physique et rhéologie des agrégats cellulaires...
TRANSCRIPT

Année 2009
THESE DE L‘UNIVERSITE DE LYON
Délivrée par
L’UNIVERSITE CLAUDE BERNARD LYON 1
ECOLE DOCTORALE
Physique et Astrophysique
DIPLOME DE DOCTORAT
(arrêté du 7 août 2006)
soutenue publiquement le
par
M. MGHARBEL Abbas
Physique et rhéologie des agrégats cellulaires
embryonnaires
Directeurs de thèse : Jean Paul Rieu
Hélène Delanoë-Ayari
JURY : M. Gillet Germain
M. Joanny Jean-François
M. Lenne Pierre-François M.Verdier Claude

Physique des agrégats cellulaires embryonnaires
2
CHAPITRE 1 10
Physique de la cellule au tissu 10 I.1. La cellule 11
I.1.1. Description générale de la cellule. 11 I.1.2. Description du cytosquelette. 13 I.1.3. Généralités sur l’adhésion cellulaire 16 I.1.4. Physique du cytosquelette et de l’adhésion. 20
I.2. Les tissus 22 I.2.1. Les tissus in vivo : l’embryogenèse 23 I.2.2. Les tissus in vitro : l’affinité cellulaire 27 I.2.3. L’hypothèse d’adhésion différentielle (HAD). 30 I.2.4 Rhéologie 42
CHAPITRE 2 47
Materials and Methods 47 II.1.Strategy, Cell lines and Aggregate preparation method 48
II.1.1. CHOVE / CHOVEtr cells 48 II.1.2. Mouse embryonic carcinoma F9 cell 49 II.1.3. Nocodazole Effect 51 II.1.4. Cell aggregates formation protocol 52 II.1.5. Storage 53
II.2. Dissected embryonic cells 54 II.2.1. Embryonic zebrafish cells 54 II.2.2. Embryonic chicken cell retina 55
II.3. Imaging cells through aggregates 55 II.3.1. Sections through fixed aggregates 55 II.3.2. Two-photon confocal microscopy 3D imaging 56
II.4. Segregation assays 59 II.4.1. Cell-sorting assay 59 II.4.2. Tissue-envelopment assay 59
II.5. Fusion and rounding up assay 59 II.5.1. Fusion of two symmetric cell aggregates 60 II.5.2. Rounding up of a free elliptical aggregate 60
II.6. Liquid interfacial tension and apparent tissue surface tension 61 II.6.1. Description of the final system: 62 II.6.2. Resolution 64
CHAPITRE 3 65
Measuring accurately tissue surface tension 65 III. 1. Introduction: TST measurements, for which purpose? 66 III. 2. The different methods to measure TST 67
III. 2.1. A few lines on capillarity 67 III. 2.2. The CA (Circular Arc) method 69 III. 2.3. The ELP (Exact Laplace Profile) method 70 III. 2.4. The LPF (Local Polynomial Fit) method 71

Physique des agrégats cellulaires embryonnaires
3
III.3. Critical comparison of existing methods using experimental data and numerical tests 72
III. 3.1. Experiments with pure liquid droplets or bubbles 72 III. 3.2. Experiments with embryonic cell aggregates 74 III. 3.3. Numerical tests 76
III. 4. Discussion 77 III. 4.1. Examination of literature data 78 III. 4.2. Recommendations for correct measurements 79
CHAPITRE 4 80
Physical properties of F9 tissues Role of adhesion and Cytoskeleton 80 IV.1. Surface tensions driven phenomena 81
IV.1.1. Aggregate rounding up 81 IV.1.2. Aggregate fusion 82 IV.1.3. Behaviours of F9/ F9(α-/-) pairs (spreading, sorting) 85
IV.2. Measurements of tissue surface tension 87 IV.2.1. Surface tension of F9 cell aggregates 87 IV.2.2. Surface tension of F9(α-/-) cell aggregates 88 IV.2.3. Effect of nocodazole on tissue surface tensions 89
IV.3. Discussion 91 IV.3.1. Surface tension depends on cytoskeleton contractility: DAH is contradicted 91 IV.3.2. Tissue viscosity does not depend directly on cell cohesion: an adhesion-protrusion competition hypothesis 92
CHAPITRE 5 95
Cell-cell rearrangements within aggregates: Experiments & Modelling 95 V.1. Elastic and/or viscous behaviours: previews studies 96 V.2. Experimental evidence of cell rearrangements 101
V.2.1. Direct visualization of stress induced cell rearrangements 101 V.2.2. Shape and stress relaxation experiments 103
V.3. Cell rearrangement: fluctuations and stress 106 V.3.1. Global Description of the Aggregate. 106 V.3.2. Local Description at the Cell Level. 106 V.3.3. Mechanical Behavior of the Aggregate. 107 V.3.4. Two Regimes 108 V.3.5. Compressions: Predictions. 109 V.3.6. Decompressions: Predictions. 110
V.4. Discussion 111 V.4.1. Energy Barriers and Non-released Elasticity. 111 V.4.2. Time Scales. 111 V.4.3. Validity of the plasticity measurement 112 V.4.4. Link between compression-decompression experiment and fusion? 113 V.4.5. Effect on Measured Surface Tension 114
Conclusions et Perspectives 115

Physique des agrégats cellulaires embryonnaires
4

Physique des agrégats cellulaires embryonnaires
5
Préface
Le développement embryonnaire, c’est à dire la croissance d’un organisme
multicellulaire complexe à partir d’un œuf fertilisé, est un processus qui a fasciné les
hommes et a toujours constitué un domaine actif de recherche. Il semble que ce sont
les anciens grecs qui aient lancé le premier débat autour de ce thème [1]. Aristote,
dès le quatrième siècle avant J.C., a proposé que le développement d’un embryon
comporte plusieurs stades, durant lesquels les différentes parties d’un embryon se
développent et prennent leurs places (Epigenèse), au contraire des idées du courant
préformationniste qui voulaient que l’embryon soit préformé dans l’œuf ou dans le
spermatozoïde (Figure A ci dessous) [1]. Ce n’est qu’après l’émergence de la théorie
cellulaire au 19ème siècle que le débat fut tranché en faveur de l’Epigenèse :
L’embryon n’était pas préformé ; il était développé à partir d’un ovule fertilisé (Figure
B).
Figure : Le développement embryonnaire (A) tel qu’il était imaginé par les préformationnistes [Essai de dioptrique (Paris, 1694, page 230), et téléchargé depuis wikipedia], et (B) tel qu’il était décrit par les théories cellulaires du 19ème siècle. Cette image montre un ovule fécondé (cellule initiale) à gauche ainsi que la première division cellulaire à l’intérieur de l’ovule à droite.
Les questions en biologie de développement ne se sont pas arrêtées suite à
cette découverte au 19ème siècle. Au contraire, elles se sont multipliées grâce au

Physique des agrégats cellulaires embryonnaires
6
progrès des connaissances scientifiques et des technologies innovantes permettant
maintenant de répondre aux nouvelles interrogations sur la morphogénèse, la
migration, la différentiation, la croissance et la mort cellulaire. Ces derniers mots clés
de la biologie constituent des thèmes de recherches fascinants, à notre époque, pour
les biologistes aussi bien que pour les physiciens.
La recherche menée sur ces thèmes a conduit, ces dernières années, au
développement d’une interface entre la biologie et les autres sciences dures :
physique, chimie, mécanique, mathématiques. L’interface physique-biologie qui met
les lois et les techniques de mesure Physique au profit des Biologistes et vice-versa,
permet de mieux investir une question d'intérêt biologique par de nouvelles
approches expérimentales. Pendant que la biologie du développement cherche à
identifier le ou les acteur(s) impliqué(s) dans l’évolution des tissus et leurs mise en
place sous forme d’organes, à investir comment le codage de l’ADN intervient dans
ces processus afin d’expliquer les mécanismes du développement, la physique
présente une vision différente de ces processus tout en complétant les observations
qualitatives par des mesures quantitatives physiquement pertinentes afin de tracer
les chemins du développement cellulaire et tissulaire par des lois universelles.
Comment les cellules, bien qu’issues d’une seule et même cellule initiale,
peuvent élaborer des structures complexes variées et hyper spécialisées tels que les
muscles, les yeux, la peau ou le cerveau ?
Cette question reste en tête de la liste des interrogations qu’on retrouve dans la
plupart des ouvrages de biophysique. Après l’identification d’un comportement
propre aux cellules ou aux tissus, les biologistes s’intéressent à déterminer les
composants clés (protéines d’adhésion cellule/cellule et cellule/ECM1, cytosquelette,
etc...) et à trouver leur mécanisme d’interaction permettant d’influer sur le système
en question voir sur l’organisme entier. Les physiciens s’intéressent à ces mêmes
composants et leurs interactions en essayant de les quantifier et de leur attribuer
une observable physique tels que (force, énergie, tension de surface, viscosité,
plasticité,…) afin d’arriver à modéliser les comportements par des lois physiques. Par
la suite, le changement d’une telle observable devrait refléter soit une évolution des
composants sous-jacents, soit un changement de mode d’interaction.
Cette caractérisation physique des comportements cellulaires et tissulaires
nous donne des informations très importantes pour la compréhension des processus
d’organisation des tissus dans les domaines de l’embryologie, de la cancérologie, et
de l’ingénierie tissulaire.
1 Extra cellular Matrix

Physique des agrégats cellulaires embryonnaires
7
Par exemple, l’embryogenèse fait intervenir des processus morphogénétiques
qu’on peut résumer ici par la prolifération et les mouvements des cellules
embryonnaires, contrôlées par l’expression des gènes du développement, afin de
déterminer la forme de l’embryon. Il a été montré que ces mouvements
morphogénétiques sont intimement liés aux forces mécaniques ressenties par les
cellules impliquées dans la formation des tissues [2]. Des études récentes montrent
que les forces mécaniques dans l’embryon en cours de morphogenèse peuvent
moduler l’expression de certains gènes du développement mécaniquement sensibles
[2]. Le génome est capable, grâce à ces rétroactions mécaniques, de détecter si la
forme qu'il est en charge de développer est atteinte et peut donc déclencher les
gènes suivants de la cascade qui orchestre la formation du tissu [2].
D’autre part, une meilleure compréhension des mécanismes impliqués dans la
formation des organes du vivant, accompagnée de technologies de
micromanipulations, ont permis des avancées significatives dans le domaine de
l’ingénierie tissulaire. Des techniques innovantes cherchent à reproduire en
laboratoire les processus morphogénétiques afin de fabriquer des organes de
remplacement pour la médecine régénérative [4]. Ce but deviendra peut être un jour
accessible maintenant qu’il est possible d’obtenir de structures mimant celles
d’organes à partir de cellules embryonnaires dissociés-réaggrégées et en particulier
grâce à la physique de l’auto-assemblage des cellules et agrégats biologiques. Ainsi
les propriétés liquides de certains tissus ont pu être mises à profit dans la maîtrise de
l’auto-assemblage des structures tridimensionnelles cellulaires vivantes [5,6, 7,8].
Ce travail de thèse s’inscrit en amont de ces applications. L’objectif de ce travail
a été, en premier temps, de développer des outils de mesure physique pour arriver à
quantifier la dynamique cellulaire à l’intérieur des tissus, et pour essayer d’identifier
les lois physiques régissant les comportements de la matière cellulaire à différentes
échelles du temps. Ensuite nous avons cherché à identifier quels étaient les
composants biologiques qui pourraient être à l’origine de cette dynamique et leur
influence sur les comportements collectifs des cellules.
Ce manuscrit se décompose en cinq chapitres :
Le premier présente un résumé sur les comportements liquides de certains
tissus biologiques modèles, les acteurs à l’échelle subcellulaire responsables de
cette dynamique, ainsi que leur rôle dans le fonctionnement d’une cellule et d’un
tissu. De plus, une brève présentation des différentes théories ou hypothèses
mettant en évidence les caractères liquides de la matière cellulaire et ses propriétés
viscoélastiques, situera le contexte biophysique et biomécanique dans lequel s’inscrit
notre étude. Les notions de tension de surface et de viscosité y sont rappelées.

Physique des agrégats cellulaires embryonnaires
8
Dans le deuxième chapitre nous présentons les dispositifs et méthodes
expérimentaux que nous avons mis en œuvre pour cette étude. Nous détaillons
aussi les protocoles biologiques utilisés sur les cellules, ainsi que la méthode de
fabrication de nos tissus modèles : les agrégats cellulaires.
Le troisième chapitre est consacré entièrement à la mesure de la tension de
surface avec le tensiomètre des tissus développé durant la thèse. Nous insisterons
sur les lois de la capillarité, l’analyse du profil des agrégats comprimés et le rôle des
paramètres géométriques dans la mesure, en particulier l’angle de contact des
agrégats avec les plaques de compression.
Dans le quatrième chapitre, nous exposons les résultats de mesure de la
viscosité et de la tension de surface tissulaire que nous avons obtenus sur différents
types cellulaires testés. L’influence de la liaison entre l’actine et les cadhérines (via
les caténines) sur ces grandeurs physiques était l’une des questions centrales de
cette thèse. Nos résultats montrent que cette liaison influence très significativement
la tension de surface tissulaire remettant en cause l’un des postulats de l’hypothèse
d’adhésion différentielle de M. S. Steinberg, mais par contre elle ne semble pas
affecter outre mesure la viscosité tissulaire.
Enfin, dans le cinquième chapitre, nous présentons les résultats des relaxations
de forme des agrégats cellulaires. Ces mesures ajoutent une nouvelle dimension (le
temps) à la discussion sur la nature complexe de cette matière cellulaire. Nous
avons couplé ces résultats avec des observations de la forme et des réarrangements
des cellules, prises au « microscope deux photons » à l’intérieur d’un agrégat
comprimé. Cette étude a permis de valider un modèle visco-élasto-plastique basé
sur des ingrédients microscopiques bien identifiés et réalisé en collaboration avec P.
Marmottant (LSP Grenoble). Ce modèle en retour nous a lancé de multiples pistes
de travail pour des expériences futures qui sont détaillées dans les conclusions et
perspectives.
Quatre annexes viennent compléter ce manuscrit.

Physique des agrégats cellulaires embryonnaires
9
[1] L. Wolpert, R. Beddington, T. Jessell, P. Lawrence, E. Meyerowitz, and J. Smith. Principles of Development. Oxford University Press, 2002.
[2] Nicolas Desprat, Willy Supatto, Philippe-Alexandre Pouille, Emmanuel Beaurepaire, and Emmanuel Farge. Tissue Deformation Modulates Twist Expression to Determine Anterior Midgut Differentiation in Drosophila Embryos. Developmental Cell, 15:470 (2008).
[3] Joanne Whitehead, Danijela Vignjevic, Claus Fütterer, Emmanuel Beaurepaire, Sylvie Robine, Emmanuel Farge. Mechanical factors activate b-catenin-dependent oncogene expression in APC1638N/+mouse colon. Human Science Frontier Journal, 1er octobre 2008
[4] Cyrille norotte, From developmental biology to tissue-engineering: printing blood vessels. Thèse (2009)
[5] Jakab. K, Neagu. A, Mironov. V, Markwald. RR, and Forgacs. G. Engineering biological structures of prescribed shape using selfassembling multicellular systems. Proc. Natl. Acad. Sci. U.S.A., 101, 2864–2869 (2004)
[6] Jakab. K, Damon. B, Marga. F, Doaga. O, Mironov. V, Kosztin. I, Markwald. R, and Forgacs. G. Relating cell and tissue mechanics: implications and applications. Dev. Dyn., 237, 2438–2449 (2008a).
[7] Jakab. K, Norotte. C, Damon. B, Marga. F, Neagu. A, Besch-Williford. CL, Kachurin. A, Church. KH, Park. H, Mironov. V, Markwald. RR, Vunjak-Novakovic. G, and Forgacs. G. Tissue engineering by self-assembly of cells printed into topologically defined structures. Tissue Eng., 14, 413–421 (2008b).
[8] Marga. F, Neagu. A, Kosztin. I, and Forgacs. G. Developmental biology and tissue engineering. Birth Defects Res. C 81,320–328 (2007).

Physique des agrégats cellulaires embryonnaires
10
Chapitre 1 Physique de la cellule au tissu
Dans ce premier chapitre, nous présenterons d’abord un aperçu rapide de
l’architecture et des fonctions biologiques de la matière vivante depuis l’échelle
cellulaire jusqu’à l’échelle des tissus. Nous espérons qu’il pourra constituer une
introduction utile au lecteur qui n’est pas du domaine de la biologie mais aussi aux
biologistes qui pourront y trouver une revue non exhaustive des travaux de
biophysiciens travaillant dans les domaines de la biomécanique cellulaire, de
l’adhésion, de la motilité cellulaire et ou de la biologie du développement. Comme
cette thèse s’intéresse tout particulièrement aux mouvements cellulaires à l’intérieur
de tissus embryonnaires modèles (agrégats cellulaires), lors de notre description de
la cellule, nous insisterons tout particulièrement sur la description du cytosquelette,
de l’adhésion cellulaire, de leur rôle respectif dans les mouvements et de leur
couplage éventuel. En passant à l’échelle tissulaire, nous essayerons de présenter
notamment à travers l’exemple du poisson zèbre l’enjeu et les apports éventuels de
la physique à l’embryogénèse. Nous suivrons la chronologie depuis le début du 20ème
siècle des travaux sur le tri et les réorganisations cellulaires en systèmes in vitro
ayant conduit à l’utilisation des agrégats cellulaires et aux prédictions de l’hypothèse
d’adhésion différentielle de Malcom Steinberg. Enfin, nous présenterons une brève
introduction générale sur la rhéologie.

Physique des agrégats cellulaires embryonnaires
11
I.1. La cellule
I.1.1. Description générale de la cellule.
La cellule constitue, à la manière de l’atome pour la matière, la brique
élémentaire de la vie d’un organisme, il s’agit de la plus petite entité de matière
vivante qui puisse vivre, et elle contient toutes les informations nécessaires à sa
reproduction. Tout d’abord, il faut faire la distinction entre les cellules procaryotes,
sans noyau (algues, bactéries, etc.), et les cellules eucaryotes, qui disposent d’un
véritable noyau (Fig.1.1) qui contient l’information génétique de la cellule, codée
dans les filaments d’ADN, présents sous forme de chromosomes ou de chromatine.
C’est ce dernier type de cellules qui nous intéresse dans notre étude.
Fig. 1.1: Schéma de cellule animale [D’après http://www.astrosurf.com/luxorion/Bio/cellule-legende.png ]
L’intérieur de la cellule est extrêmement complexe et remarquablement
organisé. Il est difficile d’imaginer et de modéliser le comportement global de
l’ensemble des organites intracellulaires (Réticulum endoplasmique, mitochondrie,
appareil de golgi, etc.,Fig. 1.1). L’organisation spatiale ainsi que le mouvement de
ces organites sont assurés grâce à un réseau complexe et dense de bio-polymères,

Physique des agrégats cellulaires embryonnaires
12
le cytosquelette. Ce réseau, qui accomplit des fonctions essentielles tout au long de
la vie d’une cellule, joue un rôle capital dans la régulation du trafic intracellulaire.
La cellule est active, elle produit, stocke, et consomme de l’énergie. L’énergie
produite par les mitochondries sera accumulée sous forme de liaisons chimiques
fortes. La monnaie courante d’échange du travail cellulaire est l’ATP (adénosine
triphosphate). Elle est consommée dans les réactions enzymatiques ayant lieu à tous
les niveaux de la vie cellulaire. Ainsi la réaction de base ayant lieu pour fournir de
l’énergie nécessaire à un processus biochimique est :
ATP → ADP + P (1.1)
avec libération de 30,5 kJ/mol. Bien qu’il existe d’autres moyens de transférer
de l’énergie (GTP : guanosine triphosphate, …), l’hydrolyse de l’ATP reste la
carburant principal de la cellule. L’inhibition de la formation d’ATP apparaît comme
un outil standard pour examiner le rôle des processus actifs à l’intérieur de la cellule.
Fig. 1.2: Schéma de membrane cellulaire, avec ses composants lipidiques et les protéines
qui y sont enchâssées. La bicouche a une épaisseur de l’ordre de 5 nm. [D’après G. Geibel,
http://sun.menloschool.org].
La cellule est entourée d’une bicouche lipidique : la membrane plasmique (Fig.
1.2. Cette membrane est globalement imperméable, et permet donc de maintenir de
forts gradients de concentration de différents types de molécules entre le liquide
extracellulaire et le liquide intracellulaire, ou cytosol. La concentration intracellulaire
en ions Ca2+ par exemple, est 10−4 fois moins élevée que celle du milieu
extracellulaire. Des protéines sont enchâssées dans la membrane et assurent des
fonctions de reconnaissance de signaux biochimiques, et d’activation d’autres

Physique des agrégats cellulaires embryonnaires
13
molécules en réponse à ces signaux. Certaines d’entre elles, les protéines
transmembranaires, traversent la membrane de part en part. Elles peuvent donc, sur
leur domaine extracellulaire, lier certaines molécules spécifiques, ce qui active en
général leur domaine cytoplasmique pour agir sur des protéines intracellulaires
cibles. Cette « transduction du signal » permet aux cellules de répondre à divers
types de sollicitations de leur environnement.
Les cellules présentent des caractéristiques mécaniques qui semblent
cohérentes avec leur structure aussi bien qu’avec leur rôle à l’intérieur de
l’organisme. Les caractéristiques mécaniques des cellules uniques ont été beaucoup
étudiées [1-8]. Ces études ont mis en évidence l’importance de l’adhésion et du
cytosquelette dans l’établissement des propriétés mécaniques des cellules. Les rôles
de l’adhésion [9-13] et du cytosquelette [14-17] ont été les sujets de nombreuses
études biomécaniques détaillées. Ces deux paramètres interviennent dans la
mécano-transduction intercellulaire (c.f.[18] pour une revue). Nous allons maintenant
nous intéresser plus particulièrement au cytosquelette et à l’adhésion.
I.1.2. Description du cytosquelette.
Les cellules animales ont leur forme fixée par l’analogue d’un squelette qui
parcourt le cytoplasme, le cytosquelette. Ce dernier est une structure
tridimensionnelle complexe issue de l’assemblage de bio-polymères filamenteux
interconnectés. Il est constitué de trois grandes familles de filaments protéiques: les
microtubules, les filaments intermédiaires et les micro-filaments (Fig. 1.3).
Fig. 1.3: À gauche, cellule dont les polymères du cytosquelette sont rendus fluorescents
grâce aux techniques de l’ingénierie moléculaire. Les microtubules entourant le noyau en
bleu sont colorées en vert (marquées au Bodipy) alors que l’actine apparaît en rouge
(marquée à la X-Phalloïdine). [D’après le catalogue Invitrogen - Molecular Probes]. À
droite schéma de la répartition des 3 types de filaments du cytosquelette dans une cellule
épithéliale [19].

Physique des agrégats cellulaires embryonnaires
14
Les microtubules sont les polymères les plus rigides du cytosquelette. Ils sont
formés à partir des dimères de tubuline qui s’associent sous forme de tube creux.
Ces tubes ont un diamètre de 25 nm. Leur longueur de persistance vaut lp~1 mm et
leur module d’Young E∼1 GPa, grandeurs déterminées dans des expériences in vitro
en observant les fluctuations de courbure des filaments [20].
Le réseau de microtubules est une structure hautement dynamique en
constante réorganisation. D’une part, il est lié au centrosome près du noyau [21;22],
d’autre part il exerce des pressions aux extrémités de la cellule [23;24]. Le réseau
rayonne donc en aster (Fig. 1.3) autour du centrosome, tant que les cellules n’entrent
pas en division tandis qu’il s’organise sous forme de fuseau mitotique lors d’une
division cellulaire, de façon à positionner et séparer les chromosomes.
De nombreuses protéines peuvent se lier aux microtubules (les Microtubule
Associated Proteins ou MAP) ou aux monomères de tubuline, et modifier la stabilité
des filaments [25]. Certaines de ces molécules sont présentes dans les cellules et
sont activées ou non, selon les signaux reçus par la cellule ou selon les phases du
cycle cellulaire. D’autres sont utilisées comme drogues, afin de contrôler l’état du
squelette de microtubules : par exemple, le nocodazole a un effet dépolymérisant,
contrairement au taxol qui stabilise les microtubules.
Ensuite, on trouve les micro-filaments ou F-actine qui sont les filaments les plus
fins : ce sont des doubles hélices de diamètre 5 à 9 nm. On rencontre encore dans
les cellules des monomères d’actine, la G-actine. La longueur de persistance des
micro-filaments vaut environ 10-15µm [26], et leur module d’Young 0,5 GPa, est du
même ordre que celui des microtubules. Cependant, contrairement aux microtubules,
l’actine n’est pas présente dans la cellule sous forme de filaments isolés, mais sous
forme de réseaux (Fig. 1.3). La description d’un tel réseau de filaments enchevêtrés,
réticulés et pouvant même coulisser les uns par rapport aux autres n’est évidemment
pas simple. Des modèles théoriques [27;28] ainsi que des expériences d’assemblage
de gels d’actine in vitro [29-32], visant à comprendre les propriétés mécaniques de
tels gels sont en plein développement.
La dernière grande classe de polymères du cytosquelette, les filaments
intermédiaires, est plus hétérogène. Ils sont formés à partir de molécules allongées
qui s’enroulent d’abord sous forme de dimères, qui eux-mêmes s’assemblent de
façon anti-parallèle en tétramères, et finalement donnent des fibres de 8 à 12 nm de
diamètre. Ces fibres sont donc apolaires, contrairement aux filaments d’actine et aux
microtubules, et on en distingue quatre types principaux : les lamines, les kératines,
la vimentine, et les neurofilaments. Les différents types sont présents à différents
Physique des agrégats cellulaires embryonnaires
15
endroits de la cellule selon le type cellulaire. Par exemple les kératines, de divers
types, sont plus abondantes dans les cellules épithéliales.
Ces fibres sont très résistantes, ce qui leur donne un rôle essentiel dans le
maintien de l’intégrité mécanique des cellules. Les filaments de kératine, eux, sont
densément engagés dans des jonctions entre cellules épithéliales (desmosomes), et
assurent donc la résistance globale du tissu.
Contrairement à un squelette osseux, le cytosquelette est un échafaudage
relativement rigide mais dynamique qui constitue l’un des éléments clefs intervenant
au niveau du remodelage permanent d’une cellule. La dynamique de
polymérisation/dépolymérisation s’effectue à des vitesses de 1 à 10 µm/min [33], elle
est régulée de façon biochimique et mécanique et permet à la cellule de se déformer,
d’adapter sa forme en fonction des forces mécaniques environnementales ou de se
déplacer. Une fonction du cytosquelette qui nous intéressera particulièrement ici est
son aptitude à exercer des forces : le cytosquelette est à la fois le squelette et le
muscle de la cellule.
Comment ces fibres exercent-elles des forces?
En premier lieu, grâce à la polymérisation. Si un filament est attaché à une
extrémité, l’ajout d’un monomère à son autre extrémité se traduit par une force de
poussée pouvant atteindre 10 pN [34].
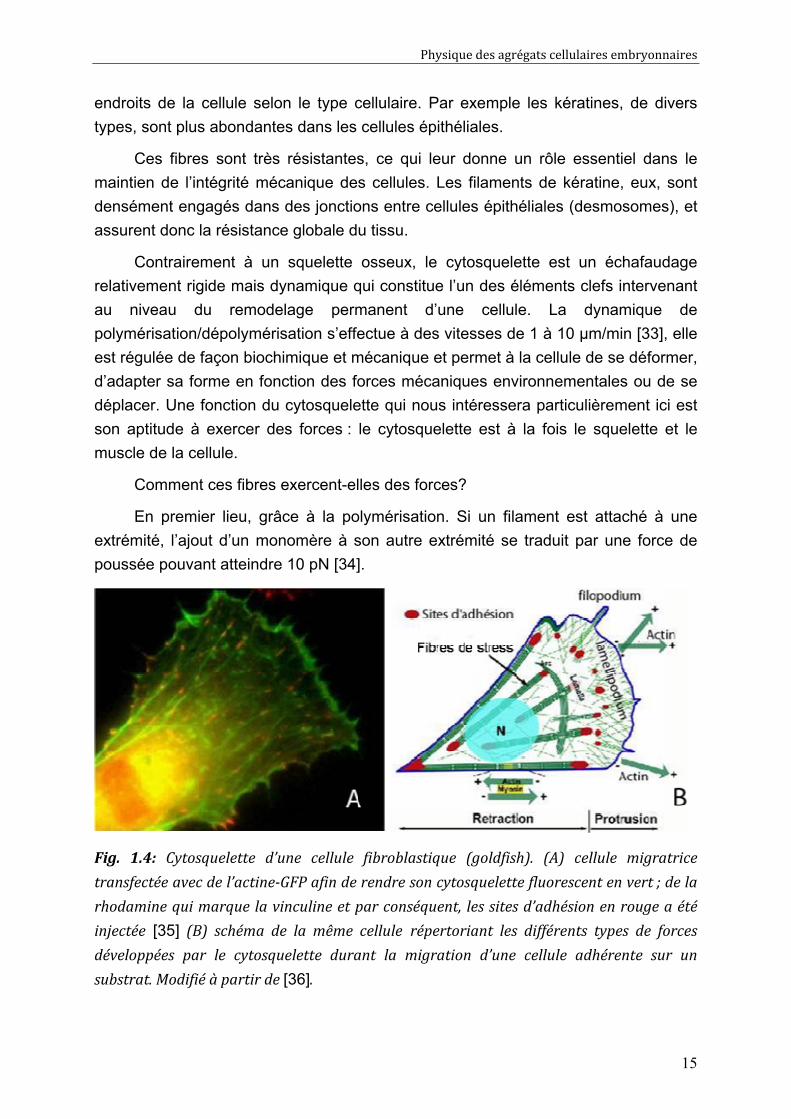
Fig. 1.4: Cytosquelette d’une cellule fibroblastique (goldfish). (A) cellule migratrice
transfectée avec de l’actine-GFP afin de rendre son cytosquelette fluorescent en vert ; de la
rhodamine qui marque la vinculine et par conséquent, les sites d’adhésion en rouge a été
injectée [35] (B) schéma de la même cellule répertoriant les différents types de forces
développées par le cytosquelette durant la migration d’une cellule adhérente sur un
substrat. Modifié à partir de [36].

Physique des agrégats cellulaires embryonnaires
16
En second lieu, des protéines qui agissent comme des moteurs moléculaires,
les myosines, sont capables de faire coulisser une fibre par rapport à l’autre, comme
dans un muscle. Ce mécanisme permet de développer des forces de l’ordre de 1.5
pN par moteur [34].
L’exemple d’une cellule fibroblastique en migration sur un substrat, (Fig. 1.4) montre
que la polymérisation de l’actine à l’avant de la cellule (dans le lamellipode),
s’accompagne de la formation de faisceaux radials (bundles) de fibres d’actine qui
poussent sur la membrane formant ainsi des pointes (filopodes). De plus, à l’intérieur
de la cellule, les fibres d’acto-myosine contribuent au développement des forces de
rétraction de la partie postérieure.
Le cytosquelette d’actine joue un rôle-clé dans l’adhésion cellulaire. Les
adhésions focales, qui sont des adhésions stables observées lorsque la cellule est
déposée sur un substrat rigide (sites rouges sur la Fig. 1.4), y sont connectées. Par
le biais de moteurs moléculaires, le cytosquelette peut exercer des forces de traction
sur les sites adhésifs, comme schématisé sur la Fig. 1.4. Cette force de traction
semble être assimilable à une contrainte constante, de l’ordre de 5 kPa : plus l’aire
d’adhésion est grande, plus la force totale exercée par la cellule est élevée. Chaque
site adhésif exerce ainsi une force comprise entre 10 et 30 nN sur le substrat [37].
C’est cette capacité à tirer sur le substrat, qui permet aux cellules de sonder les
propriétés mécaniques de leur environnement. En effet, les cellules dont l’activité de
moteurs moléculaires ciblés est inhibée (et qui donc n’exercent plus de forces sur
leurs adhésions) ne sont plus capables de former des adhésions focales [38]. Il ne
reste plus que des adhésions extrêmement transitoires, ce qui proscrit la mise en
marche de la machinerie de la division cellulaire ou de la différentiation.
Il existe d’autres types d’adhésion cellule/substrat ou cellule/cellule qui
permettent au cytosquelette d’accomplir un rôle dynamique et de développer des
forces. La partie suivante offre une présentation non exhaustive de différentes
structures d’adhésion cellulaire.
I.1.3. Généralités sur l’adhésion cellulaire
In vivo, mis à part quelques catégories de cellules pouvant "vivre" en
suspension (cellules circulantes du sang, macrophages), la plupart des cellules sont
adhérentes. Les mécanismes de l’adhésion jouent un rôle crucial pour la cellule. Ce
n’est que lorsqu’elles adhèrent qu’elles peuvent ensuite répliquer leur ADN, se
diviser, ou bien se différencier. Une cellule saine (non cancéreuse) qui se trouve
dans l’impossibilité d’adhérer se donne la mort dans de brefs délais. Ce mécanisme
d’auto-régulation évite que des cellules défaillantes ne se répandent et dégradent le

Physique des agrégats cellulaires embryonnaires
17
fonctionnement de l’organisme entier. L’adhésion est ainsi une étape-clé dans le
contrôle de la prolifération et de la différenciation.
Nombre de cellules adhèrent à un substrat de filaments protéiques enchevêtrés
(la matrice extracellulaire : extracellular matrix ou ecm), via des protéines
enchâssées dans leur membrane, les intégrines [39]. In vivo, on rencontre cette
adhésion cellule-ecm pour les cellules dans ou au contact du tissu conjonctif. In
vitro, on la rencontre lorsqu’on met des cellules en contact avec un substrat couvert
de filaments protéiques soit déposés par l’expérimentateur soit synthétisés par les
cellules elle mêmes. L’importance de cette adhésion dans la vie d’une cellule vient
du fait qu’elle assure le lien avec le milieu extérieur; adhésion et signalisation étant
étroitement couplées, les cellules sondent leur environnement par le biais de leurs
adhésions et adaptent en réponse leur activité chimique. L’adhésion implique la
formation de liaisons entre des protéines membranaires et leurs récepteurs
extracellulaires. Les protéines membranaires se répartissent en « cluster » (amas), si
bien que l’adhésion se fait par des points discrets, ainsi que la montre la figure Fig.
1.5. Si la formation de ces liaisons transmembranaires nécessite une compatibilité
chimique entre la cellule et son environnement, une compatibilité « mécanique » est
aussi requise : suivant les propriétés mécaniques de l’environnement de la cellule, et
en particulier de sa souplesse ou sa rigidité, des adhésions transitoires ou des
jonctions adhésives se forment avec une architecture protéique particulière, qui
donne lieu à un comportement cellulaire précis. On rencontre aussi ce type
d’adhésion cellule-ecm dans les agrégats de cellules fibroblastiques et dans ce cas,
l’ecm envahit probablement l’espace intercellulaire [40].
Nous n’avons pas travaillé véritablement sur ce type d’adhésion dans le cadre
de cette étude même si il est possible que la cohésion de certains de nos agrégats
(formés à partir de cellules d’Ovaire de Hamster Chinois modifiées génétiquement
(CHO VE voir chap2)) soit au moins en partie causée par leurs intégrines.
Néanmoins, une connaissance des interactions cellule-substrat reste indispensable
ne serait-ce que pour des raisons expérimentales : cela permet d’optimiser la
manipulation des cellules en culture ou en tissus (formation des agrégats), ou
d’éliminer des interactions nuisibles avec les surfaces pendant certaines expériences
(passivation).
Les cellules à l’intérieur des tissus présentent différents types d’adhésion
mutuelle. Via ces liaisons, elles sondent les propriétés mécaniques de leur
environnement cellulaire et adaptent leur activité mécanique et chimique en
conséquence. Les macromolécules qui appartiennent à la famille des cadhérines
assurent en grande partie les liaisons cellule/cellule (voir les principaux types de
contact cellulaire ci-dessous). Ces liaisons d'adhésion dépendent du calcium [41;42]
Physique des agrégats cellulaires embryonnaires
18
Ad
hé
sio
ns
Ce
llu
le/C
ell
ule
Ce
llu
le/E
CM
ADHESIONS JONCTIONNELLES ADHESIONS NON-JONCTIONNELLES
Jonctions serrées
Ceinture d’adhérence
Desmosome
Jonctions communicantes
Adhésion focal
Cadhérines
IgG-like CAMs
SélectinesIntégrines
Hémi-Desmosome Integral membrane
proteoglycan
Ad
hé
sio
ns
Ce
llu
le/C
ell
ule
Ce
llu
le/E
CM
ADHESIONS JONCTIONNELLES ADHESIONS NON-JONCTIONNELLES
Jonctions serrées
Ceinture d’adhérence
Desmosome
Jonctions communicantes
Adhésion focal
Cadhérines
IgG-like CAMs
SélectinesIntégrines
Hémi-Desmosome Integral membrane
proteoglycan
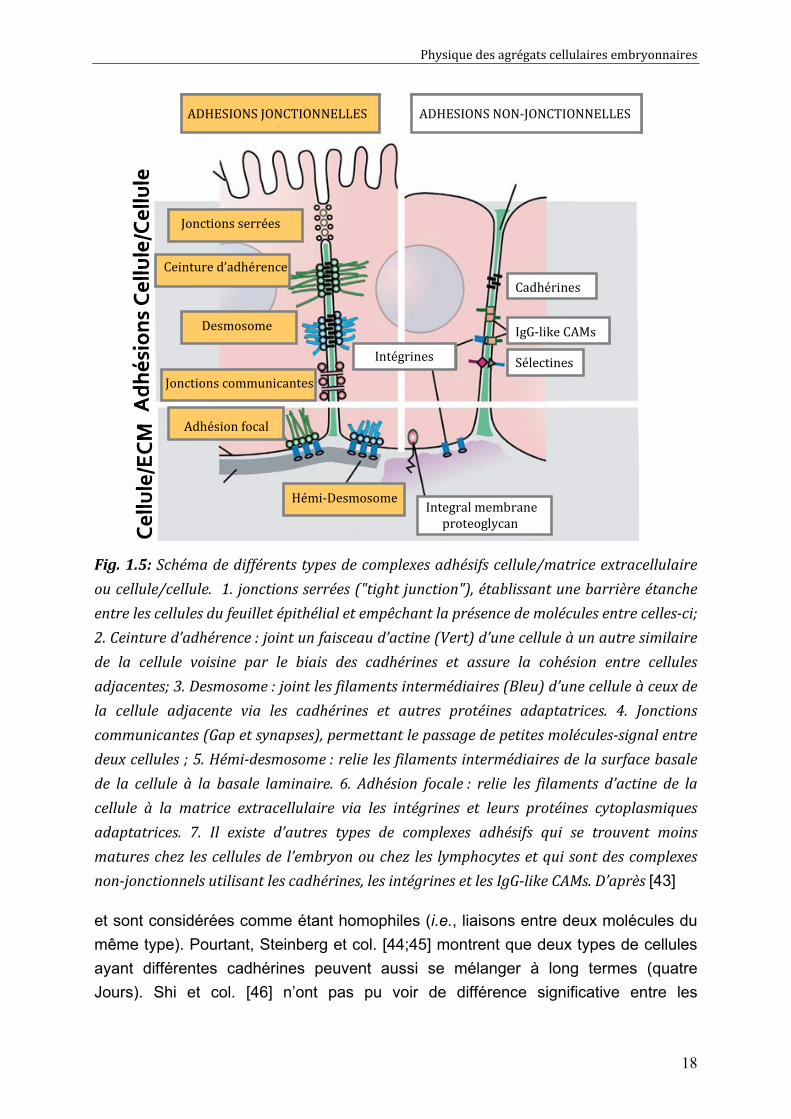
Fig. 1.5: Schéma de différents types de complexes adhésifs cellule/matrice extracellulaire
ou cellule/cellule. 1. jonctions serrées ("tight junction"), établissant une barrière étanche
entre les cellules du feuillet épithélial et empêchant la présence de molécules entre celles-ci;
2. Ceinture d’adhérence : joint un faisceau d’actine (Vert) d’une cellule à un autre similaire
de la cellule voisine par le biais des cadhérines et assure la cohésion entre cellules
adjacentes; 3. Desmosome : joint les filaments intermédiaires (Bleu) d’une cellule à ceux de
la cellule adjacente via les cadhérines et autres protéines adaptatrices. 4. Jonctions
communicantes (Gap et synapses), permettant le passage de petites molécules-signal entre
deux cellules ; 5. Hémi-desmosome : relie les filaments intermédiaires de la surface basale
de la cellule à la basale laminaire. 6. Adhésion focale : relie les filaments d’actine de la
cellule à la matrice extracellulaire via les intégrines et leurs protéines cytoplasmiques
adaptatrices. 7. Il existe d’autres types de complexes adhésifs qui se trouvent moins
matures chez les cellules de l’embryon ou chez les lymphocytes et qui sont des complexes
non-jonctionnels utilisant les cadhérines, les intégrines et les IgG-like CAMs. D’après [43]
et sont considérées comme étant homophiles (i.e., liaisons entre deux molécules du
même type). Pourtant, Steinberg et col. [44;45] montrent que deux types de cellules
ayant différentes cadhérines peuvent aussi se mélanger à long termes (quatre
Jours). Shi et col. [46] n’ont pas pu voir de différence significative entre les

Physique des agrégats cellulaires embryonnaires
19
interactions homophiles et hétérophiles à l’échelle d’un seul pont adhésif.
Récemment, Cavey et col. [47] ont montré qu’il existe deux types différents de
mécanisme de stabilisation et d’immobilisation des cadhérines. En outre, il existe
plusieurs classes de cadhérines (voir [48;49] pour une revue).
Selon la structure de la liaison ou de la nature de son association avec le
cytosquelette derrière, on peut distinguer plusieurs types de contacts cellule/cellule
(Fig. 1.5)
- Les jonctions serrées (tight junctions) sont des jonctions très étanches qui
définissent une barrière physiologique entre les compartiments extérieur et intérieur
de l'organisme.
- Les jonctions communicantes (gap junctions) permettent une communication
directe entre les cytoplasmes des cellules adjacentes. Au niveau des jonctions
communicantes, les cellules adjacentes sont unies entre elles par des petits canaux
intercellulaires tubulaires. Chaque canal intercellulaire est formé de l'aboutement de
2 hémi-canaux (ou connexions), chacun faisant partie de la membrane de chacune
des 2 cellules adjacentes.
- Les desmosomes sont des structures en forme de disque d'environ 0.1 à 0.5 µm
de diamètre et 100 nm d'épaisseur. Les desmosomes, sur lesquels s’ancrent les
filaments intermédiaires via des protéines cytoplasmiques, assurent les liaisons
intercellulaires par des molécules transmembranaires de la superfamille des
cadhérines (desmogléines et desmocollines). Ces molécules sont en relation avec la
plaque desmosomale qui contient en particulier de la plakoglobine et des
desmoplakines.
- Les jonctions adhérentes (adhesion belt) : Ces jonctions d'ancrage se forment par
l'intermédiaire des cadhérines. Bien que l'adhérence de ces molécules dépende de
leur domaine extra-cellulaire, celle-ci est modulée par des molécules
cytoplasmiques, les caténines (α-caténine, β-caténine et γ-caténine) qui se lient
d'une part au domaine cytoplasmique des cadhérines et d'autre part - par
l'intermédiaire de nombreuses protéines cytoplasmiques telle l’Eplin - aux filaments
d'actine (Fig. 1.6). La formation des jonctions adhérentes commence par l'adhésion
entre les molécules de cadhérines, puis viennent s’associer les caténines (α-
caténine, β-caténine) pour permettre de lier le domaine intracellulaire de la cadhérine
au cytosquelette. La liaison cadhérine-caténine ne semble pas être une simple
liaison physique direct [50]. Ainsi, les schémas simplificateurs tels que ceux
présentés en Fig. 1.6. peuvent donner une idée assez fausse et beaucoup trop
simplificatrice des caténines, les présentant comme de simples briques dans la

