la croissance des v égétaux -...
TRANSCRIPT

La croissance des vLa croissance des vééggéétauxtaux
Préparation à l’agrégation interne
Novembre Décembre 2008
• Françoise Saintpierre
I/ Organisation et fonctionnement des apex racinaire et I/ Organisation et fonctionnement des apex racinaire et caulinairecaulinaire
A/ Les phA/ Les phéénomnomèènes assocines associéés s àà la croissance en longueurla croissance en longueur1/ D1/ Déétermination des zones responsables de la termination des zones responsables de la
croissance en longueurcroissance en longueur
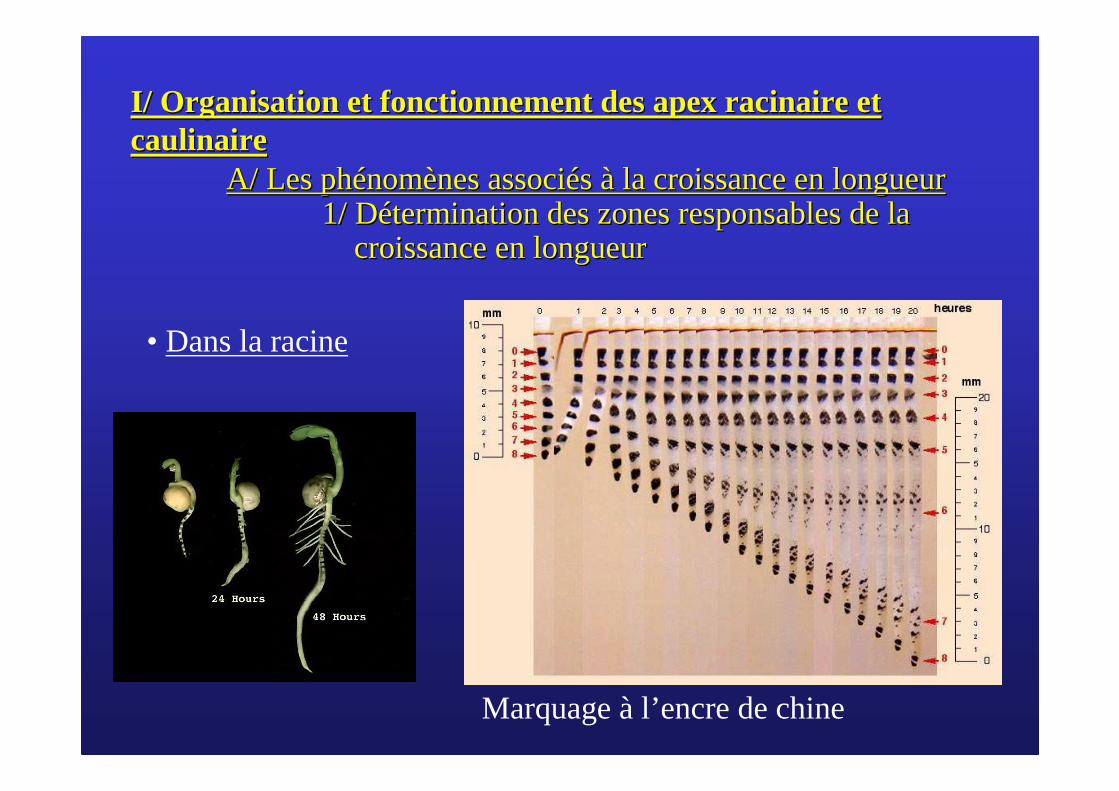
• Dans la racine
Marquage à l’encre de chine
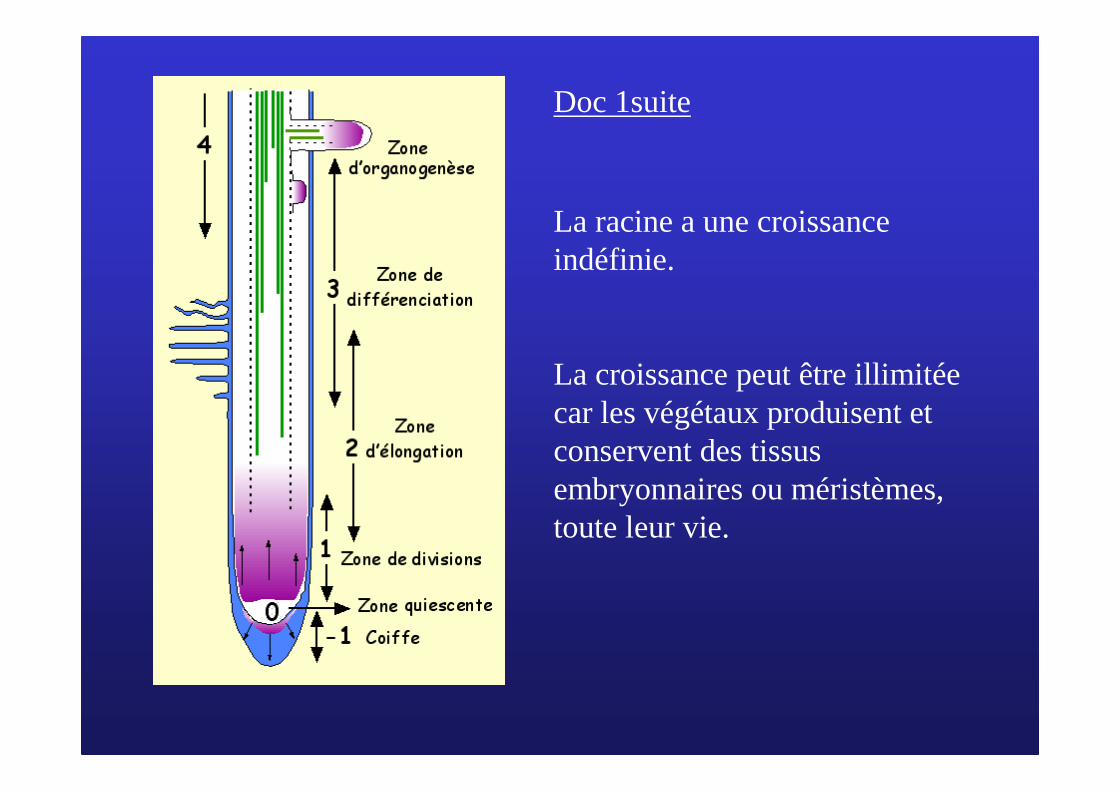
Doc 1suite
La racine a une croissance indéfinie.
La croissance peut être illimitée car les végétaux produisent et conservent des tissus embryonnaires ou méristèmes, toute leur vie.

• Dans la tige Marquage d’une tige de haricot
Coupe longitudinale d’un chou de Bruxelles
Développement d’un bourgeon de Marronnier
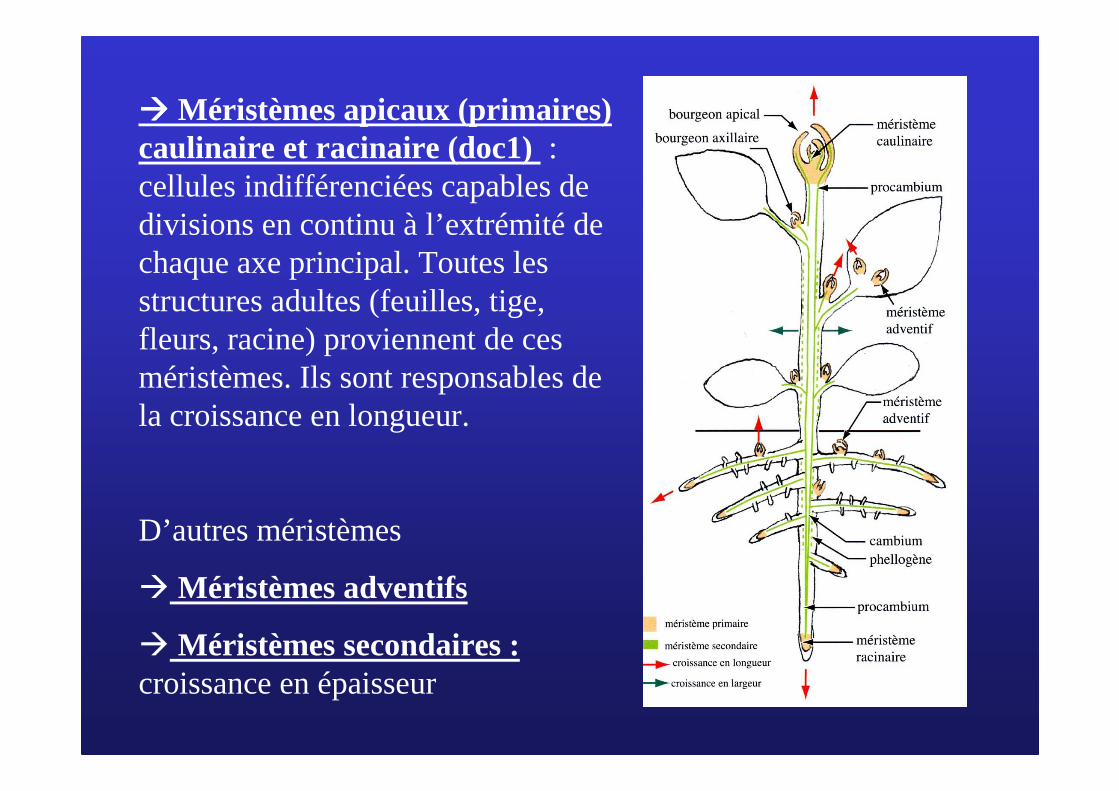
���� Méristèmes apicaux (primaires) caulinaire et racinaire (doc1) : cellules indifférenciées capables de divisions en continu à l’extrémité de chaque axe principal. Toutes les structures adultes (feuilles, tige, fleurs, racine) proviennent de ces méristèmes. Ils sont responsables de la croissance en longueur.
D’autres méristèmes
� Méristèmes adventifs
� Méristèmes secondaires : croissance en épaisseur
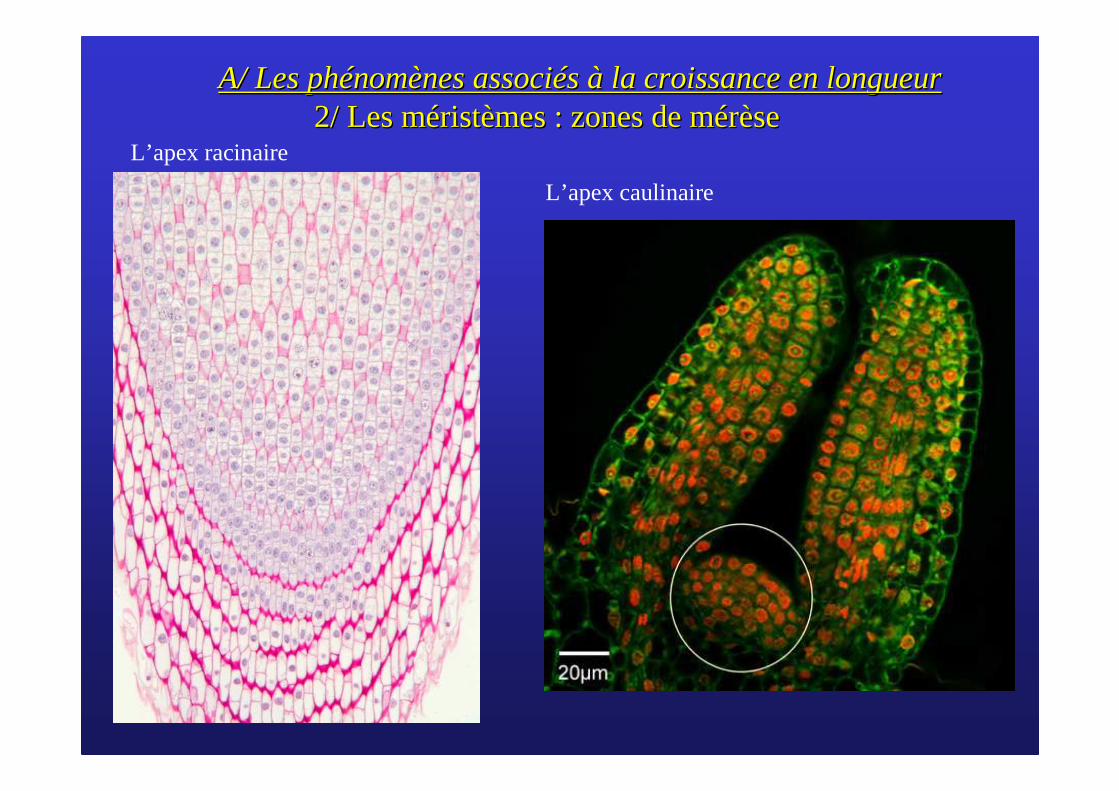
A/ Les phA/ Les phéénomnomèènes assocines associéés s àà la croissance en longueurla croissance en longueur2/ Les m2/ Les mééristristèèmes : zones de mes : zones de mméérrèèsese
L’apex racinaire
L’apex caulinaire
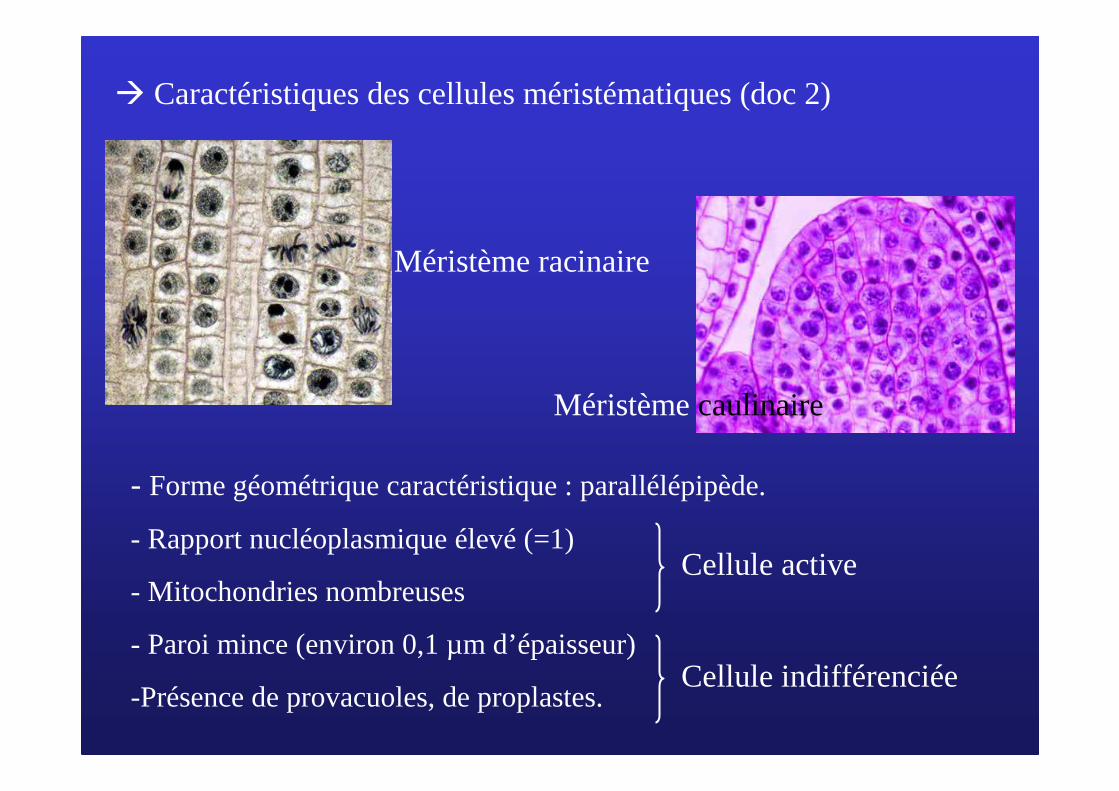
� Caractéristiques des cellules méristématiques (doc 2)
Méristème racinaire
Méristème caulinaire
- Forme géométrique caractéristique : parallélépipède.
- Rapport nucléoplasmique élevé (=1)
- Mitochondries nombreuses
- Paroi mince (environ 0,1 µm d’épaisseur)
-Présence de provacuoles, de proplastes.
Cellule active
Cellule indifférenciée
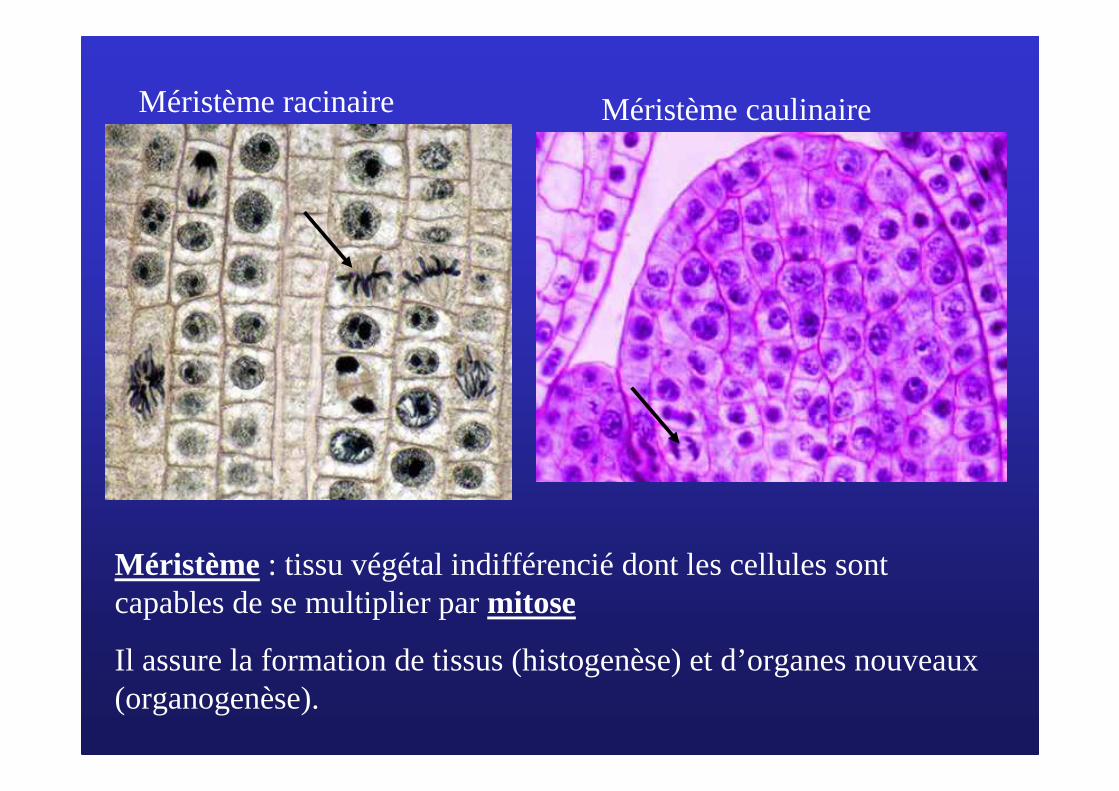
Méristème racinaire Méristème caulinaire
Méristème : tissu végétal indifférencié dont les cellules sont capables de se multiplier par mitose
Il assure la formation de tissus (histogenèse) et d’organes nouveaux (organogenèse).
