tahina razanadraibe andrianarison traitement …hydrologie.org/the/andrianarison.pdf · libres,...
TRANSCRIPT

1
UNIVERSITE MONTPELLIER II SCIENCES ET TECHNIQUES DU LANGUEDOC
THESE
pour obtenir le grade de
DOCTEUR DE L'UNIVERSITE MONTPELLIER 2
Discipline : Mécanique, Génie mécanique, Génie civi l Formation Doctorale : Eau et environnement.
Ecole Doctorale : .Terre, Eau, Espace.
présentée et soutenue publiquement le 20 octobre 2006 par
Tahina Razanadraibe ANDRIANARISON
Titre :
Traitement d’effluents urbains dans un système de 1 1 lagunes. Décontamination microbienne et élimination de l’azo te.
devant le jury composé de Miquel SALGOT DE MARÇAY , Professeur, Université de Barcelone Rapporteur Jean Luc VASEL , Professeur, Université de Liège Rapporteur Bernadette PICOT , Professeur, Université Montpellier 1 Directeur de Thèse François BRISSAUD , Professeur, Université Montpellier 2 Examinateur Alain LIENARD , Ingénieur de Recherche, CEMAGREF Examinateur

2

3
REMERCIEMENTS Ce travail a été conduit dans le laboratoire Hydrosciences de l'Université Montpellier 2 dont je remercie le directeur M. Eric SERVAT pour son accueil et son écoute. J'adresse mes sincères remerciements à Mme le Professeur Bernadette PICOT pour avoir accepté d'être le directeur de cette thèse malgré un contexte compliqué. J'exprime ma profonde reconnaissance à M. le Professeur François BRISSAUD pour avoir accepté d'être le codirecteur de cette thèse et dirigé mon travail avec une grande disponibilité. Son appui scientifique m'a beaucoup enrichie. Ses encouragements m'ont énormément soutenue. Je remercie vivement M. le Professeur Miquel SALGOT DE MARÇAY de l'Université de Barcelone ainsi que M. le Professeur Jean Luc VASEL de l'Université de Liège pour l'honneur qu'ils me font en acceptant de juger ce travail. J'adresse également mes vifs remerciements à M. Alain LIENARD du CEMAGREF qui me fait l'honneur de participer au jury de cette thèse. Mes vifs remerciements s'adressent également à M. Christian DRAKIDES du CNRS pour ses conseils et ses encouragements. Je remercie également le CCNBT pour avoir donné son accord à la réalisation de cette étude sur la station de lagunage de Mèze ainsi que pour son soutien financier. Je remercie également le laboratoire de l'Ecosite de Mèze qui a effectué les analyses bactériologiques et physico-chimiques indispensables à l'accomplissement de ce travail. Je tiens également à remercier les doctorants qui ont participé à ce travail dont Kenza Najib pour son aide précieuse dans l'utilisation du logiciel Maple ainsi que Fabiola BADROT-NICO pour son implication inestimable dans la réalisation de ma soutenance et aussi pour son amitié qui m'a été d'un grand soutien tout au long de ces années.

4

5
TABLE DES MATIERES INTRODUCTION ................................................................................................................ 11
Chapitre 1. MATERIELS ET METHODES....................................................................... 19
1.1. La station de lagunage de Mèze-Loupian ............................................................................21
1.2. Mesures hydrauliques, météorologiques et contrôles physico-chimiques in situ ................24
1.3. Echantillonnage et méthodes d’analyses..............................................................................24
1.4. Devenir de l’azote dans les lagunes de maturation ..............................................................25
1.5. Modèle de fonctionnement hydraulique...............................................................................26
Chapitre 2. PERFORMANCES DU SYSTEME DE LAGUNAGE AVA NT ET APRES SON AGRANDISSEMENT .................................................................................................. 31
2.1. Comparaison des performances en début de fonctionnement, avant et après
l’agrandissement..........................................................................................................................33
2.2. Les performances globales de la station rénovée .................................................................35
2.3. Rôle et performances de chaque étage de traitement ...........................................................37
2.3.1 . Premier étage : les lagunes anaérobies ................................................................... 38
2.3.2 . Deuxième étage : L’étage facultatif à forte charge (FFC)......................................39
2.3.3 . Troisième étage : les bassins de maturation........................................................... 40
2.3.4 . Quatrième étage : les bassins de finition ................................................................ 46
Chapitre 3. MODELISATION DE LA DECONTAMINATION BACT ERIENNE........ 47
1.1. Mécanismes de la décontamination microbienne en lagunage ............................................53
1.1.1 . La décontamination par les rayons ultra violets ..................................................... 53

6
3.1.1 . La décontamination par photooxydation ................................................................ 55
3.1.2 . Remarques .............................................................................................................. 55
3.2. Modélisation de la décontamination ....................................................................................56
3.2.1 . Méthodes ................................................................................................................ 56
3.2.2 . Application à la recherche de relations entre les cinétiques de décontamination et
les variables météorologiques............................................................................................ 59
3.2.3 . Application du modèle à l’optimisation de la gestion ............................................ 71
1.1.4 . Les limites d'une cinétique de l'abattement bactérien fonction de la température et
de l'intensité du rayonnement solaire ................................................................................ 73
1.1.5 . Influence du modèle de comportement hydraulique. ............................................. 78
Chapitre 4. ELIMINATION DE L'AZOTE................. ....................................................... 85
4.1. Le modèle de l’élimination de l’azote de Reed (1985) ........................................................90
4.1.1 . Conception.............................................................................................................. 90
4.1.2 . Application à l’étage de maturation de la station de lagunage de Mèze ............... 92
4.2. Le modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) ........93
4.2.1 . Principes de la volatilisation................................................................................... 93
4.2.2 . Modèle de la volatilisation de Pano et Middlebrooks (1982) :.............................. 95
4.2.3 . Modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) :
........................................................................................................................................... 96
4.2.4 . Application du modèle d’élimination de l’azote ammoniacal de Pano et
Middlebrooks à l’étage de maturation de Mèze ................................................................ 98
4.2.5 . Modèle de Pano et Middlebrooks (1982) et volatilisation ................................... 100
4.3. Evaluation de la volatilisation. Cinétiques de Stratton ......................................................100
4.3.1 . Cas des cours d’eau : ............................................................................................ 100

7
4.3.2 . Cas des retenues d’eau relativement abritées du vent ..........................................104
4.3.3 . Application. Volatilisation dans un lac et dans une lagune :................................ 106
4.3.4 . Volatilisation dans des retenues d’eau soumises à l’agitation du vent................. 108
4.4. Etude des pertes en azote ammoniacal par Zimmo et al. (2003) .......................................111
4.4.1 . Matériels et méthodes........................................................................................... 111
4.4.2 . Résultats ............................................................................................................... 113
4.4.3 . Conclusion............................................................................................................ 114
4.5. Application : Rôle de la volatilisation selon Stratton (1969) dans la disparition de l’azote
ammoniacal dans le dernier bassin de maturation de la station de lagunage de Mèze .............115
4.6. Nitrification et dénitrification.............................................................................................116
4.6.1 . Microbiologie et stoechiométrie........................................................................... 116
4.6.2 . Paramètres influents de la nitrification................................................................. 118
4.6.3 . Les paramètres influents de la dénitrification....................................................... 120
4.6.4 . Cinétique............................................................................................................... 120
4.7. Localisation de la nitrification et de la dénitrification dans les systèmes de lagunage par
Zimmo (2003) ...........................................................................................................................123
4.7.1 . Matériels et méthodes........................................................................................... 124
4.7.2 . Résultats ............................................................................................................... 125
4.7.3 . Interprétations....................................................................................................... 128
4.8. Modélisation du cycle de l’azote........................................................................................129
4.8.1 . Modèle du cycle de l’azote : Principe et expression ............................................ 133
4.8.2 . Méthodes d’application ........................................................................................ 134
4.8.3 . Application à l’étage de maturation de la station de lagunage de Mèze assimilé à
un bassin unique .............................................................................................................. 135

8
4.8.4 . Application du modèle du cycle de l'azote développé en 2003 à l’étage de
maturation de la station de lagunage de Mèze, bassin par bassin.................................... 142
4.8.5 . Application au dernier bassin de maturation de la station de lagunage de Mèze. 147
CONCLUSION ….……………………………………………...………………………… 153
BIBLIOGRAPHIE……………………………………………………… …………………159
ANNEXES…………………………………………………………………………………..165
Annexe 1: Calcul de l'évaporation…………………..………………………………167
Annexe 2: Rappel sur la cinétique des réactions chimiques…………………..…….169
Annexe 3: Publications………………………………………………………………172

9

10

11
INTRODUCTION

12

13
Le lagunage est la technique naturelle d’épuration la plus répandue sur les cinq continents. Un lagunage, dans sa forme traditionnelle, est constitué de plusieurs bassins étanches fonctionnant en série. L’épuration de la matière organique repose sur l’activité de bactéries libres, essentiellement aérobies, et sur de longs temps de séjour de l’eau dans les bassins successifs (Commission Européenne, 2001). Le lagunage est un procédé rustique d’épuration des eaux usées. Il ne nécessite pas ou peu d’apport d’énergie, ne comporte pas ou peu d’équipement électromécanique ; l’entretien des installations peut être assuré par des personnels peu qualifiés. Ces caractéristiques, associées à des coûts de fonctionnement et de maintenance faibles, font du lagunage une technique souvent choisie pour équiper les collectivités petites et moyennes. En France, le nombre des installations de lagunage est situé entre 2500 et 3000 (Racault et Boutin, 2005). Le principal défaut de cette technique d’épuration est la place qu’elle nécessite, estimée à 12 m2 de bassins par équivalent habitant. Aussi est-elle considérée comme destinée en priorité aux collectivités rurales. Toutefois, cette tendance n’est pas une règle universelle puisque, en Nouvelle Zélande par exemple, le lagunage est et continuera à être la technique d’épuration des eaux usées urbaines la plus fréquemment mise en oeuvre, y compris pour des agglomérations de plusieurs dizaines de milliers d’habitants (Archer et Mara, 2003). Le lagunage a d’autres propriétés qui ont concouru à son développement. A cause de l’importance du volume de ses bassins, il a une capacité remarquable à accepter les pointes de charges organique ou hydraulique, résultant par exemple soit de déversements d’effluents d’activités agro-alimentaires soit d’évènements pluvieux. D’autre part, il présente une incontestable efficacité dans la décontamination bactérienne, même si celle-ci est souvent largement fonction des conditions climatiques. Cette dernière propriété, essentielle pour la protection des eaux de baignade et des activités conchylicoles, explique en partie le développement du lagunage le long des côtes méditerranéennes du Languedoc-Roussillon. Ce procédé peut aussi contribuer à l’élimination de l’azote et du phosphore, phénomène souvent observé en région Méditerranéenne pendant la saison chaude dans des installations non surchargées. Le lagunage, qui originellement n’était pas conçu pour un tel usage, doit être capable de satisfaire à des normes qui évoluent vers de plus en plus de rigueur pour mieux protéger les milieux sensibles aux risques d’eutrophisation. La plupart des lagunages construits il y a plus de vingt ans sont maintenant surchargés. D’autre part, les normes applicables à leurs rejets ont été ou sont susceptibles d’être renforcées. Il est donc nécessaire d’augmenter les capacités et les performances de ces installations. Il arrive aussi souvent que la réhabilitation des installations d’épuration soit compliquée par l’expansion urbaine qui a rapproché considérablement les zones bâties des sites des stations d’épuration. La simple augmentation de la surface des lagunes est de moins en moins souvent la solution. En particulier, la limitation de l’espace disponible amène les concepteurs à proposer des procédés qui permettent d’intensifier le traitement des eaux usées, en recourant par exemple à des aérations par des turbines flottantes, en faisant précéder le lagunage par des traitements biologiques intensifs (Copin et al., 2004) ou en restructurant les bassins existants (Archer et Donaldson, 2003). Bien qu’un véritable stéréotype de lagunage ait longtemps prévalu dans certains pays, en France notamment, on assiste à une diversification croissante de ce procédé pour l’adapter aux nouvelles contraintes auxquelles son développement est soumis. Les lagunages sont constitués soit par une seule série de lagunes ou par plusieurs séries en parallèle. Les séries les plus courantes dans le bassin méditerranéen comprennent une lagune facultative et deux

14
lagunes de maturation – combinaison typique de la France - ou une lagune anaérobie, une lagune facultative et un ou plusieurs bassins de maturation – solution répandue notamment en Israël. D’autre types de lagunes ont été construits dans la région, à l’échelle pilote ou à l’échelle opérationnelle : lagunes aérées, lagunes haut rendement, lagunes à macrophytes et réservoirs de stabilisation plus ou moins profonds (Copin et al., 2004). La station de lagunage de Mèze a été construite en 1980 pour protéger la qualité de l’eau de l’étang de Thau, en remplacement d’une unité d’épuration à boues activées. En effet, un épisode de « malaïgue » intervenu en 1975, dû à un excès de matières organiques et à un manque d’oxygène avait d’abord remis en cause le dimensionnement de la station existante puis le type même du procédé de traitement. Le lagunage a été choisi car il présente l’avantage de garantir une bonne qualité bactériologique des eaux. De surcroît, il permet de préserver l’alimentation en sels nutritifs nécessaire à la productivité de l’Etang. Il a été choisi pour équiper l’ensemble des communes du nord du bassin de Thau (Picot et Bondon, 2002). La première station de lagunage de Mèze a été conçue selon le modèle traditionnel français ; elle était composée de trois lagunes, L1, L2, L3 en série (Photographie1). Sa superficie était alors de 8 ha pour une capacité nominale de 8000 équivalents habitants, correspondant à une charge surfacique de 50 kg DBO ha-1j-1.
Photographie1. La station de lagunage de Mèze avant son extension
Durant les deux premières années consécutives à sa mise en eau, la station recevait environ 33 kgDBOha-1j-1, soit 66% de la charge nominale, sauf en automne quand des effluents vinicoles venaient s’ajouter aux eaux usées de la ville de Mèze. Cette sous-exploitation avait permis le raccordement des eaux usées de la commune de Loupian à la station de lagunage, raccordement réalisé en 1985. Mais en décembre 1990, suite à de forts évènements pluvieux, une présence de salmonelles fut détectée dans les huîtres de l’Etang. Plusieurs actions se déroulant sur plusieurs années ont

15
alors été menées dans le but de diminuer la pression exercée par les effluents de la station de Mèze-Loupian sur l’Etang. La première d’entre elles fut, en 1991, la déconnexion des eaux résiduaires de la Cave Coopérative Vinicole de Mèze qui s’était équipée d’une unité de traitement autonome par évaporation forcée. Puis vint le curage partiel de la lagune de tête, L1, à l’automne 1994 et, en 1996, la construction de 2 lagunes de maturation supplémentaires, F1 et F2, placées en série avec les 3 premières, afin d’augmenter le temps de séjour des eaux dans la station. De 1996 à 1998, dans le cadre du Contrat de Baie de l’Etang de Thau, des travaux de réaménagement et d’extension de la station de Mèze-Loupian ont été exécutés de manière à augmenter sa capacité de traitement et faire ainsi face à l’augmentation de la population raccordée. Les principales caractéristiques de ce réaménagement furent (Photographie2):
- la construction d’un étage anaérobie en tête du système, constitué des lagunes A1 et A2,
- la construction d’un deuxième étage constitué d’un ensemble de 4 lagunes facultatives à fortes charges avec alimentation étagée et recirculation (R1 à R4), (Figure1);
- la reconversion des 3 lagunes initiales, L1 à L3, en un étage tertiaire de maturation, - le réaménagement des lagunes F1 et F2 pour constituer un quatrième étage de finition
et permettre le stockage des apports de temps de pluie.
Photographie2. La nouvelle station de Mèze-Loupian
Cette nouvelle station totalise une surface de 14,4 ha et traite actuellement une charge de 1.120 kgDBOj-1 correspondant à 19000 EH.

16
A1 A2
R1
R2
R3
R4
L1
L2
L3
F1
F2
Entréeenlet
Circuit normalflow path
Sortieet
Autres circuits
Figure 1. Schéma de la nouvelle station de Mèze-Loupian
Dès sa mise en fonctionnement, le système de lagunage de Mèze-Loupian a été l’objet de suivis réguliers de l’évolution de la qualité de l’eau au fil des bassins successifs, avec une attention particulière portée au devenir des bactéries témoins de contamination fécale (Baleux et Trousselier, 1983, Legendre et al., 1984, Trousselier et al., 1986, Mezrioui, 1987). L’installation d’une unité pilote de lagunage haut rendement a donné lieu à une série de travaux consacrés aux performances de cette technique alors nouvelle en Europe (Bahlaoui, 1990, Picot et al., 1991, Moutin et al., 1992, El Halouani et al., 1993, Mesplé et al., 1995) et, en même temps, d’effectuer des comparaisons avec le lagunage traditionnel (Picot et al., 1992, Gómez et al., 1995). En dépit de résultats très prometteurs, ce procédé n’a pas été retenu pour la réhabilitation de la station. En revanche, la construction d’un étage de deux bassins anaérobies a permis d’y étudier l’accumulation des boues, l’activité méthanogène et la production de H2S (Paing et al., 2000 et 2003, Picot et al., 2001 et 2003). De même, on a recherché comment optimiser le fonctionnement de l’ensemble des 4 bassins R1 à R4 constituant le deuxième étage de la station (Sambuco et al., 2002). Ainsi, le nombre d’études qui y ont été réalisées au cours des 25 dernières années fait du lagunage de Mèze-Loupian un site d’un intérêt remarquable ; diverses techniques de lagunage y sont mises en œuvre pour satisfaire à l’augmentation de la population desservie et à des critères de rejet renforcés. Le principal objectif de cette étude, était d’évaluer les performances de l’installation dans sa configuration actuelle, en s’attachant à quantifier la contribution de chacun des étages au résultat global. Les critères physico-chimiques de l’épuration et les critères microbiologiques courants ont été considérés. Ce travail s’appuie plus particulièrement sur les suivis réalisés entre juillet 2003 et août 2004. On a tenté de relier l’abattement des bactéries témoins de contamination fécale aux conditions météorologiques et au temps de séjour de l’eau dans les bassins. Cette approche, même si elle peut être considérée comme prometteuse, se heurte à la complexité des mécanismes qui

17
interviennent ou interfèrent dans l’élimination des microorganismes dans les systèmes naturels. Enfin, le suivi de l’installation ayant fait apparaître des épisodes de nitrification précédant ou suivant la raréfaction estivale de l’azote dans les rejets; étant donné la controverse qui entoure encore les mécanismes de l’élimination de l’azote dans les lagunages, il est apparu utile d’éclairer, à travers l’exploitation de la littérature et des suivis expérimentaux réalisés à Mèze et au laboratoire, le devenir de l’azote en lagunage. Ce travail s’appuie sur, outre le suivi des années 2003 et 2004, un suivi centré sur les lagunes L1, L2 et L3 effectué d’avril à juillet 2005 et sur une modélisation du cycle de l’azote dans les lagunes de maturation. La première partie de ce rapport est consacrée à la présentation du site, aux méthodes employées dans les suivis expérimentaux et à la modélisation du fonctionnement hydraulique du lagunage. L’évaluation des performances des différents étages du lagunage dans l’épuration de la matière organique et l’élimination des nutriments fait l’objet de la deuxième partie. La troisième partie est consacrée à l’étude de la décontamination microbienne. Dans la quatrième partie, on développe l’étude de l’élimination de l’azote dans les lagunes de maturation.

18

19
Chapitre 1. MATERIELS ET METHODES

20

21
1.1. La station de lagunage de Mèze-Loupian La station rénovée dispose donc de 2 lagunes de tête A1 et A2 en parallèle, d’un étage facultatif à fortes charges (FFC) composé de 4 lagunes facultatives R1, R2, R3 et R4 en série avec alimentation étagée et recirculation, de 3 lagunes de maturation en série, L1, L2, L3, et de 2 lagunes de finition, F1 et F2 (Figure 1.1).
A1 A2
R1
R2
R3
R4
L1
L2
L3
F1
F2
Entréeenlet
Circuit normalflow path
Sortieet
Autres circuits
Figure 1.1. Schéma de la nouvelle station de Mèze-Loupian
Tableau 1.1. Caractéristiques des bassins de la station de lagunage de Mèze
Bassins Type de lagunes Surface (ha)
Profondeur (m)
Début de mise en route
Lagunes anaérobies
1998
A1, A2 Lagunes anaérobies aérées en surface
2 x 0.23 3.1 2001
R1, R2, R3, R4 Lagunes facultatives à forte charge avec recirculation (FFC)
4 x 0.67 1.8 1999
Lagunes facultatives 1980
L1, L2, L3 Lagunes de maturation
4 + 2 + 2 1.4 – 1.7 1999
F1, F2
Lagunes de finition 1.9 + 1.2 0.8 - 1.3 1996

22
Le débit moyen rentrant dans la station était en 2003 supérieur à 3100 m3j-1. Après un prétraitement, l’eau usée se répartit entre les 2 lagunes de tête, d’une surface unitaire de 0,23 ha et profondes de 3,1 m. En 2001, afin d’éviter les nuisances olfactives, elles ont été équipées d’aérateurs de surface (Photographie 1-1), la puissance installée étant égale à 4,4 Wm-3.
Photographie 1-1. Les aérateurs de surface d'un bassin anaérobie
L’étage facultatif est constitué de 4 lagunes en série avec possibilité de recirculation. Toutefois le débit d’entrée de cet étage, Q, n’est pas introduit en totalité dans la lagune de tête R1 ; au contraire, ce débit est réparti entre les bassins de l’étage. En effet, selon son mode d’alimentation, cet étage peut avoir un fonctionnement de type algal, bactérien ou mixte. Une équi-répartition de la charge organique reçue par chaque bassin, de R1 à R4, favorise un fonctionnement de type algal ; on y parvient en répartissant le débit entrant comme suit: 50% dans le bassin R1, 30% dans R2 et 20% dans R3 (Sambuco et al., 2002). C’est ce que l’on a appelé une distribution étagée. Le débit Q accepté dans l’ensemble de l’étage facultatif correspond à une fraction seulement, évaluée en moyenne à 2300 m3j-1, du débit issu des lagunes anaérobies ; la fraction restante est acheminée directement vers L1 (Figure 1.1). L’étage facultatif fonctionne ainsi sous une charge surfacique autour de 200 kgDBOha-1j-1. Chacun de ces bassins est doté d’un aérateur de 11 kW. L’optimisation du fonctionnement de l’étage est définie par un taux de recirculation de R4 vers R1 égal à 1 Q. Les lagunes de maturation sont constituées par les 3 anciennes lagunes facultatives, L1, L2 et L3, d’une surface totale de 8 ha, qui fonctionnent maintenant sous une charge surfacique sensiblement réduite. Les lagunes de finition, F1 et F2, forment un étage de sécurité qui est destiné à protéger un milieu récepteur très sensible. Leur volume permet, en particulier, de stocker les excédents de temps de pluie, d’éviter ainsi une trop grande diminution des temps de séjour dans le lagunage et, par conséquent, une détérioration excessive de la qualité microbiologique des rejets. Cet étage peut recevoir directement, notamment en période de forts débits, des effluents venant de l’étage facultatif ; en effet, une communication directe entre R4 et F1 est périodiquement mise en service.

23
Photographie 1-2. L'aérateur d'un bassin FFC
Toujours dans le but de réguler les temps de séjour en profitant des capacités de stockage du système de lagunage, l’exploitant joue sur le niveau de l’eau dans les bassins qu’il fait varier en modifiant la hauteur des déversoirs, au moyen de l’ajout ou du retranchement de réglettes en aluminium (Photographie 1-3).
Photographie 1-3. Déversoir équipé de réglettes en aluminium

24
1.2. Mesures hydrauliques, météorologiques et contrôles physico-chimiques in situ Avant 2000, année à partir de laquelle la station a été dotée de débitmètres électromagnétiques, peu de valeurs des débits d’entrée et de sortie de la station sont disponibles. Depuis l’année 2000, les débits d’entrée et de sortie sont enregistrés et restitués sous la forme d’un cumul journalier. Les hauteurs d’eau dans les bassins sont relevées toutes les semaines. La température de l’eau, le pH et la teneur en oxygène dissous sont mesurées au même pas de temps à la sortie de chacun des étages. Une station météorologique est installée sur le site (Photographie 1-4); elle dispose d’un pluviomètre à augets basculeurs et fournit la température de l’air, l’humidité, la vitesse du vent et le rayonnement solaire au pas de 5 minutes. Quand les données météorologiques de Mèze font défaut, celles relevées à Sète, ville située sur l’autre rive de l’étang de Thau, sont utilisées.
Photographie 1-4. La station météorologique implantée sur le site de la station de lagunage de Mèze
1.3. Echantillonnage et méthodes d’analyses De 1980 à 1992, le suivi physico-chimique a été effectué, à l’entrée et à la sortie de chaque bassin, tous les 15 jours dans un premier temps puis tous les mois. A partir de 2000, il a été réalisé à l’entrée et à la sortie de la nouvelle station tous les 15 jours. De juillet 2003 à août 2004, des prélèvements supplémentaires sont effectués à la sortie des anaérobies, des bassins FFC et de maturation. De 1980 à 1992, le suivi bactériologique à l’entrée et à la sortie de chaque bassin a été plus épisodique. Il consistait en des suivis bimensuels de juin 1980 à août 1982 (Trousselier et al.,1986) puis mensuels de novembre 1984 à juin 1986 (Mezrioui, 1987) et d’octobre 1988 à août 1990 (Picot et al., 1992). D’août 1994 à février 1995, avant, pendant et après le curage de la lagune L1, des suivis ont été faits à l’entrée et à la sortie de la station selon un pas de temps variant de 1 à 3 semaines (Crabos et al., 1996). A partir de 2000, les points de prélèvement ainsi que les fréquences de suivi sont les mêmes que pour les analyses physico-

25
chimiques. De juillet 2003 à août 2004, outre les effluents à l’entrée de chaque étage et à la sortie de la station, ceux à l’entrée des bassins L2, L3 et F2 sont aussi analysés. Les analyses sont effectuées sur des échantillons moyens de 24 heures collectés à l'aide de préleveurs automatiques (Photographie 1-5). Des méthodes standards sont utilisées pour mesurer les matières en suspension (MES), la demande chimique en oxygène (DCO), la DCO filtrée (Filtres Whatman GF/C), l’azote Kjeldhal (NTK), l’azote ammoniacal (N-NH4), les nitrates (N-NO3), les nitrites (N-NO2), le phosphore total (PT) et les orthophosphates (P-PO4). La demande biochimique en oxygène (DBO) et la DBO filtrée sont analysées à l’aide d’un respiromètre Oxytop WTW. A partir de l’année 2000, les teneurs en E.Coli et Entérocoques fécaux sont déterminées par la méthode des microplaques. A partir de 2000, les analyses ont été effectuées par le laboratoire de l'Ecosite de Mèze.
Photographie 1-5. Préleveur automatique
1.4. Devenir de l’azote dans les lagunes de maturation Une étude a été réalisée entre le 7 avril et le 12 juillet 2005 afin de clarifier les mécanismes de l’élimination de l’azote dans les lagunes de maturation. Un protocole d’échantillonnage spécifiquement centré sur cette problématique a été mis en œuvre. Pendant toute cette durée, une sonde a enregistré en continu la température de l’eau, le pH et la teneur en oxygène dissous à mi-profondeur près de la sortie du bassin L3. Du 4 avril au 12 juillet, on a suivi systématiquement – avec plusieurs analyses par semaine - les différentes formes de l’azote à la sortie des lagunes L2 et L3. A la sortie des lagunes L1 et R4, nitrites et nitrates ont été suivis à partir du 22 avril, l’azote ammoniacal à partir du 17 mai et l’azote Kjeldahl à partir du 17 mai à la sortie de L1 et du 7 juin à la sortie de R4. A la sortie des lagunes anaérobies, l’azote ammoniacal et Kjeldahl sont contrôlés dès le 7 juin et les nitrites à partir du 21 juin. Pendant toute la période, les mesures de routine réalisées par l’exploitant se sont poursuivies.

26
1.5. Modèle de fonctionnement hydraulique Seuls les débits d’entrée et de sortie de la station d’épuration font l’objet d’un contrôle continu. Or, pour mener à bien plusieurs des volets de cette étude, la connaissance des flux entre les différents bassins est indispensable. Ces volets sont les suivants : - l’évaluation des rendements et des contributions de chaque étage à l’épuration globale. En effet, l’évaluation des rendements à partir des seules concentrations, soit
entréeionConcentrat
sortieionConcentratentréeionConcentrat −
est très contestable dans la mesure où elle ne rend pas véritablement compte des quantités éliminées. Une estimation plus correcte des rendements prend en compte les flux de polluants, soit
Entrée
SortieEntrée
DébitionConcentrat
DébitionConcentratDébitionConcentrat
)(
)()(
∗∗−∗
,
d’où la nécessité de connaître les débits échangés entre les différents étages du système de lagunage. - la détermination des constantes cinétiques de décontamination microbienne nécessite, elles aussi, la connaissance des débits à l’entrée et à la sortie des différents bassins ; - ces cinétiques seront ensuite utilisées pour une modélisation des performances sanitaires du système de lagunage, ce qui, une nouvelle fois, exige la connaissance des débits.
Figure 1.2. Bilan hydrique d’une lagune
La connaissance de ces débits était, de la même manière, indispensable à la modélisation du devenir de l’azote dans les lagunes de maturation. Pour chaque lagune, le modèle hydraulique calcule, au pas de temps journalier, le débit de sortie, jiQ , , du jour j à partir du débit d’entrée, jiQ ,1− , du même jour et de l’équation de bilan
hydrique ci-dessous:
( ) ( ) ijijiijjjjijiji ShhSpEPPQQQ ,1,*,,1, −+−−++= −− (Équation 1.1)
où iS est la surface (m2) de la lagune i, jih , la hauteur d’eau (m) dans la lagune i le jour j, *, jiQ représente la somme des débits entrants dans la lagune i le jour j autres que celui
provenant du bassin ( )1−i , jP les précipitations (mm) et jp les pertes par infiltration (mm)
le jour j.
Précipitations P (m/j) (m) (m)
Evaporation EP (m/j)
Débit entrant Qi-1 (m3/j) Débit sortant Qi (m
3/j)
Pertes par infiltration p (m/j)
Lagune i hi

27
L'évaporation jEP (mm) est calculée à partir de la formule de Penmann, comme développée
dans l’Annexe 1. Entre juillet 2003 et août 2004, elle est évaluée en moyenne à 7 mm.j-1 l'été, 2mm.j-1 l'automne et l'hiver et 6 mm.j-1 le printemps. Le bilan hydrique global de la station (Equation 1.2) permet de faire une estimation des pertes par infiltration.
( ) ShSpEPPQQ jjjjjEjS ⋅∆+−−+= (Équation 1.2)
où
jSQ est le débit à la sortie de la station le jour j
jEQ le débit à l'entrée de la station le jour j
jh∆ le terme relatif à l'accumulation ou le déstockage d'effluents dans la station le jour j
Comme sur le long terme∑ →∆j
jh 0 , pour la période allant de juillet 2003 à août 2004,
l'équation 1.2 s'écrit :
( )SpEPPQQj
jjjj
jEj
jS ∑∑∑ −−+= (Équation 1.3)
Les débits à l'entrée et à la sortie ainsi que les précipitations étant mesurés, une fois déterminées les pertes par évaporation, l'équation 1.3 permet d'accéder aux pertes par infiltration. Il en résulte une valeur calculée moyenne négative de celles-ci, égale à - 0,4mm.j-
1. En conséquence, dans les applications qui suivent, les pertes par infiltration sont considérées négligeables par rapport à l’évaporation. Cette valeur négative paraît être due à l’imprécision des mesures des débits ; elle est raisonnable dans la mesure où elle correspond à une erreur relative de 1.7% par rapport au débit d’entrée. Les valeurs Si sont des données, celles de P et d’EP sont déduites des observations. Aussi, si seules la pluie et l’évaporation expliquaient les écarts entre les débits d’entrée et de sortie de chaque lagune, les débits des bassins successifs seraient déduits du débit d’entrée par un calcul trivial. Tel n’est pas le cas pour deux raisons. D’une part, les niveaux de l’eau dans les bassins, hi, ne sont relevés que tous les 15 jours environ et avec une imprécision sensible dès lors que les plans d’eau sont agités. Entre deux relevés, il n’est pas rare que le niveau des déversoirs ait été modifié, entraînant d’importantes rétentions d’eau ou mouvements d’eau d’un bassin à l’autre. D’autre part, à côté du cheminement normal de l’eau d’un bassin à l’autre, d’autres circuits sont mis en jeu. Certains sont bien connus, comme l’alimentation de la lagune L1 par une partie des effluents des lagunes anaérobies ; pour ce cas, une formulation théorique
13204592141 , −⋅=→ AARR LnQQ (Équation 1.5)
41211 ,*
RRAAL QQQ →−= (Équation 1.6)
dans laquelle les débits de sortie de l’étage anaérobie, 21,AAQ et d’alimentation de l’étage
facultatif, 41 RRQ → , sont exprimés en m3j-1, permet d’évaluer le débit correspondant, sans
toutefois que sa validité ait été précisément examinée. L’existence d’une possibilité d’apport direct de la lagune R4 à la lagune F1 est aussi bien connue ; ce circuit peut être mis en route en période de fortes précipitations ; toutefois, l’expérience a montré que l’importance de ce court-circuit est quelquefois très sous estimée. Dans la mesure où cette installation a fait
28
l’objet de plusieurs restructurations, il n’est pas impossible que des circuits censés avoir été supprimés ou obturés soient encore le lieu de faibles circulations. Des incertitudes pèsent donc sur les valeurs de ih et de *
iQ . Le modèle sert donc à les reconstituer. Le modèle calcule successivement les débitsjiQ , , depuis l’étage anaérobie jusqu’à la sortie du
lagunage,2FQ . Les valeurs de jih , , les jours où ces hauteurs ne sont pas mesurées, et celles de
*1LQ et de *
1FQ ont été ajustées manuellement de manière à reproduire au mieux les valeurs
observées du débit2FQ . Le nombre relativement élevé de grandeurs à ajuster et l’absence d’un
protocole clairement formalisé de gestion des niveaux d’eau ont été d’incontestables sources d’incertitude. La détermination de *
1FQ , qui représente le passage direct d’une partie de l’eau
de la lagune R4 dans la lagune F1, s’est révélée particulièrement délicate. Ce débit a été mis en évidence pendant la période d’octobre 2003 à fin avril 2004 par des incohérences apparues dans les résultats des contrôles microbiologiques, dont une sous estimation de *
1FQ s’est
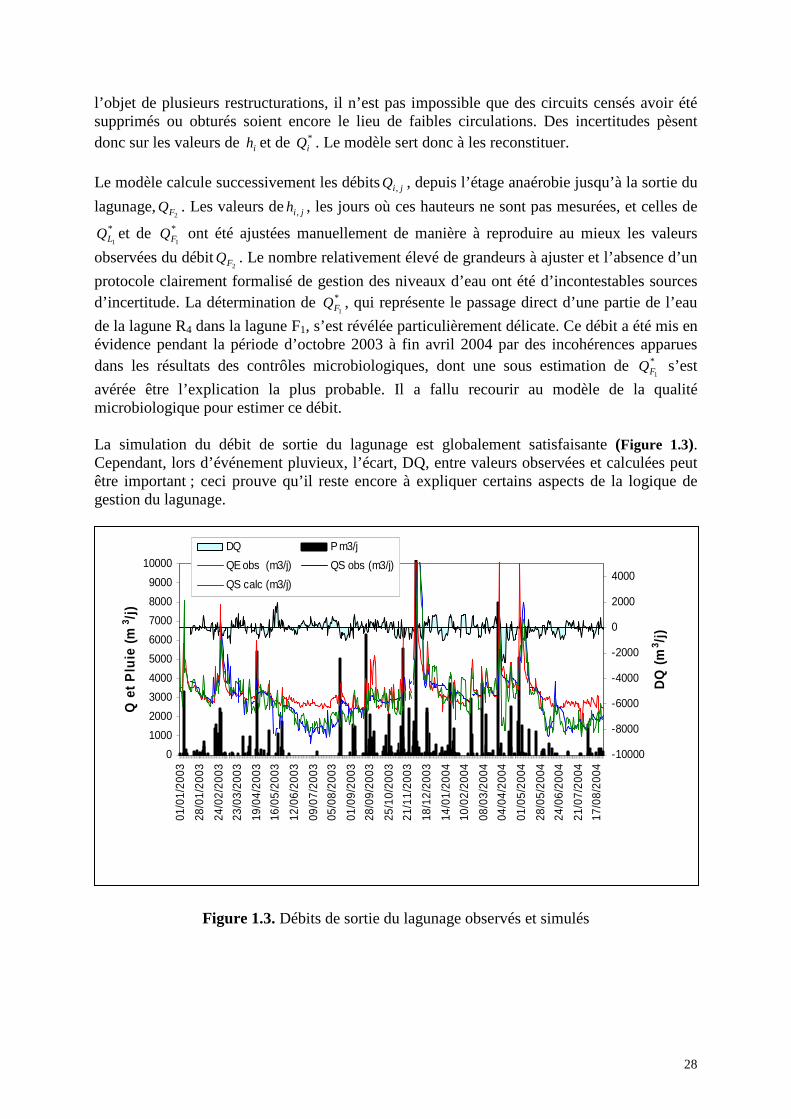
avérée être l’explication la plus probable. Il a fallu recourir au modèle de la qualité microbiologique pour estimer ce débit. La simulation du débit de sortie du lagunage est globalement satisfaisante (Figure 1.3). Cependant, lors d’événement pluvieux, l’écart, DQ, entre valeurs observées et calculées peut être important ; ceci prouve qu’il reste encore à expliquer certains aspects de la logique de gestion du lagunage.
Figure 1.3. Débits de sortie du lagunage observés et simulés
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
01/0
1/20
03
28/0
1/20
03
24/0
2/20
03
23/0
3/20
03
19/0
4/20
03
16/0
5/20
03
12/0
6/20
03
09/0
7/20
03
05/0
8/20
03
01/0
9/20
03
28/0
9/20
03
25/1
0/20
03
21/1
1/20
03
18/1
2/20
03
14/0
1/20
04
10/0
2/20
04
08/0
3/20
04
04/0
4/20
04
01/0
5/20
04
28/0
5/20
04
24/0
6/20
04
21/0
7/20
04
17/0
8/20
04
Q e
t Plu
ie (
m3 /j)
-10000
-8000
-6000
-4000
-2000
0
2000
4000
DQ
(m
3 /j)
DQ P m3/j
QE obs (m3/j) QS obs (m3/j)
QS calc (m3/j)

29

30

31
Chapitre 2. PERFORMANCES DU SYSTEME DE LAGUNAGE AVANT ET APRES SON AGRANDISSEMENT

32

33
Grâce aux suivis effectués á différentes périodes depuis la construction du lagunage, il a été possible d’évaluer les effets de la charge appliquée et ceux de la restructuration du lagunage sur ses performances aussi bien physico-chimiques que bactériologiques. 2.1. Comparaison des performances en début de fonctionnement, avant et après l’agrandissement Les comparaisons faisant intervenir les premières années de fonctionnement sont à prendre avec précaution car les suivis de 1981-1982 présentent plusieurs particularités qui les distinguent des travaux ultérieurs. L’échantillonnage à l’entrée du lagunage a été réalisé à l’intérieur du premier bassin á proximité de l’arrivée des effluents ; les valeurs des teneurs en MES et de la DCO à l’entrée sont des moyennes de 3 bilans de 24h ; les teneurs en sels nutritifs ont été déterminées par la méthode HACH. Durant les 2 premières années, la station recevait une charge surfacique relativement faible, égale en moyenne à 33 kg DBOha-1j-1. Dans ces conditions, l’installation a montré de bonnes performances physico-chimiques : 73% de la DCO, 66% de la DBO et 55% de N-NH4 étaient éliminés, les teneurs respectives à la sortie étaient elles mêmes assez faibles Tableau 2.1. L’élimination du phosphore était plus modeste, avec une teneur en sortie de 10 mgP-PO4.L
-1. L’abattement des bactéries témoins de contamination fécale était seulement de 2,9 unités logarithmiques (u.log) pour les coliformes fécaux et 2,6 u.log pour les entérocoques. Par ailleurs, les performances bactériologiques variaient fortement avec les saisons, avec des écarts des concentrations de sortie des coliformes fécaux allant jusqu’à 3 ou 4 u.log entre l’été et l’hiver (Figure 2.1). Cette variation saisonnière, toujours aussi accentuée dans les années 1984-1986, parait quelquefois moins nette pour ce qui concerne les entérocoques. Il faut aussi noter que les seuils de détection, le plus souvent fixés à 100 UFC/100 mL, conduisent à minimiser les abattements en période estivale. Il reste que, si la réglementation actuelle fixant les teneurs limites en E. Coli et entérocoques à 1000 UFC/100mL en été et à 10.000 UFC/100mL en hiver avait été appliquée, la station n’aurait pas été conforme en hiver. A cette époque là, la station de lagunage constituait une protection efficace de la qualité des eaux de baignade ; elle ne protégeait pas aussi bien les installations conchylicoles, sachant que le pic de la commercialisation des coquillages coïncide avec les fêtes de fin d’année. Après 9 ans de fonctionnement, l’augmentation de la population a entraîné celle de la charge organique (65 kgDBOha-1j-1) qui a doublé. Débits et concentrations d’entrée ont augmenté, à l’exception des phosphates. La diminution de l’usage du phosphore dans les lessives commençait à se faire sentir. Ce doublement de la charge organique s’est traduit par une nette diminution des rendements d’élimination, notamment pour l’azote ammoniacal (22% et la DCO brute (60%). Azote et phosphore étaient mal éliminés et, comme en témoignent les écarts respectifs en sortie du lagunage entre DCO et DCO filtrée, NTK et N-NH4 et Phosphore total et P-PO4, une forte concentration algale était évacuée vers le milieu récepteur. Cette dégradation des performances a affecté aussi l’élimination des coliformes fécaux, dont l’abattement n’était plus que de 2.3 u.log ; la décontamination était sensiblement moins efficace en été comme en hiver (Figure 2.1). L’effet de l’accroissement de la charge a vraisemblablement été amplifié par l’accumulation de boues dans le premier bassin, accumulation qui a réduit le temps de séjour hydraulique. Le curage en 1994 de la partie envasée du bassin L1 n’a fait que reculer l’échéance de la restructuration et de l’accroissement de la capacité de traitement de l’installation ; les travaux correspondants se sont échelonnés entre 1996 et 1998.
34
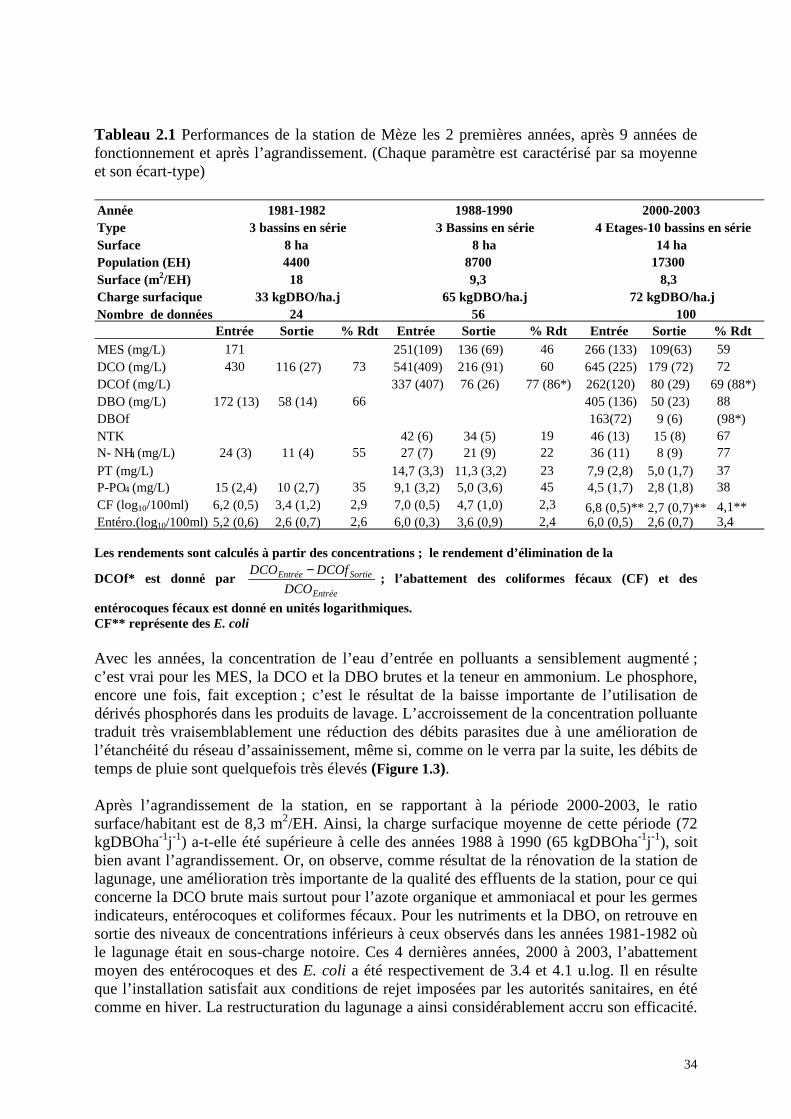
Tableau 2.1 Performances de la station de Mèze les 2 premières années, après 9 années de fonctionnement et après l’agrandissement. (Chaque paramètre est caractérisé par sa moyenne et son écart-type)
Les rendements sont calculés à partir des concentrations ; le rendement d’élimination de la
DCOf* est donné par Entrée
SortieEntrée
DCO
DCOfDCO − ; l’abattement des coliformes fécaux (CF) et des
entérocoques fécaux est donné en unités logarithmiques. CF** représente des E. coli Avec les années, la concentration de l’eau d’entrée en polluants a sensiblement augmenté ; c’est vrai pour les MES, la DCO et la DBO brutes et la teneur en ammonium. Le phosphore, encore une fois, fait exception ; c’est le résultat de la baisse importante de l’utilisation de dérivés phosphorés dans les produits de lavage. L’accroissement de la concentration polluante traduit très vraisemblablement une réduction des débits parasites due à une amélioration de l’étanchéité du réseau d’assainissement, même si, comme on le verra par la suite, les débits de temps de pluie sont quelquefois très élevés (Figure 1.3). Après l’agrandissement de la station, en se rapportant à la période 2000-2003, le ratio surface/habitant est de 8,3 m2/EH. Ainsi, la charge surfacique moyenne de cette période (72 kgDBOha-1j-1) a-t-elle été supérieure à celle des années 1988 à 1990 (65 kgDBOha-1j-1), soit bien avant l’agrandissement. Or, on observe, comme résultat de la rénovation de la station de lagunage, une amélioration très importante de la qualité des effluents de la station, pour ce qui concerne la DCO brute mais surtout pour l’azote organique et ammoniacal et pour les germes indicateurs, entérocoques et coliformes fécaux. Pour les nutriments et la DBO, on retrouve en sortie des niveaux de concentrations inférieurs à ceux observés dans les années 1981-1982 où le lagunage était en sous-charge notoire. Ces 4 dernières années, 2000 à 2003, l’abattement moyen des entérocoques et des E. coli a été respectivement de 3.4 et 4.1 u.log. Il en résulte que l’installation satisfait aux conditions de rejet imposées par les autorités sanitaires, en été comme en hiver. La restructuration du lagunage a ainsi considérablement accru son efficacité.
Année Type Surface Population (EH) 4400 8700 17300 Surface (m2/EH) 18 9,3 8,3Charge surfacique Nombre de données 24 56 100
Entrée Sortie % Rdt Entrée Sortie % Rdt Entrée Sortie % Rdt MES (mg/L) 171 251(109) 136 (69) 46 266 (133) 109(63) 59DCO (mg/L) 430 116 (27) 73 541(409) 216 (91) 60 645 (225) 179 (72) 72DCOf (mg/L) 337 (407) 76 (26) 77 (86*) 262(120) 80 (29) 69 (88*) DBO (mg/L) 172 (13) 58 (14) 66 405 (136) 50 (23) 88DBOf 163(72) 9 (6) (98*) NTK 42 (6) 34 (5) 19 46 (13) 15 (8) 67N- NH 4 (mg/L) 24 (3) 11 (4) 55 27 (7) 21 (9) 22 36 (11) 8 (9) 77PT (mg/L) 14,7 (3,3) 11,3 (3,2) 23 7,9 (2,8) 5,0 (1,7) 37P-PO 4 (mg/L) 15 (2,4) 10 (2,7) 35 9,1 (3,2) 5,0 (3,6) 45 4,5 (1,7) 2,8 (1,8) 38CF (log10/100ml) 6,2 (0,5) 3,4 (1,2) 2,9 7,0 (0,5) 4,7 (1,0) 2,3 6,8 (0,5)** 2,7 (0,7)** 4,1** Entéro.(log10/100ml) 5,2 (0,6) 2,6 (0,7) 2,6 6,0 (0,3) 3,6 (0,9) 2,4 6,0 (0,5) 2,6 (0,7) 3,4
1988-19903 Bassins en série
8 ha
65 kgDBO/ha.j
2000-20034 Etages-10 bassins en série
14 ha
72 kgDBO/ha.j
1981-1982
33 kgDBO/ha.j
3 bassins en série 8 ha

35
Cette amélioration doit beaucoup à la combinaison de plusieurs dispositions : la multiplication des bassins, l’apport d’énergie électrique pour aérer les deux premiers étages et la diversification des types de lagunage (bassins anaérobies, bassins facultatifs à forte charge et recirculation, bassins de maturation et de finition}, comme aux possibilités offertes par l’installation rénovée pour amortir les effets des évènements pluvieux. Une analyse des données recueillies au cours des années 2000-2004 et un examen détaillé, étage par étage, permettent de mettre en évidence les contributions respectives de ces dispositions.
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
Juin
Juill
et
Aoû
t
Sep
tem
bre
Oct
obre
Nov
embr
e
Déc
embr
e
Janv
ier
Fév
rier
Mar
s
Avr
il
Mai
Juin
Juill
et
Aoû
t
Sep
tem
bre
Oct
obre
Nov
embr
e
Déc
embr
e
Janv
ier
Fév
rier
Mar
s
Avr
il
Mai
Juin
Juill
et
Aoû
t
Col
iform
es fé
caux
(ou
E.c
oli)/
100
mL
1980-1982
1984-1986
1988-1990
2002-2004
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
Juin
Juill
et
Aoû
t
Sep
tem
bre
Oct
obre
Nov
embr
e
Déc
embr
e
Janv
ier
Fév
rier
Mar
s
Avr
il
Mai
Juin
Juill
et
Aoû
t
Sep
tem
bre
Oct
obre
Nov
embr
e
Déc
embr
e
Janv
ier
Fév
rier
Mar
s
Avr
il
Mai
Juin
Juill
et
Aoû
t
Ent
éroc
oque
s/10
0 m
L
1980-1982
1988-1990
2002-2004
Figure 2.1. Evolution de la décontamination microbienne
2.2. Les performances globales de la station rénovée De 2000 à 2003, période pendant laquelle la population raccordée est passée de 14 000 à 18.800 habitants, le débit journalier moyen reçu par la station a augmenté de 1875 à 3127 m3j-1. L’augmentation de la population raccordée est insuffisante à expliquer cette variation. Les précipitations plus importantes sur les bassins, passant en moyenne de 126 m3j-1 en 2000 à 254 m3j-1 en 2003, participent à l’augmentation des débits mais ne sont pas comptabilisées dans les débits entrés. L’explication est plutôt à rechercher du côté d’un accroissement des débits parasites acceptés par la station par temps de pluie (Figures 1.3 et 2.2). Pendant les périodes de forte pluie, les débits à l’entrée peuvent dépasser 10 000 m3j-1.