Physique des agrégats cellulaires embryonnaires
20
formation de la liaison cadhérine-cytosquelette. Néanmoins, les caténines sont
souvent nécessaires pour une adhésion effective [51]. En effet la mobilité d’un amas
dense de cadhérines est régulée par la liaison actine-cadhérine via l’α-caténine. Les
caténines ont d’autres fonctions allant de la régulation de la dynamique de l’actine
[52], jusqu’à l’interaction avec plusieurs voies de signalisation [48]. Enfin, il faut
mentionner qu’il existe d’autre types de liaisons entre les cadhérines et le
cytosquelette [53;54].
Me
mb
ran
e p
lasm
iqu
e
Ep
linE
plin
Me
mb
ran
e p
lasm
iqu
e
Ep
linE
plin
Fig. 1.6: Schéma de liaison entre le domaine cytoplasmique des cadhérines et le
cytosquelette. Cette liaison fait appel à des protéines cytoplasmiques adaptatrices tel que
les α-caténines, les β-caténines, p120, et d’autres protéines récemment identifiées sous le
nom d’EPLIN.
I.1.4. Physique du cytosquelette et de l’adhésion.
De par le large nombre de molécules en interaction, comprendre la contribution
du cytosquelette et de ses mécanismes moléculaires sur les propriétés physiques
des cellules reste encore un immense challenge expérimental et théorique. Janmey
et col. [17] ont étudié en détails les propriétés de gels de microtubules, de filaments
d’actine, et de filaments intermédiaires de vimentine in vitro. Selon les résultats de
cette étude, les réseaux de filaments d’actines sont les plus rigides (i.e., un gel de
filaments d’actine est moins déformable pour le même taux de stress appliqué qu’un
gel de microtubules ou de filaments intermédiaires). Les microtubules se
déstabilisent plus facilement que les filaments d’actine, ils contribuent aux propriétés
mécaniques des cellules d’autant qu’ils sont liés aux autres filaments du

Physique des agrégats cellulaires embryonnaires
21
cytosquelette. Les filaments intermédiaires sont faciles à déformer à des petits taux
de stress, mais sous une grande déformation ils deviennent plus rigides.
Depuis, une large variété de techniques et de mesures biophysiques ont été
appliquées afin d’étudier les propriétés du cytosquelette, in vitro et in vivo : pinces
optiques et magnétiques, microscopie à force atomique, méthodes rhéologiques
(cisaillement, oscillation), micro-rhéologie passive (diffusion des molécules). En
particulier, les effets d’organisation spatiale et la régulation d’interaction ont pu être
élucidés par des études in vivo [55-57]. D’ailleurs, le cytosquelette influence la
rigidité ainsi que l’organisation spatiale des jonctions adhérentes, et vice versa, les
jonctions adhérentes influencent la composition et l’organisation du cytosquelette,
d’une manière qui commence à être élucidée [54].
De même, l’adhésion est une propriété incontournable quant on parle de la
physique à l’échelle de la cellule vivante. Physiquement, l’adhésion est
l’établissement d’un pont entre les molécules, il pourra être facilement décrit par une
force attractive, ou une énergie potentielle qui diminue avec un contact. Bell. et al.
[58;59] ont montré que l’action physique des molécules adhésives dépend de leurs
dynamique chimique et que les sites d’adhésion mobiles peuvent se concentrer dans
des régions adhésives. Evans [60;61] a montré qu’il existe une transition entre une
forme continue et une forme discrète de l’adhésion le long de la membrane
cytoplasmique. L’adhésion continue n’est correcte que lorsque les molécules
d’adhésion sont suffisamment concentrées sur la membrane. Plus tard, la
spectroscopie de la molécule unique a permis de montrer que l’établissement et la
rupture d’un pont d’adhésion entre les macromolécules, ainsi que la force associé à
ces événements dépendent de la vitesse d’approche ou de séparation [10;11;62].
Dans le cas d’une cellule unique, modéliser l’adhésion par une attraction
continue [8;63], ou le cytosquelette par un gel d’actine ou par des fils et des cordes
interconnectés [64] ont amenés des avancés importantes sur notre compréhension
de la biomécanique cellulaire. Mais comment les propriétés physiques et chimiques
de l’adhésion et/ou du cytosquelette agissent sur les propriétés mécaniques aux
échelles tissulaires reste encore une question très ouverte. Des mécanismes
complexes et collectifs d'adhésion et de mouvements cellulaires interviennent lors de
la formation, de la cohésion et de l’organisation des tissus [65;66]. Cependant, établir
des relations ou des formules mathématiques reliant les comportements ou la forme
des tissus à des paramètres physiques mesurables d’une part, et à la mécanique
des cellules et leurs propriétés d’adhésion d’autre part, n’est toujours pas évident.
À l’intérieur des tissus, les cellules sont entourées par d’autres cellules
mutuellement adhérentes, et les molécules d’adhésion intercellulaires sont recrutées

Physique des agrégats cellulaires embryonnaires
22
dans des jonctions qui interagissent avec le cytosquelette. C’est une structure
tellement complexe et difficile à étudier expérimentalement ou théoriquement, qu’à
notre connaissance, une étude complète et cohérente qui prend en compte les
différentes échelles temporelles ou spatiales allant des cellules aux tissus n’existe
pas encore. Néanmoins, il est bien admis qu’il existe une relation directe entre les
comportements tissulaires et la mécanique des cellules qui les constituent. Et bien
sûr, les propriétés mécaniques du cytosquelette des cellules, ainsi que l’adhésion
cellulaire ont une influence primordiale sur la mécanique tissulaire, sur la forme et la
structure des tissus [67-70].
Dans cette thèse, nous avons essayé de mieux comprendre les
comportements collectifs des cellules dans des tissus modèles (agrégats cellulaires).
Nous avons développé des méthodes expérimentales originales issues de la
physique pour les caractériser. En particulier, nous souhaitions étudier les
écoulements et la réponse mécanique des cellules soumises à une force extérieure,
mesurer les quantités « liquides » associées (tension de surface, viscosité,
plasticité…) et essayer de les relier aux propriétés du cytosquelette et de l’adhésion.
I.2. Les tissus
Un tissu est un groupement plus ou moins homogène de cellules qui ont la
même origine et la même fonction. Les tissus constituent un niveau d'organisation
intermédiaire entre les cellules et les organes. Souvent, différents types de tissus
coexistent pour construire un organe. Par exemple, l’œil composé de l’iris, de la
rétine ganglionnaire juxtaposée à la rétine pigmenté, et des couches plus externes,
la sclérotique, etc … constitue l’organe de la vision chez l’être vivant. Tous ces tissus
accomplissent leurs missions spécifiques et transforment les vibrations
électromagnétiques de la lumière en un flux nerveux qui sera transmis au cerveau.
Un tissu apparaît dans le corps d’un organisme dès le stade précoce de son
développement embryonnaire, il se développe et se manifeste sous différentes
formes jusqu’à finir par prendre une place précise sous une conformation définie. La
mise en place d’un tissu qui est le domaine d’étude central de la biologie du
développement, fait intervenir toute une cascade de processus aussi bien
biologiques, chimiques que physiques et mécaniques. En partant d’une cellule, la
formation d’un tissu commence par la division cellulaire, qui donne lieu à un amas de
cellules ayant les mêmes informations génétiques. Ensuite, cet amas subit de
nombreuses étapes de réorganisation tissulaires qui font appel aux interactions
biochimiques, aux forces mécaniques et à des mouvements cellulaires collectifs
actifs (écoulements).

Physique des agrégats cellulaires embryonnaires
23
Dans la suite de cette partie, nous allons détailler les comportements des tissus
à partir de leur formation ainsi que leur mise en place tout en essayant d’expliquer
l’impact des forces mécaniques sur leur organisation. Dans une deuxième partie,
nous nous intéresserons aux processus de réorganisation cellulaire in vitro dans des
agrégats de cellules réagrégées puis à la caractérisation physique de ces matériaux
cellulaires modèles et enfin à leur description rhéologique.
I.2.1. Les tissus in vivo : l’embryogenèse
Le développement embryonnaire démarre généralement après la fécondation
d’un ovule par un spermatozoïde. Ensuite, la cellule fécondée se divise rapidement.
Ce stade de division cellulaire intense est suivi par une phase de réarrangement
massif au niveau cellulaire et tissulaire, la gastrulation [71]. Ces réarrangements
conduisent à la formation des trois feuillets primordiaux de l’embryon : l’ectoderme,
l’endoderme, et le mésoderme.
L’origine de tous les tissus de l’embryon peut être associée à l’un de ces
feuillets. Ces trois feuillets se rencontrent chez tous les animaux à l’exclusion de
quelques classes d’organismes très primitifs tels que les diblastiques (coraux,
méduses, anémones de mer) ou les spongiaires. Le feuillet le plus superficiel,
l’ectoderme, est à l’origine de l’épiderme, des structures tégumentaires et du
système nerveux. Le feuillet le plus interne, l’endoderme, fournit le tube digestif et les
glandes associées telles les poumons, le foie ou le pancréas. Du feuillet
intermédiaire, le mésoderme, dérive la charpente de l’organisme constitué du
squelette et des muscles.
La morphogenèse est la phase durant laquelle les tissus et les organes seront
mis en place afin de donner à l’organisme sa forme finale. Afin de mieux illustrer les
différentes phases du développement embryonnaire, on va considérer l’exemple du
développement du poisson zèbre. De plus, il s’agit de l’un des modèles cellulaires
que nous avons étudié pendant cette thèse en collaboration avec C.P. Heisenberg et
G. Krens de l’institut MaxPlanck de Dresden.
Le poisson zèbre ou Danio (Danio rerio) est une espèce de poisson ayant une
taille moyenne de 3-4 cm (Fig. 1.7). Son habitat naturel est en Asie [72]. Le poisson
zèbre est l'un des organismes modèles les plus courants en biologie du
développement. En plus de sa petite taille, de la facilité de son entretien, de sa
reproduction rapide, il présente un certain nombre d’avantages : il permet une
analyse génétique (grande collection de mutants, stratégies d’analyse du génome en
plein essor) ainsi qu'une approche embryologique classique facilitée par la
transparence des embryons et leur développement très rapide (gastrulation après 5h

Physique des agrégats cellulaires embryonnaires
24
de développement). Son développement à l’extérieur de la mère permet un suivi
détaillé de ses mouvements morphogénétiques durant les différents stades de
développement.
Fig. 1.7: image d’un poisson zèbre adulte de longueur de 3-4cm.
[http://fr.wikipedia.org/wiki/Poisson_z%C3%A8bre]
Le développement embryonnaire commence à partir d’un œuf fertilisé. Après
fécondation, la première cellule, située au sommet d'une grande réserve vitelline, va
subir une succession de divisions synchrones, tout en diminuant leurs tailles,
conduisant à un ensemble de cellules identiques et totipotentes. Jusqu'au stade de
transition miblastuléenne (1024 cellules), le développement s'effectue sous contrôle
de l'information maternelle [73]. Ce n'est qu'à partir de ce stade que le génome
zygotique commence à être transcrit et que les cellules deviennent mobiles. Les
cellules, situées au pôle animal2, vont commencer à se diviser de manière
asynchrone puis vont progressivement recouvrir la réserve vitelline3 (Fig. 1.8) au
cours du phénomène d’épibolie4. Lorsque 50 % du vitellus est recouvert par les
cellules embryonnaires, la gastrulation commence afin de transformer le blastula non
structuré en un embryon ayant 3 feuillets définis : l’ectoderme, l’endoderme et le
mésoderme.
Durant la gastrulation, trois mouvements, nécessaires à l’établissement de ces
trois feuillets primordiaux, sont à considérer : le mouvement d'épibolie qui se poursuit
jusqu'à ce que toute la réserve vitelline soit recouverte par les cellules
embryonnaires, le mouvement d'invagination des cellules de la marge (Fig. 1.9) qui
va permettre la formation des deux feuillets embryonnaires profonds, l’endoderme et
le mésoderme, et le mouvement de convergence qui conduit les cellules ventrales
2 Partie la plus antérieure de l’embryon (tête). 3 Jaune d’œuf. 4 Mouvement conduisant au recouvrement de la réserve vitelline par les cellules embryonnaires situées au pôle
animal.

Physique des agrégats cellulaires embryonnaires
25
vers la face dorsale de l’embryon où s'allonge l'axe embryonnaire [74]. C'est
immédiatement après la transition mi-blastuléenne qu'une cascade d’événements
moléculaires va progressivement conduire les cellules de l'embryon à adopter un
devenir particulier tel qu'on peut le définir sur la carte des territoires présomptifs
établie à la fin du stade blastula (Fig. 1.9.B). À ce stade, les cellules embryonnaires
forment une couche unique recouvrant l'hémisphère animal du vitellus. Par analogie
Fig. 1.8: (A-B): Deux vues d’un œuf fertilisé. (A) avec chorion protecteur, et (B) après
retrait du chorion, sur le pôle animal on repère la première cellule en haut de l’image. Au
milieu: Exemple d’une image d’embryon après les premières divisions cellulaires. (C) au
stade de 2 cellules –(D) au stade de 16 cellules. La taille d’une cellule diminue à chaque
division avec l’augmentation du nombre de cellules embryonnaires. À droite, embryons au
stade de Blastula (E) entre “high and oblong stage” et (F) entre “oblong and sphere stage”.
Barre 250 μm. Image modifiée à partir de [75].
avec le système de coordonnées utilisé pour définir une position sur le globe
terrestre on peut caractériser le devenir d'une cellule donnée en fin de stade blastula
en fonction de deux coordonnées : sa longitude, qui correspond à la position de cette
cellule de long de l’axe dorso-ventral (D/V, avec pour méridien d'origine l'axe
embryonnaire qui se forme dorsalement) et sa latitude qui correspond à la position
de cette même cellule le long de l’axe margino-animal (Fig. 1.9.B).

Physique des agrégats cellulaires embryonnaires
26
Biologiquement, en utilisant les méthodologies de perturbation de la formation
des axes embryonnaires (par la perte de fonction d’éléments requis pour la formation
de ces axes, [76;77] ou bien en inhibant la fonction des gènes d’intérêt par la
technique de morpholino knock-down [78], ou au contraire en réalisant des
expériences de surexpression d’un facteur par injection d’ARN dans des embryons à
des stades très précoces de développement), de nombreuses études (voir [73] pour
une revue) ont montré que la coordonnée «longitude» d'une cellule, est définie par
l’action morphogénique des gènes BMP5 (voir [79] pour une revue).
Outre l'information D/V, pour définir son identité spatiale au sein de la blastula,
une cellule donnée requiert une coordonnée de latitude, c'est-à-dire une information
moléculaire sur sa position le long de l'axe margino-animal. Cette coordonnée
permet de définir sa position le long de l'axe antéro-postérieur (A/P) en fin
5 bone morphogenetic proteins
Fig. 1.9 : Carte des territoires présomptifs d’un embryon de poisson zèbre au stade de
blastula tardive. A. Les cellules embryonnaires sont disposées sur l'hémisphère animal de la
réserve vitelline. Le mouvement d’épibolie conduit les cellules de la marge jusqu'au niveau
de l’équateur. PA : pôle animal ; PV : pôle végétal ; D : dorsal ; V : ventral. B. Carte des
territoires présomptifs de l’embryon au même stade qu'en A (5h de développement, d’après
[54, 55]). La région marginale comprend des cellules endodermiques, les cellules
mésodermiques incluant la plaque préchordale (pp), la notochorde (Not), les muscles et le
territoire sanguin. Au niveau de l’ectoderme, on distingue la moelle épinière ou corde
spinale (C. sp), le rhombencéphale (Rhomb), le mésencéphale (Més), le diencéphale (Di), le
télencéphale (Tél) et l’épiderme (Épi). Le devenir de chaque cellule peut être défini selon
ses coordonnées (flèches rouges) : sa longitude (le long de l’ave D/V) et sa latitude (le long
de l’axe marge/pôle animal ou antéro-postérieur, A/P). C. Structures dérivant des
territoires présomptifs décrits en B. Même code couleur. La glande d'éclosion (Gl) dérive de
la plaque préchordale [74]

Physique des agrégats cellulaires embryonnaires
27
d’embryogenèse. En effet, l'observation de la carte des territoires présomptifs (Fig.
1.9.B) à comparer avec (Fig. 1.9.C) montre que les territoires situés au pôle animal
vont conduire à la formation des yeux et du cerveau antérieur (télencéphale et
diencéphale) alors que des territoires situés dans des régions progressivement plus
végétales se différencient en structures neurales plus postérieures telles que le
cerveau moyen (mésencéphale) puis le cerveau postérieur (rhombencéphale) et
enfin la moelle épinière. En revanche, dans la région marginale, les cellules vont
ensuite migrer sous l’ectoderme en direction du pôle animal en formant les feuillets
internes (mésoderme et endoderme). Selon des études biochimiques, l’organisation
établie selon l’axe A/P est le résultat des activités de différents facteurs moléculaires
(voir [80-83] pour détails).
Ainsi, durant la morphogenèse, le mouvement cellulaire est gouverné par des
informations génétiques sécrétant des facteurs moléculaires, et implique
l’intervention de signaux mécaniques [84] et biochimiques. Ces signaux permettent
aux cellules d’investir leur environnement et de répondre aux ordres afin de
« s’écouler » vers leur destination à l’intérieur de l’organisme. Si les biologistes du
développement ont fourni des efforts énormes afin d’identifier les facteurs qui dirigent
l’organisation des tissus, les effets de chaque gène durant la cascade du
développement, l’intervention de scientifiques de différents domaines (physiciens,
mathématiciens, et chimistes, etc.) a laissé des contributions marquantes pour la
biologie du développement. Aujourd’hui encore, leur intervention est nécessaire pour
caractériser les tissus par des techniques originales ou expliquer les mécanismes
complexes de la morphogenèse [85] et plus généralement de la migration cellulaire.
Les parties suivantes décrivent les réponses et les hypothèses issues de la physique
depuis un peu plus d’un siècle vis-à-vis des questions du développement.
I.2.2. Les tissus in vitro : l’affinité cellulaire
Dès le début du 20ème siècle, de nombreux chercheurs ont essayé de percer
les mystères de l’organisation tissulaire. Parmi eux, Edmund B. Wilson, en 1907, a
découvert que les cellules ou tissus dissociés à partir d’une éponge marine peuvent
se réunir à nouveau dans un seul corps cellulaire, l’agrégat. De plus cet agrégat
évolue jusqu’à la reconstruction d’une éponge fonctionnelle [86].
Ces travaux furent ensuite poursuivis plus systématiquement par Johannes
Holtfreter, qui mélangea in vitro des cellules embryonnaires d’amphibiens provenant
d’espèces et de tissus distincts. Une deuxième découverte d’importance majeure a
lieu en 1936 : Townes et Holtfreter établissent que les cellules ainsi recombinées
subissent une ségrégation spatiale (Fig. 1.9), en ce sens qu’au lieu d’adopter une

Physique des agrégats cellulaires embryonnaires
28
disposition aléatoire à l’intérieur d’un amas, les cellules s’associent par types. « Qui
se ressemble s’assemble », dit le proverbe : les cellules épidermiques migrent
ensemble vers la périphérie de l’amas, et les cellules mésenchymateuses vers
l’intérieur, les deuxièmes étant le plus souvent complètement entourées par les
premières.
Fig. 1.9: Réassociation des cellules de neurulas de batraciens. Deux suspensions de cellules,
ont été mélangées, l’une provenant de l’ectoderme présomptif d’un embryon fortement
pigmenté, et l’autre de la future plaque neurale d’un embryon non pigmenté. Ces cellules se
rassemblent et se trient de manière telle que l’épiderme présomptif recouvre l’autre tissu.
(D’après Townes & Holtfreter [87])
En 1955, Townes et Holtfreter complètent ces résultats et publient une
impressionnante série d’expériences de tri cellulaire [87]. Ils notent que la position
finale des cellules dans les amas recombinés reflète leurs positions normales dans
l’embryon, c'est-à-dire que le mésoderme est plus central que l’ectoderme, à la
surface interne duquel il adhère fermement (Fig. 1.10). Le mésoderme adopte
également une position centrale par rapport à l’endoderme (Fig. 1.10). Toutefois
lorsqu’on mélange des suspensions cellulaires préparées à partir des trois feuillets,
l’endoderme se place au centre de l’amas ; il est alors recouvert par le mésoderme,
lui-même situé sous l’ectoderme qui finalement occupe la périphérie (Fig. 1.10). Des
agrégats provenant de la recombinaison de cellules de l’épiderme et de la plaque
neurale sont également capables de reproduire la situation normale dans l’embryon
(Fig. 1.10D et Fig. 1.10E). Enfin, lorsque la suspension de cellules comporte des
cellules présomptives axiales (notochrode) en plus des cellules présomptives
épidermiques et neurales, l’amas recombiné présente une couche épidermique
superficielle, du tissu nerveux au centre, et du tissu mésodermique entre les deux
Physique des agrégats cellulaires embryonnaires
29
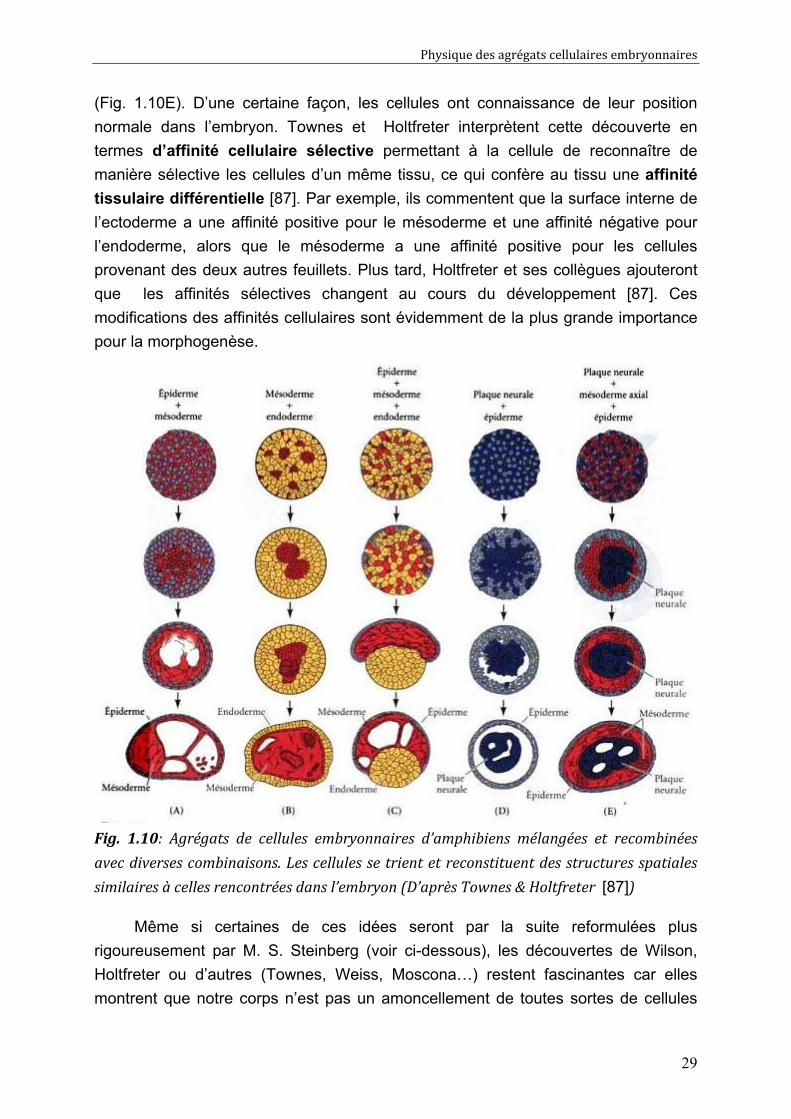
(Fig. 1.10E). D’une certaine façon, les cellules ont connaissance de leur position
normale dans l’embryon. Townes et Holtfreter interprètent cette découverte en
termes d’affinité cellulaire sélective permettant à la cellule de reconnaître de
manière sélective les cellules d’un même tissu, ce qui confère au tissu une affinité
tissulaire différentielle [87]. Par exemple, ils commentent que la surface interne de
l’ectoderme a une affinité positive pour le mésoderme et une affinité négative pour
l’endoderme, alors que le mésoderme a une affinité positive pour les cellules
provenant des deux autres feuillets. Plus tard, Holtfreter et ses collègues ajouteront
que les affinités sélectives changent au cours du développement [87]. Ces
modifications des affinités cellulaires sont évidemment de la plus grande importance
pour la morphogenèse.
Fig. 1.10: Agrégats de cellules embryonnaires d’amphibiens mélangées et recombinées
avec diverses combinaisons. Les cellules se trient et reconstituent des structures spatiales
similaires à celles rencontrées dans l’embryon (D’après Townes & Holtfreter [87])
Même si certaines de ces idées seront par la suite reformulées plus
rigoureusement par M. S. Steinberg (voir ci-dessous), les découvertes de Wilson,
Holtfreter ou d’autres (Townes, Weiss, Moscona…) restent fascinantes car elles
montrent que notre corps n’est pas un amoncellement de toutes sortes de cellules

Physique des agrégats cellulaires embryonnaires
30
assemblées au hasard, que les agrégats de cellules dissociées-réagrégées sont un
système modèle pertinent de tissus. La biologie moléculaire va pouvoir s’en servir
avec ses nouveaux outils extrêmement puissants [44;85;88;89].
On peut pour finir cette partie résumer ci-dessous les découvertes majeures
réalisées durant la première moitié du 20ème siècle sur la reconstruction in vitro des
tissus à partir de cellules dissociées :
(i) Quand les cellules des différents tissus embryonnaires sont dissociées puis
mélangées ensemble, elles sont capables de rétablir des liaisons de contact entre
elles afin de former un agrégat commun.
(ii) Lorsqu’un agrégat est formé de plusieurs types des cellules, chacun de ces
types se regroupe et se trie comme dans les différents tissus d’origines.
(iii) Les tissus se reconstruisent dans des positions définies à l’intérieur de l’agrégat.
(iv) Dans le cas où les tissus proviennent d’un même complexe chez l’embryon, la
structure de ce complexe sera bien reflétée par leur reconstruction in vitro.
I.2.3. L’hypothèse d’adhésion différentielle (HAD).
Les cellules ne s’associent donc pas au hasard, mais sont capables de se
déplacer et de se reconnaître pour s’organiser en tissus complexes. Plusieurs
théories ou hypothèses ont été proposés afin d’expliquer ces « écoulements actifs »
et d’identifier les forces mises en jeu durant la morphogenèse. L’Hypothèse
d’Adhésion Différentielle (HAD) développée par Steinberg dans les années 60,
postule une analogie intéressante entre les liquides et les tissus et en particulier
comme on le verra, l’existence d’une tension de surface tissulaire. Cette hypothèse
permet d’attribuer un sens physique à l’affinité cellulaire puis de quantifier à partir
des lois de la thermodynamique et de l’hydrodynamique le comportement des
agrégats cellulaires comme l’arrondissement ou le tri-cellulaire.
I.2.3.1 Le contexte de l’HAD et l’analogie liquides/tissus.
Durant le développement normal, on rencontre fréquemment la situation où un
tissu A s’écoule de sa position initiale afin de couvrir la surface d’un autre tissu B
(voir par exemple le phénomène d’épibolie du poisson zèbre, Fig. 1.9). D’autre part,
dans un mélange de ces mêmes deux types de cellules dissociées et réagrégées
dans un agrégat cellulaire in vitro, on peut aboutir à une configuration finale mimant
celle du développement embryonnaire. Tirant parti de cette découverte, Steinberg
postule que les fonctions responsables de cette organisation sont en liaison directe
avec les propriétés des cellules. Dans le but d’identifier ces propriétés, il réalisa lui
Physique des agrégats cellulaires embryonnaires
31
aussi au début des années 60 des expériences sur le tri (ségrégation) in vitro de
différents types de tissus dissociés d’embryons de poulet (rétine ganglionnaire et
pigmenté, cœur, etc.) [65;90]. Suite à ces expériences, il postule : (i) que les cellules
durant le tri ont un mouvement actif dirigé ainsi qu’une adhésion mutuelle sélective,
(ii) que le tri entre deux tissus mélangés ne peut avoir lieu que si les deux tissus
présentent une adhésion différentielle entre leurs cellules motiles. Une combinaison
de ces deux propriétés a été proposée par plusieurs auteurs à l’époque (voir [65]
pour une revue) même avant que Steinberg développe son hypothèse HAD en 1963.
Cependant, le mérite de Steinberg est de repérer puis de formaliser les
comportements liquides chez de tel matériaux cellulaires. Examinons d’où provient
cette idée de similarité entre tissus et liquides.
Les gaz sont constitués d’unités (atomes ou molécules) de très grande mobilité
avec des interactions négligeables. Les solides ont des forces de cohésion énormes
qui empêchent le mouvement à distance de leurs unités. Entre ces deux états de la
matière, se trouvent les liquides où les unités présentent à la fois une grande mobilité
et des forces de cohésion.
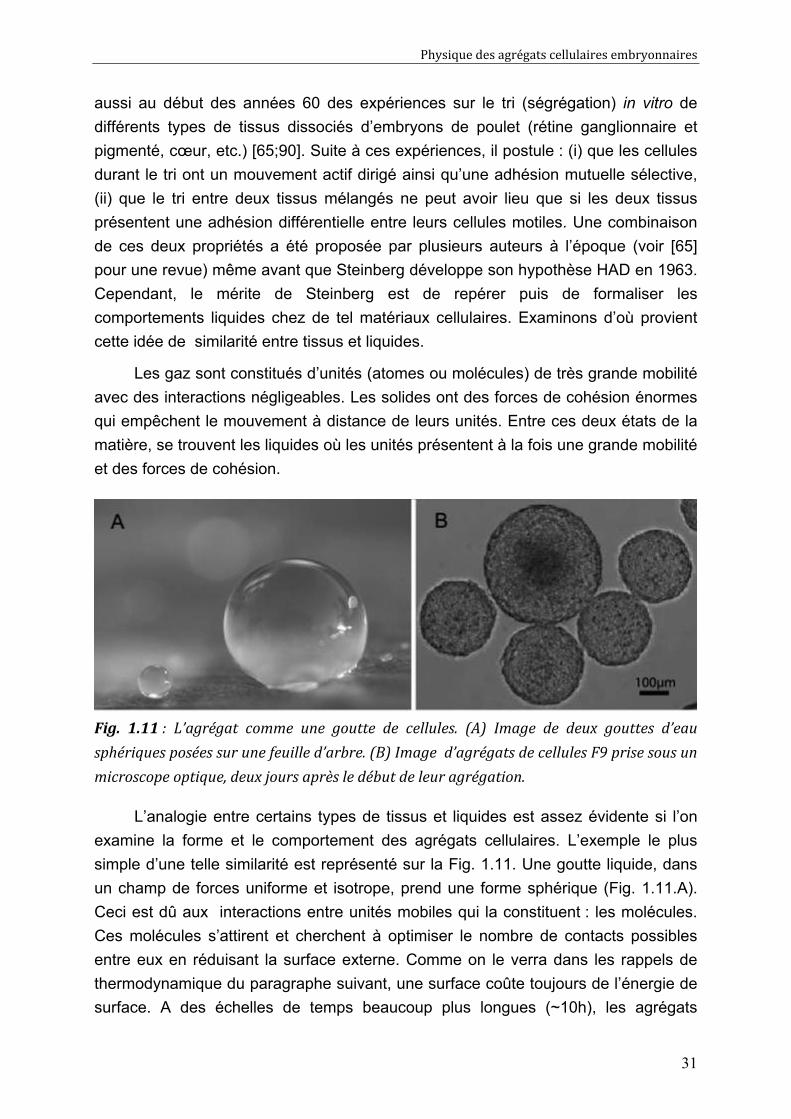
Fig. 1.11 : L’agrégat comme une goutte de cellules. (A) Image de deux gouttes d’eau
sphériques posées sur une feuille d’arbre. (B) Image d’agrégats de cellules F9 prise sous un
microscope optique, deux jours après le début de leur agrégation.
L’analogie entre certains types de tissus et liquides est assez évidente si l’on
examine la forme et le comportement des agrégats cellulaires. L’exemple le plus
simple d’une telle similarité est représenté sur la Fig. 1.11. Une goutte liquide, dans
un champ de forces uniforme et isotrope, prend une forme sphérique (Fig. 1.11.A).
Ceci est dû aux interactions entre unités mobiles qui la constituent : les molécules.
Ces molécules s’attirent et cherchent à optimiser le nombre de contacts possibles
entre eux en réduisant la surface externe. Comme on le verra dans les rappels de
thermodynamique du paragraphe suivant, une surface coûte toujours de l’énergie de
surface. A des échelles de temps beaucoup plus longues (~10h), les agrégats

Physique des agrégats cellulaires embryonnaires
32
cellulaires profitent de la motilité de leurs unités (cellules) afin d’évoluer vers une
situation similaire de contact maximum entre cellules : l’agrégat prend une forme
sphérique (Fig. 1.11.B). Steinberg sera le premier à postuler que ces agrégats
cellulaires possèdent aussi une tension de surface tissulaire apparente.
Les autres exemples, qui ont mis en évidence cette analogie liquides/tissus,
sont les phénomènes de tri-cellulaire (Fig. 1.12.A) similaires à la démixtion de deux
liquides immiscibles, ou encore la fusion entre deux agrégats simulant la
coalescence entre deux gouttes de liquide (Fig. 1.12.B).
Fig. 1.12 : Comportements liquides des agrégats cellulaires. (A) tri cellulaire et
arrondissement des agrégats formés, en haut, d’un mélange de cellules neurales et
pigmentés de rétine de poulet [91], et en bas de cellules d’ectoderme et d’endoderme de
l’hydre [92]. (B) Coalescence de deux agrégats identiques de cellules embryonnaires de
souris (F9), voir chapitre 4.
Au-delà de l’aspect qualitatif, l’intérêt de cette analogie est de fournir un point
de départ vers une interprétation thermodynamique et hydrodynamique de l’évolution
des tissus. Avant de décrire le modèle thermodynamique original de Steinberg
décrivant le tri cellulaire, il est utile de rappeler quelques notions élémentaires de
physique des liquides et des surfaces.
I.2.3.2 Notions de physique des liquides et des surfaces.
Un des principes fondamentaux de la thermodynamique prédit qu’un système
tend toujours vers l’état le plus stable, ce qui se mesure par une décroissance de son
potentiel thermodynamique. Ce principe thermodynamique peut être appliqué à une
goutte mais aussi à des agrégats de certains types de cellules. L’état d’équilibre
d’une goutte de liquide pur correspond au minimum de son énergie libre qui est, à
volume constant, la potentielle thermodynamique à minimiser. À température T,
nombre de molécules N et volume V constants, ce potentiel n’a qu’une contribution
surfacique. L’énergie libre de surface est le résultat des liaisons pendantes à la
surface de la goutte. Alors que les molécules à l’intérieur de la goutte ont un certain
nombre de molécules voisines qui les entourent de façon isotrope, les molécules de
surface présentent une anisotropie qui se reflète par un déficit de molécules vers
l’extérieur ou thermodynamiquement, par un excès d’énergie de surface. Cette

Physique des agrégats cellulaires embryonnaires
33
énergie est proportionnelle à l’aire de la surface et les variations infinitésimales de
l’énergie libre par unité de surface seront donné par :
.dF dAσ=
avec σ la tension de surface (ou tension superficielle) définie comme étant
l’énergie libre par unité de surface (à T, V, N constants), et A l’aire de la surface. Par
conséquent l’énergie libre F atteint un minimum pour dA=0. L’état d’équilibre
correspond donc à une surface minimale, par exemple une goutte sphérique si celle-
ci est libre en suspension. Dans les cas où deux phases liquides a et b immiscibles
sont en contact, la tension résultante est appelée tension interfaciale notée σab [93].
Fig. 1.13 : Expérience démontrant que la tension de surface est une force par unité de
longueur (A) la surface tire sur la barrière mobile d’une cuve de Langmuir (A), ou sur le fil
de laine souple refermant un film de savon sur un cadre (B).
Le travail qu’il faut dépenser pour accroître la surface d’un liquide de largeur
constante L à T, V, N constants est ∆W=σ∆A=σL∆x=F∆x (Fig. 1.13). Donc
σ s'interprète aussi comme une force par unité de longueur (F=σ L) d’où le nom de tension de surface.
La différence de pression entre l’intérieur et l’extérieur d’une interface fluide
courbe dépend des deux rayons de courbure et de la tension de surface selon la loi
de Laplace [93]:
int
1 2
1 1( )extP P PR R
σ∆ = − = +
Cette équation montre que des petites gouttes de liquide ont des pressions
internes plus grandes que des grandes. Nous le verrons dans le chapitre 3, lorsque
l’on comprime une goutte entre deux plaques, deux termes de force de surface sont
transmis aux plaques : l’un dû à la surpression de Laplace, l’autre dû à la force le
long de la ligne de contact (goutte/plaque/milieu extérieur comme sur la Fig. 1.14) .

Physique des agrégats cellulaires embryonnaires
34
Fig. 1.14 : Goutte de liquide (A) posée sur un autre liquide et (B) contre un solide. Il existe
une relation d’équilibre (somme vectorielle) entre les tensions de surface et les angles
(relation d’Young-Dupré).
Quand une goutte de liquide est déposée contre un solide ou un autre liquide,
soit elle s’étale et mouille complètement la surface, soit elle ne s’étale pas
complètement. Dans ce cas du mouillage partiel, la géométrie atteinte résulte de
l’équilibre entre les tensions interfaciales et superficielles aux lignes solide-liquide-
gaz avec un angle de contact π−θ bien défini (nous avons ici utilisé la notation
généralement rencontrée dans les articles de tensiométrie des tissus où θ est le
complémentaire de l’angle de contact). Pour la goutte « a » sur la surface solide
« s » (Fig. 1.14B), cette relation de Yong-Dupré est la suivante :
( )coss as aσ σ σ π= + − Θ
où σa, σa, σas sont respectivement les tension de surface du solide avec la phase
gazeuse, du liquide avec la phase gazeuse et de l’interface a/s. Quelques autres
quantités sont intéressantes à définir comme le travail de cohésion Wa qui est le
travail nécessaire pour séparer une unité d’aire du milieu a, du contact à l’infini (Fig.
1.15A) , ou le travail d’adhésion Wab qui est le travail nécessaire pour séparer une
unité d’aire d’une interface entre un milieu a et un milieu b, du contact à l’infini (Fig.
1.14B) :
( )( )2 et + - = 1+cos - a a ab a b ab aW Wσ σ σ σ σ π θ= =
où la dernière égalité à droite a été obtenu en utilisant la loi d’Young (cas du
mouillage partiel).
Toutes ces considérations sont vraies pour deux liquides a et b immiscibles c'est-à-
dire pour lesquels σab>0. [94]. La miscibilité de deux liquides (σab≤0) n’est pas
prédictible à partir des valeurs des tensions de surface σa et σb. Les molécules d’eau
par exemple sont tenues ensemble par de fortes liaisons hydrogène (liaison H
résultant d’interactions entre charges des dipôles de l’eau) qui lui confèrent une forte

Physique des agrégats cellulaires embryonnaires
35
Fig. 1.15 : (A) Travail de cohésion et (B) Travail d’adhésion
tension de surface de 72mN/m à 25°C [93;95]. L’éthanol CH3CH2OH est aussi un
liquide polaire ayant une tension de surface de 22,3 mN/m [95]. Il est miscible dans
l’eau. L’huile d’olive est un liquide non polaire (lipophile) ayant une tension de
surface de 33 mN/m à 25°C [96] donc proche de l’éthanol. Mais, elle ne se mélange
pas à l’eau : l’huile et l’eau démixent en deux phases séparées, l’huile formant par
exemple un film à la surface de l’eau. La raison est que l’huile contrairement à
l’éthanol n’est pas polaire et ne peut ainsi former de liaisons H avec l’eau. Il faut
retenir de cet exemple simple que la prédiction de l’état final d’un mélange de deux
liquides nécessite la connaissance de leurs interactions, c'est-à-dire de σab.
L’autre propriété importante d’un liquide qui nous intéresse ici est sa viscosité
dynamiqueη qui représente la résistance du liquide à l’écoulement. Lorsque deux
couches d'un fluide séparées d’une distance dz sont animées d’une vitesse relative
dv l’une par rapport à l’autre et dirigée suivant x, sous l'effet de la viscosité, une force
F sera transmise d’une couche (de surface S) à l’autre. La viscosité η est alors
définie par la relation suivante :
dz
dSF
νη ..= (1.3)
La viscosité dynamique η se mesure en pascal-seconde (Pa.s). On trouve
encore parfois l’unité du système CGS, le poise (Po) : 1 Pa.s = 10 Po.
I.2.3.3 Modèle thermodynamique de la démixtion et du tri-cellulaire
Considérons le cas d’un mélange de deux types cellulaires différents a et b
sans interaction avec d’autres surfaces. Comme pour les liquides, les différents
niveaux d’adhésion et de cohésion pourront être quantifiés par les travaux

Physique des agrégats cellulaires embryonnaires
36
nécessaires à établir une liaison cohésive ou adhésive par unité de surface Wa, Wb,
et W ab. Ce raisonnement emprunté à la physique des surfaces a été d’abord
employé dans le contexte des agrégats cellulaires par Steinberg [65] puis complété
par Graner [97].
Comme dans l’exemple de la goutte, les cellules se réarrangent dans le but de
minimiser l’énergie libre du système entier. Cette énergie est minimale quand la
somme totale des travaux accomplis à l’intérieur du système (travaux de cohésion et
d’adhésion) est maximale. Selon les valeurs de ces travaux, il y a plusieurs cas à
distinguer :
(i) Le travail d’adhésion Wab est supérieur ou égal à la moyenne des travaux de
cohésion : 2
baab
WWW
+≥ soit σab < 0.
Dans un tel cas, le travail maximum que peut accomplir le système, correspond à un
mélange des unités a et b d’une façon alternée, les deux phases sont combinées
(motif en échiquier ou « checkerboard »). C’est le cas 1 de la Fig. 1.16.
(ii) Au contraire si Wab est inférieure à la moyenne des travaux de cohésion,
2
baab
WWW
+< , soit σab > 0
le travail est maximum quand les deux types d’unités sont complètement ségrégées.
Cependant, il y a différents cas de ségrégations thermodynamiquement possibles.
Pour simplifier les cas symétriques, supposons que le travail de cohésion Wa est
supérieur au travail de cohésion Wb.
(ii-1) Si les unités b sont plus adhésives aux unités a que cohésives entre elles,
on résume alors la situation par l’inégalité suivante :
2
baabb
WWWW
+<≤ , soit σa > σab + σb
Ces conditions ne sont respectées que lorsque aba WW > . Sachant que
l’énergie libre du système est proportionnelle à la surface, sa minimisation doit
se manifester par la formation d’une sphère composite contenant à sa
périphérie le type cellulaire le moins cohésif (b) et en son centre le plus cohésif
(a). De plus l’anisotropie à l’interface entre les deux types a et b fait que cette
interface contribue à l’énergie libre du système entier. Une condition requise
afin de minimiser l’énergie libre est de minimiser l’interface a/b. En conclusion
la phase a assumera aussi une forme sphérique, entouré d’une couronne
concentrique contenant la phase b. C’est le deuxième cas du schéma de la Fig.
1.16.

