bcm 6225 enzymologie moléculaire et...
TRANSCRIPT

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
BCM 6225 Enzymologie moléculaire et mécanistique
L’évolution dirigée des enzymes

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Avantages des enzymes • leurs sites actif peuvent être chimio-,
régio- et stéréosélectifs • les taux réactionnels atteints sont
souvent 1010 fois plus rapides que les réactions non-catalysées

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Désavantages des enzymes • solubilité limitée dans les solvants organiques • dépendance des co-facteurs et co-enzymes • spécificité trop élevée pour permettre une
application large • instabilité :
– dénaturation ou mauvais repliement (insolubilité) – oxydation – hydrolyse

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Modification des enzymes • pour redresser leurs lacunes
– modification chimique : réaction des résidus (typiquement Cys) avec des réactifs électrophiles
– sémi-synthèse : synthèse d’un grand fragment peptidique, incorporant des acides aminés non-naturels, et la ligaturation de ce fragment dans sa place dans la protéine entière
• couplage peptidique est souvent problématique • ligaturation spontanée est rarement possible (P450) • ligaturation de Kent est aussi possible seulement entre un
thiolester C-terminal et un résidu Cys N-terminal – expression des protéines recombinantes : modification
du gène qui code pour une protéine, suivie par la biosynthèse de la protéine modifiée

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La mutagenèse dirigée • développée par Michael Smith, à UBC, (Prix Nobel 1993) • un gène qui code pour une protéine est modifié (reconstruit)
pour contenir une mutation dans sa séquence qui a l’effet de changer un codon pour qu’il code pour un différent acide aminé
• la surexpression de ce gène muté donne la protéine mutée contenant la mutation ponctuelle désirée
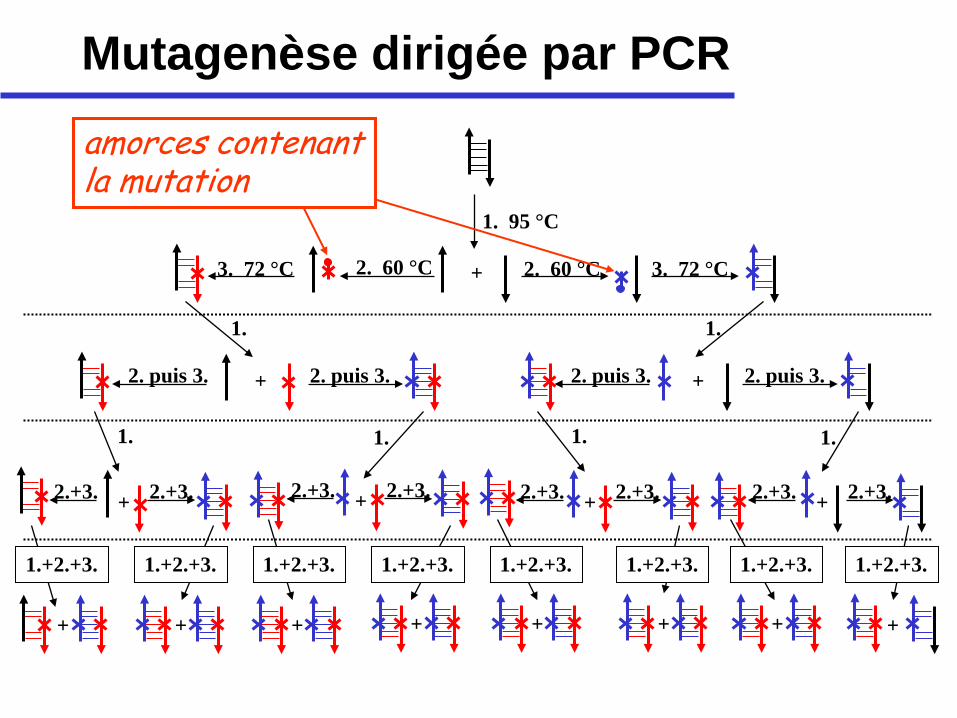
• actuellement, on fait la mutagenèse dirigée souvent à l’aide du PCR, en utilisant des amorces qui contiennent le codon (et anti-codon) modifié
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
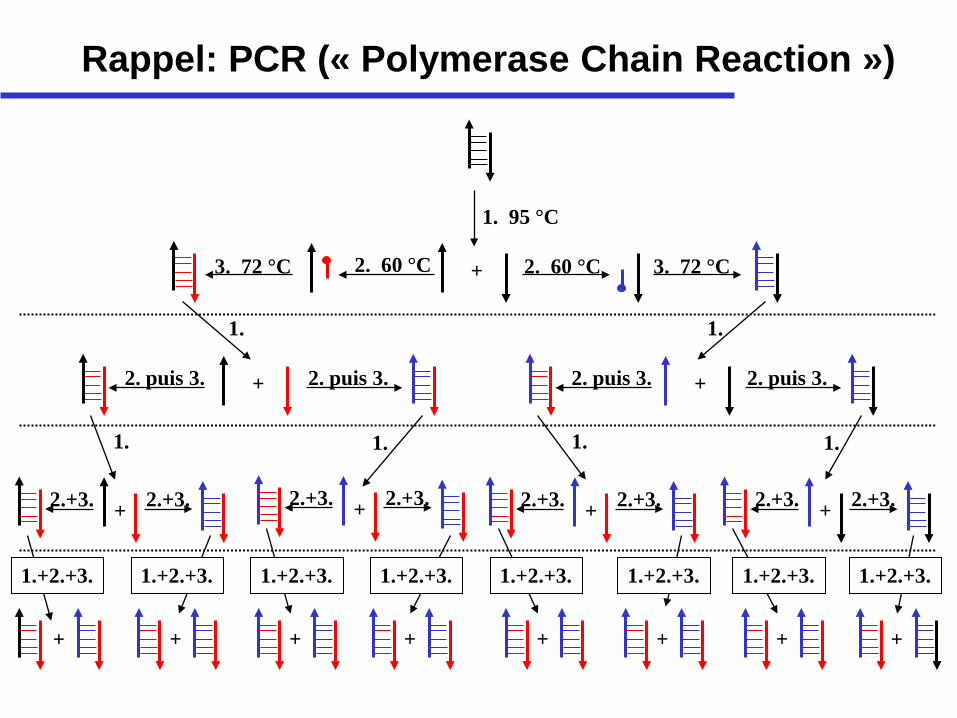
Rappel: PCR (« Polymerase Chain Reaction »)
1. 95 °C
+ 2. 60 °C 2. 60 °C 3. 72 °C 3. 72 °C
+
1. 1.
+ 2. puis 3. 2. puis 3. 2. puis 3. 2. puis 3.
2.+3. 2.+3. 2.+3. 2.+3. 2.+3. 2.+3. 2.+3. 2.+3. + + + +
1. 1. 1. 1.
+ + + + + + + +
1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3.
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Mutagenèse dirigée par PCR
1. 95 °C
+
1. 1.
1. 1. 1. 1.
1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3. 1.+2.+3.
2. 60 °C 2. 60 °C 3. 72 °C 3. 72 °C
+ 2. puis 3. 2. puis 3. 2. puis 3. 2. puis 3. +
+ 2.+3. 2.+3. + 2.+3. 2.+3. + 2.+3. 2.+3. + 2.+3. 2.+3.
+ + + + + + + +
amorces contenant la mutation

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique • modification rationnelle de la spécificité de la
trypsine par mutagenèse dirigée – Biochemistry 1987, 26, 2616-2623
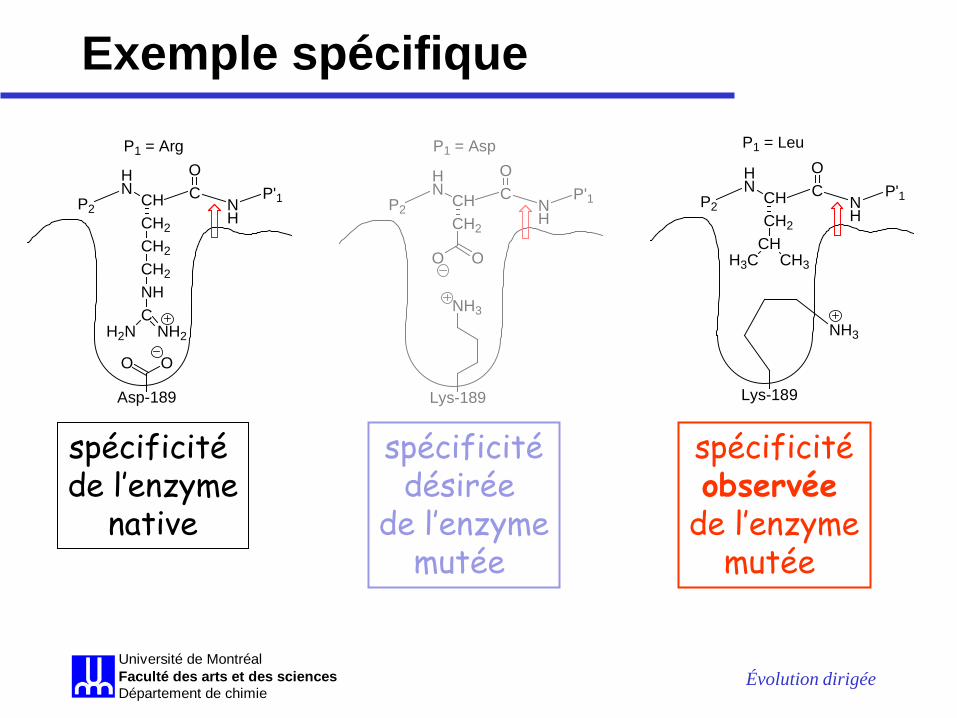
• la trypsine catalyse le clivage des liens peptidiques « après » (en C-terminal) des résidus Arg et Lys (résidus P1) – la charge négative du carboxylate du Asp189 donne la
spécificité pour des substrats ayant des chaînes latérales positivement chargées (Lys et Arg)
• le Asp-189 a été muté en Lys et la spécificité de l’enzyme mutée D189K a été caractérisée

