ministere de l’enseignement superieur et de la...
TRANSCRIPT

N° Ordre........../FSI/UMBB/2015
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE
SCIENTIFIQUE
UNIVERSITE M’HAMED BOUGARA-BOUMERDES
Faculté des sciences de l’ingénieur
Thèse de Doctorat
Présentée par :
Mme
TALANTIKITE - KELLIL SOUAD
En vue de l’obtention du diplôme de DOCTORATen :
Filière : TECHNOLOGIE ALIMENTAIRE.
Option : GENIE ALIMENTAIRE ET BIOTECHNOLOGIE.
TITRE :
Purification et caractérisation d’une enzyme coagulante
d’origine microbienne pour application en fromagerie
Devant le jury composé de :
Mr BENAMARA Salem Pr. UMBB Président
Mme FAZOUANE Fethia Pr. UMBB Directrice de thèse
Mme MECHAKRA Aicha Pr. U. Constantine Examinatrice
Mr BELLAL Mohamed Mouloud Pr. ENSA El Harrach Examinateur
MrAMIALI Malek MCA ENSA El Harrach Examinateur
Mr NOUANI Abdelouahab MCA. UMBB Invité
Année Universitaire 2014/2015

A la mémoire de mes parents
A mon époux
A mes enfants Mourad, Sid-Ahmed et Maya.
A mes belles filles,
A mes petits enfants Anis, Hella et Lina
A mes frères et sœurs.

Remerciements
J’adresse mes vifs remerciements à Madame Fazouane-NaimiFethia
professeur à l’université de Boumerdes qui a dirigé ce travail, je lui
adresse toute ma reconnaissance pour sa disponibilité, qu’elle trouve
ici l’expression de ma profonde gratitude.
Je tiens à remercier les membres du Jury particulièrement Monsieur
Benamara Salem professeur à l’UMBB qui me fait l’honneur de
présider ce jury, Madame Mechakra-Maza Aicha professeur à
l’université Mentouri de Constantine d’avoir accepté d’évaluer ce
travail. Mes remerciements s’adressent aussi à monsieurBellal
Mohamed Mouloud professeur à l’ENSA d’El Harrach ainsi que
monsieurAmiali Malek maitre de conférence (A)qui me font l’honneur
de juger ce travail.
Mes remerciements s’adressent également à monsieur
NouaniAbdelouahab maitre de conférences (A) à l’UMBB pour ses
encouragements et ses conseils durant la réalisation de ce travail.
Je tiens à remercier également Madame Louisa Gillmann Maitre de
conférences à l’I.U.T d’Angers de m’avoir accueillie dans son
laboratoire.
Je remercie chaleureusement Madame Monique Saunier Maitre de
conférences à l’université d’Angers pour son aide tout au long de mes
travaux au sein du laboratoire LASBA.
Ces remerciements s’adressent aussi au personnel de l’I.U.T d’Angers
en particulier Blandine Bregeon,PascalTulik ,LaetitiaAugeraud ainsi
qu’à Laurence Graindorge avec qui j’ai partagé de bons moments, ce
qui a permis de rendre mon séjour plus agréable.
Je voudrais remercier aussi Madame Odile Rolet-Repecaud de l’INRA
de Poligny pour m’avoir fourni gracieusement la poudre de lait
écrémé « lowheat » (substrat standard de Berridge)

Je remercie également Monsieur Arnaud-Dudoit responsable du
service micropolluants du laboratoire d’hydrologie et d’hygiène
d’Angers pour son aide dans l’analyse statistique lors de la réalisation
des plans d’expériences.
Enfin je remercie le personnel du département de technologie
alimentaire de la faculté des sciences de l’ingénieur pour leur soutien
au cours de l’élaboration de ce travail.

Résumés

i
Titre :Purification et caractérisation d’une enzyme coagulante
d’origine microbienne pour application en fromagerie
Résumé
L’objectif de notre travail consiste dans un premier temps à isoler des microorganismes
provenant de la terre prélevée à proximité d’une laiterie fromagerie située à BOUDOUAOU
(Boumerdes –Algérie), le milieu utilisé est le P.C.A, contenant 10% de lait écrémé stérile
auquel on a ajouté 2% de l’actidione.
Les bactéries sont cultivées dans un milieu contenant 5% de son de blé, 0,3% d’extrait de
levure, 0,4% de glucose, 0,2% de caséine et 0,3% d’Na2HPO4. C’est un milieu qui a été
adopté dans des travaux précédents pour la culture de Bacillus.
L’incubation a lieu à 30°c pendant 24h sous agitation 110 rpm. L’extraction de l’enzyme
s’effectue après filtration du milieu sur un tissu pour fromagerie. Le filtrat subit une
centrifugation à 4°c pendant 30 minutes à 4200g. Le surnageant est maintenue à basse T° pour
le test d’activité.
Après avoir testé une soixantaine de souches nous avons sélectionné trois d’entre-elles qui
paraissent différentes (I33M, FK6A et P47M), elles ont été identifié par biologie moléculaire.
Les résultats du BLAST donnent : I33M : Bacillus mojavensis, P47M : Bacillus subtilis,
FK6A: Bacillus amyloliquefaciens.
La souche I33M a été sélectionnée par rapport à son activité coagulante très satisfaisante.
Nous avons réalisé deux plans d’expériences type composite centré, pour le premier la
composition du milieu de production est de : 5% de son, 0,3% d’extrait de levure, 0,3%
d’NA2HPO4, 0,05% de glucose, 1,01% de caséine et à T° 39°c. L’agitation est à 200 rpm…
Nous pouvons conclure que le glucose n’étant pas significatif. Un deuxième plan nous a
donné : 7% de Son, 0 ,094% de caséine, 0,3% d’extrait de levure, 0,3% d’Na2HPO4, la
température est de 39°c et l’agitation est de 150 rpm. L’activité coagulante est de
2,18UP.L’activité protéolytique est de 19,302 µgtyr/ml/min. Lors de la fermentation, la
croissance est suivie par dénombrement des colonies en milieu PCA après des dilutions
successives, puis l’activité coagulante est mesurée.
La production de l’enzyme a lieu en fin de phase exponentielle et augmente avec la phase
stationnaire.
Pour ce qui est des moisissures, la souche sélectionnée a été identifiée comme étant Mucor
circinelloides. Un plan statistique (composite centré) a été employé afin d’optimiser les
conditions de production de la coagulase par le mucor sélectionné. L’utilisation de la
fermentation sur milieu solide contenant : 30gr son de blé, 1gr de caséine, 60ml de sulfate
d'ammonium (0,01%) à 30°c pendant 5 jours ont donné une activité maximale de 2,667
UP/ml. La caractérisation de la protéase a révélé des conditions optimales : température
d’activité coagulante de 55°c, un pH de 5,5 et une concentration de CaCl2 de 0,03 M.
Mots clés : Mucor circinelloides ,Bacillus mojavensis, enzyme coagulante, optimisation.

ii
Title:Purification and characterization of a milk clotting enzyme of
microbial origin for use in cheese
Summary
The objective of our work for the first step is to isolate microorganisms from soil. They were
collected near a cheese dairy, located at Boudouaou (Boumerdes-Algeria). The medium used
is the PCA, with 10% sterile skim milk and which we’veadded 2% of the actidione.
The bacteriaare cultured in a medium containing 5% of wheatbran, 0.3% of yeast extract,
0.4% of glucose, 0.2% of casein and 0.3% of Na2HPO4. The mediumis generally adopted in
precedent work for theBacillus’s culture.
The incubation was carried out at 30 ° C for 24 h with stirring 110 rpm. The enzyme’s
extraction is performed after the medium is filtered on a cheese cloth. Thefiltrate was
centrifuged at 4 ° C for 30 minutes at 4200g, and then the supernatant was kept at low temp
for activity test.
After testing sixty souches we have selected three of them that seem different (I33M, FK6A
and P47M). Theywere identificated by molecular biology. Theresults of BLAST gives:
I33M: Bacillus mojavensis, P47M: Bacillus subtilis, FK6A: Bacillus velezensis.
The I33M strain was selected from its very satisfactory clotting activity.
We conducted two experimental plans, for the first composition of the culture medium is: 5%
wheat bran, 0.3% of yeast extract, 0.3% of Na2HPO4, 0.05% of glucose, 1.01% of casein,
under T 39 ° C and 200 rpm agitation .... We can conclude that, the glucose is not significant.
A second plane has given us:7% of His, 0, 094% of casein, 0.3% of yeast extract, 0.3% of
Na2HPO4, under temperature about 39 ° C and agitation 150 rpm The clotting activity is 2.18
UP.L, proteolytic activity is 19.302 μgtyr / ml / min. Under fermentation, growth was
followed by counting the colonies in PCA medium after successive dilutions and clotting
activity is measured.
The enzyme’s production takes place at the end of the exponential phase and increases with
the stationary phase.
As for molds, the selected strain was identified as Mucor circinelloides. A statistically
significant (centered composite) was used to optimize the conditions of production of
coagulase by the selected Mucor. Use of the solid fermentation medium containing: 30g
wheat bran, 1gr casein, 60 ml ammonium sulfate (0.01%) at 30 ° C for 5 days, gave maximal
activity of 2.667 UP / ml. The characterization of the protease revealed optimal conditions:
temperature of clotting activity of 55 ° C, pH 5.5 and a concentration of 0.03M CaCl2.
Keywords : Mucor circinelloides, Bacillus mojavensis, milk clotting enzyme, optimization.

iii
ميكروبيةلالستخدامفيالجبنأصلذو تنقيةوتوصيفانزيمتخثر: العنوان
:ملخص
التي عزاللكائنات الدقيقةمنالتربةالتي تم جمعهابالقرب مناأللبانوالجبنبالخطوة األولىفيالهدفمنعملنا
الحليب الخالي من ٪ 01أضافactidioneعلىPCAهي(، والوسيلة المستخدمة الجزائر/ بومرداسبودواو)بتقع
الدسممعقمة.
٪ 1.0٪الجلوكوزالكازينو 1.0٪، 1.0مستخلصالخميرة٪ 5حتوي علىنخالةالقمحيمثقففيوسط الالبكتيريا
1.0٪Na2HPO4العصويةثقافة سابقة للدراسات فيعليها عتماداالتم طهو وسBacillu..
دورة في الدقيقة. يتم تنفيذ استخراج انزيم بعد 113ساعة مع التحريك 42درجة مئوية لمدة 03تم تنفيذ حضانة بها في
طاف تم 2433gدقيقة في 03درجة مئوية لمدة 2اللراشح كان طرد في تصفية المتوسطة على قطعة قماش الجبن
االحتفاظ بها في درجة الحرارة المنخفضة الختبار النشاط
إلى ت(، أرسلP47MوI33M ،FK6Aالتييبدومختلفا) منها لثالثنااخترسالالت بعد اختبارستين
، العصويةالرقيقة: mojavensis ،P47Mالعصوية: I33M: انفجارإعطاء، ونتائج المختبرلتحديدالبيولوجيا
FK6A :العصويةvelezensis.
مننشاطهاتخثرمرضية للغاية.I33Mوقد تم اختيارساللة
٪ 1.31٪ 3.35 الجلوكوزNa2HPO4 0.3٪٪ 3.0 ،خالصةالخميرة٪5مكون من وسطمنىولاألأجريناخطتينتجريبية
الكازين٪ 7 نخالة :الثانيةلناالخطة ،أعطتغير هامنستنتجأنالجلوكوز.... دورةفيالدقيقةالتحريض 433 و T 39 ° C الكازين
4.12 دورةفيالدقيقةواالنفعاالتوالنشاطتجلطهي Na2HPO4 39 ° C 150 درجةحرارة٪ 3.0 الخميرةمستخلص ،392٪
UP.L 19.034 النشاطهوبروتين μgtyr / مل / min.lors الوسط التخمير،وأعقبالنموعنطريقعدالمستعمراتفي
PCAبعدالتخفيفاتالمتتاليةويتمقياسالنشاطتجلط.
ت.ابثالمرحلة ترتفع فيالواآلسيةفينهاية المرحلة يحدثإنتاجإنزيمإن
( مركزمركب) ذاتداللةإحصائية A .العفنة circinelloides ،تمالتعرفعلىساللةمختارةكمابالفطرياتفيما يتعلق
نخالةالقمح،من غ 03: استخداموسيلةالتخميرالصلبةالتيتحتويعلى. كانتتستخدملتحسينشروطإنتاجالمخثرةقباللعفنةالمحدد
1GR ،4.007 أيامأعطىالنشاطالقصوىمن 5 درجةمئويةلمدة 03 عند٪( 3.31) ملكبريتاتاألمونيوم 03 الكازين UP / مل .
3.30M وتركيز 5.5 درجةمئوية،ودرجةالحموضة 55 درجةحرارةالنشاطتجلطالدممن: توصيفالبروتينيكشفالظروفالمثلى
CaCl2
،تخثراالنزيمmojavensis العفنة،العصوية circinelloides :لمفتاحية:الكلماتا

iv
ABREVIATIONS
AA : Acidesaminés
AC : Activité coagulante
AP :Activité protéolytique
BSA: Bovine sérum albumine
BYNB: :Buffered Yeast Nitrogen Base
CMP: Caseinomacro peptide
CCD : Central composite design
DFP : Diisopropylfluorophosphate
ECC : Expérience composite centrale
EEB : Extrait enzymatique brut
EEP : Extrait enzymatique purifié
E.C. :Enzyme Commission numbers
EFT: Expérience factorielle totale
FDA: Food drug administration
G.R.A.S: Generally Recognized As Safe
IMCU: International milk clotting unite
PCA: Plate count agar
PCC: Phosphate de calcium colloïdal
PCR: Polymerase Chain Reaction
RSM : Méthode de réponse surface
SEL : Son Extrait de Levure
Rpm :Rotation par minute
TCA : Acide trichloro-acetique
UHT : Ultra haute température
UP : Unité présure

v
Liste des figures
Figures Titres des figures Pages
Figure 1 : Structure primaire de la caséine -CN B-1P 9
Figure 2 : Modèle de micelle de caséine avec sous-unités 10
Figure 3 : Les différentes étapes de la fabrication d’un fromage 37
Figure 4: Phase de la coagulation enzymatique du lait 39
Figure 5 : Classification des mycètes 48
Figure 6: La place de Mucor sp dans la classification des champignons 52
Figure 7 : Surface de réponse pour un domaine expérimental 61
Figure 8 : Photo de la caséolyse 66
Figure 9 : Diagramme de la fabrication d’un fromage type « EDAM » à L.F.B 87
Figure 10: Photos des observations microscopiques 99
Figure 11 : Galerie API 50CH pour la souche I33M 104
Figure 12: Galerie API 50CH pour la souche P47γ 104
Figure 13: Galerie API 50CH pour la souche FK6A 104
Figure 14 : Blast de la souche I33M 107
Figure 15 : Dendrogramme de la souche I33M (Bacillus mojavensis) 108
Figure 16 : Surface de réponse de l’activité coagulante de la protéase produite par Bacillus
mojavensis, avec effet de la température et de l’agitation 111
Figure 17 : Graphique de Pareto obtenu avec les résultats des essais du plan 111
Figure 18 : Evolution de la croissance bactérienne en fonction du temps d’incubation 116
Figure 19 : Détermination du temps de génération de B.mojavensis 117

vi
Figure 20 : Effet de la variation du pH du milieu sur la croissance bactérienne(I33M) 117
Figure 21 : Evolution de l’activité coagulante de l’enzyme en fonction du temps
d’incubation de la souche I33M 118
Figure 22 : Variation du taux de protéines totales de l’extrait enzymatique brute en
fonction du temps d’incubation de la souche I33M 118
Figure 23 : Variation de l’activité protéolytique de l’enzyme en fonction du
temps d’incubation de la souche I33M 119
Figure 24:Profil chromatographique de l’extrait enzymatique produit par I33M filtré
sur gel Séphadex G-75
122
Figure 25 : Influence de la température du lait sur L’activité coagulante de l’EEB (I33M) 123
Figure 26 : Influence de la température du lait sur L’activité coagulante de l’EEP (I33M) 124
Figure 27: Influence du pH du lait sur l’activité coagulante de l’EEB de B.mojavensis 125
Figure 28: Influence du pH du lait sur l’activité coagulante de l’EEP de B.mojavensis 125
Figure 29: Influence de la concentration en CaCl2 du lait sur L’activité coagulante
de l’EEB de B.mojavensis 127
Figure 30: Influence de la concentration en CaCl2 du lait sur L’activité coagulante
de l’EEP de B.mojavensis 127
Figure 31: Photos des aspects macroscopiques de certaines moisissures isolées à
partir du sol 130
Figure 32 : Photos des aspects microscopiques des moisissures isolées 131
Figure 33 : Blast de la souche « 4D » 134
Figure 34 : Dendrogramme de Mucor circinelloides
135

vii
Figure 35 : Surface de réponse de l’activité coagulante des protéases produites par
Mucor circinelloides en fonction des facteurs choisis 140
Figure 36 : Profil d’élution sur Séphadex G75 de l’extrait enzymatique brut de
Mucor circinelloides 142
Figure 37 : Profil d’élution sur Séphadex G75 de l’extrait enzymatique précipité de
Mucor circinelloides 143
Figure 38 : Influence de la température du lait sur l’activité coagulante de l’EEB et l’EEP de
Mucor circinelloides 145
Figure 39: Influence de pH du lait sur l’activité coagulante de l’EEB et l’EEP de Mucor
circinelloides 146
Figure 40. : Influence de CaCl2 du laitsur l’activité coagulante de l’EEB et l’EEP de Mucor
circinelloides 147

viii
Liste des tableaux
Tableaux Titre des tableaux Pages
Tableau 1 : La composition moyenne du lait de vache 4
Tableau 2 : Les matières azotées totales 6
Tableau 3 : Composition moyenne de la micelle de caséine en g/100g 7
Tableau 4 : Caractéristiques physico-chimiques des caséines 11
Tableau 5 : Origine de différentes enzymes utilisées pour coaguler le lait 26
Tableau 6 : Plantes locales d’Algérie pour la coagulation du lait 28
Tableau 7 : Spécificité de quelques succédanés d’origine animale 30
Tableau 8 : Préparations commerciales 33
Tableau 9 : Impact de la température sur la coagulation du lait par la présure 41
Tableau 10 : Les paramètres d’une expérience composée centrale rotatable 64
Tableau 11 : Récapitulatif des souches bactériennes isolées à partir du sol 68-71
Tableau 12 : Caractères étudiés des bactéries sélectionnées ainsi que leur
technique d'ensemencement
76
Tableau 13 : Variation des bornes 80
Tableau 14 : Volumes d’EEB utilisés dans les essais de fabrication de
l’EDAM
84
Tableau 15 : Niveaux expérimentaux des trois facteurs utilisés dans le plan
composite centré
91
Tableau 16 : Matrice expérimentale utilisée pour mettre en place les
expériences
91
Tableau 17: Bactéries possédant une enzyme coagulante 96
Tableau 18 : Observation macroscopique des différentes souches 98

ix
Tableau 19 : Résultats des tests en galerie pour les différentes souches 100
Tableau 20 : Résultats des tests et interprétation des Hugh et Leifson 101
Tableau 21 : Résultats des tests en galerie pour les différentes souches 102
Tableau 22 : Comparaison Résultats du plan d’expérience 103
Tableau 23 : Résultats du plan d’expérience 109
Tableau 24 : L’analyse de la variance de l’activité coagulante 112
Tableau 25 : Effets estimés de l’activité coagulante 112
Tableau 26 : Coefficient de régression de l’activité coagulante 112
Tableau 27 : Résultats du suivi de la fermentation 115-116
Tableau 28 : Activité coagulante des éluâts en fonction du numéro du tube 121
Tableau 29 : Activité coagulante de l’extrait enzymatique brut 123
Tableau 30 : Description macroscopique de la collection des moisissures
isolées du sol
129
Tableau 31 : Souches possédant une enzyme coagulante et caractéristiques 132
Tableau 32 : Matrice expérimentale et activité coagulante des extraits bruts
obtenus
137
Tableau 33 : Activité de précipitation de l’extrait enzymatique brut au sulfate
d’ammonium
141
Tableau 34 : Bilan de purification de l’extrait coagulant de Mucor
circinelloides
144

SOMMAIRE

i
Sommaire
Résumés …………………………………………………………………………………….. i
Abréviations ……………………………………………………………………………….. iv
Liste des figures ……………………………………………………………………………. v
Liste des tableaux viii
Sommaire x
Introduction ………………………………………………………………………………… 1
CHAPITRE 1- SYNTHESE BIBLIOGRAPHIQUE
I. Généralités sur le lait 4
I.1 Définition légale du lait............................................................................................ ……….. 4
I.2 La composition du lait ........................................................................................................ .. 4
I.2.1 L’eau : .................................................................................................................................. 5
I.2.2 Lactose : ............................................................................................................................... 5
I.2.3 Matière grasse : .................................................................................................................... 5
II. Généralités sur les proteases ………………………………………………………….. 13
II.1 Définition ............................................................................................................................... 13
II.2 Classification ......................................................................................................................... 13
II.2.1 Selon le mode d’attaque de la chaine polypeptidique ........................................................ 14
II.2.2 Selon le pH d’activité ......................................................................................................... 14
II.2.3 Selon la nature de résidu impliqué dans le site actif ........................................................... 14
II.3 Applications alimentaires ...................................................................................................... 16
II.3.1 Industrie laitière .................................................................................................................. 16
II.3.2 Industrie de panification ..................................................................................................... 17
II.3.3 Synthèse d’aspartame ......................................................................................................... 17
II.3.4 Préparation des produits à base de soja .............................................................................. 17
II.4 Applications médicales et pharmaceutiques .......................................................................... 18
II.5 Autres applications ................................................................................................................ 18
II.5.1 Détergents ........................................................................................................................... 18
II.5.2 Tanneries ............................................................................................................................ 18
II.5.3 Traitement de divers rejets.................................................................................................. 18
II.5.4 Législation .......................................................................................................................... 18
III. Les enzymes coagulant le lait .............................................................................................. 20
III.1 La présure ............................................................................................................................. 20

ii
III.1.1 Origine et dénomination .................................................................................................... 20
III.1.2 Propriétés et mode d’action ............................................................................................... 20
III.1.3 Les composants de la présure ............................................................................................ 20
III.1.4 Obtention de la présure commerciale ......................................... 23
III.1.5 Propriétés spécifiques exigées des succédanés de présure ................................................ 23
III.2 Autres enzymes coagulant le lait .......................................................................................... 24
III.2.1 Les enzymes coagulantes d’origine végétale .................................................................... 27
III.2.2 Les enzymes coagulantes d’origine animale ..................................................................... 29
III.2.3 Les enzymes coagulantes d’origine microbienne .............................................................. 30
III.2.4 La chymosine produite par génie génétique ...................................................................... 33
III.3Action des enzymes coagulantes sur les caséines du lait ...................................................... 34
IV. Le fromage ………………………………………………………......... .............................. 36
IV.1 Généralités ........................................................................................................................... 36
IV-2 Différentes étapes de la transformation du lait en fromage ................................................. 37
IV.3 Coagulation .......................................................................................................................... 38
IV.3.1 Coagulation acide .............................................................................................................. 38
IV.3.2Coagulation par voie enzymatique ..................................................................................... 38
IV.3.3 Coagulation mixte ............................................................................................................. 40
IV.4 Egouttage ............................................................................................................................. 42
IV.5 Salage ................................................................................................................................... 42
IV.6 Affinage ............................................................................................................................... 42
V. Coagulases d’origine bacterienne ……………………………………………………… 42
V.1. Le genre Bacillus .................................................................................................................. 44
V.2. Intérêt industriel ................................................................................................................... 45
V.3 Les protéases de Bacillus....................................................................................................... 45
VI. Coagulases d’origine fongique ............................................................................................ 46
VI.1Generalités sur les moisissures ............................................................................................. 46
VI.2 Structure .............................................................................................................................. 47
VI.3 Aperçu de la classification .................................................................................................. 47
VI.4 Cycle de vie des moisissures ................................................................................................ 48
VI.5 Conditions de croissance des moisissures ........................................................................... 49
VI.5.1 Éléments nutritifs ............................................................................................................. 50
VI.5.2 Activité de l’eau ................................................................................................................ 50

iii
VI.5.3 Température ...................................................................................................................... 50
VI.5.4 pH ...................................................................................................................................... 50
VI.5.5 Oxygène ............................................................................................................................ 50
VI.6 Importance économique des moisissures dans la production d’enzymes ............................ 51
VI.7 Les especes exploitées industriellement............................................................................... 51
VI.7.1 Définition, caractéristiques et position taxonomique du genre Mucor ............................. 51
VI.7.2 Les protéases du genre Mucor et leur application en fromagerie ..................................... 53
VII. La fermentation industrielle ............................................................................................. 53
VII.1 Buts de la fermentation industrielle .................................................................................... 53
VII .2 Types de fermentation ....................................................................................................... 53
VII .2.1 Fermentation sur milieu liquide ...................................................................................... 55
VII .2.2 La fermentation solide ou semi-solide ............................................................................ 55
VII. 3 Production d’une enzyme industrielle d’origine microbienne ........................................... 58
VII.4 Sélection d’une souche microbienne .................................................................................. 58
VII.5 Production industrielle ........................................................................................................ 59
VII.6 Extraction et purification .................................................................................................... 59
VIII. Planification experimentale ............................................................................................. 59
VIII.1 L’optimisation du milieu de culture .................................................................................. 59
VIII.2 Plan d’expérience .............................................................................................................. 60
VIII.3 Notion d'espace expérimental ............................................................................................ 60
VIII.4 Notion de surface de réponse ............................................................................................ 60
VIII.5 Expérience factorielle de 2éme
ordre ................................................................................. 61
VIII.6 L’expérience composite centrée ........................................................................................ 62
VIII.7 Types d’expériences composées centrales ........................................................................ 63
VIII.8 Etape de planification, de réalisation et de traitement des résultats de l’expérience
composite centrée .........................................................................................................................
64
CHAPITRE 2. MATERIEL ET METHODES
I.Methodes d’isolement des microorganismes d’intérêt ......................................................... 65
II.Protease d’origine bacterienne .............................................................................................. 66
II.1. Isolement des souches bactériennes...................................................................................... 66
II.1.1. Choix et purification des souches ...................................................................................... 66
II.1.2. Observations macroscopiques et microscopiques ............................................................. 66
II.1.3.Conservation des souches ................................................................................................... 67

iv
II.2. Sélection des souches à activité coagulante ......................................................................... 71
II.2.1 Les milieux de culture : étude préalable ............................................................................. 72
II.3 Détermination des activités enzymatiques............................................................................. 73
II.3.1Obtention de l’extrait enzymatique brut .............................................................................. 73
II.3.2 Méthode de détermination de l’activité coagulante ............................................................ 73
II.3.3 Méthode de détermination de l’activité protéolytique ........................................................ 74
II.3.4 Dosage de la quantité protéique .......................................................................................... 75
II.4 Identification de souches à activité coagulante par voie biochimique .................................. 75
II.4.1 Tests d'identification en tubes............................................................................................. 75
II.4.2 Galerie miniaturisée API 20E ............................................................................................. 77
II.4.3 Galerie miniaturisée API 50CH .......................................................................................... 77
II.5 Identification par biologie moléculaire .................................................................................. 77
II.5.1 Les différentes étapes de l’identification moléculaire ........................................................ 78
II.5.2 Cas de l’identification de la souche I33M .......................................................................... 78
II.6 Optimisation par un plan d’expérience de la production d’enzyme ...................................... 79
II.6.1Méthodologie des surfaces de réponses ............................................................................... 79
II.7 Suivi de la croissance de la souche I33M en fermenteur....................................................... 81
II.7.1. Préparation du milieu, conditions de culture et inoculum ................................................. 81
II.7.2 Dénombrement et activités ................................................................................................. 81
II.10 Essai de fabrication de fromage ........................................................................................... 84
III. Protease d’origine fongique ................................................................................................ 88
III.1 Isolement des souches ......................................................................................................... 88
III.1.1 Méthode d’isolement ........................................................................................................ 88
III.1.2 Choix et purification des souches ..................................................................................... 88
III.1.3 Observations macroscopiques et microscopiques ............................................................ 88
III.2 Sélection des souches fongiques coagulantes ...................................................................... 88
III.2.1 Les milieux de culture ....................................................................................................... 88
III.2.2 La préparation de l’inoculum et l’ensemencement des milieux ........................................ 89
III.3Determination des activités enzymatiques ........................................................................... 89
III.3.1 Obtention de l’extrait enzymatique brut ............................................................................ 89
III.3.2 Méthode de détermination de l’activité coagulante .......................................................... 89
III.3.3 Méthode de détermination de l’activité protéolytique ...................................................... 89
III.3.4. Dosage de la quantité protéique ....................................................................................... 90

v
III.4 Sélection des souches fongiques .......................................................................................... 90
III.5 Identification des moisissures par biologie moleculaire ...................................................... 90
III.6 Optimisation de la production de la protéase ....................................................................... 90
III.6.1 Optimisation des conditions opératoires ........................................................................... 90
III.6.2 Ensemencement des milieux de culture ............................................................................ 92
III.6.3 Extraction de l’enzyme ...................................................................................................... 92
III.6.4 Evaluation statistique des composants du milieu de culture par le plan composite
centré ............................................................................................................................................
92
III.7 Essai de purification ............................................................................................................. 93
III.7.1Précipitation au sulfate d’ammonium ................................................................................ 93
III.7.2 Dessalage de l’extrait enzymatique par dialyse ................................................................ 93
III.8 Caractérisation de l’extrait enzymatique purifié ................................................................. 94
III.8.1 Influence de la température ............................................................................................... 94
III.8.2 Influence du pH ................................................................................................................. 95
III.8.3 Influence de la concentration en CaCl2 ............................................................................. 95
III.9 Essai de fabrication de fromage à pâte pressée non cuite type « EDAM » à partir de
l’extrait enzymatique brut (EEB) de Mucor circinelloides ..........................................................
95
CHAPITRE 3. RESULTATS ET DISCUSSION
I. Protease d’origine bacterienne .............................................................................................. 96
I.1 Sélection des souches possédant une enzyme coagulante ...................................................... 96
I.2 Observations macroscopique et microscopique ...................................................................... 98
I.3 Tests d'identification en tubes ................................................................................................. 99
I.4 Test d’identification en galerie pour les différentes souches .................................................. 101
I.4.1 Galerie Api 20 E .................................................................................................................. 101
I.4.2 Galerie API 50 CH ............................................................................................................... 103
I.5 Résultats de l’identification bactérienne ................................................................................. 105
I.6 Résultats de l’optimisationdu milieu de production ............................................................... 109
I.7 Résultats de la croissance durant lafermentation de la souche I33 M .................................... 115
I.7.1Analyse des résultats ............................................................................................................. 119
I.7.2.Discussion ............................................................................................................................ 120
I.8. Essai de purification de l’extrait enzymatique brut de Bacillus mojavensis .......................... 121
I.9 Caractérisation de l’enzyme coagulante de Bacillus mojavensis ............................................ 122
I.9.1 Détermination de l’activité coagulante ................................................................................ 122

vi
I.9.2.Influence de la température du lait ....................................................................................... 123
I.9.3 Influence de pH du lait ......................................................................................................... 125
I.9.4Influence de concentration en CaCl2 ..................................................................................... 126
II.Protease d’origine fongique ................................................................................................... 128
II.1Collection de souches ............................................................................................................. 128
II.2 Observations macroscopiques ............................................................................................... 128
II.3 Observations microscopiques ................................................................................................ 130
II.4 Détermination de l’activité enzymatique des extraits bruts des moisissures isolées. ............ 132
II.4.1Activité coagulante .............................................................................................................. 132
II.4.2L’activité protéolytique ....................................................................................................... 132
II.5 Identification des moisissures par biologie moléculaire ........................................................ 133
II.5.1 Séquences d’ADNr-18S obtenues ...................................................................................... 134
II.5.2Identification de la souche 4D ............................................................................................. 134
II.6 Optimisation de la production de l’enzyme coagulante ........................................................ 136
II.6.1Evaluation statistique des composants du milieu de culture par le plan composite
centré ......................................................................................................................................... ..
136
II.6.2L’analyse de l’influence de la composition du milieu de culture sur l’activité
coagulante des enzymes ...............................................................................................................
136
II.6.3 Essais de purification de l’extrait coagulant brut ............................................................... 140
II.7 Caractérisation de l’extrait enzymatique brut ........................................................................ 144
II.7.1 Influence de la température du lait sur l’activité coagulante .............................................. 144
II.7.2 Influence du pH de lait ....................................................................................................... 145
II.7.3 Influence de la concentration de CaCl2 .............................................................................. 146
Conclusion…………………………………………………………………………...…..…. 148
Références Bibliographiques ……………………….…………………………………….... 154
Annexes 167

Introduction

Introduction
1
La première étape de la transformation du lait en fromage nécessite traditionnellement
l’emploi d’un agent coagulant, appelé présure.
La coagulation joue un rôle déterminant dans la technologie fromagère. Elle est réalisée en
vue d’exploiter une propriété particulière des gels lactés qui est de s’égoutter spontanément.
Cette évolution se traduit par la séparation progressive de la majorité de l’eau constitutive du
lait sous forme de lactosérum et d’un substrat semi-solide ou caillé primitif.
Le lait coagulé constitue le début de la fabrication du fromage. Au final, l’action conjuguée et
variable d’enzymes et de microorganismes (affinage ou maturation) permettront l’obtention
d’un produit aux saveurs spécifiques, le fromage maturé. Ce dernier peut être caractérisé
comme la conversion d’un aliment périssable à grande teneur d’humidité (le lait) en un
aliment compact de longue conservation et de haute valeur nutritive. Il occupe une place de
choix comme aliment de bonne conservation, riche en protéines, matière grasse, calcium,
phosphore et autres éléments nutritifs (DAVIS,1976).Ainsi, les fromages sont par excellence
des aliments hautement digestibles, capables de satisfaire le besoin nutritionnel élémentaire et
intéressants pour tous les groupes d’âges.
L’origine du fromage remonte à la plus haute antiquité, des textes vieux de plus de 4000ans
en faisait déjà mention. Il est établi que des techniques fromagères étaient bien connues en
Mésopotamie il y a 5000ans. Différents textes bibliques prouvent par ailleurs que le fromage
était un aliment recherché dans le Proche Orient.
Il est probable que la production fromagère soit le résultat d’une recherche fortuite.En effet, la
légende l’attribue à un marchand d’Arabie qui, partant en voyage à travers le désert, aurait
rempli de lait une outre faite de l’estomac d’un animal. A la chaleur du soleil, les enzymes
des parois stomacales auraient fait coaguler le lait et avec les mouvements dus au transport,
cela a entrainé la séparation du caillé et du petit lait. Le processus de coagulation est
provoqué par l’action d’un coagulant ajouté à un taux bien défini au lait de fabrication, lui-
même utilisé à une température et un pH précis. Le coagulant traditionnel de l’industrie
laitière est la présure ; elle est extraite de l’estomac du jeune veau nourri au lait.
En ce qui concerne l’Algérie, le secteur de l’industrie laitière connait beaucoup de problèmes
du fait de l’insuffisance de la quantité de lait produite localement. En effet, la consommation
de lait est estimé à 143l/an /habitant : elle est la plus importante du Maghreb. De ce fait, nous
importons plus d’un milliard de litres de lait par an (Ministère de l’agriculture, Mars 2014).

Introduction
2
En outre, notre pays est totalement dépendant des firmes étrangères quant à
l’approvisionnement en agents coagulants.
Concernant les agents coagulants, la quantité de présure de veau disponible est devenue
insuffisante du fait de l’augmentation de la production mondiale de fromage :le besoin de
coagulants de remplacement s’est fait sentir dès le début des années 1960.De nombreux
substituts d’origine variable (animale, végétale, ou microbienne) ont été proposés.
(Ernesrom, 1997).
Cependant, les recherches continuent dans le but de trouver des enzymes de remplacement de
la présure. Depuis une quarantaine d’années, les industries de fermentations se sont
considérablement développées dans le monde et produisent des molécules variées dont un
grand nombre d’enzymes.
En particulier, un travail considérable a été réalisé dans la sélection de microorganismes
capables de produire des enzymes coagulantes.
Les plus intéressants sont des coagulants microbiens extraits des champignons
microscopiques (Rhizomucor pusillus, Rhizomucor miehei, Cryphonectria parasitica),ainsi
que la chymosine elle-même, obtenue par fermentation à partir d’un microorganisme cloné.
Plusieurs raisons sont à l’origine de cet intérêt.D’une part, ces enzymes microbiennes peuvent
être produites en quantité pratiquement illimitée sans qu’il y ait dépendance d’un
approvisionnement en matière première. En effet, le microorganisme utile est cultivé sur un
milieu spécifique de composition optimisée et son cycle de développement, limité à quelques
jours, est rapide comparativement à celui nécessaire à la génération d’un animal ou d’une
plante. Les enzymes sont extraites à partir du milieu réactionnel, puis purifiées selon des
techniques bien élaborées qui garantissent une composition constante.
D’autre part, les microorganismes producteurs d’enzymes exo cellulaires sont relativement
simples à cultiver et de ce fait les prix de revient sont satisfaisants. Enfin, l’usage des
préparations coagulantes d’origine microbienne est universel : il peut être envisagé dans les
pays où des règles philosophique ou religieuse peuvent limiter l’emploi d’autres catégories de
coagulants (Cuvellier, 1993)
De multiples souches de bactéries, moisissures et levures ont été étudiées en vue de la
production de protéases coagulantes. Les enzymes d’origine fongique ont été les plus

Introduction
3
largement explorées et développées (Scriban, 1999). Elles ont donné des résultats souvent
comparables et parfois supérieurs à ceux obtenus avec la présure. Aussi, ont-elles été
rapidement validées en fabrication industrielle. Différentes préparations sont commercialisées
sur le marché international par des laboratoires spécialisés et sont employées à plus ou moins
grande échelle selon le pays (Yadav et Mishra, 1994 ; Goursaud, 1999)
L’objectif de notre travail est donc d’isoler à partir du sol algérien, une souche microbienne
capable de fournir une protéase de bonne activité coagulante et de faible activité
protéolytique, d’optimiser les conditions de production par le choix d’un milieu de culture
adéquat ainsi que de purifier et de caractériser l’enzyme obtenue.
L’ensemble de ce travail est reparti en plusieurs volets :
La première partie comporte une synthèse bibliographique traitant des protéases en général et
des enzymes coagulantes en particulier, de la fabrication du fromage, des microorganismes
impliqués, du processus de fermentation et des plans d’expériences. Une deuxième partie
consacrée aux méthodes employées afin d’atteindre les objectifs (optimisation de la
production d’enzymes, extraction, caractérisation, et un essai de fabrication de fromage),
respectivement pour les bactéries et pour les moisissures.
Une autre partie comporte la purification des enzymes puis les résultats seront présentés
discutés dans un nouveau chapitre.

Chapitre 1
Synthèse
bibliographique

Chapitre 1. Synthèse bibliographique
4
I. Généralités sur le lait
I.1 Définition légale du lait
Le lait est le produit de sécrétion des glandes mammaires des mammifères, comme la vache,
la chèvre et la brebis, destiné à l’alimentation du jeune animal naissant. Du point du vue
physicochimique, le lait est un produit très complexe. Une connaissance approfondie de sa
composition, de sa structure et de ses propriétés physiques et chimiques est indispensable à la
compréhension des transformations du lait et des produits obtenus lors des différents
traitements industriels(Amiot et al., 2002).
I.2 La composition du lait
Tableau 1 : Composition moyenne du lait de vache (Alais et al., 1984).
Eléments Composition (g/l)
Eau 905
Glucides 49
Lipides :
Matière grasse :
Lécithine (phospholipide)
Partie insaponifiable (stérols, carotène)
35
34
0,5
0,5
Matières azotées :
Caséines
Protéines solubles (globulines, albumines)
Substances azotées non protéiques
34
27
5,5
1,5
Sels 9
Constituants divers (vitamines,
enzymes, gaz dissous)
Traces

Chapitre 1. Synthèse bibliographique
5
I.2.1 L’eau :
Le lait contient en moyenne 875 g l-1
d’eau. Cette eau se trouve sous deux états :
- l’eau extramicellaire représente environ 90% de l’eau totale, et contient la quasi-totalité du
lactose, des sels minéraux solubles, de l’azote soluble, etc... Une petite partie de cette eau est
liée aux éléments hydrosolubles, dont les protéines solubles.
- l’eau intermicellaire représente environ 10% de l’eau totale ; une fraction de cette eau est
liée aux caséines et l’autre conserve des propriétés solvants, mais les transferts de cette eau
dans les opérations de déshydratation et hydratation seraient beaucoup plus lents (Mahaut et
al., 2003).
I.2.2 Lactose :
Diholoside parfois appelé « sucre de lait », il est synthétisé par la glande mammaire à partir
du glucose sanguin (Boudier et Luquet, 1981). Présent à une dose de 47 à 52 g/l, c’est le
constituant le plus abondant du lait de vache. Il joue un rôle important, lié notamment à sa
valeur nutritionnelle et à sa fermentescibilité qui commande l’élaboration de divers produits
laitiers. C’est un disaccharide réducteur qui existe sous deux formes isomères α et ß, formes
se distinguant par certains caractères physiques (Mahaut et al., 2003).
Le lactose est un disaccahride constitué d’une unité galactose et d’une unité glucose, le β-D-
galactopyranosyl (1-4) D-glucopyranose, α ou β (Jeantet et al., 2007).
I.2.3 Matière grasse :
La matière grasse du lait se compose principalement de triglycérides, de phospholipides et
d’une fraction insaponifiable, constituée en grande partie de cholestérol et de ß-carotène.
La matière grasse du lait se présente sous la forme de petits globules sphériques qui sont
invisibles à l’œil nu (Amiot et al., 2002).
La teneur en matière grasse du lait est standardisée pour les laits de consommation :
- 36g/l pour le lait entier
- de 14,45 au minimum à 18,15 au maximum g/l pour le lait ½ écrémé.

Chapitre 1. Synthèse bibliographique
6
- au maximum 3,09g/l pour le lait écrémé (Luquet, et al., 1986).
I. 2.4 Matières azotées :
On peut distinguer 2 groupes de matières azotées dans le lait : les protéines et les matières
azotées non protéiques (ANP) qui représentent respectivement 95 % et 5 % de l’azote minéral
du lait. Ces 2 groupes sont sans aucune comparaison, ni sur le plan quantitatif ni sur le plan
qualitatif (Luquet et al., 1985).
Tableau 2 :Matières azotées totales du lait (Mathieu, 1998).
Groupes de constituants Teneur moyenne exprimée en
gramme par litre
Substances azotées totales
Substances azotées non protéiques
Substances azotées protéiques
Protéines solubles
Caséines
33,6
1,6
32
6
26
I .2 .4.1 Protéines :
Ce sont des composés organiques azotés macromoléculaires à l’état colloïdal, donnant par
hydrolyse des acides aminés. Constituants essentiels des tissus des êtres vivants, poids
moléculaire compris entre 15 et 500 kDa (Boudier et Luquet, 1981).
Elles comprennent les caséines (insolubles à pH 4,6) et les protéines du lactosérum (solubles à
pH 4,6) (Guillou et al., 1986
Les caséines du lait :
La caséine entière représente 80% des protéines du lait de vache et se présente sous une forme
micellaire. La micelle est formée par l’association des caséines αs1, αs2, β, κ et de composants
salins dont les deux principaux sont le calcium et le phosphate. Les proportions moyennes des

Chapitre 1. Synthèse bibliographique
7
différents constituants de la micelle sont données dans le tableau 3. Toutes les micelles n’ont
pas les mêmes dimensions, ni la même composition. Les grosses micelles ont une charge
minérale plus élevée et des proportions relatives de caséines β et κ plus faibles que les petites
(Brule et al., 1997).
Le diamètre moyen généralement admis est voisin de 180 nm avec une distribution entre 100
et 500nm (Cayot et Lorient, 1998). On estime que la masse micellaire doit être comprise
entre 0,5 et 1 × 109 Da. La forme est considérée comme sphérique mais avec une surface
granuleuse (Schmidt, 1982 ; Walstra, 1999).
Tableau 3 : Composition moyenne de la micelle de caséine en g/100g (Brule et al., 1997)
Caséines Composants salins
αѕ1………………………………………………………………..33
αѕ2…………………………………….........11
β…………………………………………....33
κ…………………………………………....33
γ…………………………………………….4
total caséines………………………………92
Calcium………………………………….2,9
Magnésium……………………………...0,2
Phosphate inorganique…………………4,3
Citrate …………………………………..0,5
Total……………………………………..8,0
1. Les caséines αS :
La caséine αS1 :
C’est la protéine la plus importante en masse, elle possède 199 AA et un poids moléculaire de
23 614 g/mol. Cette caséine est très sensible au calcium au pH normal du lait (=6,7) : quelle
que soit la température et en présence de calcium, on constate une formation de flocons.

Chapitre 1. Synthèse bibliographique
8
La caséine αS2 :
Elle représente 8 à 11% de la micelle de caséine, possède 207 AA et 13 à 10 phosphates (il
s’agit deαS2 ou αS3 ou αS4ou αS6selon le nombre de phosphates) et son poids moléculaire
estimé varie de 25150 à 25390 g/mol. Grâce à la présence des 2 résidus cystéine, les
molécules peuvent s’associer en dimères qui s’agrègent entre eux par interactions
électrostatiques pour former des polymères (αS5 dimère de αS3 et αS4).
Par sa richesse en phosphate, elle est très sensible au calcium, et comme pour αS1, la
caséineαS2 semble ne pas être en surface de la micelle.
2 . Les caséines β et γ :
La caséine ß :
Représentant 25 à 35% de la micelle, avec ses 209 AA et ses 5 groupements phosphates, elle
possède beaucoup d’analogie avec la caséine αS1.
Elle est sensible au calcium à température ambiante mais après déphosphorylation, la
molécule perd cette sensibilité et devient capable d’empêcher la précipitation de la caséine
αS1 par le calcium.
La caséine γ :
Il s’agit des fragments C-terminaux résultant de la protéolyse de la caséine ß par la plasmine.
3. La caséine κ :
Une grande majorité de cette caséine se trouve à la surface de la micelle, accessible à la
présure. Il s’agit d’une protéine de 169 AA, phosphorylée (Serine 149) comportant 2 variants
génétiques A et B. Elle comporte un constituant majeur non glycosylé (Fig.1) et des
constituants mineurs glycosylés dont la structure précise est élucidée (Eigelet al., 1984).
Cette caséine est insensible au calcium et stabilise les autres caséines phoshorylées vis à vis
de ce cation. La coagulation du lait se fait suite à la protéolyse de cette caséine par la présure
(ou chymosine : enzyme naturelle de la caillette du jeune bovin pré-ruminant) qui scinde la

Chapitre 1. Synthèse bibliographique
9
molécule en deux parties : la partie N-terminale (1-105)ou paracaséine et le fragment C-
terminal (106-169) ou caséinomacropeptide (CMP) aux propriétés très contrastées.
Dans le caillé, seules sont récupérées les caséines αS1, αS1etß et la paracaséine k tandis que le
CMP se retrouve dans le lactosérum. Il est à noter que le CMP contient tous les glucides,
quand ils existent, sur les Thréonine 131, 133, 135 et 136 (variant A uniquement).
PyroGlu-Glu-Gln-Asn-Gln-Glu-Gln-Pro-Ile-Arg(10)-Cys-Glu-Lys-Asp-Glu-Arg-Phe-
Phe-Ser- Asp(20)
Lys-Ile-Ala-Lys-Tyr-Ile-Pro-Ile-Gln-Tyr(30)-Val-Leu-Ser-Arg-Tyr-Pro-Ser-Tyr-
Gly-Leu(40)- Asn-Tyr-Tyr-Gln-Gln-Lys-Pro-Val-Ala-Leu (50)-Ile-Asn-Asn-Gln-
Phe-Leu-Pro-Tyr-Pro-Tyr (60)- Tyr-Ala-Lys-Pro-Ala-Ala-Val-Arg-Ser-Pro (70)-Ala-
Gln-Ile-Leu-Gln-Trp-Gln-Val-Leu-Ser (80)- Asp-Thr-Val-Pro-Ala-Lys-Ser-Cys-Gln-
Ala(90)-Gln-Pro-Thr-Thr-Met-Ala-Arg-His-Pro-His (100)- Pro-His-Leu-Ser-Phe-
Met-Ala-Ile-Pro-Pro (110)-Lys-Lys-Asn-Gln-Asp-Lys-Thr-Glu-Ile-Pro (120)- Thr-
Ile-Asn-Thr-Ile-Ala-Ser-Gly-Glu-Pro (130)-Thr-Ser-Thr-Pro-Thr-Ile*-Glu-Ala-Val-
Glu (140)- Ser-Thr-Val-Ala-Thr-Leu-Glu-Ala*-Ser-Pro (150)-Glu-Val-Ile-Glu-Ser-
Pro-Pro-Glu-Ile-Asn (160)-
PThr-Val-Gln-Val-Thr-Ser-Thr-Ala-Val-OH (169)
Glu (1)…..Phe (105) …..Met (106)-…. Val(169) (Action de la chymosine)
Figure 1 : Structure primaire de la caséine -CN B-1P (Eigel et al ; 1984).
Structure
La grande majorité des caséines sont présentes sous la forme de particules colloïdales
nommées micelles de caséine qui sont passablement stables. Les quatre principales façons
d’induire l’agrégation sont : l’utilisation d’enzymes protéolytiques, les conditions acides, les
traitements par la chaleur.
Modèle avec sous-unités
Le modèle avec sous-unités, présenté à la Figure 2, suggère que les sous-unités n’étaient pas
toutes de même composition mais que certaines étaient plus riches en κ-CN et étaient situées

Chapitre 1. Synthèse bibliographique
10
en surface de la micelle. Finalement, Schmidt (1982) précisa que les sous-micelles étaient
reliées par des agrégats de Ca9 (PO4)6.
Dans le modèle actuel, les micelles sont en fait composées de sous-unités comprenant de 10 à
100 molécules de caséines et nommées sous-micelles. Les sous-micelles sont reliées entre
elles par des ponts phosphate de calcium. La composition des sous-micelles au centre et en
périphérie est différente. En effet, les caséines β et αs1 sont plus présentes au centre de la
micelle et forment le cœur hydrophobe alors que la partie externe, davantage hydrophile, est
formée de caséine αs1, αs2 et κ (Amiot et al., 2002).
Figure 2 : Modèle de micelle de caséine avec sous-unités (Amiot et al., 2002).
I.2.4.1.2 Propriétés des caséines :
pHi et charge électrique :
Les groupements acides libres des résidus glutamyle, aspartyle et phosphoryle en nombre
supérieur aux groupements basiques libres –NH des lysines et autres acides aminés diaminés,

Chapitre 1. Synthèse bibliographique
11
confèrent à la caséine entière un pHi de 4.65, une charge négative et des propriétés acides
(réaction avec les métaux alcalino-terreux).
Propriétés associatives des caséines :
A pH = 7, lorsqu’on élève la température, les caséines ß donnent des polymères d’une
vingtaine à une trentaine d’unités, les différentes molécules étant unies par des liaisons
hydrophobes. De plus, des polymères résultent de liaisons disulfures S-S intermoléculaires.
Le Ca+2
complexe les molécules αs1, αs2, ß et diminue ainsi leur charge, leur hydrophilie et les
insolubilise (Rattray et al., 1997).
Propriétés des différentes fractions caséiniques de la micelle
Les caséines ont un certain nombre de caractères communs, la présence de phosphore sous la
forme de groupements phosphoséryls, la forte proportion de résidus apolaires. Elles se
distinguent les unes des autres par le nombre de groupements phosphoséryls, la présence ou
non de cystéine, la présence ou non de glucides, leur caractère plus ou moins hydrophobe.
La présence des groupements phosphoséryls confère aux caséines une très grande affinité vis-
à-vis du calcium, du magnésium et des oligoéléments. Dans le cas des caséines αs1, αs2, et β,
qui sont hautement phosphorylées, les sites phosphorylés sont en majeure partie groupés, ce
qui a pour conséquence de créer dans la chaîne peptidique des zones à caractère acide et
hydrophile très marqué.
Tableau 4 : Caractéristiques physico-chimiques des caséines (Creamer, 2002).
Caractéristiques Caséine-αѕ1 Caséine-αѕ2 Caséine-β Caséine-κ
Poids moléculaire (Da) 236172 25230,0 23984,8 19007,0
Conc. g/l de lait 10,25 2,75 9,62 3,5
Nombre d’acides aminés 199 207 209 169
Présence de glucides - - - +
Sensibilité au Ca++
++ +++ + -
Les différentes formes d’un type de caséine se distinguent par leur composition, en particulier,
le nombre de groupements phosphate (Mathieu, 1998 ; Creamer, 2002).