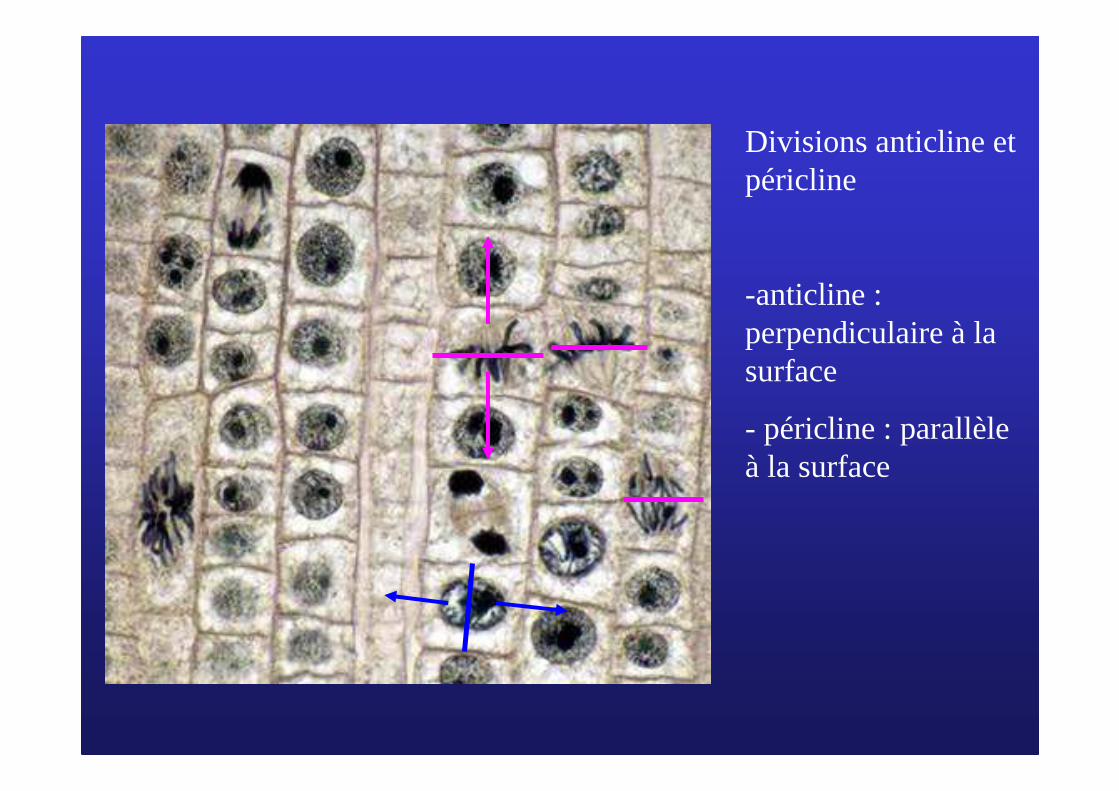
Divisions anticline et péricline
-anticline : perpendiculaire à la surface
- péricline : parallèle à la surface
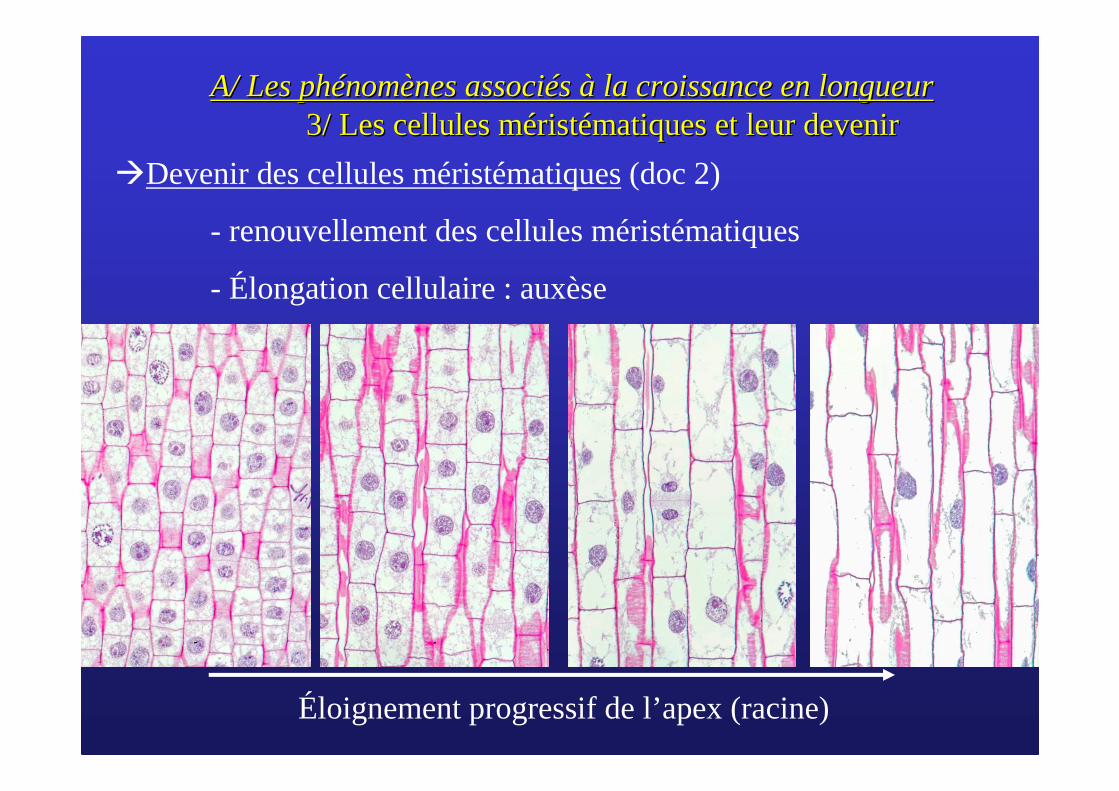
�Devenir des cellules méristématiques(doc 2)
- renouvellement des cellules méristématiques
- Élongation cellulaire : auxèse
Éloignement progressif de l’apex (racine)
A/ Les phA/ Les phéénomnomèènes assocines associéés s àà la croissance en longueurla croissance en longueur3/ Les cellules 3/ Les cellules mmééristristéématiques matiques et leur deveniret leur devenir
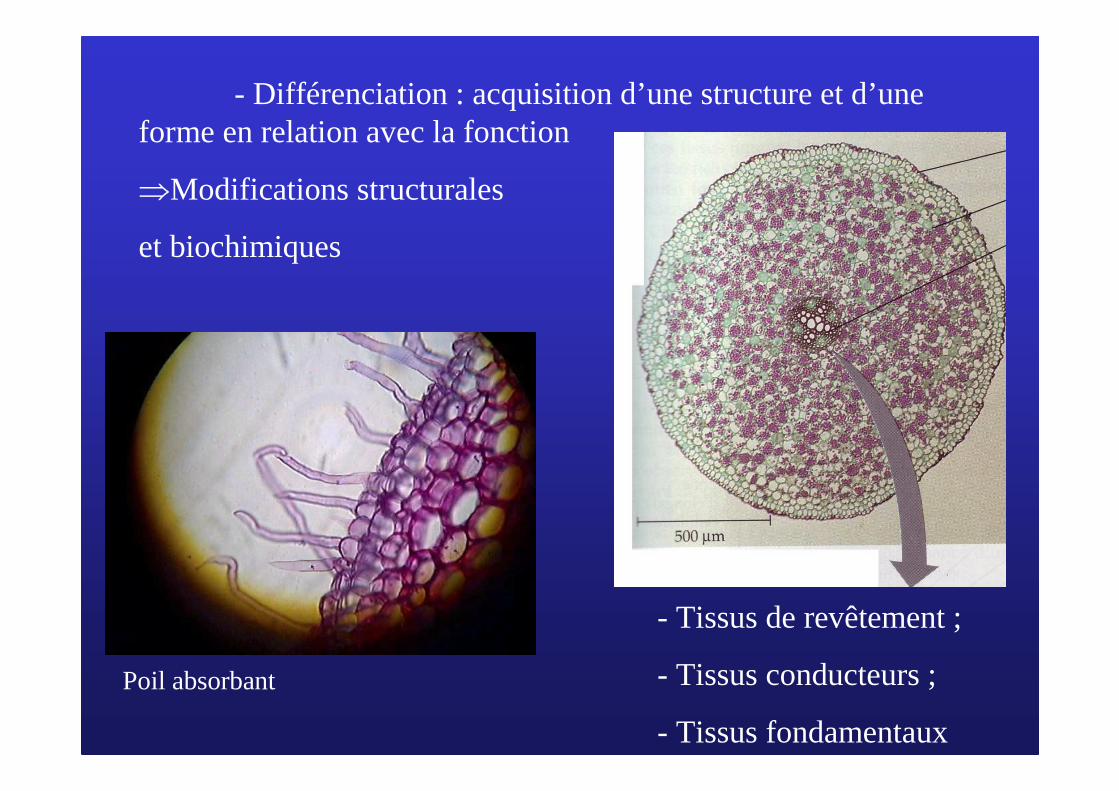
- Différenciation : acquisition d’une structure et d’une forme en relation avec la fonction
⇒Modifications structurales
et biochimiques
Poil absorbant
- Tissus de revêtement ;
- Tissus conducteurs ;
- Tissus fondamentaux
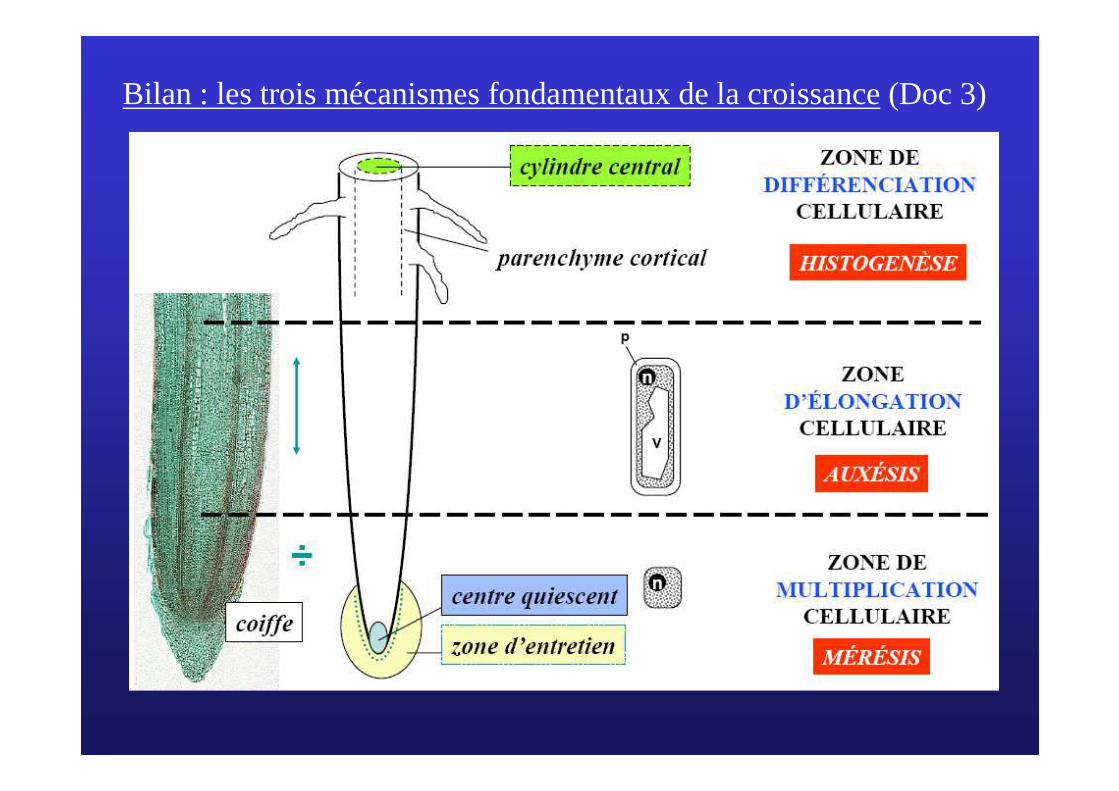
Bilan : les trois mécanismes fondamentaux de la croissance(Doc 3)
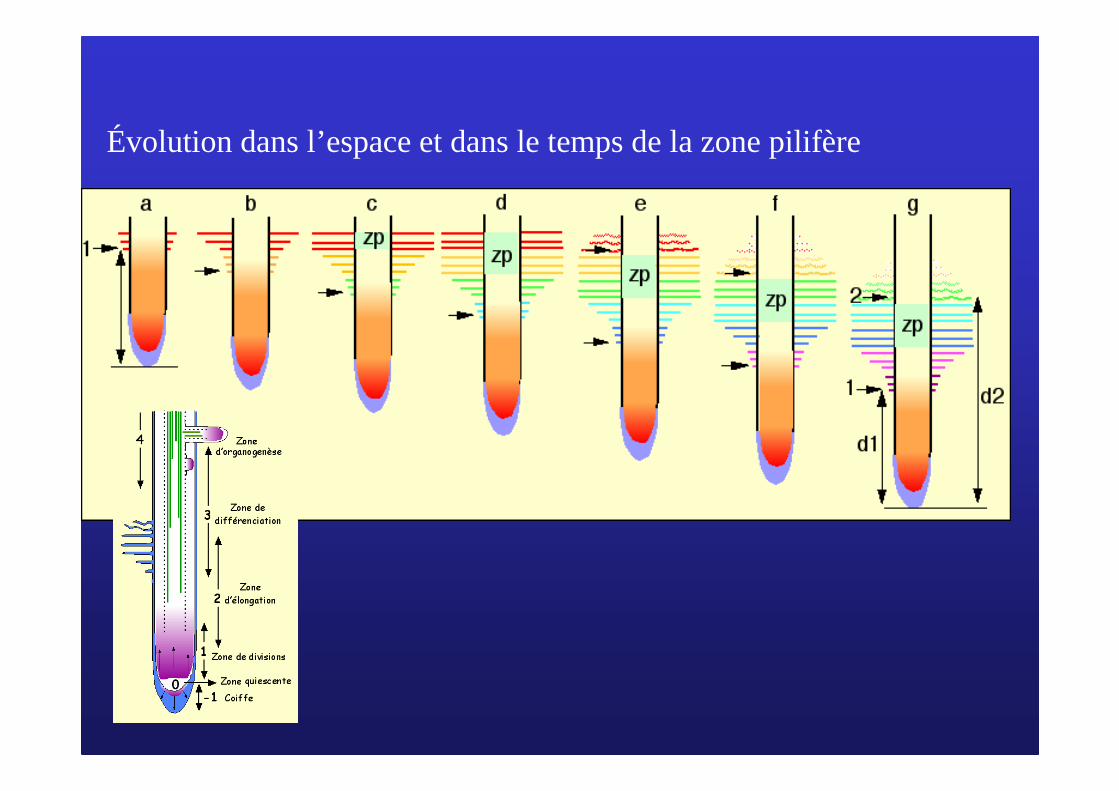
Évolution dans l’espace et dans le temps de la zone pilifère
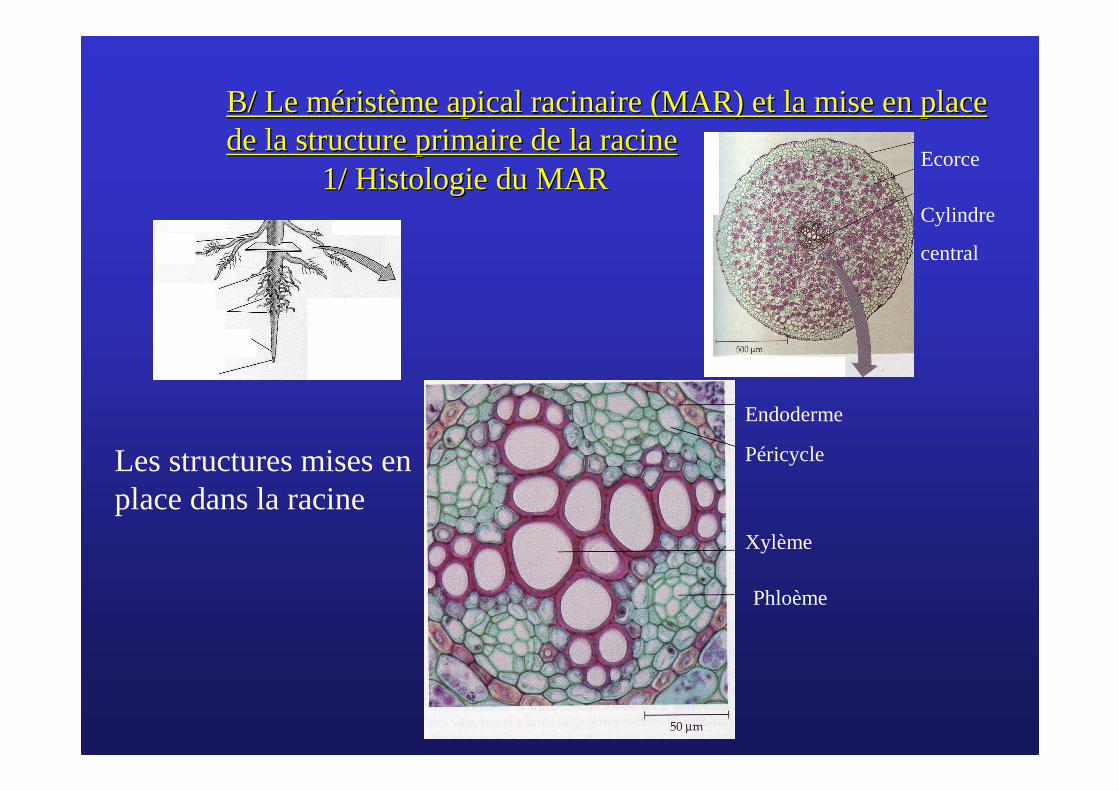
B/ Le mB/ Le mééristristèème apical racinaire (MAR) et la mise en place me apical racinaire (MAR) et la mise en place de la structure primaire de la racinede la structure primaire de la racine
1/ Histologie du MAR1/ Histologie du MAR
Endoderme
Péricycle
Xylème
Phloème
Ecorce
Cylindre
central
Les structures mises en place dans la racine

Arabidopsis thaliana
- Arabette des dames ;
- Cycle de développement court (environ 2 mois en conditions optimales)
- Plante prolifique (plusieurs milliers de graines par individu)
- le plus petit génome végétal connu et entièrement séquencé(25498 gènes)
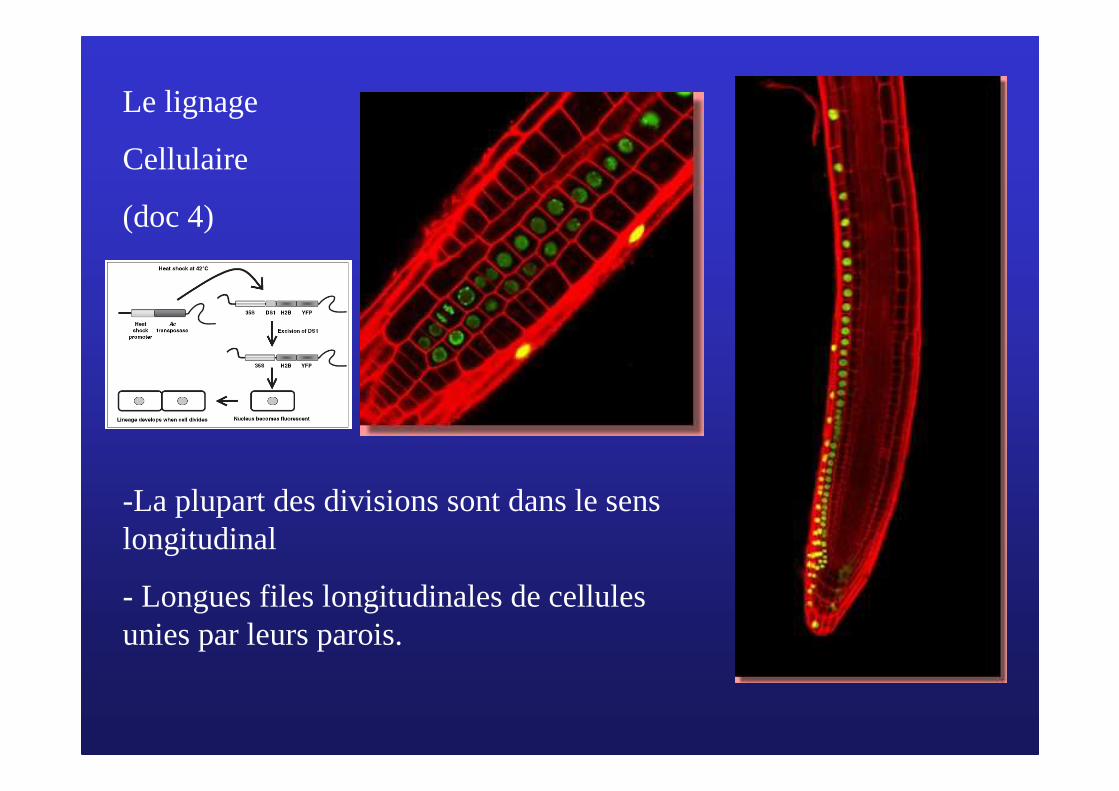
Le lignage
Cellulaire
(doc 4)
-La plupart des divisions sont dans le sens longitudinal
- Longues files longitudinales de cellules unies par leurs parois.
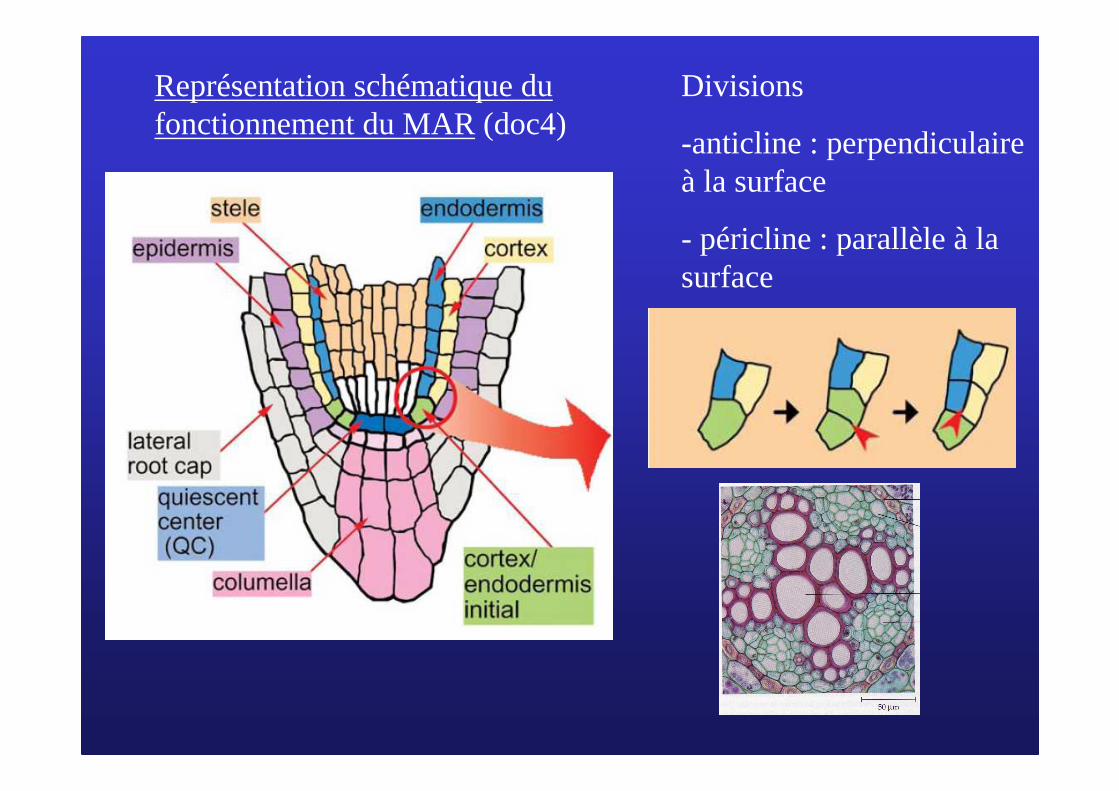
Divisions
-anticline : perpendiculaire à la surface
- péricline : parallèle à la surface
Représentation schématique du fonctionnement du MAR(doc4)
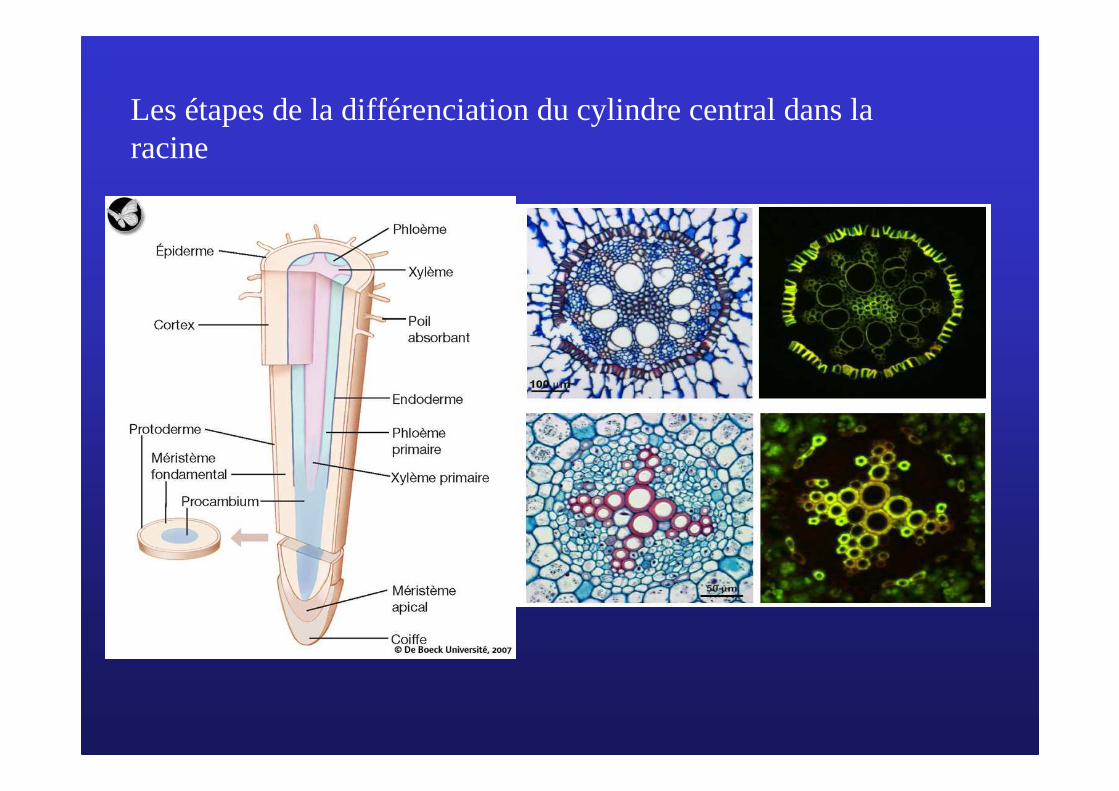
Les étapes de la différenciation du cylindre central dans la racine
B/ Le mB/ Le mééristristèème apical racinaire (MAR) et la mise en place me apical racinaire (MAR) et la mise en place de la structure primaire de la racinede la structure primaire de la racine
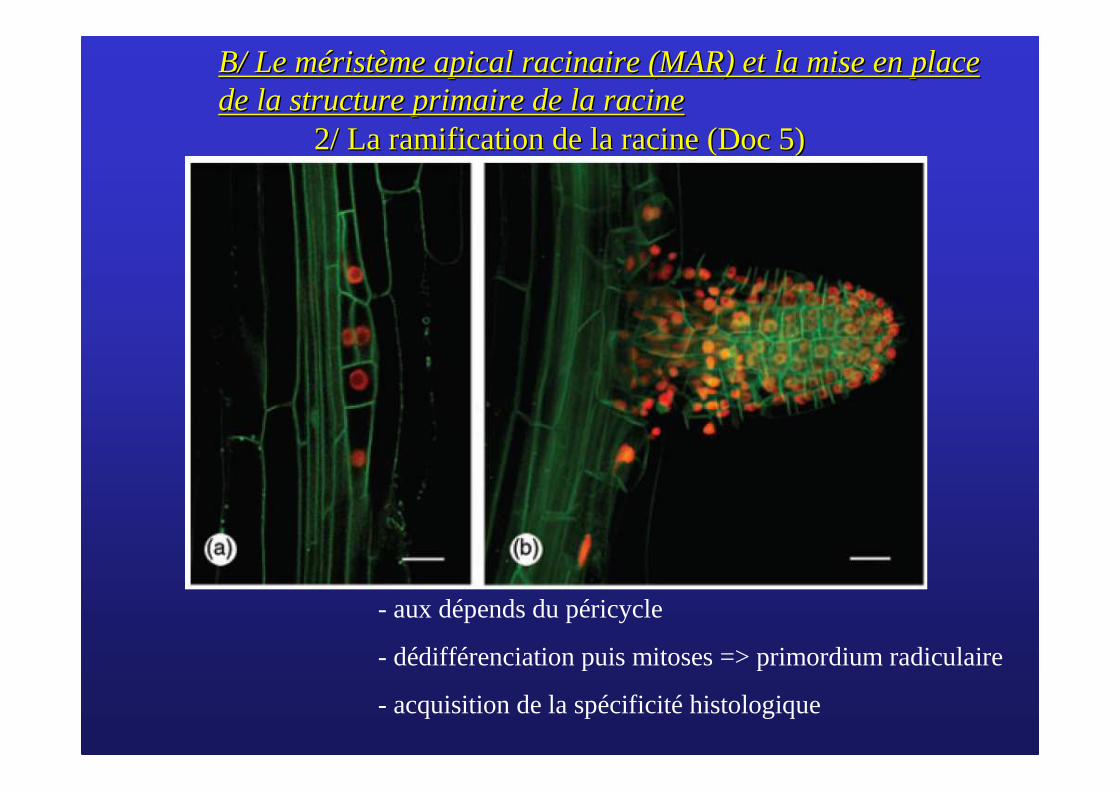
2/ La ramification de la racine (Doc 5)2/ La ramification de la racine (Doc 5)
- aux dépends du péricycle
- dédifférenciation puis mitoses => primordium radiculaire
