toft et al
TRANSCRIPT

746Q 2003 Estuarine Research Federation
Estuaries Vol. 26, No. 3, p. 746–758 June 2003
The Effects of Introduced Water Hyacinth on Habitat Structure,
Invertebrate Assemblages, and Fish Diets
JASON D. TOFT1,*, CHARLES A. SIMENSTAD1, JEFFERY R. CORDELL1, and LENNY F. GRIMALDO2
1 School of Aquatic and Fishery Sciences, University of Washington, P. O. Box 355020, Seattle,Washington 98195-5020
2 California Department of Water Resources, 3251 S Street, Sacramento, California 95816
ABSTRACT: The South American floating aquatic plant water hyacinth (Eichhornia crassipes) has a history of worldwideinvasions, including a 1904 introduction into the Sacramento-San Joaquin Delta, California. The native pennywort (Hy-drocotyle umbellata) occupies similar habitats in the Delta and is extensively used by resident invertebrates and fish. Wesought to discover if an invader would be functionally equivalent to the native plant, by asking whether the encroachinghyacinth modified the invertebrate assemblage structure and fish-invertebrate food web relative to pennywort. We sam-pled epiphytic, epibenthic, and benthic invertebrates, and plant canopy insects in patches of hyacinth and pennywort,and analyzed fish diets at three sites in the Delta during 1998. We also measured habitat structure (leaf density, rootbiomass, and surface area). In 1999, following control and absence of hyacinth, we again measured epiphytic invertebratesin pennywort. We found differences between hyacinth and pennywort in structure, associated invertebrates, and fishdiets. Most measurements inferred functional non-equivalency between hyacinth and pennywort, although some func-tional equivalency and natural variation existed. Leaf and insect densities were significantly higher in pennywort andthere were also significant differences in insect assemblage compositions. Hyacinth roots in the water column had sig-nificantly more surface area. Densities of epibenthic and benthic aquatic invertebrates were typically greater in pennywortand taxonomic compositions of aquatic invertebrate assemblages showed significant differences. Amphipods and isopodsliving epiphytically in the root masses were particularly abundant, including several newly discovered introduced species:the amphipod Crangonyx floridanus and the isopods Caecidotea racovitzai and Asellus hilgendorfii. The native amphipodHyalella azteca was more abundant in pennywort and heavily preyed upon by fish, while the non-indigenous C. floridanuswas more abundant in hyacinth and not prevalent in fish diets. The introduction of hyacinth to the Delta has causedsignificant ecological alterations in the surrounding community, due to hyacinth being functionally different from nativepatches of pennywort.
Introduction
Ecological theory predicts that non-indigenousspecies that will have the most impact are thosethat represent something fundamentally new to agiven system (Simberloff 1991; Ruesink et al. 1995;Parker et al. 1999). Although this appears to bemost often the case, we sought to illuminate wheth-er an invader that was similar in appearance to acommon native could be non-equivalent in itsfunction. Floating aquatic vegetation (FAV) cano-pies of the non-indigenous water hyacinth (Eichhor-nia crassipes) in the Sacramento-San Joaquin Deltaof California are fundamentally similar to those ofthe native pennywort (Hydrocotyle umbellata), asboth form canopies of floating vegetation. Al-though hyacinth and pennywort form similarpatches of floating vegetation, it is a questionwhether or not they are functional equivalents—providing similar functions to the surrounding bi-otic community. We compared the habitat struc-
* Corresponding author: tele: 206/221-5460; fax: 206/685-7471; e-mail: [email protected].
ture, invertebrate assemblages, and diets of fish as-sociated with hyacinth and pennywort in order todetermine whether or not they were functionalequivalents.
Comparative studies of functional equivalencyare often used to test how human-altered systemscompare to natural states, such as in restored wet-lands (Simenstad and Thom 1996) or transplantedseagrass beds (Williams and Davis 1996). Specifictests of the functional role of non-indigenous spe-cies relative to natives are rare. One such test in aterrestrial system (Stromberg 1998) has shown thatthere can be some functional redundancy (West-man 1990), while other tests show significant alter-ations (Vitousek 1986). Filling such gaps in ourknowledge is important, as invasive species are of-ten difficult if not impossible to keep under con-trol (Drake and Mooney 1989), and the congruentecological impacts of specific invaders are not wellknown (Parker et al. 1999). If we can uncover suchecological effects, we can better manage and pre-dict the fates of current and future biological in-vasions.
Hyacinth is native to Brazil and has become

The Effects of Introduced Water Hyacinth 747
widespread on a global scale, first being intro-duced to the Sacramento River in California by or-namental pond enthusiasts in 1904 (Finlayson1983; Gopal 1987). Once the plants are introducedthey rapidly increase in coverage, as hyacinth hasthe highest growth rate of any saltwater, freshwater,or terrestrial vascular macrophyte (Wolverton andMcDonald 1978). In the Delta, hyacinth covered506 ha or 22% of the waterways by 1981 (Finlayson1983). Such prevalence sparked an extensivechemical control program headed by the Califor-nia Department of Boating and Waterways (CDBW1998), which actively controls coverage of hyacinthby spraying the chemical 2,4-D. As a result of theseefforts, the extensive mats found especially in trop-ical climates (Gopal 1987) were controlled duringthe course of our study, and patches of hyacinthand pennywort were similar in coverage.
In many systems hyacinth is an ecosystem engi-neer ( Jones et al. 1997; Crooks and Khim 1999),modifying the surrounding habitat by providing astructurally complex canopy. In addition to form-ing a dense floating mat, roots hanging in the wa-ter column and leaves projecting above the watersurface provide complex structures that can be in-habited by other species. Organic deposition fromthe canopy can cover the underlying benthic zone,leading to low dissolved oxygen levels (Gopal1987). Effects of these characteristics on commu-nity dynamics when hyacinth coexists with nativeFAV counterparts are unknown, and are often asubject of curiosity among investigators (Gopal1987; Masifwa et al. 2001).
FAV can be beneficial as a nursery habitat forjuvenile fishes and invertebrates (Werner and Hall1979; Gopal 1987; Schramm and Jirka 1989; Dibbleet al. 1996). Roots hanging down into the watercolumn are important as habitat for epiphytic in-vertebrates, especially amphipods (Schramm et al.1987). Epiphytic invertebrates are typically muchmore abundant than benthic and epibenthic in-vertebrates, and their densities are positively cor-related to the amount of FAV surface area availablefor colonization (Crowder and Cooper 1982;Schramm et al. 1987). Fish such as bluegills (Le-pomis macrochirus) selectively feed on epiphytic in-vertebrates over other sources of prey (Werner andHall 1979; Schramm and Jirka 1989). The effectsof hyacinth on the fish-invertebrate food web inthe Delta could be pervasive due to the promi-nence of hyacinth in shallow water areas.
Pennywort is the predominant native FAV in theDelta. Although pennywort and hyacinth bothform floating canopies and tend to occupy similarhabitats, we hypothesize that a hyacinth invasioncould alter key habitat structure components, lead-ing to surrounding community alterations. Such
effects may not be as readily apparent comparedto invaders that either occupy vacant niches andsubstantially modify the existing environment,such as introductions of fish predators when nativepiscivores are absent (Brown and Moyle 1991), ormussel invasions into mudflats previously uninhab-ited with mussel mats (Crooks and Khim 1999).Our research sought to illuminate any communityalterations caused by the hyacinth invasion.
MethodsSTUDY AREA
Our three study sites were located in the centraland western Sacramento-San Joaquin Delta of Cal-ifornia: Site A (Mandeville Tip, 38.0849N,1218329W), Site B (Brown’s Island, 38.0829N,1218529W), and Site C (Mildred Island, 37.08599N,1218319W). These sites were chosen because theycontained discrete patches of both hyacinth andpennywort. The Delta is heavily influenced by hu-man activity, including agricultural, recreational,and industrial activities (Nichols et al. 1986). His-torically, the Delta was almost all wetlands charac-terized by tule vegetation (Scirpus spp.) with somenatural levees and riparian habitat (woody vegeta-tion, mostly Salix spp.; TBI 1998). Approximately95% of these wetlands have been leveed anddrained predominantly for agricultural purposes,and the majority of the Delta’s remaining historicchannels are rip-rapped and constrained (TBI1998). Although salt water historically entered theDelta, current water management attempts to pre-vent this, mainly through controlled water flowsthat increase freshwater inflow during summermonths to maintain freshwater available for irri-gation and drinking purposes (Nichols et al. 1986).Benthic sediments in the Delta are an unconsoli-dated mixture of sand, silt, and clay (Hymanson etal. 1994). Tidal influence ranges between 1 to 2 m(CDWR 1993).
