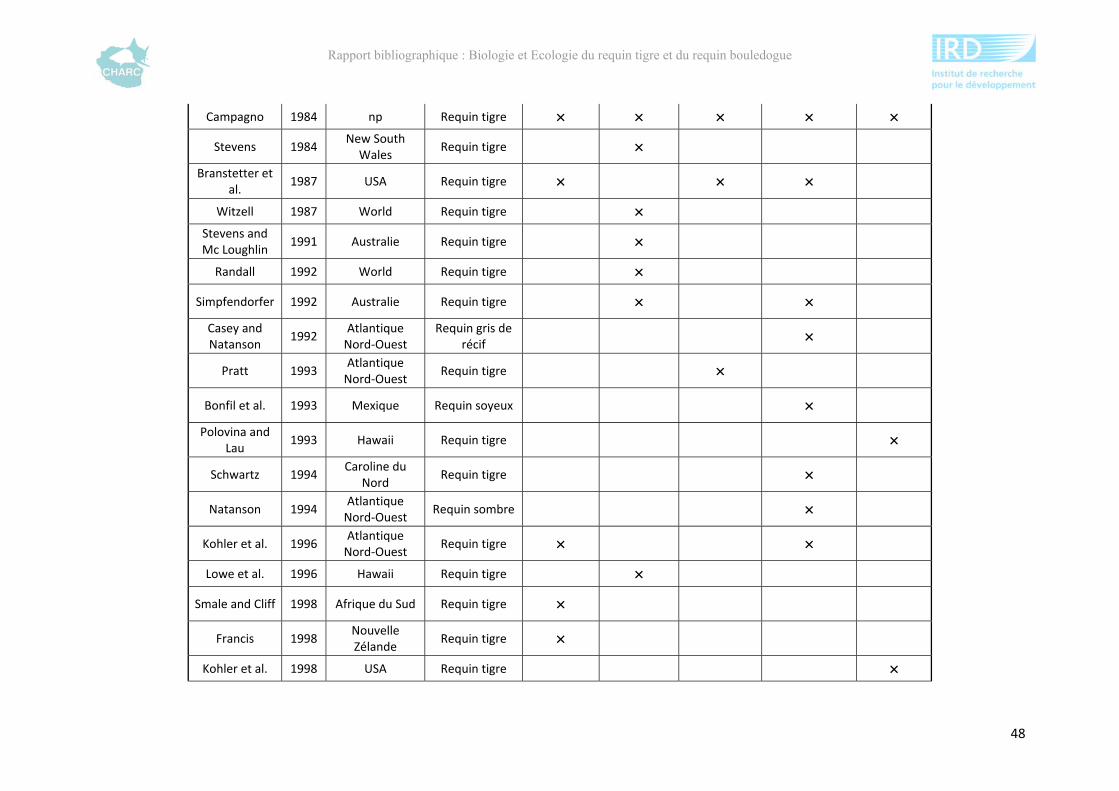
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
48
Campagno 1984 np Requin tigre × × × × ×
Stevens 1984 New South Wales
Requin tigre ×
Branstetter et al.
1987 USA Requin tigre × × ×
Witzell 1987 World Requin tigre × Stevens and Mc Loughlin
1991 Australie Requin tigre ×
Randall 1992 World Requin tigre ×
Simpfendorfer 1992 Australie Requin tigre × ×
Casey and Natanson
1992 Atlantique Nord‐Ouest
Requin gris de récif ×
Pratt 1993 Atlantique Nord‐Ouest
Requin tigre ×
Bonfil et al. 1993 Mexique Requin soyeux ×
Polovina and Lau
1993 Hawaii Requin tigre ×
Schwartz 1994 Caroline du
Nord Requin tigre ×
Natanson 1994 Atlantique Nord‐Ouest
Requin sombre ×
Kohler et al. 1996 Atlantique Nord‐Ouest
Requin tigre × ×
Lowe et al. 1996 Hawaii Requin tigre ×
Smale and Cliff 1998 Afrique du Sud Requin tigre ×
Francis 1998 Nouvelle Zélande
Requin tigre ×
Kohler et al. 1998 USA Requin tigre ×
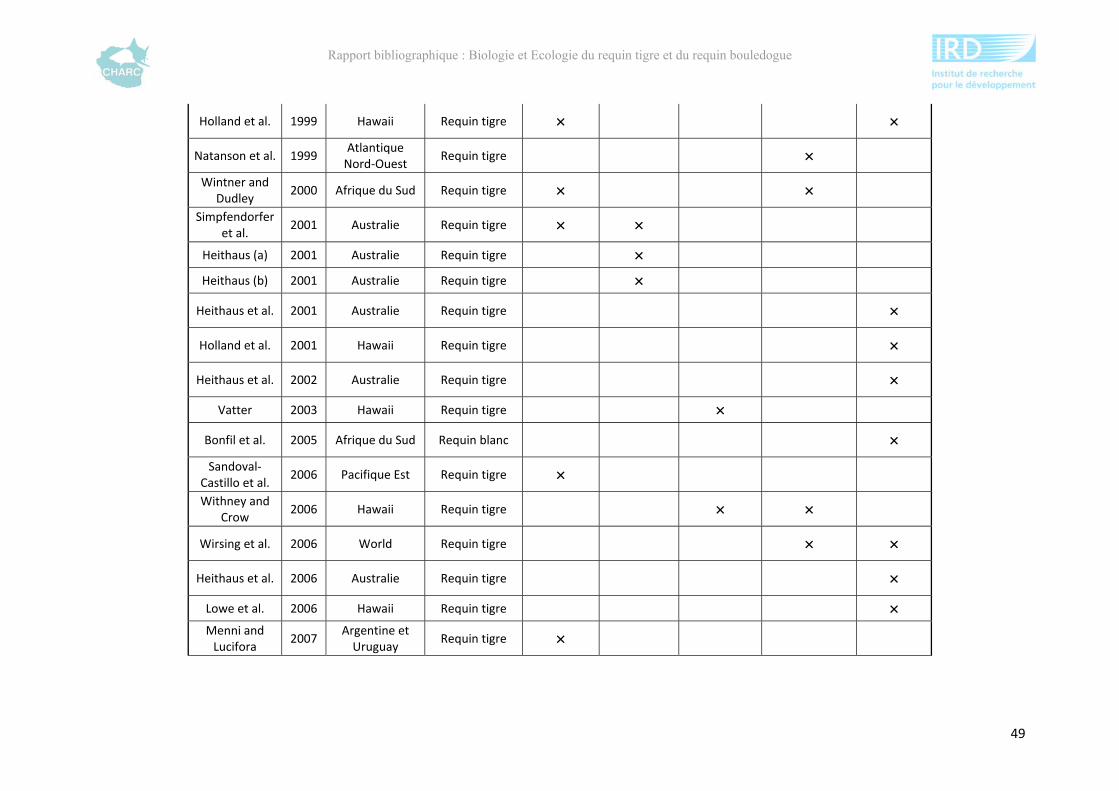
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
49
Holland et al. 1999 Hawaii Requin tigre × ×
Natanson et al. 1999 Atlantique Nord‐Ouest
Requin tigre ×
Wintner and Dudley
2000 Afrique du Sud Requin tigre × ×
Simpfendorfer et al.