- acquisition de la spécificité histologique
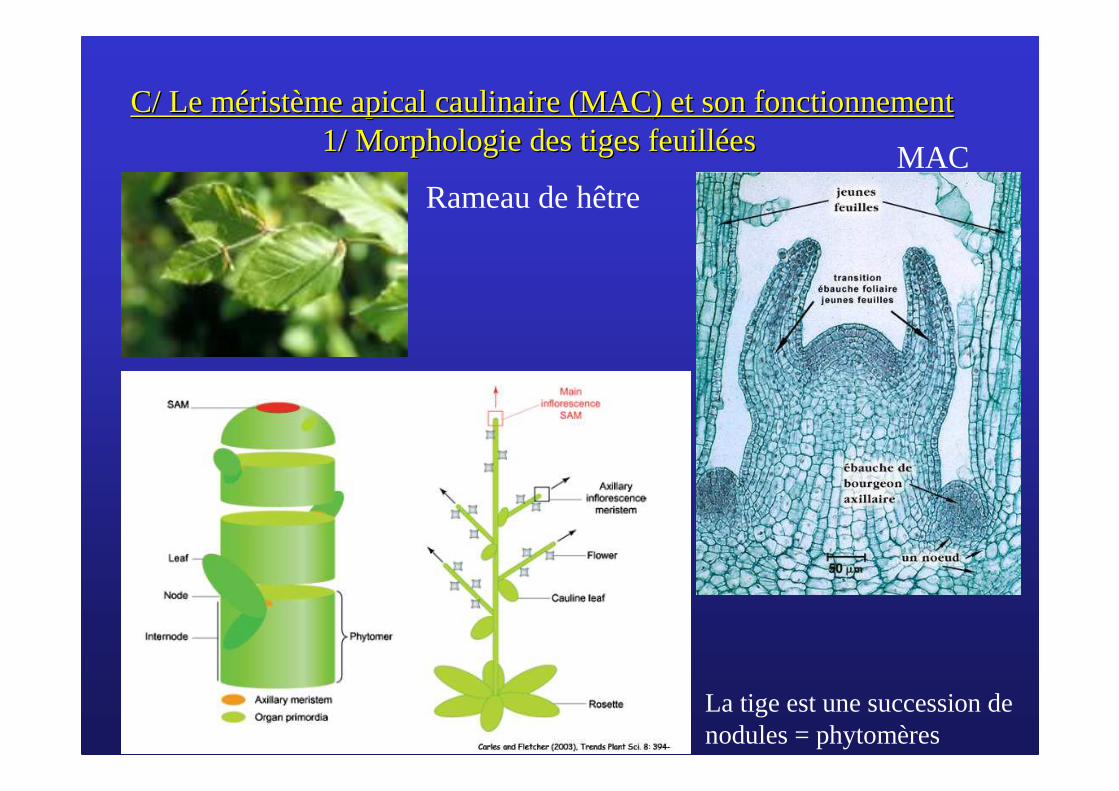
C/ Le mC/ Le mééristristèème apical caulinaire (MAC) et son fonctionnementme apical caulinaire (MAC) et son fonctionnement1/ Morphologie des tiges feuill1/ Morphologie des tiges feuillééeses
Rameau de hêtre
La tige est une succession de nodules = phytomères
MAC
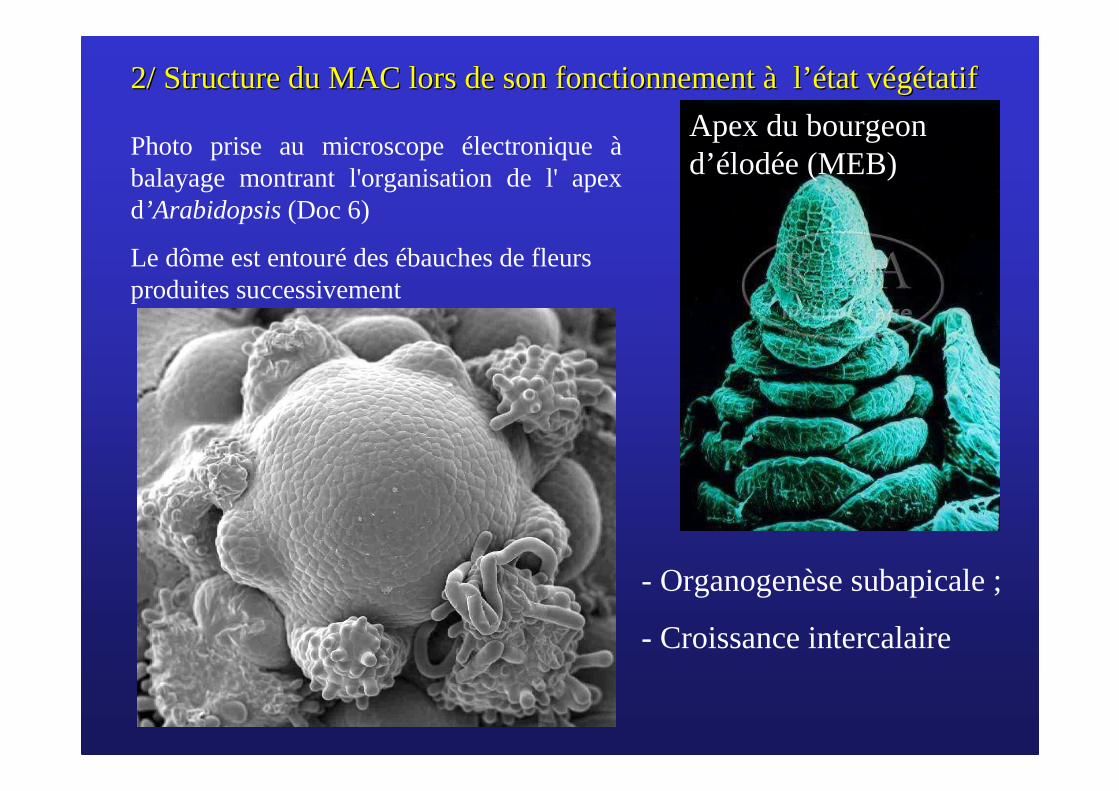
2/ Structure du MAC lors de son fonctionnement 2/ Structure du MAC lors de son fonctionnement àà ll ’é’état vtat vééggéétatiftatif
Photo prise au microscope électronique àbalayage montrant l'organisation de l' apex d’Arabidopsis (Doc 6)
Le dôme est entouré des ébauches de fleurs produites successivement
Apex du bourgeon d’élodée (MEB)
- Organogenèse subapicale ;
- Croissance intercalaire
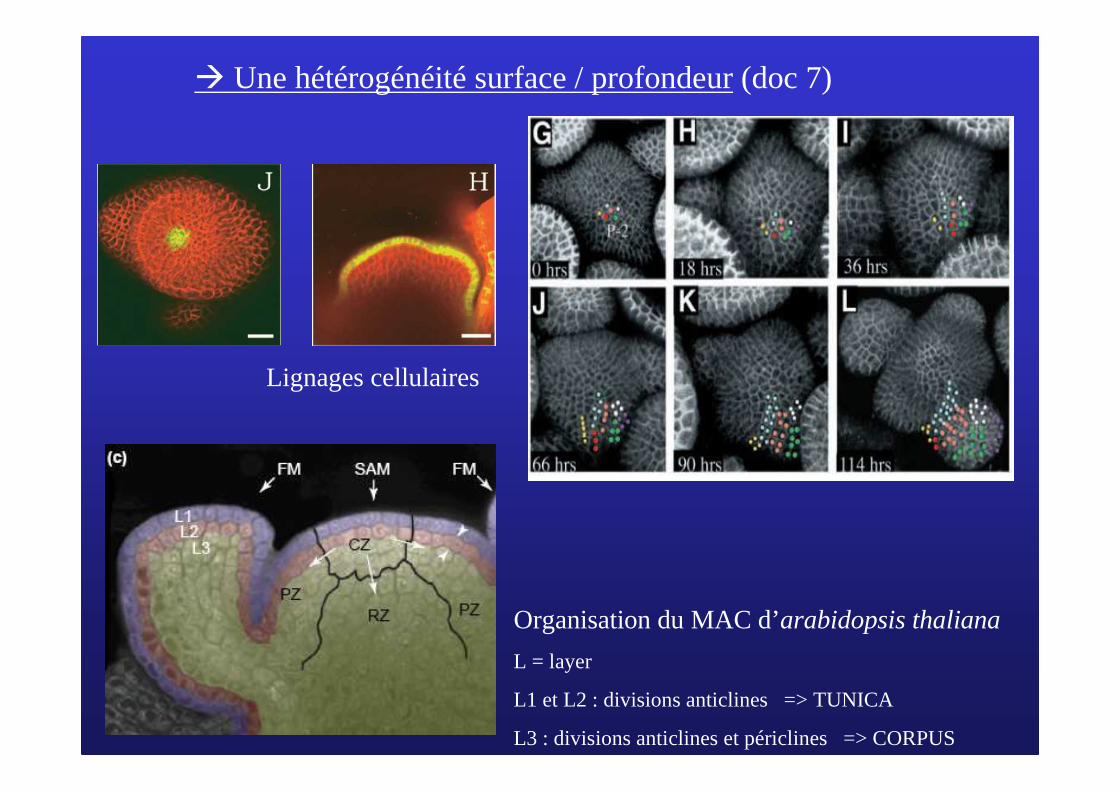
� Une hétérogénéité surface / profondeur(doc 7)
Organisation du MAC d’arabidopsis thaliana
L = layer
L1 et L2 : divisions anticlines => TUNICA
L3 : divisions anticlines et périclines => CORPUS
Lignages cellulaires

� Une zonation cytologique(doc 7)
Organisation du MAC d’Arabidopsis
ZC : zone centrale: sommet inactif, maintien des cellules méristématiques ;
ZP : zone périphérique: activité mitotique maximale, fonctionnement plastochronique et mise en place des feuilles ;
ZM : zone médullaire: histogène, mise en place des tissus de la tige ;
P : primordium
Utiliser l’index mitotique => activitémitotique différentielle
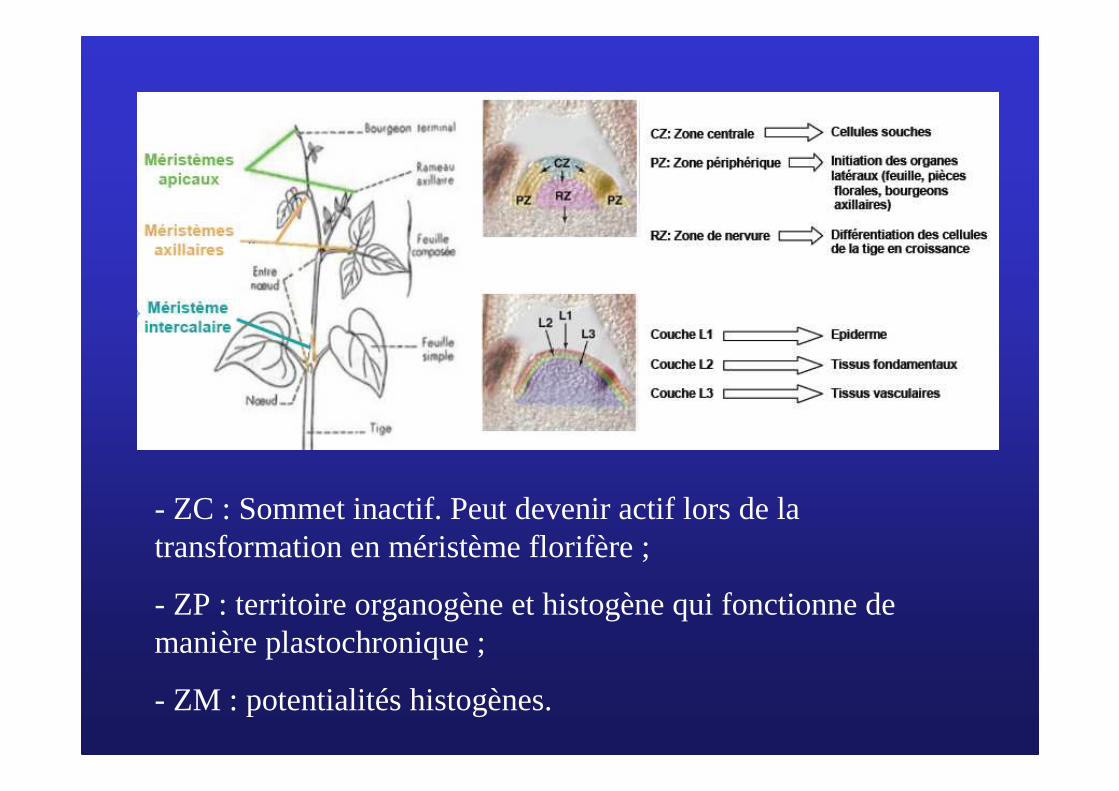
- ZC : Sommet inactif. Peut devenir actif lors de la transformation en méristème florifère ;
- ZP : territoire organogène et histogène qui fonctionne de manière plastochronique ;
- ZM : potentialités histogènes.
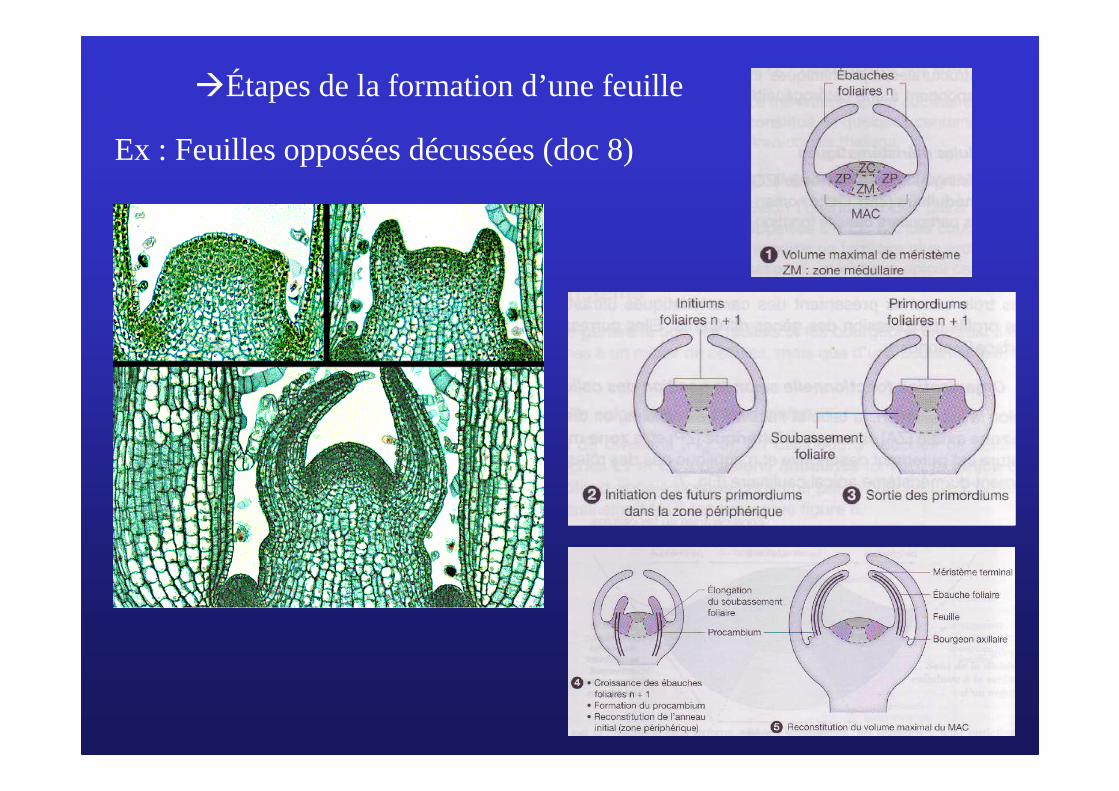
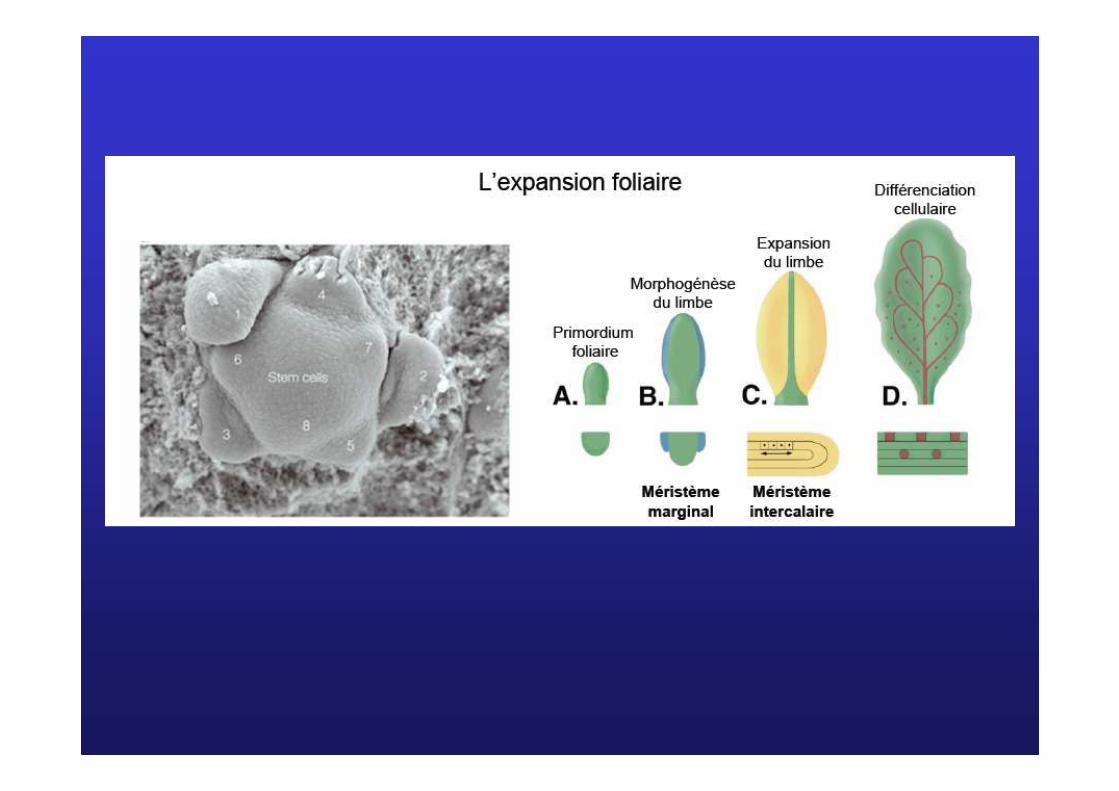
�Étapes de la formation d’une feuille
Ex : Feuilles opposées décussées (doc 8)

C/ Le mC/ Le mééristristèème apical caulinaire (MAC) et son fonctionnementme apical caulinaire (MAC) et son fonctionnement3/ Le contrôle g3/ Le contrôle géénnéétique du fonctionnement du MACtique du fonctionnement du MAC
�Utilisation des mutantsd’Arabidopsis thaliana
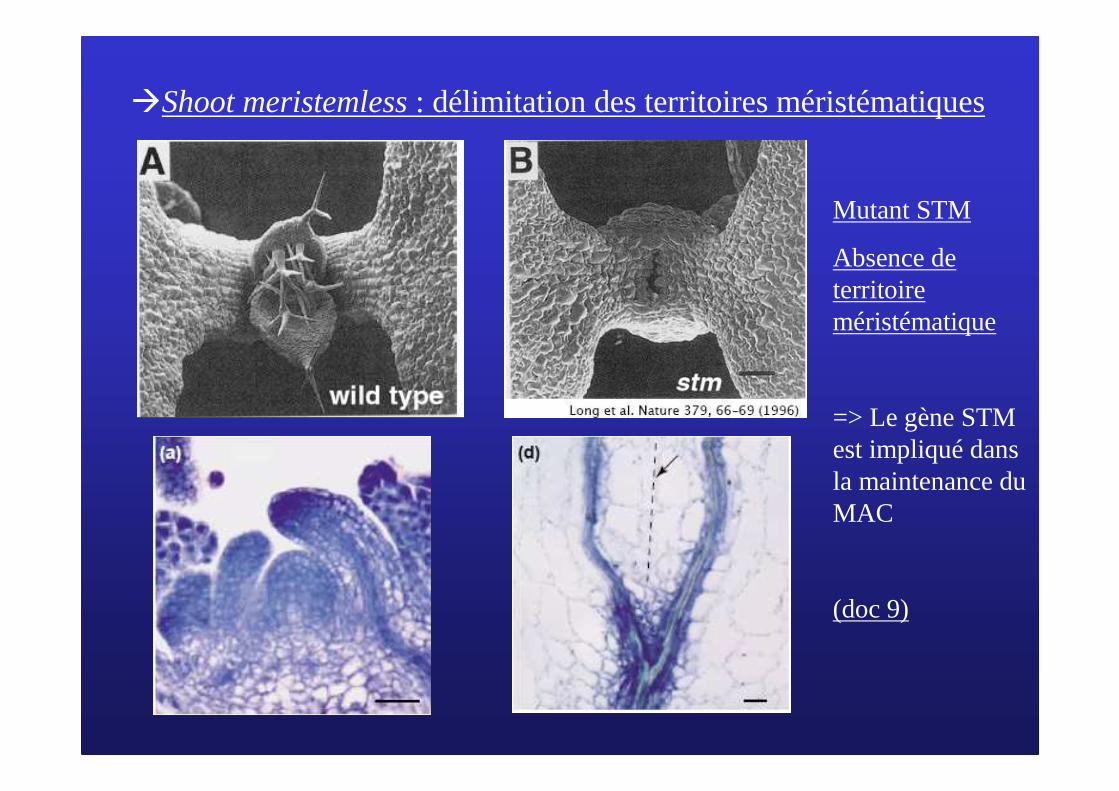
�Shoot meristemless: délimitation des territoires méristématiques
Mutant STM
Absence de territoire méristématique
=> Le gène STM est impliqué dans la maintenance du MAC
(doc 9)
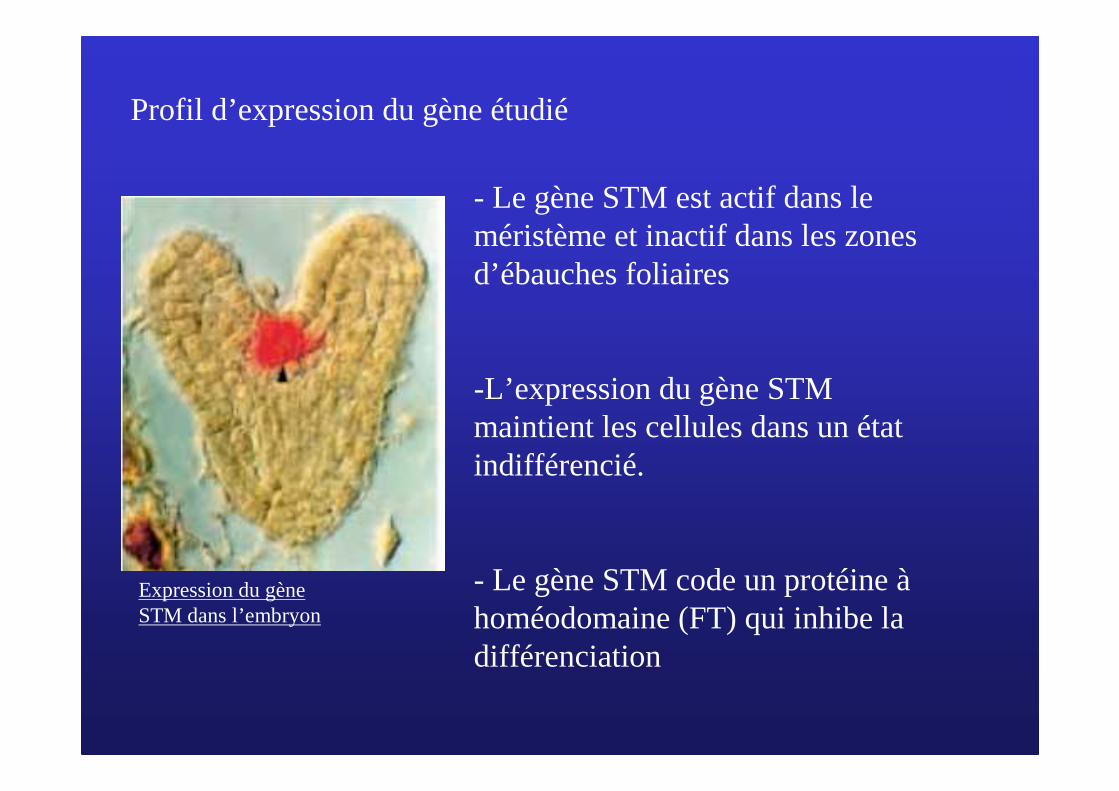
Expression du gène STM dans l’embryon
- Le gène STM est actif dans le méristème et inactif dans les zones d’ébauches foliaires
-L’expression du gène STM maintient les cellules dans un état indifférencié.