STUDY DESIGN
Healthy FAV patches not chemically controlledwith 2,4-D were sampled in areas that bordered amarsh. Habitat structure measurements and inver-tebrate samples were collected in 5 patches of bothhyacinth and pennywort at each site. Patches werechosen by a random number generator corre-sponding to the patch number along a transectdrawn parallel to shore (Fig. 1). These patches oc-cupied indentations in the marsh and were gen-erally separated by clumps of tule vegetation.Patches were fairly small and discrete (30.96 m2
average surface area, 3.92 SE) and were used onlyif they formed a fringe along the marsh, not if theyoccupied an entire channel. Since the overall sizeand shape of the patches could not be controlled

748 J. D. Toft et al.
Fig. 1. Illustration of a typical sampling design at a study site.Shallow water is light gray, tule marsh (Scirpus spp.) is hatched,H 5 hyacinth patches, and P 5 pennywort patches. Circledpatches are those randomly selected for sampling along a tran-sect south to north (patches 1, 3, 6, 7, 8 for hyacinth; 1, 4, 5,8, 10 for pennywort). Inset is the specific location of samplecollection, 1 m in from the water’s edge at the center of eachpatch.
due to the study design of sampling natural patch-es, we minimalized the natural variation by desig-nating sampling to the center of each patch, 1 min from the water’s edge (Fig. 1). We focused onthe outer edge as other studies have documentedchanges in invertebrate communities between theedge and interior of patches (Bailey and Litterick1993; Masifwa et al. 2001). Patch edges also offeredhigher association with surrounding fish and easeof access for sampling by boat. Patches of hyacinthand pennywort were not intermixed, but occasion-ally bordered each other. All patches were sampledin approximately 1 m of water, with the roots hang-ing freely halfway into the water column.
Description of the habitat structure and inver-tebrate sampling frequency is summarized in Table1. In 1998, habitat structure and epiphytic inver-tebrates were sampled in June at Site A, and in
August at Sites B and C. Epibenthic-benthic inver-tebrates and terrestrial canopy insects were sam-pled in April, June, and July at Site A and Augustat Site B. Sampling was not conducted at Site Aafter July as hyacinth patches were exterminatedwith chemical control by CDBW.
Hyacinth was absent at all three study sites in1999, although residual patches remained in inte-rior marsh channels and boat marinas in the sur-rounding area. The lack of hyacinth in 1999 wasattributed to intensive chemical spraying by CDBWduring fall and winter 1998 and spring 1999, sev-eral cold freezes during the winter that causedhigh mortality of overwintering plant material, andhigh winter outflows that flushed clumps of hya-cinth out of the Delta into San Francisco Bay. Wetook advantage of this unforeseen hyacinth remov-al experiment by sampling epiphytic invertebratesin pennywort at all three sites in June 1999. Sucha plan allowed us to measure interannual variationand to determine if the invertebrate communitychanged with the absence of hyacinth. Althoughthis was an originally unplanned segment of ourresearch, the issue of the effect of the removal ofnon-indigenous species is a vital question in bothresearch and management (Westman 1990).
PHYSICAL SAMPLING
Intensive sampling of hyacinth and pennyworthabitat structure was conducted in June 1998 atSite A and in August 1998 at Sites B and C, con-gruent with epiphytic invertebrate sampling (Table1). Entire patch surface area was estimated by mea-suring the maximum length and width of eachpatch. Leaf density was measured at the locationof sampling by counting the number of leaves in a0.5 m2 quadrat (n 5 5 site21). Roots collected forepiphytic invertebrate sampling (described below)were measured for biomass and surface area. Rootwet biomass was measured by blotting the roots drywith a towel, allowing to air dry for 10 min, andweighing to the nearest milligram. Root surfacearea was measured using the surfactant techniqueoriginally developed by Harrod and Hall (1962). Asoapy solution consisting of 60 ml of Liquinox soapin 6 l of water was mixed in a bucket and left over-night so that the bubbles dispersed. For each rootsample, five 1-g subsamples were dipped in thesoapy solution. The excess soap was shaken off sothat a monolayer of soapy solution was retainedaround the root surface. Each subsample was thenreweighed. The surface area of the root sampleswas determined by regressing the differences inweight to those measured from the same processfor known surface areas of tinfoil (25, 100, 225,400, 625, and 900 cm2; n 5 5).

The Effects of Introduced Water Hyacinth 749
TABLE 1. Invertebrate and habitat structure sampling design. Numbers are sample size, blanks indicate no sampling conducted. H5 hyacinth, P 5 pennywort. Patches of hyacinth were exterminated with chemical control at Site A (Mandeville Tip) in August 1998,after which sampling was shifted to Sites B and C (Brown’s Island and Mildred Island, respectively). Patches of hyacinth were absentat all study sites in 1999 (see text).
1998 1999
April
H P
June
H P
July
H P
August
H P
June
H P
Site A, Mandeville TipEpiphyticEpibenthic-BenthicInsectHabitat Structure
55
55
5555
5555
55
55
5
Site B, Brown’s IslandEpiphyticEpibenthic-BenthicInsectHabitat Structure
5555
5555
5
Site C, Mildred IslandEpiphyticHabitat Structure
55
55
5
BIOLOGICAL SAMPLING
Epiphytic InvertebratesEpiphytic invertebrates living in association with
the root masses of hyacinth and pennywort werecollected in June 1998 at Site A and in August 1998at Sites B and C, and in pennywort at all three sitesin June 1999 (n 5 5 site21; Table 1). Epiphytic in-vertebrates were sampled by manually collecting 1–2 plants (# 0.5 m2 quadrat described above) andimmediately placing them in a bucket (Schrammet al. 1987; Schramm and Jirka 1989). The bucketcontained a 10% isopropyl alcohol solution, caus-ing the invertebrates to detach from the root mas-ses by vigorously shaking each sample. This wasthen sieved at 0.5 mm to collect only the macro-invertebrates and fixed in a 10% buffered formal-dehyde solution. These samples were later trans-ferred to 70% isopropanol in the laboratory. Theroots from each sample were retained separately in70% isopropanol and brought to the laboratory formeasurements of surface area and biomass (de-scribed above). Any additional invertebrates thatdid not detach from the roots in the alcohol so-lution were later separated in the laboratory. Thistechnique effectively sampled invertebrates livingin direct association with the roots, as Schrammand Jirka (1989) found that most invertebrates re-mained on the roots when disturbed.
We calculated densities of epiphytic inverte-brates (number above m2 bottom; Schramm andJirka 1989) by correlating the number of leaves ineach epiphytic plant sample (# 0.5 m2) to theleaves in a 0.5 m2 quadrat (described above). In-vertebrates were counted and identified to the low-est practical taxonomic level with light microscopy.
Standing stock of invertebrates was estimated bymeasuring preserved wet biomass. Each taxonomicgroup was blotted dry and weighed to the nearest0.1 mg. Numbers and standing stock of inverte-brates were standardized to number above 1 m2
bottom, allowing for comparisons between strataand with epibenthic-benthic and insect sampling asdensity (number m22 bottom) and standing stock(g m22 bottom; Schramm and Jirka 1989). All es-timates are based on the same unit definition ofarea. Taxa richness (number of taxa) and theShannon-Wiener diversity index were also calculat-ed:
n
H9 5 2 p log pO i ii51
where pi 5 the proportion of species i in the com-munity (the # of individuals of species i/the total# of individuals in the community; Zar 1996).
Epibenthic-Benthic InvertebratesA 1-m long corer was used to sample epibenthic
and benthic invertebrates beneath hyacinth andpennywort canopies at Site A in April, June, andJuly 1998 and at Site B in August 1998 (n 5 5site21; Table 1). Cores with an internal core areaof 0.0024 m2 were taken to a 10 cm depth beneaththe sediment surface. The core was inserted intoeach patch at approximately 1 m water depth, sam-pling both epibenthic invertebrates at the water-sediment interface and benthic invertebrates inthe sediment. Samples were immediately fixed in10% buffered formaldehyde solution containingrose bengal dye, and later transferred to 70% iso-propanol in the laboratory. Invertebrates were

750 J. D. Toft et al.
counted and identified to the lowest practical tax-onomic level with light microscopy. Numbers of in-vertebrates were standardized to number m22 bot-tom, and taxa richness and the Shannon-Wienerdiversity index calculated (as above for epiphyticinvertebrates).
