micro-anatomie des tissus lymphoÏdes...
TRANSCRIPT

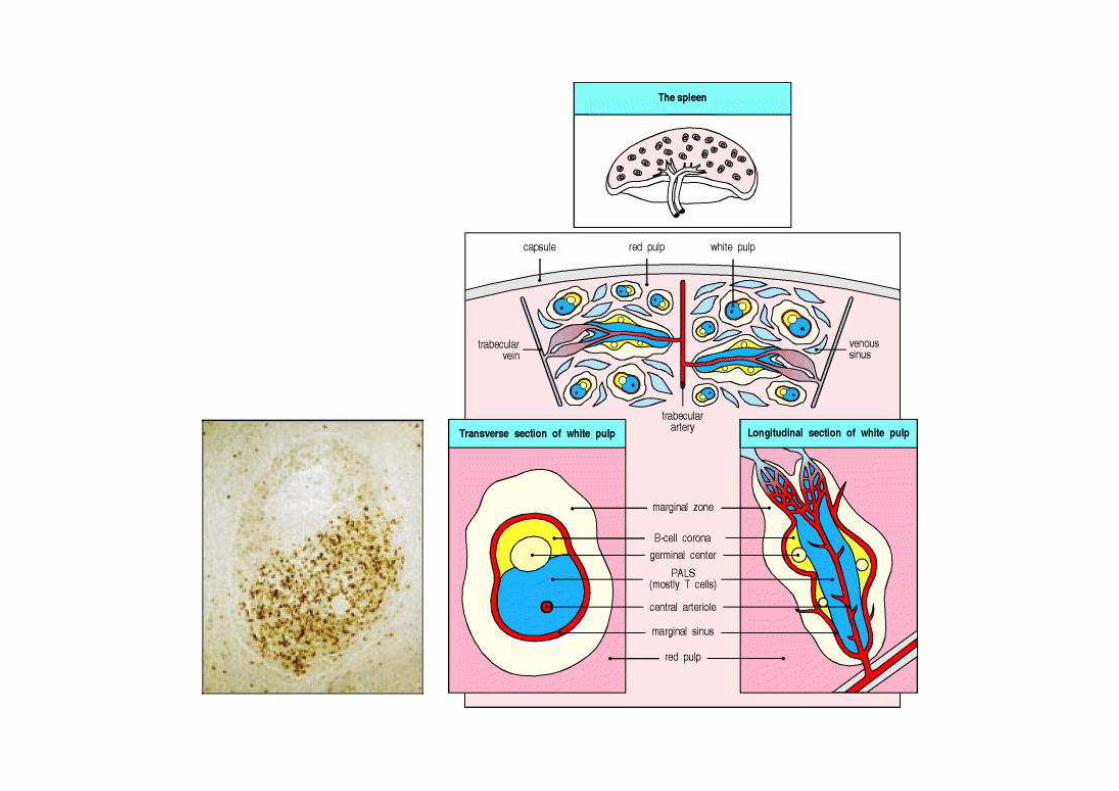
MICRO-ANATOMIE DES TISSUS LYMPHOÏDES SECONDAIRES

LES DEUX PHASES DU DÉVELOPPEMENT LYMPHOCYTAIRE B
Ag
VH-DH-JH
LYMPHOPOÏÈSE(tissus lymphoïdes foetaux et moelle osseuse )
VL-JL
cellule souche
pro-B pre-B
VH-DHJH
VL-JL
LB immature
VHDHJH
VL-JL
µ+ ΨΨΨΨCL IgM
LB mature naïf
IgD IgM
VHDHJH
VLJL
VHDHJH
VLJL
IMMUNOPOÏÈSE(tissus lymphoïdes secondaires)
centre germinatif
centroblaste centrocyte
LB à mémoire(Igs mutées)
plasmocyte(Igs mutées)
plasmocyte(Igs non mutées)

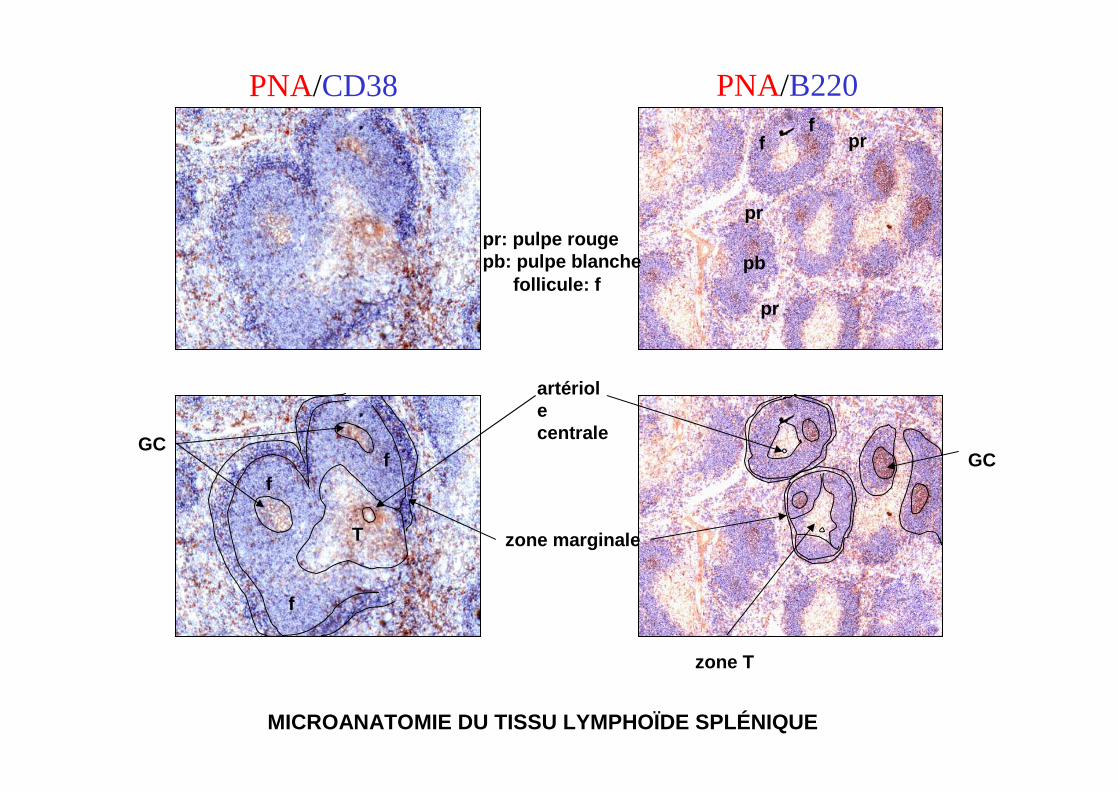
MICROANATOMIE DU TISSU LYMPHOÏDE SPLÉNIQUE
follicule: fpr
PNA/CD38 PNA/B220
pr
prpr: pulpe rougepb: pulpe blanche
zone marginale
ff
zone T
T
pb
artériolecentrale
f
f
f GCGC
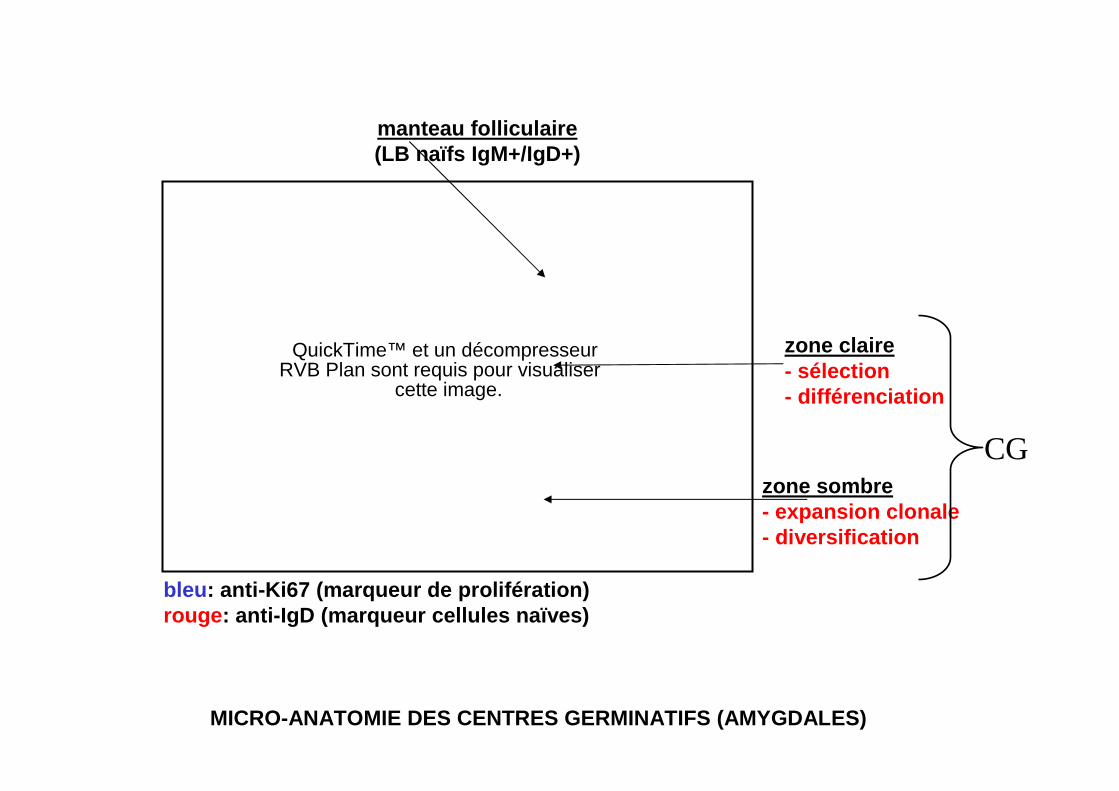
QuickTime™ et un décompresseurRVB Plan sont requis pour visualiser
cette image.
zone sombre- expansion clonale- diversification
zone claire- sélection- différenciation
manteau folliculaire(LB naïfs IgM+/IgD+)
bleu : anti-Ki67 (marqueur de prolifération)rouge : anti-IgD (marqueur cellules naïves)
MICRO-ANATOMIE DES CENTRES GERMINATIFS (AMYGDALES)
CG

RÉCEPTEURS MEMBRANAIRES IMPLIQUÉS DANS L ’ACTIVATION DES LYMPHOCYTES B
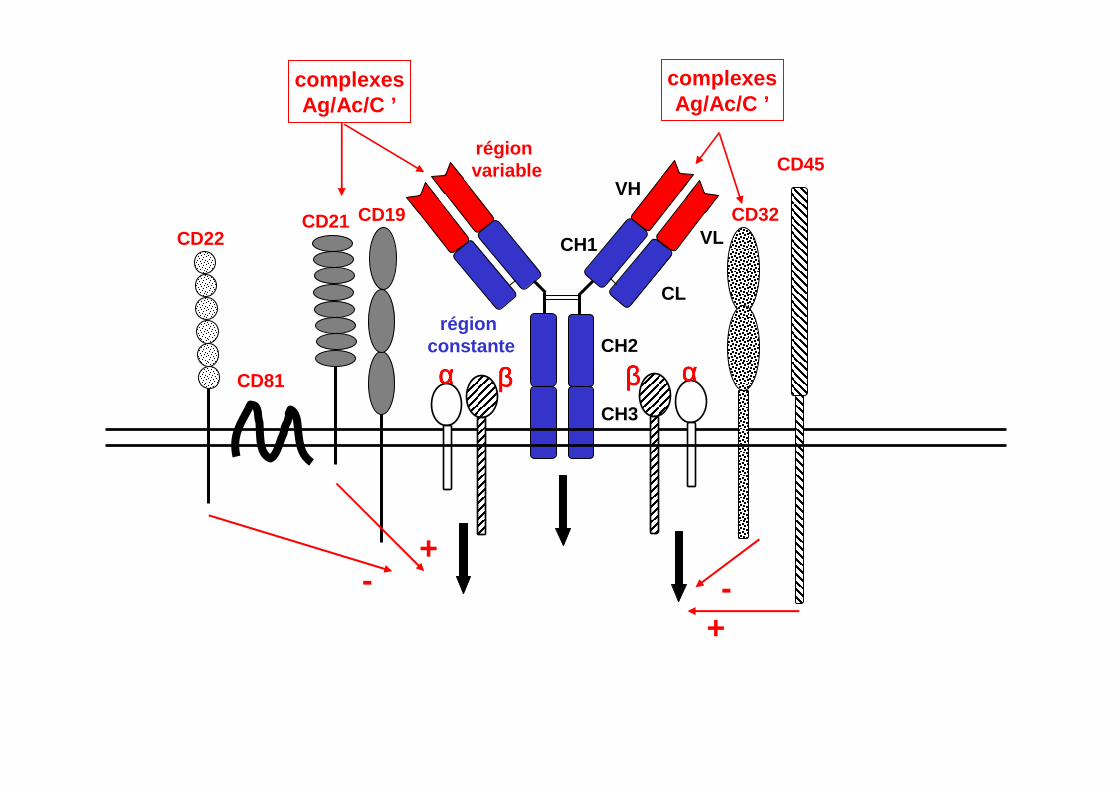
CH3
CH2région
constante
VH
VLCH1
CL
région variable
αααα ββββ ααααββββ
CD45
CD19
CD81
CD21 CD32CD22
complexesAg/Ac/C ’
complexesAg/Ac/C ’
-+
+
-

LA SUPERFAMILLE DES RÉCEPTEURS DU TNF
TNFααααLTαααα
LTαααα2/ββββ1
TWEAKTL1A
CD95/FasTNF-R1 DR3 TRAIL-R1
TRAIL-R2DR6
CD178
TRAIL
?
TACI
BCMABAFF-R
RÉCEPTEURS A DOMAINE DE MORT
BAFF/Blys
APRIL
RÉCEPTEURSDE SIGNAUX DE
SURVIE
CD30-L
TNF-R2CD27 CD30 NGF-R
OX-40CD40
CD70
NGF OX-40-LCD154
TNFααααLTαααα
LTαααα2/ββββ1
RÉCEPTEURSDE SIGNAUX
D ’ACTIVATION

Les TLR et leurs ligands
?LPS Taxol (Plant)
Diacyllipopeptides(MALP-2)
ssARN etligands synthétiques (Imidazoquinoline,
R848)ds ARN
(poly I:C)
CpG ADN
Triacyllipopeptides(PAM3CSK4)
Flagelline
CD-14
MD-2
TLR1 TLR2 TLR6 TLR2 TLR4 TLR5 TLR7 TLR8TLR3 TLR9
Bactéries Gram + Mycobactéries
LevuresBactéries Gram -
TLR11
Bactéries uropathogéniques

LA RÉPONSE AUX Ag THYMO-DÉPENDANTS

LBfolliculaires
- Maturation d ’affinité de la réponse Ac
- Formation de centres germinatifs
- Production de LB à mémoire (rate)
- Production de plasmocytes à longuedurée de vie (moelle)
Ag TD Ag TI
++++
-+/---
- Réponse Ac chez des souris athymiques - +
LB de la ZM

LES LIGNAGES LYMPHOCYTAIRES B
2 lignages B (LB1 et LB2) forment le compartiment d es LB naïfs. Ce compartimentse subdivise en 4 sous-populations:1. les LB folliculaires (FO)2. les LB de la zone marginale (ZM)3. les LB1a (CD5+)4. les LB1b (CD5-)Les LB1 et les LB ZM sont « spécialisés » dans la rép onse aux Ag TI
moelleosseuse
foie fœtalrate
LB1a (CD5+)
LB1b (CD5-)Ag TI
Ag TDLB
folliculaires(FO)
OLS et circulation
LB de la zone marginale
(ZM)
rate
-cavités séreuses-OLS-circulation
LB 1
LB 2