- Le gène STM code un protéine àhoméodomaine (FT) qui inhibe la différenciation
Profil d’expression du gène étudié
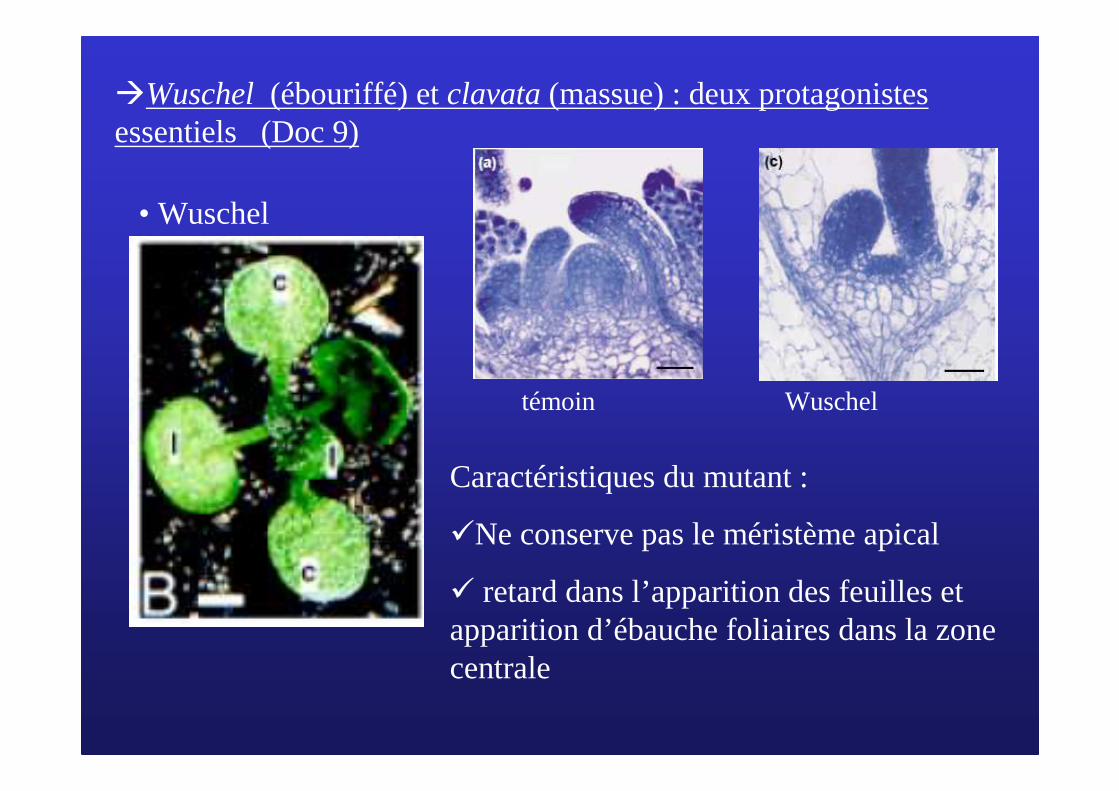
�Wuschel(ébouriffé) et clavata (massue) : deux protagonistes essentiels (Doc 9)
• Wuschel
Caractéristiques du mutant :
�Ne conserve pas le méristème apical
� retard dans l’apparition des feuilles et apparition d’ébauche foliaires dans la zone centrale
Wuscheltémoin
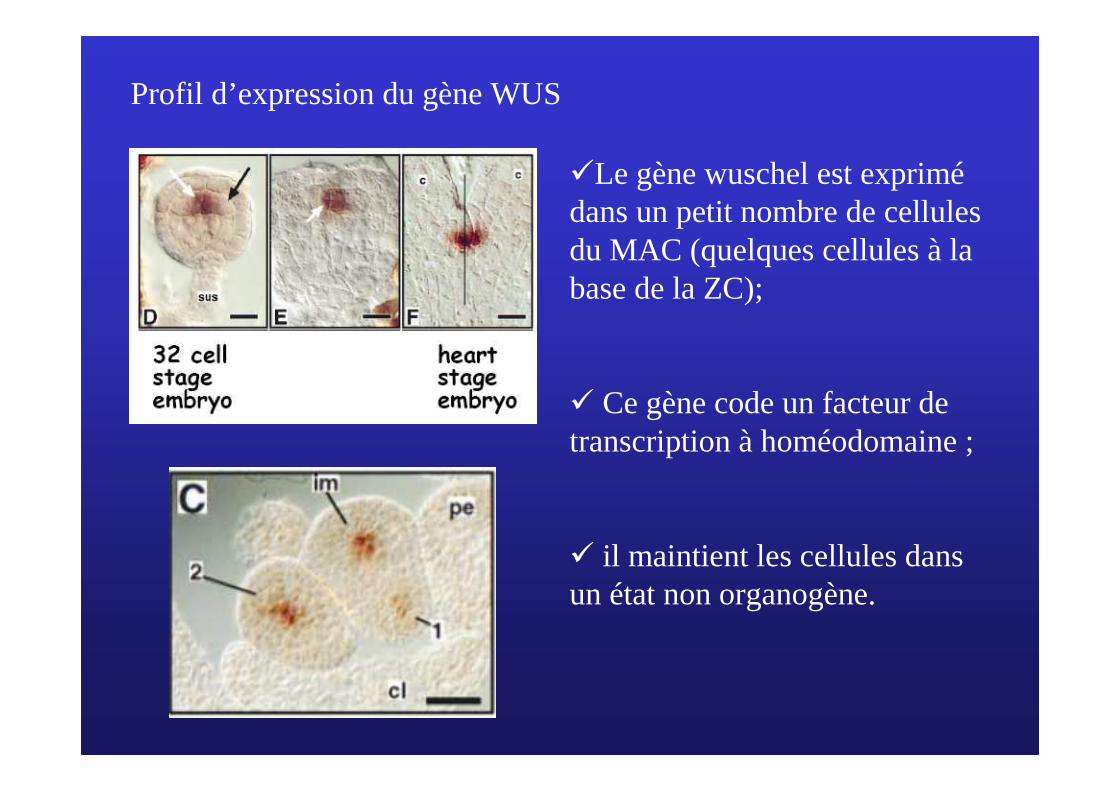
Profil d’expression du gène WUS
�Le gène wuschel est exprimédans un petit nombre de cellules du MAC (quelques cellules à la base de la ZC);
� Ce gène code un facteur de transcription à homéodomaine ;
� il maintient les cellules dans un état non organogène.
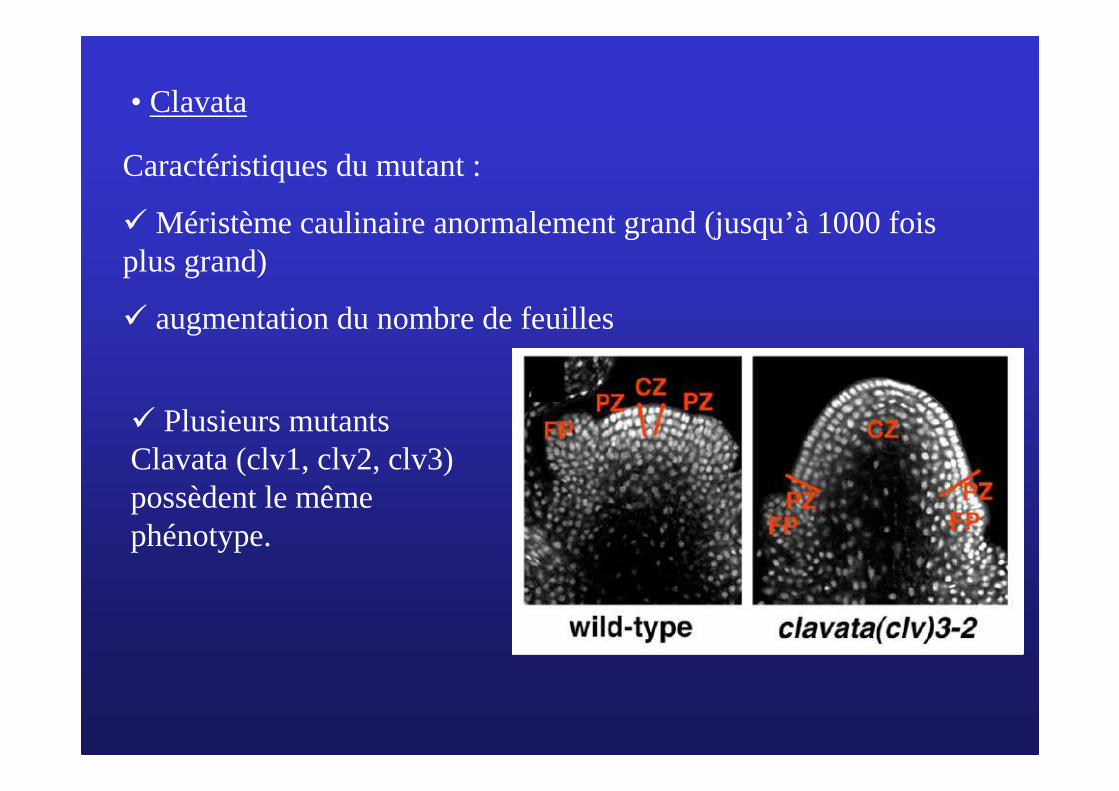
• Clavata
Caractéristiques du mutant :
� Méristème caulinaire anormalement grand (jusqu’à 1000 fois plus grand)
� augmentation du nombre de feuilles
� Plusieurs mutants Clavata (clv1, clv2, clv3) possèdent le même phénotype.

�Les gènes clv1 et clv2 code un récepteur kinase associé à un complexe protéique
� Le gène clv3 code une petite protéine extracellulaire qui peut se lier au récepteur clv1/clv2
� La fixation de clv3 entraine une cascade de réaction qui inhibe l’expression du gène WUS
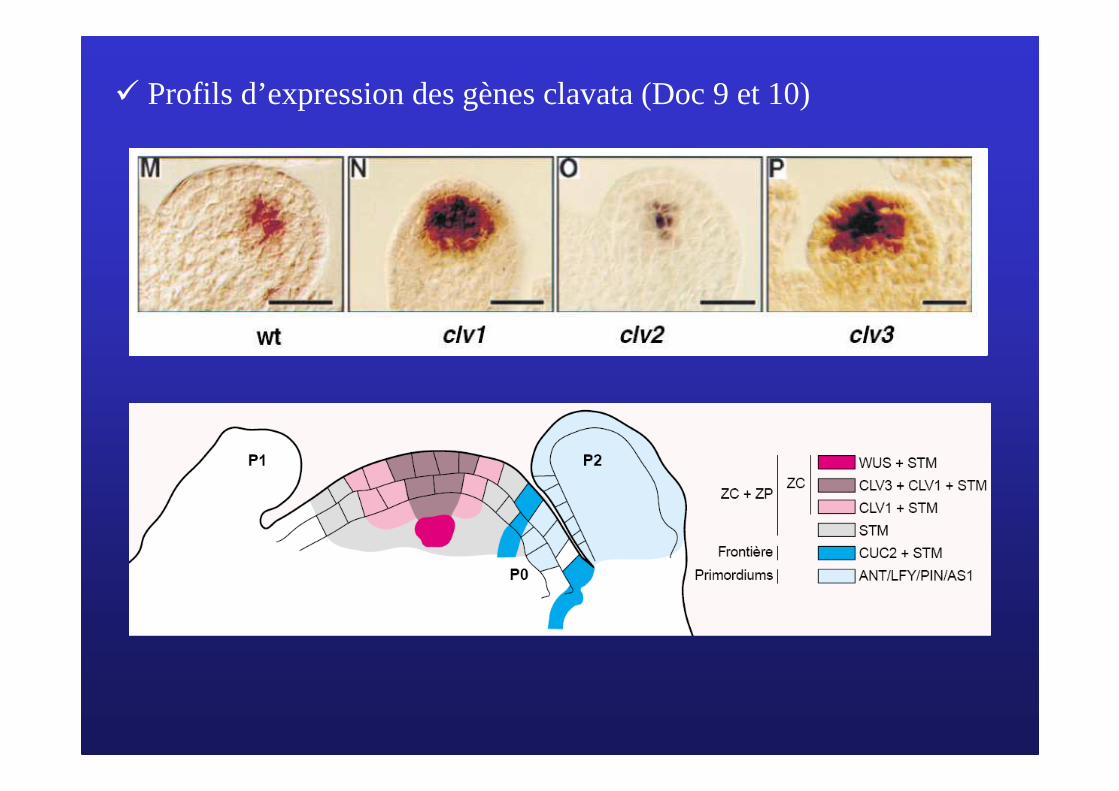
� Profils d’expression des gènes clavata (Doc 9 et 10)
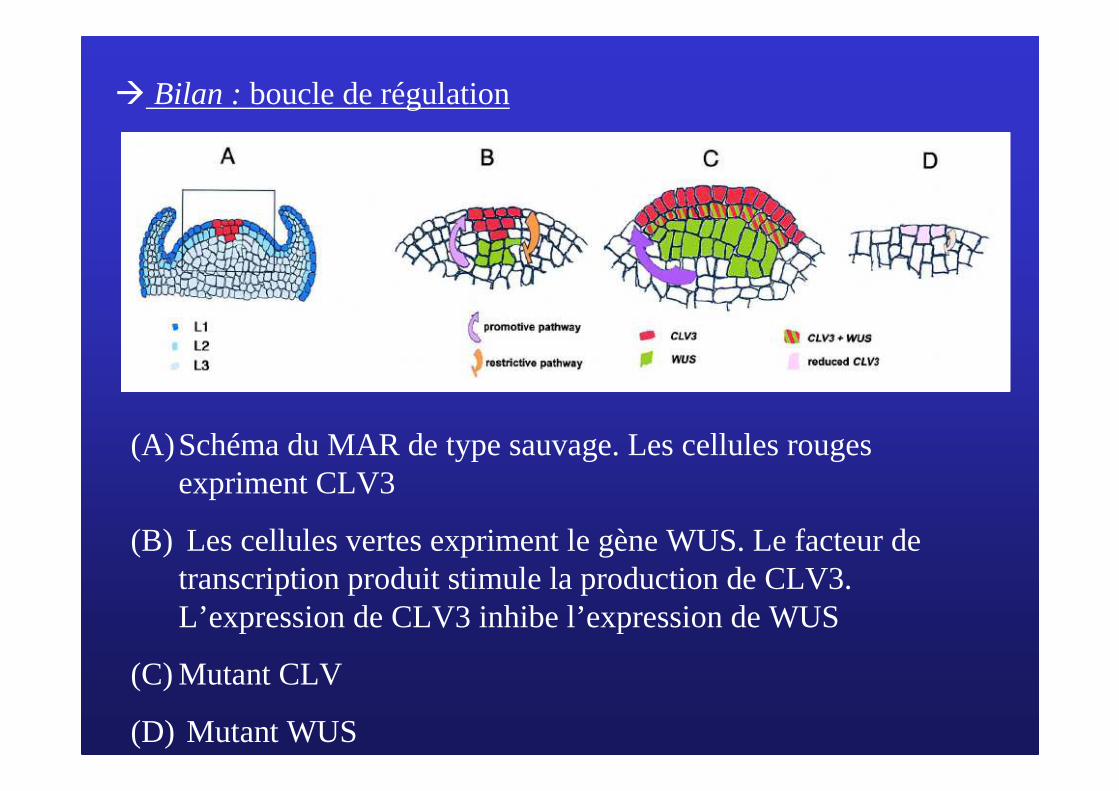
� Bilan : boucle de régulation
(A)Schéma du MAR de type sauvage. Les cellules rouges expriment CLV3
(B) Les cellules vertes expriment le gène WUS. Le facteur de transcription produit stimule la production de CLV3. L’expression de CLV3 inhibe l’expression de WUS
(C) Mutant CLV
(D) Mutant WUS
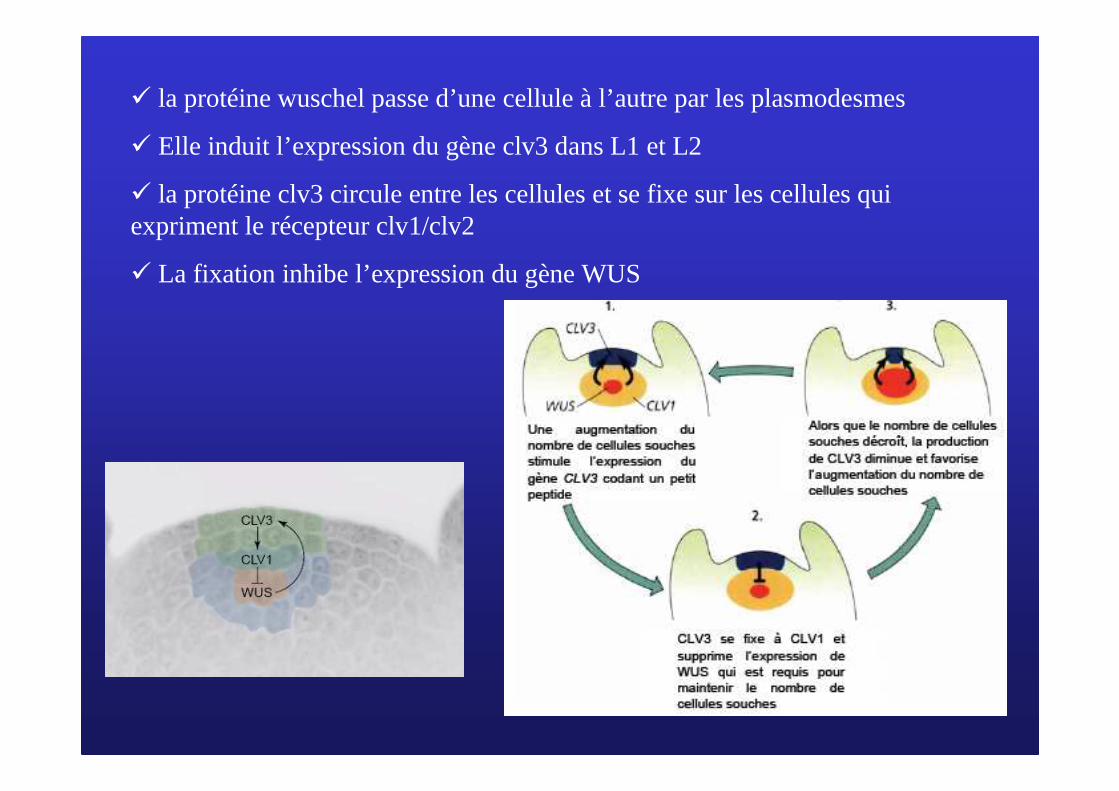
� la protéine wuschel passe d’une cellule à l’autre par les plasmodesmes
� Elle induit l’expression du gène clv3 dans L1 et L2
� la protéine clv3 circule entre les cellules et se fixe sur les cellules qui expriment le récepteur clv1/clv2
� La fixation inhibe l’expression du gène WUS
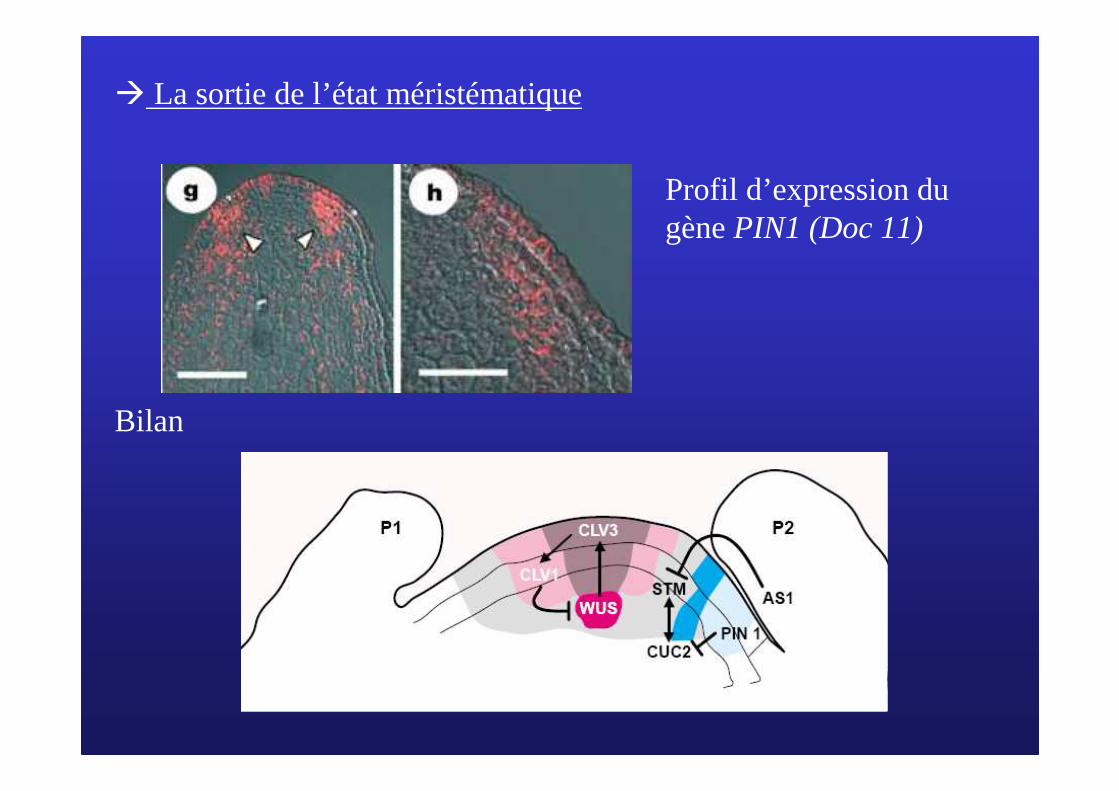
� La sortie de l’état méristématique
Profil d’expression du gène PIN1 (Doc 11)
Bilan
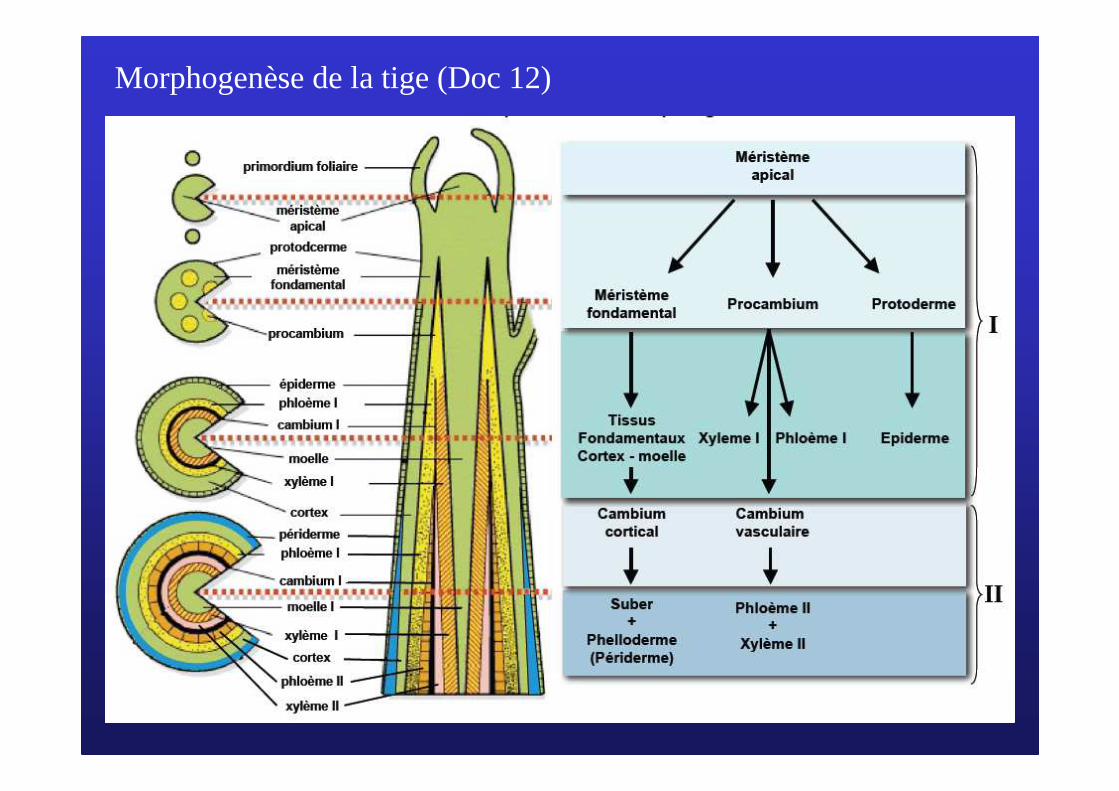
Morphogenèse de la tige (Doc 12)

II/ Les phII/ Les phéénomnomèènes cellulaires de la croissance : grandissement nes cellulaires de la croissance : grandissement cellulaire et histogencellulaire et histogenèèsese
A/ MA/ Méécanisme cellulaire de canisme cellulaire de ll ’’ auxauxèèsese1/ Caract1/ Caractééristiques cellulairesristiques cellulaires
Les petites vacuoles fusionnent en une grosse vacuole qui occupe pratiquement tout le cytoplasme
Éloignement progressif de l’apex (racine)
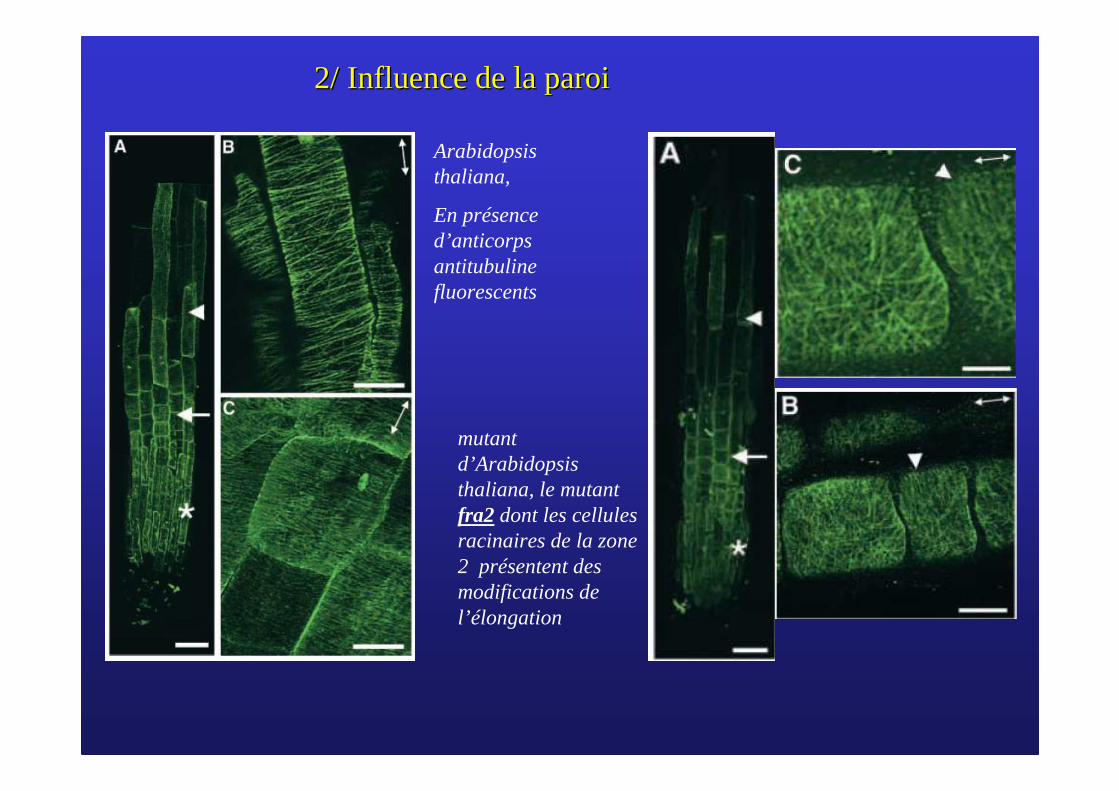
2/ Influence de la paroi2/ Influence de la paroi
mutantd’Arabidopsis thaliana, le mutant fra2 dont les cellulesracinaires de la zone 2 présentent des modifications de l’élongation
Arabidopsis thaliana,
En présence d’anticorps antitubuline fluorescents

Pourcentage de microfibrilles de cellulose par rapport au nombre total pour chacun des angles. La valeur 0 correspond arbitrairement aux microfibrillesperpendiculaires à l’axe de croissance.
