thèse de doctorat 3 cycle par hicham khattabi intérêts de l'étude
TRANSCRIPT

INTRODUCTION GENERALE
6
TThhèèssee ddee DDooccttoorraatt 33èèmm CCyyccllee
Présentée à l’UFR des Sciences et TechniquesPour l’obtention de titre de DOCTEUR DE L’UNIVERSITEA L’Institut des Sciences de l’EnvironnementSpécialité: Sciences de l’eau et de l’environnement
Par Hicham KHATTABI
Intérêts de l’étude des paramètres hydrogéologiques et
hydrobiologiques pour la compréhension du
fonctionnement de la station de traitement des
lixiviats de la décharge d’ordures ménagères
d’Etueffont (Belfort, France)
Soutenu le 25-02-2002
Devant la commission d’examen:Mr Erick CARLIER (Professeur, UNIVERSITE D'ARTOIS) Rapporteur
Mr Jean Louis MOREL (Professeur, ENSAIA de NANCY) Rapporteur
Mr L. ALEYA (Professeur, Université de Franche-Comté) Examinateur
Mr Jacky MANIA (Professeur, Ecole Universitaire D'Ingénieurs de Lille), Examinateur
Mr Jacques MUDRY (Professeur, Université de Franche-Comté) Examinateur
Mr François REMY (Professeur honoraire, Université de Franche-Comté) Examinateur
Mr Hervé GRISEY (PAST, Vice président du SICTOM) Examinateur
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
7
bbssttrraacctt:: TToo ffaacciilliittaattee aaeerroobbiicc ddeeggrraaddaattiioonn,, tthhee EEttuueeffffoonntt mmuunniicciippaall llaannddffiillll wwaassttee ((NNoorrtthheeaasstt ooff
BBeellffoorrtt,, FFrraannccee)) ppoosssseesssseess aa mmooddee ooff eexxppllooiittaattiioonn bbyy ggrriinnddiinngg wwaasstteess wwiitthhoouutt ccoommppaaccttiioonn.. TThhiiss tteecchhnniiqquuee lleeaaddss
ttoo aa pprroodduuccttiioonn ooff lleeaacchhaattee.. TThhee lleeaacchhaattee wwaass ttrreeaatteedd bbyy nnaattuurraall llaaggoooonnss ccoonnssiissttiinngg ooff 44 ddeeccaannttaattiioonn bbaassiinnss
ffoolllloowwiinngg rreelleeaassee iinnttoo tthhee MMoonntt BBoonnnneett BBrrooookk.. HHoouurrllyy ((11999999)),, mmoonntthhllyy ((11999988--11999999)) aanndd aannnnuuaall ((11999933 ttoo 11999988))
ssaammpplliinnggss ooff ddoommeessttiicc llaannddffiillll lleeaacchhaattee wweerree ttaakkeenn iinn aann aatttteemmpptt ttoo eexxaammiinnee ddiiffffeerreenncceess iinn lleeaacchhaattee ccoommppoossiittiioonn
bbeettwweeeenn oolldd aanndd nneeww aarreeaass ooff tthhee ffiillll aanndd tthheeiirr sseeaassoonnaall eevvoolluuttiioonn.. VVaarriiaattiioonnss iinn ffllooww rraattee,, rraaiinnffaallll,, tteemmppeerraattuurree,,
eelleeccttrriiccaall ccoonndduuccttiivviittyy,, OO22,, ppHH,, CCll,, SSOO44,, ZZnn,, CCuu,, FFee,, NNii,, HHCCOO33,, NNOO33,, NNHH44,, BBOODD aanndd CCOODD wweerree mmoonniittoorreedd.. TThhee
rreessuullttss ffrroomm mmoonntthhllyy ssaammpplliinnggss sshhoowweedd,, cclloossee ccoouupplliinngg bbeettwweeeenn ffllooww aanndd ccoonncceennttrraattiioonnss ooff aallll ppaarraammeetteerrss iinn tthhee
lleeaacchhaattee.. IInn ffaacctt,, tthhee llaatttteerr ddeeccrreeaasseedd wwiitthh iinnccrreeaassiinngg ffllooww rraattee.. AAllssoo,, mmaassss eemmiissssiioonnss ppeerr uunniitt wwaassttee mmaassss
ddeeccrreeaasseedd wwiitthh iinnccrreeaassiinngg wwaassttee aaggee.. TThhee hhoouurrllyy mmoonniittoorriinngg ppooiinntteedd ttoo aann iinnccrreeaassee iinn eelleeccttrriiccaall ccoonndduuccttiivviittyy,,
cchhlloorriiddee aanndd CCOODD ccoonncceennttrraattiioonnss,, ppeeaakkiinngg aatt aarroouunndd mmiiddddaayy.. TThhiiss wwaass mmoosstt lliikkeelleeyy dduuee ttoo tteemmppeerraattuurree
fflluuccttuuaattiioonnss.. IInn oorrddeerr ttoo eevvaalluuaattee ggrroouunnddwwaatteerr qquuaalliittyy sseevveerraall pphhyyssiiccoo--cchheemmiiccaall ppaarraammeetteerrss wweerree ssttuuddiieedd iinn
eeiigghhtt wweellllss llooccaatteedd nneeaarr ttoo tthhee EEttuueeffffoonntt llaannddffiillll.. TThhee rreessuullttss sshhoowweedd hhiigghh ccoonncceennttrraattiioonnss ooff mmoosstt ppaarraammeetteerrss iinn
tthhee wweellll PPZZ3300 aanndd PPZZ33,, ssiittuuaatteedd nneeaarr tthhee llaannddffiillll.. TThhee ssppaattiioo--tteemmppoorraall ddiissttrriibbuuttiioonn ooff tthhee pphhyyssiiccoo--cchheemmiiccaall
ppaarraammeetteerrss iinn tthhee 88 wweellllss wwaass ggoovveerrnneedd bbyy ggeeoollooggiiccaall,, hhyyddrrooggeeooggiiccaall,, pphhyyssiiccaall,, cchheemmiiccaall,, aanndd cclliimmaattiicc ffaaccttoorrss..
IInn aaddddiittiioonn,, tthhee ccoonncceennttrraattiioonnss ooff ccoonnttaammiinnaannttss ddeeccrreeaasseedd iinn tthhee llaasstt wweellll ((11 KKmm ffrroomm tthhee llaannddffiillll)),, iinnddiiccaattiinngg aa
ppuurriiffiiccaattiioonn aanndd ddiilluuttiioonn eeffffeecctt.. NNeevveerrtthheelleessss,, wwee ssuussppeecctt sshhoorrtt--tteerrmm ddeeggrraaddaattiioonn ooff tthhee ggrroouunnddwwaatteerr iinn tthhee
EEttuueeffffoonntt ssiittee.. TToo eevvaalluuaattee tthhee llaannddffiillll lleeaacchhaattee iimmppaacctt oonn tthhee ssttrreeaamm wwaatteerr qquuaalliittyy,, wwee aasssseesssseedd tthhee ssppaattiiaall
eevvoolluuttiioonn ooff sseevveerraall aabbiioottiicc aanndd bbiioottiicc ppaarraammeetteerrss.. SSaammpplleess wweerree ttaakkeenn iinn 1133 JJuunnee 11999999 wwiitthh aa VVaann--DDoorrnn bboottttllee
ffrroomm uuppssttrreeaamm ttoo ddoowwnnssttrreeaamm.. TThhee ssttuuddyy sshhoowweedd tthhaatt lleeaacchhaattee rruunnooffff ccaauusseedd aann iinnccrreeaassee iinn ppHH,, NNOO33,, CCll aanndd
CCOODD ccoonncceennttrraattiioonnss iinn tthhee ffiirrsstt mmeetteerr.. IInn ddoowwnnssttrreeaamm,, hhoowweevveerr tthhee rreevveerrssee wwaass qquuootteedd.. BBaacctteerriiaall aabbuunnddaannccee
iinnccrreeaasseedd ffrroomm uuppssttrreeaamm ttoo ddoowwnnssttrreeaamm wwiitthh nnoo rreellaattiioonnsshhiipp bbeettwweeeenn tthhiiss bbiioollooggiiccaall ccoommppaarrttmmeenntt aanndd lleeaacchhaattee
rruunnooffff.. FFiinnaallllyy,, iitt sseeeemmss lliikkeellyy tthhaatt tthhee aabbiilliittyy ooff sseellff rreegguullaattiioonn ooff tthhee bbrrooookk wwaass hhiigghheerr iinn dduurriinngg lleeaacchhaattee--
iinndduucceedd ppoolllluuttiioonn.. TThhee ssppaattiioo--tteemmppoorraall eevvoolluuttiioonn ooff RRoottiiffeerraa aabbuunnddaannccee wwaass ssttuuddiieedd ffrroomm MMaayy 11999988 ttoo MMaayy
11999999 iinn tthhee ssttaabbiilliizzaattiioonn ppoonnddss ooff tthhee EEttuueeffffoonntt llaannddffiillll lleeaacchhaattee.. TThhee RRoottiiffeerraa wweerree ccoolllleecctteedd mmoonntthhllyy bbyy aa
ppllaannkkttoonn nneett ((ppoorroossiittyy == 8855 µµmm)).. TThhee rreessuullttss sshhoowweedd tthhee pprreesscceennccee ooff aa mmaaxxiimmaa ooff rroottiiffeerraass dduurriinngg ssuummmmeerr 11999988
((110055 cceellllss.. ll--11)),, ccooiinncciiddiinngg wwiitthh tthhee ddeevveellooppeemmeenntt ooff aallggaaee ((PPhhaaccuuss sspp.. aanndd EEuugglleennaa sspp..)).. TThhee RRoottiiffeerraa wwaass
ddoommiinnaatteedd bbyy BBrraacchhiioonnuuss ccaallyycciifflloorruuss aanndd BBrraacchhiioonnuuss rruubbeennss ((110000%% iinn bbaassiinnss 11,, 22 aanndd 9988 %% iinn 33 aanndd 44)).. IInn
wwiinntteerr,, aabbuunnddaannccee ooff RRoottiiffeerraannss ddeeccrreeaasseedd ttoo 110022 cceellllss.. ll--11,, dduuee ttoo llooww tteemmppeerraattuurree aanndd tthhee ccoollllaappssee ooff
bbiiooaavvaaiillaabbllee pphhyyttooppllaannkkttoonn wwhhiicchh wwaass ddoommiinnaatteedd bbyy tthhee ddiiaattoomm SStteepphhaannooddiissccuuss sspp.. IInn aaddddiittiioonn,, aabbuunnddaannccee ooff
iinnddiivviidduuaallss iinnccrreeaasseedd ffrroomm tthhee ffiirrsstt bbaassiinn ttoo tthhee llaasstt ffoolllloowwiinngg tthhee ddeeccrreeaassee iinn aammmmoonniiaa ccoonncceennttrraattiioonnss ((115500 mmgg..
ll--11 iinn bbaassiinn 11 ttoo 5500 mmgg.. ll--11 iinn bbaassiinn 44)) aanndd tthhee aabbuunnddaannccee ooff tthhee CCllaaddoocceerraann MMooiinnaa sspp.. EEffffiicciieennccyy ooff tthhee eeppuurraattiioonn
wwaass eessttiimmaatteedd bbyy ccaallccuullaattiioonn tthhee nneett bbuuddggeett ooff eelleemmeennttss ((==IInnppuutt--OOuuttppuutt)).. TThhee rreessuullttss sshhoowweedd tthhaatt llaaggoooonnss
eelliimmiinnaatteedd tthhee qquuaassii--ttoottaalliittyy ooff oorrggaanniicc mmaatttteerr ((aabboouutt 8800 %% ooff BBOODD)) aanndd ddiissssoollvveedd ssuubbssttaanncceess ((aabboouutt 5500 ttoo 9900 %%
)).. NNeevveerrtthheelleessss,, aa ddeeccrreeaassee ooff eeppuurraattiioonn eeffffiicciieennccyy tthhaatt hhaass bbeeeenn oobbsseerrvveedd iinn wwiinntteerr ooff aallll eelleemmeennttss ((tteemmppeerraattuurree,,
eelleeccttrriicc ccoonndduuccttiivviittyy,, ppHH,, OO22,, EEhh,, CCll,, FFee,, MMnn,, NNii,, ZZnn,, ttoottaall nniittrrooggeenn,, ttoottaall pphhoosspphhoorruuss,, NNHH44,, BBOODD,, CCOODD)),, wwaass
dduuee ttoo tthhee ddeeccrreeaassee ooff ppHH ((ppHH == 44,,55)).. HHoowweevveerr,, iinn tthhee bbeeggiinniinngg ooff ssuummmmeerr oonnllyy CCuu22++ aanndd FFee22++ ddeeccrreeaasseedd.. TThhee
ccoollllaappssee ooff BBOODD rreemmoovvaall ((--88 %% oobbsseerrvveedd iinn SSeepptteemmbbeerr 11999988)) wwaass lliinnkkeedd ttoo tthhee eexxcceeddeennttaarryy ssttoocckk ooff oorrggaanniicc
mmaatttteerr pprroodduuccttss iinn tthhee bbaassiinnss dduurriinngg tthhee ssuummmmeerr.. TThhee aannaallyyssiiss ooff tthhee mmoonntthhllyy eevvoolluuttiioonn ooff tthhee eeffffiicciieennccyy
AA
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
8
eeppuurraattiioonn ooff NNHH44,, ttoottaall nniittrrooggeenn ((TTNN)) aanndd ttoottaall pphhoosspphhoorruuss ((TTPP)) iinnddiiccaattee ssiimmiillaarr ffeeaattuurreess.. TThhee nneett bbuuddggeett
fflluuccttuuaatteess bbeettwweeeenn 5500 %% iinn wwiinntteerr aanndd 7700 %% iinn ssuummmmeerr,, wwiitthh mmiinniimmaall vvaalluueess nnootteedd iinn JJaannuuaarryy 11999999 ((1199 %% ffoorr
NNHH44,, 1133 %% ffoorr TTNN aanndd 1144 %% ffoorr TTPP)).. TThhiiss wwoorrkk sshhoowweedd tthhaatt tthhee nnaattuurraall llaaggoooonniinngg iiss iinneeffffeeccttiivvee ffoorr tthhiiss ttyyppee ooff
llaannddffiillll lleeaacchhaatteess..
Key-words: Landfill, leachates, water pollution, lagooning, metals, BOD, COD and plankton.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
9
I. LES ORDURES MENAGERES
I. 1. Définition
Les ordures ménagères «stricto sensu» correspondent aux déchets de l’activité
domestique quotidienne des ménages.
I. 2. Classification des décharges d’ordures ménagères
La classification des décharges d’ordures ménagères est basée sur l’aptitude d’un
sol à accueillir une décharge c’est à dire sur la valeur du coefficient de perméabilité (ks) ou
coefficient de Darcy, qui définit la vitesse de percolation des eaux dans un sous-sol. Les trois
grands types de sites qui découlent de la loi n°75-633 du 15 juillet 1975 sont représentés dans
le tableau 1 et sont classés comme suit:
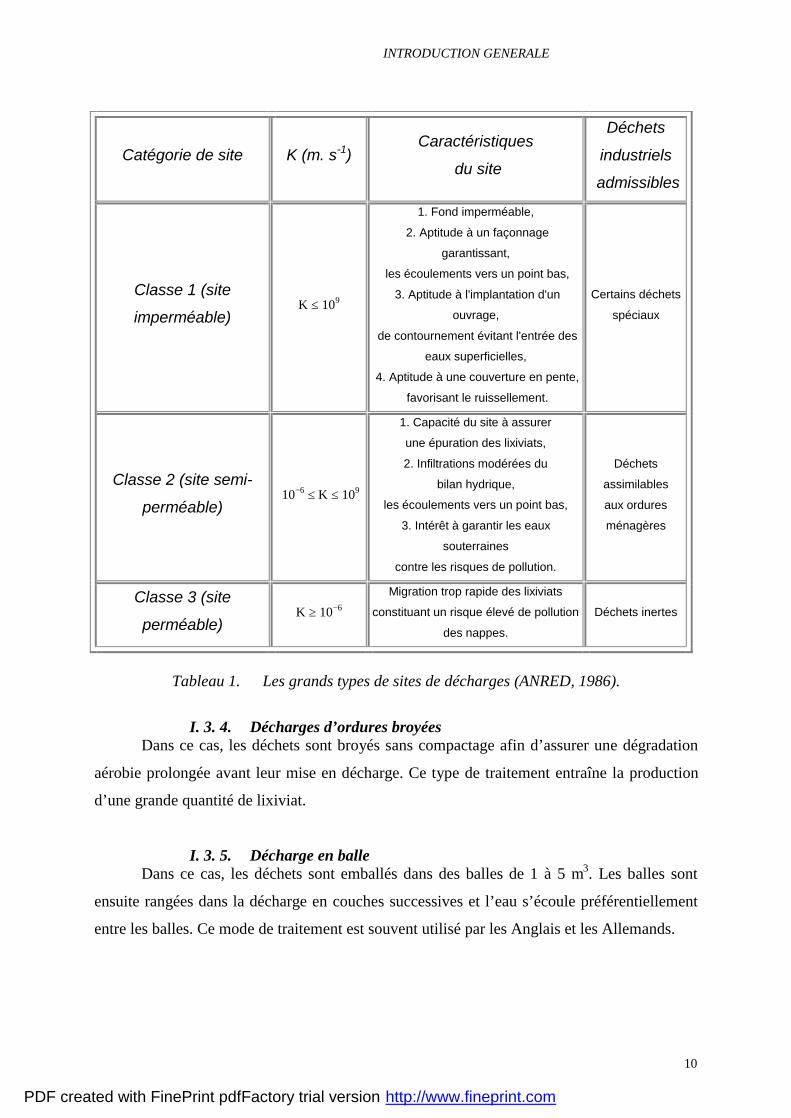
Les sites de classe 3, très perméables ne sont destinés à recevoir que les déchets
inertes, les sites de classe 2, peu perméables peuvent recevoir des déchets ménagers ou
assimilés, alors que les sites de classe 1 pratiquement étanches peuvent recevoir les déchets
dangereux.
I. 3. Les divers modes de mise en décharge d’ordures ménagères
On dénombre en France cinq modes principaux de mise en décharge des orduresménagères.
I. 3. 1. Décharge contrôlée traditionnelleElle se base sur un étalement des déchets en fines couches (0,5 à 2 m) d’épaisseur,
puis à la fin de chaque journée on couvre les déchets par une couche de matériaux inertes de
10 à 30 cm d’épaisseur.
I. 3. 2. Décharge contrôlée compactéeDans ce cas les déchets sont compactés avant leur mise en décharge. Le compactage
augmente la densité des déchets en chassant l’air contenu dans les déchets, aboutissant ainsi à
la formation d’un ensemble peu perméable.
I. 3. 3. Décharge aérobieElle consiste à traiter séparément les déchets fermentescibles des déchets inertes qui
subissent un traitement différent. Les déchets fermentescibles sont broyés et mixés à l’aide
d’engins spécialisés. Ce procédé se base essentiellement sur une dégradation aérobie.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
INTRODUCTION GENERALE
10
Catégorie de site K (m. s-1)Caractéristiques
du site
Déchets
industriels
admissibles
Classe 1 (site
imperméable)Κ ≤ 109
1. Fond imperméable,
2. Aptitude à un façonnage
garantissant,
les écoulements vers un point bas,
3. Aptitude à l'implantation d'un
ouvrage,
de contournement évitant l'entrée des
eaux superficielles,
4. Aptitude à une couverture en pente,
favorisant le ruissellement.
Certains déchets
spéciaux
Classe 2 (site semi-
perméable) 10−6 ≤ Κ ≤ 109
1. Capacité du site à assurer
une épuration des lixiviats,
2. Infiltrations modérées du
bilan hydrique,
les écoulements vers un point bas,
3. Intérêt à garantir les eaux
souterraines
contre les risques de pollution.
Déchets
assimilables
aux ordures
ménagères
Classe 3 (site
perméable)Κ ≥ 10−6
Migration trop rapide des lixiviats
constituant un risque élevé de pollution
des nappes.
Déchets inertes
Tableau 1. Les grands types de sites de décharges (ANRED, 1986).
I. 3. 4. Décharges d’ordures broyéesDans ce cas, les déchets sont broyés sans compactage afin d’assurer une dégradation
aérobie prolongée avant leur mise en décharge. Ce type de traitement entraîne la production
d’une grande quantité de lixiviat.
I. 3. 5. Décharge en balleDans ce cas, les déchets sont emballés dans des balles de 1 à 5 m3. Les balles sont
ensuite rangées dans la décharge en couches successives et l’eau s’écoule préférentiellement
entre les balles. Ce mode de traitement est souvent utilisé par les Anglais et les Allemands.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
11
I. 4. Evolution des déchets dans une décharge d’ordures ménagères
Une fois déposés dans la décharge, les déchets subissent une dégradation aérobie
favorisée par l’oxygène en place. Durant cette phase aérobie, ce sont les micro-organismes
hydrolytiques qui assurent la dégradation de la matière organique solide présente dans les
déchets par l’action d’enzymes spécifiques (protéolytiques, cellulolytiques, lipoprotéiques). Il
résulte de ces dégradations une transformation des grosses molécules organiques insolubles
en molécules plus petites qui pourront ultérieurement servir de substrats pour d’autres micro-
organismes. Ce métabolisme aérobie peut continuer jusqu’à une minéralisation complète des
substrats biodégradables et conduit à des métabolites finaux (CO2, H2O, CO32-, HCO3
-, NO3-,
PO43- et SO4
2-). Après c’est la dégradation anaérobie qui prend le relais produisant ainsi du
CO2 et CH4 (Gordon, 1988; Joly, 1988) (fig. 1). Enfin, Chian (1985) et Pohland (1985) ont
défini 5 phases de digestion anaérobie des déchets:
I. 4. 1. Phase de latence
Durant laquelle on assiste à un remplissage initial, augmentation de l’humidité et un
début de tassement des alvéoles.
I. 4. 2. Phase de transition
Cette phase est caractérisée par la première production des lixiviats et un début de la
dominance de la phase anaérobie qui se manifeste par une substitution de l’oxygène par les
nitrates ou les sulfates comme accepteur final d’électron. De plus, durant cette étape, on
assiste à une hydrolyse bactérienne aérobie ou anaérobie des complexes organiques
aboutissant à la formation de sucres simples, d’acides gras et d’acides aminés. Notons enfin
les premières formations des AGV (acides gras volatils).
I. 4. 3. Phase acidogène
C’est la phase des AGV qui peuvent constituer jusqu’à 95 % du carbone organique
total (COT) (Harmsen, 1983). Ceci est dû au développement d’une autre flore bactérienne
capable de transformer les produits issus de l’hydrolyse en acide acétique, en acétates (avec
formation d’H2 et CO2), en alcool ou en AGV. L’ensemble de ces composés conduit à la
production de lixiviats acides.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
12
I. 4. 4. Phase de fermentation méthanique
Au cours de cette étape, les AGV, l’acide acétique et le méthanol formés dans la phase
acétogène sont métabolisés. Les résidus de cette métabolisation sont des gaz: CH4 (45 à 60
%), CO2 (35 à 50 %) et de grosses molécules stabilisées (acides humiques). Cette étape est
favorisée par des bactéries strictement anaérobies. De plus, lors de cette étape, on enregistre
souvent une élévation du pH donnant des valeurs proches de la neutralité. Enfin la portion
organique des lixiviats diminue et il s’y adjoint un phénomène de complexation et de
précipitation des métaux.
I. 4. 5. Phase de mâturation finale
Cette phase est caractérisée par une diminution des teneurs en nutriments et une
diminution de biogaz.
I. 5. Les lixiviats de décharges
I. 5. 1. DéfinitionLe terme lixiviat désigne les eaux météoriques qui en percolant à travers la décharge
se chargent d’éléments polluants minéraux et organiques. Cependant, la lixiviation est la
migration des substances solubles, et le lessivage correspond à la migration des particules en
suspension.
I. 5. 2. CompositionLa composition des lixiviats de décharges est très aléatoire est varie en fonction de la
nature des déchets, l’âge de la décharge, la technique d’exploitation et les conditions
climatiques. Toutefois, Christensen et al. (2001), en étudiant plusieurs sites ont défini les
fourchettes de fluctuations de la composition physico-chimique des lixiviats d’ordures
ménagères (tab. 2). La fraction minérale des lixiviats de décharge est constituée
essentiellement de chlorures, sulfates, bicarbonates, potassium, sodium et ammonium, alors
que la partie organique est très liée à l’âge des déchets. Si les jeunes décharges sont
caractérisées par la dominance des acides gras volatiles (AGV) (surtout les acides acétiques,
propioniques et butyriques), ce sont des substances assimilées aux composés fulviques et
humiques qui sont prépondérantes dans les lixiviats âgés.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
13
I. 5. 3. Principaux facteurs influençant la production des lixiviatsLe lixiviat provient de l’eau du déchet, des précipitations météoriques et des eaux
provenant de la nappe phréatique (Farquhar, 1989). Leur intensité gouverne le volume du
lixiviat produit. La composition qualitative et quantitative du lixiviat dépend des pluies, de la
couverture de la décharge, de la topographie du site, du taux de compactage, de la teneur en
eau des déchets ou capacité au champ, du mode de comblement et de la température
(Deunevy, 1987). Cependant, selon Kmet (1982), 20 % des précipitations conduisent à la
production des lixiviats. Alors que Stegmann (1980) a trouvé des pourcentages différents en
fonction de l’état de compaction des déchets, 15 à 25 % des pluies forment du lixiviat pour les
décharges compactées et 25 à 50 % dans le cas d’une décharge non compactée. Enfin, on
notera que généralement l’étude qualitative et quantitative d’une décharge est très délicate et
diffère d’une décharge à une autre suite à la présence de plusieurs mécanismes biologiques et
physico-chimiques qui gèrent la production des lixiviats au sein d’une décharge.
II. LAGUNAGE NATUREL
II. 1. Définition Le lagunage naturel est souvent défini par plusieurs termes: "oxidation pond", "sewage
oxydation pond", "sewage lagoon", "waste stabilization-ponds", "étang de mâturation", "étang
de stabilisation". C’est un procédé biologique de traitement des eaux usées. Il consiste à
laisser l’eau se reposer dans des bassins ouverts peu profonds de 1 à 5 m de profondeur
pendant une durée variant de 30 à 60 jours. Il aboutit d’une part à l’abattement de la charge
polluante et d’autre part à la stabilisation des boues produites, sous l’action des organismes se
développant dans le milieu.
II. 2. HistoriqueLa première technique d’épuration utilisée date du milieu du siècle dernier, elle
consistait à une aspersion des champs par l’eau usée. Les micro-organismes naturels étaient
censés suffire à l’épuration. Le lagunage naturel est connu depuis le début du 20e siècle. Les
premières applications pratiques de l’épuration par l’action oxydante naturelle des micro-
organismes ont eu lieu au Texas en 1901.
Mais ce n’est qu’à partir des années 1920 que le lagunage naturel a pris son essor sous
divers climats aussi bien aux U.S.A qu’au Canada, en Suède et en Australie (Glonya, 1972).
Le bassin de stabilisation le plus ancien en Europe est celui de la ville de Munchen en
Allemagne construit en 1920. Du fait de sa facilité de mise en œuvre, son efficacité de
traitement, sa bonne intégration au milieu rural et de son coût raisonnable vis à vis des autres
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
14
traitements, le lagunage naturel a connu une expansion considérable dans le monde entier. Le
procédé de lagunage était presque ignoré en France jusqu’à une époque très récente, du fait
d’un certain préjugé à de possibles inconvénients. Pourtant il y a plusieurs siècles déjà que les
étangs naturels ont servi de réceptacles aux eaux vannes, longtemps avant que soit reconnue
la possibilité d’épurer les eaux par un tel moyen. Le premier lagunage naturel en France est
celui du Grau-du Roi construit en 1964, puis d’autres suivent en 1970 et ceci après certaines
années d’attente.
Tableau 2. Données bibliographiques en fourchettes de variation de la composition
physico-chimique du lixiviat de décharge d'ordures ménagères d'après Christensen et al.
2001. Les teneurs sont exprimées en mg. l-1.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
15
Fig. 1. Etapes de la méthanisation selon Bryant (1979).
II. 3. Processus d’épurationLe lagunage naturel repose sur une décantation directe au fond des bassins des
matières en suspension, et indirecte des substances solubles introduites ou remises en solution
par les sédiments après leur absorption par les microphytes. Ce procédé est proche de l’auto-
épuration. Il est basé sur un cycle biologique, où les bactéries assurent l’élimination des
matières organiques en présence d’O2 fourni par les algues (fig. 2).
En fonction de la nature et de l’importance des végétaux nous distinguerons: (i) le
lagunage à microphytes (caractérisé par une profondeur importante, afin de limiter le
développement des macrophytes), (ii) le lagunage à macrophytes (pour lequel la tranche d’eau
est évidemment plus faible par rapport au premier) et (iii) le lagunage composite: composé
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
16
par l’association des deux procédés cités précédemment. Dans les bassins de lagunage naturel
on retrouve généralement les trois zones suivantes:
Zone 1: appelée aussi zone de surface, très ensoleillée favorisant ainsi des conditions
idéales pour le développement des espèces végétales. L’activité photosynthétique des
végétaux enrichit la lagune en oxygène indispensable à la croissance des bactéries aérobies.
Ces micro-organismes assurent la dégradation de la quasi-totalité de la charge organique de
l’effluent. L’oxygénation de la lagune s’effectue par ailleurs directement par l’atmosphère en
liaison avec l’agitation de l’eau superficielle par le vent.
Zone 2: C’est une zone insuffisamment éclairée pour assurer une productivité algale
assez importante, et dont l’échange est faible avec l’atmosphère à cause de son éloignement
de la surface. Elle engendre ainsi un environnement pauvre en oxygène, favorisant ainsi le
développement de bactéries anaérobies. Ces bactéries produisent des gaz malodorants qui
peuvent constituer une nuisance sérieuse pour le voisinage. Deux cas sont à envisager :
* Si l’effluent est faiblement chargé le bassin bascule vers l’aérobiose,
* Si l’effluent est fortement chargé le bassin bascule vers l’anaérobiose.
Rocques (1980) a relaté les travaux de Marias et Shan publiés en 1961. Ils ont mis en place
une expression mathématique qui relie l’aérobiose et l’anaérobiose et ceci en se reposant sur
des données de plusieurs lagunes en Afrique du sud et aux Etats Unis. L’expression est
définie comme suit:
X = 1000/2H+8
X : concentration en substrat dans la lagune exprimée en mg. l-1 de DBO
H : profondeur du bassin exprimée en m
La zone au-dessus de la courbe (fig. 3) désigne la partie aérobie, alors que la zone au-
dessous représente la partie anaérobie du bassin.
Zone 3 ou zone de fond est la partie du bassin où s’accumulent les matières en
suspension de l’effluent résultant des cellules animales et végétales mortes. Elle est pauvre en
oxygène et assure la minéralisation de la matière organique avec dégagement de gaz: CH4 et
H2S. L’application de nos données à ce modèle indique que les quatre bassins de la station
d'Etueffont fonctionnent en aérobiose du fait que la profondeur des bassins ne dépasse pas 1,5
m et que la charge organique exprimée en terme de DBO n'atteint jamais les 100 mg. l-1 d'O2.
Les bassins de lagunage sont souvent colonisés par une multitude d’espèces qui
varient en fonction: des conditions climatiques, de la nature de l’effluent traité, de la
profondeur de la lagune et de la charge de l’effluent.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
17
Fig. 2. Cycles dans un lagunage naturel (d’après Champiat 1988).
Fig. 3. Limite entre aérobiose et anaérobiose dans les bassins de traitements des eaux usées(Rocques, 1980).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
18
Les micro-organismes assurent toujours la part prépondérante, voire la totalité de la
dégradation de la matière organique quel que soit le procédé biologique mis en œuvre. Pour
cela, les installateurs de bassins de stabilisation ont recours à la création d’une biomasse
microbienne la plus importante possible. Ces micro-organismes épurateurs proviennent le
plus souvent de l’eau brute. Fréquemment pour les bactéries une compétition d’ordre
trophique prend naissance entre les différentes espèces bactériennes. Les espèces les mieux
adaptées prennent le pas sur les autres espèces en raison de leur vitesse de multiplication
assez élevée. Deux grandes classes de micro-organismes sont à distinguer en fonction du type
de métabolisme mis en jeu:
* Métabolisme aérobie où les micro-organismes ne peuvent vivre qu’en présence
d’oxygène, soit moléculaire, soit exceptionnellement incorporé à des éléments minéraux (cas
du nitrate). Ces germes sont capables de garantir la transformation de la matière organique
dissoute en matière minérale (nutriments et gaz).
* Métabolisme anaérobie où peuvent se développer des micro-organismes méthagènes
(produisant le méthane). Ces germes peuvent tolérer des concentrations assez faibles en
oxygène et assurent la minéralisation de la matière organique au niveau des sédiments.
En milieu aérobie les micro-organismes les plus rencontrés sont les bactéries, les
levures et les champignons, tandis qu’en milieu anaérobie il s’agit exclusivement de bactéries.
Afin d’assurer les besoins énergétiques nécessaires à la multiplication, la
locomotion,..., les micro-organismes, en particulier les bactéries sont nourris par les matières
organiques et éliminent les polluants par différents processus:
q par absorption des particules sur les floques bactériens,
q par conversion en matière cellulaire: croissance bactérienne et des micro-
organismes associés,
q par oxydation en CO2 et H2O qui produit l’énergie nécessaire au
fonctionnement et à la croissance bactérienne.
Les matières organiques sont soumises tout d’abord à l’action d’enzymes extra-
cellulaires (cellulolytiques, lipolytiques…) sécrétés par les micro-organisme qui dégradent les
grosses molécules insolubles en molécules de petite taille solubles, pouvant ultérieurement
servir de substrats à ces micro-organismes. Selon l’utilisation du substrat, les bactéries
émettent dans l’eau soit des métabolites intermédiaires lorsqu’il s’agit d’un métabolisme
incomplet, tel que les polypeptides pour un substrat protéique ou les acides gras pour des
substrats graisseux. Ces métabolites peuvent être d’une grande importance dans le piégeage
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
19
des métaux par complexation. Soit des métabolites finaux (CO2, H2O, NO3-…) pour le cas
d’une minéralisation complète du substrat. Ces métabolites influent directement sur le pH de
l’eau qui contrôle la mobilité des ions métalliques. Le métabolisme de la matière organique
peut s’écrire comme suit:
H2O + Pollution organique + Micro-organismes + Oxygène H2O + Micro organismes en excès + CO2
Mais ces lagunes peuvent être aussi un milieu idéal pour le développement de micro-
organismes défavorables tel que les salmonelles (agents responsable de la fièvre typhoïde).
Ces espèces sont souvent présentes en faible quantité. Les chercheurs préfèrent utiliser la
méthode du test de contamination fécale, qui correspond à un comptage des indicateurs de
pollution fécale (coliformes ou streptocoques). L’ampleur de ces indicateurs renseigne sur la
présence probable des germes pathogènes. Enfin, il faut noter que le lagunage assure une
réduction assez importante des germes afin d’atteindre les paramètres microbiologiques requis
pour une eau de baignade (103. 100-1 ml CF, CF: coliformes fécaux). Plusieurs facteurs
interviennent dans l’abattement des micro-organismes dans les lagunes (chute de température,
temps de séjour assez élevé, ensoleillement).
Au deuxième rang on cite les microphytes qui peuvent être soit planctoniques
(dispersés dans la masse d’eau), soit périphytiques (fixés sur des support immergés) ou
épipéliques (déposés à la surface des sédiments). Les groupes les plus rencontrés dans le
lagunage naturel sont les cyanobactéries, chlorophycées, chrysophycées et les euglénophytes.
Les microphytes sont caractérisés par leur grande diversité, environ 15000 espèces, mais
celles qui caractérisent les bassins de lagunage sont relativement peu nombreuses (Gloyna
1972). Cependant Palmer (1974) n’a recensé que 75 genres d’algues dans 72 installations de
lagunage à travers 18 états dans U.S.A, dont 50 % d’algues vertes, 25 % de flagellées, 15 %
de cyanobactéries et 10 % de diatomées.
III. PRESENTATION DU TRAVAIL PROPOSE
Nous avons choisi de diviser ce travail pluridisciplinaire en trois parties distinctes. La
première partie du travail présenté dans ce mémoire est destinée à l'étude bibliographique des
lixiviats de décharges d'ordures ménagères et du lagunage naturel ainsi que les protocoles
expérimentaux pour les différents éléments choisis.
La seconde partie du travail aborde:
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

INTRODUCTION GENERALE
20
L'étude des lixiviats produits par la décharge d'Etueffont: Après un rappel concernant
les théories citées en littérature, nous abordons le coté pratique de ce travail, c'est à dire les
applications que nous avons entreprises. La première de ces applications constitue l'étude de
l'évolution temporelle des lixiviats avec des pas d’échantillonnage allant de l'échelle horaire à
l'échelle inter-annuelle. La deuxième est une étude d’impact des lixiviats sur l'environnement
et plus précisément sur les eaux souterraines et superficielles. Afin d'évaluer l'impact sur les
eaux souterraines, nous avons procédé à un suivi de certains polluants organiques et
inorganiques dans des piézomètres installés en amont et en aval de la décharge. Alors que
l'impact sur les eaux superficielles a été évalué par un suivi de certains paramètres en aval des
bassins de traitement.
L'évaluation d'un lagunage naturel pour le traitement: Nous soulignons ici et à notre
connaissance que ce procédé n'a jamais été appliqué pour le traitement des lixiviats de
décharges d’ordures ménagères. Cette efficacité a été mise en évidence par le calcul du
rendement épuratoire pour la majorité des éléments chimiques.
La troisième partie du travail aura trait à l’étude de l'évolution saisonnière des
paramètres abiotiques et de la dynamique des compartiments bactérioplanctoniques,
phytoplanctoniques et zooplanctoniques.
Enfin, en guise de conclusion, nous faisons une évaluation globale du travail réalisé et
soulevons certains des nombreux points non encore traités. A partir de cette constatation nous
faisons quelques propositions pour la poursuite de la recherche dans ce domaine.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
21
I. DESCRIPTION DU SITE
I. 1. Situation géographiqueLe site étudié est la décharge d’ordure ménagère située à Etueffont (NE de Belfort).
Cette décharge est gérée par le SICTOM du pays sous Vosgien (Syndicat Intercommunal de
Traitement des Ordures Ménagères). Le SICTOM a été créé en 1972. Le Broyage des déchets
et la collecte ont commencé en mai 1976.
I. 2. Caractéristiques de la décharge d'EtueffontLa décharge reçoit les déchets de ménages de 66 communes, 42 du territoire de
Belfort, 15 du Haut-Rhin et 9 de la Haute-Saône, soit un total de 47650 habitants. La
décharge d’ordures ménagères du Pays Sous-Vosgien occupe une surface de 40000 m2,
comporte 200.000 tonnes de déchets et haute de 15 m. Elle a été fonctionnelle à partir de 1976
jusqu'à 2002 et présente un mode d’exploitation à ciel ouvert par broyage des déchets sans
compactage. Cette gestion entraîne la production d’un lixiviat collecté en aval de la décharge,
et traité par lagunage naturel dans quatre bassins installés en série. L'installation de traitement
des lixiviats de la décharge d'Etueffont par lagunage naturel est située en aval de la décharge.
Elle est composée par l'association en série de 4 bassins (fig. 4). Elle a été fonctionnelle
depuis 1994. Le tableau 3 regroupe les caractéristiques morphométriques des quatre bassins
de la station d'Etueffont.
Tableau 3. Caractéristiques morphométriques des quatre bassins de la station d'Etueffont.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
22
Fig. 4. Schéma et caractéristiques des bassins de lagunage naturel.
Localisation des stations de prélèvement.
Sens d'écoulement.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
23
I. 3. TempératureLe climat de la région est de type tempéré humide, ainsi les thermomètres installés au
niveau de l’usine de broyage ont mis en évidence des températures ambiantes très élevées en
été avec un maximum de 35 °C observé le 20 août 1998 et des températures très basses en
automne et en hiver, avec un minimum de –12 °C enregistré le 24 novembre 1998,
provoquant la gelée prématurée des bassins dès le mois de novembre 1998. Les températures
estivales sont les plus élevées, au contraire l’hiver fut plus doux (fig. 5).
Fig. 5. Evolution de la température (°C) ambiante dans la station d’Etueffont.
I. 4. PrécipitationsLe suivi des précipitations à une échelle de temps plus fine à l'aide d'un pluviomètre
installé au voisinage de la décharge montre une présence des précipitations tout au long de
l'année. Cependant le régime pluviométrique montre deux pics majeurs de pluie; l'un en
octobre – novembre 1998 et l'autre en mars – avril 1999 avec quelques orages en été
(fig. 6).
Fig. 6. Evolution des pluies (mm) dans la station d’Etueffont.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
24
I. 5. GéologieL’étude géologique de la station d’étude (décharge et lagunes) montre que la station
d’Etueffont se situe dans un contexte schisteux du Dévono-dinantien (Primaire). Les sondages
ont montré la présence d’une zone d’altération très poussée, caractérisée par une domination
de la fraction argileuse et une variation de couleur en profondeur généralement au-delà de
10 m. Les schistes sont limités à l’Ouest par des grés du Permien par l’intermédiaire d’un
contact faillé. On note que les plans de schistosité sont perpendiculaires au plan de la faille, ce
qui nous laisse penser à l’effet écran de cette faille comme barrière géologique contre toute
pollution éventuelle par la station (annexe 10).
I. 6. HydrogéologieLes forages effectués par le bureau d’étude ont montré la présence de deux arrivées
d’eau dans les schistes, une dans la zone d’altération (entre 3 et 4 m) caractérisant la nappe de
surface et l’autre dans les schistes sains caractérisant la nappe profonde (12 à 15 m). Ces
essais montrent aussi que la nappe profonde est en charge avec de faibles variations des
niveaux d’eau. La nappe profonde semble guider les écoulements et éventuellement tend à
repousser les polluants vers le haut. Dans les grés du Permien on note la présence d’une seule
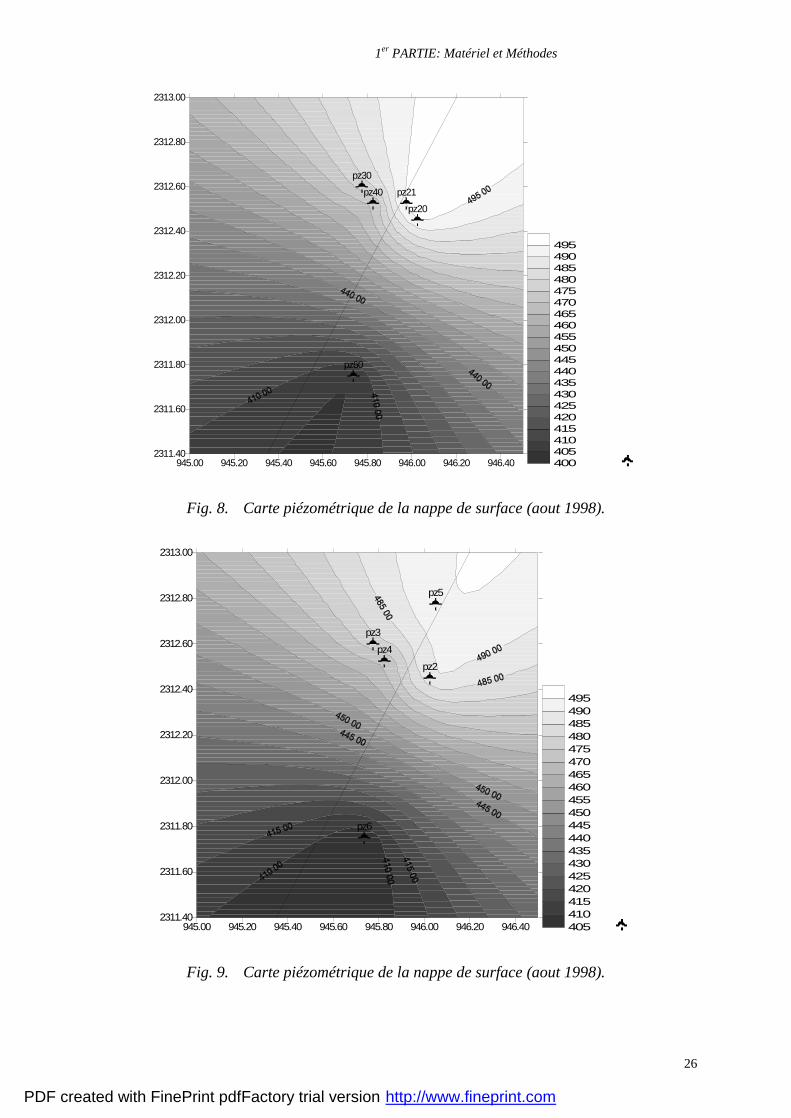
arrivée d’eau dans la zone profonde (12 à 15 m). Les cartes piézométriques relevées entre
1998 et 1999 présentées sur les figures 8 et 9, montrent un sens d’écoulement de la nappe des
grés vers la nappe des schistes prenant une direction NE-SW, ce qui empêche toute pollution
éventuelle de la nappe des grés. Enfin, la piézomètre de la nappe des grés présente des
fluctuations très importantes qui nous indiquent que cette nappe est très dépendante des
précipitations en surface si on la compare à la nappe des schistes (fig. 7).