Physique des agrégats cellulaires embryonnaires
37
(ii-2) Si l’adhésion a/b est légèrement inférieure à la cohésion entre une
paire d’unité de la phase b, mais si aucune tension de surface n’est supérieure
à la somme des deux autres, c’est à dire :
abab WWW ≤< soit σa < σab + σb, et σab < σa + σb
la phase b recouvre partiellement la surface de la phase a. C’est le troisième
cas schématisé sur la figure (Fig. 1.16).
(ii-3) Enfin si abab WWW ≤< (σa < σab + σb), mais σab > σa + σb
La condition de Young ne peut être vérifiée car σab est supérieur à la somme
des deux autres tensions de surface. Deux phases isolées se forment. À
l’équilibre, chaque phase forme un sphéroïde et l’interface a/b disparaît. C’est
l’autre troisième cas (du bas) schématisé sur la figure (Fig. 1.16).
Après avoir développé cette description thermodynamique cohérente et validée
par ses expériences, Steinberg pousse naturellement un peu plus loin l’analogie
entre liquides simples et tissus biologiques en postulant que ces derniers ont aussi
une tension de surface résultant de l’énergie de cohésion entre leurs cellules due
Fig. 1.16 : Types de distribution de phase, à l’équilibre thermodyna-mique, d’une population constituée de deux types d’unités mobiles cohésives entre elles et adhésives les unes aux autres. Le travail de cohésion Wb est arbitrairement choisi égal à 1. En fonction du rapport des travaux de cohésion, et d’adhésion, ce schéma, basé sur un modèle thermodyna-mique, peut prédire la configuration finale adoptée par les phases mobiles constituant le système (d’après [65]).

Physique des agrégats cellulaires embryonnaires
38
aux molécules d’adhésion. La preuve expérimentale et la mesure de cette tension
de surface tissulaire devient dès la fin des années 1960 pour Steinberg et ses
collaborateurs un important défi scientifique qui nécessitera une vingtaine d’année
encore pour aboutir.
I.2.3.4 Preuves expérimentales de l’HAD : expériences de tri cellulaire et
mesures de tension de surface tissulaire
On a vu que l’arrondissement d’un fragment de tissu peut donner un agrégat
sphérique (Fig. 1.11B), que deux agrégats d’un même tissu, mis en contact,
fusionnent vers un gros agrégat sphérique (Fig. 1.12B), qu’un mélange de cellules
dissociées de plusieurs types de tissus redonne après une période de tri cellulaire
des domaines homotypiques (Fig. 1.12A). Bien que les cellules qui constituent ces
agrégats ne soient pas à l’équilibre thermodynamique, tous ces comportements sont
analogues au retour à l’équilibre d’un liquide simple piloté par sa tension de surface.
Le bilan d’équilibre des forces existe pour un système hors d’équilibre. La tension de
surface étant une force par unité de longueur tout au long de la surface peut donc
être introduite pour les agrégats cellulaires. Le terme tension de surface tissulaire
sera utilisé pour décrire les forces agissantes tout au long de l’interface entre un tissu
et un milieu de culture, et tension tissulaire interfaciale pour caractériser les
mêmes forces agissant à l’interface de deux tissus différents en contact.
La première méthode à laquelle on peut penser pour déterminer
expérimentalement la cohésion d’un tissu est de mesurer les forces d’adhésion entre
paires de cellules le constituant [9;58]. Mais ceci s’avère vite une expérience très
délicate dépendant de multiples paramètres expérimentaux comme la vitesse de
rétraction lors du détachement des deux cellules [59]. La tension de surface tissulaire
par contre est une mesure macroscopique sur un ensemble de cellules, elle apporte
une quantification directe de la cohésion tissulaire sans aucune hypothèse sur les
interactions microscopiques entre cellules.
Avant de devenir quantitative et absolue, la mesure de tension de surface est
longtemps restée qualitative et relative. Phillips et Steinberg comparent la forme
d’agrégats cellulaires de même volume et soumis à des forces de centrifugation de
même amplitude afin de classer les différentes types de tissus embryonnaires de
poulets vis-à-vis de leur tension de surface. Ils ont démontré que les fragments de
tissus ainsi que les agrégats déjà arrondis aboutissent à la même configuration finale
[98;99]. Pendant ces expériences la force centrifuge qui tend à aplatir les agrégats
est équilibrée par la tension de surface tissulaire qui tend à arrondir l’agrégat et ces
deux paramètres déterminent la forme de l’agrégat. Les différences relatives de
tension de surface évaluées prédisent précisément la hiérarchie de leurs
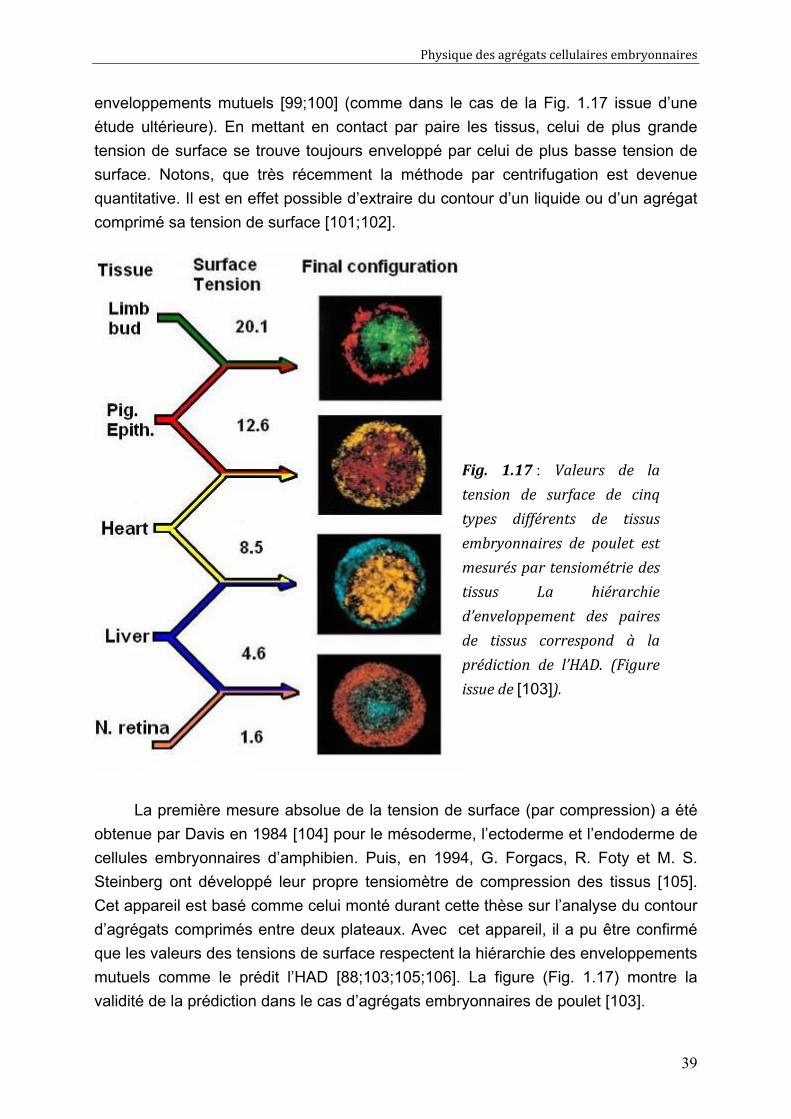
Physique des agrégats cellulaires embryonnaires
39
enveloppements mutuels [99;100] (comme dans le cas de la Fig. 1.17 issue d’une
étude ultérieure). En mettant en contact par paire les tissus, celui de plus grande
tension de surface se trouve toujours enveloppé par celui de plus basse tension de
surface. Notons, que très récemment la méthode par centrifugation est devenue
quantitative. Il est en effet possible d’extraire du contour d’un liquide ou d’un agrégat
comprimé sa tension de surface [101;102].
Fig. 1.17 : Valeurs de la
tension de surface de cinq
types différents de tissus
embryonnaires de poulet est
mesurés par tensiométrie des
tissus La hiérarchie
d’enveloppement des paires
de tissus correspond à la
prédiction de l’HAD. (Figure
issue de [103]).
La première mesure absolue de la tension de surface (par compression) a été
obtenue par Davis en 1984 [104] pour le mésoderme, l’ectoderme et l’endoderme de
cellules embryonnaires d’amphibien. Puis, en 1994, G. Forgacs, R. Foty et M. S.
Steinberg ont développé leur propre tensiomètre de compression des tissus [105].
Cet appareil est basé comme celui monté durant cette thèse sur l’analyse du contour
d’agrégats comprimés entre deux plateaux. Avec cet appareil, il a pu être confirmé
que les valeurs des tensions de surface respectent la hiérarchie des enveloppements
mutuels comme le prédit l’HAD [88;103;105;106]. La figure (Fig. 1.17) montre la
validité de la prédiction dans le cas d’agrégats embryonnaires de poulet [103].
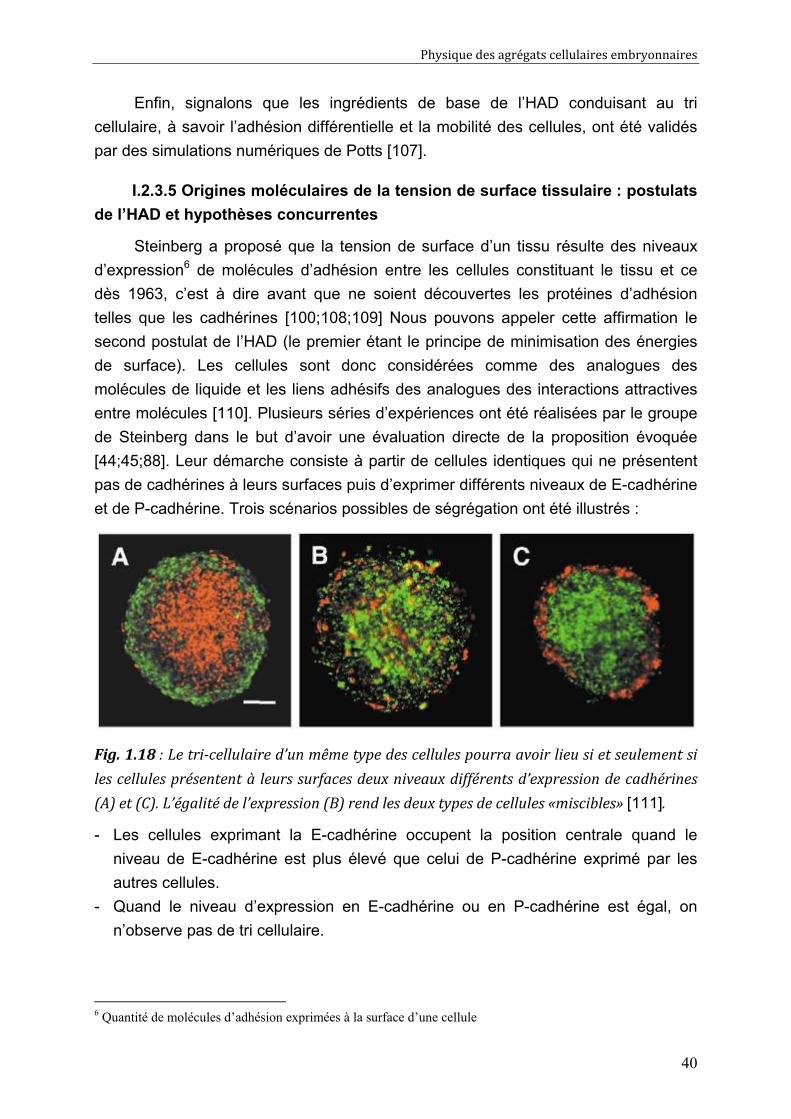
Physique des agrégats cellulaires embryonnaires
40
Enfin, signalons que les ingrédients de base de l’HAD conduisant au tri
cellulaire, à savoir l’adhésion différentielle et la mobilité des cellules, ont été validés
par des simulations numériques de Potts [107].
I.2.3.5 Origines moléculaires de la tension de surface tissulaire : postulats
de l’HAD et hypothèses concurrentes
Steinberg a proposé que la tension de surface d’un tissu résulte des niveaux
d’expression6 de molécules d’adhésion entre les cellules constituant le tissu et ce
dès 1963, c’est à dire avant que ne soient découvertes les protéines d’adhésion
telles que les cadhérines [100;108;109] Nous pouvons appeler cette affirmation le
second postulat de l’HAD (le premier étant le principe de minimisation des énergies
de surface). Les cellules sont donc considérées comme des analogues des
molécules de liquide et les liens adhésifs des analogues des interactions attractives
entre molécules [110]. Plusieurs séries d’expériences ont été réalisées par le groupe
de Steinberg dans le but d’avoir une évaluation directe de la proposition évoquée
[44;45;88]. Leur démarche consiste à partir de cellules identiques qui ne présentent
pas de cadhérines à leurs surfaces puis d’exprimer différents niveaux de E-cadhérine
et de P-cadhérine. Trois scénarios possibles de ségrégation ont été illustrés :
Fig. 1.18 : Le tri-cellulaire d’un même type des cellules pourra avoir lieu si et seulement si
les cellules présentent à leurs surfaces deux niveaux différents d’expression de cadhérines
(A) et (C). L’égalité de l’expression (B) rend les deux types de cellules «miscibles» [111].
- Les cellules exprimant la E-cadhérine occupent la position centrale quand le
niveau de E-cadhérine est plus élevé que celui de P-cadhérine exprimé par les
autres cellules.
- Quand le niveau d’expression en E-cadhérine ou en P-cadhérine est égal, on
n’observe pas de tri cellulaire.
6 Quantité de molécules d’adhésion exprimées à la surface d’une cellule

Physique des agrégats cellulaires embryonnaires
41
- L’augmentation du niveau d’expression de la P-cadhérine au delà de celui de
cellules exprimant la E-cadhérine aboutit à l’inversion de la configuration finale des
cellules.
La figureFig. 1.18 illustre les trois cas présentés ci-dessus. Cette expérience
montre clairement que le tri-cellulaire n’est pas forcément causé par la différence
dans la nature des molécules d’adhésion de différents types de tissus. Seule la
différence du niveau exprimé de la molécule est suffisant pour le tri-cellulaire. La Fig.
1.18B montre aussi que cette dernière différence est nécessaire pour que le tri
cellulaire puisse avoir lieu.
Finalement en 2005, Foty et Steinberg ont montré que la tension de surface
d’agrégats de L-cells transfectées avec des cadhérines est quasi-directement
proportionnelle au nombre de molécules d’adhésion exprimées par ces cellules
[112]. Les cellules étaient transfectées de façon à exprimer différents niveaux de
cadhérines. Le graphe de la Fig. 1.19 montre la dépendance linéaire de la tension de
surface en fonction du nombre de cadhérines exprimées par chaque cellule à
l’intérieur de l’agrégat.
Ces résultats ont été interprétés par le groupe de Steinberg comme une
indication définitive sur la dépendance exclusive de la tension de surface en fonction
de l’expression des molécules d’adhésion. La contractilité des cellules (tension
corticale) n’aurait aucun rôle sur le tri-cellulaire. Cependant, un changement des
propriétés mécaniques des cellules transfectées avec les cadhérines comparées aux
cellules originales est très probable étant donné que ces molécules d’adhésions sont
en interaction continue avec le cytosquelette. Dans le cas du poisson zèbre, il a été
montré que la différence quantitative de la tension de surface dirige la ségrégation
des tissus [113] mais que la tension corticale joue un rôle dans ce tri-cellulaire et
dans la gastrulation [85]. Ces mesures ont été obtenues en combinant des mesures
de résistance mécanique et d’adhésion par AFM sur cellules uniques, ainsi que des
expériences de tri in vitro et in vivo des trois feuillets primordiaux (ectoderme,
endoderme, mésoderme), en régulant la tension corticale par le facteur de
croissance « Nodal/ TGFβ-signalling ». De plus, des expériences avec des drogues
affectant le cytosquelette ont montré des variations de tension de surface
mesurables [114;115]. Il nous parait donc très difficile de séparer complètement
comme le fait le second postulat de l’HAD la mécanique cellulaire de l’adhésion
cellule-cellule. On peut raisonnablement penser que ces deux facteurs contribuent
au devenir des cellules dans les tissus.
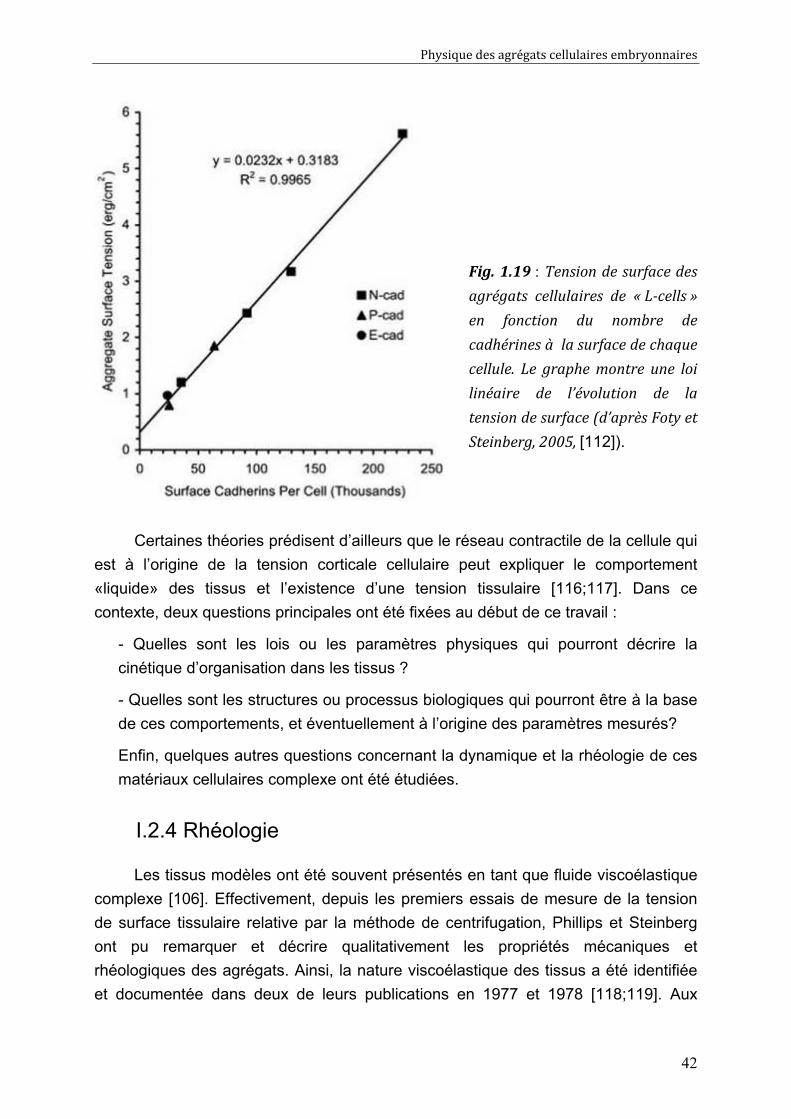
Physique des agrégats cellulaires embryonnaires
42
Fig. 1.19 : Tension de surface des
agrégats cellulaires de « L-cells »
en fonction du nombre de
cadhérines à la surface de chaque
cellule. Le graphe montre une loi
linéaire de l’évolution de la
tension de surface (d’après Foty et
Steinberg, 2005, [112]).
Certaines théories prédisent d’ailleurs que le réseau contractile de la cellule qui
est à l’origine de la tension corticale cellulaire peut expliquer le comportement
«liquide» des tissus et l’existence d’une tension tissulaire [116;117]. Dans ce
contexte, deux questions principales ont été fixées au début de ce travail :
- Quelles sont les lois ou les paramètres physiques qui pourront décrire la
cinétique d’organisation dans les tissus ?
- Quelles sont les structures ou processus biologiques qui pourront être à la base
de ces comportements, et éventuellement à l’origine des paramètres mesurés?
Enfin, quelques autres questions concernant la dynamique et la rhéologie de ces
matériaux cellulaires complexe ont été étudiées.
I.2.4 Rhéologie
Les tissus modèles ont été souvent présentés en tant que fluide viscoélastique
complexe [106]. Effectivement, depuis les premiers essais de mesure de la tension
de surface tissulaire relative par la méthode de centrifugation, Phillips et Steinberg
ont pu remarquer et décrire qualitativement les propriétés mécaniques et
rhéologiques des agrégats. Ainsi, la nature viscoélastique des tissus a été identifiée
et documentée dans deux de leurs publications en 1977 et 1978 [118;119]. Aux

Physique des agrégats cellulaires embryonnaires
43
échelles de temps long (heures), ils présentent des comportements similaires à ceux
d’un liquide visqueux. Aux temps courts, ils répondent plus comme des solides
viscoélastiques. Les comportements sont donc intimement dépendants de l’échelle
de temps considérée. Cependant, ces comportements sont susceptibles d’être
modifiés dès qu’on applique une force extérieure [120]. Ainsi, une étude physique
des tissus embryonnaires ne pourra être complète et pertinente que si elle prend en
compte toute la richesse de ces comportements en ayant recours à la rhéologie des
tissus. Le « challenge » est donc d’établir un modèle rhéologique des tissus basé sur
des paramètres physiques qui reflètent la structure ainsi que les processus ayant
place à l’intérieur de ce matériau vivant.
Aujourd’hui, l’étude rhéologique est une branche active de la physique des
tissus. Les premières mesures de rhéologie datent de plus de 10 ans [121]. Elles ont
été réalisées sur des tissus de taille macroscopique placés dans un rhéomètre. Dans
la suite de cette sous-partie, nous allons présenter une introduction générale à la
rhéologie ainsi que les modèles mécaniques les plus répandus. Dans l’introduction
du chapitre 5, nous détaillerons les modèles viscoélastiques plus complexes des
tissus vivants et les expériences qui ont permis de les établir.
I.2.4.1 Comportement viscoélastique
Un fluide newtonien a une viscosité η bien définie. Cette viscosité reste intacte
et indépendante des contraintes que subit ce fluide de la vitesse d’écoulement, de la
déformation, ainsi que de la vitesse à laquelle on le déforme. Toute déviation de ces
règles est le signe d’un comportement non-newtonien.
Sous une déformation, un matériau viscoélastique présente un comportement
complexe. Sa réponse à une déformation présente à la fois un aspect élastique
(contrainte proportionnelle à la déformation) et un aspect visqueux (contrainte
proportionnelle à la vitesse de déformation).
Un exemple particulièrement spectaculaire de fluide viscoélastique est la pâte
de silicone connue sous le nom commercial de ”silly-putty”. Une boule de cette pâte
rebondit sur le sol comme une balle élastique; pourtant, si on pose cette boule sur
une surface horizontale et si on attend quelques minutes, on voit la pâte s’étaler
comme un fluide visqueux.
Le même matériau réagit donc d’une manière très différente lorsqu’il est
soumis à une sollicitation très rapide (en le faisant rebondir sur le sol) ou lorsque la
contrainte est appliquée pendant un temps très long. Dans le premier cas, le temps
de sollicitation est inférieur à un temps caractéristique du matériau, les composants
élémentaires n’ont pas le temps de se déformer de manière importante et on observe

Physique des agrégats cellulaires embryonnaires
44
une réponse élastique. En revanche, lorsque le temps de sollicitation est plus grand
que le temps caractéristique, on observe une réponse de type visqueux.
Cette réponse complexe d’un matériau soumis aux contraintes peut être
reconstruite sous forme d’un système mécanique modèle, composé des éléments
élastiques et visqueux. La réponse élastique du matériau est modélisée par des
ressorts et celle visqueuse par des amortisseurs ou pistons [122;123]. Le ressort
réagit instantanément en encaissant une déformation proportionnelle au stress
appliqué. À l’échelle des petites déformations, une relation linéaire force-déformation
s’applique en accord avec la loi de Hooke :
σélastique(t) = E ε(t) (1.4)
où σ et ε représentent respectivement le stress appliqué et la déformation encaissée
[122] Le facteur de proportionnalité E est le module d'élasticité ou le module de
Young du matériau. Il est mesuré en Pascal (Pa) (1Pa = 1N.m-2). Le piston se
comporte comme un liquide newtonien, i.e. la viscosité modélisée est indépendante
de la vitesse de déformation ε’. Le piston présente une réponse en retard par
rapport à l’application du stress. Il se déforme à vitesse constante pour répondre au
stress qu’il subit, et sa relation stress-déformation est donnée par :
σvisqueux(t)=η ε’(t) (1.5)
où η est la viscosité mesurée en poise (P) ou en Pa.s. Avec 1Pa.s = 10 P.
I.2.4.2 Modèles mécaniques.
Le modèle le plus simple de matériau viscoélastique consiste à additionner les
contraintes d’origine élastique et les contraintes d’origine visqueuse :
σ(t)=σélastique(t)+σvisqueux(t)=E ε(t)+η ε’ (t) (1.6)
Une représentation graphique de ce modèle, dit solide de Kelvin-Voigt (Fig.
1.20), est l’association en parallèle d’un ressort et d’un piston. On peut également
associer en série un ressort et un piston (modèle du liquide de Maxwell) (Fig. 1.20),
dans ce cas, les déformations élastique et visqueuse s’additionnent et les contraintes
sont identiques, soit :
ε’(t)=η
σ
dt
dσ
Edt
dε
dt
dε+=+ 1visqueuxélastique
(1.7)

Physique des agrégats cellulaires embryonnaires
45
Fig. 1.20 : (À gauche) Modèle mécanique d’un solide simple dit de Kelvin Voigt représenté
par une association en parallèle d’un ressort élastique et d’un piston dissipatif. (À droite)
modèle de Maxwell pour un fluide viscoélastique simple.
I.2.4.3 Loi de comportement sous stress : Fluage.
Dans une expérience de fluage, le matériau est soumis, à partir d’un instant
donné, à une contrainte constante σσσσ0. La déformation εεεε(t) est reliée à la contrainte
par la fonction fluage f(t) :
εεεε (t) = f(t) σσσσ0 (1.8)
La fonction fluage a la dimension de l’inverse d’une contrainte. Pour le solide
de Kelvin-Voigt, la fonction fluage a pour expression :
(1.9)
où J = 1/E est la complaisance élastique et ττττ est le temps de relaxation défini ci-
dessus. Le solide de Kelvin-Voigt a un comportement élastique aux temps longs
(t>>ττττ), après un régime transitoire dont la durée est définie par ττττ. Il s’agit d’une
élasticité retardée par la présence de l’élément visqueux et inversement
proportionnelle au module élastique E (Fig. 1.21).
Pour le liquide de Maxwell, la fonction de fluage est :
(1.10)
Aux temps longs, le comportement est celui d’un fluide newtonien avec une
viscosité ηηηη : la déformation augmente linéairement avec le temps. La différence avec
un fluide newtonien correspond à la présence d’une élasticité instantanée aux temps
courts (t <<ττττ) (Fig. 1.21).

Physique des agrégats cellulaires embryonnaires
46
Fig. 1.21 : (à gauche) Fonction fluage d’un solide de Kelvin-
Voigt (à droite) Fonction fluage d’un liquide de Maxwell.
Ces deux modèles constituent une description assez simple de la mécanique
de la matière dans ses deux états, solide et liquide. Par rapport à cette échelle de
classification, les tissus présentent des comportements combinés. Ces modèles
simples ne pourront donc pas être à même d’expliquer et de décrire complètement la
complexité des tissus. Ainsi, certains tissus se comportent comme des solides
viscoélastiques aux courtes échelles de temps et comme des liquides visqueux aux
longues échelles de temps. Les comportements liquides aux temps longs ont permis
la mesure de certains paramètres physiques tels que la tension de surface (Chapitre
3) et la viscosité (Chapitre 4) et ensuite de prédire les comportements individuels et
d’enveloppement mutuel des tissus ou des mélanges de cellules. Le chapitre 5 sera
l’occasion de s’intéresser à l’énergie élastique de ces « solides » aux temps courts,
et à la façon dont cette énergie sera dissipée et mènera à des écoulements aux
temps longs.

Physique des agrégats cellulaires embryonnaires
47
Chapitre 2
Materials and Methods
When embryonic tissue fragments or cell lines from near confluent plates are
dissociated into single cells using trypsin treatment and subsequently reaggregated,
they form cell aggregates or spheroids. These cell aggregates constitute an
interesting model system of tissues for different studies in embryology, cancer
research or tissue engineering. The aim of this PhD thesis was to quantify the
physical properties of cell aggregates and to study the role of cytoskeleton/cadherin
link in these properties. For that purpose, we needed various experimental
developments summarized in this chapter:
- The installation of a culture room well equipped to ensure the handling of cells
in good physiological conditions necessary for their survival
- The establishment of experimental protocols for the formation and manipulation
of aggregates, adapted to each cell type and each type of experiments (fusion
rounding up, shape relaxation…).
- The development of a compression plate apparatus (tensiometer) recording the
shape of a compressed aggregate and the forces acting on it in order to measure its
surface tension.

Physique des agrégats cellulaires embryonnaires
48
II.1.Strategy, Cell lines and Aggregate preparation method
As we wanted to test the DAH hypothesis, and especially wanted to see if the
cytoskeleton was able to affect tissue cohesiveness, we searched in the literature for
a cell line which has been modified by biomolecular engineering so as to express the
same cadherin level as the original cell line but whose cadherin-cytoskeleton link has
been impaired. Our first try was with Chinese Hamster Ovary cell lines CHO,
transfected to express cadherin or truncated cadherin (i.e., cadherin, with no
intracellular domain). As we have encountered problems with this cell line, we have
used another type of cells which will appear to have the same interest as the CHO
cells (i.e., same cadherin level of expression and for one of the derivative clones the
link between the cadherin and the cytoskeleton impaired). These were F9 cells line.
II.1.1. CHOVE / CHOVEtr cells
Chinese Hamster Ovary (CHO-K1) cells have been transfected to express
Vascular Endothelium-cadherin GFP or a truncated form of this cadherin (whose
cytoplasmic part had been completely deleted) [66]. They were grown at 37°C with
5% CO2 in Dulbecco's Modified Eagle Medium (41965-039, GIBCO) supplemented
with 10% Foetal Bovine Serum (2902 P-241021, Biotech GmbH), 1% antibiotics
(penicillin streptomycin, GIBCO Invitrogen corporation, Cat. no. 15140-122) and
400µg/ml geneticin (GIBCO Invitrogen corporation, Cat. no. 10131-027).
In order to check the stability of the transfections, we have performed (thanks
to karine Ferri and G. Gillet, IBCP Lyon) a western blot analysis which showed that
the mother CHO-K1 clone doesn’t express VE cadherin in contrast to CHOVE and
CHOVEtr which expressed the same amount of cadherin. However, this global level
of expression reflect a much more diverse reality. The level of expression was really
inhomogeneous among cells on a petri culture dish, especially after several days in
culture, as if cells were progressively losing their capacity to express cadherin, even
if some selective medium has always been used. However this progressive loss did
not change enormously their cohesiveness properties they always were able to form
aggregates. In the same way, non transfected CHO-K1 cells were as well able to
aggregate. Consequently, we thought that the cohesiveness of the different CHO
tissues was at least partially maintained by integrins/extra cellular matrix adhesion
between cells. Indeed, CHO-K1 cells natively express α5β1 integrin, which was
showed to be able to mediate strong adhesion in a cell aggregate [40]. Our suspicion
was reinforced by some other experimental facts; formed aggregates present a rapid

Physique des agrégats cellulaires embryonnaires
49
and enormous adhesion to untreated glass substrates; aggregate time of formation
was much longer than for some other cell types: more than 4 days in hanging drop.
In addition, we suspect that CHO aggregates presented some deviation from liquid
behaviours, even at very long time scale, making it impossible to seriously test DAH
using this cell line (see some preliminary results in annex 2).
II.1.2. Mouse embryonic carcinoma F9 cell
Mouse embryonic carcinoma F9 cell line and their derivatives were a generous
gift from A. Nagafuchi (Institute of molecular embryology and genetics, Kumamoto
univ., Japan) [109]. Cells were maintained in DMEM (41965-039, GIBCO),
supplemented with 10% foetal bovine serum (2902 P-241021, Biotech GmbH) and
1% antibiotics (penicillin streptomycin, GIBCO Invitrogen corporation, Cat. no. 15140-
122), in plastic tissue culture dishes pre-coated for 15 min with gelatin (2%, Sigma
G1393) diluted to 0.2% in PBS.
II.1.2.1 F9 cell vs F9(αααα−/−−/−−/−−/−)
As was shown in the introduction, the link between cadherin and actin is
mediated mainly through catenins [124;125]. α-catenin, associates with the carboxy-
terminal region of the cadherin cytoplasmic domain via β-catenin to form a functional
cadherin-catenin cell adhesion complex [126] (see chapter1, Fig. 1.6). This Cadherin-
cytoskeleton interaction via catenins is essential for cadherin to act as cell adhesion
receptors [125;127;128]. Nagafuchi’s lab who was interested in the biological role of
this catenin in cell adhesion and differentiation produced F9 knocked out cells on the
two alleles of the alpha catenin genes. We will note this cell line F9(α−/−). Interestingly,
the loss of α-catenin does not change the level of cadherin expressed by F9 cells
[129] as shown by the western blotting and Immunoprecipitation analysis of Fig.
2.1A. Not only was the total expression of cadherin in these two cells lines the same,
but also their level of expression at the plasma membrane [129]; it is almost
impossible from the staining images presented in Fig. 2.1E-F to distinguish one cell
line from the other.
The F9(α-/-) mutation does not prevent in particular the same organisation of
cadherins at cell-cell junction.
This was completely different from what happened in the case of the CHOVEtr
cell line where the total cytoplasmic part of the cadherin has been deleted. In this cell
line, cadherins are present at cell-cell junction but are not uniformly distributed, and
form large clusters on the cell membrane [66]. To prevent such a huge aggregation,
a link to the cytoskeleton has to be maintained in a minimal way, as it seems to be
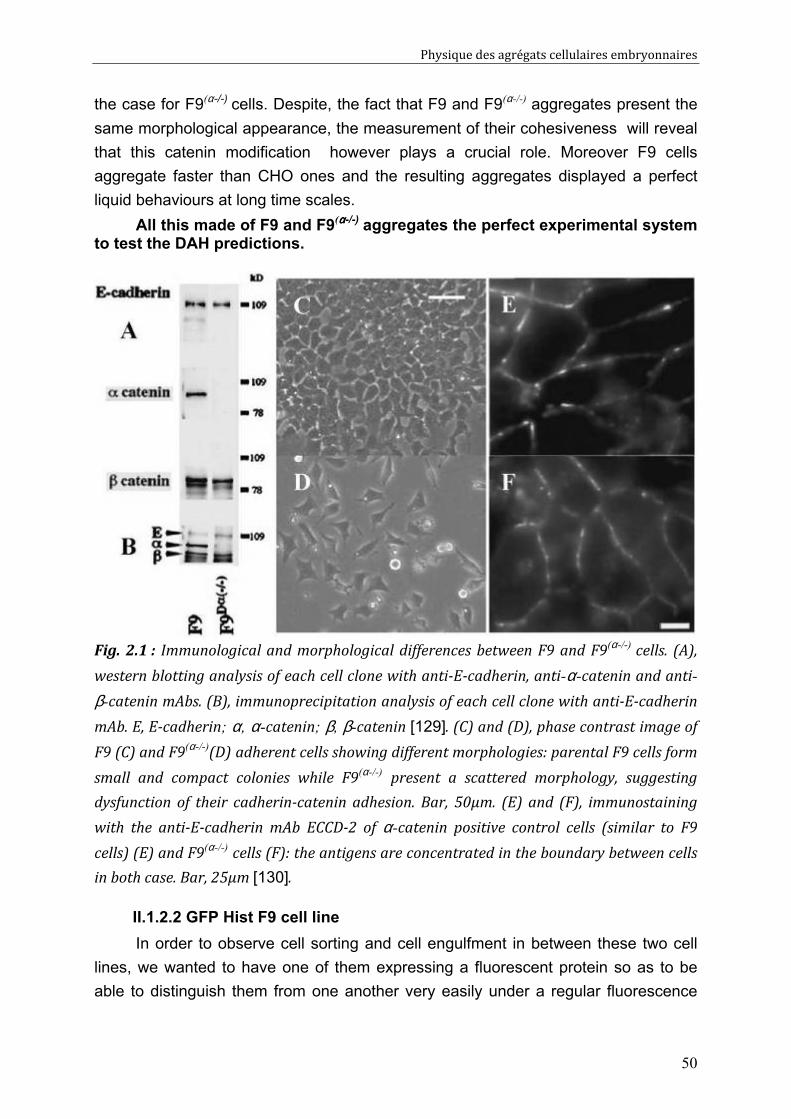
Physique des agrégats cellulaires embryonnaires
50
the case for F9(α-/-) cells. Despite, the fact that F9 and F9(α-/-) aggregates present the
same morphological appearance, the measurement of their cohesiveness will reveal
that this catenin modification however plays a crucial role. Moreover F9 cells
aggregate faster than CHO ones and the resulting aggregates displayed a perfect
liquid behaviours at long time scales.
All this made of F9 and F9(αααα-/-) aggregates the perfect experimental system to test the DAH predictions.
Fig. 2.1 : Immunological and morphological differences between F9 and F9(α-/-) cells. (A),
western blotting analysis of each cell clone with anti-E-cadherin, anti-α-catenin and anti-
β-catenin mAbs. (B), immunoprecipitation analysis of each cell clone with anti-E-cadherin
mAb. E, E-cadherin; α, α-catenin; β, β-catenin [129]. (C) and (D), phase contrast image of
F9 (C) and F9(α-/-)(D) adherent cells showing different morphologies: parental F9 cells form
small and compact colonies while F9(α-/-) present a scattered morphology, suggesting
dysfunction of their cadherin-catenin adhesion. Bar, 50µm. (E) and (F), immunostaining
with the anti-E-cadherin mAb ECCD-2 of α-catenin positive control cells (similar to F9
cells) (E) and F9(α-/-) cells (F): the antigens are concentrated in the boundary between cells
in both case. Bar, 25µm [130].
II.1.2.2 GFP Hist F9 cell line
In order to observe cell sorting and cell engulfment in between these two cell
lines, we wanted to have one of them expressing a fluorescent protein so as to be
able to distinguish them from one another very easily under a regular fluorescence

Physique des agrégats cellulaires embryonnaires
51
microscope. Histone GFP DNA was again a generous gift from A. Nagafuchi.
Detailed on this vector can be found in [131].
For the transfection step, we followed the same protocol as described in [131];
expression vectors were transfected using the lipofectamine 2000 system (GIBCO).
We used a 24-well culture plate for transfection. One day after transfection, the
transfected cells were transferred to two 10-cm culture dishes: one dish contained
1/2 of the transfected cells and another 1/20. As transfection was successful, we
found many colonies even on the 1/20 dish. Two days after transfection, culture
medium was exchanged for one which contained antibiotics for selection. In this
case, 400 microgram/milliliter G418 was used. Culture medium was changed
everyday for a week. And then, medium change was done every two or three days.
Two or three weeks after transfection, we were able to isolate colonies (Fig. 2.2).
In Culture
F9aggregate
50 µm
100 µm
In Culture
F9aggregate
50 µm
100 µm
Fig. 2.2 : Phase contrast
(left) and fluorescence
images (right) of
transfected F9 cells (F9).
(Top) an isolated clone of
adherent F9 cells in
culture dish. (Bottom) an
aggregate of F9 cells.
The cell line obtained from one of the Hist-GFP F9 clone which was mainly
used for our experiments will be further denoted as F9.
II.1.3. Nocodazole Effect
Another way of testing the hypothesis according to which the cytoskeleton may
have a role to play in cell cohesiveness as measured by surface tension was to play
on cell contractility using drugs. This work is still under progress, and we will present
in the context of this thesis only preliminary results obtained using nocodazole. This
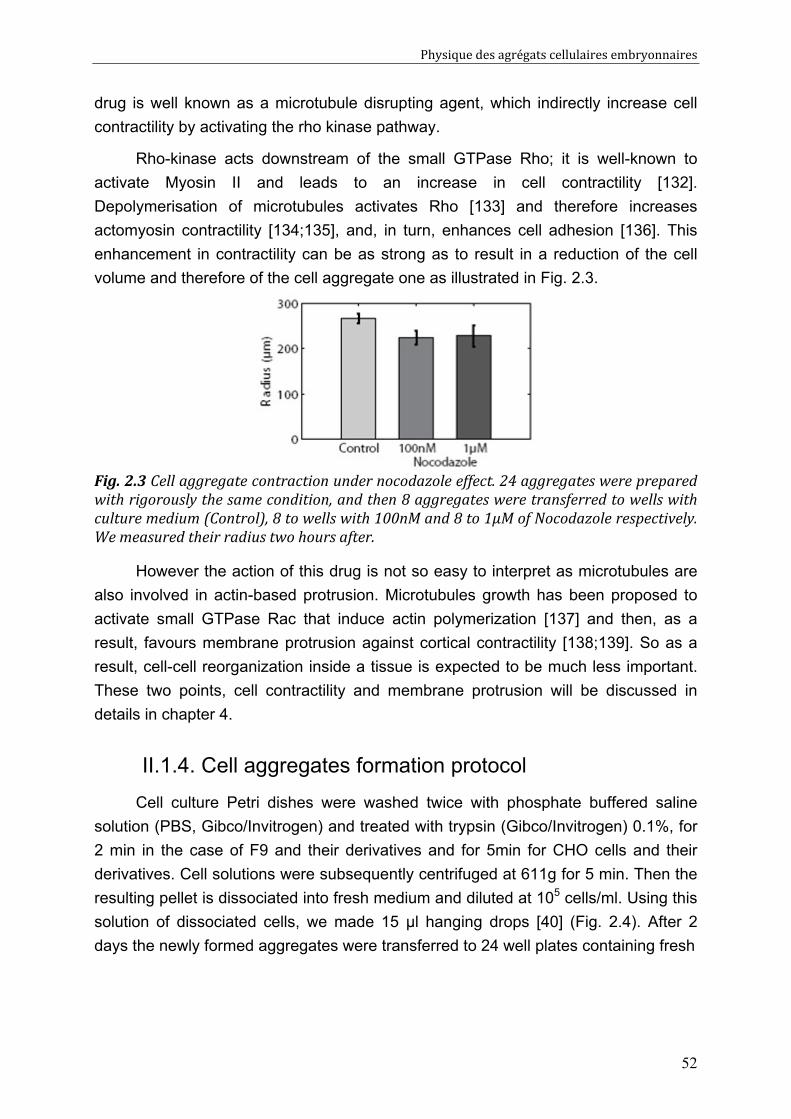
Physique des agrégats cellulaires embryonnaires
52
drug is well known as a microtubule disrupting agent, which indirectly increase cell
contractility by activating the rho kinase pathway.
Rho-kinase acts downstream of the small GTPase Rho; it is well-known to
activate Myosin II and leads to an increase in cell contractility [132].
Depolymerisation of microtubules activates Rho [133] and therefore increases
actomyosin contractility [134;135], and, in turn, enhances cell adhesion [136]. This
enhancement in contractility can be as strong as to result in a reduction of the cell
volume and therefore of the cell aggregate one as illustrated in Fig. 2.3.
Fig. 2.3 Cell aggregate contraction under nocodazole effect. 24 aggregates were prepared with rigorously the same condition, and then 8 aggregates were transferred to wells with culture medium (Control), 8 to wells with 100nM and 8 to 1µM of Nocodazole respectively. We measured their radius two hours after.
However the action of this drug is not so easy to interpret as microtubules are
also involved in actin-based protrusion. Microtubules growth has been proposed to
activate small GTPase Rac that induce actin polymerization [137] and then, as a
result, favours membrane protrusion against cortical contractility [138;139]. So as a
result, cell-cell reorganization inside a tissue is expected to be much less important.
These two points, cell contractility and membrane protrusion will be discussed in
details in chapter 4.
II.1.4. Cell aggregates formation protocol
Cell culture Petri dishes were washed twice with phosphate buffered saline
solution (PBS, Gibco/Invitrogen) and treated with trypsin (Gibco/Invitrogen) 0.1%, for
2 min in the case of F9 and their derivatives and for 5min for CHO cells and their
derivatives. Cell solutions were subsequently centrifuged at 611g for 5 min. Then the
resulting pellet is dissociated into fresh medium and diluted at 105 cells/ml. Using this
solution of dissociated cells, we made 15 µl hanging drops [40] (Fig. 2.4). After 2
days the newly formed aggregates were transferred to 24 well plates containing fresh

Physique des agrégats cellulaires embryonnaires
53
Fig. 2.4 : Hanging drops used for the preparation of cell aggregates. Inset, scheme of the
hanging drop with cells (in white) aggregating under gravity at the bottom of the red
drops. Left, top view of the petri-dish with drops on the cover. Right, side view with the
drop and its image on the cover in dark and light pink respectively.
medium and then incubated on a gyratory shaker at 120 RPM (yellow line 4mm
diameter) or on an agar bed (see section II.4) with 5% CO2 at 37°C for 24 to 36h in
the case of F9 and their derivatives and for 3 to 4 days in the case of CHO cells and
their derivatives. This protocol reproducibly resulted in three-dimensional multicellular
spheroids of similar sizes. For experiments outside the incubator, the cell aggregates
were transferred to CO2 independent medium (18045-054, GIBCO) at 37°C.
II.1.5. Storage
All cells were cryopreserved at early passages and were stored until needed for
experimental procedures. Following trypsinization, cells were pelleted via
centrifugation and resuspended in a freezing medium containing 10% dimethyl
sulfoxide (SIGMA, D8779), 10% DMEM (41965-039, GIBCO) and 90% FBS serum
(2902 P-241021, Biotech GmbH). Cells were transferred to cryogenic screw cap vials
(Fischer Scientific, PA) and incubated in at -20°C freezer for 24 hours. After 24
hours, vials were transferred to a -80°C freezer or to a liquid nitrogen tank for long
term storage.
For studies on the role of surface tension in development, dissected embryonic
cells were used. As we only had time for preliminaries studies, all the results
concerning these primary cells will be presented in Annexe1.