36
Figure 2.2. Evolution des teneurs en MES, DCO, DBO, NTK, N-NH4, PT et phosphate des eaux à l’entrée et à la sortie de la station de 2000 à 2004
0
2
4
6
8
10
12
14
16
mar
s-00
juil.
-00
nov.
-00
mar
s-01
juil.
-01
nov.
-01
mar
s-02
juil.
-02
nov.
-02
mar
s-03
juil.
-03
nov.
-03
mar
s-04
juil.
-04
PT mg/l Entrée
PT mg/l Sort ie
0
50
100
150 NTK mg/l Entrée
NTK mg/l Sortie
N-NH4 mg/l Entrée
N-NH4 mg/l Sortie
0
200
400
600
800
1000 DBO mg/l Entrée
DBO mg/l Sortie
0
400
800
1200
1600 DCO mg/l Entrée
DCO mg/l Sortie
DCO mg/l Effluent filtré
0
200
400
600
800
1000 MES mg/l Entrée
MES mg/l Sortie
0
2000
4000
6000
8000
10000
12000 0
20
40
60
80
100
120
Pluie (mm/j)
Entrée (m3/j)
Sortie (m3/j)

37
Hors des périodes estivales, les débits entrants et sortants restent globalement proches, avec au moment des pluies un amortissement des pics des débits d’entrée. L’été, au contraire, l’évaporation peut induire des écarts très importants puisque le débit sortant peut être réduit de moitié (Figure 1.3). D’où l’importance d’évaluer les performances du système de traitement à partir des flux de polluants. On distingue des évolutions saisonnières plus ou moins bien marquées des paramètres qui caractérisent la qualité des influents et des effluents (Figure 2.2). Les pics de la DCO et de la DBO des influents enregistrés en automne proviennent d’effluents de caves vinicoles privées fortement chargés en DCO soluble. Ils ne se traduisent pas par une détérioration de la qualité des effluents de la station. Pas plus dans les mois qui suivent que dans le reste de l’année, la DCO filtrée et la DBO des effluents du lagunage n’apparaissent véritablement dépendre de la saison. Cela est vraisemblablement dû grâce à un temps de séjour assez long (95 j), mais aussi à l’aération artificielle des premiers étages. En été, une importante production d’algues fait augmenter la teneur en MES, en DCO et DBO particulaires ; de plus, la forte évaporation contribue à l’accroissement de la DCO et de la DBO brutes. Les augmentations des teneurs en NTK et en ammonium à l’entrée apparaissent liées à la saison touristique estivale ; par contre, à la même saison, les teneurs en phosphate et surtout en ammonium diminuent dans les effluents ; plusieurs mois par an, l’élimination de l’azote est quasi totale, à l’exception de l’azote particulaire exporté avec les algues. Contrairement à ce qui a été observé dans les périodes antérieures à l’agrandissement, le caractère saisonnier de la décontamination microbienne n’est pas particulièrement évident. Si on observe encore une nette augmentation des teneurs en bactéries témoins de contamination fécale pendant l’hiver 2002-2003, par la suite, les évolutions de ces mêmes teneurs n’ont plus rien de saisonnier. En effet, la décontamination s’est beaucoup améliorée en hiver (Figure 2.1). Les raisons en sont pour une bonne part la multiplication des bassins. Une analyse plus précise est rendue malaisée par la gestion des débits de temps de pluie, avec des actions de stockage ou de déstockage d’eau pouvant être appliquées tout au long de la filière et particulièrement dans les deux derniers étages en vue de minimiser l’impact des évènements pluvieux, et aussi par les courts-circuits hydrauliques plus ou moins maîtrisés. 2.3. Rôle et performances de chaque étage de traitement Tableau 2.2. Concentrations moyennes annuelles (écart-type) à chaque étage de traitement ; rendements globaux (en concentrations et en flux) et abattements globaux pour les témoins de contamination fécale Entrée Bassins anaérobies FFC
FP Bassins de maturation Sortie
% concentration % flux MES (mg/l) 256 (85) 158 (99) 81 (47) 54 (39) 70 (62) 73 79 DCO (mg/lL) 557 (208) 369 (169) 183 (72) 107 (46) 117 (66) 79 81 DCOf (mg/l) 237 (94) 161 (70) 86 (35) 51 (17) 55 (20) 77 77 DBO (mg/l) 347 (133) 187 (82) 63 (22) 32 (26) 26 (24) 92 94 NTK (mg/l) 51 (16) 43 (10) 39 (7) 20 (10) 16 (9) 69 68 N-NH4 (mg/l) 36 (9) 37 (7) 31 (5) 15 (13) 13 (11) 68 73 PT (mg/l) 7.1 (1.7) 6.2 (1.5) 5.6 (1.2) 5.3 (0.7) 4.3 (1.2) 40 39 P-PO4 (mg/l) 4.3 (1.4) 3.5 (1.0) 3.0 (1.8) 4.0 (1.5) 3.0 (1.5) 33 36 E. Coli (log/100ml) 6.8 (0.4) 6.4 (0.6) 5.1 (0.8) 2.5 (0.7) 2.2 (0.7) 4.6 4.7 Entero.(log/100ml) 6.3 (0.4) 6.0 (0.3) 4.5 (0.9) 2.4 (0.8) 2.1 (0.6) 4.1 4.2
Rendement global

38
La détermination du rôle et des performances de chaque étage de traitement se fait à partir du suivi de 2003–2004. La station élimine globalement plus de 90% de la DBO, plus de 75% de la DCO et des MES, autour de 65% des composés azotés, autour de 40% des composés phosphorés. En matière de décontamination, l’abattement est supérieur à 4 unités logarithmiques (Tableau 2.2). 2.3.1. Premier étage : les lagunes anaérobies Depuis 2001, les deux bassins anaérobies sont équipés d’aérateurs de surface développant une puissance de 4,4 Wm-3.dans le but de résoudre les problèmes de nuisances olfactives. Durant la période 2003-2004, ces bassins fonctionnent sous une charge volumique moyenne de 105 g DBO m-3.j-1 et un temps de séjour moyen de 3,5 jours. Cet étage est le plus performant de la station en matière de réduction des MES (39%), de la DCO (34%) et de la DBO (47%) (Figure 2.3). C’est en automne que sa contribution à l’élimination de la matière organique est la plus importante (Figure 2.7).
39 3447
16 12
22 24
26
6 7
21 23
19
36
6
-3 -1
2
7
14
21 206
35
61
-20%
0%
20%
40%
60%
80%
100%
MES DCO DBO NT PT
Effluent
F12
L123
FFC
A12
Figure 2.3. Contribution de chaque étage au rendement global calculé avec les flux
Comparés aux résultats obtenus de 1998 à 2000 (élimination de 55% des MES et 30% de la DBO), période pendant laquelle l’étage fonctionnait sans aérateur et recevait une charge volumique de 86 g DBO m-3.j-1 avec un temps de séjour de 4,6 jours (Paing et al., 2003), on peut conclure que la mise en service des aérateurs a entraîné une diminution de l’abattement des MES et a amélioré par contre l’élimination de la DBO avec une consommation d’énergie inférieure à 0,5 kWh par kg de DBO éliminée. Cet étage a une efficacité non négligeable concernant l’élimination des nutriments (16% N et 12% P). Il contribue faiblement à la désinfection en abattant seulement de 0,42 u.log les E. Coli et de 0,45 u.log les Entérocoques. L’utilisation de bassins anaérobies aérés en surface en début de filière pour remédier aux problèmes générés par une surcharge de cet étage semble être une excellente solution, notamment quand on cherche à réduire l’emprise au sol des stations d’épuration.

39
2.3.2. Deuxième étage : L’étage facultatif à forte charge (FFC)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
A1
2
FF
C
L1
L2
L3 F1
F2
Réd
uctio
n (U
nité
s lo
g)
E.coli
Entérocoque
Figure 2.4. Contribution de chaque étage à l’abattement global des E. Coli et Entérocoques
calculée à partir des flux Le temps de séjour moyen dans cet étage est de 29,5 jours. Les bassins FFC éliminent 22% des MES entrant dans la station, 24% de la DCO, 33% de la DBO dissoute et 26% de la DBO reçues par la station. Les FFC sont très performants car ces chiffres représentent 50% d’abattement des MES, 50% de la DCO, 78% de la DBO dissoute et 66% de la DBO brute reçues en entrée de l’étage. A la sortie de cet étage l’effluent est en conformité avec la directive européenne de 1991 vis à vis de la DCO et de la DBO. La contribution de cet étage est par contre minime concernant l’élimination des nutriments (6% de N et 7% de P). Pour la décontamination microbienne, son rôle est important puisqu’il permet une réduction de 1,3 unités logarithmiques des E. Coli et 1,4 u.log des entérocoques Figure 2.4. L’effet saisonnier sur l’élimination des deux germes est peu marqué, avec toutefois des abattements un peu plus élevés en période estivale (Tableaux 2.3 et 2.4). Il faut remarquer que la recirculation, en injectant de la biomasse algale active dans le premier bassin, y contribue au maintien des conditions aérobies et accroît son efficacité et sa capacité ainsi que celle de l’ensemble de l’étage. Ainsi les FFC sont très performants malgré une charge surfacique appliquée (166 kgDBOha-1j-1) double de celle généralement en usage dans les lagunes facultatives non aérées. Il n’est alors pas étonnant si pendant un an (2001), cet étage a pu rester très efficace alors que les bassins anaérobies de tête n’étaient pas en fonctionnement. L’inconvénient de ce schéma est l’accumulation plus rapide des boues dans le premier bassin. Pour s’assurer du maintien des conditions aérobies surtout la nuit, l’étage est équipé de quatre aérateurs. La consommation d’énergie correspondante est de 0,5 kWh par kg de DBO éliminée. Dans une station de lagunage, l’étage FFC peut remplacer un bassin facultatif et produire un effluent de bonne qualité avec une occupation du sol et, sans doute, un coût moindres.

40
2.3.3. Troisième étage : les bassins de maturation Les bassins de maturation sont caractérisés par une charge surfacique de 38 kg DBOha-1j-1 et un temps de séjour moyen de 48 jours. Cet étage a une performance comparable à celle de l’étage FFC en ce qui concerne les MES (21% de l’abattement total) et la DCO (23% de l’abattement total). Il est moins efficace pour éliminer la DBO (19% de l’abattement total) mais celle-ci a déjà été considérablement réduite dans les étages précédents. Par rapport aux charges reçues en entrée de L1, l’étage de maturation élimine 56% des MES, 57% de la DCO et 72% de la DBO.
0
5
10
15
20
25
30
35
40
09/0
7/03
07/0
8/03
07/0
9/03
06/1
0/03
04/1
1/03
03/1
2/03
09/0
1/04
07/0
2/04
08/0
3/04
06/0
4/04
05/0
5/04
03/0
6/04
14/0
7/04
09/0
8/04
Tem
pér
atur
e, N
H4
n
0
1
2
3
4
5
6
7
8
9
NO
2 , N
O 3
T (°C)
N-NH4 (mg/L)
N-NO2 (mg/L)
N-NO3 (mg/L)
Figure 2.5. Evolution en fonction de la température des teneurs en N-NH4, N-NO2 et N-NO3
dans le bassin de maturation L3
Il faut noter que concernant les composés azotés, les effluents de L3 se singularisent par d’assez fortes concentrations en azote nitrique en octobre 2003 (5 mgL-1) et en juin 2004 (8 mgL-l) (Figure 2.5). Une nitrification suivie d’une dénitrification comme nous le montrerons dans le dernier chapitre pourrait expliquer la faible concentration en azote ammoniacal en été (0,7 mgL-1 en moyenne) des effluents de L3. En octobre, quand la température baisse, la diminution en intensité de la dénitrification entraîne l’augmentation de la concentration en nitrate. En novembre, la température de l’eau étant inférieure à 13°C, la nitrification ne pouvant plus avoir lieu, la teneur en N-NO3 diminue tandis que celle de N-NH4 augmente. La concentration en N-NO3 est inférieure à 0.5 mgL-1 en hiver et au printemps. Elle augmente de nouveau en juin quand la température atteint 24°C. Cet étage tient le rôle le plus important dans l’élimination de NTK avec un rendement de 38%. Cette élimination est fortement influencée par les saisons (60% en été et 20% en hiver) (Figures 2.8 et 2.9). On reviendra plus en détails sur l’élimination de l’azote dans le troisième chapitre de ce rapport.

4
1
0 50
100
150
200
250
300
350
400
450
500
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
MES (mg/l)
Entrée
Sortie A
12
Sortie FFC
Sortie L3
Sortie
0
200
400
600
800
1000
1200
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
DCO (mg/l)
Entrée
Sortie A
12
Sortie FFC
Sortie L3
Sortie
0
100
200
300
400
500
600
700
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
DBO (mg/l)
Entrée
Sortie A
12
Sortie FFC
Sortie L3
Sortie
F
igure 2.6. Contribution de chaqu
e étage de traite
ment à l’élim
ination des ME
S, de la D
CO
et de la D
BO
42
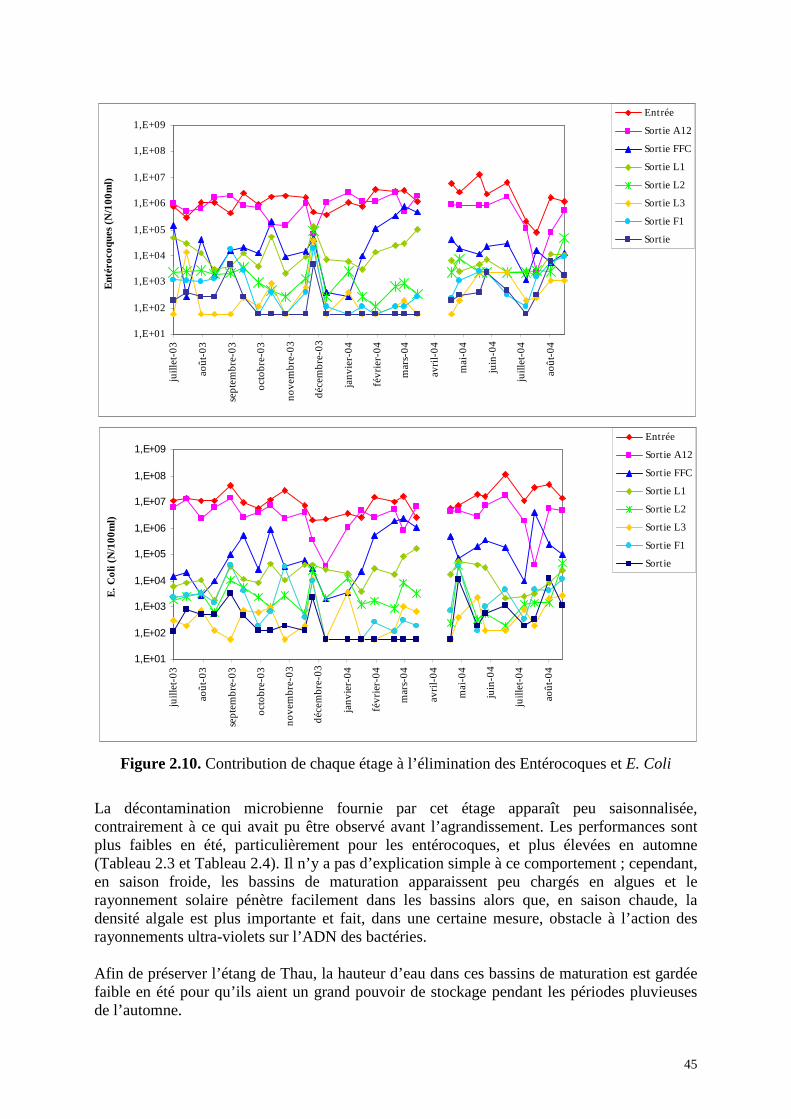
L’étage de maturation joue un rôle essentiel dans la décontamination microbienne puisque, à lui seul, il fournit un abattement (exprimé en unités logarithmiques) des bactéries témoins de contamination fécale équivalent à celui de tous les autres étages réunis. D’après la Figure 2.10 dans laquelle sont reportées les concentrations mesurées à la sortie des différents bassins, le premier bassin de maturation, L1, semble contribuer faiblement à la décontamination microbienne. Ceci est contredit par la Figure 2.4 qui indique que L1 est plus performant que l’étage FFC dans la mesure où il réduit de 1,49 u.log les flux d’E. Coli et de 1,43 u.log les flux d’entérocoques. Cette contradiction s’explique par le fait que rentrent dans L1 les effluents de R4, autrement dit de l’étage FFC, et aussi des effluents de l’étage anaérobie, ces derniers étant beaucoup plus chargés en germes de contamination fécale, ce qui est pris en compte dans le calcul des abattements.
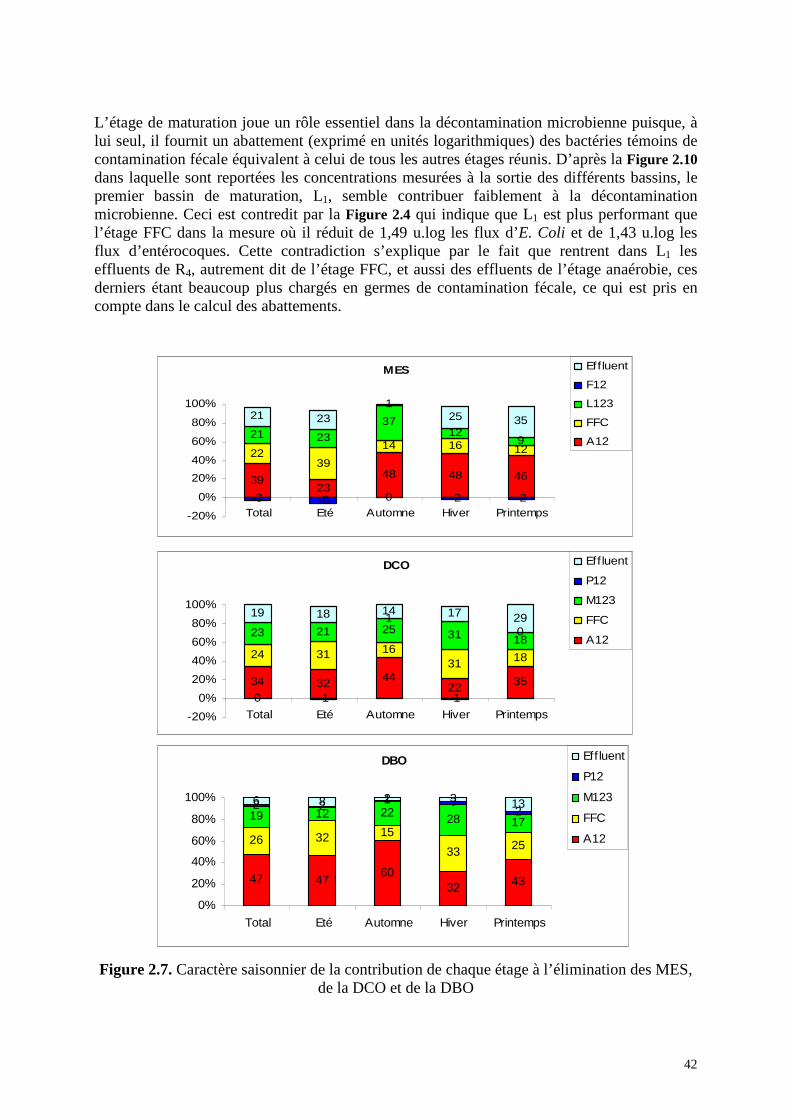
Figure 2.7. Caractère saisonnier de la contribution de chaque étage à l’élimination des MES, de la DCO et de la DBO
MES
3923
48 48 46
2239
14 16 1221 23
3712
9
-3 -8 0 -2 -2
21 231
25 35
-20%
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Effluent
F12
L123
FFC
A12
DCO
34 32 4422 35
24 31 1631 18
23 21 25 31 18
0 -1
1
-1
0
19 18 14 17 29
-20%
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Eff luent
P12
M123
FFC
A12
DBO
47 4760
32 43
26 32 15
33 25
19 12 22 28 17
2 2 1 32
6 8 2 3 13
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Eff luent
P12
M123
FFC
A12

4
3
0 20
40
60
80
10
0
12
0
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
NTK (mg/l)
En
trée
So
rtie A
12
So
rtie F
FC
So
rtie L
3
So
rtie
0 10 20 30 40 50 60 70 80
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
N-NH4 (mg/l)
Entrée
Sortie A
12
Sortie FFC
Sortie L3
Sortie
0 2 4 6 8 10 12
juil.-03
août-03
sept.-03
oct.-03
nov.-03
déc.-03
janv.-04
févr.-04
mars-04
avr.-04
mai-04
juin-04
juil.-04
août-04
PT (mg/l)
Entrée
Sortie A
12
Sortie FFC
Sortie L3
Sortie
Figure 2.8. C
ontribution de chaque éta
ge de traitem
ent à l’élimination des nutrim
ents

44
NTK
16 9 20 9 236 14
-23
838
6049
2010
8
610
69
3211
23
62 50
-20%
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Effluent
F12
L123
FFC
A12
PT
12 13 16 11 87 9
-29 13
619
6 6
-11
15
22
22 92
6137
58 6688
-20%
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Effluent
F12
L123
FFC
A12
NH4
12 16 214 -3
38
-4 -315
41
72
48
14 7
-3
4
11
10 6
47
224
75 76
-20%
0%
20%
40%
60%
80%
100%
Total Eté Automne Hiver Printemps
Effluent
F12
L123
FFC
A12
Figure 2.9. Contribution par saison de chaque étage à l’élimination des nutriments
Le bassin L2, qui comme L3, est deux fois moins grand que L1, a un rôle non négligeable dans l’élimination de E. Coli. Il est moins efficace par rapport aux entérocoques. Des 3 bassins de maturations, L3 est apparemment le moins performant des bassins de maturation mais son action est suffisante pour que les teneurs en E. Coli et entérocoques dans son eau de sortie soient inférieures à 1000 cfu/100ml avec, même, des valeurs inférieures à la limite de détection, ce qui, par ailleurs, induit à sous-estimer l’efficacité de ce bassin (Figure 2.10).
45
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
1,E+09
juill
et-
03
ao
ût-
03
sep
tem
bre
-03
oct
ob
re-0
3
no
vem
bre
-03
dé
cem
bre
-03
jan
vie
r-0
4
févr
ier-
04
mar
s-0
4
avr
il-0
4
ma
i-0
4
juin
-04
juill
et-
04
ao
ût-
04
E.
Co
li (N
/10
0m
l)
Entrée
Sortie A12
Sortie FFC
Sortie L1
Sortie L2
Sortie L3
Sortie F1
Sortie
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
1,E+09
juill
et-
03
ao
ût-
03
sep
tem
bre
-03
oct
ob
re-0
3
no
vem
bre
-03
dé
cem
bre
-03
jan
vie
r-0
4
févr
ier-
04
mar
s-0
4
avr
il-0
4
ma
i-0
4
juin
-04
juill
et-
04
ao
ût-
04
Ent
éro
coqu
es (
N/1
00
ml)
Entrée
Sortie A12
Sortie FFC
Sortie L1
Sortie L2
Sortie L3
Sortie F1
Sortie
Figure 2.10. Contribution de chaque étage à l’élimination des Entérocoques et E. Coli
La décontamination microbienne fournie par cet étage apparaît peu saisonnalisée, contrairement à ce qui avait pu être observé avant l’agrandissement. Les performances sont plus faibles en été, particulièrement pour les entérocoques, et plus élevées en automne (Tableau 2.3 et Tableau 2.4). Il n’y a pas d’explication simple à ce comportement ; cependant, en saison froide, les bassins de maturation apparaissent peu chargés en algues et le rayonnement solaire pénètre facilement dans les bassins alors que, en saison chaude, la densité algale est plus importante et fait, dans une certaine mesure, obstacle à l’action des rayonnements ultra-violets sur l’ADN des bactéries. Afin de préserver l’étang de Thau, la hauteur d’eau dans ces bassins de maturation est gardée faible en été pour qu’ils aient un grand pouvoir de stockage pendant les périodes pluvieuses de l’automne.

46
Tableau 2.3. Effet de la saison sur la contribution de chaque étage à la réduction des entérocoques calculée à partir du flux
Abattement des entérocoques (unités log)
A12 FFC L1 L2 L3 F1 F2 Total
E té 0,57 1,33 1,16 0,69 0,59 0,80 0,46 5,60
Automne 0,09 1,42 1,66 1,57 0,66 1,05 0,28 6,73
Hiver 0,65 1,42 1,47 0,49 0,91 0,13 0,31 5,38
Printemps 0,54 1,44 1,59 0,61 0,80 0,12 0,44 5,54
Tableau 2.4. Effet de la saison sur la contribution de chaque étage à la réduction des E .Coli calculée à partir du flux
Abattement des E. Coli (unités log)
A12 FFC L1 L2 L3 F1 F2 Total
E té 0,73 1,68 1,35 0,98 0,78 0,20 0,98 6,70
Automne 0,37 1,05 1,75 1,05 0,98 1,53 0,38 7,11
Hiver 0,26 1,19 1,45 1,51 0,62 0,48 0,44 5,95
Printemps 0,22 1,22 1,57 1,48 0,62 0,51 0,44 6,06
2.3.4. Quatrième étage : les bassins de finition La première attribution des deux bassins de finition est le stockage de grandes quantités d’eau lors des pluies d’automne afin de limiter leur impact sur l’étang de Thau. La charge surfacique moyenne qui y est appliquée est de 26 kg DBOha-1j-1 avec un temps de séjour moyen de 20 jours. Sa production d’algues conduit même à une augmentation des teneurs en MES tout au long de l’année et de la DCO en été ; ce phénomène est observé sur d’autres lagunes tertiaires de la région. Les performances de cet étage sont faibles pour tous les paramètres à l’exception du phosphore. Cependant ces bassins participent, de manière modeste mais significative, à l’élimination de l’azote sous sa forme ammoniacale (Figure 2.8). Ceci a lieu hors de la saison estivale, quand la nitrification n’a pas éliminé tout le N-NH4 dans les bassins de maturation. Quinze pour cent du phosphore rentrant dans la station est éliminé, essentiellement en été et en automne, dans l’étage de finition (Figure 2.9). Celui-ci contribue donc à hauteur de 40% à la performance de la station pour ce nutriment. L’examen des concentrations en indicateurs de contamination fécale montre que, pendant la durée de l’étude, la qualité de l’effluent se dégrade dans le premier bassin de finition F1. Deux explications sont invoquées pour expliquer cette évolution. La première est une contamination par les fientes d’oiseaux qui, à certaines saisons, s’y trouvent en abondance et beaucoup plus nombreux qu’ailleurs en raison de la présence dans ce bassin d’une digue immergée. La deuxième est l’existence d’une fuite dans le by-pass qui relie le bassin R4 au bassin F1. Cette dernière hypothèse semble la plus probable, au moins pour une bonne partie de la durée de l’étude, de mi-octobre 2003 à fin avril 2004. La calibration du modèle numérique de décontamination microbienne suggère que, pendant cette période, les flux bactériens provenant de L3 et de R4 sont d'ordre de grandeur comparables. L’abattement dans la lagune F2 compense la contamination dans la lagune F1.

47
Chapitre 3. MODELISATION DE LA DECONTAMINATION BACTERIENNE

48

49
L'élimination bactérienne dans les bassins de lagunage est généralement reconnue excellente et bien supérieure à celle obtenue dans les procédés biologiques conventionnels. C’est ce qui a conduit à choisir cette technique pour épurer les eaux usées du versant nord de l’étang de Thau. La restructuration du lagunage n’a pas seulement permis de maintenir les performances des premières années de fonctionnement ; elle les a améliorées et désaisonnalisées. Cet effet, spectaculaire et assez inattendu, a trouvé quelques explications dans l’analyse de la contribution de chaque étage (deuxième partie). On peut toutefois tenter d’aller plus loin dans la compréhension des performances de décontamination microbienne à travers une procédure de modélisation. Cette procédure inclue le pari de relier, pour chaque bassin, la décontamination et plus précisément ses constantes cinétiques à des variables liées à la météorologie (température de l’eau et rayonnement solaire) et à la teneur en matières en suspension. En cas de succès, le modèle peut constituer un outil permettant d’améliorer la gestion du lagunage en vue de meilleures performances ; il pourrait aussi être utilisé dans l’élaboration des systèmes de lagunage quand la décontamination microbienne fait partie du cahier des charges. Comme pour les autres techniques d’épuration des eaux usées, la décontamination microbienne est généralement étudiée à travers le dénombrement d'organismes indicateurs de contamination fécale tels que Escherichia coli, coliformes totaux, coliformes fécaux et entérocoques, qui sont caractérisés par des procédures rapides et peu coûteuses d'identification et de dénombrement. En effet, la grande diversité des organismes pathogènes, les difficultés et les coûts de leur identification et de leur dénombrement rendent impossible leur étude systématique. Heureusement, même si ils sont trop peu nombreux, quelques travaux ont été réalisés sur des organismes autres que les germes indicateurs de contamination fécale. Les exemples évoqués ci-après donnent un aperçu des difficultés de l’évaluation des performances sanitaires dans le cas particulier des systèmes de lagunage. Emparanza-Knörr et Torrella (1995) se sont intéressés au devenir des coliformes totaux, des coliformes fécaux, des salmonelles (Salmonella spp) et des coliphages somatiques dans le système de lagunage de Guardamar de Segura dans la province d'Alicante en Espagne. Ce système comprend un étage anaérobie, composé de 3 bassins en parallèle, et un étage facultatif, constitué de 2 bassins fonctionnant aussi en parallèle. En raison de la surcharge de l’installation, l’étage facultatif était anoxique. Les résultats des suivis bactériologiques à l'entrée et à la sortie de chacun des bassins semblent montrer une différence de comportement par rapport au milieu environnemental entre d'une part, les coliformes totaux et les coliformes fécaux et d'autre part, les salmonelles. Le comportement des coliphages somatiques se différenciait aussi bien de celui des coliformes que de celui des salmonelles. Ces spécificités se manifestant par des différences dans la contribution de chaque étage à l’élimination des différents germes (Tableau 3.1). En effet, les réductions totales des coliformes totaux et des coliformes fécaux ont été évaluées respectivement à 99,18 et 97,86 %, dont 94,0 et 76,1% dans l'étage anaérobie. L'élimination des salmonelles est assez variable mais relativement élevée en moyenne avec une contribution presque exclusive de l'étage facultatif, de l'ordre de 99,8%. L'étage facultatif apparaît aussi responsable de 67,4% de la disparition totale des coliphages somatiques, chiffrée à seulement 89,7%.

50
Tableau 3.1. Abattements moyens, en unités logarithmiques, des microorganismes dans le lagunage de Guardamar de Segura (d’après Emparanza-Knörr et Torrella ,1995) Etage anaérobie Etage facultatif Total Coliformes totaux 1,22 0,86 2,08 Coliformes fécaux 0,62 1,05 1,67 Salmonellae spp ε 2,7 2,7 Coliphages somatiques 0,11 0,87 0,99 Arridge et al. (1995) ont étudié les éliminations de Vibrio cholerae et des salmonelles avec celle des coliformes fécaux, des entérocoques et du Clostridium perfringens dans un système de lagunage à 5 étages dans le nord-est du Brésil. Le premier étage comprend 2 bassins anaérobies en parallèle ; le second, 5 bassins facultatifs en parallèle ; le troisième, un bassin de maturation primaire ; le quatrième, 5 bassins de maturation secondaire en parallèle et le cinquième, 4 bassins de maturation tertiaire en parallèle. Les résultats des suivis bactériologiques réalisés à l’entrée et à la sortie de chacun des bassins sont réunis dans le Tableau 3.2. Quelque soit le type de bactérie considéré, le rendement global d’élimination est supérieur à 99,7% ou 2,5 U.log. Le Tableau 3.3 donne la contribution de chaque étage, obtenue en faisant le
rapport :( )
SE
xSxE
NN
NN
−− 100
où
xEN est la teneur de l’organisme considéré à l’entrée de l’étagex
xSN la teneur à la sortie de l’étagex
EN la teneur à l’entrée de la station
SN la teneur à la sortie de la station
Ces travaux de Arridge et al. (1995), conduits dans un contexte très différent, confirment en partie ceux de Emparanza-Knörr et Torrella (1995). Dans l’étage anaérobie, les salmonelles sont moins bien éliminées que les bactéries témoins de contamination fécale. Dans l’étage facultatif, elles enregistrent une disparition ou plus forte ou comparable à celle des bactéries témoins de contamination fécale, selon le mode d’évaluation retenu (Tableau 3.3 ou Tableau 3.4). L’abattement des salmonelles reste significatif dans le premier étage de maturation tandis qu’il est faible dans les deux étages suivants où la teneur en salmonelles ne dépasse pas quelques unités par 100 mL. Les entérocoques et les coliformes fécaux présentent par contre une grande similitude de comportement, à l’exception de l’étage de maturation tertiaire. Cette étape ultime du lagunage n’affecte guère les teneurs en entérocoques alors que l’abondance des coliformes fécaux continue à décroître au même rythme que dans les étages précédents (Tableau 3.4). Dans les 3 premiers étages, l’abattement de Clostridium perfringens obéit aux mêmes tendances que celui des germes témoins de contamination fécale. Mais les deux derniers étages de maturation n’apportent qu’une contribution très modeste à l’élimination de cette bactérie. L’étage anaérobie constitue aussi l’étage le plus performant pour le Vibrio cholerae, il est vrai moins abondant que les autres bactéries, disparaît dès la sortie de l’étage de maturation primaire. Selon Arridge et al. (1995), la bonne performance de l'étage anaérobie dans la disparition du V. cholerae est due essentiellement à la forte teneur en sulfure, 11mg.L-1, des eaux de ce lagunage.
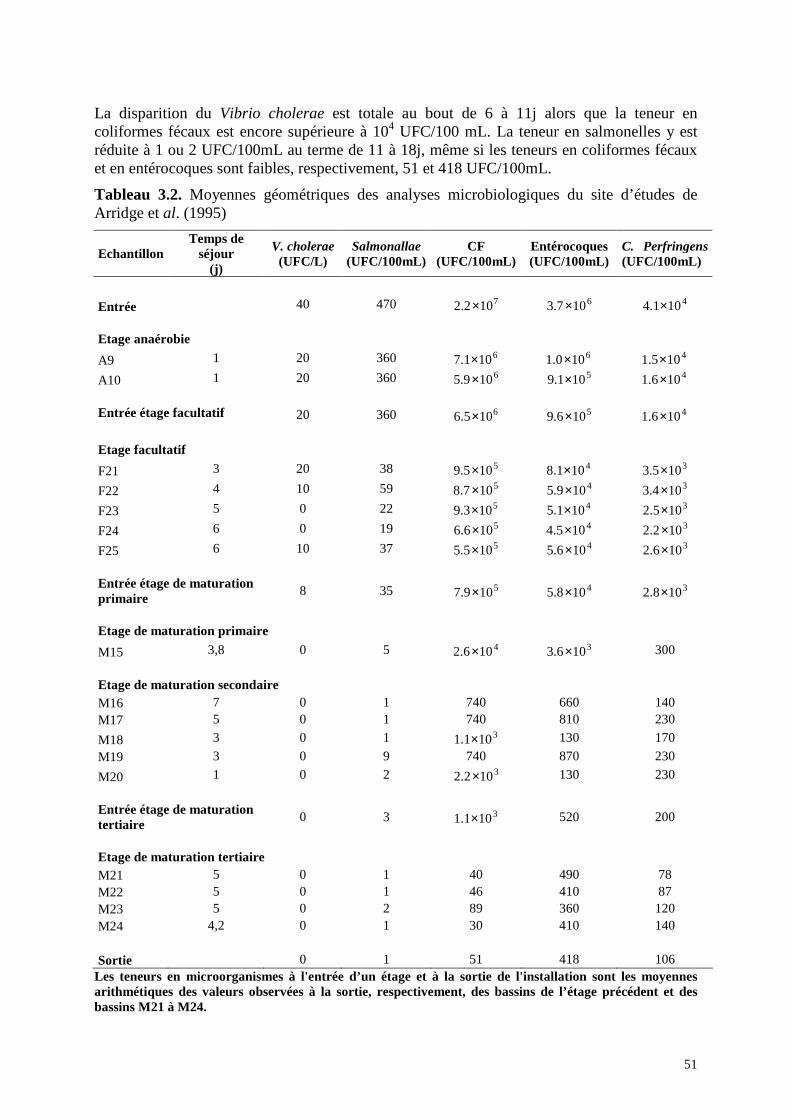
51
La disparition du Vibrio cholerae est totale au bout de 6 à 11j alors que la teneur en coliformes fécaux est encore supérieure à 104 UFC/100 mL. La teneur en salmonelles y est réduite à 1 ou 2 UFC/100mL au terme de 11 à 18j, même si les teneurs en coliformes fécaux et en entérocoques sont faibles, respectivement, 51 et 418 UFC/100mL.
Tableau 3.2. Moyennes géométriques des analyses microbiologiques du site d’études de Arridge et al. (1995)
Echantillon
Temps de séjour
(j)
V. cholerae (UFC/L)
Salmonallae (UFC/100mL)
CF (UFC/100mL)
Entérocoques (UFC/100mL)
C. Perfringens (UFC/100mL)
Entrée 40 470 7102.2 × 6107.3 × 4101.4 × Etage anaérobie
A9 1 20 360 6101.7 × 6100.1 × 4105.1 ×
A10 1 20 360 6109.5 × 5101.9 × 4106.1 × Entrée étage facultatif 20 360 6105.6 × 5106.9 × 4106.1 × Etage facultatif
F21 3 20 38 5105.9 × 4101.8 × 3105.3 ×
F22 4 10 59 5107.8 × 4109.5 × 3104.3 ×
F23 5 0 22 5103.9 × 4101.5 × 3105.2 ×
F24 6 0 19 5106.6 × 4105.4 × 3102.2 ×
F25 6 10 37 5105.5 × 4106.5 × 3106.2 × Entrée étage de maturation primaire
8 35 5109.7 × 4108.5 × 3108.2 ×
Etage de maturation primaire
M15 3,8 0 5 4106.2 × 3106.3 × 300
Etage de maturation secondaire M16 7 0 1 740 660 140 M17 5 0 1 740 810 230
M18 3 0 1 3101.1 × 130 170
M19 3 0 9 740 870 230
M20 1 0 2 3102.2 × 130 230
Entrée étage de maturation tertiaire
0 3 3101.1 × 520 200
Etage de maturation tertiaire M21 5 0 1 40 490 78 M22 5 0 1 46 410 87 M23 5 0 2 89 360 120 M24 4,2 0 1 30 410 140 Sortie 0 1 51 418 106 Les teneurs en microorganismes à l'entrée d’un étage et à la sortie de l'installation sont les moyennes arithmétiques des valeurs observées à la sortie, respectivement, des bassins de l’étage précédent et des bassins M21 à M24.

52
Tableau 3.3. Contribution, exprimée en pourcents du rendement global, de chaque étage du site d’études de Arridge et al. (1995) à la disparition de Vibrio cholerae, des Salmonallae, de C. Perfringens, des coliformes fécaux et des entérocoques
Rendement d’élimination (%) V. cholerae Salmonallae CF Entérocoques C. perfringens Etage anaérobie 50,0 23,5 70,5 74,2 62,4 Etage facultatif 30,0 69,3 25,9 24,2 31,0 Etage de maturation primaire 20,0 6,4 3,5 1,5 6,2
Etage de maturation secondaire 0,5 0,1 0,1 0,2 Etage de maturation tertiaire 0,3 0,0 0,0 0,2 Tableau 3.4. Abattement, exprimé en unités logarithmiques, de Vibrio cholerae, des Salmonallae, de C. Perfringens, des coliformes fécaux et des entérocoques dans chaque étage du site d’études de Arridge et al. (1995).
V.cholerae Salmonellae CF Entérocoques C. perfringens (Unités logarihmiques) Etage anaérobie 0,3 0,12 0,53 0,59 0,42 Etage facultatif 0,4 1,01 0,91 1,21 0,74 Etage de maturation primaire NQ 0,85 1,48 1,21 0,98
Etage de maturation secondaire NQ 0,25 1,37 0,84 0,18
Etage de maturation tertiaire NQ 0,35 1,33 0,10 0,27
NQ : non quantifiable Davies-Colley et al. (2005) ont comparé l'élimination des phages somatiques avec celle des E. coli dans une unité pilote de lagunage du type Advanced Pond Systems (APS) de Ruakara, en Nouvelle-Zélande. Le système est composé d'une unité anaérobie en série avec un bassin à haut rendement, un bassin de décantation algale et un bassin de maturation. Les suivis bactériologiques ont été réalisés pendant l'été (du 4 au 27 février) et pendant l'hiver (du 3 juin au 21 juillet) à l'entrée de la station et à la sortie de chaque unité. Contrairement à la disparition des E. coli, celle des phages présente un caractère fortement saisonnier (Figure 3.1). L’abattement des E. coli est en effet de 4,0 U.log en été et 4,1 U.log en hiver. L'élimination des phages somatiques est moins importante, soit 2,3 U.log en été, et beaucoup plus faible encore en hiver, avec 0,45 U.log seulement.

53
Figure 3.1. Décontamination dans chaque étage de la station pilote de lagunage "APS" de Ruakura, en Nouvelle-Zélande selon Davies-Coley et al. (2005)
Ces différents travaux montrent que les différents microorganismes étudiés se différencient entre eux par leur comportement dans l'environnement des lagunes. Ils illustrent la difficulté à prévoir, à partir des témoins de contamination fécale habituellement utilisés dans les contrôles et les évaluations de performance sanitaire, le degré d’élimination des pathogènes. La compréhension et la modélisation des processus d’élimination des organismes indicateurs ne présument pas de celles des pathogènes mais constituent toutefois une étape importante dans la caractérisation de la décontamination microbienne dans les lagunages. 3.1. Mécanismes de la décontamination microbienne en lagunage On s’accorde généralement à considérer que le rayonnement solaire joue un rôle prépondérant la décontamination microbienne en lagunage suivant 2 principaux processus, l'irradiation par les rayonnements ultraviolets (UV) et la photooxydation en milieu alcalin. 3.1.1. La décontamination par les rayons ultra violets Les rayonnements UV sont des radiations génotoxiques (Metcalf et Eddy, 1991). Ils regroupent les rayonnements électromagnétiques dont les longueurs d'onde sont comprises entre 100 et 400 nm. Selon leur mode d'action sur les cellules, ils sont subdivisés en 3 groupes :
- Les UVA, de 315 à 400 nm, représentent 95 % des rayonnements UV solaires qui arrivent à la surface de la terre et attaquent l'ADN par l’intermédiaire des radicaux libres.
- Les UVB, de 280 à 315 nm, composent 5 % des rayonnements UV solaires, agissent directement sur l'ADN.
- Les UVC, de 100 à 280 nm, constituent la gamme la plus agressive ; ils sont normalement absorbés en totalité dans l'atmosphère. Ce sont les rayonnements utilisés dans les systèmes conventionnels de décontamination. La longueur d'ondes la plus bactéricide est autour de 250nm. Aussi, les lampes qui produisent artificiellement des UV émettent des rayonnements de longueurs d’onde voisines de 254nm.

54
L'action abiotique des radiations UV est d'autant plus efficace que la structure de l'être vivant se rapproche de la structure monocellulaire. Les microbes, virus, bactéries, sont donc particulièrement sensibles aux rayonnements UV. La désinfection par UV augmente avec la transmittance de l'eau. Plus le milieu est turbide, plus il adsorbe le rayonnement solaire et moins grande est l’élimination des microorganismes par irradiation. La désinfection par UV est une fonction croissante du temps d'exposition.
Figure 3.2. Comparaison des variations des taux d’inactivation des phages somatiques et des phages ARN-F suivant la profondeur avec le gradient des rayons UVB en fonction de la
profondeur dans la couche superficielle du bassin de décantation algale de l’unité pilote de lagunage "APS".
Moeller and Calkins (1980) affirment que les rayons UVB entre 280 et 320nm sont responsables de la disparition des coliformes fécaux en lagunage. Selon Curtis et al. (1992) leur pénétration dans les bassins de lagunage est trop improbable pour qu’ils puissent agir sur les bactéries. En se référant aux travaux de Davies-Coley et al. (2005), il semblerait en effet que les rayons UVB soient décelés uniquement dans la tranche d’eau superficielle épaisse de 5cm (Figure 3.2). Toutefois, les gradients verticaux du taux d’inactivation des phages ARN-F et des phages somatiques apparaissent bien en relation avec la pénétration des rayonnements UVA et UVB, ce qui suggère, pour ces microorganismes au moins, un rôle primordial de ces rayonnements dans leur inactivation. Selon Curtis et al. (1992), la seule action de la lumière solaire ne peut avoir d'effet sur les coliformes fécaux présents dans les lagunages. Les résultats des études de la décontamination le long de transects verticaux effectués par Pearson et al. (1987) dans plusieurs bassins d’un lagunage au Portugal semblent aller dans ce sens. Ces derniers ont en effet constaté que la tranche d’eau ayant la plus faible teneur en coliformes fécaux n’est pas systématiquement la couche superficielle. Tel est le cas de celle de la première lagune facultative de leur site d’études, dont le pH égal à 8,4, n’aurait pas été assez élevé pour permettre un abattement des coliformes fécaux. Par ailleurs, les mêmes auteurs ont aussi remarqué que de 2 bassins montés en parallèle, de configurations et de charges surfaciques identiques, le plus turbide, mais de pH élevé, peut être le plus performant en matière de décontamination. Curtis et al. (1992) déduisent de cette étude et de la littérature que le pH joue un rôle décisif, ce qui suggère pour le moins que d’autres mécanismes que l’action directe des rayonnements UV doivent être pris en considération. La photooxydation en fait évidemment partie.

55
3.1.1. La décontamination par photooxydation La photooxydation est décrite par Curtis et al. (1992) comme étant un mécanisme en 3 étapes. La première consiste en une absorption des longueurs d’onde de la lumière solaire comprises entre 280 et 700 nm – rayonnements UV inclus - par les substances humiques. Ces substances humiques étant des produits de la décomposition de toute matière organique, d'origine animale, végétale ou bactérienne, sont omniprésentes dans les bassins de lagunage. La seconde étape est le retour des substances humiques à leur état fondamental. L'énergie ainsi libérée conduit généralement à la formation de formes toxiques de l’oxygène tels que le superoxyde, le radical hydroxyl, le peroxyde d’hydrogène ou l'oxygène à l'état singulet. La dernière étape est l’oxydation de la membrane interne des bactéries, qui joue un rôle important dans le maintien du pH interne nécessaire à leur survie. Curtis et al. (1992) soutiennent que pour les coliformes fécaux dont le pH interne est très faiblement basique, entre 7.6 et 7.8 (Booth, 1985), la photooxydation agit en synergie avec les pH élevés. Curtis et al. (1992) affirment que les longueurs d’onde dans le visible supérieures à 440 nm, qui sont les plus pénétrantes, ne peuvent nuire aux CF que si la photosynthèse élève le pH au moins à 8. Ils affirment aussi par ailleurs, que le pH sans la lumière ne présenterait de danger pour les bactéries qu’au delà de 9,3. Les matières organiques et les nutriments sont des facteurs inhibant de la photooxydation. En fonction de leurs concentrations, leurs réactions de dégradation peuvent conduire à un appauvrissement du milieu en oxygène, élément indispensable à la photooxydation. En outre, il semblerait que la disponibilité en nutriments permette aux bactéries de mieux résister aux pH élevés (Booth, 1985). Des teneurs distinctes en nutriments pourraient expliquer les différences dans l’évaluation de la valeur minimale du pH permettant l’action de décontamination du rayonnement solaire qui serait de 8 pour Curtis et al. (1992) et 9 pour Pearson et al. (1987). 3.1.2. Remarques Si l’irradiation par les rayonnements UV et la photo oxydation sont bien les mécanismes principaux de la décontamination, distinguer leurs contributions respectives paraît bien délicat. La principale difficulté vient de ce que ces deux processus dépendent tous deux du rayonnement solaire. Cette dépendance est directe pour les rayonnements UV, mais l’irradiation dépend de la turbidité, donc de la teneur en algues qui, elle-même, dépend du rayonnement solaire et d’autres facteurs parmi lesquels, très vraisemblablement, la température de l’eau. La photo oxydation dépend du rayonnement solaire et de la teneur en oxygène dissous, laquelle dépend de l’activité de la photosynthèse, donc encore une fois du rayonnement solaire et de la teneur en algues. L’action de la photo oxydation dépend du pH, qui dépend de la photosynthèse. L’approche mécaniste de la décontamination microbienne s’avère donc particulièrement délicate, et cela d’autant plus que d’autres processus que les rayonnements UV et la photo oxydation interviennent. Ce sont essentiellement la sédimentation, capable de jouer un rôle significatif dans les bassins de tête, la prédation quand les populations de protozoaires abondent ou encore, mais vraisemblablement beaucoup plus rare, la rareté des substrats nutritifs. Très peu d'études ont été consacrées à de tels mécanismes malgré leurs rôles potentiellement non négligeables, notamment dans les régions faiblement ensoleillées ou

56
pendant les saisons froides. La décontamination par sédimentation en particulier, peu dépendante du climat, peut se révéler prépondérante dans les bassins fortement chargés, aux teneurs élevées en matières en suspension. Elle est probablement à l'origine des fortes activités de décontamination enregistrées dans certains bassins anaérobies. Dans ces conditions, les démarches empiriques restent plus réalistes et mieux capables de répondre aux besoins de l’ingénierie. Parmi les travaux les plus intéressants, il faut citer ceux de Troussellier et al. (1986) relatifs à une étude au moyen d’une méthode de régressions multiples (path analysis) de la décontamination bactérienne dans le lagunage de Mèze. Cette étude montre bien l’interdépendance des variables susceptibles d’expliquer l’élimination des coliformes fécaux. Les principales variables explicatives sont le rayonnement transmis, le pH et la teneur en chlorophylle a. La plupart des modèles empiriques sont eux aussi issus d’une analyse statistique de résultats expérimentaux. Ils font intervenir pluieurs des variables suivantes : le rayonnement solaire, la DBO, le pH, l’oxygène dissous, voire la profondeur des bassins (Sarikaya et al.,1987; Saqqar and Pescod, 1992; Curtis et al., 1992; Mayo, 1995, Von Sperling et al. 2005). Cependant, nombreux sont les auteurs qui privilégient le rôle de la température (Nascimento, 1987; Mills et al., 1992; Saqqar and Pescod, 1992; Auer and Niehaus, 1993; Johansson et al., 1996), suivant en cela la très classique formulation de la cinétique de décontamination établie par Marais (1974):
( )2020
−×= TT kk θ (Équation 3.1)
où
20k est le coefficient cinétique de la dégradation bactérienne à 20°C, θ un coefficient et T la
température en °C. Même si cette relation reste une solide référence utilisée dans des travaux récents (Banda et al. 2005), les valeurs très variables de 20k et de θ données par la littérature
impliquent la nécessité d’introduire d’autres termes que la température dans la prévision de la décontamination. 3.2. Modélisation de la décontamination 3.2.1. Méthodes D’une manière générale, la modélisation de la décontamination microbienne résulte de la combinaison d’une cinétique de décontamination et d’une distribution des temps de séjour de l’eau dans chaque lagune considérée. Les cinétiques les plus couramment admises sont des cinétiques du premier ordre telles que :
kNt
N −=∂∂
(Équation 3.2)
où N est la teneur en bactéries de l'effluent, t le temps en j et k le coefficient cinétique de la décontamination en j-1. La distribution des temps de séjour de l’eau résulte soit d’un modèle – c’est le cas le plus courant - soit de l’exploitation de traçages effectués dans la lagune. Ces modèles sont simples. On peut soit considérer qu’une lagune est assimilable à un réacteur parfaitement mélangé, soit admettre que le déplacement de l’eau est purement convectif (déplacement piston) ou convectif dispersif. Le choix du modèle n’est pas aisé car le régime hydrodynamique des lagunes peut considérablement varier d’un jour à l’autre ou d’une heure à l’autre (Brissaud et al. 2003). Il peut dépendre des objectifs correspondants ; on n’adoptera pas nécessairement la

57
même attitude s’il s’agit d’un projet d’ingénierie ou d’un suivi expérimental. Le choix du modèle de distribution des temps de séjour n’est pas sans conséquence sur les évaluations des coefficients cinétiques de décontamination qui consistent le plus souvent à interpréter des concentrations d’entrée et de sortie d’une lagune à travers une distribution supposée des temps de séjour de l’eau dans cette lagune. Dans cette étude, nous avons choisi de considérer que les lagunes sont des réacteurs parfaitement mélangés. Cette hypothèse va de soi pour les lagunes de tête qui disposent d’aérateurs. En ce qui concerne les autre lagunes, des travaux récents ont montré que pendant 65% du temps en été et plus de 80% du temps au printemps, les températures de l’eau sont uniformes dans la lagune, verticalement et horizontalement, ce qui constitue un bon indice de mélange. Par ailleurs, nous avons adopté l’expression empirique du coefficient cinétique suggérée par Xu et al., (2002), qui est fonction de la température et du rayonnement solaire:
( ) mIT ek χβα ⋅⋅= −20 (Équation 3.3)
où T est la température (°C),mI l’intensité lumineuse moyenne reçue (Jcm-2j-1) et α, β et χ
sont des constantes.
mI est obtenue en intégrant, sur la hauteur de l’eau h de la lagune, la relation de Beer-
Lambert : Kx
x eII −= 0 (Équation 3.4)
où
xI est l’intensité lumineuse (Jcm-2j-1) à la profondeur x (m)
0I l’intensité lumineuse reçue à la surface de la lagune (Jcm-2j-1)
K le coefficient d’extinction (m-1)
Kh
eII
Kh
m
−−= 10 (Équation 3.5)
Selon Xu et al., (2002),K est relié à la teneur en matières en suspension par la formule empirique :
09.2469.0 += MESK (Équation 3.6)
dans laquelle MES représente la teneur en matières en suspension (mg/l) Cette formulation du coefficient cinétique fait intervenir le rayonnement solaire et la température de l’eau ; cette dernière variable peut jouer un rôle important en Europe. Associée à l’hypothèse de mélange parfait, elle a été appliquée avec un succès relatif par Xu et al. (2002) et Barbagallo et al. (2003). Le modèle de décontamination consiste en un bilan bactérien. C’est un modèle en régime transitoire qui permet d’établir pour chaque lagune une relation entre les flux bactériens et la cinétique de décontamination. En supposant que chaque bassin est parfaitement mélangé et que l’abattement des germes suit une cinétique du premier ordre, le bilan bactérien s’écrit :
( )( ) ( ) ( ) ( ) ( ) ( ) ijiijijijiji
iji
jii VNkNQNQ
t
VN
t
NV ,,,,1,1,
, −−=∆
∆+
∆∆
−− (Équation 3.7)

58
où i est la lagune i et j le jour j
iV le volume d’eau dans le bassin i (m3)
( ) ( ) ( )1,,, −−=∆ jijiji NNN
t∆ est le pas de temps, 1=∆t j
( )jiN ,1− et ( )jiN , sont les concentrations en microorganismes à l’entrée et à la sortie de la
lagune i le jour j (UFC/ 100ml)
( )jiQ ,1− et ( )jiQ , les débits à l’entrée et à la sortie de la lagune i le jour j (m3/j)
ik est la constante de la cinétique d’élimination dans la lagune i (j-1)
D’autres flux entrants de différentes concentrations peuvent être pris en compte en introduisant les termes correspondants dans l’Equation 3-7. Le modèle a été utilisé de plusieurs manières. - D’une part, connaissant les débits et les concentrations à l’entrée et à la sortie d’une lagune, on peut en déduire la valeur de la constante cinétique et en déduire l’évolution de cette constante pendant différentes périodes. On appellera expérimentales les valeurs de la constante ainsi calculées. - D’autre part, nous avons utilisé le modèle pour tenter de relier les cinétiques de décontamination aux conditions météorologiques, autrement dit pour déterminer les valeurs des paramètresα , β etχ . Dans l’Equation 3-7, la constante cinétique k était remplacée par son expression donnée par l’Equation 3-3. Une procédure d’optimisation mathématique a été mise en œuvre ; elle consiste à rechercher les valeurs de ces paramètres qui permettent le mieux d’ajuster les concentrations calculées ( )jiN , aux concentrations observées ( )
', jiN à la
sortie des lagunes, connaissant, pour chaque lagune, les débits d’entrée et de sortie, les volumes d’eau, les concentrations d’entrée, la température de l’eau, le rayonnement solaire et la teneur en MES. L’optimisation consiste à rechercher le minimum de la fonction critère :
2'),(
1),( ))()(( yji
n
yyji NLogNLog −∑
=
où n est le nombre de mesures. L’utilisation de cette méthode suppose que l’on dispose d’un nombre de mesures strictement supérieur au nombre de paramètres à caler et que ces mesures soient réparties sur la totalité de leur domaine de variation. La première condition est largement satisfaite puisque pour 3 constantes à caler plus d’une vingtaine de mesures sont disponibles. De meilleurs résultats auraient par contre pu être obtenus avec une méthode de mesures plus sensible. Les débits et les volumes sont donnés par le modèle hydraulique ; les imperfections de la reconstitution des débits notées dans le paragraphe 1.5 se répercutent inévitablement sur le calcul des paramètres à optimiser. Les concentrations bactériennes sont observées tous les 15 jours en moyenne. Le modèle fonctionnant au pas de temps journalier, il a fallu procéder à des interpolations linéaires des concentrations entre les observations. - La dernière application du modèle est son usage en prévision. Dès lors que les valeurs des paramètres α , β etχ sont connues, autrement dit dès que l’on a établi les relations entre les constantes cinétiques de décontamination et les variables environnementales que sont la température de l’eau et le rayonnement solaire – sans oublier les teneurs en MES – on peut se servir du modèle pour prévoir l’effet de différents scénarios de gestion sur la décontamination bactérienne. Ces simulations supposent connues les débits d’entrée dans le lagunage et les chroniques de valeurs météorologiques concomitantes.

