rotavirus viroplasm proteins interact with the cellular
TRANSCRIPT

Rotavirus Viroplasm Proteins Interact with the Cellular SUMOylationSystem: Implications for Viroplasm-Like Structure Formation
Michela Campagna,a Laura Marcos-Villar,a Francesca Arnoldi,b Carlos F. de la Cruz-Herrera,a Pedro Gallego,a
José González-Santamaría,a Dolores González,a Fernando Lopitz-Otsoa,c Manuel S. Rodriguez,c,d Oscar R. Burrone,e Carmen Rivasa
Centro Nacional de Biotecnología, CSIC, Madrid, Spaina; Dipartimento Universitario Clinico di Scienze Mediche, Chirurgiche e della Salute, University of Trieste, Trieste,Italyb; Proteomics Unit, CIC bioGUNE, CIBERehd, Derio, Spainc; Ubiquitylation and Cancer Molecular Biology laboratory, Inbiomed, San Sebastian-Donostia, Gipuzkoa,Spaind; International Centre for Genetic Engineering and Biotechnology, Trieste, Italye
Posttranslational modification by SUMO provides functional flexibility to target proteins. Viruses interact extensively with thecellular SUMO modification system in order to improve their replication, and there are numerous examples of viral proteinsthat are SUMOylated. However, thus far the relevance of SUMOylation for rotavirus replication remains unexplored. In thisstudy, we report that SUMOylation positively regulates rotavirus replication and viral protein production. We show that SUMOcan be covalently conjugated to the viroplasm proteins VP1, VP2, NSP2, VP6, and NSP5. In addition, VP1, VP2, and NSP2 canalso interact with SUMO in a noncovalent manner. We observed that an NSP5 SUMOylation mutant protein retains most of itsactivities, such as its interaction with VP1 and NSP2, the formation of viroplasm-like structures after the coexpression withNSP2, and the ability to complement in trans the lack of NSP5 in infected cells. However, this mutant is characterized by a highdegree of phosphorylation and is impaired in the formation of viroplasm-like structures when coexpressed with VP2. These re-sults reveal for the first time a positive role for SUMO modification in rotavirus replication, describe the SUMOylation of severalviroplasm resident rotavirus proteins, and demonstrate a requirement for NSP5 SUMOylation in the production of viroplasm-like structures.
Rotavirus, a member of the Reoviridae family, is the majoretiological cause of severe gastroenteritis of viral origin in
infants and young children. The infective virion consists of anonenveloped triple-layered particle (TLP). Inside the innerlayer, composed by pentamers of the structural protein VP2,are contained the 11 double-stranded RNA (dsRNA) segmentsof the viral genome, the RNA-dependent RNA polymeraseVP1, and the RNA capping enzyme VP3, altogether formingthe core of the virus. Around the core is present a second in-termediate layer, composed by the structural protein VP6,forming a double-layered particle (DLP) that is surrounded bythe third outermost layer composed by the proteins VP7 andVP4 forming the fully assembled infectious TLP.
Upon virus entry in the host cell, the outermost layer of thevirus is lost and DLPs become active in transcribing the viralmRNA from the dsRNA genome, acting VP1 also as a transcrip-tase. Even though it has been shown in vitro that the minimalrequirement for viral replication is represented by VP1 and VP2(1, 2), in vivo replication and packaging occur in viral factories,called viroplasms (3). These structures are formed, apart fromVP1 and VP2, also by the other structural proteins necessary forthe formation of the DLPs, VP3 and VP6, and two nonstructuralproteins, NSP2 and NSP5. Both nonstructural proteins are essen-tial for viroplasm formation and virus replication (4–6), but whileNSP2 has been proposed to be the molecular motor responsible ofthe packaging of rotavirus genome in newly synthesized cores (7,8), the role for NSP5 is less clear. The NSP5 protein, synthesized bythe smallest segment of rotavirus genome, has a molecular mass of26 kDa, a very high content of serine and threonine (25%), and alarge number of lysines at its C terminus. NSP5 is posttranslation-ally modified by O-GlcNAc glycosylation (9) and by extensivephosphorylation that causes, in infected cells, the appearance of asmear of bands that span up to 34 kDa (10, 11). NSP5 is able to
interact with the polymerase VP1 and NSP2 both in infected cellsand in cotransfection experiments (10, 12). In addition, cotrans-fection of mammalian cells with NSP5, together with NSP2 orVP2, causes the formation of spherical structures, resembling vi-roplasms, called viroplasm-like structures (VLS) known as VLS-NSP2i when NSP2-induced and VLS-VP2i when VP2-induced,respectively (13, 14). A recent study has shown that NSP5 is theonly viral protein necessary for the formation of VLS and therecruitment of all other viroplasmic proteins to these structures,suggesting a fundamental role for NSP5 in viroplasms formation(13). However, the mechanism through which NSP5 induces theformation of viroplasms has still to be clearly elucidated.
The small ubiquitin-like modifier (SUMO) is a molecule of11.5 kDa that is covalently bound to lysine residues of target pro-teins. Usually the target lysine is located in the consensus sequence�KxE (where � is a hydrophobic residue, and x is any residue) (15,16). However, SUMO can be also conjugated to lysine residueslocated in nonconsensus sequences. To date, four SUMO iso-forms have been discovered in mammals: SUMO1, the most sim-ilar to the yeast Smt3; SUMO2 and SUMO3, very similar to oneanother and characterized by an internal SUMOylation site thatallows the formation of SUMO chains; and SUMO4, which hasbeen correlated to diabetes (17–19). SUMOylation regulates awide range of processes, such as protein stability or nucleocyto-
Received 21 June 2012 Accepted 25 October 2012
Published ahead of print 31 October 2012
Address correspondence to Michela Campagna, [email protected], orCarmen Rivas, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01578-12
January 2013 Volume 87 Number 2 Journal of Virology p. 807–817 jvi.asm.org 807
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.

plasm transport, but its main function is to regulate protein-pro-tein interactions (20). In addition, an increasing number ofSUMOylated proteins can also interact with SUMO in a noncova-lent manner, through a SUMO-interacting-motif (SIM) (21).
Viral proteins were among the first substrates shown to bemodified by SUMO and SUMOylation seems to facilitate viralinfection in cells (22, 23). Although the list of viruses able to ex-ploit the SUMOylation machinery has considerably increased inthe last years, the role of SUMO in the replication of members ofthe Reoviridae family has not been reported thus far. We show herethat a change in the levels of SUMOylation machinery compo-nents in the cells alters both rotavirus replication and rotavirusprotein production. In addition, we demonstrate that rotavirusproteins that localize in viroplasms are SUMOylated and interactwith SUMO in a noncovalent manner. Finally, we show that theexpression of NSP5 mutated in the SUMOylation sites abolishesthe formation of VLS-VP2i. To our knowledge, this is the firstdemonstration of exploitation of the cellular SUMOylation ma-chinery by a member of the Reoviridae family.
MATERIALS AND METHODSCell lines, transfections, and virus. MA104, HeLa, COS7, and HEK-293cells were maintained in Dulbecco modified Eagle medium supplementedwith 10% fetal calf serum (Gibco), 5 mmol of L-glutamine (Invitrogen)/liter, and penicillin-streptomycin (Invitrogen). HEK-293 cells were trans-fected using FuGene (Roche), and COS7, HeLa, and MA104 cells weretransfected with Lipofectamine 2000, according to the manufacturer’sinstructions. When stated, cells were infected with T7-recombinant vac-cinia virus (strain vTF7.3) at an multiplicity of infection (MOI) of 5 and 1h later transfected with plasmids DNA for 16 h, as previously described(24). Infection with T7-recombinant vaccinia virus was used to increasethe expression level of proteins encoded by the transfected plasmids.
The OSU strain of rotavirus was propagated and titrated in MA104
cells as described previously (25). Rotaviruses were activated by incuba-tion with 10 �g of trypsin per ml at 37°C for 30 min.
Plasmids, siRNAs, and reagents. Plasmids pcDNA3-NSP5wt, pcDNA3-NSP2, pcDNA3-SV5-VP1, and pcDNA3-VP2 were previously described (12,14). Lysine-to-arginine mutations were carried out using the QuikChangePCR-based site-directed mutagenesis kit (Stratagene, La Jolla, CA) accordingto the manufacturer’s instructions, pcDNA3-NSP5 plasmid DNA as a tem-plate, and the oligonucleotides listed in Table 1. Plasmids pcDNA-His6-SUMO1, pcDNA-His6-SUMO2, and pcDNA-Ubc9 were previously de-scribed (26, 27). siNSP5 has been previously described (4). Smart-pool smallinterfering RNAs (siRNAs) against Ubc9 (siUbc9) and scramble siRNA(siIRR) were purchased from Dharmacon. GST-SENP1 was purchased fromBiomol.
