immunité adaptative. programme 1a. antigènes –définition –antigénicité et immunogénicité...
TRANSCRIPT

Immunité adaptative

Programme
• 1a. Antigènes– Définition– Antigénicité et immunogénicité– Structure moléculaire des antigènes– Propriétés qui influencent l’immunogénicité des
antigènes• Liées à l’antigène lui-même• Liées à l’hôte• Liées aux « circonstances » de la rencontre
– Les adjuvants

Programme
• 1b. Epitopes– Définition– Epitopes B et épitopes T– Epitopes séquentiels et non séquentiels– Interaction des épitopes et des CDR– Compétition des épitopes
• Épitopes dominants et épitopes cryptiques
– Epitopes T et agrétopes
• 1c. Haptènes– Définition et exemples– Implications cliniques

Programme
• 2a. Organisation des gènes d’immunoglobulines– Introduction sur la structure des immunoglobulines
• Chaînes lourdes et chaînes légères
• Classes et sous-classes

Programme
• 2b. Génération de la diversité– Tonegawa et l’illustration de variations somatiques du
DNA des cellules de l’immunité adaptative
– Concept de familles multigéniques
– Choix de segments géniques dans chaque famille : rôle des recombinases
– Implication clinique : les scid
– Encore plus de diversité : diversité jonctionnelle et TdT

Programme
• 2b. Génération de la diversité– Réarrangements productifs et non productifs– Exclusion allélique– Encore plus de diversité : l’hypermutation
somatique : rôle de l’AID
• 2c. Commutation isotypique– Mécanismes de la commutation isotypique– Epissage alternatif et sécrétion des Ig

Programme
• Epitopes– Définition
– Epitopes B et épitopes T
– Epitopes séquentiels et non séquentiels
– Compétition des épitopes• Épitopes dominants et épitopes cryptiques
• Organisation des gènes d’immunoglobulines• Génération de la diversité

Antigènes : molécules reconnues par des immunorécepteurs (BCR,
immunoglobulines, TCR)

Antigène
• Concept minimaliste, qualitatif
• Une substance sera dite antigénique si au moins dans certaines conditions, et au moins chez certains sujets, elle est capable d’induire une réponse en se liant de façon spécifique aux immunorécepteurs (immunoglobulines ou TCR)

Antigène
• Donc on peut parler de l’injection d’un antigène qui pourtant ne provoquera dans les conditions et chez l’individu considérés aucune réponse immunitaire

Immunogénicité
• Définition plus quantitative, plus circonstancielle
• Pouvoir d’un antigène à induire une réponse immunitaire chez un individu donné et dans des conditions données
Certains antigènes sont très immunogènes, d’autres le sont peu

Antigènes
• Protéines
• Polysaccharides
• Acides nucléiques
• Lipides et glycolipides

Propriétés des antigènes qui influencent leur immunogénicité

Le degré d’altérité
• Plus un antigène provient d’un organisme distant dans l’évolution, plus il sera immunogène

Le poids moléculaire
• Poids moléculaire idéal : 100 kD
• Les protéines de poids moléculaire < 5 kD sont parfois antigéniques mais généralement peu immunogènes

Capacité à être apprêté et présenté par une molécule du
CMH
• phagocytose par une cellule présentatrice d’antigène– Grosses protéines insolubles, agrégats
• affinité pour une molécule du CMH– Les molécules CMH ne sont pas les mêmes
chez tous les individus (chez toutes les espèces)– La nature des molécules CMH présentes
détermine l’immunogénicité de l’antigène considéré

Facteurs génétiques
• L’immunogénicité d’un antigène n’est pas la même chez tous les individus d’une même espèce– Molécules du CMH différentes chez tous les
individus– Autres facteurs

Concentration de l’antigène (dosage)
• Une concentration optimale est requise pour une bonne immunogénicité
• Si l’antigène est trop ou trop peu concentré, l’immunogénicité sera mauvaise. Une tolérance peut même s’installer

Voie d’entrée (ou d’administration)
• Selon la porte d’entrée de l’antigène, il interagira avec différentes cellules présentatrices d’antigènes et induira des réponses immunitaires d’intensité différentes
• La voie orale est généralement (mais pas toujours) peu immunogène voire génératrice de tolérance
• La voie la plus efficace : sous-cutanée ou intramusculaire

Adjuvants
• Substances utilisées dans les vaccins et qui permettent d’augmenter l’immunogénicité des antigènes utilisés

Les adjuvants
• Deux types d’adjuvants avec des fonctions différentes– Favoriser et prolonger la durée de l’interaction entre
l’antigène et le système immunitaire• Ex. hydroxyde d’alumine
– Recruter et activer des cellules de l’immunité n naturelle pour qu’elles induisent la réponse adaptative
• Les PAMP sont souvent de bons adjuvants
• Certains molécules sont à la fois des antigènes et des PAMP

Epitope ou déterminant antigénique
• Le terme d’antigène désigne généralement la molécule antigénique dans son ensemble même si seule une partie de celle-ci est reconnue par l’immunorécepteur
• On réserve le terme d’épitope (= déterminant antigénique) à la partie de l’antigène qui interagit réellement avec l’immunorécepteur
• Un même antigène peut avoir de multiples épitopes
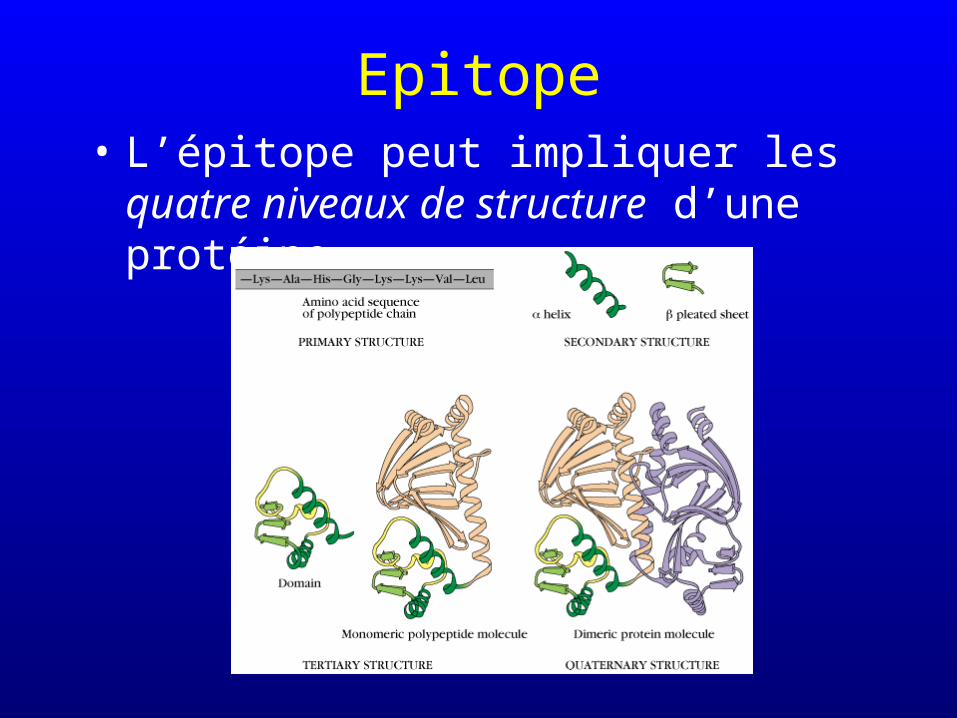
Epitope• L’épitope peut impliquer les quatre niveaux
de structure d’une protéine

Epitope T / épitope B• Les épitopes reconnus par les lymphocytes
T sont distincts de ceux reconnus par les lymphocytes B