L’orientation des microfibrilles de cellulose détermine le sens d’élongation
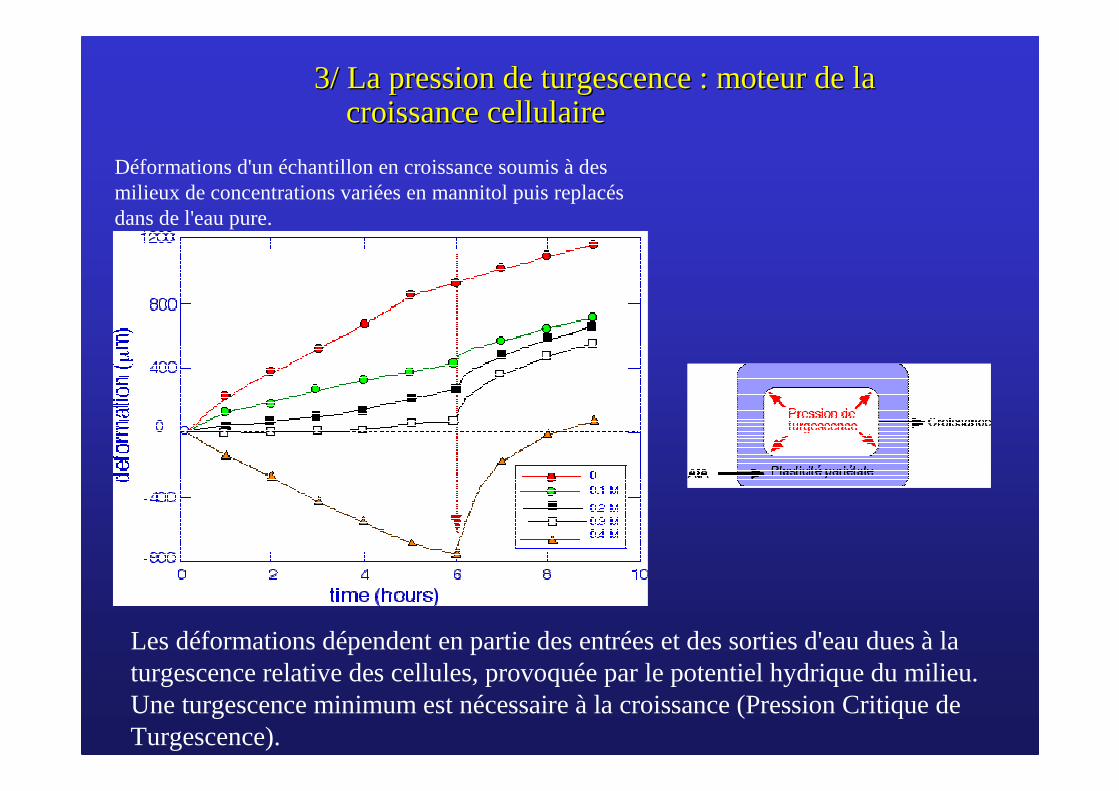
3/ La pression de turgescence : moteur de la 3/ La pression de turgescence : moteur de la croissance cellulairecroissance cellulaire
Déformations d'un échantillon en croissance soumis à des milieux de concentrations variées en mannitol puis replacés dans de l'eau pure.
Les déformations dépendent en partie des entrées et des sorties d'eau dues à la turgescence relative des cellules, provoquée par le potentiel hydrique du milieu. Une turgescence minimum est nécessaire à la croissance (Pression Critique de Turgescence).
II/ Les phII/ Les phéénomnomèènes cellulaires de la croissance : grandissement nes cellulaires de la croissance : grandissement cellulaire et histogencellulaire et histogenèèsese
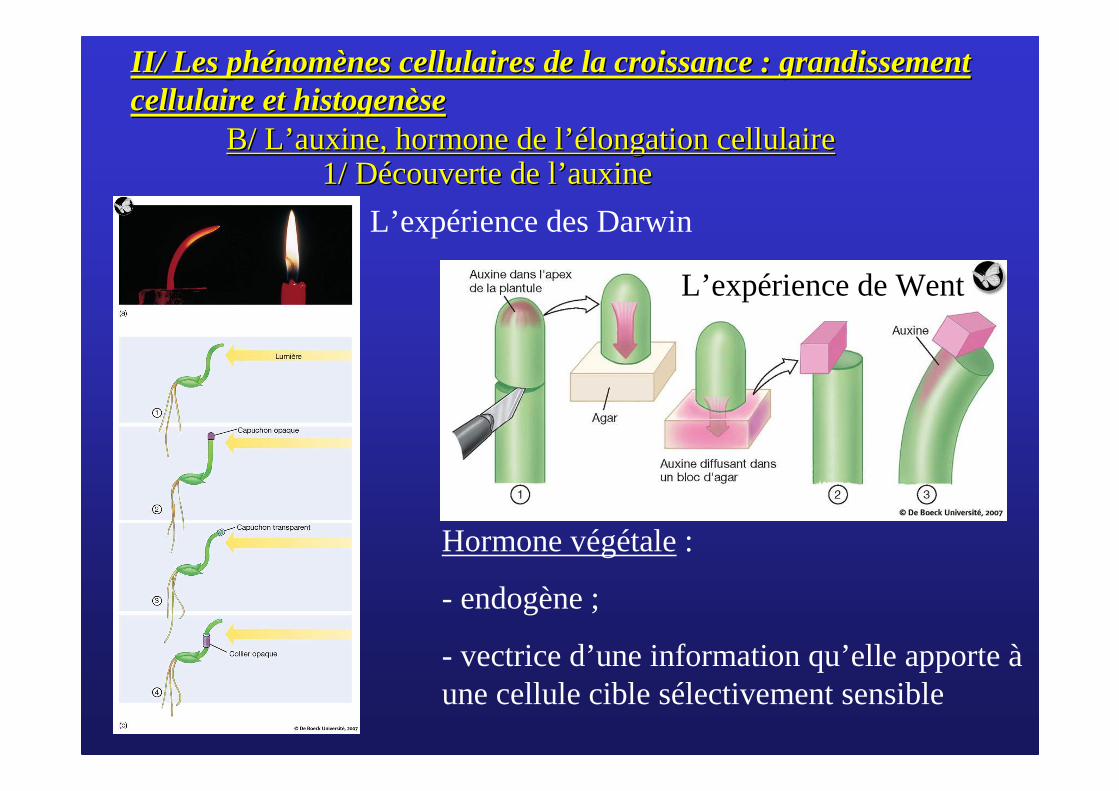
B/ LB/ L’’ auxine, hormone de lauxine, hormone de l’é’élongation cellulairelongation cellulaire1/ D1/ Déécouverte de lcouverte de l’’ auxineauxine
L’expérience des Darwin
Hormone végétale:
- endogène ;
- vectrice d’une information qu’elle apporte àune cellule cible sélectivement sensible
L’expérience de Went
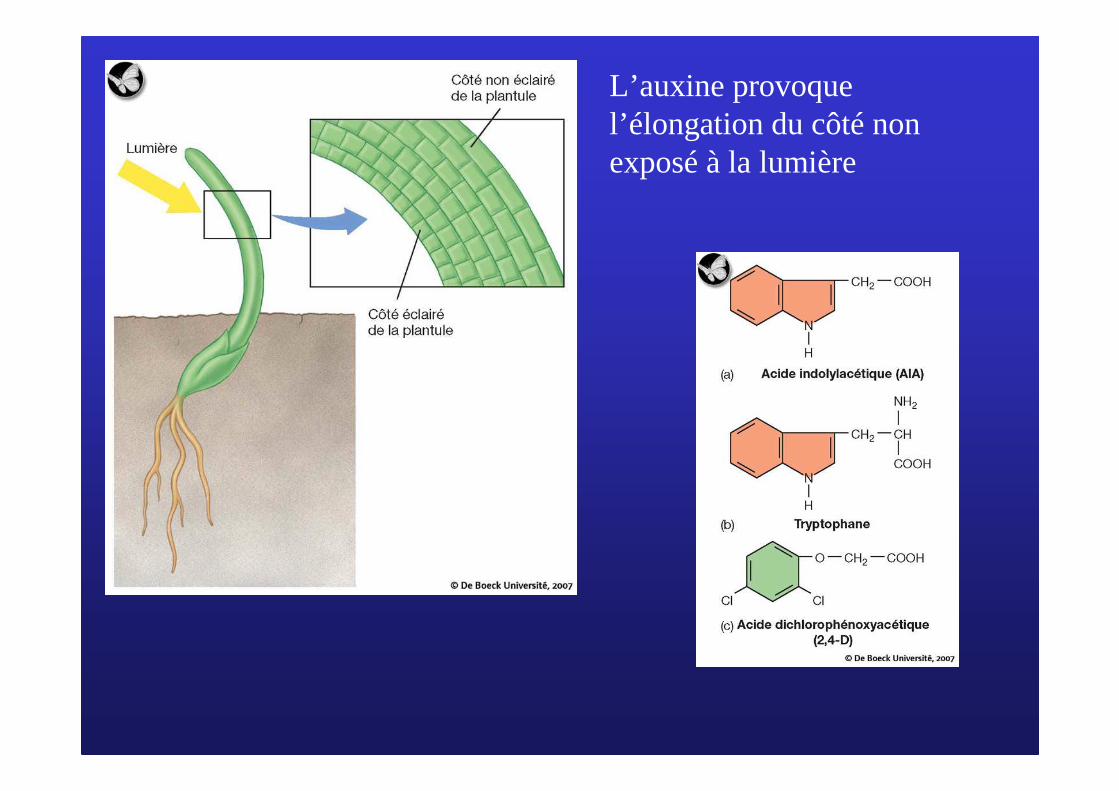
L’auxine provoque l’élongation du côté non exposé à la lumière
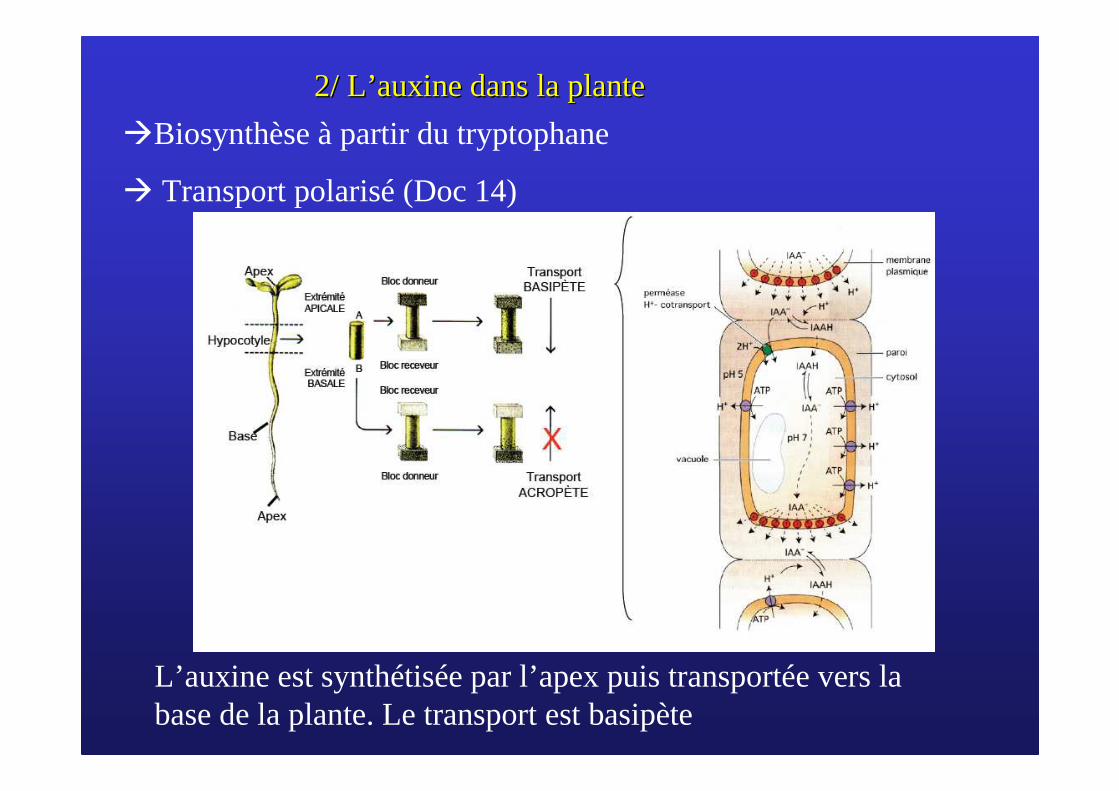
2/ L2/ L’’ auxine dans la planteauxine dans la plante
�Biosynthèse à partir du tryptophane
� Transport polarisé (Doc 14)
L’auxine est synthétisée par l’apex puis transportée vers la base de la plante. Le transport est basipète
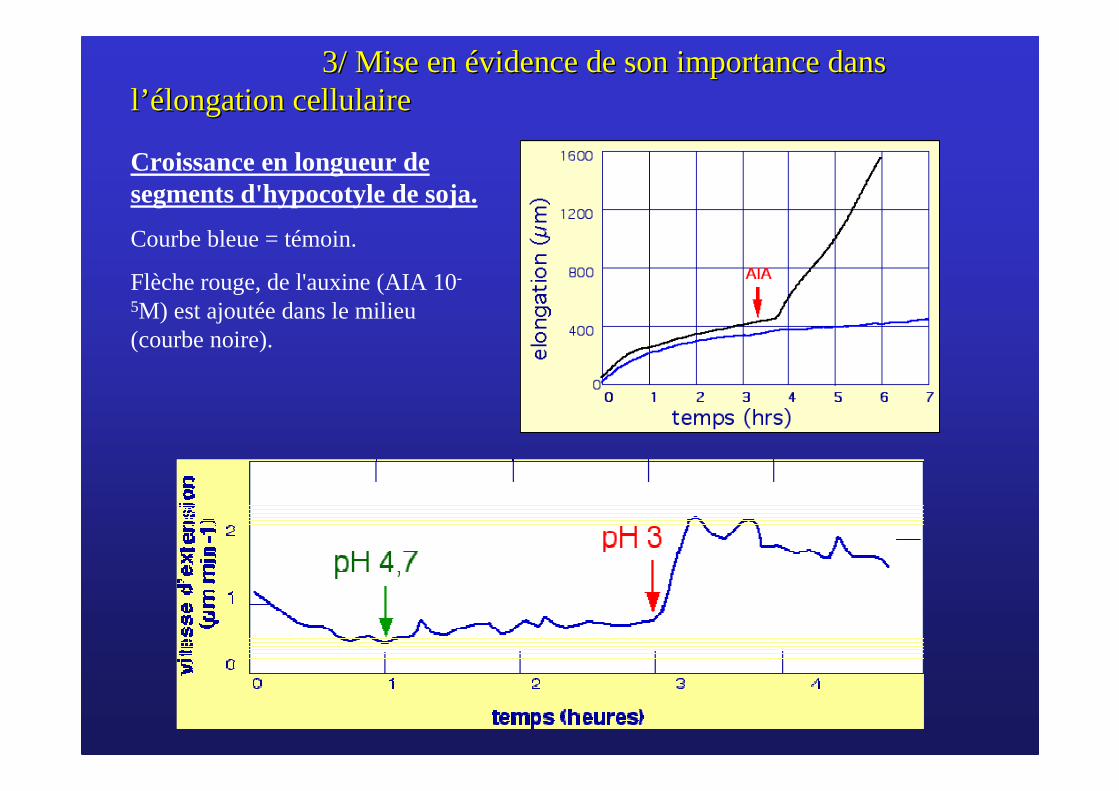
3/ Mise en 3/ Mise en éévidence de son importance dans vidence de son importance dans ll ’é’élongation cellulairelongation cellulaire
Croissance en longueur de segments d'hypocotyle de soja.
Courbe bleue = témoin.
Flèche rouge, de l'auxine (AIA 10-
5M) est ajoutée dans le milieu (courbe noire).
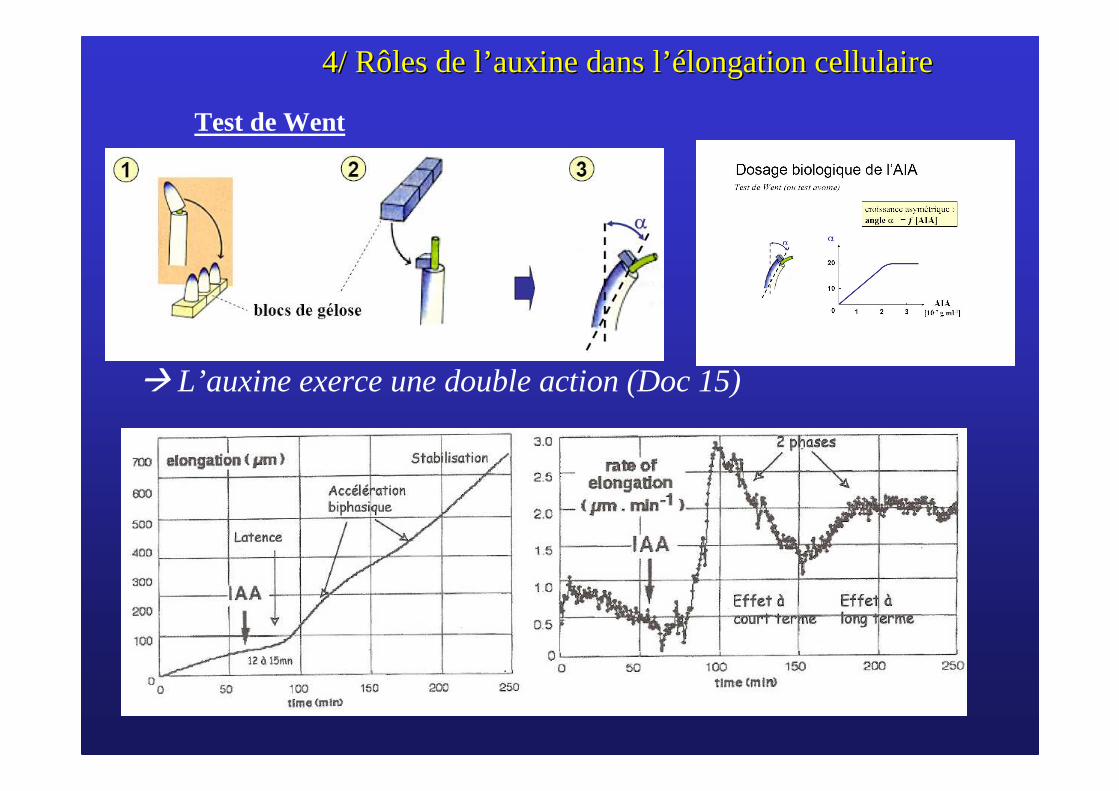
4/ Rôles de l4/ Rôles de l’’ auxine dans lauxine dans l’é’élongation cellulairelongation cellulaire
� L’auxine exerce une double action (Doc 15)
Test de Went
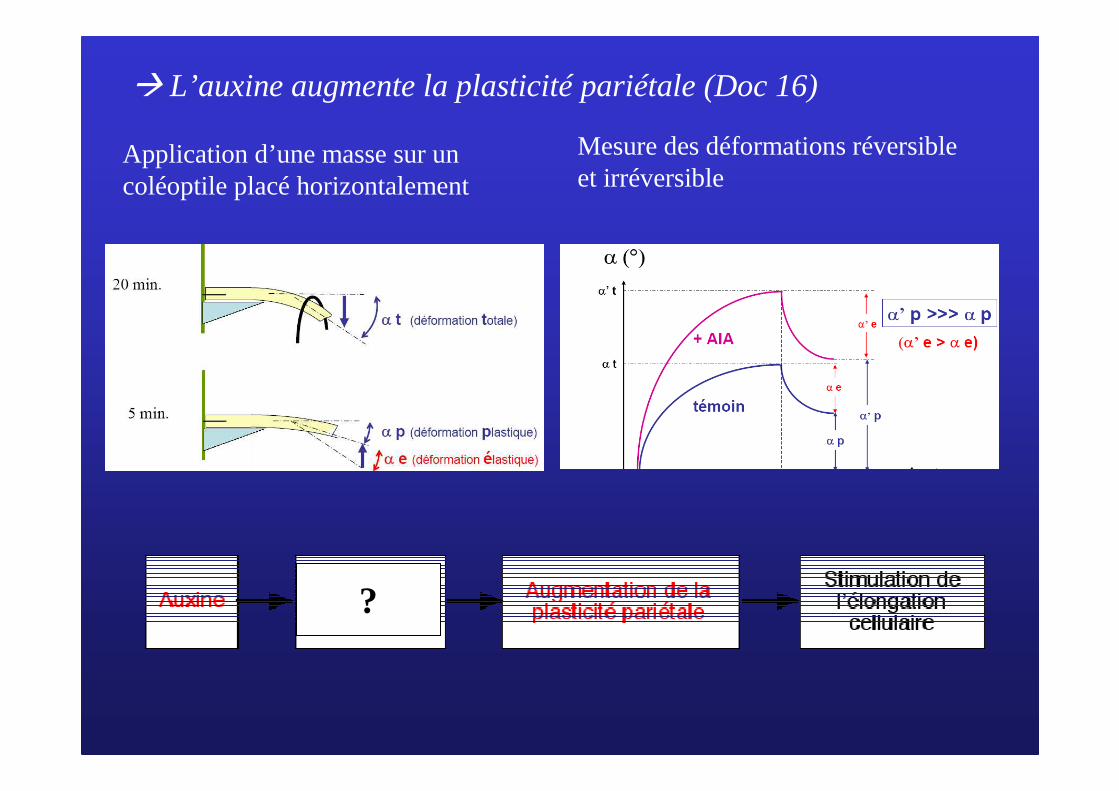
� L’auxine augmente la plasticité pariétale (Doc 16)
Application d’une masse sur un coléoptile placé horizontalement
Mesure des déformations réversible et irréversible
?
