licence de biologie cellulairecl.riviere.free.fr/haploids-y2k.pdf · faculté des sciences de...
TRANSCRIPT

FACULTÉ DES SCIENCES DE LUMINY
—————————
Licence de Biologie Cellulaire
Génétique formelledes organismes haploïdes
Transparents et problèmes
—————————
par
Claude Rivière, Dr. Sc.Maître de Conférence des Universités
2000-2001
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 2/24
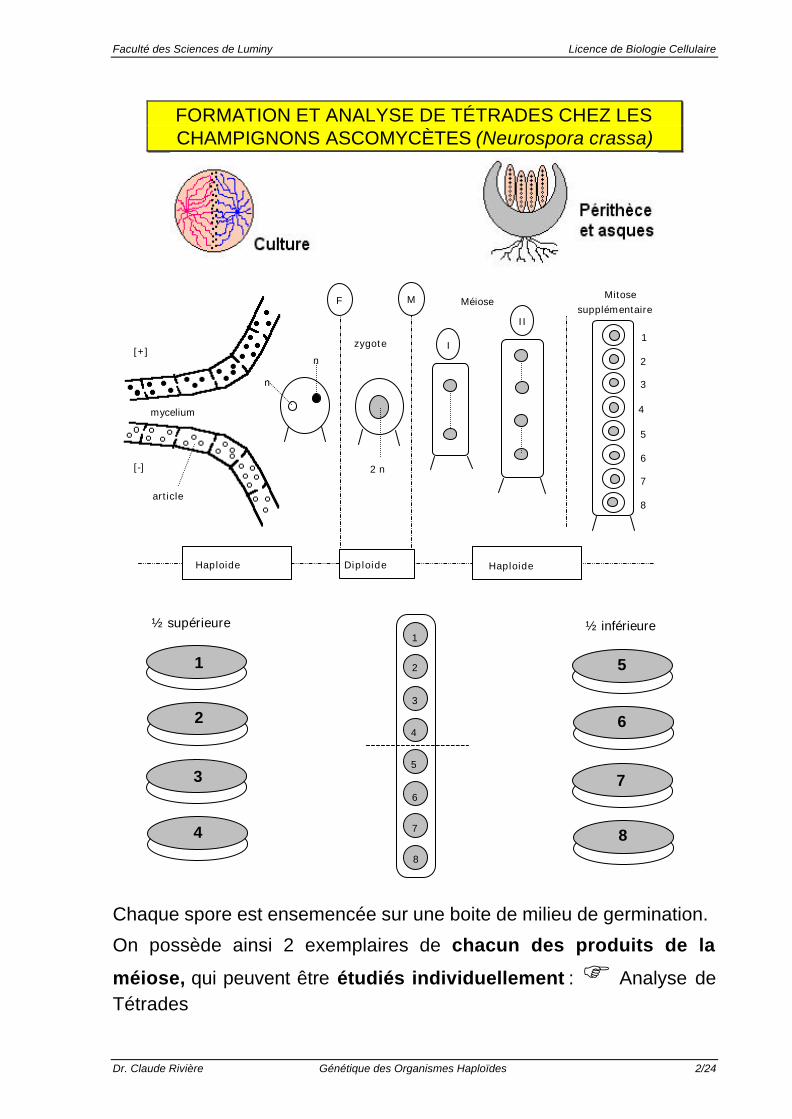
FORMATION ET ANALYSE DE TÉTRADES CHEZ LESCHAMPIGNONS ASCOMYCÈTES (Neurospora crassa)
F M
zygote
2 n
n
n[+]
[-]
Méiose Mitosesupplémentaire
1
2
3
4
5
6
7
8
Diploide Haploide
I
II
Haploide
article
mycelium
1
2
3
4
5
6
7
8
½ supérieure ½ inférieure
1
2
3
4
5
6
7
8
Chaque spore est ensemencée sur une boite de milieu de germination.
On possède ainsi 2 exemplaires de chacun des produits de la
méiose, qui peuvent être étudiés individuellement : F Analyse deTétrades

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 3/24
SÉGRÉGATION D'UN COUPLE DE GÈNES ALLÈLES
I. Rappel organismes diploïdes
• Croisement de deux races pures:
P : AA
× aa
→ F1 : Aa
• Back-Cross de l'hybride F1 par le parent récessif
A
a ×
aa
:
A a
a [A] [a]
50 % 50 %
+ La ségrégation des allèles s'effectue au moment de laformation des gamètes de l'hybride F1, mais ne peut êtreconstatée qu'à travers l'observation des phénotypes de laF2.
+ L’égalité de fréquences entre les deux sortes de gamètes del'hybride F1 est appelée «ségrégation mendélienne».
+ La phase haploïde (= les gamètes) n'est pas observable.
II. Chez les organismes haploïdes
Les gamètes, ce sont les ascospores et les mycelium haploïdes issusde ces dernières. Leurs caractères morphologiques ou biochimiquessont directement observables:
+ La phase haploïde (ascospores + mycelium) est observablegrâce à la technique de l'analyse de tétrades qui permet deconnaître les caractères de chacun des quatre gamètes(tétrade) issus d'une méiose individuelle.
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 4/24
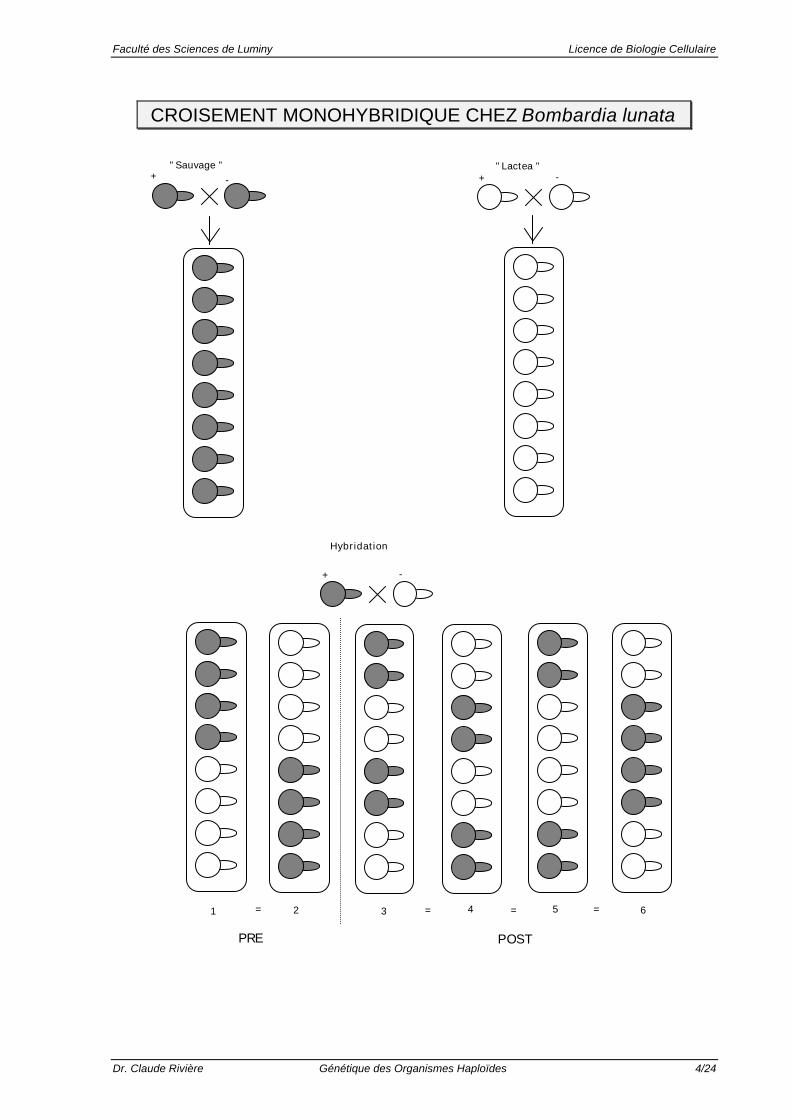
CROISEMENT MONOHYBRIDIQUE CHEZ Bombardia lunata
" Sauvage "+ -
" Lactea "+ -
+ -
Hybridation
PRE POST
= = = =1 2 3 4 5 6