In vitro SUMO conjugation assay. In vitro SUMO conjugation assayswere performed on [35S]methionine-labeled in vitro-transcribed/trans-lated proteins as described previously (28). Briefly, [35S]methionine-la-beled proteins were incubated with E1 in a 10-�l reaction including anATP regenerating system (50 mM Tris [pH 7.6], 5 mM MgCl2, 2 mM ATP,10 mM creatine phosphate, 3.5 U of creatine kinase/ml, and 0.6 U ofinorganic pyrophosphatase/ml), 10 �g of SUMO1 or SUMO2, and 600 ngof Ubc9. The reactions were incubated at 30°C for 45 min. After termi-nating the reactions with sodium dodecyl sulfate (SDS) sample buffercontaining �-mercaptoethanol, the reaction products were fractionatedby SDS-PAGE and detected by fluorography. The in vitro transcription/translation of proteins was performed by using 1 �g of plasmid DNA anda rabbit reticulocyte-coupled transcription/translation system accordingto the instructions provided by the manufacturer (Promega).
In vitro deSUMOylation assay. NSP5-SUMO1 obtained in an in vitroSUMOylation reaction as described above was incubated with 2 �g ofGST-SENP1 (Biomol) in 30 �l of reaction buffer containing 50 mM Tris(pH 7.5), 2 mM MgCl2, and 5 mM �-mercaptoethanol. Reactions wereincubated at 37°C for 1 h and terminated with SDS sample buffer contain-ing mercaptoethanol. Reactions products were then fractionated on a12% SDS-polyacrylamide gel, dried for 1 h, and exposed to X-ray film.
TABLE 1 Oligonucleotides used in site-directed mutagenesis
Amino acid position(s)
Oligonucleotide
Orientationa Sequence (5=–3=)24 (NSP5-wt siRES) F TAAAAATGAATCGTCTTCAACAACGTCAACTCTTTCTGG
R CCAGAAAGAGTTGACGTTGTTGAAGACGATTCATTTTTA
24 (NSP5-SUMOmut siRES) F TAGAAATGAATCGTCTTCAACAACGTCAACTCTTTCTGGR CCAGAAAGAGTTGACGTTGTTGAAGACGATTCATTTCTA
19 F CTTCTAGTATCTTTAGAAATGAATCGTCTTCTACAACGR CGTTGTAGAAGACGATTCATTTCTAAAGATACTAGAAG
82 F CGAATGCAGTTAGGACAAATGCAGACGCR GCGTCTGCATTTGTCCTAACTGCATTCG
133 F CAATCTCAACTGATAATAGAAAGGAGAAATCCAAGR CTTGGATTTCTCCTTTCTATTATCAGTTGAGATTG
138 F GGAGAAATCCAGGAAAGATAAAAGTAGGR CCTACTTTTATCTTTCCTGGATTTCTCC
134, 136 F CAATCTCAACTGATAATAGAAGGGAGAGATCCAGGR CCTGGATCTCTCCCTTCTATTATCAGTTGAGATTG
139, 141, 144 F CCAGGAGAGATAGAAGTAGGAGACACTACCCGR CGGGTAGTGTCTCCTACTTCTATCTCTCCTGG
a F, forward; R, reverse.
Campagna et al.
808 jvi.asm.org Journal of Virology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.

Immunoprecipitation assay. Cells were lysed in TNN buffer (100mM Tris-HCl [pH 8], 250 mM NaCl, 0.5% NP-40) at 4°C, centrifuged at15,800 � g for 5 min, and immunoprecipitated overnight at 4°C afteraddition of 1 �l of the specified antibody and 50 �l of 50% protein A-Sepharose CL4B beads (GE Healthcare). The beads were then washed fourtimes with TNN buffer and resuspended in 30 �l of SDS-PAGE loadingbuffer.
Lambda phosphatase treatment. [35S]methionine-labeled in vitro tran-scribed/translated NSP5 SUMOmut was incubated with 1 �l of �-phospha-tase (BioLabs) in buffer for lambda-phosphatase treatment (50 mM Tris-HCl[pH 7.5], 100 mM NaCl, 0.1 mM EGTA, 2 mM dithiothreitol, 0.01% Brij 35;New England BioLabs) supplemented with 2 mM MnCl2. The reaction wasincubated for 30 min at 30°C and was stopped with SDS-PAGE loadingbuffer.
Western blot analysis and antibodies. Cells were washed in phos-phate-buffered saline (PBS), scraped into SDS-PAGE loading buffer, andboiled for 5 min. Proteins of total extracts were separated by SDS-PAGEand transferred onto a nitrocellulose membrane. The membranes wereincubated with the following antibodies: anti-NSP2 mouse serum (1:3,000), anti-NSP5 guinea pig serum (1:10,000), anti-VP2 guinea pig se-rum (1:5,000), and anti-SV5 mouse monoclonal antibody (1:10,000). Sig-nals were detected by using chemiluminescence. Quantification of bandintensities was performed by using ImageJ software and normalized by theactin densitometry values.
Immunofluorescence staining. Cells cultured on coverslips were in-fected with T7-recombinant vaccinia virus and 1 h later transfected withthe indicated plasmids. At 16 h posttransfection, the cells were fixed with2% paraformaldehyde, permeabilized with 0.2% Triton X-100 in PBS,and blocked with 2% bovine serum albumin (BSA). Cells were incubatedwith primary antibodies overnight in a moist chamber at 4°C. Coverslipswere washed extensively with PBS and further incubated with appropriateAlexa-conjugated secondary antibodies for 1 h at room temperature.Nuclei were stained with DAPI (4=,6=-diamidino-2-phenylindole), andcoverslips were mounted with ProLong (Molecular Probes). Mouse anti-NSP5 antibody was used at a dilution of 1:3,000, and anti-VP2 guinea pigserum was used a dilution of 1:3,000. Rabbit anti-SUMO2 antibody (1:200) was obtained from Zymed Laboratories. Secondary Alexa 488-con-jugated, Alexa 594-conjugated, and Alexa 555-conjugated antibodies wereobtained from Molecular Probes. Analysis of the samples was carried outon a Leica TCS SP5 confocal laser microscope using simultaneous scans toavoid shift between the optical channels. Colocalization analysis was doneby calculating the overlap coefficient using the LAS-AF software version2.0.2 (Leica). Particle size analysis was done by using the ImageJ software.Images were exported by use of Adobe Photoshop version 9.0.2.
Purification of His-tagged conjugates. The purification of His-taggedconjugates using Ni2�-NTA-agarose beads allowing the purification ofproteins that are covalently conjugated to SUMO was performed as de-scribed previously (29). Briefly, cells were lysed in 4 ml of 6 M guani-dinium-HCl, 0.1 M Na2HPO4/NaH2PO4, and 0.01 M Tris-HCl (pH 8.0),plus 5 mM imidazole and 10 mM �-mercaptoethanol per 75-cm3 flask.Then, lysates were mixed with 50 �l of Ni2�-nitrilotriacetic acid-agarosebeads prewashed with lysis buffer and incubated for 2 h at room temper-ature. Beads were successively washed with the following: 6 M guani-dinium-HCl, 0.1 M Na2HPO4/NaH2PO4, and 0.01 M Tris-HCl (pH 8.0),plus 10 mM �-mercaptoethanol; 8 M urea, 0.1 M Na2HPO4/NaH2PO4,0.01 M Tris-HCl (pH 8.0), 10 mM �-mercaptoethanol; 8 M urea, 0.1 MNa2HPO4/NaH2PO4, 0.01 M Tris-HCl (pH 6.3), and 10 mM �-mercap-toethanol (buffer A) plus 0.2% Triton X-100; buffer A; and finally buffer Awith 0.1% Triton X-100. After the last wash with buffer A, the beads wereeluted with 200 mM imidazole in sample buffer. The eluates were sub-jected to SDS-PAGE and Western blotting as indicated above.
GST pulldown. Glutathione S-transferase (GST) pulldown experi-ments, allowing the identification of noncovalent interaction between thetarget proteins and SUMO, were performed using [35S]methionine-la-beled in vitro-transcribed/translated rotavirus proteins and GST or
GST-SUMO1 as described previously (29). Briefly, in vitro-translated[35S]methionine-labeled rotavirus proteins were incubated with GST orGST-SUMO1 bound onto glutathione-Sepharose 4B overnight at 4°C inbinding buffer containing 50 mM Tris-HCl (pH 7.8), 150 mM NaCl, 0.5mM EDTA, 0.1% (vol/vol) Triton X-100, 0.1% (vol/vol) Nonidet P-40, 5mM MgCl2, 10% (vol/vol) glycerol, 50 �M ZnCl2, and protease inhibitorcocktail. The resin was then washed four times with 1 ml of binding buffer,and bound proteins were eluted with sample buffer and heated at 95°C for5 min. Proteins were separated on SDS-PAGE gels and detected by fluo-rography.