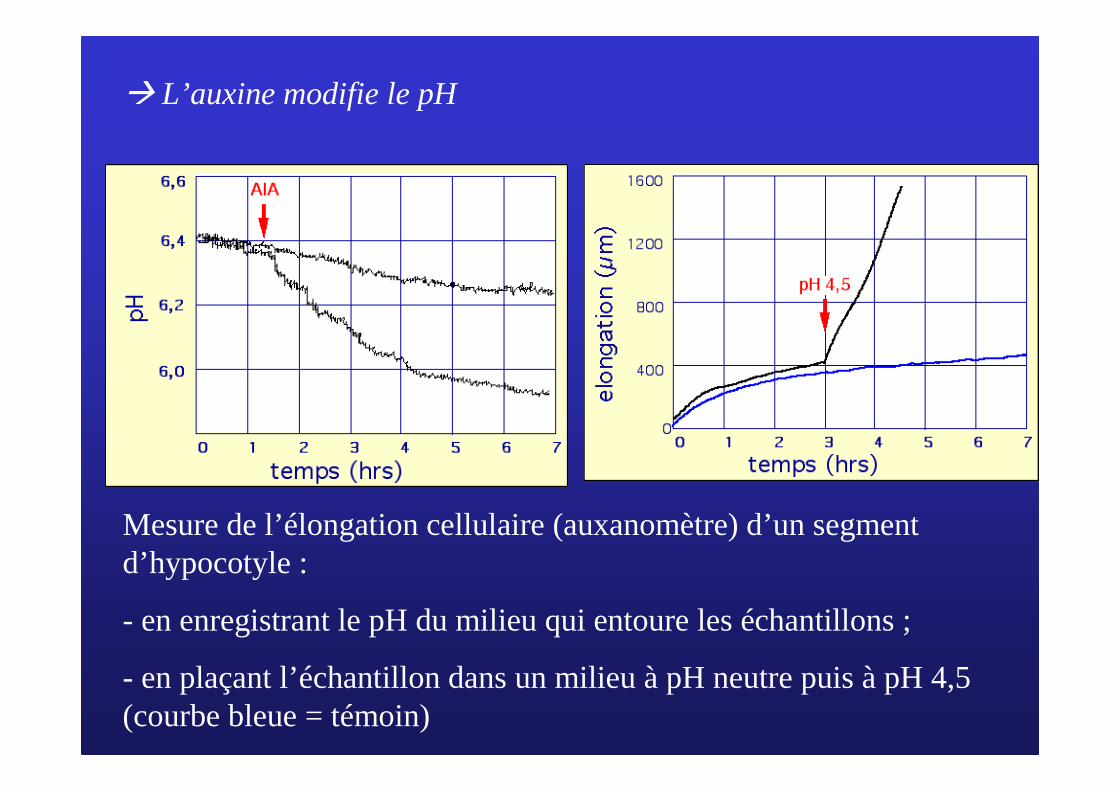
� L’auxine modifie le pH
Mesure de l’élongation cellulaire (auxanomètre) d’un segment d’hypocotyle :
- en enregistrant le pH du milieu qui entoure les échantillons ;
- en plaçant l’échantillon dans un milieu à pH neutre puis à pH 4,5 (courbe bleue = témoin)
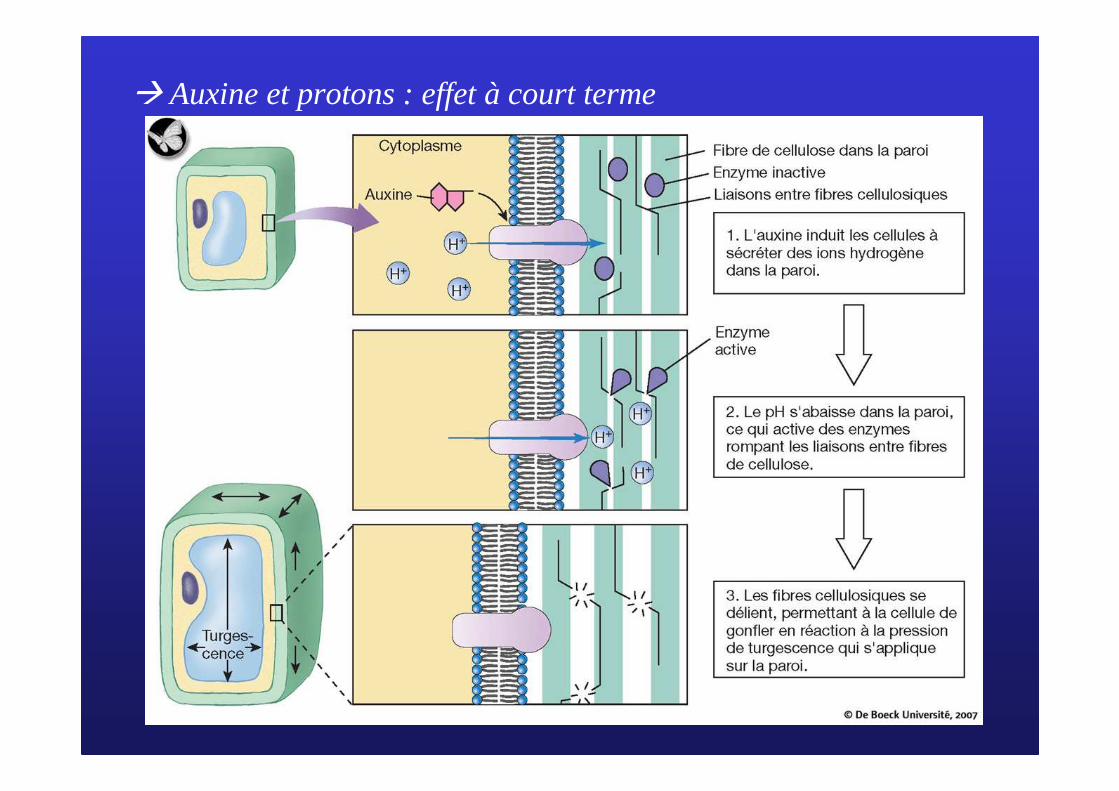
� Auxine et protons : effet à court terme
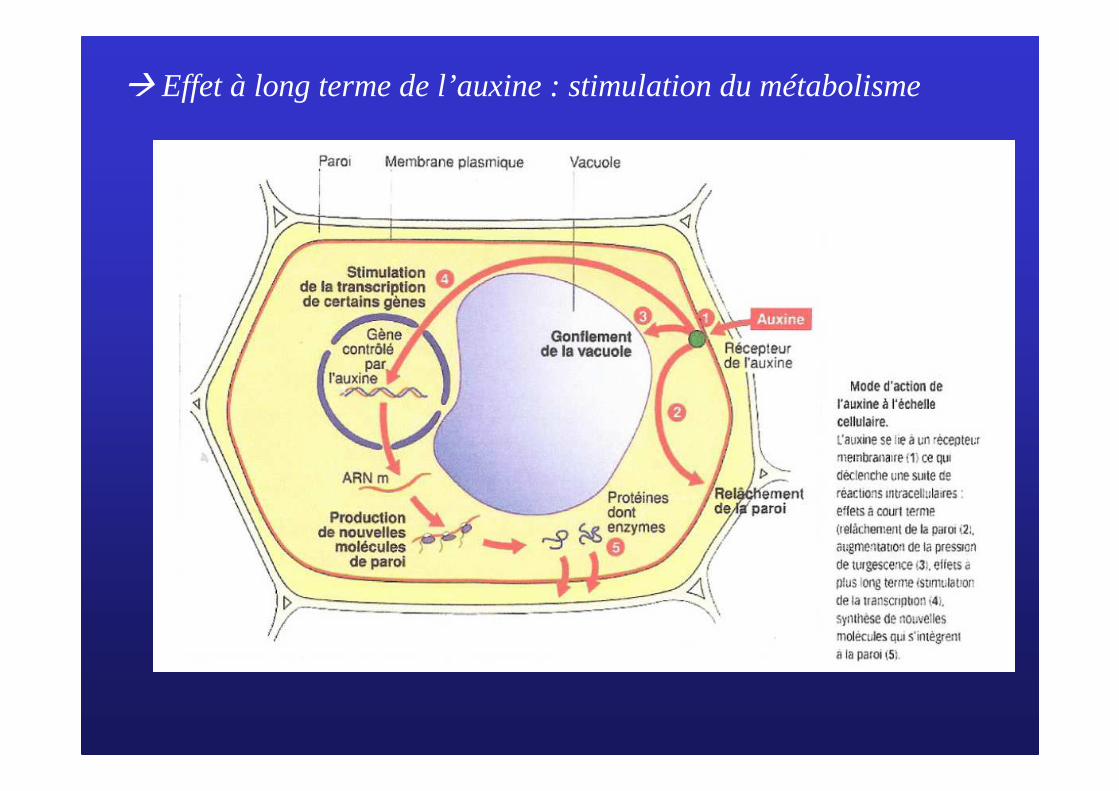
� Effet à long terme de l’auxine : stimulation du métabolisme
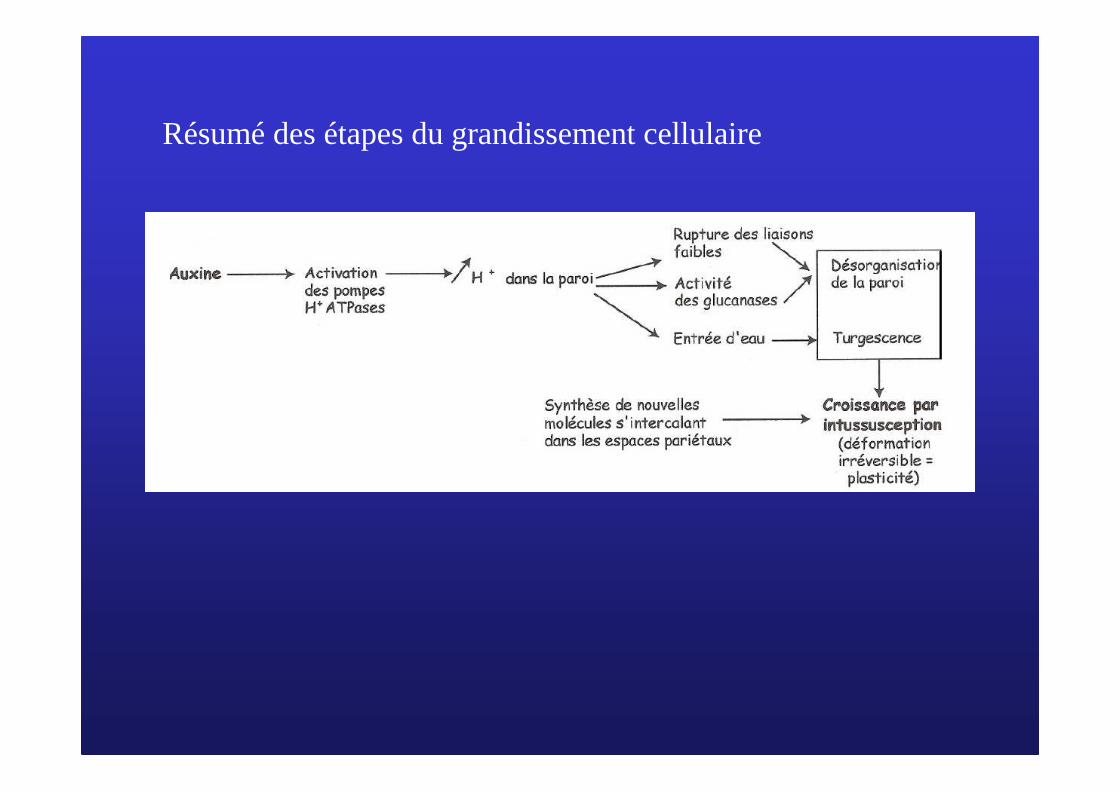
Résumé des étapes du grandissement cellulaire
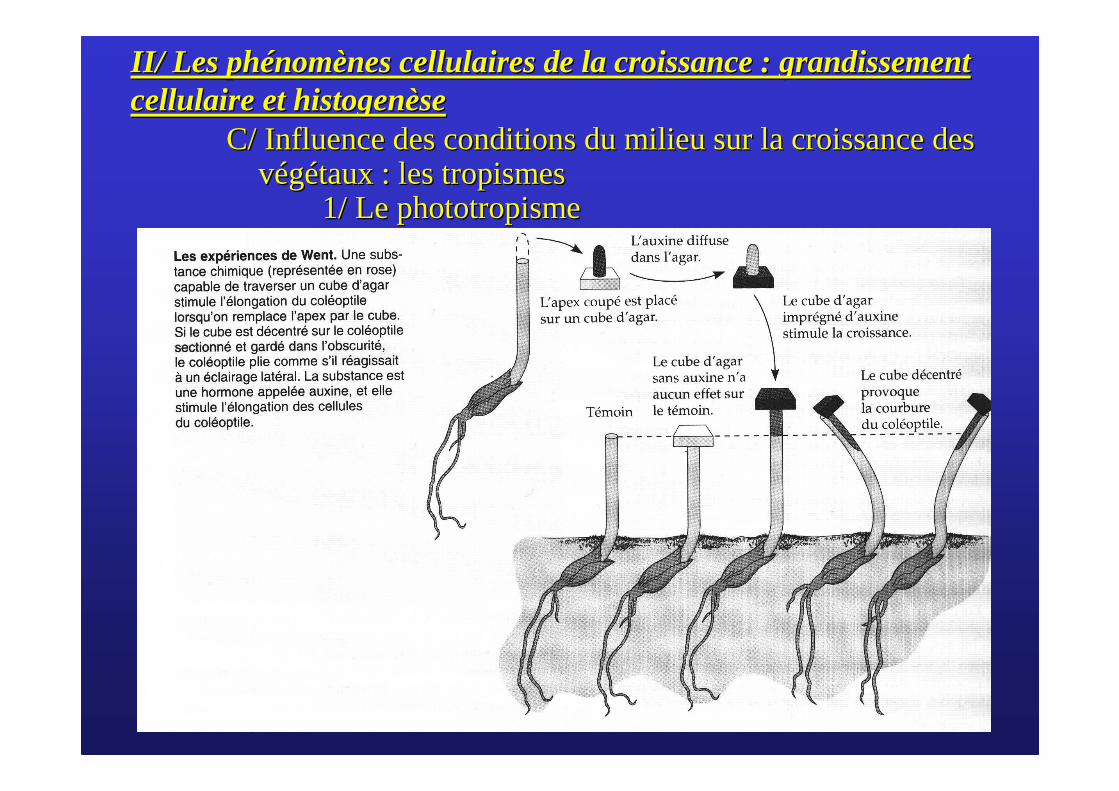
II/ Les phII/ Les phéénomnomèènes cellulaires de la croissance : grandissement nes cellulaires de la croissance : grandissement cellulaire et histogencellulaire et histogenèèsese
C/ Influence des conditions du milieu sur la croissance des C/ Influence des conditions du milieu sur la croissance des vvééggéétaux : les tropismestaux : les tropismes
1/ Le phototropisme1/ Le phototropisme
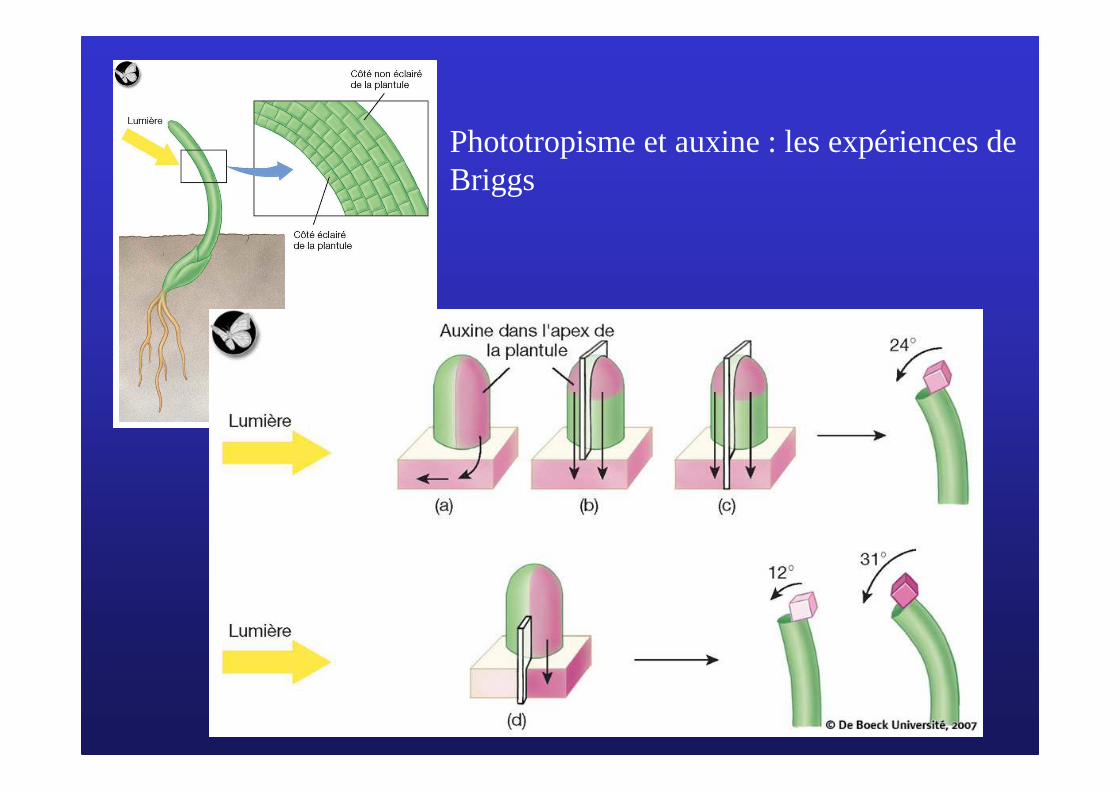
Phototropisme et auxine : les expériences de Briggs
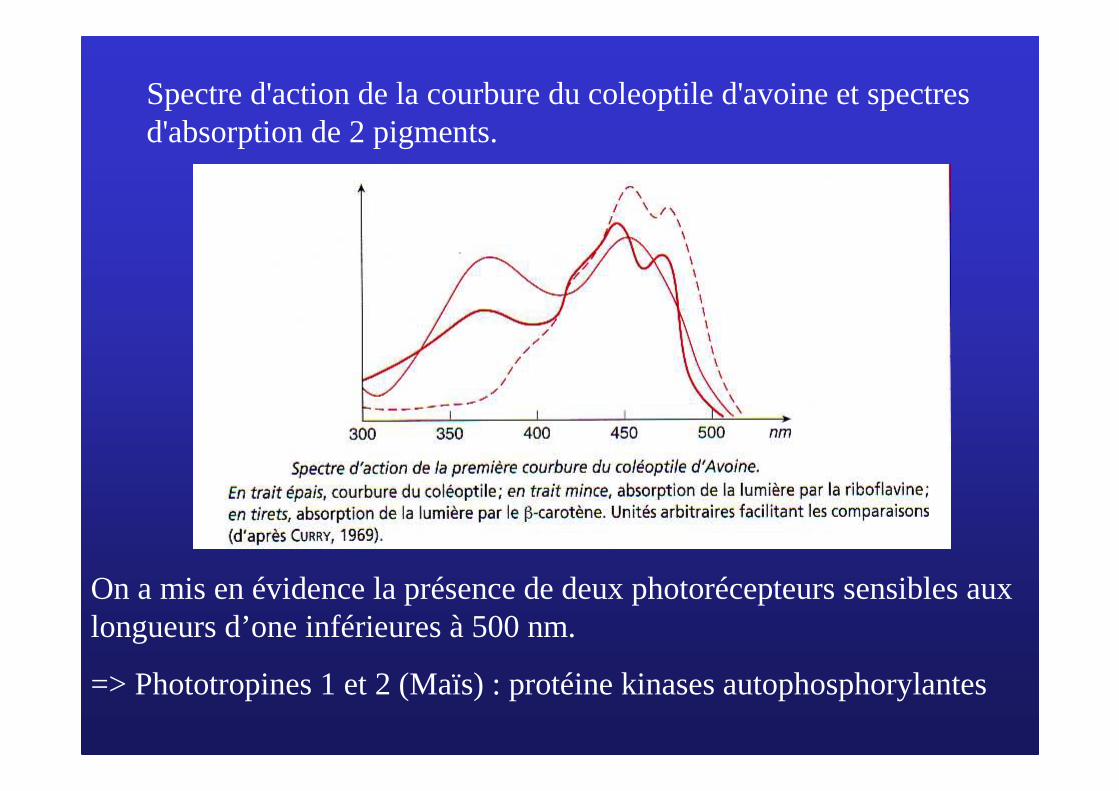
Spectre d'action de la courbure du coleoptile d'avoine et spectres d'absorption de 2 pigments.
On a mis en évidence la présence de deux photorécepteurs sensibles aux longueurs d’one inférieures à 500 nm.
=> Phototropines 1 et 2 (Maïs) : protéine kinases autophosphorylantes
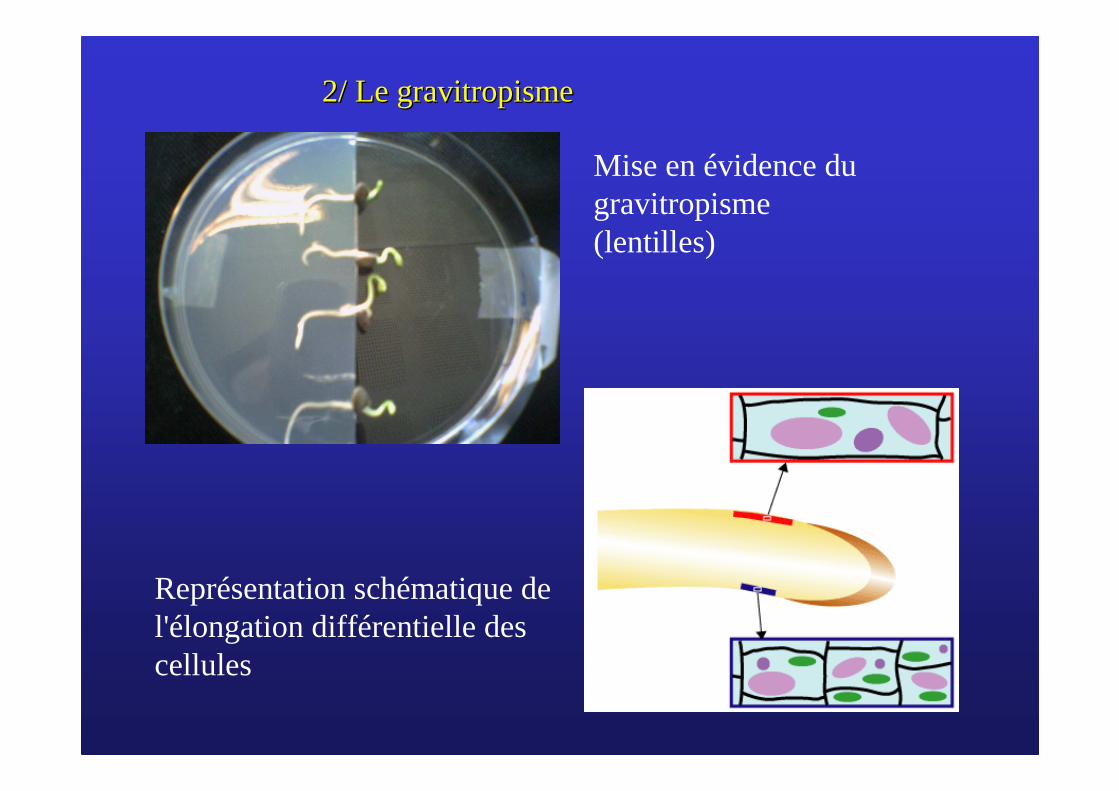
2/ Le 2/ Le gravitropismegravitropisme
Mise en évidence du gravitropisme (lentilles)
Représentation schématique de l'élongation différentielle des cellules
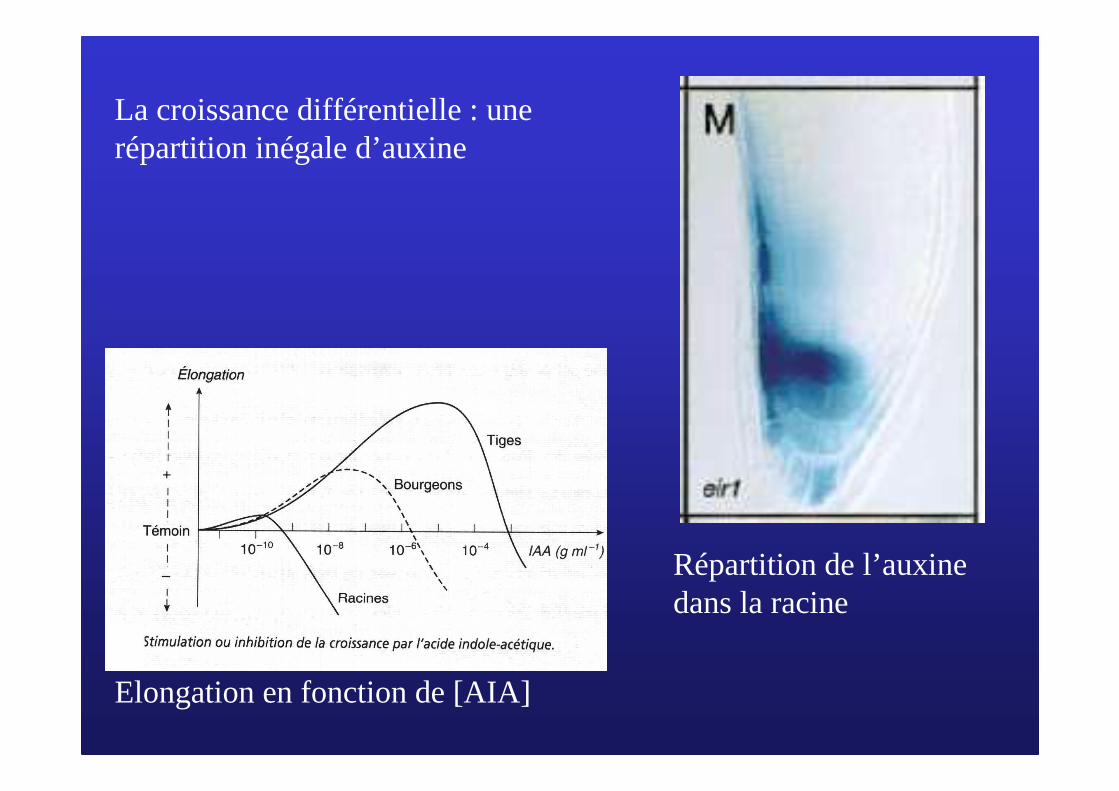
Elongation en fonction de [AIA]
Répartition de l’auxine dans la racine
