production et utilisation du carbone organique dissous...
TRANSCRIPT

Production et utilisation
du carbone organique
dissous dans une lagune tropicale
Jean PA&S* et Lionel LEMASSON*
L’utilisation du W’-U-glucose esf déferminèe par le faux de décroissance de sa concentraiion dans le milieu, k > ef par le faux d’incorporation du fraceur dans les particules, k’. Ces deux faux sont proportionnels à la biomasse plancfonique mesurée par Cv, P,, ch1 et ATP. Le rapport k/C, et le pourcentage de glucose respire, R %, sonf fonctions inverses des quotients CINIP de la matière organique dissoute (MOD), f 1 ui expriment la dégradabilité de la MOD. R :7/0 et k sonf influencés par la lumière.
L’excrétion de carbone organique dissous (COD) par le phyfoplancfon est dèferminée par la méthode au 14C. Elle est liée à la production parficulaire; l’excrétion relative est fonction inverse de la biomasse et de l’èclairemenf.
Les concentrafions in sit,u de COD sonf stables sur la journée. Leur correlation avec les formes minérules de N et P indique l’exisfence d’une part réfractaire de la MOD. Un bilan de la production ef de l’utilisation du COD esf fenfè, ainsi qu’une evaluafion de la production héférofrophe.
MOTS-CLÉS : Gl~ose - Excrétion - Carbone organique dissous.
PRODUCTION AND UTILIZATION OF DISSOLVED ORGANIC CARBON IN A TROPICAL LAGO~N
%2-U-glucose ufilizafiotz is measured by fhe rate of decrease of ifs cotzcet~ft~afiot~ in fhe mediutn, k, and fhe rate of fracer upfakr by fhe microorganisms, 14. ‘Bofh rates are posifively correlafed tvifh total biomass evaluafed by Cv, P,, ch1 and ATP. The rafio k/C, is negafively correlafed rvifh fhe C/P ratio of dissolved organic maffer (DOM). The percenfage of respired glucose, R 70, correlates negafively rvifh fhe CINIl’ ratio of DOM. k and R y(, are influ- enced by lighf.
Dissolved organic carbon (DOC) excrefion by phyfoplankfon is defermined by fhe 14C mrfhod. Excrefion is correlafed mifh parficulafe production; relative excrefion is negafively correlafed rvifh biomass and irradiante.
In situ DOC concenfrafions do tzof uary fhrough ouf fhe day. Their correlation tvifh N and P inorganic /ortns hinfs ut fhe existence of a yefracfory pavf of fhe total DOM. We fry fo esfablish fhe DOC budget a.nd fo evaluafe fhe hefet~ofrophic production.
KEY WORD~: Glucose - Excretion - Dissolved Organic Carbon.
l Océanographes O.R.S.T.O.M., C.R.O., B.P. V 18, Abidjan (République de Côte d’ivoire). Adresse actuelle: Antenne O.R.S.T.O.M.. Station INRA, Avenue de Corzent, 74203 Thonon-les-Bains (France).
Rev. Hydrobiol. trop. 14 (2): 83-101 (1981).

1. INTHODUCTTON
Le c.arbone organique dissous (COD) joue un rOle important dans les milieux aquatiques, oil sa concen- kkion est de l’ordre de 10 fois celle du carbone «r~i~Ilicpl~ particulaire (OP). On peui. remarquer que c*ett,e di&inc.tion est assez arbitraire puisque fonction de Iii tec,hnique d’analyse; nous considkrons comme dissous ce qui n’est pas retenu sur un filtre de porosité donnée, souvent. 0,s prn, alors que le spect.re des diamktres est continu (O~LJRA, 1970; SHARP, 1!17:?). Depuis les premiers travaux sur l’exsudation de matibre organique par le ph+~- planckon [Foco, 1952; Foc~ et WESTLAKE, 1955), ce processus a GtC! souvent considéré comme un artefact expérimental (SHARP, 1978); pour les rnacrophytes, il semble certain que cett.e excrétion est physiologique (FANKBONEH et de BURGH, 1977); pour le phytoplancton, il est admis qu’un minimum de prPc:rutlons puisse éviter tout dommage mka- nique aux cellules (SMITH et WIEBE, 1976; HARRIS et PININTN, 1977; ~ARONSON, 1978). La production de CUD par le phytoplancton vivant n’esl, pas sa seule source; la dégradation de COP dbtrique, par les baotéries ou par autolyse, peut contribuer A l’apparition de COD (WETZEL rf al., 19X?), de même que le broyage des cellules par broutage (LAMPEI~T, 1978).
On peu1 considérer le COD comme une forme de stockage rxtracellulaire pour une c.ommunauté; sa r6utili&ion par les bactéries peut continuer jusqu’a la synthke de COP utilisable par des prédateurs, encore que la production de COP puisse 6tre aussi un processus abiotique (PAR~ONS et STRICKLAND, l!XQ). La production de COP par les bactéries a été considkée par certains auteurs comme la source principale de COP (SOROKIN, 1971); cette opinion a t;t:é diwutée, tant en ce qui concerne 1’importanc.e relative de la production hétérokophe (BANSE, 1974; SIIXUIRTH et al., 1978) que sur la fraction du COD qui est c.atabolisée et. donc perdue pour la communauté (SMITH et Cd., 1977).
En plus dr son rcile indirect comme substrat pour les hétérotrophes, avec utilisation simultanée d’oxy- gène, le COD a un rOle chimique dire&. La survie de bactéries telluriques dans l’eau est accrue par de fortes concentrations de COD (JAXNASCH, 1968). Le COD peut former des c,omplexes avec des mAaux (SCHINVLER et al., 1972; GONPE ei ~ARPENTER, 1974; GILLES~IE et VACCARO, 1978; VAN DEN BERG et al., 1979); cet.te action c.omplexante est k la base de l’effet favorabk du COD sur la croissance du phyt.o- plancton (BARBER et KYTHER, 1969).
La. nature chimique du COD est, assez mal c.onnue (DEGENS, 1970); mème pour les quelques familles chimiques les mieux étudiées, les techniques emplo-
RPZI. llydr«hiul. trop. Id (2): 83-107 (1981).
yées peuvent biaiser les rkulhts ( DAWSCJN et GOCKE, 1978). La plupart des études sur les flux de COD ont utilisé des traceurs, déterminant ainsi soit un taux de renouvellement (PAR~ONS et STRICKLAND, 1962), soit, des constantes cinktiques (WRII;HT et HOBBIE, 1965; WILLIAMS et ASKEW, 1%8); dans tous les c.as, l’identité, du traceur avec le substrat naturel est admise.
Au cours d’une @tude gknérale du cycle de la matière organique dan, r; une lagune saumàtre tro- picale, la lagune Ebrié, il nous a semblé nécessaire de c.onsidérer la production et. l’utilisation du COD parall~lernent aux études de production primaire (PAGÈS et L~~hssoN, iri r~rel).). Nous avons utilisé pour cela des traceurs radioactifs, glucose et carbo- nate marqués au IX.
La lagune Ebrit est une lagune saumàtre, d’une surface approximative de 650 km2, d’une profondeur
Lista des symboles rt zznités
A : production photosynthtWyue particu- laire. . . . . . . . . . . . . . . . . . . . . . . gatg Cl-*
ch1 : chlorophylle (1.. . . . . . . . . . . . . . . . . pg.l-’ CID : carbone minéral dissous. . . . . . . . . . . patg Cl-1
l CID : act.ivit& du rarhone minéral dissous.. . . c.p.m. COD : carbone organique dissous. . . . . . [Late Cl-’
CII : carbone particulaire.. . . . . . . . . . . . Fatg C.l-l ‘CO : activite du carbone organique dissous. . c.p.m. ‘C, : activité des particules (pr«duct.ic.m pri-
maire) . . . . . . . . . . . . . . . . . . . . c.p.m. E : excrétion jminimuzni.. . . . . . . . . patg C.1 -l E’ : excrtition (masimuml. . . . . . . . . . . . patg Cl-l Est :
E/t. : ‘F : Ic : Ii’ : NID : NOD : NTD : N, : POD : PO, :
PTD : P, : R s{, : r s :
D s, :
lEa :
escrbtion d’npz+s les expc;riencrs en dia- lyse ............................... pat.g (1.h. j-l
escr6tion par unit.6 de temps ........... &atg (I.h.)-* acfi\-itt des particules (glucose). ........ c..p.m. IAUX de disparition du glucose .......... h-l taux d’incorporation du glucose. ....... h-l
.. azote mzneral tlzssous ................... pat,v N.l-’ azote organique dissous ................ patg N.l-l azot.e total dissous .................... p.atg N.l-l azote particulaire ..................... yatg N.l-l phosphore organique dissous. .......... patg P.l-1 phosphore nlirléra1 (ph0Sphat.P r+actif)
dissous.. .......................... patg P.l-1 phosphore total dissous ................ palg P.I-1 phosphore particulaire. ................ patg P.l-1 pourcentage de glucose rrspirB. coefficient de corrélation. concent.ration du substrat. !glucose) dans
le milieu ........................... patg (2.1-l 6cart type. somme de l’rscrétiou minimum rt de la
production primaire : (2, = ASE). .. katg Cl-l somme de l’escr6tion maximum et. de la
production particulaire (2, = A+E’). patg C.l-1