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 5/24
SÉGRÉGATION D'UN COUPLE DE GÈNESCHEZ Bombardia lunata: Principaux résultats
+ Règle de pureté des gamètes : Dans chaque asque, on netrouve que 2 types de spores, identiques soit à l'un, soit àl'autre des parents.
+ Ségrégation mendélienne (4:4) : Dans chaque asque, ontrouve autant de spores d'un type parental que de l'autre (4blanches : 4 noires).
+ 6 types d'asques différents (correspondant à 6répartitions internes différentes des ascospores) composentla descendance :
• Types �� et �� : ségrégation (= séparation) des 2 allèlesà la première division de la méiose: Pré-réduction.
• Types �� , �� , �� et �� : ségrégation des 2 allèles à ladeuxième division de la méiose: Post-réduction. Leurfréquence est, pour le cas étudié, de 66 %.
Notez que ces deux dernières notions font abstraction desévénements chromosomiques à la méiose.
+ Répartition des divers types d'asques:
• Les deux types pré-réduits (�� et �� ) ont mêmefréquence.
• Les quatre types post-réduits (�� , �� , �� et �� ) ontmême fréquence.
• La fréquence de post-réduction est constante pour lecouple de gènes considéré (mais elle varie selon lescouples de gènes).
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 6/24
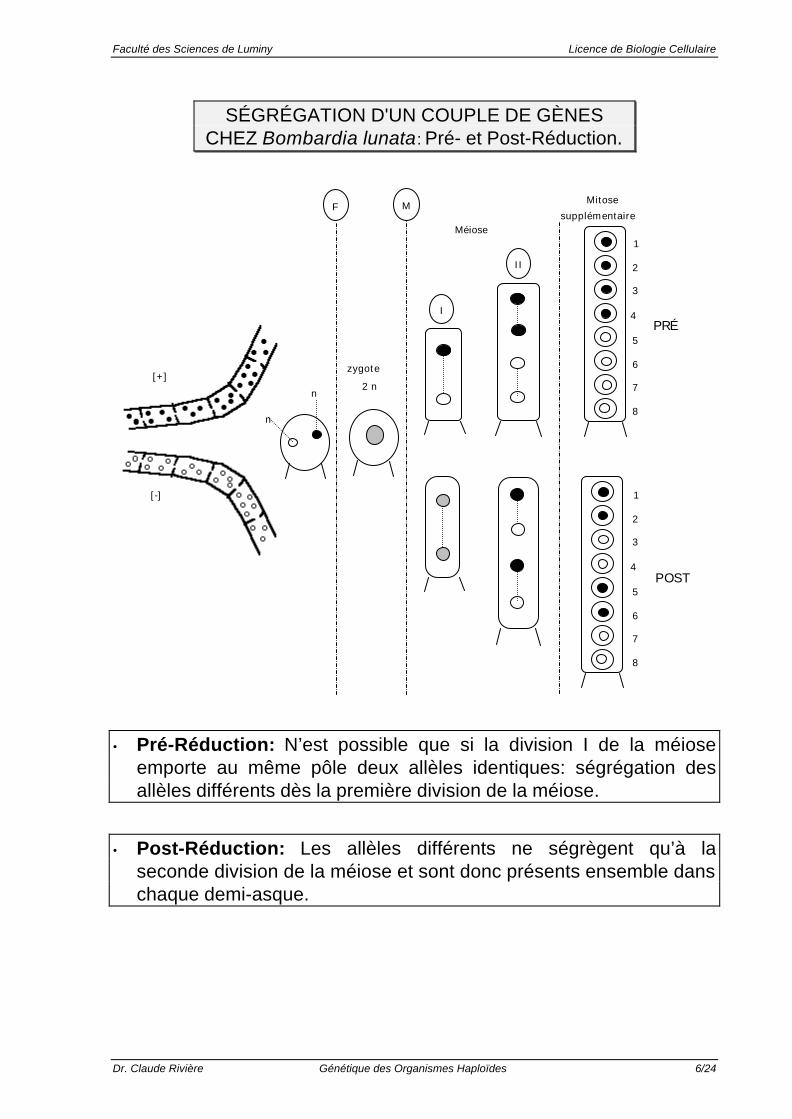
SÉGRÉGATION D'UN COUPLE DE GÈNESCHEZ Bombardia lunata: Pré- et Post-Réduction.
F M
zygote
n
n[+]
[-]
Méiose
Mitosesupplémentaire
I
II
1
2
3
5
6
7
8
4
1
2
3
5
6
7
8
4
PRÉ
POST
2 n
• Pré-Réduction: N’est possible que si la division I de la méioseemporte au même pôle deux allèles identiques: ségrégation desallèles différents dès la première division de la méiose.
• Post-Réduction: Les allèles différents ne ségrègent qu’à laseconde division de la méiose et sont donc présents ensemble danschaque demi-asque.