Physique des agrégats cellulaires embryonnaires
54
II.2. Dissected embryonic cells
II.2.1. Embryonic zebrafish cells
The experiments were conducted in collaboration with G. Krens from C.P.
Heisenberg Lab (Max-Planck-Institute for Molecular Cell Biology and Genetics,
Dresden) and V. Laudet (IGFL, ENS Lyon).
II.2.1.1. Fish stocks
Wild-type embryos were obtained from TL, and AB zebrafish lines. Maternal-
zygotic one-eyed-pinhead (Mzoep) mutant fish were used to obtain embryos
consisting almost exclusively of ectodermal cells [140]. Embryos were grown at 31 °
C in Danieaus medium and staged according to [75].
II.2.1.2. Injections of mRNA and morpholino oligonuclotides
To generate mesoderm progenitors, one-cell-stage wild-type zebrafish embryos
were injected with cyclops (cyc) mRNA (100 pg) to induce mesendodermal cell faith
and 2 ng casanova (cas)antisence morpholino oligonucleotides (MO) (GeneTools) to
inhibit endodermal cell faith [141;142]. For ectoderm progenitors, embryos were
injected with 100pg lefty1 (lft1) mRNA or Materna Zygotic (MZ) one eye pinhead
(oep) mutant zebrafish [143]. The specific identity of germ-layer progenitors was
previously confirmed by in situ staining of the injected embryos [85]. These injections
allowed efficient induction of mesoderm and ectoderm progenitor types.
II.2.1.3. Aggregates formation
For all experiments embryos were manually dechorionated during cleavage
stages (0,5-2,5 hours post fertilization (hpf)) and kept in 1x Danieaus medium buffer
at 31°C. At high to oblong stage (3-3,5 hpf), the animal poles of the embryos were
dissected manually from the yolk sac, using watchmaker’s forceps and the resulting
tissue fragments were transferred into 2%-agarose-coated petridishes with fresh 1x
Daniaus buffer to round up. Rounding of the tissue fragments (explants / aggregates)
usually took 30 minutes, where they were transferred to 2% agar coated 96 well
plates containing 150 ml CO2-independent DMEM/F-12 1:1 buffered in 15 mM Hepes
and supplemented antibiotics (Penicillin (100 U ml-1) and Streptomycin (0.1 mg ml-1)).
One explant/aggregate was placed per well. Fetal Calf Serum (FCS) was added to
the DMEM/F12 medium in concentrations varying between 1- 10%, as indicated in
the text. The explants ranged in diameter between 0.25 - 0.35 mm and were kept at
31 degrees until used for compression analysis.
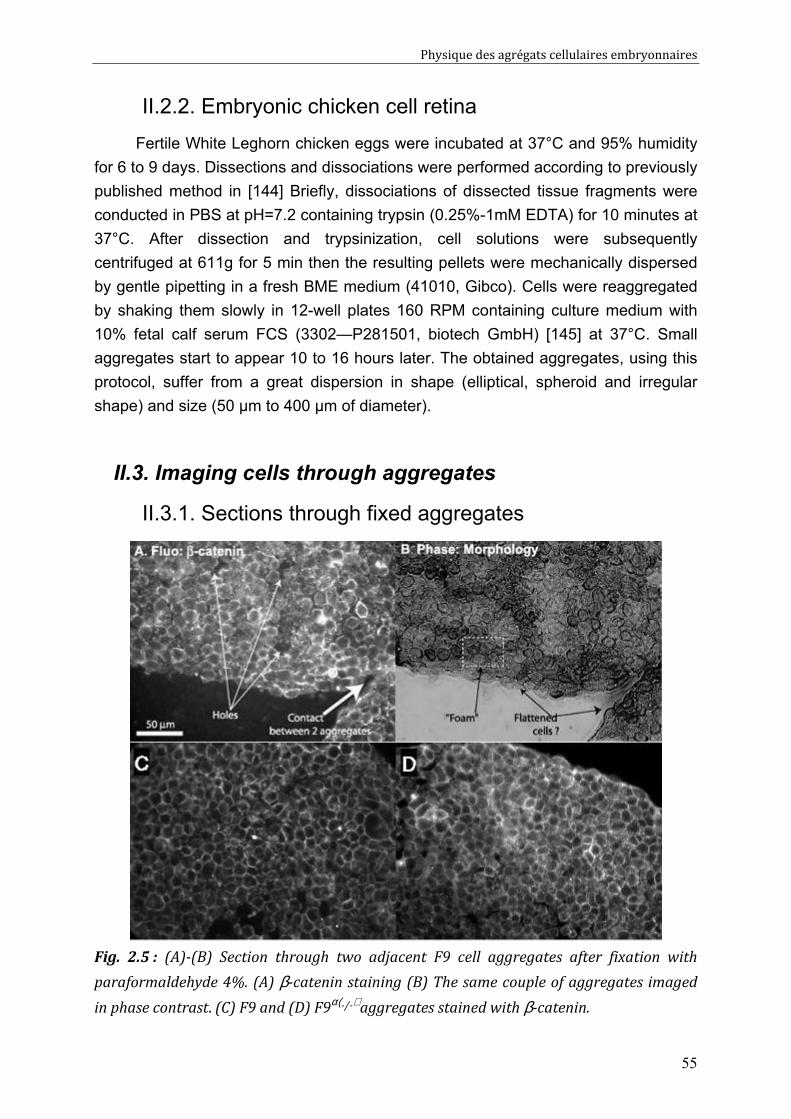
Physique des agrégats cellulaires embryonnaires
55
II.2.2. Embryonic chicken cell retina
Fertile White Leghorn chicken eggs were incubated at 37°C and 95% humidity
for 6 to 9 days. Dissections and dissociations were performed according to previously
published method in [144] Briefly, dissociations of dissected tissue fragments were
conducted in PBS at pH=7.2 containing trypsin (0.25%-1mM EDTA) for 10 minutes at
37°C. After dissection and trypsinization, cell solutions were subsequently
centrifuged at 611g for 5 min then the resulting pellets were mechanically dispersed
by gentle pipetting in a fresh BME medium (41010, Gibco). Cells were reaggregated
by shaking them slowly in 12-well plates 160 RPM containing culture medium with
10% fetal calf serum FCS (3302—P281501, biotech GmbH) [145] at 37°C. Small
aggregates start to appear 10 to 16 hours later. The obtained aggregates, using this
protocol, suffer from a great dispersion in shape (elliptical, spheroid and irregular
shape) and size (50 µm to 400 µm of diameter).
II.3. Imaging cells through aggregates
II.3.1. Sections through fixed aggregates
Fig. 2.5 : (A)-(B) Section through two adjacent F9 cell aggregates after fixation with
paraformaldehyde 4%. (A) β-catenin staining (B) The same couple of aggregates imaged
in phase contrast. (C) F9 and (D) F9α(-/-) aggregates stained with β-catenin.

Physique des agrégats cellulaires embryonnaires
56
Making sections through aggregates allows to investigate internal
morphogenetic of cell structures and to prepare fixed cell monolayers for histological
staining. These sections were realized by S. Bodennec (postdoc at LPMCN) in
collaboration with H. Mertani (Centre Léon Bérard, Lyon). The detailed protocol
(fixation, paraffin embedding and subsequent removal, immunofluorescence) is
presented in annex 3. Fig. 2.5A-B shows a section through two adjacent aggregates.
Cells are densely packed with only a few holes. In the aggregate periphery, cells
seems more flattened than inside where they have polyhedral shapes and are
sometimes organized with a foam structure. Fig. 2.5A,C-D illustrate the kind of
staining we could obtain in order to identify and localize the proteins implicated in
adhesion between cells. We checked here for instance, that the expression of β-
catenin was identical at the plasma membrane in F9 and F9(α−/−).
II.3.2. Two-photon confocal microscopy 3D imaging
Two-photon laser scanning microscopy was performed in collaboration with J.
C. Vial and B. van der Sanden (LSP, Grenoble). We used an Olympus BX50WI
upright microscope coupled to a Biorad MRC 1024 confocal scanhead with a
nondescanned detector to increase the observation depth. A 800-nm excitation beam
from a Tsunami femtosecond Ti:Saphire laser (Millenia V, Spectra-Physics) was
focused on the surface of the spheroids using a 20× water-immersion objective (0.95
NA, Xlum Plan Fl Olympus). The beam scanned the xy plane to acquire 512×512
pixel images in 0.9 s. Variation of the observation depth (z-scan) was realized by
moving the motorized objective vertically.
Our objective with these experiments was to visualize cell contours deeply
inside live aggregates. For that, cells possessing a homogeneous internal
fluorescence as our F9 fluorescent cell line are not suitable. On the other hand, we
demonstrated that the soluble dye sulforhodamine B (SRB) injected at a
concentration of 8 µg/ml in the culture medium quickly diffuses inside aggregates and
stains the inter-cellular space with intensity comparable to the SRB present outside of
the aggregates (sides of Fig. 2.6, Fig. 2.7). In addition, the cell membrane of dead
cells becomes permeable to SRB [146], as confirmed by experiments showing that
photodamaged cells submitted to a high laser power (700 mW) are completely filled
with the dye (central zone of Fig. 2.6). This demonstrates that SRB provides in the
same time a cell viability assay.
In order to evaluate the damage due to compression of aggregates between
two plates, we have developed a small compression apparatus, designed to be

Physique des agrégats cellulaires embryonnaires
57
mounted on the stage of the two-photon microscope with a thermostated circulating
water bath to control temperature at 37°C (Fig. 2.7A-B, Fig. 2.8).
Fig. 2.6 : The contrast between alive cells and photo-damaged cells by application of a
high laser power (700w) on a restricted zone of a compressed aggregate. Bar 25µm.
Fig. 2.7 : Two-photon microscopy assay. (A) and (B) shows the different components of
the compression apparatus mounted on the stage of a two-photon microscope. (C), (D)
and (E) Two-photon microscopic images of the middle plane of a F9 aggregate
compressed between parallel plates at successive compression rates of 0% (A), 27% (B)
and 57% (C) respectively. Images were acquired 2 min after each compression. The
extracellular space is stained with the freely diffusible sulforhodamine B dye. Bar 100μm.
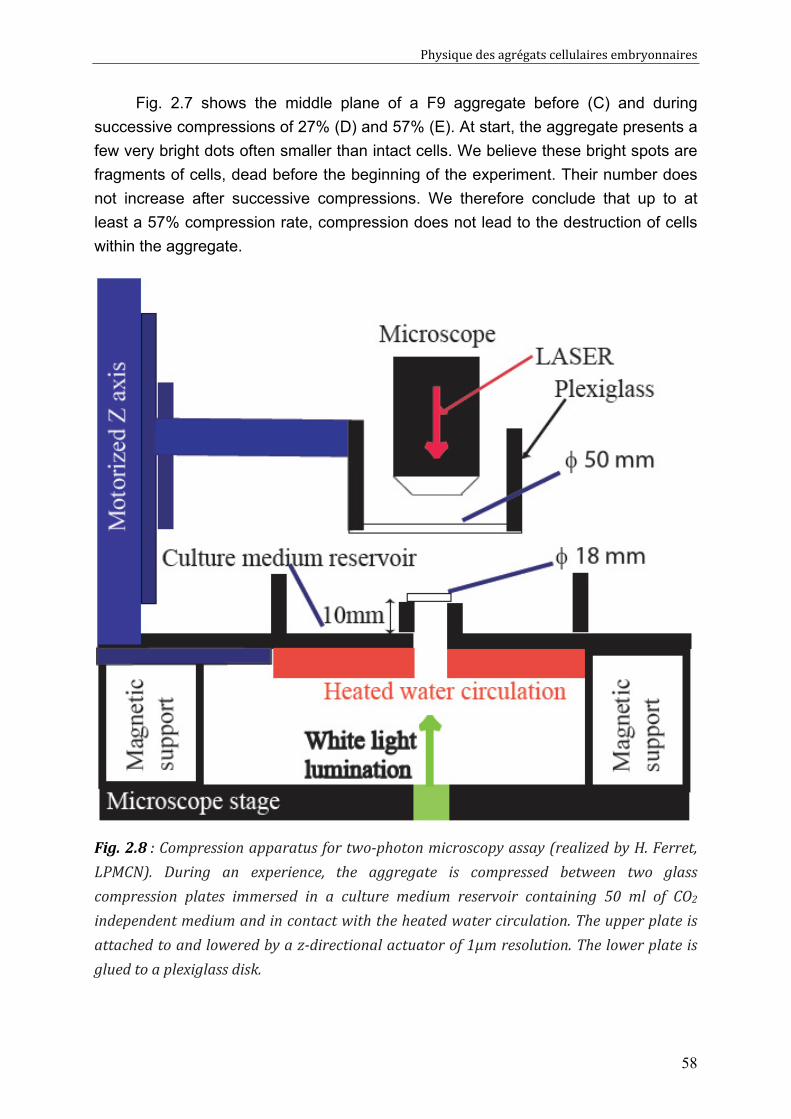
Physique des agrégats cellulaires embryonnaires
58
Fig. 2.7 shows the middle plane of a F9 aggregate before (C) and during
successive compressions of 27% (D) and 57% (E). At start, the aggregate presents a
few very bright dots often smaller than intact cells. We believe these bright spots are
fragments of cells, dead before the beginning of the experiment. Their number does
not increase after successive compressions. We therefore conclude that up to at
least a 57% compression rate, compression does not lead to the destruction of cells
within the aggregate.
Fig. 2.8 : Compression apparatus for two-photon microscopy assay (realized by H. Ferret,
LPMCN). During an experience, the aggregate is compressed between two glass
compression plates immersed in a culture medium reservoir containing 50 ml of CO2
independent medium and in contact with the heated water circulation. The upper plate is
attached to and lowered by a z-directional actuator of 1µm resolution. The lower plate is
glued to a plexiglass disk.

Physique des agrégats cellulaires embryonnaires
59
II.4. Segregation assays
II.4.1. Cell-sorting assay
The sorting pattern of mixed F9 cell types was assessed within a unique
multicellular spheroid containing two different clones of dissociated cells. Briefly,
300µm spheroids were prepared from mixtures of F9/ F9(α−/−) and F9/F9 (as a control
to assess that the transfection did not change the physical properties (surface
tension, viscosity) of the aggregate. They were then incubated at 37°C with 5% CO2
for 24 to 36 hours on a gyratory shaker (120 RPM) or in static conditions (i.e., on an
agar bed, SIGMA-A12967, to prevent aggregate/surface adhesion) . Cell-sorting was
evaluated every 12 hours by observing the cells at the surface of the aggregate on a
Nikon Eclipse 2000 fluorescence microscope.
II.4.2. Tissue-envelopment assay
The ability of F9 and their mutant F9(α-/-) cell types to spread upon one another
was assessed by bringing into contact two different types of aggregates: one only
composed of F9, the other one made uniquely of F9(α−/−). The two spheroids were
placed side-by-side on an agar bed (in 96-well plates). Spreading of one spheroid
over the other was evaluated using the Nikon Eclipse 2000 fluorescence microscope
every several hours.
II.5. Fusion and rounding up assay
For viscous liquids, various simple geometries are available to estimate from
the analysis of aggregate shape relaxation kinetics the ratio of the surface tension σ
to the viscosity η. Fusion and rounding up assay are two methods we used for this
purpose. Using a motorized microscope (Nikon Eclipse TE 2000 E), we recorded,
with a period of 10min, the fusion of several couples of symmetric aggregate
prepared under the same physiological and experimental conditions. We also used
the same imaging method for recording the shape relaxation of an elliptical
aggregate towards a spherical one with a period of 30 min between two recorded
loops.
For these experiments, aggregates were transferred into 48 well plates,
precoated with 1% of agar to prevent adhesion, and filled with a CO2 independent
medium (Gibco/invitrongen). In order to assure the required temperature during
7 The protocol of SIGMA was respected in the preparation and deposition of a 1% agar gel.

Physique des agrégats cellulaires embryonnaires
60
experiments, the microscope was enclosed in a home-made polystyrene chamber,
heated thanks to a radiator controlled by a circulating water bath thermostated at
37°C. We waited several hours for the thermal equilibrium to be achieved before
starting the experiment.
II.5.1. Fusion of two symmetric cell aggregates
When two identical viscous drops of radius R0 are placed in contact they slowly
fuse together (Fig. 2.9). The initial evolution of the radius X of the disc of contact
between drops may be described by the following equation as a function of time t:
ησ
0
2 tRX = (2.1)
where σ is the surface tension of the drop with the external medium and η the drop
viscosity. This equation is based on the balance of the work of surface tension and
the viscous dissipation. All other forces, including gravity, are neglected. In the
original work by Frenkel [34], there was a mistake about incompressibility, detected
by Eshelby (page 806 of the discussion of ref. [35]) who removed the extra prefactor
2/3 on the right hand side. We also found a calculus error on the line above his
equation 7 and removed an extra pre-factor 1/π. Our corrected formula therefore
does not contain any numerical pre-factor. This is consistent with a more extensive
calculation of the fusion process at large times [36].
Fig. 2.9 : Schematic view of the initial regime of fusion of two identical cell aggregates.
II.5.2. Rounding up of a free elliptical aggregate
As predicted from hydrodynamics and capillary laws of viscous liquids, the
elliptical deformation d (Fig. 2.10) during the rounding up of an elliptical droplet
follows a decreasing exponential law with a characteristic liquid time TLiq [144;147]
given by the following equation:
−=
LiqT
tdd exp0 and FLiq RT
σηβ= (2.2)
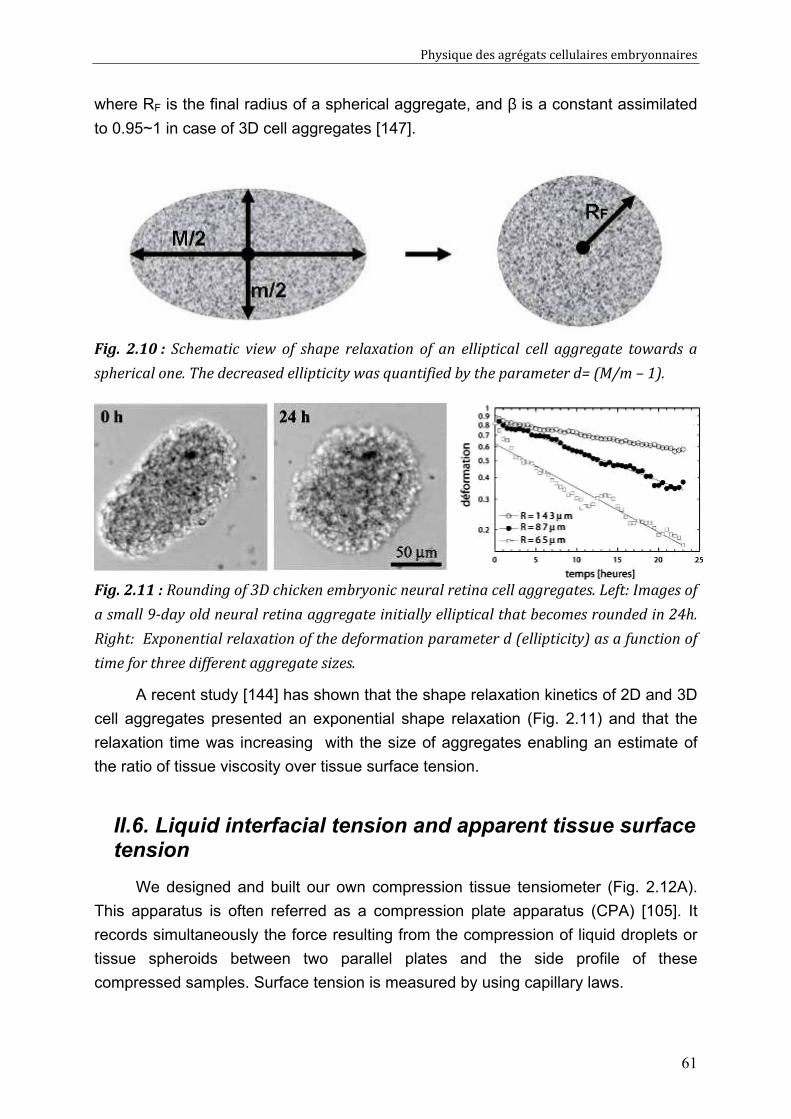
Physique des agrégats cellulaires embryonnaires
61
where RF is the final radius of a spherical aggregate, and β is a constant assimilated
to 0.95~1 in case of 3D cell aggregates [147].
Fig. 2.10 : Schematic view of shape relaxation of an elliptical cell aggregate towards a
spherical one. The decreased ellipticity was quantified by the parameter d= (M/m – 1).
Fig. 2.11 : Rounding of 3D chicken embryonic neural retina cell aggregates. Left: Images of
a small 9-day old neural retina aggregate initially elliptical that becomes rounded in 24h.
Right: Exponential relaxation of the deformation parameter d (ellipticity) as a function of
time for three different aggregate sizes.
A recent study [144] has shown that the shape relaxation kinetics of 2D and 3D
cell aggregates presented an exponential shape relaxation (Fig. 2.11) and that the
relaxation time was increasing with the size of aggregates enabling an estimate of
the ratio of tissue viscosity over tissue surface tension.
II.6. Liquid interfacial tension and apparent tissue surface tension
We designed and built our own compression tissue tensiometer (Fig. 2.12A).
This apparatus is often referred as a compression plate apparatus (CPA) [105]. It
records simultaneously the force resulting from the compression of liquid droplets or
tissue spheroids between two parallel plates and the side profile of these
compressed samples. Surface tension is measured by using capillary laws.
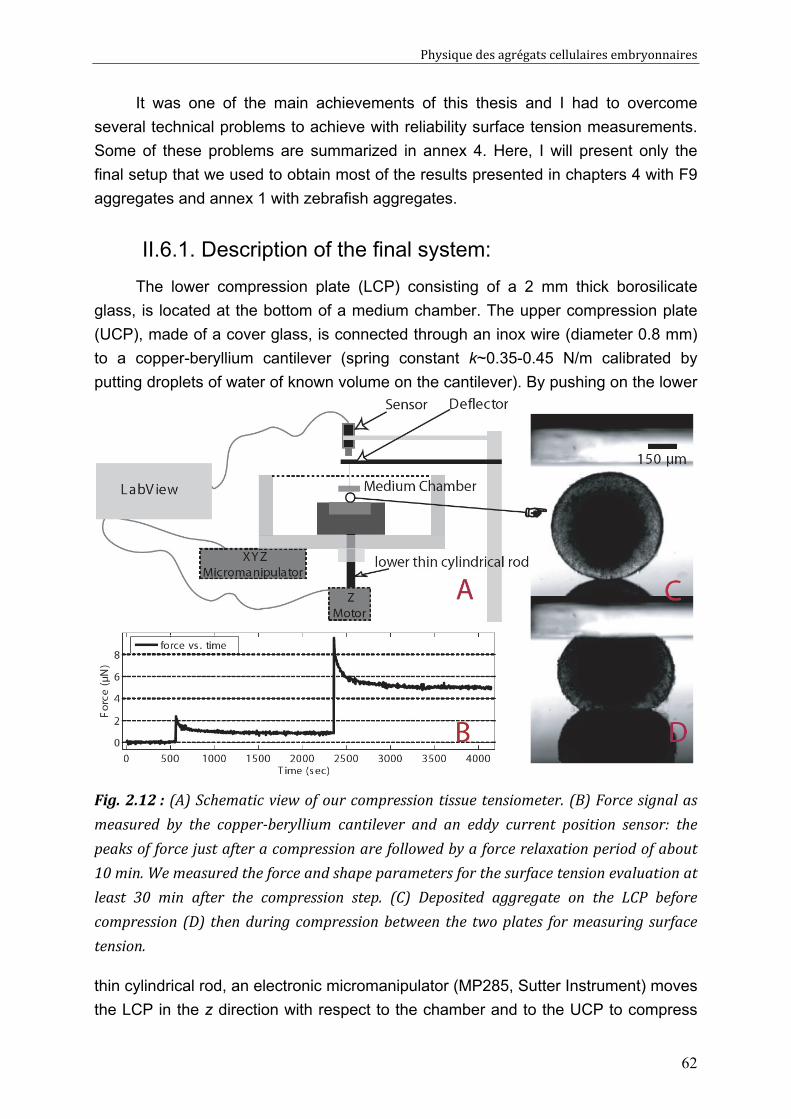
Physique des agrégats cellulaires embryonnaires
62
It was one of the main achievements of this thesis and I had to overcome
several technical problems to achieve with reliability surface tension measurements.
Some of these problems are summarized in annex 4. Here, I will present only the
final setup that we used to obtain most of the results presented in chapters 4 with F9
aggregates and annex 1 with zebrafish aggregates.
II.6.1. Description of the final system:
The lower compression plate (LCP) consisting of a 2 mm thick borosilicate
glass, is located at the bottom of a medium chamber. The upper compression plate
(UCP), made of a cover glass, is connected through an inox wire (diameter 0.8 mm)
to a copper-beryllium cantilever (spring constant k~0.35-0.45 N/m calibrated by
putting droplets of water of known volume on the cantilever). By pushing on the lower
Fig. 2.12 : (A) Schematic view of our compression tissue tensiometer. (B) Force signal as
measured by the copper-beryllium cantilever and an eddy current position sensor: the
peaks of force just after a compression are followed by a force relaxation period of about
10 min. We measured the force and shape parameters for the surface tension evaluation at
least 30 min after the compression step. (C) Deposited aggregate on the LCP before
compression (D) then during compression between the two plates for measuring surface
tension.
thin cylindrical rod, an electronic micromanipulator (MP285, Sutter Instrument) moves
the LCP in the z direction with respect to the chamber and to the UCP to compress

Physique des agrégats cellulaires embryonnaires
63
aggregates. The cantilever deflection is measured with a noncontact eddy current
position measurement apparatus (DT 3701-U1-A-C3, micro-epsilon). The medium
chamber is made in Teflon-inox with a pair of slide glasses glued on the sides to
enable optical visualization. It is filled with liquid (i.e., culture medium for aggregates)
and it has a hole that fits perfectly to the lower thin cylindrical rod supporting the
UCP. It can be moved in the x,y,z directions to center the sample on the optical axis
by the same micromanipulator. The complete setup is mounted in a thermally
isolated external chamber to maintain the desired temperature (37°C for aggregates)
using a thermal resistance, which is controlled by a Lakeshore 331 apparatus.
Aggregate profile is recorded using a binocular (MZ16, Leica) and a digital
camera (A 686M, Pixelink) (Fig. 2.12C-D). The lightning is adjusted by a KL 1500
LCD cold light source (Schott) through “flexible tubes.” The whole setup is controlled
with Labview and image analysis is performed with Matlab. The medium and external
chambers in which samples are deposited contain openings to facilitate
manipulations. In order to determine the interfacial tension between two liquids, the
inner chamber containing the LCP was first filled with the liquid of lower density (e.g.,
oil) before deposition of one liquid droplets (sizes ranged from 500 µm to 1000 µm in
diameter) of higher density (e.g., water) on the LCP.
For cell aggregates (300µm to 600µm in diameter), the medium chamber was
filled with CO2 independent medium (Gibco/Invitrogen), the free surface is covered
with a thick mineral oil layer to prevent evaporation. Glasses surfaces are carefully
cleaned with soap and pure water (sonicated 30 min with 2% Microson detergent,
Fisher Bioblock, France). They are first made hydrophobic by silanization with
perfluorosilane (ABCR, F06179) then incubated in 10 mg/ml Pluronic F-127 (Sigma)
for 5 min and finally rinsed briefly with water and dried. This treatment ensures a
minimum of aggregates adhesiveness.
The experiment starts by raising the LCP and compressing the droplet between
the two plates (Fig. 2.12D). Each droplet or aggregate was subjected to at least four
compressions. For tissues, we waited 30 min between two compression steps in
order for the aggregate to reach force and shape equilibrium. In (Fig. 2.12B), we
present the force signal as measured by our cantilever and position sensor: the peak
of force just after a compression is followed by a force relaxation period of about
10min. Chapter 3 is devoted to a detailed description of the capillary laws used to
analyze experiments and of the imaging analysis methods.
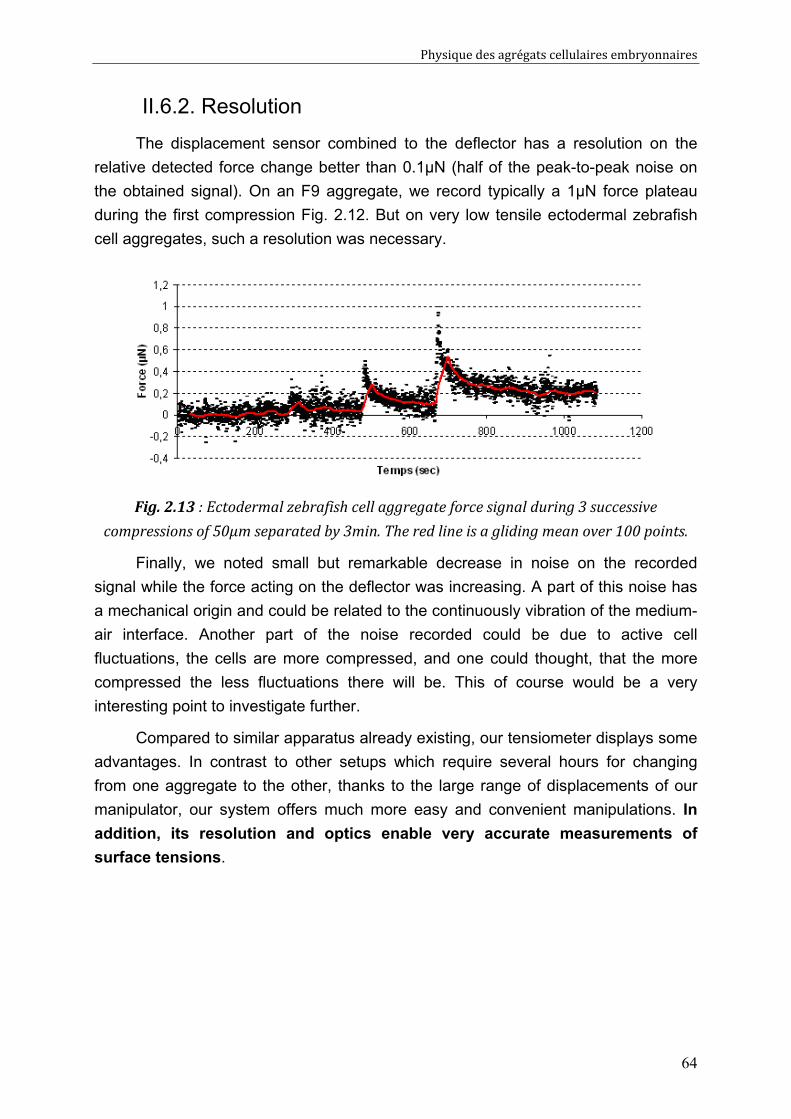
Physique des agrégats cellulaires embryonnaires
64
II.6.2. Resolution
The displacement sensor combined to the deflector has a resolution on the
relative detected force change better than 0.1µN (half of the peak-to-peak noise on
the obtained signal). On an F9 aggregate, we record typically a 1µN force plateau
during the first compression Fig. 2.12. But on very low tensile ectodermal zebrafish
cell aggregates, such a resolution was necessary.
Fig. 2.13 : Ectodermal zebrafish cell aggregate force signal during 3 successive
compressions of 50µm separated by 3min. The red line is a gliding mean over 100 points.
Finally, we noted small but remarkable decrease in noise on the recorded
signal while the force acting on the deflector was increasing. A part of this noise has
a mechanical origin and could be related to the continuously vibration of the medium-
air interface. Another part of the noise recorded could be due to active cell
fluctuations, the cells are more compressed, and one could thought, that the more
compressed the less fluctuations there will be. This of course would be a very
interesting point to investigate further.
Compared to similar apparatus already existing, our tensiometer displays some
advantages. In contrast to other setups which require several hours for changing
from one aggregate to the other, thanks to the large range of displacements of our
manipulator, our system offers much more easy and convenient manipulations. In
addition, its resolution and optics enable very accurate measurements of
surface tensions.

Physique des agrégats cellulaires embryonnaires
65
Chapitre 3
Measuring accurately tissue surface tension
Tissue surface tension allows the quantification of cell-cell cohesion and was
reported to be a powerful indicator for the cellular rearrangements that take place
during embryonic development or for cancer progression. The measurement is
realized with a parallel compression plate tensiometer using the capillary laws.
Although it was introduced more than a decade ago, it is based on various
geometrical or physical approximations. Surprisingly, these approximations have
never been tested. Using our own tissue tensiometer, we compare the two currently
used methods to measure tissue surface tension and propose a third one, based on
a local polynomial fit (LPF) of the profile of compressed droplets or cell aggregates.
Several compression experiments of water drops in mineral oil (W-O), air bubbles in
culture medium (A-M) and embryonic cell aggregates in culture medium (C-M) are
presented. We show the importance of measuring the contact angle between the
plate and the drop/aggregate to obtain real accurate measurement of surface tension
when applying existing methods. We can suspect that many reported values of
surface tension are greatly affected because of not handling this parameter properly.
We show then the benefit of using the newly introduced LPF method, which is not
dependent on this parameter. These findings are confirmed by generating
numerically compressed droplet profiles and testing the robustness and the
sensitivity to errors of the different methods.

Physique des agrégats cellulaires embryonnaires
66
III. 1. Introduction: TST measurements, for which purpose?
As discussed in chapter 1, at long time scales (hours or days depending on the
cell type), certain embryonic tissues mimic the behavior of highly viscous liquid
droplets [110]. They can round up into spherical shapes, fuse when they are brought
into mutual contact [147], sort out … All these processes are driven by surface
tension σ and resisted by viscosity η [147]. Interestingly enough, these two
quantities are accessible experimentally. Tissue surface tension (TST) can be
measured using a compression plate tensiometer [105] while the apparent viscosity
follows from the analysis of aggregate shape relaxation kinetics [92;144;147] (see
chapter 4).
For biological applications, tissue surface tensiometry is a new technology to
explore fundamental issues regarding cell-cell and cell-substratum interaction in (i)
morphogenesis, (ii) cancer progression and (iii) tissue engineering. Foty and
Steinberg [148], Marga et al. [149] have recently reviewed these issues, which we
briefly summarize here.
(i) The analysis and sorting/envelopment behaviour of germ layer progenitor
cells in amphibians [150] and zebrafish [113] suggests that TST (measured in these
studies) is involved in guiding germ layer morphogenesis during gastrulation.
Downregulation of E-cadherin levels in the later study leads to a decrease in the
measured surface tension and a corresponding reversal of germ layer positioning in
cell sorting experiments.
(ii) It is generally assumed that the invasiveness of cancer cells largely
depends on a loss of cell cohesion. Cadherins have been linked with transition to
malignancy for a variety of tumors. In particular, the expression of E-cadherin often,
but not always, inversely correlates with tumor aggressiveness [148]. However, for
brain tumors, it was shown that dexamethasone mediated decreased invasiveness
correlates with increased TST (i.e., cohesivity) but not with N-cadherin expression
[151]. Surface tension represents a global property of a tissue that may depend from
other interactions than the only cadherin-cadherin interactions. αααα5ββββ1 integrin-
fibronectin interactions can indeed mediate strong cohesion [40]. The interactions of
a cell with the surrounding matrix are also, of course very important to control the
invasive properties of tumors [152].
(iii) Tissue engineering aims at reproducing morphogenesis in the laboratory,
i.e., to fabricate replacement organs for regenerative medicine. It has been shown
that the liquid properties of some tissues, in particular the capability to fuse or

Physique des agrégats cellulaires embryonnaires
67
reaggregate, may be used to self-assemble cellular aggregates into 3D living
structures [149;153;154]. TST and viscosity are parameters that can be used both
experimentally and theoretically to control and predict this self- or re-assembly. In
addition, surface tension can also strongly influence the ability of tissues to interact
with other biomaterials [155].
III. 2. The different methods to measure TST
III. 2.1. A few lines on capillarity
Although a new method based on centrifugation followed by axisymmetric drop
shape analysis has been recently described [101], the most widely used quantitative
method to measure TST (σ) is by compression plate tensiometry [105;152]. At long
time scales, once elastic forces are relaxed, σ is measured assuming that cell
aggregates verify the same physical laws of capillarity as liquid droplets.
When a droplet is compressed between two identical plates (i.e., with identical
surface properties), it has a rotational symmetry around the z-axis and a reflection
symmetry with respect to its equatorial plane, in which it has the two principal radii of
curvatures R1 and R2 shown in Fig. 3.1. R3 is the radius of the droplet’s circular area
of contact with the compression plates. The compression force F applied to the upper
(or lower) plate is balanced by two capillary forces each proportional to σ. The first
one is due to the excess pressure inside the drop due to curvature given by the
Laplace formula:
∆p =σ(1/ R1 +1/ R2).
When the radii of curvature are positive as it is generally the case with cellular
aggregates, this first term is always positive (i.e., repulsion between the two plates).
The second term is proportional to the drop perimeter and is always negative (i.e.,
attraction between the two plates). For an arbitrary horizontal plane, at mechanical
equilibrium, the equilibrium condition when evaluated along the vertical axis, implies
F =∆p.A− σ.P.sinφ. Here A and P represent the cross-sectional area and the
perimeter of the liquid drop in this plane respectively, and φ is the angle between the
horizontal and the tangent to the profile of liquid drop at the plane.
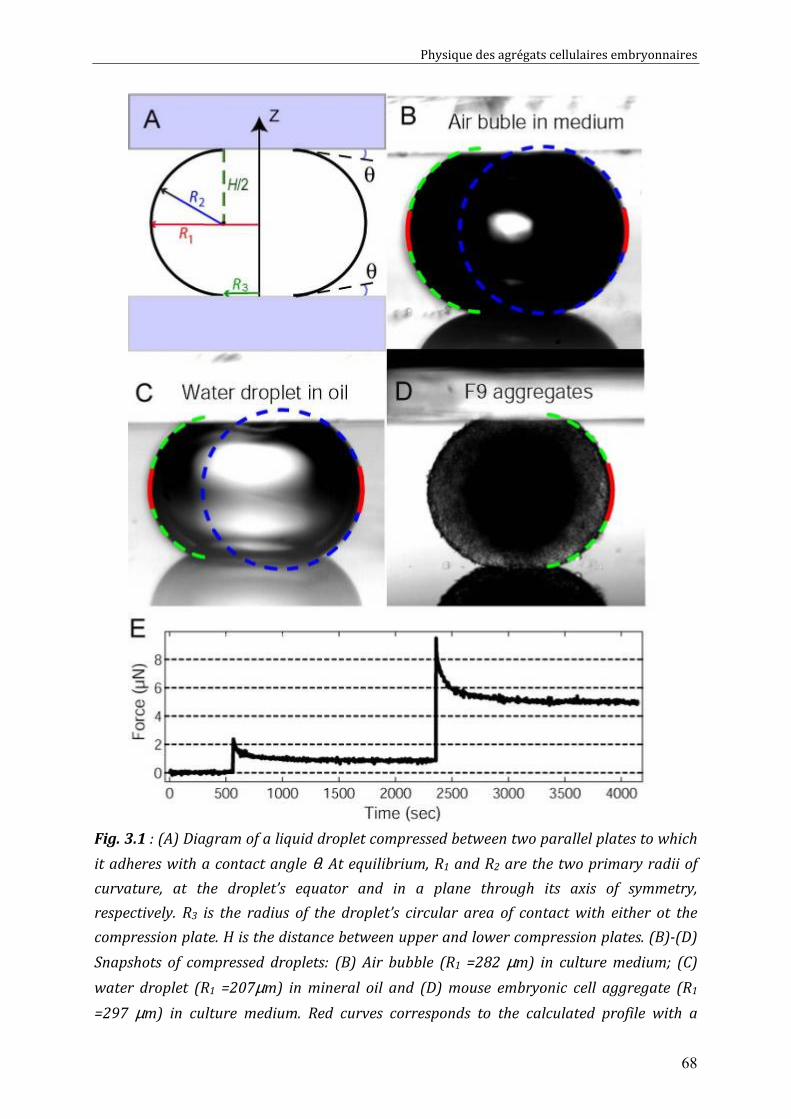
Physique des agrégats cellulaires embryonnaires
68
Fig. 3.1 : (A) Diagram of a liquid droplet compressed between two parallel plates to which
it adheres with a contact angle θ. At equilibrium, R1 and R2 are the two primary radii of
curvature, at the droplet’s equator and in a plane through its axis of symmetry,
respectively. R3 is the radius of the droplet’s circular area of contact with either ot the
compression plate. H is the distance between upper and lower compression plates. (B)-(D)
Snapshots of compressed droplets: (B) Air bubble (R1 =282 µm) in culture medium; (C)
water droplet (R1 =207µm) in mineral oil and (D) mouse embryonic cell aggregate (R1
=297 µm) in culture medium. Red curves corresponds to the calculated profile with a

Physique des agrégats cellulaires embryonnaires
69
second order polynomial fit (LPF method), dotted lines correspond to the calculated
profiles with the exact Laplace profile method (ELP, green) and the circular arc
approximation (blue). The latter is not represented in order to appreciate the embryonic
aggregate roughness in (D). (E) Force signal as measured by our cantilever and eddy
current position sensor: the peaks of force just after a compression are followed by a force
relaxation period of about 10 min. After [156]
For the horizontal boundary plane located just underneath the upper plate, A= πR3
2,
P=2πR3 and φ =θθθθ8. Thus, by using the Laplace formula for ∆p, one obtains:
2
3 3
1 2
1 1 -2 sin PF R R L
R Rπ π θ σ σ
= + =
(3.1)
where LP is therefore just a geometrical parameter depending on R1, R2, R3 and θθθθ. Similarly, for the median plane of the compressed drop at H/2, where H is the
compressed drop height, one has A= π R12, P=2π R1 and φ =π/2. The force depends
on the geometrical parameter LM:
11
2
1 M
RF R L
Rπ σ σ
= − =
(3.2)
These two equations (3.1) and (3.2) are of course equivalent as the force in
each droplet horizontal plane is conserved along the vertical axis. Let us examine
how to use them efficiently to get the more accurate and reliable TST measurements.
III. 2.2. The CA (Circular Arc) method
While R1 and F can be accurately measured, the determination of σ from either
one of these two equations requires also the measured values for R2 and/or θθθθ and R3
that can only be obtained with large errors. In earlier published studies this problem
has been circumvented by making the approximation that the lateral profile of the
drop is a portion of circular arc [103;113]:
2 2
3 1 2 2 - - ( /2)R R R R H= + (3.3)
In order to even simplify the analysis, plates are generally treated to prevent
aggregate/plate adhesion and it is assumed that θ=0°, R2=H/2 and R3 = R1 −R2 [150].
8 Note that the angle θ defined here is the complementary of the contact angle in capillarity. We use here the
convention used in previous TST papers.