Chapitre 1. Synthèse bibliographique
12
La caséine k, bien que non majoritaire dans la micelle est la protéine laitière de loin la plus
étudiée car elle détient le rôle clef dans la coagulation du lait par la présure. Le variant
génétique A est le plus fréquemment rencontrée dans le lait (75 à 85 % des cas). La liaison
peptidique 105-106 entre le résidu phénylalanyl et le résidu méthionyl est la liaison
hydrolysée spécifiquement par la chymosine après emprésurage du lait. La partie qui
correspond au glycomacropeptide (106-169) est de nature très hydrophile et donne un
caractère très amphiphile à la protéine.
La partie N-terminale (1-105) est légèrement cationique et très hydrophobe. Lors de
l’hydrolyse par la chymosine, la para-caséine k (1-105) reste accrochée à la micelle ; la
micelle en perdant son pôle hydrophile stabilisant, précipite. De toutes les caséines, c’est la
seule qui soit glycosylée. Les sites possibles de glycosylation ont été déterminés comme étant
situés sur les résidus de thréonine 131, 133, 135, et 136 ( Creamer, 2002).
Le calcium et le phosphate sont les constituants inorganiques majeurs de la micelle. Le
calcium est en partie (environ 1/3) directement fixé sur les caséines, essentiellement sur les
groupements esters phosphoriques (Brule et al., 1997). Le phosphore lié aux résidus sérine,
appelé phosphore organique, représente environ 40% du phosphore de la micelle. L’autre
fraction du calcium est à l’état de phosphate de calcium colloïdal (PCC) présent en petits
amas sphériques d’un diamètre proche de 1nm et de composition Ca9(PO4)6 (Schmidt,1982).
I.2.4.1.3 Protéines sériques
La fraction des protéines sériques englobe toutes les protéines solubles à pH 4,6. La ß-
lactoglobuline, l’α-lactalbumine, la bovine sérum albumine, les immunoglobulines, la
lactoferrine représentent plus de 90% des protéines sériques totales.
Ce sont majoritairement des protéines globulaires présentant une grande sensibilité aux
traitements thermiques. Elles sont globalement riches en acides aminés soufrés et possèdent
des résidus tryptophane leur conférant une excellente valeur nutritionnelle (Jeantet et al.,
2007).
I.2.4.2 Matières azotées non protéiques
De nombreuses substances azotées restent en solution dans les conditions de précipitation des
protéines du lait : acidification, élévation de la température ou addition de présure ; leurs

Chapitre 1. Synthèse bibliographique
13
molécules ne s’agrègent pas mais demeurent séparées par l’eau. Elles ne précipitent pas sous
l’action de l’acide trichloracétique (TCA) dans les proportions de 12 g pour 100g. Bien que
contenant de l’azote, ce ne sont pas des protéines ; elles sont dites non protéiques (Mathieu,
1998).
II. Généralités sur les protéases
II.1 Définition
Les protéases ou peptidases sont des enzymes protéolytiques qui catalysent l´hydrolyse de
liaisons peptidiques. Dans certains cas, les enzymes sont hautement spécifiques et hydrolysent
une unique liaison peptidique d´une protéine donnée. Dans d´autres cas, les peptidases
hydrolysent plusieurs liaisons peptidiques qui présentent une séquence ou conformation
déterminée (Hartely, 1960).
Les protéases sont nombreuses, représentant environ 2% de tous les produits géniques
(Rawlings et Barrett, 1994) et environ 10% des enzymes réunies dans la liste « EC ».
Les protéases sont la seule classe d’enzyme qui occupe une position centrale par rapport à leur
application dans les domaines physiologique et commercial. Les progrès des techniques
analytiques ont démontré que les protéases induisent des modifications hautement spécifiques
et sélectives des protéines : comme l’activation de la forme zymogène par protéolyse, la lyse
des caillots des fibrines (coagulation du sang), la transformation et le transport des protéines
secrétées à travers les membranes (Rao et al., 1998).
II.2 Classification
Selon la nomenclature de l’Union internationale de Biochimie et Biologie moléculaire, les
protéases sont classées dans le sous-groupe 4 du groupe 3 des hydrolases. Cependant elles ne
se soumettent pas facilement à ce système de classification à cause de la complexité de leur
structure et leurs mécanismes d’action. Récemment, les protéases sont classées sur la base de
trois critères majeurs : le type de réaction à catalyser, la nature chimique du site catalytique et
l’évolution du site de référence.

Chapitre 1. Synthèse bibliographique
14
II.2.1 Selon le mode d’attaque de la chaine polypeptidique
En fonction de leur mode d’attaque, les protéases sont divisées en deux groupes :
Les exopeptidases qui réalisent une hydrolyse à partir des extrémités N-terminale, ou
C-terminale des peptides et libèrent soit un unique résidu d’acide aminé, soit un
dipeptide ou un tripeptide (Scriban, 1999 ; Rao, 1998).
Les Endopéptidases qui hydrolysent une liaison peptidique interne.
II.2.2 Selon le pH d’activité
Selon ce paramètre, les enzymes protéolytiques de différentes origines sont classées en trois
groupes: les protéases acide, neutre et alcaline (Sumantha et al., 2006).
II.2.3 Selon la nature de résidu impliqué dans le site actif
Certains chercheurs ont développé une nouvelle approche de classification des endopeptidases
en 5 groupes distincts, sur la base de leur site actif et leur sensibilité à différents inhibiteurs. Il
existe les endopeptidases à serine, à cystéine, acide aspartique, à métal et à thréonine.
II.2.3.1 Protéases à serine
Les protéases à serine sont très répandues dans la nature aussi bien chez les eucaryotes, que
chez les bactéries et virus. Leur mécanisme catalytique implique la présence de groupement
serine dans leur site actif, ce dernier agit comme un nucléophile qui attaque alors une liaison
peptidique. Les protéases à serine sont classées en 2 superfamilles ; chymotrypsine et
subtilisine (Barrett, 1994).
Les protéases appartenant à la famille S1 (chymotrypsine), sont rencontrées chez les micro-
organismes procaryotes et eucaryotes, les plantes et les animaux vertébrés tandis que celles
appartenant à la famille S8 (subtilisine) sont uniquement rencontrés chez les bactéries. La
structure tridimensionnelle est différente pour ces deux familles mais elles possèdent la même
géométrie du site actif et le mécanisme catalytique se déroule de la même manière (Barrett et
Rawlings, 1995).
Les protéases à serine sont actives à pH neutre et alcalin avec un optimum entre 7 et 11, leur
poids moléculaire varie entre 18 et 35 kDa. Les pH isoélectriques des protéases à serine sont
généralement entre 4 et 6.

Chapitre 1. Synthèse bibliographique
15
Les protéases à serine sont inhibées irréversiblement par le diisopropylfluorophosphate
(DFP).
II.2.3.2 Protéases à cystéine
Les protéases à cystéine sont présentes chez les microorganismes, les plantes et les animaux.
Leur activité dépend du site catalytique qui est composé de cystéine et d’histidine dont l’ordre
diffère entre les familles. Elles sont généralement actives seulement en présence d’agents
réducteurs comme le cyanure et la cystéine. Selon la spécificité de leurs chaines, les protéases
à cystéines sont divisées en : papaïne, trypsine, acide glutamique spécifique et autres
(Barrett, 1994).
La majorité des protéases à cystéine présente un pH optimum à la neutralité et peu d’entre
elles sont actives à pH acide. Elles sont sensibles aux agents sulfidryl, mais ne sont pas
affectées par le DFP, et le chélateur métallique (Rao et al., 1998).
II.2.3.3 Protéases à acide aspartique
Elles sont communément connues comme des protéases acides, ce sont des endopeptidases
qui dépendent de la présence d’un résidu aspartyl pour leurs activités catalytiques. Les
protéases acides sont regroupées en trois familles nommées : pepsine, rétropepsine et les
enzymes de rétrovirus (Hartley, 1960; Barrett, 1995).
La majorité des protéases à acide aspartique montre une activité maximale à faible pH (3 à 4)
avec un pH isoélectrique dans l’intervalle est de 3 à 4,5. Leurs masses moléculaires varient de
30 à 45 kDa.
Les protéases à acide aspartique sont inhibées par la pepstatine.
La protéase à aspartyl d’origine microbienne peut être divisée en deux groupes :
La pepsine comme les enzyme produite par Aspergillus, Penicillium, Rhizopus et
Neurospora (Kumar et al., 2005).
La rénine comme les enzymes produites par Endothia et Mucor sp (Sardinas, 1968 ;
Escobar et Barnett, 1993).

Chapitre 1. Synthèse bibliographique
16
II.2.3.4 Les Métalloprotéases
Les métalloprotéases sont le groupe le plus diversifié. Il se caractérise par la présence d’ions
métalliques divalents (généralement le zinc) dont le rôle est important pour leurs activités.
Elles incluent des enzymes de différentes origines dont les plus importantes sont : les
collagénases des organismes supérieurs, les toxines hémorragiques du venin de serpent et la
thermolysine (Barrett et al., 1995).
II.2.3.5 Protéases à thréonine
Ce nouveau type catalytique n´a été découvert que très récemment grâce aux recherches
effectuées sur les protéasomes. Un protéasome est, en effet, un complexe de protéases
impliqué dans la dégradation protéique. Il est d´une grande importance pour la croissance et la
viabilité de la cellule en raison de ses fonctions dans le cycle mitotique et dans le contrôle de
la demi-vie de la plupart des protéines cellulaires.
La recherche chez les procaryotes de particules ressemblant aux protéasomes eucaryote a
conduit à la découverte d´un complexe protéolytique chez Thermoplasma acidophilum.
II.3 Applications alimentaires
II.3.1 Industrie laitière
L’application principale des protéases dans l’industrie laitière est la préparation du fromage.
Les enzymes coagulant le lait sont classées dans trois principaux groupes : la présure animale,
les coagulants d’origine microbienne et la chymosine génétiquement modifiée. Les deux
premiers groupes proviennent de la classe des protéases acide à aspartique dont le poids
moléculaire varie de 30 à 45 kDa (Foltman, 1971; Fox, 1995).
Des recherches intensives dans ce domaine visent à produire des enzymes qui sont
complètement inactivées à la température de pasteurisation et contenant un faible taux de
protéases non spécifiques. Les protéases produites : GRAS (Generally Recognized As Safe)
par Mucor miehei, Bacillus subtilis et Endothia parasitica, remplacent graduellement la
chymosine dans la production de fromage (Sardinas, 1968 ; Fernandez-Lahore et al., 1998;
Escobar et Barnett, 1993 ; Shieh et al., 2009).

Chapitre 1. Synthèse bibliographique
17
II.3.2 Industrie de panification
La farine de blé est le composant principal dans l’industrie de panification. Il contient des
protéines insolubles appelées gluten qui détermine les propriétés du pain. Les endos et les
exopeptidases d’Aspergillus oryzae sontutilisées pour modifier par protéolyse, le gluten de la
farine du blé (Bourgeois et al., 1989).
Le traitement enzymatique du pain par les protéases améliore son extensibilité, réduit le temps
de mélange et augmente son volume.
II.3.3 Synthèse d’aspartame
L’utilisation d’aspartame comme un édulcorant artificiel hypocalorique est approuvée par la
F.D.A
L’aspartame est un dipeptide composé d’acide aspartique et de l’ester méthyle de L-
phénylalanine. La configuration L des deux acides aminés est responsable du goût sucré de
l’aspartame, mais la maintenance de cette configuration reste un processus couteux. Les
protéases sont des enzymes hydrolytiques, qui catalysent la réaction reverse sous certaines
conditions contrôlées. L’immobilisation de la thermolysine de Bacillus thermoproteolyticus
est utilisée pour la synthèse enzymatique d’aspartame. Toya Soda(Japon) et DSM (The
Netherland) sont les principaux producteurs industriels d’aspartame.
II.3.4 Préparation des produits à base de soja
Le soja est une source d’aliment riche du fait de sa contenance en protéines de bonne qualité.
Les protéases sont utilisées depuis longtemps pour la préparation de la sauce de soja et
d’autres produits à base de soja. Les protéases neutres et alcalines d’origine fongique jouent
un rôle important dans la préparation de la sauce de soja. Les modifications protéolytiques des
protéines de soja conduisent à l’amélioration de leurs propriétés fonctionnelles. Du traitement
des protéines de soja avec les protéases alcalines à pH8 résultent un bon rendement en
protéines, des hydrolysats très solubles et faible amertume (Bourgeois et al., 1989 ; Botton et
al., 1990 ).

Chapitre 1. Synthèse bibliographique
18
II.4 Applications médicales et pharmaceutiques
La grande diversité et spécificité des protéases a été exploitée pour le développement d’agents
thérapeutiques. L’administration orale de protéases d’Aspergillus oryzae améliore la digestion
suite à la déficience de certaines enzymes lytiques. L’utilisation de la subtilisine en
combinaison avec les antibiotiques pour le traitement des brûlures et des blessures.
L’asparaginase isolée d’Escherichia coli est utilisée pour éliminer l’asparagine du sang
leucémique. Et la protéase alcaline de Conidiobolus coronatus est utilisée pour remplacer la
trypsine dans les cultures de cellules animales (Chiplunkar et al., 1985)
II.5 Autres applications
II.5.1 Détergents
A l’heure actuelle, les protéases sont ajoutées comme des ingrédients clé dans la formulation
des détergents pour usage domestique (détergents à lessive, détergents à vaisselle), les
produits de nettoyage pour usage industriel et les produits de nettoyage pour les lentilles
cornéennes et les appareils dentaires. Cependant, le marché le plus important au niveau des
détergents est de loin celui des détergents à lessive. Une protéase détergente idéale doit avoir
une large spécificité de substrat et demeurer stable dans l’environnement hostile de la
machine à laver (température élevée et pH alcalin) (Rao et al., 1998). Bien que la pepsine soit
utilisée depuis 1913, la plupart des protéases ajoutées dans les détergents sont produites par
des souches de Bacillus (Gupta et al., 2002).Clear-lens Pro®, une marque de Novozyme
Danemark est utilisée pour enlever les dépôts à base de protéines ainsi que les films
protéiniques présentent dans les lentilles cornéennes (Sumantha et al., 2006).
II.5.2 Tanneries
Les protéases sont utilisées en tannerie depuis le début du siècle dernier pour leurs capacités à
libérer les poils et la laine. Les trois traitements de la peau à tanner (le reverdissage, le
délainage et le confitage) sont susceptibles de solutions enzymatiques de protéases produites
par B. licheniformis, A. oryzae, B. amylolyquefasciens. Jusqu'à présent, l’usage des protéases
a été limité car leur emploi est souvent plus coûteux que l’utilisation de produits chimiques.
Par contre, l’emploi de produits chimiques comporte plusieurs inconvénients (Rao et al.,
1998; Gupta et al., 2002).

Chapitre 1. Synthèse bibliographique
19
II.5.3 Traitement de divers rejets
Les protéases sont considérées aussi comme moyen efficace pour le traitement des rejets
riches en protéines. La protéase neutre de B. subtilis peut être également utilisée pour le
décreusage de la soie naturelle. Les protéases sont employées aussi avec des mélanges
d’enzymes hydrolytiques pour dégrader les polymères constitutifs de la matière végétale
servant pour l’alimentation animale (Aviron-Violet et al., 1982). Une autre utilisation des
protéases neutres est la récupération d’argent à partir des films photographiques par hydrolyse
de la gélatine (Sumantha et al., 2006).
II.5.4 Législation
D’un point de vue réglementaire, comme tout additif l’emploi des enzymes doit être soumis à
une règlementation stricte qui permet d’éviter tout risque de toxicité (Noor-Develliet et al.,
1983).
Le milieu de culture utilisé dans le cas des enzymes microbiennes ne doit laisser aucun résidu
nocif ou toxique ou de substances cancérigènes. Les préparations enzymatiques
commercialisées ne doivent pas contenir de contaminants microbiens. Les agents de
stabilisation et de dilution doivent être dans un état d’innocuité. En ce qui concerne les
chymosines de fermentation elles suivent l’arrêté du 28 avril 1998 qui concernent Escherichia
coli k12 renfermant un gène de prochymosine A de veau, Aspergillus niger var. awamoriet
Kluyveromyces lactis renfermant un gène de prochymosine V de veau. Elles sont utilisables
pour la fabrication des fromages à l’exception des fromages bénéficiant d’une appellation
d’origine.
III. Les enzymes coagulant le lait
III.1 La présure
III.1.1 Origine et dénomination
La présure de veau est la préparation coagulante traditionnelle la plus utilisée pour la
coagulation du lait (Alais, 1984 ; Wigley, 1996). De moindres quantités sont obtenues à partir
de l’estomac de chevreau et d’agneau. La dénomination présure est réservée à l’extrait
coagulant provenant de la troisième poche de l’estomac appelée abomasum ou caillette. Elle

Chapitre 1. Synthèse bibliographique
20
renferme deux enzymes actives. La chymosine est la protéase majeure responsable d’au moins
85% de l’activité coagulante totale ; le complément est apporté par la pepsine. On observe les
plus fortes teneurs en chymosine chez les animaux non sevrés ; dès que la ration alimentaire
renferme des aliments solides et que le jeune animal commence à brouter, la proportion de
chymosine chute très fortement ; à l’inverse, la pepsine devient dominante et caractérise la
sécrétion stomacale du mammifère adulte (Cogitore, 1982).
III.1.2 Propriétés et mode d’action
La chymosine et la pepsine sont des holoprotéines dont le poids moléculaire est voisin de
30 000 Da. Ce sont des enzymes qui ont une double action sur la caséine.Elles ont une
activité élevée et spécifique sur la caséine Kappa qui conduit à la libération du
caseinomacropeptide et à la déstabilisation micellaire.Elles ont une activité faible de
protéolyse générale sur les autres fractions de la caséine. Celle-ci intervient dès la mise en
coagulation et se poursuit pendant l’affinage du fromage.Comme pour toutes les préparations
enzymatiques, l’activité protéolytique de la présure est fortement influencée par les facteurs
de milieux en particulier le pH et par la température.
- Influence du pH :
Le pH optimum d’activité coagulante de la présure sur le lait est voisin de 5,5. Au pH du lait
frais (pH6,65) l’activité est modérée. En fabrication fromagère, on a intérêt à acidifier
légèrement le lait pour accroitre l’activité enzymatique.
- Influence de la température :
La température optimum d’activité de la présure se situe à 40-42°C. En dessous de 20°C,
l’activité devient faible. L’inactivation thermique totale de l’enzyme se produit à 65°C.
En fromagerie classique les températures du lait au moment de l’emprésurage se situent dans
la fourchette 20-40°C.
- Influence de la concentration en présure :
Il existe une règle approximative de proportionnalité entre la dose de présure et l’inverse du
temps de floculation : plus la dose est forte plus le temps est court.

Chapitre 1. Synthèse bibliographique
21
- Influence de la concentration en calcium :
La présence d’ions Ca++
est indispensable au déroulement de la phase secondaire. Toute cause
susceptible de faire baisser la teneur de ces ions dans le lait entrave la coagulation. Ces causes
peuvent être naturelles.
Un traitement thermique sévère insolubilise le calcium : il est nécessaire de restaurer la charge
en ions calciques par un apport de CaCl2 lorsque ces laits sont destinés à être coagulés par la
présure.
L’apport de calcium soluble modifie les équilibres salins vers les formes insolubles, l’addition
de CaCl2 provoque en outre une légère baisse de pH favorable à l’action présure. Ce
phénomène résulte d’un échange entre ions H+ fixés sur les protéines et le calcium incorporé.
III.1.3 Les composants de la présure
III.1.3.1 La chymosine
La chymosine est la protéase majeure responsable d’au moins 85% de l’activité coagulante
totale (Ramet, 1997). Elle est synthétisée sous forme de prochymosine, activée sous l’action
du suc gastrique. Elle subit alors une conversion en chymosine active.
La chymosine est une holoprotéine, de poids moléculaire voisin de 30 kDa (Scriban, 1993).
Elle est stable entre pH 5 et 6 ; son activité est optimale à pH voisin de 5 ; elle est inactivée à
pH 7,5 et est dénaturée à pH 8. Sa température optimale d’action est voisine de 40°C.
L’inactivation thermique a lieu dès 50°C, elle est totale à 61°C (Lenoir et al, 1985 ; Scriban,
1993).
Production de la chymosine fermentaire
Grâce au développement du génie génétique, il est actuellement possible de produire de la
chymosine à partir de micro-organismes (Alais et Linden., 1997 ; Collin et al., 1997 ; Rao et
al., 1998 ; Beldarrain et al., 2000 ; Munoz et al., 2004).
La chymosine fermentaire résulte du clonage du gène responsable de la production de la
chymosine à partir de l’estomac de veau sur certains micro-organismes. Les plus utilisés sont :
Escherichia coli, Kluyveromyces et Aspergillus.

Chapitre 1. Synthèse bibliographique
22
Cette méthode constitue l’une des voies porteuses d’espoir pour la synthèse d’enzymes
utilisables dans l’industrie laitière.
III.1.3.2 La pepsine
La pepsine est le constituant mineur de la présure dont la sécrétion gastrique ne devient
prépondérante qu’après sevrage (Ramet, 1997). Elle est produite sous forme d’un précurseur
inactif, le pepsinogène. Il passe par acidification sous la forme active : la pepsine de poids
moléculaire 35 kDa (Alais, 1984).
A l’opposé de la chymosine, la pepsine possède une activité protéolytique élevée et une faible
activité coagulante. D’après (Broome et Hickey, 1990), 20% de l’activité coagulante est
assurée par la pepsine dans la fabrication fromagère (Cheddar, Emmental,…).
La pepsine est relativement stable à des pH compris entre 5 et 5,5. Son activité enzymatique
est plus élevée entre pH 1 et 4 avec un maximum vers 1,8 et varie selon la nature du substrat.
C’est une enzyme thermosensible en solution après 55°C. Elle est dénaturée à des
températures à 70°C (Graiday, 1978).
Comme pour toutes les préparations enzymatiques, l’activité protéolytique de la présure est
fortement influencée par les facteurs de milieux en particulier par le pH et par la température.
L’activité la plus élevée se situe à pH 5,5 et à la température de 42◦C. De part et d’autre de ces
valeurs, l’activité décroit rapidement ; elle devient nulle au voisinage de 0◦C et vers 65
◦C.
En fromagerie, on règle l’action de la présure en ajustant les conditions de milieu à des
valeurs plus ou moins éloignées de ces optimums et en fixant la concentration enzymatique.
Ces paramètres sont particuliers à chaque famille de fromages (Eck et Gillis ,1997).
III.1.4 Obtention de la présure commerciale
Industriellement la présure est extraite par macération de morceaux de caillettes sèches de
veaux non sevrés, dans une saumure à 10% NaCl contenant des additifs nécessaires à la
préparation (telle que le phosphate disodique), à la conservation (les plus utilisés étant les
antiseptiques et les antifongiques) et à la coloration (caramel). Le jus brut est ensuite purifié
par diverses méthodes physico-chimiques.

Chapitre 1. Synthèse bibliographique
23
Selon Goursaud (1999), il faut de 1 à 2 caillettes de 60 g pour obtenir 1L d’extrait
commercial au 10000e. Ce dernier, après ajustement avec de la saumure, contient alors 10 à
16% de NaCl. Il est de couleur jaune d’or.
Le mélange contient les deux enzymes : la chymosine et la pepsine bovine (Goursaud, 1999).
III.1.5 Propriétés spécifiques exigées des succédanés de présure
Les enzymes de remplacement de la présure doivent répondre aux critères suivants (Ramet,
1997) :
- Une bonne solubilité dans l’eau.
- Une odeur et une couleur faibles ou nulles.
- Une activité coagulante bonne et une durée de consommation raisonnable.
- Une absence de toxicité pour le consommateur et un degré de pureté élevé pour éviter tout
accident de prolifération de microorganismes indésirables.
- Les propriétés rhéologiques du coagulum doivent être évaluées après floculation de façon à
permettre le travail mécanique du gel dans les délais habituels.
- La synérèse du coagulum au cours de l’égouttage et les modalités de l’affinage devrait
permettre d’obtenir les caractéristiques usuelles des fromages dans un délai sensiblement égal
à celui de la présure.
- Les rendements fromagers doivent être très proches ou supérieurs à ceux révélés lors de
l’emploi de la présure.
III.2 Autres enzymes coagulant le lait
Les enzymes sont des catalyseurs biochimiques de nature protéique qui interviennent dans
toutes les réactions métaboliques énergétiquement possibles, qu’elles accélèrent par activation
spécifique dans des conditions douces de température et de pH.
Ce sont des outils clés de la biotechnologie et de la bio- industrie. Un grand nombre
d’enzymes protéolytiques ont la propriété de coaguler le lait (Tableau 5) (Mietton, 1991).

Chapitre 1. Synthèse bibliographique
24
Les enzymes coagulantes, la présure ou ses substituts, sont des endopeptidases appartenant au
groupe des aspartyl protéases. Ces enzymes ont une double activité : l’une très spécifique sur
la caseine k, l’autre de protéolyse générale portant sur toutes les protéines, qui sont
susceptibles de se manifester aux cours de l’affinage des fromages.
La fabrication des fromages nécessite une phase de coagulation du lait qui permettra
l’expulsion (synérèse) d’une plus ou moins grande partie de l’eau et de matière soluble(le
sérum). On obtiendra ainsi un caillé ou fromage non affiné (Lenoir et al., 1983 ; Brule et
Lenoir, 1997).
De nombreuses protéases sont capables de provoquer la coagulation du lait, mais toutes ne
sont pas pour autant aptes à la fabrication fromagère car elles ne présentent pas les propriétés
biochimiques et technologiques requises (Ramet, 1997).
Les inconvénients majeurs de l’utilisation des protéases de remplacement sont dans certains
cas, la baisse du rendement fromager (Amiot et al., 2002).
Les protéases, en plus de leur activité coagulante, interviennent dans l’affinage des fromages.
(Mahaut et al., 2003).
En industrie laitière, le caractère fluctuant de l'approvisionnement en présure a conduit à
l'emploi de préparations enzymatiques d'origines diverses, coagulant le lait de façon analogue
à la présure (Guérard, 1987).
Des préparations enzymatiques coagulant le lait dans lesquelles la chymosine est remplacée
plus ou moins totalement par d'autres enzymes à mode d'action analogue.
Dans cette optique, un certain nombre d'enzymes ont été caractérisées.
Il s'agit toujours de protéases susceptibles d'attaquer les caséines du lait et plus
particulièrement la caséine k, dont la rupture d'une liaison peptidique particulièrement labile,
la liaison Phe105-Met106 provoque la coagulation du lait (Swaisgood, 1982).
Ces enzymes appartiennent au groupe des pepsines. La pepsine bovine, qui est assez proche
de la chymosine au plan de la structure primaire (Foltmann, 1970), est déjà présente dans la
présure avec la chymosine.

Chapitre 1. Synthèse bibliographique
25
La présure est encore à l’heure actuelle la préparation la plus largement utilisée en fromagerie
(Choisy et al .,1997).
Tableau 5 : Origine de différentes enzymes utilisées pour coaguler le lait(Mietton, 1991):
ORIGINES ENZYMES
Animaux
Ruminants
-Veaux
-Agneaux
-chevreaux
-Bovins adultes
Monogastrique
-Porcs
Oiseaux
-Poulets
chymosine₊pepsine
chymosine₊pepsine
chymosine₊pepsine
chymosine+pepsine
Pepsine
Pepsine
Végétaux
-Figuier (suc)
-Ananas (tige)
-Chardon, artichaut
-Gaillet
-Courge…
Ficine
Broméline
Cardosines
Cyprosine
Moisissures -Endothiaparasitica
-Mucorpusillus
-Mucormeihei
-Aspergillus niger
Protéase
Protéase
Protéase
Protéase
Chymosine (génétique)
Levures -Kluyveromyces lactis Chymosine (génétique)
Bactéries -Echerichia coli
-Bacillus subtilis
Chymosine (génétique)
Subtiline(génétique)

Chapitre 1. Synthèse bibliographique
26
III.2.1 Les enzymes coagulantes d’origine végétale
Contrairement à la présure qui est spécifique à la κ-caséine, les aspartyl-protéases des plantes
peuvent hydrolyser les caséines α, β et κ, ce qui provoque, une amertume et des défauts de
texture des fromages et qui limite leur utilisation industrielle. (Simöes et Faro, 2004).
Plusieurs préparations coagulantes sont issues du règne végétal et sont extraites par
macération de différentes parties de plantes supérieures (Ramet, 1997). L’utilisation de la
fleur de cardon comme agent coagulant a été considérée comme l’un des facteurs
déterminants de la qualité des fromages typiques portugais au lait de brebis (Agboola, 2002,
Zhao et al., 2003) l’extrait coagulant du Cynara cardunculus a fait l’objet de nombreuses
études (Macedo et al., 1993 ;Martin et al., 1996 ; Vioque et al., 2001).
Cependant il a été conclu que cet agent coagulant se caractérise par une activité protéolytique
excessive ce qui confère un goût amer au fromage. D’autres végétaux fournissent des
coagulases telles que les ficines extraites du latex du figuier, la papaine du papayer, la
bromelaine de l’ananas (Sardinas, 1968) etc… L’activité coagulante de ces préparations
végétales est très variable car elle est fortement influencée par l’état de maturité de la plante et
par les conditions de collecte et de stockage (Lopez et al, 1996). De ce fait, l’emploi de ces
protéases coagulantes est toujours resté limité aux aires locales de production (Eck, 1987).
Généralement, les protéases végétales sont rarement utilisées en industrie fromagère en raison
de leur activité protéolytique très élevée, qui se traduit par l’apparition d’un ensemble
d’inconvénients technologiques tels que (Alais, 1984) : mauvaise fermeté des caillés obtenus,
baisse du rendement fromager, apparition d’amertume.
Ces difficultés sont liées au fait que ces préparations brutes renferment souvent des mélanges
de plusieurs enzymes dont l’activité manque de spécificité sur la caséine k (Ramet, 1997).
Malgré les inconvénients évoqués précédemment, des études récentes ont été réalisés par
différents auteurs, qui approuvent quelques avantages aux protéases extraites des végétaux
(Tab 6). Ces dernières, sont plus stables à la chaleur par rapport aux protéases d’origine
microbienne et animale (Morsli, 1996).

Chapitre 1. Synthèse bibliographique
27
Tableau 6 : Plantes locales d’Algérie utilisées pour la coagulation du lait (Morsli, 1996)
Nom scientifique Nom vulgaire
Français Anglais Algérien
Bryoniadoïcajacq Bryone White bryony Tailoula /kerma beïda
Chrysanthemum
partheniumL.pers.
matricaire Fever few Mouniat
Conium maculatum L. Ciguë Hemlock Sikrane/Guebaba
Cucurbita pépo L. Citrouille Pumpkin Takhiart/ Garâa Kabouïa
Cucumis melo L. Melon ……. Feggouce/Betikha
Cucumis sativus L. Concombre Cucumber Takhiart/ Lekhiar
Cynara cardunculus
L.
Cardon Cardoon Thaga/ khorchef
Cynara humilisL. Cardon nain …….. Tharédouit/ Fegaa
Cynara scolymusL. Artichaut Artichocke Karnoune
Dipsacus sylvestris
Mill.
Cardére Teasel Tadjahnit-guizen/ denb-es-
sebaa
Ficus carica L. Figuier Figtree Taguérourt/ Kerma
Galium verum L. Gaillet Lady’sbedstraw Fouaoua sefra
Heracleum spondylum
L.
Berce Hogweed Aréoul
Malva sylevetris L. Mauve Common Amedjir/ Khobbeïza
Ricinus communis L. Ricin Casteroilseeds Akhilouane/ Kharoua
Silybum marianum L.
gaertn.
Chardon
Marie
Milk
thistle……….
Douj-n-ilou’man/ Chouket-el-
beïda
Senecio jacobaeaL. Séneçon Ragwort Debouz-el-arab
Salanum dulcamara
L.
Douce amére ………. Aouzizi/ H’LLOUA M’RRA
Urtica dioïca L. Ortie Stingingnettle Azedouf /Bouzegdouf

Chapitre 1. Synthèse bibliographique
28
III.2.2 Les enzymes coagulantes d’origine animale
Des substituts de présure sont produits par plusieurs mammifères comme le chameau
(Elagamy, 2000 ; Siboukeur, 2005), le buffle (Mohanty et al., 2003)…et même par les
viscères de poissons (Guérard, 1987). L’étude comparative des activités coagulantes et
protéolytiques de ces enzymes a révélée qu’elles présentent des caractéristiques proches de
celles de la présure (Polaina et Mac Cabe, 2007).
De nombreuses protéases d’origine animale ont fait l’objet d’expérimentation en vue d’une
utilisation potentielle en fromagerie (Tab. 7). Cependant toutes ne sont pas pour autant aptes
à la fabrication fromagère ; seules les pepsines porcines et bovines présentent un intérêt
industriel (Eck, 1987).
Les pepsines ont une activité protéolytique assez voisine de celle de la chymosine ; leurs
sensibilités aux variations des facteurs de milieu sont semblables sauf pour l’effet pH ; elles
ont un caractère plus acide avec un optimum au voisinage de pH 1,5-2,0(Ramet, 1997).
Certains succédanés d’origine animale peuvent être considérés comme des produits de
remplacement acceptables de la présure de veau.
Plusieurs pepsines de poissons, tels que celles issues de:sardine (Noda et Murakami,1981),
morue d’Atlantique (Brewer et al, 1984) et requin (Nungaray et Legoffic, 1996) ont été
purifiées et partiellement caractérisées.
Les chercheurs sont également arrivés à extraire la pepsine à partir de la muqueuse gastrique
du phoque (Shamsuzzaman et Haard, 1985) et de l’estomac du lapin (Ramman, 1994)
L’obtention de la pepsine sous forme cristallisée à partir d’extraits de muqueuse gastrique
bovine a été réalisée pour la première fois par Northrop en 1929(Moll et Moll, 1998). La
deuxième enzyme gastrique obtenue sous forme cristallisée été la pepsine porcine et cela par
Herriot en 1938 (Banga-Mboko et al., 2002).

Chapitre 1. Synthèse bibliographique
29
Tableau 7: Spécificité de quelques succédanés d’origine animale (Boudier, 1974)
Origine Enzyme Marque commerciale
Préparation de présure de veau Chymosine-pepsine Rennet Hansen
Présure de veau pur, cristallisée Chymosine Strersen (Bengeas)
Pepsine de porc Pepsine Pantex-peptilac (Hansen)
Pepsine de bœuf Pepsine Colepsine
Mélange enzymatique animal Chymosine Metroclot (Pfizer)
III.2.3 Les enzymes coagulantes d’origine microbienne
En pratique, l’utilisation des préparations enzymatiques microbiennes a été soumise à une
stricte réglementation, imposant des contrôles hygiéniques (liés à leur production et
extraction) et toxicologiques sévères, afin d’éviter tout risque de toxicité lié à la présence
d’antibiotiques et/ou d’aflatoxines (Noor-Devillietet al., 1983).On distingue deux catégories
de protéases microbiennes : les succédanés d’origine bactérienne et les succédanés d’origine
fongique.
D’origine bactérienne
Depuis une quarantaine d’années, une puissante industrie de transformations s’est développée
dans le monde. Elle produit des substances variées dont une grande partie d’enzymes qui
trouvent de nombreuses applications dans des secteurs industriels variés et en particulier des
protéases susceptibles de coaguler le lait.
Ce sont surtout les souches du genre Bacillus : Bacillus subtilis, Bacillus cereus, Bacillus
polymixa et Bacillus coagulans, qui ont fait l’objet de plusieurs recherches pour la production
de coagulases. Leur utilisation demeure limitée par suite de réglementation stricte et leur prix
de revient. D’autre part, leur aptitude à la coagulation est meilleure que celle d’origine
végétale et moins bonne que celle des enzymes produites par les moisissures. Les caillés

Chapitre 1. Synthèse bibliographique
30
obtenus manquent de cohésion du fait d’une trop forte activité protéolytique par comparaison
à la présure animale (Alais , 1984).
Selon Puhan et Irivine (1973 a et b) , des fabrications de fromages type cheddar canadien
avec la coagulase de Bacillus subtilis, en comparaison avec un fromage témoin fait avec la
présure traditionnelle, présentaient des différences notables de point de vue rendement
fromager et qualité organoleptique due à un degré excessif de protéolyse, corrigé par
acidification du lait préalablement à l’addition de l’enzyme au lait.
Un travail considérable a été réalisé dans la sélection de microorganismes capables de
produire des enzymes coagulantes. Plusieurs raisons sont à l’origine de cet intérêt.
Les enzymes microbiennes peuvent être produites en quantité pratiquement illimitée sans
qu’il y ait dépendance d’un approvisionnement en matière première. Le microorganisme utile
est cultivé sur un milieu spécifique de composition optimisée. Son cycle de développement,
limité à quelques heures ou quelques jours, est rapide comparativement à celui nécessaire à la
génération d’un animal ou d’une plante.
A la fin du cycle de développement du microorganisme, les enzymes sont extraites et
purifiées selon des techniques bien élaborées qui garantissent une composition constante. On
préfère sélectionner des microorganismes producteurs d’enzymes exo cellulaires plus faciles à
isoler du milieu de culture que les enzymes endocellulaires qui impliquent de réaliser une
rupture préalable des structures bactériennes pour libérer le composé actif.
En Algérie, (Matoub, 2000) a mis en évidence la possibilité d’obtenir une coagulase à partir
d’une souche locale de Bacillus subtilis sélectionnée (Lc33), mais leur utilisation n’a pas
dépassé le stade expérimental en raison de leur prix de revient et de la réglementation stricte.
D’origine fongique :
Actuellement, c’est sur les protéases d’origine fongique que l’on fonde le plus d’espoir.
Contrairement aux protéases bacteriennes, les protéases fongiques ont donné des résultats
meilleurs, souvent comparables à ceux obtenus avec la présure (Luquet ,1985).
Par ailleurs, certains auteurs (Green et Stackpoole, 1975 et Jarmulet al., 1982) ont démontré
l’efficacité d’utiliser une préparation fongique "Noury ou fromase"en mélangeavec la pepsine
dans la fabrication de fromage.

Chapitre 1. Synthèse bibliographique
31
Plusieurs préparations sont déjà commercialisées sur le marché international et utilisées à plus
ou moins grande échelle selon les pays (Tab.8), elles proviennent de trois espèces de
moisissures (Alais ,1984 ; Green et Stackpoole , 1975 ; Berankovaet al., 1987) :
-Endothiaparasitica : moisissure parasite du châtaignier de laquelle, la firme américaine
"PFIZER" a extrait une préparation coagulante en poudre commercialisée sous le nom de
"sure-curd" et "suparen".
-Mucor pusillusLindt : moisissure banale mésophile du sol, de laquelle la firme
japonaise "MEITO-SANG YO" a extrait une préparation commercialisée par " NOURY".
-Mucor miehei : moisissure banale thermophile du sol dont quatre préparations
enzymatiques différentes sont exploitées (Awad et al., 1999).Il s’agit de :
Rennilase commercialisée par la firme danoise « NOVO ».
Fromase : commercialisée par la firme française « RAPIDASE ».
Marzyme commercialisée par la firme américaine « MARSHALL ».
Hannilase commercialisée par la firme danoise « HANSEN »
Ces préparations enzymatiques, ont été soumises à des contrôles hygiéniques et
toxicologiques sévères, afin de démontrer l’absence d’antibiotiques et d’aflatoxines (Noor-
Devilliet et al., 1983). Ainsi, il ressort d’après plusieurs recherches que les préparations issues
de Mucor miehei, sont celles qui se rapprochent le plus de la présure animale (Alais et
Lagrange, 1972). Cité par Scriban (1993), il confirme qu’il est parfaitement possible grâce à
l’utilisation d’enzyme extraite de Mucor miehei (solution Rennilase), de fabriquer des
fromages d’excellentes qualités en leur adoptant les processus technologiques requis.
Parmi les substituants de la chymosine,les enzymes de Mucor pusillus et de M. miehei ont
connu le plus large développement. De par ses propriétés, la protéase de M. miehei se
rapproche le plus de la présure animale, notamment pour ce qui concerne le ratio activité
coagulante/ activité protéolytique (Joyeaux, 1982).
De plus, les enzymes fongiques se caractérisent par une stabilité thermique analogue à celle
de la présure (Goursaud, 1993).

Chapitre 1. Synthèse bibliographique
32
III.2.4 La chymosine produite par génie génétique
Les méthodes modernes du génie génétique ont été récemment appliquées à la production de
chymosine par des micro-organismes clonés par le gène induisant l’expression de la
prochymosine. Actuellement, plusieurs préparations commerciales sont disponibles sur le
marché (Maxiren, Chymax, Chymogen,…).
La présure bovine recombinante est produite en utilisant comme hôtes les espèces
microbiennes suivantes : Saccharomyces cerevisiae, Kluyveromyces lactis, Aspergillus niger
et Trichoderma reesei. (Mohantyet al., 1999). Par ailleurs, la protéase recombinante
d’agneau a été produite par Rogelj et al. (2001) en utilisant E.coli.
La composition de ces préparations diffère de celle de la présure par l’absence de pepsine.
L’analyse biochimique fine montre une identité absolue entre les isomères de la chymosine
naturelle et celles produites par les micro-organismes recombinés. L’activité protéolytique
mesurée sur les différentes fractions de la caséine est également rigoureusement la même.
Sur le plan technologique, les essais nombreux et variés de fabrication de fromages réalisés
dans le monde indiquent qu’aucune différence majeure ne peut être détectée entre les produits
obtenus avec les chymosines génétiques et la présure traditionnelle en ce qui concerne les
modalités de la coagulation, de l’égouttage et de l’affinage, et les caractéristiques
organoleptiques(Ramet, 1997). Plusieurs préparations commerciales sont disponibles sur le
marché.
Tableau 8 : Préparations commerciales d’origines fongiques (Ramet, 1997).
Microorganisme recombiné Société productrice Nom de préparation
Kluyveromyceslactis
Kluyveromyceslactis
Aspergillus awamori
Gist brocardesn.v (NL)
Pfizer Inc (U.S.A)
Ch. hansen Lab (DK)
Maxiren
Chymax
chymogen

Chapitre 1. Synthèse bibliographique
33
III.3Action des enzymes coagulantes sur les caséines du lait
Le site principal pour la réaction de coagulation est la liaison Phe105-Met106 de la caséine-ĸ.
D’une manière générale, les principales protéases utilisées pour la coagulation du lait
hydrolysent la même liaison peptidique (Phe105-Met106), à l’exception de la protéase de
Cryphonectria parasitica qui hydrolyse la liaison Ser104-Phe105 (Lucey, 2002).Toutefois,
l’attaque d’autres sites dans la caséine ĸ, la caséine αs et β se produit et elle est généralement
indésirable. Cette action est souvent qualifiée d’action protéolytique générale ou non
spécifique. Les protéases acides utilisées en fromagerie bien qu’elles présentent généralement
une grande spécificité dans leur action, exercent une activité protéolytique non spécifique.
Cependant, l‘extension de la protéolyse diffère d’une enzyme à une autre. La chymosine est la
protéase qui donne le minimum de protéolyse, L’attaque de la caséine β et de la caséine αs1
par la chymosine est plus lente que l’attaque spécifique de la caséine ĸ (Dalgleish, 1982).
Ainsi une telle action se produit avec une vitesse minimale au cours de la coagulation. Les
substituts de la présure sont généralement choisis par rapport à leur activité coagulante élevée
combinée et à une activité protéolytique générale limitée.
Une activité protéolytique non-spécifique élevée au cours de la coagulation peut donner
naissance à des peptides qui seront perdus en solution provoquant la diminution du rendement
fromager. D’une manière générale la majorité des enzymes coagulantes montre une activité
protéolytique plus importante que la chymosine lorsque la caséine est prise comme substrat.
Cependant les différences sont moins évidentes lorsqu’elles sont comparées dans le lait
écrémé ou durant la fabrication du fromage. Toutefois, il a été démontré que la plupart des
enzymes coagulantes provoquent des pertes dans le rendement comparé à la présure.
(Barbano et Rasmussen, 1992).
L’activité protéolytique durant l’affinage du fromage qui est due à la quantité d’enzyme
retenue par le caillé, joue un rôle essentiel dans la dégradation des caséines. La quantité
d’enzyme retenue par le gel est influencée par la température de coupe du coagulum et le pH
au cours du drainage du lactosérum. En effet, l’abaissement du pH augmente le taux
d’adsorption de la chymosine sur les micelles de caséine. Toutefois, le taux retenu pour les
protéases fongiques ne semble pas influencé par l’abaissement du pH (Ramet, 1997).

Chapitre 1. Synthèse bibliographique
34
L’activité résiduelle est également liée à la stabilité thermique de l’enzyme coagulante
employée. La protéase de Cryphonectria parasitica, la pepsine bovine, la chymosine, les
protéases de Rhizomucor pusillus et Rhizomucor miehei montrent une meilleure stabilité
thermique .D’autre part, le taux d’hydrolyse des fractions caséiniques, au cours de l’affinage,
dépend fortement du taux d’enzyme coagulante employée
La caséine αs est hydrolysée par la chymosine au niveau de la liaison Leu192-Tyr193, alors
que la caséine αs1 est hydrolysée au niveau des liaisons Phe23- Phe24 ou bien Phe24-Val25.
L’hydrolyse de la caséine αs1 par la chymosine est beaucoup plus rapide que celle de la
caséine αs. La formation du goût amer est due à l’action successive des enzymes coagulantes
et des protéases microbiennes des cultures employées dans l’affinage, sur les caséines.
En effet, il y a formation de peptides de taille moyenne sous l’action des enzymes coagulants.
Ces peptides sont ensuite hydrolysés à leur tour par les protéases microbiennes en peptides de
bas poids moléculaire responsables d’amertume (Sousa et al., 1997).
La majorité des enzymes coagulantes employés présentent la même spécificité que la
chymosine au niveau de la caséine α et la caséine β en attaquant les mêmes liaisons
précédemment citées, cependant, avec une vitesse d’action variable.
La pepsine bovine et la protéase de Rhizomucor miehei hydrolysent la caséine β plus
lentement que la chymosine, alors que la protéase de Cryphonectria parasitica et celle de
Cyanara cardunculus montrent la plus grande activité protéolytique en attaquant plus
rapidement la caséine β (Barbano etRasmussen, 1992).
La pepsine de poulet semble attaquer plus rapidement la caséine αs comparée à la présure.
Du cheddar préparé à l’aide de pepsine de poulet a montré un taux de protéolyse et
d’amertume plus élevé que celui préparé par la présure. (Green et al., 1977). Cependant,
l’utilisation de la pepsine de poulet dans la fabrication de certains fromages à pâte cuite tels
que "Emmental" et "Kashkaval" a donné des résultats satisfaisants
Cela est due probablement au fait que la quantité d’enzyme résiduelle maintenue par le
coagulum a été détruite par le chauffage au cours de l’étape de cuisson. Ceci indique la
possibilité de l’emploi de la pepsine de poulet dans la fabrication de fromages à pâte cuite.
Son emploi pour les fromages à pâte fraîche, qui ne subissent pas d’affinage, semble

Chapitre 1. Synthèse bibliographique
35
également possible du fait que la protéolyse au cours de cette étape est évitée. (Sousa et al.,
2002).
IV. Le fromage
IV.1 Généralités
Non seulement le lait se consomme à l’état nature, il peut également subir différentes
biotransformations qui contribuent à élargir considérablement ses qualités sensorielles et
nutritionnelles. L’un des dérivés de ces transformations est le fromage (Henri et al., 2008).
Très vite, l’homme a compris que le lait produit par les animaux pouvait constituer une partie
de son alimentation. Lorsqu’il a voulu le conserver il s’est heurté à sa fragilité. En effet, de
par sa composition favorable au développement de nombreux micro-organismes qui peuvent
l’altérer et le déstabiliser, il change d’aspect. Il se transforme en gel qui laisse s’échapper un
liquide clair de couleur jaunâtre.
En ce faisant, les éléments les plus nutritifs se retrouvent sous certaines conditions en une
masse acide de plus longue conservation que le lait. Celle-ci peut évoluer vers des produits
possédant des gouts différents et variés.
L’origine exacte de la transformation du lait est incertaine. On s’entend pour dire que le
fromage serait originaire du sud-ouest asiatique et daterait d’environ 8000 ans. Les Romains
auraient stimulé le développement de nouvelles variétés durant leur invasion de l’Europe
entre 60 av.j-c.et 300 apr.j.c. (St-Gelais et Tirard-Collet, 2002).
Progressivement, des techniques de transformation se sont développées, souvent autour de
pratiques régionales guidées par la nature du lait disponible, les habitudes alimentaires et le
contexte socioculturel. L’accumulation des connaissances en science du lait et la maitrise des
procédés de transformation font qu’aujourd’hui on dénombre de très nombreuses variétés de
fromages possédant chacune sa propre originalité.
Il est d’usage, pour les comparer ou pour en faciliter le commerce, de les classer en grandes
catégories selon différents critères tels que l’espèce animale, la teneur en eau, le degré
d’affinage, le type de croutage, les moisissures apparentes, la texture, les techniques de
fabrication.

