modulation de la structure, localisation et fonction des récepteurs pour l’inositol...
TRANSCRIPT

Modulation de la structure, localisation et fonction des récepteurs pour
l’inositol trisphosphate: le rôle des interactions protéiques.
Jan B. Parys
K.U.LeuvenOrsay, 7 février 2003

(Clapham, 1995)
Intracellular Ca2+ homeostasis

Ca2+
Lumen of ER
Cytoplasm
Structure of the IP3R – the simplified view
• 4 subunits = 1 functional IP3R (homo- or heterotetrameric)
C
Lumen of ER
N
• each subunit can be divided in three parts:1. an IP3-binding domain (about 600 aa)2. a coupling domain (about 1500 aa)3. a channel domain (about 600 aa)


• Interactions of the monomers to form a tetramer
• Interactions of monomer/tetramer with associated proteins
- regulatory proteins CaM
- activatory proteins CaBP1
- kinases / phosphatases PKA, PP1
- anchor proteins homer
- determinants of the IP3R localization talin, ankyrin, 4.1N, …
- proteins regulated by the IP3R trp-channels
Protein interactions to consider:
ALL THESE INTERACTIONS MAY BE MODULATED,DEPENDING ON THE PHYSIOLOGICAL SITUATION

1) Inter- and intramolecular interactions with the N-terminal region of the IP3R(aa 1-225)
2) Dynamics concerning the intracellular localization of the IP3R

1) Inter- and intramolecular interactions with the N-terminal region of the IP3R(aa 1-225)

Why the N-terminal 225 amino acids of the IP3R1?
• Aa 1-225 act as a suppressor of IP3 binding
activity (Yoshikawa et al., 1999).
• Homer proposed to couple to aa 48-55
(Tu et al., 1998).
• CaM interacts with aa 1-159 (Adkins
et al., 2000).

What isthe
relationbetween
CaMandIP3R
?

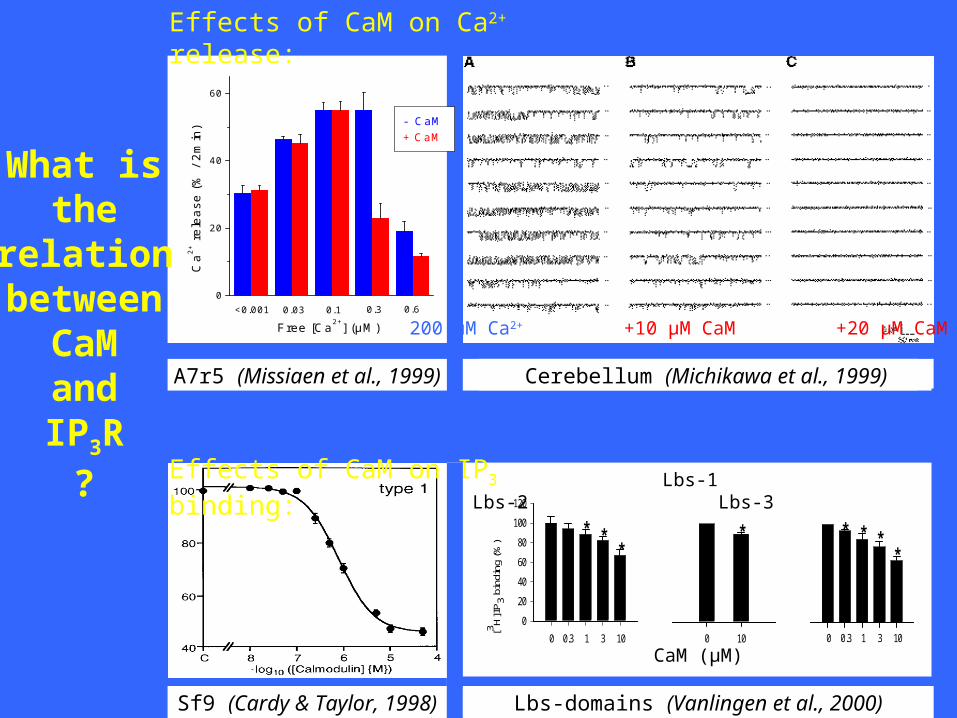
A7r5 (Missiaen et al., 1999)
0
20
40
60
- CaM + CaM
Free [Ca2+] (µM) 0.60.30.10.03<0.001
Ca2+
rel
ease
(%
/ 2m
in)
Cerebellum (Michikawa et al., 1999)
200 μM Ca2+ +10 μM CaM +20 μM CaM
Sf9 (Cardy & Taylor, 1998)
[3 H]I
P3 b
indi
ng (
%)
0
20
40
60
80
100
120
* * **
**
*
0 10
*
0 0.3 1 3 10 0 0.3 1 3 10
Effects of CaM on Ca2+ release:
Effects of CaM on IP3 binding:
CaM (μM)
Lbs-domains (Vanlingen et al., 2000)
Lbs-1 Lbs-2 Lbs-3
What isthe
relationbetween
CaMandIP3R
?
A7r5 (Missiaen et al., 1999)
0
20
40
60
- CaM + CaM
Free [Ca2+] (µM) 0.60.30.10.03<0.001
Ca2+
rel
ease
(%
/ 2m
in)
Cerebellum (Michikawa et al., 1999)
200 μM Ca2+ +10 μM CaM +20 μM CaM
Sf9 (Cardy & Taylor, 1998)
[3 H]I
P3 b
indi
ng (
%)
0
20
40
60
80
100
120
* * **
**
*
0 10
*
0 0.3 1 3 10 0 0.3 1 3 10
Effects of CaM on Ca2+ release:
Effects of CaM on IP3 binding:
CaM (μM)
Lbs-domains (Vanlingen et al., 2000)
Lbs-1 Lbs-2 Lbs-3
What isthe
relationbetween
CaMandIP3R
?