Physique des agrégats cellulaires embryonnaires
70
In all the previous reported studies except for Norotte et al. [157] and our recent work
[156], this CA method was used probably by matching manually on the screen the
complete contour of the aggregate by a printed “abacus” as no details were given on
the numerical method. We have also tested this approximation but R2 was obtained
by a fit of the recorded profile (see LPF method section below) using the function
fitellipse.m under matlab (http://www.mathworks.com/matlabcentral/fileexchange/15125). We
have calculated the surface tension using this circular arc approximation in three
different ways: (i) using Equation (3.2), where the force is expressed at median plane
(CAcm method), (ii) by combining equations (3.1) and (3.3), where force is
expressed on plates, with θ=0° (original CA method), and (iii) by combining equations
(3.1) and (3.3) using the measured θ (CAcp). R1 is obtained in the same way than for
ELP, LPF and CA methods (see description in the LPF section). When it is needed,
i.e., for the CA and CAcp methods, H is obtained as explained in the ELP method
section.
III. 2.3. The ELP (Exact Laplace Profile) method
Norotte et al. [157] have used another method based on the exact solution of
Laplace equation and on the only measurements of θθθθ, H and R1. When gravity is
neglected, a dimensionless parameter α is calculated by solving numerically the
following equation:
( )( )
1/ 21
2
2
1 ,
= ( ) 12 1
H xf dx
R xθ
β α θ
αα α
− = − + −
∫ (3.4)
where ( ) ( ) ( )( )2, sin( ) sin 4 1 2β α θ θ θ α α α= + + −
and σ is then given by
( )12 1 EF R Lπ α σ σ= − = (3.5)
They claim their method is weakly sensitive on the angle, as long as θθθθ≤20°.
They also claim that the previously existing method, based on the circular arc
approximation (CA method), fails to give consistent results in a certain range of the
compressive force or contact angle. But they did not present any quantitative
analysis on the geometrical parameter sensibility of the different methods.
We have tested these two reported methods and performed a complete
quantitative and comparative analysis. Concerning this ELP method, we have found

Physique des agrégats cellulaires embryonnaires
71
(see results) that it is not so insensitive to θθθθ which is difficult to measure accurately.
Impose θθθθ ≈0° may be sometimes difficult because cell aggregates may adhere to the
plates after prolonged compression despite the fact that a coating minimizing
adhesion is used. In our experience, θθθθ was not zero and varied during a compression
experiment. It is a difficult parameter to measure optically with high accuracy
because of a number of interfering factors, such as; the large field of focus, the
imperfect parallelism between plates, light multi-reflections or optical aberrations.
For the determination of H, it is important to use the deflection signal to
evaluate precisely the position at which the upper plate contacts the aggregate. We
found that this position can be hidden on the image due to slight inclination of upper
compression plate, or of optical axis or of illumination. With our tissue tensiometer, H
is determined with a typical resolution of about 2 µm. Finally R1 is measured
accurately on the images of compressed aggregates as explained in next section.
III. 2.4. The LPF (Local Polynomial Fit) method
Our motivation was to establish a more direct and robust method to measure
the absolute values of TST accurately and reliably independently of the contact
angle. For that purpose, we use Eq. (3.2) to calculate surface tension from the
measurement of the two radii of curvature R1 and R2 (Fig. 3.1A). This requires the
analysis of the aggregate profile r(z) along vertical axis z. It is drawn by hand on the
right and left side of the aggregate. We did not make any automated contour analysis
because of inhomogeneities and varying lightning conditions during subsequent
compression steps.
A curve can always be approximated locally by a second order polynomial
r=az2+bz+c, the local radius of curvature is then given by R2=1/2a. The narrower the
window on which the profile will be fitted, the lower the residual dr=rpredicted-rdata on the
fitted profile and the better the evaluation on the curvature will be (see Fig. 3.3). Of
course, we are limited by the image resolution on the contour and, in the case of
aggregates, by the surface roughness. This prevents us from using two small
windows. In practice, this window is adjusted between one fourth and one half of the
whole side contour. R1 is then taken to be half of the distance between points having
the highest r-coordinates on both right and left fitted contours.
For each side, the error of R1 is set to the maximum value of the residual dr
and the error on R2 is given by the 95% confidence interval given on the polynomial
fit using the curve fitting toolbox of matlab. The error on both sides is averaged and
divided by 21/2. It is about 3 µm and 10 µm for R1 and R2 respectively.

Physique des agrégats cellulaires embryonnaires
72
III.3. Critical comparison of existing methods using experimental data and numerical tests
III. 3.1. Experiments with pure liquid droplets or bubbles
We first compressed air bubbles in culture medium (CO2 independent medium,
37°C). These bubbles are easily nucleated by pouring cold medium on the
tensiometer plates already at 37°C and waiting 2 hours for temperature stabilization.
The complementary contact angle θ is small when glass is clean and hydrophilic. But
because glass surfaces are frequently re-used, θ may change from one experiment
to the other, or after successive compressions. Only for half of the compressions
investigated (n=20 compression steps corresponding to 3 different bubbles), we
could really obtained an angle 5θ ≤ ° for all four recorded glass/air/medium contact
lines (Fig. 3.1B). The angle was comprised between 10° and 25° otherwise. Fig. 3.2A
shows the plot of geometrical parameters LP, LM or LE defined in Eqs. (3.1), (3.2) and
(3.5) for each method as a function of the deflection transmitted to the upper plate
(δ=F/k where F is the force). The points in the graph correspond to three different
bubbles. This plot displays a linear relationship between δ and L for each method as
expected from capillary laws. The slope k/σ gives the surface tension σ. However,
with ELP and CA methods, points are much more dispersed and the surface tension
values (σ=56±2.6 and 39±3.1 mN/m respectively9) are significantly different from a
direct measurement using the wilhelmy plate pressure sensor of a Langmuir trough
(NIMA, England): σ=51±2 mN/m. In contrast, the LPF method provides the correct
value, with the lower error: σ=51.2±1.2 mN/m. When it is calculated at the median
plane, the circular arc approximation (CAcm) gives also a reasonable agreement
although the value is slightly lower (σ=48±1.4 mN/m). These differences are due to
the fact that some methods are independent of θ which is measured with errors but
others not.
9 Errors ∆σ are calculated from the 95% confidence interval on fitted slopes by the curve fitting toolbox of
matlab (relative errors on slopes are the same as relative error on σ as the relative error on k is negligible).
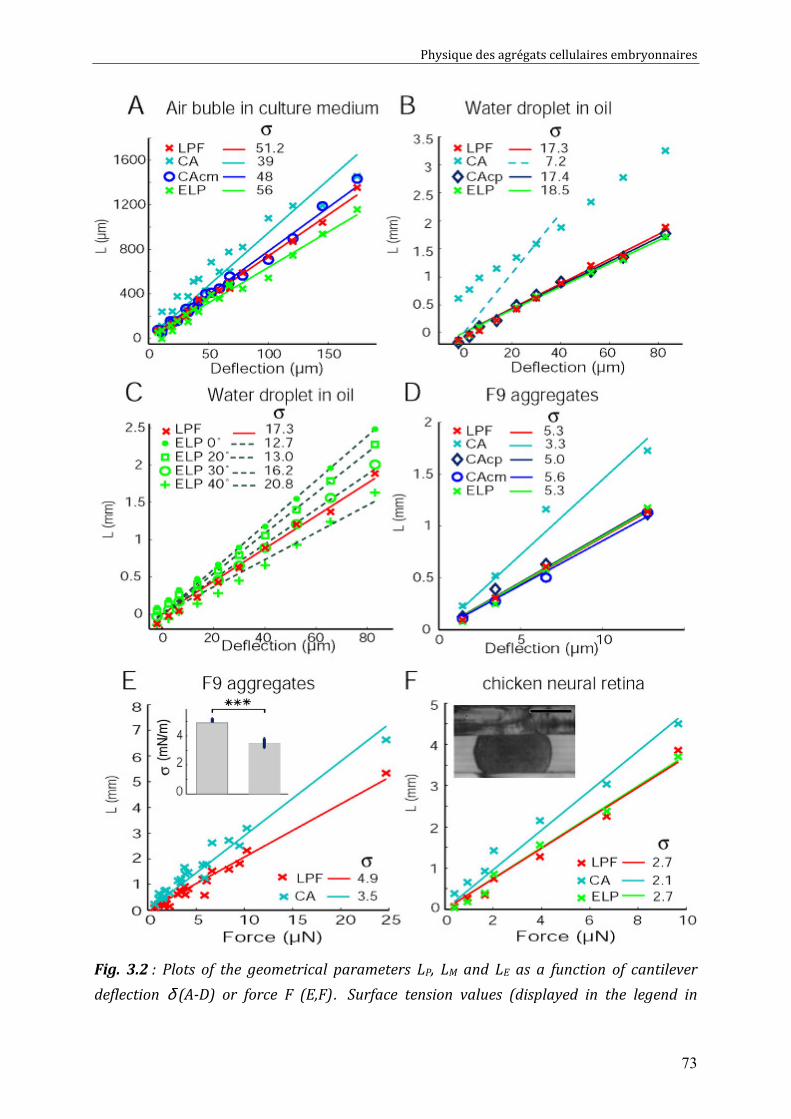
Physique des agrégats cellulaires embryonnaires
73
Fig. 3.2 : Plots of the geometrical parameters LP, LM and LE as a function of cantilever
deflection δ (A-D) or force F (E,F). Surface tension values (displayed in the legend in

Physique des agrégats cellulaires embryonnaires
74
mN/m) are obtained by taking the slope of the obtained linear fits, with LP, LM and LE
obtained as defined in Eqs. 3.1, 3.3, 3.5, and δ = F/k (with k~0.36 N/m for all experiments).
Air bubble in CO2 independent culture medium at 37°C (A), water droplet in mineral oil at
room temperature (B-C), F9 embryonic cell aggregate in CO2 independent culture medium
at 37°C (D-E) and chicken embryonic neural retina aggregate in CO2 independent culture
medium at 37°C (F). In the inset of (E), we show that the difference between the mean
measured values with each method is statistically significant. In the inset of (F), we show a
snapshot of compressed neural aggregate (bar, 200 µm). Abbreviations of the different
methods tested are as follows: ELP, Exact Laplace Profile; LPF, Local Polynomial Fit; CA:
Circular Arc; CAcm, Circular Arc with force calculated at median plane; CAcp, Circular Arc
with force calculated on plates. After [156]
We followed up our studied by compression analysis of water droplet,
immerged in mineral oil (Sigma) at room temperature. The complementary contact
angle was approximately θ=28±5 ° for the five investigated droplets, with minimal
changes from one compressions to the other. The original CA method that ignores
such a large angle (i.e., it uses Eqs. (3.1) and (3.3) with θ=0°!) gives clearly an
underestimated surface tension. The curve LP versus the deflection is not linear. But
if we use only one or two compression points as done in most reported papers
except for Norotte et al. (2008), the estimated surface tension can be more than two
times smaller than the expected one. It is for instance the case, if we use the point at
deflection 30 µm corresponding to nearly a 50% deformation (see dotted line in Fig.
3.2B). When the ELP and the CAcp (calculated on plates) are used with the angle
measured on images, they give similar results to the angle independent LPF method.
Values of surface tension are σ=18.5±1.5, 17.4±1 and 17.3±1 mN/m for ELP, CAcp
and LPF methods respectively. This is in agreement with reported values for water/oil
interfacial tension [157;158]. Fig. 3.2C shows the angle sensibility of ELP method
when angle is not properly chosen. In this case, the surface tension varies a lot,
σ=12.7±0.8, 13.0±0.8, 14.0±0.7, 16.2±0.7 and 20.8±2 mN/m for θ=0°, 10°, 20°, 30°
and 40° respectively. Note that the points are still linearly aligned even if the slopes
are very different! The error is not increasing particularly, and therefore one has to
keep in mind that a low error or a linear alignment as predicted by Laplace law is not
an insurance of correct surface tension measurement, using ELP method.
III. 3.2. Experiments with embryonic cell aggregates
Tissues display elastic behaviour at short timescales [119], which contribute to
the force signal shortly after a compression step. This elasticity is usually rapidly
dissipated and a force plateau is reached (see Fig. 3.1E). We have assumed here as

Physique des agrégats cellulaires embryonnaires
75
it was done in previous studies [106], that when this plateau is reached, the system is
at equilibrium, which enables the surface tension measurement (see a discussion
about the validity of this assumption in chapter 5)
When compressed, F9 aggregates show a complementary contact angle
between 20° and 30° on glass plates treated with Pluronic F-127 (Fig. 3.1D). We
present in Fig. 3.2D the results for one aggregate (n=4 steps of compression). With
such angle values, the non angle corrected CA method gives a much lower surface
tension value (σ=3.3±0.7 mN/m) than the angle independent methods: σ=5.3±0.7 and
5.6±0.8 mN/m for LPF and CAcm respectively (Fig. 3.2D). When the angle-
measurements from the image profiles are properly taken into account, ELP and
CAcp methods give similar values but with higher errors: σ=5.3±1.0 and 5.0±1.0
mN/m respectively. Alternatively, the choice of the correct angle may be done by an
adjustment of the compressions points of angle dependent and independent
methods. This provides a way to evaluate the complementary angle when image
profile is not clear because of very small (θ <10°) or because image is partially hidden
by for instance the upper plate, as in Fig. 3.1D.
To further show the statistical difference between the results obtained with the
usual CA method and the newly proposed LPF method we analyzed seven different
F9 aggregates, 23 compression steps (Fig. 3.2E). The mean surface value obtained
with the CA method is 3.5±0.3 mN/m compared to 4.9 ±0.3 mN/m for the LPF
method. The p-value obtained comparing the two results is inferior to 0.00110. This
indicates a very good reproducibility in the finite contact angle of these aggregates.
The seven investigated aggregates have diameters before compression in the range
of 372 to 585 µm. Their volumes are therefore comprised between 27.106 and
105.106 µm3 (i.e., a four fold increase). We can conclude that the surface tension of
these F9 aggregates is volume independent.
We finally confirmed our results on a different cell line: chicken embryonic
neural retina cells (Fig. 3.2F). The value of the contact angle was estimated to be
around 30 °. The results obtained are similar to the F9 ones. The surface tension
value with the CA method is 2.1±0.3 mN/m as previously measured [103] which is
again lower than for the ELP end LPF method (2.7±0.3 mN/m).
10 The comparison between surface tension values obtained with CA methods and the one obtained with angle
independent methods was done with a Kruskal-Wallis non parametric test. As the Kruskal-Wallis test was found
significant (P < 0.05), a Wilcoxon non parametric test was performed (R 2.6.0, R foundation for statistical
computing, Vienna, Austria). P values <0.001 were considered as significant and noted with three stars.
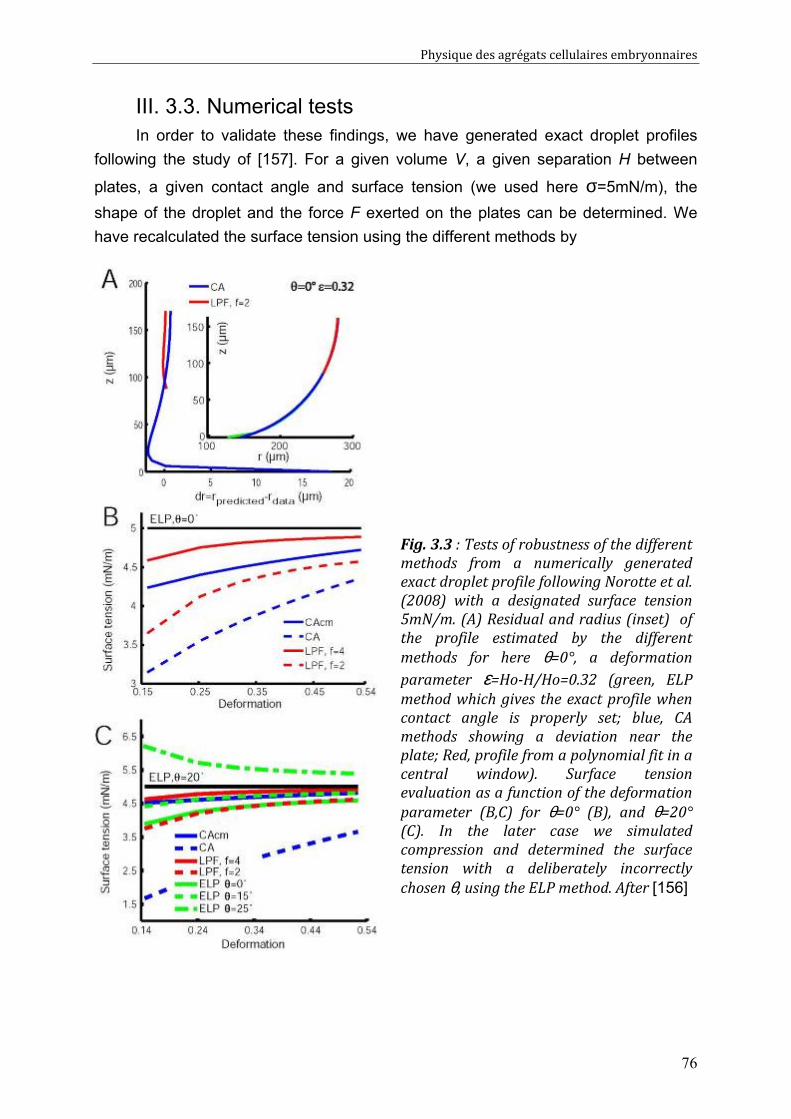
Physique des agrégats cellulaires embryonnaires
76
III. 3.3. Numerical tests In order to validate these findings, we have generated exact droplet profiles
following the study of [157]. For a given volume V, a given separation H between
plates, a given contact angle and surface tension (we used here σ=5mN/m), the
shape of the droplet and the force F exerted on the plates can be determined. We
have recalculated the surface tension using the different methods by
Fig. 3.3 : Tests of robustness of the different methods from a numerically generated exact droplet profile following Norotte et al. (2008) with a designated surface tension 5mN/m. (A) Residual and radius (inset) of the profile estimated by the different methods for here θ=0°, a deformation
parameter ε=Ho-H/Ho=0.32 (green, ELP method which gives the exact profile when contact angle is properly set; blue, CA methods showing a deviation near the plate; Red, profile from a polynomial fit in a central window). Surface tension evaluation as a function of the deformation parameter (B,C) for θ=0° (B), and θ=20° (C). In the later case we simulated compression and determined the surface tension with a deliberately incorrectly chosen θ, using the ELP method. After [156]

Physique des agrégats cellulaires embryonnaires
77
eventually introducing small errors in the input geometrical parameters (θ, see below,
and H, not shown) in order to simulate the ‘measurement inaccuracies’ of a real
experiment. Of course, the ELP method gives always the exact value when the
correct angle is introduced (black curves in Fig. 3.3B-C).
For a null complementary contact angle, one can appreciate in Fig. 3.3A the
deviation of the real profile (green curve) to the circular arc (blue curve). The
deviation is only localised in the vicinity of the plate but it results in a 15% error in the
determination of R3 used in Eq. (3.1) and thus in σ. This explains why the CA method
based on this equation has the largest error on the surface tension even when the
complementary contact angle is 0° (Fig. 3.3). When Eq. (3.2) is used, the error in σ
by using the CAcm or LPF method is introduced by the measurement of R2 itself.
Both methods underestimate σ because R2 is slightly overestimated. The deviation to
the real value (σ=5 mN/m) decreases for large compressions (i.e., large deformation
parameter ε=Ho-H/Ho where Ho is the uncompressed aggregate height). The LPF
method gives the best results when the proportion H/f of the profile around the
median plane used for the local second order polynomial fit is not too large: σ=4.88
and 4.58 mN/m for f=4 and 2 respectively for a large compression (ε=0.5). The CAcm
method gives an intermediate value 4.66 mN/m when using the full profile (f=1).
When the complementary contact angle is larger (i.e., θ=20°, Fig. 3.3C), again
the LPF method provides the best results for a narrow fitting window (f=4) followed by
the CAcm method (Fig. 3.3C). The error is even lower compared to the analysis
where θ=0°: σ=4.90 and 4.78 mN/m for the LPF (f=4) and CAcm methods
respectively for ε=0.5. Error is much larger with the ELP method when an error of
only 5° is introduced on θ: for θ=25°, σ =5.5 mN/m (i.e., 10% error). The CA methods
using θ=0° gives completely wrong results.
III. 4. Discussion
In this chapter, we have shown that when capillary laws are properly employed,
the three methods to analyze surface tension from compressed droplet (ELP, LPF
and CA) give similar results with slightly different errors. Nevertheless, each method
has a different sensibility to geometrical parameters making some of them more
robust than others.

Physique des agrégats cellulaires embryonnaires
78
III. 4.1. Examination of literature data
To date, except in the study of Norotte el al. [157], the circular arc
approximation method is the only one used to measure tissue surface tension. A
large but non-exhaustive list of papers is given in the introduction of this chapter.
Even when the deviation to the circular arc profile is evident near the plates (Fig.
3.3A), the radius of curvature R2 at median plane is correctly calculated. As a result,
the CAcm method which consists in a circular arc fit combined with the force Eq.
(3.2) at median plane provides very acceptable results (5-10% error on simulated
drops depending on contact angle and compression rate, Fig. 3.3B-C). The
simultaneous use of Eq. (3.1) with θ=0 and of Eq. (3.3) leads to some
inconsistencies when the complementary contact angle actually deviates from zero
The result is that the measured surface tension is systematically lower than the
actual tissue surface tension value, and can lead to an underestimation of more than
100% (Fig. 3.2B,D and Fig. 3.3C). One can therefore postulate that some of the
reported surface tension values are incorrect because of a finite contact angle.
Specifically, we have identified two studies where contact angles are visibly large
[106;113]. In a third one with CHO aggregates [40], there is no images of
compressed aggregates as most of the other reported studies. However, from our
own experiences, CHO aggregates are sticking and even “wetting” any surface due
to integrin-ecm interactions. A finite contact angle may explain why the authors
observed systematically a larger surface tension for the second compression than for
the first one. Indeed, in Fig. 3.2B, the slope of the dotted line intercepting the origin
(which is inversely proportional to σ) decreases when deflexion (i.e., compression)
increases. In most of these studies, polyHEMA was used to prevent cell/substrate
adhesion. We have tested this protocol and formed thick (i.e., several tenth of µm)
films of polyHEMA on glass surfaces. Aggregates indeed poorly adhere to it but they
also penetrate partially into the film making difficult a real evaluation of θ or H. For
that reason, we used Pluronic F-127 which adsorbs as a monolayer and also prevent
rather well adhesion11 but show a small but finite (complementary) contact angle.
Finally, it is important to stress that even if the errors on the surface tension appear
to be as low for the CA method as for the other angle independent methods, this
does not mean that the value obtained is correct as the capillary laws are not
properly used.
11 Aggregates do not stick to the surfaces when they are decompressed after a prolonged compression

Physique des agrégats cellulaires embryonnaires
79
III. 4.2. Recommendations for correct measurements
Inserting the correct angle in Eq. (3.1) allows a correct estimate of σ, when
using the CAcp method. However, there is no need to use Eq. (3.1) at the plane of
the plate, because this requires the input of four geometrical parameters R1, R2, θ and H, of which especially R2 and θ are usually measured with large errors. In contrast,
the ELP method, based on the exact solution of Laplace equation, is more robust
from a computational point of view. The resolution of Eqs. (3.4) and (3.5) is
numerically easy and does not necessitate a least squared fit of the radius of
curvature R2 as in case of LPF or CA methods. However, we found that an error of 5°
on θ introduces non negligible relative errors on the surface tension σ (about 10%).
Such an error is not particularly exaggerated for cell aggregates when θ is small
because cell aggregates are rough, not as regular as fluid droplets. The contact
angle may also change after several subsequent compressions. The ELP method
also requires the input of the aggregate compressed height H which may suffer
errors for instance due to optical aberrations, hidden part of the droplets. On the
other hand, if θ is well controlled or if some error on σ is acceptable, one can build a
tissue tensiometer under an inverted or upright microscope and record only H and R1
from the top view (not R2 from the side view) and use Eqs. (3.4) and (3.5). This may
enable to follow the cell reorganizations and motion during compression and to
measure σ at the equilibrium.
As a conclusion, when performing compression experiments, we strongly
recommend the use of an angle independent method that requires only the two
parameters R1 and R2. The CAcm method may have the lowest error when
aggregate is symmetric but rough. Otherwise, the proposed LPF method is robust
and provides the lowest error when the profile is smooth as the fitting window can be
narrowed. This method is a straightforward application of Laplace equation. R2 is
measured by a second order polynomial fit at median plane. While other methods
assume reflection symmetry with respect to equatorial plane, the LPF method deals
naturally with up-down asymmetries in the aggregate profile. Such asymmetries may
arise either because of a slightly titled field of view, non-perfectly parallel-aligned
compression plates, effect of gravity or because of differences in adhesion affinities
and contact angles with the top and bottom compression plates.

Physique des agrégats cellulaires embryonnaires
80
Chapitre 4 Physical properties of F9 tissues
Role of adhesion and Cytoskeleton
Several studies on embryonic tissues have attempted to characterize the
material properties of cell aggregates by a few physical parameters (surface tension
and viscoelasticity) [106;159;160]. We focus in this chapter on the comparative
physical properties of F9 cell aggregates and their α-catenin double knocked out
derivative (F9(α-/-)) in order to evaluate the effect of the cytoskeleton-cadherin bond to
the measured quantities. In the absence of external forces, F9 aggregates fuse and
round up to form spheres (see the supplementary movie M4). These processes are
classic examples of surface minimization processes, which as in ordinary liquids, are
driven by tissue surface tension (TST) [147;161]. F9(α-/-) cell aggregates have a lower
TST than F9 aggregates as measured with our compression plate apparatus
(tensiometer). The kinetics of the fusion of two symmetric aggregates is used to
evaluate the tissue viscosity. This quantity is similar for each cell line. In parallel, we
performed cell sorting and engulfment assays and showed that whatever the initial
condition F9 aggregates (with the higher TST) are enveloped by F9(α-/-).
We therefore confirm that TST is measurable and is a crucial quantity to predict
tissue mutual behaviours and organisation. However, our results contradict the
“second postulate” of the differential adhesion hypothesis (i.e., TST depends only on
the level of expression of adhesion molecules) as TST depends explicitely on the
cadherin-actin bond or on the presence of nocodazole (a contractile drug). We
believe that a differential contractility hypothesis (cytoskeleton mechanics or
actomyosin contractility) is likely to be the origin of TST.
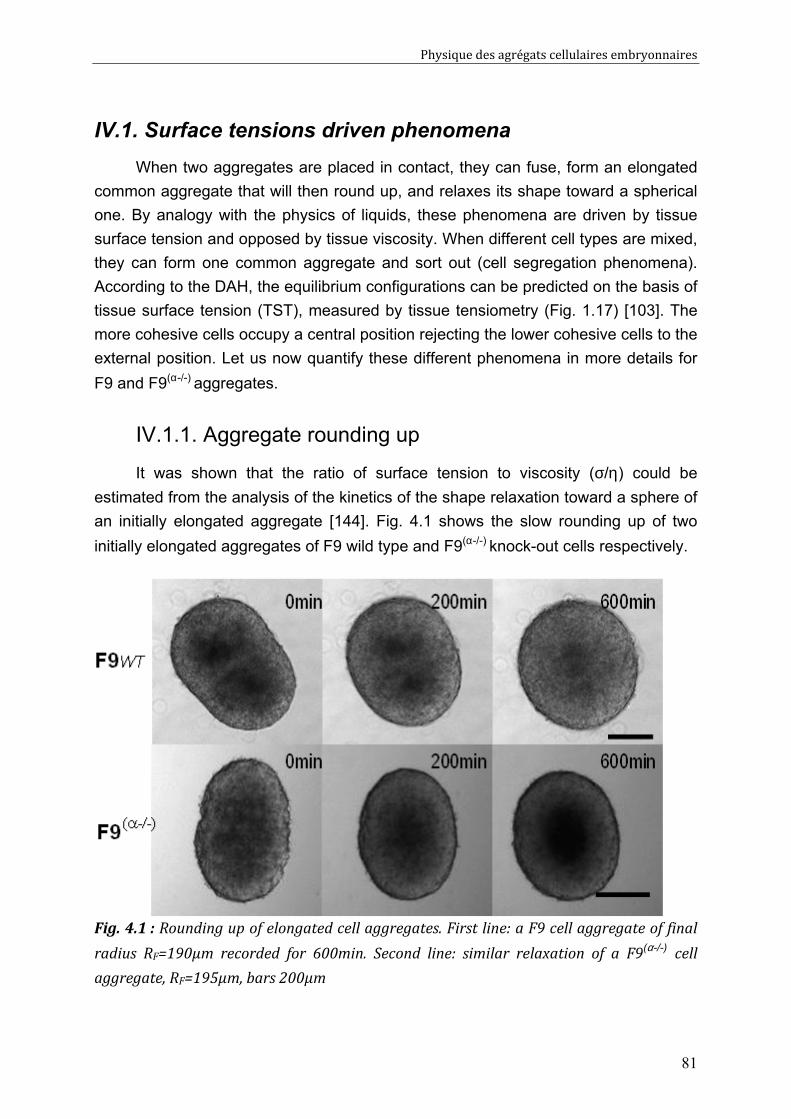
Physique des agrégats cellulaires embryonnaires
81
IV.1. Surface tensions driven phenomena
When two aggregates are placed in contact, they can fuse, form an elongated
common aggregate that will then round up, and relaxes its shape toward a spherical
one. By analogy with the physics of liquids, these phenomena are driven by tissue
surface tension and opposed by tissue viscosity. When different cell types are mixed,
they can form one common aggregate and sort out (cell segregation phenomena).
According to the DAH, the equilibrium configurations can be predicted on the basis of
tissue surface tension (TST), measured by tissue tensiometry (Fig. 1.17) [103]. The
more cohesive cells occupy a central position rejecting the lower cohesive cells to the
external position. Let us now quantify these different phenomena in more details for
F9 and F9(α-/-) aggregates.
IV.1.1. Aggregate rounding up
It was shown that the ratio of surface tension to viscosity (σ/η) could be
estimated from the analysis of the kinetics of the shape relaxation toward a sphere of
an initially elongated aggregate [144]. Fig. 4.1 shows the slow rounding up of two
initially elongated aggregates of F9 wild type and F9(α-/-) knock-out cells respectively.
Fig. 4.1 : Rounding up of elongated cell aggregates. First line: a F9 cell aggregate of final
radius RF=190µm recorded for 600min. Second line: similar relaxation of a F9(α-/-) cell
aggregate, RF=195µm, bars 200µm
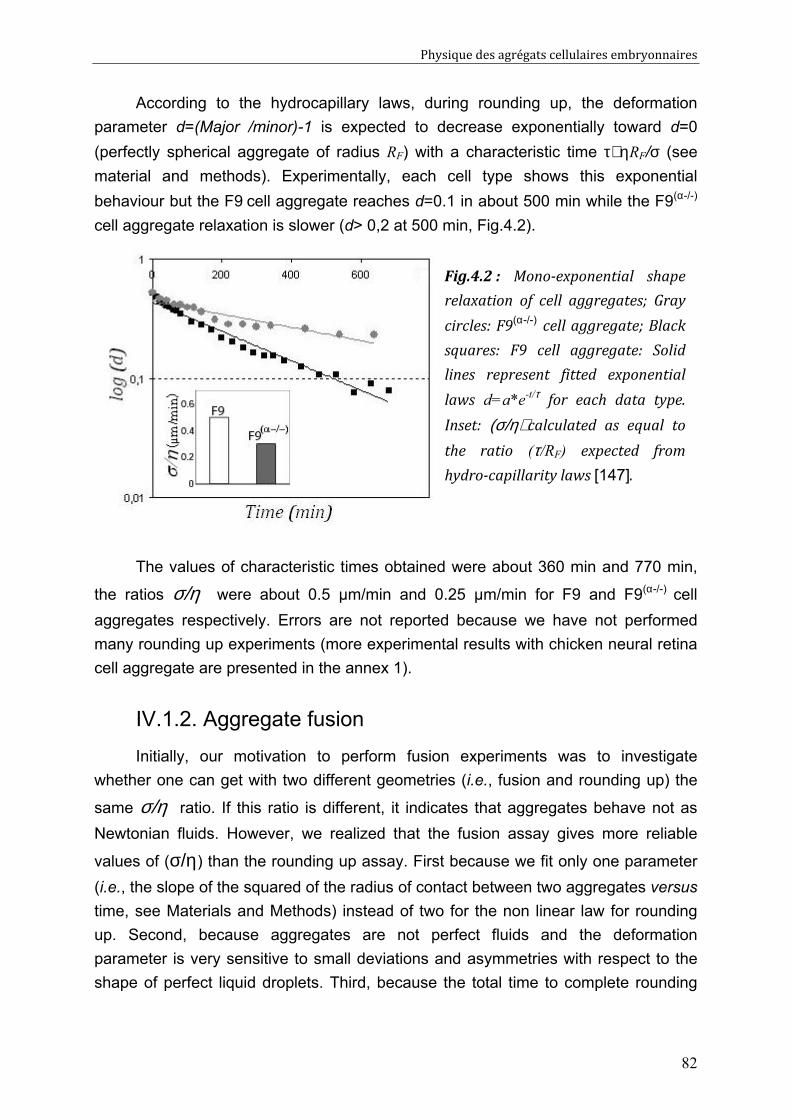
Physique des agrégats cellulaires embryonnaires
82
According to the hydrocapillary laws, during rounding up, the deformation
parameter d=(Major /minor)-1 is expected to decrease exponentially toward d=0
(perfectly spherical aggregate of radius RF) with a characteristic time τ∼ηRF/σ (see
material and methods). Experimentally, each cell type shows this exponential
behaviour but the F9 cell aggregate reaches d=0.1 in about 500 min while the F9(α-/-)
cell aggregate relaxation is slower (d> 0,2 at 500 min, Fig.4.2).
Fig.4.2 : Mono-exponential shape
relaxation of cell aggregates; Gray
circles: F9(α-/-) cell aggregate; Black
squares: F9 cell aggregate: Solid
lines represent fitted exponential
laws d=a*e-t/τ
for each data type.
Inset: (σ/η) calculated as equal to
the ratio (τ/RF) expected from
hydro-capillarity laws [147].
The values of characteristic times obtained were about 360 min and 770 min,
the ratios σ/η were about 0.5 µm/min and 0.25 µm/min for F9 and F9(α-/-) cell
aggregates respectively. Errors are not reported because we have not performed
many rounding up experiments (more experimental results with chicken neural retina
cell aggregate are presented in the annex 1).
IV.1.2. Aggregate fusion
Initially, our motivation to perform fusion experiments was to investigate
whether one can get with two different geometries (i.e., fusion and rounding up) the
same σ/η ratio. If this ratio is different, it indicates that aggregates behave not as
Newtonian fluids. However, we realized that the fusion assay gives more reliable
values of (σ/η) than the rounding up assay. First because we fit only one parameter
(i.e., the slope of the squared of the radius of contact between two aggregates versus
time, see Materials and Methods) instead of two for the non linear law for rounding
up. Second, because aggregates are not perfect fluids and the deformation
parameter is very sensitive to small deviations and asymmetries with respect to the
shape of perfect liquid droplets. Third, because the total time to complete rounding