La croissance différentielle : une répartition inégale d’auxine
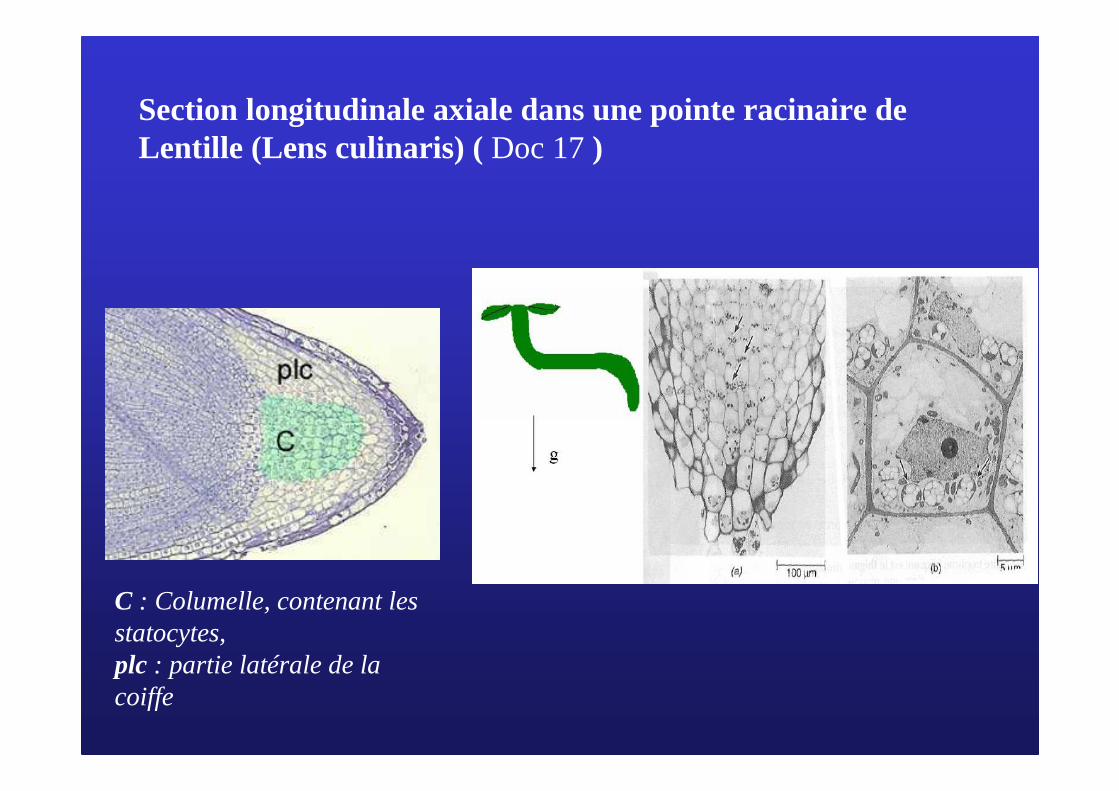
Section longitudinale axiale dans une pointe racinaire de Lentille (Lens culinaris) ( Doc 17)
C : Columelle, contenant lesstatocytes,plc : partie latérale de la coiffe
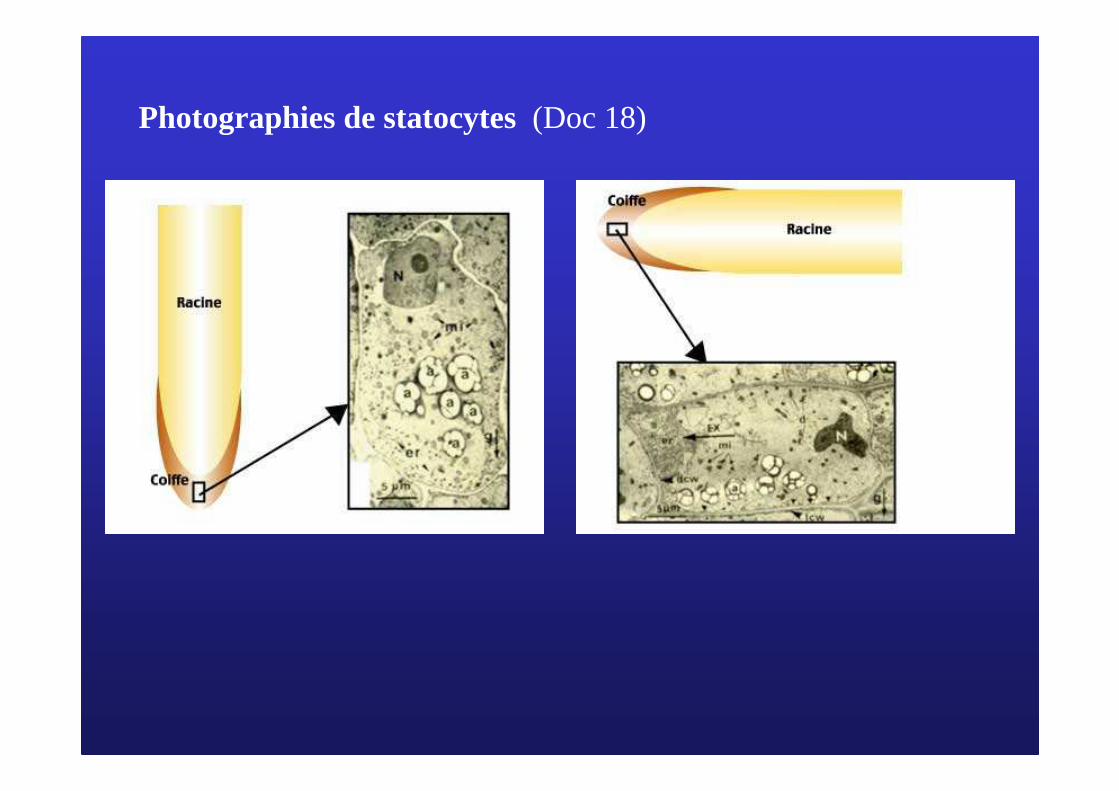
Photographies de statocytes(Doc 18)
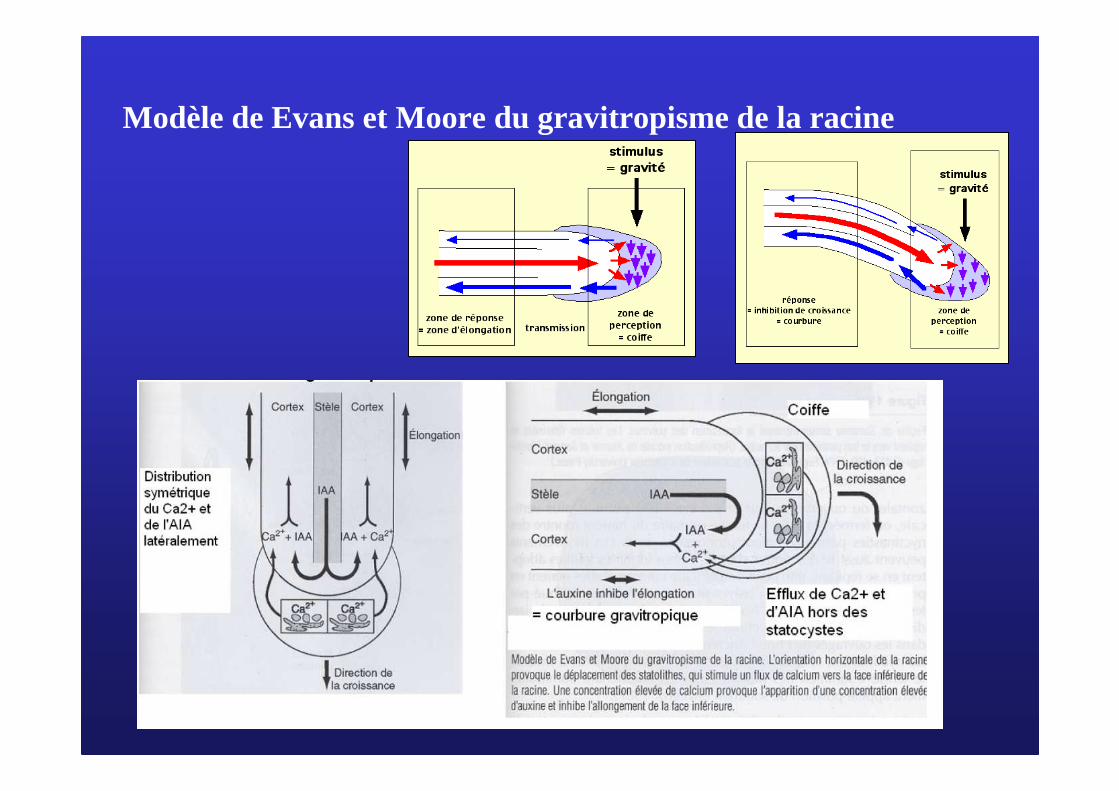
Modèle de Evans et Moore du gravitropisme de la racine

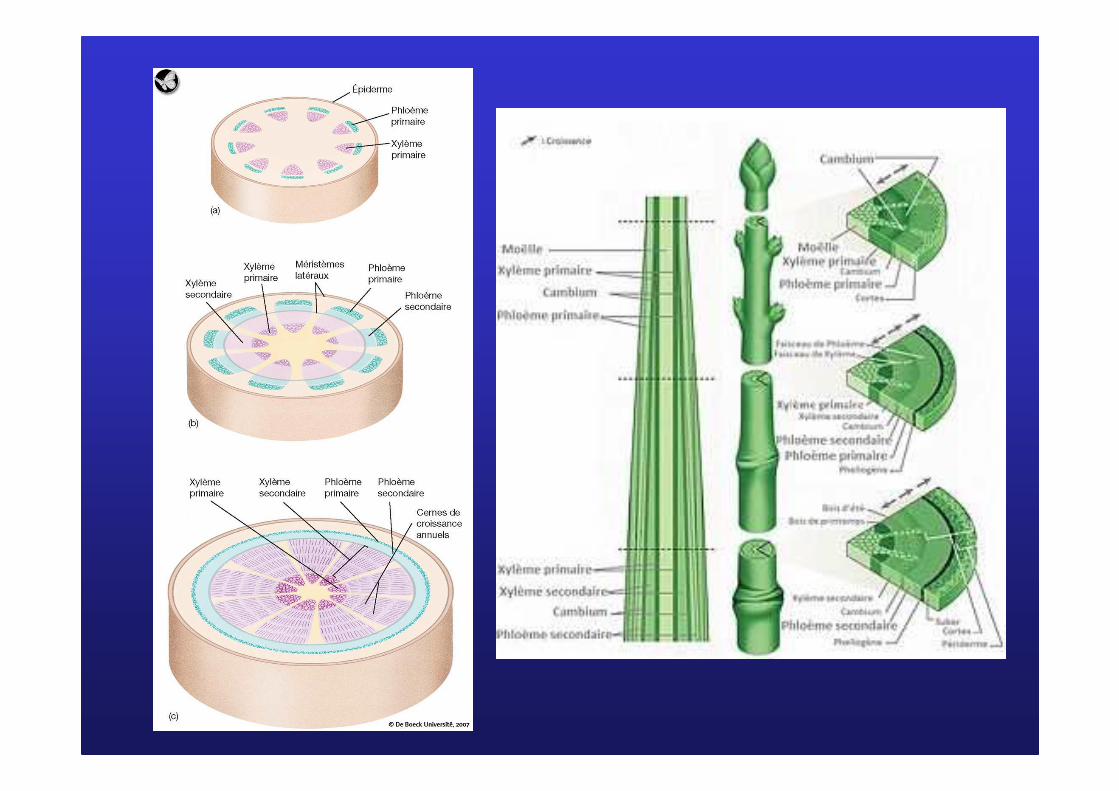
III/ MIII/ M ééristrist èèmes secondaires et croissance en mes secondaires et croissance en éépaisseurpaisseurA/ Dans la tigeA/ Dans la tige
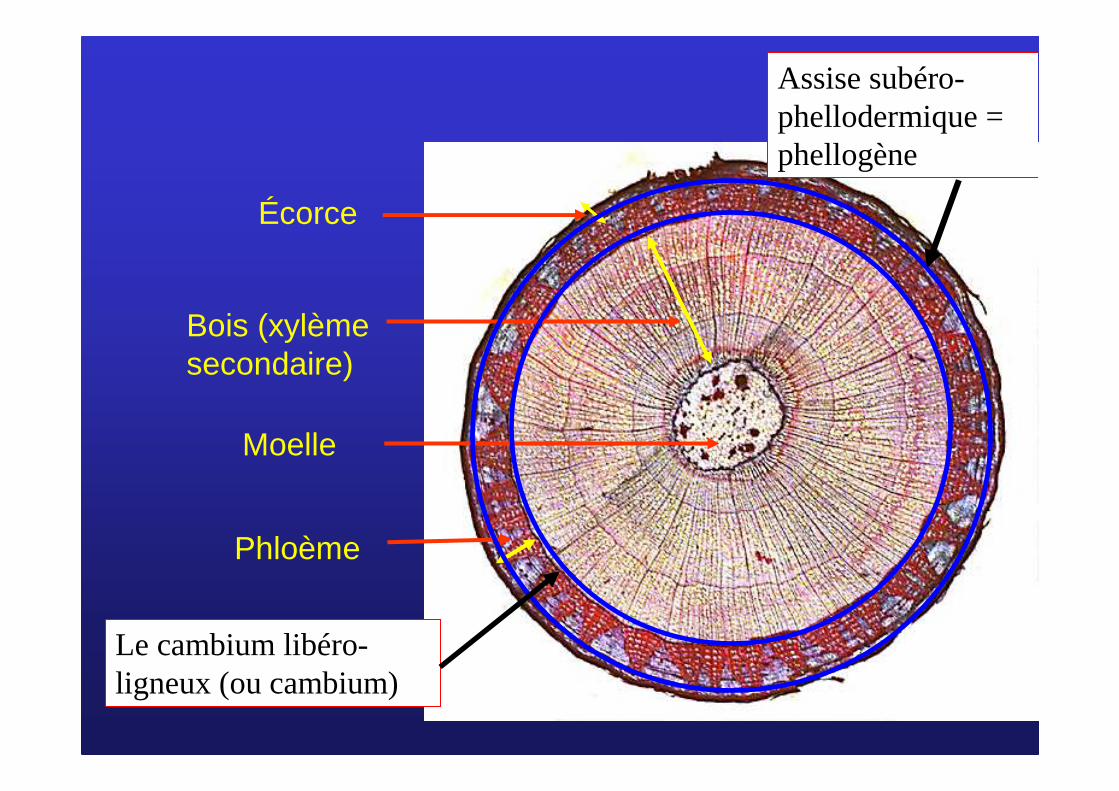
Écorce
Bois (xylème secondaire)
Moelle
Phloème
Le cambium libéro-ligneux (ou cambium)
Assise subéro-phellodermique = phellogène
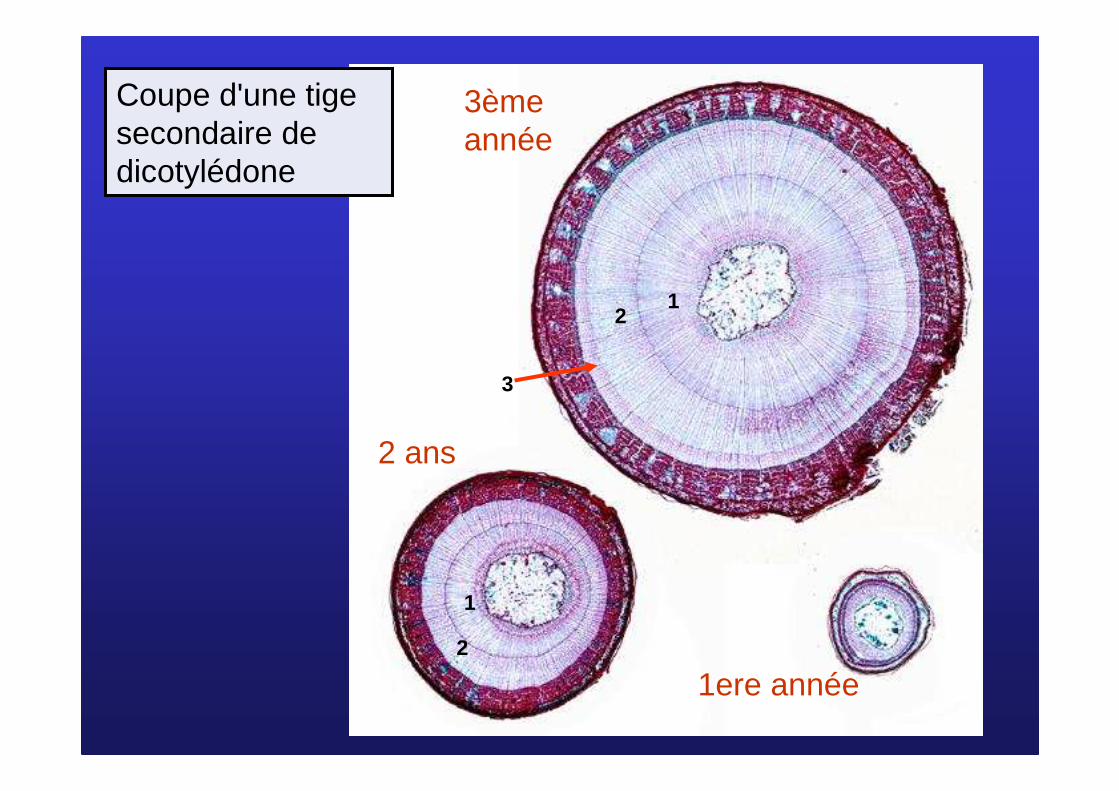
Coupe d'une tige secondaire de dicotylédone
1ere année
2 ans
3ème année
12
3
1
2
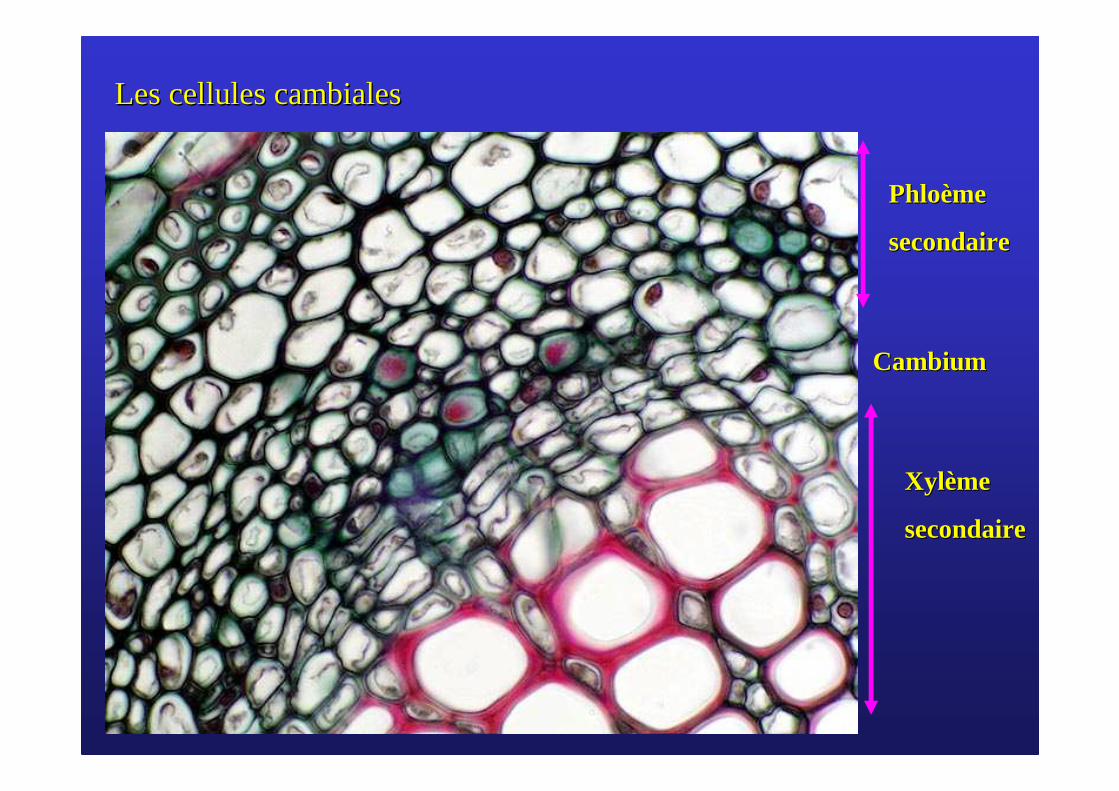
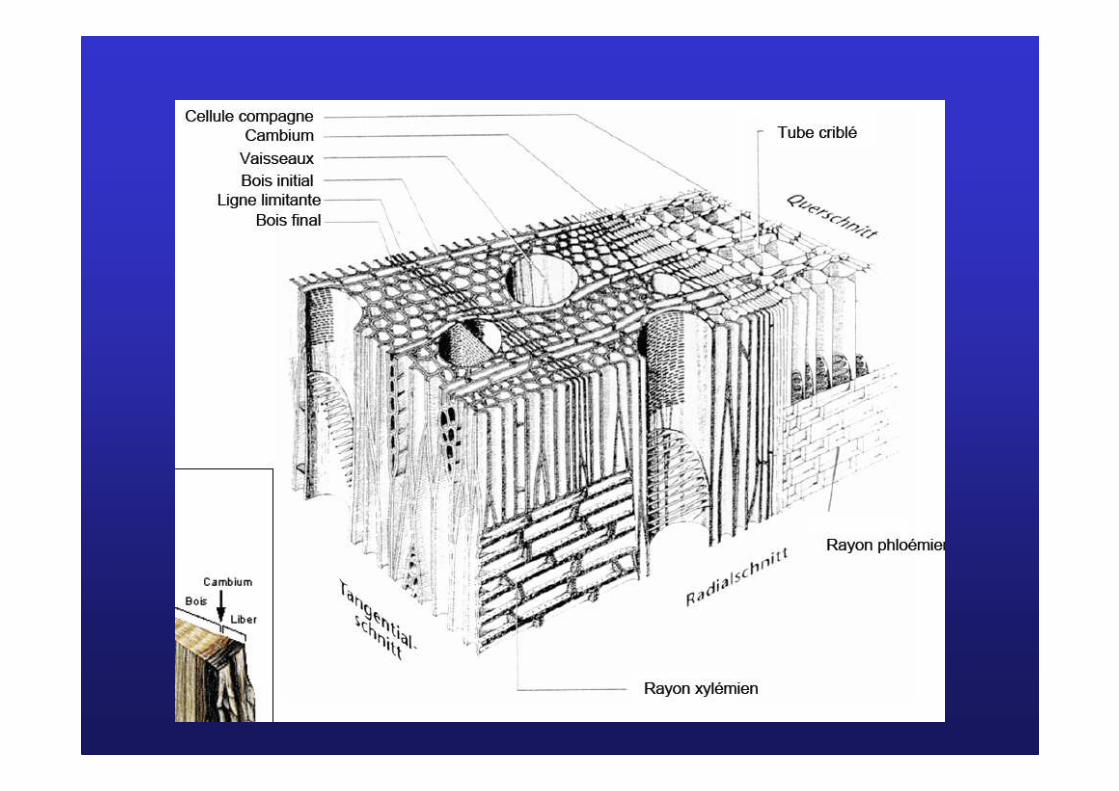
Les cellules cambialesLes cellules cambiales
PhloPhloèèmeme
secondairesecondaire
XylXyl èèmeme
secondairesecondaire
CambiumCambium
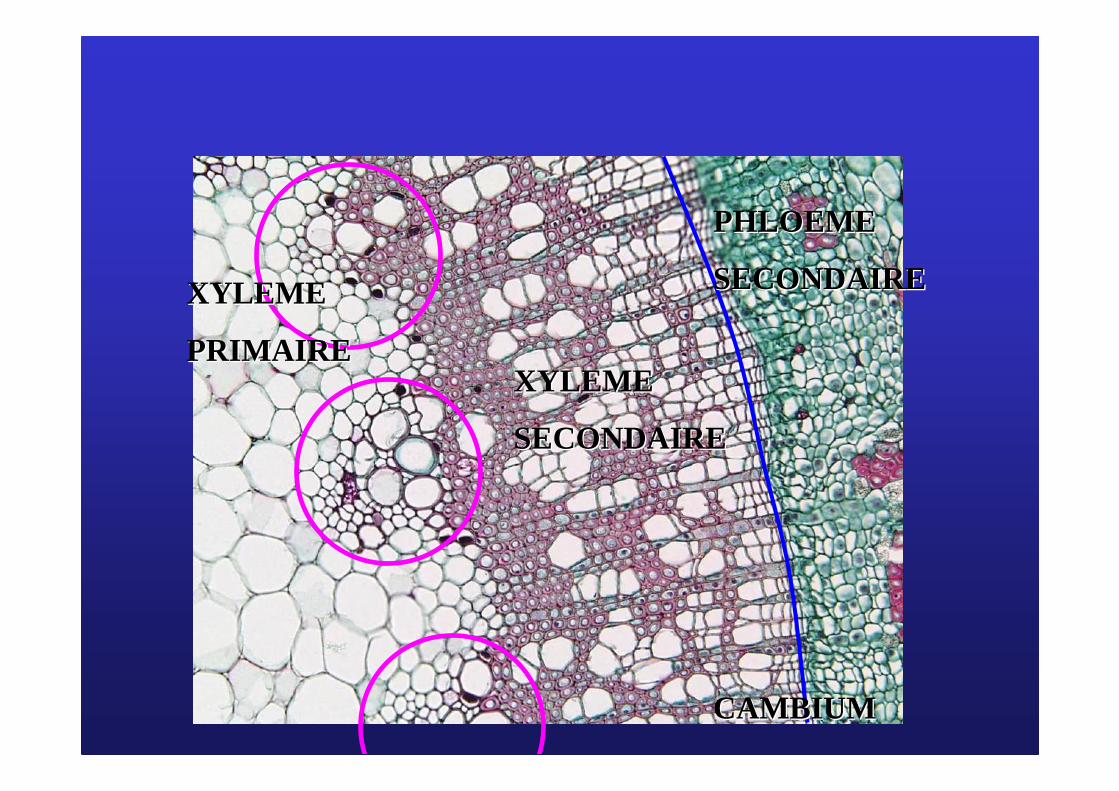
XYLEMEXYLEME
SECONDAIRESECONDAIRE
PHLOEMEPHLOEME
SECONDAIRESECONDAIRE
CAMBIUMCAMBIUM
XYLEMEXYLEME
PRIMAIREPRIMAIRE
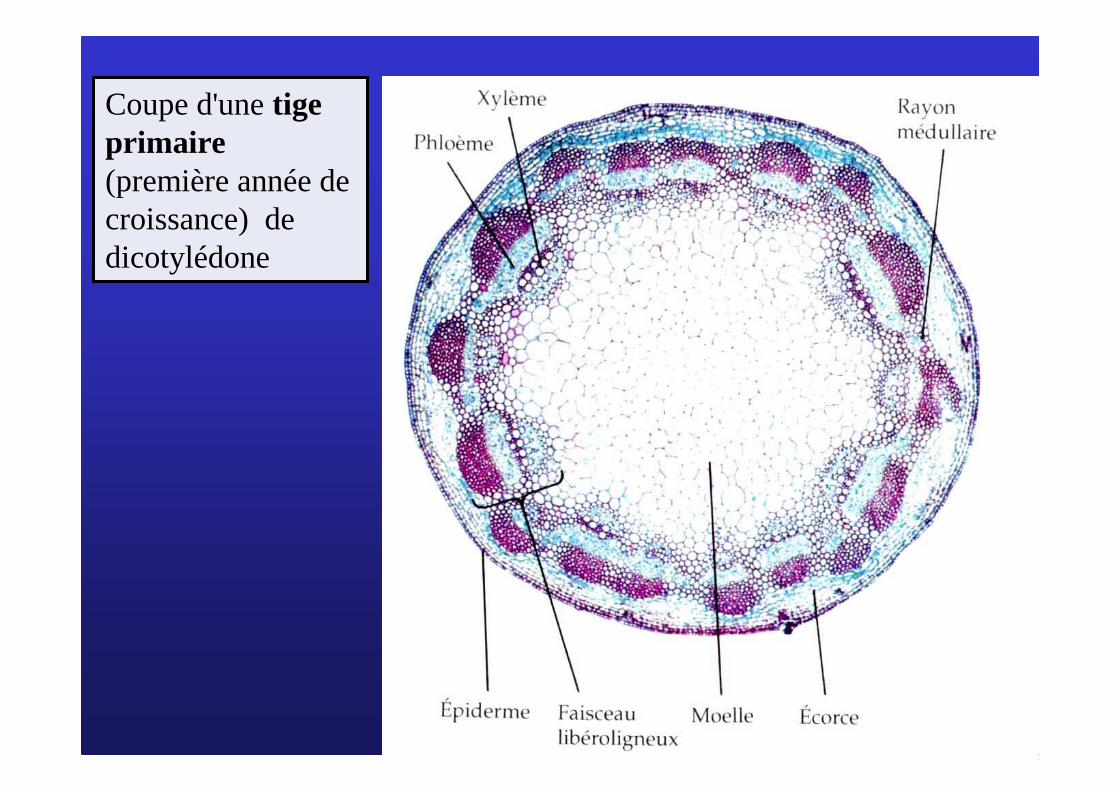
Coupe d'une tige primaire(première année de croissance) de dicotylédone
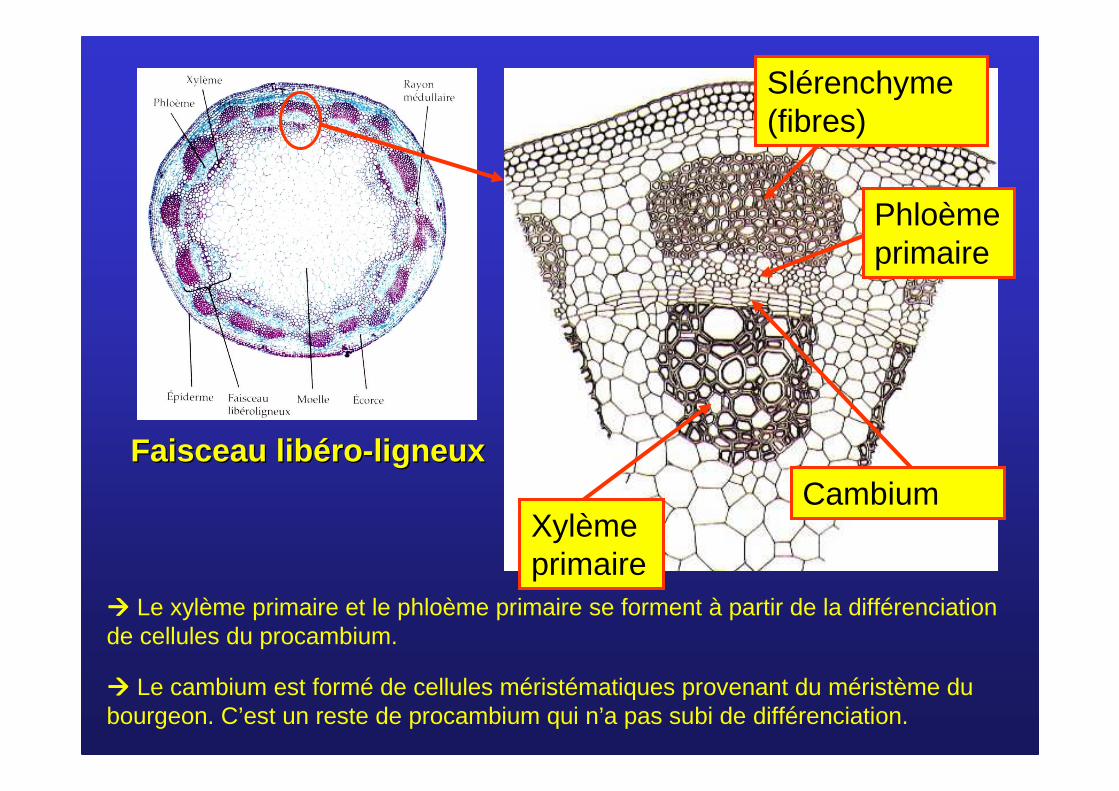
Phloèmeprimaire
Xylème primaire
CambiumFaisceau Faisceau liblib ééroro --ligneuxligneux
���� Le cambium est formé de cellules méristématiques provenant du méristème du bourgeon. C’est un reste de procambium qui n’a pas subi de différenciation.
