synthese et secretion des proteines...
TRANSCRIPT

SYNTHESE ET SECRETION
DES PROTEINES
L’information génétique est
exprimée,
maintenue,
répliquée et
quelquefois améliorée
par des processus spécifiques:
Synthèse de l’ARN et des protéines,
Réparation de l’ADN,
Réplication de l’ADN et
Recombinaison génétique

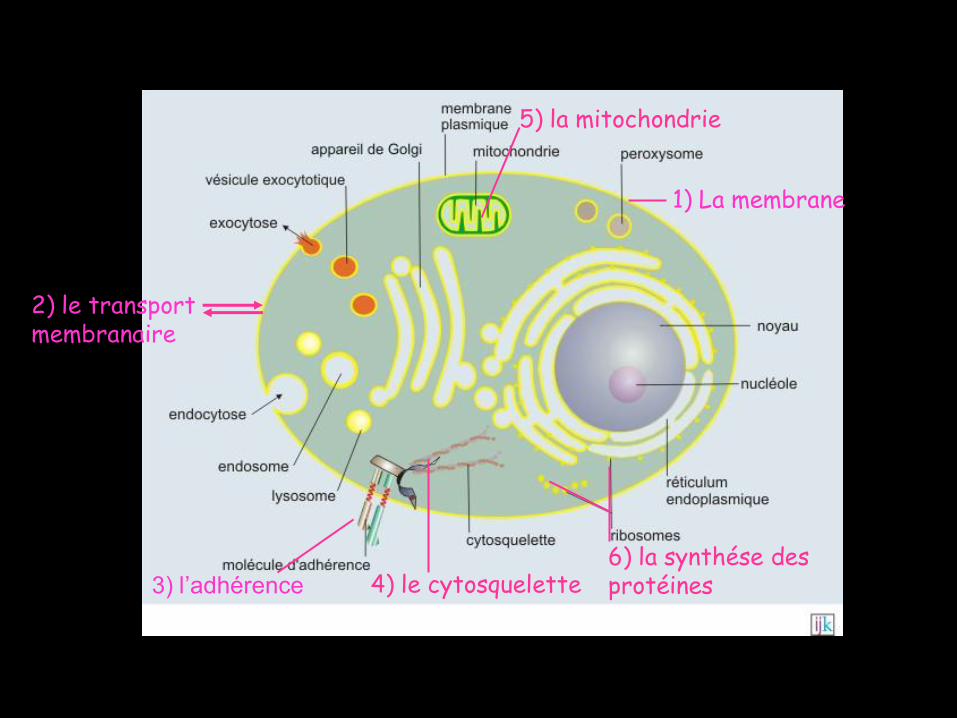
2) le transport membranaire
1) La membrane
3) l’adhérence 4) le cytosquelette
5) la mitochondrie
6) la synthése des protéines
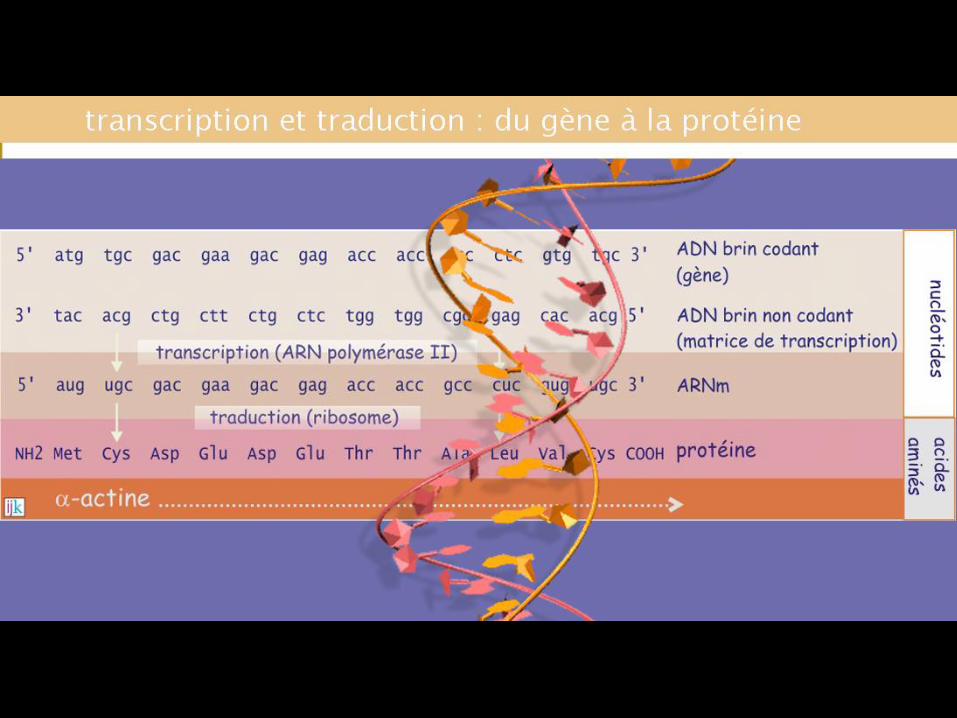


MATERIAUX REQUIS
-“briques” – aminoacides
-“transporteurs” – ARNt specifiques
-“constructeurs” – ribosomes
- “le plan”, ARNm
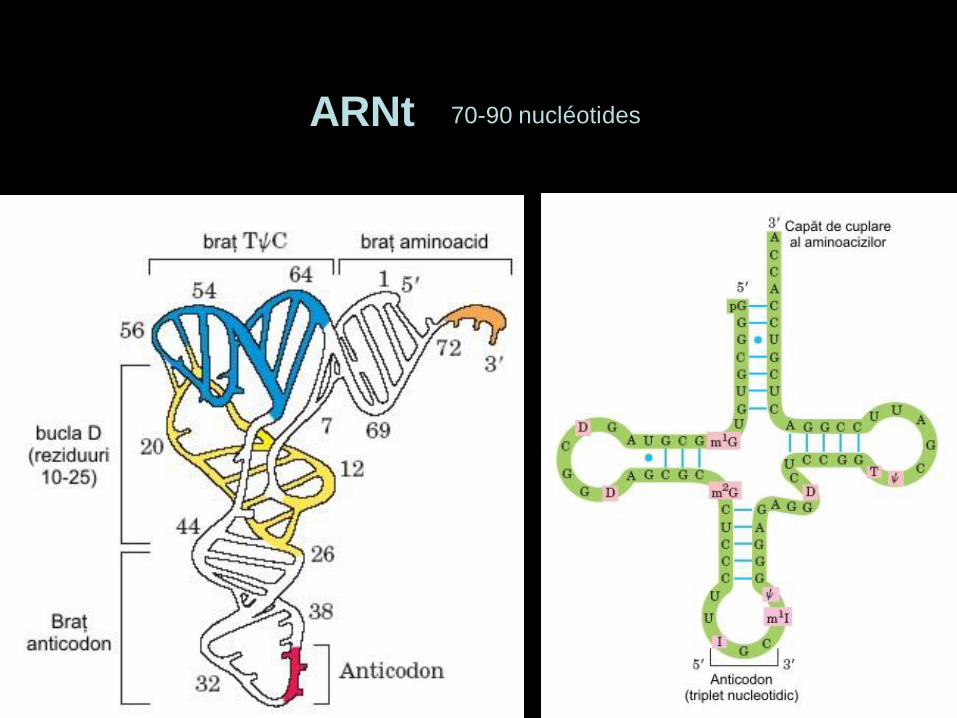
ARNt 70-90 nucléotides
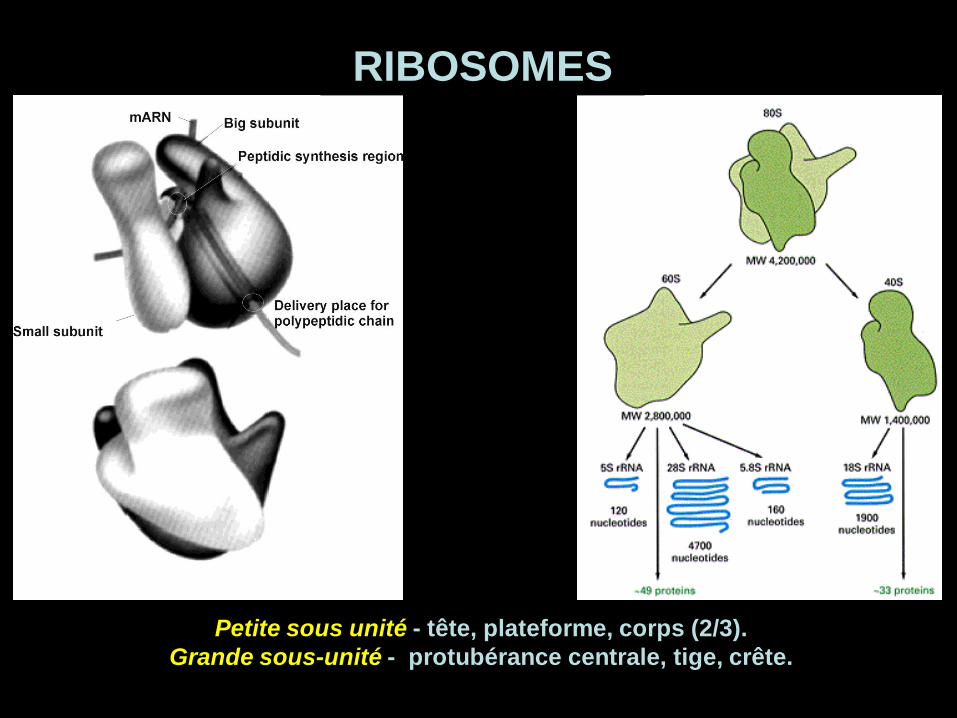
Petite sous unité - tête, plateforme, corps (2/3).