CARBONE ORGANIQUE 1)ISSOUG
Pro. 1. - Carte. de la lagune fibriti, et. position des stations
moyenne de 4,8 m, Situ&e par environ 5O N (fig. 1). Ses caractéristiques physico-chimiques générales ont ét6 décrites par ailleurs (TASTET, 1974; VARLET, 1978; Pac;Ès ef nl., 1979).
2. MATÉRIEL ET MBTHODES
2.1. Méthodes d’analyse
Les symbole.5 utilisks sont rassemblés dans le t,ableau 1.
Les stations mentionnées ic.i (fig. 1) ont été occu- pées au cours de plusieurs campagnes en 1976 et 1977 (voir les dat,es, tableau II). Le dosage des sels nutritifs a été rkalisé suivant les méthodes habitruelles (STRIK- LAND et PARSONS, 1968). Le dosage du carbone organique dissous (COD) b,tait fait en évaporant a 60OC, dans de petit,es nacelles d’aluminium, des volumes suc.cessifs de 200 1~1 d’eau préalablement filtrée sur filtres Whatman GF/C et acidifiée; l’extrait sec t%ait dos8 dans un analyseur CHN (Hewlett- Packard).
L’azote total dissous (NTD) et le phosphore total dissous (PTD) étaient, déterminés par oxydation aux U.V. d’eau filtrée, et, dosage des nitrates et des phosphates résultant. L’azote organique dissous (NOD) et le phosphore organique dissous (POD) sont déterminés par ditY8renc.e entre le t,otal dissous et la frac.tion minérale dosés parall+.lement.
2.2. Incorporation du glucose
L’utilisation du glucose a été étudiée en addition- nant une quantité const.an1.e de 14C-U-glucose (CEA., 275 PCi. @I-1) Q des Pchantillons d’eau tamisée sur 200 pm, à des doses d’environ 165 000 dpm par
Rerr. Hgdrobiol. trop. 14 (2): SS101 (1881).
65 ml d’échantillon (soit 0,3 p-; C.l-1). Les inwbations étaient. faites a température ambiant,e (,3OOC), généralement à l’obscurité; des éc.hant,illons formolés servaient de témoins. Après incubation et filtration douce (<lOO mm Hg) sur filtres What.man GF/C, les filtres 6taient séc.hés, puis comptés en scintillation liquide. Pour déterminer la quantitk de glucose non utilisé, les fikrats étaient traités immédiat.ement et conservés à - 15 OC; 5 ml de filtrat acidifié & pH 2 ktaient bullés puis comptés en scintillation liquide (Intertechnique SL 30) avec le mélange scintillant Aquasol@.
2.3. Excrétion
La production de COD par le phytoplancton a été étudiée au cours des mesures de la production pri- maire par la méthode au 14C, avec des wtivités de l’ordre de 106 cpm par 300 ml d’échantillon, égale- ment tamisés sur 200 km; les filtrats bt,aient traités comme précédemment; des knoins étaient incubés a l’obscuritk. Les expériences avec. dialyse étaient réalisées comme suit : un échant,illon de 65 ml était inoculé, incubé pendant trois heures, filtré (100 mm Hg); le filtrat acidifié et bu116 fournit le chiffre *CO,; le filtre d’une incubation en double fournit le chiffre *Cpl (dans tout ce qui suit, un astérisque indique des cpm). Le filtre était t,ransféré rapidement dans une enceinte à dialyse, de l’eau de lagune filtrée étant utilisée à l’intérieur et à l’extérieur de l’enceinte. Aprés une deuxième incubation de trois heures, le contenu de l’enceinte ét.ait repris sur un filtre, le filtrai donnant le terme *CO,.
Le carbone minéral dissous (CID) était mesuré par ac.idificat.ion d’un éc.hantillon, bullage, piégeage du CO, degagé et dosage en retour.
J. PAGÈS, L. LEMASSON
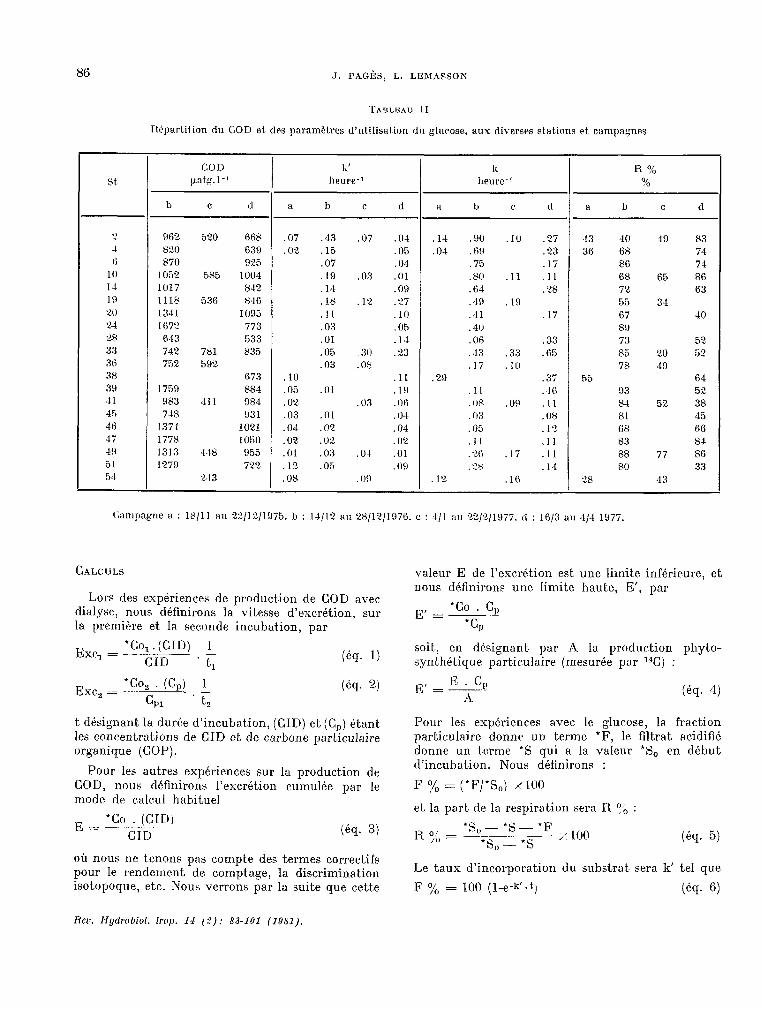
TABLEAU II
Rbpartilion du COD et des paramPt.res d’utilisation du gl~wc.~se, aux diverses stations et campagn~.s
St
CC)D Ii' ii R 90 patg.l-1 heure-’ lleure-1 0,;
h C CI a b c d a b , d a h c d
? OI?2 520 668 .07 .43 .07 .04 .14 .!lO .10 . 27 43 40 39 Y3 4 HLO 638 .02 .15 . 05 .04 60 .23 36 68 74 6 870 9% .07 . 04 .75 .17 86 74
III 1052 585 1004 .19 .0x .Ol .80 .ll .II 68 65 86 1.4 1017 842 .14 .09 .64 -28 72 63 19 1118 536 X46 .1x .12 . LT . 49 I!I . 55 34 20 1341 1095 .11 . l(i .41 .lï 67 40 24 167? 773 .03 .05 .40 P!i 28 643 533 01 . 14 .06 .33 73 52 3.7 742 7x1 835 .05 . 30 .23 . 43 .33 .65 85 20 52 36 xi2 592 .03 .or .lï .111 7x 49 38 673 10
884 :05 .11 . 29 .s 3; 55 64
39 1759 .Ul . 19 .11 .46 93 52 41 983 411 984 .02 03 . (JO OP .OH .ll P-4 52 38 45 74x 931 .03 .Ol 0-I 03 . 08 81 45 36 1.771 1021 .0-i 02 .0-i 05 1.' . - 68 66 47 1778 1050 .02 .02 02 .lI .11 83 84 49 1313 418 955 . C)l .03 O-I- .Ol .t>A .17 .ll 88 77 86 31 1279 722 1 i2 .05 09
. cx 14 80 33
51 243 .OP .09 . 12. .lR 28 43
fkmpagne il : 18/11 au ??(12/1975. ù : 14/12 au 2X/12/1976. c : 4/1 a,~ 22/2/1977. 11 : 16/3 a,~ 4/4 1977.
CALCULS
L,ors des expériencex de production de COD avec. dialyse, nous définirons la vitesse d’exwétion, sur la premiisre et la sec.onde incubat,ion! par
Exr., = *ch,. (cm) I
CID ‘t,
Exc,, = tc:y% . ((2 1 (é4. 2) u c 1’1 . t,
t. désignant la durée d’incubation, (CID) et (C,) étant. les corwentrat,ions de CID et de carbone particulaire organique (OP).
Pour les autres expériences sur la production de MD, nous dkfinirons l’exc.rét.ion cumulée par le mode de calcul habituel
E _ *ch . (Cm) CTD (éq. 3)
ou nous ne tenons pas compte des termes correctifs pour le rendement de comptage, la discrimination isotopoque, et.(:. Nous verrons par la suite que cette
valeur E de l’excrétion est une limite inférieure, et nous définirons une limite haute, E’, par
E’ = *” ’ ‘,, - .- *c,
soit., en dssignant. par A la production phyto- synthétique particulaire (mesurée par W) :
E, = E . q, -_ A
(éq. 4)
Pour les expériences avec le glucose, la fraction partiçulaire donne un terme *F, le filtrat, acidifié donne un terme *S qui a la valeur *B, en début d’incubation. Nous définirons :
F yO = (*FI*S,) x 100
et la part, de la respiration sera R 0; :
Le taux d’incorporation du substrat. sera k’ tel que F ‘+$ = 100 (1-e-k’. t) (éq* 6)
REF. Hydrobiol. trop. 14 (2): 83-191 (1981).

CARBONE ORc;ANIQUE DISSOUS 57
et le taux de disparition au subst.rat sera k, défini par ‘S = *ao _ (.+ . t (éq. T)
Dans toul ce qui suit, les çoff1cient.s de corrélation, r, sont indiqués avec leur signific.ation : x : o,c)5 xx : 0,oi ; xxx : 0,001.
3. RÉSULTATS ET DISCUSSION
3.1. Utilisation du glucose
Nous décrirons t,out d’abord l’évolution dans le temps des divers paramètres au c.ours d’une incuba- tion. Les valeurs absolues seront considérées ensuite.
3.1.1. ÉVOLUTION TEMPORELLE ET DESCRIPTION CINÉ- TIQUE
Assimilniiorz
L’utilisation d’un substrat dét,erminé peut être .%udiée par son incorporation dans les microorga- nismes. Cette voie a été employée par de nombreux auteurs depuis les premiers travaux sur la mesure de l’activité héttrotrophe (PARSONS et STRICKLAND, 1962; HOBBIE et. WRIGHT, 1965).
Certains auteurs ont observé un taux d’assirnila- tion const,ant, que ce soit sur le glwose (WRIGHT et HOBBIE, 1965; VACCARO et .JANNASCH, 1966; BELL et al., 1974; PAERL, 1974) ou sur d’aut,res substrats (MUNRO et BROCK, 1968; HOBBIE et CRAWFORD, 1969). D’autres auteurs observent une augmentation non linéaire de la radioactivité particulaire sur différent,s substrat,s ( HA~+IILTON et Am~m, 1967 ; WETZEL et WANNY, 1972; GRIFFITHY ei al., 1974; SATJNDERS, 19%; BERMAN f?t Ul., 1977; JTURRIAC;A et HOPPE, 1977; WIEBE et Shnm, 1977). Les c.onditions expériment,ales semblent jouer un rcYe assez d&er- minant; la lumière peut intervenir (-JONES et BILLINGTON, 1976); la proportion de substrat ut,ilisée en fin d’incubation doit %re faible (HOBBIE et WRIGHT, 1965; WRIG;HT et HORBIE, 1965).
Dans la plupart des cas, nos courbes expérimen- t,aIes (fig. 2) ne sont, pas linéaires. La description des courbes observées offre certaines difficultés; la théorie prévoit que l’évolution de l’activité parti- culaire, dans un système à trois compariiments, sera décrite par une somme de deux exponentielles (BERMIAN et SCHOENFELD, 1956; SMITH, 1974); en fait, l’ajustement de points expérimentaux & une telle espression exige une précision extréme aes données (ATKIN~, 1973), alors que le co%cient de variation moyen de nos chiffres de.*F est de l’ordre de 9 7: (PAGES, LEBIASYON et DUFOUR, en prépa- ration). Nous avons été amenés à utiliser la relation approchée (éq. 6) : F oie = 100 (l-e-k’ . t). Sur 47 incubations, comportant chacune de 4 A 8 points,
A 4 a
s
2 0
0 33
0 10
0 1000 mrn
FIG. 2. - Évolot.ion, au cours d’incubations ti différentes stations, de la proportion de substrat fixte dans les particules, ‘F y0 (cercles) et. de la proportion de substrat. disparu du filtrat (triangles). Le temps est exprimé en minutes, avec origine arbitraire & 06:30 (heure locale). Les symboles pleins
indiquent les incubations Si l’obscurité
seules 5 fournissent pour cette relation un c.cefficient de corrélation entre F O’ /” et t non significatif (à moins de 5 %).
A une station donnée, k’ ne montre pas de tendance particulière au cours d’une journée; dans le cas OU Ii’ varie fortement,, une inspection des autres caracté- ristiques physico-chimiques des échantillons a montré qu’il s’agissait de masses d’eau différentes. Nous avons calculé, de ce fait, la valeur moyenne de k’ pour chaque station et chaque journée d’observation (tabl. IL). Le faible nombre de données ne permet pas de définir une tendance saisonniPre ni géogra- phique.
Utilisntion du substrat
La décroissance de la conc.entrat.ion de substrat rest,ant dans le milieu, S, a été suivie par de nombreux
Heo. Hydrobiol. trop. 14 (9): Sd-101 (1981).