Chapitre 1. Synthèse bibliographique
36
La fabrication de fromage par la méthode classique comporte trois phases successives : la
coagulation, l'égouttage et l’affinage.
IV-2 Différentes étapes de la transformation du lait en fromage
Préparation
-standardisation
-maturation
Physique
Biochimique
Micro-organismes
Coagulant Coagulation = changement d’état
Formation d’un réseau
Égouttage= concentration
Elimination de lactosérum
Sortie lactosérum -décaillage
Intragranulaire -brassage
(En cuve et moule) -(chauffage)
-acidification
intergranulaire -retournements
(En moule) -(pressage)
Salage modification des constituants
Affinage
Figure 3 : Les différentes étapes de la fabrication d’un fromage
Lait
Lactosérum
Caillé
Fromage affiné
Gel

Chapitre 1. Synthèse bibliographique
37
IV.3 Coagulation
La coagulation du lait est une étape importante dans le processus de fabrication du fromage. Il
s’agit de la transformation du lait liquide en un gel, appelé aussi coagulum ou caillé qui, après
un certain nombre de transformations, deviendra un fromage.
La coagulation peut être obtenue par abaissement du pH (acidification), par action des
enzymes coagulantes ou par action mixte.
On distingue donc trois types de coagulation.
IV.3.1 Coagulation acide
L’acidification du lait peut conduire suivant les conditions, soit à un précipité de caséine, soit
à la formation d’un gel.
Elle consiste à précipiter les caséines à leur point isoélectrique (pHi =4.6) par acidification
biologique à l’aide des ferments lactiques qui transforment le lactose en acide lactique ou par
acidification chimique (injection de CO2, addition de gluconoδ-lactone ou ajout de protéines
sériques à pH acide).La voie chimique (acide organique) est uniquement utilisée en France
pour la standardisation du pH du lait avant emprésurage. L’addition d’acide minéral n’est
quant à elle pas autorisée.
Une acidification progressive obtenue par fermentation lactique, conduit à la formation d’un
gel lisse homogène qui occupe entièrement le volume initial du lait (Eck et Gillis, 1997).Ce
type de gel de par la solubilisation du phosphate de calcium colloïdal au cours de
l’acidification présente une bonne perméabilité mais une friabilité élevée ; le manque de
structuration du réseau (liaisons de faibles énergies de type hydrophobe) a pour conséquences
une élasticité et une plasticité pratiquement nulles et une faible résistance aux traitements
mécaniques (Mahaut et al., 2003).
IV.3.2Coagulation par voie enzymatique
Un grand nombre d’enzymes protéolytiques d’origine animale, végétale ou microbienne ont la
propriété de coaguler le complexe caséinique.
Mécanisme de la coagulation enzymatique :

Chapitre 1. Synthèse bibliographique
38
La coagulation comporte trois phases (Fig.4) :
L’hydrolyse enzymatique de la caséinek.
L’agrégation des micelles déstabilisées.
La réticulation et la formation du gel.
Figure 4:Phases de la coagulation enzymatique du lait (Mietton, 1994)
1ère
phase (hydrolyse enzymatique)
Elle concerne l’hydrolyse de la caséine k,au niveau de la liaison peptidique phénylalanine
(105) et méthionine (106), qui conduit à la formation de paracaséine k (1-105), et de
CMP (106-169). La libération du CMP se traduit par une réduction du potentiel de surface
des micelles et en conséquence, une diminution des répulsions électrostatiques qui à l’état
initial contribue à la stabilité du système colloïdal (Eck et Gillis, 1997).
Cette phase est indépendante des ions de Ca2+
et elle se produit à des températures variées (de
0 à 50°).

Chapitre 1. Synthèse bibliographique
39
2ème phase ou dite d’agglomération
Cette phase prend place lorsque 80% environ de la caséine k sont hydrolysés. La vitesse
d’agrégation s’accroit rapidement avec l’état d’avancement de la réaction enzymatique (Eck
et Gillis, 1997).
3ème phase ou de protéolyse générale
Les micelles agrégées subissent de profondes réorganisations : des liaisons de nature variée
s’établissent entre les micelles (électrostatiques, hydrophobes et salines) pour former un gel
constitué par un réseau lâche emprisonnant le lactosérum et la matière grasse. Dans le gel
ainsi formé, la caséine est présente sous une forme fortement minéralisée et ce degré de
minéralisation élevé confère au gel présure des caractères différents de ceux du gel lactique.
Le gel présure est souple, très cohésif, imperméable et contractile (Scriban, 1999).
IV.3.3 Coagulation mixte
Elle résulte de l’action conjuguée de la présure et de l’acidification. La multitude de
combinaisons conduisant à différents états d’équilibres spécifiques est à l’origine de la grande
diversité des fromages à pâte molle et à pâte pressée non cuite (Mahaut et al., 2003).
Dans la pratique industrielle, un gel mixte peut être obtenu selon deux techniques :
l’emprésurage d’un lait acide, l’acidification d’un gel présure (Veisseyre, 1975).
IV.3.3.1 Facteurs influençant l’activité coagulante
IV.3.3.1.1 Concentration en enzyme
Le temps de coagulation est inversement proportionnel à la concentration en enzyme, ce qui
peut se formaliser selon l’équation suivante (Mahaut et al ., 2003)
tc =K/E +ta
Avec : tc : temps de coagulation (s)
K : inverse de la constante de vitesse
E : concentration en enzyme

Chapitre 1. Synthèse bibliographique
40
ta : temps écoulé entre la fin de la réaction enzymatique et le point de coagulation (s).
IV.3.3.1.2Température
Le phénomène de coagulation est fortement dépendant de la température :
Au-dessous de 10°C, la coagulation du lait ne se produit pas.
Dans l’intervalle 10 à 20° C, la vitesse de coagulation est lente.
Au-dessus de 20°C, elle augmente progressivement jusqu'à 40 à 42°C, au-delà elle
diminue.
Au-dessus de 65°C, il n’y a plus de coagulation, l’enzyme est inactivée.
La température influe considérablement sur l’étape de l’interaction micellaire relativement à
son action sur l’étape de l’hydrolyse enzymatique (Brule et al ., 1987).
Tableau 9 : Impact de la température sur la coagulation du lait par la présure
(St-Gelais et Tirard-Collet, 2002).
Température Impact
<10°C Il n’y a pas de coagulation mais l’enzyme agit quand même.
10-20°C La coagulation est lente
30-42°C Il y a une augmentation progressive de la vitesse de coagulation.
42-55°C La vitesse de coagulation diminue.
55°C et + Il y a absence de coagulation.
IV.3.3.1.3 pH
L’abaissement du pH du lait entraine un temps de coagulation plus court. Cela résulte d’une
part d’un effet sur l’activité de l’enzyme, dont le pH optimal d’action sur la caséine kest de
5,5, d’autre part de la diminution de la stabilité des micelles, liée à la neutralisation des

Chapitre 1. Synthèse bibliographique
41
charges et de la libération d’ions calcium à partir des complexes dissous et colloïdaux
(Mahaut et al ., 2003).
IV.3.3.1.4 Teneur en CaCl2
L’addition de CaCl2 entraine une augmentation du calcium ionisé et du calcium colloïdal
ayant pour conséquence une augmentation de la taille des micelles (plus la dimension de la
micelle est grande, plus le temps de coagulation est court.)(Lenoir, 1985 ; Mahaut et al .,
2003).
IV.4 Egouttage
L’égouttage est l'étape de séparation du caillé (la phase solide) et du lactosérum ou petit lait
(la phase liquide) composée d'eau et de matières solubles telles que le lactose (75%), des
minéraux (essentiellement du calcium), des vitamines et des protéines (10%). Durant cette
phase, ce sont 80% de l'eau contenue dans le caillé qui sont éliminés.
Cette étape est importante puisque la nature du fromage fabriqué dépend de la méthode
d'égouttage pratiquée. Il faut notamment savoir que la capacité à s'égoutter d'un caillé est
directement en relation avec sa coagulation et que l'égouttage commence dès la coagulation
du lait.
Les pertes sont de deux natures :
- directes : chutes de caillé ou de résidus collant aux moules durant le pressage.
- indirectes : une irrégularité dans la répartition des grains (plus ou moins gros) au cours du
moulage ainsi qu’une irrégularité de traitement après moulage (température, hygrométrie,
pression) vont entrainer une évaporation irrégulière du sérum et accroitre les dispersions de
poids et d’extrait sec de fromages vendus à la pièce. On peut chercher à détecter ces
irrégularités en comparant les écarts types avant et après pressage, dans le cas de pates pressés
et voir ainsi la part des différents facteurs. Il est plus délicat d’évaluer les dispersions dues
respectivement au moulage et à l’égouttage des pâtes molles (Pointurier et al., 2003).
IV.5 Salage
Le salage peut s’effectuer selon deux techniques :

Chapitre 1. Synthèse bibliographique
42
salage à sec par saupoudrage superficiel, par frottage ou par incorporation dans la
masse du caillé (cantal, cheddar) ;
salage en saumure par immersion dans une solution saturée en NaCl.
L’incorporation du chlorure de sodium dans le fromage a pour objectifs :
- d’assurer un complément d’égouttage ;
- de contribuer éventuellement à la formation de la croute ;
- de régler l’activité de l’eau (aw) du fromage qui oriente et freinte les développements
microbiens et les actions enzymatiques au cours de l’affinage ;
- d’accroître le potentiel organoleptique fromage (Mahaut et al., 2003)
IV.6 Affinage
L’affinage correspond à une phase de digestion enzymatique des constituants du caillé, c’est
un processus biochimique complexe pour plusieurs raisons :
- d’une part, la matrice issue de la coagulation et de l’égouttage du lait présente une très
grande hétérogénéité physicochimique ;
- d’autre part, les enzymes intervenant dans l’affinage ont plusieurs origines.
Elles peuvent être présentes à l’origine dans le lait (plasmine, lipase, etc.), ajoutées au lait
(enzymes coagulantes, microorganismes), ou produites au cours de l’affinage par synthèses
microbiennes (bactéries, levures, moisissures).
L’affinage est dominé par trois grands phénomènes biochimiques : la fermentation du lactose,
l’hydrolyse de la matière grasse et la dégradation des protéines.
Les transformations confèrent à la pâte fromagère des caractères nouveaux. Elles la modifient
dans son aspect, dans sa composition, dans sa consistance. Simultanément, saveur, arome et
texture se développent.

Chapitre 1. Synthèse bibliographique
43
V. Coagulases d’origine bacterienne
Des études ont permis de mettre en évidence les potentialités de certaines bactéries à produire
des coagulases. Ce sont surtout les genres Bacillus (Bacillus subtilis, Bacillus coagulans,
Bacillus licheniformis) et Pseudomonasqui ont été plus particulièrement explorés (Green,
1977).
V.1 Le genre Bacillus
Le genre Bacillus englobe des bactéries à Gram positif en forme de bâtonnets droits,
généralement mobiles grâce à des cils péritriches, capables de former des endospores,
aérobies ou anaérobies facultatives. Les cultures âgées peuvent apparaitre gram négatif.
Les Bacillus sont très répandus dans la nature surtout dans le sol ; ils font partie de la flore
banale contaminant de nombreux produits alimentaires et sont souvent protéolytiques.
Le genre de Bacillus est classiquement subdivisé en fonction de la morphologie et de la
position de la spore en trois groupes :
Le groupe I : ce groupe est constitué des bacilles à Gram positif, présentant une spore
centrale ou terminale, sphérique ou ovoïde, ne déformant pas la cellule. Ce groupe est divisé
en 2 sous-groupes : le groupe IA constitué des bacilles d’un diamètre supérieur à 1 mm et
contenant des inclusions de poly-bêta-hydroxybutyrate (Bacillus anthracis, Bacillus cereus,
Bacillus megaterium, Bacillus mycoides, Bacillus pseudomycoides, Bacillus thuringensis) ; et
le groupe IB rassemblant des bacilles d’un diamètre inférieur à 1 mm et dépourvus
d’inclusions de poly-bêta-hydroxybutyrate (Bacillus coagulans, Bacillus firmus, Bacillus
licheniformis, Bacillus subtilis, Bacillus pumilus...).
Le groupe II : ce groupe est constitué des espèces à Gram variable, présentant une spore
ovoïde, centrale ou terminale, déformante (Bacillus circulans, Bacillus stearothermophilus).
Le groupe III : ce groupe est caractérisé par des bacilles à Gram variable et présentant une
spore sphérique, déformante, terminale ou sub-terminale (Bacillus globisporus, Bacillus
insolitus, ...).

Chapitre 1. Synthèse bibliographique
44
V.2 Intérêt industriel
Le développement considérable qu’a connu le genre Bacillus dans le domaine de la
production d’enzymes industrielles, revient aux caractéristiques intéressantes que présente ce
groupe :
Ces micro-organismes se cultivent facilement et s’adaptent parfaitement à un
développement industriel,
Ils présentent un fort potentiel génétique : les différentes espèces se rencontrent
dans des habitats variés et présentent de nombreux types trophiques
(psychrophiles, mésophiles, thermophiles, neutrophiles, acidophiles,
alcanophiles).
Ils excrètent à l’extérieur des cellules une quantité importante de leurs
protéines enzymatiques.
Enfin, ces enzymes exocellulaires sont des hydrolases, famille qui a connu le
plus large développement pour les applications industrielles, notamment les
amylases et les protéases (Schiff, 1992).
V.3 Les protéases de Bacillus
Les protéases de Bacillus sont des enzymes sécrétées dans le milieu extracellulaire. Parfois
leurs productions sont réprimées par l’adjonction en milieu de culture d’un hydrolysat de
protéines ou de mélange d’acides aminés (Arnaud et Guiraud, 1993).
Elles sont parmi les enzymes les plus anciennement connues et sécrétées par la plupart des
espèces de Bacillus telles que : Bacillus megaterium et Bacillus subtilis (Sastryet Mathur,
1979).
La quasi-totalité des protéases extracellulaires du genre Bacillus sont :
V.1.3.1 Les protéases alcalines (sérines protéases) :
Leur appellation est due à leur pH optimum d’action compris entre 9 et 11.
Les sérines protéases ont un poids moléculaire compris entre 25 000 et 30 000 et
sont caractérisées par la présence d’un résidu sérine au niveau de leur site actif.

Chapitre 1. Synthèse bibliographique
45
Elles sont stabilisées aux hautes températures par les ions Ca2+
.
En fonction de leurs propriétés générales, elles peuvent être classées en deux groupes :
Subtilopeptidase A : enzymes sécrétées par Bacillus licheniformis et Bacillus pumilus.
Subtilopeptidase B : enzymes sécrétées par Bacillus amyloliquefaciens et Bacillus
subtilis.
V.1.3.2 Les protéases neutres (métalloprotéases)
Elles ont un poids moléculaire variant entre 32,9 et 47,5 KDa. Les métalloprotéases
représentent le groupe des métalloendopeptidases ayant une activité maximale à un pH neutre
ou se rapprochant de la neutralité.
Les protéases neutres ne présentent aucune activité estérasique. L’insertion de l’ion de zinc au
niveau de la molécule enzymatique est primordiale pour son activité.
VI. Coagulases d’origine fongique
VI.1Généralités sur les moisissures
Les moisissures sont définies comme l’ensemble des champignons microscopiques
saprophytes présentant une végétation notable et qui ont de l’importance dans l’industrie
humaine (Leyral et Vierling, 2001).
Ce sont des eucaryotes (noyau vrai) avec des noyaux typiques entourés d’une membrane et
contenant des chromosomes (Marie, 1997).
Elles sont hétérotrophes car elles ne peuvent pas, comme les plantes vertes, synthétiser la
matière organique à partir du gaz carbonique atmosphérique (Marie, 1997).
Les plus répandues sont les Pénicilliums et les Aspergillus. Elles sont aérobies strictes.
Exemptes de chlorophylle, elles sont incapables de photosynthèse et tirent leur énergie de la
respiration et de la fermentation des matériaux organiques solubles qu’elles trouvent dans leur
environnement. Hétérotrophes et peu exigeantes, elles sont capables d’assimiler une grande
variété de substrats organiques grâce à leur potentiel élevé de sécrétion d’enzymes exo

Chapitre 1. Synthèse bibliographique
46
cellulaires leur permettant de s’attaquer à une gamme très large de composés naturels, dont
certains sont particulièrement rebelles comme la cellulose, la chitine ou la lignine. La plupart
des hydrates de carbone sont facilement assimilés. Les sources d’azote utilisées sont
nombreuses : sels d’ammonium, nitrites et nitrates, urée, acides aminés et source complexes
d’azote organique (Florent, 1993).
VI.2 Structure
Les champignons sont des organismes vivants constitués en grande partie de filaments de
cellules de structure simple et de quelques cellules plus spécialisées qui donneront naissance à
des spores. Les champignons ont un matériel génétique confiné dans un noyau au même titre
que les plantes et les animaux. Ils possèdent toutefois un certain nombre de caractéristiques
qui en font un groupe à part : parois contenant de la cellulose et de la chitine, absence de
chlorophylle et de mobilité. L’ensemble de ces caractéristiques fait en sorte que les
taxonomistes classent les champignons dans un règne distinct, soit celui des mycètes
(Kendrick, 1999 ; Malloch, 1997).
VI.3 Aperçu de la classification
La classification des moisissures, tout comme celle des autres champignons, est d’abord basée
sur le mode de reproduction sexuée ou phase téléomorphe. Ce critère définit quatre des cinq
ordres des mycètes, soit les Chytridiomycètes, les Zygomycètes, les Basidiomycètes et les
Ascomycètes (Fig.5).

Chapitre 1. Synthèse bibliographique
47
-Protozoaires
-Chromiste
---Eucaryotes -Plantes
(matériel génétique dans un noyau) -Animaux
-Mycètes Chytridiomycètes
Monde vivant (champignons) Zygomycètes
Basidiomycètes
Ascomycètes
Deutéromycètes
---Procaryotes - -Eubactéries
(matériel génétique diffus) -Archaebactéries
Figure 5 : Classification des mycètes (champignons) (Blackwell et al., 1998)
Certaines moisissures sont le plus souvent ou exclusivement rencontrées à des stades de
multiplication asexuée, dits anamorphes, et sont alors classées d’après le mode de production
des spores asexuées ou conidies. Ces espèces sont classées dans le cinquième ordre, les
Deutéromycètes, ou Fungi imperfecti.
VI.4 Cycle de vie des moisissures
Le réservoir naturel des moisissures se situe à l’extérieur, sur les végétaux, la matière
organique en décomposition, à la surface d’eau stagnante ainsi que dans le sol ou à la surface
de ce dernier (Malloch, 1997). Lorsque les conditions le permettent, les moisissures
produisent, à maturité, des spores qui peuvent être transportées par les courants d’air ou par
les humains et les animaux domestiques et se retrouver éventuellement dans les maisons et
édifices.

Chapitre 1. Synthèse bibliographique
48
Dans les climats tempérés, c’est durant la saison de croissance des végétaux et quelque temps
après celle-ci, que le nombre de spores se trouvant à l’extérieur est le plus élevé (Who, 2000).
Ces spores sont une forme latente des moisissures. La dispersion des spores peut se faire sur
de grandes distances. Des études effectuées en milieu contrôlé ont démontré qu’une petite
proportion des spores aéroportées peut se retrouver jusqu’à cent mètres de la source
d’émission bien que la grande majorité de celles-ci se retrouvent à proximité de leur lieu de
libération (Carlile et Watkinson, 1994).
En présence de conditions favorables à la sporulation, le mycélium donnera naissance à des
structures plus spécialisées, qui produiront des spores asexuées (conidies) ou, plus rarement,
des spores sexuées (Kendrick, 1999).
Chaque moisissure produit un très grand nombre de spores dont l’ensemble, appelé sporée, se
présente très souvent sous un aspect poudreux et coloré à la surface de la moisissure. La taille,
la forme et la couleur des spores de moisissures varient grandement d’une espèce à l’autre.
Par contre, en microscopie, toutes les spores d’une même espèce sont de couleur, de
dimension et de forme relativement constante ce qui, dans bien des cas, constitue un élément
d’identification taxonomique (Acgih, 1999). Le diamètre des structures fongiques de
reproduction varie entre 2 et 200 μm (Stetzenbach et Buttner, 2000), bien qu’il se situe pour
une forte proportion des spores entre 2 et 20 μm (Who, 2000). Certaines spores se retrouvent
à l’intérieur de structures fongiques spécialisées qui peuvent mesurer jusqu’à 250 μm de
diamètre. Tous les types de spores pourront dans des conditions favorables, recommencer un
cycle de vie, soit à proximité du thalle original ou même à forte distance de celui-ci, dans les
jours ou les mois suivant sa production.
VI.5 Conditions de croissance des moisissures
Dans leur milieu naturel, la plupart des moisissures sont saprophytes, tirant leur nourriture de
matières organiques mortes ou plus ou moins décomposées. Même si toute matière organique
peut constituer un substrat de croissance pour les moisissures, les conditions optimales de
croissance peuvent varier d’une espèce à l’autre, chacune d’entre elles ayant un degré
différent d’adaptation à son environnement. Certaines moisissures requièrent un taux
d’humidité très élevé pour croître tandis que d’autres préfèrent des taux beaucoup moins
élevés. Certaines peuvent croître sur des feuilles en décomposition, substance humide et
facilement pénétrable, tandis que d’autres s’attaqueront à des matières plus ligneuses, telles le

Chapitre 1. Synthèse bibliographique
49
bois ou même à des matières animales chitineuses tels les cheveux et les ongles. De plus, la
compétition inter-espèces procurera un avantage aux moisissures les mieux adaptées, référant
à la notion de niches écologiques particulières pour la croissance optimale de chaque type de
moisissure (Robbins et al., 2000 ; Malloch, 1997 ; Grant et al., 1988).
VI.5.1 Éléments nutritifs
La majorité des moisissures sont peu exigeantes quant aux éléments nutritifs nécessaires à
leur croissance. Les plus importants sont le carbone et l’azote, utilisés sous forme de
composés organiques, et des ions minéraux (Potassium, Phosphore, Magnésium…) en
quantités très faibles.
VI.5.2 Activité de l’eau
Les spores ne germent pas lorsque la teneur en eau d’un substrat est inférieure à 13% ou 14%
cependant, les exigences et la tolérance vis-à-vis de l’eau sont variables selon les groupes :
- Certaines moisissures ne se développement que sur substrat humide, c’est le cas par
exemple des mucorales ;
- D’autre peuvent proliférer sur des substrats dont l’humidité est très faible. Ce sont les
moisissures xérophiles qui rassemblent les espèces les plus osmophiles. Parmi ces espèces, les
mieux adaptées aux substrats secs sont les Aspergillus (Leyral et Vierling, 2001).
VI.5.3 Température
La végétation maximale est produite entre 20°C et 30°C, mais les moisissures tolèrent
généralement de +3°C à 40°C. Certaines espèces sont capables de continuer à croître à des
températures supérieures à 50°C. Les spores de moisissures sont détruites par un chauffage à
120°C pendant 20 à 30 min en chaleur humide, 3 heures à160°C en chaleur sèche (Leyral et
Vierling, 2001).
VI.5.4 pH
La majorité des espèces se développent dans des zones de pH assez larges, entre 3 et 8
(Florent, 1993).

Chapitre 1. Synthèse bibliographique
50
VI.5.5 Oxygène
Les moisissures se développent normalement dans les conditions aérobies (Florent, 1993).
VI.6 Importance économique des moisissures dans la production
d’enzymes
Les moisissures ou champignons filamenteux, ont une grande importance économique, en
raison à la fois de leur nocivité et de leur utilité. Leurs activités néfastes sont multiples :
altération des produits alimentaires et détérioration dans de nombreux autres domaines,
production de mycotoxines, vie parasitaire au dépond de l’homme, des animaux et des
plantes.
Les actions nocives de ces micro-organismes sont, heureusement, largement compensées par
leurs activités bénéfiques. Responsables de la destruction d’une grande partie de la matière
organique terrestre, les moisissures contribuent largement à l’accomplissement des grands
cycles biologiques naturels. Elles sont utilisées depuis fort longtemps par l’homme pour la
préparation d’aliment, intervenant notamment comme agents de fermentation dans la
fabrication de fromage. Elles synthétisent un grand nombre de substances complexes
économiquement très importantes : enzymes, acides organique, antibiotiques, etc. (Florent,
1993).
VI.7 Le genre Mucor
VI.7.1 Définition, caractéristiques et position taxonomique du genre Mucor
Selon (Gueguen, 1992), les Mucorales sont des champignons à "glucides simples", elles
compensent une faiblesse dans leur équipement enzymatique par leur rapidité foudroyante de
croissance, en consommant de petites molécules organiques. L’azote inorganique est utilisé
sous forme de sel d’ammonium. Un apport en macroéléments (P, K, Mg) et en oligo-éléments
(Zn, Fe, Mg, Cu, Mo) est nécessaire.
La plupart des Mucor vivent sur des matières végétales ou animales en décomposition mais
aussi dans le sol. Leurs spores asexuées, habituellement dispersées par le vent, se développent
dans les sporanges à l’extrémité des hyphes aériens. La reproduction sexuée produit des

Chapitre 1. Synthèse bibliographique
51
zygotes durs, entourés par une paroi épaisse et appelés zygospores ; ils restent dormants
quand l’environnement est trop rude pour la croissance du mycètes.
Une classification, proposée par Botton et al. (1990) ainsi que celle de (Yadav et Mishra,
1994), est illustrée par la figure 6. Cette classification est limitée à l’espèce utilisée dans notre
étude.
Protistes
Procaryotes Eucaryotes
Amastigomycota Mastigomycota Gymnomycota
Zygomycotina Ascomycotina Deuteromycotina Basidiomycotina
Entomophtorales Mucorales
Thamnidiaceae Mucoraceae
Thamnidium Mucor Mucor sp
Rhizopus
Absidia
Figure 6: la place de Mucor sp dans la classification des champignons (Botton et al., 1990)
VI.7.2Les espèces exploitées industriellement
Deux espèces ont été plus particulièrement exploitées au niveau industriel.

Chapitre 1. Synthèse bibliographique
52
Mucor pusillus
Il s’agit d’une moisissure banale du sol, exploitée par la firme japonaise METTOSANGYO. La
préparation enzymatique est commercialisée par Noury sous la dénomination de Noury- renn.
L’extrait est utilisé surtout aux Etats-Unis pour la fabrication du cheddar depuis de
nombreuses années. Ce fromage est semblable à celui obtenu par la présure
En ce qui concerne le fromage de type EDAM, la flaveur est moins fréquemment défectueuse
que l’EDAM fabriqué par la présure (Eck, 1987).
D’après Pien (1974), cette enzyme a permis d’obtenir des fromages de tous les types de
bonne qualité. Cependant, il a été constaté qu’une des causes possible d’une faible perte de
rendement peut être la fragilité un peu plus grande des caillés obtenus avec cette enzyme. Il
faut éviter une agitation trop grande au moment du décaillage pour que les poussières de caillé
ne partent pas dans le sérum. Toujours d’après le même auteur, on peut obtenir des fromages
un peu secs avec cette enzyme.
Mucor miehei :
Les préparations obtenues à partir de cette moisissure banale du sol, sont produites
par diverses firmes:
Française RAPIDASE sous la dénomination fromase,
Danoise nano industrie sous le nom de rennilase,
Américaine mille sous l’appellation marzyme,
Danoise Ball Hansen sous le nom hammilase.
Endothia parasitica :
C’est une moisissure parasite du châtaignier.
La préparation enzymatique est produite par la firme américaine PFIEZER et commercialisée
sous le nom sufaren.

Chapitre 1. Synthèse bibliographique
53
VI.7.3 Les protéases du genre Mucor et leur application en fromagerie
Mucor pusillus secrète, selon Somkuti et Babel (1968), une seule protéase acide active à pH
3,8 et 5,6 sur substrat hémoglobine et caséine respectivement. L’enzyme est très thermostable,
son optimum thermique est de 50°C; elle est irréversiblement dénaturée à 65°C.
Cependant (Khanet al., 1979) ont montré la production de deux protéases à la place d’une
seule ; l’une est semblable à la présure, l’autre caractérisée par une activité protéolytique
insignifiante.
D’après (Luquet, 1985), cette enzyme a permis d’obtenir des fromages de bonne qualité.
Cependant, la diminution du rendement fromager observée peut être due, selon (Houins et al.,
1973), à une production importante de peptides solubles dans l’eau. En effet selon les mêmes
auteurs, l’enzyme extrait du Mucor pusillus se caractérise par une forte concentration de gros
peptides pouvant expliquer le gout amer décelé dans certains fromages fabriqués avec cette
enzyme.
En revanche, les travaux de (Polychroniadou et Manolkidis, 1984), ont montré la bonne
aptitude que possèdent les extraits coagulants microbiens : Noury-Rennet, Rennilase et
Suparen à la fabrication de fromage de chèvre à croûte fleurie.
Par ailleurs, certains auteurs (Green et Stackpoole, 1975 ; Jarmul et al., 1982) ont démontré
l’efficacité d’utiliser une préparation fongique (Noury ou fromase) en mélange avec la
pepsine dans la fabrication de fromage.
VII. La fermentation industrielle
Le mot fermentation présente deux significations différentes pour les biochimistes et les
microbiologistes industriels. En biochimie, les fermentations sont des voies cataboliques
anaérobies au cours desquelles des composés organiques servent à la fois de donneurs et
d’accepteurs d’électrons, la synthèse d’ATP étant réalisée par phosphorylation au niveau du
substrat. En microbiologie industrielle, le terme de fermentation désigne l’opération unitaire
qui permet de produire de la biomasse ou des produits de bioconversion par la culture de
microorganismes.

Chapitre 1. Synthèse bibliographique
54
Ainsi, contrairement au sens biochimique, le terme de fermentation industrielle ne se réfère
pas au métabolisme du microorganisme. Ce terme s’applique en industrie pour des
métabolismes aérobies et anaérobies (Pascal et al., 2004).
VII.1 Buts de la fermentation industrielle
La mise en œuvre des procédés de la fermentation industrielle peut tendre vers divers buts qui
sont :
- L’obtention des micro-organismes eux-mêmes ; en quantité importante pour la
préparation des protéines, ou des autre constituants cellulaires ;
- La production de composés ou produits sécrétés naturellement par les micro-
organismes comme les enzymes, les pigments ou les toxines ;
- La production des métabolites, dans certaine condition, comme les antibiotiques, les
vitamines ou les aminoacides par les micro-organismes (Simon et Meunier, 1970).
VII .2 Types de fermentation
VII .2.1 Fermentation sur milieu liquide
Cette technique a été la première utilisée. Elle consiste à cultiver les moisissures sur la surface
d’un milieu nutritif liquide, elle nécessite des conditions aseptiques strictes. La culture en
milieu liquide s’effectue dans des récipients agités, et le plus souvent dans des Erlenmeyers,
placés sur des appareils d’agitation qui favorisent la diffusion de l’air à travers les tampons de
coton servant à les boucher.
Ce type de fermentation a été traditionnellement utilisé pour la production industrielle des
enzymes, en raison de la facilité de contrôle des différents paramètres comme le pH, la T°,
l’aération, l’oxygène dissout et l’humidité. La fermentation submergée est généralement plus
facile à exploiter de manière aseptique par rapport à la fermentation solide et peut être
appliquée plus facilement à l’échelle industrielle
VII .2.2 La fermentation solide ou semi-solide
VII .2.2.1. Définition

Chapitre 1. Synthèse bibliographique
55
La fermentation solide (fermentation de substrats solides, fermentation humide, culture solide
est généralement définie comme une croissance microbienne sur des particules solides
humides en l’absence d’eau libre (Durand, 2003 ; Gervais et al., 2003 ; Rahardjoet al.,
2006). De façon simplifiée, les microorganismes se développent dans un système à trois
phases : une matrice solide, une phase liquide qui lui est liée et une phase gazeuse prise au
piège dans les particules ou entre celles-ci.
Rahardjo et al. (2006) expliquent la diffusion des moisissures filamenteuses dans le substrat
solide humide. Selon les substrats considérés, l’apparition d’eau libre se manifeste pour des
teneurs en eau comprises entre 12 et 90 %, soit 0,65 et 0,98 d’activité d’eau (aw). (Mathot,
1996 ; Gervais et al., 2003).
VII .2.2.2 Avantages de la fermentation solide
La fermentation solide possède une réputation de technique simple à mettre en œuvre, ne
nécessitant pas obligatoirement une stérilisation énergétique du substrat de culture et requiert
un faible espace. Elle assure une forte productivité en métabolites. La faible teneur en
humidité empêche la contamination bactérienne. Cependant, cette renommée est largement
surfaite, surtout lors du développement d’applications pilotes ou en vraie grandeur. Dans ces
conditions et bien d’autres, la fermentation solide présente de nombreux inconvénients
(Assamoi et al., 2008).
VII .2.2.3 Inconvénients des fermentations solides
Les problèmes de transfert d’oxygène et de chaleur rendent difficile l’augmentation d’échelle
des procédés. En effet, la faible quantité d’eau ralentit les échanges de chaleur, pouvant créer
des problèmes de surchauffe lorsque des masses importantes sont mises en fermentation.
L’évaporation compense partiellement cet échauffement, mais en réduisant l’eau disponible.
L’évacuation des calories métaboliques peut donc poser un problème qu’il s’agit de résoudre
lors du passage de petits essais de laboratoire aux applications en vraie grandeur.
La nature solide et hétérogène des substrats utilisés complique le suivi direct des paramètres
de fermentation. Les sondes utilisées en fermentation liquide ne sont pas utilisables, même si
Bellon-Maurel et al. (2003) proposent de nouveaux types de sondes adaptées aux cultures
solides. Il est pratiquement difficile d’assurer une distribution parfaitement homogène de
substances ajoutées au substrat et donc du milieu de culture. Ce qui rend le contrôle on line

Chapitre 1. Synthèse bibliographique
56
des paramètres de culture tels que le pH, l’humidité et la concentration des nutriments, assez
aléatoire. Les microorganismes étant inséparables du substrat, l’estimation de la biomasse est
délicate. Étant donné la forte concentration des métabolites obtenus, des produits inhibiteurs
générés par les microorganismes peuvent s’accumuler en concentration élevée dans le milieu
de culture.
VII .2.2.4 Les diverses étapes suivies en fermentations solides
Ce sont respectivement la préparation du substrat ou du milieu de culture, la stérilisation du
milieu (généralement à 121 °C pendant 21 minutes) suivie par le refroidissement de celui-ci,
l’inoculation du milieu de culture, l’incubation du milieu inoculé et l’incubation en
maintenant dans la mesure du possible les conditions environnementales optimales
(température, pH, teneur en eau) (Assamoi et al., 2008).
L’isolement des microorganismes : il n’existe que deux moyens d’obtenir des
microorganismes :
- les isoler à partir des fermentations naturelles de l’eau, de l’air, du sol, ou les matériaux
organiques ou même inorganiques plus au moins décomposées.
- les obtenir déjà isolés à l’état de culture en s’adressant au collecteur des souches (Simon
et Meunier, 1970).
La préparation du substrat carboné : les substrats carbonés utilisés en fermentation
solide proviennent essentiellement des résidus agricoles et agroalimentaires. Ce sont
les substrats lignocellulosiques (sous forme de paille, de son, de bagasse, de pulpe,
etc.), les féculents (résidus de banane, de soja, les graines, etc.
La préparation du substrat carboné vise à lui faire subir divers traitements physique,
chimique ou biologique (enzymes) afin de favoriser une grande accessibilité des
constituants aux microorganismes. Le traitement physique peut se faire soit
mécaniquement (broyage, concassage, hachage, aplatissage), soit thermiquement
(chauffage, autoclavage, traitement à la vapeur), soit par irradiation ou par ultrasons.
Le traitement chimique se fait par la voie des alcalis, acides, oxydants, gaz et solvants
(Assamoi et al., 2008).
La fermentation proprement dite: après avoir amélioré la souche par culture dans
des milieux sélectifs afin de favoriser le développement rapide de la souche et après

Chapitre 1. Synthèse bibliographique
57
l’avoir purifiée, tout cela à une température adéquate, on prépare l’inoculum. Il faut
inoculer largement le milieu nutritif afin que la souche ensemencée se développe
rapidement dans le plus court délai, à une température spécifique. Cette technique est
utilisée souvent dans la production enzymatique et pour la production de divers
produits d’intérêt industriel agroalimentaire (Aguilar et al., 2008).
L’inoculation du milieu de culture : l’inoculation du milieu de culture se fait le plus
souvent à partir d’une suspension de spores (Mathot, 1996). Celles-ci restent viables
plus longtemps que du mycélium ; elles sont moins sensibles aux conditions externes
et se conservent plus facilement. La quantité optimale de spores à inoculer diffère
selon les cas. Un excès de spores peut parfois inhiber la germination. Les spores sont
cependant métaboliquement dormantes, impliquant que la dégradation du substrat ne
peut s’installer qu’après la germination. Pour minimiser cet inconvénient, une pré
germination des spores est parfois envisagée. Des inoculum spores + mycélium +
substrat de production peuvent être utilisés également (Assamoi et al., 2008).
L’optimisation de la température, de la teneur en eau, du pH et de l’aération de
la culture :à l’échelle industrielle, les contrôles de la température de culture et de
l’humidité du milieu sont très importants (Bellon-Maurel et al., 2003). La faible
conductibilité thermique des substrats utilisés et leur faible teneur en eau réduisent le
transfert de chaleur, qui lui-même dépend de la taille des particules de la couche
solide. Une élévation de la température dans la masse fermentable due à un
dégagement de chaleur métabolique peut aller jusqu’à atteindre 80°C, causant un
assèchement de la culture et une baisse de l’aw et de la disponibilité en nutriments
(Assamoi et al., 2008).
L’extraction et la purification des enzymes : la fermentation terminée, les enzymes
doivent être séparées des cellules et du milieu, et traitées de façon à obtenir une
préparation commerciale répondant aux critères de pureté et de stabilité souhaités. Ces
traitements sont des opérations unitaires simples telles que centrifugation, filtration,
évaporation, précipitation, séchage… Les préparations enzymatiques sont
commercialisées sous différentes formes : liquide, poudre, concentré, lyophilisée,
immobilisée (Scriban, 1988).
VII. 3 Production d’une enzyme industrielle d’origine microbienne
La production industrielle d’enzymes s’est orientée vers les processus fermentaires.

Chapitre 1. Synthèse bibliographique
58
Selon (Joyeaux, 1982), des fermentations en profondeur assurent une meilleure homogénéité
et un contrôle plus strict des paramètres critiques de la synthèse de l’enzyme au cours de la
fermentation.
VII.4 Sélection d’une souche microbienne
Les souches productrices d’enzymes doivent présenter un certain nombre de caractéristiques
pour être définitivement retenues :
- pousser sur milieu simple et bon marché.
- produire le moins possible des métabolites secondaires (antibiotiques,…).
- ne pas conduire à différents polluants.
- produire une enzyme exocellulaire facile à séparer et purifier.
- ne pas être pathogène ou produire des composés toxiques.
VII.5 Production industrielle
Actuellement les cultures submergées sont préférées aux cultures de surface car elles sont
mieux adaptées aux différents contrôles de température, d’aération et d’humidité, par des
méthodes modernes, et réduisent les risques de contamination.
VII.6 Extraction et purification
Une fois la fermentation terminée, les enzymes doivent être séparées des cellules et du milieu
de culture et traitées par des opérations unitaires simples telles que la centrifugation, filtration,
évaporation, précipitation, séchage.
Les préparations enzymatiques peuvent être commercialisées sous différentes formes : liquide
concentrée, poudre, lyophilisée.
VIII. Planification expérimentale
VIII.1 L’optimisation du milieu de culture
La recherche de conditions optimales de production est très importante en analyse
fondamentale pour l’industrie et l’ingénierie. En effet optimiser une production ou même un
système c’est aboutir à un rendement maximal.

Chapitre 1. Synthèse bibliographique
59
Les expériences au laboratoire ou en contexte industriel concernant généralement l’étude de
l’influence de nombreux facteurs sur les résultats d’un phénomène ou d’un procédé. Pendant
longtemps la procédure expérimentale consiste à faire varier un facteur à la fois en maintenant
fixe tous les autres facteurs. L’utilisation d’un plan d’expérience où plusieurs facteurs varient
à la fois, a permis d’alléger l’expérimentation en terme de temps et de coûts, tout en
permettant l’étude des interactions entre les facteurs et en augmentant la précision des
estimations des effets.
Ainsi, les plans d’expérience concernés sont utilisés pour l’amélioration de la qualité et de la
sécurité des produits et des procédés (Christine et al., 2006).
VIII.2 Les plans d’expériences
C’est le plan d’organisation des essais expérimentaux dans le but de connaître le
comportement du résultat (réponse) à partir de la variation des facteurs choisis. Un bon plan
permet une diminution notable du nombre d’essais tout en donnant une bonne précision dans
la détermination des résultats.
Les plans d'expériences permettent d'organiser au mieux les essais qui accompagnent une
recherche scientifique ou des études industrielles (Goupy, 2001). Ils sont applicables à de
nombreuses disciplines et à toutes les industries à partir du moment où l’on recherche le lien
qui existe entre une grandeur d’intérêt (réponse), y, et des variables(facteur), xi. Il faut penser
aux plans d'expériences si l’on s’intéresse à une fonction du type : y = f (xi)
Avec les plans d'expériences on obtient le maximum de renseignements avec le minimum
d'expériences. Pour cela, il faut suivre des règles mathématiques et adopter une démarche
rigoureuse (Box, 2005). Il existe de nombreux plans d'expériences adaptés à tous les cas
rencontrés par un expérimentateur. Les principes fondamentaux de cette science seront
indiqués et les principaux plans seront passés en revue. La compréhension de la méthode des
plans d'expériences s'appuie sur deux notions essentielles, celle d'espace expérimental et
celle de modélisation mathématique des grandeurs étudiées.

Chapitre 1. Synthèse bibliographique
60
VIII.3 Notion d'espace expérimental
Un expérimentateur qui lance une étude s'intéresse à une grandeur qu'il mesure à chaque
essai. Cette grandeur s'appelle la réponse, c'est la grandeur d'intérêt. La valeur de cette
grandeur dépend de plusieurs variables.
Au lieu du terme «variable» on utilisera le mot facteur. La réponse dépend donc de un ou de
plusieurs facteurs.
VIII.4 Notion de surface de réponse
On définit un axe orthogonal à l’espace expérimental et on l’attribue à la réponse. La
représentation géométrique de plan d’expériences et de la réponse nécessite un espace ayant
une dimension de plus que l’espace expérimental.
A chaque point du domaine d’étude correspond une réponse. À l’ensemble de tous les points
du domaine d’étude correspond un ensemble de réponses qui se localisent sur une surface
appelée la surface de réponse (Fig.7).
Les plans du second degré ou plans pour surfaces de réponse permettent d'établir des modèles
mathématiques du second degré (Box, 2005). Ils sont utilisés pour les variables continues.
Pour trois facteurs, on a :
y = α0+ α1X1+α2X2 + α3X3+ α11X2 2+α22 X2
2 +α33X3
2 + α12 X1X2+α13X1X3+α23X2X3
La représentation géométrique de plan d’expériences et de la réponse nécessite un espace
ayant une dimension de plus que l’espace expérimental.

Chapitre 1. Synthèse bibliographique
61
Figure 7 : Surface de réponse pour un domaine expérimental
VIII.5 Expérience factorielle de 2éme
ordre
L’expérience factorielle totale (EFT) est habituellement utilisée pour l’obtention d’un modèle
mathématique linéaire. Cependant, très souvent, les liaisons entre les facteurs et les
paramètres sont plus complexes. Dans ce cas leur description au moyen du modèle
mathématique de 1er
ordre est insuffisante.
VIII.6 L’expérience composite centrée
Parmi les catégories d’expériences factorielles pouvant être utilisées pour l’obtention du
modèle mathématique de 2éme
ordre, figurent les expériences dite composées centrales (ECC).
Ces plans ont été proposés pour la première fois par Box et Wilson(1951). Ils sont
construits à partir des plans factoriels complets ou fractionnaires et permettent d’estimer
efficacement un modèle du 2e degré en assignant aux facteurs plus de 2 niveaux.
L’expérience composite centrée (ECC) à K facteurs comporte trois groupes d’essais :

Chapitre 1. Synthèse bibliographique
62
Les essais dans le noyau du plan :
Ce noyau représente l’expérience factorielle totale ou partielle. Dans le premier cas, le
nombre d’essais dans ces points est égale à :
Nn=φK
Où : φ= nombre de niveau de chaque facteur =2
K= nombre de facteurs
Donc Nn=2K
Les essais dans les points étoilés ou points axiaux :
Pour chaque facteur il y a deux points étoilés. Le nombre d’essais dans ces points est égal
à : Npe=2K
Les essais dans le centre de l’expérience :
Le nombre d’essai au centre du domaine.
Remarque :
Le nombre total d’essais dans l’expérience composée centrale est égale :
N=Nn+Npe+Nce.

Chapitre 1. Synthèse bibliographique
63
Il faut cependant noter que, dans l’expérience composée centrale, chaque facteur varie sur 5
niveaux. En valeurs codées, nous avons les niveaux suivants :
Xi= -α (Niveau inférieur dans le point étoilé ; α est souvent appelée épaule étoilée
Xi= -1 (Niveau inférieur)
Xi= 0(Niveau zéro)
Xi= +1(Niveau supérieur)
Xi= + α (Niveau supérieur dans le point étoilé).
VIII.7 Types d’expériences composées centrales
En fonction de la valeur α et du nombre d’essais dans le centre de l’expérience Nc, on distingue
deux types d’expériences composées centrales :
L’expérience composée centrale orthogonale ;
L’expérience composée centrale rotatable.
A noter que le deuxième type d’expérience est le plus répandu. Les paramètres de cette expérience
sont présentés dans le tableau 10.
Tableau 10 : Les paramètres d’une expérience composée centrale rotatable.
Nombre de facteurs Nn Npe Nce N α Noyau du plan
2 4 4 5 13 1.414 EFT22
3 8 6 6 20 1.682 EFT23
4 16 8 7 31 2.000 EFT24
5 32 10 10 52 2.378 EFT25
6 64 12 15 91 2.828 EFT26

Chapitre 1. Synthèse bibliographique
64
VIII.8 Etape de planification, de réalisation et de traitement des résultats de
l’expérience composite centrée
La planification, la réalisation et le traitement des résultats de l’expérience composite centrée
comprennent les étapes suivantes :
Le choix de plan factoriel de l’expérience ;
La codification des facteurs ;
La réalisation des essais dans le noyau (toutes les expériences) ;
La réalisation des essais aux points étoilés et au centre de l’expérience ;
L’estimation de la variance de l’erreur de l’expérience;
Le calcul des coefficients du modèle mathématique de 2éme
ordre ;
La vérification de la signification statistique des coefficients du modèle
mathématique ;
La vérification de l’adéquation du modèle mathématique obtenu.

Chapitre 2
Matériel et méthodes

Chapitre 2. Matériel et Méthodes
65
I..Méthode d’isolement des microorganismes d’intérêt
Une première étape du travail a consisté à isoler des microorganismes susceptibles de
produire des enzymes coagulant le lait, à partir du sol.
La terre utilisée pour la recherche des microorganismes a été prélevée à proximité de la
laiterie fromagerie de Boudouaou, précisément aux alentours du local contenant les sacs de
poudre de lait.100 à 150 grammes de sol sont pris à des profondeurs variant entre 3 et 15cm à
l’aide d’une spatule stérile et stockés dans des contenants stériles.
Les échantillons de terre sont tamisés dans des tamis métalliques à maille de 2mm de diamètre
environ. Le produit de tamisage est recueilli sur des grandes feuilles de papier sulfurisé ; 50 à
100g sont préservés en flacon stérile. Puis de manière aseptique, 10 gr de terre tamisés sont
pesés, puis broyés le plus finement possible afin de libérer au mieux les microorganismes. La
terre est transférée dans une fiole de 100ml, complétée avec de l’eau physiologique stérile. Le
mélange est ensuite homogénéisé pendant 20 minutes sous agitation magnétique (Davet et
Rouxel, 1997). Nous obtenons ainsi une dilution au 1/10.
A partir de là nous préparons les autres dilutions (1ml de10-1
dans 9ml d’eau physiologique
stérile pour avoir 10-2
et ainsi de suite jusqu’à 10-9
).
Les milieux de culture à utiliser sont coulés en boîte de Pétri (d= 9cm), puis ensemencés (pour
les dilutions de 10-3
à 10-9)
par étalement de 0,1 ml de chaque dilution à l’aide d’un râteau à la
surface des milieux
En ce qui concerne l’isolement des bactéries, le milieu employé est le milieu Plate Count
Agar (P.C.A.), additionné de 10% lait écrémé UHT et d’actidione (cycloheximide) à2%.
L’incubation s’effectue à 30°C pendant 48h à 72h.
Dans le cas des moisissures, le milieu utilisé est le Sabouraud-chloramphénicol, dans lequel
est ajouté 10% lait écrémé UHT. L’incubation a lieu à 25° pendant 3 à 4j.
Dans un premier temps, nous ferons référence aux méthodes utilisées lors de l’étude portant
sur les bactéries. Celles qui sont relatives aux moisissures, seront évoquées dans un deuxième
temps.

