exercice 1 - poly-prepas.com td biologie 4.pdf · la souris a reçoit au jour j0 une injection...
TRANSCRIPT

EXERCICE 1 :
Première expérienceOn expérimente sur deux souris de même souche (doc. 1).La souris A reçoit au jour J0 une injection péritonéale d'hématies de mouton. La souris B témoinne subit aucune intervention.Six jours après, on sacrifie les deux animaux A et B, on prélève leur rate (la rate est un organevolumineux impliqué dans la fabrication et la destruction des cellules du sang). On en dissociedes cellules dans 10 ml de liquide physiologique isotonique et on introduit dans ce liquidephysiologique une suspension d'hématies de mouton.
Une goutte de chaque milieu est, après incubation, observée au fort grossissement d'unmicroscope optique. Le milieu A montre des corps en rosette formés de cellules de la rate ayantagglutiné les hématies de mouton. Il n'y a pas de corps en rosette dans le milieu B.
Deuxième expérienceUne deuxième série d'expériences semblables est réalisée. Mais on ajoute du sérum de cobayetrèsriche en complément (système enzymatique complexe).Autour des cellules spléniques (= cellules de la rate) de l'animal A, on observe une région sanshématies appelée plage de lyse.
Expliquez ces phénomènes :1. Pourquoi l'apparition de corps en rosette ne s'observe qu'à partir des cellules de la rate del'animal A ? (Première série d'expériences).2. Quel est le mécanisme qui provoque la formation de ces corps en rosette ?3. Quel type de cellules immunitaires est responsable de cette réaction ?4. Nommez l'élément responsable de ces corps en rosette. Donnez-en un schéma légende.5. Pourquoi n'observe-t-on pas de lyse au cours de la première expérience ?6. Rappelez brièvement le mécanisme mis en jeu lors de la deuxième série d'expériences.

EXERCICE 2 :
La communication nerveuse
Document 1
Le document 1 représente une structure en coupe transversale, intervenant dans la communicationnerveuse chez un organisme vertébré.
1. À quel niveau une telle structure peut-elle être observée ? L'encéphale, la moelle épinière, unnerf rachidien, un muscle ?2. Les éléments observés sur le grossissement sont-ils : des cellules, des parties de cellules, desorganites, des nerfs ?3. Ces éléments sont-ils doués : d'intégration d'informations, de conduction de messages,d'excitabilité, de sécrétions ?4. Ces éléments possèdent des propriétés électriques. Sont-elles par exemple : l'existence d'unepolarité naturelle, la possibilité de conduire des potentiels d'action, la faculté de répondre à desstimulations par la loi du tout ou rien ?5. Ces éléments ont-ils : tous le même diamètre, tous la même fonction, tous le même rôle, tous lemême seuil de dépolarisation, tous la même vitesse de conduction des messages, tous la mêmelongueur ?
EXERCICE 3 :
À partir des conclusions tirées de l'analyse des documents 1 et 2, montrez que la différencephenotypique entre les deux souches parentales croisées doit être attribuée à l'expressiond'au moins deux gènes situés sur le même chromosome.
Document 1Les yeux des Drosophiles de type sauvage ont une couleur rouge sombre. Cette couleur est due,en fait, à la présence de deux pigments, l'un brun et l'autre rouge vif. Lorsqu'une Drosophile nepeut fabriquer le pigment brun, ses yeux sont rouge vif. La chaîne de biosynthèse du pigmentbrun comprend plusieurs étapes, chacune d'entre elles faisant intervenir une enzyme dont lasynthèse dépend d'un gène situé sur un chromosome déterminé. Le blocage de n'importe laquelledes étapes de cette chaîne de biosynthèse ou de plusieurs d'entre elles entraîne l'absence dupigment brun et, en conséquence, l'apparition du phenotype « yeux rouge vif ». Le gène V estporté par le chromosome 1, Cn par le chromosome 2, Cd et St par le chromosome 3.

Document 2On croise deux Drosophiles de race pure, l'une à yeux rouge sombre (Rs) et l'autre à yeux rougevif (rv). Tous les individus de première génération (F1) ont pour phenotype (Rs).• On effectue un test-cross entre une femelle F1 et un mâle de type parental aux yeux rougevif. Les résultats du test-cross sont, pour une population de 212 descendants :- 87 mouches de phenotype « yeux rouge sombre » ;- 125 mouches de phenotype « yeux rouge vif ».
