les gehringiidae : famille relictuelle de coléoptères...
TRANSCRIPT

L ’espèce Gehringia olympica Darlington (1933),découverte en 1927, est l’un des Coléoptères
Adephaga les plus surprenants et parmi les plus problé-matiques qui soient. En effet, ses caractéristiques morpho-logiques l’éloignaient jusqu’à présent de toutes les autresespèces connues et elle avait été placée dès sa découvertedans une «nouvelle tribu de Carabidae», les GehringiiniDarlington (1933), ce qui revient à une sous-famille ouà une famille à part entière, selon le rang utilisé aujour-d’hui pour les principales subdivisions des Caraboidea.
Gehringia olympica a d’abord été trouvée dans leMontana et dans l’Etat de Washington, puis dans d’autresstations du nord des Montagnes Rocheuses, toujours àhaute altitude, dans l’Oregon, la Colombie Britannique,les confins des Territoires du Nord-Ouest, etc. CeCarabique est au premier abord remarquable par sa trèspetite taille – 1,7 à 1,8 mm –, ce qui est exceptionnelpour une espèce épigée d’Adephaga. Des caractères anato-miques particuliers l’isolent par ailleurs au sein desAdephaga terrestres. De plus, cet insecte se déplaceraittrès lentement, à la façon d’un Clavicorne plutôt qued’un Carabique, rendant difficile la recherche et la recon-naissance des imagos sur le terrain (Lindroth 1960).
Résumé. Des descriptions morphologiques détaillées sont données pour deux espèces réunies dans lafamille des Gehringiidae (Coleoptera, Adephaga) : Gehringia olympica Darlington, 1933, des MontagnesRocheuses septentrionales, et Helenaea torretassoi Schatzmayr & Koch, 1934, du désert du Sinaï (cetteseconde espèce identifiée comme Gehringiidae par R. Sciaky en 1997). Plusieurs autapomorphies condui-sent à réunir ces deux taxons. Néanmoins, la position phylogénétique des Gehringiidae au sein desAdephaga demeure problématique. Plusieurs hypothèses sont discutées qui rapprochent cette famillerespectivement des Trachypachidae, des Trechidae, des Psydridae et des Melaenidae. Finalement, denombreux caractères sous un état plésiomorphe donnent à cette famille une position très basale dans laphylogénie des Coléoptères Adephaga (en particulier le large processus intermétacoxal des ventritesabdominaux), mais de bons arguments isolent les Gehringiidae, notamment l’abdomen de type nébridienavec les glandes défensives associées au tergite VIII. Cette dernière association d’états de caractèrespourrait rapprocher les Gehringiidae du genre Cymbionotum Baudi, 1864 (Melaenidae), en accord avecde récents résultats d’analyses moléculaires, mais ce serait à l’encontre de la quasi-totalité des autrescaractères morphologiques.
Abstract. Gehringiidae: a relictual family of Coleoptera Adephaga, alticolous in the Rocky Mountainsand deserticolous in Sinai. Detailed morphological descriptions are given for two species assembledwithin the Gehringiidae family (Coleoptera, Adephaga): Gehringia olympica Darlington, 1933, from thenorthern Rocky Mountains, and Helenaea torretassoi Schatzmayr & Koch, 1934, from the Sinai Desert(this last species identified as belonging to the Gehringiidae by R. Sciaky in 1997). Several autapomor-phies lead to assemble both taxa together. Nevertheless, the phylogenetic placement of the Gehringiidaewithin the Adephaga remains problematic. Several hypotheses are discussed, which bring this familycloser to the Trachypachidae, the Trechidae or the Psydridae respectively. Finally, many plesiotypic statesof characters give for this family a very basal position in the phylogeny of adephagan Coleoptera (in partic-ular the wide intermetacoxal process of abdominal ventrites), but some good arguments isolate theGehringiidae, in particular the nebridian-type abdomen with defensive glands linked to tergite VIII. Thislast association of characters states could put the Gehringiidae closer to the genus Cymbionotum Baudi,1864 (Melaenidae), in accordance with some recent molecular analyses results, but this would be incontradiction with almost all other morphological data.
Les Gehringiidae : famille relictuelle de Coléoptères Adephaga, alticole dans les Montagnes Rocheuses et déserticole dans le Sinaï
Thierry DEUVE
Muséum National d’Histoire NaturelleDépartement de Systématique & Évolution, USM 601
Entomologie, Case Postale 50F-75231 Paris cedex 05, France
Ann. Soc. entomol. Fr. (n.s.), 2005, 41 (1) : 69-80. ARTICLE
69
E-mail : [email protected]é le 22/04/2005

Si la singularité de Gehringia olympica fait encore l’ob-jet d’interrogations sur sa position phylogénétique, sonhabitat et ses caractéristiques morphologiques semblentindiquer qu’il s’agit d’un témoin relictuel de peuplementstrès anciens, probablement du Mésozoïque, aujourd’huidisparus. L’intérêt de cette espèce est donc tout à faitexceptionnel et n’a pas échappé aux auteurs qui ont abordéla question (Darlington 1933; Jeannel 1941, 1942, 1946;Lindroth 1960 ; Ball 1960 ; Bell 1964, 1966 ; Hammond1979 ; Maddison 1985 ; Deuve, 1993 ; Liebherr & Will1998 ; Ball & Bousquet 2001). C’est d’ailleurs un dessinde cette espèce par David R. Maddison qui a été choisipour illustrer la couverture et le frontispice de l’ouvragequi, à sa mort, a été dédié en hommage à la mémoire deP. J. Darlington (Ball 1985). D’après Lindroth (1960),la découverte de Gehringia olympica en 1927 constitue-rait l’un des plus grands événements de l’histoire de lataxonomie des Carabiques.
Mais tout aussi étonnante est sans doute la décou-verte toute récente effectuée par R. Sciaky (1997, inlitteris), qui a étudié les collections du Muséumd’Histoire naturelle de Milan. Il a pu en effet constaterque l’espèce Helenaea torretassoi Schatzmayr et Koch(1934), décrite sur un spécimen unique comme unTrechidae Bembidiinae proche du genre Limnastis, étaiten réalité lui aussi un Gehringiidae, partageant d’im-portants caractères avec Gehringia olympica. Il s’agit aussid’une espèce de taille exceptionnellement petite– 1,1 mm ! –, surprenante surtout par sa localisationgéographique et son habitat : Helenaea torretassoi a étédécouvert en 1933 près d’une source sulfureuse enÉgypte, près d’El Tor, à basse altitude dans le Sinaï.
Ainsi donc, les Gehringiidae seraient connus par seule-ment deux espèces minuscules découvertes respective-ment en 1927 et en 1933 : l’une vivant en altitude dansle nord des Montagnes Rocheuses, la seconde au borddu golfe de Suez près d’une source dans un désert chaud.
L’objet de cette note est de donner des descriptionsanatomiques complémentaires de ces deux espèces, ensoulignant leurs évidentes affinités, mais aussi de discu-ter leur position phylogénétique au sein des ColéptèresAdephaga.
Descriptions des espèces
Gehringia olympica Darlington (fig. 1)
Gehringia olympica : DARLINGTON 1933 : 111.
Série typique. HOLOTYPE : 1 femelle, États-Unisd’Amérique, État de Washington, Monts Olympic, prèsdes sources chaudes de Sol Duc (P. J. Darlington, 5 août
1927), in coll. Museum of Comparative Zoology,Cambridge (Massachusetts). Allotype : 1 mâle de lamême provenance. Paratypes : 6 exemplaires, mâles etfemelles, de la même provenance, et 6 autres exem-plaires, mâles et femelles, États-Unis, État du Montana,Glacier Park (E. Mank, 30 juin et 8 juillet 1927).
Espèce très petite (longueur : 1,7-1,8 mm), ailée. Coloris entiè-rement noir luisant, les appendices et les élytres avec une teintede fond brun sombre, le dernier article des palpes testacé pâle.