InsectsInsect fallout traps (Sutherland 1996) were used
to sample insects living in association with hyacinthand pennywort canopies at Site A in April, June,and July 1998 and at Site B in August 1998 (n 55 site21; Table 1). These traps consisted of a rect-angular tray (0.0782 m2) filled with approximately3 cm of soapy water. The trays were nestled intoeach canopy, so that they floated on the water sur-face with the leaves of the canopy surrounding thetray. Trays were tethered to PVC poles at each spe-cific site, allowing vertical movement with the tides.The trays were deployed for 24 h, after which thecontents were sieved at 0.106 mm and the insectspreserved in 70% isopropanol. Insects were count-ed and identified to the lowest practical taxonomiclevel with light microscopy. Numbers of insectswere standardized to number m22 bottom, andtaxa richness and the Shannon-Wiener diversity in-dex were calculated (as above for epiphytic andepibenthic-benthic invertebrates).
FishFish were sampled directly adjacent to hyacinth
and pennywort patches at Site A during June andJuly 1998 in order to see if fish surrounding FAVincorporate FAV prey into their diet. Rectangularblock-net enclosures (range 30–49 m2) were usedto collect fishes within intertidal and nearshoresubtidal areas less than 1.3 m wading depth. En-closure methods are recommended for collectingsmall and juvenile fishes because they have highcatch efficiencies and provide quantitative datauseful for comparing fish densities between habi-tats and sites (Rozas and Minello 1997). Sampleareas were delineated with 4 perimeter stakes atleast 24 h in advance to minimize disturbance ofthe area at the time of sampling. The following daythe sampling area was quickly enclosed with block-nets secured to the perimeter stakes. A minimumof 4 passes with a beach seine (7.6 3 1.2 m, 3.2mm mesh) was taken within each enclosure to col-lect the fish.
A subsample of collected fishes were preservedin 10% buffered formaldehyde solution and trans-ferred to the laboratory for diet analysis. We fo-cused on non-indigenous bluegills (L. macrochirus)as an indicator species for examining potential ef-fects of hyacinth on food web dynamics becausethey are omnivorous fish that have been shown to
feed opportunistically on invertebrates in FAV hab-itats (Werner and Hall 1979; Schramm and Jirka1989), and they accounted for 55% of the numer-ical fish catch in preliminary sampling directly un-derneath hyacinth (n 5 5 patches sampled) andwere one of the most abundant fish collected inall fish sampling.
Fish were also sampled in many areas surround-ing the study sites using the same methods asabove. Additional fish species from these areaswere saved for diet analysis to see if prey itemsfound in FAV habitats were incorporated at a larg-er scale into the Delta food web.
Fish saved for diet analysis were measured (forklength) and weighed (preserved wet weight). Thestomachs were then dissected and the gut contentsremoved. Overall gut contents were blotted dryand weighed. Taxa were then separated and iden-tified to the lowest possible taxonomic level withlight microscopy. Each taxonomic group was thencounted, blotted dry, and weighed. Levels of stom-ach content digestion and stomach fullness wereranked using a standardized system (Terry 1977).Prey items were then ranked based on modifiedIndex of Relative Importance values (IRI; Pinkas etal. 1971; Simenstad et al. 1991):
IRI 5 % frequency of occurrence
3 [% numerical composition
1 % gravimetric composition]
Diet overlap with sampled aquatic prey resourceswas calculated using a modified Percent SimilarityIndex (PSI; Hurlbert 1978):
n
PSI 5 minimum(p , p )O xi yii51
where pxi 5 percentage of prey i in predator x andpyi 5 percentage of prey i in the sampled aquaticprey resources y. Ivlev’s electivity index was used tocompare differences in fish selectivity on the majoramphipod taxa present in the sampled prey re-sources (Ivlev 1961). The Ivlev electivity index waschosen as it is easy to interpret and gives similarresults to other indices (Lechowicz 1982; Kline andWood 1996).
DATA INTERPRETATION AND STATISTICAL TESTS
Parametric two-sample t-tests were used to statis-tically compare means for biological and physicalsampling between hyacinth and pennywort at eachstudy site (a 5 0.05). For all measurements be-tween hyacinth and pennywort Ho: m1 5 m2 and Ha:m1 ± m2. The parametric t-test is a robust statistic,meaning it can withstand considerable departuresfrom its underlying assumptions of normality and
The Effects of Introduced Water Hyacinth 751
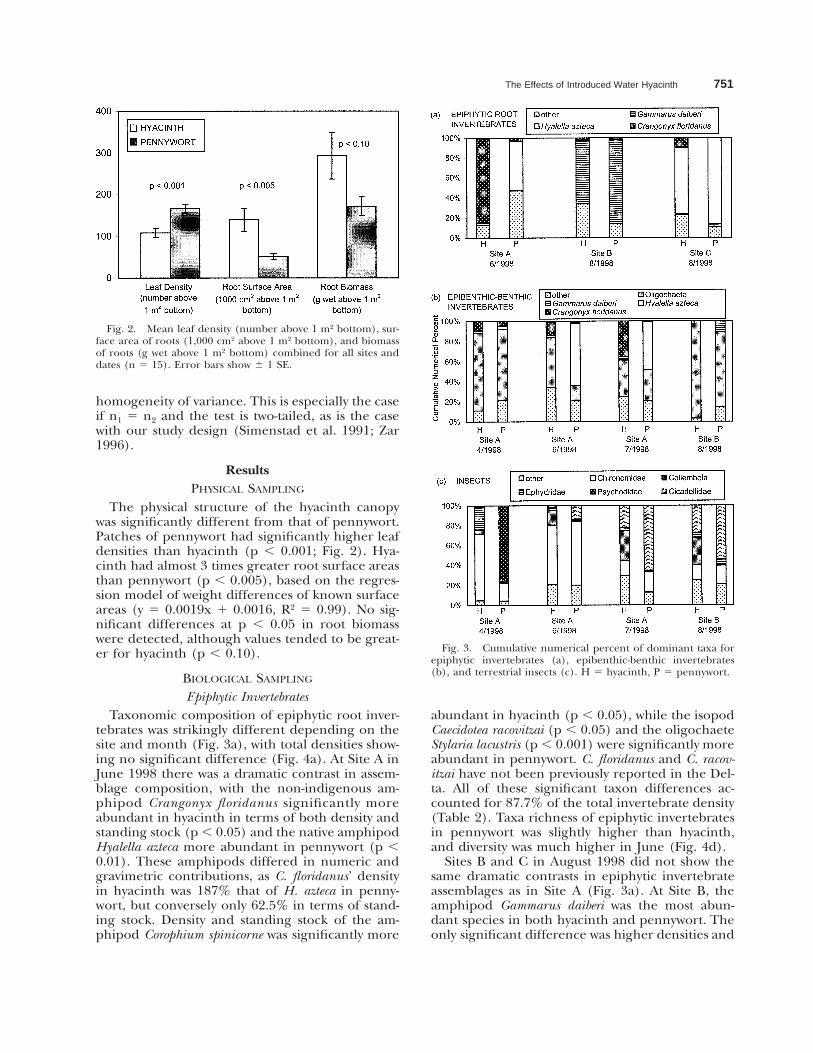
Fig. 2. Mean leaf density (number above 1 m2 bottom), sur-face area of roots (1,000 cm2 above 1 m2 bottom), and biomassof roots (g wet above 1 m2 bottom) combined for all sites anddates (n 5 15). Error bars show 6 1 SE.
