perception de la proximité génétique chez la souris ... · nicolas busquet perception de la...
TRANSCRIPT

UNIVERSITE PARIS XIII ________________________________________
Ecole Doctorale: Vivant et Société
THESE
Présentée pour obtenir le grade de
DOCTEUR DE L'UNIVERSITE PARIS XIII
Biologie des organismes (Mention: Biologie du comportement)
par
Nicolas BUSQUET
Perception de la proximité génétique chez la souris
glaneuse Mus spicilegus : mécanismes et fonctions.
soutenue le 06 décembre 2004 devant le jury composé de:
P. JAISSON Professeur (Université Paris XIII)
Président
F. S. DOBSON Professeur (Auburn University, USA))
Rapporteur
R. PORTER Directeur de Recherches CNRS (UMR PRC 6073, INRA/CNRS, Nouzilly)
Rapporteur
F. BONHOMME Directeur de Recherches CNRS (UMR 5171 Université Montpellier II )
Examinateur
M. HAUSBERGER Directeur de Recherches CNRS (UMR 6552 Université Rennes I)
Examinatrice
C. BAUDOIN Professeur (Université Paris XIII)
Examinateur (Directeur)
Laboratoire d’Ethologie Expérimentale et Comparée (CNRS FRE 2413)

Sommaire
SOMMAIRE
Introduction générale.................................................................................................................. 1
Pourquoi étudier la reconnaissance de parentèle ?................................................................. 2
La théorie de l’évolution .................................................................................................... 2
Le gène égoïste ? ................................................................................................................ 5
Le vocabulaire de la biologie ............................................................................................. 7
Fonctions, adaptations, effets ............................................................................................. 7
Proximité génétique et apparentement ............................................................................. 10
La reconnaissance de parentèle : pourquoi ?.................................................................... 11
Evitement de l'inceste et "hybridation optimale" ............................................................. 12
La sélection de parentèle .................................................................................................. 14
Mus spicilegus...................................................................................................................... 17
Objectifs et organisation de la thèse..................................................................................... 20
Chapitre I : Odeurs et Génome................................................................................................ 21
1.1. Introduction .............................................................................................................. 22
1.1.1 Odeurs et MHC ................................................................................................ 22
1.1.2 Odeurs et MUPs ............................................................................................... 26
1.2. Odeurs et parentèle chez Mus Spicilegus................................................................. 29
1.3. Odeurs et phylogénie................................................................................................ 48
1.4. Empreinte génomique et odeurs............................................................................... 56
1.4.1 Matériels et méthodes....................................................................................... 58
1.4.2 Résultats ........................................................................................................... 60
1.4.3 Discussion ........................................................................................................ 61
Chapitre II : Mécanismes d'évaluation de la proximité génétique .......................................... 63
2.1. Introduction .............................................................................................................. 64
Les Rongeurs reconnaissent-ils leurs apparentés ? .......................................................... 67
Familiarité et reconnaissance de parentèle....................................................................... 69
Assortiment phénotypique auto référencé, effet "aisselles" et reconnaissance de parentèle
.......................................................................................................................................... 70
2.2. Préférences olfactives pour des apparentés non familiers........................................ 72
2.2.1 Matériel et méthodes ........................................................................................ 72

Sommaire
2.2.2 Résultats ........................................................................................................... 73
2.2.3 Discussion ........................................................................................................ 74
2.3. Rôle de l'expérience post-natale dans l'évaluation de la proximité génétique :
adoptions croisées interspécifiques ...................................................................................... 79
2.4. Rôle de l'expérience pré- et périnatale dans l'évaluation de la proximité génétique:
préférences olfactives de nouveau-nés ................................................................................. 90
Discussion générale : l'évaluation de la proximité génétique ........................................ 105
Chapitre III : Odeurs, facteurs d’état et proximité génétique................................................ 108
3.1. Introduction ............................................................................................................ 109
3.2. Reconnaissance de parentèle et régime alimentaire ............................................... 110
3.2.1 Matériels et méthodes..................................................................................... 112
3.2.2 Résultats ......................................................................................................... 114
3.2.3 Discussion ...................................................................................................... 116
3.3. Parentèle et facteurs d'état liés à la fitness : l'âge................................................... 117
3.3.1. Matériel et méthodes ...................................................................................... 119
3.3.2. Résultats ......................................................................................................... 120
3.3.3. Discussion ...................................................................................................... 122
3.4. Facteurs d'état et parentèle : discussion générale ................................................... 124
Chapitre IV : Aspects fonctionnels de la reconnaissance de parentèle ................................. 129
4.1. Introduction ............................................................................................................ 130
Le choix du partenaire sexuel......................................................................................... 131
Le choix de partenaire chez Mus spicilegus................................................................... 132
Rappels sur la campagne de septembre 1999 à Gyöngyös (Hongrie)............................ 133
4.2. Préférences de femelles en fonction de la provenance géographique du mâle ...... 133
4.2.1 Matériel et méthodes ...................................................................................... 134
4.2.2 Résultats ......................................................................................................... 134
4.2.3 Discussion ...................................................................................................... 135
4.3. Rôle de l'apparentement dans le choix du partenaire sexuel en situation de trios . 137
4.3.1 Matériels et méthodes..................................................................................... 138
4.3.2 Résultats ......................................................................................................... 139
4.3.3 Discussion ...................................................................................................... 140
4.4. Discussion générale : choix de partenaire et apparentement.................................. 141
Discussion générale................................................................................................................ 143

Sommaire
Particularités des signaux olfactifs ..................................................................................... 144
Perception de la proximité génétique ................................................................................. 147
Pressions de sélections et reconnaissance de parentèle...................................................... 149
La reconnaissance de parentèle : un seul mécanisme ?...................................................... 151
Evolution de la reconnaissance de la proximité génétique ................................................ 152
Capacités cognitives et reconnaissance de parentèle ......................................................... 153
Y a t’il réellement reconnaissance de parentèle chez Mus spicilegus ?............................. 154
Bibliographie.......................................................................................................................... 156

Introduction générale
Introduction générale
1

Introduction générale
Pourquoi étudier la reconnaissance de parentèle ?
La reconnaissance de parentèle est la capacité à reconnaître les apparentés. Cette définition
apparemment limpide recèle malgré tout de nombreuses ambiguïtés. En effet, elle ne précise
pas la nature de la reconnaissance. Que reconnaît-on ? Les membres de la famille,
l’apparentement lui-même, les individus partageant des gènes ? Mais quels gènes ? Quel est
l'intérêt de cette reconnaissance pour ceux qui en sont capables ? Avant d’aborder en détails
ces questions, il convient de revenir sur les raisons qui font de la reconnaissance de parentèle
une des problématiques majeures de l’étude des comportements dans une perspective
évolutive.
La théorie de l’évolution
Depuis les travaux de Darwin (1809-1882), la théorie de l’évolution occupe une place centrale
dans l’édifice de la biologie moderne. En dépit de résistances et de remaniements fréquents,
elle est à ce jour la seule théorie unificatrice en biologie (les fondements de cette théorie sont
présentés succinctement dans l’encadré 1). Les principes fondamentaux de cette théorie sont
que les êtres vivants ont évolué et évoluent, et que les espèces actuelles sont le résultat d’une
compétition, la "loi du plus fort" ("survival of the fittest" selon l’expression de Spencer, plus
tard réutilisée par Darwin) : dès lors qu’il accède à l’existence, un être vivant est soumis à de
nombreuses contraintes environnementales susceptibles de mettre en péril sa survie. La notion
fondamentale de la théorie de l’évolution est que la survie ou non d’un individu n’est pas un
phénomène totalement aléatoire : certains sont mieux armés que d’autres pour survivre. Cela
ne veut en aucun cas dire que le hasard n’a pas sa place, et c’est même là un autre aspect
primordial de la théorie de l’évolution : elle n’est pas téléologique, c’est-à-dire qu’il n’y a pas
de but ultime, déterminé à l’avance, vers lequel tendent les organismes (O'Grady, 1984).
Depuis l’avènement de la génétique, on sait que le support moléculaire de l’hérédité, voire de
l’identité, est l’ADN (acide désoxyribonucléique), présent identiquement dans toutes les
cellules d’un organisme, à l’exception des cellules sexuelles. Bien sûr, un individu ne se
résume pas à son ADN, non seulement parce que l’expression des gènes est sujette à
variations, mais aussi parce que chaque individu est la résultante d’une histoire individuelle
(qui a influencé son développement). Les gènes ne s’expriment pas tous, pas toujours, pas de
manière identique d’un tissu à l’autre, et peuvent être régulés les uns par les autres, etc.
Cependant le génotype (l’ensemble des gènes portés par un individu) est bien caractéristique
2

Introduction générale
d’un organisme puisqu’il correspond au "champ des possibles", à l’éventail des potentialités
d’un individu. Autrement dit, s’il n’est pas sûr qu’un gène porté par un individu s’exprime, un
individu ne peut exprimer que ce qu’il
porte. L’ensemble des caractéristiques
d’un individu est donc la résultante d’une
interaction du génome avec
l’environnement. Ce phénotype peut se
définir comme "l’ensemble des
manifestations physiques de l’organisme",
c'est-à-dire la somme des atomes,
molécules, macromolécules, cellules,
structures, tissus, organes ainsi que le
métabolisme, les échanges d’énergie, les
réflexes et les comportements : tout ce qui
fait partie de la structure observable, du
fonctionnement ou des comportements
d’un organisme vivant.
Le principe de sélection (naturelle ou
sexuelle) qui est le fondement de la théorie
de l’évolution, suppose que les individus
ayant le phénotype le plus performant sont
les plus à mêmes de transmettre tout ou
partie de leur génotype. Cette
"performance" est une notion très
complexe. En effet, les pressions de
sélection que subit un organisme sont multiples. Dans le cas d’un petit Mammifère
(insectivore ou Rongeur par exemple), elles comprennent la prédation, le parasitisme, les
contraintes environnementales (température, humidité, etc.), la compétition pour l’accès à la
nourriture, pour l’accès à la reproduction et d’autres encore. Il est très difficile d’établir une
liste exhaustive des pressions de sélection pesant sur un organisme, donc également très
difficile de jauger les performances relatives des individus. Ce que les Anglo-saxons appellent
la fitness, que l’on peut traduire par "adéquation adaptative", c'est-à-dire la compétitivité d’un
phénotype, n’est pas une donnée absolue mais relative, notamment dans sa dimension
Encadré 1 : Quelques principes de base de lapensée darwinienne (Cartwright, 2000). • Les individus peuvent être regroupés en espèces
sur la base de caractéristiques communes deforme, anatomie, physiologie, comportement etc.En cas de reproduction sexuée, cesregroupements sont validés par le fait que sesmembres peuvent s’accoupler les uns avec lesautres et avoir une descendance fertile.
• Au sein d’une espèce, tous les individus ne sontpas identiques du point de vue physique et/oucomportemental.
• Certaines de ces différences sont héritées etpeuvent être transmises à la génération suivante.
• Les variations sont enrichies par l’occurrencealéatoire et spontanée de nouveautés. Uncaractère inédit peut apparaître, ou bien uncaractère déjà présent à la générationprécédente peut exister à un degré inédit.
• Les ressources requises par un organisme poursurvivre et se reproduire sont finies. Il existedonc une compétition pour ces ressources, etcertains organismes auront moins dedescendance que d’autres.
• Certaines variations entre individus confèrent unavantage à leur possesseur en terme d’accès àces ressources et donc en terme de descendance.
• Les caractères conférant un avantage tendent àêtre préservés et à devenir progressivement lanorme. Si le changement ou la somme dechangements est suffisamment radical parrapport à l’origine des individus, il peut y avoirspéciation, ou apparition d’une nouvelle espèce.
• La sélection naturelle a pour conséquencel’adaptation des organismes à leurenvironnement, c’est-à-dire une capacité à senourrir, à éviter la prédation, à trouver unpartenaire sexuel etc.
3

Introduction générale
temporelle. A un instant t, un organisme peut être plus compétitif que les autres, par exemple
s’il s’octroie un accès privilégié à la reproduction. Mais ce phénotype compétitif peut l’être
moins dans un autre contexte. Non seulement le phénotype est par essence changeant, puisque
résultant de l’interaction du génome et d'un environnement variable, mais les avantages qu’il
procure sont dépendants du contexte. Un exemple criant de cette ambiguïté du caractère
avantageux d’un trait phénotypique est la queue des paons, qui est chez les mâles un caractère
sexuel secondaire important pour l’accès à la reproduction, mais qui constitue également un
handicap vis-à-vis des prédateurs. Selon le point de vue adopté, un paon mâle présentant une
queue plus longue et ornée que les autres sera avantagé par un meilleur accès à la
reproduction ou désavantagé par une vulnérabilité accrue face aux prédateurs (Wiens, 2001).
Le processus de sélection sur lequel est basée la théorie de l’évolution est ainsi souvent
décomposé en sélection naturelle (liée à la résistance aux pressions environnementales,
biotiques ou abiotiques) et en sélection sexuelle (liée au succès reproducteur et donc à des
interactions intra-spécifiques, à l’environnement social). En poursuivant l’exemple du paon, il
est possible d’essayer de prendre en compte ces deux types de sélection : supposons que les
mâles à longue queue soient avantagés au niveau reproducteur mais désavantagés du point de
vue de la survie (Houle & Kondrashov, 2002). Ces mâles se reproduisent donc mieux mais
pendant moins longtemps. Le critère permettant de réellement évaluer la fitness d’un génotype
(c'est-à-dire, chez les organismes à reproduction sexuée, d’un individu) est le succès
reproducteur global, traditionnellement assimilé au nombre de descendants (Crow & Kimura,
1970). Un individu dont le phénotype se sera révélé plus performant que les autres aura donc
la possibilité de transmettre ses gènes à la génération suivante. Dans le cas de la reproduction
sexuée, cette transmission n’est pas totale, puisqu’un seul allèle de chaque gène est transmis
via les cellules sexuelles haploïdes. Cette réduction chromatique, obtenue par méiose,
implique que le matériel génétique transmis 1) varie d’une cellule sexuelle à l’autre 2) n’est
pas équivalent à la transmission du phénotype parental. De plus, il y a nécessité d’hybridation
avec une autre cellule haploïde, celle du partenaire sexuel, pour "reconstituer" un nouvel
organisme diploïde. La transmission des gènes par reproduction sexuée est donc partielle,
puisque seule la moitié du patrimoine génétique est transmis. Les implications de cette
reproduction sexuée qui reforment des combinaisons inédites à chaque génération sont
nombreuses et il est fondamental de bien les comprendre avant de s’intéresser aux choix socio
sexuels des animaux.
4

Introduction générale
Le gène égoïste ?
Le génotype d’un individu, qui détermine sa fitness dans un environnement donné à travers la
compétitivité du phénotype résultant, n’est pas une entité indivisible. Il apparaît plus simple à
bien des égards de se placer au niveau du gène lui-même, en le considérant comme l’unité
fondamentale de la sélection naturelle (Dawkins, 1976). Chaque allèle est alors considéré
comme étant en compétition avec les autres
formes alléliques portées par les autres
individus. Les allèles les plus compétitifs sont
répliqués en plus grand nombre, en vertu de
l’avantage qu’ils confèrent à leur porteur.
Cette façon de présenter l’évolution est à la
fois imparable - puisque du point de vue du
gène, sa seule présence est la preuve de son
adéquation à l’environnement et de sa
"réussite" - et trompeuse. En effet, il est
difficile pour un biologiste de faire abstraction
de l’individu, qui est la manifestation la plus
évidente de l’expression des gènes mais aussi
l’entité la mieux observable, en particulier
pour les comportementalistes. Or les
paramètres régissant la survie d’un gène ne
sont pas nécessairement les mêmes que ceux
régissant la survie d’un individu. Autrement
dit, il est abusif de considérer qu’un gène
mieux (ou plus) représenté que les autres
corresponde à un avantage pour l’individu
porteur, puisqu’il faut pour cela accepter que
la "qualité" du génome entier soit la somme
des qualités intrinsèques de chaque gène. D’aut
définie, comme en témoignent les évolutions de s
A la question "Qu’est-ce qu’un gène ?", il y a qu
interrogées, certaines accordant une importance
au caractère héritable en passant par des
5
Encadré 2 : Qu’est-ce qu’un gène ?
Le gène a commencé son histoire comme uneentité indéterminée porteuse de l’hérédité avecMendel, à la fin du XIXème siècle. Johannsen, en1909, parle d’une abstraction sans substratmatériel ni qualité physiologique mais présentedans la cellule germinale et responsable del’hérédité, ou plus exactement de la transmissiondes caractères. La découverte de l’ADN le réduità un segment de molécule codant pour uneprotéine. Il ne correspond plus à un caractèrehéritable, tant il est rare que ces caractèressoient dus à une seule protéine. Les caractèreshéritables sont le produit d’interactionscomplexes entre les gènes. La découverte desintrons – séquences non codantes intercaléesentre les séquences codant pour une protéine –fait perdre au gène son unité topographique. Legène devient (redevient ?) une unitéfonctionnelle, avant que l’on ne découvre qu’unmême gène peut intervenir dans plusieurs voiesmétaboliques et être transcrit en plusieursprotéines. Le rôle des séquences régulatricesaugmente encore la complexité de la notion degène, puisqu’elles peuvent agir sur plusieursrégions codantes. Encore aujourd’hui, bien quela plupart des biologistes connaissent etcomprennent de façon intuitive ce qu’est ungène, sa définition reste problématique (Lambert,2003). Selon les auteurs, le gène peut êtrecomme un segment d’ADN corréléstatistiquement à un caractère ou comme unemétaphore d’une unité d’hérédité, ce quirappelle l’intuition de Mendel…. Pourtant sonexistence en tant qu’entité indépendante etmatérielle est le postulat de base de nombreuxraisonnements théoriques, notamment pour lesévolutionnistes.
re part le gène est lui-même une entité mal
a définition (voir encadré 2 et Pichot, 1999).
asiment autant de réponses que de personnes
primordiale à la nature moléculaire, d’autres
définitions philosophiques ou juridiques

Introduction générale
(Chevassus au-Louis, 2001). Les gènes sont bien les entités permettant la transmission de
l’information d’une génération à l’autre (les "réplicateurs"), mais le schéma classique "un
gène – une protéine" semble plus qu’approximatif. Curieusement, parler des gènes est
beaucoup plus aisé que de parler du gène, puisqu’on garde les propriétés fondamentales
(codage pour des protéines, support de l’hérédité) sans avoir à définir les limites de la
catégorie "gène". Le gène égoïste de Dawkins est bel et bien une métaphore : il paraît en effet
difficile de parler des gènes égoïstes au pluriel sans amoindrir leur caractère égoïste.
L’existence même d’unités de sélection (composées d’un ou de plusieurs gènes) est contestée,
et le terme d’unités héritables ("units of inheritance") est peut-être plus heuristique (Weber,
2000). En effet, la propriété fondamentale du matériel génétique est sa transmission
d’individu à individu, même si cette transmission peut être imparfaite et partielle dans le cas
de la reproduction sexuée. La question du fonctionnement de l’ADN dans un environnement
particulier se situe sur un plan différent : la sélection agit sur les caractères, sur le phénotype
d’un individu. Les individus performants, de part leur meilleur accès à la reproduction, vont
transmettre tout ou partie de leurs gènes. Statistiquement, la possession de ces gènes est
corrélée à une fitness supérieure de l’individu. En comprenant cette relation statistique entre
fragment d’ADN et sélection naturelle, il est possible de réconcilier la vision "héréditariste"
du gène, issue de travaux de Mendel, De Vries et Morgan et la vision moléculaire moderne
liée à la découverte de l’ADN. Les gènes "importants" au regard de l'évolution sont ceux qui
présentent plusieurs allèles. Ce sont ces différentes formes alléliques qui sont responsables
des variations phénotypiques héritables. En d'autres termes les différences de compétitivité
entre individus induites par la possession d'une forme allélique particulière vont être
déterminantes pour la pérennité de cette forme allélique parmi les formes existantes. D'une
manière générale, dans la littérature évolutionniste, le terme de gène est utilisé pour désigner
un emplacement du génome (un locus) qui se présente selon les individus sous différentes
formes (allèles) dont la fréquence relative est liée aux phénotypes résultants. Cette définition
tacite exclue de la catégorie "gène" les portions d'ADN fixées ainsi que les portions variables
sans corrélats sélectifs.
Les difficultés de définition de termes usuels en biologie comme la fitness ou les gènes ne
doivent pas pour autant discréditer les fondements de la théorie de l’évolution. Au fil des
générations, il y a bien une modification des bases de la molécule d’ADN qui est liée au
phénotype des individus (voir Arber (2003) pour un aperçu des mécanismes moléculaires de
l’évolution). Au sein d’une espèce à reproduction sexuée, les différents phénotypes
6

Introduction générale
individuels peuvent interagir et les conséquences de ces interactions comprennent l’apparition
de nouveaux phénotypes, via la transmission et l’hybridation du matériel génétique. Ce sont
ces interactions entre phénotypes que l’éthologue observe.
Le vocabulaire de la biologie
A travers les exemples du gène et de la fitness, il paraît clair que le vocabulaire employé en
biologie n’est pas anodin. Une explication de cette ambiguïté récurrente des termes réside
dans le fait que, contrairement aux autres sciences, la biologie s’intéresse aux fonctions de ses
objets d’étude (Ruse, 2000). Cette différence est
fondamentale et n’est pas intrinsèquement liée à la
théorie de l’évolution mais correspond à une
tendance des êtres humains à organiser le vivant
selon des différences fonctionnelles. Les jeunes
enfants accordent une importance primordiale aux
fonctions et classifient les objets, êtres vivants ou
parties de ceux-ci selon leur "utilité" (par
exemple : "les nuages servent à faire de la pluie").
Cette classification téléologique (qui attribue un
but aux choses) se restreint chez les adultes au
monde vivant (Kelemen, 1999). Il n’est pas rare
que la première réaction face à une particularité
anatomique ou comportementale chez un animal
soit "A quoi ça sert ?". Cette idée que les structures et les comportements des animaux sont là
"pour quelque chose" a de nombreuses conséquences en biologie évolutive. L'encadré 3
présente quelques définitions de termes associés à la téléologie, comme le finalisme.
Encadré 3 : Quelques définitions tirées dudictionnaire Larousse. Téléologie n. f. - Doctrine qui considère le monde comme unsystème de rapports entre moyens et fins. Finalité n. f. - 1. Caractère de ce qui tend à un but ; le faitde tendre à ce but, par adaptation de moyensà des fins. - 2. Didact. Adaptation des parties au tout(principe de la beauté dans certaines théoriesesthétiques). - 3. Adaptation des êtres vivants, des organesà quelque fin ou quelque plan déterminant. Finalisme n. m. Didact. Philosophie finaliste. Finaliste n. et adj. - Philos. Qui croit à l'action des causes finaleset, en général, à la finalité comme explicationde l'univers.
Fonctions, adaptations, effets
Parmi les objectifs de l’éthologie, la recherche des fonctions des comportements est la plus
liée aux approches évolutives. Dans le cadre de la présente thèse, par exemple, une des
fonctions supposées de la reconnaissance de parentèle est liée à l’évitement de l’inceste, lui-
même lié au caractère délétère de la consanguinité. Avant de détailler ces analyses évolutives
des comportements liés à l’apparentement, il convient de préciser ce qu’est une fonction. Là
encore, le biologiste est confronté à une notion complexe et mal définie qui pourtant est
7

Introduction générale
utilisée de façon courante. Il est possible de distinguer trois acceptions du mot "fonction" en
biologie (Bekoff & Allen, 1995) :
• Utilité actuelle : le trait phénotypique (physiologique ou comportemental) T remplit la
fonction F si F est un effet de T qui a un impact positif sur la fitness de l’individu. Cette
acception permet d’expliquer le maintien d’un trait, mais pas son origine.
• Fonction historique : le trait T remplit la fonction F si F était un effet de T chez les ancêtres
de l’organisme qui a contribué à la transmission de T à l’organisme lui-même, via le
processus de sélection naturelle. Autrement dit, un trait possède une fonction primaire
(historique) qui a présidé à son apparition, mais qui n’est plus nécessairement utilisé. Dans
cette acception, il est possible d’associer à un trait phénotypique une fonction que l’animal
n’est pas en mesure de réaliser (par exemple la fonction des ailes des poules est le vol,
même si elles en sont généralement incapables).
• Propriété : le trait T remplit la fonction F si F est un effet de T qui contribue à une
propriété de l’individu. La fonction est indépendante, dans cette définition, de sa valeur
adaptative.
Le terme de fonction a donc plusieurs significations (certains auteurs distinguent même quatre
notions distinctes au lieu des trois présentées ci-dessus (Wouters, 2003)), et renvoie à la
notion, importante en évolution, d’adaptation. Une adaptation est un changement
phénotypique sous l’effet d’une pression de sélection. C’est la connaissance de la fonction
historique d’un trait qui permet de lui attribuer le statut d’adaptation. L’efficacité actuelle
d’un trait par rapport aux autres ne signifie pas que l’animal qui le présente soit "adapté" ni
même que ce trait ait été selectionné. S. J. Gould définit trois processus possibles pour un trait
considéré comme avantageux pour l’individu le présentant (Gould & Vrba, 1998) :
• La sélection naturelle façonne un trait pour son usage actuel : c’est une adaptation.
• Un trait façonné par la sélection naturelle pour une fonction (une adaptation) trouve un
nouvel usage : c’est une cooptation.
• Un trait dont l’origine ne peut être attribuée à la sélection naturelle (une nonaptation)
trouve un nouvel usage.
8

Introduction générale
Selon Gould, seule la première origine permet d’attribuer à un trait le terme d’adaptation. Les
deux autres processus définissent une exaptation. Le trait présente bien un avantage sélectif
actuel pour l’individu, mais il n’a pas la fonction historique de cet avantage. Dans les cas des
exaptations l’avantage est un effet, une conséquence d’un trait dont l’origine est attribuable
soit à la sélection naturelle pour un autre usage soit à la dérive génétique ou à des contraintes
structurelles. Gould et Lewontin, dans un article célèbre, critiquent le programme
adaptationniste qui consiste à voir en chaque trait d’un organisme une adaptation (Gould &
Lewontin, 1979). Cette vision "Panglossienne" fait de la sélection naturelle une force
omniprésente résultant dans l’optimisation de tous les traits observés. Toujours d’après ces
deux auteurs, cette vision est dangereuse car elle peut conduire à formuler des hypothèses
orientées en contradiction avec les mécanismes de l’évolution tels qu'ils sont aujourd'hui
compris. Les pressions de sélection ne sont pas directement accessibles à l'observation, et
supposer qu'un comportement répond à une de ces pressions reste souvent spéculatif. Il peut
s'ensuivre une certaine récursivité de la pensée dont un exemple sera exposé dans le chapitre
1, à propos des signaux permettant la reconnaissance de parentèle. Une notion fortement liée à
ce "Panglossianisme" est celle de progrès, qui suppose que les organismes vont
inexorablement vers une amélioration de leurs capacités et une optimisation de leurs
stratégies. La vision "Gouldienne" de l'évolution réfute la notion de progrès, présentée comme
un concept exclusivement culturel, non testable et non opérationnel. Dawkins, quant à lui,
défend une vision où le progrès est au contraire un des attributs majeurs de l'évolution,
indissociable de la complexité et de la diversité du monde vivant. Une analyse du débat
Gould/Dawkins autour de la notion de progrès en évolution, au niveau conceptuel et
méthodologique est présentée par Shanahan (2001).
La sensibilité aux dangers de la téléologie est donc variable selon les auteurs (Ruse, 1989).
Pour des raisons de clarté, il semble raisonnable d’utiliser un vocabulaire courant dans la suite
de l’exposé. Par essence, ce vocabulaire est téléologique mais présente l’avantage d’être
reconnu, admis par l’ensemble des biologistes qui pour la plupart sont conscients de ses
limitations : ce vocabulaire peut être perçu comme un outil méthodologique permettant de
guider les recherches (Ratcliffe, 2000). Cependant la vigilance est de mise, spécialement dans
l'étude de la sélection et de la reconnaissance de parentèle, un de thèmes majeurs de la
biologie évolutive.
9

Introduction générale
Proximité génétique et apparentement
L'apparentement entre deux individus correspond au pourcentage d'allèles partagés par
descendance. Bien qu'on voit mal comment ces allèles pourraient être partagés autrement, la
notion de descendance est fondamentale à la compréhension de ce qu'est la parentèle. Au sein
d'une espèce, la plupart des allèles sont fixés : ils correspondent au patrimoine commun
codant pour le phénotype de l'espèce. Les croisements entre conspécifiques n'affectent pas ces
allèles. Certains gènes présentent en revanche des formes alléliques variables selon les
individus. C'est le partage ou non de ces éléments variables du génome qui détermine
l'apparentement. Le coefficient d'apparentement r permet de rendre compte du pourcentage de
la partie variable du génome qui est commune à deux individus selon les règles de l'hérédité
mendélienne. Ainsi un père et un fils partagent 50 % de la partie variable du génome, puisque
la moitié des gènes sont hérités du père. Le coefficient r est donc de 0,5. Le coefficient r
traduit l'apparentement moyen, puisqu'il est défini par les liens de parenté et non par une
connaissance des allèles partagés réellement. Le postulat sous-tendant le calcul d'un
apparentement est que le partage d'allèles est le seul fait de la relation "généalogique" étudiée.
Ainsi le coefficient d'apparentement de deux frères est toujours de 0,5 quelle que soit la
population étudiée. De plus, en l'absence de connaissance exacte de la généalogie, on
considère souvent par défaut que les individus se sont toujours accouplés avec des individus
non-apparentés, ce qui n'est pas nécessairement vrai. La notion même d'individus non-
apparentés est équivoque, puisqu'il existe toujours un ancêtre commun. En fonction de la
population étudiée, un même coefficient d'apparentement ne correspondra pas forcément au
même nombre d'allèles partagés. Ce point est crucial pour l'étude de la reconnaissance de
parentèle. Il existe une confusion latente entre apparentement et proximité génétique. Cette
dernière peut être définie comme la proportion de matériel génétique commune à deux
individus. Contrairement à l'apparentement, le mode d'acquisition de cette similarité importe
peu. Un exemple s'impose: imaginons l'arrivée sur une île d'un couple de Rongeurs. Leur
espèce n'étant pas présente sur cette île, leurs petits n'auront d'autre choix que de s'accoupler
les uns avec les autres. Ce goulot d'étranglement génétique va réduire considérablement la
proportion variable du génome. Après la colonisation de l'île par cette nouvelle espèce, un
observateur naïf calculant les coefficients d'apparentement considérera les liens de parenté
observables au sein de cette nouvelle population, et passera sous silence le fait que la
proximité génétique globale y est supérieure à celle de la population d'origine. C'est donc la
connaissance de la généalogie qui permet de calculer le coefficient d'apparentement moyen r.
10

Introduction générale
La valeur de cet indice n'est pas directement comparable à la proximité génétique. Pourtant ce
raccourci est fréquemment emprunté en
biologie évolutive, ou l'apparentement est
parfois défini comme "une mesure de la
similarité génétique" (Griffin & West, 2002).
Cette confusion me semble poser un problème
théorique important, exposé dans l'encadré 4.
En effet, deux types d'apparentement peuvent
être distingués : l'apparentement absolu, qui
correspond à la similarité allélique réelle (par
descendance), et l'apparentement relatif, qui
compare la similarité allélique de deux
individus à la similarité moyenne au sein de la po
dernier qui doit être utilisé pour émettre des p
compte les potentialités d'optimisation des comp
entre deux individus peut se concevoir comme
totale (r = 1) et la "dissimilarité" maximale parm
définition opérationnelle serait donc liée à l'optim
environnement social. La connaissance de la gé
calculer r. Il convient maintenant de présenter l
l'apparentement un élément crucial des stratégies
La reconnaissance de parentèle : pour
L'apparentement, comme exposé ci-dessus, est av
généalogiques ont des conséquences sur les com
parenté peuvent être également perçus comme
certaines adaptations, notamment comporteme
l'apparentement peuvent être catégorisées selon d
la sélection de parentèle.
11
Encadré 4 : Que signifie un apparentement nul?
Le coefficient d'apparentement varie de 0 à 1. Lavaleur '1' correspond au partage de tous lesallèles, tel qu'on peut le constater chez desjumeaux vrais. La valeur '0', par contre, necorrespond pas à une réalité accessible. Desindividus non apparentés sont différents à tous lesloci variables de… leur espèce ? Leurpopulation ? Les non-apparentés sont simplementmoins apparentés, ont moins de gènes en communque les apparentés (Todrank & Heth, 2003) .Lecoefficient d'apparentement est donc borné parune valeur qui n'est pas objectivable. Pourtant cecoefficient est utilisé comme une valeurmathématique dans de nombreuses équations,permettant notamment de prévoir lescomportements altruistes (Hamilton, 1964).
pulation (Crozier & Pamilo, 1996). C'est ce
rédictions évolutives, puisque il prend en
ortements. Autrement dit, l'apparentement r
un indice borné par la similarité génétique
i les partenaires potentiels (r = 0). Une telle
isation des stratégies de l'individu dans son
néalogie apparaît bien le meilleur moyen de
es raisons qui font de la prise en compte de
individuelles.
quoi ?
ant tout une notion généalogique. Ces liens
portements des animaux. Mais les liens de
des causes évolutives du développement de
ntales. Les pressions évolutives liées à
eux grands axes : l'évitement de l'inceste et

Introduction générale
Evitement de l'inceste et "hybridation optimale"
Pour que les organismes aient développé des stratégies conduisant à éviter l'hybridation avec
des individus trop proches, il doit exister une pression de sélection correspondante, autrement
dit un désavantage lié à la consanguinité. Cette "dépression de consanguinité" a été très
amplement étudiée chez les végétaux (Barrett & Harder, 1996), notamment dans le cadre de
recherches agronomiques. Chez les animaux, la détection de ce coût de la consanguinité est
plus difficile en nature. Cependant, l'étude de réintroductions d'animaux consanguins en
nature ou des effets de la réduction de diversité génétique due à un isolement de population
permettent de déceler cette dépression de consanguinité (Pusey & Wolf, 1996). Les
désavantages liés à l'hybridation avec un génotype trop proche sont principalement liés à une
augmentation de l'homozygotie. L'homozygotie croissante augmente le risque d'expression
d'allèles délétères récessifs. Outre ce risque, la perte de l'hétérosis est en elle-même
désavantageuse, puisque le fait de disposer de versions différentes du même gène peut être
une garantie d'adaptabilité face à des conditions environnementales changeantes. Enfin, la
descendance produite avec un apparenté n'exprimera qu'une faible variabilité phénotypique et
présentera alors vraisemblablement une même vulnérabilité face aux pressions biotiques et
abiotiques. Un brassage constant du matériel génétique permet de présenter de nouvelles
formules à chaque génération, et ainsi de ne pas perdre de terrain dans la course aux
armements entre organismes, qui est l'une des bases de l'évolution (théorie de la reine rouge
(Van Valen, 1973)). La dépression de consanguinité a été décrite entre autres chez les
Cervidés (Slate et al., 2000), les Rats-Taupes (Clarke & Faulkes, 1999), les Chauves-Souris
(Rossiter et al., 2001), les Perdrix (Hoglund et al., 2002)…
Mais il peut également exister une baisse de la fitness quand le partenaire est "trop peu
apparenté", ou trop distant génétiquement. Cette baisse s'explique principalement par le risque
de "casser" des complexes de gènes co-adaptés et par la perte d'adaptations locales (Pusey &
Wolf, 1996). Cependant cette "dépression d'hybridation excessive" est plus souvent théorisée
que décrite. Des exemples existent, mais ils sont souvent liés à des croisements forcés par
l'homme dans le cadre de pratiques d'élevage (par exemple chez les saumons (Gharrett et al.,
1999)) ou de gestion d'animaux en captivité (par exemple chez les rhinocéros (Zschotte &
Baur, 2002)). Le problème principal dans la démonstration de ce type de dépression est celui
de la délimitation de la population où elle peut s'exercer. Il convient de considérer les choix
possibles pour l'animal et envisager les éventuels coûts liés au choix d'un individu trop peu
apparenté. Cependant, il existe un exemple chez les Souris où la réalisation de croisements
12

Introduction générale
entre non-apparentés lors de deux générations consécutives résulte en des petits avec une plus
faible croissance pondérale que ceux provenant de croisements entre cousins, mais cet effet
est faible et n'est plus significatif quand les effets de la taille de la portée sont contrôlés
(Keane, 1990). Si les exemples de dépression d'hybridation excessive sont rares pour des
populations naturelles, elle a été largement décrite d'un point de vue théorique, comme le
pendant de la dépression de consanguinité. La résultante de ces deux forces sélectives est
l'"optimal outbreeding", c'est-à-dire l'existence d'un optimum de distance génétique
permettant de maximiser la fitness (Bateson, 1983). Cet optimum est très difficile à objectiver,
notamment parce que l'hybridation procure à la fois des avantages (augmentation de
l'hétérozygotie) et des inconvénients (rupture de complexes géniques co-adaptés) et que la
saillance de ces effets et leur dynamique varient énormément selon les taxons (Edmands,
2002). D'une manière générale, il semble que les conséquences néfastes liées à l'hybridation
excessive apparaissent clairement lorsque les parents sont issus de populations isolées depuis
longtemps, voire de sous-espèces ou d'espèces différentes. Les effets néfastes de l'hybridation
au sein de populations naturelles semblent quasi-inexistants, et de toute façon restreints à la
première génération, les générations suivantes semblant au contraire bénéficier d'un léger
avantage (Edmands, 2002).
Compte tenu de ces incertitudes, il semble raisonnable de considérer qu'en conditions de
laboratoire, les choix des animaux ainsi que les conséquences en terme de fitness de ces choix
seront liés principalement à l'évitement de la consanguinité. En effet, les études en laboratoire
impliquent souvent l'utilisation de populations où, comparativement à une population
naturelle, la diversité génétique est moins forte ou en passe de le devenir.
Il existe un autre argument permettant de minimiser, voire d'écarter les pressions de sélection
liées à l'hybridation excessive : celui des probabilités de rencontres avec un partenaire
potentiel trop différent génétiquement. Dans les populations "visqueuses", c'est-à-dire les
populations où l'apparentement est corrélé avec la distance entre individus, la possibilité de
rencontrer un partenaire "trop peu apparenté" est faible. La combinaison de cette faible
probabilité avec les coûts peu évidents d'une hybridation excessive rend l'existence de
pressions de sélection correspondantes peu crédible. Bien sûr, cette conclusion ne s'applique
qu'à des études intra spécifiques : il peut en effet exister des pressions de sélection relatives à
l'hybridation entre espèces ou sous-espèces. La question de savoir si ces individus peuvent
être considérés comme des apparentés trop éloignés est complexe, et nécessiterait un exposé
13

Introduction générale
approfondi du concept d'espèce et de ses limites. Cette problématique, au demeurant
passionnante, dépasse le cadre de cette thèse par sa complexité et son apparente insolubilité
(Hey, 2001). Par ailleurs, il semble que ce concept d'espèce n'est pas généralisable à tous les
êtres vivants et ne correspond pas nécessairement à une entité signifiante dans l'étude des
processus évolutifs (de Meeus et al., 2003).
En guise de conclusion sur le sujet de l'évitement de l’inbreeding et de l’outbreeding, il faut
noter que si la consanguinité dans les populations naturelles est en général faible, elle n'est pas
nulle et ne semble pas avoir d'effet délétère sur la fitness (Marshall et al., 2002), ce qui peut
s'interpréter comme un équilibre entre évitement de la consanguinité et évitement de
l'hybridation excessive (Bateson, 1983), mais semble plus parcimonieusement expliquée par
un évitement "imparfait" de la consanguinité, c'est-à-dire par une tolérance d'un faible degré
de consanguinité, d'autant plus acceptable que ses conséquences sont faibles sur la fitness des
individus.
La sélection de parentèle
Les individus apparentés sont, par définition, des individus qui possèdent en commun un
grand nombre de gènes du fait de leur ascendance commune. Ces unités héritables peuvent
donc être transmises directement ou indirectement, via la reproduction des apparentés. Les
individus apparentés sont génétiquement proches et une bonne partie de ce qui peut être
transmis à la génération suivante par un individu l'est aussi par ses apparentés. A la notion de
fitness, qui traduit l'adéquation adaptative d'un individu à son environnement, estimée par la
transmission de ses unités héritables que sont les gènes, se substitue la notion d'inclusive
fitness. Cette adéquation adaptative globale se conçoit comme la transmission des gènes
portés par l'individu lui-même et par les autres porteurs de ses gènes (Hamilton, 1964). Dans
ce raisonnement, on postule (souvent de façon tacite) que les gènes concernés ne sont que
ceux qui sont variables entre les individus interagissant potentiellement. Les gènes fixés dans
une population ou une espèce ne rentrent pas en ligne de compte, comme ils ne rentraient pas
en compte dans le calcul de l'apparentement (cf. plus haut). C'est d'ailleurs cet apparentement
qui apparaît le meilleur indicateur du partage de ces gènes, et donc le meilleur guide pour
estimer la fitness indirecte. Cette théorie de l'inclusive fitness a permis de dresser des
hypothèses pertinentes concernant l'émergence des comportements altruistes et l'apparition de
14

Introduction générale
la socialité. L'hypothèse fondatrice de ce qui deviendra la sociobiologie est la règle de
Hamilton, formalisée sous la forme de l'inéquation :
Cx < Rx-y × By
Où Cx est le coût de l'acte altruiste de x envers y (coût en terme de diminution de la fitness
propre), Rx-y est l'apparentement entre x et y et By le bénéfice de y lié à cet acte
(augmentation de la fitness propre).
Cette règle prévoit que les comportements altruistes surviennent lorsque les bénéfices
indirects de l'aide aux apparentés surpassent les coûts de ces comportements. L'un des apports
majeurs de cette règle est la création d'un modèle permettant d'expliquer l'existence de castes
stériles chez les hyménoptères sociaux.
D'une façon plus générale, le raisonnement tenu est applicable aux relations sociales entre
apparentés : la sélection de parentèle peut permettre d'augmenter l'inclusive fitness (Queller &
Strassmann, 2002). En aidant les apparentés à se reproduire, l'individu propage indirectement
ses propres gènes via les copies portées par ces apparentés. Cette aide n'est pas
nécessairement assimilable à de l'altruisme, puisqu'il n'y a pas nécessairement de coût associé
pour l'individu aidant. Au contraire, il peut exister des bénéfices réciproques à une entraide
entre individus. On parle alors de mutualisme, de coopération. Le bénéfice d'une entraide
entre apparentés est amplifié : aux gains propres s'ajoutent les bénéfices indirects de
l'augmentation de fitness des apparentés. On parle parfois de népotisme (Hauber & Sherman,
1998), qui se définit comme une aide accordée préférentiellement aux apparentés. Il peut être
mis en évidence lorsqu'un choix est observé et que ce sont les congénères les plus apparentés
qui sont aidés. Il suppose donc une perception de l'apparentement, voire une catégorisation
des individus en fonction de cet apparentement (voir chapitre II pour une étude des
mécanismes de reconnaissance de parentèle).
D'une manière générale, deux approches se distinguent quant à l'explication de la coopération
chez les animaux. La première, dont nous avons vu un aperçu plus haut, suppose que la
coopération apparaît préférentiellement entre apparentés. La deuxième se base sur la
prédictibilité des interactions entre individus, et postule donc que la coopération peut
apparaître même entre individus non apparentés (altruisme réciproque) (Trivers, 1971). Cette
approche est héritée de la théorie des jeux (Axelrod & Hamilton, 1981). Si les deux approches
15

Introduction générale
sont souvent séparées, elles peuvent être conciliées (Creel, 1993). Cependant, des travaux
théoriques récents semblent indiquer que la sélection de parentèle serait un frein au
développement de l'altruisme réciproque (Marshall & Rowe, 2003).
La sélection de parentèle est un phénomène éminemment difficile à objectiver. La simple
mise en évidence d'un apparentement fort entre les "coopérants" ne suffit pas (Griffin & West,
2002). D'une part cet apparentement peut-être le résultat de traits d'histoire de vie : par
exemple les soins parentaux sont normalement dirigés vers des proches apparentés, mais on
ne peut pas en déduire que c'est l'apparentement élevé qui est le déclencheur de ces
comportements, en l'absence de groupe témoin ou la progéniture ne serait pas apparentée…
D'autre part, la distinction des bénéfices propres et indirects est problématique. Chez les
marmottes Marmota flaviventris, dont les cris d'alarme sont traditionnellement interprétés
comme un exemple d'altruisme (le guetteur qui avertit les autres d'un danger augmente la
fitness des membres du groupe, puisqu'ils se réfugient et échappent au prédateur, mais décroît
la sienne puisque les vocalisations et la position de guetteur le rendent plus vulnérable), il a
été montré que les femelles ayant mis bas augmentent leurs comportements de guetteurs
(Blumstein et al., 1997). Cette augmentation d'un comportement "à risque" semble indiquer
des bénéfices directs : la femelle augmenterait sa fitness propre (en protégeant sa progéniture)
plutôt que d'augmenter celle des autres apparentés. Néanmoins ces résultats ne sont pas
nécessairement la démonstration d'une plus grande importance de la fitness propre. Si la mise-
bas est le déclencheur d'une augmentation des comportements de surveillance, les autres
membres du groupe, souvent apparentés, sont également bénéficiaires de ce comportement.
La question de la distinction entre altruisme et égoïsme semble alors vaine, puisque ce
comportement bénéficie toujours à des apparentés (Hauber & Sherman, 1998).
La prise en compte du facteur "apparentement" peut intervenir au niveau du choix du
partenaire sexuel comme au niveau des préférences sociales. Il est temps à présent de
présenter l'espèce qui a servi de modèle à cette thèse, et les particularités qui en font un
modèle particulièrement intéressant pour l'étude de la reconnaissance de parentèle.
16

Introduction générale
Mus spicilegus
L’espèce étudiée dans le cadre de cette thèse est la souris des steppes ou souris glaneuse Mus
spicilegus (Sokolov et al., 1998). Son appartenance taxonomique est explicitée dans l’encadré
0.5. La phylogénie du genre Mus est bien connue, notamment grâce à la biologie moléculaire
(Bonhomme et al., 1984 ; Sage et al., 1993 ;
Boursot et al., 1993). Mus spicilegus est
phylogénétiquement proche de Mus macedonicus, et
plus éloignée des autres espèces du genre,
notamment des deux sous espèces Mus musculus
domesticus et M. m. musculus. Elle est sympatrique
avec Mus musculus musculus et parapatrique avec
Mus macedonicus mais présente un isolement
reproducteur qui se traduit dans ses comportements
vis-à-vis de partenaires potentiels de ces deux
espèces (Ivantcheva & Cassaing, 1999). Son aire de répartition est représentée dans la figure
0.1. Cette espèce présente des particularités qui en font un sujet d’étude très original. Parmi
ces particularités, la plus frappante est la présence de tumulus édifiés en automne au sein
desquels les jeunes animaux passent la période hivernale (Naumov, 1940). Ces tumulus sont
constitués d’un amas de graines recouvert de terre qui surmonte un réseau de galeries. C’est
d’ailleurs le comportement de déplacement et d’entassement des graines et épis qui a donné
son nom latin à l’espèce. L’avantage évolutif procuré par cette construction de tumulus n’est
pas encore très bien appréhendé, mais il semble que, plus que la réserve de nourriture offerte
par les graines stockées, la chaleur dégagée par la fermentation du matériel végétal soit
l’élément le plus important, puisqu’il permettrait une régulation de la température interne du
tumulus et ainsi une protection contre le gel en hiver.
Encadré 0.5 : position taxonomique deMus spicilegus. Super règne : Eucaryotes Règne : animal Sous règne Métazoaires Phylum Chordés Embranchement Vertébrés Sous embranchement Gnathostomes Super classe Tétrapodes Classe Mammifères Sous classe Euthériens Ordre Rongeurs Famille Muridés Sous famille Murinés
Quel que soit l’avantage procuré par l’édification de tumulus, ce comportement très
particulier présente des conséquences drastiques sur le cycle biologique de Mus spicilegus. En
effet, il semble que deux périodes très distinctes se suivent au cours de l’année. Dès le début
de l’automne et jusqu’au printemps, les animaux se regroupent dans les tumulus et ne se
reproduisent pas. Dès le printemps, les souris vivent en petites unités familiales hors des
tumuli qui restent abandonnés (Naumov, 1940 ; Orsini et al., 1983). Lors de cette période, il
17