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 7/24
SÉGRÉGATION D'UN COUPLE DE GÈNESCHEZ Bombardia lunata: Signification des résultats
Règle de pureté des gamètes:
• Vérifiable par les croisements impliquant des races différant par descaractères observables sur la phase haploïde.
• Signifie que les allèles apportés par chaque parent se réunissentprovisoirement dans l'hétérozygote, puis se séparent sans mélangelors de la formation des gamètes, après un doublement (ð 4produits de la méiose = tétrade).
La ségrégation mendélienne (4:4):
• Directement décelable par l'analyse de tétrades (observation desgamètes).
• Montre que les deux souches croisées ne diffèrent que par un seulcouple de gènes allèles.
Post-réductions:
• Détectables sur les tétrades ordonnées linéairement. Ce qui ne veutpas dire qu'elles n'existent pas dans les autres, mais on ne les«voit» pas.
• Leur existence prouve que, après la 1ère division de la méiose etavant la 2ème division de la méiose, il existe un stade où chaquedemi-asque contient les deux allèles différents.
• La réplication s'est donc produite avant l'anaphase I, sans doute aucours de la prophase I.
• Le pourcentage de post-réduction est constant, pour un coupled'allèles donné.
Pré-réductions:
• Détectables sur les tétrades ordonnées linéairement. Ce qui ne veutpas dire qu'elles n'existent pas dans les autres, mais on ne les«voit» pas.
• Leur existence implique que la mitose I emporte au même pôle deuxallèles identiques (chaque demi-asque est homogène).

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 8/24
SÉGRÉGATION DE DEUX COUPLES DE GÈNES INDÉPENDANTSCHEZ Neurospora crassa (Tétrades ordonnées)
Croisement Fluffy + × fluffy –: [F + × f –]
• F/f: mycelium aérien abondant / non-abondant• +/–: types conjugaux compatibles
Résultats:
1. Pour les deux couples de caractères, on constate:
• Dans chaque asque, [4 F : 4 f], [4 + : 4 – ]
+ ségrégation mendélienne 4:4.
• 2 sortes d’asques pré-réduits, 4 sortes d’asques post-réduits.
2. Les F sont soit (+), soit (–), de même les f:
+ aussi bien des associations PAR. (F +, f –) que REC. (F –, f +).
+ Les 2 couples de gènes sont vraiment distincts (≠ pléiotropie).
3. Associations parentales et recombinées ont même fréquence:
+ indépendance.
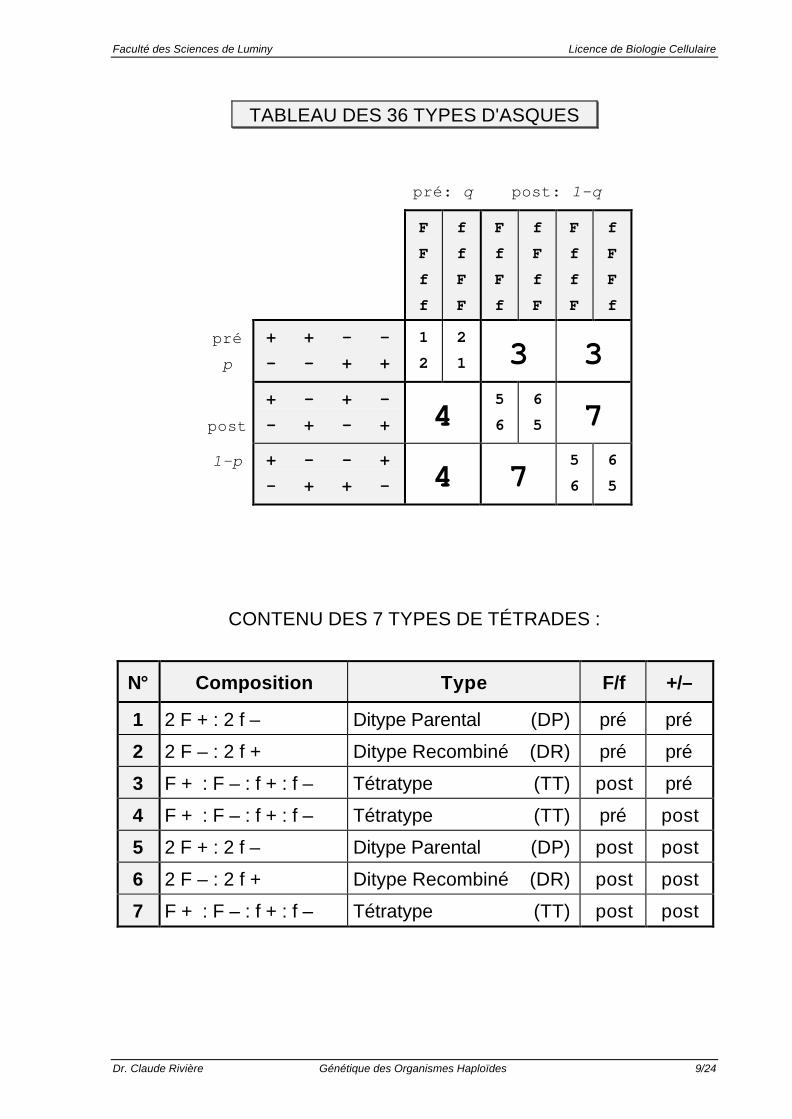
Le Tableau suivant permet de prévoir tous les types d’asques (62 = 36)qui seront obtenus dans la descendance.
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 9/24
TABLEAU DES 36 TYPES D'ASQUES
pré: q post: 1-q
F
F
f
f
f
f
F
F
F
f
F
f
f
F
f
F
F
f
f
F
f
F
F
f
pré
p
+ + - -
- - + +
1
2
2
1 3 3
post
+ - + -
- + - + 45
6
6
5 71-p + - - +
- + + - 4 75
6
6
5
CONTENU DES 7 TYPES DE TÉTRADES :
N° Composition Type F/f +/–
1 2 F + : 2 f – Ditype Parental (DP) pré pré
2 2 F – : 2 f + Ditype Recombiné (DR) pré pré
3 F + : F – : f + : f – Tétratype (TT) post pré
4 F + : F – : f + : f – Tétratype (TT) pré post
5 2 F + : 2 f – Ditype Parental (DP) post post
6 2 F – : 2 f + Ditype Recombiné (DR) post post
7 F + : F – : f + : f – Tétratype (TT) post post

