isolement et caractérisation de bactéries à fort potentiel probiotique
TRANSCRIPT

NOUR BEN ABDALLAH
ISOLEMENT ET CARACTERISATION DE BACTERIES A FORT
POTENTIEL PROBIOTIQUE À PARTIR DU TRACTUS
GASTROINTESTINAL DE VOLAILLE
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en Microbiologie agroalimentaire pour l'obtention du grade de maître es sciences (M.Sc)
DEPARTEMENT DE SCIENCE ET TECHNOLOGIE DES ALIMENTS FACULTÉ DES SCIENCES DE L'AGRICULTURE ET DE L'ALIMENTATION
UNIVERSITÉ LAVAL QUÉBEC
2010
© Nour Ben Abdallah, 2010
.

À mes parents Majid et Raja

RESUME
Les souches bactériennes à Gram négatif sont répertoriées comme étant celles qui causent
le plus de toxi-infections alimentaire au Canada. Parmi les aliments incriminés, la viande
de volailles demeure le plus connu des véhicules de transmission de ces pathogènes. Le
contrôle de la flore pathogène chez la volaille consiste non seulement de réduire le taux
de mortalité en élevage mais également d'assurer une meilleure qualité microbiologique à
l'arrivé aux abattoirs et par conséquent de produire des denrées salubres sans danger
pour le consommateur.
L'antibiothérapie et « l'antibioprévention » sont actuellement les seuls moyens utilisés
pour contrer les problèmes sanitaire et économique liés aux pathogènes aviaires.
Cependant le recours aux antibiotiques a connue ses limites, en raison de l'émergence de
nouvelles souches pathogènes multi-résistantes causée par l'utilisation abusive de ces
composés dans le secteur avicole. Récemment, de nouvelles stratégies de prévention ont
été proposées comme alternatives aux antibiotiques pour réduire l'incidence des
pathogènes entériques chez la volaille. Parmi ces stratégie, le recours aux probiotiques
semble offrir les résultats les plus prometteurs.
L'objectif général de ce projet de recherche était d'isoler et de caractériser à partir du
microbiote colique de la volaille des souches ayant un fort potentiel d'utilisation comme
probiotique en aviculture. Plusieurs isolats provenant de contenus intestinaux de volailles
ont été isolées et caractérisées sur le plan microbiologique et biologique. Parmi ces
isolats, vingt quatre isolats ayant démontré une activité inhibitrice contre des pathogènes
Gram négatifs ont été utilisées. Sept de ces souches ont été retenues par la suite en raison
de leur forte activité inhibitrice. Une caractérisation préliminaire des substances
inhibitrices produites par ces souches a permis de retenir trois isolats produisant des
composés de nature protéique, thermostable et de faible poids moléculaire très apparentés
à des bactériocines.

I I
AVANT-PROPOS
Ce mémoire comporte une introduction générale et trois chapitres. Dans l'introduction, le
contexte, la problématique de recherche, l'hypothèse de même que les objectifs
spécifiques poursuivis ont été présentés. Le premier chapitre correspond à une revue de la
littérature récente en rapport avec la problématique des pathogènes entérique en élevage
avicole, les moyens couramment utilisés pour réduire leur incidence de même que les
nouvelles alternative dont les probiotiques. Le second, rédigé sous forme d'article
scientifique, porte sur le criblage et la caractérisation de l'activité inhibitrice de souches
intestinales d'origine aviaire vis-à-vis des pathogènes entériques à Gram négatifs. Le
dernier chapitre, rédigé également sous forme d'articles scientifiques a permis
l'identification et la caractérisation des souches intestinales qui ont présenté la meilleure
activité inhibitrice face aux pathogènes entériques incluant Escherichia coli et
Salmonella. Finalement, une conclusion générale incluant des perspectives d'avenir est
présentée.

Ill
REMERCIMENTS
J'aimerai tout d'abord remercier très sincèrement, mon directeur de recherche, le Dr
Ismail Fliss pour la confiance et l'aide très précieuse, qu'il a su m'apporter sous bien des
formes tout au long de ce projet. L'intérêt qu'il a démontré pour l'avancement de mes
recherches et pour tous les conseils et orientations judicieuses prodiguées, qui ont
développé mon esprit scientifique et élargi mes connaissances.
Je voudrais dire un merci tout spécial à Céline Paquin à qui je dois beaucoup pour ses
nombreux avis, sa disponibilité en tout temps, son aide de toute sorte et ses
encouragements très précieux.
Toute ma gratitude va aussi à Nassra Dabour pour son aide et son savoir faire dans la
partie moléculaire.
Je remercie toute ma formidable équipe pour leur soutien et plus spécialement Nadia,
Riadh, Hajer, Benoit et Christophe sans qui le travail dans le laboratoire et mon passage
au Québec n'aurait pas été le même.
Je ne pourrais jamais passer outre cette formidable famille, qui sans le savoir, ma donnée
une chance mon oncle et mon grand frère Lotfi, cette femme formidable Lilia et mes
deux frères que j'aime énormément Hakim et Karim.
Finalement, j'adresse toute ma gratitude et tout mon amour à mes parents, qui m'ont
soutenu tout le long de mes études et qui m'ont toujours encouragé à aller jusqu'au bout
de mes rêves. Je vous serai éternellement reconnaissante pour votre soutien, votre
confiance et votre fierté. Malgré la distance, vous êtes présents à chaque fois que j'ai
besoin de vous. Je remercie aussi ma chère sœur et mon petit frère pour leur soutien, et
leur encouragement. J'ai la chance de vous avoir tous les quatre, je vous aime tant.

IV
TABLE DES MATIERES
RESUME i
AVANT-PROPOS ii
REMERCIMENTS iii
TABLE DES MATIÈRES iv
LISTE DES TABLEAUX vi
LISTE DES FIGURES vii
INTRODUCTION GÉNÉRALE 1
CHAPITRE 1. REVUE DE LITTÉRATURE 4
1.1. Les empoisonnements alimentaires 5
1.2. Le microbiote intestinal chez la volaille 5
1.2.1. Généralités 5
1.2.2. Description du tube digestif chez la volaille 6
1.2.3. Composition et évolution du microbiote colique endogène chez la volaille9
1.2.4. Les pathogènes entériques de la volaille 13
1.2.5. Contrôle des infections entériques chez la volaille 18
1.3. Les probiotiques 25
1.3.1. Définition 25
1.3.2. Critères de sélection 26
1.3.3. Les effets santé associés aux probiotiques 27
1.3.4. Activité antimicrobienne des probiotiques 28
1.3.5. Les bactériocines 29
1.3.6. Les probiotiques en aviculture 35
Hypothèse et objectifs du mémoire 38
CHAPITRE 2. Criblage et caractérisation de l'activité inhibitrice de souches intestinales vis-à-vis des pathogènes entériques gram négatif 39
2.1. Résumé 40
2.2. Introduction 41
2.3. Matériel et méthode 43

2.3.1. Souches bactériennes et conditions de culture 43
2.3.2. Mise en évidence de l'activité inhibitrice 45
2.4. Résultats et discussions 47
2.4.1. Criblage des souches 47
2.4.2. Test de microdilution 53
CHAPITRE 3. Identification et caractérisation de souches à fort potentiel probiotique isolées du tube digestif de volailles 63
3.1. Résumé 64
3.2. Introduction 65
3.3. Matériel et méthode 67
3.3.1. Souches bactériennes et conditions de culture 67
3.3.2. Tests de sérotypage 67
3.3.3. Méthodes moléculaires 67
3.4. Résultats et discusion 70
3.4.1. Analyse microscopique, coloration Gram et galerie API20 70
3.4.2. Tests de sérotypage 70
3.4.3. Analyses moléculaires 71
Conclusion générale 77
Références bibliographique 79
Annexes 88

VI
LISTE DES TABLEAUX
Tableau 1.1 : Composition du microbiote digestif du poulet déterminée par
dénombrement bactériens 10
Tableau 1.2 : Composition du microbiote digestif du poulet déterminée par méthodes
moléculaires 12
Tableau 1.3 : Principaux critères de sélection des probiotiques 26
Tableau 1.4 : Classification des colicines et leurs cibles cellulaires 33
Tableau 1.5 : Les dix microcines découvertes 34
Tableau 2.1 : Souches à fort potentiel probiotique 43
Tableau 2.2 : Souches sensibles 43
Tableau 2.3 : Activité inhibitrice contre Escherichia coli MC4100 du surnageant de
culture de la souche Escherichia coli LR05 cultivée à 37 °C dans le milieu
M63 pendant 18h sous agitation 47
Tableau 2.4 : Évaluation de l'activité inhibitrice (diamètre d'inhibition) contre des
pathogènes à gram négatif des surnageants de culture des 24 souches
intestinales tel que déterminée par le test de diffusion sur gélose optimisé
49
Tableau 2.5 : Évaluation de l'activité inhibitrice des surnageants de culture des souches
après un traitement thermique à 100°C pendant 5 min et/ou une filtration sur
membrane de 0,22nm 50
Tableau 3.1 : Sérotypage des souches 11004,11006 et 11026 69
Tableau 3.2 : Résultats des homologies de séquences entre les souches isolées et les
souches de références répertoriées dans Gen Bank 72
Tableau 3.3 : Les gènes codant pour la septicémie détectés chez les 3 souches 74

Vil
LISTE DES FIGURES
Figure 1.1: Schéma illustrant les caractéristiques physiologiques du tractus digestif chez
la volaille 8
Figure 1.2: Pathogénie de la souche entérohémorragique Escherichia coli 0157: H7
chez l'humain 16
Figure 1.3 : Les principaux effets bénéfiques sur la santé attribués aux
probiotiques 28
Figure 1.4: Mécanismes d'actions proposés des microorganismes probiotiques dans le
traitement des infections entériques 29
Figure 2.1 : Schéma illustrant le principe de test de diffusion sur gélose 44
Figure 2.2 : Suivie de la croissance microbienne sur microplaque 45
Figure 2.3 : Activité du surnageant de la souche Escherichia coli LR05 contre la souche
sensible Escherichia coli MC4100. A : Croissance 8h à 37°C. B : Croissance
18hà37°C 47
Figure 2.4 : Test de diffusion sur gélose montrant l'activité inhibitrice des surnageant de
culture des souches 11006 (A), 11004 (B) et 11026 (C) contre la souche
sensible Escherichia coli MC4100 suite aux différents traitements 51
Figure 2.5 : Courbes de croissance d'Escherichia coli MC4100 en absence et en présence
des surnageants brut des différentes souches intestinales 53
Figure 2.6 : Croissance d'Escherichia coli MC4100 en présences des surnageants des
cultures des souches intestinales dilué 1/2 (A), 1/4(B), 1/8 (C) et 1/16 (D).54
Figure 2.7 : Croissance d'Escherichia coli 0157 :H7 en absence et en présence des
surnageants de culture des différentes souches intestinales 56

vm
Figure 2.8 : Croissance d'Escherichia coli 0157 :H7 en présences des surnageants des
cultures des souches intestinales dilué 1/2 (A) et 1/4 (B) 57
Figure 2.9 : Croissance de la souche Salmonella enteritica en absence et en présence des
surnageants des souches bactériocinogènes 59
Figure 2.10 : La croissance de Salmonella enteritica en présences des surnageants des
cultures des souches intestinales dilué 1/2 (A) et 1/4 (B) 60
Figure 3.1 : Analyse du produit d'amplification du gène d'ARN 16S par électrophorèse
sur gel d'agarose 16S 70

INTRODUCTION GENERALE
Selon le Centre Québécois d'Inspection des Aliments et de Santé Animale (CQIASA,
2008), E. coli 0157.H7 (34%), Salmonelles (28%) et Campylobacter (17%) ont été
identifiées comme étant les agents pathogènes les plus souvent mis en cause dans des
toxi-infections alimentaires rapportées entre le 1er avril 2007 et le 31 mars 2008. De
plus, une augmentation des cas attribuables à Listeria monocytogenes a été signalée en
2009. La filière « Viandes et volailles » a été incriminée dans plus de 40,4 % des cas de
toxi-infections alimentaires rapportés. En ce qui concerne les espèces animales
impliquées, mentionnons les volailles (19,2%), le bœuf (13,1%), le porc (1,5%),
l'agneau (0,5%) et des viandes diverses sans différenciation d'espèce (6,3 %). Ces
résultats sont comparables à ceux de 2006-2007 (CAAAQ 2008).
Selon l'Organisation des Nations Unies pour l'alimentation et l'agriculture, la
production de poulet a enregistré une croissance régulière dans le monde depuis le
début des années 90. De 1985 à 2005, la production mondiale a augmenté de 158 %
alors que la production canadienne s'est accrue de 77 %. La majorité de cette
production (60,1%) se situe en Ontario et au Québec (statistiques Canada). Selon
Statistique Canada la consommation de poulet par habitant a bondi de 136 % au Canada
au cours des 30 dernières années, passant de 13 kg en 1975 à 30,7 kg en 2005 alors que
la consommation de bœuf et de porc a diminué de 35 % et 4 % respectivement durant la
même période. Les volailles sont prédisposées à de nombreuses infections en particulier
celles des systèmes gastro-intestinal et respiratoire. Malgré une amélioration importante
des conditions d'hygiène et de salubrité dans le domaine de la production et de
l'élevage de volailles, les infections entériques chez cette espèce persistent ce qui
causent des pertes économiques importantes pour l'éleveur. Les animaux atteints de
maladies gastro-intestinales manifestent un gain de poids de moins en moins important
ainsi que des signes de faiblesse. Ceux sérieusement atteints peuvent mourir peu de
temps après l'observation des premiers signes cliniques.

Jusqu'à présent la microflore intestinale des oiseaux et ses variations étaient contrôlées
par les antibiotiques qui sont utilisés de façon systématique comme facteurs de
croissance en aviculture. Les antibiotiques sont définis comme des substances d'origine
microbienne, produites par des bactéries du sol et certains champignons et qui, à très
petites doses, agissent sur d'autres microorganismes. Une de leurs caractéristiques est
de ne pas être toxique pour les cellules d'eucaryotes. Cette propriété, associée à la
grande efficacité des molécules, a fait des antibiotiques le remède « magique » du
20lème siècle. Chez la volaille, l'administration quotidienne de faibles doses
d'antibiotiques a permit d'obtenir une croissance accélérée et une consommation
moindre d'aliments en modifiant la flore intestinale et la digestibilité des aliments.
Cependant, avec le temps, leur administration répétée et parfois inappropriée chez
l'homme comme chez l'animal a contribué à l'augmentation de la résistance des agents
pathogènes vis-à-vis des antibiotiques. Face à l'apparition croissante de la résistance
aux antibiotiques conventionnels, à l'émergence de nouveaux agents pathogènes tant
chez l'homme que chez l'animal, à l'augmentation des coûts des traitements, à la baisse
de leur efficacité, les antibiotiques ne garantissent plus une réponse favorable. Avec
leur suppression annoncée en Europe en 2006, des alternatives comme les probiotiques
devront être développées pour mieux maîtriser le microbiote digestif chez la volaille.
Les probiotiques ont récemment été définies par la FAO comme étant des
microorganismes vivants qui lorsqu'ils sont administrés en quantité adéquate, procurent
un effet bénéfique pour la santé de l'hôte (FAO/WHO 2002). Ds sont utilisés au niveau
industriel depuis les années 1960 dans l'alimentation destinée aux élevages d'animaux
et depuis les années 1980 dans certains aliments ou en tant que compléments
alimentaires pour l'homme. Leur efficacité au niveau intestinal est maintenant bien
reconnue. Les probiotiques représentent une approche naturelle d'enrichissement de la
flore intestinale et d'exclusion compétitive pour lutter contre les bactéries pathogènes.
En renforçant l'écosystème microbien des volailles, les probiotiques contribuent à la
défense immunitaire et protègent les poulets contre les conséquences de stress tels que
la vaccination et les changements de températures (Patterson and Burkholder 2003).
Des améliorations en terme de gain de poids et de l'indice de consommation ont ainsi
été observée suite à la consommation de probiotiques (Jin, Ho et al. 1998); (Simon,

Jadamus et al. 2001). Les probiotiques peuvent se comparer à de véritables usines
actives capables de véhiculer des principes actifs qu'ils contiennent comme les enzymes
qui aident à la digestion des fibres, des substances antibactériennes comme les acides
organiques et les bactériocines pour combattre plusieurs micro-organismes pathogènes
(Sreekumar and Hosono 1998); (Fooks, Fuller et al. 1999).
Les bactériocines sont des protéines produites par certaines bactéries et qui sont capable
d'inhiber d'autres bactéries en se fixant sur des récepteurs spécifiques (Joerger 2003).
Parmi celles produites par les bactéries de l'écosystème intestinal, les microcines
représentent une nouvelle catégorie encore peu étudiée et présentant un intérêt majeur.
Ces dernières se caractérisent par leur faible poids moléculaire, leur résistance à la
température et à de nombreuses proteases, et surtout par leur spectre d'action
extrêmement restreint avec une activité dirigé surtout contre les bactéries
phylogénétiquement proches des souches productrices. Les mécanismes de régulation
de la synthèse des microcines sont encore peu connus mais généralement, un milieu
nutritionnel pauvre favorise leur production. Cependant, très peu de travaux ont
clairement démontré la production de bactériocines dans les conditions physiologiques
du tube digestif et l'implication de ces molécules comme mécanisme d'action des
probiotiques contre la flore entérique pathogène.
Le but de cette recherche est d'identifier des souches intestinales de volaille
productrices de substances inhibitrices actives contre des pathogènes entériques
incluant Escherichia coli et Salmonella et de les caractériser clairement pour
d'éventuelle utilisation comme souches potentiellement probiotique pour le contrôle
des microorganismes pathogènes en aviculture.

CHAPITRE 1. REVUE DE LITTERATURE

1.1. LES EMPOISONNEMENTS ALIMENTAIRES
Les empoisonnements alimentaires sont des maladies liées à la consommation
d'aliments contaminés par des germes pathogènes. Leur durée, leur sévérité, ainsi que
leur évolution dépendent beaucoup de l'agent pathogène impliqué. Santé Canada estime
que chaque année environ deux millions de canadiens sont victimes de maladies
d'origine alimentaire et une trentaine d'entre eux en meurent. Selon le bilan annuel du
Centre Québécois d'Inspection des Aliments et de Santé Animale (CQIASA 2008), les
empoisonnements alimentaires les plus répandus sont les salmonelloses, les toxi-
infections à Staphylocoque, la listériose ainsi que le botulisme. Parmi les
empoisonnements alimentaires signalés au MAPAQ en 2007-2008, 47,8 % sont
survenues après la consommation d'aliments à la maison, alors que dans 45,5 % des
signalements, les symptômes sont apparus à la suite d'un repas au restaurant, dans
d'autres catégories d'établissements (4,4 %) ou dans des institutions de restauration
collective (2,3 %).
La filière « Viandes et volailles » a été la plus souvent visée par les déclarations
d'empoisonnement alimentaires (40,4 %). Les agents pathogènes les plus souvent
signalés en 2007-2008 ont été E. coli 0157.H7, Salmonella et Campylobacter. De plus,
une augmentation de cas attribuables à Listeria monocytogenes a été signalée cette
année. Pour l'ensemble des cas rapportés, 25,9 % des signalements, ont été confirmés
par un diagnostic médical, par l'isolement de l'agent causal dans les aliments ou par
une enquête épidémiologique, alors que dans 26,5 % des cas, aucun lien avec
l'alimentation n'a été établi. Sur tout les cas rapportés confirmés ou probables, 91,8 %
sont d'origine microbiologique. Cette donnée est comparable aux résultats des années
antérieures (CQIASA 2008).
1.2. LE MICROBIOTE INTESTINAL CHEZ LA VOLAILLE
1.2.1. Généralités
Selon la définition d'Isolauri et ses collaborateurs, le microbiote intestinale normale est
un consortium complexe et en équilibre de microorganismes qui habitent normalement

le tractus gastro-intestinal et qui remplissent un rôle dans la nutrition, la physiologie et
le fonctionnement du système immunitaire de l'hôte (Isolauri, Sutas et al. 2001). La
composition de ce microbiote intestinal est en équilibre relativement stable dans le tube
digestif. Cet équilibre peut être rompu avec l'âge, les conditions d'hygiène, le stress ou
à la suite d'une agression extérieure comme lors de l'utilisation d'antibiotiques, de
facteurs de croissance (Gabriel, Mallet et al. 2003). Ainsi, on note des populations
microbiennes plus élevées chez des animaux élevés au sol sur litière propre ou litière
contaminée par une bande précédente par rapport à des animaux élevés en cage
individuelle (Gabriel, Mallet et al. 2003). Selon les conditions d'élevage,
l'augmentation de la densité d'élevage ou les stress thermiques semblent globalement
augmenter les bactéries néfastes au détriment des bactéries bénéfiques (Gabriel, Mallet
et al. 2005). Outre ces conditions, la présence de parasites intestinaux comme les
coccidies, peut entraîner la dégradation de la muqueuse intestinale et la production de
nouveaux substrats pour la microflore, modifiant ainsi sa composition (Kimura,
Shiosaka et al. 1976). La flore est modifiée aussi par l'alimentation. Ainsi, le type de
céréales en particulier la présence de polysaccharides non amylacés hydrosolubles
(Mathlouthi, Mallet et al. 2002) ou leur mode de présentation (Gabriel, Mallet et al.
2003) entraînent des changements de la flore. De même, les matières grasses, ou le type
d'amidon peuvent avoir un effet sur la composition du microbiote (Weurding, Enting et
al. 2003).
7.2.2. Description du tube digestif chez la volaille
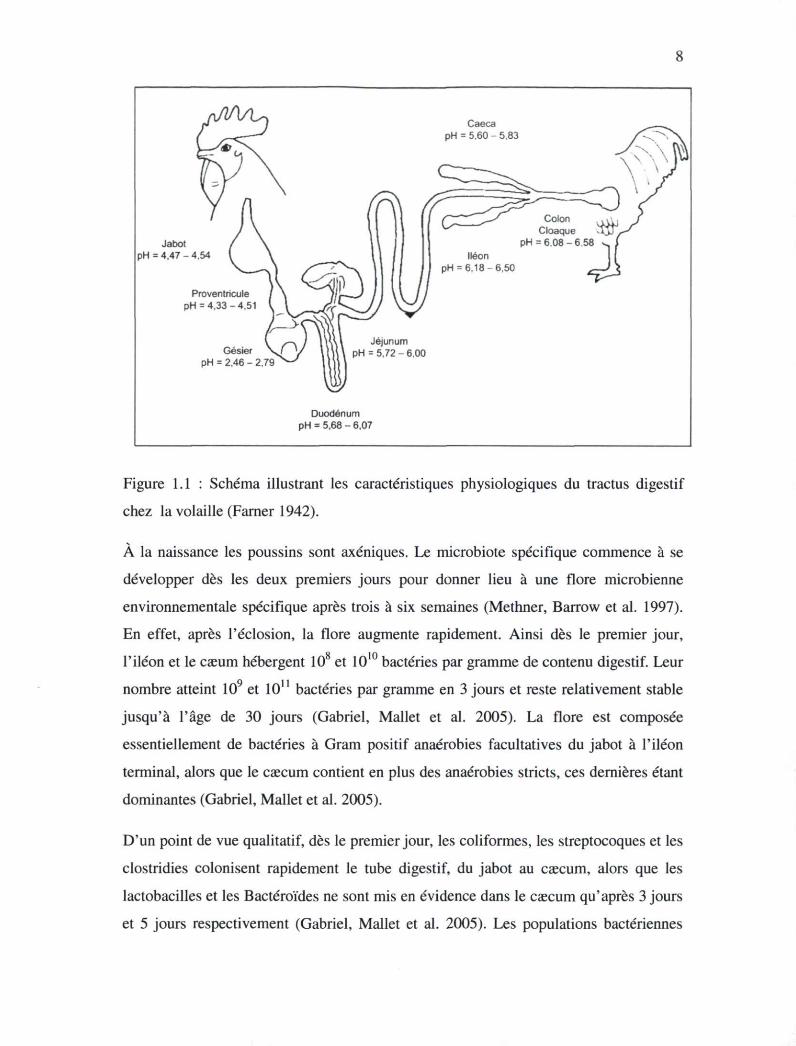
Le tractus gastro-intestinal présente quelques particularités anatomiques (Figure 1.1).
On distingue différents compartiments ; la cavité buccale ne comprend ni lèvres ni
dents, mais un bec corné qui permet la préhension et une certaine fragmentation des
aliments. Les glandes salivaires sont peu développées. D n'y a ni voile de palais, ni
épiglotte, si bien que la déglutition est un phénomène uniquement mécanique par
redressement de la tête. Dans la bouche, les aliments sont peu fragmentés et
grossièrement insalivès (M. Larbier and Leclercq 1992). L'œsophage contient un
renflement dont l'épithélium est riche en glandes à mucus : le jabot. Cet organe de pH
variant entre 4,47 et 4,54 (Farner 1942) peut entreposer des aliments qui s'y humectent

et s'y ramollissent, il fonctionne chez le poulet alimenté à volonté. Il est le lieu d'une
digestion microbienne et comporte essentiellement des Lactobacilles, d'une partie de
l'amidon (hydrolyse avec formation d'acides lactique) et de formation d'acide gras
volatiles (M. Larbier and Leclercq 1992). Le proventricule est riche en glandes
sécrétoires (acide chlorhydrique et pepsinogène précurseur de la pepsine) et permettant
la digestion chimique : c'est l'estomac chimique. La protéolyse y débute à pH de 3 à
4,5. Dans le gésier et le proventricule, le faible pH fait chuter la population bactérienne
(Farner 1942). Le gésier, estomac mécanique est caractérisé par une couche
superficielle très dure entourée de muscles puissants. Il y règne un pH très bas (2 à 3,5)
et il peut contenir de petits graviers qui sont nécessaires aux animaux consommant des
grains intacts. C'est donc au niveau du gésier que se produit véritablement la protéolyse
sous l'action de la pepsine (Gabriel, Mallet et al. 2005). Dans l'intestin,
l'environnement devient plus favorable à la croissance bactérienne en raison de la plus
faible pression d'oxygène et de la faible concentration en enzyme et en sels biliaires et
d'un pH variant dans le duodénum entre 5,68 et 6,07, dans le jéjunum entre 5,72 et 6,
dans le caecum entre 5,6 et 5,83, l'iléon entre 6,18 et 6,50 et dans le colon entre 6,08 et
6,58 (Farner 1942).
8
Jabot pH = 4,47 - 4.54
Proventricule pH = 4,33 -4,51
Gésier \ ^ f \ pH = 2,46 - 2,79
Duodénum pH = 5,68 - 6,07
Figure 1.1 : Schéma illustrant les caractéristiques physiologiques du tractus digestif
chez la volaille (Farner 1942).
À la naissance les poussins sont axéniques. Le microbiote spécifique commence à se
développer dès les deux premiers jours pour donner lieu à une flore microbienne
environnementale spécifique après trois à six semaines (Methner, Barrow et al. 1997).
En effet, après l'éclosion, la flore augmente rapidement. Ainsi dès le premier jour,
l'iléon et le caeum hébergent 108 et 1010 bactéries par gramme de contenu digestif. Leur
nombre atteint 109 et 10" bactéries par gramme en 3 jours et reste relativement stable
jusqu'à l'âge de 30 jours (Gabriel, Mallet et al. 2005). La flore est composée
essentiellement de bactéries à Gram positif anaérobies facultatives du jabot à l'iléon
terminal, alors que le caecum contient en plus des anaérobies stricts, ces dernières étant
dominantes (Gabriel, Mallet et al. 2005).
D'un point de vue qualitatif, dès le premier jour, les conformes, les streptocoques et les
clostridies colonisent rapidement le tube digestif, du jabot au caecum, alors que les
lactobacilles et les Bactéroïdes ne sont mis en évidence dans le caecum qu'après 3 jours
et 5 jours respectivement (Gabriel, Mallet et al. 2005). Les populations bactériennes