I. 7. PerméabilitéLes essais par injection d’eau effectués dans les différents forages montrent que la
perméabilité est plus importante dans la zone altérée des grés (10-5 m. s-1) que dans celle des
schistes (10-6 m. s-1).
I. 8. Caractéristiques physico-chimiques des formationsLes analyses chimiques effectuées par le bureau Géotechnique Est sur des échantillons
de sédiments récupérés à différentes profondeurs des deux massifs (schisteux et gréseux) sont
présentées dans le tableau 4. Les analyses montrent une différence nette entre les deux entités
lithologiques. Les grés semblent plus basiques (pH compris entre 8,4 à 9,6) que les schistes
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
25
(8,3 à 8,9). les schistes présentent des concentrations en arsenic plus élevées que celles du
grés (2 à 3 fois plus élevées). La teneur en baryum est plus élevée dans les grés que dans les
schistes (5 à 40 fois plus élevée). Les teneurs en fer sont élevées dans les deux entités avec
des concentrations plus fortes dans le PZ2 par rapport aux autres piézomètres, alors que les
teneurs en nitrites et nitrates sont quasi-identiques pour la totalité des forages. Enfin, de fortes
fluctuations de la teneur en manganèse dans les deux formations qui oscillent entre 78 à 1422
ppm dans le compartiment schisteux et de 142 à 1225 ppm dans le compartiment gréseux.
Fig. 7. Fluctuation du niveau piézométrique des deux nappes d’eau.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
1er PARTIE: Matériel et Méthodes
26
pz30pz40
pz60
pz20pz21
945.00 945.20 945.40 945.60 945.80 946.00 946.20 946.402311.40
2311.60
2311.80
2312.00
2312.20
2312.40
2312.60
2312.80
2313.00
400405410415420425430435440445450455460465470475480485490495
Fig. 8. Carte piézométrique de la nappe de surface (aout 1998).
pz2
pz3pz4
pz5
pz6
945.00 945.20 945.40 945.60 945.80 946.00 946.20 946.402311.40
2311.60
2311.80
2312.00
2312.20
2312.40
2312.60
2312.80
2313.00
405410415420425430435440445450455460465470475480485490495
Fig. 9. Carte piézométrique de la nappe de surface (aout 1998).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
27
Tableau 4. Composition chimique des schistes et des grés.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
28
II. STRATEGIE D'ECHANTILLONNAGE
Des prélèvements d’eau destinés aux analyses physico-chimiques et biologiques ont
été réalisés de mai 1998 à mai 1999, selon un pas de temps mensuel. De plus, et afin d'évaluer
les fluctuations journalières du lixiviat brut, un suivi horaire a été effectué à la buse de sortie
de la décharge. Les prélèvements dans les bassins ont été réalisés entre 13 heures et 15 heures,
c'est à dire au moment où l'activité photosynthétique atteint son maximum.
III. PARAMETRES ABIOTIQUES
III. 1. Fixation et conservation des échantillons L'eau prélevée dans chaque bassin est conservée dans des flacons en verre d'une
capacité de 250 ml. Les flacons sont conservés à basse température. Les échantillons d'eau
destinés à des analyses de cations et d'ions métalliques sont acidifiés à l'acide nitrique
concentré 65 % (Rodier, 1984).
III. 2. . Méthodes d'analyseIII. 2. 1. Température, pH, potentiel Redox et Conductivité
Ces paramètres ont été mesurés in situ à l'aide d'une sonde multiparamètres de marque
WTW (Multiline P3 PH/LF-SET).
III. 2. 2. Oxygène dissous
La teneur en oxygène dissous est déterminé sur place par une sonde de type WTW
(Oxi330/SET).
III. 2. 3. Matières en suspension (MES)
La quantité de MES contenue dans l'eau a été déterminée par la méthode de filtration
(Rodier, 1984).
III. 2. 4. Ammonium
L'ion NH4+ est dosé par colorimétrie suite à une catalyse en milieu alcalin par une
solution de nutriprussiate de sodium. La longueur d'onde utilisée est de 630 nm.
III. 2. 5. Chlorures, fluor, sulfate, nitrite et nitrate
Ces anions sont dosés par chromatographie ionique (Dionex DX-100).
III. 2. 6. Azote total
Cet élément a été dosé par distillation (Rodier, 1984).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
29
III. 2. 7. Calcium, Sodium, Potassium et Magnésium
Ces cations majeurs ont été dosés par adsorption atomique à l'aide d'un
spectrophotomètre à flamme de type Perkin-Elmer (1103).
III. 2. 8. Fer, Manganèse, phosphore total et la teneur en métaux
lourds (Cr, Ni, Zn, Pb, Cu, Sn et Cd)
Le fer, le manganèse, le phosphore total et les métaux traces sont dosés par
colorimétrie sur un spectrophotomètre de type WTW (Photolab spektral, exactitude de 10 %).
III. 2. 9. DBO et DCO
La Demande Biologique en Oxygène (DBO) et la Demande Chimique en Oxygène
(DCO) ont été mesurées par photométrie à l'aide d'un spectrophotomètre de type WTW
(Photolab Spektral).
III. 2. 10. Les bicarbonates
Les bicarbonates ont été dosés par potentiométrie (Rodier, 1984).
IV. PARAMETRES BIOTIQUES
IV. 1. Phytoplancton
IV. 1. 1. Analyse qualitative
La collecte des échantillons destinés à l’identification du phytoplancton a été réalisée
en traînant à la surface de chaque bassin un filet à plancton dont le vide des mailles est de 40
µm. La détermination des espèces algales a été faite en utilisant les ouvrages suivants:
Bourrelly (1972, 1981 et 1985) et Germain (1981).
IV. 1. 2. Analyse quantitative
Les échantillons d’eau (250 ml) préfiltrés sur un nylon de 160 µm de vide des mailles
sont fixés au Lugol, ce fixateur permet la coloration des réserves amylacées et la conservation
du matériel cellulaire. Au laboratoire, 20 ml d’échantillon d'eau fixée sont placés dans une
cuve à sédimentation. La durée de sédimentation des échantillons n’a jamais été inférieure à
48 heures, afin de s’assurer de la récolte des phytoflagellés. Les comptages algaux ont été
réalisés à l’aide d’un microscope inversé en contraste de phase de marque (Leica DM IRB).
Généralement un comptage de 30 champs optiques (50 à 200 individus) est suffisant pour
minimiser les erreurs, car selon Lund et al. (1958), les erreurs liées au dénombrement
montrent qu’il suffit de dénombrer 16 individus dans le volume d’observation pour avoir une
précision de 50 %. Le nombre de cellules phytoplanctoniques a été calculé en utilisant la
formule suivante:
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
30
N = X/n * Rs * 1/v
Avec,
N: nombre de cellules par ml, X: nombre de cellules comptées, n: nombre de champs
optiques, Rs: rapport de la surface de la cuve sur la surface d'un champ optique et v: volume
d'eau sédimenté.
IV. 1. 3. Chlorophylle a
Les pigments chlorophylliens (Chl a) ont été extraits selon le protocole expérimental
proposé par le groupe de travail SCOR UNESCO (1966). L'eau prélevée est préalablement
stockée à l’abri de la lumière et à basse température. Les algues sont concentrées sur des
filtres en fibre de verre (Whatman GF/C de porosité 0,45 µm) par filtration de l’eau sous vide
avec addition de MgCO3. Les filtres sont ensuite broyés dans quelques ml d’acétone à 90 %,
puis placés à 4 °C dans l’obscurité. Après 6 heures, l'échantillon d'eau est centrifugé et
complété à 10 ml par l’extrait acétonique. Les mesures ont été faites sur un spectrophotomètre
BECKMAN. Le calcul des teneurs en pigments a été réalisé en utilisant les équations
proposées par SCOR-UNESCO (1966).
IV. 1. 4. Estimation de la biomasse phytoplanctonique
La biomasse phytoplanctonique a été estimée selon le protocole de Lohman (1908) qui
consiste à évaluer le volume cellulaire de chaque espèce en rapportant celle-ci à une où
plusieurs formes géométriques calculables à partir des dimensions moyennes. Le facteur de
conversion des biovolumes en biomasses est: 1 µm3 = 0,12 10-6 µgC.
Algues biovolume (µm3)Chlamydomonas sp. 314Coelastrum sp. 78,5Chaetoptilis sp. 153,86Kircheneilleria sp. 153,86Sphaeryocystis sp. 314Phacus sp. 706,5Euglena sp. 1256Trachylomonas sp. 1256Stephanodiscus sp. 113,04
Tableau 5. Biovolumes spécifiques calculés pour les espèces algales les plus abondantes
dans les bassins d’Etueffont.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
31
IV. 1. 5. Composition biochimique du matériel particulaire (0,45-
160µm)
A partir d'un échantillon d'eau (250 ml) préfiltrée sur un tamis de 160 µm de porosité
de maille et recueillis sur des filtres Whatman GF/C, préalablement decontaminé pendant 6
heures à 500 °C. Les concentrations en protéines et glucides ont été déterminées (fig. 10).
Le dosage des glucides est réalisé selon la méthode de Dubois et al. (1956) et
développé par Moal et al. (1985), le standard utilisé étant le glucose. La concentration en
protéines est déterminée par la méthode de Lowry (1951), le standard utilisé étant l'albumine
(BSA).
Fig. 10. Dosage de la composition biochimique du matériel particulaire (Moal, 1985).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
32
IV. 2. Zooplancton
IV. 2. 1. Analyse qualitative
Les prélèvements de zooplancton ont été réalisés par des traits de filet horizontaux et
des traits verticaux ascendants du fond jusqu’à la surface. Le filet à plancton utilisé est
constitué d’un tissu synthétique de 30 cm de largeur, de 1 m de long et de 85 µm de vide des
mailles. La détermination des espèces a été faite en se référant aux clés de détermination
d'Amoros (1984) pour les Cladocères; Pourriot, Francez (1986) et Kost (1978) pour les
Rotifères.
IV. 2. 2. Analyse quantitative
Plusieurs études portant sur le zooplancton soulignent que les méthodes de
prélèvement posent beaucoup de problèmes. La récolte du zooplancton a été faite par
filtration de 10 l d’eau à l’aide d’un filet à plancton (le vide des mailles est de 85 µm). Le
zooplancton récupéré dans le collecteur est immédiatement fixé au formaldéhyde (à 5 % de
concentration) puis conservé jusqu’au moment du comptage. La quantité de zooplancton a été
déterminée par l’observation de 20 à 30 ml d’échantillon placé dans une boîte de pétri à fond
quadrillé à l’aide d’un microscope inversé (Leica DM IRB, objectif 40×).
IV. 2. 3. Estimation des biomasses
IV. 2. 3. 1. Les Rotifères
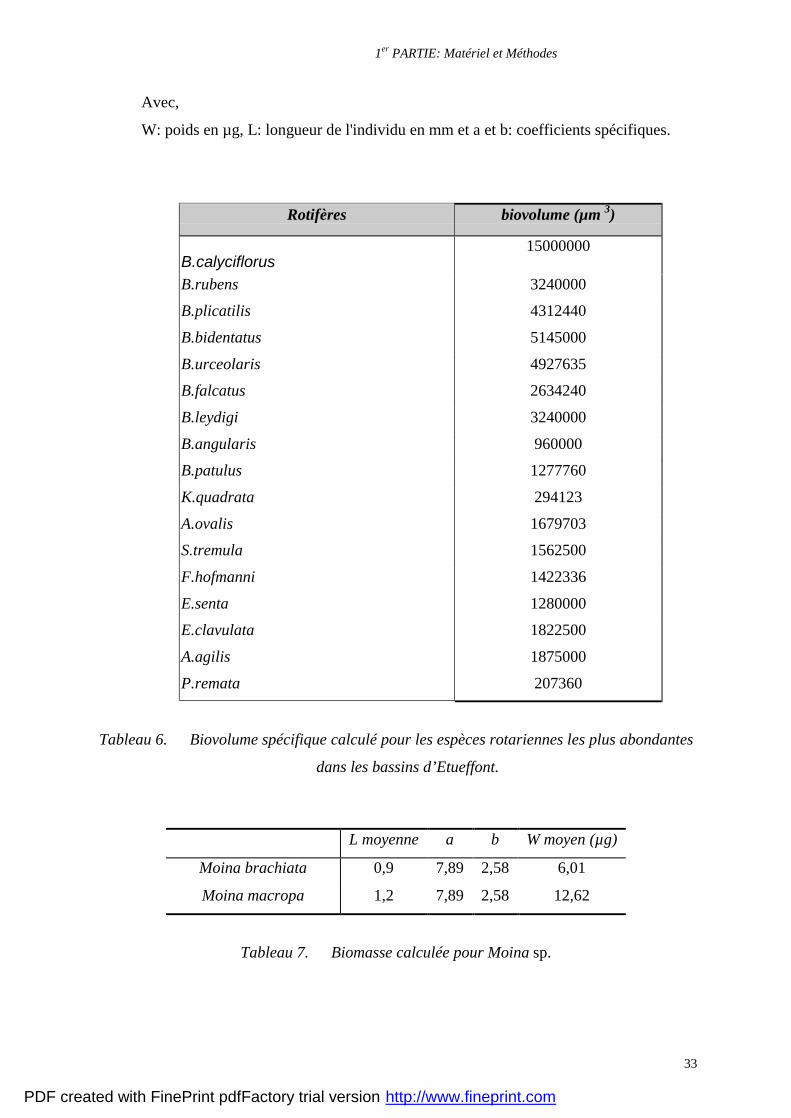
Les biomasses des Rotifères ont été calculées (± 10 %) à partir de biovolumes de
chaque espèce, en assimilant ces organismes à des formes géométriques générales de volume
aisément calculable (Botttrel et al., 1976). Le biovolume calculé est converti en poids par
conversion d'unité (1µg = 106 µm3) du fait que le zooplancton a une densité voisine de l'eau.
IV. 2. 3. 2. Les Cladocères
L'évaluation de la biomasse des Cladocères peut s'effectuer selon plusieurs procédés
(Osmera, 1966; Edmondson et Winberg, 1971; Howmiller, 1972; Dumont et al., 1975 et
Bottrel et al., 1976), la biomasse des Moinidae a été calculée par la méthode indirecte
développée par Bottrel et al. (1976) qui se base sur une équation mathématique reliant la
longueur de l'individu à son poids. La formule utilisée est la suivante:
W = a.Lb
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
1er PARTIE: Matériel et Méthodes
33
Avec,
W: poids en µg, L: longueur de l'individu en mm et a et b: coefficients spécifiques.
Rotifères biovolume (µm 3)
B.calyciflorus15000000
B.rubens 3240000
B.plicatilis 4312440
B.bidentatus 5145000
B.urceolaris 4927635
B.falcatus 2634240
B.leydigi 3240000
B.angularis 960000
B.patulus 1277760
K.quadrata 294123
A.ovalis 1679703
S.tremula 1562500
F.hofmanni 1422336
E.senta 1280000
E.clavulata 1822500
A.agilis 1875000
P.remata 207360
Tableau 6. Biovolume spécifique calculé pour les espèces rotariennes les plus abondantes
dans les bassins d’Etueffont.
L moyenne a b W moyen (µg)
Moina brachiata 0,9 7,89 2,58 6,01
Moina macropa 1,2 7,89 2,58 12,62
Tableau 7. Biomasse calculée pour Moina sp.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
34
IV. 3. Bacterioplancton
IV. 3. 1. Analyse qualitative
Un échantillon d’eau (1 l) a été prélevé dans chaque bassin à l’aide d’une bouteille
stérile. Après homogénéisation de l’échantillon, une prise d’essai de 1 ml de l’échantillon pur
et sa dilution (10-1) sont incorporées dans environ 15 ml de gélose nutritive, maintenue en
surfusion à environ 44 °C (± 2°C). Les 2 boites ensemencées (échantillon pur et dilué à 10-1)
sont mises à incuber à 22 °C ± 1 °C.
Les résultats sont exprimés en nombre moyen d’unités formant colonies par ml
d’échantillon (UFC/mL) à 3 et 10 jours.
IV. 3. 2. Analyse quantitative
Une fraction d’aliquote (50 ml) de chaque prélèvement a été préfiltrée sur une toile en
nylon de 160 µm de vide de maille, puis fixée au formaldéhyde (concentration finale ± 2 %).
Au laboratoire 5 ml de chaque échantillon ont été colorés au DAPI (4,6 diamino 2
phénylindol) et filtrés sur une membrane millipore noire de porosité 0,2 µm. Afin d’avoir une
dispersion homogène des cellules, un filtre noir en acétate de cellulose (Sartorius, porosité 1.2
µm) est placé au-dessous du filtre noir. Après une incubation de 15 mn, le filtre est monté
entre lame et lamelle et immédiatement congelé à –25 °C. le dénombrement bactérien a été
effectué à l’aide d’un microscope inversé à épifluorescence (Leica DM IRB) en ultra violet
(Porter et Feig, 1980). Le filtre utilisé est de type: UG1- DM400-L420 permettant
l'observation en UV des bactéries hétérotrophes qui émettent une intense fluorescence bleue
brillante alors que le matériel détritique est coloré en jaune pâle.
Les dénombrements bactériens ont toujours porté sur 500 à 800 bactéries (± 10 % de
précision). Le nombre de cellules compté est ensuite transformé en nombre de cellules par ml
à l'aide de la formule suivante:
N = n.S/s.V
Avec,
n: nombre de cellules dénombrées,
N: nombre de bactéries par ml,
s: surface observée en µm2,
S: surface filtrante du filtre en µm2,
V: Volume d'eau filtrée en ml.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

1er PARTIE: Matériel et Méthodes
35
IV. 3. 3. Estimation de la biomasse bactérienne
Pour chaque communauté bactérienne, on a estimé un bio-volume moyen en mesurant
au minimum une centaine de cellules à l'aide d'un micromètre oculaire. Le bio-volume
cellulaire moyen a été calculé à partir de la formule proposée par Jassby (1975) (cité par
Carrias 1996) (Biov. cell. = π/4 * l2 * (L-(l/3))). Pour les formes cocci, L = 1. Les biomasses
sont estimées en utilisant le facteur de conversion suivant : 0,56 10-6 µgC/ µm3 (Bratback
1985, Simon et Azam 1989). Le bio-volume bactérien calculé est de 0,39 µm3 ce qui est
supérieur à celui enregistré pour les milieux oligotrophes et mésotrophes (Jordan et Likens,
1980: Scavia et Laird, 1987; Nagata, 1988; Ochs et al., 1995; Krstulovic et al., 1995 et
Carrias, 1996) et proche des valeurs données dans la littérature pour les cas extrêmes (0,02 à
0,80 µm3) (Johansen, 1983; Andersson, 1983; Nagata, 1988; et Amblard et al., 1994).
IV. 3. 4. Fiabilité de la méthode à épifluorescence
Le dénombrement par le biais de la microscopie à épifluorescence après une coloration
au DAPI permet la prise en compte de bactéries alors que la méthode classique sur milieu
gélosé les occulte. C’est le cas de bactéries ayant des exigences de croissance et un
métabolisme particulier ou étant stressées, conservent des fonctions physiologiques actives, et
sont donc viables, mais non cultivables. Toutefois, un dénombrement parallèle en milieu
solide des bactéries pour quelques échantillons nous a permis de confirmer les travaux publiés
par plusieurs auteurs (Haulidier et al., 1988; Servais et al., 1992) selon lesquels le
dénombrement des bactéries sur gélose conduit à une sous-estimation considérable qui peut
atteindre un facteur 1000. En effet au cours de nos essais, la mise en culture en milieu solide a
abouti à des densités bactériennes plus faibles que celles enregistrées par la méthode à
épifluorescence et on enregistre souvent une très grande hétérogénéité des résultats entre les
deux méthodes (écart - type souvent supérieur à 50).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
36
I. INTRODUCTIONLa mise en décharge des déchets issus de l’activité anthropique a été pratiquée depuis
des siècles. De mise en œuvre aisée, cette technique autorise une élimination efficace et peu
coûteuse des résidus urbains. Cette gestion conduit toutefois à l’émanation d’odeurs
nauséabondes, à la production de biogaz et surtout de lixiviat. Plus précisément, le jus de
décharge une fois injecté dans les milieux aquatiques, peut induire des phases de
dysfonctionnement de ces écosystèmes d’autant plus dramatiques qu’elles sont aggravées par
l’intensification croissante des activités urbaines. Conséquence directe, plusieurs pays dont la
France ont adopté des lois et règlements (ISO 14000) stipulant l’interdiction de la mise en
décharge des déchets à partir du 1er Juillet 2002.
Par ailleurs, les travaux ayant trait à la composition des lixiviats réalisés tant en
laboratoire (Bookter et Ham, 1982; Blaky, 1992) qu’in situ (Jasper et al., 1985) ont permis de
mettre en évidence une relation étroite entre le climat, l’hydrologie, l’origine et la technique
d’enfouissement des déchets d’une part et la composition chimique du lixiviat d’autre part. De
plus, certaines études, du fait de leur grande hétérogénéité ont abouti à la formulation
d’hypothèses souvent contradictoires. Ainsi, si Chian et Dewalle (1976), Navarro et al. (1985)
et Blanchard et al. (1989) rapportent un impact direct de l’âge de la décharge sur la nature des
lixiviats, la division royale de traitement des déchets en Irlande indique une absence totale de
relation entre ces deux paramètres (KCWS, 1992). L’effet des conditions climatiques et en
particulier des pluies sur la régénération des lixiviats n’a pas échappé à cette controverse. De
fait, alors que des études sur plusieurs décharges en Allemagne indiquent que ces deux
paramètres étaient indépendants (Ehrig, 1978), d’autres comme Mattias et Nilsson (1997)
rapportent des résultats totalement opposés.
En conséquence, nous nous sommes proposés dans ce chapitre d’étudier l’évolution
temporelle de divers paramètres caractérisant les lixiviats d’une décharge à ciel ouvert
(Territoire de Belfort, France) et d’identifier leurs interactions avec les précipitations et l’âge
de ce site. Afin de détecter les variations à très court terme des divers paramètres étudiés, un
échantillonnage à pas de temps horaire a été réalisé le 13 juin 1998. Enfin, et afin de
confirmer l'effet de l'âge des déchets dans la caractérisation du lixiviat la nouvelle alvéole
installée à coté de la décharge nous a servi de station pilote.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
37
II. CHOIX DES POINTS DE PRELEVEMENTLes prélèvements sont effectués par immersion dans la zone euphotique des bassins
(B1, B2, B3 et B4), du lixiviat brut (E), à la sortie du quatrième bassin (S), à l’amont (avant
rejet) (A) puis enfin à l’aval du ruisseau (V) où se déverse le lixiviat traité. L’échantillonnage
est effectué d’une façon manuelle à l’aide d’un flacon en plastique de 5 l à large ouverture.
III. CARACTERISTIQUES DE LA STATION PILOTELa station pilote a une surface de 40 sur 15 m et une profondeur de 6 m. Une
membrane etanche en PEHD de 2mm a été installée au fond de l'alvéole encadrée de deux
nappes de géotextile. Enfin, la couche de geotextile a été couverte par une couche de graviers
(0,4 m) équipée de drains PEHD. Le fond de l'alvéole est légèrement incliné (1:100)
permettant le drainage du lixiviat produit à travers les tuyaux de drainage. Le lixiviat produit
rejoint les bassins de traitement. En 2000, l’épaisseur de déchets enregistrée dans la station
pilote est de 2 m.
IV. RESULTATS IV. 1. Pluies
Les valeurs pluviométriques enregistrées au cours de notre étude varient de 60 mm en
juin 1998 à 360 mm en janvier 1999 (moyenne ± écartype, m ± δ = 165 ± 64 mm) (fig. 11a).
Ces résultats indiquent que le site d’Etueffont est bien arrosé.
IV. 2. Débits
Les résultats des débits mesurés dans le jus de décharge évoluent de 0,06 à 2,5 l. s-1 (m
± δ = 0,45 ± 0,45 l. s-1) (fig. 11a). L’examen comparé de l’évolution des débits avec ceux de
la pluie met en évidence deux pics (1,12 l. s-1 en septembre 1998 et 2,5 l. s-1 en février 1999)
qui ne correspondent pas aux valeurs maximales de pluviosité.
IV. 3. Température
Les températures enregistrées dans les lixiviats varient de 7,4 à 19,5 °C (m ± δ = 14 ±
5 °C) (fig. 11b). Les valeurs maximales sont obtenues au cours de la saison estivale
parallèlement à une faible pluviosité et un débit réduit. Durant le suivi horaire les
températures évoluent de 12,9 à 28,5 °C (m ± δ = 22,4 ± 5,46 °C). Les valeurs maximales de
températures ont été enregistrées entre 12 heures et 17 heures (fig. 14a).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
38
IV. 4. Oxygène dissous
Les valeurs d'oxygène dissous mesurées à la sortie de décharge varient de 0,5 à 11
mg. l-1 (m ± δ = 3,64 ± 3,09 mg. l-1) (fig. 11c). L'évolution saisonnière de ce paramètre est
caractérisée par de faibles valeurs en été et de fortes valeurs en hiver. Sur un pas horaire, il
apparaît que l'oxygène dissous fluctue entre 5 et 9 mg. l-1 (m ± δ = 6,72 ± 2,94 mg. l-1) (fig.
14b). Il semble d'après ce suivi journalier que l'oxygène dissous diminue au cours de la
journée et atteint un minimum entre 11 heures et 18 heures. Enfin, l'évolution du potentiel
redox montre une évolution mensuelle et horaire similaire à celle de l'oxygène dissous.
IV. 5. Conductivité électrique
Au cours du suivi mensuel, les valeurs de la conductivité électrique enregistrées dans
les lixiviats oscillent entre 2100 à 3700 µS. cm-1 (m ± δ = 4374 ± 1416 µS. cm-1) (fig. 11d).
Les valeurs les plus élevées ont été enregistrées en été (6600 µS. cm-1 en juin 1999, 6800 µS.
cm-1 en juillet 1998 et 7300 µS. cm-1 en août 1998). C'est en hiver, par contre, qu'ont été
mesurées les valeurs les plus faibles de conductivité (2400 µS. cm-1 en janvier 1999 et 2100
µS. cm-1 en février 1999). Au cours de la saison automnale, on obtient une conductivité
électrique de 5080 µS. cm-1 (novembre 1998). Pendant le suivi horaire la conductivité
électrique diminue légèrement au fil de la journée en passant de 3060 µS. cm-1 le matin (8
heures) à 2850 µS. cm-1 le soir (20 heures) (fig. 14d). Les variations inter-annuelles de ce
paramètre indiquent une augmentation des valeurs en fonction du vieillissement de la
décharge (de 5000 µS. cm-1 en 1993 à 7000 µS. cm-1 en 1998) (tab. 9).
IV. 6. pH
Les valeurs de pH relevées dans le lixiviat varient de 5,1 à 8,2 (m ± δ = 7,09 ± 1,08)
(fig. 11b). En été, les valeurs de pH sont voisines de 8. En hiver, les lixiviats produits sont
légèrement acides (pH = 5 en janvier 1999). Au printemps et dès le mois d'avril 1999, le
lixiviat reprend légèrement sa basicité estivale (pH = 7,4) (fig. 11e). Le suivi journalier de pH
indique que ce dernier fluctue entre 8,14 et 8,23 (m ± δ = 8,17 ± 0,03). Les valeurs maximales
de pH ont été enregistrées entre 10 heures et 16 heures, ces valeurs coïncident avec les fortes
températures enregistrées dans le lixiviat (fig. 14a). L’évolution annuelle de cet élément
présente une légère augmentation passant de 8 en 1993 à 8,21 en 1998 (tab. 9).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
39
IV. 7. Eléments inorganiques
Les teneurs en chlorures évoluent de 150 à 839 mg. l-1 (m ± δ = 480,95 ± 263,73
mg. l-1) (fig. 11f). Les plus fortes valeurs sont observées en été (810 mg. l-1 en juin 1998). La
teneur maximale en chlorures du lixiviat a été enregistrée en novembre 1998. Les fortes
valeurs de chlorures coïncident avec les fortes valeurs de conductivités enregistrées dans le
lixiviat. L'évolution horaire montre que les teneurs du jus de décharge en ions chlorures
varient de 265 à 295 mg. l-1 (m ± δ = 281,34 ± 10,84 mg. l-1) (fig. 14c). Les concentrations
maximales de cet élément ont été enregistrées entre 10 heures et 17 heures. Les teneurs en
sulfates mesurées dans le lixiviat varient de 93 à 233 mg. l-1 (m ± δ = 185,47 ± 77,72 mg. l-1)
(fig. 11g). Il semble que cet élément ne présente pas d'évolution saisonnière nette. Les valeurs
maximales ont été enregistrées d'août 1998 en janvier 1999. En effet l'évolution horaire
montre un enrichissement du lixiviat en sulfate de 8 heures à 20 heures passant de 213 mg. l-
1à 8 heures à 239 mg. l-1 à 16 heures. A partir de 16 heures les teneurs en sulfates diminuent
légèrement et atteignent 224 mg. l-1 à 20 heures (fig. 14c). Les concentrations de fer dans le
lixiviat oscillent entre 0,2 et 10,3 mg. l-1 (m ± δ = 4,37 ± 3,63 mg. l-1). L'évolution saisonnière
de cet élément est très marquée avec des teneurs élevées en été et faibles en hiver (fig. 12b).
IV. 8. Teneurs en éléments métalliques
Durant cette étude, les 3 éléments métalliques les plus abondants dans le lixiviat de la
décharge d'Etueffont et les plus cités en littérature ont été suivis (Cu, Zn et Ni). Les valeurs
relatives à la teneur du jus de décharge en zinc oscillent entre 0,3 et 1,85 mg. l-1 (m ± δ= 0,99
± 0,45 mg. l-1) (fig. 11h). Les concentrations maximales en zinc correspondent aux maxima de
pluviosité observées au cours de notre suivi. Celles relatives au cuivre évoluent entre 0 et 1,77
mg. l-1 (m ± δ= 0,46 ± 0,52 mg. l-1). La figure 12a montre que les maxima en cuivre
correspondent aux minima de débits enregistrés pour les lixiviats. Enfin le nickel a une
évolution similaire a celle observée pour le cuivre avec des minima en périodes pluvieuses
(fig. 12c). Les teneurs en nickel dans le lixiviat varient de 0,17 à 8,64 mg/l (m ± δ = 2,08 ±
2,86 mg/l).
IV. 9. Teneurs en éléments nutritifs
Les teneurs en nitrate ont varié entre 0,16 et 135 mg. l-1 (m ± δ = 23.27 ± 43.63
mg. l-1) (fig. 12e). Les concentrations en azote total (NT) du lixiviat varient de 76 à 273 mg. l-1
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
40
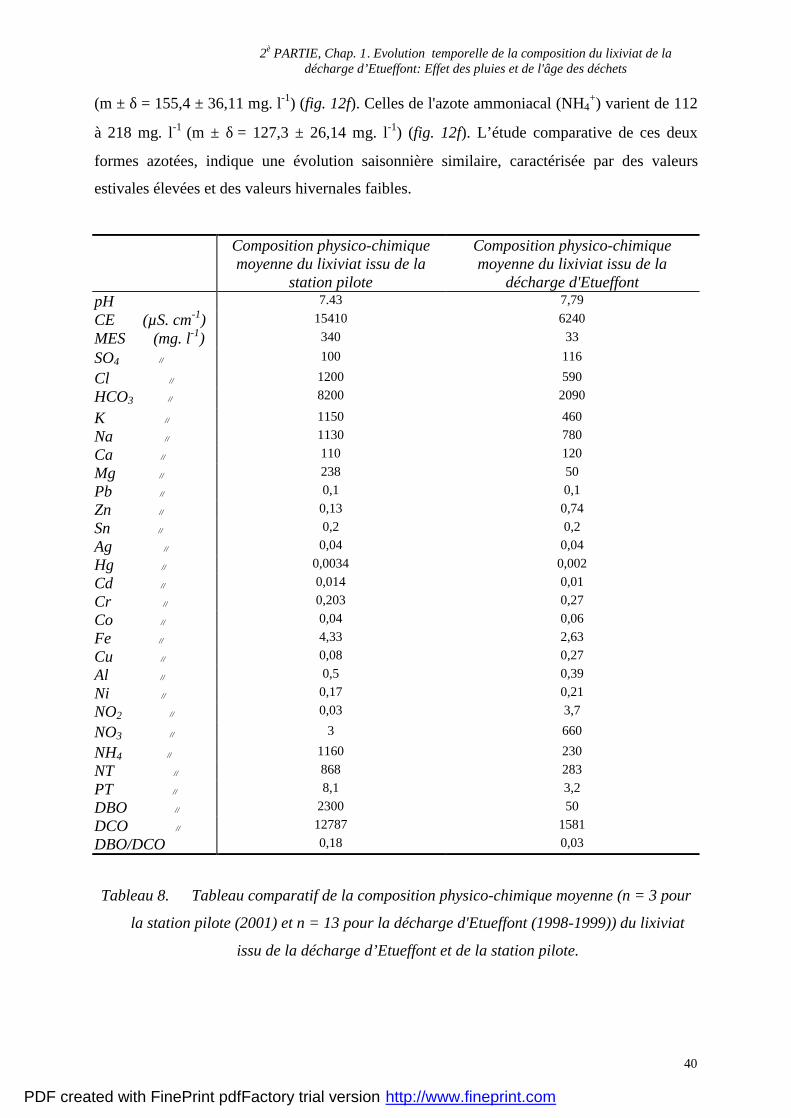
(m ± δ = 155,4 ± 36,11 mg. l-1) (fig. 12f). Celles de l'azote ammoniacal (NH4+) varient de 112
à 218 mg. l-1 (m ± δ = 127,3 ± 26,14 mg. l-1) (fig. 12f). L’étude comparative de ces deux
formes azotées, indique une évolution saisonnière similaire, caractérisée par des valeurs
estivales élevées et des valeurs hivernales faibles.
Composition physico-chimiquemoyenne du lixiviat issu de la
station pilote
Composition physico-chimiquemoyenne du lixiviat issu de la
décharge d'EtueffontpH 7.43 7,79CE (µS. cm-1) 15410 6240MES (mg. l-1) 340 33
SO4 // 100 116
Cl // 1200 590HCO3 // 8200 2090
K // 1150 460Na // 1130 780Ca // 110 120Mg // 238 50Pb // 0,1 0,1Zn // 0,13 0,74Sn
// 0,2 0,2Ag // 0,04 0,04Hg // 0,0034 0,002Cd // 0,014 0,01Cr // 0,203 0,27Co // 0,04 0,06Fe // 4,33 2,63Cu // 0,08 0,27Al // 0,5 0,39Ni // 0,17 0,21NO2 // 0,03 3,7
NO3
// 3 660
NH4
// 1160 230NT // 868 283PT // 8,1 3,2DBO // 2300 50DCO // 12787 1581DBO/DCO 0,18 0,03
Tableau 8. Tableau comparatif de la composition physico-chimique moyenne (n = 3 pour
la station pilote (2001) et n = 13 pour la décharge d'Etueffont (1998-1999)) du lixiviat
issu de la décharge d’Etueffont et de la station pilote.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
41
En novembre 1998, on assiste à une augmentation brutale de la concentration de ces
deux nutriments, vraisemblablement due à un apport excessif d'azote par la décharge, puis les
teneurs diminuent en hiver atteignant des minima de 76 mg. l-1 de NT et 65 mg. l-1 de NH4+
(janvier 1999). A partir de mars 1999, les teneurs en NH4+ et en NT augmentent avec
respectivement 140 et 126 mg. l-1.
Toutefois les teneurs en orthophosphates sont quasi nulles alors que les teneurs en
phosphate total ont une évolution similaire à celle de l'azote total avec des lixiviats plus riches
en phosphate en été et en automne et des lixiviats peu phosphorés en hiver en relation avec
l'effet de dilution. Toutefois, les teneurs en PT ont varié entre 0,3 à 11 mg. l-1 (m ± δ = 0,41 ±
0,86 mg. l-1) (fig. 11b).
IV. 10. Matières en suspension
Les valeurs de matières en suspension varient de 17 à 340 mg. l-1 (m ± δ = 81,41 ±
94,36 mg. l-1). Les concentrations maximales de ce paramètre ont été enregistrées en
septembre et avril 1999 avec respectivement 225 et 380 mg. l-1 (fig. 13). Ces fortes valeurs des
matières en suspension coïncident avec les maxima de débit enregistrés dans les lixiviats.
IV. 11. Bicarbonates
Les teneurs en bicarbonates du jus de décharge fluctuent entre 900 et 2997 mg. l-1 (m
± δ = 2012 ± 696 mg. l-1). En été, on enregistre des valeurs élevées de bicarbonates de l'ordre
de 2900 mg. l-1. En hiver, on assiste à une diminution des teneurs en bicarbonates du lixiviat
(fig. 12d).
IV. 12. Teneurs en matières organiques
Les valeurs de la teneur en matières organiques oxydables (DBO) importées par la
décharge varient de 25 à 187 mg. l-1 (m ± δ = 62 ± 44,5 mg. l-1) (fig. 12g). Les valeurs les plus
élevées ont été enregistrées en novembre 1998, avec 187 mg. l-1. En été, on observe une
légère diminution des valeurs du flux de DBO de l'ordre de 25 mg. l-1 qui coïncident avec les
maxima de températures ambiantes enregistrées sur le site. En automne (octobre et novembre
1998), on assiste à une augmentation des teneurs en DBO dans le jus de décharge. Celles-ci
commencent à diminuer avec l’arrivée de l’hiver tout en demeurant élevées. L’évolution
annuelle de ce paramètre montre une décroissance des teneurs en fonction de l’âge de la
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
42
décharge, passant de 135 mg. l-1 en 1995 à 35 mg. l-1 en 1998 (tab. 9). En terme de flux les
apport en DBO par la décharge en unité de surface varient de 5 à 25 Kg. j-1. (m2)-1. Les
concentrations de la DCO relevées dans le jus de décharge fluctuent entre 313 et 1550 mg. l-1
(m ± δ = 868 ± 482 mg. l-1) (fig. 12h). L'évolution saisonnière de ce paramètre, semble être
opposée à celle de la DBO en été. De plus, sur une échelle interannuelle, la DCO augmente de
881 mg. l-1 en 1995 à 1550 mg. l-1 en 1998 (tab. 9). Le flux de DCO importé par la décharge
varie de 6 à 331 Kg. J-1. (m2)-1. L'évolution horaire des teneurs en DCO à la mi-journée
semble être liée à l'élévation des températures. Le rapport DBO/DCO varie de 0,01 à 0,17 (m
± δ = 0,096 ± 0,061). En été, on enregistre des valeurs faibles (0,01 en mai, juin et juillet
1998). En hiver, les valeurs sont plus élevées (0,16 en janvier 1999 et 0,17 en février 1999) en
relation avec l'élévation hivernale de la DBO. L'évolution inter-annuelle montre une
diminution du rapport de 1995 (0,15) à 1998 (0,02).
IV. 13. Bactéries hétérotrophes
Le peuplement bactérien est essentiellement représenté par des cocci (77 % du
peuplement) avec une dominance des formes libres à grande taille (≥ 0,5 µm pour les cocci et
≥ 0,9 pour les bacilles). La plus forte abondance des formes bacilles a été observée en
décembre 1998 (42 % du peuplement). L’effectif bactérien dans le lixiviat brut oscille entre
0,04.106 bactéries. ml-1 observé en avril 1999 et 0,34.106 bactéries. ml-1 enregistré en octobre
1998 (m ± σ = 0,132 ± 0,122) (fig. 13). Les maxima d’abondance ont été enregistrés en
période estivale. La biomasse bactérienne varie de 8,78 à 77,51 µgC. l-1, (m ± σ = 28,94 ±
26,79 µgC. l-1).
V. DISCUSSIONV. 1. Relation pluies et débit du lixiviat
Pour déterminer le rôle des précipitations atmosphériques dans la génération des
lixiviats en hiver, nous avons recherché s’il existait d'éventuelles relations entre les
précipitations et divers paramètres abiotiques. Il apparaît une absence de relation
statistiquement significative entre ces paramètres (r = - 0,3). En effet, l’évolution temporelle
des valeurs des débits et des précipitations met en évidence l’existence d’un faible décalage
entre les pics observés pour ces deux paramètres (fig. 11a). La décharge ne répond pas
immédiatement aux pluies mais accuse un retard en raison d’une part de la faible teneur en
eau des déchets et d'autre part de la décharge. De plus, cette absence de corrélation s’explique
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
43
partiellement par l’existence d’apports hydriques collatéraux superficiels, provenant
essentiellement du bassin versant.
Fig. 11. Evolution temporelle des pluies (a), de la température (b), de la conductivitéélectrique (d), des teneurs en PT (b), de O2 (c), du pH (e), des Cl (f), des SO4 (g) et du Zn(h) en fonction du débit dans le lixiviat (toutes les concentrations sont exprimées en mg/l
le débit en l/s et les pluies en mm).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
44
Fig. 12. Evolution des teneurs en Cu (a), en Fe (b), en Ni (c), en HCO3 (d), en NO3 (e), enNH4 (f), en NT (f) de la DBO (g) et de la DCO (h) en fonction du débit dans le lixiviat(Toutes les concentrations sont exprimées en mg/l, le débit en l/s et les pluies en mm).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
45
V. 2. Evolution des substances organiques produites par la décharge
La teneur en matières organiques exprimée en DBO et DCO contenus dans le lixiviat
de la décharge d’Etueffont sont comparables à ceux trouvés par Ehrig (1983) pour les
décharge en phase de méthagenèse. Toutefois, plusieurs auteurs (Chian et DeWalle, 1977;
Harmsen, 1987) ont monté que les matières organiques contenues dans le lixiviat de
décharges d’ordures ménagères sont dominés par les acides gras volatiles (AGV). Au vu de
nos résultats, il apparaît que c’est en période estivale alors que la température du jus de
décharge est la plus élevée que le flux de matières organiques biodégradables (DBO) sortant
de la décharge présente ses valeurs maximales (fig. 12g). Cette augmentation de la charge
organique est associée au maximum du développement de l'abondance bactérienne (fig. 13) et
une diminution des teneurs en oxygène succède à la consommation de ce dernier par les
décomposeurs. Cette évolution simultanée semble indiquer que le réchauffement des eaux a
été à l’origine d’une intensification de l’activité hétérotrophe bactérienne. Notre hypothèse est
corroborée par divers travaux effectués dans le cas des eaux douces et attribuant un rôle
prépondérant de la température des eaux dont l’élévation stimulerait les activités
enzymatiques bactériennes (Kato, 1985, Marvalin et al., 1989). De plus, l’absence probable
de protistes photosynthétiques dans le jus de décharge écarterait toute possibilité d’offrir aux
bactéries du carbone organique dissous nécessaire à leur développement tant par leurs
excrétions que par la lyse des cellules sénescentes (Meffert et Overbeck, 1979; Jones et al.,
1983; Feuillade et al., 1988).
Enfin, des travaux réalisés sur d’autres sites de décharge ont permis de mettre en
évidence une augmentation de l’activité reminéralisatrice bactérienne à la suite du
réchauffement des lixiviats (Straub et Lynch, 1982; Mattias et Nilsson, 1997). Ainsi, dans la
mesure où la décharge d’Etueffont est une décharge à ciel ouvert donc sensible aux variations
climatiques, il semblerait qu’en période de faible débit, c’est vraisemblablement l’activité
hétérotrophe bactérienne qui module la composition physico-chimique des lixiviats.
