le broutement de la rÉgÉnÉration par le cerf de ... · sapin baumier. ce projet vise à évaluer...
TRANSCRIPT

CHRISTINE CASABON LE BROUTEMENT DE LA RÉGÉNÉRATION PAR LE CERF DE VIRGINIE (ODOCOILEUS VIRGINIANUS)
DANS LES GRANDES COUPES AVEC PROTECTION DE LA RÉGÉNÉRATION ET DES SOLS (CPRS) DE
L’ÎLE D’ANTICOSTI
Mémoire présenté à la Faculté des études supérieures de l’Université Laval
dans le cadre du programme de maîtrise en sciences forestières pour l’obtention du grade de maître ès sciences (M.Sc.)
FACULTÉ DE FORESTERIE
UNIVERSITÉ LAVAL QUÉBEC
2006 © Christine Casabon, 2006

Résumé Le broutement par le cerf de Virginie sur l’île d’Anticosti rend difficile la régénération du
sapin baumier. Ce projet vise à évaluer l’impact de coupes avec protection de la
régénération et des sols (CPRS) de grandes dimensions sur la régénération du sapin
baumier, des espèces compagnes et sur la diversité végétale et ce, en présence d’une
population abondante de cerf. Les résultats montrent que la distance de la lisière boisée
n’influence pas la distribution, le nombre et la hauteur des semis. La distribution, le nombre
de semis/hectare et la croissance en hauteur du sapin et du bouleau à papier sont diminués
par le broutement. Les débris ligneux peuvent soustraire temporairement les semis au
broutement. Le cerf n’a pas d’effet global sur le nombre d’espèces végétales, mais il a un
impact négatif marqué sur le recouvrement total des espèces et il entraîne une modification
de la composition en espèces. Les grandes CPRS ne permettent pas de reconstituer les
sapinières originales.

ii
Abstract Browsing by white-tailed deer on Anticosti Island makes balsam fir regeneration difficult.
This project aims at evaluating the impact of large clear-cuts on balsam fir regeneration,
associated species and vegetation diversity when they are submitted to high deer densities.
Results show that distance from forest edge does not affect the stocking, the density and the
height of seedlings. Stocking, number of seedlings/hectare and height growth of fir and
white birch are reduced by browsing. Woody debris can protect temporarily seedlings from
browsing. In general, deer does not affect the number of plant species, but produces an
important negative impact on total cover of species and causes a modification in the species
composition. Large clear-cuts fail to restore balsam fir forests.

Résumé long Le broutement effectué par la forte population de cerf de Virginie (Odocoileus virginianus
Zimmermann) sur l’île d’Anticosti rend difficile la régénération du sapin baumier qui
constitue à la fois sa nourriture et son abri de prédilection. Les sapinières originales de l’île
sont graduellement remplacées par des pessières blanches, l’épinette blanche étant très peu
broutée par le cerf. Il est donc nécessaire de trouver des moyens permettant de régénérer les
sapinières afin de conserver les écosystèmes initiaux et de fournir de la nourriture au cerf
pour assurer la pérennité de la chasse, qui représente l’activité économique principale de
l’île d’Anticosti. Par ailleurs, plusieurs études ont établi que sur le continent, le centre des
grandes coupes forestières est généralement épargné par le broutement du cerf. Cela
pourrait potentiellement constituer une solution pour régénérer les sapinières de l’île
d’Anticosti.
Ce projet vise donc à évaluer l’impact de coupes avec protection de la régénération et des
sols (CPRS) de grandes dimensions sur la régénération du sapin baumier et de ses espèces
compagnes, et sur la diversité végétale en fonction de la distance de la lisière boisée à l’île
d’Anticosti. Des surfaces clôturées adjacentes à des parcelles non clôturées ont donc été
mises en place en 1995 et 1996 dans de grandes CPRS pour étudier ce phénomène. La
régénération et la diversité végétale sont les deux volets traités dans l’étude. Pour le volet
régénération, les hypothèses à vérifier sont que le nombre de semis de sapin baumier et des
espèces compagnes augmente proportionnellement à la distance des lisières boisées, que la
croissance en hauteur de ces semis est diminuée par le broutement du cerf, que le cerf
réduit le nombre et la distribution des semis et que la présence de débris de coupe peut
soustraire temporairement les semis au broutement. Les hypothèses concernant la diversité
végétale sont que la richesse en espèces et le recouvrement total des espèces augmentent
parallèlement à la distance des lisières boisées, que le cerf réduit le nombre d’espèces et le
pourcentage de recouvrement total de celles-ci, et qu’il entraîne une modification de la
composition en espèces en faveur des espèces non broutées ou tolérantes au broutement.

iv
Les résultats démontrent que le coefficient de distribution, le nombre de semis à l’hectare et
la croissance en hauteur du sapin et de ses espèces compagnes sont diminués par le
broutement et ce, peu importe la distance de la lisière boisée. Nous avons observé que les
débris de coupe peuvent soustraire temporairement les semis au broutement. En ce qui
concerne la diversité végétale, le cerf n’a pas d’effet global sur le nombre d’espèces.
Cependant, il a un impact négatif marqué sur le pourcentage de recouvrement total des
espèces végétales. De plus, une modification de la composition en espèces est observée en
présence du cerf. Il apparaît donc que les CPRS de grandes dimensions ne permettront pas
une régénération suffisante du sapin baumier pour reconstituer les sapinières de l’île
d’Anticosti.

Avant-Propos En premier lieu, je tiens à remercier mon directeur David Pothier qui est toujours
disponible, même quand je lui demandais in extremis de m’écrire une lettre de référence
pour le lendemain! Donc un gros merci David d’être toujours là quand j’ai besoin de toi et
aussi pour ta simplicité qui nous met à l’aise de discuter avec toi. Je veux aussi sincèrement
remercier Sonia De Bellefeuille qui a aussi toujours été disponible. Merci pour tes
nombreux conseils, pour la traduction de mes textes et présentations, et pour ta gentillesse.
Je veux dire un merci spécial aussi à mon conjoint, Jérôme Rioux, qui m’a toujours donné
son avis et qui m’a soutenue tout au long de ma maîtrise. De plus, je tiens à remercier
Robert Weladgi et Sophie Baillargeon pour leur aide statistique. Merci aussi à Nelson
Thiffault et Vincent Roy qui ont évalué mon mémoire. Ensuite, c’est merveilleux d’avoir
l’île d’Anticosti comme aire d’étude mais ce qui fait la différence entre un super été terrain
et un été ennuyant, c’est les personnes avec qui on travaille. Alors merci à Annabelle
Goupil de m’avoir non seulement aidée dans la prise de données sans ne jamais se plaindre,
mais aussi pour m’avoir ouverte à la pensée « biologiste grano »! Il me semble que les
journées passaient plus vite avec nos discussions. Merci aussi à Jérôme et à Marie-Soleil
Germain-Tessier pour l’aide terrain. Différentes personnes sont intervenues à un moment
ou à un autre au cours de mon projet et je les remercie beaucoup, je parle ici de Françoise
Jenniss, Gaétan Laprise, Steeve Côté, Denis Duteault et Marie-Hélène Rousseau. Il faut
aussi souligner l’aide des partenaires de la Chaire de recherche CRSNG-Produits forestiers
Anticosti sans qui tout cela n’aurait pas été possible, c'est-à-dire les Produits forestiers
Anticosti inc., le CRSNG, le Centre d’études nordiques, le ministère des ressources
naturelles et de la faune du Québec et l’Université Laval.
Le quotidien à la maîtrise nous amène à discuter et à connaître davantage les étudiants
gradués des laboratoires dont nous faisons partie. Je retiendrai de bons moments au sein des
deux labos, puisque étant une forestière avec un projet dont la Chaire de recherche est en
biologie, j’ai eu la chance de côtoyer deux groupes d’étudiants dynamiques et dont
sympathiques. Je pense donc au laboratoire de sylviculture et aux gens du « Love labo ».
Merci à vous tous pour vos conseils et pour les discussions qui m’ont ouvert l’esprit.

vi
Finalement, tout cela n’aurait jamais été possible sans ma famille… mes parents qui m’ont
amené dans le bois dès ma naissance. C’est grâce à eux que j’ai une passion aussi forte
pour la forêt et la faune et que j’ai décidé d’en faire mon métier. La famille c’est la chose la
plus importante dans ma vie et je vous dis un énorme MERCI!

Table des matières RÉSUMÉ I ABSTRACT II RÉSUMÉ LONG III AVANT-PROPOS V TABLE DES MATIÈRES VII LISTE DES TABLEAUX IX LISTE DES FIGURES X
INTRODUCTION 1
MATÉRIEL ET MÉTHODES 4
AIRES D’ÉTUDE 4 DISPOSITIF EXPÉRIMENTAL 6 ANALYSES STATISTIQUES 10
RÉSULTATS 14
RÉGÉNÉRATION FORESTIÈRE 14 CONDITIONS AVANT LA COUPE 14 EFFET DES LISIÈRES BOISÉES 15 COEFFICIENT DE DISTRIBUTION ET NOMBRE DE SEMIS À L’HECTARE DE SAPIN BAUMIER ET DES ESPÈCES COMPAGNES 16 HAUTEUR MOYENNE DES SEMIS DE SAPIN BAUMIER ET DES ESPÈCES COMPAGNES 22 IMPACT DES DÉBRIS LIGNEUX 24 DIVERSITÉ VÉGÉTALE 27 EFFET DES LISIÈRES BOISÉES SUR LA DIVERSITÉ VÉGÉTALE 27 IMPACT DU CERF SUR LA RICHESSE EN ESPÈCES VÉGÉTALES 27 IMPACT DU CERF SUR LE POURCENTAGE DE RECOUVREMENT TOTAL DES ESPÈCES VÉGÉTALES 28 MODIFICATION DE LA COMPOSITION VÉGÉTALE PAR LE CERF 29
DISCUSSION 33
RÉGÉNÉRATION FORESTIÈRE 33 CONDITIONS AVANT LA COUPE 33 EFFET DES LISIÈRES BOISÉES 33 COEFFICIENT DE DISTRIBUTION ET NOMBRE DE SEMIS À L’HECTARE DE SAPIN BAUMIER ET DES ESPÈCES COMPAGNES 35 HAUTEUR MOYENNE DES SEMIS DE SAPIN BAUMIER ET DES ESPÈCES COMPAGNES 38 IMPACT DES DÉBRIS LIGNEUX 40 DIVERSITÉ VÉGÉTALE 41 EFFET DES LISIÈRES BOISÉES SUR LA DIVERSITÉ VÉGÉTALE 41 IMPACT DU CERF SUR LA RICHESSE EN ESPÈCES VÉGÉTALES 42 IMPACT DU CERF SUR LE POURCENTAGE DE RECOUVREMENT TOTAL DES ESPÈCES VÉGÉTALES 44

viii
MODIFICATION DE LA COMPOSITION VÉGÉTALE PAR LE CERF 45 RÉVERSIBILITÉ ET EXTINCTION LOCALE DES ESPÈCES VÉGÉTALES 47
CONCLUSION 49
PROPOSITIONS D’AMÉNAGEMENT 50
BIBLIOGRAPHIE 52

Liste des tableaux TABLEAU 1: NIVEAU DE SIGNIFICATION (P) DES CONDITIONS AVANT LA COUPE SUR LES
VARIABLES MESURÉES POUR LES DIFFÉRENTES ESSENCES, OBTENU À PARTIR DE LA PROCÉDURE MIXED DE SAS. ........................................................................................................... 14
TABLEAU 2: RÉSULTATS DE L’ANALYSE DU MODÈLE AVEC LA PROCÉDURE MIXED DE SAS TESTANT LE CDR DU SAPIN BAUMIER, AVANT LA RÉDUCTION DES TERMES NON SIGNIFICATIFS INCLUANT LA VARIABLE À EFFET FIXE (LISIÈRE)........................................ 15
TABLEAU 3: NIVEAU DE SIGNIFICATION (P) DE LA DISTANCE DE LA LISIÈRE BOISÉE SUR LES VARIABLES MESURÉES POUR LES DIFFÉRENTES ESSENCES TEL QUE CALCULÉ PAR LA PROCÉDURE MIXED DE SAS. ........................................................................................................... 15
TABLEAU 4: NIVEAU DE SIGNIFICATION (P) DU CLÔTURAGE (ZONE CLÔTURÉE ET ZONE NON CLÔTURÉE) SELON LES DIFFÉRENTES ANNÉES POUR LE COEFFICIENT DE DISTRIBUTION ET LE NOMBRE DE SEMIS À L’HECTARE POUR LE BOULEAU À PAPIER. CES RÉSULTATS ONT ÉTÉ OBTENUS AVEC LA PROCÉDURE MIXED DE SAS. ..................... 20
TABLEAU 5: NIVEAU DE SIGNIFICATION (P) DU CLÔTURAGE (ZONE CLÔTURÉE ET ZONE NON CLÔTURÉE) SELON LES DIFFÉRENTES ANNÉES POUR LA HAUTEUR MOYENNE POUR L’ÉPINETTE BLANCHE ET LE BOULEAU À PAPIER, TEL QU’OBTENU À L’AIDE DE LA PROCÉDURE MIXED DE SAS. ........................................................................................................... 24
TABLEAU 6: NIVEAU DE SIGNIFICATION (P) DU CLÔTURAGE (ZONE CLÔTURÉE ET ZONE NON CLÔTURÉE) SELON LES DIFFÉRENTES ANNÉES POUR LE NOMBRE D’ESPÈCES VÉGÉTALES OBTENU À L’AIDE DE LA PROCÉDURE MIXED DE SAS..................................... 27
TABLEAU 7: NIVEAU DE SIGNIFICATION (P) DU CLÔTURAGE (ZONE CLÔTURÉE ET ZONE NON CLÔTURÉE) SELON LES DIFFÉRENTES ANNÉES POUR LE RECOUVREMENT TOTAL DES ESPÈCES VÉGÉTALES. CES RÉSULTATS ONT ÉTÉ OBTENUS À L’AIDE DE LA PROCÉDURE MIXED DE SAS. .................................................................................................................................... 28
TABLEAU 8: POURCENTAGE DE RECOUVREMENT MOYEN DES ESPÈCES COUVRANT 0,5 % ET PLUS EN MOYENNE DANS LES ZONES CLÔTURÉES OU NON CLÔTURÉES. ......................... 32