• On réalise alors le test-cross inverse : mâle F1 aux yeux rouge sombre croisé avec unefemelle de type parental, donc homozygote, aux yeux rouge vif. (Il est établi, caractèreexceptionnel, qu'il n'y a jamais de crossing-over lors des méioses chez la Drosophile mâle.)Les résultats de ce test-cross sont les suivants pour une population de 402 descendants :- 199 mouches de phenotype « yeux rouge sombre » ;- 203 mouches de phenotype « yeux rouge vif ».
EXERCICE 4 :
L'Homme de Néanderthal a coexisté avec Homo sapiens en Europe, dans l'Ouest de l'Asie et auMoyen-Orient. Les débats actuels portent sur la question de savoir s'il s'agit de deux espècesdifférentes ou d'une seule ce qui implique une hybridation possible.
Vous montrerez en quoi les documents 1 et 2 confirment l'arbre phylogénétique dudocument de référence et conclurez en indiquant quelle est l'hypothèse qui semble validée.
Document de référence. Arbre phylogénétique reconstitué à l'aide du Logiciel « Phylogène »(INRP) à partir de séquences D'ADN mitochondrial (nean- : Néanderthalien, cro- : homme deCro-Magnon, PAN- : Chimpanzé)


EXERCICE 5 :
Au moment de sa découverte, l'AMH a été surnommée hormone antiféminine par le chercheurl'ayant mis en évidence. Discutez de la validité de ce surnom à partir des informations tirées desdocuments 1 à 3.
Document 1 Un cas cliniqueUn garçon de trois ans (de caryotype XY) est amené à la consultation d'un hôpital, car bien queses organes génitaux externes soient naturellement masculinisés, il est cryptorchide (testiculesnon en place dans les bourses). De plus, il présente une hernie inguinale (à l'aine) de côté droit.L'opération réalisée pour supprimer cette hernie, révèle la présence d'une gonade surmontée d'unetrompe conduisant à un utérus. À gauche, on découvre la présence d'une autre gonade. Lechirurgien pratique une biopsie de ces gonades qui révèle la présence de nombreux tubesséminifères. Une analyse sanguine révèle que la concentration de l'hormone AMH chez cet enfantest nulle, alors qu'elle est en moyenne de 40 ng.mL-1 à 3 ans.
Document 2 Expériences de transgenèseDes chercheurs ont injecté dans des œufs de souris mâles et femelles, le gène codant pour l'AMHhumaine. La technique utilisée fait que ce gène s'exprime dans tous les tissus et dès la vie fœtale.Souris femelles (organes génitaux externes femelles)
Souris mâles (organes génitaux externes mâles) : la majorité des mâles transgéniques se sontdéveloppés normalement mais deux lots où l'AMH est sécrétée en très grande quantité, montrentune féminisation des organes génitaux externes et ces mâles XY possédaient de petits testiculesnon descendus dans les bourses, pas d'épididyme ni de vésicules séminales.
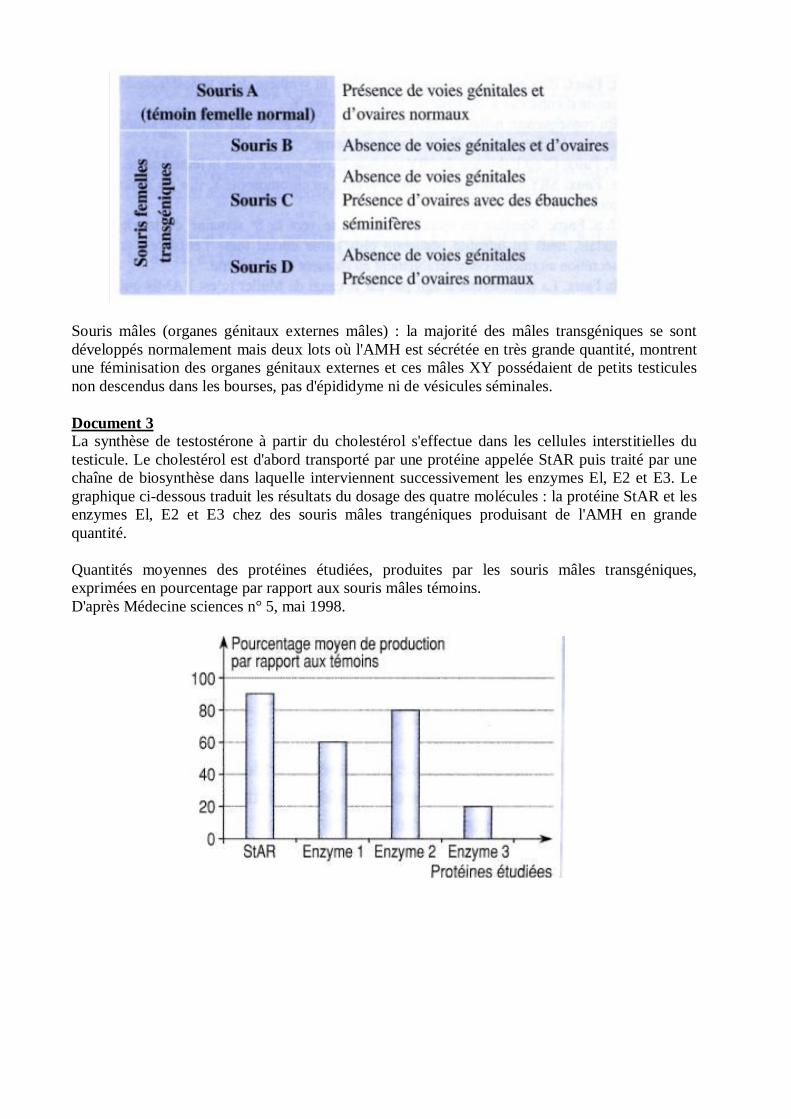