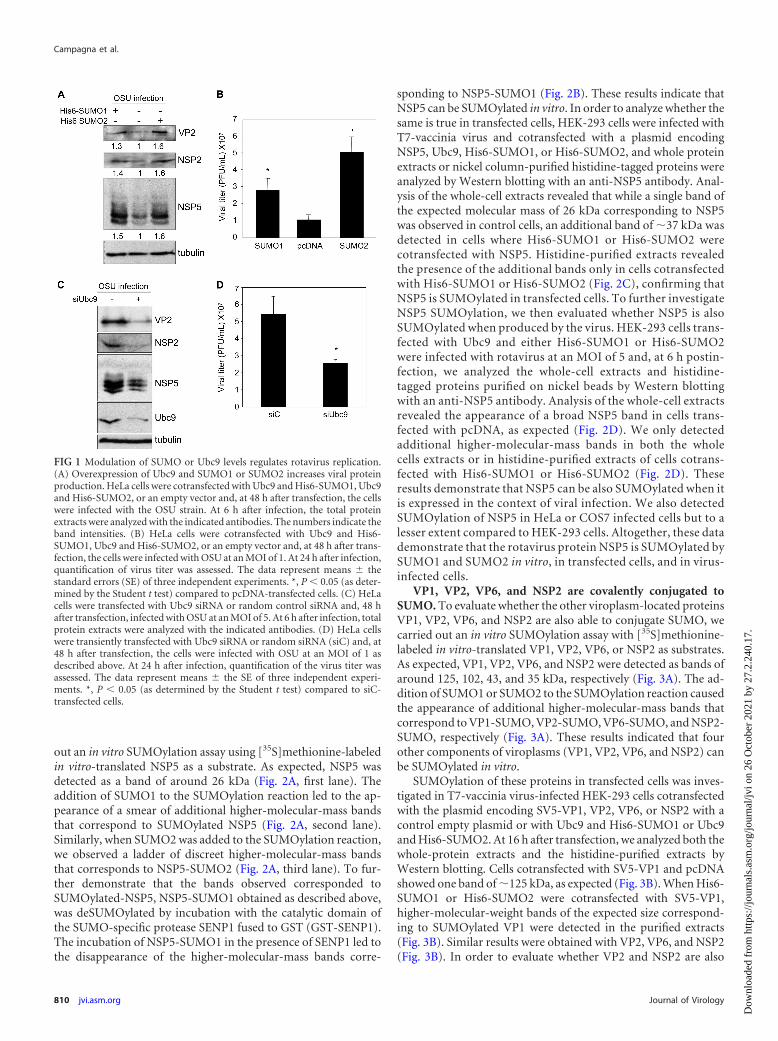
RESULTSModulation of SUMO or Ubc9 levels regulates rotavirus repli-cation. In order to evaluate whether SUMOylation influencedrotavirus replication, as observed for other viruses, we analyzedthe effect of Ubc9 and SUMO1 or Ubc9 and SUMO2 overexpres-sion on rotavirus protein synthesis. HeLa cells were cotransfectedwith plasmids encoding Ubc9 and His6-SUMO1, Ubc9, and His6-SUMO2 or a control empty vector. At 48 h after transfection cellswere infected with OSU strain at an MOI of 5 and 6 h after infec-tion, protein extracts were analyzed by Western blotting with theindicated antibodies. As shown in Fig. 1A, overexpression ofSUMO1 or SUMO2 caused an increase in the production of therotavirus proteins NSP5, NSP2, and VP2. Similar results were ob-served in COS7 or HEK-293 cells. We then decided to evaluate ifoverexpression of SUMO had also an effect in rotavirus replica-tion. HeLa cells were transfected with Ubc9 and His6-SUMO1,Ubc9, and His6-SUMO2 or a control vector as described aboveand, at 48 h after transfection, the cells were infected with OSU atan MOI of 1. At 24 h after infection, the production of infectivevirus was determined. Overexpression of SUMO1 or SUMO2caused an increase in the virus production, indicating that SUMOhas a positive role in rotavirus replication (Fig. 1B). An increase invirus production in COS7 cells transfected with Ubc9 andSUMO2 was also detected. A positive effect of SUMOylation ma-chinery components on rotavirus replication was indeed con-firmed, downmodulating the levels of the E2 SUMO conjugatingenzyme Ubc9 using an Ubc9 specific siRNA. HeLa cells weretransfected with siRNAs against Ubc9; at 48 h after transfection,the cells were infected with OSU at an MOI of 5 and, at 6 h afterinfection, the protein extracts were analyzed by Western blottingwith the indicated antibodies. Transfection of siRNAs againstUbc9 (siUbc9) caused a clear decrease in the Ubc9 levels, as ex-pected (Fig. 1C). In addition, we observed a concomitant decreasein the production of the structural and nonstructural viral pro-teins NSP5, NSP2, and VP2 (Fig. 1C). A decrease in rotavirusprotein synthesis in cells with reduced Ubc9 levels was also ob-served in COS7 cells. Moreover, we also evaluated the productionof infective particles in HeLa cells transfected with siUbc9 at 24 hafter infection. Downmodulation of the Ubc9 levels produced asignificant decrease in the yield of infective particles, indicating apositive role for Ubc9 in rotavirus replication (Fig. 1D). Takentogether, these results indicate that SUMOylation increases thereplication of rotavirus.
NSP5 is covalently modified by SUMO. Rotavirus replicationtakes place in viroplasms. It has been previously demonstratedthat NSP5 is essential for the formation of viroplasms in rotavirus-infected cells, and it is the only protein necessary for the formationof VLS in cotransfection experiments (4, 5, 13). For this reason, toevaluate whether SUMOylation has a role in viroplasm formation,we first investigated whether NSP5 was SUMOylated. We carried
Modulation of Rotavirus Replication by SUMOylation
January 2013 Volume 87 Number 2 jvi.asm.org 809
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.
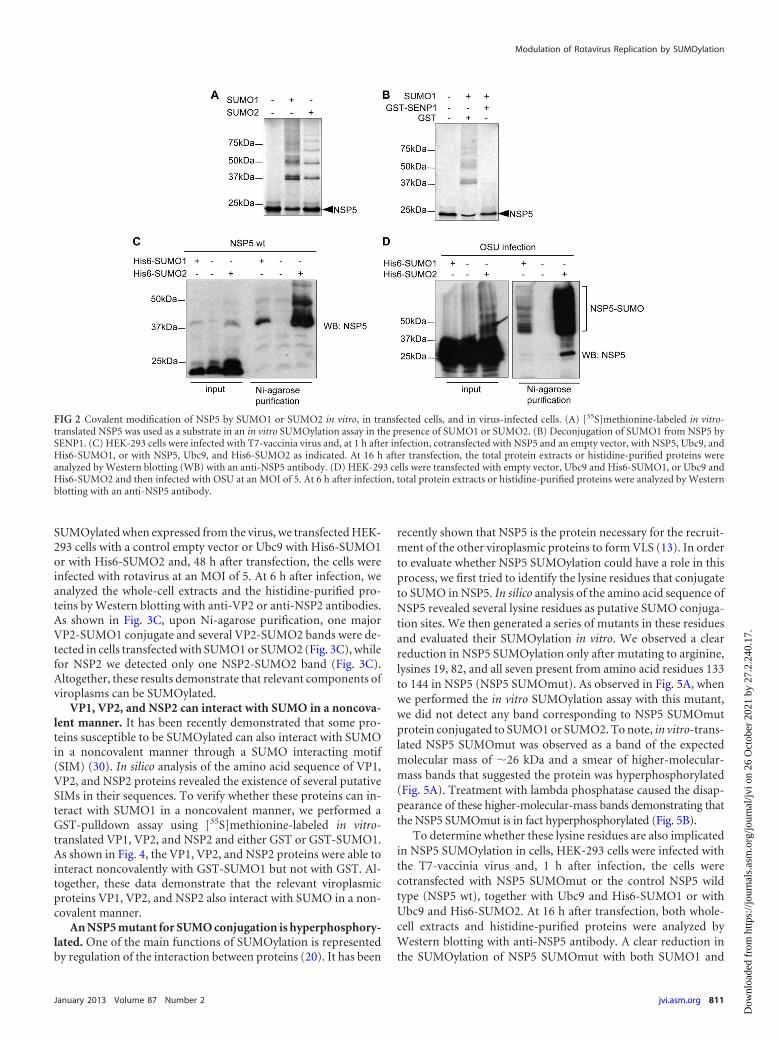
out an in vitro SUMOylation assay using [35S]methionine-labeledin vitro-translated NSP5 as a substrate. As expected, NSP5 wasdetected as a band of around 26 kDa (Fig. 2A, first lane). Theaddition of SUMO1 to the SUMOylation reaction led to the ap-pearance of a smear of additional higher-molecular-mass bandsthat correspond to SUMOylated NSP5 (Fig. 2A, second lane).Similarly, when SUMO2 was added to the SUMOylation reaction,we observed a ladder of discreet higher-molecular-mass bandsthat corresponds to NSP5-SUMO2 (Fig. 2A, third lane). To fur-ther demonstrate that the bands observed corresponded toSUMOylated-NSP5, NSP5-SUMO1 obtained as described above,was deSUMOylated by incubation with the catalytic domain ofthe SUMO-specific protease SENP1 fused to GST (GST-SENP1).The incubation of NSP5-SUMO1 in the presence of SENP1 led tothe disappearance of the higher-molecular-mass bands corre-
sponding to NSP5-SUMO1 (Fig. 2B). These results indicate thatNSP5 can be SUMOylated in vitro. In order to analyze whether thesame is true in transfected cells, HEK-293 cells were infected withT7-vaccinia virus and cotransfected with a plasmid encodingNSP5, Ubc9, His6-SUMO1, or His6-SUMO2, and whole proteinextracts or nickel column-purified histidine-tagged proteins wereanalyzed by Western blotting with an anti-NSP5 antibody. Anal-ysis of the whole-cell extracts revealed that while a single band ofthe expected molecular mass of 26 kDa corresponding to NSP5was observed in control cells, an additional band of �37 kDa wasdetected in cells where His6-SUMO1 or His6-SUMO2 werecotransfected with NSP5. Histidine-purified extracts revealedthe presence of the additional bands only in cells cotransfectedwith His6-SUMO1 or His6-SUMO2 (Fig. 2C), confirming thatNSP5 is SUMOylated in transfected cells. To further investigateNSP5 SUMOylation, we then evaluated whether NSP5 is alsoSUMOylated when produced by the virus. HEK-293 cells trans-fected with Ubc9 and either His6-SUMO1 or His6-SUMO2were infected with rotavirus at an MOI of 5 and, at 6 h postin-fection, we analyzed the whole-cell extracts and histidine-tagged proteins purified on nickel beads by Western blottingwith an anti-NSP5 antibody. Analysis of the whole-cell extractsrevealed the appearance of a broad NSP5 band in cells trans-fected with pcDNA, as expected (Fig. 2D). We only detectedadditional higher-molecular-mass bands in both the wholecells extracts or in histidine-purified extracts of cells cotrans-fected with His6-SUMO1 or His6-SUMO2 (Fig. 2D). Theseresults demonstrate that NSP5 can be also SUMOylated when itis expressed in the context of viral infection. We also detectedSUMOylation of NSP5 in HeLa or COS7 infected cells but to alesser extent compared to HEK-293 cells. Altogether, these datademonstrate that the rotavirus protein NSP5 is SUMOylated bySUMO1 and SUMO2 in vitro, in transfected cells, and in virus-infected cells.