Liste des figures FIGURE 1: LOCALISATION DE L'ÎLE D'ANTICOSTI SUR LE TERRITOIRE QUÉBÉCOIS.................... 5 FIGURE 2: SEPT COUPES DE 3 KM2 (CPRS 1995 ET 1996) ET 52 AIRES D’ÉCHANTILLONNAGE
(POINTS) SITUÉES À DIFFÉRENTES DISTANCES DE LA LISIÈRE BOISÉE. ............................... 7 FIGURE 3: ZONES CLÔTURÉE ET NON CLÔTURÉE D’UN RAYON DE 5 M QUI CONTIENNENT 10
MICRO-PLACETTES DE 4 M2 (1,13 M DE RAYON). LES ZONES SONT ESPACÉES DE 15 À 45 M. .............................................................................................................................................................. 8
FIGURE 4: ÉVOLUTION A) DU COEFFICIENT DE DISTRIBUTION (MOYENNE DE PRÉSENCE DANS LES 10 MICRO-PLACETTES) ET B) DU NOMBRE DE SEMIS À L’HECTARE DE SAPIN BAUMIER DANS LES ZONES CLÔTURÉES ET DANS LES ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR-TYPE. ........................................................ 17
FIGURE 5: ÉVOLUTION A) DU COEFFICIENT DE DISTRIBUTION (MOYENNE DE PRÉSENCE DANS LES 10 MICRO-PLACETTES) ET B) DU NOMBRE DE SEMIS À L’HECTARE DE L’ÉPINETTE BLANCHE DANS LES ZONES CLÔTURÉES ET DANS LES ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR-TYPE. ....................... 19
FIGURE 6: ÉVOLUTION A) DU COEFFICIENT DE DISTRIBUTION (MOYENNE DE PRÉSENCE DANS LES 10 MICRO-PLACETTES) ET B) DU NOMBRE DE SEMIS À L’HECTARE DU BOULEAU À PAPIER DANS LES ZONES CLÔTURÉES ET DANS LES ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR-TYPE. ....................... 21
FIGURE 7: ÉVOLUTION DE LA HAUTEUR A) DU SAPIN BAUMIER ET B) DU BOULEAU À PAPIER DANS LES ZONES CLÔTURÉES ET DANS LES ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR-TYPE.......................................................................... 23
FIGURE 8: A) COEFFICIENT DE DISTRIBUTION ET B) NOMBRE DE SEMIS À L'HECTARE DE SAPIN BAUMIER EN FONCTION DU REGROUPEMENT DES CLASSES DE RECOUVREMENT DE BRANCHES AU SOL UN AN APRÈS LA COUPE. CES GRAPHIQUES PRÉSENTENT LA DISTRIBUTION DE LA MOYENNE DES 10 MICRO-PLACETTES DE CHACUNE DES 52 ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR TYPE. .............. 25
FIGURE 9: NOMBRE DE SEMIS À L'HECTARE DE SAPIN BAUMIER EN FONCTION DU REGROUPEMENT DES CLASSES DE RECOUVREMENT D'OBSTACLES AU SOL 8,5 ANS APRÈS LA COUPE. CE GRAPHIQUE DÉMONTRE LA DISTRIBUTION DE LA MOYENNE DES 10 MICRO-PLACETTES DES 52 ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR TYPE. .................................................................................................. 26
FIGURE 10: ÉVOLUTION DU POURCENTAGE DE RECOUVREMENT TOTAL DES ESPÈCES DANS LES ZONES CLÔTURÉES ET DANS LES ZONES NON CLÔTURÉES. LES BARRES VERTICALES REPRÉSENTENT L’ERREUR-TYPE.......................................................................... 29
FIGURE 11: RÉPARTITION DES ESPÈCES AYANT UN RECOUVREMENT MOYEN D’AU MOINS 0,5 % EN LIEN AVEC LES ZONES CLÔTURÉES ET LES ZONES NON CLÔTURÉES 8,5 ANS APRÈS LA COUPE, OBTENUE PAR L’ANALYSE EN COMPOSANTE PRINCIPALE DU LOGICIEL CANOCO (VOIR CORRESPONDANCE DES CODES AU TABLEAU 8). L’AXE 1 REPRÉSENTE L’AXE HORIZONTAL ALORS QUE L’AXE 2 EST L’AXE VERTICAL. POUR L’APPELLATION DES SITES, CHAQUE BLOC (COUPES A, B, C, D, E, F ET H) EST SUIVI DE C S’IL S’AGIT D’UNE ZONE CLÔTURÉE (TRIANGLE) ET DE NC SI C’EST UNE ZONE NON CLÔTURÉE (CERCLE). ON RETROUVE UNE ZONE CLÔTURÉE ET UNE ZONE NON CLÔTURÉE PAR BLOC, IL Y A 7 BLOCS AU TOTAL ET DONC 14 POINTS REPRÉSENTANT LES ZONES. LE POSITIONNEMENT DE LA MOYENNE DES ZONES CLÔTURÉES ET DES ZONES NON CLÔTURÉES EST AUSSI IDENTIFIÉ.......................................................................... 30
FIGURE 12: ZONE CLÔTURÉE À L’INTÉRIEUR D’UNE GRANDE COUPE PRÉSENTANT UNE STRUCTURE DE PRAIRIE................................................................................................................... 38
FIGURE 13: SEMIS DE SAPIN BAUMIER PRÉSENTANT UN FAIBLE ESPACEMENT ENTRE LES DIFFÉRENTS VERTICILLES DANS UNE ZONE NON CLÔTURÉE. .............................................. 39

Introduction
Les populations de cerf de Virginie (Odocoileus virginianus Zimmermann) sont en
augmentation en Amérique du Nord depuis le siècle dernier et elles atteignent présentement
des niveaux jamais observés depuis plusieurs centaines d’années (Rooney, 2001). La forte
pression de broutement effectuée par une densité élevée de cerf peut grandement modifier
un milieu en inhibant la croissance de la régénération naturelle ou artificielle (Stoeckeler et
al., 1957; Bellingham et Allan, 2003), en modifiant la composition (Anderson et Loucks,
1979; Tilghman, 1989; Veblen et al., 1989; Anderson et Katz, 1993) et la structure des
peuplements forestiers (Stewart et Burrows, 1989; Alverson et Waller, 1997; Healy, 1997;
Rooney, 2001; Horsley et al., 2003) et même en causant l’extinction localisée d’espèces
végétales (Case et McCullough, 1987; Augustine et Frelich, 1998; Cornett et al., 2000;
Russell et Fowler, 2004). Ces effets du broutement du cerf peuvent avoir un impact majeur
sur l’économie et sur l’industrie forestière d’une région donnée (Tilghman, 1989 ; Conover,
1997).
Le cerf de Virginie a été introduit sur l’île d’Anticosti à la fin du 19e siècle et la population
avoisine aujourd’hui les 125 000 cerfs, la densité moyenne estimée étant supérieure à 15
cerfs/km2 (Rochette et al., 2003 dans Potvin et Poirier, 2004) et pouvant atteindre jusqu’à
30 cerfs/km2 en certains endroits. À ces densités, le cerf a un impact marqué sur le milieu
en limitant le développement de la régénération et en modifiant les écosystèmes (Potvin et
al., 2003a; Potvin et Poirier, 2004). Le broutement excessif du sapin baumier (Abies
balsamea (L.) Mill.) par le cerf pourrait entraîner une conversion des sapinières originales
de l’île en pessières blanches, l’épinette blanche (Picea glauca (Moench) Voss) étant une
espèce peu broutée (Potvin et al., 2003a). Conséquemment, la population de cerf de l’île est
appelée à diminuer fortement en raison du déclin prononcé des superficies couvertes par
des sapinières sur un horizon de 40 à 50 ans (Potvin et al., 2003a). Puisque l’île d’Anticosti
représente une destination de chasse très populaire qui constitue son activité économique
principale, il devient nécessaire de trouver un moyen de régénérer les sapinières de l’île

2
afin d’assurer un maintien d’un habitat de qualité pour le cerf. L’objectif de préserver les
écosystèmes initiaux est aussi de première importance. Présentement, la seule méthode
efficace qui est utilisée pour régénérer les sapinières est le clôturage systématique des aires
de coupe suivi d’une plantation de sapin baumier. Ces clôtures seront conservées jusqu’à ce
que les sapins aient atteint une hauteur suffisante pour que leur flèche terminale soit hors de
portée du cerf. Cependant, l’installation de telles clôtures représente des coûts très élevés et
il y a donc avantage à trouver une ou des méthodes sylvicoles alternatives pour régénérer
les sapinières.
Au Québec et ailleurs dans le monde, des études ont montré que le centre de grandes
coupes pouvait être épargné par le broutement du cerf en raison de la grande distance de la
lisière boisée et de l’augmentation parallèle du risque de prédation (Drolet, 1978; Welch et
al., 1991; Kay, 1993; Schmitz, 2005). L’île d’Anticosti est caractérisée par l’absence de
prédateurs, mais l’instinct du cerf de même que la longue saison de chasse qui s’étend sur
près de quatre mois pourraient entraîner les mêmes comportements. La présente étude vise
donc à tester l’effet de grandes coupes avec protection de la régénération et des sols
(CPRS) sur la régénération de sapin baumier et de ses espèces compagnes à de fortes
densités de cerf. Deux volets sont traités : la régénération forestière et la diversité végétale.
Les hypothèses à vérifier pour le volet de la régénération sont que 1) le nombre de semis de
sapin baumier et des espèces compagnes augmente parallèlement à la distance par rapport
aux lisières boisées; 2) la présence de débris de coupe peut temporairement soustraire les
semis au broutement du cerf tout en conservant les conditions nécessaires à leur survie; 3)
le cerf réduit le nombre et la distribution des semis de sapin et des espèces compagnes; 4) la
croissance moyenne en hauteur des semis de sapin et des espèces compagnes est fortement
diminuée par le broutement du cerf. Pour le volet de la diversité végétale, les hypothèses à
vérifier sont que 1) la richesse en espèces et le recouvrement total des espèces augmentent
parallèlement à la distance par rapport aux lisières boisées; 2) le cerf réduit la richesse et le
recouvrement total des espèces végétales; 3) une modification de la composition des
espèces végétales est engendrée par la pression de broutement du cerf sur le milieu. Ces
hypothèses pourront être testées à l’aide d’un dispositif expérimental comprenant des

3
grands blocs de coupe à l’intérieur desquels des placettes de régénération ont été établies à
l’intérieur et à l’extérieur de superficies clôturées.

Matériel et méthodes
Aires d’étude L’île d’Anticosti est située dans le Golfe du Saint-Laurent (49° 28’ N, 63° 00’ O) dans la
partie est du Québec (Figure 1). L’île s’étend sur une longueur de 222 km et couvre une
superficie de 7 943 km2. La topographie est peu accidentée et l’altitude moyenne est de 126
m. Les dépôts de l’île sont d’origine marine post glaciaire dans la partie basse et côtière
alors que le centre est recouvert de dépôts morainiques. De vastes étendues de dépôts
organiques formant des tourbières se retrouvent principalement dans l’est de l’île (Potvin et
Poirier, 2004). Les précipitations de neige sont abondantes et excèdent d’environ 20 % les
précipitations mesurées dans l’aire de répartition du cerf de Virginie ailleurs au Québec
(Huot, 1982). Le climat correspond à celui de la zone bioclimatique maritime sub-boréale
qui est caractérisée par des étés frais et des hivers relativement doux. La température
moyenne est d’environ -11 °C en janvier et 16 °C en juillet (Environnement Canada, 2006).

