introduction à l'étude du métabolisme cellulaire ... · la bioénergétique, un domaine de...
TRANSCRIPT

1
Introduction à l'étude du métabolisme cellulaire : quelques notions de thermodynamique, principaux types de réactions, importance des coenzymes et de l'ATP
Introduction : Un organisme vivant fonctionne en réalisant un ensemble de réactions chimiques intégrées qui constituent son métabolisme. Ainsi, plus d'un millier de réactions chimiques s'effectuent dans un organisme aussi simple que la bactérie Escherichia coli. On peut schématiquement classer les réactions métaboliques en deux grandes catégories : - les réactions cataboliques qui correspondent aux réactions de dégradation permettant de convertir l'énergie de molécules source d'énergie en des formes biologiquement utilisables; - les réactions anaboliques qui regroupent les réactions de synthèse nécessitant un apport d'énergie pour pouvoir s'effectuer. Au cours de ces réactions, il y a donc conversion d'une forme d'énergie en une autre. De telles conversions d'énergie sont décrites par une branche de la physique qui s'appelle la thermodynamique. Le 1er principe de la thermodynamique par exemple stipule que la quantité d'énergie de l'univers reste constante, et que, par conséquent, au cours de n'importe quelle transformation physique ou chimique, l'énergie ne peut être ni créée ni détruite : elle ne peut qu'être convertie d'une forme en une autre. La bioénergétique, un domaine de la biochimie étudiant les transformations et les utilisations de l’énergie par les êtres vivants, s’appuie donc sur les principes de la thermodynamique. Quelles sont les lois de la thermodynamique régissant la réalisation des réactions du métabolisme cellulaire? Quelles sont les principales réactions impliquées dans le métabolisme cellulaire? Quelles sont les sources d'énergie utilisées par les cellules? En quelle forme utilisable par les cellules cette énergie est-elle convertie? Comment s'effectue cette conversion? L'étude du métabolisme montre que, malgré le très grand nombre de réactions impliquées, on peut tout de même dégager un schéma général incluant un certain nombre de points communs à toutes ces réactions : seulement quelques grands types de réactions, l'utilisation d'une unité universelle d'énergie, l'ATP, et l'apparition d'un nombre limité d'intermédiaires activés, les coenzymes. Dans la 1ère partie, nous donnerons donc les quelques notions de thermodynamique permettant d'appréhender ces phénomènes de conversion énergétique, et nous verrons comment ils s'appliquent aux principaux types de réactions métaboliques. Puis nous montrerons l'organisation de ces réactions métaboliques au sein d'une cellule en insistant sur le rôle central de l'ATP et des coenzymes.
I. CONDITIONS THERMODYNAMIQUES DE REALISATION DES REACTIONS METABOLIQUES
A. L'enthalpie libre et l'évolution d'un système thermodynamique
1) Les cellules, des systèmes thermodynamiques ouverts
• Un système thermodynamique est une région de l’espace dans laquelle se trouve une certaine quantité de matière et où s’effectuent des transformations (par exemple une cellule ou un réacteur dans un labo de chimie). Ce qui est extérieur au système constitue son environnement. • Les transformations qui s'effectuent dans un système peuvent ou non donner lieu à des échanges avec l'environnement. On distingue : - les systèmes ouverts qui échangent de l'énergie et de la matière avec leur environnement; - les systèmes fermés qui échangent uniquement de l'énergie avec leur environnement; - les systèmes isolés qui n'échangent ni énergie ni matière avec leur environnement. • L'énergie d'un système est sa capacité à fournir un travail. Elle s'exprime en J (ou en cal). 1cal=4,18J ou 1J=0,24cal. (1 cal = quantité d'énergie nécessaire pour augmenter la T° d'1g d'eau de 1°C à 25°C) L'énergie peut être : - de l'énergie potentielle = énergie qui peut être libérée lors de la survenue de certains processus (par exemple un déplacement, une réaction entre deux molécules....); - de l'énergie cinétique = énergie associée à un mouvement;

2
- de la chaleur, qui est une manifestation de l'énergie cinétique liée au mouvement des particules d'un système. [1] p 69 Energie potentielle et énergie cinétique sont des formes d'énergie "utiles" pouvant donner naissance à un travail, contrairement à la chaleur qui est une forme d'énergie "désordonnée" non utilisable par les systèmes thermodynamiques (sauf les machines thermiques). Ainsi, les systèmes biologiques ne peuvent pas utiliser l'énergie calorique pour réaliser un travail (toutefois, celle-ci augmente
l'agitation moléculaire ce qui facilite les réactions métaboliques ce n'est pas tout à fait de l'énergie "perdue"…). • Une cellule prélève dans son environnement de la matière et de l'énergie utile grâce à laquelle elle effectue ses travaux, et elle rend à son environnement des déchets (matière qui ne lui est pas utile), et des formes dégradées d'énergie (énergie thermique) : c’est un système thermodynamique ouvert. Les lois de la thermodynamique s’appliquent normalement aux systèmes fermés mais on va tout de même pouvoir les transposer aux cellules. [1] p 67
2) La description des systèmes thermodynamiques
Un système thermodynamique peut être décrit par ses variables d’état : Volume, Masse, Pression, Température… mais également par ses fonctions d'état = grandeurs thermodynamiques dont la valeur dépend des variables d'état : énergie interne U, enthalpie H, enthalpie libre G et entropie S.
a) Energie interne et premier principe de la thermodynamique • L'énergie interne d'un système est définie par U = Q + W où Q est la quantité d'énergie calorique échangée avec l'extérieur, W le travail fourni. • Le 1er principe de la thermodynamique stipule que l'énergie interne U de l'univers est constante. Donc lorsqu'un système subit des transformations, (cad passe d'un état initial i à un état final f), toute variation du contenu énergétique du système est contrebalancée par une variation symétrique du contenu énergétique de son environnement.

3
Si une réaction chimique a lieu à P constante (ce qui est le cas dans les cellules), alors Q équivaut à l’enthalpie H. A P constante, le travail des forces de pression vaut - PΔV. La variation d'énergie interne d'un système est alors ΔU = ΔH - PΔV. Dans les cellules, les réactions se font en général à volume constant, et donc ΔU = ΔH. Dans les systèmes biologiques, la variation d'enthalpie décrit la variation d'énergie interne du système. Si ΔH>0, le système absorbe de l'énergie (sous forme de chaleur) et la transformation est endergonique. Si ΔH<0, le système libère de l'énergie (sous forme de chaleur) et la transformation est exergonique. Cependant, l'étude de la variation d’enthalpie et le 1er principe ne permettent pas de prévoir le sens de l’évolution d’un système : une réaction spontanée peut être exothermique (ex: combustion de l’essence), ou, plus rarement, endothermique (ex: dissolution du NaCl)
b) Entropie et deuxième principe de la thermodynamique • Le 2è principe de la thermodynamique stipule que l'univers se modifie constamment de façon à devenir plus désordonné. • L’entropie S mesure le degré d’organisation ou la probabilité de l’existence d’un système :
- plus la distribution des éléments d’un système est aléatoire, plus sa probabilité d'existence est grande et plus son entropie est élevée; - au contraire, plus les éléments sont organisés, plus leur probabilité d’existence est petite et plus leur entropie est faible.
Le niveau 0 de l’entropie correspond à un cristal parfait à la T de 0K. Le 2è principe, qui signifie que le désordre est plus probable que l'ordre, peut donc s'exprimer de la façon suivante : l’entropie totale de l’univers augmente (augmentation du degré de désordre) OU la somme des variations d’entropie d’un système et de son nvironnement est positive : ΔSsystème + ΔSenvironnement > 0. Un système aura donc tendance à évoluer spontanément vers un état "désordonné" qui va aboutir à une uniformisation de ses paramètres, c'est-à dire à un état d'équilibre. Dans cet état d'équilibre, il n'y a plus de travail possible, et donc plus de possibilité de retour spontané à l'état ordonné. L'énergie interne du système est alors nulle. • Dans une cellule, les processus cellulaires vont contribuer à augmenter l'ordre du système (ΔS<0, ce qui semble contradictoire avec le 2è principe), mais cette diminution de l'entropie est contrebalancée par une augmentation encore plus importante de l'entropie de l'environnement par libération de chaleur (ΔS>0). On aura donc bien ΔSsystème + ΔSenvironnement > 0. [1] p 68 L'étude de l'entropie d'un système, et plus particulièrement d'une nouvelle fonction dépendant de S, l'enthalpie libre G, va donc permettre de prévoir son évolution.