Physique des agrégats cellulaires embryonnaires
83
up is about 10 hours, while fusion is completed in only a couple of hours for the same
F9 cell line (Fig. 4.3A).
The ratios σ/η in the fusion experiments were calculated according to modified
Frenkel’s model [161-163] as described in chapter 2. Briefly, the squared of the
contact radius X in the neck region is expected to be linear as a function of time, the
slope allows an evaluation of σ/η (Fig. 4.3C). We limited our measurement to
XLimit=(2/3)R0, the expected boundary for application of capillary law in case of two
liquid droplets [162], where R0 is the mean initial radius of two symmetric aggregates.
Fig. 4.3 : Fusion of two pairs of cell aggregates. A (first line): pair of F9 cell aggregates; B
(second line): pair of F9(α-/-) cell aggregates; Bars 100µm. Note that F9(α-/-) cell aggregates
appear to be rougher on surface than F9 cell aggregates. C: Linear evolution of the squared
of the contact radius as a function of time x R0 for three different pairs of F9 aggregates.
IV.1.2.1 Fusion of F9 and F9(αααα-/-) in culture medium
Figures Fig. 4.3A-B show images at different times of the fusion process for
pairs of F9 and F9(α-/-) aggregates in culture medium. The fusion of F9 cell
aggregates is obviously faster than the fusion of F9(α-/-). This result is confirmed by
Fig. 4.4 that summarizes the mean values of σ/η obtained for different experiments.
The ratio 0,48±0.10 µm/min for F9 cell aggregates is more than twice larger than the
ratio 0.22±0.02 µm/min for F9(α-/-) ones. These values are fully consistent with the
values obtained from the rounding up experiments. Therefore these aggregates do
not display a non Newtonian behaviour at least from the presented experiments.
Tissue viscosity values will be calculated independently and discussed together with
TST measurements at the end of this chapter.
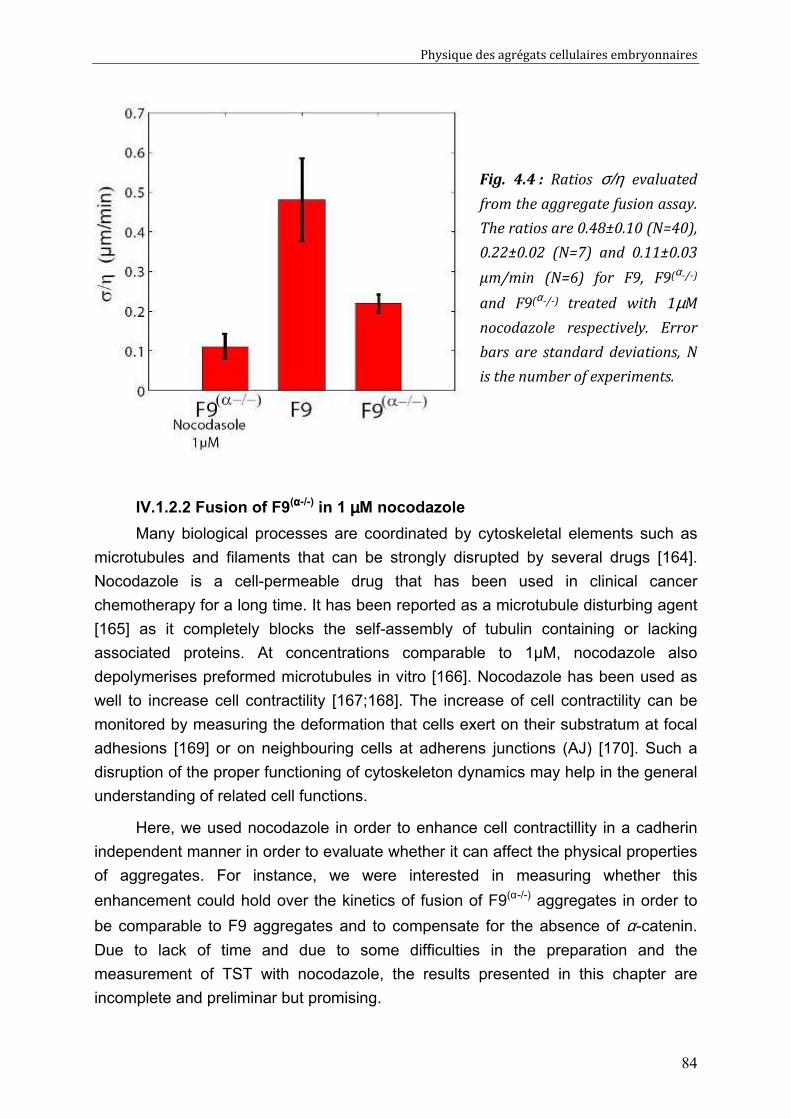
Physique des agrégats cellulaires embryonnaires
84
Fig. 4.4 : Ratios σ/η evaluated
from the aggregate fusion assay.
The ratios are 0.48±0.10 (N=40),
0.22±0.02 (N=7) and 0.11±0.03
µm/min (N=6) for F9, F9(α-/-)
and F9(α-/-) treated with 1µM
nocodazole respectively. Error
bars are standard deviations, N
is the number of experiments.
IV.1.2.2 Fusion of F9(αααα-/-) in 1 µµµµM nocodazole
Many biological processes are coordinated by cytoskeletal elements such as
microtubules and filaments that can be strongly disrupted by several drugs [164].
Nocodazole is a cell-permeable drug that has been used in clinical cancer
chemotherapy for a long time. It has been reported as a microtubule disturbing agent
[165] as it completely blocks the self-assembly of tubulin containing or lacking
associated proteins. At concentrations comparable to 1µM, nocodazole also
depolymerises preformed microtubules in vitro [166]. Nocodazole has been used as
well to increase cell contractility [167;168]. The increase of cell contractility can be
monitored by measuring the deformation that cells exert on their substratum at focal
adhesions [169] or on neighbouring cells at adherens junctions (AJ) [170]. Such a
disruption of the proper functioning of cytoskeleton dynamics may help in the general
understanding of related cell functions.
Here, we used nocodazole in order to enhance cell contractillity in a cadherin
independent manner in order to evaluate whether it can affect the physical properties
of aggregates. For instance, we were interested in measuring whether this
enhancement could hold over the kinetics of fusion of F9(α-/-) aggregates in order to
be comparable to F9 aggregates and to compensate for the absence of α-catenin.
Due to lack of time and due to some difficulties in the preparation and the
measurement of TST with nocodazole, the results presented in this chapter are
incomplete and preliminar but promising.
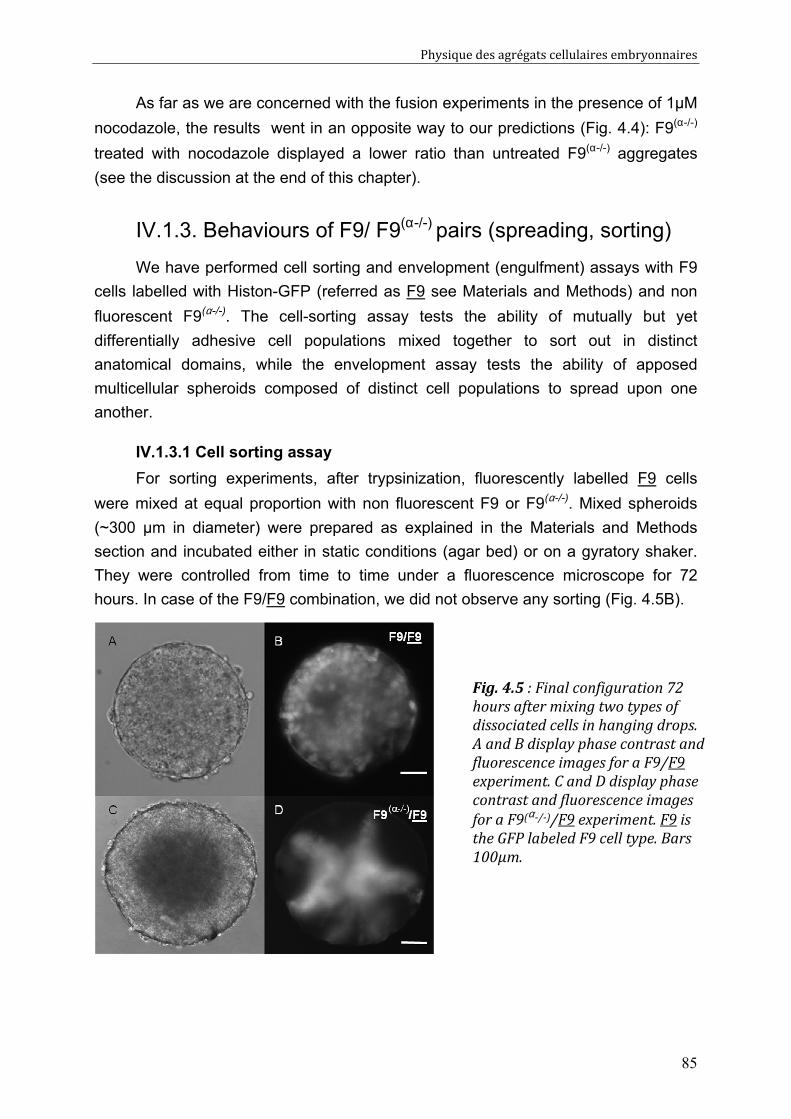
Physique des agrégats cellulaires embryonnaires
85
As far as we are concerned with the fusion experiments in the presence of 1µM
nocodazole, the results went in an opposite way to our predictions (Fig. 4.4): F9(α-/-)
treated with nocodazole displayed a lower ratio than untreated F9(α-/-) aggregates
(see the discussion at the end of this chapter).
IV.1.3. Behaviours of F9/ F9(α-/-) pairs (spreading, sorting)
We have performed cell sorting and envelopment (engulfment) assays with F9
cells labelled with Histon-GFP (referred as F9 see Materials and Methods) and non
fluorescent F9(α-/-). The cell-sorting assay tests the ability of mutually but yet
differentially adhesive cell populations mixed together to sort out in distinct
anatomical domains, while the envelopment assay tests the ability of apposed
multicellular spheroids composed of distinct cell populations to spread upon one
another.
IV.1.3.1 Cell sorting assay
For sorting experiments, after trypsinization, fluorescently labelled F9 cells
were mixed at equal proportion with non fluorescent F9 or F9(α-/-). Mixed spheroids
(~300 µm in diameter) were prepared as explained in the Materials and Methods
section and incubated either in static conditions (agar bed) or on a gyratory shaker.
They were controlled from time to time under a fluorescence microscope for 72
hours. In case of the F9/F9 combination, we did not observe any sorting (Fig. 4.5B).
Fig. 4.5 : Final configuration 72 hours after mixing two types of dissociated cells in hanging drops. A and B display phase contrast and fluorescence images for a F9/F9 experiment. C and D display phase contrast and fluorescence images for a F9(α-/-)/F9 experiment. F9 is the GFP labeled F9 cell type. Bars 100µm.
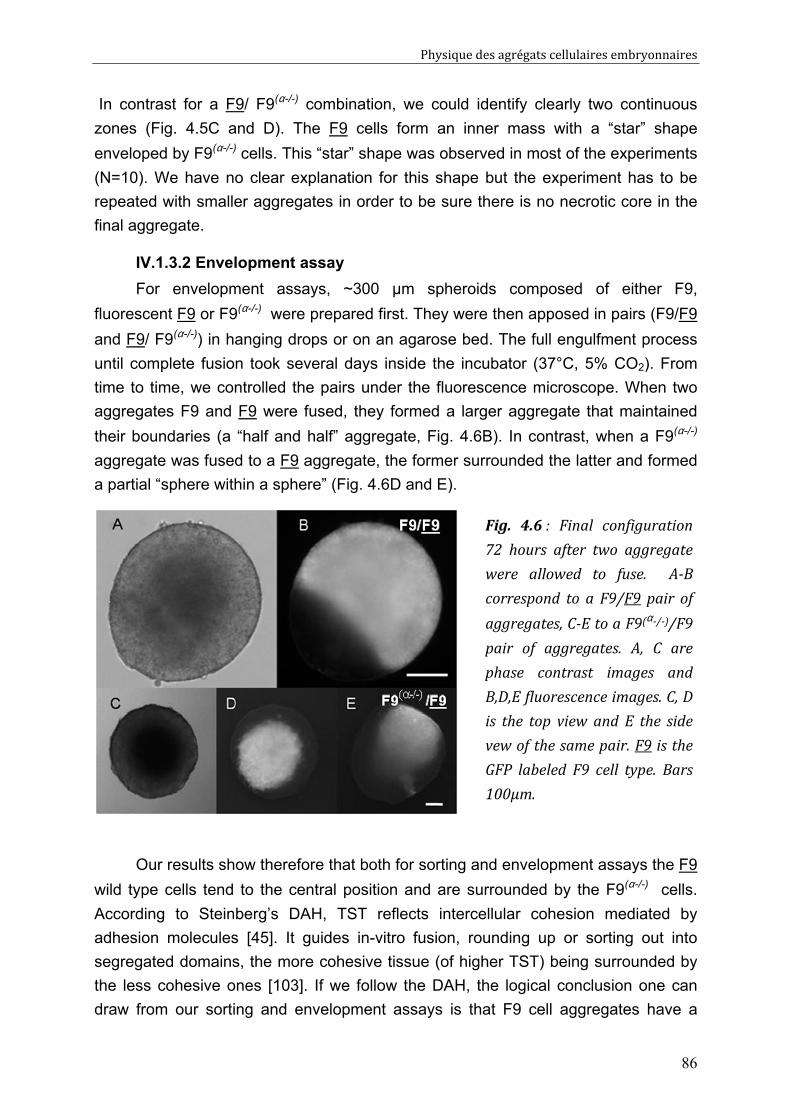
Physique des agrégats cellulaires embryonnaires
86
In contrast for a F9/ F9(α-/-) combination, we could identify clearly two continuous
zones (Fig. 4.5C and D). The F9 cells form an inner mass with a “star” shape
enveloped by F9(α-/-) cells. This “star” shape was observed in most of the experiments
(N=10). We have no clear explanation for this shape but the experiment has to be
repeated with smaller aggregates in order to be sure there is no necrotic core in the
final aggregate.
IV.1.3.2 Envelopment assay
For envelopment assays, ~300 µm spheroids composed of either F9,
fluorescent F9 or F9(α-/-) were prepared first. They were then apposed in pairs (F9/F9
and F9/ F9(α-/-)) in hanging drops or on an agarose bed. The full engulfment process
until complete fusion took several days inside the incubator (37°C, 5% CO2). From
time to time, we controlled the pairs under the fluorescence microscope. When two
aggregates F9 and F9 were fused, they formed a larger aggregate that maintained
their boundaries (a “half and half” aggregate, Fig. 4.6B). In contrast, when a F9(α-/-)
aggregate was fused to a F9 aggregate, the former surrounded the latter and formed
a partial “sphere within a sphere” (Fig. 4.6D and E).
Fig. 4.6 : Final configuration
72 hours after two aggregate
were allowed to fuse. A-B
correspond to a F9/F9 pair of
aggregates, C-E to a F9(α-/-)/F9
pair of aggregates. A, C are
phase contrast images and
B,D,E fluorescence images. C, D
is the top view and E the side
vew of the same pair. F9 is the
GFP labeled F9 cell type. Bars
100µm.
Our results show therefore that both for sorting and envelopment assays the F9
wild type cells tend to the central position and are surrounded by the F9(α-/-) cells.
According to Steinberg’s DAH, TST reflects intercellular cohesion mediated by
adhesion molecules [45]. It guides in-vitro fusion, rounding up or sorting out into
segregated domains, the more cohesive tissue (of higher TST) being surrounded by
the less cohesive ones [103]. If we follow the DAH, the logical conclusion one can
draw from our sorting and envelopment assays is that F9 cell aggregates have a
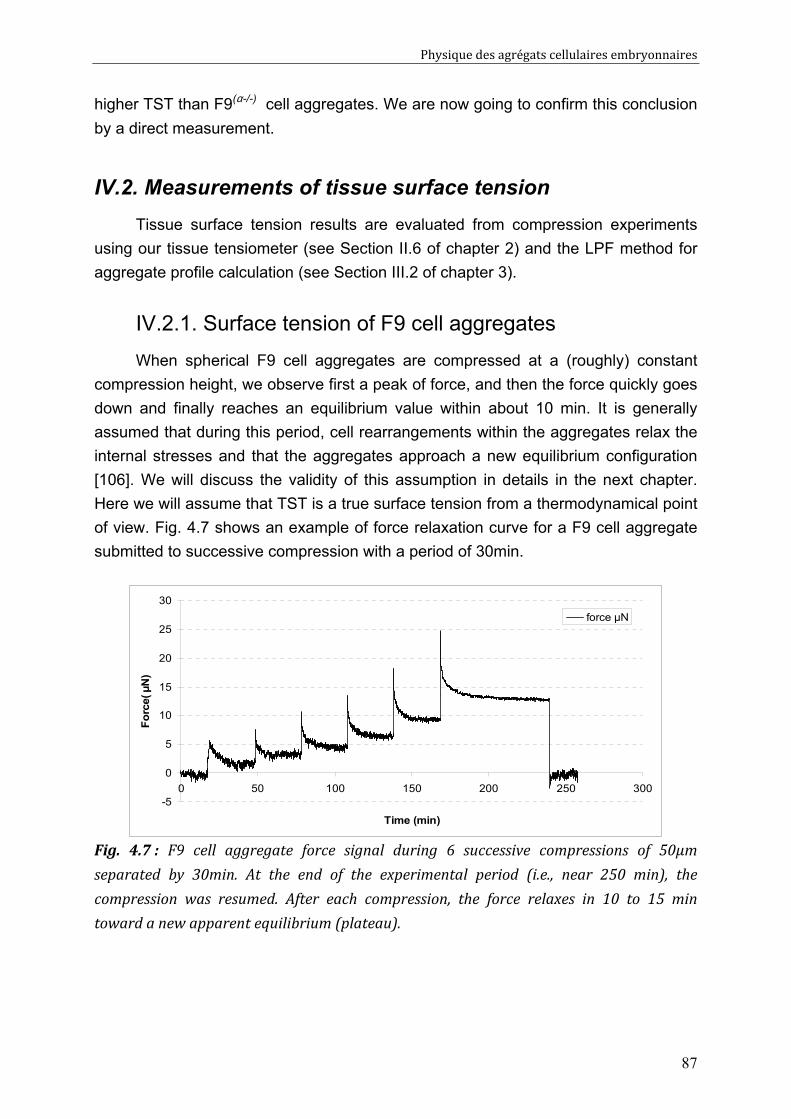
Physique des agrégats cellulaires embryonnaires
87
higher TST than F9(α-/-) cell aggregates. We are now going to confirm this conclusion
by a direct measurement.
IV.2. Measurements of tissue surface tension
Tissue surface tension results are evaluated from compression experiments
using our tissue tensiometer (see Section II.6 of chapter 2) and the LPF method for
aggregate profile calculation (see Section III.2 of chapter 3).
IV.2.1. Surface tension of F9 cell aggregates
When spherical F9 cell aggregates are compressed at a (roughly) constant
compression height, we observe first a peak of force, and then the force quickly goes
down and finally reaches an equilibrium value within about 10 min. It is generally
assumed that during this period, cell rearrangements within the aggregates relax the
internal stresses and that the aggregates approach a new equilibrium configuration
[106]. We will discuss the validity of this assumption in details in the next chapter.
Here we will assume that TST is a true surface tension from a thermodynamical point
of view. Fig. 4.7 shows an example of force relaxation curve for a F9 cell aggregate
submitted to successive compression with a period of 30min.
-5
0
5
10
15
20
25
30
0 50 100 150 200 250 300
Time (min)
Force( µN)
force µN
Fig. 4.7 : F9 cell aggregate force signal during 6 successive compressions of 50µm
separated by 30min. At the end of the experimental period (i.e., near 250 min), the
compression was resumed. After each compression, the force relaxes in 10 to 15 min
toward a new apparent equilibrium (plateau).
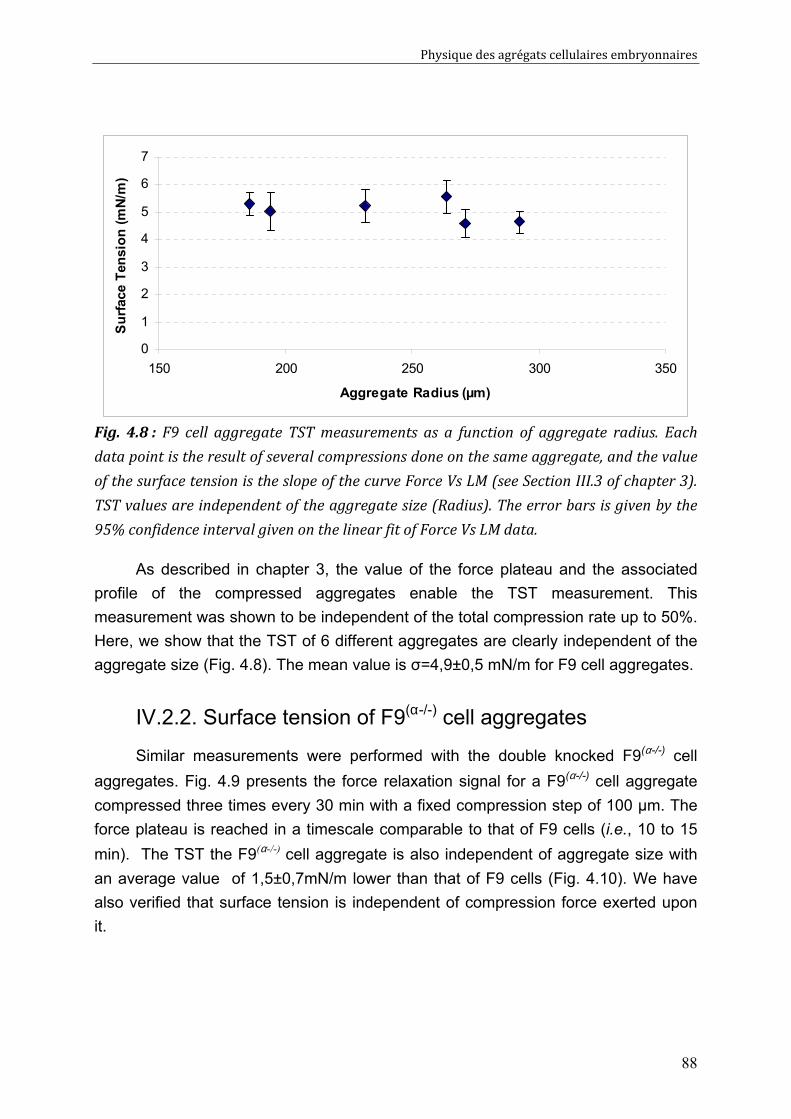
Physique des agrégats cellulaires embryonnaires
88
0
1
2
3
4
5
6
7
150 200 250 300 350
Aggregate Radius (µm)
Surface Tension (mN/m
)
Fig. 4.8 : F9 cell aggregate TST measurements as a function of aggregate radius. Each
data point is the result of several compressions done on the same aggregate, and the value
of the surface tension is the slope of the curve Force Vs LM (see Section III.3 of chapter 3).
TST values are independent of the aggregate size (Radius). The error bars is given by the
95% confidence interval given on the linear fit of Force Vs LM data.
As described in chapter 3, the value of the force plateau and the associated
profile of the compressed aggregates enable the TST measurement. This
measurement was shown to be independent of the total compression rate up to 50%.
Here, we show that the TST of 6 different aggregates are clearly independent of the
aggregate size (Fig. 4.8). The mean value is σ=4,9±0,5 mN/m for F9 cell aggregates.
IV.2.2. Surface tension of F9(α-/-) cell aggregates
Similar measurements were performed with the double knocked F9(α-/-) cell
aggregates. Fig. 4.9 presents the force relaxation signal for a F9(α-/-) cell aggregate
compressed three times every 30 min with a fixed compression step of 100 µm. The
force plateau is reached in a timescale comparable to that of F9 cells (i.e., 10 to 15
min). The TST the F9(α-/-) cell aggregate is also independent of aggregate size with
an average value of 1,5±0,7mN/m lower than that of F9 cells (Fig. 4.10). We have
also verified that surface tension is independent of compression force exerted upon
it.
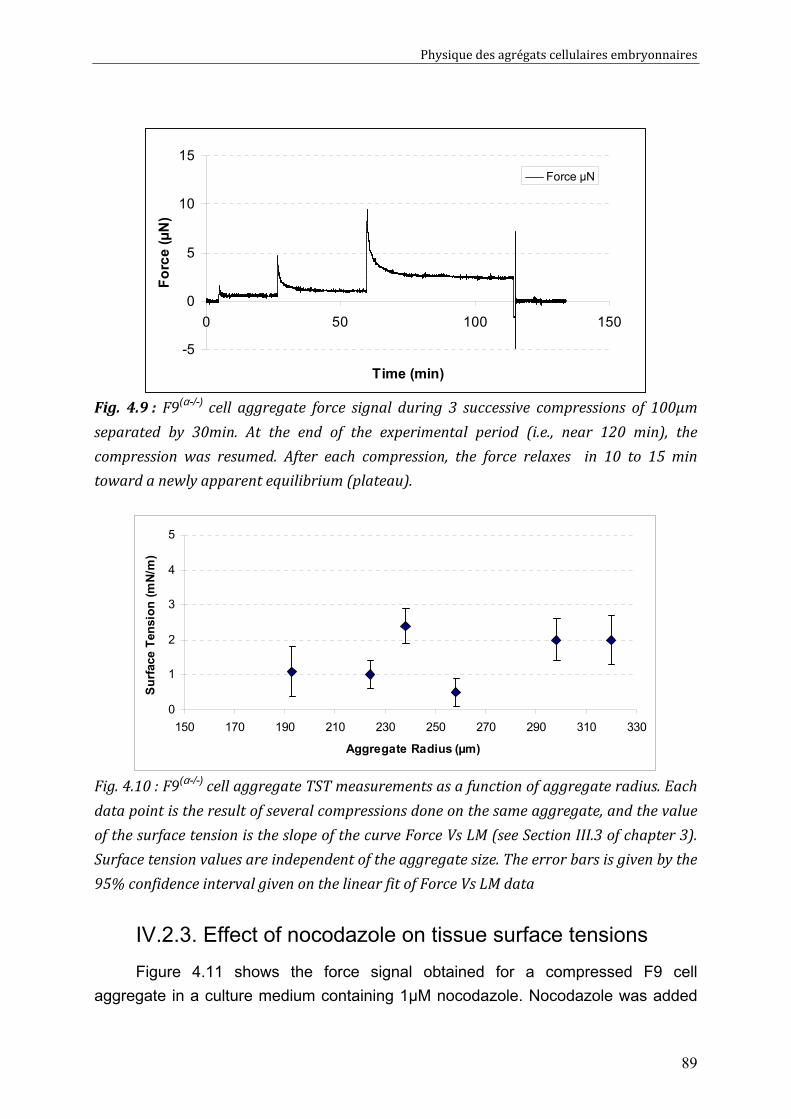
Physique des agrégats cellulaires embryonnaires
89
-5
0
5
10
15
0 50 100 150
Time (min)
Force (µN)
Force µN
Fig. 4.9 : F9(α-/-) cell aggregate force signal during 3 successive compressions of 100µm
separated by 30min. At the end of the experimental period (i.e., near 120 min), the
compression was resumed. After each compression, the force relaxes in 10 to 15 min
toward a newly apparent equilibrium (plateau).
0
1
2
3
4
5
150 170 190 210 230 250 270 290 310 330
Aggregate Radius (µm)
Surface Tension (mN/m
)
Fig. 4.10 : F9(α-/-) cell aggregate TST measurements as a function of aggregate radius. Each
data point is the result of several compressions done on the same aggregate, and the value
of the surface tension is the slope of the curve Force Vs LM (see Section III.3 of chapter 3).
Surface tension values are independent of the aggregate size. The error bars is given by the
95% confidence interval given on the linear fit of Force Vs LM data
IV.2.3. Effect of nocodazole on tissue surface tensions
Figure 4.11 shows the force signal obtained for a compressed F9 cell
aggregate in a culture medium containing 1µM nocodazole. Nocodazole was added
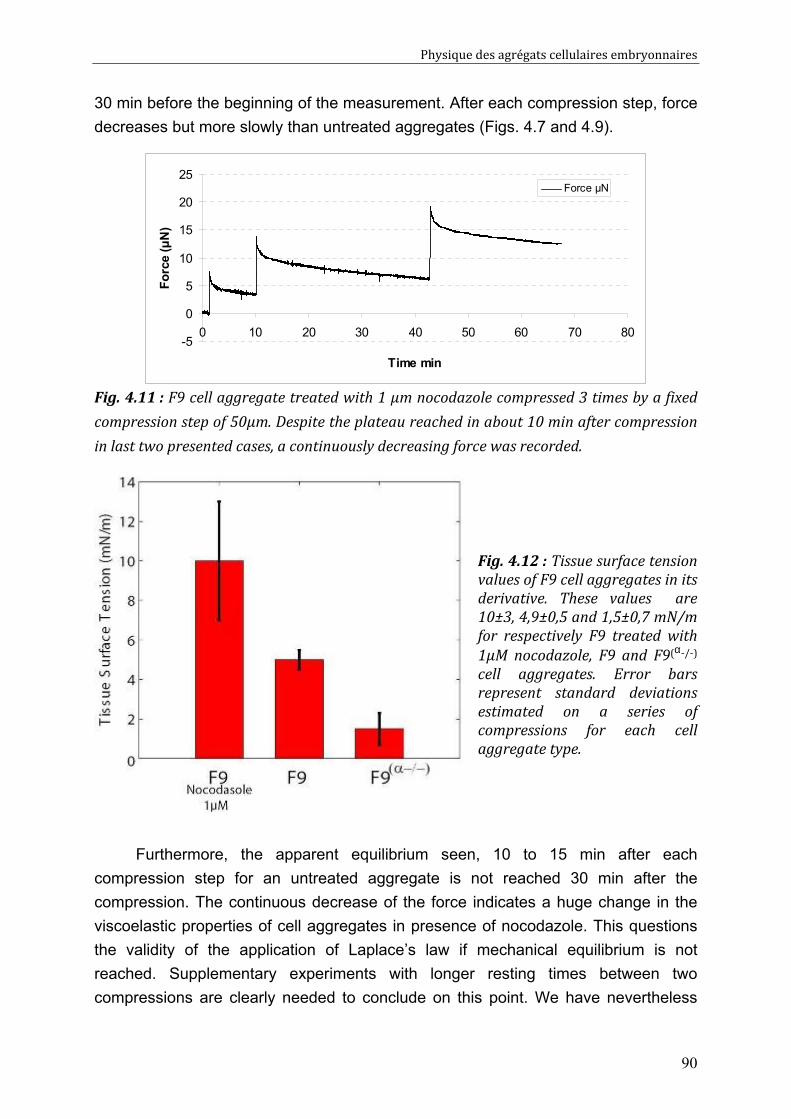
Physique des agrégats cellulaires embryonnaires
90
30 min before the beginning of the measurement. After each compression step, force
decreases but more slowly than untreated aggregates (Figs. 4.7 and 4.9).
-5
0
5
10
15
20
25
0 10 20 30 40 50 60 70 80
Time min
Force (µN)
Force µN
Fig. 4.11 : F9 cell aggregate treated with 1 µm nocodazole compressed 3 times by a fixed
compression step of 50µm. Despite the plateau reached in about 10 min after compression
in last two presented cases, a continuously decreasing force was recorded.
Fig. 4.12 : Tissue surface tension values of F9 cell aggregates in its derivative. These values are 10±3, 4,9±0,5 and 1,5±0,7 mN/m for respectively F9 treated with 1µM nocodazole, F9 and F9(α-/-)
cell aggregates. Error bars represent standard deviations estimated on a series of compressions for each cell aggregate type.
Furthermore, the apparent equilibrium seen, 10 to 15 min after each
compression step for an untreated aggregate is not reached 30 min after the
compression. The continuous decrease of the force indicates a huge change in the
viscoelastic properties of cell aggregates in presence of nocodazole. This questions
the validity of the application of Laplace’s law if mechanical equilibrium is not
reached. Supplementary experiments with longer resting times between two
compressions are clearly needed to conclude on this point. We have nevertheless

Physique des agrégats cellulaires embryonnaires
91
estimated an apparent tissue surface tension value of about 10mN/m for F9 cell
aggregates in 1µM nocodazole by taking the force value 30 min after each
compression. This value is twice larger than that of the same F9 cell aggregates in
culture medium and more than four times larger than F9(α-/-) cell aggregates (Fig.
4.12).
IV.3. Discussion
IV.3.1. Surface tension depends on cytoskeleton contractility: DAH is contradicted
Among its goals, developmental biology aims to understand the mechanisms or
factors that direct tissue organisation during development. It is now well admitted that
some morphogenetic tissue organisation and the sorting out of embryonic cells have
a common explanation [44;65;103;148;171]. The differential adhesion hypothesis
(DAH) proposed that segregation phenomena of differentially adhesive cells should
lead to an equilibrium configuration of minimal surface energy (equivalent to a
maximal binding energy) where the most cohesive cells (of higher surface tension)
are surrounded by the less cohesive cells (of lower surface tension) like do two
immiscible liquids when mixed together. If energetically favoured, this equilibrium
configuration should eventually be reached from any initial starting configuration and
reveal the operation of a strict rule of transitivity [160].
Our own TST measurements (summarized in Fig. 4.12) reinforce this surface
energy minimisation principle (that we can call the “first postulate of DAH”) for an
embryonic cell line never investigated to date. The cell-sorting and envelopment
assays (Fig. 4.5 and Fig. 4.6) show that F9 with the higher surface tension are
always surrounded by F9(α-/-) cells. However, this segregation and this difference in
value of TST occur at constant level of expression of E-cadherin. According to the
“second postulate of DAH”, TST depends only on the expression of adhesion
molecules [45;148]. As as a consequence, our two cell lines should possess the
same TST and the resulting final aggregate after an envelopment assay should be
“half-and-half” as in Fig. 4.6B. Another experimental evidende that TST depends on
cytoskeleton contractility is the fact that in presence of 1 µm nocodazole (a
cytoskeleton contractile drug), the TST of F9 aggregates seems to increase as
compared to control aggregates in culture medium.
The “second postulate of DAH” is therefore strictly wrong. Our results show that
a differential cytoskeleton’s mechanics, or cytoskeleton contractility, affect directly the
value of TST and can predict the mutual envelopment behaviour of cell aggregates

Physique des agrégats cellulaires embryonnaires
92
(Figs.Fig. 4.5, Fig. 4.6, Fig. 4.12). That means that aggregate cohesivity is a
cytoskeleton dependent property.
It should be emphasised that we are not the first to claim that conclusion or to
report experimental data supporting it. Harris or Brodland have already postulated
that TST derives not from the strengths of cell-cell adhesions but rather from
contractile forces [85;116;117]. Jakab et al. [115] have measured a direct relationship
between TST and cell contractility with CHO cells. They have shown that TST
decreases linearly with the concentration of Latrunculin A (a drug that is known to
reduce cell contractility by F-actin destruction, [164]). The presence of this drug has a
second noticeable effect: it shortens the relaxation time of force curves during
compression (i.e., aggregates with a lower contractility become more fluid).
Curiously, the authors do not comment about the second postulate of DAH. They just
aknowledge that “latrunculin renders cells less adhesive (presumably by both
destabilizing the cytoskeletal attachment of CAMs12 and the cytoskeleton itself)”. But
the logical conclusion of this sentence is that cell cytoskeleton itself controls the TST
whatever the initial expression of CAMs.
Finally, one can briefly comment the argumentation of Steinberg’s group about
this second postulate of DAH. First they showed in sorting out assays using L cell
aggregates transfected to express N-cadherins in various amounts, that the cells with
the higher N-cad expression segregate internally to cells with a lower expression
[44]. Latter, they measured the TST of L cell aggregates transfected with not only E-
but also, P- or E-cadherin in varied, measured amounts A direct, linear correlation
was observed between TST and cadherin expression level [45;112;152]. But, it
could be that modifying cadherin expression modifies the mechanics of cytoskeleton
by recruitment of actin into adhesion sites as described in [127;172]. And one can
anticipate that modifying cadherin expression will also modify cell contractility as
myosin is a well known component of cell-cell junction [173]. So this experiment can
not be taken alone as a direct test of the DAH.
IV.3.2. Tissue viscosity does not depend directly on cell cohesion: an adhesion-protrusion competition hypothesis
TST measurements have been performed as a function of the age and tissue
type for dissected embryos [103;113;150], of the level of cadherin expression [45] or
more recently as function of drug concentration [115]. Other physical properties like
tissue viscosity were not so systematically investigated. Here, we combined TST
measurements and tissue surface tension driven phenomena (fusion, rounding up) to
evaluate an effective tissue viscosity for each clone of the F9 cell line. The calculated
12 Cell adhesion molecules

Physique des agrégats cellulaires embryonnaires
93
viscosity were respectively about (6,1±2)×105Pa.s and (4±2)×105Pa.s for F9 and
F9(α−/−) tissues. Despite the clear difference in TST, the effective viscosity of F9(α−/−)
aggregates is only slightly lower than that of F9 tissues. The difference is not
significant due to the large error resulting from two independent measurements of σ
and σ/η.
In the literature, two studies at 24 years interval reported independent
measurements of the fusion kinetics and of TST for the same chicken embryo tissues
[103;147]. We have therefore evaluated the following set of tissue viscosity
values: 1.47×106, 1.3×106 and 1.43×106Pa.s for 6-day heart, 6-day liver, and 7-day
neural retinal13 tissues respectively. Again, these values lie almost in the same
range while the corresponding values of surface tensions were respectively 8.5, 4.6
and 1.6mN/m (i.e., a factor of 5 between the two extreme values). Another recent
study measured both a TST of 16 mN/m (tensiometer) and a tissue viscosity of 2×106
Pa.s for cardiac cushion tissue from 6-day chick embryos [115]. Of course, tissue
viscosity of F9 cells, is about three times smaller than the one of chicken embryo
tissues, it may depend on the age of the embryos as we measured a significant
dependence of σ/η as a function of this parameter for neural retina14. But it seems
very unlikely that the tissue independent viscosity of chick embryos is fortuitous.
Perhaps the age at which all these chick embryos were used was a compromise
between large dissected tissues and still fluid aggregates. By aging, cells may form
more irreversible adhesive bonds or simply be less healthy in the aggregates.
Whatever this issue, the independence of tissue viscosity on the actin-cadherin
attachment for F9 and F9(α−/−) tissues is rather a surprising result. First because, in a
monolayer assay, it was shown that F9(α−/−) are much less motile than F9 cells [129].
For the less motile F9(α−/−) cells, one could therefore expect a larger viscosity (which
is the inverse of the mobility in colloidal physics). From another point of view, our
result shows that tissue viscosity is not proportional to cell cohesion (TST). In Potts
model simulations, Käfer measured a TST of 2D aggregates proportional to cell-cell
adhesion energy [174], and tissue viscosity was recently simulated by Mombach et
al. using the fusion geometry. Viscosity is well proportional to surface tension (J.C.
Mombach, personal communication).
However, we can propose the following explanation. It is reasonable to assume
that tissue viscosity has two main origins: (i) cell cohesivity and (ii) cell protrusion
13 We used, for the neural retina tissue, our own measurement of (σ/η) evaluated from the rounding up assay. 14 Unfortunately we could not properly measure TST for different ages of neural retina aggregates but in the
literature TST values are relatively constant or increasing by less than a factor of two after 3 to 4 days in culture
(Foty et al., 1996) (see annex 1)

Physique des agrégats cellulaires embryonnaires
94
activity. The larger is the cell cohesion, the more difficult it is for cells to break
adhesive bonds, to change neighbours and past over each other during the large
reorganization events that occur for instance during fusion: viscosity is therefore
proportional to cell cohesion. On the other hand, the larger is the protrusion activity
the easier and faster the cells can move relative to each other: viscosity is therefore
inversely proportional to cell protrusion activity.
Let us now examine what happens in our experiments. Impairment of
cytoplasmic domain of cadherin linked to cytoskeleton was reported to reduce
dramatically cell movement [14;125;175] and we have measured here that it
decreases cohesivity. Consequently, we think there is a kind of compensation
between the losses in cell protrusion activity and the weakness of the cohesivity for
F9(α−/−) cells which leads to keep the effective viscosity unchanged.
Interestingly, this hypothesis appears to explain also the twice lower value of
σ/η for F9(α−/−) aggregates in presence of 1µM nocodazole than in culture medium
(Fig. 4.4). Although, we have not yet measured σ for such an aggregate, we expect
a larger value than in culture medium as observed for F9 aggregates in the presence
or not of nocodazole (Fig. 4.12). Therefore, the viscosity of F9(α−/−) aggregates in
presence of 1 µm nocodazole in much larger than in culture medium. To explain this
fact, we briefly review the assumed role of microtubule and its polymerisation
inhibition by nocodazole.
We have shown that aggregate cohesivity depends on actin-cadherin
attachment and actomyosin cell contractility (F9 Vs. F9 with nocodazole TST
measurement), and probably as well on other intracellular biochemical signalling
pathways. As was discussed in chapter 2, it seems that in case of F9(α−/−)
aggregates the second effect is not compensated by the first. Preliminary tests done
using two photons microscopy showed that the dynamics of shape relaxation in a
compressed aggregate treated with nocodazole is slower than the relaxation of
untreated one arguing in favour of a lower protrusion activity in treated aggregates.
Notice that the two origins of tissue viscosity (cell cohesiveness and cell
protrusion activity) appear in the model of complex cellular fluid developed in the next
chapter (Eq. 5.11). Cell cohesiveness is related to an energy barriers ∆εT1, and cell
protrusion activity has two different features: an attempt frequency f and a fluctuation
energy ξ respectively. This double effect compensation hypothesis will be the subject
of next experimental work in the future.

Physique des agrégats cellulaires embryonnaires
95
Chapitre 5 Cell-cell rearrangements within
aggregates: Experiments & Modelling
We have seen that embryonic tissues do behave as viscous liquids at long
timescales (chap. 4) and that surface tension was a measurable parameter and a
relevant one to explain cell sorting (chaps. 1, 3 and 4).
However, even if differences in surface tension contribute to the driving force
for movement during morphogenesis, by themselves they reveal little about the time
course of such processes. The kinetics of cell sorting and tissue envelopment
depends upon the dynamical properties of cell populations that flow as complex
(probably non-Newtonian) fluids [106;120]. Their behaviour depends on the time
scale and duration of the measurements.
Existing models usually involve a phenomenological viscoelastic description
[106;118;119;176]. This is not surprising considering that major components of
individual cells themselves display viscoelastic properties [177-181]. However,
aggregates of cells might have richer behaviours than single cells. Under stress, they
could be deformed like single cells but they will as well be able to rearrange like liquid
foam [182]. Therefore, cell aggregates belong to the class of cellular materials [183].
As such, they can display viscous, elastic and plastic behaviours [184].
This chapter is reserved to present and discuss the complex fluid aspect of cell
aggregates. We start by a review of previous studies on viscous and elastic
behaviours, and then we suggest an experimental method to quantify plasticity of cell
aggregates. An analytical model based on the cellular structure of cell aggregates
has been developed in collaboration with P. Marmottant (LSP Grenoble) [185] This
model captures the complexity of their triple mechanical behaviour. Combined to our
experiments, it allows evaluating several relevant rheological parameters of cell
aggregates such as elasticity, characteristic stress, characteristic time and effective
viscosity under stress.

Physique des agrégats cellulaires embryonnaires
96
V.1. Elastic and/or viscous behaviours: previews studies
Viscoelastic characteristics of tissues have been observed in early
experiments performed by Phillips and Steinberg [118;119].They have mechanically
deformed cell aggregates obtained from several embryonic chick tissues by
centrifuging them against solid substrata. Elastic-solid and viscous-liquid properties
of cell aggregates have been characterized at different time scales.
For example, Leghorn embryo cell aggregates subjected to brief centrifugation
periods (8–16 min) flatten quickly (cell shapes flatten too), and round up quickly (in
<2 min) once centrifugal forces are removed, reaching a shape very similar to the
initial one [119]. Such brief centrifugation experiments provide some support for a
simple elastic-solid model in which aggregate shape changes are accompanied by
cell deformations rather than cell redistributions. In particular, since cell migration
tends to occur quite slowly, the very rapid aggregate flattening in response to
external force presumably requires cell stretching, these aggregates exhibit
considerable elasticity that presumably reflects the swift relaxation of cell stretching
as the centrifugal force is removed (Fig. 5.1).
In contrast, during prolonged centrifugation (1 day), Leghorn embryo cell
aggregates continue to flatten more slowly (while cells gradually regain their original
shapes) [118;186]; once centrifugal forces are removed, these aggregates round up
slowly (in ~1 day), like a viscous liquid (Fig. 5.1B). Furthermore, by applying
prolonged centrifugation, it was shown that an initially very flat aggregate (freshly cut
from an embryo) and an initially round aggregate (obtained after incubating and
shaking overnight the flat piece of embryo) adopted the same final intermediate
shape [119]. In this long term experiments, aggregate behaviour is difficult to explain
on the basis of purely viscoelastic-solid models (as the one of Fig. 5.2A) but can be
readily explained by viscous-liquid models where cells are able to flow and change
neighbours at long time scales (Fig. 5.2B).
Following their centrifugation experiments, Phillips and Steinberg showed that
embryonic cell aggregates behave like deformable solids during brief experimental
manipulations, but like viscous liquids in long-term organ cultures. Then, they
suggest explaining these material properties by modelling solid and liquid behaviours
separately using simple mechanical elements (See Fig. 5.2, symbols used: spring15
15 The spring has an immediat and linear response to stress. It tends to return to its not deformed initial configuration.
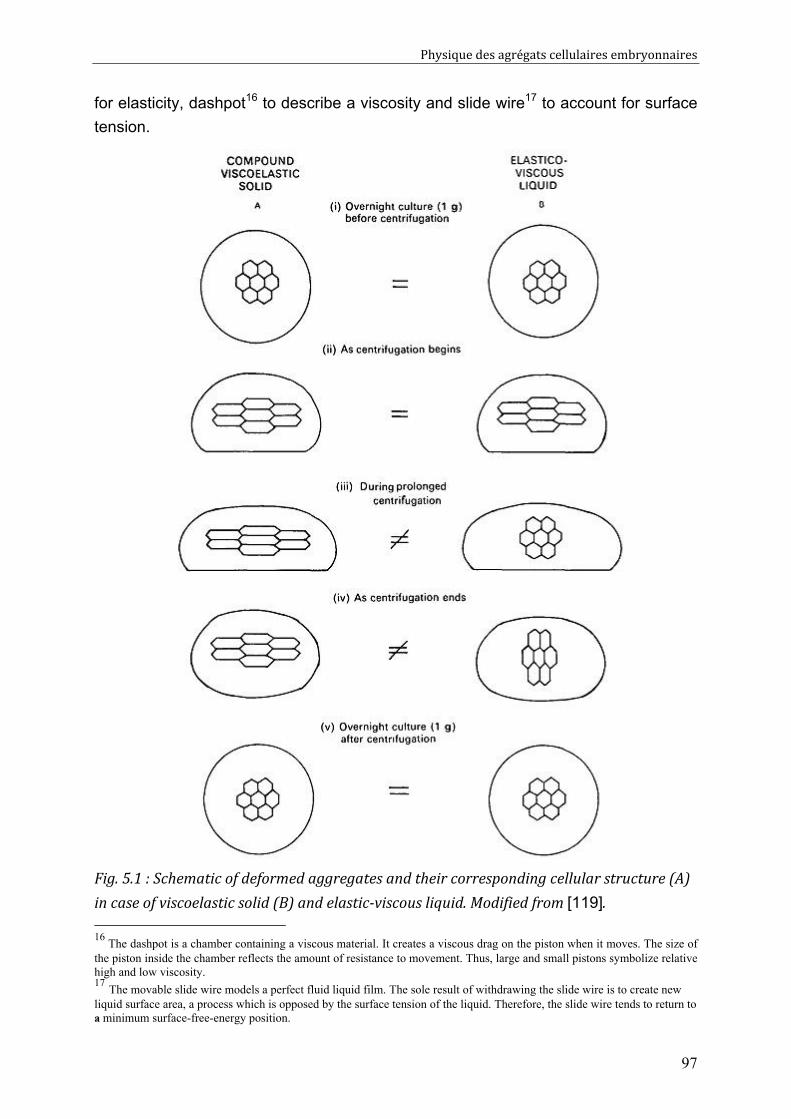
Physique des agrégats cellulaires embryonnaires
97
for elasticity, dashpot16 to describe a viscosity and slide wire17 to account for surface
tension.
Fig. 5.1 : Schematic of deformed aggregates and their corresponding cellular structure (A)
in case of viscoelastic solid (B) and elastic-viscous liquid. Modified from [119]. 16 The dashpot is a chamber containing a viscous material. It creates a viscous drag on the piston when it moves. The size of
the piston inside the chamber reflects the amount of resistance to movement. Thus, large and small pistons symbolize relative
high and low viscosity. 17 The movable slide wire models a perfect fluid liquid film. The sole result of withdrawing the slide wire is to create new
liquid surface area, a process which is opposed by the surface tension of the liquid. Therefore, the slide wire tends to return to
a minimum surface-free-energy position.
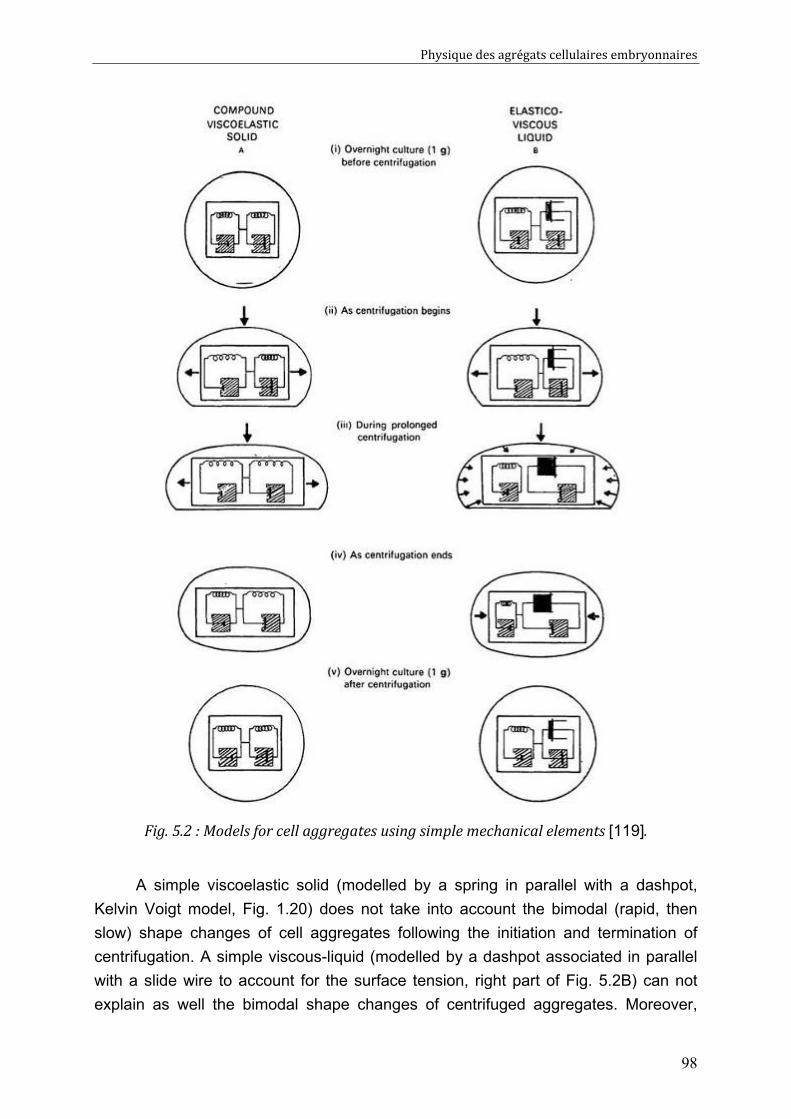
Physique des agrégats cellulaires embryonnaires
98
Fig. 5.2 : Models for cell aggregates using simple mechanical elements [119].
A simple viscoelastic solid (modelled by a spring in parallel with a dashpot,
Kelvin Voigt model, Fig. 1.20) does not take into account the bimodal (rapid, then
slow) shape changes of cell aggregates following the initiation and termination of
centrifugation. A simple viscous-liquid (modelled by a dashpot associated in parallel
with a slide wire to account for the surface tension, right part of Fig. 5.2B) can not
explain as well the bimodal shape changes of centrifuged aggregates. Moreover,

Physique des agrégats cellulaires embryonnaires
99
surface tension could hardly be expected to govern the abrupt aggregate shape
changes after sudden changes in applied force. At this time scale, the response is
dominated by elasticity.
Phillips and Steinberg thought about compound models. They associated the
fast-responding (low viscosity) viscoelastic solid in series with a slow-responding
(high viscosity) viscoelastic solid (Fig. 5.2A) or with a viscous liquid (Fig. 5.2B),
diagrammed in the right half of each model.
The compound viscoelastic solid model describes the bimodal character of
centrifuged aggregate shape changes by assuming that cells are compound
viscoelastic solids, possessing both quickly stretchable (less viscous) and slowly
stretchable (more viscous) elastic components (Fig. 5.2A). However, the tissue flows
- e.g. during the rounding-up of initially flat aggregates - must be accompanied by
rearrangements of component cells. Then an elasticoviscous liquid model was
established by replacing the slowly stretchable elastic-solid components postulated in
that compound solid model by slowly rearranging, viscous-liquid components.
The utility of these mechanical models is that they dissect the complex
behavior of cell aggregates into separable components with a minimal number of
mechanical elements. Here, the elasticoviscous-liquid model is favored to describe
tissues.
Other rheological studies were conducted by Steinberg and his collaborators
[106] in order to analyze the behaviors of living tissues in terms of standard models
of viscoelasticity and then get some quantitative details about tissue rheology. In a
first experiment, cell aggregates subjected to a very brief compression spring back
almost to their original shapes when released, whereas aggregates compressed for a
longer time do not [103], they display viscoelastic characters similar to that shown by
[119]. Then the authors suggest a method to separate viscous from elastic
contribution and obtain the purely viscous effect by recording the force relaxation of a
compressed aggregate, whose elasticity would have been completely dissipated by a
number of previous compression-decompression cycle. On the other hand,
aggregates subjected to uninterrupted force show combined viscous liquid and
elastic effects. The relaxation process is analyzed in terms of a generalized Kelvin-
body model of viscoelasticity including surface tension as shown in Fig. 5.3.
The system presents two distinct relaxation times. The force or stress
relaxation can be summarized as follows: the initial sudden compression produces
an instantaneous deformation of the slide wire element, u0, and of the two springs, u1
and u2, whereas the initial displacement of the dashpots (s1(0) and s2(0)) is zero.