Grande sous-unité - protubérance centrale, tige, crête.
RIBOSOMES
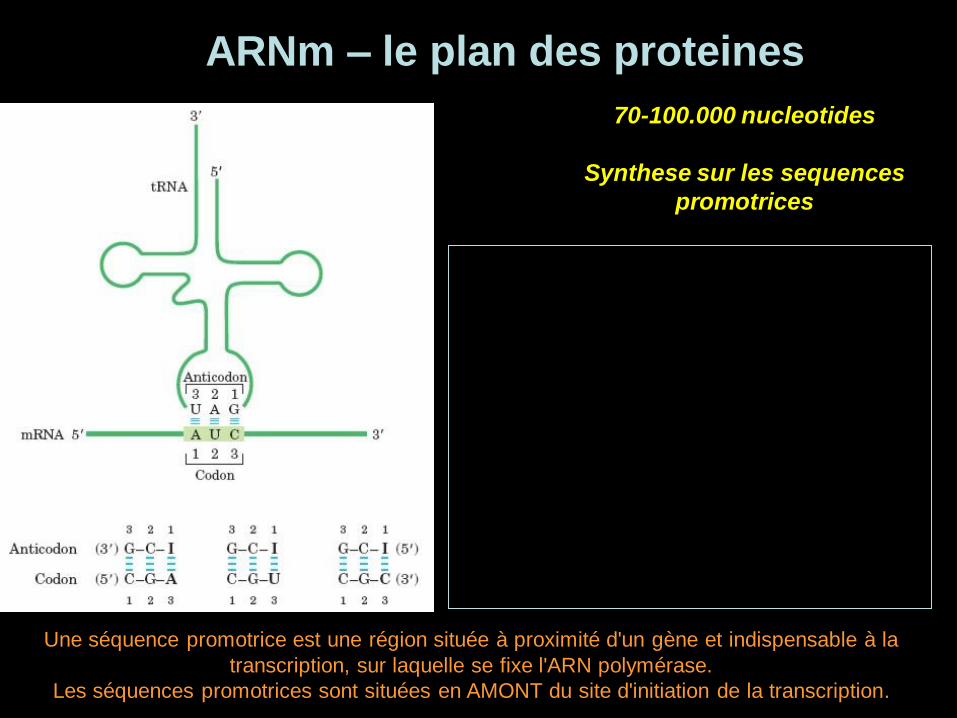
70-100.000 nucleotides
Synthese sur les sequences
promotrices
ARNm – le plan des proteines
Une séquence promotrice est une région située à proximité d'un gène et indispensable à la
transcription, sur laquelle se fixe l'ARN polymérase.
Les séquences promotrices sont situées en AMONT du site d'initiation de la transcription.
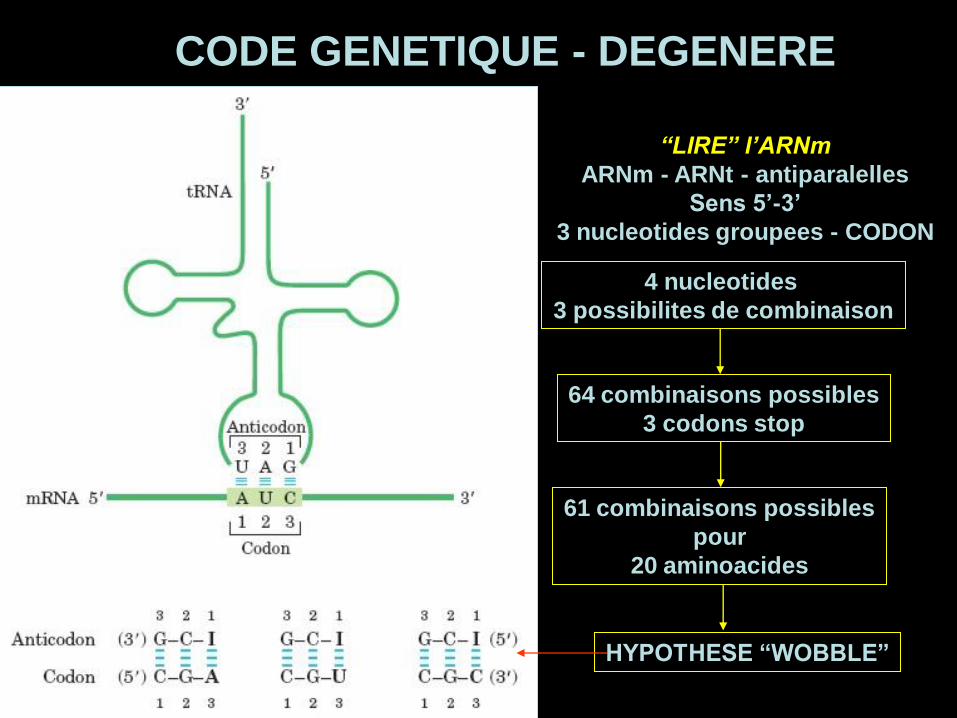
“LIRE” l’ARNm
ARNm - ARNt - antiparalelles
Sens 5’-3’
3 nucleotides groupees - CODON
CODE GENETIQUE - DEGENERE
4 nucleotides
3 possibilites de combinaison
64 combinaisons possibles
3 codons stop
61 combinaisons possibles
pour
20 aminoacides
HYPOTHESE “WOBBLE”

•Le principe de la biologie cellulaire: ADN → ARNm → AA → PROTEINES
•Aminoacides – spécifiés par les codons sur l’ARNm = triplets de nucléotides.
•Traduction - molécule adaptrice = ARNt – anticodons – qui corresponds au
codons sur l’ARNm. ARNt vont insérer les aminoacides dans leur positions
spécifiques dans la chaine peptidique.
•AUG – START - UAA, UAG et UGA - STOP.
•Le code génétique standard est dégénéré: il comprend plusieurs mots code
pour presque chaque aminoacide.
•Les mot code sont universels pour toutes les espèces, exception pour les
mitochondries et quelques unicellulaires.
•La troisième position du codon est moins spécifique.