59
3.2.2. Application à la recherche de relations entre les cinétiques de décontamination et les variables météorologiques On a procédé, bassin par bassin et selon la procédure d’optimisation indiquée ci-dessus, à la détermination des paramètres α, β et χ pour les entérocoques et les E. coli à partir des données collectées entre juillet 2003 et août 2004. Les résultats correspondants sont donnés dans le Tableau 3.5.
Tableau 3.5. Valeurs ajustées des paramètres α, β et χ avec les données enregistrées de juillet 2003 à août 2004
( ) mIT ek ⋅− ⋅⋅= χβα 20
ENTEROCOQUES E. COLI
α β χ α β χ
A12 0.033 0.990 0.393 0.255 1.030 0.140 FFC 0.358 1.008 0.002 0.410 1.056 0.000 L1 0.662 0.955 0.011 3.035 1.097 0.000 L2 0.066 0.824 0.082 0.123 0.926 0.082 L3 0.001 1.049 0.290 0.020 0.952 0.141 F1 0.090 0.948 0.000 0.109 0.960 0.046 F2 0.033 1.009 0.083 0.137 1.001 0.053 Nous avons reporté dans les Figures 3.3, 3.5, 3.7, 3.9, 3.11, 3.13 et 3.15 les concentrations en entérocoques et en E. Coli calculées par le modèle à la sortie des bassins à partir des concentrations d’entrée observées respectives, des conditions météorologiques et en utilisant les valeurs des paramètres α, β et χ issues de la procédure d’optimisation. Les constantes cinétiques calculées dans la même procédure peuvent être comparées aux valeurs expérimentales, déterminées comme indiqué précédemment, sur les Figures 3.4, 3.6, 3.8, 3.10, 3.12, 3.14 et 3.16. Les principales questions que soulèvent ces simulations sont les suivantes :
1. est-ce que la tentative de lier les cinétiques de décontamination au rayonnement solaire, à la température de l’eau et à la teneur en MES permet une représentation correcte des abattements ?
2. Est-ce que les constantes cinétiques qui en découlent (ou constantes calculées) reflètent les constantes expérimentales ?
3. Quels degrés de cohérence peut-on trouver entre les valeurs des constantes calculées et les conditions physiques et environnementales des lagunes ? Y a-t-il une rationnalité perceptible derrière leurs variations ?
4. Une dernière question que l’on pourrait poser mais qui ne trouvera pas de réponse dans le cadre de ce travail est celle de savoir si on n’aurait pas dû essayer de relier les cinétiques de décontamination à d’autres variables. Des tentatives auraient sans doute pu être faites en utilisant le pH ou la DBO ; l’utilisation de la teneur en oxygène dissous aurait été beaucoup plus problématique car elle connaît des variations considérables avec la localisation et l’heure de la mesure.

60
La consultation des Figures 3.3, 3.5, 3.7, 3.9, 3.11, 3.13 et 3.15 permet d’apporter une réponse globalement satisfaisante à la première question. Toutefois, on note des difficultés qui n’ont pas été résolues. La procédure mathématique d’ajustement a buté sur des valeurs observées singulières pas toujours faciles à interpréter ; on a même rencontré des concentrations d’entrée inférieures aux concentrations de sortie d’une même lagune. La représentativité de certaines mesures ou des erreurs analytiques peuvent être invoquées. Par ailleurs, le pas de temps de l’échantillonnage, fixé à 2 semaines, n’a pas toujours été adapté au fonctionnement de l’installation, en particulier quand de grandes masses d’eau passent brutalement d’un bassin au suivant sous l’effet des variations des niveaux d’eau imposés par l’exploitation du lagunage. Enfin, la limite de détection des concentrations bactériennes, située à environ 58 UFC/100 ml a posé de sérieux problèmes à la détermination des paramètres des constantes cinétiques dans les derniers bassins.

61
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
04/0
8/20
03
31/0
8/20
03
27/0
9/20
03
24/1
0/20
03
20/1
1/20
03
17/1
2/20
03
13/0
1/20
04
09/0
2/20
04
07/0
3/20
04
03/0
4/20
04
30/0
4/20
04
27/0
5/20
04
23/0
6/20
04
20/0
7/20
04
16/0
8/20
04
Q m
3/j
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Ent
éroc
oque
s (N
/100
ml)
P m3/j
Qe
Qs calc
BactEntrée A12
BactSortie A12Exp
BactSortie A12 Calc
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
09/1
0/20
03
09/1
1/20
03
10/1
2/20
03
10/0
1/20
04
10/0
2/20
04
12/0
3/20
04
12/0
4/20
04
13/0
5/20
04
13/0
6/20
04
14/0
7/20
04
14/0
8/20
04
Q m
3/j
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E.
Col
i (N
/100
ml)
P m3/jQe
Qs calcBactEntrée A12
BactSortie A12 Exp
BactSortie A12 Calc
Figure 3.3. Concentrations en entérocoques et E. coli calculées et observées à la sortie des bassins anaérobies
0,001
0,01
0,1
1
10
100
09/0
7/03
09/0
8/03
09/0
9/03
09/1
0/03
09/1
1/03
09/1
2/03
09/0
1/04
09/0
2/04
09/0
3/04
09/0
4/04
09/0
5/04
09/0
6/04
09/0
7/04
09/0
8/04
k Ent
éro (j-1
)
Calc
Exp
0,001
0,01
0,1
1
10
100
09/0
7/03
09/0
8/03
09/0
9/03
09/1
0/03
09/1
1/03
09/1
2/03
09/0
1/04
09/0
2/04
09/0
3/04
09/0
4/04
09/0
5/04
09/0
6/04
09/0
7/04
09/0
8/04
k E. C
oli (j-1
)
Calc
Exp
Figure 3.4. Contantes cinétiques d’élimination des Entérocoques et E. coli observées et calculées dans les bassins anaérobies
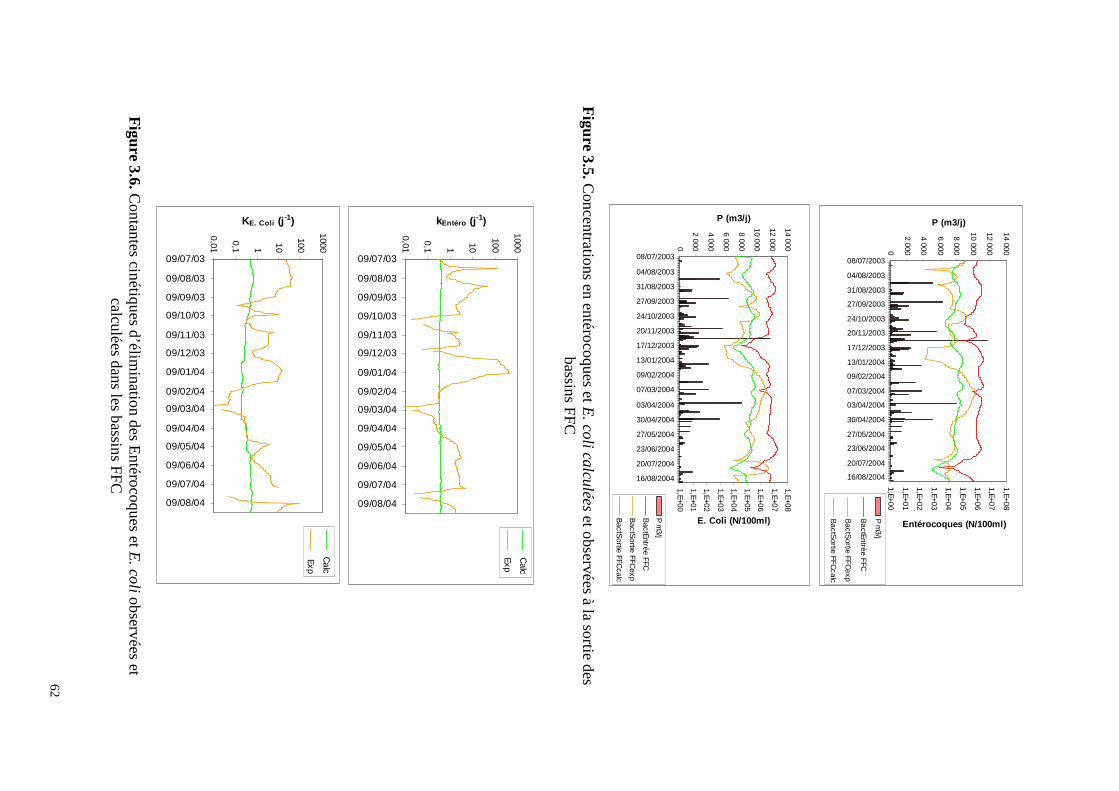
6
2
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)
P m3/j
BactEntrée FFC
BactS
ortie FFCexp
BactS
ortie FFCcalc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)
P m3/j
BactEntrée FFC
BactS
ortie FFCexp
BactS
ortie FFCcalc
Figure 3.5. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
es b
assins F
FC
0,01
0,1 1 10
100
1000
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kEntéro (j -1)
Calc
Exp
0,01
0,1 1 10
100
1000
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
KE. Coli (j -1)
Calc
Exp
Figure 3.6. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s les bassin
s FF
C

6
3
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)P m3/j
BactEntrée L1
BactS
ortie L1 Exp
BactS
ortie L1 Calc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)P m3/j
BactEntrée L1
BactS
ortie L1 Exp
BactS
ortie L1 Calc
Figure 3.7. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
u
bassin
L1
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kEntéro (j-1)
Calc
Exp
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kE. Coli (j-1)
Calc
Exp
Figure 3.8. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s le bassin
L1

6
4
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)P m3/j
BactEntrée F1
BactS
ortie F1 Exp
BactS
ortie F1 Calc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)
P m3/j
BactEntrée F1
BactS
ortie F1 Exp
BactS
ortie F1 Calc
Figure 3.9. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
u
bassin
L2
0,0
01
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kEntéro (j -1)
Calc
Exp
0,0
01
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kE. Coli (j -1)
Calc
Exp
Figure 3.10. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s le bassin
L2
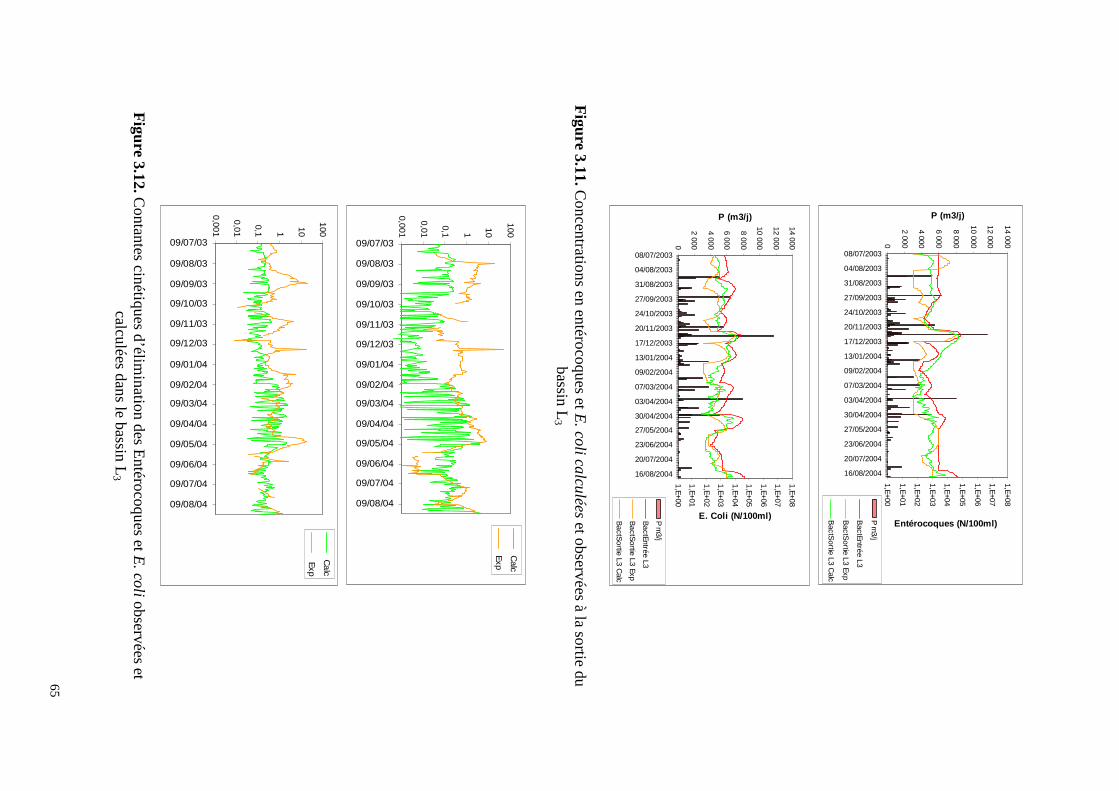
6
5
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)
P m3/j
BactEntrée L3
BactS
ortie L3 Exp
BactS
ortie L3 Calc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)P m3/j
BactEntrée L3
BactS
ortie L3 Exp
BactS
ortie L3 Calc
Figure 3.11. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
u
bassin
L3
0,001
0,01
0,1 1 10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
Calc
Exp
0,001
0,01
0,1 1 10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
Calc
Exp
Figure 3.12. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s le bassin
L3
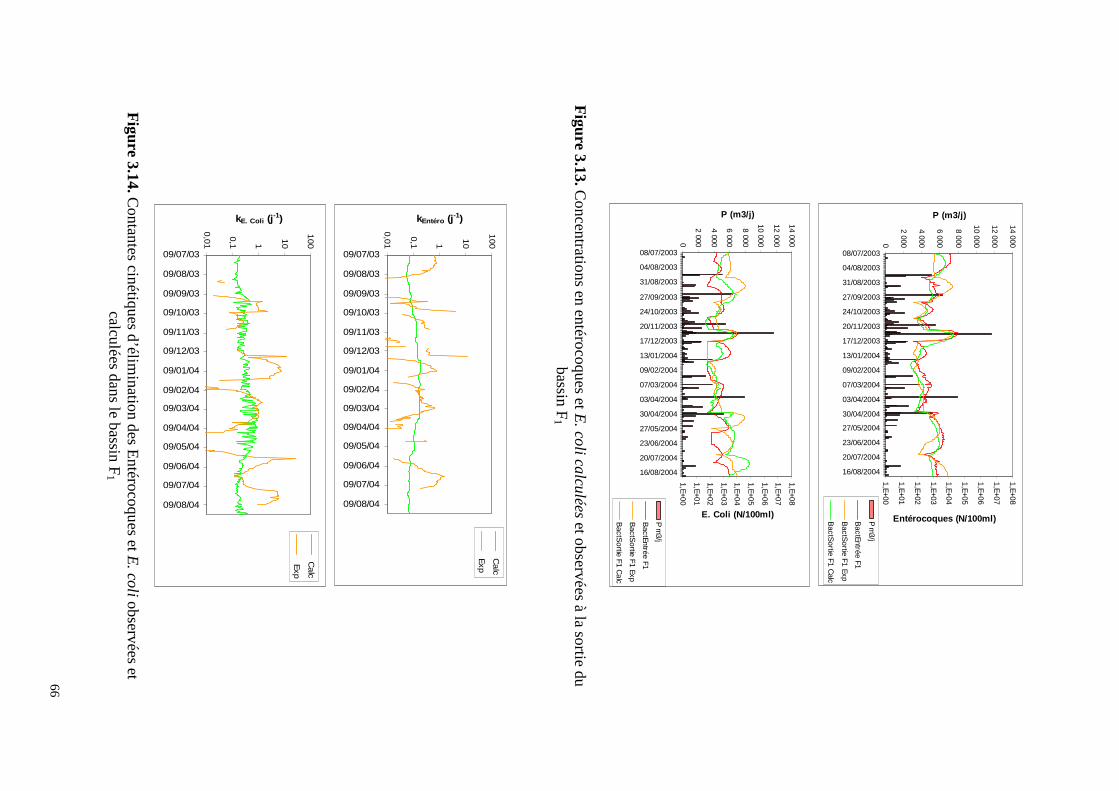
6
6
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)P m3/j
BactEntrée F1
BactS
ortie F1 Exp
BactS
ortie F1 Calc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)
P m3/j
BactEntrée F1
BactS
ortie F1 Exp
BactS
ortie F1 Calc
Figure 3.13. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
u
bassin
F1
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kEntéro (j-1)
Calc
Exp
0,0
1
0,1 1
10
100
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kE. Coli (j-1)
Calc
Exp
Figure 3.14. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s le bassin
F1
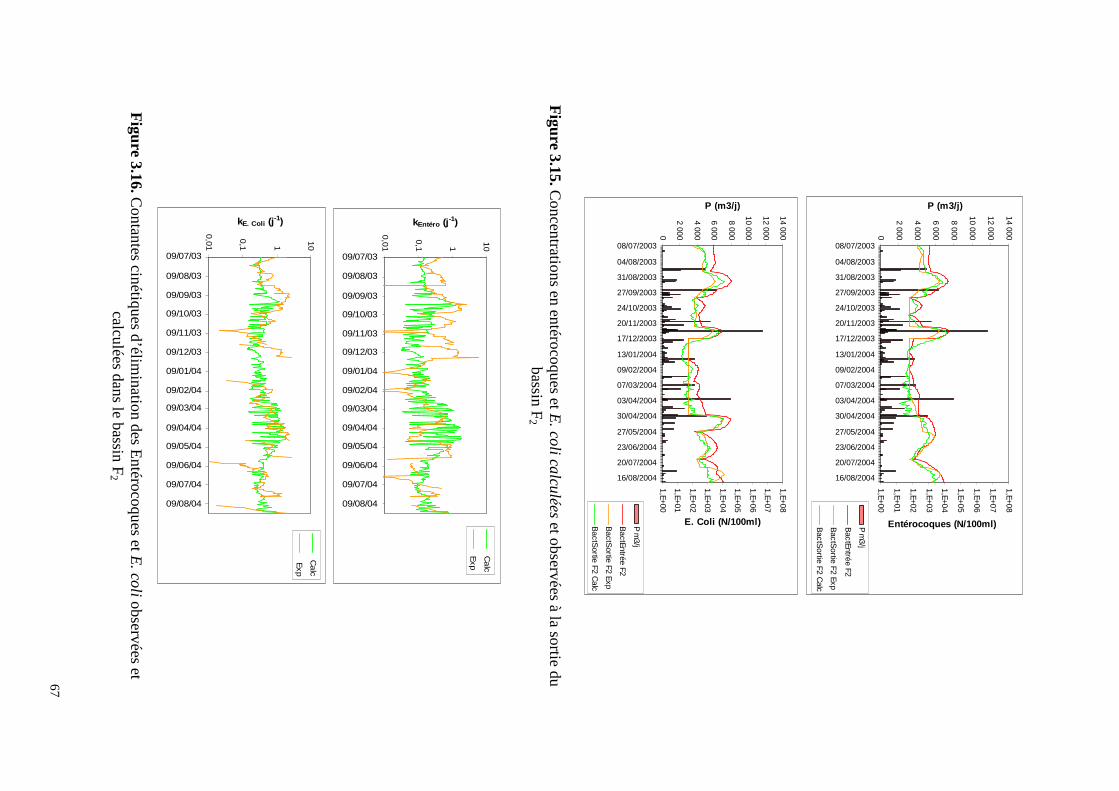
6
7
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Entérocoques (N/100ml)P m3/j
BactEntrée F2
BactS
ortie F2 Exp
BactS
ortie F2 Calc
0
2 000
4 000
6 000
8 000
10 000
12 000
14 000
08/07/2003
04/08/2003
31/08/2003
27/09/2003
24/10/2003
20/11/2003
17/12/2003
13/01/2004
09/02/2004
07/03/2004
03/04/2004
30/04/2004
27/05/2004
23/06/2004
20/07/2004
16/08/2004
P (m3/j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. Coli (N/100ml)P m3/j
BactEntrée F2
BactS
ortie F2 Exp
BactS
ortie F2 Calc
Figure 3.15. C
on
centratio
ns en
entéro
coq
ues et
E. co
li calcu
lées et o
bservé
es à la sortie d
u
bassin
F2
0,0
1
0,1 1
10
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kEntéro (j -1)
Calc
Exp
0,0
1
0,1 1
10
09/07/03
09/08/03
09/09/03
09/10/03
09/11/03
09/12/03
09/01/04
09/02/04
09/03/04
09/04/04
09/05/04
09/06/04
09/07/04
09/08/04
kE. Coli (j-1)
Calc
Exp
Figure 3.16. C
on
tantes cin
étiqu
es d’élim
inatio
n d
es En
téroco
qu
es et E
. coli o
bservées et
calculé
es dan
s le bassin
F2

68
La simulation de l’abattement des entérocoques n’est pas sensiblement moins bonne que celle des E. coli.
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
03/0
8/20
03
29/0
8/20
03
24/0
9/20
03
20/1
0/20
03
15/1
1/20
03
11/1
2/20
03
06/0
1/20
04
01/0
2/20
04
27/0
2/20
04
24/0
3/20
04
19/0
4/20
04
15/0
5/20
04
10/0
6/20
04
06/0
7/20
04
01/0
8/20
04
P (
m3/
j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Ent
éroc
oqu
es (
N/1
00m
l)
P m3/j
BactEntrée
BactSortie Exp
BactSortie Calc
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
04/0
8/20
03
31/0
8/20
03
27/0
9/20
03
24/1
0/20
03
20/1
1/20
03
17/1
2/20
03
13/0
1/20
04
09/0
2/20
04
07/0
3/20
04
03/0
4/20
04
30/0
4/20
04
27/0
5/20
04
23/0
6/20
04
20/0
7/20
04
16/0
8/20
04
P (
m3/
j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E. C
oli (
N/1
00m
l)
P m3/j
BactEntrée
BactSortie Exp
BactSortie Calc
Figure 3.17. Modélisation des teneurs en E. Coli et entérocoques de l’effluent de la station de lagunage de Mèze
On a aussi simulé les concentrations de sortie du lagunage à partir des seules concentrations mesurées à l’entrée de la station – autrement dit, sans tenir compte des mesures effectuées à la sortie des différents étages -, des débits déterminés par le modèle hydraulique et des données de rayonnement, de température de l’eau et des teneurs en MES. Le résultat est assez satisfaisant, en particulier pour ce qui concerne les Escherichia coli (Figure 3.17). On peut penser que si les seuils de détection des dénombrements microbiologiques avaient été moins élevés, on aurait noté moins d’écarts au printemps 2004 entre les valeurs observées et les valeurs calculées. Il semble donc que le modèle ainsi calibré soit capable de fournir une représentation très convenable des variations des teneurs en bactéries témoins de contamination fécale à partir des débits d’entrée dans la station, du rayonne ment solaire, de la température de l’eau et d’une estimation des teneurs en MES. Dans ces conditions, il peut constituer pour ce lagunage particulier un outil d’aide à la gestion. En effet, il existe à Mèze des degrés de liberté que les opérateurs peuvent utiliser pour optimiser les performances de l’installation. Toutefois, il convient de nuancer ce propos car les simulations ont été effectuées pour la période qui a servi au calibrage. Il n’a pas été possible de valider le modèle ; en effet, il aurait fallu pour cela disposer au moins d’une année de suivi supplémentaire.
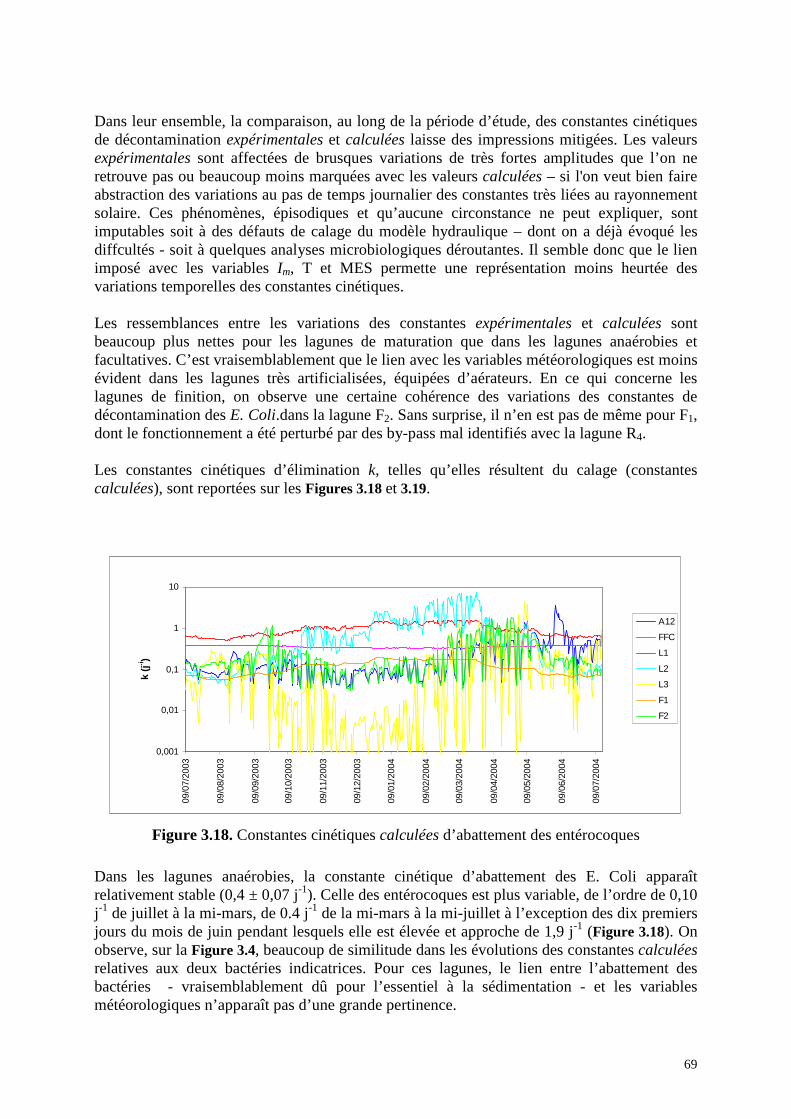
69
Dans leur ensemble, la comparaison, au long de la période d’étude, des constantes cinétiques de décontamination expérimentales et calculées laisse des impressions mitigées. Les valeurs expérimentales sont affectées de brusques variations de très fortes amplitudes que l’on ne retrouve pas ou beaucoup moins marquées avec les valeurs calculées – si l'on veut bien faire abstraction des variations au pas de temps journalier des constantes très liées au rayonnement solaire. Ces phénomènes, épisodiques et qu’aucune circonstance ne peut expliquer, sont imputables soit à des défauts de calage du modèle hydraulique – dont on a déjà évoqué les diffcultés - soit à quelques analyses microbiologiques déroutantes. Il semble donc que le lien imposé avec les variables Im, T et MES permette une représentation moins heurtée des variations temporelles des constantes cinétiques. Les ressemblances entre les variations des constantes expérimentales et calculées sont beaucoup plus nettes pour les lagunes de maturation que dans les lagunes anaérobies et facultatives. C’est vraisemblablement que le lien avec les variables météorologiques est moins évident dans les lagunes très artificialisées, équipées d’aérateurs. En ce qui concerne les lagunes de finition, on observe une certaine cohérence des variations des constantes de décontamination des E. Coli.dans la lagune F2. Sans surprise, il n’en est pas de même pour F1, dont le fonctionnement a été perturbé par des by-pass mal identifiés avec la lagune R4. Les constantes cinétiques d’élimination k, telles qu’elles résultent du calage (constantes calculées), sont reportées sur les Figures 3.18 et 3.19.
0,001
0,01
0,1
1
10
09/0
7/20
03
09/0
8/20
03
09/0
9/20
03
09/1
0/20
03
09/1
1/20
03
09/1
2/20
03
09/0
1/20
04
09/0
2/20
04
09/0
3/20
04
09/0
4/20
04
09/0
5/20
04
09/0
6/20
04
09/0
7/20
04
k (j
-i)
A12
FFC
L1
L2
L3
F1
F2
Figure 3.18. Constantes cinétiques calculées d’abattement des entérocoques
Dans les lagunes anaérobies, la constante cinétique d’abattement des E. Coli apparaît relativement stable (0,4 ± 0,07 j-1). Celle des entérocoques est plus variable, de l’ordre de 0,10 j-1 de juillet à la mi-mars, de 0.4 j-1 de la mi-mars à la mi-juillet à l’exception des dix premiers jours du mois de juin pendant lesquels elle est élevée et approche de 1,9 j-1 (Figure 3.18). On observe, sur la Figure 3.4, beaucoup de similitude dans les évolutions des constantes calculées relatives aux deux bactéries indicatrices. Pour ces lagunes, le lien entre l’abattement des bactéries - vraisemblablement dû pour l’essentiel à la sédimentation - et les variables météorologiques n’apparaît pas d’une grande pertinence.

70
0,001
0,01
0,1
1
1009
/07/
03
09/0
8/03
09/0
9/03
09/1
0/03
09/1
1/03
09/1
2/03
09/0
1/04
09/0
2/04
09/0
3/04
09/0
4/04
09/0
5/04
09/0
6/04
09/0
7/04
A12
FFC
L1
L2
L3
F1
F2
Figure 3.19. Constantes d’abattement calculées des E. Coli
Les constantes d’abattement dans les FFC semblent subir des variations saisonnières plus ou moins nettes, bien que le traitement des données ne fasse pas apparaître de dépendance du rayonnement solaire. Pour les E. Coli, l’effet saisonnier se manifeste clairement : De 1,1 j-1 en juillet et août, k diminue jusqu’à 0,17 j-1 à la fin de janvier puis augmente à la fin d’avril pour atteindre la valeur de 0.8 j-1 à la mi- juillet. Pour les entérocoques k varie peu, et l’été apparaît légèrement meilleur pour la désinfection: 0,5 j-1 de juillet à début octobre, 0,3 j-1 d’octobre à fin mars et 0.4 j-1 de fin mars à mi-juillet. Le lien avec les variables météorologiques semble forcer le caractère saisonnier de l’évolution de k pour les E. Coli. La variation de la constante d’abattement des E. Coli dans le premier bassin de maturation L1 faut aussi apparaître de nettes tendances saisonnières: En juillet et août les valeurs de k varient autour de 5,2 j-1, diminuent jusqu’à atteindre un minimum de 0,87 j-1 à la fin de décembre puis augmentent de 0,98 à 4,2 j-1 de fin mars à mi-juillet. Dans ce cas, le lien avec les variables météorologiques ou, plus exactement, la procédure d’optimisation, permet de lisser les valeurs expérimentales, en laissant à la seule température le soin d’expliquer les variations de la constante. Pour les entérocoques, k calculé varie dans de moindres proportions: la constante est de l’ordre de 0,9 j-1 de juillet à début octobre et de mi- mai à mi-juillet ; elle est proche 1,1 j-1 de début octobre à mi-mai. Elle paraît, curieusement, être un peu plus élevée en hiver. Les variations erratiques de la constante expérimentale ne permettent pas à la procédure d’optimisation de justifier les variations de cette cinétique. Dans le second bassin de maturation, la constante d’abattement des E. Coli varie autour de 0.5 j-1 sauf du début février à la mi-mai où elle est de l’ordre de 1,44 j-1. Cet accroissement en fin d’hiver est assez surprenant. Pour les entérocoques, k fluctue beaucoup : de l’ordre de 0,2 j-1 de juillet à mi-octobre, 0,7 j-1 de mi-octobre à fin décembre, 1,54 j-1 de fin décembre à début février, 3,55 j-1 de début février à fin mars et 0,5 j-1 de fin mars à mi-juillet. Ces fluctuations conduisent à considérer que la désinfection est plus faible l’été et l’automne qu’en hiver (Figure 3.9). Les singularités dans les valeurs observées à l’entrée de la lagune L2 apparaissent introduire d’importantes difficultés dans la procédure d’optimisation. Les variations des constantes d’élimination avec le climat, à l’opposé des tendances habituelles, accentuées par la procédure d’optimisation, ne sont pas faciles à justifier. On peut toutefois invoquer un rôle

71
important du rayonnement dans des bassins à une période où les algues, dans ce bassin, sont encore rares. La constante d’abattement des E. Coli calculée dans le dernier bassin de maturation est généralement de l’ordre 0,25 j-1 sauf du début février au début mai (0,8 j-1). La constante d’abattement des entérocoques présente la même tendance : de l’ordre de 0,1 j-1 sauf du début février à la mi-mai (0,5 j-1). On retrouve donc une certaine similitude avec l’évolution des constantes calculées dans L2 avec des valeurs élevées à la fin de l’hiver et au début du printemps. Les variations des constantes d’abattement des E. Coli et des entérocoques présentent la même allure dans la première lagune de finition : de faibles valeurs en été et de mi-mai à mi-juin, de l’ordre de 0,2 j-1, une augmentation sensible pendant une période intermédiaire entre fin décembre à fin mars, plus marquée pour les E. Coli que pour les entérocoques. Ces variations surprenantes conduisent à une représentation perturbée de la décontamination de mi-novembre à fin janvier aussi bien pour les E. Coli que pour les entérocoques (Figure 3.13). Pour ce bassin des remarques analogues à celles émises pour L2 peuvent être formulées. Là aussi la décontamination plus faible en été pourrait être expliquée par un développement algal important limitant l’action du rayonnement solaire sur les bactéries. La procédure d’optimisation accentue le caractère saisonnier de la décontamination calculée dans le dernier bassin de finition, avec des valeurs moyennes de k respectivement égales à 0,64 et 0,30 j-1 de juillet à septembre, à 0,28 et 0,04 j-1 d’octobre à février et de 0,78 à 0,3 de mars à juin pour les E. Coli et les entérocoques. On retrouve encore des fortes valeurs, cette fois un peu plus tard, de mars à mai. Au total, le lien imposé par la procédure d’optimisation accentue - ou force - les caractères saisonniers de l’abattement des bactéries témoins de contamination. Des observations plus précises mériteraient d’être effectuées pour vérifier la pertinence des hypothèses d’interprétation que l’on a avancées. Les calculs indiquent que, pour les E. coli, la lagune L1 offre les plus fortes valeurs de k pendant l’essentiel de la période d’étude (Figure 3.19). Cela est aussi vrai pour les entérocoques mais avec des valeurs de k nettement plus faibles (Figure 3.18). La lagune L1, qui reçoit une partie des effluents des bassins anaérobies, joue donc un rôle très important dans la décontamination. Mais, en fin d’hiver, les valeurs de k sont plus fortes dans le bassin L2, voire dans L3 ou même F2 en début de printemps. C’est, d’ailleurs, pour cette période que les simulations donnent les meilleures performances en décontamination. 3.2.3. Application du modèle à l’optimisation de la gestion On sait que la gestion du lagunage, telle qu’elle est mise en oeuvre actuellement, implique :
- qu’une partie des effluents des bassins anaérobies est directement dirigée vers le bassin L1 sans passer par les bassins facultatifs,
- que des variations des niveaux d’eau dans les bassins sont imposées dans le but d’amortir l’effet des accroissements de débit d’entrée dans la station par temps de pluie.
D’autre part, de début octobre 2003 à fin avril 2004, une partie des effluents de R4 est passée directement dans la lagune de finition F1, soit comme résultat d’une gestion par temps de pluie, soit parce que des fuites n’ont pas été arrêtées.

72
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
09/0
8/20
03
10/0
9/20
03
12/1
0/20
03
13/1
1/20
03
15/1
2/20
03
16/0
1/20
04
17/0
2/20
04
20/0
3/20
04
21/0
4/20
04
23/0
5/20
04
24/0
6/20
04
26/0
7/20
04
Q m
3/j
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
Ent
éroc
oque
s (N
/100
ml)
P m3/j
BactEntrée
BactSortie Avec By-Pass
BactSortie Ss by-Pass
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/20
03
04/0
8/20
03
31/0
8/20
03
27/0
9/20
03
24/1
0/20
03
20/1
1/20
03
17/1
2/20
03
13/0
1/20
04
09/0
2/20
04
07/0
3/20
04
03/0
4/20
04
30/0
4/20
04
27/0
5/20
04
23/0
6/20
04
20/0
7/20
04
16/0
8/20
04
Q m
3/j
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
E.
Col
i (N
/100
ml)
P m3/j
BactEntrée
BactSortie Avec By-Pass
BactSortie Ss by-Pass
Figure 3.20. Décontamination microbienne avec et sans by-pass
Nous avons supposé qu’une meilleure décontamination pourrait résulter d’un autre mode de gestion, plus simple que le précédent. Il consisterait à supprimer les deux by-pass, respectivement de la sortie des bassins anaérobies à la première lagune de maturation L1 et de la lagune R4 à la première lagune de finition F1. Ainsi, tous les bassins fonctionnent strictement en série. La distribution étagée dans les lagunes facultatives est conservée. On ne suppose pas de recirculation. On a pris un niveau d’eau moyen dans les bassins. Les constantes cinétiques ont les valeurs calculées précédemment, ce qui revient à admettre que le nouveau mode de gestion ne modifie pas les cinétiques de décontamination. Les concentrations en indicateurs de contamination fécale, montre que, pour l’essentiel de la période, ce fonctionnement devrait bien améliorer la décontamination. Cela est particulièrement vrai pour les E. coli, pour lesquels, pendant les périodes chaudes, les concentrations seraient diminuées de plus de deux U.log. Pour les entérocoques, ce gain n’excèderait guère une unité logarithmique. Les gains sont moins sensibles pendant la période froide. On peut même observer l’absence de gains de qualité au moment des forts épisodes pluvieux.

73
Les gains de performance prévus par la simulation pendant la plus grande partie de la période considérée sont dus à la suppression des by-pass qui polluent les effluents de l’étage facultatif à l’entrée de L1 et les effluents de l’étage de maturation à l’entrée de F1. La diminution des gains voire des pertes de performance apparaissent dans les prévisions pour les périodes pluvieuses vraisemblablement parce que nous avons choisi de ne pas stocker d’eau les apports en excès en faisant varier les niveaux de l’eau dans les bassins. Ces simulations montrent, d’une part que la gestion des apports de temps de pluie actuellement mise en œuvre est plutôt un succès mais que, d’autre part, pendant la plus grande partie de l’année, des performances bien supérieures pourraient être obtenues en supprimant les by-pass. Des scénarios combinant suppression des by-pass et variation des volumes dans les bassins mériteraient d’être testés. 3.1.4. Les limites d'une cinétique de l'abattement bactérien fonction de la température et de l'intensité du rayonnement solaire Il est rappelé que les applications précédentes sont basées sur une constante cinétique de l'élimination bactérienne exprimée en fonction de la température et de l'intensité du rayonnement solaire selon l'Equation 3-3. Les variations des valeurs des coefficients α, β et χ d’un bassin à l’autre tendent à montrer que ces paramètres à eux seuls ne suffisent pas pour expliquer l'abattement bactérien. Afin de mieux définir les limites d'une telle cinétique, une modélisation de l’élimination des coliformes fécaux en 1986 et de 1988 à 1990 (Figure 3.21) ainsi que des entérocoques de 1988 à1990 (Figure 3.22) ont été effectuées pour étudier la reproductibilité des constantes cinétiques de l'abattement des coliformes fécaux ou E. coli et des streptocoques dans les 2 dernières lagunes de maturation. Ayant fonctionné en tant que système facultatif avant l’agrandissement, la première lagune de maturation n’est pas incluse dans cette étude. Par contre, les lagunes L2 et L3 sont des lagunes de maturation avant et après la rénovation de la station. On pourrait donc s’attendre à trouver une relative stationnarité de la relation entre leurs cinétiques de décontamination et les variables explicatives, Im et T. Les Figures 3.21 et 3.22 montrent de très satisfaisantes simulations de la décontamination dans les lagunes L2 et L3. Les valeurs des coefficients α, β et χ correspondantes sont présentées dans le Tableau 3.6. On y observe des différences importantes des valeurs de α et des différences significatives mais moins importantes de β et χ. La stationnarité espérée n’est pas au rendez-vous. Cela est mieux illustré par les Figures 3.23 à 3.30. Les Figures 3.23 et 3.25 représentent la reconstitution de l’évolution des teneurs en E. coli à la sortie des lagunes L2 et L3 pour la période 2003-2004 en utilisant les paramétrages de l'Equation 3-3 effectués grâce aux données des 3 périodes 1986, 1988-1990 et 2003-2004. Les évolutions correspondantes des constantes cinétiques de décontamination sont données dans les Figures 3.24 et 3.26. Le même travail a été effectué pour les entérocoques, avec les concentrations représentées sur les Figures 3.27 et 3.29 et les constantes cinétiques sur les Figures 3.28 et 3.30.

74
0
1 000
2 000
3 000
4 000
5 000
6 000
7 000
01/0
1/86
22/0
1/86
12/0
2/86
05/0
3/86
26/0
3/86
16/0
4/86
07/0
5/86
28/0
5/86
18/0
6/86
Q (m
3 /j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
N/1
00m
l n
P m3/j
Qe
Qs calc
CF Entrée L2 Exp
CF Sortie L2 Exp
CF Sortie L2 calc
0
1 000
2 000
3 000
4 000
5 000
6 000
7 000
01/0
1/86
22/0
1/86
12/0
2/86
05/0
3/86
26/0
3/86
16/0
4/86
07/0
5/86
28/0
5/86
18/0
6/86
Q (m
3 /j)
1,00E+00
1,00E+01
1,00E+02
1,00E+03
1,00E+04
1,00E+05
1,00E+06
N/1
00m
l n
P m3/jQe
Qs calcCF Entrée L3 Exp
CF Sortie L3 ExpCF Sortie L3 Calc
0
1 000
2 000
3 000
4 000
5 000
6 000
01/1
0/19
88
26/1
1/19
88
21/0
1/19
89
18/0
3/19
89
13/0
5/19
89
08/0
7/19
89
02/0
9/19
89
28/1
0/19
89
23/1
2/19
89
Q (m
3 /j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
N/1
00m
l n
P m3/j
Qe
Qs calc
CF Entrée L2 Exp
CF Sortie L2 Exp
CF Sortie L2 calc
0
1 000
2 000
3 000
4 000
5 000
6 000
01/1
0/19
88
27/1
1/19
88
23/0
1/19
89
21/0
3/19
89
17/0
5/19
89
13/0
7/19
89
08/0
9/19
89
04/1
1/19
89
31/1
2/19
89
Q (m 3
/j) n
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
N/1
00m
l n
P m3/jQe
Qs calcCF Entrée L3
CF Sortie L3 ExpCF Sortie L3 Calc
Figure 3.21. Teneurs en coliformes fécaux observées et calculées dans les 2 derniers bassins de maturation en 1986 et de 1988 à 1990
0
1 000
2 000
3 000
4 000
5 000
6 000
01/1
0/19
88
27/1
1/19
88
23/0
1/19
89
21/0
3/19
89
17/0
5/19
89
13/0
7/19
89
08/0
9/19
89
04/1
1/19
89
31/1
2/19
89
Q (m
3 /j)
m
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
N/1
00m
l
p
P m3/jQe
Qs calcEntéro Entrée L2 Exp
Entéro Sortie L2 ExpEntéro Sortie L2 Calc
0
1 000
2 000
3 000
4 000
5 000
6 000
01/1
0/19
88
29/1
1/19
88
27/0
1/19
89
27/0
3/19
89
25/0
5/19
89
23/0
7/19
89
20/0
9/19
89
18/1
1/19
89
16/0
1/19
90
Q (m
3 /j)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
N/1
00m
l
m
P m3/jQeQs calcEntéro Entrée L3Entéro Sortie L3 ExpEntéro Sortie L3 Calc
Figure 3.22. Teneurs en streptocoques fécaux observées et calculées dans le dernier bassin de maturation de 1988 à 1990

75
Tableau 3.6. Coefficients cinétiques ajustés avec les données de 1986, de 1988 à 1990 et de 2003 à 2004 de l'abattement des coliformes fécaux et des entérocoques dans les 2 derniers bassins de maturation
Coliformes fécaux Entérocoques α β χ α β χ L 2 1986 0.726 1.127 0.000 1988-1990 0.052 0.987 0.227 1.065 1.079 0.000 2003-2004 0.123* 0.926* 0.082* 0.066 0.824 0.082 L 3 1986 0.735 0.992 0.031 1988-1990 0.976 1.199 0.000 0.436 0.994 0.000 2003-2004 0.020* 0.952* 0.141* 0.001 1.049 0.290
*coefficients cinétiques de l'abattement des E. coli
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
E. c
oli (
N/1
00m
l)
Entrée L2 Obs
Sortie L2 Obs
Sortie L2 Calc/ k_2003-2004
Sortie L2 Calc/ k_1988-1990
Sortie L2 Calc/ k_1986
Figure 3.23. Teneurs observées et simulées en E. coli à la sortie de L2 de juillet 2003 à août 2004
1,E-02
1,E-01
1,E+00
1,E+01
1,E+02
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
k L2
(j-1)
Obs
2003-2004
1988-1990
1986
Figure 3.24. Constantes cinétiques observées et simulées de l’abattement des E. coli dans L2 de juillet 2003 à août 2004

76
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+0508
/07/
2003
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
E. c
oli (
N/1
00m
l) Sortie L2 Obs
Sortie L3 Obs
Sortie L3 Calc/ k_2003-2004
Sortie L3 Calc/ k_1988-1990
Sortie L3 Calc/ k_1986
Figure 3.25. Teneurs observées et simulées en E. coli à la sortie de L3 de juillet 2003 à août 2004
1,E-02
1,E-01
1,E+00
1,E+01
1,E+02
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
k L3
(j-1)
Obs
2003-2004
1988-1990
1986
Figure 3.26. Constantes cinétiques observées et simulées de l’abattement des E. coli dans L3 de juillet 2003 à août 2004

77
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+0611
/08/
2003
15/0
9/20
03
20/1
0/20
03
24/1
1/20
03
29/1
2/20
03
02/0
2/20
04
08/0
3/20
04
12/0
4/20
04
17/0
5/20
04
21/0
6/20
04
26/0
7/20
04
Ent
éroc
oque
s (N
/100
ml) Entrée L2 Obs
Sortie L2 Obs
Sortie L2 Calc/ k_2003-2004
Sortie L2 Calc/ k_1988-1990
Figure 3.27. Teneurs observées et simulées en entérocoques à la sortie de L2 de juillet 2003 à août 2004
0,001
0,01
0,1
1
10
100
04/0
8/20
03
01/0
9/20
03
29/0
9/20
03
27/1
0/20
03
24/1
1/20
03
22/1
2/20
03
19/0
1/20
04
16/0
2/20
04
15/0
3/20
04
12/0
4/20
04
10/0
5/20
04
07/0
6/20
04
05/0
7/20
04
02/0
8/20
04
k L2 (
j-1) Obs
2003-2004
1988-1990
Figure 3.28. Constantes cinétiques observées et simulées de l’abattement des entérocoques dans L2 de juillet 2003 à août 2004

78
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
12/0
8/20
03
17/0
9/20
03
23/1
0/20
03
28/1
1/20
03
03/0
1/20
04
08/0
2/20
04
15/0
3/20
04
20/0
4/20
04
26/0
5/20
04
01/0
7/20
04
06/0
8/20
04
Ent
éroc
oque
s (N
/100
ml)
Sortie L2 Obs
Sortie L3 Obs
Sortie L3 Calc/ k_2003-2004
Sortie L3 Calc/ k_1988-1990
Figure 3.29. Teneurs observées et simulées en entérocoques à la sortie de L3 de juillet 2003 à août 2004
1,E-03
1,E-02
1,E-01
1,E+00
1,E+01
1,E+02
04/0
8/20
03
01/0
9/20
03
29/0
9/20
03
27/1
0/20
03
24/1
1/20
03
22/1
2/20
03
19/0
1/20
04
16/0
2/20
04
15/0
3/20
04
12/0
4/20
04
10/0
5/20
04
07/0
6/20
04
05/0
7/20
04
02/0
8/20
04
k L3 (
j-1) Obs
2003-2004
1988-1990
Figure 3.30. Constantes cinétiques observées et simulées de l’abattement des entérocoques dans L3 de juillet 2003 à août 2004
3.1.5. Influence du modèle de comportement hydraulique. Le comportement hydraulique d'un bassin est un facteur important de l'élimination microbienne puisqu'il détermine la distribution des temps de séjour. Les équations 3.8 à 3.10 représentent les expressions respectives de la teneur en bactéries à la sortie d’une lagune pour un écoulement piston, une lagune se comportant comme un mélangeur parfait et un écoulement convectif dispersif.
τpistonkeNN −⋅= 0 (Équation 3.8)
τmélangek
NN
+=
10 (Équation 3.9)

79
( ) ( ) d
a
d
a
d
eaea
aeNN
2222
2
1
0
11
4−
−−+= (Équation 3.10)
où 0N est la teneur en coliformes à l'entrée, dka dispersifτ41+= , d le nombre de dispersion
et τ le temps de séjour de l’eau défini comme entre le volume et le débit transitant dans la lagune considérée.
Tableau 3.7. Formules empiriques du nombre de dispersion
d Référence
H
L
l
L ⋅+⋅ 055.031.0
1
Namèche et Vasel (1998)
( )
⋅+−−
⋅⋅
+ l
H
l
H
L
H
LlH
Hl385.1981.0410.0
4
23102.0
τν
Agunwamba et al. (1992)
l
L
l
L
l
L⋅
++−2
014.1254.0261.0
1 Yanez (1993)
l
L1
Von Sperling (2005)
Notations : L longueur (m) l largeur (m) H hauteur (m) τ temps de séjour moyen (j) ν viscosité cinématique de l'eau (m2/j) avec 450.0325.0 −⋅= Tν d'après Von Sperling (1999) Plusieurs formules empiriques du nombre de dispersion ont été élaborées, dont quelques unes sont présentées dans le Tableau 3.7. Sans des essais de traçage, il est impossible de définir avec précision le comportement hydraulique d'un système. Par ailleurs, selon Drakides et Djajadiningrat (1981), les travaux entrepris dans ce sens font généralement ressortir le fait que l'hydraulique dans les bassins de lagunage oscillerait essentiellement en fonction des conditions météorologiques, entre un écoulement avec court-circuit et un écoulement en piston, en passant par un comportement de mélangeur parfait. Ils affirment aussi que ces travaux mettent en outre communément en évidence l'existence de zones mortes. La géométrie des bassins constitue avec les conditions météorologiques, les paramètres fondamentaux de la détermination du modèle de comportement hydraulique. Von Sperling

80
(1999) affirme que si le rapport eurL
Longueur
argn'excède pas l'unité de façon significative, le
modèle décrivant le mieux l'hydraulique serait le mélange parfait, sinon le modèle convectif dispersif devrait être privilégié. Les résultats d’essais de traçage sur des bassins de lagunage ont amené Namèche et Vasel (1998) à déduire que la valeur du nombre de Peclet Pedéfini par l’Equation 3-11 détermine le modèle de comportement hydraulique.
dPe
1= (Équation 3.11)
où d est le nombre de dispersion. Le mélangeur parfait serait caractérisé par 2≤Pe , l'écoulement en piston par 4≥Pe . Si
42 << Pe , le système serait partiellement mélangé. Namèche et Vasel ont en outre conclu à partir de données fournies par la littérature et de celles dont ils disposaient que la valeur maximale du nombre de Peclet est de 2,70 pour les systèmes dont le rapport L/l ≤ 8 et que l'erreur induite sur la prévision de leur performance en appliquant le modèle du mélangeur parfait pour l'hydraulique serait alors inférieure à 13%.
Tableau 3.8. Nombres de Peclet des bassins de maturation et de finition de la station de lagunage de Mèze
Bassins H* (m) Longueur (m) Largeur (m) l
L Pe**
M1 2.20 557 70 7.96 16.39 M2 1.90 351 57 6.16 12.07 M3 2.20 300 65 4.62 8.93 F1 3.60 220 90 2.44 4.12 F2 2.83 200 77 2.60 4.73
* profondeur du bassin et non de l'eau ** calculs effectués à partir de l’expression selon Namèche et Vasel (1998) du nombre de dispersion (Tableau 3.7) D’une part, Namèche et Vasel (1998) ont montré qu’un bassin dont le rapport L/l excède largement l’unité peut avoir un nombre de Peclet inférieur ou égal à 2 et donc un comportement hydraulique se rapprochant d’un mélange parfait. D’autre part, un système dont le rapport L/l est inférieur ou égal à 8 peut avoir un nombre de Peclet largement supérieur à 2, impliquant un écoulement en piston pour l’hydraulique. Tel est le cas des bassins de maturation et de finition de la station de lagunage de Mèze. Chacun de ces bassins est en effet caractérisé par un rapport L/l ≤ 8 et un nombre de Peclet supérieur à 4 (Tableau 3.8). Le calcul de k dépend du modèle de comportement hydraulique retenu. Von Sperling (2002) a établi des correspondances entre les valeurs issues du choix des trois modèles ; il montre que, pour un système donné, si Cpistonk °20, , Cmélangek °20, et Cdispersifk °20, sont les coefficients cinétiques
de l'élimination bactérienne si le modèle de l’hydraulique adopté est celui respectivement de l’écoulement en piston, du mélange parfait et de l’écoulement convectif dispersif, alors :

81
Si 4.01.0 ≤≤ d et , 100 20, ≤⋅≤ ° τCdispersifk , avec τ en jours et d = l/L,
( ) 4145.10137.320,
20,
20, 0020.00.1 −°
°
° ⋅+= dkk
kCdispersif
Cdispersif
Cmélange τ (Équation 3.12)
( ) 1880.04157.020,
20,
20, 2414.00.1 dkk
kCdispersif
Cdispersif
Cpiston τ⋅−= °°
° (Équation 3.13)
Si 0.11.0 ≤≤ d et 50 20, ≤⋅≤ ° τCdispersifk ,
( ) 8426.08166.120,
20,
20, 0540.00.1 −°
°
° ⋅+= dkk
kCdispersif
Cdispersif
Cmélange τ (Équation 3.14)
( ) 3415.05351.020,
20,
20, 2425.00.1 dkk
kCdispersif
Cdispersif
Cpiston τ⋅−= °°
° (Équation 3.15)
Les études de la cinétique de l'élimination des coliformes fécaux (ou E. coli) effectuées sur 186 bassins de lagunage facultatif et de maturation dont 156 implantés au Brésil et 30 répartis dans 9 autres pays (l'Argentine, la Colombie, le Chili, le Venezuela, le Mexique, l'Espagne, la Belgique, le Maroc et la Palestine), ont amené Von Sperling (2005) à rechercher l'existence d'une relation entre la hauteur de l'eau dans les bassins et la constante cinétique à 20°C. La constante k est supposée varier avec la température selon l’expression
)20(20 07,1. −= Tkk (Équation 3.16)
Von Sperling calcule les constantes k en supposant que le comportement hydraulique des lagunes est de type convectif dispersif ; le nombre de dispersion étant la forme donnée dans l’expression (E305d). A l’aide d’analyses de régression, il obtient effectivement une relation entre constante cinétique et hauteur d’eau :
456.120, 549.0 −
° = Hk Cdispersif (Équation 3.17)
Le coefficient de détermination de Cdispersifk °20, est alors de 0,571 tandis que celui de la
prévision du logarithme décimal de la teneur en bactéries à la sortie des lagunes est de 0,845. En utilisant les Equations 3.18 à 3.21, on peut relier les valeurs de Cmélangek °20, et Cpistonk °20, à
la hauteur d’eau par les équations suivantes : Si 4.01.0 ≤≤ d et , 100 20, ≤⋅≤ ° τCdispersifk ,
4145.10137.38439.5456.120, 0002.0549.0 −−−
° += dHHk Cmélange τ (Équation 3.18) 1880.04157.00613.2456.1
20, 1033.0549.0 dHHk Cpiston τ−−° −= (Équation 3.19)
Si 0.11.0 ≤≤ d et 50 20, ≤⋅≤ ° τCdispersifk ,
8426.08166.11010.4456.120, 0100.0549.0 −−−
° += dHHk Cmélange τ (Équation 3.20) 3415.05351.02351.2456.1
20, 0966.0549.0 dHHk Cpiston τ−−° −= (Équation 3.21)