9
présentes dans le tractus digestif représentent une large gamme de types métaboliques
et morphologiques. Leur nombre total est plus important que le nombre de cellules
eucaryotes constituant le corps de l'hôte. On distingue les bactéries dominantes (>106
UFC /g contenu), sous-dominantes (105 à 103 UFC / g contenu), et résiduelles (<103
UFC / g contenu). Chez le poulet, les sites principaux d'activité bactérienne sont le
jabot, le caecum et, dans une moindre mesure, l'intestin grêle (Cole and Fuller 1984).
Ainsi, dans le caecum et l'iléon, on trouve respectivement 1011 et 109 bactéries par g de
contenu (Apajalahti, Kettunen et al. 2004). Les études effectuées sur le microbiote des
oiseaux ont concerné principalement le caecum (Gabriel, Mallet et al. 2005).
1.2.3. Composition et évolution du microbiote colique endogène chez la
volaille
La flore digestive des oiseaux a été très étudiée, et s'avère différente de celle des
mammifères (Smith 1965), probablement du fait de différences anatomiques et
physiologiques. En particulier, les mammifères ont un côlon très développé par rapport
aux oiseaux. Le tube digestif des oiseaux, comme celui des mammifères renferme une
population microbienne extrêmement riche et diversifiée, composés de nombreux
microorganismes différents (Andrieu 1995; Pascual, Hugas et al. 1999).
La colonisation initiale des poussins par des agents inoffensifs prévient l'intrusion
d'agents pathogènes ou indésirables. Ces flores naturelles ne sont pas entièrement
connues, aussi bien en terme de composition bactérienns qu'en terme d'effet induits.
On parle alors de microbiote indéfini. Des nombreuses études effectuées sur le
microbiote digestif des oiseaux depuis les années 1950 ont fait appel aux cultures de
bactéries sur milieu sélectif. Or une proportion très élevée de bactéries, jusqu'à 90%
selon les estimations, n'est pas cultivable (Lan, Hayashi et al. 2002). Pour résoudre ce
problème, des techniques de biologie moléculaire ont été développées. Elles permettent
de mettre en évidence, grâce à leur ADN ribosomal 16 S, les microorganismes quelles
que soient leurs conditions de viabilité (Gabriel, Mallet et al. 2005).
10
1.2.3.1. Données de microbiologie classique
Les méthodes classiques d'identification bactérienne reposent sur la capacité des
cellules de pousser et de former des colonies visibles sur des milieux solides (boites de
Pétri). Ces méthodes montrent que la flore, constituée principalement de bactéries à
Gram positif, est composée essentiellement d'anaérobies facultatives du jabot à l'iléon
terminal, alors que le caecum contiennent en plus des anaérobies strictes, ces dernières
étant dominantes ((Cole and Fuller 1984), Tableau 1.1). Dans le jabot, on trouve
principalement des Lactobacilles qui sont attachés à l'épithélium et forment presque
une couche continue. On trouve aussi des streptocoques, des coliformes et des levures.
Dans le gésier et le pro ventricule, le faible pH (Figure 1.1) fait chuter la population
bactérienne (Gabriel, Mallet et al. 2005). Dans le duodénum, les conditions ne sont pas
propices au développement de la flore : présence de nombreuses enzymes, forte
pression en oxygène, présence de fortes concentrations de composés antimicrobiens tels
que les sels biliaires et mouvements de reflux du jéjunum au gésier entraînant une
modification rapide des conditions de milieu (Gabriel, Mallet et al. 2005). Plus loin
dans l'intestin, l'environnement devient plus favorable à la croissance bactérienne en
raison de la plus faible pression d'oxygène et de la faible concentration en enzymes et
en sels biliaires (réabsorbés par l'hôte et dégradés en partie par la microflore) (Gabriel,
Mallet et al. 2005).
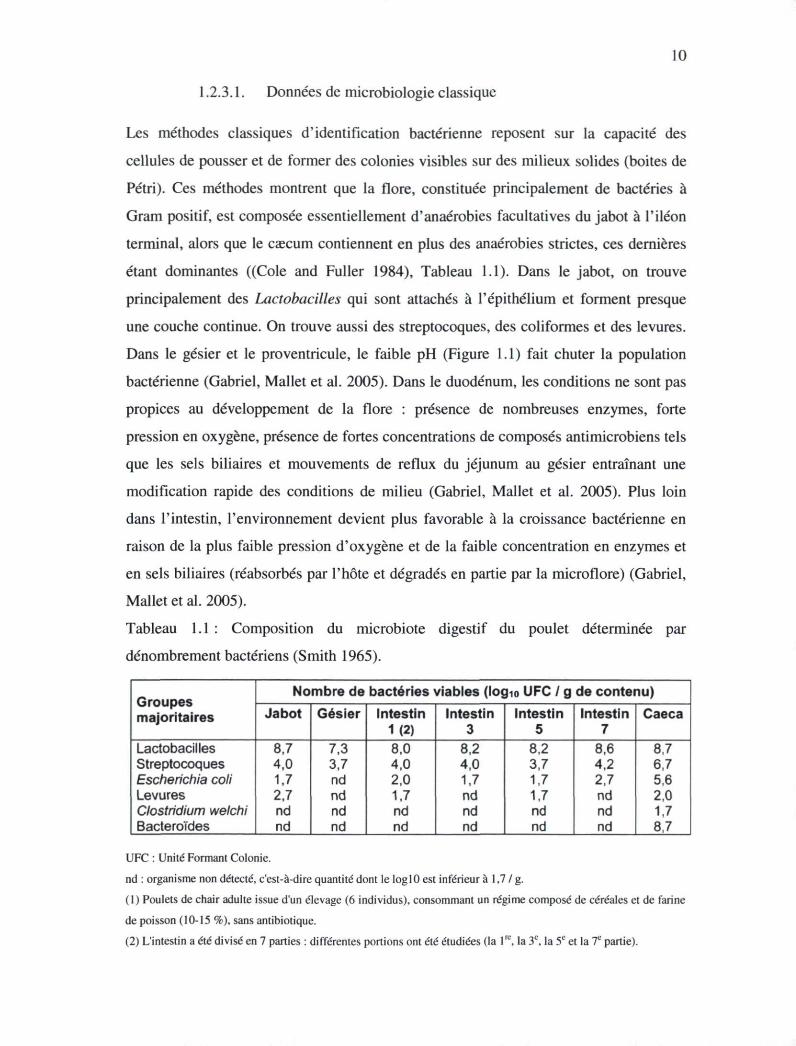
Tableau 1.1 : Composition du microbiote digestif du poulet déterminée par
dénombrement bactériens (Smith 1965).
Groupes majoritaires
Nombre de bactéries viables (log™ UFC / g de contenu) Groupes majoritaires Jabot Gésier Intestin
1(2) Intestin
3 Intestin
5 Intestin
7 Caeca
Lactobacilles Streptocoques Escherichia coli Levures Clostridium welchi Bacteroïdes
8,7 4,0 1,7 2,7 nd nd
7,3 3,7 nd nd nd nd
8,0 4,0 2,0 1.7 nd nd
8,2 4,0 1,7 nd nd nd
8,2 3,7 1,7 1,7 nd nd
8,6 4,2 2,7 nd nd nd
8,7 6,7 5,6 2,0 1,7 8,7
UFC : Unité Formant Colonie.
nd : organisme non détecté, c'est-à-dire quantité dont le log 10 est inférieur à 1,7 /g .
(1) Poulets de chair adulte issue d'un élevage (6 individus), consommant un régime composé de céréales et de farine
de poisson (10-15 %), sans antibiotique.
(2) L'intestin a été divisé en 7 parties : différentes portions ont été étudiées (la lre, la 3e, la 5e et la 7e partie).

11
Les méthodes de cultures conventionnelles ont conduit à l'identification chez le poulet
de 29 genres bactériens, chaque genre étant représenté par 3 à 4 espèces, et chaque
espèce par 3 à 4 types métaboliques différents, ce qui ferait plus de 200 souches
différentes (Mead, Norris et al. 1989). D'autres miroorganismes dont l'activité
métabolique a été mise en évidence n'ont pas pu être isolés et caractérisés du fait de
leur besoin d'anaérobiose stricte ou de l'ignorance des composants nécessaires à leur
croissance (Mead, Norris et al. 1989). Ainsi, seulement 25 % des souches seraient
identifiées.
1.2.3.2. Données de microbiologie moléculaire
Au niveau moléculaire, de nombreuses méthodes ont fait l'objet d'expérimentations, en
particulier à l'aide de méthodes à base de PCR et plus récemment les puces à ADN (Ye,
Wang et al. 2001) ainsi, que les sondes PNA (Stender, Fiandaca et al. 2002). La
majorité des méthodes utilisées actuellement repose sur une amplification pour
augmenter la sensibilité. La PCR est très généralement utilisée (Hellyer, DesJardin et
al. 1999; McKillip, Jaykus et al. 1999) mais on trouve également des méthodes
d'hybridation directe (Meijer, Roholl et al. 2000). Les données présentent après des
tests moléculaires ont confirmé certains résultats obtenus par les méthodes de culture
conventionnelle. Ainsi, la présence majoritaire des bactéries à Gram positif dans le tube
digestif et des Lactobacilles au niveau de l'intestin grêle, ainsi que la diversité plus
importante des populations bactériennes au niveau du caecum sont confirmées par
(Gong, Forster et al. 2002) et (Lu et al. 2003), tel que présenté dans le tableau 2. Les
méthodes moléculaires font aussi apparaître des différences. Ainsi les Clostridiaceae
seraient beaucoup plus importantes quantitativement surtout dans les caecum (Lu, Idris
et al. 2003), tableau 1.2).
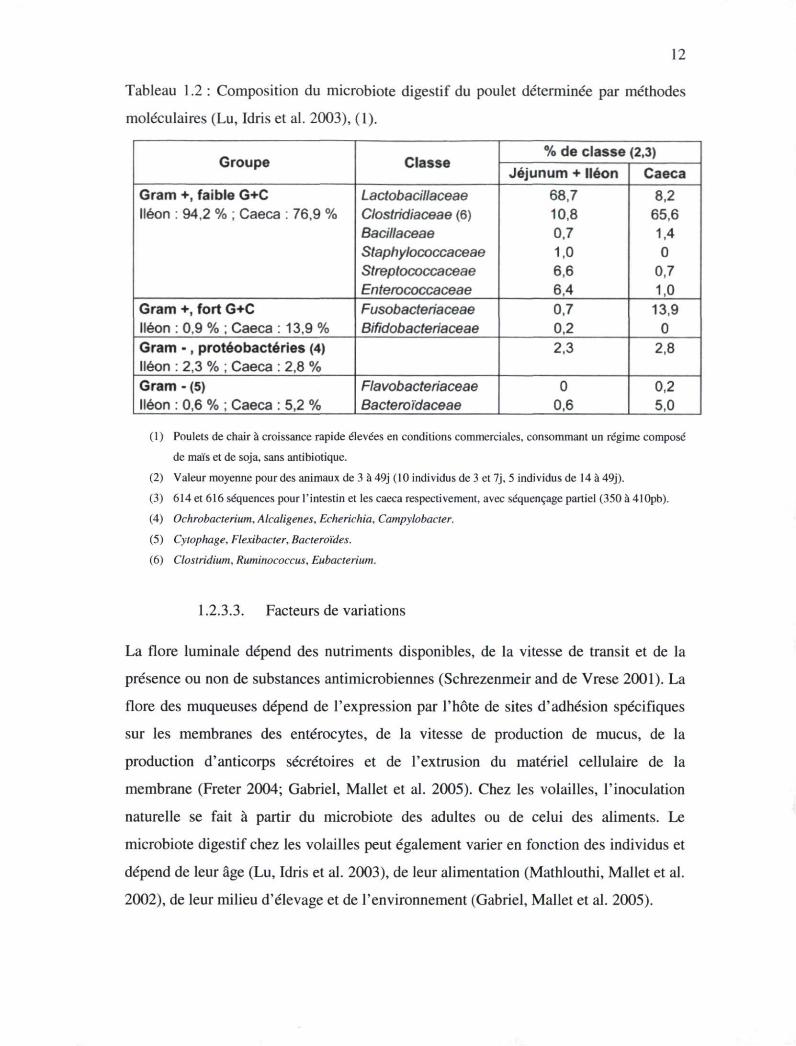
12
Tableau 1.2 : Composition du microbiote digestif du poulet déterminée par méthodes
moléculaires (Lu, Idris et al. 2003), (1).
Groupe Classe % de classe (2,3)
Groupe Classe Jéjunum + Iléon Caeca
Gram +, faible G+C Iléon : 94,2 % ; Caeca : 76,9 %
Lactobacillaceae Clostridiaceae (6) Bacillaceae Staphylococcaceae Streptococcaceae Enterococcaceae
68,7 10,8 0.7 1,0 6,6 6,4
8,2 65,6 1,4 0
0,7 1,0
Gram +, fort G+C Iléon : 0,9 % ; Caeca : 13,9 %
Fusobacteriaceae Bifidobacteriaceae
0,7 0,2
13,9 0
Gram - , protéobactéries (4) Iléon : 2,3 % ; Caeca : 2,8 %
2,3 2,8
Gram - (5) Iléon : 0,6 % ; Caeca : 5.2 %
Flavobacteriaceae Bacteroïdaceae
0 0,6
0,2 5,0
(1) Poulets de chair à croissance rapide élevées en conditions commerciales, consommant un régime composé
de maïs et de soja, sans antibiotique.
(2) Valeur moyenne pour des animaux de 3 à 49j (10 individus de 3 et 7j, 5 individus de 14 à 49j).
(3) 614 et 616 séquences pour l'intestin et les caeca respectivement, avec séquençage partiel (350 à 410pb).
(4) Ochrobacterium, Alcaligenes, Echerichia, Campylobacter.
(5) Cytophage, Flexibacter, Bacteroïdes.
(6) Clostridium, Ruminococcus, Eubacterium.
1.2.3.3. Facteurs de variations
La flore luminale dépend des nutriments disponibles, de la vitesse de transit et de la
présence ou non de substances antimicrobiennes (Schrezenmeir and de Vrese 2001). La
flore des muqueuses dépend de l'expression par l'hôte de sites d'adhésion spécifiques
sur les membranes des entérocytes, de la vitesse de production de mucus, de la
production d'anticorps sécrétoires et de l'extrusion du matériel cellulaire de la
membrane (Fréter 2004; Gabriel, Mallet et al. 2005). Chez les volailles, l'inoculation
naturelle se fait à partir du microbiote des adultes ou de celui des aliments. Le
microbiote digestif chez les volailles peut également varier en fonction des individus et
dépend de leur âge (Lu, Idris et al. 2003), de leur alimentation (Mathlouthi, Mallet et al.
2002), de leur milieu d'élevage et de l'environnement (Gabriel, Mallet et al. 2005).

13
1.2.3.4. Fonction du microbiote intestinal
La microflore intestinale exerce de nombreuses fonctions physiologiques dont les
répercussions sur l'hôte sont, pour la plupart, bénéfiques. Parmi les grandes fonctions
du microbiote est sa capacité à convertir une grande variété de substrats (incluant
glucides, protéines et lipides) en substances nutritives. Ces substances génèrent une
diversité d'effets bénéfiques sur la santé de l'hôte, sur la fermentation des substrats
disponibles au niveau du côlon (Gérard and Bernalier-Donadille 2007) et sur le
développement d'une barrière microbiologique contre la colonisation par les micro
organismes pathogènes. La microflore permet également le développement et la
maturation du système immunitaire intestinal. Cette microflore interagit avec les
cellules épithéliales pour le maintien de la santé de l'hôte (Gérard and Bernalier-
Donadille 2007). La microflore intestinale doit ainsi être considérée dans son contexte
environnemental, incluant l'hôte et l'aliment. Les interrelations entre ces différents
constituants assurent l'homéostasie de l'écosystème microbien digestif. Toute rupture
de l'équilibre entre ces constituants est susceptible de perturber le fonctionnement de
l'écosystème et d'être à l'origine de pathologies digestives (fonctionnelles,
inflammatoires, infectieuses, etc) (Patterson and Burkholder 2003).
1.2.4. Les pathogènes entériques de la volaille
1.2.4.1. Maladies entériques chez la volaille
Les volailles sont prédisposées à de nombreuses infections parasitaires, bactériennes,
mycoplasmiques et virales en particulier celles des systèmes respiratoire et gastro
intestinal. Les maladies entériques à forte prévalence comme la cryptosporidiose ou la
paratuberculose, conduisent à des pertes économiques élevées dans les élevages. De
nombreuses espèces de Salmonella peuvent attaquer les oiseaux adultes et causer des
taux élevés de mortalité et de morbidité. Une autre problématique liée à la présence de
ces pathogènes dans le tube digestif est la contamination des denrées alimentaires
provenant de la volaille. Les infections par le Welchia sont responsables de l'entérite
nécrosante. Les infections infracliniques résultent dans des performances altérées et des
lésions du foie. La maladie clinique aiguë entraîne une mortalité accrue (Chafai 2006).

14
Le traitement de choix suppose l'utilisation d'antibiotiques, mais ne prévient pas la
récurrence de la maladie après arrêt de la médication. Les mesures de contrôle
consistent en une pratique d'hygiène adéquate dans la chaîne de production et au niveau
du consommateur.
1.2.4.2. Escherichia coli
Escherichia coli est une bactérie fréquente du tube digestif de l'homme et des animaux
à sang chaud. La plupart des souches d'E. coli sont sans danger. Certaines souches,
cependant, comme les souches entérohémorragiques (ECEH), peuvent être à l'origine
de toxi-infections alimentaires (TIA) graves. Son importance pour la santé publique est
apparue en 1982, à la suite d'une flambée de TIA aux États-Unis. ECEH fabrique des
toxines, connues sous le nom de verotoxines ou de toxines de type Shiga en raison de
leur ressemblance avec les toxines élaborées par Shigella dysenteriae.
Les Escherichia coli aviaires, bien que considérés par beaucoup comme pathogènes
secondaires, représentent à l'heure actuelle l'une des plus importantes causes de pertes
économiques dans le secteur avicole. Les Escherichia coli sont des hôtes commensaux
du tractus digestif de la volaille et la plupart des souches ne sont pas pathogènes.
Cependant, un certain nombre de celles-ci appelées « Avian Pathogenic E. coli » ou
APEC et appartenant à des serotypes bien particuliers sont associées au syndrome de la
colibacillose (Stordeur and Mainil 2002). La voie d'entrée principale de l'agent
pathogène est le tractus respiratoire, via l'inhalation de particules de poussières
contaminées par les E. coli excrétées du tractus digestif d'animaux sains. Les intestins
sont, en effet, le réservoir le plus important des E. coli pathogènes aviaires ou APEC.
Après une première multiplication au niveau du tractus respiratoire supérieur, les
bactéries colonisent les voies respiratoires profondes à savoir les sacs aériens et les
poumons. Dans une troisième étape, la bactérie atteint le sang et colonise les organes
internes comme le cœur, le foie et la rate (Jordan and Pattison 1996).
Les colibacilloses sont sans doute les infections bactériennes les plus fréquentes et les
plus importantes en pathologie aviaire. Elles peuvent entrainer de la mortalité, des
baisses de performances et des saisies à l'abattoir (Stordeur and Mainil 2002).

15
Contrairement aux infections des mammifères, les colibacilloses aviaires prennent des
formes générales, avec une voie d'entrée respiratoire ou génitale (Stordeur and Mainil
2002). L'agent étiologique de la colibacillose est la bactérie Escherichia coli. Elle est
caractérisée par les antigènes O (somatique), H (flagellaire), F (pilus) et K (capsulaire),
qui permettent d'identifier plusieurs serotypes. Chez les oiseaux, les serotypes
considérés comme pathogènes sont 01 Kl, 02K1 et O78K80. De nouveaux serotypes
pathogènes (non typables) sont en émergence. Plusieurs facteurs de virulence potentiels
sont identifiés chez les E. coli aviaires : adhésines de fimbriae, protéine à activité
hémagglutinante, système aérobactine de captation du fer, antigène capsulaire
polysaccharidique, résistance au pouvoir bactéricide du sérum, toxines et cytotoxines
(Stordeur and Mainil 2002; Mainil 2003; Mainil 2003; Mainil and Van Bost 2004).
Le jeune âge, le stress, un taux élevé d'ammoniac, une baisse de la température, des
infections concomitantes, favorisent la colibacillose. La contamination est
essentiellement par voie aérienne via des aérosols. Les bactéries sont inhalées et
contaminent les sacs aériens. Ceux-ci peuvent prolonger l'infection aux organes
génitaux par contact. Certains Escherichia coli intestinaux provoquent des infections
après entérite. La transmission verticale est possible mais rare.
Escherichia coli 0157:H7 est un entéropathogène non-invasif, décrite pour la première
fois en 1982, les infections d'Escherichia coli 0157.H7 sont rapidement devenues une
cause majeure de diarrhées et de troubles rénaux aigus chez l'humain (Lamps 2007). Il
possède des facteurs de virulence lui permettant de s'attacher aux cellules intestinales et
de libérer des toxines, connues sous le nom de vérotoxines ou toxines de type Shiga en
raison de leur ressemblance avec les toxines élaborées par Shigella dysenteriae (Figure
1.2). Les vérotoxines jouent un rôle important dans l'apparition des symptômes gastro
intestinaux mais l'étape cruciale dans la pathogenèse d'E. coli 0157.H7 est
l'attachement aux cellules de l'hôte (LeBlanc 2003). Suite à l'infection, des
hémorragies, de l'oedème et une infiltration de neutrophiles situés au niveau de la
lamina propria peuvent se produire au niveau des cellules intestinales (Nataro and
Kaper 1998).
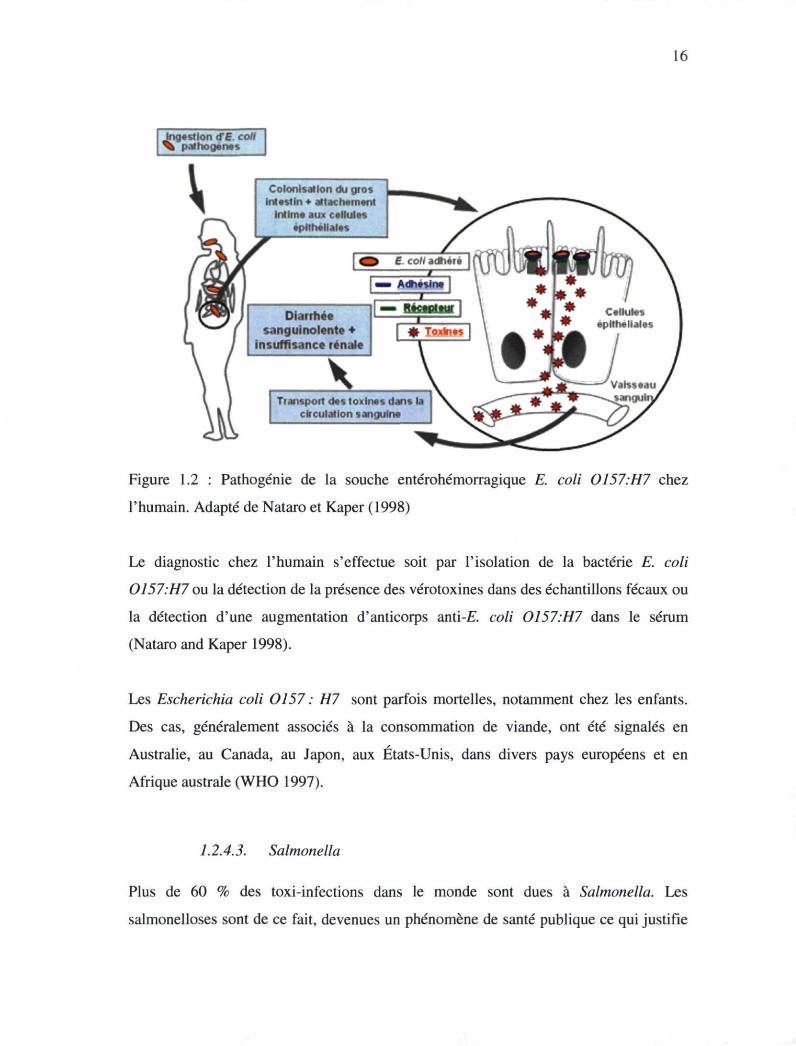
16
<? estlon <T£. coli pathogènes
Colonisation du gros intestin + attachement
intime aux cellules epltbellales
Diarrhée sanguinolente ♦
insuffisance rénale
Transport des toxines dans la circulation sanguin*
Figure 1.2 : Pathogénie de la souche entérohémorragique E. coli 0157.H7 chez
l'humain. Adapté de Nataro et Kaper (1998)
Le diagnostic chez l'humain s'effectue soit par l'isolation de la bactérie E. coli
0157.H7 ou la détection de la présence des vérotoxines dans des échantillons fécaux ou
la détection d'une augmentation d'anticorps anti-£. coli 0157:H7 dans le sérum
(Nataro and Kaper 1998).
Les Escherichia coli 0157 : H7 sont parfois mortelles, notamment chez les enfants.
Des cas, généralement associés à la consommation de viande, ont été signalés en
Australie, au Canada, au Japon, aux États-Unis, dans divers pays européens et en
Afrique australe (WHO 1997).
1.2.4.3. Salmonella
Plus de 60 % des toxi-infections dans le monde sont dues à Salmonella. Les
salmonelloses sont de ce fait, devenues un phénomène de santé publique ce qui justifie