RESUME
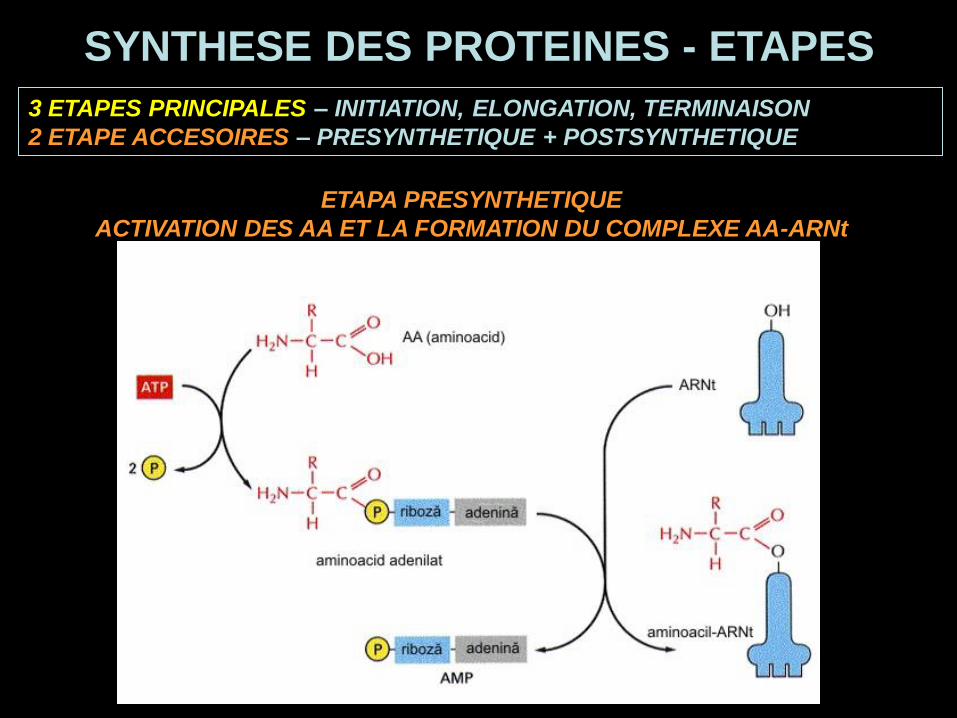
3 ETAPES PRINCIPALES – INITIATION, ELONGATION, TERMINAISON
2 ETAPE ACCESOIRES – PRESYNTHETIQUE + POSTSYNTHETIQUE
SYNTHESE DES PROTEINES - ETAPES
ETAPA PRESYNTHETIQUE
ACTIVATION DES AA ET LA FORMATION DU COMPLEXE AA-ARNt
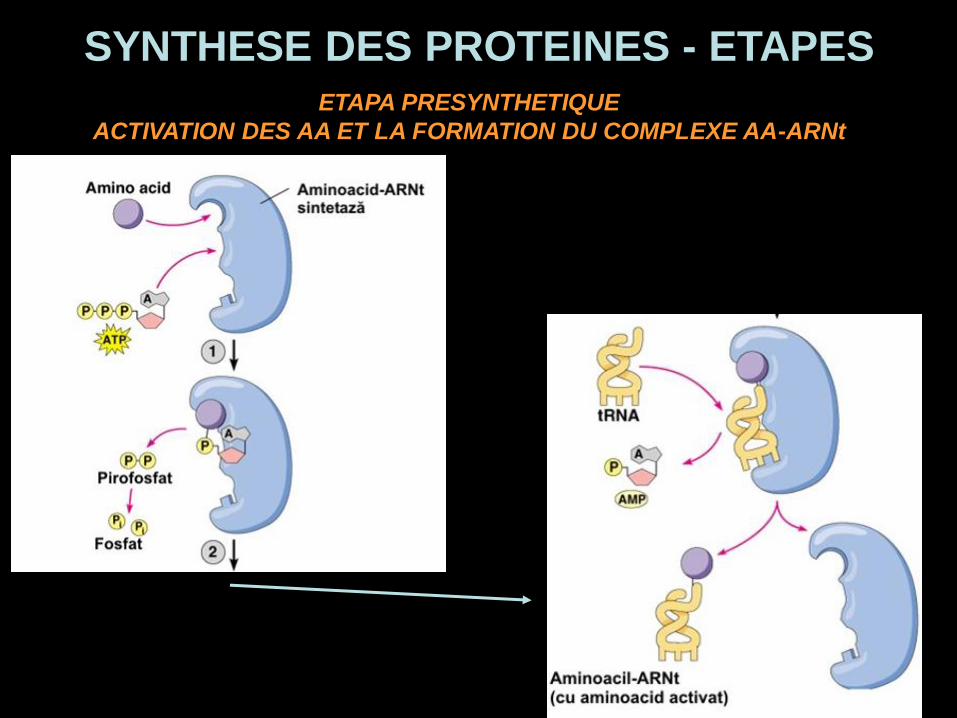
SYNTHESE DES PROTEINES - ETAPESETAPA PRESYNTHETIQUE
ACTIVATION DES AA ET LA FORMATION DU COMPLEXE AA-ARNt
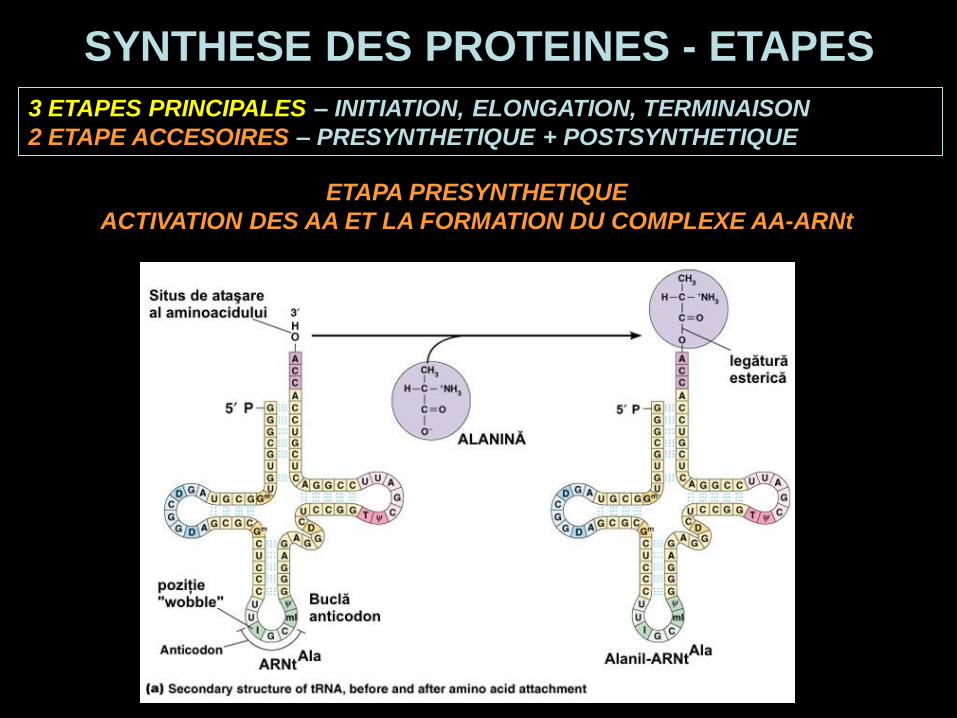
ETAPA PRESYNTHETIQUE
ACTIVATION DES AA ET LA FORMATION DU COMPLEXE AA-ARNt
3 ETAPES PRINCIPALES – INITIATION, ELONGATION, TERMINAISON
2 ETAPE ACCESOIRES – PRESYNTHETIQUE + POSTSYNTHETIQUE
SYNTHESE DES PROTEINES - ETAPES

ETAPE D’INITIATION
3 ETAPES PRINCIPALES – INITIATION, ELONGATION, TERMINAISON