82
Nous avons calculé les valeurs des constantes cinétiques dans les bassins de maturation et de finition de Mèze en utilisant l’équation 3.17 afin de voir si une approche reposant sur la seule considération de la géométrie des lagunes, de la température de l’eau et du temps de séjour moyen permet d’approcher correctement la décontamination observée. La comparaison a été faite en ramenant les valeurs de kdispersif telles que calculés par l’équation 3.17 à des valeurs de kmélange en utilisant les relations 3.18 et 3.20. Connaissant les valeurs de kmélange, on a calculé les concentrations en E. coli mais aussi en entérocoques à la sortie des lagunes de maturation et de finition à partir des concentrations observées à l’entrée des bassins respectifs. Ces concentrations ont été comparées avec les concentrations observées et calculées à l’aide des constantes cinétiques issues de la procédure d’optimisation. Il se trouve que la solution de Von Sperling restitue aussi bien les concentrations en E. coli. à la sortie de la lagune L1 que ne l’a fait notre modèle. Les résultats sont moins convaincants pour ce qui concerne les entérocoques ... mais la solution de Von Sperling a été établie pour les coliformes.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
08/
07/2
00
3
08/
09/2
00
3
08/
11/2
00
3
08/
01/2
00
4
08/
03/2
00
4
08/
05/2
00
4
08/
07/2
00
4
Ent
éroc
oque
s (N
/100
ml)
SL1 calc
SL1 Exp
SL1 V. S.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
08/
07/2
00
3
08/
09/2
00
3
08/
11/2
00
3
08/
01/2
00
4
08/
03/2
00
4
08/
05/2
00
4
08/
07/2
00
4
E.
Col
i (N
/100
ml)
SL1calc
SL1 Exp
SL1 V. S.
Figure 3.31. Concentrations en entérocoques et E. coli observées (Exp) et calculées à la sortie de la lagune L1 en utilisant les cinétiques de Von Sperling (V.S.) et celles déduites de la procédure d’optimisation (calc)
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
En
téro
coqu
es (
N/1
00m
l)
SL2 calc
SL2 Exp
SL2 V. S.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
E. C
oli (
N/1
00m
l)
SL2 calc
SL2 Exp
SL2 V. S.
Figure 3.32. Concentrations en entérocoques et E. coli observées (Exp) et calculées à la sortie de la lagune L2. en utilisant les cinétiques de Von Sperling (V.S.) et celles déduites de la procédure d’optimisation (calc)

83
La représentation des concentrations en E. coli à la sortie de L2 est, moins satisfaisante que celle fournie par notre propre modèle pendant les périodes chaudes. Il apparaît donc que la loi de variation des cinétiques avec la température surestime les cinétiques pour les températures élevées. La représentation des concentrations en entérocoques est, comme pour L1, moins bonne.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
Ent
éroc
oque
s (N
/100
ml)
SL3 calc
SL3 Exp
SL3 V. S.1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
E. C
oli (
N/1
00m
l)
SL3 calc
SL3 Exp
SL3 V. S.
Figure 3.33. Concentrations en entérocoques et E. coli observées (Exp) et calculées à la sortie de la lagune L3. en utilisant les cinétiques de Von Sperling (V.S.) et celles déduites de la
procédure d’optimisation (calc)
Dans le bassin L3, la solution de Von Sperling conduit à des représentations contrastées de l’évolution des concentrations en indicateurs de contamination ; pendant la plus grande partie du temps, les valeurs ainsi calculées sont proches des valeurs observées. Cependant les abattements sont surestimés pendant l’été 2004.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
Ent
éroc
oque
s (N
/100
ml)
SF1 calc
SF1 Exp
SF1 V. S. 1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08
/07
/20
03
08
/09
/20
03
08
/11
/20
03
08
/01
/20
04
08
/03
/20
04
08
/05
/20
04
08
/07
/20
04
E.
Col
i (N
/100
ml)
SF1 calc
SF1 Exp
SF1 V. S.
Figure 3.34. Concentrations en entérocoques et E. coli observées (Exp) et calculées à la sortie de la lagune F1 en utilisant les cinétiques de Von Sperling (V.S.) et celles déduites de la
procédure d’optimisation (calc)

84
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
Ent
éroc
oque
s (N
/100
ml)
SF2 calc
SF2 Exp
SF2 V. S.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
08/0
7/20
03
08/0
9/20
03
08/1
1/20
03
08/0
1/20
04
08/0
3/20
04
08/0
5/20
04
08/0
7/20
04
E. C
oli (
N/1
00m
l)
SF2 calc
SF2 Exp
SF2 V. S.
Figure 3.35. Concentrations en entérocoques et E. coli observées (Exp) et calculées à la sortie de la lagune F2 en utilisant les cinétiques de Von Sperling (V.S.) et celles déduites de la
procédure d’optimisation (calc)

85
Chapitre 4. ELIMINATION DE L'AZOTE

86

87
L’azote est l’un des principaux nutriments contenus dans les effluents de station d’épuration. C’est un composé essentiel à l’activité biologique dans l’eau. Toutefois, sa présence dans les rejets des stations d’épuration entraîne un risque d’eutrophisation des milieux aquatiques, particulièrement quand ceux-ci se renouvellent lentement ou pas du tout, tels que les étangs, les lacs et les rivières lentes. L’eutrophisation peut se manifester par une accélération de la croissance des algues et d’autres plantes aquatiques. Outre les problèmes esthétique et olfactif qu’elles engendrent, les algues et les plantes aquatiques peuvent affecter l’utilisation des ressources en eau. Elles constituent un supplément de matières organiques à traiter pour produire de l’eau potable. La sédimentation et la décomposition des plantes mortes limitent les activités de pêche par déplétion de l’oxygène en profondeur. La présence d’algues constitue aussi une source de perturbation pour la pratique d’activités de loisirs telles que la baignade et la plongée. Des concentrations importantes en azote dans les effluents peuvent également avoir d’autres effets négatifs en raison de la toxicité des formes oxydées et des propriétés de la forme ammoniacale. L’ammoniac présente une grande toxicité pour plusieurs organismes aquatiques. Une teneur faible en NH3 de 1 mg/l est déjà toxique pour certaines espèces de poissons (Hemens, 1966; Minocha et Prabhakar, 1988). De plus l’ammoniac peut conduire à un appauvrissement du milieu en oxygène. Par ailleurs, la contamination des ressources en eau par l’azote ammoniacal diminue l’efficacité des opérations de désinfection par chloration. Le chlore devient alors moins disponible en se combinant avec l’ammoniac pour former des chloramines, moins aseptisantes. Aussi, dans de nombreux pays, la législation environnementale exige d’ores et déjà que les stations d’épuration éliminent l’azote. Depuis un certain temps, on reconnaît cette capacité au lagunage, tout en sachant que son efficacité varie selon la nature des bassins et les conditions environnementales. Les principaux mécanismes d’élimination mis en jeu (Figure 4.1) sont en effet fortement dépendants des conditions météorologiques, chimiques et biologiques qui interfèrent avec le cycle biologique de l’azote. La variabilité de ces conditions rend l’élimination de l’azote difficilement maîtrisable en lagunage. D’autre part, faute d’études suffisamment approfondies, les avis des spécialistes divergent sur le(s) mécanisme(s) prépondérant(s) dans l’élimination de l’azote. Reed (1985) mentionnait déjà que « les articles et la littérature technique publiés au cours des deux dernières décades font apparaître des contradictions quant aux voies et aux mécanismes responsables des pertes d’azote. ». Cet état de fait constitue un handicap préoccupant dès lors qu’il s’agit de trouver des solutions pour améliorer les performances des lagunages afin de mieux protéger les milieux récepteurs sensibles et de répondre aux nouvelles exigences réglementaires. Les voies d’élimination de l’azote dans les lagunages telles que énumérées par Reed (1985), sont l’assimilation dans les algues, la volatilisation de l’ammoniac (gaz NH3), la nitrification-dénitrification, la rétention dans les boues accumulées au fond des bassins et l’adsorption de l’ammonium, cette dernière voie étant rarement mentionnée. L’assimilation dans les algues ou les plantes peut s’apprécier diversement. Dans le cas des lagunes à macrophytes, l’assimilation dans les plantes signifie une élimination de l’azote dès lors que ces plantes sont récoltées. Dans les lagunes à microphytes ou quand les macrophytes ne sont pas récoltées, l’azote assimilé est, soit exporté dans les effluents soit, après la mort et la décomposition des plantes, sédimenté dans le fond des bassins et, de là, recyclé dans la colonne d’eau ou stocké dans les boues non minéralisées. King (1978) avait chiffré à 4% la part de la récolte des algues (macrophytes) dans l’élimination de l’azote dans le premier bassin d’un système de lagunage alimenté par des effluents domestiques secondaires et à 9% cette même part pour l’ensemble du système qui comptait 4 bassins. La récolte des lentilles

88
d’eau dans un système de 4 bassins pilotes en série alimentés par des eaux usées domestiques représentait de 10 à 27% de l’élimination de l’azote, cette proportion variant avec les saisons (Zimmo et al, 2004). Les résultats du suivi du devenir de l’azote réalisé par Reedy (1983) à l’aide de l’isotope +
415NH paraissent attribuer à l’assimilation algale un rôle marginal puisque
5% seulement de l’azote était assimilé par les algues alors que 21% de l’azote était allé dans les sédiments, 21% retrouvé dans la colonne d’eau et la perte par d’autres voies était estimée à 53%. Il est cependant vraisemblable qu’une partie significative des sédiments provenait de la décomposition des algues et, donc, indirectement de l’assimilation. En adoptant la théorie, qui prévalait dans la littérature de l’époque, selon laquelle le lagunage n’est pas un contexte particulièrement favorable au développement du processus de nitrification-déntrification - théorie apparemment corroborée par l’observation que l’élimination de l’azote croit avec le pH, ce qui favorise la volatilisation de NH3, et avec les fortes teneurs en oxygène dissous, elles même peu favorables à la dénitrification - , King (1978) déduit que les pertes en azote proviennent essentiellement d’émissions de gaz NH3 à partir des bassins vers l’atmosphère ou, autrement dit, de la volatilisation. Pano et Middlebrooks (1982), d’une part, et Ferrara et Avci d’autre part (1982), ont étudié en parallèle les mêmes stations de lagunage et, notamment, celle de Corinne-Utah. Pourtant, leurs travaux, même s’ils partagent certains postulats, aboutissent naturellement à des conclusions différentes. Pour Pano et Middlebrooks (1982), le fait que les teneurs en nitrite et nitrate soient toujours faibles dans les bassins de lagunage, aussi bien en été qu’en hiver, signifie l’absence de nitrification et, donc, d’une possible élimination de l’azote par dénitrification. Il leur reste alors à expliquer l’élimination de l’ammonium par la volatilisation et l’assimilation dans la biomasse algale. Ils affirment la prépondérance de la volatilisation pendant les saisons froides parce que les activités biologiques y sont fortement ralenties. En périodes chaudes, en plus de la volatilisation, ils disent prendre en compte, d’une manière indirecte, l’assimilation algale et le relargage d’ammonium à partir des sédiments dans l’estimation des teneurs en azote ammoniacal. La modélisation qu’ils proposent est empirique mais sa forme emprunte largement à la théorie de la volatilisation. Ferrara et Avci (1982) élaborent un modèle dynamique qui prend en compte l’essentiel des processus du cycle de l’azote à ceci près qu’ils font aussi l’hypothèse de la quasi-inexistence de la nitrification. La cinétique de la volatilisation y est représentée conformément à la théorie à base physique établie par Stratton en 1968 et 1969. Or, comme on le verra par la suite, cette théorie et les mesures directes d’émission de gaz NH3 (Zimmo et al., 2003) montrent que la volatilisation ne prend, le plus souvent, qu’une part modeste à l’élimination de l’azote dans les lagunes. Dès lors, la modélisation de Ferrara et Avci aboutit à la conclusion que l’élimination de l’azote se fait essentiellement par sédimentation de la forme organique dans les boues. Dans le lagunage à haut rendement, l’absence de nitrates ou la faible teneur observée en azote nitreux et en azote nitrique ont aussi conduit plusieurs auteurs dont Nurdogan (1988) et El Halouani et al. (1993) à conclure à l’inexistence ou du moins à l’insignifiance de la nitrification. Le bilan moyen entrée-sortie de l’azote sous toutes ses formes autres que nitreuse et nitrique établi alors par El Halouani et al. (1993) suggérait que l’assimilation algale et la volatilisation sont les principaux mécanismes d’élimination. Dans un bilan de l’azote dressé pour un lagunage haut rendement faisant suite à deux étages de traitement anaérobie, El Hafiane et El Hamouri (2005) ont implicitement postulé l’absence de nitrification, d’où l’absence de contrôle des concentrations des formes oxydées de l’azote. Dès lors, ils ont attribué le déficit du bilan entrée-sortie du lagunage à haut rendement à la volatilisation de NH3. Quinze pour cent de l’azote entrant restaient dans les bassins sous

89
forme dissoute, 39% étaient éliminés par assimilation algale et 46% étaient supposés perdus par stripping de l’ammoniac. Le pH élevé généralement enregistré dans ce genre de système peut justifier l’importance de la volatilisation mais les fortes valeurs de l’oxygène dissous observées, y compris la nuit, suggèrent une nitrification significative. Les exemples précédents montrent une large diversité des points de vue sur l’importance relative des différentes voies d’élimination de l’azote ; cette diversité est due au postulat que la nitrification est négligeable, à l’absence de référence à des mesures directes de la volatilisation de NH3, à la non quantification directe de l’accumulation de l’azote dans les boues et, quelquefois aussi, à des ambiguïtés dans la prise en compte de l’assimilation algale.
Figure 4.1. Cycle de l’azote dans les bassins de lagunage
La principale raison qui a poussé de nombreux auteurs à négliger l’importance de la nitrification est que des concentrations significatives des formes oxydées de l’azote sont assez rarement observées dans les lagunages. Cependant Reed (1985) avançait que de faibles teneurs en nitrite et nitrate ne prouvent pas obligatoirement l’inexistence de la nitrification puisqu’elles peut résulter du caractère évanescent des formes oxydées de l’azote sous l’effet de la dénitrification. Mais, selon lui, la nitrification-dénitrification n’est probablement pas pour autant le principal mode d’élimination de l’azote en lagunage classique. Reed pensait que l’environnement anaérobie requis par la dénitrification est restreint au voisinage des boues déposées dans le fond des bassins de lagunage. Il soutenait que, dans cet environnement la production de nitrates est improbable et que, donc, la disponibilité en nitrates pour la dénitrification ne peut qu’être très limitée. Il supposait aussi que la majeure partie des nitrates formés dans la partie aqueuse doit probablement y rester. Aussi Reed estimait qu’en lagunage les principaux modes d’élimination de l’azote sont la volatilisation - laquelle, sous des conditions optimales de température et de pH peut être importante -, et la sédimentation en raison des longs temps de séjour. Or, cette vision de l’hydrodynamique des bassins de lagunage et de la distribution des teneurs en oxygène dissous considérées comme obéissant à
Ammonification N-Organique
Sédimentation
Assimilation par les microorganismes
N-Ammoniacal Nitrification
N-Nitreux et N-Nitrique
Dénitrification
Régénération benthique
Volatilisation
Assimilation par les microorganismes

90
des régimes permanents est loin de correspondre à la réalité (Brissaud et al., 2003). Curieusement, dix ans plus tard, Reed, Crites et Middlebrooks (1995) proposent de dimensionner des lagunes à macrophytes chargées d’éliminer l’azote en se fondant sur les seuls mécanismes de la nitrification et de la dénitrification. Plusieurs travaux récents ont mis en évidence l’existence d’une nitrification substantielle et montré que la combinaison nitrification/dénitrification n’est pas une voie mineure d’élimination de l’azote dans les lagunages. Les travaux de Zimmo et al (2004) ont permis de mesurer la part prise par la nitrification/dénitrification dans l’élimination de l’azote dans des pilotes de lagunage ; cette part variait entre 25 et 33 %. Dans cette étude, les sédiments étaient régulièrement retirés des bassins et la sédimentation comptait pour 52 à 61 % de l’élimination de l’azote. Picot et al (2005), en se fondant sur les variations saisonnières des teneurs en ammonium et en formes oxydées de l’azote dans les bassins de maturation du lagunage de Mèze, estiment que les faibles teneurs en azote observées en été dans les effluents sont très vraisemblablement imputables à la nitrification/dénitrification. Archer et O’Brien (2005) mettent en avant le rôle essentiel de la nitrification/dénitrification dans l’élimination de l’azote en soulignant l’importance des surfaces susceptibles de supporter le développement d’un biofilm, qu’il s’agisse des berges des lagunes ou des digues de graviers dont plusieurs lagunages sont équipés en Nouvelle Zélande. Evans et al. (2005) ont montré que nitrification et dénitrification peuvent conduire à une élimination très importante de l’azote dans les lagunes à haut rendement traitant des effluents d’abattoir. Ainsi, au fil des ans, l’hypothèse du rôle essentiel de la nitrification/dénitrification dans l’élimination de l’azote a-t-elle pris de plus en plus de consistance. Cela est d’autant plus vrai que, parallèlement, les mesures directes de la volatilisation (Zimmo et al., 2003) ou la faible élimination de l’azote observée dans des circonstances a priori très favorables à la volatilisation (Archer et O’Brien, 2005) montrent que l’importance de ce processus a été généralement très surévaluée. A cet égard, les résultats les plus éclairants sont les mesures directes des émissions de gaz NH3 effectuées par Zimmo et al. (2003) sur des lagunes pilotes, qui leur ont permis de conclure que les pertes en azote par volatilisation représentaient au maximum 1.5% de l’élimination totale de l’azote. On verra toutefois par la suite que ces mesures ne sont pas sans défaut. Il apparaît donc opportun de revisiter les théories et les modèles relatifs au devenir de l’azote dans les lagunages, d’en situer les hypothèses et les limites. Dans cette analyse, une place particulière sera faite aux expérimentations permettant la mesure de voies élémentaires d’élimination de l’azote. 4.1. Le modèle de l’élimination de l’azote de Reed (1985) Le modèle de Reed est un cas particulier intéressant car, bien que son auteur soit persuadé de la large prépondérance de la volatilisation, il ne suppose aucune hypothèse sur la nature de la ou des voie(s) principale(s) d’élimination de l’azote. 4.1.1. Conception Deux hypothèses sont à la base de l’élaboration de ce modèle : - l’élimination de l’azote total suit une cinétique du premier ordre ;

91
- un écoulement piston traduit mieux le comportement hydraulique des lagunages naturels aux temps de séjour généralement longs, à bassins multiples et surtout non aérés mécaniquement. Le modèle de Reed prend alors la forme générale :
),,( τpHTfk
E
S eN
N ⋅−= (Équation 4.1)
où
SN est la concentration en azote total à la sortie du lagunage (mg/l),
EN la concentration en azote total à l’entrée (mg/l), k la constante cinétique de l’élimination de l ’azote total exprimée en j-1,
),,( τpHTf une fonction de la température T, du pH et du temps de séjour τ exprimé en jours.
Conformément à l’hypothèse de l’écoulement piston, les valeurs des concentrations EN et SN
prises en compte sont décalées d’un nombre de jours égal à τ. Pour la détermination dek et de la fonctionf , Reed s’est servi des suivis d’un cycle complet d’un an effectués sur 4 stations étudiées par l’EPA et situées à Peterborough dans le New Hampshire (3 bassins en série), Kilmichael dans le Mississipi (3 bassins en série), Eudora au Kansas (3 bassins en série) et Corinne dans l’Utah (7 bassins en série). Il a alors comparé les teneurs en azote à l’entrée avec celles observées à la sortie, respectivement, du premier bassin, des 2 premiers bassins en série et des 3 premiers bassins en série de chaque site. Les 12 systèmes ainsi définis avaient des temps de séjour compris entre 27 et 231 jours. Pendant l’étude, les températures et le pH enregistrés allaient de 1.2 à 26.3°C et de 6.4 à 9.5 respectivement. Des analyses mathématiques de l’influence de chacune des variables T, pH et τ ont été faites en utilisant des périodes durant lesquelles 2 des 3 variables étaient restées relativement constantes, donnant au modèle de Reed l’expression finale :
( )[ ]6.66.60 −+−= pHk
E
S TeN
N τ (Équation 4.2)
où ( )20
20−⋅= T
T kk θ (j-1)
avec 0064.020 =k (j-1)
039.1=θ
T (°C) La totalité des données n’ayant pas été utilisée pour l’élaboration du modèle, la prévision globalement satisfaisante des concentrations moyennes mensuelles à la sortie des lagunes sur l’ensemble de la période fut considérée comme une première validation du modèle. Une deuxième validation fut obtenue, avec des résultats très convaincants, à partir des données relatives à 5 autres stations situées dans le Mississipi et au Canada. Il était donc tentant de voir dans quelle mesure ce modèle pouvait rendre compte des observations effectuées sur le lagunage de Mèze.
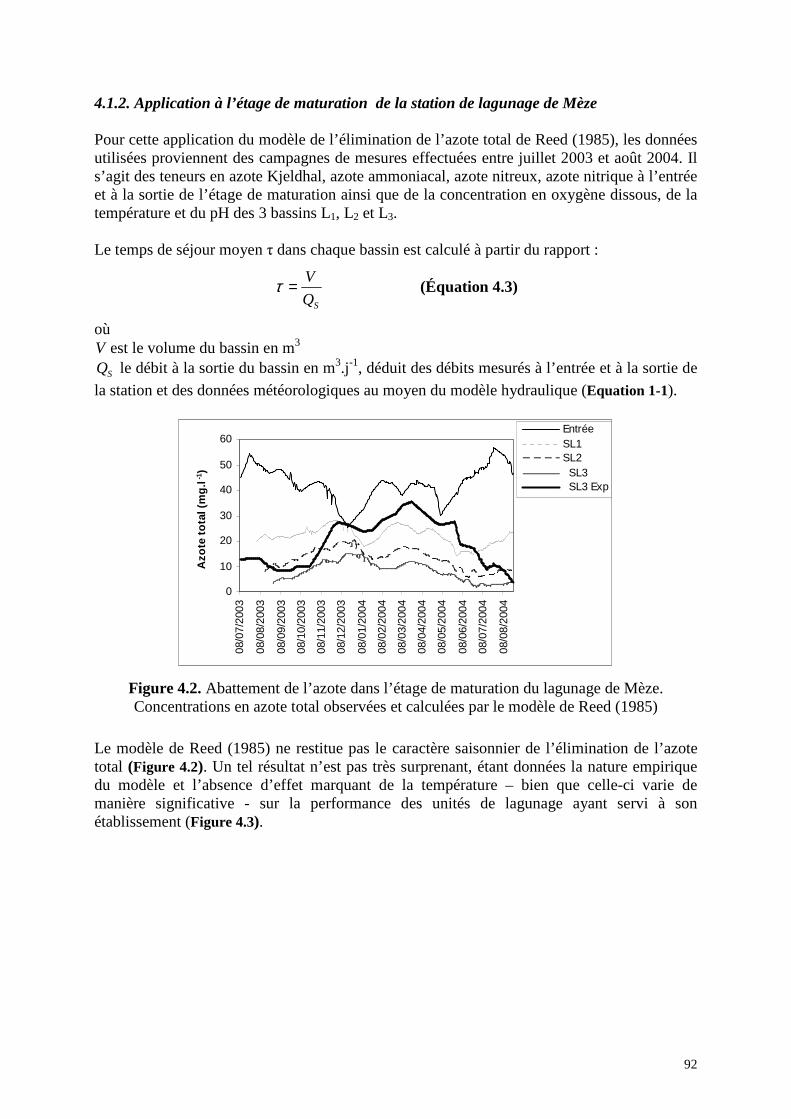
92
4.1.2. Application à l’étage de maturation de la station de lagunage de Mèze Pour cette application du modèle de l’élimination de l’azote total de Reed (1985), les données utilisées proviennent des campagnes de mesures effectuées entre juillet 2003 et août 2004. Il s’agit des teneurs en azote Kjeldhal, azote ammoniacal, azote nitreux, azote nitrique à l’entrée et à la sortie de l’étage de maturation ainsi que de la concentration en oxygène dissous, de la température et du pH des 3 bassins L1, L2 et L3. Le temps de séjour moyen τ dans chaque bassin est calculé à partir du rapport :
SQ
V=τ (Équation 4.3)
où V est le volume du bassin en m3
SQ le débit à la sortie du bassin en m3.j-1, déduit des débits mesurés à l’entrée et à la sortie de
la station et des données météorologiques au moyen du modèle hydraulique (Equation 1-1).
0
10
20
30
40
50
60
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
Azo
te to
tal (
mg.
l-1)
EntréeSL1SL2 SL3 SL3 Exp
Figure 4.2. Abattement de l’azote dans l’étage de maturation du lagunage de Mèze. Concentrations en azote total observées et calculées par le modèle de Reed (1985)
Le modèle de Reed (1985) ne restitue pas le caractère saisonnier de l’élimination de l’azote total (Figure 4.2). Un tel résultat n’est pas très surprenant, étant données la nature empirique du modèle et l’absence d’effet marquant de la température – bien que celle-ci varie de manière significative - sur la performance des unités de lagunage ayant servi à son établissement (Figure 4.3).

93
aK
Peterborough, New Hampshire
0
10
20
30
40
50
60
70
Janv
Fév
r
Mar
s
Avr Mai
Juin
Juil
Aoû
t
Sep
t
Oct
Nov
Déc
(mg.l -1)
0
5
10
15
20
25 (°C)
NT Sortie
NT Entrée
Température
Corinne, Utah
0
10
20
30
40
50
60
70
Janv
Fév
r
Mar
s
Avr Mai
Juin
Juil
Aoû
t
Sep
t
Oct
Nov
Déc
(mg.l -1)
0
5
10
15
20
25 (°C)
Eudora, Kansas.
0
10
20
30
40
50
60
70
Janv
Fév
r
Mar
s
Avr Mai
Juin
Juil
Aoû
t
Sep
t
Oct
Nov
Déc
(mg.l -1)
0
5
10
15
20
25 (°C)
Kilmichael, Mississipi.
0
10
20
30
40
50
60
70
Janv
Fév
r
Mar
s
Avr Mai
Juin
Juil
Aoû
t
Sep
t
Oct
Nov
Déc
(mg.l -1)
0
5
10
15
20
25 (°C)
Figure 4.3. Teneurs en azote total à l’entrée et à la sortie du troisième bassin de chacune des stations de lagunage de Peterborough, d’Eudora, de Corinne et de Kilmichael (Reed, 1985)
4.2. Le modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) Selon Pano et Middlebrooks, les processus pouvant intervenir dans l’élimination de l’azote ammoniacal sont la volatilisation, l’assimilation algale et le relargage d’ammonium par les sédiments. Pour décrire l’ensemble de ces processus, ils ont opté pour un modèle de l’élimination de l’azote ammoniacal qui est largement empirique mais dont la forme générale est similaire à celle qu’aurait l’équation du bilan de masse dans le cas où seule la volatilisation aurait un effet significatif (ou modèle de la volatilisation). 4.2.1. Principes de la volatilisation Dans les solutions aqueuses, l’azote ammoniacal existe sous des formes ioniques ( +
4NH ) et
moléculaires ( )(3 AqNH ) selon l’équilibre :
+4NH ++ HNH Aq)(3 (Réaction 4-1)
La constante d’acidité aK a comme expression :
[ ] [ ][ ]+
+
=4
)(3
NH
HNHK Aq
a (Équation 4.4)
où

94
( )[ ]AqNH3 , [ ]+4NH et[ ]+H sont les concentrations respectives en ammoniac dissous, en
ammonium et en protons exprimées en mol.l-1 D’après Reynolds (1977) :
TpKa 032.005.10 −= (Équation 4.5)
où T est la température en °C. Comme :
AA KpK log−=
[ ]( )+−= HpH log ; alors :
[ ] [ ]apKpH
AqNHNH −
+ =10
)(34 (Équation 4.6)
Comme tout composé volatile, l’ammoniac peut faire l’objet d’échanges à l’interface entre l’eau et l’atmosphère: HK
)(3 gNH )(3 AqNH (Réaction 4-2)
La distribution à l’équilibre de l’ammoniac entre les 2 phases gazeuse et aqueuse est donnée par la loi de Henry :
( )[ ]H
NH
Aq Kp
NH=
3
3 (Équation 4.7)
où
3NHp est la pression partielle de l’ammoniac dans l’atmosphère en atmosphères ;
HK la constante de Henry pour l’ammoniac en mole.l-1.atm-1 Dans l’air atmosphérique au volume illimité, si l’ammoniac se comporte comme un gaz parfait, sa pression partielle est uniquement dépendante de la température suivant la relation :
( )[ ]gNH NHRTp 33= (Équation 4.8)
où R la constante des gaz parfaits (0.082 l.atm.K-1.mol-1) T la température de l’air en °K
( )[ ]gNH3 est la concentration en ammoniac de l’air atmosphérique, égale à 10102 −× mol.l-1
selon Stumm et Morgan (1995) Dans le système ouvert que forment les bassins de lagunage avec l’air atmosphérique, la concentration en ammoniac dans la phase gazeuse est supposée ne pas être modifiée par les éventuels échanges à l’interface. La solubilité de l’ammoniac dans l’eau est alors donnée par l’équation 4-9 déduite des Equations 4-6 et 4-7 :
[ ] [ ]34)(3 10 NHH
pKpHAq pKNHNH a == +− (Équation 4.9)

95
DK
avec :
T
K H
180882.7log −=
La volatilisation est le processus de transfert d’ammoniac d’un plan d’eau vers l’atmosphère par évaporation afin de tendre vers l’état idéal d’équilibre caractérisé par les égalités de l’ équation 4-9. 4.2.2. Modèle de la volatilisation de Pano et Middlebrooks (1982) : Si le régime hydraulique est permanent, si le débit est conservé entre l’entrée et la sortie, s’il n’y a pas d’autre source d’azote ammoniacal que le flux entrant et si la volatilisation est l’unique voie d’élimination de l’azote ammoniacal dans un bassin et si la volatilisation suit une cinétique du premier ordre, l’équation du bilan de masse s’écrit :
[ ] [ ]( ) ( )[ ]AqNSE NHSkNHNHQdt
dCV 34343 −−= −− (Équation 4.10)
Q est le débit (m3/j) [ ]ENH 43− la concentration à l’entrée en azote ammoniacal (mg/l de N)
[ ]SNH 43− la concentration à la sortie en azote ammoniacal (mg/l de N)
[ ]43−NH la concentration moyenne en azote ammoniacal (mg/l de N) V le volume du bassin (m3) S la surface du bassin (m2) t le temps (j)
( )[ ]AqNH3 la concentration d’ammoniac dissous (mg/l de N)
Nk le coefficient de transfert de masse (m/j).
Pour des bassins aérés naturellement, Nk est une fonction croissante de la vitesse du vent.
En régime permanent par rapport aux concentrations, l’équation du bilan de masse peut s’écrire :
[ ] [ ] ( )[ ]AqNSE NHSkNHQNHQ 34343 += −− (Équation 4.11)
La coexistence des 2 formes de l’azote ammoniacal dans l’eau peut aussi être décrite à l’aide de la réaction de dissociation :
−+ + OHNH4 ( ) OHNH Aq 23 + (Réaction 4-3)
[ ][ ]( )[ ]Aq
D NH
OHNHK
3
4−+
= (Équation 4.12)
où
[ ]+4NH est la concentration de l’ammonium (mg/l de N)
[ ]−OH la concentration d’hydroxyde (mg/l)

96
( )[ ]AqNH3 la concentration d’ammoniac dissous (mg/l de N)
Par ailleurs, la constante de dissociation de l’eau étant :
[ ] [ ]−+= OHHKeau (Équation 4.13)
Les équations 4-12 et 4-13 impliquent que :
[ ] [ ][ ] D
eauAq
KH
KNHNH +
+
= 4)(3 (Équation 4.14)
Comme
eaupK = -log eauK
DpK = -log DK pH = -log ([H+])
on peut avoir l’expression de la concentration en ammoniac dissous en fonction du pH :
( )[ ] [ ]Deau pKpHpKAq
NHNH −−
+
=10
43 (Équation 4.15)
La prédominance de l’une ou l’autre des 2 formes ammoniacales dépend du pH et de la température. L’influence de la température se manifeste par l’intermédiaire des constantes de dissociation. Pour des bassins parfaitement mélangés[ ] [ ]SNHNH 4343 −− = , l’équation 4-15 peut aussi
s’écrire :
( )[ ] [ ]pHpKpK
SAq
Deau
NHNH −−
−
+=
10143
3 (Équation 4.16)
Les équations 4-11 et 4-16 conduisent à celle de la volatilisation de Pano et Middlebrooks établie pour un régime permanent:
[ ][ ]
++
=
−−−
−
pHpKpKNE
S
Deauk
Q
SNH
NH
101
11
1
43
43 (Équation 4.17)
4.2.3. Modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) : Le modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks a été obtenu en
remplaçant Nk par NK et le terme pHDeau pKpK −−+101
1 par ( )pHf dans l’équation 4-17 :
[ ][ ] ( )pHfK
Q
SNH
NH
NE
S
.1
1
43
43
+=
−
− (Équation 4.18)
où

97
NK est le coefficient d’élimination de l’azote ammoniacal (m/j)
Comme Nk , NK est fonction de la température et des conditions de brassage des bassins.
De même, la fonction( )pHf dépend du pH et de la température comme le terme qu’il remplace. Pano et Middlebrooks se sont appuyés sur les performances moyennes mensuelles du premier bassin de chacune des 3 stations de lagunage suivies par l’EPA implantées à Peterborough dans le New Hampshire, Eudora au Kansas et Corinne dans l’Utah, pour définir les formes et expressions de NK et ( )pHf . Le choix de ces bassins en particulier se justifiait par le fait que
la majeure partie de l’élimination de l’azote ammoniacal y avait lieu. Les analyses statistiques de leurs résultats expérimentaux ont induit Pano et Middlebrooks à choisir une fonction exponentielle pour( )pHf et l’équation 4-18 devenait:
[ ][ ] pHa
NE
S
eKQ
SNH
NH
.43
43
.1
1
+=
−
− (Équation 4.19)
où a est fonction de la température Pour traduire l’absence d’élimination d’azote ammoniacal qu’ils avaient constatée pour des valeurs de pH inférieures ou égales à 6.6, l’équation 4-19 était devenue :
[ ][ ] )6.6('43
43
.1
1
−−
−
+=
pHaN
E
S
eKQ
ANH
NH (Équation 4.20)
Ils ont aussi relié le coefficient'NK à la température par une fonction exponentielle du type de
l’équation de Van’t Hoff-Arrhenius comme tel est le cas pour la plupart des constantes cinétiques de réaction. Il en est ressorti que l’élimination de l’azote ammoniacal devait être traduite par deux formules : une première, l’équation 4-21, pour le cas où la température de l’eau ne dépassait pas 20 °C et une deuxième, l’équation 4-22, pour une eau dont la température est comprise entre 21 et 25 °C.
( ) ( ) ( )[ ]6.6044.0041.1exp000134.00038.01
1
−+++=
pHTTQ
AC
C
E
S (Équation 4.21)
( )[ ]6.6540.1exp10035.51
1
3 −×+=
− pHQ
AC
C
E
S (Équation 4.22)
L’effet de brassage du vent n’apparaît pas dans ces 2 équations car Pano et Middlebrooks estimaient qu’en choisissant des bassins ayant la même configuration et implantés dans des endroits soumis aux mêmes conditions climatiques, comme tel est le cas pour les 3 villes citées auparavant, NK peut être considéré comme uniquement dépendant de la température.

98
Par ailleurs, Pano et Middlebrooks ont émis des limites quant à l’utilisation de ces équations : elles ne peuvent être appliquées que pour des bassins dont la profondeur est comprise entre 1.22 et 1.52m et qui sont implantés dans des endroits au climat similaire à celui des 3 régions de leurs études. A Peterborough, Eudora et Corinne, l’hiver, les températures de l’eau sont comprises entre 1 et 5 °C et il arrive que les bassins soient recouverts de glace ; l’été, les températures de l’eau varient autour de 20 °C et dépassent rarement 25 °C. La validité du modèle semblait confirmée par de bonnes prévisions de la concentration en azote ammoniacal à la sortie d’autres installations ainsi qu’à celles des seconds et troisièmes bassins en série des stations de Peterborough, Eudora et Corinne. 4.2.4. Application du modèle d’élimination de l’azote ammoniacal de Pano et Middlebrooks à l’étage de maturation de Mèze Le modèle d’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) a été appliqué à la simulation des performances de l’étage de maturation du lagunage de Mèze, pendant la période juillet 2003 à août 2004. En régime transitoire, si les 3 bassins de maturation sont des mélangeurs parfaits et si l’élimination de l’azote ammoniacal suit la cinétique de Pano et Middlebrooks, le bilan de masse est donné par la relation :
[ ]( ) [ ] [ ] ( )[ ] [ ] jijiNijijijkk
jkjiji
NHpHfKSNHQNHQdt
NHVd,43,,43,,43,
,43,−−−
− ⋅⋅⋅−−=∑ (Équation 4.23)
où
jiV , est le volume en m3 du bassin i le jour j
[ ] jiNH ,43− la teneur en azote ammoniacal en mg.l-1 du bassin i le jour j
jkQ , le débit en m3.j-1 de l’effluent k à l’entrée du bassin i le jour j
[ ] jkNH ,43− la teneur en azote ammoniacal de l’effluent k le jour j
jiQ , le débit en m3.j-1 à la sortie du bassin i le jour j
iS la surface en m2 du bassin i
( )pHfK N ⋅ la constante de la cinétique de l’élimination de l’azote ammoniacal de Pano et
Middlebrooks définie d’après les équations 4-18, 4-21 et 4-22 pour le bassin i le jour j par :
( )[ ]( ) ( )( )[ ]
( )[ ] ( )
≤°≤−×
°≤−++=⋅
− 2521,6.6540.1exp10035.5
20,6.6044.0041.1exp000134.00038.0
,,3
;,,,
; CTpH
CTpHTTpHfK
jiji
jijijiji
jiN (Équation 4.24)
jiT , et jipH , sont la température en °C et le pH du bassin i le jour la teneur en azote
ammoniacal à la sortie du bassin i le jour j est donné par :
[ ][ ] [ ]
( )[ ] tpHfKStQVV
tNHQNHVNH
jiNjijijiji
kjkjkjiji
ji ∆⋅⋅⋅+∆⋅+−
∆⋅+=
−
⋅−−−
−
∑
;,,1,,
,43,1,43,
,43 2 (Équation 4.25)
Les évolutions selon Pano et Middlebrooks (1982) des concentrations en azote ammoniacal à la sortie des 3 bassins composant l’étage de maturation sont représentées dans la Figure 4.4.
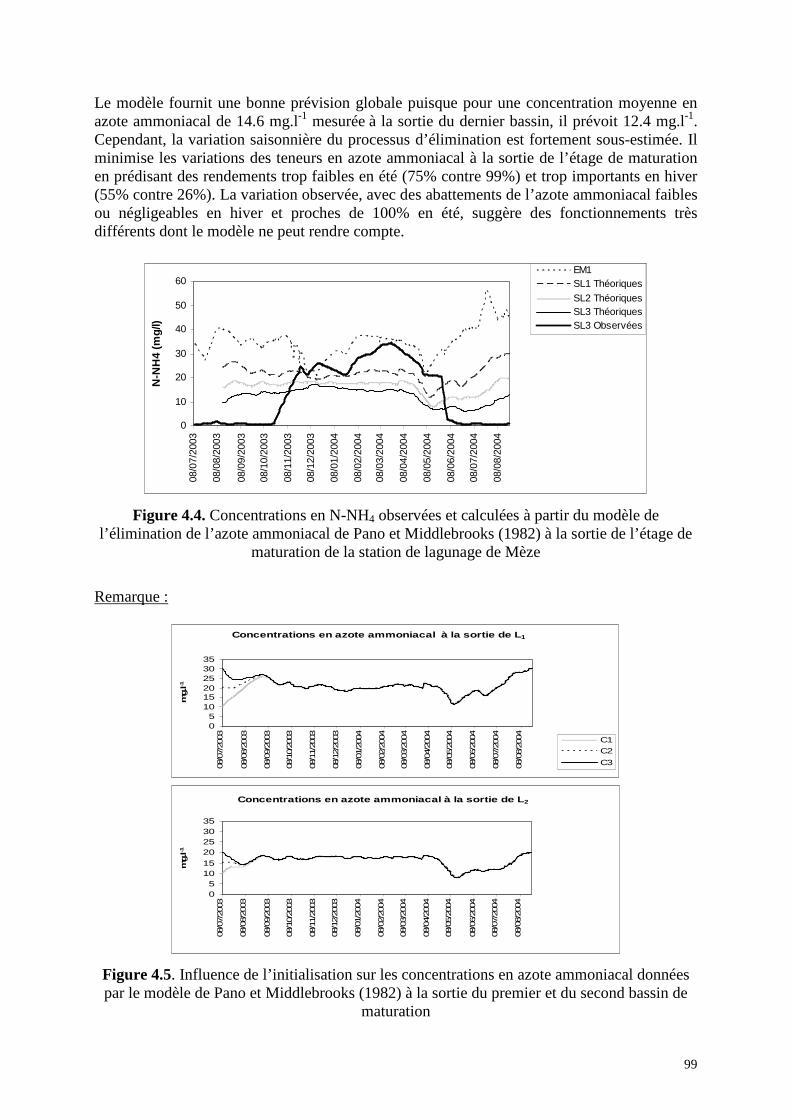
99
Le modèle fournit une bonne prévision globale puisque pour une concentration moyenne en azote ammoniacal de 14.6 mg.l-1 mesurée à la sortie du dernier bassin, il prévoit 12.4 mg.l-1. Cependant, la variation saisonnière du processus d’élimination est fortement sous-estimée. Il minimise les variations des teneurs en azote ammoniacal à la sortie de l’étage de maturation en prédisant des rendements trop faibles en été (75% contre 99%) et trop importants en hiver (55% contre 26%). La variation observée, avec des abattements de l’azote ammoniacal faibles ou négligeables en hiver et proches de 100% en été, suggère des fonctionnements très différents dont le modèle ne peut rendre compte.
0
10
20
30
40
50
60
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
N-N
H4
(mg/
l)
EM1SL1 Théoriques
SL2 ThéoriquesSL3 ThéoriquesSL3 Observées
Figure 4.4. Concentrations en N-NH4 observées et calculées à partir du modèle de l’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) à la sortie de l’étage de
maturation de la station de lagunage de Mèze
Remarque :
Concentrations en azote ammoniacal à la sortie de L1
05
101520253035
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
mg.
l-1
C1
C2
C3
Concentrations en azote ammoniacal à la sortie de L 2
05
1015
20253035
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
mg.
l-1
Figure 4.5. Influence de l’initialisation sur les concentrations en azote ammoniacal données par le modèle de Pano et Middlebrooks (1982) à la sortie du premier et du second bassin de
maturation

100
Les concentrations en azote ammoniacal à la sortie du premier ainsi que du second bassin n’ayant pas été mesurées, leur détermination à partir de l’équation 4-25 a nécessité des initialisations. Il s’est révélé que la durée de la simulation est suffisamment longue pour que le choix des valeurs initiales n’influe pas sur les résultats du calcul pendant la plus grande partie de la durée considérée. En effet, seuls les résultats des premiers 45jours environ en dépendent (Figure 4.5). 4.2.5. Modèle de Pano et Middlebrooks (1982) et volatilisation Le système formé par les équations 4-21 et 4-22 est reconnu généralement comme étant le modèle de Pano et Middlebrooks (1982) ; il est souvent assimilé à tort au modèle de la volatilisation (Silva et al. 1995, Soares et al. 1996, Zimmo et al. 2003, Archer et O’Brien, 2005). Ce dernier est en réalité représenté par l’équation 4-17. D’ailleurs, faute de mesure des émissions de gaz NH3, la constante cinétique Nk n’a pas fait l’objet de détermination directe
et l’équation 4-17 n’est pas exploitable pour évaluer la volatilisation. La confusion à propos du modèle de Pano et Middlebrooks (1982) est due essentiellement au fait que sa formulation est dérivée de celle du modèle de la volatilisation et à des ambiguïtés de formulation de la part de ses concepteurs. En effet, d’une part ils ont affirmé que la volatilisation était traduite par l’équation 4-17 et qu’ils avaient dû y apporter des modifications pour développer un modèle qui prendrait en compte l’intervention possible de processus biologiques d’élimination ou de régénération de l’ammonium ; d’autre part, Middlebrooks et al. (1982) ont désigné l’équation 4-18 comme étant celle de la volatilisation. De plus, ils ont expliqué l’absence d’élimination aux pH inférieurs ou égaux à 6.6 par le blocage de la volatilisation de l’azote ammoniacal, ce qui revient implicitement à négliger les autres formes de l’azote et les activités biologiques inhibées par les pH faibles et susceptibles de contribuer à l’élimination de l’azote. Etant donné que d’autres processus étaient pris en compte par le modèle de disparition de l’azote ammoniacal de Pano et Middlebrooks (1982), l’assimiler à celui de la volatilisation revient à surévaluer celle-ci. C’est pourtant ce que nombre d’auteurs ont fait. Les mesures directes de la volatilisation de l’azote ammoniacal montrent pourtant que cette voie d’élimination de l’azote est loin d’être prépondérante (Zimmo, 2003). 4.3. Evaluation de la volatilisation. Cinétiques de Stratton Stratton a réalisé deux études fondamentales sur la volatilisation publiées en 1968 et 1969. Après Ferrara et Avci (1982), il a fallu attendre une vingtaine d’années et les recherches de Zimmo et al. (2003) pour les voir à nouveau prises en compte dans les travaux sur le lagunage. La première étude est consacrée à la volatilisation de l’ammoniac dans les cours d’eau (Stratton, 1968), la deuxième s’intéresse au même processus dans les retenues d’eau (Stratton, 1969). 4.3.1. Cas des cours d’eau : Matériels. Pour sa première étude de la cinétique de la volatilisation, Stratton (1968) a simulé en laboratoire un cours d’eau à l’aide d’eau distillée additionnée de sulfate d’ammonium et de solution tampon de phosphate de potassium circulant dans un canal équipé d’un système de recirculation (Figure 4.6). Des conditions de turbulence ont été imposées au cours d’eau

101
artificiel par des aspérités carrées collées sur le fond du canal. Un pivot permet d’ajuster la pente du canal, fixée à 1% durant l’ensemble de l’expérience. Le système est caractérisé par une surface libre d’environ 1m2 et une profondeur constante de 0.03m. Un autre dispositif composé uniquement du système de recirculation a été construit, de manière à permettre de déterminer la part de la volatilisation imputable au seul écoulement dans le canal.
Figure 4.6. Schéma du principal dispositif de simulation de cours d’eau de Stratton (1968)
Suivis physico-chimiques. Les 2 dispositifs fonctionnaient en parallèle, dans des conditions identiques de température, de pH et de contrôle. Une série d’expériences est caractérisée par :
- un suivi permanent de la température et son ajustement éventuel, - une mesure régulière du pH et un ajustement au besoin de celui-ci à l’aide de NaOH
ou de H2SO4, - une analyse périodique de la teneur en azote ammoniacal, - une analyse des NO2
- et NO3-au début et à la fin de chaque série.
Les mesures in situ et échantillonnages sont effectuées dans le puisard (Figure 4.6). Théorie générale. Stratton (1968) a écrit l’équation de la cinétique de la volatilisation du cours d’eau artificiel en supposant : - l’absence de tout processus d’élimination de l’azote autre que la volatilisation,
- le régime d’écoulement permanent, - le brassage vertical du cours d’eau assez important par rapport à la volatilisation pour
que la concentration en NH3 soit homogène dans la colonne d’eau, - la cinétique de la volatilisation du premier ordre par rapport à la concentration en
azote ammoniacal dans l’eau
[ ]43'
'
−= NHkdt
dM (Équation 4.26)
où 'M est la masse de N-NH3 volatilisé en mg,
'k la constante cinétique de la perte en azote ammoniacal en l.h-1, [ ]43−NH la concentration en azote ammoniacal dans l’eau en mg.l-1,
t le temps.
Pompe
Pivot
Puisard
Thermomètre

102
Un départ de NH3 déplace l’équilibre de la réaction 1 acide-base du couple
( )( )AqNHNH 34 ;+ dans le sens de sa formation pour que la proportionnalité définie par
l’ équation 4-4 reste vérifiée. La perte est alors partagée entre les 2 formes. En conséquence, dans un plan d’eau où la perte en azote ammoniacal provient uniquement de la volatilisation, la masse 'M de N-NH3 émis correspond à la diminution de la masseM d’azote ammoniacal dissous indépendamment de sa forme :
dMdM −=' (Équation 4.27)
Comme :
[ ]433.10. −= NHdVdM (Équation 4.28)
où V est le volume du plan d’eau exprimé en m3 Très souvent, la volatilisation est exprimée en masse d’azote ammoniacal émis par unité de surface. La cinétique correspondante a comme expression :
[ ]43' . −= NHk
dt
dMS
S (Équation 4.29)
où
S
kkS
'' = exprimé en l.h-1.m-2
Sétant la surface libre du plan d’eau en m2
SM la masse de N-NH3 émis par unité de surface
Or [ ]43
310.−−= NHd
S
VdMS (Équation 4.30)
La cinétique de la volatilisation par unité de surface peut être formulée comme suit :
[ ] [ ]43
3'43 10.
−
−− −= NH
H
k
dt
NHd S (Équation 4.31)
où H est la profondeur du cours d’eau (ou du plan d’eau) en m
En posant : H
kk S
3' 10. −
= (Équation 4.32)
l’ équation 4-31 devient :
[ ] [ ]4343
−− −= NHk
dt
NHd (Équation 4.33)
où k est exprimé en h-1 Effets de la température et du pH sur la constante cinétique de volatilisation. Pour les 2 dispositifs, des courbes semi-logarithmiques de la variation de la concentration en azote ammoniacal en fonction du temps, sous différentes conditions de température (15, 20, 25 et 30 °C) et de pH (7.5, 8.0, 8.5, 9.0, et 9.5) ont été tracées dans le but d’en déterminer les pentes

103
qui représentaient les constantes cinétiquesk d’après l’équation 4-33. Celles correspondant au canal sont calculées avec la relation :
( )2
121 V
Vkkk −= (Équation 4.34)
où, sous chaque condition donnée de température et de pH,
1k est la constante cinétique de la volatilisation du dispositif combiné représenté par la Figure 4.6.
2k la constante cinétique de la volatilisation dans le système de recirculation
1V le volume d’eau ayant transité dans le dispositif combiné durant le temps t de l’expérience
2V le volume d’eau ayant transité dans le système de recirculation durant le même temps t Résultats. L’hypothèse de l’absence d’activité biologique significative pouvant conduire à une élimination d’azote ammoniacal par nitrification-dénitrification au sein du cours d’eau a été vérifiée par des analyses négatives de la teneur en azote nitreux et en azote nitrique au début et à la fin de chaque série d’expériences. La linéarité des variations du logarithme de la teneur en azote ammoniacal en fonction du temps confirment l’hypothèse d’une cinétique de la volatilisation d’ordre 1. Des analyses de régression faites sur les valeurs des pentes ont conduit Stratton à relier la constante cinétique de la volatilisation k à la température et au pH par les équations :
( )
( )
°==
==−
−
CTek
pHekpH
T
20,080.0
5.8,048.05.857.1
2013.0
(Équation 4.35)
Tableau 4.1. Valeurs expérimentales de la constante cinétique d’élimination k de l’azote par volatilisation en fonction de la température et du pH selon Stratton (1968)
Expérience Température (°C) pH k (h-1) 1 15 8.5 0.018 2 20 8.5 0.076 3 25 8.5 0.089 4 30 8.5 0.154 5 20 7.5 0.012 6 20 8.0 0.032 7 20 8.5 0.076 8 20 9.0 0.137 9 20 9.5 0.295
Remarque : La cinétique de la volatilisation composée par le système d’équations 4-35 définie par Stratton (1968) donne 2 valeurs différentes dek , 0.048 et 0.080 h-1, pour les mêmes conditions ; soit 20°C et pH 8.5. Une autre formulation résoudrait ce problème :
( ) ( )5.857.12013.0. −−= pHT eeAk (Équation 4.36)

104
où 31034.55 −×=A est déterminé par le minimum de la fonction( ) ( )( )29
1∑
=
−i
Expicalci kk
k étant dépendant de la hauteur du cours d’eau simulé (ici 3 cm), une cinétique plus générale est formulée en terme de 'Sk exprimé en l.m-2.h-1, calculé à partir de l’équation 4-32 :
( ) ( )5.857.12013.0' 66.1 −−= pHTS eek (Équation 4.37)
ou en l.m-2.j-1 : ( ) ( )5.857.12013.0' 84.39 −−= pHT
S eek (Équation 4.38)
4.3.2. Cas des retenues d’eau relativement abritées du vent Dans le but de déterminer les pertes en azote par émission de NH3 d’une retenue d’eau de concentration donnée en azote ammoniacal, sous des conditions bien définies de température et de pH, Stratton (1969) a réalisé des expériences similaires à celles relatives au cas du cours d’eau, toujours à l’échelle de laboratoire mais réalisées en extérieur et sur des pilotes de retenues d’eau relativement protégés du vent. Matériels. Les pilotes, des récipients remplis de 6 L de solution sur une profondeur de 0,17 m, contenaient 4 solutions d’azote ammoniacal constituées de sulfate d’ammonium dissous dans de l’eau dé-ionisée et tamponnées respectivement à pH 7.5, 8.0, 8.5 et 9.0. Suivis physico-chimiques. Pour chaque série de tests de cette seconde expérience, les variations des concentrations en azote ammoniacal en fonction du temps ont été suivies tous les 2 jours, pendant une vingtaine de jours environ, au bout desquels l’absence de nitrification-dénitrification a été vérifiée par des analyses. Les variations de concentration dues à l’évaporation ont été corrigées par ajout d’un volume adéquat d’eau dé-ionisée suivi d’une légère agitation. Durant la période de l’étude, de juin à août 1968, la station météorologique la plus proche avait relevé des moyennes journalières de température de 19.6°C et de vitesse du vent de 2m/s Résultats. La constante cinétique la plus pratiquement accessible est k . En effet, k est la pente de la représentation en coordonnées semi-logarithmiques de la variation des concentrations en azote ammoniacal en fonction du temps. En opérant à 19°6C et pH 8.5, on a ( )5.8,6.19 Ck ° .
En supposant que la concentration en azote ammoniacal reste homogène dans tout le volume
d’eau, la détermination de ( )5.8,6.19 Ck ° permet celle det
M CS
∆∆ ° )5.8,6.19( pour différentes
concentrations à partir d’une formule déduite des équations 4-29 et 4-32 :
( ) [ ]433
5.8,6.19)5.8,6.19( 10 −°
° ⋅=∆
∆NHkH
t
MC
CS (Équation 4.39)
Les résultats obtenus par Stratton peuvent se traduire par l’expression suivante :

105
( ) [ ]43,6.19 22 −
° =∆
∆NH
t
M pHCS (Équation 4.40)
Figure 4.7. Courbes de la variation de la concentration en azote ammoniacal en fonction du temps des retenues d’eau simulées par Stratton (1969)
La perte d’azote par volatilisation d’une retenue d’eau de concentrationC en azote ammoniacal, dans des conditions données de température et de pH, peut être calculée en faisant le produit de la perte trouvée à 19,6°C et pH 8.5 et des fonctions exponentielles des variations de la constante cinétique avec la température et le pH déterminées par Stratton dans ses essais sur maquette de cours d’eau:
( ) ( ) ( ) ( )
t
Mee
t
M CSpHTpHTS
∆∆
⋅⋅=∆
∆ °−− 5.8,6.195.857.12013.0, (Équation 4.41)
ou
( ) ( ) ( ) [ ]435.857.12013.0, 22 −
−− ⋅⋅⋅=∆
∆NHee
t
MpHTpHTS (Équation 4.42)
Selon Stratton (1969), le domaine de validité de cette cinétique est défini par 5.95.7 << pH . Au-delà d’un pH 9.5, la cinétique de la volatilisation ne serait plus du 1er ordre. Les évolutions des concentrations en azote ammoniacal sont bien représentées en coordonnées semi-logarithmiques par des droites, malgré des écarts attribués aux variations de température (Figure 4.7). L’importance de l’influence du pH est manifeste : une élévation de 0.5 unité fait augmenter nettementk . Les variations de k avec le pH vérifient la relation 4-42.