Introduction générale
semble que les souris deviennent très territoriales et relativement intolérantes à la présence de
congénères (Dobson & Baudoin, 2002 ; Patris et al., 2002). Dans ce contexte, il apparaît
essentiel de s’interroger sur la structuration génétique des groupements à l’intérieur des
tumulus et surtout sur l’influence de cette structuration sur les choix ultérieurs de partenaires
sexuels. En effet, parmi les individus rentrant dans les tumulus, seuls les jeunes survivent. Ces
jeunes sont issus de reproduction en été, et ne sont donc pas sexuellement matures au moment
de l'entrée dans les tumulus. En laboratoire, ces souris peuvent se reproduire dès l'âge de deux
mois, mais la première reproduction ne se produit pas avant l'âge de six mois en nature, c'est-
à-dire à la sortie des tumulus. En revanche, même si la reproduction est rare au sein des
tumulus, le fait de débuter sa vie par une période longue de cohabitation "forcée" peut avoir
des implications fortes sur les préférences sexuelles des souris (voir le chapitre 2 pour une
description du rôle de la familiarité chez les Rongeurs). Garza et al. (1997) ont étudié la
structuration génétique au sein de quelques tumulus, et il semble que les individus présents
soient le plus souvent apparentés par la mère. Les juvéniles passent donc l'hiver au sein d'un
groupe social comprenant des apparentés de deux types : des frères et sœurs et des cousins.
Figure 0.1 : Aire de répartition des différentes espèces du genre Mus en Europe (d'après Bonhomme, 1992).
Une autre particularité comportementale des Souris glaneuses est son système d'appariement.
En effet, par rapport à Mus musculus domesticus qui est polygame, Mus spicilegus présente
des traits associés à la monogamie. Au moment de l'œstrus, les femelles montrent en effet une
préférence pour un mâle familier (qu'il y ait eu accouplement ou non) alors que les femelles
18

Introduction générale
M. m. domesticus ne montrent aucune préférence. Ceci est l'indication d'un lien fort entre
partenaires chez M. spicilegus (Patris & Baudoin, 1998). De plus, les mâles de cette espèce
présente des comportements paternels importants (Patris & Baudoin, 2000), ce qui est un
autre trait lié à la monogamie (Kleiman, 1977). Lors d'expériences en enclos semi-naturels, on
observe des associations stables mâle-femelle compatibles avec un système monogame
(Dobson & Baudoin, 2002 ; Baudoin et al., sous presse). Cependant les données de terrain
indiqueraient un système d'appariement relativement flexible et influencé, au moins au
printemps, par la cohabitation hivernale au sein des tumuli (Gouat et al., 2003).
La reconnaissance de parentèle serait une capacité intéressante pour les Souris glaneuses –
comme elle l'est pour les autres animaux (cf. plus haut) – mais aussi en raison de leurs
particularités comportementales. En particulier, l'évitement de l'inbreeding revêt une
importance capitale pour une espèce même partiellement monogame. En effet, une femelle
d'une espèce polygame qui s'accouplerait "accidentellement" avec un mâle apparenté risque
d'avoir une portée moins adaptée, du fait du coût de la consanguinité. Une femelle monogame
commettant la même "erreur" verrait la fitness de toute sa descendance diminuer. De la même
façon, le coût de l'inbreeding pour le mâle chez une espèce polygame est faible puisqu'il ne
s'investit pas dans les soins parentaux et que la prochaine reproduction lui permettra
vraisemblablement de s'accoupler avec une femelle moins apparentée. Un mâle Mus
spicilegus qui forme une association stable avec une femelle va consacrer un temps important
à s'occuper des petits. Cet investissement fort doit être "payant".
19

Introduction générale
Objectifs et organisation de la thèse
Les différents points soulevés dans cette introduction générale ont guidé une approche
"prudente" de l'étude de la reconnaissance de parentèle chez Mus spicilegus. La première
étape dans cette étude est la recherche du lien entre proximité génétique et odeurs chez cette
espèce. Le terme de proximité génétique est ici préféré à celui d'apparentement car la
recherche de similarités odorantes ne s'est pas restreinte aux relations de parenté mais a été
examinée en considérant des entités plus larges (populations et espèces). Le chapitre I
comprend également une étude visant à examiner les rôles respectifs du père et de la mère
dans la transmission du phénotype odorant (effet "parent d'origine" et empreinte génomique).
Dans le chapitre II, après une revue de la littérature sur les mécanismes permettant la
reconnaissance de parentèle, ce sont les processus de perception de la proximité génétique qui
sont étudiés, en laissant délibérément de côté le rôle de la familiarité. Les préférences des
Souris pour des odeurs d'individus apparentés ou non sont ainsi examinées. Dans un deuxième
temps, la réalisation d'adoptions croisées interspécifiques permet d'apporter de précieux
éléments de réponse dans la recherche du mécanisme sous-tendant l'évaluation de la proximité
génétique chez ces rongeurs. Enfin l'étude des préférences olfactives de nouveau-nés permet
d'aborder l'ontogénèse de ce mécanisme.
Le chapitre III vise à éprouver les hypothèses issues des deux chapitres précédents en
s'interrogeant sur le rôle des facteurs d'état, liés à l'expérience de l'individu, dans la réalisation
d'une reconnaissance efficace de la proximité génétique. Sont examinés un facteur
alimentaire, considéré comme sélectivement neutre, et un facteur théoriquement fortement lié
à la fitness, l'âge.
Le chapitre IV considère les aspects fonctionnels de la reconnaissance de parentèle, et
s'interroge sur les stratégies déployées par les individus pour prendre en compte le facteur
"apparentement" dans leurs choix socio sexuels.
Enfin, la discussion générale fait le bilan des connaissances acquises lors de ce doctorat et
propose une vision de la reconnaissance de parentèle tenant compte des critiques
conceptuelles exposées dans l'introduction générale et les différents chapitres.
20

Chapitre 1 : Odeurs et Génome
Chapitre I :
Odeurs et Génome
21

Chapitre 1 : Odeurs et Génome
1.1. Introduction
Les Rongeurs utilisent de façon prédominante le canal olfactif pour communiquer. Cette
affirmation est reflétée dans la très grande quantité de travaux portant sur cette
communication olfactive. De par leur nature, les odeurs individuelles renferment de
nombreuses informations (Beauchamp & Yamazaki, 2003). Parmi ces informations, un grand
nombre reflète des facteurs d’état, c’est-à-dire des facteurs liés à l’environnement et/ou à
l’histoire individuelle de l’animal. Ces facteurs d’état incluent le régime alimentaire (Brown
et al., 1996 ; Ferkin et al., 1997), le statut de dominance (Drickamer, 1992), l’état de santé
face à une charge virale (Penn et al., 1998) ou parasitaire (Willis & Poulin, 2000). D’autre
part, il semble exister une variabilité inter-individuelle suffisamment importante pour
permettre une reconnaissance individuelle (Bowers & Alexander, 1967). Cette unicité de
l’odeur individuelle est liée au génome, lui aussi unique, de l’individu. Une partie de ce
génome a été particulièrement étudiée, le Complexe Majeur d'Histocompatibilité, ou MHC.
1.1.1 Odeurs et MHC
A partir d’un mélange d’odeurs individuelles (destiné à homogénéiser l’action des facteurs
environnementaux) de souris génétiquement identiques appartenant à deux lignées différant à
un locus du Complexe Majeur d’Histocompatibilité (Major Histocompatibility Complex ou
MHC), (Penn & Potts, 1998b) ont démontré une capacité à discriminer les odeurs des deux
lignées, capacité qui ne peut s’expliquer que par cette différence à un seul locus du MHC.
L’analyse chimique des composés volatiles présents dans l’urine révèle qu’une telle
différence à un seul locus du MHC se traduit par des variations dans les proportions de
8 composés (Singer et al., 1997). La "popularité" du MHC comme vecteur potentiel des
informations liées à la reconnaissance de parentèle est due à sa fonction primaire de
reconnaissance du soi et du non-soi au niveau cellulaire (voir encadré 1.1 pour une
présentation succincte du Complexe Majeur d’Histocompatibilité). Cette fonction
immunologique primaire s’accompagne de propriétés qui se sont révélées intéressantes non
seulement pour les immunologistes, mais aussi pour les évolutionnistes. La plus importante de
ces propriétés est un très grand polymorphisme (Zavazava & Eggert, 1997). Chez la Souris, le
nombre d’allèles à chaque locus du MHC est estimé à 100 (Tregenza & Wedell, 2000). Le
maintien de cette diversité est classiquement expliqué par la nécessité de présenter une
formule immunitaire efficace et sans cesse renouvelée, face à des agents pathogènes évoluant
22

Chapitre 1 : Odeurs et Génome
eux aussi (mais voir Hedrick, 1999 pour l'ensemble des hypothèses et la suite du texte pour
une discussion sur ce point). Cependant certains auteurs considèrent que la fonction
immunitaire actuelle serait une exaptation : la fonction primaire serait de signaler
l'individualité génétique (Singh, 2001). Une telle vision des choses est à mettre en relation
avec les nombreuses études ayant dressé un parallèle entre MHC et comportement (Zavazava
& Eggert, 1997).
Puisque le MHC est traditionnellement associé à la reconnaissance de parentèle, le principal
comportement avec lequel il a été relié est le choix de partenaire, dans le but de démontrer
son rôle dans l'évitement de l'inceste
(cf. introduction générale). Deux modèles
prédominent dans la littérature, les Souris et
l'être humain. Chez les Souris, il a été montré
une corrélation entre choix de partenaire et
MHC dans des populations semi-naturelles
(Potts et al., 1991). Les préférences sociales
semblent également être corrélées au MHC, les
femelles préférant fonder une structure de
crèche (élevage en commun des petits) avec
des femelles de MHC similaire (Manning et
al., 1992). De plus, la reconnaissance des petits
par la mère peut être influencée par le MHC
quand celui-ci est la seule partie variable du génome : les femelles préfèrent ainsi ramener au
nid des petits de même haplotype que le leur (Yamazaki et al., 2000). Il est à noter que les
préférences sexuelles comme sociales liées au MHC semblent apprises, puisque la réalisation
d’adoptions peut renverser ces préférences (Penn & Potts, 1998a ; Yamazaki et al., 2000).
Encadré 1.1 : le MHC
Le Complexe Majeur d’Histocompatibilité estune ensemble de gènes impliqués dans la réponseimmunitaire. Ces gènes sont multi-alléliques,c’est-à-dire qu’il existe de très nombreusesvariantes entre individus, très étroitement liés, cequi implique qu’ils sont transmis ensemble etenfin ils sont codominants (tous les allèless’expriment également). L’association deplusieurs gènes du MHC sur un chromosomeforme un haplotype. Le MHC code pour des glycoprotéines qui fixentdes peptides intra (gènes du MHC de classe I) ouextra-cellulaire (classe II) et les présentent auxlymphocytes T, qui sont les effecteurs de laréponse immunitaire. Ces lymphocytes possèdentdes récepteurs membranaires (TCR : T-CellReceptor) qui reconnaissent le complexe protéinedu MHC + peptide. Il y a activation de laréponse immunitaire quand le peptide OU laprotéine est différent du soi.
En ce qui concerne la transcription en odeurs du MHC, il semble qu'elle ne s'effectue pas
exclusivement via la réponse immunitaire, puisque des souches "stériles" (élevées sans
contact avec des pathogènes) de MHC différents produisent des odeurs discriminables pour
des congénères (Yamazaki et al., 1990). Ceci ne signifie en rien que le MHC soit seul à
l'origine de la production de l'odeur individuelle. En effet, les autres régions du génome et les
facteurs environnementaux influencent également les productions odorantes (Brown, 1995).
De plus, la composition de l'urine est aussi corrélée au reste du génome (Eggert et al., 1996).
23

Chapitre 1 : Odeurs et Génome
Enfin, les différences olfactives liées au MHC sont relativement faibles, comparativement aux
différences liées au régime alimentaire (Brown et al., 1996). L'environnement joue un rôle
prépondérant dans l'expression des différences au niveau du MHC. L'élevage en milieu stérile
peut rendre des souches ne différant qu'au niveau du MHC non-discriminables (Schellinck et
al., 1995). Ces résultats contradictoires avec Yamazaki et al. (1990) (voir plus haut) peuvent
s'expliquer par le fait que la discrimination de souches différant au niveau du MHC n'est pas
universelle, et semble ne concerner que les différences reflétées dans la partie fixant les
peptides (Carroll et al., 2002). D'une manière générale, le processus par lequel le MHC est
transcrit en odeurs reste mal connu (Singh, 1999 ; Penn, 2002). Les glandes préputiales
seraient impliquées chez la Souris, puisque leur ablation inhibe la préférence des femelles
pour un mâle de MHC différent (Ninomiya & Brown, 1995). Même s'il peut exister une
transcription directe du MHC en odeurs en laboratoire (Yamazaki et al., 1999 ; Montag et al.,
2001), il est difficile d'estimer l'importance de cette transcription directe par rapport à la
contribution des autres parties du génome, d'autant que les souches de souris classiquement
utilisées ont un génome identique hormis pour les locus du MHC. L'action du MHC sur la
flore bactérienne (Toivanen et al., 2001) plaide pour une action principalement indirecte, via
les produits de digestion. Chez les Rats élevés dans un environnement stérile, la
discrimination individuelle sur la base des urines semble même impossible : elle est
cependant rétablie une fois les animaux transférés en milieu non stérile, c'est à dire une fois
que la flore bactérienne a pu recoloniser le tractus digestif (Singh et al., 1990). De façon
similaire, le retrait d'une partie de la flore bactérienne altère l'odeur individuelle des rats : les
odeurs de rats connus ainsi traités ne sont pas plus explorées que celles de rats inconnus
traités identiquement (Schellinck & Brown, 2000). Cependant la discrimination individuelle
après traitement entre deux rats connus est plus facile qu'entre deux rats inconnus, ce qui
indique qu'il subsiste dans l'odeur des indices individuels malgré l'élimination d'une partie de
la flore bactérienne (Schellinck & Brown, 2000). Il semble que les molécules du MHC, elles-
mêmes non volatiles, fixent les molécules volatiles produites par la flore bactérienne et les
transportent dans l'urine. L'injection à un rat de glycoprotéines du MHC différentes de celles
qu'il produit entraîne une modification drastique de l'odeur de l'urine, bien que ces molécules
ne soient pas elles-mêmes perçues. Il y aurait donc un transport sélectif des volatiles dans
l'urine sous contrôle du MHC (Pearse-Pratt et al., 1999).
La diversité du MHC est un phénomène qui a intrigué (et intrigue encore) de nombreux
chercheurs (Edwards & Hedrick, 1998). Les divers travaux montrant des choix
24

Chapitre 1 : Odeurs et Génome
comportementaux corrélés au MHC, chez la Souris (Arcaro & Eklund, 1999) mais également
chez l'homme (Lenington, 1994 ; Wedekind & Furi, 1997 ; Milinski & Wedekind, 2001 ;
Wedekind & Penn, 2000) ont amené certains auteurs à s'interroger sur le rôle du
comportement dans le maintien de cette diversité. La tendance générale à préférer un
partenaire sexuel différent au niveau du MHC permettrait de conserver sa diversité allélique
(Jordan & Bruford, 1998 ; Eggert et al., 1999). Le "rôle" du MHC dans le signalement de
l'identité génétique pourrait même être le moteur de sa diversité, préalablement à son rôle
immunitaire (Singh, 2001). Ce raisonnement me semble un exemple marquant de la confusion
qui règne autour du MHC et de ses fonctions supposées. En effet, l'historique des travaux sur
le sujet peut se résumer ainsi : 1) le MHC permet la reconnaissance cellulaire donc peut-être
la reconnaissance de parentèle 2) la reconnaissance de parentèle est importante pour
l'évitement de l'inceste 3) puisque le MHC permet cette reconnaissance, il a du être
sélectionné pour cela. La faille de ce raisonnement est que les pressions de sélection qui
rendent importante la reconnaissance de parentèle ne sont pas prises en compte. L'évitement
de l'inceste est important notamment parce qu'il permet à la progéniture d'hériter d'une
formule immunitaire efficace et nouvelle (le fait d'avoir des parents de MHC dissimilaire
augmente la fitness des descendants). Il y a confusion entre les pressions de sélection, liées
entre autres au MHC, et les mécanismes par lesquels ce MHC est maintenu variable. Supposer
que la fonction historique du MHC est le signalement de l'identité génétique permettant la
reconnaissance de parentèle implique l'existence de pressions de sélection n'incluant pas la
compétence immunitaire (puisque l'apparition du système immunitaire serait ultérieure). Or
la compétence immunitaire est indéniablement une des forces de sélection les plus
universellement reconnues, y compris pour l'apparition de la reproduction sexuée (Hamilton
et al., 1990 ; Ravary, 2003). Le raisonnement ci-dessus est donc téléologique (voir
l'introduction générale). Cela ne signifie pas qu'il soit nécessairement faux, mais il est peu
plausible et la littérature fournit des arguments contraires. Par exemple, dans des populations
naturelles d'ovins, la diversité du MHC est corrélée à la survie des jeunes et à la résistance à
des parasites intestinaux (Paterson et al., 1998) mais les choix de partenaires ne suivent en
rien le MHC (Paterson & Pemberton, 1997). Même chez la Souris, qui est pourtant le modèle
de référence dans l'étude du MHC, le choix de partenaire n'est pas toujours basé sur le MHC
(Eklund, 1999 ; Ehman & Scott, 2001). L’hétérozygotie au niveau des loci du MHC est
pourtant bien associée à une meilleure fitness, puisque les souris hétérozygotes résistent
mieux à des infections bactériennes (Penn et al., 2002). Toujours chez les Souris de
25

Chapitre 1 : Odeurs et Génome
laboratoire, la résistance aux parasites est également influencée par le MHC (McClelland et
al., 2003a). Dans les cas d’infections multiples, les hétérozygotes montrent une supériorité par
rapport aux homozygotes (McClelland et al., 2003b). Le moteur de la diversité du MHC
semble bien être la résistance aux maladies (Jeffery & Bangham, 2000). Dans cette vision des
choses, la résistance aux pathogènes est à l'origine des choix basés sur l'apparentement, qu'ils
soient sexuels ou sociaux (Lewis, 1998). Il est important de comprendre que si les pressions
de sélections semblent bien liées à la fonction immunitaire, le choix du partenaire peut être un
des mécanismes permettant de maintenir la diversité du MHC (Hedrick, 1999; Penn & Potts,
1999). Chez les Souris, ce choix de partenaire a été attesté par des études proposant un choix
entre un partenaire similaire ou différent au niveau du MHC. Or cette stratégie n'est pas la
seule permettant de maintenir la diversité allélique du MHC. Chez les épinoches, les femelles
choisissent les mâles les plus hétérozygotes au niveau du MHC, indépendamment de leur
proximité génétique (Reusch et al., 2001). Cette hypothèse n'a jamais été testée chez les
Souris, même si une telle stratégie serait moins adaptative chez une espèce où le risque
d'inbreeding est plus important que chez les épinoches (Bakker & Zbinden, 2001). Les études
sur le MHC, sa diversité et son rôle dans les stratégies socio sexuelles des animaux sont
majoritairement réalisées sur des Mammifères, ce qui semble conduire à une trop grande
focalisation sur un seul mécanisme de sélection (le choix d'un partenaire de MHC
dissimilaire) (Bernatchez & Landry, 2003). Parmi les mécanismes encore méconnus
permettant d'expliquer le maintien de la diversité du MHC, il en est un qui semble
particulièrement intéressant : les influences parentales sur l'haplotype de la progéniture. Des
souris infectées par un virus produisent une descendance au MHC plus hétérozygote que celle
des souris témoins : l'existence de pathogènes favoriserait la sélection parmi les œufs de ceux
présentant les combinaisons les plus efficaces face à ces pathogènes, c'est-à-dire les plus
hétérozygotes (Rulicke et al., 1998). En extrapolant à partir de cette découverte, on est amené
à considérer que le degré d'hétérozygotie du MHC d'une population naturelle n'est pas
nécessairement le produit du choix de partenaire.
1.1.2 Odeurs et MUPs
Les Protéines Urinaires Majeures, ou MUPs, sont des composés importants de l'urine des
Mammifères. Ces protéines encapsulantes de la famille des lipocalines ont en effet la capacité
de lier de plus petits composés volatiles (Flower, 1996). Cette liaison protège les volatiles de
la dégradation et prolonge ainsi leur action, puisqu'ils peuvent être re-libérés à partir d'urine
26

Chapitre 1 : Odeurs et Génome
séchée (Hurst et al., 1998). Les MUPs convoient ainsi des molécules impliquées dans la
communication phéromonale (Novotny et al., 1999 ; Dulac & Torello, 2003). A l'instar du
MHC, elles présentent une grande variabilité interindividuelle (Robertson et al., 1996) chez
les souris domestiques. En vertu de cette variabilité, il a été proposé récemment que ces
MUPs soient impliquées dans la reconnaissance individuelle (Hurst et al., 2001). Dans ces
travaux, le comportement de souris mâles Mus musculus domesticus est examiné, et plusieurs
situations leur sont présentées : leur propre urine, l'urine d'un individu non-apparenté, l'urine
d'un frère présentant un profil de MUPs différent et l'urine d'un frère de même profil. Les
temps d'investigation révèlent que seules les urines dont le profil de MUPs diffère de celui du
sujet sont statistiquement plus explorées que l'urine du sujet. Les comportements de marquage
sont également plus nombreux en présence de l'odeur du frère au profil de MUPs différent et
du mâle non apparenté qu'en présence du même profil MUPs, qu'il provienne de l'urine d'un
frère ou du sujet lui même. Une seconde expérience présente aux sujets leur propre urine
modifiée par ajout de MUP recombinante. Cette modification ne modifie pas le temps
d'investigation mais induit une augmentation du comportement de marquage. La conclusion
de ces travaux est que les MUPs seraient le support de l'identité chimique des Souris. L'un des
problèmes de cette étude est que les auteurs se basent sur la non significativité des tests
statistiques pour conclure à un traitement équivalent. Or un résultat non significatif ne
démontre pas l'égalité des deux catégories, mais exprime l'impossibilité de conclure à une
différence. L'affirmation selon laquelle l'odeur d'un frère avec le même profil de MUPs est
confondue avec l'odeur propre n'est donc pas exacte, d'autant qu'aucune situation de choix
direct n'est proposée. L'autre problème majeur de cette étude est théorique. Les frères de
même profil de MUPs sont issus de couples formés avec des parents de deux populations
sauvages géographiquement distantes. Cette progéniture devrait donc présenter une variabilité
phénotypique plus importante que celle existante au sein d'une seule population sauvage.
Malgré cela, Hurst et al. (2001) ont pu trouver, apparemment facilement, des mâles
présentant la même combinaison de MUPs. Ce doit donc être encore plus fréquent dans les
populations naturelles. Si les MUPs sont le support de l'identité olfactive, doit-on en conclure
que, dans de nombreux cas, les frères peuvent confondre l'odeur de leur frère avec leur propre
odeur et donc ne pas se discriminer mutuellement ? Supposer qu'une des fonctions des MUPs
soit le signalement de l'individualité semble donc excessif et peu plausible. Cependant les
MUPs sont des molécules intéressantes à plus d'un titre, parce qu'elles présentent une
variabilité qui semble les distinguer de "simples" protéines encapsulantes (Beynon & Hurst,
27

Chapitre 1 : Odeurs et Génome
2003). Les activités de sur-marquage (marquage sur une marque laissée par un congénère)
sont influencées par les MUPs (Humphries et al., 1999). Il semble que l'information contenue
dans l'urine déposée lors d'un marquage puisse être décomposée en deux fractions : la fraction
volatile de l'odeur attire à distance et suscite l'intérêt et l'investigation ; la partie non volatile
(contenant les MUPs) détermine le déclenchement de l'activité de sur-marquage (Nevison et
al., 2003). Cette dualité du traitement de l'information est à mettre en rapport avec l'existence
de deux systèmes de l'olfaction chez les Mammifères : le système olfactif principal et l'organe
voméronasal. C'est ce dernier qui traiterait l'information portée par les MUPs. En effet, des
femelles à qui on a enlevé l'organe voméronasal présentent une capacité de discrimination
inchangée mais des comportements de marquage perturbés par rapport à des femelles intactes
(Petrulis et al., 1999). La reconnaissance du sexe du donneur d'odeur semble notamment
affectée, et serait une des fonctions de l'organe voméronasal (Keverne, 2002). Chez les
campagnols monogames Microtus ochrogaster, une lésion de l'organe voméronasal empêche
la formation du lien entre partenaires (Curtis et al., 2001). D'une manière générale, cet organe
est considéré comme l'opérateur de fonctions phéromonales, liées notamment à la régulation
de la physiologie de la reproduction (Halpern & Martinez-Marcos, 2003). Quoi qu'il en soit,
l'action des MUPs via l'organe voméronasal, si elle semble importante, semble plus liée à la
motivation à effectuer les comportements qu'au signalement de l'individualité. Ainsi les souris
mâles confrontées à des marques odorantes d'autres mâles protégés d'une grille interdisant
l'accès à la fraction non-volatile montrent une diminution de l'activité de marquage par
rapport à une situation ou toute la marque est disponible, mais pas de diminution des
comportements d'investigation (Nevison et al., 2003). Curieusement quelques auteurs
persistent à vouloir attribuer aux MUPs une fonction de signalement de l'individualité
génétique, ce qui me semble une démarche téléologique assez similaire à celle qui attribue
cette même fonction au MHC.
En l'absence de démonstration flagrante de l'existence d'une famille de molécules spécialisée
dans le signalement de l'identité génétique, les expériences réalisées dans cette thèse ont
systématiquement utilisé les odeurs globales, c'est-à-dire des odeurs les plus complexes
possible (souvent obtenues en frottant une coupelle sur la zone uro-génitale, de façon à
recueillir à la fois les parties volatiles et non volatiles provenant de l'urine mais aussi
d'éventuelles secrétions directes).
28

Chapitre 1 : Odeurs et Génome
Dans les expériences décrites ci-dessous, c'est la corrélation entre l'odeur individuelle et la
proximité génétique qui est explorée, indépendamment de l'utilisation que peuvent en faire les
souris. Cette investigation est d'abord concentrée sur les relations de parentés puis vers les
relations intra et interspécifiques.
1.2. Odeurs et parentèle chez Mus Spicilegus
L’objectif des expériences décrites dans l’article ci-joint est de rechercher les similarités
d’odeurs chez Mus spicilegus, puisque ce sont ces similarités d’odeurs qui fournissent le
substrat d’une reconnaissance de parentèle par comparaison soit avec l’odeur propre soit avec
un modèle de référence mémorisé (voir le chapitre 2 pour une synthèse des différentes
théories sur les mécanismes permettant la reconnaissance de parentèle). Il est important de
noter à nouveau que l’existence de telles similarités proportionnelles à l’apparentement ne
présume en rien de leur utilisation par l’animal dans un contexte de reconnaissance de
parentèle. Cependant, ces similarités sont un pré-requis indispensable à l’établissement d’une
réelle reconnaissance de parentèle par le canal olfactif.
Les résultats exposés dans cet article peuvent se résumer ainsi : les odeurs uro-génitales de
souris mâles apparentées sont perçues comme similaires par des congénères (eux-mêmes
systématiquement non apparentés aux donneurs d'odeur). Ainsi, les odeurs de deux frères
(r = 0.5), de deux doubles cousins (r = 0.25) ou de deux cousins (r = 0.125) présentent entre
elles des similarités par rapport à l'odeur d'un individu non-apparenté. Pour tester l'hypothèse
d'un signal familial partagé par toutes ces classes d'apparentement, le même type de test est
réalisé avec cette fois des donneurs d'odeur dont l'apparentement est plus ou moins grand par
rapport au donneur d'odeur d'habituation. L'odeur d'un double cousin est considérée comme
semblable à l'odeur d'habituation quand elle est comparée à celle d'un cousin. En revanche,
l'odeur d'un "super demi-frère" (r = 0.375) ne suscite pas une investigation plus importante
que l'odeur d'un frère. Ces résultats suggèrent que la relation entre odeurs et gènes est
graduelle, puisque plus d'apparentement se traduit par plus de similarité olfactive perçue.
Cette relation graduelle est démontrée pour des taux d'apparentement faible mais ne peut être
établie pour de forts apparentements.
Un autre résultat important de cette étude est plus "implicite" : il s'agit de la capacité
démontrée par les Souris à estimer la proximité olfactive entre deux individus qui ne leur sont
29

Chapitre 1 : Odeurs et Génome
pas apparentés. Chez Mus spicilegus, cette capacité pourrait se traduire par une
reconnaissance des individus appartenant à la même portée voire au même tumulus (puisque
cette capacité s'étend jusqu'aux cousins et que les tumuli sont souvent habités par une
combinaison de frères et de cousins (Garza et al., 1997)). En début de période automnale,
cette capacité pourrait également favoriser l'expression de comportements de coopération
entre juvéniles : le comportement adopté face à un individu non familier pourrait être différent
selon qu'il appartienne ou non à la même fratrie qu'un individu familier.
La relation forte entre odeurs et proximité génétique ne se limite pas aux relations de parenté,
comme nous le verrons par la suite.
30

Chapitre 1 : Odeurs et Génome
Odour similarities as a basis for discriminating degrees of kinship in rodents: evidence
from Mus spicilegus.
Running headline: Busquet & Baudoin: Kinship odours in mice.
Keywords: mound-building mice, kin recognition, odours, habituation-generalisation.
Nicolas BUSQUET, Claude BAUDOIN - Université Paris 13, Laboratoire d'Ethologie
Expérimentale et Comparée, 93430 Villetaneuse, France
Corresponding author: Nicolas Busquet
Université Paris 13, Laboratoire d'Ethologie Expérimentale et Comparée
99 avenue Jean-Baptiste Clément - 93430 Villetaneuse, France
Email: [email protected]
Word count: 5412
Abstract:
In order to achieve odour-based kin recognition, rodents need olfactory cues that indicate
kinship. Odour-genes covariance is a phenomenon by which closer genetic relationships
between individuals are reflected in greater similarity in their odours. Here odour-genes
covariance was tested among kin groups in Mus spicilegus. Using a habituation –
generalisation procedure designed to assess responses to similarities, males (unrelated to any
of the odour donors) recognized similarities in the odours of brothers (r = 0.5) when
compared to an unrelated male. With the same protocol, similarities were also found between
the odours of double cousins (r = 0.25) and cousins (r = 0.125). This demonstrates that
kinship from siblings to cousins is reflected in odour similarity. To investigate the precision
of animals’ responses to odour similarities based on relatedness, similar tests were conducted
using odours from males of different relatedness to the habituation odour donor. Mice
responded to the similarities between odours of double cousins compared to cousins. This is a
clear demonstration of the subtlety of odour-genes covariance among close kin. Mice did not,
however, treat odours of brothers as similar compared to those of “super half-brothers”
(r = 0.375). Implications of these findings for differential responses based on degrees of
relatedness, including kin recognition, are discussed.
31

Chapitre 1 : Odeurs et Génome
Kin recognition or discrimination, the ability to respond differentially to conspecific
individuals depending on their genetic relatedness, is a widespread phenomenon across the
animal kingdom. Animals that help close kin but do not mate with them are thought to be
favoured by evolution, because they improve their inclusive fitness (Hamilton 1964) and
decrease the costs of inbreeding (Pusey & Wolf 1996). Whatever the putative underlying
mechanisms (Blaustein 1983, Hepper 1991, Tang-Martinez 2001), kin recognition can be
treated as a communication issue (Beecher 1991). Prior to acting differentially towards kin
and non-kin, the receiver must be able to perceive the relevant signals and distinguish
appropriately between them. This is only possible if the sender provides cues that vary with
relatedness. The study of those cues and the amount of information they provide is a useful
approach that can help to understand kin biased behaviour (or absence of it) in animals
(Beecher 1989). In the present study we investigated olfactory signals of mound-building
mice (Mus spicilegus) and whether they can reliably indicate degrees of kinship.
Olfactory signals are extremely important in rodent communication (Bowers &
Alexander 1967; Brown, 1985a & b, Beauchamp & Yamazaki 2003). Rodents can make
subtle distinctions among odours and adapt their behaviour accordingly. Odours can reflect
changing biological states, such as diet (Brown et al. 1996; Ferkin et al. 1997), social status
(Drickamer 1992) or health (Penn et al. 1998; Willis & Poulin 2000). More importantly for
this context, odours can also reflect unchanging traits such as genetic relatedness (Todrank et
al. 1998, Mateo 2002, Todrank & Heth 2003). This is especially useful as it could allow
discriminative responses to kin and non-kin, an ability already demonstrated in several
species of rodents (house mice (Hayashi & Kimura 1983), rats (Hepper 1987), ground
squirrels (Holmes 1986), golden hamsters (Heth et al. 1998), beavers (Sun & Muller-
Schwarze 1997)) and has strong fitness consequences (Barnard & Fitzsimons 1989, Barnard
et al. 1991). Individual odours are complex signals: for example, slight differences in genes of
the Major Histocompatibility Complex (MHC) are reflected in the proportion of volatiles that
contribute to the odour of the urine (Singer et al. 1997). But the olfactory signals produced by
an individual are not exclusively MHC-derived products (Eggert et al. 1996). Indeed, major
urinary proteins (MUPs) provide about the same level of diversity between individuals as
MHC-products and are found in higher concentrations in the urine (Hurst et al. 2001). The
existence of a set of odorous compounds specialized for kin recognition still being
controversial, it seems reasonable to investigate animal’s perception and categorization of
complex individual odours. To the extent that an individual's odour reflects aspects of its
32

Chapitre 1 : Odeurs et Génome
genotype, genetically closer individuals should smell more similar than less genetically
similar individuals. This phenomenon, termed “odour-genes covariance”, has been
demonstrated in mole-rats (Heth & Todrank 2000) and in mice (Heth et al. 2001). In Mus
spicilegus, for example, odours of males from the same population were treated as similar
when compared to a male from another population (Heth et al. 2001). Odour-genes covariance
findings raised the possibility that odour similarities could be used to assess genetic
relatedness. This was demonstrated in mice at the species and population levels (Heth et al.
2003). If odour-genes covariance can be demonstrated at various levels among kin groups, it
would provide a basis for mice to assess subtle differences in genetic relatedness, enabling
graded, fine-tuned kin discrimination through the same mechanism that was found for graded
species and population discrimination (Heth et al. 2003). Odour-genes covariance could also
enable fine tuned responses to slight differences in genetic relatedness, documented in turkish
and golden hamsters (Heth et al. 1999), in ground squirrels (Mateo 2002) and in oldfield mice
(Koeninger Ryan & Lacy 2003).
In the present paper, we investigated this relationship between odours and genes
among kin groups of Mus spicilegus. Our prediction is that closer kinship between individuals
should be reflected in greater similarities in their odours. Odour-genes covariance has
previously been demonstrated in a wide spectrum of rodent species and at various levels of
genetic relatedness (see Todrank & Heth, 2003 for a review), but has not been investigated for
mice sharing various degrees of relatedness. In previous studies of odour similarities among
close kin, the dissimilar odour was from an unrelated individual (e.g. Todrank et al. 1998,
1999). In some tests of the current study we challenged the subjects to make much more
subtle distinctions among odours of individuals that were all close relatives.
Mus spicilegus, the mound-building mouse, is a feral species that is characterised by
some life-history traits which make it a good model for studying kin discrimination (see
Sokolov et al. 1998). Gathering within the mounds is kin biased (Garza et al. 1997) and
related juveniles spend winter inside mounds without reproducing (Gouat et al. 2003). In
spring, mound-building mice leave the mounds and start the breeding season. When compared
to the polygamous mouse Mus musculus domesticus, the mound-building mouse exhibits
some behavioural traits linked with monogamy: a strong pair-bond (Patris & Baudoin, 1998;
Dobson & Baudoin 2002, Baudoin et al. In Press) and a great involvement of males in
paternal care (Patris & Baudoin, 2000).
33

Chapitre 1 : Odeurs et Génome
We investigated odour similarities among close kin, using the habituation-
generalisation technique (Todrank et al. 1998, 1999). Three degrees of kinship were tested
(coefficient of average relatedness r = 0.5, 0.25 and 0.125 compared to r = 0). The same
procedure was then used to assess perceived similarities when comparing odours of
individuals of varying relatedness (r = 0.25 vs 0.125 and r = 0.5 vs 0.375). Behavioural
responses of male mice in these tests were expected to reveal odour-genes covariance among
close kin.
METHODS
Animals:
Mus spicilegus males (N = 192) were used in this study. They had been kept in cages with
same-sex littermates since they were 35 days old (weaning occurs at 28 days and litters are
divided into same-sex groups at 35 days) and had never engaged in reproduction or been used
in experiments. These males, aged 3 to 6 months, were used only once, either as odour donors
or subjects. All the animals were 3rd generation mice from a population originally collected
in Gyöngyös, Hungary, in October 1999. From the time of their collection, genealogy of
every individual was known and all the reproductive couples had been formed in a way to
avoid inbreeding (the initial couples were formed with partners captured in distant sites).
Animals were isolated in standard polycarbonate cages (26 x 16 x 14 cm) a week before
testing to avoid physiological consequences of any cage-mate hierarchies that could be
reflected in the odours (deCantanzaro & Gorzalka 1979; Drickamer 1992). They were kept on
a shifted 14:10 light: dark cycle in constant temperature (21 ± 1°C) and humidity
(approximately 50 %). Water and food (mice pellets, M20 type, SDS, Witham, England) were
supplied ad libitum, and cotton balls were provided for nesting material. To conform to
French legislation, animals were not regrouped or returned to the breeding stock after the
experiments, and euthanasia was carried out by singly introducing animals into a CO2
saturated atmosphere.
Habituation Generalisation Procedure:
In this series of experiments, the hypothesised odour similarity under investigation was
among close kin. The two test odours were from individuals that were differentially related to
34

Chapitre 1 : Odeurs et Génome
the habituation odour donor. We used a variation on a standard habituation-discrimination
technique (Halpin 1986) described by Heth et al. (2001) to assess odour similarities. After
habituation to one individual’s odour for 10 minutes (odour A), odours of two different
individuals are presented for 5 minutes (odour A’ and B). The donor of odour A’ is more
closely related to the habituation odour donor than the donor of odour B. If the subject
perceives odour A’ as more similar in quality to the habituation odour, he should spend more
time investigating the more different odour (B).
Protocol:
The subject was transferred into a clean plastic cage (28 × 9 × 8 cm, Iffa Credo, Lyon)
covered with a Perspex top, and left alone for 5 minutes for familiarisation with the testing
apparatus. In the meantime, the odour of the habituation odour donor was sampled by gently
rubbing a small Petri dish (3 cm diameter) on the ano-genital area for a few seconds. This area
is investigated a lot during encounters between mice and is likely to contain information from
different sources, including urine and preputial glands. This Petri dish was placed in the
middle ofthe test cage for 10 minutes (habituation phase). The subject was observed during
that time to ensure that contact with the Petri dish had occurred and investigation of the
stimulus was underway. Before the end of the habituation phase, the two test stimuli were
prepared in the same way, starting randomly with one or the other. After removal of the
habituation stimulus, those two Petri dishes were introduced simultaneously into the cage, and
the subject was observed for 5 min (test phase). The test stimuli were placed at opposite ends
of the cage and their location counter-balanced across tests. Time spent investigating both
odours was recorded with stopwatches by an experimenter unaware of the nature of each
stimulus. The subject was considered to be investigating the stimulus when its nose was
directly above the Petri dish.
Four degrees of relatedness were used in the present study: brothers, “super half-
brothers”, “double cousins” and cousins. On average, half of the genes received from the two
parents is common in brothers, which is expressed by an average coefficient of relatedness of
r = 0.5. “Super half-brothers” are males that have the same father and whose mothers are
sisters. The associated coefficient of relatedness is r = 0.375. “Double cousins” are
individuals that are related through both their father and their mother. Among the two sets of
parents whose offspring are double cousins, parents are not related to one another, but each
parent in one couple is the sibling of one parent in the other couple. Such offspring can be
35

Chapitre 1 : Odeurs et Génome
obtained, for example, by mating two brothers with two sisters from a different family.
Double cousins have a coefficient of relatedness of r = 0.25. For cousins, average relatedness
is r = 0.125. Unrelated individuals are individuals that do not have a common ancestor since
genealogy is known. Since the animals used are from the 3rd generation in the lab and the first
couples were formed with animals captured as juveniles at distant sites (therefore unlikely to
be related), the theoretical maximum relatedness is 0.0625 (for the very unlikely case where
the 8 great grandparents of one male were siblings to the 8 great grandparents of the other
male). The actual relatedness is probably much lower than that, and for reasons of clarity, we
will state that their relatedness is r = 0.
Two experiments were conducted, corresponding to two aims in the investigation of
odour-genes covariance. The first experiment, divided into three tests, is designed to answer
the following question: “Do kin related mice have perceptually similar odours ?” The
prediction was that similarities among close kin should be perceived when compared to
unrelated individuals. We used three degrees of relatedness to answer that question: brothers
(test 1a), double cousins (test 1b) and cousins (test 1c), dividing the coefficient of relatedness
by two in each step (0.5, 0.25, and 0.125).
The second experiment was designed to assess the accuracy of odour-genes covariance
by presenting in the test phase odours of two related individuals varying in their degree of
relatedness. After habituation to the scent of a male, we presented to the subject the odours of
a male double cousin vs. a male cousin (experiment 2a) or of a brother vs. a super-half-
brother of the habituation odour donor (experiment 2b). In those two tests, the predicted
outcome was that more relatedness would be reflected in greater similarities between the
individual odours, and thus that the scent of the less related donor would be investigated
more.
In every trial, the two test odour donors were the same age ± one week. In the test
involving brothers, the habituation odour donor was from a previous litter, and the age
difference between the two related donors was always bigger than the age difference between
the two unrelated or less related ones, so that any effects of the donors' age could only act
against our hypothesis.
Statistical treatment:
For each test, times spent on the two test stimuli were compared using a two-tailed Wilcoxon
matched paired test. Because of the nature of the test (simultaneous presentation of two
36

Chapitre 1 : Odeurs et Génome
stimuli), no conclusions can be drawn from comparison with the habituation phase, nor can
data of one test be compared with data of another one.
RESULTS
In all experiments, the subjects were active and investigated all the presented odours.
Although a lot of time was devoted to other activities, such as self-grooming and investigation
of the apparatus, there was no sign of avoidance of any of the stimuli. Investigation of the
stimuli was seldom interrupted by those activities. Mice were never seen urinating on or in the
close vicinity of the stimulus. All subjects were unrelated and unfamiliar to any of the odour
donors, and no behavioural differences was observed between the groups.
Experiment 1:
In the three tests, subjects spent a longer time investigating the odour of the donor that was
unrelated to the habituation odour donor. Results of those three tests are presented in figure 1.
In experiment 1a, after the period of habituation to the odour of the first male (male A),
subjects spent significantly less time investigating the odour of A’ (brother of A) than B
(unrelated to A), thus demonstrating perception of a similarity between A and A’ (Wilcoxon
matched paired test N = 8, Z = 2.52, p = 0.012). The odours of brothers were treated as
similar by an unrelated individual. In experiment 2b where A’ was a double cousin of A
(r = 0.25), the odour of B (the unrelated odour donor) was again investigated significantly
longer (N =1 0, Z = 2.5, p = 0.013). In experiment 2c, A’ was a cousin of the habituation
odour donor A (r = 0.125). Again, subjects investigated the stimulus from odour donor B
significantly more than A’ (N = 10, Z = 2.19, p = 0.028). When compared to the odour of an
unrelated individual, odours from close kin (brothers, double cousins and cousins) were
treated as similar by a naïve subject.
Experiment 2:
In experiments 2a and 2b, the two test odours were from individuals that were both related to
the habituation odour donor A, with one of them (A’) being more closely related than the
other (A’’). Odour-genes covariance would suggest that subjects should investigate the odour
of the less related individual more, because of its greater overall difference from the
37

Chapitre 1 : Odeurs et Génome
habituation odour. In experiment a, A’ was a double cousin (r = 0.25) whereas A’’ was a
cousin (r = 0.125) of the habituation odour donor. The odour A’’ was investigated
significantly more than A’, providing evidence that the subjects responded to the greater
similarity between the odours of double cousins than cousins (N = 10, Z = 2.50, p = 0.013).
In experiment b, A’ was a brother of A (r = 0.5) and A’’ a “super-half-brother” (r = 0.375).
There was no significant difference in the time spent investigating A’ and A’’ (N = 10,
Z = 0.25, p = 0.799), and therefore no evidence that subjects treated odours of super half
brothers as different from odours of brothers.
DISCUSSION
Male Mus spicilegus mice treated odours of close kin (brothers, double cousins and cousins)
as having similar qualities, i.e., after habituation to the odour of one individual, subjects in the
test trial spent less time investigating the odour of a relative of the habituation odour donor
than the odour of an unrelated individual (experiment 1). Thus genetic relatedness between
two mice is reflected in similarities in their individual odours. Moreover, when both test
odours were from individuals related to the habituation odour donor (experiment 2), the
responses were consistent with detecting the greater similarity in the qualities of odours of the
more closely related individuals when the comparison was between double cousins as
opposed to cousins. This provides evidence of odour-genes covariance at a much subtler level
than has been tested previously in mice (Heth et al. 2001). The differences between odours of
brothers and super half brothers were not salient enough, however, for subjects to respond to
this small degree of closer relatedness (r = 0.5 as opposed to r = 0.375).
Although male mice from the same kin group have similar odours, because closer
kinship is reflected in greater odour similarity (experiment 2b), it cannot be said that mice
from the same kin group share a "kinship odour". Indeed, such an odour would divide animals
into two groups: those who have it and those who do not. For instance if double cousins were
genetically close enough to be part of that category and cousins not close enough, there would
be no explanation for the perceived similarity between cousins when compared to an
unrelated individual (cousins vs. unrelated test, experiment 1c). The greater detected
similarity between odours of double cousins compared to cousins indicates a graded and
continuous relationship between odours and genes within kin groups, rather than a discrete
38

Chapitre 1 : Odeurs et Génome
one. These results are consistent with previous studies demonstrating odour-genes covariance
for mice belonging to broader genetic entities (populations of Mus spicilegus and species of
the Mus genus (Heth et al. 2001)). The present demonstration of odour-genes covariance
among close kin in Mus spicilegus echoes, but also importantly extends previous results on
different rodent species (reviewed in Todrank & Heth 2003) suggesting, although it has not
yet been thoroughly tested, that such a relationship between odours and genes is likely to be
found in most rodent species. Likewise, although results of the present study were obtained
with male subjects and odour donors, there is no reason to think this relationship between
odours and genes would not be found in females.
Given that, the result of the brother vs. super half brother (experiment 2b) seems puzzling. In
the absence of a consistent pattern, it is impossible to tell whether the odours did not convey
enough information of if the animals did not respond to the available information. The testing
situation is not appropriate to assess the subject's ability to discriminate between odours of
brothers and super half brothers. It is possible that odours of animals sharing r = or > 0.375
with the habituation odour donor were all treated as similar, because none of them is
"sufficiently novel". Furthermore, the comparison between r = 0.5 and r = 0.375 may at first
glance seem to be the same difference as between 0.25 and 0.125, but it is not: after
habituation to the individual odour A, the subject is presented with A’ (the brother of A) and
A’’ (“super half-brother” of A) that by descent share respectively 50 % and 37.5 % of their
genes with A. If we assume a continuous covariance between odours and genes, A’ is only
marginally closer to A than A’’: r(A-A’) = (4/3)r(A-A’’) (0.5 = 4/3*0.375). In the double
cousin vs. cousin experiment (2a), A’ (the double cousin) was twice as close to A compared
with A’’ (the cousin): r(A-A’) = 2r(A-A’’) (0.25 = 2*0.125). Thus the detection of the
difference between cousins as opposed to double cousins was probably more obvious than the
differences between odours of brothers and super half brothers. In experiment 2b, neither
odour was consistently treated as similar to the habituation odour. Overall, the results of the
present study demonstrate a high level of precision in the behavioural responses of mice to
odours of individuals of varying degrees of close genetic relatedness. Olfactory signals of
mound-building mice convey sufficient information about genetic relatedness and are
perceived in ways that could enable kin discrimination.
It is remarkable that mice are able to treat the odours of cousins as similar (experiment 1c)
while treating them as different when a more similar odour was present (experiment 2a). This
indicates more subtle discriminative and categorization abilities than previously documented
39