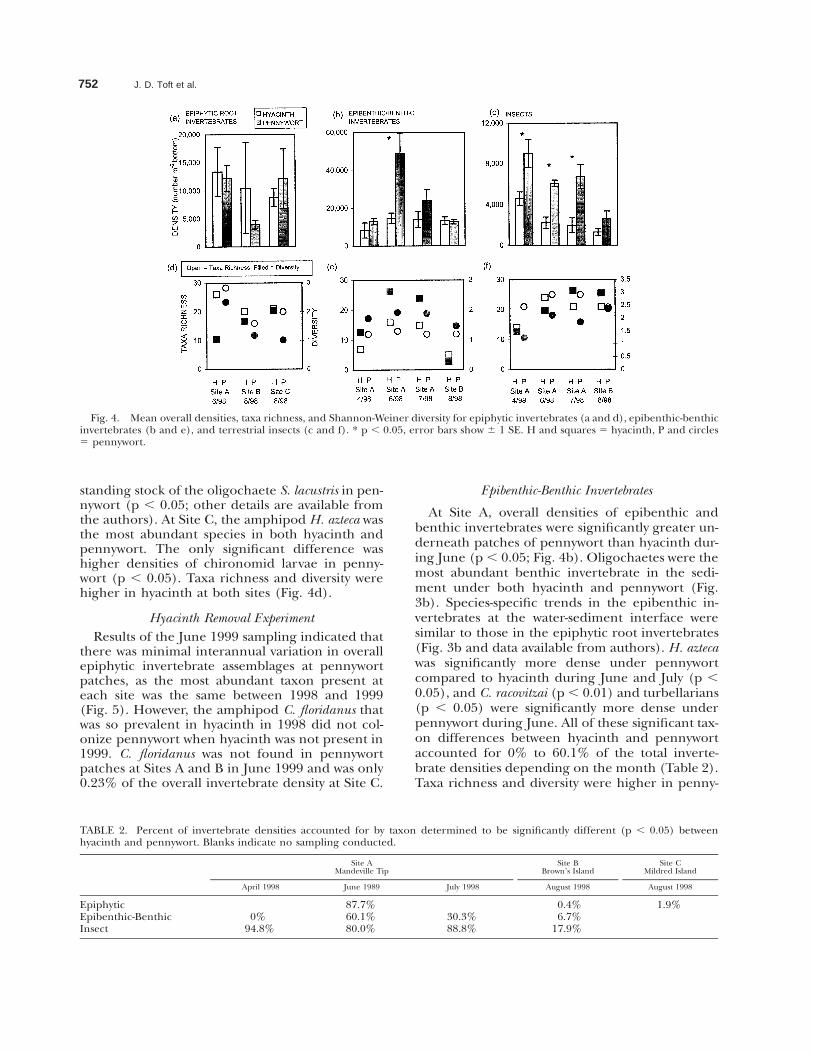
Fig. 3. Cumulative numerical percent of dominant taxa forepiphytic invertebrates (a), epibenthic-benthic invertebrates(b), and terrestrial insects (c). H 5 hyacinth, P 5 pennywort.
homogeneity of variance. This is especially the caseif n1 5 n2 and the test is two-tailed, as is the casewith our study design (Simenstad et al. 1991; Zar1996).
ResultsPHYSICAL SAMPLING
The physical structure of the hyacinth canopywas significantly different from that of pennywort.Patches of pennywort had significantly higher leafdensities than hyacinth (p , 0.001; Fig. 2). Hya-cinth had almost 3 times greater root surface areasthan pennywort (p , 0.005), based on the regres-sion model of weight differences of known surfaceareas (y 5 0.0019x 1 0.0016, R2 5 0.99). No sig-nificant differences at p , 0.05 in root biomasswere detected, although values tended to be great-er for hyacinth (p , 0.10).
BIOLOGICAL SAMPLING
Epiphytic InvertebratesTaxonomic composition of epiphytic root inver-
tebrates was strikingly different depending on thesite and month (Fig. 3a), with total densities show-ing no significant difference (Fig. 4a). At Site A inJune 1998 there was a dramatic contrast in assem-blage composition, with the non-indigenous am-phipod Crangonyx floridanus significantly moreabundant in hyacinth in terms of both density andstanding stock (p , 0.05) and the native amphipodHyalella azteca more abundant in pennywort (p ,0.01). These amphipods differed in numeric andgravimetric contributions, as C. floridanus’ densityin hyacinth was 187% that of H. azteca in penny-wort, but conversely only 62.5% in terms of stand-ing stock. Density and standing stock of the am-phipod Corophium spinicorne was significantly more
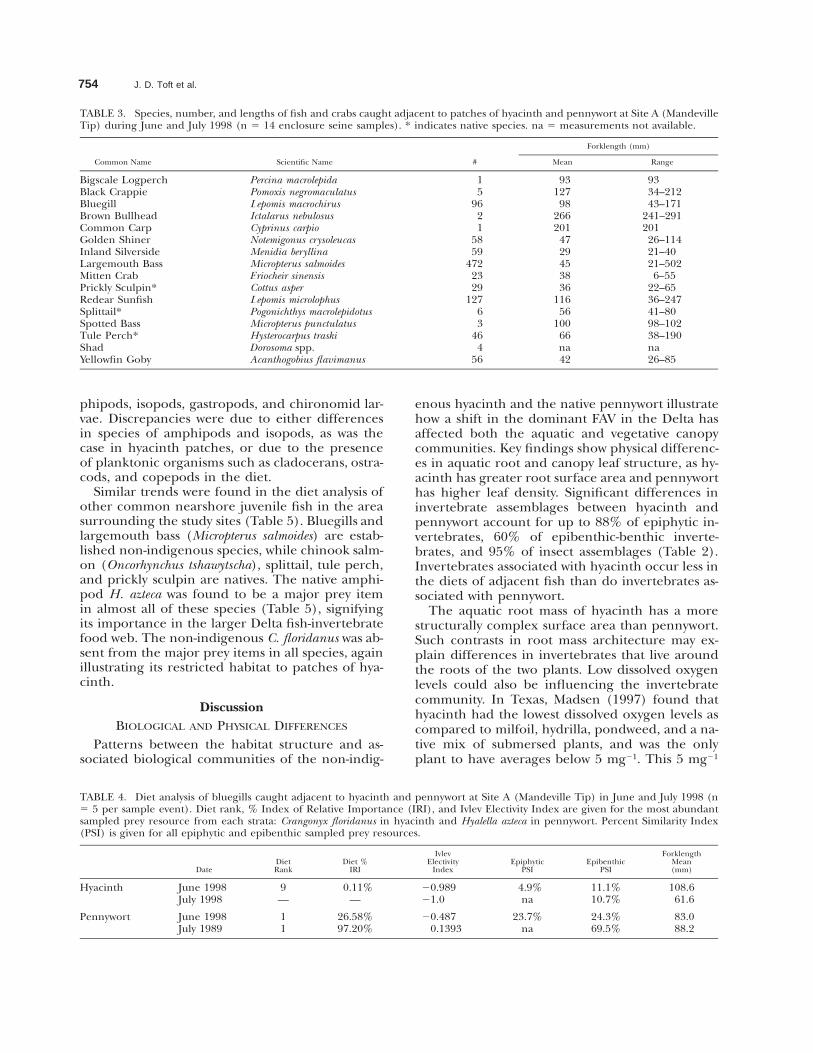
abundant in hyacinth (p , 0.05), while the isopodCaecidotea racovitzai (p , 0.05) and the oligochaeteStylaria lacustris (p , 0.001) were significantly moreabundant in pennywort. C. floridanus and C. racov-itzai have not been previously reported in the Del-ta. All of these significant taxon differences ac-counted for 87.7% of the total invertebrate density(Table 2). Taxa richness of epiphytic invertebratesin pennywort was slightly higher than hyacinth,and diversity was much higher in June (Fig. 4d).
Sites B and C in August 1998 did not show thesame dramatic contrasts in epiphytic invertebrateassemblages as in Site A (Fig. 3a). At Site B, theamphipod Gammarus daiberi was the most abun-dant species in both hyacinth and pennywort. Theonly significant difference was higher densities and
752 J. D. Toft et al.
Fig. 4. Mean overall densities, taxa richness, and Shannon-Weiner diversity for epiphytic invertebrates (a and d), epibenthic-benthicinvertebrates (b and e), and terrestrial insects (c and f). * p , 0.05, error bars show 6 1 SE. H and squares 5 hyacinth, P and circles5 pennywort.
TABLE 2. Percent of invertebrate densities accounted for by taxon determined to be significantly different (p , 0.05) betweenhyacinth and pennywort. Blanks indicate no sampling conducted.
Site AMandeville Tip
April 1998 June 1989 July 1998
Site BBrown’s Island
August 1998
Site CMildred Island
August 1998
EpiphyticEpibenthic-BenthicInsect
0%94.8%
87.7%60.1%80.0%
30.3%88.8%
0.4%6.7%
17.9%
1.9%
standing stock of the oligochaete S. lacustris in pen-nywort (p , 0.05; other details are available fromthe authors). At Site C, the amphipod H. azteca wasthe most abundant species in both hyacinth andpennywort. The only significant difference washigher densities of chironomid larvae in penny-wort (p , 0.05). Taxa richness and diversity werehigher in hyacinth at both sites (Fig. 4d).