A7r5 (Missiaen et al., 1999)
0
20
40
60
- CaM + CaM
Free [Ca2+] (µM) 0.60.30.10.03<0.001
Ca2+
rel
ease
(%
/ 2m
in)
Cerebellum (Michikawa et al., 1999)
200 μM Ca2+ +10 μM CaM +20 μM CaM
Sf9 (Cardy & Taylor, 1998)
[3 H]I
P3 b
indi
ng (
%)
0
20
40
60
80
100
120
* * **
**
*
0 10
*
0 0.3 1 3 10 0 0.3 1 3 10
CaM on Ca2+ release: INHIBITORY, Ca2+-DEPENDENT
CaM on IP3 binding: INHIBITORY, Ca2+-INDEPENDENT
CaM (μM)
Lbs-domains (Vanlingen et al., 2000)
Lbs-1 Lbs-2 Lbs-3
What isthe
relationbetween
CaMandIP3R
?

control ca cam ca cam cam1234 ca cam12340
20
40
60
80
100
Ca
2+ C
aM12
34
CaM
1234
Ca
2+ C
aM
Apo
CaM
Ca
2+
Con
trol
[3H
]IP3 b
indi
ng
(%
)
0.1 1 100
20
40
60
80
100
[3H
]IP3 b
indi
ng
(%
)
Lbs-1
Lbs-1 1-225
1
581226
581+HIS
+HIS
Effects of CaMon IP3 binding
CaM1234 (μM)

control ca cam ca cam cam1234 ca cam12340
20
40
60
80
100
Ca
2+ C
aM12
34
CaM
1234
Ca
2+ C
aM
Apo
CaM
Ca
2+
Con
trol
[3H
]IP3 b
indi
ng
(%
)
0.1 1 100
20
40
60
80
100
[3H
]IP3 b
indi
ng
(%
)
Lbs-1
Lbs-1 1-225
1
581226
581+HIS
+HIS
Effects of CaMon IP3 binding
CaM1234 (μM)
IC50 ~ 2 μM

Cyt1 Cyt2
Lbs-1
Lbs-1 1-225
1
581226
581
1 159
154 309
+HIS
+HIS
+GST
Ca2+
EGTA
Localisation of the N-terminal CaM-binding site
CaM1234

Cyt1 Cyt2
Lbs-1
Lbs-1 1-225
1
581226
581
1 159
154
+HIS
+HIS
+GST
A
B
C
E
D
F
1-5-10 1-5-10
1-5-8-14 70% IQ 76% IQ
53% IQ
A B C D E F-0.2
0.0
0.2
0.4
0.6
0.8
1.0
200 µM free Ca 2+
1 mM EGTA
309
• Band-shift experiments on non-denaturing gels
• Interaction with dansyl-CaM
Localisation of the N-terminal CaM-binding site

Cyt1 Cyt2
Lbs-1
Lbs-1 1-225
1
581226
581
1 159
154
+HIS
+HIS
+GST
A
B
C
E
D
F
1-5-10 1-5-10
1-5-8-14 70% IQ 76% IQ
53% IQ
A B C D E F-0.2
0.0
0.2
0.4
0.6
0.8
1.0
200 µM Ca2+
1mM EGTA
309
Inte
nsi
ty lo
ss (
1-B
/Bo)
Kd 0.1 μM Kd 1 μM
Localisation of the N-terminal CaM-binding site

CaM or CaBP1 ?
Inhibitory
Activatory ?(Yang et al., 2002)

CaBP1
GSTGST
1-604GST
1-225GST
226-604
Binding of (activatory?) CaBP1to the same site?
A
B
C
E
D
F
1CaM CaM
159

CaBP1
GSTGST
1-604GST
1-225GST
226-604
Binding of (activatory?) CaBP1to the same site
A
B
C
E
D
F
1CaM CaM
159
CaBP1
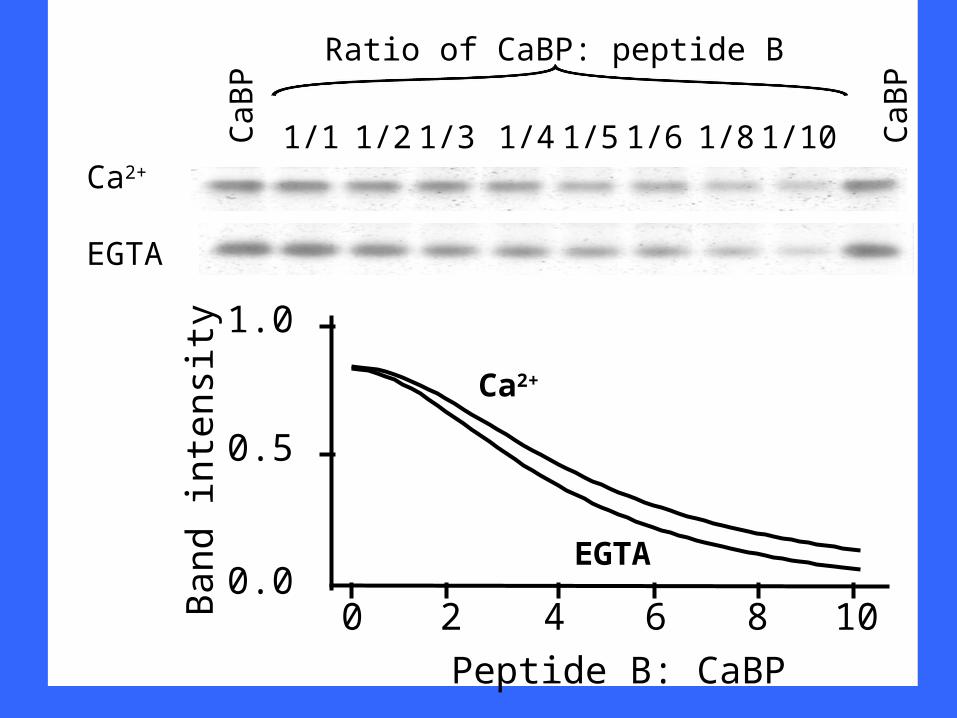
CaB
P
1/1 1/2 1/4 1/5 1/6 1/81/3 CaB
P
Ratio of CaBP: peptide B
0 2 4 6 8 100.0
0.5
1.0
Band
in
ten
sity
Peptide B: CaBP
Ca2+
EGTA
1/10
Ca2+
EGTA