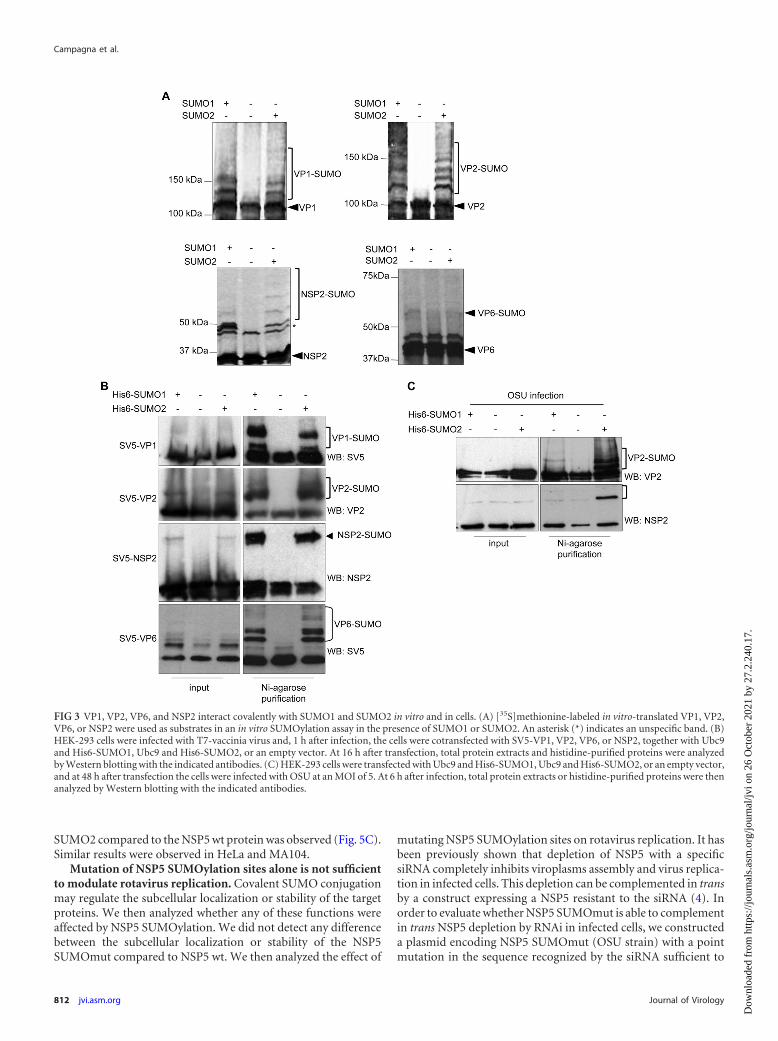
VP1, VP2, VP6, and NSP2 are covalently conjugated toSUMO. To evaluate whether the other viroplasm-located proteinsVP1, VP2, VP6, and NSP2 are also able to conjugate SUMO, wecarried out an in vitro SUMOylation assay with [35S]methionine-labeled in vitro-translated VP1, VP2, VP6, or NSP2 as substrates.As expected, VP1, VP2, VP6, and NSP2 were detected as bands ofaround 125, 102, 43, and 35 kDa, respectively (Fig. 3A). The ad-dition of SUMO1 or SUMO2 to the SUMOylation reaction causedthe appearance of additional higher-molecular-mass bands thatcorrespond to VP1-SUMO, VP2-SUMO, VP6-SUMO, and NSP2-SUMO, respectively (Fig. 3A). These results indicated that fourother components of viroplasms (VP1, VP2, VP6, and NSP2) canbe SUMOylated in vitro.
SUMOylation of these proteins in transfected cells was inves-tigated in T7-vaccinia virus-infected HEK-293 cells cotransfectedwith the plasmid encoding SV5-VP1, VP2, VP6, or NSP2 with acontrol empty plasmid or with Ubc9 and His6-SUMO1 or Ubc9and His6-SUMO2. At 16 h after transfection, we analyzed both thewhole-protein extracts and the histidine-purified extracts byWestern blotting. Cells cotransfected with SV5-VP1 and pcDNAshowed one band of �125 kDa, as expected (Fig. 3B). When His6-SUMO1 or His6-SUMO2 were cotransfected with SV5-VP1,higher-molecular-weight bands of the expected size correspond-ing to SUMOylated VP1 were detected in the purified extracts(Fig. 3B). Similar results were obtained with VP2, VP6, and NSP2(Fig. 3B). In order to evaluate whether VP2 and NSP2 are also
FIG 1 Modulation of SUMO or Ubc9 levels regulates rotavirus replication.(A) Overexpression of Ubc9 and SUMO1 or SUMO2 increases viral proteinproduction. HeLa cells were cotransfected with Ubc9 and His6-SUMO1, Ubc9and His6-SUMO2, or an empty vector and, at 48 h after transfection, the cellswere infected with the OSU strain. At 6 h after infection, the total proteinextracts were analyzed with the indicated antibodies. The numbers indicate theband intensities. (B) HeLa cells were cotransfected with Ubc9 and His6-SUMO1, Ubc9 and His6-SUMO2, or an empty vector and, at 48 h after trans-fection, the cells were infected with OSU at an MOI of 1. At 24 h after infection,quantification of virus titer was assessed. The data represent means � thestandard errors (SE) of three independent experiments. *, P 0.05 (as deter-mined by the Student t test) compared to pcDNA-transfected cells. (C) HeLacells were transfected with Ubc9 siRNA or random control siRNA and, 48 hafter transfection, infected with OSU at an MOI of 5. At 6 h after infection, totalprotein extracts were analyzed with the indicated antibodies. (D) HeLa cellswere transiently transfected with Ubc9 siRNA or random siRNA (siC) and, at48 h after transfection, the cells were infected with OSU at an MOI of 1 asdescribed above. At 24 h after infection, quantification of the virus titer wasassessed. The data represent means � the SE of three independent experi-ments. *, P 0.05 (as determined by the Student t test) compared to siC-transfected cells.
Campagna et al.
810 jvi.asm.org Journal of Virology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.
SUMOylated when expressed from the virus, we transfected HEK-293 cells with a control empty vector or Ubc9 with His6-SUMO1or with His6-SUMO2 and, 48 h after transfection, the cells wereinfected with rotavirus at an MOI of 5. At 6 h after infection, weanalyzed the whole-cell extracts and the histidine-purified pro-teins by Western blotting with anti-VP2 or anti-NSP2 antibodies.As shown in Fig. 3C, upon Ni-agarose purification, one majorVP2-SUMO1 conjugate and several VP2-SUMO2 bands were de-tected in cells transfected with SUMO1 or SUMO2 (Fig. 3C), whilefor NSP2 we detected only one NSP2-SUMO2 band (Fig. 3C).Altogether, these results demonstrate that relevant components ofviroplasms can be SUMOylated.