Hyacinth Removal ExperimentResults of the June 1999 sampling indicated that
there was minimal interannual variation in overallepiphytic invertebrate assemblages at pennywortpatches, as the most abundant taxon present ateach site was the same between 1998 and 1999(Fig. 5). However, the amphipod C. floridanus thatwas so prevalent in hyacinth in 1998 did not col-onize pennywort when hyacinth was not present in1999. C. floridanus was not found in pennywortpatches at Sites A and B in June 1999 and was only0.23% of the overall invertebrate density at Site C.
Epibenthic-Benthic Invertebrates
At Site A, overall densities of epibenthic andbenthic invertebrates were significantly greater un-derneath patches of pennywort than hyacinth dur-ing June (p , 0.05; Fig. 4b). Oligochaetes were themost abundant benthic invertebrate in the sedi-ment under both hyacinth and pennywort (Fig.3b). Species-specific trends in the epibenthic in-vertebrates at the water-sediment interface weresimilar to those in the epiphytic root invertebrates(Fig. 3b and data available from authors). H. aztecawas significantly more dense under pennywortcompared to hyacinth during June and July (p ,0.05), and C. racovitzai (p , 0.01) and turbellarians(p , 0.05) were significantly more dense underpennywort during June. All of these significant tax-on differences between hyacinth and pennywortaccounted for 0% to 60.1% of the total inverte-brate densities depending on the month (Table 2).Taxa richness and diversity were higher in penny-

The Effects of Introduced Water Hyacinth 753
Fig. 5. Cumulative numerical percent of dominant epiphyticinvertebrates at pennywort in 1998 and 1999.
wort in April, but higher in hyacinth in June andJuly (Fig. 4e).
At Site B overall densities were almost equal inhyacinth and pennywort during August 1998 (Fig.4b). Oligochaetes were once again the most abun-dant benthic invertebrate in the sediment (Fig.3b). The most striking difference was that therewere no amphipods or isopods underneath hya-cinth patches at all, so overall densities of amphi-pods and isopods were significantly higher underpennywort (p , 0.015; Fig. 3b). As with the epi-phytic invertebrates, G. daiberi was the major epi-benthic taxon under pennywort. Taxa richness anddiversity was much higher in pennywort due to hy-acinth being devoid of amphipods and isopods(Fig. 4e).
Insects
Overall densities of canopy insects were signifi-cantly greater in pennywort than hyacinth at SiteA in April (p , 0.05), June (p , 0.0005), and July(p , 0.05; Fig. 4c). Taxonomic composition wasmarkedly different as well (Fig. 3c and data avail-able from authors). In April, Ephydridae were sig-nificantly more abundant in hyacinth than penny-wort (p , 0.05), and Psychodidae were signifi-cantly more abundant in pennywort (p , 0.005).These insects both decreased in later months,when Collembola became significantly more abun-dant in hyacinth in July (p , 0.05) and Cicadelli-dae significantly more abundant in pennywort inJune (p , 0.001) and July (p , 0.0005). Chiron-omidae were significantly more abundant in hya-cinth in April (p , 0.01), but were significantlymore abundant in pennywort in June (p , 0.001)and July (p , 0.0005). Eleven less abundant taxa
also had significant density differences between hy-acinth and pennywort (data available from au-thors). The combined significant taxa differencesbetween hyacinth and pennywort accounted for80% to 94.8% of the total insect densities depend-ing on the month (Table 2). Taxa richness washigher in pennywort, while diversity was higher inhyacinth throughout all months (Fig. 4f).
At Site B, overall insect densities were not statis-tically greater in pennywort than hyacinth in Au-gust 1998 (p , 0.15; Fig. 4c). Taxonomic assem-blages were similar to those found at Site A in July1998 (Fig. 3c), although the only significant differ-ences were greater densities of Chironomidae (p, 0.05) and Araneae (p , 0.05) in pennywort thanhyacinth (data available from authors). Taxa rich-ness and diversity were both higher in hyacinth(Fig. 4f).
Fish Assemblages and Diet AnalysisMost of the fish captured adjacent to patches of
hyacinth and pennywort at Site A in June and July1998 were juveniles and non-indigenous to the Del-ta (Table 3). The native splittail (Pogonichthys ma-crolepidotus), tule perch (Hysterocarpus traski), andprickly sculpin (Cottus asper) accounted for only8.2% of the numerical catch. Also captured in lowabundance was the non-indigenous mitten crab(Eriocheir sinensis).
Bluegill feeding differed between fish caught ad-jacent to hyacinth and pennywort (Table 4). Basedon percent total importance (IRI), the predomi-nant prey item for bluegills adjacent to pennywortin both June and July was the native amphipod H.azteca, which was also the most common epiphyticand epibenthic invertebrate found in those pen-nywort patches. This was not the case in hyacinth,where the dominant fish prey items were not thesame as the most abundant aquatic invertebrates.In hyacinth, the non-indigenous amphipod C. flor-idanus was the major epiphytic and epibenthic in-vertebrate, but was only the ninth ranked prey itemin June and was not in the prey items at all in July(Table 4). Lower Ivlev electivity index values for C.floridanus in hyacinth compared to H. azteca in pen-nywort also reflect differences in preference forthese two major prey resources (Table 4).
Similarity (PSI) between fish prey items and sam-pled invertebrates was also higher in pennywort(Table 4). Similarity in pennywort was 23.7% forepiphytic and 24.3% for epibenthic invertebratesduring June, and 69.5% for epibenthic inverte-brates in July. Similarity in hyacinth was 4.9% forepiphytic and 11.1% for epibenthic invertebratesduring June, and 10.7% for epibenthic inverte-brates in July. Most of the overlap between inver-tebrate prey items and resources was due to am-
754 J. D. Toft et al.
TABLE 3. Species, number, and lengths of fish and crabs caught adjacent to patches of hyacinth and pennywort at Site A (MandevilleTip) during June and July 1998 (n 5 14 enclosure seine samples). * indicates native species. na 5 measurements not available.
Common Name Scientific Name #
Forklength (mm)
Mean Range
Bigscale LogperchBlack CrappieBluegillBrown BullheadCommon CarpGolden ShinerInland Silverside
Percina macrolepidaPomoxis negromaculatusLepomis macrochirusIctalarus nebulosusCyprinus carpioNotemigonus crysoleucasMenidia beryllina
15
9621
5859
9312798
2662014729
9334–21243–171
241–29120126–11421–40
Largemouth BassMitten CrabPrickly Sculpin*Redear SunfishSplittail*Spotted BassTule Perch*ShadYellowfin Goby
Micropterus salmoidesEriocheir sinensisCottus asperLepomis microlophusPogonichthys macrolepidotusMicropterus punctulatusHysterocarpus traskiDorosoma spp.Acanthogobius flavimanus
4722329
12763
464
56
453836
11656
10066na42
21–5026–55
22–6536–24741–8098–10238–190na26–85
TABLE 4. Diet analysis of bluegills caught adjacent to hyacinth and pennywort at Site A (Mandeville Tip) in June and July 1998 (n5 5 per sample event). Diet rank, % Index of Relative Importance (IRI), and Ivlev Electivity Index are given for the most abundantsampled prey resource from each strata: Crangonyx floridanus in hyacinth and Hyalella azteca in pennywort. Percent Similarity Index(PSI) is given for all epiphytic and epibenthic sampled prey resources.
DateDietRank
Diet %IRI
IvlevElectivity
IndexEpiphytic
PSIEpibenthic
PSI
ForklengthMean(mm)
Hyacinth June 1998July 1998
9—
0.11%—
20.98921.0
4.9%na
11.1%10.7%
108.661.6
Pennywort June 1998July 1989
11
26.58%97.20%
20.4870.1393
23.7%na
24.3%69.5%
83.088.2
phipods, isopods, gastropods, and chironomid lar-vae. Discrepancies were due to either differencesin species of amphipods and isopods, as was thecase in hyacinth patches, or due to the presenceof planktonic organisms such as cladocerans, ostra-cods, and copepods in the diet.
Similar trends were found in the diet analysis ofother common nearshore juvenile fish in the areasurrounding the study sites (Table 5). Bluegills andlargemouth bass (Micropterus salmoides) are estab-lished non-indigenous species, while chinook salm-on (Oncorhynchus tshawytscha), splittail, tule perch,and prickly sculpin are natives. The native amphi-pod H. azteca was found to be a major prey itemin almost all of these species (Table 5), signifyingits importance in the larger Delta fish-invertebratefood web. The non-indigenous C. floridanus was ab-sent from the major prey items in all species, againillustrating its restricted habitat to patches of hya-cinth.