2 ETAPE ACCESOIRES – PRESYNTHETIQUE + POSTSYNTHETIQUE
SYNTHESE DES PROTEINES - ETAPES
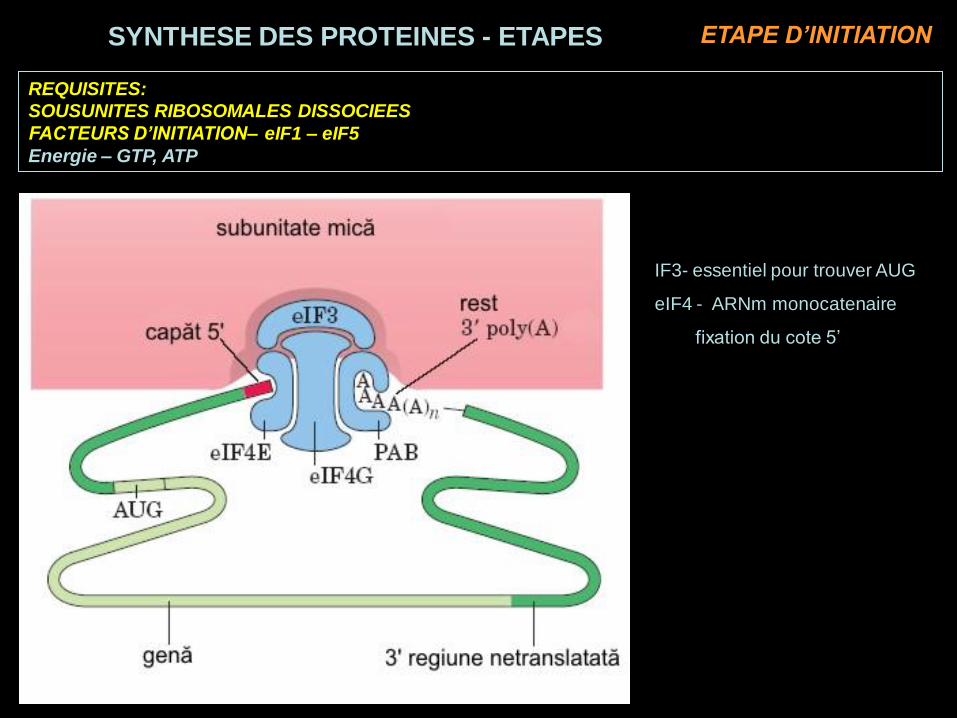
REQUISITES:
SOUSUNITES RIBOSOMALES DISSOCIEES
FACTEURS D’INITIATION– eIF1 – eIF5
Energie – GTP, ATP
IF3- essentiel pour trouver AUG
eIF4 - ARNm monocatenaire
fixation du cote 5’
SYNTHESE DES PROTEINES - ETAPES ETAPE D’INITIATION

REQUISITES:
SOUSUNITES RIBOSOMALES DISSOCIEES
FACTEURS D’INITIATION– eIF1 – eIF5
Energie – GTP, ATP
SYNTHESE DES PROTEINES - ETAPES ETAPE D’INITIATION
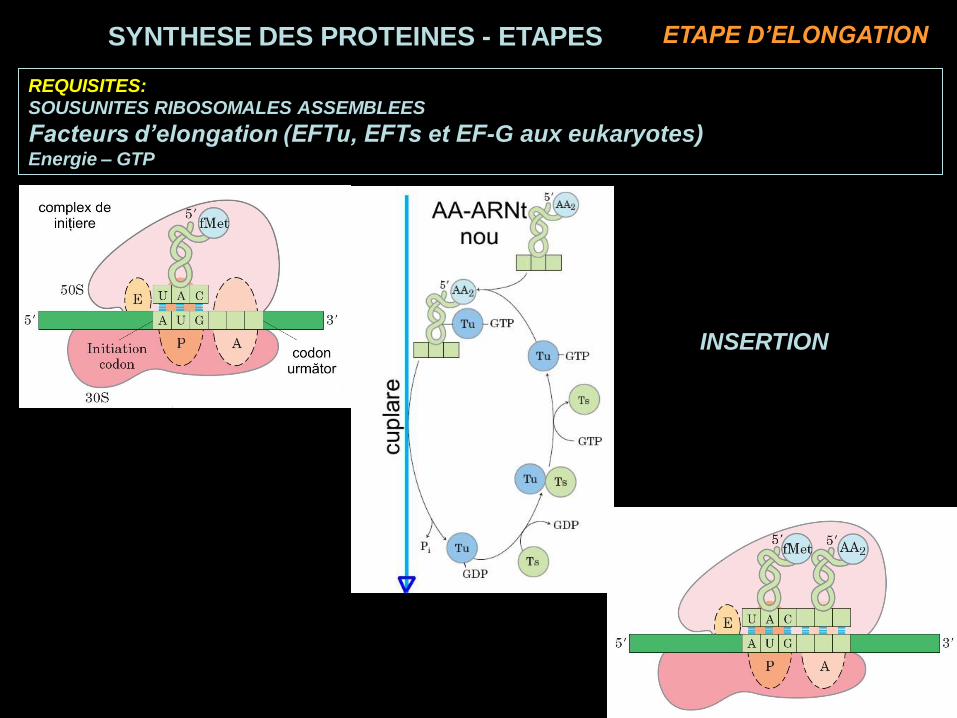
REQUISITES:
SOUSUNITES RIBOSOMALES ASSEMBLEES
Facteurs d’elongation (EFTu, EFTs et EF-G aux eukaryotes)Energie – GTP
INSERTION
SYNTHESE DES PROTEINES - ETAPES ETAPE D’ELONGATION
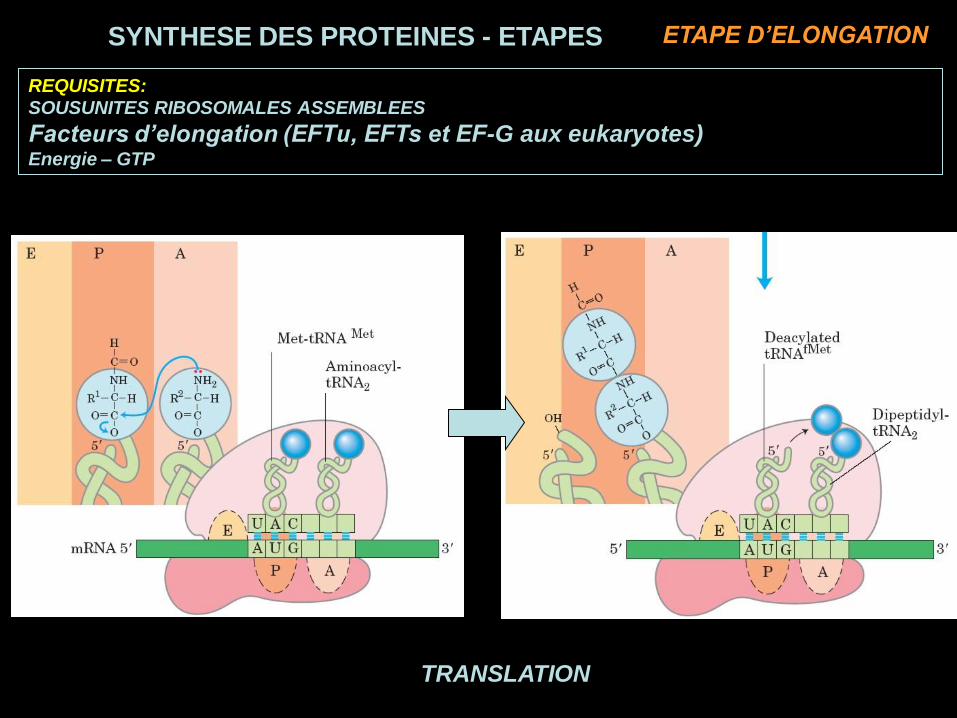