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 10/24
FRÉQUENCES RELATIVESDES DIFFÉRENTS TYPES DE GAMÈTES
Si l’on ne tient pas compte de l’ordre des spores dans les asques (casdes tétrades non-ordonnées), les 7 types de tétrades se réduisent à 3:
• Les DP (� + �) ne contiennent que des gamètes parentaux (4P)• Les DR (� + �) ne contiennent que des gamètes recombinés (4R)• Les TT (� + � + �) contiennent 4 types de gamètes (2 P + 2 R)
On constate d’autre part que:
• Les DP et les DR pré-réduits pour les deux couples de gènes ontmême fréquence (� = �).
• Les DP et les DR post-réduits pour les deux couples de gènes ontmême fréquence (� = �).
Or DP = � + � et DR = � + � , donc DP = DR
+ Nb gamètes PAR. = Nb gamètes REC. → INDÉPENDANCE
N.B. Cette égalité entre les quatre types de gamètes dans le cas del’indépendance de 2 couples de gènes se retrouve chez les diploïdesdans le cas du test-cross d’un dihybride F1 qui produit 4 types degamètes à fréquences égales:
Aa
Bb
× aa bb
Tableau de gamètes:
A B a b A b a B
a b [A B] [a b] [A b] [a B]
¼ ¼ ¼ ¼
P R

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 11/24
CALCUL DES FRÉQUENCESDES TROIS TYPES DE TÉTRADES
Ces fréquences sont fonctions des % de pré- et post-réduction dechacun des deux couples de gènes.
Soit:
q = % pré-réduction de F/f; 1-q = post-réduction de F/fp = % pré-réduction de +/–; 1-p = post-réduction de +/–
La fréquence des différents types est donnée par les équationssuivantes:
DP (� + �) = p × q
2 +
(1-q) (1-p)4
DR (� + �) = p × q
2 +
(1-q) (1-p)4
TT (� + � + �) = p (1-q) + q (1-p) + (1-q) (1-p)
2

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 12/24
SÉGRÉGATION DE DEUX COUPLES DE GÈNES LIÉSCHEZ UN HAPLOÏDE À TÉTRADES NON-ORDONNÉES
Organisme: Chlamydomonas rheinardii (algue verte unicellulaire d’eaudouce). Cycle haploïde similaire à celui des Ascomycètes:
+
-
n
n2n
zygote
multiplicationsvégétatives
méïose
Tétradenon
ordonnée
4 spores haploïdes
Croisements entre mutants métaboliques: arg– pab+ × arg+ pab–
La descendance analysée comprend trois types de tétrades (Σ = 191):
• DP (2 arg+ pab– : 2 arg– pab+) = 119• DR (2 arg– pab– : 2 arg+ pab+) = 1• TT (arg+ pab– : arg– pab+ : arg– pab– : arg+ pab+) = 71
On observe que DP > DR, donc gamètes PAR. > gamètes REC.
+ Ségrégation non-indépendante = «LINKAGE»
N.B. Situation identique au résultat du test-cross d’un double
hétérozygote pour 2 gènes liés × le double récessif (→ excès deparentaux sur les recombinés).
Linkage = existence d’une liaison physique entre 2 couples de gènes,liaison qui se rompt avec une certaine fréquence (toujours < 50%).
Valeur du linkage = pourcentage de recombinaison (pourcentage degamètes recombinés par rapport aux gamètes totaux) :
% REC = DR × 4 + TT × 2
Σ × 4 =
DR + TT / 2Σ
% REC = 1 + 71 / 2
191 = 0.191 = 19.1% ou 19.1 cM
Le pourcentage de recombinaison ou «distance génétique» entre deuxgènes (= deux couples de gènes allèles) est reproductible.
+ La recombinaison entre deux gènes liés est dûe au C.O.

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 13/24
INTERPRÉTATION CHROMOSOMIQUE DESPHÉNOMÈNES DE POST-RÉDUCTION
Il est bien connu que le crossing-over se produit dès le début de laprophase de la première division méiotique, lors de la réplication deschromosomes homologues appariés.
Le crossing-over permet d’interpréter la post-réduction:
A
APas de
Crossing-over
a
a
A
A
a
A
Aa
a
PRÉ
A
ACrossing-over
a
a
A
A
a
A
a
a
POSTa
A
a
+ Post-réduction : dûe à 1 C.O. entre le centromère et le locus.
La fréquence du C.O. sur une certaine distance étant proportionnelle àla longueur de celle-ci, le pourcentage de post-réduction d’un gène serafonction de la distance entre le centromère et le locus de ce gène.
Exemple: 20 % de post-réduction. Pour 100 méioses:
20 méioses post-réduites 2 chromatides PAR × 20 = 40 PAR
2 chromatides REC × 20 = 40 REC
80 méioses pré-réduites 4 chromatides PAR × 80 = 320 PAR
Nombre total de chromatides: 400 dont 40 remaniées, recombinées,soit 10% de recombinaison.
Une post-réduction de 20% est due à la survenue d’un crossing-overdans 10% des cas, c’est-à-dire touchant 10% des chromatides.
+ Distance gène-centromère = ½ pourcentage de post-réduction.
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 14/24
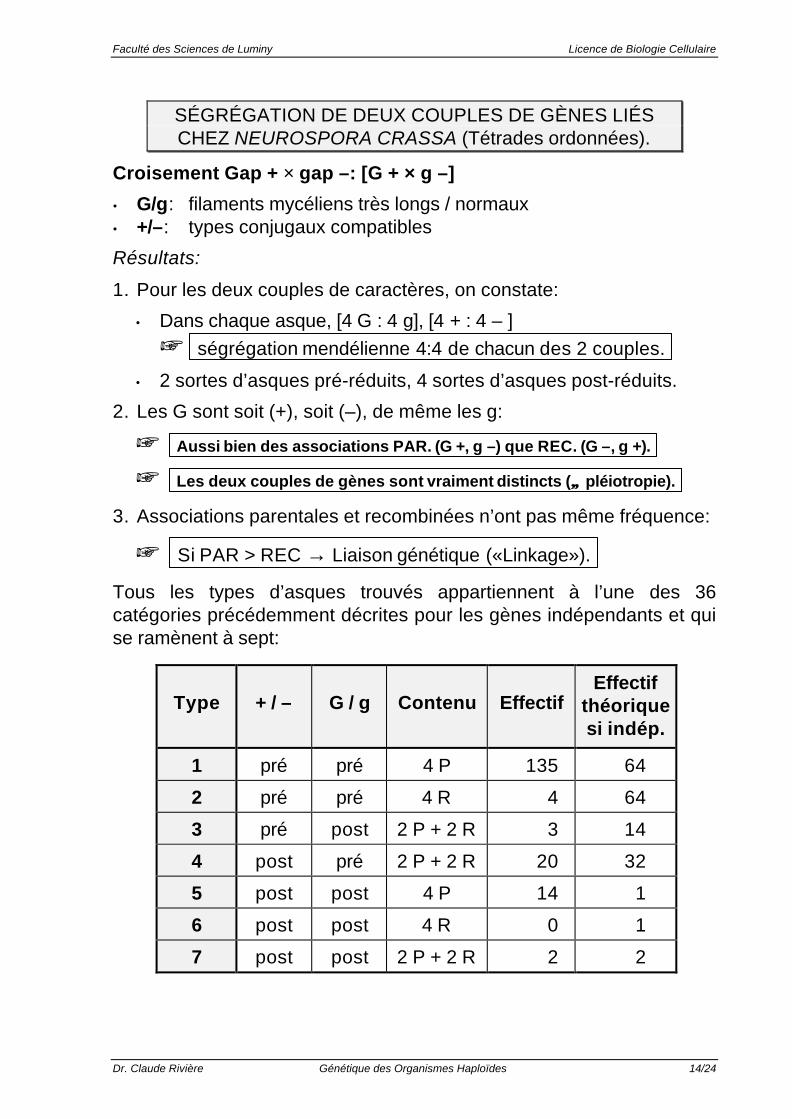
SÉGRÉGATION DE DEUX COUPLES DE GÈNES LIÉSCHEZ NEUROSPORA CRASSA (Tétrades ordonnées).
Croisement Gap + × gap –: [G + × g –]
• G/g: filaments mycéliens très longs / normaux• +/–: types conjugaux compatibles
Résultats:
1. Pour les deux couples de caractères, on constate:
• Dans chaque asque, [4 G : 4 g], [4 + : 4 – ]
+ ségrégation mendélienne 4:4 de chacun des 2 couples.
• 2 sortes d’asques pré-réduits, 4 sortes d’asques post-réduits.
2. Les G sont soit (+), soit (–), de même les g:
+ Aussi bien des associations PAR. (G +, g –) que REC. (G –, g +).
+ Les deux couples de gènes sont vraiment distincts (≠≠ pléiotropie).
3. Associations parentales et recombinées n’ont pas même fréquence:
+ Si PAR > REC → Liaison génétique («Linkage»).
Tous les types d’asques trouvés appartiennent à l’une des 36catégories précédemment décrites pour les gènes indépendants et quise ramènent à sept:
Type + / – G / g Contenu EffectifEffectif
théoriquesi indép.
1 pré pré 4 P 135 64
2 pré pré 4 R 4 64
3 pré post 2 P + 2 R 3 14
4 post pré 2 P + 2 R 20 32
5 post post 4 P 14 1
6 post post 4 R 0 1
7 post post 2 P + 2 R 2 2