17
l'implication de l'Organisation Mondiale de la Santé (OMS) dans la lutte contre les
salmonelloses (Salm-Surv 2005).
Les salmonelles appartiennent à la famille des entérobactéries, ce sont des bacilles
mobiles à Gram négatif, aéro-anaérobies facultatifs, mésophiles qui se cultivent
facilement. À nos jours plus de 2300 serotypes différents de salmonelles sont identifiés,
sur le plan épidémiologique et sont classées en fonction de leur potentiel pathogène
pour l'homme ou l'animal. La virulence des salmonelles est une notion complexe,
résultant de nombreux facteurs encore largement étudiés, tant au niveau biochimique
que génétique. Les principaux facteurs de virulence sont la mobilité (reposant sur les
flagelles), l'adhésion par les pili et les fimbriae (phénomène actif de reconnaissance
spécifique entre une adhésine bactérienne et un ligand présent à la surface d'une cellule
hôte), l'invasion (par endocytose pour les entérocytes, par phagocytose pour les
macrophages), la formation de phagosomes spacieux et la fusion avec les lysosomes
(Feuillet 2007).
Actuellement plus de 200 serotypes de salmonelles sont connus chez la volaille.
Salmonella Enteritidis qui infecte les organes profonds (foie, rate, ovaire) est à l'origine
d'infection durable au niveau des troupeaux alors que Salmonella Typhimurium est
actuellement le serotype le plus incriminé dans les salmonelloses aviaires provoquant
les formes cliniques les plus graves, surtout chez les jeunes poussins prenant une allure
septicémique, avec une mortalité brutale dans les jours qui suivent l'éclosion. On
observe également de nombreuses mortalités en coquille. La première étape de
colonisation par les salmonelles est une étape de colonisation intestinale. La bactérie
arrive dans l'intestin grêle, où elle se multiplie en adhérant à l'épithélium, elle pénètre
par un phénomène d'endocytose dans les cellules épithéliales iléales et caecales,
notamment les tissus lymphoïdes incluant les plaques de Peyer, les amygdales caecales
et dans les cellules M. Dans le cas des salmonelles provoquant des maladies
systémiques, le site d'attachement préférentiel se situe au niveau des plaques de Peyer
(Feuillet 2007). L'infection est strictement limitée à la sphère digestive et peut
correspondre à un portage latent avec élimination épisodique des Salmonelles dans les
fèces. Les porteurs latents sont cliniquement et anatomiquement indécelables. Us

18
peuvent excréter des salmonelles de façon continue ou intermittente (Takase,
Nakayama et al. 1999). Ces porteurs sains sont le plus fréquemment rencontrés. Dans
ce cas, les salmonelles sont retrouvées dans différentes portions de l'intestin par culture
sur milieux sélectifs. Les symptômes sont observés essentiellement sur les poussins de
moins de 15 jours et sont rares sur les volailles de plus de 4 semaines (Takase,
Nakayama et al. 1999).
Avant l'abattage, la peau du poulet est contaminée par une flore microbienne mésophile
variée provenant de l'environnement (sol, eau, des abreuvoirs, litières, excrément,
cage...). À l'usine les sources de contaminations peuvent être les volailles elles
mêmes, c'est-à-dire les plumes, la peau, les pattes et l'intestin. Cela peut être dû aux
cages de transport ainsi qu'aux ustensiles et l'équipements de transformation ainsi qu'à
l'environnement générale de l'usine incluant l'eau, les poussières et les aérosols, le
personnel ou encore à des contaminations croisées par contact avec les produits finis ou
au cours de transformation avec les eaux usées, les viscères et autres parties éliminées.
1.2.5. Contrôle des infections entériques chez la volaille
Le recours aux antibiotiques demeure l'approche la plus préconisée pour lutter contre
ces infections. L'utilisation des antibiotiques en alimentation animale s'est progressivement
développée dès le début des années 50 et a permis d'améliorer les conditions sanitaires des
animaux et d'accroître la productivité des élevages en réduisant les coûts de production.
Cependant, l'utilisation abusive de ces antibiotiques qui possèdent beaucoup de
similitudes avec ceux utilisés en médecine humaine a entraîné une augmentation
inquiétante du nombre de souches pathogènes multi-résistantes. En effet, actuellement on
connait une vingtaine de familles d'antibiotiques. Six modes d'actions sont connus à
savoir, l'inhibition de la synthèse des protéines (Gentamicine), l'inhibition de la
synthèse du peptidoglycane (groupe de la pénicilline G), coupure des brins d'ADN et
déroulement de l'ADN (Metronidazole), l'inhibition de la synthèse de l'ADN
(Furazolidone), le blocage de la synthèse de l'acide dihydrofolique (Sulfadiazone) ou
encore le blocage de la synthèse des ARN messagers (Ripampicine).

19
Dès 1945, la pénicilline G a été utilisée pour contrer les infections à Staphylocoques, en
produisaient une enzyme capable de dégrader la pénicilline. En 1980 est apparu le
phénomène de la 1947, on repérait déjà les premières souches de Staphylocoques
résistants. Ces dernières sont multi-résistantes. Un microorganisme peut devenir multi-
résistant lorsqu'il fait l'acquisition de plusieurs gènes codant pour la résistance. Parmi
ces organismes, on compte Streptococcus pneumoniae, Staphylococcus aureus,
Mycobacterium tuberculosis, Salmonella sp., Campylobacter sp. et Escherichia coli.
Pour contrer l'action des antibiotiques, les bactéries utilisent plusieurs types de
mécanismes. De nos jours, quatre mécanismes de résistances aux antibiotiques sont
connus ; le premier connu sous le nom de « brouillage » dans lequel la bactérie est
capable de produire des enzymes capables d'inactiver les antibiotiques, ces enzymes
sont capables de détruire des liens chimiques nécessaires à l'intégrité fonctionnelle du
médicament. Les bêta-lactames sont un exemple d'enzymes produites par la bactérie
inactivant les p-lactamines telles les pénicillines et les céphalosporines. Le second
connu sous le nom du « blindage et efflux », les microorganismes sont capables dans ce
cas de se rendre imperméable à la pénétration de l'antibiotique ou le rejeter, en effet ce
mécanisme résiste dans le fait de modifié le nombre des porines et/ou la spécificité de
celle-ci. Il s'agit d'un mécanisme de résistance spécifique aux bactéries Gram négatif
puis que sur la membrane de ces bactéries on trouve les porines qui sont une sorte de
protéines qui forment des canaux permettant le passage de plusieurs types de
molécules, dont tire également profit la pénicilline. Le troisième mécanisme de
résistance des bactéries est le « camouflage » qui consiste à modifier la structure des
cibles cellulaires des antibiotiques, puisque pour être efficace, un antibiotique doit se
fixer à une cible cellulaire. Si la bactérie remplace ou modifie cette cible, l'action de
l'antibiotique sera réduite ou totalement inhibé puisqu'il ne pourra plus s'y fixer. Le
dernier mécanisme est « Esquive » ou stratégie de contoumement dans ce cas
l'antibiotique atteint sa cible. Cependant, la bactérie est capable d'utiliser d'autres voies
métaboliques pour exécuter le même travail. Les activités inhibées par l'antibiotique
sont donc remplacées.
Cette situation a conduit l'Union européenne à décréter, en 2006, l'interdiction complète
des antibiotiques à titre de facteurs de croissance dans les aliments pour animaux dont

20
les premières espèces animales touchées par cette interdiction sont les porcs et les
volailles, principales productions à recourir à ce type d'additifs. Au Canada,
l'utilisation des antibiotiques comme facteur de croissance n'est pas formellement
interdit, mais il est déconseillé. Cependant, le Québec jouit d'une réglementation
unique au Canada puisque tous les médicaments destinés aux animaux ne sont
disponibles que sur ordonnance (CAAAQ 2008). De surcroît, aucun producteur ne peut
détenir ou administrer un médicament à un animal de consommation à moins qu'il n'ait
été prescrit par un médecin vétérinaire (selon l'Ordre des médecins vétérinaires du
Québec).
Il est donc clair que la recherche de solution alternative susceptible d'assurer une
succession satisfaisante aux antibiotiques en termes d'effets zootechniques devient une
urgence.
1.2.5.1. Les huiles essentielles
Les huiles essentielles sont des substances odorantes concentrées, obtenues à partir de
plantes par entraînement à la vapeur d'eau, hydrodistillation. Le terme "huile
essentielle" a été inventé au I6lème siècle par le médecin suisse Parascelsus von
Hohenheim pour désigner le composé actif d'un remède naturel. Il existe aujourd'hui
approximativement 3000 huiles, dont environ 300 sont réellement commercialisées,
destinées principalement à l'industrie des arômes et des parfums. La tendance actuelle
des consommateurs à rechercher une alimentation plus naturelle, a entraîné un regain
d'intérêt des scientifiques pour ces substances. Depuis deux décennies, des études ont
été menées sur le développement de nouvelles applications et l'exploitation des
propriétés naturelles des huiles essentielles dans le domaine alimentaire. Les huiles
essentielles et leurs composants, actuellement employés comme arômes alimentaires
sont également connus pour posséder des activités antimicrobiennes et pourraient donc
servir d'agents de conservation alimentaire, et ce d'autant plus qu'ils sont pour la
plupart classés "généralement reconnus comme sains" (Generally Recognized As Safe
GRAS), ou approuvés comme additifs alimentaires par la Food and Drug
Administration. Ils n'ont, par conséquent, pas besoin d'autorisation d'emploi dans les

21
aliments, cependant des études préalables sont nécessaires afin de mieux cerner leur
activité antimicrobienne. Leur activité antimicrobienne varie fonction de leur
composition chimique, et en particulier de la nature de leurs composés volatils majeurs
(Pellecuer, Jacob et al. 1979). Elles agissent en empêchant la multiplication des
bactéries, leur sporulation et la synthèse de leurs toxines (Hulin, Mathot et al. 1998).
Les huiles essentielles possèdent plusieurs modes d'action sur les différentes souches
de bactéries, mais d'une manière générale leur action se déroulent en trois phases : en
premier lieu ils attaquent la paroi bactérienne par l'huile essentielle, provoquant une
augmentation de la perméabilité puis la perte des constituants cellulaires, ensuite
l'acidification de l'intérieur de la cellule, bloquant la production de l'énergie cellulaire
et la synthèse des composants de structure et finalement la destruction du matériel
génétique, conduisant à la mort de la bactérie (Oussalah, Caillet et al. 2006).
Bozkur et ces collaborateurs ont effectué une étude dans laquelle ils ont supplémenté
l'alimentation des volailles avec des huiles essentielles, les résultats de cette étude ont
montré qu'il y avait une amélioration de la fertilité ainsi qu'une augmentation du poids
(Bozkurt, Alcicek et al. 2009), cependant, aucun effet sur l'action antimicrobienne
contre les microorganismes indésirable n'a été observé. Oral et ces collaborateurs qu'on
a eux ont introduit dans l'emballage du poulet frais Y origanum onites qui a permis de
prolongé la durée de conservation (Oral, Vatansever et al. 2009). Les huiles essentielles
ont était utilisé également afin d'inhiber Clostridium perfringens chez la volaille.
Mitsch et ces collaborateurs ont montré qu'un mélange d'huiles essentielles peut
contrôler la colonisation et la prolifération de Clostridium perfringens dans l'intestin
des poulets de chair. (Mitsch, Zitterl-Eglseer et al. 2004).
Certes, les huiles essentielles ont présenté un effet bénéfique pour la santé de volailles.
Cependant, peu d'étude ont visé l'utilisation des huiles essentielles pour contrer les
pathogènes humains présents dans la viande de volaille. Leur action inhibitrice vis-à-vis
à des pathogènes alimentaires reste encore très peu étudiée.

22
1.2.5.2. Les enzymes
Les enzymes existent pratiquement partout et sont essentielles pour tous les organismes
vivants car elles favorisent les réactions chimiques nécessaires à la vie. Un peu plus de
3000 enzymes ont été identifiées et caractérisées. L'enzymologie remonte à la fin du
19ème siècle, époque à laquelle une préparation enzymatique extraite de l'estomac du
veau, appelée présure, a été employée à grande échelle pour la fabrication du fromage.
Depuis lors, cette technologie n'a cessé de croître. Actuellement les enzymes sont
employées dans plusieurs domaines. En alimentation animale, l'usage des enzymes est
par contre beaucoup plus récent et date tout au plus de 20 ans (Boguhn and
Rodehutscord 2010; Jia and Slominski 2010). La raison essentielle de l'utilisation des
enzymes en alimentation animale est d'accroître la valeur alimentaire des aliments en
augmentant l'efficacité de la digestion (vitesse et/ou ampleur) dans le tube digestif des
animaux. En effet, l'efficacité de la digestion affecte grandement les frais
d'alimentation des animaux (Vandeplas, Dauphin et al. 2009).
Vandeplas et ses collaborateurs on ajouté à un régime riche en froment de préparations
enzymatiques à activité xylanasique principale des Lactobacillus plantarum peut
conduire à une augmentation du poids final des animaux consécutivement à une
augmentation de l'ingestion du régime et de la croissance quotidienne. Les ajouts
enzymatiques réalisés dans les régimes riches en froment chez la volaille entraînent une
augmentation de la digestibilité des matières grasses, des protéines et de la cellulose
chez le jeune poulet, avec pour conséquence une augmentation des quantités de
nutriments nécessaires à la croissance de l'animal. L'ajout des Lactobacillus plantarum
quand à lui a permis d'inhiber les Salmonella Typhimurium (Vandeplas, Dauphin et al.
2009). Donc l'effet inhibiteur revient principalement à la présence de probiotiques, les
enzymes ont permis une meilleure digestion.
Les enzymes ajoutées aux aliments des volailles doivent être assimilées à une
amplification du système enzymatique endogène de leur tube digestif. Choisies de
manière pertinente, elles permettent d'inhiber les facteurs antinutritionnels présents
dans les aliments et d'améliorer la disponibilité des nutriments pour l'animal, ainsi que

23
l'amélioration la digestibilité. (Boguhn and Rodehutscord 2010; Jia and Slominski
2010)
Certains résultats ont également mis en évidence l'intérêt des enzymes pour contrôler
la flore entérique des porcs et des volailles ce qui constitue un autre avantage de ces
molécules (Yves Beckers and Piron 2009). La première revendication pour l'utilisation
d'enzymes chez la volaille est une amélioration de la digestibilité, qui se traduit par une
amélioration de l'efficacité alimentaire en relation avec une baisse de la consommation
ou une augmentation de la masse d'œufs produite.
Mori et ces collaborateurs ont rapporté que l'ajout d'enzyme améliore la production
d'œufs et l'efficacité alimentaire chez des poules recevant des régimes à base d'orge,
de blé, de maïs et même de sorgho. Sur l'ensemble des essais présentés, le taux de
ponte a augmenté de près de 2% et l'indice de consommation a été réduit en moyenne
de 3,6% (Mori agnes 2007).
En effet les rôles principaux des enzymes ajoutés aux aliments vont casser les structures
et faire qu'à la fin de l'intestin grêle les substrats pour la flore intestinale seront
différents. Lorsqu'on met des enzymes dans un aliment pour animal, on change les
substrats et donc l'équilibre de la flore (Yves Beckers and Piron 2009).
Une amélioration de la digestibilité est induite à l'ajout des enzymes dans
l'alimentation des volailles cependant peu d'étude ont ciblé l'inhibition des pathogènes
à l'aide des enzymes.
1.2.5.3. Les extraits de plantes
Depuis l'Antiquité, l'homme a utilisé les plantes pour leurs vertus bienfaisantes.
Aujourd'hui, leurs principes actifs sont à la base de nombreux médicaments. Parmi les
principales vertus des plantes, on peut citer la stimulation de la digestion, un effet anti-
inflammatoire, antalgique, antidiabétique, anti-cholestérol, tonique, antiparasitaire,
vermifuge, antiseptique : antibactérien, antifongique, antiviral (Guardia 2009).
Les principes actifs peuvent être regroupés en plusieurs 'familles' chimiques à savoir
les composés soufrés : thiosulfonates (allicine de l'ail), les aldéhydes (cinnamaldéhyde

24
de la cannelle, cuminaldéhyde du cumin), les phénols (thymol, menthol, carvacrol,
eugenol), les terpènes (pinène, cymène) et les tannins. Ces substances agissent en
bloquant la multiplication du microorganisme ou en détruisant sa paroi (Rhayour,
Bouchikhi et al. 2003).
Guardia et ses collaborateurs ont étudié l'effet d'une combinaison d'extraits végétaux
naturels (EXVa) à action anti microbienne, utilisée seule ou procédée d'une
combinaison (EXVb) à action anti-oxydante et antibactérienne, sur la performance de la
croissance des volailles. Cette étude a permis de déduire que l'utilisation seule
d'extraits de végétaux à action antimicrobienne n'a pas induit à une amélioration de la
croissance des volailles. Cependant, la combinaison (EXVb) ayant des propriétés anti
oxydantes puis antibactérienne, permet d'améliorer les performances de croissance
(Guardia 2009).
L'ail représente l'une des meilleures plantes utilisé vue ses effets bénéfiques Dès que la
gousse est hachée, l'alliin est transformée par une enzyme en allicin qui est le principe
actif majeur. Celui-ci est relativement instable puisque la moitié de l'allicin est détruite
en 2 à 4 heures à température ambiante. On attribue à l'ail de nombreuses vertus
notamment des activités antibactériennes, anti-cholestérol, anti-tumoral, etc. L'huile
d'ail obtenue par distillation à la vapeur conserve la propriété anti-tumorale et anti
oxydante mais perd une partie de l'activité antibactérienne et anti-thrombose
(Srivastava, Bordia et al. 1995).
La plupart des études ont montré que les extraits végétaux ont une plus grande activité
contre bactéries à Gram positif par rapport aux bactéries à Gram négatif (Shelef 1984;
Zaika 1988; Smith-Palmer, Stewart et al. 1998; Ceylan and Fung 2004).
Si aujourd'hui les différentes propriétés des phyto-molécules observables in vitro sont
reconnues (action antibactérienne, immuno-modulatrice, anti-oxydante) leur action in
vivo sur la croissance des animaux, notamment du poulet, montre des effets variables
(Ceylan and Fung 2004; Windisch, Schedle et al. 2008).
Aucune de ces molécules n'a encore démontré un potentiel réel pour substituer les
antibiotiques. Les enzymes, par exemple, n'ont montré qu'une action trop faible sur les

25
performances des animaux. Les huiles essentielles quant à elles, possèdent un pouvoir
antimicrobien tout en activant l'appétit et les sécrétions digestives. Néanmoins, leur
action antimicrobienne, comme leurs effets bénéfiques sur les performances
zootechniques, restent inférieurs comparés aux antibiotiques.
Une des plus récentes alternatives proposées est le recours au probiotiques qui sont des
microorganismes vivant qui, lorsqu'il est intégré en quantité suffisante, exerce un effet
positif sur la santé. Selon plusieurs chercheurs, les probiotiques sont bien placés pour
prendre la relève des additifs antibiotiques en raison de leurs aptitudes nutritionnelles et
antimicrobiennes fort intéressantes.
1.3. LES PROBIOTIQUES
1.3.1. Définition
Le terme "probiotique" est un mot relativement nouveau qui signifie "en faveur de la
vie". Le concept probiotique est né de la théorie de la longévité de Metchnikoff en
1907. Il fut le premier à proposer l'utilisation des Lactobacilles des yaourts pour la
restauration du microbiote dans le tractus gastro-intestinal. Les probiotiques ont
d'abord été développés dans les années 1960 pour les élevages d'animaux afin de
prévenir les infections et stimuler le gain de poids. La première définition officielle a
été proposée par Fuller en 1989 qui définit un probiotique comme étant « un
supplément alimentaire microbien vivant qui affecte positivement la santé de l'animal
en améliorant sa balance microbienne intestinale ». Cette définition a été révisée
plusieurs fois, notamment par la FAO (Food and Agriculture Organization of the
United Nations) et la WHO (World Health Organization). En 2001, leur nouvelle
définition s'énonce comme suit : « Les probiotiques sont des microorganismes vivants
qui lorsqu'ils sont administrés en quantité adéquate, produisent un effet bénéfique pour
la santé de l'hôte ».

26
1.3.2. Critères de sélection
Les micro-organismes doivent posséder diverses propriétés de survie pour répondre à la
définition des probiotiques (Gagnon 2007). Ils doivent présenter une activité positive et
persister durant leur passage dans le tractus digestif. Ces propriétés sont propres à
chaque souche et ne peuvent pas être extrapolables d'une souche à l'autre même au sein
d'une même espèce (Dunne, O'Mahony et al. 2001). Plusieurs critères majeurs de
sélection ont été établis par différents auteurs dans le but de sélectionner les souches
potentiellement probiotiques. Ces critères, résumés dans le tableau 1.3, sont réparties
en trois catégories à savoir les critères de sécurité, fonctionnels et technologiques.
Tableau 1.3 : Principaux critères de sélection des probiotiques. Adapté de (Klaenhammer and Kullen 1999; Saarela, Mogensen et al. 2000; Ouwehand, Salminen et al. 2002; Gueimonde and Salminen 2006).
Critères de sécurité
Critères fonctionnels
Critères technologiques
Identification taxonomique précise.
Souche caractérisée par des techniques phénotypiques et
génotypiques.
Historique de non pathogénicité et non-invasion de l'épithélium
intestinal.
Pas de transmission possible de gènes de résistance aux
antibiotiques.
Tolérance de l'acidité à la bile et aux enzymes digestives.
Adhésion aux cellules intestinales et persistance dans le tractus
intestinal.
Production de substances antimicrobiennes (bactériocines, acides
organiques, peroxyde d'hydrogène ou autres composés inhibiteurs)
et antagonisme envers les pathogènes.
Immunomodulation.
Aptitude à produire des effets bénéfiques sur la santé de l'hôte.
Stabilité au cours des procédés de fabrication et dans le produit fini.
Conservation des propriétés probiotiques après production.
Non modification des qualités organoleptiques du produit fini.
Parmi les critères reliés à la sécurité, l'identification taxonomique de la souche est une
étape importante dans l'établissement de nouvelles souches potentiellement

27
probiotiques. Chaque souche doit être identifiée par des techniques moléculaires fiables
et confrontée à une nomenclature actualisée (FAO/WHO 2002). Actuellement, le
séquençage de la région 16S est la méthode moléculaire de référence pour identifier
l'espèce d'une souche, mais cette méthode est longue et requiert une large collection de
souches de référence (FAO/WHO 2002). Le séquençage de l'ARN 16S est une
méthode très fiable couramment utilisée pour l'identification des souches probiotiques
Dans ce dernier cas, il est recommandé que la technique soit combinée avec des tests
biochimiques et phénotypiques pour s'assurer de la conformité de la souche. L'origine
de la souche est également une condition importante car l'interaction spécifique avec
l'hôte est maximisée lorsqu'elle provient du même habitat (Alvarez-Olmos and
Oberhelman2001).
Parmi l'ensemble des critères présentés au Tableau 1.3, l'aptitude à produire des effets
bénéfiques sur la santé demeure encore délicat à évaluer dû notamment au fait que les
modes d'action par lesquels les probiotiques exercent un rôle fonctionnel in vivo sont
méconnus (Klaenhammer and Kullen 1999). La compréhension des mécanismes
d'action représente un des défis scientifiques majeurs dans le domaine des probiotiques.
D'un point de vue technologique, les souches probiotiques doivent posséder plusieurs
critères telles que la facilité à être cultivée tout en conservant leurs propriétés
biologiques et leur stabilité au cours des procédés de production et d'entreposage
(Champagne, Gardner et al. 2005).
1.3.3. Les effets santé associés aux probiotiques
Plusieurs effets bénéfiques sur la santé ont été associés à la consommation des
probiotiques. La Figure 1.3 illustre la diversité des effets santé documentés et rapportés
dans la littérature.