DiscussionBIOLOGICAL AND PHYSICAL DIFFERENCES
Patterns between the habitat structure and as-sociated biological communities of the non-indig-
enous hyacinth and the native pennywort illustratehow a shift in the dominant FAV in the Delta hasaffected both the aquatic and vegetative canopycommunities. Key findings show physical differenc-es in aquatic root and canopy leaf structure, as hy-acinth has greater root surface area and pennyworthas higher leaf density. Significant differences ininvertebrate assemblages between hyacinth andpennywort account for up to 88% of epiphytic in-vertebrates, 60% of epibenthic-benthic inverte-brates, and 95% of insect assemblages (Table 2).Invertebrates associated with hyacinth occur less inthe diets of adjacent fish than do invertebrates as-sociated with pennywort.
The aquatic root mass of hyacinth has a morestructurally complex surface area than pennywort.Such contrasts in root mass architecture may ex-plain differences in invertebrates that live aroundthe roots of the two plants. Low dissolved oxygenlevels could also be influencing the invertebratecommunity. In Texas, Madsen (1997) found thathyacinth had the lowest dissolved oxygen levels ascompared to milfoil, hydrilla, pondweed, and a na-tive mix of submersed plants, and was the onlyplant to have averages below 5 mg21. This 5 mg21

The Effects of Introduced Water Hyacinth 755
TABLE 5. Percent Index of Relative Importance (IRI) of Hy-alella azteca and Crangonyx floridanus in diets of common near-shore juvenile fish in the area surrounding the study sites.
Species% IRI ofH. azteca
% IRI ofC.
floridanusSample
Size
ForklengthMean(mm)
BluegillChinook SalmonLargemouth BassPrickly SculpinSplittailTule Perch
27.8515.0419.3867.135.60
22.47
0.015—
0.024———
902950162130
78.646.196.340.251.960.2
separation is notable because it represents the levelat which many fish start to experience oxygenstress (Madsen 1997). Lower levels of dissolved ox-ygen were likely the reason that we found no epi-benthic amphipods and isopods beneath the hya-cinth canopy at Site B in August 1998. These hya-cinth patches did have an abundance of amphi-pods and isopods living epiphytically amongst theroots that suggests that these invertebrates couldpotentially be taking refuge in the root mass fromunderlying hypoxia (Gopal 1987). Pennywort atthe same site did have amphipods and isopods liv-ing both epibenthically underneath the canopiesand epiphytically among the roots. Overall densi-ties of epibenthic and benthic invertebrates weregenerally greater in pennywort than hyacinththroughout all sampling.
Patterns of taxa richness and diversity for allaquatic invertebrates tended to follow a seasonaltrend. Both taxa richness and Shannon-Weiner di-versity indices were higher in pennywort duringthe first month of sampling ( June for epiphytic,April for epibenthic-benthic), but were higher inhyacinth for subsequent months (August for epi-phytic, June and July for epibenthic-benthic; Fig.4d,e). The exception was higher richness and di-versity for epibenthic-benthic invertebrates in pen-nywort at Site B in August, but as discussed abovethis is likely due to hyacinth being almost devoidof epibenthic invertebrates, presumably caused bylow dissolved oxygen levels. Higher richness anddiversity in hyacinth later in the season could berelated to its extensive growth rate, providing morecolonizable substrate later in the year (Gopal1987). In Florida, Jantrarotai (1990) found thatoverall dry biomass of hyacinth was 161% greaterthan pennywort (Hydrocotyle ranunculodies), andmaximum root length 164% greater, maximized inlate summer.
Pennywort patches have more leaves per unitarea than hyacinth. The structurally dense canopymay explain the greater overall density of insectsin pennywort as well as taxonomic differences ofthe insect assemblages. Pennywort was also richest
in taxa throughout all sampling except for Augustat Site B, and was lower in diversity throughout allmonths (Fig. 4f).
Amphipods clearly dominate aquatic inverte-brate assemblages in FAV and are important fishprey. Numerous studies have shown that amphi-pods such as H. azteca are vulnerable to bluegillpredation (Crowder and Cooper 1982; Schrammand Jirka 1989). Amphipods were found to be pro-portionally more abundant in the FAV canopiesthan in the fish diets, presumably due to the refugeprovided by the root mass structure. This agreeswith results from Florida lakes, where amphipodswere most abundant epiphytically and less abun-dant benthically and in fish diets (Schramm andJirka 1989). The same study also found that hya-cinth roots provided a refuge from fish predationfor H. azteca, with H. azteca accounting for 69% to86% of invertebrate densities (Schramm et al.1987).
The major difference in amphipods between hy-acinth and pennywort was the prevalence of thenon-indigenous C. floridanus in hyacinth. C. flori-danus was reduced to barely detectable levels withthe absence of hyacinth in 1999, lending evenmore credence that C. floridanus is preferentiallyassociated with hyacinth. This also suggests thatonce an invader is removed from a system, aspectsof the community can return to a more naturalpre-invasion state (Westman 1990).
IRI and Ivlev electivity index values indicate thatC. floridanus is not abundant in fish diets (Tables3 and 4). This is in contrast to the native H. azteca,which was significantly more abundant in penny-wort at Site A and a common prey in fish diets. C.floridanus may not be abundant in fish diets for avariety of reasons, including refuge function of hy-acinth roots from fish predation, low caloric valueand small size of C. floridanus, and poor taste of C.floridanus. Refuge function is the most likely con-tribution to its low abundance in fish diets, givenresearch in Florida showing that hyacinth roots canprovide a refuge for invertebrates from fish pre-dation (Schramm and Jirka 1989). We consider lowcaloric value to also be influential, as the standingstock of one C. floridanus is 0.343 mg, and that ofone H. azteca is 1.056 mg (calculated from valuesin authors’ data). C. floridanus has 33.43% lessgravimetric value than H. azteca. We consider poortaste not to be much of an influence, as it is notlikely that C. floridanus is unpalatable to fish. Workhas not been done specific to C. floridanus, but re-search has shown that a close congener Crangonyxrichmondensis is eaten by chum, chinook, and sock-eye salmon fry in freshwater tidal creeks of the low-er Fraser River in British Columbia (Levings et al.1995).

756 J. D. Toft et al.
INTRODUCED SPECIES THEORY: FUNCTIONAL
EQUIVALENCY
For a variety of reasons, it is often difficult todetermine ecological effects of non-indigenousspecies on native ecosystems (Drake and Mooney1989; Parker et al. 1999). Sufficient monitoring isusually not available to document changes causedby a invading organism. By comparing the com-munity dominated by an exotic organism to thatof a native counterpart, it is possible to illuminatechanges that may have arisen due to the establish-ment of that exotic organism. This is especially use-ful in many cases such as ours when pre-invasiondata are not available.
Our research has shown that hyacinth can becharacterized by a distinctly different invertebrateassemblage and fish-invertebrate food web as com-pared to its native counterpart pennywort. Al-though floating canopies of hyacinth do not rep-resent a fundamentally new habitat type in the Del-ta, they have proven on most counts not to be func-tionally equivalent to native floating canopies ofpennywort. Some functional redundancy in the in-vertebrate assemblages does exist (Westman 1990;Stromberg 1998), notably in the epiphytic inver-tebrates at Sites B and C (Table 2). The 87.7%significant taxon difference in epiphytic assemblag-es at Site A was also reflected into the surroundingfish-invertebrate food web (Tables 3 and 4). Dif-ferences in insect assemblages were consistentthroughout monthly sampling, ranging between80.0–94.8% in the April, June, and July samplingsat Site A, while epibenthic-benthic assemblageswere more variable, ranging between 0.4–60.1%(Table 2). Although most measurements signifynon-functional equivalency between hyacinth andpennywort, some redundancy and natural variationdoes exist as there is some overlap in measuredvalues.
Detrimental impacts are much more obviouswhen an invader either occupies a vacant niche,such as introduced pikeminnow (Ptychocheilus gran-dis) filling a top fish predator role when native pis-civores are absent (Brown and Moyle 1991), or insome way modifies the existing habitat, such asMusculista senhousia converting mudflats to musselbeds (Simberloff 1991; Ruesink et al. 1995; Crooksand Khim 1999; Parker et al. 1999). Our researchhas shown that invaders can have significant eco-logical impacts even when there are similar pre-existing native species. In the case of hyacinth, cer-tain structural elements and associated inverte-brates and fish-feeding proved to be different fromthe native pennywort. It remains to be seen wheth-er this holds true only for invaders such as hya-
cinth that provide structurally complex substrateon which other organisms can inhabit.