GS
T
GS
T-H
omer
1a
The 1225 suppressor domain interacts withHomer1a
1225
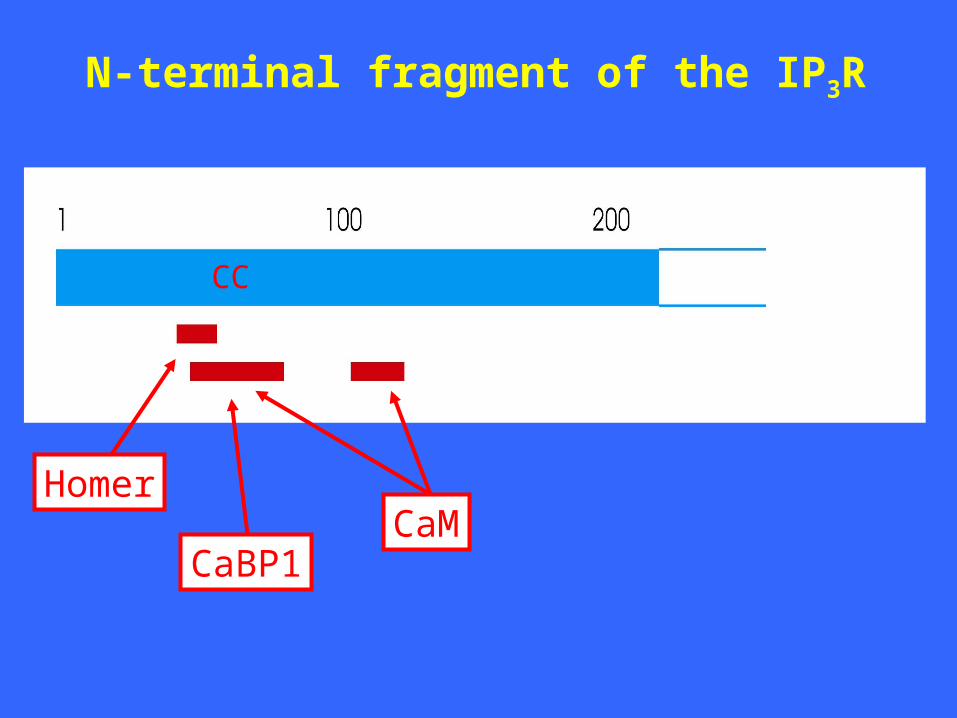
N-terminal fragment of the IP3R
CaMCaBP1
Homer
CC
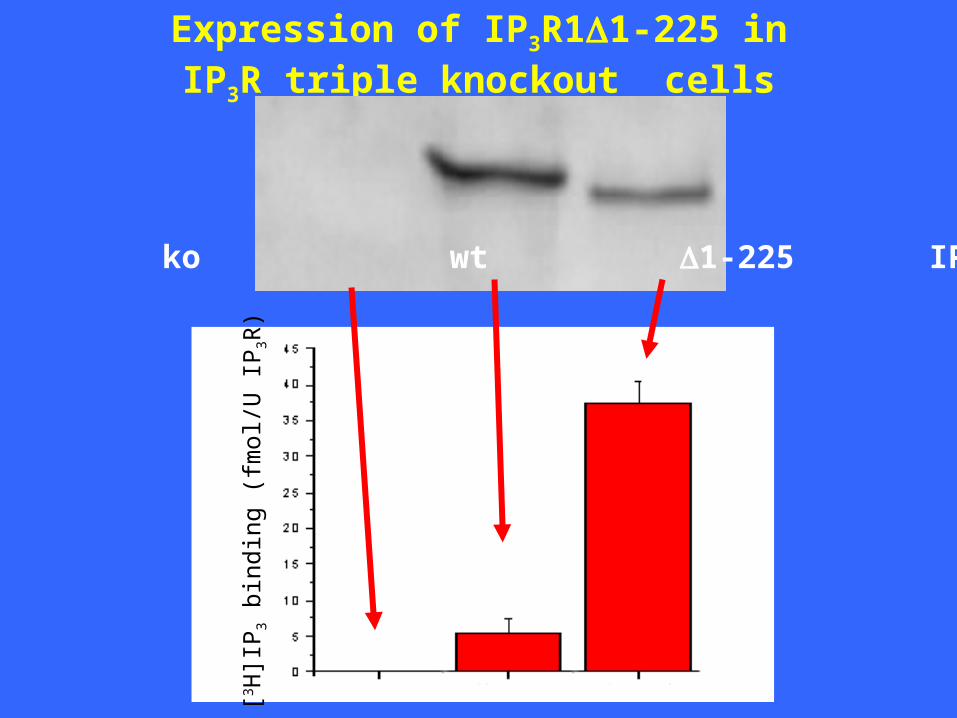
Expression of IP3R11-225 inIP3R triple knockout cells
ko wt 1-225 IP3R1[3 H
]IP
3 bi
ndin
g (f
mol
/U I
P3R
)