V. 3. Evolution mensuelle des substances inorganiques produites par la décharge
Les teneurs en substances inorganiques dans les lixiviats sont très dépendantes de
l’état de la décharge et de la teneur en substances organiques dissoutes (Christensen et al.,
2001). Toutefois, les teneurs en Mg, Ca et Na et enregistrés dans le lixiviat d’Etueffont sont
faibles et leurs évolutions saisonnières montrent bien des fortes valeurs en été,
vraisemblablement liées à la faible teneur en matière organique qui complexe ces cations.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
46
Fig. 13. Evolution saisonnière des MES et de l'abondance bactérienne dans le lixiviat.
Les teneurs en sulfates sont plus faibles en mai, juin et juillet 1998, en raison de leur
réduction en sulfures par les bactéries. Par ailleurs, l’élévation des débits induit un effet de
dilution de certains paramètres, comme le confirme la présence de relations statistiquement
significatives entre tous les éléments chimiques suivis d’une part et les débits d’autre part.
Seul le zinc est statistiquement lié aux débits (r = 0,42; pour 11 ddl) et aux précipitations (r =
0,74; pour 11 ddl). L’explication de l’existence de telles relations réside dans l’hétérogénéité
de distribution des dépôts de zinc dans la décharge nécessitant de fortes pluviosités pour
optimiser la lixiviation de ce métal. Durant tout le suivi on enregistre des teneurs en Zn2+ dans
le lixiviat similaires à celles trouvées par Park et al. (1999), alors que les teneurs en Fe2+ sont
élevées et peuvent atteindre jusqu'à 10 mg. l-1 (juillet 1998). Ces teneurs élevées en Fe2+ sont
probablement dues à la forte présence des matières organiques comme l'a suggéré Buffle
(1977). De plus, la distribution de la conductivité et des concentrations en chlorures du
lixiviat apparaît semblable, ce qui se traduit par l’existence d’une corrélation linéaire
statistiquement significative entre les deux paramètres (r = 0,93; pour 11 ddl). Si une telle
corrélation ne peut prétendre établir de liens de causalité entre processus, ce résultat semble
indiquer que les fortes teneurs en chlorures influent sur la minéralisation du lixiviat. Dans le
même ordre d’idées, l’évolution semblable des concentrations en azote total et en ammonium
(r = 0,81; pour 11 ddl) suggèrerait que l’azote lixivié par la décharge l’est majoritairement
sous forme ammoniacale.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
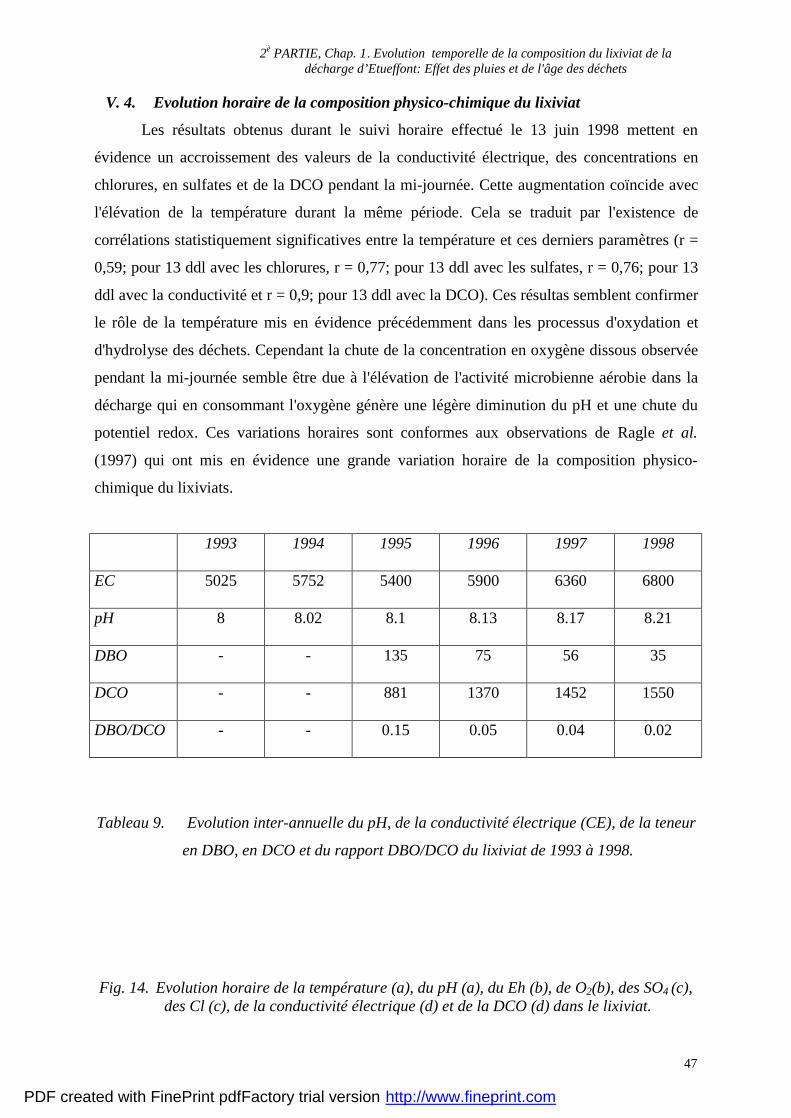
2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
47
V. 4. Evolution horaire de la composition physico-chimique du lixiviat
Les résultats obtenus durant le suivi horaire effectué le 13 juin 1998 mettent en
évidence un accroissement des valeurs de la conductivité électrique, des concentrations en
chlorures, en sulfates et de la DCO pendant la mi-journée. Cette augmentation coïncide avec
l'élévation de la température durant la même période. Cela se traduit par l'existence de
corrélations statistiquement significatives entre la température et ces derniers paramètres (r =
0,59; pour 13 ddl avec les chlorures, r = 0,77; pour 13 ddl avec les sulfates, r = 0,76; pour 13
ddl avec la conductivité et r = 0,9; pour 13 ddl avec la DCO). Ces résultas semblent confirmer
le rôle de la température mis en évidence précédemment dans les processus d'oxydation et
d'hydrolyse des déchets. Cependant la chute de la concentration en oxygène dissous observée
pendant la mi-journée semble être due à l'élévation de l'activité microbienne aérobie dans la
décharge qui en consommant l'oxygène génère une légère diminution du pH et une chute du
potentiel redox. Ces variations horaires sont conformes aux observations de Ragle et al.
(1997) qui ont mis en évidence une grande variation horaire de la composition physico-
chimique du lixiviats.
1993 1994 1995 1996 1997 1998
EC 5025 5752 5400 5900 6360 6800
pH 8 8.02 8.1 8.13 8.17 8.21
DBO - - 135 75 56 35
DCO - - 881 1370 1452 1550
DBO/DCO - - 0.15 0.05 0.04 0.02
Tableau 9. Evolution inter-annuelle du pH, de la conductivité électrique (CE), de la teneur
en DBO, en DCO et du rapport DBO/DCO du lixiviat de 1993 à 1998.
Fig. 14. Evolution horaire de la température (a), du pH (a), du Eh (b), de O2(b), des SO4 (c),des Cl (c), de la conductivité électrique (d) et de la DCO (d) dans le lixiviat.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
48
V. 5. Evolution inter-annuelle de la composition physico-chimique du lixiviat
L’évolution temporelle des valeurs de la conductivité électrique met en évidence une
faible élévation de 1994 à 1998 (5000 à 6000 µS. cm-1, tab. 9). Deux types de causes sont à
invoquer pour interpréter une telle évolution :
Ø La formation de nouveaux résaux d’écoulement au fil du temps sous l’effet des
pluies,
Ø La décharge d'Etueffont est peu stabilisée.
Théoriquement les processus de dégradation par voie aérobie et anaérobie sont
présents dans une décharge. En effet le processus par voie anaérobie se développe en même
temps que les possibilités d'accès à l'oxygène gazeux d'origine atmosphérique diminuent.
Dans le cas de la décharge étudiée les analyses effectuées entre mai 1998 et mai 1999,
montrent d'une manière assez significative la présence de deux comportements distincts de la
décharge au cours de l'année. Un comportement estival aéro-anaérobie et un comportement
hivernal aérobie. Cependant, en été, ce sont les bactéries aérobies qui en dégradant les
substrats issus de l'hydrolyse de la matière organique entraînent une diminution de l'O2 (O2 =
0,5 à 2 mg. l-1). Cet état d'anoxie entraîne le développement d'une autre faune bactérienne de
type anaérobie. Cette dernière est le siège des phénomènes de méthagenisation. Ces deux
processus aboutissent souvent à la production de méthane (CH4), du gaz carbonique (CO2) et
de l'eau (H2O) (Virpin et Pensoye, 1985). En hiver, les pluies permanentes et la diminution de
l'activité bactérienne assurent une oxygénation forte (6 mg. l-1) et continue de la décharge.
Cette oxygénation quasi permanente, nous laisse supposer que durant cette période la faune
bactérienne est dominée par des décomposeurs aérobies. Cette période hivernale est
caractérisée par une baisse des valeurs de pH (pH =5) semblablement due aux fortes teneurs
en CO2 cumulées durant les deux périodes, qui par ces équilibres calco-carboniques
engendrent une acidité au milieu.
La diminution de la charge organique exprimée par la DBO observée de 1995 à 1998
(tab. 9) est vraisemblablement attribuable à la dégradation quasi complète de la matière
organique présente dans la décharge pendant les premières années de fonctionnement.
L’augmentation observée à la même période de la matière organique chimiquement oxydable
exprimée par la DCO (tab. 9) semble être imputable d’une part à l’état de vieillissement de la
matière organique qui devient moins biodégradable et d’autre part aux composés issus d’une
biodégradation incomplète par les bactéries. Plus précisément, les auteurs ont cherché à mieux
caractériser l’activité biologique des lixiviats par le calcul du rapport DBO/DCO (Kerbachi et
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
49
Belkacemi, 1994). Ainsi, des valeurs élevées de ce rapport, de l’ordre de 0,83 indiqueraient
que les lixiviats sont générés par une décharge récente au sein de laquelle l’activité biologique
correspond à la phase acide de dégradation anaérobie de la matière organique. A l’opposé,
lorsque le rapport DBO/DCO se rapproche de zéro, les décharges sont anciennes et sont
souvent engagées dans un processus de fermentation ultime aboutissant à la production de
lixiviats basiques (Ehrig, 1982; Barkowski et al., 1987) et de fortes teneurs en ammonium
(Forgie, 1988). Il semble que c’est le cas de la décharge d’Etueffont qui a produit en 1998-
1999 des lixiviats présentant des valeurs de pH autour de 8 et des rapports DBO/DCO de
0,02.
Afin de vérifier davantage le bien fondé de notre diagnostic, nous nous sommes
appuyés sur les travaux de Kerbachi et Belkacemi (1994) qui ont établi une courbe
d’évolution des valeurs du rapport DBO/DCO en fonction de l’âge de la décharge. Il apparaît
en effet, que ce rapport établi pour une décharge de 16 ans est de 0,05. Cette valeur calculée
est proche de la valeur mesurée dans notre étude (0,02) confirmant l’existence d’une phase de
méthanisation avancée sur le site d’Etueffont.
Afin de mieux comprendre l'effet de l'âge des déchets sur la composition du lixiviats,
on a installé une alvéole étanche à côté de l'ancienne. Ce pilote à été rempli par de nouveaux
déchets. Le lixiviat produit est collecté par des conduits qui rejoignent ensuite les anciens
lixiviats. L'analyse comparative des lixiviats produits par les deux décharges (tab. 8) semble
mettre en évidence une grande hétérogénéité de la composition physico-chimique. D'après nos
résultats on constate que la conductivité est plus élevée pour la décharge récente (15410 µS.
cm-1), cette augmentation semble être liée à l’apparition de nouveaux chenaux. Ce qui
entraîne un maximum de contact entre l'eau et les déchets. Cette forte conductivité se traduit
par des teneurs élevées en Cl- (1200 mg. l-1), en HCO3- (8200 mg. l-1) et en cations majeurs.
La charge organique estimée en terme de DBO et DCO est très importante dans le lixiviat
jeune avec un rapport DBO/DCO de l'ordre de 0,18. Ceci vient confirmer notre hypothèse,
selon laquelle la DBO diminue avec l'âge de la décharge. Cependant cette diminution de la
DBO par biodégradation bactérienne entraîne une production importante en NT et en NH4+.
Le lixiviat jeune apparaît moins chargé en NO3- et en SO4
2-. Ces faibles valeurs du nouveau
lixiviat en nitrate et en sulfate sont vraisemblablement dues aux fortes conditions réductrices
qui règnent au sein de la station pilote suite aux phénomènes de biodégradations très intenses.
Enfin les teneurs en éléments métalliques apparaissent faibles et semblables pour les deux
lixiviats.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2è PARTIE, Chap. 1. Evolution temporelle de la composition du lixiviat de ladécharge d’Etueffont: Effet des pluies et de l'âge des déchets
50
VI. CONCLUSIONAu terme de ce chapitre, il convient de tirer un certain nombre de conclusions quant
aux processus gouvernant la composition qualitative des lixiviats de la décharge d’Etueffont.
En période de faibles précipitations et avec le réchauffement des eaux (été), le jus de décharge
produit est faiblement chargé en matières organiques biodégradables laissant suggérer un rôle
majeur de l’intensification de l’activité hétérotrophe bactérienne dans l’obtention d’une telle
qualité de lixiviat. En hiver, le lixiviat est plus chargé en matière organique en raison de
l’effet conjugué d’une augmentation des précipitations et d’une baisse des températures
inhibant le développement des décomposeurs.
Enfin, l’âge de la décharge semble influencer directement la composition du lixiviat,
sa charge organique biodégradable diminuant régulièrement avec le vieillissement du site. Je
vais maintenant m’intéresser à l’impact des productions de lixiviats sur les eaux souterraines.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
51
I. INTRODUCTION
L'eau qui s'infiltre dans le sous-sol est le vecteur de transfert d'éléments chimiques
vers les eaux souterraines. Dans les compartiments d'un système aquifère (le sol, la zone non
saturée (ZNS) et la zone saturée (ZS) de l'aquifère), de nombreux processus interviennent
pour limiter le transfert de ces éléments chimiques (Abraham et Hafner, 1987; Vaute, 1997).
Dans chaque compartiment, les mécanismes de dissolution et de précipitation chimique sont
étroitement liés aux conditions physico-chimiques, biologiques et hydrodynamiques. Ces
dernières réglant l'atténuation, le délai de transfert, le niveau de propagation et la rétention
définitive du produit polluant. Le risque de pollution des eaux souterraines par les lixiviats de
décharges a fait l'objet de plusieurs études. Depuis 1960, certains auteurs comme Waterton et
al., (1969) et Matter et Young (1979), ont montré en travaillant sur diverses couches
lithologiques que la pollution de la nappe par la décharge est quasiment indétectable. D'autres
Gray, Mather et Harrison (1974), Ragle et al., (1995), Howard et al., (1996), ont montré la
présence d'un réel danger sur de nombreuses décharges étudiées au Pays De Galles et au
Canada. La mobilité des éléments chimiques dans le milieu naturel est essentiellement réglée
par des phénomènes de sorption (La Force et al., 1998). Cependant, ces mécanismes sont
encore mal connus au niveau de l'évolution des polluants organiques et inorganiques issus de
la décharge sur les eaux souterraines ainsi que la nature des minéraux impliqués dans ces
mécanismes (Ludvigsen et al., 1998). L’objet de ce travail est d’étudier l'impact d'une
décharge sur les eaux souterraines du site d'Etueffont (Belfort, France) qui est un sous-bassin
d'une région étendue où les eaux souterraines sont exploitées à des fins d'adduction publique.
Dans ce cadre, nous nous attachons dans ce deuxième chapitre à l’utilisation de certains
éléments chimiques comme indicateurs de pollution organique et inorganique dans les eaux
souterraines voisines de la décharge et à caractériser les mécanismes intervenant dans la
mobilité des éléments chimiques dans le système lixiviats-nappe d'eau souterraine.
II. EQUIPEMENT DU SITE
L'étude de la contamination de la nappe concernée passe par la mise en place d'un
réseau de contrôle de 6 couples de piézomètres (PZ). Chaque couple comprend un piézomètre
court captant la zone superficielle (crépiné de 0 à 5 m de profondeur (PZ30, PZ40 et PZ60)) et
l'autre profond captant la zone saturée (crepiné de 10 à 25 m (PZ2, PZ3, PZ4, PZ5 et PZ6)).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
52
III. CHOIX DES PARAMETRES ETUDIES
Le choix des piézomètres de surveillance tient compte de leur position vis à vis de la
décharge, des bassins, du ruisseau drainant ces derniers et du sens de l'écoulement des eaux de
la nappe. L'étude à caractère physico-chimique, a permis de suivre les variations de certains
paramètres éléments indicateurs de la pollution (NO3-, Eh, O2 et métaux lourds) et l'ion
chlorure utilisé comme traceur conservatif (Appelo et Potsma, 1993). Cet ion n'est pas affecté
par les phénomènes d'adsorption ou d'échanges d'ion, et n'intervient pas dans les équilibres
acido-basiques ou d'oxydoréduction. De plus, il n'est pas retenu par les complexes argilo-
humiques des sols. Par sa grande mobilité et sa teneur importante dans les déchets (Reitzel,
1990), il peut être utilisé comme indicateur de l’impact du lixiviat sur les eaux de la nappe et
l’effet de la pluie sur sa teneur et sa vitesse de transit. De même un suivi de la conductivité
liée à la minéralisation totale des eaux a été réalisé.
IV. ECHANTILLONNAGE
Les prélèvements d’eau ont été effectués à l'aide d'un échantillonneur à pas de
prélèvement mensuel (de mai 1998 à mai 1999) dans tous les piézomètres. A des fins de
comparaison entre les valeurs mesurées dans la nappe et les mesures effectuées sur un
échantillon prélevé en surface. Toutefois, la figure 15 illustre une forte hétérogénéité des
valeurs pour le Eh qui se manifeste par une absence de corrélation positive entre les deux
mesures (r2 = 0,04), cette hétérogénéité qui se présente par une augmentation des valeurs pour
les mesures effectuées en surface n’est que le résultat d’une oxygénation par agitation de
l’échantillon d’eau lors du prélèvement. Cette hétérogénéité tend à diminuer pour les
paramètres pH et température qui sont très influencés par le potentiel redox et on retrouve des
corrélations linéaires positives de l’ordre de r2 = 0,5 pour la température et r2 = 0,6 pour le
pH. Ces différences des valeurs entre les deux mesures s’expliquent par l’échange thermique
avec l’air pour la température, alors que pour le pH ce n’est que le résultat des fluctuations du
Eh et de la température. Enfin, l’essai effectué pour la conductivité électrique ne nous montre
pas de telles variations entre les deux mesures et on enregistre une corrélation positive linéaire
très élevée (r2 = 0,93). Cet essai nous a permis de mettre en valeur la non-fiabilité des mesures
effectuées en surface après le prélèvement pour certains paramètres susceptibles d’être
influencés par le milieu extérieur.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
53
V. RESULTATS
L'étude des variations saisonnières des éléments étudiés, a été réalisée pour l'ensemble
des points d'eau suivis.
V. 1. Conductivité électrique (CE)
Les valeurs de la conductivité électrique enregistrées au cours du suivi montrent des
fluctuations très élevées pour l’ensemble des piézomètres, allant de 25 µS. cm-1 à 450
µS. cm-1 (tab. 10). Les valeurs les plus fortes sont observées dans les PZ30 (616 µS. cm-1 en
octobre 1998) et PZ6 (388 µS. cm-1 en mai 1999) situés respectivement à proximité des
bassins de traitement et à l’aval de la zone étudiée (exutoire du site). Les faibles valeurs sont
observées dans les PZ2 (32 µS. cm-1 en mars 1999) (situé à l’amont) et PZ4 (37 µS. cm-1 en
mars 1999) (situé près de la décharge mais en dehors du sens d’écoulement provenant de
celle-ci).
V. 2. Potentiel d'oxydoréduction (Eh)
L'évolution saisonnière de ce paramètre montre des fluctuations de 100 à 150 mV (fig.
16b) selon la proximité ou l’éloignement de la décharge ou des bassins de décantation.
Cependant les valeurs les plus faibles de Eh sont enregistrées dans les PZ3 et 30 (-197 mV en
octobre 1998 pour le PZ3 et -30 mV en octobre 1998 pour le PZ30)
V. 3. Oxygène dissous (O2)
Les valeurs de l’oxygène dissous varient d'un piézomètre à un autre. La variation
saisonnière est très marquée pour les piézomètres courts (2 mg. l-1en été et 5 mg. l-1en hiver).
Ce paramètre semble présenter des variations similaires à celles de l’Eh (fig. 16c).
V. 4. Chlorure (Cl-)
Les teneurs élevées en chlorures dans le lixiviat brut durant les premiers mois sont
reproduites sur cinq piézomètres (fig. 17c), cependant sur 3 autres piézomètres PZ6, PZ40 et
PZ2 on observe des valeurs faibles de chlorures (m = 2,61 mg. l-1 pour le PZ40, m = 5,36 mg.
l-1 pour le PZ6 et m = 3,41 mg. l-1 pour le PZ2), le PZ6 est loin de la décharge et les PZ2 et
PZ40 sont à l'amont des écoulements.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
54
Fig. 15. Corrélation entre les mesures effectuées à l'aide de la sonde SEBA au niveau despiézomètres et celles effectuées à l'aide de la sonde multiparamètres après
échantillonnage.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
55
L’amplitude des variations est importante pour le PZ30 (m = 48,32 mg. l-1 ) et PZ3 (m
= 19,20 mg. l-1). Il semble que ces derniers piézomètres soient sensibles aux variations de
teneurs en chlorure dans le lixiviat brut. Par contre, le pic de concentration du lixiviat brut au
mois de novembre 1998 n'a pas été répercuté sur les eaux de la nappe.
V. 5. Nitrates (NO3-)
La figure 17b montre que seuls les piézomètres courts et plus spécialement le PZ30 (m
= 19,3 mg. l-1) présentent des fluctuations importantes par rapport aux autres piézomètres. Le
PZ40 enregistre des teneurs non négligeables mais d'une ampleur relativement faible, tandis
que le PZ60 est moins influencé par cette contamination azotée.
V. 6. Les éléments métalliques
Le suivi mensuel de quelques métaux significativement présents dans les eaux de la
nappe étudiée, montre des variations saisonnières considérables de 0 à 3 mg. l-1 pour le zinc,
de 0 à 1 mg. l-1 pour le nickel et le cuivre (fig. 18 et tab. 10). Pour la plupart des piézomètres,
les teneurs faibles observées en étiage notamment celles du Cu2+, Ni2+, Mn2+, Zn2+ et le fer
total montrent une bonne dépendance avec les concentrations maximales enregistrées dans le
lixiviat brut.
VI. DISCUSSION
VI. 1 Analyse en composantes principales (ACP)
Sur l'ensemble des paramètres, une analyse en composantes principales (ACP) a été
réalisée pour l'ensemble des piézomètres suivis. L'analyse a été effectuée sur les 18
variables physico-chimiques des prélèvements d’eau. La matrice de corrélation montre que
la conductivité électrique des eaux de la nappe est influencée par tous les éléments majeurs
(r > 0,7, n = 91, hautement significatif). Par ailleurs, d'autres éléments montrent une
corrélation communément rapportée par la littérature (Na, SO4 et Cl) indiquant une origine
commune. L'analyse des variables montre que l'axe F1 exprime 61 % de la variance. On
obtient une bonne représentation (85 % d’inertie) avec les deux premiers axes seulement.
Les représentations des variables et des individus sont portées sur la figure 19.
L’axe 1 ou axe de pollution permanente est défini positivement par ces 8 variables
très regroupées Fe, CE, HCO3, Mg, K, Mn, Cl et SO4. Les individus caractérisant cet axe
du coté positif, proviennent surtout du PZ30 proche de la décharge. Les concentrations
sont élevées, surtout en bicarbonate et en chlorures. Par ailleurs, ces individus sont définis
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
56
par leur faible potentiel redox et par leur forte minéralisation. Du coté té négatif de l’axe,
on rencontre surtout les piézomètres PZ2, PZ4, PZ40, PZ6 et PZ60, donc les piézomètres
situés en amont des écoulements souterrains et en aval de la décharge et les PZ 2 et 6 dans
le compartiment des grés. Ces individus sont caractérisés par le forte oxygénation et leur
faible minéralisation.
L’axe 2, moins important (24 % d’inertie) ou axe de pollution spécifique
occasionnelle, représentée essentiellement par des individus provenant des prélèvements
effectués en septembre 1998 pour les piézomètres PZ3, PZ30, PZ6 et PZ60, ces individus
sont caractérisés par des eaux à température élevée et à forte teneur en cuivre. D’après
cette analyse statistique, on peut conclure que la distribution des valeurs correspond à une
répartition spatiale des points d’échantillonnage par rapport à la décharge. En effet, le
PZ30 est plus souvent attiré par le pôle de minéralisation et montre de fortes dégradations
de la qualité de l’eau, car il est le plus proche de la décharge. Par contre, le PZ40 captant
aussi la même zone superficielle et proche de la décharge, est moins contaminé par le
lixiviat à cause de sa situation légèrement excentrée de la zone d’influence. Les variations
observées entre les piézomètres peuvent s'expliquer par le rôle que peut jouer la lithologie
du réservoir aquifère, où sont signalées des variations verticales et latérales de faciès
(alternance d'argile, de grès et de schiste).
VI. 2 Evolution des conditions oxydantes des eaux souterraines au voisinage de la
décharge
Afin d’aller plus loin dans l’explication de l’impact des lixiviats de la décharge
d’Etueffont, un suivi de l’évolution spatiale de certains paramètres physico-chimique a été
effectué dans tous les piézomètres. Le suivi du potentiel redox dans les 8 piézomètres
étudiés met en évidence une augmentation des valeurs en s’éloignant de la décharge. Tous
les piézomètres éloignés de la décharge présentent un Eh relativement élevé, (de + 150 à +
300 mV), c'est à dire que l’on assiste à une certaine zonation du potentiel redox autour de
la décharge. Cette évolution du pouvoir oxydant est vraisemblablement attribuable d’une
part à l’oxydation des matières organiques dans les eaux souterraines avoisinant la
décharge, et d’autre part à la dilution par les eaux très oxygénées provenant de la nappe du
grès. Ces observations sont similaires à celles rapportées par d’autres auteurs sur d'autres
sites (Lyngkilde et Christensen, 1992; Rûge et al., 1995). Toutefois, les faibles valeurs du
potentiel redox observées occasionnellement en amont de la décharge (PZ5) résultent d’
un écoulement superficiel du lixiviat très chargé en matières organiques pendant les
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
57
périodes pluvieuses. La forte oxygénation des eaux, enregistrée dans les PZ2 (dans les
grès) et PZ4 (à proximité de la faille), est liée aux apports pluviométriques qui les
affectent directement.
Fig. 16. Variations saisonnières (mai 1998 à mai 1999) des précipitation mensuelles (PC,mm) (a) dans la station d'Etueffont, du potentiel redox (Eh, mV) (b) et de l'oxygène
dissous (O2, mg. l-1) (c) dans les eaux de tous les piézomètres.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
58
Fig. 17. Variations saisonnières (mai 1998 à mai 1999) des teneurs en sulfate (SO42-,
mg. l-1) (a), en nitrate (NO3-, mg. l-1) (b) et en chlorures (Cl-, mg. l-1) (c) dans les eaux de
tous les piézomètres
VI. 3 Evolution des teneurs en chlorures
Dans le cas des polluants inorganiques anioniques, tels que les chlorures, il apparaît
que le PZ30 proche de la décharge est souvent très chargé en chlorures suite à un apport quasi
permanent par la décharge, alors que l’atténuation aux autres piézomètres et plus
particulièrement le PZ3 et le PZ5 est plutôt occasionnelle. Cependant, l’évolution des
chlorures met en évidence deux indices de pollutions majeures en juin et en août 1998. Le
premier indice de pollution se manifeste sur la majorité des piézomètres et coïncide avec les
fortes concentrations dans le signal d’entrée. Cette pollution est ressentie en aval seulement
pour la nappe superficielle (PZ60). Toutefois, la dilution par les eaux provenant de la nappe
du grès contrebalance cette pollution d’où l’absence d’un tel impact en aval de la nappe
profonde (PZ6). L’élévation de la teneur en chlorures enregistrée en PZ2 est due à
l’infiltration provoquée par un ruissellement de surface provenant de la décharge. La
deuxième pollution enregistrée en août est ponctuelle, elle a été recensée tout d’abord en PZ3
du fait de la remonté du niveau piézométrique de la nappe profonde. Cette pollution est
observée plus en aval dans le PZ6, avec des teneurs plus faibles. Le suivi des teneurs en
chlorures met en évidence la présence d'une pollution imprévisible, très dépendante de l’état
de la décharge, qui peut, en période de forte activité, souvent en été (forte activité
bactérienne), produire des lixiviats très chargés qui peuvent atteindre la nappe profonde, ce
phénomène est favorisé par la diminution de la dilution par la pluie.
La même évolution a été observée dans le cas des sulfates (fig. 17a), ce qui mettra en
évidence la fiabilité de ces deux ions inorganiques comme indicateurs de pollution des eaux
souterraines par les décharges d’ordures ménagères. Par comparaison avec le signal d'entrée
en NO3 (lixiviat brut), on constate que, pour les piézomètres profonds, seul le PZ4 suit les
variations de l'entrée en montrant des pics importants en période de hautes eaux, ce qui
montre bien un "effet de chasse" instantané du flux entrant vers ce piézomètre (proche de la
décharge), alors que ce phénomène est enregistré avec un certain retard dans le PZ6 (à l'aval
de la décharge). Enfin les PZ30 et PZ40 sont les plus influencés par les nitrates, en raison de
leur proximité de la décharge. Le PZ30 montre des pics importants pour la quasi-totalité des
éléments; ce qui confirme sa grande vulnérabilité à la pollution, facilitée par sa position près
de la décharge et sa lithologie relativement plus perméable (zone superficielle altérée). On
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
59
peut expliquer une telle augmentation par une infiltration verticale non influencée par
l'écoulement latéral.
VI. 4 Evolution des polluants métalliques
L'importance de l’amplitude de variation de certains éléments chimiques (Zn, Mn
et Fe) est caractérisée par une élévation des teneurs en période estivale, associée aux fortes
teneurs contenues dans le lixiviat brut. Toutefois, des teneurs importantes ont été
enregistrées en période hiverno-printanière indépendamment des teneurs dans le lixiviat
brut, ce qui est vraisemblablement lié aux phénomènes de désorption de ces éléments dans
la zone non-saturée lors des crues ou par un apport par la décharge, notamment pour le Fe,
le Zn et le Mn. Cette remise en solution des ions métalliques est vraisemblablement
attribuée aux faibles valeurs de pH. De plus, un gradient croissant de ce dernier a été
observé en s'éloignant de la décharge, conformément aux observations rapportées par
(Chauve et al., 1986). Aucune tendance n'a été enregistrée pour le cuivre et son évolution
est caractérisée par des maxima et des minima dans l'ensemble des piézomètres. La
variabilité spatio-temporelle des teneurs est également gouvernée par les conditions
d'oxydo-réduction, notamment pour le Fe et le Mn (Chauve et al., 1986; Nimmo et al.,
1995; Tisted et Breslin, 1995). Il faut noter que la migration des métaux dans un milieu
perméable est complexe sur le plan réactionnel (réactions physico-chimiques mis en jeu)
(La force et al., 1998; Galabov, 1989; El –fedel et al., 1997; Gommy, 1997). De même la
présence des lentilles argileuses et des schistes peut constituer un facteur de rétention de
ces éléments-traces, ce qui peut se traduire par leur piégeage dans la partie superficielle.
Certains polluants métalliques illustrent bien un "phénomène de chasse" du signal d'entrée
soit instantanément, soit avec un certain décalage. Ce dernier est variable car il dépend de
l'intensité des pluies, la nature de l'élément chimique, de l'éloignement du point d'eau de la
décharge et de la position de celui-ci par rapport au sens de l'écoulement dans la nappe.
Le PZ60 et le PZ6 sont les moins touchés par cette contamination métallique, du fait de
leur situation à un kilomètre en aval du site. Ceci peut être attribué aux phénomènes de
dilution et/ou d'auto-épuration.
Tableau 10. Composition moyenne des teneurs en cuivre (Cu, mg. l-1), de la température (T,
°C) et de la conductivité électrique (CE, µS. cm-1) dans les eaux des 8 piézomètres.
VII. CONCLUSION
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
60
Enfin, la figure 20 schématise le fonctionnement global du site. La nappe peut être
contaminée par l'intermédiaire de points d'infiltration riches en polluants, représentés par
des zones plus altérées et plus perméables (cas du PZ30 et PZ40) ou par l'infiltration
diffuse d'eau de pluie sur toute la zone concernée. Les écoulements se produisent
parallèlement au ruisseau drainant la station, dans la zone non saturée en période
pluvieuse ou dans la zone saturée en période d'étiage. Au cours de l'élévation du niveau
piézométrique, l'écoulement des matières lessivées demeure toujours dans la zone
inférieure d'écoulement préférentiel. Lorsque le niveau piézométrique est le plus élevé, les
matières lessivées s'écoulent dans la zone supérieure avec un rinçage de l'ensemble par
l'arrivée d'eau moins minéralisée. La pollution la plus élevée est observée dans les lieux
les plus proches de la décharge, où prédomine l'écoulement vertical à travers la zone
altérée donc perméable (K = 10-5 m. s-1), confirmant ainsi l'importance des facteurs
hydrogéologiques dans la migration des matières lessivées. De plus, il semble que la
lithologie du site étudié (grès argileux et schistes) puisse influencer le taux des
contaminants dans la nappe.
Au terme de cette étude, il convient de tirer un certain nombre de conclusions
quant aux processus gouvernant la pollution des eaux souterraines par les lixiviats de la
décharge d’Etueffont:
Ø La pollution des eaux souterraines avoisinant la décharge d’Etueffont est
souvent ressentie dans le piézomètre PZ30, captant la nappe de surface et situé à
proximité de la décharge.
Ø La grande fiabilité des chlorures et du potentiel redox comme indicateurs
d’une telle pollution.
Ø L'amplitude et la rapidité des variations des paramètres physico-chimiques,
observées pour les éléments mesurés dans la nappe dépendent de: (i) la position
lithologique du point de prélèvement (zone superficielle ou profonde), (ii) la position par
rapport à la décharge et aux bassins de traitement, (iii) la situation piézométrique (par
rapport au sens de l'écoulement des eaux souterraines).
L'impact de la décharge en terme de qualité physico-chimique est un phénomène
complexe en raison de la mise en jeu et de l’interférence de plusieurs processus d'ordre
géologique, hydrogéologique, physico-chimique et climatique.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

2er PARTIE: Chap. 2. Impact de la décharge d’Etueffont sur les eaux souterraines
61
Fig. 18. Variations saisonnières (mai 1998 à mai 1999) des teneurs en manganèse (Mn2+,mg. l-1) (a), en fer total (Fe total, mg. l-1) (b), en zinc (Zn2+, mg. l-1) (c) et en nickel (Ni2+,
mg. l-1) (d) dans les eaux de tous les piézomètres
Fig. 19. Analyse en composantes principales...
Fig. 20. Schématisation des phénomènes de contamination des eaux souterraines dans le sited’Etueffont.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
62
I. PARAMETRES ABIOTIQUES
I. 1. Résultats I. 1. 1. Température
Les valeurs de la température du lixiviat enregistrées dans les 4 bassins d’Etueffont
varient de 3,7 à 26,2 °C (m ± δ = 14,5 ± 7,15 °C) en B1, de 1,7 à 26,2 °C (m ± δ = 13,4 ±
8 °C) en B2, de 0,9 à 23 °C (m ± δ = 12,5 ± 8 °C) en B3 et de 0 à 23,7°C (m ± δ = 12,8 ±
8,3 °C) en B4 (fig. 21a). Ces résultats mettent en évidence une évolution saisonnière similaire
dans les quatre bassins avec des lixiviats plus chauds en été et plus froids en hiver. Toutefois
une légère diminution des valeurs a été observée en passant du premier au dernier bassin.
I. 1. 2. Conductivité électriqueLes valeurs de la conductivité électrique varient de 1508 à 4567 µS. cm-1 (m ± δ = 2927
± 922 µS. cm-1) en B1, de 1130 à 3856 µS. cm-1 (m ± δ = 2243 ± 734 µS. cm-1) en B2, de
1167 à 2616 µS. cm-1 (m ± δ = 1922 ± 439 µS. cm-1) en B3 et de 1263 à 2086 µS.cm-1 (m ± δ
= 1714± 281 µS. cm-1) en B4 (fig. 21b). Les valeurs de la conductivité électriques enregistrées
durant notre étude montrent une forte minéralisation du lixivait dans le premier bassin.
Toutefois, l’évolution saisonnière de ce paramètre est faiblement ressenti à l’exception du
premier bassin où on enregistre des fortes valeurs estivales.
I. 1. 3. Oxygène dissousLes teneurs en oxygène dissous relevées dans les lixiviats des 4 bassins d’Etueffont
oscillent entre 0,17 et 12,5 mg. l-1 (m ± δ = 3,56 ± 3,8 mg. l-1) en B1, entre 0,25 et 13,6 mg. l-1
(m ± δ = 5,4 ± 3,6 mg. l-1) en B2, entre 1,5 et 17,1 mg. l-1 (m ± δ = 6,1 ± 4,3 mg. l-1) en B3 et
entre 1,5 et 16,1 mg. l-1 (m ± δ = 7,9 ± 4,5 mg. l-1) en B4 (fig. 21c). Le suivi de ce paramètre
met en évidence la présence de deux pics de fortes teneurs en oxygène, le premier, très
marqué, enregistré en été et le deuxième, avec des valeurs moins importantes, observé en
hiver. Les valeurs les plus faibles ont été observées dans le premier bassin.
I. 1. 4. Potentiel redoxLes valeurs du potentiel redox enregistrées dans le lixiviat des 4 bassins d’Etueffont
oscillent entre -90 et 94 mV (m ± δ = 0,5 ± 63 mV) en B1, entre 19 et 123 mV (m ± δ = 70 ±
34 mV) en B2, entre 67 et 217 mV (m ± δ = 113 ± 48 mV) en B3 et entre -141 et 183 mV (m
± δ = 88 ± 77 mV) en B4 (fig. 21d). Le suivi spatio-temporel de ce paramètre met en évidence
des fluctuations plus au moins importantes durant toute l’année. Toutefois, une nette
augmentation du pouvoir oxydant de l’eau a été enregistrée dans l’ordre croissant des bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
63
I. 1. 5. pHLes valeurs de pH du lixiviat enregistrées dans les 4 bassins d’Etueffont varient de 4,02
à 8,08 (m ± δ = 6,91 ± 1,24) en B1, de 4,62 à 8,21 (m ± δ = 6,88 ± 1,27) en B2, de 4,74 à 8,39
(m ± δ = 7,23 ± 1,47) en B3 et de 4,55 à 8,86 (m ± δ = 7,23 ± 1,47) en B4 (fig. 21e). Ce
paramètre présente une évolution saisonnière bien marquée, des pH basiques en été et en
automne et des pH acides en hiver et au printemps.
I. 1. 6. MESLes teneurs en matieres en suspension (MES) mesurées dans les lixiviats des 4 bassins
d’Etueffont oscillent entre 14 et 73 mg. l-1 (m ± δ = 31 ± 16 mg. l-1) en B1, entre 6 et 98 mg. l-
1 (m ± δ = 25 ± 24 mg. l-1) en B2, entre 5 et 84 mg. l-1 (m ± δ = 30 ± 25 mg. l-1) en B3 et entre
2 et 97 mg. l-1 (m ± δ = 23 ± 24 mg. l-1) en B4 (fig. 21f). Le suivi des teneurs en MES laisse
apparaître une évolution saisonnière marquée par une diminution des teneurs en hiver et au
printemps et une élévation des valeurs en été. De plus, en été, on enregistre deux maxima en
juillet et août 1998.
I. 1. 7. AzoteLes teneurs en nitrites sont quasi-nulles durant tout le suivi. L'azote total varie de 57 à
153 mg. l-1 (m ± δ = 114 ± 32 mg. l-1) dans le premier bassin, de 48 à 113 mg. l-1 (m ± δ = 74
± 20 mg. l-1) dans le deuxième bassin, de 48 à 73 mg. l-1 (m ± δ = 63 ± 10 mg. l-1) dans le
troisième bassin et de 47 à 100 mg. l-1 (m ± δ = 61 ± 11 mg. l-1) dans le dernier bassin (fig.
22a). Les teneurs en NH4+ oscillent entre 46 à 120 mg. l-1 (m ± δ = 93 ± 23 mg. l-1) dans le
premier bassin, de 36 à 88 mg. l-1 (m ± δ = 59 ± 16 mg. l-1) dans le deuxième bassin, de 36 à
94 mg. l-1 (m ± δ = 52 ± 16 mg. l-1) dans le troisième bassin et de 37 à 70 mg. l-1 (m ± δ = 49 ±
10 mg. l-1) dans le dernier bassin (fig. 23d). . Les teneurs en nitrates fluctuent entre 50 à 120
mg. l-1 (m ± δ = 51 ± 70 mg. l-1) dans le premier bassin, de 36 à 88 mg. l-1 (m ± δ = 73 ± 91
mg. l-1) dans le deuxième bassin, de 36 à 94 mg. l-1 (m ± δ = 66 ± 92 mg. l-1) dans le troisième
bassin et de 37 à 70 mg. l-1 (m ± δ = 82 ± 88 mg. l-1) dans le dernier bassin (fig. 23f).
L'évolution saisonnière de l’azote ammoniacal et celle de l'azote total sont similaires. Elle est
caractérisée par une légère diminution des teneurs en période estivale dans les trois derniers
bassins (B2 à B4), alors qu'on enregistre une évolution inverse dans le premier bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
64
Fig. 21. Evolution spatio-temporelle de la température, du pH, du O2, de la CE, du Eh et desMES dans les quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
65
I. 1. 8. PhosphoreLes teneurs en phosphore total enregistrées dans le lixiviat des 4 bassins d’Etueffont
varient de 0,15 à 3,18 mg. l-1 (m ± δ = 0,7 ± 0,8 mg. l-1) en B1, de 0,16 à 0,64 mg. l-1 (m ± δ =
0,31 ± 0,14 mg. l-1) en B2, de 0,08 à 0,45 mg. l-1 (m ± δ = 0,24 ± 0,1 mg. l-1) en B3 et de 0,09
à 0,43 mg. l-1 (m ± δ = 0,27 ± 0,08 mg. l-1) en B4 (fig. 22b). Il semble que les teneurs en
phosphore sont très faibles à l’exception du premier bassin où on enregistre des fortes teneurs
estivales.
I. 1. 9. DBO et DCOLes valeurs de DBO varient de 14 à 100 mg. l-1 d’O2
(m ± δ = 26 ± 46 mg. l-1 d’O2) en
B1, de 5 à 90 mg. l-1 d’O2 (m ± δ = 24 ± 23 mg. l-1 d’O2) en B2, de 2 à 34 mg. l-1 d’O2
(m ± δ
= 17 ± 8 mg. l-1 d’O2) en B3 et de 2 à 50 mg. l-1 d’O2 (m ± δ = 20 ± 13 mg. l-1 d’O2) en B4
(fig. 22d). Celles de la DCO oscillent entre 180 et 735 mg. l-1 d’O2 (m ± δ = 436 ± 188 mg. l-1
d’O2) en B1, entre 187 et 602 mg. l-1 d’O2 (m ± δ = 320 ± 114 mg. l-1 d’O2) en B2, entre 166
et 430 mg. l-1 d’O2 (m ± δ = 271 ± 81 mg. l-1 d’O2) en B3 et entre 153 et 379 mg. l-1 d’O2 (m ±
δ = 260 ± 74 mg. l-1 d’O2) en B4 (fig. 22e). Ces résultats mettent en évidence une évolution
spatio-temporelle bien marquée avec une diminution de la charge organique du premier au
dernier bassin et de l’été à l’hiver.
I. 2. Discussion I. 2. 1. Température
La température de l'eau est un élément essentiel dans le fonctionnement des
écosystèmes aquatiques, elle dépend de l’ensoleillement et des échanges avec l'atmosphère.