5
Figure 1: Localisation de l'île d'Anticosti sur le territoire québécois.
Du point de vue forestier, l’île d’Anticosti se situe dans le domaine bioclimatique de la
sapinière à bouleau blanc et dans le sous-domaine de l’est de la région boréale inférieure
(Grondin et al., 1996). Les principales espèces forestières sont l’épinette blanche, le sapin
baumier et l’épinette noire (Picea mariana (Mill.) B.S.P.). Des espèces telles le peuplier
faux-tremble (Populus tremuloides Michx), le peuplier baumier (Populus balsamifera L.),
le bouleau à papier (Betula papyrifera Marsh.) et le mélèze laricin (Larix laricina (Du Roi)
K. Koch) sont présentes sporadiquement. La flore herbacée est riche et diversifiée et est
dominée par la clintonie boréale (Clintonia borealis (Ait.) Raf.), la maïanthème du Canada
(Maianthemum canadense Desf.), le cornouiller du Canada (Cornus canadensis L.), la
listère cordée (Listera cordata (L.) R. Br.), la dryoptéride spinuleuse (Dryopteris spinulosa
(O.F. Muell.) Watt) et l’oxalide de montagne (Oxalis montana Raf.) (Marie-Victorin et al.,
1995). La perturbation naturelle majeure à l’île est causée par l’arpenteuse de la pruche
(Lambdina fiscellaria fiscellaria (Guen.)). Une épidémie très importante est survenue de
1931 à 1936 et la dernière infestation, de moindre envergure, s’est manifestée de 1971 à

6
1973 (Dorais et al., 1996). La tordeuse des bourgeons de l’épinette (Choristoneura
fumiferana (Clem.)) a sévi pour la première fois en 1973 (Blais, 1983 dans Potvin, 1992).
Le chablis constitue aussi une perturbation importante alors que le feu est d’une importance
secondaire.
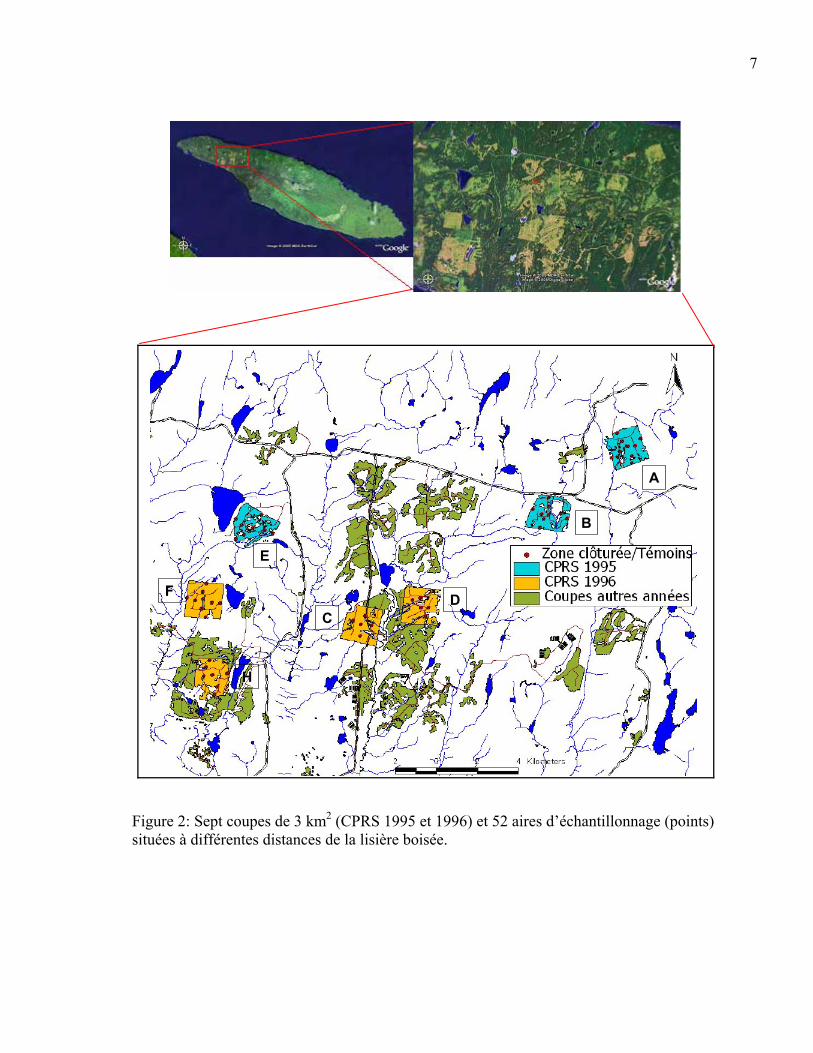
Dispositif expérimental En 1995 et 1996, sept grandes CPRS (Blocs A, B, C, D, E, F, H) d’environ 3 km2 ont été
effectuées (Figure 2) par le procédé de récolte par tronc entier à l’aide d’une abatteuse
multifonctionnelle. L’ébranchage des tiges s’est donc fait sur place, les débris de coupe
étant laissés sur le terrain. Un effort spécifique a été déployé afin que les débris de coupe
soient étalés sur le terrain et ne soient pas concentrés dans des amas. Une débardeuse à
câble a été utilisée pour amener les tiges en bordure de chemin. La machinerie était
restreinte au niveau de la circulation sur les aires de coupe pour protéger la régénération
préétablie. Ainsi, les sentiers étaient espacés d’environ 15 mètres. De 5 à 9 aires
d’échantillonnage par bloc ont été établies avant la coupe à différentes distances de la
lisière boisée pour un total de 52 aires d’échantillonnage (Figure 2). Celles-ci se composent
d’une combinaison d’une zone clôturée (zone d’exclusion du cerf) et d’une zone non
clôturée (zone de libre accès au cerf) qui représentent chacune une superficie circulaire
d’environ 80 m2 (parcelle). Les clôtures de 3 m de hauteur ont été installées immédiatement
après la coupe, soit en 1995 (Blocs A, B et E) et en 1996 (Blocs C, D, F et H). Chacune de
ces zones contient 10 micro-placettes circulaires de 4 m2 (Figure 3). En résumé, chacune
des zones contient 10 micro-placettes et il y a deux zones (une clôturée et une non clôturée)
par aires d’échantillonnage. De 5 à 9 aires d’échantillonage par bloc sont présentes et il y a
7 blocs, pour un total de 1040 micro-placettes. À la suite d’un chablis important survenu en
1996, les blocs C et D ont été fusionnés pour atteindre une superficie de près de 11 km2. Le
bloc H a aussi été fortement agrandi pour atteindre une superficie d’environ 10 km2 (Potvin
et al., 2003b).
7
A
B
DC
E
F
H
Figure 2: Sept coupes de 3 km2 (CPRS 1995 et 1996) et 52 aires d’échantillonnage (points) situées à différentes distances de la lisière boisée.

8
Figure 3: Zones clôturée et non clôturée d’un rayon de 5 m qui contiennent 10 micro-placettes de 4 m2 (1,13 m de rayon). Les zones sont espacées de 15 à 45 m.
Les placettes ont été inventoriées à 6 reprises, c’est-à-dire avant la coupe et 1, 2, 3, 5,5 et
8,5 ans après la coupe. En fait, les inventaires effectués en 2001 et en 2004 représentait
respectivement 5 ou 6 ans, et 8 ou 9 ans après la coupe, selon l’année de la coupe. Ces
années ont été regroupées puisqu’il y avait un trop grand nombre de données manquantes
lorsque les coupes étaient analysées séparément. C’est donc l’année mitoyenne qui a été
considérée, c’est-à-dire 5,5 et 8,5 ans après la coupe. Les données de régénération ont été
récoltées pour toutes ces années, alors qu’en raison de diverses contraintes, les données de
diversité végétale ont été recensées seulement après la coupe (an 0) ainsi que 5,5 et 8,5 ans
après la coupe. L’inventaire de régénération était effectué dès le début du mois de juin,
alors que l’inventaire de la végétation était effectué à partir de la mi-juillet, au moment de
la pleine feuillaison des plantes.
Pour l’inventaire de la régénération, le dénombrement des essences ligneuses commerciales
par classe de hauteur a été effectué, les classes étant < 5 cm, 5-30 cm, 31-60 cm, 61-100
cm, 101-200 cm, 201-300 et >301 cm. Trois variables ont été calculées à la suite de cette
prise de données : le coefficient de distribution, le nombre de semis à l’hectare et la hauteur
moyenne des semis par parcelle (regroupement des 10 micro-placettes). Le coefficient de
distribution de la régénération (CDR) pour chaque espèce a été calculé à partir d’une

9
donnée de présence (1) ou d’absence (0) de l’essence dans la micro-placette. Le CDR est le
pourcentage de placettes dans lesquelles au moins une tige de l’espèce ou du groupe
d’espèces désirées est retrouvé (Pominville et Ruel, 1995). La moyenne des 10 micro-
placettes produit donc un chiffre se situant entre 0 et 1, et représente le CDR de la parcelle.
Le nombre de semis des 10 micro-placettes a été utilisé pour calculer le nombre de
semis/ha de chaque parcelle. De plus, la moyenne de la hauteur des semis par essence pour
chaque micro-placette a été calculée en utilisant la valeur centrale de chaque classe de
hauteur. Par exemple, pour la classe de 5-30 cm, la valeur de hauteur considérée était de
17,5 cm. La hauteur moyenne des semis d’une micro-placette était donc représentée par la
valeur centrale de chaque classe de hauteur pondérée par le nombre de semis. La moyenne
des 10 micro-placettes a aussi été utilisée au moment de l’analyse statistique pour obtenir
une valeur moyenne par parcelle. Un inventaire du pourcentage de recouvrement
d’obstacles et de branches, ainsi que l’épaisseur et la hauteur de l’amas de branches par
rapport au sol a été effectué conjointement à l’inventaire de régénération, donc dès le début
du mois de juin. Le pourcentage de recouvrement d’obstacles au sol tenait compte de la
superficie couverte par les souches, les grosses roches et les chicots tombés au sol. Pour les
branches, le pourcentage de recouvrement a aussi été évalué dans chaque micro-placette.
L’épaisseur et la hauteur de l’amoncellement de branche ont été mesurées à l’aide d’une
règle graduée. L’épaisseur de l’amoncellement de branches correspond à l’épaisseur totale
moyenne du couvert de branches au sol dans la micro-placette, ce qui exclut l’espace vide
parfois observé entre le sol et le bas de l’amoncellement. La hauteur de l’amoncellement de
branches correspond à la distance entre le sol et le haut de l’amoncellement. Pour toutes les
variables de débris ligneux, la moyenne des 10 micro-placettes effectuée lors de l’analyse
permettait d’obtenir une valeur moyenne par parcelle.
L’inventaire de la diversité végétale consistait à noter le pourcentage de recouvrement au
sol occupé par chaque espèce ou groupe d’espèces lorsque les genres seulement étaient
considérés. Les espèces qui ont été inventoriées au niveau du genre seulement sont
amelanchier sp., aster sp., carex sp., cerastium sp., cirsium sp., equisetum sp., fragaria sp.,
galium sp., les graminées, hieracium sp., les hypnes, les lichens, les mousses, polygonum

10
sp., prenanthes sp., pyrola sp., ribes sp., senecio sp., solidago sp., trifolium sp., urtica sp.,
viola sp. et vaccinium sp.. Les classes de recouvrement utilisées étaient 1 % (< 1 %), 5 % (1
à 5 %), 10 % (6 à 15 %), 20 % (16 à 25 %), 30 % (26 à 35 %), 40% (36 à 45 %), 50 % (46
à 55 %), 60 % (56 à 65 %), 70 % (66 à 75 %), 80 % (76 à 85 %), 90 % (86 à 95 %) et 100
% (96 à 100%). Le pourcentage total pour chaque micro-placette pouvait excéder 100 %
puisqu’il s’agit d’une somme de recouvrement d’espèces de différentes hauteurs qui
peuvent donc se chevaucher. Les données recueillies ont permis de calculer trois variables.
D’abord, le nombre total d’espèces retrouvées dans chaque micro-placette, peu importe le
recouvrement et l’espèce, correspond à la richesse en espèces. Ensuite, le recouvrement
total de toutes les espèces confondues dans chaque micro-placette était aussi comptabilisé.
Le pourcentage de recouvrement de chaque espèce était aussi considéré dans le but de
vérifier la modification de la composition en espèces en présence du cerf. Au moment des
analyses statistiques, la moyenne des 10 micro-placettes a été utilisée.
Analyses statistiques Une analyse de covariance pour un plan en mesures répétées a été effectuée avec la
procédure MIXED du logiciel SAS (Milliken et Johnson, 2002). Le dispositif étudié est un
plan en blocs complets aléatoires. Les effets aléatoires retenus pour l’analyse sont le bloc,
l’aire (nichée dans le bloc) et la zone (en interaction avec l’aire). L’effet fixe de l’analyse
est la lisière. Le plan en mesures répétées est utilisé lorsqu’une variable, le temps dans le
présent cas, ne peut être aléatoire. En effet, l’année 3 ne peut venir avant l’année 1, et
l’année 1 est plus fortement liée à l’année 2 qu’à l’année 8,5 par exemple. Il est donc
important, dans le cas de mesures répétées, de sélectionner une structure de corrélation
adéquate (Milliken et Johnson, 2002). La meilleure structure de variance-covariance était
obtenue par la sélection du critère d’information d’Akaike (AIC) ayant la valeur la plus
faible. La valeur de l’AIC est dérivée du ratio de vraisemblance. Les principales structures
de covariance retenues lors de l’analyse sont ante-dependance, spatial power et
unstructured. À la suite de la sélection de la meilleure structure de variance-covariance, la
partie fixe du modèle (toutes les interactions contenant la variable lisière) était réduite
lorsque non significative pour permettre d’obtenir le meilleur modèle pour expliquer les