4
3) Les variations d’enthalpie libre et la spontanéité des réactions métaboliques
a) La variation d'enthalpie libre d'un système • On vient de voir que les transformations subies par un système biologique s'accompagnent en général d'une libération de chaleur vers son environnement, et donc d'une variation de l'entropie de cet environnement. Cette variation d'entropie sera plus importante lorsque de la chaleur sera ajoutée à un environnement froid, et moins importante lorsqu'elle sera ajoutée à un environnement plus chaud. ΔSenvironnement sera donc proportionnelle à la quantité de chaleur libérée par le système - ΔHsystème, et inversement proportionnelle à Tenvironnement. Soit : ΔSenvironnement = - ΔHsystème/T (1) On sait que : ΔSunivers = ΔSsystème + ΔSenvironnement En utilisant l'équation (1), on a ΔSunivers = ΔSsystème - ΔHsystème/T soit : -T.ΔSunivers = ΔHsystème - T.ΔSsystème -T.ΔSunivers est désignée comme la variation d'énergie libre ou variation d'enthalpie libre (G car elle a été définie par Gibbs...) : ΔG = ΔH - TΔS Elle correspond à la fraction de l’énergie récupérable pour la réalisation d’un travail (alors que TΔS représente la part de l’énergie non disponible pour la réalisation d’un travail). • Le calcul de la variation d'enthalpie libre d'un système ΔG = Gf-Gi permet donc de prévoir son évolution. En effet, si les transformations subies par un système sont spontanées, elles vont contribuer à l'augmentation de l'entropie de l'univers, soit ΔSunivers>0, et donc ΔG<0. Le système évoluera alors spontanément vers son état d'équilibre qui sera atteint lorsque ΔG=0.
- Si les transformations s'effectuent sans échanges de chaleur, ΔH = 0 et ΔG = -TΔS est bien <0 si l'entropie du système est augmentée (ΔSsystème>0). - Si les transformations sont exothermiques, ΔH < 0 et ΔG<0. - Si les transformations sont endothermiques, ΔH > 0 et, en général, ΔG>0 : les transformations ne sont pas
spontanées. Mais, dans certains cas, TΔS > ΔH et donc ΔG<0 : la transformation est spontanée tout en étant endothermique. C’est le cas de la dissolution du NaCl dans l’eau, qui s'accompagne d’une forte augmentation d’entropie du fait du passage de l’état cristallin ordonné à une répartition spatiale aléatoire des particules en solution.
b) La variation d'enthalpie libre d'une réaction • Pour étudier les réactions du métabolisme cellulaire, on étudie la variation d’enthalpie libre d’une réaction (et pas celle d'un système complet) qui permet de prévoir le sens de cette réaction lorsqu'on est à P et T constantes, ce qui est le cas dans une cellule. ΔrG = Grf-Gri (mesurée en kJ/mol) Cette variation dépend uniquement de l’état final et de l’état initial, indépendamment du chemin suivi entre ces deux états. Mais, même si le ΔrG d'une réaction est <0, le passage de l'état initial à l'état final peut nécessiter un apport d'énergie : c'est ce que l'on appelle l'énergie d'activation de la réaction (qui peut être fournie par une augmentation de température par exemple). Les catalyseurs comme les enzymes ne modifient pas le ΔrG de la réaction (ils ne peuvent pas rendre spontanée une réaction qui ne l'est pas), mais ils abaissent l'énergie d'activation nécessaire à sa réalisation, ce qui augmente la vitesse de la réaction. [1] p 73
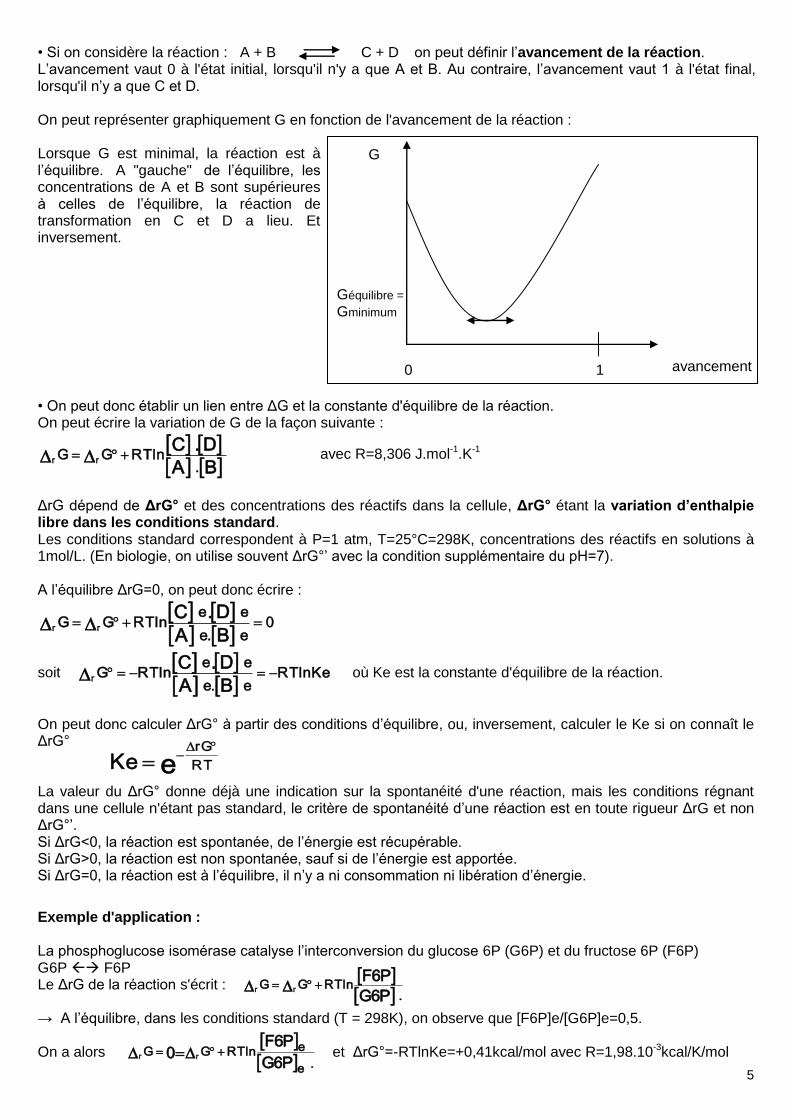
5
• Si on considère la réaction : A + B C + D on peut définir l’avancement de la réaction. L’avancement vaut 0 à l'état initial, lorsqu'il n'y a que A et B. Au contraire, l’avancement vaut 1 à l'état final, lorsqu'il n’y a que C et D. On peut représenter graphiquement G en fonction de l'avancement de la réaction : Lorsque G est minimal, la réaction est à l’équilibre. A "gauche" de l’équilibre, les concentrations de A et B sont supérieures à celles de l’équilibre, la réaction de transformation en C et D a lieu. Et inversement. • On peut donc établir un lien entre ΔG et la constante d'équilibre de la réaction. On peut écrire la variation de G de la façon suivante :
BA
D.C
.lnRTGG rr avec R=8,306 J.mol-1.K-1
ΔrG dépend de ΔrG° et des concentrations des réactifs dans la cellule, ΔrG° étant la variation d’enthalpie libre dans les conditions standard. Les conditions standard correspondent à P=1 atm, T=25°C=298K, concentrations des réactifs en solutions à 1mol/L. (En biologie, on utilise souvent ΔrG°’ avec la condition supplémentaire du pH=7). A l’équilibre ΔrG=0, on peut donc écrire :
0e.e
eelnRTGG
BA
D.Crr
soit
KelnRTe.e
eelnRTG
BA
D.Cr où Ke est la constante d'équilibre de la réaction.
On peut donc calculer ΔrG° à partir des conditions d’équilibre, ou, inversement, calculer le Ke si on connaît le ΔrG° La valeur du ΔrG° donne déjà une indication sur la spontanéité d'une réaction, mais les conditions régnant dans une cellule n'étant pas standard, le critère de spontanéité d’une réaction est en toute rigueur ΔrG et non ΔrG°’. Si ΔrG<0, la réaction est spontanée, de l’énergie est récupérable. Si ΔrG>0, la réaction est non spontanée, sauf si de l’énergie est apportée. Si ΔrG=0, la réaction est à l’équilibre, il n’y a ni consommation ni libération d’énergie.
Exemple d'application : La phosphoglucose isomérase catalyse l’interconversion du glucose 6P (G6P) et du fructose 6P (F6P) G6P F6P Le ΔrG de la réaction s'écrit : → A l’équilibre, dans les conditions standard (T = 298K), on observe que [F6P]e/[G6P]e=0,5. On a alors et ΔrG°=-RTlnKe=+0,41kcal/mol avec R=1,98.10-3kcal/K/mol
G
avancement 0 1
Géquilibre = Gminimum
e RT
rG
Ke
.e
elnRTGGP6G
P6F0 rr
.
lnRTGGP6G
P6Frr