2001 Australie Requin tigre × ×
Heithaus (a) 2001 Australie Requin tigre ×
Heithaus (b) 2001 Australie Requin tigre ×
Heithaus et al. 2001 Australie Requin tigre ×
Holland et al. 2001 Hawaii Requin tigre ×
Heithaus et al. 2002 Australie Requin tigre ×
Vatter 2003 Hawaii Requin tigre ×
Bonfil et al. 2005 Afrique du Sud Requin blanc ×
Sandoval‐Castillo et al.
2006 Pacifique Est Requin tigre ×
Withney and Crow
2006 Hawaii Requin tigre × ×
Wirsing et al. 2006 World Requin tigre × ×
Heithaus et al. 2006 Australie Requin tigre ×
Lowe et al. 2006 Hawaii Requin tigre × Menni and Lucifora
2007 Argentine et Uruguay
Requin tigre ×
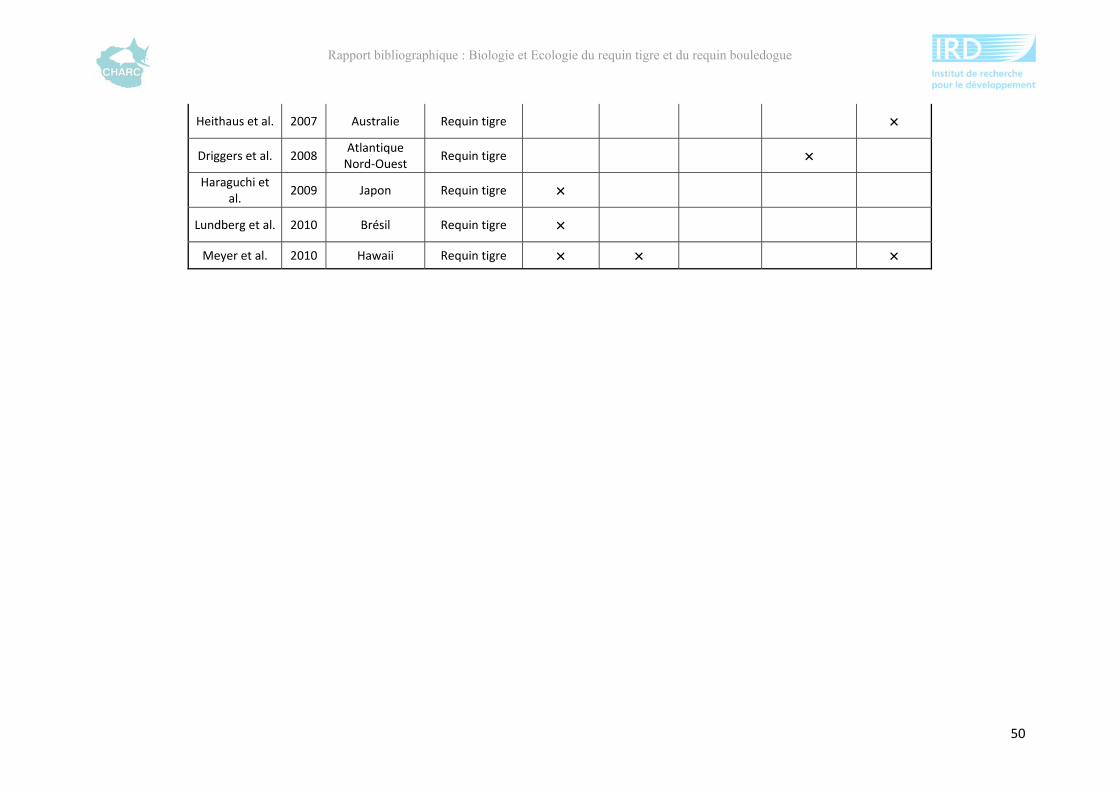
Rapport bibliographique : Biologie et Ecologie du requin tigre et du requin bouledogue
50
Heithaus et al. 2007 Australie Requin tigre ×
Driggers et al. 2008 Atlantique Nord‐Ouest
Requin tigre ×
Haraguchi et al.
2009 Japon Requin tigre ×
Lundberg et al. 2010 Brésil Requin tigre ×
Meyer et al. 2010 Hawaii Requin tigre × × ×