11
variables réponses. Le seuil alpha retenu pour déclarer un effet significatif était de 1 % pour
la variable réponse CDR puisque, étant donné le grand nombre de degrés de liberté à
l’erreur, des différences statistiquement significatives étaient détectées alors qu’elles étaient
biologiquement peu importantes à un seuil de 5 %. Pour les autres analyses, le seuil alpha
considéré pour conclure à un effet significatif était de 5 %. Ainsi, les zones clôturées et les
zones non clôturées ont été comparées dans le temps en considérant l’éloignement de la
lisière boisée.
Pour la régénération, les variables analysées étaient le coefficient de distribution, le nombre
de semis à l’hectare et la hauteur moyenne des semis. Pour assurer la validité des tests
statistiques, la compilation du nombre de semis à l’hectare et de la hauteur moyenne était
conditionnelle à la présence de l’essence. En effet, étant donné la grande quantité de valeurs
nulles (0) qui contrastaient fortement avec des valeurs élevées de semis à l’hectare et de
hauteur, les hypothèses de base de l’analyse n’étaient pas respectées. En fait, l’importante
quantité de valeurs zéro rendait impossible la normalisation des variables du nombre de
semis à l’hectare et de la hauteur moyenne. Le coefficient de distribution était donc d’abord
analysé. Ensuite, l’analyse des deux autres variables réponses (nombre de tiges/ha et
hauteur) a pu être effectuée à la condition qu’il y ait la présence d’au moins un semis dans
la micro-placette, et les micro-placettes présentant la valeur 0 (aucun semis) étaient
supprimées de l’analyse. L’inventaire de régénération effectué avant la coupe a été
considéré dans les analyses pour refléter les conditions initiales des stations. Cet inventaire
a donc été utilisé pour en extraire des covariables introduites dans des analyses de
covariance. Seuls le sapin baumier, l’épinette blanche et le bouleau à papier ont pu être
analysés puisque l’épinette noire et les peupliers faux-tremble et baumier n’étaient pas en
assez grande quantité pour assurer la validité des tests statistiques. En ce qui a trait à
l’analyse de l’effet des débris de coupe, les données ont été soumises au même test
statistique afin de déterminer l’effet du pourcentage d’obstacles, ainsi que du pourcentage,
de la hauteur et de l’épaisseur du couvert de branches sur le coefficient de distribution et le
nombre de semis. Comme pour l’analyse de la régénération, l’étude du nombre de semis
était conditionnelle à la présence de sapin dans la parcelle. La covariable dans ce cas était la

12
distance de la lisière boisée. Les débris ligneux n’ont pas été recensés avant la coupe
puisque l’objectif était de tester l’effet des déchets de coupe laissés par l’intervention. Les
variables analysées pour le volet de la diversité végétale étaient le nombre total d’espèces et
le pourcentage de recouvrement total des espèces. La covariable de l’analyse était la
distance de la lisière boisée. Aussi, des transformations de variables ont dû été réalisées
pour la plupart des analyses afin de respecter les hypothèses de base de l’analyse de
covariance. Les valeurs du CDR ont donc été transformées par l’arc sinus de la racine
carrée pour les trois espèces. Le nombre de tiges à l’hectare du sapin baumier et du bouleau
à papier a été transformé par le logarithme, alors que la racine carrée a été utilisée dans le
cas de l’épinette blanche. La hauteur moyenne a été transformée par la racine carrée pour le
sapin baumier et l’épinette blanche, et par le logarithme pour le bouleau à papier. Le
recouvrement total des espèces a été transformé par la racine carrée. Les données
présentées dans la section résultats sont les valeurs dé-transformées. Les analyses des
débris ligneux et du nombre d’espèces n’ont pas nécessité de transformation de variables.
D’autre part, afin de tester la similarité floristique entres les zones clôturées et les zones
non clôturées, un indice de similarité de Sørensen (IS) (Mueller-Dombois et Ellenberg,
1974; Magurran, 2004) a été calculé à partir de la variable du nombre d’espèces 8,5 ans
après la coupe. Cet indice permet de comparer les zones clôturées et les zones non clôturées
entre elles en termes du nombre d’espèces présentes à l’aide de la formule suivante :
IS = (2c / A+B) x 100
où c est le nombre d’espèces communes aux deux zones, alors que A et B représentent le
nombre d’espèces dans chacune des zones respectives. Une valeur de 100 % signifierait que
toutes les espèces sont présentes dans les deux zones alors qu’une valeur de 0 % indiquerait
qu’il n’y a aucune espèce commune aux deux zones.
Finalement, une analyse en composante principale a été réalisée avec le logiciel CANOCO
pour Windows 4.5 (Legendre et Legendre, 1984). Cette analyse multivariée permet de tirer

13
des conclusions sur les préférences de chaque espèce d’une population donnée en lien avec
les variables environnementales qui sont, dans notre analyse, l’absence ou la présence de
broutement par le cerf (zones clôturées et non clôturées) (Lepš et Šmilauer, 1999). Cette
analyse visait à vérifier si le cerf entraînait une modification de la composition en espèces.
Pour ce faire, une comparaison des zones clôturées et des zones non clôturées a été
effectuée avec les données 8,5 ans après la coupe. La variable analysée était le pourcentage
de recouvrement de chacune des espèces ayant un recouvrement d’au moins 0,5 % dans la
parcelle (moyenne des 10 micro-placettes). Les espèces ayant moins de 0,5 % de
recouvrement moyen ont été éliminées de l’analyse puisqu’elles étaient considérées comme
trop rares. Pour l’analyse, les corrélations entre espèces ont été effectuées et la valeur des
espèces était divisée par l’écart type. Aucune transformation des données n’a été effectuée.
La centralisation était appliquée par espèce. Cette analyse a permis de situer graphiquement
les espèces par rapport à la zone clôturée et à la zone non clôturée, et faire ainsi ressortir les
espèces qui sont plus fortement associées à l’absence ou à la présence du cerf.
Résultats
Régénération forestière
Conditions avant la coupe Pour toutes les variables mesurées et pour toutes les essences, les conditions observées
avant la coupe étaient significativement reliées aux données observées après la coupe
(Tableau 1). Ces variables ont donc pu être utilisées efficacement dans les analyses de
covariance. Le tableau 2 expose à titre indicatif les résultats du modèle complet analysé
pour le coefficient de distribution du sapin baumier avant la réduction de la partie fixe du
modèle.
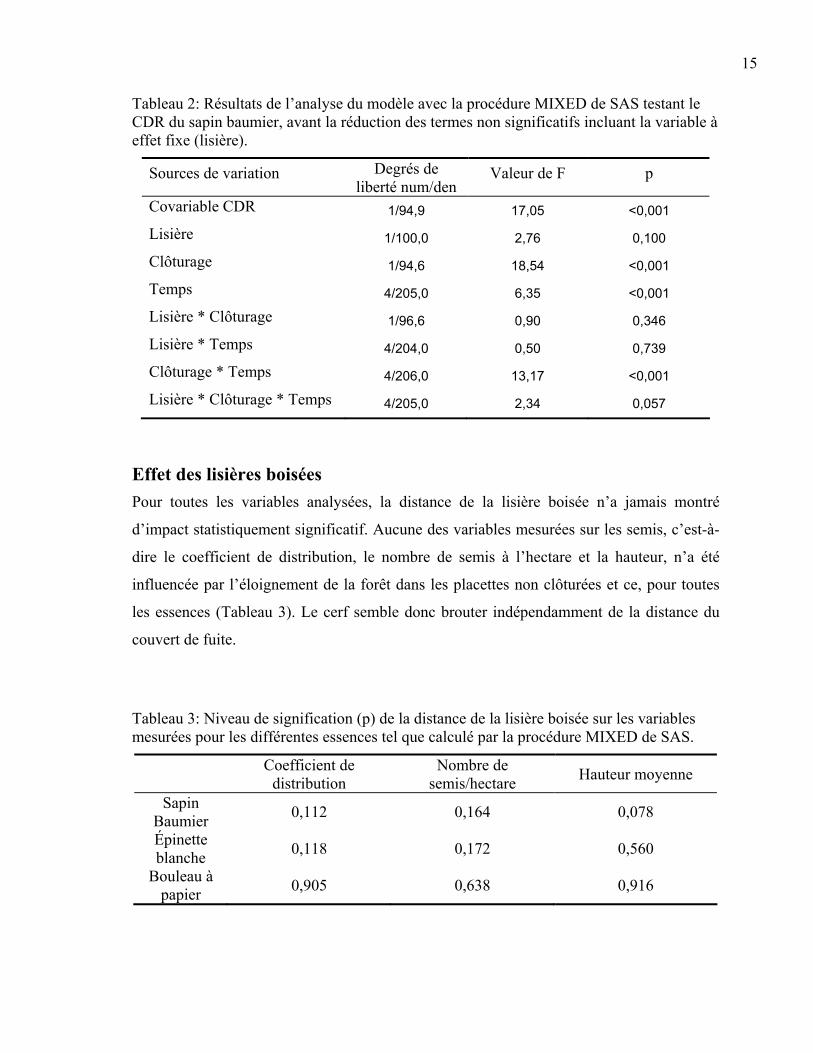
Tableau 1: Niveau de signification (p) des conditions avant la coupe sur les variables mesurées pour les différentes essences, obtenu à partir de la procédure MIXED de SAS.
Coefficient de distribution
Nombre de semis/hectare Hauteur moyenne
Sapin Baumier <0,001 <0,001 0,014
Épinette blanche <0,001 <0,001 <0,001
Bouleau à papier <0,001 0,033 <0,001
15
Tableau 2: Résultats de l’analyse du modèle avec la procédure MIXED de SAS testant le CDR du sapin baumier, avant la réduction des termes non significatifs incluant la variable à effet fixe (lisière).
Sources de variation Degrés de liberté num/den
Valeur de F p
Covariable CDR 1/94,9 17,05 <0,001
Lisière 1/100,0 2,76 0,100
Clôturage 1/94,6 18,54 <0,001
Temps 4/205,0 6,35 <0,001
Lisière * Clôturage 1/96,6 0,90 0,346
Lisière * Temps 4/204,0 0,50 0,739
Clôturage * Temps 4/206,0 13,17 <0,001
Lisière * Clôturage * Temps 4/205,0 2,34 0,057
Effet des lisières boisées Pour toutes les variables analysées, la distance de la lisière boisée n’a jamais montré
d’impact statistiquement significatif. Aucune des variables mesurées sur les semis, c’est-à-
dire le coefficient de distribution, le nombre de semis à l’hectare et la hauteur, n’a été
influencée par l’éloignement de la forêt dans les placettes non clôturées et ce, pour toutes
les essences (Tableau 3). Le cerf semble donc brouter indépendamment de la distance du
couvert de fuite.
Tableau 3: Niveau de signification (p) de la distance de la lisière boisée sur les variables mesurées pour les différentes essences tel que calculé par la procédure MIXED de SAS.
Coefficient de distribution
Nombre de semis/hectare Hauteur moyenne
Sapin Baumier 0,112 0,164 0,078
Épinette blanche 0,118 0,172 0,560
Bouleau à papier 0,905 0,638 0,916

16
Par ailleurs, l’absence d’interaction indique que les caractéristiques de la régénération des
zones clôturée et non clôturée n’ont pas été modifiées par la distance de la lisière boisée.
Cela démontre notamment qu’il n’y a pas eu d’ensemencement notable dans les placettes
clôturées près des semenciers en bordure de coupe, ce qui aurait pu représenter une cause
d’erreur de l’étude.
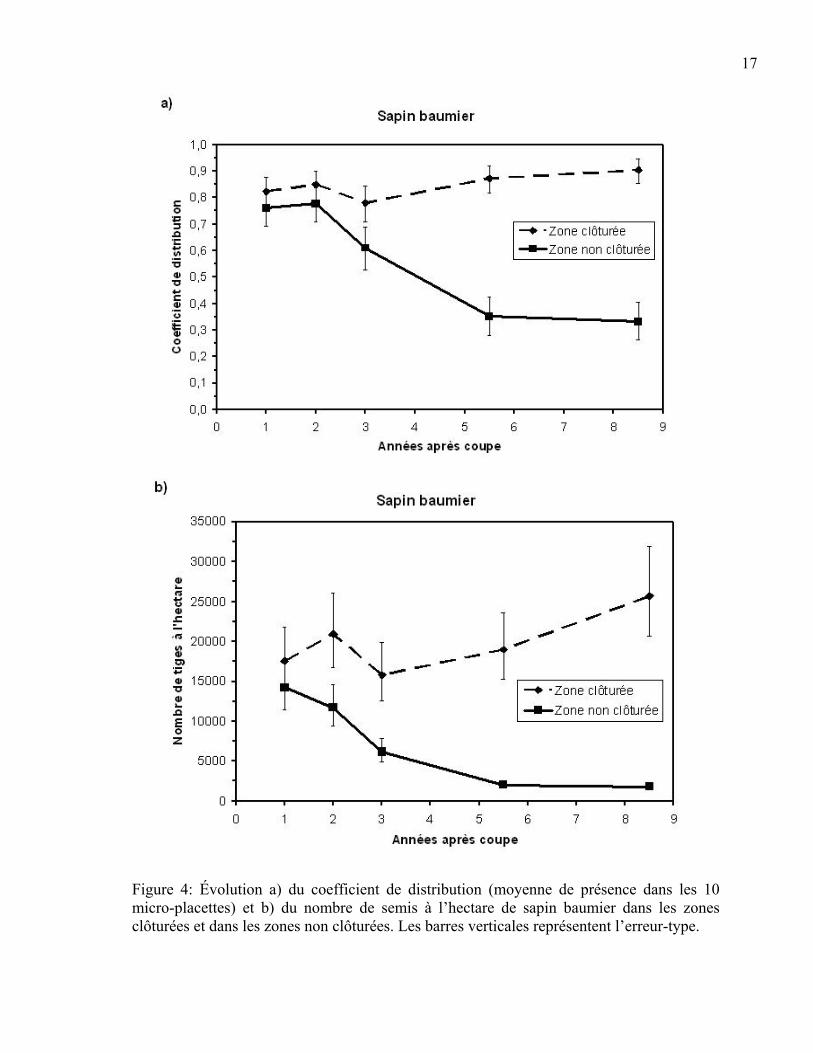
Coefficient de distribution et nombre de semis à l’hectare de sapin baumier et des espèces compagnes Le coefficient de distribution et le nombre de semis à l’hectare de sapin baumier sont
significativement différents entre les zones clôturées et les zones non clôturées dans le
temps (p<0,001). Ce résultat signifie que des différences sont observées entre les placettes
clôturées et non clôturées, mais pas pour l’ensemble des années d’observation. Ainsi, la
figure 4a montre que le coefficient de distribution du sapin est significativement plus élevé
dans les zones clôturées que dans les zones non clôturées à partir de 5,5 ans après la coupe
(p<0,001). De son côté, le nombre de semis à l’hectare de sapin est plus élevé dans les
exclusions du cerf que dans les aires non clôturées dès la deuxième année après la coupe
(an 2 p=0,003, an 3, 5,5 et 8,5 p<0,001) et l’écart entre les deux zones semble se creuser
dans le temps (Figure 4b).
17
Figure 4: Évolution a) du coefficient de distribution (moyenne de présence dans les 10 micro-placettes) et b) du nombre de semis à l’hectare de sapin baumier dans les zones clôturées et dans les zones non clôturées. Les barres verticales représentent l’erreur-type.