Tête moyenne, assez grosse relativement au pronotum,prognathe, la capsule céphalique aplanie dorso-ventralement.Vertex modérément convexe, lisse, faiblement marqué par unemicrosculpture composée de mailles transversales peu visibles.Front avec des fossettes longitudinales peu profondes et un réseaude mailles isodiamétriques assez fortes qui s’étendent aussi surle clypéus. Le front et le vertex pourvus d’une seule paire de soiesfrontales (postérieure), mais aussi d’une douzaine de soies éparsesplus petites. Clypéus trapézoïdal, bordé par quatre soies anté-rieures, le sillon épistomien peu visible (fig. 13). Genae avec une
70
T. Deuve
Figures 1-3Famille des Gehringiidae, morphologie générale et habitat. 1, Gehringiaolympica Darlington, de Linham Creek, face dorsale. 2, Helenaea torretassoiSchatzmayr et Koch, holotype, idem. 3, Distribution géographique deGehringia (a) et d’Helenaea (b).

pubescence éparse. Yeux plutôt grands mais modérément saillants,toutefois bien plus convexes que les tempes, faiblement pubes-cents, composés d’ommatidies peu nombreuses mais grandes,chacune individuellement convexe, donnant à l’oeil son aspectgranuleux. Antennes courtes, atteignant en arrière la base dupronotum, tous les articles pubescents, les quatre premiers fili-formes, les suivants moniliformes. Labre en languette libre hexa-chète, le bord antérieur faiblement incurvé (fig. 13). Piècesbuccales courtes (fig. 4, 14). Mandibules petites, falciformes, lescrobe unisétulé, le rétinacle en protubérance molaire non bifide,la dent prémolaire bien individualisée, saillante. Palpes maxil-laires et labiaux caractérisés par l’avant-dernier article dilaté etplus ou moins pubescent, tandis que le dernier article est aucontraire réduit en courte baguette étroite et achète. Le cardoglabre, le stipe avec trois soies, dont l’une sur le palpigère.Mentum libre, bien séparé du submentum par un sillon, la dentmédiane deux fois plus courte que les lobes latéraux, son sommetfaiblement bifide, sa base avec une paire de soies principales,tandis qu’une pubescence de quelques soies secondaires est visibleprès de la marge postérieure. Submentum partiellement fusionnéaux genae, avec une pubescence éparse et une paire de soies prin-cipales. Gula large, dilatée vers l’arrière.
Pronotum assez petit, 1,20 fois plus large que long, la plusgrande largeur vers le tiers antérieur, puis rétréci en arrière, lescôtés distinctement sinués avant les angles postérieurs qui sontdroits et vifs, un peu dirigés vers l’extérieur. Le bord basal faible-ment saillant. Disque modérément convexe, avec une pubes-cence éparse, le sillon médian très fin, peu visible, les margeslatérales très finement mais très régulièrement rebordées, sansgouttière ni explanation. Base marquée par des fossettes latéralespetites et arrondies, et par des ridulations longitudinales plusmédianes. Deux paires de soies marginales, médiane et basale.
Prosternum avec une faible pubescence éparse (fig. 15).Propleures lisses et glabres, la suture notopleurale bien distincte.Cavités procoxales ouvertes : le lobe propleural n’est pas raccordéà l’apophyse prosternale (fig. 6, 15).
Mésothorax glabre, relativement petit, rétréci vers l’avant(fig. 5, 6, 16), de type Disjuncta (Sloane 1923) : le mésépimé-rite atteint la cavité mésocoxale (fig. 17). Mésépisterne et mésos-ternum fusionnés. Elytres jointifs, plus longs que larges, mais àextrémité apicale tronquée, la base nullement rebordée, l’épi-pleure amenuisé vers l’arrière jusqu’à la troncature, marqué surtoute sa longueur par une strie longitudinale continue (fig. 1).Épaules étroites mais accusées, l’angle huméral aigu, le repliépipleural très distinct, avec une très étroite gouttière marginale.Disque peu convexe, lisse, seule la 1re strie profondémentmarquée, parallèle à la suture, imponctuée, continue depuis labase jusqu’à l’apex. Les 2e et 3e stries à peine perceptibles souscertaines incidences d’observation. De nombreux points séti-gères régulièrement espacés, alignés en séries parallèles situéessur les emplacements présumés des interstries. Chétotaxie margi-nale composée d’un groupe huméral de quatre soies équidis-tantes et d’une soie isolée dans la moitié postérieure de la gout-tière. Aussi une soie latéro-discale préapicale plus développéeque la pubescence ordinaire.
Métathorax assez volumineux, le métasternum de grandesdimensions (fig. 5, 7, 16), sa surface avec une pubescence éparse,la saillie postérieure intermétacoxale particulièrement large, en
rapport avec l’écartement des hanches, à extrémité bilobée, lesillon transversal paramétacoxal court et superficiel, pus oumoins distinct, pas de sillon longitudinal médian antérieur.Métépisterne grand, subrectangulaire, le métépimérite étroitdans sa partie postérieure, avec un lobe réduit qui recouvre àpeine la marge du ventrite abdominal (fig. 17). Ailes métatho-raciques membraneuses, développées, la nervation cependantrudimentaire, la bordure frangée de soies fines (cf. illustrationin Darlington 1933).
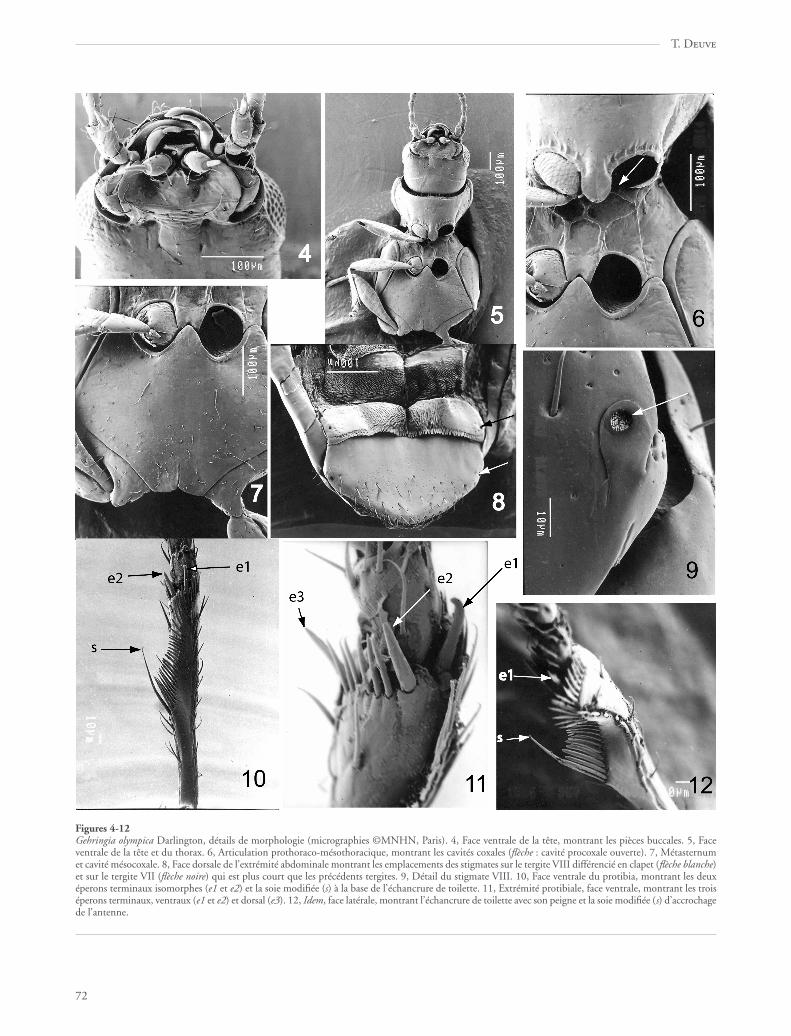
Pattes courtes mais très fines, pubescentes (fig. 5). Les coxaeproportionnellement petites par rapport au corps de l’insecte, sil’on compare avec les autres Caraboidea. Procoxae globulaires,nullement subconiques ni saillantes. Protibias échancrés en unorgane de toilette préapical (fig. 12), l’une des soies du peigne,située au talon de l’échancrure, plus longue que les autres, formantsystème d’accrochage de l’antenne (fig. 10, 12, 18, « s »). Troiséperons (ou soies modifiées ?) à l’extrémité apicale du protibia :deux ventraux (type isochète) et un dorsal (fig. 10, 11, 18, « e1,e2, e3 »). Les deux premiers articles du protarse un peu épaissischez le mâle, avec des phanères adhésifs (fig. 18).