Les températures de lixiviat enregistrées dans les quatre bassins sont très voisines et varient
entre 18 à 25 °C en été et de 0 à 8 °C en hiver (fig. 21a). Les bassins ont subi un gel au mois
de novembre 1998. Le lixiviat du premier bassin est souvent plus chaud que celui des autres
bassins du fait de la faible profondeur (0,7 m) et du réchauffement probable par les lixiviats
bruts. En outre, il est à craindre que durant la saison chaude, le lixiviat soit légèrement plus
frais et ne se soit pas écoulé longtemps avant de se mélanger aux lixiviats du premier bassin
ayant une certaine maturation, ce qui peut influencer le temps de séjour. Au cours de cette
campagne la stratification thermique est peu marquée, l'écart maximal de température entre le
fond et la surface est de 6 °C enregistré au mois de mai 1999. En fait, pendant la période d'été,
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
66
l'eau de surface s'échauffe et devient plus chaude que les eaux du fond. Le phénomène
s’inverse en hiver. Il apparaît donc une thermocline à courte durée de vie.
I. 2. 2. Conductivité électrique
Les valeurs montrent que les eaux des quatre bassins sont fortement minéralisées. La
conductivité de l'eau diminue logiquement vers l’aval. Ceci traduit l'importance des
phénomènes de décantation dans le premier bassin. L'évolution saisonnière de ce paramètre
est caractérisée par de fortes valeurs de conductivité électrique en été et de faibles valeurs en
hiver. Cette diminution hivernale semble être due aux phénomènes de dilution sous l'effet des
pluies, très fréquentes, durant cette période. Entre le fond et la surface des bassins, les écarts
de conductivité électrique sont toujours supérieurs à 100 µS. cm-1, à l'exception du mois de
janvier 1999, où l’on enregistre des valeurs identiques entre la surface et le fond. Selon
Ryding et Rast (1994), un écart inférieur à 60 µS. cm-1 est peu significatif et permet à un seul
point de refléter la qualité de l'eau. Cette stratification conductimétrique quasi-continue dans
le premier bassin ne peut être due qu'aux phénomènes de minéralisation très poussés sur le
fond du bassin 1. Alors que dans les autres bassins, on dépasse cette valeur qu'en fin d'été et
en hiver, marquant ainsi une stratification estivale et hivernale, déjà observée pour l'oxygène
et la température.
I. 2. 3. Oxygène dissous
Les teneurs des lixiviats en oxygène dissous dépendent de l'activité biologique du
milieu, c'est à dire de l'équilibre photosynthèse-respiration, des vents, de la salinité et de la
température. La teneur en oxygène est donc la résultante d'un grand nombre de facteurs
biotiques et abiotiques. Ceci explique l'interprétation complexe de ce paramètre. Les valeurs
d'oxygène dissous enregistrées dans les quatre bassins, montrent la présence d'une différence
notable entre bassins, entre le fond et la surface et d'une saison à une autre. Le cycle annuel
laisse apparaître deux séquences principales (fig. 21c). La première en été avec des valeurs
élevées de 3 à 12 mg. l-1 dans la zone euphotique liée aux blooms phytoplanctoniques
estivaux. La deuxième en hiver avec des valeurs plus faibles, 0 et 4 mg. l-1, liée à la fois à la
respiration des plantes, des animaux, des micro-organismes et à l'oxydation des organismes
sénescents et leur dégradation par les bactéries hétérotrophes. Toutefois, au mois de janvier
1999, on enregistre une forte oxygénation du milieu, vraisemblablement due à la poussée
hivernale algale et aux agitations mécaniques provoquées par les vents. L'évolution spatiale
de cet élément se traduit par une augmentation des teneurs en passant du premier au dernier
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
67
bassin. Ce qui montre bien une amélioration qualitative de lixiviat lors de son cheminement
d'une lagune à une autre. L'étude de la stratification verticale de ce paramètre, indique une
différence des teneurs bien marquée entre le fond et la surface, avec des différences qui
peuvent atteindre jusqu'à 10 mg. l-1 en été. Cette stratification estivale semble être due au
peuplement algal très important dans cette tranche superficielle d’eau.
I. 2. 4. Eh
L’évolution saisonnière des valeurs du potentiel redox est très marquée dans les quatre
bassins de la station d'Etueffont avec des conditions oxydantes en été et réductrices en hiver, à
l'exception de janvier 1999, où l’on enregistre une élévation du Eh. Cette augmentation du
pouvoir oxydant de l'eau en janvier 1999 confirme la forte oxygénation enregistrée pendant ce
mois. Cette évolution saisonnière paraît semblable à celle de l'oxygène dissous. Le premier
bassin présente des eaux à tendance réductrice à cause de l’influence directe par la décharge.
Les valeurs de Eh fluctuent entre -90 et 94 mV au bassin 1, entre 26 et 123 mV en B2, entre
70 et 216 mV en B3, entre -141 et 182 mV en B4 et entre -86 et 164 mV à la sortie (fig. 21d).
L'évolution spatiale dans l'ordre successif des bassins (de B1 à B4) de ce paramètre montre
une substitution des eaux à aspect réducteur (provenant de la décharge) par des eaux plus
oxygénées.
I. 2. 5. pH
Le pH est un paramètre important dans l'étude des milieux aquatiques. Il est très
dépendant des mécanismes chimiques et biologiques. Parmi les facteurs qui influent sur les
valeurs de pH, on cite la température, l'activité photosynthétique et la salinité. En été on
enregistre alors, des pH élevés de l'ordre de 8, alors qu'en hiver les valeurs de pH diminuent et
on enregistre des pH acides de 4 (fig. 21e). Cette tendance acide hivernale de l'eau semble être
due d'une part au ralentissement de l'activité photosynthétique consommatrice du proton H+ et
à l'apport par la décharge de lixiviats acides. L'évolution verticale de ce paramètre ne semble
pas être influencée par la profondeur de l'eau. Cependant l'écart des valeurs de pH enregistré
durant les douze mois de suivi ne dépasse pas l'unité entre la surface et le fond pour
l'ensemble des points de prélèvement.
I. 2. 6. MES
La décroissance des concentrations en MES dans l'ordre successif des bassins (de B1 à
B4) jusqu'à la sortie est très nette (fig. 21f) est vraisemblablement liée à l'augmentation de
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
68
l'activité photosynthétique et aux apports par la décharge de lixiviats très minéralisés.
Toutefois, les deux pics de MES observés en juillet et septembre 1998 dans les trois premiers
bassins coïncident avec l'élévation des MES dans le jus de décharge.
I. 2. 7. Azote
Il apparaît que l'azote ammoniacal domine quantitativement les formes azotées
(représente les 90 % de l'azote total). La diminution estivale des teneurs en azote ammoniacal
(fig. 23f) est vraisemblablement liée à une assimilation par les algues, très présentes durant
cette période, car les algues absorbent préférentiellement le NH4+. Cependant l'évolution
opposée observée pour le premier bassin (teneurs élevées en été), ne peut être expliquée que
par l'influence directe de la décharge qui alimente le premier bassin par des lixiviats très
chargés en ammonium pendant l'été suite à l'activité bactérienne très poussée (voir la 2è
partie) et qui dépasse l'assimilation estivale par le phytoplancton. L'évolution spatiale de l'ion
ammonium montre bien une diminution des teneurs d'un bassin à un autre du fait d'une
minéralisation de l'azote ammoniacal en nitrate par une action bactérienne (Nitosomonas et
Nitrobacter) (Laurent, 1972). L'étude comparative de l'évolution saisonnière des nitrates et de
l'ion ammonium, met en évidence une évolution opposée de ces deux paramètres, ce qui
indique bien une dénitrification.
I. 2. 8. Phosphore
Le phosphore est parmi les nutriments les plus importants pour la croissance algale
(Golterman, 1984). Les teneurs en PO43- s'annulent à plusieurs reprises, c'est la raison pour
laquelle nous n’avons pas représenté graphiquement ces variations spatio-temporelles (fig.
22b). Ces faibles teneurs sont donc la résultante de l'assimilation rapide de cet élément par les
bactéries et les algues (Caplancq, 1990) et/ou sa complexation avec le calcium sous forme
d'apatite (Alaoui et al., 1994). Cependant nos résultats aux rayons X, effectués pour les
matières en suspension piégées dans les quatre bassins ne mettent évidence l’absence du
phosphore qui précipite généralement sous forme hydroxyl-apatite (Ca5(PO4)OH). Cette
absence de précipitation du phosphore ne peut pas être attribuée à une absence des ligands
cationiques (Fe2+, Ca2+ et Mn2+) qui sont très présents dans le lixiviat avec des concentrations
importantes, mais il paraît que c’est le pH, pour lequel on enregistre rarement des valeurs
supérieures 8,20 dans le lixiviat qui soit le responsable direct de la non-précipitation du
phosphore (Stumm et Morgan, 1981). L’absence du phosphore dans les MES évite aussi toute
adsorption physique par des liaisons Vanderwals ou chimique par un échange de ligand OH-
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
69
avec les complexes hydrométalliques du phosphore avec les MES (Lijklema et al., 1980;
Bostrom et al., 1988). Les concentrations du phosphore total varient de 0,1 à 3,2 mg. l-1 dans
le premier bassin, de 0,15 à 0,6 mg. l-1 dans le deuxième bassin, de 0,1 à 0,4 mg. l-1 dans le
troisième bassin et de 0,09 à 0,4 mg. l-1 dans le dernier bassin. L'évolution saisonnière de ce
paramètre est marquée par une légère diminution estivale, vraisemblablement liée à
l'assimilation par les algues. Cependant l'évolution des teneurs en phosphore total d'un bassin
à un autre met en évidence une chute des teneurs en allant du premier bassin (B1) au dernier
bassin (B4). Cette diminution spatiale est liée à une précipitation du phosphore pendant le
cheminement du lixiviat d'un bassin à un autre. Enfin, les fluctuations saisonnières montrent
une élévation des teneurs en phosphore dans les bassins 2, 3 et 4 seulement (janvier 1999),
alors que dans le premier bassin on n'enregistre pas d’augmentation ce qui écarte l’hypothèse
de tout apport possible de phosphore par la décharge. Cette augmentation hivernale
vraisemblablement liée à un relargage par le sédiment sous l’effet d’une diminution de pH
pendant ce mois, peut aussi être attribuée à une régénération du phosphore par des voies
biologiques:
• libération par les algues à l’obscurité (Rocques, 1980; Florentz et al., 1984;
Somiya et al., 1988),
• par autolyse des algues mortes (Golterman, 1975),
• excrétions zooplanctoniques (Premazzi et al., 1988).
I. 2. 9. N/P
Les valeurs du rapport N/P, indicateur d'état d'eutrophisation de l'eau (Seip, 1994)
nous renseigne sur la présence probable des algues fixatrices d'azote atmosphérique pour des
valeurs inférieures à 29 (Dowing et Caule, 1992). En effet, le rapport N/P (fig. 22c), calculé
durant tout le suivi est largement supérieur à 29 ce qui confirme l'absence de Cyanobactéries
dans nos échantillons. Il oscille alors, entre 30 et 820 dans le premier bassin, de 130 et 460
dans le deuxième bassin, de 140 à 740 dans le troisième bassin et de 140 à 770 dans le dernier
bassin. Il apparaît que c'est le phosphore qui joue le rôle de facteur limitant dans les quatre
bassins, confirmé par une absence des orthophosphates durant la bonne saison. A l'exception
du mois de novembre 1998, où l’on enregistre une valeur très importante dans le premier
bassin, liée à un apport élevé d'azote par le lixiviat brut, le rapport N/P, augmente dans les
deux derniers bassins, durant les douze mois de suivi. Cette augmentation est
vraisemblablement attribuée aux phénomènes de piégeage du phosphore total (PT) dans les
boues des deux premiers bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
70
I. 2. 10. DBO et DCO
D’importantes variations de la DBO et de la DCO sont enregistrées au cours de la
saison, portant à la fois sur les niveaux de concentration des bassins et sur leur étagement.
Nos résultats mettent en évidence une chute des teneurs en matières organiques
contenues dans le lixiviats durant son cheminement d’un bassin à un autre. Cette décroissance
des teneurs est vraisemblablement liée aux phénomènes de décantation et à la dégradation de
la matière organique oxydable par les micro-organismes.
I. 2. 11. Autres paramètres physico-chimiques
Les éléments majeurs (Ca, Mg, K et Na) sont fortement présents dans les quatre
bassins, mais toujours avec une chute des teneurs en passant d'un bassin à un autre (tab. 11).
La teneur en calcium augmente dans les quatre bassins pendant la mauvaise saison sous l'effet
de la diminution de la photosynthèse. Les teneurs passent de 100 à 200 mg. l-1 en B1, de 100
à 140 mg. l-1 en B2, de 70 à 130 mg. l-1 en B3 et de 60 à 100 mg. l-1 en B4. Alors que pour
les autres cations on n’enregistre pas d’effet saisonnier bien marqué. L'évolution spatiale des
anions majeurs (Cl, F, HCO3 et SO4) dans les quatre bassins, met en évidence une évolution
similaire à celle observée pour les cations. Cependant, l'évolution saisonnière montre bien une
augmentation des teneurs en été, vraisemblablement liée à l'apport par la décharge de ces
éléments très présents dans le jus de décha
rge. Cependant, pour les chlorures, la légère augmentation en février est liée à l'arrivée
des eaux de fonte de neige. Enfin, la tendance estivale, trouvée pour la majorité des
paramètres est faiblement ressentie pour les ions métalliques. Ceci est vraisemblablement
attribuable à leur forte dépendance vis à vis des conditions physico-chimiques (tab. 11).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
71
Fig. 22. Evolution spatio-temporelle de NT (a), PT (b), DBO (c), DCO (d) et des rapportsDBO/DCO (e) et N/P (f) dans les quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è PARTIE, Chap. 1. Physico-chimie du lixiviat des 4 bassins.
72
Tableau 11. Composition physico-chimique moyenne de l'eau des quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
73
II. PARAMETRES BIOTIQUES
II. 1. Résultats II. 1. 1. Teneurs en chlorophylle a
Les teneurs en chlorophylle a, varient de 22,7 à 108,3 µg. l-1 (m ± δ = 55,36 ± 28
µg. l-1) dans le premier bassin, de 2,3 à 712 µg. l-1 (m ± δ = 147 ± 222 µg. l-1) dans le
deuxième bassin, de 0,7 à 1959 µg. l-1 (m ± δ = 201 ± 530 µg. l-1) dans le troisième bassin
et de 10,8 à 926 µg. l-1 (m ± δ = 206 ± 266 µg. l-1) dans le dernier bassin (fig. 23e). Les
valeurs les plus élevées ont été observées dans les bassins 3 et 4 et plus précisément en
été, vraisemblablement liées à l’élévation de la température et à la diminution de la
charge polluante.
II. 1. 2. Composition biochimique du materiel particulaireII. 1. 2. 1. Protèines
L’évolution spatiale des teneurs en protéines particulaires dans le lixiviat des
quatre bassins varient de 391 à 1680 µg. l-1 (m ± δ = 946 ± 467 µg. l-1) dans le premier
bassin, de 284 à 1669 µg. l-1 (m ± δ = 935 ± 452 µg. l-1) dans le deuxième bassin, de 109
à 1468 µg. l-1 (m ± δ = 701 ± 413 µg. l-1) dans le troisième bassin et de 330 à 1804 µg. l-1
(m ± δ = 797 ± 493 µg. l-1) dans le dernier bassin (fig. 23a). L’évolution saisonnière de
cet élément permet d’observer que les fortes valeurs en protéines particulaires sont notées
en période estivale, lors du développement algal.
II. 1. 2. 2. Glucides
Les teneurs en glucides particulaires varient de 8 à 324 µg. l-1 (m ± δ = 133 ± 100
µg. l-1) dans le premier bassin, de 50 à 412 µg. l-1 (m ± δ = 160 ± 126 µg. l-1) dans le
deuxième bassin, de 37 à 265 µg. l-1 (m ± δ = 125 ± 37 µg. l-1) dans le troisième bassin et
de 60 à 425 µg. l-1 (m ± δ = 190 ± 125 µg. l-1) dans le dernier bassin (fig. 23b). De façon
tout à fait identique à la teneur en protéines, les valeurs des teneurs en glucides présentent
une nette diminution des teneurs en hiver.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
74
Fig. 23. Evolution spatio-temporelle de NH4+, NO3
-, Chl a, et de la compositionbiochimique particulaire (P et G) dans les quatre bassins .
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
75
II. 2. Discussion II. 2. 1. Chlorophylle a
Les teneurs en chlorophylle a globale (Coulson, 1977) et fractionnée en classe de
taille (Aleya et Dévaux, 1988) sont souvent utilisées comme indice de productivité algale
(Carlson, 1977). Les teneurs les plus élevées en chlorophylle a, ont été enregistrées en été
dans les deux derniers bassins (B3 et B4), elles sont associées au développement de
Chlamydomonas sp. en B3 en août 1998 et au développement de Coelastrum sp. et
Chaetoptilis sp. dans le dernier bassin. Il faut noter que même dans des eaux polluées, les
teneurs en chlorophylle a restent importantes. Toutefois, des teneurs faibles en oxygène
dissous ont été observées en été dans le premier bassin et ceci malgré les fortes teneurs en
Chl a, en raison de la consommation immédiate d’oxygène par les organismes aquatiques
qui dépasse la production par le phytoplancton et l’apport atmosphérique. Une autre
explication serait que la production d'oxygène par photosynthèse est bloquée au profit
d'autres voies métaboliques. Les courbes d’évolution de la chlorophylle a et de la
biomasse algale représentées sur la figure tendent à se confondre dans les deux derniers
bassins, mettant en évidence la forte présence des chlorophycées dans ces bassins. En
confrontant les résultats des biomasses algales et celles des matières en suspension, on
observe, un chevauchement des pics des deux paramètres dans le dernier bassin. Cette
évolution similaire est le résultat de l’augmentation des MES biogènes engendrés par
l’activité algale dans ce dernier au dépend des MES allochtones.
II. 2. 2. Composition biochimique du matériel particulaire (0,45-160
µm)
L’évolution spatiale des teneurs en protéines et glucides particulaires contenues
dans le lixiviat montre que la concentration augmente du premier au dernier bassin,
vraisemblablement liée au développement algal. De plus, et sur un plan saisonnier on
retrouve la même augmentation en passant d’une saison à une autre (de l’hiver à l’été).
L’examen des valeurs obtenues à partir des mesures mensuelles dans les quatre
bassins, mettent en évidence des pics estivaux observés pour les teneurs en protéines et
glucides particulaires, vraisemblablement liés au peuplement algal très présent durant
cette période. D'un point de vue spatial, une légère augmentation des teneurs apparaît
dans le premier bassin, plus spécialement pour les teneurs en glucides, mettant en cause
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
76
l'apport par la décharge de jus de décharge riche en protéines et glucides d'origine
allochtone. La valeur du rapport Protéines/Glucides (P/G) est comprise entre 2,34 et
123,72 dans le premier bassin, de 2,4 et 309,23 dans le deuxième bassin, de 2,92 et
522,12 dans le troisième bassin et de 1,84 et 17,29 dans le dernier bassin (fig. 23c). Il
apparaît, d’après nos résultats une augmentation du rapport P/G a été observée en été,
toutefois, c’est l’apport par la décharge (très active) durant cette période et la diminution
des pluies qui soient à l’origine de tel évolution, car c’est en été où les irradiations
lumineuses sont importantes favorisant ainsi une forte accumulation des glucides au
dépend de la vitesse de synthèse des protéines (Morris et Skea, 1978; Ganf et al., 1986).
De plus une nette diminution du rapport, souvent en été, a été mise en évidence en
passant du premier au dernier bassin, vraisemblablement liée d’une part à l’éloignement
de la décharge et d’autre part à l’amélioration de la qualité de l’eau en favorisant ainsi
une faible consommation par les algues de leurs réserves glucidiques.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
77
I. BACTERIES HETEROTROPHES I. 1. IntroductionLes bactéries constituent une part prépondérante des communautés
picoplanctoniques aquatiques. Elles sont essentiellement composées par des cellules de
petite taille (0,5 à 0,9 µm) de diamètre et leur abondance dans les milieux aquatiques est
très dépendante de la température, de la disponibilité des substrats et de la prédation par
les bactérivores (Carrias, 1996). Elles constituent le groupe planctonique le plus
important des systèmes de traitement biologique des eaux usées (Tomlinson, 1956).
Toutefois, la grande variété rencontrée dans les groupes permet la dégradation de la
plupart des composés organiques rencontrés dans les déchets municipaux, industriels ou
agricoles. Les bactéries hétérotrophes sont considérées comme les principaux
décomposeurs de la matière organique dans les écosystèmes aquatiques (Fuhrman et
Azam,1982; Ducklow, 1983; Sieburth, 1996). De plus, les bactéries planctoniques
peuvent minéraliser jusqu’à 50 % de la production primaire (Cole et al., 1988; Del
Giorgio et al., 1997). Leurs effectifs varient de 105 à 107 bactéries par ml dans les stations
de traitement des eaux usées (Drakides, 1987) et sont fortement liés aux biomasses
phytoplanctoniques (Porter et Feig, 1980; Bird et Kalff, 1986; Scavia et al., 1986;
Schorter et Stockner, 1986; Gude, 1989). La majorité de ces bactéries est libre (Hodson et
al., 1981; Hope, 1984). Les bactéries présentent souvent une faible variation saisonnière
suite à l’action régulatrice par le zooplancton (Azam et al., 1983; Pomeroy et wiebe,
1988; Pedrsos-Alio, 1989) et/ou à la lyse virale (Proctor et Fuhrman, 1990 et 1992;
Murray et Eldridge, 1994). Par ailleurs, c’est plutôt les bactéries hétérotrophes Gram–
(Achromobacter, Pseudomonas, Flavobacterium, Alcaligenes.) qui caractérisent les
bassins de traitement biologique (James, 1964; Cook, 1967). Si l'étude de ce maillon a
pris une apogée dans le cas des eaux douces, eau salées et eaux usées elle est à notre
connaissance peu étudiée pour les lixiviats de décharge.
I. 2. Abondances et biomasses I. 2. 1. Résultats
Bassin 1: Le peuplement bactérien est dominé par des formes libres
essentiellement représenté par des cocci (91 % du peuplement). La plus forte abondance
des formes bacilles a été observée en octobre 1998 (36 % du peuplement). Comme dans
le lixiviat brut, les bactéries sont caractérisées par leurs grandes tailles (≥ 0,5 µm pour les
cocci et ≥ 0,9 pour les bacilles). L’abondance bactérienne varie de 0,008.106 à 0,15.106
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
78
bactéries. ml-1 (m ± σ = 0,067.106 bactéries. ml-1). Les valeurs maximales enregistrées
dans ce bassin coïncident avec une élévation des teneurs en substances nutritives dans le
lixiviat brut et dans le bassin (fig. 24). L’évolution saisonnière montre des abondances
bactériennes faibles en hiver et fortes en été. La biomasse bactérienne varie de 0,2 à 30
µgC. l-1 (m ± σ = 11,98 ± 12,04 µgC. l-1).
Bassin 2: L’analyse qualitative du peuplement bactérien dans le deuxième bassin
met en évidence une forte dominance des formes cocci au dépend des formes bacilles (96
% du peuplement bactérien). Toutefois, les bactéries observées dans ce bassin sont
caractérisées par leur petite taille (≤ 0,5 µm pour les cocci et ≤ 0,9 pour les bacilles), à
l’exception du mois de mai 1999 où l'on enregistre des cocci de grande taille dont le
diamètre est compris entre 0,5 et 0,8 µm. La densité bactérienne varie de 0,015.106 à
0,19.106 bactéries. ml-1 (m ± σ = 0,069.106 ± 0,062.106 bactéries. ml-1) (fig. 24). Dans ce
bassin on a enregistré la valeur maximale observée dans les quatre bassins durant tout le
suivi (juin 1998), l’évolution saisonnière des densités bactériennes est similaire à celle du
premier bassin, celle-ci diminue en hiver et augmente en été. La biomasse bactérienne
varie de 1,74 à 42,68 µgC. l-1 (m ± σ = 13,80 ± 14,58 µgC. l-1).
Bassin 3: Comme dans le bassin 2, les cocci dominent, le peuplement bactérien
avec 97 % au dépend des bacilles qui ne présentent que 3 % du peuplement. Toutefois,
on enregistre dans ce bassin une domination des cocci de petite taille (diamètre ≤ 0,5 µm)
très présentes en fin d’été. De plus c’est dans ce bassin qu’on observe les premières
formes coloniales avec des streptobacilles en juin 1998. Les abondances bactériennes
enregistrées dans le troisième bassin oscillent entre 0,02.106 et 0,14.106 bactéries. ml-1 (m
± σ = 0,066.106 ± 0,0199.106 bactéries. ml-1) (fig. 24). Le peuplement bactérien présente
la même évolution que celle enregistrée précédemment, mais avec des densités plus
faibles. La biomasse bactérienne varie de 2,23 à 30 µgC. l-1 (m ± σ = 12,29 ± 9,51
µgC. l-1).
Bassin 4: Dans ce dernier bassin, on enregistre une présence bactérienne
bacillaire similaire à celle observée dans le premier bassin (4 % du peuplement
bactérien). L’évolution des diamètres bactériens montre une évolution saisonnière très
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
79
nette chez les cocci, avec des cocci de petite taille en fin d’été. Les formes coloniales sont
de plus en plus présentes avec des streptocoques en avril et mai 1999, des staphylocoques
en mai 1999 et des streptobacilles en juin 1999. La densité bactérienne varie de 0,007.106
à 0,10.106 bactéries. ml-1 (m ± σ = 0,055.106 ± 0,038.106 bactéries. ml-1) (fig. 24). Ce
bassin est marqué par une évolution saisonnière bactérienne opposée à celles des autres
bassins. Cette opposition se manifeste par des densités faibles en été et fortes en hiver.
Toutefois, les densités bactériennes observées dans ce dernier bassin sont plus faibles et
suivent généralement la même tendance observée en passant du premier au dernier bassin
(diminution des densités dans le sens croissant des bassins). La biomasse bactérienne
varie de 1,52 à 23,3 µgC. l-1 (m ± σ = 8,39 ± 6,21 µgC. l-1).
I. 2. 2. Discussion
I. 2. 2. 1. Abondances et biomasses
Durant tout notre suivi, on observe une nette dominance des morphotypes cocci
(95 % de la population microbienne), par rapport aux morphotypes bacilles (4 % de la
population microbienne) et 1 % pour les autres morphotypes. Les formes bacilles sont
surtout présentes dans le premier bassin en fin d'été (septembre et octobre 1998),
vraisemblablement liée à l’élévation de la température de l’eau, à l’absence des
bactérivores et aux apports de substrats allochtones par le lixiviat brut dans le premier
bassin. Toutefois, les tailles des cellules bactériennes enregistrées dans les quatre bassins
semblent être supérieures à celles observées par certains auteurs dans les milieux
eutrophes (Sommaruga, 1995; Sommaruga et Robarts, 1997). Ces diamètres assez
importants sont attribuables à la forte charge organique quasi-permanente dans les quatre
bassins. Les densités bactériennes enregistrées dans les bassins de la station d’Etueffont
sont très aléatoires et sont inférieures à celles décrites par Davies-Colley (1998),
Bahlaoui et al. (1997) dans les eaux usées domestiques, Nagata (1988), Bennette et al.
(1990), Straskrabova (1992), Chrzanowski et Simek (1993), Pick et Hamilton (1994),
Carrias (1996) et Druvietis et al. (1998) dans les eaux douces et Ochs et al. (1995) dans
les eaux marines. Ces faibles abondances sont vraisemblablement liées à l’effet toxique
des lixiviats (Deneuvy, 1987; Devare et Bahadir, 1994; Clement et al., 1996). Il apparaît
d’après nos résultats que les densités bactériennes sont quasi-similaires dans les quatre
bassins, ceci est confirmé par l’absence de différence statistique spécifique (p = 0,88; test
Annova). Toutefois, l’étude statistique dans le premier bassin met en évidence une
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
80
corrélation positive significative entre l’effectif bactérien et le rapport DBO/DCO (r =
0,6; 11 ddl) mettant en évidence le rôle prépondérant de la matière organique oxydable
importée par la décharge dans le développement du bactérioplancton. Dans le deuxième
bassin, c’est plutôt le pH qui semblent gouverner l’abondance bactérienne avec une
corrélation positive hautement significative (r = 0,7, pour 11 ddl). Il apparaît que les
effectifs bactériens augmentent avec l’élévation de pH et des teneurs en oxygène dissous.
Ce résultat est corroboré avec les observations de Druvietis et al. (1998) qui ont trouvé
une corrélation positive (r = 0,8) dans les milieux eutrophes. En effet, le pH fait partie des
facteurs qui concourent à la disparition bactérienne dans de tels écosystèmes aquatiques.
L’influence des variations de pH dans les installations de traitement des eaux usées par
lagunage sur la disparition bactérienne a été étudiée par plusieurs auteurs. Si Pearson et
al. (1987) ont montré que les Coliformes fécaux sont affectés par les pH alcalins. A
l’opposé, ces pH alcalins favorisent le développement et la survie de certaines espèces
comme Vibrio Cholerae (Oufdou et Mzrioui, 1994). Dans le troisième bassin, l’étude
statistique entre l’abondance bactérienne et les paramètres abiotiques du milieu remet en
évidence le rôle de la matière organique biodégradable, vraisemblablement autochtone,
dans la prolifération bactérienne (r = 0,6, pour 11 ddl). Enfin, dans le dernier bassin et
suite au développement algal important qui favorise un ensemencement permanent du
milieu en matière organique, c’est le pH qui semble influencer le peuplement bactérien (r
= 0,7, pour 11 ddl).
Les faibles abondances se répercutent nettement sur les biomasses bactériennes
produites. Toutefois, ces valeurs de biomasses sont nettement inférieures à celles
trouvées dans la littérature (Furtado et al., 2001). La diminution de la biomasse
bactérienne produite en passant du premier au dernier bassin est vraisemblablement
attribuable à la diminution de la charge organique.
I. 2. 2. 2. Dynamique saisonnière
L’évolution temporelle du bactérioplancton dans les quatre bassins de la station
d’Etueffont met en évidence une évolution variable d’un bassin à un autre.
En été, on enregistre un développement bactérien important dans les trois
premiers bassins, favorisé par l’élévation de la température (Pirt, 1971; Novac, 1974;
Aloice, 1996; Jaspers et al., 2001), alors que dans le dernier bassin, le peuplement
bactérien enregistre ces faibles abondances.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
81
En automne, l’apport par la décharge de lixiviats fortement chargés en substances
nutritives et très peuplés en bactéries (3,5.106 cell ml-1 ) se répercute fortement dans le
premier bassin (octobre 1998) et en novembre 1998 dans les autres bassins (2, 3 et 4).
En hiver, on enregistre une diminution de la densité bactérienne dans l’ensemble
des bassins, cela est attribuable à la baisse de la température et à la diminution de la
charge organique sous l’effet de la dilution.
Au printemps, l’élévation de la température et le développement algal, favorisent
à nouveau le développement des bactéries hétérotrophes. La dynamique saisonnière
enregistrée dans les 3 premiers bassins est quasi-similaire à celle décrite par Bahlaoui et
al. (1996), Legendre et al. (1984) et Monfort et Baleux (1990) avec un développement
hivernal peu accentué, vraisemblablement lié aux faibles températures des bassins.
Fig. 24. Distribution des densités bactériennes dans les quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
82
II. PHYTOPLANCTON II. 1. IntroductionLes algues jouent un rôle multiple et complexe dans les processus d’épuration des
eaux usées. Par leur mécanisme de photosynthèse, elles assurent une oxygénation des
eaux à la belle saison, favorisant ainsi l’activité des décomposeurs et une élévation de pH
par consommation de CO2. Les peuplements phytoplanctoniques des bassins de lagunage
naturel sont caractérisés par leur faible richesse spécifique (Barbe, 1981; Drakides, 1987
et Belbahri, 1991). Toutefois, les algues les plus courantes en lagunage naturel sont:
Euglena, Chlamydomonas, Chlorogonium, Micractinium, Ankistrodesmus, Scenedesmus,
Chlorella Oscillatoria, Anabaena, Carteria, Phormidium, Navicula, Closterium et
Anacystis (EPA, Enviromental Protection Agency, 1971).
II. 2. Inventaire systématique des espèces rencontrées dans les quatre bassinsDe mai 1998 à mai 1999, quinze taxons ont été inventoriés. Nous constatons que
cette liste est relativement courte, confirmant, ainsi de la faible richesse spécifique des
bassins. La composition qualitative du peuplement algal rencontré dans la station
d’Etueffont rappelle celle inventoriée en littérature par certains chercheurs (Drakides,
1980 et Belbahri, 1989) dans les bassins de traitement des eaux usées domestiques.
Toutefois, la liste des espèces rencontrées s’établit comme suit:
E: CHLOROPHYTES
Cl: EUCHLOROPHYCÉES
O: Volvocales
F: Chlamydomonadacées
Chlamydomonas sp.
O: Tétrasporales
F: Tetrasporacées
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
83
Chaetoptilis sp.
Sphaerellocystis sp.
Oocystis lacustris
O: Chlorococcales
F: Scénédesmacées
Coelostrum sp.
F: Oocystacées
Kirchneriella sp.
E: EUGLÉNOPHYTES
Cl: EUGLÉNOPHYCÉES
O: Euglénales
F: Euglénacées
Euglena ehrenbergii
Euglena viridis
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
84
Phacus orbicularis
Trachelomonas curta
F: Pétalomonadacées
Petalomonas africana
Cl: BACILLARIOPHICÉES = DIATOMOPHYCÉES
O: Coscinodiscales
F: Coscinodiscacées
Stephanodiscus dubius.
Stephanodiscus hantzschii
Thalassiosira weissflogii
O: Diatomales
F: Tabellaiaridae
Tabellaria fenestrata.
F: Diatomacées
Fragilaria crotonensis
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
85
Synedra tabulata
O: Naviculales
F: Naviculacées
Navicula cuspidata
F: Tryblionellae
Nitzschia tryblionella
II. 3. Evolution spatio-temporelle du phytoplancton II. 3. 1. Résultats
Bassin 1: Les effectifs du peuplement phytoplanctonique fluctuent entre 0,016.106
et 2,9.106 cellules. l-1, (m ± σ = 0,5.106 ± 0,8.106 cellules. l-1) (fig. 25). La biomasse
algale varie de 2 à 1308 µgC. l-1 (m ± σ = 172 ± 386 µgC. l-1). La biomasse
phytoplanctonique annuelle produite est de 4,6 mgC. l-1, dont les valeurs les plus fortes
en été enregistrées en été 1998 (mai-juin-juillet-août). Le pic de production algale (1,3
mgC. l-1) enregistré en août 1998 est essentiellement associé au développement important
de Chlamydomonas sp.
Bassin 2: Les densités phytoplanctoniques enregistrées dans le deuxième bassin
fluctuent entre 0,031.106 et 4,5.106 cellules. l-1 (m ± σ = 0,8.106 ± 1,3.106 cellules. l-1)
(fig. 25). La biomasse phytoplanctonique annuelle produite est de 5,9 mgC. l-1. La
biomasse algale varie de 15 à 3769 µgC. l-1 (m ± σ = 456 ± 1052 µgC. l-1). La valeur
maximale (3,76 mgC. l-1) enregistrée dans ce bassin (août 1998) coïncide avec le
développement de Phacus sp.
Bassin 3: Les effectifs phytoplanctoniques observés dans le troisième bassin
oscillent entre 0,032 106 et 14,5.106 cellules. l-1, (m ± σ = 1,6.106 ± 4.106 cellules. l-1)
(fig. 25). La biomasse algale varie de 3,47 à 5558 µgC. l-1 (m ± σ = 791,8 ± 1777
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
86
µgC. l-1). La biomasse phytoplanctonique annuelle produite est de 10,29 mgC. l-1. La
valeur maximale (5,56 mgC. l-1) en biomasse enregistrée dans ce bassin (mai 1999) est
attribuée au le développement de Coelastrum sp. (49 % de la biomasse annuelle).
Bassin 4: Les densités phytoplanctoniques varient de 0,048.106 à 29.106 cellules.
l-1, (m ± σ = 5,1.106 ± 10,2.106 cellules. l-1) (fig. 25). En terme de biomasse, la biomasse
algale varie de 9 à 20685 µgC. l-1 (m ± σ = 3244 ± 6936 µgC. l-1). La biomasse
phytoplanctonique annuelle produite est de 42 mgC. l-1. Le maximum de production
algale a été enregistré au moi mai 1998 associé à la croissance exubérante de Coelastrum
sp. Euglena sp.
II. 3. 2. Discussion
L’étude des peuplements phytoplanctoniques est très informative
quant à la détermination du niveau de la structuration des communautés et à l’évaluation
de l’efficacité du fonctionnement d’un écosystème aquatique (Reynolds 1982 et 1991,
Aleya 1991 et 1994). L’abondance de Stephanodiscus sp. et Coelastrum sp. dans
l’ensemble des bassins, nous renseigne sur un état hypereutrophe (Reynolds 1984) dans
l’ensemble des bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
87
Fig. 25. Evolution spatio-temporelle de l’abondance phytoplanctonique.
Le suivi qualitatif saisonnier du phytoplancton dans la station d’Etueffont montre
que les Euchlorophycées représentent 10 % du peuplement algal dans le bassin 1, 13 %
dans le deuxième bassin, 39 % dans le troisième bassin et 19 % dans le dernier bassin.
Les Eugleneuphycées représentent 60 % du peuplement algal dans le premier bassin, 77
% dans le deuxième bassin, 58 % dans le troisième bassin et 76 % dans le dernier bassin.
Enfin les diatomées représentent 30 % du peuplement algal dans le premier bassin, 10 %
dans le deuxième bassin et 3 % dans le troisième bassin et 5 % dans le dernier bassin (fig.
26).
Fig. 26. Pourcentage relatif des différents groupes phytoplanctoniques dans lesdifferents bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
88
Dans le premier bassin, on enregistre une alternance de maxima et de minima
durant toute l'année, vraisemblablement liée à l’interaction de plusieurs facteurs:
ü La variation des facteurs climatiques tel que la température et l’ensoleillement,
ü L’état de la décharge, qui peut ensemencer le bassin par une charge organique et
minérale souvent nuisible au développement algal,
ü Le zooplancton (Moinidae et Rotifères) responsable du déclin des communautés en
été. En outre, la biomasse la plus faible est associée au développement simultané des
Moinidae et des Brachionidae, attribuant ainsi un rôle complémentaire l’un à l’autre
dans l’élimination du phytoplancton,
ü Sa morphométrie différente à celle des autres bassins (plus petit et moins profond).
Cet écosystème particulier et assez imprévisible entraîne la production d’une
faible biomasse phytoplanctonique et un peuplement dominé par des espèces polluo-
résistantes (Stephanodiscus sp. et Euglena sp.). Toutefois, la période des eaux claires
observée en été (mai, juin et juillet 1998) semble importante, ceci malgré l’apport
important de substances nutritives en été. Ceci est vraisemblablement lié à l’action
prédatrice exercée par le zooplancton sur le phytoplancton. L’augmentation de la
biomasse phytoplanctonique observée en hiver 1999 (janvier 1999 avec 87,68 µgC. l-1)
est vraisemblablement liée à la diminution de la charge organique et minérale (DBO
passant de 45 à 17 mg. l-1) sous l’effet de la dilution par les pluies et à l’absence du
zooplancton phytophage (Moinidae et Rotifères). Dans ce bassin, on enregistre souvent
un peuplement monospécifique avec une faible richesse spécifique (richesse spécifique =
4), or, et selon le concept de Margalef (1958, 1961), Odum (1960) et Sevrin-Ryessac
(1998) qui ont établi une relation entre la diversité spécifique et le degré de stabilité d'une
communauté, une diversité spécifique importante traduit généralement une grande
stabilité au sein de la population, du fait que dans ce cas les variations d'une espèce
influent peu sur la population. A l'opposé, une diversité faible, cas du premier bassin,
aboutit souvent à une grande instabilité du peuplement algal, de ce fait les populations de
cette communauté seront souvent caractérisées par une alternance de blooms
phytoplanctoniques et de mortalités massives.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
89
Fig. 27. Dynamique saisonnière du phytoplancton dans le premier bassin.
La succession saisonnière du plancton végétal dans les autres bassins est
caractérisée par deux croissances phytoplanctoniques communément connues dans tous
les milieux aquatiques, une croissance estivale suivie d'une croissance printanière. En été
1998, l’élévation de la température, de la photopériode, de l’intensité lumineuse
(Amblard, 1986) et la présence de substances nutritives, favorisent un développement
algal plus diversifié et plus important. Toutefois ce développement algal est
contrebalancé par le développement exubérant des Rotifères et des Moinidae qui
contribuent à l’éclaircissement des eaux du bassin par broutage des algues par le
zooplancton. Cependant, en août 1998 on enregistre la plus forte production algale
associée à la chute des effectifs zooplanctoniques. En automne, la diminution de la
température, de la radiation lumineuse et l’accumulation de la matière organique
d’origine allochtone et autochtone accentué par le brassage des eaux sont à l’origine de la
chute progressive du phytoplancton. Cependant les espèces dominantes sont des
diatomées (Stephanodiscus sp.). En hiver, ces stress physiques s’accentuent et on
enregistre la même tendance à l’exception du mois de février 1999 où l’on enregistre une
légère augmentation du phytoplancton associée à l’augmentation de la température, de
l’irradiation lumineuse, et à la diminution du broutage par le zooplancton (Sterner, 1989).
Toutefois, cette élévation phytoplanctonique enregistrée en février 1999 n’a pas été
observée dans le dernier bassin, en raison de la présence de Keratella quadrata. Cette
amélioration des conditions abiotiques continue au printemps et on enregistre la présence
d’un peuplement algal très diversifié, avec des Chlorophycées et des Euglenophycées qui
colonisent le milieu au dépens des diatomées. La diminution des densités algales en mai
1999 dans le dernier bassin semble être liée au développement important des Rotifères
stimulés par les blooms phytoplanctoniques.
II. 4. Etude de la dynamique saisonnière des espèces les plus fréquentesØ Phacus orbicularis
Durant le suivi Phacus orbicularis présente une seule phase de croissance estivale
dans le premier bassin (juin et juillet 1998 en B1) avec un maximum d'effectif de 3,9.104
ind. l-1 enregistré en juillet 1998 (figs. 27 à 30). Alors que dans les bassins 2, 3 et 4, il
apparaît que Phacus orbicularis présente deux périodes de croissance en été 1998 et au
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
90
printemps 1999. Toutefois, sa croissance s’étale en B2 de juin à août 1998 et de février à
mars 1999 (avec un maxima de 24,8 104 ind. l-1 enregistré en juillet 1998), en B3 de mai
à août 1998 et de mars à mai 1999 (avec un maxima de 16 104 ind. l-1 enregistré en mai
1999) et en B4 de mai à août 1998 avec des densités comparables à celles observées dans
les bassins 1 et 2 (la valeur maximale dans ce bassin a été observée en juillet 1998; 26,6
104 ind. l-1). Il apparaît que la population de Phacus orbicularis diminue en densité avec
une croissance plus étendue en allant du premier au dernier bassin. Cette évolution de
Phacus orbicularis de B1 à B4 coïncide avec la diminution des effectifs de Moinidae et
avec l'amélioration de la qualité physico-chimique des lixiviats. Il semble donc que la
croissance de Phacus orbicularis est gouvernée à la fois par la qualité physico-chimique
de l'eau et par l'action directe du zooplancton.
Ø Euglena sp.