6
→ Dans une cellule à T = 37°C =310K, on observe que [F6P]/[G6P]=0,19. On a alors Soit ΔrG= -0,6kcal/mol La réaction est spontanée.
B. La réalisation des principaux types de réactions métaboliques
Le métabolisme cellulaire comprend des milliers de réactions chimiques, qui peuvent toutefois être classées en six grands types catalysés par des enzymes spécifiques :
Type de réaction Description Exemple
Oxydoréduction Transfert d'électrons Succinate + FAD ↔ Fumarate + FADH2 (dans le cycle de l'acide citrique)
Ligation nécessitant le clivage de l'ATP
Formation de liaisons covalentes (par exemple liaisons C-C)
Pyruvate + CO2 + ATP + H2O ↔ Oxaloacétate + ADP + Pi + H
+
(synthèse d'acides aminés)
Isomérisation Réarrangements d'atomes pour former des isomères
Citrate ↔ Isocitrate (dans le cycle de l'acide citrique)
Transfert de groupe Transfert d'un groupe fonctionnel d'une molécule à une autre
Glucose + ATP ↔ Glucose-6P + ADP (entrée du glucose dans la glycolyse)
Hydrolytique Clivage de liaisons par addition d'eau
Amidon + H2O ↔ n Maltoses
Addition ou élimination de groupes fonctionnels
Addition de groupes fonctionnels à des doubles liaisons ou élimination de groupes fonctionnels pour former des doubles liaisons
2-Phosphoglycérate ↔ Phosphoénolpyruvate + H2O (dans la glycolyse)
[2] p 425-427 A ces 6 catégories peuvent également être ajoutées les réactions acido-basiques qui ne nécessitent pas la présence d'enzymes et qui sont au centre de nombreux processus biochimiques. Ces réactions se produisent spontanément si elles ont énergétiquement favorables, mais, dans le cas contraire, elles peuvent être couplées à d’autres réactions plus favorables. Nous allons nous intéresser plus particulièrement à deux types de réactions très importantes : les réactions d’oxydoréduction et les réactions acido-basiques.
1) Les réactions d’oxydoréduction Les réactions d’oxydoréduction sont des réactions au cours desquelles se produisent des échanges d'électrons.
a) Principe de l'oxydoréduction • Une réduction est un gain d’électrons ; une oxydation est une perte d’électrons : oxydation forme réduite forme oxydée + électron agent réducteur réduction agent oxydant (donneur d’électron) (accepteur d’électrons)
• Un couple réducteur-oxydant forme un système redox. Soient 2 systèmes redox :
Cu+ Cu2+ + e-1 et Fe2+ Fe3+ + e-1
Mis en présence l’un de l’autre, le couple Cu2+/ Cu+ est plus réducteur que le couple Fe3+/ Fe2+, c’est-à-dire que la réaction Cu+ + Fe3+ Fe2+ + Cu2+ évolue spontanément vers la droite.
Le couple Cu2+/ Cu+ cède ses électrons au couple Fe3+/ Fe2+.
)19,0ln(RT41,0.
lnRTGGP6G
P6Frr

7
• Un couple redox est caractérisé par son potentiel d’oxydoréduction E° (en V) qui est la différence de potentiel aux bornes d’une pile formée par le couple considéré (demi pile) et le couple H+/H2 (demi pile), dans les conditions standard. E°’ est le potentiel standard d’oxydoréduction à pH 7. (E°’= E° - 0,420). Un potentiel rédox >0 correspond à un couple avec un oxydant. Un potentiel rédox <0 correspond à un couple avec un réducteur. [2] p 507
b) Réaction d’oxydoréduction et variation d’enthalpie libre
• Les réactions d’oxydoréduction obéissent aux lois de la thermodynamique : le potentiel standard d’oxydoréduction permet de prévoir le sens du transfert des électrons.
En effet : rG°’ = -n . F . E°’ où n = nombre d’électrons transférés, F = constante de Faraday = 96,1 kJ.V-1.mol-1 = 23,06 kcal. V-1.mol-1
E°’ = différence des E°’ des 2 systèmes en présence = E°’2 – E°’1 = E°’ accepteur (+ fort oxydant) - E°’ donneur (+ fort réducteur) Plus la valeur de E°’ est faible plus le réducteur est fort. Plus la valeur de E°’ est forte plus l’oxydant est fort.
E°2>E°1 ΔrG°=-nF(E°2-E°1)<0 La réaction spontanée est : Ox2 + Red1 Ox1 + Red2 (flèche ) • Exemple avec des réactifs biologiques : NADH,H+ NAD+ + 2H+ + 2e- E°’1 = -0,32V ½ O2 + 2H+ + 2e- H2O E°’2 = +0,82V
E°’ +
-
Force des oxydants
Force des réducteurs
Ox2 Réd2
Ox1 Réd1
Potentiels d'oxydoréduction standard de quelques couples rédox.