18
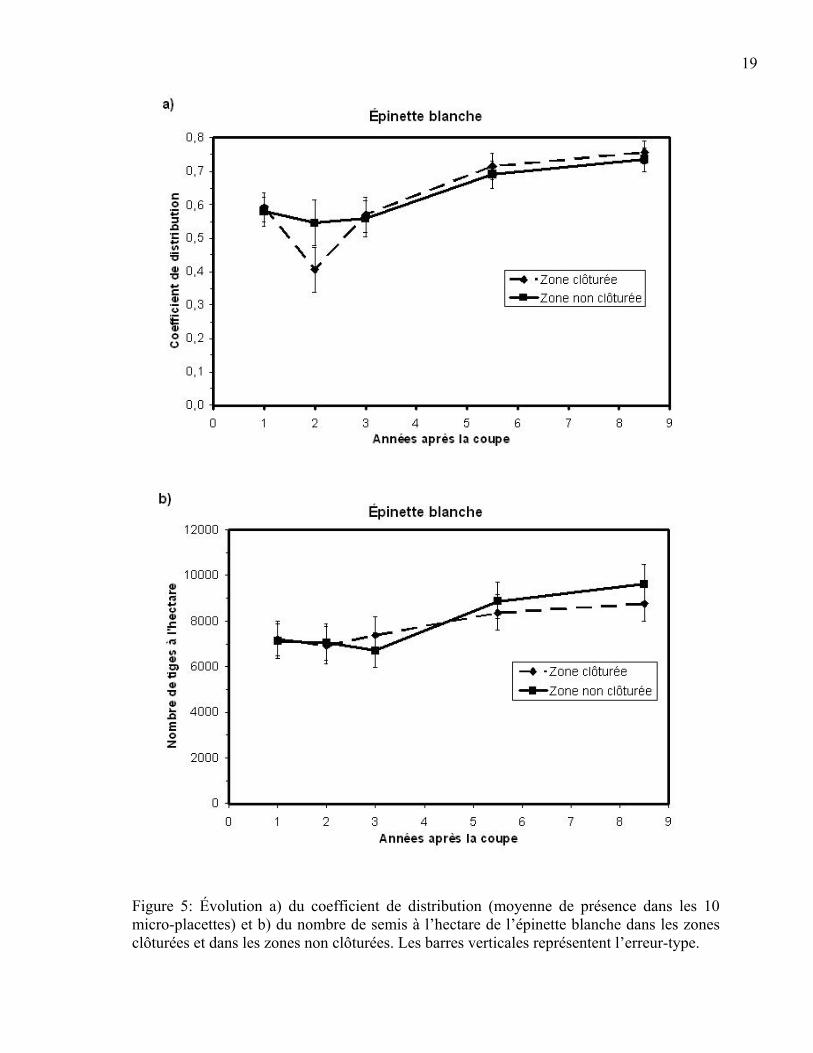
En ce qui a trait à l’épinette blanche, à la fois pour le coefficient de distribution et le
nombre de semis à l’hectare, seule l’année était fortement significative (p<0,001). La
présence de clôtures n’avait donc pas d’effet dans le temps pour cette essence (CDR :
p=0,527; Nombre de tiges/ha : p=0,487). La valeur positive du paramètre obtenu pour
chacune des années indique que ces variables augmentaient dans le temps et ce, de façon
égale pour les placettes clôturées et non clôturées (Figure 5 a et b). Le coefficient de
distribution est passé de 60 % un an après la coupe pour atteindre 75 % 8,5 ans après la
coupe dans les zones clôturées et non clôturées (Figure 5a). Le nombre de semis à l’hectare
était légèrement supérieur à 7000 un an après la coupe et s’élevait jusqu’à environ 8 500
dans les zones clôturées et 9 500 dans les zones non clôturées 8,5 ans après la coupe
(Figure 5b).
19
Figure 5: Évolution a) du coefficient de distribution (moyenne de présence dans les 10 micro-placettes) et b) du nombre de semis à l’hectare de l’épinette blanche dans les zones clôturées et dans les zones non clôturées. Les barres verticales représentent l’erreur-type.

20
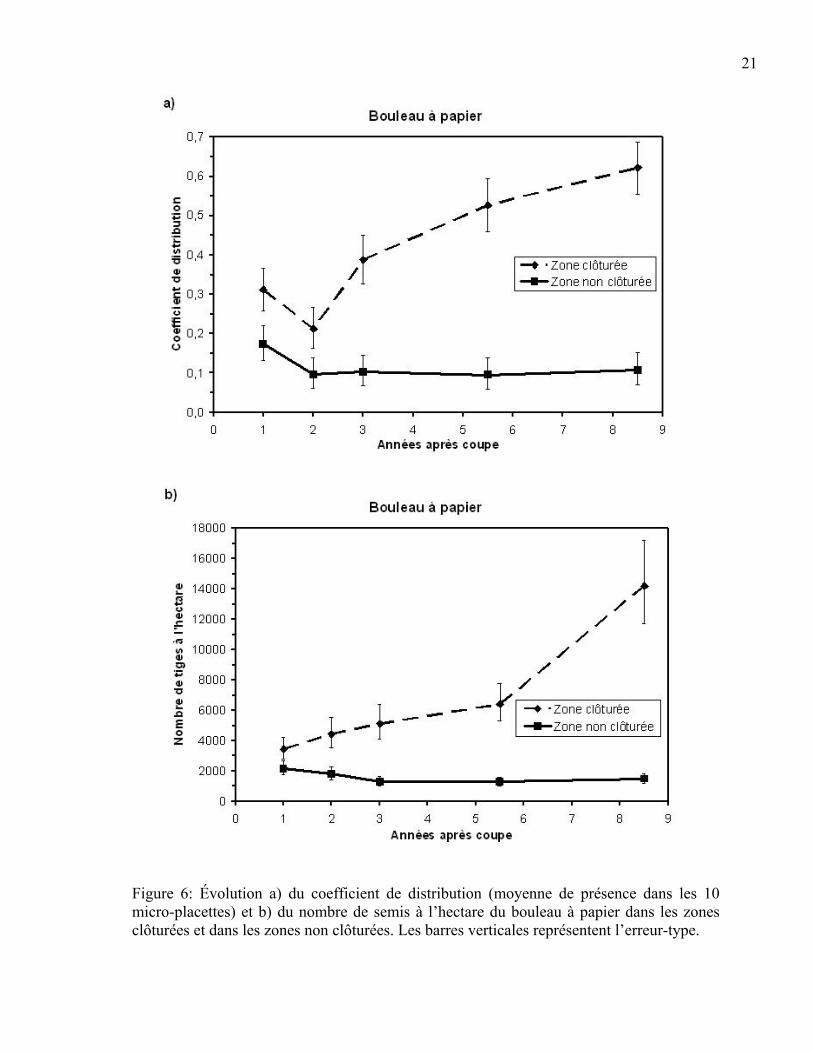
Le coefficient de distribution et le nombre de semis à l’hectare de bouleau à papier
variaient très significativement selon l’absence ou la présence du cerf (zone clôturée et non
clôturée) et l’année (p<0,001). Cet effet était vrai pour toutes les années de mesures, sauf
pour le nombre de semis à l’hectare un an après la coupe (Tableau 4). Ainsi, le bouleau est
davantage présent dans les exclusions du cerf que dans les zones non clôturées après la
coupe. Le nombre de semis à l’hectare du bouleau entre les zones clôturée et non clôturée
est différent à partir de la deuxième année suivant la coupe et l’écart entre les deux zones
augmente dans le temps par la suite (Figures 6 a et b).
Tableau 4: Niveau de signification (p) du clôturage (zone clôturée et zone non clôturée) selon les différentes années pour le coefficient de distribution et le nombre de semis à l’hectare pour le bouleau à papier. Ces résultats ont été obtenus avec la procédure MIXED de SAS.
Années après la coupe Coefficient de distribution Nombre de semis/hectare
1 0,001 0,068
2 0,009 0,002
3 <0,001 <0,001
5,5 <0,001 <0,001
8,5 <0,001 <0,001
21
Figure 6: Évolution a) du coefficient de distribution (moyenne de présence dans les 10 micro-placettes) et b) du nombre de semis à l’hectare du bouleau à papier dans les zones clôturées et dans les zones non clôturées. Les barres verticales représentent l’erreur-type.