Slérenchyme(fibres)
���� Le xylème primaire et le phloème primaire se forment à partir de la différenciation de cellules du procambium.
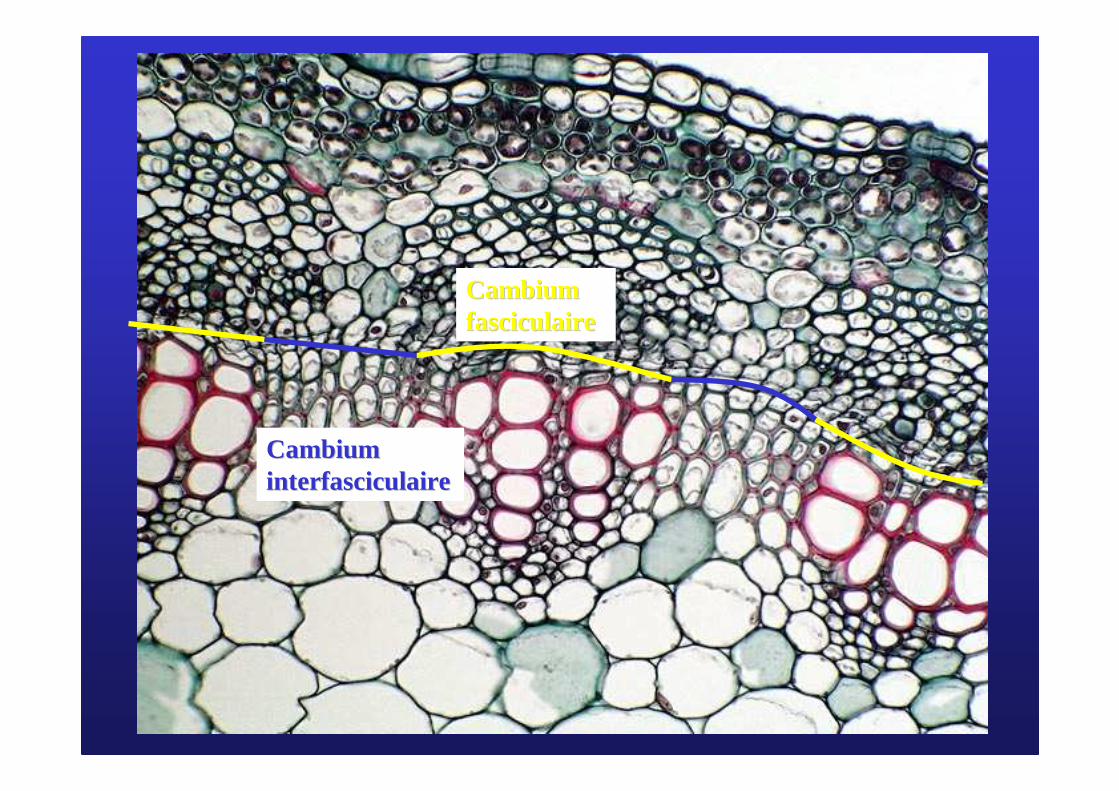
Cambium Cambium interfasciculaireinterfasciculaire
Cambium Cambium fasciculairefasciculaire
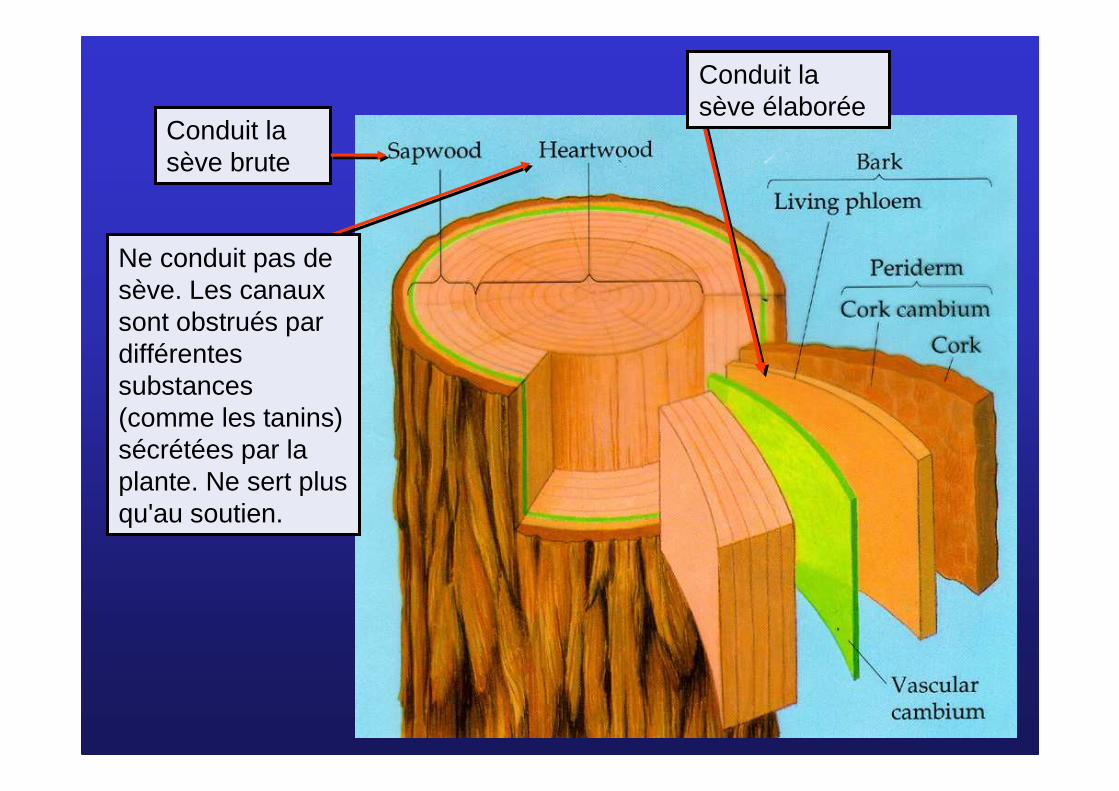
Conduit la sève brute
Conduit la sève élaborée
Ne conduit pas de sève. Les canaux sont obstrués par différentes substances (comme les tanins) sécrétées par la plante. Ne sert plus qu'au soutien.
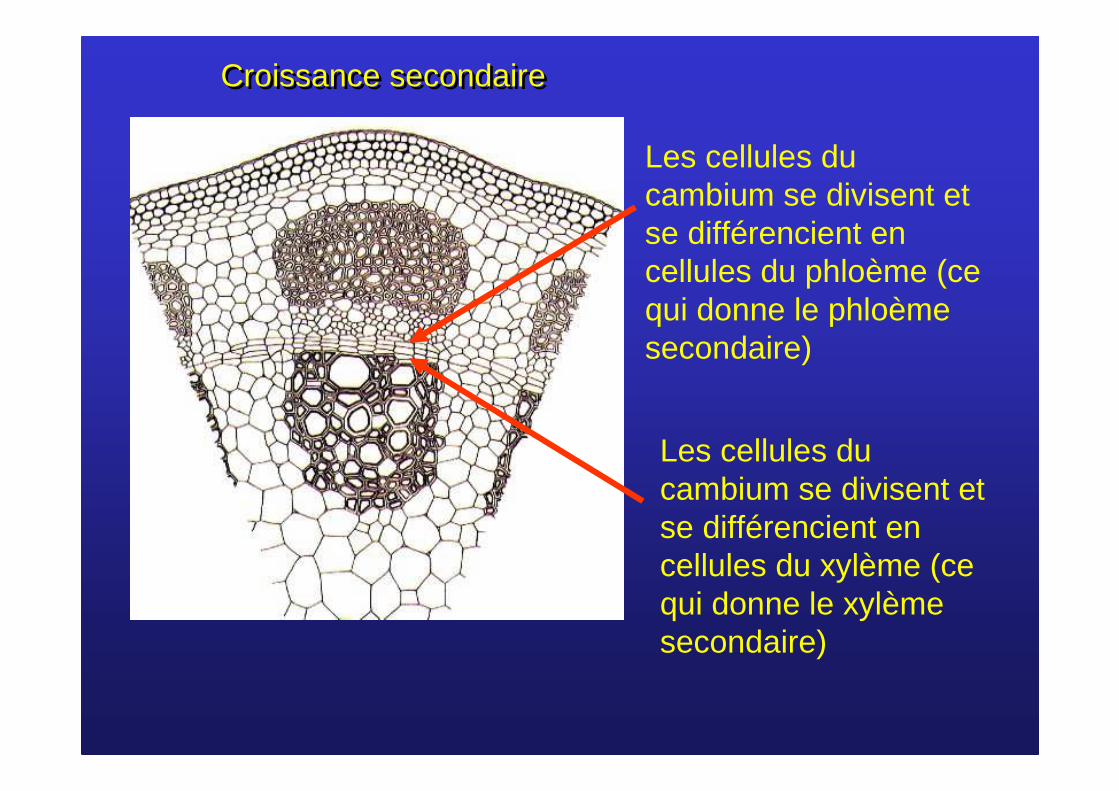
Les cellules du cambium se divisent et se différencient en cellules du phloème (ce qui donne le phloèmesecondaire)
Les cellules du cambium se divisent et se différencient en cellules du xylème (ce qui donne le xylème secondaire)
Croissance secondaireCroissance secondaire
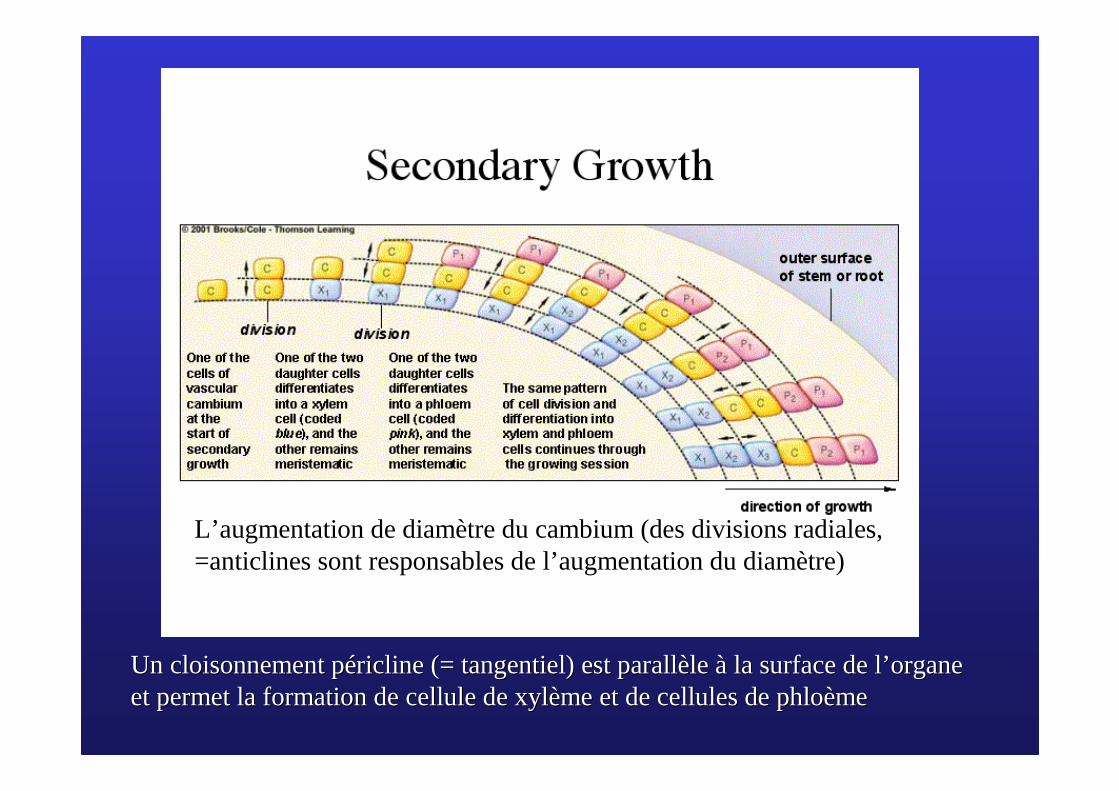
L’augmentation de diamètre du cambium (des divisions radiales, =anticlines sont responsables de l’augmentation du diamètre)
Un cloisonnement Un cloisonnement ppééricline ricline (= tangentiel) est parall(= tangentiel) est parallèèle le àà la surface de lla surface de l’’ organe organe et permet la formation de cellule de xylet permet la formation de cellule de xylèème et de cellules de me et de cellules de phlophloèèmeme
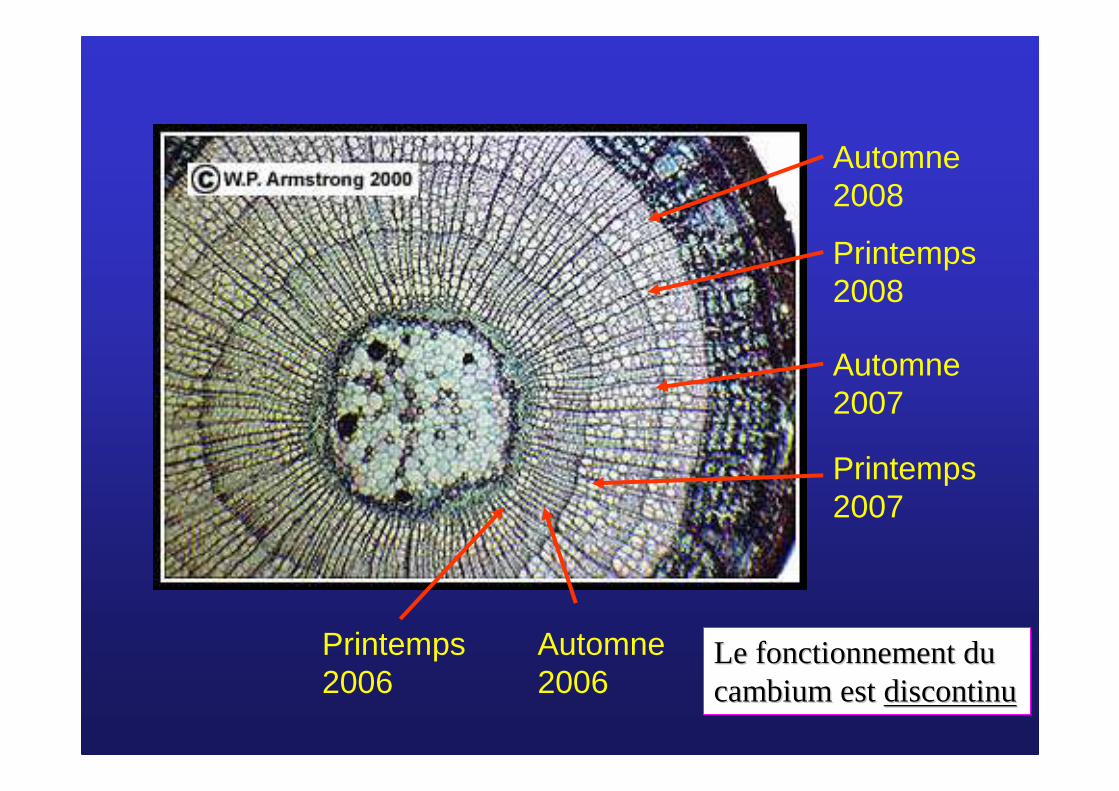
Automne 2008
Printemps 2008
Automne 2007
Printemps 2007
Automne 2006
Printemps 2006
Le fonctionnement du Le fonctionnement du cambium est cambium est discontinudiscontinu
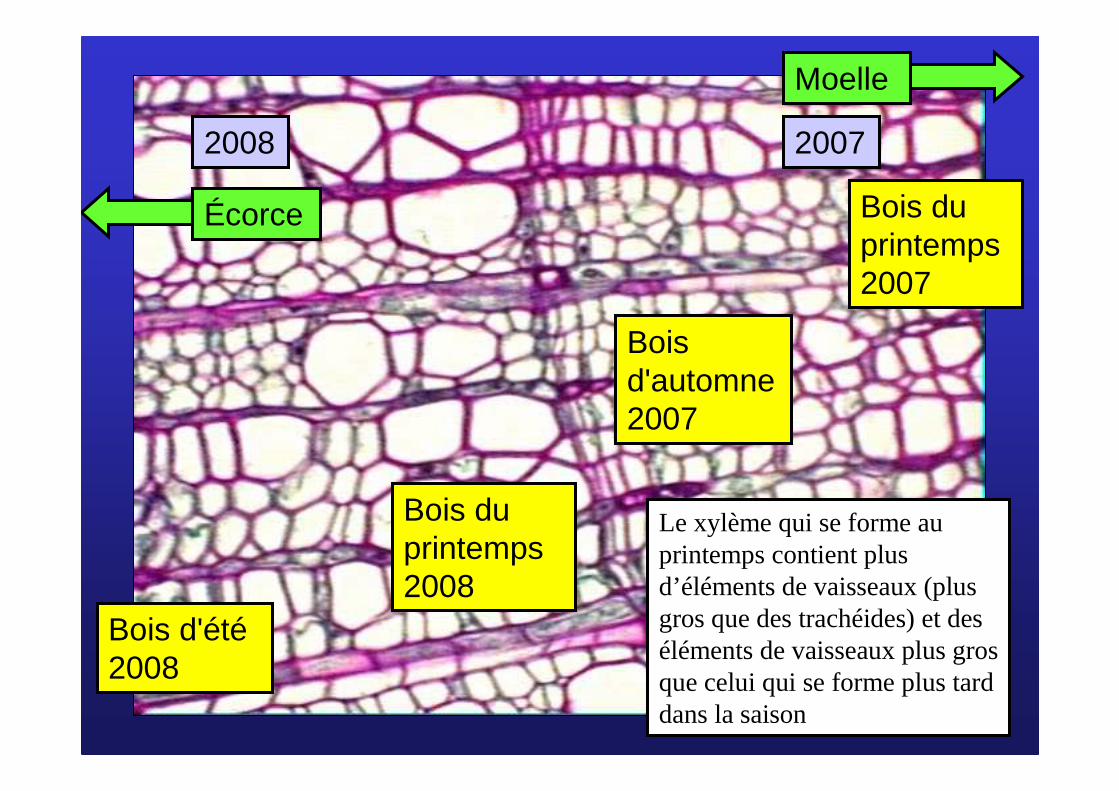
Bois du printemps 2008
Bois d'automne 2007
20072008
Bois d'été2008
Bois du printemps 2007
Moelle
Écorce
Le xylème qui se forme au printemps contient plus d’éléments de vaisseaux (plus gros que des trachéides) et des éléments de vaisseaux plus gros que celui qui se forme plus tard dans la saison

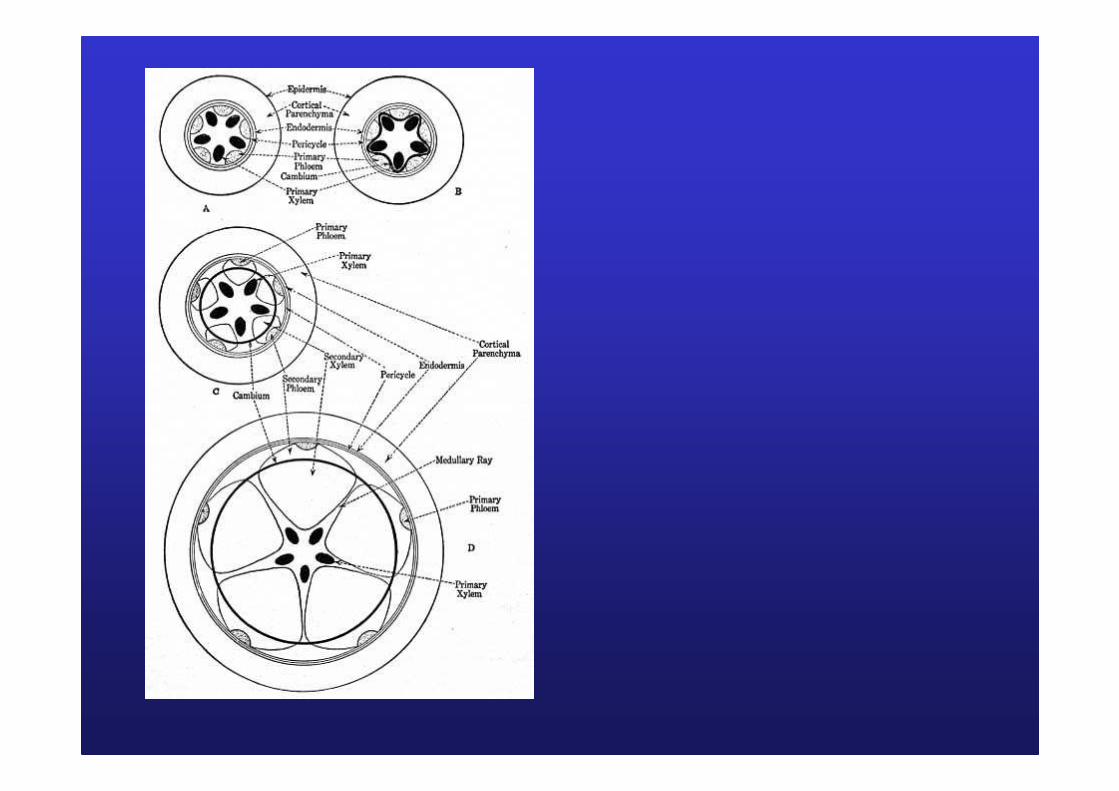
III/ MIII/ M ééristrist èèmes secondaires et croissance en mes secondaires et croissance en éépaisseurpaisseurB/ Dans la racineB/ Dans la racine

Mise en place du cambium racinaireMise en place du cambium racinaire