Epitopes B
• L’anticorps se lie à l’épitope par des interactions non covalentes, de faible affinité– Proximité directe des structures moléculaires
qui interagissent– La taille de l’épitope B ne peut être supérieure à
celle du site de liaison sur l’anticorps
Notion de complémentarité : la structure du site de liaison de l’anticorps est complémentaire de celle de l’épitope considéré
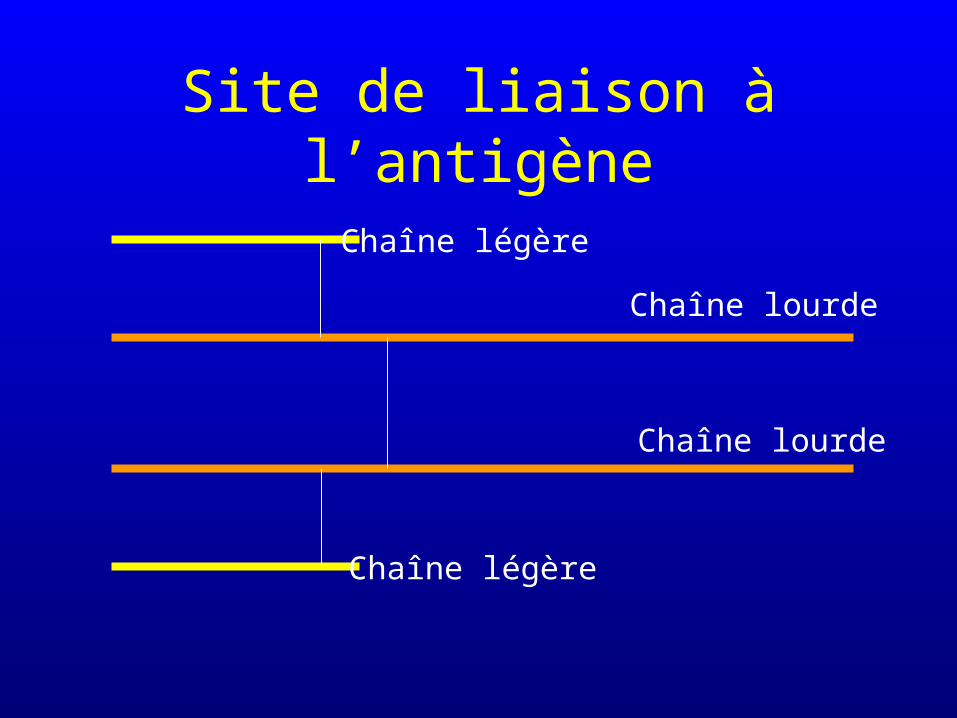
Site de liaison à l’antigène
Chaîne légère
Chaîne légère
Chaîne lourde
Chaîne lourde
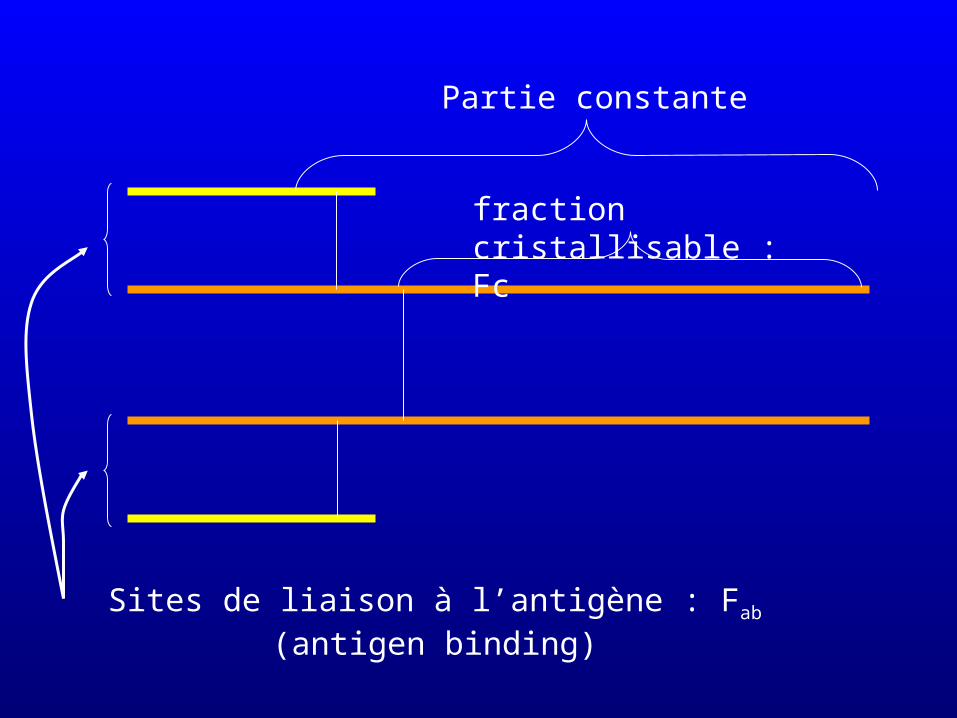
Partie constante
Sites de liaison à l’antigène : Fab (antigen binding)
fraction cristallisable : Fc

Deux sites de liaison par immunoglobuline (Fab2)
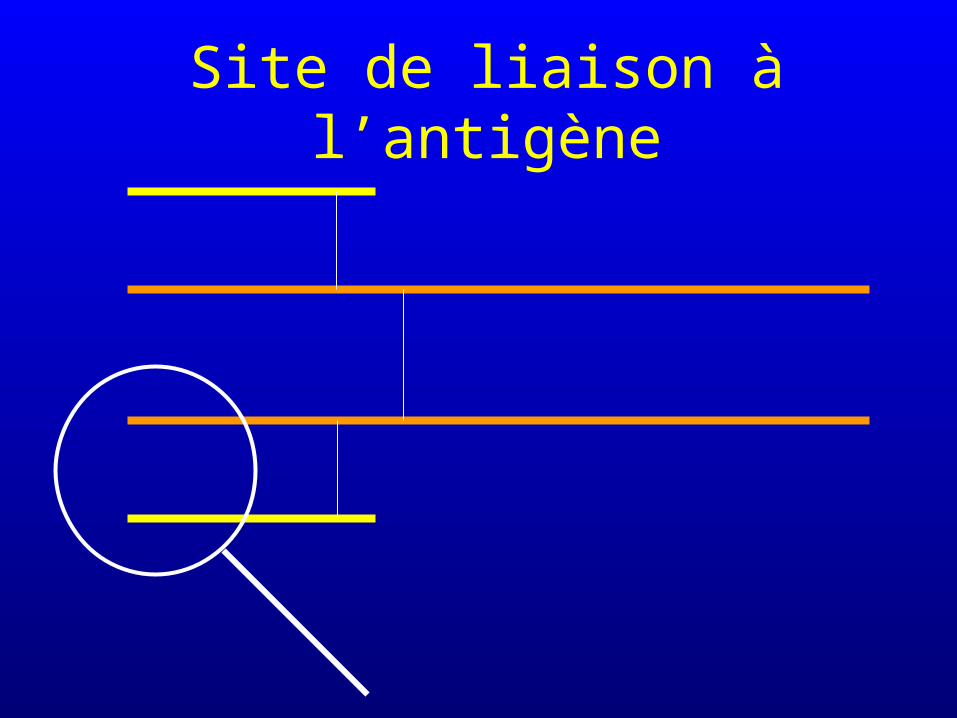
Site de liaison à l’antigène

Sites de liaison à l’antigène

Sites de liaison sur l’anticorps
• Deux types de site de liaison– « poche » dans le Fab
• petites molécules de structure compacte (peptides, oligonucléotides, sucres, autres agents chimiques : haptènes)
– surface du Fab dans son ensemble• grosses protéines