Chapitre 1 : Odeurs et Génome
in ground squirrels using a different procedure (Mateo 2002). In Mus spicilegus, this ability to
categorize individuals among an unfamiliar group into subgroups corresponding to different
degrees of relatedness would allow recognition of individuals belonging to the same litter
within a mound inhabited by animals from several litters (Garza et al. 1997). The stable
relationship between odours and genes may also be used to assess relatedness of unfamiliar
individuals to familiar ones.
Kin recognition mechanisms were not the focus of this research but the findings have
implications on future works investigating such mechanisms. The subtle odour-genes
covariance among kin groups demonstrated in the present study provides a basis for
postulating a process that relies on evaluating odour phenotypes to assess degrees of genetic
relatedness. Genetic relatedness assessment through individual odour similarities (G-ratios)
has been demonstrated in mice, at the level of populations and species, to be a graded self-
referencing process that capitalizes on odour-genes covariance (Heth et al. 2003). The
findings reported here confirm that mice perceive similarities and differences in individual
odours based on genetic relatedness at the level of kin groups. The extent to which mice use
G-ratios for subtle, graded discriminative responses toward close and less close relatives
awaits demonstration in future studies.
It is important to note that thus far, there is little knowledge about the interaction of
odour-genes covariance with changing biological states, i.e. olfactory indicators of the
individual’s experience (social status, age…) and environment (diet, parasites…). Phenotypic
similarity could be enhanced or diminished by sharing or not sharing environmental factors.
The interaction between genetics and environment in the production of individual odours and
the way this interaction affects the process of using odour-genes covariance as a basis of
genetic relatedness assessment will be the focus of subsequent studies.
The covariance between varying phenotypes and individual genotypes is not restricted
to odours nor is the process of assessing genetic relatedness by evaluating similarities in
phenotypes. Examples of other sensory modalities in which the phenotypic similarity could be
a good cue for assessing genetic relatedness include acoustic and visual communication
(vocalizations in seals (Page et al. 2001), visual recognition in swans and quails (Bateson et
al. 1980, Bateson 1983)). Further study of the phenotype-genotype covariance phenomenon
and its use in assessing genetic relatedness will be helpful in understanding kin, population,
and species discrimination across multiple taxa and sensory modalities.
40

Chapitre 1 : Odeurs et Génome
ACKNOWLEDGEMENTS
This work was carried out under a grant to NB from the French Educational Ministery of
Research and Technology. The authors would like to thank Simone Demouron for her care of
the animals, five anonymous referees for constructive comments on the manuscript, Giora
Heth and Josephine Todrank for helpful discussion. The authors have adhered to the European
recommendations (decret 87-848) for the care and use of laboratory animals adopted by the
French government in October 1987. Experiments were carried out in the Laboratoire
d’Ethologie Expérimentale et Comparée (habilitation n° 02-2651) under CB’s licence
n 04663.
REFERENCES
Barnard, C. J. & Fitzsimons, J. 1989. Kin recognition and mate choice in mice: fitness
consequences of mating with kin. Animal Behaviour 38, 35-40.
Barnard, C. J., Hurst, J. L., & Aldhous, P. 1991. Of mice and kin: The functional
significance of kin bias in social behaviour. Biological Review 66, 379-430.
Bateson, P. 1983. Optimal outbreeding. In: Mate choice (Ed. by P. Bateson), pp 257-277:
Cambridge university Press.
Bateson, P., Lotwick, W. & Scott, D. K. 1980. Similarities between the faces of parents
and offspring in Bewicks Swans and the differences between mates. Journal of Zoology,
London 191, 61-74.
Baudoin, C., Busquet, N., Dobson, F. S., Gheusi, G., Féron, C., Durand, J-L., Heth, G.,
Patris, B. & Todrank, J. In press. Male-Female Associations and Female Olfactory
Neurogenesis with Pair Bonding in Mound-building Mice. Biological Journal of the Linnean
Society.
Beauchamp, G. K. & Yamazaki, K. 2003. Chemical signalling in mice. Biochemical Society
Transactions 31, 147-150.
41

Chapitre 1 : Odeurs et Génome
Beecher, M. D. 1989. Signalling systems for individual recognition: an information theory
approach. Animal Behaviour, 38, 248-261.
Beecher, M. D. 1991. Successes and failures of parent-offspring recognition in animals.
In: Kin Recognition (Ed. By P. G. Hepper). pp 94-124: Cambridge University Press.
Blaustein, A. R. 1983. Kin recognition mechanisms: Phenotypic matching or recognition
alleles ? American Naturalist, 121, 749-754.
Bowers, J. M. & Alexander B. K. 1967. Mice: individual recognition by olfactory cues.
Science 158, 1208-1210.
Brown, R. E. 1985a. The rodents I: effects of odours on reproductive physiology (primer
effects).In: Social odours in mammals. (Ed by R. E. Brown and D. W. MacDonald), pp 245-
346: Oxford Science Publications.
Brown, R. E. 1985b. The rodents II: suborder Myomorpha. In: Social odours in mammals.
(Ed by R. E. Brown and D. W. MacDonald), pp 345-457: Oxford Science Publications.
Brown, R. E., MacIntosh Schellinck, H. & West, A. M. 1996. The influence of dietary
and genetic cues on the ability of rats to discriminate between the urinary odors of MHC-
congenic mice. Physiology and Behavior, 60, 365-372.
deCatanzaro, D. & Gorzalka, B. B. 1979. Isolation-induced facilitation of male sexual
behavior in mice. Journal of Comparative Physiology and Psychology, 93, 211-222.
Dobson F.S., & Baudoin C. 2002. Experimental tests of spatial association and kinship
in monogamous mice (Mus spicilegus) and polygynous mice (Mus musculus domesticus).
Canadian Journal of Zoology, 80, 980-986.
Drickamer, L. C. 1992. Oestrous female house mice discriminate dominant from subordinate
males and sons of dominant from sons of subordinate males by odours cues. Animal
Behaviour, 43, 868-870.
Eggert, F., Holler, C., Luszyk, D., Muller-Ruchholtz, W. & Ferstl, R. 1996.
MHC-associated and MHC-independent urinary chemosignals in mice. Physiology
and Behavior, 59, 57-62.
42

Chapitre 1 : Odeurs et Génome
Ferkin, M. H., Sorokin, E. S., Johnston, R. E. & Lee, C. J. 1997. Attractiveness of scents
varies with protein content of the diet in meadow voles. Animal Behaviour, 53, 133-141.
Garza, J. C., Dallas, J., Duryadi, D., Gerasimov, S., Croset, H. & Boursot, P. 1997. Social
structure of the mound-building mouse Mus spicilegus revealed by genetic analysis with
microsatellites. Molecular Ecology, 6, 1009-1017.
Gouat, P., Féron, C. & Demouron, S. 2003. Seasonal reproduction and delayed sexual
maturity in mound-building mice Mus spicilegus. Reproduction, Fertility and Development,
15, 187-195.
Halpin, Z. T. 1986. Individual odors among mammals: origins and functions. Advances in the
Study of Behaviour, 16, 40–70.
Hamilton, W. D. 1964. The genetical evolution of social behavior. Journal of Theoretical
Biology, 7, 1-52.
Hayashi, S. & Kimura, T. 1983. Degree of kinship as a factor regulating preferences among
conspecifics in mice. Animal Behaviour, 31, 81-85.
Hepper, P. G. 1987. The discrimination of different degrees of relatedness in the rat:
evidence for a genetic identifier. Animal Behaviour, 35, 549-554.
Hepper, P. G. 1991. Kin recognition. Cambridge: Cambridge University Press.
Heth, G. & Todrank, J. 2000. Individual odour similarities across species parallel
phylogenetic relationships in the S. ehrenbergi superspecies of mole-rats. Animal Behaviour,
60, 789-795.
Heth, G., Todrank, J., Busquet, N. & Baudoin, C. 2001. Odour-genes covariance
and differential investigation of individual odours in the Mus species complex. Biological
Journal of the Linnean Society, 73, 213-220.
Heth, G., Todrank, J., Busquet, N. & Baudoin, C. 2003. Genetic relatedness assessment
through individual odour similarities in mice. Biological Journal of the Linnean Society, 78,
595-603.
43

Chapitre 1 : Odeurs et Génome
Heth, G., Todrank, J. & Johnston, R. E. 1998. Kin recognition in golden hamsters:
evidence for phenotype matching. Animal Behaviour, 56, 409-417.
Heth, G., Todrank, J. & Johnston, R. E. 1999. Similarity in the qualities of individual odors
among kin and species in Turkish (Mesocricetus brandti) and golden (Mesocricetus auratus)
hamsters. Journal of Comparative Psychology, 113, 321-6.
Holmes, W. G. 1986. Identification of paternal half siblings by captive Belding’s ground
squirrels. Animal Behaviour, 34, 321-327.
Hurst, J. L., Payne, C. E., Nevison, C. M., Marie, A. D., Humphries, R. E., Robertson, D.
H. L., Cavaggioni, A. & Beynon, R. J. 2001. Individual recognition in mice mediated
by major urinary proteins. Nature, 414, 631-634.
Koeninger Ryan, K. & Lacy, R. C. 2003. Monogamous male mice bias behaviour towards
females according to very small differences in kinship. Animal Behaviour 65, 379-384.
Mateo, J. M. 2002. Kin-recognition abilities and nepotism as a function of sociality.
Proceedings of the Royal Society of London Series B Biological Sciences, 269, 721-727.
Page, B., Goldsworthy, S. D. & Hindell, M. A. 2001. Vocal traits of hybrid fur seals:
intermediate to their parental species. Animal Behaviour 61, 959-967.
Patris, B. & Baudoin, C. 1998. Female sexual preferences differ in Mus spicilegus and Mus
musculus domesticus: the role of familiarization and sexual experience. Animal Behaviour,
56, 1465-1470.
Patris, B. & Baudoin, C. 2000. A comparative study of parental care between two rodent
species: implications for the mating system of the mound-building mouse Mus spicilegus.
Behavioural Processes, 51, 35-43.
Penn, D., Schneider, G., White, K., Slev, P. & Potts, W. 1998. Influenza infection
neutralizes the attractiveness of male odour to female mice (Mus musculus). Ethology, 104,
685-694.
Pusey, A. & Wolf, M. 1996. Inbreeding avoidance in animals. Trends in Ecology
and Evolution, 11, 201-206.
44

Chapitre 1 : Odeurs et Génome
Singer, A. G., Beauchamp, G. K. & Yamazaki, K. 1997. Volatile signals of the major
histocompatibility complex in male mouse urine. Proceedings of the National Academy
of Sciences USA, 94, 2210-2214.
Sokolov, V. E., Kotenkova, E. V., Michailenko, A. G. 1998. Mus spicilegus. Mammalian
Species, 592, 1-6.
Sun, L. & Muller-Schwarze, D. 1997. Sibling recognition in the beaver: a field test
for phenotype matching. Animal Behaviour, 54, 493-502.
Tang-Martinez, Z. 2001. The mechanisms of kin discrimination and the evolution of kin
recognition in vertebrates: a critical re-evaluation. Behavioural Processes, 53, 21-40.
Todrank J., & Heth G. 2003. Odor-genes covariance and genetic relatedness assessments:
rethinking odor-based “recognition” mechanisms in rodents. Advances in the study
of behavior, 32, 77-130.
Todrank, J., Heth, G. & Johnston, R. E. 1998. Kin recognition in golden hamsters:
evidence for kinship odours. Animal Behaviour, 55, 377-86.
Todrank, J., Heth, G. & Johnston, R. E. 1999. Social interaction is necessary
for discrimination between and memory for odours of close relatives in golden hamsters.
Ethology, 105, 771-782.
Willis, C. & Poulin, R. 2000. Preference of female rats for the odours of non-parasited
males: the smell of good genes? Folia parasitologica, 47, 6-10.
45
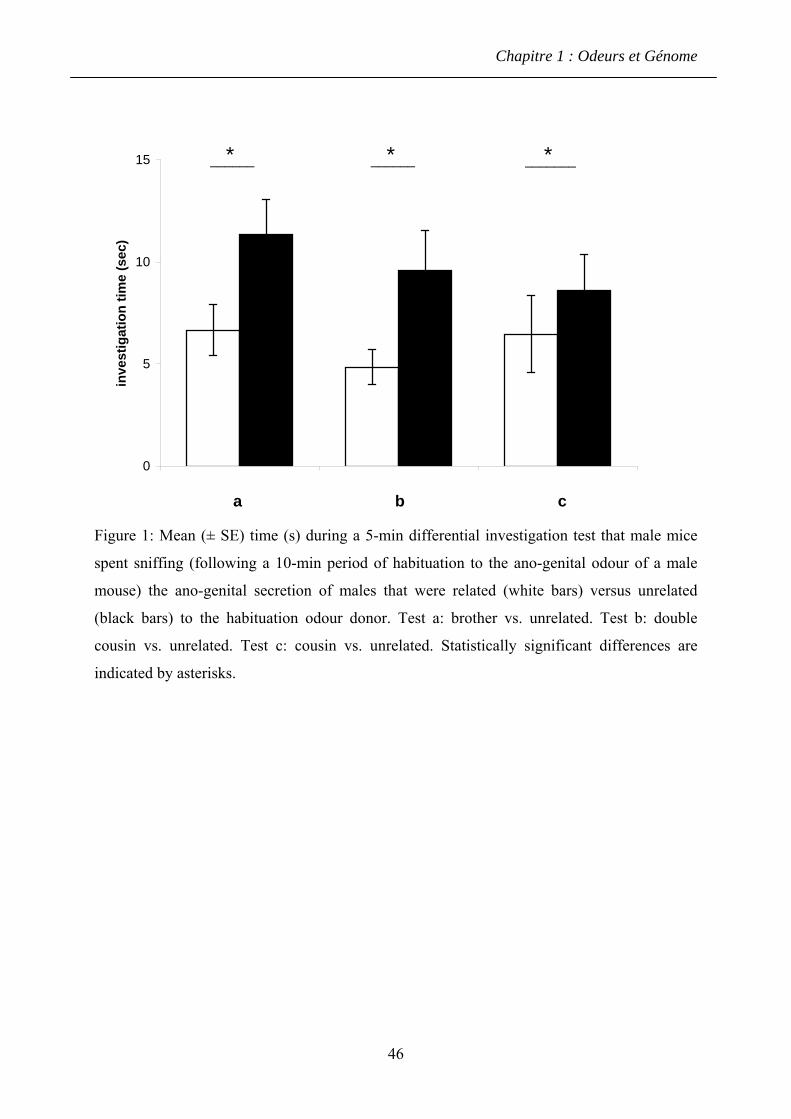
Chapitre 1 : Odeurs et Génome
0
5
10
15
inve
stig
atio
n tim
e (s
ec)
a cb
______* _______*______*
Figure 1: Mean (± SE) time (s) during a 5-min differential investigation test that male mice
spent sniffing (following a 10-min period of habituation to the ano-genital odour of a male
mouse) the ano-genital secretion of males that were related (white bars) versus unrelated
(black bars) to the habituation odour donor. Test a: brother vs. unrelated. Test b: double
cousin vs. unrelated. Test c: cousin vs. unrelated. Statistically significant differences are
indicated by asterisks.
46
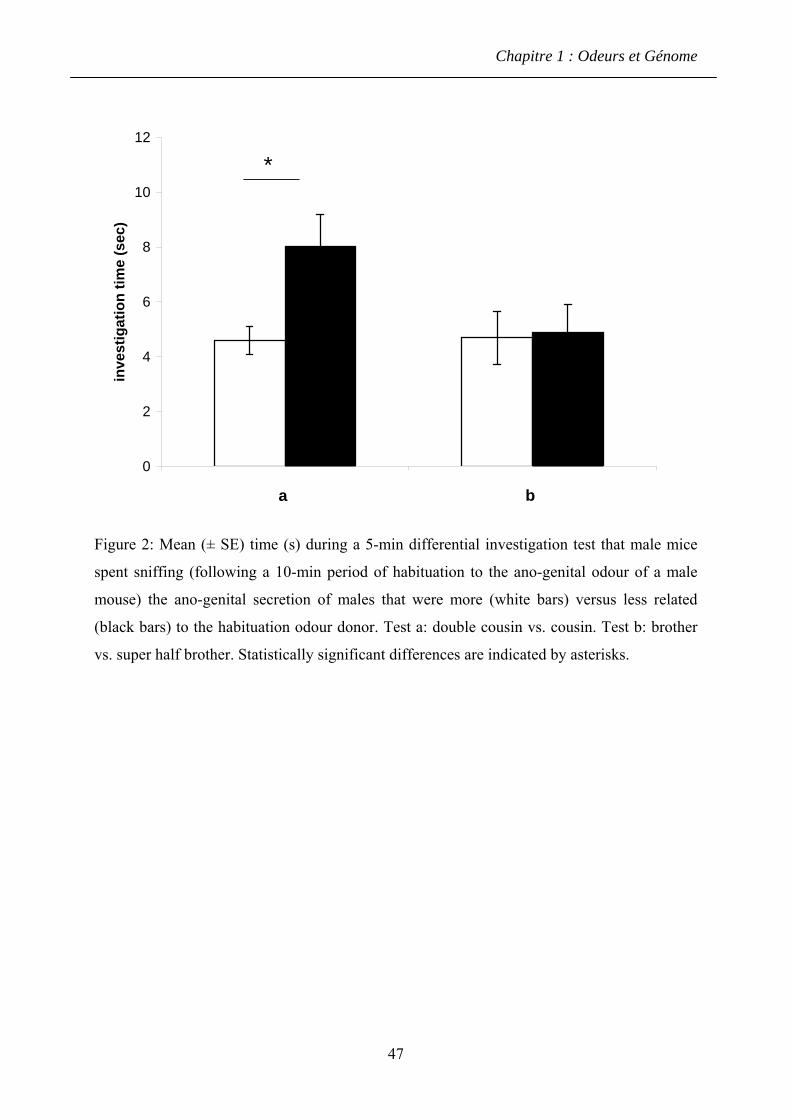
Chapitre 1 : Odeurs et Génome
0
2
4
6
8
10
12in
vest
igat
ion
time
(sec
)
_______*
a b
Figure 2: Mean (± SE) time (s) during a 5-min differential investigation test that male mice
spent sniffing (following a 10-min period of habituation to the ano-genital odour of a male
mouse) the ano-genital secretion of males that were more (white bars) versus less related
(black bars) to the habituation odour donor. Test a: double cousin vs. cousin. Test b: brother
vs. super half brother. Statistically significant differences are indicated by asterisks.
47

Chapitre 1 : Odeurs et Génome
1.3. Odeurs et phylogénie
L'article suivant rapporte les résultats d'une étude, menée en collaboration, qui recherche des
similarités olfactives au sein du genre Mus, avec un protocole semblable à celui utilisé dans
l'article précédent. Quatre espèces ont été utilisées pour cette expérience : Mus musculus, Mus
domesticus, Mus spicilegus et Mus macedonicus. M. musculus et M. domesticus sont proches
phylogénétiquement, et sont même parfois considérées comme des sous-espèces (Boursot et
al., 1993). De leur côté, M. spicilegus et M. macedonicus sont également proches entre elles,
et plus éloignées du premier couple d’espèces. Ainsi des similarités devraient exister entre les
odeurs individuelles de souris appartenant à chacun de ses "couples d'espèces". Les résultats
indiquent que ces similarités existent bien. De plus, chez Mus spicilegus, les odeurs
individuelles de mâles issus de la population capturée en octobre 1999 à Gyöngyös (Hongrie)
présentent des similarités quand on les compare à l'odeur d'une souris glaneuse d'une autre
population, plus ancienne au laboratoire (de 15 à 20 générations) et originaire de Pančevo
(Yougoslavie). La covariance odeur-gènes existe donc également au niveau des populations.
En d'autres termes la structuration génétique inhérente à chaque population se traduit par une
homogénéité des odeurs individuelles.
Enfin, une troisième expérience montre que les mâles des quatre espèces testées passent plus
de temps à explorer l'odeur d'une femelle d'une espèce proche que celle d'une femelle d'une
espèce plus éloignée phylogénétiquement. Ce résultat suggère la capacité à évaluer la
proximité génétique par rapport à soi-même, et fournit des pistes sur le mécanisme utilisé
pour effectuer cette évaluation. Ces pistes seront explorées dans le chapitre II.
48

Chapitre 1 : Odeurs et Génome
49

Chapitre 1 : Odeurs et Génome
50

Chapitre 1 : Odeurs et Génome
51

Chapitre 1 : Odeurs et Génome
52

Chapitre 1 : Odeurs et Génome
53

Chapitre 1 : Odeurs et Génome
54

Chapitre 1 : Odeurs et Génome
55

Chapitre 1 : Odeurs et Génome
1.4. Empreinte génomique et odeurs
Comme exposé dans les deux articles ci-dessus, le génome semble être reflété dans les odeurs
individuelles, puisque les similarités olfactives augmentent avec la proximité génétique. Dans
un contexte intra spécifique, si l'odeur globale des individus traduit en moyenne la proximité
génétique, alors le coefficient d'apparentement r entre deux individus pourrait fournir une
bonne estimation de la similarité olfactive entre ces individus. Cependant ce coefficient r
indique l'apparentement moyen. Il est donc possible que parmi trois individus également
apparentés, deux soient olfactivement plus similaires. Postuler que la transcription en odeurs
n'est pas liée à une seule famille de gènes comme le MHC ou les MUPs ne signifie pas pour
autant que tous les gènes influencent cette odeur. Le phénotype olfactif dépend des
interactions entre les allèles hérités du père et de la mère. Classiquement, ces interactions sont
mendéliennes, c'est-à-dire que certains allèles sont dominants, co-dominants, récessifs par
nature. Ceci peut expliquer que pour des degrés de parenté élevés, la similarité olfactive
détectée par les animaux ne suive pas exactement la proximité génétique globale. Mais
certains travaux récents permettent de supposer que le phénotype olfactif ne soit pas hérité de
façon exclusivement mendélienne.
L'empreinte génomique, ou "effet parent d'origine", est un processus par lequel l'expression
d'un allèle ne dépend pas de sa nature mais de son origine (Bartolomei & Tilghman 1997). Il
existe donc un contrôle épigénétique de l'expression de certains gènes chez les Mammifères
(dont les Souris) qui fait que c'est l'allèle hérité du père ou de la mère qui est exprimé. Les
mécanismes moléculaires permettant cet effet "parent d'origine" sont multiples (Ferguson-
Smith 2000, Ferguson-Smith & Surani 2001). L'empreinte génomique semble très importante
au cours du développement intra-utérin (Haig 1996) et dans les relations mère-jeune en
général (le "maternalisme" ; (Keverne 2001)). Puisque les gènes d'empreinte régulent entre
autres les échanges de nutriments entre le fœtus et la mère (Reik & Walter 2001), l'empreinte
génomique apparaît comme révélatrice d'un conflit entre les sexes, le père ayant intérêt à
"forcer" la femelle à allouer plus de ressources à sa progéniture, puisqu'il n'a pas à subir le
coût du développement de cette progéniture (Moore & Haig, 1991). Les exemples d'effets
parentaux sur l'expression des gènes étant de plus en plus nombreux, la notion même
d'apparentement a été réévaluée par Haig (2000). Il s'agit de décomposer le coefficient
d'apparentement moyen r en apparentement matrilinéaire m et patrilinéaire p. Dans
cette optique, r = (m + p) / 2. L'apparentement peut alors être classifié comme étant
56

Chapitre 1 : Odeurs et Génome
symétrique (m = p) ou asymétrique (m ≠ p). Les apparentés "symétriques" comprennent la
fratrie et les descendants. Les autres apparentés, parents, grands-parents, oncles, tantes,
cousins sont des apparentés "asymétriques". L'intérêt de cette distinction réside dans
l'extension de la théorie de l'inclusive fitness, en considérant l'empreinte génomique comme
un moyen de répondre à cette asymétrie d'apparentement dans les lignées paternelles ou
maternelles. Un exemple théorique du rôle de l'empreinte génomique au niveau
comportemental, dans le cadre des interactions au sein des fratries, est présenté dans Haig &
Wilkins (2000). Cette modélisation suggère que les gènes d'empreinte paternels "poussent" les
petits à la consommation des ressources maternelles, tandis que les gènes maternels freinent
cet appétit. Cette tendance trouverait son utilité notamment dans les cas de paternité multiples,
ou chacun des pères favoriserait sa propre descendance tandis que la mère essaierait
d'homogénéiser l'apport de nourriture pour toute sa progéniture. Bien évidemment, cette
approche, en dépit de sa complexité mathématique, est basée sur un scénario simple voire
simpliste où le comportement alimentaire est sous contrôle d’un ou de quelques gènes. Il
n’existe pas à ce jour de démonstration expérimentale de ces théories.
Bien que les mécanismes de l'empreinte génomique soient de mieux en mieux connus, la
question d'une asymétrie de reconnaissance liée à de tels gènes n'a été posée que récemment.
Pourtant, une éventuelle asymétrie phénotypique entre lignées paternelles et maternelles
pourrait avoir des conséquences importantes sur l'organisation sociale des Mammifères. Isles
et al. (2001) ont montré que des souris issues de croisements réciproques entre deux souches
de laboratoire préféraient l'odeur d'une troisième souche à l'odeur de la souche maternelle,
mais pas à l'odeur de la souche paternelle. Cet "évitement" de l'odeur de la souche maternelle
semble bien lié à un effet "parent d'origine" puisque les souris testées provenaient de
transferts d'embryons dans une femelle non apparentée, ce qui élimine les effets de
l'expérience pré- et post-natale. En revanche les auteurs excluent l'idée d'une traduction
olfactive de l'empreinte génomique, sur la base de résultats montrant que les deux croisements
réciproques ne sont pas discriminables dans un test d'habituation-déshabituation. Les
différences de préférence entre les deux croisements réciproques seraient dues à des
différences au niveau de la perception ou du traitement de l'information olfactive, mais pas au
niveau de la production odorante. Les mêmes résultats ont été obtenus avec deux autres
croisements entre souches de laboratoire (Isles et al. 2002). Là encore, le fait que des souris de
souche Balb / C soient incapables de discriminer les urines des deux croisements réciproques
57

Chapitre 1 : Odeurs et Génome
est présenté comme la démonstration de l'absence de différences au niveau du phénotype
odorant, d'autant que ces mêmes Balb / C discriminent parfaitement les deux souches
parentales. Cette réfutation de la transcription olfactive de l'empreinte génomique est
surprenante, car elle dissocie très (trop ?) clairement les composantes productives et
perceptives de la reconnaissance. Qu'une variation de l'expression phénotypique – à génome
égal – touche de façon drastique la perception olfactive (par exemple si les gènes "imprimés"
codent pour des récepteurs olfactifs) mais n'ait aucune conséquence sur la production
odorante ne peut être affirmé par la méthode décrite ci-dessus.
La méthode d'habituation-généralisation, largement présentée dans les pages précédentes,
permet de déduire la reconnaissance de similarité globale entre l'odeur d'habituation et l'une
ou l'autre des deux odeurs de test. C'est cette idée qui a été testée, en utilisant des souches de
souris de laboratoire, afin de vérifier si les hybrides ont en moyenne un phénotype odorant
également proche de chacune des deux souches parentales, comme le veut le modèle
mendélien, ou s'il existe une hérédité asymétrique de l'odeur individuelle. La raison de
l'utilisation de souches de parentèle est que ces souches sont considérées comme homozygotes
à l'ensemble de leurs loci. Chez une espèce sauvage où la variabilité génétique est importante,
l'effet "parent d'origine" serait en compétition avec la variabilité inter-individuelle dans les
productions olfactives. Puisque la question posée concerne la recherche d'une éventuelle
asymétrie de similarité d'odeurs entre lignées paternelles et maternelles, des souris mâles ont
été utilisées dans les tests décrits ci-dessous. En effet ceux-ci ont un chromosome X qui
provient de leur mère et qui est donc susceptible d'apporter un surcroît de similarité entre ces
mâles et la souche maternelle, si les gènes portés par le chromosome X influencent l'odeur.
1.4.1 Matériels et méthodes
Trois souches de souris de laboratoire ont été utilisées : Balb / C, C57 et DBA, obtenues
auprès de l'entreprise Charles Rivers / Iffa Credo. A partir de ces souches, les six croisements
possibles ont été réalisés : Balb / C-C57, C57-Balb/C, C57-DBA, DBA-C57, Balb / C-DBA,
DBA-Balb / C (la souche paternelle est indiquée en premier). Les souris mâles issues de ces
croisements sont utilisées comme donneuses d'odeurs d'habituation (N = 36, soit 6 mâles par
croisement), tandis que des mâles de même génération issus de croisements homologues
(Balb / C-Balb / C etc.) sont utilisés comme donneurs d'odeurs de test (représentant les
58

Chapitre 1 : Odeurs et Génome
souches paternelles et maternelles, N = 72) ou comme sujet (souche non apparentée aux
donneurs d'odeurs, N = 36). L'ensemble des tests est présenté dans le tableau 1.3.1.
sujet habituationsouche
paternelle
souche
maternelle
DD BC BB CC
DD CB CC BB
CC BD BB DD
CC DB DD BB
BB CD CC DD
BB DC DD CC
Tableau 1.3.1 : Présentation schématique des tests d'habituation-généralisation réalisés dans le cadre de l'étude sur l'empreinte génomique. Chaque catégorie est codée par deux lettres, la première correspondant à la souche paternelle, la deuxième à la souche maternelle. B = Balb / C ; C = C57 ; D = DBA.
L'habituation consiste en une présentation de 8 minutes de l'odeur hybride. Durant cette
habituation, le temps d'investigation de l'odeur est mesuré pendant les 2 premières minutes
puis les 2 minutes suivantes. Les quatre minutes restantes sont mises à profit pour préparer les
odeurs de tests (obtenus par frottement d'une boîte de Pétri sur la zone uro-génitale), ce qui
permet d'avoir des stimuli "frais". Ces deux odeurs sont ensuite présentées simultanément
pendant deux minutes, après retrait de l'odeur d'habituation.
59
Chapitre 1 : Odeurs et Génome
1.4.2 Résultats
Les résultats sont présentés dans le tableau 1.4.2. Seuls quatre sujets ne montrent pas de
diminution du temps d'investigation entre les deux premières minutes et les deux suivantes.
Ces quatre sujets sont répartis dans les quatre derniers groupes, ce qui fait que seuls les deux
premiers – où le sujet est un mâle Balb / C – montrent une habituation significative
(probabilité exacte p = 0,03125). Malgré tout l'habituation est indéniable, en vertu des temps
importants d'investigation de l'odeur par chacun des sujets (min : 8'96'' (sujet C57) ;
max : 54'57'' (sujet Balb / C) lors des deux premières minutes).
TPmPL
0-2 min 2-4 min score temps d'investigation score temps
d'investigation
BB CD 24,03 ± 4,6 15,58 ± 9,95 2/6 14,03 ± 5,16 4/6 17,27 ± 11,68
BB DC 27,93 ± 13,87 13,68 ± 6,23 1/6 11,02 ± 4,41 5/6 13,69 ± 5,18
CC BD 17,36 ± 6,91 11,83 ± 4,23 3/6 8,79 ± 2,27 3/6 8,82 ± 2,26
CC DB 13,88 ± 5,48 9,45 ± 2,97 2/6 9,33 ± 3,65 4/6 11,35 ± 4,14
DD BC 23,77 ± 12,49 12,37 ± 4,44 3/6 11,38 ± 4,43 3/6 11,73 ± 2,98
DD CB 21,22 ± 7,11 14 ± 9,52 3/6 13,8 ± 5,67 3/6 13,38 ± 3,78
Tous toutes 21,37 ± 9,65 12,82 ± 6,56 14/36 11,39 ± 4,49 22/36 12,71 ± 6,06
sujet
odeur de la souche maternelle
odeur de la souche paternelle
temps d'investigation de l'odeur d'habituation en secondes
(moyenne ± écart-type)origine du
donneur de l'odeur
d'habituation
Habituation Test
ableau 1.4.2 : résultats des tests d'habituation-généralisation. hase d'habituation : temps passé par les souris mâles à sentir l'odeur d'habituation pendant les deux premièresinutes et les deux minutes suivantes. Cette odeur est celle d'un mâle hybride entre deux souches de laboratoire. hase de test : temps passé par les souris à explorer chacune des deux odeurs de tests pendant deux minutes.'une de ces odeurs est celle d'un mâle de la lignée maternelle, l'autre provient d'un mâle de la lignée paternelle.
L'odeur d'un mâle issu de la souche paternelle du donneur d'odeur d'habituation est
globalement explorée plus longtemps que l'odeur d'un mâle de la lignée maternelle. Une
analyse globale ne permet pas toutefois de montrer une significativité de ce résultat (test de
Wilcoxon pour échantillons appariés, N = 36, p = 0,08). Une telle analyse globale est
d'ailleurs statistiquement incorrecte, puisqu'elle considère tous les tests comme équivalents.
60

Chapitre 1 : Odeurs et Génome
Or les sujets sont différents d'un groupe à l'autre : les souris Balb / C semblent être les seules à
reconnaître une plus grande similarité entre les hybrides C57-DBA (et vice-versa) et la souche
maternelle. Ces sujets sont en grande partie responsables du biais global (non significatif) en
faveur de la souche paternelle (22 sujets sur 36).
1.4.3 Discussion
Il n'a pas été possible de détecter de similarité entre l'odeur des hybrides et l'une ou l'autre des
souches parentales. Ce résultat peut être du à des propriétés attractives supérieures de
certaines souches, voire à des différences comportementales inter souches. Néanmoins, au vu
de la tendance, il est possible qu'en augmentant les effectifs une similarité plus grande puisse
être perçue entre la souche maternelle et les hybrides. L'intérêt d'une telle recherche semble
cependant limité : si un effet parent d'origine ne peut être mis en évidence facilement avec des
souches consanguines où le génome est fixé, alors il est peu plausible que cet effet soit fort
chez une espèce sauvage comme Mus spicilegus. L'empreinte génomique peut avoir un rôle
sur les comportements des Souris, notamment dans les conflits familiaux (Hager & Johnstone
2003), mais il semble qu'elle n'ait qu'un effet mineur sur les productions odorantes, si tant est
qu'elle ait un effet. Le rôle de l'empreinte génomique sur les préférences sexuelles, qui
favoriserait l'évitement de l'inceste et la dispersion via l'évitement de la souche maternelle
(Isles et al. 2002), est probablement lié à la composante perceptive de la reconnaissance. A ce
jour, aucune étude n'a montré d'effet "parent d'origine" sur une espèce sauvage. C'est pourtant
dans ce type de population que les apparentements asymétriques ont pu être sélectionnés et
maintenus. En nature, les influences maternelles et paternelles "directes" (via la familiarité) et
le biais entre celles-ci (l'environnement intra-utérin fournit une surexposition au phénotype
maternel) paraissent intuitivement beaucoup plus susceptibles d'orienter les comportements de
la progéniture. L'extrapolation à partir de résultats obtenus sur des souches de laboratoire, en
l'absence d'une connaissance exhaustive des mécanismes moléculaires, n'est que modérément
heuristique. Par exemple, les tentatives de démonstration d'une variation mesurable de la
fitness entre croisements réciproques de souches de laboratoire (Hager & Johnstone, 2003) ne
permettent pas de dresser d'hypothèses claires sur d'éventuels biais en nature, puisqu'une
population naturelle n'est pas composée de deux groupes d'individus aussi délimités que le
sont deux souches de laboratoire (voir également Smith et al. (1994) pour d'autres limites
liées à la comparaison entre souches de laboratoire et souches sauvages). En revanche les
travaux sur l'effet "parent d'origine" sur les préférences et les implications évolutives de
61

Chapitre 1 : Odeurs et Génome
l'empreinte génomique ont certainement leur place dans l'étude des zones de contact entre
espèces, particulièrement dans les cas où les hybridations sont possibles. Les interactions
entre les deux sous-espèces Mus musculus domesticus et Mus musculus musculus dans leur
zone d'hybridation paraissent un modèle idéal pour étudier la validité de ces hypothèses.
Odeurs et génome : conclusions et discussion
La proximité génétique entre deux souris se traduit par une similarité de leurs productions
odorantes, en condition de laboratoire. Cette relation entre odeurs et gènes semble exister
aussi bien au niveau interspécifique qu'au niveau intra populationnel. Il existe une covariance
entre odeurs et gènes : deux souris d'espèces proches présentent des odeurs similaires par
rapport à une troisième espèce phylogénétiquement plus éloignée, deux souris de la même
population présentent une plus grande similarité entre elles que le simple "fonds commun" de
l'espèce, et cette similarité augmente encore avec le degré d'apparentement. Bien que cela n'ait
pas été testé chez Mus spicilegus, les expériences sur les souris de laboratoire semblent
indiquer qu'il n'y a pas d'effet "parent d'origine" sur ces similarités, c'est-à-dire qu'un
apparentement par la mère n'a pas plus de corrélats olfactifs qu'un apparentement par le père.
Plusieurs conclusions importantes peuvent être tirées de ces résultats. La première, la plus
évidente, est qu'il existe un substrat olfactif pour permettre la reconnaissance de la proximité
génétique. Les Souris sont capables de percevoir les similarités d'odeurs corrélées à la
proximité génétique entre deux autres individus. Les Souris se sont révélées également
capable de distinguer l'odeur d'une espèce proche phylogénétiquement de celle d'une espèce
plus éloignée (Heth et al., 2001). Ceci est le premier indice d'une capacité à reconnaître la
proximité odorante d'une souris par rapport à soi-même. Bien sûr, il est impossible de déduire
de l'existence d'une covariance odeurs-gènes l'utilisation de celle-ci pour la reconnaissance de
parentèle. Cependant les signaux olfactifs des Souris glaneuses (et probablement des autres
Rongeurs) renferment l'information suffisante pour effectuer une reconnaissance de parentèle
efficace par assortiment phénotypique. La description des différents mécanismes théoriques
de reconnaissance de parentèle ainsi que la recherche de celui ou ceux utilisés par les Souris
glaneuses sont l'objet du chapitre suivant.
62

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Chapitre II :
Mécanismes d'évaluation
de la proximité génétique
63

Chapitre II: Mécanismes d'évaluation de la proximité génétique
2.1. Introduction
D'un point de vue évolutionniste, la reconnaissance de parentèle est intéressante parce qu'elle
procure un avantage pour les individus possédant cette capacité. Diverses pressions de
sélection peuvent favoriser les individus capables d'éviter les croisements consanguins, ou
d'aider leurs apparentés, quel que soit le ou les
mécanismes employés. Mais l'absence de
consanguinité et l'existence d'une entraide entre
apparentés ne préjugent pas de l'existence de la
reconnaissance de parentèle. Il est parfaitement
envisageable que des animaux aient développé
des stratégies ayant les mêmes conséquences que
la reconnaissance de parentèle sans y avoir
recours (la dispersion des jeunes avant leur
première reproduction, par exemple, réduit les
risques d'accouplement avec un individu
apparenté). Cependant la pression de sélection
relative à l’évitement de la consanguinité et, d’une
manière plus générale, à la prise en compte du
facteur apparentement dans les phénomènes
sociaux est bien réelle (Hamilton, 1964). La
reconnaissance de la progéniture par les parents
est un des phénomènes sociaux pour lesquels la
reconnaissance de parentèle semble la plus
profitable. En effet, chez les espèces pratiquant les
soins parentaux, il est avantageux de prodiguer
ces soins à des individus apparentés, de façon à maximiser la survie de la progéniture.
L’investissement parental parfois important (notamment chez les espèces nidicoles où le
développement des jeunes est plus long) est supposé s’accompagner de capacités de
reconnaissance élevées permettant théoriquement aux parents de ne pas "gaspiller" leur
énergie dans l’élevage de jeunes non apparentés. Curieusement, selon les taxons, les animaux
montrent des capacités de reconnaissance variables et peuvent se révéler extrêmement
efficaces ou particulièrement médiocres (Beecher, 1991). Chez les oiseaux, par exemple, la
Encadré 2.1 : Qu'est-ce que lareconnaissance ?
D'après le dictionnaire Larousse, lareconnaissance se définit entre autres comme :
• L'action de reconnaître. Reconnaître, c'est"se remettre dans l'esprit l'image d'unechose, d'une personne qu'on revoit." Cetteacceptation du terme implique l'existenced'une trace mnésique : il s'agit de"connaître à nouveau".
• Examen détaillé, vérification. Cettedéfinition implique une rechercheorientée de certains caractères.
• Une opération visant à obtenir desrenseignements sur l'ennemi dans unezone donnée. Mis à part le terme"ennemi", cette définition militaire peutcurieusement s'appliquer à lareconnaissance chez les animaux, qui viseà obtenir des informations concernant lestatut et l'identité d'un congénère.
Ces trois définitions n'éclairent pas beaucoupsur ce qu'est la reconnaissance de parentèle,mais permettent de poser des questions quantau mécanisme permettant le traitementdifférentiel des apparentés : - peut-on parler de reconnaissance en l'absenced'expérience préliminaire ? (définition 1) - la reconnaissance implique t’elle une certaineforme d'intentionnalité ? (définition 2) - la reconnaissance de parentèle doit-elle êtreconsidérée comme un sous-ensemble d'unprocessus plus général de reconnaissance ?(définition 3)
64

Chapitre II: Mécanismes d'évaluation de la proximité génétique
reconnaissance de la progéniture peut être suffisamment précise pour qu’un manchot retrouve
son petit au sein d'une colonie de milliers d’individus (Jouventin & Aubin, 2002), mais les
exemples d’espèces se laissant abuser jusqu’à élever les petits d’espèces parasites sont légion
(Smith, 1999). Comme il a été exposé dans l’introduction générale, la prise en compte du
facteur apparentement peut être également intéressante lors du choix de partenaire (Bateson,
1983), avec là aussi de grandes disparités entre les espèces. Cette variabilité importante dans
l’efficacité de la reconnaissance de parentèle est à mettre en relation avec les mécanismes
utilisés pour l’effectuer. Quatre mécanismes théoriques permettraient la reconnaissance de
parentèle (Blaustein, 1983 ; Hepper, 1991b). Ces mécanismes sont ici présentés sans exemple,
en raison de difficultés de démonstration qui seront détaillées plus bas :
1. Reconnaissance spatiale : la position spatiale des individus est un bon indicateur de
l’apparentement et les actes altruistes se dirigent préférentiellement vers les individus
occupant un lieu particulier.
2. Familiarité et apprentissage social : dans ce modèle, la détection de l’apparentement
est liée à une communauté d’évènements sociaux. Si dans les circonstances
habituelles, les individus apparentés coexistent sur une durée suffisamment longue
(par exemple lors de la période d’élevage des jeunes), alors la reconnaissance des
individus familiers est un bon moyen de reconnaître les apparentés. Ce mécanisme
inclut également la familiarité indirecte : l’association de deux individus dont l’un est
reconnu comme familier peut permettre de considérer également l’autre comme
apparenté.
3. Assortiment phénotypique (Phenotype matching) : les individus mémorisent leur
propre phénotype et/ou celui des apparentés. En se basant sur la corrélation entre
similarité phénotypique et similarité génétique, il est alors possible d’estimer
l’apparentement d’individus non familiers en comparant leur phénotype avec ce
phénotype appris (template ou modèle de référence).
4. Allèles de reconnaissance : cette dernière catégorie de mécanisme permet une
reconnaissance de parentèle indépendamment d’un quelconque apprentissage par le
biais d’allèles de reconnaissance. Il existe une base génétique fournissant à la fois le
marqueur d’apparentement et le système de reconnaissance de ce marqueur.
65

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Cette nomenclature des mécanismes de reconnaissance de parentèle est à la fois consensuelle,
au sens où elle est abondamment utilisée dans la littérature, et très critiquée (Barnard, 1999 ;
Tang-Martinez, 2001). Ces critiques peuvent s’articuler selon deux axes :
- Le premier se rapporte au fait que ces catégories ne sont pas indépendantes : non seulement
les quatre types de mécanisme ne sont pas exclusifs, mais leurs limites semblent floues. Ceci
est particulièrement vrai pour les mécanismes 2 et 3. En effet, un animal qui apprend à
reconnaître ses apparentés familiers (familiarité) ne semble pas cognitivement très éloigné de
celui qui apprend les caractéristiques phénotypiques de ses apparentés (phenotype matching).
Il est même assez probable qu’un animal capable de l’un soit capable de l’autre. (Tang-
Martinez, 2001) suggère d’ailleurs que la distinction entre ces deux mécanismes est
artificielle, puisqu’ils sont tous deux basés sur un processus sous-jacent d’apprentissage.
L'éventuelle distinction entre ces deux mécanismes est basée sur la nature des signaux appris.
- Le second s’appuie sur une objection sémantique au terme de reconnaissance de parentèle.
Dans un article intitulé "Do animals really recognize kin ?", Grafen dénonce la confusion
entre d'une part la discrimination comportementale des apparentés et d'autre part la
reconnaissance de parentèle, définie comme la reconnaissance de marqueurs ayant
spécifiquement évolué en réponse à une pression de sélection pour reconnaître les apparentés
(Grafen, 1990). Autrement dit la reconnaissance de parentèle "vraie" n’a de raison d’exister
que si aucun autre mécanisme préexistant ne permet de moduler le comportement en fonction
de l’apparentement. Dans ce cas de figure, un mécanisme dédié spécifiquement à cette
évaluation de l'apparentement peut être sélectionné. D’après lui, très peu d’études montrent
une réelle reconnaissance de parentèle, et le traitement différentiel des apparentés peut même
souvent s’expliquer par une conséquence secondaire de la reconnaissance spécifique.
En analysant les données de la littérature, il apparaît effectivement que la distinction de quatre
grands mécanismes ne soit pas aussi heuristique qu’elle semble l’être au premier abord.
Comme expliqué ci-dessus, l’argument utilisé par Grafen (1990) pour mettre en doute
l’existence d’une reconnaissance de parentèle "vraie" autre que marginale, s'appuie sur la
question de sa fonction adaptative. Puisque l’altruisme envers les apparentés et l’évitement de
l’inceste peuvent être efficacement assurés par d’autres mécanismes (notamment la
reconnaissance spécifique, toujours d’après Grafen), il n’y a pas ou peu de pressions
évolutives pour l’établissement d’une reconnaissance de parentèle per se (un système dont
l'utilité et la fonction sont d'estimer l'apparentement des conspécifiques). Par exemple, le fait
66

Chapitre II: Mécanismes d'évaluation de la proximité génétique
de mémoriser le phénotype des animaux familiers (souvent apparentés) comme étant le
"prototype" de l'espèce va conduire l'animal à traiter différentiellement les conspécifiques
différant grandement de ce phénotype, c'est-à-dire les non-apparentés. Dans ce cas, la
discrimination entre apparentés et non-apparentés serait un sous-produit de la reconnaissance
spécifique, et ne correspond pas à une "vraie" reconnaissance de parentèle. Un écueil
important à cette hypothèse est celui de "l’optimal outbreeding", c'est-à-dire la tendance à
rechercher un partenaire qui ne soit ni trop apparenté ni trop éloigné (voir l'introduction
générale). Ce phénomène, connu chez les Cailles (Bateson, 1982) et les Souris (Barnard &
Fitzsimons, 1988), semble difficilement explicable par la seule reconnaissance spécifique
(Blaustein et al., 1991). D'autres contre arguments à la définition restrictive de Grafen peuvent
être apportés, notamment concernant le rôle du développement, la part trop restreinte
accordée à l'environnement dans l'établissement du phénotype (Byers & Bekoff, 1991) et
l'aspect fonctionnel de la reconnaissance de parentèle (Stuart, 1991). Ce dernier point est
crucial car la valeur adaptative du traitement différentiel des apparentés est mal connue
(Blaustein et al., 1991). Afin de savoir si le processus guidant la discrimination des apparentés
relève de la reconnaissance de parentèle ou de la reconnaissance spécifique, il faut identifier
les pressions de sélection, tâche éminemment ardue (cf. introduction générale). C'est
l'existence de pressions de sélection pour l'établissement d'une reconnaissance de
l'apparentement qui permettrait de trancher (Grafen, 1991b). La reconnaissance de parentèle
"vraie" peut être perçue comme "la reconnaissance des apparentés par un mécanisme différent
de celui par lequel sont reconnus les individus familiers, les voisins et les autres organismes
ou objets" (Grafen, 1991a). En dépit des désaccords sémantiques sur le terme lui-même, il
semble exister un consensus sur deux points. Le premier est qu'il existe un traitement
différentiel des apparentés largement répandu dans le règne animal (Fletcher & Michener,
1987 ; Hepper, 1991a), et le second est qu'il existe des capacités de reconnaissance permettant
aux animaux d'effectuer ce traitement différentiel, quelle que soit la dénomination choisie.
Les Rongeurs reconnaissent-ils leurs apparentés ?
Avant même que ne soient recherchés les mécanismes permettant la reconnaissance de
parentèle, un traitement différentiel des apparentés était connu chez de nombreuses espèces de
vertébrés (Blaustein et al., 1987). Parmi les Mammifères, l’ordre des Rongeurs est
probablement celui qui a été le plus étudié dans le cadre de cette thématique. Des
comportements gradués selon l’apparentement ont été mis en évidence chez les Souris (Gilder
67