Document 3La synthèse de testostérone à partir du cholestérol s'effectue dans les cellules interstitielles dutesticule. Le cholestérol est d'abord transporté par une protéine appelée StAR puis traité par unechaîne de biosynthèse dans laquelle interviennent successivement les enzymes El, E2 et E3. Legraphique ci-dessous traduit les résultats du dosage des quatre molécules : la protéine StAR et lesenzymes El, E2 et E3 chez des souris mâles trangéniques produisant de l'AMH en grandequantité.
Quantités moyennes des protéines étudiées, produites par les souris mâles transgéniques,exprimées en pourcentage par rapport aux souris mâles témoins.D'après Médecine sciences n° 5, mai 1998.

EXERCICE 6 :
L'éthylène est une substance qui intervient dans la croissance des organes végétaux et lamaturation des fruits. On réalise deux expériences afin d'étudier son rôle dans la croissance.
Première expérience : des graines de soja vert sont mises à germer à l'obscurité. Les graines sontréparties en trois lots déposés dans trois récipients sur du coton. Au bout de 4 jours, leshypocotyles (portions de tige situées entre les racines et les premières feuilles appeléescotylédons) mesurent 1 à 3 cm. On place un premier lot de plantes tels quel ; un second lot estplacé dans un sac plastique fermé ; un troisième lot est placé dans un sac plastique fermé avecune pomme mûre coupée en morceaux. On signale que la pomme est une source d'éthylène.Les trois lots sont replacés à l'obscurité pendant trois jours. Puis on mesure la hauteur des plantes.
Deuxième expérience : sur des plantules de soja vert âgées de trois jours, on vaporise unesubstance proche de l'éthylène et ayant la même action. Le lendemain, on prélève des cellules del'hypocotyle et on mesure leur longueur. On effectue également la mesure sur des plantules nontraitées. Les cellules des plantules traitées sont trois fois moins longues que celles des plantulesnon traitées.
1. Justifiez le protocole expérimental mis en œuvre dans la première expérience.2. Montrez que les effets de l'éthylène au niveau cellulaire expliquent bien la croissanceobservée au niveau des organes.

CORRECTION
EXERCICE 1 :
1. La souris A a reçu, contrairement à la souris B, une injection d'hématies de mouton. La sourisA est donc la seule à être sensibilisée aux hématies de mouton (HM).L'absence de corps en rosette dans le liquide B nous permet de déduire que la rate de la souris B,non immunisée contre les HM, ne contient pas ou que peu de cellules à l'origine de la formationdes corps en rosette.La sensibilisation aux HM doit donc expliquer la formation des corps en rosette dans le liquide A.Les hématies de mouton doivent posséder sur leurs membranes des antigènes qui sont reconnuscomme non-soi par l'organisme de la souris A. Ce dernier met en place une réponse immunitairedirigée contre cet antigène.Cette réponse doit consister à produire en quantité importante des cellules qui s'accumulent dansla rate. Ces cellules forment in vitro avec les HM des corps en rosette.