Les épitopes B doivent être directement accessibles à la
surface de la protéine
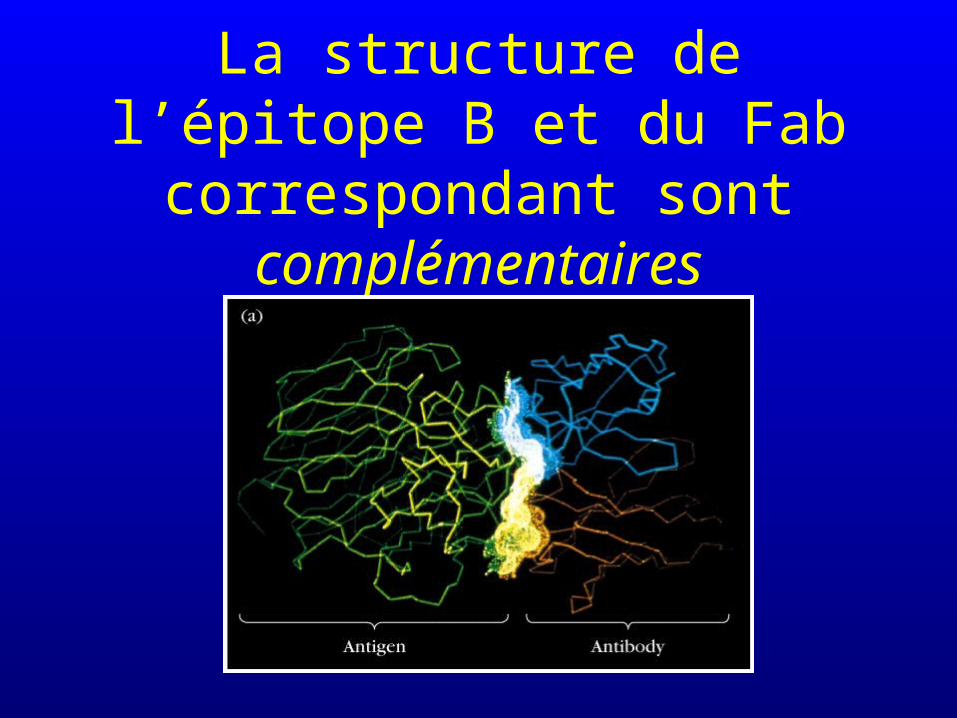
La structure de l’épitope B et du Fab correspondant sont
complémentaires
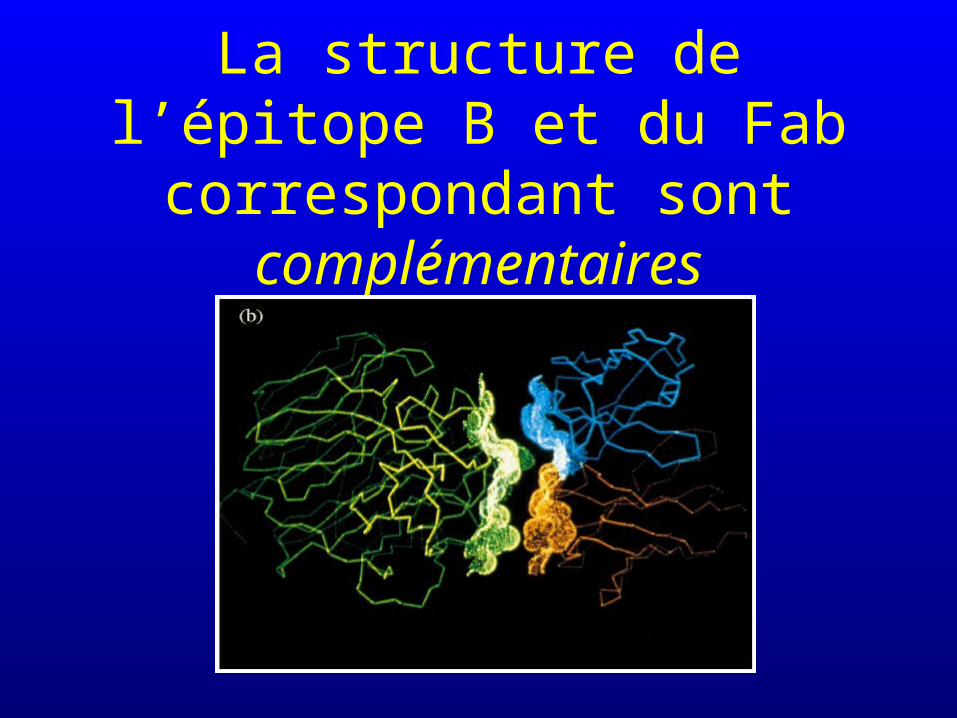
La structure de l’épitope B et du Fab correspondant sont
complémentaires

Les CDR (complementarity determining regions)
• Il existe dans la partie variable des chaînes légères et des chaînes lourdes des régions particulièrement impliquées dans la liaison à l’antigène, dans la complémentarité à la structure de celui-ci : ce sont les régions déterminant la complémentarité ou CDR (CDR1, CDR2, CDR3)

Epitopes séquentiels et non séquentiels
• Epitopes séquentiels : reconnaissance d’une séquence d’acides aminés (qui se suivent sur la protéine), indépendemment de la structure tertiaire du segment considéré– les anticorps dirigés contre des épitopes
séquentiels les reconnaîtront même si la protéine native est dénaturée
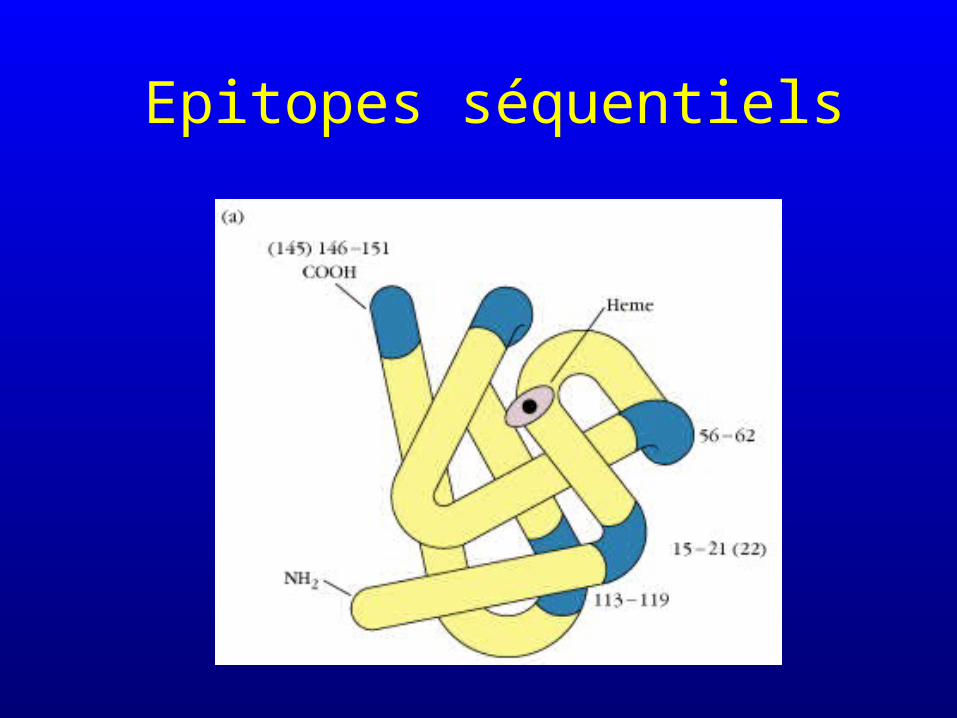
Epitopes séquentiels