REPONSES B PRIMAIRES ET SECONDAIRES AUX Ag TD: ASPECTS QUANTITATIFS
1o Ag 2o Ag
D a y s A f t e r I m m u n i z a t i o n
A b
T i
t e r
• amplitude de la réponse
– 1o - faible
– 2o - forte
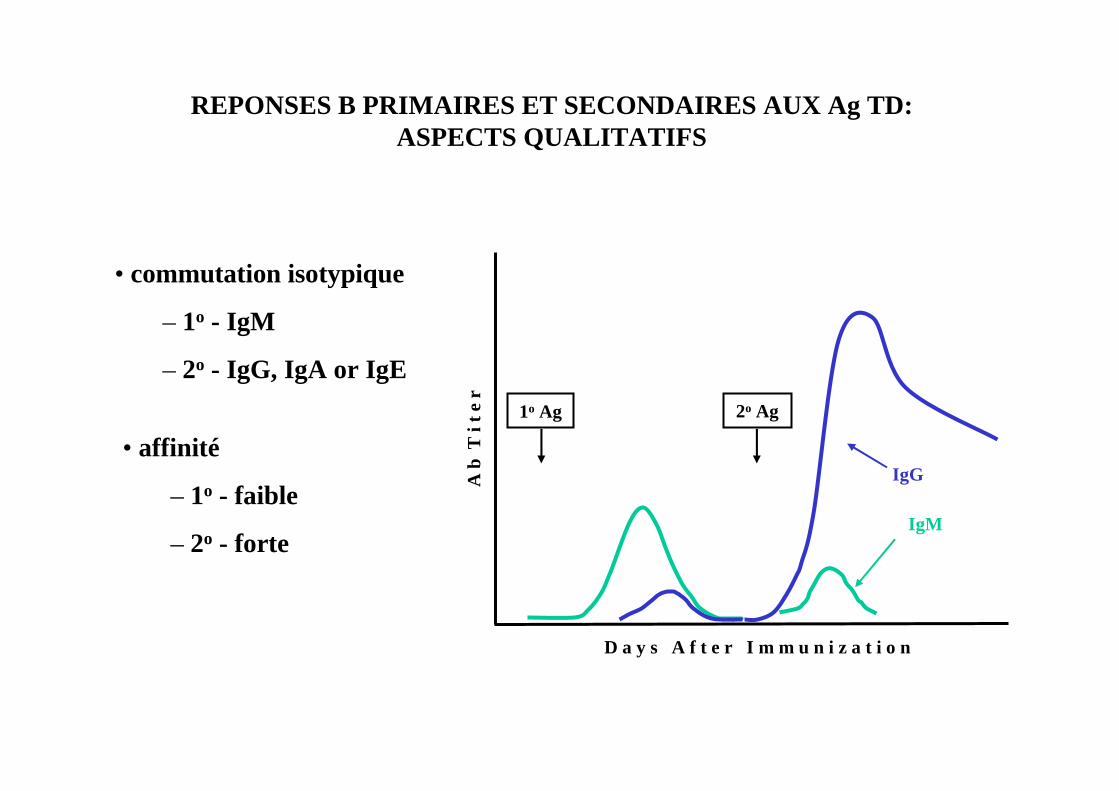
• commutation isotypique
– 1o - IgM
– 2o - IgG, IgA or IgE
1o Ag 2o Ag
D a y s A f t e r I m m u n i z a t i o n
A b
T i
t e r
IgG
IgM
REPONSES B PRIMAIRES ET SECONDAIRES AUX Ag TD: ASPECTS QUALITATIFS
• affinité
– 1o - faible
– 2o - forte
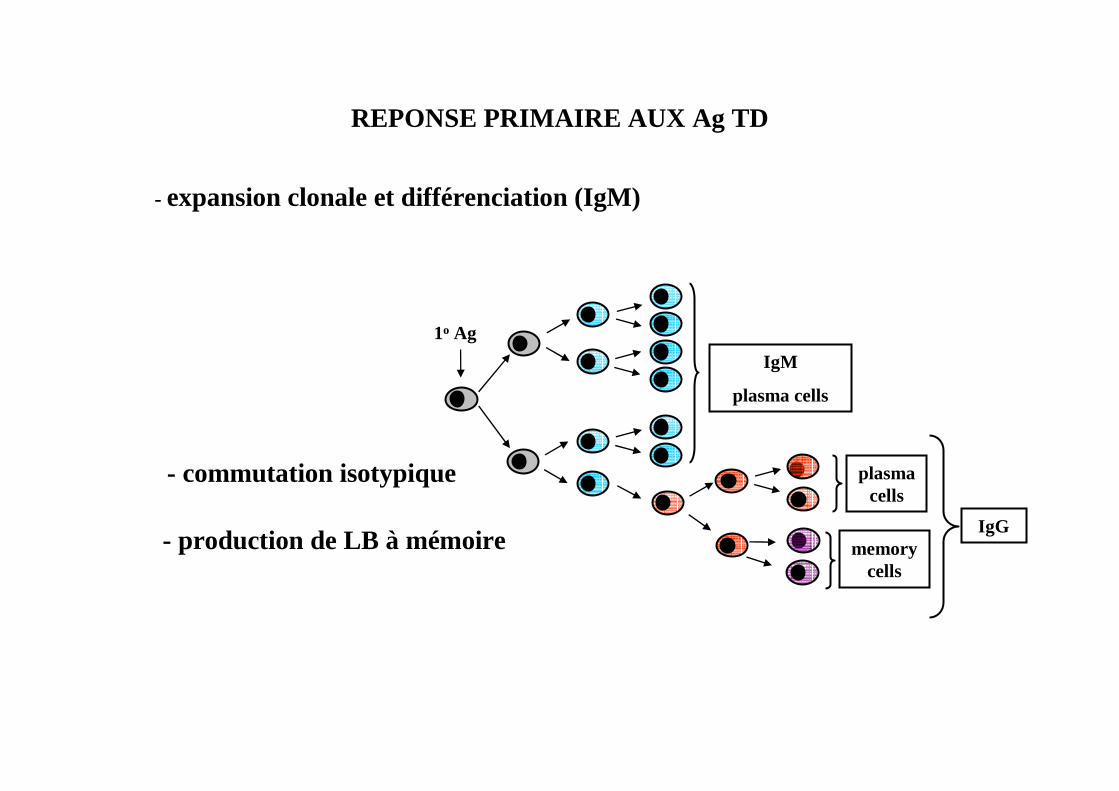
REPONSE PRIMAIRE AUX Ag TD
1o Ag
IgM
plasma cells
- expansion clonale et différenciation (IgM)
plasmacells
IgG
- commutation isotypique
memorycells
- production de LB à mémoire

Memory Pool
Virgin B cell
REPONSE SECONDAIRE AUX Ag TD
IgG Memory
Cells
IgG PC
IgG
PC
IgG Memory
Cells
-expansion et différenciationmassive des LB à mémoire
IgM
-expansion et différenciation modeste des LB naïfs
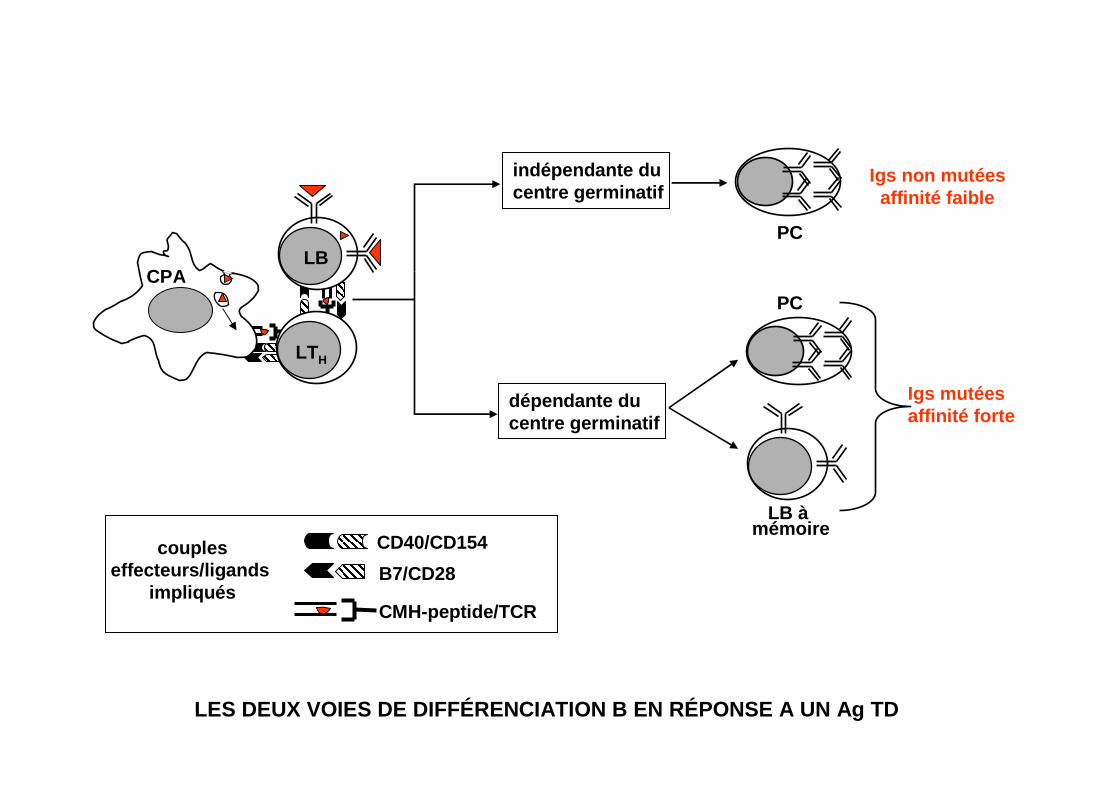
LES DEUX VOIES DE DIFFÉRENCIATION B EN RÉPONSE A UN Ag TD
CPA
LTH
LB
indépendante ducentre germinatif
dépendante ducentre germinatif
Igs non mutéesaffinité faible
Igs mutéesaffinité forte
PC
LB à mémoire
PC
coupleseffecteurs/ligands
impliqués
CD40/CD154
B7/CD28
CMH-peptide/TCR

1o Ag
D a y s A f t e r I m m u n i z a t i o n
A f
f I n
I t y 2o Ag
IgG AbIgM Ab
REPONSES B PRIMAIRES ET SECONDAIRES AUX Ag TD: LA MATURATION D’AFFINITE
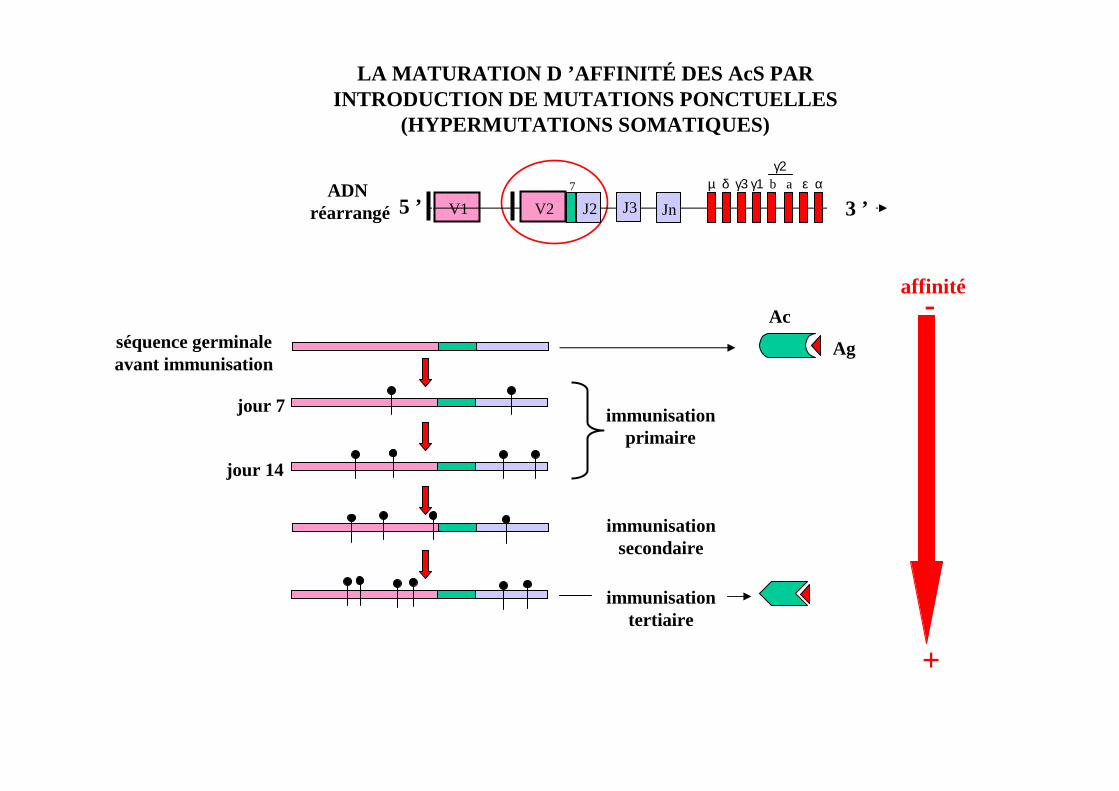
LA MATURATION D ’AFFINITÉ DES AcS PARINTRODUCTION DE MUTATIONS PONCTUELLES
(HYPERMUTATIONS SOMATIQUES)
V15 ’7
J2 J3 Jn 3 ’µ δ γ3 γ1 b a ε α
γ2
V2ADN
réarrangé
séquence germinaleavant immunisation
jour 7 immunisationprimaire
jour 14
immunisationsecondaire
immunisationtertiaire
Ac
Ag
affinité-
+

CAPTURE DE L’ANTIGENEPAR LES LYMPHOCYTES B
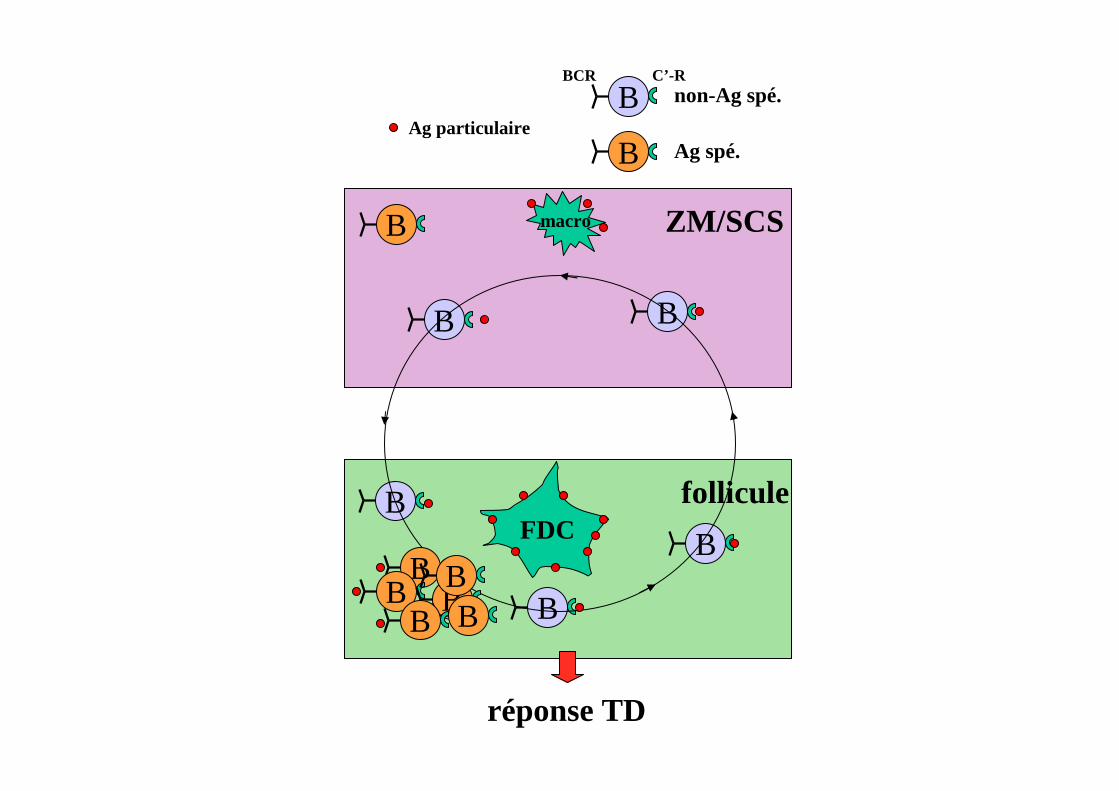
B
ZM/SCS
follicule
B
B
B
BFDC
macroB
BB BBB
B
Ag particulaireB
BCR C’-R
B
non-Ag spé.
Ag spé.
réponse TD