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique • l’enzyme mutée n’est plus capable de
catalyser l’hydrolyse des substrats contenant des résidus Arg ou Lys en P1
• MAIS, elle n’est pas capable de catalyser l’hydrolyse des substrats contenant des résidus Asp ou Glu en P1 comme désiré
• l’enzyme mutée catalyse, aussi bien que l’enzyme native, l’hydrolyse d’un peptide avec Phe ou Tyr en P1
• l’enzyme mutée catalyse, mieux que l’enzyme native, l’hydrolyse d’un peptide avec Leu en P1
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique
Asp-189
OO
HN
CH C
CH2
NH
O
CH2CH2NHC
H2N NH2
P2P'1
P1 = Arg
Lys-189
HN
CH C
CH2
NH
O
P2P'1
NH3
OO
P1 = Asp
spécificité de l’enzyme
native
spécificité désirée
de l’enzyme mutée
Lys-189
HN
CH C
CH2
NH
O
P2P'1
NH3
CHCH3H3C
P1 = Leu
spécificité observée
de l’enzyme mutée

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique • la chaîne latérale du résidu substitué (Lys) adopte
probablement une conformation où la charge positive est dirigée vers l’extérieur de la cavité, en laissant sa chaîne alkyle (hydrophobe) interagir avec le substrat, au fonds de la cavité
• on ne peut pas toujours prévoir précisément comment une mutation ponctuelle va affecter l’activité d’une enzyme
• alors, pourquoi ne pas créer une banque de protéines mutées aléatoirement et sélectionner dans cette banque pour un mutant qui a l’activité désirée? – approche combinatoire vs conception rationnelle

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Quelques chercheurs renommés • Frances Arnold, Caltech (une pionnière ) • Alan Fersht, University of London • David Baker, University of Washington • Daniel Tawfik, Weitzmann Institute of Science • Chaitan Khosla, Stanford University • Manfred Reetz, Max Planck Institut • Willem Stemmer, Maxygen, Inc. • Christopher Walsh, Harvard Medical School

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La théorie de l’évolution naturelle • les mutations génétiques sont aléatoires – sans
direction ni but – provoquées par l’irradiation UV de l’ADN, modification
chimique, erreurs de transcription, etc – peuvent avoir lieu au niveau d’une base erronée sur
1000, mais les polymérases d’ADN comprennent aussi des fonctions d’édition donc le taux d’erreur est très faible (1 erreur sur 109 bases)
• chez les êtres unicellulaires, ces mutations seront transmises à la prochaine génération
• chez les êtres plus complexes, la mutation doit être comprise dans les cellules reproductrices (ovule, spermatozoïde) pour être transmise

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La théorie de l’évolution naturelle • les phénotypes résultants peuvent conférer à un
individu des avantages ou des désavantages qui peuvent être visibles ou non (dits neutres), dépendant des conditions actuelles
• les phénotypes avantageux qui permettent à l’individu de se reproduire, aux dépens des individus qui ne possèdent pas ces phénotypes, augmentent la chance que les génotypes responsables soient retenus dans la population générale (processus de sélection)

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
L’évolution (moléculaire) dirigée • techniques expérimentales qui miment, de
façon accélérée et in vitro, le processus d’adaptation évolutionnaire
• les mutations génétiques sont introduites à grande échelle lors de la réplication d’un gène selon deux approches générales :
1.mutagenèse aléatoire par PCR de basse fidélité (à haut taux d’erreurs)
2. recombinaison aléatoire des fragments de gène (« DNA shuffling »)

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
L’évolution (moléculaire) dirigée • les copies du gène, aléatoirement mutées, sont
introduites dans des organismes hôtes (typiquement des bactéries) de sorte qu’un organisme ne reçoive en moyenne qu’une copie mutée; suivra l’expression des protéines recombinantes
• on cherche ensuite l’activité spécifique désirée dans la banque de protéines mutées aléatoirement, selon deux approches générales :
1. sélection – où l’activité désirée est reliée à la survie cellulaire 2. criblage – où l’activité désirée n’est pas reliée à la survie
cellulaire et donc, doit être évaluée différemment • on prend les gènes qui codent pour les protéines
possédant l’activité désirée et on les soumet aux rondes additionnelles de mutation et d’amélioration
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
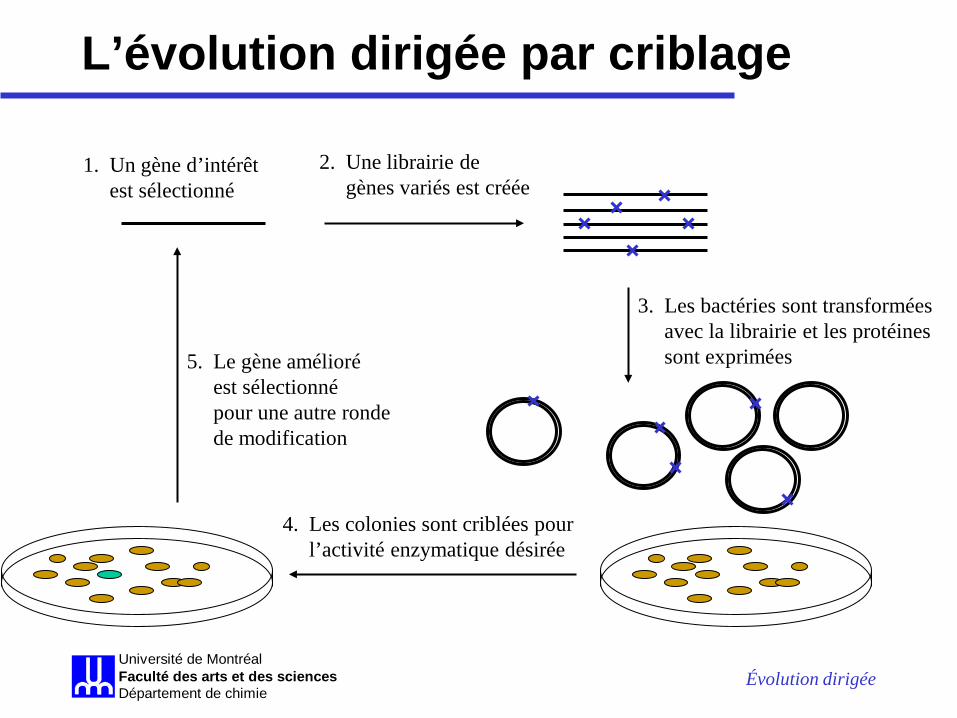
L’évolution dirigée par criblage
1. Un gène d’intérêt est sélectionné
2. Une librairie de gènes variés est créée
3. Les bactéries sont transformées avec la librairie et les protéines sont exprimées
4. Les colonies sont criblées pour l’activité enzymatique désirée
5. Le gène amélioré est sélectionné pour une autre ronde de modification
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
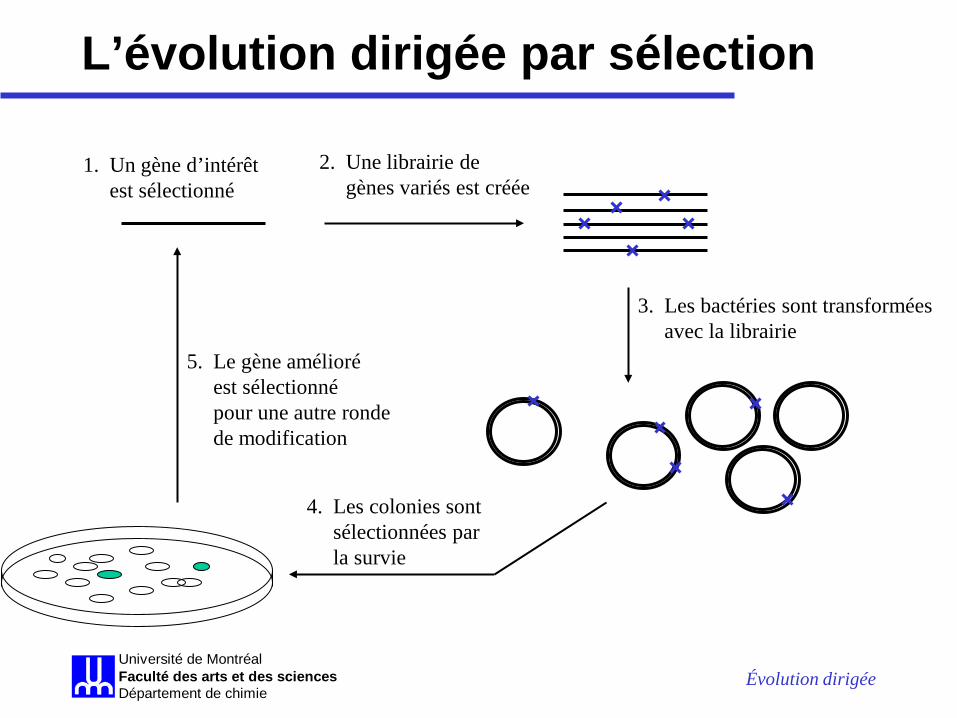
L’évolution dirigée par sélection
1. Un gène d’intérêt est sélectionné
2. Une librairie de gènes variés est créée
3. Les bactéries sont transformées avec la librairie
5. Le gène amélioré est sélectionné pour une autre ronde de modification
4. Les colonies sont sélectionnées par la survie