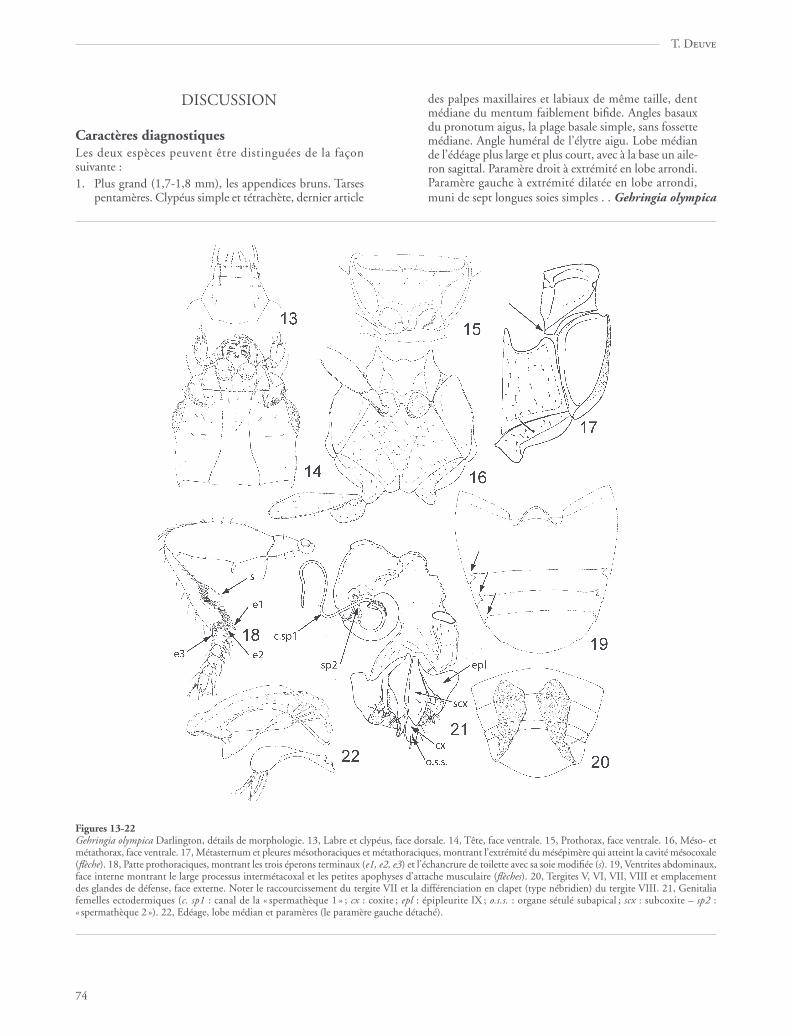
Abdomen typique des Adephaga (Jeannel & Paulian, 1944),mais les ventrites II, III et IV non pas soudés mais fusionnés(fig. 19), les ventrites V, VI et VII libres, le VIIe faiblement dilaté,« portant » le postabdomen qui est télescopé. Tous les ventriteslisses, mais avec une pubescence éparse. Pas d’épipleurites distinctssur les segments II à VII comme il s’en observe chez lesTrachypachidae (Deuve 1994), les zones péritrémales bienmembraneuses entre les tergites et les ventrites. Cavités méta-coxales très petites, les ventrites II et III formant un « triangleintermétacoxal» particulièrement large, bien développé, en rapportavec l’écartement des hanches (fig. 19). En vue interne, les extré-mités latérales de la marge antérieure, costale, des ventrites V, VIet VII sont caractérisées par la différenciation d’une petite apophyseen épine aiguë (fig. 19, flèches). Abdomen proche du type nébri-dien (Deuve, 1988a, 1993), c’est-à-dire le tergite VIII développéen un fort clapet non télescopable, sans apophyses d’attachemusculaire, qui représente le dernier segment apparent visible envue dorsale lorsque le postabdomen est rétracté (fig. 8, 20).Cependant, ce tergite est de forme non pas semi-discoïdale maisplutôt pentagonale et les glandes défensives «pygidiales » débou-chent contre le tergite VIII et non pas au niveau du IXe segment.Le tergite VII est curieusement plus court que les tergites précé-dents. Glandes défensives avec le canal collecteur débouchant versle sommet du canal efférent, celui-ci large et membraneux (fig. 20).
Genitalia femelles (fig. 21) épitopiques, à gonopore secon-daire ouvert en apparence en arrière du segment IX. EpipleuritesVIII (= « latérotergites » sensu Deuve 1993, cf. Deuve 1994)contigus. Epipleurites IX remarquables par la présence sur chacund’eux d’un prolongement appendiculiforme, à extrémité pluri-sétulée, jouxtant les coxostyles. Gonopodes IX dimères.Gonosubcoxite allongé, trois fois plus long que large, avecquelques soies éparses sur sa face externe. Gonocoxite en ongletà peine plus long que large, avec un organe bisétulé subapical.Ni gonopodes VIII ni ventrite X visibles. Oviducte ventral, coudéà sa base. Vagin dilaté en poche assez volumineuse, aminci versson extrémité antérieure et prolongé par une formation bulbeuseallongée et courbée en demi-cercle. Conduit de la spermathèquelong, fin et tubulaire. Cette description des genitalia femelles
71
Les Coléoptères Gehringiidae
72
T. Deuve
Figures 4-12Gehringia olympica Darlington, détails de morphologie (micrographies ©MNHN, Paris). 4, Face ventrale de la tête, montrant les pièces buccales. 5, Faceventrale de la tête et du thorax. 6, Articulation prothoraco-mésothoracique, montrant les cavités coxales (flèche : cavité procoxale ouverte). 7, Métasternumet cavité mésocoxale. 8, Face dorsale de l’extrémité abdominale montrant les emplacements des stigmates sur le tergite VIII différencié en clapet (flèche blanche)et sur le tergite VII (flèche noire) qui est plus court que les précédents tergites. 9, Détail du stigmate VIII. 10, Face ventrale du protibia, montrant les deuxéperons terminaux isomorphes (e1 et e2) et la soie modifiée (s) à la base de l’échancrure de toilette. 11, Extrémité protibiale, face ventrale, montrant les troiséperons terminaux, ventraux (e1 et e2) et dorsal (e3). 12, Idem, face latérale, montrant l’échancrure de toilette avec son peigne et la soie modifiée (s) d’accrochagede l’antenne.

internes (Deuve 1993) a été complétée par Liebherr & Will(1998) qui ont indiqué et illustré la présence d’une « deuxièmespermathèque ». Une formation tubuliforme petite et entortilléesur elle-même est effectivement visible à l’extrémité antérieurede la bourse copulatrice (fig. 21, sp2). En revanche, à l’opposéde ces auteurs je n’observe pas de sclérite helminthoïde maissimplement un épaississement pariétal de la formation bulbeuse.
Genitalia mâles (fig. 22) avec l’édéage couché à plat (rota-tion anatomique de 90°) en position intra-abdominale. «Anneaugénital » (formation composée des aires épipleurales IX gaucheet droite raccordées, cf. Deuve 1988b et 1994) dissymétrique,ne formant pas spiculum gastrale vers l’avant, seule la régioncostale est sclérifiée, les épipleurites proprement dits étant l’unet l’autre non visibles. Lobe médian en forme de tube incurvé,sans bulbe basal mais muni d’un robuste mais petit aileron sagit-tal. Paramères dissemblables, le droit au sommet un peu plusdilaté, arrondi, les deux munis de six ou sept soies apicales.Endophallus sans pièces copulatrices sclérifiées.
Habitat – L’espèce est connue des MontagnesRocheuses, en altitude depuis l’Oregon et le Montana,jusqu’à la Colombie Britannique, l’Alberta, et lesTerritoires du Nord-Ouest (Ball & Bousquet 2001)(fig. 3). Elle vit dans de très fins gravillons ou dans dessables grossiers, le long des torrents froids à eau claire(Lindroth 1960).
