approches moléculaires de la diversité microbienne de deux
TRANSCRIPT

Approches moleculaires de la diversite microbienne de
deux environnements extremes : les sources
hydrothermales profondes et les reservoirs petroliers
Erwan Corre
To cite this version:
Erwan Corre. Approches moleculaires de la diversite microbienne de deux environnementsextremes : les sources hydrothermales profondes et les reservoirs petroliers. Biologiemoleculaire. Universite de Bretagne Occidentale, 2000. Francais. <tel-01115381>
HAL Id: tel-01115381
http://hal.upmc.fr/tel-01115381
Submitted on 12 Feb 2015
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinee au depot et a la diffusion de documentsscientifiques de niveau recherche, publies ou non,emanant des etablissements d’enseignement et derecherche francais ou etrangers, des laboratoirespublics ou prives.

Avertissement
Au vu de la législation sur les droits d'auteur, ce travail de thèse demeure lapropriété de son auteur, et toute reproduction de cette oeuvre doit faire l'objetd'une autorisation de l'auteur. (cf Loi n°92-597; 1/07/1992. Journal Officiel,2/07/1992)

THESE
présentée pour l'obtention
du grade de Docteurde l'Université de Bretagne Occidentale
(Mention Microbiologie)
Erwan CORRE
Approches moléculaires de la diversitémicrobienne de deux environnements extrêmes :
les sources hydrothermales profondes et lesréservoirs pétroliers
Soutenue le Mercredi 2 Février 2000 devant le jury composé de :
MM. P. CAUMETTE, Professeur, Université de Pau ExaminateurM. MAGOT, Responsable de Laboratoire, Sanofi, Labège RapporteurP. NORMAND, Directeur de Recherche CNRS, Université Lyon I RapporteurB. PICARD, Professeur, Université de Bretagne Occidentale PrésidentD. PRIEUR, Professeur, Université de Bretagne Occidentale ExaminateurA.L. REYSENBACH, Professeur, Université de Portland, USA Examinatrice

Remerciements
Le travail présenté dans ce mémoire a été réalisé avec le soutien financier de la Communauté Urbaine de Brest
Je remercie tout d'abord Daniel Prieur pour m'avoir accueilli dans son équipe de recherche, d'abords en DEA puis en thèse et
de m'avoir fait confiance tout au long de ces années.
Mes remerciements s'adressent tout particulièrement à Christian Jeanthon pour son soutien, sa patience et sa disponibilité.
Je tiens à lui exprimer ici ma reconnaissance pour toutes les discussions enrichissantes, les conseils avisés et les directives salutaires
qu'il a su m'indiquer tout au long de ce travail. Merci profondément pour le temps consacré à la correction des articles.
Je remercie également Anna-Louise Reysenbach pour m'avoir accueilli dans son laboratoire à Rutgers et de m'avoir permis
de travailler sur les échantillons de Vent Cap. Son expérience en biologie moléculaire, ses conseils en phylogénie m'ont été
grandement utiles. Je la remercie également de m'avoir fait partager durant mon séjour à Rutgers sa passion pour les chevaux et
pour le trafic de fromages et de vins français.
Je remercie également les membres du jury : Pierre Caumette, Philippe Normand, Michel Magot et Bertrand Picard d'avoir
accepté de juger mon travail de thèse.
Je tiens à adresser mes plus profonds remerciements à l'ensemble des étudiants avec qui j'ai eu la joie de travailler : Cédric
Hobel (qui a essuyé les plâtres), la brillante Anne Postec, l'inénarrable mais néanmoins charmante Catherine Charpentier et enfin
le précieux Laurent Toffin avec qui j'ai passé une année de travail formidable. Une grande partie des résultats présentés dans ce
manuscrit sont le fruit de leur acharnement et de leur persévérance.
J'adresse un remerciement tout particulier à Tristan Barbeyron pour m'avoir formé aux techniques moléculaires, pour sa
disponibilité et son savoir intemporel sur tout ce qui à trait la biologie moléculaire, la microbiologie, la radioactivité, Serge Lama et
les 24 heures du Mans. Merci également à Hermie Harmsen pour ses conseils avisés à la paillasse comme à l'e-mail.
Je remercie tout particulièrement Olivier Collin, Stéphan Jouannic et Jean Ranc pour les heures qu'ils ont accepté de passer
à lire et corriger ce manuscrit. Spéciale dédicace à Stéphane et Olivier pour leur travail de pionniers dans la jungle des fautes
d'orthographes et les marais fangeux des accords mal à propos.
Je remercie également tous les membres du laboratoire. En particulier Stéphane et Edmond pour leur amitié, les foulées et la
sueur que nous avons partagé (enfin surtout eux) sur de nombreux kilomètres. Jenny pour m'avoir supporté et pour nous être
épaulé mutuellement dans la dernière (longue, très longue) ligne droite. Marc, Patrick, Olivier, Jean-Louis, Laurent, Viggo, Soizick
et Valérie pour avoir survécu dans les bureaux que nous avons partagés. Enfin je remercie les membres de la Station Biologique de
Roscoff pour leur accueil, leurs compétences et la formidable ambiance qu'ils savent entretenir dans leur laboratoire et dans les
couloirs de la station.
Merci à Odile, Claire, Catherine's, Jeff's, Didier, Vincent et Lolo, à la famille Collin à 2 jambes ou 4 pattes pour leur soutient
moral et véliplanchesque ainsi que pour leur accueil tout au long de mon travail.
Merci à l'ensemble de ma famille, à Juliette de m'avoir aiguillé vers la Station Biologique, à mes parents, "beaux"-parents et
grands-parents de leur soutien.
Merci enfin à Anne d'avoir su supporter au quotidien mes sautes d'humeurs, mes absences et d'avoir finalement, plus qu'aucune
autre personne, contribué à ce que je mène à bien ce travail.

ii
Introduction générale
La microbiologie de l'environnement est une discipline jeune, qui a pour vocation d'étudier les
relations qu'entretiennent les micro-organismes avec leur biotope. Si l'adage populaire n'accorde que
peu d'influence à ce qui est invisible à l'oeil et si la présence de micro-organismes est souvent
associée à une nuisance plutôt qu'à un bienfait, le rôle des micro-organismes est pourtant
prépondérant dans le fonctionnement de nombreux écosystèmes naturels et anthropiques. L'étude
des micro-organismes a longtemps été liée à la capacité des microbiologistes à recréer in vitro des
conditions favorables à la croissance des communautés microbiennes. Or, des écosystèmes
complexes, soumis à des contraintes physico-chimiques extrêmes telles que, de fortes températures,
de fortes pressions ou l'absence d'oxygène, ou à des fluctuations de sources de nutriments ou
d'énergie, sont particulièrement difficiles à simuler au laboratoire. Aussi, dans ces écosystèmes
,comme dans d'autres étudiés déjà depuis fort longtemps tels que le sol où l'eau de mer, un grand
nombre de micro-organismes ont échappé à toute méthode de culture. Le développement des
techniques de biologie moléculaire et l'utilisation de la molécule d'ARN ribosomique comme marqueur
évolutif a permis de s'affranchir des étapes de culture et a permis d'asseoir sur des bases
phylogénétiques les relations taxinomiques s'établissant entre les différents organismes vivants.
Le travail présenté dans ce mémoire a pour objet l'étude de la diversité microbienne de deux
écosystèmes considérés comme extrêmes : les sources hydrothermales profondes et les réservoirs
pétroliers. Les techniques mises en oeuvre sont fondées soit sur l'analyse des ARN ribosomiques
(ARNr) sans étape préalable de culture, par séquençage ou hybridation in situ, soit sur le
positionnement phylogénétique de souches isolées de ces environnements.
Avant de présenter les résultats de ce travail sous la forme d'articles en préparation, soumis ou
publiés, je me propose :
- de présenter une revue des techniques d'écologie microbienne classiques et moléculaires
couramment utilisées pour accéder à la diversité des écosystèmes microbiens, de présenter des
moyens d'accéder à la diversité fonctionnelle de ces écosystèmes et d'insister sur les biais inhérents à
chacune de ces techniques,
- d'effectuer un tour d'horizon des différents habitats microbiens qui ont été étudiés ces dernières
années et de présenter pour chacun de ces habitats les apports des techniques moléculaires dans la
description de la diversité et la découverte de nouveaux groupes phylogénétiques,
et enfin
- de présenter et de discuter les différentes techniques que nous avons mises en oeuvre pour étudier
la diversité microbienne d'un écosystème pétrolier continental et d'un site hydrothermal profond.

iii
Abréviations
A : Adénosine
ADN: Acide désoxyribonucléique
ADNc : ADN cloné
ADNr 16S : ADN codant pour la sous unité 16S de l'ARN
ribosomal
ADNr : ADN codant pout l'ARN ribosomal
AFLP : Amplified Fragment Length Polymorphism
APS : Ammonium Persulfate
ARDRA : Amplified rDNA Restriction Analysis.
ARN : Acide ribonucléique
ARNm : ARN messagé
ARNr 16S : sous unité 16S de l'ARN ribosomal
ARNr : ARN ribosomale
ATP : Adénosine tri-phosphate
BrdU : bromodéoxyuridine.
BSA : Bovine Serum Albumin
BSR: Bactéries sulfato-réductrices
C : Cytosine
C.F.B. : Cytophaga-Flavobacterium-Bacteroides
CDGE : Constant Denaturant Gel electrophoresis
CMF : Cytométrie en flux
comm. pers. : communication personelle
CTAB : Hexadecyltrimethyl ammonium bromide
CTC : 5-cyano-2,3-dotolyl tetrazolium chloride
Da : Dalton
DAPI: Acide diamino-pimélique
DGGE : Denaturing Gradient Gel Electrophoresis
DH : Deshydrogénases
DMSO : Di methyl sulfoxyde
dNTP : deoxynucléotide triphosphate
dsADN : ADN double brin (double strand)
EDTA : Ethylenediaminetetraacetic acid
EPR : East Pacific Ride
FAME : Fatty acid Methyl Ester
FISH : Fluorescent In Situ Hybridization
G : Guanidine
GES (tampon) : GIT/EDTA/SDS
GIT : Guanosine isothiocyanate
GNS : Bactéries vertes non sulfureuses
HPLC : High Presure Liquid Chromatography
INT : 2 (p-iodo-pheny)-3-(p-nitropheny)-5-phenyl tetrazolium
chloride
IPTG : isopropylthio-ß-D-galactoside
ISRT : In Situ Reverse Transcription
Knuc : distance phylogénétique
Kpb : Kilo paires de bases
LB (milieu liquide ou gélose) : Luria-Bertani
LFRFA : Low Frequency Restriction Fragment Analysis
LMW RNA: ARN de faible poids moléculaire
LPS : Lipopolysaccharides
LUCA : Last Unversal Common Ancestor
MAR : Middle Atlantic Ride
MPN : Most probable number
NPP : Nombre le plus probable
OTU : Operational Taxonomic Unit
p/v : (masse à volume)
pb : paire de base
PCB : PolyChlorinated Biphenyl
PCR : Polymerase Chain Reaction
PFLA : analyse des profils lipidiques
PGFE : Pulsed-Field Gel Electrophoresis
RAPD : Randomly Amplified Polymorphic DNA
RDP : Ribosomal Database Project
RFLP : Restriction Length Fragment Polymorphism
RPM : Rotation par minute
RT-PCR : Reverse Transcriptase-PCR
S : Svedberg
SDS : Sodium dodecyl sulfate
SET (tampon) : Sucrose/EDTA/Tris
ssADN : ADN simple brin (single strand)
T : Thymine
T-RFLP : Terminal Restriction Length Fragment
Polymorphism
TAE (tampon) : Tris/Amonium/EDTA
TBE (tampon) : Tris/Borate/EDTA
Td : Température de dissociation
TE (tampon) : Tris/EDTA
TEMED : N,N,N',N' Tétraméthyléthylènediamine
TGGE : Temperature Gradient Gel Electrophoresis
Tm : Température de fusion
TNE (tampon) : Tris/NaCl/EDTA
UFC : Unité formant colonie
v/v : volume à volume
X-Gal : 5-bromo-4-chloro-3 indolyl-ß-D-galactopyranoside

iv
Sommaire
Remerciements i
Introduction générale ii
Abréviations iii
Sommaire iv
Partie 1 : Les techniques d'écologie microbienne
I Notions d'écologie microbienne 1
II Les méthodes classiques d'écologie microbienne 3
2.1 Les cultures d'enrichissement et l'isolement 3
2.1.1 L'enrichissement 3
2.1.2 L'isolement et la taxinomie 4
2.2 Quantification et mesure d'activité microbienne 7
2.2.1 Quantification 7
2.2.2 Mesure d'activité microbienne 12
2.3 Une vue restreinte de la diversité 14
III Les acides nucléiques en écologie microbienne 15
3.1 Le développement des techniques moléculaires 15
3.1.1 La phylogénie moléculaire 16
3.1.2 L'ARN ribosomique 16
3.1.3 Le choix de l'ARN 16S 17
3.2 L'analyse phylogénétique 18
3.2.1 Les méthodes de phylogénie 18
3.2.2 L'informatique au service de la phylogénie 23
3.3 Les méthodes moléculaires 27
3.3.1 Le clonage et le séquençage des ARNr 27
3.3.2 Les sondes nucléiques 32
3.3.3 La DGGE 34
3.3.4 Les autres techniques 36
3.4 De la diversité phylogénétique à la diversité physiologique 37
3.5 Les biais des techniques moléculaires 39
3.5.1 Echantillonnage et conservation des échantillons 40
3.5.2 Les techniques de PCR 40
3.5.3 Les techniques d'hybridation 42
3.5.4 Les techniques de DGGE 43
3.5.5 Une solution : l'approche pluridisciplinaire 43
Partie 2 : Les habitats microbiens
I Les origines de la vie 45
1.1 Les premières traces de vie 45

v
1.2 Une origine de la vie controversée 46
II. L'atmosphère 49
III. Les habitats terrestres 49
3.1 Les couches superficielles de la lithosphère 49
3.1.1 Description de l'habitat 49
3.1.2 La rhizosphère 50
3.1.3 L'apport des techniques moléculaires 50
3.2 La biosphère souterraine 52
3.2.1 Description de l'habitat 52
3.2.2 Le concept de biosphère souterraine 53
3.2.3 La communauté microbienne souterraine 54
3.2.4 Des profondeurs aux astres 55
3.3 Les réservoirs pétroliers 56
3.3.1. Formation et production du pétrole 56
3.3.2. Composition physico-chimique du biotope 57
3.3.3. La diversité microbienne des puits de pétrole 58
3.3.4 Activité microbienne liée aux hydrocarbures 62
IV. Habitats aquatiques 63
4.1 L'habitat d'eau douce 64
4.2 L'habitat maritime 65
4.2.1 Les micro-organismes marins isolés 65
4.2.2 L'utilisation des techniques moléculaires 65
4.2.3 Les Fosses océaniques 69
4.3 Les zones sédimentaires marines et lacustres 69
4.3.1 Description du biotope 69
4.3.2 Les techniques moléculaires 70
4.4 Les sources hydrothermales océaniques profondes 72
4.4.1 Nature et structure des émissions hydrothermales profondes 73
4.4.2 Faune associée à l'écosystème hydrothermal 76
4.4.3 Les micro-organismes des sources hydrothermales 78
4.5 Les sites hydrothermaux continentaux et côtiers 89
4.5.1 Description des biotopes 89
4.5.2 Les micro-organismes des sites côtiers et continentaux 91
V La flore microbienne liée à l'Homme, aux animaux et à l'activité humaine 97
5.1 L'Homme comme écosystème microbien 97
5.1.1. La flore de la cavité buccale 97
5.1.2. Les micro-organismes du tractus digestif 98
5.1.3 L'apport des techniques moléculaires en microbiologie humaine 99
5.2 La flore microbienne associée aux autres animaux 100

vi
5.3 Les micro-organismes associés aux activités humaines 103
5.3.1 L'extraction de minerai 103
5.3.2. Les boues de stations d'épuration 104
5.3.3 Les autres milieux 105
VI Conclusion et présentation du projet d'étude 106
Partie 3 : Matériels et méthodes
I Echantillonnage et conservation de l'échantillon 109
1.1 Echantillonnage des puits de pétrole 109
1.2 Echantillonnage des sources hydrothermales profondes 109
1.2.1 Echantillons de cheminée 110
1.2.2 Echantillons de fluides 111
1.3 Echantillonnage des sources chaudes terrestres 111
II Analyse de l'ADNr 16S 111
2.1 Extraction d'ADN 111
2.1.1 Extraction de l'ADN sur filtre ou sur culot de centrifugation 112
2.1.2 Extraction de l'ADN chromosomique par la méthode de Sambrook et al. (1989) 112
2.1.3 Méthode d'extraction au micro-ondes 113
2.1.4 Extraction de l'ADN chromosomique par la méthode de Pitcher et al. (1989) 113
2.1.5 Extraction de l'ADN au "Bead-Beater" 113
2.2 Amplification de l'ADNr 16S 114
2.2.1 Réactions d'amplification classiques 114
2.2.2 Techniques d'optimisation de la réaction 114
2.2.3 Choix des amorces d'amplification 115
2.3 Clonage des produits de PCR 116
2.3.1 Utilisation du kit TOPO TA Cloning 116
2.3.2 Transformation des cellules par électroporation 117
2.4 Analyse des clones par RFLP : Restriction Fragment Length Polymorphism 117
2.5 Séquençage 118
2.5.1 Réactions de séquence 118
2.5.2 Amorces de séquençage 118
2.5.3 Migration des réactions de séquences 119
2.6 Analyse des séquences 120
III Hybridation in situ 120
3.1 Construction des sondes 120
3.2 Mise au point des sondes - Test de spécificité 121
3.2.1 Hybridation sur membrane 121
3.2.2 Hybridation sur culture pure 123
3.3 Hybridation in situ 123
3.3.1 L'enrobage des lames d'observation 123

vii
3.3.2 Fixation des cellules 123
3.3.3 Extraction des cellules de l'échantillon 124
3.3.4 Hybridation in situ 124
IV Observation et quantification des cellules 125
Partie 4 : Articles
Article 1 129
Article 2 154
Article 3 174
Article 4 193
Article 5 202
Article 6 206
Conclusion générale 209
Réferences bibliographiques 214
Annexes
Structure secodaire de l'ADNr 16S de E. coli
Arbre phylogénétique des Bacteria et des Archaea

Les techniques d'écologie microbienne

Les techniques d'écologie microbienne
1
La microbiologie est fondée depuis le 19ème siècle sur l'observation et la culture des micro-
organismes. Or on s'est rendu compte que les techniques de mise en culture donnent une image
faussée, puisque obligatoirement sélective, du rôle et de l'importance des micro-organismes dans la
nature. Aussi, aux techniques classiques de microbiologie (techniques de culture, de comptage,
microscopie électronique, utilisation de marqueurs radioactifs) se sont ajoutées des techniques de
biologie moléculaire (analyses du génome, sondes nucléiques, séquençage) qui permettent
dorénavant d'obtenir une image plus précise des relations des micro-organismes avec leur
environnement. Dans ce chapitre je me propose, après un bref rappel de notions d'écologie
microbienne, de présenter les techniques classiquement utilisées en écologie puis, plus
spécifiquement, les techniques fondées sur l'utilisation des acides nucléiques. Une attention
particulière sera portée aux biais inhérents à ces technologies.
I Notions d'écologie microbienne
Définitions
Un écosystème est un ensemble dynamique au sein duquel interagissent à divers degrés des
facteurs biotiques (individu, population, communauté) et abiotiques (facteurs physico-chimiques) (cf.
figure 1.1) (Begon et al., 1996).
IndividuIndividuIndividuIndividuIndividuIndividu
Individu Ecosystème
Effet des facteurs abiotiques (environnement)
Complexité de l'interaction croissante
PopulationPopulationPopulationPopulationPopulation CommunautéCommunautéCommunauté
Guilde
pH, T°, 02, NaCl, Lumière, H20, Nutriments
Figure1.1. Représentation schématique des relations au sein d'un écosystème
Individu : Unité distincte qui évolue séparément des autres, qui se reproduit dans un groupe et a un rôle (une
niche) défini.
Population : Groupe d'individus qui vivent sur une zone définie.
Guilde : Plusieurs populations distinctes réalisant des tâches similaires ou exploitant les mêmes ressources
environnementales dans un but similaire.
Communauté : Assemblage de populations dans un même espace et sur une même période de temps.
Environnement : Se réfère à tout ce qui entoure un organisme vivant. Correspond aussi bien à des facteurs
physiques, chimiques ou biologiques qu'aux forces qui agissent sur l'organisme.

Les techniques d'écologie microbienne
2
Les interactions entre les composants de l'écosystème
La structure d'une communauté microbienne va donc être soumise à des fluctuations
temporelles et spatiales qui vont correspondre à des phases de colonisation de nouveaux milieux ou
de remplacement d'une communauté par une autre plus adaptée (comme c'est le cas dans le tractus
digestif au cours de la croissance d'un individu). Nous savons qu'un grand nombre de facteurs
peuvent modifier la composition de la communauté microbienne : la disponibilité en sources de
carbone, d'énergie et en accepteurs d'électrons, la température, le pH, l'O2, la teneur en eau ou
encore la lumière. Aussi pour survivre et se développer les micro-organismes ont développé des
adaptations structurales et nutritionnelles qui correspondent à différentes stratégies en fonction des
conditions physico-chimiques auxquelles ils sont exposés (par exemple stratégies r et K, cf. tableau
1.1).Tableau 1.1. Comparaison des stratégies r et K d'après Mac Arthur (1962)
stratégies r stratégies K
Ø Taux de reproduction élevé Ø Taux de reproduction faible
Ø La forte teneur en nutriments permet une croissance importante au
détriment de cellules moins bien adaptées
Ø La faible teneur en nutriments limite la croissance de tous
les organismes.
Ø Très forte population microbienne Ø Population plus réduite
Ø Grosses variations de population lorsque la teneur en nutriments
varie
Ø Communauté pérenne et stable
Ø Ex: "Blooms" cyanobactériens dus à un apport de phosphates, ou
"blooms"" de Pseudomonas dus a un apport de sources carbonées
Ø Ex: Spirillum et Vibrio dans l'eau de mer, bactéries dans les
lac oligotrophiques
D'autres part, aux facteurs extérieurs s'ajoutent de fortes interactions qui peuvent se produire au
sein d'une population (coopération ou compétition) ou entre populations (synergie, symbiose et
antagonisme).
L'habitat naturel
L'étendue de l'habitat microbien est à mettre en rapport avec la taille du micro-organisme1. Même
si les cellules seules n'existent pas ou très peu dans la nature (on parle plutôt de micro-colonies) des
micro-environnements de l'ordre du millimètre peuvent être le siège de très fortes variations physico-
chimiques (c'est le cas notamment dans les particules de sol). De plus les cellules colonisent souvent
les surfaces sous la forme d'un biofilm. Cette structure complexe permet aux micro-organismes
d'accéder plus facilement aux nutriments nécessaires à leur croissance et de les stocker.
L'étude des populations microbiennes dans leur environnement devra donc répondre aux
questions de la composition, de la structure et de la fonction de cette population. Pour répondre à ces
questions les premiers microbiologistes disposaient de méthodes fondées sur leur capacité à recréer
le plus fidèlement possible les conditions de croissance in situ . Par la suite, le développement des
techniques moléculaires a permis de s'affranchir partiellement des étapes de mise en culture.
1 Pour une cellule de 3µm une distance de 3 mm correspond à l'équivalent d'une distance de 2 km pour l'homme.

Les techniques d'écologie microbienne
3
II Les méthodes classiques d'écologie microbienne
2.1 Les cultures d'enrichissement et l'isolement
2.1.1 L'enrichissement
La plupart de nos connaissances sur les micro-organismes proviennent d'études réalisées sur
des cultures pures isolées de l'environnement. Cet isolement est précédé par une phase
d'enrichissement qui va favoriser la croissance d'un type donné de micro-organisme, sélectionné en
fonction des conditions physiques et nutritionnelles du milieu.
On distingue deux grands types de milieux, les milieux électifs où les souches les plus adaptées
aux conditions de culture vont se développer et les milieux sélectifs où on intervient physiquement ou
chimiquement pour favoriser la croissance d'un type donné de micro-organismes2. On favorise dans
tous les cas les micro-organismes ayant les vitesses de croissance des plus élevées dans des
conditions données.
Il existe plusieurs moyens classiques de modifier la sélectivité d'un milieu :
Ø Ajouter un composé utilisé par un organisme comme source nutritive.
Ø Supprimer un composé utilisé uniquement par les organismes que l'on ne souhaite pas isoler
(supprimer toutes les sources d'azote autres que l'azote gazeux (N2) pour isoler les bactéries
fixatrices d'azote).
Ø Ajouter des antibiotiques (pénicilline pour inhiber les bactéries Gram + et certaines bactéries
Gram -, néomycine et streptomycine pour inhiber les bactéries en général, actidone et nystatine
pour inhiber les champignons).
Ø Modifier les propriétés physico-chimiques du milieu (pH, potentiel redox, etc.).
Ø Modifier les conditions d'incubation (température, teneur en eau, pression osmotique, lumière,
etc.).
Un panel de milieux d'enrichissement, présentant l'ensemble des conditions physico-chimiques
rencontrées dans le biotope d'étude, devra être choisi pour permettre la croissance du plus grand
nombre possible de types métaboliques.
Compte tenu de la complexité de certains écosystèmes, il est parfois nécessaire de mettre en
oeuvre des systèmes élaborés de culture afin de pouvoir assurer le développement de certains
organismes.
C'est le cas notamment des milieux à fort gradient (physico-chimiques, nutritionnels, etc.) pour
lesquels les systèmes suivants ont été mis au point :
Ø La colonne de Winogradsky (cf. figure 1.2).
Ø la chambre de culture en flux continu de Huber et al. (1998b) pour isoler Thermocrinis ruber, une
bactérie hyperthermophile pléomorphe formant, une fois fixée, de longs filaments roses dans les
2 Les milieux électifs sont en fait des milieux peu sélectifs.

Les techniques d'écologie microbienne
4
sources chaudes d'Octopus Spring à Yellowstone (USA). Les systèmes de culture en continu
permettent de sélectionner des micro-organismes ayant de fortes affinités avec des substrats en
faibles concentrations. Les micro-organismes n'ayant pas d'affinité suffisante sont lessivés du milieu.
Ø La "Benthic Gradient Chamber" mise au point pour reproduire les gradients sédimentaires et étudier
les phénomènes de compétitions entre bactéries pourpres et vertes (Caumette et al., 1998).
Ø Les chémostats pour maintenir en culture les bactéries formant des enchevêtrements ("mats")
filamenteux de soufre (Taylor et al., 1999).
Ø Les co-cultures pour la croissance de bactéries syntrophes : Syntrophomonas avec des bactéries
sulfato-réductrices (BSR) (Harmsen et al., 1993) ou de Proteobacteria epsilon soufre oxydantes avec
des BSR (Thiovulum sp. pour Wirsen et Jannasch, 1978 ou Arcobacter sp. pour Teske et al., 1996b).
Dans ce cas le développement du micro-organisme est conditionné par la consommation d'un
des produits de son métabolisme par un second micro-organisme.
Couvercle
Algues et cyanobactéries
Bactéries soufréespourpres
Bactéries soufréesvertes
Décompositionanaérobie et sulfato-réduction
Bouesenrichies enmatièresorganiques eten CaSO4
Eau de lacou demarais
Figure 1.2. Principe de fonctionnement d'une colonne de
Winogradsky. La phase supérieure reproduit les
conditions retrouvées dans le milieu liquide (luminosité,
aération) alors que la phase inférieure reproduit les
conditions des premiers centimètres de sédiments
(obscurité, augmentation du potentiel réducteur). Au cours
du temps, l'établissement de la flore microbienne
s'effectue en fonction de leur préférendum au sein des
gradients (d'après Madigan, 1996).
2.1.2 L'isolement et la taxinomie
A partir de l'enrichissement il est possible d'isoler des espèces pures. Pour ce faire on dispose
de trois grands types de techniques : l'isolement sur boite, la dilution successive en milieu liquide ou
dans l'agar. Pour chacune des méthodes plusieurs repiquages successifs des souches isolées seront
nécessaires pour s'assurer de la pureté de l'isolat.
Une fois la souche isolée il convient de la caractériser, c'est à dire de définir les caractères
phénotypiques et génotypiques qui permettront de la comparer avec d'autres micro-organismes
préalablement isolés. On réalise alors une taxinomie. Une partie de ces caractères est regroupée dans
le tableau 1.2.
La taxinomie microbienne a longtemps été considérée comme une discipline ingrate. A l'instar
de la taxinomie des organismes supérieurs elle était essentiellement fondée sur l'utilisation de clés
dichotomiques successives. Or ces clés n'étaient pas d'un emploi simple et étaient de plus sources

Les techniques d'écologie microbienne
5
d'erreurs. Le développement de la taxinomie numérique3 et des approches polyphasiques dans les
années 70 (Colwell, 1970; Sneath et Sokal, 1973; Vandamme et al., 1996) mais surtout l'apparition de
la taxinomie moléculaire (en particulier l'analyse de l'ARN ribosomique ou des gènes codant pour
l'ARNr (ADNr)) ont permis de redonner un véritable élan à cette discipline.
Tableau 1.2. Liste non -exhaustive des caractères phénotypiques et génotypiques déterminés pour une souche isolée.
Morphologie :
Forme de la cellule
Taille de la cellule (diamètre, longueur)
Mobilité
Flagelle
Type de division cellulaire
Différenciation cellulaire
Structures internes et externes (endospores, vésicules
gazeuses, etc.).
Coloration de Gram
Ultrastructure
Composition chimique et analyse molécul aire :
Couleur des suspensions de cellules
Pigments
Composition en base de l’ADN
Séquence de l’ARNr 16S
Réassociation ADN/ADN entre espèces proches
Acides gras totaux.
Physiologie :
Milieu de culture
Optimum et gamme de température
Optimum et gamme de pH
Croissance phototrophique ou lithotrophique
Besoins en vitamines
Sources de carbone nécessaire à la croissance
Sources d’azote nécessaire à la croissance
Relation vis à vis de l’oxygène
Modes de production d’énergie :
- donneurs d’électrons : organiques ou inorganiques
- accepteurs d’électrons : oxygène, nitrate, sulfate, CO2,
oxydes de fer, etc.
Enzymes extracellulaires :
- amylases, lipases, gélatinases, cellulases, xylanases
Autres enzymes :
- oxydase, catalase
Fermentation du glucose
Dénitrification
La notion d'espèce est le concept clé et l'unité de base de la taxinomie. Elle résulte des
premiers travaux de hiérarchisation du monde vivant au 18ème siècle par le naturaliste suédois Carl Von
Linné. Selon la définition devenue classique proposée par Ernst Mayr (1982), une espèce est un
ensemble d'individus féconds entre eux et seulement entre eux, donc isolés des autres groupes
d'êtres vivants. Or le concept de sexualité chez les procaryotes est très différent de celui des
eucaryotes puisque la reproduction est végétative et asexuée. De plus on sait que des phénomènes
de parasexualité existent chez les procaryotes (conjugaison, transformation et transduction) et que
ceux-ci passent la barrière des espèces (cf. revue de Taddei et al., 1996). A la différence des
eucaryotes, les caractères morphologiques des procaryotes considérés seuls n'ont toujours qu'une
faible signification dans la classification des bactéries car la grande majorité des micro-organismes ont
des formes trop simples pour que l'on puisse les utiliser pour la taxinomie. Le développement des
techniques moléculaires a permis de donner une définition très rigoureuse de l'espèce.
Cette définition est fondée sur les propriétés de dénaturation et de renaturation du double brin
de l'ADN génomique bactérien en fonction de la température. Elle permet de définir une espèce
3 Des caractères phénotypiques (tels que ceux présentés dans le tableau 1.2) peuvent servir de base à la comparaison de plusieurs
souches. Il faut s'assurer que les caractères comparés sont informatifs et notamment qu'ils ne correspondent pas à un caractère
dérivé. Il convient dans un premier temps de discrétiser ces caractères (sous forme de 0 et de 1) puis de les présenter sous la forme
d'une matrice. Les similitudes entre organismes vont être exprimées sous forme de dendrogrammes. Des valeurs d'hybridations
ADN-ADN ou ARN-ADN pourront être aussi utilisées comme caractères dans une taxinomie. Toutefois les hybridations ADN-ADN ne
vont être informatives que dans le cas d'espèces proches et ne pourront être utilisées entre des espèces éloignées.
Les techniques d'écologie microbienne
6
procaryotique comme constituée de souches qui présentent des valeurs d'hybridation ADN-ADN
supérieures ou égales à 70% et une valeur ∆Tm4 inférieure ou égale à 5°C (Wayne et al., 1987). Par la
suite la généralisation de l'analyse de l'ADNr a incité les auteurs à corréler les pourcentages d'identité
entre ARNr 16S et les pourcentages de réassociation ADN/ADN (Stackebrandt et Göebel, 1994). Ainsi
les séquences d'ADNr ayant des identités inférieures à 97% correspondent généralement à des
espèces différentes. Amann et al. (1995) abaissent par précaution cette limite à 95%. Selon certains
auteurs les critères d'hybridation ADN-ADN et de similarité d'ADNr 16S ne suffisent pas pour distinguer
des populations écologiquement distinctes. Il faut leur associer des mesures de similarité de gènes
codants pour des protéines du métabolisme (plus impliquées dans les mécanismes écologiques)
(Palys et al., 1997).
Le positionnement taxonomique d'une souche ne saurait toutefois se faire uniquement sur des
critères génétiques et l'utilisation de l'ensemble des caractères phénétiques et génétiques
permettent d'ancrer plus solidement l'organisme au sein d'une famille, d'un genre ou d'une espèce
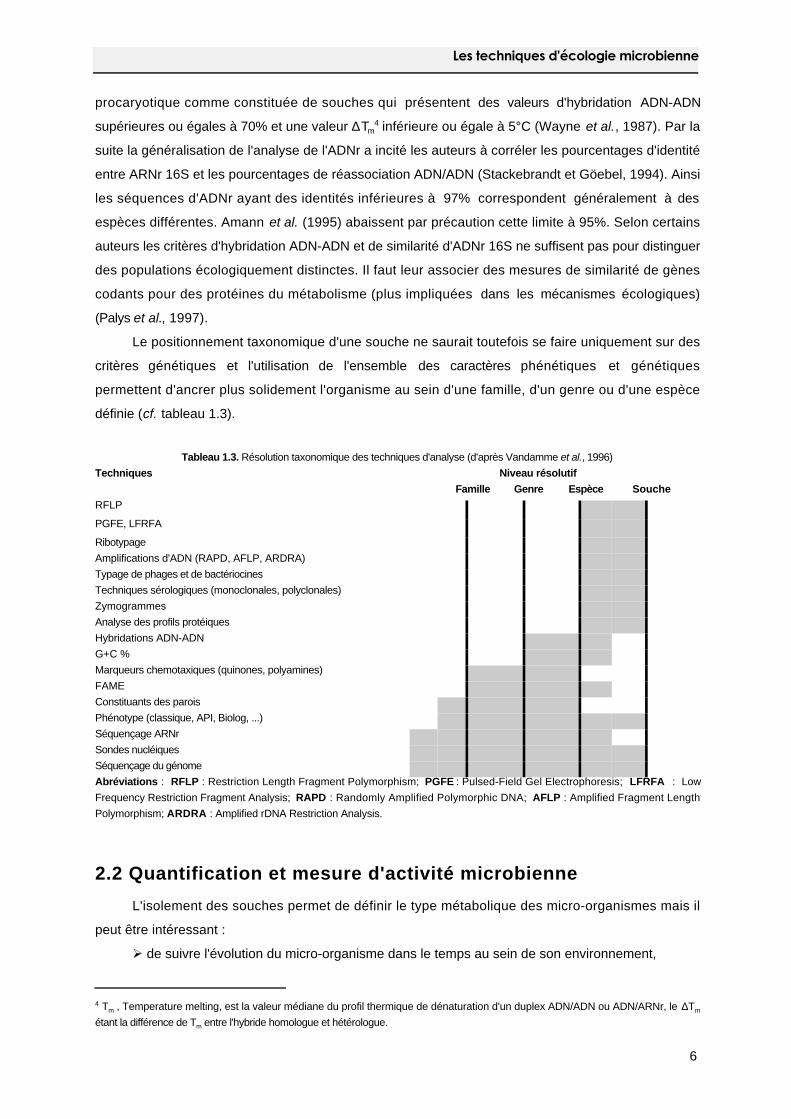
définie (cf. tableau 1.3).
Tableau 1.3. Résolution taxonomique des techniques d'analyse (d'après Vandamme et al., 1996)
Techniques Niveau résolutif
Famille Genre Espèce Souche
RFLP
PGFE, LFRFA
Ribotypage
Amplifications d'ADN (RAPD, AFLP, ARDRA)
Typage de phages et de bactériocines
Techniques sérologiques (monoclonales, polyclonales)
Zymogrammes
Analyse des profils protéiques
Hybridations ADN-ADN
G+C %
Marqueurs chemotaxiques (quinones, polyamines)
FAME
Constituants des parois
Phénotype (classique, API, Biolog, ...)
Séquençage ARNr
Sondes nucléiques
Séquençage du génome
Abréviations : RFLP : Restriction Length Fragment Polymorphism; PGFE : Pulsed-Field Gel Electrophoresis; LFRFA : Low
Frequency Restriction Fragment Analysis; RAPD : Randomly Amplified Polymorphic DNA; AFLP : Amplified Fragment Length
Polymorphism; ARDRA : Amplified rDNA Restriction Analysis.
2.2 Quantification et mesure d'activité microbienne
L'isolement des souches permet de définir le type métabolique des micro-organismes mais il
peut être intéressant :
Ø de suivre l'évolution du micro-organisme dans le temps au sein de son environnement,
4 Tm , Temperature melting, est la valeur médiane du profil thermique de dénaturation d'un duplex ADN/ADN ou ADN/ARNr, le ∆Tm
étant la différence de Tm entre l'hybride homologue et hétérologue.

Les techniques d'écologie microbienne
7
Ø de définir sur quelle distance s'étend sa niche écologique et sa zone de densité optimum,
Ø de connaître la vitesse à laquelle il dégrade les nutriments qu'il utilise,
Ø de comprendre comment les éléments qu'il métabolise s'insèrent dans les grands cycles
géochimiques et comment ils sont éventuellement incorporés dans le métabolisme d'autres
populations de micro-organismes.
2.2.1 Quantification
Les méthodes pour quantifier les micro-organismes présents dans un échantillon naturel
dérivent de celles utilisées pour quantifier les micro-organismes en culture pure. On distingue trois
grand types de méthodes : les techniques de mise en culture, les techniques de comptage direct et
les techniques d'analyse chimique de composants cellulaires.
2.2.1.1 Les techniques culturales
A l'aide de dilutions successives sur milieu liquide ou solide on détermine la dilution maximale à
laquelle il est possible d'obtenir une croissance de micro-organismes (technique du NPP: Nombre le
Plus Probable sur milieu liquide et de UFC : Unité Formant Colonie sur milieu solide). Ces techniques
sont grandement dépendantes du milieu et des conditions de culture utilisés. Elles ne permettent
d'accéder qu'à un nombre restreint de micro-organismes présents dans l'échantillon (cf. §. 2.3)
Ces techniques peuvent toutefois être utilisées conjointement avec d'autres méthodes pour détecter
des populations au type métabolique défini (par exemple les BSR, les méthanogènes, les autotrophes
soufre réducteur).
2.2.1.2 Les techniques de comptage direct
Cellules de com ptage
L'échantillon sous une forme liquide est disposé dans des cellules de comptage (cellules de
Mallassez et de Petroff-Hausser). Les cellules sont comptées sous le microscope dans un volume bien
déterminé. Cette technique n'est malheureusement applicable que dans des échantillons où les
cellules sont individualisées. Or dans la nature les cellules sont très fréquemment associées de
manière très cohésive ou associées à des surfaces. Pour simplifier le comptage et pour ne compter
que des cellules viables il est parfois possible de faire croître les cellules dans un milieu contenant un
inhibiteur de la synthèse d'ADN comme l'acide nalidixique et de l'extrait de levure avant de les
dénombrer (Kogura et Simidu, 1979). Les cellules viables vont alors former des cellules géantes (la
croissance de paroi n'étant pas stoppée). Toutefois la limite de détection de cette technique est de
l'ordre de 5.106 à 107 cellules par ml, et cette valeur est bien supérieure à la teneur en micro-
organismes que l'on peut retrouver dans l'eau de mer de surface (5.105 cellules/ml) ou de profondeur
(5.104 cellules/ml) par exemple.

Les techniques d'écologie microbienne
8
Comptage par épifluorescence
Pour ce type de comptage on utilise la propriété qu'ont certains chromophores lorsqu'ils sont
excités à une longueur d'onde (λ1), de réémettre l'énergie fournie sous forme lumineuse à une
longueur d'onde supérieure (λ2) (Zimmermann et Meyer-Reil, 1974). La fluorescence réémise a une
durée de vie plus ou moins longue (fading). Selon les colorations c'est le chromophore qui va se fixer
directement sur la cible (DAPI, SYBR Green) alors que dans d'autres cas il sera relié à une molécule
non fluorescente qui jouera le rôle d'agent de spécificité (anticorps, sondes nucléiques). La coloration
va être réalisée directement sur les cellules. Pour des environnements pauvres en micro-organismes il
est souvent nécessaire de concentrer les échantillons par filtration. On utilise couramment un filtre
noirci (Hobbie et al., 1977) sur lequel il est possible de colorer les cellules avec des marqueurs
fluorescents tels que :
Ø le DAPI (diaminodino-phenylindole) colorant l'ADN,
Ø l'acridine orange qui colore l'ADN et l'ARN,
Ø la fluorescéine isothiocianate (FITC fluorescence verte) et la rhodamine (fluorescence
rouge) qui colorent les protéines en réagissant avec les groupes sulfydryles,
Ø le SYPRO qui colore les protéines et qui a été utilisé récemment pour quantifier la biomasse
planctonique (Zubkov et al., 1999),
Ø le Calcofluor qui réagit avec la cellulose, la chitine et les polysaccharides similaires,
Ø le DANSYL chloride et le "8-anilino-1-naphthalene sulfonic acid salts" (Mg-ANS et Na-
ANS) qui réagissent (fluorescent) en entrant en contact avec les lipides microbiens.
Les deux premiers colorants ont une forte propension à se fixer sur des particules non
biologiques et sont souvent inutilisables dans des échantillons naturels. Il est donc nécessaire de
prévoir une extraction spécifique des cellules avant leur coloration. Il est aussi possible d'utiliser des
colorants beaucoup plus spécifiques des acides nucléiques tels que le SYBR Green I et II, le YOYO-1,
le POPO-1 ou le YO-PRO-1 (cf. tableau 1.4) (Noble et Fuhrman, 1998; Weinbauer et al., 1998; Marie et
al., 1999).
Certains colorants vont permettre de distinguer les cellules mortes des cellules vivantes soit en
ne pénétrant que dans les cellules vivantes (on parle de colorants métaboliques), soit en ne pénétrant
que dans les cellules mortes. De plus les colorants métaboliques permettent de détecter le niveau
d'activité des cellules (par exemple mesure du potentiel d'oxydo-réduction, Rodriguez et al., 1992).
Enfin il est aussi possible d'utiliser des anticorps fluorescents à la spécificité plus ou moins large
pour compter certaines populations de micro-organismes (en particulier dans le domaine médical et
dans l'alimentaire pour la détection et le comptage de certains sérotypes de souches pathogènes).

Les techniques d'écologie microbienne
9
Tableau 1.4. Liste non-exhaustive des fluorochromes spécifiques des acides nucléiques et utilisables pour le comptage cellulaire
(d'après le site MolecularProbes http://www.molecularprobes.com).
Colorants des acides nucléiques Ex/Em£ Propriétés et usages
4',6-Diamidino-2-phenylindole (DAPI) 358/461 Semi-perméable
Sélectif des bases AT
Etude du cycle cellulaire, détection des mycoplasmes
Colorant cellulaire et chromosomique
7-Aminoactinomycin D (7-AAD) 546/647 Peu permanent
Sélectif des bases GC
CMF (cytométrie en flux)
Acridine orange 500/526 (ADN)
460/650 (ARN)
Perméable
Mesures du taux ADN/ARN
Marquage des lysozomes, CMF
Bromure d'éthidium 518/605 Imperméable
Intercalant du dsADN (double brin)
Coloration des cellules mortes
Colorant chromosomique
Electrophorèses, CMF
Excitable par un laser Argon
Hoechst 33342 350/461 Perméable
Sélectif des bases AT
Coloration des cellules vivantes
Etude du cycle cellulaire
Colorant cellulaire et chromosomique
POPO-1 434/456 Imperméable
Forte affinité avec l'ADN
Coloration des cellules mortes
Colorant cellulaire et chromosomique
Pré-colorant d'électrophorèse
SYBR Green I 494/521 Forte sensibilité à l'ADN et aux oligonucléotides
Colorant post électrophorèse
Colorant cellulaire
SYBR Green II 492/513 Forte sensibilité à l'ARN et aux ssADN (simple brin)
Colorant post électrophorèse
Colorant cellulaire
YO-PRO-1 491/509 Imperméable
Coloration des cellules mortes
Electrophorèses
Excitable par un laser Argon
YOYO-1 491/509 Imperméable
Forte sensibilité à l'ADN
Coloration des cellules mortes
Coloration chromosomique
Pré-colorant d'électrophorèse£Longueur d'onde d'excitation et d'émission (nm/nm)
Des tentatives d'automatisation des comptages ont été entreprises pour faciliter certaines
études comparatives. L'analyse d'image assistée par ordinateur permet de reconnaître différents
paramètres morphométriques (longueur, largeur, surface, périmètre), de quantifier les cellules et de
déterminer le volume cellulaire (Van Wambeke, 1995). Mise au point sur des cultures pures, cette
technologie n'est encore pas applicable pour le comptage d'échantillons environnementaux; en effet
la diversité des morphotypes, la taille des cellules (moins importante que pour les cultures pures), le
bruit de fond important lié aux particules (sédiments, sol, matière organique) rendent l'analyse très
fastidieuse et très aléatoire. L'amélioration des technologies d'acquisition d'image (microscopie

Les techniques d'écologie microbienne
10
confocale, caméra CCD), l'accélération de l'acquisition et du traitement des données (cartes
d'acquisition et microprocesseur) devraient permettre dans les années à venir de généraliser l'analyse
d'image associée à l'épifluorescence. Récemment Blackburn et al., (1998) proposaient d'utiliser des
algorithmes de calcul fondés sur les réseaux neuronaux pour faciliter la quantification et l'analyse de
l'activité bactérienne d'échantillons de bactérioplancton prélevés en mer Baltique.
La cytométrie en flux (CMF)
Cette technique apparue dans les années 1980 pour détecter des anomalies dans les cellules
animales et analyser le cycle cellulaire est dorénavant couramment utilisée pour quantifier les micro-
organismes en particulier dans les environnements liquides comme l'eau de mer (Joux et Lebaron,
1995; Monfort et al., 1995). Différents signaux optiques permettent de mesurer différents paramètres
cellulaires (cf. tableau 1.5) :
Tableau 1.5. Paramètres mesurés en CMF en fonction des signaux optiques analysés
Signaux optiques Paramètre mesuré
Diffusion à petit angle Taille cellulaire
Diffusion à grand angle Réfringence cellulaire
Fluorescence Contenu en ADN, flux ioniques, potentiel
transmembranaire
La taille des cellules, le contenu en ADN, l'emploi de colorants particuliers ou plus récemment
l'utilisation de sondes nucléaires ou d'anticorps spécifiques permettent de distinguer et de séparer les
différentes populations contenues dans l'échantillon. Utilisée pour le comptage direct des cellules,
cette technique très puissante et très rapide (une fois mise au point) est applicable pour suivre le
métabolisme cellulaire (potentiel membranaire, activité respiratoire, activité enzymatique par exemple)
et effectuer un tri cellulaire (et donc un isolement puisque la technique n'est pas destructive). La CMF
atteint toutefois ses limites pour des échantillons où la répartition des cellules n'est pas homogène, où
on retrouve des morphotypes non réguliers tels que des filaments ou des structures en chaînettes, où
de nombreuses particules (que l'on peut confondre avec des cellules) sont présentes et où les
cellules sont associées fortement aux particules (particules de sols ou de cheminée).
2.2.1.3 Les mesures de constituants cellulaires
Ces techniques ont pour objectif la quantification d'un constituant cellulaire et permettent
d'extrapoler la biomasse microbienne en fonction de la biomasse de ce constituant (Servais, 1995). Si
certaines de ces techniques sont très précises en terme de mesure (par exemple mesure de l'ATP),
elles sont par contre extrêmement dépendantes de la qualité de l'extrapolation.
La mesure de l'ATP
L'adénosine-triphosphate peut être utilisée comme marqueur de biomasse active en écologie
microbienne et peut être détectée avec une très forte sensibilité à l'aide de techniques enzymatiques

Les techniques d'écologie microbienne
11
de bioluminescence (Holm-Hansen et Booth, 1966; Hodson et al., 1976). Cette technique est
toutefois sujette à des sources d'erreur liées : au stress de la cellule lors du prélèvement et du
traitement de l'échantillon, à la méthode d'extraction (rendement plus ou moins grand selon les
solvants utilisés), à la destruction de l'ATP lors de l'extraction, à l'inactivation des réactifs de dosages
par l'échantillon (notamment dans le cas des sédiments) ou à la présence d'ATP sous une forme libre
dans l'échantillon (par exemple liée aux argiles). De plus le taux d'ATP n'est pas directement corrélé
aux cellules vivantes et l'extrapolation "biomasse d'ATP à biomasse cellulaire" est très dépendante de
l'état physiologique des cellules (Poulicek et Danckers, 1995).
Mesure de l'Azote et du Carbone cellulaires
Par spectrométrie de masse il est possible de définir la quantité de N et C présents dans
l'échantillon et d'extrapoler cette quantité à celle contenue dans les cellules. Cette technique est très
approximative et nécessite de travailler sur des échantillons très propres.
Mesure des constituants de la paroi microbienne
Les acides muramiques contenus dans la paroi bactérienne peuvent être extraits d'échantillons
environnementaux et analysés par HPLC (High Pressure Liquid Chromatography) (Moriarty, 1977). La
quantité d'acide muramique dans la cellule est fonction du type de paroi (Gram négative ou Gram
positive : 12 µg.mg -1 en moyenne chez les Gram- et 44 µg.mg-1 chez les Gram+). Cette technique n'est
pas applicable si on ne connaît pas la proportion de Gram- et de Gram+ dans l'échantillon. Il faut de plus
tenir compte du fait que les spores contiennent 4 fois plus d'acide muramique que les cellules
végétatives.
Lipopolysaccharides (LPS)
Les LPS protéobactériens agissant comme endotoxines peuvent être détectés à de très faibles
doses (10-20 pg/mL) en analysant leur effet sur les amaebocytes de Limule (Limulus polyphemus)
(Watson et al., 1977). Ce test est utilisé en pharmacologie ou en microbiologie clinique pour détecter
les micro-organismes présents dans les fluides cérébro-spinaux, le sérum ou l'eau potable. Toutefois
la sensibilité de la méthode la rend très dépendante des contaminants et sa spécificité la limite aux
milieux colonisés par des entérobactéries.
2.2.1.4 L'analyse lipidique et les autres marqueurs moléculaires
La technique FAME (Fatty Acid Methyl Ester) est fondée sur l'analyse des profils d'acides gras
obtenus par chromatographie en phase gazeuse. Chaque profil est spécifique d'une souche et peut-
être comparé avec d'autres profils contenus dans une banque de données (Drucker et Owen, 1971;
Moss et al., 1974; Guezennec, 1995; Böttiger, 1996). En plus de l'établissement de profils lipidiques,
il est possible de rechercher un ou des lipides marqueurs spécifiques dont la présence sera
caractéristique d'un groupe de micro-organismes (par exemple étude des micro-organismes colonisant

Les techniques d'écologie microbienne
12
différents coupons en milieu hydrothermal, Guezennec et al., 1998). Les rapports entre certains
marqueurs spécifiques pourront donner une indication sur l'état physiologique de la communauté.
Cette technique peut donc être utilisée sur des souches isolées ou sur des environnements peu
diversifiés afin de suivre l'évolution d'une souche particulière. Par exemple la recherche des Nocardia
dans les boues actives (Cha et al., 1999) ou l'étude de souches isolées d'environnements naturels
(des sols (Ka et al., 1994) ou des sédiments pollués (Wagner-Döbler et al., 1998)). L’analyse des
profils lipidiques est toutefois limitée par un manque de banques de données uniformisées, par un
nombre relativement faible de micro-organismes type étudiés et par l’influence de l’état physiologique
cellulaire sur les profils observables.
En dehors des lipides, d'autres marqueurs moléculaires peuvent être utilisés pour accéder à la
diversité microbienne d'un échantillon (cf. tableau 1.6).
Tableau 1.6. Principaux marqueurs bactériens (d'après Guezennec, 1995)
Biomarqueurs Micro-organismes Remarques ou Exemples de molécules
Phospholipides (Acides Gras)
Acides ß Hydroxylés (LPS)
Toutes cellules
Bactéries Gram -
Mesure de la biomasse viable
ß OH C14:0
Acides Gras Toutes cellules
Sulfato-réductrices
Thio-oxydantes
C16:1ω7, C16:0
iC17: 1ω7c, 10MEC16:0
OH monoinsaturés, 10-11MeOC18:0
Plasmalogènes Bactéries anaérobies essentiellement
Quinones Bactéries aérobies
Bactéries anaérobies
Anaérobies facultatives
Ubiquinones
Ménaquinones
Ubiquinones et Ménaquinones
Ethers de Glycérol Archaea
Acide Muramique Bacteria
Acide diaminopimélique Bactéries Gram - et quelques Gram +
Acides aminés (D) Bacteria D-alanine, D-glutamine
Hopanes Quelques Bacteria
Phytopigments Phototrophes
2.2.2 Mesure d'activité microbienne
2.2.2.1 Incorporation des radio-isotopes
Ils permettent de suivre la dégradation, la production ou la transformation d'un composé, d'en
estimer la vitesse ou la localisation. Pour ce faire on va suivre l'incorporation d'isotopes radioactifs 14C,3H, 35S. L'incorporation du 14CO2 permettra de suivre les mécanismes de photosynthèse, ou la
production de 14CH4 permettra de suivre les mécanismes de méthanogenèse. La sulfato-réduction
pourra être suivie par mesure de la conversion de 35SO42- en H2
35S, alors que la méthylotrophie ou la
méthanogenèse acétoclastique pourra être suivie en mesurant la conversion du méthanol ou de
l'acétate marqué au 14C en 14CH4 (Zehnder et al., 1979). Les mécanismes de chimioorganotrophie
pourront être suivis par mesure de l'incorporation de sucres et d'acides aminés marqués au 14C. La
synthèse de l'ARN ou de l'ADN peut être suivie par mesure de l'incorporation de la 3H-adénine (Karl,
1979, 1981). L'activité des cellules bactériennes peut être suivie en mesurant l'incorporation de la 3H-

Les techniques d'écologie microbienne
13
thymidine dans l'ADN chez la majeure partie des hétérotrophes (Fuhrman et Azam, 1980, 1982;
Pollard, 1998), du 35SO42- (Monheimer, 1972) ou de la 3H-leucine (Kirchman et al., 1985) dans les
protéines pour l'ensemble du monde bactérien. Enfin les bactéries nitrifiantes ou dénitrifiantes sont
analysées par mesure de l'isotope 15N (Goering et Dungale, 1966). Ces mesures peuvent être
effectuées dans la nature ou sur cultures isolées. On parle alors de microautoradiographie.
2.2.2.2 Mesure des ratios isotopiques
Dans la nature chaque élément chimique existe sous la forme de plusieurs isotopes de stabilité
variable. Les ratios entre ces isotopes sont fixes (par exemple le ratio 12C/13C=19/1), or les mécanismes
de transformation cellulaire vont favoriser l'incorporation d'un isotope (en général le plus léger) et les
composants synthétisés vont donc avoir un ratio isotopique différent du ratio naturel. De plus les
mécanismes métaboliques vont incorporer différemment chaque isotope et pour une molécule
donnée un ratio isotopique pourra être attribué à chaque métabolisme.
2.2.2.3 Mesures d'activités spécifiques
L'emploi d'analogues structuraux fluorescents permet de détecter des activités enzymatiques
extra-cellulaires spécifiques comme celles des lipases, des phosphatases, des aminopeptidases.
L'incorporation de bromodéoxyuridine (BrdU), un analogue de la thymine chez les bactéries actives
(synthétisant de l'ADN) peut être suivie par méthode immunochimique. L'ADN ainsi marqué peut être
extrait et étudié (Urbach et al., 1999)
L'emploi du CTC (5-cyano-2,3-dotolyl tetrazolium chloride) ou de l'INT (2 (p-iodo-phenyl)-3-(p-
nitrophenyl)-5-phenyl tetrazolium chloride) permet de suivre l'activité respiratoire par l'apparition
(CTCàCTC-formazan) ou la disparition (INT à INT-formazan) d'un cristal fluorescent.
L'emploi du système Biolog directement à partir d'échantillon environnemental a été proposé
par Garland (1997) puis par Smalla et al. (1998) en conjonction avec une étude en TGGE. Le
métabolisme du carbone dans l'échantillon peut être ainsi appréhendé mais sans identifier les micro-
organismes qui en sont responsables.
2.2.2.4 Les microélectrodes
Si l'utilisation des microélectrodes était une chose commune en neurophysiologie et en biologie
animale, ce n'est que très récemment (ces 15 dernières années) que sous l'impulsion de Jørgensen et
de Revsbech elles ont été utilisées en écologie microbienne (de Wit , 1995; Amann et Kuhl, 1998;
Cooper, 1999). On dispose actuellement de microélectrodes qui permettent de mesurer O2, pH, S2-,
NO2, CO2, NO3-, NH4
+ et potentiels redox. D'un diamètre de 2 à 100 µm, voire 2 à 3 µm pour les
électrodes à O2, elles ont une très forte résolution spatiale qui permet de suivre des microvariations
dans des environnements où de nombreuses populations microbiennes se succèdent comme par
exemple: les sédiments ou les biofilms (Santegoeds et al., 1998 a, b; Okabe et al., 1999).

Les techniques d'écologie microbienne
14
2.3 Une vue restreinte de la diversité
L'ensemble des techniques préalablement décrites pour analyser la diversité microbiologique
d'un échantillon présente l'inconvénient de ne donner qu'une vue restreinte ou une approximation
grossière de la diversité naturelle. Les techniques qui permettent d'accéder individuellement à chaque
organisme sont notamment entachées d'erreurs inhérentes aux méthodes culturales. Celles ci sont
connues depuis fort longtemps et ont été décrites sous le terme de "la grande anomalie du comptage
sur boite" (Staley et Konopka, 1985).
Selon les auteurs entre 0,001% et 15% des bactéries présentes dans un échantillon sont
cultivables à l'aide des techniques classiques (cf. tableau 1.7)
Tableau 1.7. Cultivabilité déterminée comme le pourcentage des bactéries cultivables par rapport aux cellules totales déterminées par
comptage direct. (D'après Amann et al., 1995)
Habitat Cultivabilité (%)a
Eau de mer 0,001-0,1
Eau douce 0,25
Lac mésotrophique 0,1-1
Eaux d'estuaires non polluées 0,1-3
Boues actives 1-15
Sédiments 0,25
Sol 0,3a Bactéries cultivables mesurées en tant qu'unités formant des colonies (UFC)
Ce décalage est à attribuer à différentes causes (Amann et al., 1995; Joux et Lebaron, 1995;
Ward et al., 1997):
Ø Le milieu de culture choisi ne peut être universel et l'ensemble des conditions de
l'environnement ne peuvent être reproduites sur un seul et même milieu de culture en
particulier pour les environnements à forts gradients physico-chimiques.
Ø Certaines cellules hors de leur environnement peuvent entrer dans un état viable mais non-
cultivable (cf. tableau 1.8 pour des études réalisées sur le milieu aquatique). Cet état cellulaire
correspond à une adaptation aux conditions de stress, notamment nutritionnel. Il
s'accompagne de modifications physiologiques et structurales dont le rétablissement n'est
pas simplement possible sur milieu synthétique. Cet état peut être maintenu de plusieurs jours
à plusieurs années.
Ø Des populations inférieures numériquement peuvent supplanter des populations majoritaires
moins adaptées au milieu de culture.
Ø Des populations enrichies en liquide peuvent ne pas se développer sur milieu solide.
Ø La quantité d'inoculum choisie pour l'enrichissement influence le type de cellules enrichies.
Ø Des micro-organismes différents ayant des conditions de croissance similaires seront
difficilement distinguables.

Les techniques d'écologie microbienne
15
Ø Certaines cellules sont impossibles à obtenir en culture pure comme c'est le cas des micro-
organismes symbiotes et syntrophes.
Tableau 1.8. Pourcentages de bactéries actives par rapport au nombre total de bactéries (comptées en épifluorescence), cité
dans Servais (1995)
Méthodes Environnements Pourcentages
Autoradiographies3H acides aminés Marin (Kiel Bay)
Marin (Chesapeake Bay)
41%
45 à 76 %; 17 à 82 %3H glucose Marin (Kiel Bay) 2,3 à 56,2 %3H acétate Marin (Chesapeake Bay) 49 à 59 %; 6 à 51 %3H thymidine Marin (Chesapeake Bay)
Marin (S. California Bight)
Marin (Côte du Danemark)
15 à 94 %
28 à 50 %
20 à 80 %
Réduction d'INT Marin (mer Baltique)
Marin (Chesapeake Bay)
Eau potable (France)
Lac Léman
8 à 97%
61 %; 25 à 94 %
10 à 20 %
0,6 à 26 %
Acide nalidixique Marin (Chesapeake Bay)
Marin (Porto Rico)
7 à 15 %
0,5 à 6 %
Des techniques permettant d'accéder individuellement aux micro-organismes présents dans un
échantillon en s'affranchissant des techniques culturales ont été proposées à la fin des années 1980.
Ces techniques sont fondées essentiellement sur l'analyse, non plus de la cellule, mais des acides
nucléiques qu'elle contient sous la forme d'ADN génomique ou d'ARN ribosomaux. Dans le chapitre
suivant je me propose de décrire ces techniques après avoir présenté les molécules analysées (les
ARN ribosomaux) et les principes de phylogénie. Les apports des techniques moléculaires dans
l'analyse de différents habitats microbiens seront abordés dans la seconde partie de mon introduction.
III Les acides nucléiques en écologie microbienne
3.1 Le développement des techniques moléculaires
L'émergence des technologies moléculaires en écologie microbienne est liée au
développement de la phylogénie moléculaire à la fin des années soixante (Zuckerland et Pauling,
1965) ainsi qu'au choix de l'ARN ribosomique comme marqueur évolutif à la fin des années soixante-
dix (Woese et Fox, 1977). Les premiers travaux d'écologie microbienne moléculaire provinrent de
l'équipe de Pace qui utilisa les résultats de Woese pour accéder à la diversité d'échantillons
environnementaux (Olsen et al., 1986; Pace et al., 1986a et b).
Un grand nombre de publications se sont proposées de décrire sous forme de revue les
techniques moléculaires employées en écologie microbienne. Parmi les plus anciennes on peut citer,
en plus des deux publications précédentes : Turner et al., 1989; Olson, 1991; Pickup, 1991; Atlas et

Les techniques d'écologie microbienne
16
al., 1992; Liesack et Stackebrandt, 1992; Olson et Tsai, 1992; Ward et al., 1992. Si nombre de ces
revues décrivent avec enthousiasme les apports de ces techniques certains auteurs s'interrogent déjà
sur les nouvelles limitations qu'elles impliquent.
3.1.1 La phylogénie moléculaire
Le concept de phylogénie moléculaire est né des travaux de Zuckerland et Pauling (1965) qui
les premiers considérèrent les molécules comme des marqueurs de l'histoire évolutive et
développèrent le concept d'horloge moléculaire. Cette notion est liée à la théorie neutraliste de
l'évolution (Kimura, 1968). Les mutations s'accumulent au cours du temps dans le génome et chez
des organismes différents. Le taux d'accumulation est dicté par l'intensité de la pression de sélection
et il est du même ordre de grandeur dans des domaines homologues du génome (région soumise à la
même pression de sélection). Ainsi les gènes à mutations rapides vont pouvoir refléter des
événements évolutifs récents alors que les gènes très conservés seront les témoins d'un passé très
éloigné. Sans pour autant remettre en cause la validité de la phylogénie moléculaire, la théorie
neutraliste de l'évolution est fortement contestée (Goodman, 1981) voire niée (Li, 1993). La
différence de taux d'évolution est alors attribuée soit aux différences de temps de génération soit aux
différences d'activité du métabolisme (Li, 1993).
Près de dix ans après les travaux de Zuckerland et Pauling, Woese et ses collaborateurs (Fox et
al., 1977; Woese et Fox, 1977) appliquent cette théorie à l'étude de la phylogénie bactérienne en
utilisant l'ARN ribosomique comme marqueur de l'évolution. Les premières études portent sur
l'analyse de fragments de séquences et définissent des signatures spécifiques qui scindent le monde
du vivant en trois grands domaines. La dichotomie du monde vivant en terme de Procaryote et
Eucaryote était abandonnée au profit d'une division tripartite constituée d'Eucarya, de Bacteria et
d'Archaea. Les études menées par la suite sur des séquences entières confirmèrent ces résultats
(Woese, 1987; Woese et al., 1990).
3.1.2 L'ARN ribosomique
Les ARN ribosomiques (ARNr) sont formés de sous unités pouvant se grouper en polysomes.
Ces éléments sont caractérisés par leur constante de sédimentation exprimée en Svedberg (S). Chez
les organismes procaryotes, il existe des ribosomes 70S (de poids moléculaire 2,5.106 Da (1 Dalton= 1
g/mol)) formés de 2 sous-unités. La petite unité 30S de poids moléculaire 0,93.106 Da est composée
de 21 protéines et d'ARNr 16S (1541 nucléotides chez E. coli; 0,56.106 Da) (Moore, 1998). Lorsque
des régions sur sa séquence sont complémentaires (appariement AU et GC), le brin d'ARN se replie
sur lui même pour former une structure en "épingle à cheveux". Dans le ribosome les molécules
d'ARNr, repliées en une structure secondaire, peuvent être représentées à partir d'études
comparatives des séquences primaires (Gutell et al., 1994; Noller, 1984; Noller et Woese, 1981;
Woese et al., 1975). Une modification de la séquence primaire de l'ARNr va influencer directement sa

Les techniques d'écologie microbienne
17
structure secondaire. Pour maintenir la fonctionnalité de la molécule, la structure secondaire de l'ARNr
doit être conservée dans l'ensemble du monde vivant par le biais de mutations compensatoires
(Hancock et al., 1988; Wheeler et Honeycutt, 1988; Otsuka et al., 1999).
3.1.3 Le choix de l'ARN 16S
Le choix de Woese et de ses collaborateurs s'est porté sur la petite sous-unité de l'ARNr (16S
chez les Procaryotes et 18S chez les Eucaryotes) pour plusieurs raisons (Pace et al., 1985; Ludwig et
Schleifer, 1994; Ludwig et al., 1998) :
Ø Sa présence est universelle et il y accomplit le même rôle chez tous les organismes.
Ø Sa séquence est une alternance de domaines dont les vitesses d'évolution varient, permettant
de comparer des espèces très proches sur des domaines hypervariables et des espèces très
éloignées sur des domaines très conservés.
Ø Il a évolué lentement.
Ø Il n'est pas le résultat de transferts latéraux5.
Ø Il est relativement facile à isoler en raison de son abondance dans les cellules
Ø Sa séquence est facilement obtenue par des méthodes standard d'extraction et de
séquençage. Les techniques d'extraction (Dorsch et Stackebrandt, 1992) et de séquençage ont
été simplifiées et optimisées au fil des ans par l'introduction du séquençage des ADNr (Böttger,
1989) plutôt que des ARNr (Lane et al., 1985c), et l'utilisation du "cycle-sequencing" à l'aide
d'amorces fluorescentes (Hiraishi, 1992; Ruano et al., 1993)
Ø Il est préféré au 5S et au 23S en raison de sa taille moyenne et de sa structure secondaire moins
marquée. Sa séquence est suffisamment longue pour réaliser des comparaisons statistiquement
cohérentes (cf. § 3.2.2.2 calcul de la déviation standard).
L'ADN codant pour l'ARNr (ADNr) est organisé en opéron. Au sein d'une souche, cet opéron
peut être en copies multiples (de 1 à 14 selon les souches) et ces copies sont généralement
identiques ou très proches en raison des fortes pressions évolutives gouvernant le maintien de la
structure secondaire de l'ARNr (Hillis et al., 1991). Il a toutefois été observé au sein d'un même
organisme d'importantes variations entre les différentes copies de l'ADNr 16S (Clayton et al., 1995).
D'autre part des séquences d'ADNr 16S correspondant à des souches phénotypiquement très
distinctes pouvaient s'avérer quasiment identiques (Fox et al., 1992). Récemment Ueda et al. (1999)
attribuent ces différences entre copies d'un même opéron à deux mécanismes : d'une part une
mauvaise incorporation durant la réplication de l'ADN et d'autre part des transferts horizontaux.
Le choix de l'ARNr 16S est parfois contesté (cf. Partie 2 §1.2), il serait donc préférable d'établir
les phylogénies sur plusieurs gènes afin de ne conserver que l'arbre consensus à toutes ces
phylogénies. Des tentatives ont été entreprises (Gupta, 1998a; Huang, 1996). Mais le travail est
actuellement très laborieux. Gageons que les améliorations des techniques de séquençage :
5 Une étude récente de complémentation de E. coli (dont l'ensemble des opérons rrn (7) avaient été inactivés) par des ARNr d'une
autre espèce semble indiquer que la possibilité de transferts horizontaux de l'ARNr n'est pas à exclure (Asai et al., 1999a et b).

Les techniques d'écologie microbienne
18
augmentation du nombre d'échantillons traités simultanément, accélération des migrations dans des
microcapillaires, utilisation de puces de séquençage ou de séquençage par spectrométrie de masse,
permettront d'ici quelques années d'accéder à l'ensemble du génome des micro-organismes comme
nous accédons actuellement à l'ARNr 16S.
3.2 L'analyse phylogénétique
Une étude de diversité microbienne est fondée sur l'identification des micro-organismes
présents dans un échantillon et donc sur la comparaison de ces micro-organismes avec d'autres
préalablement isolés. Selon que l'on dispose uniquement de séquences (ADNr ou gènes du
métabolismes clonés) ou de souches isolées, les comparaisons porteront uniquement sur ces
séquences ou sur l'ensemble de l'ADN génomique et sur des caractères phénotypiques. Dans tous
les cas il convient de définir des homologies entre les organismes à comparer puis de réaliser une
phylogénie à partir des caractères homologues.
On définit par homologues des caractères dérivant d'un même caractère ancestral. Ces caractères
sont orthologues s'ils ont conservé la même fonction et paralogues si la fonction est différente.
Les caractères analysés lors d'une phylogénie moléculaire sont des séquences d'acides aminés
ou des séquences d'acides nucléiques. Ce sont des caractères discontinus qui peuvent donc être
étudiés comme des caractères discrets (absence ou présence). L'extrémité des branches d'arbres
phylogénétiques correspond aux taxons. Ces taxons constitueront des OTU (Unité Taxinomique
Opérationnelle) sur la base de caractères phénétiques (Sokal et Sneath, 1963). C'est cette unité que
choisit Moyer et al. (1994) pour définir la structure et la diversité de la communauté d'un tapis microbien
hydrothermal sur un site du Pacifique. D'autres auteurs leur préfèrent la définition de phylotype (ou
type phylogénétique) qui s'applique plus à une analyse fondée sur des caractères moléculaire
(Haddad et al., 1994; Poltz and Cavanaugh, 1995).
3.2.1 Les méthodes de phylogénie
Il n'existe pas de techniques qui permettent de réaliser l'arbre phylogénétique vrai. On ne
dispose que de moyens statistiques pour construire l'arbre "le moins faux possible".
Trois méthodes permettent de construire les arbres : l'analyse des distances (Fitch et
Margoliash, 1967), l'analyse de la parcimonie (Fitch, 1971), l'analyse du maximum de vraisemblance
(Felsenstein, 1981).
La première est une méthode phénétique c'est à dire fondée non pas sur un critère
généalogique mais sur un critère de similarité : on utilise alors des distances et il n'y a pas de tentative
pour établir des relations d'ancestralité entre des séquences différentes. Les deux autres méthodes
sont des méthodes cladistiques reposant sur des caractères discrets; on cherche par ces méthodes à
établir des relations généalogiques entre les individus étudiés. On détermine alors le nucléotide le
plus probable pour une position donnée sur la séquence ancestrale. La méthode de parcimonie est

Les techniques d'écologie microbienne
19
fondée sur un nombre minimal de mutations pour établir les relations entre les espèces et la méthode
du maximum de vraisemblance est une méthode statistique.
On considère qu'une phylogénie est solide lorsque les arbres obtenus par les trois méthodes
sont concordants.
Toutefois différentes sources d'erreurs doivent être prises en compte :
Ø Dans les zones hypervariables, l'alignement de certains nucléotides est hasardeux car la
présence d'une même base sur deux séquences différentes n'est pas liée obligatoirement à une
homologie mais à une homoplasie (cf. définition §3.2.1.1).
Ø Les différences entre deux séquences sont censées représenter un événement dans une
lignée, or un même site peut avoir changé plusieurs fois dans une lignée ou avoir changé une fois
dans deux lignées différentes.
Ø Des "gaps" doivent parfois être introduits dans des séquences qui correspondent soit à des
insertions soit à des délétions. Ces modifications peuvent correspondre à un ou plusieurs
événements.
3.2.1.1 L'alignement
L'alignement est l'étape cruciale de l'analyse phylogénétique. Il consiste à mettre en regard des
nucléotides à des positions homologues. Au sein de zones très conservées, les zones homologues
sont facilement détectables mais lorsque des événements d'insertion et de délétion (fréquents dans
l'ARNr 16S) surviennent, il convient de les répercuter sur les alignements.
Au sein de zones hypervariables, des nucléotides peuvent s'avérer identiques alors que ceux-ci
ne sont pas homologues6 mais ont subi des phénomènes de réversion, de parallélisme ou de
convergence. Il convient alors, si ces séquences sont impliquées dans une structure secondaire, de
vérifier la présence de mutation compensatoire assurant le maintien de cette structure. Des banques
de structures secondaires d'ARNr ainsi que des logiciels de détermination de structures secondaires
d'ARN sont disponibles à cet effet sur le Web (Neefs et al., 1993; Van de Peer et al., 1997, 1999).
Une fois l'alignement obtenu il faut choisir quelles positions sur la séquence seront informatives
pour l'étude. Ce choix va être dicté par la proximité des séquences prises en compte durant l'étude. Il
n'est par exemple pas informatif de choisir des portions de séquences hypervariables sur un ensemble
de souches correspondant à plusieurs genres voire plusieurs familles. Toutefois ces zones
hypervariables pourront n'être que faiblement variables voire partiellement conservées si l'étude est
restreinte aux individus d'un même genre (les zones deviennent homologues). Il va donc falloir créer
des masques qui spécifieront pour les programmes de phylogénie les zones à analyser.
6 Homoplasie : similarité qui ne dérive pas d'un caractère ancestral.
Il existe trois sortes de phénomènes qui conduisent à une homoplasie: réversion : changement dans une séquence annulé par un
second changement (A → C → A), parallélisme : changements parallèles dans deux séquences. convergence : des bases différentes
dans deux séquences ancestrales peuvent être semblables dans des séquences contemporaines.

Les techniques d'écologie microbienne
20
3.2.1.2 L'analyse des distances
Ce sont des méthodes rapides fondées sur le nombre de nucléotides différents entre deux
séquences. Les méthodes différeront en fonction de la façon dont les nucléotides seront pondérés,
dont les changements de bases (transition, transversion)7 seront traités, et des algorithmes utilisés
pour construire les arbres.
Ê Le nombre de différences observées entre deux séquences est toujours inférieur au véritable
nombre de différences en raison de phénomènes de mutation réverses et de mutations
multiples. Cette différence véritable ou distance évolutive peut être estimée par une loi de
Poisson. Si P est la proportion calculée de nucléotides différents entre deux séquences, alors
le taux de substitution selon Jukes et Cantor (1969) :
Knuc = −3
4Ln 1 −
4
3P
Cette méthode sous-entend que toutes les positions sur une molécule changent à la même
vitesse, que les transitions et les transversions7 ont la même probabilité et que chaque site
évolue de manière indépendante. Ce qui n'est pas nécessairement le cas.
Ë Selon Kimura (1980) le poids des transversions7 (proportion Q) est double par rapport à celui des
transitions (proportion P). Alors le taux de substitution
Knuc = −1
2Ln 1 − 2P − Q( ) 1− 2Q( )
1
2
Ì Van de Peer et al. (1994) proposent quant à eux de tenir compte des "gaps" qui peuvent être
introduits au cours de l'alignement. Pour P : proportion calculée de nucléotides différents entre
deux séquences, G : nombre de "gaps" dans une séquence par rapport à une autre, T : nombre
total de nucléotides pris en compte dans l'analyse. Une série de "gaps" adjacents, quel que soit
le nombre de "gap", est traitée comme un "gap". Alors
Knuc = −3
4Ln 1−
4
3P
1 −
G
T
+
G
T
Les valeurs Knuc sont alors soumises à des algorithmes produisant des arbres dont la longueur
des branches est minimisée en fonction de la proximité des séquences. Les méthodes principalement
utilisées sont celles :
Ø de UPGMA. Méthode agglomérative rapide mais peu précise. On groupe les deux espèces
les plus proches puis on construit une nouvelle matrice avec ce doublet, etc. (Sneath et Sokal, 1973).
Développée à l'origine pour réaliser de la taxinomie numérique, cette méthode a tendance à raccourcir
les branches des arbres lorsque l'on réalise une phylogénie (distances égales des branches de
chaque cotés du noeud).
Ø de Fitch et Margoliash (1967) fondée sur le principe d'additivité (la distance entre deux
taxons est égale à la somme des longueurs de branche les reliant). C'est une méthode précise mais
7 Transition : passage d'une purine à une purine (A ↔ G) ou d'une pyrimidine à une pyrimidine (T ↔ C)
Transversion : passage d'une purine à une pyrimidine et inversement.

Les techniques d'écologie microbienne
21
qui nécessite un temps de calcul élevé. Elle permet de corriger l'effet "d'écrasement" obtenu par la
méthode UPGMA en acceptant des longueurs de branches inégales de chaque coté d'un noeud.
Ø de Neighbor-joining (Saitou et Nei, 1987) la plus couramment utilisée car elle permet
d'atténuer la méthode d'additivité en tenant compte des phénomènes d'homoplasie. Cette méthode
est rapide et précise.
Dans une analyse de séquence il faut aussi tenir compte du nombre de nucléotides analysés.
Plus celui ci sera important plus l'erreur ou la déviation standard sera faible. Cette déviation standard
dépend du nombre de résidus (N), de la proportion de nucléotides différents (P), de la proportion de
nucléotides semblables (Q=1-P).
Alors ∆ =3
4.
P.QL
Q −1
4
.
Par exemple pour 50% de différence sur 1000 nucléotides ∆=0,05 soit ±10% et pour 100 nuc.
∆=0,15 soit ± 30%.
On considère que les méthodes de distance sont faciles et rapides à réaliser. Elles conviennent
à une comparaison de séquences proches, ayant un haut score de similarité. Toutefois ces méthodes
perdent de l'information (les séquences sont réduites à des distances) et ne conviennent pas à la
comparaison de séquences éloignées.
3.2.1.3 Le maximum de parcimonie
Cette méthode est fondée sur le principe de l'économie : l'arbre le plus parcimonieux sera celui
présentant le moins de mutations pour passer d'une séquence à une autre. Le noeud qui relie deux
séquences est une séquence commune ou ancestrale (Fitch, 1971).
A partir de l'alignement total, un nombre réduit de sites informatifs est utilisé pour construire des
arbres. On élimine les sites qui ne varient pas (et parfois les sites qui diffèrent par transition dans le cas
d'une parcimonie de transversion). En effet on ne considère comme informatifs que les sites qui
favorisent une topologie d'arbre parmi toutes celles possibles. Les différentes topologies d'arbres
sont testées et l'arbre le plus parcimonieux est conservé. Pour des comparaisons s'appliquant à plus
de 8 séquences les techniques de choix d'arbres par des recherches exhaustives peuvent être
difficiles et très longues, aussi on utilise une technique dite heuristique (plus rapide mais moins
précise, par exemple le "branch swaping") afin de définir l'arbre le plus parcimonieux. Cette méthode
suppose que l'horloge évolutive est constante or ce n'est pas toujours le cas et les résultats obtenus
peuvent parfois être erronés. De plus il est fréquent de générer des arbres aussi parcimonieux les uns
que les autres et le choix de l'arbre consensus est alors difficile.
3.2.1.4 Le maximum de vraisemblance
C'est une méthode statistique où les changements évolutifs dans la séquence nucléique
suivent des modèles probabilistes. On recherche l'arbre possédant la plus forte probabilité pour qu'un

Les techniques d'écologie microbienne
22
ensemble de données soit le résultat de l'évolution décrite par cet arbre. Les éventualités de
mutations sont calculées pour chaque site en suivant le modèle de Kimura dans le programme DNAML
développé par Felsenstein (1973, 1981). D'une manière similaire à la parcimonie la vraisemblance
assigne une séquence ancestrale à chaque noeud, mais elle ne suppose pas que l'évolution ait été
parcimonieuse. Pour chaque site et à chaque noeud tous les nucléotides possibles sont testés en
calculant dans tous les cas la probabilité pour que la séquence observée soit produite avec une
longueur de branche donnée. La première version du programme DNAML ne permettait pas de traiter
beaucoup de séquences à la fois, aussi Olsen a amélioré la technique en rendant possible le
traitement de plusieurs séquences (FastDNAML) (Olsen et al., 1994a).
Pour mémoire on peut citer une quatrième méthode de phylogénie qui tient compte de taux
d'évolution variables au sein des différentes branches d'un arbre. Il s'agit de la méthode des invariants
développée par Lake (1987).
3.2.1.5 Estimation de la robustesse des arbres
Ø Des méthodes différentes de phylogénie peuvent donner des arbres de topologies
différentes, aussi ne sont conservés, pour la construction de l'arbre final, que les branchements
retrouvés dans chacune des trois méthodes (Huelsenbeck et Hillis, 1993; Kim, 1993).
Ø Mais il existe aussi des méthodes statistiques qui permettent de vérifier la stabilité
(robustesse) des arbres : le "jackknife" et le "bootstrap". Cette dernière méthode est la plus
couramment utilisée. Elle consiste en la construction d'une nouvelle matrice de même taille que la
matrice initiale en réalisant des tirages avec remise des différentes colonnes de la première matrice
(Felsenstein, 1985). Aussi certaines colonnes de l'alignement pourront être utilisées plusieurs fois
alors que d'autres ne seront pas du tout utilisées pour construire la nouvelle matrice. Un arbre
phylogénétique est construit et cette opération est reproduite 50, 100 ou 1000 fois. Un arbre
consensus est calculé, portant sur chaque noeud de branches le pourcentage d'arbres où ce noeud
est retrouvé. Plus le pourcentage est élevé plus le noeud est considéré comme robuste. Dans la
pratique un noeud est considéré comme robuste lorsque le pourcentage est supérieur ou égal à 90%.
Les noeuds ayant un pourcentage inférieur à 50% ne sont pas pris en considération.
Ø Les séquences incluses dans une analyse peuvent influer sur la topologie d'un arbre. On
valide donc une topologie en incluant de nouvelles séquences et en supprimant d'anciennes.
Ø Enfin il est possible de vérifier la topologie en comparant les associations entre les séquences
sur la base de signatures spécifiques comme des nucléotides spécifiques à une position donnée ou la
présence (ou l'absence) de structures secondaires (par exemple des boucles, la longueur de structure
en épingle) à une position donnée.
Il reste toutefois difficile de définir "à coup sûr" si l'on est parvenu à réaliser une bonne
phylogénie et il est souvent nécessaire de vérifier la congruence de la phylogénie obtenue avec des
phylogénies obtenues au cours d'autres études ou à l'aide d'autres caractères.