Epitopes non séquentiels
• Reconnaissance d’acides aminés non contigus dans la structure primaire mais que la structure tertiaire de la protéine fait entrer en interaction étroite avec le site Fab de l’anticorps : on parle aussi d’épitopes conformationnels– les anticorps dirigés contre des épitopes non
séquentiels (conformationnels) ne reconnaissent plus ces derniers si la protéine est dénaturée

Epitopes non séquentiels
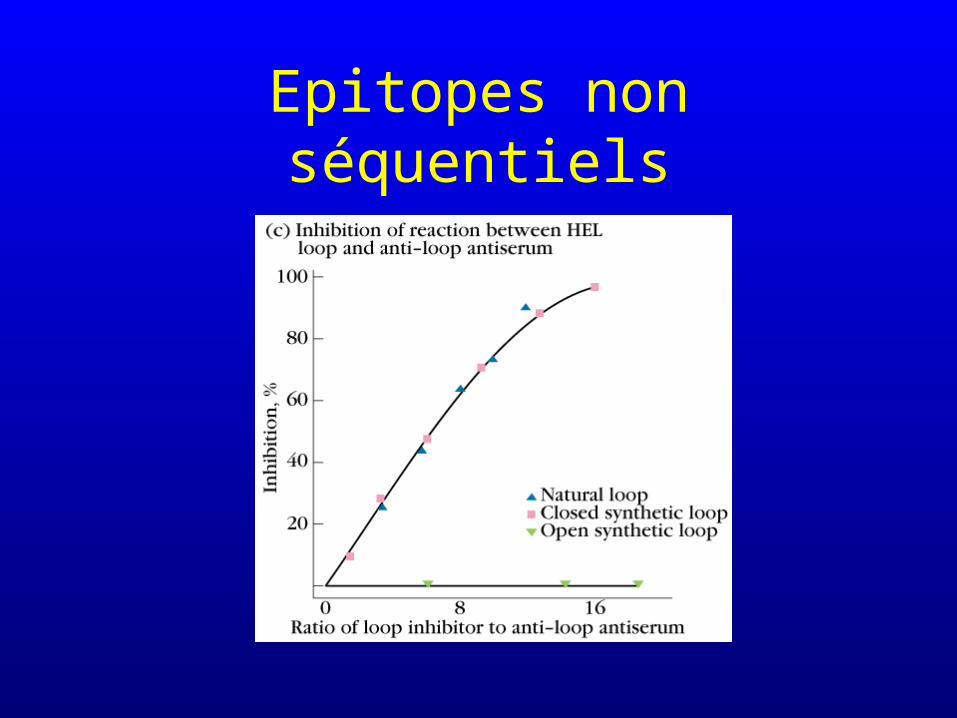
Epitopes non séquentiels

Multiplicité des épitopes et immunodominance
• les protéines possèdent généralement de nombreux épitopes potentiels dont certains peuvent se chevaucher ou entrer en compétition lors d’une réponse immunitaire
• un individu (ou un animal donné) ne développent cependant pas d’anticorps contre tous ces épitopes potentiels : il y a dominance de certains épitopes par rapport à d’autres

Multiples mécanismes de l’immunodominance
• Compétition entre différents épitopes lors des différentes étapes de l’induction d’une réponse immunitaire
• En vaccinologie, les épitopes les plus « intéressants » ne sont pas nécessairement immunodominants

Epitopes T
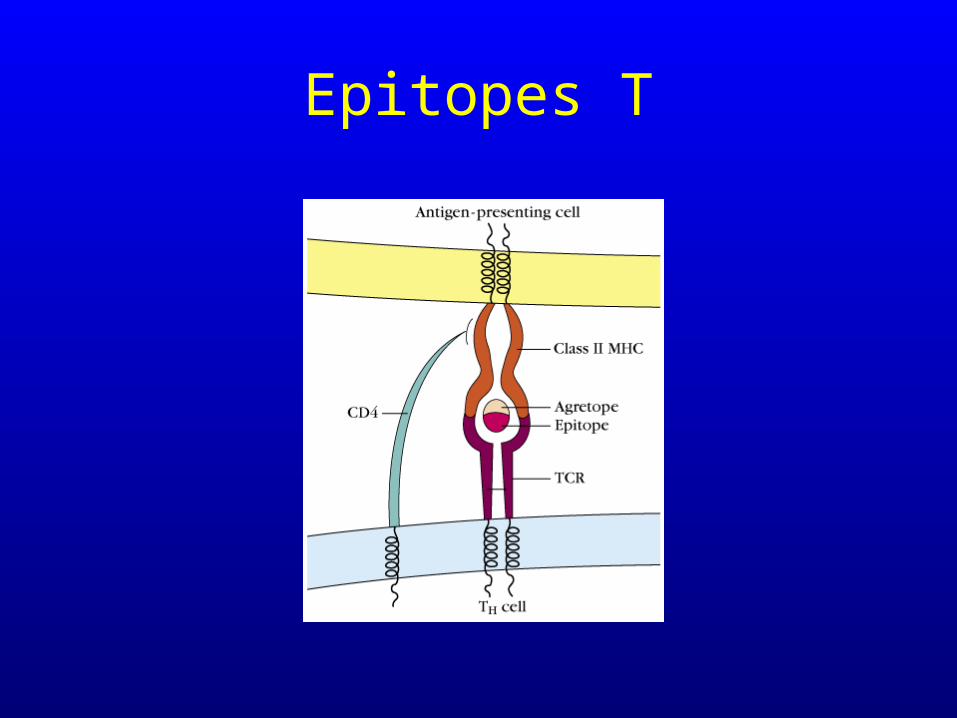
Epitopes T

Epitope et agrétope
• les antigènes reconnus par les lymphocytes doivent avoir un site de liaison pour le TCR (épitope) et un site de liaison pour la molécule CMH (agrétope)

Epitopes T
• Epitopes présentés par molécules CMH I : séquences de neuf acides aminés (nonamères)
• Epitopes présentés par molécules CMH II : séquences de 11 à 25 acides aminés
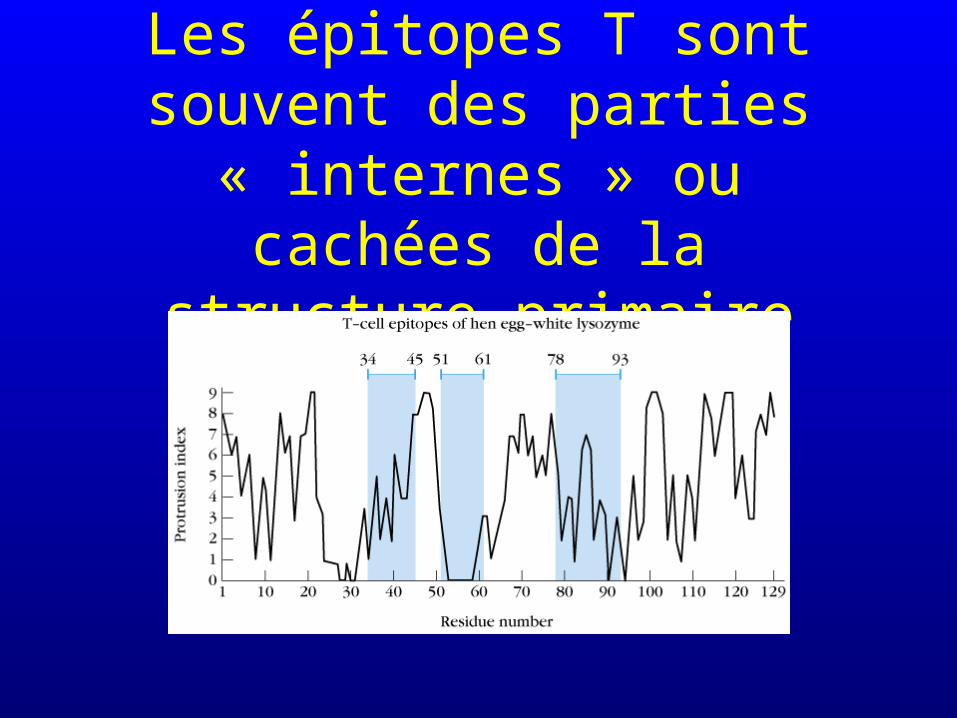
Les épitopes T sont souvent des parties « internes » ou cachées de
la structure primaire

Pas d’épitopes non séquentiels pour les épitopes T!