Helenaea torretassoi Schatzmayr & Koch 1934(fig. 2)
Helenaëa Torre-Tassoi : SCHATZMAYR & KOCH 1934 : 25.
Série typique. Holotype : 1 mâle, Égypte, Sinaï, sourcessulfureuses d’El Tor (25 février 1933), in coll. Muséumd’Histoire Naturelle, Milan.
Espèce minuscule (longueur : 1,1 mm) et ailée. Noir de poix, lelabre, les mandibules, les palpes, les antennes et les pattes jaunâtretestacé.
Tête volumineuse, large, avec de grands yeux convexes àfacettes. Front et vertex lisses et modérément convexes, avec troispaires de soies frontales à peu près alignées en agencement longi-tudinal et quelques soies plus petites en position supra-orbitaire.Sillon clypéo-frontal indistinct, le clypéus mal délimité en arrière,son emplacement marqué par deux rehaussement costulaires longi-tudinaux qui font saillie en avant au-dessus des marges latéralesdu labre (fig. 24). Labre en languette mobile à marge antérieurefaiblement incurvée, porteur de huit soies : six soies principalesdont les deux paires externes sont les plus longues, et une pairesecondaire de soies plus courtes situées au milieu (fig. 24).Mandibules courtes et robustes, sans soie scrobiculaire. Palpesspécialisés (fig. 23), le dernier article réduit en petite tigette presqueaussi longue que l’article précédent au palpe labial, mais bien pluscourte au palpe maxillaire. Le pénultième article des palpes maxil-laires épaissi et faiblement pubescent. Mentum libre, nullementfusionné au submentum, avec quatre soies principales mais aussiune dizaine de soies secondaires plus petites ; la dent médiane trèslarge et arrondie, plus de deux fois plus courte que les lobes laté-
raux. Submentum petit, avec seulement quelques soies de dispo-sition irrégulière ; la gula très large. Plusieurs soies génales.Antennes moniliformes, les articles moyens et distaux subsphé-riques, le dernier article plus long et en forme d’ogive ; le 2e articleplus court que le scape mais nettement plus long que le 3e.
Pronotum peu transverse, 1,18 fois plus large que long, laplus grande largeur au quart ou au cinquième antérieur, puisrétréci en arrière, les côtés peu sinués avant les angles postérieurs,qui sont obtus. Disque convexe, lisse, avec quelques courtes soiesdisséminées, le sillon médian distinct, fin, interrompu en arrièrepar une petite mais très profonde cavité oblongue qui cependantn’atteint pas le bord basal ; les fossettes postérieures petites maistrès bien délimitées et profondes. Ni gouttières ni rebords laté-raux bien marqués. Base régulièrement curviligne, un peusaillante. De chaque côté, une soie à l’angle antérieur, une oudeux soies marginales médianes, et une soie à l’angle basal.
Suture notopleurale bien distincte. Apophyse prosternale enlobe arrondi. Les cavités procoxales ouvertes.
Mésothorax relativement petit, rétréci vers l’avant, de typeDisjuncta. Elytres un peu allongés, d’allure générale fusiformemais franchement tronqués à l’apex, la plus grande largeur versle milieu, les épaules anguleuses et même un peu saillante versl’avant. Gouttières et rebords marginaux très étroits. Sculpturetotalement effacée à l’exception de la 1re strie qui est bien distincte,profonde surtout en avant, estompée et interrompue en arrièreavant d’atteindre le sommet de l’élytre (fig. 2). Pas de striolejuxtascutellaire. Disque lisse, marqué par des alignements de 8à 15 soies courtes le long des emplacements respectifs des anciensinterstries. Les soies marginales régulièrement disposées le longde la gouttière, mais quelques unes plus longues dans les régionshumérale et latéro-apicale.
Métathorax volumineux, le métasternum très vaste (fig. 25),sous la forme d’une grande surface lisse avec quelques soieséparses, sans sillon médian ; le sillon transversal paracoxal distinct,mais déporté en arrière, proche des articulations coxales. Lescoxae écartées l’une de l’autre. Ailes membraneuses pubescenteset frangées de longues soies très fines.
Ventrites abdominaux à surface faiblement pubescente. Leprocessus intercoxal (zone médiane du ventrite II ou III) parti-culièrement large pour un représentant des Adephaga (fig. 26).Les ventrites II, III et IV fusionnés, sans lignes de suture visibles.
Pattes courtes, tous les tarses tétramères (fig. 27). Les onglessimples, non pectinés. Protibias avec les trois éperons distauxprésents, deux ventraux (« e1 » et « e2 », l’interne un peut pluslong que l’externe) et un dorsal (« e3 »). Protarse du mâle avec le1er article plus large que les deux suivants.
Édéage (fig. 28) sans phallobase ; le lobe médian long et étroit,tubulaire, simple, la base non dilatée en bulbe et sans apodèmesen languettes latérales, avec seulement une expansion apophy-saire pour l’attache des paramères. Endophallus inerme. Paramèressétigères ; le droit plus court, l’extrémité distale dilatée en triangle ;le paramère gauche plus long, étiré et incurvé, son extrémitédistale avec des soies normales mais aussi en bordure une rangéede cinq sensilles courtes et robustes.
Habitat – Source sulfureuse près d’El Tor, dans le Sinaï(Égypte) (fig. 3). Seul est connu l’holotype, récolté en1933.
73
Les Coléoptères Gehringiidae
DISCUSSION
Caractères diagnostiquesLes deux espèces peuvent être distinguées de la façonsuivante :1. Plus grand (1,7-1,8 mm), les appendices bruns. Tarses
pentamères. Clypéus simple et tétrachète, dernier article
des palpes maxillaires et labiaux de même taille, dentmédiane du mentum faiblement bifide. Angles basauxdu pronotum aigus, la plage basale simple, sans fossettemédiane. Angle huméral de l’élytre aigu. Lobe médiande l’édéage plus large et plus court, avec à la base un aile-ron sagittal. Paramère droit à extrémité en lobe arrondi.Paramère gauche à extrémité dilatée en lobe arrondi, muni de sept longues soies simples . . Gehringia olympica
74
T. Deuve
Figures 13-22Gehringia olympica Darlington, détails de morphologie. 13, Labre et clypéus, face dorsale. 14, Tête, face ventrale. 15, Prothorax, face ventrale. 16, Méso- etmétathorax, face ventrale. 17, Métasternum et pleures mésothoraciques et métathoraciques, montrant l’extrémité du mésépimère qui atteint la cavité mésocoxale(flèche). 18, Patte prothoraciques, montrant les trois éperons terminaux (e1, e2, e3) et l’échancrure de toilette avec sa soie modifiée (s). 19, Ventrites abdominaux,face interne montrant le large processus intermétacoxal et les petites apophyses d’attache musculaire (flèches). 20, Tergites V, VI, VII, VIII et emplacementdes glandes de défense, face externe. Noter le raccourcissement du tergite VII et la différenciation en clapet (type nébridien) du tergite VIII. 21, Genitaliafemelles ectodermiques (c. sp1 : canal de la « spermathèque 1 » ; cx : coxite ; epl : épipleurite IX ; o.s.s. : organe sétulé subapical ; scx : subcoxite – sp2 :« spermathèque 2 »). 22, Edéage, lobe médian et paramères (le paramère gauche détaché).

– Plus petit (1,1 mm), les appendices jaunes. Tarses tétra-mères. Clypéus dichète, prolongé en avant par deuxcarène saillantes qui surplombent partiellement le labre,dernier article des palpes labiaux trois fois plus long quele dernier article des palpes maxillaires, dent médiane dumentum à extrémité largement arrondie. Angles basauxdu pronotum obtus, une fossette profonde à l’extrémitébasale du sillon médian discal. Angle huméral de l’élytresaillant mais arrondi. Lobe médian plus étroit et pluslong, sans aileron sagittal. Paramère droit dilaté en triangleà extrémité émoussée mais aiguë. Paramère gauche àextrémité distale faiblement dilatée en massue, avec unepubescence éparse et un alignement marginal de cinq sensilles spécialisées courtes mais fortes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Helenaea torretassoi
On ne peut que noter la relative « faiblesse » des carac-tères qui séparent les genres Gehringia et Helenaea desorte que leur même appartenance aux Gehringiidae nefait aucun doute. Les plus significatifs sont la différencede morphologie des palpes et de l’édéage, les uns et les
autres restant cependant de même type. Il ne semblepas qu’il y ait matière à classer les deux genres dans destribus ou sous-tribus distinctes sur la base de ces seulesdifférences, malgré l’éloignement géographique et ladisparité des conditions écologiques de l’habitat.