Packages généraux
- PHYLIP- PAUP*- MEGA- VOSTORG- Fitch programs- Phylo_win- Zharkikh Unix programs- ARB- DAMBE
Construction, dessin, manipulationd'arbres
- PHYLIP- PAUP*- TreeTool- TreeView- Fitch programs- NJplot- DendroMaker- Tree Draw Deck- Phylodendron- ARB- unrooted- DAMBE- MacClade- ClaDOS- PDAP- TreeTool
Calcul des distances
- PHYLIP - PAUP*- RAPDistance - MULTICOMP- MARKOV - RSVP- Microsat - DIPLOMO- OSA - DISPAN- RESTSITE - NTSYSpc- PUZZLE - Hadtree, Prepare andTrees- Wisconsin Sequence Analysis Package (GCG)- AMP - DISTREE- GCUA - DERANGE2- calcdist - POPGENE- TFPGA - REAP- MVSP - SOTA- RSTCALC - Genetix- BIOSYS-2 - RAPD-PCR package- DISTANCE - Darwin- sendbs - K2WuLi- GeneStrut - Arlequin- DAMBE - DnaSP- PAML - puzzleboot - MATRIX
Arbre consensus et comparaison d'arbres
- COMPONENT- TREEMAP- NTSYSpc- PHYLIP- PAUP*- REDCON- TAXEQ2- TreeCons
Manipulation et soumission des séquences
- PARBOOT- Random Cladistics- Tree Gardener- GDE- MUST- DNA Stacks- SeqPup- Tonex- ARB
Maximum de vraisemblance etméthodes associées
- PHYLIP- PAUP*- fastDNAml- MOLPHY- PAML- Spectrum- SplitsTree- PLATO- SPOT- PUZZLE- Hadtree, Prepare and Trees- SeqPup- Phylo_win- PASSML- ARB- Darwin- BAMBE- DAMBE- Modeltest- TreeCons- VeryfastDNAml
Maximum de Parcimonie
- PAUP*- Hennig86- MEGA- Tree Gardener- RA- Pee-Wee and Noname- PHYLIP- PHYSYS- TurboTree- Freqpars- Fitch programs- CAFCA- Phylo_win- sog- gmaes- LVB- GeneTree- TAAR- ARB- DAMBE
Méthodes des matrices de distance
- PHYLIP - PAUP*- MEGA - MacT- ODEN - Fitch programs- SINCAIDEN - ABLE- TREECON - DISPAN- RESTSITE- NTSYSpc- METREE- TreePack- TreeTree- GDA- Hadtree, Prepare and Trees- Wisconsin Sequence Analysis Package (GCG)- SeqPup- PHYLTEST- Lintre- WET- Phylo_win- njbafd- gmaes- GTREE, ADDTREE, and EXTREE- DENDRON- Molecular Analyst Fingerprinting
Bootstrap et autres tests de validation
- PHYLIP- PAUP*- Zharkikh bootstrapping programs- PARBOOT- ABLE- Random Cladistics- AutoDecay- TreeRot- RASA- DNA Stacks- OSA- DISPAN- TreeTree- PHYLTEST- Lintre- sog- njbafd- PICA95- TAXEQ2- BIOSYS-2- RAPD-PCR package- TreeCons- BAMBE- DAMBE- puzzleboot
Tableau 1.9. Logiciels dédiés aux différentes méthodes de phylogénie

Les techniques d'écologie microbienne
23
L'essor de l'écologie moléculaire bactérienne est indubitablement associé à l'essor des outils
d'analyse phylogénétique et des banques de données. La généralisation de l'outil informatique dans
les laboratoires a permis de démocratiser l'emploi de programmes de phylogénie longtemps confinés
dans des équipes disposant de centres de calculs surpuissants. Je me propose dans le chapitre
suivant de présenter différents logiciels et sites internet dédiés aux analyses de diversité.
3.2.2 L'informatique au service de la phylogénie
Les ordinateurs le plus souvent utilisés pour réaliser des calculs de phylogénie sont des stations
de travail fonctionnant sous environnement UNIX. En complément à ces machines il est désormais
possible à tout utilisateur disposant d'un ordinateur personnel de type PC ou Macintosh de faire
fonctionner sous le système d’exploitation Linux8 (http://www.linux.org) des logiciels réservés
jusqu'alors à une station de travail UNIX.
D'autre part un grand nombre de programmes de phylogénie ont été développés pour
fonctionner sur Macintosh ou sur PC sous les environnements Mac Os, DOS ou Windows.
3.2.2.1 Les programmes de phylogénie
Un grand nombre de logiciels sont disponibles pour réaliser des phylogénies à partir
d'alignements d'acides nucléiques ou d'acides aminés (cf. tableau 1.9). Je m'appliquerai ici à réaliser
une liste non exhaustive des logiciels disponibles sur Macintosh et sur Station SUN. Une liste plus
complète est disponible sur le site de l'institut Pasteur (http://www.pasteur.fr/outils.html).
Les logiciels d'alignements
La première étape d'une étude phylogénétique est celle de l'alignement. Ce travail peut être
effectué "à la main" ou à l'aide d'algorithmes regroupés dans des programmes [Clustal V ou clustal W
(GDE), Fast align sequence, aligner v1.0 ou v2.0 (ARB), pileup (GCG)]. Les résultats obtenus par ces
algorithmes ne sont malheureusement pas suffisamment fiables. Ils nécessitent toujours une
vérification de la part de l'utilisateur notamment dans le cas de l'ADNr afin de tenir compte des
structures secondaires de la molécule.
Sous environnement de type UNIX l'un des logiciels les plus ergonomiques est GDE (Genetic
Data Environment par Steven Smith). Il nécessite cependant une mémoire vive et vidéo importante
pour être utilisé sans trop de désagréments. Ce logiciel peut être utilisé seul ou dans le package ARB
(voir plus loin).
8 LINUX est un environnement gratuit de type UNIX qui a été développé à l'origine par Linus Torvalds puis qui a été amélioré par des
programmeurs du monde entier. C'est un environnement ouvert dont les sources sont disponibles gratuitement dans le cadre du projet
GNU General Public License (http://www.linux.org/info/gnu.html).

Les techniques d'écologie microbienne
24
Seaview développé par Manolo Gouy (Galtier et al., 1996) est un logiciel d'alignement multiple
très convivial. Il traite aussi bien les séquences d'acides nucléiques que les séquences d'acides
aminés (http://pbil.univ-lyon1.fr/software/seaview.html). Développé récemment, il intègre les
principales formes d'acquisitions de fichiers et un alignement manuel ou automatique sur la base des
algorithmes Clustal et de Dot Plot. Ce programme est destiné à fonctionner en aval du logiciel
d'analyse de séquences Phylo_Win (Galtier et al., 1996).
Le package ARB (voir plus loin) propose à lui seul 4 logiciels d'alignement : GDE2.2., ALE,
ARB4 et ABRNT. Certains de ces logiciels (ARB4 et ABRNT) permettent de réaliser l’alignement en
tenant compte de la structure secondaire propre à l’ADNr 16S.
Pour finir sur UNIX, TkDCSE est la version pour X-Window de DCSE (Dedicated Comparative
sequence editor)(http://indigo2.uia.ac.be/~peter/dcse).
Sur Macintosh il est aussi possible de réaliser des alignements de séquences grâce au logiciel
Seqpup . Il n'existe pas de mise à jour récente du logiciel.
Les Packages
PHYLIP (Phylogeny Inference Package) Version 3.57c est historiquement l'un des premiers
ensembles de logiciels développés pour l'analyse phylogénétique des séquences. Mis au point en
1986 par Joseph Felsenstein et ses collaborateurs de l'université de Washington, ce package a
évolué avec la mise au point de nouvelles techniques de phylogénie (Felsenstein, 1989). Toutefois
son interface uniquement en mode texte n'est pas conviviale et on lui préférera des programmes
comme ARB (voir plus loin) ayant intégré l'ensemble des méthodes de calcul de Phylip au sein d'une
interface beaucoup plus ergonomique.
GCG (Genetic Computer Group) (http://www.gcg.com) est un ensemble de logiciels très
complet mais aussi très coûteux. L'ensemble des programmes peut tourner en mode texte, mais il est
aussi possible de les faire fonctionner sous une interface graphique (SeqLab) à partir d'un Terminal X,
d'une station de travail ou à partir de n'importe quel ordinateur grâce à une "interface www" nommée
SeqWeb. L'intérêt de ce package réside dans la mise à jour régulière des logiciels et des banques de
données fournies avec ce programme. Toutefois cette dernière possibilité a été rendue caduque par
la disponibilité des banques sur le réseau.
Phylo_Win est un logiciel d'analyse phylogénétique de séquences (http://pbil.univ-
lyon1.fr/software/phylowin.html) (Galtier et al., 1996). Il accepte des alignements au format MASE
produits par Seaview. Il permet de réaliser des analyses par les méthodes du "Neighbor-joining", du
maximum de parcimonie et du maximum de vraisemblance en permettant de réaliser des
réechantillonages sur chacune de ces méthodes. Un grand nombre de facteurs de distance sont
disponibles (Jukes et Cantor, Kimura, Tajima et Nei, HKY, Galtier et Gouy (Galtier et Gouy, 1995),
LogDet pour les séquences nucléiques, la correction de Poisson pour les séquences protéiques). La
gestion des masques est très facilement effectuée à la souris et les arbres construits sont aisément
exploitables par d'autres logiciels.

Les techniques d'écologie microbienne
25
ARB est un package développé par Olivier Strunk et Wolfgang Ludwig au sein de l'université
technique de Munich (TUM) et qui est avant tout destiné à l’analyse de l’ARN ribosomique 16S. Il
fonctionne sous environnement UNIX mais une version a été récemment développée pour LINUX. Il
intègre un grand nombre de programmes déjà existants tant en alignement (GDE) qu’en phylogénie
(Phylip). ARB possède sa propre base de données de séquences. Il s'agit d’ADNr 16S préalablement
alignés par les membres de la TUM. Cette base de données contient de plus toutes les informations
disponibles sous le format GenBank (dépositaire de la séquence, publication de référence, groupe
phylogénétique de la séquence, etc.). Elle n’est malheureusement pas mise à jour suffisamment
régulièrement (dernière mise à jour en mars 1997). Il est donc préalablement nécessaire lors de la
construction d’arbres de vérifier le dépôt de nouvelles séquences au près du RDP ou auprès de
GenBank. Il est possible grâce à ce logiciel de réaliser des arbres à l’aide des trois méthodes
phénétique et cladistiques. L’arbre consensus peut être par la suite édité à l’aide du logiciel Xfig fourni
avec le package puis importé sur Macintosh ou PC sous un format poscript (type PS ou EPS).
Mais l’un des atouts majeurs de ce logiciel est le programme très ergonomique de construction
de sondes nucléiques. En choisissant dans la banque de données d’ARB les séquences cibles, il est
possible de construire puis de vérifier la validité des sondes. L’orientation "procaryotes-ADNr 16S" fait
d'ARB un outil idéal pour les microbiologistes.
3.2.2.3 Les outils d’analyse en ligne et les bases de données
Les outils d'analyse en ligne et les banques de données se sont considérablement développés
ces 5 dernières années avec la généralisation de l'accès à Internet9 au sein des laboratoires.
Dans un premier temps cantonnées à un échange de données uniquement en mode texte via
le courrier électronique (e-mail) ou ftp (file transfer protocol), les analyses sont désormais possibles via
des sites http (hypertext transfert protocol) avec une interface beaucoup plus ergonomique basée sur
le html (hypertext markup language). L'information étant disponible à tous il devient alors possible de
centraliser la source d'information, de mettre à jour plus facilement les banques, de stocker les
données sur une seule machine (The Integration of Microbial Databases (site web), 1995; Burks,
1999; FitzGerald et Blacke, 1998; Nierlich, 1996).
En France le site de l'institut Pasteur (http://www.pasteur.fr) fournit une liste des logiciels
disponibles pour la comparaison et l'alignement de séquences, les analyses phylogénétiques, la
recherche de gènes et de régions codantes, la recherche et l'extraction de motifs, les HMM (Hidden
Markov Model), les analyses de structures secondaires et tertiaires des acides nucléiques et des
acides aminés (par exemple pour l'ARNr la Mfold home page (Zuker et Jacobson, 1998)). Le
9 Internet est né il y a un peu plus de 20 ans de la volonté du département de la défense américaine de mettre en réseau l'ensemble de
ses services de recherche militaire afin de répondre le plus rapidement possible à d'éventuelles attaques nucléaires, ce premier réseau
s'appelait ARPAnet. Un protocole de communication est alors développé afin de permettre la communication entre les ordinateurs
(quelle que soit leur marque) : le protocole IP (Internet Protocol). Dix ans plus tard le développement des réseaux internes (Ethernet
local area network, ou LAN) et la pression du marché (de la part des université américaines notamment) permettent l'accès de
n'importe quel utilisateur équipé d'un ordinateur et d'un modem à ce réseau devenu un réseau de réseaux ou World Wide Web.

Les techniques d'écologie microbienne
26
BioCatalog de l'EBI (European Bioinformatics Institute) référence quant à lui la plus grande partie des
logiciels pour la biologie (UNIX, Macintosh, PC) (http://www.ebi.ac.uk/biocat).
Dans le cadre d'une analyse de séquence d'ARN ribosomique deux sites sont indiqués :
Ø Le site du NCBI (National Center for Biotechnology Information) va permettre d'accéder à
l'ensemble des séquences publiques déposées dans GenBank (Benson et al., 1999) et dans l'EMBL
(Stoesser et al., 1999). En décembre 1999 plus de 4,9.106 séquences et 3,8.109 bases étaient
disponibles dans cette banque de données (http://www.ncbi.nlm.nih.gov). Il est possible via ce site de
récupérer des séquences (Entrez), de soumettre de nouvelles séquences (BankIt et Seqin) ou de
comparer des séquences avec celles présentes dans GenBank (BLAST Basic Local Alignment Search
Tool, Altshul et al., 1997; Tatusova et Madden, 1999). Ce site permet de plus d'avoir accès au projet
PubMed gérant la banque de données développée par la NLM (National Library of Medecine)
regroupant les 9.106 références bibiographiques MEDLINE et PreMEDLINE.
Ø Le site du RDP ou Ribosomal Database Project (http://www.cme.msu.edu/RDP/html/
index.html) (Maidak et al., 1999) est une base de données et un ensemble de logiciels d'analyse en
ligne essentiellement destinés à l'analyse de l'ADNr 16 et 18S. Cette banque, gérée par le CME
(Center for Microbial Ecology) est régulièrement mise à jour à partir des séquences soumises à
GenBank mais aussi par des séquences (très souvent des clones environnementaux) soumises
directement par les utilisateurs. En plus de la banque de séquences d'ARNr, le RDP met à la
disposition des scientifiques un ensemble de programmes permettant :
Ø de comparer une ou plusieurs séquences avec celles de la banque de données,
Ø de réaliser un premier alignement sommaire,
Ø de placer des séquences dans un arbre phylogénétique rudimentaire,
Ø de vérifier la nature chimérique des séquences analysées,
Ø de vérifier la validité d'une sonde
Ø d'accéder aux banques de données de séquences d'ADNr 16S et 23S10.
Il existe aussi des banques de données concernant les micro-organismes cultivés. Ces banques
sont disponibles sur le site de l'ATCC (http://www.atcc.org) et de DSMZ (http://www.dsmz.de/). Le site
du Bergey's Manual Trust (http://www.cme.msu.edu/bergeys) donne aussi accès à la liste des
souches disponibles, aux espèces, genres, familles reconnus ou incertains, ainsi qu'à un arbre
phylogénétique type regroupant les souches isolées et les séquences provenant d'ARNr clonés à
partir d'échantillons environnementaux. Un projet de normalisation de cette liste sous la forme d'une
base de données plus aboutie est en cours sous le nom du PDP Phenotypic Database Project.
10 En complément du site du RDP deux banques inventorient les structures secondaires des petites sous-unités de l'ARNr (Van de
Peer et al., 1999) et des grandes sous-unités de l'ARNr (de Rijk et al., 1999)
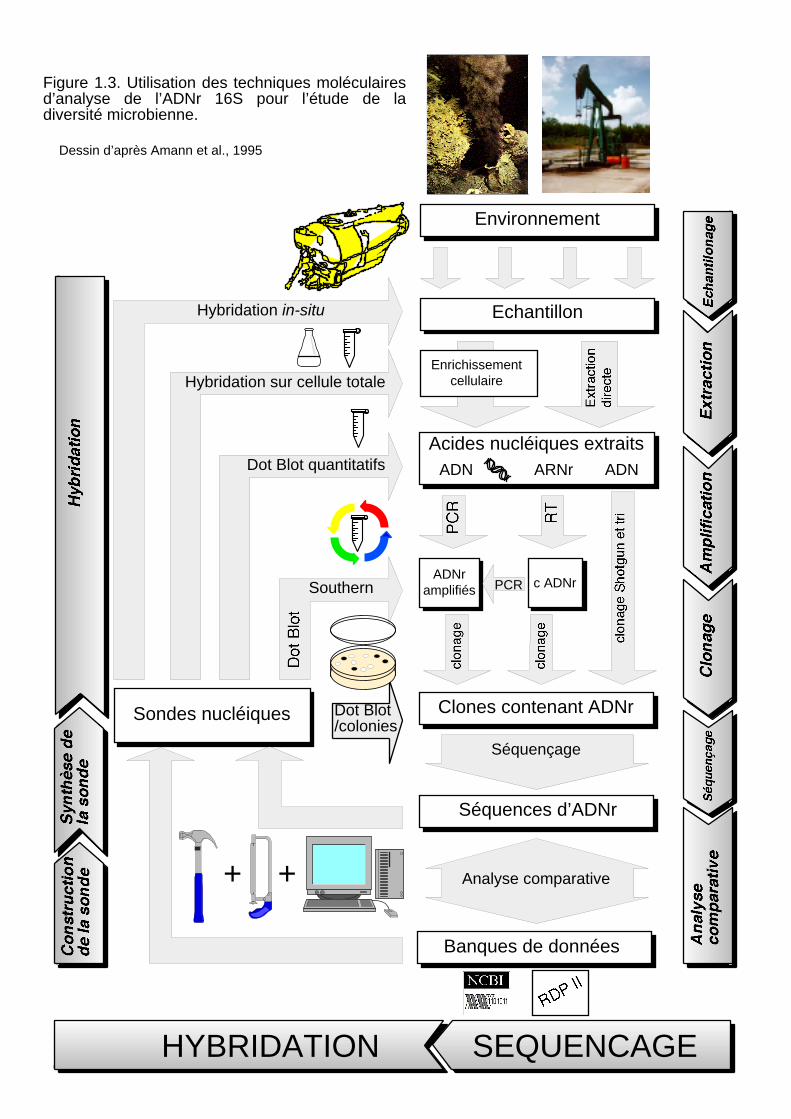
Echantillon
Acides nucléiques extraitsADN ARNr
PCR
Clones contenant ADNr
Séquences d’ADNr
Banques de données
Environnement
ADNramplifiés c ADNr
Séquençage
Analyse comparative
Enrichissementcellulaire
Sondes nucléiques
Hybridation in-situ
Hybridation sur cellule totale
Dot Blot quantitatifs
Southern
Dot Blot /colonies
ADN
SEQUENCAGEHYBRIDATION
+ +
Figure 1.3. Utilisation des techniques moléculairesd’analyse de l’ADNr 16S pour l’étude de ladiversité microbienne.
Dessin d’après Amann et al., 1995

Les techniques d'écologie microbienne
27
Après m'être intéressé au choix des acides nucléiques comme marqueurs de la diversité
microbienne et à l'analyse des séquences d'acides nucléiques, je me propose de décrire les
différentes méthodes moléculaires couramment utilisées en écologie microbienne.
3.3 Les méthodes moléculaires
Les techniques moléculaires peuvent en principe s'appliquer à tous les gènes pour peu que
ceux-ci répondent aux principes d'universalité et d'évolution que nous avons évoqués
précédemment. Toutefois nous nous attacherons dans cette partie à décrire les principales
techniques mettant en jeu l'ARNr 16S. Les hybridations ADN/ADN et la mesure du G+C% d'un ADN
génomique sont elles aussi des méthodes moléculaires, mais elles s'appliqueront plus à la
caractérisation d'une souche isolée qu'à l'étude directe d'un environnement (à l'exception de
l'approche présentée dans le §3.3.4.1).
3.3.1 Le clonage et le séquençage des ARNr
Cette technique est l'une des premières à avoir été employée pour s'affranchir de toute étape
de culture. Les différentes étapes de cette technique sont regroupées dans la partie droite de la figure
1.3. Elles consistent : à extraire l'ADN ou l'ARN total de l'échantillon environnemental, à amplifier
spécifiquement les gènes codant pour l'ARN ribosomique, puis à cloner le produit d'amplification et
enfin, à séquencer et à analyser les clones de la banque.
3.3.1.1 Historique de la méthodologie
Le résultat des premières études portant sur l'analyse de l'ARN ribosomique est reporté dans la
seconde partie de ce manuscrit. Ces études consistaient à analyser la communauté microbienne
d'échantillons naturels par séquençage des ARNr 5S directement extraits de l'échantillon. Elle
portaient sur l'analyse : de la communauté symbiotique de vers hydrothermaux considérés comme
chémo-autotrophiques (Stahl et al., 1984), de bactéries colonisant une source chaude terrestre (Stahl
et al., 1985) et des micro-organismes colonisant les bassins de biolixiviation du cuivre (Lane et al.,
1985b). Le manque de pouvoir résolutif de l'ARNr 5S (seulement 120 pb) incita les chercheurs à
analyser les ARNr 16S.
La première stratégie a été d'extraire l'ADN génomique total et de le cloner dans une banque
phagique. Les phages contenant des fragments d'ARNr ont été par la suite criblés à l'aide d'une sonde
spécifique et les clones ont été séquencés. Il a été possible à l'aide de cette technique d'analyser une
communauté picoplanctonique marine (Schmidt et al., 1991). Cette technique a été reprise par Stein
et al. (1996) pour accéder à des gènes du métabolisme à partir du même type de banque phagique.
Afin de réduire l'étape de criblage Giovannoni et al. (1990b), proposèrent à l'aide d'amorces
spécifiques d'amplifier les gènes codant pour l'ARNr 16S (ADNr 16S) puis de cloner le produit
d'amplification. Afin d'accéder à la population réellement active de l'échantillon Weller et Ward (1989)

Les techniques d'écologie microbienne
28
proposèrent de travailler sur les ADNc transcrits à l'aide d'une réverse transcriptase à partir du pool
d'ARNr 16S. Amann et al. (1992a) modifièrent cette technique en ajoutant une étape de PCR après
l'utilisation de la reverse transcriptase afin de travailler directement sur des ADNr (cf. figure 1.3).
L'approche proposée par Giovannoni et al. (1990b) est actuellement la plus utilisée. Les
modifications qui y ont été apportées s'attachent à réduire les biais inhérents à cette méthodologie en
optimisant les techniques d'extraction d'ADN, d'amplification (jeu d'amorces, cycle, etc..) et d'analyse
de la banque (RFLP, séquençage total).
Nous reviendrons en détail dans la troisième partie de ce manuscrit sur les différentes étapes de
cette méthode et le chapitre 3.4 s'attachera à décrire les biais qui peuvent entacher une étude de
diversité.
3.3.1.2 L'échantillonnage et l'extraction d'ADN
Il faut considérer que l'étude reflétera la diversité de l'échantillon avant de refléter, par
extrapolation, celle de l'environnement tout entier. Aussi l'échantillon devra représenter le plus
fidèlement possible l'environnement étudié.
Un grand nombre de dispositifs d'échantillonnage sont disponibles selon que l'on travaille sur
des échantillons solides ou liquides, profonds ou de surface. L'accessibilité du site d'échantillonnage
est la première donnée du problème. Les sources hydrothermales profondes ne sont par exemple
accessibles qu'à un nombre réduit de submersibles scientifiques. Pour les échantillons des sols, il est
possible d'effectuer des prélèvements par carottage à partir de la surface ou d'utiliser des puits pour
accéder à des échantillons profonds (Pedersen, 1996). Pour des échantillons liquides il est possible
de réaliser des prélèvements ponctuels (seringue titane, bouteille NIO, bouteille Niskin) ou de travailler
sur de grands volumes pour accéder à une diversité plus importante, notamment aux souches rares. Il
est alors nécessaire de concentrer par filtration les échantillons (Bej et al., 1991; Giovannoni et al.,
1990a; Sommerville et al., 1989).
Le mode de conservation de l'échantillon pourra lui aussi influer sur la diversité observée. Il
faudra conserver les cellules intactes, c'est à dire les fixer si on souhaite réaliser des hybridations in situ
ou les maintenir dans des conditions proches de celles où elles se développent (pH, potentiel redox).
Afin de limiter la dégradation des acides nucléiques durant le stockage, il faudra limiter l'action des
DNAses ou des RNAses qui sont libérées dans le milieu dès que les cellules sont altérées. Pour ce
faire il faudra conserver les cellules au froid, dans l'éthanol ou dans des tampons bloquant les DNAses
et les RNAses (comme le Guanidide Iso Thiocyanate). La solution idéale reste de pouvoir extraire l'ADN
des échantillons dès que ceux ci sont collectés afin de réduire au minimum le temps de stockage de
l'échantillon.
La méthode d'extraction des acides nucléiques devra lyser de manière efficace l'ensemble des
parois microbiennes, sachant que la paroi des Gram + est plus résistante que celle des Gram- et que les
Archaea ne possèdent pas de peptidoglycane et sont donc insensibles à l'action du lysozyme. De
plus, des composants présents dans l'échantillon peuvent réduire l'efficacité de l'extraction (matière
organique, métaux, etc.) en se fixant sur l'ADN libéré des cellules. Il est souvent nécessaire de

Les techniques d'écologie microbienne
29
combiner des méthodes de lyse physique (pilon et mortier, cycles de congélation-décongélation,
etc.), de lyse chimique (détergents, phénol, etc.) et de lyse enzymatique (lysozyme, protéinase K,
etc.) afin d'extraire le plus efficacement possible l'ADN contenu dans l'échantillon.
Une technique particulière de purification des ADN bactériens a été proposée par Neimark et al.
(1998) dans le cadre de l'étude de l'agent (non cultivable) du poumon gris (GLA: Grey Lung Agent).
Après extraction des ADN, ceux-ci sont séparés par électrophorèse en champ pulsé. Il est ainsi
possible de travailler sur des chromosomes entiers.
Deux autres techniques ont été proposées dans le cas d'études de milieux peu complexes pour
isoler spécifiquement les micro-organismes sans utiliser de méthodes culturales. Ces techniques
reposent sur la micromanipulation des micro-organismes ou sur leur extraction du milieu d'étude. Dans
le cadre de l'étude des bactéries symbiotes du poisson chirurgien (famille des Acanthuridae), Angert
et al. (1993) ont isolé le symbiote par micromanipulation. La taille de la bactérie (80 x 600 nm) a permis
d'utiliser cette technique habituellement appliquée aux Eucarya. Hubert et al. (1995) ont utilisé quant à
eux les pinces optiques pour isoler une Archaea hyperthermophile préalablement détectée par
extraction et séquençage des ADNr 16S à partir d'échantillons de la source chaude d'Obsidian Pool
(Barns et al., 1994). Le micro-organisme était minoritaire dans les cultures d'enrichissement et
échappait à toutes les tentatives d'isolement (Hubert et al., 1995). La technique d'isolement utilisée
est fondée sur la détection du morphotype à l'aide de sondes nucléotidiques spécifiques, puis sur la
recherche du morphotype dans des cultures d'enrichissement. Les cellules sont alors prélevées dans
le milieux à l'aide d'un LASER (Ashkin et Dziedzic, 1987)11. La seconde méthode, employée pour
étudier les symbiotes des vers hydrothermaux, consistait à isoler les cellules bactériennes des cellules
hôtes sur un gradient de Ficoll après centrifugation (Lane et al., 1985a).
3.3.1.3 L'amplification et le clonage
Chaque étape de la PCR devra être prise en compte afin d'optimiser les conditions
d'amplification (voir pour revue Arnheim et Erlich, 1992; Steffan et Atlas, 1991).
L'utilisation de l'ADNr 16S permet de concevoir des amorces de PCR aux extrémités 5' et 3' de la
molécule avec des spécificités plus ou moins importantes (Brunk et al., 1996; Marchesi et al., 1998).
On utilise généralement plusieurs jeux d'amorces universelles afin de réaliser des banques de
séquences de Bacteria, d'Archaea et parfois d'Eucarya (Hugenholtz et al., 1998a par exemple). Dans
certaines études il a été possible d'amplifier spécifiquement des séquences d'ADNr 16S
correspondant à un type métabolique défini (les BSR, Devereux et Mundfrom, 1994; Devereux et al.,
1996). En jouant sur les conditions de PCR (enzyme, nombre de cycles, température, etc.) il est
possible par la suite de modifier la spécificité de la réaction et de diminuer les risques de création de
11 L'utilisation des pinces optiques touche de nombreux domaines de la biologie. Ces pinces sont utilisées pour travailler notamment
sur les chromosomes (séparation des chromosomes lors de la division cellulaire), les mitochondries ou les microfilaments. Il est
également possible d'utiliser le LASER à la fois comme pince (maintien de la cellule) et comme ciseaux (découpage précis de la
cellule) (Berns, 1998). Récemment Arai et al. (1999) sont parvenus à l'aide de pinces optiques à faire des noeuds dans des filaments
d'actine et d'ADN et ont prouvé que l'ADN était plus solide que l'actine.

Les techniques d'écologie microbienne
30
chimères (voir §3.4.2.2). Les biais inhérents à la PCR rendent difficile une approche quantitative même
si de nombreuses techniques ont été proposées (Ferré et al., 1994 pour revue; Lee et al., 1996; Poltz
et Cavanaugh, 1997). Le développement des techniques de PCR quantitative en temps réel permet
désormais de suivre l'évolution des produits d'amplification à l'aide d'amorces marquées12 (Patel et al.,
1998; Pahl et al., 1999).
Le clonage des produits de PCR est l'étape préalable au tri. Dans la mesure où les fragments
amplifiés sont de petite taille (1,5 Kpb pour l'ADNr 16S) il est possible de les cloner dans des vecteurs
plasmidiques (généralement dérivés du plasmide pBluescript). Il est par contre nécessaire de réaliser
des banques phagiques lorsque l'on souhaite cloner l'ensemble de l'ADN génomique d'un échantillon
pour accéder à des gènes du métabolisme.
Le clonage peut être réalisé directement sur produits de PCR en utilisant le principe du TA-
cloning (Mead et al., 1991) ou après digestion du produit de PCR et d'un vecteur afin de réaliser des
clonages en bouts francs et ou en bouts cohésifs. L'avantage de la dernière méthode est de connaître
l'orientation du fragment inséré, mais le traitement du produit de PCR par des enzymes peut entraîner
une digestion partielle de ce dernier. Il n'est alors pas possible d'accéder à la séquence du gène dans
son intégrité.
3.3.1.5 Le criblage et le séquençage
Une fois les fragments clonés il est nécessaire de vérifier la nature de l'insert et sa taille. Il est
alors possible d'hybrider directement les colonies à l'aide de sondes spécifiques des ARN
ribosomiques ou plus simplement extraire le plasmide ou de réamplifier directement sur colonies
l'insert à l'aide d'amorces spécifiques du plasmide flanquant le site de clonage (T3, T7, M13, reverse,
universelle par exemple).
Afin de réduire le coût de séquençage de l'étude il peut être intéressant de réaliser un premier
tri parmi les clones de la banque. Des classes, ou OTU (Moyer et al., 1994), ou phylotypes (Ward et al.,
1998) peuvent être définis par comparaison des profils de restriction des inserts coupés à l'aide
d'enzymes tétramériques (RFLP, AFLP, ARDRA). Le choix des enzymes de restriction va conditionner
la qualité du tri (Moyer et al., 1996). Il est possible de coupler la RFLP à un marquage fluorescent. On
observe la migration des fragments de restriction marqués sur un gel de séquençage: T-RFLP (Liu et
al., 1997; Pukall et al., 1998; Chin et al., 1999; Marsh, 1999). On peut aussi analyser les profils
obtenus après séquençage des différents clones sur une seule base (Ward et al., 1990b). Suzuki et
al. (1998) analysent quant à eux le polymorphisme de taille de fragments d'ADNr 16S amplifiés.
Récemment la diminution des coûts ainsi que la simplification des techniques de séquençage a
permis de généraliser l'analyse de la totalité de la banque par séquençage. On séquence alors les
12 Les entreprises BioRad et Perkin Elmer proposent par exemple des systèmes de PCR quantitative clé en main (iCycler et Gene
Amp, TaqMan respectivement).

Les techniques d'écologie microbienne
31
clones à l'aide d'une seule amorce dans un sens (par exemple l'amorce 515F (nos études) ou l'amorce
533F (Hugenholtz et al., 1998a).
L'utilisation de séquenceurs automatiques ou le recours à des entreprises réalisant le
séquençage "à façon" a permis de grandement faciliter l'étape de séquençage dans une analyse de
diversité. L'utilisation de jeu d'amorces marquées réparties sur l'ensemble de l'ADNr 16S tous les 200
à 400 pb (cf. figure 1.4) permet de couvrir l'ensemble de la molécule en lecture double ou triple
(Edwards et al., 1989).
Figure 1.4. Exemple de jeu d'amorces permettant de couvrir l'ensemble du gène de l'ADNr 16S
L'analyse de la séquence est réalisée à l'aide des techniques de phylogénie classique
(méthodes de distance, de maximum de vraisemblance et de maximum de parcimonie) en comparant la
séquence du clone avec les séquences des micro-organismes déposées dans les banques de
données (Maidak et al., 1999).
3.3.2 Les sondes nucléiques
A la différence des techniques d'analyse par extraction et séquençage de l'ADNr 16S,
l'utilisation des sondes nucléiques permet d'accéder individuellement à chaque cellule. Cette
technique implique la formation d'un duplex entre une sonde nucléique et un ARNr cible. Comme
l'indique la figure 1.3, les sondes peuvent être utilisées aussi bien sur de l'ADN fixé sur membrane (Dot
ou Slot Blot après Southern), directement sur les colonies de clones (Grunstein et Hogness, 1975),
sur des cultures d'enrichissement ou sur souche pure (Whole-cell hybridation) ou sur des échantillons
naturels (hybridation in situ). Pour marquer la sonde on utilise soit des radioéléments (ATP-35S ou ATP-32P) (Giovannoni et al., 1988) soit des enzymes (Amann et al., 1992b) soit des marqueurs fluorescents
(fluorescéine, rhodamine, CY3, etc.); on parle alors de FISH (Fluorescent In-Situ Hybridization) (Delong
et al., 1989; Amann et al., 1990 a et b; Stahl et Amann, 1991). On retiendra deux publications
décrivant en détail les principes et les procédures d'hybridation sur membranes et cellules (Stahl et
Amann, 1991; Johansson, 1996).
3.3.2.1 La construction des sondes nucléiques
La succession de domaines, de conservés à très variables, sur la molécule d'ADNr 16S permet
de disposer de sondes au spectre plus ou moins large (de l'espèce au domaine cf. tableau 1.10). La
spécificité de la sonde s'accroît à mesure qu'elle s'hybride dans un domaine de plus en plus variable.

Les techniques d'écologie microbienne
32
Tableau 1.10. Exemple de différentes sondes ayant des niveaux de spécificité différents.
Nom de la sonde Spécificité Référence
S-*-Univ-1390-a-A-18 universelle Zheng et al., 1996
S-D-Bact-0338-a-A-18 domaine : Bacteria Amann et al., 1990a
S-D-Arch-0915-a-A-20 domaine : Archaea Amann et al., 1990b
S-K-Cren-0499-a-A-18 royaume : Crenarchaeota Burgraff et al., 1994
S-Sc-aProt-0019-a-A-17 sous classe : Proteobacteria α Manz et al., 1992.
S-G-Dsbb-0660-a-A-20 genre : Desulfobulbus Deveureux et al., 1992
S-S-Thip-0829-a-A-21 espèces : Thioplaca araucae, T. chileae Teske et al., 1995
Une grande partie des sondes ont été dessinées pour être spécifiques de l'ARNr 16S et un
grand nombre de publications décrivent la construction de ces sondes13, par exemple : sondes
générales (Amann et al., 1990b), sondes pour méthanogènes (Raskin et al., 1994a), sondes pour les
différentes sous-classes des Proteobacteria (Manz et al., 1992) ou pour les Proteobacteria δ sulfato-
réductrices (Devereux et al., 1992). Des auteurs ont ciblé d'une manière similaire l'ARNr 23S (Ludwig
et al., 1994; Trebesius et al., 1994) ou les gènes codant pour des activités métaboliques spécifiques
(Ogram et Sayler, 1988). Une technique alternative de construction de sondes a été proposée par
Fani et al. (1993). Ces derniers ont utilisé comme sondes des fragments spécifiques produits par
RAPD. Ces fragments pouvaient alors cibler l'ensemble du génome. Une technique de construction
de sondes par hybridation soustractive a été proposée par Mau et Timmis (1998) pour obtenir des
sondes très spécifiques (d'un clone par exemple) en limitant ainsi le nombre de séquences identiques
à analyser.
L'accessibilité de la sonde à l'ARN est le second facteur qui va influer sur la qualité de la sonde.
La forte structure secondaire de la molécule d'ARNr (Gutell et al., 1994) impose des contraintes
structurales d'hybridation qui peuvent réduire considérablement l'intensité du signal (Fuchs et al.,
1998).
L'un des paramètres importants de l'hybridation est la température de fusion (Tm). Cette dernière
définit pour un oligonucléotide la température d'équilibre à laquelle la moitié du duplex sonde-cible est
dissociée. La Tm est dépendante de la concentration en oligonucléotides mais est indépendante du
temps (situation d'équilibre). La Td ou température de dissociation est la température pour laquelle
50% du duplex reste intact pour une durée de lavage définie dans des conditions de non-équilibre
(donc Td peut être différent de Tm). La Td est la variable expérimentale de référence, elle définit la
température de lavage de posthybridation pour laquelle 100% des hybridations non spécifiques sont
dissociées (cf. tableau 1.11).
Une fois la sonde construite il est nécessaire de la valider en vérifiant sa spécificité dans un
premier temps sur Dot blot contre des ARN ou des ADN de souches s'hybridant ou non avec la sonde
13 Une banque de séquence de sondes (OPD: Oligonucleotide Probe Database) a été créée et est disponible sur le réseau
(http://www.cme.msu.edu/opd/). Elle n'a malheureusement pas été remise à jour récemment (Alm et al., 1996).

Les techniques d'écologie microbienne
33
puis en testant la sonde sur culture pure ou mélange de culture pure (voir par exemple Harmsen et al.,
1997b).
Tableau 1.11. Caractéristiques de la température de dissociation (Td)
La Td est déterminée par : La Td est dépendante de :
Ø le nombre de liaisons hydrogène entre bases
complémentaires,
Ø le degré de "stacking" (échange d'électrons) des
bases adjacentes sur une même chaîne,
Ø les interactions entre les phosphates (chargés
négativement) les cations en solution.
Ø la structure du duplex,
Ø la séquence de la chaîne,
Ø la longueur de la chaîne,
Ø le nombre, le type et la position des mésappariements,
Ø la présence de bases désappariées en fin de chaîne,
Ø des conditions de lavage et d'hybridation.
3.3.2.2 L'utilisation des sondes en dot blot
Les sondes marquées peuvent être utilisées en dot blot pour qualifier voire quantifier un type
de micro-organismes (s'hybridant avec la sonde) par rapport à une population totale dont l'ADN serait
fixé sur membrane. La présence ou l'absence du micro-organisme sera liée à l'hybridation ou non de la
sonde. L'intensité de l'hybridation par rapport à une sonde plus générale (par exemple une sonde
universelle ou domaine spécifique) pourra refléter la quantité de micro-organismes présents dans
l'échantillon. Cette intensité est toutefois fortement liée au nombre de ribosomes dans la cellule
(variant de 103 à 105 selon les organismes et l'état cellulaire) et à la qualité d'extraction d'ADN.
3.3.2.3 Les sondes en hybridation in situ
A la différence des hybridations sur colonies ou sur membrane, les hybridations sur cellules
entières permettent d'accéder directement à l'ARNr sans endommager la structure pariétale de la
cellule et donc d'accéder en plus du caractère phylogénétique ou métabolique lié à la sonde à un
caractère morphologique. Olsen et al., (1986) proposent les premiers d'utiliser les sondes marquées
pour identifier et compter les cellules dans des échantillons environnementaux. Le premier essai est
réalisé à l'aide de sondes radioactives (Giovannoni et al., 1988). L'hybridation consiste en une
première phase de perméabilisation de la cellule et de fixation des acides nucléiques à l'aide de
paraformaldehyde le plus souvent. Puis les cellules sont fixées sur une lame enduite de gélatine. Elles
sont incubées sur lame en présence de la sonde marquée dans des conditions de stringence qui sont
définies par la température d'hybridation ou la quantité de formamide présente dans le tampon. Après
lavage, les cellules peuvent être observées sous microscope à épifluorescence. Il est possible de
combiner l'hybridation de plusieurs sondes (ayant des spécificités différentes) au sein d'un même
échantillon par l'utilisation de différents fluorochromes (Amann et Kuhl, 1998). Il est aussi possible
d'utiliser plusieurs sondes ayant la même spécificité mais des sites d'hybridation différents afin de
détecter des micro-organismes en très faible quantité (Lee et Kemp, 1994).
Des comparaisons des différentes méthodes de quantification des cellules ont été entreprises
par Karner et Furhman (1997). Les techniques les mieux corrélées sont les sondes nucléiques et les

Les techniques d'écologie microbienne
34
techniques d'autoradiographie. Ces deux techniques détectent toutefois moins de cellules que le
comptage par DAPI.
Une technique de transcription reverse in situ (ISRT) a été développée afin de pouvoir détecter
plus facilement les bactéries actives d'un échantillon (Chen et al., 1997). Les hybridations, sur des
ADNc, à l'aide de sondes spécifiques des ARNr ou de certains ARNm, permettent d'obtenir des
signaux plus puissants (sur les bactéries actives) que par simple FISH.
3.3.3 La DGGE
3.3.3.1 Principe
Le principe de la DGGE (Denaturing Gradient Gel Electrophoresis) est fondé sur la séparation
d'un double brin d'ADN sous l'action de la chaleur ou d'un dénaturant (Fischer et Lerman, 1979). La
température à laquelle le duplex est détruit est conditionnée par deux facteurs :
Ø Le nombre de liaisons hydrogène entre les bases complémentaires. Ainsi une liaison GC (3
liaisons hydrogène) est détruite à une température plus élevée qu'une liaison AT (2 liaisons
hydrogène)
Ø L'attraction entre des bases voisines sur un même brin ou "stacking".
La molécule d'ADN aura donc une Tm (ou température de fusion) qui variera selon les domaines
et qui sera fonction de sa séquence nucléique. Deux molécules d'ADN proches, différant d'un seul
nucléotide dans une zone à faible température de fusion, auront deux températures de fusion
différentes. Lors d'une migration électrophorétique dans un gradient croissant de dénaturant
(habituellement formamide ou urée), la mobilité de la molécule d'ADN décroît alors que les zones à
faible température de fusion se dissocient. La structure de l'ADN, alors en simple brin, stoppe la
migration de la molécule. Pour éviter la dénaturation totale du double brin, une structure à très haute
température en fusion (très riche en GC) est associée lors de l'amplification. Une PCR est alors réalisée
à l'aide d'une amorce portant en 5' une succession de 40 GC : le GC clamp.
La mise au point de la technique réside essentiellement (Michaelides et al., 1995; Cotton, 1997) :
Ø dans le choix des amorces. Elles permettent d'amplifier de 100 à 400 pb dans une zone à faible
point de fusion. Le "GC clamp" doit être proche de l'extrémité amplifiée ayant le plus fort point de
fusion. Il est donc nécessaire de connaître avant l'amplification les zones qui auront une
température de fusion plus ou moins élevée. Dans le cas de l'ADNr 16S ces zones peuvent être
connues en raison de la forte stabilité de la structure secondaire de la molécule.
Ø dans la préparation du gel (nature et établissement du gradient, tension et durée
d'électrophorèse).
Dans les deux cas des outils informatiques permettent d'assister le scientifique pour déterminer
le type de gradient à réaliser dans le gel et la durée de migration estimée de l'échantillon.
En résumé les avantages et les désavantages de la méthode peuvent être présentés dans le
tableau 1.12.

Les techniques d'écologie microbienne
35
Tableau 1.12. Avantages et désavantages de la technique de DGGE
AVANTAGES DESAVANTAGES
1. Méthode très sensible
2. En routine, la mise en oeuvre est simple et ne requiert
pas l'emploi de radio éléments.
3. Les fragments de PCR après migration peuvent être
récupérés et séquencés.
1. La phase de mise au point peut être longue
2. Il est préférable d'acheter un appareillage spécifique
3. Le séquençage des fragments est limité à 400 pb.
Problématique pour réaliser des phylogénies par la suite
4. Les gènes ou régions très riches en GC sont
difficilement analysables par DGGE.14
3.3.3.2 La DGGE en écologie microbienne
Initialement développée dans le domaine médical pour détecter des mutations impliquées dans
des maladies génétiques, cette technique a été utilisée pour distinguer les différentes populations de
micro-organismes dans des échantillons naturels après amplification de leur ADNr 16S (Muyzer et al.,
1993; Muyzer, 1999).
Cette technique permet :
Ø la comparaison d'échantillons différents : évolution d'une communauté dans le temps (Santegoeds
et al., 1998b), action de différents nutriments et de l'inoculum dans des cultures en "batch" (Jackson
et al., 1998a) ou action des facteurs environnementaux dans un milieu défini (Santegoeds et al., 1996;
Ward et al. 1997),
Ø une identification des souches présentes dans un échantillon par comparaison du profil de migration
avec des profils obtenus sur des cultures pures, par séquençage des bandes après excision (Ferris et
al., 1996a) ou par hybridation sur le gel d'électrophorèse à l'aide de sondes spécifiques (Muyzer et al.,
1995).
3.3.3.3 Les variantes de la DGGE
Il existe différentes variantes de la DGGE selon le type de gradient utilisé (TGGE ou CDGE).
D'autres techniques (SSCP, HMA) sont elles aussi fondées sur les migrations de structures
secondaires d'ADNr différents.
Ø TGGE : Temperature Gradient Gel Electrophoresis. Le gradient dénaturant chimique est remplacé
par un gradient de température qui augmente au cours de la migration. (Zoetendal et al., 1998; Eichner
et al., 1999)
Ø CDGE : Constant Denaturant Gel Electrophoresis. La concentration d'agent dénaturant est
constante dans le gel et équivalente à la température de la plus base zone de fusion. Cette technique
utilisée lorsque l'on ne connaît pas la localisation des zones de mutations requiert l'utilisation de
plusieurs gels reproduisant chacun différentes conditions de dénaturation.
Ø La technique de SSCP (Single Strand Conformation Polymorphism) est fondée sur la différence de
structure secondaire et donc de migration au sein d'un gel non dénaturant entre deux fragments
14 Wu et al. (1999) ont proposé une méthode combinant la CDGE et la DGGE pour l'analyse des régions très riche en GC.

Les techniques d'écologie microbienne
36
d'ADN simples brins ayant des séquences différentes (Lee et al., 1996; Schwieger et Tebbe, 1998).
Plus facile à mettre en oeuvre que la DGGE, la SSCP nécessite l'utilisation d'ADN témoin pour identifier
les micro-organismes auxquels correspondent les bandes (Widjojoatmodjo et al., 1994). Elle peut
aussi être utilisée pour distinguer des espèces entre elles (Borin et al., 1997).
Ø La technique HMA (Heteroduplex Mobility Assay) consiste elle aussi à faire migrer des fragments
d'ADNr 16S dénaturés puis renaturés de différentes espèces. Au cours de la renaturation des
hétéroduplex se forment dont la mobilité est fonction des structures secondaires recréées dans les
régions non-appariées. La migration a lieu sur gel de polyacrylamide mais elle permet surtout de
distinguer des populations peu complexes (Espejo et al., 1998)15.
3.3.4 Les autres techniques
3.3.4.1 Les renaturations d'ADN totaux
La technique proposée par Torsvik et al. (1990a) repose sur la mesure de la vitesse de
renaturation d'un mélange d'ADN préalablement dénaturé. Plus l'échantillon est constitué d'une
population complexe plus il mettra de temps à se renaturer. Il ne s'agit pas d'une identification des
micro-organismes impliqués dans la diversité de la population mais de la mesure d'un indice de
diversité.
3.3.4.2 L'analyse des fragments de restriction.
Des endonucléases de restriction sont utilisées pour générer des fragments de différentes
tailles sur de l'ADN génomique ou sur de l'ADNr 16S amplifié. La comparaison des profils de restriction
sur gel de polyacrylamide permet d'accéder simplement et d'une manière peu onéreuse à la diversité
de l'échantillon (Moyer et al., 1994; Martinez-Murcia et al., 1995). Moyer et al. (1996), sur la base d'une
simulation réalisée à l'aide de différentes enzymes tétramériques et des séquences disponibles sur le
RDP, a proposé une liste d'enzymes particulièrement résolutives pour des RFLP sur ADNr 16S. S'il est
possible de comparer les communautés entre elles par cette technique, il n'est pas possible
d'identifier les éléments de ces communautés. Une étape de séquençage est alors indispensable.
3.4 De la diversité phylogénétique à la diversité physiologique
Si les techniques d'écologie microbienne moléculaire fondées sur l'étude de l'ADNr 16S
permettent d'accéder et de quantifier la diversité phylogénétique associée à un milieu, ces techniques
ne permettent malheureusement pas d'accéder à la diversité métabolique. En effet à l'exception de
quelques grands groupes phylogénétiques correspondant à une type métabolique donné, tels que
les Archaea méthanogènes ou les cyanobactéries, le rôle fonctionnel d'un micro-organisme détecté
15 Les mêmes auteurs proposent de corréler les profils de migration aux pourcentages de similitudes sur l'ADNr 16S entre souches.

Les techniques d'écologie microbienne
37
"moléculairement" reste dans bien des cas non défini. De plus l'isolement et la caractérisation des
micro-organismes ne nous informent que sur leur capacité métabolique dans des conditions définies.
Aussi pour accéder aux diverses contributions métaboliques au sein de l'environnement différentes
approches ont été proposées, elles consistent soit à étudier les gènes du métabolisme, soit à étudier
les ARNm.
Comme nous l'évoquerons dans la partie suivante (les habitats microbiens), à l'instar des études
réalisées sur les gènes codants pour l'ARNr 16S, des stratégies fondées sur l'emploi de sondes
nucléiques et sur l'amplification des gènes du métabolisme à l'aide d'amorces spécifiques ont déjà été
employées. Bien que le degré de conservation des gènes codants pour des fonctions métaboliques
soit moins important que celui des gènes d'ARNr, un certains nombre de gènes sont suffisamment
conservés pour pouvoir déterminer des sondes ou des amorces spécifiques. Ainsi il est possible
d'amplifier la bisulfite réductase, l'APS réductase ou l'hydrogénase impliquées dans des mécanismes
de sulfato-réduction, la méthane monooxygénase soluble (sMMO) ou particulaire (pMMO ou pmoA)
impliquées dans les phénomènes de méthanotrophie, l'ammoniaque monooxygénase (amoA)
impliquée dans la nitrification, les gènes de fixation de l'azote atmosphérique (opéron nif), la sous-
unité alpha de la méthyle coenzyme-M réductase (mer) impliquée dans la méthanogénèse ainsi que
des gènes impliqués dans des résistances aux métaux (cf. chapitre 2 §3.1.3 et §5.3.2). L'utilisation de
sondes pour des hybridations in situ est rendue aléatoire par le fait qu'à la différence des ARNr, les
gènes codant pour des fonctions métaboliques sont présents en bien moins grande quantité. Pour
circonvenir cette limitation une technique d'amplification in situ a été mise au point (Hodson et al.,
1995). Cette technique permettrait d'amplifier des gènes, des ARNm et des ARNr, mais elle n'a jusqu'à
présent été utilisée que pour étudier des cellules isolées ou des communautés peu complexes
(Tolker-Nielsen et al., 1997; Holmstrøm et al., 1999; Tani et al., 1998). L'approche la plus utilisée
consiste à amplifier directement les gènes à partir d'un pool d'ADN génomique directement extrait de
l'échantillon, puis d'une manière analogue à l'ADNr 16S, à réaliser des phylogénies en comparant les
séquences des gènes clonés avec celles de micro-organismes isolés. Les biais inhérents aux
techniques d'extraction de l'ADN, d'amplification et d'analyse phylogénétique (cf. §3.5) peuvent
malgré tout entacher cette analyse. De plus l'amplification d'un gène n'implique pas obligatoirement
que celui-ci soit transcrit. Il est donc préférable d'utiliser des techniques de PCR compétitive pour
quantifier la présence du gène dans une population (Lee et al., 1996b; Kowalchuk et al., 1999). L'une
des solutions les plus élégantes consiste donc à analyser les ARNm dont la présence dans le
cytoplasme procaryotique est directement corrélée à l'expression du gène par le micro-organisme.
Cette technique fondée sur l'extraction des ARNm suivie d'une RT-PCR à permis d'étudier
l'expression des gènes de résistance au mercure dans des échantillons d'eau (Nazaret et al., 1994),
les gènes de la manganèse péroxydase dans le sol (Bogan et al., 1996), de la naphtalène
dioxygénase dans des eaux souterraines (Wilson et al., 1999), de la fixation de l'azote chez la
communauté symbiotique du termite Neotermes kokunensis (Noda et al., 1999). La quantification de
l'expression des ARNm peut être effectuée par PCR compétitive (Tsai et Wiltbank, 1996). La
technique de "Differential Display" employée couramment dans l'étude des ARNm d'Eucarya est aussi

Les techniques d'écologie microbienne
38
applicable à partir d'ARNm procaryotique (Fleming et al., 1998), il n'est alors pas nécessaire de
connaître la séquence des ARNm dont on souhaite suivre l'induction. Une dernière technique
consiste à rechercher une fonction métabolique dans un environnement par clonage d'une banque
d'ADN génomique dans Esherichia coli, puis par criblage de la banque en suivant l'expression des
gènes clonés. Il a été possible à l'aide cette technique, à partir de différents échantillons de sol, de
détecter la présence de gènes codants pour l'utilisation du 4-hydroxybutyrate (Henne et al., 1999).
Cette technique est toutefois très laborieuse puisqu'il a fallu cribler 930000 souches d'E. coli pour
détecter 36 clones positifs dont seuls 5 possédaient un plasmide recombinant. Une étude similaire,
passant toutefois par une étape préalable clonage dans une banque de phage, a été entreprise pour
rechercher des chitinases à partir d'échantillons d'eau de mer (Cottrell et al., 1999)
L'analyse d'un gène codant pour une fonction du métabolisme ne nous renseigne pas sur la
position phylogénétique du micro-organisme qui exprime ce gène. Seules les études de
communautés très simples telles que les associations symbiotiques n'impliquant généralement qu'un
hôte et un symbiote permettent d'attribuer à un micro-organisme défini, à la fois une position
phylogénétique et une fonction métabolique. Ainsi, il a été possible d'attribuer les gènes d'une
nouvelle famille de bisulfite réductase dissimilatoire aux épibiotes de l'annélide polychète Alvinella
pompejana (Cottrell et Cary, 1999) et les gènes codants pour une flagelline au symbiote de Riftia
pachyptila (Millikan et al., 1999). Une autre méthode consiste à cloner des grands fragments d'ADN
génomique à partir d'une extraction directe d'ADN et à rechercher dans les banques des fragments
portant les gènes codant pour l'ADNr 16S (Stein et al., 1996). Il est alors possible d'étudier les gènes
adjacents disposés sur le même "contig". Comme dans le cas précédent il n'est pas garanti que les
gènes détectés soient exprimés, de plus l'effort d'échantillonnage à fournir pour détecter les gènes
d'ADNr 16S peut parfois être démesuré. Cette technique employée dans l'étude d'une communauté
simple à permis de caractériser l'ADNr 16S et les gènes de l'ADN polymérase de l'Archaea
Crenarchaeum symbiosum associée à l'éponge Axinella mexicana (Preston et al., 1996; Schleper et
al., 1997b).
Les imposants projets de séquençage génomique humain et procaryotique ont favorisé le
développement des nanotechnologies. Nombre de ces développements ne sont encore qu'à l'état
de pré-projet mais la demande en systèmes d'analyse rapides et bon marché de la part des industriels
les imposeront probablement très rapidement au sein des laboratoires. L'effort de recherche semble
s'être porté sur différentes technologies de puces à ADN fondées sur la détection du signal par
fluorescence (Marschall et Hodgson, 1998; Schena et al., 1998; Gerhold et al., 1999) ou par
spectrométrie de masse (Tang et al., 1999) ainsi que sur la miniaturisation des dispositifs de migration
d'ADN utilisant des systèmes électrophorètiques en microcapillaires de silicone (Chou et al., 1999) ou
des "crochets" animés de mouvements browniens (Bader et al., 1999). L'écologie microbienne n'a
pour l'instant que très peu bénéficié du développement de ces techniques, seule une étude décrit la
mise au point (Guschin et al., 1997b) puis l'emploi (Guschin et al., 1997a) de puces à ADN pour
détecter différentes communautés de bactéries nitrifiantes. Les programmes de séquençage
génomique ont quant à eux fourni de nombreuses informations sur l'organisation génomique, sur la

Les techniques d'écologie microbienne
39
fonctionnalité génétique et sur les mécanismes évolutifs impliqués dans les micro-organismes
séquencés (Olsen et Woese, 1996; Karp et al., 1998, 1999; Ge et Taylor, 1999). Ils permettent
désormais de comparer les micro-organismes non sur la base de caractères phénotypiques ou
phylogénétiques mais sur l'ensemble des informations contenues dans leur génome (Koonin et
Galperin, 1997; Rastan et Beeley, 1997).
3.5 Les biais des techniques moléculaires
A l'instar des techniques culturales, les techniques moléculaires sont elles aussi soumises à de
nombreux biais qui peuvent lourdement entacher l'analyse de la diversité microbienne d'un
échantillon. Ces biais peuvent intervenir à différents stades de l'analyse : lors de l'échantillonnage, de
la conservation de l'échantillon, de la lyse cellulaire, de l'amplification de l'ADNr 16S, de l'analyse des
séquences. Dans le cadre des études par hybridation in situ la construction et la spécificité de la
sonde, l'état cellulaire et la perméabilité des membranes ainsi que la sensibilité des méthodes
d'analyse peuvent influencer les résultats de l'analyse. Trois publications récentes font le point sur les
principaux bais de ces techniques. La première (Amann et al., 1995) s'attache plus particulièrement à
décrire les biais des techniques d'hybridation in situ et propose des techniques pour améliorer la
sensibilité de la méthode. La seconde (Wintzingerode et al., 1997) fait un inventaire très complet des
biais majeurs rencontrés dans les études par amplification et séquençage. La troisième (Head et al.,
1998) fait le point sur dix années d'études moléculaires ainsi que sur les biais de l'ensemble de ces
techniques.
3.5.1 Echantillonnage et conservation des échantillons
En dehors de très rares milieux où la diversité microbienne est très restreinte (par exemple
symbiose réduite à un micro-organisme) l'échantillon prélevé dans le milieu devra rendre compte le
plus fidèlement possible de l'ensemble des espèces présentes dans l'environnement. Il est donc
nécessaire de connaître les conditions physico-chimiques de chaque échantillon.
Dans la mesure où il n'est pas possible de traiter (lyse, extraction) l'échantillon dès son
prélèvement, il est nécessaire de le conserver dans les conditions qui permettront de conserver
l'intégrité des acides nucléiques. Une étude a comparé les effets de différentes techniques de
conservation sur la composition de banques de clones d'ADNr 16S (Rochelle et al., 1994). Il a été
observé que les échantillons congelés rapidement et maintenus sous atmosphère anaérobie (non-
oxydante) présentent une diversité bien plus importante que les échantillons conservés en aérobiose
ou congelés plus tardivement. La congélation tardive des échantillons est source selon les auteurs
d'enrichissement de certains phylotypes.

Les techniques d'écologie microbienne
40
3.5.2 Les techniques de PCR
3.5.2.1 La lyse cellulaire et l'extraction des acides nucléiques
La lyse cellulaire est une étape cruciale de l'analyse de la diversité. Elle devra détruire les parois
et membranes des micro-organismes en endommageant le moins possible les acides nucléiques.
L'efficacité de la technique de lyse cellulaire sur un échantillon environnemental est d'autant plus
difficile à déterminer que l'on ne connaît pas le type et la quantité de cellules à lyser. On sait toutefois
que les techniques de lyse bactérienne doivent être modifiées pour lyser des Archaea, que les
bactéries Gram + sont plus difficiles à lyser que les Gram -, que les spores et les petites cellules
résistent mieux à la lyse.Tableau 1.13. Les techniques de lyse cellulaire
Techniques physiques Techniques chimiques
congélation- décongélation
Azote liquide
pilon et mortier
bead beating
Presse de French
sonication - micro-onde
chauffage à 100°C
Résine d'échange d'ions (Chelex)
solvants
détergents (SDS)
enzymes
lysozyme
protéinase K
N-acetylmuramidase
Lysostaphin
Achromopeptidase
Les techniques de lyse les plus efficaces sont celles qui combinent à la fois les traitements
physiques et chimiques (cf. tableau 1.13). Toutefois elles peuvent conduire à une forte fragmentation
de l'ADN et favoriser la production d'amplifiats chimériques. Aussi est-il parfois nécessaire de simplifier
la technique d'extraction afin d'avoir un ADN de meilleure qualité. On est alors confronté à la présence
de contaminants tels que les acides humiques (pour des échantillons de sols) ou les métaux lourds qui
peuvent inhiber l'action de certaines ADNpolymérases.
La solution idéale consisterait à utiliser plusieurs techniques de lyse et à rassembler le résultat
des différentes extractions pour réaliser l'amplification.
3.5.2.2 L'amplification par PCR
L'amplification est l'une des sources majeures de biais dans l'étude de la diversité. Ces biais sont
introduits pour différentes raisons :
Ø inhibition de la PCR en raison de la présence de contaminants chimiques ou organiques (Wilson,
1997)
Ø l'état physiologique de la cellule va influer sur la qualité de l'amplification : les ADN des cellules à l'état
quiescent ou létal vont être plus difficilement amplifiables (Silva et Batt, 1995)

Les techniques d'écologie microbienne
41
Ø amplification préférentielle de certaines molécules : celle-ci est liée au nombre de copies de l'opéron
rrn16 (de 1 à 14) (Farrelly et al., 1995), au choix des amorces (Reysenbach et al., 1992; Brunk et Eis,
1998), à l'échantillon (Suzuki et Giovannoni, 1996), à la composition en GC de la séquence (Dutton et
al., 1993), aux proportions initiales de séquences dans l'échantillon (Suzuki et Giovannoni, 1996), aux
séquences adjacentes de la zone à amplifier (Hansen et al., 1998)
Ø formation de molécules chimériques : lors de l'appariement les simples brins d'ADN vont s'associer
entre eux plutôt qu'avec les amorces, en particulier lorsque l'ADN a été fragmenté (Pääbo et al., 1990;
Liesack et al., 1991; Kopczynski et al., 1994; Wang et Wang, 1996)
Ø formation de mutant de délétion : la structure secondaire de l'ADNr 16S va persister lors de la
dénaturation et la polymérase va "sauter" certaines structures résistantes
Ø mauvaise incorporation de la polymérase : selon les enzymes utilisées et les conditions
d'amplification le taux d'erreur variera d'une toutes les 300 bases à une toutes les 5000 bases.
Ø variation des séquences des différentes copies dans l'opéron rrn au sein d'une même souche (voir
§ 3.1.3 et Ninet et al., 1996; Reischl et al., 1998).
Ø présence de séquences de contaminants de la réaction : les tampons de réaction, l'eau ou les tubes
peuvent être source de contaminants qui passent alors pour faire partie de la population indigène de
l'échantillon (Tanner et al., 1998). L'une des solutions est d'exposer le mélange d'amplification aux
U.V. (Goldenberger et Altwegg, 1995)
3.5.2.3 Le clonage et l'analyse des séquences
Il semblerait que la technique de clonage influence la composition de la banque de clones. En
particulier pour les clonages en bouts cohésifs où il est nécessaire d'utiliser des enzymes de restriction
qui peuvent couper le fragment d'ADNr 16S (Rainey et al., 1994b). L'orientation du fragment cloné
peut elle aussi générer des différences de profils lors de l'analyse par RFLP de la banque en cas de
clonage par TA-cloning. Pitule et Pace (1999) ont proposé un vecteur spécialement construit pour le
clonage de l'ADNr 16S dans une orientation donnée.
Le criblage de la banque doit être effectué en tenant compte des espèces minoritaires. Ainsi,
dans un échantillon contenant 108 cellules.g-1 et une population présente à 102 ou 104 individus.g-1, il
sera nécessaire de cribler 104 à 106 clones pour statistiquement détecter cette population17.
La détection des chimères est une étape essentielle lors de l'analyse des séquences. Si
différents auteurs ont proposé des techniques pour réduire le nombre de chimères lors de l'étape de
PCR (voir § 3.4.2.2 et Wintzingerode et al., 1997), les outils pour détecter ces séquences sont assez
réduits. Il s'agira d'utiliser le programme CHIMERA sur le site du RDP (Robinson-Cox et al., 1995) ou de
comparer la topologie d'arbres phylogénétiques en n'utilisant que certaines parties de l'alignement
(par exemple de la partie 5' à la base 500, de 500 à 1000 et de 1000 à la partie 3'). Ces séquences
16 Ce nombre de copies peut être variable au sein d'une même espèce. Prüß et al. (1999) détectent des séquences signatures
spécifiques d'une part de souches psychrotolérantes et d'autre part de souches mésophiles sur différentes copies de l'ADNr 16S de
Bacillus cereus dont le nombre varie de 6 à 10 selon les souches.17 Lors de l'amplification, les ratios cellulaires ne sont pas maintenus, ce nombre de clones à analyser est donc théorique.

Les techniques d'écologie microbienne
42
seront d'autant plus difficiles à détecter qu'elles sont formées entre espèces proches ou entre copies
d'un même opéron (Wang et Wang, 1997).
3.5.3 Les techniques d'hybridation
Les principaux biais des techniques d'hybridation sont liés d'une part à la perméation des
cellules, et d'autre part à l'état cellulaire et à la construction des sondes.
Ø Pour la fixation des cellules et leur perméation l'utilisation d'aldéhyde comme le
paraformaldéhyde (agent de pontage) va être plus efficace sur les bactéries Gram - alors qu'un
dénaturant comme l'éthanol fixera mieux les Gram +. Pour les Gram + il est parfois nécessaire d'utiliser
du lysozyme pour faciliter la pénétration de la sonde.
Ø Des cellules actives dans l'échantillon, donc riches en ribosomes, seront plus facilement
détectées que des cellules en dormance. En effet le taux de croissance est corrélé dans la plupart des
cas avec le nombre de ribosomes par cellule. Le nombre de ribosomes n'est toutefois pas un facteur
limitant de la croissance et de la synthèse protéique chez les bactéries oligotrophes à la différence des
hétérotrophes (Fegatella et al., 1998).
Ø La construction de la sonde devra tenir compte de la spécificité de la séquence. Ainsi pour un
nucléotide de 18 bases, il y a 0,25-18 chance pour détecter une séquence non recherchée. Mais dans
la pratique des hybridations non spécifiques peuvent se produire avec quelques mésappariements
dans la sonde. Il est donc nécessaire de vérifier la validité de la sonde en la confrontant avec les
banques de séquences et par hybridation sur membrane ou sur culture pure.
Ø En jouant sur les conditions d'hybridation, Td ou pourcentage de formamide on modifie la
spécificité de la sonde. Zheng et al. (1996) ont quantifié avec plus de précision des organismes des
trois domaines (Eucarya, Bacteria et Archaea) en diminuant la Td pour différentes sondes universelles.
Ø Le type de marquage de la sonde influera sur la qualité de détection, aussi des nouveaux
marqueurs et des nouvelles techniques de détection ont été développés (Glöckner et al., 1996;
Schönhuber et al., 1997, 1999)
Ø La sonde devra "accéder" à son site d'hybridation sur la molécule d'ARNr 16S. La forte
structure secondaire de l'ARNr ainsi que les interactions avec des protéines peuvent rendre certains
sites inaccessibles. L'utilisation de formamide permet alors de réduire les liaisons hydrogène et
d'accroître l'accessibilité du site. Et en utilisant une sonde dans les régions déjà connues pour donner
des hybridations efficaces il est possible de limiter ce biais (Frischer et al., 1996). Récemment l'équipe
de Amann a établi sur la base de l'ARNr 16S d'E. coli une carte d'accessibilité de la molécule en
comparant par cytométrie en flux l'hybridation de l'ensemble des sondes couvrant les 1500
nucléotides de la séquence avec la valeur de la sonde EUB 338 donnée pour 100% d'hybridation
(Fuchs et al., 1998).

Les techniques d'écologie microbienne
43
3.5.4 Les techniques de DGGE
Les biais de la technique DGGE sont semblables à ceux rencontrés dans les techniques
d'amplification et de séquençage. Dans la mesure où il est nécessaire d'effectuer une amplification de
fragments d'ADNr 16S, l'ensemble des biais d'échantillonnage, d'extraction et d'amplification sont
présents. Seules les bandes de fortes intensités, correspondant aux populations majoritaires ou
majoritairement amplifiées pourront être détectées.
3.5.5 Une solution : l'approche pluridisciplinaire
L'engouement "excessif" du début des années 1990 vis à vis des techniques moléculaires
(Olsen et Woese, 1993; Olsen et al., 1994b; Woese, 1994 a et b) a désormais fait place à une vision
plus pondérée des différentes méthodologies d'étude de la diversité microbienne (Palleroni, 1997;
Amann et Kühl, 1998; Head et al., 1998). Si les biais des techniques culturales sont depuis longtemps
pris en compte, les résultats obtenus par des techniques d'analyse des ARNr peuvent parfois être
surprenants. D'une part :
Ø La grande majorité des séquences obtenues ne sont pas similaires à celles de souches cultivées.
Ø Un grand nombre de séquences bactériennes et archaéennes correspondent à de nouveaux
phylums.
Ø Les séquences de souches isolées de sites d'où provient une collection de clones d'ADNr 16S
sont rarement identiques à celles des clones.
Ø Des séquences de clones obtenus de sites très éloignés peuvent être très similaires.
Ø Des organismes très peu ou pas cultivés peuvent présenter une très grande variabilité
phylogénétique et être ubiquitairement présents dans la nature.
D'autre part :
Ø Les données "de base" d'écologie ne peuvent être obtenues par ces techniques; notamment en
raison des problèmes méthodologiques il est difficile de quantifier les taxons présents dans le milieu.
Ø La qualité des amplifications est dépendante de conditions physiologiques et génomiques
cellulaires que l'on ne peut prévoir.
Ø Il est difficile de définir les propriétés physiologiques, métaboliques et le rôle écologique des
organismes détectés à partir de leur position phylogénétique d'autant plus si ceux-ci forment de
nouveaux phylums.
Ø Les amorces dites "universelles" n'ont été construites qu'en utilisant une très faible fraction des
organismes connus.
Ø La présence d'un grand nombre de séquences similaires est difficile à interpréter : s'agit-il de
microhétérogénéités au sein de l'opéron rrn ou de souches très proches?
Ø La détection et la quantification des cellules par hybridation in situ est actuellement soumise à de
nombreux biais notamment lors de la détection du signal et de la perméation cellulaire.

Les techniques d'écologie microbienne
44
Ø Par amplification (séquençage et DGGE) ou par hybridation il est encore très difficile de détecter les
espèces minoritaires.
Ø Le traitement des banques de clones devrait être soumis à plus de rigueur statistique.
Ø Le pouvoir résolutif de l'ADNr 16S au niveau de l'espèce (qui est l'unité de base en écologie) est
limité. Des valeurs de 95% d'identité peuvent distinguer des espèces différentes, contre 70% de
similitude en hybridation ADN/ADN.
Si les techniques moléculaires ont permis de considérablement accroître le champ des
connaissances en écologie microbienne (cf. partie suivante), il est désormais indéniable que seul
l'emploi combiné de techniques culturales et non-culturales permettra de lier les informations
concernant la diversité et la fonction des communautés microbiennes dans leur environnement. Au
même titre que la taxinomie bactérienne a su intégrer aux techniques classiques d'identification et de
taxinomie numérique les techniques moléculaires et la phylogénie moléculaire, l'écologie microbienne
devra elle aussi intégrer l'ensemble des techniques mises à sa disposition. Cette approche
polyphasique peut s'envisager par l'utilisation conjointe des sondes moléculaires et des
microélectrodes (Schramm et al., 1996), des sondes moléculaires et des mesures d'incorporation
spécifiques (Lee et al., 1999; Ouverney et Fuhrman, 1999), le clonage de grands fragments d'ADN
génomique (Stein et al., 1996), l'utilisation des microscopes confocaux (Manz et al., 1995, 1999;
Harmsen et al., 1996b; Rocheleau et al., 1999) et des pinces optiques (Huber et al., 1995). La
biomasse procaryotique, cette "majorité invisible" telle que la décrivent Whitman et al. (1998), ne
devrait nous être accessible qu'au terme de ces efforts.

Les habitats microbiens

Les habitats microbiens
45
La Terre est un écosystème extraordinairement complexe où l'on peut distinguer trois habitats
majeurs : l'atmosphère, le sol et l'eau. Aux habitats naturels se sont adjoints de nouvelles niches
écologiques liées à la présence et aux activités humaines. L'étude de tous ces habitats a longtemps
été limitée aux grandes formes de vie et nombre de textes d'écologie articulaient leur réflexion
uniquement autour de ce qui était visible à l'œil nu. L'utilisation de techniques culturales en
microbiologie, relayé ces dernières années par l'emploi de techniques moléculaires, nous a permis
d'observer à quel point la diversité et le rôle des procaryotes pouvait être prépondérant dans l'équilibre
de la biosphère. Je me propose dans cette seconde partie introductive, après une description
sommaire des différents écosystèmes et des micro-organismes qui y ont été isolés, de présenter une
revue des apports des techniques moléculaires dans l'étude de la diversité de ces milieux. Les
écosystèmes pétroliers et hydrothermaux seront plus particulièrement approfondis. Un arbre
phylogénétique présenté en annexe permettra au lecteur de repérer les différents groupes
phylogénétiques cités dans cette seconde partie.
I Les origines de la vie
1.1 Les premières traces de vie
Aller à la recherche des premières formes de vie sur terre c'est tenter de répondre à ces trois
questions : qu'est ce que la vie? Quand cela a-t-il commencé? Et à quoi ressemblait la Terre à cette
époque?
La première question peut revêtir des aspects tant métaphysiques que physiologiques.
Norman Pace (Pace, 1991; et site web de l'équipe de Norman Pace http://pacelab.berkeley.edu)
répond qu'une forme est considérée comme vivante lorsqu'elle est autoréplicative et réalise une
chimie organique sous une forme libre.
Pour répondre à la seconde et à la troisième question nous disposons d'enregistrements
astronomiques (formation des systèmes solaires et des planètes), d'enregistrements géologiques
(fossiles, mesure d'isotopes, étude des roches, etc.) et d'enregistrements biologiques (la phylogénie
moléculaire).
On considère que la Terre est apparue il y a 4,6 milliards d'années, mais il faut attendre encore
800 millions d'années avant que l'eau liquide apparaisse sur la terre créant ainsi des conditions
compatibles avec la vie telle que nous l'imaginons. Les premières traces fossiles de micro-organismes
datent de 3,6 milliards d'années1. Ce sont des tapis microbiens fossilisés connus sous le nom de
stromatolites. On en retrouve encore en activité en Australie en Amérique et en Afrique dans des
lagunes côtières où dans des sources chaudes. A la différence des stromatolites actuels colonisés par
des cyanobactéries (aérobies) les premiers stromatolites devaient être constitués de phototrophes
1 Il existerait une trace plus ancienne de vie dans des roches prélevées au Groenland. Prélevées dans des sédiments vieux de 3,8 à 3,9
milliards d'années, des roches Isua présentent des rapport isotopiques 13C/12C laissant supposer une origine biologique.

Les habitats microbiens
46
anaérobies, l'oxygène sous une forme libre étant apparu il y a 2 à 2,5 milliards d'années. Cette période
correspond à une augmentation de la diversité morphologique des fossiles microbiens laissant
supposer un accroissement de la diversité phylogénétique.
Lorsque la vie est apparue, la Terre présentait des conditions qui nous sembleraient
particulièrement inhospitalières. C'était un environnement très réducteur où les gaz principaux étaient
le méthane, le dioxyde de carbone et l'azote. On retrouvait des traces de sulfures sous la forme H2S et
FeS. Lors du premier milliard d'années la température était alors très élevée (supérieure à 100°C). Ce
n'est que lorsque la Terre a commencé à se refroidir (4 milliards d'années) que la vie a pu se
développer (cf. figure 2.1).
400
300
200
100
°C
4,5 4 3,5 Temps en milliards d’années
“océan magmatique”
Condensation de l’eau
Synthèses abiotiques
Synthèses biotiques
Figure 2.1. Représentation schématique de l'évolution de la température à la surface de la Terre depuis sa formation.
On considère que les premières synthèses de macromolécules comme les acides nucléiques
et les protéines se sont déroulées de manière abiotique sur des surfaces de roches ayant le rôle de
catalyseur. La pyrite (FeS2) a probablement joué un rôle majeur dans ces synthèses où le rayonnement
U.V. était la source principale d'énergie (très importante à l'époque du fait de la quasi absence
d'atmosphère) (Wächtershäuser, 1988).
1.2 Une origine de la vie controversée
L'ensemble de la communauté scientifique semble s'accorder sur le type métabolique des
premiers organismes vivant sur Terre. Il s'agirait d'organismes anaérobies chimiolithotrophes tirant leur
énergie de la réduction du soufre (Pace, 1991; Stetter et al., 1996) ou plus probablement du fer (FeS
ou FeCO3) (Slobodkin and Wiegel, 1997, Vargas et al., 1998), la source de carbone provenant de
molécules organiques abiotiques2 ou du CO2 atmosphérique. La température à laquelle la vie est
apparue est quant à elle sujette à discussion.
Les similitudes observées entre les sources hydrothermales et les conditions retrouvées sur
Terre il y a 4 milliards d'années : milieu très réducteur et anoxique, forte teneur en soufre, en méthane
et métaux lourds, températures élevées, ont conduit les scientifiques, au début des années 70, à
2 La voie d'assimilation du carbone par autotrophie étant un mécanisme complexe il est peu probable qu'il ait précédé l'hétérotrophie.

Les habitats microbiens
47
considérer ces écosystèmes comme des lieux possibles d'émergence de la vie sur Terre (Corliss et al.,
1981; Baross et Hoffman, 1985)
A la même époque Woese propose une division tripartite du monde vivant en créant le
domaine des Archaebactéries (Woese, 1987) (devenant par la suite Archaea, Woese et al., 1990) qui
ne regroupait alors que des micro-organismes aux préférendums vitaux extrêmes (très forte
température, très forte salinité, production de méthane, réduction du soufre). Les branches basses de
l'arbre du vivant étaient courtes (donc ayant peu évolué) et essentiellement occupées par des micro-
organismes thermophiles (Bacteria: Aquificales, Thermotogales; Archaea: Crenarchaeota soufre-
dépendantes, Thermococcales et méthanogènes thermophiles) (Stetter et al., 1996). L'émergence
d'un concept de biosphère microbienne souterraine thermophile quelques années plus tard (Gold,
1992) abonda dans le sens d'une origine de la vie thermophile. Or cette proposition va à l'encontre de
la théorie d'un monde à ARN.
On considère que l'apparition de l'ARN a précédé celle de l'ADN (les ribonucléides avant les
desoxyribonucléides). L'ARN jouait un rôle de stockage d'information et un rôle enzymatique. Il
possède encore des capacités catalytiques qui lui permettent d'associer les acides aminés pour former
les protéines (ribosyme). De plus s'il a été prouvé que l'ADN (pour peu qu'il soit super-enroulé) pouvait
résister à de très fortes températures (Forterre, 1996; Forterre et al., 1996), l'ARN quant à lui, est très
rapidement dénaturé à mesure que la température augmente. Il est donc peu probable que les
premiers organismes (à ARN) aient pu supporter les très fortes températures que supportent les
hyperthermophiles actuels3.
De plus les phylogénies réalisées sur les ARNr ont été récemment remises en cause4.
Ø La généralisation des séquençages génomiques permet d'accéder à de nombreuses séquences
protéiques pouvant, au même titre que l'ARNr, servir de base à des phylogénies. Or si la majeure
partie des arbres obtenus sont en accord avec une division tripartite du monde vivant (33 protéines
sur 40), la monophylie de ces domaines est mise en cause dans 17 phylogénies sur 40 (Williams et
Embley, 1996). Ce résultat soulève le problème de la représentation d'un arbre universel reflétant
soit la majorité des arbres obtenus à partir de toutes les molécules, soit le résultat d'une
phylogénie sur un ou plusieurs gènes clés (Doolittle, 1996; Doolittle et al., 1996).
Ø Des phylogénies récentes placent Aquifex (la bactérie branchant le plus profondément sur l'arbre à
ARNr) non plus à la base des Bacteria mais à proximité de Bacillus ou au sein des Archaea (Klenk,
1998; Pennisi, 1998).
3 L'instabilité de l'ARN est en partie liée à la présence de cytidine , qui représente le moins stable des 3 acides nucléiques. Il a donc été
proposé que cet acide aminé pouvait être absent des premiers matériaux génétique (Levy et Miller, 1998), des liaisons A:U, G:U
pouvant être suffisamment fortes pour maintenir la structure secondaire de la molécule. Il a de plus été démontré que l'activité d'un
ribozyme était accrue lorsque celui ci était dépourvu de cytidine (Rogers et Joyce, 1999). Cet ARN primaire, stabilisé, dépourvu de
cytidine pourrait être alors en accord avec une origine de la vie dans des conditions thermophiles.4 La division du monde vivant en trois domaines telle que l'avait proposé Woese, a même été remise en cause récemment par Ernst
Mayr (1998). Cet article a conduit à un échange très vif entre les deux protagonistes par journal interposé (Woese, 1998b). Pour sa part
Gupta (1998 a, b et c) s'interroge sur les similitudes entre les Archaea et les Firmicutes et propose une nouvelle classification du
monde vivant.

Les habitats microbiens
48
Ø Il semblerait que la position d'un micro-organisme sur un arbre soit très fortement liée à la vitesse à
laquelle la molécule ayant servi à construire l'arbre a évolué. Cette fréquence mutationnelle varie au
sein de la molécule et entre les organismes induisant des phénomènes d'attraction des branches
longues5. Ces phénomènes perturbent particulièrement les analyses d'organismes "branchant
profondément" sur les arbres phylogénétiques et qui sont sensés être les plus proches de l'état
ancestral (Philippe et Laurent, 1998).
Ø On a observé que les ARNr d'hyperthermophiles étaient riches en bases G et C. Or ces dernières
forment des structures stables qui pourraient ralentir les mutations, raccourcir les branches et donc
rapprocher les branches courtes (Galtier et Gouy, 1998; Galtier et al., 1999).
Ø Enfin la détection de séquences de Crenarchaeota et d'Euryarchaeota (très probablement non
thermophiles) au sein de nombreux environnements (sols, sédiments, eaux douce et eau de mer,
symbiose avec des animaux, voir Olsen, 1994; Schleper, 1999) vient faire vaciller le dogme
d'Archaea exclusivement cantonnées dans des environnements extrêmes.
L'ensemble de ces informations présente la thermophilie comme un caractère dérivé, une adaptation à
de nouvelles niches écologiques et remet en cause une origine thermophile de la vie sur terre.
La nature de la première forme de vie sur terre est elle aussi discutée. Les phylogénies
réalisées sur les ARNr ont permis de présenter un arbre de la vie non raciné, mais des études ont été
entreprises afin d'enraciner cet arbre, à partir de travaux effectués sur les protéines codant pour les
facteurs d'élongation, les ATPases et les ADN et ARN polymérases. Ces travaux présentent la racine
de l'arbre entre la branche des Bacteria et celle des Eucarya-Archaea. Différents noms ont été
proposés pour nommer l'organisme qui serait à la base de l'arbre : progénote, cenancestor, LUCA
(Last Universal Common Ancestor). Les deux premiers termes ont le désavantage de donner une idée
primitive de la biologie de cette entité6. Or il est tout à fait probable que cet organisme ait eu un
métabolisme assez complexe, qui ait évolué en se simplifiant au cours des générations et de la
conquête de nouveaux milieux. De plus, raisonner en terme de dernier organisme commun ne doit
pas faire oublier qu'à l'origine les échanges génétiques et les fréquences de mutations devaient être
très élevés, favorisant l'apparition de gènes paralogues et xénologues (Woese, 1998a).
Le séquençage du génome de nombreux micro-organismes devrait fournir de nouveaux
matériaux pour établir des relations phylogénétiques plus solides et avancer de nouvelles hypothèses
sur la nature des premiers organismes ayant colonisé le globe.
Des trois compartiments de la biosphère, l'atmosphère constitue l'environnement le moins
propice au développement d'une vie endémique. Il semble toutefois que les micro-organismes soient
parvenus à coloniser ce milieu.
5 Les positions sur deux séquences ayant des fréquences de mutation très élevées peuvent s'avérer similaires par convergence sans
que cette similitude reflète un caractère d'ancestralité (Felsentein, 1978).6 Les termes employés pour la dénomination des domaines peuvent conduire à de faux raisonnements. Ainsi les Archaea ne sont pas
essentiellement confinés dans des environnements extrêmes (proches des conditions ancestrales) et les "pro"caryotes possèdent des
mécanismes aussi évolués que ceux des eucaryotes.