22
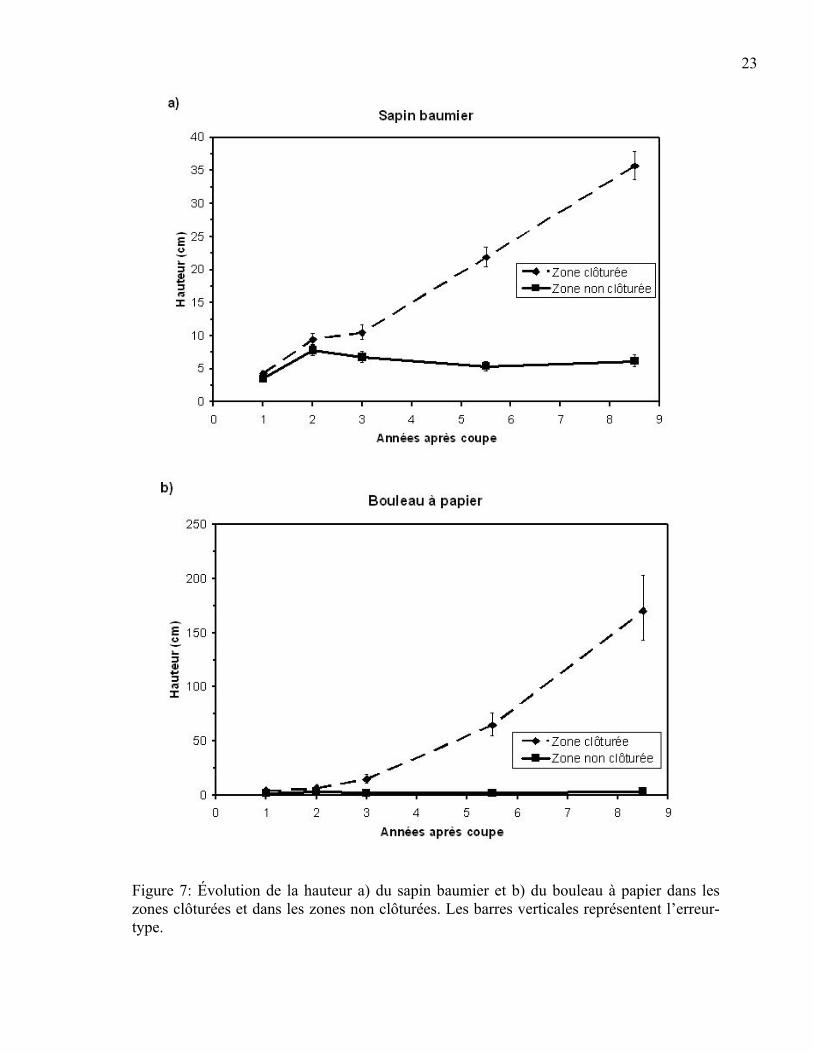
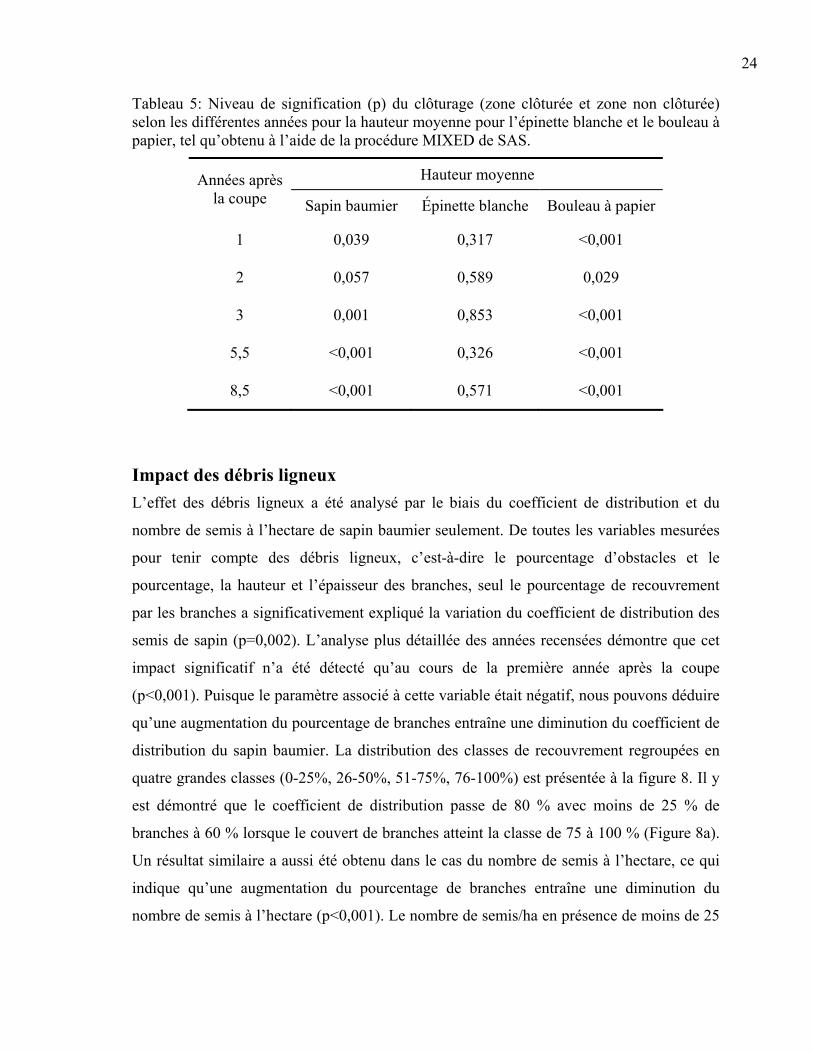
Hauteur moyenne des semis de sapin baumier et des espèces compagnes Les semis de sapin baumier avaient une hauteur moyenne plus élevée dans les zones
clôturées par rapport à celles non clôturées pour toutes les années après la coupe, sauf pour
la deuxième année (Tableau 5). Cependant, d’un point de vue biologique, la différence est
marquée à partir de la troisième année suivant la coupe (Figure 7a). La figure 7a montre
que les semis présents dans les zones non clôturées ont une hauteur moyenne stagnante à 5
cm, alors que la hauteur augmente avec le temps à l’intérieur des zones clôturées.
La différence de hauteur moyenne des semis d’épinette blanche entre les exclusions du cerf
et les zones non clôturées n’est ressortie significative pour aucune des années (Tableau 5).
Pour les zones clôturées et non clôturées, la hauteur moyenne des semis était de 10 cm un
an après la coupe pour atteindre environ 60 cm 8,5 ans après la récolte.
En ce qui concerne le bouleau à papier, il existait une différence hautement significative de
la hauteur moyenne des semis selon la présence ou l’absence du cerf et l’année (p<0,001).
Cette différence est significative pour tous les ans (Tableau 5) et est illustrée à la figure 7b.
Cette fois-ci, la hauteur moyenne dans les zones non clôturées n’est que de 2,5 cm environ.
23
Figure 7: Évolution de la hauteur a) du sapin baumier et b) du bouleau à papier dans les zones clôturées et dans les zones non clôturées. Les barres verticales représentent l’erreur-type.
24
Tableau 5: Niveau de signification (p) du clôturage (zone clôturée et zone non clôturée) selon les différentes années pour la hauteur moyenne pour l’épinette blanche et le bouleau à papier, tel qu’obtenu à l’aide de la procédure MIXED de SAS.
Hauteur moyenne Années après la coupe Sapin baumier Épinette blanche Bouleau à papier
1 0,039 0,317 <0,001
2 0,057 0,589 0,029
3 0,001 0,853 <0,001
5,5 <0,001 0,326 <0,001
8,5 <0,001 0,571 <0,001
Impact des débris ligneux L’effet des débris ligneux a été analysé par le biais du coefficient de distribution et du
nombre de semis à l’hectare de sapin baumier seulement. De toutes les variables mesurées
pour tenir compte des débris ligneux, c’est-à-dire le pourcentage d’obstacles et le
pourcentage, la hauteur et l’épaisseur des branches, seul le pourcentage de recouvrement
par les branches a significativement expliqué la variation du coefficient de distribution des
semis de sapin (p=0,002). L’analyse plus détaillée des années recensées démontre que cet
impact significatif n’a été détecté qu’au cours de la première année après la coupe
(p<0,001). Puisque le paramètre associé à cette variable était négatif, nous pouvons déduire
qu’une augmentation du pourcentage de branches entraîne une diminution du coefficient de
distribution du sapin baumier. La distribution des classes de recouvrement regroupées en
quatre grandes classes (0-25%, 26-50%, 51-75%, 76-100%) est présentée à la figure 8. Il y
est démontré que le coefficient de distribution passe de 80 % avec moins de 25 % de
branches à 60 % lorsque le couvert de branches atteint la classe de 75 à 100 % (Figure 8a).
Un résultat similaire a aussi été obtenu dans le cas du nombre de semis à l’hectare, ce qui
indique qu’une augmentation du pourcentage de branches entraîne une diminution du
nombre de semis à l’hectare (p<0,001). Le nombre de semis/ha en présence de moins de 25
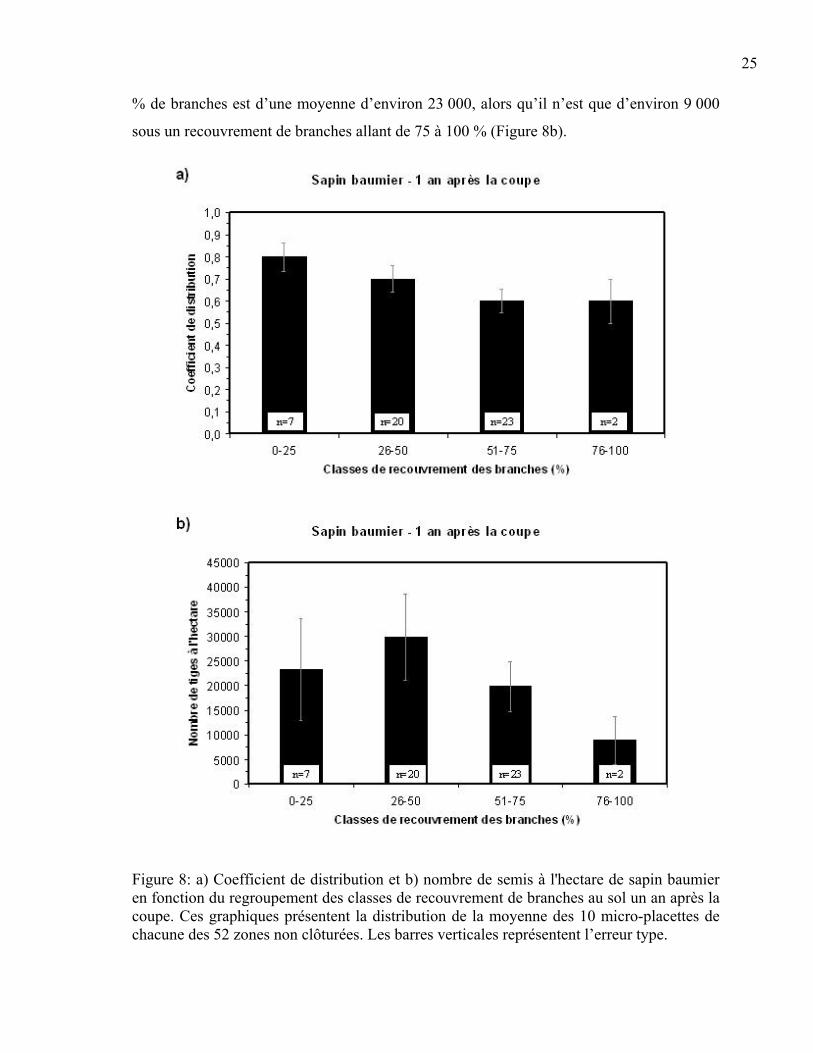
25
% de branches est d’une moyenne d’environ 23 000, alors qu’il n’est que d’environ 9 000
sous un recouvrement de branches allant de 75 à 100 % (Figure 8b).
Figure 8: a) Coefficient de distribution et b) nombre de semis à l'hectare de sapin baumier en fonction du regroupement des classes de recouvrement de branches au sol un an après la coupe. Ces graphiques présentent la distribution de la moyenne des 10 micro-placettes de chacune des 52 zones non clôturées. Les barres verticales représentent l’erreur type.
26
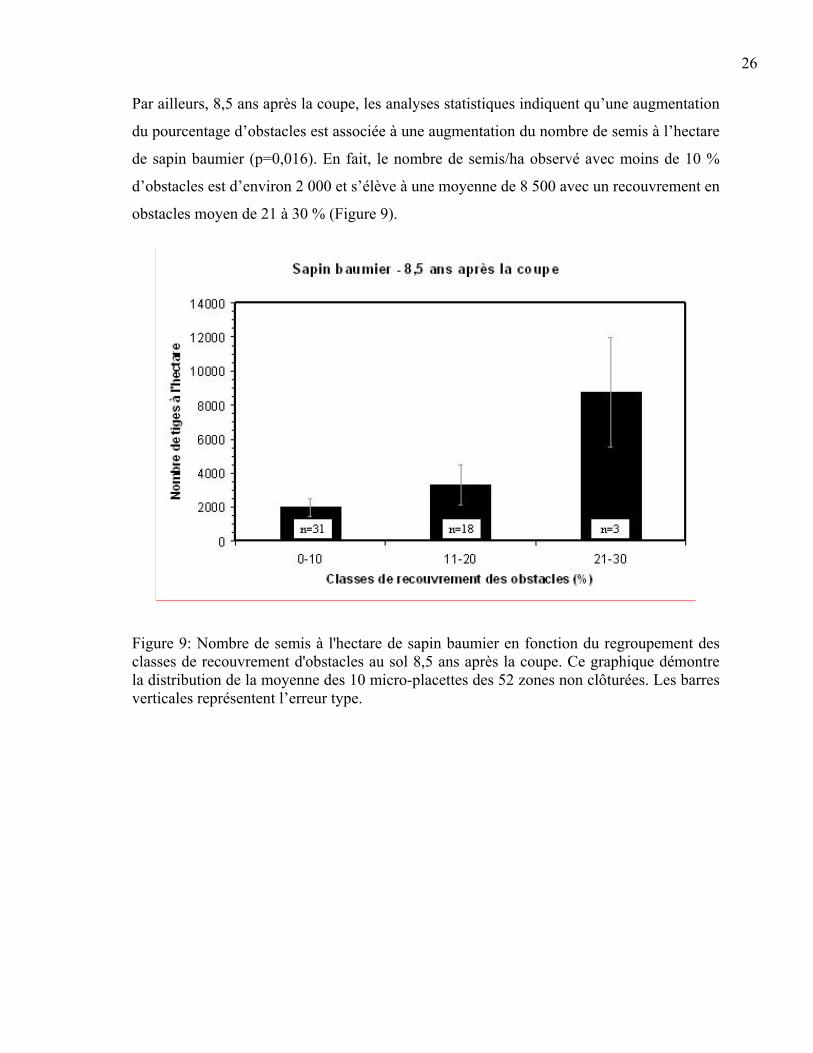
Par ailleurs, 8,5 ans après la coupe, les analyses statistiques indiquent qu’une augmentation
du pourcentage d’obstacles est associée à une augmentation du nombre de semis à l’hectare
de sapin baumier (p=0,016). En fait, le nombre de semis/ha observé avec moins de 10 %
d’obstacles est d’environ 2 000 et s’élève à une moyenne de 8 500 avec un recouvrement en
obstacles moyen de 21 à 30 % (Figure 9).
Figure 9: Nombre de semis à l'hectare de sapin baumier en fonction du regroupement des classes de recouvrement d'obstacles au sol 8,5 ans après la coupe. Ce graphique démontre la distribution de la moyenne des 10 micro-placettes des 52 zones non clôturées. Les barres verticales représentent l’erreur type.