IP3R11-225 is not functionally active
ko wt 1-225 IP3R1
0 200 400 600 800 1000 1200 1400
0
200
400
600
800
1000
1200
A2
31
87
1 µ
M I
P3
Dig
ito
nin
[Ca
2+]
(nM
)
Time (sec)
0 200 400 600 800 1000 1200 14000
200
400
600
800
25
µM
IP
3
A2
31
87
10
µM
IP
3
10
µM
IP
3
1 µ
M I
P3
1 µ
M I
P3
Dig
ito
nin
[Ca
2+]
(nM
)
Time (sec)
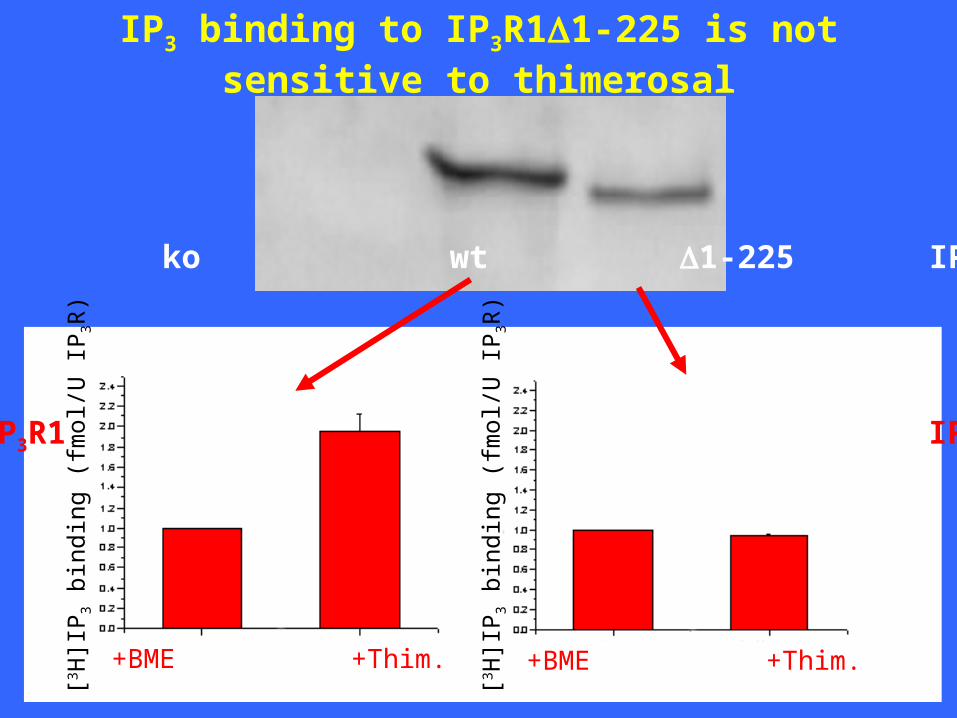
IP3 binding to IP3R11-225 is not sensitive to thimerosal
ko wt 1-225 IP3R1
[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
+BME +Thim. +BME +Thim.
wt IP3R1 IP3R1 1-225
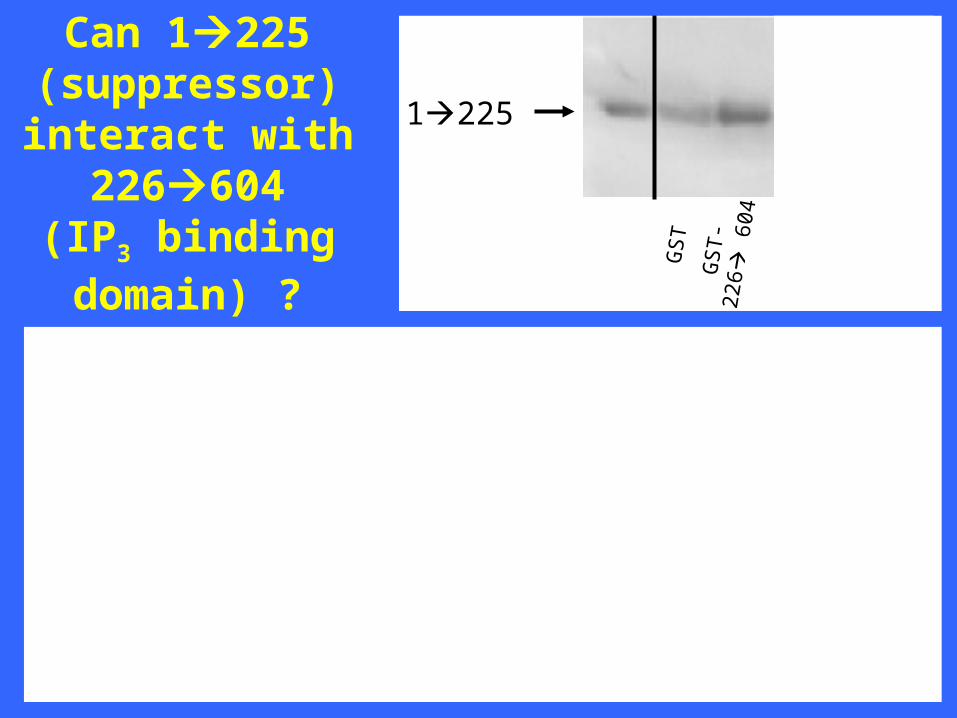
1225
GS
T
GS
T-
226
604
GS
T
GS
T-
226
604
Can 1225(suppressor)interact with
226604(IP3 binding domain) ?