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La mutagenèse aléatoire • on peut introduire de plusieurs mutations en faisant
plusieurs rondes de PCR sur un même gène à l’aide d’une polymérase qui permet l’apparition de quelques mutations et qui n’a pas d’activité exonucléique de correction des erreurs (par exemple, la polymérase de Taq)
• il faut typiquement modifier une ou plusieurs conditions réactionnelles, par rapport à celles de la PCR normale; par exemple: – Mn2+ est utilisé à la place du Mg2+ comme cofacteur – des concentrations biaisées de dNTPs sont utilisées – des analogues de dNTPs sont utilisés – des co-solvants organiques (e.g. isopropanol) diminuent
aussi la fiabilité de la polymérase

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Biais de mutations • on crée par le PCR de basse fidélité une librairie
aléatoire du gène, qui se retrouve muté non-spécifiquement et sans biais un peu partout sur toute sa longueur
• le seul biais est la faible probabilité de muter un acide aminé donné en tous les autres, étant donné le nombre de mutations nécessaire pour changer les trois bases d’un codon – par exemple, il faudrait muter 2-3 bases d’un codon de
Val pour avoir un résidu Tyr à sa place • en réalité, les mutations de A ou T vers G ou C
sont plus fréquentes que l’inverse = biais
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
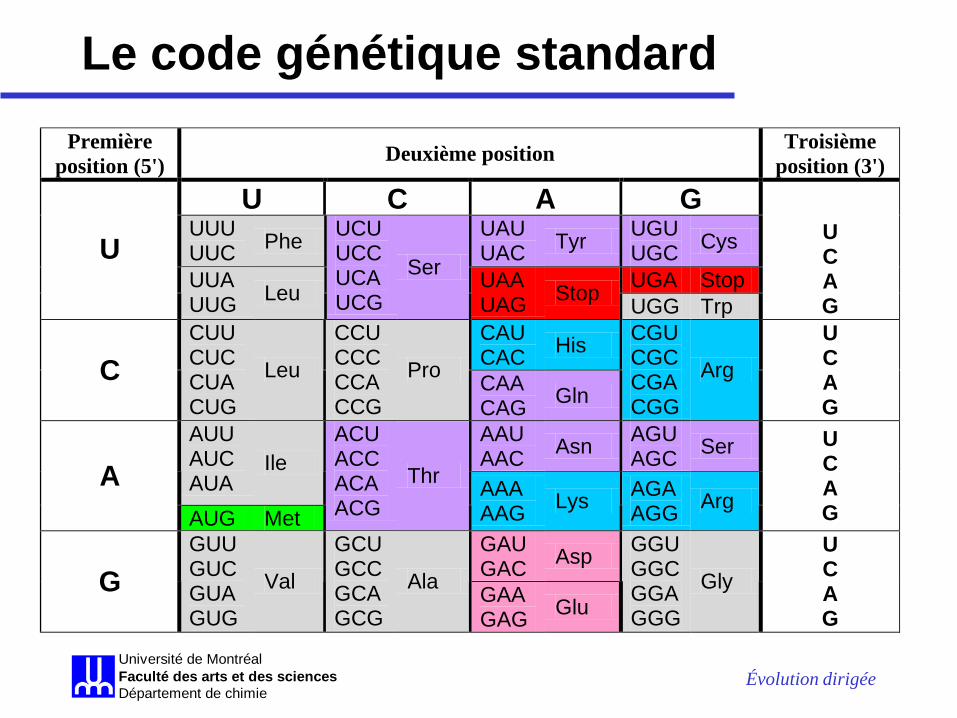
Le code génétique standard Première
position (5') Deuxième position Troisième position (3')
U C A G UUU UUC Phe UAU
UAC Tyr UGU UGC Cys
UGA Stop U
UUA UUG Leu
UCU UCC UCA UCG
Ser UAA UAG Stop UGG Trp
U C A G
CAU CAC His
C CUU CUC CUA CUG
Leu
CCU CCC CCA CCG
Pro CAA CAG Gln
CGU CGC CGA CGG
Arg
U C A G
AAU AAC Asn AGU
AGC Ser AUU AUC AUA
Ile A AUG Met
ACU ACC ACA ACG
Thr AAA AAG Lys AGA
AGG Arg
U C A G
GAU GAC Asp
G GUU GUC GUA GUG
Val
GCU GCC GCA GCG
Ala GAA GAG Glu
GGU GGC GGA GGG
Gly
U C A G

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La recombinaison aléatoire • aussi appelée le « DNA shuffling » • la mutagenèse aléatoire n’est pas une
méthode très combinatoire, puisque le point de départ pour l’introduction des mutations ponctuelles est un gène sélectionné
• dans les processus de recombinaison aléatoire, on commence avec une banque de gènes, on les coupe en fragments et on les recombine aléatoirement, ce qui mène à des réarrangements qui confèrent un plus haut degré de diversité de séquence

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
L’évolution dirigée par « DNA shuffling »
2. Les gènes sont coupés aléatoirement par la DNAse I
4. Sélection ou criblage des protéines améliorées
5. Les gènes améliorés sont sélectionnés pour une autre ronde de modification
1. Une famille de gènes similaires est sélectionnée
3. Les fragments des gènes sont recombinés aléatoirement par PCR sans amorce
Avantage: exploitation de la diversité naturelle Désavantage: le clivage et la recombinaison ne sont pas réellement aléatoires: les produits sont biaisés.

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• le point de départ est un mélange d’une série de gènes homologues d’une même famille ou bien du même gène ayant différentes mutations
• on coupe tous les gènes dans ce mélange de façon non spécifique et aléatoire à l’aide de la DNAse I
• les fragments sont ensuite remontés par hasard en faisant le PCR sans amorce – permet les fragments de se fixer les uns sur les autres au
hasard selon leur homologie de séquence
• une étape finale de PCR est entreprise à l’aide de deux amorces qui correspondent aux extrémités 3' et 5' du gène, pour amplifier spécifiquement des fragments qui ont la longueur entière du gène
L’évolution dirigée par « DNA shuffling »

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Une variante: l’évolution dirigée par assemblage de fragments synthétiques de gènes
1. Des fragments de gènes sont synthétisés (on les commande chez notre fournisseur préféré). Ils peuvent encoder autant de mutations que l’on veut, imitant les séquences naturelles, ou non.
3. Sélection ou criblage des protéines améliorées
4. Les gènes améliorés sont sélectionnés. Selon les mutations retenues, on commande de nouveaux fragments de gènes à intégrer dans une autre ronde.
2. Les fragments des gènes sont recombinés aléatoirement par PCR sans amorce
Avantage: exploitation d’une diversité illimitée Désavantage: $ € $ € $ € …mais le coût diminue chaque année.
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
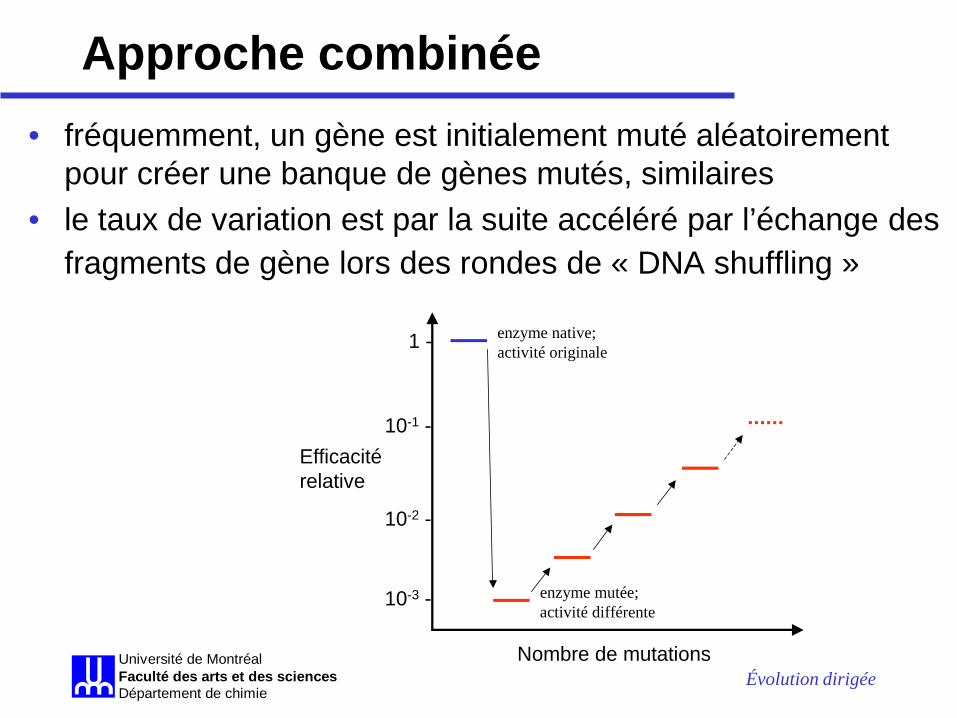
Approche combinée • fréquemment, un gène est initialement muté aléatoirement
pour créer une banque de gènes mutés, similaires • le taux de variation est par la suite accéléré par l’échange des
fragments de gène lors des rondes de « DNA shuffling »
Efficacitérelative
1 -
10-1 -
10-2 -
10-3 -
Nombre de mutations
enzyme native;activité originale
enzyme mutée;activité différente