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 15/24
On peut calculer - à partir des fréquences de PRÉ et POST de chacundes deux couples de gènes - les effectifs théoriques que donnerait leurhypothétique indépendance.
Soit p la fréquence de PRÉ de (+/–) et q celle de (G/g):
p = 135 + 4 +3
178 ≅ 0.8 et q =
135 + 4 +20178
≅ 0.9
� = � = p × q
2 =
0.8 × 0.92
= 0.36; 0.36 × 178 = 64
� = p (1 - q) = 0.8 × 0.1 = 0.08; 0.08 × 178 = 14
� = q (1 - p) = 0.9 × 0.2 = 0.18; 0.18 × 178 = 32
� = � = (1 - p) (1 - q)
4 =
0.1 × 0.24
= 0.005 0.005 × 178 = 1
� = (1 - p) (1 - q)
2 =
0.1 × 0.22
= 0.01 0.01 × 178 = 2
En multipliant les fréquences obtenues par l’effectif total, on obtient leseffectifs théoriques que donneraient une hypothétique indépendance.
La comparaison des effectifs réels et des effectifs de l’indépendancefait ressortir les points suivants:
• excès important des PAR. � et �;• déficit de REC. � et même absence de �
+ Confirmation: Les deux couples de gènes sont liés.

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 16/24
Valeur de ce linkage. Rappelons que:
% REC = DR × 4 + TT × 2
Σ × 4 =
DR + TT / 2Σ
% REC = (4 × 4) + (3 + 20 +2) 2
178 × 4 =
66712
= 0.0927 = 9.27 cM
Les deux loci sont donc distants de 9.27 cM.
D’autres précisions sur la localisation relative des gènespeuvent être obtenues. Rappelons que:
Distance gène-centromère = ½ pourcentage de post-réduction.
• POST RÉDUCTION couple (+/–) : 20 + 14 +2
178 = 0.202 = 20.2%
• POST RÉDUCTION couple (G/g) : 3 + 14 +2
178 = 0.107 = 10.7%
+ Distances des deux couples de gènes au centromère commun:
• (+/–) : 10.1 cM• (G/g) : 5.35 cM
Récapitulation des résultats obtenus sur une carte génétique:
5.35 9.27G/g +/–
—————u———————l—————————————l———10.1