[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
+BME +Thim +BME +Thim
1225
GS
T
GS
T-
226
604
GS
T
GS
T-
226
604
wt IP3R1 IP3R1 1-225
Can 1225(suppressor)interact with
226604(IP3 binding domain) ?
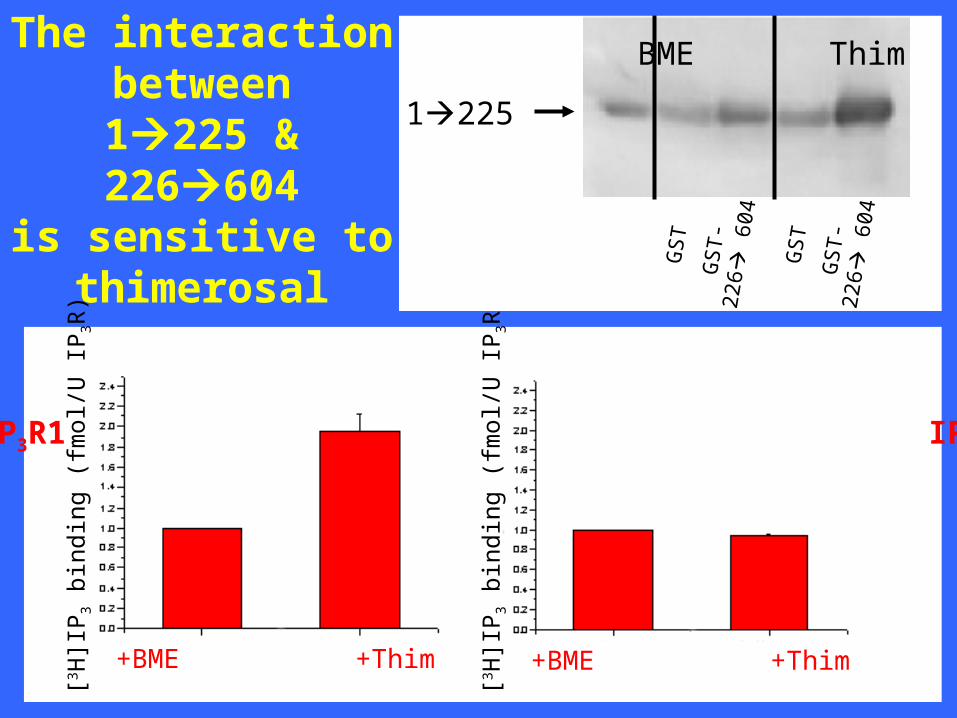
[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
[3 H]I
P3
bind
ing
(fm
ol/U
IP
3R)
+BME +Thim +BME +Thim
1225
BME Thim
GS
T
GS
T-
226
604
GS
T
GS
T-
226
604
wt IP3R1 IP3R1 1-225
The interactionbetween1225 &226604
is sensitive tothimerosal
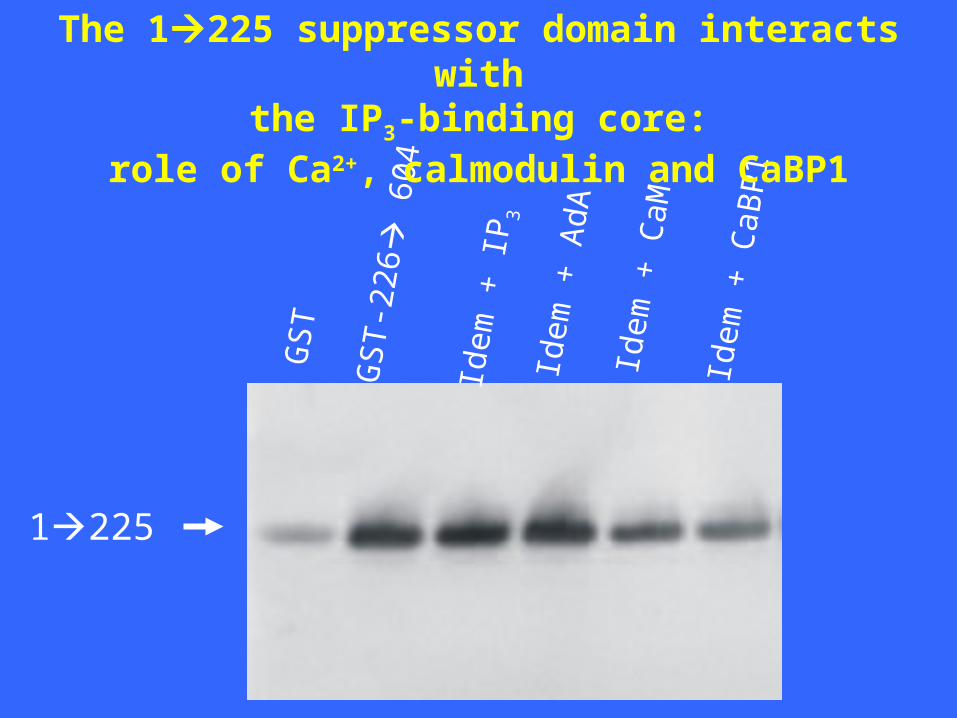
1225
GS
T
Idem
+ IP
3
GS
T-2
26
604
Idem
+ A
dA
Idem
+ C
aM
Idem
+ C
aBP
1
The 1225 suppressor domain interacts withthe IP3-binding core:
role of Ca2+, calmodulin and CaBP1

1225226604
• CaM• CaBP1• Homer• Ca2+
• Thim.
• IP3
• Ca2+
?
N
C
Interactions between N- and C-termini of IP3R1 ?

N
C
Flag3 c-myc
GST 1225
GST 226604
GST 1604
GST
= affinity matrix

N
C
GST 1225
GST 226604
GST 1604
GST
Flag3 c-myc
Flag1 2170-2749 ~60 kDa
Flag2 2253-2749 ~54 kDa
Flag3 2170-2427 ~30 kDa
Flag3 2170-2407 ~28 kDa
Flag4 2407-2749 ~41 kDa
Flag4 2560-2749 ~26 kDa
TM1-6
TM1-4
TM(5)-6
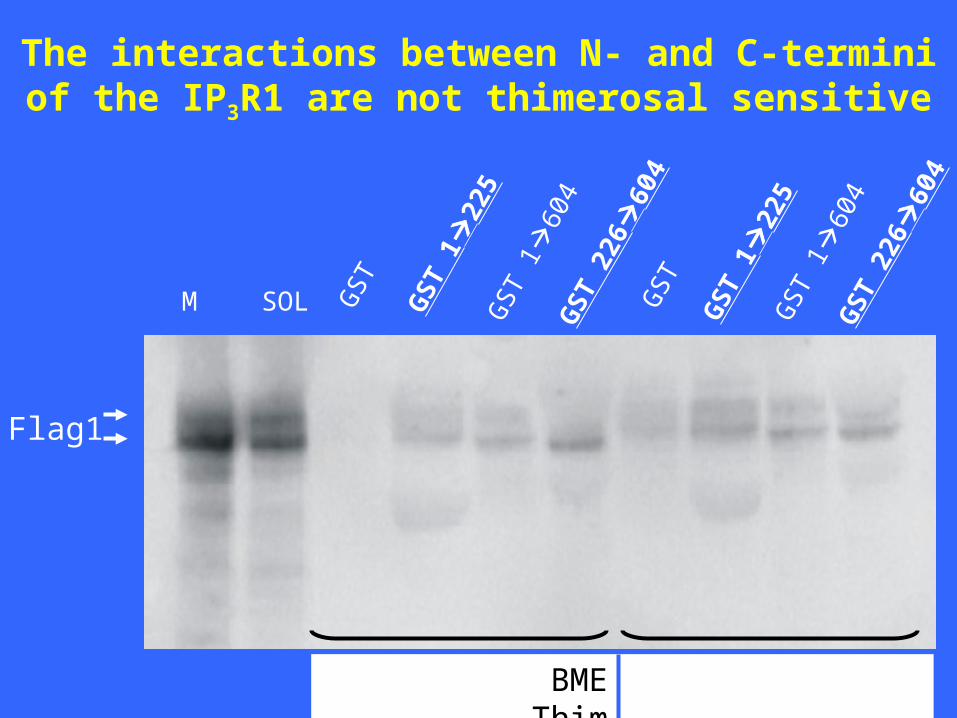
M SOL GS
T
GS
T 1
225
GS
T
GS
T 1
225
GS
T 1
604
GS
T 22
660
4
GS
T 1
604
GS
T 22
660
4
Flag1
BME Thim
The interactions between N- and C-termini of the IP3R1 are not thimerosal sensitive