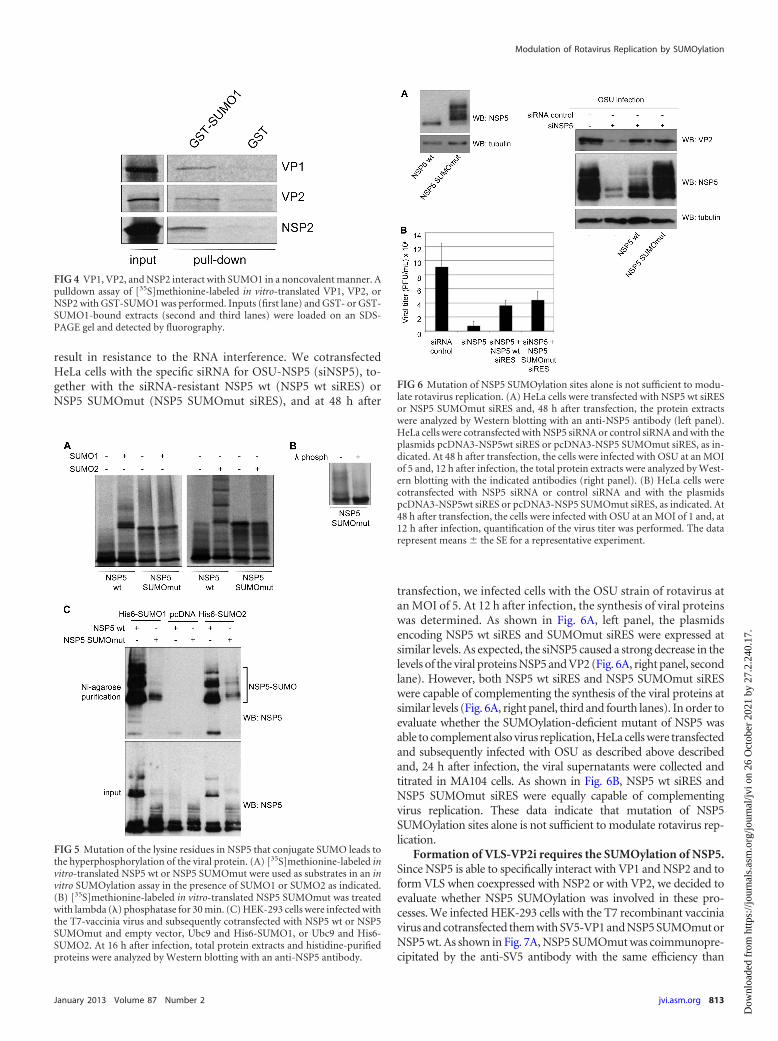
VP1, VP2, and NSP2 can interact with SUMO in a noncova-lent manner. It has been recently demonstrated that some pro-teins susceptible to be SUMOylated can also interact with SUMOin a noncovalent manner through a SUMO interacting motif(SIM) (30). In silico analysis of the amino acid sequence of VP1,VP2, and NSP2 proteins revealed the existence of several putativeSIMs in their sequences. To verify whether these proteins can in-teract with SUMO1 in a noncovalent manner, we performed aGST-pulldown assay using [35S]methionine-labeled in vitro-translated VP1, VP2, and NSP2 and either GST or GST-SUMO1.As shown in Fig. 4, the VP1, VP2, and NSP2 proteins were able tointeract noncovalently with GST-SUMO1 but not with GST. Al-together, these data demonstrate that the relevant viroplasmicproteins VP1, VP2, and NSP2 also interact with SUMO in a non-covalent manner.
An NSP5 mutant for SUMO conjugation is hyperphosphory-lated. One of the main functions of SUMOylation is representedby regulation of the interaction between proteins (20). It has been
recently shown that NSP5 is the protein necessary for the recruit-ment of the other viroplasmic proteins to form VLS (13). In orderto evaluate whether NSP5 SUMOylation could have a role in thisprocess, we first tried to identify the lysine residues that conjugateto SUMO in NSP5. In silico analysis of the amino acid sequence ofNSP5 revealed several lysine residues as putative SUMO conjuga-tion sites. We then generated a series of mutants in these residuesand evaluated their SUMOylation in vitro. We observed a clearreduction in NSP5 SUMOylation only after mutating to arginine,lysines 19, 82, and all seven present from amino acid residues 133to 144 in NSP5 (NSP5 SUMOmut). As observed in Fig. 5A, whenwe performed the in vitro SUMOylation assay with this mutant,we did not detect any band corresponding to NSP5 SUMOmutprotein conjugated to SUMO1 or SUMO2. To note, in vitro-trans-lated NSP5 SUMOmut was observed as a band of the expectedmolecular mass of �26 kDa and a smear of higher-molecular-mass bands that suggested the protein was hyperphosphorylated(Fig. 5A). Treatment with lambda phosphatase caused the disap-pearance of these higher-molecular-mass bands demonstrating thatthe NSP5 SUMOmut is in fact hyperphosphorylated (Fig. 5B).
To determine whether these lysine residues are also implicatedin NSP5 SUMOylation in cells, HEK-293 cells were infected withthe T7-vaccinia virus and, 1 h after infection, the cells werecotransfected with NSP5 SUMOmut or the control NSP5 wildtype (NSP5 wt), together with Ubc9 and His6-SUMO1 or withUbc9 and His6-SUMO2. At 16 h after transfection, both whole-cell extracts and histidine-purified proteins were analyzed byWestern blotting with anti-NSP5 antibody. A clear reduction inthe SUMOylation of NSP5 SUMOmut with both SUMO1 and
FIG 2 Covalent modification of NSP5 by SUMO1 or SUMO2 in vitro, in transfected cells, and in virus-infected cells. (A) [35S]methionine-labeled in vitro-translated NSP5 was used as a substrate in an in vitro SUMOylation assay in the presence of SUMO1 or SUMO2. (B) Deconjugation of SUMO1 from NSP5 bySENP1. (C) HEK-293 cells were infected with T7-vaccinia virus and, at 1 h after infection, cotransfected with NSP5 and an empty vector, with NSP5, Ubc9, andHis6-SUMO1, or with NSP5, Ubc9, and His6-SUMO2 as indicated. At 16 h after transfection, the total protein extracts or histidine-purified proteins wereanalyzed by Western blotting (WB) with an anti-NSP5 antibody. (D) HEK-293 cells were transfected with empty vector, Ubc9 and His6-SUMO1, or Ubc9 andHis6-SUMO2 and then infected with OSU at an MOI of 5. At 6 h after infection, total protein extracts or histidine-purified proteins were analyzed by Westernblotting with an anti-NSP5 antibody.
Modulation of Rotavirus Replication by SUMOylation
January 2013 Volume 87 Number 2 jvi.asm.org 811
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.
SUMO2 compared to the NSP5 wt protein was observed (Fig. 5C).Similar results were observed in HeLa and MA104.
Mutation of NSP5 SUMOylation sites alone is not sufficientto modulate rotavirus replication. Covalent SUMO conjugationmay regulate the subcellular localization or stability of the targetproteins. We then analyzed whether any of these functions wereaffected by NSP5 SUMOylation. We did not detect any differencebetween the subcellular localization or stability of the NSP5SUMOmut compared to NSP5 wt. We then analyzed the effect of
mutating NSP5 SUMOylation sites on rotavirus replication. It hasbeen previously shown that depletion of NSP5 with a specificsiRNA completely inhibits viroplasms assembly and virus replica-tion in infected cells. This depletion can be complemented in transby a construct expressing a NSP5 resistant to the siRNA (4). Inorder to evaluate whether NSP5 SUMOmut is able to complementin trans NSP5 depletion by RNAi in infected cells, we constructeda plasmid encoding NSP5 SUMOmut (OSU strain) with a pointmutation in the sequence recognized by the siRNA sufficient to
FIG 3 VP1, VP2, VP6, and NSP2 interact covalently with SUMO1 and SUMO2 in vitro and in cells. (A) [35S]methionine-labeled in vitro-translated VP1, VP2,VP6, or NSP2 were used as substrates in an in vitro SUMOylation assay in the presence of SUMO1 or SUMO2. An asterisk (*) indicates an unspecific band. (B)HEK-293 cells were infected with T7-vaccinia virus and, 1 h after infection, the cells were cotransfected with SV5-VP1, VP2, VP6, or NSP2, together with Ubc9and His6-SUMO1, Ubc9 and His6-SUMO2, or an empty vector. At 16 h after transfection, total protein extracts and histidine-purified proteins were analyzedby Western blotting with the indicated antibodies. (C) HEK-293 cells were transfected with Ubc9 and His6-SUMO1, Ubc9 and His6-SUMO2, or an empty vector,and at 48 h after transfection the cells were infected with OSU at an MOI of 5. At 6 h after infection, total protein extracts or histidine-purified proteins were thenanalyzed by Western blotting with the indicated antibodies.
Campagna et al.
812 jvi.asm.org Journal of Virology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.
result in resistance to the RNA interference. We cotransfectedHeLa cells with the specific siRNA for OSU-NSP5 (siNSP5), to-gether with the siRNA-resistant NSP5 wt (NSP5 wt siRES) orNSP5 SUMOmut (NSP5 SUMOmut siRES), and at 48 h after
transfection, we infected cells with the OSU strain of rotavirus atan MOI of 5. At 12 h after infection, the synthesis of viral proteinswas determined. As shown in Fig. 6A, left panel, the plasmidsencoding NSP5 wt siRES and SUMOmut siRES were expressed atsimilar levels. As expected, the siNSP5 caused a strong decrease in thelevels of the viral proteins NSP5 and VP2 (Fig. 6A, right panel, secondlane). However, both NSP5 wt siRES and NSP5 SUMOmut siRESwere capable of complementing the synthesis of the viral proteins atsimilar levels (Fig. 6A, right panel, third and fourth lanes). In order toevaluate whether the SUMOylation-deficient mutant of NSP5 wasable to complement also virus replication, HeLa cells were transfectedand subsequently infected with OSU as described above describedand, 24 h after infection, the viral supernatants were collected andtitrated in MA104 cells. As shown in Fig. 6B, NSP5 wt siRES andNSP5 SUMOmut siRES were equally capable of complementingvirus replication. These data indicate that mutation of NSP5SUMOylation sites alone is not sufficient to modulate rotavirus rep-lication.