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
L’expression de protéines mutées • tout d’abord on effectue le sous-clonage
d’une banque de gènes mutés dans des vecteurs d’expression (sous forme de plasmides)
• des bactéries sont transformées avec ces plasmides, sous conditions de haute dilution pour assurer que chaque bactérie n’en reçoive qu’un seul
• chaque bactérie se propage en colonie qui surexprime la protéine correspondant au gène d’intérêt

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La sélection • si l’activité désirée de l’enzyme d’intérêt est rendue
essentielle à la survie de la cellule transformée, il est généralement simple d’identifier les mutations bénéfiques en entreprenant une sélection par survie cellulaire – la cellule survit si, et seulement si, l’enzyme démontre
l’activité désirée – on est assuré que les colonies bactériennes qui se
propagent expriment des protéines ayant l’activité désirée; seulement les résultats positifs se voient
– on peut donc sélectionner à partir de très grandes banques de mutants (108 - 109)
• par contre, pas toutes les activités enzymatiques sont essentielles à la survie cellulaire

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Le criblage • solution de rechange dans l’éventualité où l’on ne
peut utiliser la méthode de sélection • nécessite de tester tous les clones obtenus un à un
pour l’activité désirée • normalement entrepris in vitro à l’aide d’appareillage
et/ou d’une réaction de dosage d’activité qui permettent de tester un très grand nombre de clones rapidement : – criblage à haute vitesse (« HTS, High Throughput
Screening ») – cytométrie fluorescente (« FACS, Fluorescence Assisted
Cell Sorting ») – détection de colonies fluorescentes ou colorées

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Le criblage • nécessite plus de temps et pose des limitations au
niveau du nombre de clones pouvant être criblés • par contre, le criblage est beaucoup plus flexible; on
peut cribler pour toutes sortes d’activités, sans égard à la survie cellulaire
• avertissement : c’est le criblage qui détermine ce qu’on obtient – (« you get what you screen for », Frances Arnold, Caltech) – si la méthode de criblage ne suscite pas précisément
l’activité désirée, on risque d’obtenir des ‘faux positifs’ • on peut découvrir une enzyme qui est modifiée et capable de
catalyser la réaction de criblage, tout en étant incapable de catalyser la réaction cible désirée

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Les acides aminés non-naturels • But: l’augmentation du répertoire chimique des protéines • Citons deux exemples de méthodes développées pour
incorporer des acides aminés non-naturels dans une protéine : – acides aminés non-naturels additionnels (P. Schultz)
• on utilise un ARN de transfert artificiel qui reconnaît le codon stop UAG mais il est chargé avec l’acide aminé non-naturel
• des centaines d’acides aminés ont été développés pour cette application mais un seul peut être incorporé à la fois
– acides aminés non-naturels substituants (D. Tirrell) • des tRNA synthétases sont modifiées pour avoir plus d’affinité pour
des acides aminés non-naturels • un acide aminé non-naturel est incorporé dans une protéine à la place
d’un acide aminé naturel à chaque position où se trouvait ce dernier
• En pratique, c’est peu mis en oeuvre: techniquement ardu.

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique 1: PNAS 1998, 95, 12809 • évolution dirigée d’une esterase thermostable
(Frances Arnold) • les estérases sont utilisées pour plusieurs
applications industrielles, en incluant la résolution stéréochimique
• la plupart des enzymes sont relativement instables par rapport à la température, et conçues pour être très efficaces dans les conditions cellulaires de l’organisme en question : – mésophiles : 30 – 50 C – thermophiles : 50 – 80 C – extrémophiles : > 80 C

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• les enzymes qui sont plus stables à hautes températures sont typiquement moins actives à basses températures
• le but de cette recherche est de modifier une enzyme pour qu’elle soit stable à haute température, sans perdre son activité à basse température – criblage pour les deux propriétés à la fois – Application type: lessive!
Exemple spécifique 1: PNAS 1998, 95, 12809

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• le gène qui code pour une enzyme bactérienne appelée la p-nitrobenzyl estérase (pNBE ) a été amplifié dans les conditions de PCR qui menait à un faible taux de mutagenèse – 10 fois plus de dCTP et dTTP que de dATP et dGTP – pas besoin d’ajouter de Mn2+ à la place de Mg2+
• chaque banque de gènes modifiés a été transformée dans des cellules bactériennes, propagées sur un Pétri et ensuite dans les puits d’une plaque à 96 puits
• après induction de l’expression des protéines mutées, les plaques ont été criblées pour l’activité estérase avant et après chauffage à la température de dépliement (Tm) de l’enzyme parente
Exemple spécifique 1: PNAS 1998, 95, 12809
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
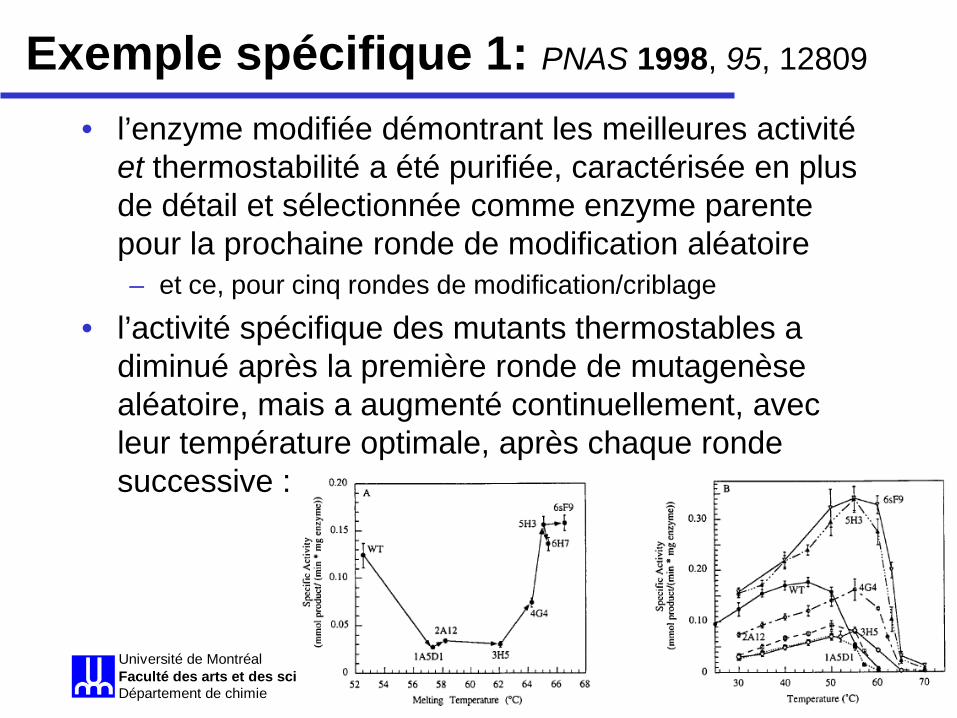
• l’enzyme modifiée démontrant les meilleures activité et thermostabilité a été purifiée, caractérisée en plus de détail et sélectionnée comme enzyme parente pour la prochaine ronde de modification aléatoire – et ce, pour cinq rondes de modification/criblage
• l’activité spécifique des mutants thermostables a diminué après la première ronde de mutagenèse aléatoire, mais a augmenté continuellement, avec leur température optimale, après chaque ronde successive :
Exemple spécifique 1: PNAS 1998, 95, 12809