Agrétope

Agrétope
• la liaison de l’antigène à la molécule CMH (via l’agrétope) est sélective mais n’a pas du tout la spécificité fine de la liaison de l’épitope sur le TCR– une même molécule du CMH peut fixer un
grand nombre de peptides différents

Epitope et agrétope
• Tous les acides aminés du peptide participent à la constitution de l’épitope, par contre seuls une partie de ceux-ci contribuent à la liaison à la molécule CMH, à l’agrétope

Dominance et CMH
• une même protéine comporte de multiples épitopes T potentiels
• seuls les peptides capables de se lier aux molécules CMH de l’individu ou de l’animal considéré pourront provoquer une réponse immunitaire T
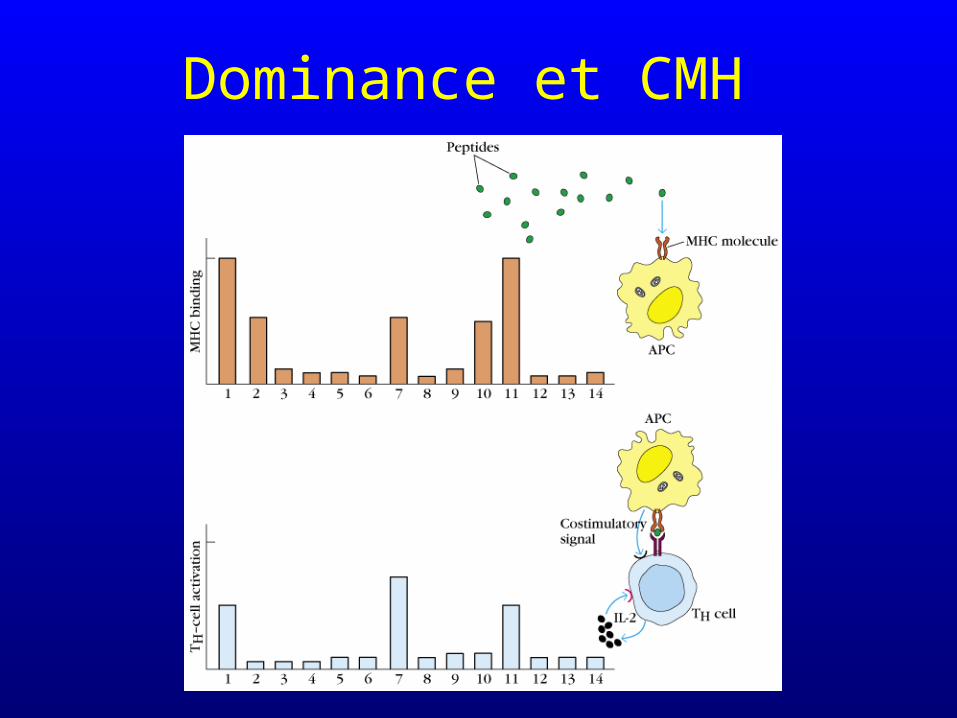
Dominance et CMH

Haptènes et porteurs
• Certaines molécules de faible poids moléculaire peuvent être reconnues par des anticorps mais ne peuvent pourtant induire la synthèse d’anticorps spécifiques
Ce sont des haptènes (haptein : nouer)

Haptènes et porteurs
• Pour qu’une réponse soit induite, il faut que l’haptène soit lié à une grosse protéine porteuse

Haptènes et porteurs

Exemples de haptènes
• Hormones stéroïdiennes– Dosages d’hormones par anticorps
• Médicaments– Base des allergies médicamenteuses
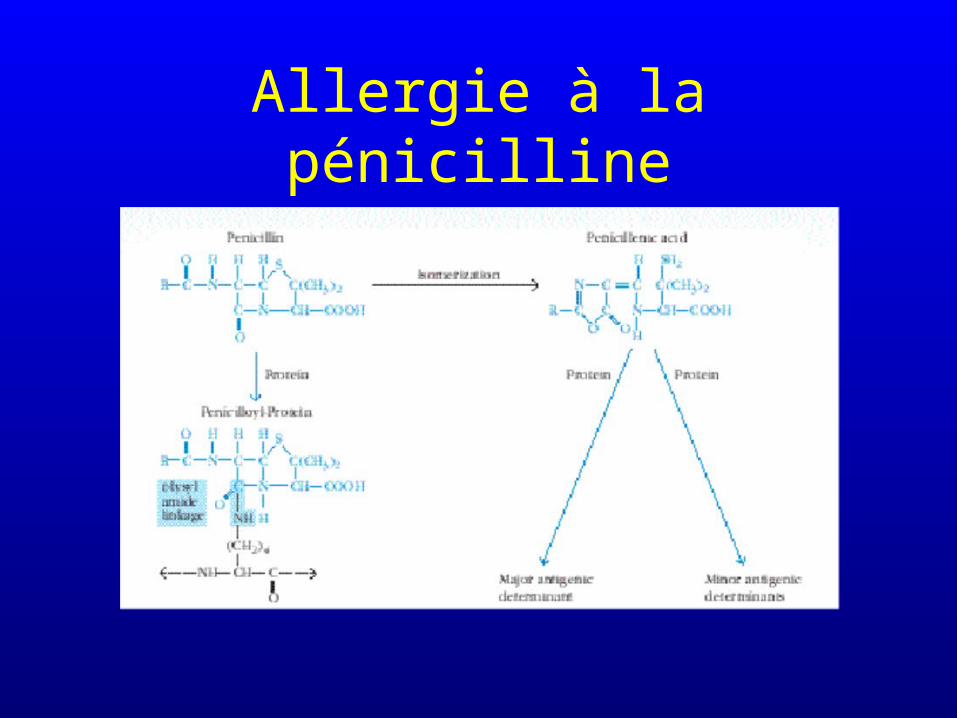
Allergie à la pénicilline

Allergies médicamenteuses
• Les médicaments qui forment des conjugués covalents avec les protéines ont un risque important de générer des allergies
• Les médicaments qui ne forment pas de conjugués covalents avec les protéines ont un risque faible de générer des allergies

Organisation et expression des gènes d’immunoglobulines

Deux chaînes lourdes et deux chaînes légères
Chaîne légère : deux types possibles (l’un ou l’autre) : ou

Au sein d’une même immunoglobuline, les deux chaînes lourdes et les deux
chaînes légères sont identiques

Deux types de chaînes légères : kappa ou lambda
Une immunoglobuline donnée utilise l’un ou l’autre type de chaîne légère

Tant les chaînes légères (qu’elles soient ou ) que les chaînes
lourdes ont une portion variable et une portion constante

La portion constante des chaînes lourdes est essentielle pour déterminer les propriétés
fonctionnelles de l’immunoglobuline
Cette portion constante des chaînes lourdes détermine l’isotype de
l’immunoglobuline
• IgM, IgD, IgG, IgE, IgA• Propriétés différentes de liaison aux récepteurs
FcR, au C1q etc.