Les habitats microbiens
49
II. L'atmosphère
La présence de micro-organismes dans l'air a été mise en évidence par les expériences de
Pasteur en 1864, mettant ainsi fin au concept de génération spontanée. Au sein de cet habitat
inhospitalier les micro-organismes ne sont qu'en transit. En haute altitude, l'absence de nutriments,
d'eau et les fortes irradiations (rayonnements U.V.) ne permettent pas le développement des micro-
organismes7. Les organismes sont donc sous la forme de spores. Au fur et à mesure que l'on s'élève
dans l'atmosphère la concentration en micro-organismes diminue.
A l'échelle humaine les micro-organismes présents dans l'air ambiant, sous forme d'aérosols,
sont responsables d'un grand nombre d'infections humaines, en particulier dans des environnements
anthropiques clos, chauffés et humides comme ceux des hôpitaux. Ces infections peuvent aussi
toucher d'autres organismes supérieurs tels que les plantes et les animaux domestiques. La teneur de
l'air en micro-organismes, qu'ils soient sous une forme viable, viable mais non cultivable ou mort, est
fonction de facteurs climatologiques (humidité, pluie, température) comme de facteurs liés à
l'ensoleillement (saison, cycle nycthéméral) (Lighthart, 1997). Les techniques de culture classiques
(Heidelberg et al., 1997; Shaffer et Lighthart, 1997; Reponen et al., 1998), comme les techniques de
biologie moléculaire (Alvarez et al., 1995; Palmer et al., 1995; Pillai et al., 1996; Bartlett et al., 1997;
Stärk et al., 1998) ont été mises à profit à de nombreuses reprises pour détecter les micro-organismes
en suspension dans l'air. Il n'a s'agit toutefois que de détecter des micro-organismes généralement
impliqués dans des infections humaines, et aucune étude ne s'est encore intéressée à la diversité
totale des micro-organismes "aériens". Ce type d'étude est beaucoup plus commun au sein des
habitats terrestres.
III. Les habitats terrestres
On distingue trois compartiments majeurs parmi les habitats terrestres : la couche superficielle
du sol qui s'étend de la surface à quelques mètres de profondeur, la biosphère souterraine qui s'étend
sur toute la lithosphère, et le cas particulier des réservoirs pétroliers. Chacune de ces niches abrite une
communauté particulière de micro-organismes.
3.1 Les couches superficielles de la lithosphère
3.1.1 Description de l'habitat
Le sol abrite de fortes quantités de micro-organismes qui interviennent activement dans
différents cycles géochimiques (C, N, S et métaux). La diversité microbienne y est fonction du type de
sols étudiés (par exemple : elle est plus abondante dans la rhizosphère que dans des sols nus) mais
7 Certains chercheurs auraient proposé l'hypothèse que des micro-organismes pourraient se développer dans les nuages, là où la
teneur en eau est plus élevée. Source : site du Microbe Zoo, Digital Learning Center for Microbial Ecology
(http://commtechlab.msu.edu/sites/dlc-me). De plus des bactéries actives ont été détectées dans des gouttes d'eau de nuage à 3000 m
d'altitude (Psenner et Sattler, 1998).

Les habitats microbiens
50
elle s'étend dans toutes les niches où la vie est thermodynamiquement possible. La proportion de
micro-organismes s’accroît à mesure que l'on s'éloigne de la surface du sol. A 10 cm de profondeur on
estime retrouver 75 à 95% de la population totale microbienne du sol. Les micro-organismes
procaryotes y côtoient des Eucarya. Par gramme de sol on retrouve 107 à 108 procaryotes, 105
champignons, 5.104 algues et 3.104 protozoaires (Madigan, 1996; Withman et al., 1998).
3.1.2 La rhizosphère
Au sein de la rhizosphère la diversité et la richesse microbienne sont très importantes. Les
populations peuvent atteindre jusqu'à 109 individus par gramme de sol. Cette richesse est à attribuer
aux relations qui s'établissent entre les plantes et les micro-organismes (Bally et al., 1999).
Ø La plante excrète des sucres, des acides aminés et des vitamines qui permettent la croissance des
micro-organismes alors que ces derniers produisent des hormones favorisant la croissance de la
plante (auxine, indole, acide acétique ou des molécules "gibbereline-like").
Ø Certains micro-organismes de la rhizosphère permettent de lutter contre les pathogènes (bactéries
et champignons) de la plante.
Ø Les polysaccharides bactériens favorisent quant à eux l'agrégation du sol autour des racines (Amellal
et al., 1998).
Ø Enfin d'importants phénomènes symbiotiques sont observables, notamment chez les
légumineuses où, au sein de bactériocystes, des Rhizobium permettent la fixation de l'azote
atmosphérique par le végétal (Ferrera-Cerrato, 1980).
Les méthodes de culture classiques ont permis d'isoler essentiellement des micro-organismes
associés aux genres Bacillus, Arthrobacter et Actinomycetes. Or les milieux de culture utilisés ne
peuvent que difficilement recréer l'ensemble des interactions que les micro-organismes entretiennent
avec le sol et les plantes. De plus les micro-organismes retrouvés en culture diffèrent de ceux
observés in situ. Ces derniers semblent avoir une paroi plus complexe et une taille plus petite (ce qui
les protégerait contre les contraintes hydriques et alimentaires).
3.1.3 L'apport des techniques moléculaires
Compte tenu de l'importance des micro-organismes tant au niveau des cycles géochimiques
que de l'augmentation des rendements de cultures, de la production d'antibiotiques, et des
mécanismes d'infection (par exemple Clostridium tetani ou Bacillus anthrax), de nombreuses études
utilisant les techniques moléculaires ont été entreprises (Liesack et al., 1997; van Elsas et al., 1998;
O'Donnel et Görres, 1999). Ces dernières ont révélé une diversité microbienne bien plus abondante
que celle obtenue par les méthodes de cultures classiques et dont les représentants se répartissent
dans toutes les branches (mésophiles) de l'arbre des bactéries (Torsvik et al., 1990a; Borneman et al.,
1996).

Les habitats microbiens
51
La première étape à franchir dans le cadre de telles études, est l'extraction d'ADN à partir
d'échantillons riches en matière humique et en substances susceptibles de fixer l'ADN telles que les
argiles (Vettori et al., 1996). Un grand nombre d'articles décrivent et comparent l'efficacité de
différentes méthodes d'extraction à partir de tels échantillons (Holben et al., 1988; Steffan et al., 1988;
Pillai et al., 1991; Tsai et Olson, 1992 a et b; Moran et al., 1993; Ogram et al., 1995; Felske et al., 1996;
Zhou et al., 1996; Jackson et al., 1997; Porteous et al., 1997; Abu Al-Soud et Rådström, 1998; Kuske
et al., 1998). Les plus efficaces de ces techniques sont décrites dans la troisième partie de ce
manuscrit.
Une fois résolus les problèmes liés à l'extraction de l'ADN, la communauté microbienne au sein
de différents types de sols a été analysée soit par séquençage direct des ADNr 16S, soit par
hybridation à l'aide d'amorce domaine-spécifique. Le tableau 2.1 présente une liste non-exhaustive
des différentes études réalisées sur les sols; chacune de ces études à contribué à la découverte de
nouvelles séquences de micro-organismes proches de micro-organismes déjà isolés mais le plus
souvent correspondant à de nouveaux phylums.Tableau 2.1. Etudes de la diversité microbienne des sols par des approches moléculaires
Sols cultivés Autres types de sols
Ø mise en évidence d'une nouvelle lignée de Crenarchaeota (Bintrin et al.,
1997; Buckley et al., 1998),
Ø recherche des bactéries actives dans le sol (Christensen et al., 1999),
Ø étude sur des sols isolés de l’archipel Hawaiien (Nüsslein et Tiedje,
1998),
Ø approche de la diversité bactérienne très complète (séquençage,
hybridation, DGGE) et quantification des espèces actives par RT-PCR
(Felske et al., 1998a; Felske and. Akkermans, 1998a),
Ø communauté des Archaea (Großkopf et al., 1998a et b) et des bactéries
oxydant le méthane (Henckel et al., 1999) se développant sur les racines
de riz,
Ø une des premières études mettant en évidence la présence de
nouvelles lignées de Planctomycetales (Liesack et Stackebrandt, 1992)
suivit des travaux de Stackebrandt et al. (1993),
Ø communauté bactérienne des pâtures (Borneman et al., 1996; Lloyd-
Jones et Lau, 1998; McCaig et al., 1999)
Ø étude des méthanotrophes et des méthylotrophes dans des sols
anoxiques ou "fluchés" en CH4 (Øvreås et al., 1998), après culture
d'enrichissement (Jensen et al., 1998),
Ø approche générale (Picard et al., 1992; Ueda et al., 1995; Lee et al.,
1996b),
Ø approche de la diversité par analyse du G+C% et la réassociation de
l'ADN génomique et par les méthodes de cultures classiques (Torsvik et
al., 1990 a et b).
Ø communautés rhizobiales (Khbaya et al., 1998;
Lafay et Burdon, 1998),
Ø sols traités par des herbicides (elFantroussi et al.,
1999),
Ø sols pollués par des hydrocarbures par la
méthode d'hybridation génomique reverse (Shen et
al., 1998),
Ø sols forestiers (Borneman et Triplett, 1997;
Jurgens et al., 1997; Widmer et al., 1999),
Ø tourbières (Hales et al., 1996; Lloyd et al., 1998),
Ø sols gelés, permafrosts (Shi et al. 1997), ou sols
de toundra (Zhou et al., 1997),
Ø sols arides (Kuske et al., 1997; Dunbar et al.,
1999),
Ø sols hypersalés : Mer Morte (Arahal et al., 1996),
marais salants et marécages (Martínez-Murcia et
al., 1995; Nübel et al., 1999)
Ø fumier et compost (Blanc et al., 1997; Kowalchuk
et al., 1999).
Ø sols pollués par le plomb et bactéries résistantes
associées (Konopka et Zakharova, 1999; Konopka et
al., 1999)
D'autre part des recherches de plasmides susceptibles, par transfert horizontal, d'être vecteurs
de résistance aux antibiotiques ont été réalisées en utilisant des techniques d'extraction directes sur
différents échantillons de sols (Götz et al., 1996). Des techniques similaires ont été utilisées pour
détecter des entérovirus dans des sols amendés par des boues actives (Straub et al., 1995).
D'autres techniques s'attachent à amplifier ou hybrider spécifiquement certains types de micro-
organismes à l'aide d'amorces et de sondes espèces- ou sous espèces- spécifiques, ou à rechercher
des métabolismes ou des gènes impliqués dans des mécanismes de résistance (cf. tableau 2.2).

Les habitats microbiens
52
Tableau 2.2. Etudes de micro-organismes ou de métabolismes spécifiques
Recherche d'une espèce spécifique Recherche d'un métabolisme ou d'une résistance
Ø une espèce de Mycobacterium (Briglia et al., 1996),
Ø un Bacillus incultivable (Felske et al., 1998b; Felske, 1999) et
une souche associée aux Verucomicrobiales (Felske et
Akkermans, 1998b) détectés en grande quantité par une étude de
diversité générale,
Ø des Acidobacterium (Barns et al., 1999),
Ø des Pseudomonas par une approche traditionnelle de culture et
d'amplification (Johnsen et al., 1999),
Ø des Leptothrix et des Sphaerotilus impliqués dans les processus
d'oxydation du fer et du manganèse (Siering et Ghiorse, 1997),
Ø des Sphingomonas hybridés à l'aide d'une sonde spécifique et
quantifiés par cytométrie en flux (Thomas et al., 1997).,
Ø des Proteobacteria β oxydant l'ammoniaque genres
Nitrosomonas et Nitrospira dans des sols cultivés (Stephen et
al., 1996, 1998) ou par DGGE dans des sables dunaires
(Kowalchuk et al., 1997) ou dans des sols cultivés (Bruns et al.,
1999),
Ø des bactéries sulfato-réductrices dans le sol (Telang et al.,
1994; Devereux et al., 1996) ou au niveau de la rhizosphère
d'une plante colonisant les marais salants (Hines et al., 1999).
On notera en particulier les études réalisées pour étudier les
bactéries magnétotactiques dont la présence dans les sédiments
fossiles est le témoin des variations du champ
électromagnétique terrestre (Spring et al., 1992, 1998; Thornhill
et al., 1995).
Ø la méthane monooxygénase (gènes mmo) et la méthanol
deshydrogénase (gènes mxaF) des micro-organismes
méthanotrophes (McDonald et al., 1995; Henckel et al., 1999),
Ø le gène mcr des méthanogènes dans des échantillons de
tourbières (Nercessian et al, 1999),
Ø la nitrogénase réductase (gène nifH) (Widmer et al., 1999),
Ø sous unité α du gène codant pour la coenzyme M réductase
(Hales et al., 1996),
Ø l'ammoniaque monooxygénase (gène amoA) (Rotthauwe et al.,
1997),
Ø la résistance au mercure (gène mer) (Bruce et al., 1992),
Ø la résistance au cadmium au cobalt et au zinc (Diels et
Mergeay, 1990),
Ø dégradation des PCB (polychlorinated biphenyl) (Walia et al.,
1990).
Si les micro-organismes colonisant les couches superficielles du sol ont fait l'objet de
nombreuses études et ont même conduit à la création d'une discipline à part entière (la microbiologie
des sols), les couches stratigraphiques plus profondes sont moins bien connues et les micro-
organismes qui s'y développent ne sont étudiés que depuis récemment.
3.2 La biosphère souterraine
3.2.1 Description de l'habitat
L'habitat souterrain est défini comme celui compris au dessous de 8 mètres pour les habitats
terrestres et en dessous de 10 cm pour les sédiments marins. En terme de volume c'est le plus gros
habitat colonisable sur le globe. Il est constitué de roches magmatiques (basaltes, granites) et de
roches sédimentaires (les argiles par exemple). La température et la pression y croissent à mesure que
la profondeur augmente. On considère que la limite inférieure de colonisation de la lithosphère ne
s'étend pas en dessous de 4 km où la température atteint 125°C et avoisine la température limite de la
vie procaryotique (Whitman et al., 1998).

Les habitats microbiens
53
3.2.2 Le concept de biosphère souterraine
La notion de biosphère souterraine est née au début des années 90, en particulier grâce aux
réflexions d'un astronome, Thomas Gold, qui le premier supposa qu'il pouvait exister dans la croûte
terrestre (ou dans l'équivalent extra-terrestre) des conditions physico-chimiques pouvant permettre la
vie de micro-organismes tout à fait indépendamment de la surface (Gold, 1992). On considérait
jusqu'alors que les micro-organismes souterrains étaient essentiellement des chimioorganotrophes se
développant très lentement en métabolisant les restes de matière organique qui s'infiltraient dans les
couches stratigraphiques profondes (Arrage et al., 1993; Paul et al., 1995). Les études réalisées par la
suite par des microbiologistes dans les environnements tels que les sources hydrothermales (Deming
et Baross, 1993), les sédiments profonds (Parkes et al., 1994; Wellsbury et al., 1997) ou les puits de
pétrole (Bernard et al., 1992, L'Haridon et al., 1995), ainsi que les travaux de Yayanos et al. (1981;
Yayanos, 1995) sur les souches barophiles des fosses océaniques, permirent de confirmer en partie
les hypothèses de Gold. D'une part les souches thermophiles isolées en profondeur avaient une
pression optimale de croissance supérieure à la pression à laquelle elles avaient été récoltées (ce qui
laissait supposer une origine plus profonde), d'autre part on retrouvait dans des environnements très
éloignés géographiquement, mais tous proches de la lithosphère (réservoirs pétroliers et sources
hydrothermales), les mêmes espèces de procaryotes, enfin les prélèvements effectués dans des
sédiments marins profonds indiquaient la présence de micro-organismes jusqu'à 750 mètres de
profondeur à des teneurs encore élevées (1,8.106 bactéries/g). Par ailleurs l'étude d'aquifères
basaltiques profonds par des méthodes moléculaires et des méthodes de cultures classiques a
indiqué la présence de micro-organismes chimiolithotrophes anaérobies (BSR, méthanogènes,
homoacétogènes) (Kotelnikova et Pedersen, 1997; Kotelnikova et al., 1998, Motamedi et Pedersen,
1998) ou thermophiles (Szewzyk et al., 1994; Boone et al., 1995) dont le métabolisme (source de
carbone et d'énergie) pourrait entièrement dériver de composés libérés par la lithosphère (Stevens et
McKinley, 1995). Ainsi H2 ne dériverait pas de la décomposition anoxique de la matière organique mais
de la réaction de l'eau sur les basaltes et pourrait ainsi servir de donneur d'électrons dans des réactions
où interviendraient en tant qu'accepteurs le CO2 ou le SO42- présents sous une forme inorganique
dans les couches profondes (cf. réactions ci dessous).
Production de H2 inorganique : 2FeO (basaltes) + H2O Õ H2 + Fe2O3
Méthanogenèse : 4H2 + CO2 Õ CH4 + 2H2O
Acétogenèse : 4H2 + 2HCO3- + H+ Õ CH3COO- + 4H2O
Réduction des sulfates : 4H2 + SO42- + H+ Õ HS- + 4H2O
Cette hypothèse a été réfutée récemment par Anderson et al. (1998) car selon eux la
production de H2 au pH in situ (pH≈8) ne peut être suffisante pour pouvoir soutenir la croissance
chimiolithotrophique de toute la biomasse souterraine. Ils attribuent la croissance des micro-
organismes à la matière organique emprisonnée dans les sédiments, ou apportée par l'eau des
aquifères. Ils n'excluent toutefois pas la possibilité que cette croissance soit liée aux gaz réduits

Les habitats microbiens
54
provenant de couches stratigraphiques inférieures et percolant au travers de la lithosphère, comme
l'avait proposé Thomas Gold.
3.2.3 La communauté microbienne souterraine
Les micro-organismes se développant dans cette biosphère ne sont étudiés que depuis une
quinzaine d'années. L'intérêt pour l'étude de la biosphère souterraine s'est accru dès lors que
l'homme a dû trouver de nouveaux espaces pour enterrer des déchets. Il s'agissait particulièrement de
se défaire des déchets ultimes radioactifs des centrales nucléaires. Cet enfouissement était réalisé
dans des zones granitiques ou argileuses où les déchets pouvaient entrer en contact avec de l'eau
d'aquifère. Ces eaux chargées en micro-organismes pouvaient accélérer la corrosion des contenants à
déchets et favoriser la dispersion des radionucléides (Pedersen, 1996a; Santo Domingo et al., 1998a;
White et al., 1998). D'autre part l'étude de la flore souterraine permettait de suivre la dégradation de
polluants tels que les hydrocarbures ou les solvants chlorés (Dojka et al., 1998; Klier et al. 1999). De
plus, on suppose que les micro-organismes interagissent avec les roches dans des processus de
formation de minéraux (formation de silicates, de carbonates ou initiation de sites de nucléation des
minéraux) comme dans des processus de dégradation (lixiviation) (Torsvik, 1997; Douglas et
Beveridge, 1998; Fisk et al, 1998). L'intérêt pour ces micro-organismes s'est traduit par la mise en
place de sections spécialisées dans l'étude de micro-organismes au sein des programmes de forages
océaniques (Ocean Drilling P roject) et par la constitution (par le Departement o f Energy des Etats-
Unis) d'une collection de souches isolées spécifiquement d'environnements souterrains (Stim, 1994;
Reeves et al., 1996; Balkwill et al., 1997). Cette collection ayant été réalisée à partir de techniques de
cultures traditionnelles, mettant en jeu des milieux de culture classiques pour hétérotrophes aérobies,
les souches isolées ne pouvaient que partiellement rendre compte de l'entière diversité du milieu.
Les études réalisées notamment par Pedersen sur des forages d'aquifères profonds ont
quant à elles mis en évidence des séquences très diverses pouvant correspondre aussi bien à des
souches aérobies qu'anaérobies. Elles se répartissent dans 11 branches du domaine bactérien et 5
nouvelles branches (Ekendahl et Pedersen., 1994; Pedersen et al., 1996 a, b, 1997). D'autre part,
des représentants des Proteobacteria α, β, γ , δ et ε, des Firmicutes à haut et bas G+C%, des
Cytophagales (ex-C.F.B) ont été retrouvés à de nombreuses reprises dans différents gisements
(Boivin-Jahns et al., 1996; Barns et Nierzwicki-Bauer, 1997; Chandler et al. 1997, 1998; Fry et al.,
1997; Stroes-Gascoyne et al., 1997). Récemment Crozier et al. (1999) ont étudié les indices de
biodiversité à partir des différentes séquences collectées sur le site Oklo par l'équipe de Pedersen. 5
sites ont été comparés et les résultats semblent indiquer que, dans aucune des études, l'ensemble de
la population n'a pu être détecté (par clonage et séquençage). Les similitudes de population entre
sites de prélèvement sont discutées en fonction de leur caractéristiques physiques.

Les habitats microbiens
55
Une étude a permis de comparer l'effet d'une pollution par des carburants sur la flore
microbienne dans un aquifère à l'aide de sondes spécifiques (domaine, classe, phylum, etc.) et de
réitérer cette expérience au laboratoire en fermenteur alimenté en nitrates (Shi et al., 1999)8.
La majeure partie des études réalisées dans de tels environnements a porté sur l'analyse de la
diversité à l'aide du gène de l'ADNr 16S, mais quelques études ont porté sur l'analyse de la naphtalène
dioxygénase (gène nahAc) (Herrick et al., 1993; Wilson et al., 1999).
L'utilisation des techniques moléculaires a permis d'accroître considérablement nos
connaissances sur la flore souterraine mais, comme dans tous les milieux où ces techniques ont été
utilisées, un gros effort reste à réaliser pour améliorer les techniques d'échantillonnage (asepsie,
pression), en particulier pour les sites profonds, ainsi que les techniques de culture (prise en compte
des gaz disponibles sur le lieu d'échantillonnage, techniques d'enrichissement in situ).
3.2.4 Des profondeurs aux astres
Le concept de biosphère souterraine intéresse particulièrement les exobiologistes. Les
observations de chimie microbienne à base de H2S, CO2, Fe3+ (Vargas et al., 1998), de minéraux
réducteurs, et en quasi absence d'eau liquide, permettent d'extrapoler cette chimie prébiotique à
d'autres planètes du système solaire (ou à leurs satellites) où ces composés sont présents en grande
quantité (Brack, 1999; Brack et Raulin, 1999).
Il y a 20 ans alors que les premières sources hydrothermales étaient découvertes, la sonde
Voyager mettait en évidence une activité volcanique sur deux satellites de Jupiter, Io et Gaymède. A la
surface d'Europe (un autre satellite de Jupiter) il a été observé des plaques (de nature inconnue) qui
se déplaceraient sur un fluide (liquide ou gazeux). Il y a de fortes présomptions pour que ces plaques
soient des glaces d'eau qui se déplacent sur un océan d'eau liquide. Les mouvements des plaques
sont à attribuer à des phénomènes de courants liés à une marée due à la force gravitationnelle de
Jupiter ou à la présence d'un noyau chaud au centre du satellite. Si ce noyau existe il est probable que
des phénomènes d'hydrothermalisme soient observables sur Europe. Aujourd'hui un programme
spatial a été initié afin de prélever des échantillons sur ce satellite et rechercher des traces de vie à
proximité de sites hydrothermaux (Delaney, 1998).
La découverte (McKay et al., 1996) (contestée par Mc Coy, 1997) de formes associées à des
micro-organismes dans la météorite martienne ALH84001, (bien que ces formes n'atteignent que 380
nm de long !?) a permis de relancer, en plus de nouveaux crédits pour la NASA, le concept d'une vie
extraterrestre martienne (Carr, 1996; Farmer, 1996; McKay, 1997). Cette découverte ravive de plus
l'idée qu'un fort bombardement météorique aurait pu initier la chimie prébiotique terrestre, c'est le
concept de panspermie (Brack et Pillinger, 1998) ou au contraire supprimer toute forme de vie et
induire la division tripartite du monde vivant (Gogarten-Boekels et al., 1995).
8 Une étude sur un environnement similaire a été réalisée par Bekins et al. (1999) en utilisant une approche culturale : NPP (MPN) et
enrichissement de métabolismes spécifiques (méthanogènes, flore aérobie, flore fermentative, etc.). Rooney-Varga et al. (1999) ont
analysé par amplification et séquençage la flore anaérobie associée à la dégradation des benzènes dans un aquifère contaminé.

1-Sédimentation de la matière organique 2-Recouvrement de la matière organique par des couches sableuses (jaune) et imperméables (vert)
3-Phénomènes géologiques de mouvements du terrain (tectonique)
4-Phénomènes géologiques de transformation de la matière organique sous l’effet de la pression et de la température (métamorphisme)
5 et 6 Migration de l’huile (vert clair) du gaz (rouge) et de l’eau (bleu cyan) dans la roche réservoir (jaune) jusqu’à atteindre la roche couverture (vert)
1 2 3
4 5 6
Figure 2.2. Mécanismes de formation du pétrole
images d’après le site Total (http://www.total.com)

Les habitats microbiens
56
3.3 Les réservoirs pétroliers
3.3.1. Formation et production du pétrole
3.3.1.1 La formation du pétrole
Le pétrole résulte de la transformation de la matière organique en hydrocarbures. Les débris
végétaux ou animaux sont théoriquement dégradés par les micro-organismes, mais certains
échappent à cette dégradation. Ils se déposent au fond des mers fermées, des lagunes, des lacs, des
deltas ou d'autres milieux aquatiques pauvres en oxygène. Protégés de l'action des bactéries, les
matières organiques se mélangent à des sédiments (sable, argile, sel...) et s'accumulent au fond des
océans où ils sont recouverts par la suite d'autres sédiments plus jeunes. Par leur propre masse, ces
couches sédimentaires s'enfoncent naturellement. Les mouvements tectoniques qui agitent la
lithosphère les cassent, et les entraînent plus profondément dans l'écorce terrestre.
Lors de cet enfouissement, pression et température s'accroissent jusqu'au point où ces débris
se transforment en hydrocarbures et sont expulsés de leur matrice originelle. L'azote et l'oxygène sont
éliminés pour ne laisser que des molécules formées de carbone et d'hydrogène qui constituent les
hydrocarbures liquides et gazeux qui se retrouvent au sein d'une roche, appelée roche mère. Moins
denses que l'eau, les hydrocarbures vont migrer vers la surface profitant de microcavités et de fissures
du sol. Si rien ne les arrête, ils s'échappent et suintent à la surface ou bien se solidifient en bitume en
perdant leurs constituants volatils. Mais si au cours de leur migration, les hydrocarbures rencontrent
une couche imperméable qu'on nomme la couverture, ils seront bloqués par cette barrière rocheuse
étanche. Piégés dans les interstices microscopiques et les fissures d'une roche dite roche réservoir,
ils vont constituer le réservoir de pétrole (cf. figure 2.2).
3.3.1.2 L'exploitation et la production du pétrole
Les premières traces d'utilisation du pétrole par d'homme datent de la haute antiquité, en
Mésopotamie. Ramassé en surface, le pétrole servait alors dans des préparations médicinales, comme
combustible d'éclairage ou pour étancher les bateaux (calfatage). Il faut attendre 1859 pour qu'en
Pennsylvanie le "colonel" Drake initie les premiers forages pétroliers qui étaient alors de quelques
mètres de profondeur (cf. figures 2.3 et 2.4).
Figure 2.3. Premier
puits commercial
d'Edwin Drake,
Titusville,
Pennsylvanie, 1859
Figure 2.4. Deux ans
plus tard exploitation à
Run Creeks,
Pennsylvanie
Images site du D.O.E. agence "fossile energy" : http://www.fe.doe.gov/
Actuellement les forages peuvent atteindre des profondeurs de plusieurs milliers de mètres.
La première phase d'une exploitation passe par l'exploration du site. A l'aide de techniques sismiques,
gravimétriques ou magnétométriques, des géologues et des géophysiciens définissent les propriétés

Les habitats microbiens
57
physiques du sous sol et confirment la présence d'hydrocarbures. La mise en évidence directe
s'opère par forage.
La production consistera à récupérer les hydrocarbures. Ils remontent à la surface sous l'effet
naturel de la pression dans le gisement. Cinq à 20% du pétrole est ainsi extrait. Afin d'accroître le taux
de production des gisements, des techniques de récupération secondaire sont mises en place. L'une
d'entre elle consiste à injecter de l'eau ou du gaz au fond des puits afin d'y exercer une pression. La
récupération peut alors atteindre 35%. Pour récupérer le maximum des 65% restants, des techniques
d'extraction tertiaires ont été mises au point. Elles consistent en des procédés complexes qui
améliorent le drainage du pétrole : injection de vapeur, injection d'eau additionnée de produits
chimiques, injection de gaz miscible, combustion in situ. (Sources : TOTAL http://www.total.com et
ELF http://www.elf.fr).
Lorsque l'extraction du pétrole devient moins rentable, les grosses sociétés pétrolières (ELF,
TOTAL, EXXON) cèdent leur place à de petites sociétés qui se chargent de tarir la nappe en injectant
plus d'eau lors de l'extraction.
3.3.2. Composition physico-chimique du biotope
Les micro-organismes que l'on peut s'attendre à trouver dans la nappe de pétrole se trouvent
en réalité dans la phase aqueuse piégée avec l'huile et les gaz sous la roche couverture.
Dans cette eau, appelée de production, les conditions physico-chimiques sont variables selon
les puits analysés, mais elles sont généralement anoxiques et légèrement acides du fait de la
dissolution du CO2 ou de l'H2S (pH entre 3 et 7). La température du puits est fonction de sa profondeur
: à raison d'une augmentation de la température de 3°C tous les 100 m, la température de certains puits
peut atteindre 130 à 150°C (généralement de 60 à 130°C, Beeder et al., 1994). Selon la profondeur
du puits la pression au sein de la nappe va varier entre 150 et 400 bars. En outre la salinité des puits
peut être très élevée. Celle-ci dépend de la salinité de la roche mère au sein de laquelle la nappe va
être emprisonnée et peut varier entre 0 et 300 g.l-1 de sels totaux (NaCl, MgCl2, KCl essentiellement)
(pour revue Magot et al., 1999; Ravot, 1996).
L'analyse des accepteurs et donneurs d'électrons potentiels dans la nappe de pétrole souffre
d'un cruel manque de données. En effet les industriels ne collectent pas de façon routinière les
données concernant par exemple le phosphore ou les nitrates. On estime toutefois que le potentiel
redox des eaux de production est relativement bas et que des accepteurs comme l'oxygène, les
nitrates et le fer ferrique sont généralement absents. Par contre la présence de sulfates et de
carbonates laisse supposer que les métabolismes que l'on peut espérer rencontrer sont la sulfato-
réduction, la méthanogenèse, l'acétogenèse et les fermentations. Les donneurs d'électrons sont
quant à eux l'H2 d'origine biotique et abiotique ainsi que des molécules organiques. Les acides
organiques comme l'acétate, le formate, le propionate et le butyrate sont souvent détectés dans les
puits de pétrole, mais on détecte aussi des acides plus complexes. Si la dégradation aérobie de
nombreuses molécules organiques présentes dans le pétrole a souvent été démontrée (n-alcanes et

Les habitats microbiens
58
alkylbenzènes), ce n'est que très récemment que des preuves d'une potentielle dégradation
anaérobie de ces composés ont été apportées (Ruetter et al., 1994; Connan et al., 1996; Heider et al.,
1999; Zengler et al., 1999). De plus des composés comme les résines et les asphaltènes, structures
riches en soufre, azote et oxygène, présentes en grandes quantités dans les huiles, pourraient servir
de sources de carbone et d'énergie à des micro-organismes anaérobies (Magot, communication
personnelle). Enfin la production anaérobie de l'hydrogène lors de la réaction de l'eau avec du basalte
pourrait servir de donneur à des hydrogénotrophes tels que les micro-organismes méthanogènes, les
sulfatoréducteurs et homoacétogènes (Stevens et McKinley, 1995) (voir §3.2.2).
3.3.3. La diversité microbienne des puits de pétrole
Même si la présence des micro-organismes dans les puits de pétrole a été établie depuis
longtemps, c'est seulement depuis une dizaine d'années que les industries pétrolières ont pris en
compte le rôle de ces micro-organismes dans la formation des huiles et les problèmes liés à la
dégradation des systèmes d'extraction. La première étude de la flore microbienne associée aux puits
de pétrole a mis en évidence des bactéries sulfato-réductrices (Bastin, 1926), et dès cette époque
l'auteur s'est interrogé sur la nature endémique des souches étudiées. La microflore des gisements
pétroliers est relativement peu diversifiée (Ravot, 1996). On retrouve principalement des micro-
organismes anaérobies qui appartiennent aux deux règnes des Procaryotes : Archaea et Bacteria. On
a cependant mis en évidence la présence d’organismes aérobies dont l'origine semble être exogène.
3.3.3.1 La colonisation exogène des gisements de pétrole
L'étude de la flore des nappes de pétrole pose le délicat problème de l'origine des micro-
organismes présents dans l'échantillon étudié. En effet, le mécanisme d'extraction du pétrole,
notamment les techniques d'extraction secondaires et tertiaires, sont des sources non négligeables
de contamination de la nappe de pétrole.
¬ L'eau injectée dans la nappe peut avoir diverses origines :
Ø eau d'un réseau interne à l'entreprise de forage : injectée, récupérée, traitée et réinjectée,
Ø eau de rivière ou de mer dans le cas d'exploitation "offshore",
Ø eau de nappe phréatique adjacente à la nappe de pétrole,
Ø eau de consommation courante lorsque qu'aucune nappe souterraine n'est proche du puits ou
lorsque ce puits est trop éloigné du site de production pour être alimenté par un réseau d'eau
interne.
La contamination peut aussi provenir du matériel utilisé. Les têtes de forage et les canalisations
peuvent perturber la composition initiale de la nappe. La nature de ces dernières favorise
particulièrement le développement de bactéries sulfato-réductrices (BSR) qui en se multipliant en
biofilm autour des canalisations provoquent une dégradation accélérée des installations connue sous
le terme de "pitting".

Les habitats microbiens
59
® Enfin, les micro-organismes présents dans les différentes couches stratigraphiques qui surmontent
la nappe peuvent eux aussi contaminer l'eau de production. Dans les couches superficielles on
retrouve des micro-organismes chimio-organotrophes se développant sur la matière organique
présente en grande quantité dans le sol. Il s'agit de souches aérobies ou anaérobies qui interviennent
dans les cycle de l'azote, du soufre et des métaux. Dans les couches plus profondes on retrouve une
flore plus spécifique (voir § 3.2.).
Des études menées sur différents gisements ont indiqué que certains puits de pétrole étaient
stériles. Ces gisements pouvaient être classés en trois catégories différentes :
Ø Les puits dont la température dépassait 82°C, ce qui semble correspondre à la température pour
laquelle aucune biodégradation des huiles n'a été observée (Philipi, 1977). Il faut toutefois noter
qu'il a été possible d'isoler des micro-organismes thermophiles et hyperthermophiles ayant des
optima de croissance supérieurs à 85°C dans les puits de pétrole terrestres et marins (Stetter et al.,
1993; Jeanthon et al., 1995; L'Haridon et al., 1995; Ravot et al., 1995b). La température élevée
des puits n'est donc pas un critère d'absence de micro-organismes. On estime toutefois que les
gisements dont la température dépasse les 130 à 150°C (température fréquemment observée
dans des puits profonds) sont stériles. Les processus indispensables à la "machinerie cellulaire" ne
peuvent se dérouler convenablement à de telles températures.
Ø Les puits dont la température est supérieure à 50°C mais dont la salinité était supérieure à celle de
l'eau de mer (Bernard et al., 1992). Cette dernière donnée corrobore l'incapacité actuelle des
microbiologistes à isoler des souches à la fois halophiles et thermophiles.
Ø Enfin des gisements dont les caractéristiques physico-chimiques les rendraient favorables au
développement de micro-organismes, mais dont le parcours géologique a porté ces gisements à
des températures plus élevées que la température actuelle ce qui a eu pour effet de "stériliser" la
nappe de pétrole (Bernard et al., 1992).
3.3.3.2 Les micro-organismes des gisements pétroliers
Les techniques moléculaires n'ont été que très peu utilisées dans les réservoirs pétroliers
comparativement à d'autres environnements. La majeure partie des études a été initiée par Gerrit
Voordouw. Elles consistent en une étude par hybridation génomique inverse sur des cultures
d'enrichissement et par séquençage des ADNr 16S (Voordouw et al., 1991, 1992, 1996; Leu et al.,
1998). Les travaux de Voordouw et de Leu ont permis de détecter, en plus des BSR, la présence de
micro-organismes aérobies ou anaérobies facultatifs semblables à ceux trouvés dans les eaux de
surface (genres Bacillus, Micrococcus, Pseudomonas). Mais il a été également possible de détecter
(amplification de l'ADNr 16S) des micro-organismes associés aux genres microaérophiles :
Oceanospirillum, Thiomicrospira et Campylobacter. Une souche nommée CVO associée aux
Campylobacter a d'ailleurs été isolée par Voordouw et al. (1996) à partir de cultures d'enrichissement
sur milieu pour Thiomicrospira denitrificans. Les Campylobacter et les Thiomicrospira pourraient trouver
une niche de développement au sein de la nappe de pétrole dans des cavités micro-aérées de la

Les habitats microbiens
60
roche mère et pourraient intervenir dans la réoxydation des sulfures (après réduction des sulfates par
les BSR) en utilisant les nitrates ou les nitrites comme accepteurs d'électrons (Voordouw et al., 1996).
Si ces micro-organismes peuvent être anaérobies facultatifs, leur caractère indigène peut être
contestable : aussi considère-t-on plus sûrement comme indigènes des micro-organismes anaérobies.
Parmi ceux-ci les micro-organismes sulfato-réducteurs, fermentaires ou homoacétogènes sont les plus
fréquemment retrouvés dans les études de diversité par une approche classique de culture.
Les micro-organismes sulfato-réducteurs
Les micro-organismes sulfato-réducteurs représentent la grande majorité des micro-
organismes endogènes aux gisements de pétrole. Ces organismes anaérobies stricts (du domaine
des Bacteria comme des Archaea) ont la particularité d'utiliser les ions sulfate SO42- comme accepteurs
d'électrons dans des oxydations génératrices d'ATP. Les ions SO42- sont réduits en plusieurs étapes
métaboliques en soufre élémentaire (ions sulfure S2- en général) qui, lorsqu'il est libéré réagit avec les
molécules d'H2O pour donner du sulfure d'hydrogène H2S en conditions acides, d'où la présence de
ce gaz en très grandes quantités dans les réservoirs.
Il a été prouvé également que ces bactéries ont la propriété de réguler leur environnement,
c'est-à-dire le biofilm, par une diminution ou une augmentation de la consommation d'ions H+. On
distingue parmi ces micro-organismes les mésophiles, impliqués dans la précipitation de sulfures de fer
ou pitting (fer provenant des systèmes d'extraction) et les thermophiles impliqués dans les
phénomènes d'acidification du réservoir.
Le tableau 2.3 énumère les souches et genres sulfato-réducteurs couramment isolés dans les
gisements pétroliers (d'après Magot et al. 1999).
Tableau 2.3. Micro-organismes sulfato-réducteurs isolés de réservoirs pétroliers.
Bacteria Archaea
Sulfato réducteur mésophile Sulfato réducteur thermophile Sulfato réducteur hyperthermophile
Genre Desulfovibrio Genre Desulfotomaculum Genre Archaeoglobus
Desulfotomaculum halophilum Desulfacinum infernum
Desulfomicrobium aspheronum Thermodesulforabdus novegicus
Genre Desulfobacter Thermodesulfobacterium mobile
Les méthanogènes
Ces micro-organismes appartiennent exclusivement au domaine des Archaea . Ce sont des
anaérobies stricts qui tirent leur énergie de la réduction du CO2 en oxydant H2 tout en produisant du
méthane (CH4). La méthanogenèse n'a été que récemment décrite au sein des gisements pétroliers.
Elle est souvent associée à des puits riches en sels. Jusqu'à présent seules des souches mésophiles
ou thermophiles modérées ont été isolées de gisements pétroliers; aucune souche hyperthermophile
n'a pu être isolée de gisements en raison de la teneur en sel trop élevée de ces derniers. Certaines
souches méthanogènes sont capables de se développer sur d'autres composés en C1 plus réduits
que le CO2, ce sont des méthanogènes méthyloclastiques. Elles se développent sur méthylamine
(CH3NH2), méthanol (CH3OH), diméthyl sulfures ((CH3)2S) (Madigan, 1996; Thauer, 1998). L'acétate

Les habitats microbiens
61
(CH3COO-) présent à de très fortes concentrations dans certains puits pourrait lui aussi servir de
substrat à certaines méthanogènes acétoclastiques soit directement, soit en syntrophie avec d'autres
organismes acétoclastiques ou acétogènes.
Parmi les souches méthanogènes isolées d'environnements pétroliers on retrouve des micro-
organismes appartenant à l'ordre des Methanobacteriales, des Methanoccoccales, des
Methanomicrobiales, des Methanosarcinales.
Les micro-organismes fermentaires
Les micro-organismes fermentaires sont des Archaea ou des Bacteria mésophiles,
thermophiles ou hyperthermophiles hétérotrophes, c'est-à-dire qu'ils utilisent de la matière organique,
comme les sucres ou les peptides, en tant que substrat énergétique en l'absence d'accepteur
d'électron externe.
Les bactéries mésophiles sont le plus souvent des halophiles modérées anaérobies et
appartiennent au genre Haloanaerobium. Certaines de ces souches présentent un intérêt pour les
industriels car en produisant des acides, des solvants et des gaz à partir des sucres elles pourraient
participer à une augmentation du rendement d'extraction du pétrole (MEOR : microbial enhancement
oil recovery). Magot et al. (1999) rapportent l'isolement de deux bactéries halophiles modérées
appartenant aux genres Spirochaeta et Dethiosulfovibrio. Les techniques fondées sur l'étude des
ADNr 16S par amplification sur des enrichissements ont quant à elles permis de détecter la présence
de micro-organismes potentiellement anaérobies stricts ou facultatifs associés aux genres Clostridium,
Eubacterium et Synergistes (Voordouw et al., 1996).
Les bactéries thermophiles appartiennent généralement à l'ordre des Thermotogales
pour lequel on retrouve dans les gisements, les genres Thermotoga, Geotoga et Petrotoga. Au sein
de cet ordre le genre Thermotoga regroupe les seuls micro-organismes du domaine des Bacteria
considérés comme hyperthermophiles. Il constitue une des branches phylogénétiques les plus
ancestrales de ce domaine. Thermotoga, isolée et identifiée dans les gisements pétroliers, pourrait
jouer un rôle dans les phénomènes de corrosion, en réduisant le thiosulfate en sulfures (Jeanthon et
al., 1995; Ravot et al., 1995a). En effet la réduction du soufre interviendrait en tant que mécanisme de
détoxication en oxydant H2 produit lors de la fermentation. D'autres familles ont également été
identifiées dans des puits pétroliers comme les Thermoanaerobiaceae capables de réduire le
thiosulfate en soufre (Thermoanaerobacterium) ou en sulfures (Thermoanaerobacter) (Grassia et al.,
1996) et la souche Anaerobaculum thermoterrenum (Rees et al. 1997) capable de réduire le soufre et
le thiosulfate.
Des Archaea fermentaires ont, elles aussi, été identifiées au sein des gisements de pétrole
(Stetter et al., 1993; L'Haridon et al., 1995). Elle appartiennent au genre Thermococcus et
Pyrococcus. Leur caractère endémique a été discuté par Stetter (1993) mais leur capacité à réduire le
soufre et le fer à température élevée en fait des candidats potentiels pour des phénomènes de
biocorrosion (Magot et al. 1994; Grassia et al. 1996; Magot, 1996).

Les habitats microbiens
62
Les bactérie s homoacétogènes et les bactéries réductrices des métaux
Les bactéries homoacétogènes se caractérisent par une production exclusive d'acétate à partir
de CO2 et de H2. En dehors des composés gazeux habituels H2 et CO2, elles peuvent réduire le CO2 à
partir d'autres composés en C1 ou de sucres. A ce jour, peu de bactéries homoacétogènes ont été
mises en évidence en milieu pétrolier (Acetobacterium romashkowii par Davydova-Charakhch'yan et
al., 1992).
Magot et al. (1999) rapportent dans leur revue l'isolement au sein de gisements pétroliers de
micro-organismes bactériens capables de réduire le fer (Shewanella putrefaciens) ou le fer et le
manganèse (Deferribacter thermophilus, Green et al., 1997). Le manque de données concernant le
cycle de ces composés au sein du gisement ne nous permet pas de déterminer le rôle de ces micro-
organismes au sein de la nappe. Il semble toutefois que les processus de réduction du Fe (III) soient
très largement répartis dans le monde procaryotique en particulier au sein des genres thermophiles.
Aussi de nombreux genres initialement décrits comme fermentatifs pourraient sous certaines
conditions respirer en anaérobiose le fer ferrique (Slobodkin et Wiegel, 1997; Vargas et al., 1998;
Slobodkin et al., 1999).
3.3.4 Activité microbienne liée aux hydrocarbures
Les techniques d'exploration et de production d'hydrocarbures bénéficient de nouvelles
informations fournies par les études microbiologiques : les propriétés physiologiques des bactéries
identifiées pourraient renseigner sur la température du milieu. Ces études peuvent aussi être un
indice pour montrer la correspondance entre deux gisements, apparemment distincts, si la flore
microbienne est similaire dans les deux puits. Un suivi régulier de la flore peut permettre de s'assurer
de l'effet de certains biocides que les producteurs injectent dans les puits lors de l'extraction des
huiles pour se défaire des BSR, ou dans les tanks pour limiter le développement des souches oxydant
les hydrocarbures en présence d'O2.
La détection de certaines bactéries peut également annoncer un risque majeur pendant la
phase de production : la biocorrosion. Les BSR constituent le groupe majeur impliqué dans ce
phénomène. La corrosion provoque des dégâts considérables sur le matériel, en particulier sur les
pipelines.
Mais la dégradation des hydrocarbures par des micro-organismes peut être aussi mise à profit
dans le cas de catastrophes écologiques majeures telles que l'échouage de pétrolier ou la rupture de
canalisations (Button et al., 1992). La décomposition des hydrocarbures est alors assurée par des
bactéries aérobies, des algues ou des champignons (Swanell et al., 1996). Les hydrocarbures
aliphatiques n'étant pas fermentables, leur dégradation se produit en présence d'O2 sous l'action de
bactéries telles que des Pseudomonas, des Corynebacterium ou des Mycobactéries. Pour accélérer
le développement de ces micro-organismes il est parfois nécessaire d'apporter des nutriments (azote
et phosphore) sous forme de "spray" en surface de la zone à nettoyer. Les micro-organismes peuvent
aussi produire des surfactants qui peuvent accélérer la dispersion des hydrocarbures dans le cas de

Les habitats microbiens
63
pollution par des nappes de pétrole (Desai et Banat, 1997). Il est possible de suivre le développement
et la présence de ces souches en amplifiant ou en hybridant spécifiquement les gènes impliqués dans
le métabolisme de dégradation des hydrocarbures (Guo et al., 1997): xylE : cathécol 2,3-dioxygénase,
tod : toluène dioxgénase, alkB : alcane hydroxylase, tmo : toluène monooxygénase, nahAcd :
composant de la naphtalène dioxygénase.
McNaughton et al. (1999) ont suivi, par mesure des profils lipidiques (PFLA) et DGGE (sur
ADNr 16S), l'évolution des populations eucaryotiques et procaryotiques lors d'une pollution simulée
aux hydrocarbures en eau de mer. Les deux techniques permettent de suivre la disparition des
eucaryotes au profit de bactéries Gram - (Proteobacteria α et Cytophagales).
Il est à noter que certains micro-organismes eucaryotes produisent des hydrocarbures en C30 jusqu'à
C36. Cette "production microbienne" de pétrole est réalisée par l'algue verte Botryococcus braunii
(Dennis et Kolattukudy, 1991; Madigan, 1996).
Si les premières études moléculaires des communautés microbiennes des sols s'attachaient
surtout à décrire la formidable diversité procaryotique qui s'y développe, l'effort s'est désormais porté
sur l'analyse de l'activité et la taille des différentes populations. L'ensemble de ces informations devrait
permettre de mieux définir l'action de l'Homme dans cet environnement et notamment d'améliorer les
pratiques agriculturales et les processus de dégradation des déchets.
IV. Habitats aquatiques
Au même titre que les habitats terrestres, on distingue trois habitats aquatiques physiquement
et chimiquement différents : l'eau douce (rivière, lac), l'eau de mer et les estuaires qui constituent la
frontière physique entre l'eau douce et l'eau de mer. Au sein de chaque habitat, de fortes variations
physiques et chimiques vont avoir lieu entre la surface de l'eau, la colonne d'eau et le fond. Ces
variations vont influer fortement sur la disponibilité en nutriments et donc sur la composition de la
communauté microbienne. D'autre part l'analyse de cette communauté est confrontée au délicat
problème de la nature endémique des souches isolées ou détectées. Au même titre que les études
réalisées dans des environnements terrestres, les premières analyses effectuées sur des échantillons
d'eau douce et d'eau de mer ont essentiellement permis d'isoler la flore aérobie, hétérotrophe,
correspondant majoritairement à des bactéries Gram - (Bertrand et Larsen, 1988). Ces bactéries étaient
en tout point semblables à celles obtenues à partir d'environnements terrestres et cette diversité
apparemment limitée comparativement à la multitude de formes observées en comptage direct laissait
présager d'un biais énorme dans les méthodes d'échantillonnage et de culture employées.
L'utilisation des méthodes moléculaires ainsi que l'introduction de techniques plus fines d'observation
(microscopie, cytométrie en flux) ont permis de révéler à quel point les milieux aquatiques pouvaient
être plus riches et diversifiés que ce que les premières études culturales pouvaient laisser présager.

Les habitats microbiens
64
4.1 L'habitat d'eau douce
Au même titre que l'eau de mer, la composition microbiologique de l'eau douce des lacs et des
rivières sera fonction de la disponibilité du milieu en oxygène et en matière organique. Dans les rivières
et dans les lacs en hiver où on peut observer de très forts courants de fond, toute la masse d'eau va
être brassée, oxygénée et alimentée en matière organique. Par contre au cours de l'été les couches
d'eau superficielles chauffées par le soleil et donc moins denses vont se disposer au dessus des eaux
plus froides et plus denses créant ainsi une stratification au sein de laquelle les micro-organismes
aérobies vont coloniser préférentiellement les couches supérieures en consommant activement la
matière organique. Les couches inférieures appauvries en oxygène vont être colonisées par les micro-
organismes micro-aérophiles ou anaérobies.
Au sein des rivières (même avec un fort courant) un apport massif de matière organique (rejet
d'industrie, boues d'épuration) peut entraîner des conditions ponctuelles d'anaérobiose qui peuvent
alors profondément perturber la composition de la communauté microbienne indigène.
Différentes méthodologies moléculaires ont été mises à profit afin d'étudier la population totale
microbienne :
Øpar extraction des ARN à bas poids moléculaire (LMW RNA), électrophorèse et séquençage des
bandes majoritaires à partir d'échantillons d'eau d'un lac eutrophe (Höfle et al., 1999). La
comparaison des profils LMW RNA permet de suivre l'évolution des populations au cours de l'année.
Ø Par amplification et séquençage de l'ADNr 16S d'échantillons d'eau d'un lac de montagne (Hiorns et
al., 1997),
Ø par amplification et DGGE sur des échantillons de différentes profondeurs d'un lac méromictique
(Øvreås et al., 1997) et d'un fjord (Teske et al., 1996a),
Ø par utilisation de sondes domaines- et phylum- spécifiques sur des échantillons de lac de montagne
(Alfreider et al., 1996; Pernthaler et al, 1998; Tonolla et al., 1998), d'eau de Fjord (Ramsing et al.,
1996), d'eau de rivière (Kenzaka et al., 1998),
Il a été aussi possible de détecter des micro-organismes spécifiques tels que des bactéries
sulfureuses pourpres associées aux Chromatiaceae dans la chémocline d'un lac suisse (Tonolla et al.,
1999), et des bactéries ammonium oxydantes (Proteobacteria β) participant activement au cycle de
l'azote dans les sédiments lacustres ou dans les eaux de surface. La présence de ces Proteobacteria
était connue depuis longtemps mais elles étaient difficilement détectables par les techniques de
culture classiques (Voytek et Ward, 1995; Hastings et al., 1998; Speksnijder et al., 1998).
Une étude très complète a été réalisée par Priscu et al. (1998) afin d'analyser la diversité
biologique au sein de lacs gelés en Antarctique. Au sein de cet environnement la température ne
dépasse pas 0°C en été. Il est toutefois possible d'y observer une activité microbienne. Durant l'été,
des poches d'eau liquide se créent à l'intérieur de la glace et des sédiments parviennent à atteindre
ces poches d'eau. Ces "oasis" permettent alors la croissance de bactéries photoautotrophes
(cyanobactéries) et hétérotrophes ainsi que de certaines algues (Priscu et al., 1998; Psenner et
Sattler, 1999).

Les habitats microbiens
65
4.2 L'habitat maritime
4.2.1 Les micro-organismes marins isolés
Les écosystèmes marins peuvent être considérés comme des environnements stressants en
raison de la salinité (3,5% soit 0,6M NaCl), de la pression (1 atm tous les 10 mètres; on considère que
90% des océans correspondent à des profondeurs supérieures à 1000 mètres), de la température
(90% des océans sont à une température inférieure à 5°C).
On distingue communément dans cet habitat deux zones principales : les océans et le littoral.
Les océans sont pauvres en matière organique (oligotrophe) et étaient donc considérés jusqu'il y a
peu comme pauvres en producteurs primaires, sauf lorsque certains courants permettent à des
nutriments de profondeur de passer en surface comme au large du Pérou et de la Californie. Les côtes
océaniques quant à elles sont des régions riches en matières organiques (eutrophe), donc riches en
producteurs primaires et riches en poissons et en coquillages. Les premières études réalisées à l'aide
de techniques de culture classiques confirmèrent ces hypothèses. Les bactéries isolées en haute mer
(Oceanospirillum, Vibrio, Photobacterium et Alteromonas) correspondaient à des micro-organismes
hétérotrophes présents en très faibles quantités (102 à 105 organismes/l). L'amélioration des
techniques de microscopie a permis au début des années 80 de mettre en évidence au milieu des
océans une diversité microbienne jusque là insoupçonnée. Dans un premier temps des équipes
américaines ont mis en évidence une abondante colonisation des eaux de haute mer par des
Cyanobactéries associées au genre Synechococcus. Par la suite les techniques de cytométrie en flux
permirent d'isoler le plus petit et le plus abondant des micro-organismes photosynthétiques connus :
Prochlorococcus. Ces deux micro-organismes cohabitent dans les océans en colonisant des zones
concomitantes. Alors que les Synechococcus colonisent les eaux de surface bien éclairées et sont
présents dans tous les océans aussi bien oligotrophes qu'eutrophes, les Prochlorococcus colonisent
des zones plus oligotrophes et sont retrouvés à des profondeurs plus importantes (pour revue
Madigan, 1996; Blanchot et al., 1998; Partensky et al, 1999).
4.2.2 L'utilisation des techniques moléculaires
4.2.2.1 Le bactérioplancton océanique et côtier
Les techniques moléculaires basées sur l'identification directe des gènes codants pour l'ARNr
16S furent appliquées très rapidement aux échantillons d'eau de haute mer. L'une des premières
études a consisté à comparer plusieurs échantillons d'eau de mer par hybridation de l'ADN génomique
total extrait (Lee et Fuhrrman, 1990). Chaque échantillon servait successivement de sonde et de cible.
Il était alors possible de déterminer des similitudes entre les échantillons mais la diversité de ces
derniers était encore inaccessible. Après la mise au point d'une technique d'échantillonnage
permettant de filtrer de très grandes quantités d'eau de mer (Giovanonni et al., 1990a), les travaux de
l'équipe de Giovanonni et de DeLong (Giovanonni et al., 1990b; Britschgi et Giovannini, 1991;
Schmidt et al., 1991) permirent de confirmer la présence d'une part (pour la moitié de la banque) de

Les habitats microbiens
66
séquences associées aux Cyanobactéries (Synechococcus et Prochlorococcus, ces derniers
n'étaient alors pas décrits) et d'autre part de séquences associées aux Proteobacteria communément
isolées (Alteromonas , Vibrio, Pseudomonas pour les Proteobacteria γ et Caulobacter pour les
Proteobacteria α), ou ne correspondant à aucun genre connu (clade SAR11). Par la suite des travaux
réalisés sur de nouveaux sites permirent de confirmer l'ubiquité des séquences précédemment
décrites comme le clade SAR11 mais aussi la présence de séquences associées aux Firmicutes à haut
G+C%, aux Flavobactéries (Fuhrman et al., 1993), aux bactéries vertes non-sulfureuses (Giovannoni et
al., 1996). Récemment une étude plus approfondie des séquences proches des Actinobacteria a
confirmé l'existence d'un clade de séquences marines provenant d'échantillons côtiers et hauturiers
sur les océans Pacifique et Atlantique (Rappé et al., 1999). Des études similaires ont été réalisées à
partir des séquences proches des Proteobacteria α associées au clade SAR11 (Fields et al., 1997), de
la sous-classe 3 des Proteobacteria α9 (Gonzalez et Moran, 1997) et des bactéries sulfureuses vertes
associées au clade SAR 406 (Gordon et Giovannoni, 1996). Des études menées par une équipe
espagnole en Méditerranée permirent dans un premier temps de suivre les variations de la diversité
microbienne de différents échantillons (formes libres et attachées, variations de profondeur, variations
temporales) ceci par une étude uniquement fondée sur l'analyse des profils RFLP (Acinas et al, 1997)
puis par séquençage des profils majeurs associés aux formes libres et attachées (Acinas et al, 1999).
L'approche suivie par Giuliano et al. (1999) sur des échantillons de surface et profonds de
Méditerranée consiste en une première étape d'isolement par dilutions successives suivie d'une
analyse des séquences d'ARNr 16S présentes dans la dernière dilution. Cette technique conduit à
l'identification de séquences proches des Proteobacteria γ et α et des Cytophagales mais elle ne
permet l'analyse que des micro-organismes majoritaires de l'échantillon.
Des études similaires ont été réalisées sur les micro-organismes associés aux neiges marines
révélant là encore une grande diversité de séquences se répartissant au sein de nombreux groupes
bactériens mais apparemment différents de ceux trouvés sous une forme libre dans la colonne d'eau
environnante (DeLong et al., 1993). On a retrouvé des séquences associées aux Proteobacteria (α, γ ,
δ, ε), aux Cytophagales , aux Planctomycetales, aux Verucomicrobiales (DeLong et al., 1993; Rath et
al., 1998). Une comparaison de la diversité bactérienne existant sous une forme libre ou sous une
forme associée avec des particules a été réalisée par extraction de l'ADN et séquençage de l'ARNr 16S
le long de la rivière Columbia (Crump et al., 1999). Les auteurs ont comparé des prélèvements dans la
rivière, dans l'estuaire et sur la côte et ont observé une plus grande diversité phylogénétique au
niveau de l'estuaire.
Une des premières études réalisées sur des sites côtiers a été effectuée dans deux lagunes
françaises en baie d'Arcachon et dans une zone saumâtre près de Palavas (Benlloch et al., 1995).
Cette étude complète (comptage, mesure du métabolisme et séquençage) mettait en évidence une
diversité bactérienne plus importante dans l'échantillon eutrophe (Méditerranée) et une grande
diversité de micro-organismes majoritairement associés aux Cytophagales et aux Proteobacteria α. Les
9 Il a été possible d'isoler des représentants de ce clade préalablement détectés par séquençage.

Les habitats microbiens
67
analyses phylogéniques de cette étude souffrent toutefois de la faible longueur d'ADNr 16S
séquencé (210 pb). L'étude effectuée sur des échantillons provenant de la côte Est des Etats-Unis
révèle encore une fois la prédominance des séquences associées aux Proteobacteria α et γ (75% de
la banque bactérienne de séquences) (Rappé et al., 1997). Par contre les eaux côtières se distinguent
des eaux du large par la présence de séquences associées aux Proteobacteria β (notamment
associées au méthylotrophes), l'absence de cyanobactéries et l'abondance de séquences plastidiales
(expliquée par l'absence de filtration de l'échantillon et l'abondante biomasse d'eucaryotes dans les
environnements côtiers). On note aussi la présence de Cytophagales, d'Actinobacteria et de
Planctomyces comme dans les études précédentes.
L'emploi de la DGGE a permis de comparer la composition du bactérioplancton sur deux
estuaires de la côte Californienne (Murray et al., 1996). Il a été ainsi possible de relier la variation de
types métaboliques détectés par des techniques classiques d'incorporation de radioisotopes à des
variations de phylotypes.
L'étude réalisée par Suzuki et al. (1997) est particulièrement intéressante car elle compare les
résultats d'une analyse directe des gènes codants pour l'ARNr 16S par séquençage et RFLP à ceux
obtenus après culture d'enrichissement sur milieu solide pour hétérotrophes marins. Si les
Proteobacteria α et γ sont bien les phylums les plus représentés dans les deux banques de
séquences, seules 4 séquences d'isolats (sur 127) sont retrouvées dans la banque
environnementale. De plus la diversité des séquences est plus grande dans la banque de clones
environnementaux que dans la banque d'isolats. Cette étude nous permet d'observer à la fois le biais
engendré par les techniques de culture classiques (la plupart des micro-organismes ne sont pas
présents sous une forme cultivable) comme celui engendré par les techniques d'amplification
(l'ensemble des micro-organismes présents dans l'échantillon n'a pas été détecté, soit il n'a pas été
amplifié, soit l'effort d'échantillonnage était trop important pour le détecter).
Enfin on peut citer l'étude réalisée sur un mésocosme constitué d'eau de mer
méditerranéenne rendue eutrophe par addition d'azote et de phosphore (Pukall et al., 1999). Les
auteurs comparent les résultats obtenus par extraction directe de l'ADN et séquençage des ADNr 16S
à ceux obtenus par isolement après dilutions successives. Parmi les 739 clones analysés (96 clones
séquencés - le reste est discriminé par dot-blot) et les 128 souches isolées, les séquences associées
à Alteromonas macleodii prédominent largement dans les deux banques. Les auteurs s'interrogent
alors sur la très forte micro-variabilité de séquence qu'ils n'attribuent pas uniquement à un grand
nombre d'opérons rrn dans cette souche mais à un possible mécanisme adaptatif.
Les études moléculaires des communautés bactériennes de l'eau mer ont confirmé que les
techniques culturales n'avaient permis d'accéder qu'à une très faible part de la diversité. Si on retrouve
toujours les même grands groupes bactériens (Proteobacteria α et γ , Cytophagales) de nouveaux
phyllums sont apparus à proximité ou au sein de ces groupes. On peut considérer toutefois que l'un
des apports majeurs de l'approche moléculaire dans l'étude des communautés microbiennes marines
a été la découverte, en grande quantité et de manière ubiquitaire, de deux grands groupes d'Archaea

Les habitats microbiens
68
rattachés aux Euryarchaeota et aux Crenarchaeota visiblement non associées à des conditions
physico-chimiques extrêmes.
4.2.2.1 L'archaeoplancton océanique et côtier
En 1992, à quelques mois d'intervalle, Fuhrman et al. (1992) et DeLong (1992) détectent dans
des eaux prélevées à 100 et 500 m de profondeur dans l'océan Pacifique (Fuhrman) et dans les eaux
côtières d'Amérique du Nord (DeLong) des séquences de micro-organismes branchant profondément
à la base des Crenarchaeota (Marine Group I - DeLong, 1992; Fuhrman et al., 1992; Fuhrman et al.,
1993) et des séquences associées aux Euryarchaeota mais formant là encore un clade bien distinct
(Marine Group II - DeLong, 1992; Fuhrman et al., 1993). Leur abondance semble indiquer qu'elles
interviennent activement dans les cycles chimiques océaniques et ne correspondraient donc pas à
des micro-organismes au métabolisme thermophile ou halophile comme le sont leur plus proches
"voisins" isolés. De plus le G+C% de leur ADNr (I : 0,51, II : 0,55-0,57) est inférieur à celui observé pour
des souches thermophiles (entre 0,60 et 0,69). DeLong propose alors que ces Archaea pourraient
correspondre soit à des méthanogènes soit à des Archaea mésophiles, aérobies10. Par la suite
l'analyse d'échantillons provenant d'eau de surface de l'Antarctique (DeLong et al., 1994) a confirmé
l'abondance des Archaea dans le picoplancton marin (jusque 34% de la biomasse procaryotique).
Ensuite des séquences associées au groupe I furent détectées dans l'estomac d'une Holoturie (Mc
Inerney et al., 1995), parmi les symbiotes d'une éponge (Preston et al., 1996) et parmi d'autres
échantillons de bactérioplancton (Stein et al., 1996; McInerney, 1997). Des séquences associées au
groupe II ont été détectées dans l'estomac de poissons (van der Maarel et al., 1998). L'approche
utilisée par Stein est particulièrement séduisante quoique fastidieuse. Elle consiste à cloner non pas
un produit d'amplification mais des fragments génomiques extraits de l'environnement. On dispose
donc à la fois de l'information phylogénétique en recherchant dans la banque de fosmides les gènes
codants pour l'ARNr 16S11 et d'informations sur le métabolisme en analysant les autres gènes
contenus dans la banque. Il faut toutefois garder à l'esprit que les gènes clonés ne sont pas forcément
exprimés. Enfin l'étude réalisée par l'équipe de DeLong à l'aide de sondes nucléiques spécifiques des
groupes I et II semble indiquer une zonation dans la colonisation de la colonne d'eau. Le groupe I serait
dominant en profondeur alors que le groupe II proliférerait en surface (Massana et al., 1997).
La détection de nouvelles Crenarchaeota dans des échantillons de sol (Bintrim et al., 1997),
d'un champ de soja (Ueda et al., 1995), de sources chaudes terrestres (Barns et al., 1994) et marines
(Takai et Sako, 1999) et de sédiments lacustres (MacGregor et al., 1997), ainsi que les séquences
d'Archaea associées au groupe II détectées avec d'autres Euryarchaeota (proches des
10 Une étude réalisée sur les éthers lipidiques présents dans la colonne d'eau associe ces derniers à des Archaea mais leur nature les
distingue clairement des méthanogènes (Hoefs et al., 1997 dans Vetriani et al, 1998). Les Crenarchaeota non thermophiles restent la
source principale de tétraéthers lipidiques dans les échantillons de plancton marin (DeLong et al., 1998). Des Archaea méthanogènes
ont toutefois été détectées dans des échantillons d'eau de mer, associées à des particules ou dans l'estomac de poissons (van der
Maarel et al., 1999)11 Un "screening" plus complet de banque fosmidique a permis par la suite de détecter des séquences de clones associées aux
Panctonomycetales (Vergin et al., 1998).

Les habitats microbiens
69
Methanosarcinales et des Halophiles) dans des sédiments de marais salants (Munson et al., 1997) et
sur des racines de riz (Großkopf et al., 1998b) nous amène à considérer ces "nouveaux" micro-
organismes comme les Archaea les plus représentées à la surface du globe (Olsen, 1994; Schleper,
1999)12.
4.2.3 Les fosses océaniques
Les fosses océaniques constituent des environnements particulièrement atypiques. En plus
de supporter des températures basses (2°C) les micro-organismes (bien souvent psychrophiles) se
développant dans de tels environnements (jusqu'à 10000 mètres de profondeur) doivent supporter
de très fortes pressions. On retrouve donc jusqu'à 4000 mètres de profondeur des micro-organismes
barotolérants, entre 5000 et 6000 mètres de profondeur des micro-organismes barophiles et des
barophiles stricts à des profondeurs avoisinants les 10000 mètres, nécessitant pour certains 700 bars
de pression minimale pour se développer (Yayanos et al., 1981; Nogi et Kato, 1999).
Très peu d'études à ce jour ont mis à profit les méthodes moléculaires pour analyser les
échantillons provenant de fosses océaniques. Seuls les travaux de Kato et al. (1997) et de
Yanagibayashi et al. (1999) associant les extractions directes et les techniques de culture en pression
et sans pression suivies d'extraction font référence. Récemment Li et al. (1999) ont comparé après
extraction de l'ADN total et séquençage la composition de sédiments abyssaux prélevés à différentes
profondeurs (entre 1159 m et 6482 m de profondeur). Ils n'ont pas observé de différences majeures
entre les échantillons dominés, pour la plupart, par des séquences proches de Pseudomonas, du
symbiote de Solemya velum ou des Proteobacteria ε.
4.3 Les zones sédimentaires marines et lacustres
4.3.1 Description du biotope
Interfaces entre le milieux terrestre et le milieu aquatique, entre des zones aérobies et
anaérobies, les sédiments sont le siège d'importants processus biogéochimiques impliquant les
principaux cycles de matière (N, C, S, O). La disponibilité de ces différents constituants dans la couche
de sédiments ainsi que les échanges de matières réalisés avec l'eau vont entraîner une forte zonation
des micro-organismes présents dans les sédiments en fonction de leur type métabolique et de leur
préférendum (cf. figure 2.5).
12 Des études similaires à celles effectuées sur les Procaryotes ont été entreprises sur les picoeucaryotes océaniques. En effet la taille
de ces organismes ainsi que la difficulté à les obtenir en culture ou à maintenir les cultures pures ont favorisé l'emploi de
méthodologies moléculaires pour accéder à la diversité eucaryotique de certains échantillons (Lim, 1996; Guillou, 1999). L'une des
techniques consiste à analyser les ADNr 16S plastidiaux contenus dans les banques de séquences (Rappé et al., 1998). Les ADNr
plastidiaux constituent dans cette étude près de 47% de la banque bactérienne.

Les habitats microbiens
70
En dehors des sédiments abyssaux qui sont aérés sur plusieurs mètres, les sédiments
présentent une couche oxygénée très fine et donc une succession de types respiratoires microbiens
très rapide. On a donc affaire à des micro-organismes réalisant des respirations aérobies sur les toutes
premières couches de sédiments puis plus profondément des bactéries fermentaires ou des bactéries
réalisant des respirations anaérobies en utilisant les nitrates (bactéries dénitrifiantes), les sulfates
(bactéries sulfato-réductrices) ou les carbonates (bactéries méthanogènes). De la même façon un
gradient s'établit entre les bactéries aérobies, anaérobies facultatives, microaérophiles et anaérobies
strictes. La disponibilité de la matière organique va conditionner quant à elle la présence de micro-
organismes hétérotrophes et chimio-autotrophes alors que le rayonnement lumineux à la surface du
sédiment va favoriser la présence de photoautotrophes.
Les techniques de culture classiques
ont conduit à l'isolement de nombreux genres
bactériens associés aux différents types
métaboliques décrits précédemment. Parmi
les plus spectaculaires on peut citer
Thiomargarita namibiensis, une bactérie sulfo-
oxydante accumulant les nitrates dans une
grande vésicule qui permet à cet organisme
d'atteindre 750 µm de diamètre (Schlulz et al.,
1999). C'est à ce jour le plus gros Procaryote
connu. Dans ces zones où l'oxygène et l'H2S
coexistent sous forme de gradient (stables ou
versatiles en raison des courants, de la marée,
des pluies qui remettent les sédiments en
suspension) on peut observer des micro-organismes accumulant le soufre sous forme de granules
internes Thioplaca ou Beggiatoa (Jørgensen et Gallardo, 1999; Teske et al., 1999) ou sous forme de
très longs filaments externes créant de véritables réseaux "mycéliens" dans les gradients d'O2 et d'H2S
(Taylor et Wirsen, 1997; Taylor et al., 1999).
4.3.2 Les techniques moléculaires
L'application des techniques moléculaires a contribué à accroître le nombre d'espèces
détectées dans cet environnement. Comme pour les études de sols, le problème principal a été
l'extraction d'ADN des échantillons de sédiments. Dès 1992 Rochelle et al. proposent des techniques
d'extraction couplées à des amplifications à partir d'échantillons de différentes profondeurs. Par la
suite, les travaux d'extraction d'ADN à partir d'échantillons de sols (cités § 3.1.3) servirent de
référence.
AIR
EA
US
ED
IME
NT
ZO
NE
AN
OX
iQU
EZ
ON
E A
ER
OB
IEA
ccep
teur
s te
rmin
aux
d’él
ectr
ons
O2
NO3-
SO42-
HCO3-
Respiration aérobie
Fermentation
Respiration nitrate
Respiration sulfate
Methanogénèse
Bactéries aérobies
Bactéries fermentatives
Bactéries dénitrifiantes
Bactéries sulfato-réductrices
Bactéries méthanogènes
O2
H2O
NO3-
N2
SO42-
H2S
HCO3-
CH4
Bactéries sulfo-oxydantes+
H2S
S
Figure 2.5. Représentation schématique des populations
bactériennes dans les couches de sédiments (d'après Marty et al.,
1988).

Les habitats microbiens
71
Compte tenu du rôle des bactéries sulfato-réductrices dans les mécanismes de production
d'H2S dans les sédiments, ces dernières ont été couramment étudiées: soit après extraction de l'ADN
total de l'échantillon et amplification à l'aide d'amorce spécifique des BSR affiliées aux Proteobacteria δ
(Devereux et Mundfrom, 1994), soit après extraction et détection à l'aide de sondes spécifiques de
ces mêmes BSR (Trimmer et al., 1997). Une étude récente réalisée sur des échantillons de sédiments
lacustres côtiers et profonds s'est attachée à étudier les BSR après isolement, séquençage de l'ADNr
16S et empreinte génomique ("genomic fingerprints") (Sass et al., 1998). Les auteurs ont ainsi
observé que même si leur souches correspondaient majoritairement (70%) au même genre
(Desulfovibrio) et présentaient de très fortes similarités d'ADNr 16S, leur contenu génomique, et donc
leur capacité physiologique, étaient très variables. Ce résultat permettrait d'attribuer l'hypervariabilité
de séquences de phénotypes proches à des souches ayant des rôles métaboliques distincts.
Des études s'intéressant à la diversité globale de l'échantillon ont été aussi réalisées :
Ø sur des échantillons de sédiments côtiers près de Seattle (Etats-Unis) où malgré le peu de clones
analysés (22 essentiellement bactériens) les auteurs observent une grande diversité
phylogénétique dans leur échantillon (Gray et Herwig, 1996). L'étude réalisée par Wise et al. (1997)
sur des échantillons prélevés dans une baie en Caroline (Etats-Unis) avec peu de clones (35 clones
bactériens) démontre encore cette très grande diversité.
Ø sur des échantillons marins provenant de sédiments sableux et boueux (Mer de Wadden en
Allemagne) hybridés à l'aide d'un grand nombre de sondes sur différentes profondeurs (Llobet-
Brossa et al., 1998)13. Un des points marquants de cette étude est la détection de séquences
associées aux Proteobacteria ε proches du genre Arcobacter.
Ø sur des sédiments coquilliers continentaux où Vetriani et al. (1998) détectent pour la première fois
dans ce biotope des séquences associées aux Archaea pélagiques proches du groupe I et d'autres
proches du groupe II. Puis sur des sédiments profonds où ces séquences sont de nouveau
détectées (Vetriani et al., 1999)
Ø des séquences similaires sont détectées (Crenarchaeota groupe II et méthanogènes) dans des
sédiments d'un lac du Michigan (Etats-Unis) (MacGregor et al., 1997; Schleper et al., 1997a). Les
séquences crearchaéennes semblent être plus abondantes dans la zone oxique du sédiment où
elles n'interviennent toutefois que pour 10% de l'ensemble des ARN archaéens.
Ø sur des sédiments lacustres d'origine postglaciaire sur lesquels une étude complète a été entreprise
(comptage total, comptage de bactéries viables, culture, extraction et séquençage) (Miskin et al.,
1998). Les 14 clones bactériens se répartissent parmi les Firmicutes à haut et bas G+C% et parmi les
Proteobacteria β et δ, et 9 clones archaéens sont quant à eux associés aux méthanogènes.
Ø enfin sur des sédiments lacustres datés de plus de 9000 ans où des séquences associées aux
bactéries pourpres sulfureuses sont détectées (à l'aide d'amorces genre-spécifique : Chromatium) et
analysées par DGGE et séquençage (Coolen et Overmann, 1998).
13 Cette étude a été suivie d'une analyse plus poussée concernant les Proteobacteria γ soufre oxydantes du genre Thiomicrospira, par
comptage, culture, amplification et DGGE (Brinkhoff et al., 1998).

Les habitats microbiens
72
D'une manière similaire aux études menées à partir d'échantillons de sols, la diversité
microbienne associée au sédiments marins ou lacustres s'avère être très importante. La part que
prennent chacun des groupes phylogénétiques détectés dans les processus géochimiques reste
malgré tout encore à définir. Si extraordinaire que semble être la découverte de nouveaux phylums
archaéens ou bactériens, il ne semble pas que ceux-ci interviennent de manière prépondérante dans
les communautés. Les études sont donc à poursuivre en s'attachant a définir les rôles que chaque
micro-organisme détecté peut tenir dans le biotope étudié.
4.4 Les sources hydrothermales océaniques profondes
Les mouvements de la croûte terrestre (ou tectonique des plaques) ont généré à la surface du
globe des zones d'anfractuosités où de l'eau (de mer ou douce) entre parfois en contact avec le
magma profond (cf. figure 2.6).
Figure 2.6. Représentation des zones d'accrétion et de subduction.
De droite à gauche : création de la lithosphère au niveau de la dorsale médio océanique. La surface du bassin océanique s'accroît alors
que sur la gauche la plaque plus ancienne glisse sous une marge passive continentale. Cette subduction est accompagnée de la
création d'une faille profonde et l'accroissement de température sur cette zone de friction crée un arc volcanique (par exemple les îles
japonaises) et une nouvelle zone d'accrétion dans le bassin arrière arc (d'après Press, F. et Siever, R. (1978) Earth (second edition).
W.H. Freeman, San Francisco).
Ces zones sont le siège de phénomènes hydrothermaux. Chargée en métaux, l'eau émerge
soit au niveau de sites océaniques profonds, de sites marins côtiers ou de sites terrestres. Ces deux
derniers sont connus depuis fort longtemps car ils sont le siège de phénomènes spectaculaires
comme les geysers (Parc de Yellowstone (USA), site de Geysir (Islande) pour les plus connus mais
aussi Russie, Nouvelle Zélande, Japon, Chili et Açores), les solfatares (Solfatara (Italie), Açores) ou les
émergences côtières (Vulcano (Italie), Ile de Santorin (Grèce), Iles Kouriles (Russie), Afrique). Si la
présence des sites hydrothermaux profonds était connue (ou supposée) déjà depuis plusieurs

Les habitats microbiens
73
années, il faut attendre la fin des années 70 pour que les premières plongées profondes mettent à
jour la formidable vie associée à ces sites.
4.4.1 Nature et structure des émissions hydrothermales profondes
4.4.1.1 Localisation des sources
La carte (figure 2.7) présente une liste non exhaustive des sites hydrothermaux étudiés
jusqu'à présent, où une faune associée a été observée (d'après Chevaldonné, 1996).
On retrouve des sites hydrothermaux profonds dans des zones à forte activité tectonique :
- Soit à proximité des zones d'accrétion, le long des dorsales médio-océaniques.
(Middle Atlantic Ride (MAR), East Pacific Ride (EPR), légende ____).
- Soit à proximité de zones de subduction, au niveau des bassins arrière arc (Bassin
de Lau, Bassin Nord Fidjien, légende tt).
Figure 2.7. Emplacements des sites hydrothermaux marins sur le globe
4.4.1.2 Composition du fluide hydrothermal
Les sources étudiées jusqu'à présent sont situées entre 700 et 4000 mètres de profondeur.
Dans ces zones où la croûte océanique a été fragilisée, l'eau de mer froide et dense s'infiltre. Elle est
alors portée à très haute température, très forte pression et peut atteindre le stade de fluide
supercritique. Sous cet état sa fluidité augmente ainsi que son pouvoir solvant. Elle remonte donc en
surface, et en percolant dans la lithosphère des échanges chimiques importants sont réalisés avec la
roche crustale. Les sulfates réagissent avec le fer contenu dans la roche pour produire du sulfure
d'hydrogène (H2S) ou des oxydes de fer, le magnésium réagit avec les hydroxydes et la silice. Le fluide
hydrothermal se charge en gaz dissous (H2S, CH4, CO, CO2, H2) et en métaux (Si, Mn, Fe, Zn). Ce fluide
est anoxique, acide, de salinité variable et ne contient pas de magnésium, de sulfates, de nitrates et de
phosphates. Sa composition contraste donc très fortement avec celle de l'eau de mer.

Les habitats microbiens
74
La température, le débit et la composition du fluide hydrothermal sont variables selon le site
étudié et même selon les points d'émission sur un même site. Ces données vont conditionner la forme
sous laquelle le fluide va être émis et les types de faunes associées.
4.4.1.3 Edification des cheminées hydrothermales
(d'après Erauso, 1994; Chevaldonné, 1996)
Les fumeurs noirs
Si le fluide en remontant dans la lithosphère ne subit pas de dilution avec l'eau de mer, il est
émis en surface à de très fortes températures (jusqu'à 400°C) et à un débit important (0,7-2,4 m.s-1)
(Converse et al. 1984). Il entre alors en contact avec l'eau de mer et forme une matrice poreuse de
sulfates (barytine ou anhydrite). La paroi de l'édifice croît alors verticalement et des grains de sulfure de
cuivre, de zinc et de fer se déposent sur la croûte externe (cf.
figure 2.8). Par la suite, la forte température du fluide induit le
dépôt d'une couche interne de sulfures de cuivre et de fer en
formant un conduit. La perméabilité de l'édifice diminue alors que
de nouveaux sulfures se déposent à la place de l'anhydrite. Les
minéraux s'arrangent de manière concentrique, stabilisant la
structure de la cheminée et créant un gradient de température
horizontal dans l'édifice. La communauté microbienne associée à
ce type de cheminée est essentiellement de type thermophile ou
hyperthermophile. On observe très peu d'Eucarya à proximité de
la zone d'émission de ces cheminées. La colonisation par des
bivalves ou des gastéropodes et les vers est plus éloignée de la
cheminée.
Les fumeurs blancs ou diffuseurs
Lorsque le fluide sort à des températures moins
élevées (<280°C), des cheminées riches en zinc peuvent alors
se former. Elles sont qualifiées de diffuseur car le conduit
central de la cheminée est absent ou très étroit et le fluide
diffuse à très faible vitesse (0,5 m.s-1) au travers d'un réseau de
petits canaux dans la cheminée. La cheminée va alors croître
dans toutes les directions car les dépôts de minéraux vont se
faire très lentement (cf. figure 2.9)
Ces diffuseurs peuvent être surmontés d'une structure en nid d'abeille au sein de laquelle des
échanges importants entre l'eau de mer et le fluide ont lieu. Ces structures très poreuses sont idéales
pour étudier les zonations de peuplement qui s'établissent en fonction des gradients de nutriments et
Figure 2.8. Fumeur noir sur le site du
Bassin Nord Fidjien
Figure 2.9. Diffuseur sur le site du Snake Pit
(MAR)

Les habitats microbiens
75
de température. On y retrouve aussi bien des micro-organismes mésophiles que thermophiles. Ces
structures sont toutefois très difficiles à échantillonner car elles sont très fragiles.
Les autres structures
En fonction de la composition du fluide et de sa célérité, d'autres types d'édifices vont être
rencontrés. Certaines cheminées sont dépourvues de sulfures et sont essentiellement constituées
d'anhydrite ("La Dame Blanche" dans le bassin nord Fidjien). Pour d'autres, le fluide a traversé une
couche de sédiments au cours de sa remontée. Le conduit de la cheminée est alors constitué de
dépôts argileux. Enfin des structures horizontales sont parfois observées. Elles se constituent
progressivement lorsque le fluide émis le long d'une surface verticale rencontre un obstacle qui initie
le dépôt des minéraux. Le fluide retenu sous ces constructions s'en échappe par les bords ou en
percolant au travers de la structure. Il se crée alors un gradient de température dans l'épaisseur de la
structure qui favorise la colonisation par une grande diversité de micro-organismes (Delaney , 1998).
4.4.1.4 Autres types d'émission
L'émission des fluides hydrothermaux peut ne pas conduire à la formation d'édifices, c'est le
cas par exemple dans le bassin de Guaymas ou au fond du lac Tanganyika. En plus des structures
classiques en cheminée, le fluide percole au travers de sédiments d'origine planctonique très riches
en matière organique et en sulfates. Ses sédiments sont le siège d'une forte activité sulfato-réductrice
se produisant à différentes profondeurs (donc à différentes températures) dans les sédiments ( pour
revue Elsgaard, 1995). Ces biotopes abritent une communauté bactérienne très riche qui se nourrit de
la matière organique présente en grandes quantités dans ces bassins.
Des suintements froids, ou à faible température, observés à proximité des marges passives
(Golfe de Californie, Golfe du Mexique), sont colonisés par une faune similaire à celle des sites
hydrothermaux profonds. La communauté microbienne est quant à elle essentiellement mésophile.
Récemment Hinrichs et al. (1999) ont analysé, par amplification et séquençage de l'ADNr 16S, la
communauté microbienne se développant dans les sédiments à proximité des suintements de
méthane émanant des réserves de méthane solidifié ("methane hydrates"). En plus de la flore
anaérobie de sulfato-réducteurs et de Gram +, les auteurs détectent des séquences associées aux
méthanogènes mésophiles (Methanomicrobiales et Methanosarcinales). L'analyse du contenu
lipidique de leur échantillon et le suivi du 13C leur permet de supposer que parmi les séquences
proches des méthanogènes certaines souches utiliseraient le méthane solidifié comme source de
carbone. La présence dans ces échantillons de sulfato-réducteurs laisserait supposer que ces
nouvelles souches réaliseraient une oxydation anaérobie du méthane en utilisant les sulfates comme
accepteurs d'électrons (Hinrichs et al., 1999; Summons, 1999).