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• si on ne crible pas pour les deux propriétés désirées, on peut facilement retenir une propriété en perdant l’autre
• N.B.: l’amélioration d’une propriété désirée, avec la détérioration d’une propriété non-désirée, ne signifie pas que les propriétés sont exclusives – ceci reflète plutôt que les conditions de sélection
n’exigent pas la rétention des deux propriétés
Exemple spécifique 1: PNAS 1998, 95, 12809
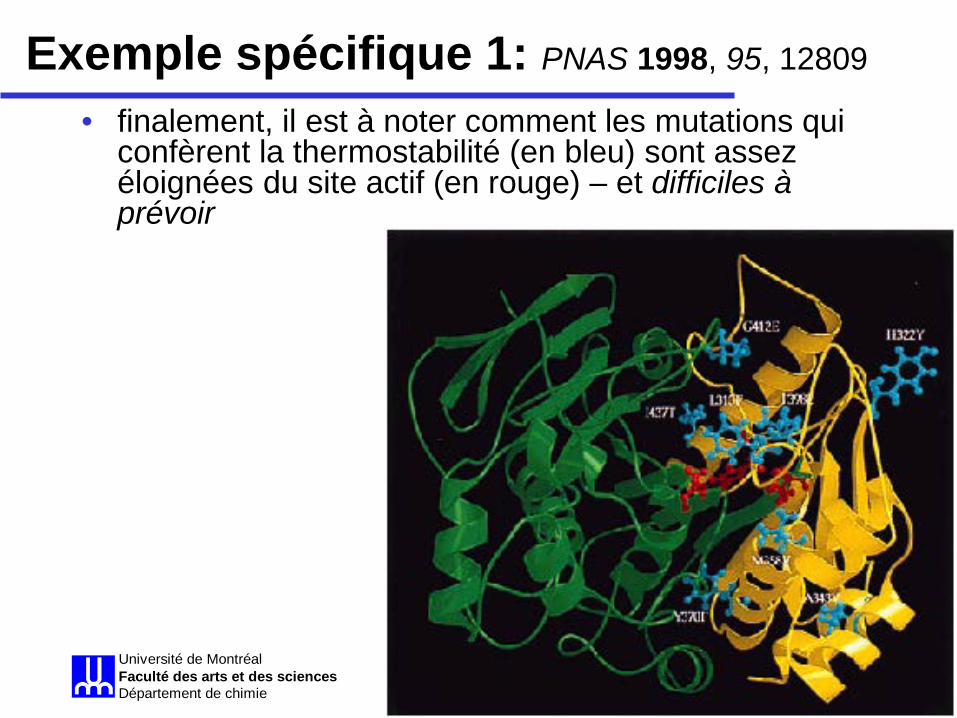
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• finalement, il est à noter comment les mutations qui confèrent la thermostabilité (en bleu) sont assez éloignées du site actif (en rouge) – et difficiles à prévoir
Exemple spécifique 1: PNAS 1998, 95, 12809

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple spécifique 2 : JACS 2004, 126, 15051
• évolution dirigée d’une glycosynthase (Virginia Cornish) • la synthèse des oligosaccharides représentent toujours
un défi et les ‘glycosynthases’ seraient très utiles – l’application des glycosidases pour la synthèse est problématique
dans l’eau – l’utilisation des glycosyltransférases nécessite des substrats
nucléotides diphosphate qui coûtent très cher
• le mutant E197A de la glycosidase Cel7B a été montré d’être une glycosynthase

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• Cornish a adapté la méthode l’approche ‘3-hybride’ pour développer une méthode de sélection pour une activité enzymatique qui n’est pas intrinsèquement essentielle à la survie cellulaire : – l’activateur de transcription DBD-AD (« DNA Binding Domain » et
« Activation Domain ») contrôle, dans ce cas-ci, l’activation du gène LEU2, responsable pour l’expression d’une protéine qui permet la survie en absence de la leucine
– le DBD-AD est clivé en deux parties et les deux fragments sont greffés à deux autres protéines pour donner les chimères DBD-DHFR et AD-GR
– les ligands à haute affinité de la DHFR (soit la méthotrexate, Mtx) et du GR (soit le dexamethasone, Dex) peuvent, une fois greffés ensemble, reconstituer le DBD-AD pour permettre la survie cellulaire
Exemple spécifique 2 : JACS 2004, 126, 15051

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• dans ce travail, le Mtx est greffé à un disaccharide donneur (Lac-F) et le Dex à un dissacharide accepteur (Cel) pour donner les substrats qui doivent être attachés l’un à l’autre par une nouvelle glycosythase pour permettre la survie – le gène LEU2 sera activé lors de la formation du :
« DBD-DHFR•Mtx-Lac-Cel-Dex•GR-AD »
Exemple spécifique 2 : JACS 2004, 126, 15051
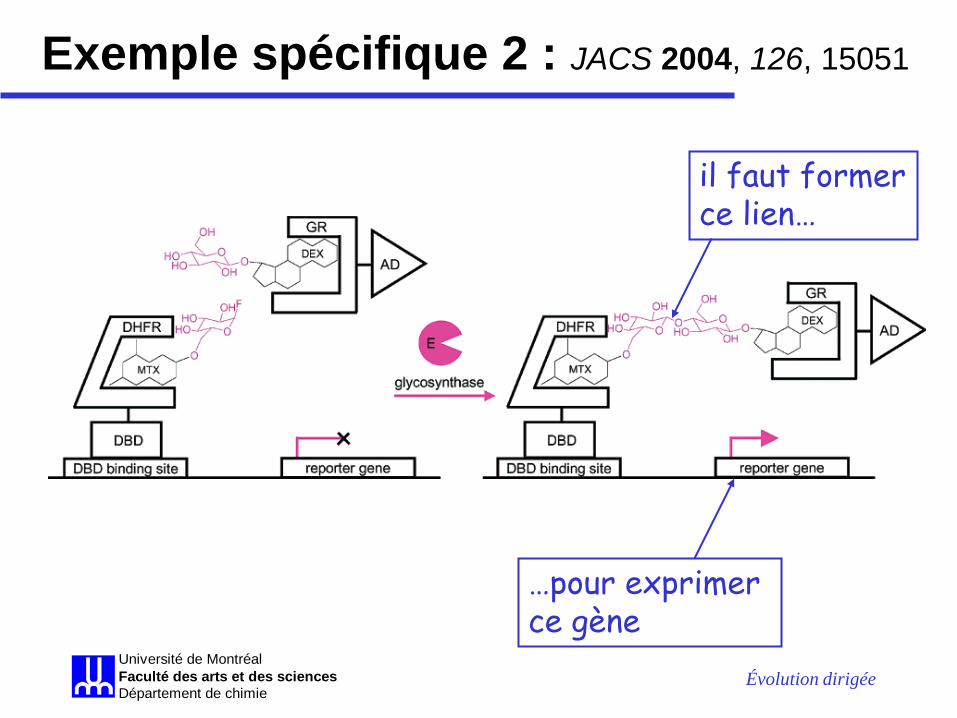
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
il faut former ce lien…
…pour exprimer ce gène
Exemple spécifique 2 : JACS 2004, 126, 15051