DC
DC
TZone T
B
Ag chargéen périphérie
T
macro BZM
B
GglionsSCS
FDC
rate
follicule
B
Ag capturé- sur les macros (gglions)
-sur les LB ZM ou FDC (rate)

LA MATURATION D ’AFFINITÉ DE LA RÉPONSEANTICORPS DANS LES CENTRES GERMINATIFS

QuickTime™ et un décompresseurRVB Plan sont requis pour visualiser
cette image.
Bcl-2 DRC1 IgD
Apoptose FDC lymphocytes B naïfs
CD45 RO(X5)
CD45 R0(X10)
IgD/Ki67
lymphocytes T prolifération
MARQUAGES IMMUNOHISTOLOGIQUES SUR COUPES D ’AMYGDALE S

GÉNÉRALITÉS SUR LES CENTRES GERMINATIFS
I. Le CG et le siège de deux processus cruciaux pou r la réponse immune humorale:1. la maturation d ’affinité des Acs, 2. la génèse de s LB à mémoire.
La plupart des mutations aboutissant à la disparitio n des CG s ’accompagnent d ’un déficit de différenciation des LB à mémoire et de production d ’Acs mutésde forte affinité.
Mais:1. il existe au moins 2 situations où les gènes V H et VL des LB sont hypermutésen l ’absence de formation des CG: a) les patients h yper IgM déficients pour l ’expression du CD40L, b)les souris KO pour Lt αααα....
2. il existe des LB présentant les caractéristiques fonctionnelles des LB à mémoire chez les souris KO pour le répresseur de tr anscription bcl-6 ne formant pas de CG
II. Le CG abrite plusieurs lignages cellulaires :
1. des cellules hématopoïétiques: LB, LT CD4+, DC e t macrophages2. des cellules non-hématopoïétiques : les cellules dendritiques folliculaires (FDC)

PHASES DE LA RÉACTION FOLLICULAIRE
FOLLICULE SECONDAIRE
(phase III: 21 j. à plusieurs mois après
immunisation)
FOLLICULE SECONDAIRE
(phase II: 4 à 21 j. après immunisation)
manteau folliculaire(LB naïfs)
FOLLICULE SECONDAIRE
(phase I: 1 à 4 j. après immunisation)
centregerminatif
blastes Bissus de la réaction
extra-folliculaire
zone sombre
zone claire
FOLLICULE PRIMAIRE
LB naïfs etFDC
réseau de FDCrésiduel

B
DIVERSIFICATION ET SÉLECTION DU REPERTOIRE B DANS LES CENTRES GERMINATIFS
expansion clonale
hypermutation desgènes V H et VL
faibleaffinité
CFD
différenciation PCLBM
sélection
forteaffinité
CFDT
signal 1BCR
CD40
Ag
CFD
CD40Lsignal 2
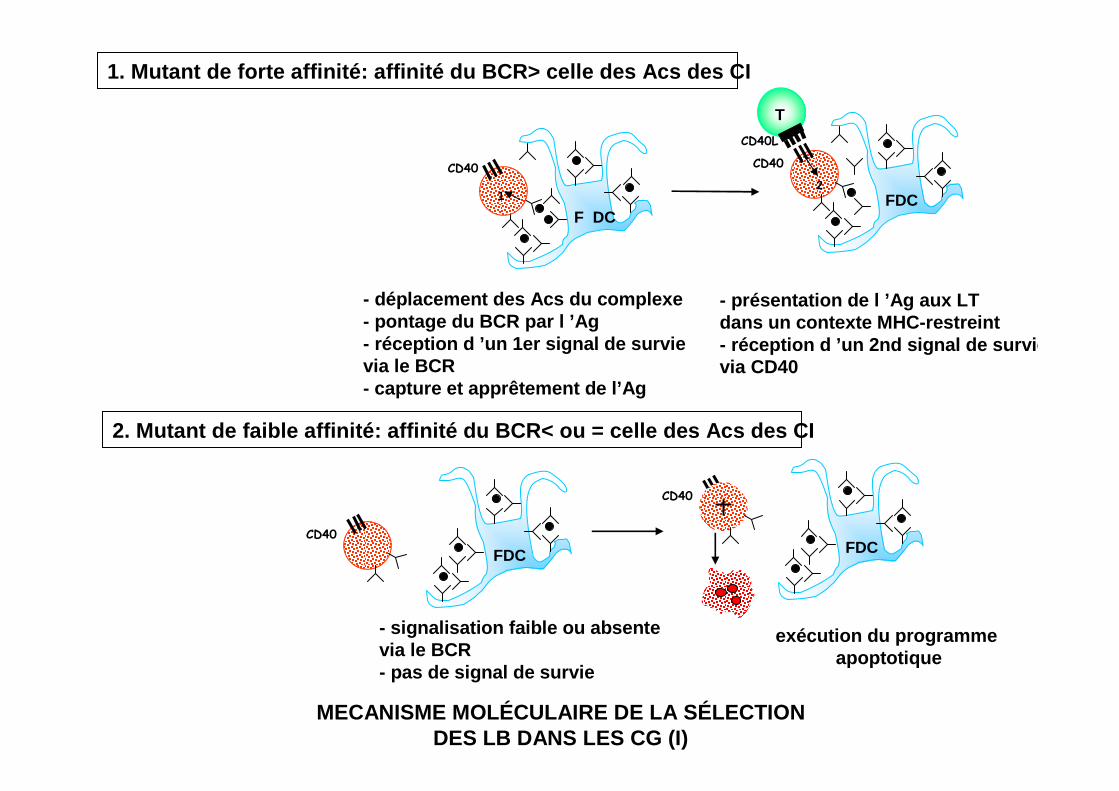
- présentation de l ’Ag aux LT dans un contexte MHC-restreint- réception d ’un 2nd signal de survie via CD40
CD40
CD40
FDC
CD40L
T
2
2. Mutant de faible affinité: affinité du BCR< ou = celle des Acs des CI
FDCCD40
- signalisation faible ou absente via le BCR- pas de signal de survie
exécution du programme apoptotique
- déplacement des Acs du complexe- pontage du BCR par l ’Ag- réception d ’un 1er signal de survie via le BCR- capture et apprêtement de l’Ag
F DC
CD40
1
FDC
1. Mutant de forte affinité: affinité du BCR> celle des Acs des CI
MECANISME MOLÉCULAIRE DE LA SÉLECTIONDES LB DANS LES CG (I)

PR
AC FOT
FO ZM
LB
LT
LB à mémoireplasmocyte
DC
LB apoptotique
FDC
LB muté
CG

LES CELLULES B À MÉMOIRE

CRITÉRES DE DÉFINITION DES LB A MÉMOIRE
2. FONCTIONNELS: - capacité à générer une réponse Ac accélérée et de f orte amplitude(susceptible de conférer une protection) après une injection de rappel-cellule ayant proliféré à l’Ag et persistant à l’éta t quiescent dans les tissus lymphoïdes
3. MOLÉCULAIRES: gènes V H et VL mutés
1. PHÉNOTYPIQUES: perte d ’expression des IgD
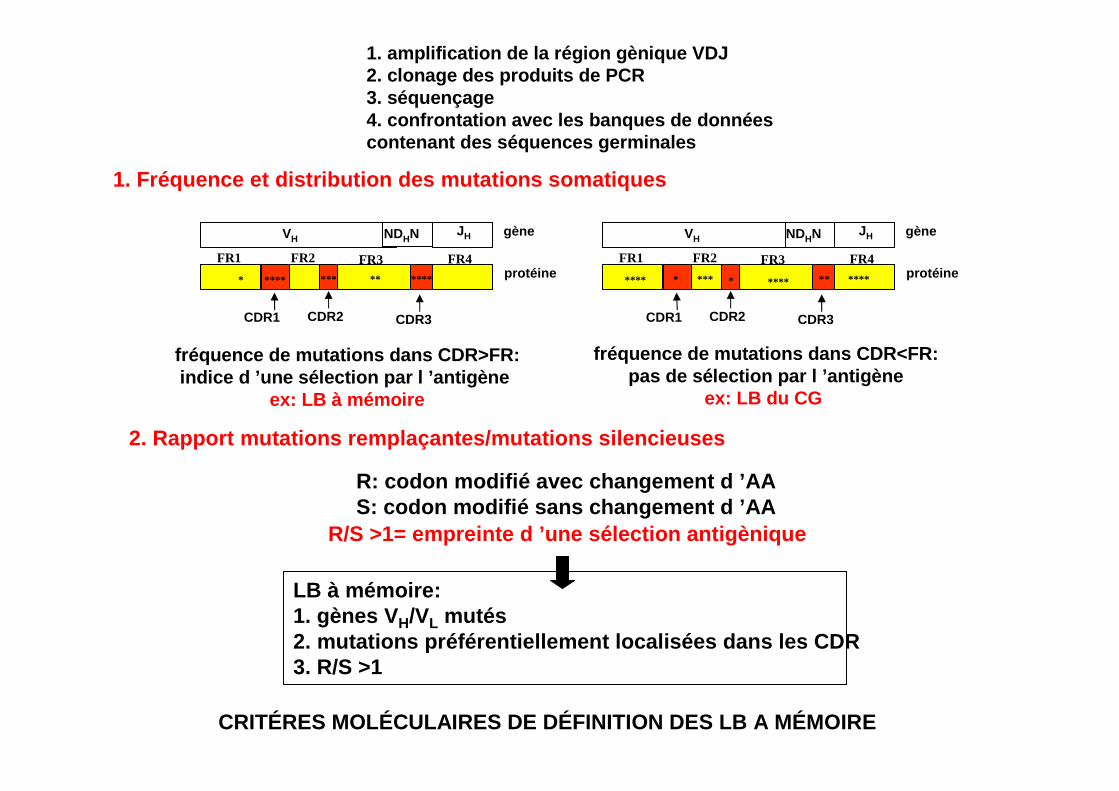
CRITÉRES MOLÉCULAIRES DE DÉFINITION DES LB A MÉMOIR E
1. amplification de la région gènique VDJ2. clonage des produits de PCR3. séquençage4. confrontation avec les banques de données contenant des séquences germinales
1. Fréquence et distribution des mutations somatiqu es
CDR1 CDR2 CDR3
FR1 FR2 FR3 FR4
VH NDHN JH gène
protéine**** *** ***** **
fréquence de mutations dans CDR>FR:indice d ’une sélection par l ’antigène
ex: LB à mémoire
2. Rapport mutations remplaçantes/mutations silenci euses
CDR1 CDR2 CDR3
FR1 FR2 FR3 FR4
VH NDHN JH gène
protéine**** *** ***** ***
fréquence de mutations dans CDR<FR:pas de sélection par l ’antigène
ex: LB du CG
****
R: codon modifié avec changement d ’AAS: codon modifié sans changement d ’AA
R/S >1= empreinte d ’une sélection antigènique
LB à mémoire:1. gènes V H/VL mutés2. mutations préférentiellement localisées dans les CDR3. R/S >1


IgD
CD
27CD27+
CD27-
mutées
nonmutées
M+D+ 60%
M+D- 10%
M+D+ 15%
M-D+ < 1%
M-D- (G+/A+) 15%
1. Tous les LB CD27+ sont mutés2. Il existe des LB mutés non switchés M+D+3. mutations somatiques et commutation isotypique dissociées
Le compartiment B à mémoire est hétérogène:- fonctions biologiques différentes?- induites en réponse à des Ag distincts?

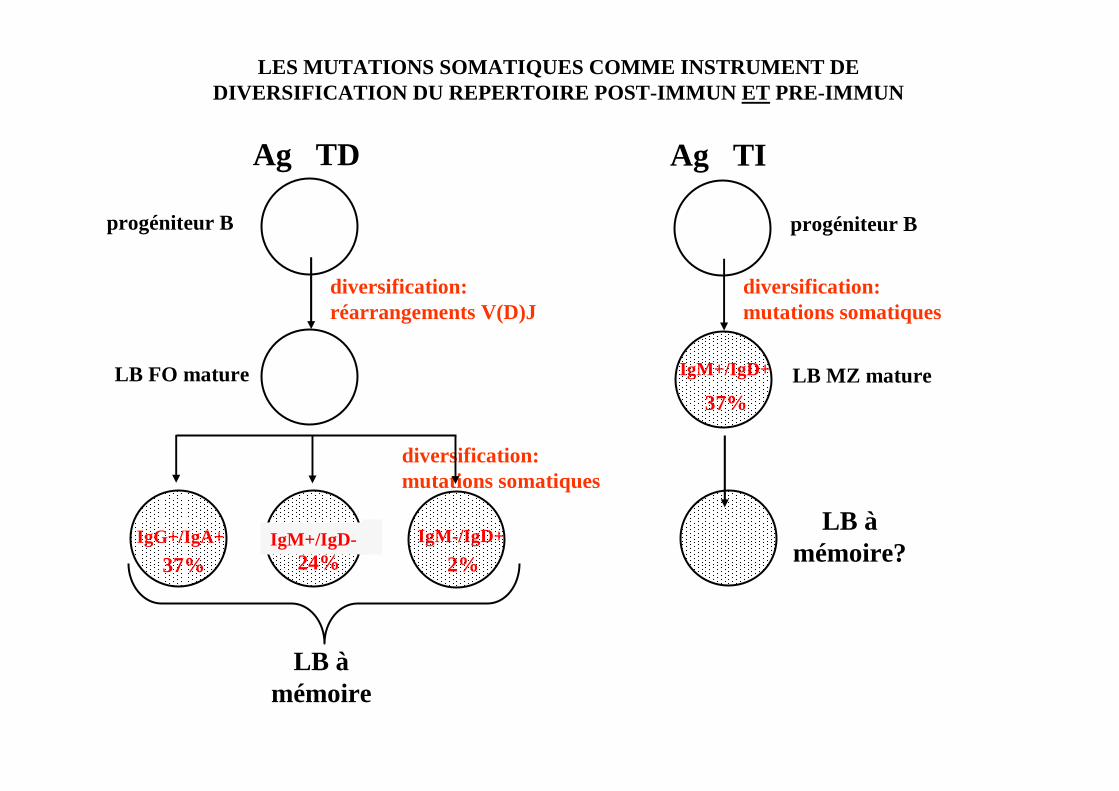
LB àmémoire
LES MUTATIONS SOMATIQUES COMME INSTRUMENT DE DIVERSIFICATION DU REPERTOIRE POST-IMMUN ET PRE-IMMUN
diversification:mutations somatiques
IgG+/IgA+ IgM+/IgD- IgM-/IgD+
progéniteur B
LB FO mature
24% 2%37%
diversification:réarrangements V(D)J
Ag TD
progéniteur B
LB MZ mature
diversification:mutations somatiques
IgM+/IgD+
37%
LB àmémoire?
Ag TI

CRITÉRES DE DÉFINITION DES LB A MÉMOIRE
2. FONCTIONNELS: capacité à générer une réponse Ac accélérée et de fo rte amplitude(susceptible de conférer une protection) après une injection de rappel
nécessite l ’isolement et le transfert adoptif de so us-populations B
3. MOLÉCULAIRES: gènes V H et VL mutés
le processus de mutations somatiques peut être util isé pour la diversification du répertoire B préimmun des LB de la ZM chezl’Homme
CD27 est aussi exprimé sur les LB naïfs de la ZM che z l’Homme
1. PHÉNOTYPIQUES:
a) perte d ’expression des IgD et expression du marque ur CD27b) perte d’expression d’un transporteur ABCB1/efflux de drogues hor s de la cellule
pas applicable chez la souris