Les habitats microbiens
76
4.4.2 Faune associée à l'écosystème hydrothermal
L'une des caractéristiques les plus remarquables des sources hydrothermales est la faune que
l'on peut y observer. Jusqu'en février 1977 (date des premières plongées réalisées par John Corliss et
John Edmond) les fonds sous-marins étaient considérés comme des espaces désertiques. Ces oasis
de vie que représentent ces nouveaux biotopes ont alors considérablement bouleversé la biologie,
remettant en question le dogme d'une vie impossible sans lumière (Corliss et Ballard, 1977; Corliss et
al., 1979).
Les premiers prélèvements réalisés sur des sites hydrothermaux entre les Iles Galápagos et
l'Equateur permirent de collecter les premiers spécimens de Calyptogena magnifica et de
Bathymodiolus thermophilus, deux mollusques se développant à proximité des fumeurs noirs. Par la
suite, de nombreux autres organismes ont été isolés de cet écosystème. Parmi ceux-ci les
vestimentifères : Riftia pachyptila et Tevnia jerichonana, les polychètes appartenant aux genres
Alvinella (pompejana et caudata) et Paralvinella. Les crevettes Rimicaris exoculata sont quant à elles
uniquement présentes sur les sites de la dorsale médio-Atlantique.
Cette faune est caractérisée par (d'après Laubier 1989; Tunnicliffe, 1991, 1992 cités dans
Cornec, 1995):
¬ Une dépendance vis à vis du système hydrothermal.
Une biomasse très importante répartie sur des surfaces limitées de l'ordre de 10 kg.m-2 (contre
quelques grammes par m2 dans les zones intra-plaques).
® Un fort taux d'endémisme : 97% de la faune observée sur ces sites est taxonomiquement distincte
de celle observée à des profondeurs équivalentes.
Un nombre important d'espèces nouvelles de grande taille appartenant à des taxons nouveaux (du
genre jusqu'à l'embranchement).
Mais l'une des caractéristiques les plus importantes de cette faune est son développement
inféodé aux symbiotes dont elle tire son énergie (voir plus loin) (Wirsen et al. 1993)
Les Riftia, les Alvinella et les
Rimicaris sont probablement les
organismes les plus surprenants de ce
biotope.
Les premiers (cf. figure 2.10)
sont de long vers pouvant atteindre 2
mètres de long pour 4-5 cm de diamètre.
Ils se développent en véritables
buissons pouvant atteindre une densité
de 100 à 200 individus au m2. Ces vers
dépourvus de système digestif tirent leur
énergie des producteurs primaires qu'ils
abritent dans leur trophosome, des
Figure 2.10. Colonie de Riftia pachyptila (photo site University of
Washington)

Les habitats microbiens
77
bactéries sulfo-oxydantes endosymbiotiques. Ces bactéries sont alimentées en H2S par
l'hémoglobine des Riftia qui possède la capacité de fixer aussi bien l'O2, le CO2 que l'H2S. Une fois
l'échange réalisé au niveau du trophosome l'hémoglobine transporte l'H+ à l'extérieur du ver.
L'hémoglobine de Riftia assure l'alimentation des symbiotes et la protection du vers vis à vis de l'H2S
hautement toxique en le transportant sous une forme stable (pour revue récente voir Zal, 1999).
Les seconds (cf. figure 2.11) sont des Alvinellidés
vivant dans des tubes construits en surface des
cheminées, mais ils peuvent aussi se déplacer librement
sur le site hydrothermal pour se nourrir de bactéries.
L'une des caractéristiques majeures de ces vers est la
présence d'une importante communauté épibiotique de
bactéries filamenteuses (voir plus loin) (pour revue
Desbruyères et al., 1998; et pour analyse moléculaire
Haddad et al., 1995; Cary et al. 1997). Si la majeure partie
de la faune se développant dans les sites hydrothermaux
se trouve dans des zones "froides" (20-30°C), les
Alvinellidés sont quant à eux fréquemment rencontrés à
proximité d'émissions à très haute température. A de
nombreuses reprises il a été observé des vers traversant le panache de fluide à la sortie d'un fumeur
où la température dépassait les 100°C (Chevaldonné et al., 1992). La température à l'intérieur des
tubes dans lesquels ces vers se réfugient correspond à un très fort gradient thermique de 60°C (entre
22°C à la sortie du tube et 81°C au fond du tube) (Cary et al., 1998).
La troisième communauté (cf. figure
2.12) très fréquemment rencontrée sur les
sites hydrothermaux de la MAR est
constituée par les crevettes du genre
Rimicaris. Rimicaris exoculata constitue une
biomasse très importante sur ces sites de 25
à 50000 individus.m -2 (Segonzac et al.,
1993; Van Dover, 1994). Elles ingèrent des
petites particules de soufre recouvertes de
bactéries et sont elles-mêmes recouvertes
de bactéries filamenteuses sur leur carapace
et autour de leur orifice buccal (Polz et
Cavanaugh, 1995).
Ces crevettes considérées auparavant
Figure 2.11. A. pompejana. Photo C. Cary. Taille 6 cm.
On observe à la surface du vers les filaments bactériens
qui dominent la communauté épibiotique.
Figure 2.12. Des essaims de Rimicaris exoculata broutant à la surface
d'une cheminée. On observe la tache blanche à la surface de la crevette
correspondant à l'oeil. Photo E. Gaten.

Les habitats microbiens
78
comme aveugles possèdent à la surface de leur céphalo-thorax un oeil fonctionnel dont le caractère
ancestral a été discuté (Gaten, 1998; Gaten et al. 1998a et b; Hering et al., 1999).
4.4.3 Les micro-organismes des sources hydrothermales
La découverte des sources hydrothermales, où la vie en l'absence de lumière s'était
développée, a incité de nombreux scientifiques à rechercher l'activité microbienne source de la
production primaire chimiolithotrophique. C'est en 1979 que Jannasch et Wirsen (entre autres) ont mis
en évidence cette présence pour la première fois en indiquant le rôle prépondérant dans cet
écosystème des bactéries sulfo-oxydantes sous une forme libre, associées symbiotiquement à la
macrofaune ou dans des consortiums microbiens (Jannasch et Wirsen, 1979). Par la suite, de
nombreuses études (comptages, techniques de culture, mesure de l'activité du métabolisme) ont
permis de mettre à jour une flore microbienne aux métabolismes et aux préférendums très diversifiés.
Si ces dernières années l'accent a été mis préférentiellement sur l'étude des micro-organismes
thermophiles, peu de choses sont en fait connues des micro-organismes mésophiles (Baross et
Deming, 1995). L'emploi récent des techniques moléculaires devrait nous permettre d'appréhender
l'écosystème hydrothermal dans sa globalité.
4.4.3.1 Habitats et types métaboliques microbiens
On considère actuellement que les communautés microbiennes s'établissent au sein des
écosystèmes hydrothermaux océaniques dans les habitats suivants (Karl, 1995) (cf. figure 2.13) :
¬ Les micro-organismes vivant sous une forme libre dans le fluide hydrothermal. Ces micro-
organismes se développeraient dans une biosphère souterraine sous-jacente aux cheminées et
seraient transportés par le fluide. Ce concept de biosphère souterraine (cf. § 3.2.2) expliquerait
l'ubiquité de certains micro-organismes dans des zones où la croûte terrestre profonde entrerait en
contact avec des milieux plus superficiels. C'est le cas par exemple des Thermoccoccales, des
Méthanogènes des Archaeoglobales retrouvées sur la quasi totalité des sites hydrothermaux
profonds et côtiers et au sein des réservoirs pétroliers (Prieur et al., 1995).
Les micro-organismes se développant à la surface des cheminées ou des roches proches des
émissions et qui seraient alimentés en nutriments par le fluide. On distingue les micro-organismes
se développant dans les cheminées (essentiellement thermophiles) des micro-organismes se
développant sous forme de tapis à proximité des cheminées (essentiellement mésophiles). La
présence de tapis bactériens très abondants est l'une des caractéristiques majeures de certains
sites hydrothermaux (Jannasch et Wirsen, 1981; Prieur, 1992). Ces enchevêtrements constituent
des zones frontières à la fois en contact avec des zones aérobies (l'eau de mer) et anaérobies (le
fluide). Le type de cheminée, sa porosité principalement, va influer sur la profondeur de
colonisation des micro-organismes. Afin d'étudier la communauté capable de se développer à la
surface des cheminées, différentes expériences ont été menées. Elles consistaient soit à prélever
directement le tapis microbien afin de réaliser des cultures d'enrichissement et des isolements, soit

Les habitats microbiens
79
à mettre à proximité du fluide des coupons de différentes matières (Téflon, acier, porcelaine,
métaux, plastiques) durant des durées d'expositions variables (Prieur et Ferra, résultats non
publiés dans Prieur, 1992; Guezennec et al., 1998)
® Les micro-organismes se développant en ecto- ou endo-symbiose avec les macro-organismes
colonisant les cheminées : vers polychètes, vestimentifères, mollusques. L'étude des symbiotes
associés aux mollusques des sources hydrothermales constitue la plus ancienne référence à
l'utilisation des techniques moléculaires pour accéder à la diversité microbienne d'un échantillon
(Stahl et al., 1984).
Les micro-organismes retrouvés dans les panaches ("plume") de fluides hydrothermaux. Ces
derniers correspondent à de l'eau de mer enrichie par les minéraux communément retrouvés dans
le fluide. Ces panaches peuvent atteindre des tailles considérables (700m d'épaisseur pour 20 km
de diamètre) (Winn et al., 1995).
° Enfin les micro-organismes se développant dans les sédiments où le fluide peut percoler comme
c'est le cas dans le bassin de Guaymas.
Oxydation/précipitation par mélange avec l'eau de mer froide environnante
FeS + O2
Mn + O2
Fe(O)OH
MnO2
Communauté Thermophiles/Mésophiles
Sous forme libre
SO réduit en S2-
4
2-Lessivage de Ca2+, Mn2+, Mg2+,Fe2+ et Cu2+
BASALTE
350°C
350°C
Eau de mer chaufféeenrichie en minéraux
350°C
Zone de mélange des eauxTempérature moyenne
20 à 40°C
Fort débitTempérature élevée
1-3 km
Percolation de l'eau de mer froide
Communauté MésophilesSous forme libre/
Sous forme de mats
Macr aune
Communauté bactériennemésophile symbionte
Communauté bactériennedans les panaches
100°C
200°C
300°C
400°C
50 à 100°C
Figure 2.13. Schéma de fonctionnement d'une source hydrothermale océanique
(d'après Jannasch et Mottl, 1985).
Si les bactéries sulfo-oxydantes et méthanotrophes semblent constituer le premier maillon de
la chaîne alimentaire de l'écosystème hydrothermal, la chimiolithoautotrophie ne constitue pas le seul
type métabolique et énergétique rencontré au sein des sources hydrothermales. De nombreux
auteurs ont proposé une liste des métabolismes bactériens observés ou potentiellement observables
à proximité des émissions de fluide (cf. tableau 2.4 d'après Prieur et al., 1987; Jannasch, 1995; Karl,
1995).
Les habitats microbiens
80
Tableau 2.4. Métabolismes microbiens observés d'isolats de sources hydrothermales profondes
Donneur Accepteur Source Température Type Processus
d'électron d'électron de carbone de réaction respiratoire métabolique
S2- O2 CO2 méso, hyper Aérobie Oxydation de sulfures
S° O2 CO2 méso, hyper Aérobie Oxydation du soufre
S2O32- O2 CO2 méso, hyper Aérobie Oxydation du thiosulfate
Fe2+ O2 CO2 méso Aérobie Oxydation du fer
Mn2+ O2 [CH2O] méso Aérobie Oxydation du manganèse
NH4+, NO2- O2 CO2 méso Aérobie Nitrification
CH4, CO O2 CH4, CO2, CO méso Aérobie Oxydation du méthane
et des composés en C1
[CH2O] O2 [CH2O] méso, hyper Aérobie Hétérotrophie aérobie
H2 O2 CO2 méso, hyper Aérobie Oxydation de l'hydrogène
H2 NO3- CO2 méso, hyper Anaérobie Dénitrification
H2 S° CO2 hyper Anaérobie Sulfo-réduction
H2 SO42- CO2 méso, hyper Anaérobie Sulfato-réduction
H2 CO2 CO2 méso, hyper Anaérobie Méthanogénèse
S2-, S°, S2O32- NO3
- CO2 méso Anaérobie Dénitrification, Sulfo-oxydation
[CH2O] S° [CH2O] hyper Anaérobie Hétérotrophie soufre réducteur
[CH2O] SO42- [CH2O] méso, hyper Anaérobie Hétérotrophie sulfatoréductrice
[CH2O] NO3- [CH2O] méso Anaérobie Dénitrification
[CH2O] [CH2O] [CH2O] méso, hyper Anaérobie Fermentation
méso : mésophile; hyper : hyperthermophile
4.4.3.2 Abondance des micro-organismes
Les comptages réalisés sur des échantillons provenant de différents sites donnent des
résultats très variables en fonction de la technique de comptage utilisée et du type d'échantillon
analysé (cf. tableau 2.5).
Tableau 2.5. Densité microbienne sur les sites hydrothermaux
Echantillon Température Densité Références (°C) $ (Cellules.ml-1); £ (Cellules.g-1);
& (Cellules.cm-2)
Fluide à basse température <50 5-10 106 $ Jannasch et Wirsen, 1979
<50 <2 106 $ Karl, 1987; Straube et al., 1990
<50 108-109 $ Corliss et al., 1979
Fluide à haute température 304 5 105 $ Baross et al., 1984
174-357 <3 105 $ Straube et al., 1990
Cheminée 65-80 2 105-5 107 £ Harmsen et al., 1997a
104-1010 £ Chevaldonné et Godfroy, 1997
Flange 20-100 et+ 106->108 Baross et Deming, 1995
Colonisation sur coupon 2 104-5 105 & (3 jours) Prieur et Ferra (com. perso.)
3 105-5 107 & (10 jours) " "
Colonisation sur Vent Cap 20 -80 4 108-2 109 $ (2-5 jours) Ce manuscrit (Article 3)
Sédiments 80-104 2 107-2 108 £ Hoaki et al., 1995
Tubes A. pompejana face externe 2-3 106 & Chevaldonné, 1989
Tubes A. pompejana face interne 0,4-2 106 & " "
Endobiote (R. pachyptila) 3 109 £ trophosome (humide)14 Cavanaugh et al., 1981
Ectobiote (R. exoculata) 8,5 106 (cellules/individu) Poltz et Cavanaugh, 1995
14 Le trophosome correspond à 40-60% du poids total du ver, ce qui fait des vers des animaux chimioautotrophes (Felbeck et
Childress, 1988)

Les habitats microbiens
81
Les trois méthodes principales de numération sont les comptages sur boites, la méthode par
dilution (NPP) et la microscopie. Les résultats obtenus par les deux premières méthodes sont
entachés des erreurs inhérentes à toute technique culturale. Les méthodes microscopiques sont
quant à elles entachées de l'erreur "humaine". Il est en effet difficile de distinguer au microscope
certaines cellules des granules de soufre qui présentent parfois une très forte autofluorescence. Je
reviendrai dans la troisième partie sur les techniques mises au point pour limiter les erreurs de
comptage en microscopie.
4.4.3.3 Les micro-organismes isolés de tapis microbiens et d'échantillons d'eau de
mer
Les micro-organismes mésophiles
Les premières observations réalisées sur les tapis microbiens du bassin de Guaymas ont
indiqué la présence de bactéries soufre-oxydantes se présentant sous la forme de longs filaments et
associées aux genres Beggiatoa et Thiothrix (Jannasch et Wirsen, 1981; Jannasch et al., 1989). Ces
micro-organismes autotrophes oxydent l'H2S en utilisant les nitrates accumulés dans une large
vacuole. Ces enchevêtrements peuvent atteindre des épaisseurs de 10 ou 20 cm et sont aussi
observables à proximité des suintements froids (Ahmad et al., 1999; Nealson, 1999). Une souche
sulfo-oxydante, autotrophe associée au genre Arcobacter a été retrouvée dans les sédiments chauds
du bassin de Guaymas. Cette souche préalablement observée dans des sédiments côtiers produit
d'importants filaments de soufre (de 0,5 à 5 µm de diamètre pour 20 à 500 µm de long) qui pourraient
servir de facteur d'initiation pour la colonisation des Alvinella (Taylor et Wirsen, 1997; Taylor et al.,
1999).
La plupart des isolats obtenus à partir d'échantillons de fluides ou de surfaces exposées aux
fluides correspondent à des bactéries soufre-oxydantes autotrophes ou mixotrophes associées aux
genres Thiomicrospira, Thiobacillus (Durand et al., 1993; Jannasch et al., 1985; Ruby et al., 1981). Des
études récentes effectuées sur un isolat de Thiomicrospira prélevé sur la dorsale médio-Atlantique,
indiquent que ce dernier correspondrait à une Thiomicrospira crunogena préalablement isolée dans le
Pacifique (Wirsen et al., 1998). Cette espèce semble donc présenter une répartition ubiquitaire. Mais
un grand nombre d'autres bactéries soufre-oxydantes hétérotrophes ou mixotrophes ont été isolées
de milieux hydrothermaux. Elles sont associées aux genres Pseudomonas, Acinetobacter et Vibrio
(Durand et al., 1994). Des bactéries sulfato-réductrices associées au genre Desulfovibrio ont été
isolées à partir d'échantillons provenant de la Ride Est-Pacifique (Elsgaard et al., 1991). Jannasch dans
sa revue (1995) rapporte l'isolement de bactéries oxydant le méthane du genre Methylococus et de
bactéries oxydant l'hydrogène du type des Hydrogenomonas. Des bactéries mangano-oxydantes et
des bactéries résistantes à de fortes concentrations en métaux lourds ont été isolées de coupons
colonisés et d'échantillons de moules et d'Alvinellidés (Ehrlich et al., 1983; Durand et al., 1990;
Jeanthon et Prieur, 1990). Elles sont associées aux genres Pseudomonas, Aeromonas et Vibrio. La
recherche de bactéries productrices d'exopolysaccharides à partir de divers échantillons

Les habitats microbiens
82
hydrothermaux a conduit à l'isolement de souches apparentées aux Vibrio et aux Alteromonas
(Vincent, 1993; Raguenes et al., 1997 a et b). Enfin des bactéries nitrifiantes, hétérotrophes, aérobies
ont été isolées à partir d'échantillons de cheminées actives sur le site 13°N, EPR et dans le bassin de
Guaymas (Mevel et al., 1996). Ces bactéries correspondent majoritairement à des Pseudomonas et
des Alcaligenes . Récemment une souche aérobie anoxygénique photosynthétique a été isolée à
2000m de profondeur dans un panache hydrothermal (Yurkov et Beatty, 1998). Cette présence
atypique (à de telles profondeurs) mais significative (près de 10% des enrichissements) n'est pas
expliquée par les auteurs.
Les micro-organismes mésophiles associés à la macrofaune
La majeure partie de la faune observée au niveau des sites hydrothermaux présente de fortes
associations avec des micro-organismes sulfo-oxydants15. Ces symbioses correspondent à des
associations hôtes-bactéries très spécifiques qui ont été étudiées dans le cas des vestimentifères
Riftia pachyptila, des mollusques bivalves Bathymodiolus thermophilus et Calyptogena magnifica, des
gastéropodes Alviniconcha hessleri et Neomphalus fretterae, des polychètes annélidés Alvinella
pompejana et A. caudata, des crustacés Rimicaris exoculata et R. chacei (Prieur, 1992; Nealson et
Fisher, 1995). L'ensemble de ces études souffre de l'impossibilité d'isoler les symbiotes de leur hôte.
Seules des techniques de mesure d'incorporation d'isotopes stables de carbone et d'azote, des
recherches d'enzymes impliquées dans le cycle de Calvin et des observations des tissus par
microscopie électronique à balayage et à transmission avaient permis, avant l'emploi des techniques
de séquençage et de marquage moléculaire, d'établir les relations trophiques s'établissant entre l'hôte
et les bactéries (Prieur, 1992). Si les relations nutritionnelles entre les ectobiotes d'A. pompejana et de
R. exoculata ne sont pas encore complètement établies16 (Poltz et Cavanaugh, 1995; Cary et al.,
1997; Desbruyères et al., 1998), celles entre les endobiotes et les mollusques ou les vestimentifères
des sources hydrothermales ont été plus étudiées et l'emploi des techniques moléculaires a permis
d'accroître considérablement le champ des connaissances sur les mécanismes relationnels hôtes-
symbiotes (Nealson, 1999; cf. § 4.4.3.4).
Les micro-organisme s thermophiles
Définition de la thermophilie
La température est l'un des facteurs qui conditionnent la vie sur terre17. On peut alors diviser le
monde vivant en trois groupes principaux, en fonction des températures optimales de croissance : les
psychrophiles se développant à de faibles températures, les mésophiles se développant à des
températures moyennes et les thermophiles se développant à des températures élevées.
15 Il a été aussi observé des symbioses avec des micro-organismes méthanotrophes chez Bathymodiolus (Cavanaugh et al., 1992;
Distel et al., 1995; Nealson et Fisher, 1995).16 Une étude a été menée par Poltz et al. (1998) afin d'établir (par assimilation de carbone marqué, techniques moléculaires de
marquage, etc.) les relations que R. exoculata entretenait avec ses ectobiotes.17 La température influe sur la structure des molécules (notamment des protéines et de l'ADN) et sur la disponibilité de l'eau par
exemple.

Les habitats microbiens
83
Les Archaea et les Bacteria thermophiles peuvent être divisées en trois sous-groupes (cf. figure 2.14):
Thermophiles modéres
Thermophiles extrêmes
Hyperthermophiles
Vit
esse
de
crois
sanc
e
Température (°C)
Frontière de la thermophilie (50 à 60°C)
Figure 2.14. Répartition des micro-organismes thermophiles (d'après Brock,
1986)
Ø les thermophiles modérés, dont
la température maximale de croissance
(Tmax) ne dépasse pas 70°C,
Ø les thermophiles extrêmes ,
capables de se développer au delà de
85°C,
Ø les hyperthermophiles, dont la
température optimale de croissance
(Topt) est supérieure à 90°C.
La notion de frontière de thermophilie, d'organismes mésophiles et thermophiles modérés ne
fait pas l'unanimité. On peut toutefois définir pour chaque organisme trois points cardinaux : la
température minimale, Tmin, à partir de laquelle la croissance est possible; le taux de croissance
augmente ensuite presque linéairement jusqu'à la Topt, puis généralement chute brutalement dans les
5 à 10°C qui suivent jusqu'à Tmax18.
Isolement des micro-organismes thermophiles
Les fortes températures relevées au sein des sites hydrothermaux profonds ont conduit à
rechercher des micro-organismes thermophiles et hyperthermophiles. Le tableau 2.6 présente les
principaux genres thermophiles isolés au sein des sources hydrothermales profondes.Tableau 2.6. Micro-organismes thermophiles isolés de sources hydrothermales profondes (d'après la revue de Stetter, 1996).
Domaine Genres Optimum Métabolisme Références
(°C) énergétique
BACTERIA Thermus 60-70 Hétérotrophe, aérobie Marteinsson et al., 1995
Bacillus 60-80 Hétérotrophe, aérobie Marteinsson et al., 1996
Thermotoga 80 Fermentaire, anaérobie Marteinsson et al., 1997
Thermosipho 70 Fermentaire, anaérobie Antoine et al., 1997
Desulfurobacterium 70 Autotrophe, anaérobie L'Haridon et al., 1998a et b
ARCHAEA
Euryarchaeota Thermococcus 80-90 Fermentaire, anaérobie Kobayashi et al., 1994
Pyrococcus 95-100 Fermentaire, anaérobie Erauso et al., 1993
"Palaeococcus" Fermentaire, anaérobie Takai et al., 1998 (resul. non pub.)
Archeaoglobus 83 Mixotrophe, anaérobie Burggraf et al., 1990
Methanococcus 85 Methanogène, anaérobie Jones et al., 1983;*
Methanopyrus 98 Methanogène, anaérobie Kurr et al., 1991
Crenarchaeota Staphylothermus 92 Hétérotrophe, anaérobie Fiala et al., 1986
Pyrodictium 97 Hétérotrophe, anaérobie Pley et al., 1991
Pyrolobus Autotrophe, aérobie Blöchl et al., 1997
Desulfurococcus 85-90 Fermentaire, anaérobie Jannasch et al., 1988
* Jeanthon et al., 1998, 1999 a et b
18 D'une manière analogue aux courbes d'efficacité enzymatique, les courbes de croissance des micro-organismes en fonction de la
température sont les résultantes de la combinaison de la loi d'Arrhenius jusqu'à la Topt et de la dénaturation du micro-organisme jusqu'à
la Tmax.

Les habitats microbiens
84
La plupart de ces micro-organismes sont des anaérobies stricts (hétérotrophes fermentaires,
méthanogènes ou sulfato-réducteurs) appartenant au domaine des Archaea . Mais il a été possible
d'isoler certaines Bacteria thermophiles (§ tableau 2.6). Il s'agit généralement de souches appartenant
à des genres déjà décrits au niveau des sources côtières ou terrestres. Ce phénomène pourrait être
expliqué soit par l'ubiquité de ces souches au sein des différents points chauds du globe, soit par un
biais lié aux techniques de cultures toutes issues d'études des environnements côtiers et ne tenant
pas compte de l'un des principaux paramètres: la pression (Prieur, 1997).
Les applications industrielles des souches thermophiles
Ces dix dernières années les études portant sur les Archaea et les Bacteria ont amené les
chercheurs à s'intéresser aux propriétés de ces micro-organismes pouvant se développer dans des
conditions extrêmes de pH, de température et de salinité. Leur capacité à produire des enzymes très
stables a très rapidement été étudiée par les industriels. Toutefois si certains de ces organismes
étaient déjà employés depuis longtemps (Archaea méthanogènes) d'autres, bien qu'offrant des
perspectives fort intéressantes, entraient en concurrence avec des bactéries déjà sur le marché et
beaucoup plus simples à produire en grande quantité. D'autre part les conditions optimales de
croissance (température, pression, anaérobiose) rendent complexe la mise en culture de telles
souches(Cowan, 1992).
Les Archaea thermophiles ont surtout été pressenties par les industriels pour les
caractéristiques de leurs enzymes. Les avantages apportés par l'utilisation d'enzymes thermophiles et
thermostables par rapport aux enzymes classiques sont :
Ø un gain de temps (pas de refroidissement) Ø une diminution des coûts de refroidissement
Ø l'amélioration de la stérilité Ø la possibilité d'extraction de composés volatils
Ø la réduction de la viscosité
La thermostabilité d'une protéine implique le plus souvent une meilleure résistance aux agents
dénaturants et à la protéolyse. Il faut garder à l'esprit que des enzymes thermostables ne réagiront pas
plus rapidement que des enzymes classiques. La vitesse de réaction d'une enzyme ayant son
optimum de température à 90°C ne sera pas 32 fois supérieure à celle de l'enzyme ayant une Topt de
37°C (la loi d'Arrhenius prévoit en effet un doublement de la vitesse de réaction pour une
augmentation de 10°C). Ceci est dû à des relations structure-fonction qui accroissent certes la stabilité
mais réduisent la flexibilité de l'enzyme aux faibles températures (Cowan, 1992). On peut augmenter la
vitesse d'une réaction en utilisant une enzyme thermostable, lorsqu'à des températures élevées le
substrat est plus susceptible d'être catalysé. C'est le cas des mécanismes de dégradation des
polymères protéiques et glucidiques.
Le tableau 2.7 donne un aperçu des applications actuelles des enzymes extraites des micro-
organismes extrêmophiles.

Les habitats microbiens
85
Tableau 2.7. Applications biotechnologiques des extrêmophiles
Organismes extrêmophiles Enzymes / Composé organique Applications Produits
Thermophiles 50 - 110°C Amylases Glucose,Fructose , Edulcorants
Xylanases Blanchiment du papier
Protéases Acides aminés produits de la kératine, détergents,
agro-alimentaire : cuisson, brasserie.
ADN-polymérases, Intéines Biologie moléculaire
Psychrophiles 0 - 20°C Protéases neutres Production laitière et fromagère
Protéases, Amylases, Lipases Additifs de dégradation dans les détergents
Acides gras polyinsaturés Industrie pharmaceutique
Déshydrogénases Biocapteurs
Acidophiles pH < 2 Oxydation des sulfures Désulfuration des minerais et du charbon
Alcaliphiles pH > 9 Cellulases, Protéases Additifs de dégradation dans les détergents
Amylases, Lipases
Cyclodextrines Stabilisation de substances volatiles
Antibiotiques Industrie pharmaceutique
Halophiles 3 - 20% sel Carotène Colorants alimentaires
Glycérol et additifs solubles Industrie pharmaceutique
Membranes Surfactants pour l'industrie pharmaceutique
Barophiles Amylase, Protéases Agro-alimentaire, Industrie pétrolière
(d'après Querellou et al., 1999; site programme européen (voir note bas de page n°19))
L'un des domaines d'application courant de telles enzymes est celui de la biologie moléculaire.
La PCR a ouvert le champ à l'utilisation de nombreuses enzymes thermostables. Dans un premier
temps la Taq polymérase extraite de la Bacteria Thermus aquaticus a remplacé le fragment de Klenow
qui exigeait des manipulations longues et était thermosensible, puis de nouvelles polymérases plus
fidèles ont été extraites d'organismes hyperthermophiles (voir pour revue récente Quérellou, 1999).
D'autres domaines d'applications de la biologie moléculaire peuvent être envisagés:
Applications ind ustrielles (König, 1989; Cowan, 1992; Querellou et al., 1999) : 75% des enzymes
utilisées dans le monde le sont dans les industries des détergents, de l'amidon et du lait. Si des
enzymes thermostables pouvaient logiquement intervenir dans de telles industries, il faudrait toutefois
que cette substitution aux enzymes classiques soit commercialement intéressante. Il faut rappeler
notamment que :
Ê Certains procédés industriels ne sont pas compatibles avec des températures élevées (ex :
industrie fromagère).
Ë Certains procédés fonctionnent de préférence à faible température (ex : détergents).
Ì Des enzymes suffisamment thermostables sont déjà utilisées (ex : saccharification de l'amidon).
Í Parfois il n'y a pas d'intérêt à augmenter la température de réaction (ex : certaines réactions de
chimie fine sur les acides aminés ou les pénicillines).
Î L'augmentation de productivité à haute température peut ne pas couvrir l'augmentation des
coûts de chauffage.
Par conséquent l'utilisation d'une enzyme thermostable doit être dictée par les règles suivantes :
À L'enzyme doit être disponible dans des quantités équivalentes à une enzyme classique.

Les habitats microbiens
86
Á Son prix par unité d'activité doit être plus faible.
 Ses performances doivent être significativement supérieures à l'enzyme classique.
à Son utilisation ne doit pas entraîner de modifications majeures de l'équipement existant.
Il s'avère donc difficile de remplacer par des enzymes extraites d'Archaea thermophiles les
enzymes déjà présentes sur le marché. L'emploi d'enzymes thermostables devra donc plutôt
s'orienter vers de nouvelles niches d'utilisation (cf. tableau 2.8).Tableau 2.8. Domaines d'applications des enzymes issues de micro-organismes thermophiles
Enzymes Applications
A.D.N. ligases recherche d'enzymes plus thermostables, plus processives et plus fidèles que la Taq polymérase
classique.
Alcool-déshydrogénases des alcools secondaires DH thermostables pourraient avoir de nombreuses applications en
chimie pour la production de molécules chirales. Leur résistance aux produits de réaction (alcool,
aldéhydes et cétones) rendent les DH de Sulfolobus solfataricus et de Hyperthermus butylicus
intéressantes.
Estérases permettant des réactions en milieu non liquide.
Enzymologie en phase gazeuse à l'aide de l'alcool DH de S. Solfataricus. Cette technique permettrait de minimiser l'emploi de
solvant.
Biolixiviation jusqu'à présent des Bacteria mésophiles étaient utilisées à très grande échelle (500 tonnes par an)
pour le traitement du minerai d'uranium. L'emploi d'Archaea sulfo- et métalo-oxydantes
thermophiles se limitait à la récupération de métaux précieux. Les Archaea acidophiles
Sulfolobus, Acidianus et Metallosphaera pourraient être d'un usage intéressant dans le cas de bio-
lixiviation thermodépendante. Le problème réside dans la sensibilité parfois trop forte aux métaux
de ces bactéries dans les réacteurs.
Actuellement le marché des enzymes et des autres composés issus de micro-organismes
extrêmophiles correspondrait à 17 milliards de dollars. Les programmes européens du type
"Extrêmophiles, usines cellulaires"19 ainsi que le séquençage de génomes totaux de micro-
organismes hyperthermophiles20 témoignent de l'intérêt grandissant des industriels pour ces
souches. Toutefois une exploitation à grande échelle de ces enzymes tarde encore à venir.
4.4.3.4 L'apport des techniques moléculaires
Les environnements hydrothermaux ont constitué l'un des premiers champs d'essai des
techniques moléculaires pour accéder à la diversité microbienne en s'affranchissant de toute étape de
culture (Stahl et al., 1984; Lane et al., 1985a). Les premiers échantillons analysés provenaient des
symbiotes de R. pachyptila, de C. magnifica et de Soleyama velum. Ces études confirment par analyse
des ARNr 5S l'affiliation de ces symbiotes avec les bactéries soufre-oxydantes correspondant aux
Proteobacteria γ (appelées alors bactéries pourpres). L'engouement suscité par l'emploi de telles
méthodes a conduit à l'analyse d'échantillons (coupons colonisés) provenant de sites hydrothermaux
terrestres (Octopus Spring dans le Parc de Yellowstone, Stahl et al., 1985) et à la publication de
nombreuses revues dans les années qui ont suivi (Pace et al., 1985, 1986 a et b; Olsen et al., 1986;
Turner et al., 1989). Par la suite les limites de détermination des souches liées à la petite taille du 5S
19 Extremophiles as cell factories http://www.tu-harburg.de/tv/ecf/ecf0_1.htm.20 Sur 17 génomes microbiens séquencés , 5 correspondent à des micro-organismes hyperthermophiles.

Les habitats microbiens
87
entraînèrent les scientifiques vers l'analyse des gènes codants pour l'ARNr 16S (1500 nucléotides
contre 120 pour le 5S). Après la mise au point d'une technique rapide et efficace de séquençage
(Lane et al., 1985b), les premières études de diversité microbienne fondées sur l'analyse des gènes
codants pour l'ARNr 16S ne sont appliquées qu'en 1990 en milieu hydrothermal continental (Ward et
al., 1990 a et b) et en 1994 et 1995 par Moyer et al. en milieu profond.
Jeanthon (1999) propose une revue très complète sur l'emploi des techniques moléculaires
en milieu hydrothermal profond. Ces dernières ont permis d'étudier à la fois les bactéries associées à la
macro-faune comme celles présentes sous une forme libre;
ce sont :
Ê les travaux de Poltz et Cavanaugh sur les ectobiotes de R. exoculata (Poltz et Cavanaugh, 1995),
Ë les travaux de l'équipe de Cary sur les symbiotes d'A. pompejana (Haddad et al., 1995; Cary et al.,
1997),
Ì les travaux de Distel sur les symbiotes de Riftia par étude du gène de l'ADNr 16S (Distel et al.,
1988, 1994),
Í les travaux de l'équipe de Vrijenhoek sur la co-évolution des symbiotes et de leur hôte chez
Calyptogena (Peek et al., 1998)et chez Riftia (Feldman et al., 1997),
Î les travaux de Moyer sur des tapis microbiens d'un site hydrothermal du Pacifique (Moyer et al.,
1994, 1995, 1998)
Ï les travaux de Harmsen sur la quantification à l'aide de sondes nucléiques des micro-organismes
de cheminées hydrothermales d'un site Atlantique (Harmsen et al., 1997 a et b)
Ð les travaux de Riley sur les communautés méthanogènes des cheminées (Riley, 1997, 1999)
Ñ les travaux de Takai sur des sources côtières et profondes au large du Japon (Takai et Horikoshi,
1999; Takai et Sako, 1999),
Des travaux Ê et Ë on retiendra la prédominance dans les banques d'ADNr 16S de phylotypes
bactériens associés aux Proteobacteria ε parmi les épibiotes de R. exoculata et A. pompejana. Ces
épibiotes correspondent dans les deux cas à de longs filaments situés sur l'épiderme dorsal des
annélides et sur des excroissances buccales des crustacés. Cette prédominance a été confirmée par
des expériences d'hybridation in situ ou de Dot-blot sur des échantillons d'animaux21. De plus il
semblerait que l'association bactérie-hôte ne soit pas exclusive. En effet il a été détecté dans
l'environnement proche des animaux (tubes externes des annélides, surfaces des roches) des
phylotypes similaires à ceux observés à la surface des animaux (jusque 60% des bactéries détectées à
la surfaces des roches dans le cas du symbiote de R. exoculata). Les épibiotes de R. exoculata
pourraient de plus correspondre à des bactéries pléomorphes comme l'indiquent les expériences
d'hybridation menées sur 3 morphotypes différents à l'aide d'une sonde spécifique du symbiote.
Les travaux Ì et Í confirmèrent les premières études portant sur l'ADNr 5S, sur le
positionnement phylogénétique des symbiotes de Riftia. Par contre il a été possible de définir deux
21 La présence de séquences associées à d'autres Proteobacteria ε a été aussi détectée dans le trophosome de vestimentifères
proches de suintements froids (Naganuma et al., 1997)

Les habitats microbiens
88
modes d'acquisition probables du symbiote pour les mollusques et les vestimentifères. Alors que les
symbiotes de Riftia n'ont jamais été détectés dans les oeufs ou les tissus juvéniles des vers et que le
même symbiote est retrouvé sur des individus d'origines géographiques distinctes ou des individus
de genre proche (Ridgea piscesae) (laissant supposer un mode d'acquisition horizontal du symbiote),
les symbiotes thioautotrophes de Calyptogena étaient quant à eux retrouvés dans les oocytes du
mollusque (mode d'acquisition vertical). La forme libre du symbiote de Riftia n'a jamais été détectée
dans l'environnement proche des vers, mais le gène codant pour une flagelline a été détecté dans
l'endosymbiote (Millikan et al., 1999). La forme libre du symbiote pourrait donc être mobile.
Les travaux Î effectués sur des échantillons de tapis microbien confirment eux aussi la
prédominance dans les banques d'ADNr 16S de phylotypes bactériens associés aux Proteobacteria ε
(différents des précédents). Ces phylotypes ne correspondent à aucun micro-organisme isolé de
source hydrothermale. La séquence bactérienne la plus proche est celle de Thiomicrospira
denitrificans22 une bactérie sulfo-oxydante chimiolithoautotrophe pouvant dénitrifier et isolée de
sédiments marins côtiers. Ces micro-organismes pourraient revêtir un rôle dans le cycle du soufre au
sein des écosystèmes hydrothermaux. Le second phylotype le plus abondant correspond à une
Proteobacteria γ proche de Xanthomonas sp. récemment isolée d'eau douce, oxydant l'ion Fe2+
(Emmerson et Moyer, 1997). En ce qui concerne les séquences archaéennes, Moyer (1998) détecte
2 groupes principaux de séquences répartis parmi les Archaea marines du groupe I (Crenarchaeota) et
les Archaea marines du groupe II (Euryarchaeota). Ces dernières ont été aussi détectées dans les
échantillons provenant de la dorsale médio-Atlantique (Reysenbach et al., 1999, résultats non
publiés).
Sur des échantillons de cheminée provenant du même site, Harmsen (Ï) détecte une forte
proportion (40%) de bactéries hybridant à la fois avec la sonde bactérienne et avec la sonde ciblant les
Aquificales. Ces micro-organismes s'avéreront correspondre aux Desulfobacteriaceae isolées des
mêmes échantillons (L'Haridon et al., 1998). Malgré l'isolement de Thermus, de Bacillus et de
Thermotoga de ces échantillons, seul ce dernier genre a pu être détecté par hybridation in situ . Ces
études confirment de plus que la colonisation microbienne est plus abondante à l'extérieur qu'à
l'intérieur des cheminées (avec de plus en plus de Bacteria par rapport aux Archaea à mesure que l'on
se rapproche du centre de la cheminée).
L'un des problèmes majeurs lors des études moléculaires d'échantillons hydrothermaux est rencontré
durant l'extraction d'ADN 23.
Riley (Ð) a probablement solutionné ce problème en extrayant directement l'ADN à bord du
bateau à l'aide d'un Ribolyser (Hybaid). Il lui a ainsi été possible d'obtenir de l'ADN génomique
amplifiable à l'aide d'amorces ribosomales. Il n'a toutefois pas pu détecter de méthanogènes comme il
l'aurait désiré.
22 Le genre Thiomicrospira est mal attribué car il ne s'agit pas ici d'une Proteobacteria γ comme les autres Thiomicrospira isolées
d'environnements hydrothermaux (Muyzer et al., 1995)23 Ce problème a été reporté d'ailleurs sur des échantillons hydrothermaux par Reysenbach et al., 1998.

Les habitats microbiens
89
Takai (Ñ) est quant à lui parvenu à extraire de l'ADN procaryotique d'échantillons de fumeurs
profonds. Une exploitation rapide des échantillons ainsi qu'une technique efficace de broyage et de
filtration des échantillons lui a permis de disposer de suffisamment d'ADN pour réaliser des
amplifications et des clonages. Il a observé une très forte diversité de séquences archaéennes se
répartissant parmi des phylums déjà détectés dans les sources profondes (Archaeoglobales,
Methanoccoccales) mais aussi parmi de nombreux nouveaux phylums dont certains sont associés aux
Euryarchaeota, aux Crenarchaeota et aux Korarchaeota. De manière très surprenante aucune
Thermoccoccales n'a été détectée durant ces études.
Les conditions extrêmes de température, de pression ainsi que les forts gradients physico-
chimiques observés dans des écosystèmes hydrothermaux profonds font de ces sites des candidats
idéaux pour les études moléculaires de la diversité. Jusqu'à présent les résultats obtenus laissent
augurer une diversité très abondante qui ne dément pas le qualificatif "d'oasis" qui avait été attribué
aux sources il y a 20 ans.
4.5 Les sites hydrothermaux continentaux et côtiers
4.5.1 Description des biotopes
4.5.1.1 Les sites terrestres
D'une manière analogue aux systèmes profonds, les systèmes continentaux et côtiers sont
liés à la rencontre d'eau d'infiltration avec le magma profond. Le parcours de cette eau dans la croûte
terrestre va conditionner sa composition chimique et sa forme d'émergence (fumerolles, geysers,
mares de boues acides ou basiques).
S'il s'infiltre une faible quantité d'eau, celle ci va s'échapper sous la forme de fumerolles comme c'est le
cas à Pozzuoli en Italie. Mais souvent ces émissions sont associées à une activité hydrothermale plus
diversifiée. Ainsi, aux fumerolles placées en haut des collines sont souvent associés des geysers et
des sources chaudes situées quant à elles dans la partie basse du bassin.
Les fumerolles et les geysers
(cf. figure 2.15) correspondent à
l'émission d'un fluide très minéralisé
qui a traversé profondément la croûte
terrestre et qui ressort violemment à
100°C dans le cas des geysers ou qui
recircule sur de longues distances
dans la lithosphère pour ressortir sous
formes de vapeurs (fumerolles).
Il existe deux formes de sources chaudes , en fonction du type de
Photos ParkVision.
Figure 2.15. Old Faithfull Geyser (à droite) et
fumerolles (en haut) au parc de Yellowstone (USA).

Les habitats microbiens
90
minéraux dont se sera chargé le fluide en traversant la lithosphère. S'il est chargé en acide sulfurique le
pH sera de 1 à 2,5. S'il est par contre chargé en carbonate ou en bicarbonate il sera de l'ordre de 6,3 à
10,2.
Dans les sources acides (cf. figure 2.16), on relève
une forte concentration en soufre et en fer mais une très
faible minéralisation (l'eau n'a pas pénétré profondément
dans le sol). Les gaz volcaniques qui s'échappent de la
source sont chargés en H2, CO2, H2, H2S, CO et NH3. En
arrivant au contact de l'air les sulfures sont oxydés en S°.
Cette oxydation est poursuivie par les bactéries soufre-
oxydantes qui produisent alors de l'acide sulfurique
H2SO4. Il peut se former de manière concomitante des
mares de boues acides. Elles seront très acides en
surface (pH 0,5 à 4) et anoxiques et plus basiques en profondeur. Elles ne constituent pas par contre
un biotope stable car les eaux de pluie modifient très régulièrement leur pH.
Les sources alcalines correspondent à des eaux
ayant circulé en profondeur dans la croûte terrestre (cf.
figure 2.17). Très chargées en minéraux qui vont se
solidifier au contact de l'air, elles constituent un biotope
très stable car les eaux de profondeur les alimentent
régulièrement et tamponnent les effets liés aux eaux de
pluie. Ces écosystèmes sont caractérisés par la
présence d'amas bactériens très stratifiés. Les sources
du parc de Yellowstone ont fait l'objet d'études très
poussées et de suivis réguliers dont des synthèses ont
été proposées récemment par D. Ward et al. (1998) et
par Reysenbach (en préparation).
4.5.1.2 Les sites côtiers
On observe des sites hydrothermaux à faible profondeur
sur les côtes de l'île Vulcano en Italie et à proximité des côtes
Islandaises (cf. figure 2.18). Comme dans les sites profonds on peut
retrouver des petites cheminées émettant un fluide chargé en H2S
mais on peut aussi observer le fluide qui percole parmi les
sédiments ou le sable. Les températures sont alors proches de
100°C mais ne peuvent atteindre de très fortes valeurs du fait de la
faible pression hydrostatique.Figure 2.18. Emissions côtières sur le site de Vulcano.
Source site web de la municipalité de Vulcano.
Il existe aussi un hydrothermalisme lacustre. Certains lacs situés sur les failles actives sont le
Figure 2.16. Bassin de boues acides dans le Upper
Geyser Bassin. Photos ParkVision. Yellowstone (USA)
Figure 2.17. Fountain Paint Pot Photo Park Vision.
Yellowstone (USA)

Les habitats microbiens
91
siège de phénomènes d'hydrothermalisme. Le fluide hydrothermal peut percoler aux travers de
sédiments à faible température ou sortir à des températures plus élevées en édifiant de petites
cheminées.
4.5.2 Les micro-organismes des sites côtiers et continentaux
Les sites hydrothermaux côtiers et terrestres ont fait l'objet de nombreuses études. En effet
leur facilité d'accès (par rapport aux sites océaniques profonds)24 permet de réaliser des suivis et des
collectes d'échantillons d'une manière beaucoup plus simple donc beaucoup plus fréquente. Ceci
explique en partie la grande diversité de souches isolées aussi bien de milieux marins que terrestres.
De plus les isolements réalisés en surface ou proches de la surface permettent d'accéder à une
communauté aérobie ou micro-aérophile sûrement moins abondante dans les gisements profonds. Il
ne semble pas qu'il y ait des genres particulièrement inféodés aux milieux terrestres et d'autres aux
milieux marins. En effet, de nombreuses souches ont été retrouvées de manière ubiquitaire dans des
sources côtières après avoir été décrites dans des sources continentales. Seules certaines
sulfothermophiles acidophiles n'ont pu être isolées de milieu marin.
4.5.2.1 Les souches mésophiles marines et continentales
Les souches mésophiles observées dans les écosystèmes marins côtiers sont les mêmes que
celles observées en milieux profonds. On retrouve notamment des bactéries sulfo-oxydantes
associées aux genres Beggiatoa et Thioplaca (Hoaki et al., 1994, 1995) et de nombreuses
hétérotrophes (Gugliandolo et Maugeri, 1998). Dans certains sédiments on retrouve la communauté
microbienne associée aux sédiments froids (BSR, méthanogènes, dénitrifiantes, voir § 4.3.1).
En milieu terrestre le facteur de dilution observé en milieu marin n'a pas cours. Les
écosystèmes sont beaucoup plus stables et sont aussi beaucoup moins étendus. Les écosystèmes
terrestres chauds ne vont donc être colonisés que par de pures thermophiles (modérées à
hyperthermophiles).
4.5.2.2 Les souches thermophiles marines et continentales.
Les écosystèmes terrestres tels que ceux observés au Parc de Yellowstone aux USA
constituent les premiers milieux d'étude des micro-organismes thermophiles. En 1964 Thomas Brock
se rend pour la première fois au Parc de Yellowstone afin d'isoler des représentants du genre Thiotrix.
Les relevés de température qu'il réalise l'incitent à rechercher des micro-organismes capables de se
développer aux températures observées in situ. En 1965 il observe et tente de cultiver de longs
filaments roses, particulièrement abondants à la sortie du fluide et dans un bassin qui deviendra par la
suite un bassin de référence pour les études microbiologiques : l'Octopus Spring25 (Brock, 1995). Si la
24 Les sites côtiers ne nécessitent pas de submersibles et sont souvent accessibles par de simples plongeurs.25 Cette source fait la couverture de la 8éme édition de "Biology of Microorganisms" de T. Brock.

Les habitats microbiens
92
position phylogénétique de ces filaments a été définie en 1994 (Reysenbach et al., 1994) ces
derniers vont rester incultivables jusqu'aux travaux de l'équipe de Stetter (Huber et al., 1998b) qui va
mettre au point une chambre de culture recréant les conditions de croissance in situ , notamment en
recréant les surfaces permettant la fixation des filaments exposés à un flux chaud de nutriments, et va
utiliser des pinces optiques pour isoler spécifiquement les micro-organismes qui seront remis en
culture par la suite.
Mais l'une des découvertes majeures réalisée à Yellowstone est l'isolement en 1966 de
Thermus aquaticus qui deviendra en 1980 la source de l'enzyme clé de la biologie moléculaire la Taq
polymérase (Brock et Freeze, 1969 puis Saiki et al., 1985, 1989).
Le tableau 2.9 présente l'ensemble des bactéries thermophiles isolées de sites terrestres et côtiers.
Tableau 2.9. Bacteria isolées de sites hydrothermaux terrestres et côtiers
Domaine Genres Optimum Origine Type Références
BACTERIA (°C) Respiratoire
Acidothermus sp. 55 t aérobie Mohagheghi et al., 1986
Aquifex sp. 85 m microaérobie Huber et al. , 1992
Alicyclobacillus (=Bacillus) 60-80 t/m aérobie (Darland et Brock 1971) Wisotzkey et al., 1992.
Calderobacterium 75 t aérobie Kryukov et al. , 1983
Chloroflexus 50 t aérobie Pierson et Castenholz,1974
Fervidobacterium 70 t anaérobie Patel et al., 1985
Hydrogenobacter 70-75 t aérobie Kawasumi et al. , 1984
Miothermus 60-70 t aérobie Chung et al. , 1997
Moorella 60 t ananérobie Slobodkin et al., 1997a
Rhodothermus 65 m aérobie Alfredsson et al. , 1988
Synechococcus 65 t aérobie
Thermoanaerobacter ium 60 t anaérobie Schink et Zeikus, 1983
Thermoanaerobacter 65 t anaérobie Zeikus et al. , 1979
Thermobrachium 55 t anaérobie Engle et al., 1996
Thermocrinis 80 t aérobie Huber et al. , 1998b
Thermodesulfobacterium 70 t anaérobie Zeikus et al., 1983
Thermodesulfovibrio 60 t anaérobie Henry et al., 1994
Thermoleophilum 60 t aérobie Zarilla et Perry, 1986
Thermomicrobium 73 t aérobie Jackson et al., 1973
Thermonema 60 t aérobie Hudson et al., 1989
Thermosipho 75 m anaérobie Huber et al., 1989b
Thermosyntropha 60 t anaérobie Svetlitshnyi et al., 1996
Thermoterrabacterium 65 t anaérobie Slobodkin et al. , 1997b
Thermothrix 72 t anaérobie Caldwell et al. , 1976
Thermotoga 80 m anaérobie Huber et al. , 1986
Thermus 60-70 t aérobie Brock et Freeze, 1969
t: terrestre; m: marine cotière, Acido: acidophile
D'une manière analogue le tableau 2.10 présente l'ensemble des souches d'Archaea
thermophiles isolées d'environnements hydrothermaux terrestres et côtiers.
Les habitats microbiens
93
Tableau 2.10. Archaea isolées de sites hydrothermaux terrestres et côtiers.
Domaine Genres Optimum Origine Type Références
ARCHAEA (°C) Respiratoire
Euryarchaeota
Thermococcales Thermococcus 80-90 t,m anaérobie Zillig et al. , 1983b
Pyrococcus 95-100 m anaérobie Fiala et Stetter , 1986.
"Aphrodite" m anaérobie Arab,1999 (result. non pub.)
Archaeoglobales Archeaoglobus 83 t,m anaérobie Stetter, 1988
Ferroglobus 85 m anaérobie Hafenbradl et al. , 1996
Méthanococcales Methanococcus 65 m anaérobie Huber et al. , 1982
Méthanobactériales Methanobacterium 55-65 t anaérobie Winter et al. , 1984
Methanothermus 83-88 t anaérobie Stetter et al. , 1981
Thermoplasmales Thermoplasma 59 t,m Acido, aérobie Darland et al. , 1970
Picrophilus 60 t Acido, aérobie Scleper et al. , 1995
Crenarchaeota
Thermoprotéales Thermoproteus 88-90 t anaérobie Zillig et al. , 1981
Thermofilum 80-88 t anaérobie Zillig et al. , 1983a.
Pyrobaculum 100 t, m anaérobie Huber et al. , 1987
Thermocladium 75 t anaérobie Itoh et al. , 1998
Désulfurococcales Desulfurococcus 85-90 t anaérobie Zillig et al. , 1982
Staphylothermus 92 m anaérobie Fiala et al. , 1986
Stetteria 93 m anaérobie Jochimsen et al., 1998
Sulfolobales Sulfolobus 65-87 t Acido, aérobie Zillig et al. , 1980
Sulfurisphaera 85 t Acido, aérobie Kurosawa et al., 1998
Stygiolobus 80 t Acido, aérobie Segerer et al. , 1991
Mettalosphaera 75 t Acido, aérobie Huber et al. , 1989a
Sulfurococcus 70-75 t Acido, aérobie Karavaiko et al. , 1994
Desulfurolobus 81 t Acido, aérobie = Acidianus sp.
Acidianus 70-90 t, m Acido, aérobie Segerer et al. , 1985
Pyrodictiales Pyrodictium 80 m anaérobie Stetter et al. , 1983
Hyperthermus 95-106 m anaérobie Zillig et al. , 1990
non-classés Aeropyrum 90-95 m aérobie Sako et al. , 1996
Caldococcus t Aoshima et al. , 1996
Igneococcus t Burggraf et al. , 1997
Sulfophobococcus 90 t anaérobie Hensel et al. , 1997
Thermodiscus 93 m Burggraf et al. , 1997
Thermosphaera 85 t anaérobie Huber et al. , 1998a
t: terrestre; m: marine côtière, Acido: acidophile
Au sein de nombreux écosystèmes terrestres, notamment les sources chaudes à pH neutre,
on retrouve comme producteurs primaires des thermophiles modérés bactériens réalisant la
photosynthèse. Ces bactéries appartiennent aux cyanobactéries du genre Synechococcus ou
Mastigocladus et des bactéries vertes non sulfureuses (GNS) du genre Chloroflexus.
4.5.2.3 L'apport des techniques moléculaires
Sur les sites hy drothermaux côtiers
D'une manière analogue aux sites profonds, très peu d'études ont été réalisées afin de
déterminer la diversité microbienne des sites côtiers par des méthodes moléculaires. Les sources
étudiées jusqu'à présent sont des sources situées à proximité des côtes japonaises ou près des îles
grecques. Yamamoto et al. (1998) ont étudié la diversité microbienne d'enchevrêtrements bactériens
soufrés par séquençage de l'ADNr 16S cloné et par hybridation in situ. Ils observent :

Les habitats microbiens
94
- qu'il n'est pas possible d'amplifier de l'ADN archaéen,
- que les banques de 16S diffèrent selon qu'elles sont construites à partir du lysat brut ou de l'ADN
purifié du lysat (diversité plus abondante dans le dernier cas),
- que ces séquences bactériennes sont majoritairement (40 à 68 %) associées aux Aquificales. Les
autres séquences correspondent à des Firmicutes et à des Proteobacteria (α, β et γ ).
Cette étude a été complétée récemment par une analyse des profils quinoliques indiquant que
lorsque la température du tapis bactérien augmente sa diversité décroît (Hiraishi et al., 1999).
Les travaux de Takai et Sako (1999) ont porté sur des sources terrestres et côtières en
réalisant des banques de séquences archaéennes et universelles. Dans le cas des sources marines
(fluide chaud : 128°C), l'ensemble des séquences isolées des banques universelles correspond à des
Bacteria se répartissant parmi des Pseudomonas, des bactéries fixatrices d'azote (Bartonella,
Rhizobium, Agrobacterium)26 et des hyperthermophiles (Aquifex). Les séquences archaéennes se
répartissent parmi les Euryarchaeota hyperthermophiles (Thermococcales) et les Crenarchaeota
proches de représentants isolés (Pyrolobus, Pyrodictium, Stetteria) ou proches du groupe I de
Crenarchaeota marines. Mais l'un des plus intéressants résultats est la détection en milieu marin de
séquences associées aux Korarchaeota détectées jusqu'à présent uniquement dans des sources
terrestres (voir plus loin). Les séquences provenant d'échantillons de sédiments hydrothermaux sont
réparties essentiellement autour des Archaea marines du groupe I, de Proteobacteria et des
Firmicutes à bas G+C%.
Enfin Dando et al. (1998) décrivent la composition microbienne de sédiments chauds sur le
site de la baie de Palaeochori sur l'arc volcanique grec. Ils ont réalisé des enrichissements
d'hyperthermophiles coccoïdes anaérobies correspondant à de nouveaux genres : Stetteria
(Jochimsen et al., 1997) et "Aphrodite" (Arab, 1999 result. non pub.) et de bactéries aérobies proches
des genres Bacillus, Achromatium et Thioplaca. Des amplifications ont été réalisées à l'aide d'amorces
spécifiques des Archaea et des Crenarchaeota conduisant à l'identification de séquences associées
aux genres Staphylothermus, Desulfurococcus, Thermodiscus, Thermosphaera, Stetteria et
Sulfofobococcus ainsi qu'à des séquences correspondant à de nouvelles lignées archaéennes.
Sur les sites continentaux
Une abondante littérature décrit l'utilisation des techniques moléculaires afin d'identifier la flore
microbienne associée aux sources hydrothermales et notamment aux sources du parc de
Yellowstone. Ces techniques ont permis non seulement d'étudier la composition de la communauté
microbienne mais de suivre l'évolution de cette communauté dans le temps et dans l'espace. Ward et
al. (1992, 1997, 1998) ont proposé un historique des travaux réalisés dans leur laboratoire, qui ont
26 Il attribue ces deux premiers types de micro-organismes à des contaminants non représentatifs de la communauté hydrothermale
comme l'avait suggéré Tanner et al. (1998).

Les habitats microbiens
95
porté essentiellement sur l'étude des tapis bactériens. Ces travaux ont été complétés par ceux de
l'équipe de Pace qui a travaillé sur des échantillons de sédiments prélevés dans un autre bassin.
Figure 2.19. Octopus Spring Figure 2.20. Obsidian Pool
Photos : N. Pace http://crab2.berkeley.edu/~pacelab/
La première étude a porté sur l'analyse de l'ARNr 5S (Stahl et al., 1985) sur le bassin d'Octopus
Spring (cf. figure 2.19). Les auteurs décrivent alors 3 phylotypes dominants : un premier associé aux
Crenarchaeota soufre-dépendantes et deux autres associés aux Bacteria proches du genre Thermus.
Cette étude a été suivie dans le laboratoire de Ward d'une analyse des ADNr 16S d'un tapis
cyanobactérien (Ward et al., 1990 a et b; Weller et al., 1992) en utilisant différentes techniques de
clonage moléculaire. Ces travaux ont conduit à la description de nouvelles séquences associées aux
cyanobactéries et aux GNS. Conjointement aux travaux de séquençage, des essais d'isolement furent
entrepris afin d'identifier les bactéries associées aux séquences de clones (Nold et al., 1996) ainsi que
des essais d'incorporation de 14CO2 pour étudier les mécanismes photosynthétiques en fonction du
cycle nycthéméral (Nold et Ward, 1996). La mise au point de sondes spécifiques des cyanobactéries
et des GNS cultivées et non cultivées (Ruff-Roberts et al., 1994) permet de mettre en évidence les
biais liés aux méthodes culturales (l'espèce cultivée n'est pas l'espèce la plus abondante dans le milieu
et elle est sélectionnée dans différents échantillons). Puis, par des techniques de dilutions
successives de l'inoculum avant l'enrichissement, il a été possible d'isoler pour la première fois un
micro-organisme correspondant à une séquence d'ADNr 16S préalablement clonée (Ferris et al.,
1996b). La technique de DGGE a été utilisée afin d'identifier la communauté microbienne colonisant
différents gradients de température au sein du tapis microbien (Ferris et al., 1996a). Il a ainsi été
possible d'observer une grande diversité de séquences toutes associées au cyanobactéries ou aux
GNS. Cette diversité pourrait correspondre à une adaptation de l'espèce en fonction des températures
du biotope. La DGGE a par ailleurs été utilisée afin de détecter les modifications dans la communauté
microbienne : (I) lors d'un enrichissement (Santegoeds et al., 199627; Ward et al., 199728), (II) lors des
changements de saisons29 (Ferris et Ward, 1997), (III) lors d'une modification du milieu30 (Ferris et al.,
1997).
27 Il est par ailleurs observé que de nouvelles séquences sont détectées après enrichissement alors que celles-ci n'avaient pas été
détectées ni par isolement ni par clonage direct.28 L'accent est mis cette fois ci sur le choix de la température d'isolement. A forte température, des souches de Thermus sont
préférentiellement isolées alors qu'elles ne représentent qu'une très faible proportion de la communauté microbienne présente dans
l'échantillon dans cette gamme de température.29 Il n'y a pas à proprement parler de succession de populations mais une adaptation des souches à différentes gammes de
températures. Certains phylotypes semblent accroître puis restreindre, au cours de l'année, leur gamme de température.

Les habitats microbiens
96
L'ensemble des résultats obtenus sur le site d'Octopus Spring a été synthétisé récemment
permettant aux auteurs d'apporter un regard critique sur leur approche polyphasique de la diversité
(Ward et al., 1998).
Au sein de l'équipe de Pace, Reysenbach et al. (1994) s'attachent à une description
phylogénétique de la communauté bactérienne associée aux filaments roses du bassin d'Octopus
Spring. Ils observent 3 phylotypes majeurs se répartissant au sein des Thermotogales (1) et au sein
des Aquificales (2). Barns et al. (1994) vont travailler sur la source Jim's Black Pool (appelée maintenant
Obsidian Pool, cf. figure 2.20). Après amplification des ADNr 16S archaéens ils observent une grande
diversité de séquences associées à la fois à des Crenarchaeota soufre-dépendantes et à des
Euryarchaeota. Un nouveau groupe de séquences enraciné très profondément à la base de l'arbre
archaéen est aussi détecté (Barns et al., 1996a). Il sera décrit plus tard comme un nouveau domaine :
les Korarchaeota (Barns et al., 1996b). Il n'a pas été possible jusqu'à présent d'isoler des
représentants de ce domaine. Brunk et Eis (1998) sont toutefois parvenus à maintenir un
enrichissement en activité dans un chemostat et à étudier les mécanismes d'amplification spécifique
des séquences d'ADNr 16S korarchaéennes en fonction du type d'amorces utilisées. Les travaux de
Barns ne portant que sur des amplification archaéennes, Hugenholtz et al. (1998a) présentent par la
suite les amplifications réalisées à l'aide d'amorces bactériennes et universelles. D'une manière
surprenante les banques universelles ne contiennent pas de séquences archaéennes (alors que
Barns en avait détecté précédemment) mais elles contiennent par contre une grande diversité de
séquences bactériennes. Ces dernières se répartissent, pour 70% d'entre elles, dans 14 des 24
divisions bactériennes couramment reconnues : parmi celles ci on retrouve des groupes thermophiles
dont la présence s'explique dans un tel environnement (Aquificales, Thermotogales, Thermus) mais
aussi des séquences correspondant à des divisions plus mésophiles (Proteobacteria , Firmicutes haut
G+C%, Cytophagales , etc.). Les 30% de séquences bactériennes restantes forment 12 nouvelles
divisons dans l'arbre bactérien au sein desquelles aucun représentant n'a été jusqu'à présent isolé.
Hugenholtz et al. (1998b) proposent donc une nouvelle version de l'arbre bactérien se répartissant en
36 divisions dont seulement 23 présentent des représentants isolés.
A la lumière de l'ensemble des résultats obtenus sur les sites hydrothermaux terrestres de
Yellowstone, il est évident que les études menées par des techniques culturales et non-culturales ont
considérablement accru nos connaissances de cet écosystème. Il est, dans bien des cas, possible de
répondre aux questions de la composition, de la structure, et de la fonction des différentes
populations de micro-organismes se développant dans cet environnement simple et stable. Si de
nombreuses questions restent encore en suspens concernant le type métabolique de certaines
séquences uniquement détectées par clonage et les relations que l'on peut établir entre les distances
phylogénétiques et les distances géographiques, Ward et al., (1998) nous démontrent la puissance
de l'approche polyphasique et pluridisciplinaire en écologie microbienne.
30 Cette approche a consisté à retirer les premiers centimètres de cellules à la surface d'un mat. Cette étude est complémentée par
l'utilisation de microélectrodes à O2 et par des mesures d'assimilation du 14CO2. Il est alors possible de suivre les successions de
populations lors de la colonisation du mat ainsi que leur métabolisme.

Les habitats microbiens
97
V La flore microbienne liée à l'Homme, aux animaux et à
l'activité humaine
5.1 L'Homme comme écosystème microbien
L'homme constitue un environnement favorable à la croissance microbienne31. C'est un
"milieu" constant physiquement (pH, température, pression osmotique) et riche en nutriments (matière
organique, facteurs de croissance). Les micro-organismes colonisent donc de nombreux
compartiments (la peau, les cavités buccale et vaginale, le tractus digestif, les voies respiratoires) à
l'exception du sang, des organes et des voies urinaires32. Mais c'est aussi un "milieu" hétérogène où
des compartiments de composition chimique particulière ont sélectionné certains organismes ayant
développé des mécanismes de résistance aux systèmes de défense de leur hôte (les
lactopéroxydases et les lysozymes de la salive, l'acide lactique des sécrétions cutanées, la bile et
l'acidité du tractus digestif). L'étude de la flore microbienne est importante car on peut ainsi détecter
des modifications lors d'infections, en déduire la source de contamination et mieux saisir (voir induire)
les mécanismes immunitaires de défense.
5.1.1. La flore de la cavité buccale
(d'après Marsh et Bradshaw, 1995; Madigan, 1996)
C'est un environnement complexe et hétérogène, il est constitué de surfaces lisses et
crevassées baignées dans la salive (qui contient des nutriments mais aussi des anti-bactériens). Les
dents sont constituées de structures minérales comme l'émail (cristaux de phosphate de calcium
formant une matrice enrichie en fluor qui limite les processus de dégradation) et de structures
"vivantes" comme la dentine et la pulpe.
Certains micro-organismes forment des plaques qui se développent comme des films à la
surface de la dent. Cette colonisation correspond à une succession de populations de micro-
organismes. Dans un premier temps les glycoprotéines de la salive créent un film fin de matière
organique à la surface des dents sur lequel des Streptoccocus (S. sanguii, S. sorbinus et S. miltis)
forment des microcolonies. Par la suite des bactéries filamenteuses (Fusobacterium et Spirochètes)
contribuent à l'épaississement de la plaque. Si aucun brossage n'intervient, des germes anaérobies
peuvent s'installer, principalement des Actinomycètes.
Si la plaque dentaire n'est pas détruite régulièrement, l'acide lactique produit par les bactéries
conduit généralement à la formation de caries par décalcification de l'émail. Deux micro-organismes
sont impliqués dans la formation des caries. S. mutans produit à partir des sucres contenus dans
31 Chez l'homme ont dénombre 1014 cellules bactériennes pour 1013 cellules purement humaines. 10% du poids de l'homme serait
d'origine bactérienne et 1/3 des fèces sont constituées de bactéries.32 Ceux-ci ne sont colonisés qu'en cas d'infection.

Les habitats microbiens
98
l'alimentation d'importants réseaux de polysaccharides (dextran) qui favorisent la fixation de S.
sorbinus qui lui utilise les sucres pour produire de l'acide lactique.
Enfin certains micro-organismes (Bacteroides gingivalis) peuvent former une plaque à la base
de la dent. Ils produisent une protéase qui peut conduire à une inflammation, puis une nécrose des
tissus de la gencive et à un déchaussement des dents.
5.1.2. Les micro-organismes du tractus digestif
Le tractus digestif est constitué de trois parties principales : l'estomac, le petit intestin
(duodénum, jéjunum, iléon) et le gros intestin (ou colon) (cf. figure 2.21).
Oesophage
Pricipales bactéries détectées
Organe
Estomac
Duodénum
Jéjunum
Iléon
Colon
Pet
it in
test
inG
os in
test
in
Lactobacilles
Enterococcus
Bacteroides
BifidobacteriumEubacteriumPeptonococcus
PeptonostreptococcusClostriumLactobacilles
Anus
Lactobacilles
Enterococcus faecalis
Enterobactéries
Ruminococcus
Figure 2.21. Micro-organismes du tractus digestif (d'après Madigan, 1996)
L'estomac est un milieu particulièrement agressif. Les sucs gastriques à un pH proche de 2
sont constitués d'HCl et de nombreuses enzymes digestives qui constituent une barrière efficace
contre les germes pathogènes (l'estomac est lui même protégé des sucs par une épaisse couche de
mucus). L'estomac est toutefois colonisé par de nombreux micro-organismes Gram + tels que des
Lactobacillus et des Streptococcus. Cette colonisation fluctue avec l'âge, l'alimentation et l'état de
santé de l'individu. L'un des germes pathogènes le plus fréquemment étudié est Helicobacter pylori.
Cette Proteobacteria ε micro-aérophile utilise l'urée contenue dans la salive et les sucs gastriques, la
convertit en ammoniaque et carbonate33 (base fortes) et contrecarre les effets de l'acidité stomacale. H.
pylori pénètre alors dans le mucus et produit des cytotoxines qui détruisent les cellules mucosales.
Sans mucus l'acidité naturelle de l'estomac et les pepsines contribuent à la formation des ulcères.
33 CO(NH2)2 + H+ +2H2O --uréase--> HCO3
- + 2(NH4)

Les habitats microbiens
99
Dans le petit intestin le pH et la densité microbienne vont s'accroître au fur et à mesure que l'on
s'éloigne de l'estomac. On peut atteindre des densités de 105-107 bactéries.g-1.
Dans le gros intestin la colonisation microbienne est très importante. Elle atteint 107 cellules.g-1
pour les micro-organismes aérobies jusqu'à 1010-1011 cellules.g-1 pour les anaérobies. Le tractus
digestif d'un nouveau né est stérile. Par la suite des micro-organismes pionniers colonisent cet
environnement, le modifient et permettent la mise en place d'une seconde flore qui prend la place des
pionniers.34 A la naissance cette flore est essentiellement constituée de Bifidobacterium puis la
modification du régime alimentaire entraîne l'apparition d'une nouvelle flore intestinale incluant un
grand nombre de genres fermentatifs comme Clostridium, Bacteroides, Enteroccocus et les coliformes
mais aussi des Archaea méthanogènes, des bactéries sulfato-réductrices et acétogènes (Pochart et
al., 1992; Bernalier et al., 1996; Morvan et al., 1996)
5.1.3 L'apport des techniques moléculaires en microbiologie humaine
La flore microbienne associée à l'homme a été intensivement étudiée par des techniques de
culture et jusqu'à présent très peu de travaux ont abordé l'étude de la diversité par une approche
moléculaire. On estime toutefois que pour cet environnement entre 20 et 40% des micro-organismes
ont échappé aux méthodes culturales (Suau et al., 1999). Les techniques moléculaires ont par contre
été mises à profit pour détecter spécifiquement certains germes commensaux ou le plus souvent
pathogènes. En microbiologie clinique la recherche de germe spécifiques prime souvent sur
l'écologie et de nombreuses techniques de détection par PCR à l'aide d'amorces spécifiques du
genre, de l'espèce, de la sous-espèce ou d'un biovariant ont été décrites pour étudier la flore
pathogène comme la flore indigène (Schmidt et Relman, 1994; Dzik, 1995; Whelen et Persing, 1996).
Certaines techniques ont toutefois été développées afin d'extraire l'ADN procaryotique de
tissus particuliers comme la plaque dentaire (Parrish et Greenberg, 1995) ou les gencives (Smith et al.,
1989 a et b).
Sur la flore de la cavité buccale, jusqu'à présent, une seule étude a été réalisée pour étudier la
diversité microbienne d'un abcès dentaire par amplification de l'ADNr 16S bactérien et séquençage
(Dymock et al., 1996). Ce travail a été approfondi par une étude plus précise de trois phylotypes
(préalablement détectés) dans des tissus de patients sains et malades par amplification à l'aide
d'amorces de PCR spécifiques (Harper-Owen et al., 1999). Au sein du même laboratoire une étude
similaire a porté sur l'amplification spécifique d'ADNr 16S codant pour le genre Eubacterium suivi d'un
séquençage (Spratt et al., 1999). Une tentative de quantification bactérienne par PCR compétitive de
la flore buccale a récemment été entreprise (Rupf et al., 1999). Mais la majeure partie des études
entreprises sur la flore buccale consiste à détecter par PCR des germes connus à l'aide d'amorces
34 Chez la souris Flavobacterium, Lactobacillus et entérococci sont les micro-organismes pionniers. Flavobacterium disparaît au bout
de 8 jours et Lactobacillus au bout de 18; Bacteroides (anaérobies stricts) apparaît par la suite et domine la communauté.

Les habitats microbiens
100
spécifiques définies sur l'ADNr 16S. Pour exemple : Helicobacter pylori (Nguyen et al., 1993; Wahlfors
et al., 1995), Treponena sp. (Willis et al., 1999), Spirochètes incultivables (Choi et al., 1994), Eikenella
corrodens et Actinobacillus actinomycetemcomitans (Furcht et al., 1996). Des sondes
oligonucléotidiques ont été utilisées pour détecter des Bacteroides forsythus sur des extraits d'ADN
buccaux (Moncla et al., 1991).
La diversité de la flore intestinale a été étudiée par culture et extraction de l'ADN total suivi de
l'amplification et du séquençage des gènes d'ADNr 16S bactériens par Wilson et Blitchington (1996).
Cette étude révèle à la fois la différence obtenue entre les deux méthodes (certaines séquences sont
abondantes dans la banque d'ADNr 16S mais sont absentes dans les cultures et inversement) et le
biais produit par la PCR (la diversité phylogénétique est bien plus importante après 9 cycles de PCR
qu'après 35 cycles). La majorité des séquences détectées sont associées à des genres ou des
espèces communément enrichies dans la flore intestinale (Bacteroides, Clostridium). Une étude plus
récente menée par Suau et al. (1999) obtient des résultats similaires. Quatre-vingt quinze pour cent de
la banque de clone sont associés à trois groupes phylogéniques (Bacteroides, Clostridium coccoides,
C. leptum), mais seuls 24% de la banque sont clairement associés à des espèces isolées. Wang et al.
(1996) quantifient quant à eux les principaux germes anaérobies retrouvés dans les flores intestinales
animales par des techniques de dilutions successives suivies de PCR à l'aide d'amorces spécifiques
de certaines espèces. Des approches fondées sur l'emploi d'un grand nombre de sondes
nucléotidiques (domaine spécifique et orientées vers la flore anaérobie) (Franks et al., 1998) et sur
l'emploi de la TGGE (Zoetendal et al., 1998) ont aussi été utilisées. Enfin d'une manière similaire à la
flore buccale, beaucoup de publications décrivent la détection par PCR d'espèces ou de genres
spécifiques à l'aide d'amorces ou de sondes dessinées sur l'ADNr 16S (cf. revue de Whelen et
Persing, 1996). Des articles récents ont mis à profit la puissance de la détection PCR pour analyser des
septicémies anciennes dans des tissus conservés dans le formol :
Ø infection de 1979, individus susceptibles d'avoir été infectés par des "armes biologiques"
(Bacillus anthracis; Jackson et al., 1998b),
Ø infection de 1997, dans des tissus d'animaux ayant été tués par le charbon (Patra et al.,
1998),
Ø infection datant des 16eme et 18eme siècles, dans des restes de pulpe dentaire sur des
squelettes d'individus probablement morts de la peste (Yersinia pestis; Drancourt et al., 1998),
Ø infection datant de 1918, dans des tissus de foie d'individus ayant été tués lors de l'épidémie
de grippe espagnole (Reid et al, 1918).
Des études similaires à celles réalisées sur la flore buccale et intestinale ont été effectuées sur
la flore pathogène pouvant infecter le liquide cérébro-spinal (Greisen et al., 1994) et sur les micro-
organismes impliqués dans les infections chroniques de la prostate (Tanner et al., 1999).
5.2 La flore microbienne associée aux autres animaux
Les micro-organismes entretiennent des relations avec un grand nombre d'Eucarya :
protozoaires, animaux comme végétaux. La microbiologie classique a permis de résoudre nombre des

Les habitats microbiens
101
relations qui peuvent être de nature commensale, mutualiste ou parasitaire. Mais certaines relations
mettent en jeu des mécanismes de symbiose très stricts que des techniques d'étude uniquement
culturales ne peuvent résoudre. Les techniques moléculaires ont donc permis d'accéder à la diversité
des micro-organismes impliqués dans ces relations, je me propose dans ce chapitre d'en décrire
certaines.
Mollusques, Eponges et Holoturies :
Les travaux sur la communauté symbiotique des mollusques hydrothermaux ont été décrits
précédemment (cf. § 4.4.3). On peut toutefois citer des travaux similaires effectués sur des
mollusques prélevés dans des environnements tropicaux côtiers (Distel et al., 1991; Durand et al.,
1996; Gros et al., 1996; Krueger et Cavanaugh, 1997).
Les travaux de Preston et al. (1996) sur les Eponges et ceux de Mc Innerney et al. (1995) sur
les Holoturies ont été décrits plus haut (cf. § 4.2.2.1). Les travaux de Preston ont été poursuivis par la
construction d'une banque fosmidique d'ADN génomique permettant d'accéder à des gènes
fonctionnels comme le gène de l'ADN polymérase (Schleper et al., 1997b) mais aussi aux micro-
hétérogénéités au sein de l'opéron ADNr (Schleper et al., 1998).
Insectes :
De nombreux insectes présentent de fortes relations symbiotiques avec les micro-organismes.
Ces derniers sont très souvent retrouvés dans le tractus digestif de ces animaux (Cazemier et al.,
1997). Les micro-organismes contenus dans l'estomac des termites ont été très étudiés. En effet
cette communauté conditionne chez cet Arthropode la digestion des composés à base de cellulose
et, à une échelle réduite, peut être comparée au fonctionnement du rumen chez un grand nombre de
ruminants. Jusqu'à présent très peu d'isolats ont pu être obtenus; les informations dont on dispose
proviennent d'études réalisées à l'aide de microélectrodes (Brune, 1998) ou d'amplifications des
gènes d'ADNr 16S à partir d'extrait d'estomac :
Ø du termite Reticulitermes speratus par analyse de l'ADNr 16S (Ohkuma et Kudo, 1996); de l'ADNr
16S, du gène mcr et du gène NifH (Kudo et al., 1998),
Ø du termite Cryptotermes domesticus (Ohkuma et Kudo, 1998),
Ø du termite Reticulitermes speratus par T-RFLP (Liu et al., 1997)
Ø du termite Reticulitermes speratus par analyse de l'ADNr 16S et hybridation in situ de la
communauté archaéenne (Shinzato et al., 1999),
Ø des symbioses entre les méthanogènes et R. speratus (Ohkuma et al., 1995) et chez d'autres
termites (Ohkuma et al., 1999),
Ø des termites Nasutitermes lujae (spirochètes incultivables, Paster et al., 1996), et de
Mastodermes darwiniensis (spirochètes incultivables, Brechtold et König, 1996), (flore totale par
hybridation in situ, Bertchold et al., 1999)
Ø et des protozoaires de l'estomac de R. speratus par analyse de l'ADNr 18S (Ohkuma et al., 1998)
et des facteurs d'élongation (Moriya et al., 1998).
Des études similaires ont été réalisées sur les micro-organismes intestinaux de Criquets (Santo
Domingo et al., 1998b). Il a même été possible d'étudier les bactéries associées à des insectes

Les habitats microbiens
102
emprisonnés dans l'ambre depuis 25 à 40 millions d'années. Après la mise au point d'une technique
efficace d'extraction d'ADN à partir d'échantillons anciens (Cano et Poinard, 1993), il a été possible de
détecter par amplification et séquençage des Bacillus dans des échantillons d'insectes (Proplebeia
dominicana) (Cano et al., 1994) puis de remettre en culture des spores de Bacillus provenant des
mêmes échantillons (Cano et Borucki, 1995)35 ainsi que des Staphylococcus (Lambert et al., 1998).
Récemment des isolements caractérisés par séquençage et FAME suivis d'une analyse moléculaire de
la communauté totale d'échantillons d'ambre par T-RFLP a été entreprise (Greenblatt et al., 1999). Les
résultats obtenus par ces deux méthodes sont cohérents mais il apparaît que les méthodes
moléculaires peuvent être biaisées en raison de la forte dégradation des ADN dans de très vieux
échantillons et par la présence d'inhibiteurs de la PCR.
Chordés:
Les micro-organismes du rumen ont été aussi intensivement étudiés que ceux des intestins
humains, dans un premier temps par des techniques culturales puis par l'emploi de sondes nucléiques
(Krause et Rusell, 1996a; Stahl et al., 1988). Dans un environnement stabilisé on peut observer des
micro-organismes dégradant la cellulose (Bacteroides, Ruminococcus), les protéines (Veillonella),
l'amidon (Selenomonas), et des producteurs de méthane (Methanobacterium). L'utilisation de sondes
plus spécifiques a permis de suivre l'évolution de certaines espèces en fonction du régime alimentaire
(Forster et al., 1996; Krause et Rusell, 1996b; Schofield et al., 1997). Récemment Tajima et al. (1999)
ont étudié le rumen de vaches en amplifiant spécifiquement l'ADNr 16S bactérien à partir
d'échantillons liquides et solides. Ils n'observent pas de différences avec les résultats obtenus par les
méthodes de mise en culture : les Firmicutes à haut G+C% et les Cytophagales dominent dans les
banques. Des techniques fondées sur l'analyse du G+C% de l'ADN total extrait de tractus digestifs de
poulets a permis de suivre l'évolution de la flore intestinale entre le petit intestin et le cæcum (Apajalahti
et al., 1998). Il n'est toutefois pas possible dans cette étude de définir précisément les genres des
bactéries présentes dans l'échantillon.
Angert et al. (1993) ont isolé par micromanipulation et séquençage le symbiote incultivable du
poisson chirurgien. Epulopiscium fishelsoni était alors la plus grande bactérie connue (80 x 600 nm).
Enfin la communauté intestinale du poisson (Platichthys flesus) a été étudiée après amplification et
séquençage. Il a été possible d'y détecter des séquences associées aux Archaea marines du groupe II
(van der Maarel et al., 1998), et des séquences associées aux methanogènes (van der Maarel et al.,
1999).
Autres:
Les symbiotes d'Acanthamoeba spp. ont été étudiés aussi par amplification de l'ADNr 16S et
séquençage (Fritshe et al., 1999). Ces organismes retrouvés de manière ubiquitaire dans le sol et l'eau
se nourrissent de micro-organismes et sont des facteurs de dissémination de germes pathogènes
notamment en milieu hospitalier.
35 Des études sur la flore intestinale de Mammouth ont été aussi réalisées mais elles consistaient uniquement en une mise en culture
des souches retrouvées dans des restes d'intestins (vieux de 12000 ans) suivi d'un séquençage des isolats (Rhodes et al., 1998). On ne
peut dans ce cas exclure la possibilité de contaminants.

Les habitats microbiens
103
Les micro-organismes associés aux algues ont été étudiés également par des techniques semblables
chez Halophila stipulacea (Weidner et al., 1996) et chez des algues vertes de la classe des
Charophyceae (Fisher et al., 1998).
Aux écosystèmes animaux, marins et terrestres naturellement colonisés par les micro-
organismes, les activités humaines ont contribué à adjoindre de nouveaux biotopes dérivant de
processus industriels de productions alimentaires ou chimiques (fermentation, bioconversion), de
transformation des déchets (traitements biologique) ou d'extraction des minerais (biolixiviation).
L'essor des biotechnologies, intégrant celui de la microbiologie industrielle, a contribué à faire prendre
pleinement conscience du rôle des micro-organismes dans notre économie. L'étude de ces
écosystèmes devait dépasser alors l'aspect purement écologique pour permettre de contrôler, c'est à
dire de prévoir et de modifier le comportement des différentes populations microbiennes au sein d'un
environnement confiné. L'emploi de méthodes moléculaires à grandement facilité cette tâche.
5.3 Les micro-organismes associés aux activités humaines
Deux biotopes liés à des activités anthropomorphiques ont été particulièrement étudiés à
l'aide des techniques moléculaires : les boues de station d'épuration et les bassins de biolixiviation. Le
fonctionnement de ces deux biotopes est intimement lié à la présence de micro-organismes qui dans
un cas (boues actives) vont accélérer la dégradation de la matière organique et les processus de
floculation et dans l'autre cas, faciliter la solubilisation des métaux précieux (lixiviation).
5.3.1 L'extraction de minerai
L'extraction des minerais comme le Cuivre ou l'Uranium est réalisée à grande échelle par des
procédés de solubilisation des minerais (soufrés généralement) par des solutions acides. Cette
solubilisation réalisée à l'air libre permet la colonisation des réservoirs par une communauté
bactérienne très abondante constituée essentiellement de micro-organismes soufre-oxydants,
autotrophes ou hétérotrophes. Les techniques moléculaires ont été mises à profit pour suivre les
successions de populations au sein de ces réservoirs soit par amplification et séquençage des ADNr
16S (Goebel et Stackebrandt, 1994), soit par amplification et séquençage de l'espace inter-génique
16-23S sur l'ADNr (Pizarro et al., 1996; Espejo et Romero, 1997; Vàsquez et Espejo, 1997), soit par
DGGE (Fournier et al., 1998). Une étude récente a permis de détecter, dans des extraits de
biolixiviation de cuivre des séquences archaéennes correspondant aux Thermoplasmales (la souche a
été cultivée mais n'a pas pu être obtenue en culture pure) (Vàsquez et al., 1999).