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• les auteurs ont fait les contrôles appropriés – le produit authentique Mtx-Lac-Cel-Dex permet la survie – les cellules qui n’expriment pas la glycosynthase (ou qui
expriment la glycosidase originale) ne survivent pas – les cellules qui poussent dans une culture qui ne contient
qu’un des deux substrats ne survivent pas
• les auteurs ont ensuite sélectionné, parmi une petite banque des 20 mutants à la position 197 (oligonucléotides dégénérés), ceux qui sont des glycosynthases et les ont caractérisé par la suite
• le but de l’article est de démontrer la généralité de la méthode (ne nécessite que la synthèse des différents substrats)
Exemple spécifique 2 : JACS 2004, 126, 15051
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
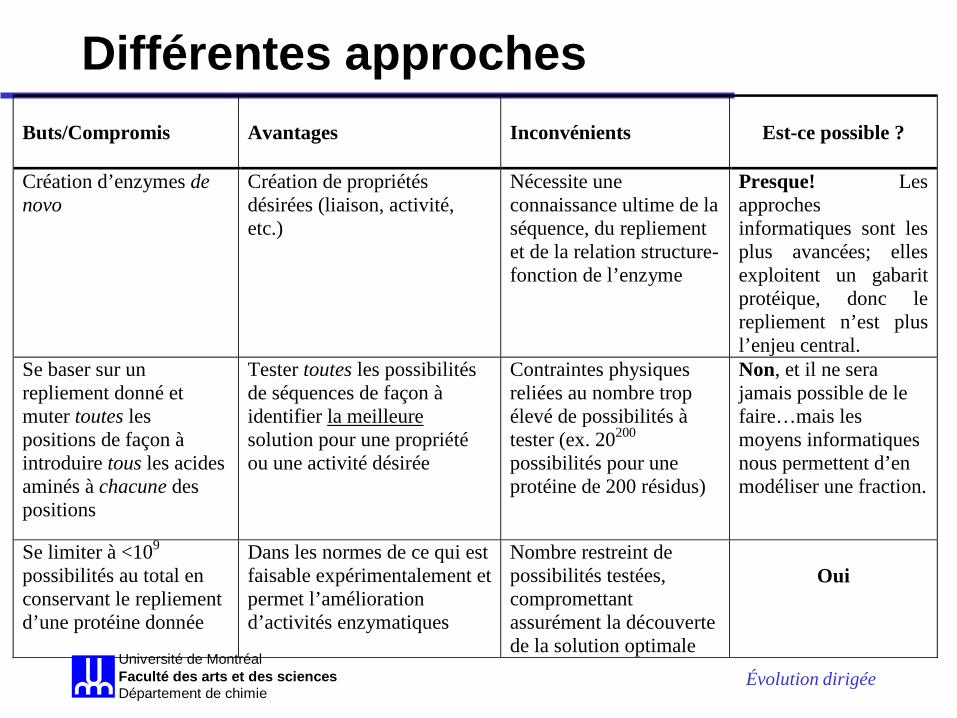
Différentes approches Buts/Compromis
Avantages
Inconvénients
Est-ce possible ?
Création d’enzymes de novo
Création de propriétés désirées (liaison, activité, etc.)
Nécessite une connaissance ultime de la séquence, du repliement et de la relation structure-fonction de l’enzyme
Presque! Les approches informatiques sont les plus avancées; elles exploitent un gabarit protéique, donc le repliement n’est plus l’enjeu central.
Se baser sur un repliement donné et muter toutes les positions de façon à introduire tous les acides aminés à chacune des positions
Tester toutes les possibilités de séquences de façon à identifier la meilleure solution pour une propriété ou une activité désirée
Contraintes physiques reliées au nombre trop élevé de possibilités à tester (ex. 20200 possibilités pour une protéine de 200 résidus)
Non, et il ne sera jamais possible de le faire…mais les moyens informatiques nous permettent d’en modéliser une fraction.
Se limiter à <109 possibilités au total en conservant le repliement d’une protéine donnée
Dans les normes de ce qui est faisable expérimentalement et permet l’amélioration d’activités enzymatiques
Nombre restreint de possibilités testées, compromettant assurément la découverte de la solution optimale
Oui
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
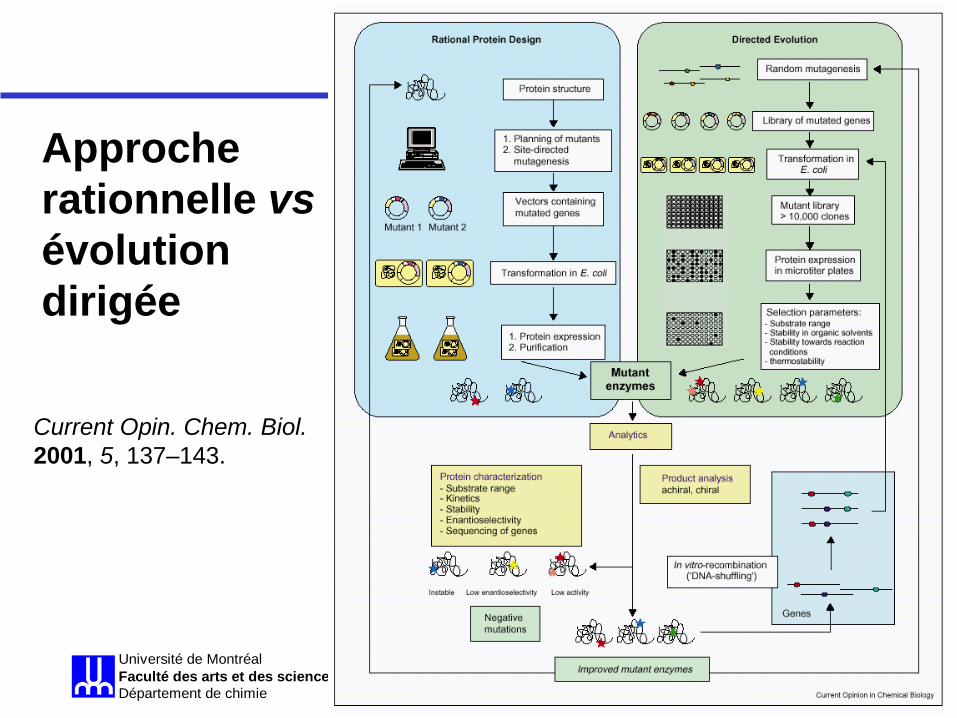
Approche rationnelle vs évolution dirigée
Current Opin. Chem. Biol. 2001, 5, 137–143.

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple 3: la modification des cytochromes P450 pour catalyser l’hydroxylation des alcanes
• Référence: JACS 2003, 125, 13442 (Frances Arnold, Caltech)
• la cytochrome P450 catalyse l’oxydation des chaînes alkyles en alcools, en utilisant l’oxygène (O2) comme oxydant – seules les P450s bactériennes sont solubles,
disponibles en grande quantité et rapides • pratiques pour des applications synthétiques
– le but ultime est de catalyser l’hydroxylation de l’éthane et du méthane en éthanol et méthanol
– modification par une approche combinée, comprenant l’évolution dirigée et la mutagenèse dirigée

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• l’activité oxydative désirée est détectée par criblage – diminution d’absorbance du NADPH à 340 nm
• le point de départ pour l’évolution dirigée: le gène d’un mutant actif, préalablement identifié, a été recombiné avec 15 autres mutants – un mutant (« J ») a été retenu pour sa capacité à accélérer
l’hydroxylation du propane et de l’octane • la mutagenèse aléatoire (par PCR de basse fidélité)
de J a produit un mutant (« 9-10A ») qui catalyse plus rapidement l’hydroxylation du propane
• ensuite, la mutagenèse dirigée a été utilisée pour diminuer le volume du site actif, afin de favoriser la reconnaissance de plus petits substrats – les mutant A328V et A82L ont démontré des meilleures
chimiosélectivités
Exemple 3: la modification des cytochromes P450 pour catalyser l’hydroxylation des alcanes