auteurs, soit, avec des substrat,s (( naturels )) (~I(;~RA et, GOTOH, 1974; HERBLAND, 1975; OGURA, ~~~~;WIEBE et, SILIITH: 1977). soit avec. des substrats chimique- ment définis ( HARIILTON et AUSTIN, 1976; FLEISC.HER, 1975 ; NALEWAJK~ et, SCHINDLER, 1967).
L’éz-olution de S A partir d’une valeur initiale Y,, est gPntralernent décrite par S = S, . e-k . t (éq. 8)
Cette forme implique, pour des valeurs de t assez grandes, que S tende vers zéro ; ceci n’est gAnérale- ment pas le cas, surtout pour des substrats naturels (GWKE, 1970). Cert.ains auteurs admettent l’exis- tence de deux substrats, et deux k, différents (C)C~RA, 1975). D’autres introduisent une valeur limite S,, correspondant SI une matière organique résiduelle ( HAL~IANN et STTLLER, 1974) ; la détermination math& matique de S,, bien que simple en théorie (CROWE et. ~ROWE, l969), est, en fait problématique à partir de points exT+rimentaux. Pour éviter la diminution de S jusqu’A zéro on peut aussi, dans les simulations num&riques du moins, introduire une re-excrétion d'une partie du substrat assimil& Un t.el processus a @té observ& (HA~LTON et AUSTIN, 1967; GOCKE, 1970; NAI.E~~A.TKO et. LEAN, 1972; FLF,ISCHGR, 1975) et peut entraîner un recyclage du t.rac.eur (BENNET et HORRIE, 1972.; MC:I<INLEIT, 1977). Ce substrat re- exc-rét.é ne peut t%re distingué du substrat original, et imit-era la part. Q réfractaire 1) observee par plusieurs auteurs (OTS~KI et HANYA, 1972; WETZEL et MANNY, 197.). Le COD exc.rét+ a souvent un poids molécu- laire plus élevé que la substanw de dé.part, ( NALE\TTA.I~ K(.I et LEAN, 1972; PAIZRL, 1974; DUNSTALL et, NALFT~.A.JKC), 197??); son ident,ification avec le COD réfractaire serait. donc possible.
Pendant, nos incubat,ions, qui durent de une heure B yuafre heures, jusqu’A 70 y$ de l’act,ivit,é. introduite peut, disparaitre du milieu (fig. 2, st. 33j. La valeur absolue de la quant.ité restant ne peut, donner d’inclice quant. A la nature chimique du COD consi- déré. dont une part au moins pourrait i%re une escr& t-inn. Une autre possibilite serait, dans notre cas au moins, que la concentration résiduelle en gluc,ose soit. devenue infPrieure A la concentration minimum observle OLI postulbe par plusieurs aut,eurs (JAN- N.~RcH, 1967, 1970; V4ccARo ef al., 1968); les concen- trat,ions de glucose ajoutées étant de l’ordre de 0,3 pg CI -l, nous serions dans le domaine des seuils indiqués. En outre, l’adsorption et le masquage du glucose par des macromolécules pourrait, aussi freiner son utilisation (STEINBERG, 1977).
Il faut. remarquer que l’équation (Sj a pour difïé- rentielle de départ l’expression dS/dt = -k. S assez fortement di&ente de la classique relation de ~Tichaelis-Ment.en
- dS/<it = V = V,,],, . S/(Ii,+S)
Heu. Hydwbiol. tmp. 14 (2): 85-101 (1981).
(éq. 9)
qui s’applique de facon tres variable a des populations naturelles (MZJNRO et BROCK, 1968; VACCARO, 1969; WILLIAMs, 1973; C;O~KE, 1974). L’intégration de l’équation (9) est difflc.ile? mais des simulations numériques montrent qu’elle fournit. des valeurs de 5 qui sont. analogues A celles données par l’equation (8). La simplicité d’emploi de cette derniére nous l’a fait retenir et 110~15 avons c.alc,ulé les cœfficients k.
De mCme que T)our k’, le faible nombre de données sur k pour chaque st.ation ne permet pas de discerner de diflërence significative entre régions et entre saisons (tabl. II). Les deux paramètres peuvent cependant fournir des renseignements, comme nous le verrons plus loin.
L’évolution du pourcentage de substrat respiré, R y{,. au cours d’une incubation, a ét.6 etudiée par quelques auteurs. HOBBIIZ et CR~WFORD (1969) et. GRIFFITHH rt al. (1974) observent un R y0 constant, respectivement. sur l’aspartate et le glut,amate, en acidifiant une culture et piégeant le CO, dégagé; au contraire, en acidifiant le fllt.rat seul, GRIFFITHS ei al. (OP. cif.) obt,iennent des valeurs plus faibles, et woissantes, de R 5;; une destruction partielle des cellules et un entraînement d’une fraction volatile du COD résultant (HARIILTON et AUSTIN, 1967; ANDERSON et ZEUTSCHEL, 1970; HANSON et WIERE, 1977) semblent probables dans le premier schéma expérimental. Dans not,re c,as, nous n’avons pu déga- ger d’évolution @néraIe de R 7; ; notre mode de calcul ut,ilise des.compt,ages sur de faibles volumes de filtrat, donc assez imprécis; il est probable que cette imprécision suffirait. A masquer une bonne part de la variat.ion réelle de R Y;. Nous avons donc,, la encore, considére la valeur moyenne pour chaque station et, chaque journée (tabl. II.). De même que pour li et k’, aucune variation significative ne peut. être distinguee sur les saisons ou les régions.
3.1.2. RELATION~ AVE(: D'A~TRES PAR~UIÈTRES
La respiration relative, R ‘TA, ne présente pas de relation avec, k, t,aux de disparition du substrat.
Nous ~VOI~S considér& un certain nombre de para- mètres c.himiques ou biologiques susc.eptibles d’avoir une influence sur les paramètres d’utilisation du gluc.ose; parmi les nombreux fac.teurs considérés (tabl. IIT), seuls quelques-uns ont fourni des relations lineaires statistiquement significatives.
Le taux de disparition du substrat, k, est de 10 A 30 y& par heure, ce qui donne des temps de demi-vie

CARBoNE ORGANIIJUE DISSOUS
TARLEAII III Ik Relations entre les paramiltres d’utilisation du glucose et des paramétres chimiqltes. (0 : pas de relation significative ;
+ : relation posilive; - : relation n@ative)
Salini LE. ........ 0 0 00 PO,. - ........... 0 0 NO,. ........... 0 0 0 NH., ............ 0 0 0 COD ........... + 0 0 N,. ............ + 0 0 PT. ............ 0 0 n c,. ............. 0 + + P,. ............ 0 + + ATP.. .......... %O + 0 ch1 ............. 0 + +
COD/PO&. ...... + 0 0 COD/NO, ....... 0 0 n COD/PTD.. .... + -- 0 COD/NTD.. .... + 0 1) C,/PO,. ........ 0 n 0 c,/ATP ......... 0 + 0 C,/P,. .......... 0 0 0
de 2 à 7 heures; dans des milieux analogues des temps de renouvellement de 0,25 a 24 heures ont, été obser- vés pour le glucose (FLE~~CHER, 1975; SAUNDERS, 1975; HANSON ~~WIEBE, 1977; C;OCKE,~~~~), encore que RAI (1979) détermine des temps de renouvelle- ment beaucoup plus longs dans des eaux douces tropicales.
Nous n’observons pas de relations entre k et la concentration de COD, au contraire des observations de ALBRIGHT (1977) ; cette absence de relation est cependant plausible car une forte quantit6 de COD peut. ne pas correspondre à une forte concentration d’analogues du glucose.
Des études de cinétique ont montré que la Vit#esse maximum d’assimilation, V,,,, est fonction de la biomasse bactérienne (WRIGHT et HOBBIE, 1965; GOCKE, 1974; OVERBEM, 1975; ROBINSON et BRANDT, 1975). Nous observons aussi une relation linéaire entre k et les évaluations de la biomasse C,, P, et. c.hl (fig. 3). Il faut remarquer que c,es para- métres évaluent la masse totale du seston; la relation btroite entre k et eh1 indiquerait que les hétérotrophes sont liés aux particules.
La liaison de k avec 1’ATP n’est significaGve qu’en met.tant B part les stations de la zone urbaine polluée, & teneur relative d’ATP forte; les stations non polluées donnent alors la relation :
Ii = 0,OOOl. ATP+O,OOts
”
L 0 20 pg. i-’
FIG. 3. - Relations entre Ir, taus de disparilion du substrat (en (heure)-l) et les paramétres de biomasse. Axes majeurs réduits : pour le carbone particulaire (en ikatg C. 1-l) : k = 0,0018 . C&+O,Ol (r = 0,78XxX) ; pour le phosphore parti- culaire (en patg P. 1-l) : lc = 0,335 P, - 0,07 (r = 0,75Y ; pour la chlorophylle (e.11 @.1-l) : lr = 0,021 ch1 - 0,03
(r = O,G9~s")
(r = 0,68”x), avec ATP en g. 1-l et k en (heure)-l. Cette séparation des stations urbaines ne peut étre due ti la seule influenc.e de la position géographique par le biais de la salinit.é, puisque celle-ci ne présente aucune relation avec. k. La relation assez faible de k avec, 1’ATP est gènante; plusieurs auteurs ont montrP, que les bactéries repré- sentent une part importante de 1’ATP de certaines populations (FERGUSON et MURDOCH, 1975; SIE- BUTRH, 1977; CAMPBELL et BAKER, 1978; COVENEY, 1975; SIEBURTH et al., 1975) mais les corrélations chlorophylle-ATP observées dans la lagune Ebrié tendraient à indiquer une faible proportion d’ATP bactérien, expliquant la faible relat,ion entre k et ATP.
Le c.aractére particulier des stations urbaines, VU avec la relation k-ATP, se retrouve dans la relation entre k et le rapport C,/ATP (fig. 4). La corrélation,
Reu. Hydrobiol. trop. 14 (2): S3-101 (1981).
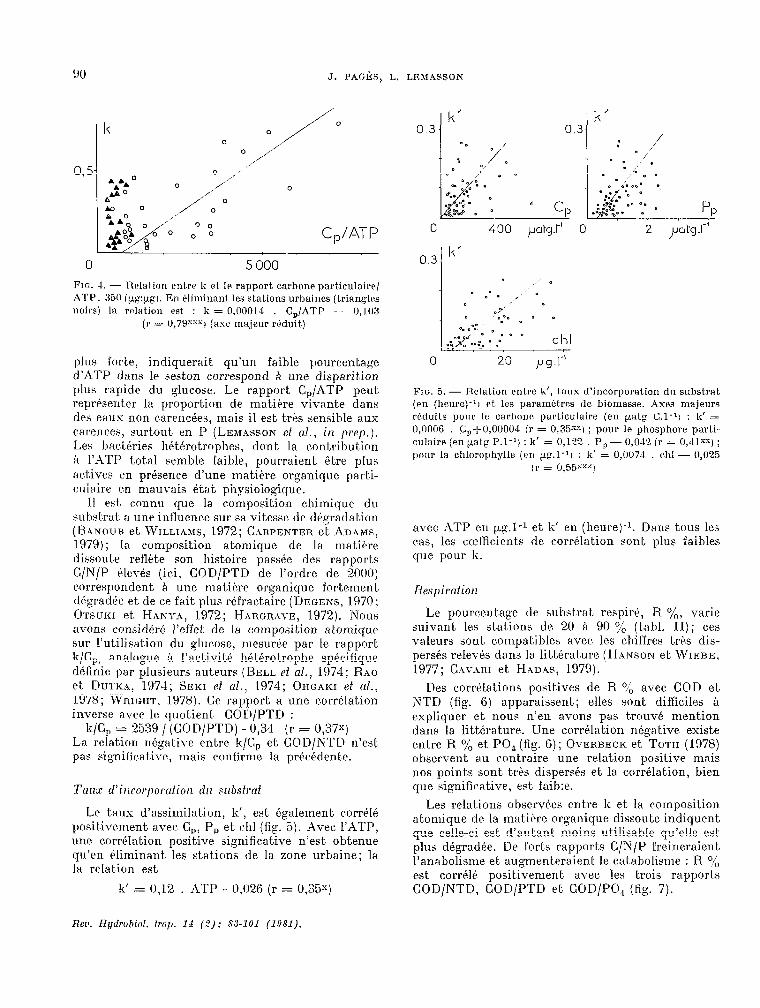
J. PAGÈS, L. LEMASSON
0.
0 5 000 FIG. 4. - Relation entre 1~ et le rapport carbone part.iculaire/ ATP. 361) ipg:pgt. En Pliminant les st.ations urbaines Itrian@les noirsj la rr~lation est : 6 = 0,00014 . C,/ATP - 0,103
(r = 0,79”“~1 (ase majeur réduit)
plus forte, indiquerait. qu’un faible pourcentage d’ATP dans 1~ seston correspond Q une disparition plus rapide du glucose. Le rapport C,/XTP peut reprf%ent..er la proportion de matihre vivante dans des eaux non c.arencées, mais il est trés sensible aus carences, surtout en P (LEn~AssoN et al., in pep.). ILes bactbries hétbrotrophes, dont la cont.ribut.ion a 1’ATP total semble faible, pourraient être plus actives en présence d’une matiére organique parfA- culaire en mauvais état physiologique.
Il est. connu que la composition chimique du subst-rat a une influence sur sa vitesse de degradation (HANfwJ3 et. WILLIAMS, 1972; ~ARPENTER et An-\~s, 1979) ; la cumpnsition atomique de la matiGre dissoute rrfléte son histoire passée des rapports G/N/P 6levés (ici, COD/PTD de l’ordre de 2000) correspondent A une matiAre organique fortement d&grad& et. de ce fait plus réfractaire (DEGENS, 1970; T)TSUKI et. HANYA, 1972; HARGRAVE, 1972). Nous avons considéré l’effet de la composition atomique sur l’ut.ilisation du glucose, mesurée par le rapport WC,, analogue A l’activité hé.térotroplie spécifique d&finie par plusieurs auteurs (BELL ef al., 1974; HA0 et W~TK~, 197-l; QEKI et al., 1974; OHC:AKI et (II., 1978; WRIGHT: 1978). Ce rapport a une wrrélation inverse avec le quotient. COD/PTD :
k/C, = 2539 / (COD/PTP) - 0,34 (r = 0,37x) La relation négative entre k/C, et COD/NTD n’est pw significatiw, mais confirme la pr6cédent.e.
Le taus d’assimilation, k’, est. également corrélé positivement avec C,, P, et ch1 (fig. 5). Avec l’ATP, une corrtYation positive signifirative n’est. obtenue qu’en 6liniinnnt. les st.ations de la zone urbaine; la la relation est
k’ = 0,12 . ATP - 0,026 (r = O,XF)
03
c! 0
0.3
I k’
FIG. 5. - Relation entre li’, taux d’incorporation du substrat (eu (lleure)-ll et. les paramèt,rss tle biomasse. Axes majeurs r&duits pour le carhont: particulaire (eu patg C.l-‘1 : 1~’ = 0,OOOG . C,+O,OOOO4 (r = 0,35xX) ; pour le phosphore parti- culaire (en palg P.l-‘1 : 1~’ = 0,lrC P, - 0,Ozl.Z (r = 0,41xX) ; pour la chlorophylle (ert kg.l-‘l : k = O,OO74 . ch1 - 0,025
avec. ATP en pg.1 -l et k’ en (heure)-l. Dans tous les cas, les ccetYicient.s de corrélation sont plus faibles que pour Ii.
Respiration
Le pouwentape de substrat respiré, R o/o, varie suivant les stations de 20 A 90 y/0 (tabl. II); ces valeurs sont, compat,ibles avec. les chiffres t&s dis- persés relevés dans la littérature (HANSON et WIEBE, 1977; CAVARI et, H4n~s, 1979).
Des corr&lations positives de R Oh avec COD et NTD (fig. 6) apparaissent,; elles sont difEc,iles à expliquer et nous n’en avons pas trouvé mention dans la 1itiSraturr. Une cor&lation négative existe entre R yo et PU, (fig. 6) ; OVERBECK et TOTH (1978) observent au contraire une relation positive mais nos point,s sont trés dispersés et la c.nrrélat~ion, bien que signific.at.ive, est faib:e.
Les relations observées enke k et la composition atomique de la mati+re organique dissoute indiquent que celle-ci est d’autant moins utilisable qu’elle est plus dégradé,e. De fcuts rapports C/N/P freineraient l’anabolisme et augmenteraient. le catabolisme : R % est, corréll positivement, avec les trois rapp0rt.s COD/NTD, COD/PTD et COD/PO, (fig. 7).