N.B. La distance la plus grande est inférieure à la somme des deuxautres. Cette observation n’est pas nouvelle et montre que sur ladistance la plus grande, des doubles crossing-over ne sont pasdétectés, si on ne considère pas un marqueur intermédiaire.
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 17/24
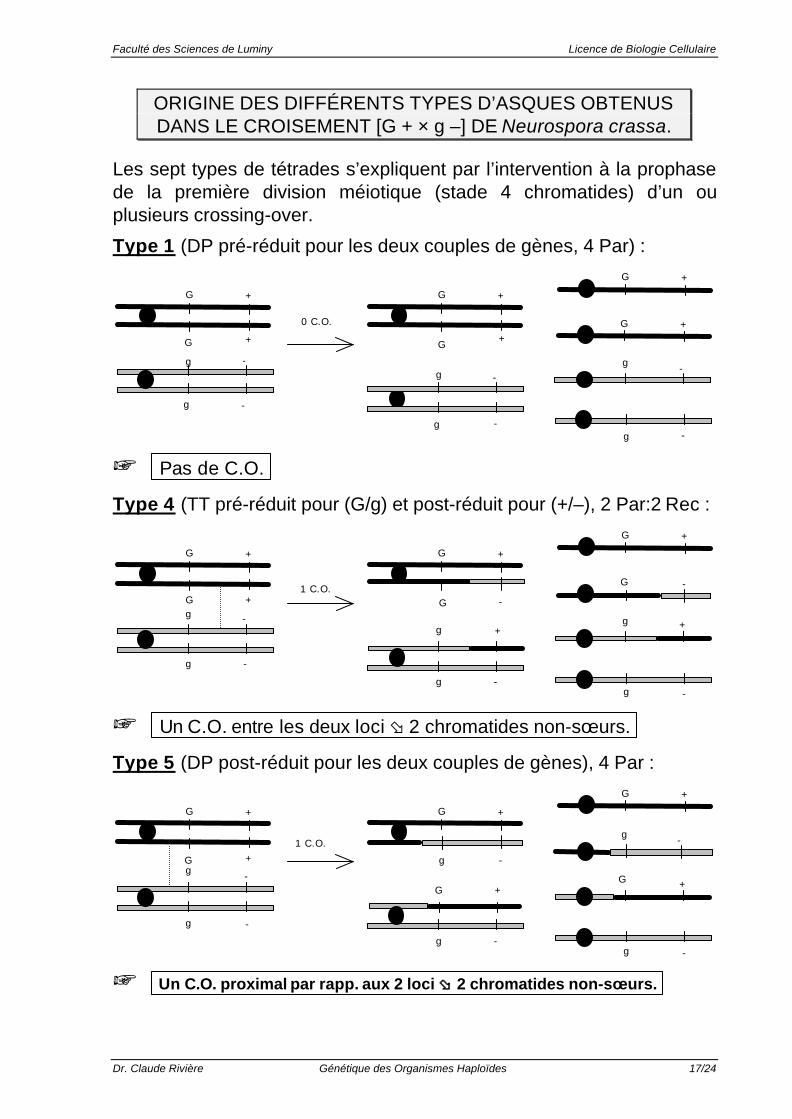
ORIGINE DES DIFFÉRENTS TYPES D’ASQUES OBTENUSDANS LE CROISEMENT [G + × g –] DE Neurospora crassa.
Les sept types de tétrades s’expliquent par l’intervention à la prophasede la première division méiotique (stade 4 chromatides) d’un ouplusieurs crossing-over.
Type 1 (DP pré-réduit pour les deux couples de gènes, 4 Par) :
G + G +G +
G +
g -
g -
g -
g -
g -
g -
G + G +0 C.O.
+ Pas de C.O.
Type 4 (TT pré-réduit pour (G/g) et post-réduit pour (+/–), 2 Par:2 Rec :
G + G +
-
G +
G
g -
g
g -
g -
g -
g
G + G
+
-
+
1 C.O.
+ Un C.O. entre les deux loci ø 2 chromatides non-sœurs.
Type 5 (DP post-réduit pour les deux couples de gènes), 4 Par :
G + G +
-
G +
G
g -
g
g -
g -
g -
gG +
G + +
-1 C.O.
+ Un C.O. proximal par rapp. aux 2 loci øø 2 chromatides non-sœurs.
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 18/24
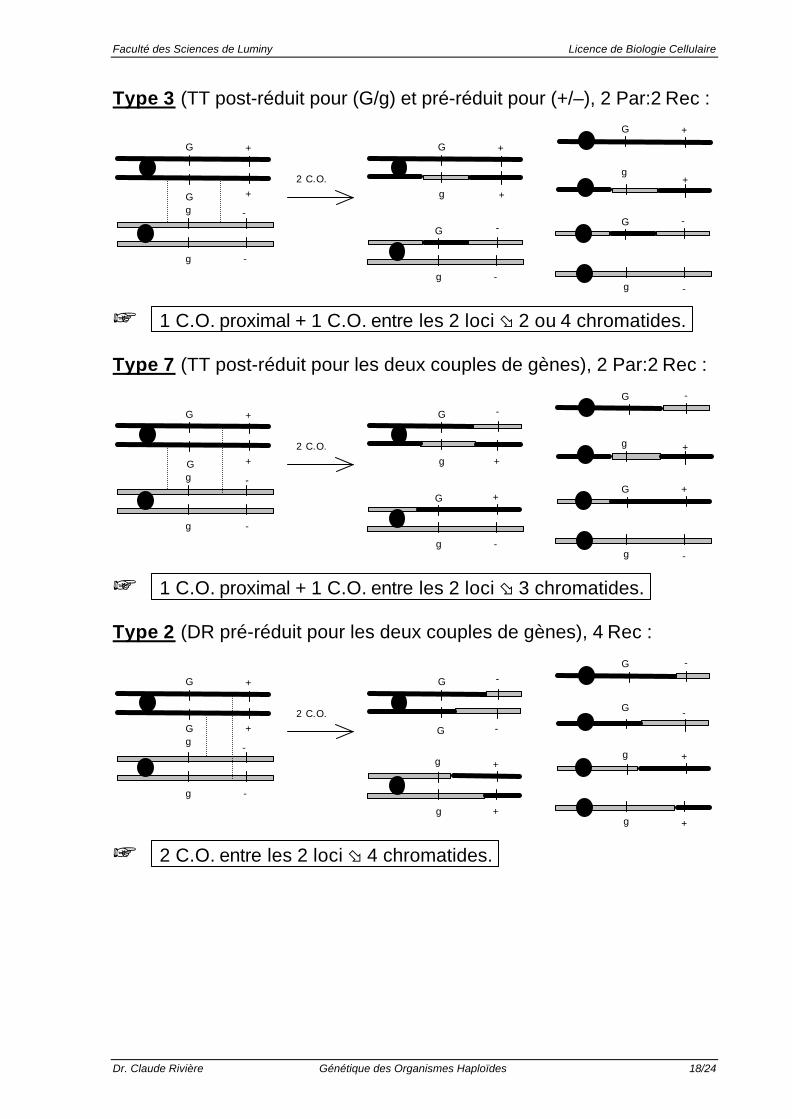
Type 3 (TT post-réduit pour (G/g) et pré-réduit pour (+/–), 2 Par:2 Rec :
G + G +
-
G +
g -
g
g -
g -
g -
G +
G
++
g
G -
2 C.O.
+ 1 C.O. proximal + 1 C.O. entre les 2 loci ø 2 ou 4 chromatides.
Type 7 (TT post-réduit pour les deux couples de gènes), 2 Par:2 Rec :
G + G
+
-G
g -
g
g -
g -
g -
G +
G
++
g
G
-
2 C.O.
+
+ 1 C.O. proximal + 1 C.O. entre les 2 loci ø 3 chromatides.
Type 2 (DR pré-réduit pour les deux couples de gènes), 4 Rec :
G + G
+
-G
g
-
g
g
-
g -
g -
G + G
++
g
G
-
2 C.O.
+
+ 2 C.O. entre les 2 loci ø 4 chromatides.