Réputée par sa grande tolérance vis à vis des eaux très polluées et plus
spécialement les eaux chargées en métaux (Hg, Cd et Pb) (Devars et al. 1998), Euglena
sp. a largement dominé les Euglenophytes. dans les quatre bassins. Cependant le suivi
mensuel montre que dans le premier bassin Euglena sp. présente des pics de croissance
brefs et intensifs en mai 1998 (480.104 ind. l-1), novembre 1998 (21,8.103 ind. l-1), et une
croissance printanière en mars et avril 1999 (respectivement 16 et 22.104 ind. l-1). Dans le
deuxième bassin et durant toute la période estivo-automnale Euglena sp. présente deux
pics de croissance brève en août 1998 où elle atteint sa densité maximale (65,6.104 ind. l-
1) et le second en novembre 1998 (0,6.104 ind. l-1). Après une absence hivernale Euglena
sp. réapparait en mars 1999 mais avec une densité plus faible que celle enregistrée en été
1998 (2,7.104 ind. l-1). Dans le troisième bassin on enregistre la même dynamique
observée en B2 avec une présence plus ou moins étendue en été (de août à octobre 1998),
le maxima d'effectif d' Euglena sp. a été enregistré en août 1998 (111.104 ind. l-1). Enfin
dans le dernier bassin sa croissance s'étale sur toute la période estivale avec une absence
en période printanière, le maxima d'effectif a été observé en août 1998 (193,4 104 ind. l-1)
(figs. 27 à 30). Il semble que ce soit la présence des Moinidae en grande quantité dans le
premier bassin qui a masqué sa présence estivale.
Ø Stephanodiscus dubius
C'est l'algue planctonique la plus abondante dans les quatre bassins du fait de son
large éventail de tolérance vis a vis des eaux polluées (Germain, 1981). Stephanodiscus
dubius est absente seulement en juillet 1998 dans le premier bassin avec un maxima
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
91
d'effectif de l'ordre de 29,2.105 ind. l-1 enregistré en août 1998. Sa dominance diminue en
B2 et on enregistre son absence au cours d'une longue période estivale (mai, juin et août
1998), son maxima d'effectif à été observé en janvier 1999 (292,8.104 ind. l-1). Dans le
troisième bassin Stephanodiscus dubius s'étale sur les treize mois de suivi, mais avec des
effectifs plus faibles que ceux observés dans les deux premiers bassins. La densité
maximale de Stephanodiscus dubius a été enregistrée en août 1998. Enfin, dans le dernier
bassin, les effectifs de Stephanodiscus dubius diminuent et on note son absence en juin et
août 1998, il présente deux maxima de développement le premier en août 1998 (266.104
ind. l-1) et le deuxième en avril 1999 (315.104 ind. l-1) (figs. 27 à 30).
Ø Chlamydomonas sp.
C'est la seule population des Euchlorophycées qui puisse se développer
communément dans les quatre bassins. Cependant sa présence n'est pas étonnante
puisqu’elle a été inventoriée souvent dans les stations de traitement des eaux usées
(Belbahri, 1989) ainsi que dans les eaux eutrophes (Prat et al., 1999). Cependant on
l’observe dans le premier bassin avec deux croissances en été et au printemps, la
croissance maximale a été enregistrée en mai 1998 (58.104 ind. l-1). Dans les autres
bassins (2, 3 et 4) Chlamydomonas sp. présente un développement plus étendu que celui
observé dans le premier bassin. Dans ces trois derniers bassins, la dynamique de
Chlamydomonas sp. est caractérisée par plusieurs développements, une croissance
estivale en mai et juin 1998 dans le bassin 2; en mai, août et septembre 1998 dans le
troisième bassin; en juin, juillet et août 1998 dans le bassin 4. L’autre croissance a été
enregistrée au printemps 1999 en février et mars dans les 3 derniers bassins. Les maxima
d’effectifs de Chlamydomonas sp. enregistrés dans ces trois derniers bassins sont de
l’ordre de 2,8.104 ind. l-1 en B2, 10,2.104 ind. l-1 en B3 et de 5,5.104 ind. l-1 dans le dernier
bassin (figs. 27 à 30).
Ø Chaetoptilis sp.
Cette espèce n’a jamais été rencontrée dans le premier bassin, sa présence dans les
autres bassins se manifeste par des pics de croissance brefs et intensifs en début d’été
avec des effectifs très importants atteignant 900.104 ind. l-1 dans le dernier bassin (figs. 27
à 30). En effet, son absence dans le premier bassin est vraisemblablement liée à sa
sensibilité aux eaux très chargées (Bourreley, 1965).
Ø Coelastrum sp.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
92
Durant toute notre étude, il apparaît que cette espèce, absente dans le premier
bassin et présente seulement en juillet 1998 dans le deuxième bassin, préfère les bassins 3
et 4 où le lixiviat est moins chargé (figs. 27 à 30).
Ø Kirchneriella sp.
Cette population n'est présente que dans le dernier bassin et son effectif peut
atteindre des densités très importantes (figs. 27 à 30) mettant en évidence sa préférence
aux eaux moins chargées en matières organiques.
Les autres populations phytoplanctoniques sont représentées toujours par de
faibles densités.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
93
Fig. 28. Dynamique saisonnière du phytoplancton dans le deuxième bassin.
Fig. 29. Dynamique saisonnière du phytoplancton dans le troisième bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
94
Fig. 30. Dynamique saisonnière du phytoplancton dans le dernier bassin.
III. ZOOPLANCTONIII. 1. Rôle du zooplancton dans les processus d’épuration
Le rôle du zooplancton dans les processus d'épuration a été souvent oublié.
Certains auteurs (Fitzegerald et al., 1958) l'ont négligé, alors que d'autres (Sampson,
1955) l'ont considéré comme nuisible. Il a fallu attendre les années 1970 pour que des
chercheurs, comme Angeli (1979), mettent en évidence le rôle primordial du zooplancton
dans l’épuration des eaux polluées. Ce rôle consiste à éliminer des bactéries, des
particules organiques et permettre la régulation de la biomasse phytoplanctonique.
Cependant, l'évolution saisonnière des peuplements zooplanctoniques est liée à la nature
des algues et aux conditions physico-chimiques qui varient d'une saison à une autre.
Toutefois, Verdy (1987) met l’accent sur la présence d’éventuels Rotifères dans ces
milieux où la forte charge organique est pourtant défavorable à leur métabolisme. Durant
tout notre suivi, 19 taxons ont été inventoriés dont 17 espèces de Rotifères et 2 espèces de
Cladocères.
III. 2. Les Rotifères
III. 2. 1. Aperçu biologique et écologie
Le terme Rotifera fut utilisé depuis 1798 par Cuvier. Ce dernier faisait référence à
la présence, en position apicale de l’animal, d’un appareil rotateur cilié donnant l’illusion
de deux roues en mouvement. Regroupant plus de 2000 espèces décrites par Kost (1978),
les Rotifères constituent une part importante des effectifs des métazoaires d’eau douce.
Leur petites tailles, de quelques centièmes de millimètres, les qualifient de
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
95
microzooplancton. Ce sont des espèces filtreuses consommatrices de microparticules.
Leurs appareils filtreurs constituent un tamis qui s’oppose à l’entrée de grosses particules
non ingérables. En effet Asplanchna sp. et Synchaeta sp. sont généralement omnivores et
consommatrices de macroparticules (Lair, 1977). Par leur capacité de reproduction
importante (Kirk, 1998; Yoshinaga et al., 1999), leur courte durée de vie ne dépassant
généralement pas deux semaines (Ruttner-Kolisko, 1974) et leur grande diversité
spécifique, les Rotifères jouent un rôle important dans la transformation de la matière
organique et dans la production des biocénoses (Lair, 1978). Leur densité est très
dépendante de l’état trophique de l’écosystème. Dans un milieu oligotrophe, leur densité
ne dépasse pas quelques dizaines d’individus par litre, alors qu’elle atteint des valeurs
voisines de 104 ind. l-1 dans les eaux eutrophes (Pourriot, 1965; Pourriot et al., 1982). La
grande majorité des Rotifères se reproduisent par parthénogenèse. Les Rotifères sont des
animaux hydrobiontes, colonisant des biotopes aquatiques ou péri-aquatiques variés, du
sol humide au milieu marin. Les Rotifères sont des métazoaires, commun, des eaux
douces où la majorité d’entre eux appartiennent aux Monogonontes. Cependant leur
densité est très faible dans les eaux marines. Leur effectif est très lié au statut trophique
des biotopes (Pourriot, 1965). Dans les lacs oligotrophes, leur densité ne dépasse
généralement pas quelques individus par litre, alors qu’elle atteint des valeurs voisines de
104 ind. l-1 dans des écosystèmes eutrophes. Les Rotifères planctoniques sont
globalement cosmopolites. Selon Pejler (1957 et 1977) les différences latitudinales
seraient plutôt liées aux ressources nutritives. Toutefois, le cosmopolitisme serait
beaucoup moins accentué chez les espèces périphytiques et benthiques, 48 % des 278
taxons montrent une distribution géographique limitée. La composition spécifique des
peuplements rotariens diffère malgré tout d’un biotope à un autre, la composition ionique
n’intervenant dans cette distribution que dans la mesure où elle conditionne la qualité et
la quantité des ressources nutritives. Enfin, les Rotifères sont aussi microphages et se
nourrissent alors de bactéries, d’algues et de détritus (mastax ramé), macrophages
carnivores (à mastax forcipé ou incudé).
III. 2. 2. Inventaire systématique des Rotifères recensées durant notre
campagne
Classe des Rotifères
Sous-classe des Monogonontes
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
96
Ordre des Pseudotroques
Famille Brachionidae
Genre Brachionus
Brachionus calyciflorus
Brachionus urceolaris
Brachionus rubens
Brachionus plicatilis
Brachionus falcatus
Brachionus leydigi
Brachionus bidentatus
Brachionus angularis
Brachionus patulis
Genre Epiphanes
Epiphanes senta
Epiphanes clavulata
Genre Keratella
Keratella quadratra
Famille Synchaetidae
Genre Synchaeta
Synchaeta tremula
Genre Polyarthra
Polyarthra remeta
Famille Gastropodidae
Genre Ascomorpha
Ascomorpha agilis
Ascomorpha ovalis
Ordre des Gnésiotroques
Sous-ordre des Monimotroques
Famille Testudinellidae
Genre Filinia
Filinia hofmanni
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
97
III. 2. 3. Dynamique saisonnière du peuplement rotarien
III. 2. 3. 1. Résultatsü Bassin 1: L’abondance rotarienne fluctue entre 0 et 43700 individus. l-1 (m ± σ =
3519 ± 12077 individus. l-1). La valeur maximale enregistrée dans ce bassin (juillet 1998)
coïncide avec une diminution de la densité des Cladocères (fig. 31). A l’exception de
janvier 1999, les Rotifères colonisent le bassin en période estivo-printannière seulement.
Les Brachionidae représentent la totalité des Rotifères recensés (100 % du peuplement) et
la richesse spécifique ne dépasse pas 2 espèces pour chaque prélèvement, toutefois, un
total de 5 espèces a été recensé dans ce bassin durant tout le suivi. La biomasse rotarienne
mensuelle varie de 0 à 223 mgC. l-1 (m ± σ = 17,6 ± 61995 µgC. l-1).
ü Bassin 2: La densité des Rotifères varie de 0 à 92000 individus. l-1 (m ± σ = 12173 ±
28997 individus. l-1) (fig. 31). Dans ce bassin on enregistre une augmentation importante
de la densité des Rotifères. Ces derniers colonisent le bassin uniquement en période
estivo-printannière. Les Brachionidae représentent 99,98 % du peuplement des Rotifères
rencontrés et on enregistre une légère augmentation de la richesse spécifique mensuelle
qui atteint 5 en mai 1998, toutefois 9 espèces ont été recensées durant tout le suivi. La
biomasse rotarienne mensuelle produite oscille entre 0 et 784 mgC. l-1 (m ± σ = 101,9 ±
248260 µgC. l-1).
ü Bassin 3: C’est le bassin où les Rotifères ont atteint des valeurs records de
développement, leurs densité varie de 0 à 100240 individus. l-1 (m ± σ = 23358 ± 59002
individus. l-1) (fig. 31). A l’exception de septembre, octobre 1998 et janvier 1999, les
Rotifères ont colonisé le bassin durant toute l’année, mais avec des abondances
importantes en été. Comme dans le deuxième bassin les Brachionidae représentent 99,98
% du peuplement des Rotifères rencontrés et la richesse spécifique mensuelle atteint 5 en
mai 1998, toutefois, 10 espèces ont été recensées dans ce bassin durant toute l’étude. La
biomasse rotarienne mensuelle produite varie de 0 à 1,07 gC. l-1 (m ± σ = 110 ± 301372
µgC. l-1).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
98
Fig. 31. Distribution des densités rotariennes dans les quatre bassins
ü Bassin 4: Dans ce bassin la densité rotarienne oscille entre 0 (février 1999) et 84300
(mai 1998) individus. l-1 (m ± σ = 10188 ± 23261 individus. l-1) (fig. 31). Toutefois, on
enregistre une augmentation de la richesse spécifique (19 espèces ont été rencontrées
durant le suivi) avec l’apparition de nouvelles espèces (Brachionus angularis,
Brachionus patulis et Polyrthra remeta). La domination des Brachionidae retrouvée dans
les trois premiers bassins semble s’affaiblir avec seulement 93 % du peuplement rotarien.
Cette augmentation de la richesse spécifique est accompagnée d'une diminution des
effectifs. L’expansion temporelle des Rotifères est semblable à celle enregistrée dans le
troisième bassin avec une absence en septembre, octobre 1998 et janvier 1999. La
biomasse rotarienne mensuelle produite fluctue entre 0 et 783 mgC. l-1 (m ± σ = 64,9 ±
215936 µgC. l-1)
III. 2. 3. 2. DiscussionD’après ces résultats, il apparaît que les abondances rotariennes dénombrées au
cours de notre suivi dépassent largement les densités rapportées dans la littérature pour
les différents milieux oligtrophes (Blavy et Druart, 1982) et eutrophes (Herzig, 1979 et
Oulad Ali, 1988). Toutefois, nos valeurs sont comparables à celles observées par
Kennedy (1995) dans des bassins de traitement des eaux issus d’une unité laitière. Le
peuplement rotarien est représenté essentiellement par le genre Brachionus
caractéristique des milieux eutrophes (Hakkari, 1978 et Pejler, 1983).
L’évolution spatio-temporelle des Rotifères montre que c’est dans le troisième
bassin que les Rotifères atteignent leur développement optimal. Cette apogée coïncide
avec un développement des Chlorophycées (39 % du peuplement algal). Toutefois, une
augmentation de la richesse spécifique a été observée en passant du premier au dernier
bassin (2 dans le premier bassin, 4 dans le deuxième bassin, 5 dans le troisième bassin et
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
99
10 dans le dernier bassin). Cette augmentation du nombre d’espèces confirme les
résultats physico-chimiques qui mettent en évidence une amélioration de la qualité des
lixiviats en passant d’un bassin à un autre. L’évolution temporelle met en évidence l'effet
de la température et de la composition qualitative du phytoplancton caractérisée par une
préférence aux euglenophycées et aux chlorophycées dans la distribution temporelle des
Rotifères. En effet, comme le soulignent Brett et Goldman (1996), la composition
spécifique du phytoplancton et du zooplancton doit être prise en compte dans
l’établissement des modèles de fonctionnement des interactions trophiques. Durant toute
notre étude, nous avons noté que Brachionus sp. a dominé quantitativement et
qualitativement le peuplement rotarien, bien qu’en période hiverno-printanière de l’année
1999 les Epiphanes sp. soient aussi importantes en densité. Quant aux autres espèces,
moins importantes en densité, elles montrent des périodes de longues absences. Leur
évolution se manifeste par des pics de croissance brefs et intensifs sur une courte période
de l’année. L’évolution saisonnière des Rotifères, laisse apparaître une dynamique
saisonnière très marquée.
En été 1998, les petits herbivores atteignent leurs maxima de développement
confirmant ainsi le rôle prépondérant de la température de l’eau dans la distribution des
peuplements rotariens (Berzin et Pejler, 1989). Cette période coïncide avec une diversité
algale très importante. Les Rotifères n'apparaissent dans le bassin 1 qu'au mois de juin
1998 avec une faible densité (400 ind. l-1). Cette absence est attribuée au développement
exubérant des Moinidae (4,6.104 ind. l-1). En juillet 1998, les Rotifères atteignent leur
apogée de croissance et on enregistre une densité de 4,3.104 ind. l-1, représentée
essentiellement par Brachionus rubens. Après, c’est en août 1998 que les effectifs
diminuent et on enregistre alors une densité comparable à celle observée en juin 1998
(300 ind. l-1). Toutefois, cette augmentation en densité et en espèces ne s’accompagne pas
d’une élévation de la richesse générique et le peuplement rotarien est dominé par
Brachionus sp. (100 % du peuplement). Le peuplement rotarien dans ce bassin s’étale sur
les quatre prélèvements estivaux (de mai à août 1998). Dans le troisième bassin, on
retrouve la même domination des Brachionidae, mais avec des effectifs encore plus
importants. Cette augmentation qualitative (Brachionus sp., Ascomorpha sp., Synchaeta
sp., Felinia sp. et Keratella sp.) et quantitative des Rotifères semble être liée à
l’amélioration qualitative et quantitative du peuplement algal avec des densités
importantes de Chlamydomonas sp. et de Phacus sp., algues préférées des Brachions
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
100
(Pourriot, 1982) et à la diminution des Moinidae (800 ind. l-1 en mai 1998 seulement).
Enfin, et dans le dernier bassin, il apparaît que le développement important de
Chlamydomonas sp. est à l’origine du développement des Rotifères. Le maximum
d’effectif enregistré dans ce bassin est de 84400 ind. l-1 (mai 1998).
En automne 1998, on enregistre une absence des Rotifères durant les mois de
septembre et octobre dans les quatre bassins. Cette absence coïncide avec l’absence
d’algues de petite taille facilement ingérables par les petits algivores (Phacus sp. et
Chlamydomonas sp.) et la présence d’un peuplement algal diatomique souvent médiocre
pour le développement des Rotifères (Devaux et Lair, 1976). De plus, il semble que la
forte désoxygénation des eaux, très fréquente durant cette période, soit à l’origine de la
disparition des Rotifères (O2 passe de 0.4 à 0.1 mg. l-1 en B1, de 8,8 à 1,7 mg. l-1 en B2 et
de 10,3 à 2,5 mg. l-1 en B3) ainsi que de l’augmentation spectaculaire des MES (qui
passent de 20 à 100 mg. l-1) dans le premier bassin. Cependant, en novembre, on assiste à
la réapparition des Rotifères avec des effectifs très faibles dans les bassins 3 et 4, 1000
ind. l-1 (Epiphane sp.) en B3 et 200 ind. l-1 (Keratella sp.). Cette réapparition montre
bien la résistance de ces deux espèces aux faibles températures qui se développent au
dépend d’un peuplement algal diatomique (Syndra sp. et Stephanodiscus sp.). Leur
présence seulement dans les bassins 3 et 4 peut être expliquée par la faible charge
organique et minérale de ces deux derniers par rapport aux bassins 1 et 2.
Le peuplement rotarien hivernal est très faible, représenté essentiellement par des
épiphanes dans le premier et le troisième bassin, respectivement 100 ind. l-1 (janvier
1999) et 1700 ind. l-1 (janvier et février). Toutefois, dans le dernier bassin on enregistre
l’apparition en février 1999 de Keratella quadrata ( 200 ind. l-1). Cette quasi-absence des
Rotifères en hiver est vraisemblablement liée à la baisse flagrante des températures de
l'eau (Pourriot et Deluzarches 1971 et Pourriot 1965), toutefois, les bassins ont été
complètement gelés en novembre 1998.
Au printemps 1999, il apparaît que la poussée phytoplanctonique printanière
(représentée par des algues assimilables Phacus sp., Chlamydomonas sp. ) soit à l’origine
de la réapparition des Rotifères représentés uniquement par Epiphane sp., caractérisé par
sa tolérance vis à vis des basses températures (Pourriot 1965). Les effectifs maximaux
observés atteignent 1200 ind. l-1 en B1, 2500 ind. l-1 en B2, 7700 ind. l-1 en B3 et 1800
ind. l-1 en B4. Dès le début de l’été 1999, on enregistre une élévation progressive des
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
101
températures de l'eau induisant ainsi un milieu favorable pour le développement de
Brachionus sp. qui a largement dominé nos bassins durant l’été précédent (1998).
III. 2. 4. Facteurs contrôlant la dynamique des Rotifères
Si la température, la disponibilité du phytoplancton et la compétition avec les
autres groupes sont les facteurs essentiels qui contrôlent le développement des Rotifères
dans la majorité des écosystèmes aquatiques, pour la station d’Etueffont d’autres facteurs
interviennent directement ou indirectement dans la distribution des rotifères parmi
lesquels:
ü La charge polluante: la diminution de cette dernière par dégradation ou par
décantation de la charge polluante le long du cheminement du lixiviat d’un bassin à un
autre favorise la colonisation des Rotifères. Toutefois, le lixiviat de chaque bassin est
caractérisé par une composition physico-chimique particulière qui impose par son
complexe paramétrique un peuplement bien précis. Chaque espèce n’arrive en effet à
coloniser et à prospérer dans un milieu que si le complexe paramétrique du bassin lui
convient. Ceci nous permettra de mettre en valeur le rôle des Brachionidae comme
indicateur des eaux très polluées,
ü Morphométrie: La différence morphométrique des quatre bassins peut intervenir
(d’autant plus importante dans les trois derniers bassin) en favorisant le développement
de biotopes supplémentaires.
III. 2. 5. Etude de la dynamique saisonnière des espèces les plus
fréquentes
Ø Brachionus calyciflorus
Brachionus calyciflorus est l’espèce la plus commune des Rotifères, elle joue un
rôle majeur dans la régulation de la biomasse algale, jusqu’à 2000 chlorelles par heure
(Kenedy, 1995, Seamann et al., 1986). C’est le Brachion le plus commun des
Brachionidae d’eau douce.
Souvent considéré comme espèce d’eaux eutrophes (Pourriot, 1965; Saksena,
1987), Brachionus calyciflorus peut se développer dans les eaux peu oxygénées (Dor et
al., 1987; Pearson et al., 1987) et dans des eaux très chargées en matières organiques (O
Brien et De Noyelles, 1972; Seamann et al., 1986 et Sladecek, 1983). Durant notre suivi
on enregistre des densités en Brachionus calyciflorus voisines de celles observées par
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
102
Kennedy (1995) dans des lagunes de traitement des dérivés laitiers. Toutefois, une nette
augmentation de l’effectif de Brachionus calyciflorus en passant du premier au dernier
bassin a été mise en évidence durant cette étude. Ceci est en accord avec les travaux de
Kennedy (1995) qui a montré que la densité de Brachionus calyciflorus augmente avec la
diminution des teneurs en NH4+. De plus, l’augmentation de la diversité spécifique
phytoplanctonique, avec une présence plus importante des chlorophycées considérés
comme nourriture préférée de Brachionus calyciflorus (Lubzens, 1987) et le
développement important des macrophytes (biotope idéal pour le développement des
Rotifères) dans les bassins 3 et 4 peuvent expliquer une telle évolution (Miracle et al.,
1988). Toutefois, on ne néglige pas l’effet probable des Moinidae comme inhibiteur du
développement de Brachionus calyciflorus qui agissent soit par leurs interférences
mécaniques (Gilbert, 1988) qui perturbent le développement de certains Rotifères soit par
une compétition trophique sur la nourriture et l’oxygène dissous contenus dans le milieu.
Dans le premier bassin on enregistre un seul développement en juillet 1998, avec
seulement 7.103 ind. l-1, vraisemblablement lié aux densités importantes des Moinidae et
de Brachionus rubens. Alors que son développement s’étale sur une longue période
estivale dans les derniers bassins avec des effectifs nettement plus élevés, les maxima
d’effectifs ont été enregistrés en mai 1998 dans les bassins 2, 3 et 4 (avec 5.104 ind. l-1 en
B2, 6,4.104 ind. l-1 en B3 et 4,3.104 ind. l-1 en B4) (figs. 32 à 35). De plus, il semble que
Brachionus calyciflorus présente sa croissance maximale pour des températures d’eau
comprises entre 17 et 18 °C, valeurs légèrement supérieures à celles citées par Kennedy
(1995). Enfin, son endémisme estival est lié à l’interaction de plusieurs facteurs
environnementaux: anoxie et absence d’algues assimilables (septembre et novembre
1998), chute de température en hiver (Berzin et Pejler, 1989) et pH acide au printemps
(Miracle et Vicente, 1983).
Ø Brachionus rubens
C’est le Brachionidae le plus représenté des quatre bassins (97 % en B1, 80 % en
B2, 7,5 % en B3 et 11 % en B4 des Rotifères) (figs. 32 à 35), sa croissance s’étale sur
une période plus au moins longue (de juin à août 1998). Toutefois, il apparaît une
substitution spatiale de Brachionus rubens par Brachionus calyciflorus mettant en
évidence un régime alimentaire commun et une tolérance plus importante du premier par
rapport au deuxième vis à vis des Moinidae et de la pollution (plus précisément la forte
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
103
teneur en ammonium). L’effectif maximal en Brachionus rubens (7,4.104 ind. l-1) a été
enregistré en juillet 1998 dans le deuxième bassin.
Ø Brachionus urceolaris
Cette espèce à été rencontrée dans les bassins 1, 2 et 4, son évolution temporelle
est marquée par des croissances brèves à faible densité. La densité maximale a été
enregistrée en mai 1999 dans le dernier bassin (476 ind. l-1) (figs. 32 à 35). Ces pics de
croissance unique, témoignent aussi d'une compétition interspécifique chez les
Brachionidae.
Ø Brachionus plicatilis
Espèce essentiellement phytophage (Pourriot, 1965), elle est souvent utilisée dans
les bassins d’aquaculture comme agent bactéricide (Markirdis 2000), sa densité maximale
a été enregistrée en juillet 1998 avec un effectif de 9600 ind. l-1 (figs. 32 à 35). Il semble
que, Brachionus plicatilis soit très sensible aux eaux chargées (absente dans le premier
bassin) et sa densité ainsi que sa pérennité augmentent légèrement du deuxième au
quatrième bassin. De plus, son développement n'est pas trop influencé par le
développement de Brachionus calyciflorus et Brachionus rubens. Enfin, son absence en
hiver est vraisemblablement attribuable à sa faible tolérance aux basses températures
(Fielder, 2000).
Ø Brachionus falcatus
Brachionus falcatus est observée dans les trois derniers bassins, en août 1998 dans
le bassin 2 (400 ind. l-1) (figs. 32 à 35), en mai et août 1998 dans les bassins 3 et 4. Son
absence en juin et juillet 1998 semble être liée à la présence des autres Rotifères et à
l'absence d’Euglena sp. De plus son absence dans le premier bassin indique sa faible
tolérance vis à vis des eaux trop chargées.
Ø Brachionus leydigii
Brachionus leydigii peut se développer dans des milieux riches en matières
organiques d'origine animale (Pourriot 1965). Son développement est caractérisé par des
croissances brèves et imprévisibles en mai 1999 dans le bassin 3 (476 ind. l-1) et en août
1998 dans le dernier bassin (100 ind. l-1) (figs. 32 à 35). Son développement semble être
lié à la nature du peuplement algal en particulier les Chlorophycées qui se sont bien
développées durant cette période et à l’amélioration de la composition physico-chimique
des lixiviats.
Ø Brachionus bidentatus
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
104
Brachionus bidentatus est apparue dans le deuxième bassin en mai 1998 (40 ind.
l-1), en mai et juin 1998 (respectivement 240 et 180 ind. l-1) dans le troisième bassin et en
mai 1998 dans le dernier bassin (320 ind. l-1) (figs. 32 à 35). Son évolution confirme aussi
le rôle de la qualité de l'eau et la grande compétition spécialement d'ordre trophique entre
cette espèce et les autres Rotifères.
Ø Brachionus angularis
Cette espèce n’a été rencontrée que dans le dernier bassin avec une densité de 238
ind. l-1 (figs. 32 à 35). Sa croissance coïncide avec le développement des Chlorophycées.
Toutefois, cette espèce semble être moins tolérante à la pollution et de plus , elle ne peut
pas se développer au dépend des peuplements eugleniens, qui dominent une longue
période le peuplement algal dans les quatre bassins, et ceci en raison de sa petite taille par
rapport aux autres brachions (Pourriot, 1965).
Ø Brachionus patulis
Cette espèce présente une évolution saisonnière similaire à celle de Brachionus
patulis et de Polyarthra remeta. Son évolution est caractérisée par un pic de croissance
unique (245 ind. l-1) enregistré en mai 1999 (figs. 32 à 35).
Ø Epiphanes sp.
Epiphane senta est apparue dans le premier bassin en janvier et avril 1999 (100 ind. l-1 et
400 ind. l-1), alors qu’Epiphane clavulata n’est présente qu’au mois d’avril 1999 (800
ind. l-1). Dans le bassin 2, les Epiphanes ont été rencontrés au printemps uniquement
(mars et avril 1999), (1500 ind. l-1 d’Epiphane clavulata, observée en avril 1999). Dans
les deux derniers bassins (3 et 4) et à l’exception de décembre 1998 (figs. 32 à 35), les
Epiphanes ont colonisé le milieu durant toute la période hiverno-printanière. Il apparaît
que le genre Epiphane ne tolère pas trop les fortes charges, de plus, Epiphane clavulata
semble sensible aux faibles températures de l'eau, la même constatation à été faite par
Oulad Ali (1988). Epiphane clavulata est observée seulement en avril et semble être
sensible aux faibles températures. Epiphanes sp. se développe avec une plus faible
densité que les autres Brachionidae et plus spécialement en l’absence de Brachionus
calyciflorus. Ce qui met en évidence la grande compétition trophique existant entre ces
deux populations.
Ø Keratella quadratra
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
105
Keratella quadratra est le microfiltreur qui ne peut se développer qu’au dépens
des petites algues phytoplanctoniques (Chlamydomonas sp., Chrysomonas sp. et
Cryptomonas sp.) des bactéries et des détritus (Pejler 1957 et Pourriot 1965). Keratella
quadratra est apparue seulement dans les deux derniers bassins, en mai 1998 et mai 1999
dans le bassin 3 et mai, novembre, décembre 1998 et mai 1999. La densité maximale de
Keratella quadratra a été observée en mai 1998 (800 ind. l-1) dans le dernier bassin (figs.
32 à 35). Son absence dans les deux derniers bassins serait probablement liée à la forte
diminution de la charge organique d'une part et à la compétition avec les autres
Brachionidae d'autre part.
Ø Synchaeta tremula
L'espèce Synchaeta tremula est observée seulement dans le dernier bassin en juin
et juillet 1998. Son maximum d'effectif à été observé en juin 1998 (4200 ind. l-1) (figs. 32
à 35). Il semble que Synchaeta tremula préfère les eaux faiblement chargées.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
106
Fig. 32. Dynamique saisonnière des Rotifères dans le premier bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
107
Fig. 33. Dynamique saisonnière des Rotifères dans le deuxième bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
108
Fig. 34. Dynamique saisonnière des Rotifères dans le troisième bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
109
Ø Polyarthra remeta
Cette espèce n’a été rencontrée qu'en mai 1999 dans le dernier bassin avec une
faible densité (238 ind. l-1) (figs. 32 à 35). Son développement coïncide avec
l'amélioration de la qualité physico-chimique de l'eau, élévation de la température, le
développement important des Chlorophycées et l'absence de Moina brachiata. Il semble
que sa croissance soit essentiellement régie par les variations de la température et la
compétition avec les Moinidae. Les mêmes effets ont été montrés par Lair (1978) et
Gilbert (1987). Toutefois Gilbert (1988) a mis en évidence que les Cladocères et plus
précisément les daphnies peuvent entraîner une suppression importante des rotifères du
milieu. De plus, l'absence de Asplancha sp., écarte toute prédation possible par ce
Rotifère qui participe souvent au déclin de cette espèce (Garnier et Lescher-Moutoué,.
1984; Fahde, 1994).
Ø Ascomorpha sp.
Ascomorpha sp. est représentée par Ascomorpha agilis au printemps 1998 (avril et
mai 1999) avec des densités respectives de 100 et 714 ind. l-1 dans le bassin B3 et
seulement en mai 1999 dans le bassin 2 (154 ind. l-1), alors que dans le dernier bassin on
enregistre l’émergence de Ascomorpha ovalis (juin 1998, 1000 ind. l-1) (figs. 32 à 35).
L’effectif maximal d’ Ascomorpha agilis à été enregistré en mai 1999 (1900 ind. l-1). En
plus de sa sensibilité vis à vis des milieux pollués (Bassin 1 et 2) et la compétition
exercée par les autres Rotifères, il apparaît la présence d’une compétition inter spécifique
entre les deux espèces qui se manifeste par des pics de croissance décalés.
Ø Filinia hofmanni
Cette espèce s'est développée uniquement dans le dernier bassin en juin-juillet
1998 (respectivement avec 4200 et 8200 ind. l-1) et mai 1999 (476 ind. l-1) (figs. 32 à 35).
Toutefois, sa présence en été, est à l’encontre des observations rapportées par Ruttner-
Kolisko (1980) qui a montré que la température limitante maximale pour le
développement Felinia sp. est de 12 °C. Sa préférence aux particules plus petites
(inférieures à 10 µm) semble être le facteur limitant pour son développement dans les
bassins de la station d’Etueffont.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
110
Fig. 35. Dynamique saisonnière des Rotifères dans le quatrième bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
111
III. 3. Cladocères
III. 3. 1. Aperçu biologique et écologieLes Cladocères constituent un compartiment important des communautés
planctoniques d’eaux douces (Lacroix, 1989). Ce sont des crustacés de petite taille (leur
longueur s’échelonne entre 0,2 et 3 mm) (Amoros, 1984). Les Cladocères colonisent tous
les types d'eau douce, à l'exception des eaux courantes, où on ne les rencontre que
lorsqu'ils sont entraînés par la dérive à partir des eaux calmes (Amoros, 1984). Malgré
leur petite taille, les Cladocères jouent un rôle primordial dans les écosystèmes
aquatiques en assurant un transfert d'énergie vers les maillons supérieurs (poisson,
insectes, etc.) (Pourriot et Francez, 1986). Grâce à leurs œufs de durée, protégés par les
éphippies, ils peuvent coloniser même les mares temporaires. Les Cladocères possèdent
un spectre alimentaire très large, ils peuvent être herbivores ou détritivores. La majorité
estt des filtreurs qui se nourrissent d’algues unicellulaires, de protozoaires, de levures, de
bactéries et de détritus suivant les cas, d’autres sont prédateurs (Leptodora, Bythotrephes,
Polyphemus) et présentent des périopodes adaptés pour capturer les proies. Les particules
sont alors rassemblées dans une gouttière thoracique ventrale puis acheminées vers la
bouche. Les espèces benthiques ou périphytiques prélèvent leur nourriture en grattant les
substrats et la filtration n’est que secondaire. Les cladocères sont soumis à la prédation
des poissons, batraciens, oiseaux, des invertébrés, des grands Rotifères, des Copépodes,
des larves de diptères planctoniques ou d’hydracariens et de certains insectes. Certaines
espèces se développent dans des eaux très chargées en matière organique, peu oxygénées
et supportent des pollutions organiques (Daphnia magna et Moina sp.). Les Cladocères
se déplacent lentement et leurs vitesses varient de 0.3 à 1.2 mm. s-1 (Pourriot et Champ P,
1982). Le mode de locomotion est en relation étroite avec leur type d’habitat. Chez les
formes planctoniques (d’eau douce ou marine) la propulsion est assurée par le
rabattement vigoureux des antennes biramées vers l’arrière du corps; elles sont alors
capables d’effectuer des migrations de 50 m à des vitesses de 0.3 cm. s-1 (ces migrations
sont provoquées par les variations d’intensité lumineuse qui interagissent avec des
gradients thermiques et chimiques, et interfèrent aussi avec l’état physiologique en
particulier les stades de développement). Les formes benthiques se déplacent grâce à des
mouvements des périopodes.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
112
III. 3. 2. Inventaire systématique des Cladocères recensés durant notrecampagne
Durant tout le suivi, un seul genre à été rencontré avec seulement deux espèces
(Moina macropa et Moina brachiata).
Embranchement des Arthropodes
Sous-embranchement des Mandibulates
Classe des Crustacés
Sous-classe des Branchiopodes
Ordre des Anomopoda
Super-famille des Chydoroïdes
Famille des Moinidae
Genre Moina
Moina macropa
Moina brachiata
III. 3. 3. Evolution spatio-temporelle de la densité du peuplementCladocérien
Le peuplement cladocérien des lagunes de la station d’Etueffont est représenté
uniquement par deux espèces de Moinidae (Moina brachiata et Moina macropa).
L'évolution saisonnière des Moinidae dans la station d'Etueffont est caractérisée par un
développement estival unique caractérisé par une présence des Moinidae en mai et juin
1998 dans le premier bassin, en mai - juin - juillet 1998 en B2, en mai et juillet en B3 et
seulement en mai 1998 (fig. 36). L’évolution spatiale met en évidence une diminution des
effectifs de Cladocères en passant du premier au dernier bassin (qui passe de 46000
ind. l-1 dans le premier bassin à 80 ind. l-1 dans le dernier bassin). Cette chute de la
densité des Moinidae en passant du premier au dernier est vraisemblablement liée au
développement important d’algues non assimilables (Coelastrum sp. très abondant) en
l’absence des prédateurs spécifiques des Cladocères (poissons). Par ailleurs, une action
prédatrice par les batraciens, très répandus dans les derniers bassins sur les Moinidae est
envisageable.
La présence d’un peuplement cladocérien monospécifique de type Moinidae
prouve bien un état de dystrophie très avancé de l’eau dans les quatre bassins, pour la
raison que les Moinidae sont les Cladocères les mieux adaptés aux fortes charges
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
113
organiques (Amoros, 1973 et Pacaud, 1939). La présence des Moinidae et en particulier
Moina macropa caractérisée par sa forte sensibilité aux éléments métalliques (Martinez-
Tabche et al., 1998) dans le premier bassin témoigne de l’absence d’une toxicité
métallique éventuelle. Si les basses températures (inférieures à 18 °C) peuvent expliquer
l'absence des Moinidae durant une longue période de l'année (octobre 1998 à avril 1999)
en provoquant un ralentissement du rythme de ponte et la rareté des ressources
phytoplanctoniques (Tifnouti et Pourriot, 1989). Un effet défavorable dû à l'élévation de
cette dernière est exclu du fait qu'on a jamais enregistré des températures supérieures à 30
°C durant tout le suivi, de plus les Moinidae sont caractérisés par une forte résistance aux
températures très élevées (Gras et Saint-Jean, 1978; Jana et Pal, 1985 et Hart, 1985 in
Tifnouti et Pourriot, 1989). La disparition des Cladocères en septembre et octobre 1998,
phénomène déjà observé chez les Rotifères met en évidence un facteur commun dans la
disparition de ces deux groupes. Cette absence coïncide avec une chute importante de la
teneur en oxygène dissous. Il est néanmoins difficile d'attribuer à la baisse de l'oxygène
dissous toute la responsabilité de cette disparition car Pacaud (1939) et Liebermann
(1970) ont montré une forte résistance des Moinidae pour des milieux faiblement
oxygénés. Il apparaît que l'oxygène n'est qu'un simple facteur qui peut accentuer
l'élimination du zooplancton dans les quatre bassins. De plus, l'effet négatif des blooms
phytoplanctoniques qui selon certains auteurs (Lefevre 1942 (cité par Angeli, 1979))
l’attribue au déclin des Moinidae peut être envisagé dans notre cas parce que les
biomasses algales enregistrées dans les quatre bassins restent néanmoins moins
importantes que celles enregistrées en littérature pour les bassins de traitement des eaux
usées. Il semble donc, que c'est plutôt la composition qualitative du phytoplancton qui
soit à l’origine du déclin des Moinidae.
Selon Benider et al. (1998) et Lynch (1989), un apport insuffisant de nourriture
(concentration en algues inférieure à 3,125 10 4 cellules. ml-1) supprime la reproduction
chez les Moinidae. De plus, durant ces deux mois (septembre et octobre 1998), on
enregistre la disparition des Chlorophycées et une dominance du genre Stephanodiscus
dubuis pour les diatomées. Toutefois, une toxicité due aux algues déjà envisagée (Fogg,
1971 et Angeli, 1979) nous paraît peu probable en raison de l'absence des Cyanobactéries
dans les quatre bassins durant tout le suivi.
Par ailleurs, il apparaît que Brachionus calyciflorus apparaît sensible et ne
coexiste pas avec les Moinidae, à l’opposé de Brachionus rubens qui apparaît peu
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
114
sensible aux fortes densités des Moinidae. Ceci est en désaccord avec les travaux de
Benider et al. (1998) qui ont mis en évidence l’absence d’une telle compétition. De plus,
la diminution de la densité des Moinidae dans l'ordre croissant des bassins est associée à
l’amélioration qualitative du peuplement rotarien. La succession taxonomique observée
par Angeli en 1979, dans laquelle il classe les groupes zooplanctoniques selon un
gradient d'oxydation de la matière organique (Ciliés - Brachionus calyciflorus ou autres
Rotifères - Daphnia magna), modifié ensuite par Tifnouti et Pourriot en 1989, qui
intercalent Moina micrura avant Daphnia magna, pourrait être définie pour le cas des
lixiviats de décharge comme suit : Ciliés - Moinidae, Brachionidae et autres Rotifères -
Daphnia magna.
Fig. 36. Distribution des densités des Cladocères dans les quatre bassins.
III. 3. 4. Etude de la dynamique saisonnière des espèces les plusfréquentes
q Moina macropa
Moina macropa à été rencontrée seulement dans les deux premiers bassins, en B1
(mai et juin 1998) et en B2 (juin et juillet 1998) (fig. 37). Malgré cette absence dans les
deux derniers bassins, Moina macropa représente 54 % des Moinidae. Son effectif
maximal a été enregistré en juillet 1998 dans le deuxième bassin avec un effectif de
l'ordre de 25000 ind. l-1.
q Moina brachiata
A l'opposé de Moina macropa, Moina brachiata, semble avoir un développement
spatial plus étendu avec une présence dans les bassins 3 et 4. Toutefois, il apparaît que,
Moina brachiata présente la même évolution saisonnière que celle observée chez Moina
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
115
macropa. Elle représente 46 % des Moinidae. L'effectif peut atteindre 3.104 ind.l-1
enregistré en juin 1998 dans le premier bassin (fig. 37). Son évolution spatio-temporelle
est marquée par un développement en mai et juin 1998 dans le premier bassin.
III. 3. 5. Facteurs responsables de l’absence de Daphniidae dans lesbassins de la station d’Etueffont
Si dans la plupart des bassins de lagunage, les chercheurs enregistrent la présence
de Daphnia sp. (Loedolf, 1965; Angeli, 1979 et Pizay-Parenty, 1985), plus eurytherme
que le genre Moina sp., nous n’avons jamais rencontré cette espèce dans les bassins de la
station d’Etueffont. Toutefois, Loedolf (1965) a montré que les Moinidae dominent les
bassins de stabilisation primaire par opposition au genre Daphnia qui préfère les bassins
moins chargés (bassins secondaires et tertiaires). La même constatation à été faite par
Angeli (1979), qui montre en étudiant les bassins de lagunage du Nord de la France un
déplacement au profit des Moinidae à forte charge polluante, à forte température et à
forte activité algale. De plus, plusieurs auteurs attribuent l’absence de Daphnia à la
présence des cynobactéries (Sterner, 1989) ou à la grande action prédatrice par les
poissons (Persson, 1988). Enfin, Ding (1973), cité par Angeli 1979, observe dans les
bassins de lagunage au Texas une substitution des Daphniidae par les Moinidae lorsque le
pH et la concentration en NH4+ deviennent élevés. Il paraît que c'est plutôt cette dernière
hypothèse qui explique l'absence des Daphnies dans la station d'Etueffont ceci en raison
de la forte concentration en NH4+ enregistrée durant tout le suivi.