Les habitats microbiens
104
5.3.2. Les boues de stations d'épuration
Les stations d'épuration constituent un système complexe où se mêlent à la fois des
techniques physiques (filtration, sédimentation, chloration, cf. figure 2.22) et des techniques
biologiques mettant en jeu une importante communauté microbienne .
La connaissance de la structure de la communauté microbienne des boues actives (traitement
aérobie de la phase soluble) et des fermenteurs (traitement anaérobie de la phase insoluble) est un
axe de recherche majeur. En effet la connaissance de cette structure permet d'influer sur sa
dynamique (en jouant sur les nutriments ou sur l'ensemencement par une flore définie) et donc sur
l'efficacité du traitement des eaux usées (cf. figure 2.22). Compte tenu de la diversité des nutriments
disponibles dans les boues, la diversité métabolique et phylogénétique est aussi très importante.
Aussi les techniques de culture, d'analyse par immunofluorescence, comme les profils quinoliques et
polyaminiques ne rendent qu'une image très restreinte de cette diversité (Snaidr et al., 1997). Les
techniques moléculaires ont donc été utilisées abondamment pour rendre compte de la diversité aussi
fidèlement que possible.
Boues solides fraction insoluble
Effluant liquidefraction soluble
Boues actives Egouttage
SéchageIncinération
Désinfection:- chloration
Effluant traité rejetté dans les rivières
SédimentationFiltrationEaux
usées
Traitement anaérobie:digestion
Traitement aérobie:oxydation
Tra
item
ent
mic
robi
olog
ique
Figure 2.22. Représentation schématique du fonctionnement d'une station d'épuration (d'après Madigan, 1996)
Les tableaux 2.11 et 2.12 se proposent d'énumérer une partie des analyses réalisées à l'aide
de techniques moléculaires sur différents échantillons de boues actives ou d'eau usées de stations
d'épuration.Tableau 2.11. Analyse de la diversité microbienne totale
Références Techniques Commentaires
Bond et al. , 1995 Amplif. 16S / Séqu. Comparaison de la population microbienne de 2 fermenteurs dont l'un élimine les
phosphates.
Bradford et al. , 1996 Micromanipulation / Isolat
Amplif. 16S / Séqu.
Sondes 16S
Etude des bactéries filamenteuses. Isolement par micromanipulation,
séquençage et localisation à l'aide de sondes spécifiques.
Eichner et al. , 1999 TGGE 16S Effet de l'addition d'un micro-organisme pouvant dégrader les phénols sur une
population microbienne lors d'un choc phénolique.
Harmsen et al. , 1996b Sondes 16S Analyse des gradients de population dans des granules des boues actives.
Analyse des successions syntrophe/méthanogènes.
Kämpfer et al. , 1996 Isolats
Sondes 16S et 23S
70% des souches isolées (Proteobacteria β) ne correspondent qu'à 5% des
bactéries détectées par hybridation in situ.
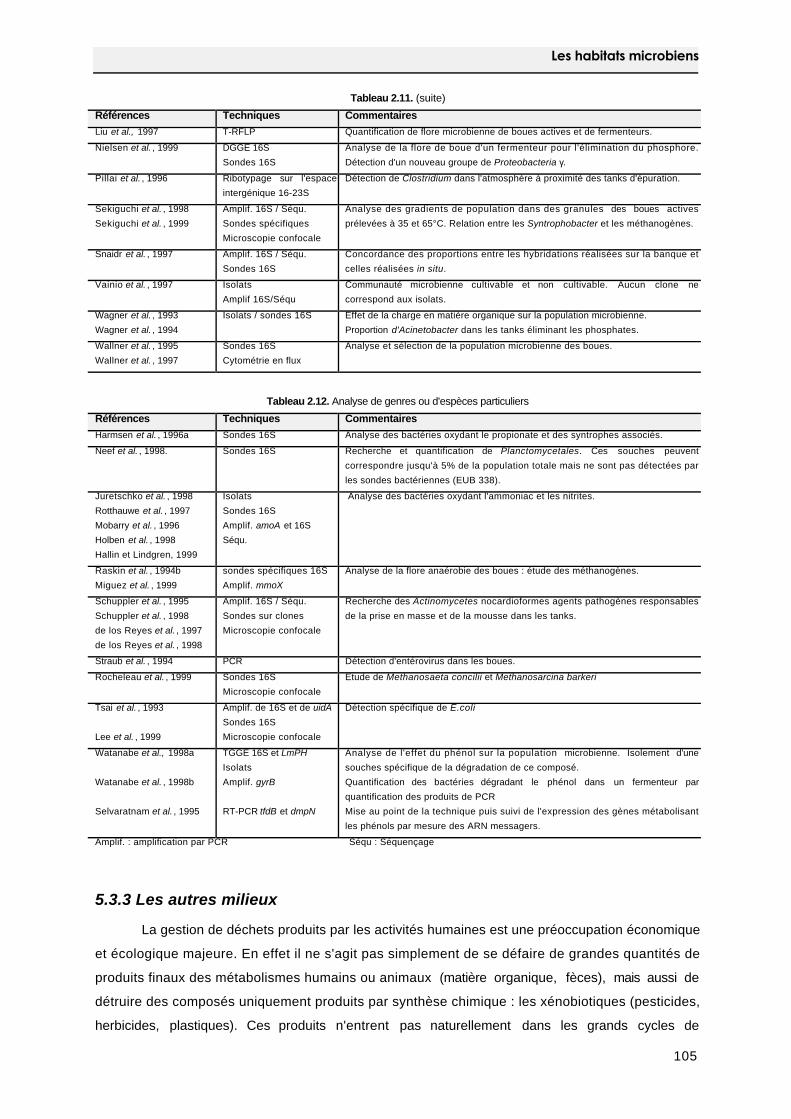
Les habitats microbiens
105
Tableau 2.11. (suite)
Références Techniques Commentaires
Liu et al., 1997 T-RFLP Quantification de flore microbienne de boues actives et de fermenteurs.
Nielsen et al. , 1999 DGGE 16S
Sondes 16S
Analyse de la flore de boue d'un fermenteur pour l'élimination du phosphore.
Détection d'un nouveau groupe de Proteobacteria γ.
Pillai et al. , 1996 Ribotypage sur l'espace
intergénique 16-23S
Détection de Clostridium dans l'atmosphère à proximité des tanks d'épuration.
Sekiguchi et al. , 1998
Sekiguchi et al. , 1999
Amplif. 16S / Séqu.
Sondes spécifiques
Microscopie confocale
Analyse des gradients de population dans des granules des boues actives
prélevées à 35 et 65°C. Relation entre les Syntrophobacter et les méthanogènes.
Snaidr et al. , 1997 Amplif. 16S / Séqu.
Sondes 16S
Concordance des proportions entre les hybridations réalisées sur la banque et
celles réalisées in situ.
Vainio et al. , 1997 Isolats
Amplif 16S/Séqu
Communauté microbienne cultivable et non cultivable. Aucun clone ne
correspond aux isolats.
Wagner et al. , 1993
Wagner et al. , 1994
Isolats / sondes 16S Effet de la charge en matière organique sur la population microbienne.
Proportion d'Acinetobacter dans les tanks éliminant les phosphates.
Wallner et al. , 1995
Wallner et al. , 1997
Sondes 16S
Cytométrie en flux
Analyse et sélection de la population microbienne des boues.
Tableau 2.12. Analyse de genres ou d'espèces particuliers
Références Techniques Commentaires
Harmsen et al. , 1996a Sondes 16S Analyse des bactéries oxydant le propionate et des syntrophes associés.
Neef et al. , 1998. Sondes 16S Recherche et quantification de Planctomycetales. Ces souches peuvent
correspondre jusqu'à 5% de la population totale mais ne sont pas détectées par
les sondes bactériennes (EUB 338).
Juretschko et al. , 1998
Rotthauwe et al. , 1997
Mobarry et al. , 1996
Holben et al. , 1998
Hallin et Lindgren, 1999
Isolats
Sondes 16S
Amplif. amoA et 16S
Séqu.
Analyse des bactéries oxydant l'ammoniac et les nitrites.
Raskin et al. , 1994b
Miguez et al. , 1999
sondes spécifiques 16S
Amplif. mmoX
Analyse de la flore anaérobie des boues : étude des méthanogènes.
Schuppler et al. , 1995
Schuppler et al. , 1998
de los Reyes et al. , 1997
de los Reyes et al. , 1998
Amplif. 16S / Séqu.
Sondes sur clones
Microscopie confocale
Recherche des Actinomycetes nocardioformes agents pathogènes responsables
de la prise en masse et de la mousse dans les tanks.
Straub et al. , 1994 PCR Détection d'entérovirus dans les boues.
Rocheleau et al. , 1999 Sondes 16S
Microscopie confocale
Etude de Methanosaeta concilii et Methanosarcina barkeri
Tsai et al. , 1993
Lee et al. , 1999
Amplif. de 16S et de uidA
Sondes 16S
Microscopie confocale
Détection spécifique de E.coli
Watanabe et al., 1998a
Watanabe et al. , 1998b
Selvaratnam et al. , 1995
TGGE 16S et LmPH
Isolats
Amplif. gyrB
RT-PCR tfdB et dmpN
Analyse de l'effet du phénol sur la population microbienne. Isolement d'une
souches spécifique de la dégradation de ce composé.
Quantification des bactéries dégradant le phénol dans un fermenteur par
quantification des produits de PCR
Mise au point de la technique puis suivi de l'expression des gènes métabolisant
les phénols par mesure des ARN messagers.
Amplif. : amplification par PCR Séqu : Séquençage
5.3.3 Les autres milieux
La gestion de déchets produits par les activités humaines est une préoccupation économique
et écologique majeure. En effet il ne s'agit pas simplement de se défaire de grandes quantités de
produits finaux des métabolismes humains ou animaux (matière organique, fèces), mais aussi de
détruire des composés uniquement produits par synthèse chimique : les xénobiotiques (pesticides,
herbicides, plastiques). Ces produits n'entrent pas naturellement dans les grands cycles de
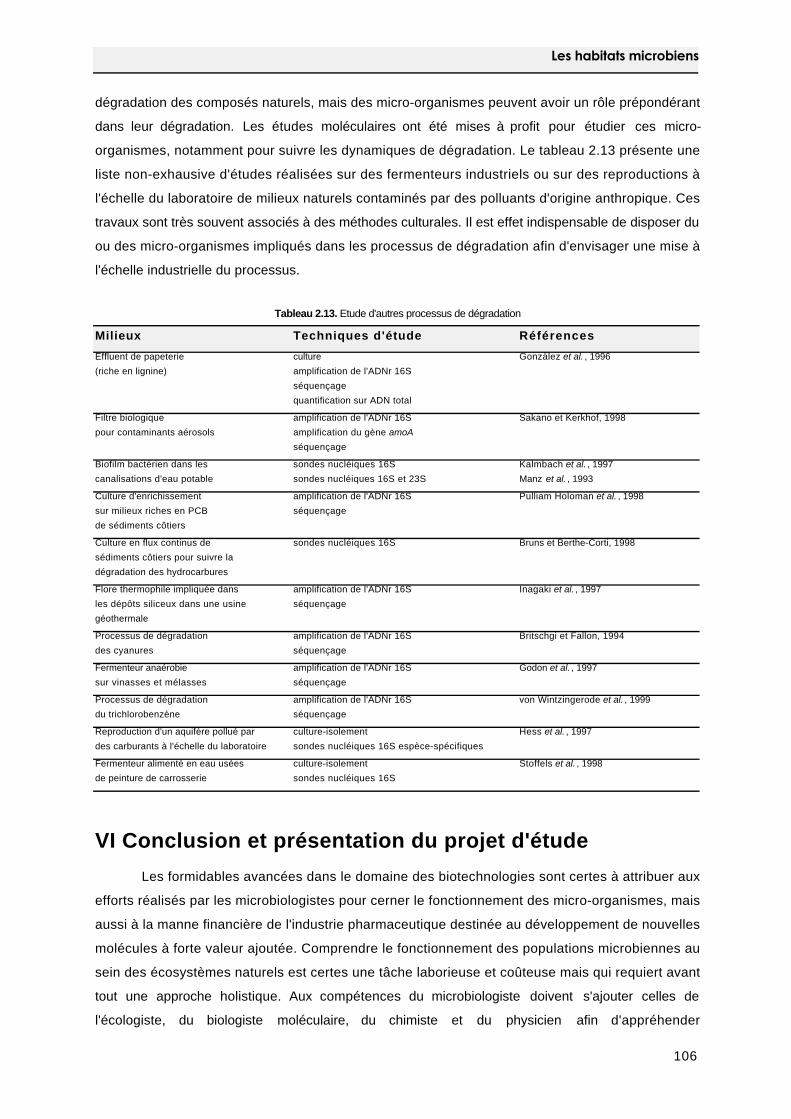
Les habitats microbiens
106
dégradation des composés naturels, mais des micro-organismes peuvent avoir un rôle prépondérant
dans leur dégradation. Les études moléculaires ont été mises à profit pour étudier ces micro-
organismes, notamment pour suivre les dynamiques de dégradation. Le tableau 2.13 présente une
liste non-exhausive d'études réalisées sur des fermenteurs industriels ou sur des reproductions à
l'échelle du laboratoire de milieux naturels contaminés par des polluants d'origine anthropique. Ces
travaux sont très souvent associés à des méthodes culturales. Il est effet indispensable de disposer du
ou des micro-organismes impliqués dans les processus de dégradation afin d'envisager une mise à
l'échelle industrielle du processus.
Tableau 2.13. Etude d'autres processus de dégradation
Milieux Techniques d'étude Références
Effluent de papeterie culture Gonzàlez et al. , 1996
(riche en lignine) amplification de l'ADNr 16S
séquençage
quantification sur ADN total
Filtre biologique amplification de l'ADNr 16S Sakano et Kerkhof, 1998
pour contaminants aérosols amplification du gène amoA
séquençage
Biofilm bactérien dans les sondes nucléiques 16S Kalmbach et al. , 1997
canalisations d'eau potable sondes nucléiques 16S et 23S Manz et al. , 1993
Culture d'enrichissement amplification de l'ADNr 16S Pulliam Holoman et al. , 1998
sur milieux riches en PCB séquençage
de sédiments côtiers
Culture en flux continus de sondes nucléiques 16S Bruns et Berthe-Corti, 1998
sédiments côtiers pour suivre la
dégradation des hydrocarbures
Flore thermophile impliquée dans amplification de l'ADNr 16S Inagaki et al. , 1997
les dépôts siliceux dans une usine séquençage
géothermale
Processus de dégradation amplification de l'ADNr 16S Britschgi et Fallon, 1994
des cyanures séquençage
Fermenteur anaérobie amplification de l'ADNr 16S Godon et al. , 1997
sur vinasses et mélasses séquençage
Processus de dégradation amplification de l'ADNr 16S von Wintzingerode et al. , 1999
du trichlorobenzène séquençage
Reproduction d'un aquifère pollué par culture-isolement Hess et al. , 1997
des carburants à l'échelle du laboratoire sondes nucléiques 16S espèce-spécifiques
Fermenteur alimenté en eau usées culture-isolement Stoffels et al. , 1998
de peinture de carrosserie sondes nucléiques 16S
VI Conclusion et présentation du projet d'étude
Les formidables avancées dans le domaine des biotechnologies sont certes à attribuer aux
efforts réalisés par les microbiologistes pour cerner le fonctionnement des micro-organismes, mais
aussi à la manne financière de l'industrie pharmaceutique destinée au développement de nouvelles
molécules à forte valeur ajoutée. Comprendre le fonctionnement des populations microbiennes au
sein des écosystèmes naturels est certes une tâche laborieuse et coûteuse mais qui requiert avant
tout une approche holistique. Aux compétences du microbiologiste doivent s'ajouter celles de
l'écologiste, du biologiste moléculaire, du chimiste et du physicien afin d'appréhender

Les habitats microbiens
107
l'environnement dans sa totalité. Un inventaire complet de la diversité microbienne terrestre n'est
certes pas envisageable mais, comme cela a été proposé par Pace (1997), il est possible de définir sur
le globe une centaine de sites (biotopes aux conditions physico-chimiques différentes) pour lesquels
une étude intensive de la diversité tant procaryotique qu'eucaryotique serait entreprise. Certains sites
naturels, comme les sources chaudes du parc de Yellowstone (Etats-Unis), font déjà l'objet de suivis
réguliers depuis de nombreuses années. D'autres sites, comme les sources hydrothermales
profondes et les réservoirs pétroliers, pourraient être des candidats potentiels à cette entreprise
d'inventaire.
Jusqu'à présent les informations concernant l'écologie de ces milieux ont été principalement
obtenues par des travaux fondés sur des techniques de cultures traditionnelles, peu d'études
moléculaires y ont été entreprises.
Le but de mon travail de thèse aura été d'utiliser des technologies moléculaires fondées sur
l'analyse des ARNr 16S (séquençage - hybridation in situ) au profit de l'étude de la diversité et de la
taxonomie microbienne, en association avec deux environnements extrêmes (un réservoir pétrolier et
des sources hydrothermales océaniques).
Après un exposé des méthodes générales utilisées, les résultats sont présentés sous forme
d'articles, soumis ou à soumettre.
Le premier article (Article 1) présenté dans ce manuscrit décrit l'analyse de la diversité
microbienne associée à un réservoir pétrolier situé dans le bassin Parisien. Si quelques travaux
décrivent l'isolement et la caractérisation de micro-organismes, mésophiles ou thermophiles, impliqués
dans des mécanismes de fermentation, de méthanogenèse ou d'acétogenèse, la majeure partie des
études réalisées sur les puits de pétrole se sont intéressées aux micro-organismes impliqués dans les
phénomènes de sulfato-réduction. L'originalité de cette étude était d'accéder à la diversité
microbienne totale d'un réservoir en s'affranchissant de toute étape de mise en culture. L'analyse a
porté sur des échantillons d'eaux de production prélevés en tête de puits et dans des collecteurs. On
observe que les communautés microbiennes s'organisent différemment selon les échantillons. Dans
certains puits, les BSR thermophiles ou mésophiles dominent les banques de clones, alors qu'elles
sont absentes d'autres échantillons provenant du même réservoir. En revanche on retrouve dans ces
échantillons des séquences associées à des souches micro-aérophiles pouvant être impliquées dans
le cycle du soufre. En introduction de cet article je présenterai une étude réalisée sur un échantillon
d'eau chaude prélevée sur un site hydrothermal terrestre en Roumanie. Les résultats obtenus sur cet
échantillon d'eau géothermale éclairent ceux qui sont présentés pour certains échantillons d'eau de
production.
Le second et le troisième articles décrivent l'utilisation conjointe de techniques
culturales (enrichissements) et moléculaires (séquençage et hybridation in situ) pour étudier la
diversité microbienne associée à des fluides hydrothermaux prélevés sur le site du "Snake Pit" sur la

Les habitats microbiens
108
dorsale médio-Atlantique durant la campagne "Microsmoke" (1995). Les fluides ont été collectés à
l'aide d'un échantillonneur original, le "Vent Cap", qui est déployé durant plusieurs jours sur le site
d'émission du fluide. Deux sites ont été échantillonnés au cours de déploiements de 2 et 5 jours.
L'article 2 décrit, par une approche d'extraction d'ADN total et séquençage des ADNr 16S, la
diversité bactérienne associée à un déploiement de 5 jours sur le "Chi". On observe que la banque de
clones est dominée par une grande diversité de micro-organismes associés ou proches des
Proteobacteria epsilon. Deux nouveaux phylums associés à la classe des Proteobacteria , mais
distincts des sous-classes décrites jusqu'à présent, sont aussi observés dans cet échantillon. Deux
sondes nucléiques spécifiques sont construites afin d'identifier les micro-organismes détectés par
séquençage.
L'article 3 (Manuscrit en préparation) présente une synthèse des résultats obtenus à partir
de l'ensemble des déploiements du "Vent Cap" lors de la campagne "Microsmoke". Il reprend les
résultats décrits dans l'article 2 sur le site "Chi" et ceux décrits par Reysenbach et al. (1999) sur le site
"Les Ruches". Il présente, de plus, des résultats originaux d'enrichissement de micro-organismes
thermophiles, de comptage de cellules totales, d'analyse par séquençage du déploiement de 2 jours
sur le site "Les Ruches" et de quantification de micro-organismes par hybridation in situ à l'aide de
sondes nucléiques spécifiques. La confrontation des résultats obtenus par ces différentes analyses
avec ceux obtenus lors de l'étude de cheminées hydrothermales nous permet de mieux cerner la
distribution des micro-organismes au sein des sources hydrothermales profondes.
Comme nous l'avons vu dans les deux premières parties de ce manuscrit, l'utilisation de
techniques moléculaires et l'emploi de méthodes culturales classiques ne sauraient s'exclure dans
l'étude de la diversité d'un écosystème. Dans le cadre de la caractérisation d'une nouvelle souche,
l'analyse phylogénétique de l'ADNr peut permettre d'asseoir une taxonomie sur une base cladistique.
Le travail que j'ai effectué pour les articles suivants constitue une contribution à la caractérisation de
souches isolées d'environnements hydrothermaux (articles 4, 5 et 6).
L'article 4 (article publié) présente la caractérisation d'une nouvelle espèce de
Methanococcales, Methanococcus vulcanius et l'identification d'une Methanococcales déjà isolée, M.
fervens.
L'article 5 (manuscrit en préparation) présente la caractérisation phylogénétique de 5
Thermococcales radiorésistantes isolées sur des sites hydrothermaux des dorsales médio-Atlantique
et est-Pacifique.
L'article 6 (manuscrit en préparation) présente la caractérisation phylogénétique d'une
souche de Bacillus thermophile isolée sur le site hydrothermal du bassin de Lau au sud-est des îles
Fidji.

Matériel et Méthodes

Paris
Ardennes
Marne
Haute-Marne
Aube
Seine-et-Marne
Val-d'Oise
Yvelines
Essonne
MONTMIRAIL
1 2
3 4
5
Légendes:1: Situation géographique de Montmirail.2: Puit de Pétrole (LMN).3: Décanteur (B01C).4: Echantillonnage en tête de puits (I50).5: Procédé de filtration de l’eau de production.
Figure 3.1. Echantillonnage des puits de pétrole. Site de Montmirail
Planche 1

Matériel et Méthodes
109
I Echantillonnage et conservation de l'échantillon
Durant ce travail de thèse, trois types de sites ont été échantillonnés :
Ø des puits de pétrole en région parisienne (échantillons d'eau de production),
Ø des sources hydrothermales profondes sur la dorsale médio Atlantique (échantillons de
fluides et de cheminées),
Ø des sources chaudes terrestres Roumaines (échantillon d'eaux).
1.1 Echantillonnage des puits de pétrole
Les échantillons proviennent des puits de pétrole du site de Montmirail, dans le Bassin Parisien
(cf. planche photos figure 3.1). La société COPAREX, spécialisée dans l'exploitation des puits en fin
de production, en assure la gestion. Les techniques d'extraction utilisées nécessitent l'envoi d'eau
pressurisée dans le réservoir afin de faire remonter l'huile. Une fois en surface, le mélange eau/huile
est envoyé par des pipelines vers des bassins de décantation, puis vers des bassins de collecte.
Les échantillons étudiés sont des mélanges eau/huile prélevés en tête de puits ou dans un
décanteur. Après décantation des échantillons, la partie aqueuse a été conservée. Afin de récupérer
un grand nombre de micro-organismes, les échantillons ont été filtrés ou centrifugés. Les extractions
d'ADN sont donc réalisées à partir d'un filtre ou d'un culot de centrifugation.
La technique de filtration est dérivée de la technique décrite par Sommerville et al., 1989. Cette
technique peut s'appliquer à de nombreux échantillons liquides. Elle permet de concentrer les
échantillons et d'effectuer directement la lyse cellulaire sur filtre. Réalisée initialement sur des filtres
Millipore Sterivex GT (2 mL), elle peut être extrapolée à des filtres de plus grande taille afin de pouvoir
filtrer de plus gros volumes d'échantillons.
Il convient de filtrer l’échantillon à l'aide d'une pompe péristaltique. Le filtre utilisé est un filtre GT
(Millipore) de 0,2 µm en polycarbonate. En fin de filtration le filtre est lavé avec 10 mL de tampon SET
stérile (Sucrose EDTA Tris, voir composition § 2.1.1) puis stocké à -20°C.
1.2 Echantillonnage des sources hydrothermales profondes
Les échantillons ont été prélevés à 3500m de profondeur durant la campagne "Microsmoke" en
1995 (chef de mission Daniel Prieur). Cette campagne a été effectuée sur le site du Snake Pit
(23°22'118N, 44°56'984W) sur la dorsale médio Atlantique (cf. planche cartes figure 3.2). Le site du
"Snake Pit" a été retenu car il était profond (donc susceptible d'être colonisé par des micro-organismes
barophiles), la faune associée était relativement simple (essentiellement des crevettes) et les
cheminées présentaient des structures minéralogiques permettant un bon échantillonnage.
Plusieurs types de cheminées sont observables sur ce site. Les cheminées dénommées "Chi"
et "les Ruches" ont retenu notre attention.
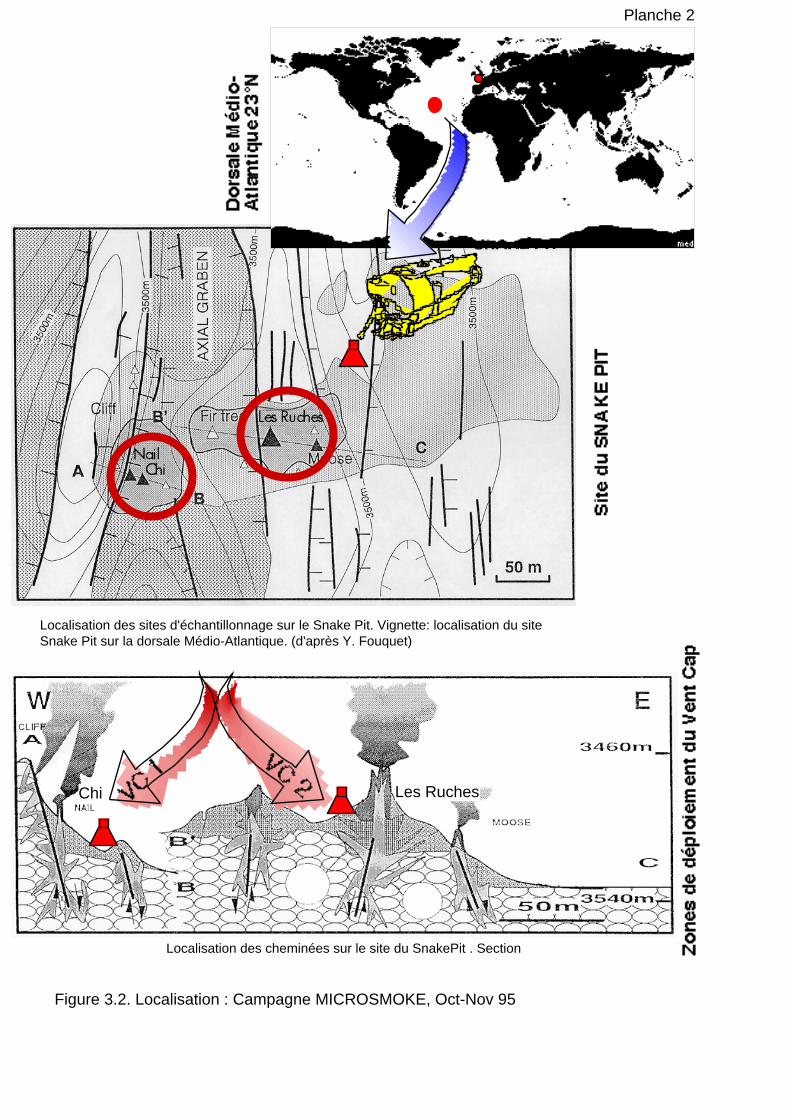
Figure 3.2. Localisation : Campagne MICROSMOKE, Oct-Nov 95
Les RuchesChi
Localisation des sites d'échantillonnage sur le Snake Pit. Vignette: localisation du site Snake Pit sur la dorsale Médio-Atlantique. (d'après Y. Fouquet)
Localisation des cheminées sur le site du SnakePit . Section
Planche 2

Matériel et Méthodes
110
Deux types d'échantillons ont été prélevés durant cette campagne : des échantillons de
cheminée (solides) et des échantillons de fluides collectés à l'aide d'un incubateur in situ , le Vent Cap.
La troisième planche de photos représente les différents échantillonneurs utilisés durant la campagne.
1.2.1 Echantillons de cheminée
Afin d'étudier la répartition des micro-organismes le long de la cheminée par des méthodes
culturales et moléculaires, les cheminées ont été sous-échantillonnées en suivant un axe longitudinal
et radial (cf. figure 3.3).
PARTIE Top
PARTIE B
PARTIE A
A
A’
B
B’
Section AA’Section BB’
12
12 3
Figure 3.3. Echantillonnage des cheminées prélevées sur le site du Snake-Pit.
Les échantillons ont été prélevés par grattage des cheminées à bord du bateau (Le Nadir) après
collecte de la cheminée à l'aide du sous marin (Le Nautile). Ils ont été conservés à -20 ou -80°C jusqu'à
extraction de l'ADN. Les échantillons conservés pour l'hybridation in situ ont été fixés par la méthode
décrite § 3.3.3.
La technique de conservation des échantillons pour l'extraction d'ADN n'a pas été optimale et a
sûrement contribué à une forte dégradation des échantillons. S'il n'est pas possible de réaliser
l'extraction d'ADN dès la remontée de la cheminée, il est préférable de conserver les échantillons dans
l'éthanol ou dans un milieu réducteur bloquant l'activité des DNAses afin d'éviter la dénaturation des
cellules et la dégradation de l'ADN. Par exemple il est possible de conserver les cellules dans un
tampon GIT (Guanidine isothiocyanate) 5 M, à raison de 1 volume de solution de GIT pour 1 volume
d'échantillon puis on stocke à 4°C.
solution de GIT 5M :
Dissoudre 60 g de GIT dans 5 mL 1 M Tris buffer pH 7,4, 5 mL 0,5 M EDTA (le GIT représente la majeure partie des 98,2 mL :
dissoudre le GIT dans le Tris l'EDTA et très peu d'eau en chauffant la solution au micro-ondes). Ajouter de l'eau MilliQ pour un volume
final de 98,2 mL. Ajouter 800 µl β-mercaptoethanol et 1 mL d’une solution de Sarkosyl stérile 10 % (p/v) (sodium N-lauroyl-
sarcosinate). Conserver la solution à température ambiante.
1.2.2 Echantillons de fluides
Le fluide a été collecté à l'aide d'un échantillonneur, le Vent Cap, qui permet, à la différence des
bouteilles Niskin ou des seringues de titane, de collecter du fluide sur plusieurs jours. Ce dispositif a
été mis au point par l'équipe de Norman Pace (Karl et al., 1988) (cf. planche photos figure 3.4).

1
2
3
4
5
Chambre
d’incubation
Jupe de
collecte
6
Légendes:
Figure 3.4. Echantillonnage des sites hydrothermaux profonds
Planche 3
Crédits photos:
1 IFREMER2,3 Laboratoire de microbiologie Roscoff4, 5, 6 A.L. Reysenbach
1 Submersible ”Le Nautile”2 Seringues de prélévement en titane3 Bouteille NIO4 Vent Cap sur site (2 jupes)5 Vent Cap sur site (1 jupe)6 Vent Cap schéma descriptif
Sonde de température

Matériel et Méthodes
111
Disposé sur le lieu d'émission du fluide (petite cheminée ou fissure), le fluide est dirigé vers une
chambre d'incubation par une jupe en Téflon. Cette chambre est ouverte et fermée par une trappe
manoeuvrable par le submersible. A l'intérieur de la chambre on peut disposer des surfaces de
différents matériaux (Téflon, acier, porcelaine, etc.) qui serviront de support au développement des
micro-organismes. Le fluide ressort de la chambre à l'extérieur de la jupe lorsque la trappe est ouverte.
Les échantillons de fluide contenus dans le Vent Cap ainsi que les coupons sont conservés dans
l'éthanol à - 20°C ou fixés pour des hybridations in situ.
1.3 Echantillonnage des sources chaudes terrestres
Un échantillon provenant d'une source chaude Roumaine a été analysé. Il provient du site de
Caciulata (Roumanie du sud) et a été prélevé (par D. Prieur et I. Gomoiu) à la sortie du puits 1003 ayant
3000 m de profondeur. La température de l'eau était de 89,6°C et le pH de 6,8-6,9. Cette eau très
riche en sel est utilisée dans un système de chauffage.
II Analyse de l'ADNr 16S
L'analyse de la diversité microbienne de nos échantillons s'est déroulée selon le schéma suivant
1. Extraction de l'ADN.
2. Amplification spécifique du gène codant pour l'ARNr 16S par PCR.
3. Clonage des fragments d'ADNr 16S dans un plasmide.
4. Analyse du polymorphisme de longueur des fragments de restriction des clones (RFLP), afin
de déterminer des classes de clones (O.T.U. ou phylotype).
5. Séquençage des différents clones.
6. Analyse des séquences à l'aide d'outils informatiques et construction d'arbres
phylogénétiques.
2.1 Extraction d'ADN
Plusieurs protocoles d'extraction d'ADN ont été utilisés et testés notamment pour extraire l'ADN
des cheminées hydrothermales (cf. figure 3.5A). Différents essais ont été entrepris pour tester
l'efficacité des méthodes d'extraction. Des cellules de E. coli on été ajoutées à un échantillon de
cheminée et l'échantillon a été soumis à différents protocoles d'extraction. Les quantités d'ADN extrait
ont été comparées par Spot -test (cf. figure 3.5B). Les codes utilisés dans cette figure correspondent
à ceux utilisés dans la figure 3.5. Les méthodes qui semblent être les plus efficaces pour les
cheminées hydrothermales sont : la technique du CTAB (2), la technique du micro-ondes (5) et la
technique de Tiedje (6). Pour les échantillons de fluide d'hydrothermaux et d'eaux de production les
protocoles les plus efficaces ont été ceux décrits ci dessous.
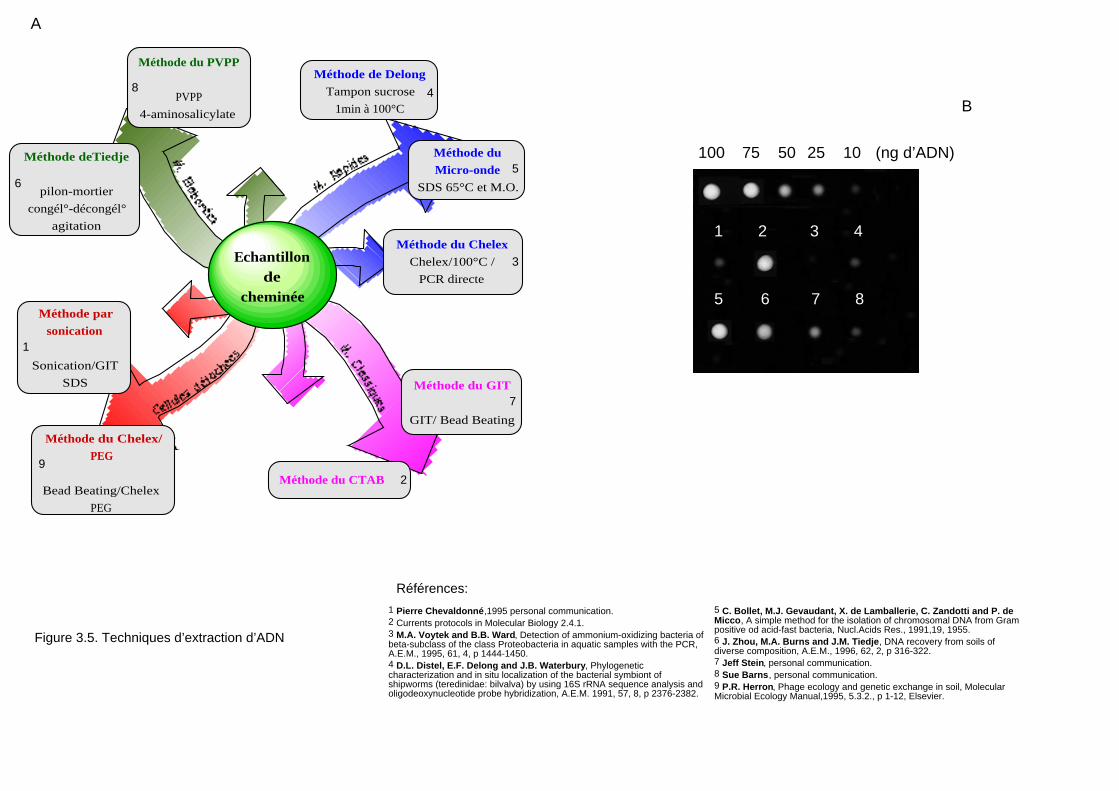
Figure 3.5. Techniques d’extraction d’ADN
1 Pierre Chevaldonné,1995 personal communication.2 Currents protocols in Molecular Biology 2.4.1.3 M.A. Voytek and B.B. Ward, Detection of ammonium-oxidizing bacteria of beta-subclass of the class Proteobacteria in aquatic samples with the PCR, A.E.M., 1995, 61, 4, p 1444-1450.4 D.L. Distel, E.F. Delong and J.B. Waterbury, Phylogenetic characterization and in situ localization of the bacterial symbiont of shipworms (teredinidae: bilvalva) by using 16S rRNA sequence analysis and oligodeoxynucleotide probe hybridization, A.E.M. 1991, 57, 8, p 2376-2382.
5 C. Bollet, M.J. Gevaudant, X. de Lamballerie, C. Zandotti and P. de Micco, A simple method for the isolation of chromosomal DNA from Gram positive od acid-fast bacteria, Nucl.Acids Res., 1991,19, 1955.6 J. Zhou, M.A. Burns and J.M. Tiedje, DNA recovery from soils of diverse composition, A.E.M., 1996, 62, 2, p 316-322.7 Jeff Stein, personal communication.8 Sue Barns, personal communication.9 P.R. Herron, Phage ecology and genetic exchange in soil, Molecular Microbial Ecology Manual,1995, 5.3.2., p 1-12, Elsevier.
Méthode du Chelex/
PEG
Bead Beating/Chelex
PEG
Méthode par
sonication
Sonication/GIT
SDS
Méthode du CTAB
Méthode du Chelex
Chelex/100°C /
PCR directe
Méthode de Delong
Tampon sucrose
1min à 100°C
Méthode du
Micro-onde
SDS 65°C et M.O.
Méthode du GIT
GIT/ Bead Beating
Méthode deTiedje
pilon-mortier
congél°-décongél°
agitation
Méthode du PVPP
PVPP
4-aminosalicylate
Echantillonde
cheminée
1
2
3
4
9
65
8
7
Références:
(ng d’ADN)10255075100
1 2 3 4
5 6 7 8
A
B

Matériel et Méthodes
112
2.1.1 Extraction de l'ADN sur filtre ou sur culot de centrifugation
(d'après Sommerville et al., 1989)
Ajouter sur le filtre ou sur le culot de centrifugation :
1,8 mL de tampon SET (voir composition plus bas) + 20 µL de lysosyme à 20 mg/mL (fragilisation
de la paroi cellulaire). Placer le mélange 15 min sur la glace. Ajouter 20 µL de SDS à 10% et 20 µL de
N-lauryl-sarcosyl à 10% (p/v) (dénaturant des protéines et des polysaccharides). Laisser 1 heure en
agitation à température ambiante. Ajouter 50 µL de protéinase K à 20 mg/mL. Laisser 2 heures à 50°C
(digestion des protéines). La solution est récupérée dans le filtre. Ajouter : 0,5 volume d'acétate
d'ammonium 7.5 M à pH 8 (précipitation des protéines et des polysaccharides). Il est possible à ce
niveau de réaliser une extraction au phénol chloroforme (extraction des protéines), mais il est aussi
possible de passer directement à l'étape suivante. Laisser 10 min à température ambiante. Centrifuger
à 12000 g à 20°C durant 10 min. Eliminer le culot et ajouter 2 volumes d'éthanol 100% au surnageant.
Laisser 20 min à -20°C (précipitation de l’ADN). Centrifuger à 10000 g à température ambiante durant
20 min. Eliminer le surnageant. Resuspendre le culot avec 1 mL d'éthanol à 70% (v/v) (lavage de l’ADN
et élimination des sels). Centrifuger à 13000 RPM à 4°C durant 30 min. Jeter le surnageant et sécher le
culot au SpeedVac.
Dissoudre le culot dans 30 µL d'eau milliQ ou du TE (voir composition ci-dessous).
Composition des tampons
SET : 20% saccharose (p/v) TE : 10 mM Tris
50 mM EDTA 1 mM EDTA
50 mM Tris pH=7,6
pH=7,6
2.1.2 Extraction de l'ADN chromosomique par la méthode de Sambrook et al. (1989)
Chaque milieu est centrifugé et les culots contenant les bactéries sont récupérés. Ces culots
sont resuspendus dans 400 µL de tampon TNE (Tris Cl /NaCl /EDTA). On ajoute 15 µL de lysozyme (5
mg/mL) et 10 µL de la protéinase K à 20 mg/mL. Ce mélange est incubé durant 1 heure à 50°C. Il est
alors possible de rajouter 0,1 volume de SDS à 10% (p/v) et 0,1 volume de L-Lauryl-Sarcosyl à 10%
(p/v) et d'incuber de nouveau pendant 2 heures à 50°C. Afin d'extraire les protéines, 1 volume de
phénol équilibré dans du Tris est ajouté. La phase aqueuse est récupérée après centrifugation de 5 à
10 min à 10000 g. Ajouter 1 volume d’un mélange contenant du phénol saturé TRIS, du chloroforme
et de l’alcool isoamylique (24:24:1) avant de centrifuger durant 10 min à 6000 g. La phase aqueuse
contenant l’ADN est récupérée et traitée au chloroforme et à l’alcool isoamylique (24:1), et centrifugée
à nouveau. L’ADN est précipité avec 0,1 volume d'acétate de sodium 3 M et 2 volumes d'éthanol
100% durant 1 heure à -20°C avant une dernière centrifugation. Le culot contenant l’ADN précipité est
séché puis repris dans de l'eau MilliQ ou dans du TE.

Matériel et Méthodes
113
2.1.3 Méthode d'extraction au micro-ondes
(d'après Bollet et al., 1991)
Laver les cellules ou l'échantillon dans du TE ou du TNE. Ajouter du SDS à une concentration
finale de 5% (p/v). Mettre 30 min à 65°C. Centrifuger et supprimer le surnageant. Mettre le culot au
micro-ondes 2 fois 1 min à 900 W (ou 3 x 1 min à 750 W). Resuspendre le culot dans du TNE et
effectuer une extraction au phénol-chloroforme suivie d'une précipitation de l'ADN à l'éthanol.
2.1.4 Extraction de l'ADN chromosomique par la méthode de Pitcher et al. (1989)
Cette méthode peut être utilisée à la place de la méthode traditionnelle décrite par Sambrook et
al., (1989) pour extraire l'ADN à partir de cultures pures. Il conviendra de rajouter du L-Lauryl-Sarcosyl
pour lyser plus efficacement les Archaea.
1,5 mL de culture en phase exponentielle (ou d'échantillon) sont centrifugés durant 5 min à
5000 g. Les cellules sont resuspendues dans 100 µL de tampon TE et lysées avec 0,5 mL de tampon
GES (5 M guanidium thiocyanate, 100 mM EDTA, 0,5% v/v SDS, pH 8). Les cellules en suspension
sont agitées brièvement et laissées à température ambiante durant 10 min. Le lysat est refroidi sur
glace et on ajoute 0,25 mL d'acétate d'ammonium 7,5 M froid. Les échantillons sont maintenus sur
glace pendant 10 min et occasionnellement agités. On ajoute 0,5 mL de chloroforme/2-pentanol
(24:1). Après agitation les phases sont séparées par 2 min de centrifugation à 2000 g. La phase
aqueuse est transférée dans un nouveau tube et 0,54 volume d'isopropanol froid est ajouté. On agite
les tubes par inversion pendant 10 min et l'ADN précipité est recueilli par centrifugation durant 1 min à
7000 g. Les culots d'ADN sont lavés à l'éthanol 70% (v/v) et séchés (Speed Vac ou air ambiant). L'ADN
est resuspendu durant une nuit dans 100 µL de tampon TE stérile (pH 8).
2.1.5 Extraction de l'ADN au "Bead-Beater"
Cette technique a été utilisée avec succès par la société Diversa et par Paul Riley (1997) pour
extraire l'ADN de sédiments ou de cheminée.
Utiliser 10 à 20 grammes de cheminée ou de sédiments. Ajouter un volume de billes pour "bead-
beating". Puis ajouter un volume (billes + échantillons) de solution de 5M GIT contenant 0.1% (p/v) de
SDS. Agiter vigoureusement (Bead-beater ou Ribolyser) durant 30 min à 1 h. Centrifuger à faible
vitesse pour éliminer les débris cellulaires et extraire l'ADN de la solution de GIT. Le culot est
resuspendu dans un tampon de lyse (TE/SDS 10% (p/v) par exemple) et traité par la protéinase K et
subit ensuite une extraction phénol/chloroforme.
2.2 Amplification de l'ADNr 16S
Afin d'amplifier l'ADNr 16S de nos échantillons nous avons utilisé une technique d'amplification
classique. Il a toutefois été nécessaire d'introduire des modifications dans la technique lorsque l'ADN
extrait était en trop faible quantité ou lorsque des contaminants (hydrocarbures, métaux) inhibaient la
réaction.
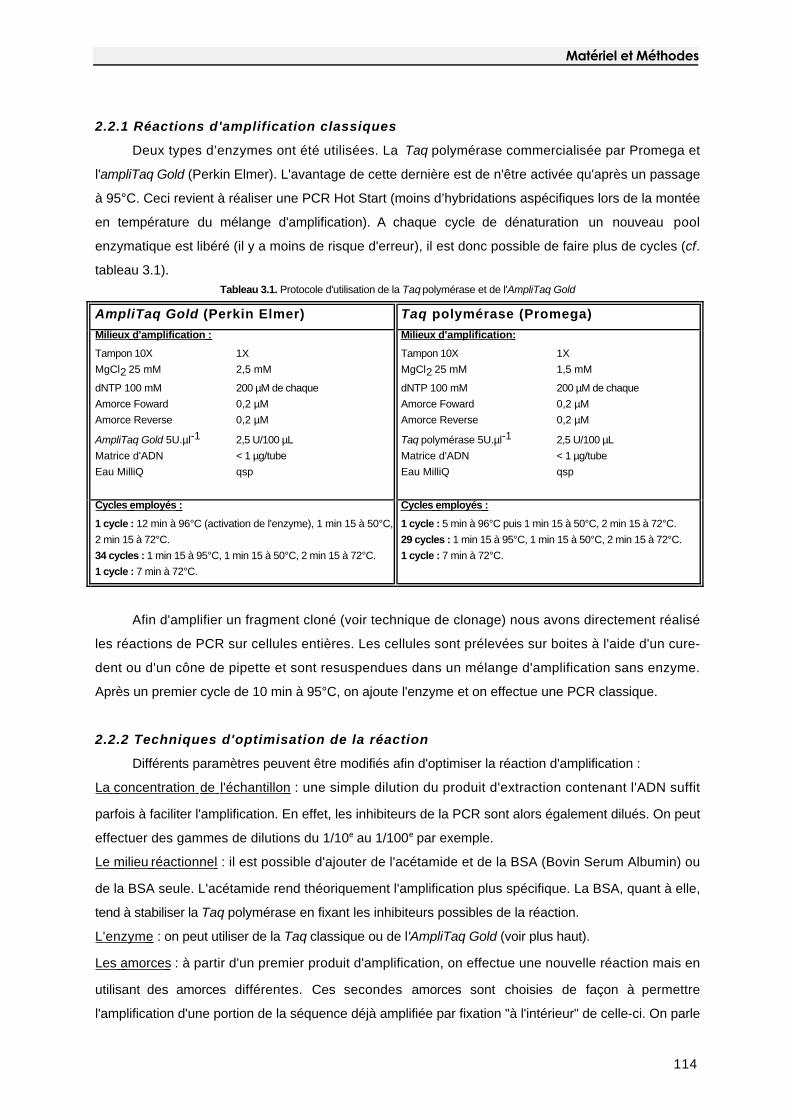
Matériel et Méthodes
114
2.2.1 Réactions d'amplification classiques
Deux types d’enzymes ont été utilisées. La Taq polymérase commercialisée par Promega et
l'ampliTaq Gold (Perkin Elmer). L'avantage de cette dernière est de n'être activée qu'après un passage
à 95°C. Ceci revient à réaliser une PCR Hot Start (moins d’hybridations aspécifiques lors de la montée
en température du mélange d'amplification). A chaque cycle de dénaturation un nouveau pool
enzymatique est libéré (il y a moins de risque d'erreur), il est donc possible de faire plus de cycles (cf.
tableau 3.1).Tableau 3.1. Protocole d'utilisation de la Taq polymérase et de l'AmpliTaq Gold
AmpliTaq Gold (Perkin Elmer) Taq polymérase (Promega)
Milieux d'amplification :
Tampon 10X 1X
MgCl2 25 mM 2,5 mM
dNTP 100 mM 200 µM de chaque
Amorce Foward 0,2 µM
Amorce Reverse 0,2 µM
AmpliTaq Gold 5U.µl-1 2,5 U/100 µL
Matrice d'ADN < 1 µg/tube
Eau MilliQ qsp
Milieux d'am plification:
Tampon 10X 1X
MgCl2 25 mM 1,5 mM
dNTP 100 mM 200 µM de chaque
Amorce Foward 0,2 µM
Amorce Reverse 0,2 µM
Taq polymérase 5U.µl-1 2,5 U/100 µL
Matrice d'ADN < 1 µg/tube
Eau MilliQ qsp
Cycles employés :
1 cycle : 12 min à 96°C (activation de l'enzyme), 1 min 15 à 50°C,
2 min 15 à 72°C.
34 cycles : 1 min 15 à 95°C, 1 min 15 à 50°C, 2 min 15 à 72°C.
1 cycle : 7 min à 72°C.
Cycles employés :
1 cycle : 5 min à 96°C puis 1 min 15 à 50°C, 2 min 15 à 72°C.
29 cycles : 1 min 15 à 95°C, 1 min 15 à 50°C, 2 min 15 à 72°C.
1 cycle : 7 min à 72°C.
Afin d'amplifier un fragment cloné (voir technique de clonage) nous avons directement réalisé
les réactions de PCR sur cellules entières. Les cellules sont prélevées sur boites à l'aide d'un cure-
dent ou d'un cône de pipette et sont resuspendues dans un mélange d'amplification sans enzyme.
Après un premier cycle de 10 min à 95°C, on ajoute l'enzyme et on effectue une PCR classique.
2.2.2 Techniques d'optimisation de la réaction
Différents paramètres peuvent être modifiés afin d'optimiser la réaction d'amplification :
La concentration de l'échantillon : une simple dilution du produit d'extraction contenant l'ADN suffit
parfois à faciliter l'amplification. En effet, les inhibiteurs de la PCR sont alors également dilués. On peut
effectuer des gammes de dilutions du 1/10e au 1/100e par exemple.
Le milieu réactionnel : il est possible d'ajouter de l'acétamide et de la BSA (Bovin Serum Albumin) ou
de la BSA seule. L'acétamide rend théoriquement l'amplification plus spécifique. La BSA, quant à elle,
tend à stabiliser la Taq polymérase en fixant les inhibiteurs possibles de la réaction.
L'enzyme : on peut utiliser de la Taq classique ou de l'AmpliTaq Gold (voir plus haut).
Les amorces : à partir d'un premier produit d'amplification, on effectue une nouvelle réaction mais en
utilisant des amorces différentes. Ces secondes amorces sont choisies de façon à permettre
l'amplification d'une portion de la séquence déjà amplifiée par fixation "à l'intérieur" de celle-ci. On parle

Matériel et Méthodes
115
de "PCR nichée". Par exemple, si le premier couple d'amorces utilisé est 8F-1525R, le second peut
être 50F-1492R. La séquence alors amplifiée sera légèrement plus courte mais le rendement de la
"PCR interne" est parfois meilleur car les amorces peuvent s'hybrider sur plus de matrice.
Le nombre de cycles effectués : sur des produits ayant subi une première réaction, il est possible de
rajouter de l'enzyme (1 ou 1,5 U) et de relancer un nouveau cycle de PCR. La première amplification
est parfois insuffisante pour permettre de visualiser une bande lors de la vérification sur gel.
Réparation préalable du matériel génétique : lors d'un cycle particulier (1 fois : 5 min à 95°C puis 10 fois
: 30 sec à 95°C, 30 sec à 54°C et 45 sec à 72°C) on met l'ADN extrait en présence d'un milieu
réactionnel classique de PCR mais sans amorce. Il se produit alors une réparation de l'ADN; c'est-à-dire
que la polymérase ajoute des dNTP aux endroits dégradés. Les petits fragments d'ADN servent en
quelque sorte d'amorces à l'enzyme. Cette technique est susceptible d'augmenter la quantité de
chimères.
2.2.3 Choix des amorces d'amplification
Les amorces suivantes (cf. tableau 3.2) sont utilisées pour les différentes réactions
d'amplification.Tableau 3.2. Spécificité et description des amorces d'amplification
Amorces Localisation Séquence Spécificité
E. coli Archaea Bacteria Eukarya
Orientation “Forward”
4F (3-20) 5'- TCCGGTTGATCCTGCCRG -3' +
21F (2-21) 5'-TTCCGGTTGATCCYGCCGGA-3' +
333F (333-348) 5'- TCCAGGCCCTACGGG -3' +
8F (8-27) 5'- AGAGTTTGATCCTGGCTCAG -3' +
50F (50-68) 5'- AACACATGCAAGTCGAACG -3' +
515F (515-533) 5'-GTGCCAGC(AC)GCCGCGGTA-3' + + +
Orientation “Reverse”
1492R (1510-1492) 5'- CGGTTACCTTGTTACGACTT -3' + + +
1525R (1541-1525) 5'- AAGGAGGTGATCCAGCC -3' + + +
Universal MCS 5'- GGAAACAGCTATGACCAT -3' hybridation avec
Reverse MCS 5'- GTAAAACGACGGCCAGTG -3' le plasmide
MCS: site multiple de clonage localisé sur le plasmide de chaque coté de l'insert.
2.3 Clonage des produits de PCR
Deux techniques de clonage ont été utilisées durant ces études. La plus fréquemment utilisée
est la technique du “TA cloning". Il a été aussi possible de transformer les cellules en utilisant un
électroporateur.
2.3.1 Utilisation du kit TOPO TA Cloning (Invitrogen )
Le protocole de clonage repose sur une erreur de la Taq polymérase qui ajoute
systématiquement un dNTP de type adénine (A) aux extrémités 3' du brin néosynthétisé. Le vecteur
utilisé possède, quant à lui, une thymine (T) au niveau du site d'insertion, à ses extrémités 3'. Une

Matériel et Méthodes
116
liaison entre ses deux bouts cohésifs est donc possible. Elle est facilitée par l'action d'une
topoisomérase qui va liguer les deux fragments d'ADN à température ambiante.
Clonage dans le plasmide pCR 2.1 et transformation:
Le protocole de clonage et de transformation est celui décrit par le fabriquant.
Analyse des clones positifs :
Couler en boîte de Pétri un milieu LB additionné de 0.1 g/L d'ampicilline et de 0.3 mL/L de tétracycline
à 50µg/mL. Appliquer sur le milieu une solution contenant 40µL de X-Gal 5% dans du DMSO (p/v) et
40µL d'IPTG 100 mM. Etaler 80 µl de la solution de transformation. Incuber au minimum 18 heures à
37°C. Puis placer les boîtes environ 2 heures à 4°C pour faire apparaître la coloration bleue. On
repique les colonies blanches sur des boîtes de Pétri contenant du milieu LB additionné d'ampicilline
et de tétracycline (même milieu que précédemment), ceci afin de les conserver. On réalise également
une PCR suivie d'une électrophorèse sur gel d'agarose 0.8% (p/v) pour vérifier la taille de l'insert. La
PCR est effectuée directement sur les souches contenues dans le milieu de réaction à l'aide des
amorces “universal et reverse“ s'hybridant sur le plasmide.
Conservation des clones :
Il est possible de conserver les clones obtenus en les cultivant sur microplaques dans du milieu LB
liquide. On ajoute par la suite 5% (final) de glycérol à la culture. La microplaque protégée (film plastique
ou couvercle) peut être conservée à - 20°C (pendant une nuit) puis - 80°C (pendant plusieurs mois).
Composition des milieux utilisés :
Milieu LB (Luria-Bertani) : 1.0% Tryptone, 0.5% d'extrait de levure (Yeast Extract), 1.0% NaCl, pH 7.0
Gélose LB : milieu LB + 15 g/L d'agar avant d'autoclaver
IPTG = isopropylthio-ß-D-galactoside
X-Gal = 5-bromo-4-chloro-3 indolyl-ß-D-galactopyranoside
2.3.2 Transformation des cellules par électroporation
Une autre technique de transformation des cellules a été utilisée. Moins rapide que la technique
du choc thermique, la technique d'électroporation est toutefois très efficace.
1- Dessaler les produits de ligation. S'il y a trop de sel dans les échantillons ou si la cuvette est trop
vieille, celle-ci peut exploser durant l'électroporation. Utiliser 2 µL de produit de ligation. Les déposer
sur un filtre 0,022 µm flottant dans une boite de Pétri remplie d'eau désionisée. Effectuer la dialyse
durant 45 min à 1 heure.
2- Décongeler les cellules compétentes (sur la glace) en évitant les chocs thermiques. Mettre la
cuvette d'électroporation dans la glace. Aliquoter dans des tubes de 5 mL, 50µL de cellules
compétentes. Conserver ces tubes au froid. Ajouter les 2 µL de produit de ligation dialysé aux cellules
compétentes. Laisser 1 min sur glace.

Matériel et Méthodes
117
3- Mettre les cellules dans la cuvette à la fin de la minute et procéder à l'électroporation.
4- Puis rapidement : Récupérer la cuvette et y ajouter directement 1 mL de milieu SOC puis
retransférer le tout dans un tube. Mettre les tubes à 37°C durant 45 min avec agitation.
5- Déposer entre 100 et 800 µL de la culture sur boite de Pétri en fonction de l'efficacité de la
transformation. Mettre les boites à 37°C durant la nuit.
2.4 Analyse des clones par RFLP : Restriction Fragment Length
Polymorphism
Afin de réaliser un premier tri parmi les clones à séquencer, nous avons analysé ces derniers par
RFLP. A chaque profil de restriction correspondait une classe, ou phylotype ou OTU. A chaque classe
correspondait des séquences différentes mais au sein d'une même classe on pouvait retrouver des
séquences différentes. Le choix des enzymes a été dicté par les résultats obtenus par Moyer et al.
(1996). MspI et HinP1I (ou HhaI son isoschyzomère) sont parmi les enzymes les plus discriminantes sur
l'ADNr 16S.
Site de coupure des enzymes de restriction :
MspI → 5’ C*CGG 3’ HinP1I → 5’ G*CGC 3’ HhaI → 5’ GCC*C 3’
3’ GGC*C 5’ 3’ CGC*G 5’ 3’ C*GCG 5’
Coupure :
Sur une microplaque, on met en présence l'ADN, le tampon et l' (les) enzyme(s) dans les
proportions suivantes (pour un puits) : 9 µL d’ADN, 1,2 µL de tampon, 0,6 µL HinP1I ou HhaI, 0,3 µl
MspI. La microplaque est placée au bain-marie à 37°C durant au moins deux heures. L'ADN matrice est
le produit de PCR ayant permis de vérifier l'efficacité du clonage.
Electrophorèse des produits de restriction:
L'électrophorèse est réalisée dans des gels résolutifs pour des fragments de (100 à 1500 pb).
Idéalement elle sera effectuée dans un gel d'agarose "NU Sieve" à 4% (p/v) dans un tampon TAE. On
pourra aussi pour des raisons d'économie faire la migration dans du gel "Sea Kem LE" à 2% (p/v) avec
un tampon TAE. Les échantillons sont chargés avec un tampon de charge contenant de l'Orange G
afin de détecter la sortie des bandes à faible poids moléculaire. Le marqueur couramment utilisé est le
"100 bp DNA ladder" (5µL par puits soit 5 U) . Ce marqueur comporte 11 fragments d’ADN double-brin
de 100 à 1500 paires de bases .
Composition des produits d'électrophorèse
Tampon TAE : 0.04 M Tris-acétate Loading Buffer : 0.25% Orange G (p/v)
0.001 M EDTA 40% saccharose dans du TE (p/v).

Matériel et Méthodes
118
2.5 Séquençage
Si une des séquences a été réalisée manuellement (souche de Bacillus SG9) la majeure partie
des séquences réalisées durant cette thèse (clones et autres souches) l'ont été à l'aide d'un
séquenceur automatique. Nous ne décrirons donc que cette méthode de séquençage.
2.5.1 Réactions de séquence
Le principe de séquençage utilisé est celui développé par Sanger : l'incorporation de
didésoxynucléotides. A ce principe de séquençage a été couplée une réaction de PCR afin d'amplifier
le signal de séquençage. Il s'agit donc de séquençage cyclique ou "cycle sequencing". L’amorce est
marquée au moyen d'un fluorochrome (Texas Red) fixé aux extrémités 3' du simple brin et l'enzyme
Thermo Séquénase, analogue de la Taq polymérase, est thermostable et très processive.
Les conditions de séquençage sont celles décrites par le fabriquant du kit de Séquençage
(Amersham). Seul le cycle de PCR est modifié et l'ADN matrice utilisé est le produit de PCR obtenu
après amplification des clones à l'aide des amorces "universal et reverse".
Cycle : 97°C - 5 min (1 cycle); 95°C - 1 min, 54°C - 1 min, 61°C - 1 min (24 fois).
2.5.2 Amorces de séquençage
Les amorces utilisées pour le séquençage (cf. tableau 3.3) sont semblables à celles utilisées pour
l'amplification de l'ADNr 16S. Il est nécessaire d'utiliser des amorces internes à l'ADNr 16S afin de
couvrir la totalité de la séquence de la molécule.
Tableau 3.3. Spécificité et description des amorces de séquençage.
Amorces Localisation Séquence Spécificité
E. coli Archaea Bacteria Eukarya
Forward
4F (3-20) 5'- TCCGGTTGATCCTGCCRG -3'TRed +
8F (8-27) 5'- AGAGTTTGATCCTGGCTCAG -3'TRed +
515F (515-533) 5'- GTGCCAGCMGCCGCGGTAA -3'TRed +/- + +
906F (906-922) 5'- GAAACTTAAAKGAATTG -3'TRed + +
Reverse
515R (533-515) 5'- TTACCGCGGCKGCTGGCAC -3'TRed +/- + +
906R (922-906) 5'- CAATTCMTTTAAGTTTC -3'TRed + +
1492R (1510-1492) 5'- CGGTTACCTTGTTACGACTT -3'TRed + + +
Universal MCS 5'- GGAAACAGCTATGACCAT -3'TRed hybridation avec
Reverse MCS 5'- GTAAAACGACGGCCAGTG -3'TRed le plasmide
MCS: site multiple de clonage localisé sur le plasmide de chaque coté de l'insert. IUB nomenclature: M=A ou C, K=G ou T.

Matériel et Méthodes
119
2.5.3 Migration des réactions de séquences
L'obtention d'une séquence nécessite donc quatre dépôts (1 par base). Le gel utilisé a un fort
pouvoir résolutif. C'est un gel dénaturant à 7% d'acrylamide-bisacrylamide. Le caractère dénaturant est
dû à la présence d'urée (6M)1.
Pour 30 mL de gel à 7% de polyacrylamide : 4,2 mL de Long Ranger 50% (mélange Acrylamide-
Bisacrylamide) TEBU-FMC Bio Products, 11g d'urée, qsp 27 mL d'eau MilliQ. Le mélange est
désionisé (aide d'une résine échangeuse d'ions) et 3 mL de TBE X10 filtrés sont ajoutés. Avant de
couler le gel on ajoute les agents polymérisants : 150µL de TEMED et 15µL d'APS 10% (p/v)2. Le gel
est coulé entre deux plaques de verre borosilicaté et polymérisé durant 45 min à 1 heure. Le gel est
placé verticalement dans le séquenceur. Un peigne à dents de requin permet de former les puits. Les
échantillons peuvent ensuite être chargés.
Principe du séquenceur
La migration de 10 réactions de séquences (40 puits) s'effectue verticalement (cf. figure 3.6).
Les brins les plus courts migrent plus rapidement; ils vont donc se retrouver les premiers en bas du
gel. 512 photodiodes sont situées en ligne au bas du gel. Un faisceau laser de longueur d'onde 594
nm traverse le gel dans sa largeur et excite les fluorochromes qu'il rencontre. Les photodiodes
enregistrent l'intensité lumineuse émise à 612 nm par les fluorochromes excités. Le signal est
numérisé et transmis à l'ordinateur qui enregistre et analyse les données.
Figure 3.6. Représentation schématique du fonctionnement d'un séquenceur automatique.
1 La teneur en urée du gel peut varier de 4 à 7 M.2 APS = Ammonium Persulfate, Long Ranger = Acrylamide et N,N'-méthylenebisacrylamide 50%, TEMED = N,N,N',N'Tétraméthyléthylènediamine

Matériel et Méthodes
120
2.6 Analyse des séquences
Les principes d'analyse des séquences ont été rappelés dans la première partie de ce
manuscrit. Après migration les séquences sont vérifiées afin d'éliminer d'éventuelles erreurs de
lecture (logiciel du séquenceur Vistra V 2.0). Puis elles sont comparées aux banques de données
(RDP et GenBank), alignées contre les séquences les plus proches et assemblées (logiciel ARB).
L'analyse phylogénétique est réalisée par la suite en comparant les résultats obtenus par les
techniques d'analyse des distances, de maximum de parcimonie et de maximum de vraisemblance
(logiciel ARB).
III Hybridation in situ
La technique d'hybridation in-situ peut elle aussi se diviser en plusieurs étapes : la construction
des sondes, la mise au point des sondes et l'hybridation in situ.
3.1 Construction des sondes
La construction des sondes est réalisée sur ordinateur. A partir d'un choix de séquences le
logiciel ARB permet de construire des sondes spécifiques des séquences à cibler. Les paramètres à
considérer sont le nombre de mésappariements, la longueur de la sonde, le Tm relatif de la sonde, le
G+C% de la sonde (plus il est important plus le signal d'hybridation sera fort).
A partir de la liste de sondes fournie par le logiciel il convient de déterminer la plus efficace , c'est à dire:
Ø la sonde qui ciblera le plus grand nombre d'organismes que l'on souhaite marquer.
Ø la sonde qui aura un nombre de mésappariements suffisant avec les micro-organismes
que l'on souhaite exclure; l'idéal est 3 mésappariements répartis sur toute la longueur
de la sonde (notamment au centre de la sonde).
Ø la sonde qui s'hybridera sur une région de l'ADNr 16S où la structure secondaire de la
molécule n'empêchera pas sa fixation (cf. la structure secondaire analysée par Fusch et
al., 1999 et voir exemple de structure secondaire en annexe).
3.2 Mise au point des sondes - Test de spécificité
Avant de réaliser les hybridations in situ il convient de tester la spécificité de la sonde sur
membrane (pour vérifier la fixation de la sonde sur l'ADN) et sur culture pure (afin de vérifier la
pénétration de la sonde dans la cellule et son hybridation sur la structure secondaire de l'ARN).
3.2.1 Hybridation sur membrane
On va déposer sur membrane l'ADN génomique, l'ARNr ou l'ADNr de souches dont on souhaite
vérifier l'hybridation avec la sonde. Il convient de choisir à la fois:
Ø des espèces qui sont spécifiquement ciblées par la sonde,

Matériel et Méthodes
121
Ø des espèces de groupes phylogénétiques proches et éloignés ne s'hybridant pas avec la sonde,
Ø des espèces non recherchées mais pour lesquelles la sonde présente peu de mésappariements.
Il est possible de fixer sur la membrane des ARNr ou de l'ADNr 16S. Les ARN sont extraits à
l'aide des techniques classiques d'extraction (cf. Harmsen et al., 1997a). Les ADNr 16S déposés sur la
membrane peuvent être le produit d'amplification de l'ADNr 16S de souches pures ou d'ADNr 16S
clonés.
Dépôt sur membrane :
On dépose sur la membrane (par puits) soit 30 ng d'ARNr soit 100 ng d'ADNr. Avant dépôt,
l'appareil est nettoyé à l'eau MilliQ et séché à l'éthanol absolu. La membrane (Zeta Probe, GT-BioRad
ou Hybond N+-Amersham) est humidifiée par flottaison dans de l'eau MilliQ stérile puis séchée sur
papier Whatman. Les ADN ou les ARN sont dénaturés durant ce temps dans 1 volume de 0,4N NaOH,
4 mM EDTA 10 min à 100°C. On ajoute 0,1 volume de solution stop (2M d'acétate d'ammonium +
Bleu). Les échantillons sont conservés sur glace avant leur dépôt. Le volume total est de 50µL/dépôt.
On dépose les matrices dénaturées sur la membrane à l'aide d'un appareil à Dot-Blot ou à Slot-Blot.
Après séchage sommaire (sur papier Whatman) et emballage dans du film alimentaire, les matrices sont
fixées sur la membrane à l'aide d'un Cross linker (2 cycles de 30 mJoules). Les membranes sont
conservées à - 20°C.
Marquage de la sonde :
La sonde est commandée non marquée. Pour les tests de spécificité la sonde est marquée à
l'[γ -32P]ATP à l'aide de la T4 polynucléotide kinase. 100ng (dans 1µL) de sonde sont marqués à
chaque réaction. On ajoute 2µL de tampon, 16µL d'eau MilliQ stérile, 1 µL de sonde et 0,5 à 1 µL de
T4-Kinase et pour finir 1 µL de [γ -32P]ATP.
Hybridation sur membrane :
Les membranes sont préhybridées dans un volume minimal de tampon d'hybridation (10 à 20
mL) préchauffé durant 15 min à 45°C. Après marquage, la sonde (20µL) est introduite dans le tube et
l'hybridation se déroule durant la nuit à 45°C. Le lendemain, le mélange d'hybridation est éliminé et la
membrane est lavée 2 ou 3 fois avec du tampon 5X SSC. 20 mL de tampon de lavage préchauffé
(45°C) sont ajoutés et la membrane est incubée 20 min à 45°C. Le tampon est renouvelé et la
membrane est incubée à Td (voir plus bas) durant 1 heure au moins. Les membranes sont emballées
(humides) dans du film alimentaire et déposées contre un écran photo-stimulable. Les écrans sont lus
après 15 min à 1 heure d'exposition sur un Fluorimager (Storm, Molecular Dynamics). Les membranes
sont stockées à -20°C jusqu'à leur lavage.
Tampon d'hybridation Tampon SSCx20 Tampon de lavage
0,5M tampon phosphate 0,15 M NaCl 1X SCC
7% SDS (p/v) 0,015M Citrate de sodium 1% SDS (p/v)
1% BSA (p/v)
1 mM EDTA pH7,2

Matériel et Méthodes
122
Mesure de la T d :
La technique de mesure de la Td est celle mise au point par Raskin et al. (1994a). Elle consiste a
effectuer des lavages successifs à des températures de lavage croissantes (avec un pas de 3 °C par
lavage par exemple).
CPM
Température de lavage
2000
4000
6000 CPM max
Td
CPM 50%
40 50 60 70 80 °C
Figure 3.7. Mesure de la température de dissociation (Td)
Dans un premier temps, la sonde est hybridée avec sa cible en duplicat ou en triplicat puis est
lavée comme précédemment 2 à 3 fois par 5X SSC. Après ce premier lavage, chaque "dot" est
découpé et déposé dans des tubes à scintillation contenant 2,5 mL de tampon de lavage préchauffé.
Le premier lavage est effectué à 38°C durant 20 min (pour supprimer l'excès de sonde). Puis un
deuxième lavage est effectué à 38°C durant 45 min à 1 heure. Après chaque lavage le "dot" est
transféré dans un nouveau tube à scintillation contenant du tampon de lavage. L'ancien tube est
rempli de 2,5 mL de liquide à scintillation et est conservé jusqu'au comptage. On procède alors à un
nouveau lavage à 41°C durant 45 min à 1 heure (puis à 44, 47, 50, etc., jusqu'à 71 ou 74°C). Lorsque
tous les lavages sont effectués, les tubes sont comptés et la courbe de valeur cumulées de
CPM=f(température de lavage) est construite. La Td est la température pour laquelle 50% de la
radioactivité aura été libérée (cf. figure 3.7).
Lavage des membranes pour réhybridation :
Les membranes peuvent être réhybridées plusieurs fois pour peu qu'elles aient été lavées.
Pour ce faire les membranes froides (-20°C) sont plongées dans une solution chaude (85°C) de 0,5%
SDS (p/v) dans de l'eau MilliQ. Elles y sont laissées durant 30 min au minimum. Puis les membranes
sont lavées avec du tampon de lavage froid, emballées dans du film alimentaire et stockées à -20°C.
3.2.2 Hybridation sur culture pure
Pour les hybridations sur culture pure, on choisit généralement deux souches. Chacune d'entre
elles s'hybride avec la sonde générale (EUB 338 par exemple pour la sonde bactérienne) et une seule
des souches s'hybride avec la sonde à analyser. Les hybridations sur souche pure permettent
d'optimiser la stringence du tampon d'hybridation en utilisant différentes concentrations de formamide.
Pour les conditions d'hybridation voir (§ 3.3).

Matériel et Méthodes
123
3.3 Hybridation in situ
3.3.1 L'enrobage des lames d'observation
Avant le dépôt des cellules, les lames sont traitées. Laver les lames dans du KOH-éthanol (10%
KOH : 10 mL 5M KOH+ 40 mL éthanol). Rincer abondamment avec de l'eau MilliQ (2X). Plonger dans
une solution chaude (70°C) de gélatine 0,1% (p/v) et CrKSO4 0,01% (p/v). Sécher à l'air et conserver à
l'abri de l'humidité.
3.3.2 Fixation des cellules
Cette technique s'applique aux cellules obtenues par culture pure ou aux échantillons.
Préparation de la solution de fixation:
Chauffer 20 mL d'eau MilliQ dans un tube de 50 mL à 60°C (micro-onde). Ajouter 2 g de
paraformaldéhyde. Ajouter 100 µl de NaOH 5N et agiter. La poudre doit se dissoudre rapidement.
Ajouter 100 µl HCl 5N. Ajouter 15 mL PBS 10x et 10 mL d'eau. Refroidir la solution à température
ambiante. Equilibrer le pH à 7,2 et ajuster à 50 mL. Stériliser par filtration à 0,2 µm.
Fixation des cellules
Centrifuger 1 mL de culture en fin de phase logarithmique dans un microtube à 4000 g durant 5 min.
Supprimer le surnageant. Resuspendre le culot dans 1 mL de PBS 3x et centrifuger à 4000 g durant 5
min. Resuspendre le culot dans 1 mL de solution de fixation (paraformaldéhyde 4% (p/v) dans du PBS
3x). Fixer les cellules durant 3 heures à 4°C (au minimum) ou 1,5 heure à température ambiante.
Centrifuger 5 min à 3000 g (attention les cellules sont fragiles). Resuspendre le culot dans 1 mL de
PBS 3x et centrifuger 3000 g (optionnel). Resuspendre le culot dans 100 µL de PBS 1x /0,1%
IGEPAL (p/v) et ajouter 100 µL d'éthanol. Stocker à -20°C.
3.3.3 Extraction des cellules de l'échantillon
Afin de limiter le bruit de fond lié à des hybridations non spécifiques des sondes sur des
morceaux de cheminée, il a été nécessaire d'extraire les cellules de l'échantillon avant de réaliser les
hybridations. Les cellules sont extraites après agitation avec un agent chélateur (le Chelex) et
migration sur un gradient de sucrose.
Déposer l'échantillon fixé dans un tube de 25 mL. Ajouter 5 mL de PBS 1x/2% PEG8000 (p/v)
et 1 g Chelex-100. Agiter vigoureusement. Puis agiter durant 1 heure à 4°C à l'aide d'un agitateur
horizontal. Centrifuger à 700 g pendant 1 min et récupérer le surnageant. Effectuer deux nouvelles
extractions sur le culot en ne rajoutant que 3 mL de PBS 1x/2% PEG8000 (p/v). Après centrifugation,
rassembler les surnageants et les centrifuger 5 min à 700 g. Le surnageant est finalement centrifugé
10 min à 4000g. Le culot (contenant les cellules) est resuspendu dans 1 mL de PBS 1x/0,1% IGEPAL
(p/v) (Sigma) et est déposé à la surface de 2 mL d'une solution de PBS 1x/0,1% IGEPAL (p/v)/ 50%
Ficoll (p/v) (Sigma). Après 1 min de centrifugation à 4000 g les cellules sont récupérées à la surface du
gradient de Ficoll et sont lavées dans du PBS 1x/0,1% IGEPAL (p/v). Elle sont resuspendues dans 60
µL de PBS 1x/ 0,1% IGEPAL (p/v) et 60µL d'éthanol 100%.

Matériel et Méthodes
124
3.3.4 Hybridation in situ
Il faut dans un premier temps fixer les cellules sur la lame puis réaliser l'hybridation. Les lames
sont ensuite lavées et conservées jusqu'à leur observation.
Fixation des cellules sur la lame :
Essuyer les côtés de la lame avec du papier Whatman. Ajouter 1 à 5 µL de la solution de cellules fixées
(1µL pour les cultures pures). Laisser sécher à l'air en agitant avec un cône de pipette. Sécher 10 min à
46°C. Pour les bactéries Gram + ajouter 10 µL de lysozyme à 2 mg/mL (5 min à température ambiante).
Laver à l'eau MilliQ et sécher à l'air. Sécher les cellules en transférant les lames dans des tubes de 50
mL contenant : 50% (v/v) d'éthanol dans l'eau MilliQ (5 min), 70% (v/v) d'éthanol dans l'eau MilliQ (5
min), 96% (v/v) ou éthanol pur (5 min). Sécher les lames à l'air comprimé.
Chambre humide :
Utiliser un petit récipient avec un couvercle. Mettre des serviettes en papier au fond et les humidifier
avec une solution de la même force ionique que le tampon d'hybridation. On peut utiliser du tampon
d'hybridation ou une vieille solution de lavage. Préchauffer la chambre à 48°C.
Hybridation :
0,5 µL de sonde (50 ng pour chacune des deux sondes) sont mélangés à 9 µL de tampon
d'hybridation (Les solution filles de sondes sont préparées à 100 ng/µL; la solution stock est
généralement à 1µg/µl). Déposer les 10 µl dans les puits de la lame. Transférer rapidement dans la
chambre humide pour éviter l'évaporation. Incuber 3h à la température d'hybridation (généralement à
46°C).
Stringence des tampon d'hybridation (% de formamide)
Pour Eub/Arch : 20% (optimum) 40% (maximum). Pour Cren/Eury : 0%.
Pour Thermus : 60%. Pour Thermus/Eub: 60%. Pour Bacillus : 40%. Pour Bacillus/Eub: 40%.
Pour Hydrogeno : 60%. Pour Hydr/Eub: 60%. Pour Toga : 20 à 40%. Pour Toga/Eub: 20 to 40%.
Pour Epsi : 40%. Pour Epsi/Eub: 40%. Pour GroupC : 40%. Pour GroupC/Eub: 40%.
Pour Cren/Arch : 30% Pour Arch : pas plus de 40%
Tampon d'hybridation Tampon de lavage
0,9M NaCl 180mM NaCl
20mM Tris pH 7,5 20mM Tris pH 7,5
0,1% SDS (p/v)
% de formamide désiré
(voir articles 2 et 3 pour la description des sondes)
Lavage des cellules :
Préchauffer 50 mL de solution de lavage dans un tube de 50 mL à la température d'hybridation plus
2°C (soit 48°C). Prévoir deux lames par tube au maximum. Rincer les lames avec de la solution de
lavage et les mettre dos à dos dans les tubes de 50 mL. Incuber de 15 minutes à 1 heure. Rincer les
lames sur les deux faces avec précaution à l'eau MilliQ pour supprimer le tampon de lavage. Sécher

Matériel et Méthodes
125
rapidement à l'air comprimé (pour éviter l'élution de la sonde par l'eau MilliQ). Conserver à 4°C ou
observer directement.
Epifluorescence :
Ajouter environ 20 µL de Citifluor (dans du PBS-glycérol) à la surface de la lame (pas directement dans
les puits) et mettre une lamelle. Laisser 15 à 30 min à température ambiante. Le Citifluor (pH 9)
équilibre le pH des cellules et limite les phénomènes de fading (affaiblissement). Visualiser les cellules
sous un microscope à épifluorescence à l'aide de filtres appropriés :
Fluorescéine : excitation à 490 nm, émission à 520 nm.
Rhodamine : excitation à 550 nm, émission à 580 nm.
IV Observation et quantification des cellules
Les échantillons fixés sont colorés au SYBR-Green I ou II pour réaliser des comptages de
cellules totales. A la différence du DAPI, le SYBR Green se fixe spécifiquement sur les acides
nucléiques. Il y a très peu de bruit de fond avec cette coloration, même sur des échantillons dans
lesquels les cellules n'ont pas été extraites.
1 à 5 µL d'échantillon ou de cellules extraites sont resuspendus dans 4,5 mL de TE stérile. Le
mélange est agité et filtré sur Anodisc 25 à 0,2 µm (Whatman). Afin d'assurer une filtration régulière un
second filtre de 0,65 µm (Millipore) est placé sous le filtre de 0,2 µm. Le filtre 0,2 µm est récupéré puis
est déposé sur une goutte (100 µL environ) de solution diluée de SYBR Green I ou II (dilution au
1/1000e de la solution mère). La coloration s'effectue par diffusion durant 15 min (au moins) à
l'obscurité. Le filtre est monté sur lame, recouvert d'un mélange d'anti-fading (100 à 150 µL) puis d'une
lamelle.
Les comptages sont effectués au grossissement X 1000. Un filtre est compté lorsque le nombre
de bactéries dans le champ est compris entre 30 et 200. On compte 20 champs par filtre et 2 filtres par
échantillon. L’oculaire de comptage permet le dénombrement soit sur toute la surface de l’oculaire soit
sur la surface quadrillée. Le filtre optique du SYBR Green est le même que celui utilisé pour
l'observation des cellules marquées à la fluorescéine.
Les comptages de cellules marquées par des sondes nucléiques sont effectués sur le même
nombre de filtres et de champs.
Solution d'anti-fading Solution Fille de SYBR Green
50% 1x PBS (v/v) dilution au 1/1000ede la solution mère dans du TE
50% Glycérol (v/v) le SYBR Green est instable dans l'eau MilliQ.
0,1% p-phénylènediamine (p/v)

Articles

127
Article 1
- Analyse moléculaire de la diversité phylogénétique de réservoirs pétroliers continentaux et d'une
source chaude terrestre.
- Article soumis à Applied and Environmental Microbiology (Janvier 2000).
Corre Erwan, Laurent Toffin,and Christian Jeanthon. Molecular Phylogenetic Analysis of the
Microbial Diversity of Continental Oil reservoirs.
Article 2
- Etude de la diversité phylogénétique bactérienne associée à un échantilloneur déployé sur un site
hydrothermal de la dorsale médio-Atlantique.
- Article à soumettre Applied and Environmental Microbiology (Janvier 2000)
Corre Erwan, Daniel Prieur and Anna Louise Reysenbach. Molecular Phylogenetic Bacterial
Diversity Associated with In Situ Growth Chambers deployed on a Vent Site on the Mid Atlantic Ridge.
Article 3
- Article en préparation :
Corre Erwan, Laurent Toffin, Julie Kirshtein, Anna-Louise Reysenbach, Daniel Prieur et Christian
Jeanthon. Analyse de la diversité procaryotique associée à des fluides hydrothermaux profonds par
séquençage des ADNr 16S, hybridation in situ et enrichissement.
Article 4
-Caractérisation d'une Archaea hyperthermophile méthanogène provenant d'un site hydrothermal
profond.
- Article publié dans International Journal of Systematic Bacteriology (1999).
Jeanthon C., S. L'Haridon, A.L. Reysenbach, E. Corre, M. Vernet, P. Messner, U.W. Sleytr and D.
Prieur. 1999. Methanococcus vulcanius sp. nov., a novel hyperthermophilic methanogen isolated
from East Pacific Rise and identification of Methanococcus spp. DSM 4213 as Methanococcus fervens
sp. nov. Int. J. Syst. Bacteriol. 49:583-589.
Article 5
- Article en préparation : Caractérisation phylogénétique d'Archaea hyperthermophiles
radiorésistantes isolées d'un site hydrothermal profond.
Edmond JOLIVET, Stéphane L'HARIDON, Erwan CORRE, Patrick FORTERRE and Daniel PRIEUR
Article 6
- Article en préparation : Caractérisation phylogénétique d’une Bacteria thermophile isolée sur le site
du Bassin de Lau.

128
Bibliographie :
Les références bibliographiques non citées dans les articles sont présentées dans la partie références
du manuscrit.