M GS
T
GS
T 1
22
5
GS
T
GS
T 1
22
5
GS
T 1
60
4G
ST
226
60
4
GS
T 1
60
4G
ST
226
60
4
SO
L
Flag1
The interactions between N- and C-termini of the IP3R1 are Ca2+ sensitive
EDTA Ca2+
1mM 25 M
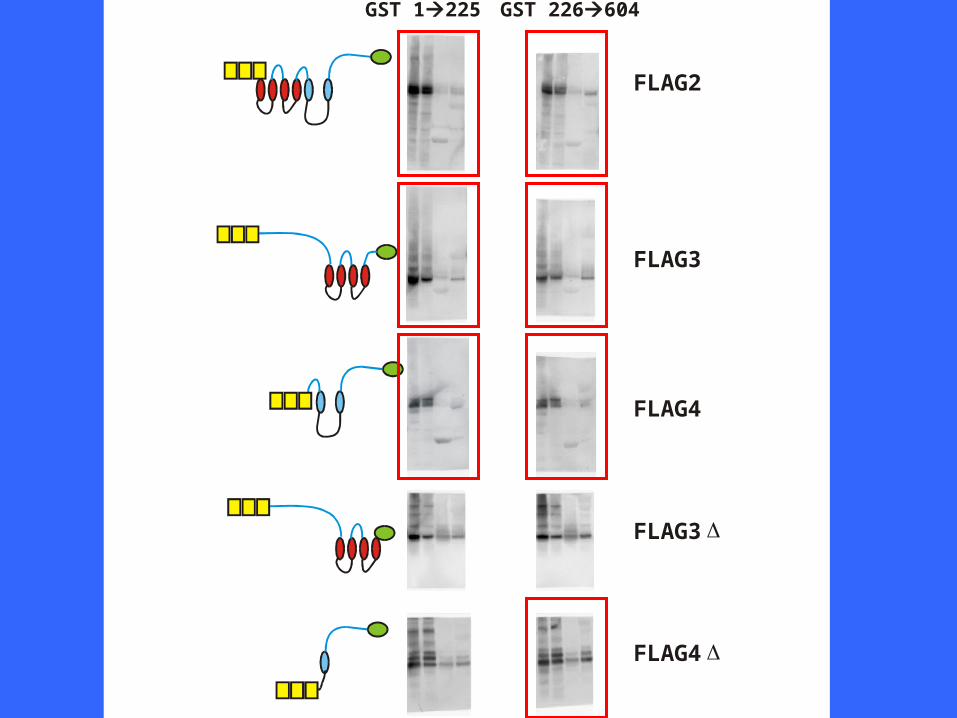
GST 1225 GST 226604
FLAG2
FLAG3
FLAG4
FLAG3
FLAG4

Multiple interactions between N- and C-termini of IP3R11225226604
• CaM• CaBP1• Homer• Ca2+
• Thim.
• IP3
• Ca2+N
C
• Ca2+

CaM/CaBP1

CC
CaM/CaBP1
CC
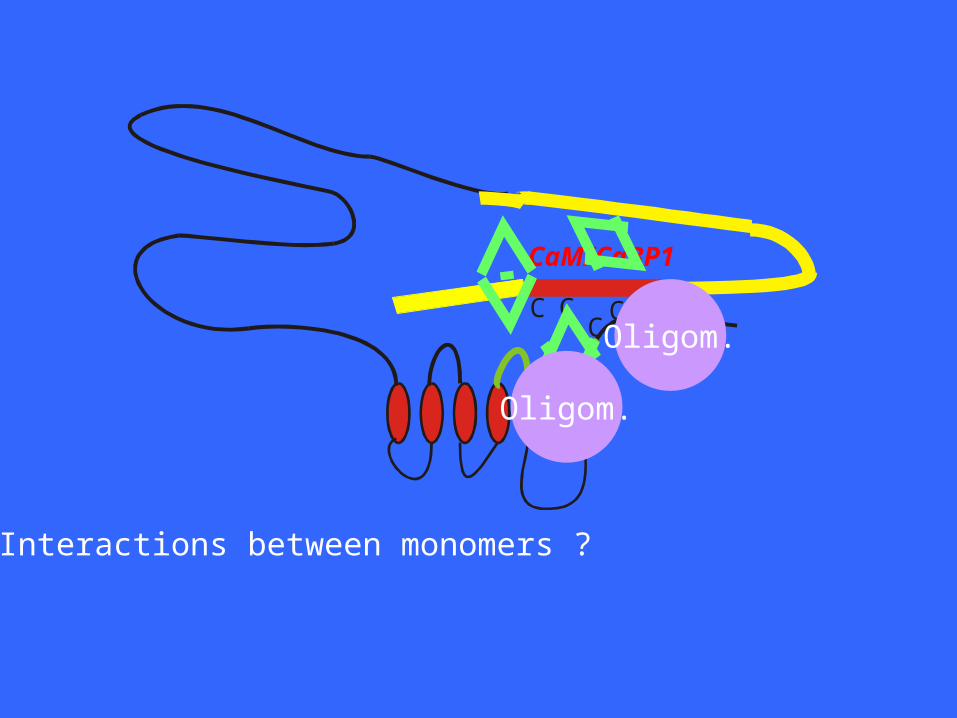
CC
CaM/CaBP1
CC
• Interactions between monomers ?
Oligom.
Oligom.