Formation of VLS-VP2i requires the SUMOylation of NSP5.Since NSP5 is able to specifically interact with VP1 and NSP2 and toform VLS when coexpressed with NSP2 or with VP2, we decided toevaluate whether NSP5 SUMOylation was involved in these pro-cesses. We infected HEK-293 cells with the T7 recombinant vacciniavirus and cotransfected them with SV5-VP1 and NSP5 SUMOmut orNSP5 wt. As shown in Fig. 7A, NSP5 SUMOmut was coimmunopre-cipitated by the anti-SV5 antibody with the same efficiency than
FIG 4 VP1, VP2, and NSP2 interact with SUMO1 in a noncovalent manner. Apulldown assay of [35S]methionine-labeled in vitro-translated VP1, VP2, orNSP2 with GST-SUMO1 was performed. Inputs (first lane) and GST- or GST-SUMO1-bound extracts (second and third lanes) were loaded on an SDS-PAGE gel and detected by fluorography.
FIG 5 Mutation of the lysine residues in NSP5 that conjugate SUMO leads tothe hyperphosphorylation of the viral protein. (A) [35S]methionine-labeled invitro-translated NSP5 wt or NSP5 SUMOmut were used as substrates in an invitro SUMOylation assay in the presence of SUMO1 or SUMO2 as indicated.(B) [35S]methionine-labeled in vitro-translated NSP5 SUMOmut was treatedwith lambda (�) phosphatase for 30 min. (C) HEK-293 cells were infected withthe T7-vaccinia virus and subsequently cotransfected with NSP5 wt or NSP5SUMOmut and empty vector, Ubc9 and His6-SUMO1, or Ubc9 and His6-SUMO2. At 16 h after infection, total protein extracts and histidine-purifiedproteins were analyzed by Western blotting with an anti-NSP5 antibody.
FIG 6 Mutation of NSP5 SUMOylation sites alone is not sufficient to modu-late rotavirus replication. (A) HeLa cells were transfected with NSP5 wt siRESor NSP5 SUMOmut siRES and, 48 h after transfection, the protein extractswere analyzed by Western blotting with an anti-NSP5 antibody (left panel).HeLa cells were cotransfected with NSP5 siRNA or control siRNA and with theplasmids pcDNA3-NSP5wt siRES or pcDNA3-NSP5 SUMOmut siRES, as in-dicated. At 48 h after transfection, the cells were infected with OSU at an MOIof 5 and, 12 h after infection, the total protein extracts were analyzed by West-ern blotting with the indicated antibodies (right panel). (B) HeLa cells werecotransfected with NSP5 siRNA or control siRNA and with the plasmidspcDNA3-NSP5wt siRES or pcDNA3-NSP5 SUMOmut siRES, as indicated. At48 h after transfection, the cells were infected with OSU at an MOI of 1 and, at12 h after infection, quantification of the virus titer was performed. The datarepresent means � the SE for a representative experiment.
Modulation of Rotavirus Replication by SUMOylation
January 2013 Volume 87 Number 2 jvi.asm.org 813
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.
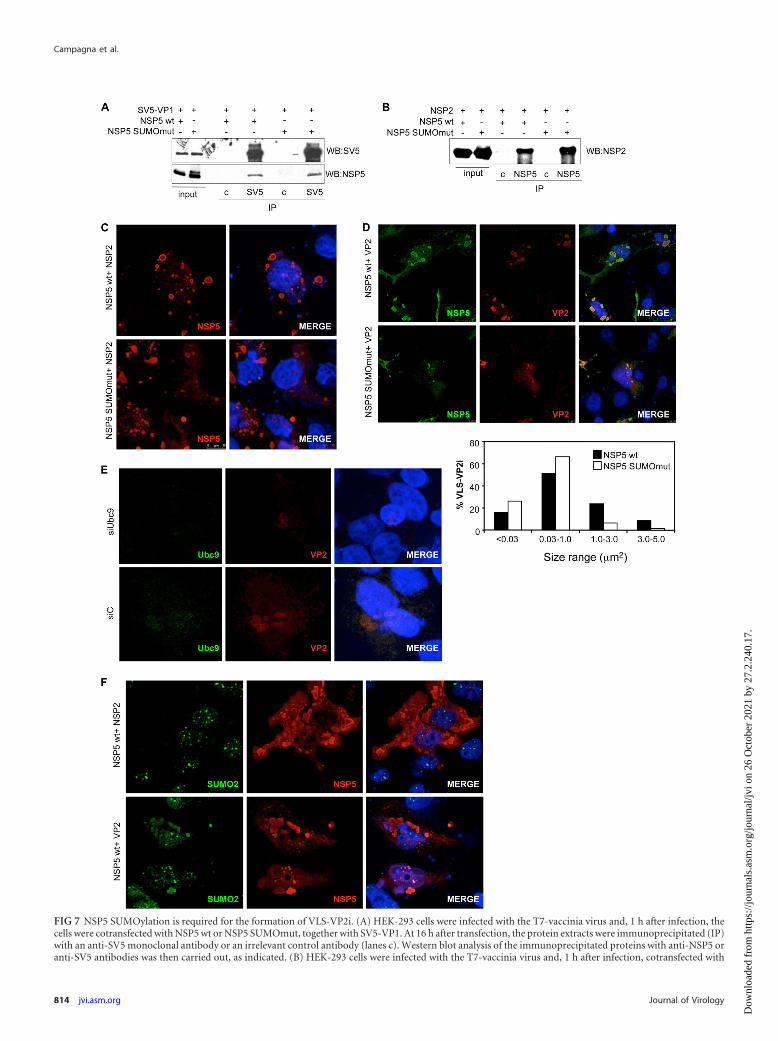
FIG 7 NSP5 SUMOylation is required for the formation of VLS-VP2i. (A) HEK-293 cells were infected with the T7-vaccinia virus and, 1 h after infection, thecells were cotransfected with NSP5 wt or NSP5 SUMOmut, together with SV5-VP1. At 16 h after transfection, the protein extracts were immunoprecipitated (IP)with an anti-SV5 monoclonal antibody or an irrelevant control antibody (lanes c). Western blot analysis of the immunoprecipitated proteins with anti-NSP5 oranti-SV5 antibodies was then carried out, as indicated. (B) HEK-293 cells were infected with the T7-vaccinia virus and, 1 h after infection, cotransfected with
Campagna et al.
814 jvi.asm.org Journal of Virology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.

NSP5 wt, suggesting that the SUMOylation status of NSP5 does notaffect its interaction with VP1. Similar results were obtained in coim-munoprecipitation experiments in cells coexpressing NSP2 withNSP5 wt of NSP5 SUMOmut (Fig. 7B). In addition, we evaluated thecapability of NSP5 SUMOmut to form VLS-NSP2i or VLS-VP2i.HeLa cells were infected with the T7-vaccinia virus and, 1 h afterinfection, cotransfected with NSP5 SUMOmut or the control NSP5wt, together with NSP2 or VP2. At 16 h after transfection, VLS for-mation was analyzed by immunostaining with the indicated antibod-ies. VLS produced after transfection of NSP2 and NSP5 SUMOmutwere similar to those observed after transfection of NSP5 wt (Fig. 7C).In contrast, the ability of NSP5 SUMOmut to form VLS when coex-pressed with VP2 (VLS-VP2i) compared to NSP5 wt was significantlyaltered, as shown by the smaller and more diffused punctuated struc-tures observed (Fig. 7D). Similar results were observed in MA104cells. Particle size analysis of at least 300 VLS-VP2i in each transfec-tion revealed that only 7% of the VLS-VP2i detected in MA104 cellstransfected with NSP5 SUMOmut had a size larger than 1 �m2 incomparison to more than 32% of the NSP5 wt-transfected cells (Fig.7D). A similar alteration in the formation of VLS-VP2i was also de-tected in cells transfected with siUbc9 in comparison to those ob-served in siC-transfected cells (Fig. 7E). In agreement with these data,immunofluorescence analysis of VLS-VP2i and VLS-NSP2i detectedin HeLa cells using anti-SUMO2 and anti-NSP5 antibodies revealed aclear colocalization between SUMO2 and NSP5 in VLS-VP2i but notin VLS-NSP2i (Fig. 7F). In addition, analysis of the colocalizationbetween SUMO1 and NSP5 in four images containing at least 80VLS-NSP2i or VLS-VP2i formed in MA104 cells as described aboverevealed that SUMO1 and NSP5 colocalized in VLS-VP2i, as shownby an overlap coefficient of 0.78 � 0.076, but not in VLS-NSP2.Taken together, these data indicate that NSP5 SUMOylation is notrequired for the interaction with VP1 and NSP2 but plays an impor-tant role in the formation of VLS-VP2i.