8
NADH,H+ est un plus fort réducteur, le dioxygène est un fort oxydant. On peut écrire la réaction complète de la façon suivante : NADH,H+ + ½ O2 H2O + NAD+ Alors ΔrG° = -nF(E°'2-E°'1) = -nF(0,82+0,32) <0. La réaction est donc spontanée. On peut même voir qu’elle libère de l’énergie, énergie qui peut être récupérée par la cellule sous forme d’ATP.
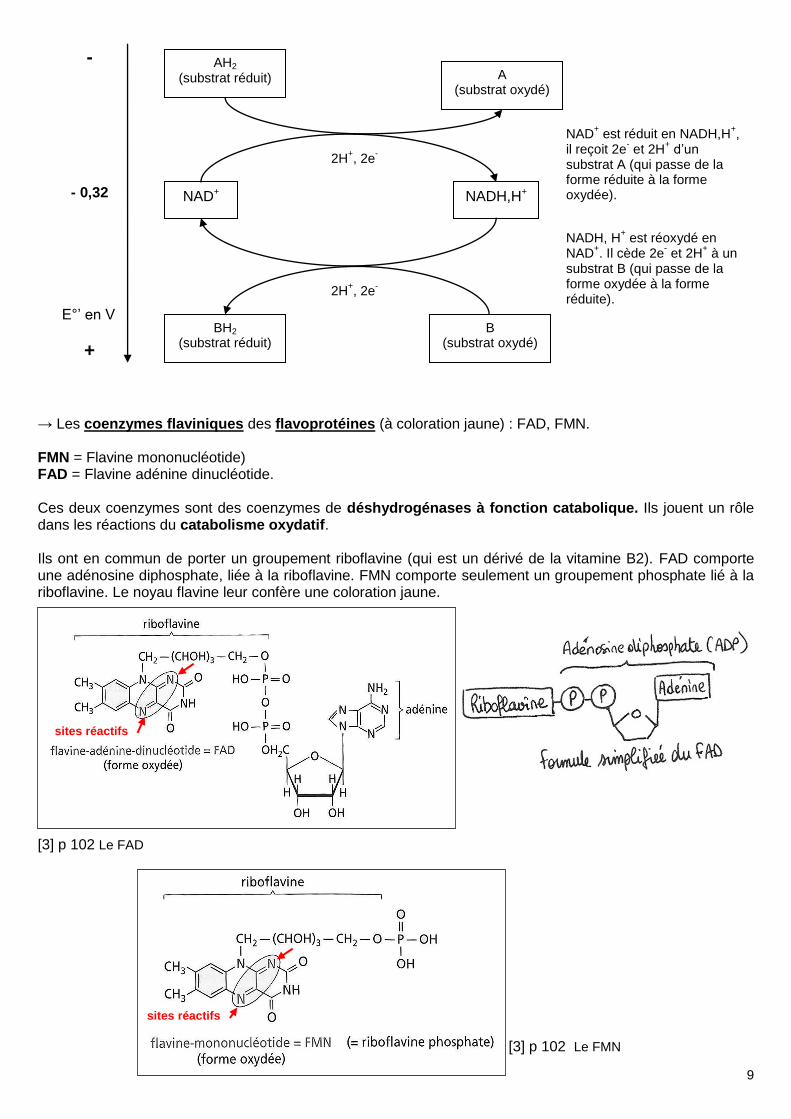
c) Rôle des coenzymes d'oxydoréduction • Les réactions d’oxydoréductions sont catalysées par des enzymes de type déshydrogénases, fonctionnant avec des coenzymes. Les coenzymes (ou cofacteurs) sont des molécules non protéiques liées à une enzyme par des liaisons covalentes ou des liaisons faibles, et nécessaires à son fonctionnement. Un certain nombre de coenzymes sont dérivés de nucléotides. • Les coenzymes des déshydrogénases (= enzymes catalysant les réactions d’oxydoréduction) sont de 2 types: → Les pyridine-nucléotides, NAD+ et NADP+. NAD+ = Nicotinamide Adénine Dinucléotide oxydé (car +) NADP+ = Nicotinamide Adénine Dinucléotide Phosphate oxydé (car +) NAD+ est le coenzyme de nombreuses déshydrogénases à fonction catabolique. Il joue un rôle central dans les réactions du catabolisme oxydatif (respiration, fermentation). NADP+ est le coenzyme de quelques enzymes du catabolisme, mais surtout de déshydrogénases à fonction anabolique. Il joue un rôle central dans les réactions de la photosynthèse. NAD comporte deux nucléotides. Un AMP est lié à un Nicotinamide-ribose phosphate. Le nicotinamide est un dérivé pyrimidique. Les deux nucléotides sont liés par liaison phosphoester. [3] p 100 Le NAD
+ NAD+ (oxydé) et NADH, H+ (réduit) forment donc un couple redox dont le potentiel rédox standard vaut -0,32V. Demi-équation rédox : NADH,H+ NAD+ + 2H+ + 2e-
site réactif (fixation de
2H+ + 2e
-)
H est remplacé par PO3
2- dans NADP
+
9
→ Les coenzymes flaviniques des flavoprotéines (à coloration jaune) : FAD, FMN. FMN = Flavine mononucléotide) FAD = Flavine adénine dinucléotide. Ces deux coenzymes sont des coenzymes de déshydrogénases à fonction catabolique. Ils jouent un rôle dans les réactions du catabolisme oxydatif. Ils ont en commun de porter un groupement riboflavine (qui est un dérivé de la vitamine B2). FAD comporte une adénosine diphosphate, liée à la riboflavine. FMN comporte seulement un groupement phosphate lié à la riboflavine. Le noyau flavine leur confère une coloration jaune. [3] p 102 Le FAD [3] p 102 Le FMN
NAD+ est réduit en NADH,H
+,
il reçoit 2e- et 2H
+ d’un
substrat A (qui passe de la forme réduite à la forme oxydée).
NADH, H+ est réoxydé en
NAD+. Il cède 2e
- et 2H
+ à un
substrat B (qui passe de la forme oxydée à la forme réduite).
AH2 (substrat réduit) A
(substrat oxydé)
NAD+ NADH,H+
2H+, 2e
-
BH2 (substrat réduit)
B (substrat oxydé)
2H+, 2e
-
- 0,32
+
E°’ en V
-
sites réactifs
sites réactifs

10
FAD (oxydé) et FADH2 (réduit) forment donc un couple redox dont le potentiel rédox standard vaut -0,22V. Demi-équation rédox : FADH2 FAD + 2H+ + 2e-
2) Les réactions acido-basiques
• Une réaction acido-basique est une réaction chimique au cours de laquelle se produit un transfert d'un ou plusieurs ions H+ en solution aqueuse. • Des échanges d'ions H+ (= protons) ont lieu spontanément dans toute solution aqueuse. Mais ces ions sont en réalité fixés aux molécules d'eau, formant des ions hydronium H3O
+. Toutefois, lorsqu'on décrit des réactions acido-basiques, on simplifie le plus souvent en désignant ces ions par H+. • La concentration des ions H+ d'une solution est exprimée par le pH : pH = -log[H+] avec [H+] exprimée en unités de molarité. Ainsi, dans l'eau pure, [H+] = 10-7M, et donc pH = 7. La constante d'équilibre de la réaction de dissociation de l'eau H2O H+ + OH- vaut : K = [H+].[OH-]/[H2O] = 1,8.10-16M. [1] p 109 Dans l'eau pure : [H2O] = 55,5M. On a donc : [H+].[OH-] = 1,8.10-16 x 55,5 = 1,0.10-14. Lorsqu'on connait le pH d'une solution, on peut donc en déduire [H+] et [OH-]. Par exemple, dans l'eau pure à pH = 7 : [H+] = 10-7M et [OH-] = 10-14/10-7 = 10-7M. Dans une solution de HCl 0,1M : [H+] = 10-1M donc pH = 1 et [OH-] = 10-14/10-1 = 10-13M. • Les réactions acido-basiques vont donc modifier le pH d'une solution en enlevant ou rajoutant des H+ à cette solution. [1] p 109 Un acide est défini comme une molécule pouvant céder un proton : AH A- + H+ Une base est définie comme une molécule pouvant accepter un proton : B + H+ B H+ AH/A- et BH+/B sont des couples acides-bases. A- est la base conjuguée de l'acide AH, et BH+ est l'acide conjugué de la base B.