106
Pompe
Remarques : On peut exprimer comme suit les constantes cinétiques '
Sk et k de la volatilisation d’une eau
sous des conditions données de température et de pH : ( ) ( )5,8.57,120.13,0' ..22 −−= pHT
S eek (Équation 4.43)
( ) ( )5,8.57,120.13,03
.1022 −−
−×= pHT eeH
k (Équation 4.44)
Cependant, le coefficient 22 est presque moitié moindre que celui de l’équation 4-38. Cet écart peut être attribué à deux raisons ; dans la maquette de cours d’eau, la colonne d’eau était toujours parfaitement mélangée, ce qui n’était pas vrai dans les pilotes de retenues ; d’autre part, la surface libre dans le cours d’eau simulée était vraisemblablement agitée. Ces deux phénomènes expliquent une plus importante volatilisation dans la maquette de cours d’eau. 4.3.3. Application. Volatilisation dans un lac et dans une lagune : Pour valider son modèle de la volatilisation dans les retenues d’eau, Stratton (1969) a mesuré et calculé les masses de N-NH3 émises par deux petits plans d’eau eutrophisés de Californie, le lac d’« Elfin Forest» et la lagune de rétention de Santee. Mesure directe de la volatilisation par Stratton (1969). La mesure directe de la volatilisation par Stratton (1969) était basée sur une collecte suivie d’un dosage de l’ammoniac volatilisé pendant un intervalle de temps d’une heure. La collecte était réalisée dans une cellule flottante qui recueillait les émissions au-dessus d’une portion de surface du plan d’eau étudié. Le contenu de la cellule était évacué de façon continue par un conduit vers une solution acide ayant pour rôle de piéger l’ammoniac (Figure 4.8). La teneur en azote ammoniacal de la solution acide était analysée par la suite.
Figure 4.8. Schéma du dispositif de collecte et de piégeage des émissions de gaz ammoniac par Stratton (1969)
Flotteur
Cellule de collecte
Solution acide

107
Un test avec un blanc constitué de l’air atmosphérique a été effectué en parallèle. La différence avec le blanc donnait la quantité d’azote ammoniacal émis par le plan d’eau. L’efficacité du piégeage acide a été évaluée à 67 % par des tests effectués en laboratoire sur de l’air contenant une quantité connue en azote ammoniacal. Des tests ont aussi été faits dans le but de vérifier que la mise en place de la cellule flottante ne perturbait pas le métabolisme des algues et n’entraînait donc pas une surévaluation de la volatilisation par une élévation du pH. Description des plans d’eau et emplacements des cellules flottantes. Le premier plan d’eau étudié était le lac d’Elfin Forest, Escondido - California, où se pratiquaient des activités de pêche et de loisirs. Le choix de ce lac se justifiait par le fait que les eaux du système d’assainissement de la municipalité et de zones d’activités agricoles représentaient une part importante de son alimentation. Sa surface est de 20.230 m2. Sa profondeur varie de 0,60 à 4,50 m. Les eaux de surface du lac avaient un pH compris entre 9.5 et 9.8, une température entre 28 et 29°C et une teneur en azote ammoniacal entre l’état de trace et 1.47 mg/l. Les mesures de pH et de teneur en azote ammoniacal ont été effectuées en 16 points. La cellule flottante avait été placée en un point où le pH était de 9.8, la température de 29 °C et la teneur en azote ammoniacal de 0,47 mg/l. Le second plan d’eau étudié était une lagune, composante du traitement tertiaire des eaux usées de Santee, California. Au moment de l’étude, elle était en fonction depuis 6 mois. Elle a une surface de 22.260 m2 et une profondeur de 1,40 m. Le pH de ses eaux de surface était assez variable en raison d’une concentration non homogène en algues. Le point de mesure de la volatilisation était situé dans une zone de forte activité algale où le pH était de 9,1, la température de 31,5 °C et la teneur en azote ammoniacal de 1,75 mg/l. Résultats et conclusions. Les pertes en azote par volatilisation calculées par Stratton (1969) ne sont pas identiques à celles déduites de l’équation 4-42. Tableau 4.2.
Plan d’eau Azote ammoniacal (mg/l)
T (°C)
pH Perte calculée (mg/m2.j)
Perte mesurée (mg/m2.j)
Elfin Forest 0.47 29.0 9.8 256 35.5
Santee 1.75 31.5 9.1 440 98.0
Tableau 4.2. Comparaison des pertes en azote ammoniacal calculées et mesurées (Stratton, 1969)
Même en faisant les corrections se rapportant à l’efficacité du mode opératoire, les pertes en azote ammoniacal mesurées restent largement inférieures à celles calculées. Ce résultat n’est pas vraiment inattendu. Ces retenues, dans des conditions de température élevées ont de grandes chances d’être très stratifiées ; c’est notamment ce que montrent les profils de pH et de teneur en azote ammoniacal du lac d’Elfin Forest où seuls les 30 premiers centimètres sous la surface de l’eau semblent mélangés (Figure 4.9). Il est vraisemblable que, sous la cloche de collecte des gaz, le milieu liquide reste stratifié durant la mesure. Dès lors, la concentration en NH4 s’épuise rapidement dans la couche d’eau la plus proche de la surface où l’effet de la

108
volatilisation de l’ammoniac, dopée par les fortes valeurs de la température et du pH, n’est pas compensé par la diffusion de NH4 dans la phase liquide. Aussi, la volatilisation chute-t-elle rapidement. Cet effet est moins sensible dans les essais effectués dans des récipients en plein air où l’agitation de l’atmosphère, les variations nychtémérales de température entre deux analyses (le pas de temps des prélèvements était de 2 jours) et les courants convectifs résultants ont vraisemblablement maintenu une relative homogénéité, sachant par ailleurs que les prélèvements s’accompagnaient aussi d’un mélange du volume d’eau.
Figure 4.9. Variation avec la profondeur du pH et de la teneur en azote ammoniacal dans le lac d’ « Elfin Forest »
Au total, on peut conclure que, sauf effet d’un vent notable, l’équation 4-42 constitue une bonne estimation de l’ordre de grandeur de la volatilisation. 4.3.4. Volatilisation dans des retenues d’eau soumises à l’agitation du vent Au cours du printemps 2006, on a repris le mode opératoire défini par Stratton (1969) pour réaliser des mesures directes de la volatilisation de l’ammoniac sur des retenues d’eau simulées soumises à l’agitation naturelle du vent et en déduire une valeur de '
Sk plus générale. Matériels et suivis physico-chimiques. Les retenues d'eau simulées sont des cuvettes de 0,12 m2, profondes de 15, 10 et 6 cm (Photographie 1-1), remplies jusqu’à ras bord de solutions de sulfate d’ammonium à 40 mgL-1 de N-NH4. Le pH, contrôlé par une solution tampon et de la soude, réajusté au moins deux fois par jour, variait avec les essais et les cuvettes entre 7,5 et 9. Ces cuvettes ont été exposées en extérieur pendant plusieurs journées consécutives à des vents variant entre 2,5 et 6,3 ms-1. Pendant cette période, la température de la solution était enregistrée en continu ; les valeurs moyennes journalières étaient comprises entre 15 et 25°C.
Pro
fon
deu
r (p
ied
s)
A

109
Les variations de concentration dues à l’évaporation ont été corrigées par ajout d’un volume adéquat d’eau dé-ionisée suivi d’une agitation.
Photographie 4-1. Retenues d'eau simulées
Résultats. Les résultats des suivis de l’évolution des teneurs en azote ammoniacal dans les retenues d’eau simulées sont reportés dans les Tableaux 4.3 à 4.5. La constante '
Sk est calculée par la relation :
[ ] [ ]( )
( ) ( ) [ ] j
pHT
jjS
NHee
HNHNHk
jj
435.857.12013.03
14343' 1
10 −−−−
+−−
⋅⋅⋅
⋅−= (Équation 4.45)
où 2
1++= jj
j
pHpHpH et [ ] [ ] [ ]
214343
43+−−
−
+= jj
j
NHNHNH
Tableau 4.3. Constante cinétique mesurée'Sk de la volatilisation avec les retenues d’eau
simulées de 6 cm de profondeur
T
(°C) pH [ ]43−NH
(mg N/L) Sk
(L.j-1.m-2) 26/04/2006 17.8 8.1 25.6 32 27/04/2006 18.1 7.3 22.9 22 28/04/2006 18.2 7.2 22.0
03/05/2006 17.8 9.0 26.9 47 04/05/2006 18.1 9.0 5.8 56 05/05/2006 18.2 9.0 0.6
10/05/2006 16.1 8.0 4.0 48 11/05/2006 17.2 8.0 27.2 22 12/05/2006 15.4 8.0 24.2

110
Tableau 4.4. Constante cinétique mesurée'Sk de la volatilisation avec les retenues d’eau
simulées de 10 cm de profondeur
T
(°C) pH [ ]43−NH
(mg N/L) Sk
(L.j-1.m-2) 26/04/2006 22.1 8.4 28.4 26 27/04/2006 20.6 7.6 24.4 11 28/04/2006 18.8 7.3 23.8
03/05/2006 18.5 9.0 37.2 40 04/05/2006 18.6 9.0 17.5 39 05/05/2006 18.6 9.0 8.4
10/05/2006 16.5 8.0 34.0 51 11/05/2006 17.6 8.0 29.8 18 12/05/2006 15.9 8.0 28.2
Tableau 4.5. Constante cinétique mesurée'Sk de la volatilisation avec les retenues d’eau
simulées de 15 cm de profondeur
T
(°C) pH [ ]43−NH
(mg N/L)
'Sk
(L.j-1.m-2) 26/04/2006 20.6 8.5 29.7 32 27/04/2006 18.8 8.0 25.1 28/04/2006 17.4 7.6 -
03/05/2006 18.9 9.0 42.7 34 04/05/2006 18.9 9.0 27.7 33 05/05/2006 19.1 9.0 18.1
10/05/2006 16.8 8.0 36.3 26 11/05/2006 17.9 8.0 34.4 10 12/05/2006 16.2 8.0 33.6
17/05/2006 23.1 7.7 34.9 4 18/05/2006 21.1 7.7 34.5 62 19/05/2006 20.9 7.7 30.1
17/05/2006 22.9 8.0 34.9 15 18/05/2006 21.0 8.0 32.6 59 19/05/2006 21.1 8.0 26.5
17/05/2006 22.64 9 25.6 25 18/05/2006 20.83 9 15.1 26 19/05/2006 21.15 9 9.8

111
'Sk varie de 4 à 62 L.m-2.j-1. Ces écarts sont explicables à la fois par les variations des vitesses
du vent et par des difficultés, plus ou moins grandes selon les tests, à maintenir le pH à sa valeur nominale. Sa valeur moyenne mesurée est de 31 L.m-2.j-1. Elle est à mi-chemin des valeurs obtenues par Stratton lors de ses mesure sur, respectivement, une maquette de cours d’eau ( '
Sk = 40 L.m-2.j-1) qui fournit une estimation maximale de la volatilisation et sur des
bacs à l’air libre assez bien abrités du vent ('Sk = 22 L.m-2.j-1).
On adoptera comme cinétique de la volatilisation l’expression suivante :
( ) ( ) ( ) [ ]435.857.12013.0, 30 −
−− ⋅⋅⋅=∆
∆NHee
t
MpHTpHTS (Équation 4.46)
4.4. Etude des pertes en azote ammoniacal par Zimmo et al. (2003) Zimmo et al. ont comparé, dans le cas d’une installation pilote de lagunage, les valeurs des pertes en azote ammoniacal mesurées directement avec celles calculées en utilisant les travaux de Stratton, celles calculées en utilisant le modèle de Pano et Middlebrooks (1982) compris comme un modèle de la volatilisation et, enfin, avec des mesures directes de volatilisation. 4.4.1. Matériels et méthodes Cette comparaison a été effectuée sur une installation pilote de traitement d’eaux usées urbaines par lagunage (Figure 4.10), située dans le campus de l’université de Birzeit en Palestine. Les 8 bassins composant le système avaient chacun une surface d’environ 3m2 et une hauteur d’eau de 0.9m. Chaque unité était alimentée par un débit constant de 0.38 m3/j à partir d'un réservoir d’eaux usées provenant, de décembre 1998 jusqu’à la mi-juillet 2000, de l’université de Birzeit, puis, de la mi-juillet 2000 à février 2001, de la ville d’Al-Bireh. Dans la première période, la teneur en azote ammoniacal était de 60 mg/l ; elle était de 110 mg/l en moyenne dans la deuxième. Cette installation comportait deux séries de bassins, la première constituant un lagunage naturel classique reposant sur l’action des microphytes (série A), la deuxième fonctionnant avec des lentilles d’eau (série B).
Figure 4.10. Schéma du système pilote de traitement par lagunage
Les pertes en azote ammoniacal observées ont été calculées avec la relation :
[ ] [ ]( )τ.
4343
S
VNHNH SE −− − (Équation 4.47)
Unité A : Bassins à microphytes
Unité B : Bassins à lentilles d’eau
B1 B2 B3 B4
A1 A2 A4 A3
Réservoir

112
où [ ]ENH 43− est la concentration en azote ammoniacal à l’entrée (mg-N/l)
[ ]SNH 43− la concentration en azote ammoniacal à la sortie (mg-N/l)
V le volume du bassin (l) S la surface (m2) τ le temps de séjour (j) Zimmo et al. (2003) ont calculé les pertes en azote dues à la volatilisation de l’ammoniac en adoptant une expression de la cinétique de la volatilisation déjà utilisée par Ferrara et Avci (1982) puis par Senzia et al. (2001). Cette expression, attribuée à Stratton par ces auteurs successifs, diffère en réalité de celle développée dans Stratton (1969). Pour ces auteurs, la cinétique de Stratton s’écrit :
[ ] [ ]Aq
Aq NHkdt
NHd3
3 −= (Équation 4.48)
où
( )20.13,031060.56 −
−×= TeH
k (Équation 4.49)
avec H la profondeur du plan d’eau en m t le temps [ ]AqNH3 est la concentration en gaz ammoniac dissous en mg N/l
Les pertes en azote ammoniacal par unité de surface sont alors calculées par :
( )[ ]AqTS NHe
t
M3
2013.060.56 −=∆
∆ (Équation 4.50)
Comme ce qui est mesurable directement en laboratoire est la teneur en azote ammoniacal global, pour déterminer la teneur en N-NH3 dissous, Zimmo et al. ont utilisé la constante
d’acidité définie par l’équation 4-4 : ( )[ ] [ ][ ]+
+
=4
3
NH
HNHK Aq
a , de l’équilibre acide-base du couple
( )( )AqNHNH 34 ;+ .
Soit α la fraction d’azote ammoniacal sous forme gazeuse dans l’eau :
( )[ ]( )[ ] [ ]++
=43
3
NHNH
NH
Aq
Aqα (Équation 4.51)
En combinant les équations 4-4 et 4-49 on a :
pHpKa −+=
101
1α (Équation 4.52)
apK est donné par l’équation 4-5

113
Soit : [ ] ( )[ ] [ ]+− += 4343 NHNHNH Aq
Une foisα et [ ]43−NH connus, la teneur en N-NH3 s’obtient par :
( )[ ] [ ]433 . −= NHNH Aq α (Équation 4.53)
Pour la mesure des émissions d’ammoniac, Zimmo et al. ont repris la procédure établie par Stratton (1969) mais avec un temps de collecte plus long, 24 h au lieu d’une heure. Ce rallongement du temps de mesure les ont conduits à ajouter à leur matériel une pompe qui envoyait de l’air à une vitesse constante de 1.1m.h-1 à travers la cellule de collecte, pour éviter qu’elle ne catalyse la volatilisation par un réchauffement de l’eau. 4.4.2. Résultats Les Tableaux 4.6 et 4.7 résument les résultats des travaux de Zimmo et al. (2003), où les teneurs en azote ammoniacal représentent les moyennes des valeurs mesurées durant la période de l’étude (octobre 1999 à février 2001); les températures et pH sont des moyennes journalières; les teneurs en N-NH3 sont calculés par les mêmes auteurs à l'aide de l'Equation 4-53. Nous y avons ajouté la perte en azote ammoniacal par volatilisation donnée par la relation 4-42 de Stratton (1969). Il ressort essentiellement de ces mesures que la présence des lentilles d’eau s’accompagne d’une moindre élévation du pH et d’une moindre perte d’ammonium, même si les écarts entre les bassins A et B ne sont pas considérables ; ces observations iraient bien a priori dans le sens d’une participation substantielle de la volatilisation à l’élimination de l’ammonium. Les pertes d’ammonium mesurées sont globalement plus importantes dans les bassins à microphytes que dans les bassins à lentilles d’eau. La différence est particulièrement importante entre les premiers bassins de chaque série.
Tableau 4.6. Valeurs moyennes des mesures effectuées par Zimmo et al (2003) [ ]43−NH (mgN/l)
T (°C)
pH α (%)
[ ]AqNH3
(mgN/l) Bassin
Entrée/Sortie Eaux des bassins Entrée 70.1
Unité A Sortie A1 46.0 59.1 16.7 8.1 3.8 2.2 Sortie A2 32.4 43.7 17.0 8.1 3.6 1.6 Sortie A3 19.4 27.3 17.3 8.0 3.0 0.8 Sortie A4 11.4 18.3 17.1 8.0 2.8 0.5
Unité B Sortie B1 58.4 67.4 16.5 8.0 3.0 2.0 Sortie B2 47.7 52.6 17.3 7.8 2.0 1.1 Sortie B3 37.0 43.4 16.9 7.8 1.7 0.8 Sortie B4 26.8 31.7 16.5 7.7 1.5 0.5

114
Tableau 4.7. Comparaison des pertes par volatilisation mesurées et calculées à partir des cinétiques inspirées de Stratton (1969) avec les pertes d’ammonium observées et déduites de Pano et Middlebrooks (1982)
Bassin Pertes en azote ammoniacal (mg.m-2.j -1) Mesures
directes Stratton * Stratton ** Pano et Middlebrooks
Mesurées par volatilisation
Unité A A1 3099 81.1 452 3607 21.7 A2 1749 60.5 347 2399 22.3 A3 1671 37.7 193 1666 19.1 A4 1029 19.8 126 1021 10.5 Unité B B1 1504 63.0 429 3199 18.1 B2 1376 41.8 271 2203 16.4 B3 1376 28.3 213 1765 14.2 B4 1311 17.7 126 1180 10.6 * calculs effectués à partir de l’équation 4-50 ** calculs effectués à partir de l’équation 4-42 Ces pertes sont bien plus importantes que les volatilisations de gaz NH3 calculées à partir des équations 4-50 et 4-42. Si l’équation 4-50 proposée par Ferrara et Avci (1982) puis par Senzia et al. (2001) et finalement adoptée par Zimmo et al. (2003) laisse penser que la volatilisation est négligeable, nos calculs effectués à partir de l’ équation 4-42 dérivée directement des essais de Stratton tendent à montrer que la volatilisation joue un rôle secondaire mais significatif dans l’élimination de l’ammonium. Les pertes en azote ammoniacal calculées à partir des équations 4-21 et 4-22 de Middlebrooks fournissent un bon ordre de grandeur de la perte d’ammonium, particulièrement dans les bassinss à microphytes. Comme dans les essais de Stratton à Elfin Forest et Santee, les mesures de la volatilisation dans une cloche flottante donnent des valeurs faibles comparées aux calculs effectués à partir de l’équation 4-43. Cela est très vraisemblablement dû au fait que, dans le volume liquide emprisonné sous la cloche, le renouvellement, dans la couche d’eau située immédiatement sous la surface, de l’ammonium perdu du fait de la volatilisation est beaucoup plus difficile que dans des eaux libres.. 4.4.3. Conclusion Le modèle d’élimination de l’azote ammoniacal de Pano et Middlebrooks (1982) fournit, d’une manière générale, d’assez bonnes estimations de la disparition de NH4. C’est vraisemblablement ce qui explique sa popularité. Toutefois, il parvient mal, comme c’est le cas à Mèze, à rendre compte des variations saisonnières des concentrations en azote ammoniacal. D’autre part, la volatilisation, telle que déterminée sur un canal de laboratoire (Stratton, 1968) et sur des petits pilotes en extérieur (Stratton, 1969 et nous-mêmes), n’est pas négligeable mais tend à jouer un rôle secondaire quand l’élimination de l’azote ammoniacal

115
est importante. Cela signifie que le modèle de Pano et Middlebrooks n’est pas un modèle de la volatilisation mais qu’il englobe tous les processus intervenant dans la disparition de l’azote ammoniacal et non uniquement la volatilisation. Il y a donc d’autres processus que la volatilisation qui ont une part substantielle dans l’élimination de l’azote. C’est notamment le cas, comme on le montrera par la suite, de la nitrification/dénitrification. 4.5. Application : Rôle de la volatilisation selon Stratton (1969) dans la disparition de l’azote ammoniacal dans le dernier bassin de maturation de la station de lagunage de Mèze Le rôle de la volatilisation dans la disparition de l’azote ammoniacal dans l’étage de maturation est déterminé en comparant les pertes globales observées dans chaque bassin entre mai et mi-juillet 2005 à celles qui seraient dues selon Stratton (1969) à l’émission de gaz ammoniac. Pour un bassin i supposé parfaitement mélangé parfaitement, l’équation du bilan de masse s’écrit :
[ ]( ) [ ] [ ] jijijijik
jkjkjiji
VPNHQNHQt
NHV,,,43,,43,
,43, −−=∆
∆−−
−∑ (Équation 4.54)
où
jiV , est le volume en m3 de l’eau dans le bassin i le jour j
[ ] jiNH ,43− la concentration en azote ammoniacal en mg.l-1 à la sortie du bassin i le jour j
[ ]( ) [ ] [ ]( ) [ ] ( )1,,,431,43,43,,43, −−−−−− −+−=∆ jijijijijijijiji VVNHNHNHVNHV
t∆ le pas de temps égal à 1j
jkQ , le débit en m3.j-1 de l’effluent k à l’entrée du bassin i le jour j
jkC , la teneur en azote ammoniacal de l’effluent k le jour j
jiQ , le débit en m3.j-1 à la sortie du bassin i le jour j
jiP, les pertes globales en azote ammoniacal par unité de volume dans le bassin i le jour j
En supposant par ailleurs que la volatilisation est régie par la cinétique de Stratton (1969) définie par les équations 4.33 et 4.44, la disparition en azote ammoniacal par volatilisation dans le bassin i entre l’intervalle de temps 1=∆t j est donnée par la relation :
( ) ( ) ( )tCee
HC ji
pHT
jiVji
jiji ∆⋅⋅×−=∆ −−−
,5.857.12013.0
,
3
,,,1098.21
(Équation 4.55)
où
jiT , la température de l’eau en °C dans le bassin i le jour j
jipH , le pH
jiH , la hauteur d’eau
Entre juin et mi-juillet 2005, la disparition en azote ammoniacal dans l'étage de maturation est en moyenne d'environ 135 kg.j-1, avec une performance croissante des bassins au fur et à mesure de la progression vers la sortie (Figure 4.11). En effet la première lagune assure 17% de l'élimination totale en azote ammoniacal de l'étage, la seconde 36% et la dernière 47%.

116
La contribution de la volatilisation est totale dans le premier bassin, de l'ordre de 24% et de 2% dans le second et le troisième bassin respectivement. Plus l’élimination globale est importante, plus la teneur en azote ammoniacal est faible et moindre est la disparition par volatilisation d’après la cinétique de Stratton (1969).
0
10
20
30
40
50
01/0
5/05
08/0
5/05
15/0
5/05
22/0
5/05
29/0
5/05
05/0
6/05
12/0
6/05
19/0
6/05
26/0
6/05
03/0
7/05
10/0
7/05
N-N
H 4 (m
g.l-1
)
Sortie L1Sortie L2
0
10
20
30
40
50
01/0
5/05
08/0
5/05
15/0
5/05
22/0
5/05
29/0
5/05
05/0
6/05
12/0
6/05
19/0
6/05
26/0
6/05
03/0
7/05
10/0
7/05
N-N
H 4 (m
g.l-1
)
Sortie L2Sortie L3
0
10
20
30
40
50
01/0
5/05
08/0
5/05
15/0
5/05
22/0
5/05
29/0
5/05
05/0
6/05
12/0
6/05
19/0
6/05
26/0
6/05
03/0
7/05
10/0
7/05
N-N
H 4 (m
g.l-1
)
Entrée L1Sortie L1
04080
120160200240280
01/0
5/20
05
08/0
5/20
05
15/0
5/20
05
22/0
5/20
05
29/0
5/20
05
05/0
6/20
05
12/0
6/20
05
19/0
6/20
05
26/0
6/20
05
03/0
7/20
05
10/0
7/20
05
N-N
H 4 (k
g)Pertes par volatilisationPertes globales
04080
120160200240280
01/0
5/20
05
08/0
5/20
05
15/0
5/20
05
22/0
5/20
05
29/0
5/20
05
05/0
6/20
05
12/0
6/20
05
19/0
6/20
05
26/0
6/20
05
03/0
7/20
05
10/0
7/20
05
N-N
H 4 (k
g)
04080
120160200240280
01/0
5/20
05
08/0
5/20
05
15/0
5/20
05
22/0
5/20
05
29/0
5/20
05
05/0
6/20
05
12/0
6/20
05
19/0
6/20
05
26/0
6/20
05
03/0
7/20
05
10/0
7/20
05
N-N
H 4 (k
g)
Figure 4.11. Rôle de la volatilisation selon Stratton (1969) dans l’élimination de l’azote
ammoniacal dans l’étage de maturation de la station de lagunage de Mèze 4.6. Nitrification et dénitrification 4.6.1. Microbiologie et stoechiométrie Nitrification . La nitrification est la transformation biochimique de l’ammonium en nitrite, puis en nitrate selon les réactions suivantes:
24 32 ONH ++ +− ++ HOHNO 422 22 (Réaction 4-4)

117
222 ONO +− −32NO (Réaction 4-5)
La réaction bilan s’écrit :
24 2ONH ++ +− ++ HOHNO 223 (Réaction 4-6)
Ces schémas sont simplificateurs dans la mesure où de nombreux stades intermédiaires interviennent. Les étapes mentionnées ici concernent les formes de l’azote présentes à des concentrations significatives dans les eaux usées. Les bactéries nitrifiantes sont principalement autotrophes. Elles utilisent le gaz carbonique, généralement sous forme d’ions bicarbonates, comme source de carbone et l’azote réduit comme source d’énergie. Selon Wong Chong et Loehr (1975), certaines bactéries hétérotrophes possèdent des capacités nitrifiantes mais avec des vitesses de réaction tellement faibles que leur contribution est insignifiante. La nitritation (Réaction R101), parfois aussi appelée nitrosation, est réalisée par des bactéries nitreuses : Nitrosomonas, Nitrosococcus, Nitrosopira, Nitrosovibrio et Nitrosolobus. Nitrosomonas est considéré comme jouant un rôle prépondérant dans ce processus. La nitratation (Réaction 4-4) est faite par des bactéries nitriques : Nitrobacter, Nitrospira, Nitrospina, Nitrococcus et Nitrocystis. De récents travaux tendent à montrer que dans la plupart des installations de traitement des eaux usées, Nitrospira et non Nitrobacter représente le genre dominant de l’oxydation des nitrites (Rittmann et McCarty, 2001). Wiesmann (1994) représente les réactions respectives de la nitritation et de la nitratation comme suit :
−+ ++ 324 1097655 HCOONH 3222275 1045754 COHOHNONOHC +++ − (Réaction 4-7)
233242 1954400 OHCOCOHNHNO ++++ −+− −++ 32275 4003 NOOHNOHC (Réaction 4-8)
Wiesmann(1994) suppose comme plusieurs autres auteurs que les bactéries de la nitratation ont besoin de l’azote ammoniacal pour leur biosynthèse. Dénitrification . La dénitrification est la réduction biochimique des nitrates ou des nitrites en azote gazeux moléculaire 2N (Réaction 4-13). Les principaux produits intermédiaires du processus sont l’oxyde nitriqueNO et l’oxyde nitreux ON2 (Réactions 4-10 et 4-11).
−+− ++ eHNO 223 OHNO 22 +− (Réaction 4-9)
−+− ++ eHNO 22 OHNO 2+ (Réaction 4-10)
−+ ++ eHNO 222 OHON 22 + (Réaction 4-11)
−+ ++ eHON 222 OHN 22 + (Réaction 4-12)
Il en est déduit que la réaction globale de la dénitrification s’écrit :

118
−+− ++ eHNO 10122 3 OHN 22 6+ (Réaction 4-13)
Rittmann et McCarty (2001) affirment que la dénitrification peut être assurée par des bactéries hétérotrophes ou autotrophes. Parmi les hétérotrophes, Metcalf et Eddy (1991) citent plusieurs genres : Achromobacter, Aerobacter, Alcaligenes, Bacillus, Brevibacterium, Flavobacterium, Lactobacillus, Micrococcus, Proteus, Pseudomonas et Spirillum. Les bactéries dénitrifiantes sont des aérobies facultatives. Pour leur respiration, elles utilisent de manière préférentielle l’oxygène dissous. Mais elles ont la faculté de se servir de l’oxygène des ions nitrates et des ions nitrites, lorsqu’elles se trouvent plongées dans des conditions d’anoxie.
Deronzier et al. (2001) ont décrit la dénitrification par la Réaction 4-14 suivante: ++− +++ OHNHNONOHC 34391918 15.439.05.461.0 OHCONNOHC 222275 3.998.527.2 +++
où 91918 NOHC et 275 NOHC représentent respectivement la matière organique nécessaire à la
réaction et le nouveau corps bactérien formé. 4.6.2. Paramètres influents de la nitrification La teneur en oxygène dissous. La réaction bilan (Réaction 4-7) implique que la nitrification d’1 mg d’azote ammoniacal requiert 4.57 mg d’oxygène. Metcalf et Eddy (1991) affirment qu’une teneur en oxygène supérieure à 1 mg.l-1 est essentielle pour que la nitrification puisse avoir lieu. Un plus faible état d’oxygénation du milieu ralentirait ou empêcherait le catabolisme des bactéries nitrifiantes. L’oxygène est un facteur limitant de la nitrification. La température et le pH. A cause de sa nature enzymatique, la nitrification est influencée par le pH et la température. Les enzymes ont en effet des domaines spécifiques d’activité optimale par rapport à ces paramètres (Figure 4.12). Les gammes données par Prescott et al. (1995) sont seulement représentatives puisque les enzymes diffèrent les unes des autres quant aux optimums et à la forme des courbes de pH et de température.
Figure 4.12. Influence du pH et de la température sur l’activité enzymatique d’après Prescott et al. (1995)
D’après Metcalf et Eddy (1991), le pH optimal pour la nitrification se situe entre 7.5 et 8.6, mais les bactéries nitrifiantes peuvent s’adapter à des valeurs en dehors de cet intervalle. D’une manière générale, la nitrification diminue avec le pH (Wong Chong et Loehr, 1975 et

119
1978). Un pH inférieur à 6.5 provoque d’une manière sensible un ralentissement de leur activité pouvant aller jusqu’à une inhibition totale en dessous de 6. De même un pH supérieur à 9.5 rend impossible la nitrification. La conversion de l’ammonium en nitrate et plus précisément l’étape de nitritation est acidifiante (Réactions 4-4 et 4-6). Sans une alcalinité suffisante qui procurerait un effet tampon sur le pH de l'eau par neutralisation des protons formés (Réaction 4-15), une nitrification poussée peut conduire à une chute de pH capable d’inhiber le processus.
+− + HHCO3 OHCO 22 + (Réaction 4-15)
La stœchiométrie stipule que la neutralisation de l’acidité produite par l’oxydation d’1mg d’azote nécessite de l’alcalinité en bicarbonate équivalent à 7.14 mg CaCO3
(Réactions 4-6 et 4-12). La charge organique. Les bactéries nitrifiantes sont présentes dans les milieux contenant des matières organiques (Rittmann et McCarty, 2001). Evans et al. (2005) ont mis en évidence l’existence d’une DCO optimale pour la nitrification. Les bactéries nitrifiantes, caractérisées par des vitesses de croissance plus lentes, ne peuvent pas entrer en compétition avec les bactéries hétérotrophes dans l’utilisation de l’oxygène dissous et des nutriments. Ainsi, une charge organique trop importante favorise les hétérotrophes au détriment des autotrophes. Mais une certaine quantité de matière organique semble nécessaire aux bactéries nitrifiantes. Ces matières organiques à travers leur dégradation s’avèrent constituer pour eux, une source essentielle de carbone inorganique. L’influence de la charge organique sur la nitrification se manifeste aussi par le rapport de la DBO et de la teneur en azote Kjeldahl (NTK). D’après Metcalf et Eddy (1991) :
Si 1<NTK
DBO <3, alors le système est juste nitrifiant. Le pourcentage en microorganismes
autotrophes y serait compris entre 21 et 8,3%.
Si NTK
DBO>5, alors le système est le siège des processus combinés de la l’oxydation du carbone
et de la nitrification. Le support. La fixation sur un support représente l’état prédominant des bactéries nitrifiantes. Dans les unités de lagunage, outre les parois latérales des bassins, le support est composé par des matériaux particulaires inorganiques ou organiques (phytoplancton, zooplancton, agrégats de bactéries. L’ammoniac dissous. Selon Surmacz-Gorska et al. (1997), l’ammoniac dissous inhibe la nitrification, même à de faibles concentrations : 10 mg.l-1 pour la nitritation et entre 0.1 et 6 mg.l-1 pour la nitratation. Le temps de séjour. Les vitesses de croissance des bactéries nitrifiantes étant faibles, un temps de séjour minimal d’environ une semaine est requis pour optimiser la nitrification (Abeliovich, 1983). L’absence de matières toxiques. Les bactéries nitrifiantes sont plus sensibles que les bactéries hétérotrophes aux composés inhibiteurs tels que les métaux lourds.

120
4.6.3. Les paramètres influents de la dénitrification La teneur en oxygène dissous. Selon Metcalf et Eddy (1991), quand la concentration en oxygène dissous augmente jusqu’à 1 mg.l-1, le taux de dénitrification décroît linéairement vers 0 mg.l-1.j-1. Rittmann et McCarty (2001) affirment que l’oxygène dissous peut agir de deux façons sur la dénitrification. En fonction de sa concentration, il peut soit favoriser le métabolisme aérobie des bactéries dénitrifiantes, soit inhiber les réactions de réduction enzymatique. Des travaux effectués sur Pseudomonas montrent qu’une teneur minimale en oxygène dissous variant de 2.5 à 5 mg.l-1 est requise pour favoriser la respiration aérobie des bactéries dénitrifiantes. Par contre, une concentration en oxygène dissous excédant 0.1 mg-l-1 suffit pour inhiber les activités de réduction des enzymes produits. Comme cette action d’inhibition tend à s’exercer en premier lieu sur les transformations des nitrites, des oxydes nitriques et des oxydes nitreux avant d’agir sur celle des nitrates, une concentration importante en oxygène dissous peut conduire à une accumulation des produits intermédiaires de la dénitrification. La charge organique. 5 électrons équivalents sont indispensables à la réduction d’une mole de nitrate en 2N gazeux (Réaction 4-12). Deronzier et al. (2001) estiment que la dénitrification est en grande partie assurée par les bactéries hétérotrophes. Ainsi les principaux donneurs d’électron du processus sont les matières organiques. Une trop faible DBO peut ralentir fortement la dénitrification ou produire une accumulation de produits intermédiaires. La température et le pH. D’une manière générale, une augmentation de température active la dénitrification. Les pH optimaux se situent entre 6.5 et 7.5. Des valeurs de pH n’appartenant pas à cet intervalle pourraient conduire à des réductions incomplètes des nitrates aboutissant aux gazNO et ON2 . La dénitrification agit aussi sur le pH. Elle assure approximativement une restitution de la moitié de l’alcalinité consommée par la nitrification. En effet, la réduction d’1g d’azote nitrique en azote moléculaire gazeux libère environ 3.29g d’équivalents alcalins exprimés en CaCO3 (Réaction 4-14). 4.6.4. Cinétique Cinétique de la nitrification. La cinétique de la nitrification est régie par celle de l’étape la plus lente, la nitritation. Soit
43/ −NHBY le rendement de bioconversion de l’azote ammoniacal en biomasse. Il est défini
par la masse de biomasse formée par unité de masse d’azote ammoniacal oxydé. En raisonnant sur une unité de volume :
[ ]( ) ionNitrificat
ANHB NHd
dY
43/ 43
−
−=−
ρ (Équation 4.56)
où

121
Aρ est la teneur en bactéries autotrophes en mg.l-1
[ ]43−NH la concentration en azote ammoniacal total en mg.l-1
Dans des limites assez larges, le rendement de bioconversion peut être supposé constant. L’équation 4-55 peut alors s’écrire :
[ ]( ) [ ]dt
NOddt
d
dt
NHddt
d
Y
A
ionNitrificat
A
NHB3243
/ 43−−
=−=−
ρρ
(Équation 4.57)
où
[ ]32−NO est la teneur en azote nitreux et en azote nitrique en mg.l-1
t le temps en j Il en est déduit que la vitesse de nitritation est directement proportionnelle à celle de la croissance de Nitrosomonas :
[ ]dt
d
Ydt
NOd A
NHB
ρ⋅=
−
−
43/
32 1 (Équation 4.58)
La vitesse de croissance spécifique des bactéries est définie par la relation de type Monod :
[ ][ ]
[ ][ ]2
2
43
43max
243
1
OK
O
NHK
NH
dt
d
ONH
A
A +⋅
+⋅=
−
−
−
µρρ
(Équation 4.59)
où [ ]2O est la teneur en oxygène dissous en mg.l-1
43−NHK la constante de demi-saturation par rapport à l’azote ammoniacal en mg.l-1
2OK la constante de demi-saturation par rapport à l’oxygène en mg.l-1
maxµ le taux maximum de croissance des bactéries autotrophes en j-1 La vitesse de la nitrification prend alors l’expression suivante :
[ ] [ ]
[ ][ ]
[ ] AONHNHB OK
O
NHK
NH
Ydt
NOd ρµ⋅
+⋅
+⋅=
−
−−
−− 2
2
43
43
/
max32
24343
(Équation 4.60)
D’après la stoechiométrie de la réaction de la nitritation (Réaction 4-7) de Wiesmann (1994) : 147.0
43/ =−NHBY mg de biomasse autotrophe formée par mg d’azote ammoniacal oxydé
Selon Fritz et al. (1979), 3.12=OK mg.l-1
L’influence de la température et du pH sur la cinétique de la nitrification s’exerce à travers le taux maximum de croissance des bactéries autotrophes et la constante de demi-saturation par rapport à l’azote ammoniacal. Pour les systèmes de lagunage, Fritz et al. (1979) les décrivent par les équations :
TpH CC ⋅⋅= µµmax (Équation 4.61)

122
58.1051.01043
−=−
TNHK (Équation 4.62)
où µ est la constante cinétique de la croissance de Nitrosomonas supposée égale à 008.0 j-1
pHC le facteur d’inhibition de la croissance des bactéries par le pH
( )
≥<−−
=2.7,1
2.7,2.78333.01
pH
pHpHC pH (Équation 4.63)
TC le coefficient de température
( )15098.0 −= TT eC (Équation 4.64)
T la température en °C L’expression du coefficient de température est typique de la loi empirique d’Arrhénius. Celle du facteur d’inhibition par le pH a été tirée des travaux de Downing (1966) sur la nitrification dans les systèmes de boues activées. Les pH basiques étudiés par Downing (1966) qui semblaient ne pas avoir d’effet notable sur la nitrification étaient compris 7.2 et 8. L’extrapolation de Fritz et al. (1979) pour de plus fortes valeurs de pH est donc à prendre avec beaucoup de précaution. Cinétique de la dénitrification. Pour des raisons analogues à celles évoquées pour la nitrification, la vitesse de dénitrification est aussi directement proportionnelle à celle de la croissance des bactéries qui en sont responsables. Wiesmann (1994) donne une cinétique différente de la croissance spécifique des bactéries de la dénitrification, supposées être composées en majeure partie par des hétérotrophes, suivant que le composé accepteur d’électron utilisé est l’azote nitrique (Equation 4-65) ou l’azote nitreux (Equation 4-66):
[ ][ ]3
3
1max1
3
1
NOK
NO
DCOK
DCO
dt
d
NODCO
H
H +⋅
+⋅= µρ
ρ (Équation 4.65)
[ ][ ]2
2
2max2
2
1
NOK
NO
DCOK
DCO
dt
d
NODCO
H
H +⋅
+⋅= µ
ρρ
(Équation 4.66)
Il en résulte 2 expressions de la cinétique de la dénitrification :
( ) [ ]
[ ] HNODCONOB NOK
NO
DCOK
DCO
Ydt
NOd ρµ⋅
+⋅
+⋅=−
3
3
1/
max13
33
(Équation 4.67)
( ) [ ]
[ ] HNODCONOB NOK
NO
DCOK
DCO
Ydt
NOd ρµ⋅
+⋅
+⋅=−
2
2
2/
max22
22
(Équation 4.68)
où 3/ NOBY et
3/ NOBY sont les rendements de bioconversion de l’azote nitrique et de l’azote
nitreux respectivement. Le
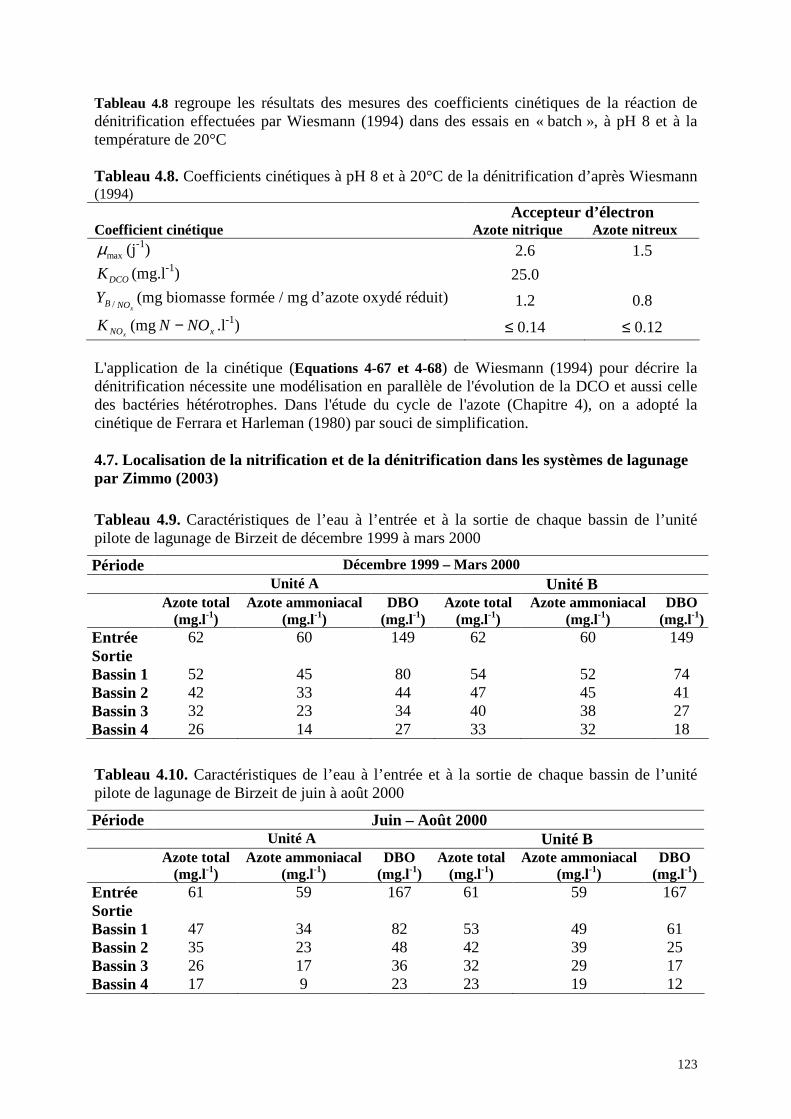
123
Tableau 4.8 regroupe les résultats des mesures des coefficients cinétiques de la réaction de dénitrification effectuées par Wiesmann (1994) dans des essais en « batch », à pH 8 et à la température de 20°C Tableau 4.8. Coefficients cinétiques à pH 8 et à 20°C de la dénitrification d’après Wiesmann (1994) Accepteur d’électron Coefficient cinétique Azote nitrique Azote nitreux
maxµ (j-1) 2.6 1.5
DCOK (mg.l-1) 25.0
xNOBY / (mg biomasse formée / mg d’azote oxydé réduit) 1.2 0.8
xNOK (mg xNON − .l-1) 14.0≤ 12.0≤
L'application de la cinétique (Equations 4-67 et 4-68) de Wiesmann (1994) pour décrire la dénitrification nécessite une modélisation en parallèle de l'évolution de la DCO et aussi celle des bactéries hétérotrophes. Dans l'étude du cycle de l'azote (Chapitre 4), on a adopté la cinétique de Ferrara et Harleman (1980) par souci de simplification. 4.7. Localisation de la nitrification et de la dénitrification dans les systèmes de lagunage par Zimmo (2003)
Tableau 4.9. Caractéristiques de l’eau à l’entrée et à la sortie de chaque bassin de l’unité pilote de lagunage de Birzeit de décembre 1999 à mars 2000
Période Décembre 1999 – Mars 2000 Unité A Unité B
Azote total (mg.l-1)
Azote ammoniacal (mg.l-1)
DBO (mg.l-1)
Azote total (mg.l-1)
Azote ammoniacal (mg.l-1)
DBO (mg.l-1)
Entrée 62 60 149 62 60 149 Sortie Bassin 1 52 45 80 54 52 74 Bassin 2 42 33 44 47 45 41 Bassin 3 32 23 34 40 38 27 Bassin 4 26 14 27 33 32 18
Tableau 4.10. Caractéristiques de l’eau à l’entrée et à la sortie de chaque bassin de l’unité pilote de lagunage de Birzeit de juin à août 2000
Période Juin – Août 2000 Unité A Unité B Azote total
(mg.l-1) Azote ammoniacal
(mg.l-1) DBO
(mg.l-1) Azote total
(mg.l-1) Azote ammoniacal
(mg.l-1) DBO
(mg.l-1) Entrée 61 59 167 61 59 167 Sortie Bassin 1 47 34 82 53 49 61 Bassin 2 35 23 48 42 39 25 Bassin 3 26 17 36 32 29 17 Bassin 4 17 9 23 23 19 12

124
Tableau 4.11. Caractéristiques de l’eau à l’entrée et à la sortie de chaque bassin de l’unité pilote de lagunage de Birzeit de mai à juin 2001
Période Mai – Juin 2001 Unité A Unité B Azote total
(mg.l-1) Azote ammoniacal
(mg.l-1) DBO
(mg.l-1) Azote total
(mg.l-1) Azote ammoniacal
(mg.l-1) DBO
(mg.l-1) Entrée 69 62 363 69 62 363 Sortie Bassin 1 55 42 203 59 51 197 Bassin 2 43 31 127 47 41 104 Bassin 3 33 24 76 38 33 68 Bassin 4 25 17 49 29 25 41 4.7.1. Matériels et méthodes Ces travaux de quantification de la nitrification et de la dénitrification ont été effectués par Zimmo (2003) sur l’installation pilote de lagunage de Birzeit (Figure 4.10) de décembre 1999 à mars 2000, de juin à août 2000 et de mai à juin 2001. Les Tableau 4.9 à Tableau 4.11 donnent les caractéristiques moyennes de l’eau à l’entrée et à la sortie de chaque bassin durant ces 3 périodes d’études. Zimmo et al ont étudié 3 couches d’eau aux profondeurs suivantes : 0 à 10 cm, 40 à 45 cm et la couche de fond entre 80 et 90cm. Des quantifications de la nitrification et de la dénitrification de chaque couche ont été réalisées sur des échantillons prélevés à ces différentes profondeurs et incubés en laboratoire. Les échantillonnages dans les 2 couches les moins profondes ont été effectués au moyen d’une pompe péristaltique. Un simple tube en verre a été utilisé pour les prélèvements dans la couche de fond où résident des sédiments. Les analyses physico-chimiques ont été faites sur la phase liquide des échantillons incubés. En parallèle, des mesures in situ ont été menées à l’aide de la mise en place dans les bassins de colonnes d’incubation, constituées par des tuyaux en PVC de 0.1m de diamètre et de 1m de long chacun. Les températures et pH d’incubation ont été ajustés à ceux des bassins au moment des prélèvements. Le rayonnement solaire a été simulé par une lampe fluorescente. Les teneurs en nitrates des échantillons étaient tellement faibles, de 1 à 2mg.l-1, que du KNO3 a été ajouté afin qu’elles atteignent 5 à mg.l-1 et qu’il en reste au terme des 24 h d’incubation. Cet apport ne devait pas interférer avec la mesure de la nitrification puisque les 2 réactions R01 et R02 étaient supposées d’ordre 0 par rapport au nitrate (Wong-Chong et Loehr, 1978). La situation d’anoxie constatée dans les couches de fond a été reproduite en laboratoire en ajoutant du sulfite de sodium. Chaque échantillon fut incubé sous 2 conditions : avec et sans inhibiteur de nitrification. Il en fut de même des incubations in situ. Les pertes par évaporation ont été corrigées par addition d’eau distillée. Les nitrites et nitrates ont été analysés au moyen des procédures HACH. La vitesse de dénitrification. La vitesse de dénitrification D est définie par la disparition par unité de temps des ions nitrites et nitrates (mg.l-1.j-1) en présence d’inhibiteur de nitrification :

125
[ ] [ ]( )t
NONOD inhibiteur
∆
−= −− 132032
(Équation 4.69)
où [ ]032−NO et [ ]132−NO sont respectivement les teneurs en azote nitreux et en azote nitrique au
début et à la fin de l’incubation exprimées en mg.l-1 t∆ la durée d’incubation (1j)
La vitesse de nitrification. Comme la nitrification et la dénitrification interviennent simultanément, la concentration en nitrate observée est le résultat net des 2 processus. La vitesse de nitrification réelleN est définie par :
[ ] [ ]( )D
t
NONON inhibiteursans +
∆
−=
−− 032132 (Équation 4.70)
N a la même unité queD La vitesse de nitrification nette représente la différence entre les taux de nitrification et de dénitrification. 4.7.2. Résultats Dans tous les bassins la couche superficielle est la plus oxygénée. La teneur en oxygène dissous décroît avec la profondeur. Des conditions d’anaérobiose ont été localisées au niveau des sédiments. Contrairement aux échantillons de fond, ceux provenant des échantillons prélevés à 0-10 et 30-40 cm de profondeur restent aérobies, avec une teneur en oxygène dissous supérieure à 1mg.l-1, du début à la fin des temps d’incubation (Figure 4.13). Les températures moyennes dans les bassins sont de 10, 20 et 21°C lors des 3 périodes d’études respectivement.
Figure 4.13. Concentrations en oxygène dissous des 3 couches, superficielle, intermédiaire ainsi que celle recouvrant les sédiments, composant la colonne d’eau dans chaque bassin durant les 3 périodes d’études de Zimmo (2003)
. Vitesses de nitrification et de dénitrification mesurées en laboratoire. Durant les incubations avec inhibiteur de nitrification, les teneurs en azote oxydé des échantillons d’eau sont restées

126
stables contrairement à celles des échantillons sédimentaires qui ont diminué sans pour autant descendre au-dessous de 4 mg.l-1. Sans inhibiteur de nitrification, les concentrations en azote oxydé ont augmenté ou sont restées constantes dans les échantillons d’eau et ont décrû dans les échantillons avec sédiments. Les vitesses de nitrification et dénitrification pour chaque bassin pendant les 3 périodes de l’étude sont donnés dans la Figure 4.14 et la Figure 4.15.
Figure 4.14.Vitesses moyennes de nitrification déterminées par incubation en laboratoire de décembre 1999 à mars 2000 (Période 1), de juin à août 2000 (Période 2) et de mai à juin 2001 (Période 3) des 8 bassins de l’unité pilote

127
Figure 4.15. Vitesses moyennes de dénitrification dans les sédiments, déterminés par incubation en laboratoire, des 8 bassins du système pilote pendant les 3 périodes de l’étude
Taux de nitrification et de dénitrification mesurés in situ. Les vitesses de nitrification et de dénitrification dans les colonnes d’eau, incluant les sédiments, isolées et incubées in situ sont présentés dans la Figure 4.16.
0
100
200
300
400
500
600
700
800
A1 A2 A3 A4
Tau
x de
nitr
ifica
tion
mg-
N.m
-2.j-
1
0
100
200
300
400
500
600
700
800
D1 D2 D3 D4
Tau
x de
nitr
ifica
tion
m
g-N
.m-2
.j-1
0
100200
300
400
500600
700
800
A1 A2 A3 A4
Taux
de
déni
trifi
catio
n
mg-
N.m
-2.j-1
0
100
200
300
400
500
600
700
800
D1 D2 D3 D4
Tau
x de
dén
itrifi
catio
n m
g-N
.m-2.j-1
-50
0
50
100
150
200
A1 A2 A3 A4
Nitr
ifica
tion
nett
e m
g-N
.m-2.j-1
Periode 1
Periode 2
Periode 3
-50
0
50
100
150
200
D1 D2 D3 D4
Nitr
ifica
tion
nett
e m
g-N
.m-2.j-1
Figure 4.16.Vitesses moyennes de nitrification et de dénitrification déterminés par incubation in situ, dans les 8 bassins pendant les 3 périodes de l’étude Les teneurs en azote oxydé dans les échantillons d’eau incubés ont diminué de 20 à 70% en présence d’inhibiteur de nitrification. Elles ont par contre augmenté, en l’absence d’inhibiteur