PROPRIETES FONCTIONNELLESDES LYMPHOCYTES B A MEMOIRE

IgG hybride M/GIgM
VL
VH
-synthèse d ’Igs-apoptose-prolifération-effectif des PC
mêmes VJ et VDJ codant pour un BCR anti-HEL de forte affinité
portions intracytoplasmiques différentes
receveur
LB-HEL-M
1.
LB-HEL-M/G
receveur
3.
receveur
LB-HEL-G
2.
HEL

receveur
LB-HEL-M1.
LB-HEL-M/G
receveur
3.
receveur
LB-HEL-G
2.
la portion cytoplasmique des IgG confère une plus gr ande capacité d ’expansion aux LB. Elle agirait en limitant l ’apop tose cellulaire
au cours des divisions succcessives
Ac sériquesanti-HEL
+
++++
++++
ASCanti-HEL
+
++++
++++
nombresde LB anti-HEL
+
++++
++++
LA PORTION CYTOPLASMIQUE DES IgG EST RESPONSABLED ’UNE PARTIE DES CARACTÉRISTIQUES FONCTIONNELLES
DES LB A MÉMOIRE

LES CELLULES B EFFECTRICES
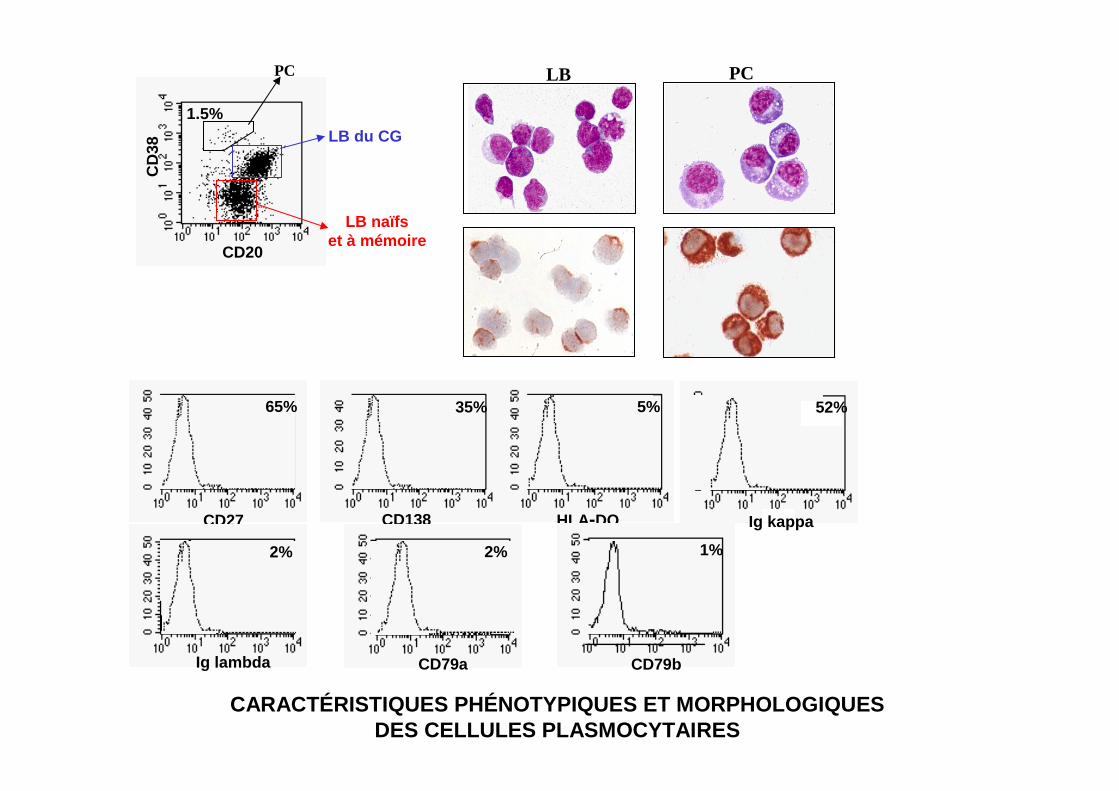
CD20
CD
38
1.5%
LB naïfset à mémoire
LB du CG
PC LB PC
CARACTÉRISTIQUES PHÉNOTYPIQUES ET MORPHOLOGIQUESDES CELLULES PLASMOCYTAIRES
65%
CD27
35%
CD138
5%
HLA-DQ κκκκ
52%52%
Ig kappa
Ig lambda
2%
CD79a
2%
CD79b
1%

PC with Russel bodies
PC Mott Cell
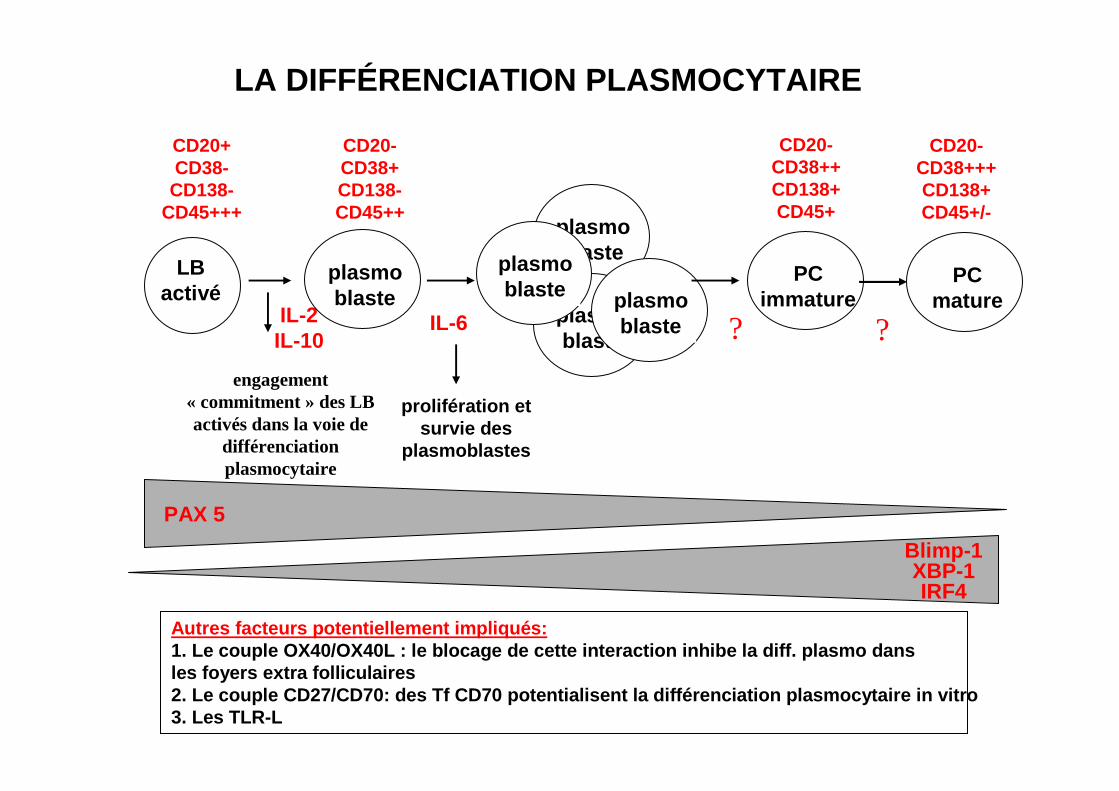
LA DIFFÉRENCIATION PLASMOCYTAIRE
LBactivé
IL-2IL-10
plasmoblaste
engagement« commitment » des LBactivés dans la voie de
différenciationplasmocytaire
plasmoblaste
plasmoblaste
plasmoblaste
plasmoblaste
IL-6
prolifération etsurvie des
plasmoblastes
PCimmature
PCmature
CD20+CD38-
CD138-CD45+++
CD20-CD38+CD138-CD45++
CD20-CD38++CD138+CD45+
CD20-CD38+++CD138+CD45+/-
? ?
Blimp-1XBP-1IRF4
PAX 5
Autres facteurs potentiellement impliqués:1. Le couple OX40/OX40L : le blocage de cette inter action inhibe la diff. plasmo dans les foyers extra folliculaires2. Le couple CD27/CD70: des Tf CD70 potentialisent la différenciation plasmocytaire in vitro3. Les TLR-L
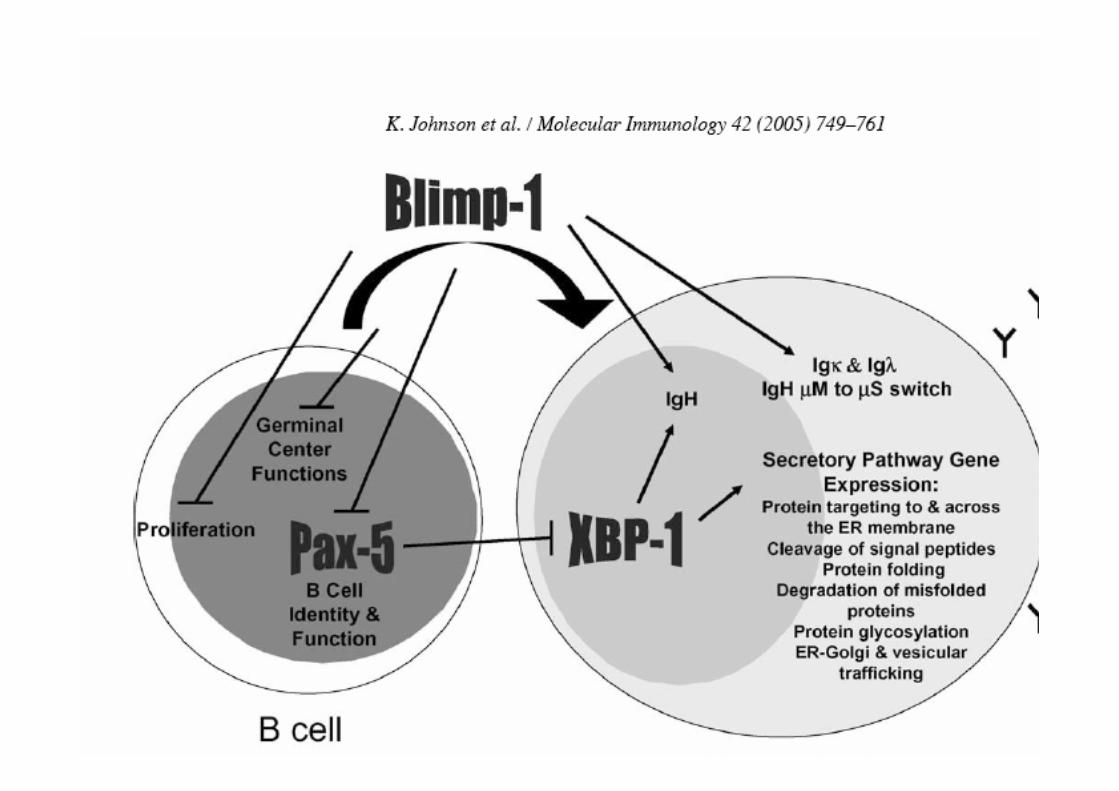

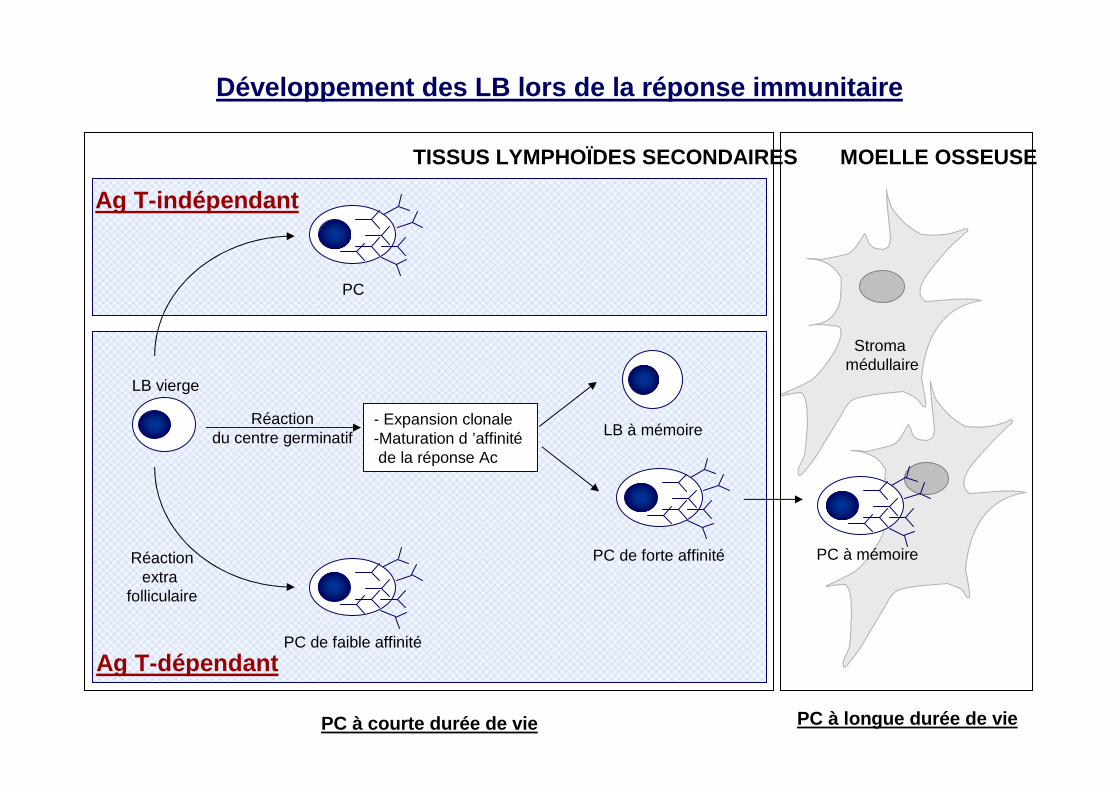
MOELLE OSSEUSE
Stroma médullaire
TISSUS LYMPHOÏDES SECONDAIRES
Réactionextra
folliculaire
LB vierge
LB à mémoire
PC de forte affinité
PC de faible affinité
Réactiondu centre germinatif
- Expansion clonale-Maturation d ’affinité de la réponse Ac
PC à mémoire
Ag T-dépendant
PC
Ag T-indépendant
PC à courte durée de vie PC à longue durée de vie
Développement des LB lors de la réponse immunitaire