Organisation et expression des gènes d’immunoglobulines
• Le système immunitaire des mammifères génère des anticorps de plus de 1010
spécificités différentes
• Si chacun de ces anticorps était codé par un gène particulier, 15% du génome seraient consacrés à cette seule fin

Variations somatiques
• Pour les lymphocytes (et seulement pour eux) :– La séquence du DNA évolue et se diversifie
considérablement au fil de la différenciation lymphocytaire
– Ce sont des variations somatiques, par opposition à la séquence initiale présente sur les cellules souches, dite séquence germinale (la même que sur toutes les autres cellules de l’organisme)

Tonegawa : prix Nobel 1987
• Les fractions constantes et variables d’une même protéine (chaîne légère d’une immunoglobuline) sont codées par des gènes qui sont initialement distincts
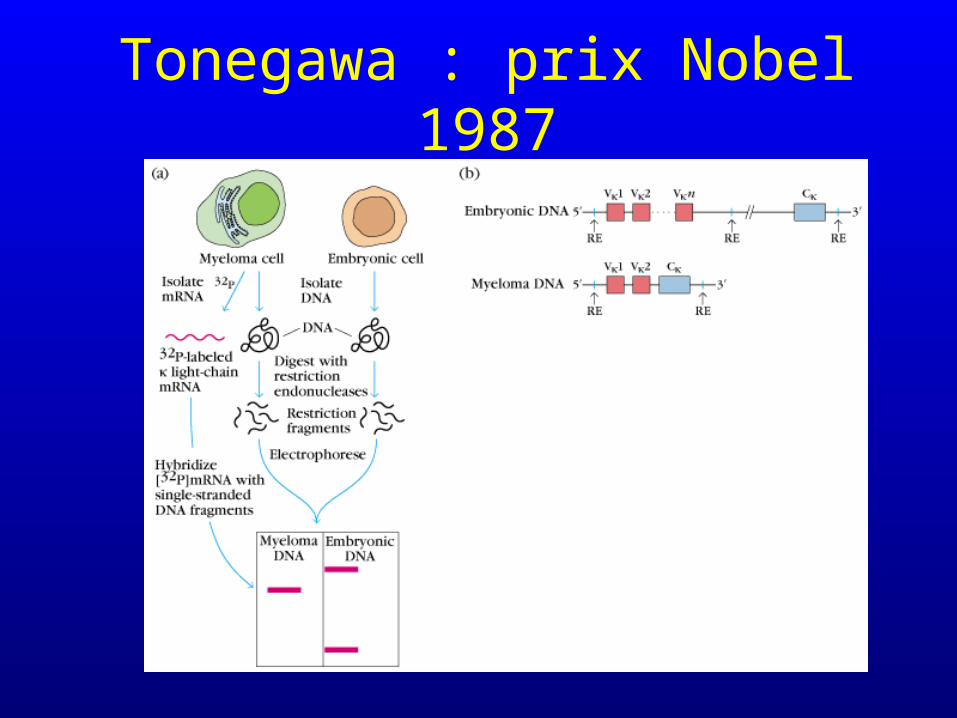
Tonegawa : prix Nobel 1987

Concepts de familles multigènes
• Existence de familles qui regroupent des « segments géniques »
• Au sein de chaque famille, un seul segment est aléatoirement choisi
• La juxtaposition de tous les segments choisis forme le gène à partir duquel le RNA de l’immunoglobuline considérée sera transcrit

Concept de familles « multigènes »
• Pour les chaînes légères de type – Deux familles responsables de la portion
variable• Famille V (V1, V2, …, V30) et famille J (J1,
J2, J3 et J4)
– Une famille responsable de la portion constante• Famille C (C1, C2, C3, et C4)
Seuls un segment V, un segment J et un segment Cseront
conservés dans la cellule mûre qui exprime une chaîne légère de type

Concept de familles « multigènes »
• Pour les chaînes légères de type – Deux familles responsables de la portion variable
• Famille V (V1,V2, …, V40) et famille J (J1, J2, J3, J4 et J5))
– Un gène responsable de la portion constante : C
Seuls un segment V, un segment J et le segment Cseront
conservés dans la cellule mûre qui exprime la chaîne légère de type
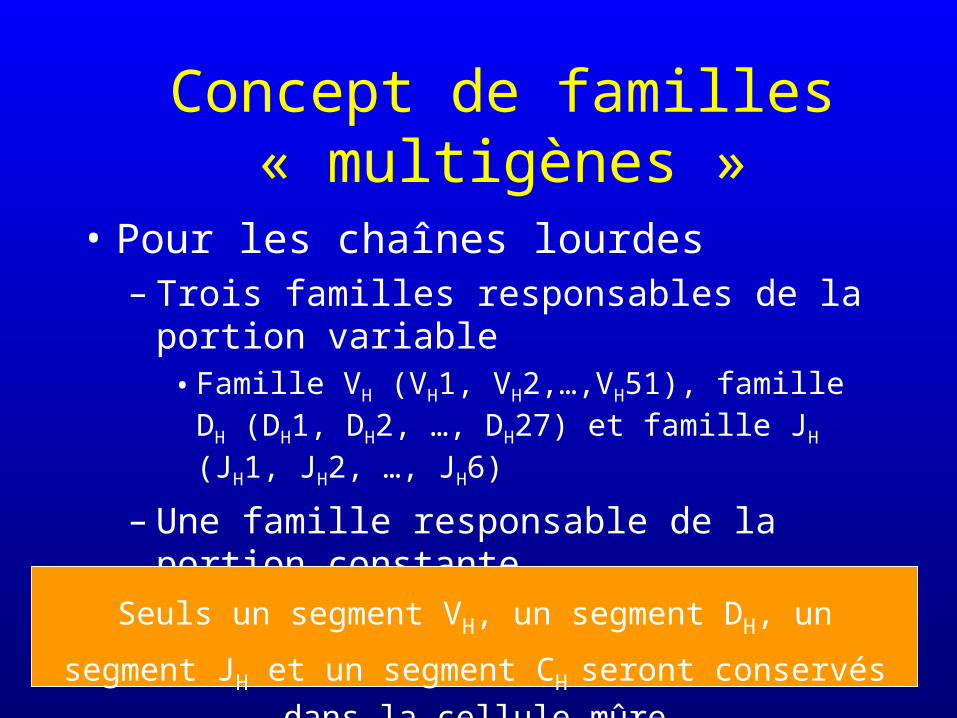
Concept de familles « multigènes »
• Pour les chaînes lourdes– Trois familles responsables de la portion
variable• Famille VH (VH1, VH2,…,VH51), famille DH (DH1,
DH2, …, DH27) et famille JH (JH1, JH2, …, JH6)
– Une famille responsable de la portion constante• Famille C (C,C, C, C et C)
Seuls un segment VH, un segment DH, un segment JH et un
segment CHseront conservés dans la cellule mûre

Réarrangement ou recombinaison du DNA pour aboutir au
rapprochement de segments aléatoirement choisis au sein de chaque famille (avec excision du
matériel superflu)

Réarrangement du DNA
• Etapes distinctes dans le réarrangement et bien coordonnées au fil de la différenciation du lymphocyte B– d’abord réarrangement gènes de chaîne lourde– puis réarrangement gènes de chaîne légère

Réarrangement du DNA
• Les enzymes en cause : les V(D)J recombinases– RAG-1 (recombination activating gene-1)
– RAG-2 (recombination activating gene-2)
– TdT : terminal deoxynucléotide transférase
• Reconnaissance de séquences signal à proximité des segments géniques à réarranger (recombination signal sequences)