28
Amélioration de la digestion du lactose (sécrétion de lactase)
Influence positive sur la flore intestinale Bonne croissance et le bien-être
Réduction des produits du catabolisme éliminés par le foie et le rein
Augmentation de la valeur nutritionnelle (bonne digestion et absorption des minéraux et vitamines
V
Prévention des infections intestinales (virus. Helicobacter pylori..) et urogénitales
Régulation de la motilité intestinale (constipation, syndrome
N d'irritation intestinale)
Prévention de : ostéoporose. cancer, hypertension et athérosclérose (réduction du j janx de cholestérol)
Modulation du système immunitaire Réduction de l'inflammation >pu des réactions allergiquesy
Figure 1.3 : Les principaux effets bénéfiques attribués aux probiotiques. Adapté de
Mercenier et al. (2003).
L'apport de probiotiques doit être régulier (Gagnon, Kheadr et al. 2004). Les
probiotiques sont souvent considérés comme des aliments fonctionnels ou des
compléments alimentaires, mais pas comme des médicaments (FAO/WHO 2002).
Malgré la diversité des allégations revendiquées, les mécanismes par lesquels les
probiotiques exercent leur effets bénéfiques sur l'hôte ne sont pas toujours connus.
1.3.4. Activité antimicrobienne des probiotiques
L'activité antimicrobienne des probiotiques est probablement l'activité la plus
documentée. Plusieurs mécanismes ont été proposés pour expliquer ces activités. Ces
mécanismes sont résumés dans la figure 1.4.

29
Mécanisme 1 : Inhibition de l'adhésion du
pathogène
Exclusion stérique
^ Adhésion au mucus
Inhibition des enzymes cytoplasmiques
Mécanisme 2 : Inhibition de la croissance du
pathogène
Lyse cellulaire
/ Acides organiques Bactériocines
Compétition stérique <
Adhésion aux cellules épithéliales
Compétition nutritionnelle
Restriction Nutriments r
Interaction avec le système , immunitaire
Mécanisme 3 : Immunomodulation
Augmentation de la défense immune
Figure 1.4 : Mécanismes d'action proposés des microorganismes probiotiques dans le
traitement des infections entériques. Adapté de Kaur, Kuhad et al. (2009) et Calder and
Kew (2002).
Un des mécanismes qui suscite de plus en plus d'intérêt en recherche dans le domaine
de probiotiques est la production de substances inhibitrices telles que les acides
organiques et les bactériocines. Paradoxalement, très peu d'études ont montré, par des
tests in vitro ou in vivo, l'implication de ces substances inhibitrices dans l'activité des
probiotiques.
1.3.5. Les bactériocines
1.3.5.1. Définition
La première définition des bactériocines date de 1953 et faisait référence aux protéines
de type colicine produites par les bactéries à Gram négatif (Jacob, Lwoff et al. 1953).
La découverte de la première bactériocine remonte à 1925. Cette dernière, isolée

30
^Escherichia coli <P, possédait une activité bactéricide envers une autre souche
d'Escherichia coli V. Elle fut nommée colicine V (Gratia 1925). La découverte de
composés similaires chez les bactéries à Gram positif, (Tagg, Dajani et al. 1976) a
élargi le terme "bactériocines" à l'ensemble des peptides antimicrobiens produits par les
bactéries, qui sont des peptides antimicrobien de faible poids moléculaire. Elles ont une
activité inhibitrice dirigée contre les bactéries proches de la souche productrice. Leur
spectre d'action est généralement étroit. Les bactériocines représentent une arme
importante dans les phénomènes de compétitions bactériennes car elles aident la
bactérie productrice à se défendre contre les autres bactéries de son environnement. À
cette époque, les bactériocines étaient encore définies comme des substances ayant un
spectre d'action limité. En 1963, Hamon and Peron (1963) ont observé que certaines
bactériocines produites par les bactéries à Gram positif pouvaient avoir un spectre
d'activité étendu. Le terme plus général de bactériocine fut proposé en 1976 pour
englober toutes les substances de nature protéique synthétisées par les bactéries et qui
ont un pouvoir antibactérien dirigé contre des bactéries taxonomiquement proches du
microorganisme producteur (Tagg, Dajani et al. 1976). Aujourd'hui, il est généralement
accepté que la majorité de ces peptides sont cationiques, hydrophobes, qu'ils sont
sensibles aux proteases et qu'ils présentent une activité maximale à pH acide (Ennahar,
Deschamps et al. 2000). La famille de bactériocine inclut une diversité des protéines en
termes de taille, cibles microbiennes, modes d'action, et mécanismes d'immunité. La
nomenclature des bactériocines est relativement simple. De la même manière que la
terminaison "ase" est associée à la dénomination d'enzymes, le suffixe «ine» est utilisé
afin de dénoter une activité bactériocinogène (Kaiser and Montville 1993). Certaines
bactériocines tirent leur nom de la souche productrice, d'autres ont été nommées en
termes généraux, par exemple les "colicines" produites par les coliformes, ou les
"lactostrepcines" produites par les streptocoques lactiques (Kozak, Bardowski et al.
1978; Klaenhammer 1988; Ecker 1992). Les bactériocines ont en effet une activité
bactéricide ou bactériostatique qui affecte, dans la plupart des cas, des bactéries
taxonomiquement proches de l'organisme producteur. Ce sont des peptides ou des
protéines synthétisées par voie ribosomale. Les bactériocines sont séparées en deux
grandes familles, celles produites par les bactéries à Gram positif et celles produites par
les bactéries à Gram négatif.

31
La famille de bactériocine inclut une diversité des protéines en termes de taille, cibles
microbiennes, modes d'action, et mécanismes d'immunité. Il faut savoir qu'un type
bactérien ou même une bactérie, peut produire plus d'une bactériocine (Gagnon et al
2007).
1.3.5.2. Utilisation
Les microorganismes produisent un grand nombre de systèmes de défenses microbiens.
Ceux-ci incluent des sous produits métaboliques tels que les acides lactiques produits
par les lactobacilles, des agents lytiques tels que les lysozymes, de nombreux types de
protéines, des exotoxines et des bactériocines qui sont employées dans plusieurs
domaines en raison de leur grand potentiel dans l'amélioration de la santé de l'être
humain et des animaux ainsi que dans des éventuelles utilisations dans le secteur
agricole (David, Oliver et al. 2007). Leur utilisation dans le domaine alimentaire est
devenue très intéressante grâce à leur potentiel d'assurer une sécurité microbienne et
une bonne qualité du produit alimentaire sans modifier la qualité organoleptique du
produit. Il s'agit donc de très bons agents conservateurs (Cutter and Siragusa 1996).
D'autres études ont montré l'utilisation des bactériocines comme agent antimicrobien
dans l'emballage des produits carnés (Ming, McDonaldMcGinn et al. 1997) et les
produits marins. Tel que déjà mentionné, l'utilisation des bactériocines n'est pas
restreinte au domaine alimentaire, mais trouvent aussi des applications dans le domaine
médical et vétérinaire. Les bactériocines pourraient servir d'alternatives aux
antibiotiques pour le traitement des maladies infectieuses tant chez l'humain que chez
l'animal. Il faut cependant, bien signaler que peu d'études sont réalisées pour la
détermination des mécanismes d'action des bactériocines. Davies, Milne et al. (1999)
ont montré qu'une augmentation en teneur des lipides réduit l'activité biologique des
bactériocines dans la matrice alimentaire. Il faut également signaler que les
bactériocines sont des peptides amphiphiles capables de s'absorber aux macromolécules
de la matrice alimentaire, mais ces propriétés peuvent limiter leur utilisation comme
agents de conservation.

32
1.3.5.3. Les bactériocines actives contre les bactéries à Gram négatifs
Les bactéries à Gram négatif produisent un certain nombre de peptides avec une activité
antibiotique. Ces bactériocines sont principalement produites par les bactéries de
l'écosystème intestinal, représentées principalement par la famille des
Enterobacteriaceae. Elles sont réparties en deux grands groupes, les colicines et les
microcines.
Les colicines
Les colicines ont été largement étudiées et une trentaine de molécules différentes ont
été identifiées. Elles sont toutes produites par Escherichia coli (Braun, Killmann et al.
2002) et possèdent un spectre d'activité restreint aux souches phylogénétiquement
proches (Gillor, Nigro et al. 2005). Les colicines possèdent en général un poids
moléculaire élevé compris entre 25 et 80 kDa (Dirix, Monsieurs et al. 2004). Ce sont
des protéines qui ne subissent pas de modifications post-traductionnelles, sauf en ce qui
concerne la methionine N-terminale qui peut être enlevée (Smajs, Smarda et al. 2003).
Elles présentent une même organisation structurale basée sur trois domaines
fonctionnels. La partie N-terminale, appelée domaine T, riche en résidus glycine,
participe à la translocation de la colicine à travers la membrane externe en interagissant
avec un système de transport. La partie centrale, appelée domaine R, représente 50 %
de la colicine et intervient dans la reconnaissance spécifique de la colicine avec son
récepteur membranaire. Enfin, la partie C-terminale, appelée domaine C, est
responsable de l'activité bactéricide de la colicine et possède une séquence de liaison
avec la protéine d'immunité (Lazdunski, Bouveret et al. 2000; Cao and Klebba 2002;
Riley and Wertz 2002).
Braun, Patzer et al. (2002)et Lazzaroni, Dubuisson et al. (2002) ont proposé une
classification des colicines selon la voie de translocation dans la cellule cible qu'elles
utilisent. Effectivement, la structure de leur domaine T leur permet d'interagir avec les
systèmes périplasmiques Toi (colicines du groupe A) ou Ton (colicines du groupe B)
(Cao and Klebba 2002; Gillor, Kirkup et al. 2004). Le groupe A comprend notamment

33
les colicines A, El à E9, K, N et DF13 tandis que le groupe B regroupe les colicines B,
D, la, Ib, M, 5 et 10 (Tableau 1.4).
Tableau 1.4 : Classification des colicines et leurs cibles cellulaires (Lazdunski,
Bouveret et al. 2000)
Groupes Colicines Cibles cellulaires
Groupe A A
DF13
HI
E2, E7, E8 et E9
E3àE6
K
N
Membrane cytoplasmique
Acides nucléiques
Membrane cytoplasmique
Acides nucléiques
Acides nucléiques
Membrane cytoplasmique
Membrane cytoplasmique
Groupe B B
D
la et Ib
M
5 et 10
Membrane cytoplasmique
Acides nucléiques
Membrane cytoplasmique
Peptidoglycane
Membrane cytoplasmique
Les microcines
Asensio et al (1976) ont été les premiers à présenter les microcines (Mec) comme une
nouvelle famille de peptides à activité antibiotique produits par les bactéries à Gram
négatif. Ils définissent les microcines en tant que peptides antibiotiques de faible poids
moléculaire, produits par des bactéries non sporulantes isolées à partir du tractus
intestinal humain, sécrétés dans le milieu de culture et agissant sur des
microorganismes phylogénétiquement proches (Asensio, Perezdiaz et al. 1976). Bien
que les microcines se rapprochent des colicines par de nombreux points, elles forment
un groupe spécifique de peptides à activité antibiotique. Elles se distinguent des
colicines par plusieurs caractéristiques dont leur petite taille (< 10 kDa) et leur
sécrétion non létale pour les bactéries productrices (David, Oliver et al. 2007).
Contrairement aux colicines, les microcines ne sont pas produites uniquement par des
souches d'Escherichia coli. Une microcine a été découverte chez une souche de K.
pneumoniae (De Lorenzo 1984). Par ailleurs, elles possèdent des points communs avec

34
les bactériocines de petite taille des bactéries à Gram positif, tels que leur nature
hydrophobe et leur stabilité à la chaleur, à certaines proteases et aux pH extrêmes
(Pons, Lanneluc et al. 2002).
Jusqu'à aujourd'hui, dix microcines ont été identifiées (Tableau 1.5), parmi lesquelles
sept sont bien caractérisées (Mec B17, C7, E492, J25, H47, L et V) et trois (Mec D93,
M et N24) sont peu connues. Des différences au niveau de leur poids moléculaire et des
modifications post-traductionnelles permettent de répartir les microcines en deux
classes (Pons, Lanneluc et al. 2002).
Tableau 1.5 : Les dix microcines découvertes.
Microcines Références
Classe I
B17 (Patzer, Baquero et al. 2003) C7 (Garcia-Bustos, Pezzi et al. 1984) D93 (Martinez, Carter-Franklin et al. 2003) J25 (Salomon and Farias 1992) E492 (Delorenzo 1984) H47 (Lavina, Gaggero et al. 1990) L (Gaillard-Gendron, Vignon et al. 2000) M (Braun, Killmann et al. 2002; Patzer, Baquero et al. 2003) N24 (O'Brien and Gibson 1970) V (Gratia 1925; Yang and Leong 1982)
Classe II
La classe I regroupe les microcines B17, C7, D93 et J25. Elles ont un très faible poids
moléculaire, inférieur à 5 kDa, sont largement modifiées post-traductionnellement et
sont toutes sécrétées par Escherichia coli (Pons, Lanneluc et al. 2002). Les microcines
de classe II comprennent les microcines E492, H47, L, M, N24 et V et sont synthétisées
sous forme de précurseurs avec des peptides signal très homologues. Elles sont de plus
grande taille (7 à 10 kDa). Les microcines de classe II sont toutes hydrophobes et non
modifiées post-traductionnellement.

35
1.3.6. Les probiotiques en aviculture
En alimentation animale de nombreux genres bactériens et fongiques incluant
Lactobacillus, Bifidobacterium, Bacillus, Streptococcus, Pediococcus, Enterococcus,
Propionibacterium, Saccharomyces, Aspergillus et Torulopsis sont utilisés, comme
probiotique (Tannock 1997). En général, les souches probiotiques sont sélectionnées
pour leurs effets bénéfiques sur la santé tout en assurant leur sécurité d'utilisation
(innocuité). L'utilisation commerciale de probiotiques en élevages industriels des
volailles est relativement nouvelle. Comme pour les autres animaux, l'utilisation des
probiotiques s'est développée à la suite des recherches effectuées sur le tractus gastro
intestinal qui ont permis une meilleure compréhension du rôle de la microflore et de
son importance sur la santé et l'hygiène digestive des animaux (Gaggia, Mattarelli et
al.). Les différentes études réalisées sur des volailles ont montré que les probiotiques
exercent des activités antibactériennes contre diverses bactéries pathogènes notamment
celles responsables d'infection chez les poulets dont Salmonella sp (Van Immerseel,
Cauwerts et al. 2002; Higgins, Higgins et al. 2008), Campylobacter et Escherichia coli
(Zacconi, Scolari et al. 1999; Ragione, Narbad et al. 2004). L'administration de
Lactobacillus salivarius A23 à des poussins a permis d'augmenter le taux des
Lactobacilles dans le jabot dès le premier jour d'administration. Par contre, aucune
augmentation significative n'a été observée au niveau du caecum. Ceci signifie que ce
probiotique colonise préférentiellement le jabot (Zacconi, Scolari et al. 1999).
L'administration d'une préparation brut de microflore caecale a permis de protéger les
animaux contre des infections à Salmonella Typhimunium et à Salmonella Enteritidis
(Andreatti, da Silva et al. 2000; Higgins, Higgins et al. 2008). Enterococcus faecium
J96, une souche probiotique isolée de l'intestin d'une poule a permis de réduire le taux
de croissance de Salmonella Pullorum, Gallinarum, Typhimurim et Enteritidis in vitro.
L'administration orale de 109 UFC de cette souche à des poussins de 30 h d'âge leur a
permis de résister à une infection à Salmonella pullorum (Audisio, Oliver et al. 2000).
Dans le même ordre d'idée, l'administration simultanée de Salmonella Enteritidis et
Lactobacillus salivarius CTC2197 par voie orale à des poussins d'un jour a permis une
élimination complète des Salmonelles (Zacconi, Scolari et al. 1999).

36
L'utilisation d'un consortium de souches probiotiques contenant Lactobacillus
salivarius et Lactobacillus Blantarum inhibent in vitro Escherichia coli et Salmonella
Typhimurium a également été proposée par Murry and Hinton (2006). De la même
façon, une suspension fécale permet de protéger les poussins contre une colonisation
par Salmonella Typhimurium, Salmonella Agona, Salmonella Infatis, Salmonella
Enteritidis (De Oliveira, Berchieri et al. 2000). Willis and Reid (2008) ont montré que
la présence de Campylobacter jejuni à diminuer chez les volailles avec un régime
alimentaire supplémenté avec 108 UFC/g d'une combinaison de Lactobacillus
acidophilus, Lactobacillus casei, Bifidobacterium thermophilus, et Escherichia
faecium.
Globalement, les études rapportées dans la littérature suggère que l'utilisation d'un
mélange de plusieurs souches probiotiques appartenant à des genres bactériens
différents a permis d'obtenir des effets plus significatifs que ceux obtenus avec des
monosouches (Jung, Houde et al. 2008; Awad, Ghareeb et al. 2009; Revolledo, Ferreira
et al. 2009; Vandeplas, Dauphin et al. 2009). Ces études ont également montré qu'il
serait possible de favoriser la colonisation du tube digestif des poussins dès l'éclosion
par l'utilisation des probiotique et que cette colonisation est plus rapide et plus efficace
pour la protection des animaux qu'une colonisation naturelle. Chez les poussins,
l'infection par des bactéries pathogènes est beaucoup plus fréquente du fait que la flore
intestinale n'est pas complètement établie. De plus, les poussins étant séparés de leur
mère dès leur éclosion dans l'élevage, n'ont pas la possibilité d'acquérir la microflore
protectrice maternelle.
Cependant, il est à noter que l'utilisation de probiotiques chez la poule pondeuse
permet d'améliorer la qualité de l'œuf (épaisseur des coquilles) mais ne protège pas le
poussin contre l'infection après l'éclosion (Panda, Reddy et al. 2003). Ainsi, pour
obtenir l'effet protecteur escompté, les probiotiques doivent être administrés en continu
dès l'éclosion.
Certaines études ont allé chercher remède pour les antibiotiques dans les probiotiques
mais cependant le mode d'action de ces derniers reste encore mal connue. D'autre part

37
peu de recherches ont utilisés des souches Gram négatifs pour contrer les pathogènes
causant le plus de mortalité en élevage et de cas de toxi-infections alimentaires.

38
HYPOTHÈSE ET OBJECTIFS DU MÉMOIRE
Hypothèse
Les souches bactériennes productrice de substances inhibitrices telles que les
bactériocines et faisant partie du microbiote normal du tube digestif de la volaille
représentent des candidates à fort potentiel probiotique d'utilisation pour la prévention
des infections entériques majeurs en aviculture
Objectifs
L'objectif général de ce travail de maîtrise est d'identifier et caractériser des souches à
fort potentiel probiotique actives contre des pathogènes à Gram négatif tel que
Salmonella et Escherichia coli de la volaille. D vise également à caractériser sur les
plans physicochimiques et biologiques les substances antimicrobiennes responsables de
cette activité biologique.

39
CHAPITRE 2. CRIBLAGE ET CARACTERISATION DE L'ACTIVITÉ INHIBITRICE DE SOUCHES INTESTINALES VIS-
À-VIS DES PATHOGÈNES ENTÉRIQUES GRAM NÉGATIF

40
2.1. RÉSUMÉ
Cette étude vise à identifier, à partir de contenus intestinaux de volailles, de nouvelles
souches probiotiques productrices de bactériocines actives contre des pathogènes à
Gram négatif incluant Escherichia coli et Salmonella. Vingt quatre souches ont été
isolées du contenu digestif de volailles en santé et ont été soumises à une première
étape de criblage visant à identifier celles présentant une activité inhibitrice
significative contre Escherichia coli 0157 : H7, Salmonella enterica et Yersinia
enterocolitica. Sept souches à savoir : 11004, 11006, 11012, 11014, 11016, 11018 et
11026 ont été alors retenues à la suite de ce premier criblage. Dans un deuxième temps,
l'activité inhibitrice des surnageants des sept cultures des souches actives a été évaluée
après un traitement thermique de 100 °C durant 5 min, une filtration sur membrane de
0,22 u.m et une combinaison des deux traitements. Cette deuxième étape nous a permis
de retenir 3 souches qui semblent produire des substances inhibitrices thermostables
très apparentées à des bactériocines et ont conservé une activité inhibitrice significative
contre Escherichia coli 0157 : H7 et Salmonella enteritica.

41
2.2. INTRODUCTION
Les maladies entériques sont une préoccupation importante en aviculture en raison des
conséquences économiques et sanitaires qu'elles engendrent. Sur le plan économique
ces maladies sont responsables de pertes importantes associées à la faible productivité
du troupeau et au taux de mortalité élevé. Sur le plan sanitaire, les animaux malades
peuvent transmettre à l'humain des microorganismes pathogènes via la consommation
de denrées contaminées.
À l'échelle mondiale, l'industrie avicole et porcine sont les deux secteurs qui
enregistrent la plus forte croissance avec des taux annuels de 2,6% et 3,7% au cours des
dix dernières années (FAO 2007). Malheureusement, les denrées alimentaires
provenant de ces deux espèces sont également les denrées les plus incriminées dans la
transmission de bactéries potentiellement pathogènes pour l'homme (Van Immerseel,
Cauwerts et al. 2002). Dans le cas de la volaille les carcasses peuvent être contaminées
par plusieurs bactéries pathogènes durant le plumage, le refroidissement, 1'evisceration,
le lavage, etc. Une des stratégies utilisée pour limiter la propagation de ces pathogènes
bactériens par les denrées de volaille est la réduction de la charge bactérienne chez
l'animal vivant. Dans ce contexte, le recours aux antibiotiques demeure l'approche la
plus préconisée pour lutter contre plusieurs infections chez la volaille. Ainsi, l'utilisation
des antibiotiques en alimentation animale a permis d'améliorer les conditions sanitaires des
animaux et d'accroître la productivité des élevages en réduisant les coûts de production.
Cependant, l'utilisation abusive de ces antibiotiques qui possèdent beaucoup de
similitudes avec ceux utilisés en médecine humaine, a entraîné une augmentation
inquiétante du nombre de souches pathogènes multi-résistantes. Cette situation a conduit
l'Union européenne à décréter, en 2006, l'interdiction complète des antibiotiques à titre
de facteurs de croissance dans les aliments pour animaux. Comme alternative aux
antibiotiques, plusieurs additifs alimentaires ont été suggérés notamment les enzymes,
les huiles essentielles et les extraits de plantes.
Certes, ces molécules ont présenté certains effets bénéfiques pour la santé de volailles,
Cependant, aucune de ces molécules n'a encore démontré un potentiel réel pour
substituer les antibiotiques. Les enzymes, par exemple, n'ont montré qu'une action trop

42
faible sur les performances des animaux ce qui ne joue pas en leur faveur pour succéder
aux additifs antibiotiques. Les huiles essentielles quant à elles, possèdent un pouvoir
antimicrobien tout en activant l'appétit et les sécrétions digestives. Néanmoins, leur
action antimicrobienne, comme leur effet bénéfique sur les performances
zootechniques, restent inférieurs comparé aux antibiotiques.
Une des plus récentes alternatives proposées est le recours au probiotiques. Les
probiotiques sont des microorganismes vivants qui, lorsqu'ils sont intégrés en quantité
suffisante, exercent un effet positif sur la santé. Selon plusieurs chercheurs, les
probiotiques sont bien placés pour prendre la relève des antibiotiques en raison de leurs
aptitudes nutritionnelles et antimicrobiennes fort intéressantes.
Les mécanismes par lesquels les probiotiques inhibent les bactéries intestinales
pathogènes incluent la concurrence pour les sites de la colonisation, la compétition pour
les nutriments, la production des composés toxiques tels que les acides organiques et
les bactériocines, ou la stimulation du système immunitaire (Calder and Kew 2002;
Kaur, Kuhad et al. 2009). Ces mécanismes ne sont pas mutuellement exclusive et
l'inhibition peut comprendre une, plusieurs ou tous ces mécanismes (Gagnon, 2007).
Bien que la production de bactériocines soit un phénomène relativement bien étudié
aussi bien chez les Gram négatifs que positifs, l'implication de ces molécules bioactives
dans l'activité antibactérienne des probiotiques n'a jamais été mise en évidence
(Gagnon, 2007).
Dans le cadre de se travail, on postule que des souches à Gram négatif d'origine
intestinale productrices de bactériocines constitueraient des souches probiotiques très
prometteuse pour la prévention des infections entériques à Escherichia coli 0157 : H7
et Salmonella chez la volaille.