Chapitre 2. Matériel et Méthodes
66
II. PROTEASE D’ORIGINE BACTERIENNE
II.1. Isolement des souches bactériennes
II.1.1. Choix et purification des souches
Au bout de 30h d’incubation, des colonies de bactéries apparaissent et nous nous intéressons
particulièrement aux colonies qui présentent un halo clair d’hydrolyse du lait ou caséolyse.
Les souches caséolytiques sont prélevées et repiquées sur des milieux P.C.A. contenant du
lait, puis purifiées éventuellement par plusieurs isolements par la méthode des quadrants
jusqu’à l’obtention de souches pures.
Au final, seules les souches isolées présentant un halo de caséolyse témoignant d’une action
sur les protéines du lait sont retenues.
II.1.2. Observations macroscopiques et microscopiques
Le pouvoir caséolytique des souches pures sélectionnées a été vérifié par un isolement sur
milieu P.C.A. additionné de lait (1ml de lait écrémé stérile pour 15ml de milieu P.C.A.).
Après incubation (30°c/48h), la présence d’un halo clair autour de la colonie caractérise le
pouvoir caséolytique : le diamètre de la zone de caséolyse mesuré (Fig.8)
Figure 8 : colonies caséolytiques après plusieurs heures d’incubation
P47 FK6
JF9

Chapitre 2. Matériel et Méthodes
67
L’étude microscopique des différentes souches est entreprise à l’aide d’une coloration Gram
et éventuellement d’une coloration de spores (Méthode Benitto), en fonction de l’observation
morphologique.
Méthode de coloration de Benito :
- déposer le frottis de bactéries sur une platine chauffante
- recouvrir largement le frottis avec du vert Malachite à 5% (5g dans 100ml d’eau
distillée)
- chauffer jusqu’à émissions de vapeur et laisser refroidir
- renouveler plusieurs fois (3) l’opération, laissée en contact 10mn en ajoutant si
nécessaire du vert Malachite pour éviter l’évaporation
- rincer à l’eau courante, puis recouvrir le frottis d’une solution de safranine à 0,25%
(0,25g dans 100ml d’eau distillée) pendant une minute
- rincer à l’eau, sécher et observer à l’objectif *100 à immersion.
- Résultats : les spores apparaissent colorées en vert, les cellules végétatives en rose
II.1.3.Conservation des souches
Au final, nous disposons d’une collection de 70 souches qui ont été codées afin de les
différencier. Elles ont été étudiées du point de vue macroscopique et microscopique en
procédant à une coloration de Gram et éventuellement une coloration de Benito (mise en
évidence de spores) et en mesurant le diamètre de la plage de caséolyse.
Ces souches sont présentées dans le tableau N°11

Chapitre 2. Matériel et Méthodes
68
Tableau N° 11 : Collection de souches bactériennes isolées à partir du sol
Code de la souche Aspect microscopique GRAM Caséolyse
en mm
FK 1 grands bâtonnets + 3
FK 2 Bacillus taille moyenne + 5
FK 3 Mélange de cocci et de bacilles courts + 1
FK 4 grands Bacillus + 5
FK 5 Mélange de bacilles + 4
FK 6 A Très longs bacilles, spore légèrement déformante + 5
FK 6 B Bacilles formant des petites chainettes, spores
terminales légèrement déformantes + 8
FK 7 petit Bacilles avec spore terminale déformante + 5
FK 8 Bacillus taille moyenne + 5
FK 10 Bacillus taille moyenne + 5
FK 11 Bacillus très granuleux plus gros que KF10 + 5
FK 12 Bacillus court + 3
FK 13 Bacillus granuleux gros trapus rectangulaire et court + 3
FK 14 Bacillus taille moyenne en voie de sporulation + 5
FK 15 Bacillus granuleux gros trapus rectangulaire et court + 2
FK 16 Bacillus granuleux gros trapus rectangulaire et court + 3
FK 17 Bacillus granuleux gros trapus rectangulaire et court + 2
A 1 Petit bacilles - 6

Chapitre 2. Matériel et Méthodes
69
A 2 Petit bacilles - 5
A 3 Cocco bacille - 5
C 1 Cocco bacille - 5
C 2 Bâtonnet assez grand + 5
C 3 Bacilles courts - 1
C 4 Bacilles courts - 5
C 5 Grands bacilles + 5
C 6 Bacilles plus courts + 4
P 43 Ressemble à JF9 en plus minces bacilles + 5
P 47 Bacilles droit extrémités carrés + 4
P 47 M Bacilles courts et fins, plus fins que FK6 + 5
P 47 A Bacilles longs et fins assez polymorphes + 6
P 47 β Petits bacilles en faisceaux, pas de spore en
apparence + 6
P 47 γ Bacilles plus longs et plus fins que P47β + 6
P 47 R Bacilles courts peu sporulés + 6
P 48 Bacilles taille moyenne plus long que P47 et plus
gros + 6
J F 2 Bacillus courts plus mince + 3
J F 3 Bacillus courts plus mince + 5
J F 5 Bacillus courts + 5
J F 6 Bacciles assez fins taille moyenne + 3

Chapitre 2. Matériel et Méthodes
70
J F 8 Bacillus taille moyenne, spores terminales
déformantes + 7
J F 9 Tres long bacillus en voie de sporulation + 5
J F 10 Bacilles courts + 5
J F 11 Bacilles courts + 5
J F 13 Bacillus granuleux assez gros, présence de spores + 6
J F 14 Bacillus taille moyenne formants des chainettes + 2
J F 15 Bacillus courts, spores terminales déformantes + 4
J F 16 Bacillus taille moyenne formants des chainettes + 2
P47 Bacillus taille moyenne + 6
F3 Bacciles taille moyenne granuleux parfois en
chainettes + 2
F4 Petit bacilles en faisceaux + 3
F5 Gros bacilles + 1
F6 long bacilles droit et fin + 1
F7 bacillus moins gros que F3 mais plus long + 2
F8 bacillus moins gros que F3 mais plus long + 3
F10 Bacilles courts trapus et fins + 2
F13 Bacilles taille moyenne granuleux parfois en
chainettes + 3
F14 Gros bacilles + 5
F15 Bacillus avec granulation fines + 6
F16 Bacillus long et fins en faisceaux + 5

Chapitre 2. Matériel et Méthodes
71
F18 Bacillus avec granulation fines F15 en plus petit + 2
I 1 Bacillus en chainette assez gros granuleux + 1
I 2 Bacilles assez long et fins + 1
I 4 Petit bacillus + 1
I 5 Bacillus assez gros granuleux spores centrale + 2
I 6 Bacillus granuleux avec spores ovoides non
déformantes + 2
I 7 Bacillus plus trapus différents de I5 et I6 + 1
I 10 Long bacilles sporulés flexueux + 3
I 11 Bacillus taille moyenne avec spores terminales
déformante + 4
I 12 Bacilles courts + 2
I 33 Bacilles courts + 1
I 33M Bacillus assez gros peu de chainettes + 6
Les souches isolées sont conservées sur gélose inclinée de milieu « conservation », formule
Institut Pasteur, et cela à température ambiante.
Les souches islées ont été testées pour leur activité coagulante, les bactéries gram- ox- ayant
été éliminées en raison du risque de pathogénicité.
II.2. Sélection des souches à activité coagulante
L’activité coagulante de 70 souches de la collection a été évaluée. Une étude préalable
concernant les milieux de production de l’enzyme a été entreprise et un nouveau milieu « au
jus de son » a été mis au point. Les extraits enzymatiques obtenus après culture ont été
caractérisés par les activités coagulante et protéolytique et un dosage de protéines.

Chapitre 2. Matériel et Méthodes
72
II.2.1Choix des milieux de culture : étude préalable
Deux milieux de culture ont été expérimentés afin de rechercher l’activité coagulante des
différentes souches. Le premier est un milieu liquide de nature synthétique (Ageito et Vallejo,
2007), le deuxième est le milieu semi-solide (S.E.L) selon Rao et Mathur, 1976.
Après avoir testé un grand nombre de souches parallèlement sur ces deux milieux, il s’est
avéré que le milieu liquide ne favorisait pas la production de l’enzyme coagulante des souches
sélectionnées. Nous avons donc opté pour des milieux semi solide : le milieu au son selon
Rao et Mathur, 1976, et un milieu au son modifié mis au point pour des raisons pratiques.
II.2.1.1 Composition des milieux de culture
Milieu liquide : en g/l (Ageito et Vallejo, 2007)
20g : Glucose, 10g : (NH4)2SO4 , 1g: KH2PO4 ; 0,4g: MgSO4; 2mg FeSO4, 2 mg : MnSO4;
50mg : NaCl, 3g : Urée, 2g : Caséine.
Semi-solide en g/l préconisé par Rao et Mathur (1976).
50g : Son de blé, 3g : Extrait de levure:4 g Glucose : 2 g caséine : 3g Na2HPO4.
II.2.1.2 Préparation du milieu au son modifié
Nous avons élaboré un milieu au son modifié, réalisé à partir du jus de son. Le jus de son est
fabriqué en mélangeant dans un flacon 125g de son et 750ml d’eau. Le mélange est autoclavé
pendant 1h à 120°C. Une fois refroidi, ce mélange est pressé manuellement dans un tissu fin,
le jus est récupéré, la matière sèche est ensuite déterminée. 5 g de jus de son (3exemplaires)
sont placés dans des petits récipients en aluminium et mis à sécher à 105°C jusqu’à poids
constant.
Le milieu est préparé en fonction de la matière sèche du jus de son. Après calcul des
pourcentages, les ingrédients sont pesés et ajoutés au son, dont nous connaissons la
composition moyenne (annexe 1)
L’avantage de ce milieu est que les substances nutritives du son sont plus disponibles pour les
microorganismes. Le milieu est plus fluide et l’étape de filtration n’est plus nécessaire.

Chapitre 2. Matériel et Méthodes
73
L’emploi de ce nouveau milieu a été validé par une étude comparative des résultats obtenus
avec ceux du milieu au son classique (Rao et Mathur, 1976).
Une fois les différents milieux préparés, ils sont répartis par 50 ml dans des erlems et sont
stérilisés par autoclavage (120°C pendant 20 minutes).
II.2.1.3 Ensemencement et incubation des milieux
L’ensemencement se fait à raison de 5 107 à 5 10
8 bactéries par millilitre de milieu. 5ml d’une
suspension bactérienne Mac Farland 3 (annexe 7) (environ 9 108/ml) sont ajoutés dans le
milieu. Le nombre des bactéries inoculées est vérifié par dénombrement sur milieux coulés en
boîte de Pétri au moment de l’ensemencement.
L’incubation a lieu à 30°C pendant 24H sous agitation à 110 rpm ((Poza et al., 2003). dans
une étuve rotative IKA® KS 4000i control shaker
II.3 Détermination des activités enzymatiques
II.3.1Obtention de l’extrait enzymatique brut
Après incubation, le milieu de culture au son de type classique est filtré sur un tissu pour
fromagerie, le filtrat est centrifugé à 4000 g pendant 30mn à 4°C dans une centrifugeuse
réfrigérée (JOUAN GR422). Le surnageant ou extrait enzymatique brut (EEB) est maintenu à
basse température pour la recherche des activités enzymatiques.
En ce qui concerne le milieu « au jus de son », l’étape de filtration est supprimée. L’E.E.B.,
est obtenu directement après centrifugation.
II.3.2 Méthode de détermination de l’activité coagulante
La détermination de l’activité coagulante du lait est faite selon la norme internationale FIL
(IDF) 157A :1997.
11 g de poudre de lait écrémé « low heat »standard de « Berridge »(substrat fourni par
l’INRA de Poligny. France) sont dissout dans 100 ml d’une solution de CaCl2 à 0,01M sous
agitation magnétique pendant 30minutes ; le pH est d’environ 6,5.Le substrat ainsi reconstitué
est reparti dans des tubes à essais à raison de 10 ml par tube. Les tubes sont placés dans un
bain marie à 32°C pendant 30 min.

Chapitre 2. Matériel et Méthodes
74
1ml de l’extrait enzymatique est alors ajouté au substrat. Le tube est agité et on note à l’aide
d’un chronomètre, l’apparition de la première floculation sur la paroi du tube.L’activité
coagulante est exprimée en unité présure (U.P.) (ml).
L’unité présure correspond à la quantité d’enzyme contenu dans un ml d’extrait enzymatique
pouvant coaguler 10 ml de substrat en 100 secondes à 32°C.
UP = 10 V / Tv
V= Volume de lait
T = temps de coagulation en secondes
v = volume de l’enzyme.
L’activité coagulante peut être également exprimée en ‘’force coagulante’’, exprimée en unité
Soxhlet et représente le nombre de volumes de lait frais coagulés par un volume de présure en
40 minutes à 35 °C.
Une unité internationale UI= 400 SU (unité soxhlet) correspondant à la quantité d’enzymes
qui coagulent les 10ml de lait en 1 minute
F = 2400.V/T. v
Il existe une relation UP=F x 0,00457 (Alais, 1984).
II.3.3 Méthode de détermination de l’activité protéolytique
La méthode utilisée est celle de Murado et al. (1993).
L’activité protéolytique est mesurée en utilisant comme substrat une solution de caseine
(Sigma) à 1% dissoute dans NaOH 0,02M.
1ml de substrat est incubé avec 1ml de la solution enzymatique dans du tampon phosphate
pH6 pendant 10 min à 35 °C. La réaction est stoppée par l’ajout de 5ml de TCA à 5%. Après
un repos de 15 min, la solution est filtrée. Puis à 1 ml de filtrat, sont ajoutés 5ml de la solution
réactive, on incube à 35°C pendant 10 min, puis on ajoute 0 ,5 ml du réactif de Folin dilué au
1/3. Après agitation, on incube à 35°C pendant 30min, puis la DO est mesuré à 660 nm.

Chapitre 2. Matériel et Méthodes
75
Une unité d’activité est définie comme la quantité d’enzyme qui libère 1µg d’acide aminé
exprimé en tyrosine par minute dans les conditions d’essais.
II.3.4 Dosage de la quantité protéique
Ce dosage est réalisé selon la méthode de Lowry (1951).
Le principe repose sur une méthode colorimétrique par référence à une courbe étalon établi à
partir d’une solution standard de protéine pure (BSA). La DO est mesuré à 660 nm.
II.4 Identification de souches à activité coagulante par voie biochimique
Un ensemble d’observations a permis de sélectionner, à partir de la collection initiale de 70
souches initiales, 8 souches dont l’activité coagulante semblait satisfaisante.
Après purification des souches et vérification du gram et du pouvoir caséolytique sur milieu
P.C.A. + lait, des tests d'identification en tubes et en galeries miniaturisées (Galerie API) ont
été réalisés pour les 8 souches sélectionnées.
Après ce travail, 3 d’entre elles ont fait l’objet d’une identification moléculaire.
II.4.1 Tests d'identification en tubes
Les manipulations ont été effectuées à partir des souches de 24h, ensemencées sur milieu
PCA. Pour chaque souche, une suspension dense (Mac Farland 2) a été réalisée dans de l'eau
distillée stérile. Elle a permis l'inoculation des milieux d'identification. Les milieux utilisés
ainsi que leur méthode d'ensemencement figure dans le tableau 12.
De plus, des milieux de Mossel +œuf (10 ml d'émulsion de jaune d'œuf /90 ml de milieu de
base) ont été utilisés. Des isolements ont été réalisés pour chaque souche afin d’en vérifier la
pureté.
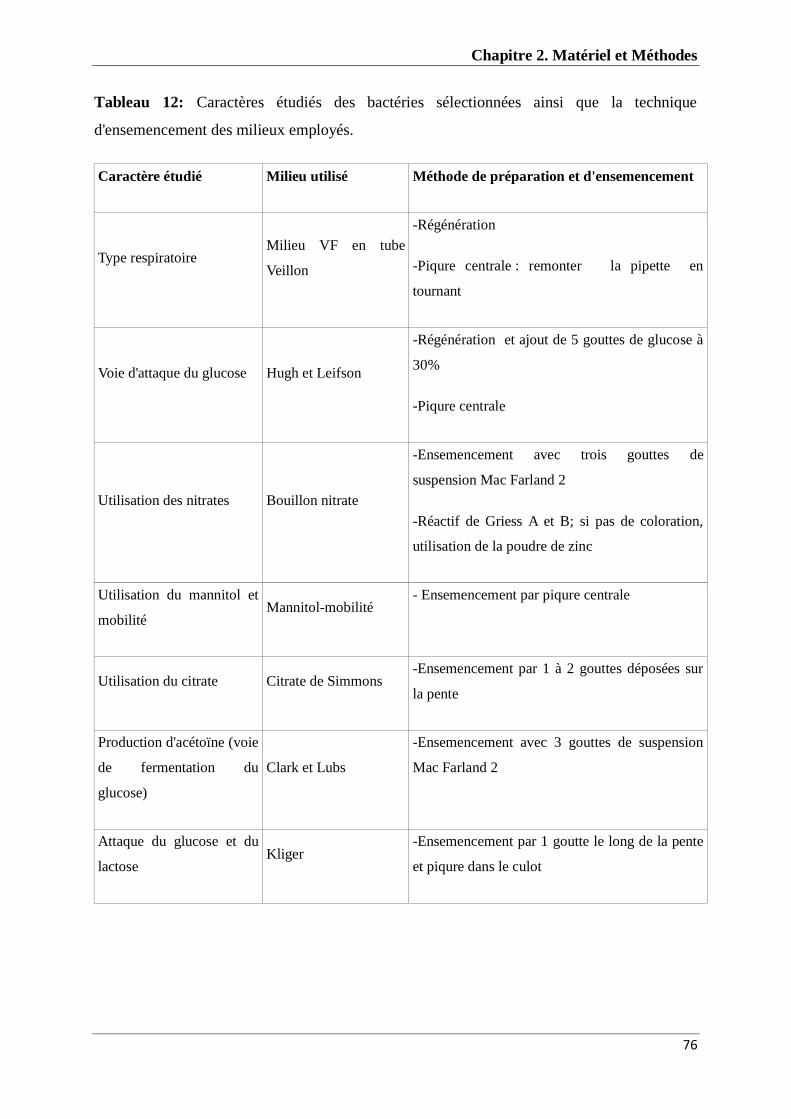
Chapitre 2. Matériel et Méthodes
76
Tableau 12: Caractères étudiés des bactéries sélectionnées ainsi que la technique
d'ensemencement des milieux employés.
Caractère étudié Milieu utilisé Méthode de préparation et d'ensemencement
Type respiratoire Milieu VF en tube
Veillon
-Régénération
-Piqure centrale : remonter la pipette en
tournant
Voie d'attaque du glucose Hugh et Leifson
-Régénération et ajout de 5 gouttes de glucose à
30%
-Piqure centrale
Utilisation des nitrates Bouillon nitrate
-Ensemencement avec trois gouttes de
suspension Mac Farland 2
-Réactif de Griess A et B; si pas de coloration,
utilisation de la poudre de zinc
Utilisation du mannitol et
mobilité Mannitol-mobilité
- Ensemencement par piqure centrale
Utilisation du citrate Citrate de Simmons -Ensemencement par 1 à 2 gouttes déposées sur
la pente
Production d'acétoïne (voie
de fermentation du
glucose)
Clark et Lubs
-Ensemencement avec 3 gouttes de suspension
Mac Farland 2
Attaque du glucose et du
lactose Kliger
-Ensemencement par 1 goutte le long de la pente
et piqure dans le culot

Chapitre 2. Matériel et Méthodes
77
II.4.2 Galerie miniaturisée API 20E
Les galeries ont été ensemencées à partir de souches de 24h cultivées sur milieu PCA. Pour
chaque souche, une colonie a été prélevée et mis en suspension dans 5 ml d'eau distillée
stérile (Mac Farland 0,5).
Une première série de galeries a été ensemencée où chaque tube de la galerie a été rempli
selon les indications du fabriquant. Certaines lectures n'étant pas très franches, des galeries
ont été recommencées en modifiant les conditions d’ensemencement : les souches s'étant
révélées aérobies strictes, nous avons proposé, en ce qui concerne les tests relatifs à
l'utilisation des sucres, d'ensemencer tubes et cupules afin d'améliorer la lisibilité des
résultats.
II.4.3 Galerie miniaturisée API 50CH
Ces galeries miniaturisées permettent d'étudier l'utilisation par les souches bactériennes de 50
substances carbonées, soit par oxydation (utilisation du milieu BYNB: Buffered Yeast
Nitrogen Base), soit par fermentation (utilisation du milieu B dans le cas des Bacillus).
Après sélection de certaines souches, au total 6 galeries ont été ensemencées, respectivement
deux pour I33M et deux pour FK6A; les deux restantes ont été utilisées pour les souches
P47M et P47γ.
La première série de galeries a été ensemencée avec du milieu BYNB à 7g/l d'agar (étude de
l'oxydation des «sucres») pour les souches P47M, I33M et FK6A.
Le deuxième série de galeries concernait les souches I33M, FK6A et P47γ, et
l'ensemencement a été fait à l'aide d'ampoules API 50CHB/E medium destiné à l'identification
des Bacillus.
II.5 Identification par biologie moléculaire
Après avoir étudié plusieurs caractères biochimiques des souches retenues, nous avons pu
conclure que certaines souches sont semblables.
Les trois d’entre elles paraissant différentes (I33M,FK6etP47M), elles ont fait l’objet d’une
identification par biologie moléculaire (Laboratoire Geno-screen, Campus de l’Institut
Pasteur de Lille, 1 rue du Professeur Calmette - 59000 Lille - France).

Chapitre 2. Matériel et Méthodes
78
II.5.1 Les différentes étapes de l’identification moléculaire
Elles ont été les suivantes :
Amplification des régions hypervariables de l’ADNr16S.
Purification et séquençage double sens des produits PCR.
Validation et correction des séquences.
Blast des séquences sur les bases de données internationales.
Rapport incluant la séquence au format fasta, l’interprétation des séquences en %
d’homologie et un arbre phylogénétique.
II.5.2 Cas de l’identification de la souche I33M
L’étude macroscopique et microscopique a été effectuée sur des boîtes de Pétri avec gélose
PCA (plate cout Agar) sur les colonies de 24 h.
Pour l'analyse de séquençage, l'ADN génomique a été extrait de la souche par la méthode
standard de l’alcool isoamyl en utilisant le kit PrepMan ultra (Applied Biosystems) et purifiés
sur des plaques Nucleo Fast (Macherey Nagel). L'amplification de l'ADNr 16S a été réalisée
par la technique de PCR, en utilisant la Taq ADN polymerase, l'ADN génomique comme
matrice, et des amorces 3’ (forward) et 5’ (reverse) universel
Les séquences de ces amorces sont les suivantes :
- F1 (5’-AGAGTTTGATCCTGGCTCAG-3’),
- R1 (5’GTATTACCGCGGCTGCTGGCAC-3’),
- F2 (5’-CTCCTACGGGAGGCAG3’)
- R2 (5’-GACACGAGCTGACGACA-3’).
Les produits PCR sont séquencés à l’aide de la technique Sanger sous des conditions
optimales réalisées par Geno screen (Laboratoire spécialisé en biologie moléculaire)
Après avoir reçu les résultats de séquençage, la séquence de nucléotides de l’ADNr 16S de la
souche a été déposée dans la GenBank, et alignée avec des séquences d’ADNr 16S
disponibles dans les bases de données publiques dans NCBI (National Center for
Biotechnology Information) (retrouvées dans les sites: http://www.ncbi.nlm.nih.gov/), et
LeBibi database (http://umr5558-sud-str1.univ-lyon1.fr/lebibi/lebibi.cgi)en utilisant le
logiciel de recherche de l’alignement local de base (BLAST).

Chapitre 2. Matériel et Méthodes
79
II.6 Optimisation du mileu de culture par un plan d’expérience de la
production d’enzyme
L’optimisation d’un milieu de production nécessite des méthodes qui consistent à faire
varier successivement chacun des facteurs qui influencent la culture, ce qui conduit à
minimiser le nombre d’expériences.
L’utilisation d’une méthodologie de recherche expérimentale ou plan d’expérience permet de
déterminer l’influence de nombreux paramètres, d’étudier leurs interactions et par la même
d’optimiser la réponse expérimentale.
Le plan choisi est un plan de surface de réponse de type composite centré (CCD).
II.6.1Méthodologie des surfaces de réponses
Cette étude est basée sur la conception composite centrale (CCD) utilisée pour obtenir des
données expérimentales, qui devraient correspondre à un modèle empirique polynomial du
second degré représentant des surfaces de réponse sur une gamme relativement large de
paramètres tel que rapporté par Duttet al. (2008).
Chaque facteur de la CCD a été étudié à 3 différents niveaux : niveau faible (-1), niveau
moyen (0) et niveau élevé (+1). Pour cela, l’approche de la surface de réponse a été utilisée et
une mise au point a été réalisée. La gamme et les niveaux des variables expérimentales
étudiées sont présentés dans le Tableau 13. Quatre différentes variables ont été utilisées pour
déterminer l’interaction entre différents facteurs (24 CCD avec 4 facteurs ont été appliqués)
incluant 6 points centrés et un ensemble de 60 expériences réalisées. Les valeurs centrales
(niveau zéro) choisies pour la conception expérimentale sont : son de blé 5% (p/v), caséïne
1.01% (p/v), température 37.5°C et agitation 150 rpm.
Les résultats ont été analysés grâce au module de conception expérimental du
STATGRAPHICS plus V5 .1 software (Statsoft, USA). Ce modèle a permis l’évaluation des
effets des conditions linéaires, quadratiques et interactives des variables indépendantes sur les
variables dépendantes. La significativité statistique des coefficients de régression a été
déterminée par le coefficient Test de T Student et le modèle d’équation du second degré a été
déterminé par le test de Fisher.
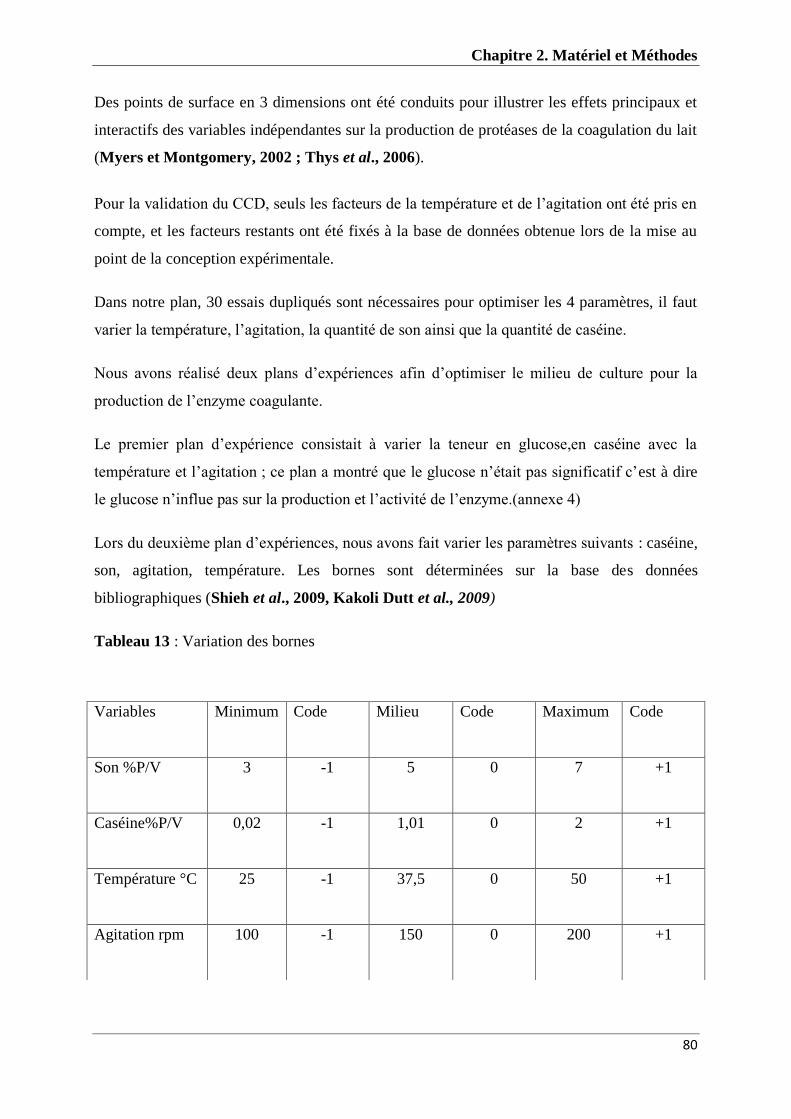
Chapitre 2. Matériel et Méthodes
80
Des points de surface en 3 dimensions ont été conduits pour illustrer les effets principaux et
interactifs des variables indépendantes sur la production de protéases de la coagulation du lait
(Myers et Montgomery, 2002 ; Thys et al., 2006).
Pour la validation du CCD, seuls les facteurs de la température et de l’agitation ont été pris en
compte, et les facteurs restants ont été fixés à la base de données obtenue lors de la mise au
point de la conception expérimentale.
Dans notre plan, 30 essais dupliqués sont nécessaires pour optimiser les 4 paramètres, il faut
varier la température, l’agitation, la quantité de son ainsi que la quantité de caséine.
Nous avons réalisé deux plans d’expériences afin d’optimiser le milieu de culture pour la
production de l’enzyme coagulante.
Le premier plan d’expérience consistait à varier la teneur en glucose,en caséine avec la
température et l’agitation ; ce plan a montré que le glucose n’était pas significatif c’est à dire
le glucose n’influe pas sur la production et l’activité de l’enzyme.(annexe 4)
Lors du deuxième plan d’expériences, nous avons fait varier les paramètres suivants : caséine,
son, agitation, température. Les bornes sont déterminées sur la base des données
bibliographiques (Shieh et al., 2009, Kakoli Dutt et al., 2009)
Tableau 13 : Variation des bornes
Variables Minimum Code Milieu Code Maximum Code
Son %P/V
3 -1 5 0 7 +1
Caséine%P/V 0,02
-1 1,01 0 2 +1
Température °C 25
-1 37,5 0 50 +1
Agitation rpm 100
-1 150 0 200 +1

Chapitre 2. Matériel et Méthodes
81
II.7 Suivi de la croissance de la souche I33M en fermenteur
Une fois les conditions optimales déterminées grâce au plan d’expériences, une croissance en
fermenteur a été effectuée. Les conditions de cette croissance correspondent aux valeurs
déterminées par le plan d’expérience précédemment décrit.
II.7.1. Préparation du milieu, conditions de culture et inoculum
La composition du milieu correspond à : 7% de son, 0 ,094% caséine, 0 ,3% Na 2, HPO4, 0,3%
extrait de levure. Nous avons préparé 1500ml de milieu stérile, le fermenteur ayant une
capacité de 2000 ml.
La culture s’effectue à une température de 39°C et sous une agitation de 150 rpm.
L’oxygénation est maintenue à 1 bar. Au cours du suivi, le pH n'est pas régulé.
L’inoculum est préparé à partir de la souche pure isolée sur boite de Pétri avec une
concentration de bactéries de 9. 108 UFC /ml (Mac Farland 3). 45ml de suspension
bactérienne sont introduites dans les 1500 ml de milieu.
II.7.2 Dénombrement et activités
Lors de la fermentation, la croissance est suivie par dénombrement des colonies sur milieu
P.C.A. après des dilutions décimales et successives. Le premier prélèvement s’effectue à To
après agitation du milieu au début de la fermentation ; les prélèvements sont ensuite effectués
toutes les 2 heures au début puis toutes les 3 h pendant 48h.
Une fois l’échantillon prélevé (5ml), son pH est mesuré, puis 1 ml est utilisé pour les
dilutions et ensemencé les boîtes de milieu de culture. Le reste de l’échantillon servira pour le
test de coagulation, le dosage des protéines et l’activité protéolytique.
II.8 Essai de purification partielle de l’extrait enzymatique brut :
L’extrait coagulant issu de Bacillus mojavensis est soumis à une chromatographie
d’exclusion moléculaire.

Chapitre 2. Matériel et Méthodes
82
Principe :
La chromatographie d’exclusion moléculaire est une Méthode fréquemment utilisée dans
la purification et la séparation des protéines en utilisant un gel (Durang et Monsan, 1974).
Les protéines sont éluées dans un ordre décroissant, les protéines de poids moléculaire
élevé sont éluées les premières, les autres de masse moléculaire moyenne et petite passent à
travers les pores du gel et sont éluées tardivement.
Les molécules quittent donc un gel Séphadex placé dans une colonne de verre dans l’ordre
des masses moléculaires décroissantes (Kamoun,1977).
Appareillage :
Colonne en verre (PHARMACIA)
Réservoir pour la solution tampon
Pompe péristaltique
Collecteur de fraction à 100 tubes
Spectrophotomètre (réglé à 280 nm)
Préparation du gel:
Le gel Séphadex G 75 est lavé avec la solution tampon (pH 7 . 0,02M), puis mis à gonfler
dans un excès de cette même solution tampon pendant 24 heures. A l’aide d’une pompe à
vide, le gel est dégazé pour éviter la formation des bulles d’air dans la colonne.
Remplissage de la colonne :
Une fois le gel prêt, il est coulé sur les parois de la colonne afin d’éviter la formation des
bulles d’air. Le gel doit être bien tassé et homogène et présenter une seule phase. Ensuite la
colonne est équilibrée avec la solution tampon.
Dépôt de l’échantillon :
Avant le dépôt de l’échantillon,on procède à un lavage du gel dans la colonne avec la
solution tampon, ensuite 1ml de la solution enzymatique brut est déposé délicatement à la
surface du gel.

Chapitre 2. Matériel et Méthodes
83
Elution :
Le débit d’élution des protéines(0,5 ml/min). Les éluats sont recueillis dans le collecteur de
fraction à raison de 2ml par tube.
La lecture de l’absorbance a été faite à une longueur d’onde de 280 nm.
La recherche de l’activité coagulante a été effectuée au niveau de chaque tube.
II.9 Caractérisation des extraits enzymatiques bruts et partiellement
purifiés :
Afin de pouvoir déterminer les conditions optimales de l’activité coagulante des protéases
extraites à partir de Bacillus mojavensis on procède à l’étude des paramètres qui influencent
sur le lait :la température, le pH, la concentration en CaCl2 .
La mesure de l’activité coagulante a été déterminée en mesurant le temps de coagulation
du lait en faisant varier les valeurs du paramètre étudié selon les conditions standards
(T=35°C, pH=6,4, concentration en CaCl2 =0,01M).
II.9.1.Influence de la température du lait :
la température optimale de coagulation du lait a été déterminée en faisant varier la
température du lait de 30 à 70°C avec un intervalle de 5°C ;les autres paramètres sont fixés
selon les conditions standards (pH=6,4, concentration en CaCl2 =0,01M ).
II.9.2.Influence du pH du lait :
Le pH optimum de coagulation du lait est déterminé en faisant varier le pH du lait de 5 à 7
par l’addition d’hydroxyde de sodium ou d’acide chlorhydrique, les autres paramètres sont
établis selon les conditions standards(T=35°C, concentration en CaCl2 =0,01M)
II.9.3.Influence de la concentration en CaCl2 :
L’activité coagulante a été déterminée en faisant varier les concentrations en CaCl2 allant de
0,005 à 0,1, les autres paramètres sont fixés selon les conditions standards (T=35°C, pH=6,4).

Chapitre 2. Matériel et Méthodes
84
II.10 Essai de fabrication de fromage
Nous avons utilisé 6 litres de lait, avec des volumes des extraits enzymatiques bruts différents
pour les quatre essais que nous avons entrepris.
L’activité protéolytique étant très élevée, nous avons diminué la quantité d’extrait
enzymatique.
Tableau 14: Volumes d’EEB utilisés dans les essais de fabrication de l’EDAM
50% EEB de Bacillus mojavensis
50% de présure
4ml
0.08 ml (dilution de 0.25g de présure dans 3 ml
d’eau physiologique
100% EEB de Bacillus mojavensis 6ml
La pasteurisation du lait se fait à température de 90° C pendant 15 secs. Ce traitement va
permettre de réduire la flore banale et d’éliminer la flore pathogène, le lait est ensuite refroidi
à 38°C pour la présure et à 45 °C pour les extraits enzymatiques bruts.
II.8.1.2 L’ensemencement
Il s’agit de rajouter au lait déjà pasteurisé :
0.3g des ferments lactiques (mésophile et thermophile)
1.5g de phosphate mono-calcique.
Quelques gouttes de colorant (Rocou).
Le melange est agité pendant 10 min.
II.8.1.3L’emprésurage
Cette étape est réalisée quand le pH est d’environ de 6.4 et à la température de 38 °C pour la
présure et à 45°C pour les extraits enzymatiques bruts (cette température reste stable jusqu’à
la fin de la fabrication dans un bain marie), et cela avec une agitation pendant 1min. La dose
des E.E.B additioné est calculée préalablement à partir de la force coagulante.

Chapitre 2. Matériel et Méthodes
85
II.8.1.4La coagulation
Le mouvement du lait doit être stoppé immédiatement afin d’évite l’obtention de caillé
feuilleté.
Le temps de coagulation des E.E.B est plus long que celui de la présure.
II.8.1.5Le décaillage
Il se fait par un couteau à une vitesse très faible pour limiter les pertes car le caillé est encore
fragile.
II.8.1.6 Le brassage
Avec une agitation et à l’aide d’un fouet dans le but dediminuer la taille des grains de caillé.
II.8.1.7Le délactosage
Un repos de 5 min est nécessaire pour le dépôt des grains au fond, puis l’évacuation du 1er
lactosérum. Ensuite un lavage des grains a été effectué, avec un volume d’eau à 38°C égal à
1/3 de la quantité de lactosérum soutiré soit 20 à 30 % du volume total du lait. Le 2ème
lactosérum povenant du 2ème
brassage est évacué après 20 min.
II.8.1.8 Le pré-préssage
Après avoir éliminé le lactosérum II, le caillé obtenu a été pré-pressé à l’aide d’une toile dans
un couscoussier.
II.8.1.9 Le moulage
Lesmoules sont couverts avec des toiles, et remplis avec le caillé.
II.8.1.10 Le préssage
Les moules remplis subissent un pressage à l’aide d’une plaquette pendant 5 heures (le
retournement se fait après 30 minutes).

Chapitre 2. Matériel et Méthodes
86
II.8.1.11 Le salage
Après le démoulage, les boules de fromage sont déposés dans un bain de saumure pendant 4
heures à température de 10 à 15°C et une concentration en NaCl de 350g/l.
II.8.1.12 L’affinage
Dans des hâloirs d’affinage, les boules de fromage ont été placées sur des planches en bois, à
une T= 10° C avec 85 à 95 % d’humidité. La durée de l’affinage est au minimum 21 jours au
bout desquels les fromages sont retournés au moins une fois.
II.8.1.13Le lavage des boules
Après l’affinage les boules sont normalement plongées dans une solution fongicide si non
dans un bain d’eau salée puis lavées avec une brosse à main.
II.8.1.14 Le grattage des boules
Les boules sont grattées à l’aide d’un couteau pour éliminer la croûte dure externe.
II.8.1.15 Le conditionnement
A la fin les fromages sont enrobés avec de la paraffine fondue, et laissés un certain temps
pour sécher et emballés sous vide dans du papier cellophane.
Le procédé de fabrication est représenté sous forme d’un diagramme
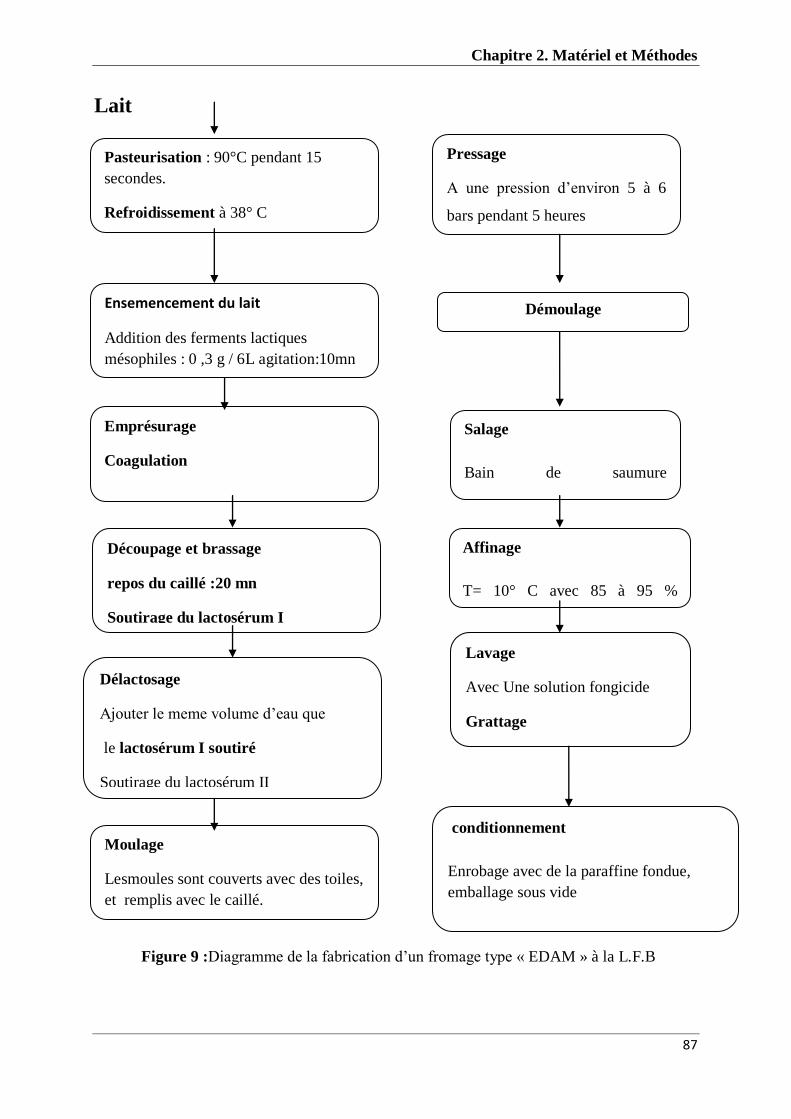
Chapitre 2. Matériel et Méthodes
87
Lait
Figure 9 :Diagramme de la fabrication d’un fromage type « EDAM » à la L.F.B
Pasteurisation : 90°C pendant 15
secondes.
Refroidissement à 38° C
Ensemencement du lait
Addition des ferments lactiques
mésophiles : 0 ,3 g / 6L agitation:10mn
Emprésurage
Coagulation
Découpage et brassage
repos du caillé :20 mn
Soutirage du lactosérum I
Délactosage
Ajouter le meme volume d’eau que
le lactosérum I soutiré
Soutirage du lactosérum II
Moulage
Lesmoules sont couverts avec des toiles,
et remplis avec le caillé.
Pressage
A une pression d’environ 5 à 6
bars pendant 5 heures
Démoulage
Salage
Bain de saumure
pendant4hhheures; T° : 10 à
15°C
Affinage
T= 10° C avec 85 à 95 %
d’humidité. La durée 21 jours
Lavage
Avec Une solution fongicide
Grattage
Pour éliminer la croûte dure
externe.
conditionnement
Enrobage avec de la paraffine fondue,
emballage sous vide

Chapitre 2. Matériel et Méthodes
88
III. PROTEASE D’ORIGINE FONGIQUE
III.1 Isolement des souches
III.1.1 Méthode d’isolement
La méthode d’isolement des souches fongiques, à partir de prélèvements de sol, est identique
à celle qui a été employée pour les bactéries. A partir de la solution mère au 1/10, on effectue
aussi des dilutions que l’on ensemence à la surface d’un milieu de culture. Il s’agit du milieu
Sabouraud-chloramphénicol, additionné de 10% de lait écrémé stérile. L’incubation s’effectue
à 27°C pendant 4 à 5 jours.
III.1.2 Choix et purification des souches
Après incubation, de nombreux thalles de moisissures apparaissent à la surface du milieu. On
procède alors aux repiquages des différents types de moisissures afin de purifier les souches.
Nous avons obtenu 24 moisissures avec des caractéristiques sensiblement différentes. Ces
souches ont été codées afin de constituer la collection de base.
III.1.3 Observations macroscopiques et microscopiques
Des photos des observations microscopiques ont été prises au laboratoire de microbiologie du
département de génie biologique de l’IUT d’ANGERS, à l’aide d’un microscope ordinaire de
marque Olympus (CH30), muni d’une tête trinoculaire permettant l’adaptation d’un appareil
photo de la même marque.
III.2 Sélection des souches fongiques coagulantes
III.2.1 Les milieux de culture
Sur la base de travaux antérieurs (Srinivasan et al.,1964 ; Tubesha et al.,2003) nous avons
choisi deux types de milieux de culture :
un milieu liquide : le milieu de CZAPEK dont la composition est en gr/l : 2,5 NaNo3 ;
1 KH2PO4 ; 0,5 MgSO4 7H2O ; 0,5KCl ; 0,01 FeSO47H2O ; 10 glucose ; 10 de poudre
de lait écrémé. Il est réparti en erlen à raison de 50ml.

Chapitre 2. Matériel et Méthodes
89
un milieu semi solide de composition suivante : 30gr de son de blé, 2,5g de poudre de
lait écrémé, additionné de 60 ml de sulfate d’ammonium à 0,01% (Levadoux, 1989),
réparti dans des erlenmeyers qui sont ensuite stérilisés à 121°C pendant 15min.
III.2.2 La préparation de l’inoculum et l’ensemencement des milieux
Pour chaque souche une fois purifiée, les spores sont récupérées à l’aide d’une solution de
tween 80 (0,01%) que l’on déverse dans la boîte de Pétri. Puis la surface est grattée et la
suspension de spores obtenue, constitue l’inoculum.
Le nombre de spores est alors déterminé par dénombrement à la cellule de Malassez. Le
volume de l’inoculum à introduire dans les erlens est calculé de manière à avoir environ 5.
106spores/ ml de milieu de culture (Guilherme Garcia da Silveira et al., 2005)
L’incubation des milieux de culture s’effectue par agitation à 27°C pendant 4 à 5 jours dans
un shaker (IKA®
KS 4000i control,) à 110 rpm.
III.3Determination des activités enzymatiques
III.3.1 Obtention de l’extrait enzymatique brut
Après fermentation, le contenu des erlens est d’abord filtré, puis à 4000g pendant
30minutes à 4°C (Jouan GR 422). Le surnageant constitue l’extrait enzymatique brut (EEB)
dont on détermine certaines activités enzymatiques et qui servira à la fabrication de fromage.
III.3.2 Méthode de détermination de l’activité coagulante
La méthode utilisée est identique à celle qui a été employée dans le cas des bactéries.
III.3.3 Méthode de détermination de l’activité protéolytique
L’activité protéolytique est mesurée selon la méthode de Murado et al. (1993)La même
méthode que celle utilisée pour les bactéries.

Chapitre 2. Matériel et Méthodes
90
III.3.4. Dosage de la quantité protéique
Le dosage des protéines de l’extrait enzymatique est réalisé selon la méthode décrite par
Lowry et al (1951).
III.4 Sélection des souches fongiques
13 souches sur 24 se sont avérées possédant des enzymes coagulantes. Leurs caractéristiques
macroscopiques et microscopiques ont été étudiées afin de les différencier. Trois d’entre elles
qui offraient certaines différences (« 4D », SF15 et « 5F »), ont été envoyées pour
identification. Une seule espèce a été retenue, pour laquelle a été entreprise la suite du travail.
III.5 Identification des moisissures par biologie moleculaire
Les moisissures ont été identifiées avec la même méthode que pour celle des bactéries dans le
même laboratoire sauf que c’est l’ADNr-18S qui est séquencé
III.6 Optimisation de la production de la protéase
III.6.1 Optimisation des conditions opératoires
L’influence de la composition du milieu et de la concentration de chaque constituant, sur
l’activité coagulante et l’activité protéolytique de certaines bactéries et moisissures, a
beaucoup été étudié (Kakoli et al., 2009 ; Sathya et al., 2009 ; Srinu Babu et al., 2007).
A cet effet, nous avons étudie l’influence de quelques facteurs sur la production des protéases
à activité coagulante.
Des cinétiques de production ont été réalisées grâce à un plan d’expérience où plusieurs
facteurs varient simultanément.
La température, la quantité de son de blé, ainsi la quantité de caséine, ont été choisis comme
étant les variables qui affectent le plus la production de protéase (Palaiswamy, 2009), alors
que les autres composants du milieu restent constants (volume et densité de l’inoculum,
volume et concentration en sulfate d’ammonium).
A cet effet 20 expériences ont été réalisées, en répétant chaque expérience deux fois avec les
trois facteurs et chaque facteur varie sur 5 niveaux. (Tab. 15 et 16).
Toutes les expériences ont été réalisées dans des ErlenMeyers de 250 ml, les cultures sont
ensuite stérilisées à 120°C pendant 20min.
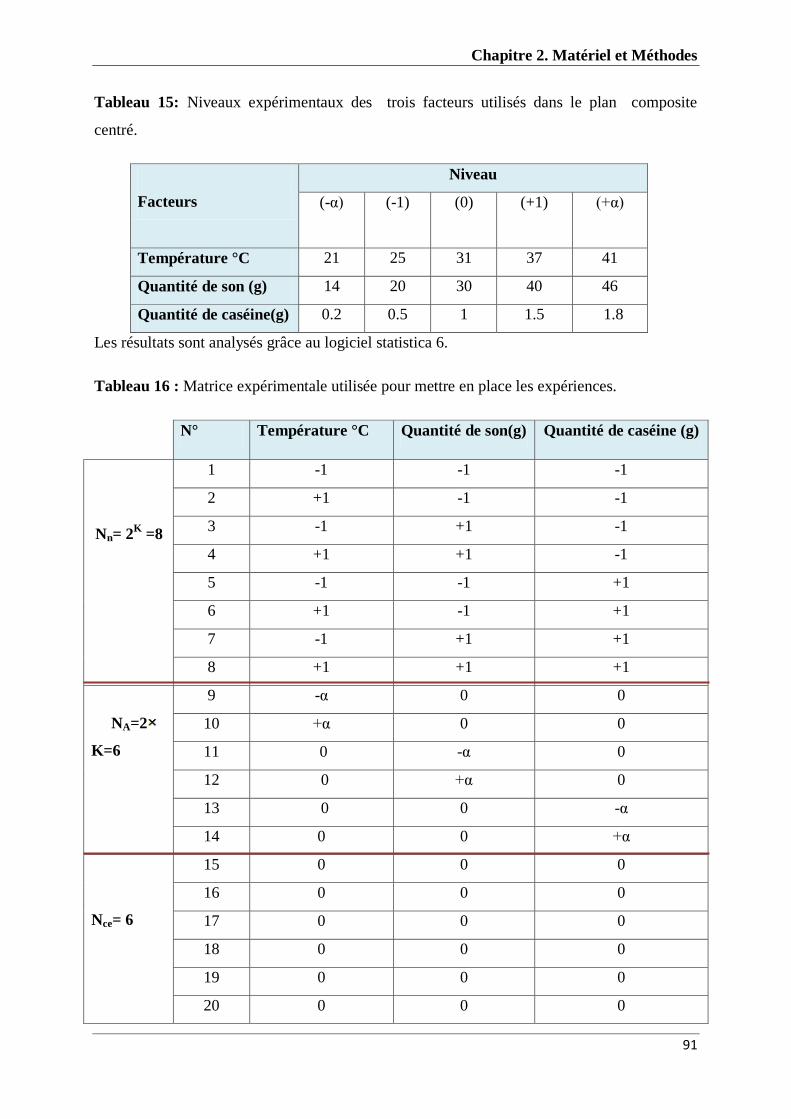
Chapitre 2. Matériel et Méthodes
91
Tableau 15: Niveaux expérimentaux des trois facteurs utilisés dans le plan composite
centré.
Facteurs
Niveau
(-α) (-1) (0)
(+1)
(+α)
Température °C 21 25 31 37 41
Quantité de son (g) 14 20 30 40 46
Quantité de caséine(g) 0.2 0.5 1 1.5 1.8
Les résultats sont analysés grâce au logiciel statistica 6.
Tableau 16 : Matrice expérimentale utilisée pour mettre en place les expériences.
N° Température °C Quantité de son(g) Quantité de caséine (g)
Nn= 2K
=8
1 -1 -1 -1
2 +1 -1 -1
3 -1 +1 -1
4 +1 +1 -1
5 -1 -1 +1
6 +1 -1 +1
7 -1 +1 +1
8 +1 +1 +1
NA=2
K=6
9 -α 0 0
10 +α 0 0
11 0 -α 0
12 0 +α 0
13 0 0 -α
14 0 0 +α
Nce= 6
15 0 0 0
16 0 0 0
17 0 0 0
18 0 0 0
19 0 0 0
20 0 0 0

Chapitre 2. Matériel et Méthodes
92
III.6.2 Ensemencement des milieux de culture
10 ml de la suspension de spore (≈3×106) (Guilherme et al ., 2005) servant à inoculer chaque
milieu de culture. Les milieux ensemencés sont ensuite incubés à cinq températures
différentes (25, 27, 31,35 et 37°C) pendant 5 jours.
III.6.3 Extraction de l’enzyme
Après l’incubation, 150 ml d’eau distillée stérile est ajoutée dans les Erlens. Après agitation
mécanique pendant 1 heure à 300 tours/min, le contenu est filtré puis centrifugé à 10 000
tours/min pendant 20 min à 4°C. Le surnageant constitue l’extrait enzymatique brut qui
servira pour le test de coagulation.
L’activité coagulante des 40 expériences est déterminée par la méthode de Berridje, (1951) et
les résultats sont interprétés par la méthode statistique selon le plan composite centré afin de
choisir les conditions optimales de la production.
III.6.4 Evaluation statistique des composants du milieu de culture par le plan
composite centré
La méthode utilisée, pour optimiser et caractériser comment les facteurs choisis affectent la
production, est celle préconisée par le plan composite centré à 5 niveaux.
Les conditions expérimentales (niveau de facteurs et réponses) figurent dans la matrice des
expériences présentées dans le tableau 15 et 16.
III.7 Essai de purification
III.7.1Précipitation au sulfate d’ammonium
Principe
Le sulfate d’ammonium provoque, dans une solution protéique, une déshydratation et une
précipitation des protéines induisant ainsi le phénomène de relargage. L’avantage du
relargage est que les protéines conservent leur conformation native et peuvent être ensuite
dissoutes habituellement sans dénaturation. Le choix s’est porté sur le sulfate d’ammonium du
fait de sa solubilité dans l’eau et de sa force ionique élevée (Kamoun, 1977).