DISCUSSION
Since its discovery in the mid-1990s, posttranslational modifica-tion by SUMO has proven to be a key regulator of protein func-tions. Viruses interact extensively with SUMO in order to regulatethe activity of either cellular or viral proteins. Although most ofthe studies have been carried out for DNA viruses, RNA viruseshave been also shown to interact at various levels with the SUMOpathway (23).
In this study we show that SUMOylation plays a positiverole in rotavirus replication. Upregulation of SUMO proteinlevels had a positive effect on the production of viral proteinsand increased rotavirus replication. Similarly, RNA interfer-ence of E2 SUMOylation enzyme Ubc9 caused a marked de-crease in the production of viral proteins and the virus titer,
demonstrating a relevant effect of the SUMOylation system onrotavirus replication. These results are similar to those ob-served with other RNA viruses where SUMO system showed tobe important for virus replication (31–33). To our knowledge,this is the first demonstration of exploitation of the cellularSUMOylation machinery by a member of the Reoviridaefamily.
Rotavirus replication occurs within highly specialized entitiescalled viroplasms, containing four structural (VP1, VP2, VP3, andVP6) and two nonstructural proteins (NSP2 ad NSP5). Bioinfor-matics analysis revealed that all of the viroplasm-resident proteinscontain one or more putative SUMOylation sites (34) and VP1,VP2, and NSP2 contain one or more putative SIMs. NSP5 hasbeen demonstrated to play a fundamental role in architecturalassembly of viroplasms and in recruitment of viroplasmic proteins(4, 5, 13). In addition, NSP5 is thus far the only rotavirus proteinthat has been shown to cause formation of VLS when coexpressedwith either NSP2 or VP2 (13, 14). Since the important role ofSUMO in protein-protein interaction, we then speculated thatSUMO might have a role in the interaction between viroplasmicresident proteins.
In the present study we demonstrated that rotavirus proteinsVP1, VP2, VP6, NSP2, and NSP5 can be SUMOylated both in vitroand in cells and that VP1, VP2, and NSP2 interact with SUMO ina noncovalent manner. Our results demonstrated that the conju-gation of SUMO1 to NSP5 led to the appearance of bands withdifferent molecular masses, indicating that NSP5 is modified bySUMO in more than one lysine residue. In silico analysis of NSP5showed one putative SUMOylation consensus site in lysine 19 andone high-score site in lysine 133. It has been described that lysineselection can be promiscuous, since the mutation of lysine to ar-ginine at preferred SUMO modification sites may result in modi-fication at secondary SUMOylation sites (35, 36). In the case ofNSP5, only after the mutation of lysines 19 and 82 and seven lysineresidues in the segment 133-144 (i.e., lysines 133, 134, 136, 138,139, 141, and 144) could we observe a clear decrease in the SUMOconjugation to NSP5 in vitro and in transfected cells.
SUMOmut protein showed a diffuse localization when ex-pressed in cells in the absence of other viral proteins, led to theformation of VLS-NSP2i after cotransfection with NSP2, inter-acted with VP1 or NSP2 in coimmunoprecipitation experiments,and it is able to complement in trans the NSP5 depletion in in-fected cells, similar to the NSP5 wt protein. Interestingly, NSP5SUMOmut showed increased phosphorylation in vitro and intransfected cells, suggesting the possible existence of an interplaybetween NSP5 SUMOylation and phosphorylation. Extensivestudies have been carried out on the function of NSP5 phosphor-
NSP2, NSP5 wt, or NSP5 SUMOmut. At 16 h after transfection, the protein extracts were cross-linked with DSP and immunoprecipitated with anti-NSP5antibody or an irrelevant control antibody (lanes c) and analyzed by Western blotting with an anti-NSP2 antibody. (C) HeLa cells were infected with theT7-vaccinia virus and, 1 h after infection, cotransfected with NSP2 and NSP5 wt or NSP5 SUMOmut. At 16 h after transfection, the cells were fixed and stainedwith anti-NSP5 antibodies and DAPI. The images were analyzed by confocal microscopy. (D) Both HeLa and MA104 cells were infected with the T7-vacciniavirus and, 1 h after infection, cotransfected with VP2 and NSP5 wt or NSP5 SUMOmut. At 16 h after transfection, the cells were fixed and stained with anti-VP2and anti-NSP5 antibodies. The images were analyzed by confocal microscopy. The top panel shows VLS-VP2i formed in HeLa cells transfected with NSP5 wt orNSP5 SUMOmut. The lower panel shows the size distribution of VLS-VP2i detected in MA104 cells enumerated by percentage of particles in each size interval,obtained after analysis of at least 300 VLS-VP2i per condition using confocal analysis and ImageJ software. (E) HeLa cells were transfected with siC or siUbc9 and,48 h after transfection, the cells were infected with the T7-vaccinia virus. At 1 h after infection, the cells were cotransfected with NSP5 wt and VP2 and, 16 h aftertransfection, the cells were fixed and stained with anti-Ubc9 and anti-VP2 antibodies. The images were analyzed by confocal microscopy. (F) HeLa cells wereinfected with the T7-vaccinia virus and, 1 h after infection, cotransfected with NSP5 wt and NSP2 or VP2. At 16 h after transfection, the cells were fixed and stainedwith anti-SUMO2 and anti-NSP5 antibodies. The images were analyzed by confocal microscopy.
Modulation of Rotavirus Replication by SUMOylation
January 2013 Volume 87 Number 2 jvi.asm.org 815
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.

ylation. NSP5 is hyperphosphorylated when produced upon virusinfection, and blocking of this phosphorylation causes the forma-tion of viroplasms with altered shape (10, 11, 37). In addition,coexpression of NSP5 with VP2 or NSP2 causes hyperphosphory-lation of the protein in the absence of virus infection (13, 14). Wedid not detect a change in the level of NSP5 phosphorylation afteroverexpression of SUMO or downmodulation of Ubc9, nor didwe detect a change in the levels of NSP5-SUMO conjugation afterthe coexpression of NSP2, VP2, or VP1. Based on these observa-tions, we cannot conclude that there is a relationship betweenthese two posttranslational modifications.
In a recent report, the polybasic region 132-146 of NSP5 hasbeen shown to be the responsible for its interaction with NSP2(38). In our study, however, NSP5 SUMOylation was not neces-sary for the NSP5-NSP2 interaction since NSP5 SUMOmut (withseveral lysines mutated in the 133-144 region) retained interactionwith NSP2. The lack of effect observed can be due to the fact thatwe did not change the polybasic nature of this region since lysineswere mutated to arginines. This result is also confirmed by the factthat no alteration in the shape and number between VLS-NSP2iinduced by NSP5 wt or NSP5 SUMOmut could be observed inimmunofluorescence.
VLS can be considered a valid model for viroplasm assembly. Ithas been shown that the viroplasm-resident proteins VP1 and VP6can be recruited to VLS-VP2i in the absence of viral infection (13).Our results demonstrate that NSP5 SUMOmut is defective in theformation of VLS-VP2i, demonstrating that SUMOylation ofNSP5 modulates the interaction with VP2. The notion that SUMOmodification plays an essential role in the formation of VLS-VP2isuggests a structural role for this protein in viroplasm assembly.However, our results indicate that SUMOylation of NSP5 is notrequired for rotavirus replication, probably because of the exten-sive SUMO interaction of the other viroplasm components. Fur-ther studies to evaluate whether inhibiting the SUMO interactionwith the other rotavirus proteins leads to a more dramatic effecton rotavirus replication are required.
In summary, these results demonstrate for the first time thatrotavirus highjack the SUMOylation machinery of the cell for ef-ficient replication probably through modification of several com-ponents of the viroplasms, structures where rotavirus replicationtake place, providing potential new therapeutic targets for rotavi-rus infection.
ACKNOWLEDGMENTS
This study was supported by BFU-2008-03784 and BFU-2011-27064.M.C. and L.M.-V. are supported by the Juan de la Cierva Programme. F.A.was supported by a FIRB grant of the Italian Ministero dell’Istruzione,dell=Università e della Ricerca. J.G.-S. is supported by an IFARHU-SENACYT predoctoral fellowship from Panama. P.G. is supported by aJAE predoctoral fellowship from CSIC. C.F.D.L.C.-H. is supported by a LaCaixa fellowship.
We are grateful to Gianluca Petris and Bartosz Muszynski (Burrone’slaboratory) for production of the mouse anti-NSP5 antibody.
REFERENCES1. Patton JT, Jones MT, Kalbach AN, He YW, Xiaobo J. 1997. Rotavirus
RNA polymerase requires the core shell protein to synthesize the double-stranded RNA genome. J. Virol. 71:9618 –9626.
2. Zeng CQ, Wentz MJ, Cohen J, Estes MK, Ramig RF. 1996. Character-ization and replicase activity of double-layered and single-layered rotavi-rus-like particles expressed from baculovirus recombinants. J. Virol. 70:2736 –2742.
3. Patton JT, Silvestri LS, Tortorici MA, Vasquez-Del Carpio R, Tarap-orewala ZF. 2006. Rotavirus genome replication and morphogenesis: roleof the viroplasm. Curr. Top. Microbiol. Immunol. 309:169 –187.