TRANSLATION
REQUISITES:
SOUSUNITES RIBOSOMALES ASSEMBLEES
Facteurs d’elongation (EFTu, EFTs et EF-G aux eukaryotes)Energie – GTP
SYNTHESE DES PROTEINES - ETAPES ETAPE D’ELONGATION
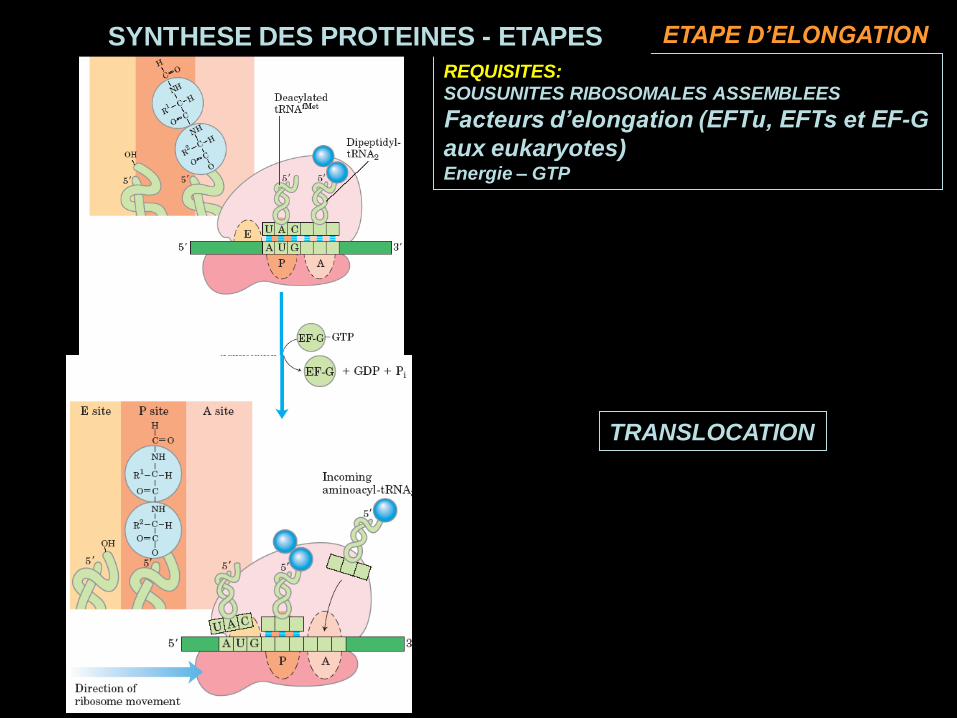
TRANSLOCATION
REQUISITES:
SOUSUNITES RIBOSOMALES ASSEMBLEES
Facteurs d’elongation (EFTu, EFTs et EF-G
aux eukaryotes)Energie – GTP
SYNTHESE DES PROTEINES - ETAPES ETAPE D’ELONGATION

REQUISITES:
SOUSUNITES RIBOSOMALES ASSEMBLEES
Facteurs d’elongation (EFTu, EFTs et EF-G aux
eukaryotes)
Energie – GTP
SYNTHESE DES PROTEINES - ETAPESTRANSLOCATION
ETAPE D’ELONGATION
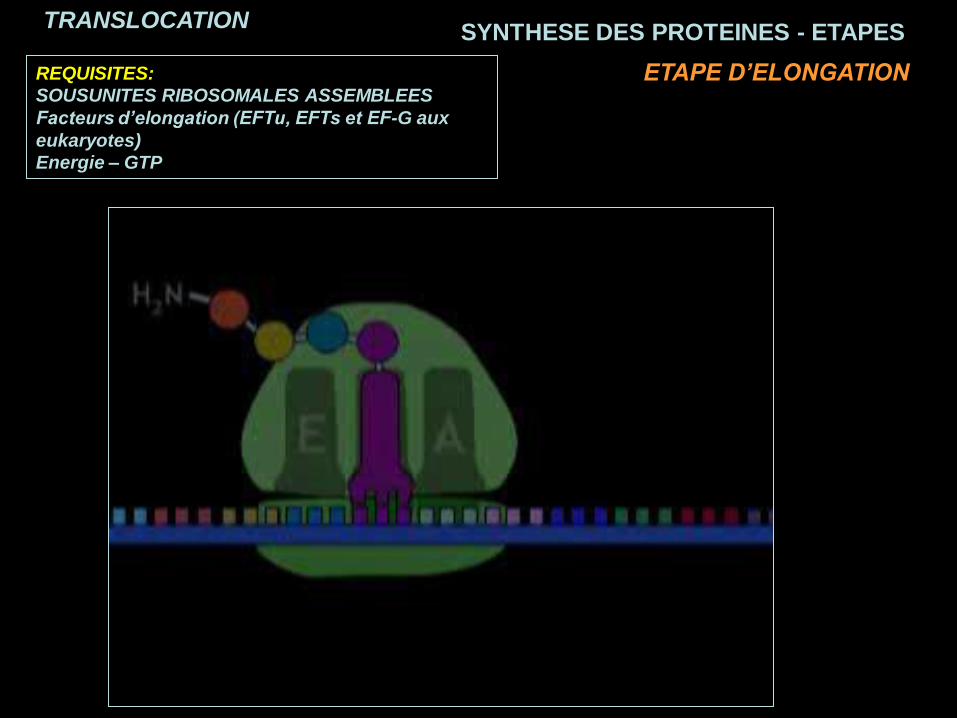
REQUISITES:
SOUSUNITES RIBOSOMALES ASSEMBLEES
Facteurs d’elongation (EFTu, EFTs et EF-G aux
eukaryotes)
Energie – GTP