La réduction du nombre d’articles des tarses chezHelenaea est liée à la miniaturisation de la taille. D’autresexemples sont connus chez les Adephaga parmi lesTrechidae de la tribu des Anillini, minuscules espècesendogées qui ont parfois les tarses tétramères, parexemple Anillotarsus tetramerus (Mateu, 1980).
Autapomorphies des GehringiidaeLe rapprochement des deux genres Gehringia etHelenaea – dont l’idée revient à R. Sciaky (1997, in litte-ris) – et leur inclusion dans la même famille desGehringiidae ne semblent faire aucun doute tant sontimportantes certaines autapomorphies significatives quileur sont communes. Les principales sont : 1) la morpho-
75
Les Coléoptères Gehringiidae
Figures 23-28Helenaea torretassoi Schatzmayr et Koch, détails de morphologie. 23, Tête, face ventrale. 24, Labre et clypéus, face dorsale. 25, Métasternum et métacoxae.26, Ventrites II, III et IV de l’abdomen fusionnés. Noter le large triangle intermétacoxal. 27, Extrémité protibiale et protarse tétramère, montrant les troiséperons terminaux (e1, e2, e3), face ventrale. 28, Edéage, lobe médian et paramères (le paramère gauche détaché).

logie abdominale, avec fusion des ventrites II, III et IV,et développement en clapet trapézoïdal du tergite VIII ;2) la morphologie identique des élytres, tronqués à leursommet, avec effacement de la sculpture élytrale maispersistance de la seule 1re strie ; 3) la dilatation du pénul-tième article des palpes et la réduction en alène dudernier article (ce caractère s’observe aussi chez lesTrechidae Bembidiinae mais, comme on le verra, il nepeut s’agir que d’une convergence) ; 4) la présence detrois éperons terminaux à l’apex des protibias, deuxventraux et un dorsal ; 5) la chétotaxie des margesélytrales avec un groupe huméral de quatre soies et ungroupe post-médian de deux soies (état de caractèrerappelant aussi certains Trechidae) ; 6) la taille minus-cule (moins de 2 mm), probablement liée à la spéciali-sation de l’habitat (état de caractère présent aussi chezquelques Trechidae Bembidiinae Tachyini et Anillini).
Plusieurs caractères pourtant importants n’ont puêtre étudiés que chez Gehringia car le seul spécimenconnu du genre Helenaea n’a pas été entièrement dissé-qué afin de ne pas le détériorer. Ainsi, nous ne savonspas si le tergite VII est plus court que les précédentsdans ce genre, à l’instar de Gehringia, ni si les glandesde défense débouchent bien à la marge du tergite VIIIcomme il est probable. Il faudra aussi connaître la femelled’Helenaea torretassoi pour en étudier les genitaliainternes et pour vérifier si les expansion digitiformes etsétigères des épipleurites IX sont présentes.
Enfin, d’autres caractères essentiels sont communs àGehringia et Helenaea mais n’ont pu être polarisés aveccertitude. Ainsi par exemple le métasternum développéen vaste plastron sans sillon médian, qui représente proba-blement une excellente autapomorphie des Gehringiidae,mais sans que l’on puisse exclure un état plésiomorphelié aux caractéristiques des insertions musculaires chezces insectes à la locomotion peu performante.
Relations phylogénétiques au sein des AdephagaLa question de la place des Gehringiidae dans la phylo-génie des Adephaga a été posée dès la découverte deGehringia olympica et n’a toujours pas été complètementrésolue. Plusieurs hypothèses ont été envisagées et cetaxon a été tour à tour rapproché principalement desTrachypachidae, des Trechidae Bembidiinae, desPsydridae et des Melaenidae.
Gehringiidae et TrachypachidaeDès sa découverte, Gehringia olympica a été placée parDarlington (1933) dans une nouvelle tribu des«Gehringiini » située par son auteur dans les Carabiquesinférieurs, c’est-à-dire la sous-famille des « Carabinae »sensu Horn (1881) ou des « Carabidae Apertae » sensuSloane (1923). Plus précisément, Darlington plaça les
Gehringiini près des «Trachypachini» et des «Cychrini».Dans cette classification, Darlington a bien situé lesGehringiidae parmi les taxons les plus basaux de la phylo-génie des Caraboidea. Quelques années plus tard, Jeannel(1941, 1942, 1946) confirma ce classement et plaçaGehringia olympica dans une famille des Gehringiidaesituée dans sa division des « Isochaeta », c’est-à-dire prèsdes « Metriidae » et des Trachypachidae. Cette positionbasale des Gehringiidae sera admise par Lindroth (1960,1961) – qui relia même précisément cette famille auxseuls Trachypachidae d’après des caractéristiques larvaires–, mais aussi par Ball (1960, 1979) et Nagel (1979).Reprenant le point de vue de Lindroth, Kryzhanovskij(1976) inclut Gehringia dans une sous-famille desTrachypachinae qu’il divisa en Trachypachini etGehringiini.
D’importants caractères rattachent Gehringia auxCarabiques inférieurs, en particulier les cavités procoxalesouvertes et les mésépimères au contact des cavités méso-coxales. Cependant, l’un des caractères les plus cruciauxa fait l’objet de discussions. En effet, contrairement auxTrachypachidae mais comme les Paussidae Metriinae etOzaeninae, Gehringia olympica présente une échancrurede toilette aux protibias. La question a été posée de savoirsi les éperons terminaux associés étaient de type isochèteou anisochète, la petite taille de l’insecte rendant malai-sée la distinction entre soies, peignes et éperons à l’ex-trémité des pattes antérieures.
Darlington (1933) décrivit Gehringia avec des proti-bias de type anisochète (« variable spur distant fromapex »), ce qui ressort aussi du dessin qu’il a publié.L’illustration donnée par Koch (1936) pour Helenaeaproduit la même impression. Jeannel (1941) a cepen-dant rangé les Gehringiidae parmi les Isochaeta et il al’année suivante publié un dessin du protibia d’où ilressort que le prétendu éperon situé au talon de l’échan-crure de toilette était en réalité une soie modifiée dupeigne, plus longue que les autres, tandis que les deuxéperons ventraux isomorphes étaient situés à l’apex, ainsid’ailleurs qu’un troisième éperon, plus dorsal (Jeannel1942, « Fig. 99e »).
Cette dernière interprétation paraît exacte : la soiemodifiée du peigne est bien visible en micrographie élec-tronique à la base de l’organe de toilette (fig. 10, 12,18, « s»), tandis que trois éperons terminaux sont présentsà l’apex du protibia, deux ventraux et un dorsal (Fig. 11,18, « e1, e2, e3 »). Une structure similaire s’observe chezHelenaea (fig. 27). Cependant, un examen plus précismontre que l’éperon ventral le plus externe semblehomologue des soies terminales qui bordent en couronnel’extrémité protibiale et il ne s’en distingue, faiblement,que par sa longueur et son épaisseur (fig. 11, « e2 »). Iln’est ainsi pas certain que cet éperon soit véritablement
76
T. Deuve

l’homologue de l’éperon de même position visible chezles autres Isochaeta. Il en est de même du troisièmeéperon à la face dorsale (fig. 18, « e3 »).