2. Le mécanisme qui provoque la formation des corps en rosette est une reconnaissance directeentre l'antigène (les hématies de mouton) et certaines cellules de la rate.
3. Les LB qui possèdent des récepteurs spécifiques des déterminants antigéniques des hématiessont responsables de cette réaction.
4. Le récepteur membranaire à l'origine de la formation de ces corps en rosette correspond à unanticorps membranaire (= immunoglobuline membranaire).
5. Le récepteur membranaire du LB neutralise uniquement l'antigène sans le détruire, ce quiexplique que la lyse ne puisse pas être observée dans la première expérience.
6. La fixation du complément sur l'anticorps associé à l'antigène permet l'activation ducomplément qui peut lyser l'hématie de mouton en formant un complexe d'attaque membranaire.

EXERCICE 2 :
1. Une telle structure peut être observée au niveau d’un nerf rachidien.
2. Les éléments observés sur le grossissement sont des parties de cellules. Les axones myélinisésdes neurones (cellules nerveuses) sont représentés en coupe transversale. On les nomme fibresnerveuses.
3. Ces fibres ne sont pas douées d'intégration. Cette fonction est assurée par le cône axonique desneurones.Elles peuvent assurer la conduction de messages, c'est-à-dire la propagation de potentielsd'action. Elles sont excitables car elles réagissent à un stimulus efficace (présence de canauxvoltage-dépendants). Les axones sécrètent des neurotransmetteurs au niveau de leur arborisationterminale.
4. Les fibres possèdent des propriétés électriques qui correspondent à l'existence d'une polariténaturelle et à la possibilité de conduire des PA.La polarité naturelle des fibres est en effet due à des différences de concentration ionique de partet d'autre de la membrane engendrant des déséquilibres de charges électriques. La possibilité deconduire des PA est liée à la création dans les milieux intra et extracellulaires de courantsélectriques nommés courants locaux.La faculté de répondre a la loi du tout ou nen n’est pas liée à des propriétés électriques mais à despropriétés physiologiques de la membrane ; les canaux voltage-dépendants qui sont tous ouvertsou tous fermés.
6.- Les axones n'ont pas tous le même diamètre.- Les fibres n'ont pas toutes la même fonction :
- certaines transmettent des influx centripètes (du récepteur au centre nerveux) et sontsensi-tives;- d'autres transmettent des messages centrifuges (du centre nerveux à l'effecteur) et sontmotrices.
- Les fibres ont toutes le même rôle : elles peuvent véhiculer des messages nerveux. Ellessécrètent également toutes de l'ACH, neurotransmetteur excitateur.- Les fibres ne possèdent pas le même seuil de potentiel, elles ne possèdent pas la mêmeexcitabilité.- Elles ne possèdent pas toutes la même vitesse de conduction qui varie avec leur diamètre.Plus le diamètre est important, moins la résistance aux courants de propagation est importante etplus la vitesse de conduction est élevée.- Elles ne possèdent pas toutes la même longueur. La longueur d'une fibre du nerf rachidiendépend du muscle à innerver.
EXERCICE 3 :
Document 1 : l'absence de pigment brun (donc le phénotype œil rouge vif) peut être due à desallèles codant une enzyme non fonctionnelle d'un des gènes contrôlant la chaîne de biosynthèsedu pigment brun. Document 2 : le phénotype « œil rouge sombre » est dominant et le phénotype «œil rouge vif » récessif.

La descendance du premier test-cross n'est pas constituée de deux phénotypes en quantités égales.En conséquence, la différence phénotypique entre les deux souches est due à plus d'un gène, aumoins deux.
La descendance du test-cross inverse comprend deux types de phénotypes en quantités égales.Envisagé seul, ce résultat conduit à penser que la différence phénotypique est due à un seul gène.La contradiction entre les conclusions tirées des deux tests-cross s'explique si on tient compte del'information fournie : absence de crossing-over durant les méioses chez les Drosophiles mâles. Ily a bien deux gènes enjeu portés par le même chromosome, mais en l'absence de crossing-overles allèles associés chez les parents le restent lorsque le F1 mâle fabrique ses spermatozoïdes.