It is important that patches of the native pen-nywort were abundant enough to provide a suit-able comparison, although it is unknown whetheror not pennywort has decreased in abundance viacompetitive exclusion since the invasion of hya-cinth. Since hyacinth remains firmly established inthe Delta despite an extensive chemical controlprogram, it will be critical to monitor abundanceof hyacinth and pennywort and their associatedcommunities in future years. Community dynamicswould undoubtedly change if hyacinth once againdeveloped extensive mats, as it does more often intropical climates (Gopal 1987).
Hyacinth in the Delta has followed a predictablepathway of plant invasion theory—once natural en-vironmental constraints in the area were lifted dueto an altered hydrological regime caused by damsand channelization, the invasive hyacinth flour-ished (Finlayson 1983; Barret 1989; Galatowitsch etal. 1999). Although disturbance is not required fora successful hyacinth invasion, such a combinationof appropriate abiotic and biotic factors often turnhyacinth into a perfect invader (Ashton and Mitch-ell 1989). Hyacinth is one of the most influentialinvaders in the Delta in terms of recent controlefforts, due to its extensive blocking of waterways.During 1998–1999, 985 ha of hyacinth were chem-ically treated with an annual budget of approxi-mately $1,000,000 (CDBW 1998). The weevils Neo-chetina bruchi and N. eichhorniae and the moth Sa-maodes albiguttalis were also imported and releasedas biological control agents in 1982 and 1983, butwithout much success (USCOE 1985). Combinedwith our ecological findings, the detrimental ef-fects of hyacinth add to those of other prominentaquatic invaders in the Delta, such as the sub-merged aquatic plant Egeria densa, the clam Corbic-ula fluminea, the Chinese mitten crab Eriocheir si-nensis, the Asian copepod Pseudodiaptomus forbesi,and numerous species of non-indigenous fish (Hy-manson et al. 1994; Cohen and Carlton 1995).
It is noteworthy that in the process of studyingone non-indigenous species, three more were dis-covered—the amphipod C. floridanus and the iso-pods C. racovitzai and Asellus hilgendorfii (Table 6;Magniez and Toft 2000; Toft et al. 2002). Such newdiscoveries of non-indigenous species are not over-ly surprising, given the extent that the San Fran-cisco Bay-Delta ecosystem has been anthropogeni-cally modified and colonized by invasive species(Nichols et al. 1986; Cohen and Carlton 1998).This sheds light on the need for more diligent re-search—for example, A. hilgendorfii has specificallybeen shown to serve as an intermediate host fornumerous species of acanthocephalans that para-

The Effects of Introduced Water Hyacinth 757
TABLE 6. Native range and non-indigenous populations of theamphipod Crangonyx floridanus and the isopods Caecidotea raco-vitzai and Asellus hilgendorfii. See Toft et al. (2002) for a moredetailed discussion of non-indigenous status.
SpeciesDescribed
Native RangeNon-Indigenous
Populations
C. floridanus Eastern and east-cen-tral U.S. (Holsinger1972; Zhang 1997)
Colorado andOregon, U.S., andJapan (Zhang1997), California(this study)
C. racovitzai Northeastern U.S.and southeasternCanada, Florida,and Georgia, U.S.(Williams 1970)
Washington andUtah, U.S. (Wil-liams 1970, Bow-man 1974), Califor-nia (this study)
A. hilgendorfii Eastern Siberia, Chi-na, and Japan(Henry and Mag-niez 1995)
California (this study;Magniez and Toft2000)
sitize salmonids and other fish in its native watersof Japan (Nagasawa et al. 1983). It is possible thathyacinth may have been the vector of introductionfor these crustaceans, and facilitated their inva-sions as a function of its habitat-providing charac-teristics. As a general test of this vector, we ordered4 hyacinth plants from an ornamental pond supplycompany, and they arrived teeming with live inver-tebrates including many amphipods (Hyalellaspp.). Hyacinth does seem like a viable vector ofintroduction especially for C. floridanus due to itsprevalence in the hyacinth community, althoughother methods of introduction are certainly avail-able (Cohen and Carlton 1998). The discovery ofthese three new species adds to the already lengthylist of 84 documented non-indigenous species inthe Delta, and keeps pace with calculations thatone new invasive species is currently established ev-ery 14 wk in this system (Cohen and Carlton 1998).Given that many of these species may already bewell established by the time they are discovered,future research should focus not only on docu-menting their occurrence but also on assessingtheir ecological roles in the surrounding commu-nity.
ACKNOWLEDGMENTS
Financial support was provided by a California Federal Bay/Delta Program Category III grant through the Metropolitan Wa-ter District of Southern California, contract number 14318.Wayne Fields, John Holsinger, Guy Magniez, Doug Smith, No-boru Nunomura, and Julian J. Lewis provided valuable inverte-brate taxonomy guidance. Special thanks to David Armstrong,Christian Grue, Daniel Schindler, Marjorie Wonham, SarahMorley, and two reviewers for commenting on previous versionsof this manuscript. Field and laboratory assistance was providedby Heather Higgins, Peter Moyle, Scott Matern, Zach Hyman-son, Rob Miller, Chris Peregrin, Eric Santos, Denise Reed, Greg
Cain, Kemper Kurowski, and other members of the WetlandEcosystem Team (University of Washington).
LITERATURE CITED
ASHTON, P. J. AND D. S. MITCHELL. 1989. Aquatic plants: Patternsand modes of invasion, attributes of invading species and as-sessment of control programmes, p. 111–154. In J. A. Drakeand H. A. Mooney (eds.), Biological Invasions: A Global Per-spective. Scope 37. John Wiley and Sons, Chichester, NewYork.
BAILEY, R. G. AND M. R. LITTERICK. 1993. The macroinvertebratefauna of water hyacinth fringes in the Sudd swamps (RiverNile, southern Sudan). Hydrobiologia 250:97–103.
BARRET, S. C. H. 1989. Waterweed invasions. Scientific American261:90–97.
BOWMAN, T. E. 1974. The Californian freshwater isopod, Asellustomalensis, rediscovered and compared with Asellus occidentalis.Hydrobiologia 44:431–441.
BROWN, L. R. AND P. B. MOYLE. 1991. Changes in habitat andmicrohabitat partitioning within an assemblage of stream fish-es in response to predation by Sacramento squawfish (Ptych-ocheilus grandis). Canadian Journal of Fisheries and Aquatic Sci-ences 48:849–856.
CALIFORNIA DEPARTMENT OF BOATING AND WATERWAYS (CDBW).1998. 1998 Summary of Operations Waterhyacinth ChemicalControl Program. CDBW, Sacramento, California.
CALIFORNIA DEPARTMENT OF WATER RESOURCES (CDWR). 1993.The Sacramento/San Joaquin Delta Atlas. CDWR, Sacramen-to, California.
COHEN, A. N. AND J. T. CARLTON. 1995. Nonindigenous aquaticspecies in a United States estuary: A case study of the biolog-ical invasions of the San Francisco Bay and Delta. A reportfor the U.S. Fish and Wildlife Service and the National SeaGrant College Program. University of California at Berkeley,Williams College-Mystic Seaport, Berkeley, California.
COHEN, A. N. AND J. T. CARLTON. 1998. Accelerating invasionrate in a highly invaded estuary. Science 279:555–558.
CROOKS, J. A. AND H. S. KHIM. 1999. Architectural vs. biologicaleffects of a habitat-altering, exotic mussel, Musculista senhou-sia. Journal of Experimental Marine Biology and Ecology 240:53–75.
CROWDER, L. B. AND W. E. COOPER. 1982. Habitat structural com-plexity and the interaction between bluegills and their prey.Ecology 63:1802–1813.
DIBBLE, E. D., K. J. KILLGORE, AND G. O. DICK. 1996. Measure-ments of plant architecture in seven aquatic plants. Journal ofFreshwater Ecology 11:311–318.
DRAKE, J. A. AND H. A. MOONEY (EDS.). 1989. Biological Inva-sions: A Global Perspective. Scope 37. John Wiley and Sons,Chichester, New York.
FINLAYSON, B. J. 1983. Water hyacinth: Threat to the Delta? Out-door California 44:10–14.
GALATOWITSCH, S. M., N. O. ANDERSON, AND P. D. ASCHER. 1999.Invasiveness in wetland plants in temperate North America.Wetlands 19:733–755.
GOPAL, B. 1987. Aquatic Plant Studies 1. Water Hyacinth. Elsev-ier Publishing, New York.
HARROD, J. J. AND R. E. HALL. 1962. A method for determiningthe surface areas of various aquatic plants. Hydrobiologia 20:173–178.