Conclusion générale

Conclusion générale
209
Le but de ce travail de thèse était d'étudier, à l'aide de techniques de phylogénie moléculaire,
la diversité microbienne inféodée aux réservoirs pétroliers et aux sources hydrothermales profondes.
A la lumière des résultats présentés dans ce manuscrit, une représentation schématique des relations
s'établissant entre les différents groupes bactériens et archaéens des écosystèmes pétroliers (cf.
figure 11.1) et hydrothermaux (cf. figure 11.2) peut être proposée.
Les processus de production primaire autotrophes au sein du réservoir pétrolier sont encore
méconnus. Il est donc probable que les micro-organismes se développant dans les réservoirs utilisent,
en absence d'oxygène, les hydrocarbures et leurs dérivés comme sources de carbone et comme
donneurs d'électrons. Les hydrocarbures peuvent soit subir une dégradation abiotique liée aux fortes
pressions, aux fortes températures et à des réactions physico-chimiques avec la roche mère, soit subir
une dégradation biotique liée aux micro-organismes présents dans l'eau de production ou dans la
roche réservoir. Il a en effet été démontré que les alcanes et un grand nombre d'alkyles benzènes
pouvaient être dégradés en anaérobiose par des micro-organismes (Heider et al., 1999 pour revue;
Zengler et al., 1999). Il s'agit de bactéries réduisant le sulfate et fer ferrique, ainsi que de bactéries
dénitrifiantes. Certains consortiums de bactéries syntrophes et méthanogènes peuvent dégrader les
hydrocarbures si l'hydrogène et l'acétate sont consommés activement par méthanogenèse
acétoclastique et hydrogénoclastique. Enfin, certaines fermentations peuvent être impliquées dans la
dégradation des hydrocarbures (Heider et al., 1999). Les composés simplifiés (chaînes d'alcanes
réduites, composés cycliques "ouverts") vont être disponibles pour des micro-organismes pratiquant
la respiration anaérobie ou la fermentation. Le méthane et le CO2 peuvent être présentés comme les
produits terminaux de cette dégradation anaérobie des hydrocarbures. Or le méthane pourrait être à
son tour consommé par d'autres micro-organismes. Il y a peu, seuls les mécanismes d'oxydation
aérobie du méthane étaient connus, mais la détection récente de micro-organismes assimilant en
anaérobiose le méthane au sein de sédiments marins (Hinrichs et al., 1999) rend cette réaction
également envisageable au sein d'un réservoir pétrolier.
Les études réalisées par des techniques moléculaires (article 1) et culturales (L'Haridon et al.,
1995; Jeanthon et al., 1995) dans le réservoir de Montmirail, nous ont permis de détecter ou de
cultiver des micro-organismes fermentaires, sulfato- ou thiosulfato-réducteurs, des bactéries
syntrophiques et des méthanogènes. Ces populations pourraient dégrader à forte température et en
absence d'oxygène les hydrocarbures et constitueraient la communauté indigène. Cette
communauté a pu être "enrichie" par des micro-organismes injectés dans le réservoir lors des
processus de récupération secondaire. Présents dans l'eau potable, l'eau de nappe phréatique ou
dans les eaux de production retraitées, ces micro-organismes, exogènes et généralement aérobies,
sont par la suite retrouvés dans les échantillons prélevés en tête de puits. Les conditions physico-
chimiques régnant au sein du réservoir ne permettent probablement pas à ces micro-organismes de
participer très activement à la dégradation des hydrocarbures. Toutefois il est probable que certaines
micro-niches, à proximité de la zone d'injection d'eau ou dans le puits, rendent cette dégradation
possible dans des conditions d'aérobiose ou de micro-aérophilie. Les canalisations des puits

CO2
CO2
NO3-
N2
Mn4+
Mn2+
Fe3+
Fe2+
SO42-
S2-
Composés en C1
CO2
CH4
MnO2(S)
NO3-
Mn4+Fe3+
SO42-
N2
Mn2+
Fe2+
S2-
Acides gras àlongue chaine
H2 CO2
Gaz d’origine géologique
CH4
Matière organique
n-Alcanes
Composéscycliques
HCO3-
Gaz
FeXOXOHX(S)
sous forme soluble
Injection d’eau
de production
Respirations anaérobies
Fermentations
Acétogenese
Méthanogenèseacétoclastique
hydrogénoclastiqueCH4
NO3- Mn4+Fe3+ SO42- S2O32-
de nappe phréatiquepotable
Bactéries du sol et souterraines
Processus indigènes thermophiles et mésophilesZone mésophile d’aérobiose
partielle
Apport de micro-organismes aérobies
contaminants
Proteobacteria epsilon(Campylobacteriaceae)
Thermococcales
Thermotogales
Propionibacteriaceae
Thermoanaerobacter
Thermodesulfobacterium
Proteobacteria delta(Desulfovibrionaceae)Peptococaccaceae(Desulfotomaculum)
Methanosarcinales
Proteobacteria delta(Desulfobulbaceae, Desulfovibrionaceae)
Archaeoglobales
Peptococaccaceae(Desulfotomaculum)
Bactéries sulfato-réductrices thermophiles
Dégradation aérobie
Acides gras àcourte chaine
Acétate
CO2
Bactéries syntrophiquesFirmicutes Proteobacteria
Alcools
Bactéries sulfato-réductrices mésophiles
Proteobacteria alpha
Proteobacteria beta
Proteobacteria gamma
Firmicutes à haut G+C%
Micro-organismes hétérotrophes ou fermentaires
CO2
Proteobacteria alpha
Proteobacteria beta
Proteobacteria gamma
Firmicutes à haut G+C%
Micro-organismes hétérotrophes ou fermentaires
Processus de sulfato-réduction etfermentaires mésophiles
induisant le “Pitting”
Fig. 11.1. Ecologie microbienne des réservoirs pétroliers

Conclusion générale
210
constituent par ailleurs des biotopes favorables au développement de micro-organismes sulfato-
réducteurs mésophiles intervenant activement dans les phénomènes de biocorrosion. La présence
de cette communauté mésophile est corrélée à un faible débit d'extraction. Plus la longueur de
canalisation est importante, plus le débit est réduit et plus il est possible aux micro-organismes de se
développer au sein des installations. Tout autour du réservoir, les communautés de micro-organismes
souterrains peuvent elles aussi interagir avec celles présentes dans le réservoir. Il a été proposé
(Voordouw et al., 1996) que des micro-organismes sulfo-oxydants (pouvant correspondre dans notre
cas à des Proteobacteria epsilon) interviendraient dans la réoxydation des composés soufrés réduits
produits par les bactéries sulfato-, thiosulfato- et soufre-réductrices du réservoir. Mais d'autres
interactions entre ces deux communautés sont envisageables notamment via les processus de
respiration anaérobie.
Les mécanismes biotiques et abiotiques se déroulant au sein des réservoirs pétroliers sont
encore mal connus en raison, notamment, des difficultés d'échantillonnage et de notre incapacité à
prévenir les contaminations par les micro-organismes des sols ou injectés dans la nappe. Aussi
considère-t-on comme endémiques des micro-organismes dont les conditions de croissance sont
compatibles avec les conditions physico-chimiques des puits. Il est toutefois difficile de connaître
l'ensemble des conditions physico-chimiques inhérentes à un réservoir s'étendant sur une surface de
plusieurs kilomètres carrés, sur plusieurs dizaines de mètres de profondeur et au travers de
différentes roches. Aussi, il est probable que des conditions de température modérée et/ou
d'aérobiose partielle puissent exister au sein du réservoir en particulier lorsque la nappe est en contact
avec la roche couverture.
Il ne nous aura pas été possible au cours de cette étude d'estimer la représentativité des
séquences détectées au sein du réservoir. Si les séquences correspondant aux micro-organismes
sulfato-réducteurs semblent dominer certaines des banques obtenues, seules des expériences
d'hybridation in situ nous permettraient de préciser la contribution de ces micro-organismes dans cet
environnement. Par ailleurs ce type d'étude n'a jamais été réalisé à partir d'échantillon de réservoirs
pétroliers.
Au sein des écosystèmes hydrothermaux profonds on distingue les micro-organismes se
développant dans la "zone chaude" à proximité des fumeurs, dans le fluide ou les parois des
cheminées (partie gauche de la figure 11.2), de ceux se développant dans la zone froide ou à
température modérée, dans l'eau de mer, sous forme libre ou en association symbiotique avec la
macro-faune des sources hydrothermales (partie droite de la figure 11.2).
Dans la zone froide, la production primaire est assurée par des bactéries chimiosynthétiques
autotrophes sulfo-oxydantes sous une forme libre ou associées à la macrofaune. Les Proteobacteria
epsilon que nous avons détectées dans les échantillons de "Vent Cap" pourraient intervenir dans ces
processus puisqu'elles semblent être associées à certaines espèces d'Annélides (Haddad et al.,
1994) et de Crustacés (Poltz et Cavanaugh, 1995) mais qu'elles sont aussi présentes sous une forme
libre dans des tapis bactériens ou sur des édifices minéraux (Moyer et al., 1995; Poltz et Cavanaugh,
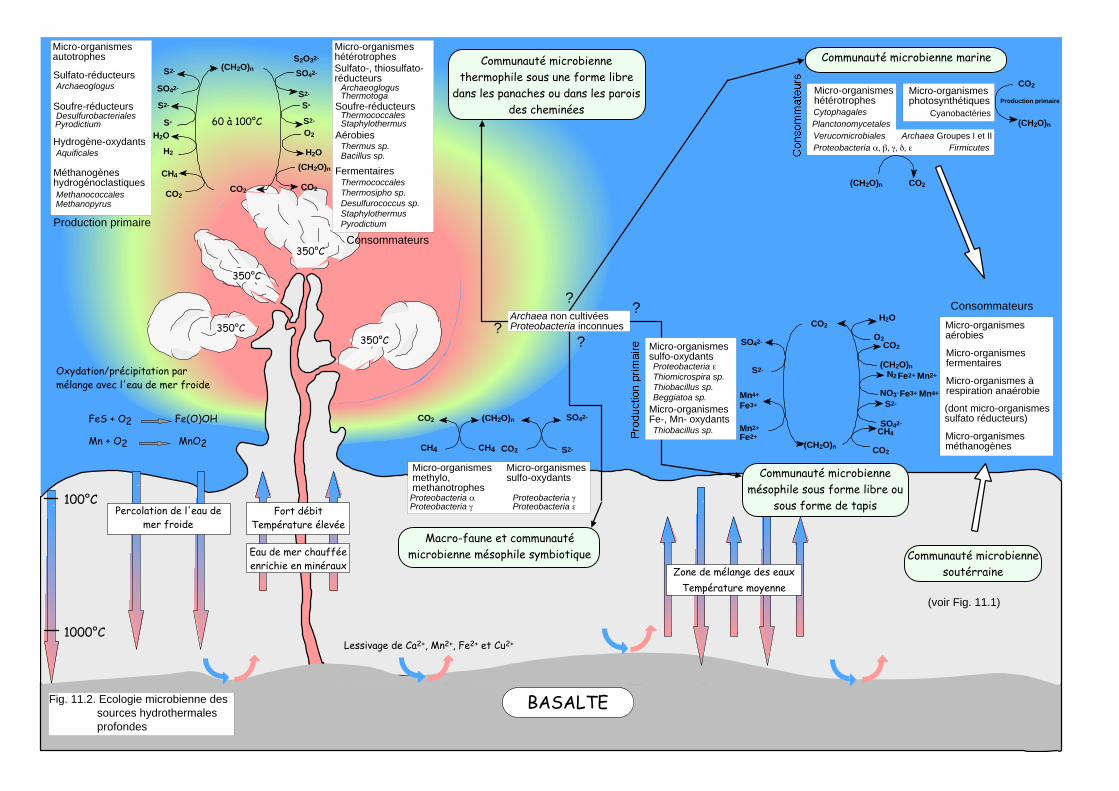
Oxydation/précipitation par mélange avec l'eau de mer froide
FeS + O2
Mn + O2
Fe(O)OH
MnO2
Lessivage de Ca2+, Mn2+, Fe2+ et Cu2+
BASALTE
Eau de mer chaufféeenrichie en minéraux
Macro-faune et communautémicrobienne mésophile symbiotique
Communauté microbiennethermophile sous une forme libre
dans les panaches ou dans les paroisdes cheminées
100°C
1000°C
Percolation de l'eau demer froide
Fort débitTempérature élevée
Communauté microbiennemésophile sous forme libre ou
sous forme de tapis
CH4 CO2
SO42-
S2-
(CH2O)n
CH4
CO2
Micro-organismes sulfo-oxydants
Micro-organismes methylo, methanotrophes
Proteobacteria Proteobacteria
Proteobacteria Proteobacteria
Communauté microbienne marine
Micro-organismes hétérotrophes
Proteobacteria
Cytophagales
Verucomicrobiales
Planctonomycetales
Firmicutes
Archaea Groupes I et II
CO2(CH2O)n
Cyanobactéries
Micro-organismes photosynthétiques
CO2
(CH2O)n
Consommateurs
Micro-organismes aérobies
Micro-organismes fermentaires
Micro-organismes à respiration anaérobie
Micro-organismes méthanogènes
(dont micro-organismes sulfato réducteurs)
Production primaire
Micro-organismes autotrophes
Sulfato-réducteurs
Soufre-réducteurs
Méthanogènes hydrogénoclastiques
Hydrogène-oxydants
Archaeoglogus
Desulfurobacteriales
Aquificales
MethanococcalesMethanopyrus
Pyrodictium 60 à 100°C
SO42-
S2-
S2-
S°
CH4
CO2
(CH2O)n
CO2
H2
H2O
S2-
S°
SO42-
S2-
CO2
(CH2O)n
O2
H2O
Micro-organismes hétérotrophesSulfato-, thiosulfato- réducteurs
Soufre-réducteurs
Fermentaires
Aérobies
Archaeoglogus
Thermococcales
Thermus sp.
Thermococcales
Bacillus sp.
Thermosipho sp.
Consommateurs
Thermotoga
S2O32-
Desulfurococcus sp.
Staphylothermus
StaphylothermusPyrodictium
Production primaire
Archaea non cultivéesProteobacteria inconnues
??
O2
H2O
SO42-
S2-
Mn4+
Fe3+
Mn2+
Fe2+
CO2
(CH2O)n
SO42-
S2-
NO3-
N2
CO2
(CH2O)n
Mn4+Fe3+
Mn2+Fe2+
CO2
CH4
??
Zone de mélange des eaux
Température moyenne
Communauté microbiennesoutérraine
Fig. 11.2. Ecologie microbienne des sources hydrothermales profondes
(voir Fig. 11.1)
Micro-organismes sulfo-oxydantsProteobacteria Thiomicrospira sp.Thiobacillus sp.Beggiatoa sp.
Micro-organismes Fe-, Mn- oxydantsThiobacillus sp.
350°C
350°C
350°C350°C

Conclusion générale
211
1995; Cary et al., 1997) ou dans le fluide (articles 2 et 3). Jusqu'à présent aucun micro-organisme
méthanotrophe autotrophe n'a été détecté sous une forme libre mais ceux-ci interviennent activement
en tant que producteurs primaires sous une forme symbiotique associée à des Mythilidés (Nealson et
Fischer, 1995). D'autre part, des activités in situ correspondant à une oxydation du méthane par des
formes libres ont déjà été enregistrées à proximité des sources hydrothermales profondes (De Angelis
et al., 1991). Des micro-organismes n'oxydant strictement que les métaux, tels que le Fe(II) et le Mn(II),
n'ont pas été isolés. Si certains Thiobacillus peuvent se développer en autotrophie en oxydant les
métaux, le fer ferreux n'est quant à lui stable que dans des conditions acidophiles qui ne sont
rencontrées dans les sources hydrothermales que dans le fluide chaud. Les consommateurs du pôle
froid sont soit les macro-organismes associés aux bactéries symbiotiques, soit d'autres micro-
organismes sous une forme libre se développant en hétérotrophie en conditions d'anaérobiose
(fermentations, respirations anaérobies) ou d'aérobiose complète ou partielle (respiration aérobie,
micro-aérophilie). On connaît peu de chose sur ces micro-organismes mésophiles en grande partie
parce que les études microbiologiques se sont très vite focalisées sur les micro-organismes
thermophiles. Il est toutefois probable qu'on puisse retrouver dans cette communauté des micro-
organismes issus des communautés microbiennes marines et souterraines. Ces micro-organismes
transportés par les courants, déposés avec les sédiments ou charriés par des émissions de fluide
hydrothermal à base température, profiteraient d'un apport abondant en matière organique pour se
développer activement et contribuer aux différents cycles géochimiques (C, N, S, métaux) au sein de
l'écosystème hydrothermal.
Dans la zone chaude, c'est à dire dans le panache du fluide hydrothermal, à proximité et dans
les parois des cheminées, la production primaire pourrait être assurée par des micro-organismes
chimiolithoautotrophes thermophiles. Il est aussi possible que la matière organique produite dans les
zones froides se retrouve dans les zones chaudes ou que des processus géochimiques de
profondeur puissent être générateurs de matière organique. Les Methanococcales réalisant une
méthanogenèse hydrogénoclastique (article 4), les Archaeoglobales sulfato-réductrices et les
Desulfurobacteriales soufre-réductrices détectées dans nos échantillons pourraient contribuer
activement à la production primaire biotique du pôle chaud (Harmsen et al., 1997b; article 3). Les
micro-organismes associés aux Aquificales détectés dans l'échantillon VC 2.1 pourraient quant à eux
intervenir dans la production primaire en oxydant l'hydrogène (article 3; Reysenbach et al., 1999). Les
micro-organismes thermophiles hétérotrophes se développant dans le pôle chaud sont mieux connus
que leurs analogues du pôle froid. Les enrichissements d'hétérotrophes thermophiles en présence
de soufre sont généralement dominés par les Thermococcales, micro-organismes fermentaires
réduisant le soufre (articles 3 et 5). Il nous a été possible, par ailleurs, de détecter des Thermotogales
et des Thermus dans de faibles proportions, pouvant eux aussi se développer, respectivement en
anaérobiose et aérobiose, au niveau du pôle chaud (article 3). Bien qu'ils n'aient pas été détectés
dans nos échantillons, des Bacillus ont déjà été isolés sur le site du Snake Pit (cf. tableau 6.1;
Marteinson et al., 1996). Ces micro-organismes sont présents de manière ubiquiste dans de
nombreux environnements terrestres ou marins, en raison notamment de leur capacité à sporuler. Ils

Conclusion générale
212
dominent généralement les enrichissements aérobies à haute température. Ces micro-organismes
peuvent prendre part activement à la consommation de la matière organique au niveau du pôle chaud
des sources hydrothermales (article 6).
Les variations de disponibilité des nutriments, liées aux caractère abrupt des gradients
physico-chimiques à la sortie des sources hydrothermales, mais aussi aux forts courants de fond,
favorisent très probablement le développement des micro-organismes mixotrophes. Mais les
techniques d'isolement généralement utilisées ne favorisent pas la détection de ce type métabolique
(Karl, 1995). La forte variabilité de séquences d'ADNr 16S observée pour de nombreux groupes
phylogénétiques (notamment pour les Proteobacteria epsilon) dans chacun de nos échantillons
(articles 2 et 3, Reysenbach et al., 1999) pourrait correspondre à une réponse adaptative corrélée aux
variabilités des conditions environnementales.
Les écosystèmes pétroliers et hydrothermaux profonds présentent de fortes similitudes tant
au niveau de leur localisation (environnements souterrains ou profonds), de leurs conditions physico-
chimiques (forte température, forte pression, anoxie) que des communautés microbiennes qu'ils
hébergent (micro-organismes thermophiles, fermentatifs, sulfato-réducteurs, soufre-oxydants). Le
soufre et les composés soufrés y sont considérés comme des éléments-clés des processus
métaboliques indigènes. Comme les micro-organismes décrits les plus intimement associés au dernier
ancêtre commun sont des hyperthermophiles anaérobies réduisant le soufre, il a été suggéré que la
réduction du soufre pourrait être l'une des premières formes de respiration microbienne (Madigan et
al., 1996). Il a cependant été récemment démontré qu'un grand nombre de micro-organismes
thermophiles isolés d'écosystèmes chauds (par exemple des méthanogènes, des fermentatifs ou des
sulfato-réducteurs) étaient également capables de réduire le fer ferrique en présence d'H2 comme
seule source d'électrons (Vargas et al., 1998; Slododkin et al., 1999). Compte tenu de l'abondance de
ce métal dans l'ensemble de la biosphère terrestre, la réduction du fer aurait pu constituer un
processus majeur sur la Terre primitive et serait encore actuellement prépondérante dans les
biosphères souterraines (Liu et al.,1997b; Lovley, 1997).
Un grand nombre de séquences, en particulier celles affiliées à des groupes phylogénétiques
où aucun isolat n'est disponible (Crenarchaeota et Euryarchaeota non cultivées, Bacteria "branchant"
profondément, nouvelles Proteobacteria ) ne peuvent clairement être associées à un biotope ou à un
processus métabolique. L'utilisation de nouvelles techniques d'isolement (comme par exemple les
pinces optiques) combinée à celle de sondes moléculaires, l'analyse phylogénétique et la
quantification de gènes impliqués dans des fonctions métabolique (via le séquençage, les puces à
ADN et l'analyse des ARNm) ainsi que la généralisation des études génomiques pourraient permettre
d'attribuer une fonction et une place dans l'écosystème à ces micro-organismes.
L'origine endémique ou cosmopolite des communautés de micro-organismes thermophiles et
mésophiles des écosystèmes hydrothermaux et pétroliers reste encore indéterminée. Si certains

Conclusion générale
213
genres, à l'instar de Thermococcus, de Bacillus, de Thermotoga, de Thermosipho ou de
Methanococcus, ont été détectés de manière ubiquiste en milieu hydrothermal côtier et profond, et
en milieu pétrolier marin et continental, d'autres semblent strictement associés à un environnement
donné, comme les Petrotoga et les Geotoga dans les réservoirs pétroliers ou les Pyrolobus et
Desulfurobacterium dans les écosystèmes hydrothermaux profonds. En poursuivant l'analyse de ces
écosystèmes, en entreprenant par exemple des études de génétique des populations, sur la base
des espaces intergéniques ou des gènes codants pour des protéines, de différencier les populations
écologiquement distinctes et d'établir leur origine biogéographique.
L'analyse de la diversité des communautés microbiennes d'environnements extrêmes peut
présenter un intérêt économique autant que fondamental. Hunter-Cevera s'interrogeait récemment
sur la valeur que pouvait présenter cette diversité (Hunter-Cevera, 1998). On peut y voir une valeur
pécuniaire au travers des processus métaboliques que l'on peut détecter, cloner et exprimer à une
échelle industrielle (Garrity et Hunter-Cevera, 1999). Mais elle présente avant tout une valeur
fondamentale. De telles études contribuent d'une part à un inventaire des espèces vivantes tel que
cela a été proposé par Pace (1997), et d'autre part à mieux définir quelles pourraient être les frontières
géographiques et physico-chimiques du vivant et la nature des premières formes de vie terrestres.

Références biobliographiques

Références bibliographiques
214
A
Abu Al-Soud W. and P. Rådström. 1998. Capacity of ninethermostable DNA polymerases to mediate DNAamplification in the presence of PCR-inhibiting samples. Appl.Environ. Microbiol. 64:3748-3753.
Acinas S.G., F. Rodriguez-Valera and C. Pedros-Alio.1997. Spatial and temporal variation in marinebacterioplankton diversity as shown by RFLP fingerprinting ofPCR amplified 16 S rDNA. FEMS Microbiol. Ecol. 24:27-40.
Acinas S.G., J. Anton and F. Rodriguez-Valera. 1999.Diversity of free-living and attached bacteria in offshoreWestern Mediterranean waters as depicted by analysis ofgenes encoding 16S rRNA. Appl. Environ. Microbiol. 65:514-522.
Ahmad A., J.P. Barry and D.C. Nelson. 1999. Phylogeneticaffinity of a wide, vacuolate, nitrate-accumulating Beggiatoasp. from Monterey Canyon, California, with Thioploca spp.Appl. Environ. Microbiol. 65:270-277.
Alfredsson G. A., J.K. Kristjansson, S. Hjorleifsdottir andK.O. Stetter. 1988. Rhodothermus marinus, gen. nov., athermophilic, halophilic bacterium from submarine hot springsin Iceland. J. Gen. Microbiol. 134:299-306.
Alfreider A., J. Pernthaler, R. Amann, B. Sattler, F.O.Glöckner, A. Wille and R. Pessner. 1996. Communityanalysis of the bacterial assemblags in the winter cover andpelagic layers of a high mountain lake by in situ hybridization.Appl. Envir. Microbiol. 62:2138-2144.
Alm E.W., D.B. Oerther, N. Larsen, D.A. Stahl and L.Raskin. 1996. The oligonucleotide probe database. Appl.Environ. Microbiol. 62:3557-3559.
Altschul S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z.Zhang, W. Miller and D.J. Lipman. 1997. Gapped BLASTand PSI-BLAST: a new generation of protein database searchprograms. Nucleic Acids Res. 25:3389-3402.
Alvarez A.J., M.P. Buttner and L.D. Stetzenbach. 1995.PCR for bioaerosol monitoring: sensitivity and environmentalinterference. Appl. Envir. Microbiol. 61:3639-3644.
Amann R. and M. Kuhl. 1998. In-situ methods forassessment of microorganisms and their activities. Curr.Opin. Microbiol. 1:352-358.
Amann R.I., B. Binder, R.I. Olson, S.W. Chisholm, R.Devereux and D.A. Stahl. 1990a. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry foranalyzing mixed microbial populations. Appl. Environ.Microbiol. 56:1919-1925.
Amann R.I., B. Zarda, D.A. Stahl and K.H. Schleifer.1992b. Identification of individual prokaryotic cells by usingenzymes-labeled, rRNA-targeted oligonucleotide probes.Appl. Envir. Microbiol. 58:3007-3011.
Amann R.I., J. Stromley, R. Deveureux, R. Key and D.A.Stahl. 1992a. Molecular and microscopic identification ofsulfate reducing bacteria in multispecies biofilm. Appl. Envir.Microbiol. 58:614-623.
Amann R.I., L. Krumholz and D.A. Stahl. 1990b.Fluorescent-oligonucleotide probing of whole cells fordeterminative, phylogenetic, and environmental studies inmicrobiology. J. Bacteriol. 172:762-770.
Amann R.I., W. Ludwig and K.H. Schleifer. 1995.phylogenetic identification and in situ detection of individualmicrobial cells without cultivation. Microbiological Reviews.59:143-169.
Amellal N, G. Burtin, F. Bartoli and T. Heulin. 1998.Colonization of wheat roots by an exopolysaccharide-producing pantoea agglomerans strain and its effect onrhizosphere soil aggregation. Appl. Environ. Microbiol.64:3740-3747.
Anderson A.W., N.R.F. Cain, G. Parrish and D. Duggan.1956. Studies on a radio-resistant Micrococcus. I. Isolation,morphology, cultural characteristics and resistance to gammaradiation. Food technology 10:575-578.
Anderson R.T., F.H. Chapelle and D.R. Lovley. 1998.Evidence against hydrogen-based microbial ecosystems inbasalt aquifers. Science 281:976-977.
Angert E.R., K.D. Clements and N.R. Pace. 1993. Thelargest bacterium. Nature 362:239-241.
Antoine E., V. Cilia., J.R. Meunier, J. Guezennec, F.Lesongeur and G. Barbier. 1997. Thermosiphomelanesiensis sp. nov., a new thermophilic anaerobicbacterium belonging to the order Thermotogales, isolated fromdeep-sea hydrothermal vents in the Southwestern Pacificocean. Int. J. Syst. Bacteriol. 47:1118-1123.
Aoshima M., Y. Nishibe, M. Hasegawa, A. Yamagishi andT. Oshima. 1996. Cloning and sequencing of a gene encoding16S ribosomal RNA from a novel hyperthermophilicarchaebacterium NC12. Gene 180:183-187.
Apajalahti J.H., L.K. Sarkilahti, B.R. Maki, J.P. Heikkinen,P.H. Nurminen and W.E. Holben. 1998. Effective recoveryof bacterial DNA and percent-guanine-plus-cytosine-basedanalysis of community structure in the gastrointestinal tract ofbroiler chickens. Appl. Environ. Microbiol. 64:4084-8408.
Arahal D.R., F.E. Dewhirst, B.J. Paster, B.E. Volcani andA. Ventosa. 1996. Phylogenetic analyses of some extremlyhalophilic archaea isolated from Dead Sea water, determinedon the basis of their 16S rRNA sequences. Appl. Environ.Microbiol. 62:3779-3786.
Arai Y., R. Yasuda, K.I. Akashi, Y. Harada, H. Miyata, K.Kinosita Jr. and H. Itoh. 1999. Tying a molecular knot withoptical tweezers. Nature 399: 446-448.
Arnheim N. and H. Erlich. 1992. Polymerase chain reactionstrategy. Annu. Rev. Biochem. 61:131-156.
Arrage A.A., T.J. Phelps, R.E. Benoit and D.C. White.1993. Survival of subsurface microorganisms exposed to UVradiation and hydrogen peroxide. Appl. Envir. Microbiol. 59:3545-3550.
Asai T., C Condon, J. Voulgaris, D. Zaporojets, B. Shen,M. Al-Omar, C. Squires and C.L. Squires. 1999a.Construction and initial characterization of Escherichia colistrains with few or no intact chromosomal rRNA operons. J.Bacteriol. 181:3803-3809.
Asai T., D. Zaporojets, C. Squires and C.L. Squires.1999b. An Escherichia coli strain with all chromosomal rRNAoperons inactivated: complete exchange of rRNA genesbetween bacteria. Proc. Natl. Acad. Sci. USA 96:1971-1976.
Askin A. and J.M. Dziedzic. 1987. Optical trapping andmanipulation of virus and bacteria. Science 235:1517-1520.
Atlas R.M., G. Sayler, R.S. Burlage and A.K. Bej. 1992.Molecular approaches for environmental monitoring ofmicroorganisms. BioTechniques 12:706-717.
B
Bader J.S., R.W. Hammond, S.A. Henck, M.W. Deem,G.A. McDermott, J.M. Bustillo, J.W. Simpson, G.T.Mulhern and J.M. Rothberg. 1999. DNA transport by amicromachined brownian ratchet device. Proc. Natl. Acad.Sci. USA 96:13165-13169.
Balkwill D.L., R.H. Reeves, G.R. Drake, J.Y. Reeves, F.H.Crocker, M.B. King and D.R. Boone. 1997. Phylogeneticcharacterization of bacteria in the subsurface microbial culturecollection. FEMS Microbiol. Rev. 20:201-216.
Bally R., T. Heulin and P. Lemanceau. 1999. Les microbeset le cultivateur. Biofutur 185:17-19.
Barns S. and S.A. Nierzwicki-Bauer. 1997. Microbialdiversity in ocean, surface and subsurface environments. fromGeomicrobiology: Interaction between microbes and minerals.J.F. Banfield and K.H. Nealson, eds. J. Revs. in Mineralogy35:35-79.
Barns S.M., C.F. Delwiche, J.D. Palmer and N.R. Pace.1996a. Perspectives on archaeal diversity, thermophily andmonophyly from environmental rRNA sequences. Proc. Natl.Acad. Sci. USA 93:9188-9193.
Barns S.M., C.F. Delwiche, J.D. Palmer, S.C. Dawson, K.L.Hershberger and N.R. Pace. 1996b. Phylogeneticperspective on microbial life in hydrothermal ecosystems,past and present. Ciba Found Symp. discussion 32-9 202:24-32.

Références bibliographiques
215
Barns S.M., R.E. Fundyga, M.W. Jeffries and N.R. Pace.1994. Remarkable archaeal diversity detected in aYellowstone National Park hot spring environment. Proc. Natl.Acad. Sci. USA 91:1609-1613.
Barns S.M., S.L. Takala and C.R. Kuske. 1999. Widedistribution and diversity of members of the bacterial kingdomAcidobacterium in the environment. Appl. Environ. Microbiol.65:1731-1737.
Baross J.A. and Deming J.W. 1995. Growth at hightemperatures: isolation and taxonomy, physiology, ecology. In:Karl DM (Ed) The Microbiology of Deep-sea HydrothermalVents (pp 169-217). CRC Press Inc, Boca Raton.
Baross J.A. and S.E. Hoffman. 1985. Submarinehydrothermal vent and associated gradients environments assites for the origin and evolution of life. Origins of life 15:327-345.
Baross J.A., J.W. Deming and R.R. Becker. 1984. Evidencefor microbial growth in high-temperature high-pressureenvironments. In: Current Perspectives in Microbial ecology(M.J. Klug and C.A. Reddy eds.), p 186-195. A.S.M.:Washington D.C.
Bartlett M.S., S.H. Vermund, R. Jacobs, P.J. Durant, M.M.Shaw, J.W. Smith, X. Tang, J.J. Lu, B. Li, S. Jin and C.H.Lee. 1997. Detection of Pneumocystis carinii DNA in airsamples: likely environmental risk to susceptible persons. J.Clin. Microbiol. 35:2511-2513.
Bastin E.S. 1926. The problem of natural reduction of sulfates.Bull. Am. Associat. Petrol. Geologists 10:1270-1299.
Beeder J., R.K. Nilsen, J.T. Rosnes, T. Torsvik and T.Lien. 1994. Achaeoglobus fulgidus isolated from hot north seaoil field waters. Appl. Environ. Microbiol. 60:1227-1231.
Begon M., Harper J.L. and C.R. Townsend. 1996. Ecology:individuals, populations and communities. 3rd Edition.Blackwell Science Eds. Oxford.
Bej A.K., M.H. Mahbubani, J.L. Dicesare, and R. Atlas.1991. Polymerase chain reaction-gene probe detection ofmicroorganisms by using filter-concentrated samples. Appl.Envir. Microbiol. 57:3529-3534.
Bekins BA, E.M. Godsy and E. Warren. 1999. Distributionof microbial physiologic types in an aquifer contaminated bycrude oil. Microb. Ecol. 37:263-275.
Benloch S., F. Rodriguez-Valera and A.J. Martinez-Murcia. 1995. Bacterial diversity in two coastal lagoonsdeduced from 16S rDNA PCR amplification and partialsequencing. FEMS Microbiol. Ecol. 18:267-280.
Benson D.A., M.S. Boguski, D.J. Lipman, J. Ostell, B.F.Ouellette, B.A. Rapp and D.L. Wheeler. 1999. GenBank.Nucleic Acids Res. 27:12-17.
Bernalier A., V. Rochet, M. Leclerc, J. Dore and P.Pochart. 1996. Diversity of H2/CO2-utilizing acetogenicbacteria from feces of non-methane-producing humans. Curr.Microbiol. 33:94-99.
Bernard F.P., J. Connan and M. Magot. 1992. Indigenousmicroorganisms in connate water of many oil fields: a newtool in exploration and production techniques, SPE 24811.Annual technical conference and exhibition of the Society ofPetroleum Engineers, Washington, D.C., October, 4-7.
Berns M. 1998. Pinces et ciseaux optiques. Pour la Science248 (Edition internet).
Bertrand C. and H. Larsen. 1988. La “bactérie marine” :mythe ou réalité, pp.3-25. In: Micro-organismes dans lesécosystèmes océaniques, M. Bianchi, D. Marty, J.C.Bertrand, P. Caumette, M. Gauthier et coll. (Eds), Masson,Paris.
Bintrin S.B., T.J. Donohue, J. Handelsman, G.P. Robertsand R.M. Goodman. 1997. Molecular phylogeny of archaeafrom soil. Proc. Natl. Acad. Sci. USA 94:227-282.
Blackburn N., Å. Hagström, J. Wikner, R. Cuadros-Hansson and P.K. Bjørnsen. 1998. Rapid determination ofbacterial abundance, biovolume, morphology, and growth byneural network-based image analysis. Appl. Envir. Microbiol.64:3246-3255.
Blanc M., L. Marilley, T. Beffa and M. Aragno. 1997. Rapididentification of heterotrophic, thermophilic, spore-forming
bacteria isolated from hot composts. Int. J. Syst. Bacteriol.47:1246-1248.
Blanchot J., L. Guillou, F. Partensky, N. Simon and D.Vaulot. 1998. Le petit peuple du grand large. Biofutur 179:22-24.
Blöchl E., R. Rachel, S. Burggraf, D. Hafenbradl, H.W.Jannasch and K.O. Stetter. 1997. Pyrolobus fumarii, gen.and sp. nov., represents a novel group of archaea, extendingthe upper temperature limit for life to 113°C. Extremophiles1:14-21.
Bogan B.W., B. Schoenike, R.T. Lamar and D. Cullen.1996. Manganese peroxidase mRNA and enzyme activitylevels during bioremediation of polycyclic aromatichydrocarbon-contaminated soil with Phanerochaetechrysosporium. Appl. Envir. Microbiol. 62:2381-2386.
Boivin-Jahns V, R Ruimy, A Bianchi, S Daumas and RChristen.1996. Bacterial diversity in a deep-subsurface clayenvironment. Appl. Environ. Microbiol. 62:3405-3412.
Bollet C., M.J. Gevaudant, X. de Lamballerie, C. Zandottiand P. de Micco. 1991. A simple method for the isolation ofchromosomal DNA from Gram positive od acid-fast bacteria,Nucl.Acids Res.19:1955.
Bond P.L., P. Hugenholtz, J. Keller and L.L. Blackall.1995. Bacterial community structures of phosphate-removingand non-phosphate-removing activated sludges fromsequencing batch reactors. Appl. Envir. Microbiol. 61:1910-1916.
Boone D.R., Y. Liu, Z.J. Zhao, D.L. Balkwill, G.R. Drake,T.O. Stevens and H.C. Aldrich. 1995. Bacillus infernus sp.nov., an Fe(III)- and Mn(IV)-reducing anaerobe from the deepterrestrial subsurface. Int. J. Syst. Bacteriol. 45:441-448.
Borin S., D. Daffonchio and C. Sorlini. 1997. Single strandconformation polymorphism analysis of PCR-tDNAfingerprinting to address the identification of Bacillus species.FEMS Microbiol. Lett. 157:87-93.
Borneman J. and E.W. Triplett. 1997. Molecular microbialdiversity in soils from eastern Amazonia: evidence for unusualmicroorganisms and microbial population shifts associatedwith deforestation. Appl. Envir. Microbiol. 63: 2647-2653.
Borneman J., P.W. Skroch, K.M. O'Sullivan, J.A. Palus,N.G. Rumjanek, J.L. Jansen, J. Nienhuis and E.W.Triplett. 1996. Molecular microbial diversity of an agriculturalsoil in Wisconsin. Appl. Envir. Microbiol. 62: 1935-1943.
Böttger E.C. 1989. Rapid determination of bacterialribosomal RNA sequences by direct sequencing ofenzymatically amplified DNA. FEMS Microbiol. Lett. 65:171-176.
Böttiger E.C. 1996. Approaches for identification ofmicroorganisms. ASM News 62:247-250.
Brack A. 1999. Vie extraterrestre : les charmes del'improbable. La Recherche 317:50-52.
Brack A. and C.T. Pillinger. 1998. Life on Mars: chemicalarguments and clues from martian meteorites. Extremophiles2:313-319.
Brack A. and F. Raulin. 1999. La vie dans le systèmesolaire. Pour la Science H.S. Les terres celestes:40-42.
Bradford D., P. Hugenholtz, E.M. Seviour, M.A.Cunningham, H. Stratton, R.J. Seviour and L.L.Blackhall. 1996. 16S rRNA analysis of isolates obtained fromgram-negative filamentous bacteria micromanipulated fromactived sludge. System. Appl. Microbiol. 19:334-343.
Brechtold M. and H. König. 1996. Phylogenetic analysisand in situ identification of uncultivated Spirochete from thehindgut of the termite Mastotermes darwiniensis. System.Appl. Microbiol. 19:66-73.
Brechtold M., A. Chatzinotas, W. Schönhuber, A. Brune,R. Amann, D. Hahn and H. König. 1999. Differentialenumeration and in situ localization of microorganisms in thehingut of the lower termite Mastodermes darwiniensis byhybridization with rRNA-targeted probes. Arch. Microbiol.172:407-416.
Briglia M., R.I.L. Eggen, W.M. de Vos, and J.D. van Elsas.1996. Rapid and sensitive method for the detection ofMycobacterium chlorophenolicum PCP-1 in soil based on 16S

Références bibliographiques
216
rRNA gene-targeted PCR. Appl. Envir. Microbiol. 62: 1478-1480.
Brinkhoff T., C.M. Santegoeds, K. Sahm, J. Kuever andG. Muyzer. 1998. A polyphasic approach to study the diversityand vertical distribution of sulfur-oxidizing Thiomicrospiraspecies in coastal sediments of the german wadden Sea. Appl.Environ. Microbiol. 64:4650-4657.
Britschgi T.B. and R.D. Fallon. 1994. PCR-amplification ofmixed 16S rRNA genes from anaerobic, cyanide-degradingconsortium. FEMS Microbiol. Ecol. 13:225-232.
Britschgi T.B. and S.J. Giovannoni. 1991. Phylogeneticanalysis of a natural marine bacterioplankton population byrRNA gene cloning and sequencing. Appl. Environ. Microbiol.57:1707-1713.
Brock T.D. 1986. Introduction: an overview of thethermophiles. In: Brock, T.D. (ed) Thermopiles: general,molecular, and applied microbiology. Wiley, New-York, p 1-16.
Brock T.D. 1995. The road to Yellowstone and beyond. Annu.Rev. Microbiol. 49:1-28.
Brock T.D. and H. Freeze. 1969. Thermus aquaticus gen.nov., a nonsporulating extreme thermophile. J. Bacteriol.98:289-297.
Bruce K.D., W.D. Hiorns, J.L. Hobman, A.M. Osborn, P.Strike and D.A. Ritchie. 1992. Amplification of DNA fromnative populations of soil bacteria by using the polymerasechain reaction. Appl. Envir. Microbiol. 58:3413-3416.
Brune A. 1998. Termite guts: the world's smallestbioreactors. Trends Biotech. 16:16-21.
Brunk C.F. and N. Eis. 1998. Quantitative measure of small-subunit rrna gene sequences of the kingdom Korarchaeota.Appl. Envir. Microbiol. 64:5064-5066.
Brunk C.F., E. Avaniss-Aghajani and C.A. Brunk. 1996. Acomputer analysis of primer and probe hybridization potentialwith bacterial small-subunit rRNA sequences. Appl. Envir.Microbiol. 62:872-879.
Bruns A. and L. Berhe-Corti. 1998. In-situ detection ofbacteria in continuous-flow cultures of seawater sedimentssuspensions with fluorescently labelled rRNA-directedoligonucleotide probes. Microbiology 144:2783-2790.
Bruns M.A., J.R. Stephen, G.A. Kowalchuk, J.I. Prosserand E.A. Paul. 1999. Comparative diversity of ammoniaoxidizer 16S rRNA gene sequences in native, tilled, andsuccessional soils. Appl. Environ. Microbiol. 65:2994-3000.
Buckley D.H., J.R. Graber and T.M. Schmidt. 1998.Phylogenetic analysis of nonthermophilic members of thekingdom Crenarchaeota and their diversity and abundance insoils. Appl. Envir. Microbiol. 64:4333-4339.
Burggraf S., H. Huber and K.O. Stetter. 1997.Reclassification of the crenarchaeal orders and families inaccordance with 16S rRNA sequence data. Int. J. Syst.Bacteriol. 47:657-660.
Burggraf S., H.W. Jannasch, B. Nicolaus and K.O.Stetter. 1990. Archaeoglobus profundus sp. nov., represents anew species within the sulfate-reducing archaeobacteria.System. Appl. Microbiol. 13:24-28.
Burggraf S., T. Mayer, R. Amann, S. Schadhauser, C.R.Woese and K.O. Stetter. 1994. Identifying members of thedomain Archaea with rrna-targeted oligonucleotide probes.Appl. Environ. Microbiol. 60:3112-3119.
Burks C. 1999. Molecular biology database list. Nucleic AcidsRes. 27:1-9.
Button D.K., B.R. Robertson, D. McIntosh and F.Jüttner. 1992. Interactions between marine bacteria anddissolved-phase beached hydrocarbons after the ExxonValdez oil spill. Appl. Environ. Microbiol. 58:243-251.
C
Caldwell D.E., S.J. Caldwell and J.P. Laycock. 1976.Thermothrix thioparus gen. et sp. nov. a facultativelyanaerobic facultative chemolithotroph living at neutral pH andhigh temperature. Can. J. Microbiol. 22:1509-1517.
Canganella F., W.J. Jones, A. Gambacorta and G.Antranikian. 1998. Thermococcus guaymasensis sp. nov.and Thermococcus aggregans sp. nov., two novelthermophilic archaea isolated from the Guaymas Basinhydrothermal vent site. Int. J. Syst. Bacteriol. 48:1181-1185.
Cano R.J. and H.N. Poinar. 1993. Rapid isolation of DNAfrom fossil and museum specimens suitable for PCR.Biotechniques 15:432-436.
Cano R.J. and M.K. Borucki. 1995. Revival andidentification of bacterial spores in 25- to 40-million-year-oldDominican amber. Science 268:1060-1064.
Cano R.J., M.K. Borucki, M. Higby-Schweitzer, H.N.Poinar, G.O. Poinar Jr and K.J. Pollard. 1994. Bacillus DNAin fossil bees: an ancient symbiosis? Appl. Environ. Microbiol.60:2164-2167.
Carr M.H. 1996. Water on early mars. Evolution ofhydrothermal ecosystems on Earth (and Mars?). Wiley,Chichester (ciba Foundation symposium 202) 249-267.
Cary C., T. Shank and J. Stein. 1998. Worms bask inextreme temperature. Nature 391:545-546.
Cary S.C., M.T. Cottrel, J.L. Stein, F. Camacho and D.Desbruyères. 1997. Molecular indentification and localizationof filamentous symbiotic bacteria associated with thehydrothermal vent annelid, Alvinella pompejana. Appl.Environ. Microbiol. 63:1124-1130.
Caumette P., M. Kühl, O. Pringault and R. De Witt. 1998.Etude expérimentale de la compétition entre bactériespourpres et bactéries vertes anoxygéniques en milieubenthique. 5ème congrès de la Société Française deMicrobiologie. 27, 28, 29 avril 1998 Lille, France.
Cavanaugh C.M., C.O. Wirsen and H.W. Jannasch. 1992.Evidence for methylotrophic symbionts in a hydrothermal ventmussel (Bivalvia: Mytilidae) from the Mid-Atlantic Ridge.Appl. Environ. Microbiol. 58:3799-3803.
Cavanaugh C.M., S.L. Gardiner, M.L. Jones, H.W.Jannasch and J.B. Waterbury. 1981. Prokaryotic cells in thehydrothermal vent tube worm, Riftia pachyptila: possiblechemoautotrophic symbionts. Science 213:340-342.
Cazemier A.E., J.H.P. Hackstein, H.J.M. Op den Camp, J.Rosenberg and C. van der Drift. 1997. Bacteria in theintestinal tract of different species of Arthropods. Microb Ecol33:189-197.
Cha D.K., J.J. Fuhrmann, D.W. Kim and C. M. Golt. 1999."Fatty acid methyl ester (FAME) analysis for monitoringNocardia levels in activated sludge." Water Research,33:1964.
Chandler D.P., F.J. Brockman, T.J. Bailey and J.K.Fredrickson. 1998. Phylogenetic diversity of Archaea andBacteria in a deep subsurface paleosol. Microb. Ecol. 36:37-50.
Chandler D.P., S.M. Li, C.M. Spadoni, G.R. Drake, D.L.Balkwill, J.K. Fredrickson and F.J. Brockman. 1997. Amolecular comparaison of culturable aerobic heterotrophicbacteria and 16S rDNA clones derived from a deepsubsurface sediment. FEMS Microbiol. Ecol. 23:131-144.
Charbonnier F., P. Forterre, G. Erauso and D. Prieur.1995. Purification of plasmids from thermophilic andhyperthermophilic Archaea, in Thermophiles, Archaea alaboratory manual, edited by F.T. Robb and A.R. Place, ColdSpring Harbor Laboratory Press.
Chen F., J.M. González, W.A. Dustman, M.A. Moran andR.E. Hodson. 1997. In situ reverse transcription, an approachto characterize genetic diversity and activities of prokaryotes.Appl. Envir. Microbiol. 63:4907-4913.
Cherry R., D. Desbruyères, M. Heyraud and C. Nolan.1992. High levels natural radioactivity in hydrothermal ventpolychetes. C. R. Acad. Sci. Paris III:21-26.
Chevaldonné P. 1989. Ecologie des Alvinellidae (annélidespolychètes des souces hydrothermales profondes) :comportement, biomasse, environnement thermique etmicroflore du tube. D.E.A d'Océanologie de l'Université d'Aix-Marseille II, 30p.
Chevaldonné P. 1996. Ecologie des cheminéeshydrothermales actives. Thèse de doctorat de l'Université dela méditérannée (Aix-Marseille II), 257p.

Références bibliographiques
217
Chevaldonné P. and A. Godfroy. 1997. Enumeration ofmicroorganisms from deep-sea hydrothermal vent chimneys.FEMS Microbiol. Lett. 146:211-216.
Chevaldonné P., D. Desbruyère and J.J. Childress. 1992.And some even hotter. Nature 359:593.
Chin K.J., T. Lukow and R. Conrad. 1999. Effect oftemperature on structure and function of the methanogenicarchaeal community in an anoxic rice field soil. Appl. Environ.Microbiol. 65:2341-2349.
Choi B.K., B.J. Paster, F.E. Dewhirst and U.B. Gobel.1994. Diversity of cultivable and uncultivable oral Spirochetesfrom a patient with severe destructive periodontitis. Infect.Immun. 62:1889-1895.
Chou H.P., C. Spence, A. Scherer and S. Quake. 1999. Amicrofabricated device for sizing and sorting DNA molecules.Proc. Natl. Acad. Sci. USA 96:11-13.
Christensen H., M. Hansen and J. Sørensen. 1999.Counting and size classification of active soil bacteria byfluorescence in situ hybridization with an rRNAoligonucleotide probe. Appl. Envir. Microbiol. 65:1753-1761.
Chung A.P., F. Rainey, M.F. Nobre, J. Burghardt andM.S. da Costa. 1997. Meiothermus cerbereus sp. nov., a newslightly thermophilic species with high levels of 3-hydroxyfatty acids. Int. J. Syst. Bacteriol. 47:1225-1230.
Clayton R.A., G. Sutton, P.S. Hinkle Jr., C. Bult and C.Fields. 1995. Intraspecific variation in small-subunit rRNAsequences in GenBank: why single sequences may notadequately represent procaryotic taxa. Int. J. Syst. Bacteriol.45:595-599.
Colwell R.R. 1970. Polyphasic taxonomy of the genus vibrio:numerical taxonomy of Vibrio cholerae, Vibrioparahaemolyticus, and related Vibrio species. J.Bacteriol.104:410-433.
Connan J., G. Lacrampe-Coulome and M. Magot. 1996.Origin of gases in reservoirs. In : Dolenc A (Ed.) Proceedingsof the 1995 International Gas Research Conference.Government Institutes Inc., Rocokville, Md.
Converse D.R., H.D. Holland and J.M. Edmond. 1984.Flow rate in the axial sprimgs of the East Pacific Rise (21°N):implications for the heat budget and the formation of massivesulfide deposits. Earth Planet. Sci. Lett. 69:159-175.
Coolen M.J. and J. Overmann.1998. Analysis of subfossilmolecular remains of purple sulfur bacteria in a lakesediment. Appl. Environ. Microbiol. 64:4513-4521.
Cooper J.M. 1999. Towards electronic Petri dishes andpicolitre-scale single-cell technologies. Trends Biotechnol.17:226-230.
Corliss J.B. and R.D. Ballard. 1977. Oases of life in the coldabyss. Natl. Geogr. 152:440-453.
Corliss J.B., J. Dymond, L.I. Gordon, J.M. Edmond, R.P.Von Herzen, R.D. Ballard, K. Green, D. Willianms, A.Bainbridge, K. Crane and T.H. van Andel. 1979.Submarine thermal springs on the galapagos Rift. Sciences203:1073-1083.
Corliss J.B., J.A. Baross and S.E. Hoffman. 1981. Ahypothesis concerning the relationship between submarine hotsprings and the orign of life on earth. Oceanol. Acta, Specialissue 59-69.
Cornec L. 1995. Mise en évidence et étude de deux enzymesthermostables : lipase et estérase, de micro-organismesthermophiles isolés d'écosystèmes hydrothermaux sous-marins. Thèse de doctorat de l'Université de Technologie deCompiègne, 205p.
Cotton R.G.H. 1997. Mutation Detection. Oxford: OxfordUniversity Press.
Cottrell M.T. and S.C. Cary. 1999. Diversity of disimilatorybisulfite reductase genes of Bacteria associated with the deep-sea hydrothermal vent polychaete annelid Alvinellapompejana. Appl. Envir. Microbiol. 65:1127-1132.
Cottrell M.T., J.A. Moore and D. Kirchman. 1999.Chitinases from uncultured marine microorganisms. Appl.Environ. Microbiol. 65:2553-2557.
Cowan D.A. 1992. Biotechnology of the Archaea. TrendsBiotechnol. 10:315-323.
Crozier R.H., P.M. Agapow and K. Pedersen. 1999.Towards complete biodiversity assesment: an evaluation ofthe subterranean bacterial communities in the Oklo region ofthe sole surviving natural nuclear reactor. FEMS Microbiol.Ecol. 28:325-334.
Crump B.C., E.V. Armbrust and J.A. Baross. 1999.Phylogenetic analysis of particle-attached and free-livingbacterial communities in the columbia river, its estuary, andthe adjacent coastal ocean. Appl. Environ. Microbiol. 65:3192-3204.
D
Dando P.R., M. Thomm, H. Arab, M. Brehmer, L.E.Hooper, B. Jochimsen, H. Schlesner, R. Stöhr, J.C.Miquel and S.W. Fowler. 1998. Microbiology of shallowhydrothermal sites of Palaeochori Bay, Milos (HellenicVolcanic Arc). Cah. Biol. Mar. 39:369-372.
Darland G. and T.D. Brock. 1971. Bacillus acidocaldariussp. nov., an acidophilic thermophilic spore-forming bacterium.J. Gen. Microbiol. 67:9-15.
Darland G., T.D. Brock, W. Samsonoff and S.F. Conti.1970. A thermophilic, acidophilic mycoplasma isolated from acoal refuse pile. Science 170:1416-1418.
Davydova-Charakhch'yan I.A., A.N. Mileeva, L.L.Mityushima and S.S. Belyaev. 1992. Acetogenic bacteriafrom oil fields of tataria and west siberia. Microbiology (Engl.Tr.) 61:306-315.
De Angelis M.A., J.A. Baross and M.D. Liley. 1991.Enhenced microbial methane oxidation in water from a deepsea hydrothermal vent field at simulated in situ hydrostaticpressures. Limmol. Oceanogr. 36:565-570.
De Bartolomeo A., F. Trotta, F. La Rosa, G.Saltalamacchia and V. Mastrandrea. 1991. Numericalanalysis and DNA base compositions of some thermophilicBacillus species. Int. J. Syst. Bacteriol. 41:502-509.
de los Reyes F.L., W. Ritter and L Raskin. 1997. Group-specific small-subunit rRNA hybridization probes tocharacterize filamentous foaming in activated sludge systems.Appl. Envir. Microbiol. 63:1107-1117.
de los Reyes M.F., F.L. de los Reyes, M. Hernandez and LRaskin. 1998. Quantification of Gordona amarae strains infoaming activated sludge and anaerobic digester systems witholigonucleotide hybridization probes. Appl. Envir. Microbiol.64:2503-2512.
De Rijk P., E. Robbrecht, S. de Hoog, A. Caers, Y. Van dePeer and R. De Wachter. 1999. Database on the structure oflarge subunit ribosomal RNA. Nucleic Acids Res. 27:174-178.
Delaney J.R. 1998. Volcanic ecology- A planetaryperspective. Thermophiles'98, International Conference, 6-11septembre, Brest France.
DeLong E.F. 1992. Archaea in coastal marine environments.Proc. Natl. Acad. Sci. USA 89:5685-5689.
DeLong E.F., D.G. Franks and A.L. Alldredge. 1993.Phylogenetic diversity of aggregate-attached vs. free-livingmarine bacterial assemblages. Limnol. Oceanogr. 38:924-934.
DeLong E.F., G.S. Wickham and N.R. Pace. 1989.Phylogenetic stains: ribosomal RNA-based probes for theidentification of single cells. Science 243:1360-1363.
DeLong E.F., K.Y. Wu, B.B. Prezelin and R.V. Jovine.1994. High abundance of Archaea in Antarctic marinepicoplankton. Nature 371:695-697.
DeLong E.F., L.L. King, R. Massana, H. Cittone, A.Murray, C. Schleper and S.G. Wakeham.1998. Dibiphytanylether lipids in nonthermophilic Crenarchaeotes. Appl. Environ.Microbiol. 64:1133-1138.
Deming J.W. and J.A. Baross. 1993. Deep-sea smokers:windows to a subsurface biosphere. Geochim. et Cosmo-chim. Acta 57:3219-3230.
Dennis M.W. and P.E. Kolattukud. 1991. Alkanebiosynthesis by decarbonylation of aldehyde catalyzed by amicrosomal preparation from Botryococcus braunii. Arch.Biochem. Biophys. 287:268-275

Références bibliographiques
218
Desai J.D. and I.M. Banat. 1997. Microbial production ofsurfactants and their commercial potential. Microbiol. Mol.Biol. Rev. 61: 47-64.
Desbruyères D., P. Chevaldonné, A.-M. Alayse, D.Jollivet, F.H. Lallier, C. Jouin-Toulmond, F. Zal, P.M.Sarradin, R. Cosson, J.C. Caprais, C. Arndt, J. O'Brien, J.Guezennec, S. Hourdez, R. Riso, F. Gaill, L. Laubier andA. Toulmond. 1998. Biology and ecology of the "Pompeiiworm" (Alvinella pompejana Desbruyères and Laubier), anormal dweller of an extreme deep-sea environment: asynthesis of current knowledge and recent developments.Deep-Sea Res. II 45:383-422.
Devereux R. and G.W. Mundfrom. 1994. A phylogenetictree of 16S rRNA sequences from sulfate-reducing bacteria ina sandy marine sediment. Appl. Environ. Microbiol. 60:3437-3439.
Devereux R., M.D. Kane, J. Winfrey and D.A. Stahl. 1992.Genus- and group- specific hybridization probes fordeterminative and environmental studies of sulfate-reducingBacteria. System. Appl. Microbiol. 15:601-609.
Devereux R., M.E. Hines and D.A. Stahl. 1996. S Cycling:characterization of natural communities of sulfates-reducingbacteria by 16S rRNA sequence comparaisons. Microb. Ecol.32:283-292.
deWit R. 1995. Measurements of sedimentary gradients ofpore water species, by use of microelectrodes. Calculations ofmicrobial metabolic processes in the sediments. Océanis287-297.
Diels L. and M. Mergeay. 1990. DNA probe-mediateddetection of resistant bacteria from soils highly polluted byheavy metals. Appl. Envir. Microbiol. 56: 1485-1491.
Dirmeier R., M. Keller, D. Hafenbradl, F.J. Braun, R.Rachel, S. Burggraf and K.O. Stetter. 1998. Thermococcusacidaminovorans sp. nov., a new hyperthermophilicalkalophilic archaeon growing on amino acids. Extremophiles2:109-114.
DiRuggiero J., N. Santangelo, Z. Nackerdien, J. Ravel andF.T. Robb. 1997. Repair of extensive ionizing-radiation DNAdamage at 95 degrees C in the hyperthermophilic archaeonPyrococcus furiosus. J. Bacteriol. 179:4643-4645.
Distel D.L., D.J. Lane, G.J. Olsen, S.J. Giovannoni, B.Pace, N.R. Pace, D.A. Stahl and H. Felbeck. 1988. Sulfur-oxidizing bacterial endosymbionts: analysis of phylogeny andspecificity by 16S rRNA sequences. J. Bacteriol. 170:2506-2510.
Distel D.L., E.F. DeLong and J.B. Waterbury. 1991.Phylogenetic characterization and in situ localization of thebacterial symbiont of shipworms (Teredinidae: Bivalvia) byusing 16S rRNA sequence analysis and oligodeoxynucleotideprobe hybridization. Appl. Environ. Microbiol. 57:2376-2382.
Distel D.L., H. Felbeck and C.M. Cavanaugh. 1994.Evidence for phylogenetic congruence among sulfur-oxidizingchemoautotrophic bacterial endosymbionts and their bivalvehosts. J. Mol. Evol. 38:533-542.
Distel D.L., H.K.W. Lee and C.M Cavanaugh. 1995.Intracellular coexistence of methano- and thioautotrophicbacteria in a hydrothermal vent mussel. Proc. Natl. Acad. Sci.USA 92:9598-9602.
Dojka M.A., P. Hugenholtz, S.K. Haack and N.R. Pace.1998. Microbial diversity in a hydrocarbon- and chlorinated-solvent-contaminated aquifer undergoing intrinsicbioremediation. Appl. Envir. Microbiol. 64:3869-3877.
Doolittle R.F., D.F. Feng, S. Tsang, G. Cho and E. Little.1996. Determining divergence times of the major kingdoms ofliving organisms with a protein clock. Science 271:470-477.
Doolittle W.F. 1996. At the core of the Archaea. Proc. Natl.Acad. Sci. USA 93:8797-8799.
Dorsch M. and E. Stackebrandt. 1992. Some modification inthe procedure of direct sequencing of PCR amplified 16SrDNA. J. Microbiol. Methods 16:271-279.
Douglas S. and T.J. Beveridge. 1998. Mineral formation bybacteria in natural microbial communities. FEMS Microbiol.Ecol. 26:79-88.
Drancourt M., G. Aboudharam, M. Signoli, O. Dutourand D. Raoult. 1998. Detection of 400-year-old Yersiniapestis DNA in human dental pulp: An approach to the
diagnosis of ancient septicemia. Proc. Natl. Acad. Sci. U.S.A.95:12637-12640.
Drucker D.B. and I. Owen. 1971. Bacterial identification bygas-liquid chromatography. J. Gen. Microbiol.69:ix-x.
Duffaud G.D., O.B. d'Hennezel, A.S. Peek, A.L.Reysenbach and R.M. Kelly. 1998. Isolation andcharacterization of Thermococcus barossii, sp. nov., ahyperthermophilic archaeon isolated from a hydrothermal ventflange formation. Syst. Appl. Microbiol. 21:40-49.
Dunbar J., S. Takala, S.M. Barns, J.A. Davis and C.R.Kuske. 1999. Levels of bacterial community diversity in fourarid soils compared by cultivation and 16S rRNA genecloning. Appl. Environ. Microbiol. 65:1662-1669.
Durand P., A.L. Reysenbach, D. Prieur and N.R. Pace.1993. Isolation and characterization of Thiobacillushydrothermalis sp. nov. a mesophilic obligatelychemolithotrophic bacterium isolated from a deep-seahydrothermal vent in Fiji Bassin. Arch. Microbiol. 159:39-44.
Durand P., D. Prieur, C. Jeanthon and E. Jacq. 1990.Présence et activité de bactéries hétérotrophes mangano-oxydantes associées aux Alvinellidés (Annélides polychètes)dans un site d'hydrothermalismes profond de la dorsale duPacifique oriental. C. R. Acad. Sci. Paris 310(III):273-278.
Durand P., E. Benyagoub and D. Prieur. 1994. Numericaltaxonomy of heterotrophic sulfur-oxidizing bacteria isolatedfrom southwestern Pacific hydrothermal vents. Can. J.Microbiol. 40:690-697.
Durand P., O. Gros, L. Frenkiel and D. Prieur. 1996.Phylogenetic characterization of sulfur-oxidizing bacterialendosymbionts in three tropical Lucinidae by 16S rDNAsequence analysis. Mol. Mar. Biol. Biotech. 5:37-42.
Dutton C.M., C. Paynton and S. Sommer. 1993. Generalmethod for amplifying regions of very high G + C content.Nucleic Acids Res. 21:2953-2954.
Dymock D., A.J. Weightman, C. Scully and W.G. Wade.1996. Molecular analysis of microflora associated withdentoalveolar abscesses. J. Clin. Microbiol. 34:537-542.
Dzik W.H. 1995. Molecular diagnostics in transfusionmedecine : the best is yet to come. Transfusion 35:183-185.
E
Edwards U., T. Rogall, H. Blöcker, M. Emde and E.C.Böttger. 1989. Isolation and direct complete nucleotidedetermination of entire genes. Characterization of a genecoding for 16S ribosomal RNA. Nucl. Acids Res. 17:7843-7853.
Eichner C.A., W.E. Rainer, K.N. Timmis and I. Wagner-Döbler. 1999 Thermal gradient gel electrophoresis analysis ofbioprotection from pollutant shocks in the activated sludgemicrobial community. Appl. Envir. Microbiol. 65:102-109.
Ekendahl S. and K. Pedersen. 1994. Carbon transformationsby attached bacterial populations in granitic groundwater fromdeep crystalline bed-rock of the Stripa research mine.Microbiology 140:1565-73.
el Fantroussi S., L. Verschuere, W. Verstraete and E.M.Top. 1999. Effect of phenylurea herbicides on soil microbialcommunities estimated by analysis of 16S rRNA genefingerprints and community-level physiological profiles. Appl.Environ. Microbiol. 65: 982-988.
Elsgaard L. 1995. Le cycle du soufre et bactéries sulfatoréductrices dans les sédiments hydrotheramux, marins etlacustres. Thèse de l'Université Paris 6. 195p.
Elsgaard L., J. Guezennec, N. Benbouzid-Rollet and D.Prieur. 1991. Fatty acid composition of sulfate-reducingbacteria isolated from deep-sea hydrothermal vent (13°N, eastPacific Rise). Kieler Meeresforsch. 8:182-187.
Emmerson D. and C. Moyer. 1997. Isolation andcharacterization of novel iron-oxidizing bacteria that grow atcircumneutral pH. Appl. Env. Microbiol. 63:4784-4792.
Engle M., Y. Li, F. Rainey, S. DeBlois, V. Mai, A. Reichert,F. Mayer, P. Messner and J. Wiegel. 1996.Thermobrachiumcelere gen. nov., sp. nov., a rapidly growing thermophilic,

Références bibliographiques
219
alkalitolerant, and proteolytic obligate anaerobe. Int. J. Syst.Bacteriol. 46:1025-1033.
Erauso G. 1994. Les Thermococcales du Bassin Nord-Fidjien, description d'une nouvelle espèce Pyrococcus abyssiet caractérisation de son plasmide. Thèse de doctorat del'Université de Bretagne Occidentale. 461 p
Erauso G., A.L. Reysenbach, A. Godfroy, J-R. Meunier,B. Crump, F. Partensky, J.A. Baross, V. Marteinsson, G.Barbier, N.R. Pace and D. Prieur. 1993. Pyrococcus abyssisp. nov., a new hyperthermophilic archaeon isolated from adeep-sea hydrothermal vent. Arch. Microbiol., 160:338-349.
Erhlich H.L. 1983. Magnanese-oxidizing bacteria fromhydrothermally active area on the Galapagos Rift. Environ.Biogeochem. Ecol. Bull. 35:357-366.
Espejo R.T. and J. Romero. 1997. Bacterial community incopper sulfide ores inoculated and leached with solution froma commercial-scale copper leaching plant. Appl. Envir.Microbiol. 63:1344-1348.
Espejo R.T., G.G. Feijóo, J. Romero and M. Vásquez.1998. PAGE analysis of the heteroduplexes formed betweenPCR-amplified 16S rRNA genes: estimation of sequencesimilarity and rDNA complexity. Microbiology 144:1611-1617.
F
Fani R., G. Damiani, C. Di Serio, E. Galllori, A. Grofoniand M. Bazzicapulo. 1993. Use of random amplifiedpolymorphic DNA (RAPD) for generating specific DNAprobes for microorganisms. Mol. Ecol. 2:243-250.
Farmer J.D. 1996. Hydrothermal systems on mars: anassesment of present evidence. Evolution of hydrothermalecosystems on Earth (and Mars?). Wiley, Chichester (cibaFoundation symposium 202) 273-299.
Farrelly V., F.A. Rainey and E. Stackebrandt. 1995. Effect ofgenome size and rrn gene copy number on PCR amplificationof 16S rRNA genes from a mixture of bacterial species. Appl.Envir. Microbiol. 61:2798-2801.
Fegatella F., J. Lim, S. Kjelleberg and R. Cavicchioli. 1998.Implications of rRNA operon copy number and ribosomecontent in the marine oligotrophic ultramicrobacteriumSphingomonas sp. strain RB2256. Appl. Envir. Microbiol.64:4433-4438.
Felbeck H. and J.J. Childress. 1988. Riftia pachyptila: ahighly integrated symbiosis. Oceanol. Acta 8:131-138.
Feldman R.A., M.B. Black, C.S. Cary, R.A. Lutz and R.C.Vrijenhoek. 1997. Molecular phylogenetics of bacterialendosymbionts and their vestimentiferan hosts. Mol. Mar.Biol. Biotechnol. 6:268-277.
Felsenstein J. 1973. Maximum-likelihood estimation ofevolutionary trees from continuous characters. Am. J. Hum.Genet. 25:471-492.
Felsenstein J. 1978. Cases in wich parsimony orcompatibility methods will be positively misleading. syst.Zool. 27:401-410.
Felsenstein J. 1981. Evolutionary trees from DNAsequences: a maximum likelihood approach. J. Mol. Evol.17:368-376.
Felsenstein J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39:783-791.
Felsenstein J. 1989. PHYLIP -- Phylogeny Inference Package(Version 3.2).Cladistics 5:164-166.
Felske A. 1999. Reviewing the DA001-files: a 16S rRNAchase on suspect #X99967, a Bacillus and Dutch undergroundactivist. J. Microbiol. Methods 36:77-93.
Felske A. and A.D.L. Akkermans. 1998a. Spatialhomogeneity of abundant bacterial 16S rRNA molecules inGrassland soils. Micro. Ecol. 36:31-36.
Felske A. and A.D.L. Akkermans. 1998b. Prominentoccurrence of ribosomes from an uncultured bacterium of theVerrucomicrobiales-cluster in grassland soils. Lett. Appl.Microbiol. 26:219-223.
Felske A., A. Wolterink, R. Van Lis and A.D.L.Akkermans. 1998a. Phylogeny of the main bacterial 16S
rRNA sequences in Drentse A grassland soils (TheNetherlands). Appl. Envir. Microbiol. 64: 871-879.
Felske A., A.D.L. Akkermans and W.M. De Vos. 1998b. Insitu detection of an uncultured predominant Bacillus in Dutchgrassland soils. Appl. Envir. Microbiol. 64: 4588-4590.
Felske A., B. Engelen, U. Nubel and H. Backhaus. 1996.Direct ribosome isolation from soil to extract bacterial rRNAfor community analysis. Appl. Envir. Microbiol. 62:4162-4167.
Ferber D.M., P.J. Haney, H. Berk, D. Lynn and J. Konisky.1997. The adenylate kinase genes of M. voltae, M.thermolithotrophicus, M. jannaschii, and M. igneus define anew family of adenylate kinases. Gene 185:239-244.
Ferré F., A. Marchese, P. Pezzoli, S. Griffin, E. Buxtonand V. Boyer. 1994. Quantitative PCR: an overview.In : ThePolymerase Chain Reaction. K.B. Mullis, F. Ferre, R.A.Gibbs, Eds. Birkhauser, Boston.
Ferreira A.C., M.F. Nobre, F.A. Rainey, M.T. Silva, R. Wait,J. Burghardt, A.P. Chung and M.S. da Costa. 1997.Deinococcus geothermalis sp. nov. and Deinococcus murrayisp. nov., two extremely radiation-resistant and slightlythermophilic species from hot springs. Int. J. Syst. Bacteriol.47:939-947.
Ferrera-Cerrato R. 1980. [Inoculation of Rhizobium phaseolionto different species of the genus Phaseolus originating inMexico]. Rev. Latinoam. Microbiol. 22:175-180.
Ferris M.J. and D.M. Ward. 1997. Seasonal distributions ofdominant 16S rRNA-defined populations in a hot springmicrobial mat examined by denaturing gradient gelelectrophoresis. Appl. Environ. Microbiol. 63:1375-1381.
Ferris M.J., A.L. Ruff-Roberts, E.D. Kopczynski, M.M.Bateson and D.M. Ward. 1996b. Enrichment culture andmicroscopy conceal diverse thermophilic Synechococcuspopulations in a single hot spring microbial mat habitat. Appl.Environ. Microbiol. 62:1045-1050.
Ferris M.J., G. Muyzer and D.M. Ward. 1996a. Denaturinggradient gel electrophoresis profiles of 16S rRNA-definedpopulations inhabiting a hot spring microbial community. Appl.Environ. Microbiol. 62:340-346
Ferris M.J., S.C. Nold, N.P. Revsbech and D.M. Ward.1997. Population structure and physiological changes within ahot spring microbial mat community following disturbance.Appl. Envir. Microbiol. 63:1367-1374.
Fiala G. and K.O. Stetter. 1986. Pyrococcus furiosus sp. nov.represents a novel genus of marine heterotrophicarchaebacteria growing optimally at 100°C. Arch. Microbiol.145:56-61.
Fiala G., K.O. Stetter, H.W. Jannasch, T.A. Langworthyand J. Madon. 1986. Staphylothermus marinus sp. nov.represent a novel genus of marine heterotrophicarchaebacteria growing optimally at 100°C. Arch. Microbiol.145:56-61.
Field K.G., D. Gordon, T. Wright, M. Rappe, E. Urback, K.Vergin and S.J. Giovannoni. 1997. Diversity and depth-specific distribution of SAR11 cluster rRNA genes frommarine planktonic bacteria. Appl. Environ. Microbiol. 63:63-70.
Fischer, S.G. and L.S. Lerman. 1979. Length-independentseparation of DNA restriction fragments in two-dimensionalgel electrophoresis. Cell 16:191-200.
Fisher M.M., L.W. Wilcox and L.E. Graham. 1998.Molecular characterization of epiphytic bacterial communitieson charophycean green algae. Appl. Environ. Microbiol.64:4384-4389.
Fisk M.R., S.J. Giovannoni and I.H. Thorseth. 1998.Alteration of oceanic volcanic glass: textural evidence ofmicrobial activity. Science 281:978-980.
Fitch W.M. 1971. Toward defining the course of evolution:minimum change for a specific tree topology. Syst. Zool.20:406-416.
Fitch W.M. and E. Margoliash. 1967. Construction ofphylogenetic trees. Science 155:279-284 .
FitzGerald L.M. and J.A. Blake. 1998. Databases for use inmolecular biology: cocepts in their design, access and use.Molecular Approaches to the Study of the Ocean. K.E.Cooksey Eds. Chapman & Hall, London pp 515-527.

Références bibliographiques
220
Fleming J.T., W.H. Yao and G.S. Sayler. 1998. Optimizationof differential display of prokaryotic mRNA: application to pureculture and soil microcosms. Appl. Envir. Microbiol. 64:3698-3706.
Forster R.J., J. Gong and R.M. Teather. 1997. Group-specific 16S rRNA hybridization probes for determinative andcommunity structure studies of Butyrivibrio fibrisolvens in therumen. Appl. Environ. Microbiol. 63:1256-1260.
Forterre P. 1996. A hot topic: the origin of hyperthermophiles.Cell. 85:789-792.
Forterre P., A. Bergerat and P. Lopez-Garcia. 1996. Theunique DNA topology and DNA topoisomerases ofhyperthermophilic Archaea. FEMS Microbiol Rev 18:237-248.
Fouquet Y., U. Von Stackelberg, J.L. Charlou, J.P.Donval, J. Erzinger, J.P. Foucher, P. Herzig, R. Mühe, S.Sokai, M. Wiedicke and H. Whitechurch. 1991.Hydrothermal activity and metallogenesis in the Lau back-arcbasin. Nature 349:778-781.
Fournier D., R. Lemieux and D. Couillard. 1998. Geneticevidence for highly diversified bacterial poulations inwastewater sludge during biological leaching of metals.Biotech. Lett. 20:27-31.
Fox G.E., J.D. Wisotzkey and P. Jurtshuk Jr. 1992. Howclose is close: 16S rRNA sequence identity may not besufficient to guarantee species identity. Int. J. Syst. Bacteriol.42:166-170.
Fox G.E., K.J. Pechman and C.R. Woese. 1977.Comparative cataloging of 16S ribosomal ribonucleic acid:molecular approach to prokaryotic systematics. Int. J. Syst.Bacteriol. 27:44-57.
Franks A.H., H.J. Harmsen, G.C. Raangs, G.J. Jansen, F.Schut and G.W. Welling. 1998. Variations of bacterialpopulations in human feces measured by fluorescent in situhybridization with group-specific 16S rRNA-targetedoligonucleotide probes. Appl. Environ. Microbiol. 64:3336-3345.
Frischer M.E., P.J. Floriani and S.A. Nierzwicki-Bauer.1996. Differential sensitivity of 16S rRNA targetedoligonucleotide probes used for fluorescence in situhybridization is a result of ribosomal higher order structure.Can. J. Microbiol. 42:1061-1071.
Fritsche T.R., M. Horn, S. Seyedirashti, R.K. Gautom,K.H. Schleifer and A. Wagner. 1999. In situ detection ofnovel bacterial endosymbionts of Acanthamoeba spp.phylogenetically related to members of the orderRickettsiales. Appl. Environ. Microbiol. 65:206-212.
Fry N.K., J.K. Fredrickson, S. Fishbain, M. Wagner andD.A. Stahl. 1997. Population structure of microbialcommunities associated with two deep, anaerobic, alkalineaquifers. Appl. Environ. Microbiol. 63:1498-1504.
Fuchs B.M., Wallner G., W. Beisker, I. Schwippl, W.Ludwig and R. Amann. 1998. Flow cytometric analysis ofthe in situ accessibility of Escherichia coli 16S rRNA forfluorescently labeled oligonucleotide probes. Appl. Environ.Microbiol. 64:4973-4982.
Fuhrman J.A. and F. Azam. 1980. Bacterioplanktonsecondary production estimates for coastal waters of BritshColumbia, Antarctica, and California. Appl. Environ. Microbiol.39:1085-1095.
Fuhrman J.A. and F. Azam. 1982. Thymidine incorporationas a mesure of heterotrophic bacterioplankton production inmarine surface waters: evaluation and field results. Mar. Biol.66:109-120.
Fuhrman J.A., K. McCallum and A.A. Davis. 1992. Novelmajor archaebacterial group from marine plankton. Nature356:148-149.
Fuhrman J.A., K. McCallum and A.A. Davis. 1993.Phylogenetic diversity of subsurface marine microbialcommunities from the Atlantic and Pacific Oceans. Appl.Environ. Microbiol. 59:1294-1302.
Furcht C., K. Eschrich and K. Merte. 1996. Detection ofEikenella corrodens and Actinobacillusactinomycetemcomitans by use of the polymerase chainreaction (PCR) in vitro and in subgingival plaque. J. Clin.Periodontol. 23:891-897.
G
Galtier N. and M. Gouy. 1995. Inferring phylogenies fromDNA sequences of unequal base compositions. Proc. Natl.Acad. Sci. U S A 92:11317-11321.
Galtier N. and M. Gouy. 1998. Inferring pattern and process:maximum-likelihood implementation of a nonhomogeneousmodel of DNA sequence evolution for phylogenetic analysis.Mol. Biol. Evol. 15:871-879.
Galtier N., M. Gouy and C. Gautier. 1996. SeaView andPhylo_win, two graphic tools for sequence alignment andmolecular phylogeny. Comput. Applic. Biosci. 12:543-548.
Galtier N., N. Tourasse and M. Gouy. 1999. Anonhyperthermophilic common ancestor to extant life forms.Science 283:220-221.
Garland J.L. 1997. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMSMicrobiol. Ecol. 24:289-300.
Garrity G. and J. Hunter-Cevera. 1999. Bioprospecting in thedeveloping world. Curr. Opin. Microbiol. 2:236-240.
Gaten E. 1998. Eye structure and phylogeny: is there aninsight? The evolution of superposition eyes in the Decapoda(Crustacea). Contributions to Zoology 67:223-235.
Gaten E., P.J. Herring, P.M.J. Shelton and M.L. Johnson.1998a. Comparative morphology of the eyes of post-larvalbresiliid shrimps obtained from the region of hydrothermalvents. Biological Bulletin. 194:267-280.
Gaten E., P.J. Herring, P.M.J. Shelton and M.L. Johnson.1998b. The development and evolution of the eyes of ventshrimps (Decapoda: Bresiliidae). Cahiers de Biologie Marine,39:287-290.
Ge Z. and D.E. Taylor. 1999. Contributions of genomesequencing to understanding the biology of Helicobacter pylori.Annu. Rev. Microbiol. 53:353-387.
Gerhold D., T. Rushmore and C.T. Caskey. 1999. DNAchips: promising toys have become powerful tools. TrendsBioSci. 24:168-173.
Giovannoni S.J., E.F. DeLong, G.J. Olsen and N.R. Pace.1988. Phylogenetic group-specific oligodeoxynucleotideprobes for identification of single microbial cells. J. Bacteriol.170:720-726.
Giovannoni S.J., E.F. DeLong, T.M. Schmidt and N.R.Pace. 1990a. Tangential flow filtration and preliminaryphylogenetic analysis of marine picoplankton. Appl. Environ.Microbiol. 56:2572-2575.
Giovannoni S.J., M.S. Rappe, K.L. Vergin and N.L. Adair.1996. 16S rRNA genes reveal stratified open oceanbacterioplankton populations related to the Green Non-Sulfurbacteria. Proc. Natl. Acad. Sci. USA 93:7979-7984.
Giovannoni S.J., T.B. Britschgi, C.L. Moyer and K.G.Field. 1990b. Genetic diversity in Sargasso Seabacterioplankton. Nature 345:60-63.
Giuliano L., M. De Domenico, E. De Domenico, M.G.Hofle and M.M. Yakimov. 1999. Identification of culturableoligotrophic bacteria within naturally occurringbacterioplankton communities of the Ligurian Sea by 16SrRNA sequencing and probing. Microb. Ecol. 37:77-85.
Glöckner F.O., R. Amann, A. Alfreider, J. Pernhaler, R.Psenner, K. Trebesius and K.H. Schleifer. 1996. An in situhybridization protocol for detection and indentification ofplanktonic bacteria. System. Appl. Microbiol. 19:403-406.
Godfroy A., F. Lesongeur, G. Raguenes, J. Querellou, E.Antoine, J.R. Meunier, J. Guezennec and G. Barbier.1997. Thermococcus hydrothermalis sp. nov., a newhyperthermophilic archaeon isolated from a deep-seahydrothermal vent. Int. J. Syst. Bacteriol. 47:622-666.
Godfroy A., J.R. Meunier, J. Guezennec, F. Lesongeur, G.Raguenes, A. Rimbault and G Barbier. 1996.Thermococcus fumicolans sp. nov., a new hyperthermophilicarchaeon isolated froma deep-sea hydrothermal vent in thenorth Fiji Basin. Int. J. Syst. Bacteriol. 46:1113-1119.