SYNTHESE DES PROTEINES - ETAPESTRANSLOCATION
ETAPE D’ELONGATION
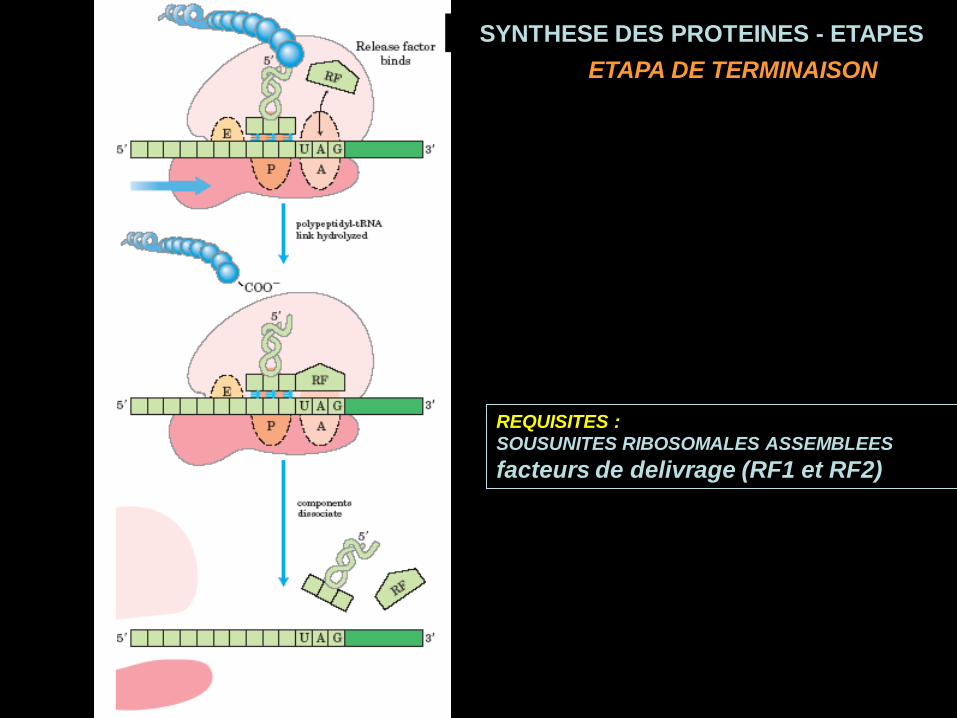
REQUISITES :
SOUSUNITES RIBOSOMALES ASSEMBLEES
facteurs de delivrage (RF1 et RF2)
ETAPA DE TERMINAISON
SYNTHESE DES PROTEINES - ETAPES
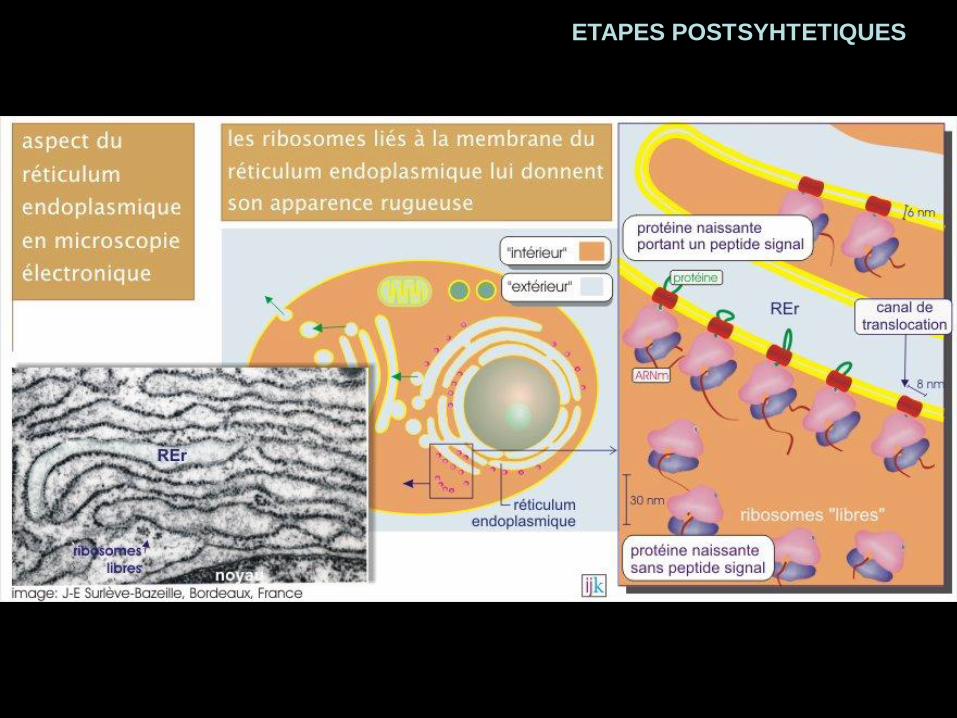
ETAPES POSTSYHTETIQUES
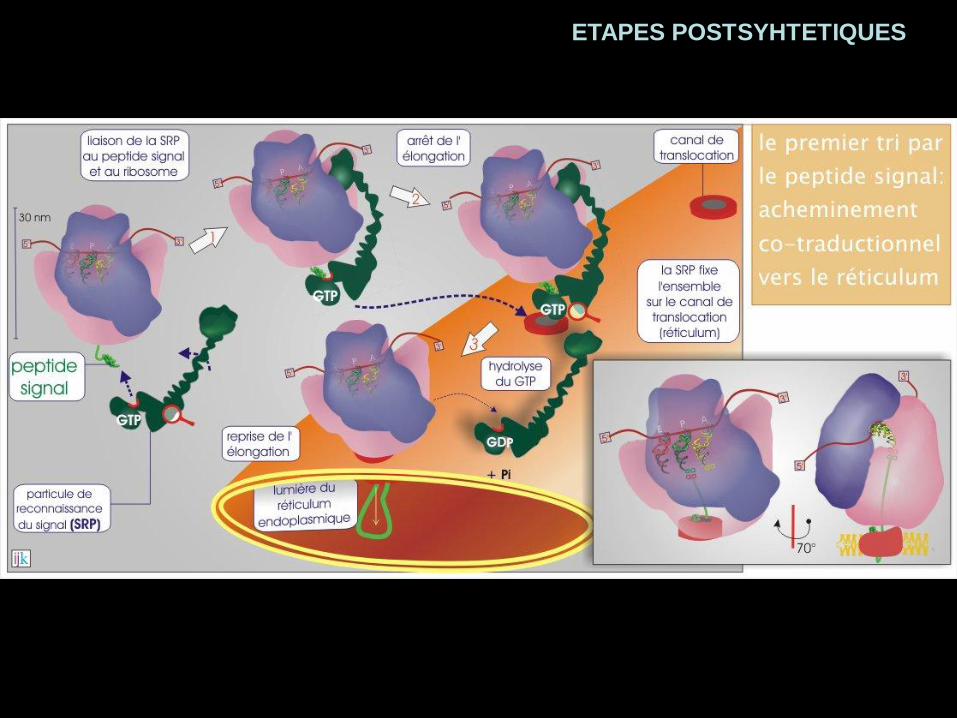
ETAPES POSTSYHTETIQUES
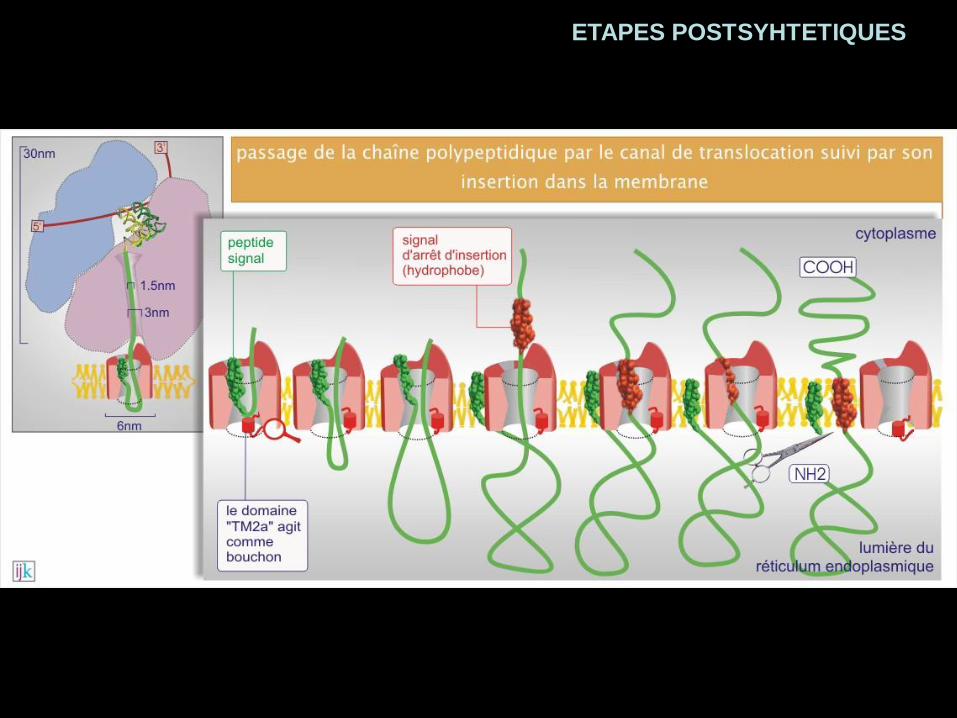
ETAPES POSTSYHTETIQUES
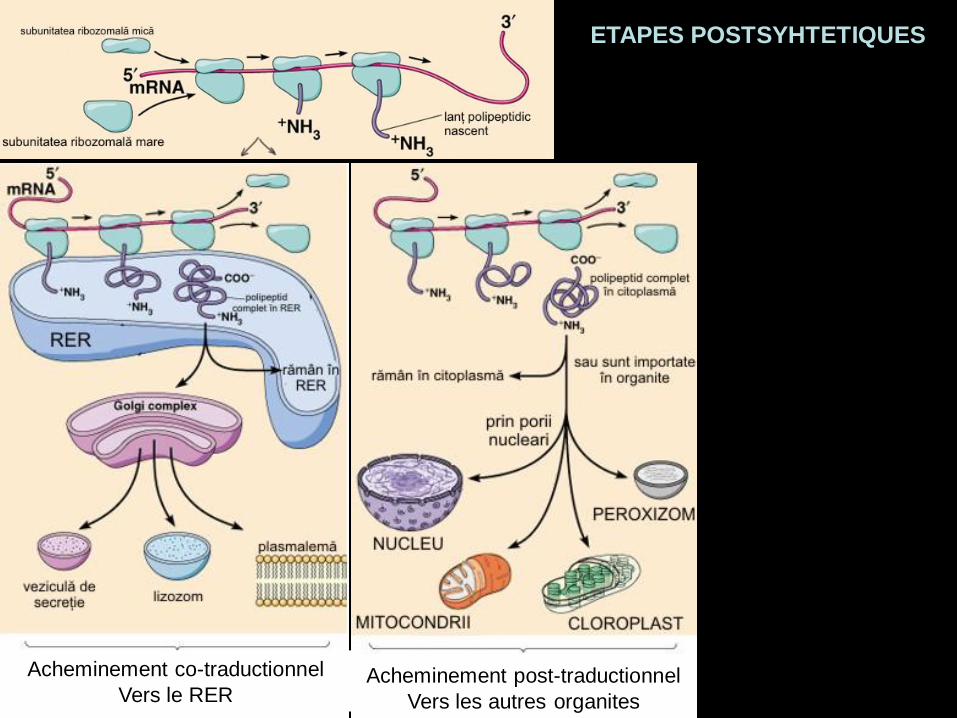
Acheminement co-traductionnel
Vers le RERAcheminement post-traductionnel
Vers les autres organites
ETAPES POSTSYHTETIQUES

RETICULUM ENDOPLASMIQUE
RUGUEUX
(RER)
MO
- formation basophile perinucleaire.
- position basale
- hépatocytes – autour du noyau - corps Berg
- neurones - corps Nissl.