Chapitre 2. Matériel et Méthodes
93
Méthode
L’extrait enzymatique brut est soumis à une précipitation au sulfate d’ammonium à trois taux
de saturation à savoir (p/v) : 40%, 60%, 80%. A 140 ml d’extrait enzymatique, on ajoute
respectivement 34,86 g, 54,6g, 78,54g de sulfate d’ammonium sous agitation douce à 4°C.
La solution enzymatique ainsi saturée est mise à décanter pendant une nuit. Après
centrifugation à 13000 tr/min pendant 30mn à 4°C, le culot obtenu est mis en suspension, lavé
deux fois avec du tampon acétate de sodium (0.1 M ; pH 5,2), puis centrifugé à 13000 tr/min
pendant 20 min à 4°C. Les culots obtenus sont remis en suspension dans un volume réduit de
tampon acétate de sodium (0,1M ; pH 5,2). L’activité enzymatique de la protéase est dosée
dans le surnageant et le culot.
III.7.2 Dessalage de l’extrait enzymatique par dialyse
Principe
La dialyse est une technique de séparation qui permet d’éliminer d’une solution
macromoléculaire des ions ou petites molécules, par leur passage à travers une membrane
semi-perméable. Les protéines, par leurs poids moléculaires élevés, ne sont pas diffusibles.
Méthode
Nous avons utilisé une membrane semi-perméable. Le boudin contenant les protéines
précipitées par le sulfate d’ammonium est fermé à ses deux extrémités et maintenu en
suspension dans la solution de dialyse. La dialyse est contre le tampon acétate de sodium
(0,1M ; pH 5,2) à 4°C et sous agitation constante pendant 24 heures. Le tampon de dialyse est
régulièrement renouvelé.
III.7.3 Chromatographie d’exclusion moléculaire
Principe
C’est une technique qui permet la séparation des molécules en fonction de leurs poids
moléculaires et de leurs tailles.
Les molécules de masse moléculaire élevée sont exclues et éluées les premières, les autres de
masse moléculaires moyenne et petite passent à travers les pores du gel et sont éluées
tardivement, la séparation est réalisée dans un ordre décroissant des poids moléculaires
(Kamoun, 1997).
Mode opératoire

Chapitre 2. Matériel et Méthodes
94
Pour la purification partielle de notre extrait enzymatique brut, notre choix s’est porté sur le
sephadex G -75 dont le domaine de fractionnement est compris entre 3 et 80 KDa.
Le gel gonflé et coulé dans une colonne de type pharmacia (1,5 30cm) est calibré par un
tampon acétate de sodium (0,1 M ; pH5, 2).
Un volume aliquote de 0,5 ml d’échantillon est élué avec le tampon acétate de sodium (0,1
M ; pH 5.2) à un débit de 0,93ml/min. Les éluâts sont recueillis dans un collecteur de fraction
à raison de 3 ml/tube et sur chaque fraction l’absorbance (280 nm) et l’activité coagulante
sont déterminées.
III.8 Caractérisation de l’extrait enzymatique purifié
Afin de déterminer l’activité coagulante optimale sur le lait, nous avons étudié l’influence de
certains facteurs.
III.8.1 Influence de la température
La température optimale de coagulation de lait a été déterminée en portant le lait à différentes
température, de 30 à 70°C avec un intervalle de 5°C. Les autres paramètres sont fixés selon
les conditions standards (pH du lait = 6,3, concentration de CaCl2= 0,01 M).
III.8.2 Influence du pH
Le pH optimal de coagulation de lait a été déterminé en mesurant le temps de coagulation du
lait à divers pH, de 5 à 7 avec un intervalle de pH de 0.5. Les autres paramètres sont fixés
selon les conditions standards (CaCl2 = 0,01, température 35°C).
III.8.3 Influence de la concentration en CaCl2
L’influence de la concentration en CaCl2 a été déterminée en faisant varier la concentration du
lait en CaCl2 de 0.005M à 0.05 M avec un intervalle de 0.005M (pH du lait = 6,3, température
35°C).

Chapitre 2. Matériel et Méthodes
95
III.9 Essai de fabrication de fromage à pâte pressée non cuite type
« EDAM » à partir de l’extrait enzymatique brut (EEB) de Mucor
circinelloides
Il s’effectue suivant les mêmes opérations de fabrication à base de présure. La dose utilisée de
l’EEB est calculée préalablement à partir de sa force coagulante.
Deux essais de fabrication de fromage type EDAM ont été effectués:
Le procédé de fabrication est le même que celui utilisé dans le cas des enzymes bactériennes.

Chapitre 3
Résultats et discussion

Chapitre 3. Résultats et discussion
97
I. PROTEASE D’ORIGINE BACTERIENNE
I.1 Sélection des souches possédant une enzyme coagulante
leSur les 70 souches isolées, seulement 20 possèdent une activité coagulante. Leurs
caractéristiques sont représentées dans tableau 17
Tableau 17 : Bactéries possédant une enzyme coagulante
Code de la
souche
Aspect
macroscopique
Aspect
microscopique
Gram Caséolyse
(largeur en
mm)
Activité
coagulante
en
UP /ml
P 47
Colonies plates très
sèches qui adhèrent
fortement à la
gélose couleur mâte
Bacillus en
faisceaux
environ 1µm
X 2/3 µm
+ 6 0, 55
FK6
Colonies d’un blanc
mât brillante
gluante b
Bacillus petits,
longs, fins
différents des
autres
+
4 0,33
F13
Colonies rondes,
sèches avec une
empreinte au centre
Gros bacilles
+ 6
0 ,16
F3
(ressemble à F13)
Colonies rondes,
sèches avec une
empreinte au centre
Bacilles + 5 0 ,270
P48
Colonies jaunes
différentes des
autres, colonies
plates relativement
sèches jaune clair.
Bacilles taille
moyenne plus
long que P47
et plus gros
+
6 0,23
JF9
Colonies rondes
avec une empreinte
au centre , pas très
sèches.
Bacilles plus
minces
+ 5 0,056
I33
Colonies
envahissantes
petites plus ou
moins brillantes
Bacilles taille
moyenne
sporulant
+ 6 0,84

Chapitre 3. Résultats et discussion
98
Code de la
souche
Aspect
macroscopique
Aspect
microscopique
Gram Caséolyse
(largeur en
mm)
Activité
coagulante
en
UP /ml
P43
Colonies
légèrement
bombées avec une
empreinte au centre
Ressemble à
JF9 en plus
minces bacilles
+ 1 0,083
F15
Souche initiale
Colonies sèches
mâtes avec un
centre
Bacillus + 4 0,27
A3
Colonies luisantes
beiges lisses
Bacilles courts - 6 0,33
C5
Colonies brillantes
lisses
Bacilles courts - 10 0,104
C6
Petites colonies
rondes, beiges
brillantes
Bacilles courts - 5 0,27
I33M Colonies plates, très
fines sèches très
étalées avec un
centre
Bacilles assez
gros amas
sporulant,
spore terminale
+ 6 1,67
FK6A Colonies larges,
muqueuses, bords
irréguliers qui
s’assèchent au bout
de 48H
Très longs
bacilles, spore
légèrement
déformante
+ 5 1,30
FK6B Larges colonies
muqueuses à la
surface, bordures
irrégulières, restent
muqueusesjusqu’à5
jours très
caseolytiques
Bacilles
formant des
petites
chainettes,
spores
terminales
légèrement
déformantes
+ 3mm 0,84
P47A Colonies sèches,
bordures
irrégulières
dentelées, surface
planes
Bacilles longs
et fins assez
polymorphes
+ 6mm 0,56
P47β Colonies sèches
mâtes, bordures
dentelées
Petits bacilles
en faisceaux,
pas de spore en
apparence
+ 6mm 0,84
P47M Colonies
irrégulières très
sèches ont
tendance à s’étaler
Bacilles courts
et fins, plus
fins que FK6
+ 6mm 1,67
P47R Ressemble à P47M Bacilles courts
peu sporulés
+ 6mm 1,67
P47 γ Ressemble à P47β Bacilles plus
longs et plus
fins que P47β
+ 6mm 1,67
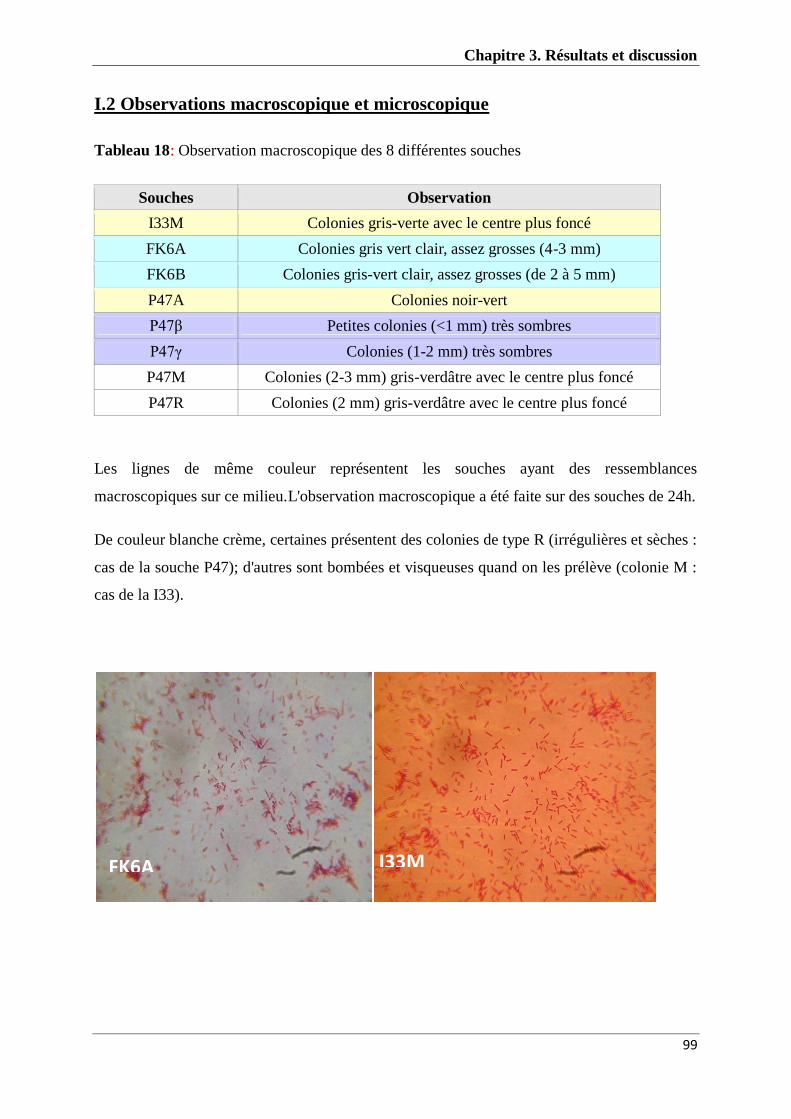
Chapitre 3. Résultats et discussion
99
I.2 Observations macroscopique et microscopique
Tableau 18: Observation macroscopique des 8 différentes souches
Souches Observation
I33M Colonies gris-verte avec le centre plus foncé
FK6A Colonies gris vert clair, assez grosses (4-3 mm)
FK6B Colonies gris-vert clair, assez grosses (de 2 à 5 mm)
P47A Colonies noir-vert
P47β Petites colonies (<1 mm) très sombres
P47γ Colonies (1-2 mm) très sombres
P47M Colonies (2-3 mm) gris-verdâtre avec le centre plus foncé
P47R Colonies (2 mm) gris-verdâtre avec le centre plus foncé
Les lignes de même couleur représentent les souches ayant des ressemblances
macroscopiques sur ce milieu.L'observation macroscopique a été faite sur des souches de 24h.
De couleur blanche crème, certaines présentent des colonies de type R (irrégulières et sèches :
cas de la souche P47); d'autres sont bombées et visqueuses quand on les prélève (colonie M :
cas de la I33).
FK6A
A A
AAA
AAA
AAA
AA
I33M

Chapitre 3. Résultats et discussion
100
Figure 10: Photos des observations microscopiques (FK6A, I33M, P47M et JF9)
Pour leur aspect microscopique, ce sont toutes des bacilles Gram+.Toutes les souches
présentent sur milieu PCA+lait, des auréoles caractéristiques de l'hydrolyse de la caséine.
Cependant les zones de caséolyse sont plus ou moins grandes suivant les souches.
I.3 Tests d'identification en tubes
Huit souches offrant les caractéristiques que nous recherchions, avaient été sélectionnées :
I33M, FK6A, FK6B, P47A, P47β, P47, P47R et P47γ
Les résultats des tests biochimiques effectués sont présentés dans les tableaux suivants (19,
20, 21, 22).
Kligler
Les résultats pour le milieu kligler ne sont pas très nets même après 48h d'incubation. On peut
cependant avoir une idée des tubes qui vont virer au jaune(Tableau 19).
P47M JF9

Chapitre 3. Résultats et discussion
101
Tableau 19: Résultats des tests sur les tubes Kligler
Souche Résultats
I33M Pousse faible, couleur initiale
FK6A Pente et culot rouge
FK6B Pente et culot rouge
P47A Pente et culot rouge
P47β Pente rouge, culot jaune
P47γ Pousse faible, couleur initiale
P47M Pente rouge, culot jaune
Mannitol mobilité
Le caractère mobilité ne peut être étudié car les souches n'ont pas poussé le long de la piqure.
Toutes les souches se sont développées en surface mise à part la P47R (aucun
développement). Quatre tubes ont viré au jaune; ce sont les tubes des souches P47A, P47β,
P47γ et P47M, elles sont donc Mannitol+.
Hugh et Leifson
Aucun changement de couleur net n'a pu être observé. Cependant, certains tubes ont
commencé à virer au jaune (Tab.20). De plus les souches se sont surtout développées en
surface.
Milieu Citrate de Simmons
Pour les souches P47R, P47β, P47A et I33M certaines zones commencent à virer au bleu,
elles sont donc citrate +, les colonies sont alors visibles et jaunes.
Pour toutes les autres, il n'y a aucun changement de l'indicateur coloré.
Tube Veillon
Ce milieu permet d'étudier le type respiratoire de la bactérie. Dans notre cas, il a démontré
que toute les souches sont aérobies strictes. Cependant pour certaines, on peut se demander si
elles ne seraient pas aussi micro-aérophiles ou si ce résultat est dû à des bulles d'air qui
auraient été introduite lors de l'ensemencement.

Chapitre 3. Résultats et discussion
102
Tableau 20 : Résultats des tests et interprétation des Hugh et Leifson
Souche Résultats Interprétation
I33M Aucun changement de
l'indicateur coloré
Pas d'oxydation du glucose
ou ensemencement
insuffisant
FK6A Virage au jaune en surface Oxydation du glucose
FK6B Aucun changement de
l'indicateur coloré
Pas d'oxydation du glucose
ou ensemencement
insuffisant
P47A Vire au jaune Oxydation du glucose
P47β Vire au jaune Oxydation du glucose
P47γ Vire au jaune Oxydation du glucose
P47M Vire au jaune Oxydation du glucose
P47R Vire au jaune Oxydation du glucose
Clark et Lubs
Après avoir vérifié que toutes les souches se sont développées dans le milieu, la voie de
Voges-Proskauer a été testée.
Il est apparu que toutes les souches étaient positives à ce test.
Bouillon nitrate
Comme dans le cas précédent, il a fallu tout d'abord vérifier que toutes les souches s'étaient
développées sur le milieu.
Au test de Griess, toutes les souches sont positives sauf I33M. Elles sont donc toutes nitrate+
sauf une.
I.4 Test d’identification en galerie pour les différentes souches
I.4.1 Galerie Api 20 E

Chapitre 3. Résultats et discussion
103
Tableau 21: Résultats des tests en galerie pour les différentes souches
Souche
ON
PG
AD
H
LD
C
OD
C
Cit
rate
H2
S
Uré
e
TD
A
Ind
ole
V.P
.
Gél
ati
na
s
e
I33M + - - - - - - - - + +
FK6A - - - - + - + - - + +
FK6B - - - - + - - - - + +
P47A + - - - + - - - - + +
P47β + - - - + - - - - + +
P47γ + - - - + - + - - + +
P47M + - - - + - + - - + +
P47R + - - - + - + - - + +
Souche
Glu
cose
Ma
nn
ito
l
Ino
sito
l
So
rbit
ol
Rh
am
no
se
Sa
cch
aro
s
e Mel
ibio
se
Am
ygd
ali
ne
Ara
bin
ose
I33M +/- + +/- + - + - + -
FK6A - +/- +/- - - - - - -
FK6B - +/- - - - - - - -
P47A + + + + - + - + -
P47β + + + + - + - + -
P47γ + + + + - + - + -
P47M + + + + - + - + -
P47R + + + + - + - + -

Chapitre 3. Résultats et discussion
104
A partir des résultats des galeries (Tab 21), on peut voir apparaître certaines différences entre
les souches. Ainsi P47A et P47β ont les mêmes caractéristiques, elles diffèrent des autres
souches P47 qui sont toutes uréase +.
Lorsque l'on compare les caractères qui ont été étudiés en galerie et en tube on peut constater
des différences.
Ainsi pour le caractère citrate, les résultats obtenus sont différents (Tab.22).
Tableau 22: Comparaison entre les tests en tubes et les galeries pour un même caractère
Souches Résultats en tube de
Simmons
Résultats de la galerie API
20 E
I33M Citrate + Citrate -
FK6A Citrate - Citrate +
FK6B Citrate - Citrate +
P47A Citrate + Citrate +
P47β Citrate + Citrate +
P47γ Citrate - Citrate +
P47M Citrate - Citrate +
P47R Citrate + Citrate +
I.4.2Galerie API 50 CH
Pour la première série de galeries ensemencées avec le milieu BYNB gélosé, les lectures se
font par estimation du trouble du à la croissance bactérienne. Bien que les souches soient
aérobies strictes, les résultats n'ont pas été très lisibles: c'est pourquoi l'étude a été reprise à
l'aide du milieu B (Bacillus) qui contient un indicateur coloré (rouge de phénol).

Chapitre 3. Résultats et discussion
105
Figure 11: Lecture à 24h et 48h de la galerie API 50CH pour la souche I33M
Figure 12:Lecture à 24h et 48h de la galerie API 50CH pour la souche P47γ
Figure 13: Lecture à 24h et 48h de la galerie API 50CH pour la souche FK6A
A partir des résultats obtenus, nous avons sélectionné trois souches, I33M, P47M et FK6A
Les tests d'identification en tubes ont fourni des résultats plutôt mitigés; en effet pour la
plupart des tests, les réponses n'étaient pas franches, ce qui n'a pas permis d'établir avec
certitude, si les souches possédaient tel ou tel caractère.

Chapitre 3. Résultats et discussion
106
Cependant, des ressemblances ont pu être notées entre les souches et il apparaissait peu
probable que les huit souches soient toutes différentes. C'est pourquoi seules certaines
souches ont été choisies pour l'étude en galeries miniaturisées Lorsque les galeries API 20E
ont été réalisées, on a pu remarquer que la I33M et les P47 avaient beaucoup plus de points
communs que les FK6.
A partir des résultats des galeries 50 CH et du tableau d’identification, Les souches ont une
forte probabilité d'être des Bacillus subtilis.
Après avoir vérifié plusieurs de leurs caractères, on peut en conclure que certaines des
souches sont semblables et qu'il y a une forte probabilité qu'elles appartiennent toutes à
l'espèce B. subtilis. Cependant, pour vérification, les souches I33M, FK6A et P47M ont été
envoyées à un laboratoire de biologie moléculaire, afin d'avoir une confirmation.
I.5 Résultats de l’identification bactérienne
Après avoir envoyé nos trois souches au laboratoire de biologie moléculaire pour
identification, nous avons reçu les résultats suivants :
FK6A :Bacillus Amyloliquefaciens
P47M :Bacillus subtilis
I33M :Bacillus mojavensis
Mais nous avons choisi la souche I33M pour la suite du travail, car c’est la bactérie qui
possède une relative faible activité protéolytique et une bonne activité coagulante

Chapitre 3. Résultats et discussion
107
Identification de la souche :
L’analyse de la séquence de gène de l’ADNr 16S a été décrite comme étant nécessaire,
principalement pour détecter certaines erreurs d’identifications du Bacillus (Ben et al., 2011).
Cette technique est également l’une des plus utilisées aujourd’hui pour l’identification
d’espèces étant donnée sa rapidité, sont efficacité et sa reproductibilité comparée aux
techniques conventionnelles utilisant des critères morphologiques et physiologiques. Pour une
meilleure caractérisation, le produit PCR de la souche I33M a été analysé par électrophorèse,
purifié, et enfin séquencé par GenoScreen (Campus de l’Institut Pasteur de Lille, France).
En plus de l’identification biochimique et morphologique, le gène de l’ADNr 16S a été
amplifié avec succès à partir de l’ADN génomique de la souche I33M (Fig.13). Cette
séquence de 999 paires de bases a une longueur de séquence Query de 195 A, 254T, 321C et
229G. La recherche BLAST de la séquence du gène de l’ADNr 16S comparée à d’autres
séquences de la base de données a montré 100% d’homologie avec la souche B. mojavensis.
Notre souche a été nommée B. mojavensis I33M et les bases de données nous ont permis
d’élaborer un dendrogramme phylogénétique pour I33M et les souches associées (Fig.14).
L’Analyse phylogénétique montre que la souche testée est proche de la souche B .subtilis
Des résultats similaires ont été reportés par Thuy et Bose (2011).
Roberts et al., 1994 ont également reporté que l’isolat a été ensuite retrouvé appartenant a un
phénotype associé au B. subtilis qui a été récemment décrit comme étant une souche B.
mojavensis.
B. mojavensis peut être distinguée uniquement par des différences dans la composition
d’acides gras de la cellule entière, dans la divergence de l’analyse de la séquence d’ADN et de
la résistance à la transformation génétique au sein du groupe B. subtilis (Bacon et Hinton,
2001)
La séquence d’ADNr 16s de la souche I33M est représentée à la figure 14

Chapitre 3. Résultats et discussion
108
CCCGAAGGGGACGTCCTATCTCTAGGATTGTCAGAGGATGTCAAGACCTGGTAAGGTTCTTCGCGTT
GCTTCGAATTAAACCACATGCTCCACCGCTTGTGCGGGCCCCCGTCAATTCCTTTGAGTTTCAGTCTT
GCGACCGTACTCCCCAGGCGGAGTGCTTAATGCGTTAGCTGCAGCACTAAGGGGCGGAAACCCCC
TAACACTTAGCACTCATCGTTTACGGCGTGGACTACCAGGGTATCTAATCCTGTTCGCTCCCCACGCT
TTCGCTCCTCAGCGTCAGTTACAGACCAGAGAGTCGCCTTCGCCACTGGTGTTCCTCCACATCTCTAC
GCATTTCACCGCTACACGTGGAATTCCACTCTCCTCTTCTGCACTCAAGTTCCCCAGTTTCCAATGAC
CCTCCCCGGTTGAGCCGGGGGCTTTCACATCAGACTTAAGGAACCGCCTGCGAGCCCTTTACGCCCA
ATAATTCCGGACAACGCTTGCCACCTACGTATTACCGCGGCTGCTGGCACGTAGTTAGCCGTGGCTT
TCTGGTTAGGTACCGTCAAGGTACCGCCCTATTCGAACGGTACTTGTTCTTCCCTAACAACAGAGCTT
TACGATCCGAAAACCTTCATCACTCACGCGGCGTTGCTCCGTCAGACTTTCGTCCATTGCGGAAGATT
CCCTACTGCTGCCTCCCGTAGGAGTCTGGGCCGTGTCTCAGTCCCAGTGTGGCCGATCACCCTCTCA
GGTCGGCTACGCATCGTTGCCTTGGTGAGCCGTTACCTCACCAACTAGCTAATGCGCCGCGGGTCCA
TCTGTAAGTGGTAGCCGAAGCCACCTTTTATGTTTGAACCATGCGGTTCAAACAAGCATCCGGTATTA
GCCCCGGTTTCCCGGAGTTATCCCAGTCTTACAGGCAGGTTACCCACGTGTTACTCACCCGTCCGCC
GCTAACATCAGGGAGCAAGCTCCCATCTGTCCGCTCGACTTGCATGTATTAGGCAC
Figure 14 : Résultat séquences D’ADNr-16S obtenues(Souche I33M)
Séquence similaire à Bacillus mojavensis à 100% de similarité sur un fragment de 999 pb
Le dendrogramme de la souche est representé à la figure 15

Chapitre 3. Résultats et discussion
109
Figure 15 : Dendrogramme de la souche I33M (Bacillus_mojavensis)
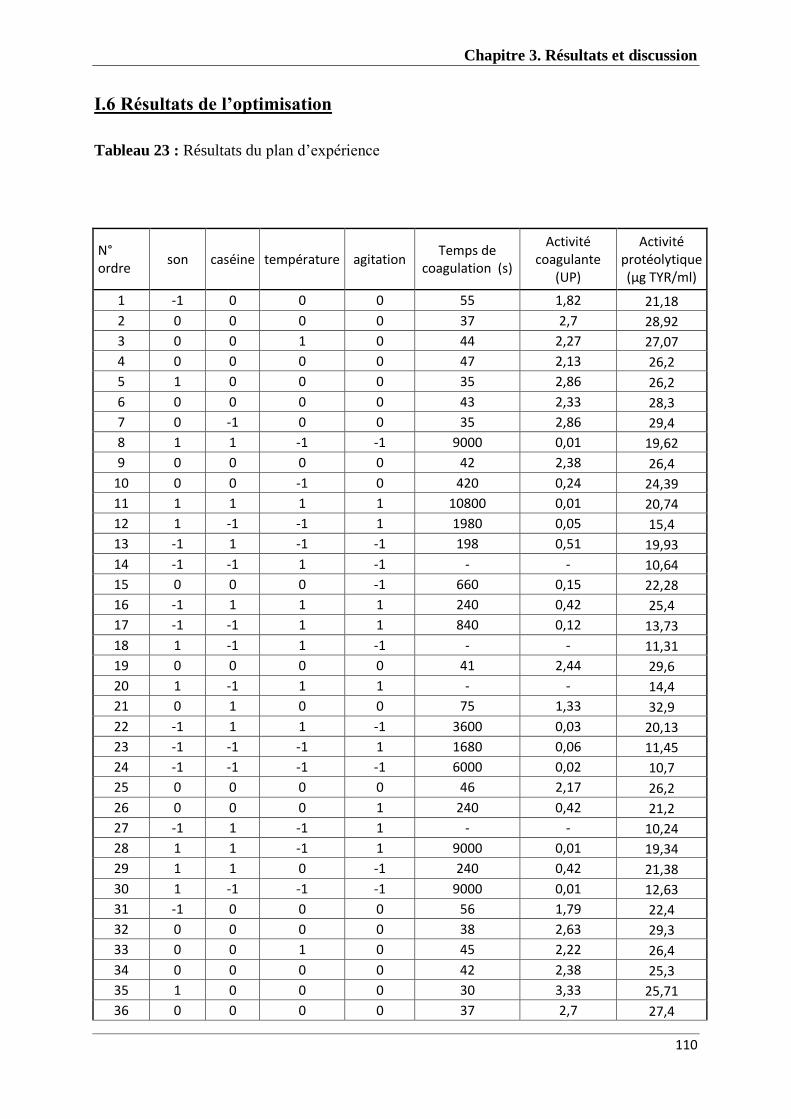
Chapitre 3. Résultats et discussion
110
I.6 Résultats de l’optimisation
Tableau 23 : Résultats du plan d’expérience
N° ordre
son caséine température agitation Temps de
coagulation (s)
Activité coagulante
(UP)
Activité protéolytique (µg TYR/ml)
1 -1 0 0 0 55 1,82 21,18
2 0 0 0 0 37 2,7 28,92
3 0 0 1 0 44 2,27 27,07
4 0 0 0 0 47 2,13 26,2
5 1 0 0 0 35 2,86 26,2
6 0 0 0 0 43 2,33 28,3
7 0 -1 0 0 35 2,86 29,4
8 1 1 -1 -1 9000 0,01 19,62
9 0 0 0 0 42 2,38 26,4
10 0 0 -1 0 420 0,24 24,39
11 1 1 1 1 10800 0,01 20,74
12 1 -1 -1 1 1980 0,05 15,4
13 -1 1 -1 -1 198 0,51 19,93
14 -1 -1 1 -1 - - 10,64
15 0 0 0 -1 660 0,15 22,28
16 -1 1 1 1 240 0,42 25,4
17 -1 -1 1 1 840 0,12 13,73
18 1 -1 1 -1 - - 11,31
19 0 0 0 0 41 2,44 29,6
20 1 -1 1 1 - - 14,4
21 0 1 0 0 75 1,33 32,9
22 -1 1 1 -1 3600 0,03 20,13
23 -1 -1 -1 1 1680 0,06 11,45
24 -1 -1 -1 -1 6000 0,02 10,7
25 0 0 0 0 46 2,17 26,2
26 0 0 0 1 240 0,42 21,2
27 -1 1 -1 1 - - 10,24
28 1 1 -1 1 9000 0,01 19,34
29 1 1 0 -1 240 0,42 21,38
30 1 -1 -1 -1 9000 0,01 12,63
31 -1 0 0 0 56 1,79 22,4
32 0 0 0 0 38 2,63 29,3
33 0 0 1 0 45 2,22 26,4
34 0 0 0 0 42 2,38 25,3
35 1 0 0 0 30 3,33 25,71
36 0 0 0 0 37 2,7 27,4

Chapitre 3. Résultats et discussion
111
37 0 -1 0 0 35 2,86 29,6
38 1 1 -1 -1 9000 0,01 15,76
39 0 0 0 0 36 2,78 25,8
40 0 0 -1 0 480 0,21 24,8
41 1 1 1 1 60 1,67 29,8
42 1 -1 -1 1 - - 12,3
43 -1 1 -1 -1 7080 0,01 19,01
44 -1 -1 1 -1 - - 10,43
45 0 0 0 -1 480 0,21 23,2
46 -1 1 1 1 10800 0,01 20,15
47 -1 -1 1 1 2700 0,04 9,89
48 1 -1 1 -1 - - 12,4
49 0 0 0 0 46 2,17 30,85
50 1 -1 1 1 300 0,33 21,84
51 0 1 0 0 72 1,39 28,48
52 -1 1 1 -1 9000 0,01 21,82
53 -1 -1 -1 1 1680 0,06 12,2
54 -1 -1 -1 -1 4800 0,02 9,14
55 0 0 0 0 47 2,13 26,8
56 0 0 0 1 600 0,17 23,82
57 -1 1 -1 1 - - 12,4
58 1 1 -1 1 7200 0,01 18,2
59 1 1 0 -1 6300 0,02 20,85
60 1 -1 -1 -1 9000 0,01 12,96
Réponse optimisée de l’activité coagulante :
Valeur optimale = 2,40661
Facteur minimum maximum Optimum
------------------------------------------------------------------------------
Son 3, 0 7, 0 7, 0
Caséine 0, 02 2,0 0,0941462
Température 25, 0 50,0 39,2587
Agitation 100, 0 200,0 150,799

Chapitre 3. Résultats et discussion
112
Surface de Réponse Estimée
son=7,0,Caseine=0,094
Temperature
Agitation
Activité
co
ag
ula
nte
Agitation
2530
3540
4550 100 120 140 160 180 200
-0,1
0,4
0,9
1,4
1,9
2,4
2,9
Figure 16 : Surface de réponse de l’activité coagulante de la protéase produite par
Bacillusmojavensis, avec effet de la température et de l’agitation.
Diagramme de Pareto pour l'Activité coagulante
Effet s tandardisé
+-
0 2 4 6 8 10
B:Caseine
D:Agitation
A:son
C:T emperature
CC
DD
Figure 17 : Graphique de Pareto obtenu avec les résultats des essais du plan. D : agitation ;
DD : effet au carré de l’agitation ; A : quantité de son ; CC : effet au carré de la température ;
AA : effet au carré de la quantité de son ; C : Température ; B : caséine.
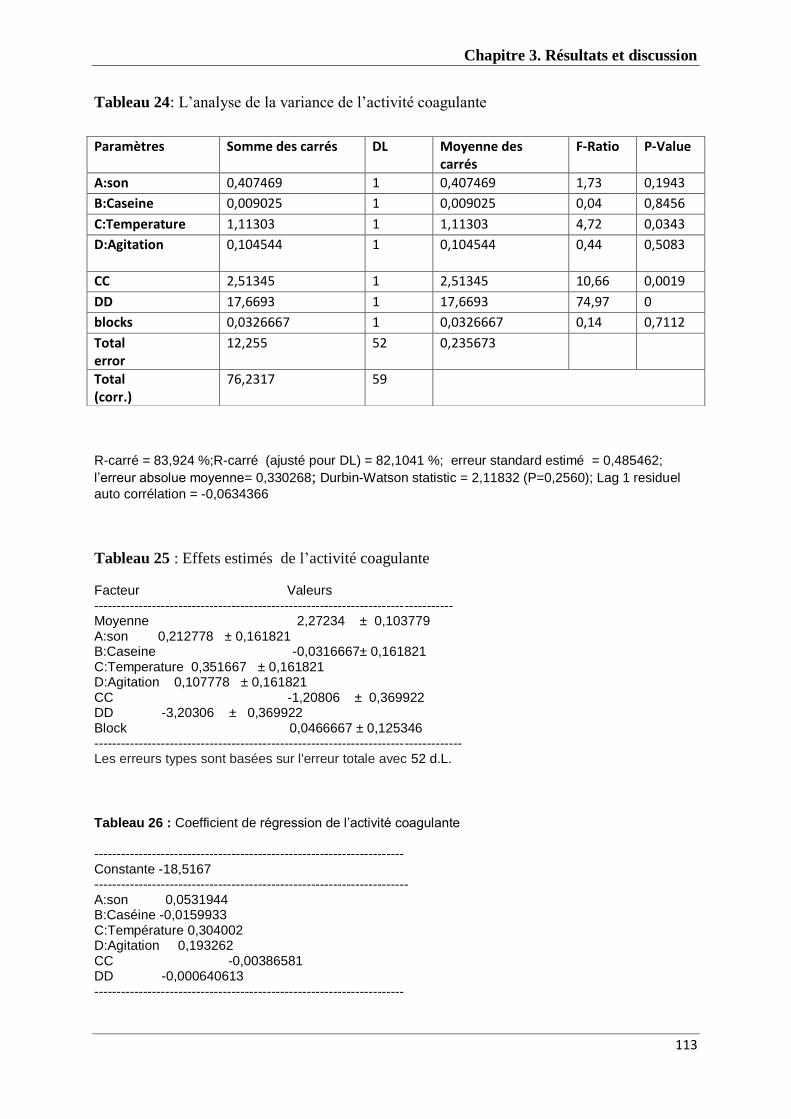
Chapitre 3. Résultats et discussion
113
Tableau 24: L’analyse de la variance de l’activité coagulante
R-carré = 83,924 %;R-carré (ajusté pour DL) = 82,1041 %; erreur standard estimé = 0,485462;
l’erreur absolue moyenne= 0,330268; Durbin-Watson statistic = 2,11832 (P=0,2560); Lag 1 residuel
auto corrélation = -0,0634366
Tableau 25 : Effets estimés de l’activité coagulante
Facteur Valeurs --------------------------------------------------------------------------------- Moyenne 2,27234 ± 0,103779 A:son 0,212778 ± 0,161821 B:Caseine -0,0316667± 0,161821 C:Temperature 0,351667 ± 0,161821 D:Agitation 0,107778 ± 0,161821 CC -1,20806 ± 0,369922 DD -3,20306 ± 0,369922 Block 0,0466667 ± 0,125346 ----------------------------------------------------------------------------------- Les erreurs types sont basées sur l'erreur totale avec 52 d.L.
Tableau 26 : Coefficient de régression de l’activité coagulante
---------------------------------------------------------------------- Constante -18,5167 ----------------------------------------------------------------------- A:son 0,0531944 B:Caséine -0,0159933 C:Température 0,304002 D:Agitation 0,193262 CC -0,00386581 DD -0,000640613 ----------------------------------------------------------------------
Paramètres Somme des carrés DL Moyenne des carrés
F-Ratio P-Value
A:son 0,407469 1 0,407469 1,73 0,1943
B:Caseine 0,009025 1 0,009025 0,04 0,8456
C:Temperature 1,11303 1 1,11303 4,72 0,0343
D:Agitation
0,104544 1 0,104544 0,44 0,5083
CC 2,51345 1 2,51345 10,66 0,0019
DD 17,6693 1 17,6693 74,97 0
blocks 0,0326667 1 0,0326667 0,14 0,7112
Total error
12,255 52 0,235673
Total (corr.)
76,2317 59

Chapitre 3. Résultats et discussion
114
I.6.1 Discussion
Optimisation par méthode de surface de réponse:
L’approche du modèle statistique utilisant la méthode de surface de réponses est utilisée pour
étudier les effets interactifs de différents facteurs chimiques et physiologiques sur la
production de la protéase coagulant le lait par B. mojavensis. L’objectif principal est de
maximiser l’activité de coagulation et de minimiser l’activité protéolytique. L’expérience a
été effectué 60fois et les données ayant le plus d’influence sur la réponse finale du système
ont été identifiées à partir de l’interaction de quatre facteurs : son de blé (source de carbone)
et caséine (source d’azote), température et agitation (facteurs physiques), ont été examinés à
travers la RSM suivant le CCD. Nous rappelons que les niveaux faibles et élevés (% p/v)
codés entre parenthèses pour ces facteurs sont définis à 3(-1) et 7(+1) pour le son de blé,
0,02(-1) et 2.0(+1) pour la caséine, 25°(-1) et 50°C(+1) pour la température, 100rpm(-1) et
200(+1) pour l’agitation.
Les résultats des expériences CCD pour l’étude de ces variables sur la production de protéases
dans l’activité de coagulation du lait (IMCU/ml) sont présentés dans le tableau 23
accompagnés des réponses observées.
L’analyse statistique de ces résultats montre que la gamme étudiée, effet quadratique de la
température et de l’agitation a un effet considérable sur la production de protéases suivi de
l’effet quadratique de l’agitation.
La température a un effet significatif (Fig.16). Le tableau 25 montre les effets estimés et les
interactions du modèle de surface de réponse du second ordre sous la forme d’une analyse de
la variance (ANOVA) et sont présentées dans le tableau 24.
Le test-F de Fisher démontre une significativité dans le modèle de régression où le coefficient
de détermination R² était de 0.8392. Le R² statistique indique que le modèle établi explique
83,924% de la variabilité de l’activité de coagulation du lait. Le R² ajusté, le plus adapté à la
comparaison des modèles avec des nombres de variables indépendantes, est de 82,1041%..
Cette équation de régression (en termes de facteurs codés) des niveaux de protéase produite
est exprimée en IMCU/ml(Y) en fonction du son de blé (A), de caséine(B), de température
(C) et de l’agitation (D).

Chapitre 3. Résultats et discussion
115
Y= -18.5167 + 0.0531944* A – 0.0159933*B + 0.304002*C + 0.193262*D –
0.00386581*CC -0.000640613*DD.
Les coefficients de régression sont listés dans le tableau 26. La magnitude des effets indique
un niveau de significativité de la variable de production de protéases. Parmi les variables
étudiées ; il semblerait que seuls les facteurs physiques (température et agitation) sont
identifiés comme les variables les plus significatives dans l’influence de la production des
enzymes coagulantes (Fig. 16) que le son de blé et de caséine.
En réalité, les résultats des réponses observées dans les expériences réalisées en utilisant le
CCD (Fig.17) montre que la température et l’agitation sont des facteurs limitants pour la
production d’enzyme quel que soit la concentration du substrat. La combinaison des niveaux
de facteurs qui maximisent l’activité de coagulation du lait a été donnée par un milieu
composé de son de blé : 7.0% ; caséine : 0,094% ; température 39,2% et agitation : 150rpm.
La valeur optimale de l’activité coagulante est de 2,4 (IMCU/ml).
La réponse de surface en 3dimensions est représentée dans la figure 16, qui montre les effets
relatifs de 2 facteurs : température et agitation qui sont en association que lorsque tous les
facteurs (concentration de son de blé et de caséine) sont maintenus à leur niveau central.
Il n’a été observé qu’une augmentation de la production de protéases à des températures
relativement élevée et une production de protéase maximale obtenue de 38 à 39° (Fig.16).
Notre modèle indique que la température a un effet majeur sur la production de protéase par la
souche B. mojavensis. Des résultats d’études RSM ont aussi montré que la température est un
facteur très important dans la production de protéase microbienne (Wu et Hang, 2000 ;Thys
et al., 2006). La méthodologie RSM a été montrée comme étant très efficace dans la
détermination des conditions optimales pour la production de protéases par Bacillus (Thys et
al., 2006)
En général, il est clairement montré qu’aucun milieu défini n’a été établi pour la production
de protéases à partir de sources microbiennes de protéases (Gupta et al., 2002). Chaque
organisme ou souche a sa propre exigence de conditions spéciales pour la production
d’enzyme maximale. Ainsi, dans notre cas, les acteurs physiques, température et agitation
sont plus significatifs que les facteurs nutritionnels.

Chapitre 3. Résultats et discussion
116
I.7 Résultats de la croissance durant lafermentation de la souche I33 M
Les dénombrements ont lieu au bout de 24h. L'activité coagulante a été mesurée ainsi que le
pH.Les résultats obtenus de la croissance sont représentés dans le tableau 27 et illustrés par
les figures 19, 20, 21, 22, 23, 24.
Un suivi de croissance et divers dosages ont permis d’évaluer les conditions de la production
de l'enzyme coagulante par la souche I33M.
Tableau 27 : Résultats du suivi de la fermentation
Temps
d'incubation
(en h)
N Ln (N) pH AC (en
UP/ml)
Protéine
µg/ml
Protéine
mg/ml
AP
µg/ml/min
0 1,42E+06 14,1648862 6,7 0 3400 3,4 9,8
1,5 1,36E+06 14,1256655 6,67 0 3200 3,2 9,4
3 2,41E+07 16,9973451 6,68 0 3357 3,4 10,2
4,5 4,36E+08 19,8939865 6,5 0 3314 3,3 9,4
6 4,91E+08 20,0117695 6,23 0,00641026 3893 3,9 12,9
7,5 7,55E+08 20,4416261 6,14 0,01754386 3871 3,9 18,1
10,5 1,25E+09 20,9500392 6,34 0,04761905 4164 4,2 21,2
13,5 1,11E+09 20,8268065 6,26 0,09803922 4257 4,3 28,4
16,5 1,09E+09 20,8102772 6,27 0,16666667 4286 4,3 28,9
19,5 1,66E+09 21,2322716 6,14 0,27777778 4071 4,1 34,0
22,5 1,43E+09 21,0790313 6,01 0,35087719 4493 4,5 37,6
25,5 1,58E+09 21,1818408 6 0,35087719 4829 4,8 31,9
28,5 1,50E+09 21,1287309 6,08 0,48780488 3936 3,9 43,1
31,5 1,81E+09 21,3160903 6,1 0,52631579 4271 4,3 41,8
34,5 2,26E+09 21,5402384 6,12 0,5 5586 5,6 47,3
37,5 1,88E+09 21,3555043 6,22 0,55555556 4350 4,4 48,2
40,5 2,09E+09 21,4608648 6,22 0,52631579 4286 4,3 48,5
43,5 1,67E+09 21,2377212 6,24 0,60606061 4343 4,3 44,9
46,5 1,84E+09 21,3310532 6,26 0,64516129 4321 4,3 49,5

Chapitre 3. Résultats et discussion
117
Figure 18 : Evolution de la croissance bactérienne en fonction du temps d’incubation
Figure 19 : Détermination du temps de génération
G = 0,36048844 G : temps de génération
21,629306721min36s taux de croissance= 1,9228
y = 1,9228x + 11,237 R² = 1
0
5
10
15
20
25
0 1 2 3 4 5
Ln (
N)
Temps d'incubation (en h)
14
15
16
17
18
19
20
21
22
0 10 20 30 40 50
Ln(N
)
Temps d'incubation (en h)
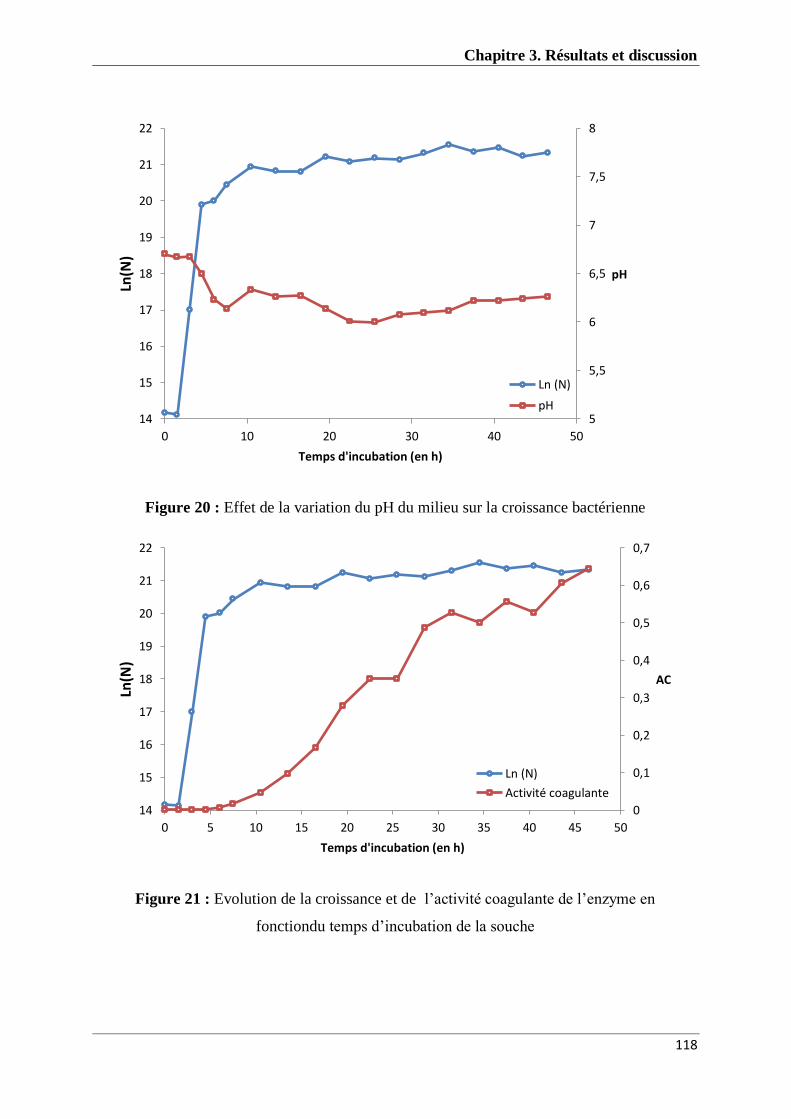
Chapitre 3. Résultats et discussion
118
Figure 20 : Effet de la variation du pH du milieu sur la croissance bactérienne
Figure 21 : Evolution de la croissance et de l’activité coagulante de l’enzyme en
fonctiondu temps d’incubation de la souche
5
5,5
6
6,5
7
7,5
8
14
15
16
17
18
19
20
21
22
0 10 20 30 40 50
pH
Ln(N
)
Temps d'incubation (en h)
Ln (N)
pH
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
14
15
16
17
18
19
20
21
22
0 5 10 15 20 25 30 35 40 45 50
AC
Ln(N
)
Temps d'incubation (en h)
Ln (N)
Activité coagulante
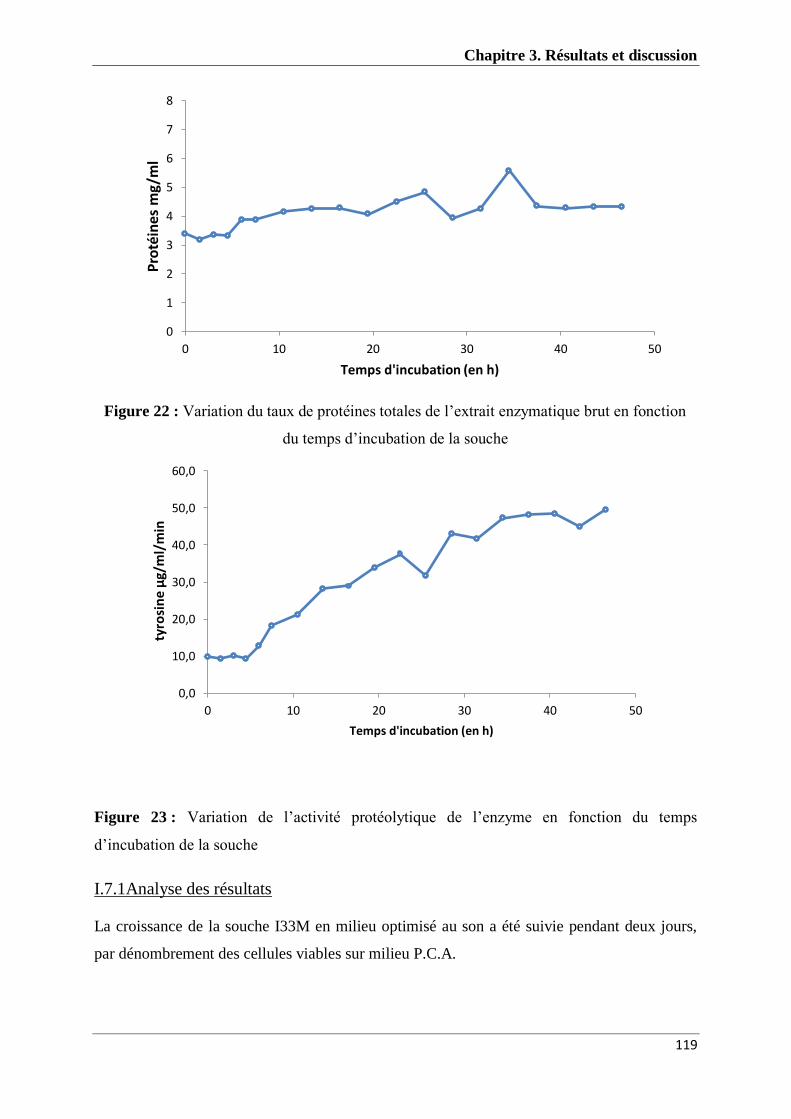
Chapitre 3. Résultats et discussion
119
Figure 22 : Variation du taux de protéines totales de l’extrait enzymatique brut en fonction
du temps d’incubation de la souche
Figure 23 : Variation de l’activité protéolytique de l’enzyme en fonction du temps
d’incubation de la souche
I.7.1Analyse des résultats
La croissance de la souche I33M en milieu optimisé au son a été suivie pendant deux jours,
par dénombrement des cellules viables sur milieu P.C.A.
0
1
2
3
4
5
6
7
8
0 10 20 30 40 50
Pro
téin
es m
g/m
l
Temps d'incubation (en h)
0,0
10,0
20,0
30,0
40,0
50,0
60,0
0 10 20 30 40 50
tyr
osi
ne
µg/
ml/
min
Temps d'incubation (en h)

Chapitre 3. Résultats et discussion
120
L’analyse de la courbe de croissance Ln (N) en f(t) (Fig.18) montre qu’après une courte
période de latence de moins de 1h, la phase exponentielle est rapide et la phase décélération
est atteinte en moins de 5h. La phase stationnaire s’est prolongée sur la durée de l’expérience,
sans que l’on puisse mettre en évidence au bout de deux jours une phase de déclin.
Le taux de croissance népérien calculé sur les 3 points de la phase exponentielle (Fig.19)
montre une vitesse de croissance extrêmement rapide puisque le temps de génération
exponentiel est évalué à 22 mn environ.
La mesure du pH en cours de croissance (Fig.20), montre que le pH voisin de 7 au départ se
stabilise à 6 dès le début de la phase stationnaire.
Concernant les activités coagulante (Fig.21) et protéolytique (Fig.23) et du surnageant, elles
augmentent progressivement dès le début de la phase stationnaire et cela jusqu’à la fin de
l’expérience. L’activité coagulante semble augmenter moins rapidement et n’apparaitre de
façon significative qu’au bout d’une dizaine d’heure de culture. La quantité totale de protéines
du surnageant augmente légèrement en parallèle de ces activités (Fig.22).
I.7.2.Discussion
La souche I33M en culture dans le milieu au son, se caractérise par un temps de génération
très faible, ce qui témoigne de son aptitude à mobiliser et dégrader les constituants du milieu
de culture employé.
La phase stationnaire se prolonge sur les deux jours de l’expérience : on peut supposer que la
stabilisation du pH contribue au maintien de la phase stationnaire. La bibliographie indique
que chez Bacillus subtilis (Dutt et al., 2008 ), elle peut durer de 3 à 8jours .
L’évolution du pH (diminution en phase exponentielle et stabilisation à pH 6) peut s’expliquer
par le fait que les bactéries consomment en premier lieu les glucides contenus dans le milieu ;
l’acidification qui en résulte est rapidement neutralisée par les produits basiques issus de
l’activité protéolytique des Bacillus.
Au bout des deux jours d’expérience, l’activité coagulante est toujours en augmentation. Il ne
nous est donc pas possible de dire si le maximum est atteint. Dutt et al. (2008) précise que
l’activité coagulante diminue au bout de 45h alors que Shieh et al. (2009) montre, dans les
conditions de l’expérience, que le maximum est atteint au bout d’1 jour ; cependant, celui-ci
signale que d’autres auteurs rapportent que l’optimum de production est atteint au bout de 3 à
8 jours.