128
de nitrification, sauf dans quelques échantillons prélevés entre mai à juin 2001 enregistrant de légères baisses. 4.7.3. Interprétations Le manque d’oxygène a empêché ou du moins a fortement affaibli la nitrification dans les sédiments et les minces couches d’eau, de 17cm environ, qui les recouvrent. L’influence de la teneur en oxygène dissous est moins manifeste dans la colonne d’eau proprement dite. En effet, les couches d’eau superficielles et intermédiaires ont des taux de nitrification comparables malgré des différences non négligeables dans leurs états d’oxygénation (Figure 4.13). Le passage de la température moyenne de l’eau de 10 à 20°C entre la première et la seconde période d’études, en intensifiant les activités des enzymes des bactéries nitrifiantes, a augmenté de 64% les taux de nitrification. Une DBO deux fois plus importante des eaux à l’entrée en dernière période a fait baisser sensiblement le taux de nitrification. Pour une augmentation de température de 10°C, Metcalf et Eddy (1991) prévoient dans les systèmes de boues activées une augmentation de la vitesse de nitrification de plus de 300%. Un milieu moins favorable au développement des bactéries nitrifiantes serait à l’origine du faible taux (64%) observé par Zimmo (2003) dans l’unité pilote de lagunage de Birzeit. La fixation sur un support représente l’état prédominant des bactéries nitrifiantes. Dans les systèmes de boues activées, le support est constitué par les bactéries elles-mêmes, hétérotrophes et autotrophes, agglomérées sous forme de flocs. Dans les unités de lagunage, outre les parois latérales des bassins, il est composé par des matériaux particulaires inorganiques ou organiques (phytoplancton, zooplancton, agrégats de bactéries). Contrairement aux boues activées qui se maintiennent en suspension, en lagunage, un nombre non négligeable de bactéries décanteraient avec leurs supports dans les sédiments. Les bactéries nitrifiantes vont y mourir ou devenir inactives par manque d’oxygène dissous. Dans l’unité A, l’augmentation de 100% de la DBO entre la première et la deuxième période n’a eu d’impact évident que dans le dernier bassin où la vitesse de dénitrification a enregistré une hausse de l’ordre de 20%. L’augmentation de 10 à 20°C de la température moyenne de l’eau de la première à la seconde période a eu sur la dénitrification le même impact que sur la nitrification : une intensification d’environ 64%. Ces travaux ont permis de vérifier la théorie de Reed (1985) sur le rôle possible de la dénitrification dans les faibles teneurs en nitrates. En effet, ils ont mis en évidence que parfois une dénitrification plus importante et non une absence de nitrification était à l’origine de la diminution de la teneur en nitrates. Ces travaux ont aussi démontré qu’outre les boues, la couche d’eau recouvrant les sédiments était généralement assez pauvre en oxygène dissous pour être le siège de la dénitrification. En conséquence, sous des conditions environnementales optimisant la nitrification et un comportement hydrodynamique permettant une bonne diffusion des ions nitreux et nitriques dans les bassins, le processus d’élimination

129
par nitrification-dénitrification était potentiellement important et pouvait atteindre 15 à 25% de l’azote total entrant. Dans le cycle de l'azote des mécanismes autres que ceux de l'élimination de l'azote, à savoir la volatilisation et le couple nitrification-dénitrification, interviennent. Il s'agit des processus d'ammonification, d'assimilation par les microorganismes, de sédimentation, de régénération de l'azote ammoniacal à partir des sédiments qui ne seront pas développés. 4.8. Modélisation du cycle de l’azote. On a montré que la volatilisation ne peut expliquer que marginalement l’élimination de l’azote dans l’étage de maturation ; la nitrification couplée avec la dénitrification semble donc être le principal moteur de la perte d’azote. Même si cette explication apparaît déjà très vraisemblable, il convient d’aller plus loin dans l’argumentation au moyen d’une modélisation mathématique du cycle de l’azote. On attend du modèle qu’il représente l’évolution des différentes formes de l’azote – azote organique, azote ammoniacal ainsi que azote nitreux et azote nitrique - et qu’il permette de quantifier les transferts relatifs aux différentes composantes du cycle. Bien évidemment, cette modélisation implique quelques hypothèses simplificatrices et la détermination d’un certain nombre de paramètres au cours du calibrage. En conséquence, une représentation correcte de l’évolution des formes de l’azote ne constituera pas une véritable démonstration mais tout de même un argument de poids dans la controverse sur le rôle des différents mécanismes dans la disparition de l’azote.
Figure 4.17. Les mécanismes composant le cycle de l’azote (Rappel)
Ammonification N-Organique
Sédimentation
Assimilation par les microorganismes
N-Ammoniacal Nitrification
N-Nitreux et N-Nitrique
Dénitrification
Régénération benthique
Volatilisation 4C
1C
2C
3C
6C
5C
7C
Assimilation par les microorganismes '3C

130
Tableau 4.12. Les cinétiques des mécanismes du cycle de l’azote
Processus Vitesse Source Ammonification [ ]OrgNTbC −⋅⋅=1 Ferrara et Avci (1982) Sédimentation [ ]OrgNdC −⋅=2 Ferrara et Halerman (1980)
Assimilation de l’azote ammoniacal par les microorganismes
( ) [ ][ ][ ]OrgNNHK
NHC
NH
T −+
⋅=−
−−
− 43'
43203
43
αµ Ferrara et Halerman (1980)
Volatilisation ( ) ( )[ ]435.857.12013.0
3
41030
−−−
−
⋅×= NHeeH
C pHT Stratton (1968), Stratton (1969)
Régénération benthique ( )H
DUC mT
R ⋅⋅= −20205 10 β Fritz et al. (1979)
Nitrification ( ) [ ]
[ ][ ]
[ ] ApHONHA
ThA C
OK
O
NHK
NH
Y
eC ρµ
⋅⋅+
⋅+
⋅⋅
=−
−−
− 2
2
43
4315
6
243
Fritz et al. (1979), Wiesmann (1994)
Dénitrification ( ) [ ]3225
7 −− ⋅⋅= NOkC Tγ Senzia et al. (2002)
Notations :
[ ]OrgN − teneur en azote organique (mg.l-1) T température (°C) [ ]43−NH teneur en azote ammoniacal (mg.l-1) H hauteur d’eau (m) [ ]32−NO teneur en azote oxydé (mg.l-1) Aρ teneur en biomasse autotrophe (mg.l-1) [ ]DCO Demande Chimique en Oxygène (mgO2 .l
-1) Hρ teneur en biomasse "hétérotrophe" (mg.l-1) [ ]2O teneur en oxygène dissous (mg.l-1) mD masse détritique azotée active par unité de surface (mg.cm-2)

131
Tableau 4.13. Les constantes des cinétiques des mécanismes du cycle de l’azote
Valeur optimisée Constante Dénomination Unité Valeur de la littérature Source
* ** b Coefficient d’ammonification °C-1.j-1 0.002-0.003 Ferrara et Harleman (1980), Senzia et al. (2002) 0.001 0.001
d Coefficient de sédimentation j-1 0.015-0.030 Ferrara et Harleman (1980), Senzia et al. (2002) 0.013 0.014 µ Taux maximal de croissance des microorganismes à 20°C j-1 0.10-0.18 Ferrara et Harleman (1980) 0.056 0.040
α Constante de température 1.02 Senzia et al. (2002) 1.363 1.07 '
43−NHK Constante de Michaelis par rapport à l’azote ammoniacal mg.l-1 18 Senzia et al. (2002) Non optimisée
Aµ Taux maximal de croissance des bactéries autotrophes à 20°C j-1 0.008 0.470
Fritz et al. (1979) Wiesmann (1994)
0.837 0.966
AY Rendement de bioconversion de 43−NH en Nitrosomonas 0.15 Wiesmann (1994) Non optimisé
h Coefficient de température 0.098 Fritz et al. (1979) 0.07 0.07
pHC Facteur d’inhibition de la croissance des bactéries par le pH 1-0.833(7.2-pH), pH<7.2
1, pH≥7.2 Downing et Knowles (1966) Non optimisée
43−NHK Constante de Michaelis par rapport à l’azote ammoniacal mg.l-1 10 58.1051.0 −T Downing et Knowles (1966), Fritz et al. (1979) Non optimisée
2OK Constante de Michaelis par rapport à l’oxygène mg.l-1 0.5 Horn et Hempel (1996) Non optimisée
20rU Taux de régénération benthique de 43−NH à 20°C j-1 0.09 Fritz et al. (1979) Non optimisée
β Coefficient de température 1.07 Fritz et al. (1979) Non optimisée
k Constante de dénitrification j-1 0.07-0.1 Ferrara et Harleman (1980), Senzia et al. (2002) 0.44 0.30 γ Coefficient de température 1.02-1.09 Senzia et al. (2002) 1.25 1.25
* valeurs optimisées à partir des suivis de 2003-2004 ** valeurs optimisées à partir des suivis de 2005
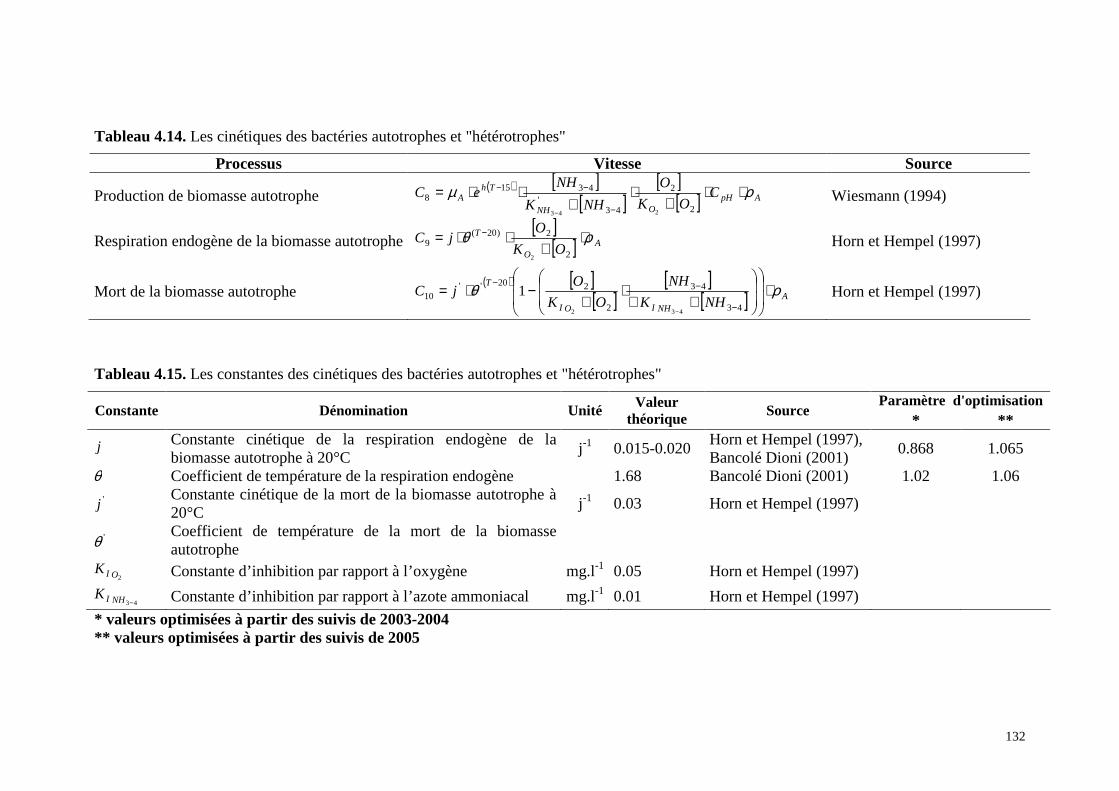
132
Tableau 4.14. Les cinétiques des bactéries autotrophes et "hétérotrophes"
Processus Vitesse Source
Production de biomasse autotrophe ( ) [ ]
[ ][ ]
[ ] ApHONH
ThA C
OK
O
NHK
NHeC ρµ ⋅⋅
+⋅
+⋅⋅=
−
−−
− 2
2
43'
43158
243
Wiesmann (1994)
Respiration endogène de la biomasse autotrophe [ ]
[ ] AO
T
OK
OjC ρθ ⋅
+⋅⋅= −
2
2)20(9
2
Horn et Hempel (1997)
Mort de la biomasse autotrophe ( ) [ ]
[ ][ ]
[ ] ANHIOI
T
NHK
NH
OK
OjC ρθ ⋅
++⋅
+−⋅=
−
−−
− 43
43
2
220''10
432
1 Horn et Hempel (1997)
Tableau 4.15. Les constantes des cinétiques des bactéries autotrophes et "hétérotrophes"
Paramètre d'optimisation Constante Dénomination Unité Valeur
théorique Source * **
j Constante cinétique de la respiration endogène de la biomasse autotrophe à 20°C
j-1 0.015-0.020 Horn et Hempel (1997), Bancolé Dioni (2001)
0.868 1.065
θ Coefficient de température de la respiration endogène 1.68 Bancolé Dioni (2001) 1.02 1.06 'j
Constante cinétique de la mort de la biomasse autotrophe à 20°C
j-1 0.03 Horn et Hempel (1997)
'θ Coefficient de température de la mort de la biomasse autotrophe
2OIK Constante d’inhibition par rapport à l’oxygène mg.l-1 0.05 Horn et Hempel (1997)
43−NHIK Constante d’inhibition par rapport à l’azote ammoniacal mg.l-1 0.01 Horn et Hempel (1997)
* valeurs optimisées à partir des suivis de 2003-2004 ** valeurs optimisées à partir des suivis de 2005

133
4.8.1. Modèle du cycle de l’azote : Principe et expression Le modèle utilisé pour décrire le cycle de l’azote est basé d’une part sur la théorie cinétique des réacteurs chimiques et d’autre part sur le principe de la conservation des masses dans un système transitoire. Les bassins sont assimilés à des réacteurs chimiques ou biochimiques parfaitement mélangés. Le modèle est défini par les bilans matières de l’azote organique, de l’azote ammoniacal et de l’azote oxydé. Ces bilans matières prennent en compte les flux entrants et sortants ainsi que la production et la disparition évaluées à l’aide de leurs cinétiques. En supposant une utilisation préférentielle de l’azote ammoniacal par les microorganismes pour leurs besoins en azote et en négligeant par conséquent l'assimilation de l'azote oxydé, les bilans de masses des différentes formes de l'azote ont comme expressions :
[ ]( ) [ ] [ ]
[ ]( ) [ ] [ ]
[ ]( ) [ ] [ ]
⋅+−+⋅−⋅=∆
∆
⋅++−−−+⋅−⋅=∆
∆
⋅+−−+−⋅−−⋅=∆−∆
−−−
−−−
∑
∑
∑
jjjjjjii
jijj
jjjjjjjjjii
jijj
jjjjjjjii
jijj
VCCNOQNOQt
NOV
VCCCCCNHQNHQt
NHV
VCCCOrgNQOrgNQt
OrgNV
)(
)(
)(
,6,732,32,32
,1,5,6,4,343,43,43
,3,2,1,,
(Équation 4.71)
où
jV est le volume d’eau en m3 le jour j
[ ] jOrgN − ,[ ] jNH 43− , [ ] jNO 32− les teneurs respectives, exprimées en mg.l-1, en azote
organique, en azote ammoniacal et en azote oxydé le jour j
jiQ , le débit de l’effluent i à l’entrée le jour j en m3.j-1
[ ] jiOrgN ,− , [ ] jiNH ,43− et [ ]ji
NO,32− sont les teneurs respectives, en mg.l-1, en azote
organique, en azote ammoniacal et en azote oxydé de l’effluent i à l’entrée jour j
jQ le débit à la sortie en m3.j-1
t∆ le pas de temps, 1=∆t j
jj CC ,7,1 ... les vitesses respectives le jour j des mécanismes de production ou de disparition
des composés azotés dont les expressions générales sont données dans le Tableau 4.12 et les constantes cinétiques dans le Tableau 4.13. La cinétique de la nitrification fait intervenir la teneur en biomasse autotrophe. Celle-ci n'ayant pas été mesurée, nécessite une initialisation ; après quoi elle est calculée à partir de l'équation de bilan de masse :
( )
jjjjAjAi
ji
AjVCCCQQ
t
Vjji
j ⋅−−+−=∆
∆∑ )( ,10,9,8, ;
ρρρ
(Équation 4.72)
où jj CC ,10,8 ... sont les vitesses le jour j des processus de production et de disparition de la
biomasse autotrophe (

134
Tableau 4.14 et Tableau 4.15) D'une manière générale, les bactéries autotrophes étant en grande partie fixées, leur flux à l'entrée est supposé négligeable. Par ailleurs, la vitesse de disparition des bactéries autotrophes par manque d'oxygène ou d'ammonium jC ,10 (Tableau 4.15) tend vers 0 puisque
2OIK et 43−NHIK sont négligeables par
rapport aux teneurs observées en oxygène dissous et en azote ammoniacal respectivement. De même, la régénération benthique met aussi à contribution un paramètre non mesuré, la masse détritique active azotée. Après initialisation, elle est calculée à partir de son bilan de masse défini par la différence entre la sédimentation azotée et la régénération benthique:
[ ] ( )j
j
mT
rjjm
DUOrgNHd
t
D⋅⋅−−
⋅=
∆
∆−20
2010β (Équation 4.73)
4.8.2. Méthodes d’application Une calibration du modèle se révèle indispensable, d'autant plus que, dans la plupart des cas, il n’y a pas de certitude a priori sur la valeur à attribuer aux paramètres entrant dans l’expression des constantes cinétiques. Un certain nombre de valeurs de paramètres trouvées dans la littérature ont été conservées (Tableau 4.13 et Tableau 4.15). Pour les autres paramètres, les valeurs issues de la littérature ont dû être modifiées pour assurer une représentation convenable des concentrations. La calibration du modèle. La calibration du modèle consiste à déterminer les valeurs à attribuer aux paramètres entrant dans l’expression des constantes cinétiques (Tableau 4.13 et Tableau 4.15) par l’ajustement mathématique des concentrations calculées en azote organique et en azote oxydé sur celles mesurées à la sortie du système étudié, à partir de la recherche des minimums des fonctions critères suivantes :
[ ]( ) [ ]( )( )2∑ −−−j
ExpjCalcj OrgNOrgN
[ ]( ) [ ]( )( )23232∑ −− −j
ExpjCalcj NONO
[ ]( )CalcjOrgN − et [ ]( )
CalcjNO 32− sont tirés des équations de leurs bilans de masse respectifs,
en donnant à[ ] jNH 43− sa valeur expérimentale.
L’expression de la volatilisation telle que formulée dans le Tableau 4.13 est considérée comme acquise. Elle a été vérifiée à travers les expériences effectuées au printemps 2006. Les teneurs calculées en azote organique, en azote ammoniacal et en azote oxydé à la sortie. Les constantes cinétiques des divers mécanismes du cycle de l'azote une fois définies, les concentrations respectives en azote organique, en azote ammoniacal et en azote oxydé que prévoit le modèle à la sortie d’une unité donnée sont obtenues en résolvant à l'aide du logiciel Maple le système formé par les équations différentielles 4-71 à 4-73.

135
Les masses d’azote journalières calculées mises en jeu par les différents mécanismes du cycle de l’azote. Une fois déterminées les constantes cinétiques ainsi que les concentrations journalières moyennes de chacune des formes de l’azote à la sortie, les masses d’azote respectives mises en jeu le jour j par les mécanismes intervenant dans le cycle de l’azote sont calculées par :
310−⋅⋅= VCM kk (Équation 4.74)
avec 7,...,1=k où
kM est la masse d’azote mise en jeu par le mécanisme k le jour j en kg N/j
kC la vitesse du mécanisme k le jour j en mgN.l-1.j-1 (Tableau 4.12)
V le volume en m3 du système étudié le jour j Les mécanismes d'élimination de l'azote et leur importance respective. D'une manière générale, le bilan de masse de l’azote total le jour j est défini par :
[ ]( ) [ ] [ ] jjjiTjjii
TjijTj
VPNQNQt
NV−⋅−⋅=
∆∆
∑ ,,
, (Équation 4.75)
où [ ] jTN représente la concentration en azote total en mg.l-1 le jour j
jP la perte en azote ammoniacal en mg.l-1.j-1 le jour j
Or la somme des équations formant le système 4-71 donne :
[ ]( ) [ ] [ ] ( ) jjiTjjii
TjijTj
VCCCCNQNQt
NV⋅+−+−⋅−⋅=
∆∆
∑ 7542,,
, (Équation 4.76)
Selon donc le modèle, le jour j, la perte en azote global du système est égale à celle due à l'élimination combinée de la volatilisation et de la dénitrification à laquelle on ajoute la perte nette par sédimentation donnée par la différence entre la quantité d'azote organique qui sédimente et celle restituée par relargage sous forme ammoniacale. 4.8.3. Application à l’étage de maturation de la station de lagunage de Mèze assimilé à un bassin unique Le modèle a été utilisé pour définir les rôles respectifs des différents mécanismes dans le cycle de l'azote entre juillet 2003 et août 2004 dans l'étage de maturation de la station de lagunage de Mèze. Durant cette période, les concentrations des composés azotés ayant été mesurées uniquement à l’entrée et à la sortie de l’étage, une étude individuelle de chacun des 3 bassins de maturation est impossible. En conséquence, pour cette première application ils sont assimilés à un unique bassin parfaitement mélangé.

136
Les cinétiques calibrées (Tableau 4.13 et Tableau 4.15). Les valeurs des coefficients des cinétiques de l’ammonification et de la sédimentation issues de la calibration ont les mêmes ordres de grandeur que celles relevées dans la littérature. Celle du taux maximal de croissance des microorganismes à 20°C relatif à l’assimilation de l’azote ammoniacal représente la moitié de celle de Ferrara et Harleman (1980). Le taux maximal de croissance des bactéries autotrophes à 20°C est par contre 2 fois supérieur à celui de Wiesmann (1994) et 100 fois à celui de Fritz et al.(1979); la valeur proposée par Fritz est une valeur supposée. Les valeurs optimisées du taux maximum de croissance et du coefficient de la respiration endogène des bactéries autotrophes sont du même ordre de grandeur (0.837 et 0.868 respectivement). Le coefficient de dénitrification déterminé par calibration est de 0.44 contre 0.07 à 0.1 selon Ferrara et Harleman (1980) et Senzia et al. (2002). Initialisation des teneurs en bactéries et des masses détritiques actives azotées surfaciques. Les incertitudes sur les concentrations calculées des différentes formes de l’azote que génère l’initialisation des teneurs en bactéries autotrophes ainsi que celle des masses détritiques actives azotées par unité de surface s'étendent sur une durée variable, entre un et quatre mois pour des teneurs en bactéries autotrophes initiales allant de 10-8 à 10 mg.l-1 et des masses détritiques actives azotées initiales comprises entre 10-3 et 5 mg.cm-2. L’influence des initialisations sur les concentrations calculées en azote organique et en azote oxydé est représentée dans la Figure 4.18.
0
5
10
15
20
25
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
mg
N/ l
Norg (ρi=10-8)
Norg (ρi=0,1)
Norg (ρi=10)
0
2
4
6
8
10
12
08/0
7/20
03
08/0
8/20
03
08/0
9/20
03
08/1
0/20
03
08/1
1/20
03
08/1
2/20
03
08/0
1/20
04
08/0
2/20
04
08/0
3/20
04
08/0
4/20
04
08/0
5/20
04
08/0
6/20
04
08/0
7/20
04
08/0
8/20
04
mg
N/ l
NO2-3 (Dmi=0,001)
NO2-3 (Dmi=0,1)
NO2-3 (Dmi=5)
Figure 4.18. Variation des teneurs calculées à la sortie en azote organique et en azote oxydé en fonction des valeurs initiales des teneurs en bactéries autotrophesiρ et des masses détritiques actives azotées par unité de surfaceimD

137
En supposant grande la probabilité pour que les caractéristiques physiques, chimiques et biologiques des bassins de l’étage de maturation soient reproduites d'une année sur l’autre, on minimise l’influence des initialisations en adoptant pour valeurs initiales celles simulées à la même date l’année suivante. Comme le montrent la Figure 4.19 et la Figure 4.20, ces valeurs ne dépendent pas de l’initialisation de cette simulation. On adopte ainsi pour le premier jour du calcul, le 8 juillet 2003, la teneur en bactéries autotrophes et la masse détritique active azotée par unité de surface prédites par la simulation pour le 08 juillet 2004, soit.
1.0=iρ mg.l-1 pour les bactéries autotrophes et 1.0=imD mg.cm-2 pour la masse détritique
active.
1,E-08
1,E-07
1,E-06
1,E-05
1,E-04
1,E-03
1,E-02
1,E-01
1,E+00
08/0
7/03
08/0
8/03
08/0
9/03
08/1
0/03
08/1
1/03
08/1
2/03
08/0
1/04
08/0
2/04
08/0
3/04
08/0
4/04
08/0
5/04
08/0
6/04
08/0
7/04
08/0
8/04
Ten
eur e
n ba
ctér
ies
auto
trop
hes
mg/
l
ρi=10-1
ρi=10-8
Figure 4.19. Variation de la teneur en bactéries autotrophes simulée en fonction de l'initialisation
0
0,2
0,4
0,6
0,8
1
08/0
7/03
08/0
8/03
08/0
9/03
08/1
0/03
08/1
1/03
08/1
2/03
08/0
1/04
08/0
2/04
08/0
3/04
08/0
4/04
08/0
5/04
08/0
6/04
08/0
7/04
08/0
8/04
Mas
se d
étrit
ique
act
ive
azot
ée
(mg/
cm2 )
Dmi=5
Dmi=0,1
Figure 4.20. Variation de la masse surfacique détritique active azotée en fonction de l'initialisation
Les teneurs calculées en azote organique, en azote ammoniacal et en azote oxydé à la sortie de l’étage de maturation. Le modèle restitue de façon satisfaisante les évolutions de l'azote ammoniacal et de l'azote organique et reproduit assez correctement les 3 pics d’azote oxydé (Figure 4.21).
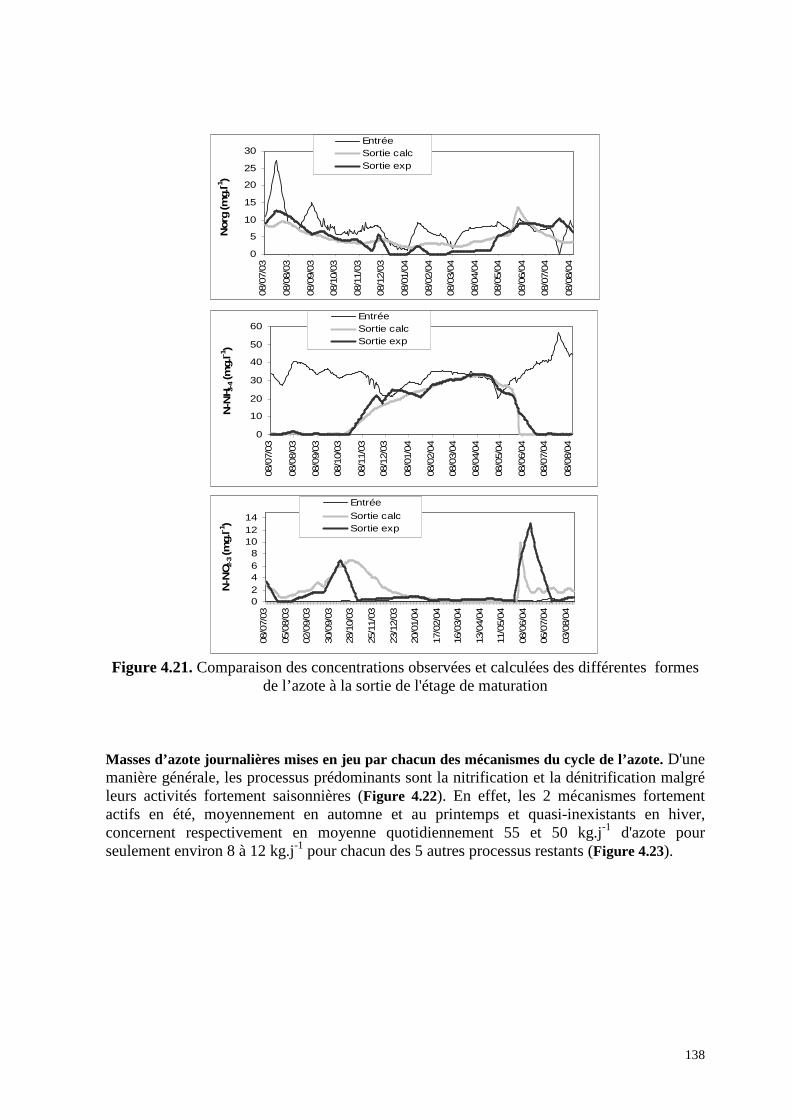
138
0
5
10
15
20
25
30
08/0
7/03
08/0
8/03
08/0
9/03
08/1
0/03
08/1
1/03
08/1
2/03
08/0
1/04
08/0
2/04
08/0
3/04
08/0
4/04
08/0
5/04
08/0
6/04
08/0
7/04
08/0
8/04
Nor
g (m
g.l-1
)
EntréeSortie calcSortie exp
0
10
20
30
40
50
60
08/0
7/03
08/0
8/03
08/0
9/03
08/1
0/03
08/1
1/03
08/1
2/03
08/0
1/04
08/0
2/04
08/0
3/04
08/0
4/04
08/0
5/04
08/0
6/04
08/0
7/04
08/0
8/04
N-N
H 3-4 (m
g.l-1
)
EntréeSortie calcSortie exp
02468
101214
08/0
7/03
05/0
8/03
02/0
9/03
30/0
9/03
28/1
0/03
25/1
1/03
23/1
2/03
20/0
1/04
17/0
2/04
16/0
3/04
13/0
4/04
11/0
5/04
08/0
6/04
06/0
7/04
03/0
8/04
N-N
O 2-3 (m
g.l-1
)
Entrée
Sortie calcSortie exp
Figure 4.21. Comparaison des concentrations observées et calculées des différentes formes
de l’azote à la sortie de l'étage de maturation Masses d’azote journalières mises en jeu par chacun des mécanismes du cycle de l’azote. D'une manière générale, les processus prédominants sont la nitrification et la dénitrification malgré leurs activités fortement saisonnières (Figure 4.22). En effet, les 2 mécanismes fortement actifs en été, moyennement en automne et au printemps et quasi-inexistants en hiver, concernent respectivement en moyenne quotidiennement 55 et 50 kg.j-1 d'azote pour seulement environ 8 à 12 kg.j-1 pour chacun des 5 autres processus restants (Figure 4.23).

139
0
100
200
300
400
500
600
700
800
900
08/0
7/20
03
26/0
7/20
03
13/0
8/20
03
31/0
8/20
03
18/0
9/20
03
06/1
0/20
03
24/1
0/20
03
11/1
1/20
03
29/1
1/20
03
17/1
2/20
03
04/0
1/20
04
22/0
1/20
04
09/0
2/20
04
27/0
2/20
04
16/0
3/20
04
03/0
4/20
04
21/0
4/20
04
09/0
5/20
04
27/0
5/20
04
14/0
6/20
04
02/0
7/20
04
20/0
7/20
04
07/0
8/20
04
kg N
/j
0
5
10
15
20
25
30
Tem
péra
ture
(°C
)
Ammonif ication
Sédimentation
Assimilation par les Microorganismes
Volatilisation
Nitrif ication
Dénitrif ication
Regénération benthique
T° Moyenne
Figure 4.22. Masses d’azote calculées journalières mises en jeu par les différents mécanismes du cycle de l'azote dans l’étage de
maturation

140
L'influence des saisons est manifeste sur la plupart des processus du cycle de l'azote. L'ammonification la plus importante a lieu en été et au printemps. Elle diminue sensiblement en automne et devient très faible l'hiver. L'évolution en fonction des saisons de la sédimentation suit celle de la concentration en azote organique, à laquelle elle est directement liée (Tableau 1). La sédimentation n'est jamais très importante. Elle est plus active en été et au printemps qu'en automne. Elle est très faible l'hiver. La régénération benthique présente un profil saisonnier semblable à celui de la sédimentation. Pour l'assimilation de l’azote ammoniacal par les microorganismes, l'influence de la température est couplée avec celle du substrat. Ainsi, ce processus enregistre sa plus forte activité au printemps, saison caractérisée par une température permettant à un processus biologique d'avoir lieu, et surtout par une grande disponibilité en substrat. La faible teneur en azote ammoniacal dans les bassins explique sa moindre importance en été. En automne et en hiver elle est négligeable. La volatilisation est négligeable l'été et l'automne en raison de la faible concentration en azote ammoniacal. Elle est faible l'hiver et enregistre sa plus forte activité au printemps.
0
20
40
60
80
100
120
140
Moyenneannuelle
Eté Automne Hiver Printemps
kg N
/ j
n
Ammonification
Sédimentation
Assimilation par les Microorganismes
Volatilisation
Nitrification
Dénitrification
Régénération benthique
Figure 4.23. Moyennes saisonnières des masses d’azote mises en jeu par les différents mécanismes du cycle de l'azote dans l’étage de maturation

141
-20
0
20
40
60
80
100
120
140
Moyenneannuelle
Eté Automne Hiver Printemps
kg N
/ j
Dénitrif ication
Volatilisation
Sédimentation -Régénération
Figure 4.24. Parts respectives de la sédimentation, de la volatilisation et de la dénitrification dans l'élimination de l'azote Les mécanismes de l'élimination de l'azote. La disparition moyenne de l’azote dans l'étage de maturation entre mi-juillet 2003 et mi-juillet 2004 est de l'ordre de 58 kg.j-1, dont 86% semble dû à la dénitrification, et 14% à la volatilisation (Figure 4.24). L’azote qui sédimente, variant entre 5 et 11 kg.j-1, serait généralement compensé par la régénération benthique. L'élimination maximale a lieu en été, évaluée à 114 kg.j-1 où elle est assurée presque totalement par la dénitrification. En automne, la diminution de la température entraîne une baisse notoire des pertes en azote, seulement 41 kg.j-1, en raison d’un fort ralentissement de la dénitrification qui en reste le principal mécanisme. L’hiver, la dénitrification est inhibée et la volatilisation est à l’origine de l’élimination minimale de 7 kg.j-1 d’azote. Les pertes journalières en azote sont de l’ordre de 68 kg au printemps dont 46 kg sont dues à la dénitrification et 22 kg à la volatilisation. Conclusions. Cette modélisation du cycle de l’azote met en évidence l’impact primordial de la nitrification et de la dénitrification dans l’élimination de l’azote dans l’étage de maturation du lagunage de Mèze. Notre évaluation expérimentale de la volatilisation, qui recoupe remarquablement les travaux de Stratton (1968 et 1969) conduit à une estimation peu contestable de l’ordre de grandeur de ce processus. Il est clair alors que la nitrification/dénitrification est le mécanisme principal de la perte d’azote. Ce mécanisme explique bien, à travers l’influence de la température sur la nitrification et la dénitrification, le caractère saisonnier de l’élimination. La simulation montre que, dans ces bassins de maturation, la sédimentation ne participe que marginalement au bilan de l’azote. Une des faiblesses de cette modélisation est l’assimilation de l’ensemble des trois bassins à un bassin unique. Cette simplification peut avoir un impact sur les valeurs des paramètres optimisés.

142
4.8.4. Application du modèle du cycle de l'azote développé en 2003 à l’étage de maturation de la station de lagunage de Mèze, bassin par bassin Des suivis bassin par bassin de l’étage de maturation ont été effectués au printemps et au début de l’été 2005, du début avril à la mi-juillet, de manière, en particulier, à vérifier si les pics de nitrate observés en 2003 et 2004 n’étaient pas accidentels. Ils nous ont donné la chance de pouvoir modéliser, cette fois, le devenir de l’azote bassin par bassin. Pour chaque bassin, les concentrations des différentes formes de l’azote sont calculées à la sortie du bassin à partir des concentrations observées à l’entrée du même bassin.
L1
01020304050
06/0
4/20
05
21/0
4/20
05
06/0
5/20
05
21/0
5/20
05
05/0
6/20
05
20/0
6/20
05
05/0
7/20
05
mg
NO
rg/l
0
10
20
30
40
50
06/0
4/20
05
21/0
4/20
05
06/0
5/20
05
21/0
5/20
05
05/0
6/20
05
20/0
6/20
05
05/0
7/20
05
mgN
-NH
3-4/
l
0
10
20
30
40
50
06/0
4/20
05
21/0
4/20
05
06/0
5/20
05
21/0
5/20
05
05/0
6/20
05
20/0
6/20
05
05/0
7/20
05
mgN
-NO
2-3/
l
EntréeSortie calcSortie Obs
L2
01020304050
16/0
5/20
05
26/0
5/20
05
05/0
6/20
05
15/0
6/20
05
25/0
6/20
05
05/0
7/20
05
mgN
Org
/l
0
10
20
30
40
50
16/0
5/20
05
26/0
5/20
05
05/0
6/20
05
15/0
6/20
05
25/0
6/20
05
05/0
7/20
05
mgN
-NH
3-4/
l
0
10
20
30
40
50
16/0
5/20
05
26/0
5/20
05
05/0
6/20
05
15/0
6/20
05
25/0
6/20
05
05/0
7/20
05
mgN
-NO
2-3/l
Figure 4.25. Comparaison des concentrations observées et calculées des différentes formes de l’azote aux sorties des 2 premiers bassins de maturation

143
0
10
20
30
40
06/0
4/05
21/0
4/05
06/0
5/05
21/0
5/05
05/0
6/05
20/0
6/05
05/0
7/05
N-N
H 3-4 (
mg/
l)
0
10
20
30
40
06/0
4/05
21/0
4/05
06/0
5/05
21/0
5/05
05/0
6/05
20/0
6/05
05/0
7/05
N-N
O 2-3 (m
g/l)
Entrée
Sortie calc
Sortie exp
L3
0
10
20
30
40
06/0
4/05
21/0
4/05
06/0
5/05
21/0
5/05
05/0
6/05
20/0
6/05
05/0
7/05
NO
rg (m
g/l)
Figure 4.26. Comparaison des concentrations observées et calculées des différentes formes de l’azote à la sortie du dernier bassin de maturation
Les composés azotés à la sortie de chacun des bassins de maturation (Figure 4.25 et Figure 4.26). Pendant le court suivi de la première lagune, du 7 juin au 12 juillet, la teneur en azote organique à la sortie est très variable, entre 8,6 et 26,4 mg.l-1. Celle de la teneur en azote ammoniacal varie peu autour de 35 mg.l-1. L'azote oxydé y est pratiquement absent, avec une concentration inférieure 0,06 mg.l-1. Entre le 17 mai et le 12 juillet, la teneur moyenne en azote organique à la sortie de la seconde lagune est de l'ordre de 13 mg.l-1. Du 17 mai au 22 juin, celle en azote ammoniacal varie peu autour de 29 mg.l-1 et celle en azote oxydé reste inférieure à 0,9 mg.l-1. Du 22 au 27 juin la concentration de l'azote ammoniacal diminue de 27 à 15 mg.l-1 tandis que celle de l'azote oxydé augmente de 0,5 à 11mgl-1. Du 27 juin au 5 juillet la teneur en azote ammoniacal reste à peu près constante de l'ordre de 14 mg.l-1 alors que celle en azote oxydé diminue de 11 à 2,3 mg.l-1. Du 5 au 12 juillet, la concentration en azote organique croît de 14 à 20 mg.l-1 quand celle en azote oxydé reste stable autour de 2.4 mg.l-1. Le suivi de la dernière lagune a été réalisé du 7 avril au 12 juillet. Dans ce bassin, durant toute cette période, la teneur en azote organique varie modérément. La concentration de l'azote ammoniacal enregistre une diminution de 28 à 3 mg.l-1 entre le 31 mai et le 9 juin accompagnée d'une augmentation de celle de l'azote oxydé de 3 à 28 mg.l-1. Suit une

144
décroissance rapide de la teneur en azote oxydé tandis que celle en azote ammoniacal reste faible et inférieure à 3 mg.l-1 jusqu'à la fin du suivi, le 12 juillet. Les teneurs observées et calculées en azote organique, en azote ammoniacal et en azote oxydé à la sortie de chacun des bassins de l’étage de maturation. Afin de pouvoir appliquer le modèle du cycle de l'azote précédemment développé, une modélisation simultanée de la teneur en biomasses autotrophes et de la masse détritique active azotée par unité de surface est indispensable. En effet ces paramètres ne sont pas mesurés. Pour chacun des bassins, on a initialisé la teneur en bactéries autotrophes et la masse détritique active en utilisant les valeurs simulées à la même époque en 2004. Le modèle prévoit pour chacune des 3 lagunes l’apparition, à la même date pour tous les bassins, d’un pic d’azote oxydé accompagné d’une diminution importante en azote ammoniacal (Figure 4.25 et Figure 4.26). Or durant la période de suivi, la première lagune se caractérise par la quasi-absence de composés oxydés et une teneur élevée en azote ammoniacal. Dans la seconde lagune le pic d’azote oxydé et la diminution de l’azote ammoniacal apparaissent environ 18 jours plus tard que prévu par le modèle ; la perte d’azote ammoniacal est moins importante que ne le voudrait la simulation. Dans la dernière lagune, de mi-mai à mi-juin, l’azote oxydé apparaît bien mais avec quelques jours de retard sur les observations et un pic sous estimé. L’azote organique est en revanche surestimé par le modèle. Les conditions ayant favorisé la nitrification et la dénitrification en 2003 se trouvant de nouveau réunies dans les bassins de maturation vers fin mai à mi-juillet en 2005, le modèle prévoit en conséquence durant cette période l'apparition d'un pic d'azote oxydé dans chacun d’eux. En effet, en se référant aux expressions des cinétiques de la nitrification et de la dénitrification (Tableau 4.13 et Tableau 4.15), la température, le pH, la teneur en oxygène dissous, les teneurs en azote ammoniacal et en azote oxydé sont les paramètres influents de la nitrification et de la dénitrification. Or le pH n'intervient que pour des valeurs inférieures à 7.2. Quant à l'oxygène dissous et à l'azote ammoniacal, ils ne sont limitants que pour des teneurs faibles comparables aux constantes de demi-saturation
43−NHK et
2OK respectivement.43−NHK qui est fonction de la température, varie entre 0.2 et 0.5 mg.l-1
quelque soit le bassin considéré pendant la période de l'étude. La valeur de2OK adoptée pour
le modèle est celle proposée par Horn et Hempel (1996) égale à 0.5 mg.l-1. Comme en 2003, vers la fin du printemps les eaux dans les bassins de maturation se caractérisent par des pH toujours supérieurs à 7.2, des teneurs en azote ammoniacal largement supérieures à
43−NHK et
des teneurs en oxygène dissous, mesurées par l’exploitant toutes les semaines, supérieures à 2 mg.l-1 (Figure 4.27). L’augmentation de la température serait alors le facteur déclenchant de la nitrification et de la dénitrification. Les profils similaires de la température dans les 3 bassins conduisent alors à des évolutions semblables des teneurs simulées en azote oxydé. Les 3 bassins de maturation se différent les uns des autres par les évolutions des 3 formes de l'azote observées à la sortie. Ces différences ne sont pas représentées par le modèle. La raison principale est vraisemblablement liée à un défaut de représentativité des mesures des teneurs en oxygène dissous. En effet, ce paramètre est extrêmement variable au cours de la journée, comme le montre la Figure 4.28- relative à un enregistrement en un point de la lagune L3 -; et dans l’espace d’un bassin. Malgré cela, le suivi du gestionnaire consiste en des mesures

145
uniques instantanées hebdomadaires. La non adéquation de la méthode de suivi se traduit vraisemblablement par une surestimation de l'oxygénation dans les 2 premiers bassins. Ce point de vue est conforté par le suivi de la matière organique en 2003-2004 ; en effet celui-ci montre qu'entre le 27 avril et le 15 juillet, de l'entrée à la sortie de l'étage de maturation, la DCO et la DBO filtrées diminuent en moyenne de 52 et 65% respectivement (Figure 4.29). Il est généralement admis que, dans les systèmes d’épuration, l’oxygène est préférentiellement consommé pour l’oxydation de la matière organique ; il est donc vraisemblable que moins d’oxygène est disponible pour l’oxydation de l’azote dans L1 que dans L3.
0
4
8
12
16
01/0
4/20
05
15/0
4/20
05
29/0
4/20
05
13/0
5/20
05
27/0
5/20
05
10/0
6/20
05
24/0
6/20
05
08/0
7/20
05
O2 d
isso
us (m
g/l)
n
L1
L2
L3
6,8
7,2
7,6
8,0
8,4
8,8
01/0
4/20
05
15/0
4/20
05
29/0
4/20
05
13/0
5/20
05
27/0
5/20
05
10/0
6/20
05
24/0
6/20
05
08/0
7/20
05
pH
L1
L2
L3
05
101520253035
01/0
4/20
05
15/0
4/20
05
29/0
4/20
05
13/0
5/20
05
27/0
5/20
05
10/0
6/20
05
24/0
6/20
05
08/0
7/20
05
Tem
péra
ture
(°C
)
n
L1
L2
L3
Figure 4.27. pH, teneur en oxygène dissous et température dans les 3 bassins de maturation
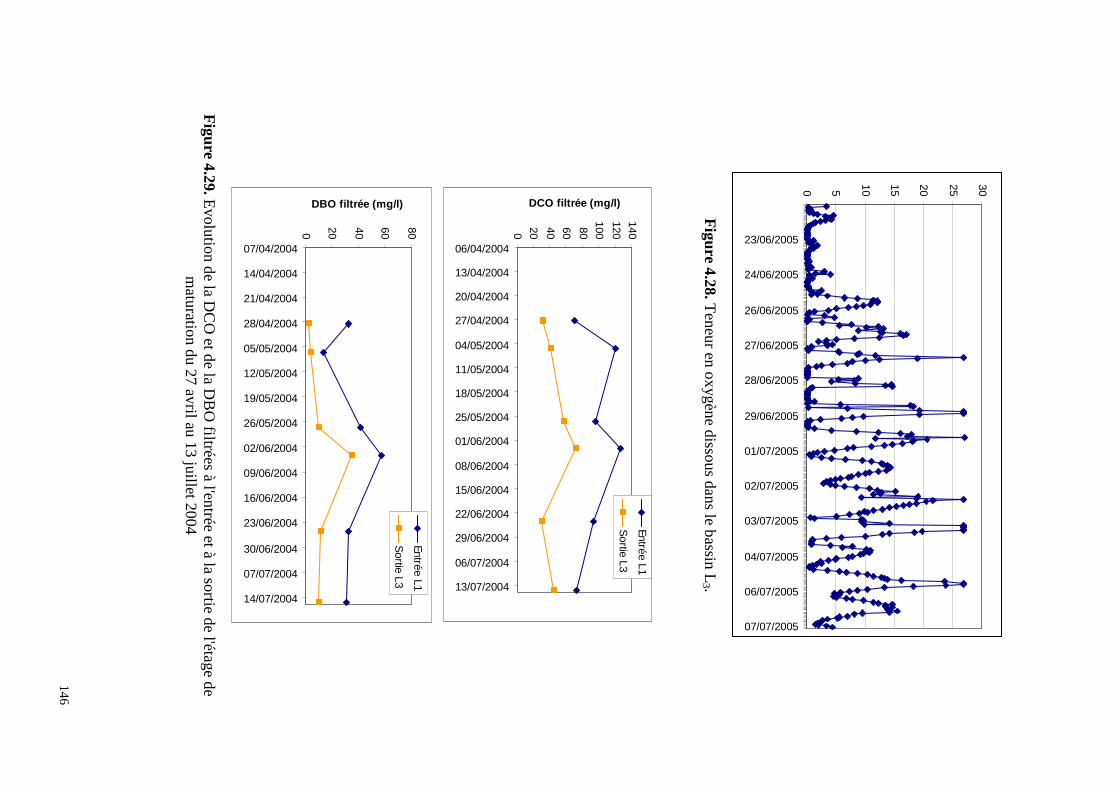
1
46
0 5 10 15 20 25 30
23/06/2005
24/06/2005
26/06/2005
27/06/2005
28/06/2005
29/06/2005
01/07/2005
02/07/2005
03/07/2005
04/07/2005
06/07/2005
07/07/2005
Figure 4.28. T
eneur en oxygèn
e dissous dans le bassin L
3 .
0 20 40 60 80
100
120
140
06/04/2004
13/04/2004
20/04/2004
27/04/2004
04/05/2004
11/05/2004
18/05/2004
25/05/2004
01/06/2004
08/06/2004
15/06/2004
22/06/2004
29/06/2004
06/07/2004
13/07/2004
DCO filtrée (mg/l)
Entrée L1
Sortie L3
0 20 40 60 80
07/04/2004
14/04/2004
21/04/2004
28/04/2004
05/05/2004
12/05/2004
19/05/2004
26/05/2004
02/06/2004
09/06/2004
16/06/2004
23/06/2004
30/06/2004
07/07/2004
14/07/2004
DBO filtrée (mg/l)
Entrée L1
Sortie L3
Figure 4.29. E
volution de la D
CO
et de la DB
O filtrées à l'entrée
et à la sortie de l'étage d
e m
aturation du 27 avril au 13 juillet 2004

147
De bonnes valeurs des teneurs en oxygène dissous des bassins n'excluent pas la nécessité d'une nouvelle optimisation. Entre autres, la constante de demi-saturation par rapport à l'oxygène donnée par Horn et Hempel (1997) peut ne plus être adaptée. D'autant plus que ces auteurs ont travaillé sur les biofilms de systèmes physiques différents des bassins de lagunage. Avec les paramètres calibrés pour représenter le devenir de l’azote entre l’entrée et la sortie de l’étage de maturation considéré comme une lagune unique au cours de l’année 2003-2004, on parvient à une simulation raisonnablement acceptable du devenir de l’azote entre l’entrée et la sortie de la seule lagune L3 en 2005. Cela tient à la forme des équations du système 4-69, dans lesquelles, quand les conditions sont réunies pour des variations importantes des concentrations des formes de l’azote, les termes ][][ NQNQ ji ij −∑ sont faibles par rapport
aux termes jn njVC∑ , ce qui fait que les vitesses de variations des concentrations dépendent
alors peu du volume du bassin. 4.8.5. Application au dernier bassin de maturation de la station de lagunage de Mèze On pouvait tenter d’améliorer le calibrage des paramètres du modèle appliqué à la lagune L3 en utilisant les données du suivi réalisé entre le 7 avril et le 12 juillet 2005. Les inconvénients de l’assimilation de 3 lagunes à une lagune unique sont ainsi contournés. Les valeurs utilisées pour l'initialisation des simulations de la teneur en bactéries autotrophes et de la masse détritique active azotée par unité de surface sont les mêmes que pour la précédente application. Les valeurs correspondantes des paramètres des cinétiques de transformations de l’azote sont présentées dans le Tableau 4.13 et le Tableau 4.15. Elles ne diffèrent pas sensiblement de celles issues de la calibration effectuée à partir des données des années 2003-2004. La Figure 4.30 montrent que le modèle ainsi calibré reproduit de façon satisfaisante les évolutions des 3 formes de l'azote, avec notamment une meilleure représentation de l’azote organique. Masses d’azote journalières mises en jeu par chacun des mécanismes du cycle de l’azote. La modélisation montre que la période de l'étude peut être divisée en deux parties définies par les intervalles de temps précédant et succédant la manifestation de la nitrification (Figure 4.31). La Figure 4.32 et la Figure 4.33 représentent les masses calculées d’azote journalières mises en jeu par chacun des mécanismes du cycle de l’azote ainsi que leurs moyennes respectives suivant la période considérée. Durant la première période, l’assimilation de l’azote par les microorganismes augmente de 6 à 16 kg.j-1 avec la température. Avec une moyenne de 13 kg N.j-1, elle est le mécanisme le plus important du cycle. L’ammonification croît aussi avec la température de 5 à 11 kg N.j-1, avec une moyenne de 10 kg N.j-1.La volatilisation met en jeu en moyenne 8.8 kg N.j-1. La sédimentation varie de 4.8 à 8.8 kg N.j-1 avec une moyenne de 7.5 kg N.j-1 et est compensée par la régénération benthique qui augmente de 6 à 10 kg N.j-1 avec une moyenne de 7.8 kg N.j-1. La nitrification et la dénitrification sont négligeables avec des moyennes respectives de 0.4 et 0.6 kg N.j-1.

148
Début juin et durant la seconde période la situation change rapidement. L’assimilation et la volatilisation décroissent jusqu’à des valeurs inférieures à 1 kgN.j-1 en raison de la forte diminution de la teneur en azote ammoniacal. Leurs valeurs moyennes sont de 3.1 et 2.5 kgN.j-1 respectivement. La sédimentation est toujours compensée par la régénération benthique avec des moyennes respectives de 4.7 et 5.8 kgN.j-1. La nitrification avec une moyenne de 76.5 kgN.j-1 et la dénitrification avec 63.5 kgN.j-1 constituent les processus les plus importants. D’une manière générale, durant la période de l’étude, la nitrification et la dénitrification sont les mécanismes prépondérants avec des moyennes respectives de 33.4 et 27.9 kgN.j-1. L’ammonification concerne quotidiennement 9.3 kg d’azote, l’assimilation par les microorganismes 8.6, la régénération benthique 7, la sédimentation 6.3 et la volatilisation 6.1
0
10
20
30
40
50
04/0
4/20
05
18/0
4/20
05
02/0
5/20
05
16/0
5/20
05
30/0
5/20
05
13/0
6/20
05
27/0
6/20
05
11/0
7/20
05
NO
rg (
mg/
l)
0
10
20
30
40
50
04/0
4/20
05
18/0
4/20
05
02/0
5/20
05
16/0
5/20
05
30/0
5/20
05
13/0
6/20
05
27/0
6/20
05
11/0
7/20
05
N-N
H 3-4
(mg/
l)
0
10
20
30
40
50
04/0
4/20
05
18/0
4/20
05
02/0
5/20
05
16/0
5/20
05
30/0
5/20
05
13/0
6/20
05
27/0
6/20
05
11/0
7/20
05
N-N
O 2-3
(mg/
l)
Entrée
Sortie calc
Sortie exp
Figure 4.30. Concentrations observées et calculées des différentes formes de l’azote à la sortie du dernier bassin de maturation

149
0
50
100
150
200
250
300
06/0
4/20
05
16/0
4/20
05
26/0
4/20
05
06/0
5/20
05
16/0
5/20
05
26/0
5/20
05
05/0
6/20
05
15/0
6/20
05
25/0
6/20
05
05/0
7/20
05
0
5
10
15
20
25
30
35
Ammonif ication
Sédimentation
Assimilation par les microorganismes
Volatilisation
Nitrif ication
Dénitrif ication
Regénération benthique
T°
Figure 4.31. Masses d’azote journalières calculées mises en jeu par les différents mécanismes du cycle de l'azote dans le dernier bassin de maturation

150
0
2
4
6
8
10
12
14
16
1806
/04/
2005
13/0
4/20
05
20/0
4/20
05
27/0
4/20
05
04/0
5/20
05
11/0
5/20
05
18/0
5/20
05
25/0
5/20
05
kgN
/j
Ammonification
Sédimentation
Assimilation par les microorganismes
Volatilisation
Nitrification
Dénitrif ication
Regénération benthique
0
50
100
150
200
250
300
01/0
6/20
05
07/0
6/20
05
13/0
6/20
05
19/0
6/20
05
25/0
6/20
05
01/0
7/20
05
07/0
7/20
05
kgN
/j
Figure 4.32. Masses d’azote journalières calculées mises en jeu par les différents mécanismes du cycle de l'azote dans le dernier bassin de maturation durant la première et la seconde parties de la période d'études
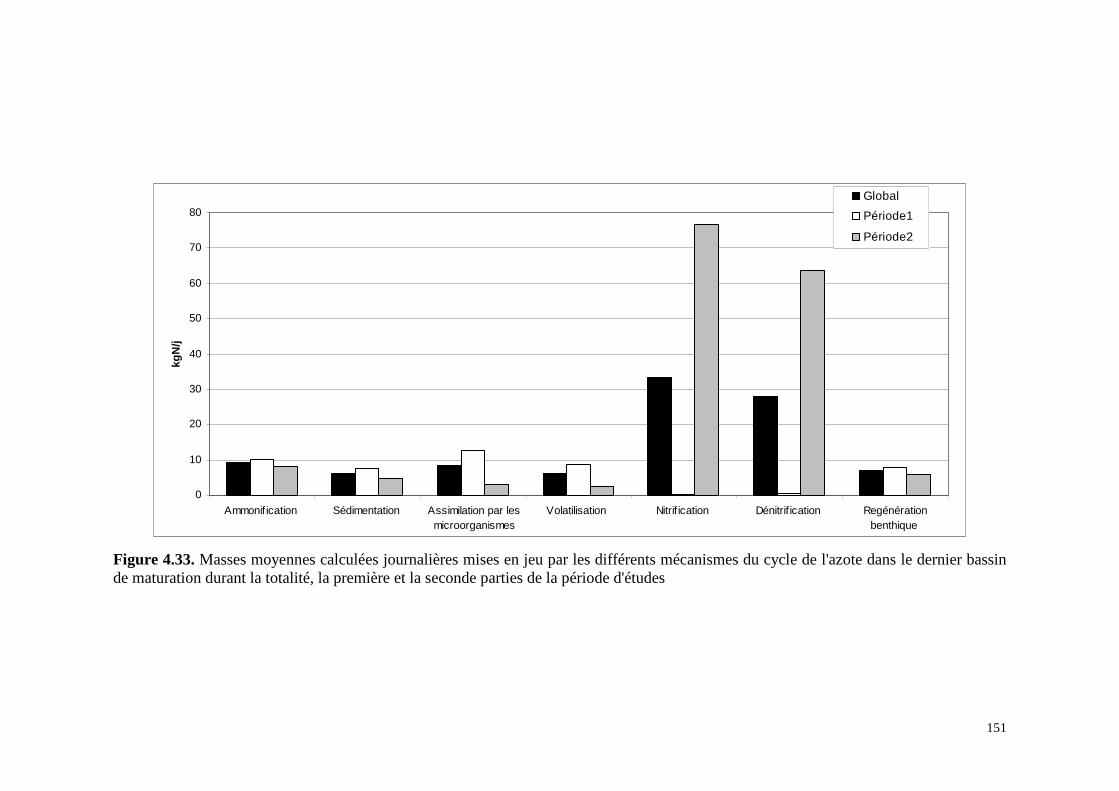
151
0
10
20
30
40
50
60
70
80
Ammonif ication Sédimentation Assimilation par lesmicroorganismes
Volatilisation Nitrif ication Dénitrif ication Regénérationbenthique
kgN
/j
Global
Période1
Période2
Figure 4.33. Masses moyennes calculées journalières mises en jeu par les différents mécanismes du cycle de l'azote dans le dernier bassin de maturation durant la totalité, la première et la seconde parties de la période d'études

152
Les mécanismes de l'élimination de l'azote. La sédimentation étant généralement compensée par la régénération benthique, seules les contributions de la volatilisation et de la dénitrification à l’élimination de l’azote sont présentées dans la Figure 4.34. La volatilisation augmente doucement de 3.8 à 13.7 du 7 avril au 12 mai puis commence à décroître avec la concentration en azote ammoniacal. Jusqu’à fin mai elle explique 97% des pertes en azote. Mais après le 3juin, elle diminue rapidement et atteint des valeurs inférieures à 3 kgN.j-1 dès le 8 juin. Les pertes en azote, dues essentiellement à la dénitrification, quant à elles, croissent jusqu’à 114 kgN.j-1 le 20 juin puis diminuent et sont comprises entre 30 et 40 kgN.j-1 dans la première semaine de juillet.
0
20
40
60
80
100
120
06/0
4/20
05
16/0
4/20
05
26/0
4/20
05
06/0
5/20
05
16/0
5/20
05
26/0
5/20
05
05/0
6/20
05
15/0
6/20
05
25/0
6/20
05
05/0
7/20
05
kg N
/ j l
0
5
10
15
20
25
30
35
Tem
péra
ture
(°C
) n
Volatilisation
Dénitrification
Pertes totales calculées
T°
Figure 4.34. Pertes simulées en azote
Conclusions. La nitrification est un processus important du cycle de l’azote. Dès que les conditions environnementales le permettent, les bactéries nitrifiantes se développent rapidement et peuvent nitrifier en peu de jours la majorité de l’azote ammoniacal présent. Après, la nitrification se limite à l’oxydation du flux d’azote ammoniacal entrant. L’augmentation de la température ainsi que la présence d’une teneur significative en DCO filtrée favorisent l’activation de la dénitrification. Toutefois, en se référant aux teneurs en oxygène dissous observées dans le suivi de l’opérateur de la station, elle n’aurait pas dû avoir lieu puisque nécessitant un milieu anoxique. Cependant, comme le montre l’enregistrement de la teneur en oxygène dissous représentée sur la Figure 4.28, l’alternance chaque jour de périodes de forts et de très faibles états d’oxygénation explique l’existence et l’importance de ces 2 mécanismes durant la seconde période de l’étude. La quantité d’azote ammoniacal oxydée à un moment de la journée se trouve réduite dans les heures suivantes. De très faibles valeurs en azote oxydé peuvent en résulter.