HENRY, J. P. AND G. MAGNIEZ. 1995. Nouvelles donnees sur lesAsellidae epiges d’Extreme-Orient (Crustacea, Isopoda, Asel-lota). Contributions to Zoology 65:101–122.
HOLSINGER, J. R. 1972. Biota of Freshwater Systems, Identifica-tion Manual No. 5. The Freshwater amphipod crustaceans(Gammaridae) of North America. Environmental ProtectionAgency, Washington, D.C.
HURLBERT, S. H. 1978. The measurement of niche overlap andsome relatives. Ecology 59:67–77.

758 J. D. Toft et al.
HYMANSON, Z., D. MAYER, AND J. STEINBECK. 1994. Long-termtrends in benthos abundance and persistence in the upperSacramento-San Joaquin estuary, summary report: 1980–1990.Technical Report 38 (FWQ/BIO/IATR/94-38). InteragencyEcological Program for the San Francisco Bay/Delta Estuary,California Department of Water Resources, Sacramento, Cal-ifornia.
IVLEV, V. S. 1961. Experimental Ecology of the Feeding of Fish-es. Translated from Russian. Yale University Press, New Ha-ven, Connecticut.
JANTRAROTAI, P. 1990. Water hyacinth (Eichhornia crassipes) andwater pennywort (Hydrocotyle ranunculodies) use in treatmentof poultry wastewater. Ph.D. Dissertation, Auburn University,Auburn, Alabama.
JONES, C. G., J. H. LAWTON, AND M. CHACHAK. 1997. Positive andnegative effects of organisms as physical ecosystem engineers.Ecology 78:1946–1957.
KLINE, J. L. AND B. M. WOOD. 1996. Food habitats and diet se-lectivity of the brown bullhead. Journal of Freshwater Ecology 11:145–151.
LECHOWICZ, M. J. 1982. The sampling characteristics of electivityindices. Oecologia 52:22–30.
LEVINGS, C. D., D. E. BOYLE, AND T. R. WHITEHOUSE. 1995. Dis-tribution and feeding of juvenile Pacific salmon in freshwatertidal creeks of the lower Fraser River, British Columbia. Fish-eries Management and Ecology 2:299–308.
MADSEN, J. D. 1997. Methods for management of nonindige-nous aquatic plants, p 145–171. In J. O. Luken and J. W.Thieret (eds.), Assessment and Management of Plant Inva-sions. Springer, New York.
MAGNIEZ, G. AND J. TOFT. 2000. Presence d’un Asellide epigeoriginaire d’Extreme-Orient en Californie. Bulletin mensuel dela Societe Linneenne de Lyon 69:127–132.
MASIFWA, W. F., T. TWONGO, AND P. DENNY. 2001. The impact ofwater hyacinth, Eichhornia crassipes (Mart) Solms on the abun-dance and diversity of aquatic macroinvertebrates along theshores of northern Lake Victoria, Uganda. Hydrobiologia 452:79–88.
NAGASAWA, K., S. EGUSA, T. HARA, AND I. YAGISAWA. 1983. Eco-logical factors influencing the infection levels of salmonids byAcanthocephalus opsariichthydis (Acanthocephala: Echinorhyn-chidae) in Lake Yunoko, Japan. Fish Pathology 18:53–60.
NICHOLS, F. H., J. E. CLOERN, S. N. LUOMA, AND D. H. PETERSON.1986. The modification of an estuary. Science 231:567–573.
PARKER, I. M., D. SIMBERLOFF, W. M. LONSDALE, K. GOODELL, M.WONHAM, P. M. KAREIVA, M. H. WILLIAMSON, B. VON HOLLE,P. B. MOYLE, J. E. BYERS, AND L. GOLDWASSER. 1999. Impact:Toward a framework for understanding the ecological effectsof invaders. Biological Invasions 1:3–19.
PINKAS, L., M. S. OLIPHANT, AND I. L. K. IVERSON. 1971. Foodhabits of albacore, bluefin tuna, and bonito in California wa-ter. California Department of Fish and Game Fish Bulletin 152:1–105.
ROZAS, L. P. AND T. J. MINELLO. 1997. Estimating densities ofsmall fishes and decapod crustaceans in shallow estuarinehabitats: A review of sampling design with focus on gear se-lection. Estuaries 20:199–213.
RUESINK, J. L., I. M. PARKER, M. J. GROOM, AND P. M. KAREIVA.1995. Reducing the risks of nonindigenous species introduc-tions: Guilty until proven innocent. Bioscience 45:465–477.
SCHRAMM, H. L. AND K. J. JIRKA. 1989. Epiphytic macroinverte-
brates as a food resource for bluegills in Florida Lakes. Trans-actions of the American Fisheries Society 118:416–426.
SCHRAMM, H. L., K. J. JIRKA, AND M. V. HOYER. 1987. Epiphyticmacroinvertebrates on dominant macrophytes in two centralFlorida Lakes. Journal of Freshwater Ecology 4:151–161.
SIMBERLOFF, D. 1991. Keystone species and community effectsof biological introductions, p. 1–19. In L. R. Ginzburg (ed.),Assessing Ecological Risks of Biotechnology. Butterworth-Hei-nemann, Boston, Massachusetts.
SIMENSTAD, C. A., C. D. TANNER, R. M. THOM, AND L. L. CON-QUEST. 1991. Estuarine habitat assessment protocol. U.S. En-vironmental Protection Agency, Seattle, Washington.
SIMENSTAD, C. A. AND R. M. THOM. 1996. Functional equivalencytrajectories of the restored Gog-Le-Hi-Te estuarine wetland.Ecological Applications 6:38–56.
STROMBERG, J. C. 1998. Functional equivalency of saltcedar (Ta-marix chinensis) and fremont cottonwood (Populus fremontii)along a free-flowing river. Wetlands 18:675–686.
SUTHERLAND, W. J. 1996. Ecological Census Techniques: AHandbook. Cambridge University Press, New York.
TERRY, C. 1977. Stomach contents methodology: Still lots ofquestions, p. 87–92. In C. A. Simenstad and S. J. Lipovsky(eds.), Fish Food Habits Studies, Proceedings 1st PacificNorthwest Technical Workshop, 13–15 October 1976, Astoria,Oregon. Washington Sea Grant WSG-WO-77-2, Seattle, Wash-ington.
THE BAY INSTITUTE (TBI). 1998. From the Sierra to the Sea: TheEcological History of the San Francisco Bay-Delta Watershed.The Bay Institute, San Francisco, California.
TOFT, J. D., J. C. CORDELL, AND W. C. FIELDS. 2002. New recordsof crustaceans (amphipoda, isopoda) in the Sacramento/SanJoaquin Delta, California and application of criteria for intro-duced species. Journal of Crustacean Biology 22:200–210.
U.S. ARMY CORPS OF ENGINEERS (USCOE). 1985. Waterhyacinth,Sacramento-San Joaquin Delta, California. State Design Mem-orandum Report. USCOE, Sacramento, California.
VITOUSEK, P. M. 1986. Biological invasions and ecosystem prop-erties, can species make a difference?, p. 163–176. In H. A.Mooney and J. A. Drake (eds.), Ecology of Biological Inva-sions of North America and Hawaii. Springer-Verlag, NewYork.
WERNER, E. E. AND D. J. HALL. 1979. Foraging efficiency andhabitat switching in competing sunfishes. Ecology 60:256–264.
WESTMAN, W. E. 1990. Park management of exotic plant species:Problems and issues. Conservation Biology 4:251–260.
WILLIAMS, S. L. AND C. A. DAVIS. 1996. Population genetic anal-yses of transplanted eelgrass (Zostera marina) beds reveal re-duced genetic diversity in Southern California. RestorationEcology 4:163–180.
WILLIAMS, W. D. 1970. A revision of North American epigeanspecies of Asellus (Crustacea:Isopoda). Smithsonian Contribu-tions to Zoology 49:1–80.
WOLVERTON, B. C. AND R. C. MCDONALD. 1978. Waterhyacinth(Eichhornia crassipes) productivity and harvesting studies. Eco-nomic Botany 33:1–10.
ZAR, J. H. 1996. Biostatistical Analysis, 3rd edition. Prentice Hall,New Jersey.
ZHANG, J. 1997. Systematics of the freshwater amphipod genusCrangonyx (Crangonyctidae) in North America. Ph.D. Disser-tation, Old Dominion University, Norfolk, Virginia.
Received for consideration, February 13, 2002Revised, July 12, 2002
Accepted for publication, July 23, 2002