CARBONE ORGANIQUE DISSOUS 91
100 100 R %
00 0 0 8 o O*
COD 1000 /Jutg.r*
I 7% 8
0 0
/* NID
100
c 0
.-
u L
.- CL
z c
%oo C!c
1
0
l-4 10030
0 2 ptg.I-’
FIG. 6. - Relations entre le pourcanta~e de substrat respire, R ‘$A, et les paramét.res du milieu. Ases majeurs reduits : pour le phosphate dissous (en patg P. 1-l) : R ‘$6 = - 38,l . PO,+86,7 (r = - 0,5Zx9 ; pour le carbone organique dissous (en patg C.l-1) : R % = 0,06.COD+13,6 (r = 0,4Sxxx) ; pour I’azobe total dissous (en patg N. 1-l) : R y0 = 4,05 . NTD -
FIG. 7. - Relations entre le pourcentage de substrat respiré, R %, et. la composition atomique de la mat.iere or,aanique dissoute. Ases majeurs réduits : pour le rapport. carbone organique/azote tot.al (at:at) : R % = 1,3 . COD/NTD+f$O (r = 0,42Xx) ; pour le rapport c.arbone/phosphore total (atxt) : R ‘+A = O,O4 . COD/PTD+-20,4 (r = 0,41x”) ; pour le rapport. carbone orjianique/phosphate dissous (103.at:at.) : R yo =
23,5 (r = 0,339 - 35,6 . PO,/COD-/-90 (r = - 0,61x9
3.1.3. EFFET DE LA LUMIÈRE
IGffërents travaux font apparaître un effet, variable de la lumiére sur I’activit,e hétérotrophe d’une population (JONES ~~BILLINGTON, 1976; MCKINLEY, 1977; WIEBE et SMITH, 1977).
Des incubations parallèles à I’obscurit,e et à la lumilre naturelle ont, été. faites a quelques stations. Le taux de disparition du subst.rat, k, est plus élevé
0 0 COD/PTD
2’000
I C OD/PO4
à l’obscurité (tabl. IV), sauf à la st. 41, où les faibles différences observées peuvent indiquer une absence d’action réelle de l’éclairement. En éliminant cet.te station, l’augmentation de k à l’obscurité est signi- ficative (test de F : 95 %). 11 semble probable que cette augmentation de l’activité hétérotrophe soit due à une fraction du phytoplanct,on qui, a I’obscu- rite, consommerait des composés organiques (PANT et, FOGG, 1976; BERMAN ef al., 1977; LEWIN et HELLEBUST, 1978).
Heu. Hydrobiol. trop. 14 (8): 83-101 (1981).

J. PA(;I?S, L. I,EMAssoN
wlrstrat rcspirc’ ; t., : heure de d+but d’incubation
. .
_
. .
. _
. .
-
Ii I H 00 I clair sombre clair sombre I
.0057 .0013 33.6 341
.on-11 . 0043 - -
1 A’ pourcentage respirr, R (‘B, dh%it significatire- ment. B la lumi+re (test de F : 90 yO), probablement par rt;-assimilation d’une partie du CO, dégage. Des mesures de production primaire, par la méthode liC1 faites simultanément, auraient. pu permetke d’estimer l’assimila’kn de carbone minéral et donc celle du MD; nos tentat.ives ont échouf5 faute d’une hypothbse satisfaisante quant à l’activitk spécifique du CC), produit, par la respirat.ion.
3.2.. Excrétion de COD par le phytoplancton
3.2.1. (:.ZLCuL DE L'EXCRÉTION
L’lypothtse de base habit,uelle pour le c.alcul de la quantitk de WD produit, E, est. que les activit.&s spéc.ifiques des divers compartiments du systkne eau-phgt(?pl;~nct.on-CC)T) sont égales. En fait, l’acti- vit6 spécifique des cellules est inférieure, ou au maximum @le, à celle du milieu extkieur (HOBWN ai al., 1976; WIEBE et SMITH, 1977; PA&S et LESHSSON, en préparation). L’activité spécifique du COD excrété dépendra, de plus, de son origine; E~LET et, SLOAN (1966) remarquent que le COD a une act,ivité spkifique plus faible que celle du COI? rkemment synt~h8tisé, ce qui implique un rkervoir s+aré pour l’excrétion (ShfITH et. WIEBE, 1970). STOHCH etA SAUNDEFS (1975 et 1978) envisagent, deux hypothPses possibles; selon celle d’un réservoir de grandes dimensions et non en équilibre isotopique, i’activitf? spécifique des protl;ines gouverne celle du
COD et, l’excrét,ion sera E”. II a étk observ0 que les macro-moléwles et les protbines accumulent une grande Pa&ie de l’activit,t; (WALLEN et. GEEN: 1971 BAULD et BROCK, 1974) mais que c.et.te awumulntion est. relativement, plus lente (.J~~TTNER et M24~usc~~~, 1978).
Nous ;1vo1is fait l’hypotlikse simplificatrice et approximative que l’akvité spécitique du COD est identique à celle du carbone particulaire, définissant ainsi not,re valeur haute E’ : E’ = *CO . C,/*C,, (éq. 3)
Cette expression sureslime l’excrétion; en effet, une partie variable, mais non négligeable, du C, est détrit,ique et ne participe donc pas aux @changes; cependant, la relation de STORCH et S~UNDERS (1975) donnant E” aboutit., par quelques transformations simples, & : E” = 1,l .E’ +1,3.E La surestimation que nous faisons par E’ sera donc plus faible que celle que nous aurions par E”. Les valeurs de E’ sont. de 2 5 10 fois plus fortes que celles de E (tabl. V); des c.onsid&ations théoriques et des simulations numériques prévoient que E reprtsente de 10 & 40 ?/A de l’exwétion réelle (PAGES et LEMAS- SON, en pr&parnt,ion).
Il est, difkile de juger de la véracité des chiffres de E et E’ en valeur absolue ou en ut,ilisant les excrétions relatives. N»U pouvons remarquer que les rapports E’/CC)D et A/C, reflétent les ac.tivit.és spécifiques du COD et. du C,, la premiére ét,ant nécessairement inférieure, ou au maximum égale, à la seconde; nous pouvons donc rejeter toutes valeurs de E’ qui ne vérifient pas cetke inégalit.6. Il est c,ertain que les deus rapports considérés sous- estiment les valeurs reelles, puisque le C, dktritique et le COU réfractaire ne participent pas aux pro- cessus; nous adinetkons que les erreurs ainsi intro- duites se r.ompensent..
3.2.2. &oLUTION DANS LE TEMPS
La quantité cur11ul6e de COII excrétt;, E, augrnent,e au cours d’une incubation. Cette augment.at.ion n’est. pas linéaire, mais plafonne A une çert-aine valeur dans la majorité de nos observat.ions (fig. 8). Plusieurs auteurs ont rnesurt; un taux d’exorktion constant (Foc;~ rt CII., 1965; C;UILI.AR» et HALLERUST, 1971 ; BAULD et, BROCK, 197-1; WILLIAhrs et YENTSCH, 1976); par cont.re, on trouve aussi des courbes de E en fonction du temps qui présentent un plateau analogue B celui que IIOUS observons. La diminution de 1’6clairement~ naturel en fin de journée a un role (NALE\VA.JKo et SCHINDLER, 1976) niais on observe aussi un plateau, en lumibre constante, en présence
CARBONE ORGANIQUE DISSOUS 93
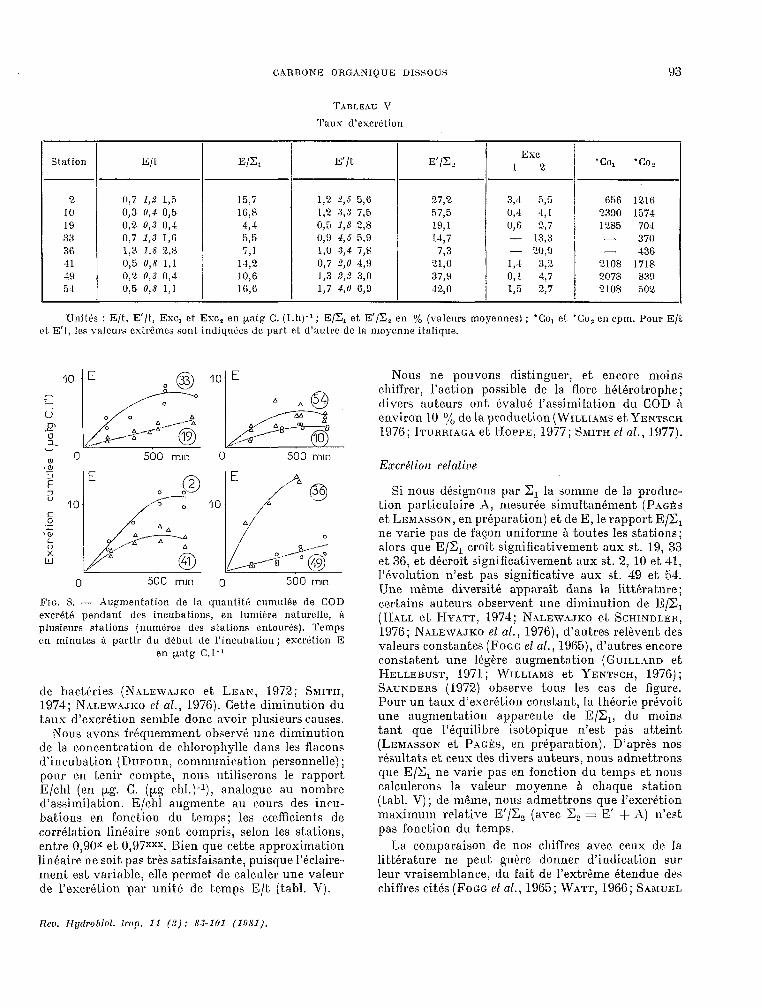
‘,?ABLEhU \’
Taux d’excrétion
Station E/t. El& E’/t E’(&
2 rl,7 1,2 1,6 15,7 1,2 2,s 5,ô 27,2 10 0,3 u,4 0,5 16,4 1,2 .3,3 7,s 57,5 19 0,2. 0,.3 0,4 a,4 0,5 1,8 2,8 19,l 33 0,ï 1,s I,6 595 lJ,Y 1,S 5.9 147 36 1,3 1,8 2,n î,l 1,o d,4 74 723 41 145 0,8 1,l 14,2 0,7 ?,O 4,!) Yl,O 19 0,2 1),.3 0,l 10,6 1,3 2,n 3,o 37,H 54 0,5 0,s 1,l I6,ü 1,7 4,O 6,O 42,o
34 5,5 656 1210 0,4 4,1 2390 1574 CI,6 2,7 , 1285 704 - 13,3 4 - 370 - 20,O - 436 14 34 2108 1718 0,l 4.7 2078 839 1,5 2,7 2108 302
Unités : E/t, E’/t, Exc, et. Exc, en palg C. (I.h)-‘; E/X, et E’/Z, en 7; (valeurs moyennes) ; ‘CO, et. ‘Co, en cpm. Pour E/t et E’I, les valeurs extrèmes sont indiquées de part et d’au& de la moyenne ilalique.
FIG. 8. - Augmentation de la quan1it.é cumulke de COD esc.rkt.6 pendant des incuba tiens, en lumiére naturelle, fi plusieurs stations (numéros des stations entourés). Temps en minutes ti partir du début, de l’incubation ; excr6lion E
en pttg C.l-1
(je bact6ries (NALEWAJKO et. LEAN, 19’72; SMITH, 1974; NALEWAJKO et al., 1976). Cette diminution du taux d’excrétion semble donc avoir plusieurs causes.
Nous avons fréquemment obaervk une diminution de la conc.entration de chlorophylle dans les flacons d’incubation (DUFOUR, communication personnelle) ; pour en tenir compte, nous utiliserons le rapport E/c.hl (en pg. C. (pg chl.)-l), analogue au nombre d’assimilation. E/chl augmente au cours des incu- bations en fonckion du temps; les cceffkients de c,orrélation linéaire sont compris, selon les stations, entre 0,90x et 0,97xXx. Bien que cette approximation linéaire ne soit pas très satisfaisante, puisque I’Cclaire- ment est variable, elle permet de calculer une valeur de l’exc&ion par unité de temps E/t (tabl. V).
Nous ne pouvons distinguer, et encore moins c.hiffrer, l’action possible de la flore héttkotrophe; divers auteurs ont évalué l’assimilation du COD A environ 10 9: de la production (WILLIAMS et, YENTSCH 1976; ITURRIAGA et, HOPPE, 1977; SMITH ei nl., 1977).
Excrèfion relative
Si nous désignons par p 1 a1 a somme de la produc- Lion particulaire A, mesurée simult,anément (PAGÈ~ et LEMASSON, en préparation) et de E, le rapport E/C, ne varie pas de facon uniforme à toutes les stations; alors que E/S, croît significativement aux st. 19, 33 et 36, et décroît significativement aux st. 2, 10 et, 41, l’évolution n’est pas significative aux st. 49 et 54. Une mème diversité apparait dans la lit,térature; certains auteurs observent. une diminution de E/X, (HALL et HYATT, 1974; NALEWAJKO et SCHINDLEK, 1976; NALEWAJKO et al., 1976), d’aut.res relèvent des valeurs c.onstantes (FOGG ef al., 1965), d’autres encore constatent une kgère augmentation (GAILLARD et HELLEBUST, 1971; WILLIAMS et. YENTSCH, 1976); SAUNDEHS (1972) observe tous les cas de figure. Pour un taux d’excrétion c.onst,ant, la théorie prévoit une augmentation apparente de E/‘C,, du moins tant que l’équilibre isotopique n’est pas atteint, (LEMASSON et PAGÈS, en préparation). D’aprés nos résult.ats et, ceu‘r des divers auteurs, nous admettrons que E/& ne varie pas en fonction du temps et nous calculerons la valeur moyenne A chaque station (tabl. V); de même, nous admettrons que l’excrbtion maximum relative E’/& (avec. X3 = E’ + A) n’est pas fonction du temps.
La comparaison de nos chiffres avec. ceux de la littérature ne peut guére donner d’indication sur leur vraisemblanc.e, du fait de l’extréme étendue des chiffres cités (FOGG et al., 1965; WATT, 1966; SAMUEL
KEV. I-lgdrobiol. frop. 14 (2): 83-101 (1.981).