Le protibia des Gehringiidae apparaît donc de typeisochète, mais selon une modalité atypique (primitive ?)où la distinction n’est pas claire entre éperons et soiesterminales. Bell (1964, 1966) a mis en doute l’appar-tenance de Gehringia aux Isochaeta pour deux raisons.D’une part, il n’a observé qu’un seul éperon à la faceventrale de l’extrémité protibiale et il a interprété l’épe-ron dorsal comme le second éperon. Il a ensuite comparécet éperon dorsal au « faux éperon » visible par exemplechez des Bembidiinae Tachyini. Son interprétation erro-née a été rectifiée par Hlavac (1971). D’autre part, Bella argué du caractère hexachète du labre de Gehringiapour l’exclure des Isochaeta, chez lesquels le labrecomporte plus de six soies. S’il est vrai que les Adephagabasaux ont généralement un labre fortement polychète,c’est aussi le cas dans quelques autres familles (parexemple les Carabidae Carabini), mais il n’est pas certainque l’état polychète soit plésiotypique. De plus, onremarque que le labre d’Helenaea comporte une pairede plus petites soies paramédianes (fig. 24).
Un second critère utilisé pour rapprocher lesGehringiidae des Trachypachidae est l’écartement desmetacoxae (fig. 16, 25). Cependant, comme je l’ai déjàindiqué, le triangle intermétacoxal du ventrite abdomi-nal est plus développé et plus large chez Gehringia qu’ilne l’est chez tous les autres Adephaga terrestres, y comprisles Trachypachidae, mais excepté les Rhysodidae (Deuve1993). Cet état de caractère n’est donc pas identiquechez les Gehringiidae et les Trachypachidae. De même,les genitalia femelles internes sont très différents. Onnotera en particulier l’absence du sclérite helminthoïdechez Gehringia.
Un autre caractère important pour séparer lesGehringiidae des Trachypachidae concerne encore lastructure abdominale. Chez les Trachypachidae, l’abdo-men est de type nébridien (Deuve 1988a) : le derniertergite apparent est le tergite VIII, qui est différencié enclapet semi-discoïdal qui « ferme » dorsalement l’extré-mité abdominal ; il n’est nullement télescopé et neprésente pas d’apophyses d’attache musculaire ; lestergites précédents, I à VII, sont indifférenciés et tousde même longueur. De plus, les glandes défensives(« pygidiales ») sont associées au segment IX. Chez lesGehringiidae, l’abdomen est aussi de type nébridien(quoique la conformation du tergite VIII soit un peudifférente : il est de forme subpentagonale et non pasvraiment semi-discoïdale), mais surtout il jouxte lesorifices des glandes défensives qui lui sont associées etne sont donc pas liées au segment IX (fig. 20).
En définitive, si de bons arguments permettent deplacer les Gehringiidae à la base de la phylogénie desAdephaga, aucun critère déterminant ne permet de lesrapprocher des Trachypachidae.
Gehringiidae et Trechidae BembidiinaeCertainement à cause de leur très petite taille, de leurhabitat ripicole, de leurs palpes spécialisés et de leuréchancrure protibiale de toilette, les Gehringiidae ontsouvent été rapprochés des Trechidae Bembidiinae. Onpeut aussi noter dans ce sens l’allure générale de la capsulecéphalique (du moins chez Gehringia), la morphologiecomparable de la denture mandibulaire, l’existence d’unéperon dorsal à l’extrémité du protibia et la présenced’un groupe huméral de quatre soies aux marges élytrales.Des antennes moniliformes pubescentes se rencontrentaussi chez de minuscules Bembidiinae Tachyini.
Dès sa description de Gehringia olympica, Darlington(1933) a évoqué certaines de ces similitudes avec lesBembidiinae, mais pour aussitôt rejeter l’hypothèsed’une proximité phylogénétique. Dans leur descriptiond’Helenaea torretassoi, Schatzmayr & Koch (1934) placè-rent au contraire leur nouvel insecte dans les Bembi-diinae, près du genre Lymnastis, mais ils exprimèrentleurs doutes en arguant de la difficulté de disséquer etde mieux examiner l’unique spécimen en leur posession(« Per non sacrificare l’unico esemplare esistente di questostranissimo insetto, dobbiamo limitarci a questa descri-zione veramente alquanto superficiale. Siamo convinti peròche un esame microscopico di questo insetto ci darebbe deirisultati inattesi. »).
Plus récemment, Bell (1967) proposa à nouveaul’hypothèse que les Gehringiidae puissent dériver desBembidiinae Tachyini, tout en reconnaissant que celaimpliquait des réversions de caractères morphologiquesimportants.
Cette hypothèse résiste mal à un premier examen,comme l’avait signalé Darlington dès 1933 : les cavitésprocoxales ouvertes, le mésothorax de type Disjuncta,les metacoxae très écartées l’une de l’autre, la réductiondu lobe métépiméral, sont autant d’états de caractèresen contradiction avec ce classement. On peut ajouter letype isochète des protibias et la morphologie de l’édéage(lobe médian et paramères), mais aussi surtout lamorphologie abdominale : l’abdomen de type nébri-dien mais avec association des glandes de défense autergite VIII ne correspond pas aux familles proches desTrechidae.
Gehringiidae et PsydridaeLes Gehringiidae ont aussi été rapprochés des Psydridaepar Erwin & Sims (1984) et Erwin (1985, 1991), «dansun groupe basal conduisant (probablement) aux Bembi-
77
Les Coléoptères Gehringiidae

diini ». Les caractères utilisés sont la sétulation des para-mères, la forme de la capsule céphalique et la spéciali-sation des palpes. Pour les mêmes raisons que cellesévoquées à l’endroit des Trechidae, cette hypothèse neme semble pas pouvoir être retenue. La morphologiedes pleures thoraciques et du processus intermétacoxaldes ventrites des Gehringiidae s’y oppose.
Gehringiidae et Nebriidae, Omophronidae et ElaphridaeUne analyse cladistique basée sur la seule morphologiedes genitalia femelles a conduit à réunir dans un mêmeclade les familles dites posséder une « seconde sperma-thèque» : Gehringiidae, Elaphridae, Nebriidae et Omo-phronidae (Liebherr & Will 1998). Cependant, lesauteurs de l’étude envisagent eux-mêmes l’hypothèse decaractères non homologues et ne retiennent pas cettephylogénie. Ils notent toutefois que les Gehringiidaesont des Anisochaeta du fait de l’état biarticulé des gono-podes femelles (subcoxa et coxa ne sont pas fusionnées).De mon point de vue, la séparation de la subcoxa et dela coxa est fondamentalement une plésiomorphie etpourrait donc sans problème être observée parmi desIsochaeta.
Une parenté des Gehringiidae avec les Nebriidae etCarabidae («Hemipleuri ») a aussi été envisagée par Bell(1967), mais comme une simple possibilité théorique.
Gehringiidae et PaussidaeUne analyse cladistique basée sur de nombreux carac-tères larvaires et imaginaux a conduit Beutel (1997) àclasser les Gehringiidae comme groupe-frère desPaussidae. Cependant, cet auteur n’a pas pris en comptede nombreux caractères importants, notamment lamorphologie abdominale et les genitalia femelles, ce quiobère l’ensemble de ses résultats. Des relations phylo-génétiques plus ou moins étroites entre les Gehringiidaeet les Paussidae Metriinae avaient déjà été évoquées parplusieurs auteurs, principalement sur la base d’une mêmeappartenance aux Isochaeta, mais les cavités procoxalesouvertes et les glandes défensives associées ausegment VIII éloignent les deux familles l’une de l’autre.
Dans un autre travail, Beutel & Haas (1996) utili-sent sensiblement les mêmes caractères imaginaux etlarvaires, mais ils aboutissent à un arbre de consensusstrict qui ne résout pas la position phylogénétique desGehringiidae.
Gehringiidae et MelaenidaeUne étude de phylogénie moléculaire portant sur uneportion de l’ADN 18S conduit au regroupement deGehringia avec le genre Cymbionotum (Maddison et al.,1999). Ce résultat inattendu a été accueilli avec inté-rêt mais aussi avec scepticisme par les auteurs du travail
car il pourrait être dû au phénomène bien connu d’at-traction des longues branches qui biaise parfois lesphylogénies moléculaires. À la fois Cymbionotum etGehringia présentent en effet de « longues branches »dans la phylogénie obtenue. Par exemple, dans la mêmeétude sont aussi parfois regroupés les Cicindelidae,Rhysodidae, Paussidae et Scaritinae (« CRPS Quartet »,Maddison et al. 1999), ce qui apparaît comme un arte-fact assez évident.