11
• La force d'un acide, c'est-à-dire sa capacité à libérer un (ou plusieurs) H+, est définie par son pKa. pKa = -log(Ka). Ka est la constante d'acidité du couple acide base. En considérant la réaction : AH A- + H+ Ka = [H+].[A-]/[AH] On peut donc exprimer pKa en fonction de pH : -logKa = -log ([H+].[A-]/[AH]) soit : pKa = pH -log[A-]/[AH] ou pH = pKa + log[A-]/[AH] Lorsque pH = pKa on aura log[A-]/[AH] = 0 et donc [A-] = [AH] . Le processus de déprotonation est à moitié complet. Lorsque pH < pKa on aura log[A-]/[AH] < 0 et donc une prédominance de AH dans la solution. Le processus de déprotonation est très incomplet. Lorsque pH > pKa on aura log[A-]/[AH] > 0 et donc une prédominance de A- dans la solution. Le processus de déprotonation est très avancé. Plus le pKa d'un acide est faible, plus la gamme de pH dans laquelle il est susceptible de libérer des H+ est importante, et plus cet acide est fort. Plus un acide possède un Ka élevé, c'est-à-dire un pKa faible, plus cet acide est fort et plus sa base conjuguée est faible. Et inversement. • De très nombreuses réactions acido-basiques se produisent donc dans les liquides intra et extracellulaires. Mais un changement significatif de pH peut modifier certaines structures moléculaires (en détruisant les liaisons faibles comme les liaisons hydrogène ou les interactions électrostatiques), et déclencher des réactions dangereuses. Par exemple, une réaction acido-basique qui augmente le pH peut entraîner une dissociation des deux brins de la double hélice d'ADN. Des variations de pH peuvent également dénaturer de nombreuses protéines en modifiant leur structure tertiaire, et leur faire perdre leur fonctionnalité. C'est pourquoi il existe dans les systèmes biologiques des tampons qui modèrent les variations de pH dues à l'ajout d'un acide ou d'une base. Une solution tampon est composée d'un couple acide/base qui va consommer les ions H+ apportés par l'ajout d'un acide à la solution (ou au contraire libérer des ions H+ qui vont se fixer sur une base ajoutée à la solution) et permettre de maintenir le pH de la solution relativement constant. Les tampons sont plus efficaces lorsque le pH est proche de leur pKa. Ainsi, dans les systèmes biologique où le pH moyen est de 7,4, les solutions tampons formées à partir d'acide phosphorique seront particulièrement efficaces. [2] p 17 Dans le sang humain, on trouve également un tampon formé par le couple H2CO3/HCO3
- qui maintient le pH sanguin entre 7,35 et 7,45.
3) Le couplage de réactions
Nous avons vu que les voies métaboliques sont formées de nombreuses réactions qui, pour pouvoir se réaliser, doivent être thermodynamiquement favorables. Mais une réaction thermodynamiquement défavorable peut être rendue possible par couplage avec une réaction favorable. L'énergie libérée par une réaction exergonique permet la réalisation d’une réaction qui ne l’est pas, à condition qu’il existe un élément de couplage entre les deux réactions. (1) A + B C + D Ke1=3.10-4 (2) D + E F + G Ke2=1,3.105 La réaction 1 est non spontanée puisque Ke1<1 donc ΔrG° >0 dans les conditions standard. La réaction 2 est par contre spontanée. L’élément de couplage est la molécule D.

12
Il est alors possible d’écrire une réaction globale : (3) A + B + E C + F + G
Ke3=Ke1.Ke2=39>1 ΔrG°3= ΔrG°1 + ΔrG°2 =-RTln (Ke1.Ke2) <0 La réaction globale est donc exergonique, elle est spontanée. D’une façon générale :
ΔrG°=ΣΔrG°i K=K1.K2….
Exemple : Glucose + Pi Glucose 6P + H2O ΔG°1= +3,3kcal/mol (1) ATP + H2O ADP + Pi ΔG°2= -7,3kcal/mol (2) ATP + Glucose Glucose 6P + ADP ΔG°3= -4kcal/mol (3) L’énergie libérée par l’hydrolyse de l’ATP permet de réaliser la phosphorylation du glucose. Les transferts d’énergie par couplage sont très importants dans la cellule, les enzymes représentant des facteurs de couplage. En particulier, les réactions faisant intervenir l’ATP sont toujours couplées. Attention !! une réaction complète d’oxydoréduction n’est pas un couplage des deux demi-réactions (celles-ci ne peuvent se dérouler de façon isolée). Quelle est la place de ces réactions dans le métabolisme cellulaire?
II. STRUCTURE GENERALE DU METABOLISME ET ROLE CENTRAL DE L'ATP
A. Les différentes sources d'énergie utilisables par la cellule
Une cellule réalise de nombreux travaux : travail chimique, travail osmotique, travail mécanique, travail électrique, travail photochimique… Ces travaux font intervenir des réactions endergoniques variées et la cellule peut utiliser différentes sources d'énergie pour les réaliser.
1) L'énergie tirée de l'établissement d'un gradient
• Lorsqu'un soluté diffuse entre deux compartiments 1 et 2 séparés par une membrane perméable à ce soluté, son sens de diffusion est déterminé par les valeurs de son potentiel électrochimique de part et d'autre de la membrane.
Le potentiel électrochimique d'un soluté est défini par :
= o + RT ln (C) + ZFV
composante liée à la
concentration
composante liée au potentiel électrique
o = potentiel électrochimique dans les conditions standard, R=constante des gaz parfaits = 2 cal.mol
-1.K
-1 = 8,31 J. mol
-1.K
-1 ,
T = température absolue (en K), C = concentration de l'ion dans la solution (en mol.l
-1)
Z = valence de l’ion considéré, F = constante de Faraday = 2,3.10
4 cal.V
-1.mol
-1 = 9,65.10
4 J. V
-1.K
-1 ,
V = potentiel électrique de l'ion dans la solution (en V).

13
Et la différence de potentiel électrochimique du soluté entre les deux compartiments Δ= 2 - 1 correspond à la quantité d'énergie consommée ou libérée pour transférer une mole de ce soluté de part et d'autre de la membrane.
Δ étant directement proportionnel à ΔrG (= différence d'enthalpie libre entre les deux compartiments), si
Δ<0, ΔrG<0 et la diffusion du soluté se fait spontanément de 1 vers 2 en libérant de l'énergie. Un soluté va donc toujours diffuser dans le sens de son potentiel électrochimique décroissant.
Δ peut s'exprimer de la façon suivante :
• Si le soluté est une molécule non chargée, Z = 0 et Δ dépend uniquement du gradient de concentration. Le soluté va diffuser passivement du milieu ou il est le plus concentré vers celui où il est le moins concentré
en libérant une quantité d'énergie proportionnelle à Δ. 1 2 ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° °
gradient de concentration • Si le soluté est une particule chargée (un ion par exemple), et qu'il existe une ddp de part et d'autre de la membrane (ce qui est le cas pour la membrane plasmique, cette ddp étant le potentiel de repos), on aura Z≠0 et V2-V1≠0. Le sens de diffusion de l'ion dépendra à la fois du gradient de concentration et du gradient électrique. Par exemple, dans le cas d'un cation (ion+) plus concentré en 1 qu'en 2, et d'une membrane dont le potentiel de repos est tel que V1-V2<0: le gradient de concentration va avoir tendance à faire passer l'ion de 1 vers 2, mais le gradient électrique va avoir tendance à le ramener de 2 vers 1. Le flux net correspondra donc à la résultante de ces deux flux. Dans l'exemple considéré, la force osmotique étant supérieure (en valeur absolue) à la force électrique, l'ion diffuse passivement de 1 vers 2 en libérant de l'énergie. 1 2 ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° °
gradient de concentration gradient électrique • Un gradient de potentiel électrochimique peut donc être utilisé comme source d’énergie potentielle : gradient de H+ dans les cellules bactériennes et végétales, de Na+ dans les cellules animales…. Ces gradients sont par exemple utilisés dans le transport actif secondaire. De tels gradients peuvent être crées grâce à l'énergie fournie par l'hydrolyse de l'ATP .
Ex 1 : la pompe ATPasique Na+/K+ de la membrane plasmique maintient les gradients ioniques responsables du potentiel de repos.
)1V2V(ZF1C
2ClnRT 1-2
Force de diffusion due au gradient de concentration = force osmotique
Force de diffusion due au gradient électrique = force électrique
Force résultante
+
+
+
-
-
-
Gradient électrochimique
de Na+
Gradient électrochimique
de K+
+ + + + +
- - - - -
Potentiel transmembranaire
de repos ≈ -70mV
Diffusion
passive