LES PC À LONGUE DURÉE DE VIE (I)
Slifka et Ahmed (Immunity, 8: 363, 1998)
LCMV
immunisation1. Le modèle expérimental
LBM: sécrétion d ’Acs après réexposition à l ’Ag in v itro
PC: sécrétion spontanée d ’Acs sans réexposition à l ’Ag
spots colorés
* * **
ELISPOT
adsorption de l’Agsur une membranede nitrocellulose
addition descellules
éliminationdes cellules
addition d ’unsystème de révélation
numération des « spots » colorésestimation de la fréquence
des cellules sécrétrices
1. MBC PC ?
2. PC ?PC
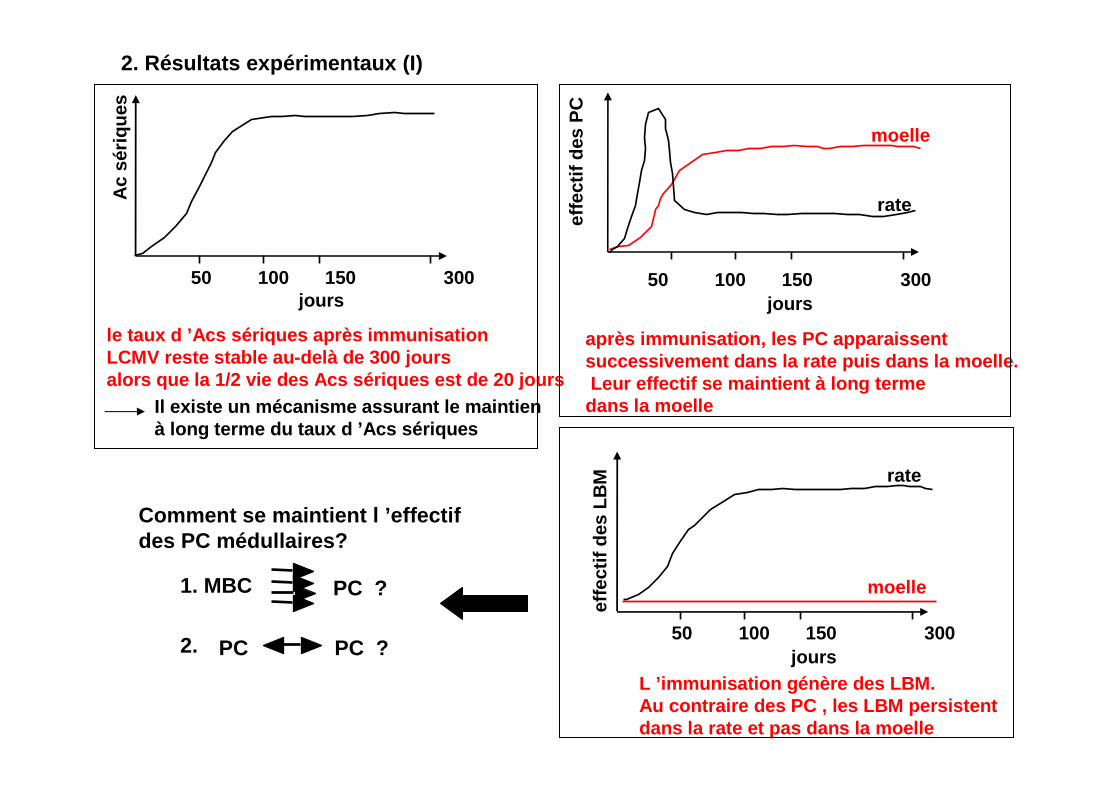
2. Résultats expérimentaux (I)
le taux d ’Acs sériques après immunisation LCMV reste stable au-delà de 300 joursalors que la 1/2 vie des Acs sériques est de 20 jou rs
jours50 100 150 300
Ac
sériq
ues
Il existe un mécanisme assurant le maintien à long terme du taux d ’Acs sériques
effe
ctif
des
PC
moelle
rate
jours50 100 150 300
après immunisation, les PC apparaissentsuccessivement dans la rate puis dans la moelle.Leur effectif se maintient à long terme
dans la moelle
jours50 100 150 300
rate
moelleef
fect
if de
s LB
M
L ’immunisation génère des LBM.Au contraire des PC , les LBM persistentdans la rate et pas dans la moelle
Comment se maintient l ’effectifdes PC médullaires?
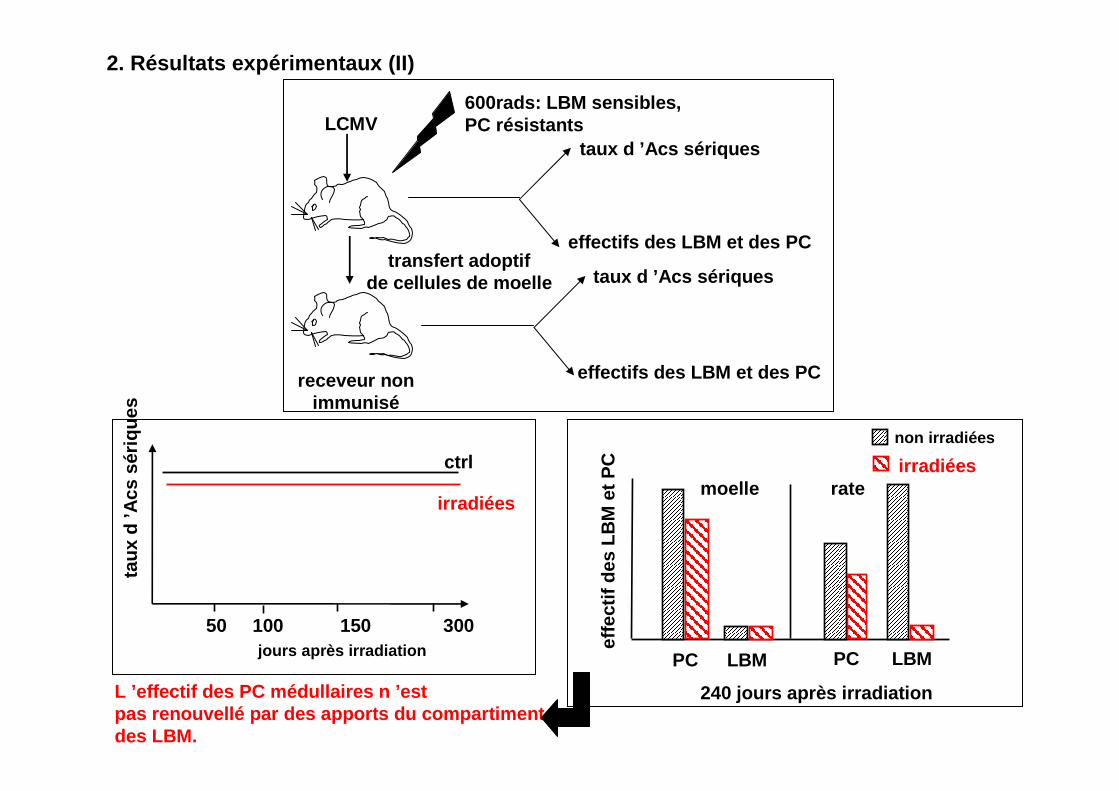
2. Résultats expérimentaux (II)
LCMV600rads: LBM sensibles, PC résistants
taux d ’Acs sériques
effectifs des LBM et des PC
receveur nonimmunisé
transfert adoptifde cellules de moelle taux d ’Acs sériques
effectifs des LBM et des PC
moelle rate
PC LBM PC LBM
non irradiées
irradiées
240 jours après irradiation
effe
ctif
des
LBM
et P
C
jours après irradiation
50 100 150 300
ctrl
irradiées
taux
d’A
cs s
ériq
ues
L ’effectif des PC médullaires n ’estpas renouvellé par des apports du compartimentdes LBM.

Les deux composantes cellulaires de la mémoire B aux Ag TD
Expression du BCR
Localisation
Production d’Acs Protecteurs
Fonction
Expression desmolécules de classe II
LB à mémoireconventionnels
+
Tissus lymphoïdessecondaires
Induite par l’antigène
Protection après réinfection
+
PC à mémoire
-
Moelle osseuse
Constitutive
Prévention de la réinfection
+/- (DR+
DQ-)
Les capacités de présentation antigènique et de réactivation par l ’Ag sont réduites chez les PC à mémoire

LA PERSISTANCE DE LA MÉMOIRE B

L’ÉVOLUTION DES CONCEPTS SUR LA PERSISTANCE DE LA M ÉMOIRE B
transfert de LB à mémoire chez des receveurs naïfs, en présence ou en l’absence de l’Ag:pas de survie à long terme des LBM si l’Ag n’est pas co-transféré
1. La longévité de la mémoire B est conditionnée par la persistance de l’AgGray D. 1988.
2. La longévité de la mémoire B n’est pas conditionnée par la persistance de l’AgRajewsky K. 2000.
3. La longévité de la mémoire B est conditionnée par des signaux non-spécifiquesd’Ag (cytokines, agents microbiens) Lanzavecchia A. 2003.
4. La longévité de la mémoire B est en partieconditionnée par la persistance de l’Ag,Weill JC & Reynaud CA. 2009.

QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.

AID-Cre-ERT2X
Rosa26-loxP-EYFP
SRBC.1 SRBC.2
J0 J30
J7 J12 J31
tamoxyfène
J200
stopx x
stopx x
EYFP
EYFP
loxP loxPCre ERT2 AID
Cre ERT2 AIDx
AID AID
Cre ERT2 AID
stopx x EYFP
AID AID
Cre ERT2 AID
EYFP+ tamoxifene
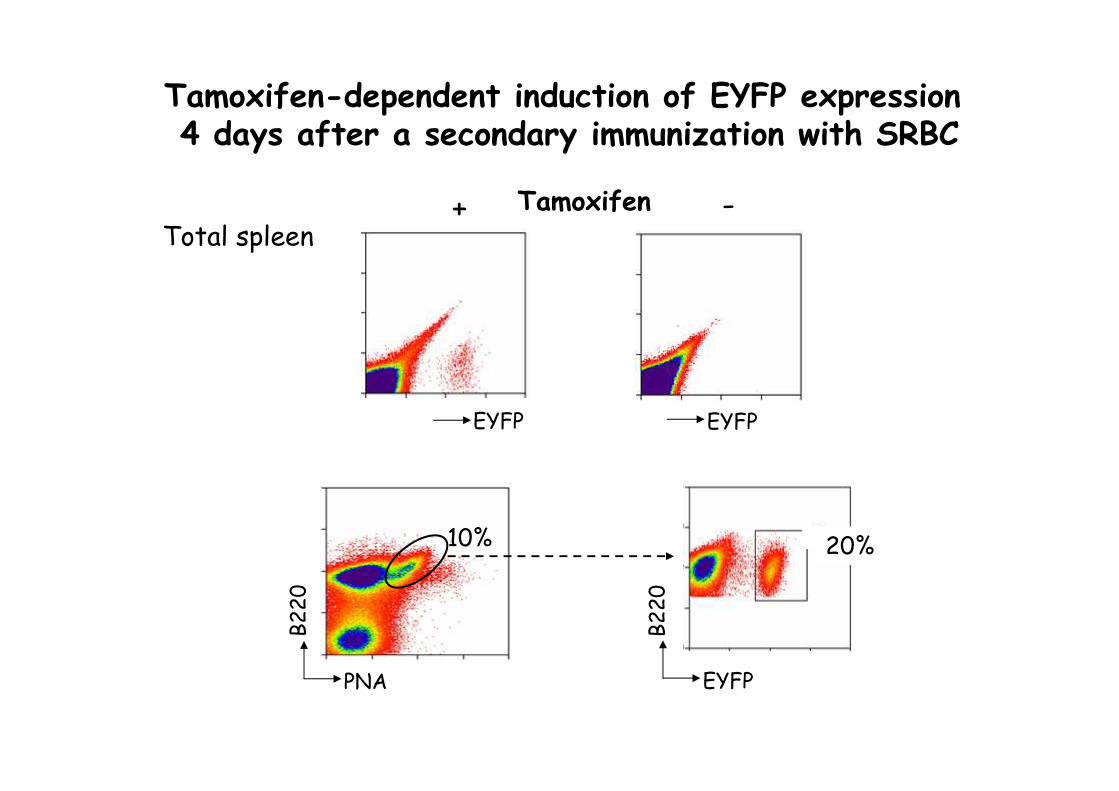
Tamoxifen-dependent induction of EYFP expression 4 days after a secondary immunization with SRBC
20%10%
PNA
B220
EYFPB220
Total spleenTamoxifen+ -
EYFPEYFP

EYFP
B220
Analysis 2 months after secondary SRBC challenge
Four EYFP+ « memory » B cell subpopulations
GL7
PNA
13%
25%
75%
87%
IgM
IgG1
20%
60%
IgG1+GL7-
IgG1+GL7+
IgM+GL7-
IgM+GL7+