Bilan : La différence phénotypique entre les deux souches est due à deux gènes intervenant dansla synthèse du pigment brun. Ces deux gènes sont situés sur le même chromosome. Il doit doncs'agir des gènes Cd et St. Désignons par Cd+ et Cd les allèles du premier gène et par St+ et Stceux du deuxième gène.
Les Drosophiles croisées étant de lignée pure les génotypes des parents sont Cd+ StV Cd+ St+ etCd St / Cd St. Le génotype des F1 est Cd+St+ / Cd St.Les mâles F1 ne produisent que des gamètes Cd+ St+ et Cd St (gamètes parentaux) en quantitéségales.
Les femelles F1 produisent 4 types de gamètes Cd+St+ et Cd St (gamètes parentaux) et Cd+St etCd St+ (gamètes recombinés). Les gamètes recombinés produits en quantités plus faiblesrésultent de méioses où il y a un crossing-over entre les deux gènes (schéma à faire). Ladescendance du premier test-cross comprend plus de Drosophiles de phénotype récessif, car seulsles gamètes Cd+ St+ sont à l'origine de Drosophiles à phénotype dominant, les trois autres typesconduisant au phénotype récessif.
EXERCICE 4 :
Analyse du document de référence
Il montre :- que Néanderthaliens, Cro-Magnon et Hommes actuels (Homo sapiens), ontun ancêtre commun qui n'est pas celui du Chimpanzé ;-que les Néanderthaliens ont un ancêtre commun qu'ils ne partagent pas avec l'Homme actuel etl'Homme de Cro-Magnon ;- que les Hommes de Cro-Magnon font partie de la même espèce que les Hommes actuels.
• Exploitation du document 1En recherchant les différences entre les séquences proposées, on constate que :- les 8 Néanderthaliens ont la même séquence pour ces 27 nucléotides ;- les Hommes de Cro-Magnon et les Hommes actuels ont la même séquence pour ces 27nucléotides et présentent 3 différences avec celles des Néanderthaliens en position 4, 14 et 26.Cela confirme que les Néanderthaliens sont plus apparentés entre eux qu'avec les Hommesactuels et de Cro-Magnon. Cela est en accord avec l'idée qu'ils ont un ancêtre commun qu'ils nepartagent pas avec les Hommes actuels et de Cro-Magnon.

D'autre part, le fait que les Hommes actuels et les Hommes de Cro-Magnon ont exactement lamême séquence de nucléotides confirme l'idée qu'ils ont un ancêtre commun exclusif et qu'ilsfont partie de la même espèce.
• Exploitation du document 2La comparaison des séquences homologues constituées de 345 nucléotides révèle que :- la séquence du Chimpanzé est celle qui présente la plus grande différence avec toutes les autreset que cette différence est du même ordre de grandeur (17-18%) ; ces valeurs sont conformes àl'idée que tous les Néanderthaliens, Hommes de Cro-Magnon et actuels possèdent un ancêtrecommun qui n'est pas celui du Chimpanzé. Le fait que la différence soit la même entre leChimpanzé et les divers Homo indique que ces données sur 1'ADN mitochondrial, sontpertinentes pour établir des relations de parenté ; - les séquences des trois Néanderthaliens ontentre 0 et 2 % de différences entre elles alors qu'elles ont une différence de l'ordre de 7 % avecles Hommes de Cro-Magnon et actuels. Cela concorde avec l'idée que les Néanderthaliens ont unancêtre commun qui n'est pas celui des deux autres ; - les Hommes de Cro-Magnon n 'ont pasplus de différences avec les Hommes actuels que les Hommes actuels entre eux : cela confirmel'idée qu'ils appartiennent à la même espèce.
ConclusionLe document 2 traduit l'existence d'un certain nombre de différences entre les séquences desHommes actuels ; les séquences des Hommes de Cro-Magnon entrent dans le domaine devariation des Hommes actuels. En revanche, celles de tous les Néanderthaliens ont trois fois plusde différences avec celles des Hommes actuels (7 à 8 %).D'autre part, leurs séquences ne sont pas plus proches des séquences des Européens que de cellesdes Russes, des Chinois... et de celles des hommes de Cro-Magnon.Ce sont ces deux arguments qui semblent indiquer que les Néanderthaliens forment un groupe àpart dont l'évolution de F ADN mitochondrial a été indépendante de celle de F ADN desHommes actuels ou de Cro-Magnon et donc qu'ils peuvent constituer une espèce différente.