Diversité végétale
Effet des lisières boisées sur la diversité végétale En aucun cas, la distance de la lisière boisée a eu un effet sur la diversité végétale. Ainsi, la
richesse en espèces (p=0,502) et le pourcentage de recouvrement total des espèces
(p=0,141) ne variaient pas avec l’éloignement de la lisière boisée. Le cerf a donc le même
impact sur la végétation, peu importe la distance de la bordure forestière.
Impact du cerf sur la richesse en espèces végétales Le nombre d’espèces végétales variait entre les zones clôturées et les zones non clôturées
dans le temps (p<0,001), mais uniquement à la même année que la coupe. En effet,
l’analyse détaillée de l’interaction entre les zones clôturées et les zones non clôturées selon
les années montre une différence significative seulement immédiatement après la coupe
(Tableau 6). À ce moment, le nombre d’espèces était plus élevé dans les zones
inaccessibles au cerf (entre 23 et 24 espèces) que dans les zones non clôturées (~ 21
espèces). Cette différence s’estompe dans le temps puisqu’il n’y a pas de différence entre
les zones clôturées et celles non clôturées 5,5 et 8,5 ans après la coupe (Tableau 6), le
nombre d’espèces étant d’environ 25. En somme, la forte densité de cerf sur l’île n’affecte
pas la richesse en espèces végétales à moyen terme.
Tableau 6: Niveau de signification (p) du clôturage (zone clôturée et zone non clôturée) selon les différentes années pour le nombre d’espèces végétales obtenu à l’aide de la procédure MIXED de SAS.
Années après la coupe Nombre d’espèces végétales
0 0,002
5,5 0,427
8,5 0,483

28
La valeur de l’indice de similarité de Sørensen obtenue est de 88,2 % 8,5 ans après la
coupe, ce qui signifie que 88,2 % des espèces sont communes aux zones clôturées et aux
zones non clôturées. À l’inverse, il est possible de déduire que 11,8 % (100 % - 88,2 %) des
espèces ne se retrouvent que dans l’une ou l’autre des zones (clôturée et non clôturée).
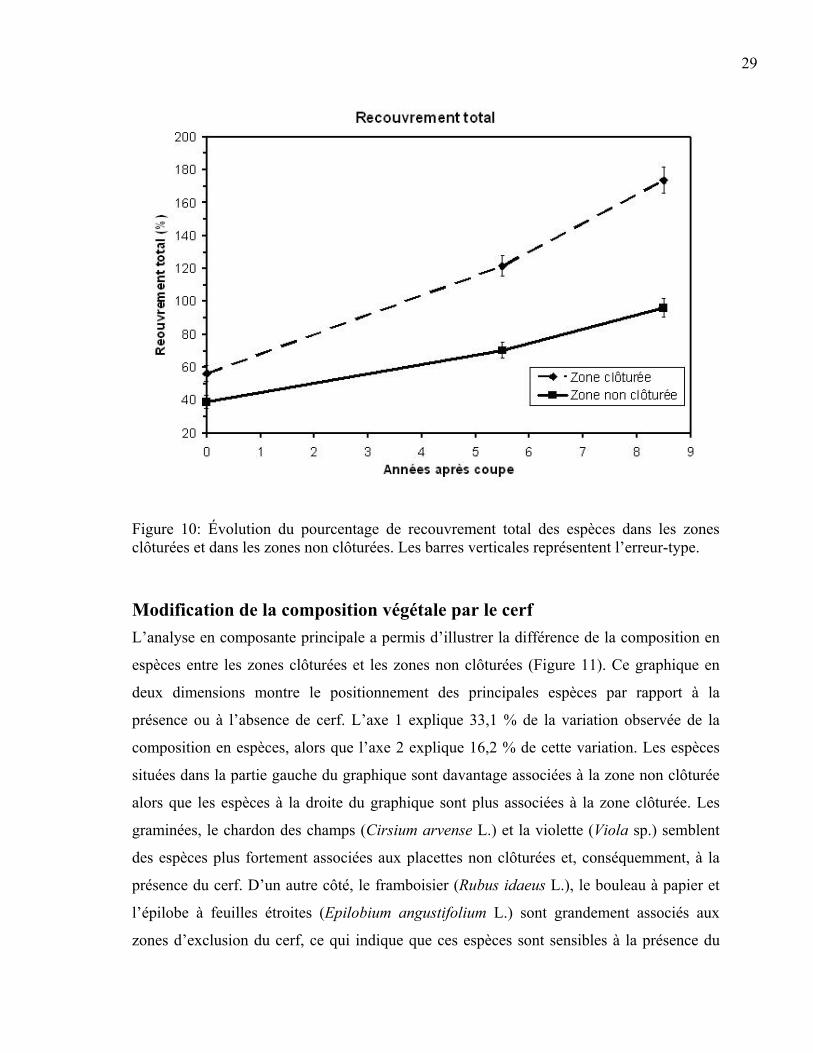
Impact du cerf sur le pourcentage de recouvrement total des espèces végétales En ce qui concerne le recouvrement total des espèces végétales, une différence hautement
significative est notée entre les zones clôturées et les zones non clôturées dans le temps
(p<0,001). Cette différence du pourcentage de recouvrement entre les espaces clôturés et
ceux non clôturés était marquée pour toutes les années analysées (Tableau 7). Le cerf a
donc eu un impact sur le pourcentage de recouvrement total des espèces et l’écart entre les
zones d’exclusion et les zones non clôturées augmente dans le temps (Figure 10). En effet,
la différence de recouvrement des espèces entre les zones avec et sans cerfs après la coupe
était d’environ 20 % pour atteindre environ 70 % 8,5 ans après la coupe.
Tableau 7: Niveau de signification (p) du clôturage (zone clôturée et zone non clôturée) selon les différentes années pour le recouvrement total des espèces végétales. Ces résultats ont été obtenus à l’aide de la procédure MIXED de SAS.
Années après la coupe
Pourcentage de recouvrement des espèces végétales
0 <0,001
5,5 <0,001
8,5 <0,001
29
Figure 10: Évolution du pourcentage de recouvrement total des espèces dans les zones clôturées et dans les zones non clôturées. Les barres verticales représentent l’erreur-type.
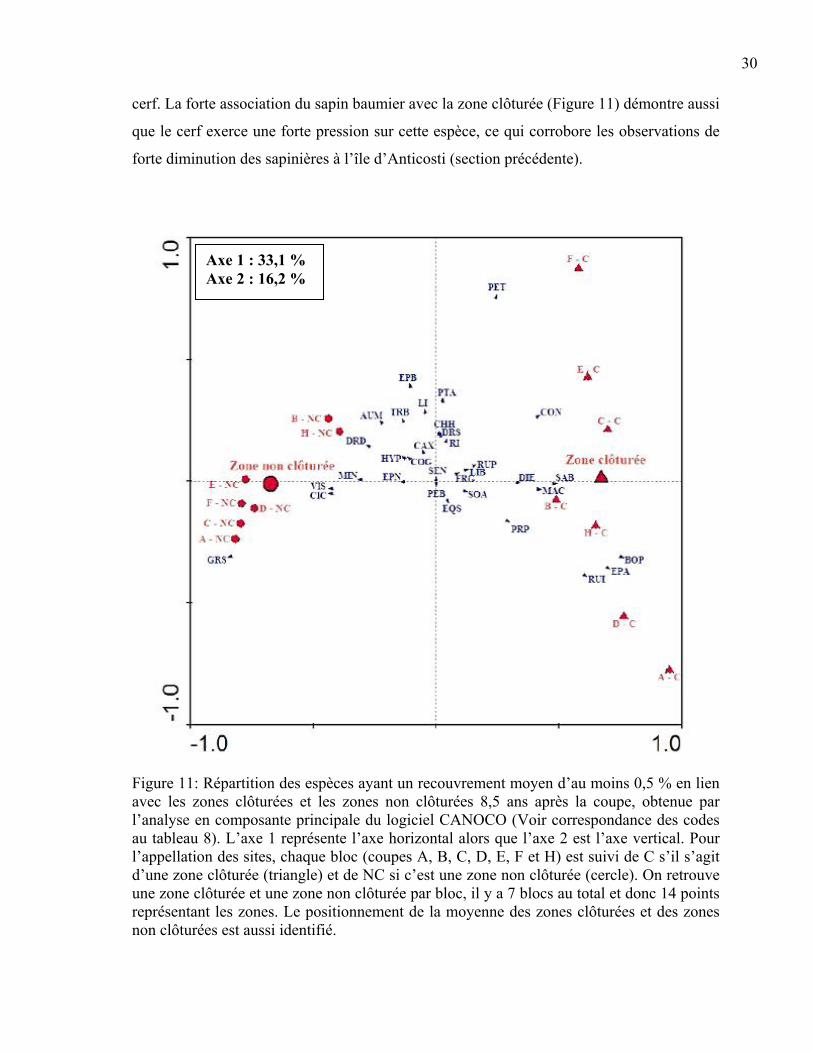
Modification de la composition végétale par le cerf L’analyse en composante principale a permis d’illustrer la différence de la composition en
espèces entre les zones clôturées et les zones non clôturées (Figure 11). Ce graphique en
deux dimensions montre le positionnement des principales espèces par rapport à la
présence ou à l’absence de cerf. L’axe 1 explique 33,1 % de la variation observée de la
composition en espèces, alors que l’axe 2 explique 16,2 % de cette variation. Les espèces
situées dans la partie gauche du graphique sont davantage associées à la zone non clôturée
alors que les espèces à la droite du graphique sont plus associées à la zone clôturée. Les
graminées, le chardon des champs (Cirsium arvense L.) et la violette (Viola sp.) semblent
des espèces plus fortement associées aux placettes non clôturées et, conséquemment, à la
présence du cerf. D’un autre côté, le framboisier (Rubus idaeus L.), le bouleau à papier et
l’épilobe à feuilles étroites (Epilobium angustifolium L.) sont grandement associés aux
zones d’exclusion du cerf, ce qui indique que ces espèces sont sensibles à la présence du
30
cerf. La forte association du sapin baumier avec la zone clôturée (Figure 11) démontre aussi
que le cerf exerce une forte pression sur cette espèce, ce qui corrobore les observations de
forte diminution des sapinières à l’île d’Anticosti (section précédente).
Axe 1 : 33,1 % Axe 2 : 16,2 %
Figure 11: Répartition des espèces ayant un recouvrement moyen d’au moins 0,5 % en lien avec les zones clôturées et les zones non clôturées 8,5 ans après la coupe, obtenue par l’analyse en composante principale du logiciel CANOCO (Voir correspondance des codes au tableau 8). L’axe 1 représente l’axe horizontal alors que l’axe 2 est l’axe vertical. Pour l’appellation des sites, chaque bloc (coupes A, B, C, D, E, F et H) est suivi de C s’il s’agit d’une zone clôturée (triangle) et de NC si c’est une zone non clôturée (cercle). On retrouve une zone clôturée et une zone non clôturée par bloc, il y a 7 blocs au total et donc 14 points représentant les zones. Le positionnement de la moyenne des zones clôturées et des zones non clôturées est aussi identifié.

31
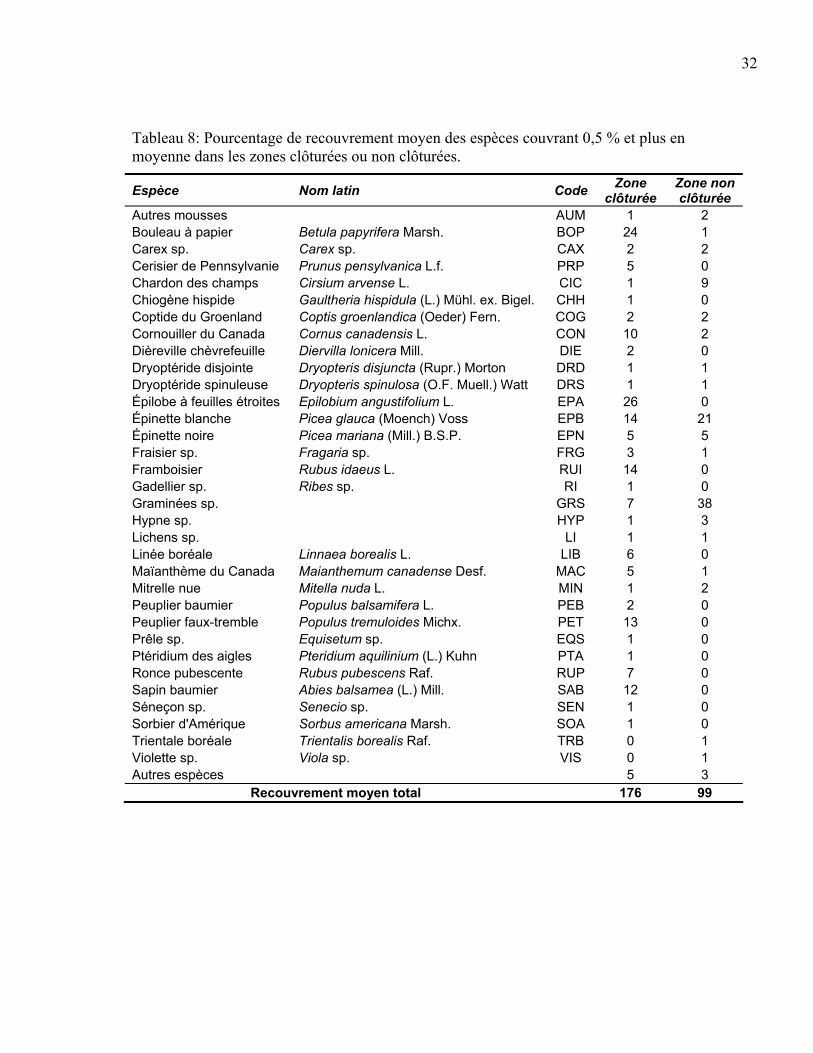
Une évaluation plus détaillée du pourcentage de recouvrement moyen pour chaque espèce
dans les zones clôturées et dans les zones non clôturées est présentée au tableau 8. Nous
pouvons remarquer que parmi les espèces ayant un plus fort recouvrement, les plus fortes
valeurs se retrouvent généralement dans les zones clôturées (BOP, PRP, CON, EPA, RUI,
LIB, MAC, PET, RUP, SAB). Des espèces ayant un fort recouvrement, les zones non
clôturées n’en comptent que trois (CIC, EPB, GRS). Le recouvrement total plus élevé dans
les exclusions du cerf tel que démontré à la figure 10 s’explique donc par ces quelques
espèces. Pour sa part la violette (VIS), qui est associée davantage à la zone non clôturée
(Figure 11), présente un très faible recouvrement dans les zones clôturées et non clôturées.
La forte association de la violette aux placettes non clôturées (Figure 11) est liée au fait que
le recouvrement est tout de même près de trois fois plus élevé dans les zones non clôturées
que dans les zones exclues du cerf.
32
Tableau 8: Pourcentage de recouvrement moyen des espèces couvrant 0,5 % et plus en moyenne dans les zones clôturées ou non clôturées.
Espèce Nom latin Code Zone clôturée
Zone non clôturée
Autres mousses AUM 1 2 Bouleau à papier Betula papyrifera Marsh. BOP 24 1 Carex sp. Carex sp. CAX 2 2 Cerisier de Pennsylvanie Prunus pensylvanica L.f. PRP 5 0 Chardon des champs Cirsium arvense L. CIC 1 9 Chiogène hispide Gaultheria hispidula (L.) Mühl. ex. Bigel. CHH 1 0 Coptide du Groenland Coptis groenlandica (Oeder) Fern. COG 2 2 Cornouiller du Canada Cornus canadensis L. CON 10 2 Dièreville chèvrefeuille Diervilla lonicera Mill. DIE 2 0 Dryoptéride disjointe Dryopteris disjuncta (Rupr.) Morton DRD 1 1 Dryoptéride spinuleuse Dryopteris spinulosa (O.F. Muell.) Watt DRS 1 1 Épilobe à feuilles étroites Epilobium angustifolium L. EPA 26 0 Épinette blanche Picea glauca (Moench) Voss EPB 14 21 Épinette noire Picea mariana (Mill.) B.S.P. EPN 5 5 Fraisier sp. Fragaria sp. FRG 3 1 Framboisier Rubus idaeus L. RUI 14 0 Gadellier sp. Ribes sp. RI 1 0 Graminées sp. GRS 7 38 Hypne sp. HYP 1 3 Lichens sp. LI 1 1 Linée boréale Linnaea borealis L. LIB 6 0 Maïanthème du Canada Maianthemum canadense Desf. MAC 5 1 Mitrelle nue Mitella nuda L. MIN 1 2 Peuplier baumier Populus balsamifera L. PEB 2 0 Peuplier faux-tremble Populus tremuloides Michx. PET 13 0 Prêle sp. Equisetum sp. EQS 1 0 Ptéridium des aigles Pteridium aquilinium (L.) Kuhn PTA 1 0 Ronce pubescente Rubus pubescens Raf. RUP 7 0 Sapin baumier Abies balsamea (L.) Mill. SAB 12 0 Séneçon sp. Senecio sp. SEN 1 0 Sorbier d'Amérique Sorbus americana Marsh. SOA 1 0 Trientale boréale Trientalis borealis Raf. TRB 0 1 Violette sp. Viola sp. VIS 0 1 Autres espèces 5 3
Recouvrement moyen total 176 99