Références bibliographiques
221
Godon J.J., E. Zumstein, P. Dabert, F. Habouzit and R.Moletta. 1997. Molecular microbial diversity of an anaerobicdigestor as determined by small-subunit rDNA sequenceanalysis. Appl. Envir. Microbiol. 63:2802-2813.
Goebel B.M. and E. Stackebrandt. 1994. Cultural andphylogenetic analysis of mixed microbial populations found innatural and commercial bioleaching environments. Appl.Envir. Microbiol. 60:1614-1621.
Goering J.J. and V.A. Dungale. 1966. Estimates of the ratesof denitrification in a subartic lake. Limnol. Oceanogr. 11:113-116.
Gogarten-Boekels M., E. Hilario and J.P. Gogarten. 1995.The effects of heavy meteorite bombardement on the earlyevolution - the emergence of the three domains of life. Originsof life and evolution of the biosphere 25:251-264.
Gold T. 1992. The deep, hot biosphere. Proc. Natl. Acad. Sci.USA 89:6045-6049.
Goldenberger D. and M. Altwegg. 1995. Eubacterial PCR:contaminating DNA in primer preparations and its eliminationby UV light. J. Microbiol. Meth. 21:27-32.
Gonzàlez J.M. and M.A. Moran. 1997. Numericaldominance of a group of marine bacteria in the alpha-subclassof the class Proteobacteria in coastal seawater. Appl. Environ.Microbiol. 63:4237-4242.
González J.M., D. Sheckells, M. Viebahn, D. Krupatkina,K.M. Borges and F.T. Robb. 1999. Thermococcuswaiotapuensis sp. nov., an extremely thermophilic archaeonisolated from a freshwater hot spring. Arch. Microbiol. 172:95-101.
Gonzàlez J.M., W.B. Withman, R.E. Hodson and M.A.Moran. 1996. Identifying numerically abundant culturablebacteria from complex communities: an example from a ligninenrichment culture. Appl. Envir. Microbiol. 62:4433-4440.
Good I.J. 1953. The population frequencies od species and theestimation of population parameters. Biometrika 40:237-264.
Goodman M. 1981. Globin evolution was apparently veryrapid in early vertebrates: a reasonable case against the rateconstancy hypothesis. J. Mol. Evol. 17:114-120.
Gordon D.A. and S.J. Giovannoni. 1996. Detection ofstratified microbial populations related to Chlorobium andFibrobacter species in the Atlantic and Pacific oceans. Appl.Environ. Microbiol. 62:1171-1177.
Gotz A., R. Pukall, E. Smit, E. Tietze, R. Prager, H.Tschape, J.D. van Elsas and K. Smalla. 1996. Detection andcharacterization of broad-host-range plasmids inenvironmental bacteria by PCR. Appl. Envir. Microbiol. 62:2621-2628.
Grassia G.S., K.M. McLean, P. Glénat, J. Bauld and A. J.Sheehy. 1996. A systematic survey for thermophilicfermentative bacteria and archaea in high temperaturepetroleum reservoirs. FEMS Microbiol. Ecol. 21:47-58.
Graves D.J. 1999. Powerful tools for genetic analysis come ofage. Trends Biotech. 17:127-134.
Gray J.P. and R.P. Herwig. 1996. Phylogenetic analysis ofthe bacterial communities in marine sediments. Appl. Environ.Microbiol. 62:4049-4059.
Greenblatt C.L., A. Davis, B.G. Clement, C.L. Kitts, T.Cox and C.J. Cano. 1999. Diversity of microorganismsisolated from amber. Micro. Ecol. 38:58-68.
Greene A.C., B.K.C. Patel and A.J. Sheehy. 1997.Deferribacter thermophilus gen. nov., sp. nov., a novelthermophilic manganese- and iron-reducing bacteriumisolated from a petroleum reservoir. Int. J. Syst. Bacteriol.47:505-509.
Greisen K., M. Loeffelholz, A. Purohit and D. Leong.1994. PCR primers and probes for the 16S rRNA gene ofmost species of pathogenic bacteria, including bacteria foundin cerebrospinal fluid. J. Clin. Microbiol. 32:335-351.
Gros O., A. Darrasse, P. Durand, L. Frenkiel and M.Moueza. 1996. Environmental transmission of a sulfur-oxidizing bacterial gill endosymbiont in the tropical lucinidbivalve Codakia orbicularis. Appl. Envir. Microbiol. 62:2324-2330.
Großkopf R., P.H. Janssen and W. Liesack. 1998a.Diversity and structure of the methanogenic community in
anoxic rice paddy soil microcosms as examined by cultivationand direct 16S rRNA gene sequence retrieval. Appl. Envir.Microbiol. 64:960-969.
Großkopf R., S. Stubner and W. Liesack. 1998b. NovelEuryarchaeotal lineages detected on rice roots and in theanoxic bulk soil of flooded rice microcosms. Appl. Envir.Microbiol. 64:4983-4989.
Grote R., L. Li, J. Tamaoka, C. Kato, K. Horikoshi and G.Antranikian. 1999. Thermococcus siculi sp. nov., a novelhyperthermophilic archaeon isolated from a deep-seahydrothermal vent at the Mid-Okinawa Trough. Extremophiles3:55-62
Grunstein M. and D. Hogness. 1975. Colony hybridization:a method for the isolation of cloned DNAs that contain aspecific gene. Proc. Natl. Acad. Sci. USA. 72:3961-3965.
Guezennec J. 1995. Approche des écosystèmes bactérienspar les lipides: force et faiblesse des marqueurs bactériens.Océanis 21:75-93.
Guezennec J., O. Ortega-Morales, G. Raguenes and G.Geesey. 1998. Bacterial colonization of artificial substrate inthe vicinity of deep-sea hydrothermal vents. FEMS Microbiol.Ecol. 26:89-99.
Gugliandolo C. and T.L. Maugeri. 1998. Temporalvariations in heterotrophic mesophilic bacteria from a marineshallow hydrothermal vent off the island of Vulcano. Microb.Ecol. 36:13-22.
Guillou L. 1999. Diversité des picoeucaryotes océaniques:description, phylogénie et écologie de nouvelles espèces.Thèse de doctorat de l'université de Paris VI. 172p.
Guo C., W. Sun, J.B. Harsh and A. Ogram. 1997.Hybridization analysis of microbial DNA from fuel oil-contaminated and noncontaminated soil. Microb. Ecol. 34:178-187.
Gupta R.S. 1998a. Protein phylogenies and signaturesequences: A reappraisal of evolutionary relationships amongarchaebacteria, eubacteria, and eukaryotes. Microbiol. Mol.Biol. Rev. 62:1435-1491.
Gupta R.S. 1998b. Life's third domain (Archaea): anestablished fact or an endangered paradigm? Theor. Popul.Biol. 54:91-104.
Gupta R.S. 1998c. What are archaebacteria: life's thirddomain or monoderm prokaryotes related to gram-positivebacteria? A new proposal for the classification of prokaryoticorganisms. Mol. Microbiol. 29:695-707.
Guschin D., B.K. Mobarry, D. Proudnikov, D.A. Stahl,B.E. Rittmann and A.D. Mirzabekov. 1997a.Oligonucleotide microchips as genosensors for determinativeand environmental studies in microbiology. Appl. Envir.Microbiol. 63:2397-2402.
Guschin D., G. Yershov, A. Zaslavsky, A. Gemmell, V.Shick, D. Proudnikov, P. Arenkov and A. Mirzabekov.1997b. Manual manufacturing of oligonucleotide, DNA, andprotein microchips. Anal. Biochem. 250:203-211.
Gutell R.R., N. Larsen and C.R. Woese. 1994. Lessonsfrom an evolving rRNA: 16S and 23S rRNA structures from acomparative perspective. Microbiol. Rev. 58:10-26.
Guyard S. 1994. Etude de la resistance aux métaux lourdschez les Eubactéries et des archaebactéries thermophiles.D.E.A., UBO. 34p.
H
Haddad A.F., P. Camacho, P. Durand and S.C. Cary. 1995.Phylogenic characterization of the epibiotic bacteriaassociated with the hydrothermal vent polychaete Alvinellapompejana. Appl. Environ. Microbiol. 61:1679-1687.
Hafenbradl D., M. Keller, R. Dirmeier, R. Rachel, P.Rossnagel, S. Burggraf, H. Huber and K.O. Stetter. 1996.Ferroglobus placidus gen. nov., sp. nov., A novelhyperthermophilic archaeum that oxidizes Fe2+ at neutral pHunder anoxic conditions. Arch. Microbiol. 166:308-314.
Hales B.A., C. Edwards, D.A. Ritchie, G. Hall, R.W.Pickupand and J.R. Saunders. 1996. Isolation andidentification of methanogen-specific DNA from blanket bog

Références bibliographiques
222
peat by PCR amplification and sequence analysis. Appl. Envir.Microbiol. 62:668-675.
Hallin S. and P.E. Lindgren. 1999. PCR detection of genesencoding nitrite reductase in denitrifying bacteria. Appl. Envir.Microbiol. 65:1652-1657.
Hancock J.M., D. Tautz and G.A. Dover. 1988. Evolution ofthe secondary structures and compensatory mutations of theribosomal RNAs of Drosophila melanogaster. Mol. Biol. Evol.5:393-414.
Hansen M.C., T. Tolker-Nielsen, M. Givskov and S. Molin.1998. Biased 16S rDNA PCR amplification caused byinference from DNA flaking the template region. FEMSMicrobiol. Ecol. 26:141-149.
Harmsen H.J., A.D. Akkermans, A.J. Stams and W.M. deVos. 1996b. Population dynamics of propionate-oxidizingbacteria under methanogenic and sulfidogenic conditions inanaerobic granular sludge. Appl. Envir. Microbiol. 62:2163-2168.
Harmsen H.J., B. Wullings, A.D. Akkermans, W. Ludwigand A.J. Stams. 1993. Phylogenetic analysis ofSyntrophobacter wolinii reveals a relationship with sulfate-reducing bacteria. Arch. Microbiol. 160:238-240.
Harmsen H.J., D. Prieur and C. Jeanthon. 1997a.Distribution of microorganisms in deep-sea hydrothermal ventchimneys investigated by whole-cell hybridization andenrichments of thermophilic subpopulations. Appl. Environ.Microbiol. 63:2876-2883.
Harmsen H.J., D. Prieur and C. Jeanthon. 1997b. Group-specific 16S rRNA-targeted oligonucleotide probes to identifythermophilic bacteria in marine hydrothermal vents. Appl.Environ. Microbiol. 63: 4061-4068.
Harmsen H.J., H.M. Kengen, A.D. Akkermans, A.J. Stamsand W.M. de Vos. 1996a. Detection and localization ofsyntrophic propionate-oxidizing bacteria in granular sludge byin situ hybridization using 16S rRNA-based oligonucleotideprobes. Appl. Envir. Microbiol. 62:1656-1663.
Harper-Owen R., D. Dymock, V. Booth, A.J. Weightmanand W.G. Wade. 1999. Detection of unculturable bacteria inperiodontal health and disease by PCR. J. Clin. Microbiol.37:1469-1473.
Hastings R.C., J.R. Saunders, G.H. Hall, R.W. Pickup andA.J. McCarthy. 1998. Application of molecular biologicaltechniques to a seasonal study of ammonia oxidation in aeutrophic freshwater lake. Appl. Environ. Microbiol. 64:3674-3682.
Head I.M., J.R.Saunders and R.W. Pickup. 1998. Microbialevolution, diversity and ecology: a decade of ribosomal RNAanalysis of uncultivated microorganisms. Micro. Ecol. 35:1-21.
Heidelberg J.F., M. Shahamat, M. Levin, I. Rahman, G.Stelma, C. Grim and R.R. Colwell. 1997. Effect ofaerosolization on culturability and viability of Gram- negativebacteria. Appl. Envir. Microbiol. 63:3585-3588.
Heider J., A.M. Spormann, H.R. Beller and F. Widdel.1999. Anaerobic bacterial metabolism of hydrocarbons. FEMSMicrobiol. Rev. 22:459-473.
Heinen U.J. and W. Heinen. 1972. Characteristics andproperties of a caldo-active bacterium producing extracellularenzymes and two related strains. Arch. Mikrobiol. 82:1-23.
Henckel T., M. Friedrich and R. Conrad. 1999. Molecularanalyses of the methane-oxidizing microbial community inrice field soil by targeting the genes of the 16S rRNA,particulate methane monooxygenase, and methanoldehydrogenase. Appl. Environ. Microbiol. 65:1980-1990.
Henne A., R. Daniel, R.A. Schmitz and G. Gottschalk.1999. Construction of environmental DNA libraries inEscherichia coli and screening for the presence of genesconferring utilization of 4-hydroxybutyrate. Appl. Envir.Microbiol. 65:3901-3907.
Henry E.A., R. Devereux, J.S. Maki, C.C. Gilmour, C.R.Woese, L. Mandelco, R. Schauder, C.C. Remsen and R.Mitchell. 1994. Thermodesulfovibrio yellowstonii, gen. nov.and sp. nov.: its phylogenetic relationship toThermodesulfobacterium commune and their origins deepwithin the bacterial domain. Arch. Microbiol. 161:62-69.
Hensel R., K. Matussek, K. Michalke, L. Tacke, B.J.Tindasll, M. Kohlhoff, B. Siebers and J. Dielenschneider.1997. Sulfophobococcus zilligii gen. nov., sp. nov., a novelhyperthermophilic archaeum isolated from hot alkalinesprings of Iceland. Syst. Appl.Microbiol. 20:102-110.
Herrick J.B., E.L. Madsen, C.A. Batt and W.C. Ghiorse.1993. Polymerase chain reaction amplification of naphthalene-catabolic and 16S rRNA gene sequences from indigenoussediment bacteria. Appl. Environ. Microbiol. 59:687-694.
Herring P.J., E. Gaten and P.M.J. Shelton. 1999. Are ventshrimps blinded by science? Nature 398:116.
Hess A., B. Zarda, D. Hahn, A. Haner, D. Stax, P. Hohenerand J. Zeyer. 1997. In situ analysis of denitrifying toluene- andm-xylene-degrading bacteria in a diesel fuel-contaminatedlaboratory aquifer column. Appl. Envir. Microbiol. 63:2136-2141.
Hillis D.M., C. Moritz, C.A. Porter and R.J. Baker. 1991.Evidence for biased gene conversion in concerted evolution ofribosomal DNA. Science 251:308-310.
Hines M.E., R.S. Evans, B.R. Sharak Genthner, S.G. Willis,S. Friedman, J.N. Rooney-Varga and R. Devereux. 1999.Molecular phylogenetic and biogeochemical studies of sulfate-reducing bacteria in the rhizosphere of Spartina alterniflora.Appl. Environ. Microbiol. 65:2209-2216.
Hinrichs K.U., J.M. Hayes, S.P. Sylva, P.G. Brewer andE.F. DeLong. 1999. Methane-consuming archaebacteria inmarine sediments. Nature 398:802-805.
Hiorns W.D., B.A. Methé, S.A. Nierzwicki-Bauer and J.P.Zehr. 1997. Bacterial diversity in Adirondack mountain lakesas revealed by 16S rRNA gene sequences. Appl. Environ.Microbiol. 63:2957-2960.
Hiraishi A. 1992. Direct automated sequencing of 16S rDNAamplified by polymerase chain reaction from bacterialcultures without DNA purification. Lett. Appl. Microbiol.15:210-213.
Hiraishi A., T. Umezawa, H. Yamamoto, K. Kato and Y.Maki. 1999. Changes in quinone profiles of hot springmicrobial mats with a thermal gradient. Appl. Environ.Microbiol. 65:198-205.
Hoaki T., M. Nishijima, H. Miyashita and T. Maruyama.1995. Dense community of hyperthermophilic sulfur-dependent heterotrophs in a geotherlly heated shallowsubmarine biotope near Kodakara-Jima island, Kagoshima,Japan. Appl. Environ. Microbiol. 61:1931-1937.
Hoaki T., M. Nishijima, M. Kato, K. Adachi, S. Mizobuchi,N. Hanzawa and T. Maruyama. 1994. Growth requirementsof hyperthermophilic sulfur-dependent heterotrophic archaeaisolated from a shallow submarine geothermal system withreference to their essential amino acids. Appl. Envir.Microbiol. 60:2898-2904.
Hobbie J.E., R.J. Dailey and S. Jasper. 1977. Use ofNucleopore filters for counting bacteria by fluorescencemicroscopy. Appl. Envir. Microbiol. 37:805-812.
Hodson R.E. O. Holm-Hannsen and F. Azam. 1976.Improved methodology for ATP determination in marineenvironments. Mar. Biol. 34:143-149.
Hodson R.E., W.A. Dustman, R.P. Garg and M.A. Moran.1995. In situ PCR for visualization of microscale distribution ofspecific genes and gene products in prokaryotic communities.Appl. Envir. Microbiol. 61:4074-4082.
Hoefs M.J.L., S. Schouten, J.W. De Leeuw, L.L. King,S.G. Wakeham and J.S. Sinninghe Damsté. 1997. Etherlipids of planktonic Archaea in marine water column. Appl.Environ. Microbiol. 63:3090-3095.
Höfle M.G., H. Haas and K. Dominik. 1999. Seasonaldynamics of bacterioplankton community structure in aeutrophic lake as determined by 5S rRNA analysis. ApplEnviron. Microbiol. 65:3164-3174.
Holben W.E., J.K. Jansson, B.K. Chelm and J.M. Tiedje.1988. DNA probe method for the detection of specificmircroorganisms in the soil bacteria community. Appl. Envir.Microbiol. 54:703-711.
Holben W.E., K. Noto, T. Sumino and Y. Suwa. 1998.Molecular analysis of bacterial communities in a three-compartment granular activated sludge system indicates

Références bibliographiques
223
community-level control by incompatible nitrificationprocesses. Appl. Envir. Microbiol. 64:2528-2532.
Holm-Hannsen O. and C.R. Booth. 1966. Themeasurement of adenosine triphosphate in the ocean and itsecological significance. Limnol. Oceanogr. 11:510-519.
Holmstrøm K., T. Tolker-Nielsen and S. Molin. 1999.Physiological states of individual Salmonella typhimuriumcells monitored by in situ reverse transcription-PCR. J.Bacteriol. 181:1733-1738.
Huang W.M. 1996. Bacterial diversity based on type II DNAtopoisomerase genes. Annu. Rev. Genet. 30:79-107.
Huber G., C. Spinnler, A. Gambacorta and K.O. Stetter.1989a. Metallosphaera sedula gen. and sp. nov. represents anew genus of aerobic, metal-mobilizing, thermoacetophilicarchaebacteria. System. Appl. Microbiol. 12:38-47.
Huber H., M. Thomm, H. König, G. Thies and K.O.Stetter. 1982. Methanococcus thermolithotrophicus, a novelthermophilic lithotrophic methanogen. Arch. Microbiol.132:47-50.
Huber R., C.R. Woese, T.A. Langworthy, H. Fricke andK.O. Stetter. 1989b Thermosipho africanus gen. nov.,represents a new genus of thermophilic eubacteria within the"Thermotogales". System. Appl. Microbiol. 12:32-37.
Huber R., D. Dyba, H. Huber, S. Burggraf and R. Rachel.1998a. Sulfur-inhibited Thermosphaera aggregans sp. nov., anew genus of hyperthermophilic archaea isolated after itsprediction from environmentally derived 16S rRNAsequences. Int. J. Syst. Bacteriol. 48:31-38.
Huber R., J.K. Kristjansson and K.O. Stetter. 1987.Pyrobaculum gen. nov., a new genus of neutrophilic, rod-shaped archaebacteria from continental solfataras growingoptimally at 100 °C. Arch. Microbiol. 149:95-101
Huber R., S. Burggraf, T. Mayer, S.M. Barns, P.Rossnagel and K.O. Stetter. 1995. Isolation of ahyperthermophilic archaeum predicted by in situ RNAanalysis. Nature 376:57-58
Huber R., T. Wilharm, D. Huber, A. Trincone, S.Burggraf, H. König, R. Rachel, I. Rockinger, H. Fricke,K.O. Stetter. 1992. Aquifex pyrophilus gen. nov. sp. nov.,represents a novel group of marine hyperthermophilichydrogen-oxidizing bacterium. System. Appl. Microbiol.15:340-351.
Huber R., T.A. Langworthy, H. Köning, M. Thomm, C.R.Woese, U.B. Sleytr and K.O. Stetter. 1986. Thermotogamaritima sp. nov. represents a new genus of unique extremelythermophilic eubacteria growing up to 90°C. Arch. Microbiol.144:324-333.
Huber R., W. Eder, S. Heldwein, G. Wanner, H. Huber, R.Rachel and K.O. Stetter. 1998b. Thermocrinis ruber gen.nov., sp. nov., a pink-filament-forming hyperthermophilicbacterium isolated from Yellowstone national park. Appl.Envir. Microbiol. 64:3576-3583.
Hudson J.A., K.M. Schofield, H.W. Morgan and R.M.Daniel. 1989. Thermonema lapsum gen. nov., sp. nov., athermophilic gliding bacterium. Int. J. Syst. Bacteriol. 39:485-487.
Huelsenbeck J.J. and D.M. Hillis. 1993. Success ofphylogenetic methods in four taxon cases. Syst. Biol. 42:247-264.
Hugenholtz P., B.M. Goebel and N.R. Pace. 1998b. Impactof culture-independent studies on the emerging phylogeneticview of bacterial diversity. J. Bacteriol. 180:4765-4774.
Hugenholtz P., C. Pitulle, K.L. Hershberger and N.R.Pace. 1998a. Novel division level bacterial diversity in aYellowstone hot spring. J. Bacteriol. 180:366-376.
Hunter-Cevera J.C. 1998. The value of microbiel diversity.Curr. Opin. Microbiol. 1:278-285.
I
Imanaka T., M. Fujii, I. Aramori and S. Aiba. 1982.Transformation of Bacillus stearothermophilus with plasmidDNA and characterization of shuttle vector plasmids between
Bacillus stearothermophilus and Bacillus subtilis. J. Bacteriol.149:824-30
Inagaki F., S. Hayashi, K. Doi, Y. Motomura E. Izawa andS. Ogata. 1997. Microbial participation in the formation ofsiliceous deposits from geothermal water and analysis of theextremly thermophilic bacterial community. FEMS Microbiol.Ecol. 24:41-48.
Itoh T., K. Suzuki and T. Nakase. 1998. Thermocladiummodestius gen. nov., sp. nov., a new genus of rod-shaped,extremely thermophilic crenarchaeote. Int. J. Syst. Bacteriol.48:879-887.
J
Jackson C.R., E.E. Roden and P.F. Churchill. 1998a.Changes in bacterial species composition in enrichmentcultures with various dilutions of inoculum as monitored bydenaturing gradient gel electrophoresis. Appl. Environ.Microbiol. 64:5046-5048.
Jackson C.R., J.P. Harper, D. Willoughby, E.E. Rodenand P.F. Churchill. 1997. A simple, efficient method for theseparation of humic substances and DNA from environmentalsamples. Appl. Envir. Microbiol. 63: 4993-4995.
Jackson P.J., M.E. Hugh-Jones, D.M. Adair, G. Green,K.K. Hill, C.R. Kuske, L.M. Grinberg, F.A. Abramova andP. Keim. 1998b. PCR analysis of tissue samples from the1979 Sverdlovsk anthrax victims: The presence of multipleBacillus anthracis strains in different victims. Proc. Natl.Acad. Sci. USA 95:1224-1229.
Jackson T.J., R.F. Ramaley and W.G. Meinschein. 1973.Int. J. Syst. Bacteriol. 23:28-36.
Jannasch H.W. 1995. Microbial interactions withhydrothermal fluids. In: Humphris SE, Zierenberg RA,Mullineaux LS & Thomson RE (Eds) Seafloor HydrothermalSystems: Physical, Chemical, Biological, and GeologicalInteractions, p 273-296. Geophysical monograph 91, AmericanGeophysical Union, Washington DC.
Jannasch H.W. and M.J. Mottl. 1985. Geomicrobiology ofdeep-sea hydrothermal vents. Science 229:717-725.
Jannasch H.W. and C.O. Wirsen. 1979. Chemosyntheticprimary production at East Pacific sea-floor spreadingcenters. Bioscience 29:592-598.
Jannasch H.W. and C.O. Wirsen. 1981. Morphologicalsurvey of microbial mats near deep sea thermal vents. Appl.Environ. Microbiol. 41:528-538.
Jannasch H.W., C.O. Wirsen ,S.J. Molyneaux and T.A.Langworthy. 1988. Extremely thermophilic fermentativearchaebacteria of the genus Desulfuroccocus from deep-seahydrothermal vents. Appl. Environ. Microbiol. 41:528-538.
Jannasch H.W., C.O. Wirsen, D.C. Nelson and L.A.Robertson. 1985. Thiomicrospira cruogena sp. nov., acolorless sulfur-oxidizing bacterium from deep-seahydrothermal vent. Int. J. Microbiol. 35:422-424.
Jannasch H.W., D.C. Nelson and C.O. Wirsen. 1989.Massive natural occurrence of unusually large bacteria(Beggiatoa sp.) at a hydrothermal vent site. Nature 342:834-836.
Jeanthon C. 1999. Molecular ecology of hydrothermal ventmicrobial communities. Antonie van Leeuwenhoek (soumis).
Jeanthon C. and D. Prieur. 1990. Suceptibility to heavymetals and characterization of heterotrophic bacteria isolatedfrom two hydrothermal vent polychete annelids, Alvinellapompejana and Alvinella caudata. Appl. Environ. Microbiol.56:3308-3314.
Jeanthon C., A-L Reysenbach, S. L'Haridon, A.Gambacorta, N. Pace, P. Glénat and D.Prieur. 1995.Thermotoga subterranea sp. nov., a new thermophilicbacterium isolated from a continental oil reservoir. Arch.Microbiol. 164:91-97.
Jeanthon C., S. L'Haridon, A.L. Reysenbach, E. Corre, M.Vernet, P. Messner, U.W. Sleytr and D. Prieur. 1999a.Methanococcus vulcanius sp. nov., a novel hyperthermophilicmethanogen isolated from East Pacific Rise and identificationof Methanococcus spp. DSM 4213 as Methanococcus fervenssp. nov. Int. J. Syst. Bacteriol. 49:583-589.

Références bibliographiques
224
Jeanthon C., S. L'Haridon, A.L. Reysenbach, M. Vernet,P. Messner, U.W. Sleytr and D. Prieur. 1998.Methanococcus infernus sp. nov., a novel hyperthermophiliclithotrophic methanogen isolated from a deep-seahydrothermal vent. Int. J. Syst. Bacteriol. 48:913-919.
Jeanthon C., S. L'Haridon, N. Pradel and D. Prieur. 1999b.Rapid identification of hyperthermophilic methanococciisolated from deep-sea hydrothermal vents. Int. J. Syst.Bacteriol. 49:591-594.
Jensen S., L. Øvreås, F.L. Daae and V. Torsvik. 1998.Diversity in methane enrichments from agricultural soilrevealed by DGGE separation of PCR amplified 16S rDNAfragments. FEMS Microbiol. Ecol. 26:17-26.
Jochimsen B., S. Peinemann-Simon, H. Völker, D.Stüben, R. Botz, P. Stoffers, P.R. Dando and M. Thomm.1997. Stetteria hydrogenophila, gen. nov. and sp. nov., a novelmixotrophic sulfur-dependent crenarchaeote isolated fromMilos, Greece. Extremophiles 1:67-73.
Johansson K.E. 1996. Oligonucleotide probescomplementary to 16S rRNA. Molecular and diagnosticProcedures in Mycoplasmology, Vol II p 29-46.
Johnsen K., Ø. Enger, C.S. Jacobsen, L. Thirup and V.Torsvik. 1999. Quantitative selective PCR of 16S ribosomalDNA correlates well with selective agar plating in describingpopulation dynamics of indigenous Pseudomonas spp. in soilhot spots. Appl. Envir. Microbiol. 65:1786-1788.
Jones W.J., J.A. Leigh, F. Mayer, C.R. Woese and R.S.Wolfe. 1983. Methanococcus jannaschii sp. nov., anextremely thermophilic methanogen from a submarinehydrothermal vent. Arch. Microbiol. 136:254-261.
Jorgensen B.B. and V.A. Gallardo. 1999. Thioplaca spp.:filamentous sulfur bacteria with nitrate vacuoles. FEMSMicrobiol. Ecol. 28:301-313.
Joux F. and P. Lebaron. 1995. Applications en écologiebactérienne des sondes oligonucléotidiques fluorescentes parles techniques d'hybridation et de cytométrie. Océanis 21:125-138.
Jukes T. H. and C.R. Cantor. 1969. Evolution of proteinmolecules, p.21-32. In H. N. Munro (ed.), Mammalian proteinmetabolism. Academic Pres, New York, N.Y.
Juretschko S., G. Timmermann, M. Schmid, K.H.Schleifer, A. Pommerening-Röser, H.P. Koops and M.Wagner. 1998. Combined molecular and conventionalanalyses of nitrifying bacterium diversity in activated sludge:Nitrosococcus mobilis and Nitrospira-like bacteria asdominant populations. Appl. Envir. Microbiol. 64:3042-3051.
Jurgens G., K. Lindstrom and A. Saano. 1997. Novel groupwithin the kingdom Crenarchaeota from boreal forest soil.Appl. Envir. Microbiol. 63: 803-805.
K
Ka J.O., W.E. Holben and J.M. Tiedje. 1994. Genetic andphenotypic diversity of 2,4-dichlorophenoxyacetic acid (2,4-D)-degrading bacteria isolated from 2,4-D-treated field soils.Appl. Envir. Microbiol. 60:1106-1115.
Kalmbach S., W. Manz and U. Szewzyk. 1997. Isolation ofnew bacterial species from drinking water biofilms and proofof their in situ dominance with highly specific 16S rRNAprobes. Appl. Envir. Microbiol. 63: 4164-4170.
Kämpfer P., R. Erhart, C. Beimfohr, J. Böhringer, M.Wagner and R. Amann. 1996. Characterization of bacterialcommunities from activated sludge: culture-dependentnumerical identification versus in situ identification usinggroup- and genus-specific rRNA-targeted oligonucleotideprobes. Microbial Ecology 32:101-121.
Karavaiko G.I., O.V. Golyshina, A.V. Troitskii, K.M.Val'ekho-Roman, R.S. Golovacheva and T.A. Pivovarova.1994. Sulfurococcus yellowstonii sp. nov/ a new species ofiron- and sulfur-oxidizing thermoacidophilicArchaeobacterium. Mikrobiologiia 63:668-682.
Karl D.M. 1979. Measurement of microbial activity andgrowth in the ocean by rates of stable ribonucleic acidsynthesis. Appl. Envir. Microbiol.38:850-860.
Karl D.M. 1981. Simultaneous rates of ribonucleic acid anddeoxyribonucleic acid syntheses for estimating growth andcell division of aquatic microbial communities. Appl. Envir.Microbiol. 42:802-810.
Karl D.M. 1987. Bacterial production at deep-seahydrothermal vent and cold seeps: evidence forchemosynthetic primary production. In: Ecology of MicrobialCommunities, Forty First Symposium of the Society forGeneral Microbiology (M. Fletcher, T.R. Gary and J.G. Joneseds.), p 319-359. Cambridge Universtity Press: Cambridge.
Karl D.M. 1995. Ecology of free-hydrothermal vent microbialcommunities. In: Karl DM (Ed) The Microbiology of Deep-seaHydrothermal Vents (p 35-124). CRC Press Inc, Boca Raton.
Karl D.M., G.T. Taylor, J.A. Novitsky, H.W. Jannasch, C.O.Wirsen, N.R. Pace, D.J. Lane, G.J. Olsen, and S.J.Giovannoni. 1988. A microbiological study of Guaymas basinhigh temperature hydrothermal vents. Deep-Sea Res. 35:777.
Karner M. and J.A. Fuhrman. 1997. Determination of activemarine bacterioplankton: a comparaison of universal 16SrRNA probes, autoradiography and nucleoid staining. Appl.Envir. Microbiol. 63:1208-1213.
Karp P.D., M. Riley, S.M. Paley, A. Pellegrini-Toole and M.Krummenacker. 1999. Eco Cyc: encyclopedia of Escherichiacoli genes and metabolism. Nucleic Acids Res. 27:55-58.
Karp P.D., M. Riley, S.M. Paley, A. Pellegrini-Toole and M.Krummenacker. 1998. Eco Cyc: encyclopedia of Escherichiacoli genes and metabolism. Nucleic Acids Res. 26:50-53.
Kato C., L. Li, J. Tamaoka and K. Horikoshi. 1997.Molecular analyses of the sediment of the 11,000-m deepMariana Trench. Extremophiles 1:117-123.
Kawasumi T., Y. Igarashi, T. Kodama and Y. Minoda.1984. Hydrogenobacter thermophilus gen. nov. sp. nov., anextremely thermophilic, aerobic, hydrogen-oxidizingbacterium. Int. J. Syst. Bacteriol. 34:5-10.
Keller M., F.J. Braun, R. Dirmeier, D. Hafenbradl, S.Burggraf, R. Rachel and K.O. Stetter. 1995. Thermococcusalcaliphilus sp. nov., a new hyperthermophilic archaeumgrowing on polysulfide at alkaline pH. Arch. Microbiol.164:390-395.
Kenzaka T., N. Yamaguchi, K. Tani and M. Nasu. 1998.rRNA-targeted fluorescent in situ hybridization analysis ofbacterial community structure in river water. Microbiology144:2085-2093.
Keswani J., S. Orkand, U. Premachandran, L. Mandelco,M.J. Franklin and W.B. Whitman. 1996. Phylogeny andtaxonomy of mesophilic Methanococcus spp. and comparisonof rRNA, DNA hybridization, and phenotypic methods. Int. J.Syst. Bacteriol. 46:727-735.
Khbaya B., M. Neyra, P. Normand, K. Zerhari and A.Filali-Maltouf. 1998. Genetic diversity and phylogeny ofrhizobia that nodulate Acacia spp. in Morocco assessed byanalysis of rRNA genes. Appl. Envir. Microbiol. 64: 4912-4917.
Kim J. 1993. Improving the accuracy of phylogeneticestimation by combining different methods. Syst. Biol. 42:331-340.
Kimura M. 1968. Evolutionary rate at the molecular level.Nature. 217:624-626.
Kimura M. 1980. A simple method for estimating evolutionaryrates of base substitutions through comparative studies ofnucleotide sequences. J. Mol. Evol. 16:111-120.
Kirchman D., F. K'Ness and R. Hodson. 1985. Leucineincorporation and its potential as a measure of proteinsynthesis by bacteria in natural aquatic systems. Appl. Envir.Microbiol. 49:599-607.
Klenk H.P. 1998. On the phylogenetic position of Aquifexpyrophilus in protein based phylogenies. Thermophiles'98,International Conference, 6-11 septembre, Brest France.
Klier N.J., R.J. West and P.A. Donberg. 1999. Aerobicbiodegradation of dichloroethylenes in surface and subsurfacesoils. Chemosphere 38:1175-1188.
Kobayashi T., Y.S. Kwak, T. Akiba, T. Kudo and K.Horikoshi. 1994. Thermococccus profundus sp. nov., a newhyperthermophilic archaeon isolated from a deep-seahydrothermal vent. System. Appl. Microbiol. 17:232-236.

Références bibliographiques
225
Kogura K. and U. Simidu. 1979. A tentative directmicroscopic method for countingliving marine bacteria. Can.J. Microbiol. 25:415-420.
König H. 1989. Archaebacteria. Biotechnology, Vol 6b,Special Microbial Processes, V.C.H. editors, 6b, p 697-728.
Konopka A. and T. Zakharova. 1999. Quantification ofbacterial lead resistance via activity assays. J. Microbiol.Methods 37:17-22.
Konopka A., T. Zakharova, M. Bischoff, L. Oliver, C.Nakatsu and R.F. Turco. 1999. Microbial biomass andactivity in lead-contaminated soil. Appl. Environ. Microbiol.65:2256-2259.
Koonin E. and M.Y. Galperin. 1997. Prokaryotic genomes:the emerging paradigm of genome-based microbiology. Curr.Opin. Genet. Dev. 7:757-763.
Kopczynski E.D., M.M. Bateson and D.M. Ward. 1994.Recognition of chimeric small-subunit ribosomal DNAscomposed of genes from uncultivated microorganisms. Appl.Environ. Microbiol. 60:746-748.
Kopylov V.M., E.A. Bonch-Osmolovskaya, V.A.Svetlichnyi, M.L. Miroshnicheko and V.S. Skobkin. 1993.Gamma-irradiation and UV-sensitivity of extremlythermophilic archaebacteria and eubacteria. Microbiologiya.62:90-95.
Kotelnikova S. and K. Petersen. 1997. Evidence formethanogenic Archaeae and homoacetogenic Bacteria in deepgranitic rock aquifers. FEMS Microbiol. Rev. 20:339-349.
Kotelnikova S., A.J. Macario and K. Pedersen. 1998.Methanobacterium subterraneum sp. nov., a new alkaliphilic,eurythermic and halotolerant methanogen isolated from deepgranitic groundwater. Int. J. Syst. Bacteriol. 48:357-367.
Kowalchuk G.A., J.R. Stephen, W. De Boer, J.I. Prosser,T.M. Embley and J.W. Woldendorp. 1997. Analysis ofammonia-oxidizing bacteria of the beta subdivision of theclass Proteobacteria in coastal sand dunes by denaturinggradient gel electrophoresis and sequencing of PCR-amplified16S ribosomal DNA fragments. Appl. Envir. Microbiol.63:1489-1497.
Kowalchuk G.A., Z.S. Naoumenko, P.J. Derikx, A. Felske,J.R. Stephen and I.A. Arkhipchenko. 1999. Molecularanalysis of ammonia-oxidizing bacteria of the beta subdivisionof the class Proteobacteria in compost and compostedmaterials. Appl. Environ. Microbiol. 65:396-403.
Krause D.O. and J.B. Russell. 1996a. How many ruminalbacteria are there? Dairy Sci. 79:1467-1475.
Krause D.O. and J.B. Russell. 1996b. An rRNA approach forassessing the role of obligate amino acid-fermenting bacteriain ruminal amino acid deamination. Appl. Environ. Microbiol.62:815-821.
Krueger D.M. and C.M. Cavanaugh. 1997. Phylogeneticdiversity of bacterial symbionts of Solemya host based oncomparative sequence analysis of 16S rRNA genes. Appl.Environ. Microbiol. 63:91-98.
Kryukov V.R., N.D. Savelyeva and M.A. Pusheva. 1983.Calderobacterium hydrogenophilum nov. gen., nov. sp., anextreme thermophilic bacterium and its hydrogenase activity.Mikrobiologiya 52:781-788.
Kudo T., M. Ohkuma, S. Moriya, S. Noda and K. Ohtoko.1998. Molecular phylogenetic identification of the intestinalanaerobic microbial community in the hindgut of the termite,Reticulitermes speratus, without cultivation. Extremophiles2:155-161.
Kurosawa N., Y.H. Itoh, T. Iwai, A. Sugai, I. Uda, N.Kimura, T. Horiuchi and T. Itoh. 1998. Sulfurisphaeraohwakuensis gen. nov., sp. nov., a novel extremelythermophilic acidophile of the order Sulfolobales. Int. J. Syst.Bacteriol. 48:451-456.
Kurr M., R. Huber, H. König, H.W. Jannasch, H. Fricke, A.Trincone, J.K. Kristjansson and K.O. Stetter. 1991.Methanopyrus kandleri, gen. and sp. nov. represents a novelgroup of hyperthermophilic methanogens, growing at 110°C.Arch. Microbiol. 156:239-247.
Kuske C.R., K.L. Banton, D.L. Adorada, P.C. Stark, K.K.Hill and P.J. Jackson. 1998. Small-scale DNA samplepreparation method for field PCR detection of microbial cellsand spores in soil. Appl. Envir. Microbiol. 64: 2463-2472.
Kuske C.R., S.M. Barns and J.D. Busch. 1997. Diverseuncultivated bacterial groups from soils of the aridsouthwestern United States that are present in manygeographic regions Appl. Envir. Microbiol. 63: 3614-3621.
L
L'Haridon S., A.-L. Reysenbach, P. Glénat, D. Prieur andC. Jeanthon. 1995. Hot subterranean biosphere in acontinental oil reservoir. Nature 377:223-224.
L’Haridon S., M. Speck, C. Vetriani, A.L. Reysenbach, D.Prieur and C. Jeanthon. 1998b. The Desulfurobacteriaceae,a new family of thermophilic bacteria from deep-seahydrothermal vents. Thermophiles 98’ InternationalConference BP 28.
L'Haridon S., V. Cilia, P. Messner, G. Raguénès, A.Gambacorta, U.W. Sleytr, D. Prieur and C. Jeanthon.1998a. Desulfurobacterium thermolithotrophum gen. nov., sp.nov., a novel autotrophic, sulfur-reducing bacterium isolatedfrom a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol.48:701-711.
Lafay B. and J.J. Burdon. 1998. Molecular diversity ofrhizobia occurring on native shrubby legumes in southeasternAustralia. Appl. Envir. Microbiol. 64: 3989-3997.
Lake J.A. 1987. A rate-independent technique for analysis ofnucleic acid sequences: evolutionary parsimony. Mol. Biol.Evol. 4:167-191.
Lambert L.H., T. Cox, K. Mitchell, R. Rossello-Mora, C.Del Cueto, D.E.Dodge, P. Orkand and R.J. Cano. 1998.Staphylococcus succinus, sp. nov., Isolated from Dominicanambert. Int. J. Syst. Bacteriol. 48:511-518.
Lane D.J., B. Pace, G.J. Olsen, D.A. Stahl, M.L. Sogin andN.R. Pace. 1985c. Rapid determination of 16S ribosomal RNAsequences for phylogenetic analyses. Proc. Natl. Acad. Sci.USA 82:6955-6959.
Lane D.J., D.A. Stahl, G.J. Olsen and N.R. Pace. 1985a.Analysis of hydrothermal vent-associated symbionts byribosomal RNA sequences. Proc. Biol. Soc. Wash. 6:389-400.
Lane D.J., D.A. Stahl, G.J. Olsen, D.J. Heller and N.R.Pace. 1985b. Phylogenetic analysis of the genera Thiobacillusand Thiomicrospira by 5S rRNA sequences. J. Bacteriol.163:75-81.
Lee D.H., Y.G. Zo and S.J. Kim. 1996a. Nonradioactivemethod to study genetic profiles of natural bacterialcommunities by PCR-single-strand-conformationpolymorphism. Appl. Envir. Microbiol. 62:3112-3120.
Lee N., P.H. Nielsen, K.H. Andreasen, S. Juretschko, J.L.Nielsen, K.H. Schleifer and M. Wagner. 1999. Combinationof fluorescent in situ hybridization and microautoradiography anew tool for structure-function analyses in microbial ecology.Appl. Envir. Microbiol. 65:1289-1297.
Lee S. and J.A. Fuhrman. 1990. DNA hybridization tocompare species compositions of natural bacterioplanktonassemblages. Appl. Environ. Microbiol. 56:739-746.
Lee S.H. and P.F. Kemp. 1994. Single-cell RNA content ofnatural marine planktonic bacteria mesured by hybridizationwith multiple 16S-rRNA targeted fluorescent probes. Limnol.Oceannogr. 39:869-879.
Lee S.Y., J. Bollinger, D. Bezdicek and A. Ogram. 1996b.Estimation of the abundance of an uncultured soil bacterialstrain by a competitive quantitative PCR method. Appl. Envir.Microbiol. 62: 3787-3793.
Leu J.Y., C.P. McGovern-Traa, A.J.R. Porter, W.J. Harrisand W.A. Hamilton. 1998. Identification and phylogeneticanalysis of thermophilic sulfate-reducing bacteria in oil fieldsamples by 16S rDNA gene cloning and sequencing.Anaerobe 4:165-174.
Levy M. and S.L. Miller. 1998. The stability of RNAbases:implications for the origin of life. Proc. Natl. Acad. Sci. USA.95:7933-7938.
Li L. C. Kato and K. Horikoshi. 1999. Bacterial diversity indeep-sea sediments from different depths. Biodiversity andConservation. 8:659-677.

Références bibliographiques
226
Li W. 1993. So, what about the molecular clock hypothesis?Curr. Opin. Gen. Dev. 3:896-901.
Liesack W. and E. Stackebrandt. 1992. Occurrence of novelgroups of the domain Bacteria as revealed by analysis ofgenetic material isolated from an Australian terrestrialenvironment. J. Bacteriol. 174: 5072-5078.
Liesack W., H. Weyland and E. Stackebrandt. 1991.Potential risk of gene amplification by PCR as determined by16S rDNA analysis of a mixed-culture of strict barophilicbacteria. Microb. Ecol. 21:191-198.
Liesack W., P.H. Janssen, F.A. Rainey, N.L. Ward-Raineyand E. Stackebrandt. 1997. Microbial diversity in soil: theneed for a combined approach using molecular and cultivationtechniques. In Modern Soil Microbiology, J.D. VanElsas, J.T.Trevors, E.M.H. Wellington Eds. Marcel Decker, New York. p375-439.
Lighthart B. 1997. The ecology of bacteria in the alfrescoatmosphere. FEMS Microbiol. Ecol. 23:263-274.
Lim E.L. 1996. Molecular identification of nanoplanktonicprotists based on small subunit ribosomal RNA genesequences for ecological studies. J. Eukaryot. Microbiol.43:101-106.
Liu W.T., T.L. Marsh, H. Cheng and L.J. Forney. 1997.Characterization of microbial diversity by determiningterminal restriction fragment length polymorphisms of genesencoding 16S rRNA. Appl. Environ. Microbiol. 63:4516-4522.
Liu S. V.,J. Zhou, C. Zhang, D.R. Cool, M. Gajdarziska-Josifovska and T.J. Phelps. 1997b. Thermophilic Fe(III)-reducing bacteria from deep subsurface: the evolutionaryimplication. Science 277:1106-1109.
Llobet-Brossa E., R. Rossello-Mora and R. Amann. 1998.Microbial community composition of Wadden sea sedimentsas revealed by fluorescence in situ hybridization. Appl.Environ. Microbiol. 64:2691-2696.
Lloyd D., K.L. Thomas, A. Hayes, B. Hill, B.A. Hales, C.Edwards, J.R. Saunders, D.A. Ritchie and M. Upton.1998. Micro-ecology of peat: minimally invasive analysisusing confocal laser scanning microscopy, membrane inletmass spectrometry and PCR amplification of methanogen-specific gene sequences. FEMS Microbiol. Ecol. 25:179-188.
Lloyd-Jones G. and P.C.K. Lau. 1998. A molecular view ofmicrobial diversity in a dynamic landfill in Québec. FEMSMicrobiol. Letters 162:219-226.
Lovley D.R. 1997. Microbial Fe(III) reduction in subsurfaceenvironments, Fems Microbiol. Rev.3-4:305-313
Ludwig W. and KH Schleifer. 1994. Bacterial phylogenybased on 16S and 23S rRNA sequence analysis. FEMSMicrobiol. Rev. 15:155-173.
Ludwig W., O. Strunk, S. Klugbauer, N. Klugbauer, M.Weizenegger, J. Neumaier, M. Bachleitner and K.H.Schleifer. 1998. Bacterial phylogeny based on comparativesequence analysis. Electrophoresis 19:554-568.
Ludwig W., S. Dorn, N. Springer, G. Kirchhof and K.H.Schleifer. 1994. PCR-based preparation of 23S rRNA-targeted group-specific polynucleotide probes. Appl. Envir.Microbiol. 60:3234-3244.
M
MacArthur R.H. 1962. Some generalized theorems of naturalselection. Proc. Natl. Acad. Sci. USA. 48:1893-1897.
MacGregor B.J., D.P. Moser, E.W. Alm, K.H. Nealson andD.A. Stahl. 1997. Crenarchaeota in Lake Michigan sediment.Appl. Environ. Microbiol. 63:1178-1181.
Madigan M.T. 1996. Brock biology of microorganisms/Michael T. Madigan, John M. Martinko, Jack Parker 8th ed.
Magot M. 1996. Similar bacteria in remote oil fields. Nature.379:681.
Magot M., J. Connan and J.L. Crolet. 1994. Les bactériesdes gisements pétroliers. La recherche. 25:936-937.
Magot M., Ollivier B. and B.K.C. Patel. 1999. Microbiologyof petroleum reservoirs. Antonie van Leeuwenhoek (soumis).
Maidak B.L., J.R. Cole, C.T. Parker Jr, G.M. Garrity, N.Larsen, B. Li, T.G. Lilburn, M.J. McCaughey, G.J. Olsen,R. Overbeek, S. Pramanik, T.M. Schmidt, J.M. Tiedje andCR Woese.1999. A new version of the RDP (RibosomalDatabase Project). Nucleic Acids Res. 27:171-173.
Manz W., K. Wendt-Potthoff, T.R. Neu, U. Szewzyk andJ.R. Lawrence. 1999. Phylogenetic composition, spatialstructure, and dynamics of lotic bacterial biofilms investigatedby fluorescent in situ hybridization and confocal laser scanningmicroscopy. Microb. Ecol. 37:225-237.
Manz W., R. Amann, R. Szewzyk, U. Szewzyk, T.A.Stenstrom, P. Hutzler and K.H. Schleifer. 1995. In situidentification of Legionellaceae using 16S rRNA-targetedoligonucleotide probes and confocal laser scanningmicroscopy. Microbiology 141:29-39.
Manz W., R. Amann, W. Ludwig, M. Wagner and K.-H.Schleifer. 1992. Phylogenetic oligodeoxynucleotide probes forthe major subclasses of Proteobacteria: problems andsolutions. Syst. Appl. Microbiol.15:593-600.
Manz W., U. Szewzyk, P. Ericsson, R. Amann, K.H.Schleifer and T.A. Stenstrom. 1993. In situ identification ofbacteria in drinking water and adjoining biofilms byhybridization with 16S and 23S rRNA-directed fluorescentoligonucleotide probes. Appl. Envir. Microbiol. 59:2293-2298.
Marchesi J.R., T. Sato, A.J. Weightman, T.A. Martin, J.C.Fry, S.J. Hiom and W.G. Wade. 1998. Design and evaluationof useful bacterium-specific PCR primers that amplify genescoding for bacterial 16S rRNA. Appl. Envir. Microbiol. 64:795-799.
Marie D., C.P.D. Brussaard, R. Thyrhaug, G. Bratbak andD. Vaulot. 1999. Enumeration of marine virus in culture andnatural samples by flow cytometry. Appl. Environ. Microbiol.65:45-52.
Marsh P.D. and D.J. Bradshaw. 1995. Dental plaque as abiofilm. J. Ind. Microbiol. 15:169-175.
Marsh T.L. 1999. Terminal restriction fragment lengthpolymorphism (T-RFLP): an emerging method forcharacterizing diversity among homologous populations ofamplification products. Curr. Opin. Microbiol. 2:323-327.
Marshall A. and J. Hodgson. 1998. DNA chips: an array ofpossibilities. Nature Biotech. 16:27-31.
Marteinsson V.T., J.-L. Birrien and D. Prieur. 1997. In situenrichment and isolation of thermophilic microorganismsfrom deep-sea hydrothermal environments. Can. J. Microbiol.43:694-697.
Marteinsson V.T., J.-L. Birrien, C. Jeanthon and D.Prieur. 1996. Numerical taxonomic study of thermophilicBacillus isolated from three geographically separated deep-sea hydrothermal vents. FEMS Microbiol. Ecol. 21:255-266.
Marteinsson V.T., J.-L. Birrien, J.K. Kristjansson and D.Prieur. 1995. First isolation of thermophilic aerobic non-sporulating heterotrophic bacteria from deep-seahydrothermal vents. FEMS Microbiol. Ecol. 18:163-174.
Marteinsson V.T., J.L. Birrien, A.L. Reysenbach, M.Vernet, D. Marie, A. Gambacorta, P. Messner, U.B. Sleytrand D. Prieur. 1999. Thermococcus barophilus sp. nov., anew barophilic and hyperthermophilic archaeon isolated underhigh hydrostatic pressure from a deep-sea hydrothermal vent.Int. J. Syst. Bacteriol. 49:351-359.
Martínez-Murcia A.J., S.G. Acinas and F. Rodriguez-Valera. 1995. Evaluation of prokaryotic diversity by restrictasedigestion of 16S rDNA directly amplified from hypersalineenvironments. FEMS Microbiol. Ecol. 17:247-256.
Marty D., J.C. Bertrand and P. Caumette. 1988. Lesmétabolismes bactériens dans les systèmes sédimentairesmarins, pp.:101-151. In: Micro-organismes dans lesécosystèmes océaniques, M. Bianchi, D. Marty, J.C.Bertrand, P. Caumette, M. Gauthier et coll. (Ed), Masson,Paris.
Massana R., A.E. Murray, C.M. Preston and E.F. DeLong.1997. Vertical distribution and phylogenetic characterization ofmarine planktonic Archaea in the Santa Barbara Channel.Appl. Environ. Microbiol. 63:50-56.
Mau M. and K.N. Timmis. 1998. Use of subtractivehybridization to design habitat-based oligonucleotide probes

Références bibliographiques
227
for investigation of natural bacterial communities. Appl.Environ. Microbiol. 64:185-191.
Mayr E. 1982. Biology is not postage stamp collecting.Science. 216:718-720.
Mayr E. 1998. Two empires or three? Proc. Natl. Acad. Sci.USA. 95:9720-9723.
McCaig A.E., L.A. Glover and J.I. Prosser. 1999. Molecularanalysis of bacterial community structure and diversity inunimproved and improved upland grass pastures. Appl. Envir.Microbiol. 65:1721-1730.
McCoy T.J. 1997. A lively debate. Nature 386:557-558.
McDonald I.R., E.M. Kenna and J.C. Murrell. 1995.Detection of methanotrophic Bacteria in environmentalsamples with the PCR. Appl. Envir. Microbiol. 61: 116-121.
McInerney J.O., M. Mullarhey, M.E. Wernecke and R.Powell. 1997. Phylogenetic analysis of group I marinearchaeal rRNA sequences emphasizes the hidden diversitywithin the primary group Archaea. Proc. R. Soc. Lond. B264:1663-1669.
McInerney J.O., M. Wilkinson, J.W. Patching, T.M.Embley and R. Powell. 1995. Recovery and phylogeneticanalysis of novel archaeal rRNA sequences from a deep-seadeposit feeder. Appl. Environ. Microbiol. 61:1646-1648.
McKay C.P. 1997. The search for life on Mars. Origins of lifeand evolution of the biosphere 27:263-289.
McKay D.S., E.K. Gibson Jr., K.L. Thomas-Keprta, H. Vali,C.S. Romanek, S.J. Clemett, X.D.F. Chillier, C.R.Maechling and R.N. Zare. 1996. Search for past life on mars:possible relic biogenic activity in martian meteoriteALH84001. Science 273:924-930.
McNaughton S.J., J.R. Stephen, A.D. Venosa, G.A. Davis,Y.J. Chang and D.C. White. 1999. Microbial populationchanges during bioremediation of an experimental oil spill.Appl. Envir. Microbiol. 65:3566-3574.
Mead D.A., N.K. Pey, C. Herrnstadt, R.A. Marcil and L.M.Smith. 1991. A universal method for the direct cloning of PCRamplified nucleic acid. Bio/Technology 9:657-663.
Mevel G., C. Fady and D. Prieur. 1996. Distribution, activity,and diversity of heterotrophic nitrifiers originating from EstPacific deep-sea hydrothermal vents. Can. J. Microbiol.42:162-171.
Michaelides K., R. Schwaab, M.R.A. Ialloz, W. Schmidtand E.G.D. Tuddenham. 1995. Mutational analysis: newmutations. In: McPherson MJ, Hames BD, Taylor, GR,editors. PCR 2 A practical approach. Oxford: IRL Press atOxford University Press. p. 255-287.
Miguez C.B., C.F. Shen, D. Bourque, S.R. Guiot and D.Groleau. 1999. Monitoring methanotrophic bacteria in hybridanaerobic-aerobic reactors with PCR and a catabolic geneprobe. Appl. Envir. Microbiol. 65:381-388.
Millikan D.S., H. Felbeck and J.L. Stein. 1999. Identificationand characterization of a flagellin gene from the endosymbiontof the hydrothermal vent tubeworm Riftia pachyptila. Appl.Environ. Microbiol. 65:3129-3133.
Miroshnichenko M.L., G.M. Gongadze, F.A. Rainey, A.S.Kostyukova, A.M. Lysenko, N.A. Chernyh and E.A.Bonch-Osmolovskaya. 1998. Thermococcus gorgonariussp. nov. and Thermococcus pacificus sp. nov.: heterotrophicextremely thermophilic archaea from New Zealandsubmarine hot vents. Int. J. Syst. Bacteriol. 48:23-29.
Miskin I., G. Rhodes, K. Lawlor, J.R. Saunders and R.W.Pickup. 1998. Bacteria in post-glacial freshwater sediments.Microbiology 144:2427-2439.
Mobarry B.K., M. Wagner, V. Urbain, B.E. Rittmann andD.A. Stahl. 1996. Phylogenetic probes for analyzingabundance and spatial organization of nitrifying bacteria. Appl.Envir. Microbiol. 62:2156-2162.
Mohagheghi A., K. Grohmann, M. Himmel, L. Leightonand D.M. Updegraff. 1986. Isolation and characterization ofAcidothermus cellulyticus gen. nov., sp. nov., a new genusofthermophilic, acidophilic, cellulolytic bacteria. Int. J. Syst.Bacteriol. 36:435-443.
Moncla B.J., S.T. Motley, P. Braham, L. Ewing, T.H.Adams and N.M. Vermeulen. 1991. Use of syntheticoligonucleotide DNA probes for identification and direct
detection of Bacteroides forsythus in plaque samples. J. Clin.Microbiol. 29:2158-2162.
Monfort P., M.H. Ratinaud, P.Got and B. Baleux. 1995.Apports de la cytométrie en flux et en image en écologiebactérienne des milieux aquatiques. Océanis 21:97-111.
Monheiner R.H. 1972. Heterotrophy by plankton in threelakes of different productivity. Nature 236:463-464.
Moore P.B. 1998. The three-dimensional structure of theribosome and its components. Annu. Rev. Biophys. Biomol.Struct. 27:35-58.
Moran M.A., V.L. Torsvik, T. Torsvik and R.E. Hodson.1993. Direct extraction and purification of rRNA for ecologicalstudies. Appl. Envir. Microbiol. 59:915-918.
Moriarty D. 1977. Improved method using muramic acid toestimate biomass of bacteria in sediments. Oecologia 26:317-323.
Moriya S., M. Ohkuma and T. Kudo. 1998. Phylogeneticposition of symbiotic protist Dinenympha [correction ofDinemympha] exilis in the hindgut of the termiteReticulitermes speratus inferred from the protein phylogeny ofelongation factor 1 alpha. Gene 210:221-227.
Morvan B., F. Bonnemoy, G. Fonty and P. Gouet. 1996.Quantitative determination of H2-utilizing acetogenic andsulfate-reducing bacteria and methanogenic archaea fromdigestive tract of different mammals. Curr. Microbiol. 32:129-133.
Moss C.W., M.A. Lambert and W.H. Merwin. 1974.Comparison of rapid methods for analysis of bacterial fattyacids. Appl. Microbiol. 28:80-55.
Motamedi M. and K. Pedersen. 1998. Desulfovibrioaespoeensis sp. nov., a mesophilic sulfate-reducing bacteriumfrom deep groundwater at Aspo hard rock laboratory, Sweden.Int. J. Syst. Bacteriol. 48:311-315.
Moyer C.L., F.C. Dobbs and D.M. Karl. 1994. Estimation ofdiversity and community structure through restrictionfragment length polymorphism distribution analysis ofbacterial 16S rRNA genes from a microbial mat at an active,hydrothermal vent system, Loihi Seamount, Hawaii. Appl.Environ. Microbiol. 60:871-879.
Moyer C.L., F.C. Dobbs and D.M. Karl. 1995. Phylogeneticdiversity of the bacterial community from a microbial mat atan active, hydrothermal vent system, Loihi Seamount, Hawaii.Appl. Environ. Microbiol. 61:1555-1562.
Moyer C.L., J.M. Tiedje, F.C. Dobbs and D.M. Karl. 1996.A computer-simulated restriction fragment lengthpolymorphism analysis of bacterial small-subunit rRNAgenes : efficacy of selected tetrameric restriction enzymes forstudies of microbial diversity in nature. Appl. Environ.Microbiol. 62:2501-2507.
Moyer C.L., J.M. Tiedje, F.C. Dobbs and D.M. Karl. 1998.Diversity of deep-sea hydrothermal vent Archaea from LoihiSeamount, Hawaii. Deep-Sea Res II 45:303-317.
Mullins T.D., T.B. Britschgi, R.L. Krest and S.L.Giovannonni. 1995. Genetic comparaisons reveal the sameunknown bacterial lineages in Atlantic and Pacificbacterioplankton communities. Limnol. Oceanogr. 40:148-158.
Munson M.A., D.B. Nedwell and T.M. Embley. 1997.Phylogenetic diversity of Archaea in sediment samples from acoastal salt marsh. Appl. Environ. Microbiol. 63:4729-4733
Murray A.E., J.T. Hollibaugh and C. Orrego. 1996.Phylogenetic compositions of bacterioplankton from twoCalifornia estuaries compared by denaturing gradient gelelectrophoresis of 16S rDNA fragments. Appl. Envir.Microbiol. 62:2676-2680.
Muyzer G. 1999. DGGE/TGGE a method for identifying genesfrom natural ecosystems. Curr. Opin. Microbiol. 2:317-322.
Muyzer G., A. Teske, C.O. Wirsen and H.W. Jannasch.1995. Phylogenetic relationships of Thiomicrospira speciesand their indentification in deep-sea hydrothermal ventsamples by denaturing gradient gel electrophoresis of 16SrRNA fragments. Arch. Microbiol. 164:165-172.
Muyzer G., E.C. de Waal, A.G. Uitterrlinden. 1993. Profilingof complex microbial populations by denaturing gradient gelelectrophoresis analysis of polymerase chain reaction-

Références bibliographiques
228
amplified genes coding for 16S rRNA. Appl. Environ.Microbiol. 59:695-700.
N
Naganuma T., C. Kato, H. Hirayama, N. Moriyama, J.Hashimoto and K. Horikoshi. 1997. Intracellular ocurrenceof ε-proteobacterial 16S rRNA sequences in theVestimentiferan Trophosome. J. Oceano. 53:193-197.
Nazaret S., W.H. Jeffrey, E. Saouter, R. Von Haven and T.Barkay. 1994. merA gene expression in aquatic environmentsmeasured by mRNA production and Hg(II) volatilization. Appl.Envir. Microbiol. 60: 4059-4065.
Nealson D.C. 1999. Recent progress in the microbiology ofdeep-sea hydrothermal vent and seeps. Cah. Biol. Mar.39:373-378.
Nealson D.C. and C.R. Fisher. 1995. Chemoautrophic andmethanotrophic endosymbiotic bacteria at deep-sea vents andseeps. In: Karl DM (Ed) The Microbiology of Deep-seaHydrothermal Vents (pp 125-167). CRC Press Inc, BocaRaton.
Neef A., R. Amann, H. schlesner and K.H. Schleifer. 1998.Monitoring a widespread bacterial group: in situ detection ofplanctomycetes with 16S rRNA-targeted probes. Microbiology144:3257-3266.
Neefs J.M., Y. Van de Peer, P. De Rijk, S. Chapelle and R.De Wachter. 1993. Compilation of small ribosomal subunitRNA structures. Nucleic Acids Res. 21:3025-3049.
Neimark H., D. Mitchelmore and R.H. Leach. 1998. Anapproach to characterizing uncultivated prokaryotes: the GreyLung agent and proposal of a Candidatus taxon for theorganism, 'Candidatus Mycoplasma ravipulmonis'. Int. J.Syst. Bacteriol. 48:389-394.
Nercessian D., M. Upton, D. Lloyd and C. Edwards. 1999.Phylogenetic analysis of peat bog methanogen populations.FEMS Microbiol. Lett. 173:425-429.
Nguyen A.M., L. Engstrand, R.M. Genta, D.Y. Grahamand F.A. el-Zaatari. 1993. Detection of Helicobacter pylori indental plaque by reverse transcription-polymerase chainreaction. J. Clin. Microbiol. 31:783-787.
Nielsen A.T., W.T. Liu, C. Filipe, L. Grady Jr, S. Molin andD.A. Stahl. 1999. Identification of a novel group of bacteria insludge from a deteriorated biological phosphorus removalreactor. Appl. Environ. Microbiol. 65:1251-1258.
Nierlich D.P. 1996. Futur directions for biomoleculardatabases. ASM News 62:251-254.
Ninet B., M. Monod, S. Emle, J. Pawlowski, C. Metral, P.Rohner, R. Auckenthaler and B. Hirschel. 1996. Twodifferent 16S rRNA genes in a mycobacterial strain. J. Clin.Microbiol. 34:2531-2536.
Noble R.T. and J. A. Fuhrman. 1998. Use of SYBR Green Ifor rapid epifluorescence counts of marine viruses andbacteria. Aquat. Microb. Ecol. 14:113-118.
Noda S., M. Ohkuma, R. Usami, K. Horikoshi and T.Kudo. 1999. Culture-independent characterization of a generesponsible for nitrogen fixation in the symbiotic microbialcommunity in the gut of the termite Neotermes koshunensis.Appl. Environ. Microbiol. 65: 4935-4942.
Nogi Y and C Kato. 1999. Taxonomic studies of extremelybarophilic bacteria isolated from the Mariana Trench anddescription of Moritella yayanosii sp. nov., a new barophilicbacterial isolate. Extremophiles 3:71-77.
Nold S.C. and D.M. Ward. 1996. Photosynthate partitioningand fermentation in hot spring microbial mat communities.Appl. Environ. Microbiol. 62:4598-4607.
Nold S.C., E.D. Kopczynski and D.M. Ward. 1996.Cultivation of aerobic chemoorganotrophic proteobacteria andGram-positive bacteria from a hot spring microbial mat. Appl.Environ. Microbiol. 62:3917-3921.
Noller H.F. 1984. Structure of ribosomal RNA. Annu. Rev.Biochem. 53:119-162.
Noller H.F.and C.R. Woese. 1981. Secondary structure of16S ribosomal RNA. Science. 212:403-411.
Nübel U., F. Garcia-Pichel, M. Kühl and G. Muyzer. 1999.Quantifying microbial diversity: morphotypes, 16S rRNAgenes, and carotenoids of oxygenic phototrophs in microbialmats. Appl. Envir. Microbiol. 65:422-430.
Nüsslein K. and J.M. Tiedje. 1998. Characterization of thedominant and rare members of a young Hawaiian soilbacterial community with small-subunit ribosomal DNAamplified from DNA fractionated on the basis of its guanineand cytosine composition. Appl. Envir. Microbiol. 64:1283-1289.
O
O'Donnell A.G. and H.E. Görres. 1999. 16S rDNA methodsin soil microbiology. Curr. Op. in Biotec. 10:225-2229.
Ogram A., W. Sun, F.J. Brockman and J.K. Fredrickson.1995. Isolation and characterization of RNA from low-biomassdeep-subsurface sediments. Appl. Envir. Microbiol. 61:763-768.
Ogram A.V. and G.S. Sayler. 1988. The use of gene probes inthe rapid analysis of natural microbial communities. J. Ind.Microbiol. 3:281-292.
Ohkuma M. and T. Kudo. 1996. Phylogenetic diversity of theintestinal bacterial community in the termite Reticulitermessperatus. Appl. Environ. Microbiol. 62:461-468.
Ohkuma M. and T. Kudo. 1998. Phylogenetic analysis of thesymbiotic intestinal microflora of the termite Cryptotermesdomesticus. FEMS Microbiol. Lett. 164:389-395.
Ohkuma M., K. Ohtoko, C. Grunau, S. Moriya and T.Kudo. 1998. Phylogenetic identification of the symbiotichypermastigote Trichonympha agilis in the hindgut of thetermite Reticulitermes speratus based on small-subunit rRNAsequence. J. Eukaryot. Microbiol. 45:439-444.
Ohkuma M., S. Noda and T. Kudo. 1999. Phylogeneticrelationships of symbiotic methanogens in diverse termites.FEMS Microbiol. Lett. 171:147-153.
Ohkuma M., S. Noda, K. Horikoshi and T. Kudo. 1995.Phylogeny of symbiotic methanogens in the gut of the termiteReticulitermes speratus. FEMS Microbiol. Lett. 134:45-50.
Okabe S., H. Satoh and Y. Watanabe. 1999. In situ analysisof nitrifying biofilms as determined by in situ hybridization andthe use of microelectrodes. Appl. Environ. Microbiol.65:3182-3191.
Olsen G.J. 1994. Archaea, archaea, everywhere. Nature371:657-658.
Olsen G.J. and C. Woese. 1996. Lessons from an Archaealgenome: what are we learning from Methanococcusjannaschii? Trends Genet. 12:377-379.
Olsen G.J. and C.R. Woese. 1993. Ribosomal RNA: a key tophylogeny. FASEB J. 7:113-123.
Olsen G.J., C.R. Woese and R. Overbeek. 1994b. The windsof (evolutionary) change: breathing new life into microbiology.J. Bacteriol. 176:1-6.
Olsen G.J., D.J. Lane, S.J. Giovannoni, D.A. Stahl andN.R. Pace. 1986. Microbial ecology and evolution: a ribosomalRNA approach. Ann. Rev. Microbiol. 40:337-365.
Olsen G.J., H. Matsuda, R. Hagstrom and R. Overbeek.1994a. fastDNAmL: a tool for construction of phylogenetictrees of DNA sequences using maximum likelihood. Comput.Appl. Biosci. 10:41-48.
Olson B.H. 1991. Tracking and using genes in theenvironment. Environ. Sci. Technol. 25:604-611.
Olson B.H. and Y.L.Tsai. 1992. Molecular approaches toenvironmental management. Environmental. Microbiology,Wiley-Liss, Inc., p 239-263.
Otsuka J., G. Terai and T. Kakano. 1999. Phylogeny oforganisms investigated by base-pair changes in the stemregions of small and large ribosomal subunit RNAs. J. Mol.Evol. 48:218-235.
Ouverney C.C. and J.A. Fuhrman. 1999. Combinedmicroautoradiography-16S rRNA probe technique fordetermination of radioisotope uptake by specific microbial celltypes in situ. Appl. Envir. Microbiol. 65:1746-1752.

Références bibliographiques
229
Øvreås L., L. Forney, F.L. Daae and V. Torsvik. 1997.Distribution of bacterioplankton in meromictic LakeSaelenvannet, as determined by denaturing gradient gelelectrophoresis of PCR-amplified gene fragments coding for16S rRNA. Appl. Environ. Microbiol. 63:3367-3373.
Øvreås L., S. Jensen, F. L. Daae and V. Torsvik. 1998.Microbial community changes in a perturbed agricultural soilinvestigated by molecular and physiological approaches. Appl.Envir. Microbiol. 64:2739-2742.
P
Pääbo S., D.M. Irwin and A.C. Wilson. 1990. DNA damagepromotes jumping between template during enzymaticamplification. J. Biol. Chem. 265:4718-4721.
Pace N.R. 1991. Origin of life-facing up to the physical setting.Cell 65:531-533.
Pace N.R. 1997. A molecular view of microbial diversity andthe biosphere. Science 276:734-740.
Pace N.R., D.A. Stahl, D.J. Lane and G.J. Olsen. 1985. Theanalysis of natural microbial populations by ribosomal RNAsequences. Am. Soc. Microbiol. News 51:4-12.
Pace N.R., D.A. Stahl, D.J. Lane and G.J. Olsen. 1986a.The analysis of natural microbial populations by ribosomalRNA sequences. Adv. Microbial Ecol. 9:1-55.
Pace N.R., D.J. Lane, G.J. Olsen and D.A. Stahl. 1986b.Phylogenetic analysis of organisms and populations usingribosomal RNA sequences. Proc. Fourth Internatl. Symp. onMicrobial Ecology, pp. 117-122.
Pahl A., U. Kühlbrandt, K. Brune, M. Röllinghoff and A.Gessner. 1999. Quantitative Detection of Borrelia burgdorferiby Real-Time PCR. J. Clin. Microbiol. 37:1958-1963.
Palleroni N.J. 1997. Procaryotic diversity and the importanceof culturing. Antonie van Leeuwenhoeck 72:3-19.
Palmer C.J., G.F. Bonilla, B. Roll, C. Paszko-Kolva, L.R.Sangermano and R.S. Fujioka. 1995. Detection of Legionellaspecies in reclaimed water and air with the EnviroAmpLegionella PCR kit and direct fluorescent antibody staining.Appl. Envir. Microbiol. 61: 407-412.
Palys T., L.K. Nakamura and F.M. Cohan. 1997. Discoveryand classification of ecological diversity in the bacterial world:the role of DNA sequence data. Int. J. Syst. Bacteriol. 47:1145-1156.
Parkes R.J., B.A. Cragg, S.J. Bala, J.M. Getiff, K.Goodman, P.A. Rochelle, J.C. Fry, A.J. Weightman andS.M. Harvey. 1994. Deep bacterial biosphere in pacific oceansediments. Nature 371:410-413.
Parrish K.D. and E.P. Greenberg. 1995. A rapid method forextraction and purification of DNA from dental plaque. Appl.Envir. Microbiol. 61:4120-4123.
Partensky F., W.R. Hess and D. Vaulot. 1999.Prochlorococcus, a marine photosynthetic prokaryote of globalsignificance. Microbiol. Mol. Biol. Rev. 63:106-127.
Paster B.J., F.E. Dewhirst, S.M. Cooke, V. Fussing, L.K.Poulsen and J.A. Breznak. 1996. Phylogeny of not-yet-cultured Spirochetes from termite guts. Appl. Environ.Microbiol. 62:347-352.
Patel B.K.C., H.W., Morgan and R.M. Daniel. 1985.Fervidobacterium nodosum gen. nov. and spec. nov., a newchemoorganotrophic, caldoactive, anaerobic bacterium. Arch.Microbiol. 141:63-69.
Patel B.K.C., M. Spanevello and B. Olivier. 1998. Microbialdiversity of the thermal waters emmited from the deepsubterrestrial aquifer, The Great Artesian Bassin of Australia.Thermophiles'98, International Conference, 6-11 septembre,Brest France.
Patra G., J. Vaissaire, M. Weber-Levy, C. Le Doujet and M.Mock. 1998. Molecular characterization of Bacillus strainsinvolved in outbreaks of Anthrax in France in 1997. J. Clin.Microbiol. 36:3412-3414.
Paul J.H., J.B. Rose, S. Jiang, C. Kellogg and E.A. Shinn.1995. Occurrence of fecal indicator bacteria in surface waters
and the subsurface aquifer in Key Largo, Florida. Appl. Envir.Microbiol. 61: 2235-2241.
Pedersen K. 1996. Investigations of subterranean bacteria indeep crystalline bedrock and their importance for the disposalof nuclear waste. Can. J. Microbiol. 42:382-391.
Pedersen K., J. Arlinger, L. Hallbeck and C. Pettersson.1996a. Diversity and distribution of subterranean bacteria ingroundwater at Oklo in Gabon, Africa, as determined by 16SrRNA gene sequencing. Mol. Ecol. 3:427-436.
Pedersen K., J. Arlinger, S. Ekendahl and L. Hallbeck.1996b. 16S rRNA gene diversity of attached and unattachedbacteria in boreholes along the acces tunnel to the Äspö hardrock laboratory, Sweden. FEMS Microbiol. Ecol. 19:249-262.
Pedersen K.,L. Hallbeck, J. Arlinger, A.-C. Erlandson andN.Jahromi. 1997. Investigation of the potential for micobialcontamination of deep granitic aquifers during drilling using16S rRNA gene sequencing and culturing methods. J.Microbiol. Meth. 30:179-192.
Peek A.S., R.A. Feldman, R.A. Lutz and R.C. Vrijenhoek.1998. Cospeciation of chemoautotrophic bacteria and deep seaclams. Proc. Natl. Acad. Sci. USA 95:9962-9966.
Pennisi E. 1998. Genome data shake tree of life. Science280:672-674.
Pernthaler J., F.O. Glockner, S. Unterholzner, A.Alfreider, R. Psenner and R. Amann. 1998. Seasonalcommunity and population dynamics of pelagic bacteria andarchaea in a high mountain lake. Appl. Environ. Microbiol.64:4299-4306.
Philippe H. and J. Laurent. 1998. How good are deepphylogenetic trees? Curr. Opin. Genet. Dev. 8:616-623.
Philippi G.T. 1977. On the depth, time, and mecanism oforigin of the heavy to medium gravity naphtenic crude oil.Geochim. Cosmochim. Acta 41:33-52.
Picard C., C. Ponsonnet, E. Paget, X. Nesme and P.Simonet. 1992. Detection and enumeration of bacteria in soilby direct DNA extraction and polymerase chain reaction. Appl.Envir. Microbiol. 58: 2717-2722.
Pickup R.W. 1991. Development of molecular methods forthe detection of specific bacteria in the environment. J. Gen.Microbiol. 137:1009-1019.
Pierson B.K. and R.W. Castenholz. 1974. A phototrophicgliding filamentous bacterium of hot springs, Chloroflexusaurantiacus, gen. and sp. nov. Arch. Mikrobiol. 100:5-24.
Pillai S.D., K.L. Josephson, R.L. Bailey. C.P. Gerba andI.L. Pepper. 1991. Rapid method for processing soil samplesfor polymerase reaction amplification of specific genesequences. Appl. Envir. Microbiol. 57: 2283-2286.
Pillai S.D., K.W. Widmer, S.E. Dowd and S.C. Ricke. 1996.Occurrence of airborne bacteria and pathogen indicatorsduring land application of sewage sludge. Appl. Envir.Microbiol. 62:296-299.
Pitcher D.G., N.A. Saunders and R.J. Owen. 1989. Rapidextraction of bacterial genomic DNA with guanidiumthiocyanate. Lett.Appl.Micro. 8:151-156
Pitulle C. and N.R. Pace. 1999. T-cloning vector for plasmid-based 16S rDNA analysis. Biotechniques 26:222-224.
Pizarro J., E. Jedlicki, O. Orellana, J. Romero and R.T.Espejo. 1996. Bacterial populations in samples of bioleachedcopper ore as revealed by analysis of DNA obtained beforeand after cultivation. Appl. Envir. Microbiol. 62:1323-1328.
Pley U., J. Schipka, A. Gambacorta, H.W. Jannasch, H.Fricke, R. Rachel and K.O. Stetter. 1991. Pyrodictiumabyssi sp. nov., represents a novel heterotrophic marinearchaeal hyperthermophile growing at 110°C. System. Appl.Microbiol. 14:245-253.
Pochart P., J. Dore, F. Lemann, I. Goderel and J.C.Rambaud. 1992. Interrelations between populations ofmethanogenic archaea and sulfate-reducing bacteria in thehuman colon. FEMS Microbiol. Lett. 77:225-228.
Pollard P.C. 1998. Estimating the growth rate of a bacterialspecies in a complex mixture by hybridization of genomicDNA. Microb. Ecol. 36:111-120.