Cependant, ce résultat ne mérite pas d’être rejetéd’emblée. La description morphologique de Gehringiaindique en effet un abdomen de type nébridien avecles glande défensives associées au tergite VIII. Or l’exa-men de la morphologie abdominale des Adephagamontre que cette association d’états de caractères estrare et s’observe seulement chez les Apotomidae et chezles Melaenidae du genre Cymbionotum (les glandesdéfensives sont associées au segment IX chez Melaenus)(Deuve 1993). Chez les Apotomidae, le type nébri-dien de l’abdomen est mal caractérisé car le tergite VIIIn’est pas plus long que les tergites précédents et il estdonc peu différencié en clapet. Chez Cymbionotum, letergite VIII est plus nettement de type nébridien,quoique très faiblement invaginé. De morphologiedifférente de celui de Gehringia, il est néanmoins demême type.
La première idée est que si une analyse d’ADNregroupe Gehringia et Cymbionotum et si, d’autre part,le type abdominal de Gehringia est exceptionnel parmiles Adephaga et ne se retrouve précisément que chezCymbionotum (et dans une mesure moindre chezApotomus), alors il ne s’agit pas d’une coïncidence.Cependant, la quasi-totalité des caractères morpholo-giques importants éloignent les Gehringiidae desMelaenidae : chez Cymbionotum les cavités procoxalessont fermées, les cavités mésocoxales sont de typeConjuncta, les métépisternes ne sont pas lobés, les proti-bias sont de type Anisochaeta, le métasternum possèdeun sillon médian, les metacoxae ne sont pas écartées, letriangle intermétacoxal du ventrite II est petit et typiquedes Caraboidea, les genitalia femelles internes sont tota-lement différents (Deuve 1993), les palpes n’ont riende commun, etc. Réunir ces deux genres conduirait àfaire intervenir des homoplasies pour tous ces carac-tères, ce qui est théoriquement possible mais difficile àconsidérer comme probable.
Il faut aussi beaucoup d’imagination pour trouverdes états de caractères morphologiques qui réunissentGehringia et Cymbionotum : citons la capsule crânienneassez volumineuse, les antennes pubescentes plus oumoins moniliformes à partir du 5e article, la formegénérale du pronotum, le métasternum relativementvaste. On conviendra que c’est insuffisant. Cependant,
78
T. Deuve

la distribution géographique de Cymbionotum enAfrique, en Asie, mais aussi en Amérique du Sud(Perrault 1994), indique qu’il s’agit d’un genre« panchronique », ancien, comme d’ailleurs le genreApotomus, et un branchement basal dans la phylogéniedes Adephaga est envisageable.
CONCLUSION
Des remarques précédentes, il ressort que lesGehringiidae présentent de nombreuses plésiomorphiesqui indiquent une position basale au sein des ColéoptèresAdephaga : les cavités procoxales ouvertes, les protibiasde type isochète (avec les réserves évoquées plus haut),le mésothorax de type Disjuncta, les métépimères faible-ment lobés, sont autant d’états de caractères primitifs.On peut ajouter l’abdomen de type nébridien qui corres-pond à l’état de la majorité des Adephaga les plus basaux.
Cependant, l’association des glandes défensives autergite VIII de type nébridien isole les Gehringiidaeparmi tous les autres Carabiques inférieurs chez lesquelsces glandes sont liées au segment IX. De ce point devue, ils occupent une place très particulière. Il n’y aguère que chez certains Melaenidae (commeCymbionotum) ou chez les Apotomidae que l’onretrouve la même association de caractères abdomi-naux. Cependant, le large triangle intermétacoxal dupremier ventrite abdominal apparent semble plus plésio-morphe chez les Gehringiidae que le petit triangle formépar le ventrite II chez tous les autres Adephaga terrestres,Rhysodidae exceptés (Deuve 1993). Ce dernier carac-tère place alors les Gehringiidae tout à fait à la base dela phylogénie des Adephaga.
Un autre caractère peu pris en compte jusqu’àprésent, est la structure du métasternum qui est dilatéen un vaste plastron indivis et faiblement convexe, sansla présence d’un sillon médian. Le sillon transversal para-coxal, lorsqu’il est présent (Helenaea) est déplacé versl’arrière, le sillon longitudinal postérieur sépare en deuxlobes distincts l’apophyse postérieure intercoxale qui estparticulièrement large. Cette morphologie particulièredu métasternum, unique parmi les Adephaga, se retrouvechez Gehringia et chez Helenaea. Associée à des parti-cularités des attaches musculaires, elle est probablementliée à la locomotion lente de ces insectes.
On peut aussi accorder une signification à l’étrangedistribution géographique des Gehringiidae. Peu defamilles d’Adephaga ont une répartition discontinuequi soit comparable. Citons les Cicindidae, qui sontaussi connus par deux genres monospécifiques, l’un en
Argentine, l’autre aux confins du Koweït et de l’Iran(Kavanaugh & Erwin 1991). Les Trachypachidae quisont distribués dans les régions circum-arctiques d’unepart et dans l’Amérique australe d’autre part. LesPromecognathidae n’ont longtemps été connus que del’Amérique du Nord-Ouest et de l’Afrique australe, avantqu’un représentant en soit récemment découvert isolédans le sud de la péninsule Ibérique (Mateu 2002 ;Mateu & Bellés 2004). Les Nebriidae, exclusifs de l’hé-misphère Nord, semblent avoir un représentant ou ungroupe-frère, les Notiokasinae (ou Notiokasidae ?), auxconfins du Brésil et de l’Uruguay (Kavanaugh & Nègre1983). Ces quelques exemples correspondent toujoursà des lignées très anciennes. Ainsi la distribution géogra-phique insolite et très discontinue des Gehringiidae est-elle un témoignage de leur ancienneté, d’autant plusqu’ils vivent chacun dans un milieu écologiquement trèscontrasté. Cette singularité biogéographique appuiel’hypothèse d’un branchement très basal (c’est-à-diretrès ancien) dans la phylogénie des Adephaga.
En définitive, si cette étude n’a pas permis de résoudreavec précision la position phylogénétique desGehringiidae au sein des Coléoptères Adephaga, elleprivilégie fortement l’hypothèse d’une position basale,sans relation particulière avec les Trachypachidae. Cettehypothèse est celle déjà exprimée par Darlington dès1933, mais aussi par Jeannel (1941, 1942, 1946), Ball(1979), Deuve (1993), Ball & Bousquet (2001) et Sciaky(1997, in litteris). L’hypothèse d’une position phylogé-nétique proche du genre Cymbionotum (Melaenidae)nécessiterait des arguments complémentaires, mais ellene peut être totalement exclue. Il ne fait en revanchepas de doute que les deux genres Gehringia et Helenaeadoivent être réunis en un même clade.
Remerciements – Je tiens à remercier très vivement leDr Riccardo Sciaky qui a eu le mérite de reconnaître en Helenaeaun Gehringiidae et qui très cordialement a bien voulu m’infor-mer en 1997 de l’état de ses recherches en me confiant alors unmanuscrit en cours de rédaction qu’il souhaitait mener à termeet publier. De mon côté je l’avais avisé de mon étude en courssur la position phylogénétique de Gehringia. Finalement, sontravail n’a pas fait l’objet d’une publication, mais je suis très rede-vable à R. Sciaky d’avoir bien voulu me mettre en contact avecle conservateur du Muséum de Milan pour l’emprunt de l’holo-type d’Helenaea torretassoi et d’avoir accepté que j’utilise l’infor-mation de sa découverte dans mon étude en cours.
Pour l’aimable communication du précieux type d’Helenaeatorretassoi, je tiens à remercier le Dr Carlo Pesarini et leDr Maurizio Pavesi, du Muséum d’Histoire Naturelle de Milan.
79
Les Coléoptères Gehringiidae

RÉFÉRENCES
Ball G. E. 1960. Carabidae : p. 55-181. In : Arnett R. H., The beetles of theUnited States. Washington D. C., Catholic University Press, 1112 p.
Ball G. E. 1979. Conspectus of carabid classification : history, holomor-phology, and higher taxa : p. 63-111. In : Erwin T. L., Ball G. E. &Whitehead D. R. (eds.), Carabid beetles : their evolution, natural history,and classification. The Hague, W. Junk, 635 p.