En conclusion on notera que les Moinidae semblent avoir un seuil de tolérance de
pollution similaire à celui des Rotifères. Les pics de croissance décalés témoignent d'un
phénomène de compétition intraspécifique et interspécifique entre les Rotifères et les
Cladocères. Enfin, indépendamment de la température et de la qualité physico-chimique
très complexe des quatre bassins qui conditionne la succession des Cladocères et des
Rotifères c'est le peuplement algal qui apparaît le facteur déterminant de la composition
zooplanctonique. L'étude de la dynamique des deux espèces de Moinidae nous montre
une tolérance de Moina macropa au dépens de Moina brachiata vis à vis de tous les
facteurs environnementaux, ce qui semble être lié à sa taille plus faible que celle de
Moina macropa, donc avec des besoins alimentaires plus faibles et une mobilité
suffisante pour diminuer l'effet de prédation possible.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
116
IV. RELATIONS ENTRE BACTERIES ET PHYTOPLANCTONA l’exception des mois de septembre et décembre 1998 pour le bassin 1, et de
novembre 1998 pour les bassin 2, 3 et 4, où l’on enregistre une dominance de la biomasse
bactérienne au dépend de la biomasse algale suite à la diminution de l’action prédatrice
par le zooplancton et à la diminution de la biomasse algale, le rapport biomasse
bactérienne sur la biomasse algale est souvent très faible, et la production
phytoplanctonique représente plus de 90 % de la production primaire. Ceci est en
désaccord avec les travaux de Hessen (1985), Simon et Tizler (1987) et Cho et Azam
(1990) qui ont mis en évidence dans les milieux lacustres une nette domination de la
biomasse bactérienne. De plus, ce rapport tend vers une valeur de 2 dans les milieux
ultraoligotrophes (Fuhrman et al., 1989 et Azam, 1990). Nos résultats montrent qu’à
l'exception des prélèvements d’avril 1999 dans le premier bassin, et ceux de janvier 1999
dans les bassins 2 et 3, les pics de biomasses bactériennes coïncident avec les pics de
biomasses phytoplanctoniques. Ces résultats confirment le rôle des algues dans la
stimulation de la croissance du bactérioplancton par l’excretion de carbone dissous
(COD). Pour tenter d’aller plus loin dans l’interprétation de nos résultats, nous avons
recherché s’il existait d’éventuelles relations statistiques entre les bactéries et le
phytoplancton. L’étude statistique entre ces deux maillons met en évidence l'existence de
corrélations exponentielles statistiquement significatives dans les bassins 2 et 3 (r = 0,67
pour le deuxième bassin et r = 0,6 pour le troisième bassin). Cela est corroboré par les
travaux d’Aleya et al. (1987), Weiss et al. (1990), Pace et al. (1990), qui ont montré que
les excrétas d'algues sont la principale source de substrat des bactéries. De plus, Feuillade
et al. (1986) ont pu montrer l'existence d'un couplage entre l'excrétion phytoplanctonique
et l'assimilation hétérotrophe bactérienne dans la zone euphotique du lac Léman.
Toutefois, l’absence d'une telle corrélation dans le premier bassin ne peut être due qu’à
l’apport par le lixiviat de matières organiques nécessaires pour le développement
bactérien. C’est vraisemblablement la décharge qui gouverne la dynamique bactérienne
par l’intermédiaire de son lixiviat qui fournit les substances organiques, en particulier le
carbone organique nécessaire à la croissance bactérienne dans le premier bassin. Ainsi,
c'est en été quand la décharge atteint son activité maximale avec la production des
lixiviats de plus en plus riches en substrats organiques, que la densité bactérienne, dans le
premier bassin, atteint sa croissance optimale. Bien que l’on n'ait pas réalisé de
spéciations de la matière organique présente dans le lixiviat de la décharge d’Etueffont, il
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
117
apparaît que les bactéries au sein de la décharge et grâce à leurs enzymes extracellulaires
semblent favoriser la tache des bactéries contenues dans le premier bassin en hydrolysant
la matière organique souvent sous la forme de biopolymères à poids moléculaire élevé.
De plus, l’utilisation du carbone organique allochtone par les bactéries est dépendant du
temps de séjour de l’eau, de l’ensoleillement et de la température (Berstrom et Jansson,
2000). Ces conditions coexistent dans le premier bassin. Enfin, Filip et al. (1998 et 2000),
ont montré que les bactéries au sein des décharges se développent essentiellement grâce
aux acides humiques. La dynamique des bactéries semble dépendante de l’apport par la
décharge de substances organiques dans le premier bassin et du peuplement
phytoplanctonique dans les bassins 2 et 3. Dans le dernier bassin et malgré les fortes
densités algales rencontrées et le faible apport de substances organiques par la décharge,
aucune relation n’a été enregistrée entre ces deux maillons. Toutefois, c’est dans ce
dernier bassin que les plus faibles densités bactériennes ont été enregistrées. Cette
absence de corrélation entre phytoplancton et bactéries dans le bassin 4 semble être
attribuée à la compétition trophique entre les deux maillons pour les substrats présents,
corroborant les observations d’Aloice et Tatsuya (1996). Ce bassin est caractérisé par une
diversité spécifique rotarienne importante. Les Rotiféres, tant par leur action prédatrice
directe sur le peuplement bactérien que par leur rôle compétitif au niveau de
l’exploitation de la matière organique détritique, sont susceptibles d’entraîner une
diminution des effectifs bactériens.
V. RELATION ENTRE BACTERIES ET ZOOPLANCTONLes microflagellés, peuvent exercer une prédation importante sur les bactéries
dans le milieu marin et dulçaquicole, en raison que 103 à 104 organismes par litre,
peuvent ingérer jusqu’à 100 % de la biomasse bactérienne par jour (Fenchel, 1986;
Nagata, 1988; Berstrom et Jansson, 2000). Les Rotifères et les Cladocères seraient
cependant susceptibles d’ingérer efficacement les bactéries regroupées en agrégats ou
fixées sur un substrat. Cette fraction est souvent très importante dans les milieux
eutrophes et peut atteindre les 50 % du peuplement bactérien (Pedros-Alios, 1989).
Cependant, les courbes d’évolution des biomasses zooplanctoniques et bactériennes
mettent en évidence une évolution identique dans les 3 premiers bassins avec des facteurs
de corrélation plus élevés dans les bassins 1 et 2, respectivement r = 0,52 et 0,48.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
118
Fig. 37. Dynamique saisonnière des Rotifères dans le quatrième bassin.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
119
Cette corrélation s’atténue dans le bassin 3 (r = 0,32) et devient négative dans le
bassin 4 (r = -0,58). L'évolution similaire des bactéries et du zooplancton dans les deux
premiers bassins semble être liée à l'absence de prédateurs donc à un développement
zooplanctonique de type phytophage qui coïncide avec le développement le plus
important des Moinidae dans ces deux bassins. La substitution spatiale des Cladocères
par des espèces de petite taille et de régime bactérivore de préférence (Rotifère) stimulé
par la présence importante dans le dernier bassin d’algues de grande taille et non
assimilables (Coelastrum sp.) et la diminution de la charge organique semblent renverser
cette tendance. Ainsi, dans le dernier bassin on enregistre une évolution opposée entre le
zooplancton et les bactéries. Toutefois, si la chute des densités bactériennes en passant du
premier au dernier bassin peut être attribuée en grande partie à une augmentation du
broutage par le zooplancton (Sterner, 1989) et à la diminution de l’état trophique du
lixiviat, l’ensoleillement par son action bactéricide semble éliminer une grande partie des
bactéries (Bahlaoui, 1990; Wei et al., 1994; Herrera Melian et al., 2000). En conclusion,
il apparaît que si le phytoplancton est la proie essentielle de ces Rotifères, l’utilisation
directe des bactéries libres ou fixées constitue un appoint de nourriture non négligeable.
Ainsi, le zooplancton peut jouer un rôle régulateur du peuplement bactérien et favoriser
ainsi comme dans le cas de la station d’Etueffont un abattement bénéfique de la charge
bactérienne.
VI. RELATIONS ENTRE ZOOPLANCTON ET PHYTOPLANCTONSelon Vedry (1987), la sélectivité du milieux peut s'exercer à différents niveaux:
toxicité organique ou inorganique, toxicité induite par la prolifération bactérienne ou,
selon Angeli (1979), par des excrétions algales et compétition intraspécifique ou
interspécifique vis a vis de l'oxygène ou de la nourriture. Les interactions trophiques
régissent en grande partie sur l’abondance de chaque population d’un écosystème
aquatique (Lazzaro et al., 1992; Miltstein et al., 1995). Toutefois, Pourriot (1968) a
effectué une synthèse bibliographique sur de nombreux travaux effectués en Europe de
l'Est (Hrbacek, 1962; Hrbacek et al., 1965; in Angeli, 1979), qui ont mis en évidence
l’efficacité des grands Cladocères filtreurs (genre Daphnia) dans la limitation des blooms
phytoplanctoniques, même si leurs effectifs sont modestes. A cet effet et dans le même
cadre Dawdowicsz (1990) souligne que c'est seulement Daphnia magna qui peut ingérer
des particules supérieures à 250 µm. Le rôle du zooplancton comme régulateur de la
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
120
production primaire phytoplanctonique a été le sujet de plusieurs travaux (Oulad ali,
1988; Norberg et al., 2000; Hernandez-Leon, 2000; Weithoff, 2000). La forte réduction
des algues sous l’effet du broutage par le zooplancton, très fréquente juste après les
poussée phytoplanctoniques printanières est connue sous le nom de phases d’eaux claires
en raison de l’augmentation de la transparence du milieu. Dans le cas des bassins de la
station d’Etueffont le phénomène d’eaux claires (bien que les profondeurs soient faibles
par rapport aux lacs) a été observé en début d’été (mai, juin et juillet 1998) et au
printemps 1999 (fig. 38). Si la disparition du phytoplancton en début de l’été peut être
attribuée à l’action conjuguée des Moinidae et des Rotifères, la disparution printanière ne
peut être liée qu’aux Rotifères. Malgré leur faible pouvoir de filtration et grâce à leur
forte densité, ce petit zooplancton apparait capable de controler la prolifération du
phytoplancton dans les bassins d’Etueffont. Ces résultats sont corroborés par les
observations d’Angeli (1979), Salomoni (1991) et Gnudi et al. (1991) dans des stations
d’épuration des eaux usées, ainsi que celles observées par Sommer et al. (1986) dans les
lacs peu profonds. L’action prédatrice par le zooplancton est accentuée par la diminution
des substances nutritives. Toutefois, et dès la fin de juillet 1998 on enregistre un
développement phytoplanctonique dominé par des formes coloniales de grande de taille
(Coelastrum sp.) difficilement ingérables par la plupart des espèces zooplanctoniques. Il
apparaît que le phytoplancton soumis à la forte pression de prédation exercée par les
Rotifères et les Moinidae au printemps et en début d’été tend à s’adapter par un
changement structural de sa communauté. Les mêmes observations ont été décrites par
Burns (1968), Gliwicz (1969 et 1980) et Sterner (1989) qui ont mis en évidence une
substitution du phytoplancton unicellulaire (de grande taille et/ou coloniale) par un
peuplement phytoplanctonique composé par de petites algues unicellulaires. L’étude du
cycle saisonnier des phytoplanctontes et des zooplanctontes (Rotifères et Cladocères)
montre une alternance entre les deux maillons, les pics de biomasses phytoplanctoniques
coïncidant avec les faibles biomasses rotariennes. Cela met en evidence le rôle primordial
de ces petits filtreurs dans la régulation du phytoplancton, comme semble l’indiquer la
présence d’une corrélation exponentielle négative statistiquement hautement significative
entre la biomasse rotarienne et la biomasse algale dans le dernier bassin (r = 0,92; n =
13). L’absence d’une telle corrélation significative dans les trois premiers bassins
n’élimine pas le rôle des Rotifères dans la régulation du phytoplancton, mais il semble
que leur action soit masquée par l’effet conjugué de la présence des grand filtreurs
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3 è Partie Chap. 2. Etude des peuplements planctoniques dans les quatrebassins
121
(Moinidae) qui exercent un broutage plus important que celui des Rotifères et à la
présence d’une autre source de nourriture représentée par de la matière organique
particulaire ou dissoute importée par la décharge.
Fig. 38. Dynamique saisonnière du zooplancton dans les quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
122
I. TRAITEMENT DES LIXIVIATS PAR LAGUNAGE NATUREL
I. 1. Introduction
Des études tant à l'échelle du laboratoire qu'à celle du pilote ou en vraie grandeur
ont été réalisées afin de sélectionner la méthode la plus fiable pour traiter ces eaux
polluées. Les premières recherches datent des années 70 (Boyle et Ham, 1974) qui se
basaient au début sur un mixage des lixiviats avec les eaux usées domestiques. La non-
crédibilité de ce procédé a poussé les chercheurs à traiter les lixiviats séparément des
eaux usées domestiques tout en utilisant les mêmes procédés appliqués à ces derniers
(lagunage aéré, les boues activées et les réacteurs en batch développés par Hosomi en
1989). En effet ces procédés ont des faiblesses de rendement pour les basses
températures. Ces faiblesses sont observées surtout dans le cas de l'azote où l’on
enregistre des taux de dénitrification très faibles (Knox, 1983 et 1985). Afin de pallier ces
inconvénients plusieurs procédés ont été proposés comme le traitement par les filtres
biologiques aérobies submergés proposés par Pedersen et Jansen (1992) et les réacteurs
biologiques (Spengel et Dzombak, 1991). Ces deux systèmes apparaissent moins
sensibles aux variations de température et aboutissent à une dénitrification significative.
D'autres comme Hollopeter et Dague (1994) ont pu obtenir un abattement de 96 % de
DCO en développant le procédé ASBR (Anaerobic Sequence Batch Reactor). Enfin
Welander et al. (1997) ont pu atteindre des taux de dénitrification de 77 % avec une
température de 5 °C en utilisant des bio-films suspendus. Dans ce chapitre on a essayé
d’évaluer l’efficacité du traitement par lagunage naturel pour ce type de lixiviat.
I. 2. Cinétique de dégradation de la matière organique
Plusieurs formules empiriques ont été proposées pour évaluer la cinétique de
dégradation de la matière organique dans les bassins de lagunage naturel (Hermann et
Gloyna, 1958; Marais et Shaw, 1961; Marais, 1966; Huang et Gloyna, 1968; Gloyna et
Espino, 1969). Toutefois et si l’on admet que toute la DBO des lixiviats est stabilisée par
des organismes facultatifs, que les eaux sont complètement mélangées et que la
dégradation se fait par une réaction du premier ordre (Gloyna, 1966). La cinétique de
dégradation peut se calculer par la formule suivante :
Kt = (C0/ Cf –1)/ Rt
Avec,
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
123
Kt: cinétique de dégradation par jour à la température T, C0: teneur en DBO (mg.
l-1) de l’eau brute, Cf: teneur en DBO (mg. l-1) de l’eau traitée et Rt: temps de rétention en
jours.
Fig. 39. Variations de la vitesse de dégradation K (j-1) de la DBO calculée en fonction dela température.
Il apparaît d’après nos résultats que les cinétiques de dégradation calculées sont
très voisines des valeurs trouvées par certains auteurs dans des bassins de stabilisation
des eaux usées (Duart, 1968; cité dans le rapport inter agence) et sont dépendantes des
températures, mettant ainsi en cause l’effet de la température dans la stimulation de
l’activité bactérienne hétérotrophe. La cinétique de dégradation la plus élevée a été
enregistrée au mois de juin 1998 qui correspond à une température de 22 °C (fig. 39),
alors qu’une température de 20 °C aboutit à une vitesse de dégradation très faible (0,066).
Ceci nous laisse penser que l’augmentation de l’activité bactérienne commence à être
sensible pour des températures supérieures à 20 °C. Les rendement épuratoires ont été
calculés par la formule suivante :
X = ((C0 - Cf) / C0) * 100
Avec:
X : Rendement épuratoire (en %), C0 : Concentration de l'élément chimique
à la sortie de la décharge et Cf: Concentration de l'élément chimique à la sortie du
quatrième bassin.
Lixiviats traité
Moy Ecar Min Max
Q (l. s-1) 2,46 2,64 0,06 8,60Cond (µS. cm-1) 1772 266 1303 2130T (°C) 12,07 7,33 1,50 20,20pH 7,12 1,49 4,09 8,77O2d (mg. l-1) 5,52 4,46 0,44 18,30Eh (mV) 94 71 -86 164MES (mg. l-1) 24 20 6 60chl a (µg. l-1) 79 50 15,5 191DBO (mg. l-1) 20 17 1 65
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
124
DCO " 240 83 114 443K+ " 148 65 17 250Mg2+ " 16 7 1 31Na+ " 212 93 20 327NH4
+ " 55 15 38 98Ca2+ " 96 59 38 260HCO3
- " 645 92 500 781F- " 0,13 0,17 0,00 0,42Cl- " 192 46 123 260NO2
- " 1,6 4,4 0,0 15,2NO3
- " 88 85 7 283SO4
2- " 119 127 51 519PT " 0,25 0,08 0,16 0,41NT " 66 12 66 66Fe2+ " 2,51 1,53 0,02 5,10Ni2+ " 0,80 1,30 0,05 4,93Zn2+ " 1,11 0,83 0,35 2,91Sn2+ " 0,19 0,34 0,00 1,04Cr3+ " 0,08 0,08 0,00 0,22Mn2+ " 1,91 1,10 0,10 3,49Cu2+ " 0,15 0,18 0,00 0,58
Tableau 12. Composition physico-chimique moyenne du lixiviat après traitement.
I. 3. Résultats et discussion
I. 3. 1. Abattement des matières en suspensions
L’étude de l’évolution du taux des MES (fig. 40a) met en évidence des
abattements qui varient de 34 à 97 % avec un maximum d’abattement enregistré en avril
1999 (97 %), cependant on constate que les lagunes arrivent toujours à éliminer une
grande partie des MES provenant du lixiviat brut ainsi que les MES engendrées par
l’activité photosynthétique. De plus, durant tous les suivis on n'a jamais dépassé les
normes concernant les eaux traitées par lagunage naturel qui préconisent des teneurs en
MES < 120 mg. l-1 (loi du 4 novembre 1980).
En effet, les deux chutes d’abattement ont été observées en août 1998 et en mai
1999 avec respectivement des rendements négatifs de –41 % et –29 % marquant ainsi le
rôle prépondérant des algues dans l’élaboration de MES biogènes.
I. 3. 2. Abattement des éléments inorganiques
Les taux d’abattement de la conductivité électrique ainsi que tous les ions
majeurs, représentés par les chlorures (fig. 40a) ont la même évolution saisonnière avec
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
125
des taux d’abattement élevés en été et faibles en hiver. Le taux d’abattement maximal de
la conductivité électrique a été enregistré en juillet 1998 avec 71 %. Toutefois, pour les
éléments métalliques on enregistre des fluctuations très importantes qui différent d’un ion
métallique à un autre. Pour le Ni le rendement calculé varie de 52 à 90 % (fig. 40e). En
effet, une baisse importante des rendements a été observée en mars 1999 pour le nickel.
Cette diminution de rendement enregistrée est vraisemblablement liée aux phénomènes
de relargage par le fond induits par la diminution importante du pH (pH compris entre 4
et 5). Pour le fer et le manganèse (figs. 40b et 40c), les courbes d'évolutions mensuelles
des rendements épuratoires sont quasi semblables avec des taux d'abattement trop élevés
qui peuvent atteindre 90 % en été, alors qu'on enregistre des valeurs négatives en début
d'été et en hiver. Il semble que le relargage de ces deux oligo-éléments par le fond est
géré à la fois par la diminution du pouvoir oxydant en été et par la diminution du pH en
hiver et début du printemps. Enfin, aucune tendance n'a été notée dans le cas du zinc, du
cuivre et de l’étain. Cependant, les rendements calculés peuvent atteindre 60 % pour le
zinc (fig. 40d), 98 % pour l’étain et 99 % pour le cuivre. On peut conclure que le
fonctionnement global de ce procédé de dépollution par lagunage naturel est très différent
entre l’été et l’hiver qui voit la diminution de la vie biologique qui influe directement ou
indirectement sur le Eh et le pH responsables des phénomènes de précipitations et de
relargages des éléments chimiques.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
126
Fig. 40. Evolution des taux d'abattement de NH4, MES, NT, DCO, DBO, Cl, PT, Zn, Ni,Mn et Fe.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
127
I. 3. 3. Abattement des substances nutritives
Le suivi de l'évolution mensuelle des rendements épuratoires des nutriments (figs.
40h, 40i et 40j) nous a permis de mettre en évidence la présence comme pour les
paramètres abiotiques des abattements importants (jusqu’à 98 %) en été et des
abattements faibles en hiver. Les abattements estivaux sont liés au temps de séjour très
important du lixiviat dans la station. Huang (2000) a trouvé des taux d’abattement de la
DBO, NT et PT de 92 % pour un temps de séjour de 85 jours. Cependant et durant la
période estivale, on enregistre les taux d’abattement les plus élevés pour l’ensemble des
paramètres liés à l'activité bactérienne intense stimulée par l'élévation de la température
(Kato, 1985; Marvalin et al., 1989), à la précipitation chimique et à l’activité algale (Tam
et Wong, 2000). Les pourcentages d’élimination du phosphore total varient de 13 à 93 %.
Les rendements est les plus faibles ont été enregistrés en hiver, vraisemblablement liés à
la faible activité algale (Ouzzani et al., 1997) et au relargage possible par le sédiment
stimulé par la forte diminution du pH (pH = 4 dans les quatre bassins). Toutefois, la
désorption hivernale provoquée par cet abaissement de pH nous renseigne sur le
phosphore qui est plutôt associé au calcium et non aux hydroxydes de fer et d’aluminium.
Cette constatation découle du fait que c’est du phosphore associé aux hydroxydes de fer
ou d’aluminium et donc que c’est plutôt une augmentation du rendement donc un
piégeage du phosphore à bas pH (Holtman et al., 1988; Fox et al., 1988). Néanmoins, on
élimine, par le relargage estival, favorisé par plusieurs facteurs tel que la température qui
provoque un ralentissement du relargage du phosphore (Saraiva-Cavalcante, 1995), la
diminution de l’oxygène dissous en fin d’été stimule le relargage du phosphore,
cependant Saraiva-Cavalcante (1995), a mis en évidence un relargage dès que la valeur
de l’oxygène dissous atteint les 2 mg. l-1 il y a une élévation de l’activité bactérienne du
sédiment qui selon Gachter et Meyer (1993) intervient dans ce processus par une
assimilation et stockage du phosphore dans les cellules, par relargage du phosphore après
leur mort, par un rôle de catalyseur des réactions de la minéralisation du phosphore
organique et un relargage de phosphore si les conditions redox ne sont pas favorables.
Toutefois cette désorption estivale du phosphore total n’influe pas sur le taux
d’abattement de ce paramètre parce qu’elle est contrebalancée par une forte assimilation
algale. Les taux d’élimination de l’azote total suivent la même évolution que celle du
phosphore total, mais avec des maxima qui ne dépassent pas les 70 % d’abattement; Les
taux d’élimination de l’azote total oscillent entre 14 et 70 %, ces valeurs sont
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
128
comparables à celles trouvées par d’autres chercheurs pour des climats semi arides
(Bouarab et al., 1994). Le faible rendement hivernal s’explique, comme pour le
phosphore, par une remise en solution par le fond associée à une mauvaise dénitrification
engendrée par l’abaissement des températures et une diminution de l’assimilation algale
(Bouarab et al., 1994). L’abattement de l’azote total ou de l’ion ammonium, du fait qu’il
présente la quasi-totalité de l’azote total, met en évidence que la station élimine mieux le
NH4+ en été, alors que ce rendement donne des valeurs critiques à basses températures.
L’augmentation des nitrates au fil des bassins témoigne de l’importance des phénomènes
de nitrification souvent considérés comme négligeables pour ces types de traitement par
plusieurs auteurs (Neel et al., 1961; Walker et Leclerc, 1973; Pujol, 1982). Cependant
d’autres comme Schleypen (1983) lui attribuent un rôle primordial, Jannes et Pritchard
(1972) cités par Golterman (1975) ont trouvé que c’est à l’entourage des MES et des
particules en aérobiose que la dénitrification s’installe.
I. 3. 4. Abattement de la charge organique
Les pourcentages d’abattement de la DBO varient de 13 à 98 % avec des maxima
d’abattement en été (fig. 40f). L’abattement de la DBO s’effectue grâce aux bactéries
hétérotrophes (Pseudomonas, Flavobacterium, Archromobacter et Alcaligenes sp.)
(Maynard et al 1999), jusqu’à 98 % en juin. Toutefois, deux chutes de rendement ont été
enregistrées pour la DBO. La première a été observée en septembre 1998 (-8 %),
vraisemblablement due aux blooms phytoplanctoniques très présents en été et qui
enrichissent le lixiviats aux cours de son transit dans les bassins. La deuxième chute de
rendement enregistrée en hiver avec seulement 13 % en janvier 1999. Cette diminution
hivernale est attribuée d'une part à la diminution de l'activité biologique et d'autre part
aux phénomènes de relargage par le fond de matières organiques sous l'effet de la forte
diminution du pH (pH compris entre 4 et 5). L’évolution du taux d’élimination des
bactéries libres montre une efficacité très importante en été (87 à 99 %), faibles en hiver
(40 à 60 %) et des valeurs intermédiaires en été. Cette efficacité estivale est
vraisemblablement liée à l’intervention de facteurs biologiques (prédation) et physiques
(ensoleillement).
La DCO présente une évolution annuelle similaire à celle de la DBO (fig. 40g).
En effet la diminution brutale du rendement en septembre 1998 est faiblement ressentie
pour la DCO. Ce qui nous renseigne sur l'aspect biodégradable des matières organiques
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
129
autochtones. Toutefois, les courbes d'évolution mensuelle des taux d'abattement du NH4,
NT et PT présentent des allures semblables à ceux de la DBO et de la DCO, témoignant
ainsi, d'un lien très fort entre ces éléments et avec les paramètres indicateurs de la matière
organique. L’abattement de la charge bactérienne atteint des valeurs très importantes de
98 % en été et 79 % au printemps. Ces maxima d’abattement coïncident avec l’élévation
de la température qui agit directement par l’action des UV et indirectement par le
développement du zooplancton bactérivore.
I. 3. 5. Conclusion
Dans ce chapitre, il ressort que le lagunage naturel pour le traitement de ces
lixiviats est très satisfaisant pour certains paramètres (DBO, NT, PT et les éléments
métalliques) durant toute la saison, alors qu’il semble être moins efficace pour la
DCO et les autres éléments chimiques à cause de leurs grandes sensibilités aux
variations des facteurs naturels non maîtrisables tel que la T, le pH et le Eh et de
leurs rendements qui dépassent rarement les 80 %.
II. IMPACT DES LIXIVIATS TRAITES SUR LES EAUX SUPERFICIELLES
II. 1. IntroductionLes lixiviats de décharges peuvent soit s’infiltrer dans le sous-sol et contaminer
les eaux souterraines, soit encore après drainage, collecte et traitement, être rejetés dans
un cours d’eau. Les études écotoxicologiques de l’impact des rejets des lixiviats de
décharges sur les écosystèmes lentisques se limitent le plus souvent à l’évaluation de la
toxicité des acides organiques présents en grande quantité dans les lixiviats et qui peuvent
former des complexes toxiques avec les éléments métalliques (Fraser et al., 2000) où à la
recherche d’indicateurs de contamination. Cependant, Cyr et al. (1987), ont montré que
les lixiviats de la décharge de Peticodiac (Canada) provoquent une augmentation de la
conductivité électrique, des chlorures, des sulfates, de l’alcalinité et de la dureté. De plus,
Keck et Jean (1990, 1991) ont mis en évidence une élévation des teneurs en chlorures, de
la conductivité électrique, de l’ammonium avec apparition des bactéries filamenteuses et
une diminution de la richesse taxonomique pour une rivière qui reçoit directement des
lixiviats d’ordures ménagères. Enfin, Nuttall (1973) indique que le rejet des lixiviats de
décharge dans une rivière peut entraîner le développement exubérant des champignons et
la domination des oligochètes chez les invertébrés. Dans le même cadre et afin d’évaluer
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
130
l’impact des lixiviats traités de la décharge d’Etueffont sur les eaux superficielles, on a
procédé à un suivi mensuel pendant 13 mois de quelques paramètres en amont (environ
50 m) et en aval du rejet de lixiviat traité (environ 100 m). De plus et afin de mettre en
évidence le pouvoir auto-épuratoire du ruisseau, on a procédé le 13 juin à un suivi de
quelques paramètres le long du ruisseau.
II. 2. Points d’échantillonnage
Les prélèvements d’eau du 13 juin 1999 ont été effectués entre 12h et 14h à la
buse de sortie du dernier bassin (S), dans le ruisseau à l’amont du rejet (A), où se déverse
le lixiviat (M), à 20 m en aval du rejet (V20), à 40 m en aval du rejet (V40), à 60 m en
aval du rejet (V60), à 100 m en aval du rejet (V100), à 200 m en aval du rejet (V200), à
300 m en aval du rejet (V300) et à 500 m en aval du rejet (V500).
II. 3. Résultats II. 3. 1. Température
Les valeurs de la température relevées mensuellement en amont du ruisseau
varient de 3,1 à 20,2 °C (m ± δ = 12,3 ± 5,5 °C), celles relevées en aval fluctuent entre
2,8 et 18,6 °C (m ± δ = 11,6 ± 5,6 °C) (tab. 13). Les valeurs de la température de l’eau
observée le 13 juin 1999 varient entre 13 °C (V500) et 17,3 °C (5M). L’évolution spatiale
de ce paramètre depuis l’amont jusqu’à 500 m en aval de la zone d’influence met en
évidence un faible réchauffement des eaux en aval du rejet (fig. 41).
II. 3. 2. Conductivité électriqueLes valeurs de la conductivité électrique mesurées en amont du ruisseau au cours
du suivi annuel varient de 40 à 4450 µS. cm-1 (m ± δ = 801 ± 1166 µS. cm-1), celles
mesurées en aval oscillent entre 700 et 9620 µS. cm-1 (m ± δ = 1880 ± 2348 µS. cm-1)
(tab. 13). Ces résultats soulignent une forte augmentation des valeurs de la conductivité
électrique en aval sous l’effet de l’apport des bassins de lixiviat fortement minéralisé.
II. 3. 3. pHLes valeurs de pH observées en amont du ruisseau mensuellement varient de 4,18
à 7,78 (m ± δ = 6,62 ± 1,08), celles relevées en aval fluctuent entre 4,26 et 7,83 (m ± δ =
6,81 ± 1,16) (tab. 13). Les valeurs de pH de l’eau observées le 13 juin 1999 varient de 7,4
à 7,8. Le pH du ruisseau est légèrement basique. Toutefois, l’injection des lixiviats traités
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
131
ayant un pH légèrement supérieur à celui du ruisseau (8,3) engendre une faible
augmentation de la basicité de l’eau du ruisseau qui passe de 7,4 (A) à 7,6 (V500) (fig.
41).
II. 3. 4. Oxygène dissousLes teneurs en oxygène dissous enregistrées mensuellement en amont du ruisseau
varient de 0,74 à 10,4 mg. l-1 (m ± δ = 5,13 ± 3,3 mg. l-1), celles enregistrées en aval
fluctuent entre 1,38 et 12,6 mg. l-1 (m ± δ = 5,11 ± 3,36 mg. l-1) (tab. 13). Les valeurs de
l’oxygène dissous mesurées le 13 juin 1999 varient de 3,1 (V20) à 6,9 mg. l-1 (V500).
L’oxygénation la plus importante enregistrée en été à 500 m en aval. L’effet du lixiviat
est très marqué par une diminution des teneurs dans les 40 m en aval du rejet (5,6 mg. l-1
en amont et 3,1 mg. l-1 a 40 m en aval) (fig. 41). Ce paramètre présente généralement des
variations similaires à celles de l’Eh.
II. 3. 5. MESLes teneurs en matières en suspension, mésurées mensuellement en amont du
ruisseau varient de 0 à 26 mg. l-1 (m ± δ = 7,7 ± 9,3 mg. l-1), celles mesurées en aval
fluctuent entre 2 et 93 mg. l-1 (m ± δ = 18 ± 23 mg. l-1) (tab. 13). Il apparaît que le rejet
du lixiviat traité entraîne une élévation des teneurs en MES dans le ruisseau.
II. 3. 6. NitratesLes teneurs en nitrates mesurées mensuellement en amont du ruisseau oscillent
entre 13 et 109 mg. l-1 (m ± δ = 36,7 ± 28,7 mg. l-1), celles observées en aval fluctuent
entre 7 et 200 mg. l-1 (m ± δ = 71 ± 58 mg. l-1) (tab. 13). Les teneurs en nitrates
enregistrées le long du ruisseau oscillent entre 30,2 et 75,4 mg. l-1. La figure 47 montre
une augmentation importante des teneurs en nitrates dans le ruisseau qui passe de 30
mg. l-1 en amont (A) à 75 mg. l-1 en aval à 20 m du rejet. Toutefois, une légère diminution
des nitrates a été observée à 500 m en aval (V500) avec 44 mg. l-1 seulement.
II. 3. 7. AmmoniumLes teneurs en NH4
+ enregistrées mensuellement en amont du ruisseau varient de
0,7 à 53 mg. l-1 (m ± δ = 14,8 ± 15 mg. l-1), celles enregistrées en aval fluctuent entre 0,8
et 55,8 mg. l-1 (m ± δ = 31,3 ± 12,6 mg. l-1) (tab. 13). Il apparaît que les fortes teneurs en
ammonium enregistrées dans le lixiviat traité se répercutent nettement en aval du
ruisseau.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
132
II. 3. 8. ChloruresLes concentrations en chlorures observées mensuellement en amont du ruisseau
varient de 0,5 à 170 mg. l-1 (m ± δ = 52,7 ± 59,9 mg. l-1), celles observées en aval varient
de 0,5 à 194 mg. l-1 (m ± δ = 112,7 ± 46 mg. l-1) (tab. 13). Les teneurs en chlorures
mésurées le long du ruisseau oscillent entre 60,7 (V300) et 144,75 mg. l-1 (M). La teneur
élevée en chlorures dans le lixiviat (165,4 mg. l-1) est largement supérieure à celle du
ruisseau (66,4 mg. l-1). Cependant cet apport allochtone de chlorures se répercute
largement en aval du rejet et on retrouve les valeurs initiales à partir de 300 m en aval
(fig. 41).
II. 3. 9. Phosphore totalLes teneurs en phosphore total mésurées mensuellement en amont du ruisseau
varient de 0,07 à 0,29 mg. l-1 (m ± δ = 0,15 ± 0,08 mg. l-1), celles mésurées en aval
fluctuent entre 0,09 et 0,3 mg. l-1 (m ± δ = 0,18 ± 0,06 mg. l-1) (tab. 13). Malgré les
faibles teneurs enregistrées dans le lixiviat traité, elle restent néanmoins supérieures à
celles observées à l’amont du ruisseau, donc une légère augmentation des teneurs en
phosphore total à l’aval du ruisseau est toujours envisageable.
II. 3. 10. DBOLes valeurs de la DBO enregistrées en amont du ruisseau au cours du suivi mensuel
varient de 0,5 à 25,6 mg. l-1 (m ± δ = 7,2 ± 7,9 mg. l-1), celles enregistrées en aval varient
de 1,6 à 179 mg. l-1 (m ± δ = 40,4 ± 49,8 mg. l-1) (tab. 13). Ces résultats mettent en
évidence une évolution spatio-temporelle bien marquée avec une diminution de la charge
organique de l’amont à l’aval et de l’été à l’hiver.
II. 3. 11. DCOLes valeurs de la DCO enregistrées en amont du ruisseau au cours du suivi
mensuel varient de 4,7 à 280 mg. l-1 (m ± δ = 66,6 ± 77 mg. l-1), celles enregistrées en
aval varient de 14,9 à 559 mg. l-1 (m ± δ = 184 ± 143 mg. l-1) (tab. 13). Les valeurs de
la DCO observées dans le ruisseau durant le suivi du 13 juin 1998 varient entre 55 mg.
l-1 (A) et 160 mg. l-1 (M). La valeur maximale de DCO a été observée sur le point (M),
c’est à dire dans la zone de brassage des eaux du ruisseau et des lixiviats, provoquant
ainsi un enrichissement en matières organiques du ruisseau (tab. 13). Toutefois,
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
133
l’éloignement de la zone d’influence montre une légère diminution de la DCO (80 mg.
l-1 à 500 m en aval), qui demeure néamoins plus élevée que la valeur enregistrée à
l’amont du ruisseau (55 mg. l-1) (fig. 41).
II. 3. 12. Bactéries hétérotrophesLes densités bactériennes observées au cours du suivi du 13 juin 1999 fluctuent
entre 0,02 et 0,04 106 bactéries. ml-1. Cependant, l’abondance bactérienne la plus faible
a été observée à l’amont du ruisseau (0,023 106 bactéries. ml-1), cette abondance reste
néanmoins comparable à celle observée dans le lixiviat (0,033 106 bactéries. ml-1). Par
ailleurs, il apparaît une légère augmentation des densités bactériennes sur les points
V300 et V500, avec des valeurs respectives de 0,04 106 bactéries. ml-1 et de 0,041 106
bactéries. ml-1 (fig. 41). Qualitativement l’abondance bactérienne s’accompagne d’une
dominance des formes bacilles (≈ 15% du peuplement bactérien) rarement présentes
dans le lixiviat (≈ 5% du peuplement bactérien). Le suivi de l’abondance bactérienne
dans le ruisseau avant et après rejet des lixiviats met en évidence une évolution inverse
à celle observée pour la DCO, mettant en évidence un lien très fort entre ces deux
paramètres.
Tableau 13. Composition physico-chimique moyenne (13 échantillons) à l’amont du
ruisseau et à 100 m en aval du rejet.
II. 4. Discussion II. 4. 1. Outil informatique utilisé
Afin de comparer les résultats obtenus dans les deux stations de mesure, on a
utilisé le test de Mann-Whitney. Le logiciel utilisé est Statistica 97.
II. 4. 2. Evolution mensuelle
Il apparaît d’après nos résultats que l’impact des lixiviats traités sur le ruisseau
récepteur varie en fonction de la saison et du paramètre chimique étudié. La température
et le pH ne montrent pas un tel impact, vraisemblablement associé à la faible profondeur
du ruisseau et notre proximité de la source. Toutefois, de légères augmentations estivales
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
134
de la basicité et de la température des eaux du ruisseau ont été enregistrées en aval. En
hiver, l’impact des lixiviats sur le ruisseau reste faible, en ne dépassant pas 1 °C pour la
température et l’unité pour le pH. L’absence d’un tel impact à été confirmé par l’absence
d’une différence significative par le biais du test Mann Whitney (p =0,6 pour la
température et p = 0,4 pour le pH).
L’apport de lixiviat fortement minéralisé dans le ruisseau s’étale sur toute l’année
à l’exception du mois de septembre 1999 où l’on enregistre des valeurs plus élevées à
l’amont qu’à l’aval (p = 0,003), vraisemblablement dues au surplus de lixiviats provenant
du premier bassin qui déversait directement dans le ruisseau sous l’effet des fortes pluies.
L’élévation de la minéralisation des eaux du ruisseau apparaît surtout en période sèche
(automne), où l’on enregistre en aval une élévation 43 fois plus forte que celle observée
en amont (la conductivité électrique passe de 40 en amont à 1725 µS. cm-1 en aval pour
un débit de 0,1 l. s-1).
Le suivi des teneurs en oxygène met en évidence l’absence de différence
significative entre les deux points de mesure (p = 0,9). Ceci est lié à la forte oxygénation
du lixiviat. De plus, en été, le lixiviat contribue à une oxygénation du ruisseau (1,2 à 2,56
mg. l-1 en août 1998) suite à une activité phytoplanctonique très élevée. Enfin, une
oxygénation due aux remous de l'eau à sa surface n’est pas négligeable.
Malgré la forte diminution des teneurs en MES du lixiviat au cours du traitement,
les teneurs restent encore plus élevées que celles du ruisseau (p = 0,05). Les teneurs
moyennes en MES passent de 8 mg. l-1 en amont à 18 mg. l-1 en aval. Cette augmentation
peut être attribuée aux remous engendrés lors du brassage du lixiviat avec les eaux du
ruisseau qui peuvent entraîner des remises en suspension de particules déjà sédimentées
au fond. Toutefois, le surplus de MES biogènes en aval n’est pas négligeable. L’apport en
substances nutritives par le lixiviat traité dans le ruisseau est très nette pour les ions
ammonium dont le traitement ne permet pas une élimination complète de cet ion (plus
précisément en hiver). Les lixiviats entraînent un enrichissement du ruisseau en
ammonium qui passe de 14 à 40 mg. l-1. La pollution nitratée semble être sempblable à
celle qui est observée pour lion ammonium, avec un impact du lixiviat très visible en
hiver. Le lixiviat rejeté dans le ruisseau entraîne aussi une élévation des teneurs en PT en
aval, favorisant ainsi un surplus de substances nutritives en aval. Les teneurs moyennes
en PT passent de 0,11 mg. l-1 en amont à 0,16 mg. l-1 en aval. Cette élévation des teneurs
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
135
est confirmée par la présence d’une différence nettement significative pour les deux
éléments entre l’amont et l’aval (p = 0,002 pour le NH4+ et p = 0,004 pour le PT).
Toutefois aucune différence significative n’a été enregistrée pour le cas des
nitrates (p = 0,004), ceci est lié aux fortes teneurs enregistrées en amont. Les chlorures,
très abondants dans le lixiviat, engendrent une forte augmentation des teneurs en aval
(112 mg. l-1), favorisée par leur faible interaction avec les autres éléments chimiques
(Appelo et Potsma, 1993).
Examinons les effets de la pollution organique du rejet dans le ruisseau
avoisinant. Le lixiviat traité, déversé contient un mélange complexe de produits, parmi
ceux-ci, une grande quantité de matières organiques biodégradables (DBO). Il apparaît
que les effets de l’apport en DBO et en DCO sont quasi-similaires (p = 0,006 pour les
deux paramètres) avec un enrichissement important en matières organiques en aval en fin
d’été, lié à la faible dégradation des matières oxydables par les bactéries en aval et aux
fortes teneurs enregistrées dans le lixiviat. Ce phénomène est accentué par l’apport de
matières organiques engendrées par photosynthèse dans le ruisseau.
II. 4. 3. Evaluation du pouvoir auto-épuratoire du ruisseau
Le suivi effectué le 13 juin 1999 le long du ruisseau met en évidence un faible
réchauffement du ruisseau par le lixiviat (élévation de 2 °C). Cette élévation tend à
disparaître à 100 m en aval du rejet et le ruisseau reprend sa température initiale (13 °C).
Pour le pH et à l’exception de la zone de mélange (M), où on assiste à une légère
augmentation des valeurs (passant de 7,4 à 8,3) suite à la forte basicité du lixiviat (8,3),
les valeurs enregistrées en aval sont quasi-identiques. Le rejet du lixiviat dans le ruisseau
entraîne une légère oxygénation du ruisseau (6,3 mg. l-1). Cette oxygénation semble être
liée au brassage mécanique provoqué lors de l’injection du lixiviat dans le ruisseau. Dès
que l’on s’éloigne de la zone du mélange, on assiste à une diminution des teneurs en
oxygène dissous (3 mg. l-1). Cette désoxygénation brutale est vraisemblablement liée à la
décomposition des matières organiques importées par le lixiviat traité qui s’étale sur 60 m
en aval. A partir de 100 m en aval, on enregistre une légère augmentation des teneurs (3,9
mg. l-1). Cette élévation des teneurs en oxygène s’accentue plus en aval (200 m), à partir
de cette distance on retrouve les valeurs initiales en oxygène dissous (jusqu'à 6,9 mg. l-1 à
500 m). Ce retour à l’état initial est lié à: (i): la dégradation de la matière organique en
amont, (ii): au développement important des algues (dominance de zones mortes et
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
136
diminution de la pollution), (iii): à l’oxygénation du ruisseau par apport d’eau du bassin
versant. Cette évolution est confirmée par une évolution similaire du potentiel Redox qui
enregistre la même tendance que celle observée pour l’oxygène dissous. Le Eh passe de –
67 mV (à 10 m du rejet) à 153 mV (à 500 m en aval). L’évolution des teneurs en nitrates
montre une augmentation des teneurs en NO3- en aval du rejet (les teneurs en nitrates
passent de 30,2 mg. l-1 en amont à 75 mg. l-1 à 20 m du rejet) par l’apport d’eau nitratée
du lixiviat, toutefois une légère diminution des teneurs en nitrates en aval (à partir de 300
m) semble être liée à l’action conjuguée de deux facteurs: (i): dilution par des eaux moins
chargées en nitrates du ruisseau et celles provenant du bassin versant, (ii) dénitrification
sous l’action microbienne (Wyer, 1988; Montuelle et al., 1996). La diminution des
teneurs en chlorures en aval de la zone d’influence pourrait être attribuée à la dilution. Le
suivi des valeurs de DCO met en évidence une chute des teneurs en DCO de l’amont à
l’aval. Les teneurs en DCO passent de 155 mg. l-1 à 10 m, à 80 mg. l-1 en aval du rejet
(500 m). La dilution et la biodégradation semblent être les principaux facteurs de cette
diminution spatiale de la charge organique. De plus, Zagorc-Konan et Omen (1999) ont
mis en évidence l’importance de l’épuration par le biais des bactéries dans les processus
d’auto-épuration des rivières polluées par des eaux industrielles. Cette diminution de la
charge organique allochtone en aval coïncide avec une légère augmentation de l’effectif
bactérien. En effet, la densité bactérienne varie de 0,023.106 bactéries. ml-1 en amont du
ruisseau à 0,041.106 bactéries. ml-1 à 500 m en aval, vraisemblablement liée à la
diminution de la charge polluante importée par le lixiviat. Toutefois, Baccella et al.