CC
CaM/CaBP1
CC
IRBITHomer
Ankyrin
4.1NPP1
• Interactions between monomers ?• Interactions with other proteins ?
Regulation of localization ?Regulation of phosphorylation ?

1) Inter- and intramolecular interactions with the N-terminal region of the IP3R(aa 1-225)
2) Dynamics concerning the intracellular localization of the IP3R

2) Dynamics concerning the intracellular localization of the IP3R

Localization of IP3R1 and IP3R3in A7r5 smooth-muscle cells
IP3R1 IP3R3
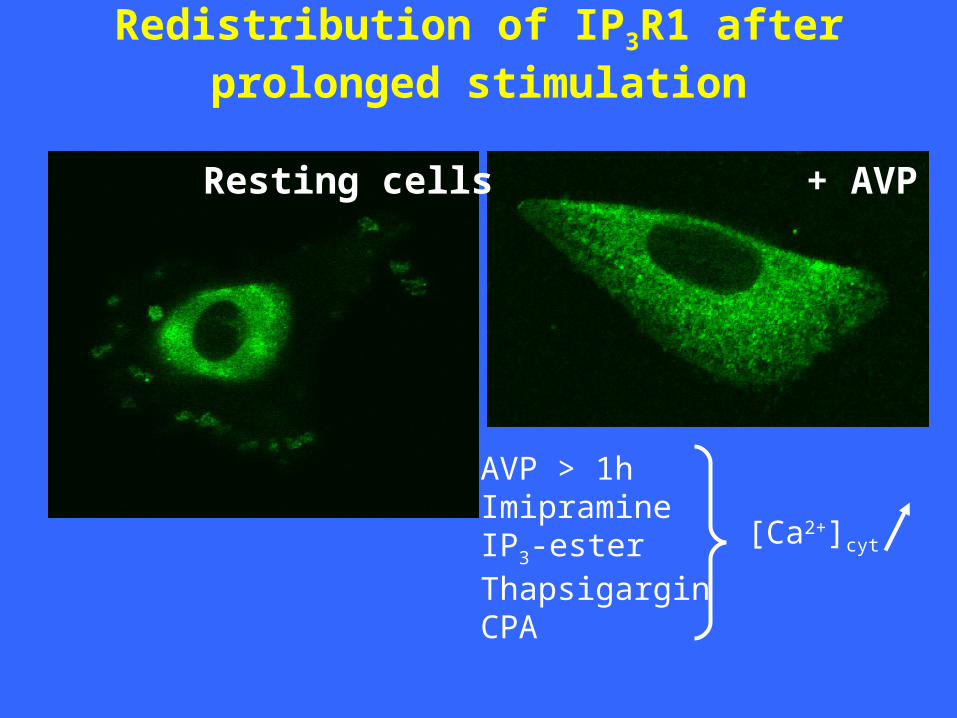
Redistribution of IP3R1 after prolonged stimulation
Resting cells + AVP
AVP > 1hImipramineIP3-ester ThapsigarginCPA
[Ca2+]cyt

Redistribution during AVP treatment:time dependence
CytoplasmicIP3R1
localization
PerinuclearIP3R1
localization
AVPtreatment
Perinuclear localizationafter AVP wash-out
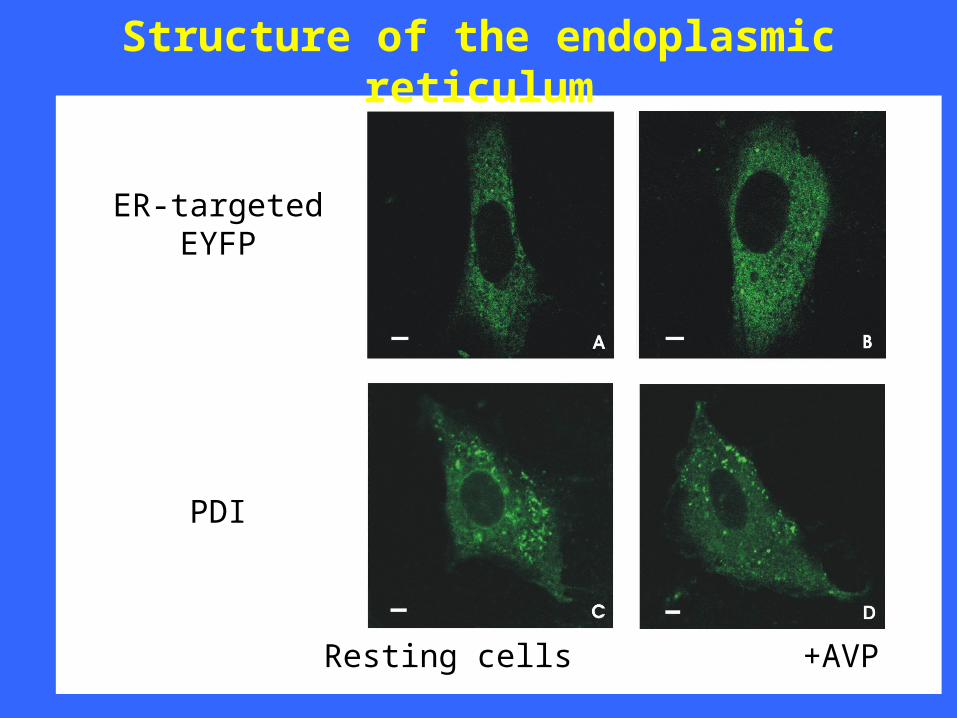
Structure of the endoplasmic reticulum
Resting cells +AVP
ER-targetedEYFP
PDI

SERCA localization and redistribution
Resting cells +AVP

0
20
40
60
80
100
Role of PKC
perinuclear IP3R1 cytoplasmic IP3R1
Percentages of cells with:C
ells
(%
)
Con
trol
Con
trol
AV
P
AV
P
AV
P+S
taur
.
AV
P+S
taur
.
TG+S
taur
.
TG+S
taur
.
AV
P+B
IM
AV
P+B
IM
OA
G
OA
G

Role of cytoskeleton
perinuclear IP3R1
Percentages of cells with:
Ce
lls (
%)
Con
trol
AV
P
AV
P+N
ocod
.
AV
P+T
axol
Noc
od.
Taxo
l
0
10
20
30
40
50
60
70
80
Resting cells
+AVP
Microtubular network
or +OAG

Role of vesicle trafficking
• Brefeldin A (2 μg/ml, 2h) complete IP3R1 redistribution
(29 ± 2 % cells with perinuclear localization)
• Cooling (15°C, 2h) inhibition of AVP-induced IP3R1 redistribution
(76 ± 3 % cells with perinuclear localization)

Conclusions (part 1)
The first 225 amino acids of IP3R1 contain important
regulatory sites, including for Ca2+-binding proteins as calmodulin and CaBP1, for homer as well as potentially for thimerosal.
Interactions occur between the N-terminal suppressor domain, the IP3-binding domain and the channel domain.
These interactions are under control of Ca2+, calmodulin/CaBP1 and thimerosal.
The role of additional interactions between monomers or with other proteins and the role of phosphorylation in this process will be investigated.