14
Ex 2 : Un gradient de protons est à l'origine de nombreux transports actifs secondaires à travers le tonoplaste des cellule végétales.
Mais ils peuvent également être utilisés pour synthétiser de l'ATP (ex : utilisation d'un gradient de H+ par les ATPsynthases de la membrane interne des mitochondries et des thylakoïdes).
2) L'énergie tirée de réactions d'oxydoréduction
Dans les réactions d'oxydoréduction, les électrons passent spontanément de la forme la plus réductrice vers la forme la plus oxydante (c'est-à-dire dans le sens des potentiels rédox croissants) et cette réaction peut libérer
rG°’ = -n . F . E°’
Avec E°’= E°’2 – E°’1 = E°’ accepteur (+ fort ox) - E°’ donneur (+ fort red). Plus E°’1 est faible, plus la réaction est facilitée. Les molécules réduites comme NADH, H+ ou NADPH, H+ représentent donc une forme d'énergie chimique permettant les réactions de réduction (ce qui est le cas de beaucoup de réactions de synthèse de molécules organiques). On dit que ces coenzymes réduits représentent un pouvoir réducteur. L'oxydation de ces coenzymes réduits peut également être utilisée pour synthétiser de l'ATP.
3) L'énergie tirée du transfert de groupements
a) Le potentiel de transfert de groupement Soit la réaction d’hydrolyse : RX + H2O XH +ROH
Son rG°’ est le potentiel d’hydrolyse. Si rG°’ > 25 kJ, la liaison rompue est dite à haut potentiel ; si
rG°’ < 25 kJ, elle est dite à bas potentiel.
Une liaison à haut potentiel est dite riche en énergie et notée (l’expression est trompeuse car une liaison ne contient pas d’énergie mais c’est sa rupture qui en libère). La rupture de ces liaisons par hydrolyse va permettre, par couplage avec une réaction endergonique, de transférer un groupement (ici R) sur une autre molécule (A).
RX + H2O XH +ROH rG°’<< 0
A + R AR rG°’>0

15
RX + A AR + X rG°’< 0 On dit que ces molécules (à haut potentiel d'hydrolyse) ont un haut potentiel de transfert de groupement. Les composés à haut potentiel d’hydrolyse qui vont pouvoir effectuer ce transfert de groupement sont des thiosters (ex : AcétylCoA) et des composés phosphorylés (ex : ATP).
Les potentiels d'hydrolyse de quelques molécules biologiques. [2] p 416
b) Le transfert de groupements acyles • Le coenzyme A est dérivé d’une adénosine 3’, 5’ triphosphate (un groupement sur le 3’ et 2 groupements sur le 5’). Le coenzyme A comporte aussi un dérivé d’acide pantothénique (vitamine B). Ce dérivé d’acide pantothénique se termine par un groupement thiol (SH) correspondant à la partie réactive : il peut fixer des radicaux acyle, en particulier acétyl (CH3)CO. Le coenzyme A et l'acétylcoenzyme A [3] p 103

16
Les groupements acyles sont d'importants constituants des molécules organiques (acides gras par exemple),
et on les retrouve dans de nombreuses réactions du métabolisme (ex : synthèse des lipides, oxydation des acides gras…). • Le coenzyme A est un coenzyme de transfert de groupement acyle (CH3)-(CH2)nCO, et plus particulièrement du groupement acétyl.
En effet, le G°’ de l'hydrolyse de l'AcétylCoA a une valeur largement négative.
AcétylCoA + H2O acétate (CH3COO-) + CoA + H+ G°’ = - 31,4 kJ/mol = -7,5 kcal/mol. On dit que l'AcétylCoA a un haut potentiel de transfert de groupement acétyle: il transporte un groupe acétyle activé qui va pouvoir être transféré sur une autre molécule par couplage de son hydrolyse avec une autre réaction.
c) Le transfert de groupements phosphoryles De nombreuses molécules phosphorylées ont un potentiel de transfert de groupement phosphoryle (par hydrolyse) plus ou moins important. L'ATP a une position intermédiaire entre : - les composé à haut potentiel qui sont formés à partir de la dégradation des molécules alimentaires et qui vont pouvoir phosphoryler l'ADP : PEP, créatine-P, 1,3DPG. - les composés à bas potentiel (contenant des liaisons peptidiques ou des liaisons osidiques) et qui peuvent être phosphorylés par l'ATP : glucose 6P, fructose 1,3biP (à peu près au même niveau que glucose 1P). Exemple : L'ATP a un rôle de plaque tournante dans le métabolisme cellulaire.
17
B. L'ATP, unité énergétique universelle des systèmes biologiques
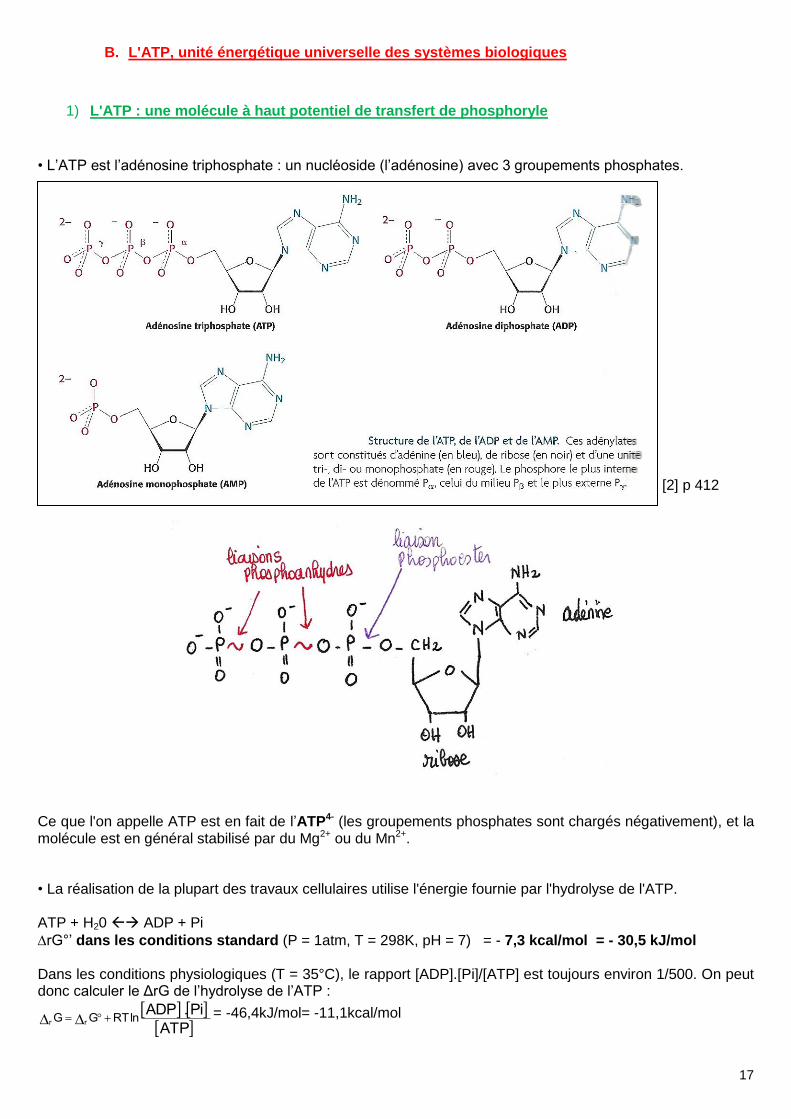
1) L'ATP : une molécule à haut potentiel de transfert de phosphoryle
• L’ATP est l’adénosine triphosphate : un nucléoside (l’adénosine) avec 3 groupements phosphates. [2] p 412 Ce que l'on appelle ATP est en fait de l’ATP4- (les groupements phosphates sont chargés négativement), et la molécule est en général stabilisé par du Mg2+ ou du Mn2+. • La réalisation de la plupart des travaux cellulaires utilise l'énergie fournie par l'hydrolyse de l'ATP. ATP + H20 ADP + Pi
rG°’ dans les conditions standard (P = 1atm, T = 298K, pH = 7) = - 7,3 kcal/mol = - 30,5 kJ/mol Dans les conditions physiologiques (T = 35°C), le rapport [ADP].[Pi]/[ATP] est toujours environ 1/500. On peut donc calculer le ΔrG de l’hydrolyse de l’ATP :
ATP
Pi.ADPlnRTGG rr = -46,4kJ/mol= -11,1kcal/mol