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• les gènes de J, 9-10A et des mutants 9-10A-A328V et 9-10A-A82L ont été soumis à une ronde de « gene shuffling » et le mutant sélectionné à partir de la banque résultante (« 1-12G ») était plus efficace, plus chimiosélective et autant énantiosélective que les enzymes parentes – par exemple, 82 % 2-OH en 39% ee (R) avec l’octane – le séquençage a confirmé que les deux mutations
ponctuelles précédentes ont été retenues dans le meilleur mutant
• voir aussi : – ChemBioChem 2005, 6, 1765 – Angew. Chem. Int. Edit. 2005, 44, 4029
Exemple 3: la modification des cytochromes P450 pour catalyser l’hydroxylation des alcanes
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
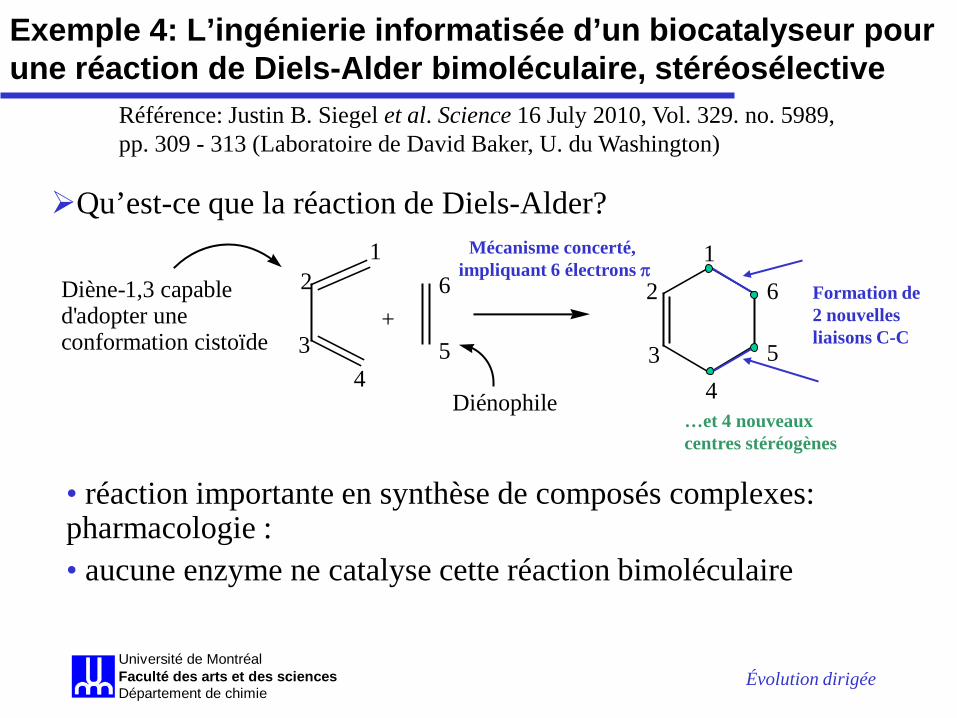
Exemple 4: L’ingénierie informatisée d’un biocatalyseur pour une réaction de Diels-Alder bimoléculaire, stéréosélective
Référence: Justin B. Siegel et al. Science 16 July 2010, Vol. 329. no. 5989, pp. 309 - 313 (Laboratoire de David Baker, U. du Washington)
+
12
34
5
61
2
34
5
6Diène-1,3 capable d'adopter une conformation cistoïde
Diénophile
Mécanisme concerté, impliquant 6 électrons π
Formation de 2 nouvelles liaisons C-C
…et 4 nouveaux centres stéréogènes
• réaction importante en synthèse de composés complexes: pharmacologie : • aucune enzyme ne catalyse cette réaction bimoléculaire
Qu’est-ce que la réaction de Diels-Alder?
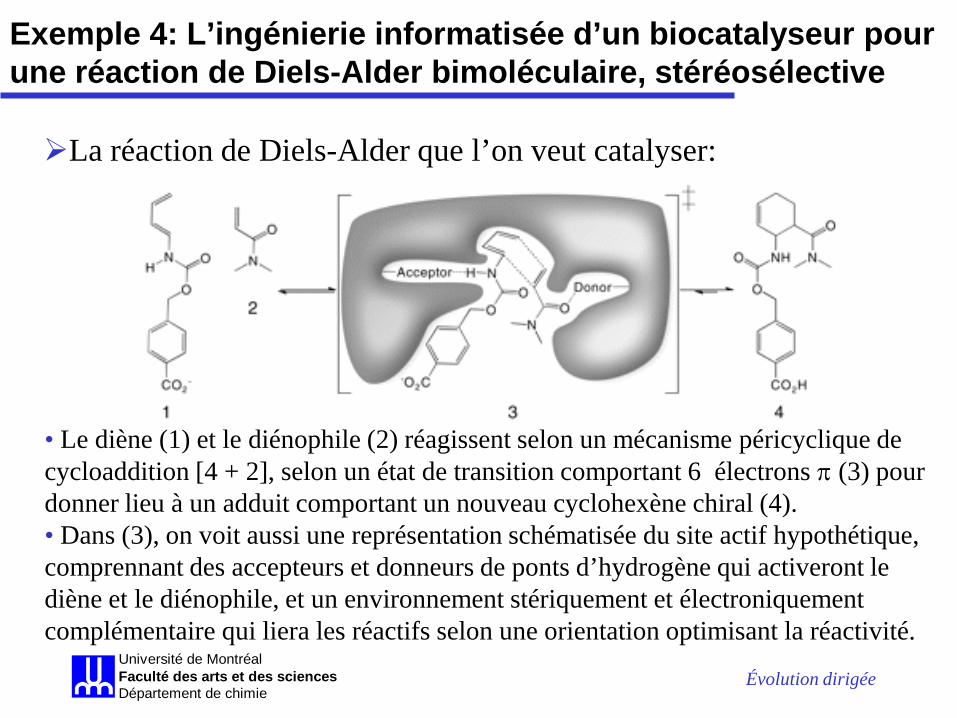
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
• Le diène (1) et le diénophile (2) réagissent selon un mécanisme péricyclique de cycloaddition [4 + 2], selon un état de transition comportant 6 électrons π (3) pour donner lieu à un adduit comportant un nouveau cyclohexène chiral (4). • Dans (3), on voit aussi une représentation schématisée du site actif hypothétique, comprennant des accepteurs et donneurs de ponts d’hydrogène qui activeront le diène et le diénophile, et un environnement stériquement et électroniquement complémentaire qui liera les réactifs selon une orientation optimisant la réactivité.
La réaction de Diels-Alder que l’on veut catalyser:
Exemple 4: L’ingénierie informatisée d’un biocatalyseur pour une réaction de Diels-Alder bimoléculaire, stéréosélective
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
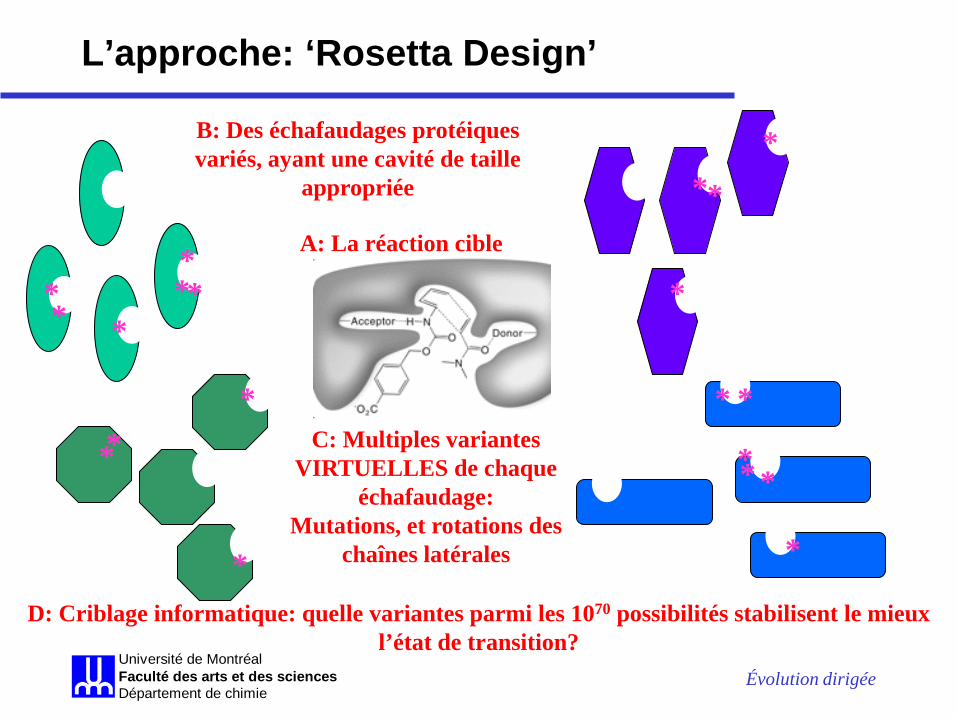
L’approche: ‘Rosetta Design’
A: La réaction cible
B: Des échafaudages protéiques variés, ayant une cavité de taille
appropriée
C: Multiples variantes VIRTUELLES de chaque
échafaudage: Mutations, et rotations des
chaînes latérales
D: Criblage informatique: quelle variantes parmi les 1070 possibilités stabilisent le mieux l’état de transition?
* * *
* * *
*
*
* *
* * *
*
* *
* *
*
*

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
L’approche: ‘Rosetta Design’
Prochaine étape: ayant identifié des variantes théorique, on les crée au laboratoire: les mutations appropriées sont apportées aux échafaudages appropriés. Puis, on teste la réactivité:
Note: knoncat pour cette réaction est de 4 x 10–4 s–1 M–1
Catalyst kcat
(hour–1) KM-diene
(mM)
KM-dienophile
(mM) kcat/KM-diene
(s–1 M–1)
kcat/KM-dienophile
(s–1 M–1)
DA_20_00 0.10 ± 0.02 3.5 ± 1.5 146.0 ± 2.5 0.008 0.0002
En créant des mutations additionnelles dans ce nouveau catalyseur, les chercheurs ont augmenté sa réactivité pour atteindre 0.455 s–1 M–1, soit une accélération de 1000 par rapport à la réaction non catalysée.