J. PAGES, L. LEMASsON
FIG. 9. - Ikolution de l’escrktion par unit.6 de chlorophylle, E/chl (en IL~ C.Ipg chlj-li, en fonction de l’intensitb moyenne 1, de I’tclairement. (en \v.m-B, Pchelle logarithmiquej en
incubations in situ
of al., 1!)71 ; HOHHIE at al., 1972; BELL~ ut al., 1973; BERM.\N et. HOLM-HANSEN, 1974; SMITH et al., 1977; PENTEXOST et HAPPEY-WOO~, 1978); ~AR~ON~ et SErtr (1970) indiquent qu’il est peu fréquent cepen- dant que l’excrétion relat.ive, notre E/T&, dépasse 30 cv,;.
Les valeurs moyennes de E/X, (tabl. V) permettent de distinguer un groupe de stations (( lagunaires o (st-. lit, SS-et A!.)) avec un E/X, de 7 76 environ, et un groupe 4 d’est-uaire 0 (St.. 2, 10, 41 et. 54) avec un EJC, d’environ 15 $,, ; la st.ation 36, eutrophe et assez polh.&, se rattacherait au premier groupe.
3.2.X EFFET IXE L'IHTENYITI? DE L'ÉCLAIREMENT
T?ri; échantillons d’eau de surface ont été inwbés irz sifu & différentes profondeurs; la température étant. uniforme dans la colonne d’eau, l’Aclairement wt le seul fac.t.eur variable; nous considérons ici l’intensiti: moyenne, I,, déduite de l’intensité moyenne de surfac.e et des profils de pénétration vertic.ale mesurks (PAGES, T,EBZASSON et. DUFOUR, en prPparation ).
E/&
50
EcIuirernenI
E /Ix, a
FI~. 10. - Variation de I’escr@tion relative, E/C, (en y&) en fonction do I’intrnsitfi moyenne de l’éclairement,, 1, (en W.m-', khelle arithm+iiyuej. Courbes tracées à l’oeil
La valeur de I’excrSon spécifique, E/chl, aug- mente avec 1, (fig. 9) a des taux variables suivant les stat.ions. L’excrétion relative, E/C,, montre une augmentation IégPre, et. souvent non significative, aux intensitks lumineuses moyennes (fig. 10) et de fortes valeurs aux faibles éclairements (1 à 10 W. m-2). Ceci est, en accord avec. les observations de BELLY rt al.,(1973) et de BAULD et BROCK (1974), selon lesquelles E/V U1 est approximativement constant entre 000 et 6 000 ft..c, soit. 20 k 250 W. m-s. D’aut>res travaux ont également montré que E/L, augmente vers le fond et en sub-surface (FOGG et al., 1965; WETZEL et al., 1972), encore que d’aukes études n’aient pu mettre en 6vidence de tendance nette (HOBBIE ef al., 1972; SMITH et al., 1977); la lumière semble agir moins par son intensité que par sa c.omposition spectrale (WALLi3v et. GEEN, 1971; BROWN et GEEN, 1974j. Une augmentation de E/& a été constatée dans toutes les conditions sub- optimales, par exemple en eau oligotrophe (ANDER- SON et ZEUTSCHEL, 1970) ou en limitation par le CO, (ROL~E, 1978); ces facteurs ne peuvent avoir agi ici, puisque tous nos échantillons provenaient, à chaque station, d’un m&me prélévement de surface.
Hezr. Hgdmbiol. frop. 13 (2): 83-101 (1951).

CARBONE ORGANIQUE DISSOUS 95
1 E-I
Fro. 11. - Helation entre I’excrcWm relative, E/X, (en %) et. les parnmttres de biomasse. Axes majeurs rtduits : pour la chlorophylle (en pg.l-‘) : E/S1 = lIl,T/chl-J-8,s (r = 0,13X) ; pour 1; carbone particulaire (en patg.l-l) : E/S, = 1&4O/C,+
6,'k (r = 11,7l"X")
3.2.4. EXPERIENCES AVE~ DI.~LYSE
Les chil’fres de c,pm obtenus lors des deux phases consécutives des expériences avec. dialyse, *CO, et *CD~, ne sont pas significativement différents (tabl. V), bien que *Goa soit généralement inférieur H *CO,; la suppression du trac.eur du milieu externe lors de la deuxi+me phase explique cette tendance. Nous voyons aussi que les différentes manipulations n’ont pas provoqué de traumatisme important. Enfin, il semble que la membrane utilisée ne gêne pas la diffusion du COU, dont la majeure par& aurait un poids moléculaire inférieur k 10.000 d’aprts les caractéristiques de la membrane; ceci est en accord avec les chiffres existant, dans la littérature, t.ant, pour la composition du COD dans les eaux que pour celle du COD produit, par diverses populations (WATT, 1966; HALL et HYAT, 1974), bien que des variat,ions import,antes aient été observées en fonc,tion de l’age et de la composition spécifique des populations (NALEWAJKO et LEAN, 1972) et en fonction de la présence de batteries (NALEWAJKO et SCHTNDLER, 1976). Nous ne pouvons pas déterminer une limite basse du poids moléculaire du COD produit ici.
Les valeurs d’excrétion, Exc,,, qui sont des
moyennes sur trois c.hiffres, sont du méme ordre que celles de $3/t. si nous exprimons les deux en patg.C (1.11)-r (t.ahl. V). D e meme, Exc, est du méme ordre de grandeur que E’/t; le mode de c,alcul de ces deux dernières excrétions est analogue, mais les conditions expérimentales sont, assez différentes, puisque I’incu- bation dont a été obt,enu Exc, s’est faite sans traceur dans le milieu extérieur.
Les valeurs de Exc, sont largement supérieures a celles de EscI, souvent d’un facteur 10, alors que nous avons vu que les chiffres de départ *Go, sont inférieurs a *GO~. La différence entre Exc, et Exc, est analogue a celle entre E et E’ et a la même origine : le mode de calcul fait intervenir le t.erme C, et surestime l’excrétion, puisque tout le C, ne participe pas aux processus, comme déja relevé plus 11aut.
3.2.5. RELATIONS AVEC D'A~TRES PARAMETRES
Le taux d’excrétion est considéré comme dépen- dant de l’état physiologique et, 1 ou nutrit,ionnel des cellules; nous avons essayé de dégager une relation ent,re les divers parannXres décrivant l’excrét.ion et les facteurs suivants : C,, chl, salinité, PO,$-P du milieu, rapport. N/P minéral du milieu, rapports atomiques C/N/P des particules et chl/ATP.
Nous observons des relations négatives entre E/& et C, d’une part, c.hl d’autre part, (fig. 11). Plusieurs travaux ont montré une diminution de l’excrét.ion relative pour une biomasse croissante, aussi bien dans des populations naturelles que dans des cultures (FOGG rf al., 1965; WATT, 1966; BROWN et GEEN, 1974); la forme mathématique de la relation entre E/G et la concentration en chlorophylle, utilisée comme mesure de la biomasse, varie suivant les auteurs. Pour nos données, une hyperbole décrit le mieux les relations négatives entre E/IZ, et les deux paramètres de biomasse C, et chl. Par contre E/Z;, n’est pas lié à ATP.
Des relat.ions negatives mais non significatives apparaissent entre E/C, et PO,P d’une part, chl/ ATP d’autre part; ces mauvaises relations, et l’absence de relat.ion avec un paramètre tel que C,/P,, qui déc.rit. l’état physiologique des organismes, sont assez surprenantes ( IGNATIADES, 1973). Aucune relation n’exist,e entre E/Ç, et C,/ATP.
Une corrélation positive existe entre E et la pro- duction part,ic.ulaire A (fig. 12); une telle relàt.ion est signalée par plusieurs auteurs, et décrite soit, par une équation log/log (ANDERSON et ZEUTSSHEL, 1970; BERMAN et HOLM-HANSEN, 1974), soit par une équat,ion arithmétique (Foc;~ et nl., 1965; SMITH ef nl., 1977) ; pour des chiffres identiques de production particulaire, nos valeurs de l’excrétion sont plus fortes que celles calculées d’après les
Rev. Hydrobiol. trop. 1-I (2): S-3-101 (1981).