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 19/24
Type 6 (DR post-réduit pour les deux couples de gènes), 4 Rec :
G + G
+
-G
g
-
g
-g -
g -
G +
G
++
g
G
-
+g3 C.O.
+ 1 C.O. proximal + 2 C.O. entre les 2 loci ø 4 chromatides.
Probabilité d’obtenir une tétrade de cette catégorie dans ladescendance:
P = 0.0535 × 0.09272 = 4.6 × 10-4 ≅ 5 × 10-4
+ Il faudrait observer environ 2 000 tétrades pour en trouver 1 du type6… À condition qu’il n’y ait pas d’interférence…
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 20/24
P R O B L È M E S
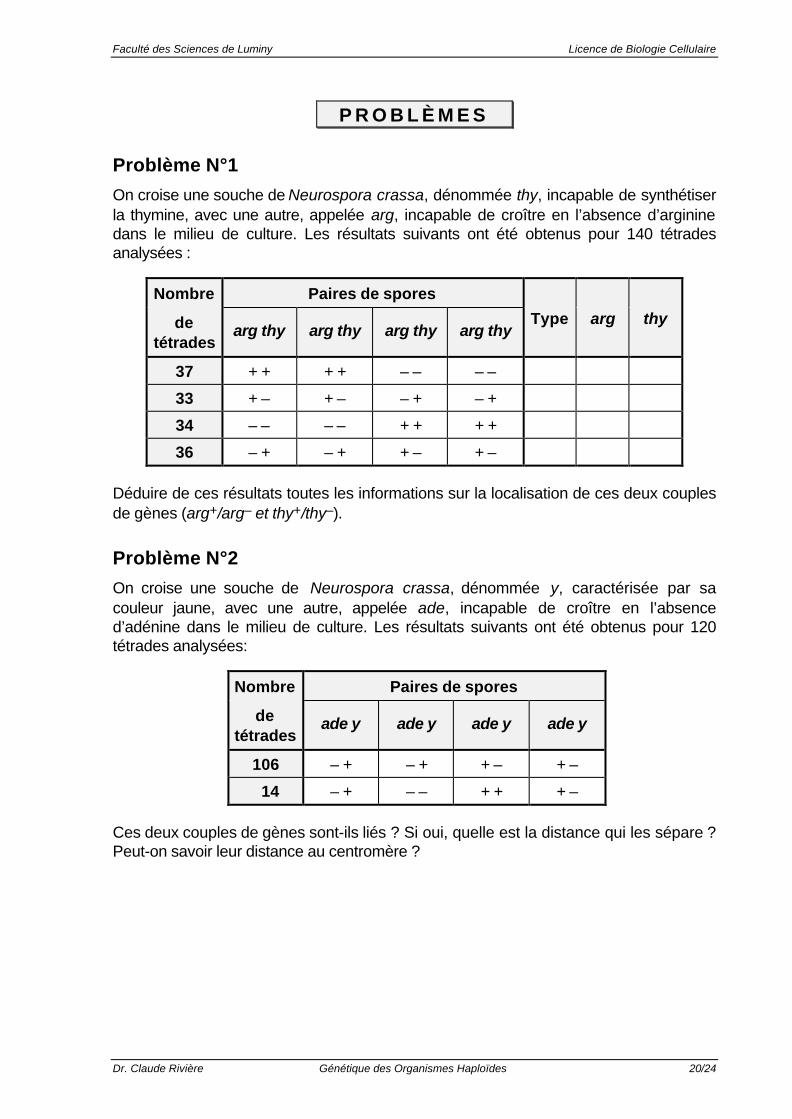
Problème N°1
On croise une souche de Neurospora crassa, dénommée thy, incapable de synthétiserla thymine, avec une autre, appelée arg, incapable de croître en l’absence d’argininedans le milieu de culture. Les résultats suivants ont été obtenus pour 140 tétradesanalysées :
Nombre Paires de spores
detétrades
arg thy arg thy arg thy arg thyType arg thy
37 + + + + – – – –
33 + – + – – + – +
34 – – – – + + + +
36 – + – + + – + –
Déduire de ces résultats toutes les informations sur la localisation de ces deux couplesde gènes (arg+/arg– et thy+/thy–).
Problème N°2
On croise une souche de Neurospora crassa, dénommée y, caractérisée par sacouleur jaune, avec une autre, appelée ade, incapable de croître en l’absenced’adénine dans le milieu de culture. Les résultats suivants ont été obtenus pour 120tétrades analysées:
Nombre Paires de spores
detétrades
ade y ade y ade y ade y
106 – + – + + – + –
14 – + – – + + + –
Ces deux couples de gènes sont-ils liés ? Si oui, quelle est la distance qui les sépare ?Peut-on savoir leur distance au centromère ?
Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 21/24
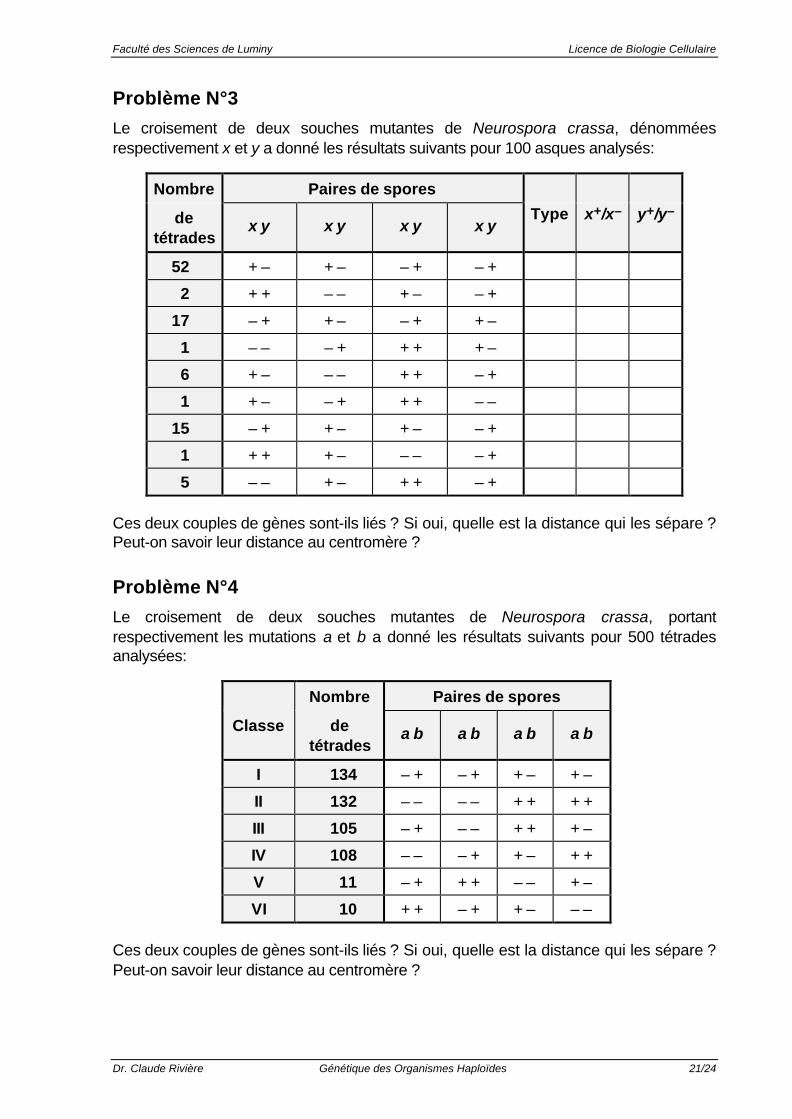
Problème N°3
Le croisement de deux souches mutantes de Neurospora crassa, dénomméesrespectivement x et y a donné les résultats suivants pour 100 asques analysés:
Nombre Paires de spores
detétrades
x y x y x y x yType x+/x– y+/y–
52 + – + – – + – +
2 + + – – + – – +
17 – + + – – + + –
1 – – – + + + + –
6 + – – – + + – +
1 + – – + + + – –
15 – + + – + – – +
1 + + + – – – – +
5 – – + – + + – +
Ces deux couples de gènes sont-ils liés ? Si oui, quelle est la distance qui les sépare ?Peut-on savoir leur distance au centromère ?
Problème N°4
Le croisement de deux souches mutantes de Neurospora crassa, portantrespectivement les mutations a et b a donné les résultats suivants pour 500 tétradesanalysées:
Nombre Paires de spores
Classe detétrades
a b a b a b a b
I 134 – + – + + – + –
II 132 – – – – + + + +
III 105 – + – – + + + –
IV 108 – – – + + – + +
V 11 – + + + – – + –
VI 10 + + – + + – – –
Ces deux couples de gènes sont-ils liés ? Si oui, quelle est la distance qui les sépare ?Peut-on savoir leur distance au centromère ?