Chapitre II: Mécanismes d'évaluation de la proximité génétique
& Slater, 1978 ; Kareem & Barnard, 1982 ; Hayashi & Kimura, 1983), les Ecureuils terrestres
(Holmes & Sherman, 1982), les Rats (Hepper, 1987), les hamsters dorés (Heth et al., 1998) ou
les Castors (Sun & Muller-Schwarze, 1997). S’il semble exister une certaine ubiquité de la
reconnaissance des apparentés chez les Rongeurs, les mécanismes proposés sont en revanche
variables d’une étude à l’autre. Parmi les quatre mécanismes "classiques" exposés plus haut,
deux ont reçu un intérêt particulier : la familiarité et l’assortiment phénotypique. Les
références au quatrième mécanisme, les allèles de reconnaissance, sont plus rares mais ne sont
pas à négliger. En effet, ce dernier mécanisme est celui qui est le plus intimement lié à la
sélection de parentèle (Hamilton, 1964). La possession d'allèles de reconnaissance
représenterait en effet la concrétisation de l'importance de cette sélection de parentèle (le
phénotype résultant de ces allèles et permettant la reconnaissance est classiquement appelé
"barbe verte" (Dawkins, 1976)). Ce mécanisme présente donc un intérêt d'un point de vue
théorique, et il est logique qu'il ait été recherché activement dans les premières études sur la
reconnaissance de parentèle utilisant des apparentés non familiers (Tang-Martinez, 2001).
Cependant une "barbe verte", un marqueur non ambigu de l'appartenance à une entité
génétique familiale, semble peu probable chez les Mammifères, et ce pour deux raisons. La
première est l'existence d'une incohérence logique : ces allèles de reconnaissance devraient
permettre de reconnaître les cousins maternels comme les cousins paternels. Ceux-ci étant non
apparentés les uns aux autres, il semble impossible qu'ils puissent porter cette "barbe verte",
puisqu’ alors ils se reconnaîtraient mutuellement comme apparentés (Todrank & Heth, 2003).
La deuxième raison de l'improbabilité de la "barbe verte" dans la reconnaissance de parentèle
est le scénario évolutif nécessaire à son apparition : il existerait une pression de sélection
pour la faire apparaître dans une population n'ayant aucun moyen de reconnaître
l'apparentement. Le premier individu développant ce signal va donc être amené à préférer les
individus… l'ayant aussi. A moins d'une apparition concomitante chez plusieurs individus, le
maintien d'un tel caractère semble improbable. Même si un tel marqueur se maintenait en
procurant un avantage à ceux le possédant, cet avantage devrait conduire rapidement à sa
propagation dans la population. Une fois propagé, ce marqueur ne présente plus d'intérêt
puisqu'il est possédé par tout le monde. Il ne permet donc plus de distinguer les apparentés
des non apparentés. En conclusion, l'effet "barbe verte" est envisageable dans le cadre de la
reconnaissance spécifique, mais semble extrêmement improbable pour la reconnaissance de
parentèle.
68

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Si les deux mécanismes "classiques" les plus étudiés chez les Rongeurs sont bien la
familiarité et l’assortiment phénotypique, nous verrons que ces deux mécanismes ne sont pas
aisément différentiables.
Familiarité et reconnaissance de parentèle
Les Rongeurs, comme tous les Mammifères, présentent la particularité d’avoir une
"cohabitation forcée" après la naissance. Cette période liée à l’allaitement implique une
familiarité avec la mère et les membres de la fratrie s’ils existent. Apprendre le phénotype de
ces individus familiers apparaît donc un bon moyen pour adapter son comportement à
l'apparentement. Les individus familiers ne
seront pas considérés comme des partenaires
sexuels potentiels, mais pourront recevoir des
comportements altruistes. La familiarité semble
effectivement jouer un rôle prépondérant chez
les Rongeurs. Les Campagnols Microtus
pennsylvanicus, par exemple, montrent une
préférence pour les individus familiers, quel que
soit l'apparentement (Ferkin & Rutka, 1990). Le
même type de résultat se retrouve chez
Peromyscus maniculatus (Dewsbury, 1988).
La familiarité semble "l'emporter" sur
l'apparentement, même si familiarité et
apparentement vont le plus souvent de pair dans
les conditions naturelles. Ce rôle prépondérant
de la familiarité a été mis en évidence par des
expériences d'adoptions croisées (Todrank &
Heth, 2001). Classiquement, un nouveau-né est
transféré dans une portée non apparentée où il se
familiarise avec les phénotypes de la mère et de la fratrie adoptives. Chez les Souris, cette
procédure induit une modification des préférences socio sexuelles à l'âge adulte, avec un rôle
prépondérant des membres de la fratrie (Kareem & Barnard, 1982 ; Kareem & Barnard,
1986). Cette modification se traduit par une préférence sexuelle pour la fratrie biologique par
rapport à la fratrie adoptive. Cependant, si les apparentés "vrais" ne sont pas reconnus à l'âge
Encadré 2.2 : Familiarité indirecte etassortiment phénotypique
Dans la description de la familiarité comme undes quatre mécanismes permettant lareconnaissance de parentèle (Blaustein, 1983),l'apprentissage du phénotype des familiers estsupposé être un bon guide pour jauger del'apparentement d'individus non familiers. Si lephénotype de ces derniers est peu différent decelui d'un individu familier, alors il estenvisageable que les mêmes règles dedécisions s'appliquent à cet individu. Chez lessouris épineuses Acomys cahirinus, des jeunesséparés pendant neuf jours se reconnaissentpourvu qu'ils aient été maintenus en contactavec un membre de la portée, suggérant que lareconnaissance s'opère par extrapolation descaractéristiques de l'individu resté familier(Porter et al., 1983). Les familiarités directeset indirectes permettant la reconnaissance deparentèle sont parfois regroupées sous leterme de "reconnaissance par association"(Halpin, 1991) On retrouve là l'ambiguïté liéeà la méconnaissance du traitement cognitif del'information : l'extrapolation à partir descaractéristiques d'un individu familier semblepeu différente de la mémorisation descaractéristiques phénotypiques des individusfamiliers, pourtant classiquement considéréecomme relevant de l'assortiment phénotypique,c'est-à-dire un mécanisme de nature trèsdifférente.
69

Chapitre II: Mécanismes d'évaluation de la proximité génétique
adulte, les préférences de juvéniles semblent influencées par le phénotype propre du sujet
(Aldhous, 1989). D'autre part, les adoptés semblent ne pas être discriminés par les adoptants,
ce qui suggère un transfert des informations olfactives mais également un rôle important de la
reconnaissance individuelle. D'une manière générale, l'objectif des adoptions croisées d'un
seul individu est de minimiser l'influence sociale des apparentés de façon à pouvoir séparer
clairement la reconnaissance "génétique" de la reconnaissance sociale. Dans le cadre des
études sur le MHC (cf. chapitre I), il a été montré que l'environnement précoce va servir de
guide à la construction des préférences sexuelles : les souris mâles transférées à la naissance
dans une portée présentant un haplotype différent du leur vont exprimer des préférences
sexuelles à l'âge adulte allant vers un évitement de l'haplotype adoptif (Beauchamp et al.,
1988) et une préférence pour leur propre haplotype en cas de choix binaire (Yamazaki et al.,
1988). Cependant ce type de protocole est basé sur un postulat d'hypothèses exclusives dont
nous avons vu plus haut qu'il est inadéquat. Il semble indéniable que la familiarité joue un
rôle prépondérant dans l'acquisition de préférences sexuelles, mais ceci ne constitue en aucun
cas une démonstration de l'inexistence d'autres mécanismes. En effet, même pour des
individus dont on a modifié l'environnement post-natal, les apparentés non familiers ne sont
pas exactement équivalents à des non-apparentés non familiers. La réunion d'individus
apparentés non familiers se traduit par des corrélats physiologiques (abaissement du seuil de
douleur comparativement à des réunions de non-apparentés non familiers), ce qui semble
traduire un certain degré de perception de cet apparentement (D'Amato, 1994).
Les mécanismes permettant cette perception de la proximité génétique indépendamment de la
familiarité supposent généralement une référence à l'odeur propre de l'individu.
Assortiment phénotypique auto référencé, effet "aisselles" et
reconnaissance de parentèle
L'assortiment phénotypique auto référencé ("self-referent phenotype matching") est un
mécanisme par lequel un individu utilise son propre phénotype comme point de comparaison
pour évaluer l'apparentement avec les individus rencontrés (Hauber & Sherman, 2001). Le
terme "d'effet aisselles" ("armpit effect"), proposé par (Dawkins, 1982) est utilisé dans le cas
des signaux olfactifs pour illustrer la théorie selon laquelle un animal est capable de comparer
l'odeur d'un congénère à son odeur propre. L'avantage d'un tel mécanisme est qu'il évite les
"erreurs" inhérentes aux systèmes de reconnaissance indirects (reconnaissance spatiale et
70

Chapitre II: Mécanismes d'évaluation de la proximité génétique
familiarité). En effet, les individus familiers ne sont pas nécessairement apparentés et les non
familiers peuvent être apparentés. Parmi les situations propices à ces erreurs, citons le cas des
animaux de mêmes parents mais de portées différentes qui peuvent se rencontrer pour la
première fois une fois adultes, celui des espèces où l'élevage des jeunes se fait en commun ou
bien celui des paternités multiples (Hauber & Sherman, 2000 ; Hauber & Sherman, 2001).
Chez les spermophiles Spermophilus beldingi, après l'hibernation, les apparentés sont toujours
discriminés des non-apparentés, alors que les individus familiers avant la période
d'hibernation de neuf mois sont traités comme des non familiers (Mateo & Johnston, 2000b).
Même quand la mémoire sociale semble épuisée, il subsiste une reconnaissance de parentèle.
Chez les hamsters Mesocricetus auratus, quand on réalise des adoptions croisées en
transférant un seul petit dans une portée non apparentée, ce jeune, une fois adulte, est capable
de discriminer ses apparentés de non-apparentés : l'environnement social post-natal n'est pas
déterminant dans la reconnaissance de parentèle chez cette espèce (Mateo & Johnston,
2000a). La conclusion qui semble s'imposer est que les hamsters, comme les spermophiles
(Mateo & Johnston, 2000b), doivent se référer à leur odeur propre. Cette démonstration est en
fait imparfaite, puisque la familiarité, quoique réduite, n'est pas totalement contrôlée (Mateo
& Johnston, 2000a), ayant réalisé les adoptions croisées de 3 à 12 heures après la naissance
(Hare et al., 2003). De plus, même si l'on passe sur cette imperfection de la méthode qui est
compréhensible compte tenu des contraintes de manipulations des animaux, la capacité de
discrimination des hamsters peut être due non pas à une référence à l'odeur propre, mais à une
préférence "innée" pour les odeurs d'apparentés. En d'autres termes, il pourrait exister un
"biais sensoriel" envers le phénotype des apparentés. Cette plus grande sensibilité aux signaux
des apparentés est un phénomène qui existe notamment dans la relation mère-embryon et qui
est interprété comme l'existence d'allèles de reconnaissance voire un effet "barbe verte" (Haig,
1996). On retrouve là encore l'ambiguïté des mécanismes proposés : la comparaison du
phénotype d'un congénère avec le phénotype propre correspond à de l'assortiment
phénotypique auto référencé, mais l'éventuelle découverte d'un biais sensoriel est considéré
comme relevant de l'existence d'allèles de reconnaissance, alors que ce n'est que l'élucidation
du mécanisme de cet assortiment auto référencé.
Les frontières entre les mécanismes "classiques" de reconnaissance de parentèle étant floues,
l'approche choisie dans cette thèse a conduit à proposer une vision nouvelle, si possible
libérée de ces ambiguïtés. Il ne faut donc pas considérer le mécanisme présenté dans la suite
71

Chapitre II: Mécanismes d'évaluation de la proximité génétique
du manuscrit comme un nouveau "concurrent" dans la compétition entre mécanismes putatifs
de reconnaissance de parentèle mais plutôt comme une tentative d'explication parcimonieuse
qui recouvre ces mécanismes.
Le chapitre I de cette thèse a apporté des éléments déterminants quant à l'information présente
dans les odeurs individuelles et le fait qu'elle révèle de façon fiable la proximité génétique. En
vertu de la proximité olfactive des individus apparentés, il est possible qu'un apparenté non
familier soit discriminé d'un non-apparenté également non familier, de la même façon que les
Souris discriminent les odeurs d'hétérospécifiques lointains ou proches (Heth et al., 2001).
2.2. Préférences olfactives pour des apparentés non familiers
Les expériences décrites ci-dessous ont pour objectif de mettre en évidence une capacité de
discrimination olfactive des souris Mus spicilegus en fonction de leur apparentement aux
donneurs d'odeurs. Ces préférences sont explorées pour un seul degré d'apparentement
(r = 0,5 soit une relation de frère et sœur) mais pour toutes les configurations: préférences de
mâles pour des odeurs de mâles et de femelles ; préférences de femelles pour des odeurs de
femelles et de mâles, tous les donneurs d'odeurs étant non familiers avec le sujet.
2.2.1 Matériel et méthodes
Les souris mâles et femelles sont issues des premiers accouplements réalisés à partir de la
population capturée à Gyöngyös (Hongrie) en septembre 1999 (génération 1). Les mâles sont
isolés une semaine avant les tests de façon à homogénéiser leur statut hormonal. Les femelles
sont gardées en groupe unisexe, telles qu'elles sont depuis le sexage à 35 jours. Ce
groupement permet d'inhiber les cycles oestriens et les ouvertures vaginales (Feron & Gheusi,
2003). Au moment du test, les animaux sont âgés de 4 à 7 mois.
Dans tous les tests, un choix est proposé entre les odeurs uro-génitales de deux individus de
même sexe et de même âge (± 7 jours). Les donneurs d'odeurs sont issus de portée différente
de celle du sujet (portée antérieure dans la moitié des tests, postérieure dans l'autre). Ils sont
donc non familiers. La différence d'âge entre le sujet et les donneurs est systématiquement
supérieure à celle entre les donneurs. Pour chacun des tests, les sujets sont issus de couples
différents de façon à éviter la pseudo-réplication.
72
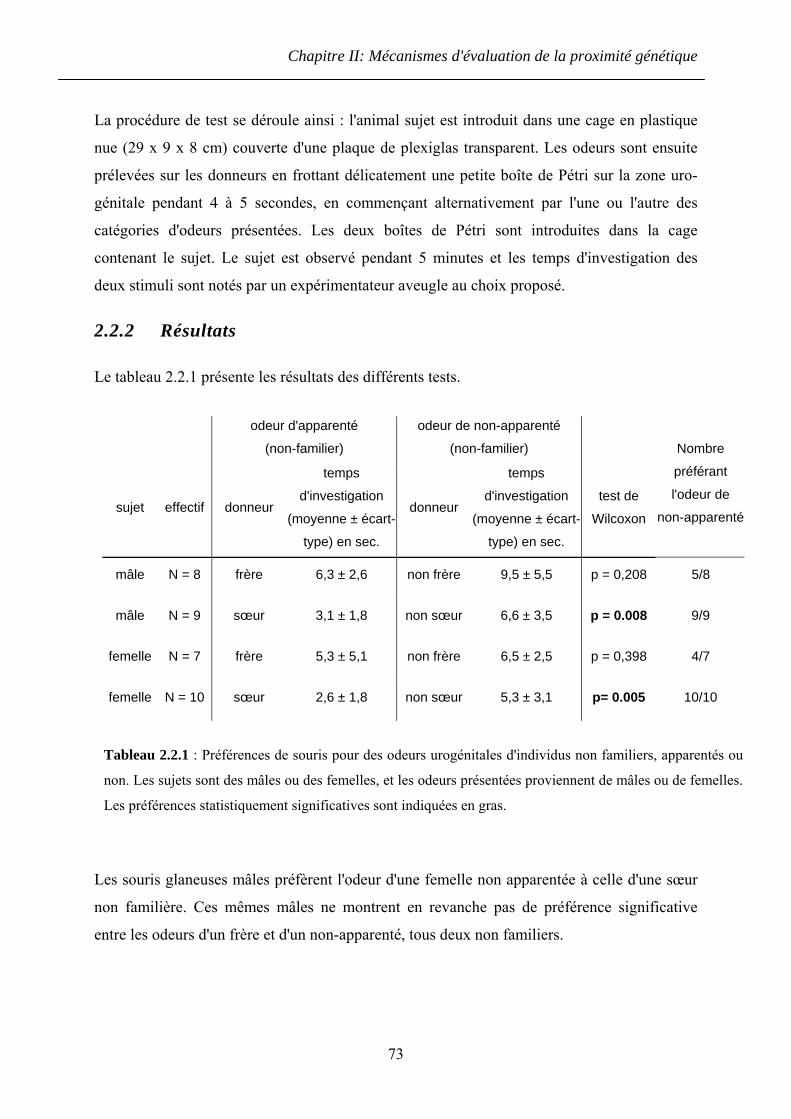
Chapitre II: Mécanismes d'évaluation de la proximité génétique
La procédure de test se déroule ainsi : l'animal sujet est introduit dans une cage en plastique
nue (29 x 9 x 8 cm) couverte d'une plaque de plexiglas transparent. Les odeurs sont ensuite
prélevées sur les donneurs en frottant délicatement une petite boîte de Pétri sur la zone uro-
génitale pendant 4 à 5 secondes, en commençant alternativement par l'une ou l'autre des
catégories d'odeurs présentées. Les deux boîtes de Pétri sont introduites dans la cage
contenant le sujet. Le sujet est observé pendant 5 minutes et les temps d'investigation des
deux stimuli sont notés par un expérimentateur aveugle au choix proposé.
2.2.2 Résultats
Le tableau 2.2.1 présente les résultats des différents tests.
s
m
m
fem
fem
Tab
non
Les
Les
non
entre
odeur d'apparenté
(non-familier)
odeur de non-apparenté
(non-familier)
ujet effectif donneur
temps
d'investigation
(moyenne ± écart-
type) en sec.
donneur
temps
d'investigation
(moyenne ± écart-
type) en sec.
test de
Wilcoxon
Nombre
préférant
l'odeur de
non-apparenté
âle N = 8 frère 6,3 ± 2,6 non frère 9,5 ± 5,5 p = 0,208 5/8
âle N = 9 sœur 3,1 ± 1,8 non sœur 6,6 ± 3,5 p = 0.008 9/9
elle N = 7 frère 5,3 ± 5,1 non frère 6,5 ± 2,5 p = 0,398 4/7
elle N = 10 sœur 2,6 ± 1,8 non sœur 5,3 ± 3,1 p= 0.005 10/10
leau 2.2.1 : Préférences de souris pour des odeurs urogénitales d'individus non familiers, apparentés ou
. Les sujets sont des mâles ou des femelles, et les odeurs présentées proviennent de mâles ou de femelles.
préférences statistiquement significatives sont indiquées en gras.
souris glaneuses mâles préfèrent l'odeur d'une femelle non apparentée à celle d'une sœur
familière. Ces mêmes mâles ne montrent en revanche pas de préférence significative
les odeurs d'un frère et d'un non-apparenté, tous deux non familiers.
73

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Les femelles passent significativement plus de temps à sentir l'odeur d'une femelle non
apparentée que celle d'une sœur non familière. Il n'existe pas de préférence claire quand le
choix est entre deux odeurs de mâles non familiers, apparentés ou non.
2.2.3 Discussion
En l'absence de familiarité avec les donneurs d'odeurs, les souris Mus spicilegus réagissent en
fonction de leur apparentement à ces donneurs d'odeurs dans certaines configurations. Les
mâles passent plus de temps à sentir l'odeur d'une femelle non apparentée que l'odeur d'une
sœur non familière. Les femelles confrontées à ce même choix montrent également une
discrimination en explorant plus longuement l'odeur d'une femelle non apparentée. Il n'a en
revanche pas été possible de montrer cette discrimination pour les mâles confrontés à des
odeurs de mâles et pour les femelles confrontées à des odeurs de mâles.
Plusieurs conclusions peuvent être tirées de ces résultats. La première est que les préférences
ne s'expriment pas dans toutes les situations. Certaines situations sont plus propices, peut-être
en raison du contexte motivationnel. Les femelles montrent une préférence pour l'odeur d'une
non-apparentée par rapport à une sœur, ce qui est délicat à interpréter. La littérature suppose
classiquement une préférence mutuelle entre apparentées, en raison de l'élevage en commun
des jeunes (Manning et al., 1992). Cet élevage en commun n'a certes jamais été démontré
chez Mus spicilegus (Baudoin et al., sous presse), mais les regroupements de femelles au
moment de l'entrée des juvéniles dans les tumuli en automne sont plausibles, en raison de
l'apparentement des portées présentes dans ces tumuli (Garza et al., 1997). La préférence pour
l'odeur d'une non-apparentée est probablement explicable par la nature "artificielle" du test : il
est peu probable que le temps supérieur passé à sentir l'odeur corresponde à une préférence
sociale pour cette femelle. En revanche, compte tenu des résultats exposés dans le chapitre I,
une explication plus parcimonieuse se fait jour : la covariance odeurs-gènes implique que les
odeurs de sœurs sont plus similaires que les odeurs de femelles non apparentées (bien que les
résultats du chapitre I ne concernent que les mâles, il n'y a pas de raison de supposer que la
covariance puisse ne pas exister chez les femelles (Busquet & Baudoin, sous presse). L'odeur
de la sœur non familière est donc plus proche de l'odeur portée par la souris testée elle-même.
Cette odeur est aussi plus proche de l'odeur portée par les sœurs de même fratrie avec qui la
femelle testée cohabite, et également d'un éventuel modèle de référence appris après la
naissance. La réaction des femelles testées, si elle correspond bien à l'apparentement, ne
74

Chapitre II: Mécanismes d'évaluation de la proximité génétique
permet pas de conclure sur le mécanisme utilisé puisque familiarité indirecte, assortiment
phénotypique (auto référencé ou non) et allèles de reconnaissance peuvent expliquer ces
résultats. En revanche, la familiarité directe, c'est-à-dire la reconnaissance individuelle des
familiers comme étant apparentés, ne peut expliquer ces résultats. En l'absence d'un contexte
motivationnel clairement défini, il convient de considérer l'hypothèse de "plus bas niveau" :
parmi les deux odeurs présentées, l'une est proche de ce à quoi l'animal testé est habitué,
l'autre est moins proche. Jusqu'au moment du transfert dans la cage de test, les récepteurs
olfactifs de la souris sont confrontés à des molécules odorantes provenant de l'animal lui-
même et de son environnement : ceci peut être perçu comme la "stimulation basale". Les
molécules odorantes d'une sœur non familière sont probablement très similaires à celles
responsables de cette stimulation basale, du fait de la covariance odeurs-gènes. L'odeur d'une
femelle non apparentée, en revanche, est susceptible de renfermer des signaux nouveaux sous
la forme de molécules différentes ou de concentrations différentes de ces molécules. Ce signal
va donc se démarquer du signal habituel et induire une stimulation "beaucoup plus différente"
de la stimulation basale que ne le fait l'odeur d'une sœur. Ces résultats sont cohérents avec
l'existence d'un biais sensoriel. Quand les femelles avaient à choisir entre deux odeurs de
mâles, dans les mêmes conditions de test, elles n'ont pas montré de préférence claire pour
l'odeur d'un non-apparenté. Par rapport aux odeurs de femelles auxquelles les souris testées
sont confrontées, les odeurs de mâles diffèrent énormément, et la stimulation des récepteurs
est d'emblée très différente, puisque les signaux présentés sont extrêmement nouveaux. Là
encore, l'interprétation est délicate : les théories classiques supposent que les femelles
choisissent le mâle, et que ce choix est le moteur de la compétition intra sexuelle et de
l’évolution des traits des mâles (Drickamer et al., 2000 ; Gavrilets et al., 2001). Or les
femelles ne semblent pas montrer de préférence pour un partenaire potentiel non apparenté
par rapport à un frère. De plus, sur les 10 femelles testées, seules 7 ont montré un intérêt pour
les odeurs, les 3 autres n’ayant exploré aucune des deux odeurs pendant le temps imparti.
L’absence de préférence des femelles pour les odeurs de mâles ne peut pas pour autant être
considérée comme une absence de choix de partenaire chez les femelles Mus spicilegus : la
nouveauté des signaux présentés (les souris testées n’ont aucune expérience des odeurs de
mâles depuis leur maturité sexuelle) est probablement responsable des réactions observées.
Pour la femelle, l’odeur d’un mâle, qu’il soit frère ou non, serait tellement différente des
odeurs sociales auxquelles elle est habituée que la différence entre les deux stimuli serait
négligeable. Pour vérifier cette interprétation, il faudrait proposer le même type de choix à des
75

Chapitre II: Mécanismes d'évaluation de la proximité génétique
femelles ayant l’expérience des odeurs de mâles (par exemple des femelles en couple). Un
autre point important dans l’analyse de ces résultats est lié à la réceptivité sexuelle des
femelles testées. En effet, celles-ci sont en anœstrus, et présentent une fermeture vaginale
induite par la cohabitation entre femelles (Féron & Gheusi, 2003). On peut donc concevoir
que les femelles ne soient pas dans l’état nécessaire à un choix de partenaire. Cependant, chez
Mus spicilegus, la réceptivité sexuelle est induite par la cohabitation avec un mâle (Féron &
Gheusi, 2003). Le choix de partenaire devrait donc être préalable à la réceptivité sexuelle. Les
comportements des mâles dans les expériences décrites ci-dessus sont particulièrement
intéressants à cet égard.
Les mâles Mus spicilegus montrent une préférence significative pour l’odeur d’une femelle
non apparentée par rapport à une sœur, toutes deux étant non familières. Ceci démontre la
capacité à réagir en fonction de l’apparentement. Mais ces résultats sont également
surprenants dans la mesure où les mâles sont classiquement perçus comme ne choisissant pas
(Bergstrom & Real, 2000 ; Drickamer et al., 2003). Dans l’expérience ci-dessus, non
seulement semblent-ils choisir, mais ils choisissent entre des odeurs de femelles non
sexuellement réceptives. Bien sûr, il est difficile d’objectiver la nature sexuelle de la
préférence olfactive, mais un argument peut tout de même être apporté pour soutenir cette
hypothèse : les femelles confrontées à des odeurs de mâles ne choisissent pas, probablement à
cause d’un "effet plafond" lié à la nouveauté des signaux présentés (voir plus haut). Les mâles
n’ont pas non plus d’expérience avec des odeurs de femelles depuis le sexage (à 35 jours). Les
signaux présentés devraient donc être "également nouveaux", comme les signaux de mâles
l’étaient pour les femelles. Le fait que ce ne soit pas le cas et qu’il y ait une préférence claire
démontrée par les mâles suggère l’existence d’une motivation à choisir, peut-être résultant de
pressions de sélection en faveur d’un choix par les mâles. Chez M. spicilegus, montrer un
intérêt pour des odeurs de femelles non sexuellement réceptives est capital, puisque celles-ci
ne peuvent devenir réceptives sans cohabitation avec un mâle (Feron & Gheusi, 2003).
L’émotivité supérieure des femelles par rapport aux mâles (Simeonovska-Nikolova, 2000) et
l’intolérance entre sexes (Patris et al., 2002) chez cette espèce suggèrent le scénario suivant :
avant tout accouplement, le mâle doit cohabiter avec la femelle pour induire la réceptivité
sexuelle. Cette cohabitation n’est pas possible directement en raison de l’agressivité des
femelles. On peut donc supposer qu’il va falloir pour le mâle passer du temps avant de se faire
accepter et de pouvoir induire la réceptivité sexuelle (ceci correspondrait bien au délai
76

Chapitre II: Mécanismes d'évaluation de la proximité génétique
observé avant la formation d'un couple stable chez cette espèce (Baudoin et al., sous presse)).
Cet "investissement" doit théoriquement être dirigé de façon optimale, c'est-à-dire que le mâle
n’a pas intérêt à passer trop de temps à "courtiser" une femelle qui n’est pas adéquate. Ce
choix de la femelle adéquate doit se faire alors même qu’elle n’est pas réceptive sexuellement.
Les résultats obtenus lors de l’expérience ci-dessus sont cohérents avec ce scénario puisque
les mâles montrent une préférence nette pour l’odeur d’une femelle qui ne leur est pas
apparentée. Ce choix par les mâles, s’il était démontré, constituerait un contre-exemple très
intéressant aux théories habituelles sur le choix de partenaire (Bergstrom & Real, 2000). En
effet celui-ci est très majoritairement attribué aux femelles et étudié, modélisé et théorisé de
ce seul point de vue (Drickamer et al., 2000 ; Jennions & Petrie, 1997 ; Jennions & Petrie,
2000 ; Houle & Kondrashov, 2002 ; Gavrilets et al., 2001 ; Roberts & Gosling, 2003 ;
Tregenza & Wedell, 2000). Bien évidemment, la préférence montrée par les mâles
M. spicilegus n’est en rien une démonstration d’un réel choix de partenaire. Il est cependant
frappant de voir que les mâles ne réagissent pas différemment à l’odeur d’un frère non
familier qu’à l’odeur d’un mâle non apparenté. Cette distinction a pourtant aisément été
montrée par les femelles dans une situation similaire. Il est probable que les résultats des
mâles confrontés à des odeurs de mâles ne soient pas le fait d’une incapacité à réagir en
fonction de la proximité génétique mais d’un problème de motivation. Les mâles sont tout à
fait capables de réagir en fonction du degré d’apparentement dans un protocole d’habituation-
généralisation (Busquet & Baudoin, sous presse) et distinguent parfaitement l’odeur d’une
sœur de celle d’une femelle non apparentée. Confrontés à des odeurs de mâles sans "mise en
condition" préalable, les sujets ont traité les deux stimuli sans se référer à l’apparentement.
Selon les sujets, l’odeur la plus explorée provenait de l’un ou l’autre des deux mâles
donneurs. L’interprétation de ces résultats en terme de compétition entre mâles est possible
mais le protocole n’ayant pas été construit pour étudier les aspects fonctionnels mais les
mécanismes de la reconnaissance de parentèle, une telle interprétation serait peu
démonstrative.
La capacité à discriminer les apparentées non familières des non-apparentées permet de
montrer que la reconnaissance individuelle n’est pas nécessaire au traitement différentiel des
apparentés. Cependant une familiarité indirecte et la mémorisation du phénotype familial
("phenotype matching") peuvent expliquer ces résultats, même si ces explications ne sont pas
les plus parcimonieuses. Il est important de comprendre, comme déjà expliqué plus haut, que
77

Chapitre II: Mécanismes d'évaluation de la proximité génétique
ces mécanismes ne sont pas des explications exclusives mais correspondent à des capacités
coexistantes. La reconnaissance individuelle n’est pas nécessaire à la reconnaissance de
parentèle, mais existe probablement chez Mus spicilegus comme elle existe chez d’autres
Rongeurs (Bowers & Alexander, 1967 ; Jacobs & Kuiper, 2000 ; Lai & Johnston, 2002 ;
Johnston, 2003). Plutôt que de chercher à déterminer lequel de ces mécanismes prend le pas
sur les autres, il semble plus intéressant de présenter aux animaux des situations dans
lesquelles ils ne peuvent plus compter sur un ou plusieurs de ces mécanismes. Dans le cas des
mâles préférant de façon unanime l’odeur d’une femelle non apparentée, ceux-ci ont été
familiers avec le phénotype d’apparentés jusqu’au moment de leur isolement une semaine
avant le test. Même s’ils avaient été isolés dès le sevrage, ils auraient toujours un biais
d’exposition aux odeurs des apparentés. La réalisation d’une adoption croisée avec transfert
d’un seul petit aurait inversé ce biais d’exposition en faveur de la famille adoptante, et se
serait probablement traduit par une préférence basée sur ce phénotype familial (Penn & Potts,
1998 ; Todrank & Heth, 2001). Ce rôle prépondérant de la familiarité ne permet évidemment
pas d’écarter les autres mécanismes. Autrement dit, le fait que la familiarité ait été le "guide"
des comportements des animaux dans un contexte expérimental donné se signifie pas qu’il en
sera toujours ainsi dans toutes les situations. En nature, il est possible de rencontrer des
individus pour lesquels la familiarité ne sera pas un outil efficace pour déterminer s’il doit être
ou non traité comme un apparenté (Todrank & Heth, 2001). C’est pourquoi, dans la procédure
d’adoption croisée présentée dans l’article qui suit, nous n’avons pas tenté d’éliminer toute
familiarité mais de la contrôler, de la niveler de façon à ce que l’exposition au phénotype
génétiquement proche soit comparable à l’exposition à l’autre phénotype proposé.
78

Chapitre II: Mécanismes d'évaluation de la proximité génétique
2.3. Rôle de l'expérience post-natale dans l'évaluation
de la proximité génétique : adoptions croisées
interspécifiques
Comme nous l'avons déjà vu, l'un des problèmes majeurs de l'objectivation de la
reconnaissance de parentèle "vraie" est celui de la superposition des phénomènes que sont la
familiarité et l'apparentement. Un individu, dans les conditions naturelles, présente un surcroît
d'expérience avec ses apparentés. L'approche développée dans l'article ci-dessus (Heth et al.,
2003) a consisté à standardiser, à contrôler cette familiarité, pour mettre en évidence une
reconnaissance de la proximité génétique "vraie". En supprimant les influences parentales
"naturelles" juste après la naissance, et en standardisant la familiarité avec les deux types de
phénotypes testés, les souris (mâles et femelles) réagissent en fonction de leur proximité
génétique avec les donneurs d'odeurs, et manifestent, une fois à l'âge adulte, une préférence
pour les odeurs des femelles génétiquement proches (membres de la même population,
conspécifiques ou hétérospécifiques proches selon les tests). Il existe donc un mécanisme
d'évaluation de la proximité génétique via la similarité des odeurs.
79

Chapitre II: Mécanismes d'évaluation de la proximité génétique
80

Chapitre II: Mécanismes d'évaluation de la proximité génétique
81

Chapitre II: Mécanismes d'évaluation de la proximité génétique
82

Chapitre II: Mécanismes d'évaluation de la proximité génétique
83

Chapitre II: Mécanismes d'évaluation de la proximité génétique
84

Chapitre II: Mécanismes d'évaluation de la proximité génétique
85

Chapitre II: Mécanismes d'évaluation de la proximité génétique
86

Chapitre II: Mécanismes d'évaluation de la proximité génétique
87

Chapitre II: Mécanismes d'évaluation de la proximité génétique
88

Chapitre II: Mécanismes d'évaluation de la proximité génétique
89

Chapitre II: Mécanismes d'évaluation de la proximité génétique
2.4. Rôle de l'expérience pré- et périnatale dans l'évaluation
de la proximité génétique: préférences olfactives de
nouveau-nés
L'article ci-dessus a permis de montrer que lorsque la familiarité est contrôlée, les souris Mus
spicilegus mais aussi les souris Mus musculus musculus, malgré une expérience post-natale
commune, montrent une capacité à réagir en fonction de leur proximité génétique aux
donneurs d'odeurs. L'éventuel modèle de référence formé à partir de l'ensemble des individus
familiers (conspécifiques et hétérospécifiques) ne permet pas de distinguer l'odeur d'un
individu génétiquement proche du sujet de l'odeur d'un individu plus éloigné mais également
proche de ce modèle de référence. Ceci ne signifie pas qu'il n'y ait pas de modèle de
référence, donc pas d'assortiment phénotypique. Quand on empêche l'utilisation de ce
mécanisme en le rendant non discriminant pour la question posée, les souris sont tout de
même capables d'évaluer la proximité génétique avec les donneurs d'odeurs. Une objection
peut néanmoins être apportée (avant d'être rejetée) : serait-il possible que les sujets de ces
expériences aient malgré tout utilisé l'assortiment phénotypique en se basant sur un modèle de
référence incluant majoritairement les caractéristiques des apparentés familiers ? Cette
hypothèse suppose qu'il y ait une sensibilité supérieure aux indices olfactifs provenant de la
fratrie biologique par rapport à la fratrie adoptive. Or cette sensibilité supérieure, ce biais
sensoriel est précisément l'essence du mécanisme d'évaluation de la proximité génétique par la
proximité des odeurs. Cette objection n'en est donc pas une.
Une des caractéristiques du mécanisme proposé est qu'il est cognitivement simple : il ne
nécessite pas d'apprentissage ni de mémorisation. L'article suivant présente des expériences
réalisées sur des nouveau-nés, et démontre une capacité de ces nouveau-nés à réagir en
fonction de la proximité génétique (y compris l'apparentement chez Mus spicilegus). De façon
à montrer que cette capacité précoce n'est pas due à un apprentissage, le test le plus révélateur
de cet article propose aux nouveau-nés un choix entre deux odeurs de femelles allaitantes non
apparentées à la mère du petit testé. Parmi ces deux femelles, l'une est cependant apparentée
au sujet lui-même, puisqu'il s'agit d'une tante paternelle. Enfin, pour réellement empêcher
l'apprentissage de ce phénotype paternel, le père est retiré de la cage au moins trois jours
avant la mise bas. Les apprentissages prénataux existent en effet mais semblent limités au tous
derniers jours de la gestation chez les Rongeurs comme le Rat (Gruest et al., 2004). Malgré
90

Chapitre II: Mécanismes d'évaluation de la proximité génétique
cette procédure, les nouveau-nés sont significativement plus attirés par l'odeur de cette tante
paternelle que par l'odeur de l'autre femelle.
91

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Preferences of newborn mice for odours indicating closer genetic relatedness:
Is experience necessary ?
Josephine Todrank1,2, Nicolas Busquet1, Claude Baudoin1 and Giora Heth1,2 1Laboratoire d'Ethologie Expérimentale et Comparée, Université Paris Nord, 99 Avenue
Jean-Baptiste Clément, F-93430 Villetaneuse, France 2Institute of Evolution, University of Haifa, Mount Carmel, Haifa 31905, Israel
Abstract
Mechanisms to discriminate between individuals on the basis of genetic relatedness
could have evolutionary significance through enabling animals to optimise reproductive
success and enhance inclusive fitness. Recently accumulated evidence about the nature of kin
recognition mechanisms indicates that in adult rodents individual recognition enables
distinctions between familiar individuals irrespective of relatedness (but including close kin)
and a separate mechanism enables discriminations based on genetic relatedness without prior
familiarity. In behavioural tests exploring the latter mechanism, adult mice could assess the
extent of their genetic relatedness to other individuals using individual odour similarities. The
ontogeny of this genetic relatedness assessment mechanism, however, had not been
investigated. Here, in two-choice tests, newborn mice differentially preferred odours of more
genetically similar lactating females (paternal aunts to unrelated conspecific and conspecific
to heterospecific) even without prior exposure to adults with the tested genotypes. The results
provide a direct demonstration of genetic relatedness assessment abilities in newborns and
show that prenatal or postnatal experience with parental odours is not necessary for genetic
relatedness distinctions.
Keywords: genetic relatedness; kin recognition; odour preferences; newborn mice
92

Chapitre II: Mécanismes d'évaluation de la proximité génétique
1. INTRODUCTION
Differential responses to other individuals based on different genetic relatedness are
thought to provide adaptive benefits, including optimal mate choice to promote
heterozygosity/genetic variability and to avoid the detrimental fitness consequences of
inbreeding and outbreeding depression (Bateson, 1983; Thornhill, 1993; Tregenza & Wedell,
2000; Edmands, 2002) and enhancing inclusive fitness (Hamilton, 1964). It is generally
assumed that kin recognition mechanisms (Fletcher & Michener, 1987; Hepper, 1991;
Sherman et al., 1997) underlying such discriminative responses depend on both some sort of
“learning” and some sort of “matching” (Tang-Martinez, 2001). In these mechanisms,
“recognition by association” involves learning characteristics of familiar individuals and
matching remembered characteristics to specific individuals on subsequent encounters, and
“phenotype matching” is thought to involve incorporating learned kin characteristics into a
composite template and matching characteristics of unfamiliar individuals against that
template (Tang-Martinez, 2001). Even self-referencing (using one’s own phenotype as the
referent) ostensibly involves learning and matching one’s own phenotypic characteristics
(Sherman et al., 1997; Hauber & Sherman, 2001). This inclination to interpret data in terms of
learning is understandable given the remarkable range of associations that rodents are able to
make. A recent review of kin recognition data (Todrank & Heth, 2003), however, raised new
hypotheses about genetic relatedness distinctions (not only between kin and non-kin) that do
not involve postnatal learning.
Although rodents can recognize familiar individuals from their odours, in several
species tested, learning these individual odours did not influence their genetic relatedness
discriminations. For example, golden hamsters (Mesocricetus auratus) raised in mixed litters
(see Todrank & Heth, 2001for an explanation of the cross-fostering procedure) recognized the
individual odours of their biological and foster siblings (Todrank et al., 1998, 1999), but they
did not incorporate these odours (or that of the female that nursed them) into a composite
categorical template for discriminating between unfamiliar kin and non-kin (Heth et al.,
1998). As a result, they responded to odours of conspecifics, whether familiar or unfamiliar,
in a graded fashion based on the degree of similarity with their own phenotype (Heth et al.,
1998). In another cross-fostering study in which pups from two species of wild mice (Mus
spicilegus and Mus musculus) were raised (by a laboratory mouse) in mixed litters from
within hours of birth, adults’ odour preferences across a range of genetic relatedness from
conspecifics through heterospecifics were graded based on similarity to their own phenotypes
93

Chapitre II: Mécanismes d'évaluation de la proximité génétique
and were not influenced by being reared with pups from another species or by a foster mother
(Heth et al., 2003). What juvenile rats reared in mixed litters (Hepper, 1983) learned about
their foster-siblings did not interfere with their genetic relatedness preferences. It was also
found, as in the studies mentioned above with hamsters and mice, that differential responses
of rats to conspecific odours were graded based on genetic relatedness (Hepper, 1987a).
Given that those rodents did not incorporate odours of the mother and littermates into a
composite template during postnatal development, they must have been using a separate
mechanism to discriminate degrees of genetic relatedness from the mechanism they used to
learn templates to recognise familiar individuals (Todrank & Heth, 2003). Evidence consistent
with separate familiarity and genetic relatedness mechanisms was also found in ground
squirrels (Spermophilus beldingi; Holmes & Sherman, 1982).
In addition, this genetic relatedness assessment mechanism enables subtle
discriminative responses across a wide range of genetic relatedness from distinctions between
siblings and half-siblings to distinctions between closer as opposed to more genetically distant
heterospecifics (reviewed in Todrank & Heth, 2003). Recent studies of the subtlety of the
differential responses (Mateo, 2002; Koeninger Ryan & Lacy, 2003; Busquet & Baudoin,
in press) also suggested that the discriminations were more fine-tuned than categorical
distinctions between kin and non-kin or conspecifics and heterospecifics. It has been found in
several rodent species and for many degrees of genetic relatedness (Todrank & Heth, 2003;
Busquet & Baudoin, in press) that similarities in the qualities of individual odours covary
with the degree of genetic relatedness between individuals, a phenomenon termed “odour-
genes covariance” (Heth & Todrank, 2000). This covariance between individual odour
phenotypes and their underlying genotypes provides a basis for assessing one’s relatedness to
another individual by comparing the degree of similarity between the other individual’s odour
and one’s own (Heth et al., 2003).
The current study was conducted to investigate the ontogeny of this genetic
relatedness assessment mechanism in two discrimination contexts: kin vs. non-kin and
conspecifics vs. heterospecifics. It has been suggested that prenatal learning primes kin
recognition because newborn rats preferred the odour of amniotic fluid from their mother to
that from another parturient female (Hepper, 1987b); however, it was not possible to
determine from that study whether familiarity with one’s own amniotic fluid was the
important factor or whether the pups were discriminating between the 2 samples on the basis
of differential genetic relatedness. The study reported here was designed to test the genetic
94

Chapitre II: Mécanismes d'évaluation de la proximité génétique
relatedness assessment abilities of newborns. It may not be possible to isolate fetal mice from
exposure to the phenotypic characteristics of their mother or other fetuses during gestation. It
is possible, however, to create an early postnatal environment that precludes the possibility of
learning associations that could facilitate genetic relatedness assessments. To this end, we
presented newborn wild mice with a choice between the ventral body odours of 2 lactating
females that were comparably unrelated to the mother but differentially related to the pups: a
paternal aunt and an unrelated female. We hypothesized that if these newborn mice (even
without exposure to the father) prefer the odour of their father’s sister to the odour of an
unrelated female, they would have to be responding to the degree of overlap with their own
phenotype. We also investigated whether newborns can discriminate between the odours of
conspecific and heterospecific lactating females.
2. METHODS
(a) Animals
The M. spicilegus mice were 5th generation born in the lab, from wild progenitors
caught in Hungary and were maintained under standard conditions (Heth et al., 2001; Busquet
& Baudoin, in press) at University of Paris XIII. The M. musculus and M. domesticus mice
were 6th generation born in the lab, from wild progenitors caught in Denmark and were
maintained under standard conditions (Smadja et al., 2004) at University of Montpellier II. In
both colonies, the genealogy was carefully monitored since the time of capture, and breeding
couples were comprised of animals selected to maintain maximal genetic diversity across the
particular population. For tests with 39 pups from 6 litters of M. musculus and M. domesticus
and 66 pups from 8 litters of M. spicilegus, the pups were from established couples that had
had several previous litters so that the time of birth was predictable. For the test with 69 pups
from 11 litters of M. spicilegus, 12 new couples were established at the same time (from a
pool of 12 pairs of brothers and sisters) in hopes of synchronising births. In some cases,
however, the paternal aunt gave birth up to 3 days later so the pups were tested between the
first and third days of life depending on when the paternal aunt gave birth. Pups tested on
their first day of life were at least 2 hours but not more than 16 hours old. For the “father-
removed” condition with 66 pups from 8 litters, M. spicilegus mothers were moved to clean
cages without the father or his odours at least 2 days before the anticipated birth, because in
contrast with M. musculus and M. domesticus, in natural conditions the father is an important
participant in parental care in M. spicilegus (Patris & Baudoin, 2000). For the conspecific-
95

Chapitre II: Mécanismes d'évaluation de la proximité génétique
heterospecific condition, 50 pups from 7 litters of M. musculus and M. domesticus were tested
on the first or second day of life. An additional 40 pups from 4 litters of M. musculus and M.
domesticus were tested in the control condition on their first day of life.
(b) Procedure
We assessed differential interest of newborn mice in two-choice tests using odours of
adult lactating females. The 2 odour donors for each test were selected based on their
genealogy to be comparably related to the pups’ mother and to have given birth as closely
together as possible, usually within 1 day but never more than 3 days apart. Odours were
collected by rubbing one edge of a small (3-cm diameter) plastic Petri dish back and forth 8
times along both sides of the mouse’s ventrum including the nipples but avoiding the ano-
genital area. The odorous edges of the 2 dishes (randomly coded as A or B) were placed 2.5-
cm apart equidistant from a line drawn on clean tissue paper. Pups were tested in warm
conditions (around 30 °C, comparable to the nest temperature). Subjects were positioned on
the line 4 cm from the odours, slid gently (anchored between the experimenter’s thumb and
index finger) up the line until the nose was in the centre between the dishes, and held until
their head moved enough that their nose touched one of the dishes, constituting the choice.
Choosing times varied between a few seconds and less than 2 min. Half of the pups from each
litter were removed to a holding cage and tested individually; then the position of the odours
was reversed and the second half of the litter was removed from the home cage and tested.
This procedure also ensures that mothers were never left without young in the nest and pups
were not left without other pups near them. The experimenters wore latex gloves during
preparation of the stimuli and testing and were blind to the position of the odours during the
test. All experiments followed the rules of French Ministries (Agriculture; Research and
Technology).
(c) Data Analysis
Each litter was treated as a statistical unit. The proportion of litters in which pups
chose the more closely related female was tested for significance with binomial tests. (No
preferential choice was shown in control tests when the odour of the same unfamiliar and
unrelated lactating female was presented on both sides: 21 left vs. 19 right.)
96