Conclusions (part 2) Various IP3R isoforms can have a different intra-
cellular distribution.
The localization of the IP3Rs is dynamic and change
with the physiological status of the cell.
This redistribution is dependent on PKC activation and on the microtubular network and is probably mediated by vesicle trafficking.
The results indicate a dynamic regulation of the structure and localization of the IP3R, which we
believe to be functionally relevant.Future work will focus on the understanding of the physiological importance of the various interactions.

Geert CALLEWAERT - Humbert DE SMEDT - Ludwig MISSIAEN - Jan B. PARYS
IP3-team (Leuven, Belgium)
Zerihun ASSEFA
Geert BULTYNCK
Patrick DE SMET
Rafael A. FISSORE
Nael NADIF KASRI
Joelle NSIMIRE CHABWINE
Ilse SIENAERT
Karolina SZLUFCIK
Kristel VAN ACKER
Sara VANLINGEN
Marijke VAN MOORHEM
Esther VENMANS
Leen VERBERT
Elke VERMASSEN

Geert CALLEWAERT - Humbert DE SMEDT - Ludwig MISSIAEN - Jan B. PARYS
IP3-team (Leuven, Belgium)
Zerihun ASSEFA
Geert BULTYNCK
Patrick DE SMET
Rafael A. FISSORE
Nael NADIF KASRI
Joelle NSIMIRE CHABWINE
Ilse SIENAERT
Karolina SZLUFCIK
Kristel VAN ACKER
Sara VANLINGEN
Marijke VAN MOORHEM
Esther VENMANS
Leen VERBERT
Elke VERMASSEN

Geert CALLEWAERT - Humbert DE SMEDT - Ludwig MISSIAEN - Jan B. PARYS
IP3-team (Leuven, Belgium)
Zerihun ASSEFA
Geert BULTYNCK
Patrick DE SMET
Rafael A. FISSORE
Nael NADIF KASRI
Joelle NSIMIRE CHABWINE
Ilse SIENAERT
Karolina SZLUFCIK
Kristel VAN ACKER
Sara VANLINGEN
Marijke VAN MOORHEM
Esther VENMANS
Leen VERBERT
Elke VERMASSEN


IP3R1IP3R2IP3R3
RyR1RyR2RyR3
Intraluminal proteins
Calreticulin
Calsequestrin
Chromogranins A and B
Annexin VI
ERmembrane proteins
Triadin
Junctin
Kinase anchor proteins
Plasma membrane proteins
Trp’s DHPR
G-proteinsCytosolic proteins
CaM CaBPs
FKBP’s
SorcinS100
IRAG
Cytoskeletal proteins
Homer
Ankyrin
TalinVinculin
Alpha-actin
Myosin II

FKBP12-binding
siteIP3
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
FKBP12
NH2
COOH
SI
SIIPORE
SIII
PP
ATP
ATP
IP3R1 VCTEGKNVYTEIKCNSLLPLDDIVRVVTHEDCIPEV
IP3R2 ACTEGKNVYTEIKCNSLLPLDDIVRVVTHDDCIPEV
IP3R3 ACAEGKNVYTEIKCTSLVPLEDVVSVVTHEDCITEV
RyR1 QAGKGEALRIRAILRSLVPlDDLVGIISLPLQIPTG RyR2 HAGKGEAIRIRSILRSLIPLGDLVGVISIAFQMPTI
RyR3 QTGKGEAIRIRSILRSLVPTEDLVGIISIPLKLPSL
Cytosol
Lumen of the store

Effects on RyR
(Rodney et al., 2001)
3614 3643
ApoCaM: Ca2+/CaM:
ACTIVATOR OF RYR1 INHIBITOR OF RYR1
Ca2+

IP3
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
CaM
CaM
NH2
COOH
SI
SII
CaM
PORE
SIII
Cytosol
Lumen of the store
PP
ATP
ATP
CaM-binding
sitesHigh-affinityCa2+-dependentTypes 1 and 2
Low-affinityCa2+-dependentNot in neuronalIP3R1 (SII+)

IP3R
A7r5 (Missiaen et al., 1999)
0
20
40
60
- CaM + CaM
Free [Ca2+] (µM) 0.60.30.10.03<0.001
Ca2+
rel
ease
(%
/ 2m
in)
Cerebellum (Michikawa et al., 1999)
200 μM Ca2+ +10 μM CaM +20 μM CaM
Sf9 (Cardy & Taylor, 1998)
[3 H]I
P3 b
indi
ng (
%)
0
20
40
60
80
100
120
* * **
**
*
0 10
*
0 0.3 1 3 10 0 0.3 1 3 10
Effects of CaM on Ca2+ release:
Effects of CaM on IP3 binding:
CaM (μM)
Lbs-domains (Vanlingen et al., 2000)
Lbs-1 Lbs-2 Lbs-3

Discontinuous (aa 49-81 and 106-128).
Low affinity.
Ca2+ independent.
Involved in inhibition of IP3 binding.
Not involved in inhibition of IP3-induced Ca2+ release.
Other possible functions:Role in conformation of the IP3R ?
Tethering of CaM ?Binding site for CaM-like proteins ? CaBP-1?Protective effect ? Proteolysis? Oxidative stress?
N-terminal CaM-binding site of the IP3R