Physique des agrégats cellulaires embryonnaires
100
Fig. 5.3 : Illustration of the
generalized Kelvin body used to
model the relaxation process [106].
µ1 and µ2 are the friction constant of
dashpots, k1 and k2 are the spring
constants. σ is the surface tension
implemented as a slide wire.
Considering the forces acting on the individual mechanical elements and
tanking in account that the overall shape of the aggregate is practically unchanged
during the force relaxation; therefore ut = u0 = const., whereas the deformations of
the springs decay while the dashpots gradually shrink. Finally, the equation
governing the relaxation of Ft takes the form
(5.1)
The solution of Eq. (5.1) is then
(5.2)
Here the two relaxation times are given by
(5.3)
As mentioned, at equilibrium all the stress is concentrated in the interfacial region.
The solution of Eq. 5.1 leads to
(5.4)
which implies that in the limit of large t, the only force which differs from zero is Fw.
In order to relate the model parameters derived from the measurements to
specific possible biological origin of relaxation process, they interpreted τ1 (~2 sec)
as the time scale characterizing the relaxation of the more or less elastic deformation

Physique des agrégats cellulaires embryonnaires
101
of each individual cell in the aggregate, together with whatever extracellular matrix
molecules might be present. This elastic stress is quickly dissipated and is followed
by a longer viscous response (relaxation time τ2~30-40 sec) in the course of which
individual cells move and the interior of the aggregate is reorganized. This motion
requires that cells break and reform their adhesive contacts with their neighbors. It is
interesting to notice that in this interpretation all the rearrangements occur in the τ2
timescale which is order of magnitudes smaller than the one obtained for the
characteristic timescales obtained for the rearrangement of cells during fusion
(chapter 4). We will discuss this point below.
Some other models have been introduced to explain typical results of
compression experiments without taking into account surface tension, and making
the assumption that cell aggregates are behaving as visco-elastic solid or as complex
fluid. [176;187].
In summary, there have been several hypotheses or theories built to explain
embryonic tissues behaviors sometimes in term of liquid and other in term of solid.
The mechanical models remain qualitative and the parameters used, could not be
easily linked to the mentioned true physical properties (cell elasticity, viscosity,
relaxation time, which could be measured independently). So no real test of the
models could be achieved. The challenge is to establish a model which account for
solid and liquid behaviors of cell aggregates and could link biological properties of
cellular materials, cell structure, geometrical tissues organization and its associated
processes with macroscopic significant and measurable physical properties. We will
describe a model that fill some of these criteria in section V.3 but before that let
present some new original and quantitative data on cell rheology within aggregates.
V.2. Experimental evidence of cell rearrangements
V.2.1. Direct visualization of stress induced cell rearrangements
Until now the only direct experimental evidence that cells could relax their
stress by changing neighbors was made by Phillips and Steinberg [118]. They
showed by electron microscopy that cells submitted to a brief centrifugation are
flattened while cells regain their original polyhedral shapes after more than one day
centrifugation (Fig. 5.4). But this experiment was done on fixed aggregates at these
only two extreme time points. No direct observation of cell displacements and
rearrangements is therefore available at intermediate time scales.
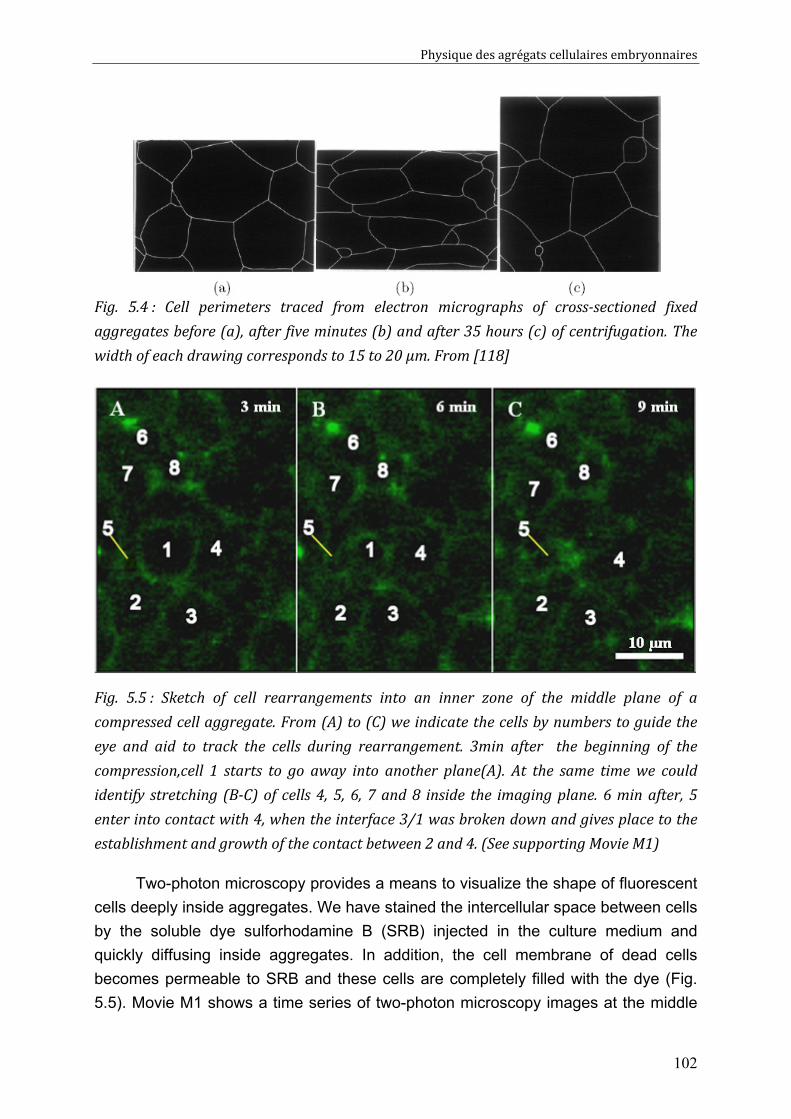
Physique des agrégats cellulaires embryonnaires
102
Fig. 5.4 : Cell perimeters traced from electron micrographs of cross-sectioned fixed
aggregates before (a), after five minutes (b) and after 35 hours (c) of centrifugation. The
width of each drawing corresponds to 15 to 20 µm. From [118]
Fig. 5.5 : Sketch of cell rearrangements into an inner zone of the middle plane of a
compressed cell aggregate. From (A) to (C) we indicate the cells by numbers to guide the
eye and aid to track the cells during rearrangement. 3min after the beginning of the
compression,cell 1 starts to go away into another plane(A). At the same time we could
identify stretching (B-C) of cells 4, 5, 6, 7 and 8 inside the imaging plane. 6 min after, 5
enter into contact with 4, when the interface 3/1 was broken down and gives place to the
establishment and growth of the contact between 2 and 4. (See supporting Movie M1)
Two-photon microscopy provides a means to visualize the shape of fluorescent
cells deeply inside aggregates. We have stained the intercellular space between cells
by the soluble dye sulforhodamine B (SRB) injected in the culture medium and
quickly diffusing inside aggregates. In addition, the cell membrane of dead cells
becomes permeable to SRB and these cells are completely filled with the dye (Fig.
5.5). Movie M1 shows a time series of two-photon microscopy images at the middle

Physique des agrégats cellulaires embryonnaires
103
plane of the same aggregate just after a 27% compression. Images are taken every
minute during 15 min, which is larger than the usual force relaxation period (Fig. 4.7,
Fig. 4.9 or Fig. 5.9). Although the aggregate’s external boundary is mostly fixed
during this relaxation period, cells inside it are clearly actively moving, changing
neighbors either in the same plane or from one plane to the other (i.e., some cells are
disappearing, others are appearing during that sequence, Fig. 5.5). These qualitative
experiments clearly validate the assumption that cells are relaxing some of their
stress by changing neighbors. A more quantitative analysis of the “amount of
rearrangements” in the course of time is needed (this is a complex task as optical
contrast is weak and rearrangements occur in 3D), but it is clear that rearrangements
start as soon as aggregates are compressed and continue during at least 15 min.
The fusion of a pair of aggregates was also recorded by confocal microscopy
and constitutes a spectacular example of cell rearrangements in the contact area
(see supplementary Movie M2).
V.2.2. Shape and stress relaxation experiments
It is possible to use the compression apparatus has a simple rheometer by
recording the shape relaxation of aggregates initially compressed to a given height
hcomp (corresponding to a deformation εcomp) during a time tcomp. To maintain a fixed
height during the compression, the elastic copper-beryllium cantilever bringing the
upper plate is replaced by a rigid pipe (See chapter 2 for more details on the set-up).
Experimentally, we quantify the aggregate deformation by its relative height
change, εtot= (hag-h)/hag. Here, hag is the aggregate’s initial height (i.e., diameter),
corresponding to the plate separation at the time of first contact; h is its current height
(h<hag). During compression, h is equal to the current plate separation hcomp; during
free decompression h increases.
For F9 mouse embryonic cell aggregates, Fig. 5.6 shows the height relaxation
after a prolonged compression (duration tcomp), followed by a sudden free
decompression (at time taken as t =0). All curves show an initial fast relaxation
(in~102s) but the aggregate does not recover completely its initial shape during a
recording time of about 30 min to 1 h (Fig. 5.6C). The compression has thus
promoted irreversible, plastic-like deformations. To recover the initial shape, it is
necessary to wait a much longer time, corresponding to the surface tension driven
rounding time (TLiq=6 h, see Section IV.1.2 of chapter 4). We will call apparent
plasticity this irreversible changes in the aggregate shape occurring at intermediate
times.
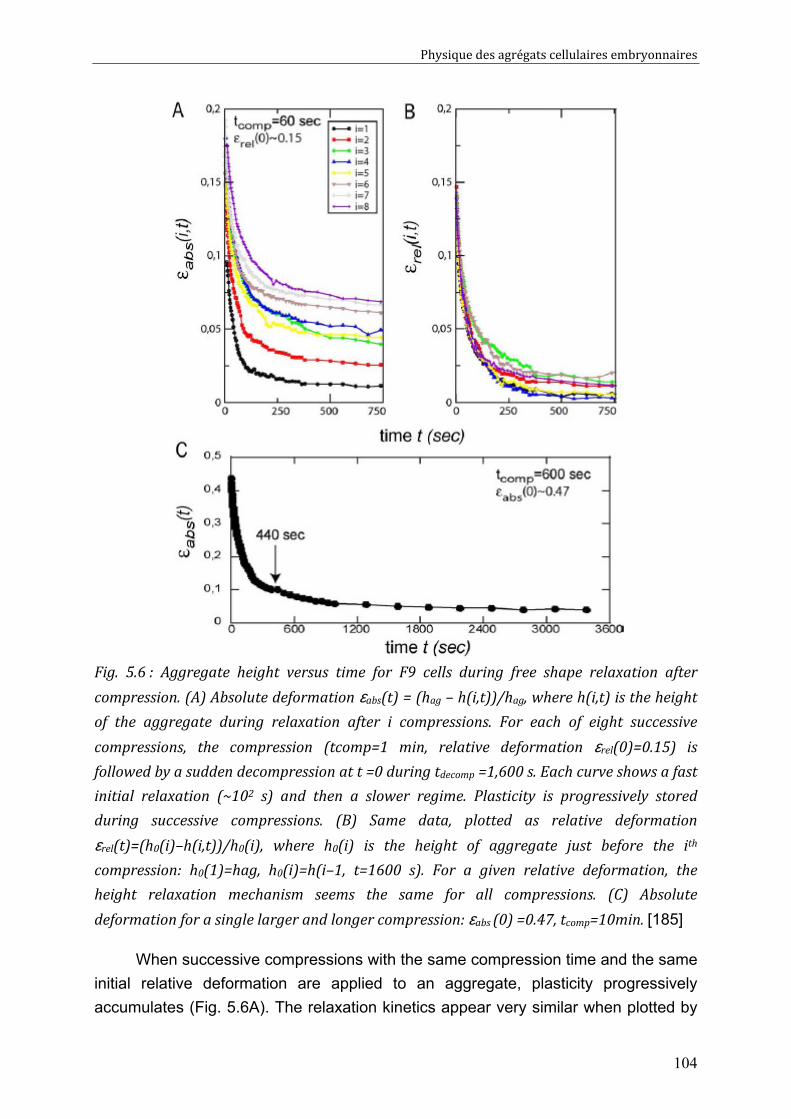
Physique des agrégats cellulaires embryonnaires
104
Fig. 5.6 : Aggregate height versus time for F9 cells during free shape relaxation after
compression. (A) Absolute deformation εabs(t) = (hag – h(i,t))/hag, where h(i,t) is the height
of the aggregate during relaxation after i compressions. For each of eight successive
compressions, the compression (tcomp=1 min, relative deformation εrel(0)=0.15) is
followed by a sudden decompression at t =0 during tdecomp =1,600 s. Each curve shows a fast
initial relaxation (~102 s) and then a slower regime. Plasticity is progressively stored
during successive compressions. (B) Same data, plotted as relative deformation
εrel(t)=(h0(i)–h(i,t))/h0(i), where h0(i) is the height of aggregate just before the ith
compression: h0(1)=hag, h0(i)=h(i–1, t=1600 s). For a given relative deformation, the
height relaxation mechanism seems the same for all compressions. (C) Absolute
deformation for a single larger and longer compression: εabs (0) =0.47, tcomp=10min. [185]
When successive compressions with the same compression time and the same
initial relative deformation are applied to an aggregate, plasticity progressively
accumulates (Fig. 5.6A). The relaxation kinetics appear very similar when plotted by
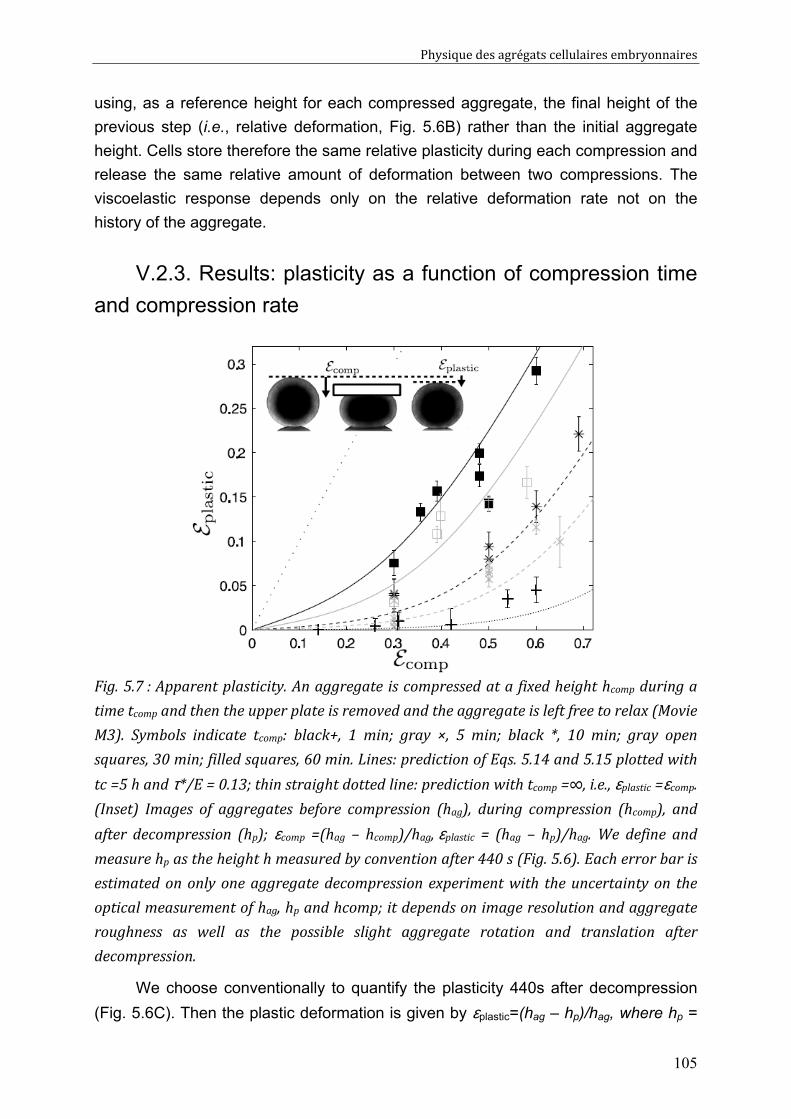
Physique des agrégats cellulaires embryonnaires
105
using, as a reference height for each compressed aggregate, the final height of the
previous step (i.e., relative deformation, Fig. 5.6B) rather than the initial aggregate
height. Cells store therefore the same relative plasticity during each compression and
release the same relative amount of deformation between two compressions. The
viscoelastic response depends only on the relative deformation rate not on the
history of the aggregate.
V.2.3. Results: plasticity as a function of compression time
and compression rate
Fig. 5.7 : Apparent plasticity. An aggregate is compressed at a fixed height hcomp during a
time tcomp and then the upper plate is removed and the aggregate is left free to relax (Movie
M3). Symbols indicate tcomp: black+, 1 min; gray ×, 5 min; black *, 10 min; gray open
squares, 30 min; filled squares, 60 min. Lines: prediction of Eqs. 5.14 and 5.15 plotted with
tc =5 h and τ*/E = 0.13; thin straight dotted line: prediction with tcomp =∞, i.e., εplastic =εcomp.
(Inset) Images of aggregates before compression (hag), during compression (hcomp), and
after decompression (hp); εcomp =(hag – hcomp)/hag, εplastic = (hag – hp)/hag. We define and
measure hp as the height h measured by convention after 440 s (Fig. 5.6). Each error bar is
estimated on only one aggregate decompression experiment with the uncertainty on the
optical measurement of hag, hp and hcomp; it depends on image resolution and aggregate
roughness as well as the possible slight aggregate rotation and translation after
decompression.
We choose conventionally to quantify the plasticity 440s after decompression
(Fig. 5.6C). Then the plastic deformation is given by εplastic=(hag – hp)/hag, where hp =

Physique des agrégats cellulaires embryonnaires
106
h (440s). Fig. 5.7 summarizes our experimental plasticity measurements as a
function of compression time as well as initial deformation of aggregates. One can
obviously notice that the higher or the longer the compression step, the larger the
remaining plasticity. We tried next to model this characteristic feature of cell
aggregate, by taking into account its visco-elastic-plastic behavior.
V.3. Cell rearrangement: fluctuations and stress
V.3.1. Global Description of the Aggregate.
At a scale much larger than the cell size, the aggregate can be characterized
by its state of deformation and stress (‘‘continuous medium’’ description). In a
parallel-plate compression experiment, we quantify the aggregate deformation by its
relative height change, εtot= (hag-h)/hag. The force exerted by the aggregate on the
plates is the cross-section area S of the aggregate through its midplane times the
internal stress of the compressed aggregate:
Fplate = (p+τ) S (5.5)
Here, p is the hydrostatic pressure and τ the elastic contribution to the stress,
which we study here. In the case of a drop made of a simple liquid (τ =0) with surface
tension σ, at equilibrium, p would be balanced by the drop mean curvature κ
[Laplace law: p = σ κ [157]].
V.3.2. Local Description at the Cell Level.
We assume the cell deformation (monitored by the relative cell size change,
εcell) and the stress are small enough to be proportional:
τ = E εcell (5.6)
where E is a stiffness (elastic modulus).
Within aggregates of inert objects such as drops (emulsions) or bubbles
(foams), a ‘‘T1’’ [182] occurs when the local stress is high enough to overcome the
energy barrier, ∆εT1 (Fig. 5.8), which depends on the interfaces tension and length. It

Physique des agrégats cellulaires embryonnaires
107
Fig. 5.8 : A neighbor swapping (‘‘T1’’ topological rearrangement [182]). (A) Sketch of
initially relaxed cells. (B) Flattened cells, with a deformation εcell due to stress. (Inset)
(Upper) Energy barrier ∆εT1 to overcome for a T1 to occur. (Inset) (Lower) Membrane
fluctuations with amplitude proportional to ξ. (C) after a T1: the cell arrangement has
changed, and the cell shapes relax, dissipating energy. [185]
marks the transition from a reversible (elastic) object shape change to an irreversible
(plastic) rearrangement, after which energy is dissipated [184] The energy barriers
are biased and easier to overcome in the direction of the stress [184]:
Rearrangements accumulate and result in a continuously increasing material
deformation, noted εrea, analogous to a flow. Because εrea does not correlate with
each object’s individual deformation, it has no direct elastic contribution to stress
(and we neglect the contribution to the stress arising from each object’s internal
viscosity).
Unlike inert objects, living cells overcome some energy barriers, with large
membrane shape fluctuations. These fluctuations are active and associated with
cytoskeleton dynamics (see movies 1 and 2), as is evidenced by studies on cells
treated with cytochalasin-B (an actin depolymerizing drug) [91;110]. An effective
temperature or, equivalently, a thermal energy scale ξ (also noted kT in the literature)
[110], describes not only the amplitude of these interface fluctuations [110;188] but
also the diffusion of a cell throughout an aggregate [189].
V.3.3. Mechanical Behavior of the Aggregate.
To capture the main physical ingredients that determine the aggregate’s liquid
behavior, we simplify its description. We consider here that all energy barriers have
the same height, that of a T1. We take the total deformation to be:
(5.7)

Physique des agrégats cellulaires embryonnaires
108
In the absence of stress, fluctuation-induced rearrangements switch back and forth,
with no flow on average. According to the Eyring model (26), the probability of
rearrangement in the direction of stress (i.e., a T1 that reduces stress) is favorable:
(5.8)
Here, f is the inverse of a time (‘‘attempt frequency’’): It is a phenomenological
quantity that characterizes how often fluctuations occur, only a fraction of which are
actually able to jump over energy barriers; τV is the work gained during a
rearrangement in the direction favored by the stress (V is of the order of a cell
volume) and δε the deformation associated with a rearrangement. In the direction
opposing stress, the probability of a rearrangement is less favorable:
(5.9)
The flow rate rea results from the difference between favored and unfavored rearrangement probabilities:
(5.10)
Here, both τ* and η* are continuous medium parameters (related to cell properties,
see Discussion). The characteristic stress τ* =ξ/V.δε is determined by the fluctuation
amplitude and cell size. The characteristic viscosity:
(5.11)
depends on τ* and energy barriers (but not on the internal viscoelasticity of cells).
V.3.4. Two Regimes
Eq. 5.10 relates the aggregate’s deformation and the applied stress; together
with Eqs. 5.6 and 5.7, it fully describes the aggregate’s mechanical response, i.e., the
time evolution of the flow for any given configuration (‘‘constitutive equation’’). The
characteristic stress τ* separates two regimes: that of fluctuation- and stress-induced
flows. If the stress τ is much smaller than τ*, the flow is due to fluctuations. The
viscous stress and the flow rate are proportional to each other, τ ≈ η*. rea (Eq.5.10).
The associated time scale over which stress disappears is:
(5.12)
If tC is smaller than or comparable with the duration of the measurement, at
large scale, the aggregate relaxes the stress like a simple (Newtonian) fluid of

Physique des agrégats cellulaires embryonnaires
109
viscosity η* (Fig. 4.3). Conversely, if tC is much larger than the duration of the
measurement, the flow is not measurable, and the aggregate seems to display a
solid (plastic) behavior.
In contrast, if the stress τ is larger than τ*, it overcomes energy barriers.
According to Eq. 5.10, the ratio of the viscous stress to the flow rate defines an
effective viscosity ηeff that is smaller than η*, by a factor:
(5.13)
The aggregate thus behaves like a ‘‘shear-thinning’’ fluid in which viscosity
decreases at higher τ.
V.3.5. Compressions: Predictions.
Eq. 5.10 applies to compressions or decompressions. We tried first to fit the
compression experiments. In this case, the distance h between plates is decreased
in a stepwise manner, then is kept constant to h = hcomp, so that εtot =εcomp, and tot=0.
This creates an initial elastic stress τ0=E.εcomp (Eqs. 5.6–5.10) then yield a first-order
differential equation. Its solution is the relaxation of stress with time t (Fig. 5.8):
(5.14)
Initially, the flow is in the stress-induced regime: The stress decreases faster
than exp(–t/tc). When the stress reaches τ*, the flow enters the fluctuation-induced
regime, where the stress decreases like exp(–t/tc).
In principle, after a sufficiently long compression time (tcomp = tc) all elastic
stresses ultimately disappear (Eq. 5.14). At this stage, the plates would only measure
the constant contribution arising from the hydrostatic pressure generated by the
aggregate surface tension. On the contrary, if tcomp is smaller (or not much larger)
than tc, the aggregate still stores some residual stress, like a plastic solid.
The shape of the relaxation curve is well fitted by presented model. The error
bar on fitted parameters is the 95% confidence interval of the nonlinear curve fit: All
fitted parameters have errors <10% of the estimated value and smaller than the data
dispersion.
During successive compressions the characteristic stress remains
approximately constant, τ* =2.4 Pa (Fig. 5.9Inset). In the curve of Fig. 5.9, the initial
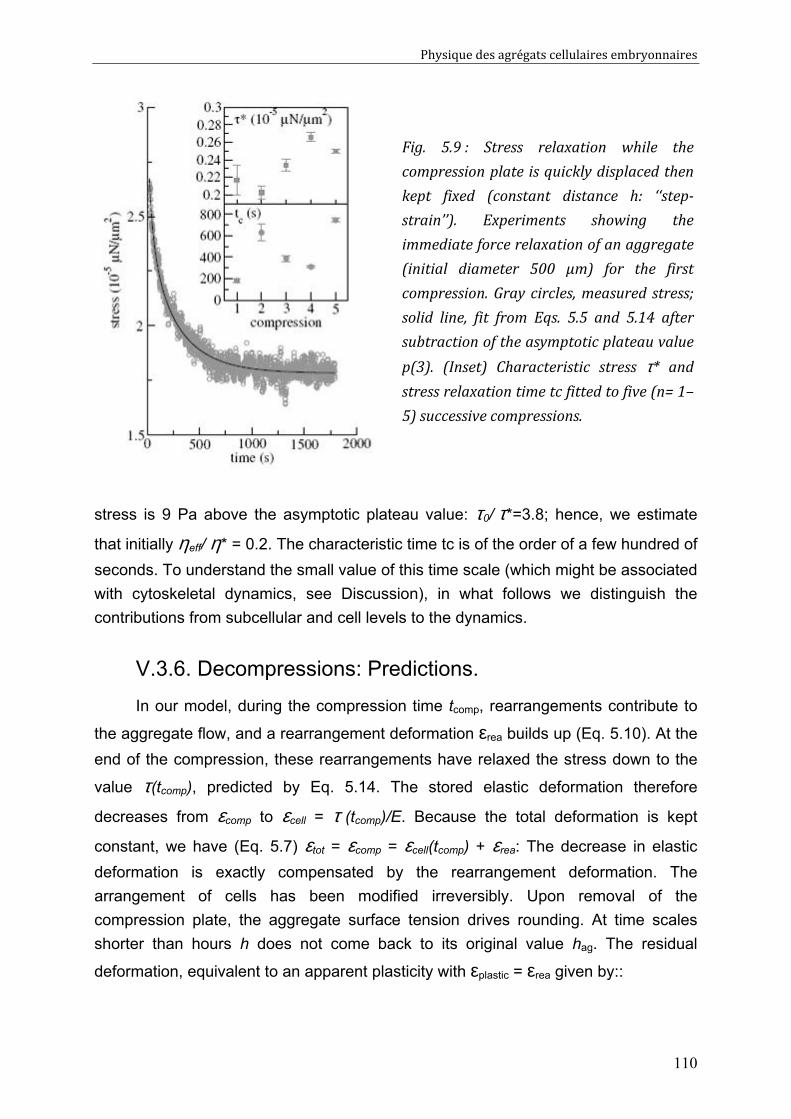
Physique des agrégats cellulaires embryonnaires
110
Fig. 5.9 : Stress relaxation while the
compression plate is quickly displaced then
kept fixed (constant distance h: ‘‘step-
strain’’). Experiments showing the
immediate force relaxation of an aggregate
(initial diameter 500 µm) for the first
compression. Gray circles, measured stress;
solid line, fit from Eqs. 5.5 and 5.14 after
subtraction of the asymptotic plateau value
p(3). (Inset) Characteristic stress τ* and
stress relaxation time tc fitted to five (n= 1–
5) successive compressions.
stress is 9 Pa above the asymptotic plateau value: τ0/ τ*=3.8; hence, we estimate
that initially ηeff/ η* = 0.2. The characteristic time tc is of the order of a few hundred of
seconds. To understand the small value of this time scale (which might be associated
with cytoskeletal dynamics, see Discussion), in what follows we distinguish the
contributions from subcellular and cell levels to the dynamics.
V.3.6. Decompressions: Predictions.
In our model, during the compression time tcomp, rearrangements contribute to
the aggregate flow, and a rearrangement deformation εrea builds up (Eq. 5.10). At the
end of the compression, these rearrangements have relaxed the stress down to the
value τ(tcomp), predicted by Eq. 5.14. The stored elastic deformation therefore
decreases from εcomp to εcell = τ (tcomp)/E. Because the total deformation is kept
constant, we have (Eq. 5.7) εtot = εcomp = εcell(tcomp) + εrea: The decrease in elastic
deformation is exactly compensated by the rearrangement deformation. The
arrangement of cells has been modified irreversibly. Upon removal of the
compression plate, the aggregate surface tension drives rounding. At time scales
shorter than hours h does not come back to its original value hag. The residual
deformation, equivalent to an apparent plasticity with εplastic = εrea given by::

Physique des agrégats cellulaires embryonnaires
111
(5.15)
For several values of εcomp and tcomp, the model agrees (Fig. 5.7) with the
experimental residual deformation with only two parameters. The relaxation time is
tC=5h. The characteristic deformation is ε*=τ*/E=0.13. It means that the characteristic
stress τ* is reached if the aggregate is quickly compressed with a deformation of at
least 0.13.
V.4. Discussion
V.4.1. Energy Barriers and Non-released Elasticity.
Elasticity and plasticity arise from contributions of cell bulk (cytoskeleton and
organelles) and surface (actomyosin-rich cortex). Adhesion and cortical contraction
contribute to create a cell–cell interfacial tension [not to be confused with the
aggregate surface tension [85;183] and, thus, energy barriers.
We study the combined effect of stress and active fluctuations to overcome
energy barriers. The model captures the main features of compression experiments,
as well as free relaxations, and distinguishes two flow regimes. Fits to the data
determine τ*, E, tC.
To interpret the values of these parameters, it is necessary to take explicitly
into account not only cell rearrangements, but also the subcellular (cytoskeleton)
dynamics. If the model were also developed for cell internal reorganization, Eq. 5.14
would be well suited to fit both types of relaxations. A future extension of our single
energy barrier model might include a more realistic distribution of energy barriers,
some of which can be passed by fluctuations, whereas others are not passed within
the experiment duration.
V.4.2. Time Scales.
The slow time scales tC =5 h obtained from the fit of our shape relaxation
experiments with the model (Fig. 5.7) can be interpreted as the relaxation time due to
cell rearrangements. Rearrangements-induced flow starts when aggregates are out
of mechanical equilibrium, e.g., compressed. Rearrangements continue over hours
until the complete equilibration of the aggregate. This interpretation agrees with the
cell shape relaxation observed after 24 h in a centrifuged aggregate [119]. It also
agrees with orders of magnitude extracted from the fusion of two F9 aggregates and

Physique des agrégats cellulaires embryonnaires
112
the rounding of the resulting aggregate, driven by surface tension, which takes 6
hours ( Fig. 4.3, Movie M4).
We observe a much shorter time scale (~102 s) in stress relaxation experiments
(Fig. 5.9), confirmed by height recovery after compression (Fig. 5.6). We believe that
it is due to internal release of cell elasticity by active cytoskeleton polymerization-
depolymerization processes, not included in the model. It is compatible with the
internal cell viscosity, probably hundreds of times smaller [190] than the tissue
viscosity η* or equivalently by lower energy barriers. Interestingly, the characteristic
stress τ* which according to the model is determined by the fluctuation amplitude and
the cell size (not the energy barrier) measured from stress relaxation curves (Fig.
5.9) is the same than the one deduced from apparent plasticity data (Fig. 5.7). To
obtain this last value, we combined the measured viscosity from fusion experiments
η*=4.4×105 Pa.s18 with the above relations τ*=0.13E and E=η*/tC. This yields
E=25Pa, then τ*=3.2 Pa. Notice that the elastic modulus is extremely low but
consistent with the measurements realized at low applied stress and the predictions
of the tensegrity model that states that the cytoskeleton prestress and architecture is
the primary determinant of the cell elastic response [191].
Aggregates are not always able to fuse and round up within the time of
experiments (CHO cell aggregates barely fuse even after 1 week [115] ). In addition,
even after relaxation, their shape sometimes displays a strong deviation from that of
a liquid drop [91;110]. This suggests that the energy barriers are too high: Both the
aggregate’s surface tension and the fluctuations are insufficient to relax the elastic
stress over the duration of the experiment. Simulations at high cell–cell tension [174]
confirm that a plateau is sometimes reached before full stress relaxation.
V.4.3. Validity of the plasticity measurement
One can question our choice of this 440s arbitrary time for plasticity
measurement. Indeed, several phenomena are occurring simultaneously during the
shape relaxation. If we want to extract one parameter characteristic of one of them,
we need a clear distinction of timescales. Let’s list the different physical origin of the
shape relaxation: (1) Individual cells may relax a stored elasticity when they are
decompressed (0-1 min). (2) They can also reorganize their cytoskeleton actively at
rather short timescales (1-5 min as explained above). At the tissue level, cell-cell
rearrangements occur induced (3) or not (4) by the stress.
18 η*=η measured by the fusion assay (see chapter 4)

Physique des agrégats cellulaires embryonnaires
113
Again our aim here was to quantify, cell rearrangement induced by the
preceding compression experiment. If during the 440s, a lot of these rearrangements
also occur due to the surface tension stress, our measurement can only be made
with a very large error.
Two arguments can be presented here to argue that this error, of course exist,
but is still very reasonable. During compression experiments under the two-photons
microscope, we could visualize these rearrangements, and even if our theory predict
a shear thinning effect we did not see a tremendous difference in terms of numbers
of rearrangement between the first 440s and for example the following ten minutes
(of course this remain to be quantify).
We can also estimate the proportion of rearrangements occurring during the
first 440 sec as a maximum value of 12%, by assuming an exponential decay in
height recovery (1-exp(-t/teff)). The effective relaxation time is lower than the
characteristic time due to shear-thinning effect 19:
c
eff
effc t
Et
t
E≤=≤= )()
55
*(
ηη
So the relative error on apparent plasticity is not too exaggerated as compared to
error bars (Fig. 5.7) and justifies a posteriori our hypothesis.
Moreover, in decompression experiment, the starting stress is of course smaller
than in the compression phase (it corresponds to the plateau value obtained during
compression), so the number of rearrangement at short timescales will of course be
smaller and so will be the error.
V.4.4. Link between compression-decompression experiment and fusion?
We have just noticed that the timescale tc obtained from compression-
decompression experiment is the same than the one obtained from fusion
experiment. However, these two timescales were derived from very different
modelling. In compression-decompression experiment, we analysed the experiment
in term of a viscous-elatic-plastic behaviour of aggregate. The surface tension does
not play a significant role it just gives the value of the plateau in force relaxation
experiment. Whereas fusion experiments were analyzed only taking into account the
fluid behaviour. Of course, the flow during fusion will necessitate the same type of
19 The ratio ηeff=η*/5 was obtained for deformations higher than 50%

Physique des agrégats cellulaires embryonnaires
114
rearrangements. We indeed visualized them under two-photon microscopy and also
observed shape deformation (see movie M2).
Is it pure coincidence that these two interpretations give the same timescales of
the same type of phenomenon (rearrangements) or is there a clear correspondence
between the two models which will then lead to the relation:
E ≈ σ /R.
This would mean that cells are able to adapt their mechanical properties to their
environmental conditions, and then to tune their elasticity in function of the pressure
they undergo. This could be an interesting point to elucidate in future work.
V.4.5. Effect on Measured Surface Tension
In compression–decompression cycle experiments, at a given relaxation time,
the longer or the stronger the compression, the larger the number of rearrangements,
and thus the stronger the apparent irreversibility, in good agreement with Eqs. 5.14
and 5.15. For the assessment of surface tension, the time between two successive
compressions is usually between 30 min and 2 h [115]. An equilibrium force seems to
be attained 2–10 min after compression. We believe that this time is large enough to
relax internal contributions; but, unlike what is assumed by [106], only a limited
number of rearrangements have taken place. Residual elastic contributions could
lead to an overestimation of surface tension. This questions the validity of the
application of Laplace’s law in past and present surface tension measurement
experiments. A likely hypothesis would explain the remaining stored elasticity in our
experiments by an active response of the cell. When compressed, cells could actively
release a part of the elasticity stored in the compressed cytoskeleton; when
decompressed, the surface tension drives the relaxation of the global aggregate
shape, and cells could actively regain part of the elasticity. This hypothesis is
compatible with the rate, 1–10 µm/min, at which single cells reorganize their
cytoskeleton [33]. It could be tested by modifying the dynamics of actin, microtubules,
or intermediate filaments using drugs (latrunculin and jasplakinolide). Two-photon
imaging of cell contours combined to simultaneous force measurements could be an
excellent assay for that investigation. If this hypothesis is correct, the elastic energy
stored in deformed cells will be less than we assume in our model, and the
overestimation of the surface tension in compression experiments will be smaller.
This will explain why published measurements of surface tension are usually robust
and do not depend on compression magnitude and duration, nor on aggregate size
[103;105;156].