18
• En fait, ce n’est pas la réaction d'hydrolyse de l’ATP qui est réalisée mais un transfert de groupement phosphoryle au cours de réactions couplées. Le groupement phosphoryle de l'ATP est transféré sur la molécule de H2O. Il vaut donc mieux parler de « potentiel de transfert de groupement phosphoryle » plutôt que de « potentiel d’hydrolyse ». Ce potentiel de transfert de groupement phosphoryle est plus élevé que celui
d'autres molécules phosphorylées (ex :Glycérol 3P, rG°’ = -9,2 kJ/mol). Inversement, le rG°’ de synthèse de l’ATP n’est pas trop haut (comparé au PEP par exemple). Il n’est donc thermodynamiquement pas si coûteux à former.
2) L'origine structurale de ce haut potentiel de transfert de phosphoryle
• Le fort potentiel de transfert de groupement phosphoryle de l’ATP provient de différences structurales entre l'ATP et ses produits d'hydrolyse. → Le groupement triphosphate de l'ATP est très instable notamment en raison de l’accumulation des charges négatives et des forces de répulsion électrostatiques ainsi générées. La densité des électrons autour de la liaison riche en énergie est anormalement élevée, ce qui s'oppose à la liaison covalente, d’où la tendance à transférer le groupement phosphoryle sur une molécule où la liaison sera plus stable.
O O O
Adénosine - O - P - O P - O P - O- répulsions électrostatiques
O- O- O- La répulsion électrostatique entre groupements chargés négativement est moins importante dans le cas de l’ADP où le nombre de charges négatives est moindre. → L’ATP est moins stabilisée par les phénomènes de résonance que le Pi ou l’ADP. L'acide phosphorique a de nombreuses formes mésomères entre lesquelles la molécule est en résonance, ce qui la stabilise.
Le groupe phosphoryle de l'ATP par contre possède un plus petit nombre de formes mésomères. → L’ATP est moins stabilisée par hydratation que l'ADP ou le Pi car l'eau se fixe plus efficacement à l'ADP et au Pi qu'à la partie phospho-anhydre de l'ATP.
Ces trois propriétés font que la molécule d’ATP4- est métastable. L’ADP3- et l’AMP2- sont des molécules plus stables. L'ATP ne peut donc pas être mis en réserve et, par conséquent, il doit être renouvelé en permanence. • D'autre part, le réaction d’hydrolyse de l'ATP en ADP + Pi , qui est spontanée et très exergonique, se déroule à une vitesse faible en solution aqueuse. Elle doit donc être accélérée par la présence d’une enzyme, ce qui facilite le transfert du groupement phosphoryle de l'ATP à une autre molécule par couplage (les enzymes étant des facteurs de couplage), et permet également une régulation de ces réactions couplées (via la régulation de l'activité enzymatique).