Chapitre 3. Résultats et discussion
121
Concernant l’enzyme coagulante produite par Bacillus mojavensis, deux hypothèses peuvent
être formulées. On peut supposer que cette protéase est très vraisemblablement un
métabolite secondaire (son taux ne cesse d’augmenter durant la phase stationnaire alors qu’un
métabolite primaire est produit durant la phase exponentielle). L’autre hypothèse est qu’il
s’agit d’une protéine intracellulaire (taux très faible en phase exponentielle) libérée
progressivement à la mort des cellules en phase stationnaire). La lyse du culot bactérien
obtenu en phase exponentielle et le dosage de l’activité coagulante permettraient de
différencier les deux hypothèses.
.8. Essai de purification de l’extrait enzymatique brut de Bacillus
mojavensis
L’avantage de la chromatographie d’exclusion moléculaire est de pouvoir être appliqué
indifféremment aux extraits bruts ou partiellement purifiés (Milliere, 1984).
Les protéines de l’extrait enzymatique brut sont séparées selon leur poids moléculaire par le
gel Séphadex G-75 (La phase stationnaire) dont la zone de fractionnement est comprise entre
3 000 et 70 000 Daltons, la phase mobile étant la solution tampon phosphate (pH 6 – 0,4M)
qui va transporter les protéines.
Le volume de l’extrait brut injecté est de 1 ml, les éluâts de la colonne sont recueillis dans le
collecteur de fraction à raison de 2 ml/tube et on détermine par la suite la densité optique de
chaque tube à 280 nm.
L’activité coagulante a été mesurée pour chaque fraction collectée. Les résultats obtenus sont
reportés sur le tableau 28.
Tableau 28: Activité coagulante des éluât en fonction du numéro du tube.
N° du tube Activité coagulante (UP)
10 ∞
11 0,081
12 0,088
13 0,094
14 0,038
15 ∞

Chapitre 3. Résultats et discussion
122
Le profil chromatographique de l’extrait coagulant de Bacillus mojavensis montre deux pics
d’adsorption bien distincts (Fig.24). L’activité coagulante a été retrouvée dans les tubes du
n°11, 12, 13 et 14 qui forment le premier pic ; ces résultats révèlent que seul le premier pic est
doué d’une activité coagulante.
Le second pic n’est pas doué d’activité coagulante et correspond aux protéines non
coagulantes. On peut dire que ce niveau de purification n’est que partiel, car il n’est pas exclu
que d’autres protéines de masses moléculaires très rapprochées peuvent être incluses dans
cette même fraction.
Figure 24: profil chromatographique de l’extrait enzymatique produit par I33M filtré sur gel
Séphadex G-75.Tampon phosphate 0,4M pH 6, debit 0,3 ml/min (2ml/tube)
1.9 Caractérisation de l’enzyme coagulante de Bacillus mojavensis
I.9.1 Détermination de l’activité coagulante
Après l’extraction, l’activité coagulante de l’extrait enzymatique brut de Bacillus mojavensis
a été déterminée par la méthode de Berridge et qui a permis d’obtenir les résultats illustrés
dans le tableau 29.
0
0,02
0,04
0,06
0,08
0,1
0,12
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
0 10 20 30 40 50 60 acti
vit
é coa
gu
lan
te (
UP
/ml)
DO
à
28
0n
m
N° de tubes
activité coagulante profil d'elution
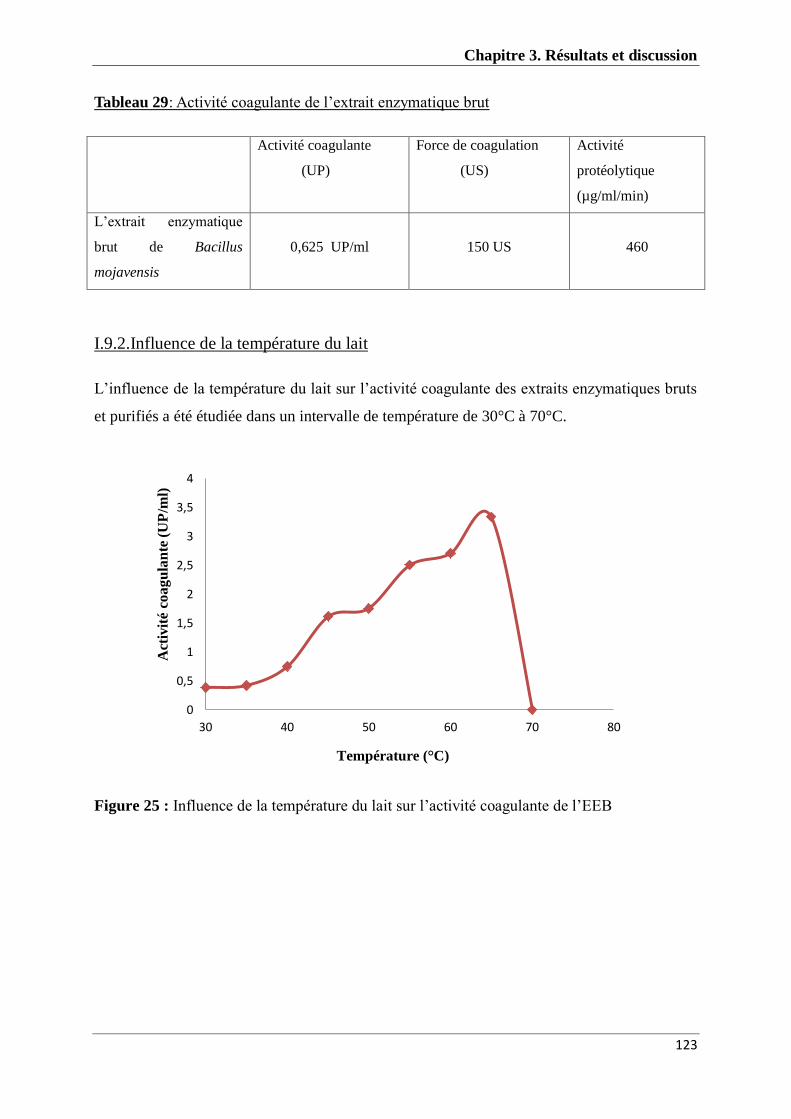
Chapitre 3. Résultats et discussion
123
Tableau 29: Activité coagulante de l’extrait enzymatique brut
Activité coagulante
(UP)
Force de coagulation
(US)
Activité
protéolytique
(µg/ml/min)
L’extrait enzymatique
brut de Bacillus
mojavensis
0,625 UP/ml
150 US
460
I.9.2.Influence de la température du lait
L’influence de la température du lait sur l’activité coagulante des extraits enzymatiques bruts
et purifiés a été étudiée dans un intervalle de température de 30°C à 70°C.
Figure 25 : Influence de la température du lait sur l’activité coagulante de l’EEB
0
0,5
1
1,5
2
2,5
3
3,5
4
30 40 50 60 70 80
Act
ivit
é co
ag
ula
nte
(U
P/m
l)
Température (°C)

Chapitre 3. Résultats et discussion
124
Figure 26 : Influence de la température du lait sur l’activité coagulante de l’EEP
D’après les résultats rapportés par les figures 26 et 27, on note que la température optimale de
l’activité coagulante de nos extraits enzymatiques est de 65°C. De 30°C à 65°C, l’activité
coagulante augmente progressivement et au delà de 65°C on note une baisse rapide de
l’activité coagulante.
Nos résultats obtenus sont similaires à ceux réalisés par Yang etal. (1987) qui ont démontré
un optimum d’activité coagulante à une température de 65°C.
Chopra et Mathur (1985)ont obtenu une coagulase purifiée de Bacillus stearothermophilus
avec une activité optimale à 70°C. Matoub (2000) a montré que l’activité optimum de
l’enzyme purifié de Bacillus subtilis est de 70°C.
Selon Garnot et Martin (1980), la présure est caractérisée par une activité optimale à une
température voisine à 42°C.D’après Morsli (1996), l’optimum d’activité obtenue pour
l’extrait enzymatique du figuier a été enregistré à 85°C, pour l’artichaut est de 72°C.
En général, les extraits enzymatiques d’origine animale ont une activité importante à des
températures modérés cependant les coagulases d’origine microbienne et végétale se montrent
plus actives aux températures élevées.
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
30 35 40 45 50 55 60 65 70
Acti
vit
é coa
gu
lan
te (
UP
/ml)
Température (°C)

Chapitre 3. Résultats et discussion
125
I.9.3 Influence de pH du lait
Nous avons exposé l’extrait enzymatique de Bacillus mojavensis à un intervalle de pH allant
de 5 à 7 afin d’optimiser l’activité exprimé en unité présure.
Figure 27:Influence du pH du lait sur l’activité coagulante de l’EEB.
Figure 28:Influence du pH du lait sur l’activité coagulante de l’EEP.
0
0,5
1
1,5
2
2,5
5 5,5 6 6,5 7 7,5
pH
Acti
vit
é coa
gu
lan
te (
UP
/ml)
0,03
0,032
0,034
0,036
0,038
0,04
0,042
0,044
0,046
5 5,5 6 6,5 7 7,5 8
Act
ivit
é co
ag
ula
nte
(U
P/m
l)
pH

Chapitre 3. Résultats et discussion
126
Les variations de l’activité coagulante des extraits enzymatiques bruts et purifiés en fonction
du pH du lait illustrées par les figures 28 et 29, montrent que l’activité coagulante augmente
progressivement de pH 5 à 6 et baisse au-delà de pH optimum (pH 6) pour s’annuler
complètement à pH 7.
Nos résultats se rapprochent de ceux de Yang et al. (1987) qui dans une étude similaire sur
une coagulase purifié de Bacillus subtilis ont rapporté un pH d’activité optimal de 5,8.
Matoub (2000) indique un pH optimum d’action de 6,2 pour une coagulase produite par une
souche locale sélectionné de Bacillus subtilis (LC33) par contre Chopra et Mathur (1985)
ont obtenu une coagulase de Bacillus stearothermophilus dont l’activité optimale est observée
à pH 8.
Selon Garnot et Martin (1980), le pH d’activité de la présure est compris entre 5,3 et 6,3.
Son optimum d’action est à pH 5,8. Elle est inactivée à pH 7,5.
Morsli (1996), indique un pH optimum de 6,5- 6,2- 6,5 pour des préparations enzymatique de
l’artichaut, du figuier et du poulet respectivement.
Scriban (1999), note une activité coagulante optimale entre pH 5,5 et 6,7 pour les enzymes
issues d’Endothia parasitica et Mucor pusillus.
En définitif, on peut conclure que le pH optimum des coagulases varie dans l’intervalle de
5,5 à 8 selon leur origine.
Les valeurs habituelles de pH utilisées en technologie fromagère sont de pH 6,5 à 6,3 ; voisin
d’un lait frais (pH 6,6) ou légèrement acidifié (pH 6,4). Dans certains cas, quand les
conditions de technologie fromagère le permettent, le pH du lait peut être abaissé au pH 6,2.
I.9.4Influence de concentration en CaCl2
D’après Lenoir (1985), l’ion calcium est un activateur des protéases, il est donc indispensable
pour la coagulation du lait en associant ainsi les micelles des caséines afin de former un gel
plus ferme. Cette coagulation se produit par l’agrégation des molécules en présence du
calcium après hydrolyse de la caséine k par la protéase coagulase.

Chapitre 3. Résultats et discussion
127
Les concentrations habituellement utilisées en technologie fromagère se situent dans
l’intervalle de 0,01 à 0,02M de CaCl2, l’extrait purifié de Bacillus mojavensis a une meilleure
activité à cette concentration, ce qui permet leur utilisation dans ces conditions. L’influence
de calcium sur l’activité coagulante de l’extrait enzymatique a été étudiée en faisant varier sa
concentration dans le lait de 0,005 à 0,1 M.
Figure 29:Influence de la concentration en CaCl2 du lait sur l’activité coagulante
de l’EEB.
Figure 30:Influence de la concentration en CaCl2 du lait sur l’activité coagulante
de l’EEP
0
0,2
0,4
0,6
0,8
1
1,2
0 0,02 0,04 0,06 0,08 0,1 0,12
Concentration en CaCl2 (M)
A
ctiv
ité
coa
gu
lan
te (
UP
/ml)
0,03
0,032
0,034
0,036
0,038
0,04
0,042
0,044
0,046
0,048
0,05
0 0,02 0,04 0,06 0,08 0,1 0,12
A
ctiv
ité
co
agu
lan
te (
UP
/ml)

Chapitre 3. Résultats et discussion
128
Les figures 30 et 31 expriment les résultats en montrant que l’activité coagulante croit lorsque
la concentration en CaCl2 augmente. On obtient ainsi un optimum d’activité pour une
concentration de 0,04M, au-delà de cette valeur l’activité diminue et ceci par un effet
inhibiteur de l’ion calcium.
Dans une étude similaire de la coagulase de Bacillus subtilis, Chemlal (1998), a obtenu une
concentration optimale de 0.05M. Par contre, Matoub (2000), a rapportée une activité
optimale de la coagulase purifiée à une concentration de 0.02M.
Par ailleurs, Morsli (1996), lors d’une étude du pouvoir coagulant de quelques plantes
locales et d’une coagulase de poulet indique une concentration du lait en CaCl2 de l’ordre de
0,02M pour l’ensemble des préparations et d’après Garnot et Martin (1980) une activité
optimale de la présure est obtenue à 0,02M de CaCl2.
En général, les extraits enzymatiques d’origine animale sont moins sensibles à la baisse
de la concentration en CaCl2 du lait, contrairement à ceux d’origine végétale qui nécessitent
une concentration plus importante.
II.PROTEASE D’ORIGINE FONGIQUE
II.1Collection de souches
24 souches isolées du sol ont constitué la collection de base (Tab.30). Le critère de
sélection a été leur pouvoir caséolytique sur milieu de culture (Sabouraud +CMP+ lait),
témoignant d’une action sur les protéines du lait. Elles ont été ensuite étudiées d’un point de
vue macroscopique puis microscopique.
II.2 Observations macroscopiques
Les résultats sont présentés dans le tableau 30 et dans la figure 31.

Chapitre 3. Résultats et discussion
129
Tableau 30: Description macroscopique de la collection des moisissures isolées du sol.
Code des
souches Aspect macroscopique des moisissures
4A souche étalée grise centre blanc
4 B halle étalé mycélium gris vert
4 C centre jaune clair pourtour jaune foncé
4 D centre jaune foncé pourtour gris-vert
5 F jaune au centre pourtour gris blanc
S 1 centre plus clair, pourtour gris foncé
S 3 centre clair pourtour foncé
S F 11 souche grise
SF 12 thalle de couleur grise
SF 19 colonies envahissante jaune foncé à la base et presque blanche
à la surface,
W centre foncé, pourtour beige foncé
SF 14 colonies jaune très envahissante,
SF 15 souches de couleur grise
4 E couche étalée pourtour gris vert,
A 2 mycélium blanc avec base jaune
I/5 colonie avec un centre claire et un pourtour vert foncé.
T7 thalle noir
Y centre jaune pourtour vert
P/M centre jaune pale pourtour gris
K/L centre duveteux blanc pourtour gris
E mycélium blanc avec spores noires
D centre jaune avec un Mycélium qui monte jusqu'au couvercle
de la boite de pétri.
A1/B1 base jaune pourtour gris vert.
A/B centre du ventre blanc pourtour gris vert.

Chapitre 3. Résultats et discussion
130
Figure 31: Aspect des colonies sur boîtes de Pétri pour quelques moisissures
Conclusion :
De nombreuses colonies isolées se caractérisent par un mycelium duveteux et très extensif.
Les couleurs varient du blanc au gris foncé ; des zones de fructification centrales sont bien
visibles et leur aspect laisse supposer que nous sommes essentiellement en présence de
Mucorales.
II.3 Observations microscopiques
La figure 32 montre l’aspect microscopique de certaines moisissures isolées. Elles sont toutes
caractérisées par un appareil de fructification caractéristique des Mucorales.
Celui-ci éclate et libère les spores à maturité. Les spores ont des formes ovoïdes ou rondes
suivant la souche isolée.
Conclusion :
Comme nous l’avons observé avec les bactéries, la microflore fongique isolée montre une
certaine homogénéité et semble essentiellement constituée de Mucorales (Rhizopus, Mucor).
Ces genres forment un très grand nombre de spores. Il est vraisemblable que dans les
prélèvements effectués, ces spores constituaient l’élément fongique majeur et sont à l’origine
des souches isolées sur milieu de culture.
C/D S2/S3 I/J
4A 5F 4C

Chapitre 3. Résultats et discussion
131
Figure 32 : aspect microscopique de quelques moisissures isolées
S1
4 C
4A
4 D
5F
SF 15

Chapitre 3. Résultats et discussion
132
II.4 Détermination de l’activité enzymatique des extraits bruts des
moisissures isolées.
La fermentation des souches sur milieu solide à base de son de blé selon Levadoux(1989) a
permis d’obtenir une quantité intéressante de protéases. La composition du milieu de
fermentation a une grande influence sur la régulation de la production de protéases par les
souches et permet d’accroitre de façon substantielle l’activité coagulante des extraits bruts.
Les résultats concernant les paramètres qui reflètent l’activité enzymatique des EEB (activités
coagulante et protéolytique) ont été déterminées pour les 24 souches de la collection.
II.4.1Activité coagulante
L’activité coagulante de la protéase issue des moisissures est exprimée en unité présure/ml.
Les résultats sont reportés dans le tableau 31.
II.4.2L’activité protéolytique
Les données du calcul de l’activité protéolytique des extraits enzymatiques bruts sont portées
dans le tableau 31.
Après avoir testé les 24 souches de moisissures, nous avons obtenu uniquement 12 souches
produisant une enzyme coagulante, elles sont présentées dans le tableau 31.
Tableau 31 : souches possédant une enzyme coagulante et caractéristiques
Code des
Souches
Aspect macroscopique ACen
UP/ml
AP en
µg/ml/min
4A Etalée, envahissante, grise, centre un peu plus clair 0.104 59.7
4C Centre clair (gris clair)
Pourtour gris vert
0.098 56.7
4D
Centre jaune foncé
Pourtour gris vert
0.208 49.06
5F
Centre jaune
Pourtour vert
0.277 62 .2
S1
Centre plus clair, pourtour gris foncé 0.098 79.34
S 2 / S3
Centre plus clair, pourtour foncé 0.208 74.28
SF14 Colonies très envahissantes/ vert clair. Centre plus clair 0.277 79.39

Chapitre 3. Résultats et discussion
133
SF15 Grise. envahissante 0.280 57. 74
I/J
Centre plus clair; pourtour vert foncé 0.128 61.8
P/M
Centre jaune ; pourtour vert. 0.119 64.5
K/L Centre clair, pourtour jaune vert. 0.277 49.89
C/D
Envahissante ; centre jaune ; pourtour gris vert. 0.084 50 .44
Conclusion
50% des souches montrent une activité coagulante, ce qui est important par rapport aux
résultats obtenus avec la microflore bactérienne. Cependant, l’activité coagulante des souches
positives est faible et varie de 0,08 à 1 avec une moyenne de 0,18 UP/ml. En revanche
l’ensemble des extraits montrent une forte activité protéolytique (moyenne = 62,2 µg/ml).
Ces résultats (fort % de souches coagulantes, faibles activités coagulantes de EEB) peuvent
s’expliquer par une certaine homogénéité des souches isolées.
II.5 Identification des moisissures par biologie moléculaire
Après avoir observé de manière minutieuse au microscope les 12 souches sélectionnées pour
produire une enzyme coagulante, il s’est avéré que seules 3 d’entre elles se différentiaient
d’un point de vue microscopique (souches 4D, 5F, SF15). Ces 3 souches ont été identifiées
par biologie moléculaire seuls les résultats de 4D sont présentés (Blast : figure 33).

Chapitre 3. Résultats et discussion
134
II.5.1 Séquences d’ADNr-18S obtenues
Souche 4D
TAATCAATAATTTTGGCTTGTCCATTATTATCTATTTACTGTGAAATGTATTATTACTTGACGCTTGAGG
GATGCTCCACTGCTATAAGGATAGGCGGTGGGGATGCTAACCGAGTCATAATCAAGCTTAGGCTTGG
TATCCTATTATTATTTACCAAAAGAATTCAGAATTAATATTGTAACATAGACCTAAAAAATCTATAAAACA
ACTTTTAACAACGGATCTCTTGGTTCTCGCATCGATGAAGAACGTAGCAAAGTGCGATAACTAATGTG
AATTGCATATTCAGTGAATCATCGAGTCTTTGAACGCAACTTGCGCTCATTGGTATTCCAATGAGCACG
CCTGTTTCAGTATCAAAACAAACCCTCTATCCAACATTTTGTTGAATAGGAATACTGAGAGTCTCTTGA
TCTATTCTGATCTCGAACCTCTTGAAATGTACAAAGGCCTGATCTTGTTTGAATGCCTGAACTTTTTTTT
AATATAAAGAGAAGCTCTTGCGGTAAACTGTGCTGGGGCCTCCCAAATAATACTTTTTTTAAATTT
GATCTGAAATCAGGCGGGATTACCCGCTGAACTTAA
Souche 4D
Séquence similaire à Mucor circinelloides à 100% de similarité sur un fragment de 586 pb.
Figure 33 : Blast de la souche « 4D »
Nous avons choisi la souche 4D pour la suite du travail car elle possède une bonne activité
coagulante et une activité protéolytique relativement basse par rapport aux 2 autres souches.
II.5.2Identification de la souche 4D
La séquence étudiée est similaire à Mucor circinelloides avec 100% de similarité sur un
fragment de 586 pb.
L’arbre phylogénétique de la souche sélectionnée est présenté dans la figure 34.

Chapitre 3. Résultats et discussion
135
Figure 34 : Position de la souche 4D dans l’arbre phylogénétique de Mucor circinelloides.

Chapitre 3. Résultats et discussion
136
II.6Optimisation de la production de l’enzyme coagulante
La production de l’enzyme coagulante a été réalisée par fermentation sur son de blé pendant 5
jours au bout desquels un bon développement du champignon fut observé donnant au milieu
une texture laineuse blanche caractéristique des mucors.
L’extraction de l’enzyme est faite après fermentation. L’activité coagulante des 40
expériences est déterminée par la méthode de Berridje (1951) et les résultats sont interprétés
par la méthode statistique selon le plan composite centré afin de choisir les conditions
optimales de la production.
II.6.1Evaluation statistique des composants du milieu de culture par le plan composite
centré
La méthode utilisée est le plan composite centré à 5 niveaux. Les conditions expérimentales
(niveau de facteurs et réponses) figurent dans la matrice des expériences présente dans le
tableau 15.
II.6.2L’analyse de l’influence de la composition du milieu de culture sur l’activité
coagulante des enzymes
Les activités coagulantes obtenues varient de 0.603 UP/ml à 2.667 UP/ml ce qui indique
l’influence des paramètres choisis sur la production (tableau 32). L’activité coagulante
maximale 2.667 UP/ml a été observée dans l’expérience N°18, où les composants du
milieu sont à un niveau moyen: température : 31°C ; quantité de son de blé : 30 g ; quantité de
caséine : 1 g.
Ce résultat est probablement du à la température choisie. En effet, selon Tubesha et Delaimy
(2003), l’enzyme de la plus forte activité a été produite à température égale à 30°C.
La faible activité coagulante 0.609 U/ml a été déterminée dans l’expérience N°6 :
température : 37°C ; quantité de son de blé : 20g ; quantité de caséine : 1,5g.
Ceci peut s’expliquer soit par la température élevée (Tubesha et Delaimy, 2003) ou la faible
quantité de son de blé.

Chapitre 3. Résultats et discussion
137
Tableau 32:Matrice expérimentale et activité coagulante des extraits bruts obtenus.
II.6.2.1Analyse statistique
L’analyse statistique des résultats est effectuée par un pro-logiciel de calcul, qui teste les
critères de l’adéquation du modèle mathématique.
Nombre
D’essais
Température
(°C)
Son de
blé
(g)
Caséine
(g)
Réponse
mesurée
Réponse prédite
statistiquement
1 25 20 0.5 1.247 1.225
2 37 20 0.5 0.715 1.444
3 25 40 0.5 1.708 1.623
4 37 40 0.5 1.442 1 .842
5 25 20 1.5 1.376 1.216
6 37 20 1.5 0.603 0.998
7 25 40 1.5 2. 488 2.069
8 37 40 1.5 1.589 1.851
9 27 30 1 1.561 1.918
10 35 30 1 2.605 1.918
11 31 25 1 1.212 0.782
12 31 35 1 1.799 1.835
13 31 30 0.6 1.732 1.498
14 31 30 1.4 1.656 1.498
15 31 30 1 2.088 2.254
16 31 30 1 2.109 2.254
17 31 30 1 2.196 2.254
18 31 30 1 2.667 2.254
19 31 30 1 2.338 2.254
20 31 30 1 2.138 2.254

Chapitre 3. Résultats et discussion
138
Vérification de l’homogénéisation des valeurs par le test de Cochran
Le test de Cochrane a été effectué sur la variable dépendante (activité coagulante) et nous a
permit de vérifier l’homogénéité de la variance (valeur de Cochrane calculée = 0.2170 est
inférieur à celle tabulée 0.3894).
Vérification de la signification statistique des coefficients du modèle
mathématique
Grâce au test de Student, les coefficients de régression non significatifs sont éliminés. Les
coefficients significatifs sont utilisés afin de construire un modèle mathématique adéquat qui
nous permet de déterminer en fonction des valeurs des variables utilisées ; l’activité
coagulante correspondante.
Le modèle mathématique est donné par l’équation :
Il a 7 coefficients significatifs :
α0 = 2.254 (la valeur de la réponse au point central du domaine expérimental).
α2 = +0.313 (effet de la quantité du son de blé).
α13= -0.109 (effet de l’interaction entre la température et la quantité de caséine).
α23= +0.114 (effet de l’interaction entre la quantité du son et la quantité de caséine).
α11= -0.119 (effet de la température).
α22= -0.334(effet de la quantité du son de blé).
α33= -0.267 (effet de la quantité de caséine).
Gc < Gt, les variances sont homogènes donc l’expérience est reproductible.
Ac = 2.254+0.313 X2 - 0.109X1X 3+0.114 X2X3-0.119 X12-0.334 X2
2-0.267 X3
2

Chapitre 3. Résultats et discussion
139
L’analyse des coefficients de régression des facteurs montre que la température exercent un
effet négatif –0.119, alors que la quantité de son exerce un effet positif +0.313.
En conclusion, plus on cultive à des températures inférieures à 37°C, meilleure est la
production ; de même, plus on augmente la quantité de son, meilleure est la réponse.
Vérification de l’adéquation de modèle mathématique obtenu
La vérification de l’adéquation du modèle obtenu permet de préciser si ce modèle décrit
fidèlement le procédé étudié. L’estimation de l’adéquation du modèle est effectuée au moyen
du critère de Fisher, (valeur de Fisher calculée = 0.118 est inférieur à celle tabulée 2.46).
Afin de déterminer les conditions optimales pour la meilleure production de coagulase, la
surface de réponse tridimensionnelle est réalisée par le logiciel Statistica 6.0 en fixant les
variables température, quantité du son et quantité de caséine sur les axes X, Y, Z
respectivement.
La figure 36 montre la réponse à l’interaction entre ces variables. L’activité coagulante
maximale prédite par le logiciel Statistica est de 2.254 UP/ml correspondant à une
température de 31°C et à une quantité du son égale à 30 grammes, alors que dans les
conditions expérimentales, cette valeur est légèrement élevée et est égale à 2.667 UP/ml. Elle
est représentée dans le graphique par la couleur marron.
La valeur déterminée expérimentalement est en corrélation avec celle déterminée par le
logiciel ce qui confirme l’adéquation de notre modèle. De ce faitMucor circinelloides produit
une protéase coagulant le lait avec une activité coagulante de 2.667 UP/ml sous les conditions
optimales choisies.
Fc < Ft, le modèle est adéquat à une probabilité de 95%.
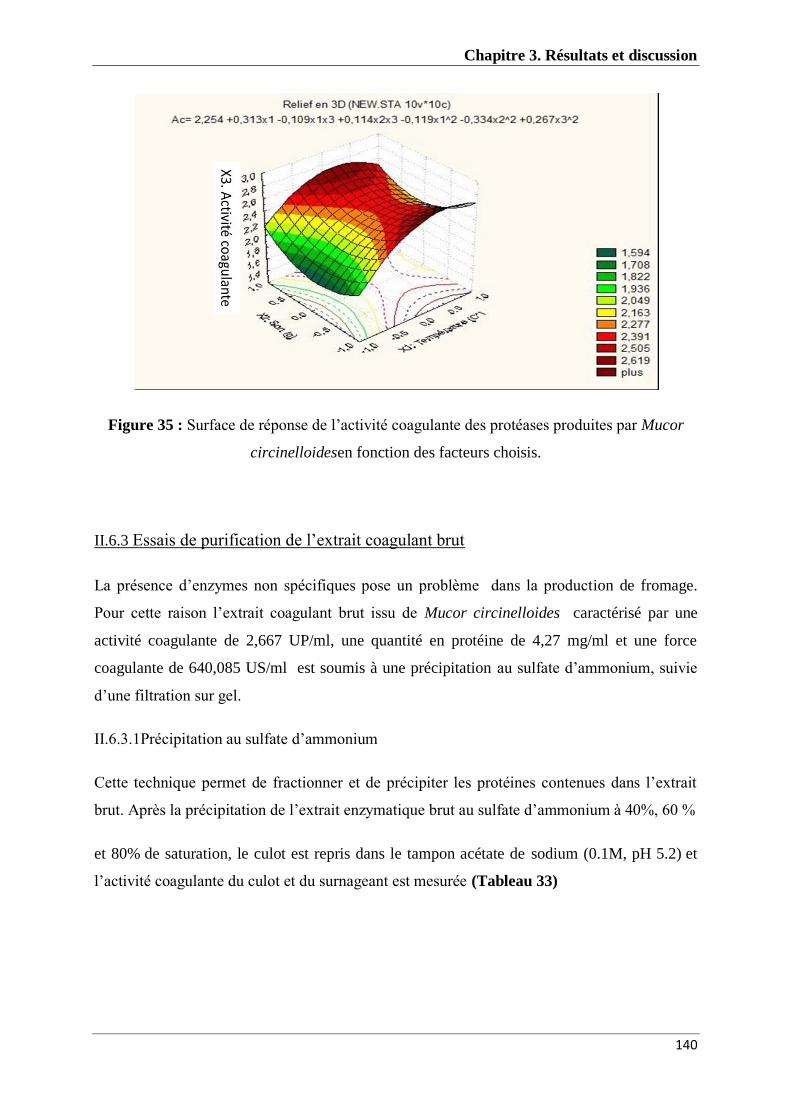
Chapitre 3. Résultats et discussion
140
Figure 35 : Surface de réponse de l’activité coagulante des protéases produites par Mucor
circinelloidesen fonction des facteurs choisis.
II.6.3 Essais de purification de l’extrait coagulant brut
La présence d’enzymes non spécifiques pose un problème dans la production de fromage.
Pour cette raison l’extrait coagulant brut issu de Mucor circinelloides caractérisé par une
activité coagulante de 2,667 UP/ml, une quantité en protéine de 4,27 mg/ml et une force
coagulante de 640,085 US/ml est soumis à une précipitation au sulfate d’ammonium, suivie
d’une filtration sur gel.
II.6.3.1Précipitation au sulfate d’ammonium
Cette technique permet de fractionner et de précipiter les protéines contenues dans l’extrait
brut. Après la précipitation de l’extrait enzymatique brut au sulfate d’ammonium à 40%, 60 %
et 80% de saturation, le culot est repris dans le tampon acétate de sodium (0.1M, pH 5.2) et
l’activité coagulante du culot et du surnageant est mesurée (Tableau 33)

Chapitre 3. Résultats et discussion
141
D’après les résultats portés dans le tableau 30, nous avons constaté que le taux de saturation
de 80% en sulfate d’ammonium a donné une activité coagulante maximale au niveau de culot
avec absence de cette activité coagulante au niveau de surnageant.
Nos résultats concordent bien avec ceux de Khan et al.,(1983) qui ont obtenu une
précipitation totale des protéines à 80% de saturation en sulfate d’ammonium. Alors que
Kasemi-Vaysari et al., (2002) rapportent que l’activité coagulante optimale de la protéase de
Mucor miehei et Rhizopus oryzae est observée pour un taux de saturation en sulfate
d’ammonium égale à 50% et à 60% respectivement.
Tableau 33 : Activité de précipitation de l’extrait enzymatique brut au sulfate d’ammonium
Pourcentage en sulfate
d’ammonium (%)
Activité coagulante
du surnageant (UP/ml)
Activité coagulante du
Culot (UP /ml)
40 0.256 0. 100
60 0.047 0.541
80 0 0.621
II.6.3.2Chromatographie d’exclusion moléculaire
La séparation des composants protéiniques de l’extrait enzymatique brut et les protéines
obtenues par précipitation à 80% de sulfate d’ammonium a été réalisée par chromatographie
sur gel Séphadex G75.Les éluats de la colonne sont recueillis dans un collecteur de fraction à
raison de 3ml/tube pour chaque fraction collectée, l’activité coagulante a été mesurée.
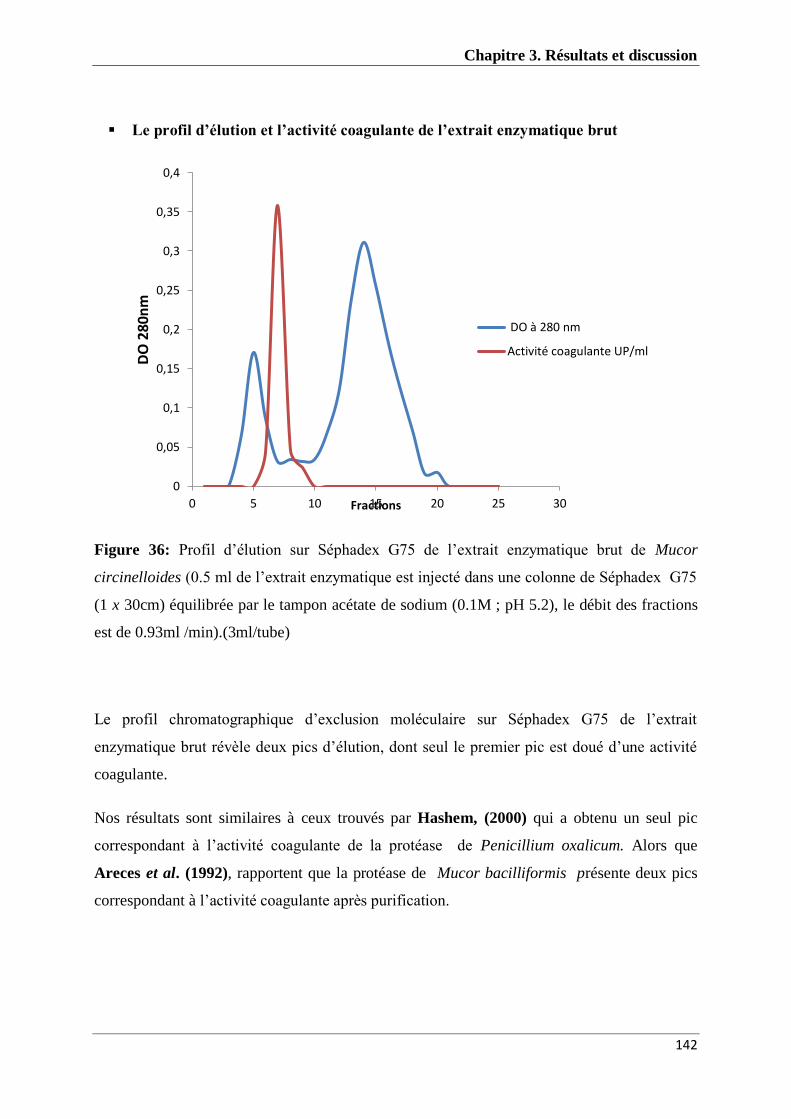
Chapitre 3. Résultats et discussion
142
Le profil d’élution et l’activité coagulante de l’extrait enzymatique brut
Figure 36: Profil d’élution sur Séphadex G75 de l’extrait enzymatique brut de Mucor
circinelloides (0.5 ml de l’extrait enzymatique est injecté dans une colonne de Séphadex G75
(1 x 30cm) équilibrée par le tampon acétate de sodium (0.1M ; pH 5.2), le débit des fractions
est de 0.93ml /min).(3ml/tube)
Le profil chromatographique d’exclusion moléculaire sur Séphadex G75 de l’extrait
enzymatique brut révèle deux pics d’élution, dont seul le premier pic est doué d’une activité
coagulante.
Nos résultats sont similaires à ceux trouvés par Hashem, (2000) qui a obtenu un seul pic
correspondant à l’activité coagulante de la protéase de Penicillium oxalicum. Alors que
Areces et al. (1992), rapportent que la protéase de Mucor bacilliformis présente deux pics
correspondant à l’activité coagulante après purification.
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 5 10 15 20 25 30
DO
280
nm
Fractions
DO à 280 nm
Activité coagulante UP/ml

Chapitre 3. Résultats et discussion
143
Le profil d’élution et l’activité coagulante de l’extrait enzymatique précipité
Figure 37 : Profil d’élution sur Séphadex G75 de l’extrait enzymatique précipité de Mucor
circinelloides (0.5 ml de l’extrait enzymatique est injecté dans une colonne de Séphadex G75
(1 x 30cm) calibrée par le tampon acétate de sodium (0.1M ; pH 5.2), le débit des fractions est
de 0.93ml /min).(3ml/tube)
Le profil chromatographique d’exclusion moléculaire sur séphadex G75 de l’extrait
enzymatique précipité révèle un seul pic d’élution, dont les fractions présentent une activité
coagulante très faible. Cela est du à la perte de l’enzyme lors de la remise du culot en
suspension dans le tampon et à la dilution de l’extrait enzymatique.
Les conditions dans lesquelles a été effectuée la filtration sur gel de l’extrait enzymatique
n’ont pas permis de déterminer le poids moléculaire de la protéase coagulante ; la résolution
de la séparation est insuffisante.
-0,01
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0 5 10 15 20 25
Do
280
nm
Fractions
Do à 280 nm
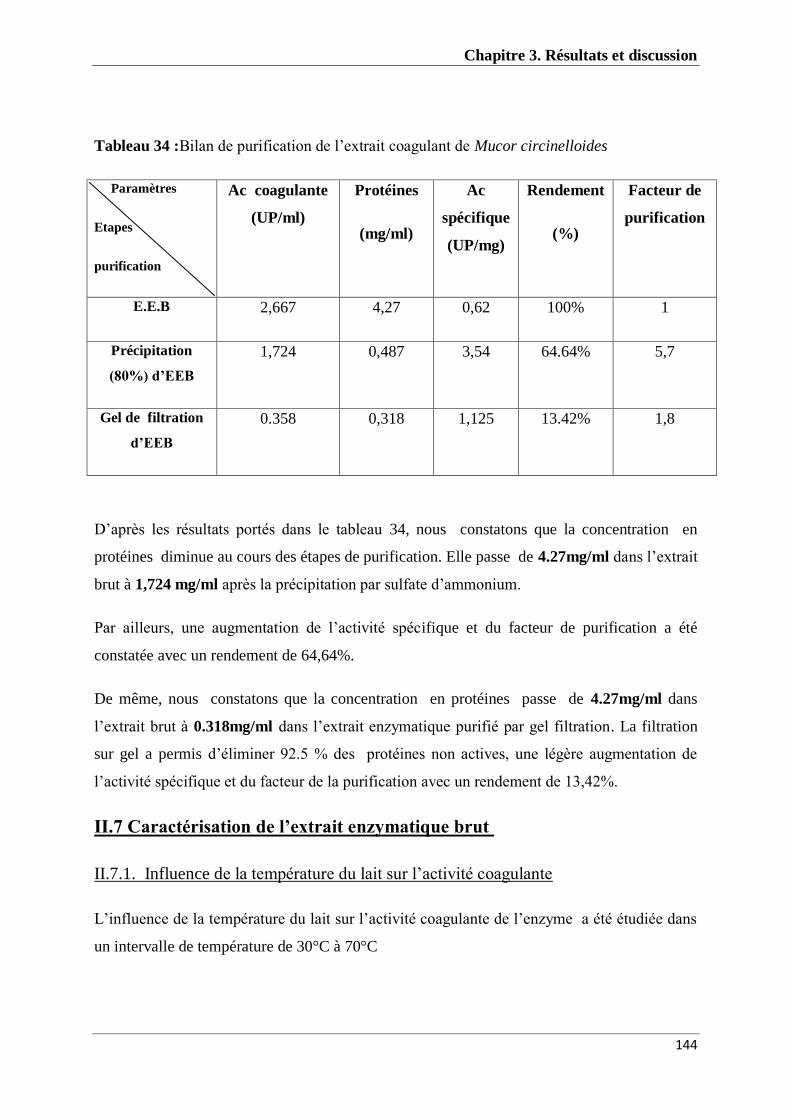
Chapitre 3. Résultats et discussion
144
Tableau 34 :Bilan de purification de l’extrait coagulant de Mucor circinelloides
Paramètres
Etapes
purification
Ac coagulante
(UP/ml)
Protéines
(mg/ml)
Ac
spécifique
(UP/mg)
Rendement
(%)
Facteur de
purification
E.E.B 2,667 4,27 0,62 100% 1
Précipitation
(80%) d’EEB
1,724 0,487 3,54 64.64% 5,7
Gel de filtration
d’EEB
0.358 0,318 1,125 13.42% 1,8
D’après les résultats portés dans le tableau 34, nous constatons que la concentration en
protéines diminue au cours des étapes de purification. Elle passe de 4.27mg/ml dans l’extrait
brut à 1,724 mg/ml après la précipitation par sulfate d’ammonium.
Par ailleurs, une augmentation de l’activité spécifique et du facteur de purification a été
constatée avec un rendement de 64,64%.
De même, nous constatons que la concentration en protéines passe de 4.27mg/ml dans
l’extrait brut à 0.318mg/ml dans l’extrait enzymatique purifié par gel filtration. La filtration
sur gel a permis d’éliminer 92.5 % des protéines non actives, une légère augmentation de
l’activité spécifique et du facteur de la purification avec un rendement de 13,42%.
II.7 Caractérisation de l’extrait enzymatique brut
II.7.1. Influence de la température du lait sur l’activité coagulante
L’influence de la température du lait sur l’activité coagulante de l’enzyme a été étudiée dans
un intervalle de température de 30°C à 70°C
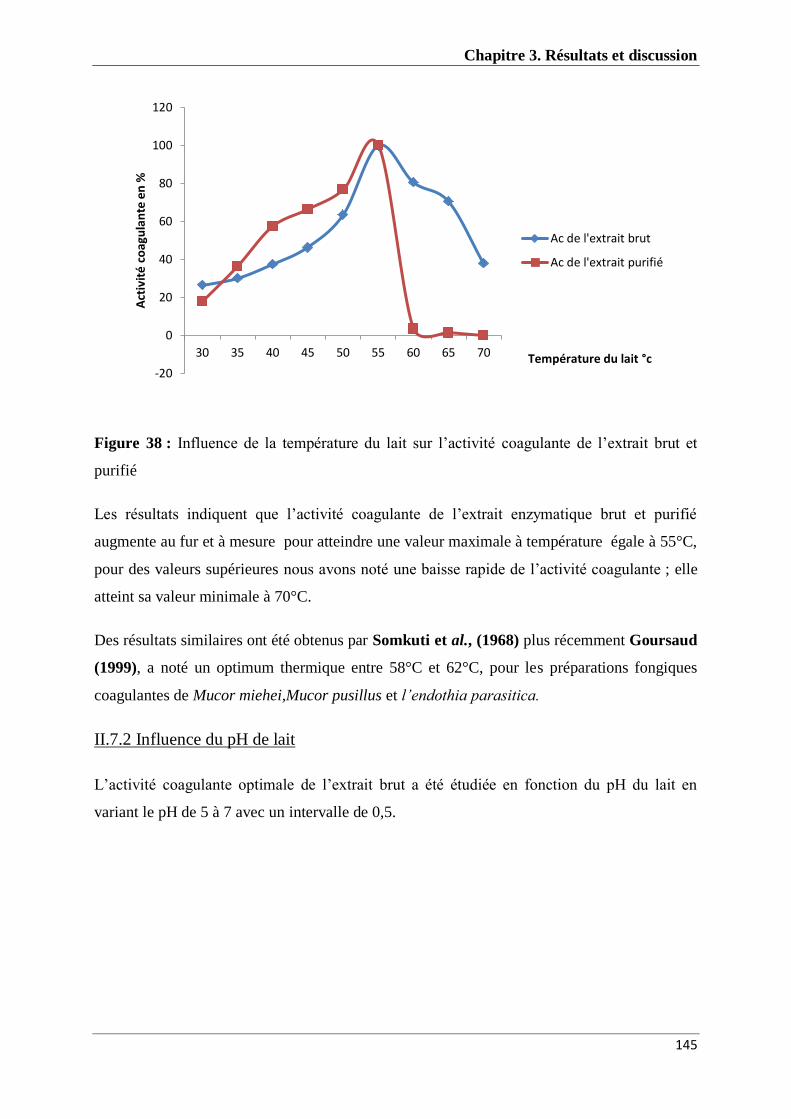
Chapitre 3. Résultats et discussion
145
Figure 38 : Influence de la température du lait sur l’activité coagulante de l’extrait brut et
purifié
Les résultats indiquent que l’activité coagulante de l’extrait enzymatique brut et purifié
augmente au fur et à mesure pour atteindre une valeur maximale à température égale à 55°C,
pour des valeurs supérieures nous avons noté une baisse rapide de l’activité coagulante ; elle
atteint sa valeur minimale à 70°C.
Des résultats similaires ont été obtenus par Somkuti et al., (1968) plus récemment Goursaud
(1999), a noté un optimum thermique entre 58°C et 62°C, pour les préparations fongiques
coagulantes de Mucor miehei,Mucor pusillus et l’endothia parasitica.
II.7.2 Influence du pH de lait
L’activité coagulante optimale de l’extrait brut a été étudiée en fonction du pH du lait en
variant le pH de 5 à 7 avec un intervalle de 0,5.
-20
0
20
40
60
80
100
120
30 35 40 45 50 55 60 65 70
Act
ivit
é co
agu
lan
te e
n %
Température du lait °c
Ac de l'extrait brut
Ac de l'extrait purifié

Chapitre 3. Résultats et discussion
146
Figure 39:Influence de pH du lait sur l’activité coagulante de l’extrait brut et purifié
A pH 5, il y a apparition des flocules avant d’ajouter l’enzyme, nous sommes dans le cas
d’une coagulation acide (pH pHi de la caséine).En effet, l’acidification du lait favorise la
réaction d’agrégation par suite de la diminution de la stabilité des micelles, liée à la
neutralisation des charges négatives et à libération d’ion calcium.
Une activité relativement importante et maximale à pH 5,5 pour l’extrait brut et purifié ; au
delà de cette valeur l’activité diminue.
Les enzymes fongiques, selon Fernandez-Lahore et al. (1998), se caractérisent par un pH
optimum entre 2.5 et 5.5.
II.7.3 Influence de la concentration de CaCl2
L’influence de la concentration en CaCl2 a été déterminée en faisant varier la concentration du
lait en ion CaCl2 de 0.005M à 0.05 M avec un intervalle de 0.005M.
0
20
40
60
80
100
120
5,5 6 6,5 7
Act
ivit
é c
oag
ula
nte
%
pH du lait
Ac de l'extrait brut
Ac de l'extrait purifié

Chapitre 3. Résultats et discussion
147
Figure 40 : Influence de CaCl2 du lait sur l’activité coagulante de l’extrait brut et purifié
Nous observons que l’activité coagulante augmente de manière proportionnelle avec la
concentration en CaCl2 pour atteindre une activité maximale à une concentration de 0,03M
pour l’extrait brut et purifié.
L’ion de calcium est un activateur des protéases .Il est indispensable pour la coagulation du
lait et joue le rôle de ciment dans la polymérisation des caséines par la formation de ponts
calciques, après hydrolyse de la caséine қ par la protéase coagulante.
0
20
40
60
80
100
120
0 0,01 0,02 0,03 0,04 0,05 0,06
Act
ivit
é co
agu
lan
te %
Ca Cl2 du lait (M)
Ac de l'extrait brut
Ac de l'extrait purifié

Conclusion

Conclusion
149
Cette étude a été entreprise dans le but de contribuer à la recherche de protéases issues de la
culture de souches microbiennes destinées à être utilisées comme succédanées de la présure.
Ce travail nous a permis d’isoler à partir d’échantillons de terre des souches microbiennes
productrices d’une enzyme coagulant le lait en vue de la fabrication de fromage. Cette
protéase doit être douée d’une bonne activité coagulante et de faible activité protéolytique.
La terre utilisée pour la recherche des microorganismes a été prélevée à proximité de la
laiterie fromagerie de Boudouaou
En ce qui concerne l’isolement des bactéries, le milieu employé est le milieu Plate Count
Agar (P.C.A.), additionné de 10% lait écrémé UHT et d’actidione (cycloheximide) à 2%.
L’incubation s’effectue à 30°C pendant 48h à 72h.
Dans le cas des moisissures, le milieu utilisé est le Sabouraud-chloramphénicol, dans lequel
est ajouté 10% lait écrémé UHT. L’incubation a lieu à 25° pendant 3 à 4j.
Dans cette première partie nous nous sommes intéressés aux bactéries qui présentent un halo
clair d’hydrolyse du lait ou caséolyse. Les souches caséolytiques sont prélevées et repiquées
sur des milieux P.C.A. contenant du lait, puis purifiées éventuellement par plusieurs
isolements par la méthode des quadrants jusqu’à l’obtention de souches pures.
Sur les 70 souches bactériennes isolées, présentant une caséolyse, uniquement 20 souches
possèdent une activité coagulante. Des observations macroscopiques et microscopiques ont
été effectuées sur l’ensemble des souches.la plupart présentaient des similitudes sur le plan
morphologique, d’où la sélection de 8 souches qui semblaient différentes. Une identification
biochimique a été réalisée pour ces dernières.
Après avoir vérifié plusieurs de leurs caractères, on a pu conclure que certaines des souches
sont semblables et qu'il y a une forte probabilité qu'elles appartiennent toutes à l'espèce B.
subtilis. Cependant, pour vérification, les souches I33M, FK6A et P47M ont été envoyées à
un laboratoire de biologie moléculaire, afin d'avoir une confirmation :nous avons obtenu
comme résultat :FK6A : Bacillus Amyloliquefaciens P47M : Bacillus subtilis et I33M :
Bacillus mojavensis

Conclusion
150
La bactérie (I33M), identifiée comme étant Bacillus mojavensis a été sélectionnée pour sa
bonne activité coagulante et sa relative faible activité protéolytique, son activité coagulante
est de 2,18 U.P et son activité protéolytique de 19,302 µg /tyr/min.
Une optimisation du milieu de culture de Bacillus mojavensis a été réalisée par les plans
d’expériences.
L’optimisation d’un milieu de production nécessite des méthodes qui consistent à faire varier
successivement chacun des facteurs qui influencent la culture, ce qui conduit à minimiser le
nombre d’expériences.
L’utilisation d’une méthodologie de recherche expérimentale ou plan d’expérience permet de
déterminer l’influence de nombreux paramètres, d’étudier leurs interactions et par la même
d’optimiser la réponse expérimentale. Le plan choisi est un plan de surface de réponse de
type composite centré (CCD). L’objectif principal est de maximiser l’activité de coagulation
et de minimiser l’activité protéolytique. Les résultats obtenus sont donnés pour un milieu
composé de : son de blé : 7% ; caséine : 0,094% ; température 39°c et agitation : 150 rpm
La valeur optimale de l’’activité coagulante est de 2,4 (IMCU/ml).
La réponse de surface en 3 dimensions montre les effets relatifs de 2 facteurs : température et
agitation qui ont un effet significatif considérable sur la production des protéases.
Une croissance en fermenteur a été effectuée pour la production de l’enzyme et parallèlement
la croissance de la souche a été suivie ainsi que l’évolution du pH, de l’activité coagulante,
protéolytique ainsi que le taux de protéines.
L’analyse de la croissance de la souche I33M en milieu optimisé au son a été suivie pendant
deux jours, par dénombrement des cellules viables sur milieu P.C.A. Elle montre qu’après une
courte période de latence de moins de 1h, la phase exponentielle est rapide et la phase de
décélération est atteinte en moins de 5h. La phase stationnaire s’est prolongée sur la durée de
l’expérience, sans que l’on puisse mettre en évidence au bout de deux jours une phase de
déclin.
Le taux de croissance montre une vitesse extrêmement rapide puisque le temps de génération
exponentiel est évalué à 22 mn environ.