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
(A) Le modèle, lié aux substrats (diène et diénophile, en violet). Les résidus catalytiques et les ponts d’hydrogène qu’ils doivent établir sont en bâtonnets.
(B) Superposition du modèle et de la structure cristalline obtenue.
L’approche: ‘Rosetta Design’
Enfin, on compare la structure du nouveau catalyseur (cristallographie) à la variante qui avait été obtenue par moyen informatiques: la réalité correspond-elle au design?
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Exemple 5: la synthèse de la sitagliptine par une approche d’évolution combinée
La sitagliptine, un composé anti-diabète (Merck)
Problème: synthèse inefficace
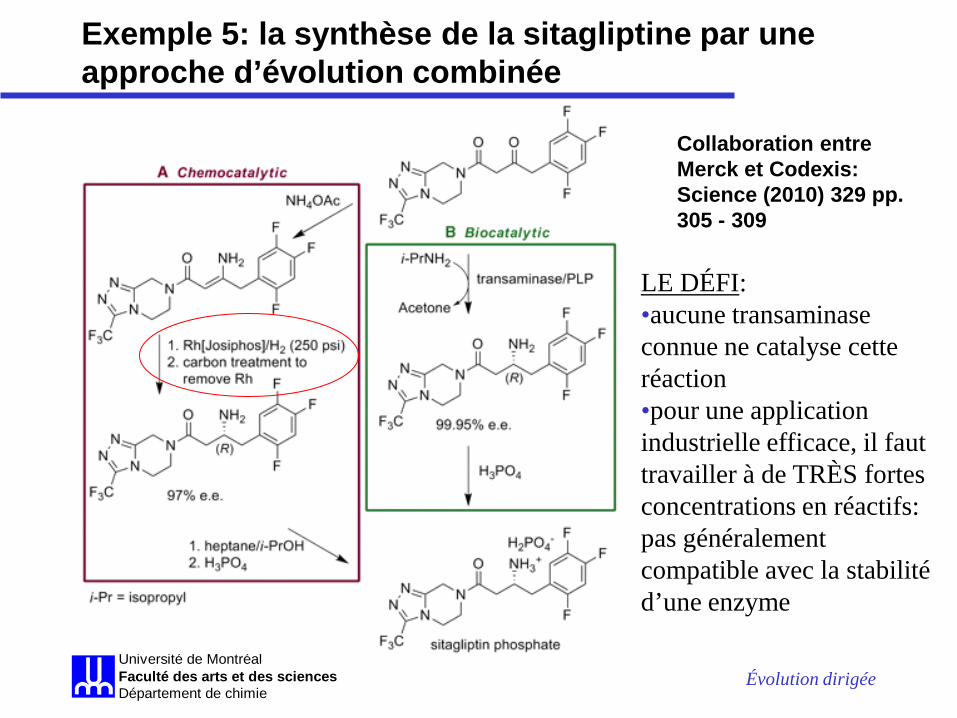
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
Collaboration entre Merck et Codexis: Science (2010) 329 pp. 305 - 309
LE DÉFI: •aucune transaminase connue ne catalyse cette réaction •pour une application industrielle efficace, il faut travailler à de TRÈS fortes concentrations en réactifs: pas généralement compatible avec la stabilité d’une enzyme
Exemple 5: la synthèse de la sitagliptine par une approche d’évolution combinée
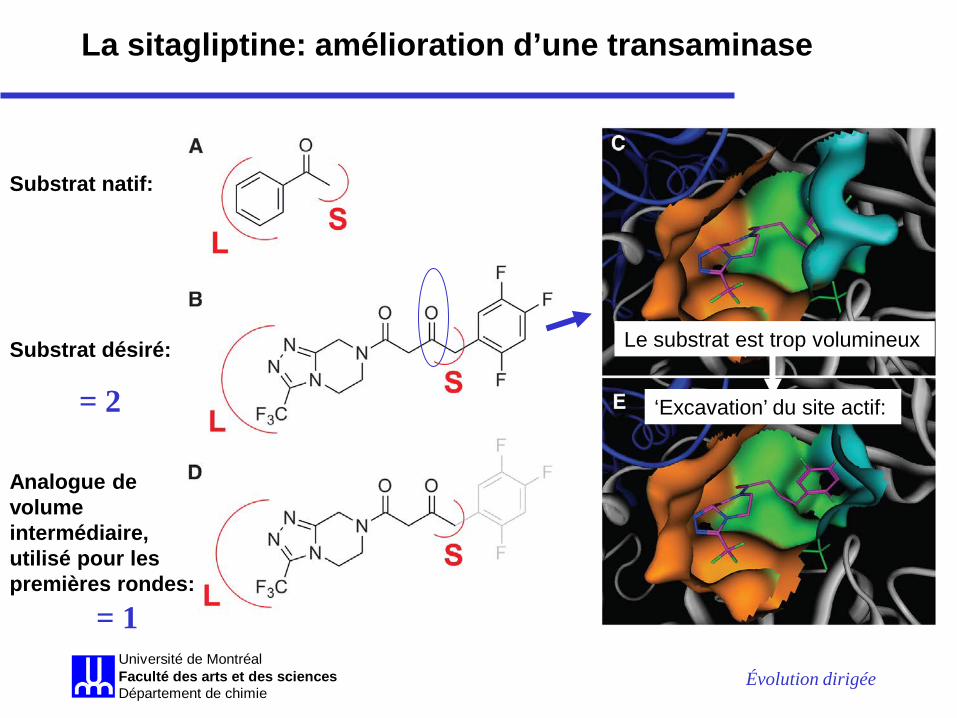
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La sitagliptine: amélioration d’une transaminase
Substrat natif:
Substrat désiré: Le substrat est trop volumineux
‘Excavation’ du site actif:
Analogue de volume intermédiaire, utilisé pour les premières rondes:
= 2
= 1
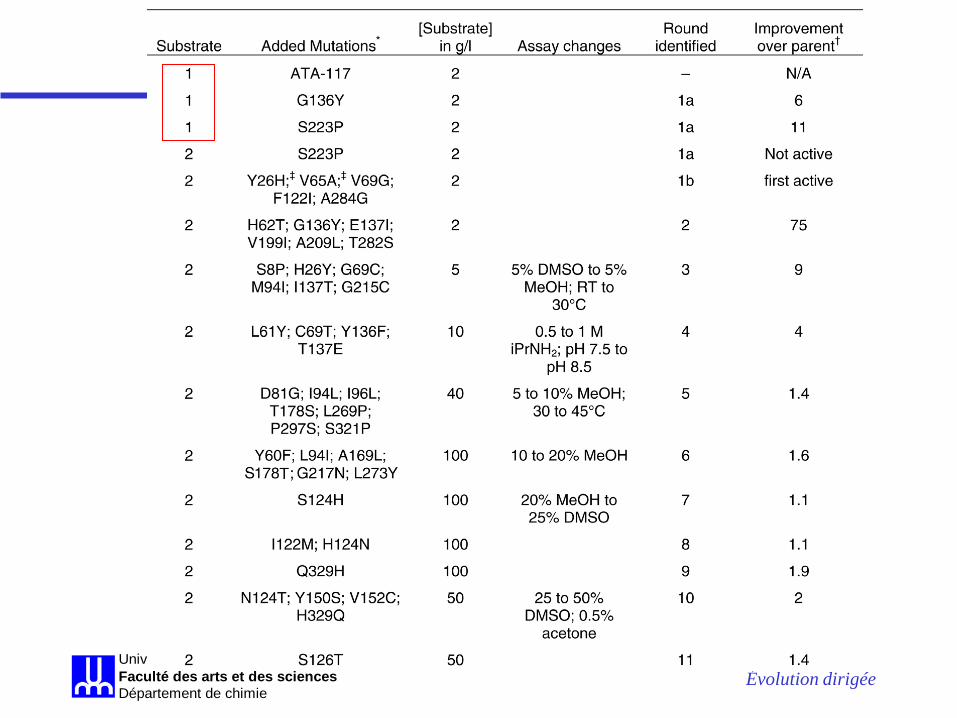
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La sitagliptine: amélioration d’une transaminase
La procédure fut-elle efficace?
•Au début: aucune transformation de 2 •À la fin: conversion de 200 g / L (!) de 2 avec un excès énantiomérique de >99.95%, en utilisant 6 g / L de transaminase en 50 % de DMSO; rendement de 92%.
Le procédé biocatalytique augmente le rendement de 10 – 13 %, avec une augmentation de productivité de 53% (donc la conversion est plus rapide = $ économisés) De plus: réduction des déchets de 19% élimination des métaux lourds aucun besoin de pression élevée donc plus besoin d’équipement spécialisé réduction des coûts de transformation
Est-ce mieux que la conversion chimique?

Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La sitagliptine: amélioration d’une transaminase
Globalement, 27 mutations furent incorporées à la transaminase améliorée: 4 par design rationnel de la petite cavité de liaison 5 suite à la mutagenèse par saturation de la grande cavité de liaison 1 suite à la mutagenèse par saturation de la petite cavité de liaison 2 suite à la mutagenèse de résidus avoisinants des cavités de liaison 10 par inclusion de mutations selon l’homologie de séquence parmi plusieurs transaminases 5 par mutagenèse aléatoire
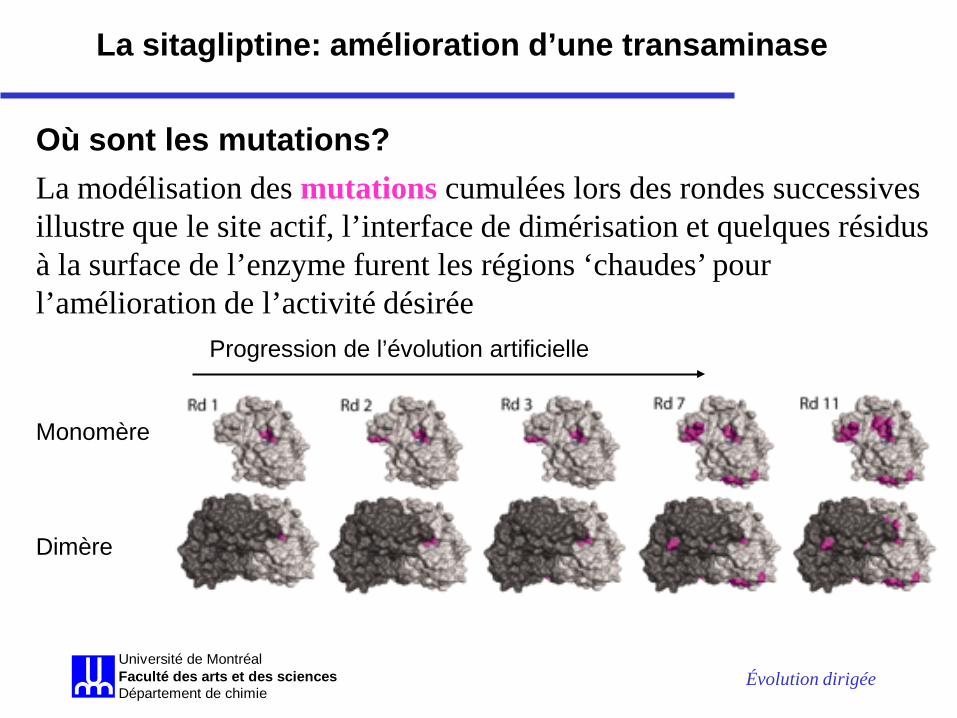
Évolution dirigée Université de MontréalFaculté des arts et des sciencesDépartement de chimie
La modélisation des mutations cumulées lors des rondes successives illustre que le site actif, l’interface de dimérisation et quelques résidus à la surface de l’enzyme furent les régions ‘chaudes’ pour l’amélioration de l’activité désirée
Monomère Dimère
Progression de l’évolution artificielle
Où sont les mutations?
La sitagliptine: amélioration d’une transaminase