153
CONCLUSION

154

155
Cette étude a tenté de tirer le meilleur parti des données recueillies au cours de suivis du lagunage de Mèze. Outre les mesures hydrauliques et les analyses fixées par le protocole de contrôle de routine de la station, un suivi plus détaillé a été effectué entre juillet 2003 et août 2004. Les analyses ont été réalisées à Mèze par le gestionnaire de la station d’épuration. Des mesures complémentaires ont été effectuées en 2005 par le laboratoire Hydrosciences. La complexité du système étudié, les fluctuations de débit liées aux évènements pluvieux et à l’évaporation et les variations de volumes stockés dans les différents bassins imposées par le gestionnaire font de la modélisation hydraulique du lagunage un outil indispensable à une évaluation quantitative des performances de la station d’épuration. Ce modèle est utilisé, d’abord dans une étape de calibrage, comme un moyen de reconstituer les données manquantes, puis pour quantifier les performances et, enfin, pour des modélisations de la décontamination microbienne et du devenir de l’azote. Le nombre élevé des données manquantes, débit des by-pass et niveau de l’eau dans les bassins au pas journalier, fait que le calage sur le débit observé en sortie de l’installation ne garantit pas que la reconstitution des données manquantes soit correcte. La reconstitution des débits de sortie s’est, elle-même, avérée délicate, notamment à la suite des épisodes pluvieux majeurs. Les limites du calibrage du modèle hydraulique ne sont pas sans impact sur les utilisations du modèle, particulièrement en ce qui concerne la modélisation des concentrations des germes témoins de contamination fécale. La comparaison des performances de la station entre les années 1988-1990 et 2000-2003, avec des charges surfaciques appliquées respectivement égales à 65 et 72 kg DBO ha-1j-1, montre que, malgré une plus grande charge surfacique, la rénovation de l’installation a permis une amélioration très nette de la qualité des effluents. Cette amélioration est significative pour la DCO brute mais très nette en ce qui concerne l’azote organique et ammoniacal et les germes témoins de contamination fécale. Cette amélioration tient à la multiplication des bassins, à la diversification des types de lagunage mais aussi, bien sûr, à l’apport d’énergie électrique dans les bassins artificiellement aérés. Les bassins anaérobies jouent un rôle primordial dans l’élimination des MES et de la matière organique. L’étage facultatif à forte charge (FFC) prend aussi une forte part à l’élimination des mêmes substances. Les bassins de maturation, grâce à un temps de séjour moyen de 48 jours, éliminent autant de MES et de DCO que l’étage FFC et, de plus, il assure l’essentiel de l’élimination de l’azote. L’étage de finition ne contribue que très marginalement à l’abattement de la matière organique et de l’azote ; il joue tout de même un rôle important dans l’élimination du phosphore mais voit les MES augmenter. Si l’on évalue l’abattement des germes témoins de contamination fécale en termes d’unités logarithmiques, les bassins anaérobies y ont une participation relativement faible, comprise entre 0,09 et 0,73 u.log, selon les bactéries considérées et les saisons, et l’étage FFC une contribution plus importante, entre 1,05 et 1,68 u.log (Tableaux 2.3 et 2.4). La contribution de ces deux étages ne dépasse pas 2 u.log. Ce sont les lagunes de maturation qui, avec des battements compris entre 2,44 et 3,89 u.log., assurent l’essentiel de la décontamination microbienne. Malgré des performances décevantes de la lagune F1 dues à des raisons que nous n’avons pas pu élucider, l’étage de finition participe tout de même de manière significative à l’élimination des entérocoques et des E. coli.

156
Au vu de ces résultats, l’utilité de l’étage de finition pourrait être discutée. Cependant, l’intérêt de cette discussion dépend de la réglementation imposée localement. Comme la sévérité de celle-ci s’accroît progressivement, l’étage de finition apparaît comme une sécurité, en particulier pour faire face aux évènements pluvieux et même si une autre stratégie de gestion de l’installation pourrait peut-être permettre de tirer un meilleur parti de ces dernières lagunes. La décontamination microbienne a été simulée en supposant que les bassins se comportent comme des mélangeurs parfaits et que les cinétiques de décontamination sont du premier ordre. On a tenté, au moyen d’une procédure d’optimisation, de relier ces cinétiques aux variables explicatives suivantes : température de l’eau, rayonnement solaire et teneur en MES des bassins. Les évolutions des constantes cinétiques ainsi obtenues (valeurs calculées) ne sont ni totalement satisfaisantes – quand on les compare aux valeurs dites expérimentales -, ni aisément interprétables. On note des ressemblances plus nettes entre les variations calculées et expérimentales pour les lagunes de maturation que pour les étages anaérobies, facultatifs et de finition. Les paramètres des relations empiriques entre constante cinétique et variables explicatives varient d’un bassin à un autre sans qu’une logique claire vienne justifier ces variations. La difficulté à obtenir, à travers la modélisation, des liaisons convaincantes entre les cinétiques de décontamination et les variables explicatives est liée pour une part aux hypothèses du modèle et à la qualité des données mais aussi à la complexité de ces liaisons dans la réalité d’une lagune. Une simple équation empirique entre les variables explicatives retenues et les constantes cinétiques ne suffit pas à en rendre compte. Cette même difficulté est illustrée par la comparaison des modélisations de la décontamination microbienne dans les lagunes de maturation L2 et L3 pour les années 1986, 1988-1990 et 2003-2004. Les valeurs des constantes cinétiques calculées pour ces différentes périodes présentent des écarts assez considérables, ce qui confirme que la seule prise en compte des variables explicatives retenues ne suffit effectivement pas à prédire la décontamination. En dépit de ses insuffisances, le modèle de la décontamination microbienne constitue un outil susceptible d’aider utilement à la réflexion relative à l’amélioration de la gestion du système de lagunage pour obtenir de meilleures performances sanitaires. Une autre voie de la modélisation de la décontamination microbienne a été explorée. La démarche de Von Sperling, qui prédit l’abattement des bactéries témoins de contamination fécale à partir de la forme des bassins, de la hauteur et de la température de l’eau, s’avère tout aussi performante que la tentative de relier les cinétiques à la température de l’eau, au rayonnement solaire et à la teneur en MES, quand cette démarche est appliquée aux lagunes de maturation. Cela confirme que la température est un facteur explicatif important. Cette approche se révèle particulièrement intéressante puisque, au moins pour les lagunes de maturation, elle est générale et n’exige pas de calibration bassin par bassin. Malgré quelques travaux récents effectués principalement dans les pays anglo-saxons, les mécanismes de la décontamination microbienne peinent à être caractérisés dans le contexte des lagunes. Un effort important dans ce domaine pourra vraisemblablement permettre à terme de développer une approche mécaniste de la décontamination microbienne.

157
L’analyse de la littérature permet de comprendre comment l’hypothèse, attribuée à Pano et Middlebrooks, selon laquelle la volatilisation de l’ammoniac est le mécanisme principal de l’élimination de l’azote a longtemps été adoptée par les spécialistes du lagunage. La littérature récente fait apparaître des contestations croissantes de ce postulat. Des expériences, semblables à celles effectuées par Stratton, nous ont permis d’affirmer que, à Mèze, la volatilisation n’a qu’une part marginale à la disparition de l’azote. Dès lors, le mécanisme principal de cette élimination ne peut être que la nitrification couplée à la dénitrification. Ce processus intervient, à partir du printemps, principalement dans l’étage de maturation et plus particulièrement dans la troisième lagune puis dans la deuxième lagune de cet étage. Il cesse durant l’automne. La sédimentation, très faible dans ces bassins, ne joue pas de rôle notable dans la disparition de l’azote. L’ensemble du cycle de l’azote dans les lagunes de maturation a fait l’objet d’une modélisation. Les valeurs des paramètres des cinétiques des différentes composantes du cycle ont été soit prises dans la littérature soit déterminées par calibrage du modèle. Celui-ci montre comment la variation de la température explique la mise en route de la nitrification puis de la dénitrification, ce qui justifie le caractère saisonnier de l’élimination de l’azote. Le calibrage du modèle, appliqué à l’ensemble de l’étage de maturation assimilé à une lagune unique, permet de représenter fidèlement l’évolution de la concentration en azote dans la lagune L3, mais pas dans les lagunes L1 et L2. Les différences observées dans le devenir de l’azote d’une lagune de maturation à une autre ne semblent pouvoir être expliquées que par des disponibilités en oxygène croissantes de L1 à L3. Ces différences n’apparaissent pas clairement dans les mesures de l’oxygène dissous utilisées pour la modélisation. Elles sont seulement suggérées par les variations de la DCO dissoute. La mesure des teneurs en oxygène dissous, qui connaissent des variations considérables dans le temps et l’espace, mériterait beaucoup plus d’attention. De meilleurs suivis de l’oxygène dissous permettraient une calibration plus pertinente du modèle. Par ailleurs, une autre représentation de la dénitrification, plus mécaniste, pourrait être alors envisagée. Cela conduirait à changer le pas de temps du modèle, qui pourrait devenir l’heure et non le jour. L’identification du principal mécanisme de l’élimination de l’azote permet d’espérer pouvoir accroître de manière significative les performances des lagunages. L’application la plus directe, déjà mise en œuvre dans plusieurs installations, consiste à augmenter les surfaces susceptibles de servir de support à la biomasse nitrifiante et dénitrifiante. Parmi les sytèmes possibles, les digues de graviers (ou rock filters) semblent les plus prometteurs. Leur installation, entre deux lagunes ou à la sortie d’un lagunage, doit permettre, entre autres objectifs, d’obtenir une élimination significative de l’azote pour des températures inférieures à celles notées au moment où nitrification et dénitrification se manifeste dans les lagunes de maturation de Mèze.

158

159
BIBLIOGRAPHIE

160

161
Abeliovich A., 1985. Nitrification of ammonia in wastewater. Field observations and laboratory studies. Water Research, 19(9), 1097-1099.
Archer, H.E. and Donaldson, S.A., 2003. Waste stabilization ponds upgrading at Blenheim and Seddon, New Zealand – case studies. Wat. Sci. Tech. 48(2), 17-23.
Archer H.E. and O’Brien B.M., 2005. Improving nitrogen reduction in waste stabilisation ponds. Wat. Sci. Tech., 51(12), 133-138.
Archer, H.E. et Mara D.D., 2003. Waste stabilisation pond developments in New Zealand. Wat. Sci. Tech., 48(2), 9-15.
Arridge H., Oragui J.I., Pearson H.W., Mara D.D. et Silva S.A., 1995.Vibrio cholerae 01 and Salmonellae removal compared with the die-off faecal indicator organisms in waste stabilization ponds in northeast Brazil. Wat. Sci. Tech., 31(12), 249-256.
Auer M. T. and Niehaus S. L., 1993. Modelling fecal coliform bacteria I: field and laboratory determination of loss kinetics. Water Research 27(4), 693-701.
Bahlaoui A., 1990. Lagunage à haut rendement expérimental. Dynamique de différents groupes bactériens et performances épuratrices sanitaires. Th. Doct., Université Montpellier 2.
Baleux B. et Trousselier M. 2003. Evolution des bactéries témoins de contamination fécale et de Pseudomonas aeruginosas et Aeromonas hydrophila. Tech. Sci. Munic. 7,33-42.
Bancolé Dioni A., 2001. L’oxydation en infiltration percolation. Thèse de l’Université de Banda C.G., Sleigh P.A. et Mara D.D., 2005. Escherichia coli removal in waste stabiliszation
ponds: a comparison of modern and classical designs. Wat. Sci. Tech. 51(12), 75-81. Barbagallo, S., Brissaud, F., Cirelli, G.L., Consoli, S. and Xu, P. (2003). Modelling of
bacterial removal in wastewater storage reservoir for irrigation purposes : a case study in Sicily, Italy. Wat. Sci. Tech./ Wat. Supply 3(4), 169-176.
Booth I.R., 1985. Regulation of cytoplasmic pH in bacteria. Microbiological reviews, 49, 379-378.
Brissaud F., Tournoud M.G., Drakidès C., Lazarova V., 2003. Mixing and its impact on faecal coliform removal in a stabilisation pond. Wat. Sci. Tech., 48(2), 75-80.
Commission Européenne, 2001. Extensive wastewater treatment processes adapted to small and middle sized communities. Office des Publications officielles des Communautés Européennes, Luxembourg.
Copin, Y., Brouillet, J.L., Rivière, Y. and Brissaud, F. (2004). Waste stabilisation ponds in the wastewater management strategy of a French department. 6th Int IWA Conf. on Waste Stabilisation Ponds, Avignon, 27 Sept – 1st Oct., 29-35.
Crabos J.L., Sambuco J.P., Bondon D., Brouillet J.L., Lebec C., Bonal. O., 1996. Lagunage de Mèze-Loupian,. Curage, deshydratation et épandage des boues. Rapport CEREMHER, IFREMER, 55 p.
Curtis T.P., Mara D.D., Silva S.A., 1992. The effect of sunlight on faecal coliforms in ponds: Implications for research and design. Wat. Sci. Tech., 26(7-8), 1729-1738.
Davies-Colley R.J., Craggs R.J., Park J., Sukias J.P.S., Nagels J.W. et Stott R., 2005. Virus removal in a pilot scale “advanced” pond system as indicated by somatic and F-RNA bacteriophages. Wat. Sci. Tech. 51(12), 107-110.
Deronzier G., Schétrite S., Racault Y., Canler J.P., Liénard A., Héduit A., Duchène P., 2001. Traitement de l'azote dans les stations d'épuration biologique des petites collectivités. Document technique FNDAE, 25. Deuxième edition, Cemagref Éditions.
Downing A.L., 1966. Population dynamics biological systems. Proceedings of Third International Conference on Water Pollution and Research, Water Pollution Control Federation, 2, 117-136.

162
Drakides C. et Djajadiningrat A., 1981. Etude de la stratification et de l'évaporation sur la caractérisation de l'écoulement en lagunage dans le cas de modèles de laboratoire. Rapport définitif. Laboratoire de Génie Chimique Appliqué aux Biotechnologies. Université des Sciences et Techniques du Languedoc.
El Hafiane F. et El Hamouri B., 2005. Anaerobic reactor/high rate pond combined technology for sewage treatment in the Mediterranean area. Wat. Sci. Tech., 51(12), 125-132.
ElHalouani H., Picot B., Casellas C., Pena G., Bontoux J., 1993. Elimination de l’azote et du phosphore dans un lagunage à haut rendement. Revue des sciences de l’eau, 6, 47-61.
Emparanza-Knörr A. et Torrella F., 1995. Microbiological performance and salmonella dynamics in a wastewater depuration pond system of southeastern spain. Wat. Sci. Tech., 31(12), 239-248.
Evans R.A., Cromar N.J. et Fallowfield H.J., 2005. Performance of a pilot-scale high rate algal pond system treating abattoir wastewater in rural South Australia: nitrification and denitrification. Wat. Sci. Tech., 51(12), 117-124.
Ferrara R.A. et Avci C.B., 1982. Nitrogen dynamics in waste stabilisation ponds. Journal Water Pollution Control Federation, 54, 361–369.
Ferrara R.A. et Harleman D.R.F., 1980. Dynamic nutrient cycle model for waste stabilisation ponds. Journal of the Environmental Engineering Division, 106(1), 37-54. Fritz J.J., Middleton A.C. et Meredith D.D., 1979. Dynamic process modeling wastewater
stabilization ponds. Water Pollution Control Federation, 51(11), 2724-2743. Hemens J., 1966. The toxicity of ammonia solutions to the mosquito fish. J. Inst. Sew. Purif.,
65(3), 265 Horn H. et Hempel D.C., 1997. Growth and decay in an auto-/heterotrophic biofilm. Water
research, 31(9), 2243-2252. King D.L., 1978. The role of ponds in land treatment of wastewater. Proceedings
International Symposium on Land Treatment of Wastewater, 191-198. Marais G.V.R., 1974. Faecal bacterial kinetics in stabilisation ponds. Journal of the
Environmental Engineering Division, American Society of Civil Engineering, 100(1), 119-139.
Metcalf et Eddy, 1991. Wastewater engineering. Treatment, disposal and reuse. Third edition, McGraw-Hill, Inc, New York. 1334p.
Middlebrooks E. J., Middlebrooks C. H., Reynolds J. H., Watters G. Z., Reed S. C., Dennis D. B., 1982. Wastewater stabilisation lagoon design, performance and upgrading. Macmillan, 356p.
Namèche Th. et Vasel J.L., 1998. Hydrodynamic studies and modelization for aerated lagoons and waste stabilisation ponds. Water research, 32(10), 3039-3045.
Pano A. et Middlebrooks E.J., 1982. Ammonia nitrogen removal in facultative wastewater stabilisation ponds. Journal Water Pollution Control Federation, 54(4), 344–351.
Prescott, Harley et Kein, 1995. Microbiologie. Seconde édition. W.C. Brown Communications, Inc. 1014p.
Racault Y. et Boutin C., 2005. Waste stabilisation ponds in France: state of the art and recent trends. Wat. Sci. Tech., 51(12), 1-9
Reddy K.R., 1983. Fate of nitrogen and phosphorus in a wastewater retention reservoir containing aquatic macrophytes. Journal of environmental Quality, 12, 137-141.
Reed S.C., 1985. Nitrogen removal wastewater stabilisation ponds. Journal Water Pollution Control Federation, 57, 39–45.

163
Rittmann B.E. et McCarty P.L., 2001. Environmental biotechnology: Principles and applications. Mc Graw Hill Companies, Inc.
Senzia M.A., Mayo A.W., Mbwette T.S.A., Katima J.H.Y. et Jorgensen S.E., 2002. Modelling nitrogen transformation and removal in primary facultative ponds. Ecological Modelling, 154, 207-215.
Silva S.A., de Oleivera R., Soares J., Mara D.D. et Pearson H.W., 1995. Nitrogen removal in pond systems with different configurations and geometries. Wat. Sci. Tech., 31(12), 321-330.
Soares J., Silva S.A., de Oleivera R., Araujo A.L.C., Mara D.D. et Pearson H.W., 1996. Ammonia removal in a pilot-scale WSP complex in northeast Brazil. Wat. Sci. Tech., 33(7), 165-171.
Stratton F.E., 1968. Ammonia Nitrogen losses from streams. Journal of the Sanitary Engineering Division, ASCE, 94, 1085-1092.
Stratton F.E., 1969. Nitrogen losses from alkaline water impoudments. Journal of the Sanitary Engineering Division, ASCE, 95, 223-231.
Stumm W. et Morgan J.J., 1995. Aquatic chemistry. Chemical equilibria and rates in natural waters. Third edition. John Wiley & Sons, Inc. 1022p.
Surmacz-Gorska J., Cichon A. et Korneliusz M., 1997. Nitrogen removal from wastewater with high ammonia nitrogen concentration via shorter nitrification and denitrification. Wat. Sci. Tech., 36(10), 73-78.
U.S. Environmental Protection Agency, 1993. Process design manual for nitrogen control. Office of Technology Transfer, Washington, D.C..
Von Sperling M., 1999. Performance evaluation and mathematical modelling of coliform die-off in tropical and subtropical waste stabilisation ponds. Wat. Res., 33(6), 1435-1448.
Von Sperling M., 2002. Relationship between first-order decay coefficients in ponds, for plug flow, CSTR and dispersed flow regimes. Wat. Sci. Tech., 45(1), 17-24.
Von Sperling M., 2005. Modelling of coliform removal in 186 facultative and maturation ponds around the world. Wat. Res., 39, 5261-5273.
Wiesmann U., 1994. Biological nitrogen removal from wastewater. Advances in biochemical engineering – Biotechnology, 51, 113-154.
Wong-Chong G.M. et Loehr R.C., 1975. Kinetics of microbial nitrification as applied to the treatment of animal wastes: Ammonium nitrogen oxidation. AIChE Symposium Series, 71(145), 70-79.
Wong-Chong G.M. and Loehr R.C., 1978. The kinetics of microbial nitrification: Nitrite-nitrogen oxidation. Water research, 12, 605-609.
Xu, P., F. Brissaud and A. Fazio, 2002. Non-steady state modelling of faecal coliform removal in deep tertiary lagoons. Wat. Research .36(12) :3074-3082.
Zimmo O.R., 2003. Nitrogen transformations and removal mechanisms in algal and duckweed waste stabilisation ponds. Ph D. University of Delft, Netherlands, 143p.
Zimmo O.R., van der Steen, Gijzen H.J., 2003. Comparaison of ammonia volatilisation rates in algae and duckweed-based waste stabilisation ponds treating domestic wastewater. Wat. Res., 37, 4587-4594.
Zimmo O.R., van der Steen, Gijzen H.J., 2004. Nitrogen mass balance across pilot-scale algae ad duckweed-based wastewater stabilisation ponds. Wat. Res., 38, 913-920.

164

165
ANNEXES

166

167
ANNEXE 1: CALCUL DE L'EVAPORATION L'évaporation EP, en mm.j-1, est calculée à partir de la formule de Penmann :
( )
( )1+∆
+∆
=
γ
γT
ERNT
EPa
(1)
▪ ( )T∆ est la pente de la courbe donnant la tension de vapeur saturante dans l’air ( )TeS ,
fonction de la température T en °C
T
TeT s
δδ )(
=∆ (2)
( )TeS est exprimée en hPa et a comme expression:
( ) T
T
s Te +×= 5.237
5.7
1011.6 (3) D'où
)²3.237(
5.75.7)3.237()10ln(1011.6)( 3.237
5.7
T
TTT T
T
+−+×××=∆ + (4)
▪γ est la constante psychrométrique. Elle est dépendante de la température. Mais dans la pratique, on adopte sa valeur à de 15°C en supposant constante la pression atmosphérique (1013 hPa):
( ) 66.015 =°Cγ hPa.°C-1 (5)
▪ aE est l'advection. Elle caractérise la quantité de vapeur d’eau journalière emmagasinable
par l’atmosphère, en mm d’eau. )54.01()(26.0 2UeeE asa +−= (6)
où
2U est la vitesse journalière moyenne du vent à 2m du sol, en m/s,
ae la pression partielle journalière moyenne de vapeur d’eau en hPa déduite des équations (3)
et (6)
s
ar e
eh = (7)
rh est l’humidité relative moyenne journalière ▪RNest le rayonnement solaire net exprimé en millimètres d’eau par jour

168
( ) ( )
+−−
+−=H
heT
H
ha
IgARN a 9.010.008.056.0
5962.018.01
594σ
(8)
où T est la température de l’air sous abris en °K, IgA la radiation solaire nette en l’absence d’atmosphère en cal/cm²/j, H et h la durée théorique du jour et la durée réelle d’ensoleillement en h σ la constante de Stefan-Boltzman, 71018.1 −×=σ cal/cm²/j/°K a l’albédo de la surface étudiée, défini par le rapport entre l’énergie réfléchie par la surface et l’énergie incidente, pour toutes les longueurs d’onde du visible, 05.0=a pour les plans d’eau avec
[ ]{ }{ })0172.0cos(033.01)²(sin)²(cos)()(cos)sin()sin(54.914 JDLDtgLtgArcDLIgA +−+⋅−⋅⋅= (9)
où L est la latitude D la déclinaison du soleil en rad J le numéro du jour dans l'année La déclinaison D a comme expression:
[ ] [ ][ ] [ ])57.17(0688115.0sin000149.0)42.21(0516086.0sin003009.0
)85.42(0344057.0sin006675.0)95.81(0172029.0sin406149.00066241.0
−+−+−+−+=
JJ
JJD (10)
H la durée théorique du jour dépend aussi de L et de D:
[ ]
−+−=
)²(sin)²(cos
01065.0)()(cos*6394.7
DLDtgLtgArcH (11)
H déterminée, la durée réelle du jour h peut être déduite de l'équation suivante:
)62.018.0(H
hIgARG += (12)
où RG est le rayonnement global en cal/cm²/j

169
ANNEXE 2: RAPPEL SUR LA CINETIQUE DES REACTIONS CHI MIQUES
A.1. Expression générale de la loi de vitesse
La vitesse v d'une réaction chimique dépend de plusieurs variables, dont les plus importantes sont les concentrations des différentes espèces chimiques (réactifs et/ou produits ,...,, CBA ) et la température T :
[ ] [ ] [ ]( ),...,...,,, TCBAfv = (13)
La fonction f a généralement comme expression :
[ ] [ ] [ ]( ) ( ) [ ] [ ] [ ] ....,...,...,...,,, Cba CBATkTCBAf = (14)
où k est la constante de vitesse de la réaction
,....,, cba sont les ordres partiels de la réaction par rapport respectivement à ,...,, CBA L’ordre global σ de la réaction est définie par:
...+++= cbaσ (15)
L’influence de la température est habituellement décrite par la loi d’Arrhenius :
( )2RT
E
T
Logk A=∂
∂
où k est la constante de vitesse en s-1 R la constante des gaz parfaits en J.mol-1.K-1 T la température en °K
AE l’énergie d’activation en J.mol-1 L’énergie d’activation est généralement déterminée expérimentalement. L’intégration de l’équation d’Arrhenius donne la fonction reliantk à la température :
RT
E
A
kLog A−=
ou RT
EA
eAk−
⋅= (16)
où A est une constante de même unité que k A.2. Cas particulier des réactions enzymatiques : Modèle de Michaelis-Menten Les réactions biochimiques enzymatiques obéissent aux mêmes lois cinétiques que les réactions chimiques. Etant souvent du second ordre, le calcul de leurs constantes de vitesse s’avère complexe et fait appel au modèle de Michaelis-Menten.

170
Ce modèle est basé sur l’hypothèse de l’existence d’un complexe enzyme-substrat E-S. Une réaction enzymatique est alors représentée par le système réactionnel :
1k 2k
SE + ES PE + (Réaction A-1)
1−k La vitesse de la réaction est définie par l’étape limitante, la plus lente, constituée par celle de la formation du produit P à partir du complexe ES:
[ ]ESkv 2= (17)
2k étant difficilement accessible mathématiquement, sa détermination peut être contournée en opérant sous certaines conditions expérimentales et en appliquant l’approximation de l’état quasi-stationnaire de Bodenstein. Dans le cas où [ ] [ ]SE T << tout l’enzyme est sous forme de complexeES, la réaction enzymatique atteint sa vitesse maximale :
[ ]Tm Ekv 2= (18)
Les équations (E022) et (E023) permettent d’éliminer 2k de l’expression de la vitesse de la réaction :
[ ][ ]Tm E
ESvv = (19)
Par ailleurs, l’approximation de Bodestein stipule qu’à l’état quasi-stationnaire le milieu réactionnel est caractérisé par :
[ ]0=
dt
ESd (20)
Ceci n’est possible que si la vitesse de formation1v du complexeESest égale à celle de sa
dissociation ou de destruction2v .
[ ][ ]SEkv 11 = (21)
( )[ ]ESkkv 212 += − (22)
Auquel cas :
[ ][ ][ ] 1
21
k
kk
ES
SE += − (23)
Ce rapport définit la constante de Michaelis mK .
Comme :

171
[ ] [ ] [ ]ESEE T += (24)
Alors :
[ ][ ][ ] [ ]SSES
EK T
m −= (25)
Les équations (E024) et (E030) permettent le calcul de la vitesse de la réaction en fonction uniquement de mK et de [ ]S selon l’équation de Michaelis:
[ ][ ]SK
Svv
Mm +
= (26)
Cette équation implique queMK est la concentration en substrat pour laquelle la vitesse est égale à la moitié de la vitesse maximale.

172
ANNEXE 2: PUBLICATIONS
Publication1. Twenty years monitoring of Mèze stabilisation ponds. I – removal of organic matter and nutrients B. Picot*, T. Andrianarison**, J.P. Gosselin*** and F. Brissaud** * Département Sciences de l'Environnement et Santé Publique, UMR 5569 Hydrosciences, MSE,- Université Montpellier I, Faculté de Pharmacie, BP 1149, 34093 Montpellier Cedex 5, France. (E-mail: picot@univ–montp1.fr) ** Hydrosciences, MSE, Université Montpellier II, 34095 Montpellier Cedex 05, France. (E-mail : [email protected], [email protected]) *** Ecosite du Pays de Thau, Parc Scientifique et Environnement, B.P.118, 34140 Mèze, France. (E-mail: [email protected]) Abstract The Mèze stabilisation pond system has been monitored over more than twenty years. Despite the enlargement of the plant, the organic load doubled between the early eighties and recent years, the removal of organic matter and nutrients has been maintained at the same level for COD and increased for BOD5, N and P. Combining anaerobic, step fed aerated and maturation ponds and multiplying the number of cells resulted in a significant improvement in the performances of the plant. Respectively 34, 24 and 23 % of the applied COD was eliminated in the anaerobic, the step fed and the first three maturation ponds, while the figures for BOD5 were 47, 26, and 19% respectively. 38% of the applied nitrogen was eliminated in the first 3 maturation ponds. Nitrification and denitrification seem to be a major process of nitrogen removal in warm periods. Most of the phosphorus removal was observed to take place in the two polishing ponds. Keywords Nitrogen, phosphorus, removal; step fed facultative pond®; upgrading; waste stabilisation pond
INTRODUCTION Waste stabilisation ponds (WSPs) systems have been widely developed during the 30 last years in France. They account for over 20% of the wastewater treatment plants. They are chosen both for small communities because of their low operation and maintenance costs, low risk of failure and low production of sludge and for bigger coastal towns because of their ability to treat variable loads in tourist zones and to achieve good removal of pathogens when the discharge is located in bathing waters or shellfish breeding areas. Very few waste stabilisation ponds have been monitored for SS, organic matter, nutrients and microbiological faecal indicators over more than twenty years. The Mèze WSP system which

173
aims to protect the Thau lagoon where more than 10 % of the French production of oysters takes place, is an outstanding exception. This paper presents the performance of the plant from the beginning of the eighties up to 2004. The impacts of pollution load, the ageing of the plant and its enlargement and retrofitting on the removal of organic matter and nutrients are discussed. The specific performance of each stage of treatment is quantified and methods of pollution removal specified. MATERIEL AND METHODS Site description
A1 A2
R1
R2
R3
R4
M1
M2
M3
P1
P2
Inlet
Normal flow path
Outlet
Ponds Type Area
(ha) Depth
(m) Start up
A1, A2 Anaerobic ponds
then deep aerated ponds 2 x 0.23 3.1
1998 2001
R1, R2, R3, R4 Step fed facultative ponds with recirculation (SFP)®
4 x 0.67 1.8 1999
M1, M2, M3 Facultative
then maturation ponds 4 + 2 + 2 1.4 -1.7
1980 1999
P1, P2 Polishing ponds 1.9 + 1.2 0.8 -1.3 1996
Figure 1. Lay-out of the current Mèze waste stabilisation pond system. The Mèze WSP plant located on the Mediterranean coast (03°35’06’’E, 43°25’10’’N) was a typical example of the French state of the art when it was constructed in 1980. Made up of three ponds with a total surface of 8ha, the plant was designed with a treatment capacity of 8,000 p.e. and a design organic load of 50kg BODha-1d-1. It received an initial average organic load of only33kg BODha-1d-1 augmented with winery wastewater, particularly in autumn. It discharged into the Thau lagoon. As the population connected to the plant increased, as sludge accumulated and the criteria applying to the treated water quality became more exacting, maintenance and enlargements works had to be undertaken. In October 1994, a 1.5ha area of the 4 ha primary pond was dredged and 800 tonnes of dried matter were extracted. In 1996 the 3 initial ponds (M1, M2, M3) were supplemented with 2 polishing cells in series (P1 and P2), in order to improve the microbiological quality of effluents. Two anaerobic ponds in parallel (A1 and A2) were added in 1998 and 4 step fed facultative ponds (SFP) in series (R1 to R4) with recirculation and aeration® in 1999 (Entech patent). Each
Other flow paths

174
pond has a surface area of 0.65ha, is 1.6-1.8 deep and equipped with an 11kW stirring aerator. Step feeding distribution (50% R1, 30% R2, 20% R3), and recirculation from R4 to R1 were based on a previous study (Sambuco et al. 2002). The total area of the ponds nowadays is 14.4 ha; the load to be treated amounts to up 1120kg BOD per day, being equivalent to 19,000p.e.. The present installation can be operated combining new and old ponds either in series or in parallel, the 2 polishing cells serving in both cases (figure 1). Actually SFP received a constant flow of about 2500m3d-1, excess of flow at the outlet of A1-2 entered directly in M1.
Sampling and analysis methods. From 1980 to 1992 the WSP was monitored at the inlet and outlet of the 3 initial facultative ponds M1, M2 and M3 fortnightly at first and then every three months. Since 2000 the inlet and the outlet have been monitored fortnightly and the inflow and outflow recorded with an electromagnetic flow meter. During the year 2003-2004 analysing the water quality fortnightly at the inlet and outlet of (i) the anaerobic ponds in parallel, (ii) the step fed ponds, (iii) the 3 first maturation ponds and (iv) the two last polishing ponds allowed to specification of the role of each type of pond in the overall performance. Composite 24 h samples were taken and analysed daily. The following parameters were measured according to Standard Methods: suspended solids (SS), total chemical oxygen demand (COD), filtered COD (Whatman GF/C filters), Kjeldhal nitrogen (TKN), ammonia (NH4-N), nitrate (NO3-N), nitrite (NO2-N), total phosphorus (TP) and orthophosphate (PO4-P). Biological oxygen demand (BOD) and filtered BOD were measured with a respirometer Oxytop WTW. E.Coli and faecal enterococci were analysed using microplate methods. Removal efficiency was calculated in % concentration (inlet concentration – outlet concentration reported to inlet concentration) or in % mass flow (inlet mass flow - outlet mass flow reported to inlet mass flow). RESULTS AND DISCUSSION Comparison of performance at the starting up and prior and after upgrading During the two first years of operation the average overall organic load on the system was 33kg BODha-1d-1 and performances were high, 73% COD and 55% N-NH4 removal, 2.9log unit faecal coliform and 2.6 log unit enterococci reductions (table 1). After 9 years of operation the organic load had increased (65kg BODha-1d-1) due to population growth. Meanwhile removal, mainly nitrogen removal, (19%), tCOD (60%) and faecal coliform (2.3 log unit removal efficiency) had decreased due to both increase in loading rates and accumulation of sludge in the first cell which reduced the hydraulic residence time. After the enlargement and retrofitting at the end of the nineties the area of the plant was 8.3m2/P.E. The overall improvements to effluent quality from the upgraded WSP have been substantial, particularly for faecal coliform and enterococci indicators (average removal during the last four years of 4.1 and 3.4 log. units respectively). Brissaud et al. (2004) investigated this improvement in the microbiological effluent quality. Despite the enlargement of the plant, the surface organic loading still doubled between the early eighties and the present time but the same level of COD has been maintained and even BOD and nutrient removals increased. While influent ammonia concentrations increased with time,

175
effluent ammonia concentrations decreased. Influent phosphorus concentrations decreased with time due to less use of phosphorus in washing products, P removal was the same but effluent concentrations were lower. It is interesting to investigate the performance of the first two stages of treatment: a deep aerated pond (AP) and step fed facultative pond (SFP). With an area of 1.7m2/p.e removal efficiencies were high for SS, COD and BOD (68, 67 and 82% respectively). These first two stages (or one of them) could be used when retrofitting to increase the capacity of overloaded WSPs. Table 1. Comparison of Mèze WSP performance during the two first years of operation, after 9 years of operation and after upgrading works for overall plant. Mean, [standard deviation], % removal and log unit reduction for faecal indicators
YearsTypeSurface areaPopulation (p.e.) 4400 8700 17300Area (m²/p.e.) 18 9.3 8.3Surface loading
In Out % In Out % In Out %TSS (mg/L) 171 251(109) 136 (69) 46 266 (133) 109(63) 59COD (mg/L) 430 116 (27) 73 541(409) 216 (91) 60 645 (225) 179 (72) 72fCOD (mg/L) 337 (407) 76 (26) 77 (86*) 262(120) 80 (29) 69 (88*)BOD (mg/L) 172 (13) 58 (14) 66 405 (136) 50 (23) 88fBOD (mg/L) 163(72) 9 (6) 94 (98*)NTK (mg/L) 42 (6) 34 (5) 19 46 (13) 15 (8) 67NH4-N (mg/L) 24 (3) 11 (4) 55 27 (7) 21 (9) 22 36 (11) 8 (9) 77TP (mg/L) 14.7 (3.3) 11.3 (3.2) 23 7.9 (2.8) 5.0 (1.7) 37PO4-P (mg/L) 15 (2.4) 10 (2.7) 35 9.1 (3.2) 5.0 (3.6) 45 4.5 (1.7) 2.8 (1.8) 38FC (log10 /100ml) 6.2 (0.5) 3.4 (1.2) 2.9 7.0 (0.5) 4.7 (1.0) 2.3 6.8 (0.5) 2.7 (0.7) 4.1Entero. (log10 /100ml) 5.2 (0.6) 2.6 (0.7) 2.6 6.0 (0.3) 3.6 (0.9) 2.4 6.0 (0.5) 2.6 (0.7) 3.4
33 kgDBO/ha.d
3 Cells in series8 ha
1981-1982 2000-20034 Stages-10 cells in series
14.4 ha
72 kgDBO/ha.d
1988-19903 Cells in series
8 ha
65 kgDBO/ha.d
% removal was calculated with concentration (in-out/in); fCOD* removal was calculated CODin-f CODout/ CODin ; FC and enterococci removals were expressed in mean(log. unit) reduction.

176
Evolution of performances of the upgraded Mèze WSP
Figure 2. Times series (2000-2004) of influent wastewater and effluent from Mèze WSP for flow, SS, COD, BOD, Kjeldahl Nitrogen (TKN), ammonia (NH4-N), total-P and
phosphate Mean daily inflow increased between 2000 and 2003 from 1875 m3 d-1 to 3127 m3d-1. In summer the outflow is halved by evaporation(Figure 2). During periods of heavy rain, the plant can have an inflow greater than 10,000 m3d-1 and the maturation ponds M1-2-3 and polishing ponds P1-2 have a reserve height of 30 and 60 cm respectively to cope with sudden increases in hydraulic charge . During the same time population increased between 14000 to 18800 p.e. Significant seasonal variations of organic matter and nutrients contents and removal were observed, due to tourist population, winery effluents and climate conditions.
0
500
1000 SS mg/L Influent
SS mg/L Effluent
0
400800
1200
1600 COD mg/L Influent
COD mg/L Effluent
COD mg/L Filtered effluent
0200400600800
1000BOD mg/L Influent
BOD mg/L Effluent
0
20
40
60
80 TKN mg/L Influent
TKN mg/L Effluent
NH4-N mg/L Influent
NH4-N mg/L Effluent
0
5000
10000
15000 0
50
100
150
Rainfall (mm/d)
Inflow (m3/d)
Outflow (m3/d)
02468
10121416
26
/03
/20
00
26
/06
/20
00
26
/09
/20
00
26
/12
/20
00
26
/03
/20
01
26
/06
/20
01
26
/09
/20
01
26
/12
/20
01
26
/03
/20
02
26
/06
/20
02
26
/09
/20
02
26
/12
/20
02
26
/03
/20
03
26
/06
/20
03
26
/09
/20
03
26
/12
/20
03
26
/03
/20
04
TP mg/L Influent
TP mg/L Effluent
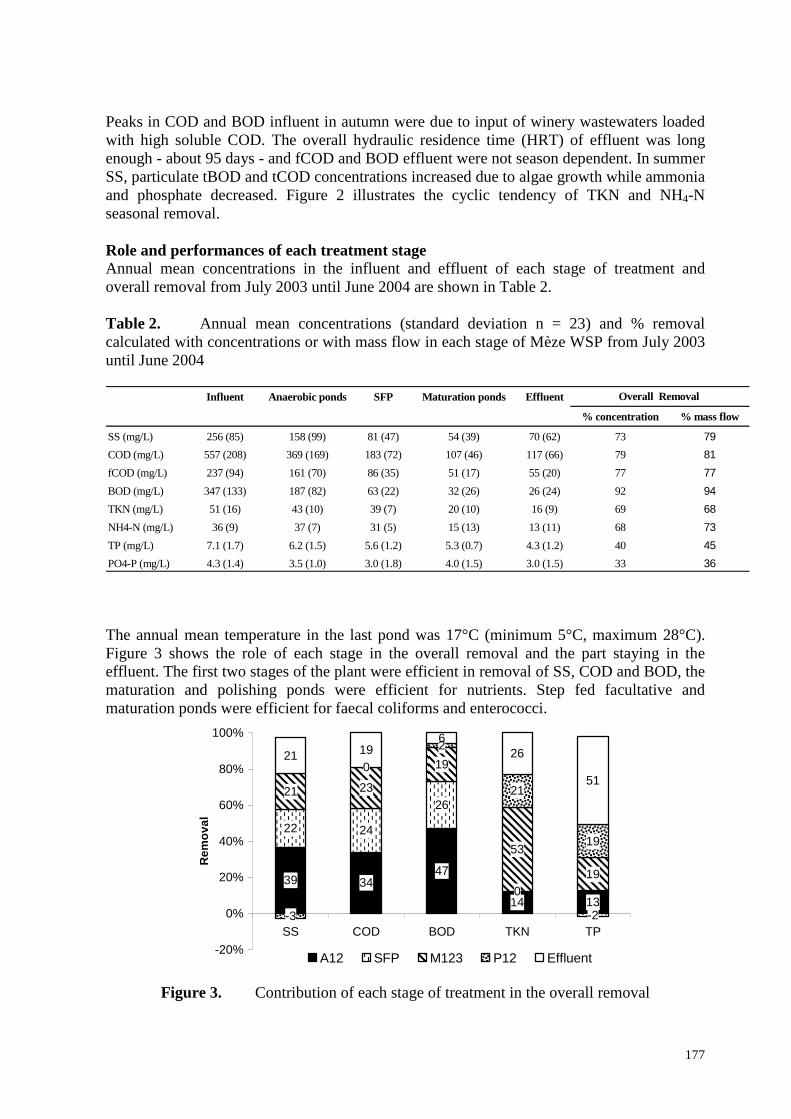
177
Peaks in COD and BOD influent in autumn were due to input of winery wastewaters loaded with high soluble COD. The overall hydraulic residence time (HRT) of effluent was long enough - about 95 days - and fCOD and BOD effluent were not season dependent. In summer SS, particulate tBOD and tCOD concentrations increased due to algae growth while ammonia and phosphate decreased. Figure 2 illustrates the cyclic tendency of TKN and NH4-N seasonal removal. Role and performances of each treatment stage Annual mean concentrations in the influent and effluent of each stage of treatment and overall removal from July 2003 until June 2004 are shown in Table 2. Table 2. Annual mean concentrations (standard deviation n = 23) and % removal calculated with concentrations or with mass flow in each stage of Mèze WSP from July 2003 until June 2004
Influent Anaerobic ponds SFP Maturation ponds Effluent
% concentration % mass flow
SS (mg/L) 256 (85) 158 (99) 81 (47) 54 (39) 70 (62) 73 79
COD (mg/L) 557 (208) 369 (169) 183 (72) 107 (46) 117 (66) 79 81
fCOD (mg/L) 237 (94) 161 (70) 86 (35) 51 (17) 55 (20) 77 77
BOD (mg/L) 347 (133) 187 (82) 63 (22) 32 (26) 26 (24) 92 94
TKN (mg/L) 51 (16) 43 (10) 39 (7) 20 (10) 16 (9) 69 68
NH4-N (mg/L) 36 (9) 37 (7) 31 (5) 15 (13) 13 (11) 68 73
TP (mg/L) 7.1 (1.7) 6.2 (1.5) 5.6 (1.2) 5.3 (0.7) 4.3 (1.2) 40 45
PO4-P (mg/L) 4.3 (1.4) 3.5 (1.0) 3.0 (1.8) 4.0 (1.5) 3.0 (1.5) 33 36
Overall Removal
The annual mean temperature in the last pond was 17°C (minimum 5°C, maximum 28°C). Figure 3 shows the role of each stage in the overall removal and the part staying in the effluent. The first two stages of the plant were efficient in removal of SS, COD and BOD, the maturation and polishing ponds were efficient for nutrients. Step fed facultative and maturation ponds were efficient for faecal coliforms and enterococci.
39 3447
14 13
22 24
26
0
-2
21 23
19
53
19
-3
0
2
21
19
21 196
26
51
-20%
0%
20%
40%
60%
80%
100%
SS COD BOD TKN TP
Rem
oval
A12 SFP M123 P12 Effluent
Figure 3. Contribution of each stage of treatment in the overall removal

178
First stage: Anaerobic ponds later transformed into deep aerated ponds The first stage of treatment is two deep cells (3.1 m) in parallel. From 1998 to 2000 these ponds were anaerobic with a mean volumetric loading of 86 g BOD/m3d and a mean HRT of 4.6 days. SS removal was 55% and BOD5 removal only 30% (Picot et al. 2003). However odour nuisance due to the emission of hydrogen sulphide (H2S) led to the addition of an aeration power of 4.4 W/m3 to each cell (Paing et al, 2003). These aerators have been in operation since 2001 and have resolved odour nuisances. Volumetric loading is now 105 g BOD/m3d and mean residence time is 3.5 days. Aeration decreases SS performance particularly in summer (39% average removal, 17% in summer) but increases BOD removal (47%) with a power consumption of less than 0.5 kWhr per kg of BOD5 eliminated. This primary step of treatment has low efficiencies for removal of nutrients (14% N and 13%P) and faecal indicators (E. coli: 0.5 log unit, Enterococci: 0.23 log unit). It is possible to use deep aerated ponds successfully at the beginning of the plant to upgrade an overloaded WSP plant with a low energetic cost. (Copin et al., 2004). Second stage: Step fed facultative ponds with recirculation and aeration® (SFP) The mean residence time in the 4 step-fed facultative ponds with recirculation and aeration (SFP) is about 25 days. The organic loading applied to the four ponds is around 166 kg BODha-1d-1 overall. A recirculation of once the inflow was applied after doing tests with ratios of 0.5, 1 and 2 (Sambuco et al , 2002.). Four aerators can work if necessary, preferably by night (0.5kWhr/kgBOD eliminated). This step had very high remove performance SS (50%), COD (50%), dissolved (77%) and total (66%) BOD. It was also efficient to remove microbiological indicators (E. coli: 1.62 log unit in summer, 1.19 log unit in winter). This stage had a minimal contribution to the elimination of N (10%) and P (14%). The SFP effluent after these two stages of treatment is below the limit set by the EU Directive 1991 for COD and BOD. Recirculation of effluent from the four ponds also has the primary advantage of seeding active algal biomass into the first pond and maintaining aerobic conditions thus increasing pond efficiency. It is therefore possible to more than double the organic load on the first pond in comparison to the usual configuration of stabilisation ponds in series (Shelef and Kanarek,1995). During one year (2001) the first stage of treatment was stopped stop but SFP still showed good efficiency even though the influent was not pre-treated in the anaerobic pond. However in this case sludge increased in the pond R1. After four years of operation (including one of raw wastewater), sludge heights were still low, 0.30, 0.21, 0.18 and 0.14m in R1, R2, R3 and R4 respectively, leading to a reduction of volume of 16%R1, 11% R2, 10%R 3 and 7% R4. The SFP is a stage which can replace the facultative pond in the waste stabilisation pond process and produces high effluent quality with decreased land requirements and low operating costs. (Copin et al, 2004). Third stage: Maturation ponds The actual average organic load on the 3 maturation ponds M123 is 38 kg BODha-1d-1 and the hydraulic residence time is about 47 days. This stage of treatment removes the majority of total N (38%) and contributes to faecal indicators removal. The water level is kept low in the three ponds during summer to allow them to stock heavy autumn rain and limit its impact on the Thau lagoon.

179
Fourth stage: Polishing ponds The two last cells P1 and P2 are polishing ponds. They were commissioned to stock heavy autumn rain and limit its impact on the Thau lagoon. Loading rate was 26kg BODha-1d-1 and the hydraulic residence time was 20 days. Removal was very low in these ponds for all parameters except phosphorus (20%). Increases in SS and particulate COD concentration were even observed due to algae in summer. Decrease of organic pollution Respectively 34, 24 and 23 % of the applied COD was eliminated in the anaerobic, the step fed and the first three maturation ponds, while the figures for BOD5 were 47, 26, and 19 % respectively. The K coefficients for degradation, calculated according to the first order kinetic equation of Marais (complete mixing) were high for the first two stages: KCOD(AP) = 0.1d-1 in summer, 0.16d-1 in winter and KBOD (AP) 0.20d-1 in summer, 0.28d-1 in winter for deep aerated pond and KCOD(SFP)= 0.04 the same in summer and winter and KBOD(SFP)= 0.08 in summer , 0.06 in winter for step fed ponds Behaviour of nitrogen and phosphorus; After enlargement and retrofitting, the Mèze WSP system provided high nitrogen removal. Mean influent nitrogen concentration was 49.5 mg/L, 75% as ammonia. The overall mean removal was 72%. Twenty one % of influent total nitrogen was discharged into the Thau lagoon in the form of ammonia, 5% as organic-N and 2% as nitrite or nitrate. The proportion of organic-N was higher in summer and the beginning of autumn, than in winter or spring when N-NH4 effluent concentrations were less than 1mg/L. The first two stages of treatment (AP and SFP) removed very little nitrogen. Maturation ponds provided the bulk of the total nitrogen removal. Mechanisms involved in N removal are stripping of NH3, nitrification/denitrification and sedimentation (of bacteria, algae and zooplankton) after biological nitrogen uptake. However scientists disagreed over predominant mechanisms. Pano and Middlebrooks (1982) argued that ammonia volatilisation largely explains total nitrogen removal from ponds. On the other hand Zimmo et al, (2003) found that ammonia volatilisation in waste stabilisation ponds accounts for only a small fraction (1.5%) of total N-removal from domestic sewage and suggested that other nitrogen removal mechanisms such as nitrification/denitrification or sedimentation may be more important in overall nitrogen removal. On the basis of the low prevailing nitrate concentrations in ponds systems, several studies concluded that nitrification does not take place and consequently denitrification does not play a major role in nitrogen removal (Reed, 1985). In the Mèze WSP, although average concentrations of the oxidised forms of nitrogen in the effluent were low (annual mean 1.6mg/L), nitrate concentration reached 5mgN/L in the M3
effluent for more than 15 days in October 2003 and 8mg/L in June 2004 even though it was not present in the SFP effluent. Nitrification followed by denitrification could explain the low N-NH4 concentration observed in summer in pond M3 effluent. In October, when temperature was decreasing, denitrification was limited and N-NO3 concentration increased; in November, water temperature was less than 13°C and nitrate decreased and ammonia increased. Nitrate was <0.5mg/L in winter and spring and increased again in June when temperatures reached 24°C. For the same pond M3 prior to upgrading, Gomez et al(1995) demonstrated correlations that were negative between temperature and ammonium and positive between

180
loss of ammonia and the oxidised forms of nitrogen and suggested the existence of a nitrification/denitrification processes. The annual total phosphorus removal was 45%, higher in summer (63%) when phosphate concentration in effluent was <1mg/L and lower in spring (2%). Most of the phosphorus removal was observed to take place in the two polishing ponds, except in summer where it took place also in maturation ponds CONCLUSION The Mèze facility is a successful example of enlargement and retrofitting of an overloaded WSP; though more sophisticated than the initial system and requiring more energy consumption, it remains reliable, easy to operate and maintain, with only a moderate increase in operation costs and improved performances. The deep aerated pond was very efficient in removing SS and organic matter so it could be used to upgrade an overloaded WSP plant with low land requirement and energetic cost. The step fed facultative pond with recirculation and aeration is an innovative design pond efficient in removing BOD and faecal indicators; its surface organic loading is twice higher than what is usually applied in primary facultative ponds. Maturation and polishing ponds efficiently removed nutrients. They may have a significant part in the mitigation of storm events. The quality of effluent discharged is in accordance to European Directives and could be improved in warmest periods when replacing polishing ponds with rock filters or constructed wetlands that would be more effective to remove SS and particulate pollution. ACNOWLEDGEMENTS The authors wish to acknowledge the supports of the CCNBT, Conseil Général de l’Hérault, Agence de l’Eau, RMC, SATESE 34, ENTECH and Ecosite de Mèze. Our thanks to Helen Burnett for her kind assistance in translation . REFERENCES Brissaud F., Andrianarison T., Brouillet J.L. and Picot B. (2004). Twenty years monitoring of Mèze
stabilisation ponds II Removal of faecal indicators. 6th Int Specialist Group Conference on Waste Stabilisation Ponds, Avignon, 28 Sept. – 1st Oct.
Copin Y. Patent N° 12024 Copin Y. Brouillet J.L., Rivière Y. and Brissaud F. (2004). Waste stabilisation ponds in the wastewater
management strategy of a French department 6th Int Specialist Group Conference on Waste Stabilisation Ponds, Avignon, 28 Sept. – 1st Oct.
Gomez E., Casellas C., Picot B. and Bontoux J. (1995). Ammonia elimination processes in stabilisation and high rate algal pond systems. Wat. Sci. and Tech., 31(12), 303-312.
Paing J., Picot B. and Sambuco J.P. (2003). Emission of H2S and mass balance of sulfur in anaerobic ponds. Wat. Sci. Tech., 48(2), 227-234
Pano A. and Middlebrooks E.J., (1982). Ammonia nitrogen removal in facultative WSP. JWPCF 54,344-351
Picot B., Bahlaoui A., Moersidik S. Baleux B. and Bontoux J. (1992). Comparison of the purifying efficiency of high rate algal pond with stabilisation pond. Wat. Sci. Tech. 25(12), 197-206.