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 22/24
Problème N°5
Chez Neurospora crassa le locus du gène a se trouve à 5 cM du centromère sur lechromosome I et celui du gène b à 10 cM du centromère sur le chromosome VII. Onréalise le croisement a– b+ × a+ b–.
1. Quels seront les pourcentages des tétrades DP, DR et TT ?
2. Quel sera le pourcentage de spores recombinantes ?
3. Si les mutations a– et b– représentent des exigences nutritionnelles pour lessubstances A et B, respectivement, quel sera le pourcentage des spores de ladescendance qui pousseront sur milieu minimum (ne contenant ni A, ni B) ?
Problème N°6
Chez Neurospora crassa, on réalise le croisement ade– y– s– × ade+ y+ s+ (ade =adénine dépendance, y = yellow, s = signe sexuel). 1 000 tétrades ont été analysées etclassées en cinq catégories dont les caractéristiques sont données dans le tableausuivant:
Classe Effectif ade y s ade y s ade y s ade y s
I 303 – – – – – – + + + + + +
II 297 – – + – – + + + – + + –
III 97 – – – + – – – + + + + +
IV 105 – – + + – + – + – + + –
V 102 – – – – + – + – + + + +
VI 96 – – + – + + + – – + + –
Déterminer les relations qualitatives et - éventuellement - quantitatives entre les troiscouples de gènes et leur localisation respective par rapport au(x) centromère(s).
Problème N°7
Chez l’algue unicellulaire à tétrades non-ordonnées Chlamydomonas rheinardii, on aanalysé 100 tétrades issues du croisement a– b– c– × a+ b+ c+ et on les a classées enquatre catégories:
a b c a b c a b c a b c
40 – – – – – – + + + + + +
42 – – + – – + + + – + + –
10 – + – + + – – – + + – +
8 – + + + + + – – – + – –
Quelles informations pouvez-vous tirer de ces résultats quant aux relations génétiquesentre ces trois couples de gènes et à leur localisation chromosomique ?

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 23/24
Problème N°8
Le croisement de deux souches de Neurospora crassa, l’une porteuse d’une mutationde dépendance à la méthionine (met—) et de type sexuel s—, l’autre porteuse d’unemutation de dépendance à l’adénine (ade—) et de type sexuel s+ a donné les résultatssuivants :
Classe Effectif met ade s met ade s met ade s met ade s
I 46 – + – – + – + – + + – +
II 6 – + – + + – – – + + – +
III 4 – + – – + + + – – + – +
IV 2 – + – + – – – + + + – +
V 2 – + – + + + – – – + – +
VI 1 – + – + + + – – + + – –
1. Tirer de ces résultats le maximum d’informations concernant la localisationrespective des loci des gènes met, ade et s.
2. Reconstituer sur des schémas les événements méiotiques ayant abouti auxdifférentes sortes de tétrades :
Problème N°9
Les loci de quatre mutations his-dépendantes obtenues indépendamment ont étélocalisés respectivement sur quatre chromosomes différents de Neurospora:
• his -1 à 1 cM de ad-3 et du centromère;• his -2 à 1 cM du centromère;• his -3 à 10 cM du centromère;• his -4 à 30 cM du centromère;
On dispose de quatre souches portant chacune l’une de ces mutations.
On isole une nouvelle souche his– qui porte par ailleurs la mutation ad-3 très liée aucentromère. On désire savoir si cette mutation est identique à l’une des quatre déjàconnues. À cette fin, on la croise par une souche sauvage ne portant aucune mutationad ou his. Les 50 tétrades analysées, sans tenir compte de l’ordre des spores, serépartissent en 10 D.P., 10 D.R. et 30 T.T.
1. Déduire de ces résultats laquelle des quatre mutations his est le plus probablementportée par cette nouvelle souche.
2. Quelle expérience complémentaire devrait-on faire pour confirmer l’hypothèseprécédemment avancée ?

Faculté des Sciences de Luminy Licence de Biologie Cellulaire
Dr. Claude Rivière Génétique des Organismes Haploïdes 24/24
Problème N° 10
Chez l’Ascomycète Sordaria fimicola, dont le cycle de reproduction est identique àcelui de Neurospora, les spores des souches sauvages sont noires. Deux mutants depigmentation des spores (résultant chacun de la mutation d’un seul gène) ont été isolésde manière indépendante :
• l’un (I) à spores incolores ou «blanches»• l’autre (II) à spores marron.
Trois croisements : a, b et c, ont été réalisés entre souches de types sexuelscompatibles. Les résultats sont regroupés dans le tableau suivant, dans lequel il n’a pasété tenu compte de l’ordre des spores :
1 2 3 4
a I × sauv. noir noir blanc blanc 100 %
b II × sauv. noir noir marron marron 100 %
marron marron blanc blanc 357
c I × II noir noir blanc blanc 3
blanc blanc noir marron 213
Interprétez les résultats de ce croisement et déterminez les relations existant entre lesdeux gènes impliqués.