43
2.3. MATÉRIEL ET MÉTHODE
2.3.1. Souches bactériennes et conditions de culture
2.3.1.1. Souches productrices
Vingt-quatre souches isolées de contenus intestinaux de volailles et ayant démontré une
activité inhibitrice contre Escherichia coli K12 et Salmonella enteritidis ont été utilisées
lors de cette étude. Ces souches sont présentées dans le tableau 2.1.
La souche Escherichia coli LR05 qui est une souche sauvage productrice des MccL,
B17, D93 et J25 et résistante à la MccV (ou ColV) et à la colicine M (Sable, Duarte et
al. 2003) a été utilisée comme témoin positif.
2.3.1.2. Souches sensibles
Les 24 souches bactériennes ont était testées contre 5 souches pathogènes ainsi que
contre une souche non pathogène de référence appelée Escherichia coli MC4100. Cette
souche, qui est un dérivé d'Escherichia coli K12, est particulièrement intéressante car
elle est sensible à toutes les microcines connues sauf, la microcine D93. Les 5 souches
pathogènes testées proviennent de la collection de l'Université Laval et de l'Université
La Rochelle et sont énumérées dans le tableau 2.2.
2.3.1.3. Propagation des souches productrices et sensibles
Toutes les souches sont repiquées deux fois dans du milieu Brain Heart Infusion (BHI;
Difco, Sparks, MD) et incubées à 37°C. Avant leur utilisation, ces souches sont
conservées à -80°C dans une solution cryoprotectrice contenant du glycerol à 20%.
Pour les tests d'évaluation de l'activité inhibitrice, les souches ont été propagées dans
un milieu minimum M63 (Miller 1972). La composition de ce milieu de même que les
conditions de culture ont été optimisées pour la production de bactériocines. Ainsi, les
effets de l'agitation, de l'ajout ou non de la thiamine et du glucose de même que le
temps d'incubation sur la production de bactériocines ont été étudiés.

44
Tableau 2.1 : Souches à fort potentiel probiotiques
11001 Activité inhibitrice vis-à-vis d'E. coli K12 et 5. enteritidis 11004 Activité inhibitrice vis-à-vis d'E. coli K12 et 5. enteritidis 11006 Activité inhibitrice vis-à-vis d'E. coli K12 et 5. enteritidis
Activité inhibitrice vis-à-vis d'E. coli K12 et S. enteritidis 11007 11010 Activité inhibitrice vis-à-vis d'E. coli Kl2 et 5. enteritidis
Activité inhibitrice vis-à-vis d'E. coli Kl2 et 5. enteritidis (faible), V+ 11012 11013 Activité inhibitrice vis-à-vis d'E. coli Kl2 11014 Activité inhibitrice vis-à-vis d'E. coli K12 11015 Activité inhibitrice vis-à-vis d'E. coli Kl2 11016 Activité inhibitrice vis-à-vis d'E. coli K12 11018 Activité inhibitrice vis-à-vis d'E. coli K12 11019 Activité inhibitrice vis-à-vis d'E. coli Kl2 11022 Activité inhibitrice vis-à-vis d'E. coli K12 et S. enteritidis (faible),
V+,B17+,J25+
11023 Activité inhibitrice vis-à-vis d'E. coli K12 et 5. enteritidis 11024 Activité inhibitrice vis-à-vis d'E. coli Kl2 et 5. enteritidis 11025 Activité inhibitrice vis-à-vis d'E. coli Kl2 et 5. enteritidis 11026 Activité inhibitrice vis-à-vis d'E. coli K12 et 5. enteritidis 11038 Activité inhibitrice vis-à-vis d'E. coli Kl2 et S. enteritidis (faible), V 11039 Activité inhibitrice vis-à-vis d'E. coli Kl2 11040 Activité inhibitrice vis-à-vis d'E. coli Kl2 11058 Activité inhibitrice vis-à-vis d'E. coli Kl2 et S. enteritidis (faible), V+
11060 Activité inhibitrice vis-à-vis d'E. coli Kl2 11061 Activité inhibitrice vis-à-vis d'E. coli K12 11062 Activité inhibitrice vis-à-vis d'E. coli Kl2 J25+ : Possède le gène de structure de la MccJ25 B17+ : Possède le gène de structure de la MccB17 V+ : Possède le gène de structure de la MccV (ColV)
Tableau 2.2 : Souches sensibles
Souche Origine E. coli MC4100 La Rochelle France
E. coli 0157 : H7 ATCC 35150 E. coli 0157 : H7 Ottawa (Canada), isolé à partir d'un patient
S. enterica ssp enterica serotype seftenberg ATCC 8400 S. enterica ssp enterica serotype Montevideo ATCC 8387
Y. enterocolitica serotype 0:3 Kapperud (Norvège), isolé à partir d'un patient

45
2.3.2. Mise en évidence de l'activité inhibitrice
2.3.2.1. Test de diffusion sur gélose
Les cultures bactériennes sont centrifugées à 4°C pendant 10 min à une vitesse de
5000xg et les surnageants de culture obtenus sont utilisés directement ou après un
traitement thermique de 100°C pendant 5min, une filtration 0,22 im et une
combinaison des deux traitements avant de tester l'activité inhibitrice pour tester leur
activité inhibitrice en utilisant le test de diffusion sur gélose tel que décrit par Tagg,
Dajani et al. (1976). Les tests ont été réalisés dans du milieu M63 gélose préalablement
enrichi en hydrolysat de caséine (Fluka biochemika) et en MgS04 (Aldrich) (Portrait
1999). La souche E. coli LR05 a été utilisée comme témoin positif.
Traitement thermique + Filtration
Filtration Témoin -
Traitement thermique Sans traitement Témoin +
Figure 2.1 : Schéma illustrant le principe de test de diffusion sur
gélose
2.3.2.2. Test de microdilution
La croissance d'E. coli 0157 : H7 et de 5. enteritica a été évaluée en absence et en
présence des surnageants des souches intestinales en utilisant la méthode de dilution
critiques sur microplaque. La figure 2.2 illustre le principe de fonctionnement de ce
test. Ainsi que la disposition des échantillons analysés.

46
Chaque microplaque permet l'évaluation de l'activité inhibitrice contre une souche
sensible. Les puits de la ligne 1 correspondent au témoin négatif et sont remplis avec
175ul du milieu BHI. Les puits de la ligne 2 correspondent au témoin positif et sont
remplis avec 125uJ de milieu BHI et inoculé avec 50ul de la souche sensible. Dans les
autres puits de la microplaque, les surnageants des souches à tester sont ajoutés en
duplicata en présence de 50|ri de la même souche sensible. Les microplaques sont
ensuite incubées à la température optimale de la souche sensible utilisée. Des mesures
d'absorbance (DO) à 650nm sont effectuée à tous les 15min durant 24h afin de suivre la
croissance des souches sensibles.
1 2 3 4 5 6 7 8 9 10 11 12
C
H
00OOOOOOOOOO 00OOOOOOOOOO ©©oooooooooo oeoo oooooooo ©©OOOOOOOOOO ©©oooooooooo ©©oooooooooo ©©oooooooooo
^s
Figure 2.2 : Suivie de la croissance microbienne sur microplaque.

47
2.4. RÉSULTATS ET DISCUSSIONS
2.4.1. Criblage des souches
Optimisation du milieu et des conditions opératoires
Plusieurs paramètres opératoires ont été modifiés pour la mise en évidence de la
production de bactériocines à savoir l'agitation, l'ajout ou non de la thiamine et/ou du
glucose ainsi que le temps d'incubation. La figure 2.3 montre l'activité du surnageant
de la souche Escherichia coli LR05 contre la souche sensible Escherichia coli MC4100
à deux temps d'incubation différents pour la production 18h ou 8h à 37°C. Le tableau
2.3 présente l'activité du surnageant de la souche Escherichia coli LR05 cultivé dans le
milieu M63 enrichi en thiamine et/ou en glucose. Le milieu minimal M63 ne contient
pas de source de fer, car la production des bactériocines et reliée au système de capture
du fer. L'ajout de glucose et de thiamine permet cependant la production d'une
biomasse plus importante. L'agitation à 150 rpm montre que la production de substance
inhibitrice par les souches intestinales retenues nécessite une bonne aération. L'étude
de ces paramètres nous a permis de définir un milieu de culture modèle de même que
des conditions opératoires optimales pour l'évaluation de l'activité inhibitrice des
surnageant de culture des souches intestinales testées. Nos résultats ont montré que la
meilleure production de biomasse est obtenue dans un milieu minimal M63 enrichi en
glucose (concentration final de 4 g/L) et en thiamine (inoculum de 0,1 % d'une solution
à 0,1% en concentration) incubé sous agitation de 150 rpm durant 18 h à une
température de 37°C.

48
Figure 2.3 : Activité du surnageant de la souche E. coli LR05 contre la souche sensible E. coli MC4100. A : Croissance de 8h à 37°C. B : Croissance de 18h à 37°C.
Tableau 2.3 : Activité inhibitrice contre E. coli MC4100.du surnageant de culture de la souche E. coli LR05 cultivée à 37°C pendant 18h sous agitation dans le milieu M63. 1 : milieu M63 supplémenté de thiamine; 2 : milieu M63 supplémenté de glucose; 3 : milieu M63 supplémenté de glucose et de thiamine.
Ajout dans le milieu M63
f 1 : Thiamine (0,1%)
f 2 : Glucose (4g/l) f 3 : Thiamine + Glucose f Le tableau 2.4 présente les diamètres d'inhibition obtenus avec les surnageants des
différentes souches intestinales contre différents pathogènes à Gram négatif. Sept des

49
24 souches testées ont été retenues pour leur forte activité inhibitrice et ont été soumise
à un deuxième essai dans le but d'identifier celles sécrétant des substances inhibitrices
thermostables. Les surnageants de culture de ces sept souches ont subit trois traitement
différents soit un chauffage à 100°C pendant 5 minutes ou une filtration avec un filtre à
0,22 [im ou bien la combinaison des deux traitements avant de procéder à l'évaluation
de leur activité antimicrobienne contre trois souches indicatrices.
Les résultats de cette étude sont présentés dans le tableau 2.5. Cette deuxième étape de
criblage a permis de retenir trois souches parmi les sept testées à savoir 11004,11006 et
11026 puisqu'elles présentent une activité inhibitrice sur les souches sensibles même
suite aux traitements thermiques et de filtration. Des exemples d'activité inhibitrice
obtenue avec les trois souches retenues lors du test de diffusion sur gélose sont
présentés à la figure 2.4.
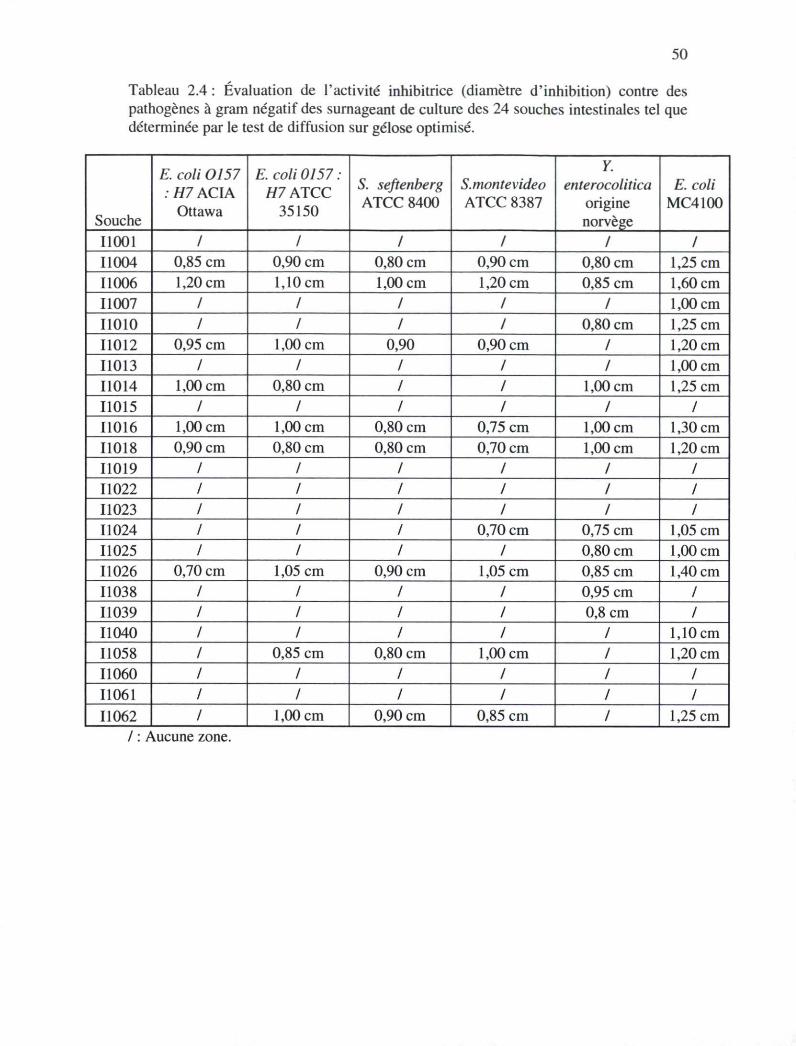
50
Tableau 2.4 : Évaluation de l'activité inhibitrice (diamètre d'inhibition) contre des pathogènes à gram négatif des surnageant de culture des 24 souches intestinales tel que déterminée par le test de diffusion sur gélose optimisé.
Souche
E. coli 0157 : H7 ACIA
Ottawa
E. coli 0157 : H7 ATCC
35150
S. seftenberg ATCC 8400
S.montevideo ATCC 8387
Y. enterocolitica
origine norvège
E. coli MC4100
11001 / / / / / / 11004 0,85 cm 0,90 cm 0,80 cm 0,90 cm 0,80 cm 1,25 cm 11006 1,20 cm 1,10cm 1,00 cm 1,20 cm 0,85 cm 1,60 cm 11007 / / / / / 1,00 cm 11010 / / / / 0,80 cm 1,25 cm 11012 0,95 cm 1,00 cm 0,90 0,90 cm / 1,20 cm 11013 / / / / / 1,00 cm 11014 1,00 cm 0,80 cm / / 1,00 cm 1,25 cm 11015 / / / / / / 11016 1,00 cm 1,00 cm 0,80 cm 0,75 cm 1,00 cm 1,30 cm 11018 0,90 cm 0,80 cm 0,80 cm 0,70 cm 1,00 cm 1,20 cm 11019 / / / / / / 11022 / / / / / / 11023 / / / / / / 11024 / / / 0,70 cm 0,75 cm 1,05 cm 11025 / / / / 0,80 cm 1,00 cm 11026 0,70 cm 1,05 cm 0,90 cm 1,05 cm 0,85 cm 1,40 cm 11038 / / / / 0,95 cm / 11039 / / / / 0,8 cm / 11040 / / / / 1,10cm 11058 / 0,85 cm 0,80 cm 1,00 cm 1,20 cm 11060 / / / / / 11061 / / / / / 11062 / 1,00 cm 0,90 cm 0,85 cm 1,25 cm
/ : Aucune zone.
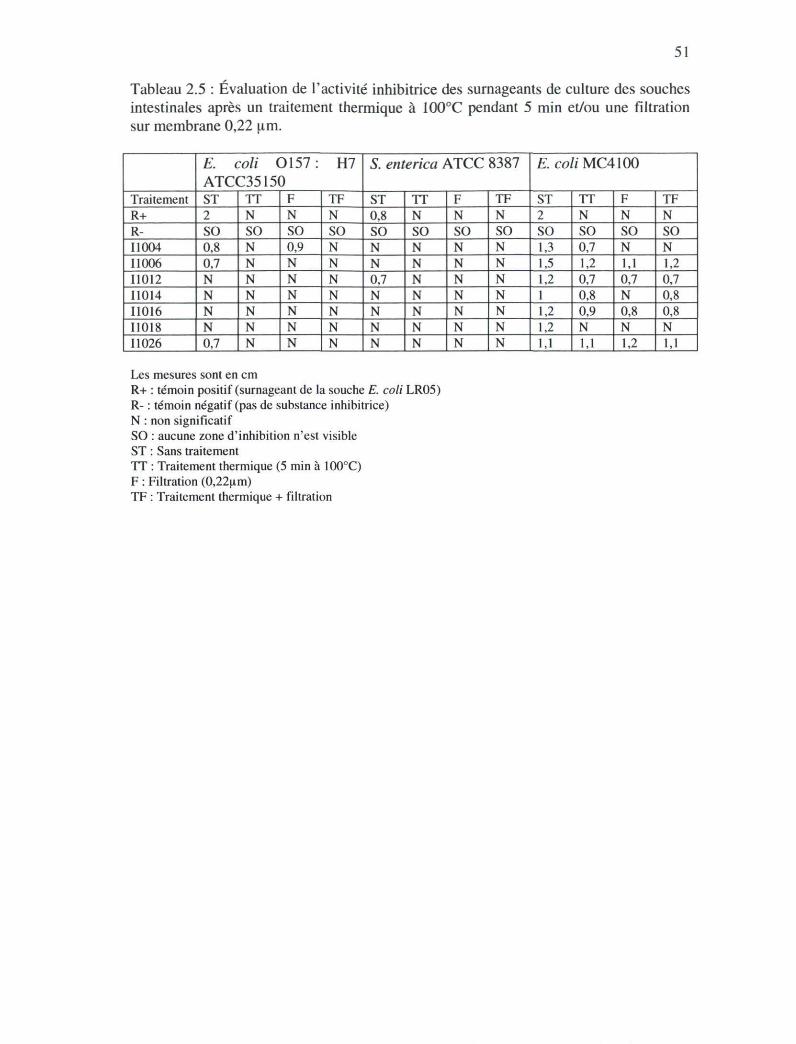
51
Tableau 2.5 : Évaluation de l'activité inhibitrice des surnageants de culture des souches intestinales après un traitement thermique à 100°C pendant 5 min et/ou une filtration sur membrane 0,22 [xm.
E. coli ( ATCC3515I
D157: )
H7 5. enterica ATCC 8387 E. coli MC4100
Traitement ST TT F TF ST TT F TF ST TT F TF R+ 2 N N N 0,8 N N N 2 N N N R- SO SO SO SO SO SO SO SO SO SO SO SO 11004 0.8 N 0,9 N N N N N 1,3 0,7 N N 11006 0.7 N N N N N N N 1,5 1,2 1,1 1,2 11012 N N N N 0,7 N N N 1,2 0,7 0,7 0,7 11014 N N N N N N N N 1 0,8 N 0,8 11016 N N N N N N N N 1,2 0,9 0,8 0,8 11018 N N N N N N N N 1,2 N N N 11026 0,7 N N N N N N N 1,1 1.1 1,2 1,1
Les mesures sont en cm R+ : témoin positif (surnageant de la souche E. coli LR05) R- : témoin négatif (pas de substance inhibitrice) N : non significatif SO : aucune zone d'inhibition n'est visible ST : Sans traitement TT : Traitement thermique (5 min à 100°C) F : Filtration (0,22u.m) TF : Traitement thermique + filtration

52
Figure 2.4 : Test de diffusion sur gélose montrant l'activité inhibitrice des surnageant de culture des souches 11006 (A), 11004(B) et 11026(C) contre la souche sensible E. coli MC4100 suite aux différents traitements.

53
2.4.2. Test de microdilution
2.4.2.1. Activité inhibitrice contre E. coli MC4100
La figure 2.5 représente la courbe de croissance de E. coli MC4100 seule ou en
présence du surnagent de culture des souches 11004, 11006 et 11026. Le surnageant de
la souche E. coli LR05 qui produit les MccL, B17, D93 et J25 a été utilisé comme
contrôle positif. Les courbes de croissances obtenues montrent une inhibition de la
croissance d'E. coli MC4100 en présence des différents surnageant de culture pendant
les 14 premières heures d'incubation. Une croissance progressive a été ensuite
observée. Cette croissance peut être expliquée par deux phénomènes possibles soit
l'épuisement de la substance antimicrobienne produite en raison de la présence de
proteases dans le milieu soit par la nature de l'activité antimicrobienne exercée qui
serait plutôt de type bactériostatique.
De plus nous avons démontré que l'activité inhibitrice observée contre E. coli MC4100
est de type dose dépendante. Tel que présenté dans la figure 2.6, l'effet inhibiteur des
différents surnageants diminue lorsque les surnageants sont dilués au 1/2, 1/4, 1/8 et
1/16.
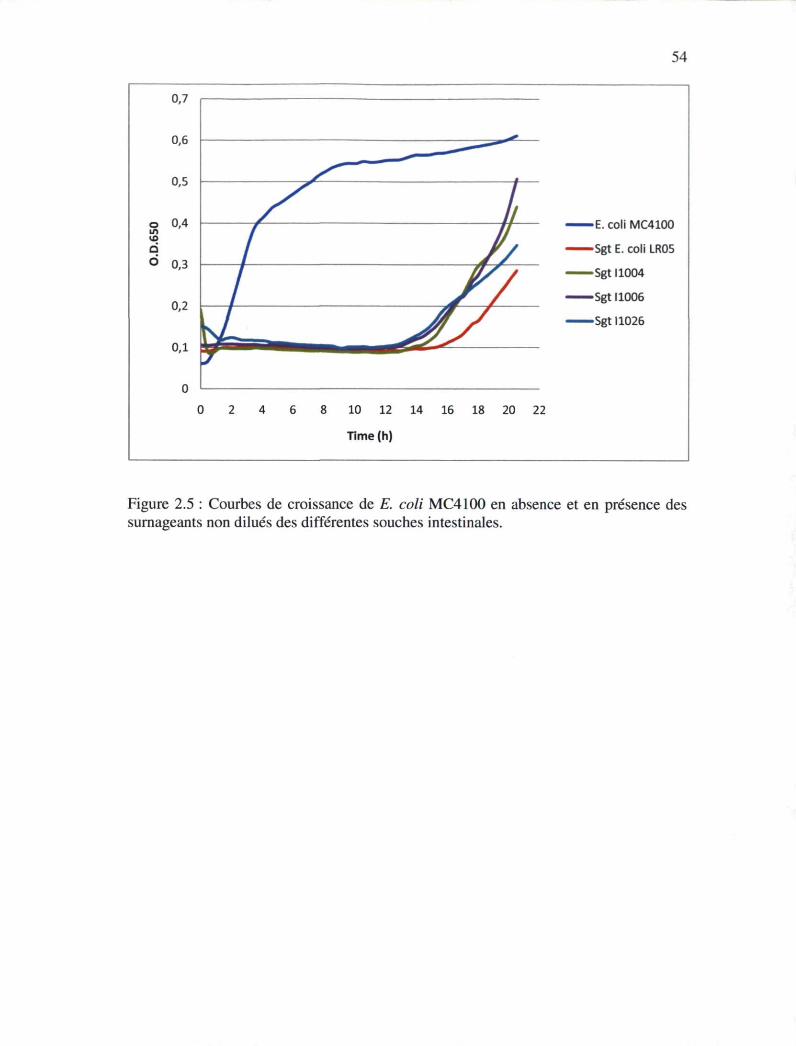
54
0,7
0,6
0,5
o in
Q
0,4
O 0,3
0,2
0,1
•E. coliMC4100
■Sgt E. coli LR05
■Sgt 11004
■Sgt 11006
■Sgt 11026
0 2 4 6 8 10 12 14 16 18 20 22
Time (h)
Figure 2.5 : Courbes de croissance de E. coli MC4100 en absence et en présence des surnageants non dilués des différentes souches intestinales.
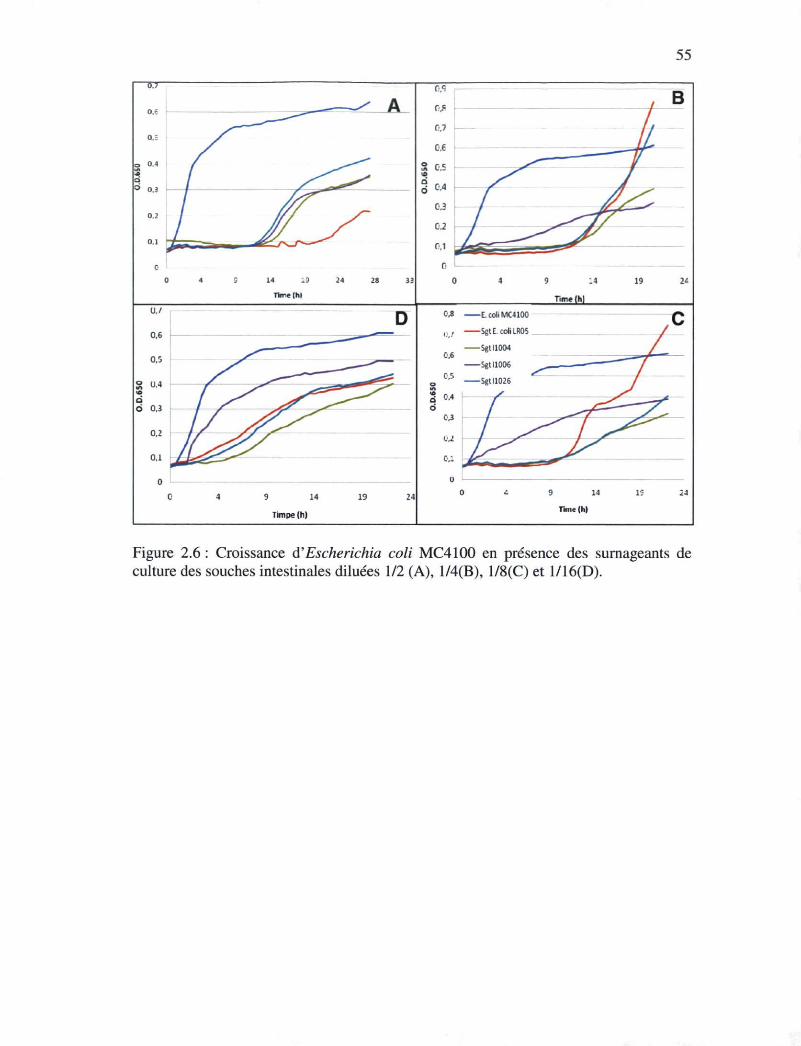
55
19 2 i
0,6
o.r.
0.4
0.3
DJ
0,1
u
E.coliMC4100
Set E. coli LR05
SgtllCKM
SgtllOOe
Sgtll026
9 14
Time (h)
IS 24
Figure 2.6 : Croissance d'Escherichia coli MC4100 en présence des surnageants de culture des souches intestinales diluées 1/2 (A), 1/4(B), 1/8(C) et 1/16(D).