4 sous-populations LB mémoire(EYFP+)
M-G1+M-G1+PNA-
M-G1+PNA+
M+G1-M+G1-PNA+
M+G1-PNA-(60%)
(20%)
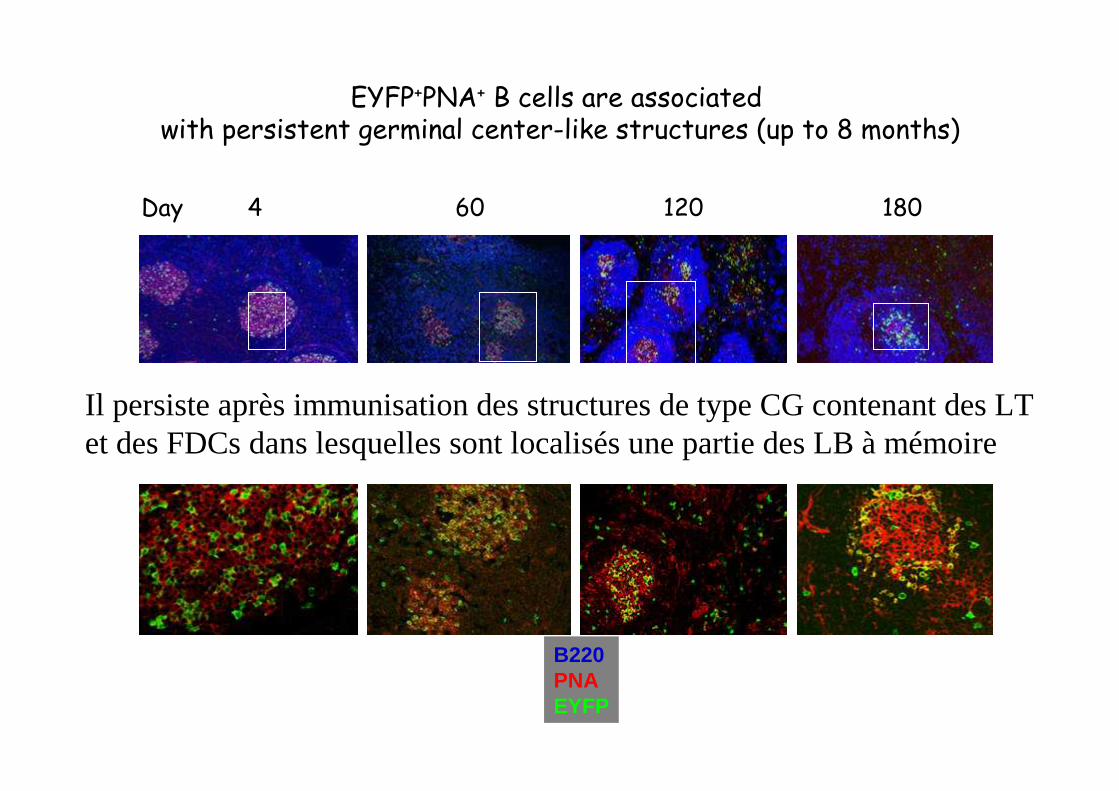
B220PNAEYFP
Day 4 60 120 180
EYFP+PNA+ B cells are associatedwith persistent germinal center-like structures (up to 8 months)
Il persiste après immunisation des structures de type CG contenant des LTet des FDCs dans lesquelles sont localisés une partie des LB à mémoire
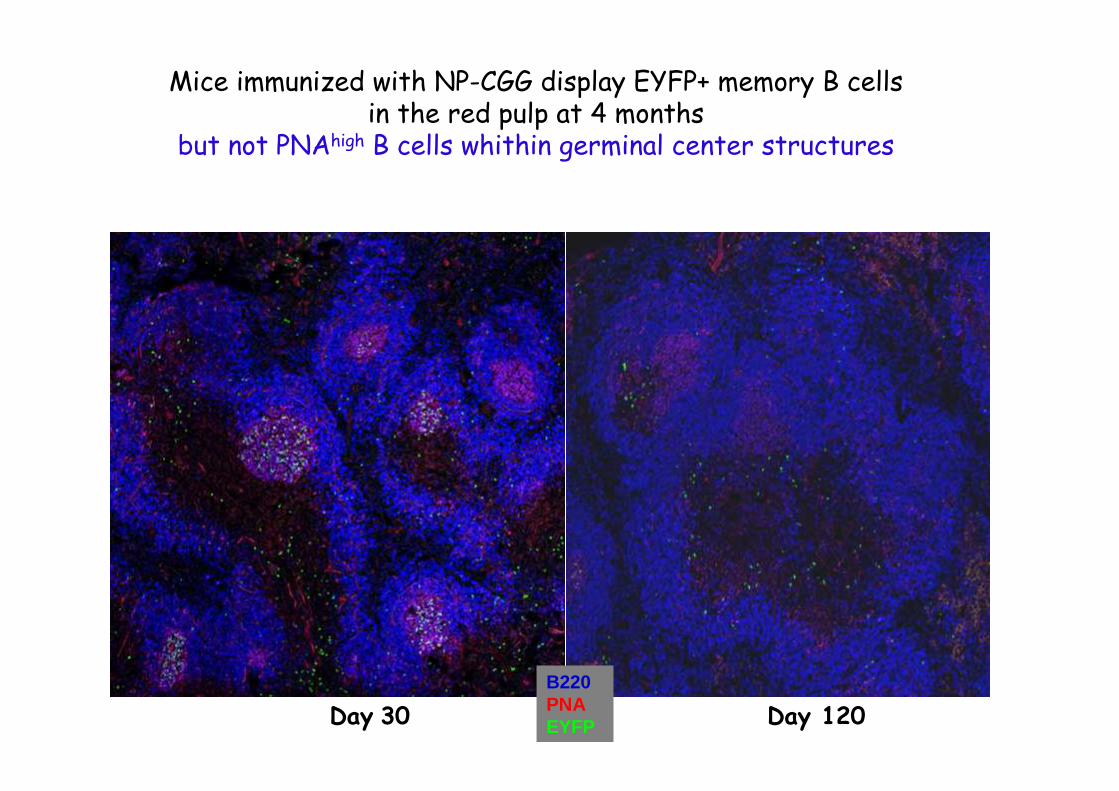
Day 30 Day 120
Mice immunized with NP-CGG display EYFP+ memory B cellsin the red pulp at 4 months
but not PNAhigh B cells whithin germinal center structures
B220PNAEYFP
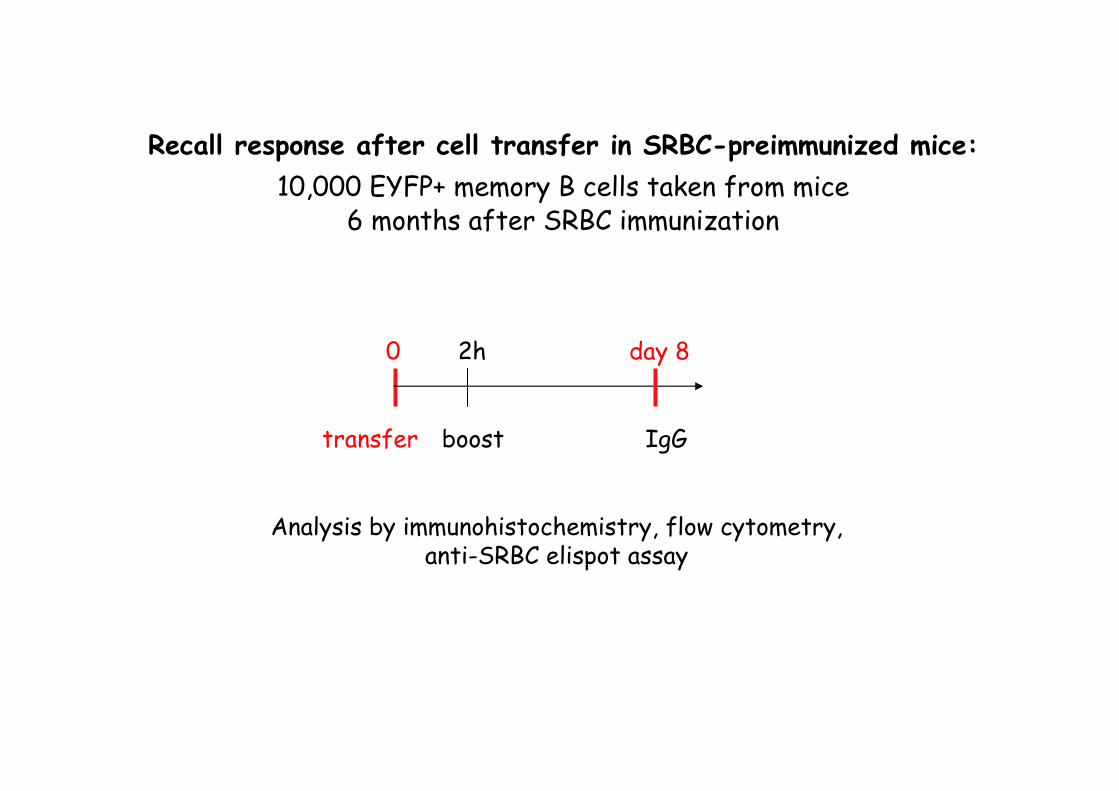
0 2h day 8
transfer boost
Analysis by immunohistochemistry, flow cytometry,anti-SRBC elispot assay
Recall response after cell transfer in SRBC-preimmunized mice:
10,000 EYFP+ memory B cells taken from mice6 months after SRBC immunization
IgG
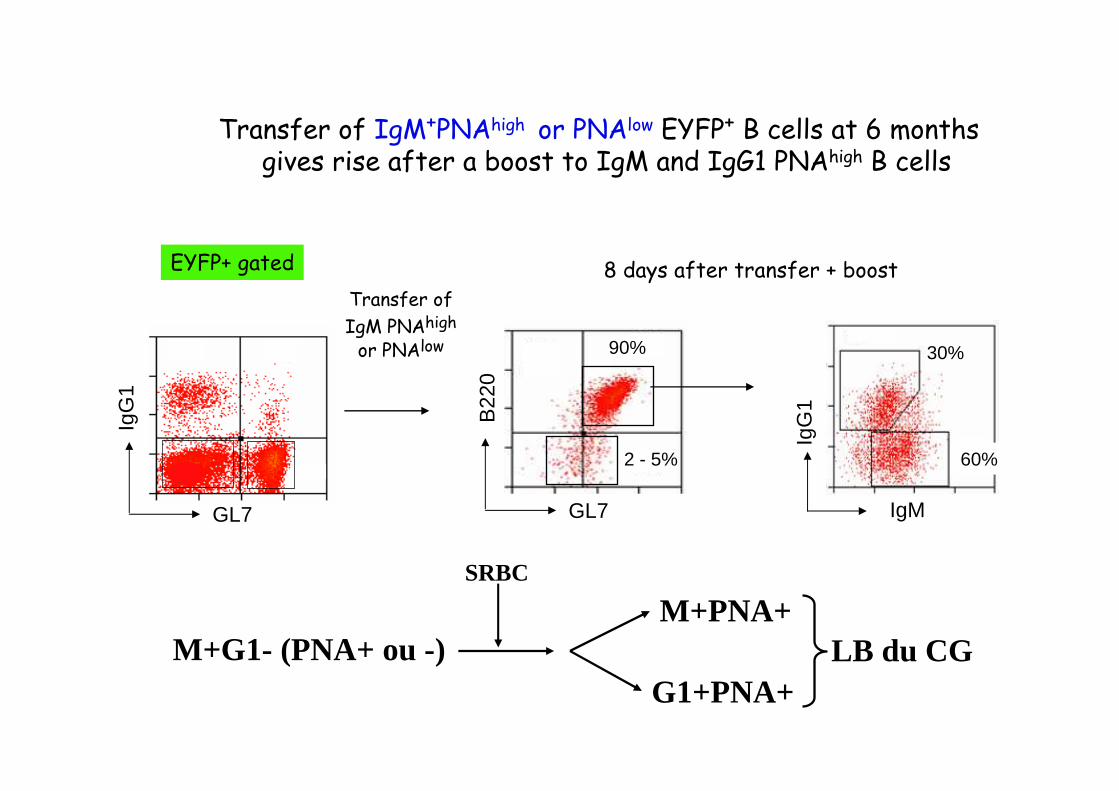
GL7
IgG
1
Transfer of
IgM PNAhigh
or PNAlow
IgM
IgG
1
30%90%
2 - 5%
8 days after transfer + boost
Transfer of IgM+PNAhigh or PNAlow EYFP+ B cells at 6 months gives rise after a boost to IgM and IgG1 PNAhigh B cells
GL7
60%
B22
0
EYFP+ gated
M+G1- (PNA+ ou -)
SRBC
M+PNA+
G1+PNA+LB du CG

B220PNAEYFP
Transfer of IgM+ PNAhigh or PNAlow EYFP+ B cells at 6 months gives rise after a boost to germinal center B cells
germinal centerEYFP+ cells

Total IgG
Transfer ofIgG+PNAlow
Elispot onB220-EYFP+ cells
Transfer of IgG+ PNAlow memory B cells at 6 months gives rise after a boost to plasma cells (anti-SRBC)
and not to germinal center B cells
GL7
IgG
90%
anti-SRBC IgG
PNAB
220
EYFP+ gated
M-G1+ (PNA+ ou -)
SRBC
LB B220-/PNA- = plasma cells

1) GC like structures containing centroblasts and FDCs can persistfor a long time after immunization with a particulate antigen(SRBC)
but not with a soluble protein + adjuvant (NP-CGG)
2) B cell memory against SRBC is composed of two long-lived populations:an « antigen-dependent subset » of centroblasts
and an « antigen-independent » subset of memory B cellsBoth subsets are composed of an IgM and an IgG component
3) Upon a boost: the IgG subset has immediate effector functions and does not self maintain :
« effector function » the IgM subset resplenishes the memory pool and produces effector B cells in
prevision of a next encounter with the antigen : « central function »
CONCLUSIONS
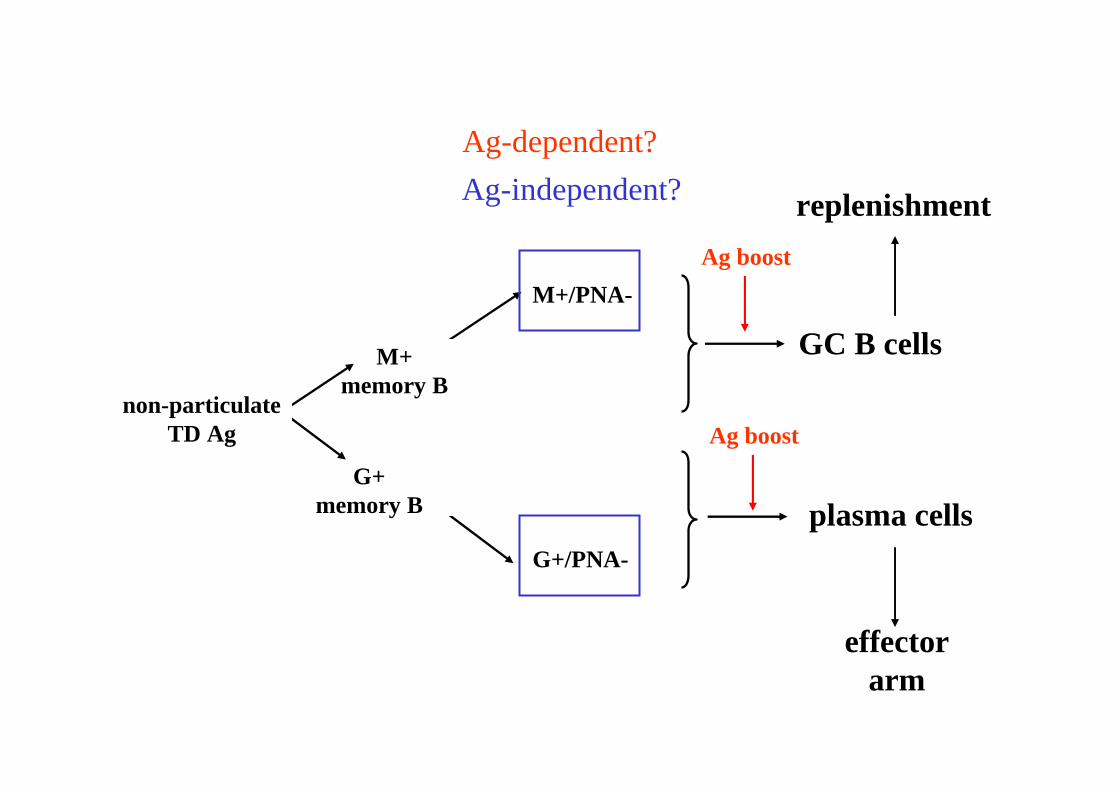
particulateTD Ag
M+memory B
G+memory B
M+/PNA+
M+/PNA-
G+/PNA-
G+/PNA+
Ag boost
Ag boost
GC B cells
plasma cells
Ag-dependent?
Ag-independent?
non-particulateTD Ag
effectorarm
replenishment

LA RÉPONSE AUX Ag THYMO-INDÉPENDANTS

Protéines
Ag TD(divalent)
Ag TI-2(multivalent)
Ag TI-1
Polysaccharides bactériensPathogen AssociatedMolecular Patterns
TLR
Reconnaissance par le récepteurd’antigène (BCR)
Reconnaissance par un récepteur
distinct du BCR
Immunité adaptative Immunité innée
LBfolliculaires
Ex: LPS
?
Follicule
LB1(CD5+)
LB ZM
Péritoine et rate
Zonemarginale