Chapitre II: Mécanismes d'évaluation de la proximité génétique
3. RESULTS AND DISCUSSION
In this investigation of odour-based genetic relatedness discrimination in newborn
mice, more pups chose the odour of the more genetically similar female in all the litters
tested. Mouse pups preferred the odour of their lactating paternal aunt to the odour of an
unrelated lactating female (Table 1; Mus spicilegus: n = 11 litters, P = 0.001, corresponding to
54 out of 69 pups; Mus musculus complex: n = 6, P = 0.031, and 31out of 39 pups), indicating
that pups discriminate between odours of adults on the basis of genetic relatedness. It was
possible in these tests, however, that the pups learned enough about their father’s odour
during the first few hours of life to facilitate the discrimination between the father’s sister and
a female that was not related to either of the parents. In a key test, however, M. spicilegus
newborns (less than 16 hours old) from litters that were born in cages from which the father
and his odours had been removed at least 2 days prior to their birth also preferred odours from
their lactating paternal aunts (Table 1; n = 8, P = 0.008, and 56 out of 66 pups). Thus the same
pattern was found whether the father was present or not. Postnatal exposure to the mother’s
lactation odour could not have helped pups to discriminate between odours of lactating
females that were comparably unrelated to their mother. These results provide the first direct
and non-confounded demonstration of the ability of newborns to discriminate between odours
of kin and non-kin without the necessity of prenatal or postnatal learning about their parents.
It is a clear demonstration of the remarkable ability of newborns to make genetic relatedness
discriminations on the basis of odours.
These odour-based genetic relatedness distinctions were not restricted to kinship
discrimination, however. In tests using odour donors that were conspecifics and
heterospecifics, newborn M. musculus and M. domesticus mice preferred the odour of a
lactating female from a different population of conspecifics to that of a heterospecific
lactating female (Table 1; n = 7, P < 0.016, and 39 out of 50 pups), demonstrating that a wide
range of genetic relatedness distinctions is possible from birth. Although using the mother’s
odour as a referent could have helped the newborns make the discrimination between
conspecifics and heterospecifics, it is interesting to note that in previous tests with adults
raised in mixed litters of M. musculus and M. spicilegus, the odour of the rearing lactating
female did not affect subjects’ preference for the odour of the more genetically similar
individual (Heth et al., 2003). It is also surprising, given the greater differences between the
donors’ odours in this test, that the effect was no stronger for discrimination between
conspecific and heterospecific than for aunt as opposed to unrelated conspecific.
97

Chapitre II: Mécanismes d'évaluation de la proximité génétique
With the experimental design used in this study, it is not possible to rule out effects of
environmental odours, including odours of littermates, but we have shown in previous tests
with mice from these species that were cross-fostered within a few hours of birth that
environmental odours, including odours of fostered littermates, did not affect their genetic
relatedness assessments (Heth et al., 2003), thus it seems unlikely that environmental
odours affected the discriminative abilities or choices of newborns.
In previous self-referencing studies (reviewed in Todrank & Heth, 2003) that involved
adult rodent subjects, the discrimination tasks used odours from other adults. In those
studies, subjects were making comparisons between odours of animals that were all the
same age. In the current study, the subject’s own odour was that of a newborn while the
test odours were from lactating adult females, probably posing a more challenging task for
the subjects. Nonetheless, newborn mice completed the discrimination task successfully.
When the findings from the odour-genes covariance studies were combined with the
results of the self-referencing studies, it became clear that the genetic relatedness assessment
mechanism being used was different from the traditionally conceived kin recognition
mechanisms (Todrank & Heth, 2003). First, because more closely related individuals produce
odours that have “relatively” more similar qualities (whether they are double cousins
compared with cousins (Busquet & Baudoin, in press) or conspecifics from different
populations compared with heterospecifics (Heth et al., 2001) or closer as opposed to more
distant heterospecifics (Heth & Todrank, 2000)), the distinctions could be quite subtle.
Second, the range across which rodents are able to make subtle genetic relatedness
distinctions precludes simple categorical distinctions such as between kin and non-kin or
conspecifics and heterospecifics. To distinguish it from self-referent phenotype matching, we
called the mechanism based on odour-genes covariance “genetic relatedness assessment
through individual odour similarities” and abbreviated it “G-ratios” (Heth et al., 2003).
Previous evidence indicates that the G-ratios mechanism in adult rodents is not affected by
learning the individual odours of nest mates (Todrank & Heth, 2003), but the ontogeny of this
process had not been explored previously. The current results extend what had previously
been demonstrated only in adult rodents to include newborns as well.
Exposure to odours during gestation is known to influence postnatal behaviour in
some rodent species (e.g., Hepper, 1987b, 1988; Beauchamp et al., 1994; Hudson et al., 1999;
Gruest et al., 2004). Determining whether exposure to littermates during gestation or
98

Chapitre II: Mécanismes d'évaluation de la proximité génétique
immediately after birth influences or is necessary for successful genetic relatedness
distinctions would require embryo-transplant experiments. Nonetheless, it is clear from the
evidence presented here that prenatal and postnatal experience with parental odours are not
necessary for newborns to make genetic relatedness discriminations at the levels of kinship
and species.
Acknowledgments:
We thank Dr. G. Ganem, Laboratoire de Génétique et Environnement, ISEM, Université
Montpellier II for providing animals and research space, P. Gouat for consultation on animal
genealogies, and S. Demouron and M. Perriat for technical assistance.
REFERENCES:
Bateson, P. (1983). Optimal outbreeding. In P. P. G. Bateson, (Ed.) Mate choice. (pp. 257-
277). Cambridge University Press, Cambridge.
Beauchamp, G. K., Yamazaki, K., Curran, M., Bard, J. & Boyse, E. A. (1994). Fetal odour
types are evident in the urine of pregnant female mice. Immunogenetics, 39, 109-113.
Busquet, N. & Baudoin, C. Odour similarities as a basis for discriminating degrees of kinship
in rodents: evidence from Mus spicilegus. Animal Behaviour (in press).
Edmands, S. (2002). Does parental divergence predict reproductive compatibility ? Trends in
Ecology & Evolution, 17, 520-527.
Fletcher, D. J. C. & Michener, C. D. (Eds.) (1987). Kin recognition in animals. Chichester:
John Wiley & Sons.
Gruest, N., Richer, P. & Hars, B. (2004). Emergence of long-term memory for conditioned
aversion in the rat fetus. Developmental Psychobiology, 44, 189-198.
Hamilton, W. D. (1964). The genetical evolution of social behaviour. I. II. Journal of
Theoretical Biology, 7, 1-52.
99

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Hauber, M. E. & Sherman, P. W. (2001). Self-referent phenotype matching: theoretical
considerations and empirical evidence. Trends in Neuroscience, 24, 609-616.
Hepper, P. G. (1983). Sibling recognition in the rat. Animal Behaviour, 31, 1177-1191.
Hepper, P. G. (1987a). The discrimination of different degrees of relatedness in the rat:
evidence for a genetic identifier ? Animal Behaviour, 35, 549-554.
Hepper, P. G. (1987b). The amniotic fluid: an important priming role in kin recognition.
Animal Behaviour, 35, 1343-1346.
Hepper, P. G. (1988). Adaptive fetal learning: Prenatal exposure to garlic affects postnatal
preferences. Animal Behaviour, 36, 935-936.
Hepper, P. G. (Ed.) (1991). Kin Recognition. Cambridge: Cambridge University Press.
Heth, G. & Todrank, J. (2000). Individual odour similarities across species parallel
phylogenetic relationships in the S. ehrenbergi superspecies of mole rats. Animal Behaviour,
60, 789-795.
Heth, G., Todrank, J. & Johnston, R. E. (1998). Kin recognition in golden hamsters: evidence
for phenotype matching. Animal Behaviour, 56, 409-417.
Heth, G., Todrank, J., Busquet, N. and Baudoin, C. (2001). Odour-genes covariance and
differential investigation of individual odours in the Mus species complex. Biological Journal
of the Linnean Society, 73, 213-220.
Heth, G., Todrank, J., Busquet, N. & Baudoin, C. (2003). Genetic relatedness assessment
through individual odour similarities (G-ratios) in mice. Biological Journal of the Linnean
Society, 78, 595-603.
Holmes, W. G. & Sherman, P. W. (1982). The ontogeny of kin recognition in female
Belding’s ground squirrels. American Zoolologist, 22, 38-47.
Hudson, R., Schaal, B. & Bilko, A. (1999). Transmission of olfactory information from
mother to young in the European rabbit. In H. O. Box & K. R. Gibson, (Eds.) Mammalian
100

Chapitre II: Mécanismes d'évaluation de la proximité génétique
social learning: Comparative and ecological perspectives (pp. 141-157). Cambridge
University Press, Cambridge.
Koeninger Ryan, K. & Lacy, R. C. (2003). Monogamous male mice bias behaviour towards
females according to very small differences in kinship. Animal Behaviour, 65, 379-384.
Mateo, J. M. (2002). Kin recognition abilities and nepotism as a function of sociality.
Proceedings of the Royal Society London B, 269, 721-727.
Patris, B. & Baudoin, C. (2000). A comparative study of parental care between two rodent
species: implications for the mating system of the mound-building mouse Mus spicilegus.
Behavioural Processes, 51, 35-43.
Sherman, P. W., Reeve, H. K. & Pfenning, D. W. (1997). Recognition Systems. In J. R. Krebs
& N. B. Davies (Eds.) Behavioural Ecology: A Behavioural Approach (pp. 69-6) Blackwell,
London.
Smadja, C., Catalan, J. & Ganem, G. (2004). Strong premating divergence in a unimodal
hybrid zone between two subspecies of the house mouse. Journal of Evolutionary Biology,
117, 165-176.
Tang-Martinez, Z. (2001). The mechanisms of kin discrimination and the evolution of kin
recognition in vertebrates: A critical re-evaluation. Behavioural Processes, 53, 21-40.
Thornhill, N.W. (Ed.) (1993). The Natural History of Inbreeding and Outbreeding. Chicago:
University of Chicago Press.
Todrank, J. & Heth, G. (2001). Re-thinking cross-fostering designs for studying kin
recognition mechanisms. Animal Behaviour, 61, 503-505.
Todrank, J. & Heth, G. (2003). Odor-genes covariance and genetic relatedness assessments:
Rethinking odor-based “recognition” mechanisms in rodents. Advances in the Study
of Behavior, 32, 77-130.
101

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Todrank, J., Heth, G. & Johnston, R. E. (1998). Kin recognition in golden hamsters: evidence
for kinship odours. Animal Behaviour, 55, 377-386.
Todrank, J., Heth, G. & Johnston, R. E. (1999). Social interaction is necessary
for discrimination and memory for odours of close relatives in golden hamsters. Ethology,
105, 771-782.
Tregenza, T. & Wedell, N. (2000). Genetic compatibility, mate choice and patterns
of parentage: Invited review. Molecular Ecology, 9, 1013-1027.
102
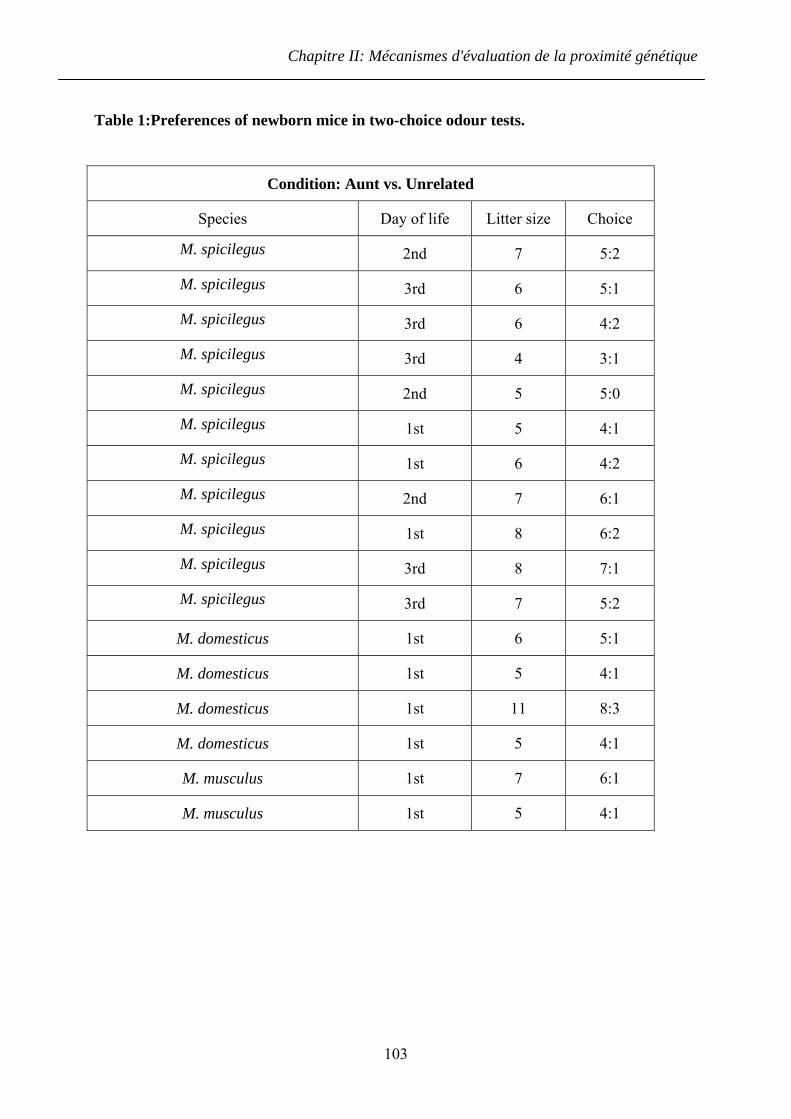
Chapitre II: Mécanismes d'évaluation de la proximité génétique
Table 1:Preferences of newborn mice in two-choice odour tests.
Condition: Aunt vs. Unrelated
Species Day of life Litter size Choice
M. spicilegus 2nd 7 5:2
M. spicilegus 3rd 6 5:1
M. spicilegus 3rd 6 4:2
M. spicilegus 3rd 4 3:1
M. spicilegus 2nd 5 5:0
M. spicilegus 1st 5 4:1
M. spicilegus 1st 6 4:2
M. spicilegus 2nd 7 6:1
M. spicilegus 1st 8 6:2
M. spicilegus 3rd 8 7:1
M. spicilegus 3rd 7 5:2
M. domesticus 1st 6 5:1
M. domesticus 1st 5 4:1
M. domesticus 1st 11 8:3
M. domesticus 1st 5 4:1
M. musculus 1st 7 6:1
M. musculus 1st 5 4:1
103
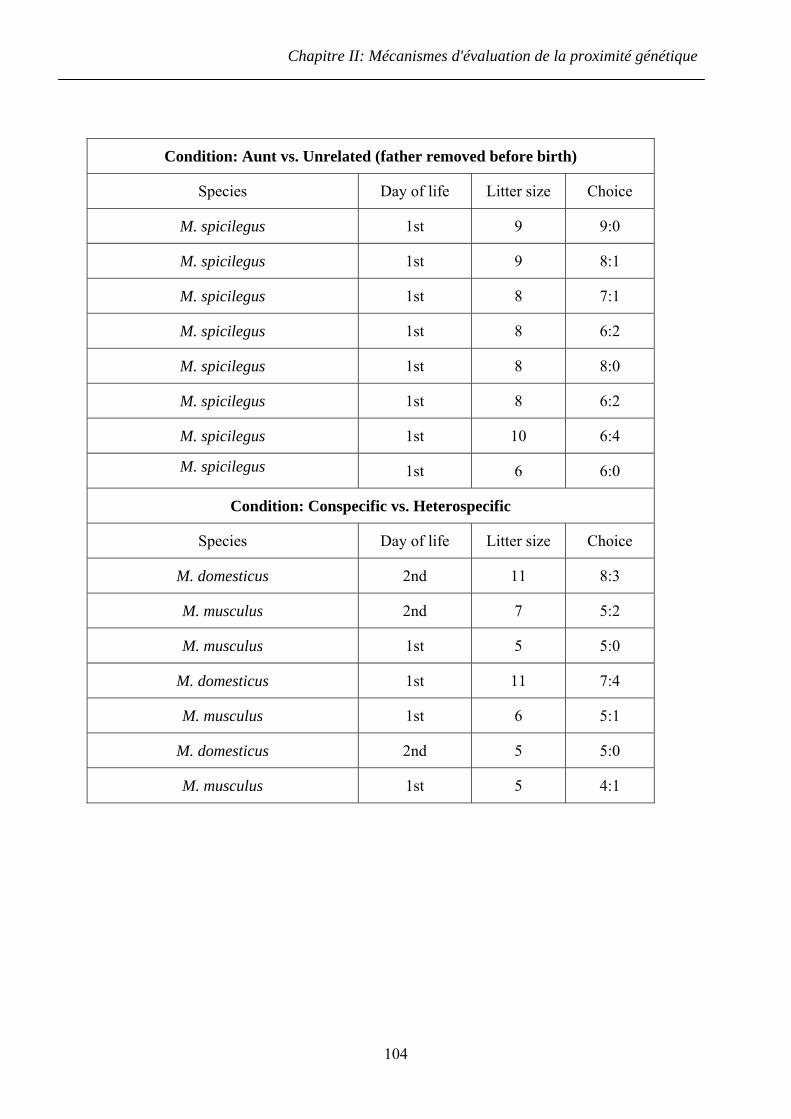
Chapitre II: Mécanismes d'évaluation de la proximité génétique
Condition: Aunt vs. Unrelated (father removed before birth)
Species Day of life Litter size Choice
M. spicilegus 1st 9 9:0
M. spicilegus 1st 9 8:1
M. spicilegus 1st 8 7:1
M. spicilegus 1st 8 6:2
M. spicilegus 1st 8 8:0
M. spicilegus 1st 8 6:2
M. spicilegus 1st 10 6:4
M. spicilegus 1st 6 6:0
Condition: Conspecific vs. Heterospecific
Species Day of life Litter size Choice
M. domesticus 2nd 11 8:3
M. musculus 2nd 7 5:2
M. musculus 1st 5 5:0
M. domesticus 1st 11 7:4
M. musculus 1st 6 5:1
M. domesticus 2nd 5 5:0
M. musculus 1st 5 4:1
104

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Discussion générale : l'évaluation de la proximité génétique
Les expériences présentées dans ce chapitre ont apporté des éléments significatifs dans la
compréhension du mécanisme d'évaluation de la proximité génétique. Tout d'abord, cette
évaluation est possible, puisque les Souris peuvent réagir par rapport à la proximité génétique
du donneur et ce, en l'absence de familiarité avec ce donneur. Il existe une reconnaissance de
parentèle qui dépasse la simple reconnaissance des apparentés familiers.
Le substrat de cette évaluation de la proximité génétique est lié à la similarité des productions
odorantes du sujet et du donneur d’odeurs (Heth et al., 2003). Cette évaluation de la proximité
génétique dépasse le cadre spécifique, ce qui rappelle l'argument de Grafen qui voyait le
traitement différentiel des apparentés non pas comme une réelle reconnaissance de parentèle
mais comme un "sur fonctionnement", une conséquence de la reconnaissance spécifique
(Grafen, 1990). En proposant cette interprétation, Grafen souhaitait dénoncer la dérive
téléologique consistant à attribuer à n'importe quel traitement différentiel des apparentés la
fonction historique (voir introduction générale) de reconnaissance de parentèle. Autrement
dit, selon Grafen, il existe des pressions de sélection pour reconnaître l'espèce qui amènent à
sélectionner un mécanisme de reconnaissance spécifique et ce mécanisme nouveau permet
également la reconnaissance de parentèle, par cooptation. Le postulat de ce raisonnement est
que la reconnaissance spécifique est une adaptation, ce qui pose problème. En effet, les
arguments de Grafen sur la reconnaissance de parentèle sont applicables à la reconnaissance
spécifique : on ne peut parler de reconnaissance spécifique "vraie" que si l'on peut montrer
qu'elle est apparue en réponse à des pressions de sélection chez des organismes à qui
manquait cette capacité. Ce point crucial sera exposé plus avant dans la discussion générale de
cette thèse. L'évaluation de la proximité génétique par la similarité des odeurs est en effet un
mécanisme qui permet la reconnaissance de parentèle et la reconnaissance spécifique mais
dont "l'origine évolutive" reste floue et soulève de nombreuses questions.
Intuitivement, l'évaluation de la proximité génétique montrée par les Souris semble être un
processus de traitement de l'information "de bas niveau". La capacité des nouveau-nés à réagir
en fonction de l'apparentement apparaît en effet difficilement conciliable avec l'utilisation de
processus cognitifs complexes tels que l'existence d'une catégorie mentale regroupant les
apparentés. Bien que rien ne permette d'éliminer ce type d'explication, il semble plus
parcimonieux d'envisager un processus plus simple.
105

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Les expériences sur les nouveau-nés ont permis de montrer que la reconnaissance de parentèle
était possible sans référence à l'odeur maternelle, puisque celle-ci est non apparentée aux deux
donneuses d'odeur. Ceci plaide pour un mécanisme auto référencé similaire à celui trouvé
chez les adultes (Heth et al., 2003) mais il subsiste une possibilité "d'alloréférence". En effet,
les petits sont en contact les uns avec les autres dans les heures qui suivent la naissance. De
plus, dans l'utérus maternel, les communications chimiques entre embryons - ou du moins les
échanges de molécules - sont possibles, même si chaque fœtus possède une poche amniotique
individuelle (C. Féron, comm. pers.). Outre le phénotype maternel, chaque nouveau-né est
donc familier avec des phénotypes d'apparentés. Cette familiarité n'est pas aisément
objectivable étant donné que le petit qui vient de naître n'est probablement pas à même de
distinguer les odeurs de ses frères et sœurs de ses propres productions odorantes. Le concept
d'habituation est peut-être plus adapté à ce contexte : les récepteurs olfactifs du sujet sont
habitués à une stimulation basale, mélange de molécules odorantes produites par l'animal lui-
même, par les frères et sœurs et par la mère. Du fait de la covariance odeurs-gènes, l'odeur de
la tante paternelle présente probablement quelques caractéristiques communes avec ce
"cocktail". Les nouveau-nés s'orienteraient alors préférentiellement vers cette odeur, qui est la
"moins nouvelle".
Les mécanismes explicatifs de cette discrimination comportementale par les nouveaux-nés
peuvent se situer à deux niveaux d'explication :
1) Au niveau sensoriel : l'appareil olfactif des nouveaux-nés est plus réceptif aux
stimulations provenant des apparentés. Les récepteurs olfactifs sont "programmés"
pour répondre préférentiellement aux signaux des apparentés.
2) Au niveau productif : l'appareil olfactif est "totipotent" mais a été exposé à de
productions odorantes d'apparentés (y compris lui-même), ce qui s'est traduit par une
habituation des récepteurs à ces signaux. Le traitement différentiel des apparentés est
une conséquence de cette habituation différentielle.
Ces deux hypothèses sont malheureusement difficilement distinguables : s'il est
théoriquement possible de soustraire une souris à l'exposition au phénotype d'apparentés, en
pratiquant le transfert d'un embryon unique dans l'utérus d'une femelle non apparentée, il
paraît en revanche difficile d'empêcher l'exposition des récepteurs olfactifs du sujet à ses
propres molécules.
106

Chapitre II: Mécanismes d'évaluation de la proximité génétique
Une autre manière d'aborder le problème est de modifier les odeurs auxquelles sont exposées
les souris en jouant sur les facteurs environnementaux. L'autre intérêt de cette manipulation de
facteurs externes (ou facteurs d'état) est d'évaluer la fiabilité du mécanisme d'évaluation de la
proximité génétique en situation de conflit avec d'autres facteurs. Cette étude de l'interaction
des facteurs génétiques (liés à l'apparentement) et des facteurs environnementaux est l'objet
du chapitre suivant.
107

Chapitre III: Odeurs, facteurs d’état et proximité génétique
Chapitre III :
Odeurs, facteurs d’état
et proximité génétique
108

Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.1. Introduction
Le chapitre I a démontré l’existence d’une covariance odeurs-gènes. Cette covariance est le
pré-requis à l’existence d’un mécanisme de reconnaissance de parentèle auto référencé
(cf. chapitre II). Cependant, il est important de garder à l’esprit que la covariance odeurs-
gènes a été démontrée en laboratoire. Les facteurs environnementaux y sont contrôlés et
standardisés. En nature, les animaux sont confrontés à des conditions environnementales
variables dans le temps mais aussi dans l’espace. Leur métabolisme s’en trouve affecté et par
conséquent leur odeur individuelle doit être modifiée. De plus, le régime alimentaire, s’il est
globalement le même, subit nécessairement des variations interindividuelles dues à la
disponibilité des ressources voire à l’expression de préférences individuelles. Enfin, un
individu, au cours de son existence, va voir son métabolisme changer en fonction de son âge,
de son statut hormonal, de sa charge parasitaire etc. Les Souris sont donc susceptibles de
rencontrer des congénères non familiers qui, outre leur apparentement, vont présenter des
caractéristiques provenant de leur histoire personnelle, proche ou même lointaine. Si
l’apparentement semble détectable en laboratoire, grâce au phénomène de la covariance
odeurs-gènes, il paraît intéressant de se demander comment les fluctuations d’origine
environnementale de l’odeur individuelle vont interagir avec ce phénomène. Il existe deux
versants à cette question. Le premier concerne les odeurs individuelles elles-mêmes et
l’éventail des variations qu’elles peuvent subir. Le second concerne les modulations des
préférences olfactives des Souris liées à ces facteurs environnementaux. C’est ce second
aspect qui a été privilégié, car à travers lui, c’est la robustesse et la fonctionnalité de la
reconnaissance de parentèle qui est examinée. De plus, dans l'étude des mécanismes de la
reconnaissance de parentèle (et de l'évaluation de la proximité génétique en général), certaines
hypothèses font état de l'utilisation de l'odeur individuelle comme référent. Cette odeur peut-
être soit l'odeur propre de l'animal (self referent phenotype matching ou armpit effect), soit
l'odeur des congénères (phenotype matching), soit un mélange des deux (cf. chapitre II). De
plus, cette odeur peut être apprise, comme c'est le cas dans l'assortiment phénotypique, mais
aussi utilisée de façon instantanée (Todrank & Heth, 2003). Un des moyens d'estimer la
proximité génétique est d'utiliser la similarité des odeurs individuelles (Heth et al., 2003). Or
ces odeurs individuelles sont susceptibles d'être influencées par les facteurs
environnementaux (Beauchamp & Yamazaki, 2003). Si le phénotype est modifié, alors le
résultat de ces comparaisons phénotypiques peut lui aussi être modifié. En revanche, la
109

Chapitre III: Odeurs, facteurs d’état et proximité génétique
reconnaissance de parentèle par l'utilisation d'allèles de reconnaissance ne doit pas être
influencée par les variations de l'état du sujet. La recherche d'éventuelles modifications des
préférences des Souris à la suite de modifications environnementales des odeurs permet donc
théoriquement d'élucider plus avant les mécanismes de reconnaissance de parentèle.
Les préférences olfactives des Souris ont donc été examinées dans le contexte de la sélection
sexuelle, en se basant sur les résultats présentés dans le chapitre II. Les souris mâles
confrontées à deux odeurs de femelles en diœstrus passent plus de temps à sentir l’odeur
d’une femelle non apparentée qu'à sentir celle d'une soeur. Cette "préférence" pour une
femelle non apparentée, si elle se traduit dans le choix réel du partenaire, aura des
conséquences bénéfiques sur la fitness de la descendance. Il paraît donc important qu’elle soit
robuste, même quand les odeurs sont affectées par des facteurs d’état. Ces facteurs d’état ne
sont pas tous équivalents : selon leur nature, ils affectent plus ou moins la qualité du
partenaire potentiel. La reconnaissance de parentèle sur la base des odeurs individuelles a
donc été étudiée en interaction avec un facteur théoriquement peu lié à la qualité du
partenaire, le régime alimentaire. Dans un second temps, un facteur plus important dans le
choix de partenaire, l’âge, a été étudié.
3.2. Reconnaissance de parentèle et régime alimentaire
Le régime alimentaire est un facteur d’état dont les variations sont réversibles à court terme :
ces conséquences en terme de fitness sont relativement faibles. D’autre part, le régime
alimentaire est perceptible chez la Souris domestique. Moles & D'Amato (2000) ont montré
que les vocalisations ultrasoniques de femelles Mus domesticus (souche NMRI) lors de la
rencontre avec un congénère sont affectées par le régime alimentaire de celui-ci et par l’état
motivationnel du sujet. Ainsi une femelle affamée vocalise en présence d’un congénère
nourri, indépendamment du type de nourriture, alors qu’une femelle rassasiée ne vocalise que
si le congénère a ingéré des aliments appétants. Ce travail permet d’inférer une capacité à
distinguer le type de régime alimentaire sur la base d’informations olfactives. Bien sûr, les
souris de cette étude interagissaient directement et recevaient donc des informations plus
complexes que la simple odeur individuelle, mais la transcription en odeur du régime
alimentaire est l’explication la plus logique et la plus parcimonieuse.
110

Chapitre III: Odeurs, facteurs d’état et proximité génétique
D’autres études se sont intéressées au régime alimentaire et aux odeurs chez les Rongeurs.
Les campagnols mâles Microtus pennsylvanicus sont plus intéressés par l’odeur d’une femelle
ayant ingéré de la nourriture à forte teneur en protéines que par l’odeur d’une femelle ayant
reçu un régime faible en protéines mais néanmoins suffisant pour maintenir une activité
reproductrice. Les femelles montrent la même préférence pour l’odeur du mâle ayant un
régime à fort taux de protéines (Ferkin et al., 1997). Cette préférence est interprétée par les
auteurs comme un choix du partenaire le plus intéressant, car en situation de compétition pour
les ressources alimentaires en nature, l’animal ayant ingéré de la nourriture très énergétique
"démontre" ainsi sa capacité à se procurer cette nourriture et signale donc sa qualité. Le
régime alimentaire peut également jouer un rôle dans un contexte différent du choix de
partenaire, la relation mère-jeune. Chez les Souris épineuses Acomys cahirinus, les jeunes
préfèrent l'odeur d'une femelle nourrie avec le régime maternel à l'odeur d'une femelle nourrie
différemment. De plus, les jeunes montrent cette préférence même si la femelle nourrie
comme la mère biologique est d'une espèce différente et assez éloignée (Mus domesticus) et
que l'autre femelle proposée (et nourrie différemment) est conspécifique (Porter & Doane,
1977). Cette expérience suggère que des variables environnementales comme l'alimentation
peuvent induire des choix préférentiels, même vis-à-vis d'une autre espèce. Il semble donc
que le régime alimentaire soit perceptible dans les productions odorantes des Rongeurs, ce qui
en fait un paramètre intéressant à étudier en interaction avec la reconnaissance de parentèle.
Comme expliqué dans le chapitre I, le MHC est considéré par de nombreux chercheurs
comme le support génétique de l’individualité et de l’apparentement. Dans ce contexte,
Brown et al. (1996) se sont intéressés aux rôles respectifs de l’alimentation et du MHC dans la
production de l’odeur individuelle. Pour ce faire, Ils ont utilisé un processus de
conditionnement opérant appliqué à des rats dont la tâche était de discriminer les odeurs de
souris de laboratoire de deux souches variant à un seul locus du MHC selon les modalités
suivantes : 1) MHC différent, même régime alimentaire ; 2) MHC différent, régime différent ;
3) MHC identique, régime identique ; 4) MHC identique, régime différent. Les rats
discriminent facilement les odeurs des situations 2) et 4), c’est-à-dire les odeurs de souris
ayant reçu une alimentation différente. L’apprentissage est plus difficile sur les tâches 1) et 3).
Pour cette dernière, les rats ne conservent pas l’apprentissage d’un jour à l’autre mais sont
capables de discriminer lors d’une même session, ce qui tend à montrer que des souris, bien
que génétiquement identiques et nourries pareillement, produisent des odeurs discriminables,
111

Chapitre III: Odeurs, facteurs d’état et proximité génétique
même par des rats. La plus grande facilité de l’apprentissage induite par la différence
d’alimentation entre les souris démontre la capacité à discriminer les odeurs d’animaux
nourris différemment mais aussi indique une plus grande saillance des informations liées au
régime alimentaire par rapport au informations liées à la différence génétique (très faible dans
cette expérience). Cette dichotomie entre les deux types d’informations - génétiques et
alimentaires - est très ambiguë. En effet, les deux phénomènes semblent liés : le MHC
détermine en partie la composition de la flore bactérienne chez les Souris (Toivanen et al.,
2001), et cette flore bactérienne joue sur les processus digestifs et donc la formation des
odeurs.
La covariance odeur-gènes démontrée chez Mus spicilegus (cf. chapitre 1) est donc
susceptible d’être modulée par l’alimentation. L’hypothèse d’une comparaison phénotypique
avec l’odeur propre de l’animal (self-referent phenotype matching) exposée au chapitre 2
permet de faire la prédiction suivante : les variations du phénotype odorant, induites par
exemple par un changement de régime alimentaire, sont de nature à modifier les préférences
olfactives des animaux ayant subi ces variations. Pour tester cette hypothèse, il est important
de choisir un facteur alimentaire n’affectant pas la qualité du partenaire proposé. Dans cette
expérience, deux variétés de pommes ont été utilisées, en partant du postulat que les
propriétés énergétiques de ces deux régimes alimentaires étaient similaires.
3.2.1 Matériels et méthodes
Les souris utilisées dans cette expérience ont été nourries avec la nourriture standard
complémentée de morceaux de pommes. Chaque animal recevait des morceaux de pomme de
la variété "Royal Gala" ou bien de la variété "Granny Smith" pendant une semaine avant le
début des tests et jusqu’à la fin de l’expérience. Les morceaux de pommes étaient renouvelés
tous les jours et il était vérifié que les morceaux précédents avaient été consommés. Durant
cette semaine, les mâles étaient isolés et les femelles laissées en petits groupes de deux ou
trois sœurs, de façon à prévenir la réceptivité sexuelle (Féron & Gheusi, 2003). Les tests sont
réalisés de façon similaire aux tests de préférences déjà exposés : les mâles sont transférés
dans une cage propre et sont mis en présence de deux odeurs de femelles recueillies sur une
boîte de Pétri frottée sur la zone uro-génitale. Pendant 5 minutes, le temps d’investigation est
chronométré par deux observateurs non informés de la nature des stimuli présentés. Trois tests
ont été réalisés :
112

Chapitre III: Odeurs, facteurs d’état et proximité génétique
- Le premier choix proposé au mâle se fait entre les odeurs de deux sœurs non
familières (non apparentées au sujet), l’une ayant reçu le même régime (la même variété de
pomme) que lui, l’autre ayant mangé l’autre variété de pomme. Les deux femelles sont issues
de la même portée séparée en deux groupes d’au moins deux femelles. Ainsi, la proximité
génétique avec le sujet est la même et la relative homogénéité du background permet de
limiter le risque d’une préférence spontanée pour l’une ou l’autre des deux femelles. Si le
choix du sujet est dépendant de son régime alimentaire, alors il est possible de conclure à une
perception du régime alimentaire par les Souris glaneuses, ainsi qu’à une discrimination très
fine de ce régime alimentaire, puisque seule la variété des pommes change.
- Le second test propose à l’animal testé le choix entre l’odeur d’une femelle
apparentée (une sœur d’une portée ultérieure) et une femelle non apparentée selon deux
modalités. Dans le 1er cas, le sujet et les donneuses d’odeurs ont reçu le même régime
alimentaire, tandis que dans le 2e cas, le régime alimentaire diffère entre le sujet et les deux
femelles. Cette deuxième modalité permet de tester l’hypothèse d’un masquage des
informations liées à l’apparentement par les informations liées au régime alimentaire.
- Le troisième test est conditionné par les résultats des deux précédents, puisqu’il
s’agit de mettre en conflit les éventuelles préférences mises en évidence dans ces tests. Si les
animaux préfèrent les femelles au régime alimentaire différent dans le 1er test et les non
apparentées dans le deuxième, alors le choix sera proposé entre l’odeur d’une femelle non
apparentée avec le même régime alimentaire que le sujet et l’odeur d’une femelle apparentée
avec un régime différent. C’est ce dernier test qui permet d’envisager l’interaction des
facteurs génétiques et alimentaires.
113
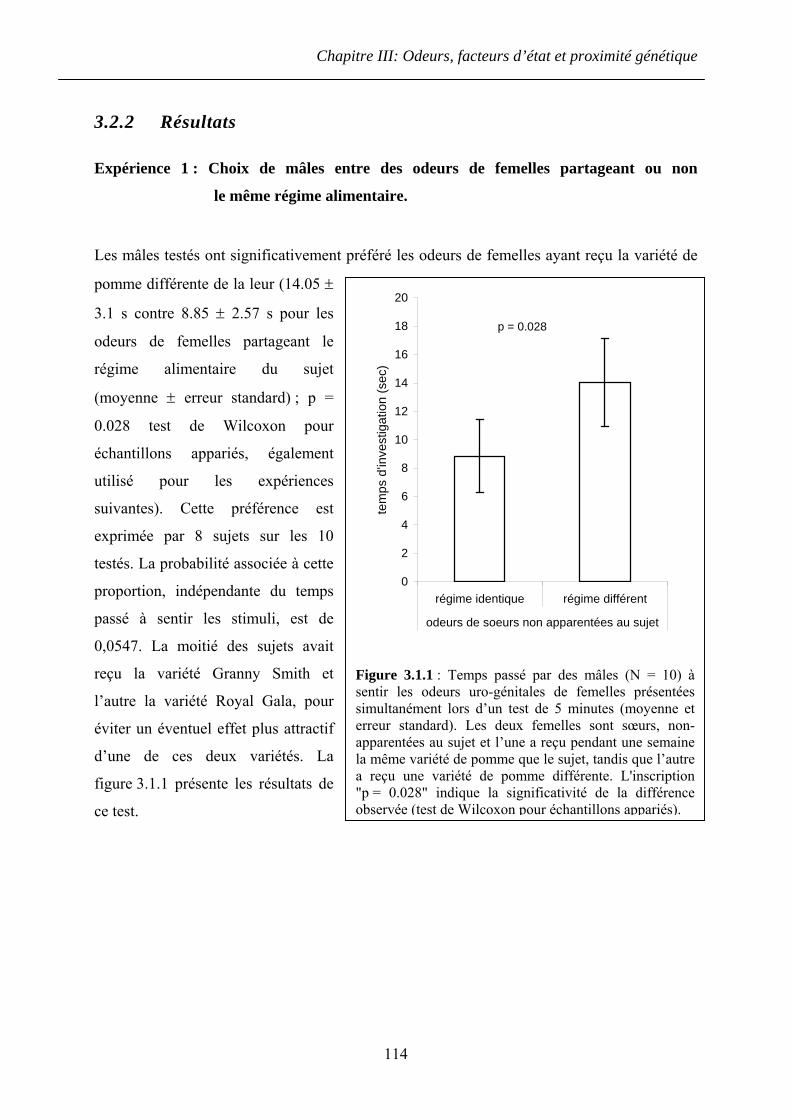
Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.2.2 Résultats
Expérience 1 : Choix de mâles entre des odeurs de femelles partageant ou non
le même régime alimentaire.
Les mâles testés ont significativement préféré les odeurs de femelles ayant reçu la variété de
pomme différente de la leur (14.05 ±
3.1 s contre 8.85 ± 2.57 s pour les
odeurs de femelles partageant le
régime alimentaire du sujet
(moyenne ± erreur standard) ; p =
0.028 test de Wilcoxon pour
échantillons appariés, également
utilisé pour les expériences
suivantes). Cette préférence est
exprimée par 8 sujets sur les 10
testés. La probabilité associée à cette
proportion, indépendante du temps
passé à sentir les stimuli, est de
0,0547. La moitié des sujets avait
reçu la variété Granny Smith et
l’autre la variété Royal Gala, pour
éviter un éventuel effet plus attractif
d’une de ces deux variétés. La
figure 3.1.1 présente les résultats de
ce test.
0
2
4
6
8
10
12
14
16
18
20
régime identique régime différent
odeurs de soeurs non apparentées au sujet
tem
ps d
'inve
stig
atio
n (s
ec)
p = 0.028
Figure 3.1.1 : Temps passé par des mâles (N = 10) à sentir les odeurs uro-génitales de femelles présentées simultanément lors d’un test de 5 minutes (moyenne eterreur standard). Les deux femelles sont sœurs, non-apparentées au sujet et l’une a reçu pendant une semaine la même variété de pomme que le sujet, tandis que l’autrea reçu une variété de pomme différente. L'inscription"p = 0.028" indique la significativité de la différence observée (test de Wilcoxon pour échantillons appariés).
114

Chapitre III: Odeurs, facteurs d’état et proximité génétique
Expérience 2a : choix de mâles entre des odeurs de femelles apparentées ou non
ayant toutes deux le même régime alimentaire que le sujet
Les mâles testés ne montrent pas de préférence claire pour l’une ou l’autre des deux
catégories de stimuli présentés. Les odeurs de sœurs sont investiguées en moyenne 7,98
secondes contre 7,67 s pour les odeurs de femelles non apparentées. Sur les 10 mâles testés,
6 passent plus de temps à sentir l’odeur de leur sœur non familière (p = 0,386).
Expérience 2b : Choix de mâles entre des odeurs de femelles apparentées ou non
ayant un régime alimentaire différent de celui du sujet
Lorsque les deux femelles donneuses d’odeurs sont nourries avec une variété de pomme
différente de celle du mâle, ceux-ci sont plus intéressés par l’odeur de la moins apparentée des
deux (8 mâles sur 10). Cette préférence n’est toutefois pas significative (p = 0,074, test de
Wilcoxon). La probabilité exacte d’avoir 8 sujets sur 10 préférant la même catégorie d’odeurs
selon l’hypothèse nulle d’attractivité égale des deux stimuli est de 0,0547. Les résultats des
expériences 2a et 2b sont présentés dans la figure 3.1.2.
0
2
4
6
8
10
12
régime similaire à celui du sujet régime différent de celui du sujet
tem
ps d
'inve
stig
atio
n (s
ec)
sœur non familière
femelle non apparentée
p = 0.386 p = 0.0742a 2b
Figure 3.1.2 : Temps passé par des mâles à sentir les odeurs uro-génitales de femelles présentées simultanément lors d’un test de 5 minutes (moyenne et erreur standard). L’une des femelles est unesœur non familière du sujet, l’autre une femelle non familière non apparentée au sujet. Dans le test 2a,les deux donneuses d’odeurs ont reçu pendant une semaine la même variété de pomme que le sujet(N = 10). Dans le test 2b, les deux femelles ont reçu le même régime alimentaire, différent de celui du sujet (N = 10). Les valeurs de probabilité données par le test de Wilcoxon pour échantillons appariéssont indiquées pour chacun des deux tests.
115
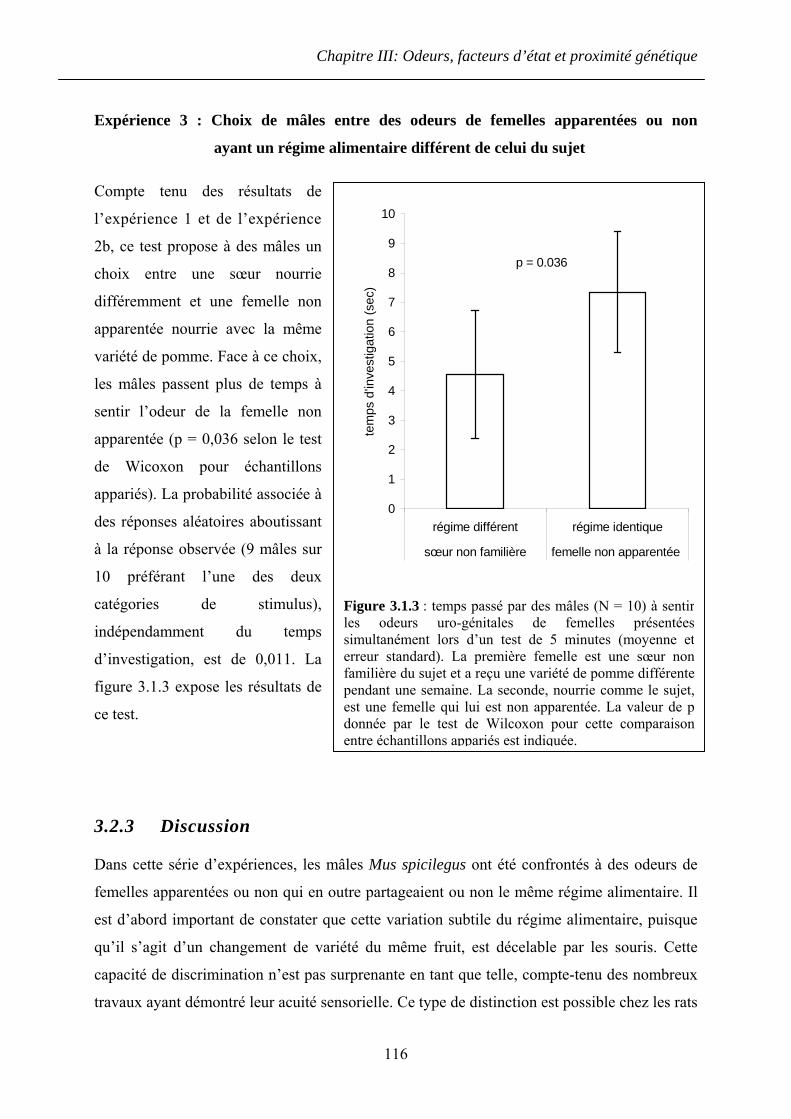
Chapitre III: Odeurs, facteurs d’état et proximité génétique
Expérience 3 : Choix de mâles entre des odeurs de femelles apparentées ou non
ayant un régime alimentaire différent de celui du sujet
Compte tenu des résultats de
l’expérience 1 et de l’expérience
2b, ce test propose à des mâles un
choix entre une sœur nourrie
différemment et une femelle non
apparentée nourrie avec la même
variété de pomme. Face à ce choix,
les mâles passent plus de temps à
sentir l’odeur de la femelle non
apparentée (p = 0,036 selon le test
de Wicoxon pour échantillons
appariés). La probabilité associée à
des réponses aléatoires aboutissant
à la réponse observée (9 mâles sur
10 préférant l’une des deux
catégories de stimulus),
indépendamment du temps
d’investigation, est de 0,011. La
figure 3.1.3 expose les résultats de
ce test.
3.2.3 Discussion
Dans cette série d’expériences, les m
femelles apparentées ou non qui en o
est d’abord important de constater qu
qu’il s’agit d’un changement de var
capacité de discrimination n’est pas s
travaux ayant démontré leur acuité sen
0
1
2
3
4
5
6
7
8
9
10
régime différent régime identique
sœur non familière femelle non apparentée
tem
ps d
'inve
stig
atio
n (s
ec)
p = 0.036
Figure 3.1.3 : temps passé par des mâles (N = 10) à sentirles odeurs uro-génitales de femelles présentéessimultanément lors d’un test de 5 minutes (moyenne eterreur standard). La première femelle est une sœur nonfamilière du sujet et a reçu une variété de pomme différentependant une semaine. La seconde, nourrie comme le sujet,est une femelle qui lui est non apparentée. La valeur de pdonnée par le test de Wilcoxon pour cette comparaisonentre échantillons appariés est indiquée.
âles Mus spicilegus ont été confrontés à des odeurs de
utre partageaient ou non le même régime alimentaire. Il
e cette variation subtile du régime alimentaire, puisque
iété du même fruit, est décelable par les souris. Cette
urprenante en tant que telle, compte-tenu des nombreux
sorielle. Ce type de distinction est possible chez les rats
116