- membranes RER se continuent a la membrane nucléaire externe et l’interieur du RER se continue avec l’espace perinucleaire
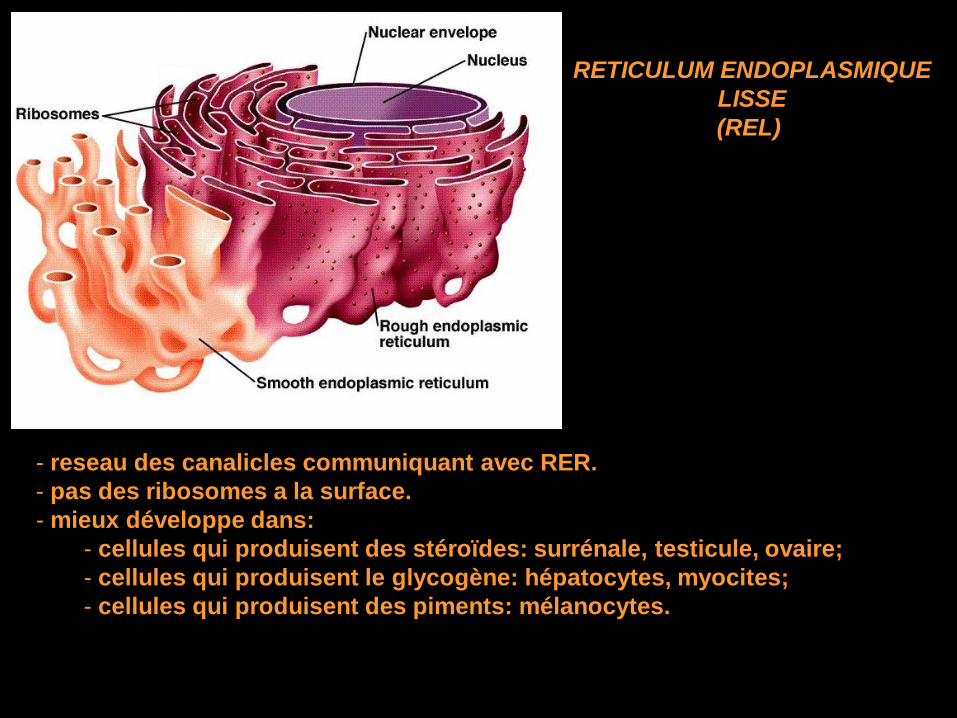
- reseau des canalicles communiquant avec RER.
- pas des ribosomes a la surface.
- mieux développe dans:
- cellules qui produisent des stéroïdes: surrénale, testicule, ovaire;
- cellules qui produisent le glycogène: hépatocytes, myocites;
- cellules qui produisent des piments: mélanocytes.
RETICULUM ENDOPLASMIQUE
LISSE
(REL)
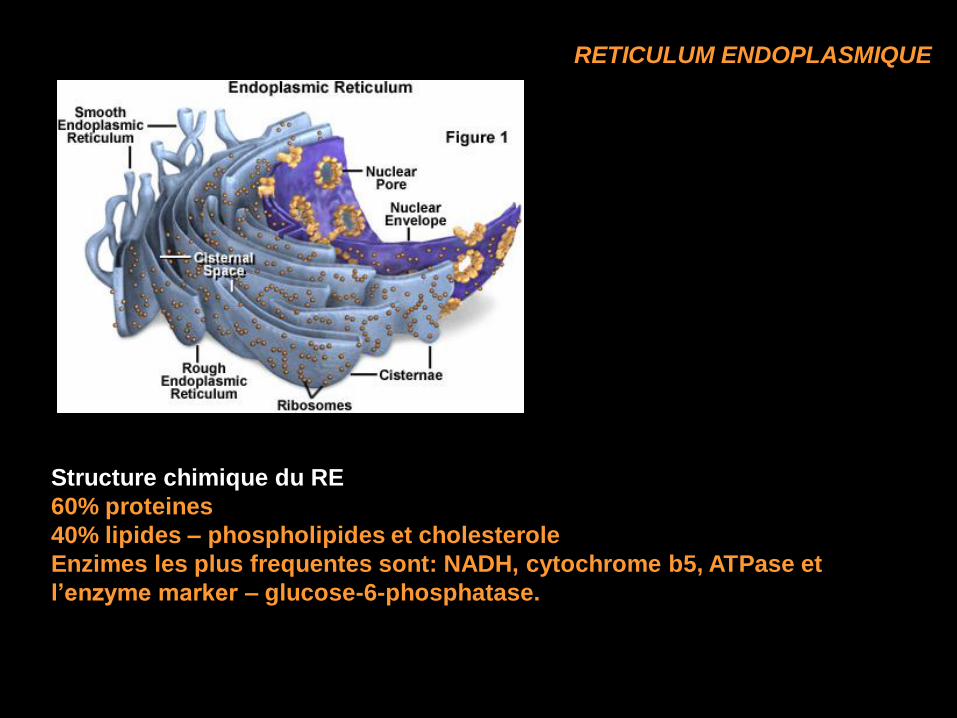
RETICULUM ENDOPLASMIQUE
Structure chimique du RE
60% proteines
40% lipides – phospholipides et cholesterole
Enzimes les plus frequentes sont: NADH, cytochrome b5, ATPase et
l’enzyme marker – glucose-6-phosphatase.

Fonctions RE
Fonctions specifiques RER
- synthese des proteines par des ribosomes attaches;
- glycosylation de la chaine polypeptidique;
- changements des chaines laterales par la formation des liaisons
disulfidiques;
Fonctions specifiques REN
- synthese des lipides, dans les gonades, intestin;
- detoxification, par des enzymes; r. de oxydation, hidrolise, reduction ou
conjugation;
- delivrer la glucose dans les hepatpcytes: enzima marker glucose-6-
phosphatase.
Fonctions communes RER et REN
- RE – systeme circulatoire intracytoplasmique
- RE - synthese des phospholipides;
- RE – support mechanique pour la membrane;
- RE – fabrique des membranes
RETICULUM ENDOPLASMIQUE
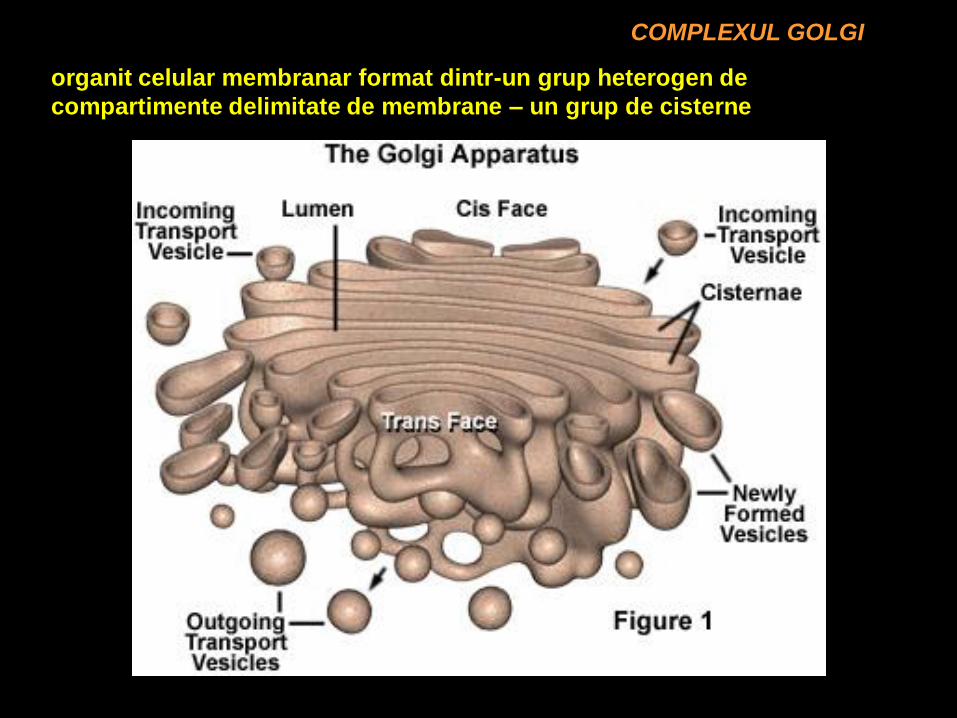
COMPLEXUL GOLGI
organit celular membranar format dintr-un grup heterogen de
compartimente delimitate de membrane – un grup de cisterne
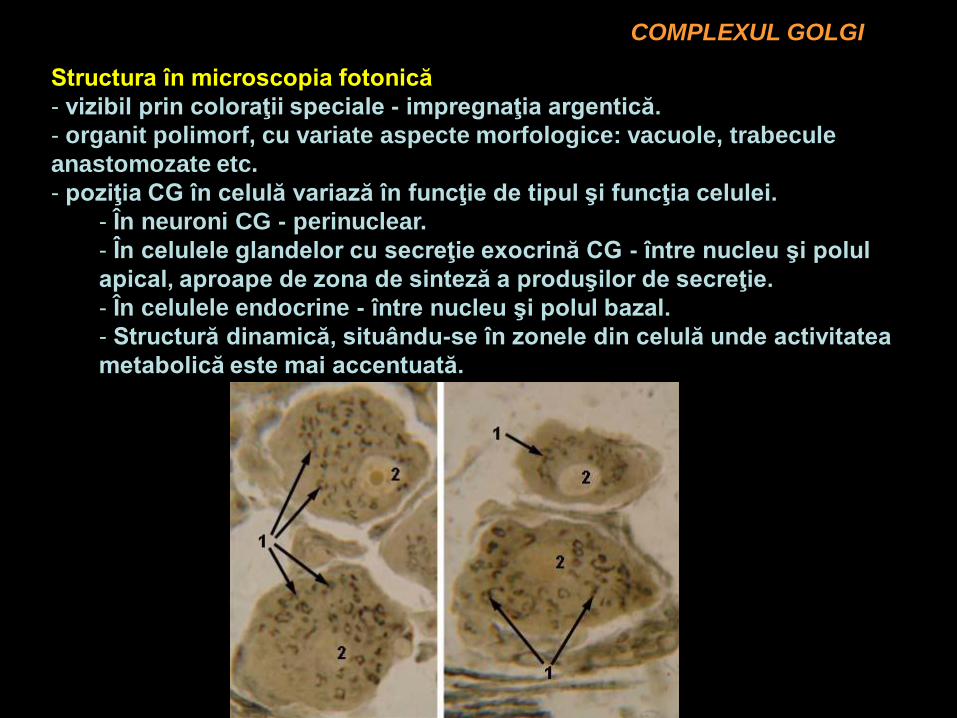
COMPLEXUL GOLGI
Structura în microscopia fotonică
- vizibil prin coloraţii speciale - impregnaţia argentică.