Ball G. E. (ed.) 1985. Taxonomy, phylogeny and zoogeography of beetles andants. A volume dedicated to the memory of Philipp Jackson Darlington, Jr.(1904-1983). Dordrecht, Boston, Lancaster, W. junk, 514 p.
Ball G. E., Bousquet Y. 2001. 6. Carabidae Latreille, 1810. In : Arnett R.H. & Thomas M. C., American beetles. Volume 1. Archostemata,Myxophaga, Adephaga, Polyphaga : Staphyliniformia. Boca Raton, London,New York, Washington D.C., CRC Press, p. 32-132.
Bell R. T. 1964. Does Gehringia belong to the Isochaeta ? (Coleoptera :Carabidae). The Coleopterists’ Bulletin, 18 : 59-61.
Bell R. T. 1966. Trachypachus and the origin of the Hydradephaga(Coleoptera). The Coleopterists’ Bulletin, 20 : 107-112.
Bell R. T. 1967. Coxal cavities and the classification of the dephaga(Coleoptera). Annals of the entomological Society of America, 60 : 101-107.
Beutel R. 1997. Ueber Phylogenese une Évolution der Coleoptera (Insecta),insbesondere der Adephaga. Abhandlungen des naturwissenschaftlichenVereins in Hamburg (N. F.), 31 : 1-164.
Beutel R. G., Haas A. 1996. Phylogenetic analysis of larval and adult charac-ters of Adephaga (Coleoptera) using cladistic computer programs.Entomologica scandinavica, 27 : 197-205.
Darlington P. J. 1933. A new tribe of Carabidae (Coleoptera) from westernUnited States. The Pan-Pacific Entomologist, 9 : 110-114.
Deuve Th. 1988a. Étude phylogénétique des Coléoptères Adephaga : redé-finition de la famille des Harpalidae, sensu novo, et position systéma-tique des Pseudomorphinae et Brachinidae. Bulletin de la Société ento-mologique de France, 92 : 161-182
Deuve Th. 1988b. Les sternites VIII et IX de l’abdomen sont-ils visibleschez les imagos des Coléoptères et des autres insectes holométaboles ?Nouvelle Revue d’Entomologie (N. S.), 5 : 21-34.
Deuve Th. 1993. L’abdomen et les genitalia des femelles de ColéoptèresAdephaga. Mémoires du Muséum national d’Histoire naturelle, 155 :1-184.
Deuve Th. 1994. Sur la présence d’un « épipleurite » dans le plan de basedu segment des Hexapodes. Bulletin de la Société entomologique de France,99 : 199-210.
Erwin T. L., Sims L. L. 1984. Carabid beetles of the West Indies (Insects :Coleoptera) : a synopsis of the genera and checklists of tribes ofCaraboidea, and of the West Indian species. Quaestiones entomologicae,20 : 351-466.
Erwin T. L. 1985. The taxon pulse : a general pattern of lineage radiationand extinction among carabid beetles : p. 437-472. In : Ball G. E. (ed.),Taxonomy, phylogeny and zoogeography of beetles and ants. A volume dedi-cated to the memory of Philipp Jackson Darlington, Jr. (1904-1983).Dordrecht, Boston, Lancaster, W. junk, 514 p.
Erwin T. L. 1991. The ground-beetles of Central America (Carabidae),Part II : Notiophilini, Loricerini, and Carabini. Smithsonian Contributionsto Zoology, 501 : 1-30.
Hammond P. M. 1979. Wing-folding mechanisms of beetles, with specialreference to investigations of adephagan phylogeny (Coleoptera) : pp.113-180. In : Erwin T. L., Ball G. E. & Whitehead D. R. (eds.),Carabid beetles : their evolution, natural history, and classification. TheHague, W. Junk : 635 p.
Hlavac T. F. 1971. Differentiation of the carabid antenna cleaner. Psyche,78 : 51-66.
Horn G. H. 1881. On the genera of Carabidae with special reference to thefauna of Boreal America. Transactions of the american entomologicalSociety, 9 : 91-196.
Jeannel R. 1941. Coléoptères Carabiques I. Faune de France, 39 : 1-571.Jeannel R. 1942. La genèse des faunes terrestres. Eléments de biogéographie.
Paris, Presses Universitaires de France, 514 p. + 8 pls h. t.Jeannel R. 1946. Coléoptères Carabiques de la région Malgache, I. Faune
de l’Empire français, 6 : 1-372.Jeannel R., Paulian R. 1944. Morphologie abdominale des Coléoptères et
systématique de l’Ordre. Revue française d’Entomologie, 11 : 65-110.Kavanaugh D. H., Erwin T. L. 1991. The tribe Cicindini Bänninger
(Coleoptera : Carabidae) : comparative morphology, classification, natu-ral history and evolution. Proceedings of the entomological Society ofWashington, 93 : 356-389.
Kavanaugh D. H., Nègre J. 1983. Notiokasini. New tribe of Carabidaefrom southeastern South America. Coleopterists’ Bulletin, 36 (1982) :549-566.
Koch C. 1936. Ueber Helenaëa Torre-Tassoi Schatzm.-Koch (Coleoptera.Carabidae). Bulletin de la Société royale entomologique d’Egypte, 20 : 298.
Kryzhanovskij O. L. 1976. Revised classification of the family Carabidae(Coleoptera). Entomological Review, 1 : 80-91.
Liebherr J. K., Will W. 1996. Inferring phylogenetic relationships withinCarabidae (Insecta, Coleoptera) from characters of the female repro-ductive tracts, p. 107-170. In : Ball G. E., Casale A., Vigna Taglianti A.(eds.), Phylogeny and classification of Caraboidea. Torino, Museo regio-nale di Scienze naturali, 543 p.
Lindroth C. H. 1960. The larvae of Trachypachus Mtsch., Gehringia Darl.,and Opisthius Kby. (Col. Carabidae). Opuscula entomologica, 25 : 30-42.
Maddison D. R. 1985. The discovery of Gehringia olympica Darlington(Coleoptera : Carabidae) : p. 35-37. In : Ball G. E., ed., Taxonomy,phylogeny and zoogeography of beetles and ants. A volume dedicated to thememory of Philipp Jackson Darlington, Jr. (1904-1983). Dordrecht,Boston, Lancaster, W. junk, 514 p.
Maddison D. R., Baker M. D., Ober K. A. 1999. Phylogeny of carabidbeetles as inferred from 18S ribosomal DNA (Coleoptera : Carabidae).Systematic Entomology, 24 : 103-138.
Mateu J. 1980. Un nouveau Anillini tétramère, gen. & sp. nov. récolté dansles grottes du Pérou. Mémoires de Biospéléologie, 7 : 201-207.
Mateu J. 2002. Sur un genre nouveau et une espèce cavernicole inéditeappartenant à une nouvelle sous-famille de Coléoptères CarabiquesPromecognathidae. Revue française d’Entomologie, (N. S.), 24 : 67.
Mateu J., Bellés X. 2004. Position systématique et remarques biogéogra-phiques sur Dalyat mirabilis Mateu, 2002 (Coleoptera : Adephaga :Promecognathidae), cavernicole du Sud-Est Ibérique. Annales de laSociété entomologique de France (N. S.), 39 (2003) : 291-303.
Nagel P. 1979. The classification of Carabidae : pp. 7-14. In : Den BoerP. J., Thiele H. U., Weber F. (eds.), On the evolution of behaviour incarabid beetles. Wageningen : H. Veenman & B. V. Zonen.
Perrault G. P. 1994. Estudios sobre los Carabidos venezolanos. II.Cymbionotum negrei n. sp. (Coleoptera : Carabidae : Cymbionotini).Boletin de Entomologia venezolana. (N. S.), 9 : 207-208.
Schatzmayr A., Koch C. 1934. Risultati scientifici della spedizione ento-mologica di S.A.S. il Principe Alessandro della Torre e Tasso in Egittoe nella penisola Sinai. Bolletino della Società adriatica di Scienze natu-rali in Trieste, 33 : 7-25.
Sloane T. G. 1923. The classification of the family Carabidae. Transactionsof the entomological Society, London, 1-2 : 234-250.
80
T. Deuve