Références bibliographiques
230
Poltz M.F. and C.M. Cavanaugh. 1995. Dominance of onebacterial phylotype at a Mid-Atlantic Ridge hydrothermal ventsite. Proc. Natl. Acad. Sci. USA 92:7232-7236.
Poltz M.F. and C.M. Cavanaugh. 1997. A simple method forquantification of uncultured microorganisms in theenvironment based on in vitro transcription of 16S rRNA. Appl.Envir. Microbiol. 63:1028-1033.
Polz M.F., J.J. Robinson, C.M. Cavanaugh and C.L. VanDover. 1998. Trophic ecology of massive shrimpaggregations at a Mid-Atlantic Ridge hydrothermal vent site.Limnol. Oceanogr. 43:1631-1638.
Porteous L.A., R.J. Seidler and L.S. Watrud. 1997. Animproved method for purifying DNA from soil for polymerasechain reaction amplification and molecular ecologyapplications. Mol. Ecol. 6:787-791.
Poulicek M. and V. Danckers. 1995. L'ATP et la chargeénergétique adénylique (ECA) en écologie microbienne desmilieux aquatiques. Océanis 21:191-223.
Preston C.M., K.Y. Wu, T.F. Molinski and E.F. DeLong.1996. A psychrophilic crenarchaeon inhabits a marine sponge:Crenarchaeum symbiosum gen. nov., sp. nov. Proc. Natl.Acad. Sci. USA 93:6241-6246.
Prieur D. 1992. Physiology and biotechnological potential ofdeep-sea bacteria, in: Molecular biology and Biotechnology ofextremophiles. R.A. Herbert and R.J. Sharp, Eds. Blackie,Glasgow & London. p 163-197.
Prieur D. 1997. Deep-sea hydrothermal vents. TIBTech.15:242-244.
Prieur D., C. Jeanthon and E. Jacq. 1987. Lescommunautés bactériennes des sources hydrothermalesprofondes du Pacifique Oriental. Vie Milieu 28:419-424.
Prieur D., C. Jeanthon and G. Erauso. 1995.Hyperthermophilic life at deep-sea hydrothermal vents.Planet. Space Sci. 43:115-122.
Priscu J.C., C.H. Fritsen, E.E. Adams, S.J. Giovannoni,H.W. Paerl, C.P. McKay, P.T. Doran, D.A. Gordon, B.D.Lanoil and J.L. Pinckney. 1998. Perennial Antarctic lake ice:an oasis for life in a polar desert. Science 280:2095-2098.
Prüß B.M., K.P. Francis, F. von Stetten and S. Scherer.1999. Correlation of 16S ribosomal DNA signature sequenceswith temperature-dependent growth rates of mesophilic andpsychrotolerant strains of the Bacillus cereus group. J.Bacteriol. 181:2624-2630.
Psenner R. and B. Sattler. 1998. Life at the freezing point.Science 280:2073-2074.
Pukall R., E. Brambilla and E. Stackebrandt. 1998.Automated fragment lenght analysis of fluorescently-labeled16S rDNA after digestion with 4-base cutting restrictionenzymes. J. Microbiol. Methods 32:55-63.
Pukall R., O. Päuker, D. Buntelfuß, G. Ulrichs, P. Lebaron,L. Bernard, T. Guindulain, J. Vives-Rego, E.Stackebrandt. 1999. High sequence versity of Alteromonasmacleodii-related cloned and cellular 16S rDNAs fromMediterranean seawater mesocosm experiment. FEMSMicrobiol. Ecol. 28:335-344.
Pulliam Holoman T.R., M.A. Elberson, L.A. Cutter, H.D.May and K.R. Sowers. 1998. Characterization of a defined2,3,5,6-tetrachlorobiphenyl-ortho-dechlorinating microbialcommunity by comparative sequence analysis of genescoding for 16s rRNA. Appl. Envir. Microbiol. 64:3359-3367.
Q
Quérellou J. 1999. La biologie moléculaire colonisée par uneenzyme. La Recherche 317:34-35.
Quérellou J., Y. Guéguen, G. Barbier and J. Dietrich. 1999.Des enzymes pour l'industrie. La Recherche 371:87-88.
R
Raguenes G.H., A. Peres, R. Ruimy, P. Pignet, R.Christen, M. Loaec, H. Rougeaux, G. Barbier and J.G.
Guezennec. 1997a. Alteromonas infernus sp. nov., a newpolysaccharide-producing bacterium isolated from a deep-seahydrothermal vent. J. Appl. Microbiol. 82:422-430.
Raguenes G.H., R. Christen, J. Guezennec, P. Pignet andG. Barbier. 1997b. Vibrio diabolicus sp. nov., a newpolysaccharide-secreting organism isolated from a deep-seahydrothermal vent polychaete annelid, Alvinella pompejana.Int. J. Syst. Bacteriol. 47:989-995.
Rainey F.A., D. Fritze and E. Stackebrandt. 1994a. Thephylogenetic diversity of thermophilic members of the genusBacillus as revealed by 16S rDNA analysis. FEMS Microbiol.Lett. 115:205-211.
Rainey F.A., N. Ward, L.I. Sly and E. Stackebrandt. 1994b.Dependence on the taxon composition of clone libraries forPCR amplified, naturally occuring 16S rDNA, on the primerpair and the cloning system used. Experientia 50:796-797.
Ramsing N.B., H. Fossing, T.G. Ferdelman, F. Andersenand B. Thamdrup. 1996. Distribution of bacterial populationsin a stratified fjord (Mariager Fjord, Denmark) quantified by insitu hybridization and related to chemical gradients in thewater column. Appl. Environ. Microbiol. 62:1391-1404.
Rappé M.S. P.F. Kemp and S.J. Giovannoni. 1997.Phylogenetic diversity of marine coastal picoplankton 16SrRNA genes cloned from the continental shelf off CapeHatteras, North Carolina. 1997. Limnol. Oceanogr. 42:811-826.
Rappé M.S., D.A. Gordon, K.L. Vergin and S.J.Giovannoni. 1999. Phylogeny of Actinobacteria small subunit(SSU) rRNA gene clones recovered from marinebacterioplankton. System. Appl. Microbiol. 22:106-112.
Rappé M.S., M.T. Suzuki, K.L. Vergin and S.J.Giovannoni. 1998. Phylogenetic diversity of ultraplanktonplastid small-subunit rRNA genes recovered in environmentalnucleic acid samples from the Pacific and Atlantic coasts ofthe United States. Appl. Environ. Microbiol. 64:294-303.
Raskin L., J.M. Stromley, B.E. Rittmann and D.A. Stahl.1994a. Group-specific 16S rRNA hybridization probes todescribe natural communities of methanogens. Appl. Envir.Microbiol. 60:1232-1240.
Raskin L., L.K. Poulsen, D.R. Noguera, B.E. Rittmannand D.A. Stahl. 1994b. Quantification of methanogenic groupsin anaerobic biological reactors by oligonucleotide probehybridization. Appl. Envir. Microbiol. 60:1241-1248.
Rastan S. and L.J. Beeley. 1997. Functional genomics: goingforwards from the database. Curr. Opin. Genet. Dev. 7:777-783.
Rath J., K.Y. Wu, G.J. Herndl and E.F. DeLong. 1998. Highphylogenetic diversity in a marine-snow-associated bacterialassemblage. Aquat. Microb. Ecol. 14:261-269.
Ravot G. 1996. Nouvelles approches microbiologiques de lathiosulfato-reduction en milieu pétrolier. Thèse de doctorat de3eme cycle Université Aix - Marseille I : 6-17.
Ravot G., B. Ollivier, M. Magot, B.K.C. Patel, J.L. Crolet,M.L. Fardeau and J.L. Garcia. 1995a. Thiosulfate reduction,an important physiological feature shared by members oforder Thermotogales. Appl. Envir. Microbiol. 61:2053-2055.
Ravot G., M. Magot, M.L. Fardeau, B.K.C. Patel, G.Prensier, A. Egan, J.L. Garcia and B. Ollivier. 1995b.Thermotoga elfii sp. nov., a novel thermophilic bacterium froman african oil-producing well. Int. J. Syst. Bacteriol. 45:308-314.
Rees G.N., B.K. Patel, G.S. Grassia and A.J. Sheehy. 1997.Anaerobaculum thermoterrenum gen. nov., sp. nov., a novel,thermophilic bacterium which ferments citrate. Int. J. Syst.Bacteriol. 47:150-154.
Reeves R.H., J.Y. Reeves and D.L. Balkwill. 1996.Strategies for phylogenetic characterization of subsurfacebacteria. J. Microbiol. Methods 21:235-251.
Reid A.H., T.G. Fanning, J.V. Hultin and J.K.Taubenberger. 1999. Origin and evolution of the 1918"Spanish" influenza virus hemagglutinin gene. Proc. Natl.Acad. Sci. USA 96:1651-1656.
Reischl U., K. Feldmann, L. Naumann, B. J. M. Gaugler,B. Ninet, B. Hirschel and S. Emler. 1998. 16S rRNAsequence diversity in Mycobacterium celatum strains caused

Références bibliographiques
231
by presence of two different copies of 16S rRNA gene. J. Clin.Microbiol. 36:1761-1764.
Reponen T.A., S.V. Gazenko, S.A. Grinshpun, K. Willekeand E.C. Cole. 1998. Characteristics of airborneActinomycete spores. Appl. Envir. Microbiol. 64:3807-3812.
Reysenbach A.L., G.S. Wickham and N.R. Pace. 1994.Phylogenetic analysis of the hyperthermophilic pink filamentcommunity in Octopus Spring, Yellowstone National Park.Appl. Environ. Microbiol. 60:2113-2119.
Reysenbach A.L., J.D. Kirshtein, D. Lane, K. Longneckerand N.R. Pace. 1999. Microbial diversity associated with insitu growth chambers deployed on a deep sea hydrothermalvent. (en préparation).
Reysenbach A.L., L.J. Giver, G.S. Wickham and N.R.Pace. 1992. Differential amplification of rRNA genes bypolymerase chain reaction. Appl. Environ. Microbiol. 58:3417-3418.
Reysenbach A.L., N.G. Holm, K. Hershberger, D. Prieurand C. Jeanthon. 1998. In search of a subsurface biosphereat a slow-spreading ridge. In: Herzig PM, Humphris SE,Miller DJ & Zierenberg RA (Eds) Proc ODP Sci Results Vol158 (pp 355-360), College Station, TX.
Rhodes A.N., J.W. Urbance, H. Youga, H. Corlew-Newman, C.A. Reddy, M.J. Klug, J.M. Tiedje and D.C.Fisher. 1998. Identification of bacterial isolates obtained fromintestinal contents associated with 12,000-year-old mastodonremains. Appl. Envir. Microbiol. 64:651-658.
Riley P. 1999. Detection of methanogen communities. Report.Amores: Azores Mid-Oceanic Ridge Ecosystem Studies.Final Detailed scientific report. Theme N°3 (1/2)
Riley P.W. 1997. The development of an alternative to 16SrDNA, the mcrA gene, as a phylogenetic tool in the descriptionof methanogen communities in the marine environment. FirstInternational Symposium on Deep-sea Hydrothermal VentBiology. p. 109.
Risatti J.B., W.C. Capman and D.A. Stahl. 1994.Community structure of a microbial mat: the phylogeneticdimension. Proc. Natl. Acad .Sci. USA 91:10173-10177.
Robison-Cox J.F., M.M. Bateson and D.M. Ward. 1995.Evaluation of nearest-neighbor methods for detection ofchimeric small-subunit rRNA sequences. Appl. Environ.Microbiol. 61:1240-1245.
Rocheleau S., C.W. Greer, J.R. Lawrence, C. Cantin, L.Laramee and S.R. Guiot. 1999. Differentiation ofMethanosaeta concilii and Methanosarcina barkeri inanaerobic mesophilic granular sludge by fluorescent in situhybridization and confocal scanning laser microscopy. Appl.Environ. Microbiol. 65:2222-2229.
Rochelle P.A., B.A. Cragg, J.C. Fry, R. J. Parkes and A.J.Weightman. 1994. Effect of sample handling on estimation ofbacterial diversity in marine sediments by 16S rRNA genesequence analysis. FEMS Microbiol. Ecol. 15:215-226.
Rochelle P.A., J.C. Fry, R.J. Parkes and A.J. Weightman.1992. DNA extraction for 16S rRNA gene analysis todetermine genetic diversity in deep sediment communities.FEMS Microbiol. Lett. 179:59-65.
Rodriguez G.G., D. Phipps, K. Ishiguro and H.F.Ridgway. 1992. Use of a fluorescent redox probe for directvisualization of actively respiring bacteria. Appl. Envir.Microbiol. 58:1801-1808.
Rogers J. and G.F. Joyce. 1999. A ribozyme that lackscytidine. Nature. 408:323-325.
Ronimus R.S., A. Reysenbach, D.R. Musgrave and H.W.Morgan. 1997. The phylogenetic position of theThermococcus isolate AN1 based on 16S rRNA genesequence analysis: a proposal that AN1 represents a newspecies, Thermococcus zilligii sp. nov. Arch. Microbiol.168:245-248.
Rooney-Varga J.N., R.T. Anderson, J.L. Fraga, D.Ringelberg and D.R. Lovley. 1999. Microbial communitiesassociated with anaerobic benzene degradation in apetroleum-contaminated aquifer. Appl. Environ. Microbiol.65:3056-3063.
Rotthauwe J.H., K.P. Witzel and W. Liesack. 1997. Theammonia monooxygenase structural gene amoA as afunctional marker: molecular fine-scale analysis of natural
ammonia-oxidizing populations. Appl. Envir. Microbiol. 63:4704-4712.
Ruano G., M.E. Lewis and R.E. Kouri. 1993. Cycled primerextension: a method for DNA amplification and labelling fromtemplates of unknown sequence. Annal. Biochem. 212:1-6.
Ruby E.G., C.O. Wirsen and H.W. Jannasch. 1981.Chemolithotrophic sulfur-oxidizing bacteria from theGalapagos Rift hydrothermal vents. Appl. Environ. Microbiol.42:317-324.
Rueter P., R. Rabus, H. Wilkes, F. Aeckersberg, F.A.Rainey, H.W. Jannasch and F. Widdel. 1994. Anaerobicoxidation of hydrocarbons in crude oil by new types ofsulphate-reducing bacteria. Nature 372:455-458.
Ruff-Roberts A.L., J.G. Kuenen and D.M. Ward. 1994.Distribution of cultivated and uncultivated cyanobacteria andChloroflexus-like bacteria in hot spring microbial mats. Appl.Environ. Microbiol.60:697-704.
Rupf S., K. Merte and K. Eschrich. 1999. Quantification ofbacteria in oral samples by competitive polymerase chainreaction. J. Dent. Res. 78:850-856.
S
Saiki R.K., D.H. Gelfand, S. Stoffel, S.J. Scharf, R.Higuchi, G.T. Horn, K.B. Mullis and H.A. Erlich. 1988.Primer-directed enzymatic amplification of DNA with athermostable DNA polymerase. Science 239:487-491.
Saiki R.K., S. Scharf, F. Faloona, K.B. Mullis, G.T. Horn,H.A. Erlich and N. Arnheim. 1985. Enzymatic amplificationof β-globin genomic sequences and restriction site analysis fordiagnostic of sickle cell anemia. Science 230:1350-1354.
Saitou N. and M. Nei. 1987. The neighbor-joining method: anew method for reconstructing phylogenetic trees. Mol. Biol.Evol. 4:406-425.
Sakano Y. and L. Kerkhof. 1998. Assessment of changes inmicrobial community structure during operation of anammonia biofilter with molecular tools. Appl. Envir. Microbiol.64:4877-4882.
Sako Y., N. Nomura, A. Uchida, Y. Ishida, H. Morii, Y.Koga, T. Hoaki and T. Maruyama. 1996. Aeropyrum pernixgen. nov., sp. nov., a novel aerobic hyperthermophilicarchaeon growing at temperatures up to 100 degrees C. Int. J.Syst. Bacteriol. 46:1070-1077.
Sambrook J., E.F. Fritsch and T. Maniatis. 1989. Molecularclonning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
San Martin R., D. Bushell, D.J. Leak and B.S. Hartley.1992. Development of a synthetic medium for continuousanaerobic growth and ethanol production with a lactatedehydrogenase mutant of Bacillus stearothermophilus. J. Gen.Microbiol.138:987-996.
Santegoeds C.M., A. Schramm and D. de Beer. 1998a.Microsensors as a tool to determine chemical microgradientsand bacterial activity in wastewater biofilms and flocs.Biodegradation 9:159-167.
Santegoeds C.M., S.C. Nold and D.M. Ward. 1996.Denaturing gradient gel electrophoresis used to monitor theenrichment culture of aerobic chemoorganotrophic bacteriafrom a hot spring cyanobacterial mat. Appl. Environ.Microbiol. 62:3922-3928.
Santegoeds C.M., T.G. Ferdelman, G. Muyzer and D. deBeer. 1998b. Structural and functional dynamics of sulfate-reducing populations in bacterial biofilms. Appl. Environ.Microbiol. 64:3731-3739.
Santo Domingo J.W., C.J. Berry, M. Summer and C.B.Fliermans. 1998a. Microbiology of spent nuclear fuel storagebassins. Curr. Microbiol. 37:387-394.
Santo Domingo J.W., M.G. Kaufman, M.J. Klug and J.M.Tiedje. 1998b. Characterization of the Cricket hindgutmicrobiota with fluorescently labeled rRNA-targetedoligonucleotide probes. Appl. Envir. Microbiol. 64:752-755.
Sass H., E. Wieringa, H. Cypionka, H.D. Babenzien and J.Overmann. 1998. High genetic and physiological diversity of

Références bibliographiques
232
sulfate-reducing bacteria isolated from an oligotrophic lakesediment. Arch. Microbiol. 170:243-251.
Schink B. and J.G. Zeikus. 1983. Clostridiumthermosulfurogenes sp. nov., a new thermophile that produceselemental sulfur from thiosulfate. J. Gen. Microbiol. 129:1149-1158 .
Schleper C. 1999. Les archaebactéries sont parmi nous. LaRecherche 317:30-33.
Schleper C., E.F. DeLong, C.M. Preston, R.A. Feldman,K.Y. Wu and R.V. Swanson. 1998. Genomic analysisreveals chromosomal variation in natural populations of theuncultured psychrophilic archaeon Cenarchaeum symbiosum.J. Bacteriol. 180:5003-5009.
Schleper C., G. Puehler, I. Holz, A. Gambacorta, D.Janekovic, U. Santarius, H.P. Klenk and W. Zillig. 1995.Picrophilus gen. nov., fam. nov.: a novel aerobic,heterotrophic, thermoacidophilic genus and family comprisingarchaea capable of growth around pH 0. J.Bacteriol. 177:7050-7059.
Schleper C., R.V. Swanson, E.J. Mathur and E.F. DeLong.1997b. Characterization of a DNA polymerase from theuncultivated psychrophilic archaeon Cenarchaeumsymbiosum. J. Bacteriol. 179:7803-7811.
Schleper C., W. Holben and H.P. Klenk. 1997a. Recoveryof crenarchaeotal ribosomal DNA sequences fromfreshwater-lake sediments. Appl. Envir. Microbiol. 63:321-323.
Schmidt T.M. and D.A. Relman. 1994. Phylogeneticidentification of uncultured pathogens using ribosomal RNAsequences. Methods Enzymol. 235:205-222.
Schmidt T.M., E.F. DeLong and N.R. Pace. 1991 Analysisof a marine picoplankton community by 16S rRNA genecloning and sequencing. J. Bacteriol. 173:4371-4378.
Schofield P., A.N. Pell and D.O. Krause. 1997. Molecularbeacons: trial of a fluorescence-based solution hybridizationtechnique for ecological studies with ruminal bacteria. Appl.Environ. Microbiol. 63:1143-1147.
Schönhuber W., B. Fuchs, S. Juretschko and R. Amann.1997. Improved sensitivity of whole-cell hybridization by thecombination of horseradish peroxidase-labeledoligonucleotides and tyramide signal amplification. Appl.Envir. Microbiol. 63:3268-3273.
Schönhuber W., B. Zarda, S. Eix, R. Rippka, M. Herdman,W. Ludwig and R. Amann. 1999. In situ identification ofcyanobacteria with horseradish peroxidase-labeled, rRNA-targeted oligonucleotide probes. Appl. Envir. Microbiol. 65:1259-1267.
Schramm A., L.H. Larsen, N.P. Revsbech, N.B. Ramsing,R. Amann and K.H. Schleifer. 1996. Structure and functionof a nitrifying biofilm as determined by in situ hybridization andthe use of microelectrodes. Appl. Environ. Microbiol. 62:4641-4647.
Schulz H.N., T. Brinkhoff, T.G. Ferdelman, M. HernandezMarine, A. Teske and B.B. Jorgensen. 1999. Densepopulations of a giant sulfur bacterium in Namibian shelfsediments. Science 284:493-495.
Schuppler M., F. Mertens, G. Schon and U.B. Gobel.1995. Molecular characterization of nocardioformactinomycetes in activated sludge by 16S rRNA analysis.Microbiology 141:513-521.
Schuppler M., M. Wagner, G. Schon and U.B. Gobel.1998. In situ identification of nocardioform actinomycetes inactivated sludge using fluorescent rRNA-targetedoligonucleotide probes. Microbiology 144:249-259.
Schwieger F. and C.C. Tebbe. 1998. A new approach toutilize PCR-single-strand-conformation polymorphism for16S rRNA gene-based microbial community analysis. Appl.Envir. Microbiol. 64:4870-4876.
Segerer A., K.O. Stetter and F. Klink. 1985. Two contrarymodes of lithotrophy in the same archaebacterium. Nature313:787-789.
Segerer A.H., A. Trincone, M. Gahrtz and K.O. Stetter.1991. Stygiolobus azoricus gen. nov., sp. nov. represents anovel genus of anaerobic, extremely thermoacidophilicarchaebacteria of the order Sulfolobales. Int. J. Syst. Bacteriol.41:495-501.
Segonzac M., M. de SaintLaurent and B. Casanova. 1993.L'énigme du comportement trophique des crevettesAlvinocarididae des sites hydrothermaux de la dorsale medio-atlantique. Cah. Biol. Mar. 34:535-571.
Sekiguchi Y., Y. Kamagata, K. Syutsubo, A. Ohashi, H.Harada and K. Nakamura. 1998. Phylogenetic diversity ofmesophilic and thermophilic granular sludges determined by16S rRNA gene analysis. Microbiology 144:2655-2665.
Sekiguchi Y., Y. Kamagata, K. Nakamura, A. Ohashi andH. Harada. 1999. Fluorescence in situ hybridization using 16SrRNA-targeted oligonucleotides reveals localization ofmethanogens and selected uncultured bacteria in mesophilicand thermophilic sludge granules. Appl. Envir. Microbiol.65:1280-1288.
Selvaratnam S., B.A. Schoedel, B.L. McFarland and C.F.Kulpa. 1995. Application of reverse transcriptase PCR formonitoring expression of the catabolic dmpN gene in a phenol-degrading sequencing batch reactor. Appl. Envir. Microbiol.61:3981-3985.
Servais P. 1995. Estimation de la biomasse et de l'activité desbactéries en milieux aquatique. Océanis 21:141-160.
Shaffer B.T. and B. Lighthart. 1997. Survey of culturableairborne bacteria at four diverse locations in oregon: urban,rural, forest and coastal. Microb. Ecol. 34:167-177.
Sharp R.J., K.J. Bown and A. Atkinson. 1980. Phenotypicand genotypic characterization of some thermophilic speciesof Bacillus. J. Gen. Microbiol. 117:201-210.
Sharp R.J., P.W. Riley and D. White. 1992. Heterotrophicthermophilic Bacilli. In: Thermophilic Bacteria (KritjanssonJ.K. Ed.) p 19-50. CRC Press, Boca Raton. Fl.
Shen Y., L.G. Stehmeier and G. Voordouw. 1998.Identification of hydrocarbon-degrading bacteria in soil byreverse sample genome probing. Appl. Envir. Microbiol.64:637-645.
Shena M., R.A. Heller, T.P. Theriault, K. Konrad, E.Lachenmeier and R.W. Davis. 1998. Microarrays:biotechnology's discovery platform for functional genomics.Trends Biotech. 16:301-306.
Shi T., R.H. Reeves, D.A. Gilichinsky and E.I. Friedmann.1997. Characterization of viable bacteria from Siberianpermafrost by 16S rDNA sequencing. Microb. Ecol. 33:169-179.
Shi Y., M.D. Zwolinski, M.E. Schreiber, J.M. Bahr, G.W.Sewell and W.J. Hickey. 1999. Molecular Analysis ofmicrobial community structures in pristine and contaminatedaquifers: field and laboratory microcosm experiments. Appl.Envir. Microbiol. 65:2143-2150.
Shinzato N., T. Matsumoto, I. Yamaoka, T. Oshima and A.Yamagishi. 1999. Phylogenetic diversity of symbioticmethanogens living in the hindgut of the lower termiteReticulitermes speratus analyzed by PCR and in situhybridization. Appl. Envir. Microbiol. 65:837-840.
Siering P.L. and W.C. Ghiorse. 1997. Development andapplication of 16S rRNA-targeted probes for detection of iron-and manganese-oxidizing sheathed bacteria in environmentalsamples. Appl. Envir. Microbiol. 63: 644-651.
Silva M.C. and C.A. Batt. 1995. Effect of cellular physiologyon PCR amplification efficiency. Mol. Ecol. 4:11-16.
Slobodkin A., A.L. Reysenbach, F. Mayer and J. Wiegel.1997a. Isolation and characterization of the homoacetogenicthermophilic bacterium Moorella glycerini sp. nov. Int. J. Syst.Bacteriol. 47:969-974.
Slobodkin A., A.L. Reysenbach, N. Strutz, M. Dreier andJ. Wiegel. 1997b. Thermoterrabacterium ferrireducens gen.nov., sp. nov., a thermophilic anaerobic dissimilatory Fe(III)-reducing bacterium from a continental hot spring. Int. J. Syst.Bacteriol. 47:541-547.
Slobodkin A., C. Jeanthon, S. L'Haridon, T. Nazina, M.Miroshnichenko and E. Bonch Osmolovskaya. 1999.Dissimilatory reduction of Fe(III) by thermophilic bacteria andarchaea in deep subsurface petroleum reservoirs of WesternSiberia. Curr. Microbiol.. 39:99-102
Slobodkin A.I. and J. Wiegel. 1997. Fe(III) as an electronacceptor for H2 oxidation in thermophilic anaerobicenrichment cultures from geothermal areas. Extremophiles1:106-109.

Références bibliographiques
233
Smalla K., U. Wachtendorf, H. Heuer, W.T. Liu and LForney. 1998 Analysis of BIOLOG GN substrate utilizationpatterns by microbial communities. Appl. Envir. Microbiol.64:1220-1225.
Smith G.L., C. Sansone and S.S. Socransky. 1989a.Comparison of two methods for the small-scale extraction ofDNA from subgingival microorganisms. Oral Microbiol.Immunol. 4:135-140.
Smith G.L., S.S. Socransky and C.M. Smith. 1989b. Rapidmethod for the purification of DNA from subgingivalmicroorganisms. Oral Microbiol. Immunol. 4:47-51.
Snaidr J., R. Amann, I. Huber, W. Ludwig and K.H.Schleifer. 1997. Phylogenetic analysis and in situ identificationof bacteria in activated sludge. Appl. Envir. Microbiol.63:2884-2896.
Sneath P.H. and R.R. Sokal. 1973. Numerical taxonomy.San Francisco: W.H. Freeman and Co.
Sokal R.R. and P.H. Sneath. 1963. Principles of numericaltaxonomy. Freeman, San Francisco.
Somerville C.C., I.T. Knight, W.L. Straube and R.R.Colwell. 1989. Simple, rapid method for direct isolation ofnucleic acids from aquatic environments. Appl. Envir.Microbiol. 55:548-554.
Speksnijder A.G., G.A. Kowalchuk, K. Roest H.J.Laanbroek. 1998. Recovery of a Nitrosomonas-like 16SrDNA sequence group from freshwater habitats. Syst. Appl.Microbiol. 21:321-330.
Spratt D.A., A.J. Weightman and W.G. Wade. 1999.Diversity of oral asaccharolytic Eubacterium species inperiodontitis--identification of novel phylotypes representinguncultivated taxa. Oral Microbiol. Immunol. 14:56-59.
Spring S., R. Amann, W. Ludwig, K.H. Schleifer and N.Petersen. 1992. Phylogenetic diversity and identification ofnonculturable magnetotactic bacteria. System. Appl.Microbiol. 15:116-122.
Spring S., U. Lins, R. Amann, K.H. Schleifer, L.C.Ferreira, D.M. Esquivel and M. Farina. 1998. Phylogeneticaffiliation and ultrastructure of uncultured magnetic bacteriawith unusually large magnetosomes. Arch. Microbiol.169:136-147.
Stackebrandt E. and B.M. Goebel. 1994. Taxonomic note: aplace for DNA-DNA reassociation and 16S rRNA sequenceanalysis in the present species definition in bacteriology. Int. J.Syst. Bacteriol. 44:846-849.
Stackebrandt E., W. Liesack and B.M. Goebel. 1993.Bacterial diversity in a soil sample from a subtropicalAustralian environment as determined by 16S rDNA analysis.FASEB. J. 7:232-236.
Stahl D.A. and R. Amman. 1991. Development andapplication of nucleic acid probes, p. 205-248. In E.Stackebrandt and E. Goodfellow (ed.), Nucleic acid techniquesin bacterial systematics, John Wiley and Sons, Chichester,England.
Stahl D.A., B. Flesher, H.R. Mansfield and L.Montgomery. 1988. Use of phylogenetically basedhybridization probes for studies of ruminal microbial ecology.Appl. Environ. Microbiol. 54:1079-1084.
Stahl D.A., D.J. Lane, G.J. Olsen and N.R. Pace. 1984.Analysis of hydrothermal vent-associated symbionts byribosomal RNA sequences. Science 224:409-411.
Stahl D.A., D.J. Lane, G.J. Olsen and N.R. Pace. 1985.Characterization of a Yellowstone hot spring microbialcommunity by 5S ribosomal RNA sequences. Appl. Environ.Microbiol. 49:1379-1384.
Staley J.T. and A. Konopka. 1985. Measurement of in situactivities of nonphotosynthetic microorganisms in aquatic andterrestrial habitats. Annu. Rev. Microbiol. 39:321-346.
Stärk K.D.C., J. Nicolet and J. Frey. 1998. Detection ofMycoplasma hyopneumoniae by air sampling with a nestedPCR assay. Appl. Envir. Microbiol. 64:543-548.
Steffan R.J. and R.M. Atlas. 1991. Polymerase chainreaction: applications in environmental microbiology. Annu.Rev. Microbiol. 45:137-161.
Steffan R.J., J. Goksøyr, A.K. Bej and R.M. Atlas. 1988.Recovery DNA from soils and sediments. Appl. Envir.Microbiol. 54: 2908-2915.
Stein J.L., T.L. Marsh, K.Y. Wu, H. Shizuya and E.F.DeLong. 1996. Characterization of uncultivated prokaryotes:isolation and analysis of a 40-kilobase-pair genome fragmentfrom a planktonic marine archaeon. J. Bacteriol. 178:591-599.
Stephen J.R., A.E. McCaig, Z. Smith, J.I. Prosser and T.M.Embley. 1996. Molecular diversity of soil and marine 16SrRNA gene sequences related to beta-subgroup ammonia-oxidizing bacteria. Appl. Envir. Microbiol. 62:4147-4154.
Stephen J.R., G.A. Kowalchuk, M.A.V. Bruns, A.E.McCaig, C.J. Phillips, T.M. Embley and J.I. Prosser. 1998.Analysis of beta-subgroup proteobacterial ammonia oxidizerpopulations in soil by denaturing gradient gel electrophoresisanalysis and hierarchical phylogenetic probing. Appl. Envir.Microbiol. 64:2958-2965.
Stetter K.O. 1988. Archaeoglobus fulgidus gen. nov., sp. nov.:a new taxon of extremely thermophilic archaebacteria.System. Appl. Microbiol. 10:172-173.
Stetter K.O. 1996. Hyperthermophilic procaryotes. FEMSMicrobiol. Rev. 18:149-158.
Stetter K.O., H. König and E. Stackebrandt. 1983.Pyrodictium, a new genus of submarine disc-shaped sulfurreducing archaebacteria growing optimally at 105°C. System.Appl. Microbiol. 4:535-551.
Stetter K.O., M. Thomm, J. Winter, G. Wildgruber, H.Huber, W. Zillig, D. Janecovic, H. König and P. Palm.1981. Methanothermus fervidus, sp. nov., a novel extremelythermophilic methanogen isolated from an Islandic hot spring.Zbl. Bakt. Hyg., I. Abt. Orig. C 2:166-178.
Stetter K.O., R. Huber, E. Blöchl, M. Kurr, R.D. Eden, M.Fielder, H. Cash and L. Vance. 1993. Hyperthermophilicarchaea are thriving in deep North Sea and Alaskan Oilreservoirs. Nature 365:743-745.
Stevens T.O. and J.P. McKinley. 1995. Lithotrophicmicrobial ecosystems in deep basalts aquifiers. Science270:450-454.
Stim K.P. 1994. A phylogenetic analysis of micro-organismsisolated from subsurface environments. Mol. Ecol. 3:1-10.
Stoesser G., M.A. Tuli, R. Lopez and P. Sterk. 1999. TheEMBL Nucleotide Sequence Database. Nucleic Acids Res.27:18-24.
Stoffels M., R. Amann, W. Ludwig, D. Hekmat and K.H.Schleifer. 1998. Bacterial community dynamics during start-up of a trickle-bed bioreactor degrading aromatic compounds.Appl. Environ. Microbiol. 64:930-939.
Straub T.M., I.L. Pepper and C.P. Gerba. 1995. Comparisonof PCR and cell culture for detection of enteroviruses insludge-amended field soils and determination of theirtransport. Appl. Envir. Microbiol. 61:2066-2068.
Straub T.M., I.L. Pepper, M. Abbaszadegan and C.P.Gerba. 1994 A method to detect enteroviruses in sewagesludge-amended soil using the PCR. Appl. Envir. Microbiol.60:1014-1017.
Straube W.L., J.W. Deming, C.C. Somerville, R.R. Colwelland J.A. Baross. 1990. Particulate DNA in smoker fluids:evidence for existence of microbial populations in hothydrothermal systems. Appl. Environ. Microbiol. 56:1440-1447.
Stroes-Gascoyne S., K. Pedersen, S.A. Haveman, K.Dekeyser, J. Arlinger, S. Daumas, S. Ekendahl, L.Hallbeck, C.J. Hamon, N. Jahromi and T.L. Delaney. 1997.Occurrence and identification of microorganisms incompacted clay-based buffer material designed for use in anuclear fuel waste disposal vault. Can. J. Microbiol. 43:1133-1146.
Studholme D.J., R.A. Jackson and D.J. Leak. 1999.Phylogenetic analysis of transformable strains of thermophilicBacillus species. FEMS Microbiol. Lett. 172:85-90.
Suau A., R. Bonnet, M. Sutren, J.J. Godon, G.R. Gibson,M.D. Collins and J. Doré. 1999. Direct analysis of genesencoding 16s rRNA from complex communities reveals manynovel molecular species within the human gut. Appl. Envir.Microbiol. 65: 4799-4807.

Références bibliographiques
234
Summit M. and J.A. Baross. 1999. Thermophilicsubseafloor microorganisms from 1996 North Gorda Ridgeeruption. Deep-Sea Res. II 45:2751-2766.
Summons R. 1999. Molecular probing of deep secrets.Nature. 398:752-753.
Suzuki M., M.S. Rappe and S.J. Giovannoni. 1998. Kineticbias in estimates of coastal picoplankton community structureobtained by measurements of small-subunit rRNA gene PCRamplicon length heterogeneity. Appl. Environ. Microbiol.64:4522-4529.
Suzuki M.T. and S.J. Giovannoni. 1996. Bias caused bytemplate annealing in the amplification of mixtures of 16SrRNA genes by PCR. Appl. Environ. Microbiol. 62:625-630.
Suzuki M.T., M.S. Rappe, Z.W. Haimberger, H. Winfield,N. Adair, J. Strobel and S.J. Giovannoni. 1997. Bacterialdiversity among small-subunit rRNA gene clones and cellularisolates from the same seawater sample. Appl. Environ.Microbiol. 63:983-989.
Svetlitshnyi V., F. Rainey and J. Wiegel. 1996.Thermosyntropha lipolytica gen. nov., sp. nov., a lipolytic,anaerobic, alkalitolerant, thermophilic bacterium utilizingshort- and long-chain fatty acids in syntrophic coculture with amethanogenic archaeum. Int. J. Syst. Bacteriol. 46:1131-1137.
Swannell R.P., K. Lee and M. McDonagh. 1996. Fieldevaluations of marine oil spill bioremediation. Microbiol. Rev.60: 342-365.
Szewzyk U., R. Szewzyk and T.A. Stenström. 1994.Thermophilic, anaerobic bacteria isolated from deep boireholein granite in Sweden. Proc. Natl. Acd. Sci. USA 91:1810-1813.
T
Tachibana Y., A. Kuramura, N. Shirasaka, Y. Suzuki, T.Yamamoto, S. Fujiwara, M. Takagi and T. Imanaka. 1999.Purification and characterization of an extremely thermostablecyclomaltodextrin glucanotransferase from a newly isolatedhyperthermophilic archaeon, a Thermococcus sp. Appl.Environ. Microbiol. 65:1991-1997.
Taddei F., I. Matic and M. Radman. 1996. Du nouveau surl'origine des espèces. La recherche. 291:52-59.
Tajima K., R.I. Aminov, T. Nagamine, K. Ogata, M.Nakamura, H. Matsui and Y. Benno. 1999. Rumen bacterialdiversity as determined by sequence analysis of 16S rDNAlibraries. FEMS Microbiol. Ecol. 29:159-169.
Takai K. and K. Horikosi. 1999. Genetic diversity of archaeain deep-sea hydrothermal vent environments. Genetics.152:1285-1297.
Takai K. and Y. Sako. 1999. A molecular view of archaealdiversity in marine and terrestrial hot water environmentsFEMS Microbiol. Ecol. 28:177-188.
Tang K., D.J. Fu, D. Julien. A. Braun, C.R. Cantor and H.Köster. 1999. Chip-based genotyping by mass spectrometry.Proc. Natl. Acad. Sci. USA. 96:10016-10020.
Tani K., K. Kurokawa and Nasu M. 1998. Development of adirect in situ PCR method for detection of specific bacteria innatural environments. Appl. Envir. Microbiol. 64:1536-1540.
Tanner M., B.M. Goebel, M.A. Dojka and N.R. Pace. 1998.Specific ribosomal sequences from diverse environmentalsettings correlate with experimental contaminants. Appl.Environ. Microbiol. 64:3110-3113.
Tanner M., D. Shoskes, A. Shahed and N.R. Pace. 1999.Prevalence of corynebacterial 16S rRNA sequences inpatients with bacterial and "nonbacterial" prostatitis. J. Clin.Microbiol. 37:1863-1870.
Tatusova T.A. and T.L. Madden. 1999. BLAST 2Sequences, a new tool for comparing protein and nucleotidesequences. FEMS Microbiol. Lett. 174:247-250.
Taylor C.D. and C.O. Wirsen. 1997. Microbiology and ecolgyof filamentous sulfur formation. Science 277:1483-1485.
Taylor C.D., C.O. Wirsen and F. Gaill. 1999. Rapidmicrobial production of filamentous sulfur mats athydrothermal vents. Appl. Environ. Microbiol. 65:2253-2255.
Telang A.J., G. Voordouw, S. Ebert, N. Sifeldeen, J.M.Foght, P.M. Fedorak and D.W. Westlake. 1994.Characterization of the diversity of sulfate-reducing bacteria insoil and mining waste water environments by nucleic acidhybridization techniques. Can. J. Microbiol. 40:955-964.
Teske A., C. Wawer, G. Muyzer and N.B. Ramsing. 1996a.Distribution of sulfate-reducing bacteria in a stratified fjord(Mariager Fjord, Denmark) as evaluated by most-probable-number counts and denaturing gradient gel electrophoresis ofPCR-amplified ribosomal DNA fragments. Appl. Envir.Microbiol. 62:1405-1415.
Teske A., M.L. Sogin, L.P. Nielsen and H.W. Jannasch.1999. Phylogenetic relationships of a large marine Beggiatoa.Syst. Appl. Microbiol. 22:39-44.
Teske A., N.B. Ramsing, J. Kuver and H. Fossing. 1995.Phylogeny of Thioploca and related filamentous sulfide-oxidizing bacteria. Syst. Appl. Microbiol. 18:517-526.
Teske A., P. Sigalevich, Y. Cohen and G. Muyzer. 1996b.Molecular identification of bacteria from a coculture bydenaturing gradient gel electrophoresis of 16S ribosomal DNAfragments as a tool for isolation in pure cultures. Appl. Envir.Microbiol. 62:4210-4215.
Thauer R.K. 1998. Biochemistry of methanogenesis : a tributeto Marjory Stephenson. Microbiology 144:2377-2406.
The Integration of Microbial Databases Report of aWorkshop held at Michigan State University, August 11-12,1995, Organized by the Center for Microbial Ecology,http://www.cme.msu.edu/index.html/PUBLICATIONS/IMD_WORKSHOP/TofC.html.
Thomas J.C., M. Desrosiers, Y. St Pierre, P. Lirette, J.G.Bisaillon, R. Beaudet and R. Villemur. 1997. Quantitativeflow cytometric detection of specific microorganisms in soilssamples using rRNA targeted fluorescent probes and ethidiumbromide. Cytometry 27:224-232.
Thornhill R.H., J.G. Burgess and T. Matsunaga. 1995.PCR for direct detection of indigenous uncultured magneticcocci in sediment and phylogenetic analysis of amplified 16Sribosomal DNA. Appl. Envir. Microbiol. 61: 495-500.
Tolker-Nielsen T., K. Holmstrøm and S. Molin. 1997.Visualization of specific gene expression in individualSalmonella typhimurium cells by in situ PCR. Appl. Envir.Microbiol. 63:4196-4203.
Toniuc M., F. Popea and I. Gomoiu. 1998. Studies on theBacteria from thermal habitats in Romania. Thermophiles'98International conference, 6-11 September, Brest, FRANCE.
Tonolla M., A. Demarta, D. Hahn and R. Peduzzi. 1998.Microscopic and molecular in situ characterization of bacterialpopulations in the meromictic lake Cadagno pp. 31-44. In:Lake Cadagno: a meromictic Alpine lake, Peduzzi, R., R.Bachofen,and M. Tonolla (Eds), Documenta Ist. ital. Idrobiol.63.
Tonolla M., A. Demarta, R. Peduzzi and D. Hahn. 1999. Insitu analysis of phototrophic sulfur bacteria in the chemoclineof meromictic lake cadagno. Appl. Environ. Microbiol.65:1325-1330.
Torsvik T. 1997. Bacteria/rock interaction in exploring thedeep sub-seafloor biosphere Workshop. Bristol 2-4 July 1997.
Torsvik V., F.L. Daae, R.A. Sandaa and L. Øvreås. 1998.Novel techniques for analysing microbial diversity in naturaland perturbed environments. J. Biotech. 64:53-62.
Torsvik V., J. Goksøyr and F.L. Daae. 1990a. High Diversityin DNA of Soil Bacteria. Appl. Envir. Microbiol. 56:782-787.
Torsvik V., K. Salte, R. Sørheim and J. Goksøyr. 1990b.Comparison of phenotypic diversity and DNA heterogeneity ina population of soil bacteria. Appl. Envir. Microbiol. 56:776-781.
Trebesius K., R. Amann, W. Ludwig, K. Mühlegger andK.H. Schleifer. 1994. Identification of whole fixed bacterialcells with nonradioactive 23S rRNA-targeted polynucleotideprobes. Appl. Envir. Microbiol. 60:3228-3235.
Trimmer M., K.J. Purdy and D.B. Nedwell. 1997. Processmeasurement and phylogenetic analysis of the sulfatereducing bacteria communities of two contrasting benthic sitesin the upper estuary of the Great Ouse, Norfolk, UK. FEMSMicrobiol. Ecol. 24:333-342.

Références bibliographiques
235
Tsai S.J. and M.C. Wiltbank. 1996. Quantification of mRNAusing competitive RT-PCR with standard curve methodology.BioTechniques. 21:862-866.
Tsai Y.L. and B.H. Olson. 1992a. Detection of low numbersof bacterial cells in soils and sediments by polymerase chainreaction. Appl. Envir. Microbiol. 58:754-757.
Tsai Y.L. and B.H. Olson. 1992b. Rapid method forseparation of bacterial DNA from humic substances insediments for polymerase chain reaction. Appl. Envir.Microbiol. 58:2292-2295.
Tsai Y.L., C.J. Palmer and L.R. Sangermano. 1993.Detection of Escherichia coli in sewage and sludge bypolymerase chain reaction. Appl. Envir. Microbiol. 59:353-357.
Turner S., E.F. DeLong, S.J. Giovannoni, G.J. Olsen andN.R. Pace. 1989. Phylogenetic analysis of microorganismsand natural populations using ribosomal RNA sequences. In,"Microbial Mats: Ecological Physiology of Benthic MicrobialCommunities," Y. Cohen and E. Rosenberg (eds.). Am. Soc.Microbiol. pp. 390-401.
U
Ueda K., T. Seki, T. Kudo, T. Yoshida and M. Kataoka.1999. Two distinct mechanisms cause heterogeneity of 16SrRNA. J. Bacteriol. 181:78-82.
Ueda T., Y. Suga and T. Matsuguchi. 1995. Molecularphylogenetic analysis of a soil microbial community in asoybean field. Eur. J. Soil Sc. 46:415-421.
Urbach E., K.L. Vergin and S.J. Giovannoni. 1999.Immunochemical detection and isolation of DNA frommetabolically active Bacteria. Appl. Envir. Microbiol. 65:1207-1213.
V
Vainio E.J., A. Moilanen, T.T. Koivula, D.H. Bamford andJ. Hantula. 1997. Comparison of partial 16S rRNA genesequences obtained from activated sludge bacteria. Appl.Microbiol. Biotech. 48:73-79.
Van de Peer Y., E. Robbrecht, S. de Hoog, A. Caers, P. DeRijk and R. De Wachter. 1999. Database on the structure ofsmall subunit ribosomal RNA. Nucleic Acids Res. 27:179-183.
Van de Peer Y., J. Jansen, P. de Rijk and R. de Wachter.1997. Database on the structure of small ribosomal subunitRNA. Nucleic Acids Res. 25:111-116.
Van de Peer Y., J.M. Neefs, P. deRijk, P de Vos and R. deWachter. 1994. About the order of divergence of the majorbacterial taxa during evolution. System. Appol. Microbiol.17:32-38.
Van der Maarel M.J., R.R. Artz, R. Haanstra and L.J.Forney. 1998. Association of marine archaea with thedigestive tracts of two marine fish species. Appl. Environ.Microbiol. 64:2894-2898.
Van der Maarel M.J.E.C., W. Sprenger, R. Haanstra andL.J. Forney. 1999. Detection of methanogenic archaea inseawater particles and the digestive tract of a marine fishspecies. FEMS Microbiol. Lett. 173:189-194.
Van Dover C.L. 1994. in Hydrothermal Vents and Processes,Special Publication 87, eds. Parson L.M., Walker, C.L. andDixon D.R. (Geol. Soc., London), 257-294.
Van Elsas J.D., G.F. Duarte, A.S. Rosado and K. Smalla.1998. Microbiological and molecular biological methods formonitoring microbial inoculants and their effects in soilenvironment. J. Microbiol. Methods 32:133-154.
Vandamme P., B. Pot, M. Gillis, P. DeVos, K. Kersters andJ. Swings. 1996. Polyphasic Taxonomy, a consensusapproach to bacterial systematics. Appl. Environ. Microbiol.60:407-438.
VanWambeke F. 1995. Numération et taille des bactériesplanctoniques au moyen de l'analyse d'image couplée àl'épifluorescence. Océanis 21:113-124.
Vargas M., K. Kashefi, E.L. Bunt-Harris and D.R. Lovley.1998. Microbiological evidence for Fe(III) reduction on earlyearth. Nature 395:65-67.
Vàsquez M. and R.T. Espejo. 1997. Chemolithotrophicbacteria in copper ores leached at high sulfuric acidconcentration. Appl. Envir. Microbiol. 63:332-334.
Vàsquez M., E.R.B. Moore and R.T. Espejo. 1999.Detection by polymerase chain reaction-amplification andsequencing of an archaeon in a commercial-scale copperbioleaching plant. FEMS Microbiol. Lett. 173:183-187.
Vergin K.L., E. Urbach, J.L. Stein, E.F. DeLong, B.D.Lanoil and S.J. Giovannoni. 1998. Screening of a fosmidlibrary of marine environmental genomic DNA fragmentsreveals four clones related to members of the orderPlanctomycetales. Appl. Environ. Microbiol. 64:3075-3078.
Vetriani C., A.L. Reysenbach and J. Dore. 1998. Recoveryand phylogenetic analysis of archaeal rRNA sequences fromcontinental shelf sediments. FEMS Microbiol. Lett. 161:83-88.
Vetriani C., H.W. Jannasch, B.J. MacGregor, D.A. Stahland A.L. Reysenbach. 1999. Population structure andphylogenetic characterization of marine benthic Archaea indeep-sea sediments. Appl. Env. Microbiol. 65:4375-4384.
Vettori C., D. Paffetti, G. Pietramellara, G. Stotzky and E.Gallori. 1996. Amplification of bacterial DNA bound on clayminerals by the random amplified polymorphic DNA (RAPD)technique. FEMS Microbiol. Ecol. 20:251-260.
Vieille C., D.S. Burdette and J.G. Zeikus. 1996.Thermozymes. Biotechnol. Annu. Rev. 2:1-83.
Vincent P. 1993. Etude d'eubactéries productricesd'exopolysaccharides, originaires d'un site hydrothermalprofond (13°N). Thèse de doctorat de l'Université de BretagneOccidentale, 368p.
von Wintzingerode F., B. Selent, W. Hegemann and U.B.Göbel. 1999. Phylogenetic analysis of an anaerobic,trichlorobenzene-transforming microbial consortium. Appl.Envir. Microbiol. 65:283-286.
Voordouw G., J.K. Voordouw, R.R. Karkhoff-Schweizer,P.M. Fedorak and D.W.S. Weslake. 1991. Reverse samplegenome probing, a new technique for identification of bacteriain environmental samples by DNA hydridization, and itsapplication to the identification of sulfate-reducing bacteria inoil field samples. Appl. Environ. Microbiol. 57:3070-3078.
Voordouw G., J.K. Voordouw, T.R. Jack, J. Foght, P.M.Fedorak and D.W.S. Weslake. 1992. Identification of distinctcommunities of sulfate-reducing bacteria in oil fields byreverse sample genome probing. Appl. Environ. Microbiol.58:3542-3552.
Voordouw G., S.M. Armstrong, M.F. Reimer, B. Fouts,A.J. Telang, Y. Shen and D. Gevertz. 1996. Characterizationof 16S rRNA genes from oil field microbial communitiesindicates the presence of a variety of sulfate-reducing,fermentative, and sulfide-oxidizing bacteria. Appl. Envir.Microbiol. 62:1623-1629.
Voytek M.A. and B.B. Ward. 1995. Detection of ammonium-oxidizing bacteria of the beta-subclass of the classProteobacteria in aquatic samples with the PCR. Appl.Environ. Microbiol. 61:1444-1450.
W
Wächtershäuser. 1988. Pyrite formation, the first energysource for life: a hypothesis. System. Appl. Microbiol. 10:207-210.
Wagner M., R. Amann, H. Lemmer and K.H. Schleifer.1993. Probing activated sludge with oligonucleotides specificfor Proteobacteria: inadequacy of culture-dependent methodsfor describing microbial community structure. Appl. Envir.Microbiol. 59:1520-1525.
Wagner M., R. Erhart, W. Manz, R. Amann, H. Lemmer,D. Wedi and K.H. Schleifer. 1994. Development of an rRNA-targeted oligonucleotide probe specific for the genusAcinetobacter and its application for in situ monitoring inactivated sludge. Appl. Envir. Microbiol. 60:792-800.
Wagner-Döbler I., A. Bennasar, M. Vancanneyt, C.Strömpl, I. Brümmer, C. Eichner, I. Grammel and E.R.B.

Références bibliographiques
236
Moore. 1998. Microcosm enrichment of biphenyl-degradingmicrobial communities from soils and sediments. Appl. Envir.Microbiol. 64:3014-3022.
Wahlfors J., J.H. Meurman, J. Toskala, A. Korhonen, P.Alakuijala, E. Janatuinen, U.M. Karkkkainen, P. Nuutinenand J. Janne. 1995. Development of a rapid PCR method foridentification of Helicobacter pylori in dental plaque and gastricbiopsy specimens. Eur. J. Clin. Microbiol. Infect. Dis. 14:780-786.
Walia S., A. Khan and N. Rosenthal. 1990. Constructionand application of DNA probes for detection of polychlorinatedbiphenyl-degrading genotytpes in toxic organic-contaminatedsoil environments. Appl. Envir. Microbiol. 56:254-259.
Wallner G., B. Fuchs, S. Spring, W. Beisker and R.Amann. 1997. Flow sorting of microorganisms for molecularanalysis. Appl. Envir. Microbiol. 63:4223-4231.
Wallner G., R. Erhart and R. Amann. 1995. Flow cytometricanalysis of activated sludge with rRNA-targeted probes. Appl.Envir. Microbiol. 61:1859-1866.
Wang G.C. and Y. Wang. 1996. The frequency of chimericmolecules as a consequence of PCR co-amplification of 16SrRNA genes from different bacterial species. Microbiology142:1107-1114.
Wang G.C.Y. and Y. Wang. 1997. Frequency of formation ofchimeric molecules as a cosequence of PCR coamplificationof 16S rRNA genes from mixed bacterial genomes. Appl.Environ. Microbiol. 63:4645-4650.
Wang R.F., W.W. Cao and C.E. Cerniglia. 1996. PCRdetection and quantitation of predominant anaerobic bacteria inhuman and animal fecal samples. Appl. Environ. Microbiol.62:1242-1247.
Ward D.M., C.M. Santegoeds, S.C. Nold, N.B. Ramsing,M.J. Ferris and M.M. Bateson. 1997. Biodiversity within hotspring microbial mat communities: molecular monitoring ofenrichment cultures. Antonie von Leeuwenhoek 71:143-150.
Ward D.M., M.J. Ferris, S.C. Nold and M.M. Bateson.1998. A natural view of microbial biodiversity within hot springcyanobacterial mat communities. Microbiol. Mol. Biol. Rev.62:1353-1370.
Ward D.M., M.M. Bateson, R. Weller and A.L. Ruff-Roberts. 1992. Ribosomal RNA analysis of microorganismsas they occur in nature. Adv. Microb. Ecol. 12:219-286.
Ward D.M., R. Weller and M. Bateson. 1990a. 16S rRNAsequences reveal numerous uncultured microorganisms in anatural community. Nature 345:63-65.
Ward D.M., R. Weller and M. Bateson. 1990b. 16S rRNAsequences reveal uncultured inhabitants of a well-studiedthermal community. FEMS Microbiol. Rev. 6:105-115.
Watanabe K., M. Teramoto, H. Futamata and S.Harayama. 1998a. Molecular detection, isolation, andphysiological characterization of functionally dominant phenol-degrading bacteria in activated sludge. Appl. Envir. Microbiol.64:4396-4402.
Watanabe K., S. Yamamoto, S. Hino and S. Harayama.1998b. Population dynamics of phenol-degrading bacteria inactivated sludge determined by gyrB-targeted quantitativePCR. Appl. Envir. Microbiol. 64:1203-1209.
Watson S.W., T.J. Novitsky, H.L. Quinby and F.W. Valois.1977. Determination of bacterial number and biomass in themarine environment. Appl. Envir. Microbiol. 33:940-946.
Wayne L.L., D.J. Brenner, R.R. Colwell, P.A.D. Grimont,O. Kandler, K.I. Krichevsky, L.H. Moore, W.E.C. Moore,R.G.E. Muray, E. Stackebrandt, M.P. Starr and H.G.Trüper. 1987. Report of the Ad Hoc committee onreconciliation of approaches to bacterial systematics. Int. J.Syst. Bacteriol. 37:463-464.
Weidner S., W. Arnold and A. Puhler. 1996. Diversity ofuncultured microorganisms associated with the seagrassHalophila stipulacea estimated by restriction fragment lengthpolymorphism analysis of PCR-amplified 16S rRNA genes.Appl. Environ. Microbiol. 62:766-771.
Weinbauer M.G., C. Beckmann and M.G. Höfle. 1998.Utility of green fluorescent nucleic acid dyes and aluminumoxide membrane filters for rapid epifluorescence enumerationof soil and sediment Bacteria. Appl. Envir. Microbiol. 64:5000-5003.
Weller R. and D.M. Ward. 1989. Selective recovery of 16SRNA sequences from natural microbial communities in theform of cDNA. Appl. Envir. Microbiol. 55:1818-1822.
Weller R., M.M. Bateson, B.K. Heimbuch, E.D.Kopczynski and D.M. Ward. 1992. Uncultivatedcyanobacteria, Chloroflexus-like inhabitants, and spirochete-like inhabitants of a hot spring microbial mat. Appl. Environ.Microbiol. 58:3964-3969.
Wellsbury P., K. Goodman, T. Barth, B.A. Cragg, S.P.Barnes and R.J. Parkes. 1997. Deep marine biospherefuelled by increasing organic matter availability during burialand heating. Nature 388:573-576.
Wheeler W.C. and R.L. Honeycutt. 1988. Paired sequencedifference in ribosomal RNAs: evolutionary and phylogeneticimplications. Mol. Biol. Evol. 5:90-96.
Whelen A.C. and D.H. Persing. 1996. The role of nucleicacid amplification and detection in the clinical microbiologylaboratory. Annu. Rev. Microbiol. 50:349-373.
White D., R.J. Sharp and F.G. Priest.1993. A polyphasictaxonomic study of thermophilic Bacilli from a widegeographical area. Antonie Van Leeuwenhoek 64:357-86
White D.C., T.J. Phelps and T.C. Onstott. 1998. What's updown there? Curr. Opin. Microbiol. 1:286-290.
Whitman W.B., D.C. Coleman and W.J. Wiebe. 1998.Prokaryotes: the unseen majority. Proc. Natl. Acad. Sci. USA95:6578-6583.
Widjojoatmodjo M.N., A.C. Fluit and J. Verhoef. 1995.Molecular identification of bacteria by fluorescence-basedPCR-single- strand conformation polymorphism analysis ofthe 16S rRNA gene. J. Clin. Microbiol. 33:2601-2606.
Widmer F., B.T. Shaffer, L.A. Porteous and R.J. Seidler.1999. Analysis of nifH gene pool complexity in soil and litter ata Douglas Fir forest site in the Oregon cascade mountainrange. Appl. Envir. Microbiol. 65:374-380.
Williams D.M. and T.M. Embley. 1996. Microbial diversity:domains and kingdoms. Annu. Rev. Syst. 27:569-595.
Willis S.G., K.S. Smith, V.L. Dunn, L.A. Gapter, K.H.Riviere and G.R. Riviere. 1999. Identification of sevenTreponema species in health- and disease-associated dentalplaque by nested PCR. J. Clin. Microbiol. 37:867-869.
Wilson I.G. 1997. Inhibition and facilitation of nucleic acidamplification. Appl. Environ. Microbiol. 63:3741-3751.
Wilson K.H. and R.B. Blitchington. 1996. Human colonicbiota studied by ribosomal DNA sequence analysis. Appl.Environ. Microbiol. 62:2273-2278.
Wilson MS., C. Bakermans and E.L. Madsen. 1999. In situ,real-time catabolic gene expression: extraction andcharacterization of naphthalene dioxygenase mRNAtranscripts from groundwater. Appl. Envir. Microbiol. 65:80-87.
Winn C.D., J.P. Cowen and D.M. Karl. 1995. Microbes indeep-sea hydrothermal plumes. In: Karl DM (Ed) TheMicrobiology of Deep-sea Hydrothermal Vents (pp 255-273).CRC Press Inc, Boca Raton.
Winter J., C. Lerp, H.P. Zabel, F.X. Wildenauer, H. Königand F. Schindler. 1984. Methanobacterium wolfei, sp. nov., anew tungsten-requiring, thermophilic, autotrophic methanogen.System. Appl. Microbiol. 5:457-466.
Wintzingerode F.V., U.B. Gobel and E. Stackebrandt.1997. Determination of microbial diversity in environmentalsamples: pitfalls of PCR-based rRNA analysis. FEMSMicrobiol. Rev. 21:213-229.
Wirsen C.O., H.W. Jannasch and S.J. Molyneaux. 1993.Chemosynthetic microbial activity at Mid-Atlantic Ridgehydrothermal vent site. J. Geophys. Res. 98:9693-9703.
Wirsen C.O., T. Brinkhoff, J. Kuever, G. Muyzer, S.Molyneaux and H.W. Jannasch. 1998. Comparison of a newThiomicrospira strain from the mid-atlantic ridge with knownhydrothermal vent isolates. Appl. Environ. Microbiol. 64:4057-4059.
Wirsen C.O. and H.W. Jannash. 1978. Physiological andmorphological observation on Thiovulum sp. J. Bacteriol.136:765-774.

Références bibliographiques
237
Wise M.G., J.V. McArthur and L.J. Shimkets. 1997.Bacterial diversity of a Carolina bay as determined by 16SrRNA gene analysis: confirmation of novel taxa. Appl. Environ.Microbiol. 63:1505-1514.
Wisotzkey J.D., P.Jr. Jurtshuk, G.E. Fox, G. Deinhard andK. Poralla. 1992. Comparative sequences analyses on the16S rRNA (rDNA) of Bacillus acidocaldarius, Bacillusacidoterrestris, and Bacillus cycloheptanicus and proposal forcreation of a new genus, Alicyclobacillus gen. nov. Int. J. Syst.Bacteriol. 42:263-269.
Woese C.R. 1987. Bacterial evolution. Microbiol. Rev.51:221-271.
Woese C.R. 1994a. There must be a prokaryote somewhere:microbiology's search for itself. Microbiol. Rev. 58:1-9
Woese C.R. 1994b. Microbiology in transition. Proc. Natl.Acad. Sci. USA. 91:1601-1603.
Woese C.R. 1998a. The universal ancestor. Proc. Natl. Acad.Sci. USA 95:6854-6859.
Woese C.R. 1998b. Default taxonomy: Ernst Mayr's view ofthe microbial world. Proc. Natl. Acad. Sci. USA 95:11043-11046.
Woese C.R. and G. Fox. 1977. Phylogenetic structure of theprokaryotic domain: The primary kingdoms. Proc. Natl. AcadSci. USA 74:5088-5090.
Woese C.R., G.E. Fox, L. Zablen, T. Uchida, L. Bonen, K.Pechman, B.J. Lewis and D. Stahl. 1975. Conservation ofprimary structure in 16S ribosomal RNA. Nature 254:83-86.
Woese C.R., O. Kandler and M.L. Wheelis. 1990. Towardsa natural system of organisms : proposal for the domainsArchae, Bacteria, Eucarya. Proc. Natl. Acad. Sci. USA87:4576-4579.
Wu Y., R.P. Stulp, P. Elfferich, J. Osinga, C.H. Buys andR.M. Hofstra. 1999. Improved mutation detection in GC-richDNA fragments by combined DGGE and CDGE. NucleicAcids Res. 27:e9.
Y
Yamamoto H., A. Hiraishi, K. Kato, H.X. Chiura, Y. Makiand A. Shimizu. 1998. Phylogenetic evidence for theexistence of novel thermophilic bacteria in hot spring sulfur-turf microbial mats in Japan. Appl. Envir. Microbiol. 64:1680-1687.
Yanagibayashi M., Y. Nogi, L. Li and C. Kato. 1999.Changes in the microbial community in Japan Trenchsediment from a depth of 6292 m during cultivation withoutdecompression. FEMS Microbiol. Lett. 170:271-279.
Yayanos A.A. 1995. Microbiology to 10,500 meters in thedeep sea. Annu. Rev. Microbiol. 49:777-805.
Yayanos A.A., A.S. Dietz and R. Van Boxtel. 1981.Obligately barophilic bacterium from the Mariana trench.Proc. Natl. Acad. Sci. USA 78:5212-5215.
Yurkov V. and J.T. Beatty. 1998. Isolation of aerobicanoxygenic photosynthetic bacteria from black smoker plumewaters of the Juan de Fuca ridge in the Pacific Ocean. Appl.Environ. Microbiol. 64:337-341.
Z
Zal F. 1999. "Riftia pachyptila" le vers géant des abysses. LaRecherche 317:90-95.
Zarilla K.A. and J.J. Perry. 1986. Thermoleophilum album,gen. nov. and sp. nov., a bacterium obligate for thermophilyand n-alkane substrates. Arch. Microbiol. 137:286-290 .
Zehnder A.J.B., B. Huser and T.D. Brock. 1979. Measuringradioactive methane with liquid scintillation counter. Appl.Envir. Microbiol. 37:897-899.
Zeikus J.G., M.A. Dawson, T.E. Thompson, K. Ingvorsenand E.C. Hatchikian. 1983. Microbial ecology of volcanicsulfidogenesis: isolation and characterization ofThermodesulfotobacterium commune gen. nov. and spec. nov.J. Gen. Microbiol. 129:1159-1169.
Zeikus J.G., P.W. Hegge and M.A. Anderson. 1979.Thermoanaerobium brockii gen. nov., sp. nov., a newchemoorganothrophic caldoactive anaerobic bacterium. Arch.Microbiol. 122:41-48.
Zengler,K., H.H. Richnow, R. Rossello-Mora, W.Michaelis and F. Widdel. 1999. Methane formation fromlong-chain alkanes by anaerobic microorganismes. Nature401:266-269.
Zheng D., E.W. Alm, D.A. Stahl and L. Raskin. 1996.Characterization of universal small-subunit rRNAhybridization probes for quantitative molecular microbialecology studies. Appl. Envir. Microbiol. 62:4504-4513.
Zhou J., M.A. Bruns and J.M. Tiedje. 1996. DNA recoveryfrom soils of diverse composition Appl. Envir. Microbiol.62:316-322.
Zhou J., M.E. Davey, J.B. Figueras, E. Rivkina, D.Gilichinsky and J.M. Tiedje. 1997. Phylogenetic diversity of abacterial community determined from Siberian tundra soilDNA. Microbiology 143:3913-3919.
Zillig W., A. Gierl, G. Schreiber, S. Wunderl, D. Janekovic,K.O. Stetter and H.P. Klenk. 1983a. The archaebacteriumThermophilum pendens represents a novel genus of thethermophilic, anaerobic sulfur respiring Thermoproteales.System. Appl. Microbiol. 4:79-87.
Zillig W., I. Holz, D. Janekovic, H.P. Klenk, E. Imsel, J.Trent, S. Wunderl, V.H. Forjaz, R. Coutinho and T.Ferreira. 1990. Hyperthermus butylicus, a hyperthermophilicsulfur-reducing archaebacterium that ferments peptides. J.Bacteriol. 172:3959-3965.
Zillig W., I. Holz, D. Janekovic, W. Schäfer and W.D.Reiter. 1983b The archaebacterium Thermococcus celerrepresents a novel genus within the thermophilic branch of thearchaebacteria. System. Appl. Microbiol. 4:88-94.
Zillig W., K.O. Stetter, D. Prangishvilli, W. Schäfer, S.Wunderl, D. Janekovic, I. Holz and P.Palm. 1982.Desulfurococcaceae, the second family of the extremelythermophilic, anaerobic, sulfur-respiring Thermoproteales.Zbl. Bakt. Hyg., I. Abt. Orig. C 3:304-317.
Zillig W., K.O. Stetter, W. Schäfer, D. Janekovic, S.Wunderl, J. Holz and P. Palm. 1981. Thermoproteales: anovel type of extremely thermoacidophilic anaerobicarchaebacteria isolated from Icelandic solfataras. Zbl. Bakt.Hyg., I. Abt. Orig. C 2:205-227.
Zillig W., K.O., Stetter, S. Wunderl, W. Schulz, H. Priessand J. Scholz. 1980. The Sulfolobus-"Caldariella" group:taxonomy on the basis of the structure of DNA-dependentRNA polymerases. Arch. Microbiol. 125:259-269.
Zimmermann R. and L.A. Meyer-Reil. 1974. A new methodfor fluorescent staining of bacterial population on membranefilters. Kiel. Meeresforsh. 30:24-27.
Zoetendal E.G., A.D. Akkermans and W.M. de Vos. 1998Temperature gradient gel electrophoresis analysis of 16SrRNA from human fecal samples reveals stable and host-specific communities of active bacteria. Appl. Environ.Microbiol. 64:3854-3859.
Zubkov M.V., B.M. Fuchs, H. Eilers, P.H. Burkill and R.Amann. 1999. Determination of total protein content ofbacterial cells by SYPRO staining and flow cytometry. Appl.Environ. Microbiol. 65:3251-3257.
Zuckerland E. and L. Pauling. 1965. Evolutionarydivergence and convergence in proteins, In Evolving Genesand Proteins, V. Bryson and H.J. Vogel (éd), academic Press,New York, pp 97-166.
Zuker M. and A.B. Jacobson. 1998. Using reliabilityinformation to annotate RNA secondary structures. RNA4:669-679.

Annexes
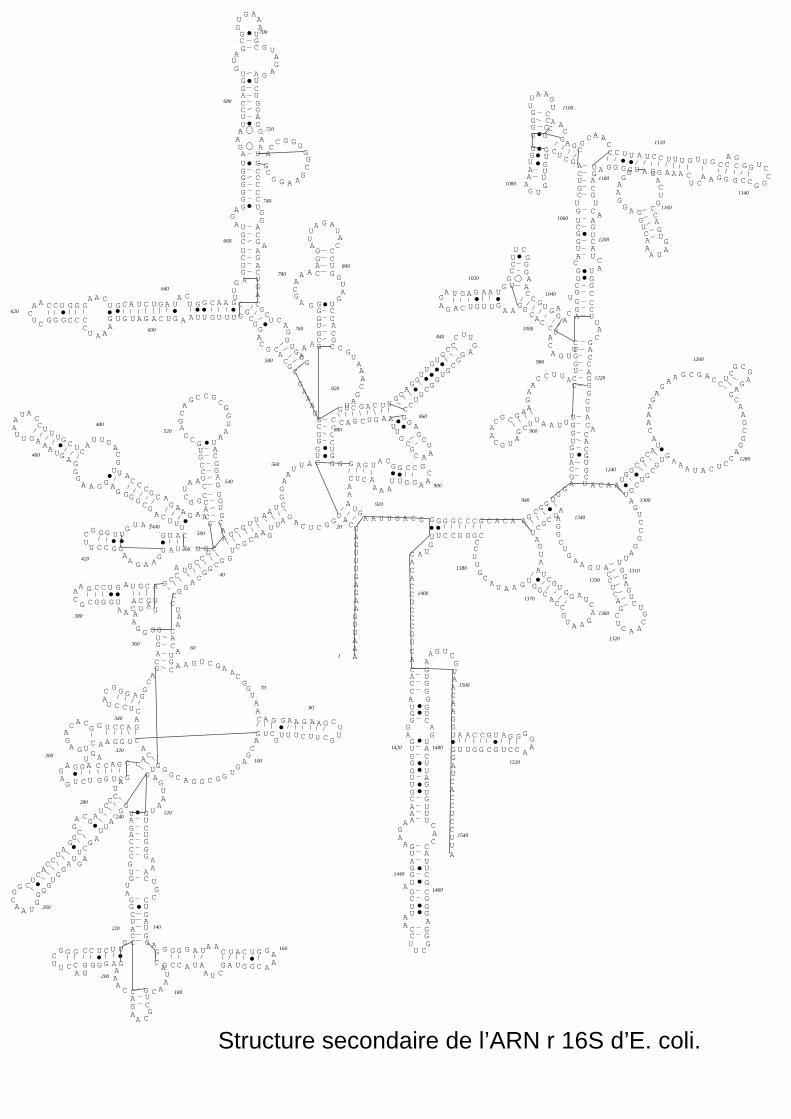
AAAUUGAAGAGUUUGAUC
AUGGCUCAGAUU
GAACGCUGGCGG
CAGGC
CUAA
CACAU
GC A
A G U C G A ACGGUAAC A G G A A G A A G C
UU
GCUUCUUUGCUG
ACGA
GUGGCGGACG
GGUG A
GUAA
UGUCUGGG AAA
C UGCC
UGAUGG AG G G G G A UA A
C U A C U G G AA
ACGGUAGCUA
AUACCGCAUA
ACGUCGCA
AGACC
AAAGAGGGG
GACCUU
CG G G C C U C U U
G CCAUCGGAUGUGCCCAGAU
GG
GA
UUAG
CUA
GU
AG
GUG
GGG
UAACGG C
UCACC
UAGGC
GA C
GAUCCCUA GCUGGUCUG
AGA G G A
UG AC C A G C C
ACA
CUGGAACUGA
GAC A C
G G U C C A GACUC
CUAC G G G A G
GCAGCAGUGGGG
AAU A U
UGCACAA
UGGGCGCAA G C C U G A U G C A G C
CA UG
CCGCGUGUAU
GAAGA
AGGCCU
UC G G G U U
G U A A AG U A C
UUUC
AGC
GGGG
AGGAAGGGA
GUAA
AGUUAAU A C
CU U UG C UCAU U G
ACGUU A
C C CG C AG AA G A A
GCACC
GGCUA A
CUCCGUGC
CAGCAG C C G
CGGUA
AUACGGAGGGUGCAA G
C GU U
AAU CGGAAU U A C
UGGGCGUAAAG
CGC
ACG
CAG
GCGGUUUGUUAAGUCAGAUGUG
AAAU
CCCCGGGCU
CA A C C U G G G
A A CU G C A U C U G A U
A CU G G C A A G C
UUG AGUCUCGUA
GA GGGGGGUAGAAUUCCAGGU
GUAGCGGUG A
AAUGC G U
AGAGA
UCUGGAGGAAU A
CC G G U
GGCG
AAGGCGG
CCCCCUGGACG AAGACUGACGCU C
AGGUGCG
AA A G
CGUGGGGA
GCAA A C
AGG
AUUA G A
UAC
CCUG G
UAG
UCCACGC C G
UAAACGAUG U C G A C U U
GGAGGUUGU
GCCC U U
GAG
GC
GUG
GCU
UCC
GG AGCUAAC
GC
GUUAAGUCGACC
GCCUGG G G A G U A C
G G C C GCAAGGUU
AAAACUC
AAAUG
A A U U G A C G G G G G C C C G C A C A AGCG
G UGGAGCAUGUGGU
UUAAUUCGA
UGCAAC G C G
AAGAAC C U U A
CCUGGUCU
UGACAUC
CAC
GGAAGUUUUCAGA
GA U G A G A A U
G UGCCUU C
GGGAACC GU G A
G AC AGGUGCUGCAUGGCUGUCGUCAG
CUCG
UGUUG
UGAAAUGU
UGGGUU A A
GU
CCC
GCA A
CGA GC
GC A A
CC C U U A U C C U U U G U U G C C
A GC G G U C
CGGC
CGGGAACU
CAAAGGA GACUG
CCAGUGA
UAAACUGGAG
GAAGG UGGGGAUG
ACGUC AAGUCAU CAU
GGCCCUUAC
GACCAGGGCUACACACGUGCU A C A A
UGG
CGCA
UACAAAGAGA A
G C G A C CUCG C
GAG
AGCAAGCGGA
CCUCAUAA
AGU
GC
GUC
GUAGUCCGGA
UUGGAGUC U
GCAAC
UCGACUCCAUGAA
GUCGGAA
UCGCUAGUAAU
CGUGG A U
CAGAAU
GCC
ACG
GUGAAUA
CGUUCCCGGGCCUU
GUAC
ACACCGCCCGUCA
CACCAUGGGAGUGGGUUGCAAAA
GAA GUAGGUAGCUUA
A CCUU C
GGGAGGGCGCUUACCAC
UUUGUGAUUCAUGA
CUGGGGUGAA G U C
GUAACAAGGU A A C C G U A G G G G
AACCUGCGGUUG
GAUCACCUCCUUA
1
20
40
60
70
80
100
120
140
160
180
200
220
240
260
280
300320
340
360
380
400
420
440
460
480
500
520
540
560
580
600
620
640
660
680
700
720
740
760
780
800
820
840
860
880
900
920940
960
980
1000
1020
1040
1060
1080
1100
1120
1140
1160
1180
1200
1220
1240
1260
1280
1300
1310
1320
1330
1340
1360
1370
1380
1400
1420
1440
1460
1480
1500
1520
1540
Structure secondaire de l’ARN r 16S d’E. coli.
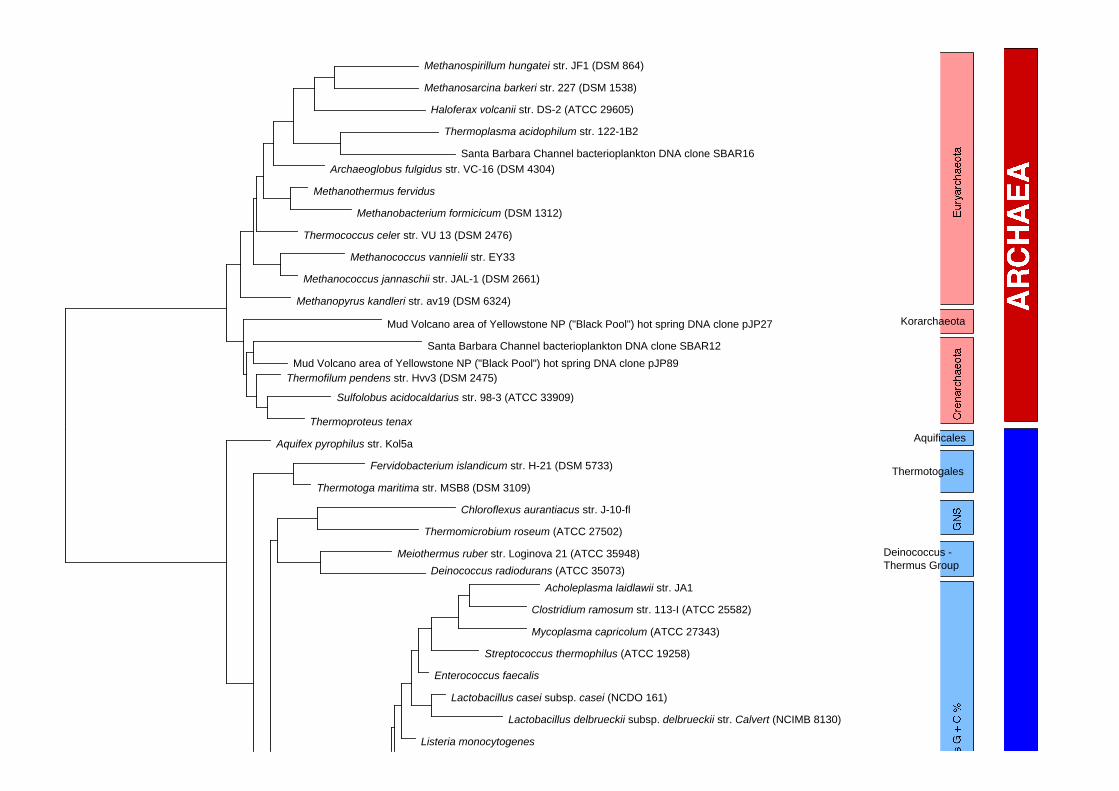
Methanospirillum hungatei str. JF1 (DSM 864)
Methanosarcina barkeri str. 227 (DSM 1538)
Haloferax volcanii str. DS-2 (ATCC 29605)
Thermoplasma acidophilum str. 122-1B2
Santa Barbara Channel bacterioplankton DNA clone SBAR16 Archaeoglobus fulgidus str. VC-16 (DSM 4304)
Methanothermus fervidus
Methanobacterium formicicum (DSM 1312)
Thermococcus celer str. VU 13 (DSM 2476)
Methanococcus vannielii str. EY33
Methanococcus jannaschii str. JAL-1 (DSM 2661)
Methanopyrus kandleri str. av19 (DSM 6324)
Mud Volcano area of Yellowstone NP ("Black Pool") hot spring DNA clone pJP27
Santa Barbara Channel bacterioplankton DNA clone SBAR12
Mud Volcano area of Yellowstone NP ("Black Pool") hot spring DNA clone pJP89 Thermofilum pendens str. Hvv3 (DSM 2475)
Sulfolobus acidocaldarius str. 98-3 (ATCC 33909)
Thermoproteus tenax
Aquifex pyrophilus str. Kol5a
Fervidobacterium islandicum str. H-21 (DSM 5733)
Thermotoga maritima str. MSB8 (DSM 3109)
Chloroflexus aurantiacus str. J-10-fl
Thermomicrobium roseum (ATCC 27502)
Meiothermus ruber str. Loginova 21 (ATCC 35948)
Deinococcus radiodurans (ATCC 35073)
Acholeplasma laidlawii str. JA1
Clostridium ramosum str. 113-I (ATCC 25582)
Mycoplasma capricolum (ATCC 27343)
Streptococcus thermophilus (ATCC 19258)
Enterococcus faecalis
Lactobacillus casei subsp. casei (NCDO 161)
Lactobacillus delbrueckii subsp. delbrueckii str. Calvert (NCIMB 8130)
Listeria monocytogenes
Korarchaeota
Aquificales
Thermotogales
Deinococcus -Thermus Group
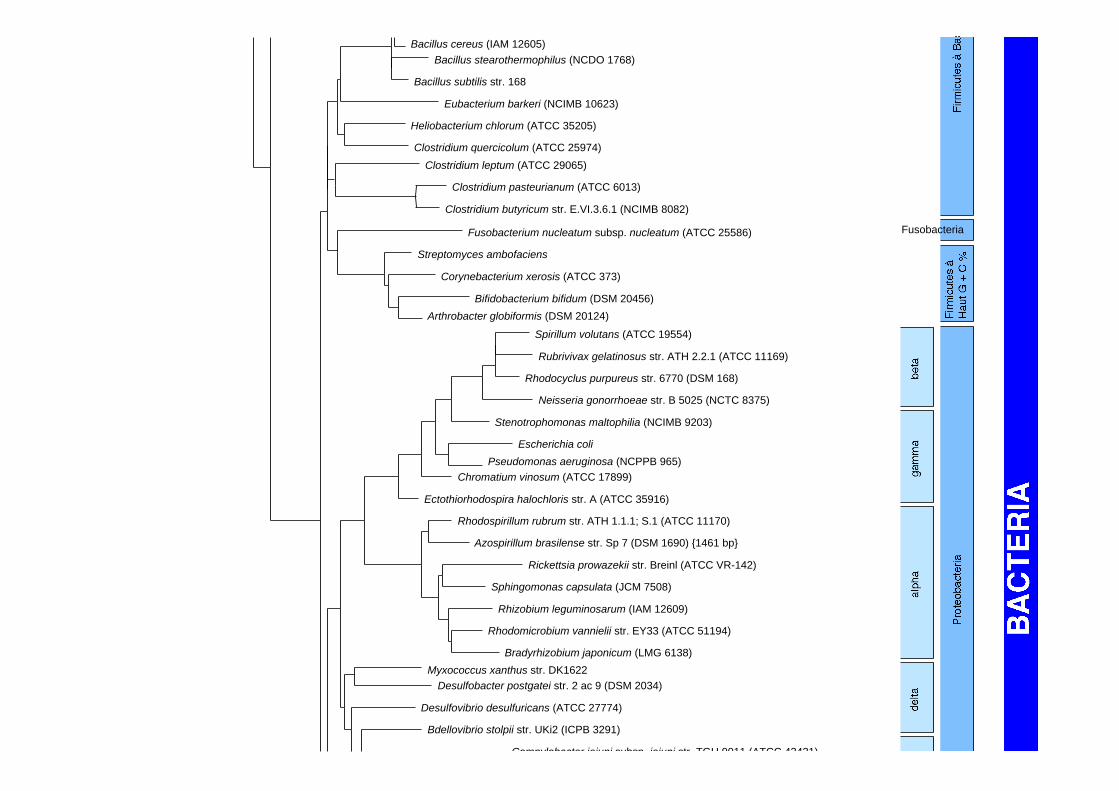
Bacillus cereus (IAM 12605)
Bacillus stearothermophilus (NCDO 1768)
Bacillus subtilis str. 168
Eubacterium barkeri (NCIMB 10623)
Heliobacterium chlorum (ATCC 35205)
Clostridium quercicolum (ATCC 25974)
Fusobacterium nucleatum subsp. nucleatum (ATCC 25586)
Streptomyces ambofaciens
Corynebacterium xerosis (ATCC 373)
Bifidobacterium bifidum (DSM 20456)
Arthrobacter globiformis (DSM 20124)
Clostridium leptum (ATCC 29065)
Clostridium pasteurianum (ATCC 6013)
Clostridium butyricum str. E.VI.3.6.1 (NCIMB 8082)
Spirillum volutans (ATCC 19554)
Rubrivivax gelatinosus str. ATH 2.2.1 (ATCC 11169)
Rhodocyclus purpureus str. 6770 (DSM 168)
Neisseria gonorrhoeae str. B 5025 (NCTC 8375)
Stenotrophomonas maltophilia (NCIMB 9203)
Escherichia coli
Pseudomonas aeruginosa (NCPPB 965)Chromatium vinosum (ATCC 17899)
Ectothiorhodospira halochloris str. A (ATCC 35916)
Rhodospirillum rubrum str. ATH 1.1.1; S.1 (ATCC 11170)
Azospirillum brasilense str. Sp 7 (DSM 1690) {1461 bp}
Rickettsia prowazekii str. Breinl (ATCC VR-142)
Sphingomonas capsulata (JCM 7508)
Rhizobium leguminosarum (IAM 12609)
Rhodomicrobium vannielii str. EY33 (ATCC 51194)
Bradyrhizobium japonicum (LMG 6138)
Myxococcus xanthus str. DK1622 Desulfobacter postgatei str. 2 ac 9 (DSM 2034)
Desulfovibrio desulfuricans (ATCC 27774)
Bdellovibrio stolpii str. UKi2 (ICPB 3291)
Campylobacter jejuni subsp. jejuni str. TGH 9011 (ATCC 43431)
Fusobacteria
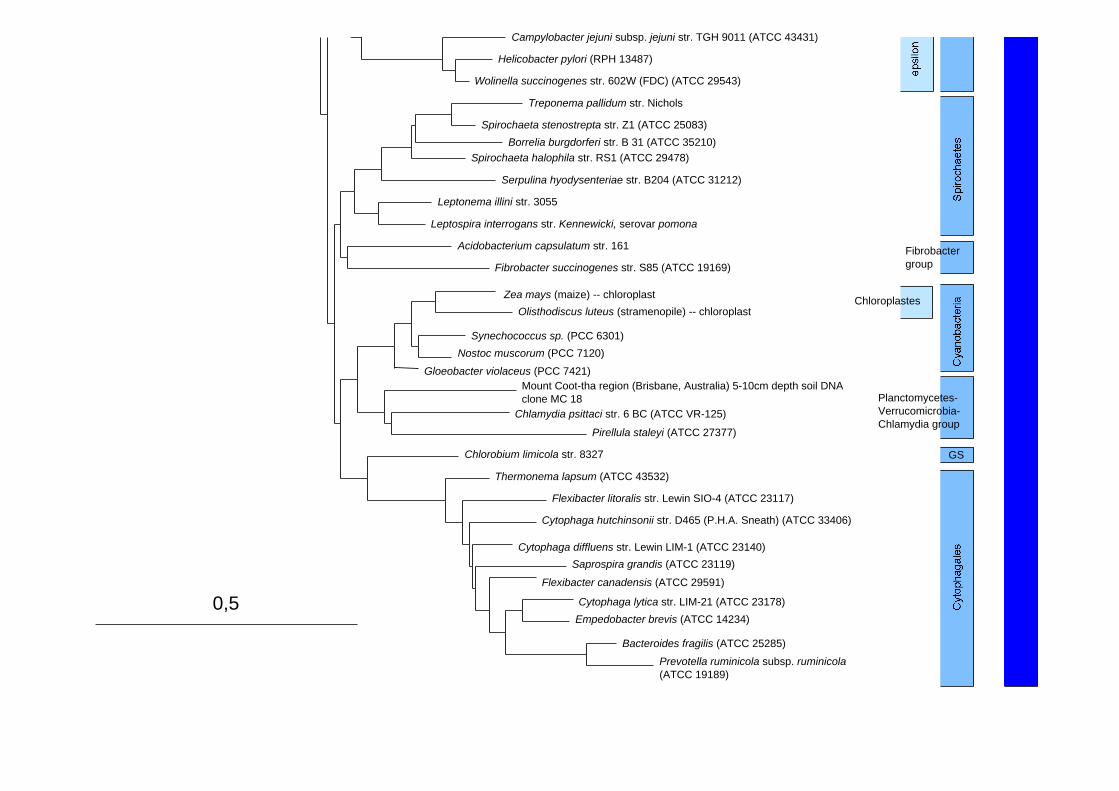
Campylobacter jejuni subsp. jejuni str. TGH 9011 (ATCC 43431)
Helicobacter pylori (RPH 13487)
Wolinella succinogenes str. 602W (FDC) (ATCC 29543)
Treponema pallidum str. Nichols
Spirochaeta stenostrepta str. Z1 (ATCC 25083)
Borrelia burgdorferi str. B 31 (ATCC 35210)Spirochaeta halophila str. RS1 (ATCC 29478)
Serpulina hyodysenteriae str. B204 (ATCC 31212)
Leptonema illini str. 3055
Leptospira interrogans str. Kennewicki, serovar pomona
Acidobacterium capsulatum str. 161
Fibrobacter succinogenes str. S85 (ATCC 19169)
Zea mays (maize) -- chloroplast
Olisthodiscus luteus (stramenopile) -- chloroplast
Synechococcus sp. (PCC 6301)
Nostoc muscorum (PCC 7120)
Gloeobacter violaceus (PCC 7421)Mount Coot-tha region (Brisbane, Australia) 5-10cm depth soil DNA clone MC 18
Chlamydia psittaci str. 6 BC (ATCC VR-125)
Pirellula staleyi (ATCC 27377)
Chlorobium limicola str. 8327
Thermonema lapsum (ATCC 43532)
Flexibacter litoralis str. Lewin SIO-4 (ATCC 23117)
Cytophaga hutchinsonii str. D465 (P.H.A. Sneath) (ATCC 33406)
Cytophaga diffluens str. Lewin LIM-1 (ATCC 23140)
Saprospira grandis (ATCC 23119)
Flexibacter canadensis (ATCC 29591)
Cytophaga lytica str. LIM-21 (ATCC 23178)
Empedobacter brevis (ATCC 14234)
Bacteroides fragilis (ATCC 25285)
Prevotella ruminicola subsp. ruminicola (ATCC 19189)
Chloroplastes
Fibrobacter group
Planctomycetes-Verrucomicrobia-Chlamydia group
GS
0,5

RESUMEL'étude des communautés de micro-organismes des sources hydrothermales profondes et des réservoirs
pétroliers a longtemps été limitée par l'utilisation exclusive de méthodes culturales ne permettant d'accéder qu'à unepart restreinte de la diversité microbienne. Afin de dépasser cette restriction, nous avons utilisé les approchesculturales en combinaison avec des approches moléculaires pour étudier les communautés microbiennes de deuxréservoirs pétroliers du bassin parisien et des fluides hydrothermaux profonds collectés sur la dorsale médio-Atlantique. Ces approches ont consisté d'une part en une analyse phylogénétique des gènes codant pour l'ARNr 16Spar amplification et hybridation in situ en s'affranchissant de toute étape de culture, d'autre part dans l'enrichissementde diférents types métaboliques à haute température, et enfin dans le positionnement phylogénétique de souchesisolées d'environnements hydrothermaux.
L'étude réalisée à partir d'échantillons d'eau de production de deux réservoirs pétroliers a conduit à ladescription de séquences d'Archaea méthanogènes, halophiles ou hyperthermophiles affiliées aux Methanosarcinalesou aux Thermococcales. Pour deux échantillons, les séquences de Bacteria étaient associées à des Thermotogales,des Proteobacteria delta et des Firmicutes à bas G+C% généralement impliqués dans des processus thermophiles etmésophiles de sulfato- ou thiosulfato-réduction. Des séquences associées au genre Arcobacter pouvant intervenirdans le cycle des composés soufrés au sein du réservoir ont été détectées dans un troisième échantillon. Enfin undernier échantillon au sein duquel les séquences associées au genre Herbaspirillum, provenant probablement d'unecontamination liée aux processus d'extraction secondaires des hydrocarbures, étaient dominantes.
Des fluides hydrothermaux ont été échantillonnés à l'aide d'un collecteur original, le "Vent Cap". Desenrichissements de différents types métaboliques (soufre-réducteurs autotrophes et hétérotrophes, sulfato- etthiosulfato-réducteurs, et méthanogènes) à 65 et 80°C ont été positifs dans la majorité de nos échantillons.L'amplification et le séquençage des ADNr 16S bactériens et archaéens a conduit à la détection d'une grande diversitéde séquences. Les Proteobacteria epsilon dominent généralement les banques de séquences bactériennes et sontdétectées avec les Desulfurobacteriales en forte proportion par hybridation in situ. Des séquences d'Archaeacommunément isolées en milieu hydrothermal profond, telles que les Thermococcales, les Methanococcales et lesArchaeoglobales ont également été détectées.
L'utilisation combinée de différentes méthodes moléculaires et de méthodes culturales nous a permis dedépasser les limitations inhérentes à chacune de ces méthodes et de proposer une image des relations s'établissantentre les différentes communautés de micro-organismes présents dans ces deux environnements.
ABSTRACTStudies of the microbial communities from deep sea hydrothermal vents and oil reservoir have been for a long
time exclusively limited to the use of cultural techniques which described only a restricted part of the diversity. Tocircumvent this limitation we used cultural techniques in conjunction with molecular methods to investigate microbialcommunities of continental oil reservoirs in the East Paris Basin and of hydrothermal fluids collected on the Mid AtlanticRidge. Our approach combined total 16S rDNA sequencing and in situ hybridization without any cultural step,enrichments of differents metabolic types at high temperature and phylogenic characterisation of strains isolated fromhydrothermal environment.
The study of oil reservoirs water production lead to the description of archaeal sequences affiliated with themethanogenic, halophilic or hyperthermophilic, Methanosarcinales and Thermococcales orders. Bacterial sequencesare mostly affiliated, in two of our samples, to thermophilic or mesophilic sulfate- or thiosulfate-reducing bacteriabelonging to the Thermotogales and δ-Proteobacteria phylum and to the Low G+C Gram-positive bacteria. Sequencesaffiliated to the genus Arcobacter, probably involved in the sulphur cycle inside/or in the vicinity of the oil reservoir, werealso detected in another sample. Finally a last sample was dominated by sequences related to the aerobic genusHerbaspirillum, that may come from contaminations occuring during the secondary oil recovery process.
Deep hydrothermal fluids were sampled by a special collector, the Vent Cap. Enrichments of heterotrophic andautotrophic sulphur-reducers, sulphate and thiosulfate reducers and methanogenes at 65 and 80°C were positive inmost of our samples. Archaeal and bacterial 16S rDNA analyses revealed a wide sequences diversity. ε-Proteobacteriadominated the bacterial clones libraries and have been detected in high proportions by in situ hibridization withDesulfurobacteriales . Archaeal sequences associated to genera commonly isolated in the deep hydrothermalenvironment, as Thermococcales, Methanococcales and Archaeoglobales were also detected.
Combined use of molecular and cultural methods allows us to overcome the limitations inherent to eachtechniques and yield to the description of the relations between the different microbial communities in theirenvironments.