Réarrangement du DNA
• Les déficits génétiques qui touchent les enzymes impliquées dans la recombinaison provoquent une absence de lymphocytes B et de lymphocytes B fonctionnels : ce sont les SCID (severe combined immunodeficiencies)

Réarrangement du DNA et diversité jonctionnelle
• La diversité intrinsèquement liée à la recombinaison est fortement accrue par le mode de jonction des segments sélectionnés
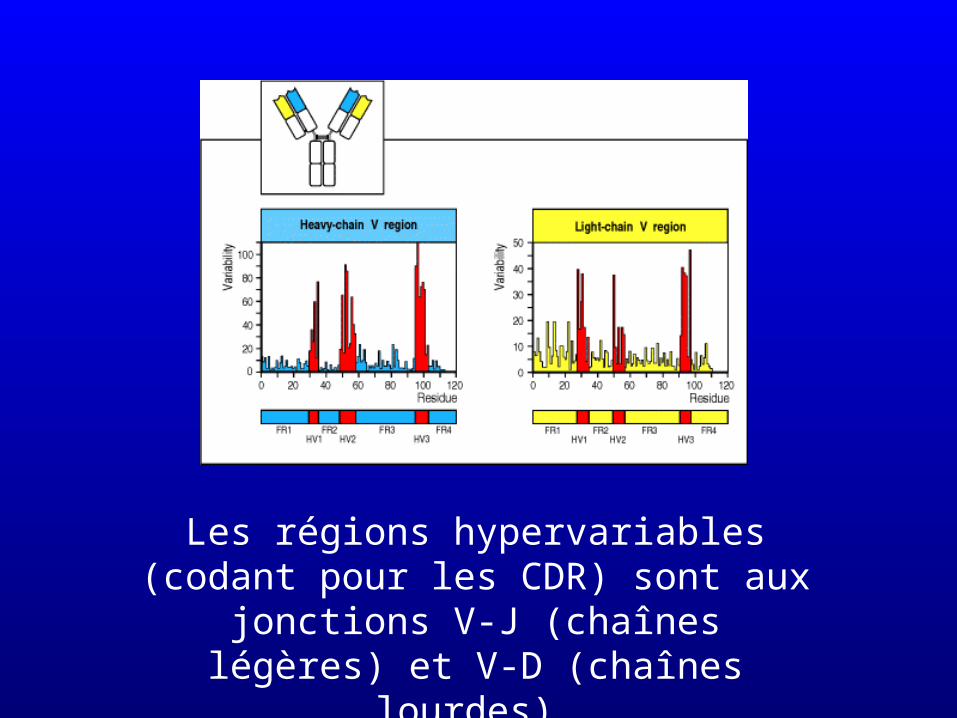
Les régions hypervariables (codant pour les CDR) sont aux jonctions V-J (chaînes
légères) et V-D (chaînes lourdes)

Diversité jonctionnelle
Imprécision de jonction des séquences codantesFlexibilité jonctionnelle

Diversité jonctionnelle
Addition de nucléotides palindromiques (P nucléotides) et de nucléotides non palindromiques (N nucléotides)

Diversité jonctionnelle
• L’addition des N-nucléotides (jusqu’à 15) est totalement aléatoire, et constitue une source importante de diversité
• Elle est catalysée par la TdT (terminal déoxynucléotide transférase)

Réarrangements productifs et non productifs

Exclusion allélique
• Les lymphocytes B sont des cellules diploïdes mais n’expriment que les gènes réarrangés d’un seul chromosome qu’il s’agisse des gènes de chaîne légère ou de ceux de la chaîne lourde : c’est l’exclusion allélique.
• Un clone de lymphocytes B donné ne possède qu’une seule spécificité (pas deux)

Exclusion allélique
• Dès qu’un réarrangement productif de chaîne lourde est obtenu, il bloque les réarrangements de chaîne lourde et enclenche les réarrangements de la chaîne légère
• Dès qu’un réarrangement productif de la chaîne est obtenu, il bloque les réarrangements de chaîne légère et les chaînes ainsi obtenues s’assemblent
• Dans le cas contraire, on passe au réarrangement de la chaîne légère

Hypermutation somatique
• Processus qui touche le produit fini du réarrangement génique
• 1mutation/1000 bases par division cellulaire soit 105 fois plus que dans les autres gènes
• Généralement substitutions (plutôt que délétions)
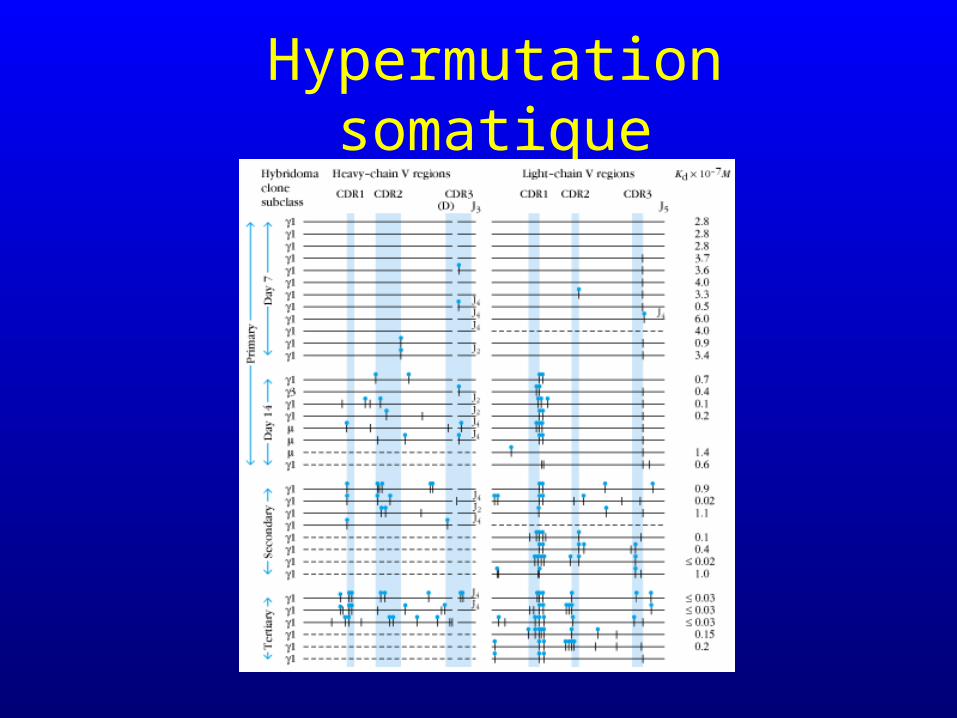
Hypermutation somatique

Hypermutation somatique
• Processus qui touche les lymphocytes B différenciés (plasmocytes) et qui intervient après la phase de réarrangement du DNA
• Processus influencé par la présence de l’antigène
• Concept de maturation d’affinité et de sélection par l’antigène

AID
• Le processus d’hypermutation somatique dépend strictement de l’activité de l’enzyme AID : activation induced cytidine deaminase
• Cette enzyme transforme les dC en dU sur le DNA• Le mismach G-U qui en résulte est le signal
nécessaire au processus d’hypermutation et à la commutation isotypique

Dernier niveau de la génération de diversité
• Association aléatoire d’une chaîne lourde et d’une chaîne légère

Diversité et CDR

Commutation isotypique
• La dernière phase de réarrangement n’accroît pas la diversité et juxtapose un multisegment VDJ déjà réarrangé à un segment CH particulier

Les lymphocytes B matures qui n’ont jamais rencontré leur antigène
expriment des IgM et des IgD