Conclusion
151
Le pH voisin de 7 au départ se stabilise à 6 dès le début de la phase stationnaire. La stabilité
du pH du milieu de fermentation, peut s’expliquer par le fait que les bactéries consomment en
premier lieu les glucides contenus dans le milieu ; l’acidification qui en résulte est rapidement
neutralisée par les produits basiques issus de l’activité protéolytique des Bacillus.
Concernant les activités coagulante et protéolytique, elles augmentent progressivement dès le
début de la phase stationnaire et cela jusqu’à la fin de l’expérience. L’activité coagulante
semble augmenter moins rapidement et n’apparaitre de façon significative qu’au bout d’une
dizaine d’heure de culture.
Concernant l’enzyme coagulante produite par Bacillus mojavensis, deux hypothèses peuvent
être formulées. On peut supposer que cette protéase est très vraisemblablement un
métabolite secondaire. L’autre hypothèse est qu’il s’agit d’une protéine intracellulaire (taux
très faible en phase exponentielle) libérée progressivement à la mort des cellules en phase
stationnaire).
Des essais de purification de l’extrait enzymatique brut ont été effectués, nous avons obtenus
deux pics, dont uniquement un est doué d’une activité coagulante.
Une caractérisation de l’extrait enzymatique a été effectuée, on note des valeurs optimales de
l’activité coagulante, qui sont 65°C pour la température, un pH de 6 et une concentration
enCaCl2de 0,04M
Plusieurs essais de fabrication de fromage affiné à pâte pressée non cuite type «EDAM » ont
été réalisés, en faisant varier le type et la concentration en extrait coagulant.
Nous avons obtenu des fromages fabriqués avec 100% présure, 100% EEB Bacillus
mojavensis, 50% EEB Bacillus mojavensis et 50% présure, Pour résoudre le problème causé
par l’activité protéolytique élevé de nos extraits enzymatiques, nous avons cuit le caillé après
l’étape de brassage, et nous avons obtenu un caillé de bonne consistance.
La deuxième partie du travail concerne les moisissures. 24 souches ont été isolées et
purifiées, la plupart de souches sont des Mucor, dont seulement 12 possèdent une activité
coagulante. Nous avons remarqué que beaucoup de souches se ressemblent après observation
au microscope. Seulement 3 souches (5F, 4D et SF15) semblaient être différentes des autres,
et elles ont été identifiées par biologie moléculaire. La souche 4D, a été retenue pour la suite
du travail, et a été identifié comme étant Mucor circinelloides.

Conclusion
152
Une optimisation du milieu de culture, de type composite centré a été réalisée comme pour les
bactéries Les meilleurs résultats concernant l’activité coagulante ont été obtenus pour un
milieu de culture composé de 30g de son de blé auxquels on ajoute 1g de caseine puis 50 ml
de sulfate d’ammonium à 0,01%. Et la température d’incubation de 31°C
L’extrait coagulant brut issu de Mucor circinelloides caractérisé par une activité coagulante
de 2,667 UP/ml, une quantité en protéine de 4,27 mg/ml et une force coagulante de 640,085
US/ml, a subit une précipitation au sulfate d’ammonium d’abord, puis une purification sur
gel. Un profil chromatographique de l’extrait enzymatique sur gel séphadex (G75) a été
effectué avec le tampon acétate de sodium (0,1M ; pH 5,4).
La caractérisation de l’enzyme fongique a donné une température optimale de 70°C, un pH de
6 et une concentration en CaCl2 de 0,03M
La fabrication de l’EDAM avec l’extrait enzymatique brut (EEB) de Mucor circinelloides,
s’effectue suivant les mêmes opérations de fabrication à base de présure. La dose utilisée de
l’EEB est calculée préalablement à partir de sa force coagulante.
Deux essais de fabrication de fromage type EDAM ont été effectués ; (1) le fromage est
préparé à partir de 50% présure et 50% de EEB, (2) le fromage est préparé à partir de 100%
EEB.
Actuellement les substituts d’origine fongique dominent le marché mondial des agents
coagulant le lait comparés aux présures de remplacement d’origine animale. Certes, le génie
génétique a développé le clonage de la Chymosine de veau et constitue aujourd’hui, avec
quelques rares restrictions règlementaires une autre voie d’application dont l’emploi en
fromagerie est en pleine croissance.
L’analyse de ces résultats indiquent que les deux souches ont une aptitude à la coagulation et
peuvent être utilisées comme des succédanées de présure dans la fabrication du fromage,
mais l’étude comparative entre les 2 souches indique que le Bacillus mojavensis a une
meilleure activité coagulante que celle de Mucor circinelloides, mais que son activité
protéolytique (64 µg/ml) reste toujours plus élevée que celle de Mucor sp (46µg/ml).
Cette étude nous a permis de contribuer aux travaux de recherches préexistants dans le
domaine de l’industrie fromagère, et de renforcer l’idée de pouvoir trouver de nouvelles
sources potentielles de succédanés de présure.

Conclusion
153
Enfin nous proposons de compléter cette étude par une suite de recherche qui portera sur :
Tester d’autres milieux de fermentation (solide, semi solide varier la
composition du milieu).
Vérification de la pureté de l’enzyme par électrophorèse SDS-PAGE.
Etudier les domaines d’application de l’enzyme et son mécanisme d’action.
Il serait intéressant d’approfondir les connaissances concernant les souches
sélectionnées afin de valoriser leurs capacités pour de meilleures applications
industrielles.

154
Références bibliographiques
Acgih., 1999. American Conference Of Governmental Industrial Hygienists Bioaerosols :
Assessment And Control. Publication 3180. Janet Macher Editor. 526 P
Agboola S., 2002. Cheesemaking from ultrafiltered milk using plant rennet. The Australian
Journal of DairyTechnology. 57, p.143.
Aikawa J., Yamashita T., Nishiyama M., Horinouchi S., and Beppu T., 1990. Effects of
glycosylation on the secretion and enzyme activity of Mucor, rennin, an aspartic proteinase of
Mucor pusillus, produced by recombinant yeast. J. Biol. Chem. 265, 13955–13959.
Ageitos J.M., Vallejo, J.A., Sestelo, A.B.F., Poza, M., and Villa T.G., 2007. Purification
and characterization of a milk-clotting protease from Bacillus licheniformis strain USC13. J.
Appl. Microbiol. 103, 2205–2213.
Alais C., and Lagrange A., 1972. Etude biochimique d’une protéase coagulante produite par
Mucor miehei . I. Activité coagulante et activité protéolytique. Le Lait 52, 407–427.
Alais C., 1984. Science du lait : Principes des techniques laitières (Société d’édition et de
promotion agro-alimentaires, industrielles et commerciales
Alais C., and Linden G., 1997. Abrégé de biochimie alimentaire (Masson).
Aguilar J.A., Díaz-Pérez C., Díaz-Pérez A.L., Rodríguez-Zavala J.S., Nikolau B.J., and
Campos-García J., 2008. Substrate specificity of the 3-methylcrotonyl coenzyme A (CoA)
and geranyl-CoA carboxylases from Pseudomonas aeruginosa. J. Bacteriol.190, 4888–4893.
Amiot J., Fournier F., Lebeuf Y., Paquin P et Simpson R., 2002. Science et technologie du
lait : transformation du lait. Presses internationales Polytechnique, Montréal, 1-73.
Aparna M ., Mala B., Mohini S., Ghatge V. 1998. Molecular and Biotechnological aspects
of microbial protease. Microbiology and Molecular Biology Reviews, 62, 3: 597-635.
Areces L. B., Biscoglio de Jimenez Bonino M. J., Parry M.A.A., Faraile E.R., Fernandez
H.M et Cascone O., 1992. Purification and characterization of milk-clotting protease from
Mucor bacilliformis. Applied Biochemistry and Biotechnology. 37:238-294.
Arnaud A., Guiraud J. P., 1993. Le métabolisme microbien. Dans Biotechnologie.
SCRIBAN R. [éd.], Technique et Documentation- Lavoisier, Londres, New York, Paris, 904
p.
Assamoi Allah Antoine, Jacqueline Destain, Philippe Thonart ., 2008. Aspects
microbiologiques de la production des protéases par fermentation solide des endo-β-1,4-
xylanases de moisissures : le cas de Penicillium canescens
Assamoi A.A., Destain J., and Thonart P., 2009. Aspects microbiologiques de la production
par fermentation solide des endo-β-1,4-xylanases de moisissures: le cas de Penicillium
canescens. Biotechnologie, agronomie, société et environnement 13.

155
Aviron-Violet P., Monsan P., and Durand G., 1982. Les Enzymes: production et
utilisations industrielles (Paris: Gauthier-Villars).
Awad S., Lüthi-Peng Q.Q., and Puhan Z., 1999. Proteolytic activities of suparen and
rennilase on buffalo, cow, and goat whole casein and beta-casein. J. Agric. Food Chem. 47,
3632–3639.
Bacon C.W., Yates I.E., Hinton D.M., and Meredith F., 2001. Biological control of
Fusarium moniliforme in maize. Environ. Health Perspect. 109 Suppl 2, 325–332.
Banga-Mboko H., Godeau J.M., Drion P.V., El Amiri B., Drion V., Perenyi Z., Sousa
N.M., Et Beckers J.F., 2002. Évaluation de l’utilisation du pepsinogène sanguin comme bio
marqueur de l’intégrité de la muqueuse gastrique. Chez le porc –Historique, physiologie des
muqueux gastriques et différents formes de pepsinogènes.
Ann. Méd. Vét. 146: 339-346.Barbano D.M., and Rasmussen R.R., 1992. Cheese Yield
Performance of Fermentation-Produced Chymosin and Other Milk Coagulants1. Journal of
Dairy Science 75, 1–12.
Baarn, Hollande Samson A., 2002. Cheese making from ultrafiltred milk using plant rennet.
The Australian journal of dairy technology, .57, 2 : 39-73PP.
Berridge. N.J., 1952. Determination of the activity of rennet.Analyst.77:57-62.
Barrett A.J., 1994. Proteolytic enzymes: serine and cysteine peptidases.Methods
enzymology.244: 1-15
Barrett A.J., and Rawlings N.D., 1995. Families and clans of serine peptidases. Arch.
Biochem. Biophys. 318, 247–250.
Beldarrain A., Acosta N., Montesions R., Mata M et Cremata J., 2000. Characterization
of Mucor pusillus rennin expressed in Pichra pastoris: enzymic, spectroscopie and
calorimetric, studies. Biotechnol. Appl. Biochem., 31, 77-84.
Bellon-Maurel V., Orliac O., & Christen P., 2003. Sensors and Measurements In Solid-
State Fermentation: A Review. Process Biochem., 38, 881-896.
Ben Maachia S, Errakhi R, Mathieu F, Chérif M, Lebrihi A 2011. Identification and
partial characterization of antifungal and antibacterial activities of two Bacillus sp. Strains
isolated from salt soil in Tunisia. Afr. J. Microbiol. Res. 5(13):1599-1608.
Berankova E., Rauck P., Kasj.,Husek V. et Paluska E., 1987. Détermination indirecte de
l’activité de coagulation du lait de Mucor miehei présente dans les mélanges de Présure. J.
Dairy Res., 54 :(3), 407-413.
Blackwell M., R. Vilgalyset J.W. Taylor., 1998. Fungi, Eumycota. In The Tree Of Life,
D.R. Maddison Et W.P. Maddison Editor, University Of
Arizona.Http://Tolweb.Org/Tree?Group=Fungi&Contgroup=Eukaryotes.
Botton B., Breton A ., Fever M., Gauthier S .,Guy P., Larpent J. P., Reymond P.,
Sanglier J J.,Vayssier Y et Veau P. 1990. Moisissures utiles et nuisibles importance
industrielle, Masson, 2éme
édition. Paris, p 512.

156
Boudier J.F., 1974. Présure et succédanés de la présure. Série de synthèses bibliographiques.
Ed. Tec et Doc., APRIA., France. 3 (3), 873.
Boudier J.F., Luquet F. M., 1981. Dictionnaire Laitier 2ed
; PP 11, 75, Tec & Doc, Lavoisier
Paris).
Bourgeois C.M., Mescle J.F., Zucca J., 1989. Microbiologie alimentaire. Aspect
microbiologique de la sécurité et de la qualité des aliments. Lavoisier.
Brewer P, Helbig N etHaard N.F. 1984. Atlantic cod pepsin. Characterization and use as a
rennet substitute. Can. Int. Food sci. and Technol. J, 17:38-43.
Brinkman D., Duiven M., 1972. Purifiaction and properties of two protease from brewer’s
yeast, Ind, Aliment, Agric, 89: 1755-1758.
Broome M.C., et Hickey M.W. 1990. Comparison of fermentation produced chymosin and
calf rennet in cheddar cheese. Australian J. Dairy. Technology., 45, 2 , 53-59.
Broome M.C., et Limsowtin G. K. Y., 1998. Milk coagulants. Australian journal of dairy
technology, .53, .3, 188-201.
Brule, G., Lenoir, J., et Remeuf, F., 1997. La micelle de caséine et la coagulation du lait in:
Le Fromage. Ed., A. Eck, 3ème Ed., Technique Et Documentation Lavoisier, Paris,
P.741,891p.
Brûlé G. et Lenoir J., 1990. Les mécanismes généraux de la transformation du lait en
fromage. In: Fromage. Coord Eck. A. Tech. Doc. Lavoisier Paris 1-21.
Box G.E.P., et Wilson K.B., 1951.On the experimental attainment of optimum conditions.
Journal of the Royal Statistical Society, B, 13(1):1-41.
Box G.E.P., Hunter W.G et Hunter J., 2005.Stuart «Statistics for Experimenters» deuxième
édition. John Wiley and Sons. New-York. 633 pages.
Carlile M.J., & Watkinson S.C., 1994. The Fungi, Academic Press, London, Boston,
SanDiego, New York, Sydney, Tokyo, 9-76, 77-139, 153-172, 191-201.
Carnot P. Martin P., 1980. Présure, composition, activité, son rôle en fromagerie, Tech,
laitière, 190, 3, 27-30 PP.
Cayot P., et Lorient D., 1998. Structures et techno-fonctions des protéines du lait. Technique
et Documentation Lavoisier, 363 p.
Chazarra S. Sidrach1l. Loopez – Molina D., et Neptuno Rodriguez- Loopez N., 2007.
Characterization of milk – clotting properties of extracts from artichoke (Cyanara
Scolymus, L) flowers. International Dairy Journal 17: 1393-1400 PP.
Chemlal B., 1998. Caractérisation et purification partielle de coagulases des souches Bacillus
subtilis et Bacillus coagulans, Mémoire, ing, U.S.T.H.B, Alger.

157
Chiplunkar J. M., Gangodkar S. V., Wagh UV., Ghadge G D., Rele M V., Srinivasan M
C., 1985. Applications of alkaline protease from Conidiobolus in animal cell culture; Biotech.
Lett. 7:65-68.
Choisy C, Desmazeaud M, Gripon J.C, Lamberet G et Lenoir J., 1997. La biochimie de
l’affinage. In : Le fromage, Tec et Doc Lavoisier, Paris, 1997.
Chopra et Mathur 1985. Purification and characterisation of heat stable proteases from
Bacillus stearothermophilusJ. App Bacteria., 15, p 355 -358.
Christine D., Hennequet C.et Schaeffer B., 2006. Plan d’experiences,Mèthodes
mathématiques pour l’amelioration de la qualité.
Collin J.C., Grappin R et Legraet Y., 1997. Etude de la méthode de mesure, selon Berridge,
du temps de coagulation du lait additionné d’une solution enzymatique. Rev. lait. Franç.,
N°335.p.p.389-394.
Creamer L. K., 2002. Casein Nomenclature, Structure and Association Properties (Ed.)
Encyclopedia of Dairy Science. Academic Press, Amsterdam, the Netherlands; 1895–1902
Cogitore A., 1982. Traité pratique de réglementation laitière, laits et produits laitiers (Antoine
Cogitore).
Cuvellier G.F., 1993 Production des enzymes in Biotechnologie. Coord Scriban R. 4ème
édition Tec. et Doc.Lavoisier PP948.
Davet P et Rouxel F., 1997.Techniques for direct isolation of microorganisms from soil.
Collection Techniques et pratiques. INRA Editions (Rennes, France).
Davis J.G., 1976. la fromagerie et les variétés de fromage du bassin méditerranéen.
Dalgleish D.G., 1982.The enzymatic coagulation of milk (in Development in dairy chemistry
1.proteins) Applied Science Publishers London and New York.
Dehove R.A., 1990. La Réglementation des Produits, Qualité et Répression des Fraudes. Ed.
Lomy. France Pp: 550-560.
Durand A., 2003. Bioreactor Designs for Solid-State Fermentation. Biochem. Eng. J., 13,
113-125.
Durand G et Monsan P.,1974.Les enzymes immobilisées.Serie de synthese bibliographique
Ed.Tech et Doc.,A.P.R.I.A.France,5.
Dutt K, Meghwashi GK., Gupta P., Saxena RK., 2008. Role of casein induction and
enhancement of production of a bacterial milk clotting protease from an indigenously isolated
Bacillus subtilis. Lett. Appl. Microbiol. 46:513-518.
Eck A., 1987. Le fromage. Paris, Lavoisier, Tec et Doc.,891p.
Eck André et Jean Claude Gillis., 1997. Les Enzymes D’origine Animale : Le Fromage De
La Science A L’assurance Qualité, 3ème
Edition, Lavoisier Tec& Doc. Paris.

158
Eigel WN, Butler JE, Ernstrom., 1984. Nomenclature of proteins of cow’s milk .fifth
revision. J. Dairy Sci., 67: 1599-1631
Elagamy E.I., 2000. Physicochemical, Molecular and Immunological Characterization Of
Camel Calf Rennet: A Comparison With Buffalo Rennet. J. Dairy Res. 67: 73-81.
Ernesrom C., 1997. Les agents de transformation du lait dans le fromage de la science de
l’assurance qualité,Edition Lavoisier technique.
Escobar J. et Barnett S.M., 1993. Effect of agitation speed on the synthesis of Mucor meihei
acid protease. Enzyme Microbiology Technology.15:1009-1013.
Fernandez-lahore H., Auday R., Fraile E., Machalinski C., 1998. Purification and
characterisation of an acido proteinase from mesophilic Mucor sp, solid-state cultures. J,
Peptide, Res. 53: 599-605 PP.
FIL .Norme Internationale 157 A:1997(Federation Internationale du Lait) ., Determination
de l’activité coagulante totale du lait., Bruxelles-Belgique
Florent J., 1993. Microbiologie industrielle –les microorganismes d’intérêt industriel, ed, Tec
et Doc, Lavoisier, Paris, 248P.
Foltman B., 1970. Prochymosin and Chymosin (Prorennin and Rennin). Methods In
Enzymology, 19: 421-436.
Foltmann B., 1971. The biochemistry of prorennin and reninn (chymosin), édition Technique
de documentation, Lavoisier, Paris : 24-28PP.
Fox P.F et Whitaker J.R., 1977. Isolation and characterization of sheep pepsin. Biochem. J.,
161: 389-398;
Fox P., Singh T., Macsweeney H.P., 1995. Biogenesis of flavour compounds in cheese,
Plenum Press, New York: 59-97.
Garnot P., ET Martin P., 1980. la présure : composition, détermination de l’activité, son
rôle en fromagerie. Technique laitière. 930, N°3, 27-30.
Gaucheron F., 2004. Minéraux Et Produits Laitiers, Tec & Doc , PP 473.
Gervais P. et Molin P., 2003. The Role of Water In Solid-State Fermentation. Biochem. Eng.
J., 13: 85-101.
Goursaud J., 1999. Réacteurs traditionnels à enzyme libre : cas de l’industrie laitière. in :
Biotechnologie. Coord. Scriban, 5ème
edition. PP 365 401.
Goupy J., 2001.Introduction aux Plans d'expériences. Dunod. Paris. 303 pages.
Goursaud J., 1993 –Coagulation enzymatique du lait In :Scriban R.(cood).Biotechnologie.
Ed.Tec et Doc. Lavoisier,Paris,4ème
éd, pp393-405.
Graiday P., 1978. Détermination de l’activité enzymatique d’extraits coagulants d’origine
animale. Revue du Technicien du lait, 83 : p.p. 5-47.

159
Grant C., C.A. Hunter B. Flannigan et A.F. Bravery., 1988. The Moisture Requirements
Of Moulds Isolated From Domestic Dwellings. Int. Biodeterioration, 25: 259-84.
Green M.L., 1977. Review of the progress of dairy science: Milk coagulants J. of Dairy
Res.44:159-188.
Green M. L. et Stackpoole A., 1975. The Preparation And Assesment of A Suitable Mucor
Pusillus Lindt Proteinase- Swine Pepsine Mixture For Cheddar Cheese-making. J. of Dairy
Res. 42:297-312.
Gueguen M., 1992. Les Moisissures. In : Les Groupes microbiens d’intérêt Laitier. Cepilinra.
325-368
Guérard F., 1987.Une utilisation des enzymes protéolytiques extraites des viscères de
poissons : La coagulation du lait. Institut pêches maritimes.49, N°.3, PP 199-203.
Gupta R., Beg QK. Lorenz P., 2002.Bacterial alkaline proteases: molecular approaches and
industrial applications. Appl. Microbiol. Biotechnol. 59:15-32.
Guilherme G.S., Gustavo M.O, Eloizio J.R., Rubens M et Jonas C., 2005. Microbial
Rennet produced by Mucor meihei in solid –state and submerged Fermentation.Brasilian
Archives of Biology and Technology 48:6 931-935
Guillou H., Pelissier J.P., et Grappin R., 1986. Méthodes de dosage des protéines du lait de
vache. Le Lait 66:143-175.
Hashem A.M., 1999 .Optimization of milk clotting enzyme productivity by penicillium
oxalicum.Bioresource Technology 70, 203-207.
Hashem AM., 2000. Purification and properties of a milk-clotting enzyme produced by
Penicillium oxalicum. Bioresource Technology ; 75: 219–22.
Hartley B.S., 1960. Proteolytic Enzymes. Ann Rev. Biochem. 29 : 45-72.
Henri Goudéranche, Bénédicte Camier-Caudron, Jean-Yves Gassi, Pierre Schuck.,
2008. Procédés De Transformation Fromagère- Partie 1, Pp 305.
Houins G., Dervanne C., et Coppens R., 1973. Etude Comparative de L’activité Coagulante
et du pouvoir protéolytique de la présure animale et de trois de ses succédanés. Le Lait 529-
530, 610-624.
Hunter, C.A., A.V. Hull, D.F. Highman et Al., 1996. Fungi and Bacteria. In Berry R.W. et
Al. Ed., Indoor Air Quality in Homes; The Building Research Establishment Indoor
Environment Study Part I, Construction Research Communications. London.
Jarmul I., Reps A., Poznanski S. et Zelazowska H., 1982. Utilisation du mélange de la
pepsine avec la préparation microbienne «Fromase» dans La fabrication des Fromages EDAM
et Kortowski. Le Lait, 62, 75 – 86.
Jeantet Romain, Thomas Croguennec, Pierre Brule., 2007. Science Des Aliments :
Biochimie, Microbiologie, Procédés, Produits. 2 : 12, 15 Tec & Doc Lavoisier. Londres-
Paris-New York.

160
Joyeaux A., 1982. les préparations industrielles d’enzymes In : les enzymes ; production et
utilisation industrielles, 3ème
trimestre, 352p.
Kasemi-Vaysari A.,Kheirolomoom A., Arjmand M et Habibollah M., 2002.Optimisation
of Mucor miehei renning production and recovery.scienta.Iranica.9(1):99-104
Kamoun., 1977 ; Appareils et méthodes in biochimie. 3ème
ed flammarian ; Paris.
Kendrick B., 1999. The Fifth Kingdom. 2nd
Edition. Mycologie Publications.
Http://Www.Mycolog.Com/Fifthtoc.Html.
Khan M. R., Blain J.A. Et Patterson J. D. E. (1979).Extracellular Proteases of Mucor
pusillus . App. Envir. Microb., 37, 4, 719, 724.
Khan M .R., Blain J.A. et Patterson J.D.E.,1983.Partiel purification of Mucor pusillus
,intracellular proteases. Applied and Environmental Microbiology.45[1]:94
Kumar S., Sharma N. S., Sharab M.R, et Singb R., 2005. Extracellular acid protease from
Rhizopus oryzae : purification and characterization.Process Biochemistry. Vol: 40. PP 1701-
1705.
Kumar V., Sharma J., Sharma A.K., Grover S et Batish K.V., 2006. Purification and
characterization of milk clotting enzyme from goat (Capra hircus), Comparative
Biochemistry and Physiology, Part B 145: 108-113 PP.
Leclerc M., 2000. Microbiologie générale, édition Technique de documentation, Lavoisier,
Paris : 156-160 PP.
Lenoir J., Lamberet G.et Schmidt., 1983. L’élaboration d’un fromage, l’exemple du
camembert. De la science,à l’Assurance qualité 69, 30-42.
Lenoir J., 1985. Les caséines du lait. Rev lait franç, 440 : 17-23.
Lenoir J, Lamberet G, Schmidt J.L. et Tourneur C., 1985. La maitrise du biocatalyseur
fromage. Biofutur., (12). 23-50.
Levadoux W., Johnson M. Et Ander G., 1989.Monitoring the degradation of commercial
microbial rennet preparations. J. Dairy Sci., 72 (3):627 – 634.
Leyral C., Vierling E., 2001. Microbiologie et Toxicologie des Aliments, Hygiène
Alimentaire, Ed, Dunod, Paris, 652p.
Lopez M.B., Jordan M.J., Hellin P. et Laencia J., 1996 : Technological suitability of
different rennets and coagulant enzymes applied in Murciano- Gradina goat milk.
International Dairy Federation.,52: 144-148
Lowry O.M., Rosebrough N.J., Farr A.L., Randall R. J., 1951. Protein measurement with
the folin phenol reagent. J. Bio. Chem. 193: 265-275.
Lucey J.K., 2002. Rennet Coagulation of Milk In: Encyclopedia of Dairy Science. Ed., H.
Royinski, J. Fuquay And P. Fox, Elsevier Science Ltd, P. 286-293.

161
Luquet F.M., 1990. Lait Et Produits Laitiers Vache, Brebis, Chèvre et les Produits laitiers.
Transformation et Technologies. Tome 2.
Luquet F M., 1985. Lait Et Produit Laitiers, Vache- Brebis Et Chèvre. Les Produits Laitiers,
Transformation et Technologie. Ed. Technique Et Documentation Lavoisier. Tome 2, 633 P.
Luquet F M., J. Keilling R., De Wilde., 1985.Laits Et Produis Laitiers ; Vache, Brebis,
Chèvre. Les Produits Laitiers, Transformation Et Technologie. Ed. Tech&Doc, Lavoisier
Paris, Tome 1. PP 37.
Luquet .F.M, J. Keilling, R. De Wilde., 1986. Laits et Produis Laitiers Vache, Brebis,
Chèvre .Les Tomes 3 ; Pp 94-96, 101-104. Tec & Doc, Lavoisier Paris.
Lynn H., Kuntz R., 1993. The rheological and facture properties of Gouda cheese, Ph. D
thesis, Wageningen Agricultural Univ : 11-23 PP.
Mahaut M., Jeantet R. et Brulé G., 2003. Initiation à la technologie fromagère. Paris,
Lavoisier, Technique Et Documentation, Lavoisier, France ; Pp 24-102.
Malloch D., 1997. Moulds: Their Isolation, Cultivation and Identification. Department of
Botany, University of Toronto.
M. Marie, Juin 1997. Les Moisissures : Nature, Biologie Et Contamination ; Marie-France
Roquebert professeur ; Muséum national d’histoire naturelle,
Martin L.A.P., Pestana de Vasconcelos M.M.et de Sousa R.B., 1996.Thistle (Cynara
cardunculus) flower as a coagulant agent for cheese making.Short characterization.
Lait,76:473-477.
Mathieu J., 1998. initiation à la physicochimie du lait, technique et documentation Lavoisier
Paris pp 12-199.
Mathot P., 1996. modélisation d’un réacteur simplifié pour la fermentation solide de produits
et sous-produits agricoles. valorisation de l’aliment fermenté par le Porc. Thèse De Doctorat :
Faculté Universitaire Des Sciences Agronomiques De Gembloux (Belgique).
Matoub L., 2000. Essai de purification et de caractérisation d’une coagulase produite par la
souche locale Bacillus subtilis sélectionnée (Lc33). Mémoire de Magister. Sciences
alimentaires. I.N.A. El-Harrach.
Macedo Q., Faro C.J et Pires E.M., 1993: Specificity and kinetics of the milk-clotting
enzyme from cardoon ( Cynara cardunculus L.) toward bovine K-casein. J. agric .food Chem.
41: 1535- 1540.
Mietton B., 1991. Methods in enzymology, 19 (1970), 446-459.
Mietton B., Dermazeaud M., Deroissart H. Et Weber F.,(1994).Transformation du lait en
fromage. In : Bactéries lactiques. T2. Coord. Deroissart H. et Luquet F.M. Ed. Lorica
Merheb-Dini C., Gomes E., Da Saliva R., 2010. Production and characterization of milk-
clotting protease in the crude enzymatic extract from the newly isolated Thermomucor
indicae- seudaticae . Food Chemistry.120: 87-93.

162
Milliere J.B., 1984. Flavobacterium, bacterie psychrotrophe du lait cru, considération
taxonomique; localisation cellulaire, purification de L. Leucrylamidase et de la protéase.
Doctorat ès-sci., Paris-Nord, 101 – 126.
Mohanty A.K., Mukhopadhyay U.K., Grover S., Batish V.K., 1999. Bovine Chymosin:
Production By R .DNA Technology and Application In Manufacture. Cheese Biotechnol. Adv.
17: Pp 205-217.
Mohanty A.K., Mukhopadhyay U.K., Kaushik J.K., Grover S., Batish V.K., 2003.
Isolation, Purification and Characterization of Chymosin from Riverine Buffalo
(Bubalbubalis). J. Dairy Res. 70: 37–43.
Mahon M.C et Brown R.T., 1985. Chymosine, présure, protéases microbiennes et pepsine.
Effet du type d’enzyme sur la coagulation du lait. J. of Dairy Sci 68(3) : 628-632.
Moll M et Moll N., 1998. Additifs alimentaires et auxiliaires technologiques. Ed. Dunod,
Paris, 218p.
Munoz R.,Garcia J.L., Carrascosa A.V. et Gonzalez R., 2004. Cloning of the authentic
bovine gene encoding pepsinogenA and its expression in microbial cells. Rev. Appl. Envir.
Microb., 70, .5, pp 2588-2595.
Murado M.A., Siso I.G., Gonzalez P., Montemayor I., Pastrana L.et Pintado J., 1993.
Characterization of microbial biomasses and amylolytic preparation obtained from mussel
processing waste treatment. Bioresource technology 43:117-125.
Myers RH., Montgomery DC., 2002. Response surface methodology: process and product
optimization using designed experiments. New York: Wiley.
Nafa N., Si Ziani S., 2002. Extraction et caractérisation de l’extrait brut coagulant le lait de
Bacillus megaterium isolé localement. Mémoire ing, Uni. T. Ouzou : PP 62 .
Noda M., et Murakami K., 1981. Studies of proteinases from the digestive organs of
sardine. II. Purification and characterization of two acid proteases from the stomach.
Biochem. Biophys. Acta., 658(1):27-34.
Noor-Develiet P.E., Gist-Brocades N.N. Et Delft N.C.D. 1983. Les Enzymes Alimentaires :
Utilisation et Innocuité. Microbiol. Alim. Nut., 1 : 15.
Nungary A et Legoffic F,(1996)-étude d’une lipase hépatique et de la pepsine d’un requin
benthique en vue de la valorisation de ses viscères .utilisation des INIST.CNRS, travaux
universitaires, université de Paris06.
Pascal C., Joël C., et Françoise G., 2004. Opérations Unitaires en Génie Biologique
(Fermentation), Biologie Et Technique.
Palaiswamy M et al., 2009. Production of milk clotting enzyme protease by a local isolate of
Mucorsp under solid stat fermentation using agro industriel wastes.biotech.
Biotech.Bioprocess.Eng. 14:788-794.
Pien., 1974. Etude de la coagulation du lait par la présure. Rev. Tech, PP. 21-29.

163
Pointurier Hubert., 2003.La gestion matières dans l’industrie laitière. Edition Technique Et
Documentation, Lavoisier ; Londres-Paris- New York. PP 292-293.
Polaina J., Maccabe A.P., 2007. Industrial Enzymes, Structure, Function and Applications.
Springer. Netherlands.
Polychronia Dou A. et Manolkidis K. S., 1984. Etude biochimique d’un fromage de chèvre
à croûte fleurie à l’aide d’extraits coagulants microbiens. lait 64, 469-483.
Poza M., Sieiro C., CarreiraL. and Barros-Velazquez J., 2003. Production and
characterization of the milk-clotting protease of Myxococcus Xanthus strain 422. J. Ind.
Microbiol. Biotechnol., 30: 691- 698.
Preetha S et Boopathy R., 1997. Purification and characterization of a milk clotting protease
from Rhizomucor meihei. World journal of Microbiolology & Biotechnology 13: 573 - 578 .
Puhan Z., Irivine D.M., 1973a. Proteolysis of caséins by protéases of Bacillus subtilis used
to make Canadian cheddar cheese. J. Dairy. Sci. 56: 317-322.
Puhan Z., Irivine D.M., 1973b. Reduction of proteolytic activity of Bacillus subtilis protease
by acidification of milk before cheddar cheese manufactures. J. Dairy. Sci. 56: 323-327.
Rahardjo Y.S.P., Tramper J. & Rinzema A., 2006.Modeling Conversion and Transport
Phenomena In Solid-State Fermentation: A Review And Perspectives. Biotechnol. Adv., 24
(2), 161-179.
Ramesh M., Girich B., Mahalingeshwra K., 2000. Thermophilic fungi their physiology and
enzymes. Microbiology and Molecular Biology reviews, 64, 3: 461- 488.
Ramet J.P., 1980. La micelle de caséine et la coagulation du lait. In: Le Fromage. Ed., A.
Eck, 3ème Ed., Tec Et Doc, Lavoisier 60, 1-13.
Ramet, J.P., 1997. Les agents de transformation du lait; la présure et les enzymes coagulantes
In: Le Fromage. Ed., A. Eck, 3ème Ed., Tec Et Doc, Lavoisier, P.101-107, 539p
Ramman A.M., 1994. Rabbit pepsin as a rennet subtitute. Minu. J. Agric. Dev. Egypt., 12:
1685 - 1690.
Rao K., Mathur D, K., 1976. Studies on the production of bacterial rennet in pilot plant
fermentor. Biotech and Bioeng. 17, 1349-1351
Rao M.B., Tanksale A.M., Ghatge M.S. et Deshpande V., 1998. Molecular and
biotechnological aspects of microbial protease. Microbiology and Molecular Biology
Reviews. 60 (3): 597-635.
Rattray W, Gallman P, Jelen P., 1997. Nutritional, sensory and physicochemical
characterization of protein standardized UHT milk. Le Lait, 77: 279-296.
Rawlings N.D., Barret A.J., 1993. Evolutionary Families of Peptidases. Biochem J. 290:
205-218

164
Rawlings N.D., and Barrett A.J. 1994. Families of serine peptidases. Meth. Enzymol. 244,
19–61.
Rawlings N.D., and Barrett A.J. 1995. Evolutionary families of metallopeptidases. Meth.
Enzymol. 248, 183–228.
Reps A., Poznanski S., Babichowski A. et Jedrychowski L., 1979. Propriétés des substituts
de presure fabriqués à partir de Mucor miehei. Le lait 580-582
Roberts MS., Nakamura LK., Cohan FM., 1994.Bacillus mojavensis., distinguishable from
Bacillus subtilis by sexual isolation, divergence in DNA sequence, and differences in fatty
acid composition. Int. J. Syst. Bacteriol. 44:256-264
Robbins C.A., L.J. Swenson M.L. Neally R.E. Gots Et B.J. Kelman ., 2000. Health Effects
of Mycotoxins in Indoor Air: A Critical Review. Appl. Occup. Environ. Hyg, 15 (10): 773-84.
Rogelj I., Perko B., Francky A., Penca V., Pungercar J., 2001. Recombinant Lamb
chymosin as an Alternative Coagulating Enzyme In Cheese Production. J. Dairy Sci. 84:
1020-1026.
Rowan N.J., C.M. Johnstone R.C. Mclean J.G. Anderson et J.A. Clarke., 1999.
Prediction of Toxigenic Fungal Growth in Buildings By Using A Novel Modelling System.
Appl Environ Microbiol 65 (11): 4814-21.
Somkuti G.A., Rabel F.J., 1968. Purification and properties of Mucor pusillus acid protéase,
journal of Bacteriology, . 96, 4: 1407-1414.
Sardinas J.L., 1968. Rennin enzyme of Endothia parasitica. Appl. Microbiol., 16, 248-253.
Sastry K., Mathure B., 1979. Studies on milk clotting enzyme from Bacillus mégaterium K-
4. 1. Effect of some nutrients on enzyme production. J. of food Sci. and techno. 16. 1., 11-15.
Sathya R., Pradeep B.V., Angayarkanni J. et Palaniswamy M., 2009.Production of Milk
Clotting Protease by a local isolate of Mucor circinelloides under SSF using Agro-industrial
Wastes.Biotechnology and Bioprocess Engineering., 14: 788-794.
Schiff H.E., 1992. Environmental production and safety measures Related to biotechnological
production of industrial enzymes at novo nordisk based upon genetically microorganisms.
Dechema, sichercheit in biotechnology. In: Biotechnologies. Cood. Scriban., R. 4eme
èd. 357-
361.
Schmidt D.G., 1982. Association of caseins and casein micelle structure. in: Developments of
Dairy Chemestry1-Proteins. Ed., P. F. Fox, Applied Science Publishers. Ltd, 61-86.
Scriban R., 1999. Biotechnologie. 5e Edition. Tech Et Doc. Lavoisier. Paris.
Shamsuzzaman K. et Haard N.F., 1985. Coagulation du lait et aptitude en technologie
fromagère d’une enzyme analogue à la chymosine extraite de la muqueuse du phoque. J.
Food. Biochem., 9, 173 N 180.
Shieh CJ., Phan Thi LA., Shih IL 2009.Milk-clotting enzymes produced by culture of
Bacillus subtilis natto. Biochem. Eng.Journal 43:85-91.

165
Shinjiro I., Takuzo G., Kei A., 1967. The enzyme production and the properties of crude
enzyme. Agr, Biol, Chem, .31, 5:546-551.
Siboukeur O., Mati A., Hessas B., 2005. Amélioration de l’aptitude à la coagulation du lait
camelien (camelus dromedarus). utilisation d’extraits enzymatiques coagulants gastrique de
dromadaire. Cahier d’études et de recherches francophones / Agriculture.14 : 473-483.
Sicart P.J., 1980. Compte rendu de la journée technologique enzymatique. Beckman, Paris.
Sicart P.J., 1985. Utilisation des enzymes comme aides technologiques dans les industries
agro-alimentaires. Biofutur, 35, 61-64.
Simöes I., Faro C., 2004. Structure and Function of Plant Aspartic Proteinases. Eur. J.
Biochem. 271: 206–207.
Simon P., Meunier R., 1970. Microbiologie industrielle et génie biochimique. Ed Masson Et
Cie. Paris. PP 567.
Somkuti G. A. et Babel F. J., 1968. Purification and Properties of Mucor pusillus acid
protease. J. Bact., 95, 4, 1407-1414.
Sousa M.J et Malcata F.X., 1997. comparison of plant and animal rennets in terms of
microbiological,chemical and proteolysis characteristics of ovine cheese j.agri.food.chem.,45,
74-81.
Srinivasan R .A.,M.K.K.Iyengar,I.J.Babbar S.C. Chakravorty A.T.Dudani.,1964. Milk –
clotting enzymes from microorganisms Applied microbiology .12 .6 .475-478
Srinu Babu G., Shiva Kiran R. R., Lokeswari N. et Jaya Raju K., 2007.Optimization of
Protease Production from Aspergillus Oryzae Sp. Using Box-Behnken Experimental Design.
4, . 2, pp 145-153.
St-Gelais D. et Patrick Tirard-Collet., 2002. In : Science Et Technologie Du Lait,
Transformation Du Lait, Canada, Pp 349.
Sterberg M., 1976: Microbial rennet. Applied Microbiology, .20: 135-153.
Stetzenbach L.D. et Buttner M.P., 2000. Airborne microorganisms and in door air quality,
pp.116-125, in encyclopedia of microbiology, vol.1, second edition, academic press.
Rockefeller university.
Sumantha A., Larroche C.,and Pandey A., 2006 .Microbiolology and industrial
biotechnology of food-grade proteases:A perspective.Food Technology Biotechnology 44 (2)
:211-220
Swaisgood H.E., 1982. Chemistry of Milk Protein. In Development Dairy Chemistry 1-
Protein: 1-60. Ed. P. F. Fox London App. Sci. Publish.
Tavares J.F.P., Babtita J.A.B., Marcone M.F., 1997. Milk-coagulating enzymes of tuna
fish waste as a rennet substitute. International journal of Food Science and Nutrition,
.48,3:169-171.

166
Taylor J.W., Bowman B., Berbee M.L., White T.J., 1993. Fungual model organisms.
phylogenetics of saccharomyces, Aspergillus and Neurospora. Systematic Biology. 42., 440-
457.
Thuy DTB., et Bose SK., 2011. Characterization of multiple extracellular proteases produced
by a Bacillus subtilis strain and identification of the strain. Int. J. Biol. 3 (1):101-110.
Thys RCS, Guzzon SO, Florencia CO, Brandelli A., 2006. Optimization of protease
production by Microbacterium sp in feather meal using response surface methodology.
Process Biochem. 41:67-73.
Tubesha Z. A and Al-Delaimy K .S., 2003. Rennin-like milk coagulant enzyme produced
by a local isolate of Mucor. International Journal of Dairy Technology. 56, 4 , 237 – 241.
Veisseyre R., 1975. Technologie du lait. Constitution, récolte, traitement et transformation du
lait. Ed .La maison rustique, 3ème
éd., Paris,714p.
Vioque M., Gomez R., Sanchez E., Mata C., Tajada L. et Fernandez-Salguero J., 2000 :
chemical and microbiological caracteristics of Ewes’milk cheese manufactured with extracts
from flowers of cynara cardunculus and cynara humilis as coagulants. J.Agric. Food
Chem.48: 451-456.
Walstra P., 1999. Casein Sub-Micelles: Do They Exist. International Dairy Journal 9: 189-
192.
Wigley R.C., 1996. Cheese and whey in industrial enzymology. Second edition. Chapitre 27.
Ed.Godfrey and Wiest., 135 N° 142.
Wilkinson M.G., 1993. Acceleration of cheese repening. In Fox. P.F. Ed. Cheese chemistry
physic and microbiology. Chapman and Hall. London. P 523-530
World Health Organization (Who), 2000. Human Exposure Assessment. Environmental
Health Criteria, No.214. 375 P.
Wu LC., Hang YD., 2000. Acid protease production from Neosartorya fischeri. Lebenm.
Wiss. Technology 33:44-47.
Yadav J. S. et Mishra N. C., 1994. Eucaryotic Cells and Expression Vechors. In: Food
Biotechnology: Micro Organisms. Coord. Houi Y. H. Khachat Ourian G. Vch. Publichers,
Inc, PP 133-181.
Yang W.P ., Kobayashi H., et Kusakabe I., 1987. Purification and characterization of
soymilk clotting- enzymes from Bacillus sp 2956 . Bio.Agri.Biochem.59.9.2343-2349.
Yu J., Iwasaki S., Tamara G., Arima K., 1968. Physical properties and amino acid
composition of Mucor rennin crystal isolated from Mucor pusillus. Agr, Biol, Chem, 9: 16-
24PP.
Zhao J., Chen S. et Agboola S.O., 2003. Effect of starter culture on the biochemical and
sensory properties of ovine cheese manufactured with a plant coagulant. Austrialian J. of
Dairy Techn., 58 (2), 219.

ANNEXES

167
Annexe 1 :
Composition Du son utilisé
Origine Moulin ‘Algérie’ Blida
- Humidité: 14. 5 %
- Matières minérales :5.6 %
- Protides (N X 5.7) : 15.7%
- Protides (N X 6.25) : 17.2%
- Lipides : 7.25%
- Amidon : 23.05%
- Acidité (g H2SO4): 0.375
- Sucres réducteurs exprimés en glucose au % de matière sèche : 0.465 %
168
Annexe 2
Resultats de l’analyse statistique pour minimiser l’activité protéolytique Multiple Response Optimization ------------------------------ Data variables: Activité coagulante Activité protéolytique Observed Response Minimum Maximum ------------------------------------------------ Activité coagulante 0,0 3,33 Activité protéolytique 9,14 32,9 ------------------------------------------------ Desirability Weights Response Low High Goal First Second Impact ------------------------------------------------------------------------------------------ Activité coagulante 0,0 3,0 Maximize 1,0 3,0 Activité protéolytique 9,0 32,0 Minimize 1,0 3,0 ------------------------------------------------------------------------------------------ Predicted Observed Row Activité coagulante Activité protéolytique Desirability Desirability ------------------------------------------------------------------ 1 1,82 21,18 0,541658 0,534226 2 2,7 28,92 0,354779 0,347162 3 2,27 27,07 0,350608 0,402728 4 2,13 26,2 0,354779 0,423135 5 2,86 26,2 0,477186 0,490312 6 2,33 28,3 0,354779 0,353471 7 2,86 29,4 0,481947 0,328281 8 0,01 19,62 0,0 0,042358 9 2,38 26,4 0,354779 0,439499 10 0,24 24,39 0,409494 0,162695 11 0,01 20,74 0,176673 0,0403966 12 0,05 15,4 0,106105 0,109677 13 0,51 19,93 0,0 0,298686 14 0,0 10,64 0,149887 0,0 15 0,15 22,28 0,290586 0,145363 16 0,42 25,4 0,152741 0,200434 17 0,12 13,73 0,226958 0,178252 18 0,0 11,31 0,288033 0,0 19 2,44 29,6 0,354779 0,291324 20 0,0 14,4 0,290431 0,0 21 1,33 32,9 0,144699 0,0 22 0,03 20,13 0,0863406 0,0718392 23 0,06 11,45 0,0 0,133677 24 0,02 10,7 0,0 0,0785743 25 2,17 26,2 0,354779 0,42709 26 0,42 21,2 0,299052 0,256396 27 0,0 10,24 0,0 0,0 28 0,01 19,34 0,0489329 0,0428344 29 0,42 21,38 0,202956 0,254251 30 0,01 12,63 0,0 0,0529835 31 1,79 22,4 0,545852 0,499043 32 2,63 29,3 0,355754 0,320801

169
33 2,22 26,4 0,352868 0,424469 34 2,38 25,3 0,355754 0,48073 35 3,33 25,71 0,479803 0,522951 36 2,7 27,4 0,355754 0,424264 37 2,86 29,6 0,484899 0,315402 38 0,01 15,76 0,0 0,0485142 39 2,78 25,8 0,355754 0,499797 40 0,21 24,8 0,414421 0,148031 41 1,67 29,8 0,186748 0,230752 42 0,0 12,3 0,152316 0,0 43 0,01 19,01 0,0 0,043389 44 0,0 10,43 0,190723 0,0 45 0,21 23,2 0,300904 0,163654 46 0,01 20,15 0,174291 0,0414414 47 0,04 9,89 0,253934 0,113214 48 0,0 12,4 0,310072 0,0 49 2,17 30,85 0,355754 0,190175 50 0,33 21,84 0,30649 0,220434 51 1,39 28,48 0,139535 0,26629 52 0,01 21,82 0,124255 0,0384104 53 0,06 12,2 0,0 0,131215 54 0,02 9,14 0,0 0,0814008 55 2,13 26,8 0,355754 0,400652 56 0,17 23,82 0,307827 0,141963 57 0,0 12,4 0,0 0,0 58 0,01 18,2 0,107782 0,0447214 59 0,02 20,85 0,220111 0,0568497 60 0,01 12,96 0,0 0,0525302 ------------------------------------------------------------------ The StatAdvisor --------------- This procedure helps determine the combination of experimental factors which simultaneously optimize several responses. It does so by maximizing a desirability function. You may set various characteristics of the desirability function through the Analysis Options dialog box. The goals for each of the responses are currently set as: Activité coagulante - maximize Activité protéolytique - minimize Optimize Desirability -------------------- Optimum value = 0,634486 Factor Low High Optimum ----------------------------------------------------------------------- son 3,0 7,0 3,0 Caseine 0,02 2,0 0,0200001 Temperature 25,0 50,0 40,2713 Agitation 100,0 200,0 153,333 Response Optimum --------------------------------- Activité coagulante 2,18755 Activité protéolytique 19,302

170
--------------------------------- The StatAdvisor --------------- This table shows the combination of factor levels which maximize the desirability function over the indicated region. It also shows the combination of the factors at which that optimum is achieved. Use the Analysis Options dialog box to indicate the region over which the optimization is to be performed.
Surface de Réponse Estiméeson= 3,0,Caseine= 0,02
Temperature
Agitation
Obje
ctif
25 30 35 40 45 50100
150
200
0
0.2
0,4
0,6
0,8

171
Annexe 3
Photos de quelques bactéries sélectionnées

172
Annexe 4
Résultats du 1er
plan d’expérience
Standardized Pareto Chart for Activité coagulante
Standardized effect
+-
0 1 2 3 4
B:Caseine
D:Agitation
A:Glucose
C:Temperature
BB
CC
Main Ef fects Plot for Activité coagulante
0,9
1,2
1,5
1,8
2,1
2,4
Activité
co
ag
ula
nte
GlucoseCaseine
TemperatureAgitation

173
Estimated Response SurfaceTemperature=40,0,Agitation=200,0
Glucose Caseine
Activité c
oagula
nte
0 1 2 3 4 50
0,40,8
1,21,6
20,9
1,2
1,5
1,8
2,1
2,4
Analysis Summary ---------------- File name: C:\Documents and Settings\automate\Bureau\Souad 260309\Souad 260309 replicate.sfxCentral composite design: 2^4 + star Comment: Plan Souad Estimated effects for Activité coagulante ---------------------------------------------------------------------- average = 2,13812 +/- 0,196957 A:Glucose = -0,0871 +/- 0,307112 B:Caseine = 0,0019 +/- 0,307112 C:Temperature = -0,1149 +/- 0,307112 D:Agitation = 0,0215667 +/- 0,307112 BB = -2,17448 +/- 0,702058 CC = -2,22848 +/- 0,702058 block = -0,0270067 +/- 0,237888 ---------------------------------------------------------------------- Standard errors are based on total error with 52 d.f. The StatAdvisor --------------- This table shows each of the estimated effects and interactions. Also shown is the standard error of each of the effects, which measures their sampling error. To plot the estimates in decreasing order of importance, select Pareto Charts from the list of Graphical Options. To test the statistical significance of the effects, select ANOVA Table from the list of Tabular Options. You can then remove insignificant effects by pressing the alternate mouse button, selecting Analysis Options, and pressing the Exclude button.