Chapitre III: Odeurs, facteurs d’état et proximité génétique
même quand les animaux donneurs d'odeurs sont génétiquement identiques (MacIntosh
Schellinck et al., 1992). Ce qui est peut-être plus surprenant, c’est que cette discrimination est
induite par le propre régime alimentaire du sujet. En effet, l’expérience 1 présente un choix
entre deux sœurs de la même portée, donc également intéressantes pour le mâle. La moitié des
sujets ayant consommé la variété Granny Smith et l’autre la Royal Gala, il est impossible
d’attribuer la discrimination observée à un plus grand intérêt pour l’une ou l’autre de ces
variétés. La préférence pour l'odeur de la femelle nourrie différemment d'eux-mêmes doit être
due à une forme de comparaison, soit avec l'odeur propre de l'animal, soit avec une trace
mnésique, un souvenir olfactif. L'odeur uro-génitale de femelles contient donc des
informations similaires à celles portées et/ou mémorisées par l'animal.
3.3. Parentèle et facteurs d'état liés à la fitness : l'âge
Pour poursuivre l'étude des interactions entre facteurs d'état et parentèle, il a semblé judicieux
de s'intéresser à un facteur important pour la fitness des animaux. En effet, dans la première
partie de ce chapitre, les préférences des mâles étaient dirigées de façon "adéquate" malgré les
variations de régime alimentaire. Le fait d'avoir consommé l'une ou l'autre des deux variétés
de pomme n'est pas associé à un gain de fitness. Certains facteurs traduits dans l'odeur
individuelle sont en revanche associés à une meilleure fitness : ainsi l'état de santé ; le statut
social et l'âge peuvent se traduire par des modifications importantes de l'odeur (Beauchamp &
Yamazaki, 2003). C'est ce dernier facteur, l'âge, qui a été choisi pour l'étude suivante.
Pourquoi est-ce un facteur fortement lié à la fitness ? Il existe plusieurs réponses qu'il
convient de présenter rapidement. Tout d'abord, en nature, atteindre un âge avancé, c'est avoir
fait la démonstration de sa résistance aux pressions de sélection, liées à la prédation, la
compétition, le parasitisme ou les conditions abiotiques. Un partenaire âgé est donc
théoriquement porteur de "bons gènes" qui lui ont permis cette survie et cette longévité
(Brooks & Kemp, 2001). En nature, une femelle qui choisirait un mâle âgé pourrait faire
bénéficier à sa progéniture de ces bons gènes. L'un des problèmes de cette théorie est qu'il est
difficile de dire si les femelles préfèrent les signaux révélateurs de l'âge parce qu'ils sont
corrélées à une meilleure fitness ou si elles préfèrent les signaux indicateurs de cette meilleure
fitness, signaux plus fréquents chez les individus âgés (Proulx et al., 2002). Une autre raison
pour une femelle de préférer s'accoupler avec un mâle âgé est leur implication plus grande
dans les soins parentaux, démontrée chez les oiseaux (Brooks & Kemp, 2001). Chez Mus
117

Chapitre III: Odeurs, facteurs d’état et proximité génétique
spicilegus où le père s'investit beaucoup dans l'élevage des jeunes (Patris & Baudoin, 2000),
l'efficacité de ces soins peut être importante (Il faut noter qu'il n'existe à ce jour aucune
indication d'une corrélation entre l'âge du père et les soins parentaux chez cette espèce).
Un partenaire âgé n'est cependant pas toujours préférable. Le potentiel reproducteur décroît
avec l'âge, chez les mâles comme chez les femelles. De plus, un individu âgé a plus de
chances d'avoir accumulé au cours de sa vie des mutations délétères (Brooks & Kemp, 2001).
Chez une espèce présentant un lien fort entre partenaires comme Mus spicilegus, une trop
grande différence d'âge entre les partenaires signifie que l'un des deux risque de mourir avant
l'autre: les reproductions successives seront interrompues, et le partenaire restant devra
retrouver un nouveau partenaire, ce qui représente un coût considérable. La femelle devra de
plus assurer seule l'élevage des jeunes si le mâle meurt peu après l’accouplement.
Avant de tester l'interaction du facteur âge avec les préférences olfactives liées à la parentèle,
il est important de vérifier que ce facteur âge est bien perceptible par les souris. Le
métabolisme devrait changer avec l'âge chez les Souris glaneuses, comme il le fait par
exemple chez les rats-taupes (O'Connor et al., 2002) et les souris de laboratoire (Osada et al.,
2003), et donc permettre une discrimination olfactive. D'autre part, il est intéressant de
prendre en compte l'âge du sujet en même temps que celui des donneurs d'odeurs. En effet, si
le phénotype varie bien avec l'âge de façon similaire pour tous les individus, alors la
préférence peut être influencée par la comparaison des phénotypes du sujet et des donneurs.
Enfin, les donneurs utilisés sont des frères et sœurs d'âge différent, obtenus en laboratoire de
couples stables s'étant reproduit pendant plus de quatre mois. Ce dernier point est capital car il
permet d'éviter la critique évoquée plus haut à la théorie de l'âge comme révélateur de bons
gènes : en utilisant des frères et sœurs comme donneurs, on s'assure qu'aucun des deux n'est
génétiquement plus intéressant pour le sujet. L'âge, via ses conséquences sur l'odeur
individuelle, devrait donc être le seul critère de choix pour les souris.
La première étape de ce travail est de vérifier s'il existe des préférences olfactives basées sur
l'âge des donneurs, et si ces préférences sont influencées par l'âge du sujet.
118

Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.3.1. Matériel et méthodes
Pour cette étude, 10 mâles âgés, 10 mâles jeunes, 10 femelles âgées et 10 femelles jeunes ont
été utilisés. Au moment du test, les souris âgées ont environ 8 mois et les jeunes sont âgés de
4 à 5 mois. Une telle différence d'âge correspond, en milieu naturel, à des cohortes très
différentes, nées au début ou à la fin de la période de reproduction. Cette différence d'âge peut
sembler faible, mais il faut garder à l'esprit que ces individus ont les mêmes parents. Il a donc
fallu sélectionner des couples ayant une activité reproductrice pendant plus de trois mois, ce
qui correspond à plus de quatre portées successives. De plus, il est également nécessaire de
disposer d'origines variées pour éviter la pseudo réplication.
Les mâles sont isolés au moins une semaine avant le début des expériences, et les femelles
sont maintenues en groupe unisexe depuis l'âge de 35 jours. Aucune de ces femelles n'a donc
eu d'expérience de la reproduction, et le regroupement inhibe les cycles oestriens (Féron &
Gheusi, 2003).
Le protocole utilisé est très similaire à celui présenté dans l'étude précédente. Le sujet est
transféré dans une cage propre et confronté à deux odeurs d'individus du sexe opposé, l'un du
même âge, l'autre d'un âge différent. Les deux donneurs d'odeurs, non familiers, sont non
apparentés au sujet. Les quatre tests sont donc :
• préférences de mâles âgés pour l'odeur d'une femelle âgée ou l'odeur d'une femelle jeune
• préférence de mâles jeunes confrontés au même choix que ci-dessus
• préférences de femelles âgées pour l'odeur d'un mâle âgé ou l'odeur d'un mâle jeune
• préférence de femelles jeunes confrontées au même choix que ci-dessus
L'éventuelle préférence est objectivée par la mesure du temps d'investigation de l'un et l'autre
des deux stimuli (des boîtes de Pétri frottées sur la zone uro-génitale des donneurs) pendant
5 minutes.
119
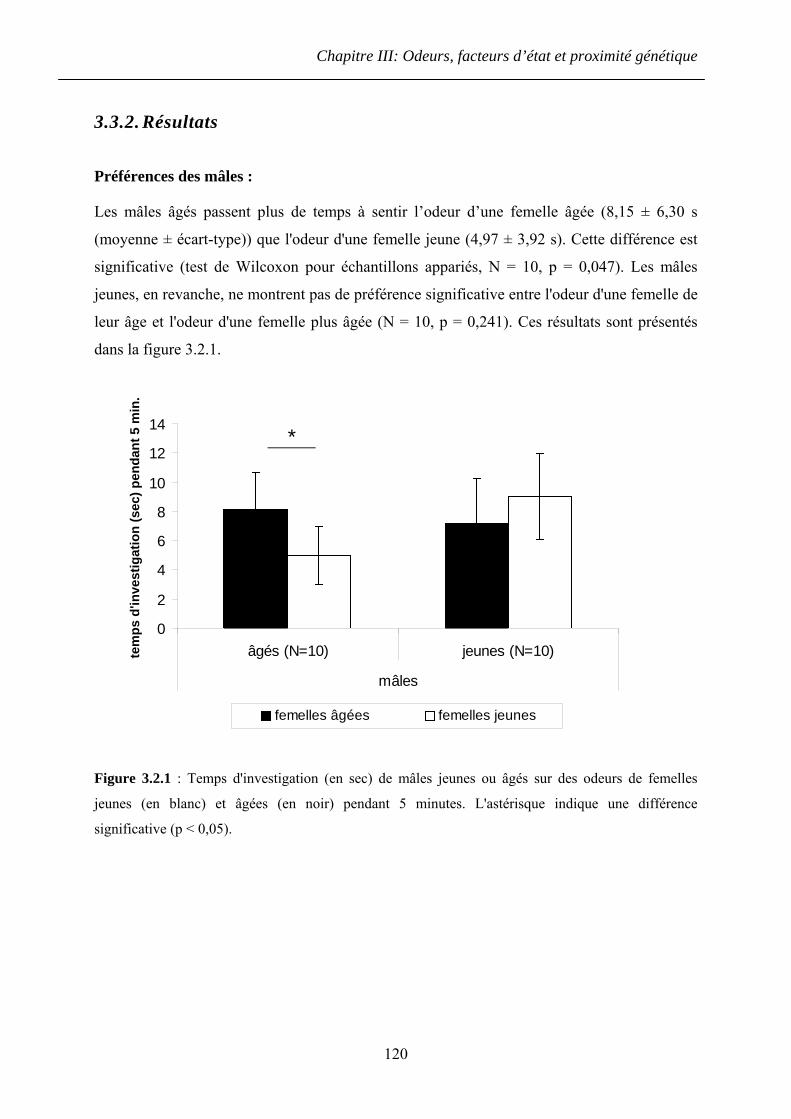
Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.3.2. Résultats
Préférences des mâles :
Les mâles âgés passent plus de temps à sentir l’odeur d’une femelle âgée (8,15 ± 6,30 s
(moyenne ± écart-type)) que l'odeur d'une femelle jeune (4,97 ± 3,92 s). Cette différence est
significative (test de Wilcoxon pour échantillons appariés, N = 10, p = 0,047). Les mâles
jeunes, en revanche, ne montrent pas de préférence significative entre l'odeur d'une femelle de
leur âge et l'odeur d'une femelle plus âgée (N = 10, p = 0,241). Ces résultats sont présentés
dans la figure 3.2.1.
0
2
4
6
8
10
12
14
âgés (N=10) jeunes (N=10)
mâles
tem
ps d
'inve
stig
atio
n (s
ec) p
enda
nt 5
min
.
femelles âgées femelles jeunes
______*
Figure 3.2.1 : Temps d'investigation (en sec) de mâles jeunes ou âgés sur des odeurs de femelles
jeunes (en blanc) et âgées (en noir) pendant 5 minutes. L'astérisque indique une différence
significative (p < 0,05).
120
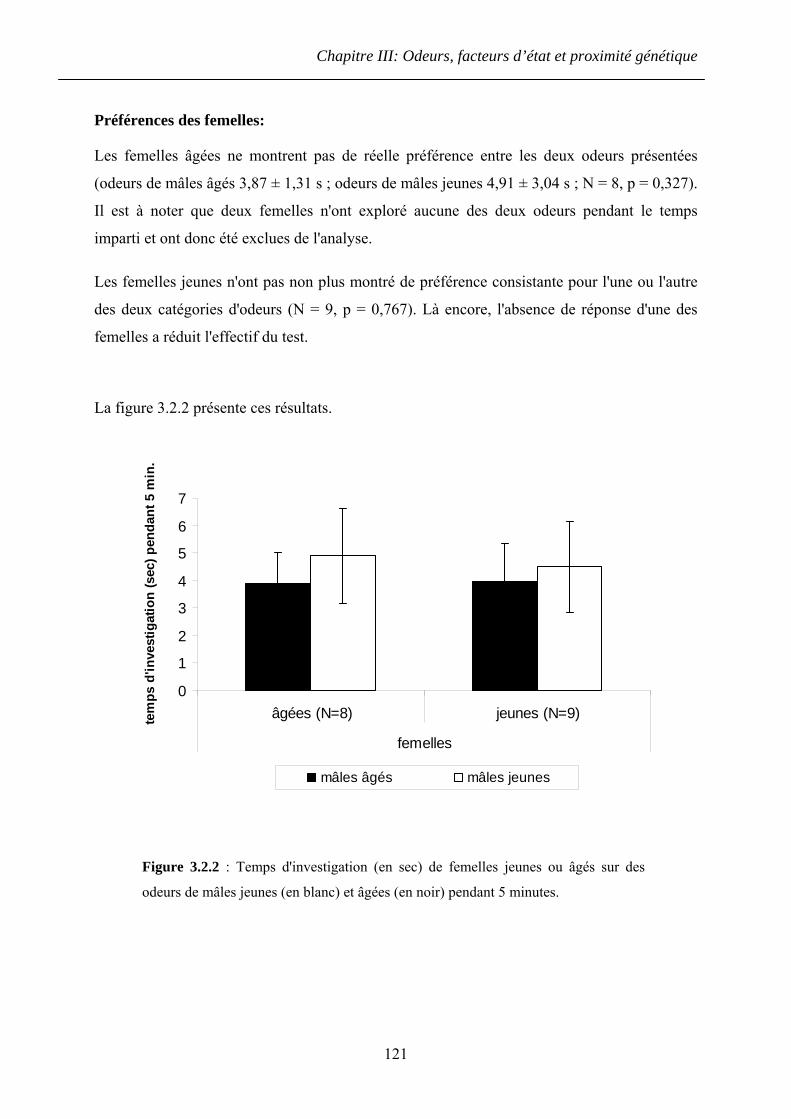
Chapitre III: Odeurs, facteurs d’état et proximité génétique
Préférences des femelles:
Les femelles âgées ne montrent pas de réelle préférence entre les deux odeurs présentées
(odeurs de mâles âgés 3,87 ± 1,31 s ; odeurs de mâles jeunes 4,91 ± 3,04 s ; N = 8, p = 0,327).
Il est à noter que deux femelles n'ont exploré aucune des deux odeurs pendant le temps
imparti et ont donc été exclues de l'analyse.
Les femelles jeunes n'ont pas non plus montré de préférence consistante pour l'une ou l'autre
des deux catégories d'odeurs (N = 9, p = 0,767). Là encore, l'absence de réponse d'une des
femelles a réduit l'effectif du test.
La figure 3.2.2 présente ces résultats.
0
1
2
3
4
5
6
7
âgées (N=8) jeunes (N=9)
femelles
tem
ps d
'inve
stig
atio
n (s
ec) p
enda
nt 5
min
.
mâles âgés mâles jeunes
Figure 3.2.2 : Temps d'investigation (en sec) de femelles jeunes ou âgés sur des
odeurs de mâles jeunes (en blanc) et âgées (en noir) pendant 5 minutes.
121

Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.3.3. Discussion
Les mâles âgés semblent bien montrer une préférence pour l'odeur d'une femelle du même âge
qu'eux par rapport à une femelle plus jeune. Ceci permet de déduire une capacité à discriminer
entre les deux catégories. Cette capacité de discrimination peut être attribuée aux
conséquences olfactives de l'âge via les changements métaboliques liés au vieillissement,
mais il n'est pas possible d'être catégorique sur ce point. En effet, les femelles utilisées font
partie de deux cohortes différentes : les femelles âgées sont nées au même moment de l'année
et faisaient partie de l'une des premières portées de leur mère. Les femelles jeunes sont
également nées au même moment, 3 à 4 mois plus tard que les femelles âgées, de mères
relativement âgées ayant déjà eu de nombreuses portées. Bien que la saisonnalité soit
contrôlée le plus possible en laboratoire, par l'application d'un éclairage artificiel, d'une
photopériode stable et d'une température constante, et qu'il n'y ait pas de référence
bibliographique traitant de changements du phénotype odorant lié à l'âge de la mère, il n'est
pas impossible que ces facteurs associés à l'âge des souris testées aient pu influencer le choix
des mâles âgés. Pour lever cette ambiguïté, il faudrait prélever l'odeur de femelles de la même
portée, congeler les prélèvements puis effectuer un deuxième prélèvement à la fin de leur vie.
De plus, cette procédure permettrait d'étudier les modifications de l'odeur individuelle avec
l'âge, puisque le même individu pourrait fournir plusieurs échantillons d'odeurs échelonnés
dans le temps, et permettrait également d'étudier finement les capacités de discrimination et
de catégorisation en fonction de l'âge. Cependant, les deux catégories de femelles sont bien
discriminables, quelle que soit l'origine de leur différence. L'hypothèse d'un changement
olfactif lié à l'âge lui-même reste la plus parcimonieuse (Osada et al., 2003).
L'interprétation de la préférence des mâles âgés est malaisée. Il est difficile d'associer ce choix
à une plus grande qualité des femelles âgées, et ce pour deux raisons : 1) le potentiel
reproducteur des femelles âgées est fatalement plus faible que celui des jeunes. De plus, l'âge
n'est pas dans ce cas le signe d'une plus grande expérience, ni d'une meilleure compétence
dans l'élevage des jeunes, puisque toutes les femelles testées sont nullipares. 2) Si les femelles
âgées étaient réellement plus intéressantes pour un mâle, alors cette même préférence devrait
se retrouver chez les mâles jeunes. Or ceux-ci montrent une tendance inverse (non
significative). Ce dernier point permet de proposer une interprétation alternative : les mâles
ont tendance à choisir la femelle de leur âge, ce qui rappelle certains travaux chez les Oiseaux
où il existe une association prédominante de partenaires d'âge similaire (Coulson & Thomas,
122

Chapitre III: Odeurs, facteurs d’état et proximité génétique
1983). Cependant les préférences pour les odeurs de femelles s'expliquent certainement de
façon plus simple. Si l'âge a des conséquences olfactives similaires chez tous les individus,
alors il est probable que les odeurs d'individus du même âge présentent des qualités similaires.
Les mâles ont alors pu être attirés par cette similarité relative à ce à quoi ils sont habitués,
c'est-à-dire leur propre odeur. La motivation à choisir ce stimulus par rapport à l'autre est,
encore une fois, difficile à analyser.
Les femelles ne montrent pas de préférence, ni même de tendance à préférer les mâles en
fonction de leur âge. Il n'y a pas non plus d'indication que leur âge dicte une préférence,
comme chez les mâles. De façon similaire aux résultats présentés dans le chapitre II, les
femelles ne semblent pas choisir entre les odeurs de mâles.
L'absence de résultats probants concernant le rôle de l'âge dans les préférences olfactives des
Souris glaneuses empêche la réalisation de l'étape suivante, l'étude de l'interaction de ce
facteur avec la reconnaissance de parentèle. En effet, si les souris avaient montré une
préférence claire pour les odeurs de l'autre sexe basée sur l'âge des donneurs, il aurait été
possible de proposer un choix opposant préférence liée à l'apparentement et préférence liée à
l'âge, comme il a été proposé un choix opposant préférence génétique et préférence lié au
régime alimentaire (voir ci-dessus). Par exemple, si les mâles - quelque soit leur âge - avaient
préféré l'odeur d'une femelle jeune, le choix aurait été entre une sœur jeune et une femelle non
apparentée plus âgée. La seule préférence significative obtenue dans ces tests a été celle des
mâles âgés pour l'odeur d'une femelle également âgée. En croisant ce facteur avec le facteur
apparentement, le choix pour les vieux mâles aurait été entre l'odeur d'une sœur du même âge
et l'odeur d'une femelle jeune et non apparentée. Or le résultat de cette comparaison est
difficilement interprétable : en effet, la proximité génétique comme le partage d'un état
commun (ici l'âge) accroissent les similarités olfactives. Dans l'hypothèse d'un mécanisme
basé sur la similarité des odeurs, une telle comparaison n'est pas heuristique, puisqu'au lieu de
présenter une situation où la similarité génétique et la similarité liée au partage de facteurs
d'état sont en conflit, le choix serait entre femelle "doublement similaire" (par l'âge et par
l'apparentement) et une femelle "doublement différente".
Il a donc été impossible d'étudier l'interaction de l'apparentement et de l'âge comme avait été
étudiée l'interaction de l'apparentement et du régime alimentaire.
123

Chapitre III: Odeurs, facteurs d’état et proximité génétique
3.4. Facteurs d'état et parentèle : discussion générale
Les résultats exposés dans ce chapitre ne permettent malheureusement pas de répondre aux
interrogations qui ont présidé à l'élaboration de ces protocoles. La raison d'être de cette étude
sur l'interaction des facteurs d'état et de la parentèle était de tester de façon indirecte la
validité de l'hypothèse présentée dans le chapitre II, c'est-à-dire l'utilisation par les Souris
glaneuses de la proximité des odeurs individuelles pour évaluer la proximité génétique. Une
modification du phénotype odorant est de nature à perturber un tel mécanisme.
Dans la première étude, l'ingestion par le sujet (mâle) d'une variété de pomme influence ses
préférences olfactives : une femelle ayant consommé l'autre variété de pomme aura tendance
a être préférée à une femelle ayant le même régime alimentaire que le sujet. Une différence
subtile de régime alimentaire est donc suffisante pour infléchir les préférences olfactives. Le
sujet choisit l'odeur la moins similaire à la sienne. Le facteur "nouveauté" paraît primordial
pour comprendre cette préférence : ce n'est pas une variété de pomme qui est préférée, c'est la
variété de pomme que le sujet ne connaît pas. Ces résultats sont tout à fait cohérents avec la
littérature existante : le changement de régime alimentaire modifie l'odeur et peut même
permettre de discriminer les odeurs d'individus qui nourris identiquement ne sont pas
discriminés (Brown & Schellinck, 1992). Ces résultats sont également cohérents avec
l'hypothèse d'une comparaison du phénotype rencontré avec le phénotype propre : la similarité
est plus grande entre l'odeur du sujet et celle du donneur nourri avec la même variété de
pomme qu'entre l'odeur du sujet et celle du donneur nourri différemment. Dans le chapitre II,
les préférences olfactives des mâles étaient dirigées vers l'odeur d'une femelle non apparentée
plutôt que vers l'odeur d'une sœur (non familière). La covariance odeurs-gènes implique en
effet une plus grande similarité entre le sujet et sa sœur qu'entre le sujet et une femelle non
apparentée. En croisant les deux facteurs, les mâles ont montré une préférence pour l'odeur
d'une femelle non apparentée pourtant nourrie de la même façon que lui-même.
L'interprétation la plus parcimonieuse est que le phénotype d'une apparentée est demeuré plus
proche, malgré le régime alimentaire différent. L'odeur de cette femelle est donc perçue
comme moins attractive que celle d'une non-apparentée. Cette expérience ne permet toutefois
pas de démontrer que c'est bien la similarité phénotypique qui guide les préférences olfactives
de mâles. En effet, l'hypothèse d'un traitement distinct des informations liées à
l'apparentement et de celles liées au régime alimentaire est tout à fait plausible. Un moyen
indirect de tester cette hypothèse est d'accroître la différence entre les odeurs en modifiant de
124

Chapitre III: Odeurs, facteurs d’état et proximité génétique
façon plus drastique le régime alimentaire. Cette procédure devrait être réalisée de la façon
suivante :
1) Nourrir les souris avec deux régimes alimentaires A et B très différents mais
n'entraînant pas de changement de leur fitness, c'est-à-dire conserver un taux de
protéines constant et une valeur énergétique constante tout en jouant sur les composés
volatiles.
2) Présenter aux sujets mâles A (nourris avec le régime A) l'odeur d'une femelle A et
celle d'une femelle B (et vice-versa), toutes deux non apparentées. Le résultat de ce
test devrait être le même qu'avec les variétés de pommes, c'est-à-dire une préférence
pour la nouveauté (l'odeur de la femelle B).
3) Proposer le choix au mâle A entre une femelle non apparentée A et une sœur B. Selon
le résultat de ce test, deux mécanismes peuvent être envisagés :
• Si la femelle non apparentée est préférée, bien qu'à apparentement égal la
femelle au régime alimentaire différent soit préférée, alors ceci constitue une
indication forte d'un traitement séparé des facteurs de parentèle et des facteurs
d'état présents dans l'odeur. Ce résultat seul restant néanmoins interprétable
comme une prédominance de l'apparentement par rapport au régime alimentaire
dans l'odeur, il faudrait pour confirmer cette interprétation mettre en œuvre une
série de tests d'habituation-généralisation similaires à ceux décrits dans les
études sur la covariance odeur-gènes (Busquet & Baudoin, sous presse ; Heth et
al., 2001). Si ces tests confirment que les odeurs du sujet et de la femelle non
apparentée soumise au même régime sont perçues comme plus similaires que
les odeurs du sujet et d'une sœur nourrie différemment, alors la préférence pour
la femelle non apparentée dans le test croisé ne peut s'expliquer par l'utilisation
de la similarité des odeurs. Le mécanisme sous-jacent serait dans ce cas proche
de l'utilisation d'allèles de reconnaissance et par là même d'une reconnaissance
de parentèle "vraie" sensu Grafen (1990).
• Si l'odeur de la sœur nourrie différemment du sujet est préférée par celui-ci,
alors l'hypothèse de l'utilisation de la similarité des odeurs se verrait confortée.
Le sujet s'est trouvé en quelque sorte "trompé" par la manipulation de son
phénotype et de celui des femelles donneuses. La plus grande similarité de son
125

Chapitre III: Odeurs, facteurs d’état et proximité génétique
odeur avec celle de la femelle non apparentée lui ferait considérer l'odeur de sa
sœur comme plus intéressante.
Un autre moyen de tester ces deux hypothèses et de jouer sur la similarité des odeurs liée à
l'apparentement, plutôt que sur la similarité liée aux facteurs d'état. Par exemple, si l'odeur
d'une non-apparentée est préférée à celle d'une double cousine (r = 0,25), ce qui n'a encore
jamais été testé, il est possible de proposer à un mâle un choix croisé entre l'odeur d'une
double cousine nourrie avec une variété de pomme différente et l'odeur d'une non apparentée
nourrie avec la même variété que le sujet. Ainsi, avec un test en tout point comparable à celui
présenté dans ce chapitre, il est possible que la préférence du mâle se porte vers l'odeur de la
double cousine. La similarité odorante "d'origine génétique" étant moindre entre doubles
cousins qu'entre frères et sœurs, la similarité "d'origine environnementale" pourrait l'avoir
emporté et ainsi renversé la préférence. L'idéal serait d'entreprendre une étude de grande
envergure où les similarités phénotypiques seraient manipulées selon les deux axes,
l'apparentement et le régime alimentaire. Une telle étude de l'interaction des facteurs de
parentèle avec un facteur d'état (l'alimentation) permettrait indirectement de mettre à l'épreuve
le mécanisme d'évaluation de la proximité génétique par la similarité des odeurs.
L'autre facteur d'état étudié dans ce chapitre était l'âge. Bien qu'il n'ait pas été possible de
mener l'étude à son terme en proposant le choix croisé entre une sœur de l'âge préféré et une
femelle non apparentée de l'autre classe d'âge, quelques enseignements intéressants peuvent
être tirés. Tout d'abord les femelles n'ont pas montré de choix basé sur l'âge des mâles
donneurs. Ceci semble moins être lié à une absence de perception du facteur âge qu'à une
probable absence de motivation à choisir entre des odeurs de mâles. En effet, les résultats
exposés dans le chapitre II faisaient déjà état d'une absence de choix des femelles entre les
odeurs de mâles (qui différaient par leur apparentement). Les mâles, quant à eux, montrent
une préférence qui semble dirigée vers les femelles de leur classe d'âge, bien que cette
tendance ne soit significative que pour les mâles âgés. Les mâles ont été attirés par le
phénotype qui semble en toute logique le plus proche du leur. Bien que ce point n'ait pas été
vérifié (les tests d'habituation-généralisation entrepris dans ce but n'ont pu être réalisés, les
animaux montrant un manque d'intérêt pour les odeurs présentées), le fait que les jeunes
mâles ne montrent pas de préférence pour ces femelles âgées mais plutôt une tendance inverse
indique que ce n'est pas le phénotype des femelles âgées qui est plus attractif mais bien la
relation entre le sujet et les donneuses d'odeurs qui influence la préférence. Il est frappant de
126

Chapitre III: Odeurs, facteurs d’état et proximité génétique
voir que les préférences des mâles pour des odeurs de femelles ne sont pas aussi facilement
interprétables que les résultats du chapitre II ne le laissaient penser. Il est à noter que d'autres
pistes ont été explorées concernant l'interaction des facteurs de parentèle avec des facteurs
d'état liés à la fitness, mais ces pistes ont toutes été abandonnées, faute de préférence
constante des souris pour l'une ou l'autre des catégories présentées (raison pour laquelle ces
pistes ne sont pas exposées dans cette thèse). La privation temporaire de nourriture des
donneurs d'odeurs, par exemple, ne se traduit pas par un évitement de l'odeur de ces donneurs,
qui sont pourtant théoriquement des partenaires potentiels moins intéressants. Dans le cadre
du choix de partenaire, il paraît primordial de trouver un facteur d'état influençant fortement
les préférences sexuelles des Souris glaneuses afin d'étudier son interaction avec les facteurs
de parentèle. L'un des candidats les plus prometteurs est le parasitisme, en vertu d'études
montrant que les individus parasités tendent à être évités lors du choix du partenaire sexuel
(Willis & Poulin, 2000 ; Ehman & Scott, 2002 ; Kavaliers et al., 2004). En évitant un
partenaire parasité, une souris évite d'une part de se voir transmettre ce parasite et d'autre part
de s'accoupler avec un partenaire dont le système immunitaire est manifestement déficient
(Penn & Potts, 1998). Le parasitisme est un facteur décelable dans les odeurs individuelles, et
qui se traduit par des préférences olfactives fortes chez les souris de laboratoire (Kavaliers
& Colwell, 1995). De plus, l'état du sujet peut influer sur les préférences pour des odeurs de
partenaires potentiels parasités ou non. En effet, la simple exposition à l'odeur d'un mâle
parasité fait chuter la préférence de femelles Mus domesticus pour l'odeur d'un mâle sain
(Kavaliers et al., 2003). Des femelles elles-mêmes parasitées réagissent beaucoup moins que
des femelles saines à l'odeur de mâles parasités, mais réagissent de façon similaire à l'odeur
d'un mâle stressé, ce qui indique que la cause de la baisse de réactivité à l'odeur d'un mâle
parasité est moins liée aux modifications comportementales induites par le parasite chez la
femelle qu'à une plus grande familiarité avec les signaux chimiques révélateurs de la charge
parasitaire (Kavaliers et al., 1998). Une étude de l'interaction du facteur parasitisme avec
l'apparentement dans les préférences socio sexuelles des Souris glaneuses permettrait de
relativiser le rôle de la proximité génétique face à un facteur extrêmement important dans le
cadre du choix de partenaire. Ce faisant, c'est l'aspect fonctionnel de la reconnaissance de
parentèle qui serait mis à l'épreuve. Cet aspect fonctionnel est absolument primordial pour
deux raisons :
- La première est liée à la compréhension des stratégies des animaux sur le terrain. Si
les facteurs d'état interviennent fortement dans le choix de partenaire, alors l'étude de la
127

Chapitre III: Odeurs, facteurs d’état et proximité génétique
structuration de ces facteurs d'état en milieu naturel devient fondamentale pour comprendre
les stratégies des animaux. Concrètement, si le fait de partager un état commun (par exemple,
un régime alimentaire identique) est important dans les préférences sexuelles, alors
l'utilisation de la similarité des odeurs comme guide des préférences socio sexuelles recevrait
un appui significatif, bien qu'empirique.
- La deuxième raison se rapporte à l'évolution des mécanismes de reconnaissance de
parentèle. Comme suggéré en introduction du chapitre I, à propos du MHC et des MUPs, la
pensée évolutionniste conduit parfois à supposer qu'en raison de l'importance des pressions de
sélection relatives à la prise en compte de l'apparentement dans les comportements, un
mécanisme a du être développé pour remplir cette fonction capitale. L'étude des facteurs
environnementaux covariants avec l'apparentement en nature permettrait de proposer un ou
des scénarios évolutifs alternatifs, où serait prise en compte la contingence de ces facteurs
(Ryan et al., 2001). La discussion générale reprendra et développera ce point crucial.
Dans ce chapitre ainsi que dans les précédents, les choix des souris portaient sur des odeurs
individuelles. Il a souvent semblé judicieux de rapprocher ces préférences olfactives de
préférences sexuelles ou sociales, sans qu'il soit possible d'en faire la démonstration. Le
prochain chapitre s'est intéressé à cet aspect fonctionnel de la reconnaissance de parentèle, en
essayant de corréler les capacités de discrimination et les préférences mises en évidence
jusqu'ici avec un réel choix de partenaire.
128

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
Chapitre IV :
Aspects fonctionnels de la
reconnaissance de parentèle
129

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
4.1. Introduction
Les fonctions théoriques de la reconnaissance de parentèle ont été largement présentées dans
l’introduction générale. Elles s’articulent autour de deux grands axes qui sont le choix du
partenaire sexuel et les comportements altruistes. Compte tenu des résultats nouveaux
exposés dans les chapitres précédents, il est apparu intéressant de rechercher chez Mus
spicilegus des comportements qui traduisent cette reconnaissance de parentèle. En effet, les
Souris sont capables de discrimination fine entre les odeurs individuelles, notamment liée à
l'apparentement. Il semble donc que les conditions soient réunies pour une discrimination
comportementale : 1) le signal olfactif contient l'information nécessaire à la traduction des
degrés d'apparentement. 2) cette traduction olfactive est perceptible par les Souris. La
troisième composante de la reconnaissance, suivant la production et la perception, est
l'action. C'est par la mise en évidence de cette troisième composante que l'existence d'une
reconnaissance de parentèle peut être validée. En effet, il est supposé exister des pressions de
sélection ayant amené les animaux à prendre en compte le facteur apparentement dans leurs
choix socio-sexuels. De plus, l'existence de pressions de sélection ayant spécifiquement
présidé à l'élaboration d'une reconnaissance de parentèle est perçue par certains auteurs
comme le critère permettant de définir la reconnaissance de parentèle "vraie" (Grafen 1990 ;
Barnard 1999 ; cf. chapitre II).
Comme exposé dans l'introduction générale, la mise en évidence de pressions de sélection en
laboratoire est une tâche ardue, puisque la seule validation réelle de la meilleure fitness d'une
catégorie d'individus est le succès reproducteur, succès qui n'est estimable que par le nombre
de descendants accédant à la reproduction. Cette mesure n'a de sens qu'en milieu naturel, où
les animaux subissent effectivement l'ensemble des pressions de sélection (prédation,
parasitisme, compétition, pressions abiotiques etc.). D'autre part, les principes de l'évolution
impliquent un certain degré d'optimisation des comportements. Si une aptitude telle que la
reconnaissance de parentèle représente effectivement un avantage majeur, alors il est probable
que l'ensemble des individus de l'espèce étudiée présente cette capacité. Autrement dit, il
n'existe probablement pas de "groupe témoin", d'individus ne manifestant pas cette capacité,
qui permettrait de quantifier l'avantage sélectif des individus pratiquant la reconnaissance de
parentèle. Or la démonstration la plus efficace (et la plus classique) en sciences est la
comparaison de deux groupes différant par un seul trait. Cette démarche ne peut pas
s'appliquer à la recherche de la fonctionnalité d'un comportement, comme elle ne pouvait pas
130

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
s'appliquer à la recherche d'un mécanisme (Hauber & Sherman, 2003). Il est cependant
possible d'imaginer des stratégies permettant d'estimer les conséquences fonctionnelles de la
reconnaissance de parentèle. Du fait du lien durable existant entre partenaires chez Mus
spicilegus qui est une des caractéristiques majeures de cette espèce, il semble logique de
rechercher dans le choix de partenaire les éventuelles conséquences fonctionnelles des
capacités de reconnaissance de parentèle.
Le choix du partenaire sexuel
L'une des fonctionnalités théoriques de la reconnaissance de parentèle est le choix d'un
partenaire sexuel qui ne soit ni trop apparenté ni trop éloigné (Bateson, 1983). Il semble donc
important de s'intéresser au choix de partenaire chez Mus spicilegus, en proposant des
partenaires potentiels différant par leur degré d'apparentement. La mise en évidence de ce
choix est difficile pour des raisons pratiques et théoriques.
Une préférence olfactive ne traduit pas nécessairement un choix sexuel. Cette préférence peut-
être guidée par un contexte motivationnel auquel l'observateur n'a pas accès. Il faut donc
pouvoir montrer une activité copulatoire traduisant la nature sexuelle de la préférence.
Deux partenaires sont impliqués dans l'accouplement. La préférence de l'un ne correspond pas
nécessairement à la préférence de l'autre, à moins qu'un seul des deux sexes ne présente une
préférence. Chez les Mammifères (comme chez de nombreux animaux), les femelles sont
considérées comme étant celles qui choisissent, en vertu de l'investissement supérieur dans la
reproduction, lié au coût de la gestation et de l'allaitement. Les explications évolutives de ce
choix par les femelles sont nombreuses et anciennes (Bateson, 1983 ; Jennions & Petrie,
1997). Certaines de ces explications supposent un avantage sélectif de ce choix (Houle &
Kondrashov, 2002). Les femelles devraient donc effectuer des choix qui permettent de
maximiser leur succès reproducteur. Chez les Souris domestiques, le respect du choix des
femelles se traduit par une meilleure survie des jeunes et des caractéristiques
comportementales liées à une meilleure fitness (Drickamer et al., 2000). Le choix en lui-
même semble donc avoir une importance, indépendamment de la "qualité" des partenaires
(cette assertion est invérifiable puisque la qualité du partenaire est multifactorielle et n'est pas
directement observable, surtout en l'absence d'hypothèses orientées présidant au choix
proposé à la femelle). Cette importance du choix n'est pas l'apanage exclusif des femelles,
puisque les mâles Mus domesticus produisent plus de petits quand ils sont accouplés à la
131

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
femelle qu'ils préfèrent (Gowaty et al., 2003). Même dans une espèce polygyne comme Mus
domesticus (Bronson, 1979), les préférences des mâles existent. Il est donc plausible que chez
une espèce comme Mus spicilegus chez laquelle existe une association pérenne entre
partenaires (Patris & Baudoin, 1998 ; Baudoin et al., sous presse) et un investissement
important du mâle dans les soins parentaux (Patris & Baudoin, 2000), le choix soit mutuel.
Malheureusement, les approches théoriques permettant d'analyser ces choix de partenaire
mutuels sont encore rares et peu développées (Bergstrom & Real, 2000). Ce choix mutuel ne
signifie pas pour autant que les deux partenaires potentiels soient équivalents. Il existe des
modèles qui expliquent le choix de la femelle non pas par la recherche de caractéristiques
attractives chez le mâle mais par une tendance évolutive à réduire le coût de l'accouplement
(Gavrilets et al., 2001). La femelle, en refusant de s'accoupler avec le premier mâle venu,
diminue le risque d'avoir été fécondée par un mâle qui lui donnerait une progéniture peu
nombreuse ou inadaptée (ce modèle n'est pas exclusif des modèles supposant une préférence
de la femelle pour certains traits (Kokko et al., 2003)). En extrapolant cette idée, il est
possible de proposer un scénario dans lequel le refus de la femelle implique un choix de la
part du mâle. Si l'accouplement nécessite un investissement de la part du mâle, alors il a
intérêt à ne pas "gaspiller" son énergie à convaincre une femelle qui n'est pas une partenaire
adéquate, soit parce qu'elle est trop apparentée, soit parce qu'elle présente des traits
négativement corrélés à sa fitness.
Le choix de partenaire chez Mus spicilegus
Au début de la saison de reproduction, les animaux sortis des tumulus se dispersent. Cette
dispersion peut être un moyen efficace d'éviter la consanguinité (Keane 1990, Ferriere et al.
2000). Bien que les données de terrain permettant d'évaluer cette dispersion chez Mus
spicilegus manquent encore, il semble plausible que ces souris migrent à distance de leur
tumulus d'origine, ne serait-ce que pour éviter les individus apparentés et familiers avec
lesquels elles ont cohabité. Ce comportement ne peut être sélectionné que s'il apporte un
avantage surpassant les coûts liés à la dispersion. Les choix de partenaire pourraient donc
suivre un gradient géographique, avec une préférence pour les animaux originaires d'un site
différent par rapport aux animaux du même site. Cette question a pu être posée aux animaux
capturés en Hongrie en octobre 1999.
132

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
Rappels sur la campagne de septembre 1999 à Gyöngyös (Hongrie)
Lors de ce travail de terrain, 83 individus furent capturés par plusieurs chercheurs de l'équipe
(Claude Baudoin, Patrick Gouat, Chantal Poteaux) afin de mettre en place un nouvel élevage.
8 individus sont morts lors du transfert, 7 jeunes d’un même tumulus et l’unique adulte
capturé (une femelle). Les 75 individus survivants ont été conservés en cage, avec une double
contrainte :
• Respect de l’origine : les animaux capturés dans un tumulus ont été groupés dans la même
cage, et aucun transfert d’animaux n’a été effectué.
• Évitement des accouplements non désirés : il est nécessaire, pour les besoins de la conduite
d’élevage, de connaître et de maîtriser la paternité. De plus, même sans fécondation, un
accouplement pourrait avoir des conséquences importantes sur le développement d’un lien
socio sexuel particulier susceptible de biaiser nos expériences. Les deux sexes ont donc
été séparés le plus rapidement possible.
La sex-ratio observée était biaisée en faveur des mâles, avec 48 d’entre eux pour seulement 27
femelles. Le démarrage de l’élevage étant une priorité absolue, il était nécessaire d’optimiser
la reproduction en "utilisant" toutes les femelles disponibles. Afin de combiner au mieux les
impératifs liés au démarrage de l’élevage et la question posée du choix de partenaire, c'est le
choix des femelles qui a été évalué.
4.2. Préférences de femelles en fonction de la provenance
géographique du mâle
L'objectif principal de cette expérience est d'évaluer la validité d'une préférence pour
objectiver un choix de partenaire. Chaque femelle se verra offrir un choix entre un mâle issu
de son propre site (mais d'un tumulus différent, de façon à minimiser le risque de présenter un
apparenté proche) et un mâle d'un autre site distant de quelques kilomètres. Cette question
écologiquement valide est utilisée pour tenter d'approcher les conséquences du choix de la
femelle. En effet, la moitié des femelles verra son choix respecté (elle sera mise en couple
avec le mâle qu'elle aura préféré) tandis que l'autre moitié sera mise en couple avec le mâle
non préféré.
133

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
4.2.1 Matériel et méthodes
25 femelles et 50 mâles capturés en octobre 1999 ont été utilisées dans cette expérience,
réalisée en mars 2000 (les sujets étaient alors âgés d'environ 7 mois). Chaque femelle est
introduite dans la zone centrale d'un terrarium (dimensions : 60 x 30 x 45 cm), et chacune des
extrémités est occupée par un mâle, l'un originaire du même site, l'autre d'un site différent. De
façon à éviter les agressions, les animaux ne peuvent pas entrer en contact direct : des
séparations en plexiglas perforé doublé de grillage permettent cependant un contact naso-
nasal et une communication olfactive. Le test dure 30 minutes, et le temps de contact naso-
nasal entre la femelle et chacun des deux mâles est mesuré. Les résultats de ce premier test
permettent de répartir les femelles en deux groupes où le choix de la femelle est respecté
(groupe CR) ou non (groupe CNR). La rencontre entre les deux partenaires est ensuite filmée
et les comportements notés, de façon à obtenir un indice agonistique permettant de confirmer
ou d'infirmer la préférence (ce travail a été réalisé par Patrick Gouat). Les couples sont ensuite
régulièrement surveillés et les naissances notées.
4.2.2 Résultats
Pendant les 30 minutes de test, les femelles passent entre 0 et 59 secondes en contact avec un
mâle. L'interaction avec les mâles du même site dure 16,52 ± 13,92 secondes (moyenne ±
écart-type) contre 17,74 ± 13,54 s pour les mâles d'un site différent. Parmi les 25 femelles,
13 passent plus de temps en interaction avec le mâle du même site contre 12 pour le mâle du
site différent. Les femelles ne montrent donc pas de préférence pour une de ces deux
catégories de mâles (p = 0,798, test pour échantillons appariés de Wilcoxon).
Lors de la rencontre des deux partenaires, aucune différence comportementale n'a pu être
relevée entre les deux groupes CR et CNR. Ni le nombre d'interactions ni l'occurrence des
comportements agonistiques ne distinguent les deux groupes (P. Gouat, comm. pers.).
Seuls 5 couples ont donné naissance à des petits pendant les 80 jours après la mise en couple,
et cet effet plancher n'a pu permettre de déceler de différence entre groupes (à la suite de ces
80 jours, les femelles nullipares ont été mises en couple avec un nouveau partenaire et ont pu
commencer à se reproduire).
134

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
4.2.3 Discussion
Dans cette étude, les femelles ne choisissent pas en fonction de l'origine géographique des
mâles. De plus, l'intérêt des femelles pour l'un ou l'autre des mâles présentés ne semble
corrélé ni à une différence de comportements lors de la rencontre, ni à un succès reproducteur
supérieur. La reproduction est d'ailleurs rare dans les couples formés. En l'absence de
nombreuses reproductions, il est impossible de tester les effets du choix (Drickamer et al,.
2000) ou les effets de la dispersion (Keane, 1990). Un point important dans la compréhension
de ces résultats est la nature du test : puisque des animaux vigiles étaient présentés aux
femelles, il est probable que le comportement de celles-ci ait été influencé par le
comportement du mâle, plus que par son identité. Les variations comportementales et/ou
physiologiques interindividuelles seraient donc responsables de l'absence de tendance dans les
préférences des femelles. La présentation d'animaux vigiles comme stimulus ne semble pas
plus heuristique que la simple présentation d'odeurs. De plus la recherche mutuelle du contact
n'est pas synonyme de préférence sexuelle.
Cette expérience fournit un exemple de la difficulté à valider un choix de partenaire chez Mus
spicilegus. Une fois le couple formé, en revanche, les femelles semblent montrer une
préférence pour le mâle familier (Patris & Baudoin, 1998), même si l'influence du
comportement du mâle sur cette préférence est mal connue. D'une manière générale, la
familiarité semble être un facteur très important chez Mus spicilegus, puisque c'est celle-ci qui
semble être responsable de l'absence de reproduction au sein des tumulus (Gouat et al., 2003).
Une autre approche de ces résultats consiste à reconsidérer les choix proposés en terme de
proximité génétique, en essayant a posteriori de corréler le comportement des femelles avec
leur proximité génétique des mâles présentés. Des échantillons ont été prélevés par Chantal
Poteaux sur l'ensemble des animaux capturés sur le terrain, donc sur l'ensemble des sujets de
l'expérience ci-dessus. L'outil génétique a permis d'attribuer à chacun des deux mâles
présentés à la femelle un coefficient de distance génétique, calculé à partir des différences et
similarités au niveau de 5 loci de microsatellites, grâce à l'outil Relatedness®. Il est à noter
que ce coefficient de distance génétique, en dépit du nom du programme, ne peut en aucun
cas être assimilé à un coefficient d'apparentement (voir introduction générale). Deux
enseignements ont pu être tirés des coefficients calculés. D'une part, le mâle issu d'un site
différent de celui de la femelle ne présente pas nécessairement un coefficient de proximité
135