181
Picot B., Paing J., Sambuco J.P., Costa R.H.R and Rambaud A. 2003. Biogas production, sludge accumulation and mass balance of carbon in anaerobic ponds. Wat. Sci. Tech., 48(2), 243-250
Reed S.C.(1985). Nitrogen removal in stabilisation ponds. JWPCF, 57(1),39-45 Sambuco J.P., Costa R.H.R., Paing J. and Picot B. 2002. Influence of load distribution and recycle rate in
step-fed facultative ponds. Wat. Sci. Tech. 45(1), 33-39. Shelef G. and Kanarek A. (1995). Stabilization ponds with recirculation. Wat. Sci. Tech., 31(12), 389-397 Silva S.A., de Oliviera R., Soares J., Mara D.D. and Pearson H.W. (1995). Nitrogen removal in pond
systems with different configurations and geometries. Wat. Sci. Tech. 35(12), 321-330 Zimmo O.R., van der Steen N.P. andGijzen H.J. (2003). Comparison of ammonia volatilisation rates in
algae and duckweed-based waste stabilisation ponds treating domestic wastewater. Water research, 37, 4587-4594.

182
Publication2. Twenty years monitoring of Mèze stabilisation ponds II – removal of faecal indicators F. Brissaud*, T. Andrianarison*, J.L. Brouillet** and B. Picot*** Hydrosciences, MSE, Université Montpellier II, 34095 Montpellier cedex 05, France. (E-mail : [email protected], [email protected]) **Conseil General de l’Hérault, DARE-DEMA, rue d’Alco 34087 Montpellier cedex France (E-mail : dema-gestionglobaleeau@cg34) ***Département Sciences de l'Environnement et Santé Publique, UMR 5569 Hydrosciences- UM1, Faculté de Pharmacie, BP 1149, 34093 Montpellier cedex 5, France. (E-mail: picot@univ –montp1.fr)
Abstract The WSP system serving Mèze and Poussan (French Mediterranean coast) was constructed in 1980 and enlarged and upgraded from 1994 to 1998. Water quality along the WSP system has been monitored over the years; thus allowing to assess the influence of enlargement and upgrading works. A significant enhancement of the average microbiological quality of the effluent was observed, with respective E. coli and streptococci average abatements of 4.1 and 3.4 log. units. Former seasonal variations of microbiological removal have vanished. The contribution of the different ponds to the disinfection performance of the WSP system was analysed. A microbiological quality model was proposed to evaluate the die-off kinetics related to the different ponds and as a tool for the design and management of WSP systems. Though the relationships between die-off coefficients and environmental factors appeared somewhat frail, this modelling is considered a promising approach for the prediction of WSP microbiological performance.
Keywords Die-off kinetics; disinfection; modelling; waste stabilisation ponds.
INTRODUCTION Twenty five years ago waste stabilisation ponds (WSPs) have been recommended for the treatment of wastewater to be reused for unrestricted irrigation (WHO, 1989). About the same years, WSP technology was chosen to protect bathing waters and shellfish breeding areas along the Mediterranean coast of Languedoc and Roussillon (Southern France). A prominent implementation of this policy was the construction of WSPs to treat the sewage of all the municipalities of the northern part of the watershed of Thau coastal lagoon. Since then, disinfection in WSP has been much studied, at the real scale, on pilot plants and at the laboratory. However, predicting disinfection performance still remains a somewhat tricky aspect of WSP state of the art. This is due to the high number of factors and inter-related processes involved. Disinfection in WSP can be seen as resulting from the combination of pond hydrodynamics, which control water retention times, and micro-organism decay processes. Both, hydrodynamics and die-off processes, are highly dependent on

183
meteorological factors, mainly temperature, solar irradiation and wind. The natural variability of these factors adds to the difficulty of disinfection investigation and prediction (Brissaud et al., 2003). Therefore, despite the valuable and promising findings of many investigators, from Racault et al. (1984) to Shilton & Harrison (2003) on hydrodynamics and from Mezrioui (1987) to Davies-Colley et al. (2003), on micro-organism die-off kinetics, there is still much to be learnt from the monitoring of full scale plants. Monitoring the impact of upgrading and enlargement works on effluent quality may be highly significant for practical purposes (Archer & Donaldson, 2003). Moreover a microbial quality model was proposed in order to evaluate die-off kinetics in the different ponds of the system and as a tool for the design and management of WSP systems. MATERIALS AND METHODS
Mèze WSP system The main characteristics of Mèze WSP system are described in Picot et al. (2004). The plant was constructed in 1980 in order to protect the shellfish breeding areas of the Thau coastal lagoon. It was a 3 cells system (M1,2,3) with a total surface of 8 ha and a treatment capacity of 8,000 p.e.. In the early eighties, the treated load corresponded to 4,400 p.e. -augmented with winery effluents in autumn (Figure 1). At that time, no microbiological criterion was set on the effluents discharged in the Thau lagoon. The plant became overloaded a few years later, due to serviced population growth as well as increasing industrial loads. Nowadays, the load to be treated amounts to up to 1120 kg BOD5 per day, being equivalent to 19,000 p.e.. Furthermore, the new consent conditions require that WSP effluent Escherichia coli and enterococci contents are less than 1000 cfu/100 mL in summer and 10,000 cfu/100 mL in winter. These requirements might be strengthened in a near future. Therefore the plant had to be retrofitted in order to cope with these new conditions. Eight new ponds were added to the 3 old ones between 1996 end 1998, the total pond surface reaching 14 ha. At first, in the early eighties, a typical French 3 ponds system, it nowadays is a sophisticated system of 11 ponds, some of them being aerated, step fed and with possible recirculation. Moreover, the management of the system has been complicated in order to optimise the performance of the step fed facultative stage and alleviate the impact of storm events on microbiological water quality. A part of the effluents of the anaerobic ponds are derived to M1 without passing through the step-fed ponds (Figure 1). Water depth is controlled in every pond through weir level adjustments. Wastewater is stored during storm events in order to increase water detention times and prevent degradation of the effluent microbiological quality. Raising the water levels entails temporary modifications in the hydraulic circuits which tend to affect the microbiological quality.
Monitoring Water quality was monitored at the inlet and outlet of ponds M1,2, 3 fortnightly from June 1980 to August 1982 (Trousselier et al., 1986), monthly from November 1984 to June 1986 (Mezrioui, 1987) and from October 1988 to January 1990 (Picot et al., 1992). Water quality

184
was observed at the inlet of M1 and outlet of M3 from August 1994 to February 1995 before, during and after desludging of pond M1, with a time step varying from 1 to 3 weeks (Crabos et al., 1996). From 1980 to 1998, inlet and outlet flow rates have not been recorded; only a few measures are available. The current facility shown on figure 1 was completed in 1999. From then, inlet and outlet flow rates were recorded and water quality analysed on a fortnight basis at the plant inlet, the outlet of the deep surface aerated ponds A1 and A2, the outlet of R4 which is the 4th pond of the step-fed facultative aerated stage, the outlet of the maturation pond M3 and the outlet of the last polishing pond P2. Additional analysis were made, from July 2003 to August 2004 at the outlets of M1, M2 and F1 in order to be able to identify the part played by each pond in the performance of the system. Meteorological data were available, recorded either on site or at Sète, on the South bank of Thau lagoon. Water temperature was measured fortnightly.
A1 A2
R1
R2
R3
R4
M1
M2
M3
P1
P2
Inlet
Normal flow path
Outlet
Other flow paths
Figure 1 : Lay-out of the current Mèze WSP system
Modelling microbial decontamination The model of microbial decontamination is based on bacterial mass balance, assuming that each lagoon is perfectly stirred and bacterial decay is a first order kinetic process. The bacterial balance is expressed for each pond as:
where i is the ith pond and j the jth time; vi is the water volume in the pond i (m3); ∆n(i, j) = n(i, j) - n(i, j-1); ∆t is the time step, ∆t =1 day; n (i-1, j) and n(i, j) are microorganism concentrations at the inlet and in lagoon i respectively in the day j (cfu/100ml); q(i-1, j) and q(i, j) are the inflow and outflow rates of lagoon i (m3/day); ki is the die-off coefficient of lagoon i (xu et al., 2002). several inlets with different concentrations can be taken into account by adding
Vij)(i,
Nj)(i,
Nj)(i,
Qj)1,(i
Nj)1,(i
Qt
Vij)N(i,
t
j)N(i,Vi ki−−−−=
∆∆+
∆∆ (1)

185
corresponding terms in equation 1. flow rates have been calculated through water balances, at each time step, from inlet flow rate in the plant, rainfall, evaporation and water levels. during the last years, water levels were measured fortnightly. Die-off coefficient, ki, is a key parameter for disinfection predictions; it is known to depend on several environmental variables, mainly temperature, solar radiation, suspend solids, pH, DO, …; its values can be calculated directly from Equation 1 provided inlet and outlet microorganism concentrations have been analysed and hydraulic parameters are known. However it is preferable for prediction purposes to link ki to the more influential variables; according to the work of Xu et al., (2002), temperature, solar radiation and SS were considered. Die-off rates were expressed as follows:
k = α* β(T-20) * e 0.χIm (2) where T is the temperature, Im, the depth averaged solar intensity – which is a function of SS - and α, β and χ are constants determined through a multiple regression process. Values of α, β and χ minimise the difference between values of N (i,j) respectively observed and calculated using Equation 1 where N(i-1,j) is an observed value and ki is expressed according to Equation 2.
RESULTS Microbiological performances Average removals of faecal coliforms (or E. coli) and streptococci were respectively only 2.9 and 2.6 log units during the first years of operation. High seasonal variations were observed; winter outlet faecal coliform contents were up to 3 log units higher than summer values, which meant a lower protection of shellfish breeding areas (Figure 2). During the following years, as the applied load increased progressively, microbiological performances slowly degraded. Measures from August 1994 to February 1995 reflect the disruptions due to desludging works.
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
June
Aug
ust
Oct
ober
Dec
embe
r
Feb
ruar
y
Apr
il
June
Aug
ust
Oct
ober
Dec
embe
r
Feb
ruar
y
Apr
il
June
Aug
ust
Fae
cal c
olifo
rms
(or
E.c
oli)/
100
mL
1980-1982
1984-1986
1988-1990
1994-1995
2002-2004
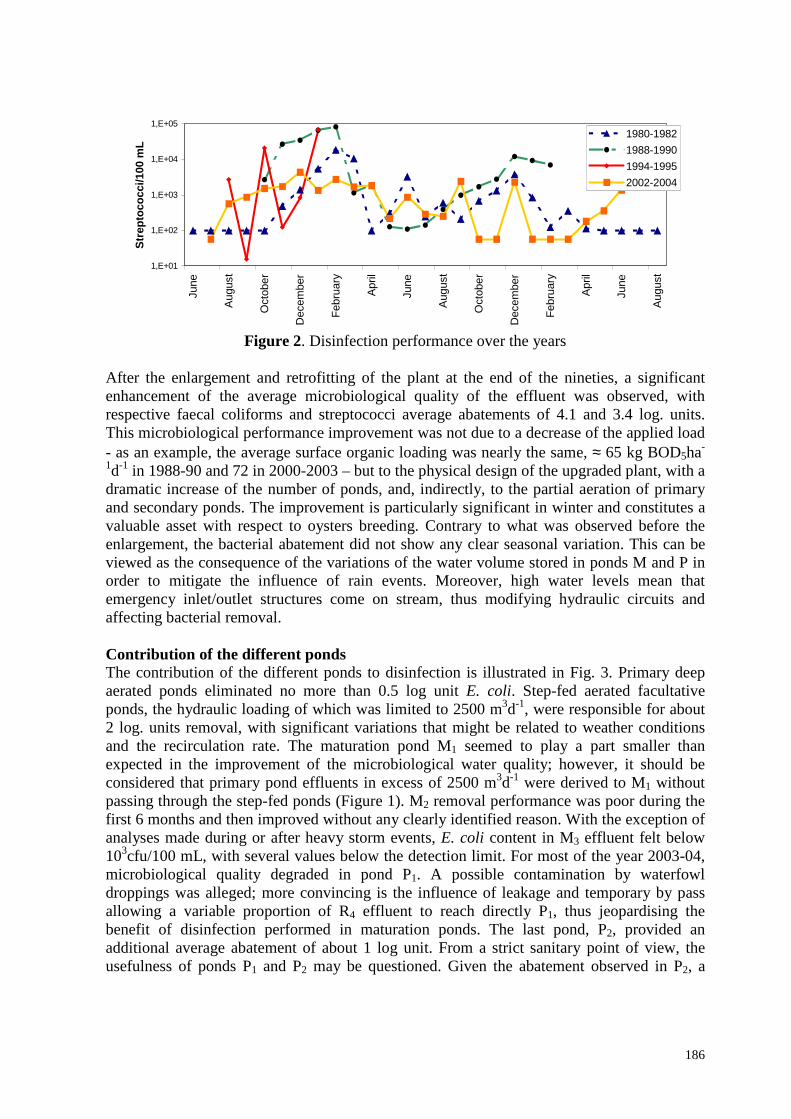
186
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
June
Aug
ust
Oct
ober
Dec
embe
r
Feb
ruar
y
Apr
il
June
Aug
ust
Oct
ober
Dec
embe
r
Feb
ruar
y
Apr
il
June
Aug
ust
Str
epto
cocc
i/100
mL
1980-1982
1988-1990
1994-1995
2002-2004
Figure 2. Disinfection performance over the years
After the enlargement and retrofitting of the plant at the end of the nineties, a significant enhancement of the average microbiological quality of the effluent was observed, with respective faecal coliforms and streptococci average abatements of 4.1 and 3.4 log. units. This microbiological performance improvement was not due to a decrease of the applied load - as an example, the average surface organic loading was nearly the same, ≈ 65 kg BOD5ha-
1d-1 in 1988-90 and 72 in 2000-2003 – but to the physical design of the upgraded plant, with a dramatic increase of the number of ponds, and, indirectly, to the partial aeration of primary and secondary ponds. The improvement is particularly significant in winter and constitutes a valuable asset with respect to oysters breeding. Contrary to what was observed before the enlargement, the bacterial abatement did not show any clear seasonal variation. This can be viewed as the consequence of the variations of the water volume stored in ponds M and P in order to mitigate the influence of rain events. Moreover, high water levels mean that emergency inlet/outlet structures come on stream, thus modifying hydraulic circuits and affecting bacterial removal. Contribution of the different ponds The contribution of the different ponds to disinfection is illustrated in Fig. 3. Primary deep aerated ponds eliminated no more than 0.5 log unit E. coli. Step-fed aerated facultative ponds, the hydraulic loading of which was limited to 2500 m3d-1, were responsible for about 2 log. units removal, with significant variations that might be related to weather conditions and the recirculation rate. The maturation pond M1 seemed to play a part smaller than expected in the improvement of the microbiological water quality; however, it should be considered that primary pond effluents in excess of 2500 m3d-1 were derived to M1 without passing through the step-fed ponds (Figure 1). M2 removal performance was poor during the first 6 months and then improved without any clearly identified reason. With the exception of analyses made during or after heavy storm events, E. coli content in M3 effluent felt below 103cfu/100 mL, with several values below the detection limit. For most of the year 2003-04, microbiological quality degraded in pond P1. A possible contamination by waterfowl droppings was alleged; more convincing is the influence of leakage and temporary by pass allowing a variable proportion of R4 effluent to reach directly P1, thus jeopardising the benefit of disinfection performed in maturation ponds. The last pond, P2, provided an additional average abatement of about 1 log unit. From a strict sanitary point of view, the usefulness of ponds P1 and P2 may be questioned. Given the abatement observed in P2, a

187
better control of flow paths between the ponds should result in a high microbiological quality effluent.
1 ,E +01
1 ,E +02
1 ,E +03
1 ,E +04
1 ,E +05
1 ,E +06
1 ,E +07
1 ,E +08
09/0
703
07/0
8/03
07/0
9/03
06/1
0/03
04/1
1/03
03/1
2/03
09/0
1/04
07/0
2/04
08/0
3/04
06/0
4/04
05/0
5/04
03/0
6/04
11/0
7/04
09/0
8/04
D ate
E. c
oli /
100
mL
Inlet
A1-A2 outlet
R 4 outlet
M1 outlet
M2 outletM3 outlet
P 1 outlet
P 2 outlet
Figure 3. Contribution of the different ponds to E. coli removal
Die-off coefficients E. coli and streptococci die-off coefficients have been calculated for ponds M1,2,3, from direct application of Equation 1 for the periods 1980-82, 1984-86 and 1988-89. Data related to E. coli are analysed in this paper. Mean E. coli die-off coefficient of pond M1, at that time a facultative pond, was found to be about only 0.13 d-1 in 1980-82 and 1984-86 and 0.46 d-1 in 1988-89; no seasonal variation appeared. Though highly variable from one year to another, seasonal effects were more easily identified in maturation ponds M2 and M3. M2 E. coli die-off coefficient, k2, was 35 d-1 from June to September 1980, 0.45 d-1 from October 1980 to March 1981, only 1.4 d-1 from June to September 1981; it dropped to less than 0.1 d-1 in December, then increased to 4 d-1 in March and April 1982 and 96 d-1 from May to August. k2 value was 0.23 d-1 in November 1984, dropped to less than 0.001 d-1 in March 1985, raised to 26 in July, was about 0.4 d-1;from September 1985 to March 1986 and raised to 1.5 d-1 from April to June. Data from 1988-89 monitoring revealed clear seasonal variations of k2 with minimum values of 0.2 and 0.13 d-1 in December 1988 and November 1989 and a maximum of 15 d-1 in May and June. Variations of M3 E. coli die-off coefficient, k3, were also shown to depend on seasons. From 1.55 d-1 in June 1980, k3 raised to 5 d-1 in September, then reached a minimum of 0.13 d-1 in January 1981 and a maximum of 13 d-1 at the end of April, a second minimum of 0.7 d-1 at the end of October before reaching 5 d-1 in April 1982 and 9 d-1 in August. In November 1984, k3 was about 0.15 d-1; it reached 170 d-1 in May 1985, decreased to 0.001 d-1 in January 1986 to reach a maximum of 16 d-1 in March and only 5 d-1 in April 1986. From 0.07 d-1 in November 1988, k3 reached a maximum of 4.5 d-1 in July 1989 and decreased to 0.25 d-1 in December. These data demonstrated that, beyond a clear seasonal effect in maturation ponds, die-off coefficient were highly variable.
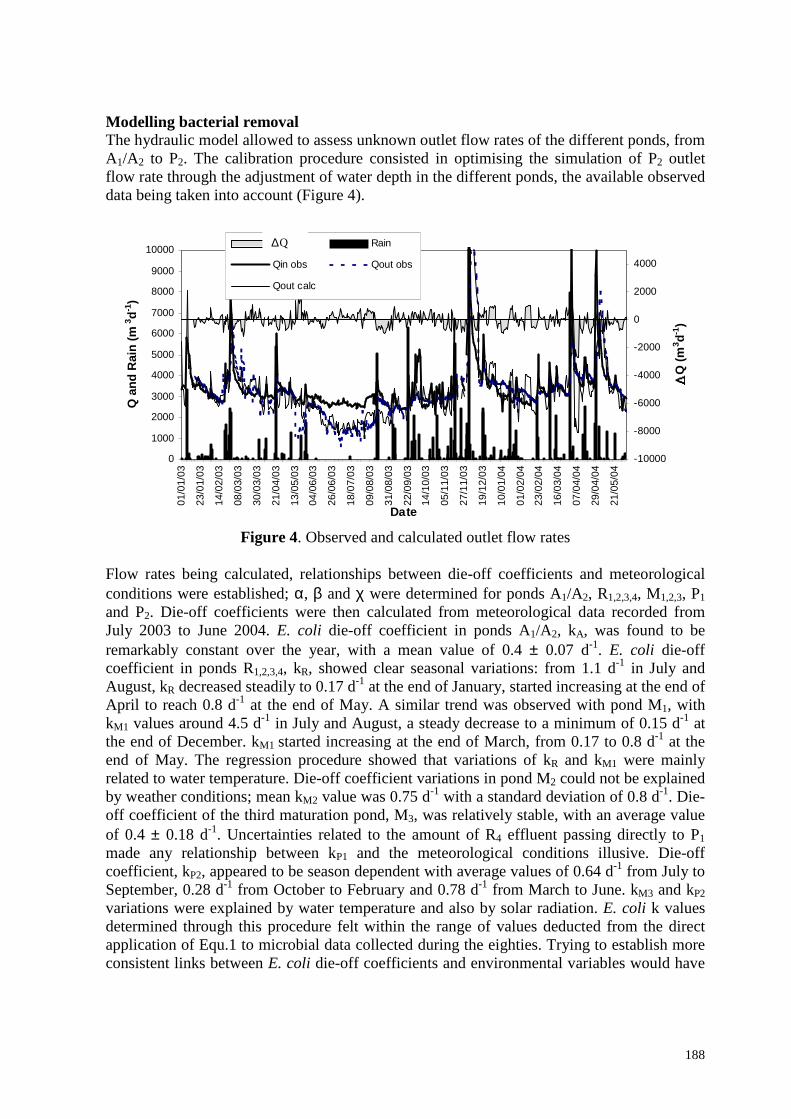
188
Modelling bacterial removal The hydraulic model allowed to assess unknown outlet flow rates of the different ponds, from A1/A2 to P2. The calibration procedure consisted in optimising the simulation of P2 outlet flow rate through the adjustment of water depth in the different ponds, the available observed data being taken into account (Figure 4).
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
01/0
1/03
23/0
1/03
14/0
2/03
08/0
3/03
30/0
3/03
21/0
4/03
13/0
5/03
04/0
6/03
26/0
6/03
18/0
7/03
09/0
8/03
31/0
8/03
22/0
9/03
14/1
0/03
05/1
1/03
27/1
1/03
19/1
2/03
10/0
1/04
01/0
2/04
23/0
2/04
16/0
3/04
07/0
4/04
29/0
4/04
21/0
5/04
Date
Q a
nd R
ain
(m3 d
-1)
-10000
-8000
-6000
-4000
-2000
0
2000
4000
Q (
m3 d
-1)
DQ Rain
Qin obs Qout obs
Qout calc
∆Q
Figure 4. Observed and calculated outlet flow rates
Flow rates being calculated, relationships between die-off coefficients and meteorological conditions were established; α, β and χ were determined for ponds A1/A2, R1,2,3,4, M1,2,3, P1 and P2. Die-off coefficients were then calculated from meteorological data recorded from July 2003 to June 2004. E. coli die-off coefficient in ponds A1/A2, kA, was found to be remarkably constant over the year, with a mean value of 0.4 ± 0.07 d-1. E. coli die-off coefficient in ponds R1,2,3,4, kR, showed clear seasonal variations: from 1.1 d-1 in July and August, kR decreased steadily to 0.17 d-1 at the end of January, started increasing at the end of April to reach 0.8 d-1 at the end of May. A similar trend was observed with pond M1, with kM1 values around 4.5 d-1 in July and August, a steady decrease to a minimum of 0.15 d-1 at the end of December. kM1 started increasing at the end of March, from 0.17 to 0.8 d-1 at the end of May. The regression procedure showed that variations of kR and kM1 were mainly related to water temperature. Die-off coefficient variations in pond M2 could not be explained by weather conditions; mean kM2 value was 0.75 d-1 with a standard deviation of 0.8 d-1. Die-off coefficient of the third maturation pond, M3, was relatively stable, with an average value of 0.4 ± 0.18 d-1. Uncertainties related to the amount of R4 effluent passing directly to P1 made any relationship between kP1 and the meteorological conditions illusive. Die-off coefficient, kP2, appeared to be season dependent with average values of 0.64 d-1 from July to September, 0.28 d-1 from October to February and 0.78 d-1 from March to June. kM3 and kP2 variations were explained by water temperature and also by solar radiation. E. coli k values determined through this procedure felt within the range of values deducted from the direct application of Equ.1 to microbial data collected during the eighties. Trying to establish more consistent links between E. coli die-off coefficients and environmental variables would have

189
required highly reliable microbial and hydraulic data. A one day time step monitoring would have been suitable. Effluent quality was calculated through the application of Equation 1 in each cell, one cell after the other, k values being derived at each time step from the relationships established between α, β and χ and water temperature, solar radiation and SS content. Microorganism contents in each cell were thus calculated from the observed microorganism content in inlet raw wastewater. Despite some discrepancies between the observed and simulated k values, particularly after the storm of early May, the simulation was considered satisfactory (Fig. 5).
0
2000
4000
6000
8000
10000
12000
14000
08/0
7/03
23/0
7/03
07/0
8/03
22/0
8/03
06/0
9/03
21/0
9/03
06/1
0/03
21/1
0/03
05/1
1/03
20/1
1/03
05/1
2/03
20/1
2/03
04/0
1/04
19/0
1/04
03/0
2/04
18/0
2/04
04/0
3/04
19/0
3/04
03/0
4/04
18/0
4/04
03/0
5/04
18/0
5/04
02/0
6/04
Date
Rai
n (m
3 d-1 )
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
CF
U/1
00m
L
Rain
Inlet obs. contentOutlet obs. content
Outlet calc. content
detection limit
Figure 5. Modelling E. coli content in the effluent of Mèze WSP system
CONCLUSION Retrofitting of Mèze WSP system resulted in a significant enhancement of the average microbiological quality of the effluent, with respective faecal coliforms and streptococci average abatements of 4.1 and 3.4 log. units. This improvement was not due to a surface loading decrease but to the subdivision of the plant into a high number of cells and to the step-fed deep aerated pond system. Former seasonal variations of microbiological removal have vanished ; this is considered the consequence of the plant management in order to mitigate the impact of storm events more than the result of opposite effects of environmental factors. The contribution of the polishing ponds to the disinfection performance is not without ambiguity. Therefore, it might be considered that, owing to the microbiological requirements of the present consent, the total pond surface could have been significantly reduced. This is considered as a beneficial effect of the design of the two first stages of the retrofitted plant. However, as they have a significant part in the mitigation of storm events, polishing ponds are not likely to be decommissioned. A microbiological quality model was proposed to evaluate the die-off kinetics in the different ponds and as a tool for the design and management of WSP systems. Though relationships between die-off coefficients and environmental factors appeared somewhat frail, this

190
modelling is considered a promising approach for the prediction of WSP microbiological performance and the design of management strategies. ACKNOWLEDGEMENTS The authors would like to thank the CCNBT, the Conseil Général de l’Hérault, the Agence de l’Eau RMC for their financial support and SATESE 34, ENTECH and Ecosite de Mèze for their kind assistance. REFERENCES Archer, H.E. and Donaldson, S.A. (2003). Waste stabilisation ponds upgrading at Blenheim and
Seddon, New Zealand – case studies Wat. Sci Tech., 48(2), 17-23. Brissaud, F., XU, P. and Auset, M. (2003). Extensive reclamation technologies for irrigation reuse.
Wat. Sci. Tech./Water Supply. 3(4), 209-216. Crabos J.L., Sambuco J.P., Bondon D., Brouillet J.L., Lebec C., Bonal. O. (1996). Lagunage de
Mèze-Loupian,. Curage, deshydratation et épandage des boues. Rapport CEREMHER, IFREMER, 55 p.
Davis-Colley, R.J., Craggs, R.J. and Nagels, J.W. (2003). Disinfection in a pilot scale “advanced” pond system (APS) for domestic sewage treatment in New Zealand. Wat. Sci Tech., 48(2), 81-88.
Lloyd, B.J., Vorkas, C.A. and Guganesharajah, R.K. (2003). Reducing hydraulic short-circuiting in maturation ponds to maximize pathogen removal using channels and wind breaks. Wat. Sci Tech., 48(2), 153-162.
Mezrioui, N.E. (1987). Etude expérimentale des effets du pH, du rayonnement et de la température sur la disparition des bactéries d’intérêt sanitaire et évaluation de la résistance aux antibiotiques d’E. coli lors de l’épuration des eaux usées. Th. Doct., Montpellier 2. 171p.
Picot B., Bahlaoui A., Moersidik S. Baleux B. and Bontoux J. (1992). Comparison of the purifying efficiency of high rate algal pond with stabilisation pond. Wat. Sci. Tech. 25(12), 197-206.
Picot, B., Andrianarison, T., Gosselin, J.P. and Brissaud, F. (2004). Twenty years monitoring of Méze stabilisation ponds. I – Removal of arganic matter and nutrients. 6th IWA Int. Conf. on Waste Stabilisation Ponds.
Racault, T., Boutin P. and Douat J. (1984). Etude par traçage du comportement hydraulique d’une lagune d’épuration: influence de la géométrie du bassin. Rev. Sci. Eau 3, 197-218.
Trousselier, M., Legendre, P. and Baleux, B. (1986). Modeling of the evolution of bacterial densities in an eutrophic ecosystem (sewage lagoon). Microb. Ecol. 12, 355-379.
Shilton, A. and Harrison, J. (2003). Development of guidelines for improved hydraulic design of waste stabilisation pond. Wat. Sci Tech., 48(2), 173-180.
Xu, P., F. Brissaud and A. Fazio, 2002. Non-steady state modelling of faecal coliform removal in deep tertiary lagoons. Wat. Research .36(12) :3074-3082.

191
Article soumis.
Modelling the fate of nitrogen in maturation ponds T. Andrianarison*, X. Wang**, J. Qiu**, F. Brissaud*and B. Picot*** *UMR 5569 Hydrosciences, MSE, Université Montpellier II, 34095 Montpellier cedex 05, France. (E-mail : [email protected], [email protected] ) ** School of Environmental Science and Engineering, Shanghai Jiaotong University, 800 Dongchuan Rd, Shanghai, 200240 China (E-mail: [email protected]; [email protected] ) ***Département Sciences de l'Environnement et Santé Publique, UMR 5569 Hydrosciences- UM1, Faculté de Pharmacie, BP 14491, 34093 Montpellier cedex 5, France. (E-mail: picot@univ–montp1.fr) Abstract Nitrogen removal in stabilization ponds has become, over the years, a matter of controversy. Some doubts existed on the main mechanisms responsible for nitrogen removal. Volatilization has long been assumed to be the most important process while more and more authors are putting forward that nitrification and denitrification are the major mechanisms involved in N content reduction. From 1st April to 13th July 2005, the influent and effluent water quality of the third 2-ha maturation pond of the Mèze wastewater treatment plant were monitored to verify the suggestion that nitrification/denitrification might be responsible for N removal. A dynamic mechanistic model was developed to simulate the fate of nitrogen. The model considers organic nitrogen, ammonia and nitrates in the liquid phase as well as the organic nitrogen accumulated in benthic sludge. The mechanisms simulated in the model include organic nitrogen mineralization and sedimentation, ammonia assimilation and volatilization, regeneration of organic N contained in benthic sludge and nitrification and denitrification. The model simulated successfully organic N, NH3-4-N and NO2-3
—
N concentrations in the pond effluent. The model showed that, during the period considered, nitrification-denitrification and volatilization were respectively responsible for 82 and 18 % of total nitrogen removal. Keywords: Denitrification; maturation ponds; modelling; nitrification; nitrogen removal; volatilization.
INTRODUCTION Waste stabilization ponds (WSP) have been widely used for wastewater treatment in small communities throughout the world during the 30 last years. This technology has often been chosen for its low operation and maintenance costs, low risk of failure and low sludge production. Nowadays, wastewater treatment systems are required to protect sensitive receiving water bodies; that is to say, to remove nutrients. While upgraded conventional treatment systems capable of removing nitrogen and phosphorus are now commonplace, not enough attention has been paid to the potential of pond systems for nutrient reduction.

192
Many mechanisms are involved in the fate of nitrogen in ponds: uptake of inorganic nitrogen by algae and bacteria, mineralization, sedimentation, volatilization of ammonia and nitrification coupled with denitrification (Mayo and Mutamba, 2005). For a long time, contradictory assumptions on the main mechanism responsible for nitrogen removal were proposed. Some researchers believed that ammonia volatilization largely explains total nitrogen removal from WSPs (Silva et al., 1995; Soares et al., 1996). However other researches showed or suggested that nitrification/denitrification is the predominant mechanism for N-removal (Zimmo et al., 2003; Picot et al., 2005). Many factors affect the nitrogen cycle, including temperature, solar radiation, mixing (Azov and Tregubova, 1995) and pH (Gomez et al., 1995). A survey of 178 French WSP systems showed no significant seasonal variations in COD, BOD5 and TSS removal but a contrasting pattern as regards nutrients (Racault et al., 1995); indeed, the average ammonia nitrogen concentration in the outlet water varied from 7.2mg/L in summer to 20.8mg/L in winter. Significant amounts of nitrate were observed at the outlet of the maturation stage of the Mèze-France WSP facility in Autumn 2003, just before the cold season rise of N-NH4 content, and in Spring 2004, simultaneously with the hot season N-NH4 drop (Picot et al., 2005). Therefore it was suggested that nitrification combined with denitrification, two temperature dependant mechanisms, might be responsible for N elimination (Mc Lean et al., 2000). The aim of this work was to determine the mechanisms responsible for nitrogen removal in the final maturation pond of the Mèze WSP system, which is located in a French Mediterranean region. Volatilization has been assessed through literature and specific experiments. A mathematical modeling of the fate of nitrogen was undertaken in order to verify whether the dramatic drop in nitrogen content observed in Spring season can be explained by nitrification combined with denitrification. MATERIALS AND METHODS
A1A2
R1
R2
R3
R4
M1
M2
M3
P1
P2
Inlet
Normal flow path
Outlet
Other flow paths
A1, A2: anaerobic ponds R1 to R2: step fed ponds M1, M2, M3: maturation ponds P1, P2 : polishing ponds
Figure 1. Lay-out of the Mèze waste stabilization pond system

193
Site description The Mèze WSP plant is located on the Mediterranean coast (03°35’06’’E, 43°25’10’’N). Enlargement and modifications have been undertaken in the years 1996-1999, including aeration of the anaerobic and step-fed ponds (Figure 1). The total area of the ponds nowadays is 14.4 ha, and the organic load amounts to 1120 kg BOD per day, being equivalent to 19,000p.e.. A more detailed description of the systems was presented in Brissaud et al. (2005) and Picot et al. (2005). The third maturation pond, M3, with an area of 19500m2 and a water depth that can vary from 1.6m to 2.0m, was chosen to study the fate of nitrogen. Sampling and analysis From 1st April to 13th July 2005, influent and effluent flows of the Mèze WSP system were monitored with an electromagnetic flowmeter; the water depth was measured manually everyday. Meteorological data were recorded on site. Dissolved oxygen (DO), water temperature and pH were monitored at mid-depth near the outlet and recorded. Water samples were taken at the inlet and outlet of pond M3 every week and analyzed for suspended solids (SS), total chemical oxygen demand (COD), filtered COD, Kjeldhal nitrogen (TKN), ammonia (NH4-N), nitrite (NO2-N) and nitrate (NO3-N), according to standard methods. Volatilization assessment Ammonia volatilization tests were conducted according to the experimental method employed in Stratton (1969). Ammonia losses were measured in outdoor jars filled with 15, 10 and 6 cm water heights, the pH varying between 7.5 and 9, the mean daily temperature between 15 and 25°C and the daily mean wind velocity between 2.5 and 6.3 m.s-1. Mathematical model The model for nitrogen transformation and removal was adapted from Fritz et al. (1979), Senzia et al. (2002) and Mayo and Mutamba (2005). Pond M3 inlet and outlet flow rates were calculated at a daily time step from the records of the plant inlet flow rate, water depth observations and rainfall and evaporation derived from local meteorological records; water losses due to leakage were considered negligible because of the high ground water level.. Pond M3 was assumed to behave like a perfectly mixed reactor. The model simulates the concentrations of ammonia (NH4-3-N), organic nitrogen (Org-N) and nitrite and nitrate nitrogen (NO2-3
--N) in the effluent, the respective inlet concentrations being observed or interpolated from observed values. Seven processes of nitrogen transformation were considered, including ammonia assimilation (ra), ammonia volatilization (rv), organic matter sedimentation (rs), benthic sludge regeneration (rr), organic matter mineralization (rm), nitrification (rn) and denitrification (rd), ra, rv, rs, rr, rm, rn and rd being the respective transformation rates in mg.L-1d-1 (Figure 2). Ammonia content was high enough to admit that NO3
--N assimilation was negligible. Expressions of the transformation rates are shown in Table 1.

194
Figure 2. Nitrogen transformation in waste stabilization ponds
Table 1. Nitrogen transformation processes equations and parameters description Process Transformation rates Reference
Ammonia assimilation
( ) [ ][ ] [ ]Norg
NNHK
NNHafr T
a −−+
−=−
−−
43
4320.
f = 0.04 (d-1), biomass maximum growth rate a = 1.07, microorganism growth temperature coefficient K = 18 mgL-1, ammonia uptake half saturation constant
Ferrara and Harleman (1980) Ferrara and Avci (1982)
Ammonia Volatilization ( ) ( ) [ ]NNHee
Hr pHTv −⋅×= −
−−−
435.857.1)2013.0
31030
H = water depth (m)
Stratton (1968 and 1969)
Organic matter sedimentation
[ ]NOrgdrs −= .
d = 0.014 (d-1), settling coefficient
Ferrara and Harleman (1980)
Benthic sludge regeneration
( )
H
Dr mTr
2007.19.010 −××=
Dm,= Org N in active benthic sludge (mg cm-2)
Fritz et al (1979)
Organic matter mineralization
[ ]NOrgTbrm −= ..
b = 0.001 (d-1oC-1), mineralization coefficient T = temperature (oC)
Ferrara and Avci (1982)
Nitrification
[ ][ ]
[ ][ ]
( )Y
eDO
DO
NNH
NNHgr Th
Tn
1
5.01015
4358.1051.0
43 ⋅+
⋅−+
−⋅= −
−−
− ρ
g = 0.966 (d-1), nitrifier growth rate at 15 oC; h = 0,07 ρ = nitrifier concentration (mg L-1); Y = 0.15, yield coefficient;
Fritz et al (1979), Wiesmann (1994)
Denitrification
( )[ ]NNOmkr Td −⋅= −
−−
3225
k = 0.3 (d-1), denitrification rate at 25 oC; m = 1.25, denitrification temperature coefficient ;
Senzia et al. (2002)
Influent Influent Influent
Benthic sludge
Organic nitrogen
Ammonia Nitrite and Nitrate
Effluent Effluent
Mineralization
Assimilation
Volatilization Denitrification
Nitrification
Effluent
Regeneratition Sedimentation

195
Mass balance equations for water, organic nitrogen, ammonia nitrogen, nitrate nitrogen, autotrophic nitrifying bacteria and benthic sludge were developed and detailed in Table 2. The modeling process was carried out using a fourth-order Runge-Kutta method, and the calculation time step varied from 1hour to 1 day. The values of several coefficients were taken from the literature, while the unknown values were determined through the calibration procedure. Table 2. Mass balance equations Substance Mathematic expression
Ammonia rvnmae
ii rrrrrNNH
V
QNNH
V
Q
t
NNH +−−+−−−−=−−−
− ][][d
][d4343
43
Organic nitrogen smse
ii rrrNOrg
V
QNOrg
V
Q
t
NOrg +−−−−−=−][][
d
][d
Benthic sludge rsm rrt
D −=d
d
Nitrate dne
ii rrNNO
V
QNNO
V
Q
t
NNO −+−−−=−−−
− ][][d
][d3232
32
Nitrifier ( ) [ ][ ] ρρρ ⋅
+⋅⋅−⋅+⋅−= −
DO
DOnbrYQ
tT
nji 5.0d
d 20,
Water balance evrie QQQQ −+=
Parameter description: Qe: effluent flow rate; Qi: influent flow rate; Qr: rainfall (water height multiplied by the pond surface); Qev: evaporation losses (evaporation multiplied by the pond surface); b: maximum decay coefficient at 20 oC
RESULTS AND DISCUSSION The evolution of N species concentrations in pond M3 effluent was characterized by moderate variations in [Org-N], a drop of ammonia from 28 to 3 mg.L-1 between May 31st and June 9th and a simultaneous increase of [NO2-3-N] from 3 to 28 mg.L-1(Figure 3). Then [NO2-3-N] decreased rapidly, [NH3-4-N] remaining at values lower than 3 mg.L-1 until the end of the survey in mid July. The [NH3-4-N] drop was soon followed by dramatic nitrogen losses which continued from mid June till the end of the survey (Figure 4). Ammonia volatilization was calculated using a mathematical expression derived from Stratton (1968 and 1969) with a coefficient 30.10-3 determined from the outdoor experiments we have carried out (Table 1). Ammonia volatilization depends on pH, water temperature and ammonia concentration. The pH varied from 7.9 in early April to a peak of 8.6 on June 20th. The temperature was less than 15°C at the beginning of the survey and rose steadily up to 20°C on May 22nd and more than 28°C on June 21st, then decreased to 23°C (Figure 4). As a result, NH3 volatilization increased gently from around 4 kg N d-1 in early April up to 13.7 kg d-1 on May 12th and then started decreasing with the ammonia concentration. Until the end of May, it could explain most of the nitrogen losses but NH3 volatilization fell rapidly after June 3rd and reached values lower than 3 kg N d-1 from June 8th to the end of the survey (Figure 4). In the meantime, nitrogen losses soared up to 116 kg N d-1 on June 20th and then decreased down to 30-40 kg N d-1 in the first week of July. This last decrease was mainly due to a lower

196
N content entering the pond after June 20th. The dramatic nitrogen losses observed in pond M3 in June and July could not be explained by ammonia volatilization. Sedimentation was unlikely to explain the nitrogen losses in hot Spring and Summer. Therefore, the only pathway that allowed explaining the hot season nitrogen losses was nitrification combined with denitrification.
0
10
20
30
40
50
04/04/05 02/05/05 30/05/05 27/06/05
N o
rg (
mg/
L)Inlet - observedOutlet - calculatedOutlet - observed
0
10
20
30
40
50
04/04/05 02/05/05 30/05/05 27/06/05
N -
NH
4 (m
g/L)
Inlet - observed
Outlet - calculated
Outlet - observed
0
10
20
30
40
50
04/04/05 02/05/05 30/05/05 27/06/05
N -
NO
x (m
g/L)
Inlet - observedOutlet - calculatedOutlet - observed
Figure3. Effluent nitrogen species : observed and simulated values

197
0
20
40
60
80
100
120
06/04/05 26/04/05 16/05/05 05/06/05 25/06/05
N tr
ansf
orm
atio
ns (
Kg
N d
-1)
0
5
10
15
20
25
30
35
Tem
pera
ture
(°C
)
Volatilisation
Denitrification
Nitrogen losses
Temperature
Figure 4. Nitrogen losses (simulated values)
[NO2-3-N] started increasing at the end of May when the temperature exceeded 22°C. The filtered COD of the inlet water was lower than 80 mg.L-1 and, though highly variable from one day to another and all along a 24 hours day (Figure 5), DO content, with average daily values around 6 mg.L-1, was sufficient to allow the development of nitrification within the pond. Temperature increase seems to have been the key factor of the nitrification development.
0
5
10
15
20
25
30
22/6/05 24/6/05 26/6/05 28/6/05 30/6/05 2/7/05 4/7/05 6/7/05 8/7/05
DO
(m
g/L)
Figure 5. Dissolved oxygen variations
After June 10th, [NO2-3-N] decreased, which was not offset by any ammonia or [Org-N] augmentation. Simultaneously, dramatic nitrogen losses were observed. The most probable explanation of these losses was denitrification. However, several conditions are required for
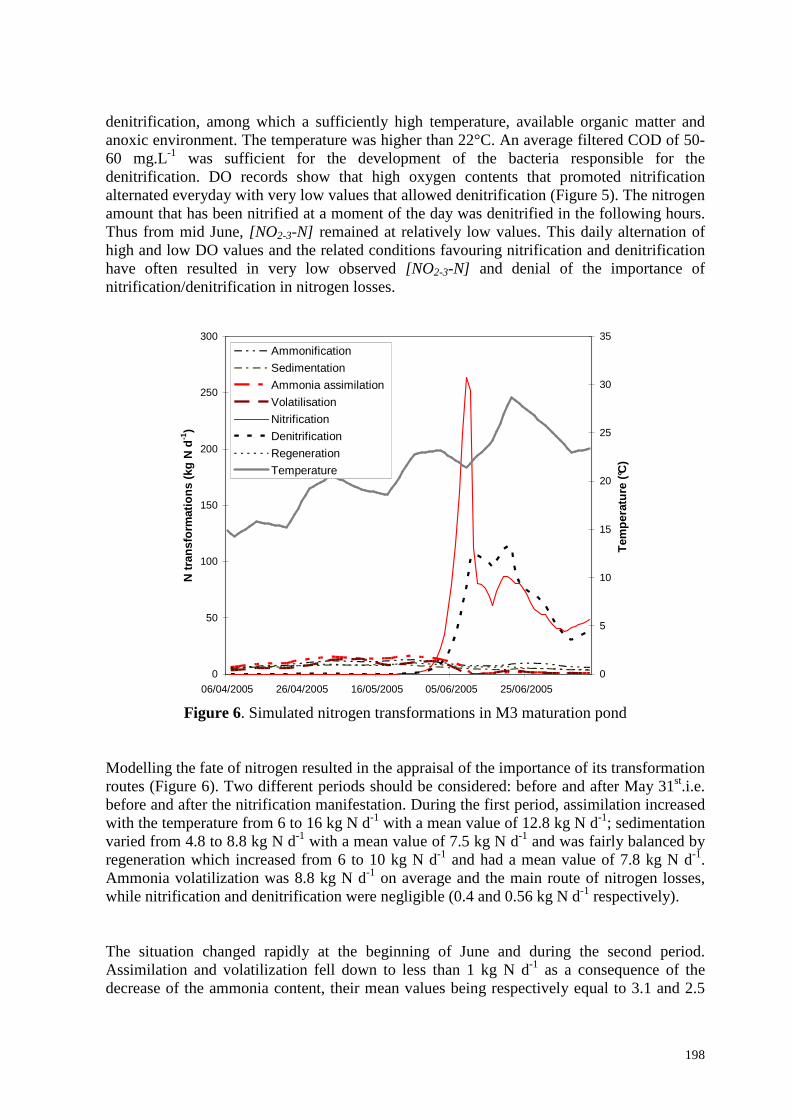
198
denitrification, among which a sufficiently high temperature, available organic matter and anoxic environment. The temperature was higher than 22°C. An average filtered COD of 50-60 mg.L-1 was sufficient for the development of the bacteria responsible for the denitrification. DO records show that high oxygen contents that promoted nitrification alternated everyday with very low values that allowed denitrification (Figure 5). The nitrogen amount that has been nitrified at a moment of the day was denitrified in the following hours. Thus from mid June, [NO2-3-N] remained at relatively low values. This daily alternation of high and low DO values and the related conditions favouring nitrification and denitrification have often resulted in very low observed [NO2-3-N] and denial of the importance of nitrification/denitrification in nitrogen losses.
0
50
100
150
200
250
300
06/04/2005 26/04/2005 16/05/2005 05/06/2005 25/06/2005
N tr
ansf
orm
atio
ns (
kg N
d-1
)
0
5
10
15
20
25
30
35
Tem
pera
ture
(°C
)
Ammonification
Sedimentation
Ammonia assimilation
Volatilisation
Nitrification
Denitrification
Regeneration
Temperature
Figure 6. Simulated nitrogen transformations in M3 maturation pond
Modelling the fate of nitrogen resulted in the appraisal of the importance of its transformation routes (Figure 6). Two different periods should be considered: before and after May 31st.i.e. before and after the nitrification manifestation. During the first period, assimilation increased with the temperature from 6 to 16 kg N d-1 with a mean value of 12.8 kg N d-1; sedimentation varied from 4.8 to 8.8 kg N d-1 with a mean value of 7.5 kg N d-1 and was fairly balanced by regeneration which increased from 6 to 10 kg N d-1 and had a mean value of 7.8 kg N d-1. Ammonia volatilization was 8.8 kg N d-1 on average and the main route of nitrogen losses, while nitrification and denitrification were negligible (0.4 and 0.56 kg N d-1 respectively). The situation changed rapidly at the beginning of June and during the second period. Assimilation and volatilization fell down to less than 1 kg N d-1 as a consequence of the decrease of the ammonia content, their mean values being respectively equal to 3.1 and 2.5

199
kg N d-1. Sedimentation was around 4.7 kg N d-1 and nearly balanced by regeneration, the mean value of which being 5.8 kg N d-1. Nitrification was the paramount nitrogen transformation; as soon as the conditions could favor its development, nitrifying bacteria rapidly multiplied and could nitrify within a few days the bulk of the ammonia present in pond M3. Afterwards, nitrification was limited to the oxidation of the ammonia entering the pond. Peak and average values of nitrification were respectively 260 and 76 kg N d-1 during the second period. Denitrication, the mean value of which was 64 kg N d-1, soon followed nitrification and eliminated most of the nitrate. CONCLUSION A dynamic mathematical model was elaborated to simulate the nitrogen transformations in a maturation pond. The model together with an experimental independent evaluation of volatilization provided consistent explanations of the nitrogen losses. In April and May, volatilization was by far the main route for nitrogen losses with a mean rate of 484 mg N.m-
2d-1, a much higher value than those measured by Zimmo et al (2003). As soon as temperature reached 22°C, nitrification combined with denitrification was the main if not the only mechanism of nitrogen removal. The mean denitrification rate over the period of June and July was 3,300 mg N.m-2d-1. About 82% of total nitrogen removal was accomplished through nitrification and denitrification in the survey period. REFERENCES Azov, Y. and Tregubova, T. (1995). Nitrification processes in stabilisation reservoirs. Wat. Sci.Tech.,
31(12), 313-319. Brissaud, F., Andrianarison, T., Brouillet, J. L. and Picot, B. (2005). Twenty years' monitoring of
Meze stabilisation ponds: part II - removal of faecal indicators. Wat. Sci. Tech., 51(12), 33-41. Ferrara R.A. and Harleman D.R.F. (1980). Dynamic nutrient cycle model for waste stabilization
ponds. J. of the Environmental Engineering Division 106(1), 37-54. Ferrara R.A. et Avci C.B., 1982. Nitrogen dynamics in waste stabilisation ponds. J. Wat. Pollut.
Control Fed., 54, 361–369. Fritz J.J., Middleton A.C. et Meredith D.D., 1979. Dynamic process modeling wastewater
stabilization ponds. J. Wat. Pollut. Control Fed. 51(11), 2724-2743. Gomez, E., Casellas, C., Picot, B. and Bontoux, J. (1995). Ammonia elimination processes in
stabilisation and high-rate algal pond systems. Wat. Sci. Tech., 31(12), 303-312. Mayo, A. W. and Mutamba, J. (2005). Modelling nitrogen removal in a coupled HRP and unplanted
horizontal flow subsurface gravel bed constructed wetland. Physics and Chemistry of the Earth, 30(11-16), 673-679.
Mclean, B. M., Baskaran, K. and Connor, M. A. (2000). The use of algal bacterial biofilms to enhance nitrification rates in lagoons: experience under laboratory and pilot-scale conditions. Wat. Sci. Tech., 42(10-11), 187-194.
Picot, B., Andrianarison, T., Gosselin, J. P. and Brissaud, F. (2005a). Twenty years' monitoring of Meze stabilisation ponds: Part I - Removal of organic matter and nutrients. Wat. Sci. Tech., 51(12), 23-31.
Picot, B., Sambuco, J. P., Brouillet, J. L. and Riviere, Y. (2005b). Wastewater stabilisation ponds: sludge accumulation, technical and financial study on desludging and sludge disposal case studies in France. Wat. Sci. Tech., 51(12), 227-234.
Racault, Y., Boutin, C. and Seguin, A. (1995). Waste stabilization ponds in France: A report on fifteen years experience. Wat. Sci. Tech., 31(12), 91-101.

200
Senzia, M. A., Mayo, A. W., Mbwette, T. S. A., Katima, J. H. Y. and Jorgensen, S. E. (2002). Modelling nitrogen transformation and removal in primary facultative ponds. Ecol. Model., 154(3), 207-215.
Silva, S. A., De Oliveira, R., Soares, J., Mara, D. D. and Pearson, H. W. (1995). Nitrogen removal in pond systems with different configurations and geometries. Wat. Sci. Tech., 31(12), 321-330.
Soares, J., Silva, S. A., De Oliveira, R., Araujo, A. L. C., Mara, D. D. and Pearson, H. W. (1996). Ammonia removal in a pilot-scale WSP complex in northeast Brazil. Wat. Sci. Tech., 33(7), 165-171.
Stratton F.E., 1968. Ammonia Nitrogen losses from streams. Journal of the Sanitary Engineering Division, ASCE, 94, 1085-1092.
Stratton F.E., 1969. Nitrogen losses from alkaline water impoudments. Journal of the Sanitary Engineering Division, ASCE, 95, 223-231.
Wiesmann U., 1994. Biological nitrogen removal from wastewater. Advances in Biochemical Engineering – Biotechnology, 51, 113-154.
Zimmo, O. R., Van Der Steen, N. P. and Gijzen, H. J. (2003). Comparison of ammonia volatilisation rates in algae and duckweed-based waste stabilisation ponds treating domestic wastewater. Wat. Res., 37(19), 4587-4594.