Kinetics of Ab Response toT-independent Ags
• IgM antibody
• No secondary response
1o Ag 2o Ag
D a y s A f t e r I m m u n i z a t i o n
A b
T i
t e r
IgM Ab
� pas de présentation via molécules du CMH
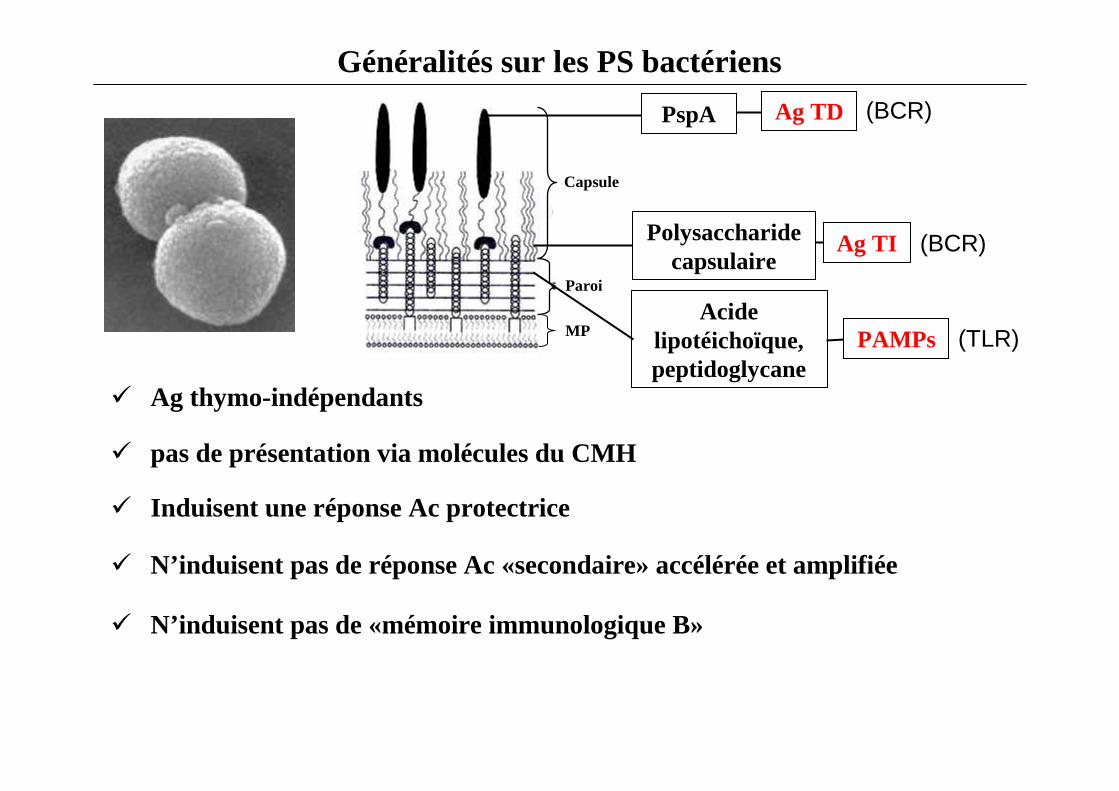
Généralités sur les PS bactériens
� Induisent une réponse Ac protectrice
� N’induisent pas de réponse Ac «secondaire» accélérée et amplifiée
� N’induisent pas de «mémoire immunologique B»
Acidelipotéichoïque,peptidoglycane
Ag TD
Ag TI
Capsule
Paroi
MP
PspA
Polysaccharidecapsulaire
(BCR)
(BCR)
PAMPs (TLR)
� Ag thymo-indépendants
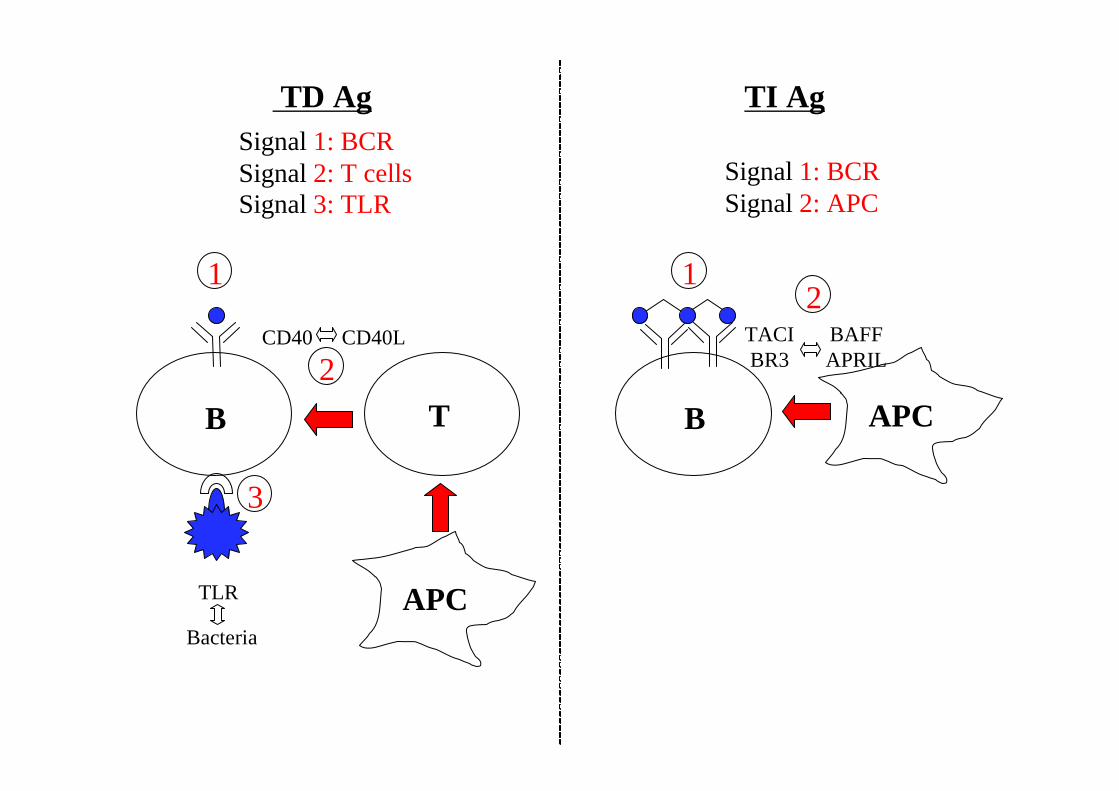
TD Ag TI Ag
B T
APC
B APC
CD40 CD40L TACIBR3
BAFFAPRIL
Signal 1: BCRSignal 2: T cellsSignal 3: TLR
1 1
2
2
TLR
Bacteria
3
Signal 1: BCRSignal 2: APC
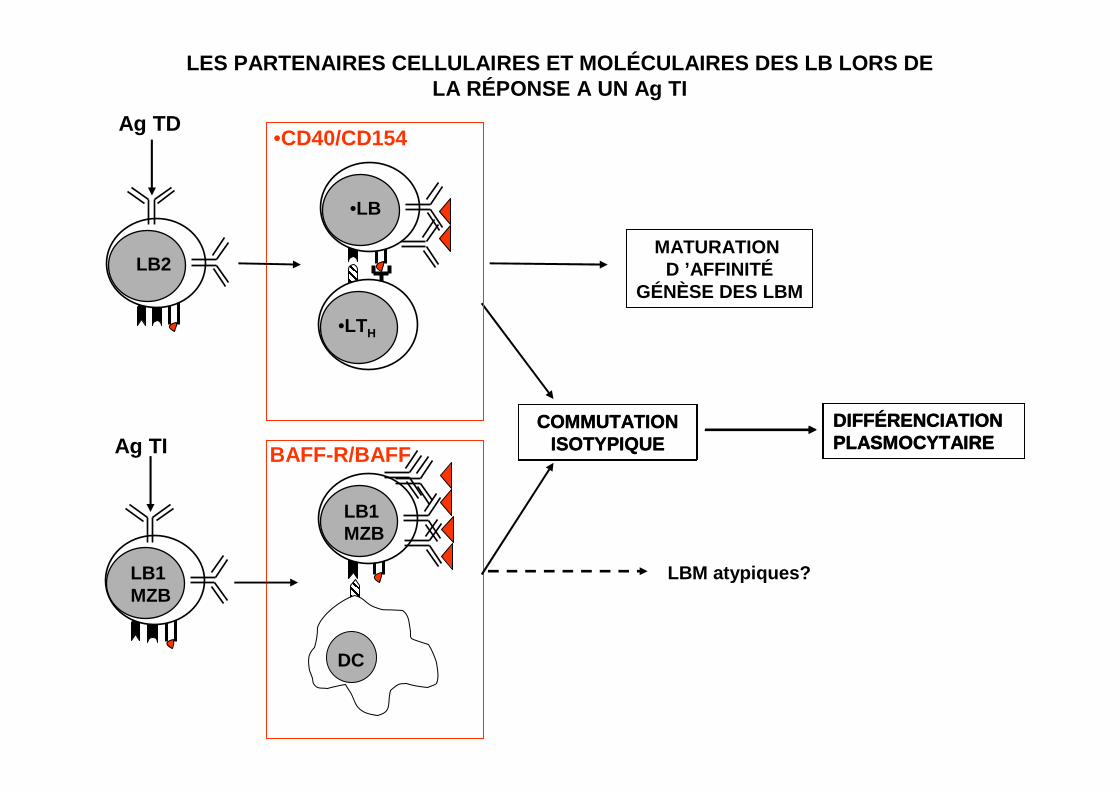
LES PARTENAIRES CELLULAIRES ET MOLÉCULAIRES DES LB LORS DELA RÉPONSE A UN Ag TI
COMMUTATIONISOTYPIQUE
DIFFÉRENCIATIONPLASMOCYTAIRE
MATURATION D ’AFFINITÉ
GÉNÈSE DES LBM
LB1MZB
DC
BAFF-R/BAFF
•LTH
•LB
•CD40/CD154
LB1MZB
Ag TI
LB2
Ag TD
COMMUTATIONISOTYPIQUE
DIFFÉRENCIATIONPLASMOCYTAIRE
LBM atypiques?

LE PARADOXE IMMUNOLOGIQUE DE LA REPONSE B AUX PS BACTERIENS
les PS bactériens n’induisent ni réponse Ac « secondaire » accélérée et amplifiée ni « mémoire immunologique B »
les vaccins polysaccharidiques purs sont protecteurs
mémoire B non conventionnelleen réponse aux Ag TI?

Les cellules B-1b sont le support de la mémoire B aux Ag TI
les LB-1b confèrent la protection après immunisatio n par la bactérie B. hermsii
les LB-1b confèrent la protection après immunisatio n par le PS3 de S. pneumoniae

LES COMPOSANTES CELLULAIRES DE LA MÉMOIRE B
- lymphocytes B à mémoire « conventionnels »
- plasmocytes à mémoire
- lymphocytes B à mémoire « atypiques »
Ag TDLB-2
Ag TILB-1b

LE SECOND PARADOXE DE LA REPONSE B AUX Ag TI
les Ag TI (PS bactériens) induisent la production de LB-1b « à mémoire »
une immunisation secondaireavec un Ag TI n’induit
pas de réponse Ac amplifiéeet/ou accélérée
par quel mécanisme les Ag TIpeuvent-ils conférer une protection?

0 5 7 14 28 30 31 33 350
5
10
15
20
25
30
PS3PBS
PS3Primaire
PS3Secondaire
Temps (j)
PS3 DOES NOT GENERATE AN AMPLIFIED SECONDARY RESPONSEIN IMMUNOCOMPETENT MICE
PS31ary
PS32ary
Time (days)
anti-
PS
3 Ig
M(µ
g/m
l)

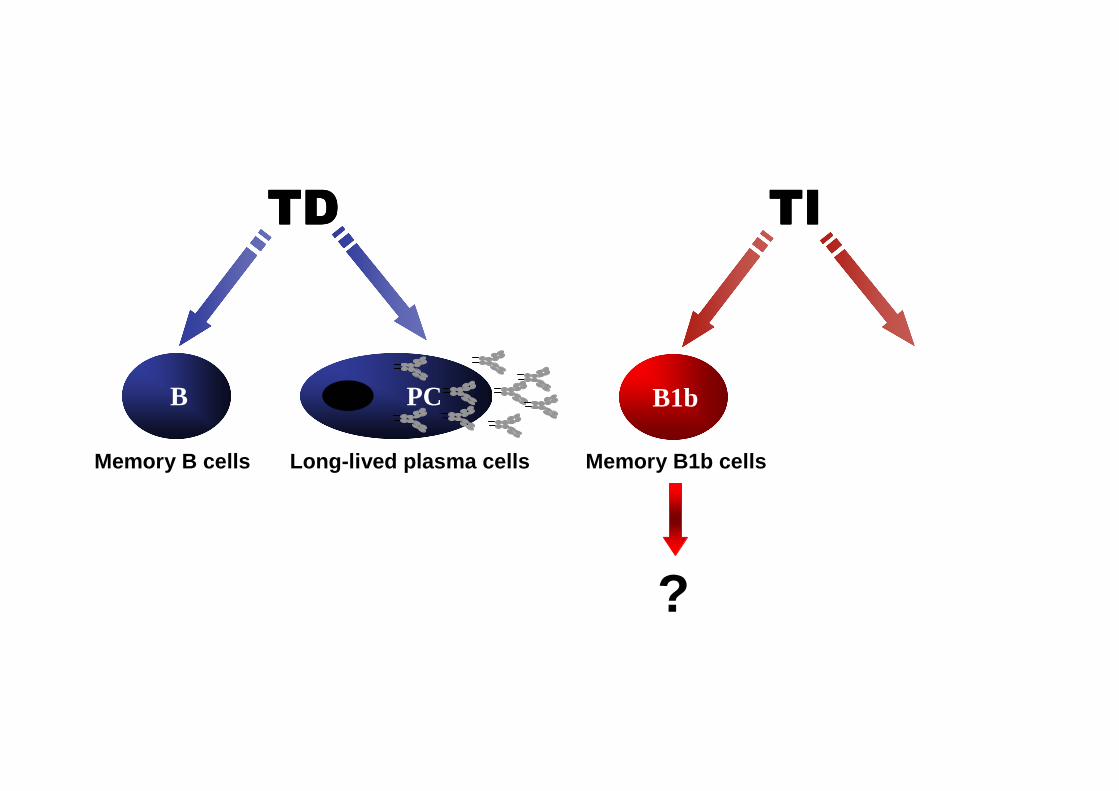
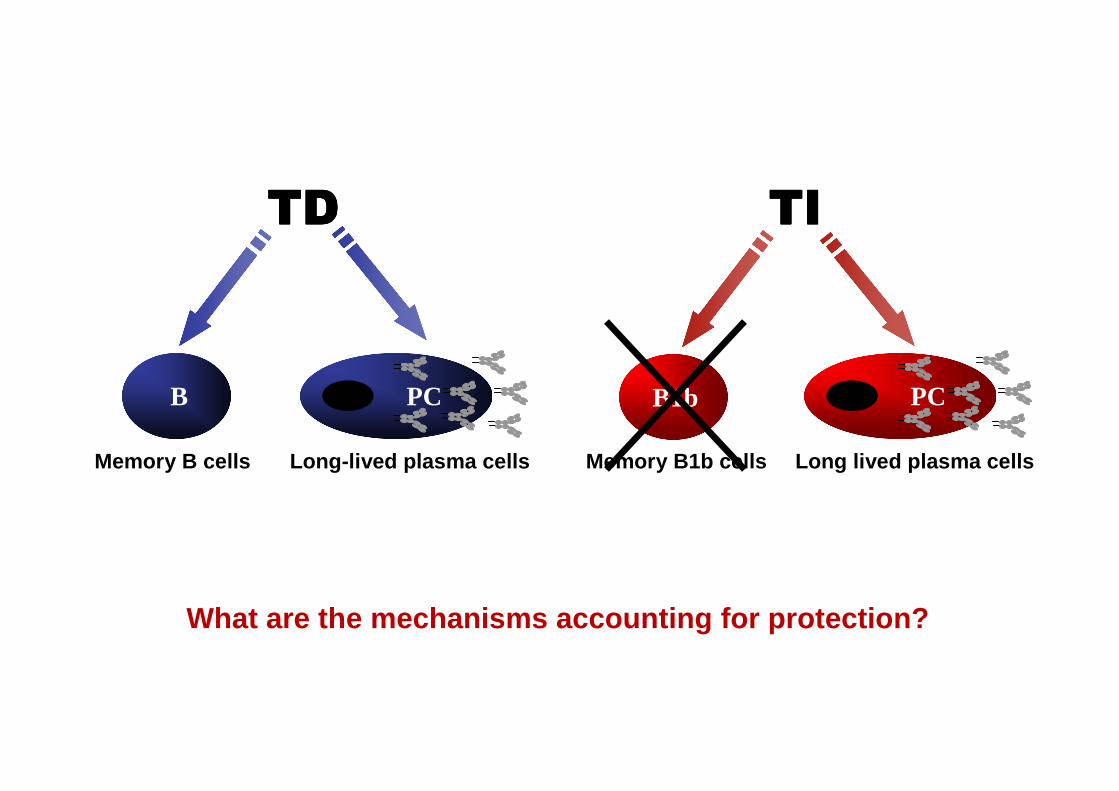
TITITITITDTDTDTD
Memory B cells
PCB
Long-lived plasma cells
????
Memory B1b cells
B1b
?
TITITITITDTDTDTD
Memory B cells
B PC
Long-lived plasma cells Memory B1b cells
B1b PC
Long lived plasma cells
What are the mechanisms accounting for protection?

QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
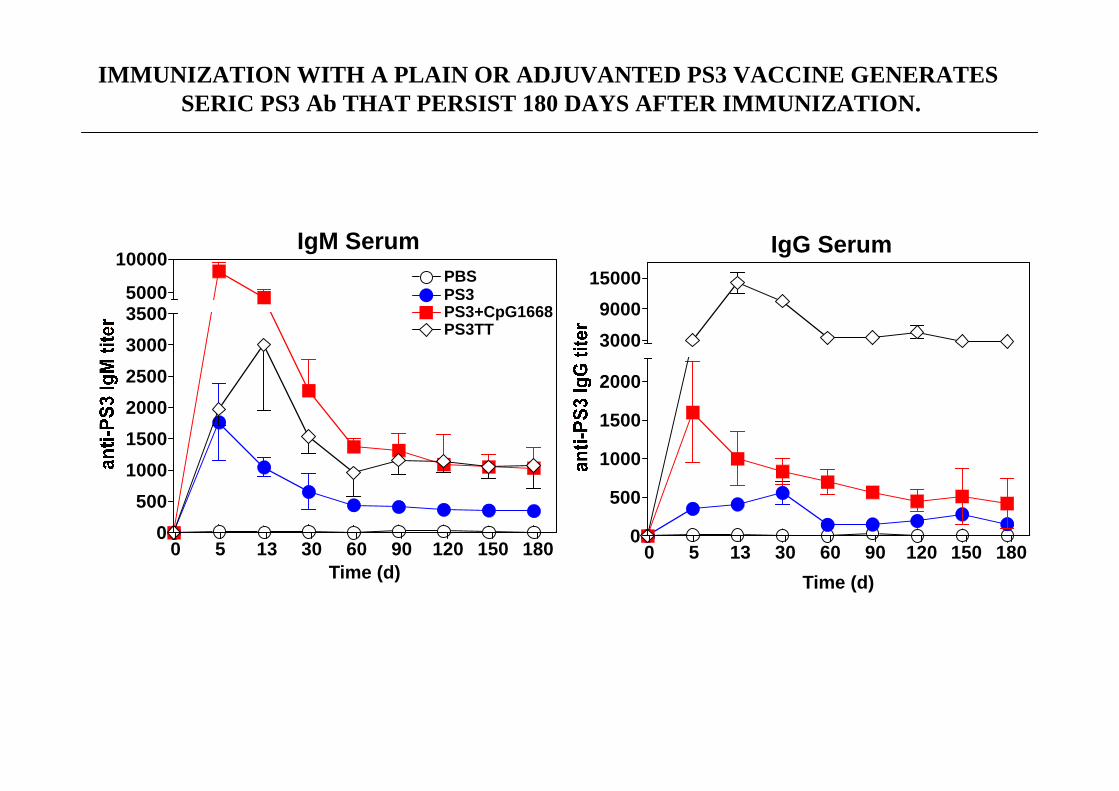
IgM Serum
0 5 13 30 60 90 120 150 1800
500
1000
1500
2000
2500
3000
3500PS3PBS
PS3+CpG1668PS3TT
5000
10000
Time (d)
IgG Serum
0 5 13 30 60 90 120 150 1800
500
1000
1500
2000
3000
9000
15000
Time (d)
IMMUNIZATION WITH A PLAIN OR ADJUVANTED PS3 VACCINE GENERATES SERIC PS3 Ab THAT PERSIST 180 DAYS AFTER IMMUNIZATIO N.
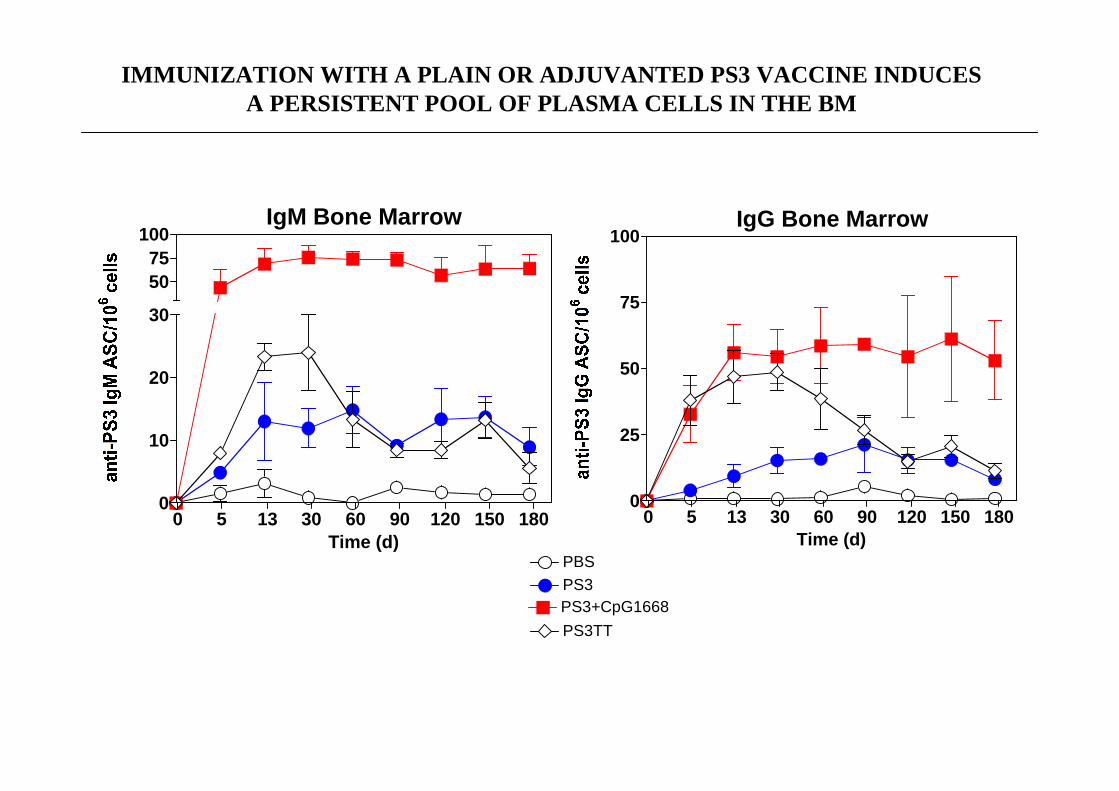
IMMUNIZATION WITH A PLAIN OR ADJUVANTED PS3 VACCINE INDUCESA PERSISTENT POOL OF PLASMA CELLS IN THE BM
PS3+CpG1668
PS3TT
PS3PBS
IgM Bone Marrow
0 5 13 30 60 90 120 150 1800
10
20
30
5075
100
Time (d)
IgG Bone Marrow
0 5 13 30 60 90 120 150 1800
25
50
75
100
Time (d)
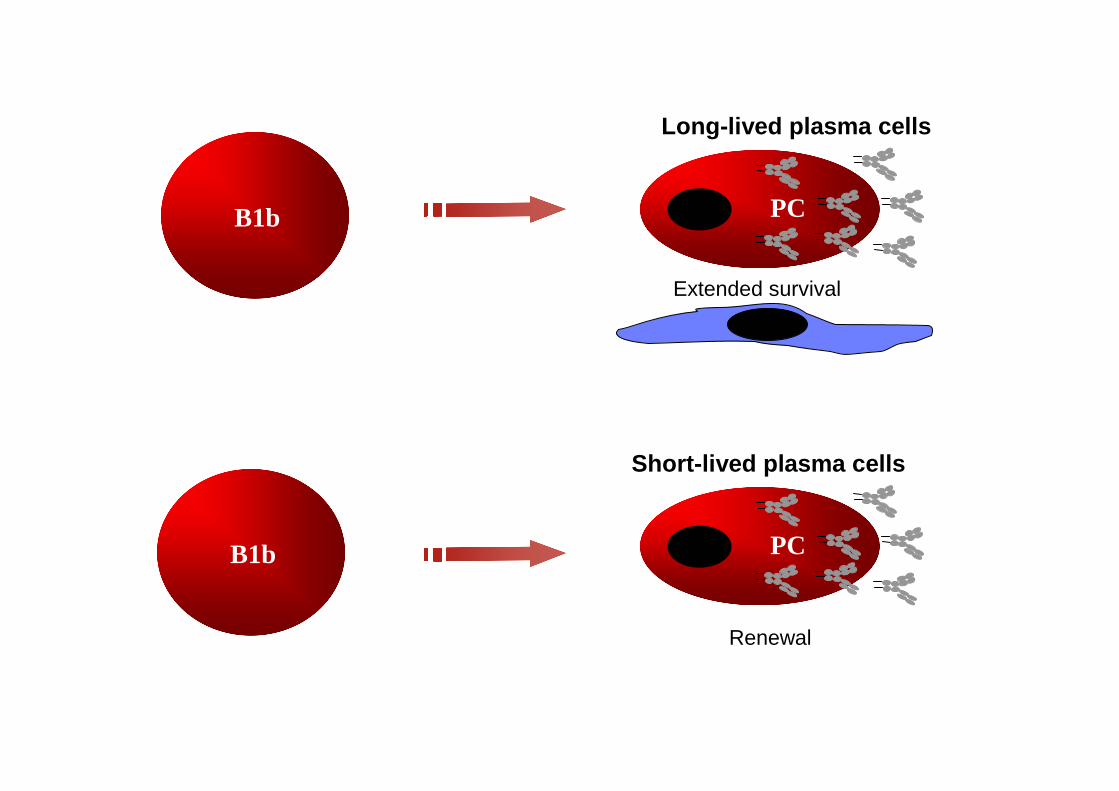
B1b
B1b
PC
Long-lived plasma cells
Extended survival
PC
Short-lived plasma cells
Renewal
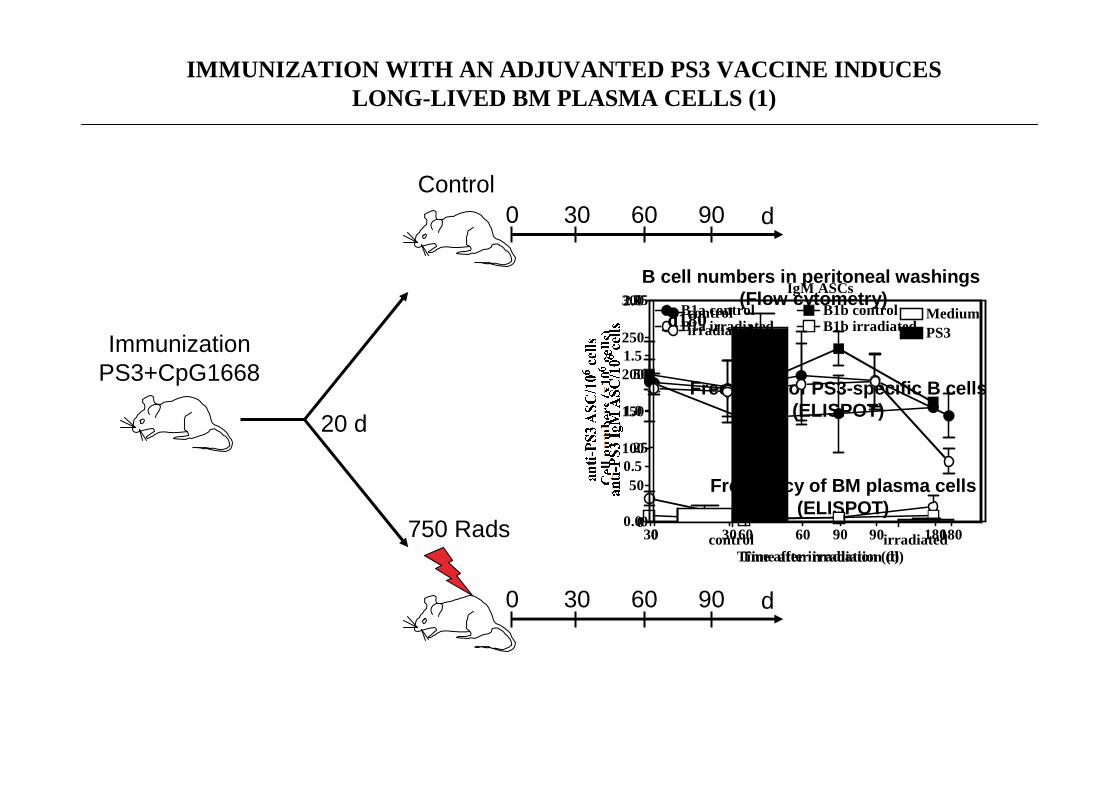
750 Rads
Control
ImmunizationPS3+CpG1668
20 d
B cell numbers in peritoneal washings(Flow cytometry)
Frequency of BM plasma cells(ELISPOT)
d30 90600
d30 90600
IMMUNIZATION WITH AN ADJUVANTED PS3 VACCINE INDUCESLONG-LIVED BM PLASMA CELLS (1)
Frequency of PS3-specific B cells(ELISPOT)
30 60 90 1800.0
0.5
1.0
1.5
2.0B1a controlB1a irradiated
B1b controlB1b irradiated
Time after irradiation (d)
IgM ASCs
0 30 60 90 1800
25
50
75controlirradiated
Time after irradiation (d)
0
50
100
150
200
250
300
control irradiated
PS3Mediumd180

PS3+
CpGPS3-TT
WT
d40
BM
µMT
anti-PS3IgM titers
d30 d45 d60

15 30 45 60 75 900
20
40
60
80PS3+CpG
PS3TTPS3+CpG CD138 depleted
PS3TT CD138 depleted
Time after transfert (d)
IMMUNIZATION WITH AN ADJUVANTED PS3 VACCINE INDUCESLONG-LIVED BM PLASMA CELLS (2)
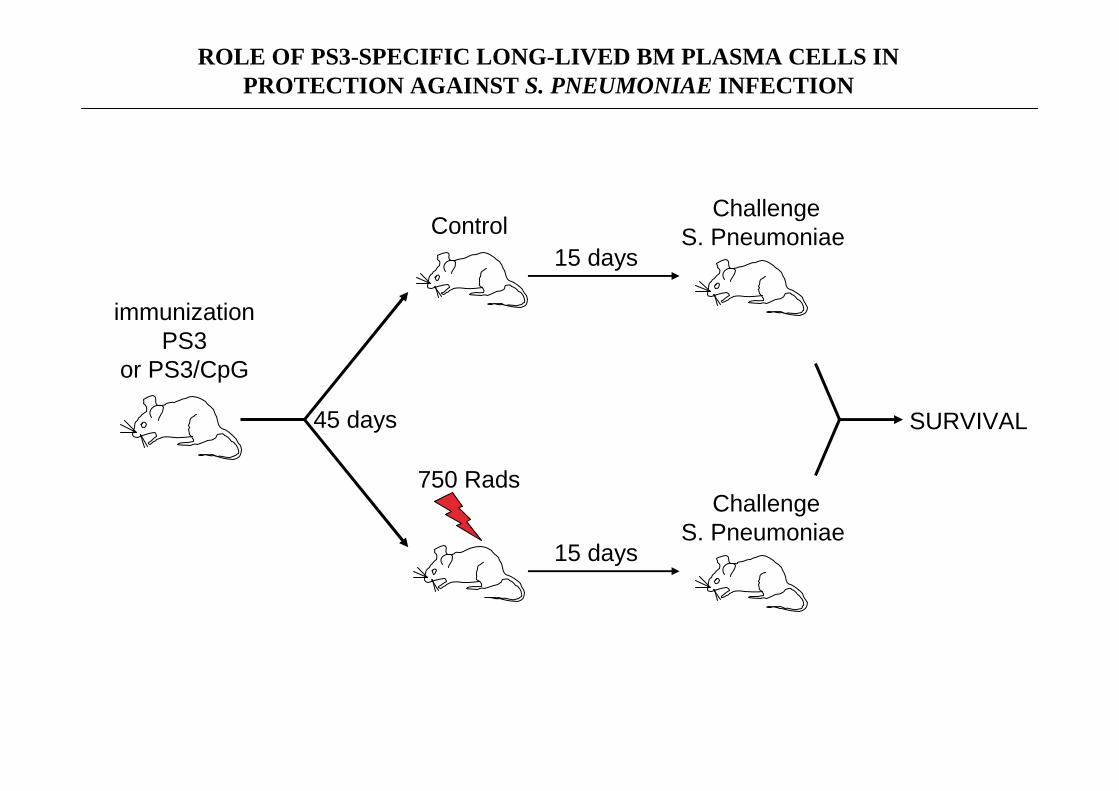
ROLE OF PS3-SPECIFIC LONG-LIVED BM PLASMA CELLS IN PROTECTION AGAINST S. PNEUMONIAE INFECTION
immunizationPS3
or PS3/CpG
45 days
750 Rads
Control15 days
ChallengeS. Pneumoniae
15 days
ChallengeS. Pneumoniae
SURVIVAL

LONG-LIVED BM PLASMA CELLS ARE RESPONSIBLE FORTHE IMMUNE PROTECTION CONFERRED BY A PS3 VACCINE
106 cfu
0 1 2 3 4 5 6 7 8 9 100
20
40
60
80
100
PS3 0 radPS3 750 rad
PS3+CpG1668 0 radPS3+CpG1668 750 rad
Time (d)
107 cfu
0 1 2 3 4 5 6 7 8 9 100
20
40
60
80
100
Time (d)

TLR-L can restore responsiveness of young mice to bacterial PS
� Young mice are unresponsive to TI Ag
Paradigms on the B cell response to TI Ag have to be reconsidered
Primed B-1 cells generate an amplified and accelerated Ab response after transfer in Rag2 recipients. TI Ag can induce a secondary Ab response in AID KO mice.
� TI Ag do not promote bona fide secondary responses
Adoptive transfer of B-1b cells from PS-immune mice protects naive mice
� TI Ag do not generate B cell memory
High numbers of PS3-specific PC can be detected in the bone marrow of mice immunized with PS3 and TLR-L
� TI Ag do not generate long-lived plasma cells