EXERCICE 5 :
• Exploitation du document 1En l'absence d'AMH (gène non fonctionnel) durant la vie fœtale, on constate qu'il y aconservation des voies génitales féminines. Cela confirme l'idée que l'AMH est responsable de ladestruction des voies génitales femelles au cours du développement et, en cela, c'est une hormoneanti-féminine.La différenciation masculine des organes génitaux externes est la preuve que les gonades del'enfant ont sécrété de la testostérone durant la vie fœtale. Cependant, il n'y a pas eu descente destesticules dans les bourses, ce qui laisse à penser que l'AMH intervient, directement ouindirectement, dans la mise en place des testicules.
• Exploitation du document 2Chez la femelle, non seulement l'AMH agit en provoquant la disparition des voies génitalesfemelles embryonnaires, mais elle peut également agir sur la différenciation des ovaires de façonplus ou moins importante. Chez le mâle, l'AMH en grande quantité a empêché la masculinisationdes organes génitaux externes qui a lieu normalement sous l'action de la testostérone, ainsi que ladifférenciation complète des voies génitales et glandes annexes qui a lieu également sous l'actionde la testostérone. On peut donc supposer :- soit que l'AMH agit directement sur les voies génitales et les organes génitaux externes ;- soit qu'elle agit indirectement en inhibant la production de testostérone par les testicules.

• Exploitation du document 3 :Les données du document 3 permettent de tester la deuxième hypothèse émise.Le document indique que l'AMH agit sur les cellules interstitielles. On constate que la productiondes protéines impliquées dans la synthèse de la testostérone par les cellules interstitielles estinférieure à celle de témoins mâles, le déficit étant particulièrement important pour l'enzyme E3(20 %). Il suffit d'un déficit important au niveau d'une étape d'une chaîne de biosynthèse pourdiminuer l'efficacité de cette chaîne.
En conséquence, on peut penser que chez les souris mâles transgéniques, l'AMH agit au stadefœtal sur les cellules interstitielles en freinant l'expression de gènes codant pour les protéinesimpliquées dans la synthèse de la testostérone ; d'où la non-masculinisation de l'appareil génitalen développement.
Bilan :Aussi bien chez l'homme que chez la souris, l'AMH humaine, lorsqu'elle est absente chez le mâle,entraîne le maintien des voies génitales femelles et, lorsqu'elle est présente à forte concentrationdans le milieu intérieur d'un foetus femelle entraîne la disparition des voies génitales femelles.Par son action sur les voies génitales, l'AMH a une action anti-féminine. À forte concentration,l'AMH tend à agir sur l'ovaire en le faisant disparaître ou en provoquant son atrophie et même samasculinisation, ce qui confirme son action anti-féminine.
À forte concentration chez le mâle, supérieure à la normale, l'AMH agit sur les cellulesinterstitielles du testicule en réprimant l'expression de gènes qui codent pour les protéinesenzymatiques intervenant dans la synthèse de la testostérone. Par là, à fortes doses, l'AMH aégalement une action antimasculine.
En définitive, l'AMH peut être qualifiée de répresseur sexuel même si dans les conditionsnaturelles normales elle a une action anti-féminine.
EXERCICE 6 :
1. Le premier lot laissé à l'air libre sans pomme sert de témoin. Le second lot sous sacplastique permet de comparer la croissance de ses plantules avec celles qui ont les morceaux depomme ; on peut ainsi savoir si les effets observés sont vraiment dus à la pomme.
2. Dans la première expérience, on observe que les plantes du lot 3 ont peu grandicontrairement aux plantes témoin et à celles sous sac plastique. Cela montre que les morceaux depomme ont un effet négatif sur la croissance des plantules. On peut supposer que cet effet négatifest dû à l'éthylène.
Dans la seconde expérience, une molécule ayant les mêmes effets que l'éthylène empêchel'élongation normale des cellules de l'hypocotyle, puisqu'elles sont trois fois moins longues quedes cellules témoin.
Ces deux expériences permettent de conclure que l'éthylène est une substance qui agit sur lescellules de la zone d'élongation en empêchant leur allongement. Par conséquent, les organes ontune croissance réduite.