(2000) ont mis en évidence une évolution inverse entre les bactéries et la DCO. Il
apparaît, d’après nos résultats que les variations de la composition physico-chimique de
l’eau du ruisseau sous l’effet du rejet du lixiviat se répercutent faiblement sur
l’abondance bactérienne. Ce qui nous laisse penser à un taux de toxines peu important
dans le lixiviat et/ou à la présence d’une charge organique à dominance non
biodégradable. De plus, une évolution morphotypique a été observée avec une présence
plus importante des formes bacilles, rarement présent dans le lixiviat et à l’amont du
ruisseau. Cette substitution des cocci par les bacilles en aval vient confirmer
l’amélioration qualitative de l’eau à l’aval du rejet.
II. 5. Conclusion
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4 è Partie Chap.1. Evaluation de l’efficacité du traitement par lagunagenaturel
137
Il ressort de nos observations qu’un impact limité des lixiviats traités de la
décharge d’Etueffont s’effectue sur le ruisseau avoisinant. Il semble que la pollution ne
dépasse pas le pouvoir auto-épuratoire du milieu récepteur en période estivale. Les
phénomènes d’auto-épuration dans le ruisseau semblent minimiser l’impact du lixiviat
traité sur le milieu récepteur (ruisseau). De plus, sur un plan strictement méthodologique,
l’évolution des teneurs en oxygène dissous donne une information préliminaire utile de
l'impact des lixiviats traités de la station d’Etueffont sur la qualité des eaux superficielles
d’un petit ruisseau. Il reste toutefois à étudier la pathogénicité des bactéries décomptées
ainsi que l’écotoxicité des lixiviats concernés.
Fig. 41. Evolution du pH, T, NO3, Cl, Eh, O2, DCO et de l'abondance bactérienne del'amont (A) jusqu'à 500 m en aval du rejet des lixiviats dans le ruisseau récepteur.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
138
I. INTRODUCTIONL'élimination par oxydation biologique est le noyau des technologies de
traitement des eaux résiduaires (Tay and Show, 1992). Le traitement biologique entraîne
l’accumulation de volumes importants de boues au fond des bassins. L’importance de la
décantation dans les stations de lagunage dépend exclusivement de la charge organique et
minérale de l’eau à traiter. De plus, la sédimentation biogène est induite par les fortes
productions internes. La sédimentation dans les bassins de traitement, d'origine détritique
et biogénique, est un phénomène dont les impacts sont fortement négatifs. Ces impacts se
manifestent par la formation de boues très chargées en matières organique et minérale,
plus précisément en métaux dont le traitement est complexe et donc coûteux. Plusieurs
auteurs se sont intéressés à l’étude des boues et différents aspects ont été abordés, et plus
spécialement les travaux ayant souvent traits à l’étude de l’effet fertilisant des boues
(Chaussod, 1977; Chaussod et al., 1981; Chaussod et al., 1985; Mench et al., 1989;
Cadot, 1993; Bousselhaj et Ouzzani, 1994). L’originalité de ce chapitre réside d’une part
dans la caractérisation des boues de lagunage des lixiviats et d’autre part dans l’utilisation
de pièges à sédiments pour la détermination du taux de sédimentation.
II. TECHNIQUES DE PIEGEAGE DE MATIERES EN SUSPENSIONS II. 1. Filtration
Cette méthode a été développée par Etcheber et al. en 1977. Elle se base sur une
aspiration du sédiment à l'aide d'une pompe péristaltique. Les particules sont récupérées
par décantation de 8 à 15 jours. Cette méthode se heurte aux problèmes de relargage des
métaux lors de l'aspiration.
II. 2. CentrifugationC'est une méthode largement utilisée (Bertrand, 1977 et 1980; Cauwet et al.,
1980). Elle consiste à filtrer un volume d'eau important en un temps très court (100 à 500
l. h-1), elle permet de recueillir une quantité de MES suffisante très rapidement. Cette
méthode a l'avantage de minimiser les risques de contamination métallique dans la
centrifugeuse (inférieurs à 2 %) par rapport à la filtration (Etcheber, 1978).
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
139
II. 3. Piège à sédimentsCe procédé consiste à une récolte des MES qui se déposent au fond des bassins
dans un réceptacle. Le piège à sédiments doit être facilement repérable, suffisamment
élevé au fond, afin d'empêcher l'effet des agitations du fond provoquées lors du
prélèvement ou par certaines espèces benthiques. Il est nécessaire que le piège reste en
place assez longtemps (généralement 15 jours à 2 mois) pour qu’une quantité appréciable
de matières en suspension se dépose à l’intérieur. Enfin, il faut souligner que les
sédiments piégés ne seront pas exactement comparables au sédiment sur place, car le
temps de dépôt long, fait que cet échantillon est sujet aux variations hydrauliques et à
l’action bactérienne.
Fig. 42. Schéma simplifié du piège à sédiments.
III. STRATEGIE D'ECHANTILLONNAGE III. 1. Boues draguées
Trois prélèvements de boues ont été effectués dans chaque bassin à l'aide d'une
drague manuelle en été et en hiver. Les prélèvements ont été effectués aux mêmes points
choisis pour l'échantillonnage d'eau. Cet échantillonnage a été effectué sur les premiers
centimètres de vases (10 cm) des boues.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
140
III. 2. Sédiments piégésA des fins de comparaison des sédiments cumulés (boues sur place) avec les boues en
néo-formation (matières en suspensions sédimentées) et afin de déterminer le taux
d'envasement dans les quatre bassins, on a procédé à un suivi spatio-temporel des
matières en suspension piégées dans les quatre bassins en utilisant des pièges à sédiment.
Le piège à sédiment développé découle du modèle décrit par Hakonson (1976), auquel on
a apporté quelques modifications, afin de l'adapter à nos bassins. Le piège à sédiments est
composé d'un récipient cylindrique de 20 cm de diamètre et de 10 cm de hauteur, une tige
métallique de 1,60 m de hauteur, deux barres métalliques de 1,5 m chacune portant des
contre-poids pour stabiliser le piège au fond du bassin (fig. 42). Enfin, on note que la
durée moyenne d'immersion des pièges dans les bassins est de l'ordre de deux mois.
III. 3. Conservation des échantillonsLes sédiments dragués et piégés sont stockés dans des sacs en plastiques et conservés
à 4 °C jusqu’au laboratoire.
III. 4. Séchage des échantillonsLe séchage est la meilleure façon pour conserver les sédiments jusqu’à ce que le
dosage soit réalisable. Ce qui évite toute évolution de la composition chimique des
échantillons. De plus les chercheurs tendent souvent à rapporter les teneurs en métaux
des sédiments par rapport au poids sec, du fait que les teneurs en eau sont très variables
d’un sédiment à un autre. Montiel (1980), a mis en évidence une hétérogénéité des
résultats plus importante avec des dispersions qui peuvent atteindrent 37 % pour le
sédiment humide et 16 % sur le même échantillon séché à 110 °C à l'étuve. L'auteur
préconise un séchage à l'étuve avec des températures allant de 40 à 110 °C (valeurs
choisies pour le dosage du mercure). Enfin, certains chercheurs (Mac Henry et al., 1977;
Sanchez et Lee, 1978 et Soldatini et al., 1976) préfèrent un séchage avec une température
ambiante. Enfin, d'autres chercheurs préconisent un séchage par lyophilisation en
déshydratant les échantillons de sédiment (Ackermann, 1980; Arnoux et al., 1980 et
Prosi, 1979). Pour notre étude Le séchage des sédiments a été effectué à l'étuve à l'aide
d'une température constante de 105 °C.
IV. DESCRIPTION DES SEDIMENTSLes sédiments dragués et piégés ont une couleur noir foncé dans le premier bassin
virant à une couleur marron noir dans le dernier bassin. L’aspect et la couleur des
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
141
sédiments témoignent d'une fermentation anaérobie très intense dans le premier bassin,
qui tendent à s'estomper dans les derniers bassins suite à la diminution de la charge
organique provenant de la décharge (tab. 14).
Couleur Aspect Odeur
Bassin 1 noire foncée homogène/liquide forte
Bassin 2 noire marron homogène/liquide moyenne
Bassin 3 noire marron homogène/liquide moyenne
Bassin 4 marron homogène/liquide moyenne
Tableau 14. Description des sédiments recueillis.
Tableau 15. Variations du taux de sédimentation dans le bassin 1.
Tableau 16. Variations du taux de sédimentation dans le bassin 2.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
142
Fig. 43. Implantation des pièges dans les quatre bassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
143
V. RESULTATS ET DISCUSSIONV. 1. Taux de sédimentation
Afin de comprendre et estimer l’évolution des taux de sédimentation dans les
bassins de la station d’Etueffont, 8 pièges ont été installés par bassin. Ceci nous permettra
de cartographier les zones de forte et de faible sédimentation dans chaque bassin (fig. 44).
Les taux de sédimentation varient de 0,08 à 4,2 dans le premier bassin, de 0,5 à 0,27 dans
le bassin 2, de 0,09 à 0,9 dans le bassin 3 et de 0,05 à 0,4 dans le dernier bassin (tabls. 15
à 18).
Tableau 17. Variation du taux de sédimentation dans le bassin 3.
Tableau 18. Variations du taux de sédimentation dans le bassin 4.
Les résultats montrent que dans le premier bassin le taux de sédimentation est très
élevé pour les points B11 (1,8 g. j-1) et B18 (4,2 g. j-1), alors qu'il est plus faible sur les
autres points variant entre 0,08 et 0,34 g. j-1. Dans le bassin 2, le taux de sédimentation
varie de 0,05 à 0,1 g. j-1 pour tous les points à l'exception du point B23 où l’on enregistre
un taux de sédimentation important atteignant une valeur de 0,35 g. j-1. Dans le troisième
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
144
bassin, les taux de sédimentation sont faibles et ne nous montrent pas de grandes
variations entre les points de mesure (0,09 à 0,5 g. j-1) avec un maximum au point B37
(0,8 g. j-1). Enfin, dans le dernier bassin, le taux de sédimentation fluctue entre 0,1 et 0,15
g. j-1. Le taux maximum de sédimentation est enregistré au point B45 avec 0,45 g. j-1.Il
apparaît d’après nos essais que les taux de matières en suspensions piégées sont plus
importants dans le premier bassin que dans les autres bassins, à l’aplomb de
l’alimentation et sur les bordures de chaque bassin. Cette diminution traduit le classique
processus de la réduction du phénomène de décantation du matériel particulaire de
l’amont et à l’aval de la station. L’élévation des valeurs de taux de sédimentation au
niveau des berges ne peut être attribuée qu'aux conditions hydrodynamiques favorables à
la sédimentation (eau peu agitée).
L’étude comparative entre les deux essais de piégeage effectués en hiver et en
été pour les quatre bassins met en évidence des taux de sédimentation élevés en été. Cette
augmentation estivale semble être (essentiellement liée à l’augmentation des matières en
suspension émises en été par la décharge dont l'activité biologique est intense. De plus, la
présence de conditions idéales à la sédimentation (absence d’agitation mécanique due
aux vents) et la présence probable de matières en suspension d'origine autochtone
élaborées par l’activité algale (Jigorel et Bertru, 1993; Jigorel et Morin, 1994; Andrieu et
al., 1997) peuvent aussi expliquer l'importance des dépôts observés. Enfin, Mayo (1996)
a montré que dans une station de traitement des eaux usées 160 à 240 % de la DBO est
due aux MES et aux biomasses particulaires.
Afin de mettre en évidence le rôle biogénique des matières en suspension en été et
plus précisément dans les derniers bassins, on a suivi l’évolution spatio-temporelle des
MES, de la DCO et de la DBO de l’eau dans les quatre bassins (fig. 45). Les résultats
montrent une alternance des pics de MES et de DBO, mettant en évidence une faible
proportion organique contenue dans ces MES. En effet, dans les autres bassins les pics de
DBO tendent à s’approchent de ceux des MES, jusqu’à un chevauchement des pics dans
les deux derniers bassins. Ce chevauchement entre des pics de DBO et de MES montre la
dominance organique des MES, vraisemblablement engendrée par la forte activité algale
plus importante dans ces derniers.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
145
Fig. 44. Evolution spatiale du taux de sédimentation dans les quatre bassins (essais dedécembre 1998 et juin 1999).
V. 2. Vitesse d'accumulationLes vitesses d'accumulation calculées pour les quatre bassins (hiver et été) (tab.
19), mettent en évidence de fortes valeurs dans les bassins 1 et 2 (2,96 et 2,65 cm. an-1),
par rapport aux bassins 3 et 4 (0,26 et 0,32 cm. an-1). Ceci est vraisemblablement
attribuable à la diminution de l'apport par la décharge de lixiviats très chargés en MES
dans les bassins 3 et 4. On enregistre une légère augmentation de la vitesse de
sédimentation dans le bassin 4 par rapport au bassin 3, ce qui confirme le rôle des
matières en suspension biogènes. En effet, les vitesses d’accumulation obtenues sont
inférieures à celles rapportées dans la littérature (Drakides, 1983; Beaumier, 1988). De
plus, ces valeurs ne correspondent pas aux épaisseurs mesurées sur place. Ceci est
vraisemblablement lié à la faible digestion des boues au sein des pièges, suite au faible
temps de piégeage (28 jours). Enfin, il apparaît que la vitesse d'accumulation ne semble
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
146
pas être influencée par la température, contrairement à ce que l’on constate lors de l'étude
du taux de sédimentation. L’absence de cet effet saisonnier ne peut être expliqué que par
une dégradation des substances sédimentées.
V. 3. Composition physico-chimiqueV. 3. 1. Boues draguées
Les résultats d’analyses des teneurs en éléments métalliques des boues effectuées
au laboratoire de chimie des eaux de Besançon, montrent qu’entre la lagune B1 qui reçoit
les jus de la décharge et la lagune B4, on constate une diminution des teneurs en métaux
(tab. 20). Les teneurs en étain passent de 852 à 426 mg. kg-1 de MS (MS = Matière
Sèche), en cuivre passent de 412 à 50 mg. kg-1 de MS, en plomb passent de 398 à moins
de 0,005 mg. kg-1 de MS, en chrome total passent de 368 à 36 mg. kg-1 de MS, en baryum
passent de 264 à 226 mg. kg-1 de MS, en nickel passent de 120 à 80 mg. kg-1 de MS et
celles relatives à l’arsenic passent de 71,2 à 16,6 mg. kg-1 de MS. Cette chute des teneurs
en éléments métalliques est vraisemblablement attribuable à l’accentuation des
phénomènes de décantation dans le premier bassin. A l’inverse, les teneurs en fer et en
manganèse tendent à augmenter dans le dernier bassin (les teneurs en fer passent de 134
en B1 à 116 mg. kg-1 de MS en B4, celles relatives aux teneurs en manganèse passent de
422 en B1 à 1530 mg. kg-1 de MS en B4). Ceci est lié à la forte oxygénation du lixiviat
dans le dernier bassin qui favorise sa précipitation chimique. Il faut noter que pour le
cadmium on a toujours enregistré des valeurs inférieures au seuil analytique (<0,005 mg.
kg-1 de MS). Les teneurs en matières organiques des boues ne montrent qu’une faible
diminution spatiale des teneurs. Le carbone organique total passe de 6,43 en B1 à 5,33
mg. kg-1 de MS en B4, l’azote kjeldahl passe de 7,34 en B1 à 7,06 mg. kg-1 de MS en B4
et les teneurs en phosphore total passent de 0,26 en B1 à 0,17 mg. kg-1 de MS en B4.
Cette faible diminution est vraisemblablement liée à l’apport de matières organiques par
le plancton qui atteint son apogée dans le dernier bassin. Il semble que l’apport organique
par le plancton contrebalance la décantation et la dégradation favorisées par l’élévation
du temps de séjour.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
147
Vitesse d'accumulation en cm. an-1
Bassin 1 2,96
Bassin 2 2,61
Bassin 3 0,26
Bassin 4 0,32
Tableau 19. Vitesse d'accumulation calculée pour les quatre bassins.
Fig. 45. Variations spatio-temporelles de la DBO, la DCO et les MES dans les quatrebassins.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4è Partie Chap.2. Etude de la partie sédiment
148
Enfin et en guise de conclusion, il apparaît que les boues prélevées dans les quatre
bassins présentent des teneurs métalliques bien plus faibles que celles définies dans les
normes exigées par l’union européenne (décret du 8 décembre 1997 concernant
l’épandage des boues). Enfin, Le rapport K (Valeur de référence/Teneur dans la boue)
défini par la norme NF U 44-041 est souvent supérieur à 2 pour tous les métaux. Ce qui
les classent dans la catégorie des boues peu chargées en métaux.
V. 3. 2. Sédiments piégésL'analyse aux Rayons X, effectuée à l’Institut des Sciences et Techniques de
l’Environnement de Montbéliard sur les sédiments piégés, montre la présence de
diffractogrammes identiques pour tous les sédiments dans les quatre bassins (annexe 10),
mais avec une présence majoritaire de silice, très marquée dans les deux derniers essais
(printemps et été). Ceci est vraisemblablement dû aux frustules diatomiques très
répandues dans les quatre bassins, et met en évidence la dominance probable de
sédiments biogènes.
L'analyse chimique détaillée des diffractogrammes de quelques éléments
chimiques présents dans le sédiment, nous montre que le fer est très présent dans le
premier bassin par rapport aux autres éléments. Il précipite souvent sous différentes
formes avec une dominance de la forme hydroxylée (apatite ferrique). Le chrome
précipite dans les quatre bassins avec des pourcentages décroissants du premier au
dernier bassin. La forme CrO4 n'est présente que dans le premier bassin (pH élevé). Le
manganèse est rencontré seulement en B2 et son absence dans le premier bassin est due à
la forte compétition du manganèse avec le fer qui forme des complexes plus stables avec
les ions hydroxydes qu’avec le manganèse. Le plomb est un métal très exigeant
nécessitant la présence suffisante des ions OH-, des pH et des Eh assez élevés pour
précipiter. En effet ces conditions ne sont présentes que dans le dernier bassin, ce qui
explique sa présence dans les boues du quatrième bassin. Les orthophosphates sont
rapidement piégés dès leur entrée en B1 sous forme de FePO4.
La comparaison entre les teneurs en métaux contenus dans les sédiments piégés et
celles enregistrées dans les boues sédimentées est très significative avec une faible
présence métallique dans le sédiment piégé. Cette différence est essentiellement liée à
l’effet cumule qui fait augmenter les teneurs en éléments métalliques au fil du temps et à
la forte adsorption des métaux sur les boues en place.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
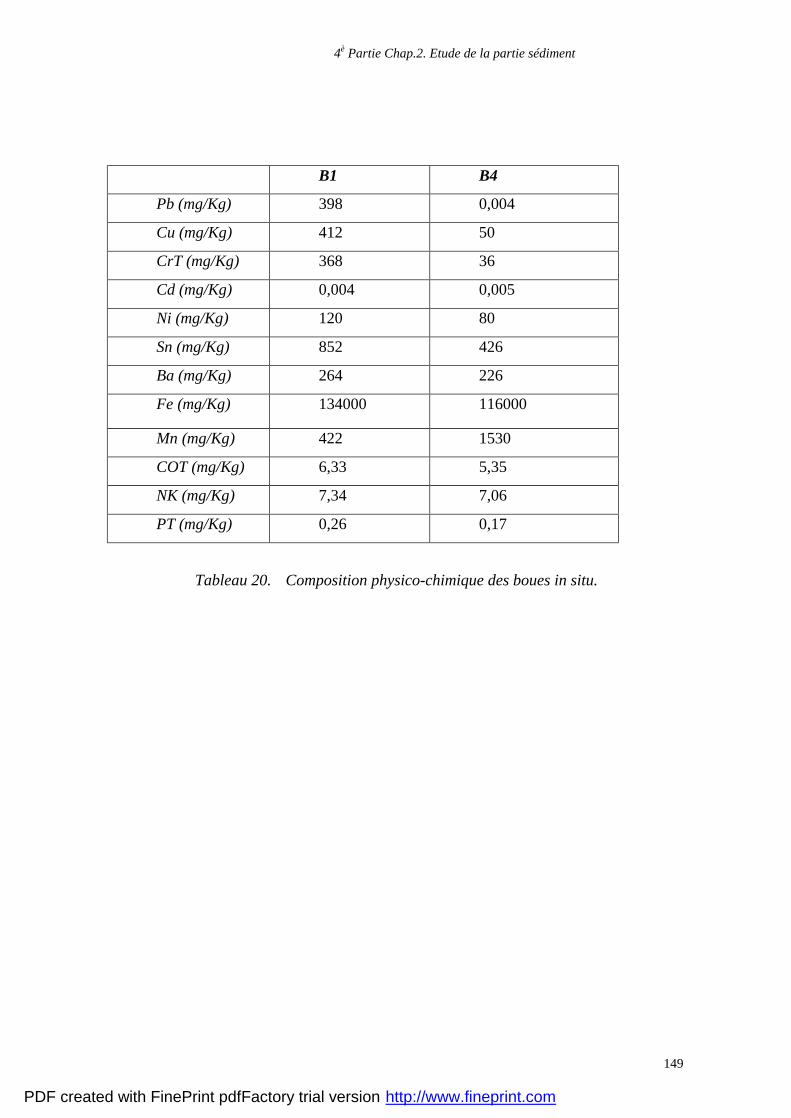
4è Partie Chap.2. Etude de la partie sédiment
149
B1 B4
Pb (mg/Kg) 398 0,004
Cu (mg/Kg) 412 50
CrT (mg/Kg) 368 36
Cd (mg/Kg) 0,004 0,005
Ni (mg/Kg) 120 80
Sn (mg/Kg) 852 426
Ba (mg/Kg) 264 226
Fe (mg/Kg) 134000 116000
Mn (mg/Kg) 422 1530
COT (mg/Kg) 6,33 5,35
NK (mg/Kg) 7,34 7,06
PT (mg/Kg) 0,26 0,17
Tableau 20. Composition physico-chimique des boues in situ.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Conclusion générale
150
CCoonncclluussiioonn GGéénnéérraalleeDans les prochaines années la surveillance des décharges et plus précisément celle
des lixiviats de décharge sera renforcée. Selon les experts, l’amélioration des outils de
traitement des lixiviats passera par une meilleure connaissance de la composition
physico-chimique des lixiviats et la supervision des décharges afin de mieux comprendre
les mécanismes complexes qui gouvernent la qualité des lixiviats. Toutefois, une
attention particulière sera portée sur l’influence des conditions climatiques.
Ce travail pluridisciplinaire repose sur l’étude de l’évolution temporelle des lixiviats
d’une décharge à ciel ouvert, la fiabilité de l’application d’un lagunage naturel pour le
traitement des lixiviats produits et l’évaluation du risque du complexe décharge unité de
traitement sur les eaux souterraines et superficielles. Pour atteindre ces objectifs, nous
avons instrumenté le site avec plusieurs équipements: (i) une petite station
météorologique (composée d’un pluviomètre et d’un thermomètre, (ii) une station pilote
qui nous permettra de comparer les lixiviats en fonction de l’âge des déchets, (iii) des
couples de piézomètres courts et profonds ont été installés au voisinage de la station afin
d’évaluer l’impact de la décharge sur les eaux souterraines avoisinantes et (iiii) enfin des
pièges à sédiment ont été implantés au sein des bassins afin d’estimer les taux de
sédimentation dans les quatre bassins de la station de lagunage.
Le bilan météorologique, les suivis qualitatifs et quantitatifs à différents pas de
temps, nous ont permis de suivre le comportement de la décharge à tout instant. Ce
comportement se résume à des variations de la composition des lixiviats pour les
différents pas de temps choisi, mettant en cause l’effet de la température et les
précipitations atmosphériques dans la génération des lixiviats. A tout cela s’ajoute
l’impact de l’âge de la décharge qui semble influencer directement la composition du
lixiviat, sa charge organique biodégradable diminuant régulièrement avec le
vieillissement du site.
Bien que la décharge d’Etueffont soit conçue avec un maximum d’étanchéité du
fait de sa position sur un substratum schisteux, le suivi de la composition des eaux
souterraines avoisinantes met en évidence une pollution occasionnelle de la nappe
profonde plus précisément en période estivale où la décharge est en pleine activité,
alors que la nappe de surface est souvent touchée par le flux polluant et surtout les
piézomètres les plus proches de la décharge. Toutefois, les phénomènes d’auto-
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Conclusion générale
151
épuration et de dilution semblent contrebalancer cette pollution en aval. De plus, et sur
le point méthodologique, le suivi des teneurs en chlorure apparaît comme un indicateur
fiable pour une telle pollution.
Le calcul des rendements épuratoires de la station d’Etueffont laisse apparaître
que le lagunage naturel appliqué pour le traitement de ces lixiviats est très satisfaisant
pour certains paramètres (DBO, NT, PT et les éléments métalliques) durant toute la
saison. Le pourcentage d’abattement de la matière organique représenté en terme de DBO
peut atteindre 98 % en été, alors que la DCO et les autres éléments chimiques présentent
des taux d’abattement nettement inférieurs du fait de leur grande sensibilité à des facteurs
naturels non maîtrisables tel que la température, le pH et le potentiel rédox. Toutefois,
deux périodes de dysfonctionnement de la station ont été observées, la première a été
observée essentiellement pour la DBO (Septembre 1998, -8 %), vraisemblablement due
aux blooms phytoplanctoniques très présents en été et qui enrichissent le lixiviats au
cours de son transit dans les bassins. La deuxième chute de rendement a été enregistrée
en hiver pour l’ensemble des éléments chimiques suivis. Cette diminution hivernale est
attribuée d'une part à la diminution de l'activité biologique et d'autre part aux
phénomènes de relargage par le fond de matières organiques sous l'effet de la forte
diminution du pH (pH compris entre 4 et 5). De plus, l’évolution du taux d’élimination
des bactéries libres montre une efficacité très importante en été (87 à 99 %), faibles en
hiver (40 à 60 %) et des valeurs intermédiaires en été. Cette efficacité estivale est
vraisemblablement liée à l’intervention de facteurs biologiques (prédation) et physiques
(ensoleillement).
Il ressort de l’étude hydrobiologique des bassins d’Etueffont que le peuplement
phytoplanctonique est dominé par des espèces polluo-résistantes (Stephanodiscus sp. et
Euglena sp.). Cependant, une nette amélioration qualitative a été enregistrée en passant
du premier au dernier bassin, caractérisée par une substitution des diatomées (très
fréquent dans le premier bassin) par les Chlorophyceae (Phacus sp.). Le zooplancton
dans ces lagunes est dominé par le genre Brachionus pour les Rotifères (80 % du
peuplement) et par le genre Moina pour les Cladocères (100 % du peuplement). De plus,
il apparaît que l’abondance des moinidae diminue du premier au quatrième bassin. A
l’opposé, les Rotifères présentent une diversité et des effectifs croissants en allant du
premier au dernier bassin. L’augmentation de la diversité spécifique, très marquée chez
les Rotifères, coïncide avec l’amélioration qualitative du lixiviat au cours du
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Conclusion générale
152
cheminement de l’eau d’un bassin à un autre. Enfin, le bacterioplancton met en évidence
une nette dominance des morphotypes cocci (95 % de la population microbienne), par
rapport aux morphtypes bacilles (4 % de la population microbienne) et 1 % pour les
autres morphotypes. Les formes bacilles sont surtout présentes dans le premier bassin en
fin d'été (septembre et octobre 1998), vraisemblablement en raison de l’élévation de la
température de l’eau, de l’absence de bactérivores et des apports de substrats allochtones
par le lixiviat brut dans le premier bassin. Enfin, l’étude quantitative du bactérioplancton
ne montre pas une telle amélioration spatiale.
Concernant l’impact des lixiviats traités sur les eaux superficielles, le suivi
mensuel et journalier du ruisseau récepteur montre que le pouvoir auto-épuratoire naturel
du ruisseau semble minimiser l’impact du lixiviat traité sur le milieu récepteur (ruisseau).
Sur un plan strictement méthodologique, l’évolution des teneurs en oxygène dissous
donne une information préliminaire utile de l'impact des rejets sur la qualité des eaux.
La partie sédiment nous a permis de mettre en évidence la fiabilité des pièges à
sédiments pour la détermination des taux de sédimentation dans les quatre bassins avec
des maxima de décantation à l’aplomb et aux bordures des bassins. De plus, les analyses
physico-chimiques effectuées sur les échantillons de boues prélevées in situ et les
sédiments piègés laissent apparaître leur faible teneur en éléments métalliques.
Enfin, il apparaît que cette étude apportera des informations nouvelles sur
l’évolution physico-chimique et biologique des lixiviats et leur traitement par lagunage
naturel, ce qui suggère une attention toute particulière des autorités municipales et
communales afin de multiplier les efforts et s’orienter vers une meilleure gestion des
lixiviats de décharge d’ordures ménagères.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
AABRAHAM, HAFNER, 1987. Heavy metal transport in aquifer material: Laboratory investigations and
parameter estimation. IAHS, Pub., 173, 103-111.
ACKERMANN F.,1980. Aprocedure for correcting the grain size effect in heavy metal analysis of estuarinr
and coastal sediments, Environmental Technology Letters, 1, 518-527.
ALAOUI M., ALEYA L., DEVAUX J., 1994. Phosphorus exchanges between sediment and water in
trophically different reservoirs. Wat. Res., 28, 1971-1980.
ALEYA L., 1991. The concept of ecological succession applied to an eutrophic lake trough the seasonal
coupling of diversity index and several parameters. Arch. Hydrobiol., 3, 327-343.
ALEYA L., DESMOLLES F., BONNET M. P., DEVAUX J., 1994. The deterministic factors of the
Microcystis aeruginosa blooms over a biyearly survey in hypereutrophic reservoir of Villerest
(Roanne, France). Arch. Hydrobiol. Suppl., 99, 1-26.
ALOICE W. M., TATSUYA N., 1996. Effects of temperature and pH on the growth of heterotrophic bacteria
in waste stabilization ponds, Water Research, 30, 447-455.
AMBLARD C., 1986. Les nucléotides adéniliques: intêret pour l’étude de la biomasse, de l’activité
métabolique et de la structure des peuplements phytoplanctoniques. Thèse de doctorat d’Etat de
l’Université de Clermont-Ferrand, 317pp.
AMBLARD C., BOURDIER G., CARRIAS J. F., MAURIN N., QUIBLIER C., 1994. Etude des réseaux
trophiques microbien du lac de Vasssivière en Limousin. Rapport scientifique. 68pp.
AMOROS C., 1973. Evolution des populations de cladocères et copépodes dans trois étangs piscicoles de la
Dombes. Annls Limnol., 9, 135-155.
AMOROS C., 1984. Introduction pratique à la systématique des organismes des eaux continentales
françaises les crustacés Cladocères N°4, Lyon, Association française de limnologie , 5-63.
AMOROS C., 1986. Introduction pratique à la systématique des organismes des eaux continentales
françaises les Rotifères n° 5, Lyon, Association française de limnologie, 6-37.
ANDRIEU J. P., CAULT J. B., BOUCHARD J. P., COMTET A., COTTIN L. GARROS-BERTHET H.,
GUILLEMOT B., GIGOREL A., LEVENQ J., LURIN P., MORIN J. P., 1997. Expérience française
récente dans le domaine de la géstion des sédiments dans les réservoirs. 19èm Congrés des Grands
Barrages, Florence, 365-382.
ANGELI N., 1979. Relations entre le plancton et la qualité de l’eau. Thèse de Doctorat de 3èm cycle,
Université des Sciences et Techniques de Lille.
APPELO C. A. J., POTSMA D., 1993. Geochemistry, groundwater and pollution. A.A. Balkema,
Rotterdam, 540 pp.
ASHBEE E., FLITCHER I., 1993. Reviewing the options for leachate treatment. Wastes Management, 8,
32-33.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
155
ARNOUX A., NIENCHEWSKI L. P., TATOSSIAN J., 1980. Comparaison de quelques méthodes d’attaque
des sédiments pour l’analyse des métaux lourds. Conf.: Les métaux à l’état de trace dans les
eaux, organisé par l’association pharmaceutique française pour l’hydrologie , Lille, 28-05-1980.
AZAM F., FENCHEL T., FIELD J. G., GRAY. J. S., MAYER L. A., THINGSTAD F., 1983. The ecological
role of water column microbes in the Sea. Mar. Ecol. Prog. Ser., 10, 257-263.
B
BACCELLA S., CERICHELLI G., CHIARINI M., ERCOLE C., FANTAUZZI E., LEPIDI A., TORO L., VEGLIO
F., 2000. Biological treatment of alkaline industrial waste waters, Process. Biochemistry, 35, 595-
602.
BAHLAOUI A., 1990. Lagunage à haut rendement experimental: dynamique de differnts groupes
bacteriens et perfermances épuratrices sanitaires. Thése de 3è cycle, Montpellier II, 186pp.
BAHLAOUI M. A., BALEUX B., TROUSSELLIER M., 1997. Dynamics of pollution-indicator and
pathogenic bacteria in high-rate oxydation wastewater treatment ponds. Wat. Res., 31, 630-
638.
BARBE J., 1981. Les peuplements planctoniques des installations de lagunage naturel en France.
Relazione presentata in occasione del convegno Intenazionale su fitodepurazione e impiego
delle biomasse. Parma, 15-16.
BARBER C., REDHEAD, 1978. Maendy landfill./ A study of possible water pollution by leachate. Note
technique WLR n° 11, département de l'environnement, Londres.
BARKOWSKI D., GUNTHER P., ROCHERT R., 1987. Atlasten. Edition C.F. Muller, Karlsruhe et al. 87-
93.
BELBAHRI S., 1991. Relations eau sédiment en épuration par lagunage naturel. Thèse de 3è Cycle,
Université Montpellier II.
BENIDER A., TIFNOUTI A., POURRIOT R., 1998. Reproduction parthenogenitique de Moina macropa
(Straus 1820) (Crustacea: Cladocera). Influence des conditions trophiques, de la densité de la
population, du groupement et de la température. Annls Limnol. 34, 387-399.
BERGSTROM A. K., JANSSON M., 2000. Bacterioplankton production in humique lake Ortrasket in
relation to input of bacterial cells and input of allochthonous organic carbon. Microbial
ecology, 39, 105-115.
BERTRAND PH., 1977. Etude expérimentale par fractionnement, de la réparttition géochimique du zinc
dans les matières en suspension estuariennes. Cas de la Gironde, J. Rech. Océanogr., 4, 39-51.
BERTRAND PH., 1980. Etude expérimentale par fractionnement, de la réparttition géochimique de
quelques métaux (Zn, Cu, Pb, Ni) dans les matières en suspension de l’estuaire de la Gironde.
Thèse de 3èm cycle, N° 1547, Univ. Bordeaux I, 149pp.
BERZIN B., PEJLER B., 1989. Rotifer occurrence in relation to température. Hydrobiologia, 175, 223-
231.
BLAKY N. C., 1992. Model prediction of landfill leachate production. Elsevier. Appl. Sci. London,
England. 17-34.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
156
BLANCHARD J. M., NAVARRO A., REVIN PH., CYRCLEWSKI J., VOVARD S., PILLARD G., 1989. Les
mâchefers d'incinération d'ordures ménagères II. Utilisation en traitement des lixiviats de
décharge. TSM. 3, 133-140.
BLAVY G., DRUART J. C., 1982. Les Rotifères. In: Le Leman, synthèse 1959-1982. Commission
Internat. Pour la protection des Eaux du Léman contre la Pollution, Lausanne, 304-313.
BOOKTER T. J. B., HAM R., 1982. Decomposition of solid waste in test lysimeters. J. Environ. Engin.
Div. ASSCE., 108, 1147-1170.
BOSTROM B., ANDERSEN J. M., FLEISCHER S., JANSSON M., 1988. Exchanges of phosphorus across the
sediment water interface. Hydrobiologia. 170, 229-238.
BOTTRELL H. H., DUNCAN A., GLIWICZ Z. M., GRYGIEREK E., HERZIG A., HILLBRICHT-ILKOWSKA
A., KURASAWA H., LARSSON P., WEGLENSKA T., 1976. Areview of some problems in
zooplankton production studies. Norw. J. Zool., 24, 419-456.
BOUARAB L., OUZZANI N., OUDRA B., DARLEY J., PICOT B., BONTOUX J., 1994. Evaluation des
formes d’azote dans la station expérimentale de lagunage naturel de Marrakech. Quatrième
conférence internationale des limnologues d’Expression française , Marrakech.
BOURRELLY P., 1972. Les algues d'eau douce. Tome I: les algues vertes. Paris, France, Boubée et Cie,
572 pp.
BOURRELLY P., 1981. Les algues d'eau douce. Tome II : les algues jaunes et brunes. Paris, France,
Boubée et Cie, 517 pp.
BOURRELLY P., 1985. Les algues d'eau douce. Tome III : les algues bleues et rouges. Paris, France,
Boubée et Cie, 606 pp.
BOUSSELHAJ K., OUAZZANI N., 1994. Etude des potentialites fertilisantes d’une boue de lagunage
anaerobie. Quatrième conférence internationale des limnologues d’Expression française ,
Marrakech.
BOYLE W. C., HAM R. K., 1974. Biological treatability of landfill leachate. Wat. Res. Poll. Cont. Fed.,
46, 860-872.
BRETT M. T., GOLDMAN C. R., 1996. A meta-analysis of the freshwater trophic cascade. Proc. Natl.
Acad. Sci. USA., 93, 7723-7726.
BRYANT M. P., 1979. Microbial methane production, Theorical aspects. Journal of Animal Science, 48,
193-201.
BUFFLE J. P., 1977. Les substances humiques et leurs interactions avec les ions minéraux. TSM., 1, 72.
BURNS C. W., 1968. The relationship between body size of filter feeding Cladocerans and the
maximum size of particles ingested. In: Limnol. Oceanogr., 13, 675-678.
C
CADOT L., 1993. Impact de facteurs pédoclimatiques sur la biodégradation des boues résiduaires de
station d’épuration dans deux sols. Modélisation et simulation. Thèse de 3èm cycle à l’INRA
DE Paris-Grignon, 175pp.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
157
CAMERON R. D., KOCH F. A., 1980. Toxicity of landfill leachates. J. Watt. Pollut., Cont. Fed., 52, 760-
769.
CAUWET G., ELBAZ F., JEANDEL C., JOUANNEAU J. M., LAPAQUELLERIE Y., MARTIN J. M., THOMAS
A., 1980. Comportement géochimique des éléments stables et radio-actifs dans l’estuaire de la
Gironde en periode de crue, Bull. Inst. Géol. Bassin d’Aquitaine, 27, 5-33.
CHAMPIAT D., LARPENT J. P., 1988. La biologie des eaux : méthodes et techniques. Paris, Masson, 374
pp.
CHAUSSODE R., GERMON J. C., CATROUX G., 1981. Académie de France Extrait du procés verbal de la
séance du 13 mai 1981, 762-771.
CHAUSSODE R., GUPTA S. K., HALL J. E., POMMEL B., WILLIAMS J. H., 1985. Nitrogen and
phosphorus value of sewage sludges. Edited by Hall J. E. and Williams J. H., XII/ENV/35/82-
REV.
CHAUVE P., MANIA J., MIGNOT C., RECOULES A., REMY. F., TRUCHE C., 1986. Paramètres physico-
chimiques, lithologiques et hydrologiques influençant l'évolution dans le temps des teneurs en
fer et en manganèse; critique des résultats obtenus sur des champs expérimentaux de la plaine
alluviale de la Saône (Franche Comté). J. Hydrology, 86, 367-390.
CHIAN E. S. K., DEWALLE F. B., 1976. Sanitary landfill leachates and their treatment. J. Environ.
Engin. Div. ASSCE. 102 (EE2), 411pp.
CHIANESE A., ROLANDO R., VERDONE N., 1999. Treatment of landfill leachate by reverse osmosis.
Wat. Res., 33, 647-652.
CHO B., AZAM F., 1990. Biogeochemical significance of bacterial biomass in the ocean’s euphotic
zone. Mar. Ecol. Prog. Ser., 63, 253-259.
CHRISTENSEN T. H., KJELDSEN P., BJERG P. L., JENSEN D. L., CHRISTENSEN J. B., BAUN A.,
ALBRECHTSEN H. J., HERON G., 2001. Biogeochemistry of landfill leachate plumes. Appl.
Geochemistry, 16, 659-718.
CLEMENT B., PERSOONE G., JANSSEN C., LE DÙ-DELPIERRE A., 1996. Estimation of the hazard of
landfills through toxicity testing of leachates. Chemosphere, 11, 2203-2320.
COLE J. J., FINDLAY S., PACE M., 1988. Bacterial production in fresh and salwater ecosystems: a
cross-system overview. Mar. Ecol. Prog. Ser., 43, 1-10.
D
DAWDOWICSZ P., LOOSE C. J., 1990. Metabolic cost during predator-induced diel vertical migration of
Daphnia. Limnol. Oceanogr, 37, 1585-1595.
DEL GIORGIO P. A., COLE J. J., CIMBLERIS A., 1997. Respiration rates in bacteria exceed
phytoplancton productio in uproductive aquatic systems. Nature, 385, 148-151.
DEVARE M., BAHADIR M., 1994. Biological monitoring of landfill leachate using plants and
luminescent bacteria. Chemosphere, 28, 261-271.
DEUNEVY J. P., 1987. Les lixiviats de décharges, Approche méthodologique de leur toxicité aigu en
fonction des différents modes de traitement. Thèse de troisième cycle INSA Lyon. 333p.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
158
DEVARS S., HERNANDEZ R., MORENO-SANCHEZ R., 1998. Enhanced heavy metal tolerance in two strains
of photosynthetic Euglena gracilis by preexposure to mercury or cadmium. Archives of
Environmental Contamination and Toxicology , 34 128-135.
DEVAUX J., LAIR N., 1976. Production primaire et biomasse des populations algales dans un petit lac
oligotrophe du massif central Français. Hydrobiologia, 50, 209-220.
Dinges W. R., 1973. Ecology of Daphnia in stabilization pods. Texas departement of Health Ressources,
Division of wastewater Technology and Surveillance , 155pp
DOR I., SCETER H., BROMLEY H. J., 1987. Limnology of hypertrophic reservoir storing wastewater
effluent for agriculture at Kibbutz Na’an. Israel. Hydrobiologia, 150, 225-241.
DRAKIDES C., GERVAIS M., 1987. Potentialités désinfectantes du lagunage par microphytes en climat
chaud. Premières observations. Inter-University Seminar on Wastewater in Waste Stabilization
Ponds and Aerated Lagoons.
DRAKIDES C., 1981. Etude de l’influence de la stratification et de l’évaporation sur la caractérisation de
l’écoulement en lagunage dans le cas de modèles de laboratoire. Rapport interne. Laboratoire de
génie chimique.
DRAKIDES C., 1983. Lagunes à charge estivale fonctionnement et conception. ABRMC, 194pp.
DUBOIS M., GILLES K. A., REBERS P. A., SMITH F., 1956. Colorimetric method for determination of
sugars and related substances. Annal. Chem., 28, 350-356.
DUMONT H. J., VAN DE VELDE I., DUMONT S., 1975. The dry weight estimate of biomass in a selection of
Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental
waters. Oceanologia, 19: 75-97.
E
EDMONDSON W. T., WINBERG G. G., 1971. A manual on methods for the assessment of secondary
productivity in fresh water. I.B.P. Handbook 17, Blackwell Scientifique publication, Oxford, 358p.
EHRIG A., 1978. Beitrag zum quantitativen und qualitativen Wasserhaushalt von Mûlldeponien.
Verôffentlichungen des institut fûr Stadtbauwesen, Technischen Universitât Braunschweig,
Germany.
EL-FADEL M., FINDIKAKIS A. N., LECKIE J. O., 1997. Modelling leachate generation and transport in
solid waste landfills. Environmental Technology, 18, 669-686.