La rencontre avec l’antigène déclenche la commutation isotypique
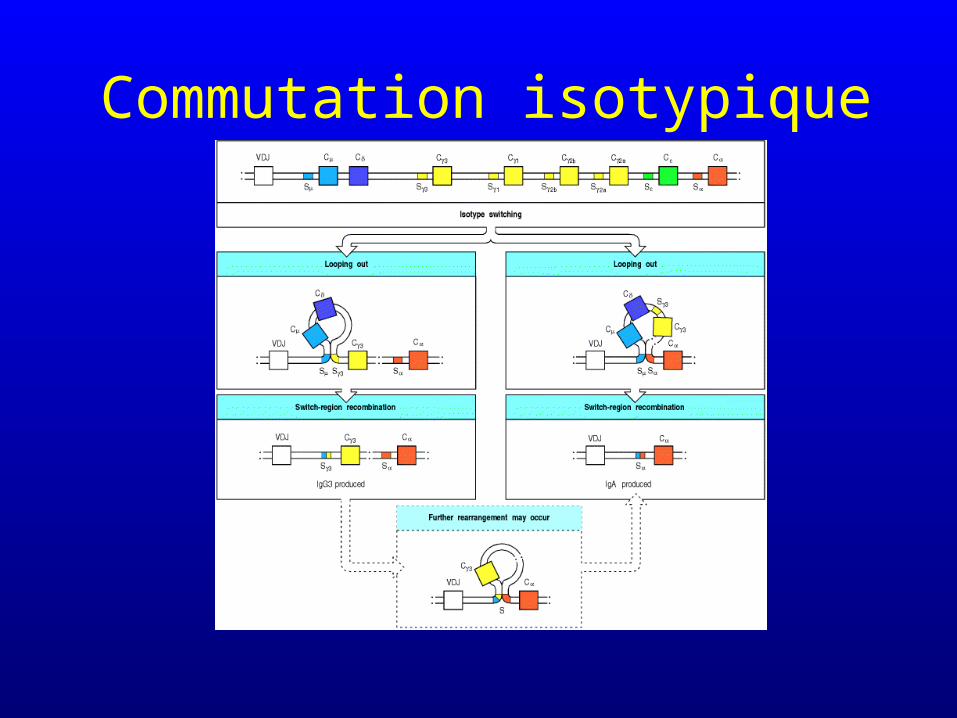
Commutation isotypique

Commutation isotypique
• Réarrangements non aléatoires selon une séquence donnée
• Influencés par la présence de l’antigène et certaines cytokines et l’aide des lymphocytes T
• Responsables de la sécrétion séquentielle d’anticorps d’isotypes différents mais de même spécificité
• Dépendance de l’AID (activation induced cytidine deaminase)

Epissage alternatif
• Epissage (splicing) : excision des introns du transcrit primaire avant polyadénylation et synthèse du mRNA
• L’épissage alternatif détermine si l’immunoglobuline sera sécrétée ou membranaire
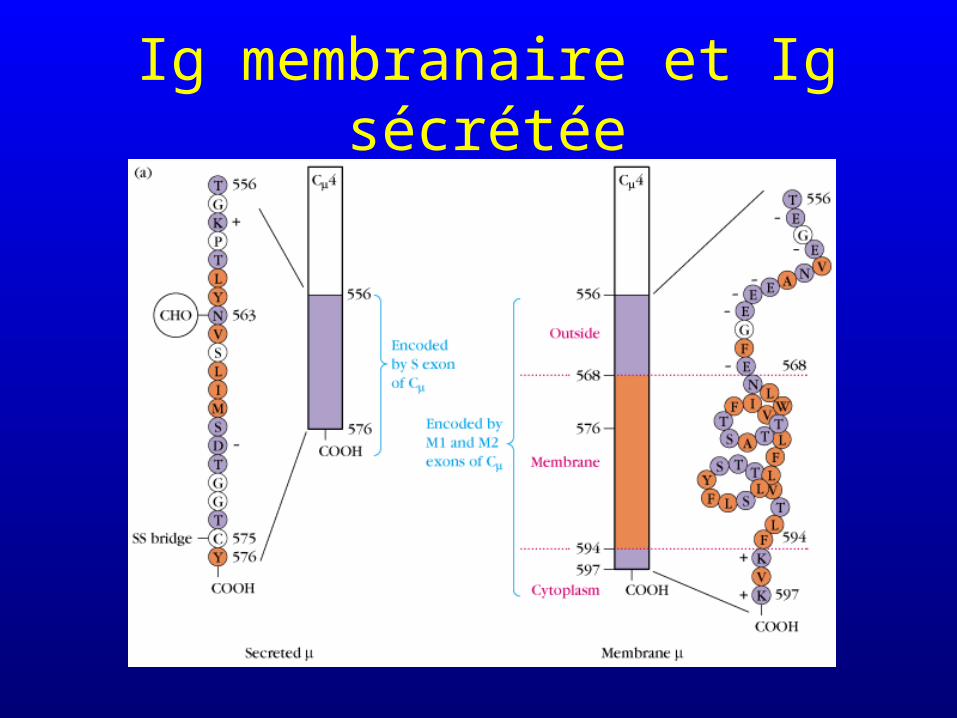
Ig membranaire et Ig sécrétée
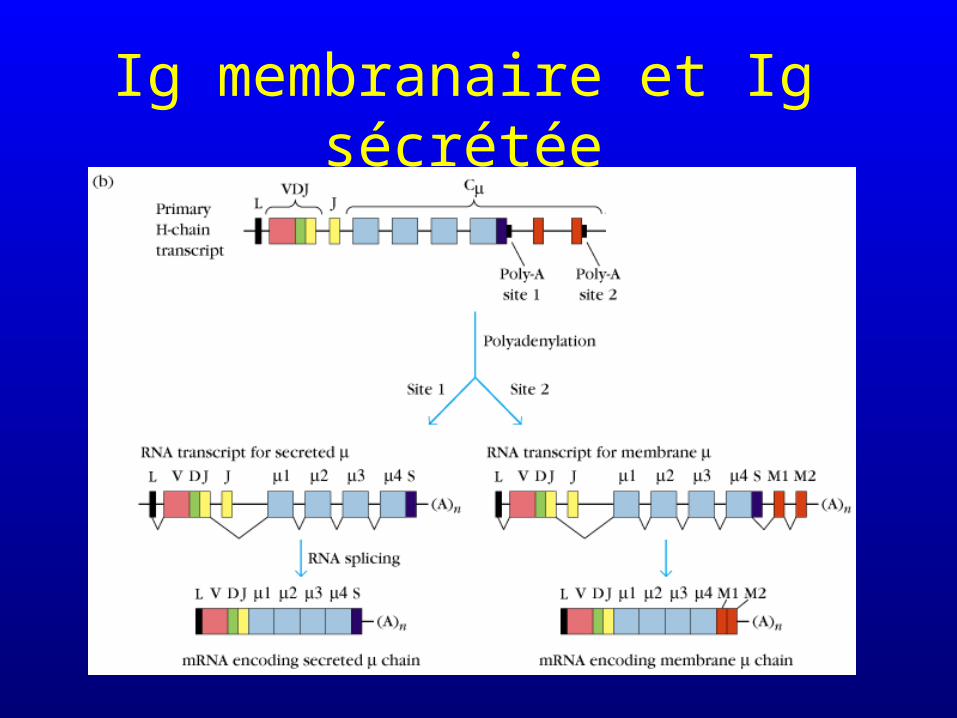
Ig membranaire et Ig sécrétée

Epissage alternatif
• Intervient dans la synthèse d’IgM et d’IgD par les mêmes cellules (en même temps)
• Intervient dans la synthèse d’immunoglobulines membranaires et d’immunoglobulines solubles (mais généralement pas par les mêmes cellules : l’épissage est influencé par la différenciation cellulaire)