J. PA&S, L. LEMASSON
0 50 Production (pUt’J.Y >
FIG. 1% - Relation entw es&lion (E) et production parli- culaire (A), tout.es tleuu en ~dlg.l-~. Ase majeur reduit :
E = I:l,?16 . A+ 1,37 ,r = 0 7WX”I 1 ,
relations clonnées par les auteurs mentionnés; il faut remarquer que, bien que la corrélation entre E et h soit. haut.ement significative, les points sont fortement dispersés.
3.3. Distribution du COD
X3.1. C:ON~:ENTH~'I.IONS
,-\u cours de stations de 24 heures en huit points, nous n’avons pu mettre en évidence de variation si@fic:ative des concentrations de COU in sitrz. IJne relative c.onstanre du COD est logique si le systéme est ?II équilibre, production et utilisation se compen- sant.. La concentration du COD peut également ne ne pas varier de facon perceptible si la fraction labile est trop faible pour que ses variations soient. mesurables.
Les wneent~rations de COD sont, suivant les st,ationx. de 500 à 1600 patg C.I-1 (tabl. II); elles sont plus élevees que la majorité des chiffres releves dans la littérafure (P~RL et GOLDMAN, 1972; GILLRPSIE et VACCARO. 1978) mais les valeurs eitees pour des environnements analogues à la lagune sont du rnéme ordre que les notres (HAPP et cd., 1977). Les concent~rations les plus faibles sont., logiquement, dans la zone d’estuaire, mais il n’existe pas de relation ent,re COD et, salinité. ilucune tendance saisonnik ne peut ét,re dist.in@e.
La composition de la matiere organique divs0ut.e est. variable. Il n’existe aucune relation si@ficative entre COD et les formes organiques NOD et POD; le COD et l’azote minera1 sont lies par la relat,ion NTD = 0,015 . COD - 6,7 (r = O,:jtix~)
1
0
20
n
NJ. D. pu tg .i-’
“. 1’000 rutg.1-’
PO4 patg,l“
FIG. 13. - Relation entre le WD et les formes minkales de N et P (tous en (&g.l-ll. Pour le Pc),, les stat.ions du
chenal central (voir texte\ sont. en noir
L’ext.rapolat.ion à teneur nulle d’azote donne un COD cc résiduel 1) de 147 pal,,- C.l-l (fig. 1.3). Pour le phosphore mintral, une minorite de stations à fort. PO,, situées dans le chenal central, doit étre separée de la majorité des stations, qui fournit la relation : PO, = 0,0007 . COD - 0,25 (r = O,ï;Oxx)
ce qui indiquerait, un COD residuel de 357 Fa& C.l-l a teneur nulle de PO, (fig. 1.3).
Le rapport atomique moyen de la matière orga- nique diss0ut.e COD/PTD est de 1022, alors qu’il est de 160 pour la matière particulaire (LEMASSON, en préparation). Cette différence illustre la degradation de la rnatiére organique dissoute.
3.3.2. %8.41 DE BILAN
Nous avons vu que le t.aux de disparition du @mose est, de 10 à 30 ?” par heure; l’excrétion de COD par le phytoplancton représenterait, suivant le mode de c,alrul et les stations, de 0,l a 1 y:, du COD par heure. Cette nette différenee d’ordre de
Rru. Hydrohiol. frop. Id (2): 83-101 (1981).

CARBONE ORGANIQUE DISSOUS
TABLEAU VI
Comparaison des productions hétérotrophe et. primaire, calculées sur 12 heures de jour
97
Station
2 10 19 33 36 41 49 54
Excrttion
ptg CI-1
E E’
14 20 5 40 4 22
16 54 22 41 10 24 4 26
10 48
z
m
6,0 1,5
10,o 2,5 %O 8,5 2,2 2,5
HtWrotrophir
mg C.m-z
min. mas.
202 432 18 144 96 528 96 324
103 192 204 490
21 137 60 288
Photosynth.
mg C.m-*
1593 326
2760 4525
651 176 258 803
Httéro./Phot.
143
min. mas.
13 27 5 44 3 1 !:, 2 7
16 29 26 63
8 53 7 36
grandeur est une premike indicat‘ion de l’impossi- bilité d’étendre les résultats obtenus pour le glucose r’r. la t,otalité du COT), comme l’ont fait remarquer plusieurs auteurs (H~BBIE et WRIGHT, 1965; OVER- BECK, 1975). Divers travaux montrent que 5 à 10 % du COD est de faible poids moléculaire (OGUR.~, 1974; DUNSTALL et NALE~AJKO, 1975; WHEELER, 1976; STABEL, 1978) et les sucres libres représentent moins de 5 oh de la mat.iére organique dissoute (STUMM - ZOLLINC;ER~~HARRIS, 1971 ;OGURA~~GOTOH,~~~~). En regard de cette faible portion du COD facilement dégradable, nous avons pu constater indirectement qu’une forte part du COD est probablement réfrac.- taire.
Nous ne pouvons donc guère obtenir de c.hiffres, meme approçht%, d’utilisation du COD à partir des observations sur le glucose.
Sources de CXID
L’excrBtion par le phytoplanc.ton vivant est, une source de GOD. Sa destruction mécanique par le zooplanckon a étk mentionnée ( LABIPERT, 1978). Plusieurs auteurs estiment cIue la dégradation du C, détritique est la source princ.ipale de COD (SAUN- DERS, 1975; GERLETTI, 1965); de nombreux travaux indiquent que O,5 A 2 y& du C, sont ainsi transformés en COD par jour (HANDA &YANA~, 1969; HANDA, 1970; GOCKE, 1978; WETZEL el al., 1972). Les c.oncentrations de C, total é,t,ant, dans la lagune, de 5 à 25 o/. de celles du COD, la dégradation du C, détrit,ique fournira moins de COD que I’excrktion phytoplanctonique. Les apports externes par les débris végktaux jouent un rOle important dans certains cas; mais, en t,enant compte de la morpho- logie de la lagune, et en utilisant des chiffres relevés
dans des c.limats a~mlogues (~(LINGE et, RODRIGUE~, 1968), nous voyons que ces apports seront négli- geables. Enfin, les apports par les riviéres sont peu quantifiables, mais il est. probable que les cours d’eau a eau fort.ement colorée apportent peu de COD dégradable. Nous devons donc. considérer que la majeure partie de la production de COD est due à l’excrétion.
Nous avons admis que l’assimilation de COD est équilibrée par l’excrétion due au phytoplancton. Le rendement molaire ((( molar growth yield n) sur des substrats simples est. de 14 à 35 yo suivant. les conditions (BAUCHOP ~~EL~DEN, 1960; SCHULTZE et LIPE, 1964; OXENBURGH et SN«WSWELL, 1965; STOUTHAMER et BETTENHAUSEN, 19%); il eSt de l’ordre de 30 yo sur des substrats complexes (GOCKE, 1970). Si nous admett,ons un rendement. de 20 y0 sur le COD, nous pouvons calculer la production hétéro- trophe à partir du COD excrkté et, la comparer à la production primaire intkgrée sur la colonne d’eau et la période d’klairement (PA&S, LEIMSSON et DUFOUR, en prkparation). Comme d&jA signalé, les divers auteurs donnent des chiffres assez variables quant à l’importance des bac.téries (SOROKIN, 1971; BANSE, 1974). Cette production hétérotrophe appa- rait comme non négligeable (tabl. VI) mème en considérant les valeurs obtenues ti partir de E; les bornes hautes, calculées sur la base de E’, donnent des rapports (production hétérotrophelproduction primaire) compris entre 10 et. 60 94, ceci ‘calculé sur 12 heures (tabl. VI) : ces chiffres augmenteraient encore en considérant une période de 24 heures.
Manuscrit reçu au Service des Édifions de f’O.R.S.T.0.M. le 16 septembre 1980.
Heu. Hgdrobiol. trop. 14 (2): 83-101 (1981).
2

J. PAGES, L. LEMASSON
BIBLIOGRAPHIE
A~~o~soiv (Si, 197% - Excretion of organic matter by phytopl~rrkton in vitro. Limnol. Oceunogr., %?/A.
ALHHIG~‘~ IL. .J.i, 1977. - Hetcrotrophic bactmial dynamics in the tower Fraser River, its est,uary ancl Georgia SIreait, British Columbia, Canada. ~11~. Biol., 39/3 : 21%211.
;INDEHSON (c:. G.!, ZEUTSCHEL (R. P.), 1970. - Release of ttissolvett orgsnic matter by marine phytoplankton in eoastal and offshore areas of the northeast Pacitlc. CJwan. Limnol. Ocennogr., I5/3 : 4024.07.
ATKINS (C:. L.), 1973. - Modeles à compartiments multiples pour les systemes biologiques. Gauthier-Villars (Parisj, pp. 1Yl.
Ba~oun (M. W.], WtLLt4hrs iP. J.1, 1972. - Measurements of microbial activily and organic material in thr western Mrditerrnnean Sea. Deep Setr Res., 19/6 : ‘4~‘~-4.4‘~. .c *
BAN~~ (Ii.), 1971. - In thr role of bacterioplanliton in thr tropical CJrean. nlar. Biol., 24/1 : 29-168.
RAHRII.R (R. T.1, RYTHER (.J. H.l, 1969. - Organic chrlators : factors affecting primary production in the Cromwell rurrrnt upwelling. J. e.cp. mur’. Biol. Ecol., 3 : 191-199.
BAIJCHOP (T.), EL~~EN (S. R.), 1960. - Thc growth of micro- organisms in relation to their energy supply. J. qen. Alicrobic~l., 13 : 457~46!).
BAULD (J.), BROC~ (T. D.), 1971. - Algal excrct.ion anti bacterial assimilation in hot. spring algat mats. .J. Phycol., lc)/l : lCIl-106.
BI?LL (W. H.I. LANG (J. M. J, J~ITC:HELL (R.), 1974. - Selective st-imulation of marine bacteria hy algal estracellular protlncts. Limnol. Ocecznogr., 19/5 : 833-839.
BELL-Y (R. T.), TANSEY (M. R.), BROCR (T. D.), 1973. - Algal excretion of W. labeled compounds and microbial interactions in C~ywzdirzm czzldarizzm mats. J. Phycol., 9 : 123-127.
BENRETT (M. E.), HOBBIE (J. E.I, 1972. - The upt.ake of glucose hy Chlamydomonas sp. J. Phycol., 8/4 : 392-398.
BEWAN (T.), HOLI\I-HANSEN (O.?, 1974. - Release of pboto- assimilat.ed carbon as dissolved organic mat.ter bg marine phytoplankton. ~Xlnr. BM., 28/1 : 305.
BERMAN (M.), SGHOENFELD (R.j, 1956. - Invariants in experimental data on lincar kinetics ami t.he formu- lation of models. J. rlppl. Phys., 27 : 1361-1370.
BERhrAiY (T.j, Hanas (CI.), KAPLAN (B.). 1977. -- Uptake and respiration of organir compounds and hetrrotrophic growth in Pedinstrzzm dzzple.c (Moyen). Fresh. Biol., 5/5 : 195~5w2.
ROL~E (A. l, 197X. - Drr Einfluss van CCJz- Versorgung, BelrurhtuIlasstWrt~e und Temperatm auf die Konzcn- tration extrazellnl8rer oryanischer Suhstanz in Syn- chronkulturen V,BIl Scenedesmrzs nczzfus. Arch. Ilydrohiol., Suppl. 511-i : 114-455.
BROWN (T. J.), C:EEX (CG. H.1, 1974. - The affect of light. quality on thc c.arbon metabolism and estracellular release of ~~hlanz~~d(lmc~nas reinizurdifii. J. Phycol., 10/?2 : 213-220.
CAMPRELL (P. G. C;.i, BARER (-1. H.), 197% - &Ieasuremrnt of sulphatr uptake in thr dark by suspended micro- organisms : applicaf iorr to running mat.ers. iverh. infernaf. Serein Limnol., ?(Jr2 : 1423-1428.
G~RPENTER is. H.1, hDAhlS jar. S.j, 1979. - Effects of nutrients ami temperature on docomposition of Myrfophyllnm spicnfzrm L. in a hard-water eutrophic lake. Limnol. Oceanogr., 2.1/3 : 520-5-X.
CAVARI (H. Z.), H.4w.s (0. I, 1979. - Hrterotrophic activity, glucose uptake and priniary productivity in Lake Kinneret. Fresh. Biol., Y/4 : 3’29-338.
C:CWENEY- (M. F.i, 1!17Y. Separation of algae and bacteria in lake water hy sise. fractionation. Verh. inferntrf. Verein Limnol., 20/2 : 1264-1269.
CROWE (A. j, ~:RO~T (A.), 1969. - ;LIat.hematics for biologists. Academic PrcSs (Lond»ni, pp. 303.
DAWSON (R.), Gocrt~ (K.), 1978. - Heterolrophic activity in COIIlpiUiSOIl tu tho free XniKlO afitl C»IICeIl~KitiOIlS
in Ballic sra mater samples. Ocannol. Acfa, l/l : 45-54.
DEGENS (E. T.), 19711. - Molccular nat.uro of nit.rogcnous coumpounds in sea water ad recent marine scdiments. In : Clrgnnic niat.t.er in nat.ural water (Symposium Univ. Alaska, 19681. Ed. by Il. W. Hootl. Insf. Mnr. Sri. Ocçasionat Publ. 110 1 : 77-106.
DUNSTALL (T. G.1, N.~L.EWA.TKO (C.1, 1975. - Extracellular releasr in planktonic bacteria. T’erh. infernuf. Iverein Limnol., 19/4 : 2643-‘26l!J.
E~PLEY (R. W.), SLCIAN (P. R.1, 1965. - Carbon balance esperiments with marine phyt oplankton. J. Fish. Res. Board Car~, 22./4 : 1 tIP3-1097.
FANICRON~R (P. ‘I-.i, BURGH de (M. E.I, 1977. - Diurnal exudation of W. labelled compounds by t.he large kelp AIacrocysfis infegriîcrlizz Bory. .J. e.cp. mar. Biol. Ecol., 28 : 151-162.
FER~LISON (R. L.j, MURDOCH (M. B. t, 1975. - Microbial ATP and organic carhon in sediments of t.he Newport River est.uary, N.C. In Estuarine Research. L. E. Cronin cd. Acati. Press. : 22%‘250.