19
Ces deux propriétés : métastabilité (l’ATP cède facilement un groupement phosphoryle) et faible vitesse d’hydrolyse (libération d’énergie nécessitant une enzyme) font de l'ATP un très bon vecteur d'énergie dans la cellule.
C. Les différentes modalités de synthèse d’ATP dans la cellule
1) La respiration cellulaire convertit l’énergie potentielle du glucose en ATP
• L’équation générale de la respiration cellulaire est : C6H12O6 + 6O2 6 CO2 + 6 H2O + énergie Il s’agit globalement d’une réaction d’oxydoréduction : la matière organique (glucose) est oxydée en CO2 et O2 (accepteur final des électrons) est réduit en H2O. Mais la respiration cellulaire est en fait constituée de nombreuses réactions intermédiaires, avec deux grandes phases : la glycolyse et les oxydations mitochondriales. • La synthèse d’ATP dans la respiration cellulaire se fait selon 2 modalités : - par couplage de réactions, dans la glycolyse, ou le cycle de Krebs : c’est la phosphorylation par couplage ou phosphorylation liée au substrat. Des composés à plus fort potentiel cèdent leur groupement phosphate à l’ADP. - en utilisant l’énergie libérée par des réactions d’oxydoréductions (réactions impliquant NAD ou FAD et le dioxygène), on parle de phosphorylation oxydative dans la mitochondrie. En effet, la réoxydation de coenzymes réduits peut libérer de l’énergie :
NADH + H+ + 1/2 O2 NAD+ + H2O rG°’ = -2 x 96,1. [(+0,82)-(-0,32)] = -219 kJ/mol
Ce rG°’ peut être comparé à celui de l'hydrolyse de l’ATP = -30,5 kJ.mol-1 . La synthèse de plusieurs molécules d’ATP peut être réalisée grâce à l’oxydation d’une seule molécule de NAD réduit. La réoxydation des coenzymes permet la synthèse d’ATP.
20
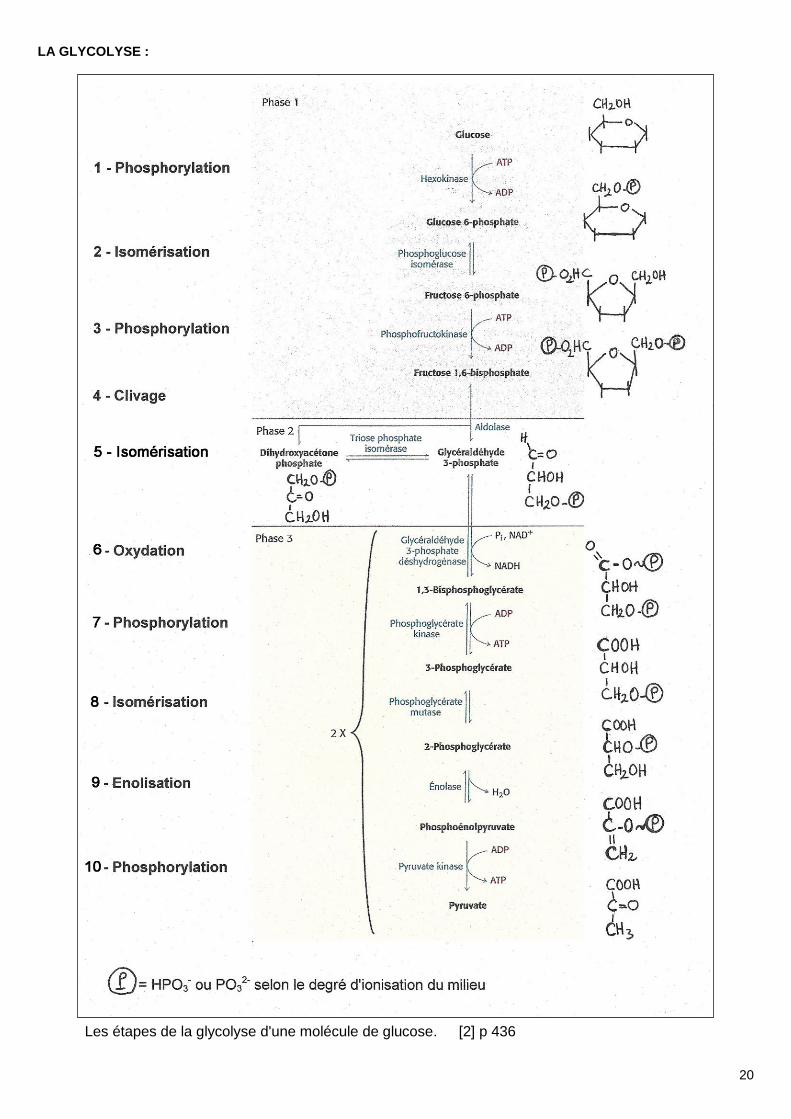
LA GLYCOLYSE :
Les étapes de la glycolyse d'une molécule de glucose. [2] p 436
21
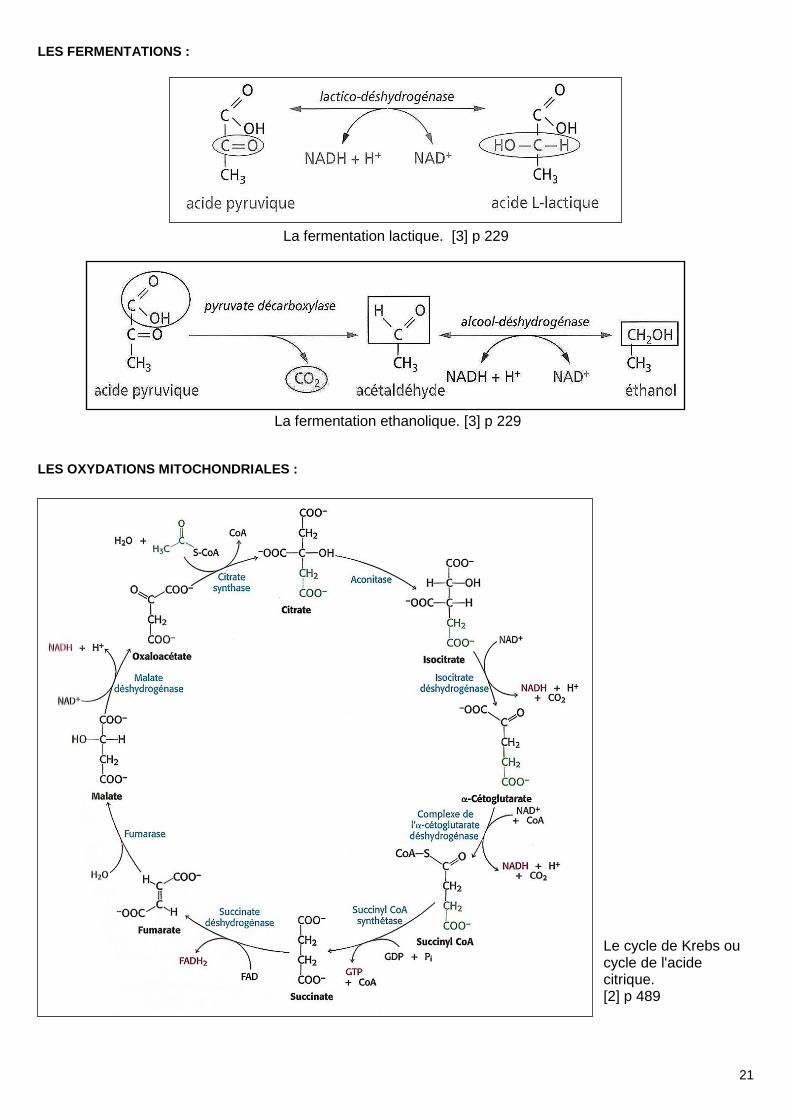
LES FERMENTATIONS :
La fermentation lactique. [3] p 229
La fermentation ethanolique. [3] p 229
LES OXYDATIONS MITOCHONDRIALES :
Le cycle de Krebs ou cycle de l'acide citrique. [2] p 489
22
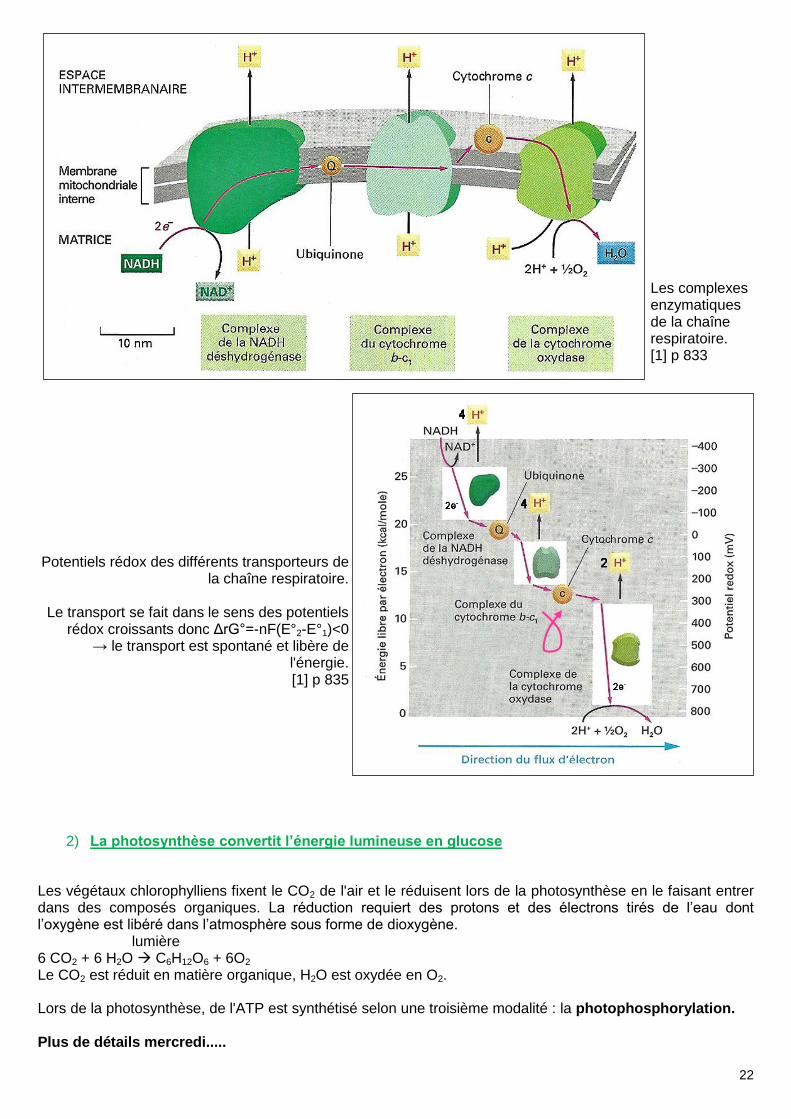
Les complexes enzymatiques de la chaîne respiratoire. [1] p 833
Potentiels rédox des différents transporteurs de la chaîne respiratoire.
Le transport se fait dans le sens des potentiels
rédox croissants donc ΔrG°=-nF(E°2-E°1)<0 → le transport est spontané et libère de
l'énergie. [1] p 835
2) La photosynthèse convertit l’énergie lumineuse en glucose
Les végétaux chlorophylliens fixent le CO2 de l'air et le réduisent lors de la photosynthèse en le faisant entrer dans des composés organiques. La réduction requiert des protons et des électrons tirés de l’eau dont l’oxygène est libéré dans l’atmosphère sous forme de dioxygène.
lumière 6 CO2 + 6 H2O C6H12O6 + 6O2 Le CO2 est réduit en matière organique, H2O est oxydée en O2. Lors de la photosynthèse, de l'ATP est synthétisé selon une troisième modalité : la photophosphorylation. Plus de détails mercredi.....

23
Conclusion : Le monde biologique réalise donc des conversions d'énergie qui s’accompagnent d’une dégradation constante de cette énergie. Soit une certaine quantité d’énergie rayonnante en provenance du Soleil (seule source d’énergie pour les êtres vivants sur la Terre) :
E h (haute énergie, c’est-à-dire susceptible d’être utilisée par les cellules pour effectuer un travail : synthèse d’ATP lors
de la phase photochimique de la photosynthèse) E chimique potentielle (moyenne E pouvant être libérée pour être utilisée par les des molécules organiques cellules, ex : synthèse d’ATP lors de la glycolyse)
E calorique (basse E, destin inéluctable de l’énergie solaire à la sortie du monde vivant : production de chaleur lors de la respiration cellulaire) A la fin de son passage à travers le monde vivant, l’énergie est devenue inutilisable : les êtres vivants « restituent » l’énergie sous forme de chaleur et contribuent ainsi à l'augmentation de l'entropie de l'Univers. Les cellules sont donc des systèmes thermodynamiques ouverts en état d’équilibre dynamique qui réalisent l'ensemble de ces conversions par le biais des réactions du métabolisme cellulaire. Ces réactions métaboliques possèdent de nombreux points communs qui trouvent leur origine dans un héritage commun partagé au cours de l'évolution. [2] p 429 Ainsi des transporteurs activés tels qu'ATP, NADH, H+, FADH2, CoA auraient évolué à partir des RNA ancestraux. Ces processus métaboliques sont contrôlés par trois mécanismes principaux : le contrôle de l'accessibilité des substrats (augmenté par la compartimentation chez les Eucaryotes), le contrôle des quantités d'enzymes et le contrôle de leur activité catalytique.
BIBLIOGRAPHIE [1] : B.ALBERTS ET AL; 2011; Biologie moléculaire de la cellule, 5è édition; Médecine-Sciences, Lavoisier. [2] : L.STRYER ET AL; 2008; Biologie moléculaire de la cellule, 6è édition; Médecine-Sciences, Flammarion. [3] : J.H. WEIL ET AL; 2001; Biochimie générale, 9è édition; Dunod.