174
Analysis of Variance for Activité coagulante - Plan Souad -------------------------------------------------------------------------------- Source Sum of Squares Df Mean Square F-Ratio P-Value -------------------------------------------------------------------------------- A:Glucose 0,0682777 1 0,0682777 0,08 0,7778 B:Caseine 0,00003249 1 0,00003249 0,00 0,9951 C:Temperature 0,118818 1 0,118818 0,14 0,7098 D:Agitation 0,00418609 1 0,00418609 0,00 0,9443 BB 8,14327 1 8,14327 9,59 0,0031 CC 8,55275 1 8,55275 10,08 0,0025 blocks 0,0109404 1 0,0109404 0,01 0,9100 Total error 44,1406 52 0,848858 -------------------------------------------------------------------------------- Total (corr.) 104,441 59 R-squared = 57,7363 percent R-squared (adjusted for d.f.) = 52,9517 percent Standard Error of Est. = 0,921335 Mean absolute error = 0,611637 Durbin-Watson statistic = 3,10235 (P=0,0000) Lag 1 residual autocorrelation = -0,600491 The StatAdvisor --------------- The ANOVA table partitions the variability in Activité coagulante into separate pieces for each of the effects. It then tests the statistical significance of each effect by comparing the mean square against an estimate of the experimental error. In this case, 2 effects have P-values less than 0,05, indicating that they are significantly different from zero at the 95,0% confidence level. The R-Squared statistic indicates that the model as fitted explains 57,7363% of the variability in Activité coagulante. The adjusted R-squared statistic, which is more suitable for comparing models with different numbers of independent variables, is 52,9517%. The standard error of the estimate shows the standard deviation of the residuals to be 0,921335. The mean absolute error (MAE) of 0,611637 is the average value of the residuals. The Durbin-Watson (DW) statistic tests the residuals to determine if there is any significant correlation based on the order in which they occur in your data file. Since the P-value is less than 0.05, there is an indication of possible serial correlation. Plot the residuals versus row order to see if there is any pattern which can be seen. Regression coeffs. for Activité coagulante - Plan Souad ---------------------------------------------------------------------- constant = -11,1569 A:Glucose = -0,017596 B:Caseine = 2,24177 C:Temperature = 0,614234 D:Agitation = 0,000143778 BB = -1,10931 CC = -0,00773777 ---------------------------------------------------------------------- The StatAdvisor

175
--------------- This pane displays the regression equation which has been fitted to the data. The equation of the fitted model is Activité coagulante = -11,1569 - 0,017596*Glucose + 2,24177*Caseine + 0,614234*Temperature + 0,000143778*Agitation - 1,10931*Caseine^2 - 0,00773777*Temperature^2 where the values of the variables are specified in their original units. To have STATGRAPHICS evaluate this function, select Predictions from the list of Tabular Options. To plot the function, select Response Plots from the list of Graphical Options. Optimize Response ----------------- Goal: maximize Activité coagulante Optimum value = 2,1932 Factor Low High Optimum ----------------------------------------------------------------------- Glucose 0,05 5,0 0,0500001 Caseine 0,02 2,0 1,00992 Temperature 28,0 52,0 39,6883 Agitation 50,0 200,0 200,0 The StatAdvisor --------------- This table shows the combination of factor levels which maximizes Activité coagulante over the indicated region. Use the Analysis Options dialog box to indicate the region over which the optimization is to be performed. You may set the value of one or more factors to a constant by setting the low and high limits to that value.
176
Annexe 5
Courbe d’étalonnage de la tyrosine et de la BSA
Figure : Courbe étalon de la tyrosine.
Figure : Courbe étalonde sérum d’albumine bovin (BSA).
y = 0,009x + 0,090 R² = 0,993
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 10 20 30 40 50 60
Ab
sorb
ance
à 6
60 n
m
μg/ml
y = 0,0014x + 0,0192 R² = 0,9946
0
0,1
0,2
0,3
0,4
0,5
0,6
0 50 100 150 200 250 300 350 400 450
Ab
sorb
ance
à 6
50 n
m
μg/mL

177
Annexe 6
Photos de quelques moisissures isolées
S1-52
4c-57
4a-61
4a-62
C/D-68
5F-71

178
Annexe 7
Echelle de Mc Farland
In microbiology, McFarland standards are used as a reference to adjust the turbidity of
bacterial suspensions so that the number of bacteria will be within a given range to
standardize microbial testing. An example of such testing is antibiotic susceptibility testing by
measurement of minimum inhibitory concentration which is routinely used in medical
microbiology and research. If a suspension used is too heavy or too dilute, an erroneous result
(either falsely resistant or falsely susceptible) for any given anti microbial agent could occur.
Original McFarland standards were mixing specified amounts of barium chloride and sulfuric
acid together. Mixing the two compounds forms a barium sulfate precipitate, which causes
turbidity in the solution. A 0.5 McFarland standard is prepared by mixing 0.05 mL of 1.175%
barium chloride dihydrate (BaCl2•2H2O), with 9.95 mL of 1% sulfuric acid (H2SO4).
Now there are McFarland standards prepared from suspensions of latex particles, which
lengthens the shelf life and stability of the suspensions. The standard can be compared
visually to a suspension of bacteria in sterile saline or nutrient broth. If the bacterial
suspension is too turbid, it can be diluted with more diluent. If the suspension is not turbid
enough, more bacteria can be added.

179
McFarland Nephelometer Standards:
McFarland Standard No. 0.5 1 2 3 4
1.0% Baruim chloride (ml) 0.05 0.1 0.2 0.3 0.4
1.0% sulruric acid (ml) 9.95 9.9 9.8 9.7 9.6
Approx. cell density (1X10^8 CFU/mL) 1.5 3.0 6.0 9.0 12.0
% Transmittance* 74.3 55.6 35.6 26.4 21.5
Absorbance* 0.08 to 0.1 0.257 0.451 0.582 0.669
*at wavelength of 600 nm
McFarland standards. No. 0.5, 1 and 2.

African Journal of Biotechnology Vol. 11(100), pp. 16594-16606, 13 December, 2012 Available online at http://www.academicjournals.org/AJB DOI: 10.5897/AJB12.1115 ISSN 1684–5315 ©2012 Academic Journals
Full Length Research Paper
Isolation and identification of bacterial strain I33M producing milk-clotting enzyme: Optimization of culture
parameters using response surface
Talantikite-Kellil, S1, Nouani, A. 1,2*, Saunier, M3, Gillmann, L3, Brahmi, A4 and Fazouane, F1
1Food Technology Laboratory, University of Boumerdes, Algeria.
2Food Technology laboratory, ENSA, Algeria.
3lasba-Sonas (UPRES - EA 921 , Antenne IUT - Université d’Angers 49000, France.
4Department of Mathematics and Statistics - Moncton University, NB, Canada.
Accepted 19 October, 2012
A strain I33M which produces a milk-clotting enzyme was screened from Algerian soil near a dairy factory. This strain was identified as Bacillus mojavensis based on morphology and internal transcription spacer sequence. Sequencing analysis of 16S rDNA gene showed 100% identity of the tested strain with the B. mojavensis in the database. Phylogenetic analysis of this strain showed that it was most closely related to Bacillus subtilis strain. The optimum levels of these significant parameters to obtain the highest milk clotting activity and the lowest proteolytic activity were determined employing the response surface methodology (RSM), which revealed these as follows: wheat bran 7%, casein 0.094%, temperature 39°C, agitation size (rpm) 150. Among the various variables screened, agitation and temperature were most significant in submerged fermentation (SmF). The optimal value of milk clotting activity (MCA) is esteemed at 2.40. Key words: Milk clotting protease, Bacillus, response surface methodology, sequencing analysis.
INTRODUCTION The proteases constitute at least 65% of the total industrial enzyme market (Rao et al., 1998) and are also known as peptidyl_/peptide hydrolases (EC 3.4.21-24 and 99) which catalyze the hydrolysis of a peptide bond in a protein molecule such as casein for cheese making production (Beg et al., 2003).
In cheese technology, these extracellular proteases are obtained especially from microbial culture and throughout the world, many strains of clotting protease microorganisms are being produced industrially mainly fungi such as Mucor miehei, Mucor pusillus, Endothia parasitica, Irpex lacteus, Aspergillus niger var. Awamori, Kluyveromyces lactis and Escherichiacoli using solid state fermentation process (Olson, 1995; Chen et al., 2011). *Corresponding author. E-mail: [email protected].
It is well known that with the increase in world cheese production and consumption, attempts were made to find substitutes for this enzyme (Neelakantan et al., 1999). So far, most efforts for calf rennet substitute focus on finding new milk clotting-enzymes that could be used in cheese production and many microbial and plant proteases were identified and have been suggested as milk coagulants. Microbial proteases, especially from Bacillus sp. have traditionally held the predominant share of the industrial enzyme market of the worldwide enzyme sales with major application such as detergent product, waste treatment, food industry (Sepahy and Jabalameli, 2011). However, few studies have demonstrated the value of proteases as a substitute for rennet. Although, Ding et al. (2011) report that only few research work on bacteria are reported, wild bacteria with a high milk-clotting activity (MCA) in submerged fermentation show a great potential, due to shorter fermentation period, larger capacity of extracellular secretion and higher material utilization ratio.

Talantikite-Kellil et al. 16595
Figure 1. Photograph of microscopic observation of the strain I33M and other Bacillus strain Gram+ (A. B. C not studied).
Protease production is an inherent capacity of all microorganisms, however, only those microbes that produce substantial amounts of extracellular enzyme are of industrial importance such as dairy industry (Beg et al., 2003). Also, it is well known that extracellular protease production by microorganisms is greatly influenced by physical factors such as pH, temperature, time and by others factors (Thys et al., 2006) where submerged fermentation (SmF) has been traditionally used. The optimization of parameters becomes obvious and the RSM is a collection of statistical techniques for designing experiments, building models, evaluating the effects of factors and searching for the optimum conditions (Chen et al., 2011).
Thus, our present study is a contribution of the achievement of a milk clotting enzyme from the isolation of a bacterial strain local. Statistical methods have been applied for optimization of protease production (Gangadharan et al., 2008). A statistical approach has been employed for identifying significant variables influencing protease production under submerged fermentation (SmF). The levels of the significant variables were further optimized using response surface methodology.
MATERIALS AND METHODS
Microorganism, culture conditions and Isolation method
Fifty grams of soil samples were collected near a dairy factory in the region of Boumerdes, Algeria. A small amount of soil was homogenized under magnetic agitation for 20 mn with 90 mL sterile physiological water to make an initial dilution (10
−1) (Davet and
Rouxel, 1997). Serial dilutions were made for each sample and 0.1 mL of the appropriate dilution to 10
−9 was spread on plate count
agar (PCA) medium containing 2% (w/v) actidione antifungal agent to prevent fungal growth and 12% (w/v) sterilised skimmed milk powder. The plates were incubated at 30°C for 48 h for the enumeration of the caseolytic strain producing milk clotting protease by the presence of a clear halo around the colony (Figures 1 and 2). Colonies with distinct morphologies differences such as color (Gram technique), shape and size were selected (Table 1) and purified by streaking using the same medium. Of the 68 strains selected, 52 were Gram positive and 16 were Gram negative. All isolates were checked for milk clotting production and examined microscopically before stock preparations. The Gram positive bacterial isolates identified as Bacillus was purified and the selected strains are those with a clotting activity and are maintained on agar conservation.
Protease production in shake flask cultures, medium composition and inoculum
According to Rao and Mathur (1976), the basal minimal medium

16596 Afr. J. Biotechnol.
Figure 2. Caseolyse area of some strains tested. Strain I33M and other Bacillus strain Gram positive (A. B. C not studied).
Table 1. Experimental range and levels of the independent variables used in RSM in terms of actual and coded factors.
Variable Range of level
Actual Coded Actual Coded Actual Coded
Wheat bran (%w/v) 3 -1 5 0 7 +1
Casein (%w/v) 0.02 -1 1.01 0 2 +1
Temperature (°C) 25 -1 37.5 0 50 +1
Agitation (RPM) 100 -1 150 0 200 +1
(pH 6.8) used for milk clotting protease production, contained (g.l
-1):
Wheat bran (50), yeast extract (3), glucose (4), casein (2), Na2HPO4 (3). The production medium (50 ml) in a 250 ml Erlenmeyer flask) was inoculated with 10
7 to 5.10
8 bacteria per
milliliter (Mac Farland3) and incubated at 30°C on a rotary shaker with agitation (110 rpm) for 24 h (Poza et al., 2003 ) in a IKA
® KS
4000i control shaker. After incubation, the culture broth was filtered and then centrifuged at 4,000 g for 30 min at 4°C in a cold centrifuge (Jouan GR 422). Total protease yield and milk clotting activity was determined in the cell-free supernatant. Strain that showed the highest milk-clotting activity and the lowest proteolytic activity was selected for further studies and identified using 16S rDNA sequencing. The selected strain is called I33M.
In the next stage, response surface methodology is used to study the interactive effects of four variables, that is, wheat bran, casein,
temperature and agitation for improving total protease production by the strain I33M
Response surface methodology
The present work was based on the central composite design (CCD) utilized to obtain the experimental data, which would fit an empirical, full second-order polynomial model representing the response surfaces over a relatively broad range of parameters as reported by Dutta et al. (2004). Each factor in the central composite design (CCD) was studied at three different levels, low (-1), medium (0) and high (+1). For this purpose, the response surface approach was used and a set of experiment was performed. The range and levels of experimental variables investigated are presented in Table 2. Four independent variables were used to approach the
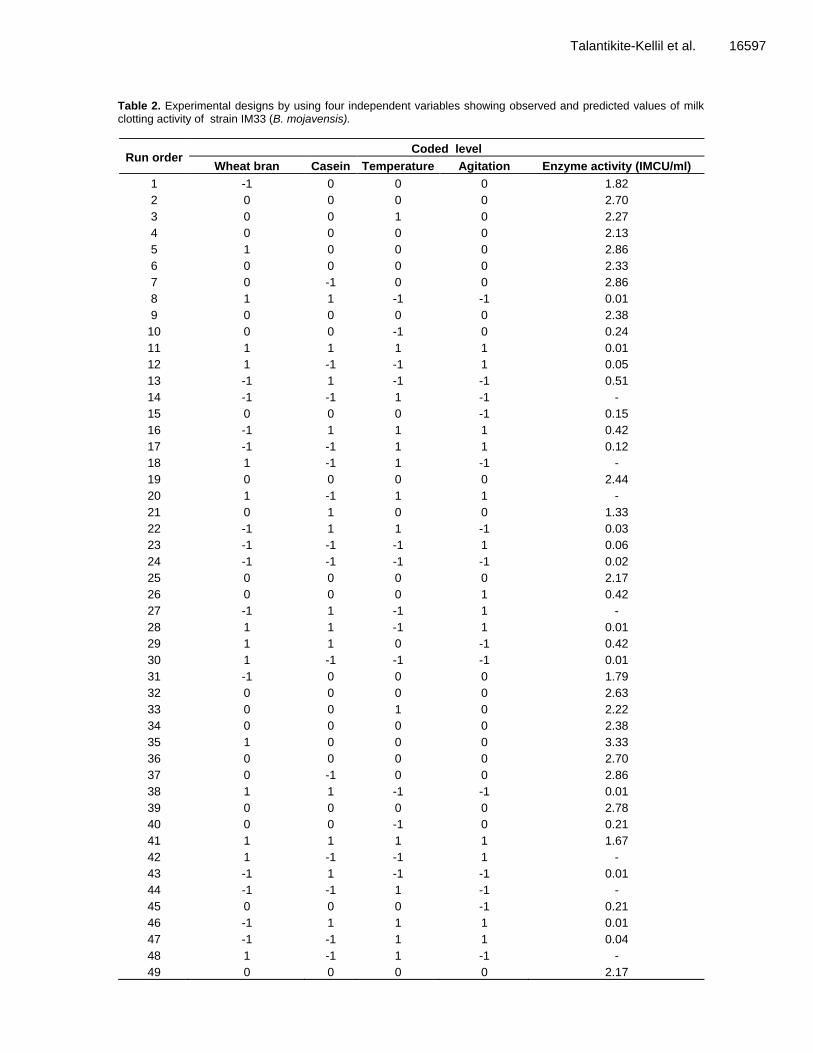
Talantikite-Kellil et al. 16597
Table 2. Experimental designs by using four independent variables showing observed and predicted values of milk clotting activity of strain IM33 (B. mojavensis).
Run order Coded level
Wheat bran Casein Temperature Agitation Enzyme activity (IMCU/ml)
1 -1 0 0 0 1.82
2 0 0 0 0 2.70
3 0 0 1 0 2.27
4 0 0 0 0 2.13
5 1 0 0 0 2.86
6 0 0 0 0 2.33
7 0 -1 0 0 2.86
8 1 1 -1 -1 0.01
9 0 0 0 0 2.38
10 0 0 -1 0 0.24
11 1 1 1 1 0.01
12 1 -1 -1 1 0.05
13 -1 1 -1 -1 0.51
14 -1 -1 1 -1 -
15 0 0 0 -1 0.15
16 -1 1 1 1 0.42
17 -1 -1 1 1 0.12
18 1 -1 1 -1 -
19 0 0 0 0 2.44
20 1 -1 1 1 -
21 0 1 0 0 1.33
22 -1 1 1 -1 0.03
23 -1 -1 -1 1 0.06
24 -1 -1 -1 -1 0.02
25 0 0 0 0 2.17
26 0 0 0 1 0.42
27 -1 1 -1 1 -
28 1 1 -1 1 0.01
29 1 1 0 -1 0.42
30 1 -1 -1 -1 0.01
31 -1 0 0 0 1.79
32 0 0 0 0 2.63
33 0 0 1 0 2.22
34 0 0 0 0 2.38
35 1 0 0 0 3.33
36 0 0 0 0 2.70
37 0 -1 0 0 2.86
38 1 1 -1 -1 0.01
39 0 0 0 0 2.78
40 0 0 -1 0 0.21
41 1 1 1 1 1.67
42 1 -1 -1 1 -
43 -1 1 -1 -1 0.01
44 -1 -1 1 -1 -
45 0 0 0 -1 0.21
46 -1 1 1 1 0.01
47 -1 -1 1 1 0.04
48 1 -1 1 -1 -
49 0 0 0 0 2.17
16598 Afr. J. Biotechnol.
Table 2. Continued.
50 1 -1 1 1 0.33
51 0 1 0 0 1.39
52 -1 1 1 -1 0.01
53 -1 -1 -1 1 0.06
54 -1 -1 -1 -1 0.02
55 0 0 0 0 2.13
56 0 0 0 1 0.17
57 -1 1 -1 1 -
58 1 1 -1 1 0.01
59 1 1 0 -1 0.02
60 1 -1 -1 -1 0.01
interaction among different factors (2
4 CCD with four factors was
applied) including 6 centre points and a set of 60 experiments was carried out. The central values (zero level) chosen for experimental design were: wheat bran 5% (w/v), casein 1.01% (w/v), temperature 37.5°C and agitation 150 rpm.
Results were analyzed by the experimental design module of the STATGRAPHICS plus V 5.1software (Statsoft, USA). The model permitted evaluation of the effects of linear, quadratic and interactive terms of the independent variables on the dependent variable. The statistical significance of the regression coefficients was determined by Student’s t-test and the second order model equation was determined by Fisher’s test. Three-dimensional surface plots were drawn to illustrate the main and interactive effects of the independent variables on milk clotting protease production (Myers and Montgomery, 2002; Thys et al., 2006).
For the validation of CCD, only two factors, temperature and agitation were taken, and remainder of the factors were fixed on the basis of the data obtained from the first set of experimental design. Analytical procedures Milk clotting activity assay Milk clotting activity (MCA) was measured according to the procedure described by international dairy federation (IDF) standard (IDF, 2002). Skim milk powder was reconstituted by dissolving 12.5 g in 100 ml of 0.5 g/L of CaCl2 solution (pH 6.5). The clotting assay was carried out by mixing 10 ml of this substrate with 1 ml of sample. Clotting activity was expressed in international milk clotting units (IMCU)/ml Proteolytic activity Proteolytic activity was determined according to the study of Murado and Siso (1993). One millilitre of the enzymatic extracts was added to 1 ml of 2% (w/v) alkali soluble casein in 0.02 M sodium-citrate buffer (pH 5.2). The reaction mixture was incubated at 35°C in a water bath for 10, 30, 60, 90 and 180 min, and the reaction was terminated by adding 5 ml of 12% (w/v) trichloroacetic acid. After 15 min, the precipitates formed were removed by filtration through Whatman no. 1 filter paper. Five millilitres of 0.5 M sodium carbonate solution was added to 1 ml of the aforementioned clear filtrate. The mixtures were incubated at 35°C for 10 min, when 0.5 ml of Folin–Ciocalteu reagent was added. The reaction was incubated for 20 min at 35°C for colour development. The optimal density at 660 nm expresses the rate of tyrosine released during hydrolysis.
Protein estimation Protein concentration was measured by the method of Lowry et al. (1951), with crystalline bovine albumin as the standard. Identification of strain I33M Macroscopy and microscopy were performed on Petri dishes with plate count agar (PCA) on colonies of 24 h. For sequencing analysis, the genomic DNA was extracted from the strain by the standard chloroform isoamyl alcohol method using PrepMan ultra sample preparation reagent kit (Applied Biosystems) and purified on plates NucleoFast (Macherey Nagel). The amplification of the 16S rDNA was performed through PCR technique, using Taq DNA polymerase, genomic DNA as a template, and 3 forward and 5reverse universal primers. The sequences of these primers used are as follows: F1 (5’-AGAGTTTGATCCTGGCTCAG-3’), R1 (5’-GTATTACCGCGGCTGCTGGCAC-3’), F2 (5’-CTCCTACGGGAGGCAG-3’) and R2 (5’-GACACGAGCTGACGACA-3’). PCR products were sequenced by Sanger technique using optimised conditions performed by Geno screen (Geno-screen Campus de l’Institut Pasteur de Lille 1, rue du Professeur Calmette – 59000 Lille – France). Receiving the sequencing results, 16S rDNA nucleotide sequence of the strain was deposited in GenBank, and aligned with the 16S rDNA sequences available in public data bases in National Center for Biotechnology Information (NCBI) (Available at: http://www.ncbi.nlm.nih.gov/), and LeBibi database (http://umr5558-sud-str1.univ-lyon1.fr/lebibi/lebibi.cgi) using basic local alignment search tool (BLAST) software. Bacterial growth and protease production in a fermentor The final validation of the statistical approach to protease production from B. mojavensis was carried out in a 2 l bioreactor (Sartorius Biostat A+, Switzerland) with a working volume of 1.5 L. The optimized medium (pH 6.8, 7% (w/v) wheat bran and 0.094% (w/v) casein) containing 0.3%(w/v) yeast extract and 0.3% (w/v) Na2HPO4 was sterilized in situ at 120°C for 30 min. The medium was inoculated aseptically with 60 ml of inoculum (Mc Farland 3) (9. 10
8 CFU / ml). The fermentation was carried out at 39°C for 50 h
with uncontrolled pH and the impeller speed was initially adjusted to 150 rpm. Samples were withdrawn periodically at an interval of 2 h and analyzed for protein and protease production (as described previously) and biomass estimation was determined using the technique described by Thys et al. (2006). The nutrient agar plates loaded by bacterial suspension were incubated for 24 h at 37°C and

Talantikite-Kellil et al. 16599
Figure 3. Sequence of the 16S rDNA of strain I33M (B. mojavensis).
counts performed on plates having between 20 and 300 colonies. Other fermentation parameters, such as pH, were continuously monitored using microprocessor-controlled probes.
RESULTS AND DISCUSSION Screening of strains producing bacterial
Milk-clotting enzyme
In this study, some strain with different milk-clotting activity (MCA) were isolated from soil and identified as bacillus Gram positive by Biochemical tests (data not shown). Among these strains, four Bacillus sp (A, B, C and I33M ) had a good milk-clotting activity and caseolytic effect on casein agar plates as shown in Figure 2 and were selected to be studied further. In the present work and in a first step, we report the results of extracellular protease activity and molecular identification and phylogenetic analysis of strain I33M which also has the best enzyme activity (Up to 3.3 IMCU/ml) followed by high proteolytic effect. It is well known that protease production is an inherent capacity of all microorganisms, however only those microbes that produce substantial amounts of extracellular enzyme are of industrial importance (Gupta et al., 2002; Kole et al., 1988). Thus, the most common and widely used bacteria for industrial proteases belong to the genus Bacillus (Ara et al., 2007). Ghaffour and Hasnain (2010) reported that the strains of B. subtilis was highly active on milk agar plates and caseinase activity was confirmed by growing the organism on casein agar plates concentration ranging from 0.3 to 1%. In light of the examined literature, no work, in our opinion, has reported on milk clotting proteases produced by B. mojavensis. So, According to the results, B. mojavensis could be a good producer of extracellular milk clotting protease. As Fungal milk-
clotting enzymes, the Bacillus species could find applications in cheese industry. Identification of the strain The 16SrDNA gene sequence analysis has been described to be necessary mainly to detect some misidentification of Bacillus and related strains (Ben et al., 2011). Also, this technique is one of the most commonly used today for the identification of species, as it is a quick, reliable, and reproducible method compared to conventional technique using morphological and physiological criteria. For further characterization, the PCR product of strain I33M was examined by electrophoresis, purified, and then sequenced by Geno-screen (Campus de l’Institut Pasteur de Lille, France). In addition to biochemical and morphological identification, the 16S rDNA gene was successfully amplified from the genomic DNA of the strain I33M (Figure 3). This sequence consists of 999 bp and the length of Query sequence is: 195 A, 254 T, 321 C and 229 G. The BLAST search of 16S rDNA gene sequence against sequences in nucleotide database has shown 100% homology with B. mojavensis strain. Therefore, our strain was named B. mojavensis I33M and the data bases allowed us to elaborate a phylogenetic dendrogram for I33M and related strains (Figure 4). The phylogenetic analysis showed that the tested strain was closest to B. subtilis. Similar results were reported by Thuy and Bose (2011). Also, Roberts et al. (1994) report that isolate was subsequently found to belong to the closely related B. subtilis-like phenotype that has been recently described as B. mojavensis. B. mojavensis can be distinguished only by differences in whole-cell fatty acid composition, divergence in DNA sequence analysis, and resistance to genetic transformation between taxa within the B. subtilis group. Moreover, Bacon and Hinton (2001) revealed
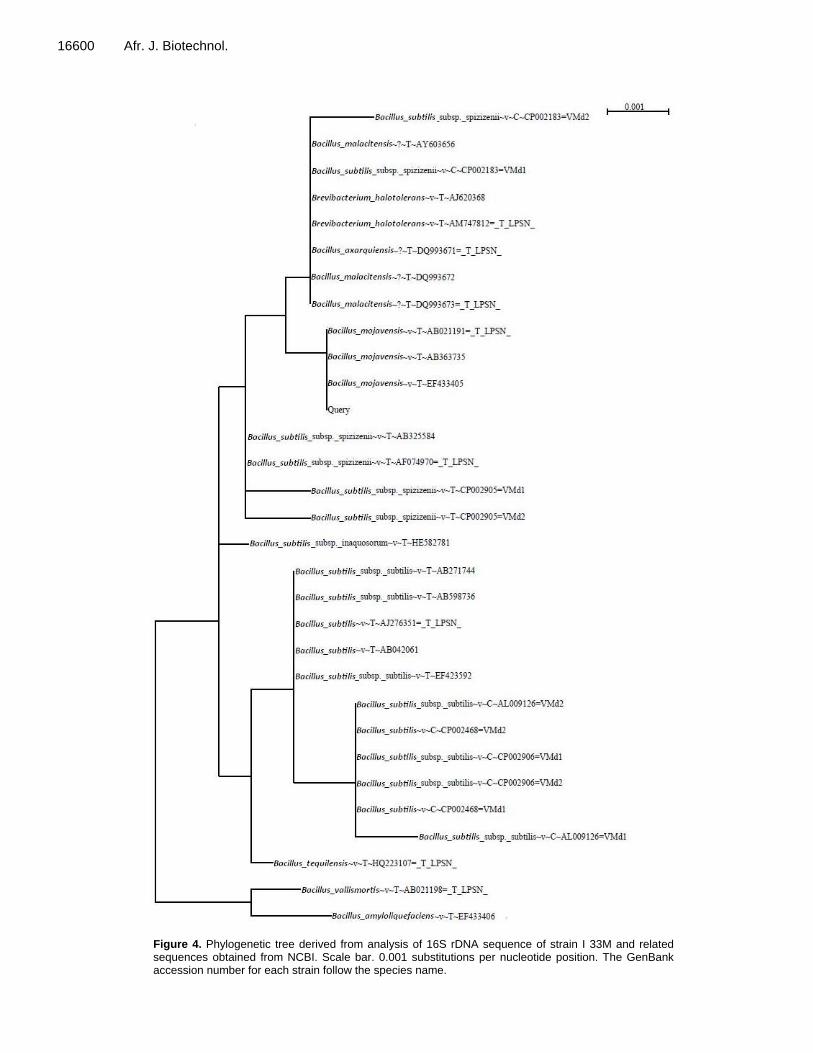
16600 Afr. J. Biotechnol.
Figure 4. Phylogenetic tree derived from analysis of 16S rDNA sequence of strain I 33M and related sequences obtained from NCBI. Scale bar. 0.001 substitutions per nucleotide position. The GenBank accession number for each strain follow the species name.

Talantikite-Kellil et al. 16601
Figure 5. Bar graph of standardized estimated effects of the different variables tested in the prospective experiment on milk clotting protease production by strain I33M (B. mojavensis). The variables tested were wheat bran. temperature. agitation and casein. The point at which the effect estimates were statistically significant (at P = 0. 05) is indicated by the vertical line.
Table 3. Estimated effects for Milk clotting activity.
Factor Value
Average 2.27234 ±0.103779
A:Wheat bran 0.212778 ± 0.161821
B:Casein -0.0316667 ± 0.161821
C:Temperature 0.351667 ± 0.161821
D:Agitation 0.107778 ± 0.161821
CC -1.20806 ± 0.369922
DD -3.20306 ± 0.369922
Block 0.0466667 ± 0.125346
Standard errors are based on total error with 52 d.f.
in their works that the identity of a patented endophytic bacterium was established by 16S rRNA sequence analysis as a strain of B. mojavensis, a recently erected species within one of the B. subtilis subgroups. Optimization by response surface methodology The statistical design approach using response surface methodology was used to study the interactive effects of various chemicals and physical factors on protease (milk clotting) production by B. mojavensis. The primary aim is to maximize the milk clotting activity and to minimize the proteolytic activity. The experiment was conducted in 60 runs and the input variables that have the maximum influence on the final response of the system were identified from the interaction of four factors namely, wheat bran (carbon source) and casein (nitrogen source)
(nutritional factors), temperature and agitation (physical factors), was examined through RSM following the CCD. We recall that the respective low and high levels (% w/v) with the coded levels in parentheses for the factors were defined as 3 (-1) and 7 (+1) for wheat bran, 0.02 (-1) and 2.0 (+1) for casein, 25°C (-1) and 50°C (+1) for temperature, 100 rpm (-1) and 200 (+1) for the agitation.
The results of CCD experiments for studying the effects of these variables on protease production in milk clotting activity (IMCU/ml) are presented in Table 2 along with the observed responses.
Statistical analysis of results showed that in the range studied, quadratic effect of temperature and agitation has a strong effect on protease production followed by the quadratic effect of agitation. The temperature have a significant effect but slightly higher than the limit of significance for 95% of significance (Figure 5). Table 3 shows the estimate effects and interactions. The results
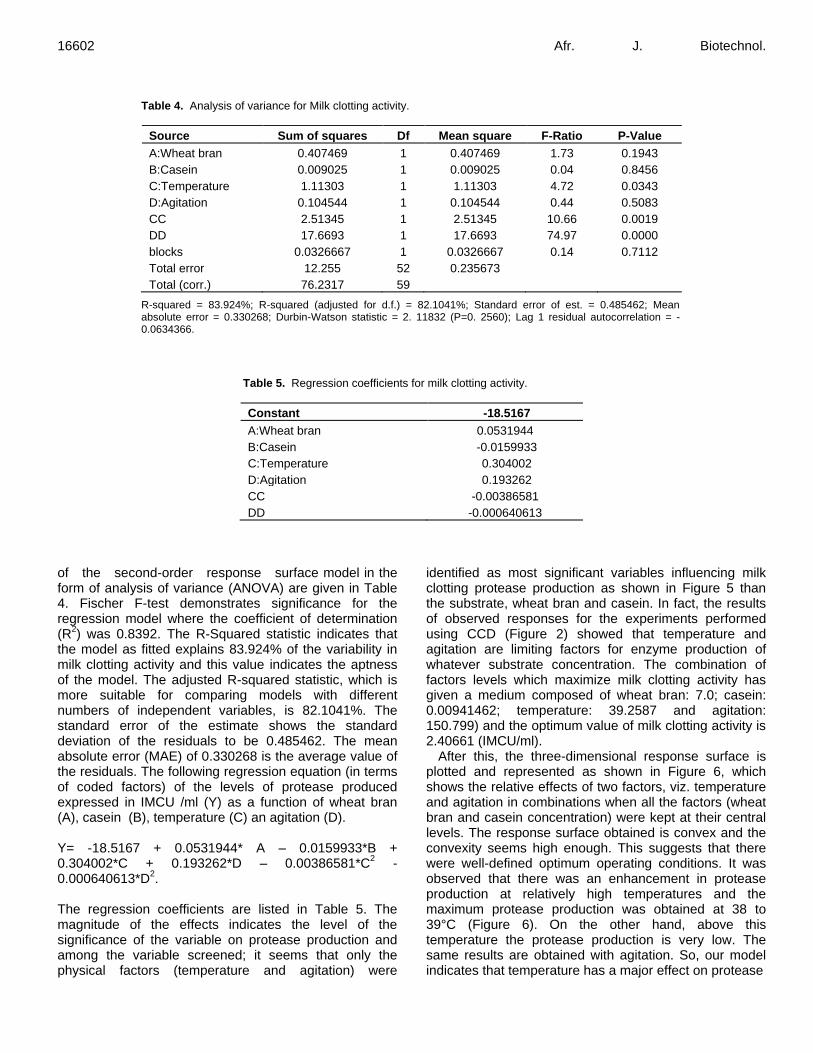
16602 Afr. J. Biotechnol.
Table 4. Analysis of variance for Milk clotting activity.
Source Sum of squares Df Mean square F-Ratio P-Value
A:Wheat bran 0.407469 1 0.407469 1.73 0.1943
B:Casein 0.009025 1 0.009025 0.04 0.8456
C:Temperature 1.11303 1 1.11303 4.72 0.0343
D:Agitation 0.104544 1 0.104544 0.44 0.5083
CC 2.51345 1 2.51345 10.66 0.0019
DD 17.6693 1 17.6693 74.97 0.0000
blocks 0.0326667 1 0.0326667 0.14 0.7112
Total error 12.255 52 0.235673
Total (corr.) 76.2317 59
R-squared = 83.924%; R-squared (adjusted for d.f.) = 82.1041%; Standard error of est. = 0.485462; Mean absolute error = 0.330268; Durbin-Watson statistic = 2. 11832 (P=0. 2560); Lag 1 residual autocorrelation = -0.0634366.
Table 5. Regression coefficients for milk clotting activity.
Constant -18.5167
A:Wheat bran 0.0531944
B:Casein -0.0159933
C:Temperature 0.304002
D:Agitation 0.193262
CC -0.00386581
DD -0.000640613
of the second-order response surface model in the form of analysis of variance (ANOVA) are given in Table 4. Fischer F-test demonstrates significance for the regression model where the coefficient of determination (R
2) was 0.8392. The R-Squared statistic indicates that
the model as fitted explains 83.924% of the variability in milk clotting activity and this value indicates the aptness of the model. The adjusted R-squared statistic, which is more suitable for comparing models with different numbers of independent variables, is 82.1041%. The standard error of the estimate shows the standard deviation of the residuals to be 0.485462. The mean absolute error (MAE) of 0.330268 is the average value of the residuals. The following regression equation (in terms of coded factors) of the levels of protease produced expressed in IMCU /ml (Y) as a function of wheat bran (A), casein (B), temperature (C) an agitation (D). Y= -18.5167 + 0.0531944* A – 0.0159933*B + 0.304002*C + 0.193262*D – 0.00386581*C
2 -
0.000640613*D2.
The regression coefficients are listed in Table 5. The magnitude of the effects indicates the level of the significance of the variable on protease production and among the variable screened; it seems that only the physical factors (temperature and agitation) were
identified as most significant variables influencing milk clotting protease production as shown in Figure 5 than the substrate, wheat bran and casein. In fact, the results of observed responses for the experiments performed using CCD (Figure 2) showed that temperature and agitation are limiting factors for enzyme production of whatever substrate concentration. The combination of factors levels which maximize milk clotting activity has given a medium composed of wheat bran: 7.0; casein: 0.00941462; temperature: 39.2587 and agitation: 150.799) and the optimum value of milk clotting activity is 2.40661 (IMCU/ml).
After this, the three-dimensional response surface is plotted and represented as shown in Figure 6, which shows the relative effects of two factors, viz. temperature and agitation in combinations when all the factors (wheat bran and casein concentration) were kept at their central levels. The response surface obtained is convex and the convexity seems high enough. This suggests that there were well-defined optimum operating conditions. It was observed that there was an enhancement in protease production at relatively high temperatures and the maximum protease production was obtained at 38 to 39°C (Figure 6). On the other hand, above this temperature the protease production is very low. The same results are obtained with agitation. So, our model indicates that temperature has a major effect on protease

Talantikite-Kellil et al. 16603
Figure 6. Response surface curve of milk clotting protease production from strain I33M (B. mojavensis) showing interaction between temperature and agitation.
production by the B. mojavensis strain. This result is reported by Thys et al. (2006) and other RSM studies also showed that temperature is a very relevant factor for microbial protease production (Wu and Hang, 2000). The RSM methodology was found to be very efficient to determine the optimal conditions for protease production by Bacillus sp. PE-11 (Thys et al., 2006). In general, it is clearly shown that no defined medium has been established for producing protease from different microbial sources (Gupta et al., 2002). Each organism or strain has its own requirement of special conditions for maximum enzyme production. So, in our case, the physical factors, temperature and agitation are more significant than nutritional factors. Growth and protease production at optimal conditions in laboratory-scale bioreactor During the period of growth, we notice that the lag phase is short and the exponential phase, after 10 h of fermentation, the stationary phase begins and continues for 48 h, the growth rate is 1.9228 while the generation time is 29 min 36 s (Figure 7). In terms of the stationary phase, there have been a number of studies conducted on B. subtilis fermentation. These studies show that it can last from 3 to 8 days. Thus, Dutt et al. (2008) report that clotting activity decreases after 45 h whereas maximum is reached after one day (Shieh et al., 2009). The pH was fairly stable throughout the fermentation
period (Figure 7). This could be explained by the fact that the bacteria consuming sugars produced acids that were quickly neutralized by the basic products from the proteolytic activity of Bacillus. The milk clotting and proteolytic activities increase over time while the protein level remains more or less telling; the two activities are linked (Figures 8 and 9).
On the clotting enzyme produced by B. mojavensis, the results show that this protease is a secondary metabolite because its rate continues to increase during stationary phase when a primary metabolite is produced during the exponential phase. Conclusion From this study, it can be concluded that milk clotting protease from B. mojavensis can be considered as a suitable alternative to the conventional rennet but it is more important to increase in the milk clotting activity (MCA) per proteolytic activity (PA) index. On the other hand, the present work show that protease production in B. mojavensis can be improved by controlling various factors simultaneously viz. agitation and temperature and this interaction could only be well understood by suitable RSM selection. In fact, this statistical method which includes factorial design and regression analysis helps in evaluating the effective factors and in building models to study interaction and select optimum conditions of variables for a desirable response. In our case, maximum
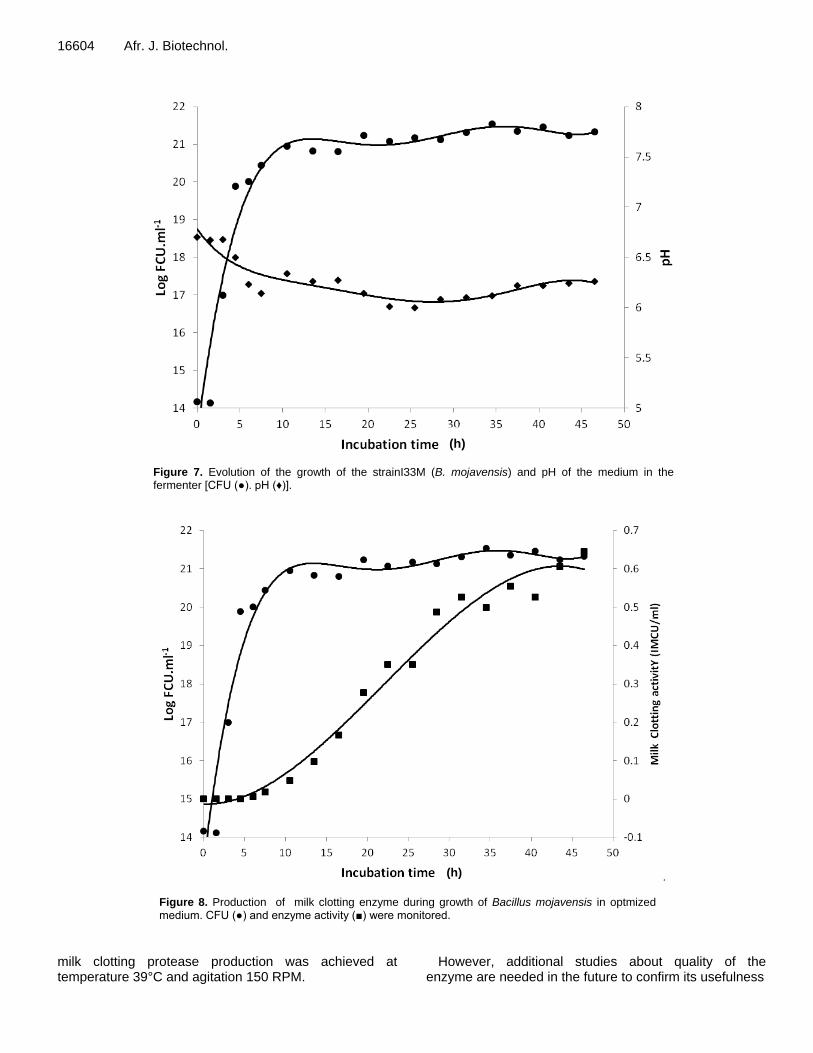
16604 Afr. J. Biotechnol.
Figure 7. Evolution of the growth of the strainI33M (B. mojavensis) and pH of the medium in the fermenter [CFU (●). pH (♦)].
.
Figure 8. Production of milk clotting enzyme during growth of Bacillus mojavensis in optmized medium. CFU (●) and enzyme activity (■) were monitored.
milk clotting protease production was achieved at temperature 39°C and agitation 150 RPM.
However, additional studies about quality of the enzyme are needed in the future to confirm its usefulness
(h)
(h)

Talantikite-Kellil et al. 16605
Figure 9. Curve of the proteolytic activity of strainI33 M (B.mojavensis) depending on the incubation time.
in the dairy processing. ACKNOWLEDGEMENTS Dr. ARNAUD DUDOIT, responsible micropollutants service in the laboratory of Hydrology and hygiene, Angers, is acknowledged for his assistance in statistical analysis. We also acknowledge the helpful collaborations of Odile ROLET-REPECAUD engineer, INRA (POLIGNY (39801, France). REFERENCES Ara K, Ozaki K, Nakamura, Yamane K, Sekiguchi J, Ogasawara N
(2007). Bacillus minimum genome factory: effective utilization of microbial genome information. Biotechnol. Appl. Biochem. 46:169-178.
Bacon CW, Hinton DM (2001). Endophytic and Biological Control Potential of Bacillus mojavensis and Related Species. Biol. Control 23:274-284.
Beg QK, Sahai V , Gupta R (2003)- Statistical media optimization and alkaline protease production from Bacillus mojavensis in a bioreactor. Process. Biochem. 39:203-209.
Ben Maachia S, Errakhi R, Mathieu F, Chérif M, Lebrihi A (2011). Identification and partial characterization of antifungal and antibacterial activities of two Bacillus sp. Strains isolated from salt soil in Tunisia. Afr. J. Microbiol. Res. 5(13):1599-1608.
Chen HQ, Chen XM, Chen TX, Xu XM, Jin ZY (2011).Optimization of solid-state medium for the production of inulinase by Aspergillus ficuum JNSP5-06 using response surface methodology. Carbohydr. Polym. 86:249-254.
Davet P, Rouxel F (1997). Techniques for direct isolation of microorganisms from soil. Collection Techniques et pratiques. INRA Editions (Rennes, France).
Ding Z, Liu S, Gu Z, Zhang L, Zhang K , Shi G (2011). Production of milk-clotting enzyme by Bacillus subtilis B1 from wheat bran. Afr. J. Biotechnol. 10(46):9370-9378.
Dutt K, Meghwashi GK, Gupta P, Saxena RK (2008). Role of casein induction and enhancement of production of a bacterial milk clotting protease from an indigenously isolated Bacillus subtilis. Lett. Appl. Microbiol. 46:513-518.
Dutta J R , Dutta P K , Banerjee R (2004). Optimization of culture parameters for extracellular protease production from a newly isolated Pseudomonas sp. using response surface and artificial neural network models. Process Biochem. 39:2193-2198
Gangadharan D, Sivaramakrishnan S, Nampoothiri K M, Sukumaran R K., Pandey AS (2008).Response surface methodology for the optimization of alpha amylase production by Bacillus amyloliquefaciens. Bioresour. Technol. 99:4597-4602
Ghaffour A, Hasnain S (2010). Purification and characterization of an extracellular protease from Bacillus subtilis EAG- 2 strain isolated from ornamental plant nursery. Pol. J. Microbiol. 59(2):107-112.
Gupta R, Beg QK, Lorenz P (2002). Bacterial alkaline proteases: molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 59:15-32.
IDF (2002). Milk and milk products—Microbial coagulants— Determination of total milk-clotting activity (2002) - IDF Standard 176. International Dairy Federation, Brussels, Belgium.
Kole MM, Draper I, Gerson DF (1988). Production of protease by Bacillus subtilis using simultaneous control of glucose and ammonium concentrations. J. Chem. Technol. Biotechnol. 41:197-206.
Lowry OH, Rosenbrough NJ, Farr AI, Randall R J (1951). Protein measurement with the Follin phenol reagent. J. Biol. Chem. 193:265-275.
Murado MA, Siso MI, Gonzalez MP, Pintado DJ (1993). Characterization of microbial biomasses and amylolytic preparations
(h)

16606 Afr. J. Biotechnol.
obtained from mussel processing waste treatment. Bioresour. Technol. 43:117-125
Myers RH, Montgomery DC (2002). Response surface methodology: process and product optimization using designed experiments. New York: Wiley.
Neelakantan S, Mohanty AK, Kaushik JK (1999).Production and use of microbial enzymes for dairy processing. Curr. Sci. 77:143-148.
Olson NF (1995). Cheese. In Enzymes, Biomass, Food and Feed, Biotechnology, Vol. 9, eds Rehm HJ & Reed G pp: 353-384. Weinheim. VCH.
Poza M, Sieiro C, Careira L, Banos- valazquez J, Villa TG (2003). Production and characterization of the milk-clotting protease of Myxococcus xanthus strain 422. J. Ind. Microbiol. Biotechnol. 30:691-698
Rao K, Mathur DK (1976).Studies on the production of bacterial rennet in pilot plant fermentor). Biotechnol. Bioeng. 17:1349-1351.
Rao MB, Tanksale AM, Ghatge MS, Deshpande VV (1998). Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 62:597-635
Roberts MS, Nakamura LK, Cohan FM (1994). Bacillus mojavensis sp. nov., distinguishable from Bacillus subtilis by sexual isolation, divergence in DNA sequence, and differences in fatty acid composition. Int. J. Syst. Bacteriol. 44:256-264.
Sepahy AA , JabalameliL (2011). Effect of culture Conditions on the
Production ofan Extracellular Protease by Bacillus sp. isolated from soil sample of lavizan jungle park. Enzyme Res. 2011:1-7.
Shieh CJ, Phan Thi LA, Shih IL (2009). Milk-clotting enzymes produced by culture of Bacillus subtilis natto. Biochem. Eng. 43:85-91.
Thuy DTB , Bose SK (2011). Characterization of multiple extracellular proteases produced by a Bacillus subtilis strain and identification of the strain. Int. J. Biol. 3(1):101-110.
Thys RCS, Guzzon SO, Florencia CO, Brandelli A (2006). Optimization of protease production by Microbacterium sp in feather meal using response surface methodology. Process Biochem. 41:67-73.
Wu LC, Hang YD (2000). Acid protease production from Neosartorya fischeri. Lebenm. Wiss. Technology 33:44-47.