CARBONE ORGANIQUE DISSOUS 99
FLEISCHER (S.), 1975. - Sugar turn-over in lake water and sediment. Verh. infcrnnf. T’erein Limnol., 19/4 : 2627- 2635.
FO(X (G. E.), 1952. - The production of estracellular nitro- genous substances by a blue-green alga. Proc. roy. Soc., ser. B, 139 : 372-397.
Foc;~ (G. E.), \VEOTLAKE (D. F.), 1955. - The importance of extracellular products of algae in freshwater. Verh. internat. Tlrrein. Limnot., 12 : 219-2.32..
Font (G. E.), NALEWA.TK» (Ci, WATT (W. D.), 1965. - Estracellular products of phytoplankton photo- synthesis. Proc. R. Soc. (Londonj, ser. B. 162 : 517-534.
GERLETTI (M.), 1965. - The soluble protein content of Lago Maggiore waters, with considerations on ot.her organic subst.ances. Mem. Ist. ital. Idrobiol., 18 : 217-239.
GILLESPIE (P. A.), VACCARo (R. F.), 1978. - A baCteria1 bio-assay for measuring the copper-c.helation capacity of seawater. Lirmol. Oceanogr., 23/3 : 543-548.
GOCKE (K.), 1970. - Untersuchungen über Abgabe uud Aufnahme von Aminosauren und Polypeptiden durch Plankt«norpanismen. Arch. Hydrobiol., 67/3 : 285-367.
GOCKE (IL), 1974. - Methodische Probleme bei Int.ersu- chungen zur mikrobiellen Stoffaufnahme in Gewassern. Kieler Meeresforschung, SO/I : 12-21.
G~CKE (K.), 1977. - Comparison of methods for determining the turnover times of dissolved orranic compounds. Mur. Biol., 1212 : 131-141. .
GOÇICE (K.), 1978. - Untersuchungen über den Jahrescyclus der heterotrophen Aktivitat. in der Kieler Forde. drch. Hlydrobiol., 82/1-4 : 123-141.
GONYE (E. R.), ~ARPENTER (E. .J.), 1974. - Production of ironbinding compounds by marine micro-organisms. Limnol. Oceanogr., 19/5 : 840-M%
GRIFFITHR (R. P.), HANUS (F. J.), MORITA (R. Y.\, 1974. - Thc effect. of various water sample treatments on lbc apparent upt,ake of glutamic acid by natural microbial p»pUkLtionS. Cm. J. Microbiol., 20 : 1261-1266.
GUILLARD (R. R. L.), HELLEBLJST (J. A.), 1971. - Growth and the production of extracellular substances by two strains of Phoeocysiis pouchefi. J. Phycol., 714 : 330-338.
HALL (Ii. J.), IIYATT (K. D.), 1974. - Marion Lake (IBP) from bacteria to flsh. .T. Fish. Hes. Board Cnrz., 31/5 : 893-911.
HALMANK (SI.), STILLER (M.), 1974. - Turn-over and uptake of dissolved phosphate in freshwater. A study in Lake Kinneret. Limnot. Oceanogr., 19/5 : 774-783.
HAMILTON (R. D.), AUBTIN (K. E.j, 1967. - Assay of relative heterotrophic potential in the sea : the use of speci- flcally labelled @ucose. Can. J. Aricrobiot., 13 : 1165 1173.
HANDA (N.1, 1970. - Dissoved and particulate carbohydrates 171 Organic matter in natural waters (Symposium Univ. Alaska 1968). Ed. by D. W. Wood. Inst. Mur. Sci. Occasional Publ. n’> 1, pp. 129.
HarvnA (N.), ~ANAGI (K.), 1969. - Studies on water-extrac- table carbohyrirxtes of the part.iculate matter from the Northwest Pacifie Ocean. Alar. Biot., 4/3 : 197-207.
Reo. Hydrobiol. trop. 14 (2): S3-101 (19X1).
HAN~OK (R. B.), WIEBE (W. J.), 1977. - Heterotrophic activity associated with particulate size fract.ions in a Spartinu aZterni/Tnrn Salt-marsh estuary, Sape10 Island, Georgia, USA, and the continental shelf waters. Alar. Biol., 42 : 321-330.
IlArP (G.), GOSSELINR [J. G.), DAY (J. W. Jr.), 1977. - The seasonal distribution of organic carbon in a Louisiana estuary. Estuar. coastal mur. Ski., 5 : 695-705.
HARGRAVE (B. T.), 1972. - Aerobic decomposition of sedi- ment and det.ritus as a function of particle surface area and organic content. Limnol. Oceanogr., 17/4 : 583-596.
HARRIS (G. P.), PICCININ (B. B.), 1977. - Photosynthesis by natural phyboplankton populations. Arch. Hydro- biol., 80/4 : 405-457.
HERBLANn (A.), 1975. - Utilisation par la flore h&@rotrophe de la matière organic naturelle dans l’eau de mer. J. erp. mur. Biol. Ecot., 19/1 : 19-31.
HOBBIE (J. E-j, CRAWFORD (C. c.). 1969. - Respiration corrections for bacterial uptake of dissolved organic compounds in natural waters. Limnot, Oceanogr., 14/4 : 528-532.
HORBIE (J. E.), WRIIXIT (R. T.), 1965. - Bioassay with bacterial uptake kinetics : glucose in freshwater. Limnot. Oceanogr., 1013 : 471-174.
Honnr~ (J. E.), HOLM-HANSEN (O.j, PAcIiAnn (T. T.), POMEROY (L. R.), SHELDON (R. W.), THOMAS (J. P.), WIEBE (W. J.), 1972. - A study of the distribution and activity of microorganisms in ocean water. Limnol. Oceanogr., 1714 : 544-555.
HOBSON (L. A.), MORRIS (W. J.), PIRQUET (K. T.j, 1976. -. Theoretical and esperimental analysis of t.he *% fech- nique and its use in st.udies of primary produc.tion. .J. Pish. Res. Bonrd Can., 33(8 : 1715-1721.
TüN.4TrAnEs (L.), 1973. - Studies on t.he fact.ors affecting the release of organic matter by Skeletonema costafum in field conditions. J. mar. biot. A~S. Lr.K., 53/4 : 923-935.
ITURRIA~A (R.), HOPPE (H. G.), 1977. - Obserwtions of heterotrophic act.ivity on photoassimilsted organic matter. Mur. Riol., 40/2 : 101-108.
JANNASCFI (H. W.), 1967. - Growth of marine bacterin at, limibing concentrations of organic carbon in seawat.er. Limnot. Ocennogr., 12/2 : 261-271.
J.\NNASCI~ (H. W.), 1968. - Competitive elimination of Enterobacteriaceae from seawater. Appl. AIicrobioZ.t 16/10 : 1616-1621.
JANNASCH (H. W.), 1970. - Threshold concentrations of carbon souces limiting bacterial growth in sea water. In Organic Matter in Natural Waters, ed. D. W. Hood. Inst. Mar. Sci., U. of Alaska, Pub. no 1 : 321-328.
JONES (A. K.), RILLIN~TOE (C. A.), 1976. - Observations on carbon dioxide fixation in the Nant-y-Mach reservoir, C,eredigion, Wales. Hydrobiot., EiO/l : 33-42.
JUTTNER (F.), MATUXHEK (T.), 1978. - The release of low molecular meight. compounds by the phytoplankton in an eutrophic lake. Water Res., 12/4 : 251-255.