4. Campagna M, Eichwald C, Vascotto F, Burrone OR. 2005. RNA inter-ference of rotavirus segment 11 mRNA reveals the essential role of NSP5 inthe virus replicative cycle. J. Gen. Virol. 86:1481–1487.
5. Lopez T, Rojas M, Ayala-Breton C, Lopez S, Arias CF. 2005. Reducedexpression of the rotavirus NSP5 gene has a pleiotropic effect on virusreplication. J. Gen. Virol. 86:1609 –1617.
6. Silvestri LS, Taraporewala ZF, Patton JT. 2004. Rotavirus replication:plus-sense templates for double-stranded RNA synthesis are made in vi-roplasms. J. Virol. 78:7763–7774.
7. Schuck P, Taraporewala Z, McPhie P, Patton JT. 2001. Rotavirus non-structural protein NSP2 self-assembles into octamers that undergo ligand-induced conformational changes. J. Biol. Chem. 276:9679 –9687.
8. Taraporewala Z, Chen D, Patton JT. 1999. Multimers formed by therotavirus nonstructural protein NSP2 bind to RNA and have nucleosidetriphosphatase activity. J. Virol. 73:9934 –9943.
9. Gonzalez SA, Burrone OR. 1991. Rotavirus NS26 is modified by additionof single O-linked residues of N-acetylglucosamine. Virology 182:8 –16.
10. Afrikanova I, Fabbretti E, Miozzo MC, Burrone OR. 1998. RotavirusNSP5 phosphorylation is upregulated by interaction with NSP2. J. Gen.Virol. 79(Pt 11):2679 –2686.
11. Afrikanova I, Miozzo MC, Giambiagi S, Burrone O. 1996. Phosphory-lation generates different forms of rotavirus NSP5. J. Gen. Virol. 77(Pt9):2059 –2065.
12. Arnoldi F, Campagna M, Eichwald C, Desselberger U, Burrone OR.2007. Interaction of rotavirus polymerase VP1 with nonstructural proteinNSP5 is stronger than that with NSP2. J. Virol. 81:2128 –2137.
13. Contin R, Arnoldi F, Campagna M, Burrone OR. 2010. Rotavirus NSP5orchestrates recruitment of viroplasmic proteins. J. Gen. Virol. 91:1782–1793.
14. Fabbretti E, Afrikanova I, Vascotto F, Burrone OR. 1999. Two non-structural rotavirus proteins, NSP2 and NSP5, form viroplasm-like struc-tures in vivo. J. Gen. Virol. 80(Pt 2):333–339.
15. Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD. 2002. Struc-tural basis for E2-mediated SUMO conjugation revealed by a complexbetween ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108:345–356.
16. Sampson DA, Wang M, Matunis MJ. 2001. The small ubiquitin-likemodifier-1 (SUMO-1) consensus sequence mediates Ubc9 binding and isessential for SUMO-1 modification. J. Biol. Chem. 276:21664 –21669.
17. Guo D, Li M, Zhang Y, Yang P, Eckenrode S, Hopkins D, Zheng W,Purohit S, Podolsky RH, Muir A, Wang J, Dong Z, Brusko T, AtkinsonM, Pozzilli P, Zeidler A, Raffel LJ, Jacob CO, Park Y, Serrano-Rios M,Larrad MT, Zhang Z, Garchon HJ, Bach JF, Rotter JI, She JX, Wang CY.2004. A functional variant of SUMO4, a new I B� modifier, is associatedwith type 1 diabetes. Nat. Genet. 36:837– 841.
18. Matic I, van Hagen M, Schimmel J, Macek B, Ogg SC, Tatham MH, HayRT, Lamond AI, Mann M, Vertegaal AC. 2008. In vivo identification ofhuman small ubiquitin-like modifier polymerization sites by high accu-racy mass spectrometry and an in vitro to in vivo strategy. Mol. Cell. Pro-teomics 7:132–144.
19. Tatham MH, Jaffray E, Vaughan OA, Desterro JM, Botting CH, Nais-mith JH, Hay RT. 2001. Polymeric chains of SUMO-2 and SUMO-3 areconjugated to protein substrates by SAE1/SAE2 and Ubc9. J. Biol. Chem.276:35368 –35374.
20. Zhao J. 2007. Sumoylation regulates diverse biological processes. Cell.Mol. Life Sci. 64:3017–3033.
21. Geiss-Friedlander R, Melchior F. 2007. Concepts in sumoylation: a de-cade on. Nat. Rev. Mol. Cell. Biol. 8:947–956.
22. Boggio R, Chiocca S. 2006. Viruses and sumoylation: recent highlights.Curr. Opin. Microbiol. 9:430 – 436.
23. Wimmer P, Schreiner S, Dobner T. 2012. Human pathogens and the hostcell SUMOylation system. J. Virol. 86:642– 654.
24. Fuerst TR, Niles EG, Studier FW, Moss B. 1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizesbacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. U. S. A. 83:8122– 8126.
25. Estes MK, Graham DY, Gerba CP, Smith EM. 1979. Simian rotavirusSA11 replication in cell cultures. J. Virol. 31:810 – 815.
26. Desterro JM, Rodriguez MS, Hay RT. 1998. SUMO-1 modification ofIB� inhibits NF-B activation. Mol. Cell 2:233–239.
Campagna et al.
816 jvi.asm.org Journal of Virology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.

27. Vertegaal AC, Andersen JS, Ogg SC, Hay RT, Mann M, Lamond AI.2006. Distinct and overlapping sets of SUMO-1 and SUMO-2 target pro-teins revealed by quantitative proteomics. Mol. Cell. Proteomics 5:2298 –2310.
28. Campagna M, Herranz D, Garcia MA, Marcos-Villar L, Gonzalez-Santamaria J, Gallego P, Gutierrez S, Collado M, Serrano M, EstebanM, Rivas C. 2011. SIRT1 stabilizes PML promoting its sumoylation. CellDeath Differ. 18:72–79.
29. Marcos-Villar L, Lopitz-Otsoa F, Gallego P, Munoz-Fontela C, Gonza-lez-Santamaria J, Campagna M, Shou-Jiang G, Rodriguez MS, Rivas C.2009. Kaposi’s sarcoma-associated herpesvirus protein LANA2 disruptsPML oncogenic domains and inhibits PML-mediated transcriptional re-pression of the survivin gene. J. Virol. 83:8849 – 8858.
30. Song J, Durrin LK, Wilkinson TA, Krontiris TG, Chen Y. 2004. Iden-tification of a SUMO-binding motif that recognizes SUMO-modified pro-teins. Proc. Natl. Acad. Sci. U. S. A. 101:14373–14378.
31. Sun D, Xu P, He B. 2011. Sumoylation of the P protein at K254 plays animportant role in growth of parainfluenza virus 5. J. Virol. 85:10261–10268.
32. Tseng CH, Cheng TS, Shu CY, Jeng KS, Lai MM. 2010. Modification ofsmall hepatitis delta virus antigen by SUMO protein. J. Virol. 84:918 –927.
33. Xu K, Klenk C, Liu B, Keiner B, Cheng J, Zheng BJ, Li L, Han Q, WangC, Li T, Chen Z, Shu Y, Liu J, Klenk HD, Sun B. 2011. Modification of
nonstructural protein 1 of influenza A virus by SUMO1. J. Virol. 85:1086 –1098.
34. Chattopadhyay S, Bagchi P, Dutta D, Mukherjee A, Kobayashi N,Chawla Sarkar M. 2010. Computational identification of posttransla-tional modification sites and functional families reveal possible moon-lighting role of rotaviral proteins. Bioinformation 4:448 – 451.
35. Eladad S, Ye TZ, Hu P, Leversha M, Beresten S, Matunis MJ, Ellis NA.2005. Intra-nuclear trafficking of the BLM helicase to DNA damage-induced foci is regulated by SUMO modification. Hum. Mol. Genet. 14:1351–1365.
36. Lin DY, Huang YS, Jeng JC, Kuo HY, Chang CC, Chao TT, Ho CC,Chen YC, Lin TP, Fang HI, Hung CC, Suen CS, Hwang MJ, Chang KS,Maul GG, Shih HM. 2006. Role of SUMO-interacting motif in DaxxSUMO modification, subnuclear localization, and repression of sumoy-lated transcription factors. Mol. Cell 24:341–354.
37. Campagna M, Budini M, Arnoldi F, Desselberger U, Allende JE, Bur-rone OR. 2007. Impaired hyperphosphorylation of rotavirus NSP5 in cellsdepleted of casein kinase 1alpha is associated with the formation of viro-plasms with altered morphology and a moderate decrease in virus repli-cation. J. Gen. Virol. 88:2800 –2810.
38. Martin D, Ouldali M, Menetrey J, Poncet D. 2011. Structural organisa-tion of the rotavirus nonstructural protein NSP5. J. Mol. Biol. 413:209 –221.
Modulation of Rotavirus Replication by SUMOylation
January 2013 Volume 87 Number 2 jvi.asm.org 817
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Oct
ober
202
1 by
27.
2.24
0.17
.