ETCHEBER H., JOUANNEAU J. M., LATOUCHE C., 1977. Teneurs en Zn, Pb, Cu, Ni des suspensions de
l’estuaire de la Gironde. Evolution amont-aval de novembre 1975 à août 1976, Bull. IGBA, 22,
123-137.
ETCHEBER H., 1978. Etude de la répartition et du comportement de quelques oligo-éléments métalliques
(Zn, Pb, Cu et Ni) dans le compexe fluvio-esturien de la Gironde, Thèse de 3èm cycle, Univ.
Bordeaux 1, n° 1455, 209pp.
EPA., 1971. Waste treatment lagoon-state of the art. Water pollution Control Research., Series: 17090,
07/71.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
159
F
FARQUHAR G. J., 1989. Leachate: production and characterization. Can. J. Eng., 16 (3), 317-325.
FEUILLADE M., DUFOUR Ph., FEUILLADE J., 1988. Organic carbon release by phytoplankton and bacterial
reassimilation. Schweiz. Z. Hydrol,. 50, 115-135.
FENCHEL T., 1986. The ecology of heterotrophic microflagellates. Adv. Microbiol. Ecol., 9, 57-97.
FIELDER D. S., PURSER G., BATTAGLENE S. C., 2000. Effect of rapid changes in temperature and salinity
on availability of the rotifers Brachionus rotundiformis and Brachionus plicatilis, Aquaculture, 189, 85-99
FITZEGERALD G. P., ROHLICH G. A., 1958. A evaluation of stabilization pond litterature. Sew. Rev.
Hydrobiol., 49, 63-131.
FLIP Z., CLAUS H., DIPPELL G., 1998. Mikrobieller Abau von Huminstoffen cine Übersicht. Z.
PflanzenernAHR. Bodendk., 161, 605-612.
FLIP Z., PECHER W., BERTHELIN J. 2000. Microbial utilization and transformation of humic acide-like
substances extracted from a mixture of municipal refuse and sewage sludge disposed of in a
landfill. Environ. Pollution., 109, 83-89.
FLORENTZ M., HARTMANN P., GILLES P., 1984. Expérimentation sur pilote à alternance de phase portant
sur l’élimination biologique du phosphore. Rev. Sci. Eau., 3: 1-17.
FRASER J. K., BUTLER C.A., TIMPERLEY .M. H., EVAN C. W., 2000. Formation of copper complexes in
landfill leachate and their toxicity to zebrafish embryos. Environmrntal toxicology and chemistry,
19, 1397-1402.
FÜHRAMAN J., A., SLEETER T. D., CARLSON C. A., PROCTOR L. M., AZAM F., 1989. Dominance of
bacterial biomass in the sargasso sea and its ecological implications. Mar. Ecol. Prog. Ser., 57,
207-217.
FURTADO A. L., CASPER P., ESTEVES F. A., 2001. Bacterioplankton abundance, biomass and production
in a brazilian coastal logoon and in two german lakes. Anais da Academia Brasileira de Ciencias,
73, 39-49.
G
GALABOV M., 1989. La migration souterraine des polluants chimiques radioactifs cas particulier d'un
milieu très peu perméable. Hydrogéologie, 1, 45-49.
GANF G. G., STONE S. J. L., OLIVER R. L., 1986. Use protein to carbohydrate ratios to analyse for nutrient
deficiency in phytoplankton. Aust. J. Mar. Freshwat. Res., 37, 183-197.
GERMAIN M., 1981. Flore des Diatomées, eaux douces et saumâtres. Paris, France, Boubée, 444 pp.
GILBERT J. J., 1988. Suppression of rotifer populations by Daphnia a review of the evidence, the
machanisms, and the effects on zooplankton community structure, Limnol. Oceanogr., 33, 1286-
1303.
GLIWICZ Z. M., 1969. The food sources of zooplankton. Ecol. Polska, B15, 221-231.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
160
GLIWICZ Z. M., 1980. Filtering rates, food size selection and feeding rates in Cladocerans-another aspect
of interspecific competition in filter feeding zooplankton. In Kerfoot, W. C. (ed.) Ecology of
Zooplankton Communities. Univ. press, New England, pp: 282-291. Ecol. Polska, B15, 221-231.
GLOYNA E. F., 1971. Waste stabilization ponds. WHO Monogr. Ser., N° 60, Genève, Suisse.
GNUDI S., GIACOMETTI N., SALOMONI C., 1991. Rotifer control in wastewater high rate algae ponds. M.
Madoi (Ed.), Biol. Appr. Sewage Trat. Proc. Current State and Perspect. Perugia. Italie, 367-371.
GOMMY C., 1997. Optimisation d'un schéma de spéciation des métaux Pb, Zn, Cd et Cu : Application à des
sols pollués du nord de la France. Thèse de doctorat de troisième cycle de l'Université de
Technologie de Compiègne, 355pp.
GORDON A. M., MACBRIDE R. A., FISKEN A. J., VORONEY R. P., 1988. Effect of landfill leachate
spraying on soil respiration and microbial biomass in a northern hardwood forset ecosystem.
Waste. Mgt. Res., 6, 141-148.
GRAS R., ILTIS A., SAINT-JEAN L., 1978. Taux de natalité et relations entre paramètres d'accroissement et
d'abondance dans une population à structures d'age stable: cas d'une population de cladocères à
reproduction par parthénogenèse. Hydrobiol., 12: 19-63.
GRAY D. A., MATHER J. D., HARRISON I. B., 1974. Review of groundwater pollution from waste disposal
site in England and Wales, with provisional guidelines for future site selection. Q. J, Engng. Geol.,
7, 181-196.
GRISEY H., 1997. Rapport d'étude hydrogéologique. Hydro-Géotechnique Est (1997).
H
HAKANSON, 1976. A bottom sediment trap for recent sedimentary deposits. Limnol. Oceagr., 21, 170–174.
HAKKARI L., 1978. Interrelationships of phytoplankton and zooplankton production in some lakes of
Central Finland. Verh. Internat. Verain. Limnol., 20, 905-909.
HALL P., 1984. Chemical fluxes at the sediment-sea water interface : insitu investigations with bentic
chambers. Ph. D. Thesis, Kemihuset, Chalmers, Tekniska.
HAKONSON L., 1976. Abottom sediment trap for recent sedimentary deposits. Limnol. Oceanogr., 21, 170-
174.
HARMSEN J., 1983. Identification of organic compounds in leachate from a waste tip. Wat. Res., 17, 699-
705.
HARPER S. C., MANOHARAN R., MAVINIC D. S., RANDALL C. W., 1996. Chronium and nickel toxicity
during the biotreatment of high ammonia landfill leachate. Wat. Environ. Res., 68, 19-24.
HART R. C., 1985. Embryonic development times of entomostracan zooplankton from lake LeRoux
(Orange river, South Africa) and their possible relationships to seasonal succession.
Hydrobiologia, 127, 17-26.
HAULIDIER K., PAQUIN J. L., FRANÇOIS T., 1988. Biofilm-growth in a drinking water network: a
priliminary industrial pilot-plant experiment. Wat. Sci; Technol., 20, 109-115.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
161
HERRERA MELIÁN J. A., DOÑA RODRÍGUEZ J. M., VIERA SUÁREZ A., TELLO RENDÓN E., VALDÉS DO
CAMPO C., ARANA J., PÉREZ PEÑA J., 2000. The photocatalytic disinfection of urban waste
waters. Chemosphere, 41, 323-327
HERNANDEZ-LEON S., ALMEIDA C., PORTILLO-HAHNEFELD A., GOMEZ M., MONTERO L., 2000.
Biomass and potential feeding, respiration and growth of zooplankton in the Bransffield Strait
(Antarctic Peninsula) during austral summer. Polar Biology, 23, 679-690.
HERZIG A., 1979. The zooplankton of the open lake, in: H. Lofler (ed), Neusiedlersee, Limology of
shallow lake in central Europe. DR. W. Junk Publ., The Hague., Monogr. Biol., 37, 281-235.
HESSSEN D. O., 1985. The relation between bacterial carbon and dissolved humiqc compounds in
oligotrophic lakes. Microbiol. Ecol., 31, 215-223.
HOLLOPETER J. A., DAGUE R., 1994. Anaeorobic sequencing batch reactor treatment of landfill leachate.
49th Purdue Industrial Waste Conference Proceedings , 277-284.
HOSOMI M., MATSUSIGE K., INAMORI Y., SUDO R., YAMADA K., YOSHINO Z., 1989. Sequencing batch
reactor activated sludje processes for the treatment of municipal landfill leachate: removal of
nitrogen and refractory organic compounds. Water. Sci. Tecnol., 21, 1651-1654.
HOWARD KEN W. F., EYLES N., LIVINGSTONE S., 1966. Municipal landfill practice and its impact on
grounwater resources in and around urban Toronto, Canada. Hydrgeôlogy Journal, 4, 64-79.
HOWMILLER R. P., 1972. Effects of preservatives on weight of som common macrobenthic invertebrates.
Trans. Amer. Fish. Soc., 101, 743-746.
HRBACEK J., DVORAKOVA M., KORINEK, PROCHASOVA L., 1961. Demonstration of the effects of the fish
stock on the species composition of zooplankton and the intensity of metabolism of the whole
plankton associations. Verh. Internat. Verein. Limnol., 14, 192-195.
HRBACEK J., 1962. Species composition and the amount of zooplankton in relation to the fish stock.
Rozpravy, CSAV, rada mat. A. prir. Ved., 72, 1-116.
HUANG Y., ZHAO Z., XU M., GAO Y., 2000. Biological approaches for disposing and reusing chemical
wastewater. Ecological Engineering. 16, 281-292.
J
JANA B. B., PAL G. P., 1985. The life history parameters of Moina micrura (KURZ) grown in different
cultiring media. Water. Res., 19: 863-868.
JASPER S. E., ATWATER J. W., MAVINIC D. S., 1985. Leachate production and characteristics as a
function of water input and landfill configuration. Wat. Poll. Res. J. Can., 20, 43-56.
JASPERS E., NAUHAUS K., CYPIONKA H., OVERMANN J., 2001. Multitude and temporal variability of
ecological niches as indicated by the diversity of cultivated bacterioplankton. FEMS Microbiology
Ecology, 36, 153-164.
JIGOREL A., BERTRU G., 1993. Endogenic development of sediments in a eutrophic lake. Hydrobiologia,
268, 45-55.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
162
JIGOREL A., MORIN J. P., 1994. Bilan de la sédimentation dans retenue eutrophisée quinz ans aprés sa
création. Proceedings Seventh International Congress International Association of Engineering
Geology. Lisbonne, 2667-2674.
Jolanta B., Michal B., Joanna G., 2001. Application of pressure-driven membranes techniques to
biological treatment of landfill leachates. Process Biochemistry. 36, 641-646.
JOLY P. L., 1988. Théorie et pratique du traitement aérobie des lixiviats de décharge en lit bactérien: Etude
cinétique et modélisation, conception et réalisation d’une unité d’épuration. Thèse de Doctorat de
l’INSA de Lyon.
JONES J. G., SIMON B. M., MITCHELL R., 1983. Estimates of bacterial growth from changes in uptake
rates and biomass. Appl. Environ. Microbiol., 44, 1296-1307.
K
KATO K., 1985. Seasonal observations of heterotrophic activity and active bacterial number in lake
constance. Arch. Hydrobiol./ suppl., 66, 3, 309-319.
KENEDY F., ROCHE., 1995. Growthe of the rotifer Brachinus calyciflorus pallas in Dairy waste
stabilization ponds. Wat. Res., 29, 2255-2260.
KERBACHI R., BELKACEMI M., 1994. Caractérisation et évolution des lixiviats de la décharge de Oued-
Smar à Alger. TSM.,11.
KING COUNTRY SOLID WASTE DIVISION (KCSWD), 1992. Cedar Hills Regional Landfill Leachate
Monitoring Data Base. Seattle.
KIRK K. L., 1998. Enrichment can stabilize population dynamics: autotoxins and density dependence.
Ecology, 79, 2456-2462.
KOST W., 1978. Rotatoria Die Radertiere Mitteleuropas II. Tafelbandi Gerbruder Borntraeger, Berlin,
Stuttgart.
KMET P., 1982. Environmental Protection Agency’s 1975, water balance method, its use and limitations.
Guidance Report, Bureau of Solide Waste Management, Wisconsin Department of Natural Ressources,
Madison.
KNOX K., 1983. Treatability studies on leachate from a co-disposal landfill. Environ. Poll. (Series B), 5,
157-174.
KNOX K., 1985. Leachate treatment with nitrification of ammonia. Water. Res., 19, 595-904.
KROM M. D., BERNER R. A., 1980. Adsorption of phosphate in anoxic marine sediments. Limnol.
Oceanogr., 25, 797-806.
L
LA FORCE M. J., FENDORF S. E., LI G. C., SCHNEIDER G. M., ROSENZWEIG R. F., 1998. A laboratory
evaluation of trace element mobility from flooding and nutrient loading of Cower d'Alene river
sediments. J. Environ. Qual., 27, 318-328.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
163
LAIR N., 1977. Regimes alimentaires et nutrition du zooplancton, le cas du copepode Cyclops abyssorum
praealpinus (Kieffer 1939). Bull. Ecol., 2, 137-182.
LAIR N., 1978. Répartition spatio-temporelle, Biomasse et reproduction des populations zooplanctoniques
du lac d’Aydat en periode estivale. Hydrobiologia, 61, 237-256.
LAZZARO X., DRENNER R. W., STEIN R. A., SMITH J. D., 1992. Planktinivores and plankton dynamics:
effects of fish biomass and planktinivores type. Can. J. Fish. Sc., 49, 1466-1473.
LEFEVRE M., 1942. L'utilisation des algues d'eau douce par les Cladocères. Bull. Biol., 76, 250-276.
LEGENDRE L., BALEUX B., TROUSSELLIER M., 1984. Dynamics of pollution inducator and heterotrophic
bacteria in sewage treatment lagoons. Appl. Environ. Microbiol., 48, 586-593.
LIEBERMANN M. E., 1970. The response of moina brachiata (Jurine) to biological oxygen demand, oxygen
and light. Hydrobiologia, 36, 6-16.
LIJKLEMA L., 1980. Interaction of orthophosphate with iron (III) and aluminium hydroxydes. Environ. Sci.
Technol. 14: 537-541.
LOEDOLF G. J., 1965. The function of Cladocera in oxydation ponds. Proce. 2d int. Water. Poll. Res.
Conf., Tokyo: 307-325.
LOHMAN, 1908. Untersuchungen zur festetellung des vollstandigen gehaltes des meeres an plankton. Wiss.
Meeresunters, Abt. Kiel N. F. 10, 132-170.
LOWRY O. H., ROSEBROUGH N. G., FAR A. L., RANDALL R. G., 1951. Protein measurement with the
Folin phenol reagent; J. Biol. Chem., 193, 265-275.
LUBZENS E., 1987. Raising rotifers for use in aquaculture. Hydrobiologia, 147, 245-255.
LUDVIGSEN L., ALBRECHTSEN H. J., HERON G., BJERG P. L., CHRISTENSEN T. H., 1998. Anaerobic
microbial rédox processs in a landfill leachate contamined aquifer (Grindsted, Danemark). J. of
Contaminant Hydrology, 33, 273-291.
LUND J. W. G., KIPLING C., LE CRENNE D., 1958. The inverted microscope methode of estimating algal
numbers and the statistical basis of estimations by counting. Hydrobiol., 11, 143-170.
LYNCH M., 1989. The life history consequences of resource depression in Daphnia pulex. Ecology, 700,
1246-1256.
LYNGKILDE J., CHRISTENSEN T. H., 1992. Rédox Zones of landfill leachate pollution plume (Vejen,
Danemark). J. of Contaminant Hydrology, 10, 273-289.
M
MAC HENRY J. R., RIICHIE J. C., 1977. Physical and chemical parametrs affecting transport of Cs137 in
arid watersheds, Water Resources Research, 6, 923-927.
Margalef R., 1958. Temporary succession and spacial heterogeneity in phytoplankton. In Buzzati
Traverso. A. A. (Ed), Perspectives in Marine Biology. Univ. California press, Berkeley, CA., 323-
347.
Margalef R., 1961. Algunas applicaciones de la informacion en el campo de biologia y concretamenta a la
ecologia y al studio de la evolucion. Scientia, ser., 55, 1-7.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
164
MARTINEZ-TABCHE L., RAMIREZ MORA B., GERMAN-FAZ C., DIAZ-PARDO E., 1998.
Acetylcholinesterase activity of the freshwater cladoceran, Moina macropa, used as an in vitro
screening tool for water quality. Aquatic Ecosystem Health and Management Society , 1, 317-332.
MARVALIN O., ALEYA L., HARTMANN H. J., LAZAREK S. 1989., Coupling of the seasonal patterns of
bacterioplankton and phytoplankton in a eutrophic lake. Can. J. Microbiol., 35, 706-712.
MATTER J. D., YOUNG C. P., 1979. Recherches actuelles sur la pollution des eaux souterraines par les
décharges. Bulletin du B.R.G.M., 3, 233-244.
MATTIAS B. A., NILSSON P., 1997. Seasonal changes of leachate production and quality from test cells. J.
Envir. Eng. Div. ASSCE. 892-900.
MAYNARD H. E., OUKI S. K., WILLIAMS S. C., 1999. Tertiary lagoons: A review of removal mechanisms
and performance. Wat. Res., 33, 1-13.
MAYO A. W., 1996. DBO5 Removel in facultative ponds: Experience in Tanzania. Wat. Sci. Technol., 34,
447-455.
MEFFERT M. E., OVERBECK J., 1979. Regulation of bacterial growth by algal release products. Arch.
Hydrobiol./ suppl., 87, 118-121.
MENCH M., CLAIRON M., SOBESKY O., NAGOU D., 1989. Dynamique en temps court de l’azote minéral
en sol ferrallitique nu après apport d’une boue urbaine. Agronomie, 9, 785-793.
MILLERIOUX G., 1976. Contribution à l’étude du lac d’Aydat (Puy de Dome). Thése de 3è Cycle,
Université Clermont Ferrand II, 107pp.
MILTSTEIN A., ALKON A., KOEHBA M., AVMMENECH Y., KARPLUS I., 1995. Combined effects of
fertilization rate manuring and feed pellets applications on fish performances and water quality in
polyculture ponds. Aquacult. Res., 26, 55-65.
MIRACLE M. R., VICENT E., 1983. Vertical distribution and rotefer concentrations in the chemocline of
meromectic lakes. Hydrobiologia, 104, 259-267.
MIRACLE M. R., SERRA M., OLTRA R., VICENT E., 1988. Differantial distributions of Brachionus species
in three coastl lagoons. Vrh. Internat. Limnol., 23, 2006-2015.
MOAL J., SAMAIN J. F., LE COZ J. F., DANIEL J. Y., 1985. Protéines, glucides et lipides particulaires:
aspects méthodologiques. Oceanis, 11, 487-502.
Moehlum T., 1995. Treatment of landfill leachate in on – site lagoons and constructed wetlands. Wat. Sci.
Tech., 32, 3, 129-135.
Morris I., Skea W., 1978. Products of photosynthesis in natural populations of marine phytoplankton from
the Gulf of Maine. Mar. Biol., 47, 303-312.
MONFORT P., BALEUX B., 1990. Dynamics of aeromonas hydrophila, aeromonas sobria and aeromonas
caviae in a sewage treatment pond. Appl. Environ. Microbiol., 56, 1999-2006.
MONTIEL A., 1980. Method de determination des micropolluants organiques et minéraux contenus dans les
sédiments, 25è Congrés de l’Association Française de Limnologie , Paris, le 19-05-1980.
MONTUELLE, B. VOLAT B., TORIO-FERNANDEZ M. M., NAVARRO E., 1996. Changes in nitrobacter
serotypes biodiversity in a river: impact of a wastewater treatment plant discharge, Wat Res, 30,
1057-1064.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
165
N
NOVAC J. T., 1974. Temperature-substrate interaction in biological treatment. J. Wat. Pollut. Control. Fed.,
46, 1984-1994.
NAGATA T., 1988. The microflagellate-picoplankton food linkage in the water column of lake Biwa.
Limnol. Oceanogr., 33, 504-517.
NAVARRO A., BERNARD D., MILLOT N., 1988. Les problèmes de pollution par les lixiviats de décharges.
TSM., 3, 541-545.
NIMMO D.R., WILLOX M. J., KARICH J. F., TESSARI J. D., CRAIG T. L., GASSER E. G., SELF R. J.,
1995. Non-availability of metal from an urban landfill in Virginia. Chemical Speciationand
Bioavailability, 7, 2, 65-72.
NORBERG J., 2000. Resource-niche complementarity and autrophic compensation determines ecosystem-
level responses to increased cladoceran species richness. Oecologia, 122, 264-272.
O
O BRIEN J., DENOYELLES F., 1972. Photosynthetically elevated pH as a factor in zooplankton mortality in
nutrient enriched ponds. Ecology., 53, 605-614.
ODUM E. P., CANTLON J. E., KORNICKER., 1960. An organizational hierarchy postulate for the
interpretation on species individual distribution, species entropy, ecosystem evolution and the
meaning of species variety index. Ecology, 41, 395-399.
OSMERA S., 1966. Zur methods der festsetzung der biomasse einiger plankton Krebstiere. Zool. Listly., 15:
79-83.
OUFDOU K., MEZRIOUI N., 1994. Effets du pHet du rayennement solaire sur la survie de Vibrio cholerae et
d’Echerichia coli au cours d’un traitement des eaux usées par lagunage. 4ème Conférence
Internationale des Limnologues d’Expression Française , Tome 2, Marrakech, 25 au 28 avril 1994.
OUALAD ALI H., 1988. Role et importance des Rotifères planctoniques dans un écosystème lacustre
Eutrophe de la zone tempérée nord: Le lac d’Aydat (France). Thése de 3è Cycle, Université Blaise
Pascal de Clermont-Ferrand.
OUZZANI N., BOUARAB L., PICOT B., LAZREK H. B., OUDRA B., BONTOUX J., 1997. Variatios
saisonnières des formes de phosphore dans une station de traitement des eaux usées par lagunage,
sous climat aride de Marrakech (Maroc). Rev. Franç. des Sci. de l’eau, 4: 527-544.
P
PACAUD A., 1939. Contribution à l'ecologie des cladocères. Bull. Biol., suppl., 25, 226p.
PACE M. L., MCMANUS G. B., FINDLAY S. E. G., 1990. Planktonic community structure determines the
fate of bacterial production in a temperate lake. Limnol. Oceanogr., 35, 795-808.
PARK S., JOE KS., HAN SH., EOM TY., KIM HS., 1999. Characteristics and distribution of metallic
elements in landfill leachates. Environmental Technology., 20, 443-448.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
166
PAVLOV M., ANDERS J. F., JORUNN S., OLAV V., 2000. Control of bacterial flora of Brachionus Plicatilis
and Artemia franciscana by incubation in bacterial suspension. Aquaculture, 185, 207-218.
PEARSON H. W., MARA D. D., MILLS S. W., SMALLMAN D. J., 1987. Factors determining algal
populations in waste stabilization ponds and the influence of algae on pond performance, Wat. Sci.
Technol., 19, 131-140.
PERSSON L., ANDERSSON S., HAMRIN F., JOHANSSON L., 1988. Predator regulation and primary
production along eutrophication gradient of temperate lake ecosystems, In: Complex interactions
in lake communities (Carpenter. S. R., ed.), 45-65. Springer Verlag.
PEDERSON B. M., JANSEN J. I. A. C., 1992. Treatment of leachate -polluted groundwater in an aerobic
biological filter Euro. Water. Poll. Control., 2, 40-45.
PEDROS-ALLIO C., 1989. Toward and autecology of bacterioplankton.b In: Sommer U. (ed), Plankton
ecology: succession in plankton communities. Brock/Springer, Berlin, 297-336.
PEJLER B., 1957. Taxonomical and ecological studies on planktonic Rotatoria from northern Swedish
Sapland. Kungl. Svenska. Handlingar, 6, 1-68.
PEJLER B., 1977. General problems on rotifer taxonomy and global distribution. Arch. Hydrobiol. Beith., 8,
212-220.
PETERS T., STANFORD P., 1993. L’osmose inverse et disc-tube modulé dans le traitement des lixiviats.
Tribune de l’eau, 556, 6, 67-72.
PIRT S. J., 1971. Pricinpales of microbes and Cell Cultivation Blackwell, London.
PIZAY-PARENTY M. D., 1985. Bassins de lagunage d'Aniche-Auberchicourt (Nord): Cinetiques
saisonnières et spatiales du zooplancton et des descripteurs de la qualité de l'eau. Thèse de 3è cycle,
Université des Sciences et Techniques de Lille, 195pp.
POHLAND F. G., HARPER S. R., 1985. Critical review and summery of leachate and gas production from
landfills. Georgia, Tech. Project, n°E-20-G01.EPA, CR 809-997, Georgia.
POURRIOT R., 1965. Recherche sur l’écologie des Rotifères. Thèse de 3è cycle, Université de Paris.
POURRIOT R., DELUZARCHES M., 1971. Recherche sur la biologie des rotifères. II. Influence de la
température sur la durée du développement embryonnaire et post-embryonnaire. Annl. Limnol., 7,
25-52.
POURRIOT R., 1982. Ecologie du plancton des eaux continentales, Paris, Masson, 198pp.
POURRIOT R., CHAMP P., 1982. Consommateurs et production secondaire, in Pourriot et coll., Ecologie du
plancton des eaux continentales Paris, Masson, 49-112.
POURRIOT R., FRANCEZ A. J., 1986. Introduction pratique à la systématique des organismes des eaux
continentales Françaises. Bulletin Mensuel Société Linéenne de Lyon , 55, 148-176.
PRAT N., TOJA J., SOLA C., BURGOS M. D., PLANS M., RIERADEVALL M., 1999. Effect of dumping and
cleaning activities on the aquatic ecosystems of the Guadiamar River following a toxic flood. The
Science of the total Environment, 242 132-248.
PREMAZZI G., MARENGO G., 1988. Sedimentation rates in Swisse-Italian lake mesured with seity Press,
384pp
PREMAZZI G., MARENGO G., 1988. Sedimentation rates in Swisse-Italian lake mesured with seity Press,
384pp
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
167
PROSI F., 1979. Biovailabilty of haevy metals in different fresh water sediments : Uptake in macrobenthos
and biomobilization in: Proceedings of the international Conference Management and Control of
heavy metals in the environment, London, Sept., Published by CEP consultants, 288-291.
R
REITZEL A., 1990. The temporal characterization of municipal solid waste leachate. Waterloo, Ontario,
University of Waterloo, Masters of applied Science Thesis, 144pp.
REYNOLDS C. S., JAWORSKI G. H. M., CMIECH H. A., LEEDALE G. F., 1981. On the annual cycle of
blue-green alga Microcystis aeruginosa. Kutz. Phil. Trans. R. Soc., London., 293, 419-477.
REYNOLDS C. S., 1984. The ecology of freshwater phytoplankton. Cambridge studies in ecology,
Cambridge University Press, 384pp.
RIISE G., VAN HEES P., LUNDSTROM U., TAU STRAND L., 2000. Mobility of different size fractions of
organic carbon, Al, Fe, Mn and Si in podzols. Geoderma, 28, 237-247.
ROBINSON H. D., 1990. On site treatment of leachates from landfilled wastes. Journal of institution of
water and environmental management , 4, 78-89, Londre England.
ROBINSON H. D., GRANTHAM G., 1988. The treatment of landfill leachates in on-site arerated lagoon
plants: Experience in Britain and ireland. Wat. Res., 22, 733-747.
ROCQUES M., 1980. Fondement théorique de traitement biologique des eaux.. vol. II, ed. Technique et
Documentation. Paris.
RODIER J., 1984. L'analyse de l'eau. 7e édition Dunod, Paris.
RÜGGE K., BJERG P.L., CHRISTENSEN T.H., 1995. Distribution of organic compounds from municipal
solid waste in the grounwater down gradient of lanfill (Grindsted, Danemark). Envir. Sc. Technol.,
39, 1395-1400.
RUTTNER-KOLISKO A., 1974. The vertical distribution of plankton rotifers in small alpine lake with a
sharp oxygen depletion (Lunzer observee). Verh. Internat. Verein. Limnol., 19, 1286-1294.
S
SALOMONI C., 1991. Biotrattamento de reflui suinicoli per la produzione di organismi aquatici. Riv.
Suinicoltura, 2, 33-37.
SAMPSON E. O., 1955. A double duty oxydation pond. Sewage. ind. waste., 27, 1440pp.
SANCHEZ I., LEE G. F., 1978. Environmental chemistry of cooper in Lake Monona, Wisconsin, Wat. Res.,
12, 899-903.
SAUVE, SEBASTIEN, HENDERSHOT, WILLIAM, ALLEN, HERBERT E., 2000. Solid-Solution partitioning of
metals in contaminated soils: dependence on pH, tolal metal burden and organic matter. Environ.
Sci. Tech., 34, 1125-1131.
SCHNETTER R. W., MIDDLEBROOK E. J., 1983. Cold region wasterwater lagoon sludge accumulation.
Water Research, 18, 1201 – 1206.
S.C.O.R.- U.N.E.S.C.O., 1966. Determination of photosynthetic pigments in sea water. Monographs on
oceanographic methodology 1. 2nd ed. Paris 9-18.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
168
SCHETRITE S., RACAULT Y., 1995. Purification by natural waste stabilization pond : influence of weather
and ageing on treatment quality and sediment thickness. Wat. Sci. Tech., 31, 191-200.
Seamann M. T., Gophen M., Cavari B. Z., Azoulay B., 1986. Brachionus calyciflorus Pallas as agent for
the removal of E. coli in sewige ponds. Hydrobiologia, 135, 56-60.
SERVAIS P., LAURENT P., RANDON G., 1992. Mesure de la biomasse et de l’activité bacterienne dans l’eau
de distribution. Rev. Sci. L’Eau., 5, 473-488.
SEVRIN-REYSSAC J., 1998. Biotreatment of swine manure by production of aquatic valuable biomasses.
Agriculture Ecosystems & Environment, 68, 177-186.
Simon M., Tilzer M. M., 1987. Bacterial reponse to seasonal changes in primary production and
phytoplankton biomass in lake Constance. J. Plankton. Res., 9, 535-552.
SLATER C., AHLERT R., UCHRIN C., 1983. Treatment of landfill leachates by reverse osmosis. Environ
Prog., 2, 251-256.
SLADECEK V., 1983. Rotifers as indicators of water quality. Hydrobiologia, 100, 169-201.
SOLDATINI G. F., RIFFALDI R., LEVI-MINZI R., 1976. Pb adsorption by soils water, air and soil poll., 6,
11-118.
SOMIYA I. TSUNO H. MATSUUMOTO M., 1984. Phosphorus release-storage reaction and organic substrate
behavior in biological phosphorus removal. Wat. Res., 22, 49-58.
SOMMARUGA R., 1995. Microbial and classical food webs: A visit to a hypertrophic lake. FEMS
Microbiology Ecology, 17, 257-270.
SOMMARUGA R., ROBARTS R. D., 1997. The signicance of autrophic and hypertrophic picoplankton in
hypertrophic ecosystems. FEMS Microbiology Ecology, 24, 187-200.
SOMMAIR U., GLIWICZ Z. M., LAMPERT W., DUNCAN A., 1986. The plankton ecology groupe (PEG)
model of seasonal succession of planktonic events in freshwaters. Arch. Hydrobiol. Beih., 106,
433-471.
SPENGEL D. B., DZOMBAK D. A., 1991. Treatment of landfill leachate with rotating biological contactors:
bebch-scale experiments. Res. J. WPFC., 63, 971-981.
STAAB K. F., EICHBERGER M, ZWICKL S., 1999. Dponiesickerwasser-Behandlung auf der
sonderabfalldeponie Billigheim. Entsorgungs Praxis, 5, 38-41.
STEGMAN R., EHRIG H. G., 1980. Operation and design of biological leachate treatment plants. Prog.
Wat. Technol., 12, 919-947.
STERNER R. W., 1988. The role of grazers in phytoplankton succession. In: Plankton Ecology (Sommer,
U., Ed), 107-170. Springer Verlag.
STRAUB W. A., LYNCH D. R., 1982. Models of landfill leaching : Organic Strength. J. Environ. Engin. Div.
ASSCE. 108, 251-268.
T
TAM N. F. Y., WONG Y. S., 2000. Effect of immobilized microalgal bead concentrations on wastewaternutrient removal. Envirnmental Pollution, 107, 145-151.
TAY J. H., SHOW K. Y., 1992. The use of lime-blended sludge for production of cementitious material.Wat. Envir. Res., 64, 6-12.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Références bibliographiques
169
TIFNOUTI A., POURRIOT R., 1989. Dynamique d'une population de Moina micrura (Crustacea, Cladocera)
dans un bassin de lagunage à Marrakech (Maroc). Rev. Hydrobiol. Trop., 22, 239-250.
TISDEL S. E., BRESLIN V. T., 1995. Charactérisationand leaching of elements from municipal solid waste
compost. J. Environ. Qual., 24, 827-845.
V
VAUTE L., 1997. Contribution à l'étude du transport des contaminants dans les aquifères karstiques. Thèse
de 3è cycle, université Montpellier II - sciences et techniques du Languedoc, France, 169 pp.
VEDRY B., 1987. L’analyse écologique des boues activées. Vicq: SEGETEC, 2è éd., 117pp.
VIRPIN F., PENSOYE I., 1985. Compte rendu des visites de décharge en Allemagne. ANRED, 40pp.
W
WEBER B., HOLTZ F., 1991. Landfill leachate treatment by reverse osmosis. In tuner MK, editor. Effective
Industrial Membrane processese : Benefits and Opportunities. London, New York: Elsevier
Applied Science, 143-154.
WATERTON T., 1969. The effects of tipped domestic refuse on groundwater quality. Wat. Treatemnt
examen, 18, 15-69.
WEI C., LIN W., ZAINAL Z., WILLIAMS N. E., ZHU K., KRUZIC A. P., SMITH R; L., RAJESHWAR K.,
1994. Bactericidal activity of TiO2 Photocatalyst in aqueous media: toward a solar-assisted water
desinfection system. Env. Sci. Technol., 28, 934-938.
WEISS T., MULLER H., PINTO-COELHO R. M., SCHWEIZER A. SPRINGMANN D., BALDRINGER G., 1990.
Response of the microbial loop to the phytoplankton spring bloom in a large prealpine lake;
Limnol. Oceanogr., 35, 781-794.
WEITHOFF G., LORKE A., WALZ N., 2000. Effects of water-column mixing on bacteria, phytoplankton,
and rotifers under different levels of herbivory in a shallow eutrophic lake. Oecologia, 125, 91-
100.
WELANDER U., HENRYSSON T., WELANDER T., 1997. Nitrification of landfill leachate using suspended-
carrier biofilm technology. Wat. Res., 31, 2351-2355.
WYER M. D., 1988. Nitrification in Ontario stream sediments, Wat. Res., 22, 287-292.
Y
YOSHINAGA T., HAGIWARA A., TSUKAMOTO K., 1999. Effect of conditioned media on the asexual
reproduction of the monogonont rotifèr Brachionus plicatilis. O.F. Muller. Hydrobiologia, 412,
103-110.
Z
ZAGORC-KONAN J.and OMEN J., 1999. A simple test for montoring biodegradable industrial pollution in
receiving stream. Wat Sci Technol, 39, 221-224 .
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Liste des tableaux
170
Tableau 1. Les grands types de sites de décharges (ANRED, 1986). ___________________________ 7Tableau 2. Données bibliographiques en fourchettes de variation de la composition physico-chimiquedu lixiviat de décharge d'ordures ménagères d'après Christensen et al. 2001. Les teneurs sont exprimées enmg. l-1. 11Tableau 3. Caractéristiques morphométriques des quatre bassins de la station d'Etueffont. ________ 18Tableau 4. Composition chimique des schistes et des grés. ________________________________ __ 24Tableau 5. Biovolumes spécifiques calculés pour les espèces algales les plus abondantes dans lesbassins d’Etueffont. ________________________________ ________________________________ ___ 27Tableau 6. Biovolume spécifique calculé pour les espèces rotarinnes les plus abondantes dans lesbassins d’Etueffont. ________________________________ ________________________________ ___ 30Tableau 7. Biomasse calculée pour Moina sp. ________________________________ ___________ 30Tableau 8. Tableau comparatif de la composition physico-chimique moyenne (n = 3 pour la stationpilote (2001) et n = 13 pour la décharge d'Etueffont (1998-1999)) du lixiviat issu de la décharged’Etueffont et de la station pilote. ________________________________ ________________________ 37Tableau 9. Evolution inter-annuelle du pH, de la conductivité électrique (CE), de la teneur en DBO, enDCO et du rapport DBO/DCO du lixiviat de 1993 à 1998.________________________________ _____ 44Tableau 10. Composition moyenne des teneurs en cuivre (Cu, mg. l -1), de la température (T, °C) et de laconductivité électrique (CE, µS. cm-1) dans les eaux des 8 piézomètres.___________________________ 58Tableau 11. Composition physico-chimique moyenne de l'eau des quatre bassins. ________________ 73Tableau 12. Composition physico-chimique moyenne du lixiviat après traitement. _______________ 125Tableau 13. Composition physico-chimique moyenne (13 échantillons) à l’amont du ruisseau et à 100 men aval du rejet. ________________________________ ________________________________ _____ 134Tableau 14. Description des sédiments recueillis. ________________________________ _________ 143Tableau 15. Variations du taux de sédimentation dans le bassin 1. ___________________________ 143Tableau 16. Variations du taux de sédimentation dans le bassin 2. ___________________________ 143Tableau 17. Variation du taux de sédimentation dans le bassin 3. ____________________________ 145Tableau 18. Variations du taux de sédimentation dans le bassin 4. ___________________________ 145Tableau 19. Vitesse d'accumulation calculée pour les quatre bassins. _________________________ 149Tableau 20. Composition physico-chimique des boues in situ. _______________________________ 151
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Liste des tableaux
171
Fig. 1. Etapes de la méthanisation selon Bryant (1979). ________________________________ ____ 12Fig. 2. Cycles dans un lagunage naturel (d’après Champiat 1988). ____________________________ 14Fig. 3. Limite entre aérobiose et anaérobiose dans les bassins de traitements des eaux usées (Rocques,1980). 14Fig. 4. Schéma et caractéristiques des bassins de lagunage naturel. ___________________________ 19Fig. 5. Evolution de la température (°C) ambiante dans la station d’Etueffont. ___________________ 20Fig. 6. Evolution des pluies (mm) dans la station d’Etueffont. ________________________________ 20Fig. 7. Fluctuation du niveau piézométrique des deux nappes d’eau. ___________________________ 22Fig. 8. Carte piézométrique de la nappe de surface (aout 1998). ______________________________ 23Fig. 9. Carte piézométrique de la nappe de surface (aout 1998). ______________________________ 23Fig. 10. Dosage de la composition biochimique du matériel particulaire (Moal, 1985). ___________ 28Fig. 11. Evolution temporelle des pluies (a), de la température (b), de la conductivité électrique (d),des teneurs en PT (b), de O2 (c), du pH (e), des Cl (f), des SO4 (g) et du Zn (h) en fonction du débit dans lelixiviat (toutes les concentrations sont exprimées en mg/l le débit en l/s et les pluies en mm). __________ 40Fig. 12. Evolution des teneurs en Cu (a), en Fe (b), en Ni (c), en HCO3 (d), en NO3 (e), en NH4 (f), enNT (f) de la DBO (g) et de la DCO (h) en fonction du débit dans le lixiviat (Toutes les concentrations sontexprimées en mg/l, le débit en l/s et les pluies en mm). ________________________________ ________ 41Fig. 13. Evolution saisonnière des MES et de l'abondance bactérienne dans le lixiviat. ___________ 43Fig. 14. Evolution horaire de la température (a), du pH (a), du Eh (b), de O 2(b), des SO4 (c), des Cl (c),de la conductivité électrique (d) et de la DCO (d) dans le lixiviat. _______________________________ 45Fig. 15. Corrélation entre les mesures effectuées à l'aide de la sonde SEBA au niveau des piézomètreset celles effectuées à l'aide de la sonde multiparamètres après échantillonnage. ____________________ 52Fig. 16. Variations saisonnières (mai 1998 à mai 1999) des précipitation mensuelles (PC, mm) (a)dans la station d'Etueffont, du potentiel redox (Eh, mV) (b) et de l'oxygène dissous (O 2, mg. l-1) (c) dans leseaux de tous les piézomètres. ________________________________ ____________________________ 55Fig. 17. Variations saisonnières (mai 1998 à mai 1999) des teneurs en sulfate (SO 4
2-, mg. l-1) (a),en nitrate (NO3
-, mg. l-1) (b) et en chlorures (Cl-, mg. l-1) (c) dans les eaux de tous les piézomètres _____ 56Fig. 18. Variations saisonnières (mai 1998 à mai 1999) des teneurs en manganèse (Mn 2+, mg. l-1) (a),en fer total (Fe total, mg. l-1) (b), en zinc (Zn2+, mg. l-1) (c) et en nickel (Ni2+, mg. l-1) (d) dans les eaux detous les piézomètres ________________________________ ________________________________ ___ 60Fig. 19. Analyse en composantes principales.________________________________ ____________ 61Fig. 20. Schématisation des phénomènes de contamination des eaux souterraines dans le sited’Etueffont. 62Fig. 21. Evolution spatio-temporelle de la température, du pH, du O 2, de la CE, du Eh et des MES dansles quatre bassins. ________________________________ ________________________________ ____ 65Fig. 22. Evolution spatio-temporelle de NT (a), PT (b), DBO (c), DCO (d) et des rapports DBO/DCO(e) et N/P (f) dans les quatre bassins. ________________________________ _____________________ 72Fig. 23. Evolution spatio-temporelle de NH4
+, NO3-, Chl a, et de la composition biochimique
particulaire (P et G) dans les quatre bassins . ________________________________ ______________ 75Fig. 24. Distribution des densités bactériennes dans les quatre bassins. _______________________ 82Fig. 25. Evolution spatio-temporelle de l’abondance phytoplanctonique. ______________________ 86Fig. 26. Pourcentage relatif des différents groupes phytoplanctoniques dans les differents bassins. __ 87Fig. 27. Dynamique saisonnière du phytoplancton dans le premier bassin. _____________________ 89Fig. 28. Dynamique saisonnière du phytoplancton dans le deuxième bassin. ___________________ 93Fig. 29. Dynamique saisonnière du phytoplancton dans le troisième bassin. ____________________ 94Fig. 30. Dynamique saisonnière du phytoplancton dans le dernier bassin. _____________________ 95Fig. 31. Distribution des densités rotariennes dans les quatre bassins_________________________ 99Fig. 32. Dynamique saisonnière des Rotifères dans le premier bassin. _______________________ 107Fig. 33. Dynamique saisonnière des Rotifères dans le deuxième bassin. ______________________ 108Fig. 34. Dynamique saisonnière des Rotifères dans le troisième bassin. ______________________ 109Fig. 35. Dynamique saisonnière des Rotifères dans le quatrième bassin. _____________________ 111Fig. 36. Distribution des densités des Cladocères dans les quatre bassins. ____________________ 115Fig. 37. Dynamique saisonnière des Rotifères dans le quatrième bassin. _____________________ 119Fig. 38. Dynamique saisonnière du zooplancton dans les quatre bassins. _____________________ 122Fig. 39. Variations de la vitesse de dégradation K (j -1) de la DBO calculée en fonction de latempérature.________________________________ ________________________________ ________ 124Fig. 40. Evolution des taux d'abattement de NH4, MES, NT, DCO, DBO, Cl, PT, Zn, Ni, Mn et Fe. _ 127
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Liste des tableaux
172
Fig. 41. Evolution du pH, T, NO3, Cl, Eh, O2, DCO et de l'abondance bactérienne de l'amont (A)jusqu'à 500 m en aval du rejet des lixiviats dans le ruisseau récepteur. __________________________ 139Fig. 42. Schéma simplifié du piège à sédiments. ________________________________ _________ 141Fig. 43. Implantation des pièges dans les quatre bassins. ________________________________ _ 144Fig. 44. Evolution spatiale du taux de sédimentation dans les quatre bassins (essais de décembre 1998et juin 1999). ________________________________ ________________________________ _______ 147Fig. 45. Variations spatio-temporelles de la DBO, la DCO et les MES dans les quatre bassins. ____ 149
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

Liste des tableaux
173
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com