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
génétique inférieur à l'autre mâle (une dizaine de situations de choix semble même proposer
un mâle d'un site différent plus proche génétiquement que le mâle issu du même site que la
femelle). D'autre part, les situations de choix, au regard des coefficients de distance génétique
calculés, sont très ambigus ; en effet, dans certains des tests, les deux mâles présentés sont
tous deux génétiquement très proches de la femelle, tandis que dans d'autres, les deux mâles
sont éloignés. Il apparaît donc que, si l'on s'en réfère au coefficient de distance génétique
calculé, les tests effectués ont été de nature très différente, donc sont difficiles à interpréter de
façon claire : dans le contexte théorique de "l'optimal outbreeding", les choix de partenaire
sexuel correspondent à une distance génétique intermédiaire entre les frères et sœurs et les
non-apparentés (mais voir l'introduction générale pour une discussion de la validité de cette
théorie). Ne sachant pas à quelle relation de parenté peuvent correspondre ces coefficients, il
serait difficile d'interpréter une tendance. Or la comparaison des résultats de l'expérience de
choix avec les données génétiques n'a pas permis de mettre en évidence une tendance. De
même, la relecture des données comportementales obtenues lors de la mise en couple (Patrick
Gouat) ne permet pas de montrer une relation entre le niveau d'agression (par exemple) et ce
coefficient de distance génétique. Cette absence de résultats probants amène une remise en
question des protocoles utilisés. Il aurait été plus rigoureux, si les données génétiques avaient
été disponibles au départ, de proposer des choix plus standardisés aux femelles, en proposant
par exemple un mâle très proche génétiquement et un mâle plus lointain.
Il est difficile de ne pas dresser un parallèle entre ces résultats et les tests de préférence
présentés dans les chapitres précédents. Dans aucun des tests réalisés il n'a été possible
d'observer une préférence claire de la part des femelles entre deux odeurs de mâles. Ceci est
probablement dû au statut hormonal de ces femelles au moment du test, c'est-à-dire un
anœstrus induit par le regroupement entre femelles (Féron & Gheusi, 2003). Les femelles
étaient par contre tout à fait à même de choisir entre les odeurs d'autres femelles et de façon
peut-être plus surprenante, les mâles ont montré un intérêt dans les odeurs de femelles en
anœstrus qui s'est traduit par l'expression de préférences nettes, qu'elles soient pour l'odeur
d'une femelle apparentée ou l'odeur d'une femelle au régime alimentaire différent
(cf. chapitre II). Malheureusement ces tests de préférence n'ont pas été suivis de mises en
couple, où les mâles auraient été accouplés soit avec la femelle préférée (la femelle non
apparentée) soit avec l'autre femelle (une sœur). Il n'est pas toutefois certain que ces mises en
couple auraient été informatives : la présence du mâle est nécessaire pour déclencher la
136

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
réceptivité sexuelle, et il est probable que les mâles se seraient accouplés avec la femelle dès
cette réceptivité sexuelle atteinte. La seule alternative à ce résultat serait une induction moins
efficace de l'oestrus par les signaux d'un mâle apparenté, ce qui n'est pas évoqué dans la
littérature.
La mise en évidence d'un choix sexuel via une préférence olfactive semble hasardeuse, même
en permettant ensuite à l'animal de s'accoupler avec le donneur de l'odeur préférée. L'idéal est
de présenter un choix sexuel simultané. C'est cette approche qui a été choisie, via la
réalisation de trios.
4.3. Rôle de l'apparentement dans le choix du partenaire
sexuel en situation de trios
Sur le terrain, les souris Mus spicilegus sont souvent trouvées en paires durant la période de
reproduction. Cependant il arrive que par piégeage, on trouve 3 animaux associés
spatialement (Gouat et al., 2003a). Cette situation se retrouve parfois en enclos semi naturels
(Dobson & Baudoin, 2002 ; Baudoin et al., sous presse). Des trios constitués de 2 sœurs et
1 mâle non apparenté ont été constitués par Gouat et al. (données non publiées). La
reproduction est possible dans ces conditions, mais elle s'accompagne d'une diminution
importante de la fitness des femelles.
Cette dernière étude a permis d'envisager la possibilité de constituer des trios avec un mâle et
deux femelles. Il est à noter que cette possibilité ne remet pas en cause le caractère monogame
de l'espèce : ce type de trios, en nature, découlerait d'une prolongation du lien social entre
individus du même tumulus. En enclos, l'initiation de la reproduction a tendance à scinder ces
trios pour aboutir à une structure sociale monogame (Baudoin et al., sous presse).
Pour mettre en évidence l'aspect fonctionnel de la reconnaissance de parentèle chez Mus
spicilegus, des trios ont donc été réalisés de la façon suivante : un mâle introduit dans la cage
de deux femelles, l'une étant une sœur non familière de ce mâle, l'autre étant non apparentée à
cette fratrie. L'hypothèse est que les animaux, en cohabitant, auront la possibilité d'exprimer
une préférence sexuelle. Le mâle peut théoriquement choisir entre les deux femelles, et
chaque femelle a la possibilité de refuser les tentatives d'accouplement du mâle. Une
préférence claire du mâle comme une reconnaissance de la part de la femelle devraient
137

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
amener un évitement de la consanguinité, objectivée par plus de reproduction entre le mâle et
la femelle non apparentée.
4.3.1 Matériels et méthodes
20 trios ont été réalisés. La première étape dans cette réalisation est la réunion des deux
femelles non apparentées. Ces femelles sont d'âge similaire, et la différence d'âge entre elles
est inférieure à la différence d'âge avec le mâle. Pour chaque duo constitué, l'une des deux
femelles est marquée par une coupe de poils du flanc, ce qui permet une identification
ultérieure. Chaque femelle est extraite de sa cage, qu'elle partage depuis le sexage avec ses
sœurs, et introduite dans une cage propre avec l'autre femelle. De la nourriture est disposée
sur la sciure couvrant le fond de la cage et du coton est fourni, ce qui permet aux souris de
construire un nid. Un seul de ces duos est réalisé par jour, et il est surveillé pendant la
première demi-heure puis inspecté toutes les demi-heures, de façon à s'assurer de l'absence
d'agression. Les deux femelles coexistent une semaine avant l'introduction du mâle.
Les 20 mâles utilisés sont isolés une semaine avant la "mise en trio". Ils sont issus d'une
portée différente (antérieure) de celle de leur sœur, qui est donc non familière.
Les origines sont contrôlées de façon à ce que les femelles non apparentées n'aient pas de
grand-parent en commun avec les frères et sœurs avec qui elle est mise en cage. Les mâles
sont originaires de 8 couples, mais pour éviter la pseudo-réplication, aucun des mâles issus du
même couple n'est mis en cage avec une femelle non apparentée de la même origine.
Une seule introduction de mâle dans la cage du duo de femelles est réalisée par jour, de façon
à permettre un suivi de la rencontre.
Les trios ainsi constitués sont suivis pendant 80 jours. Les mises bas sont notées et attribuées
à l'une ou l'autre des femelles soit quand la mise bas est directement observée, soit par
l'activité d'allaitement et l'état des tétines, révélateur de la lactation. Les données intéressantes
pour cette étude sont l'identité des femelles mettant bas (apparentées ou non), la latence de
première reproduction et le nombre de petits.
138
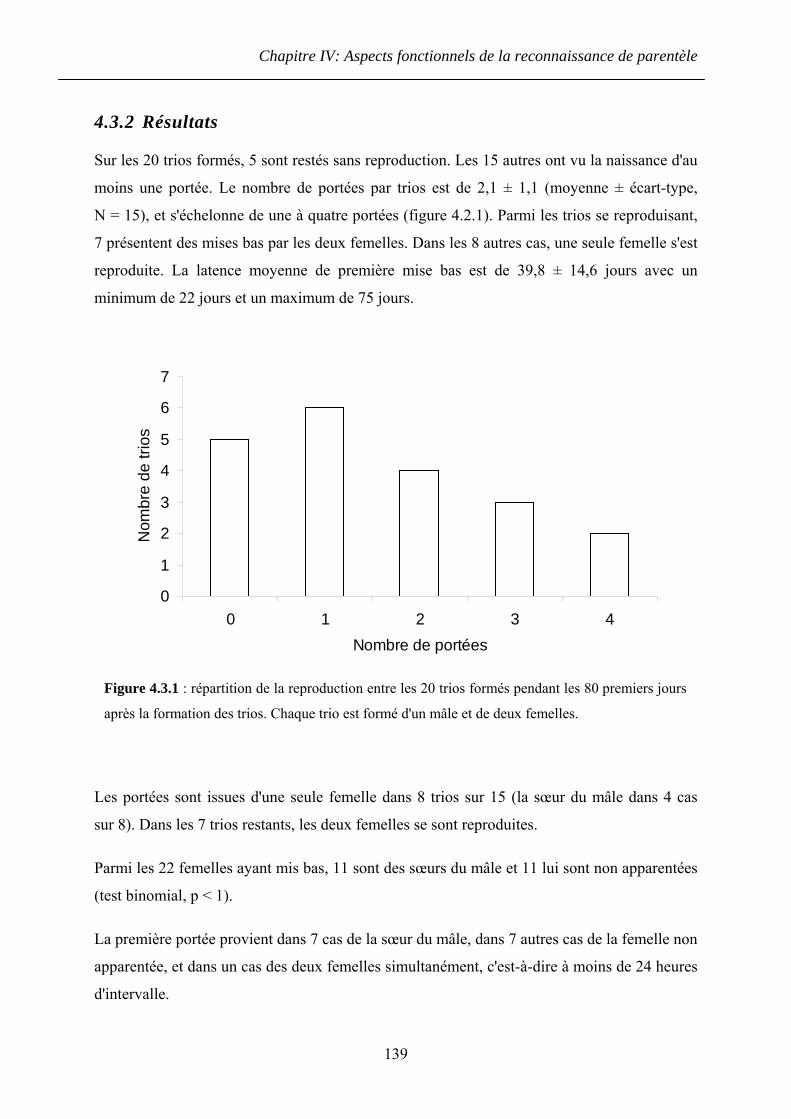
Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
4.3.2 Résultats
Sur les 20 trios formés, 5 sont restés sans reproduction. Les 15 autres ont vu la naissance d'au
moins une portée. Le nombre de portées par trios est de 2,1 ± 1,1 (moyenne ± écart-type,
N = 15), et s'échelonne de une à quatre portées (figure 4.2.1). Parmi les trios se reproduisant,
7 présentent des mises bas par les deux femelles. Dans les 8 autres cas, une seule femelle s'est
reproduite. La latence moyenne de première mise bas est de 39,8 ± 14,6 jours avec un
minimum de 22 jours et un maximum de 75 jours.
0
1
2
3
4
5
6
7
0 1 2 3 4Nombre de portées
Nom
bre
de tr
ios
Figure 4.3.1 : répartition de la reproduction entre les 20 trios formés pendant les 80 premiers jours
après la formation des trios. Chaque trio est formé d'un mâle et de deux femelles.
Les portées sont issues d'une seule femelle dans 8 trios sur 15 (la sœur du mâle dans 4 cas
sur 8). Dans les 7 trios restants, les deux femelles se sont reproduites.
Parmi les 22 femelles ayant mis bas, 11 sont des sœurs du mâle et 11 lui sont non apparentées
(test binomial, p < 1).
La première portée provient dans 7 cas de la sœur du mâle, dans 7 autres cas de la femelle non
apparentée, et dans un cas des deux femelles simultanément, c'est-à-dire à moins de 24 heures
d'intervalle.
139

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
La latence de 1ère mise bas pour les sœurs est de 40,8 ± 14,9 jours contre 41,9 ±14,2 jours
pour les femelles non apparentées. Il n'y a pas de différence significative entre ces deux types
de femelles (test de Mann-whitney, procédure exacte p = 0,739).
Le nombre de petits lors de la première mise bas ne diffère pas non plus entre les sœurs du
mâle (5,22 ± 0,97) et les femelles non apparentées (5,1 ± 2,13) (test de Mann-whitney,
procédure exacte p = 0,818). Les portées simultanées ont été exclues de l'analyse, face à
l'impossibilité d'attribuer les petits à l'une ou l'autre des femelles.
Une analyse plus globale du nombre de petits par portée ne révèle pas non plus de différence
entre les accouplements consanguins (5,38 ± 1 petits par portée) et les autres (5,71 ± 2,4
petits) (test de Mann-whitney, procédure exacte p = 0,532).
4.3.3 Discussion
La conclusion la plus évidente de ces expériences est qu'il n'y a aucune indication d'un
quelconque évitement de l'inceste dans ces conditions expérimentales. Au sein des trios, la
reproduction est enclenchée avec l'une ou l'autre des femelles, de façon indépendante de son
apparentement avec le mâle. Comme le mâle induit la réceptivité sexuelle, il est possible qu'il
se soit accouplé avec la première femelle atteignant l'oestrus. Cependant on peut être surpris
par la latence relativement importante de la première mise bas, qui s'échelonne entre 22 et 76
jours. Soit l'influence du mâle est relativement peu efficace pour induire l'oestrus, soit il existe
des oestrus qui ne donnent pas lieu à des accouplements ou à des fécondations. Si
effectivement les femelles étaient réceptives avant le premier accouplement fécondant, alors
le mâle avait la possibilité de réaliser un choix. De plus, le fait que 5 trios sur 20 soient restés
stériles indique qu'il est peu probable que le mâle ait agi uniquement en fonction de la
réceptivité des femelles. Quelque chose dans la situation sociale a rendu l'activité
reproductrice plus difficile qu'en situation de couple. A titre de comparaison, les 23 couples
formés dans le cadre de l'élevage central du laboratoire début 2004 présentent le bilan
suivant :
• Sur les 23 couples formés, 17 se sont reproduits, soit à peu près la même proportion que
dans les trios (15 / 20). Les trios infertiles sont peut-être dus à une défaillance de certains
mâles, et pas liés directement à la situation de trio.
140

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
• Le nombre de petits dans la première portée est légèrement supérieur (6 ± 1,3) à celui
observé chez les trios (5,3 ± 1,7), sans que cette différence ne soit significative (test de
Mann-whitney, procédure exacte p = 0,116).
• La latence de première mise bas est de 32,2 ± 11,9 jours, ce qui est significativement
moins qu'en situation de trios (test de Mann-whitney, procédure exacte p = 0,009).
La situation de trios semble donc perturber la reproduction chez Mus spicilegus.
Une autre conclusion importante de ces résultats est qu'il n'y a pas de coût apparent lié à
l'inceste, ni en terme de nombre de petits ni en terme de latence de reproduction.
Mais l'enseignement le plus important de ces trios est que même dans une situation de contact
direct entre partenaires, ni le mâle, qui peut théoriquement choisir avec qui s'accoupler, ni la
femelle, qui peut refuser de s'accoupler, ne montre le moindre signe d'évitement de l'inceste.
La reconnaissance de parentèle qui est pourtant possible chez cette espèce ne se traduit pas
par un choix de partenaire adéquat dans ces conditions.
4.4. Discussion générale : choix de partenaire et
apparentement
Un réel choix de partenaire est particulièrement difficile à mettre en évidence chez Mus
spicilegus. D'autres pistes ont été envisagées pour tenter de mettre en évidence la
fonctionnalité de la reconnaissance de parentèle, mais elles ont toutes du être abandonnées
faute de résultats probants lors des pré-tests. Parmi ces pistes, certaines s'adressaient au choix
d'un partenaire social. Par exemple, un dispositif a été construit où des femelles pouvaient
construire un nid commun à travers des grilles les séparant. La femelle du milieu étant
apparentée à l'une de ces voisines, l'hypothèse était qu'elle allait construire plus de ce côté. Or
ce n'est absolument pas le cas. J'ai également voulu induire chez les mâles un comportement
de marquage lorsqu'ils étaient confrontés à des odeurs de femelles, pour mimer ce qui se passe
probablement sur le terrain. La littérature sur le domaine indique qu'il s'agit là d'un aspect
important de la communication sociale chez de nombreuses espèces de Rongeurs (Thomas
& Kaczmarek, 2002 ; Humphries et al., 1999 ; Mech et al., 2003 ; Nevison et al., 2003 ;
Petrulis et al., 1999 ; Thomas, 2002). Cette piste a du être abandonnée, faute d'avoir su
trouver le protocole adéquat pour induire le marquage. La conclusion de ces tentatives
141

Chapitre IV: Aspects fonctionnels de la reconnaissance de parentèle
avortées est qu'il est extrêmement difficile de démontrer la fonctionnalité d'un mécanisme en
laboratoire. L'une des raisons pour cela est que les conditions sont contrôlées au point qu'il
n'existe peu ou pas de pressions de sélection : les animaux sont nourris, protégés du froid, ne
subissent pas la prédation, ne sont pas en compétition pour la nourriture etc. Les animaux qui
en nature pourraient être parasités, malades, affamés, dominés, ou tout autre facteur les
rendant moins intéressants, sont en laboratoire égaux aux autres. De même, la baisse de
l'immunocompétence des petits issus de croisements consanguins n'est pas observable quand
le système immunitaire n'est pas mis à l'épreuve. Ni le choix de partenaire lui-même ni les
conséquences de ce choix en termes de fitness ne peuvent être étudiés de façon convaincante
au laboratoire. Pour réellement mettre en évidence une reconnaissance de parentèle
fonctionnelle, il faudrait travailler sur le terrain ou au moins en conditions semi naturelles.
Il reste néanmoins surprenant que malgré des capacités de discrimination très fines et des
préférences olfactives marquées, aucun signe de reconnaissance de parentèle ne soit
perceptible. Les implications de ces résultats dans la compréhension de l'évolution des
mécanismes de reconnaissance de parentèle et plus généralement d'estimation de la proximité
génétique sont non négligeables, comme nous le verrons dans la discussion générale de cette
thèse.
142

Discussion générale
Discussion générale
143

Discussion générale
Les différentes expériences réalisées dans le cadre de cette thèse ont permis de proposer des
réponses à certaines questions, concernant notamment les informations liées à la proximité
génétique et le mécanisme d'évaluation de celle-ci (chapitres I et II). D'autres questions sont
restées sans réponse : l'interaction de la proximité génétique et des facteurs d'état mériterait un
approfondissement (chapitre III), tout comme les aspects fonctionnels de la perception de la
similarité génétique chez la Souris glaneuse (chapitre IV). Mais la prise en compte de la
globalité des résultats permet d'aborder un domaine plus intangible, celui de l'évolution.
Avant de présenter ma réflexion dans ce domaine, il est important de résumer les principaux
points développés dans cette thèse et de détailler leur articulation.
La reconnaissance de parentèle, comme toute reconnaissance, peut être décomposée en trois
éléments : la production d'éléments discriminants, la perception de ces éléments et l'action
consécutive (Beecher, 1989). L'action (par exemple, l'évitement de la consanguinité ou le
népotisme) n'est possible que s'il y a perception de l'apparentement dans le phénotype des
congénères. Pour qu'il y ait perception de la proximité génétique, il doit exister une
production de signaux révélateurs. Il ne faut pas pour autant penser que cette production est
dirigée, qu'elle a pour but l'évaluation de la proximité génétique. Il serait plus exact (et moins
finaliste) de dire que la perception et l'action sont inféodées à l'existence de signaux
révélateurs de l'apparentement, plutôt qu'à une production dans ce but. Ces signaux
révélateurs de l'apparentement peuvent être de nature très différente.
Particularités des signaux olfactifs
Les signaux olfactifs utilisés par les Souris glaneuses sont-ils directement comparables avec
les autres types de signaux existant dans le règne animal ? En fonction des modalités
sensorielles, les signaux semblent "plus ou moins produits". La communication acoustique est
l'exemple type d'une communication basée sur des signaux produits activement. Le signal
existe quand l'animal décide de vocaliser, striduler, frapper un support etc., l'éventuelle
information présente dans le signal peut alors traduire l'individualité, l'apparentement ou toute
autre information à destination des congénères. Chez les manchots, par exemple, les signaux
acoustiques révèlent l'individualité et permettent la reconnaissance mutuelle entre partenaires
(Aubin et al., 2000). La complexité du signal semble dépendre du contexte écologique de
l'espèce et de la structure sociale : les signaux sont d'autant plus complexes qu'il n'existe pas
de moyen alternatif pour se reconnaître (Jouventin & Aubin, 2002 ; Searby et al., 2004). Ces
144

Discussion générale
signaux acoustiques semblent donc bien être produits "pour" permettre la reconnaissance
individuelle. En termes moins téléologiques, il est plausible que l'avantage sélectif des
individus produisant des vocalisations complexes soit suffisamment fort pour que cette
complexité se soit propagée dans la population. Chez certaines espèces de manchots, la
complexification de la production acoustique est une adaptation à un environnement physique
et social. Est-il possible d'appliquer le même type de raisonnement aux signaux chimiques des
Souris ?
Tout d'abord, il faut noter que le terme de "production" ne semble pas aussi adapté à la
communication olfactive qu'il l'est à la communication acoustique. Bien sûr il existe des
productions odorantes, et le déclenchement d'une activité de marquage, par exemple, ne
semble pas très différent du déclenchement d'une vocalisation. Ce qui demeure cependant
différent c'est qu'au moins une partie du signal olfactif chez les Souris est présent tout le
temps : un oiseau peut être silencieux, mais une souris peut difficilement ne pas sentir. Le
signal est porté par l'animal en permanence.
Certains auteurs envisagent la communication olfactive comme un équivalent pour de
nombreux Mammifères de ce qu’est pour nous la communication visuelle (Beauchamp &
Yamazaki, 2003). Outre mes doutes sur l’aspect heuristique d’une telle comparaison, je ne
pense pas que les signaux olfactifs soient directement comparables avec les autres types de
signaux, qu'ils soient visuels ou acoustiques. Dans le cadre de la reconnaissance de parentèle,
les signaux visuels peuvent être de bons indicateurs de la similarité phénotypique. Chez
l’homme, la ressemblance entre apparentés est une quasi évidence et peut avoir des
conséquences comportementales fortes (la ressemblance perçue augmente la confiance portée
à un individu pourtant non familier (DeBruine, 2002)). Le visage humain présente des
caractéristiques qui ont la particularité d’être stables dans le temps : la longueur du nez,
l’écartement des yeux ou la hauteur des pommettes sont autant de traits invariants, imposés
par la forme du crâne. Bien sûr, un visage peut grossir, vieillir, être maquillé, présenter des
expressions ou des émotions différentes, mais, sauf accident, rien de tout cela ne change la
forme du crâne et l’organisation résultante des éléments du visage. Dans la communication
olfactive des Rongeurs, certaines molécules vont révéler l’état du sujet (réceptivité sexuelle,
statut de dominance, régime alimentaire, parasitisme etc.) et, en conditions de laboratoire,
l’odeur globale permet de révéler la proximité génétique entre individus (Todrank et al.,
1998 ; Heth & Todrank, 2000 ; Heth et al., 2001 ; Busquet & Baudoin, 2004). Cependant, que
145

Discussion générale
ce soit dans la littérature ou dans les résultats exposés dans cette thèse, rien ne permet
d’affirmer que l’odeur globale présente une stabilité quelles que soient les variations
environnementales ou l’état interne de l’animal. Autrement dit, il n’est pas sûr qu’il existe un
"crâne" olfactif, une partie de l’odeur individuelle qui reste constante et sur laquelle les
animaux pourraient se baser pour effectuer une reconnaissance de la proximité génétique
fiable. Même le Complexe Majeur d’Histocompatibilité (MHC), le support génétique de la
réponse immunitaire, ne fournit pas toujours des différences d’odeurs discriminables
(Schellinck et al., 1995). Si le MHC semble toujours s’exprimer, les différences olfactives
induites par un MHC différent semblent d’autant plus discriminables qu’elles sont couplées
avec d’autres différences, notamment d’alimentation (Brown et al., 1996). L'existence de
signaux stables dans l'odeur individuelle en conditions naturelles n'est pas certaine. En
conditions contrôlées, en revanche, l'odeur individuelle est bien représentative d'un individu
(par exemple chez les rats (Gheusi et al., 1997)). Cette individualité de l'odeur est-elle
conservée tout au long de la vie de l'animal, quelque soit ses conditions de vie ? A l'heure
actuelle, il n'existe pas de réponse catégorique à cette question, ni chez les Rongeurs, ni chez
les autres Mammifères. Par exemple, les marquages individuels de femelles Marmosets
Callithris jacchus en conditions contrôlées ne présentent pas de différences qualitatives (pas
de composés révélateurs de l'identité) mais des différences quantitatives : c'est la balance
entre les différents composés qui semble être propre à chaque individu (Smith et al., 2001).
Une étude sur les secrétions de la glande subcaudale des blaireaux Meles meles, prélevées sur
le terrain tout au long de l'année, montre que la composition de ces secrétions est influencée
par la saison, l'âge, le statut reproducteur et la santé de l'animal (Buesching et al., 2002a).
Malgré ces variations, les membres du même groupe tendent à avoir des secrétions proches
(Buesching et al., 2002a). Il semble d'autre part qu'il y ait peu de variation à court terme du
profil individuel, mais de grandes variations saisonnières liées à la biologie des blaireaux
(Buesching et al., 2002b). Dans ces deux exemples, les composés analysés proviennent de
secrétions glandulaires et semblent présenter une relative stabilité, au moins à court terme,
mais cette stabilité ne semble pas due à certains composés "spécialisés". Bien qu'il ne soit pas
possible d'extrapoler ces résultats aux Rongeurs, il est intéressant de constater que les signaux
chimiques d'autres Mammifères ne présentent pas le "crâne" olfactif évoqué plus haut.
Les signaux olfactifs utilisés par les Rongeurs présentent une autre particularité qui les
distingue des signaux visuels ou acoustiques : ils sont liés à des produits de dégradation. Il
146

Discussion générale
existe bien sûr des glandes spécialisées, souvent impliquées dans les comportements de
marquage (Brown, 1985), mais l'urine est un produit métabolique dont au moins l'une des
fonctions est de débarrasser l'organisme des produits de la digestion, des "déchets". Cette
urine est pourtant un support de communication très important chez les Souris (Jones &
Nowell, 1973 ; Davies & Bellamy, 1974 ; Jemiolo et al., 1992).
L'influence des facteurs environnementaux sur l'odeur de l'urine et la coexistence de la
fonction métabolique avec une implication dans la communication interindividuelle incitent à
penser que le signal utilisé par les Souris pour la reconnaissance de la proximité génétique
n'est probablement pas "honnête". Des erreurs sont possibles, et le signalement de la
proximité génétique peut être imparfait. Mais comme l'ont souligné des approches théoriques,
un signal biologique n'a pas besoin d'être honnête pour être maintenu par l’évolution, il doit
être fiable "en moyenne" (Johnstone & Grafen, 1992 ; Johnstone & Grafen, 1993). Comme
déjà évoqué, l'éventail de variations que peut subir une odeur individuelle est un domaine peu
exploré. S'il est plausible qu'une certaine stabilité existe, elle n'est pas nécessairement la
résultante de l'existence de signaux dont cette stabilité serait la fonction et qui seraient
présents en permanence.
La production des signaux olfactifs permettant la reconnaissance de parentèle est d'une nature
particulière, qu'il sera important de prendre en compte dans la compréhension de l'évolution
de la reconnaissance de la proximité génétique.
Perception de la proximité génétique
Les résultats exposés dans les chapitres I et II ont permis de proposer un mécanisme par
lequel les Souris glaneuses peuvent évaluer la proximité génétique soit entre deux individus,
soit d'un individu à elles-mêmes. La base de ce mécanisme est la similarité des odeurs. Du fait
de la covariance odeurs-gènes, la similitude entre les odeurs traduit la proximité génétique,
tout du moins en conditions de laboratoire. Mais les odeurs peuvent être proches pour d'autres
raisons que la proximité génétique, et influencer ainsi ce mécanisme. Dans le chapitre III de
cette thèse, un régime alimentaire légèrement différent (des pommes de deux variétés
distinctes) entraînait une différence olfactive perceptible, puisque les préférences de mâles
pour des odeurs de femelles s’en trouvaient modifiées. Les mâles préféraient ainsi l’odeur
d’une femelle ayant mangé une autre variété de pomme que celle qu’ils avaient eux-mêmes
147

Discussion générale
consommée. Cette préférence basée sur le régime alimentaire n'inversait toutefois pas la
préférence pour une femelle non apparentée par rapport à une sœur. Mais, hormis la variété de
pomme ajoutée à l’alimentation, toutes les autres caractéristiques des femelles étaient
contrôlées. Il serait inexact d’interpréter la préférence pour une femelle non apparentée
malgré son régime alimentaire similaire comme l’existence de signaux invariants permettant
la reconnaissance de parentèle. Dans les conditions naturelles, les odeurs individuelles sont
susceptibles d’être modifiées de façon beaucoup plus drastique que par une simple
modification de la variété de pomme consommée. Si l’odeur individuelle des Souris est en
partie liée au métabolisme, puisque constituée notamment de produits d’excrétion (voir plus
haut), alors les modifications du métabolisme vont se traduire dans l'odeur individuelle. Le
métabolisme est influencé par l’alimentation, par l’état hormonal (lui-même lié au statut
reproducteur et social) et par les conditions environnementales. Quand le régime alimentaire
présente des qualités nutritives variables (contrairement aux variétés de pomme utilisées dans
cette thèse), il peut fortement influencer les préférences olfactives : les campagnols Microtus
pennsylvanicus préfèrent les urines, fèces ou secrétions anogénitales d’individus ayant reçu un
régime riche en protéines par rapport à ceux ayant une alimentation plus pauvre (Ferkin et al.,
1997). Un régime riche en protéines est associé à une meilleure fitness du partenaire potentiel,
puisque corrélé à une maturation plus rapide, une meilleure utilisation de l’espace et de
meilleures capacités de fourragement (Bucyanayandi & Bergeron, 1990 ; Jones, 1990;
Bergeron et al., 1990). Mais ce qui est le plus intéressant dans l’étude des mécanismes de
reconnaissance de parentèle, c’est que les corrélats olfactifs du régime alimentaire semblent
pouvoir interférer avec les préférences liées à la proximité génétique : les jeunes Souris
épineuses préfèrent l'odeur d'une femelle ayant consommé un régime similaire à celui de la
mère à l'odeur d'une femelle nourrie différemment, et ce même si la femelle préférée n'est pas
de la même espèce (Porter & Doane, 1977). Cela suggère la possibilité d'une intégration d'un
facteur environnemental (ici l'alimentation) dans une prise de décision pourtant théoriquement
basée sur la proximité génétique. Cette piste a été pour le moins négligée dans les études sur
la reconnaissance de parentèle, pour des raisons bien compréhensibles. En effet la démarche
scientifique repose le plus souvent sur des expériences dans lesquelles un seul facteur varie à
la fois. Les études sur la reconnaissance de parentèle ont donc présenté aux sujets des
animaux en tout point équivalents, à l’apparentement près. Pourtant certaines hypothèses
concernant les mécanismes de reconnaissance de parentèle supposent un apprentissage d’une
odeur (soit un "template" familial, soit l’odeur propre) dont on sait qu’elle peut être influencée
148

Discussion générale
par des facteurs environnementaux. La modification de cette odeur apprise par ces facteurs
apparaît donc le meilleur moyen de tester ces hypothèses. Ainsi dans l’hypothèse d’un
mécanisme d’assortiment phénotypique, c’est-à-dire l’utilisation par l’animal d’un modèle de
référence appris socialement et utilisé comme point de comparaison pour estimer la distance
d’autres individus à ce modèle et donc indirectement à lui-même (voir Chapitre II), il doit être
possible d’élever des portées génétiquement comparables (par adoptions croisées ou en
utilisant des souches consanguines de laboratoire) et de les élever avec un régime alimentaire
différent. Une fois à l’âge adulte et malgré un retour à une alimentation standard, les
préférences olfactives pourraient se trouver infléchies par le régime alimentaire des donneurs
d’odeur : à apparentement égal, un mâle utilisant l’assortiment phénotypique préférerait
l’odeur d’une femelle ayant un régime alimentaire différent de celui qu’il a lui-même reçu au
moment de la formation du modèle de référence. En modifiant suffisamment le régime
alimentaire (voire d’autres paramètres environnementaux), il est théoriquement possible de
modifier suffisamment le phénotype appris pour inverser les préférences des animaux. Ainsi
un mâle pourrait être amené à considérer une sœur comme une meilleure partenaire sexuelle
potentielle qu’une femelle non apparentée, puisque cette dernière aura été amenée
artificiellement à avoir un phénotype proche de celui appris. L’intérêt d’une telle
expérimentation dépasse la simple démonstration de l’importance de la familiarité et de
l’apprentissage précoce, elle permet théoriquement de jauger l’importance des différents
mécanismes de reconnaissance de parentèle les uns par rapport aux autres, comme nous le
verrons par la suite.
Pressions de sélections et reconnaissance de parentèle
La reconnaissance de parentèle est importante parce qu'il existe des pressions de sélection
liées notamment à l'évitement de la consanguinité. Ce n'est pas le degré d'apparentement entre
les parents en tant que tel qui détermine la force de ces pressions de sélection (cf. Introduction
générale). Ce degré d'apparentement est une donnée généalogique inféodée à la connaissance
par l'observateur des relations familiales entre les animaux. Selon les populations, un même
degré d'apparentement ne va correspondre exactement ni au partage des mêmes gènes ni au
partage de la même proportion du génome. Par exemple, une population qui serait
homozygote à tous les locus (tous les individus présentent les mêmes allèles) n'a aucun intérêt
à éviter l'inceste puisque les accouplements d'une femelle avec un frère ou un non-apparenté
149

Discussion générale
auront comme résultat la production d'individus avec le même génome. Dans un cas moins
extrême (et plus plausible) où la population présente une forte consanguinité, l'accouplement
d'une femelle avec un cousin "combinera" les génomes paternels et maternels de la même
façon qu'une fécondation entre frère et sœur dans une population moins consanguine. En
l'absence d'une connaissance exhaustive de la population étudiée, il est impossible de dresser
une correspondance entre le coefficient d'apparentement r et la proximité génétique réelle,
c'est-à-dire le nombre d'allèles communs entre deux individus. Cette idée est illustrée dans la
figure 5.1. Cette figure essaie de schématiser à quelle proximité génétique "réelle" peut
correspondre un coefficient d'apparentement en fonction du taux de consanguinité de la
population.
1
0,5
0,25
0,12
5
0,06
25 0
0
0,5
199,0%
99,2%
99,4%
99,6%
99,8%
100,0%
prox
imité
gén
étiq
ue
Coefficient d'apparentementTaux de
consanguinité
Figure 5.1 : représentation théorique de la relation entre coefficient d'apparentement et proximité
génétique en fonction du taux de consanguinité de la population étudiée. Les valeurs numériques de la
proximité génétique réelle ne correspondent pas à une réalité. Le minimum de proximité génétique
entre conspécifiques a été fixé arbitrairement à 99 %.
150

Discussion générale
L'apparentement en lui-même n'est probablement pas un guide fiable des choix
comportementaux des animaux dans tous les cas. Ce n'est pas le coefficient d'apparentement
entre les parents qui détermine la fitness de la descendance, c'est la proximité génétique réelle
et surtout, le contexte social. La proximité génétique réelle des parents détermine par exemple
le degré d'hétérozygotie au niveau du MHC, et donc l'efficacité de la réponse immunitaire de
la progéniture. Parmi les membres d'une population, les plus avantagés seront ceux qui ont
effectué les meilleurs choix, même si ces choix ne sont pas optimaux dans l'absolu.
Il n'y a pas à proprement parler de pressions de sélection pour reconnaître l'apparentement. Il
y un avantage évolutif lié à l'accouplement avec un partenaire suffisamment éloigné
génétiquement, mais cet éloignement n'est pas traduit de façon fiable par le coefficient
d'apparentement. Ceci est à mettre en relation avec l'idée que la reconnaissance de parentèle et
la reconnaissance spécifique peuvent être opérées par le même mécanisme, en vertu de la
relation graduelle entre odeurs et gènes (voir chapitre II).
La reconnaissance de parentèle : un seul mécanisme ?
Comme exposé dans le chapitre II, les mécanismes "classiques" ayant été proposés pour
expliquer la reconnaissance de parentèle ne sont pas exclusifs les uns des autres (Tang-
Martinez, 2001 ; Todrank & Heth, 2003). Il est de fait souvent difficile de les différencier à
cause de la concomitance des conditions nécessaires à leur réalisation : par exemple, chez les
Rongeurs en conditions naturelles, les familiers sont des apparentés et l’apprentissage d’un
modèle de référence familial permet théoriquement les mêmes discriminations qu’un
mécanisme auto référencé (Hauber & Sherman, 2003). Mais il est également possible, et
même probable, que ces mécanismes coexistent. Intuitivement il semble presque impossible
que les Rongeurs ne possèdent qu’un seul de ces mécanismes. En effet les aptitudes
sensorielles ou cognitives liées à ces mécanismes sont souvent également liées à d’autres
capacités. La reconnaissance individuelle est bien connue chez les Rongeurs (Bowers &
Alexander, 1967 ; Jacobs & Kuiper, 2000 ; Johnston, 2003 ; Lai & Johnston, 2002). Or être
capable de reconnaître un individu, c’est apprendre tout ou partie de son phénotype (Beecher,
1989). La reconnaissance des familiers est nécessairement associée à une capacité
d’apprentissage de caractéristiques phénotypiques, ce qui semble bien être le pré requis pour
la réalisation de l’assortiment phénotypique : dans les deux cas, il y a comparaison d’un
phénotype rencontré avec un phénotype mémorisé.
151

Discussion générale
La reconnaissance du lieu est un autre des moyens "classiques" de reconnaître les apparentés
(Blaustein, 1983). Cette reconnaissance est intimement liée aux capacités d’orientation
spatiales des animaux, capacités primordiales pour le retour au gîte ou le fourragement.
L'équipement sensoriel et cognitif permettant la reconnaissance du lieu a d'autres utilités que
de reconnaître les apparentés.
Pour le mécanisme qui nous a intéressé durant cette thèse, l’évaluation de la proximité
génétique par la similarité des odeurs (Heth et al., 2003), les capacités requises sont elles aussi
des capacités utiles à d’autres processus.
Evolution de la reconnaissance de la proximité génétique
Reconnaître la proximité génétique est une aptitude intéressante pour les Souris d'un point de
vue théorique. Mais ce n'est pas cette proximité génétique qui est perçue, c'est la similarité des
phénotypes odorants qui l'est. Comment a pu évoluer cette capacité à percevoir les similarités
odorantes entre les odeurs globales? Il est très difficile de répondre à cette question (voir
introduction générale). Il semble toutefois clair que les capacités sensorielles permettant
l'évaluation de la proximité génétique par la similarité des odeurs sont à la base de multiples
autres comportements. L'acuité sensorielle remarquable des Rongeurs a évolué parce qu'elle
procurait un avantage sélectif. Parmi les bénéfices d'une olfaction développée, on peut citer
les discriminations alimentaires, la détection de la présence de compétiteurs ou de prédateurs,
la détection du sexe et de l'état d'un congénère, etc. sans qu'il ne soit possible d'attribuer à
aucune de ces activités la fonction historique d'une olfaction développée. Il en est de même
pour la perception de la similarité des odeurs: l'évolution des capacités perceptives
correspondantes ne peut être attribuée à cette fonction.
A ce jour, on ne connaît pas de neurones sensoriels dans les épithéliums olfactifs qui soient
apparus au cours de l'évolution pour traiter des informations liées à la proximité génétique, ce
qui est logique étant donné que l'existence de ces informations est elle-même douteuse (voir
ci-dessus). Les bases moléculaires de la perception olfactive sont trop complexes pour être
abordées ici – voir cependant (Lancet & Pace, 1987) et (Firestein, 2001) – mais il semble qu'il
n'y ait spécialisation ni des signaux olfactifs ni des récepteurs correspondant à la
reconnaissance de la proximité génétique. Se pourrait-il qu'il existe des capacités cognitives
directement liées à cette reconnaissance ?
152

Discussion générale
Capacités cognitives et reconnaissance de parentèle
Comme il a déjà été évoqué, la comparaison avec l’odeur propre – qui est la base des
mécanismes auto référencés d’estimation de la proximité génétique – n’est pas nécessairement
un processus actif tel que décrit par l’expression "effet aisselles" (voir chapitre II). Il n’est pas
nécessaire de concevoir qu’une souris en rencontrant une autre va enclencher un processus
d’auto inspection – à la suite duquel elle pourra effectuer la comparaison avec l’odeur
rencontrée – pour comprendre ces mécanismes. Une attitude parcimonieuse conduit à
envisager des mécanismes de plus bas niveau, ne faisant pas appel à un processus "conscient".
Parmi ces mécanismes, l’habituation des récepteurs olfactifs semble primordiale : les
récepteurs cellulaires soumis à une stimulation constante vont réagir de moins en moins à
cette stimulation : ils vont s’habituer. Les récepteurs olfactifs d’une souris sont en
permanence soumis à l’odeur individuelle de celle-ci. Cette stimulation n’induit pas (ou plus)
de réponse chez l’animal lui-même, du fait de l’habituation des récepteurs olfactifs. Il est
donc tout à fait possible que l’odeur d’un individu apparenté, donc similaire à l’odeur propre
en vertu de la covariance odeurs-gènes, se démarque peu de cette stimulation basale. La
réaction de l'animal peut donc être graduée selon l'apparentement sans qu'il n'y ait implication
de processus cognitifs de plus haut niveau. Par exemple, pour comprendre l'évitement de
l'inceste, il n'est pas nécessaire de supposer une représentation mentale de ce but
(Shettleworth, 2000). Autrement dit, rien ne suggère qu'il existe une catégorie mentale
"apparentés" similaire à ce qui existe chez l'homme (Jones, 2004).
En revanche, il existe bien un traitement particulier des odeurs sociales chez les Rongeurs
(Young, 2002) : l'odeur d'une autre souris ne peut pas être confondue avec une odeur non
sociale, alimentaire ou autre. Ce traitement différentiel des odeurs sociales englobe t’il
d'autres catégories ? On peut penser que les odeurs sociales peuvent être catégorisées selon le
genre (Schwanz & Lacey, 2003), selon la familiarité (Mateo & Johnston, 2000), mais on ne
sait pas si elles peuvent être catégorisées selon la proximité génétique. Les résultats de cette
thèse suggèrent une éventuelle catégorisation selon la similarité par rapport à l'odeur propre,
mais cette similarité peut être influencée par des facteurs environnementaux et n'est pas
exclusivement liée à la proximité génétique.
153

Discussion générale
Y a t’il réellement reconnaissance de parentèle chez Mus
spicilegus ?
Cette question peut sembler surprenante au vu des capacités démontrées par les Souris dès le
plus jeune âge dans les expériences de préférences (chapitre II). Il y a bien perception d'une
communauté de signaux entre individus proches génétiquement, et cette perception peut se
traduire par une préférence, comme chez les nouveau-nés. Cependant il n'a pas été possible de
mettre en évidence de corrélats fonctionnels de cette perception. Le choix de partenaire a
semblé indépendant de l'apparentement (chapitre IV). De plus, les aspects cognitifs n'ont pas
été abordés expérimentalement. La perception d'une similarité phénotypique entre le sujet et
l'individu rencontré n'est peut-être pas "intégrée". Il faudrait savoir si les Souris peuvent
former une catégorie mentale contenant les apparentées et si oui, sur quelle base. Un des
moyens de répondre à cette question est d'utiliser des tâches d'apprentissage : dans un premier
temps, il faudrait apprendre à un sujet à réagir à l'odeur d'un apparenté (par exemple pour
obtenir de la nourriture) et ensuite présenter l'odeur d'un autre apparenté. Si ce nouvel
individu est traité de la même façon que le premier, alors il y a effectivement une catégorie
mentale. Les choix de stimuli pourraient de plus permettre de préciser les frontières et les
bases de cette catégorie. Le test le plus important serait d'apprendre à l'animal à réagir à
l'odeur d'un cousin paternel puis de voir si cet apprentissage est transféré à un cousin maternel
(ou vice-versa). Les deux cousins n'étant pas apparentés l'un à l'autre, la seule règle
permettant de les "classer" ensemble serait leur apparentement au sujet. Une telle
catégorisation validerait l'hypothèse d'une catégorisation auto référencée. Les capacités des
nouveau-nés à discriminer les apparentés semblent toutefois indiquer qu'une telle
catégorisation n'est pas nécessaire.
Mais l'argument le plus important pour décider si la discrimination des apparentés mérite
l'appellation de reconnaissance de parentèle est le scénario évolutif présidant à son apparition
(Grafen, 1990). En passant en revue les trois aspects de la reconnaissance que sont la
production, la perception et l'action, il apparaît que dans le cas de la reconnaissance de
parentèle, aucun ne semble exister en réponse à une pression de sélection :
• les signaux utilisés pour l'estimation de la proximité génétique sont des conséquences de la
reproduction sexuée. La structuration génétique des populations, espèces ou groupe
154

Discussion générale
d'espèces, fournit tous les éléments nécessaires à la reconnaissance de parentèle mais aussi
à la reconnaissance d'appartenance populationnelle ou spécifique.
• la perception des signaux, elle aussi, est liée à des avantages évolutifs qui dépassent
largement le cadre de la reconnaissance de parentèle.
• l'action consécutive à cette perception n'est pas objectivement graduée selon
l'apparentement, mais selon la proximité génétique réelle.
Supposer qu'un de ces trois niveaux ait été modelé par des pressions de sélection pour la
reconnaissance de parentèle constitue une dérive téléologique. La reconnaissance de la
proximité génétique, traditionnellement scindée en reconnaissance de parentèle et
reconnaissance spécifique, présente toutes les caractéristiques d'une exaptation (Gould &
Lewontin, 1979). Tous les éléments nécessaires au traitement différentiel des apparentés (et
des hétérospécifiques) sont des conséquences de la biologie de l'espèce. A partir de
phénotypes trahissant la proximité génétique et grâce à un système sensoriel permettant de
percevoir ces phénotypes, les animaux peuvent moduler leur comportement et répondre aux
pressions de sélection liées à la prise en compte de l'apparentement. De plus, la structuration
des traits d'histoire de vie (période prénatale et postnatale de cohabitation "obligatoire" avec
les apparentés) et l'existence de stratégies minimisant les risques de consanguinité telles que
la dispersion (Gandon, 1999 ; Ferriere et al., 2000 ; Lehmann & Perrin, 2002) ou l'inhibition
de la reproduction entre familiers (chez Mus spicilegus (Gouat et al., 2003) mais aussi décrite
ou proposée chez de nombreuses espèces y compris l'homme (Weisfeld et al., 2003)) font que
les Souris, Rongeurs voire Mammifères n'ont jamais eu "besoin" de développer un système de
reconnaissance de la proximité génétique per se. Pourtant le signalement de l'identité
génétique est souvent présenté comme une "nécessité". Cette présentation téléologique des
choses a été un frein à la compréhension des mécanismes comportementaux d'évitement de la
consanguinité, en focalisant les travaux sur la recherche de "marqueurs d'apparentement"
(voir les exemples du MHC et des MUPs dans le chapitre I). De plus, en faisant de
l'apparentement une entité primordiale au regard de l'évolution, ces dérives téléologiques ont
cloisonné de façon totalement artificielle les différents champs d'investigation des
comportements socio sexuels des Rongeurs (en les calquant sur des représentations
essentiellement humaines), empêchant une compréhension globale des mécanismes de
reconnaissance olfactive et de leur évolution.
155

Bibliographie
Bibliographie
156

Bibliographie
Aldhous, P. 1989. The effects of individual cross-fostering on the development of intrasexual
kin discrimination in male laboratory mice, Mus musculus L. Animal Behaviour, 37,
741-750.
Allen, C., Bekoff, M. & Lauder, G. 1998. Nature's purposes: analyses of function and design
in biology. Cambridge, Massachusetts: the MIT press.
Arber, W. 2003. Elements for a theory of molecular evolution. Gene, 317, 3-11.
Arcaro, K. F. & Eklund, A. 1999. A review of MHC-based mate preferences and fostering
experiments in two congenic strains of mice. Genetica, 104, 241-244.
Aubin, T., Jouventin, P. & Hildebrand, C. 2000. Penguins use the two-voice system to
recognize each other. Proceedings of the Royal Society of London Series B Biological
Sciences, 267, 1081-1087.
Axelrod, R. & Hamilton, W. D. 1981. The evolution of cooperation. Science, 211, 1390-6.
Bakker, T. C. & Zbinden, M. 2001. Counting on immunity. Nature, 414, 262-263.
Barnard, C. 1999. Still having trouble with relatives. Trends in Ecology and Evolution, 14,
448.
Barnard, C. J. & Fitzsimons, J. 1988. Kin recognition and mate choice in mice : the effects of
kinship, familiarity and social interference on intersexual interaction. Animal
Behaviour, 36, 1078-1090.
Barrett, S. C. H. & Harder, L. D. 1996. Ecology and evolution of plant mating. Trends in
Ecology and Evolution, 11.
Bartolomei, M. S. & Tilghman, S. M. 1997. Genomic imprinting in mammals. Annual Review
of Genetics, 31, 493-525.
Bateson, P. 1982. Preferences for cousins in Japanese quail. Nature, 295, 236-237.
Bateson, P. 1983a. Mate choice. Cambridge University Press.
157