56
2.4.2.2. Activité inhibitrice contre Escherichia coli 0157 :H7
Une expérience similaire à la précédente a été réalisée avec la souche pathogène
Escherichia coli 0157 :H7. La figure 2,7 illustre les courbes de croissance
d'Escherichia coli 0157 :H7 en présence et en absence des surnageants des souches
intestinales. Cette figure montre un effet inhibiteur qui est moins important que celui
observé contre la souche sensible Escherichia coli MC4100. Suite à la dilution des
surnageants, l'effet inhibiteur diminue rapidement et aucune inhibition n'a été observée
à partir de la dilution 1/4 (figure 2.8). La figure 2.8 montre aussi que parmi les quatre
souches testées, la souche 11026 est celle qui montre l'activité inhibitrice la plus
marquée. Cependant, la souche utilisée comme témoin positif E. coli LR05 reste la plus
efficace de toutes les souches testées.
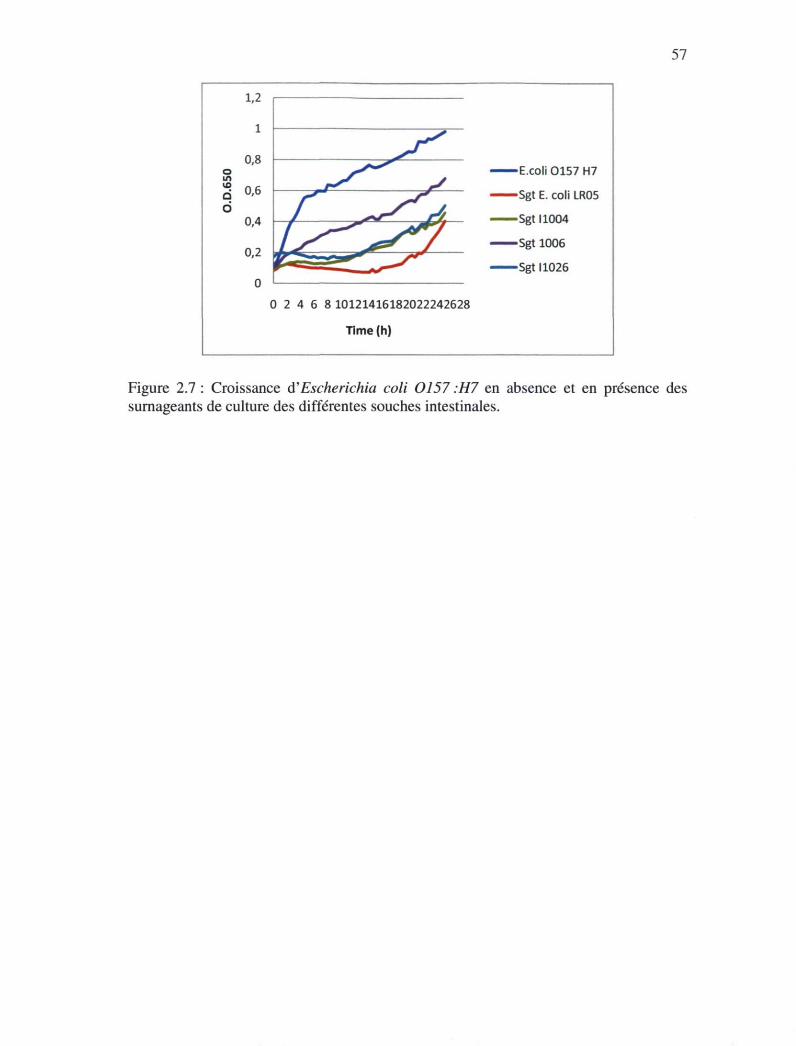
57
0 2 4 6 810121416182022242628
Time (h)
■E.coli 0157 H7
■Sgt E. coli LR05
■Sgt 11004
■Sgt 1006
■Sgt 11026
Figure 2.7 : Croissance d'Escherichia coli 0157 :H7 en absence et en présence des surnageants de culture des différentes souches intestinales.
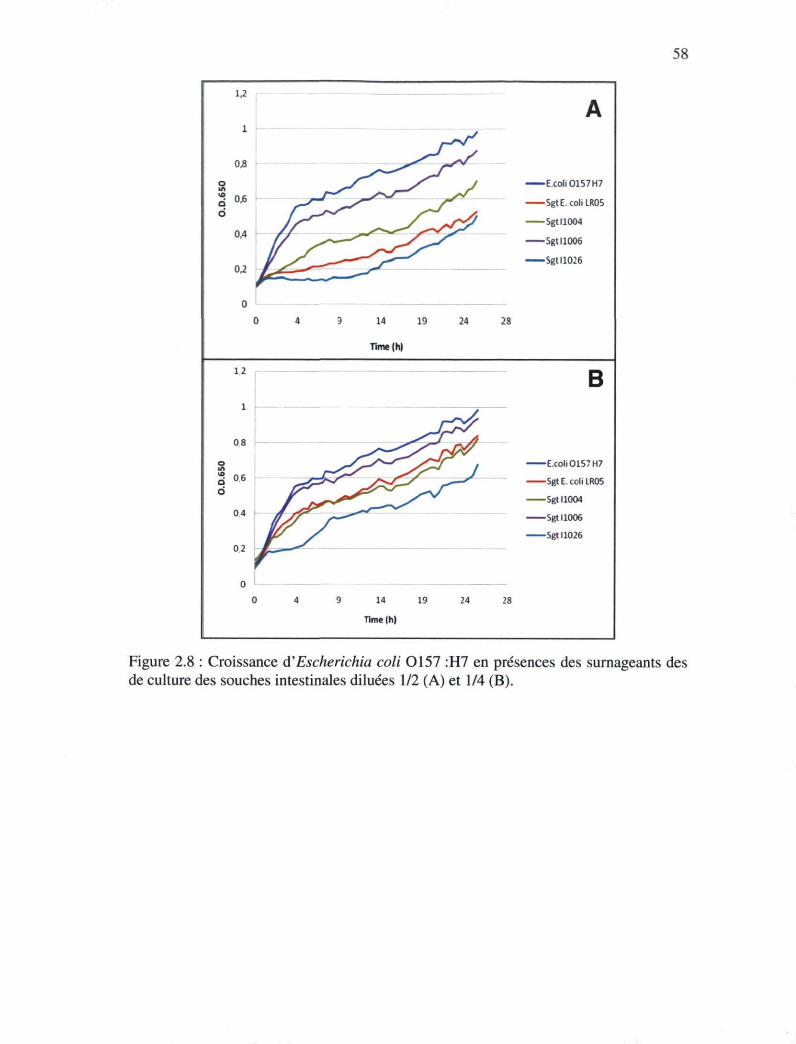
58
Q O
-Ecoli0157H7
- Sgt E. coli LROS
-Sgtll004
-Sgt 11006
-Sgt 11026
E.coli0157H7
Sgt E. coli LR05
Sgt 11004
Sgt 11006
Sgt 11026
Figure 2.8 : Croissance d'Escherichia coli 0157 :H7 en présences des surnageants des de culture des souches intestinales diluées 1/2 (A) et 1/4 (B).

59
2.4.2.3. Activité inhibitrice contre Salmonella enterica ssp enterica
serotype Montevideo
La figure 2.9 montre un effet inhibiteur des trois souches inhibitrices contre la
croissance de Salmonella enterica, suite à une expérience similaire aux deux autres.
Cependant cette figure montre un effet inhibiteur moins important que celui observé
contre la souche Escherichia coli MC4100 mais semblable à celui observé contre la
souche pathogène Escherichia coli 0157 : H7. De plus, suite à la dilution des
surnageants quatre fois l'effet inhibiteur diminue rapidement (figure 2.10) tout comme
observé pour la souche pathogène Escherichia coli 0157 : H7, avec la souche 11026
étant la plus active.
Nurmi et Rantale (1973), proposaient le concept d'exclusion compétitive en mettant en
évidence l'effet protecteur de la flore intestinale d'animaux adultes distribuée à des
poussins pour prévenir la colonisation par Salmonella Infantis. L'expérience consistait
à traiter des poussins juste après l'éclosion avec une flore caecale de poulet adultes
sains (Nurmi 1973). Ce concept a été repris plus tard par Zhang-Barber et al. (1998) qui
ont confirmé que l'administration à des poussins nouveau-nés de flores intestinales de
poulets adultes, en bonne santé, prélevées entre 37 et 80 jours d'âge permet de
développer une résistance aux infections intestinales. L'utilisation de flore caecale
normale peut ainsi réduire la colonisation caecale par les souches pathogènes,
cependant le mode d'action de cette flore reste encore mal étudié. Selon Nisbet (2002)
plusieurs modes d'actions sont à l'origine de l'activité des flores bactériennes mais les
mécanismes ne sont pas clairement identifiés. D faut noter que certains effets
inhibiteurs mis en évidence in vitro ne sont pas toujours reproductibles in vivo
(Patterson and Burkholder 2003).
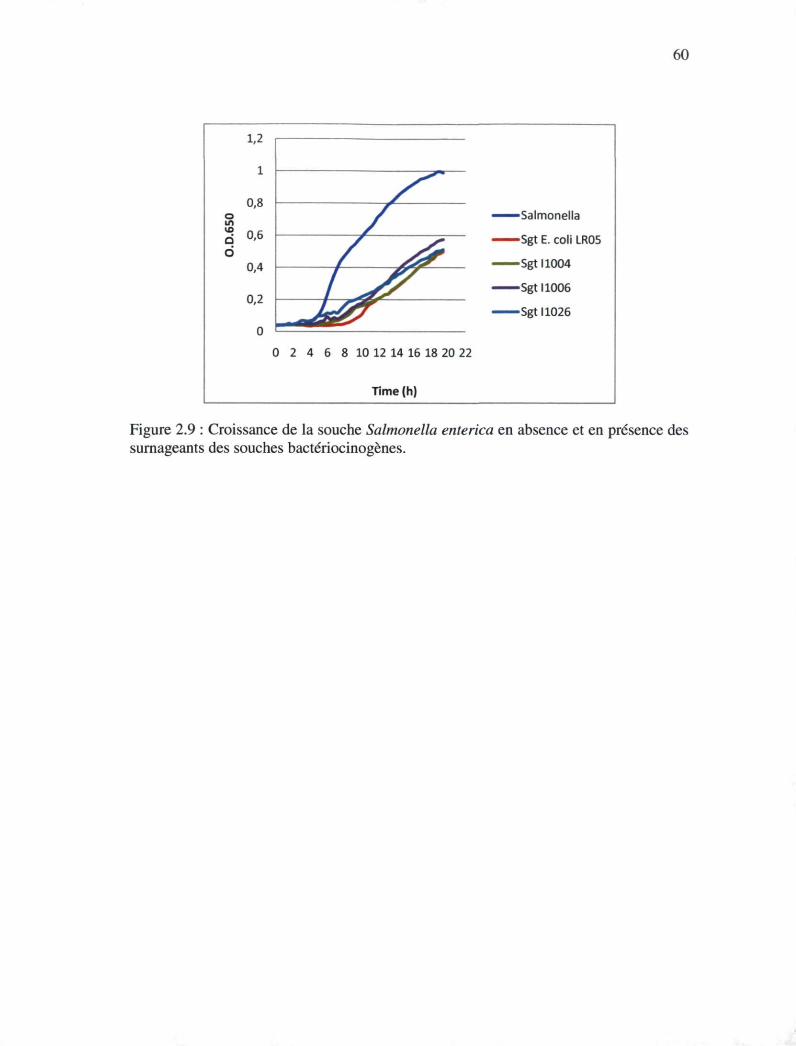
60
1,2 1,2
1
0,8 o
/ " Salmonella
0,8 o / Salmonella
2 0,6 ci
0,4
0,2
/ — S g t E. coli LR05
Sgt 11004
2 0,6 ci
0,4
0,2
/ < * — S g t E. coli LR05
Sgt 11004
2 0,6 ci
0,4
0,2 J T Sgt 11006
— S g t 11026
2 0,6 ci
0,4
0,2 Jstr Sgt 11006
— S g t 11026 0
(
t ^ t ^ ^ m ^ ^ s i ^ 0
( ) 2 4 6 8 10 12 14 16 18 20 22
Time (h)
Figure 2.9 : Croissance de la souche Salmonella enterica en absence et en présence des surnageants des souches bactériocinogènes.
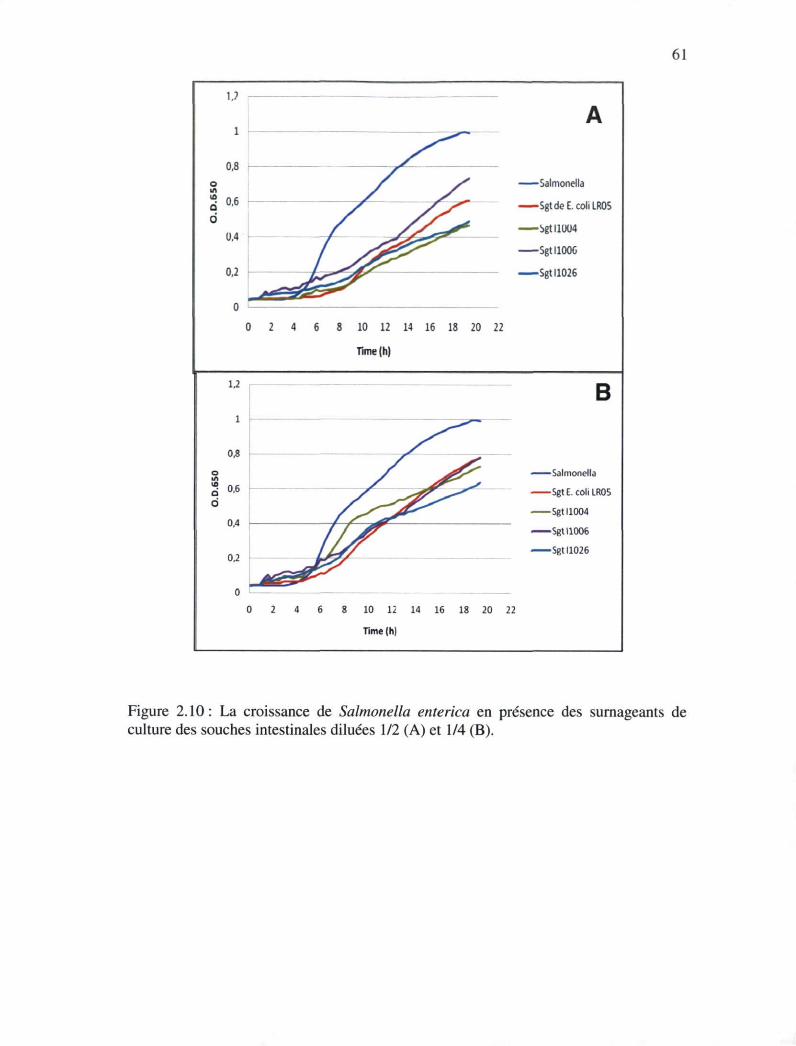
61
-Salmonella
-SgtdeE. coliLROS
-Sgt 11004
-SgtllOOG
■Sgt 11026
0 2 4 6 8 10 12 14 16 18 20 22
Time f h]
S ID d O
1.2
O.S
0,6
0.4
0,2
B
-Salmonella
-Sgt E. coliLROS
-Sgt 11004
-Sgt11006
-Sgt 11026
8 10 12 14 16 18 20 22
Time (h)
Figure 2.10: La croissance de Salmonella enterica en présence des surnageants de culture des souches intestinales diluées 1/2 (A) et 1/4 (B).

62
De manière générale, les études portant sur l'utilisation des probiotiques chez les
animaux d'élevage sont difficiles à évaluer car la plupart n'ont pas été statistiquement
analysées. De plus, les protocoles expérimentaux ne sont pas clairement définies, les
micro-organismes n'ont pas été identifiés et la viabilité des organismes n'a pas été
vérifiée (Stavric, 1992). Dans de nombreux cas, l'état de l'environnement et le stress
infligé aux volailles ne sont pas pris en considération dans la réponse vis-à-vis
l'admission de cette flore fécale.
L'utilisation des souches à Gram négatif dans l'alimentation des animaux a été
proposée pour la première fois suite à la constatation de la tolérance à l'acidité de
certaines souches d'E. coli (Bearson, Bearson et al. 1997; Ganzle, Hertel et al. 1999).
Depuis, plusieurs études ont utilisés des souches de E. coli productrices de colicin pour
réduire la charge digestive de E. coli pathogènes chez les animaux d'élevage (Tkalcic,
Zhao et al. 2003). Portrait et al. (1999) ont aussi montré l'effet inhibiteur de la
microcine J25 produite par certaines souches d'E. coli sur Salmonella enteritidis in
vitro. Toutefois, l'effet des bactériocines mis en évidence in vitro n'est pas facile ni à
mettre en évidence ni à reproduire in vivo.
Finalement, Schroeder et al. (2006) ont montré que la souche probiotique E. coli Nissle
1917 a été utilisée avec succès pour prévenir la diarrhée aiguë induite par des
pathogènes entérotoxinogènes chez les porcs (Schroeder, Duncker et al. 2006).
Les résultats obtenus dans le cadre de cette étude ont permis de sélectionner des
souches intestinales à Gram négatif productrices de substances inhibitrices apparentées
à des bactériocines. Trois de ces souches ont montré une activité inhibitrice marquée
contre des pathogènes de la volaille à savoir Salmonella (Enteritidis et Enterica) et E.
coli 0157 :H7. Ces souches présentent donc un potentiel certain d'utilisation comme
probiotique pour prévenir les infections entériques chez la volaille. Cependant et afin de
répondre aux exigences de la réglementation actuelle en termes de probiotiques
animaux, une identification poussée de ces souches de même que la démonstration de
leur innocuité demeurent nécessaire avant de permettre l'ajout de ces souches comme
supplément dans l'alimentation de la volaille. Ces deux aspects font l'objet du chapitre
3 de ce mémoire.

63
CHAPITRE 3. IDENTIFICATION ET CARACTERISATION DE SOUCHES À FORT POTENTIEL PROBIOTIQUE ISOLÉES DU
TUBE DIGESTIF DE VOLAILLES

64
3.1. RESUME
Trois souches bactériennes isolées du contenu intestinal de la volaille et baptisées
11004, 11006 et 11026 ont été identifiées et sélectionnées pour leur forte activité
inhibitrice vis-à-vis E. coli et Salmonella deux pathogènes entériques chez la volaille.
Une identification poussée utilisant des approches différentes mais complémentaires de
microscopie, biochimie et biologie moléculaire ont été utilisées pour l'identification
exacte de ces souches. Ainsi l'utilisation de la coloration Gram combinée à une
observation microscopique, une analyse du profil de fermentation des sucres par les
galeries API 20 E et une amplification de la région d'ARN16S ont permis d'identifier
les trois souches comme étant des Escherichia coli.
L'analyse du lipo-polysaccharides et des antigènes « O » ont permis de déterminer le
serotype de chacune des trois souches, Finalement et dans le but d'évaluer l'innocuité
des souches, la recherche de quatre gènes de virulence à savoir le gène codant pour
l'aérobactine (Aero), pour les facteurs cytotoxiques nécrosants (toxines CNF), pour les
adhésines fimbriaires ( P) et pour l'hémagglutinine sensible à la température (Tsh) a été
réalisée en utilisant la méthode d'amplification par PCR. Les résultats obtenus ont
montré que les trois souches possèdent les gènes de septicémie Aero et Tsh et la souche
11006 possède en plus le gène P.

65
3.2. INTRODUCTION
Les probiotiques sont des microorganismes vivants qui lorsqu'ils sont administrés en
quantité adéquate, produisent un effet bénéfique pour la santé de l'hôte (FAO/WHO.,
2001). Plusieurs effets bénéfiques sur la santé ont été associés à la consommation des
probiotiques tel que l'amélioration de la digestion, l'influence positive sur la flore
intestinale, la régulation de la motilité intestinale, la prévention de certaines maladies
(ostéoporose, cancer, hypertension...), la modulation de la réponse immunitaire et la
prévention des infections intestinales et urogénitales.
Plusieurs souches bactériennes à fort potentiel probiotiques ont été isolées et
caractérisées et sont aujourd'hui utilisées chez l'humain avec diverses allégations santé.
Cependant, l'utilisation des probiotiques chez les animaux est encore à un stage très
précoce. À titre d'exemple, l'emploi commercial de probiotiques en aviculture est
relativement nouveau. Des études récentes ont montré que l'incorporation de certaines
cultures probiotiques chez la volaille permet de renforcer l'écosystème microbien et
contribue à stimuler les mécanismes de défense immunitaire tout en protégeant les
poulets contre les conséquences de stress tels que la vaccination et les changements de
températures (Feuillet 2007; Awad, Ghareeb et al. 2009).
Selon la réglementation actuelle au Canada, l'utilisation des probiotiques en élevage
des animaux est régie par la loi T-3 159 sur les aliments pour bétail sous la rubrique
D cultures microbiennes vivantes et levures". Selon cette loi, une culture bactérienne
vivante ne peut ajoutée comme supplément à l'alimentation animale que si elle satisfait
à certaines exigences relatives à son innocuité et son efficacité. Entre autre, le
demandeur doit clairement identifier les espèces de bactéries utilisées et soumettre un
rapport d'analyse indiquant les méthodes de dénombrement et d'identification ainsi que
le profil biochimique de chacune des souches. Une description des méthodes de
confirmation utilisées doit aussi être soumise.
La grande majorité des souches actuellement approuvées et utilisées comme
probiotiques sont généralement des bactéries Gram positifs. Ces souches présentent peu
d'intérêt en aviculture puisque la majorité des souches pathogènes rencontrées chez la
volaille sont des bactéries à Gram négatif. La caractérisation et l'identification des

66
souches à fort potentiel probiotique actives contre les pathogènes à Gram négatif tels
que Salmonella et E. coli 0157 :H7 demeurent une réelle nécessité pour le secteur
avicole.
Dans le chapitre précédent et dans le but de sélectionner des souches à fort potentiel
d'utilisation comme probiotique chez le poulet, nous avons décrit les travaux relatifs à
l'identification de trois souches isolées de l'intestin de volailles en santé et qui étaient
capable d'inhiber certains pathogènes entériques de la volaille à savoir Salmonella et E.
coli. Bien que ces souches présentent déjà un certain potentiel, leur utilisation comme
probiotique en alimentation animale nécessite obligatoirement une identification
poussée et une caractérisation biochimique et moléculaire. De plus, leur innocuité doit
être démontrée sans aucun doute possible aussi bien pour les volailles que pour l'être
humain.
L'objectif général de ce chapitre est d'utiliser une approche polyphasique pour
l'identification et la caractérisation des trois isolats 11004,11006 et 11026 sélectionnées
pour leur activité inhibitrice. Il vise également à évaluer l'innocuité de ces isolats en
vue de leur utilisation comme supplément pour aliment de bétail.

67
3.3. MATERIEL ET METHODE
3.3.1. Souches bactériennes et conditions de culture
Les trois souches bactériennes 11004, 11006 et 11026 isolées du contenu intestinale de
volailles et ayant démontré une activité inhibitrice contre E. coli Kl2, S. enteritidis, S.
enteritica, E. coli 0157 : H7 et E. coli MC4100 ont été utilisées lors de cette étude.
La souche E. coli LR05 qui est une souche sauvage productrice des MccL, B17, D93 et
J25 et qui est résistante à la MccV (ou ColV) et à la colicine M (Sable, Duarte et al.
2003) a été utilisée comme témoin positif.
Toutes ces souches sont conservées à -80°C dans une solution cryo-protectrice
contenant du glycerol à 20%. Toutes les souches sont repiquées deux fois dans du
milieu BHI ; (Difco, Sparks, MD) et incubées à 37°C.
Dans un premier temps, des cultures fraîches des trois souches ont été soumises à une
observation microscopique ainsi à un test de coloration Gram. Dans un deuxième
temps, une analyse biochimique à l'aide des galeries API20E a été réalisée. Ces deux
premières étapes ont permis une identification partielle des souches.
3.3.2. Tests de sérotypage
Le sérotypage est une technique immunologique qui consiste en la mise en évidence
d'antigènes structuraux bactériens et donc de classer les souches en sérogroupes ou
sérovars. Ce test est basé la réaction spécifique entre un anticorps présent dans un
sérum test et un antigène porté par le microorganisme étudié. Cette réaction est
visualisée grâce à test d'agglutination qui est le résultat macroscopique de la réaction
antigène-anticorps. Lors de cette étude, plus de 85 antiserums ont été utilisés.
3.3.3. Méthodes moléculaires
Les méthodes moléculaires utilisées dans cette partie du projet avaient pour finalité
d'une part, d'identifier les souches isolées et d'autre part, d'évaluer leur innocuité. Cinq

68
gènes ont été alors ciblés, le gène codant pour l'ARN 16S et quatre gènes codant pour
des facteurs de virulence chez E. coli à savoir les gènes Aero, P, Tsh et CNF.
3.3.3.1. Amplification et séquençage de l'ARN 16S
Le gène ADNr 16S a été amplifié chez les trois souches en utilisant les amorces
spécifiques au domaine ADNr 16S à savoir SSU27F
S'AGAGTTTGATCMTGGCTCAG'S et SSU1492R
5'TACGGYTACCTTGTTACGACTT'3 (Lane, 1991). Le mélange réactionnel (25 [il)
était constituée de 19,25 ul d'eau «nuclease free water » (Invitrogen Life
Technologies, Carlsbed, CA, USA), 2,5 ul de 10X Taq buffer, 1 ui de dNTP (A, T, G et
C à 10 uM), (New England biolabs Ltd, ON, Canada) 1 [il d'une solution d'ADN cible
(25 ng/[il), 0,5 [il de chacun des deux amorces (10 uiM/uil) et 0,25 [il (1,25 U) de
l'enzyme Taq polymerase (New England Biolabs). L'amplification par PCR a été
réalisée dans un Tgradient (Montréal, Dorval, PQ, Canada) en utilisant 35 cycles
d'amplification. Chaque cycle comprend les étapes suivantes : une dénaturation de 30 s
à 94°C, une hybridation des amorces à 55°C durant 30 s, une elongation à 72°C pour
1,5 min et une extension de 5 min. Les différents produits d'amplification ont été
analysés par électrophorèse sur gel d'agarose à 0,8% (Invitrogen) dans un tampon TAE
IX contenant 40mM de Tris-aceatate, lmM d'EDTA (Sambrook, Fritsch, & Maniatis
1989). La coloration a été réalisée à l'aide du bromide d'éthidium et une visualisation à
l'aide des UV (Gel Doc, BioRad, Mississauga, ON, Canada) a été effectuée.
Le séquençage de l'ARDr 16S a été réalisé au service de l'Université Laval (Life and
Health Sciences Pavillon, Québec, Québec, Canada) avec l'appareil ABI Prism 3100.
L'analyse des séquences d'ADN a été effectuée à l'aide du logiciel « Genetics
Computer Group Wisconsin Package software » (version 10.2) (Devereux, Haeberli,
and Smithies, 1984). Les séquences obtenues ont été comparées à celles de la base de
données utilisée pour la recherche de similitude (Pearson and Lipman, 1988) à l'aide du
service FASTA du centre national de biotechnologie information NCBI (National
Center of Biotechnology Information) (http://www.ncbi.nlm.nih.gov).