KL.INC~E (H.), HI~IIR~~IJES (W’. A.), 1968. - Litter production in an area of Amazonian terra firma forest. Part. 1 : Litter fall, organic carhon and total nit.ropen rontents of litter. Amazoniana, l/.i : 287-302.
Lanwwr (W.), 197X. - Helease of dissolved organic carbon by grazing zooplankton. Limno/. Oceunogr., S/d : 831-834.
LEWIN (J.), HIZLLEBUST (.J. A. t, 1978. - Utilization of glutamate and ghlcose for heterotrophic growlh hy thr marine pennat e diatom Nifischiu hreui.s. ftlnr. Bic~/., 47/1 : l-8.
MCKINLF.Y (K. H.r. 1977. - [email protected] uptake of 3H-gl~lcose in a small hard-mater lake. Ecology, W/O : 1358-1365.
MUKERJI (D.j, GLVVER (H. E.), MOHRIS (I.j, 1978. - Diversity in the mechanism of carbon dioxide fixation in »unelie[ltr ferfiolecta (Chlorophyceae). .J. Phycol., 13-/1 : 137-l 12.
MUNRO [A. L. 9.1, BRIXX I,T. D.,, 196S. - Distinction bet.ween baotwial and algal utilization of soluble substances in the Sea. ,7. yt=n. rllicrobinl., 51/1 : 35-42.
NALMWA.IR(.I (Ci, LEAN (D. R. S.), 1972. - Growlh and excw- tien in planktonic algae and bacteria. .7. Phycd., 814 : 361-366.
NALEWM~O (C.1, ,C;c~rwr~~m (D. IV.), 1976. - Primary production, cstracellular release and heterotrophy in two lakes in the E.L.A., Nort.hwestern Ontario. J. Fish. HF.~. Bourti Crin., 33/2 : 219-226.
NALEWAJKO (C.), Du~~ST.~LL (T. G.), SIIRAR (H.1, 1976. - Kint‘tirs est.racellular release in axenic algae and in miked algal-bac.t,erial cult,ures : significance in esti- mation of total (gross) phyboplankton excret.ion rates. J. Pkyccd., l’L/l : I-5.
OclIRA iN.1, 1970. - On the presence of 0.1-0.5-p diSS«lWd or-anic malier in seawater. Limnol. Oceanogr., 1513 : 47647Y < .
OGURA (N.1, 1971. - Xolccular weight fractionnal.ion of dissolved organic matter in coastal seawater by ultra filtration. Mur. Biol., 2d/J: : 306-312.
~C;URA (8.1, 1976. - Further studies on decomposition of diasolved orranic matter in coasial sea-water. Mnr. &(Il., 31/1 : 101-111.
C)~URA iN.1, GOTOH (T.), 1974. - Decomposition of dissovetl carbohydrates derived from diatoms of lake \-uno-ko. Inf. Revue ges. Hytlrctbiol., 5911 : 39-47.
0HG.4ICI (S.;, HWAR’~ (S. L.), &L4TSUMoTO (J.), 1978. - Kinetics of attached microbial prowt.h in a continuous stirrpd tank reactor. Water Res., 121-t : 243-219.
Orsu~r (A.), H.4~1.A (T.j, 1972. - Production of dissolved or-anic matt.er from dead green algal cells : 1 : Aeroùic microhial dwomposition. II : Anaerobic microbial decomposition. Limnol. Octwzogr., 17/2. : 248-264.
OVERRECK (J.1, 1976. - E CO o~y of aquatie. organisms. 1 : 1 Bact.eria. Dislribution pattern of uptake kinetic responses in a stratitied wtrophic lake (Plus see eco- system shldy IV). Verh. internat. Verein Limnol., 19/4 : 2600-2615.
OVERRECIC (J.), Toru (D.), 1978. - Eintluss des Phosphat- gehalts auf dit! Glucoseaufnahme bsi Bakterien. Arch. H!ydrohiol., x1, 1/1 : 11+1X
OXENBURG~ (M, Y.), SNC)~WELL (A. M.), 1965. - TTse of molar gromth yields for t.he evaluat.ion of energy-producing pathways in Lacfc~bacillzzs plantarum. J. Ba~t., 89/3 : 913-911.
PAERL (H. W.1, 19ï-1-. - Bact.erial uptake of dissolved organic matter in .relat.ion to detrital ageregation in marine and freshwa t er sys t rms. Limnd. Oceanngr., 1916 : 966-972.
PAERL (H. W.i, Gor.onr \N (Cl. F:.,, 1972. - Heterotrophic assays in the deteCt.i»u of watrr masses at Lake Tahoe, I,alif. Lirnnol. Ocrcznoyr., 1 ï/I : 14.5-I 48.
PAI?ES (3.1, LF.M.~SSOX (L.I, DUP~UR (P.), 1979. - Éléments nutritifs et prtrduct.iori primaire dans les lagunes de Cote d’lvoirr. C;ycle annuel. Arrh. Scienf. Centre Reck. Ocennogr. -1bi$jan., 5/1 : l-60.
PANT (A.), Foc~; (G. E.i, 1976. - Uptakr of glycollate by Sh-elefctnema cosftrtiun~ in 1)acterized culture. J. ecp. mar. BM. Ecd., OY/3 : 5!?7-234.
!?~RSONS (T. B.), SEKZ (I.I.), 1970. - ImportancP and general implications of organir matter in aquatic environ- ments. In 0r;anic matier in natural waters (Univ. Alaskaj. Ed. by D. IV. Hood. Insf. Mur. 8ci. Occasional Publ. 110 1 : l-27.
P.~RSONS (7. R.), STHICKLANIJ (J. r). H.), 196% - On the production of particulate organic carbon by hetero- trophic procesbes in sea-wit.er. Dwp. .%a Research, 8 : 211-222.
PENTECOST (Ai, HaPpE>--Wnnrr ((:. M.i, lW8. - Primary pIXdUC~.iOU Stlidit:S in tW« linked ht. C»Ilt~aS~ing
Welsh lakes. Freshzzvfer Riof., 8/1 : 9-23.
Bar (H.), 1979. - Mic.robioio-y of Central Amazon Lakes. Ama~oniana, 6/i : 583-599.
Hao (s. S.j, DUTK.~ (B. J.), 1971-. - Influence of temperat.ure on lake bacterial activity. IVafer Res., 818 : 525-528.
ROBINSON (C;. G. C:.), RRANDT (L.), 1975. - The influence of temperatuw and bacterial numhws on t.he het.ero- trophic uptake of “(:-glllclJSe in West. Blue Lake, Manitoba. TTer. irzfernnf.~ T?crein Limnol., 19/1 : 2616- 2619.
SAXUEL (S.), SAAII IN. BI.,, Foc~ (G. E.), 1971. -- Liheration of extracellular products of photosynthesis by tropical phytoplankton. J. mur. bid. dssoc. C?I<., 61/4 : 793-ï98.
,~A’LINL)ERS (G. W.), 1072. - The kinetics of extracellular release of so111l~le orrduic mattcr hy plankt.on. Ver-h. internol. Vwein ungrzr,. Limnol., Ir‘/l : I-lO-146.
SAUNIIIZRB (Ci. W.), 1975. - Decomposition in fresh water. In : The rolr of terrestria and aquatic oqanisms in decomposition processrs. Ed. by J. M. Andersen and A. Macfadyen. (Blackwrll Scientifia Publications) : 311-771 * . .
SCHINDLER 1.1. E.i, ALREWS (J. .J.j, Honrw (K. H.), 1972. - A premilinary investigation of organic-inorganic associations in a stagnating system. Limnol. Oceanogr., 1716 : 952-957.
Ru*. Hytfrubid. trop. 14 (2) : 83-101 (iY~Y1).

ChRBCINE CIRMNIQUE UISSOUS 101
SC~ULZE (K. L.), LIPE (R. S.), 1964. - Relationship between substrate roncemration, growth rate and respiration rate of E. coli in cont.inuous culture. Arch. Mikrobiof., 18 : I-20.
Smir (H.), NAIX (T.), OT~BE (I-I.), 197-1. - Turnover rate of dissolved materials in the Philippine Sea at winter of 1973. Arch. Hydrobiol., 7312 : 238-2M.
SHARP (J. H.), 1973. - Size classes of organic carbon in sea water. Limnol. Ocennoyr., 18/3 : 341-1-17.
SIIAHP (J. H.), 1978. -- Reply to comment by S. Aaronson. Limnol. Oceanoyr., ?3/1 : 839-840.
SIEBURTH (.J. hfc.N.), 1977. - International Helgoland Symposium : Con\-ener’s report on the Informa1 Session on biomass and productivity of micro-organisms in planktonic ecosystems. Helgctfifnder roiss. Meere- sunfers., 30 : 697-701.
SIEBURTH (J. hic. N.), SMETACF.K (\‘.), LFXZ (J.), 1978. - Pela@ ecosystrm structure : Heterotrophic compart- me& of the plankton sud their relationship to plankton size fract.ions. Linmol. Ocennogr., 23/6 : 1256-1263.
SMITH (D. F.), 197-1. - Quantitative analysis of t.he ftmctional relationships existing between ecosystem components. 1 : Analysis of t.he linear intercomponent mass transfrrs. II : Analysis of the non-linear re1at.ionship.s. Oecoloylicl (Berl.), 16 : 97-lli.
S~IITH (D, F.), WIERE (W. J.), 1976. - Constant. relrase of photosynthate from marine phytoplankt.on. Appl. Environ. Microbiol., 32/1 : 75-79.
SMITH (W. 0. Jr.), B&REF.~ (R. T.1, HUNTBMAN (S. A.), 1971. - Primary production of the coasl of northwest. Afrira : excrrtion of dissolved organic matter and its heterotrophic uptake. Deep Se.0 Res., 21/1 : 35-47.
SOROIUN (1-u. I.), 1971. - On thu role of bacteria in tho productivity of tropical oceanic waters. Inf. R~r~zze ges. Hydrobiol., 56 : l-18.
STABEL (H.-H.), 1978. - Zur hIoleBulargewichtsverteil~~ng grliM.er organischer hfoleküle in vcrschiedenen Ober- flachengew%ssern. Arch. Hydrobiol., 8’2/1--1 : 88-97.
STEINRERF (C. J, 1977. - Schwer abbaubare, stickstoffhaltige geliiste organisc.he Substanzen im Schijsee und in Algenkulturen. Arch. Hydrobiol., Suppl. 53/1 : 48-158.
STORCH (T. A.), SAUÏXDERS (G. W.), 1975. - Estimating daily rates of estracellular dissolved organic carbon relrase by phytoplankton populations. Verh. internat. Verein. Limnol.z 19/2 : 952-958.
STORCFI (T. A.), SAUNDERS (G. W.), 1978. - Phytoplankton extracellular relrase and its relation to t,he seasonal cycle of dissolved organic c.arbon in a entrophic Mie. Limnol. Oceanoyr., 23/1 : 112-110.
STOUTI~AMER (A. H.), BETTEN~IAUSSBN (C.), 1972. - Influence of hydrogen acceptors on growth and energy production of Profeus mirabifis. dnf. I,. Leeuw., 38/1 : 81-90.
STRICKLAKD (J. 11. D.), PARSOXS (T. R.), 1968. - A practical handbook of sea-water analysis. BuIf. Fish. Res. Board Can., 167 : 311.
STUMM-ZOLLIR.FER (E.), HARKIS (R. H.), 1971. - Kinetics of biologically mediat.ed aerobic osidation of organic com1nmnds in receiving %aters and in waste i.real.ment. In : Organic compounds in aquatic environments. Ed. bp 5. D. Faust Gi-. J. V. Hunter. BI. Dekker Inc. (NY) 1971 : 555-598.
Heu. Hydrobiol. frop. II (2): SS101 (1981).
TASTET (J. P.), 197 1. - L’environnement physique du systéme lagunaire Ébrié;. Univ. Abidjan ; sér. Document. no 11, Dep. Sc.i. Terre : 25.
VACCARO (R. F.), 1969. - The response of natural microbial populations in sea-water t.o organic enrichment. Linmol. Oceunogr., 1415 : 726-735.
VACCARO (R. F.), JANNASCH (H. W.), 1966. - Studies on heterot.rophic activity in sea-water based on glucose assimilation. Limncrl. Oceanogr., 11 /-i : 596-607.
Vacca~o (R. F.), Hrcxs (S. E.), JANNASCH (H, W.), ÇAREY (F. G.), 1968. - The occurrence and role of glucose in sea-water. Limnol. Ocaanogr., 1312 : 356-360.
VAN DEN BER~; (C. ïN. G.), WONC: (P. T. S.), Cm4u (Y. K.), 1979. - Measurement of complexing mat.erials excreted from algae and their ability to ameliorate copper t.osicity. J. Fish. RES. Board Cun., 36/8 : 901-905.
VARLET (F.), 1978. - Le regime de la lagune Ébrié (Cote d’ivoire). Traits physiques essentiels. Trau. Dot. O.R.S.T.O.M., no 83 : 162.
WALLEN (D. G.), GEEN (G. H.), 1971. - The nat.ure of t.he photosynthate in natural phytoplankton populations in relation to light quality. Mar. Biol., lO/S2 : 157-168.
WATT (W. D.), 1966. - Release of dissolved organic material îrom the cells of phytoplankton populations. Proc. Roy. Soc., B, 164 : 521-551.
WETZEL (R. G.), MANNY (B. A.), 1972. - Decomposition of dissolved organic carbon and nitrogen compounds from leaves in an experimental hard-water stream. Limnol. Oceanogr., 17/6 : 927-931.
WETZEL (R. G.), RICA (P. H.), MILLER (RI. C.), ALLE~ (H. L.), 1972. - Mctabolism of dissolved and particulate detrital carbon in a temperate hard-water lake. Mem. Isf. ifal. Idrobiol., 29 Suppl. : 185-213.
WHEELER (J. R.), 1976. - Fractionation hy molccular weight. of organic substances in Georgia coastal water. Limnof. Oceanogr., 21/6 : S.i6-852.
WYIEBE (W. J.), SMITH (D. F.), 1977. - Direct measurement. of dissolved carbon release by phytoplankton and incorporation by micrnheterotrophs. AIar. Bief., 12/1 : 213-223.
WILLIAMS (P. .T. Le B.), 1973. - The validity of the applicat.ion of simple kinetic analyzis to heterogeneous microhial populations. Limnol. Oceanoqr., 18/1 : 159-165.
WILLI.&MS (P. J. Le B.), A~I<E~ (C.), 1968. - A method of measuring the mineralization by microorganisms of organic compounds in sea-water. Deep Sea Rrs., 15 : 365-375.
WILLIAMS (P. J. Le B.), YENTSCH (C, S.), 1976. -An esamina- tien of photosynthetic production, excretion of phyto- synthetic produc.ts and heterotrophic utilization of dissolved organic compounds with reference to results from a coastal subtropical sea. Alar. Bief., 35/1 : 31-10.
WILLIAMS (P. J. Le B.), BERJCAN (T.), H~LRI-HANSEN (O.)? 1976. - Amino-acid uptake and respiration by marine het.erot.rophs. Mar. Bief., 3511 : 11-47.
WRIGHT (R. T.), 1978. - Measurement and significance of specific activity in the heterotrophic bacberia of nat.ural waters. Appl. enuiron. Alirrobiol., 36/2 : 297-305.
WRIGAT (R. T.), H~~ure (J. E.), 1965. - The uptake of ’
organic solutes in lake waler. Limnol. Oceanogr., lO/l : 22-28.