- organit polimorf, cu variate aspecte morfologice: vacuole, trabecule
anastomozate etc.
- poziţia CG în celulă variază în funcţie de tipul şi funcţia celulei.
- În neuroni CG - perinuclear.
- În celulele glandelor cu secreţie exocrină CG - între nucleu şi polul
apical, aproape de zona de sinteză a produşilor de secreţie.
- În celulele endocrine - între nucleu şi polul bazal.
- Structură dinamică, situându-se în zonele din celulă unde activitatea
metabolică este mai accentuată.
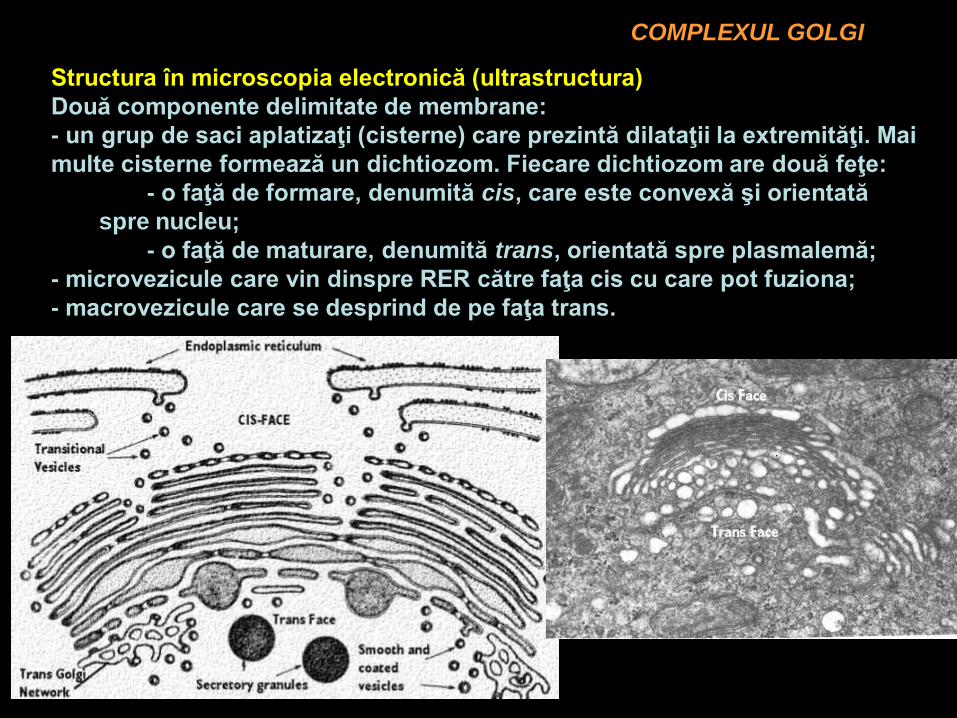
COMPLEXUL GOLGI
Structura în microscopia electronică (ultrastructura)
Două componente delimitate de membrane:
- un grup de saci aplatizaţi (cisterne) care prezintă dilataţii la extremităţi. Mai
multe cisterne formează un dichtiozom. Fiecare dichtiozom are două feţe:
- o faţă de formare, denumită cis, care este convexă şi orientată
spre nucleu;
- o faţă de maturare, denumită trans, orientată spre plasmalemă;
- microvezicule care vin dinspre RER către faţa cis cu care pot fuziona;
- macrovezicule care se desprind de pe faţa trans.

COMPLEXUL GOLGI
Fonctions CG
Fonctions în secreţia celulară
- Formarea de granule de secreţie;
- Glicozilarea terminală a proteinelor: produşii de secreţie proveniţi din RE
sunt glicozilaţi terminal în prezenţa glicozil-transferazei şi -manozidazei;
- Glicozilarea gangliozidelor şi cerebrozidelor are loc în celulele din creier şi
rinichi şi este asistată de glicoziltransferază;
- Sulfatarea produşilor proveniţi din RE, în prezenţa sulfotransferazelor: CG
are un rol important în secreţia mucopolizaharidelor;
- Concentrarea produşilor de secreţie: are loc în sacii CG;
- Maturarea produşilor de secreţie: proinsulina este transformată în insulină
-Biogeneza lizozomilor: enzimele lizozomale prezintă un marker, manoză-6-
fosfat, pentru care există receptori la nivelul zonelor dilatate din coarnele
CG. Aici enzimele sunt împachetate în vezicule care se desprind ca lizozomi
primari.
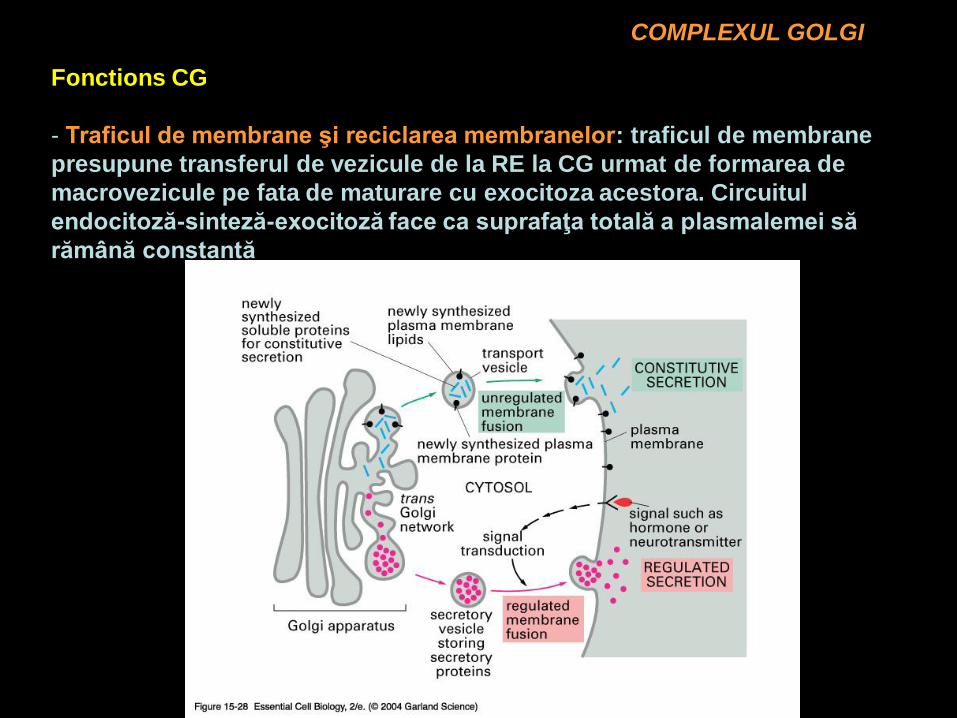
COMPLEXUL GOLGI
Fonctions CG
- Traficul de membrane şi reciclarea membranelor: traficul de membrane
presupune transferul de vezicule de la RE la CG urmat de formarea de
macrovezicule pe fata de maturare cu exocitoza acestora. Circuitul
endocitoză-sinteză-exocitoză face ca suprafaţa totală a plasmalemei să
rămână constantă