69
3.3.3.2. Recherche et amplification des gènes de virulence
Quatre gènes de virulence connus chez E. coli ont été ciblés à savoir le gène Aero
codant pour l'aérobactine, un système de captation de fer codée par un opéron
plasmidique ou chromosomique (Mainil 2003), le gène CNF codant pour un facteur
cytotoxique nécrosants (Mainil 2003; Van Bost, Jacquemin et al. 2003), le gène P
codant pour une adhésine fimbriaire (Feria, Machado et al. 2001; Mainil 2003) et
finalement le gène Tsh codant pour une hémagglutinine thermosensible (Mainil and
Van Bost, 2004). L'amplification de ces gènes a été réalisée au laboratoire de référence
pour Escherichia coli de Faculté de médecine vétérinaire, Université de Montréal,
Saint-Hyacinthe, Qc.

70
3.4. RESULTATS ET DISCUSION
3.4.1. Analyse microscopique, coloration Gram et galerie API20
Suite à la réalisation des testes biochimique à l'aide du système d'identification API
20E, nos codes d'identification des trois souches sont :
- 11004:514 4552. - 11006:504 4572. - 11026:504 0552.
Les essais d'observation en microscopie, la coloration Gram de même que les résultats
des tests biochimiques obtenus avec les galeries API 20E indiquent sans aucun doute
possible que les souches 11004,11006 et 11026 appartiennent l'espèce Escherichia coli.
3.4.2. Tests de sérotypage
Selon la littérature, les sérogroupes d'E. coli les plus incriminés dans les cas de toxi-
infections alimentaires ou dans les maladies entériques qui causent des mortalités en
aviculture sont les serotypes Oi, O2, O35 et O78 représentant de 15 à 61 % des souches
isolées bien que d'autres soient aussi présents. Les autres serotypes représentés de
manière significative sont : 08 , O15, Oig, O35, Og8, O109, O115 et OHÔ (Stordeur and
Mainil, 2002)

71
Tableau 3.1 : Résultats du test de sérotypage des souches 11004,11006 et 11026
■ ■ J - -
Echantillons Sérotypie1
11004 018-023
11006 ONT
11026 Ol
ONT : non typé
'Liste des antiserums testés pour l'antigène O :
O,, 0 2 , 0 4 , 0 5 , 0 6 , Ov, Og, 0 9 , O,o, O n , 01 5 , 0, 6 , 0 1 7 , 0 1 8 , O20, 0 2 1 , 02 2 , 0 2 3 , 02 5 , 02 6 , 027, O34, O35, O36, O45, O49, O53, O54, O55, Oô0, 064, Oô9, O71, O73, O75, O76, Û78, 0 8 2 , 083, Os4, 086, 08 8 , O91, Oc2, 098, O100, Oioi, O]03, Oifj8, O109, Oui, O112, O113, O114, O115, On6, O117, On», O119» Oi2o, O121, Oi23, Oi25, Oi26, Oi28, O131, Oo2 , O137, 0|38 , O141, O143, O145, O146, O147, Oi48, O149, O153, O157, O159, Oi63, Ono, Oni, Oi72, O173.
Le tableau 3.1 nous permet de voir que la souche 11026 appartient à l'un des serotype le
plus incriminé dans les toxi-infections alimentaires ainsi que dans les maladies
entériques causant un taux élevé de mortalité en élevage de volailles. Dans le cas de la
souche 11004, une agglutination significative a été observée avec les antiserums à 018
et 023 ce qui classerait cette souche dans les serotypes les plus incriminés dans les
maladies entériques chez l'humain et les cas de mortalité dans l'élevage des volailles.
Enfin, la souche 11006 n'a agglutiné en présence d'aucuns des 85 antiserums utilisés.
3.4.3. Analyses moléculaires
La séquence de l'ARNr 16S (environ 1500 nucleotides) est spécifique de chaque
microorganisme et de son évolution depuis un ancêtre commun. De plus, le grand
nombre de séquences actuellement disponible (plus de 6000 séquences de procaryotes)
permet de dessiner un vaste arbre phylogénétique de microorganismes et d'y placer
chaque nouvelle séquence. À partir des alignements de séquences, il est possible
d'identifier les souches. Dans certains cas, des espèces sont si proches qu'elles ne
possèdent que très peu de différences dans la séquence 16S. Une région plus divergente
sera alors utilisée. Le pouvoir pathogène quand à lui est lié à la présence de gènes de
virulence qu'il faudra cibler.

72
Figure 3.1 : Analyse du produit d'amplification du gène d'ARN 16S par
d'électrophorèse sur gel d'agarose
La figure 3.1 nous présente le gel d'électrophorèse suite à l'amplification de la région
codant pour l'ADNr 16S chez les trois souches intestinales.
Le tableau 3.2 illustre les homologies des séquences obtenues avec celles des souches
déjà séquencées et disponibles dans GenBank. L'ADNr 16S de la souche 11004 montre
un grand degré de similitude (99%) avec des gènes d'E. coli S88 et E. coli 0127 : H6I
(GenBank Accession number CU928161.2 et FM180568.1). Une homologie de 99%
a également été observée entre la souche 11006 et d'E. coli SYW003 et BEBJ3
(GenBank Accession number EF620924.1 and EF560787.1). Finalement, le séquençage
de la région 16S de l'ADNr de la souche 11026 révèle une homologie de 99% avec la
souche E. coli IAI1, SE 11 et E. coli 0157.H7 EC4115 (GenBank Accession number
CU928160.2, AP009240.1 et CP001164.1). Ces résultats confirment que les trois
souches isolées lors de cette étude sont identifiées comme étant des souches d'E. coli
pathogènes.
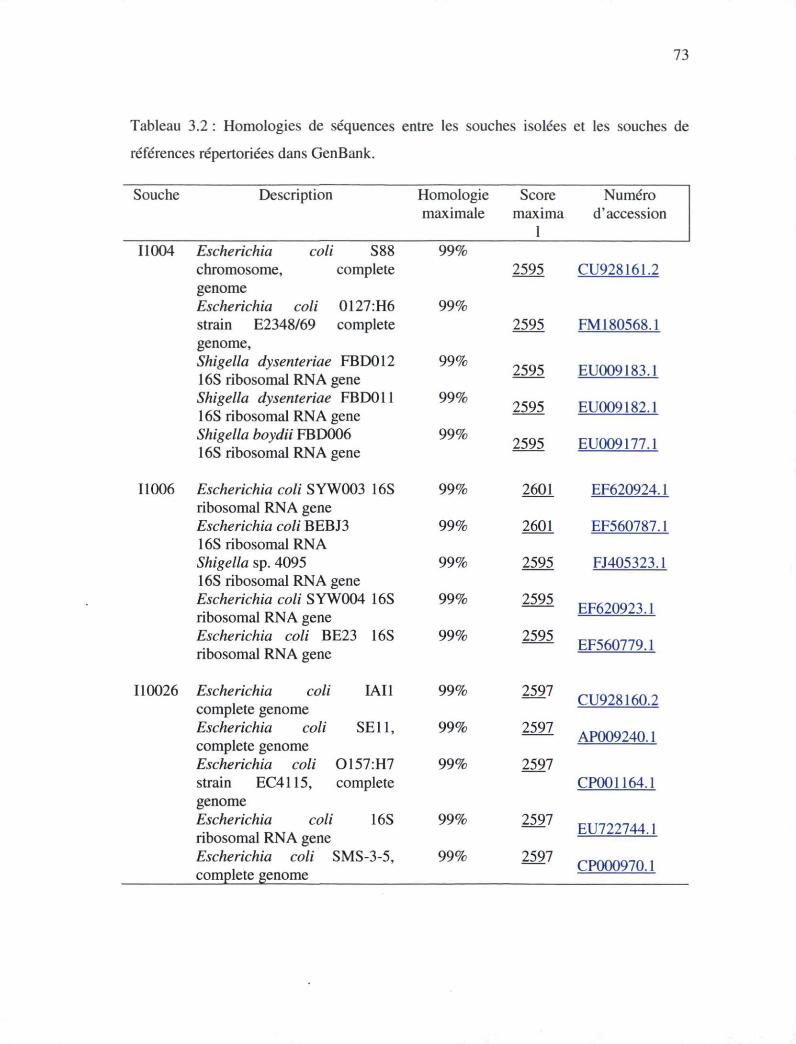
73
Tableau 3.2 : Homologies de séquences entre les souches isolées et les souches de
références répertoriées dans GenBank.
Souche Description Homologie Score Numéro maximale maxima
1 d'accession
11004 Escherichia coli S88 99% chromosome, complete 2595 CU928161.2 genome Escherichia coli 0127 : H6 99% strain E2348/69 complete 2595 FM 180568.1 genome, Shigella dysenteriae FBD012 16S ribosomal RNA gene
99% 2595 EU009183.1
Shigella dysenteriae FBD011 16S ribosomal RNA gene
99% 2595 EU009182.1
Shigella boydii FBD006 16S ribosomal RNA gene
99% 2595 EU009177.1
11006 Escherichia coli SYW003 16S 99% 2601 EF620924.1 ribosomal RNA gene Escherichia coli BEBJ3 99% 2601 EF560787.1 16S ribosomal RNA Shigella sp. 4095 99% 2595 FJ405323.1 16S ribosomal RNA gene Escherichia coli SYW004 16S ribosomal RNA gene
99% 2595 EF620923.1
Escherichia coli BE23 16S ribosomal RNA gene
99% 2595 EF560779.1
110026 Escherichia coli IAI1 complete genome
99% 2597 CU928160.2
Escherichia coli SE11, complete genome
99% 2597 AP009240.1
Escherichia coli 0157 : H7 99% 2597 strain EC4115, complete CP001164.1 genome Escherichia coli 16S ribosomal RNA gene
99% 2597 EU722744.1
Escherichia coli SMS-3-5, complete genome
99% 2597 CP000970.1

74
L'espèce E. coli comprend de nombreuses souches pathogènes pour l'homme et les
animaux basés sur la possession de nombreux facteurs de pathogènicité. On distingue
alors les souches à tropisme intestinal (entérotoxinogènes, entéropathogènes,
entérohémorragiques, vérotoxinogènes et entéro-invasives) des souches à tropisme
extra-intestinal (uropathogènes et invasives). Si les propriétés et les facteurs spécifiques
de virulence des souches à tropisme intestinal sont relativement bien connus et
caractérisés, ceux des souches à tropisme extra-intestinal le sont beaucoup moins,
surtout chez les animaux (Mainil 2003).
Plusieurs articles de synthèse ont fait le point sur les connaissances actuelles en rapport
avec les facteurs de virulence et les propriétés spécifiques des souches invasives d'E.
coli. Chez les « Avian Pathogenic E. coli » (APEC), le pouvoir pathogène est liée à la
production d'aérobactine, la résistance au complément fimbriae P et/ou d'adhésines
(Mainil 2003; Mainil 2003; Mainil and Van Bost 2004). Plusieurs études ont également
montré que la plupart des souches APEC (73-98%) possède le système d'acquisition du
fer appelé aérobactine, alors que les souches non pathogènes le produisent moins
fréquemment (Stordeur and Mainil 2002). Comme le montre le tableau 3.3 les trois
souches analysées possèdent le gène codant pour l'aérobactine.
La toxine CNF a été découverte dans les années 80 provenant d'une souche d'E. coli
issue de nourrissons atteints de diarrhée. La toxine CNF induisait la formation de
cellules HeLa géantes multi-nuclées. Son rôle principal serait de permettre aux
bactéries de pouvoir se multiplier dans le sang ou les organes autres que l'intestin
(Stordeur and Mainil 2002). Les toxines CNF semblent agir en modifiant une petite
protéine G de 21kDa appelée Rho qui joue un rôle majeur dans la régulation du
cytosquelette des cellules eucaryotes et donc leur division et que cette altération
provoque une polymérisation des filaments d'actine du cytosquelette entraînant le
blocage de la motilité cellulaire et de la cytokinèse. De plus, il a été montré que ce
phénomène pourrait permettre l'entrée de bactéries normalement non invasives dans les
cellules épithéliales de l'hôte. Les résultats obtenus lors de notre étude ont montré que
les trois souches intestinales étudiées ne possèdent pas le gène CNF (Tableau 3.3) ce
qui est en parfaite concordance avec les travaux de Mainil (2003) et de Van Bost,
Jacquemin et al. (2003) qui ont montré que la majorité des souches d'E. coli provenant

75
de l'espèce aviaire aucune ou très peu de souches APEC sont positives pour les toxines
CNF.
Les adhésines fimbriaires de la famille P se définissent sur la base de leurs propriétés
d'agglutination des erythrocytes humains du groupe sanguin O. Des souches exprimant
des fimbriae P, ou en possédant les gènes, d'origine urinaire, systémique ou intestinale
ont aussi été isolées chez les volailles (Feria, Machado et al. 2001; Mainil 2003). La
présence du gène P chez la souche 11006 suggère un pouvoir septicémique très
important de cette souche puisque ces adhésines permettent à la bactérie de résister aux
mécanismes de défense de l'organisme et, ce faisant, de se multiplier sur place,
provoquant la formation de microcolonies.
D'autre part, il a été montré que le gène Tsh isolé d'une souche APEC de poulet est
associé préférentiellement à des souches pathogènes et ne se retrouve pas dans des
fèces d'animaux sains (Stordeur and Mainil 2002; Mainil and Van Bost 2004) ce qui
laisserai conclure que sa présence est un indicateur de pathologie. Tel qu'indiqué dans
le tableau 3.3 les trois souches d'E. coli isolées sont porteurs du gène Tsh ce qui
renforce l'hypothèse de ces trois souches soient potentiellement pathogènes.
Tableau 3.3 : Les gènes de septicémies détectées chez les trois souches.
Echantillons Gènes détectés dans
l'échantillon
11004 Aero, Tsh
11006 P, Aero, Tsh
11026 Aero, Tsh
En conclusion à ce dernier chapitre, il est clair que E. coli est une espèce bactérienne
qui est très présente parmi le microbiote colique normale de la volaille. Plusieurs
serotypes ont été déjà identifiés comme pathogène chez la volaille mais plusieurs autres
demeurent encore inconnues.
Lors de cette étude, la souche 11026 a été identifiée comme étant une E. coli 01 , ce qui
l'associe directement aux serotypes le plus incriminés dans les toxi-infections

76
alimentaire ainsi que dans les maladies entériques causant un taux élevé de mortalité en
aviculture. La présence des gènes codant pour l'aérobactine et l'hémagglutinine
confirme cette affirmation. En ce qui concerne la souche 11004 et bien que le serotype
exact n'a pas été déterminé en raison de l'agglutination des deux antiserums
différents 018 et 023, sont caractère pathogène a été confirmé par la présence des
gènes Aero et Tsh. Finalement pour la souche 11006 et bien qu'aucun serotype n'a été
obtenu (absence d'agglutination avec les 85 antiserums testés), la présence de trois des
quatre gènes de virulence suggère également un fort potentiel de pathogénicité chez
cette souche. Malgré l'activité inhibitrice intéressante observée, les trois souches
sélectionnées lors de cette étude ne peuvent être utilisées comme probiotiques en
alimentation animale.

77
CONCLUSION GENERALE
À l'échelle mondiale, les élevages de porcs et de volailles sont les sous-secteurs qui
enregistrent la plus forte croissance avec des taux annuels de 2,6% et 3,7% au cours des
dix dernières années (FAO, 2007). Plus de 40,4 % des cas de toxi-infections
alimentaires proviennent de la filière « Viandes et volailles », dont 19,2% représentés
par les volailles occupant ainsi la première place (CQIASA, 2008). Les pathogènes
alimentaires associés à la chaire de volailles sont surtout des souches à Gram négatif. À
l'heure actuelle, la prévention de ces maladies repose encore essentiellement sur
l'antibiothérapie. Cependant, les antibiotiques connaissent actuellement leurs limites
d'utilisation vue l'émergence de souches résistantes.
Dans le présent travail, vingt quatre isolats de contenus intestinaux de volailles ont été
criblées pour leur activité inhibitrice contre des souches pathogènes à Gram négatif en
vue de leur utilisation comme probiotique en aviculture.
Parmi les vingt quatre isolats étudiés, sept ont présenté une activité importante contre
Salmonella, Escherichia coli et Yersinia enterocolitica. L'analyse poussée des
surnageants de culture de ces souches a permis d'identifier trois (11004,11006 et 11026)
qui ont préservé leur activité inhibitrice à la suite d'un traitement thermique de 5
minutes à 100°C et d'une filtration sur une membrane de 0.22 [im. Le suivi de la
croissance des souches pathogènes entériques en présence des surnageants des trois
souches a révélé une forte activité inhibitrice pendant les 10 à 14 premières heures
d'incubation et une prolongation de la phase de latence. Une croissance progressive a
été ensuite observée. Cette croissance peut être expliquée par deux phénomènes
possibles soit l'épuisement de la substance antimicrobienne produite en raison de la
présence de proteases dans le milieu ou soit par la nature de l'activité antimicrobienne
exercée qui serait plutôt de type bactériostatique.
L'utilisation d'une approche multidisciplinaire combinant des méthodes biochimiques
et moléculaire a permis de conclure que nos trois souches sont des Escherichia coli.

78
Finalement, l'analyse de l'innocuité de ces souches a permis d'identifier la présence de
plusieurs gènes de pathogénicité indiquant le caractère pathogène de ces souches.
À la lumière de l'ensemble des résultats obtenus et en tenant compte de l'aspect
réglementaire, les souches intestinales ayant fait l'objet de cette étude ne peuvent pas
être utilisées comme probiotique en alimentation avicole.
Ce mémoire représentait une phase préliminaire d'un projet d'envergure et laisse
entrevoir une possibilité à l'avenir de trouver des souches présentant des
caractéristiques intéressantes et répondant aux normes de la réglementation.

79
REFERENCES BIBLIOGRAPHIQUE
Alvarez-Olmos, M. I. and R. A. Oberhelman (2001). "Probiotic agents and infectious diseases: A modem perspective on a traditional therapy." Clinical Infectious Diseases 32(11): 1567-1576.
Andreatti, R. L., E. N. da Silva, et al. (2000). "Use of anaerobic cecal microflora, lactose and acetic acid for the protection of broiler chicks against experimental infection with Salmonella Typhimurium and Salmonella Enteritidis." Brazilian Journal of Microbiology 31(2): 107-112.
Andrieu, V. (1995). Intérêt des probiotiquesdans le gavage du canard. Application à la région des landes, faculté de médecine. Université de Nantes.
Apajalahti, J., A. Kettunen, et al. (2004). "Characteristics of the gastrointestinal microbial communities, with special reference to the chicken." Worlds Poultry Science Journal 60(2): 223-232.
Asensio, C , J. C. Perezdiaz, et al. (1976). "New Family of Low-Molecular Weight Antibiotics from Enterobacteria." Biochemical and Biophysical Research Communications 69(1): 7-14.
Audisio, M. C , G. Oliver, et al. (2000). "Protective effect of Enterococcus faecium J96, a potential probiotic strain, on chicks infected with Salmonella pullorum." Journal of Food Protection 63(10): 1333-1337.
Awad, W. A., K. Ghareeb, et al. (2009). "Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens." Poultry Science 88(1): 49-56.
Bearson, S., B. Bearson, et al. (1997). "Acid stress responses in enterobacteria." Ferns Microbiology Letters 147(2): 173-180.
Boguhn, J. and M. Rodehutscord (2010). "Effects of nonstarch polysaccharide-hydrolyzing enzymes on performance and amino acid digestibility in turkeys." Poultry Science 89(3): 505-513.
Bozkurt, M., A. Alcicek, et al. (2009). "Effect of an herbal essential oil mixture on growth, laying traits, and egg hatching characteristics of broiler breeders." Poultry Science 88(11): 2368-2374.
Braun, M., H. Killmann, et al. (2002). "Diffusion through channel derivatives of the Escherichia coli FhuA transport protein." European Journal of Biochemistry 269(20): 4948-4959.
Braun, V., S. I. Patzer, et al. (2002). "Ton-dependent colicins and microcins: modular design and evolution." Biochimie 84(5-6): 365-380.
CAAAQ (2008). Rapport de la commission sur l'avenir de l'agriculture et de l'agroalimentaire québécois: Agriculture et agroalimentaire: assurer et bâtir l'avenir, propositions pour une agriculture durable et en santé.
Calder, P. C. and S. Kew (2002). "The immune system: a target for functional foods?" British Journal of Nutrition 88: S165-S176.
Cao, Z. and P. E. Klebba (2002). "Mechanisms of colicin binding and transport through outer membrane porins." Biochimie 84(5-6): 399-412.

80
Ceylan, E. and D. Y. C. Fung (2004). "Antimicrobial activity of spices." Journal of Rapid Methods and Automation in Microbiology 12(1): 1-55.
Chafai, S. (2006). Effet de l'addition des probiotiques dans les régimes alimentaires sur les performances zootechnique du poulet de chair.
Champagne, C. P., N. J. Gardner, et al. (2005). "Challenges in the addition of probiotic cultures to foods." Critical Reviews in Food Science and Nutrition 45(1): 61-84.
Cole, C. B. and R. Fuller (1984). "Bile acid deconjugation and attachment of chicken gut bacteria: Their possible role in growth depression." British Poultry Science 25(2): 227-231.
CQIASA (2008). Bilan annuel Toxi-infection alimentaires et plaintes requérent des prélèvements alimentaires. Québec: 29.MAPAQ
Cutter, C. N. and G. R. Siragusa (1996). "Reductions of Listeria innocua and Brochothrix thermosphacta on beef following nisin spray treatments and vacuum packaging." Food Microbiology 13(1): 23-33.
David, M. G., E. Oliver, et al. (2007). The diversity of bacteriocins in Gram-negative bacteria. Bacteriocins: Ecology and Evolution. M. A. Riley and M. A. Chavan.
Davies, E. A., C. F. Milne, et al. (1999). "Effective use of nisin to control lactic acid bacterial spoilage in vacuum-packed bologna-type sausage." Journal of Food Protection 62(9): 1004-1010.
De lorenzo, V. (1984). "Isolation and Characterization of Microcin E-492 from Klebsiella-Pneumoniae." Archives of Microbiology 139(1): 72-75.
De Oliveira, G. H., A. Berchieri, et al. (2000). "Prevention of Salmonella infection by contact using intestinal flora of adult birds and/or a mixture of organic acids." Brazilian Journal of Microbiology 31(2): 116-120.
Dirix, G., P. Monsieurs, et al. (2004). "Peptide signal molecules and bacteriocins in Gram-negative bacteria: a genome-wide in silico screening for peptides containing a double-glycine leader sequence and their cognate transporters." Peptides 25(9): 1425-1440.
Dunne, C , L. O'Mahony, et al. (2001). "In vitro selection criteria for probiotic bacteria of human origin: correlation with in vivo findings." American Journal of Clinical Nutrition 73(2): 386s-392s.
Ecker, K. F. (1992). "Bacteriocin and food applications." Dairy Food And Environmental Sanitation (12): 204-209.
Ennahar, S., N. Deschamps, et al. (2000). "Natural variation in susceptibility of Listeria strains to class Ha bacteriocins." Current Microbiology 41(1): 1-4.
Ennahar, S., T. Sashihara, et al. (2000). "Class Ha bacteriocins: biosynthesis, structure and activity." Ferns Microbiology Reviews 24(1): 85-106.
FAO (2007). Les changements profonds de la production mondiale de viande risquent d'accroître les maladies
FAO salle de presse. Rome. FAO/WHO (2002). Joint working group report on guidelines for the evaluation of
probiotics in food. London, Ontario, Canada. Farner, D. S. (1942). "The hydrogen ion concentration in avian digestive tracts."
Poultry Science 21: 445-450. Feria, C , J. Machado, et al. (2001). "Virulence genes and P fimbriae PapA subunit
diversity in canine and feline uropathogenic Escherichia coli." Veterinary Microbiology 82(1): 81-89.