Bibliographie
Bateson, P. 1983b. Optimal outbreeding. In: Mate choice (Ed. by Bateson, P.), pp. 257-277:
Cambridge university Press.
Beauchamp, G. K., Yamazaki, K., Bard, J. & Boyse, E. A. 1988. Preweaning experience in
the control of mating preferences by genes in the major histocompatibility complex of
the mouse. Behavior Genetics, 18, 537-547.
Baudoin, C., Busquet, N., Dobson, F. S., Gheusi, G., Feron, C., Durand, J.-L., Heth, G.,
Patris, B. & Todrank, J. sous presse. Male-Female Associations and Female Olfactory
Neurogenesis with Pair Bonding in Mound-building Mice. Biological Journal of the
Linnean Society.
Beauchamp, G. K. & Yamazaki, K. 2003. Chemical signalling in mice. Biochemical Society
Transactions, 31, 147-150.
Beecher, M. D. 1989. Signalling systems for individual recognition: an information theory
approach. Animal Behaviour, 38, 248-261.
Beecher, M. D. 1991. Successes and failures of parent-offspring recognition in animals. In:
Kin Recognition (Ed. by Hepper, P. G.), pp. 94-124. Cambridge: Cambridge
University Press.
Bekoff, M. & Allen, C. 1995. Teleology, function, design and the evolution of animal
behaviour. Trends in Ecology and Evolution, 10, 253-255.
Bergeron, J. M., Brunet, R. & Jodin, L. 1990. Is space management of female voles (Microtus
pennsylvanicus) related to nutritive quality of plants ? Oecologia, 82, 531-536.
Bergstrom, C. T. & Real, L. A. 2000. Towards a theory of mutual mate choice: Lessons from
two-sided matching. Evolutionary Ecology Research, 2, 493–508.
Bernatchez, L. & Landry, C. 2003. MHC studies in nonmodel vertebrates: what have we
learned about natural selection in 15 years ? Journal of Evolutionary Biology, 16,
363-377.
Beynon, R. J. & Hurst, J. L. 2003. Multiple roles of major urinary proteins in the house
mouse, Mus domesticus. Biochemical Society Transactions, 31, 142-146.
158

Bibliographie
Blaustein, A. R. 1983. Kin recognition mechanisms: Phenotypic matching or recognition
alleles ? American Naturalist, 121, 749-754.
Blaustein, A. R., Bekoff, M. & Daniels, T. J. 1987. Kin recognition in vertebrates (excluding
primates): Empirical evidence. In: Kin recognition in animals (Ed. by Fletcher, D. J. &
Michener, C. D.), pp. 287-331. New York: John Wiley.
Blaustein, A. R., Bekoff, M., Byers, J. A. & Daniels, T. J. 1991. Kin recognition in
vertebrates: what do we really know about adaptative value ?. Animal Behaviour, 41,
1079-1083.
Blumstein, D. T., Steinmetz, J., Armitage, K. B. & Daniel, J. C. 1997. Alarm calling in
yellow-bellied marmots: II. The importance of direct fitness. Animal Behaviour, 53,
173-184.
Bonhomme, F., Catalan, J., Britton-Davidian, J., Chapman, V. M., Moriwaki, K., Nevo, E. &
Thaler, L. 1984. Biochemical diversity and evolution in the genus Mus. Biochemical
Genetics, 22, 275-303.
Bonhomme, F. 1992. Genetic diversity and evolution in the genus Mus. In: Techniques for the
Genetic Analysis of Brain and Behaviour : Focus on the Mouse. (Ed. by Goldowitz,
D., Whalsten, D. & Wimer, R. E.), pp. 41-56. Amsterdam: Elsevier Science
Publishers.
Boursot, P., Auffray, J.-C., Britton-Davidian, J. & Bonhomme, F. 1993. The evolution of
house mice. Annual Review of Ecology and Systematics, 24, 119-152.
Bowers, J. M. & Alexander, B. K. 1967. Mice: individual recognition by olfactory cues.
Science, 158, 1208-1210.
Brennan, P. A. 2004. The nose knows who's who: chemosensory individuality and mate
recognition in mice. Hormones and Behavior, in press.
Bronson, F. H. 1979. The reproductive ecology of the house mouse. Quarterly Review of
Biology, 54, 265–299.
159

Bibliographie
Brooks, R. & Kemp, D. J. 2001. Can older males deliver the good genes ? Trends in Ecology
and Evolution, 16, 308-313.
Brown, R. E. 1985. The rodents II: suborder Myomorpha. In: Social odours in mammals
(Ed. by Brown, R. E. & MacDonald, D. W.), pp. 345-457: Oxford Science
Publications.
Brown, R. E. 1995. What is the role of the immune system in determining individually
distinct body odours ? International Journal of Immunopharmacology, 17, 655-661.
Brown, R. E. & Schellinck, H. M. 1992. Interactions among the MHC, diet and bacteria in the
production of social odors in rodents. In: Chemical Signals in Vertebrates VI (Ed. by
Doty, R. L. & Muller-Schwarze, D.), pp. 175-181. New York: Plenum Press.
Brown, R. E., Schellinck, H. M. & West, A. M. 1996. The influence of dietary and genetic
cues on the ability of rats to discriminate between the urinary odors of MHC-congenic
mice. Physiology and Behavior, 60, 365-372.
Bucyanayandi, J. D. & Bergeron, J. M. 1990. Effects of food quality on feeding patterns of
meadow voles (Microtus pennsylvanicus) along a community gradient. Journal of
Mammalogy, 71, 390-396.
Buesching, C. D., Waterhouse, J. S. & Macdonald, D. W. 2002a. Gas-chromatographic
analyses of the subcaudal gland secretion of the European badger (Meles meles) part I:
chemical differences related to individual parameters. Journal of Chemical Ecology,
28, 41-56.
Buesching, C. D., Waterhouse, J. S. & Macdonald, D. W. 2002b. Gas-chromatographic
analyses of the subcaudal gland secretion of the European badger (Meles meles)
part II: time-related variation in the individual-specific composition. Journal of
Chemical Ecology, 28, 57-69.
Busquet, N. & Baudoin, C. in press. Odour similarities as a basis for discriminating degrees
of kinship in rodents: evidence from Mus spicilegus. Animal Behaviour.
Byers, J. A. & Bekoff, M. 1991. Development, the conveniently forgotten variable in "true
kin recognition". Animal Behaviour, 41, 1088-1090.
160

Bibliographie
Carroll, L. S., Penn, D. J. & Potts, W. K. 2002. Discrimination of MHC-derived odors by
untrained mice is consistent with divergence in peptide-binding region residues.
Proceedings of the National Academy of Sciences U S A, 99, 2187-2192.
Cartwright, J. 2000. Evolution and Human Behaviour. Palgrave.
Chevassus-au-Louis, N. 2001. Qu'est-ce qu'un gène? La Recherche, 348, 50-56.
Clarke, F. M. & Faulkes, C. G. 1999. Kin discrimination and female mate choice in the naked
mole-rat Heterocephalus glaber. Proceedings of the Royal Society of London Series B
Biological Sciences, 266, 1995-2002.
Coulson, J. C. & Thomas, C. S. 1983. Mate choice in the kittiwake gull. In: Mate choice (Ed.
by Bateson, P.), pp. 361-376: Cambridge University Press.
Creel, S. 1993. Why cooperate? Game theory and kin selection. Trends in Ecology and
Evolution, 8, 71-72.
Crow, J. & Kimura, M. 1970. Introduction to Population Genetics Theory. New York: Harper
and Row.
Crozier, R. H. & Pamilo, P. 1996. Evolution of social insect colonies. Oxford: Oxford
University Press.
Curtis, J. T., Liu, Y. & Wang, Z. 2001. Lesions of the vomeronasal organ disrupt mating-
induced pair bonding in female prairie voles (Microtus ochrogaster). Brain Research,
901, 167-174.
D'Amato, F. R. 1994. Physiological evidence for genetically mediated sibling recognition in
mice. Behavior Genetics, 24, 493-496.
Davies, V. J. & Bellamy, D. 1974. Effects of female urine on social investigation in male
mice. Animal Behaviour, 22, 239-241.
Dawkins, R. 1976. The Selfish Gene. Oxford: Oxford University Press.
Dawkins, R. 1982. The extended phenotype. San Francisco: W.H. Freeman.
161

Bibliographie
de Meeus, T., Durand, P. & Renaud, F. 2003. Species concepts: what for ? Trends in
Parasitology, 19, 425-427.
DeBruine, L. M. 2002. Facial resemblance enhances trust. Proceedings of the Royal Society of
London Series B Biological Sciences, 269, 1307-1312.
Dewsbury, D. A. 1988. Kinship, familiarity, aggression, and dominance in deer mice
(Peromyscus maniculatus) in seminatural enclosures. Journal of Comparative
Psychology, 102, 124-128.
Dobson, F. S. & Baudoin, C. 2002. Experimental tests of spatial association and kinship in
monogamous mice (Mus spicilegus) and polygynous mice (Mus musculus
domesticus). Canadian Journal of Zoology, 80, 980-986.
Drickamer, L. C. 1992. Oestrous female house mice discriminate dominant from subordinate
males and sons of dominant from sons of subordinate males by odours cues. Animal
Behaviour, 43, 868-870.
Drickamer, L. C., Gowaty, P. A. & Holmes, C. M. 2000. Free female mate choice in house
mice affects reproductive success and offspring viability and performance. Animal
Behaviour, 59, 371-378.
Drickamer, L. C., Gowaty, P. A. & Wagner, D. M. 2003. Free mutual mate preferences in
house mice affect reproductive success and offspring performance. Animal Behaviour,
65, 105-114.
Dulac, C. & Torello, A. T. 2003. Molecular detection of pheromone signals in mammals:
from genes to behaviour. Nature Reviews Neurosciences, 4, 551-562.
Edmands, S. 2002. Does parental divergence predict reproductive compatibility ? Trends in
Ecology and Evolution, 17, 520-527.
Edwards, S. V. & Hedrick, P. W. 1998. Evolution and ecology of MHC molecules: from
genomics to sexual selection. Trends in Ecology and Evolution, 13, 305-310.
162

Bibliographie
Eggert, F., Holler, C., Luszyk, D., Muller-Ruchholtz, W. & Ferstl, R. 1996. MHC-associated
and MHC-independent urinary chemosignals in mice. Physiology and Behavior, 59,
57-62.
Eggert, F., Muller-ruchholtz, W. & Ferstl, R. 1999. Olfactory cues associated with the major
histocompatibility complex. Genetica, 104, 191-197.
Ehman, K. D. & Scott, M. E. 2001. Urinary odour preferences of MHC congenic female mice,
Mus domesticus: implications for kin recognition and detection of parasitized males.
Animal Behaviour, 62, 781-789.
Ehman, K. D. & Scott, M. E. 2002. Female mice mate preferentially with non-parasitized
males. Parasitology, 125, 461-466.
Eklund, A. C. 1999. Use of the MHC for mate choice in wild house mice (Mus domesticus).
Genetica, 104, 245-248.
Ferguson-Smith, A. C. 2000. Genetic imprinting: silencing elements have their say. Current
Biology, 10, 872-875.
Ferguson-Smith, A. C. & Surani, M. A. 2001. Imprinting and the epigenetic asymmetry
between parental genomes. Science, 293, 1086-1089.
Ferkin, M. H. & Rutka, T. F. 1990. Mechanisms of sibling recognition in meadow voles.
Canadian Journal of Zoology, 68, 609-613.
Ferkin, M. H., Sorokin, E. S., Johnston, R. E. & Lee, C. J. 1997. Attractiveness of scents
varies with protein content of the diet in meadow voles. Animal Behaviour, 53,
133-141.
Feron, C. & Gheusi, G. 2003. Social regulation of reproduction in the female mound-builder
mouse (Mus spicilegus). Physiology and Behavior, 78, 717-722.
Ferriere, I. I., Belthoff, J. R., Olivieri, I. I. & Krackow, I. I. 2000. Evolving dispersal: where
to go next ? Trends in Ecology and Evolution, 15, 5-7.
Firestein, S. 2001. How the olfactory system makes sense of scents. Nature, 413, 211-218.
163

Bibliographie
Fletcher, D. J. & Michener, C. D. 1987. Kin recognition in animals. New York: John Wiley.
Flower, D. R. 1996. The lipocalin protein family: structure and function. Biochemical
Journal, 318 (part 1), 1-14.
Gandon, S. 1999. Kin competition, the cost of inbreeding and the evolution of dispersal.
Journal of Theoretical Biology, 200, 345-364.
Garza, J. C., Dallas, J., Duryadi, D., Gerasimov, S., Croset, H. & Boursot, P. 1997. Social
structure of the mound-building mouse Mus spicilegus revealed by genetic analysis
with microsatellites. Molecular Ecology, 6, 1009-1017.
Gavrilets, S., Arnqvist, G. & Friberg, U. 2001. The evolution of female mate choice by sexual
conflict. Proceedings of the Royal Society of London Series B Biological Sciences,
268, 531-539.
Gharrett, A. J., Smoker, W. W., Reisenbichler, R. R. & Taylor, S. G. 1999. Outbreeding
depression in hybrids between odd- and even-broodyear pink salmon. Aquaculture,
173, 117-129.
Gheusi, G., Goodall, G. & Dantzer, R. 1997. Individually distinctive odours represent
individual conspecifics in rats. Animal Behaviour, 53, 935–944.
Gilder, P. M. & Slater, P. J. 1978. Interest of mice in conspecific male odours is influenced by
degree of kinship. Nature, 274, 364-365.
Gouat, P., Féron, C. & Demouron, S. 2003a. Seasonnal reproduction and delayed sexual
maturity in mound-building mice Mus spicilegus. Reproduction, Fertility and
Development, 15, 187-195.
Gouat, P., Katona, K. & Poteaux, C. 2003b. Is the socio-spatial distribution of mound-
building mice, Mus spicilegus, compatible with a monogamous mating system?
Mammalia, 67, 15-24.
Gould, S. J. & Lewontin, R. C. 1979. The spandrels of San Marco and the Panglossian
paradigm: a critique of the adaptationist programme. Proceedings of the Royal Society
of London Series B Biological Sciences, 205, 581-598.
164

Bibliographie
Gould, S. J. & Vrba, E. S. 1998. Exaptation - A missing term in the science of form. In:
Nature's purposes: analyses of function and design in biology (Ed. by Allen, C.,
Bekoff, M. & Lauder, G.), pp. 519-540: MIT press.
Gowaty, P. A., Drickamer, L. C. & Schmid-Holmes, S. 2003. Male house mice produce fewer
offspring with lower viability and poorer performance when mated with females they
do not prefer. Animal Behaviour, 65, 95–103.
Grafen, A. 1990. Do animals really recognize kin? Animal Behaviour, 39, 42-54.
Grafen, A. 1991. A reply to Byers & Bekoff. Animal Behaviour, 41, 1091-1092.
Grafen, A. 1991. Kin vision?: a reply to Stuart. Animal Behaviour, 41, 1095-1096.
Griffin, A. S. & West, S. A. 2002. Kin selection: fact and fiction. Trends in Ecology and
Evolution, 17, 15-21.
Gruest, N., Richer, P. & Hars, B. 2004. Emergence of long-term memory for conditioned
aversion in the rat fetus. Developmental Psychobiology, 44, 189-198.
Hager, R. & Johnstone, R. A. 2003. The genetic basis of family conflict resolution in mice.
Nature, 421, 533-535.
Haig, D. 1996. Gestational drive and the green-bearded placenta. Proceedings of the National
Academy of Sciences U S A, 93, 6547-6551.
Haig, D. 2000. The kinship theory of genomic imprinting. Annual Review of Ecology and
Systematics, 31, 9-32.
Haig, D. & Wilkins, J. F. 2000. Genomic imprinting, sibling solidarity and the logic of
collective action. Philosophical Transactions of the Royal Society of London series B
Biological Sciences, 355, 1593-1597.
Halpern, M. & Martinez-Marcos, A. 2003. Structure and function of the vomeronasal system:
an update. Progress in Neurobiology, 70, 245-318.
Halpin, Z. T. 1991. Kin recognition cues of vertebrates. In: Kin recognition (Ed. by Hepper,
P. G.), pp. 220-258. Cambridge: Cambridge University Press.
165

Bibliographie
Hamilton, W. D. 1964. The genetical evolution of social behavior. Journal of Theoretical
Biology, 7, 1-52.
Hamilton, W. D., Axelrod, R. & Tanese, R. 1990. Sexual Reproduction as an Adaptation to
Resist Parasites (A Review). Proceedings of the National Academy of Sciences U S A,
87, 3566-3573.
Hare, J. F., Sealy, S. G., Underwood, T. J., Ellison, K. S. & Stewart, R. L. M. 2003. Evidence
of self-referent phenotype matching revisited: airing out the armpit effect. Animal
Cognition, 6, 65-68.
Hauber, M. E. & Sherman, P. W. 1998. Nepotism and marmot alarm calling. Animal
Behaviour, 56, 1049-1052.
Hauber, M. E. & Sherman, P. W. 2000. The armpit effect in hamster kin recognition. Trends
in Ecology and Evolution, 15, 349-350.
Hauber, M. E. & Sherman, P. W. 2001. Self-referent phenotype matching: theoretical
considerations and empirical evidence. Trends in Neurosciences, 24, 609-616.
Hauber, M. E. & Sherman, P. W. 2003. Designing and interpreting experimental tests of self-
referent phenotype matching. Animal Cognition, 6, 69-71.
Hayashi, S. & Kimura, T. 1983. Degree of kinship as a factor regulating preferences among
conspecifics in mice. Animal Behaviour, 31, 81-85.
Hedrick, P. W. 1999. Balancing selection and MHC. Genetica, 104, 207-214.
Hepper, P. G. 1987. The discrimination of different degrees of relatedness in the rat : evidence
for a genetic identifier. Animal Behaviour, 35, 549-554.
Hepper, P. G. 1991a. Kin recognition. Cambridge: Cambridge University Press.
Hepper, P. G. 1991b. Recognizing kin: ontogeny and classification. In: Kin recognition
(Ed. by Hepper, P. G.), pp. 259-288. Cambridge: Cambridge University Press.
Heth, G., Todrank, J. & Johnston, R. E. 1998. Kin recognition in golden hamsters: evidence
for phenotype matching. Animal Behaviour, 56, 409-417.
166

Bibliographie
Heth, G. & Todrank, J. 2000. Individual odour similarities across species parallel
phylogenetic relationships in the S. ehrenbergi superspecies of mole-rats. Animal
Behaviour, 60, 789-795.
Heth, G., Todrank, J., Busquet, N. & Baudoin, C. 2001. Odour-genes covariance and
differential investigation of individual odours in the Mus species complex. Biological
Journal of the Linnean Society, 73, 213-220.
Heth, G., Todrank, J., Busquet, N. & Baudoin, C. 2003. Genetic relatedness assessment
through individual odour similarities in mice. Biological Journal of the Linnean
Society, 78, 595-603.
Hey, J. 2001. The mind of the species problem. Trends in Ecology & Evolution, 16, 326-329.
Hoglund, J., Piertney, S. B., Alatalo, R. V., Lindell, J., Lundberg, A. & Rintamaki, P. T. 2002.
Inbreeding depression and male fitness in black grouse. Proceedings of the Royal
Society of London Series B Biological Sciences, 269, 711-715.
Holmes, W. G. & Sherman, P. W. 1982. The ontogeny of kin recognition in two species of
ground squirrels. American Zoologist, 22, 491–517.
Houle, D. & Kondrashov, A. S. 2002. Coevolution of costly mate choice and condition-
dependent display of good genes. Proceedings of the Royal Society of London Series B
Biological Sciences, 269, 97-104.
Humphries, R. E., Robertson, D. H., Beynon, R. J. & Hurst, J. L. 1999. Unravelling the
chemical basis of competitive scent marking in house mice. Animal Behaviour, 58,
1177-1190.
Hurst, J. L., Payne, C. E., Nevison, C. M., Marie, A. D., Humphries, R. E., Robertson, D. H.,
Cavaggioni, A. & Beynon, R. J. 2001. Individual recognition in mice mediated by
major urinary proteins. Nature, 414, 631-634.
Hurst, J. L., Robertson, D. H. L., Tolladay, U. & Beynon, R. J. 1998. Proteins in urine scent
marks of male house mice extend the longevity of olfactory signals. Animal
Behaviour, 55, 1289-1297.
167

Bibliographie
Isles, A. R., Baum, M. J., Ma, D., Keverne, E. B. & Allen, N. D. 2001. Urinary odour
preferences in mice. Nature, 409, 783-784.
Isles, A. R., Baum, M. J., Ma. D, Szeto, A., Keverne, E. B. & Allen, N. D. 2002. A possible
role for imprinted genes in inbreeding avoidance and dispersal from the natal area in
mice. Proceedings of the Royal Society of London Series B Biological Sciences, 269,
665-670.
Ivantcheva, A. & Cassaing, J. 1999. Male-female interactions and socio-sexual isolation of
Mus spicilegus toward two other species of east-European mice, M. macedonicus and
M. m. musculus. Comptes Rendus de l'Academie des Sciences, Serie III-Sciences de la
Vie, 322, 597-605.
Jacobs, D. S. & Kuiper, S. 2000. Individual recognition in the Damaraland mole-rat,
Cryptomys damarensis (rodentia: Bathyergidae). Journal of Zoology, London, 251,
411-415.
Jeffery, K. J. & Bangham, C. R. 2000. Do infectious diseases drive MHC diversity ?
Microbes and Infection, 2, 1335-1341.
Jemiolo, B., Xie, T. M. & Novotny, M. 1992. Urine marking in male mice: responses
to natural and synthetic chemosignals. Physiology and Behavior, 52, 521-526.
Jennions, M. D. & Petrie, M. 1997. Variation in mate choice and mating preferences: a review
of causes and consequences. Biological Reviews of the Cambridge Philosophical
Society, 72, 283-327.
Jennions, M. D. & Petrie, M. 2000. Why do females mate multiply ? A review of the genetic
benefits. Biological Reviews of the Cambridge Philosophical Society, 75, 21-64.
Johnston, R. E. 2003. Chemical communication in rodents: from pheromones to individual
recognition. Journal of Mammalogy, 84, 1141-1162.
Johnstone, R. A. & Grafen, A. 1992. Error-prone signalling. Proceedings of the Royal Society
of London Series B Biological Sciences, 248, 229-233.
168

Bibliographie
Johnstone, R. A. & Grafen, A. 1993. Dishonesty and the handicap principle. Animal
Behaviour, 46, 759-764.
Jones, D. 2004. The universal psychology of kinship: evidence from language. Trends in
Cognitive Sciences, 8, 211-215.
Jones, E. N. 1990. The effect of forage availability on the home range and population density
of Microtus pennsylvanicus. Journal of Mammalogy, 71, 382-389.
Jones, R. B. & Nowell, N. W. 1973. Aversive effects of the urine of a male mouse upon the
investigatory behaviour of its defeated opponent. Animal Behaviour, 21, 707-710.
Jordan, W. C. & Bruford, M. W. 1998. New perpectives on mate choice and the MHC.
Heredity, 81, 127-133.
Jouventin, P. & Aubin, T. 2002. Acoustic systems are adapted to breeding ecologies:
individual recognition in nesting penguins. Animal Behaviour, 64, 747-757.
Kareem, A. M. & Barnard, C. J. 1982. The importance of kinship and familiarity in social
interaction between mice. Animal Behaviour, 30, 594-601.
Kareem, A. M. & Barnard, C. J. 1986. Kin recognition in mice: age, sex and parental effects.
Animal Behaviour, 34, 1814-1824.
Kavaliers, M., Choleris, E., Agmo, A. & Pfaff, D. W. 2004. Olfactory-mediated parasite
recognition and avoidance: linking genes to behavior. Hormones and Behavior, 46,
272-283.
Kavaliers, M. & Colwell, D. D. 1995. Discrimination by female mice between the odours
of parasitized and non-parasitized males. Proceedings of the Royal Society of London
Series B Biological Sciences, 261, 31-35.
Kavaliers, M., Colwell, D. D., Braun, W. J. & Choleris, E. 2003. Brief exposure to the odour
of a parasitized male alters the subsequent mate odour responses of female mice.
Animal Behaviour, 65, 59-68.
169

Bibliographie
Kavaliers, M., Colwell, D. D. & Choleris, E. 1998. Parasitized female mice display reduced
aversive responses to the odours of infected males. Proceedings of the Royal Society
of London Series B Biological Sciences, 265, 1111-1118.
Keane, B. 1990. The effect of relatedness on reproductive success and mate choice in the
white-footed mouse. Animal Behaviour, 39, 264-273.
Kelemen, D. 1999. Function, goals and intention: children's teleological reasoning about
objects. Trends in Cognitive Sciences, 3, 461-468.
Kokko, H. 2001. Fisherian and "good genes" benefits of mate choice: how (not) to distinguish
between them. Ecology Letters, 4, 322-326.
Keverne, E. B. 2001. Genomic Imprinting, Maternal Care, and Brain Evolution. Hormones
and Behavior, 40, 146–155.
Keverne, E. B. 2002. Mammalian pheromones: from genes to behaviour. Current Biology, 12,
807-809.
Kleiman, D. G. 1977. Monogamy in mammals. Quarterly Review of Biology, 52, 39-69.
Lai, W. S. & Johnston, R. E. 2002. Individual recognition after fighting by golden hamsters:
a new method. Physiology and Behavior, 76, 225-239.
Lambert, G. 2003. La légende des gènes : Anatomie d'un mythe modern e. Paris: Dunod.
Lancet, D. & Pace, U. 1987. The molecular basis of odor recognition. Trends in Brain
Science, 12, 63-64.
Lehmann, L. & Perrin, N. 2002. Altruism, Dispersal, and Phenotype-Matching Kin
Recognition. American Naturalist, 159, 451-458.
Lenington, S. 1994. Of mice, men and the MHC. Trends in Ecology and Evolution, 9,
455-456.
Lewis, K. 1998. Pathogen resistance as the origin of kin altruism. Journal of Theoretical
Biology, 193, 359-363.
170

Bibliographie
MacIntosh Schellinck, H., West, A. M. & Brown, R. E. 1992. Rats can discriminate the urine
odors of genetically identical mice maintained on different diets. Physiology and
Behavior, 51, 1079-1082.
Manning, C. J., Wakeland, E. K. & Potts, W. K. 1992. Communal nesting patterns in mice
implicate MHC genes in kin recognition. Nature, 360, 581-583.
Marshall, J. A. & Rowe, J. E. 2003. Kin selection may inhibit the evolution of reciprocation.
Journal of Theoretical Biology, 222, 331-335.
Marshall, T. C., Coltman, D. W., Pemberton, J. M., Slate, J., Spalton, J. A., Guinness, F. E.,
Smith, J. A., Pilkington, J. G. & Clutton-Brock, T. H. 2002. Estimating the prevalence
of inbreeding from incomplete pedigrees. Proceedings of the Royal Society of London
Series B Biological Sciences, 269, 1533-1539.
Mateo, J. M. & Johnston, R. E. 2000a. Kin recognition and the “armpit effect”: evidence of
self-referent phenotype matching. Proceedings of the Royal Society of London Series
B Biological Sciences, 267, 695-700.
Mateo, J. M. & Johnston, R. E. 2000b. Retention of social recognition after hibernation
in Belding's ground squirrels. Animal Behaviour, 59, 491-499.
Mateo, J. M. & Johnston, R. E. 2003. Kin recognition by self-referent phenotype matching:
weighing the evidence. Animal Cognition, 6, 73-76.
McClelland, E. E., Granger, D. L. & Potts, W. K. 2003. Major histocompatibility complex-
dependent susceptibility to Cryptococcus neoformans in mice. Infection and Immunity,
71, 4815-4817.
McClelland, E. E., Penn, D. J. & Potts, W. K. 2003. Major histocompatibility complex
heterozygote superiority during coinfection. Infection and Immunity, 71, 2079-2086.
Mech, S. G., Dunlap, A. S. & Wolff, J. O. 2003. Female prairie voles do not choose males
based on their frequency of scent marking. Behavioural Processes, 61, 101-108.
Milinski, M. & Wedekind, C. 2001. Evidence for MHC correlated perfume preferences
in humans. Behavioral Ecology, 12, 140-149.
171

Bibliographie
Moles, A. & D'Amato F, R. 2000. Ultrasonic vocalization by female mice in the presence
of a conspecific carrying food cues. Animal Behaviour, 60, 689-694.
Montag, S., Frank, M., Ulmer, H., Wernet, D., Gopel, W. & Rammensee, H. G. 2001.
"Electronic nose" detects major histocompatibility complex-dependent prerenal and
postrenal odor components. Proceedings of the National Academy of Sciences USA,
98, 9249-9254.
Naumov NP. 1940. Ecology of the mound-builder mouse Mus musculus hortulanus. Works
Institute Evolution Morphology USSR 3: 33-76.
Nevison, C. M., Armstrong, S., Beynon, R. J., Humphries, R. E. & Hurst, J. L. 2003. The
ownership signature in mouse scent marks is involatile. Proceedings of the Royal
Society of London Series B Biological Sciences, 270, 1957-1963.
Ninomiya, K. & Brown, R. E. 1995. Removal of the preputial glands alters the individual
odors of male MHC-congenic mice and the preferences of females for these odors.
Physiology and Behavior, 58, 191-194.
Novotny, M. V., Jemiolo, B., Wiesler, D., Ma, W., Harvey, S., Xu, F., Xie, T. M. & Carmack,
M. 1999. A unique urinary constituent, 6-hydroxy-6-methyl-3-heptanone, is a
pheromone that accelerates puberty in female mice. Chemistry and Biology, 6, 377-
383.
O'Connor, T. P., Lee, A., Jarvis, J. U. & Buffenstein, R. 2002. Prolonged longevity in naked
mole-rats: age-related changes in metabolism, body composition and gastrointestinal
function. Comparative Biochemistry and Physiology Part A, 133, 835-842.
O'Grady, R. T. 1984. Evolutionary theory and teleology. Journal of Theoretical Biology, 107,
563-578.
Orsini, P., Bonhomme, F., Britton-Davidian, J., Croset, H., Gerasimov, S. & Thaler, L. 1983.
Le complexe d'epèces du genre Mus en Europe Centrale et Orientale. II. Critères
d'identification, répartition et caractéristiques écologiques. Zeitshrift für
Säugetierkunde, 48, 86-95.
172

Bibliographie
Osada, K., Yamazaki, K., Curran, M., Bard, J., Smith, B. P. C. & Beauchamp, G. K. 2003.
The scent of age. Proceedings of the Royal Society of London Series B Biological
Sciences, 270, 929-933.
Paterson, S. & Pemberton, J. M. 1997. No evidence for major histocompatibility complex-
dependent mating patterns in a free-living ruminant population. Proceedings of the
Royal Society of London Series B Biological Sciences, 264, 1813-1819.
Paterson, S., Wilson, K. & Pemberton, J. M. 1998. Major histocompatibility complex
variation associated with juvenile survival and parasite resistance in a large
unmanaged ungulate population. Proceedings of the National Academy of Sciences
USA, 95, 3714-3719.
Patris, B. & Baudoin, C. 1998. Female sexual preferences differ in Mus spicilegus and Mus
musculus domesticus: the role of familiarization and sexual experience. Animal
Behaviour, 56, 1465-1470.
Patris, B. & Baudoin, C. 2000. A comparative study of parental care between two rodent
species: implications for the mating system of the mound-building mouse Mus
spicilegus. Behavioural Processes, 51, 35-43.
Patris, B., Gouat, P., Jacquot, C., Christophe, N. & Baudoin, C. 2002. Agonistic and sociable
behaviors in the mound-building mice, Mus spicilegus: a comparative study with Mus
musculus domesticus. Aggressive Behavior, 28, 75-84.
Pearse-Pratt, R., Schellinck, H., Brown, R., Singh, P. B. & Roser, B. 1999. Soluble MHC
antigens and olfactory recognition of genetic individuality: the mechanism. Genetica,
104, 223-230.
Penn, D. 2002. The scent of genetic compatibility: sexual selection and the Major
Histocompatibility Complex. Ethology, 108, 1-21.
Penn, D. & Potts, W. 1998a. Chemical signals and parasite-mediated sexual selection. Trends
in Ecology and Evolution, 13, 391-396.
173

Bibliographie
Penn, D. & Potts, W. 1998b. MHC-disassortative mating preferences reversed by cross-
fostering. Proceedings of the Royal Society of London Series B Biological Sciences,
265, 1299-1306.
Penn, D. & Potts, W. 1999. The Evolution of Mating Preferences and Major
Histocompatibility Complex Genes. American Naturalist, 153, 145-164.
Penn, D. & Potts, W. K. 1998c. Untrained mice discriminate MHC-determined odors.
Physiology and Behavior, 64, 235-243.
Penn, D., Schneider, G., White, K., Slev, P. & Potts, W. 1998. Influenza infection neutralizes
the attractiveness of male odour to female mice (Mus musculus). Ethology, 104,
685-694.
Penn, D. J., Damjanovich, K. & Potts, W. K. 2002. MHC heterozygosity confers a selective
advantage against multiple-strain infections. Proceedings of the National Academy of
Sciences U S A, 99, 11260-11264.
Petrulis, A., Peng, M. & Johnston, R. E. 1999. Effects of vomeronasal organ removal on
individual odor discrimination, sex-odor preference, and scent marking by female
hamsters. Physiology and Behavior, 66, 73-83.
Pichot, A. 1999. Histoire de la notion de gène. Paris : Flammarion.
Porter, R. H., Deni, R. & Doane, H. M. 1977. Responses of Acomys cahirinus pups to
chemical cues produced by a foster species. Behavioral Biology, 20, 244-251.
Porter, R. H. & Doane, H. M. 1977. Dietary-dependent cross-species similarities in maternal
chemical cues. Physiology and Behavior, 19, 129-131.
Porter, R. H., Matochik, J. A. & Makin, J. W. 1983. Evidence for phenotype matching in
spiny mice (Acomys cahirinus). Animal Behaviour, 31, 978-984.
Potts, W. K., Manning, C. J. & Wakeland, E. K. 1991. Mating patterns in seminatural
populations of mice influenced by MHC genotype. Nature, 352, 619-621.
Proulx, S. R., Day, T. & Rowe, L. 2002. Older males signal more reliably. Proceedings of the
Royal Society of London Series B Biological Sciences, 269, 2291-2299.
174

Bibliographie
Pusey, A. & Wolf, M. 1996. Inbreeding avoidance in animals. Trends in Ecology and
Evolution, 11, 201-206.
Queller, D. C. & Strassmann, J. E. 2002. Kin selection. Current Biology, 12, 832.
Ratcliffe, M. 2000. The function of function. Studies in History and Philosophy of Biological
and Biomedical Sciences, 31, 113-133.
Ravary, F. 2003. Comportement et organisation sociale de la fourmi parthénogénétique
Cerapachys biroi (Forel). Thèse de doctorat. pp. 202 : Université Paris 13 -
Villetaneuse.
Reik, W. & Walter, J. 2001. Genomic imprinting: parental influence on the genome. Nature
Reviews Genetics, 2, 21-32.
Reusch, T. B. H., Haberli, M. A., Aeschlimann, P. B. & Milinski, M. 2001. Female
sticklebacks count alleles in a strategy of sexual selection explaining MHC
polymorphism. Nature, 414, 300-302.
Roberts, S. C. & Gosling, L. M. 2003. Genetic similarity and quality interact in mate choice
decisions by female mice. Nature Genetics, 35, 103-106.
Robertson, D. H., Cox, K. A., Gaskell, S. J., Evershed, R. P. & Beynon, R. J. 1996. Molecular
heterogeneity in the Major Urinary Proteins of the house mouse Mus musculus.
Biochemical Journal, 316 ( Pt 1), 265-272.
Rossiter, S. J., Jones, G., Ransome, R. D. & Barratt, E. M. 2001. Outbreeding increases
offspring survival in wild greater horseshoe bats (Rhinolophus ferrumequinum).
Proceedings of the Royal Society of London Series B Biological Sciences, 268,
1055-1061.
Rulicke, T., Chapuisat, M., Homberger, F. R., Macas, E. & Wedekind, C. 1998. MHC-
genotype of progeny influenced by parental infection. Proceedings of the Royal
Society of London Series B Biological Sciences, 265, 711-716.
Ruse, M. 1989. Teleology in biology: is it a cause for concern ? Trends in Ecology and
Evolution, 4, 51-54.
175

Bibliographie
Ruse, M. 2000. Teleology: Yesterday, Today, and Tomorrow ? Studies in History and
Philosophy of Biological and Biomedical Sciences, 31, 213-232.
Ryan, M. J., Phelps, S. M. & Rand, A. S. 2001. How evolutionary history shapes recognition
mechanisms. Trends in Cognitive Sciences, 5, 143-148.
Sage, R. D., Atchley, W. R. & Capanna, E. 1993. House mice as models in systematic
biology. Systematic Biology, 42, 523-561.
Schellinck, H. M. & Brown, R. E. 2000. Selective depletion of bacteria alters but does not
eliminate odors of individuality in Rattus norvegicus. Physiology and Behavior, 70,
261-270.
Schellinck, H. M., Rooney, E. & Brown, R. E. 1995. Odors of individuality of germfree mice
are not discriminated by rats in a habituation-dishabituation procedure. Physiology and
Behavior, 57, 1005-1008.
Schwanz, L. E. & Lacey, E. A. 2003. Olfactory discrimination of gender by colonial tuco-
tucos (Ctenomys sociabilis). Mammalian Biology, 68, 53-60.
Searby, A., Jouventin, P. & Aubin, T. 2004. Acoustic recognition in macaroni penguins: an
original signature system. Animal Behaviour, 67, 615-625.
Shanahan, T. 2001. Methodological and Contextual Factors in the Dawkins/Gould Dispute
Over Evolutionary Progress. Studies in History and Philosophy of Biological and
Biomedical Sciences, 32, 127-151.
Shettleworth, S. J. 2000. Animal cognition and animal behaviour. Animal Behaviour, 61,
277-286.
Simeonovska-Nikolova, D. M. 2000. Strategies in open field behaviour of Mus spicilegus and
Mus musculus musculus. Belgian Journal of Zoology, 130, 115-120.
Singer, A. G., Beauchamp, G. K. & Yamazaki, K. 1997. Volatile signals of the major
histocompatibility complex in male mouse urine. Proceedings of the National
Academy of Sciences U S A, 94, 2210-2214.
176

Bibliographie
Singh, P. B. 1999. The present status of the 'carrier hypothesis' for chemosensory recognition
of genetic individuality. Genetica, 104, 231-233.
Singh, P. B. 2001. Chemosensation and genetic individuality. Reproduction, 121, 529-539.
Singh, P. B., Herbert, J., Roser, B., Arnott, L., Tucker, D. K. & Brown, R. E. 1990. Rearing
rats in a germ-free environment eliminates their odors of individuality. Journal of
Chemical Ecology, 16, 1667-1681.
Slate, J., Kruuk, L. E., Marshall, T. C., Pemberton, J. M. & Clutton-Brock, T. H. 2000.
Inbreeding depression influences lifetime breeding success in a wild population of red
deer (Cervus elaphus). Proceedings of the Royal Society of London Series B
Biological Sciences, 267, 1657-1662.
Smith, J., Hurst, J. L. & Barnard, C. J. 1994. Comparing behaviour in wild and laboratory
strains of the house mouse: Levels of comparison and functional inference.
Behavioural Processes, 32, 79-86.
Smith, J. 1999. The cuckoo’s contested history. Trends in Ecology and Evolution, 14, 415.
Smith, T. E., Tomlinson, A. J., Mlotkiewicz, J. A. & Abbott, D. H. 2001. Female marmoset
monkeys (Callithrix jacchus) can be identified from the chemical composition of their
scent marks. Chemical Senses, 26, 449-458.
Sokolov, V. E., Kotenkova, E. V. & Michailenko, A. G. 1998. Mus spicilegus. Mammalian
Species, 592, 1-6.
Stuart, R. J. 1991. Kin recognition as a functional concept. Animal Behaviour, 41, 1093-1094.
Sun, L. & Muller-Schwarze, D. 1997. Sibling recognition in the beaver: a field test for
phenotype matching. Animal Behaviour, 54, 493–502.
Tang-Martinez, Z. 2001. The mechanisms of kin discrimination and the evolution of kin
recognition in vertebrates: a critical re-evaluation. Behavioural Processes, 53, 21-40.
Thomas, S. A. 2002. Scent marking and mate choice in the prairie vole, Microtus ochrogaster.
Animal Behaviour, 63, 1121-1127.
177

Bibliographie
Thomas, S. A. & Kaczmarek, B. K. 2002. Scent-marking behaviour by male prairie voles,
Microtus ochrogaster, in response to the scent of opposite- and same-sex conspecifics.
Behavioural Processes, 60, 27-33.
Todrank, J. & Heth, G. 2001. Rethinking cross-fostering designs for studying kin recognition
mechanisms. Animal Behaviour, 61, 503-505.
Todrank, J. & Heth, G. 2003. Odor-Genes Covariance and Genetic Relatedness Assessments:
Rethinking Odor-Based "Recognition" Mechanisms in Rodents. Advances in the study
of behavior, 52, 77-130.
Todrank, J., Heth, G. & Johnston, R. E. 1998. Kin recognition in golden hamsters: evidence
for kinship odours. Animal Behaviour, 55, 377-386.
Toivanen, P., Vaahtovuo, J. & Eerola, E. 2001. Influence of major histocompatibility complex
on bacterial composition of fecal flora. Infection and Immunity, 69, 2372-2377.
Tregenza, T. & Wedell, N. 2000. Genetic compatibility, mate choice and patterns of
parentage: invited review. Molecular Ecology, 9, 1013-1027.
Trivers, R. L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology, 46,
35-47.
Van Valen, L. 1973. A new evolutionary law. Evolutionary Theory, 1, 1-30.
Weber, T. P. 2000. Biological objects, units of selection and character decomposition. Trends
in Ecology and Evolution, 15, 304-305.
Wedekind, C. & Furi, S. 1997. Body odour preferences in men and women: do they aim for
specific MHC combinations or simply heterozygosity ? Proceedings of the Royal
Society of London Series B Biological Sciences, 264, 1471-1479.
Wedekind, C. & Penn, D. 2000. MHC genes, body odours, and odour preferences.
Nephrology Dialysis Transplantation, 15, 1269-1271.
Weisfeld, G. E., Czilli, T., Phillips, K. A., Gall, J. A. & Lichtman, C. M. 2003. Possible
olfaction-based mechanisms in human kin recognition and inbreeding avoidance.
Journal of Experimental Child Psychology, 85, 279-295.
178

Bibliographie
Wiens, J. J. 2001. Widespread loss of sexually selected traits: how the peacock lost its spots.
Trends in Ecology & Evolution, 16, 517-523.
Willis, C. & Poulin, R. 2000. Preference of female rats for the odours of non-parasitised
males: the smell of good genes ? Folia Parasitologica (Praha), 47, 6-10.
Wouters, A. G. 2003. Four notions of biological function. Studies in History and Philosophy
of Biological and Biomedical Sciences, 34, 633-668.
Yamazaki, K., Beauchamp, G. K., Kupniewski, D., Bard, J., Thomas, L. & Boyse, E. A. 1988.
Familial imprinting determines H-2 selective mating preferences. Science, 240,
1331-1332.
Yamazaki, K., Beauchamp, G. K., Curran, M., Bard, J. & Boyse, E. A. 2000. Parent-progeny
recognition as a function of MHC odortype identity. Proceedings of the National
Academy of Sciences USA, 97, 10500-10502.
Yamazaki, K., Beauchamp, G. K., Imai, Y., Bard, J., Phelan, S. P., Thomas, L. & Boyse, E.
A. 1990. Odor types determined by the major histocompatibility complex in germfree
mice. Proceedings of the National Academy of Sciences U S A, 87, 8413-8416.
Yamazaki, K., Singer, A. G. & Beauchamp, G. K. 1999. Origin, functions and chemistry
of H-2 regulated odorants. Genetica, 104, 235-240.
Young, L. J. 2002. The neurobiology of social recognition, approach, and avoidance.
Biological Psychiatry, 51, 18-26.
Zavazava, N. & Eggert, F. 1997. MHC and behavior. Trends in Immunology, 18, 8-10.
Zschotte, S. & Baur, B. 2002. Inbreeding, outbreeding, infant growth, and size dimorphism
in captive Indian rhinoceros (Rhinoceros unicornis). Canadian Journal of Zoology,
80, 2014-2023.
179

Perception de la proximité génétique chez la souris glaneuse Mus spicilegus : mécanismes et fonctions Résumé: La distance génétique entre individus est un paramètre fondamental dans l'élaboration des théories en biologie évolutive. Elle est en effet un bon indicateur des pressions de sélection liées au choix socio-sexuels des animaux. Au niveau spécifique la proximité génétique est souvent assimilée à l'apparentement. Les animaux qui aident leurs proches apparentés mais ne s'accouplent pas avec sont favorisés, d'une part parce qu'ils augmentent leur inclusive fitness, d'autre part parce que leur descendance ne souffre pas des conséquences de la consanguinité. Cette capacité à traiter les congénères différemment en fonction de leur proximité génétique est très répandue et peut être la résultante de multiples stratégies. Chez la souris glaneuse Mus spicilegus, comme chez tous les Rongeurs, l'olfaction est un canal de communication privilégié. Les expériences présentées dans cette thèse démontrent l'existence d'une relation graduelle entre proximité génétique et similarité des odeurs individuelles. Cette covariance odeurs-gènes existe aussi bien au niveau des relations de parenté qu'au niveau phylogénétique. Lors de tests de choix, les préférences des souris sont guidées par la proximité génétique entre le sujet et les donneurs d'odeurs. Cette capacité à discriminer les odeurs d'apparentés est présente dès la naissance et semble ne pas nécessiter d'apprentissage des caractéristiques familiales. Le rôle de facteurs d'état sur cette évaluation de la proximité génétique par la similarité des odeurs est ensuite étudié, avant d'envisager les conséquences fonctionnelles de cette capacité. Parallèlement, cette thèse s'articule autour d'une réflexion sur l'évolution de la reconnaissance de la proximité génétique. Ni les signaux révélateurs de la proximité génétique ni la capacité à les percevoir ne semblent avoir été sélectionnés pour permettre le traitement différentiel des apparentés ou même des conspécifiques. La confrontation de cette réflexion aux résultats expérimentaux permet d'envisager la perception de la proximité génétique chez les Rongeurs comme une exaptation. Perception of genetic relatedness in the mound-building mouse Mus spicilegus: mechanisms and functions Abstract: In evolutionary biology, genetic relationships are supposed to modulate interactions between individuals, because they are linked with selective pressures on social and sexual behaviours. Animals that help close relatives but do not mate with them increase their inclusive fitness while avoiding the cost of inbreeding. The ability to discriminate relatives from non-relatives is widespread across the animal kingdom, and can result from many different behavioural strategies. Mound-building mice Mus spicilegus, like all rodents, use olfaction as a primary means of communication. The experiments presented in this dissertation demonstrate a continuous relationship between individual odour similarities and genetic relatedness. This odour-genes covariance is found at the kinship level as well as across species. In choice tests, mice respond based on their relatedness with the odour donors. This ability, present in adult and newborn mice, does not seem to rely on learning because it is demonstrated in experimental designs precluding the use of a learned familial template. Interactions of state factors with genetic relatedness assessment are considered, and the use of this ability in mate choice experiments is investigated. This dissertation also addresses theoretical concerns about the evolution of kin recognition. This ability does not seem to result from direct selection on the olfactory signals or the sensory apparatus to perceive them, and can hardly be considered an adaptation. Instead, evidence reported in this PhD and in the literature suggests that the perception of genetic relatedness in rodents is more accurately described as an exaptation. Discipline: Biologie du comportement Mots-clés: Mus spicilegus, Rongeurs, souris sauvages, comportement, odeurs, apparentement, reconnaissance de parentèle, fonction, adaptation, téléologie. Laboratoire d'Ethologie Expérimentale et Comparée – Université Paris-XIII – 93430 Villetaneuse