Physique des agrégats cellulaires embryonnaires
115
Conclusions et Perspectives
Nous avons réalisé une caractérisation physique et rhéologique des agrégats
cellulaires embryonnaires. Ces agrégats formés à partir d’auto-assemblages de
cellules dissociées constituent un système modèle des tissus. Ils permettent
d’étudier les réorganisations cellulaires qui interviennent dans les domaines de
l’embryologie, la cancérologie et l’ingénierie tissulaire. C’est grâce à ces
réorganisations que les cellules peuvent, se reconnaitre, se trier, s’adapter à leur
environnement afin de générer une structure complexe, possédant une forme définie,
le tissu. Nous avons utilisé plusieurs moyens physique, cohérents entre eux, afin
d’apporter des réponses sur l’origine, la cinétique et la dynamique des
réorganisations des cellules dans les tissus.
La tension de surface influe sur la forme d’un liquide. Qu’il soit en suspension,
posé sur une surface, ou mélangé avec un autre liquide, le liquide doit définir ses
interfaces de façon à obéir à la contrainte de sa tension de surface et donc de
manière à minimiser son énergie totale. L’établissement d’un tel équilibre engendre
toujours des écoulements dans les liquides, la viscosité, quant à elle, va s’opposer
aux déplacements relatifs des unités du liquide. Chez certains agrégats cellulaires
embryonnaires, des phénomènes qui semblent guidés par la tension de surface
apparente et freinés par une viscosité effective ont été bien identifiés. Ceux-ci sont,
en effet, capables de s’arrondir, de fusionner ou de se trier à des échelles de temps
très longues comparées à celles des liquides visqueux usuels.
La compréhension des comportements cellulaires en tissus fait partie des
questions ouvertes de la biologie de développement. Il a été montré que la tension
de surface permettait de prédire les comportements mutuels d’enveloppement des
tissus. C’est pourquoi, mesurer proprement la tension de surface dans un premier
temps puis identifier son origine structurale à l’intérieur des tissus ont été les idées
directrices de notre travail. Un tensiomètre des tissus ainsi qu’une méthode
d’analyse adéquate (indépendante de l’angle de contact) ont été développés à cet
effet. En parallèle, le rôle du cytosquelette dans la cohésion du tissu a été étudié en
quantifiant sa contribution à la tension de surface. La suppression de l’expression de
l’α-caténine indispensable pour la liaison cytosquelette/ cadhérine, dans des cellules
exprimant un même niveau de cadhérine, fait baisser la tension de surface alors
qu’elle est toujours capable de prédire les comportements d’enveloppement mutuels
et de tri cellulaire dans les expériences de ségrégations. Ce nouveau résultat remet
en cause l’origine proposée pour la tension de surface par l’hypothèse d’adhésion
différentielle HAD (i.e., le niveau d’expression des molécules d’adhésion seul

Physique des agrégats cellulaires embryonnaires
116
conditionne la tension de surface tissulaire), tout en renforçant son idée de base : le
tri cellulaire et la morphogenèse sont gouvernés par la tension de surface des tissus.
Tirant parti de l’analogie entre tissus et liquides, nous avons pu quantifier
d’autres propriétés physiques de plusieurs lignées de cellules telles que la viscosité.
Nous avons mesurés une viscosité similaire pour deux types cellulaires n’ayant pas
la même cohésion. Des résultats similaires ont été trouvés dans la littérature. Des
mesures préliminaires en présence de nocodazole révèlent une augmentation de la
viscosité des tissus. Suite à ces résultats, nous proposons que la viscosité tissulaire
est fixée par une compétition entre les phénomènes protrusifs d’une part et la
cohésion d’autre part.
En parallèle, nous avons réalisé des expériences rhéologiques de relaxation de
force ou de relaxation de forme d’un agrégat initialement compressé. Des
comportements visco-élasto-plastiques ont été rencontrés. Afin de capturer la
complexité de la réponse des agrégats sous stress, une modélisation analytique des
réarrangements cellulaires à l’intérieur de l’agrégat a été menée. La combinaison
expériences-modèle nous a permis d’évaluer certains paramètres rhéologiques
significatifs des tissus embryonnaires (E=25 Pa, τ*=2.4 Pa, ηeff=0,2.η∗, tc=5h) et de
classer les comportements des agrégats sous stress τ selon deux régimes : régime
dominé par les fluctuations τ < τ* ou régime rhéofluidifiant τ > τ*.
Ce travail offre de multiples perspectives.
Dans un avenir proche, grâce en particulier à l’imagerie deux photons, on
pourra espérer quantifier les réarrangements et déformations car ce type d’imagerie
permet d’avoir accès aux champs de vitesse et de déformation à l’intérieur de
l’agrégat. Outre la géométrie de compression, il parait intéressant de travailler avec
des agrégats que l’on force à s’écouler dans des microcanaux de diamètre inférieur à
celui des agrégats par aspiration afin possiblement d’obtenir une estimation de la
viscosité tissulaire indépendante de la tension de surface. Ces données pourront être
confrontées à des simulations numériques par éléments finis de modèles purement
visqueux ou visco-élasto-plastiques de ces agrégats. Le rôle de l’élasticité pourra
ainsi être quantifié puis mesuré indépendamment sur le tensiomètre en y
implémentant un mode dynamique (rhéomètre en compression) . Il parait également
très prometteur de mettre en évidence les composants du cytosquelette à l’origine de
l’élasticité en jouant sur différentes drogues contractiles car ce travail a été tout juste
commencé durant cette thèse. Les mesures effectuées au chapitre 5 sur les cellules
F9 pourront aussi être effectuées sur des cellules F9(α-/-) afin de déterminer le rôle du
cytosquelette sur les différents paramètres mesurés.

Physique des agrégats cellulaires embryonnaires
117
Enfin, de nombreuses expériences en collaboration avec des biologistes,
pourront être effectuées afin de mettre en évidence les différents chemins de
signalisation jouant un rôle sur les paramètres physiques que nous sommes
maintenant facilement capable de mesurer.

Physique des agrégats cellulaires embryonnaires
118
Bibliography [1] Evans E.A. & Waugh R. Osmotic correction to elastic area compressibility measurements on red cell membrane. Biophys. J. (1977) 20(3): p. 307–313. [2] Thoumine O., Cardoso O., & Meister J.J. Changes in the mechanical properties of fibroblasts during spreading: a micromanipulation study. Eur. Biophys. J. (1999) 28(3): p. 222–234. [3] Guilak F., Erickson G.R., & Ting-Beall H. P. The effects of osmotic stress on the viscoelastic and physical properties of articular chondrocytes. Biophys. J. (2002) 82(2): p. 720–727. [4] Baaijens F.P., Trickey W.R., Laursen T.A., & Guilak F. Large deformation finite element analysis of micropipette aspiration to determine the mechanical properties of the chondrocyte. Ann. Biomed.Eng. (2005) 33(4): p. 494–501. [5] Peeters E.A., Oomens C.W., Bouten C.V., Bader D.L., & Baaijens F. P. Viscoelastic properties of single attached cells under compression. J. Biomech. Eng. (2005) 127(2): p. 237–243. [6] Trickey W.R., Baaijens F.P., Laursen T.A., Alexopoulos L.G., & Guilak. Determination of the poisson’s ratio of the cell: recovery properties of chondrocytes after release from complete micropipette aspiration. J. Biomech (2006) 39(1): p. 78–87. [7] Rosenbluth M.J., Lam W.A., & Fletcher D.A. Force microscopy of nonadherent cells: a comparison of leukemia cell deformability. Biophys. J. (2006) 90(8): p. 2994–3003. [8] Cuvelier D., Théry M., Chu Y.S., Dufour S., Thiéry J.P., Bornens M., Nassoy P.,& Mahadevan L. The universal dynamics of cell spreading. Curr. Biol. (2007) 17(8): p. 694–699. [9] Sato-Maeda M., Uchida M., Graner F., & Tashiro H. Quantitative evaluation of tissue-specific cell adhesion at the level of a single cell pair. Dev. Biol. (1994) 162(1): p. 77–84. [10] Merkel R., Nassoy P., Leung A., Ritchie K., & Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature (1999) 397(6714): p. 50–53. [11] Zhu B., Chappuis-Flament S., Wong E., Jensen I. E., Gumbiner B. M., & Leckband D. Functional analysis of the structural basis of homophilic cadherin adhesion. Biophys. J. (2003) 84(6): p. 4033–4042. [12] Perez T.D., Nelson W.J., Boxer S.G., & Kam L. E-cadherin tethered to micropatterned supported lipid bilayers as a model for cell adhesion. Langmuir. (2005) 21(25): p. 11963–11968. [13] Leckband D. & Prakasam A. Mechanism and dynamics of cadherin adhesion. Annu. Rev. Biomed. Eng. (2006) 8: p. 259–287. [14] Janmey P. A. Mechanical properties of cytoskeletal polymers. Curr.Opin. Cell Biol. (1991) 3(1): p. 4–11. [15] Brangwynne C.P., MacKintosh F.C., Kumar S., Geisse N.A., Talbot J., Mahadevan L., Parker K.K., Ingber D.E., & Weitz D.A. Microtubules can bear enhanced compressive loads in living cells because of lateral reinforcement. J. Cell Biol. (2006) 173(5): p. 733–741. [16] Lenormand G., Henon S., Richert A., Simeon J., & Gallet F. Direct measurement of the area expansion and shear moduli of the human red blood cell membrane skeleton. Biophys. J. (2001) 81(1): p. 43–56. [17] Janmey P.A., Euteneuer U., Traub P., & Schliwa M. Viscoelastic properties of vimentin compared with other filamentous biopolymer networks. J. Cell Biol. (1991) 113(1): p. 155–160. [18] Janmey P.A. & McCulloch C.A. Cell mechanics: integrating cell responses to mechanical stimuli. Annu. Rev. Biomed. Eng. (2007) 9: p. 1–34. [19] Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, & James D.Watson. Molecular biology of the cell. Flammarion Médecine- & Sciences pour l’édition française. Third Edition, 1995.

Physique des agrégats cellulaires embryonnaires
119
[20] Gittes F., Mickey B., Nettleton J., & Howard J. Flexural rigidity of microtubules and actin filaments measured from thermal fluactuations in shape. J. Cell Biol. (1993) 120: pp. 923-934. [21] Théry M. Contrôle de la polarité des cellules adhérentes. Université Paris 7-Denis Diderot. 2006. [22] Holy T.E. LS. Dynamic instability of microtubules as an efficient way to search in space. Proc.Natl Acad. Sci. USA (1994) 91: pp. 5682-5685. [23] Tolić-Nørrelykke I.M. Push-me-pull-you: how microtubules organize the cell interior. Eur Biophys J. (2008) 37(7): pp. 1271-1278. [24] Wühr M., Dumont S., Groen A.C., Needleman D.J., Mitchison T.J. How does a millimeter-sized cell find its center? Cell Cycle (2009) 8(8): pp. 1115-1121. [25] Desai A &MT. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. (1997) 13: pp. 83-117. [26] Isambert H., Venier P., Maggs A.C., Fattoum A., Kassab R., Pantaloni D., & Carlier M.F. Flexibility of actin filaments derived from thermal fluctuations. effect of bound nucleotide, phalloidin, and muscle regulatory proteins. J. Biol.Chem. (1995) 270: pp. 11437-11444. [27] MacKintosh F.C. KJ&JP. Elasticity of semiflexible biopolymer networks. Phys. Rev. Lett. (1995) 75(24): pp. 4425-4428. [28] Kruse K., Joanny J.F., Jülicher F., Prost J. & Sekimoto K. Generic theory of active polar gels : a paradigm for cytoskeletal dynamics. Eur.Phys. J. E (2005) 16: pp. 5-16. [29] Gardel M.L., Nakamura F., Hartwig J.H., Crocker J.C., Stossel T.P., & Weitz D.A. Prestressed f-actin networks cross-linked by hinged filamins replicate mechanical properties of cells. Proc. Natl. Acad. Sci. USA (2006) 103: pp. 1762-1767. [30] Bugyi B., Le Clainche C., Romet-Lemonne G.& Carlier MF. How do in vitro reconstituted actin-based motility assays provide insight into in vivo behavior? FEBS Lett. (2008) 582(14): pp. 2086-2092. [31] Tharmann R., Claessens M. M. A. E., & Bausch A. R. Viscoleasticity of isotropipcally cross-linked actin network. Phys. Rev.Lett. (2007) 98: p. 088103. [32] Le Goff L., Amblard F., & Furst E.M. Motor-driven dynamics in actin-myosin networks. Phys. Rev. Lett. (2002) 88: p. 018101. [33] Watanabe N. &MT. Single-molecule speckle analysis of actin filament turnover in lamellipodia. Science (2002) 295(5557): p. 1083–6. [34] Alice Nicolas. L’adhésion cellulaire, une sonde de l’environnement mécanique dans les tissus. CNRS, 2007. [35] Small J.V.,Stradalb T.,Vignala E.& Rottnerb K. The lamellipodium: where motility begins. Nat Rev. Mol. Cell Biol. (2002) 12(3): pp. 112-120. [36] Kaverina I.,Krylyshkina O.,Beningo K.,Anderson K., Wang Y.L. & Small J.V. Tensile stress stimulates microtubule outgrowth in living cells. Journal of Cell Science (2002) 115: pp. 2283-2291. [37] Balaban N.Q., Schwarz U.S., Riveline D., Goichberg P., Tzur G., Sabanay I., Mahalu D., Safran S., Bershadsky A., Addadi L.& Geiger B. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. (2001) 3: pp. 466-472. [38] Folsom T. D. SDS. Disruption of actin-myosin interactions results in the inhibition of focal adhesion assembly in xenopus xr1 glial cells. GLIAEJ (1999) 26(3): pp. 245-259 . [39] Bongrand P. Adhesion of cells in structure and dynamics of membranes. R. Lipowsky & E. Sackmann (Ed.). Elsevier, Amsterdam, Vol. IB, pp 755-803, 1995. [40] Robinson E.E., Zazzali K.M., Corbett S.A., & Foty R.A. Alpha5beta1 integrin mediates strong tissue cohesion. J Cell Sci (2003) 116: pp. 377-386. [41] Hyafil F., Morello D., Babinet C. & Jacob F. A cell surface glycoprotein involved in the compaction of embryonal carcinoma cells and cleavage stage embryos. Cell (1980) 21: pp. 927-934. [42] Ogou S., Okada T. S., & Takeichi M. Cleavage stage mouse embryos share a common cell adhesion system with teratocarcinoma cells. Dev.Biol. (1982) 92: p. 521.

Physique des agrégats cellulaires embryonnaires
120
[43] Alberts B., Johnson A., Lewis J., Raff M., Roberts K., & Walter P. Molecular biology of the cell. Garland Science NY, 2002. [44] Duguay D., Foty R.A., & Steinberg M.S. Cadherin-mediated cell adhesion and tissue segregation: qualitative and quantitative determinants. Dev. Biol. (2003) 253(2): p. 309–323. [45] Foty R.A. & Steinberg M.S. The differential adhesion hypothesis: a direct evaluation. Dev. Biol. (2005) 278(1): p. 255–263. [46] Shi Q., Chien Y.H., & Leckband D. Biophysical properties of cadherin bonds do not predict cell sorting. J. Biol. Chem. (2008) 283(42): pp. 28454-28463. [47] Cavey M., Rauzi M., Lenne P.F., & Lecuit T. A two-tiered mechanism for stabilization and immobilization of e-cadherin. Nature (2008) 453(7196): p. 751–756. [48] Gumbiner B.M. Regulation of cadherin-mediated adhesion in morphogenesis. Nat. Rev. Mol. Cell Biol. (2005) 6(8): p. 622–634. [49] Halbleib J.M. & Nelson W.J. Cadherins in development: cell adhesion, sorting, and tissue morphogenesis. Genes Dev. (2006) 20(23): p. 3199–3214. [50] Yamada S., Pokutta S., Drees F., Weis W.I., & Nelson W.J. Deconstructing the cadherin-catenin-actin complex. Cell (2005) 123(5): p. 889–901. [51] Gates J. & Peifer M. Can 1000 reviews be wrong? actin, alpha-catenin, and adherens junctions. Cell (2005) 123(5): p. 769–772. [52] Drees F., Pokutta S., Yamada S., Nelson W.J., & Weis W.I. Alpha-catenin is a molecular switch that binds e-cadherin-beta-catenin and regulates actin-filament assembly. Cell (2005) 123(5): p. 903–915. [53] Mège R.M., Gavard J., & Lambert M. Regulation of cell-cell junctions by the cytoskeleton. Curr. Opin. Cell Biol. (2006) 18(5): p. 541–548. [54] Kovacs E.M. &YA. Cell-cell contact: cooperating clusters of actin and cadherin. Curr. Biol. (2008) 18(15): p. R667–R669. [55] Kerssemakers J.W., Munteanu E.L., Laan L., Noetzel T.L., Janson M.E.& Dogterom M. Assembly dynamics of microtubules at molecular resolution. Nature (2006) 442(7103): pp. 709-712. [56] Biener G., Vrotsos E., Sugaya K.& Dogariu A. Optical torques guiding cell motility. Opt Express. (2009) 17(12): pp. 9724-9732. [57] Grishchuk E.L., Molodtsov M.I., Ataullakhanov F.I.& McIntosh J.R. Force production by disassembling microtubules. Nature (2005) 438(7066): pp. 384-388. [58] Bell G.I. Models for the specific adhesion of cells to cells. Science (1978) 200(4342): p. 618–627. [59] Bell G.I., Dembo M., & Bongrand, P. Cell adhesion. competition between nonspecific repulsion and specific bonding. Biophys. J. (1984) 45(6): p. 1051–1064. [60] Evans E. Detailed mechanics of membrane-membrane adhesion and separation. i. continuum of molecular cross-bridges. Biophys. J. (1985a) 48(1): p. 175–183.. [61] Evans E. Detailed mechanics of membrane-membrane adhesion and separation. ii. discrete kinetically trapped molecular cross-bridges. Biophys. J. (1985b) 48(1): p. 185–192. [62] Evans E.A. & Calderwood D.A. Forces and bond dynamics in cell adhesion. Science (2007) 316(5828): p. 1148–1153. [63] Frisch T. & Thoumine O. Predicting the kinetics of cell spreading. J. Biomech. (2002) 35(8): p. 1137–1141. [64] Cañadas P., Laurent V. M., Oddou C., Isabey D., Wendling S. A cellular tensegrity model to analyse the structural viscoelasticity of the cytoskeleton. J. Theor. Biol. (2002) 218: pp. 155-173 . [65] Steinberg M.S. Reconstruction of tissues by dissociated cells. some morphogenetic tissue movements and the sorting out of embryonic cells may have a common explanation. Science (1963) 141: p. 401–408. [66] Hélène Delanoe-Ayari. Nucléation et croissance de contacts entre cellules biologiques. Université Joseph Fourier Grenoble I. 2003. [67] Lecuit T. & Lenne P.F. Cell surface mechanics and the control of cell shape, tissue patterns and morphogenesis. Nat. Rev. Mol. Cell Biol. (2007) 8(8): p. 633–644.

Physique des agrégats cellulaires embryonnaires
121
[68] Pilot F. & Lecuit T. Compartmentalized morphogenesis in epithelia: from cell to tissue shape. Dev. Dyn. (2005) 232(3): p. 685–694. [69] Classen A.K., Anderson K.I., Marois E., & Eaton S. Hexagonal packing of drosophila wing epithelial cells by the planar cell polarity pathway. Dev. Cell (2005) 9(6): p. 805–817. [70] Carthew R.W. Adhesion proteins and the control of cell shape. Curr. Opin. Genet. Dev. (2005) 15(4): p. 358–363. [71] Olivier Pourquié. Biologie du développement. Hermann (Ed.). , 2002. [72]http://www.fishbase.org. Danio rerio. [73] Thisse B. TC. Établissement des axes embryonnaires au cours du développement du poisson zèbre. Medecine Sciences (2002) 18: pp. 193-204. [74] Kimmel C.B. WR. Tissue-specific cell lineages originate in the gastrula of the zebrafish. Science (1986) 231: pp. 365-368. [75] Kimmel C., Ballard W., Kimmel S., Ullman B., & Schilling T. Stages of embryonic development of the zebrafish. Developmental Dynamics (1995) 203: pp. 253-310 . [76] Driever W, Solnica-Krezel L, Schier AF, Neuhauss SC, Malicki J, Stemple DL, Stainier DY, Zwartkruis F, Abdelilah S, Rangini Z, Belak J, & Boggs C. A genetic screen for mutations affecting embryogenesis in zebrafish. Development (1996) 123: p. 37-46.. [77] Haffter P, Granato M, Brand M, Mullins MC, Hammerschmidt M, Kane DA, Odenthal J, van Eeden FJ, Jiang YJ, Heisenberg CP, Kelsh RN, Furutani-Seiki M, Vogelsang E, Beuchle D, Schach U, Fabian C, & Nüsslein-Volhard C. The identification of genes with unique and essential functions in the development of the zebrafish, danio rerio. Development (1996) 123: pp. 1-36. [78] Nasevicius A., & Ekker S. Effective targeted gene knockdown in zebrafish. Nat. Genet. (2000) 26: pp. 216-220. [79] DeRobertis E.M., Larrain J., Oelgeschlâger M., & Wessely O. The establishment of spemann’s organizer and patterning of the vertebrate embryo. Nat Genet (2000) 1: pp. 171-181. [80] Meno C., Saijoh Y., Fujii H., Ikeda M., Yokoyama T., Yokoyama M., Toyoda Y., & Hamada H. Left-right asymmetric expression of the tgfb family member lefty in mouse embryos. Nature (1996) 381: pp. 151-155. [81] Oulad-Abdelghani M., Chazaud C., Biullet P., Matteri M.G., Dollé P., & Chambon P. Stra 3/lefty, a retinoid acidinducible novel member of the transforming growth factor b family. Int J Dev Biol (1998) 42: pp. 23-32. [82] Daopin S., Piez K.A., Ogawa Y., Davies D.R. Crystal structure of transforming growth factor-beta 2: an unsual fold for the superfamily. Science (1992) 257: pp. 369-373. [83] Schlunegger M.P. &GM. An unusual feature revealed by the crystal structure at 2.2 a resolution of human transforming growth factorbeta 2. Nature (1992) 395: pp. 185-189. [84] Pouille P.A., Ahmadi P., Brunet A.C., Farge E. Mechanical signals trigger myosin ii redistribution and mesoderm invagination in drosophila embryos. Sci Signal (2009) 2(66): p. ra16. [85] Krieg M., Arboleda-Estudillo Y., Puech P.H., Käfer J., Graner F., Müller D.J., & HeisenbergC.P. Tensile forces govern germ-layer organization in zebrafish. Nat. Cell Biol. (2008) 10: pp. 429-436. [86] Wilson H.V. One some phenomena of coalescence and regeneration in sponges. J. Exp. Zool. (1907) 5: pp. 53-120. [87] Townes P.L. &HJ. Directed movements and selective adhesion of embryonic amphibians cells. J Exp Zool (1955) 128: pp. 53-120. [88] Steinberg M.S. & Takeichi M. Experimental specification of cell sorting, tissue spreading, and specific spatial patterning by quantitative differences in cadherin expression. Proc. Natl. Acad. Sci. U.S.A. (1994) 91(1): p. 206–9. [89] Godt D. TU. Drosophila oocyte localization is mediated by differential cadherin-based adhesion. Nature (1998) 395: pp. 387-391. [90] Steinberg M.S. On the mechanism of tissue reconstruction by dissociated cells. iii: free energy relations and the reorganization of fused, heteronomic tissue fragments. Proc. Natl. Acad. Sci. U.S.A. (1962) 48: p. 1769–1776.

Physique des agrégats cellulaires embryonnaires
122
[91] Mombach J. C.,Glazier J. A. ,Raphael R. C.& Zajac M. Quantitative comparison between differential adhesion models and cell sorting in the presence and absence of fluctuations. Phys. Rev. Lett. (1995) 75: p. 2244 –2247. [92] Rieu J.P. SY. Hydrodynamics and cell motion during the rounding of 2d hydra cell aggregates. Eur. Phys. J. B (2002) 27: p. 167–172. [93] P. de Gennes, F. Brochard-Wyart, D. Quéré. Gouttes, bulles, perles et ondes. Belin, 2005. [94] Isenberg C. The science of soap bubbles. New York: Dover Inc. (Ed.). , 1992. [95] Glaser R.. Biophysics. Springer (Ed.). , 1999. [96] C. Gianino. Measurement of surface tension by the dripping of a needle. Physics Eductaion (2006) 41: p. 440–444. [97] Graner F. & Sawada Y. Can surface-adhesion drive cell rearrangement.2. a geometrical model. J. theor. Biol. (1993) 164(4): p. 477–506. [98] Phillips H. Equilibrium measurements of embryonic cell adhesiveness: physical formulation and testing of the differential adhesion hypothesis. John Hopkins University. 1969. [99] Phillips H. & Steinberg M. Equilibrium measurements of embryonic cell adhesiveness: i. shape equilibrium in centrifugal fields. PNAS (1969) 64: p. 121–127. [100] Steinberg M. S. Adhesion in development: an historical overview. Dev. Biol. (1996) 180: p. 377–88. [101] Kalantarian A., Ninomiya H., Saad S. M. I., David R., Winklbauer R., Neumann A. W. Axisymmetric drop shape analysis for estimating the surface tension of cell aggregates by centrifugation. Biophysical Journal (2009) 96: p. 1606–1616. [102] David R., Ninomiya H., Winklbauer R. , Neumann A.W. Tissue surface tension measurement by rigorous axisymmetric drop shape analysis. Colloids and Surfaces B: Biointerfaces (2009) 72: pp. 236-240 . [103] Foty R.A., Pfleger C.M., Forgacs G., & Steinberg. M.S. Surface tensions of embryonic tissues predict their mutual envelopment behavior. Development (1996) 122: pp. 1611-1996. [104] Davis G.S. Migration-directing liquid properties of embryonic amphibian tissues. Am. Zool. (1984) 24: p. 649655. [105] Foty R.A., Forgacs G., Pfleger C. M., & Steinberg M. S. Liquid properties of embryonic tissues: measurement of interfacial tensions. Physical Review Letters (1994) 72(14): p. 2298–2301. [106] Forgacs G., Foty R.A., Shafrir Y., & Steinberg M.S. Viscoelastic properties of living embryonic tissues: a quantitative study. Biophys. J. (1998) 74: pp. 2227-2234. [107] Graner F. GJ. Simulation of biological cell sorting using a two-dimensional extended potts model. Phys Rev Lett. (1992) 69(13): pp. 2013-2016. [108] Becam I. HJ. Régulation génétique de l’adhérence intercellulaire ou comment les cadhérines sculptent la drosophile. Medecine Sciences (2007) 23: pp. 285-290. [109] Nagafuchi A., Shirayoshi Y., Okazaki K., Yasuda K., & Takeichi M. Transformation of cell adhesion properties by exogenously introduced e-cadherin cdna. Nature (1987) 329: pp. 341-343. [110] Beysens D.A., Forgacs G., & Glazier J.A. Cell sorting is analogous to phase ordering in fluids. Proc. Natl. Acad. Sci. USA (2000) 97: p. 9467–9471. [111] Duguay D., Foty R.A., & Steinberg M.S.. Cadherin-mediated cell adhesion and tissue segregation: qualitative and quantitative determinants. Dev. Biol. (2003) 253(2): p. 309–23. [112] Foty R.A. & Steinberg M. S. The differential adhesion hypothesis: a direct evaluation.. Dev. Biol. (2005) 278(1): p. 255–63. [113] Schötz E., Burdine R.D., Jülicher F., Steinberg M.S., Heisenberg C.P., & Foty R.A. Quantitative differences in tissue surface tension influence zebrafish germ layer positioning. HFSP J. (2008) 2: pp. 42-56. [114] Eva-Maria Schotz. Dynamics and mechanics of zebrafish embryonic tissues. 2007.

Physique des agrégats cellulaires embryonnaires
123
[115] Jakab K., Damon B., Marga F., Doaga O., Mironov V., Kosztin I., Markwald R.& Forgacs G. Relating cell and tissue mechanics: implications and applications. Dev. Dyn. (2008) 237(9): pp. 2438-2449. [116] Brodland G. The differential interfacial tension hypothesis (dith): a comprehensive theory for the self-rearrangement of embryonic cells and tissues. J. Biomech. Engineer. (2002) 124: pp. 188-197. [117] Harris A.K. Is cell sorting caused by differences in the work of intercellular adhesion? a critique of the steinberg hypothesis. Journal of Theoretical Biology (1975) 61(2): p. 267–285. [118] Phillips H. & Steinberg M. Embryonic tissues as elasticoviscous liquids. ii. direct morphological evidence for cell slippage in centrifuged aggregates. Dev.Biol (1977) 59: p. 124–134. [119] Phillips H. & Steinberg M. Embryonic tissues as elaticoviscous liquids. i. rapid and slow shape changes in centrifuged aggregates. J.Cell.Sci. (1978) 30: p. 1–20. [120] Brodland G.W., Chen D.I., & Veldhuis J.H. A cell-based constitutive model for embryonic epithelia and other planar aggregates of biological cells. Int. J. Plast. (2006) 22: p. 965–995. [121] Fredberg et al. Friction in airway smooth muscle : mechanism, latch and implications in asthma. J. Appl. Physiol. (1996) 81: pp. 2703-2712. [122] Fung Y. Mechanical properties of living tissues. Springer (Ed.). , 1993.. [123] Glaser R. Biophysics. Springer, 1999. [124] Kemler R. From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. (1993) 9: p. 317–321. [125] Nagafuchi A.,Ishihara S.,& Tsukita S. The roles of catenins in the cadherin-mediated cell adhesion: functional analysis of e-cadherin-alpha catenin fusion molecules. The Journal of Cell Biology (1994) 127(1): pp. 235-245. [126] Nagafuchi A. TSTM. Transmembrane control of cadherin-mediated cell-cell adhesion. Semin. Cell Biol. (1993) 4: p. 175–181. [127] Adams C.L., Nelson W.J., & Smith S.J. Quantitative analysis of cadherin-catenin-actin reorganization during development of cell–cell adhesion. J. Cell Biol. (1996) 135: p. 1899–1911. [128] Yap A.S., Brieher W.M., Pruschy M. , & Gumbiner B.M. Lateral clustering of the adhesive ectodomain: a fundamental determinant of cadherin function. Curr. Biol. (1997) 7: pp. 308-315. [129] Maeno Y.,Moroi S.,Nagashima H.,Noda T.,Shiozaki H.,Monden M.,Tsukita S.,& Nagafuchi A. Alfa-catenin-deficient f9 cells differentiate into signet ring cells. American Journal of Pathology (1999) 154(5): p. 1323:1328. [130] Maeno Y., Moroi S., Nagashima H., Noda T., Shiozaki H., Monden M., Tsukita S. & Nagafuchi A. Alpha-catenin-deficient f9 cells differentiate into signet ring cells. Am J Pathol. (1999) 154(5): pp. 1323-1328. [131] Fukunaga Y., Liu H., Shimizu M., Komiya S., Kawasuji M., Nagafuchi A. Defining the roles of beta-catenin and plakoglobin in cell-cell adhesion: isolation of beta-catenin/plakoglobin-deficient f9 cells. Cell Struct Funct. (2005) 30(2): pp. 25-34. [132] Kimura K., Ito M., Amano M., Chihara K., Fukata Y., Nakafuku M., Yamamori B., Feng J., Nakano T., Okawa K., Iwamatsu A. & Kaibuchi K. Regulation of myosin phosphatase by rho and rho-associated kinase (rho-kinase). Science (1996) 273: p. 245–248. [133] Krendel M. ZF&BG. Nucleotide exchange factor gef-h1 mediates cross-talk between microtubules and the actin cytoskeleton. Nat. Cell Biol. (2002) 4: p. 294–301. [134] Danowski B.A. Fibroblast contractility and actin organization are stimulated by microtubule inhibitors. J. Cell Sci. (1989) 93: p. 255–266. [135] Pletjushkina O.J., Belkin A.M., Ivanova O.J., Oliver T., Vasiliev J.M., Jacobson K.. Maturation of cell–substratum focal adhesions induced by depolymerization of microtubules is mediated by increased cortical tension. Cell Adhes. Commun. (1998) 5: p. 121–135. [136] Kadi A., Berthet V., Pichard V.,Abadie B.,Rognoni J-B.,Marvaldi J. & Luis J. Involvement of fak, pi3-k and pkc in cell adhesion induced by microtubule isruption. bulletin de Cancer (2002) 89.

Physique des agrégats cellulaires embryonnaires
124
[137] Rohatgi R, Henri Ho H & Kirschnera M. Mechanism of n-wasp activation by cdc42 and phosphatidylinositol 4,5-bisphosphate. J. Cell Biol. (2000) 150: p. 1299–1310. [138] Waterman-Storer C.M., Worthylake R.A., Liu B.P., Burridge K. & Salmon E.D. Microtubule growth activates rac1 to promote lamellipodial protrusion in fibroblasts. Nat. Cell Biol. (1999) 1: p. 45–50. [139] Bershadsky A. D. VEA&VJM. Pseudopodial activity at the active edge of migrating fibroblast is decreased after drug-induced microtubule depolymerization. Cell Motil. Cytoskeleton (1991) 19: p. 152–158. [140] Gritsman K., Zhang J., Cheng S., Heckscher E., Talbot W.S., & Schier A.F. The egf-cfc protein one-eyed pinhead is essential for nodal signaling. Cell (1999) 97(1): pp. 121-132. [141] Aoki T.O., David N.B., Minchiotti G., Saint-Etienne L., Dickmeis T., Persico G.M., Strähle U., Mourrain P. & Rosa F.M. Molecular integration of casanova in the nodal signalling pathway controlling endoderm formation. Development (2002) 129: p. 275–286. [142] Carmany-Rampey A. &SAF. Single-cell internalization during zebrafish gastrulation. Curr. Biol. (2001) 11: p. 1261–1265. [143] Gritsman K., Zhang J., Cheng S., Heckscher E., Talbot W.S., Schier A.F. The egf–cfc protein one-eyed pinhead is essential for nodal signaling. Cell (1999) 97: p. 121–132. [144] Mombach JC.M., Robert D., Graner F., Gillet G., Thomas G.L., Idiart M., & Rieu J.P. Rounding of aggregates of biological cells: experiments and simulations. Physica A (2005) 352: pp. 525-534. [145] Gillet G., Michel D., Crisanti P., Guerin M., Herault Y., Pessac B., Calothy G., Brun G., Volovitch M. Serum factors and v-src control two complementary mitogenic pathways in quail neuroretinal cells in culture. Oncogene (1993) 8: p. 565. [146] Ricard C., Vial J.C., Douady J., & van derSanden B. In vivo imaging of elastic fibers using sulforhodamine b. J. Biomed. Opt. (2007) 12: p. 064017. [147] Gordon R., Goel N.S., Steinberg M.S., & Wiseman L.L. A rheological mechanism sufficient to explain the kinetics of cell sorting. J Theor Biol. (1972) 37: p. 43–73. [148] Foty R.A. &SM. Cadherin-mediated cell-cell adhesion and tissue segregation in relation to malignancy. Int. J. Dev. Biol. (2004) 48: p. 397–409. [149] Marga F., Neagu A., Kosztin I., & Forgacs G. Developmental biology and tissue engineering. Birth Defects Res. C (2007) 81: p. 320–328. [150] Davis G.S., Phillips H.M., & Steinberg M.S. Germ-layer surface tensions and "tissue affinities" in rana pipiens gastrulae:quantitative measurements. Dev. Biol. (1997) 192: p. 630–644. [151] Winters B.S., Shepard S.R., & Foty R.A. Biophysical measurement of brain tumor cohesion. Int. J. Cancer (114) 2005: p. 371–379. [152] Hegedüs B., Marga F., Jakab K., Sharpe-Timms K.L., & Forgacs G. The interplay of cell-cell and cell-matrix interactions in the invasive properties of brain tumors. Biophys. J. (2006) 91: p. 2708–2716. [153] Jakab K., Neagu A., Mironov V., Markwald R.R. & Forgacs, G. Engineering biological structures of prescribed shape using self-assembling multicellular systems. Proc. Natl. Acad. Sci. USA (2004) 101: pp. 2864-2869. [154] Jakab K., Norotte C., Damon B., Marga F., Neagu A., Besch-Williford C.L., Kachurin A., Church K.H., Park H., Mironov V., Markwald R., Vunjak-Novakovic G.& Forgacs, G. Tissue engineering by self-assembly of cells printed into topologically defined structures. Tissue Engineering (2008) 14: pp. 413-421. [155] Ryan P.L., Foty R.A., Kohn J. & Steinberg M.S. Tissue spreading on implantable substrates is a competitive outcome of cellcell vs. cell-substratum adhesivity. Proc. Natl. Acad. Sci. U.S.A. (2001) 98: p. 4323–4327. [156] Mgharbel A., Delanoe-Ayari H., & Rieu J-P. Measuring accurately liquid and tissue surface tension with a compression plate tensiometer. HFSP J. (2009) 3(3): pp. 213-221. [157] Norotte C., Marga F., Neagu A., Kosztin I., & Forgacs G. Experimental evaluation of apparent tissue surface tension based on the exact solution of the laplace equation. EPL (2008) 81: p. 46003.

Physique des agrégats cellulaires embryonnaires
125
[158] du Noüy P. L. An interfacial tensiometer for universal use. J. Gen. Physiol. (1925) 7: p. 625. [159] Schotz E.M. Dynamics and mechanics of zebrafish embryonic tissues. Der Fakultat fur Physik der Technischen Universitat Dresden eingereichte. 2007. [160] Norotte C. From developmental biology to tissue-engineering:printing blood vessels. the Faculty of the Graduate School at the University of Missouri-Columbia. 2009. [161] Frenkel J. Viscous flow of crystalline bodies. J. Phys. (1945) 9: p. 385–391. [162] Pokluda O., Bellehumeur C.T., & Machopoulos J. Modification of frenkel’s model for sintering. AIChE J. (1997) 43: p. 3253–3256. [163] Shaler A.J. Seminar on the kinetics of sintering. Metals Trans. (1949) 185: p. 796–813. [164] Delphine Icard – Arcizet. Modifications mécaniques et biologiques induites dans des cellules en culture par application locale d’une force contrôlée. Université Paris 7. 2007. [165] de Brabander M.J., van de Veire R.M., Aerts F.E., Borgers M. & Janssen P.A. The effects of methyl [5-(2-thienylcarbonyl)-1h-benzimidazole-2yl ] carbamate (r 17934; nsc 238159), a new synthetic antitumoral drug interfering with microtubules, on ammalian cells cultured in vitro. Cancer Res. (1976) 36: p. 905–916. [166] Samson F, Donoso JA, Heller-Bettinger I, Watson D, Himes RH. Nocodazole action on tubulin assembly, axonal ultrastructure and fast axoplasmic transport. J Pharmacol Exp Ther. (1979) 208(3): p. 411:417. [167] Bershadsky A., Chausovsky A., Becker E., Lyubimova A. ,& Geiger B. Cytoskeleton, contractility and adhesion: control of morphogenetic behavior. Curr. Biol. (1996) 6: p. 1279. [168] Liu B.P., Chrzanowska-Wodnicka M., & Burridge K. Microtubule depolymerization induces stress fibers, focal adhesions, and dna synthesis via the gtp-binding protein rho. Cell Adhes. Commun (1998) 5: p. 249. [169] Danowski B.A. Fibroblast contractility and actin organization are stimulated by microtubule inhibitors. J. Cell. Sci. (1989) 93: pp. 255-266. [170] Brevier J., Vallade M., & Riveline D. Force-extension relationship of cell-cell contacts. PRL (2007) 98: p. 268101. [171] Steinberg M.S. Does differential adhesion govern self-assembly processes in histogenesis? equilibrium configurations and the emergence of a hierarchy among populations of embryonic cells. J. Exp. Zool. (1970) 173: p. 395–434. [172] Yonemura S., Itoh M., Nagafuchi A. , & Tsukita S. Cell-to-cell adherens junction formation and actin filament organization: similarities and differences between non-polarized fibroblasts and polarized epithelial cells. J. Cell Sci. (1995) 108: p. 127–142. [173] Bischofs I.B., Klein F., Lehnert D., Bastmeyer M., Schwarz U.S. Filamentous network mechanics and active contractility determine cell and tissue shape. Biophys J. (2008) 95(7): pp. 3488-3496. [174] Jos Käfer. Des cellules aux tissus : modélisation physique du comportement collectif des cellules embryonnaires. Université Grenoble I - Joseph Fourier. 2008. [175] Pacquelet A. RP. Regulatory mechanisms required for de-cadherin function in cell migration and other types of adhesion. J. Cell Biol. (2005) 170: pp. 803-812. [176] Malik W.A., Prasad S.C., Rajagopalk R., Preziosi L. On the modeling of the viscoelastic response of embryonic tissues. Mathematics and Mechanics of Solids (2008) 13(1): pp. 81-91. [177] Valberg P. A., & Albertini, D.F. Cytoplasmic motions, rheology and structure probed by a novel magnetic particle method. J. Cell Biol. (1985) 101: p. 130 –140. [178] Sakanishi TY. Effects of viscoelasticity of cytoplasm on the complex viscosity of red blood cell suspensions. Biorheology (1988) 25: p. 123–128. [179] Stamenović D. Rheological behavior of mammalian cells. Cell Mol Life Sci. (2008) 65(22): pp. 3592-3605. [180] Balland M., Desprat N., Icard D., Féréol S., Asnacios A., Browaeys J., Hénon S., Gallet F. Power laws in microrheology experiments on living cells: comparative analysis and modeling. Phys Rev E Stat Nonlin Soft Matter Phys. (2006) 74(2 Pt 1): p. 021911. [181] Ragsdale G. K., Phelps J., & Luby-Phelps K. Viscoelastic response of fibroblasts to tension transmitted through adherens junctions. Biophys. J. (1997) 73: p. 2798 –2808.

Physique des agrégats cellulaires embryonnaires
126
[182] Weaire D., & Rivier, N. Soap, cells and statistics—random patterns in two dimensions. Contemp Phys (1984) 25: p. 59–99. [183] Graner F. Can surface-adhesion drive cell-rearrangement. 1. biological cellsorting. J. Theor. Biol. (1993) 164: p. 455–476. [184] Marmottant P., Raufaste C., Graner F. Discrete rearranging disordered patterns, part ii: 2d plasticity, elasticity and flow of a foam. Eur Phys J E (2008) 25: p. 371–384. [185] Marmottant P., Mgharbel A., Käfera J., Audren B., Rieu J-P.,Viala J-C.,van der Sanden B., Marée A.F.M.,Graner F.,& Ayari H.D. The role of fluctuations and stress on the effective viscosity of cell aggregates. PNAS, Published online, (2009) doi: p. 10.1073/pnas.0902085106 . [186] Yang J. & Brodland G.W. Estimating interfacial tension from the shape histories of cells in compressed aggregates: a computational study. Ann Biomed Eng (2009) 37: p. 1019–1027. [187] Preziosi L. , Ambrosi D.,Verdier C. An elasto-visco-plastic model of cell aggregates. J. Theor. Biol. (2009) doi: p. 10.1016/j.jtbi.2009.08.023. [188] Rieu J-P., Upadhyaya A., Glazier J.A., Ouchi N.B., Sawada Y. Diffusion and deformations of single hydra cells in cellular aggregates. Biophys. J. (2000) 79: p. 1903–1914. [189] Mombach J.C. GJ. Single cell motion in aggregates of embryonic cells. Phys. Rev. Lett. (1996) 76: p. 3032–3035. [190] Yamada S., Wirtz D., Kuo S.C. Mechanics of living cells measured by laser tracking microrheology. Biophys. J. (2000) 78: p. 1736–1747. [191] Stamenovic D., & Coughlin M.F. The role of prestress and architecture of the cytoskeleton and deformability of cytoskeletal filaments in mechanics of adherent cells: a quantitative analysis. J. theor. Biol. (1999) 201: pp. 63-74.