81
Feuillet, L. (2007). Étude comparative des vaccins et des flores bactériennes dans la lutte contre les Salmonelles en élevage de poules pondeuses. Médecine vétérinaire. Alfort, École Nationale vétérinaire d'Alfort: 17-21.
Fooks, L. J., R. Fuller, et al. (1999). "Prebiotics, probiotics and human gut microbiology." International Dairy Journal 9(1): 53-61.
Fréter, R. (2004). Factors affecting the gut by lactobacilli and other bacteria. department of microbiology and immunology. University of Michigan.
Gabriel, I., S. Mallet, et al. (2003). "Differences in the digestive tract characteristics of broiler chickens fed on complete pelleted diet or on whole wheat added to pelleted protein concentrate." British Poultry Science 44(2): 283-290.
Gabriel, I., S. Mallet, et al. (2005). "Digestive microflora of bird: factors of variation and consequences on bird." Productions Animales 18(5): 309-322.
Gaggîa, F., P. Mattarelli, et al. "Probiotics and prebiotics in animal feeding for safe food production." International Journal of Food Microbiology In Press, Corrected Proof.
Gagnon, M. (2007). Rôle des probiotiques lors d'infection entériques d'origine bactérienne et virale : anlyses in vitre et études in vivo chez des modèles murins. Département des sciences des aliments et de nutrition Québec. Université Laval. Ph.D: 155.
Gagnon, M., E. E. Kheadr, et al. (2004). "In vitro inhibition of Escherichia coli 0157 : H7 by bifidobacterial strains of human origin." International Journal of Food Microbiology 92(1): 69-78.
Gaillard-Gendron, S., D. Vignon, et al. (2000). "Isolation, purification and partial amino acid sequence of a highly hydrophobic new microcin named microcin L produced by Escherichia coli." FEMS Microbiology Letters 193(1): 95-8.
Ganzle, M. G., C. Hertel, et al. (1999). "Effect of bacteriocin-producing lactobacilli on the survival of Escherichia coli and Listeria in a dynamic model of the stomach and the small intestine." International Journal of Food Microbiology 48(1): 21-35.
Garcia-Bustos, J. F., N. Pezzi, et al. (1984). "Microcin 7: purification and properties." Biochemical and Biophysical Research Communications 119(2): 779-85.
Gérard, P. and A. Bernalier-Donadille (2007). "Les fonctions majeures du microbiote intestinal." Cahiers de Nutrition et de Diététique 42(Supplement 2): 28-36.
Gill, H. S., K. J. Rutherfurd, et al. (2000). "Enhancement of natural and acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019)." British Journal of Nutrition 83(2): 167-176.
Gillor, O., B. C. Kirkup, et al. (2004). "Colicins and microcins: the next generation antimicrobials." Advances in Applied Microbiology 54: 129-46.
Gillor, O., L. M. Nigro, et al. (2005). "Genetically engineered bacteriocins and their potential as the next generation of antimicrobials." Current Pharmaceutical Design 11(8): 1067-1075.
Gong, J. H., R. J. Forster, et al. (2002). "Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum." Ferns Microbiology Ecology 41(3): 171-179.
Gratia, A. (1925). "Sur un remarquable exemple d'antagonisme entre deux souches de Colibacille." C. R. Soc. Biol. 93: 1040-1041.

82
Gratia, A. (1925). "Sur un remarquable exemple d'antagonisme entre deux souches de Colibacille." C. R. Soc. biol.(93): 1040-1041.
Guardia, S. S., Recoquillay, F,Juin, Hervé H, Lessire, Michel M, Leconte, Maryse M, Guillot, J.F, Gabriel, Irène I (2009). Effet de l'utilisation de combinaisons d'extraits végétaux naturels dans l'alimentation du poulet de chair selon la densité d'élevage sur les performances de croissance. 8èmes Journées de la Recherche Avicole, St Malo (FRA).
Gueimonde, M. and S. Salminen (2006). "New methods for selecting and evaluating probiotics." Digestive and Liver Disease(38): S242-S247.
Hamon, Y. and Y. Peron (1963). "Bactériologie - Quelques Remarques Sur Les Bactériocines Produites Par Les Microbes Gram-Positifs." Comptes Rendus Hebdomadaires Des Seances De L Académie Des Sciences 257(5): 1191-&.
Hellyer, T. J., L. E. DesJardin, et al. (1999). "Quantitative analysis of mRNA as a marker for viability of Mycobacterium tuberculosis." Journal of Clinical Microbiology 37(2): 290-295.
Higgins, S. E., J. P. Higgins, et al. (2008). "Evaluation of a Lactobacillus-Based Probiotic Culture for the Reduction of Salmonella Enteritidis in Neonatal Broiler Chicks." Poultry Science 87(1): 27-31.
Hulin, V., A. G. Mathot, et al. (1998). "Antimicrobial properties of essential oils and flavour compounds." Sciences Pes Aliments 18(6): 563-582.
Isolauri, E., Y. Sutas, et al. (2001). "Probiotics: effects on immunity." American Journal of Clinical Nutrition 73(2): 444S-450.
Jacob, F., A. Lwoff, et al. (1953). "Definition of some terms relative to lysogeny." ann. inst. Pasteur (Paris)(84): 222-224.
Jia, W. and B. A. Slominski (2010). "Means to improve the nutritive value of flaxseed for broiler chickens: The effect of particle size, enzyme addition, and feed pelleting." Poultry Science 89(2): 261-269.
Jin, L. Z., Y. W. Ho, et al. (1998). "Effects of adherent Lactobacillus cultures on growth, weight of organs and intestinal microflora and volatile fatty acids in broilers." Animal Feed Science and Technology 70(3): 197-209.
Joerger, R. (2003). "Alternatives to antibiotics: bacteriocins, antimicrobial peptides and bacteriophages." Poultry Science 82(4): 640-647.
Jordan, F. T. W. and M. Pattison (1996). Poultry diseases. S. company. London: 38-43. Jung, S. J., R. Houde, et al. (2008). "Effects of Galacto-Oligosaccharides and a
Bifidobacteria lactis-Based Probiotic Strain on the Growth Performance and Fecal Microflora of Broiler Chickens." Poultry Science 87(9): 1694-1699.
Kaiser, A. L. and T. J. Montville (1993). "The Influence of Ph and Growth-Rate on Production of the Bacteriocin, Bavarian Mn, in Batch and Continuous Fermentations." Journal of Applied Bacteriology 75(6): 536-540.
Kaur, I. P., A. Kuhad, et al. (2009). "Probiotics: Delineation of Prophylactic and Therapeutic Benefits." Journal of Medicinal Food 12(2): 219-235.
Kimura, T., S. Shiosaka, et al. (1976). "Effect of Dietary Amino-Acids and Food-Intake on Intestinal Sucrase and Leucineaminopeptidase Activities of Rats." Nutrition Reports International 14(6): 657-670.
Klaenhammer, T. R. (1988). "Bacteriocins of Lactic-Acid Bacteria." Biochimie 70(3): 337-349.

83
Klaenhammer, T. R. and M. J. Kullen (1999). "Selection and design of probiotics." International Journal of Food Microbiology 50( 1 -2): 45-57.
Kozak, W., J. Bardowski, et al. (1978). "Lactostrepcins - Acid Bacteriocins Produced by Lactic Streptococci." Journal of Dairy Research 45(2): 247-257.
Lamps, L. W. (2007). "Infective disorders of the gastrointestinal tract." Histopathology 50(1): 55-63.
Lan, P. T. N., H. Hayashi, et al. (2002). "Phylogenetic analysis of cecal microbiota in chicken by the use of 16S rDNA clone libraries." Microbiology and Immunology 46(6): 371-382.
Lavina, M., C. Gaggero, et al. (1990). "Microcin H47, a chromosome-encoded microcin antibiotic of Escherichia coli." Journal of Bacteriology 172(11): 6585-8.
Lazdunski, C , E. Bouveret, et al. (2000). "Colicin import into Escherichia coli cells requires the proximity of the inner and outer membranes and other factors." International Journal of Medical Microbiology 290(4-5): 337-344.
Lazzaroni, J. C , J. F. Dubuisson, et al. (2002). "The Toi proteins of Escherichia coli and their involvement in the translocation of group A colicins." Biochimie 84(5-6): 391-397.
LeBlanc, J. J. (2003). "Implication of virulence factors in Escherichia coli 0157 : H7 pathogenesis." Critical Reviews in Microbiology 29(4): 277-296.
Lu, J. R., U. Idris, et al. (2003). "Diversity and succession of the intestinal bacterial community of the maturing broiler chicken." Applied and Environmental Microbiology 69(11): 6816-6824.
M. Larbier and B. Leclercq (1992). Nutrition et alimentation des volailles. Paris. Mainil, J. (2003). "Virulence factors and specific properties of invasive Escherichia
coli: I) Adhesins and colonisation factors." Annales De Médecine Vétérinaire 147(2): 105-+.
Mainil, J. (2003). "Virulence factors and specific properties of invasive Escherichia coli: n. Mucosal transfer and invasive properties." Annales De Médecine Vétérinaire 147(3): 159-171.
Mainil, J. and S. Van Bost (2004). "Virulence factors and specific properties of invasive Escherichia coli : IV) Necrotoxigenic strains." Annales De Médecine Vétérinaire 148(3): 121-132.
Martinez, J. S., J. N. Carter-Franklin, et al. (2003). "Structure and membrane affinity of a suite of amphiphilic siderophores produced by a marine bacterium." Proceedings of the National Academy of Sciencees of the United States of America 100(7): 3754-9.
Mathlouthi, N., S. Mallet, et al. (2002). "Effects of xylanase and $\beta$-glucanase addition on performance, nutrient digestibility, and physico-chemical conditions in the small intestine contents and caecal microflora of broiler chickens fed a wheat and barley-based diet." Animal Research 51(05): 395-406.
McKillip, J. L., L. A. Jaykus, et al. (1999). "Nucleic acid persistence in heat-killed Escherichia coli 0157 : H7 from contaminated skim milk." Journal of Food Protection 62(8): 839-844.
Mead, G. C, A. P. Norris, et al. (1989). "Differentiation of Staphylococcus-Aureus from Freshly Slaughtered Poultry and Strains Endemic to Processing Plants by

84
Biochemical and Physiological Tests." Journal of Applied Bacteriology 66(2): 153-159.
Meijer, A., P. J. M. Roholl, et al. (2000). "Chlamydia pneumoniae in vitro and in vivo: a critical evaluation of in situ detection methods." Journal of Clinical Pathology 53(12): 904-910.
Mercenier, A., S. Pavan, et al. (2003). "Probiotics as biotherapeutic agents: Present knowledge and future prospects." Current Pharmaceutical Design 9(2): 175-191.
Methner, U., P. A. Barrow, et al. (1997). "Comparative study of the protective effect against Salmonella colonisation in newly hatched SPF chickens using live, attenuated Salmonella vaccine strains, wild-type Salmonella strains or a competitive exclusion product." International Journal of Food Microbiology 35(3): 223-230.
Miller, J. H. (1972). "Experiments in molecular Genetics." (Cold Spring harbor laboratory, Cold Spring harbor laboratory, NY): 218-220.
Ming, J. E., D. M. McDonaldMcGinn, et al. (1997). "Heterotaxia in a fetus with campomelia, cervical lymphocele, polysplenia, and multicystic dysplastic kidneys: Expanding the phénotype of Cumming syndrome." American Journal of Medical Genetics 73(4): 419-424.
Mitsch, P., K. Zitterl-Eglseer, et al. (2004). "The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestines of broiler chickens." Poultry Science 83(4): 669-675.
Mori agnes, F. m., Gautron Joël, Geraert Pierre-André (2007). Les enzymes chez la pondeuse : des benefices au-delà de la valeur nutritionnelle. Septième journées de la recheche Avicole. Tours.
Murry, A. and A. Hinton (2006). "Comparison of in vitro inhibition of growth of Salmonella Typhimurium and Escherichia coli on broiler feed media by two sources of Bacillus subtilis." Poultry Science 85: 192-193.
Nataro, J. P. and J. B. Kaper (1998). "Diarrheagenic Escherichia coli." Clinical Microbiology Reviews 11(1): 142.
Nisbet, D. (2002). "Defined competitive exclusion cultures in the prevention of enteropathogen colonisation in poultry and swine." Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 81(1-4): 481-486.
Nurmi, E., Rantale, M (1973). "New aspects of Salmonella infection in broiler production." Nature 241: 210-211.
O'Brien, I. G. and F. Gibson (1970). "The structure of enterochelin and related 2,3-dihydroxy-N-benzoylserine conjugates from Escherichia coli." Biochimica et Biophvsica Acta 215(2): 393-402.
Oral, N., L. Vatansever, et al. (2009). "Effect of absorbent pads containing oregano essential oil on the shelf life extension of overwrap packed chicken drumsticks stored at four degrees Celsius." Poultry Science 88(7): 1459-1465.
Oussalah, M., S. Caillet, et al. (2006). "Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli 0157 : H7 and Listeria monocytogenes." Journal of Food Protection 69(5): 1046-1055.
Ouwehand, A. C , S. Salminen, et al. (2002). "Probiotics: an overview of beneficial effects." Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 82(1-4): 279-289.

85
Panda, A. K., M. R. Reddy, et al. (2003). "Production performance, serum/yolk cholesterol and immune competence of white leghorn layers as influenced by dietary supplementation with probiotic." Tropical Animal Health and Production 35(1): 85-94.
Pascual, M., M. Hugas, et al. (1999). "Lactobacillus salivarius CTC2197 prevents Salmonella enteritidis colonization in chickens." Applied and Environmental Microbiology 65(11): 4981-4986.
Patterson, J. A. and K. M. Burkholder (2003). "Application of prebiotics and probiotics in poultry production." Poultry Science 82(4): 627-631.
Patzer, S. L, M. R. Baquero, et al. (2003). "The colicin G, H and X determinants encode microcins M and H47, which might utilize the catecholate siderophore receptors FepA, Cir, Fiu and IroN." Microbiology 149(Pt 9): 2557-70.
Pellecuer, J., M. Jacob, et al. (1979). "Therapeutic Value of the Essential Oil of Cultivated Satureia Montana." Planta Medica 36(3): 256-257.
Pons, A. M., I. Lanneluc, et al. (2002). "New developments in non-post translationally modified microcins." Biochimie 84(5-6): 531-7.
Pons, A. M., I. Lanneluc, et al. (2002). "New developments in non-post translationally modified microcins." Biochimie 84(5-6): 531-537.
Portrait, V. G.-G., S. Cottenceau, G. and Pons, A.M. (1999). "Inhibition of pathogenic Salmonella enteritidis growth mediated by Escherichia coli microcin J25 producing strains." Candian Journal of Microbiology 45: 988-994.
Ragione, R. M. L., A. Narbad, et al. (2004). "<i>In vivo</i> characterization of <i>Lactobacillus johnsonii</i> FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry." Letters in Applied Microbiology 38(3): 197-205.
Revolledo, L., C. S. A. Ferreira, et al. (2009). "Prevention of Salmonella Typhimurium colonization and organ invasion by combination treatment in broiler chicks." Poultry Science 88(4): 734-743.
Rhayour, K., T. Bouchikhi, et al. (2003). "The mechanism of bactericidal action of oregano and clove essential oils and of their phenolic major components on Escherichia coli and Bacillus subtilis." Journal of Essential Oil Research 15(5): 356-362.
Riley, M. A. and J. E. Wertz (2002). "Bacteriocin diversity: ecological and evolutionary perspectives." Biochimie 84(5-6): 357-364.
Saarela, M., G. Mogensen, et al. (2000). "Probiotic bacteria: safety, functional and technological properties." Journal of Biotechnology 84(3): 197-215.
Sable, S., M. Duarte, et al. (2003). "Wild-type Escherichia coli producing microcins B17, D93, J25, and L; cloning of genes for microcin L production and immunity." Canadian Journal of Microbiology 49(5): 357-361.
Salm-Surv, G. (2005). un reseau de l'OMS pour la surveillance des maladies d'origine alimentaire, OMS.
Salomon, R. A. and R. N. Farias (1992). "Microcin 25, a novel antimicrobial peptide produced by Escherichia coli." Journal of Bacteriology 174(22): 7428-35.
Schrezenmeir, J. and M. de Vrese (2001). "Probiotics, prebiotics, and synbiotics -approaching a definition." American Journal of Clinical Nutrition 73(2): 361s-364s.

86
Schroeder, B., S. Duncker, et al. (2006). "Preventive effects of the probiotic Escherichia coli strain nissle 1917 on acute secretory diarrhea in a pig model of intestinal infection." Digestive Diseases and Sciences 51(4): 724-731.
Shelef, L. A. (1984). "Antimicrobial Effects of Spices." Journal of Food Safety 6(1): 29-44.
Simon, O., A. Jadamus, et al. (2001). "Probiotic feed additives - effectiveness and expected modes of action." Journal of Animal and Feed Sciences 10: 51-67.
Smajs, D., J. Smarda, et al. (2003). "The Escherichia fergusonii iucABCD iutA genes are located within a larger chromosomal region similar to pathogenicity islands." Folia Microbiologica 48(2): 139-147.
Smith-Palmer, A., J. Stewart, et al. (1998). "Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens." Letters in Applied Microbiology 26(2): 118-122.
Smith, H. W. (1965). "Observations on Flora of Alimentary Tract of Animals and Factors Affecting Its Composition." Journal of Pathology and Bacteriology 89(1): 95-&.
Sreekumar, O. and A. Hosono (1998). "Antimutagenicity and the influence of physical factors in binding Lactobacillus gasseri and Bifidobacterium longum cells to amino acid pyrolysates." Journal of Dairy Science 81(6): 1508-1516.
Srivastava, K. C , A. Bordia, et al. (1995). "Garlic (Allium-Sativum) for Disease Prevention." South African Journal of Science 91(2): 68-77.
Stavric, S. (1992). "Defined Cultures and Prospects." International Journal of Food Microbiology 15(3-4): 245-263.
Stender, H., M. Fiandaca, et al. (2002). "PNA for rapid microbiology." Journal of Microbiological Methods 48(1): 1-17.
Stordeur, P. and J. Mainil (2002). "Colibacillosis in poultry." Annales De Médecine Vétérinaire 146(1): 11-18.
Tagg, J. R., A. S. Dajani, et al. (1976). "Bacteriocins of Gram-Positive Bacteria." Bacteriological Reviews 40(3): 722-756.
Takase, K., T. Nakayama, et al. (1999). "Growth of Salmonella typhimurium and Salmonella enteritidis in egg yolks from highly immunized hens." Journal of Veterinary Medical Science 61(8): 959-960.
Tannock, G. W. (1997). "Probiotic properties of lactic-acid bacteria: Plenty of scope for fundamental R&D." Trends in Biotechnology 15(7): 270-274.
Tkalcic, S., T. Zhao, et al. (2003). "Fecal shedding of enterohemorrhagic Escherichia coli in weaned calves following treatment with probiotic Escherichia coli." Journal of Food Protection 66(7): 1184-1189.
Van Bost, S., E. Jacquemin, et al. (2003). "Multiplex PCRs for identification of necrotoxigenic Escherichia coli." Journal of Clinical Microbiology 41(9): 4480-4482.
Van Immersed, F., K. Cauwerts, et al. (2002). "Feed additives to control Salmonella in poultry." Worlds Poultry Science Journal 58(4): 501-513.
Vandeplas, S., R. D. Dauphin, et al. (2009). "Efficiency of a Lactobacillus plantarum-xylanase combination on growth performances, microflora populations, and nutrient digestibilities of broilers infected with Salmonella Typhimurium." Poultry Science 88(8): 1643-1654.

87
Vandeplas, S., R. D. Dauphin, et al. (2009). "Efficiency of a Lactobacillus plantarum-xylanase combination on growth performances, microflora populations, and nutrient digestibilities of broilers infected with Salmonella Typhimurium." Poultry Science 88(8): 1643-1654.
Weurding, R., H. Enting, et al. (2003). "The relation between starch digestion rate and amino acid level for broiler chickens." Poultry Science 82(2): 279-284.
Willis, W. L. and L. Reid (2008). "Investigating the Effects of Dietary Probiotic Feeding Regimens on Broiler Chicken Production and Campylobacter jejuni Presence." Poult Sci 87(4): 606-611.
Windisch, W., K. Schedle, et al. (2008). "Use of phytogenic products as feed additives for swine and poultry." Journal of Animal Science. 86(14_suppl): El40-148.
Yang, C. C. and J. Leong (1982). "Production of deferriferrioxamines B and E from a ferroverdin-producing Streptomyces species." Journal of Bacteriology 149(1): 381-3.
Ye, R. W., T. Wang, et al. (2001). "Applications of DNA microarrays in microbial systems." Journal of Microbiological Methods 47(3): 257-272.
Yves Beckers and F. Piron (2009). Utilisation des enzymes exogènes en alimentation porcine et avicole. Belgique, Unité de Zootechnie, Gembloux Agro-bio tech, université de Liège.
Zacconi, C , G. Scolari, et al. (1999). "Colonisation of chicken intestinal tract by Lactobacillus salivarius A23 strain." Annali Di Microbiologia Ed Enzimologia 49: 103-115.
Zacconi, C , G. Scolari, et al. (1999). "Effect of administration of Lactobacillus salivarius and lactic microflora in chick digestive tract." Annali Di Microbiologia Ed Enzimologia 49: 117-123.
Zaika, L. L. (1988). "Spices and Herbs - Their Antimicrobial Activity and Its Determination." Journal of Food Safety 9(2): 97-118.
Zhang-Barber, L., A. K. Turner, et al. (1998). "Protection of chickens against experimental fowl typhoid using a nuoG mutant of Salmonella serotype Gallinarum." Vaccine 16(9-10): 899-903.

88
ANNEXES
Annex 1: Séquences des gènes d'ADNr 16S des différentes souches intestinales
11004 (1416 base)
ACACATGCAGTCGAACGGTAACAGGAAGCAGCTTGCTGCTTTGCTGACGAGTGGCGGACGGGTGAGTAAT GTCTGGGAAACTGCCYGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAG ACCAAAGAGGGGGACCTTMGGGCCTCTTGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGT AACGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACG GTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCC GCGTGTATGAAGAAGGCCTTCGGGTTGTAAAGTACTTTCAGCGGGGAGGAAGGGAGTAAAGTTAATACCT TTGCTCATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGG GTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATC CCCGGGCTCAACCTGGGAACTGCATCTGATACTGGCAAGCTTGAGTCTCGTAGAGGGGGGTAGAATTCCA GGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGACT GACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATG TCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGT ACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATT CGATGCAACGCGAAGAACCTTACCTGGTCTTGACATCCACGGAAGTTTTCAGAGATGAGAATGTGCCTTC GGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCA ACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTCCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAAA CTGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGACCAGGGCTACACACGTGCTACAATG GCGCATACAAAGAGAAGCGACCTCGCGAGAGCAAGCGGACCTCATAAAGTGCGTCGTAGTCCGGATTGGA GTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTGGATCAGAATGCCACGGTGAATACGTT CCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTANCCTT CGGGAGGGCGCTACCA
11006 (1419 base)
ACACATGCAGTCGAACGGTAACAGGAANCAGCTTGCTGTTTNGCTGACGAGTGGCGGACGGGTGAGTAAT GTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAAYGTCGCAAG ACCAAAGAGGGGGACCTTCGGGCCTCTTGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGT AACGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACG GTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCC GCGTGTATGAAGAAGGCCTTCGGGTTGTAAAGTACTTTCAGCGGGGAGGAAGGGAGTAAAGTTAATACCT TTGCTCATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGG GTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATC CCCGGGCTCAACCTGGGAACTGCATCTGATACTGGCAAGCTTGAGTCTCGTAGAGGGGGGTAGAATTCCA GGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGACT GACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATG TCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGT ACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATT CGATGCAACGCGAAGAACCTTACCTGGTCTTGACATCCACAGAACCTTGTAGAGATACGAGGGTGCCTTC GGGAACTGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCA ACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTCCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAAA

89
CTGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGACCAGGGCTACACACGTGCTACAATG GCGCATACAAAGAGAAGCGACCTCGCGAGAGCAAGCGGACCTCATAAAGTGCGTCGTAGTCCGGATTGGA GTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTGGATCAGAATGCCACGGTGAATACGTT CCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTT CGGGAGGGCGCTACCACTT
11026 (1411 base)
ACACATGCAAGTCGAACGGTAACAGGAAGAAGCTTGCTTCTTTGCTGACGAGTGGCGGACGGGTGAGTAA TGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAA GACCAAAGAGGGGGACCTTCGGGCCTCTTGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGG TAAAGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACAC GGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGC CGCGTGTATGAAGAAGGCCTTCGGGTTGTAAAGTACTTTCAGCGGGGAGGAAGGGAGTAAAGTTAATACC TTTGCTCATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAG GGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAAT CCCCGGGCTCAACCTGGGAACTGCATCTGATACTGGCAAGCTTGAGTCTCGTAGAGGGGGGTAGAATTCC AGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGAC TGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGAT GTCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAG TACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAAT TCGATGCAACGCGAAGAACCTTACCTGGTCTTGACATCCACAGAACTTTCCAGAGATGGATTGGTGCCTT CGGGAACTGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGC AACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTCCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAA ACTGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGACCAGGGCTACACACGTGCTACAAT GGCGCATACAAAGAGAAGCGACCTCGCGAGAGCAAGCGGACCTCATAAAGTGCGTCGTAGTCCGGATTGG AGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTGGATCAGAATGCCACGGTGAATACGT TCCCGGGCCTTGTACACACCGCCCGTCACACCANGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCT TCGGGAGGGCG