Discussion
Régénération forestière
Conditions avant la coupe Les données récoltées avant la coupe étaient liées à celles récoltées après la coupe pour
toutes les variables mesurées. À cet effet, Pominville et Ruel (1995) ont démontré que le
nombre de semis observé jusqu’à cinq ans après une coupe à blanc était proportionnel à la
quantité de régénération présente avant la coupe, cet effet étant encore plus évident lorsque
seul le sapin était considéré. Aussi, Parent et Ruel (2002) ont démontré qu'il y avait une
relation étroite entre le taux d'accroissement des semis de sapin baumier à l'ouverture du
couvert par la récolte forestière et le taux d'accroissement avant la coupe. Ces résultats
indiquent qu’il est important de tenir compte des conditions avant la coupe puisqu’elles
peuvent influencer les observations après la coupe. L’introduction des variables avant la
coupe comme covariables rend donc plus précise la validation de nos hypothèses. Cela
permet de mettre les différentes aires échantillonnées sur une base commune et comparable,
ce qui pourrait éviter d’attribuer les résultats aux traitements alors qu’ils seraient plutôt
attribuables aux conditions avant la coupe.
Effet des lisières boisées La distance d’accès au couvert de fuite n’a pas influencé le comportement de broutement
du cerf de Virginie qui semble occuper la totalité de la superficie des grandes coupes.
L’hypothèse de départ voulant que le centre de grandes coupes soit épargné du broutement
du cerf est donc invalidée. Cette hypothèse était fondée sur plusieurs études ayant démontré
une diminution de l’utilisation du brout avec l’éloignement de la lisière boisée dans les
coupes à blanc (Drolet, 1978; Welch et al., 1991; Schmitz, 2005). De même, Kay (1993) a
constaté une réduction des dommages avec l’augmentation de la grandeur de la coupe au
cours d’expériences effectuées en présence de chevreuil (Capreolus capreolus L.). Par
contre, toutes ces études démontrant un effet de la distance de la bordure forestière ont été
effectuées dans des milieux où des prédateurs étaient présents contrairement aux conditions

34
retrouvées sur l’île d’Anticosti. À cette hypothèse s’ajoute la possibilité que les populations
d’ongulés étaient moins nombreuses dans ces études. L’impact de la présence des
prédateurs a d’ailleurs été souligné par Bergquist et Örlander (1998b). Pour expliquer leurs
résultats, ils soulèvent la possibilité que l’exposition visuelle face aux prédateurs étant
beaucoup plus élevée dans les jeunes coupes, il devient probablement très coûteux en
termes d'énergie et d'augmentation du risque de prédation pour un cerf d'utiliser ces stations
pour se nourrir.
Par ailleurs, d’autres auteurs ont obtenu des résultats similaires à ceux trouvés dans la
présente étude. D’abord, Williamson et Hirth (1985) ont noté que la plupart des espèces
sont broutées près de la bordure forestière, mais que les espèces préférées et abondantes au
centre des coupes sont aussi consommées même si le risque de prédation est augmenté par
l’éloignement du couvert de fuite. En Nouvelle-Écosse, des coupes ont été inventoriées
plusieurs années après leur réalisation et la fréquence de broutement augmentait ou était
stable avec la distance de la lisière boisée dans les coupes effectuées depuis 1 et 4 ans
respectivement, alors que la fréquence diminuait avec la distance de la bordure forestière
cinq ans après coupe (Reyes et Vasseur, 2003). Dans une étude effectuée dans l’état de
New-York, Cadenasso et Pickett (2000) démontraient que dans les zones coupées à blanc,
les semis étaient endommagés de façon constante par le broutement du cerf à différentes
distances de la lisière boisée. Une autre expérience effectuée en Suède en présence d’une
forte densité d’orignaux (Alces alces L.) présentait la même tendance que celle obtenue
dans la présente étude, c’est-à-dire que la pression de broutement n’était pas
significativement différente à diverses distances de la lisière boisée (Andrén et Angelstam,
1993). Pour expliquer ces résultats contraires à ceux démontrant un effet de l’éloignement
de la bordure, ces derniers auteurs ont émis l’hypothèse que le rapport nourriture/orignal
était plus élevé que celui des autres études. Conséquemment, en s’appuyant sur la théorie
de la sélection d’habitat dépendante de la densité [Traduction libre : density-dependant
habitat selection theory] (Fretwell et Lucas, 1970), certains individus faisant partie d’une
population à forte densité sont forcés d’aller se nourrir dans des habitats plus contraignants,
ce qui inclut les endroits éloignés des lisières boisées. Borlowski (2000) avait aussi noté

35
qu’en présence de fortes densités de cerf, ces derniers utilisent davantage les habitats de
moindre qualité. Une autre hypothèse énoncée stipulait que les orignaux n’étant pas
confrontés à un niveau de prédation élevé, ils ont pu modifier leur comportement en
réduisant leur tendance à demeurer près du couvert de fuite pour se nourrir (Andrén et
Angelstam, 1993). Reyes et Vasseur (2003) ont repris cette hypothèse pour expliquer
l’absence d’effet de la distance de la bordure forestière étant donné la faible pression de
prédation historiquement retrouvée en Nouvelle-Écosse.
L’île d’Anticosti présente ces mêmes caractéristiques de densité élevée d’ongulés et
d’absence de prédation. L’idée que le cerf ait pu modifier son comportement face à
l’absence de prédateur est plausible, mais va à l’encontre de l’hypothèse des fantômes des
prédateurs passés [Traduction libre : ghosts of predators past] (Byers, 1997). Cette théorie
soutient que le cerf craint les prédateurs instinctivement même s’ils sont absents.
Néanmoins, l’hypothèse qui nous apparaît plus probable est celle s’appuyant sur la théorie
de sélection d’habitat dépendante de la densité. En effet, le manque de nourriture associé à
la forte densité de cerf obligerait ces derniers à s’éloigner de la couverture forestière pour
se nourrir, même si ces stations présentent théoriquement plus de risques que celles situées
près de la forêt.
Coefficient de distribution et nombre de semis à l’hectare de sapin baumier et des espèces compagnes Les résultats obtenus montrent que le cerf de Virginie a un impact marqué à court ou
moyen terme sur le coefficient de distribution et le nombre de semis/ha du sapin baumier et
du bouleau à papier. Le coefficient de distribution et le nombre de semis/ha des placettes
clôturées tendent à augmenter progressivement alors qu’une diminution ou une stagnation
peut être observée dans les placettes non clôturées (Figures 4 et 5). L’impact du cerf est
donc de plus en plus marqué avec les années.

36
Des résultats similaires ont déjà été observés au terme d’une étude réalisée dans des coupes
à blanc où la régénération de thuya géant (Thuja plicata Donn ex D. Don) était fortement
réduite en présence du cerf à queue noire de Sitka (Odocoileus hemionus sitchensis
Merriam) (Martin et Baltzinger, 2002). Dans une autre étude avec des coupes à blanc,
Tilghman (1989) notait que le nombre de tiges d'essences non commerciales était
dramatiquement réduit lorsque la densité de cerf était forte. De plus, il y avait
substantiellement moins de semis d'espèces sensibles au broutement dans les classes de
hauteur les plus grandes à des densités de cerf très élevées. La même tendance a été
observée par Case et McCullough (1987) sur l’île North Manitou où les cerfs ont été
introduits. Sous couvert, les essences favorites du cerf étaient davantage présentes aux
endroits où le cerf n’avait pas accès alors que l’essence la moins broutée, le hêtre, était
davantage présente dans les zones accessibles au cerf. En sous étage, il a aussi été démontré
que le broutement augmente significativement le taux de mortalité des semis de pruche
(Tsuga canadensis (L.) Carrière) (Frelich et Lorimer, 1985 ; Alverson et Waller, 1997) et
prévient le recrutement de semis de thuya de l’est (Thuja occidentalis L.) (Rooney, 2001).
À cet effet, Cornett et al. (2000) ont démontré que le thuya de l'est et le sapin baumier ont
un pourcentage de recouvrement substantiellement plus grand dans la classe de hauteur 0,5-
2 m à l'intérieur qu'à l'extérieur des zones inaccessibles au cerf. L’importance du sapin
baumier dans les forêts de l’île Royale a été diminuée par le broutement de l’orignal (Alces
alces andersoni Peterson) (Brandner et al., 1990). De plus, les fortes densités de cerf
préviennent la régénération et diminuent la densité de semis de nombreuses essences
feuillues telles le chêne (Quercus sp.) (Healy, 1997 ; Rooney et Waller, 2003 ; Russell et
Fowler, 2004), le frêne d’Amérique (Fraxinus americana L.) (Cross, 1998) et plusieurs
autres espèces (Veblen et al., 1989 ; Witmer et deCalesta, 1991).
Par ailleurs, l’absence de différence significative entre les exclusions du cerf et les aires
non clôturées en ce qui concerne l’épinette blanche indique que le cerf évite de brouter
cette espèce. En fait, les fortes densités de cerf n’affectent pas le coefficient de distribution
et le nombre de semis à l’hectare de cette espèce dans les grandes coupes. L’épinette
blanche présente un bon coefficient de distribution (75 %) et un nombre de semis/ha, toutes

37
hauteurs confondues, assez élevé (entre 8 500 et 9 500 semis/ha) 8,5 ans après la coupe. Si
cette tendance se maintenait, les sapinières d’Anticosti seraient prédisposées à être
graduellement remplacées par des pessières blanches, ce qui avait déjà été évoqué par
Potvin (1992), Potvin et al. (2000) et Potvin et al. (2003a). Ce remplacement du sapin
baumier au profit de l’épinette blanche avait aussi été observé sur l’île Royale en présence
de l’orignal (McInnes et al., 1992). L’épinette noire, qui est également évitée par le cerf,
pourrait accompagner l’épinette blanche dans ces futurs types forestiers. Aussi longtemps
que la densité de cerf ne sera pas réduite sur l’île, il est probable que la coupe des sapinières
favorise l’établissement de pessières.
Par ailleurs, à certains endroits où l’épinette n’a pu se régénérer, nous avons observé que
les coupes présentent davantage une structure de prairie avec des tiges d’épinette blanche
éparses (Figure 12). Plusieurs auteurs ont noté cette tendance de passage d’une forêt à une
prairie ou une savane, les essences initiales étant souvent remplacées par des herbacées ou
des arbustes non broutés par le cerf (Jane, 1994; Healy, 1997; Putman et Moore, 1998;
Russell et Fowler, 2004). Cette transformation pourrait donc survenir après la coupe de
certaines stations dans l’île d’Anticosti si la densité de population de cerf reste aussi élevée
dans l’avenir.

38
Figure 12: Zone clôturée à l’intérieur d’une grande coupe présentant une structure de prairie.
Hauteur moyenne des semis de sapin baumier et des espèces compagnes Selon les résultats obtenus, la hauteur moyenne des semis de sapin baumier et de bouleau à
papier est significativement inférieure dans les placettes non clôturées par rapport aux
placettes clôturées, sauf pour la deuxième année suivant la coupe dans le cas du sapin. Le
cerf de Virginie a donc un impact marqué sur l’accroissement en hauteur des semis. Les
semis des zones clôturées, à l’abri du broutement par le cerf, augmentent en hauteur dans le
temps alors qu’une stagnation est observée dans les espaces non clôturés. Dans ces derniers
endroits, le broutement empêche les semis de croître au-delà d’environ cinq cm dans le cas
du sapin et de 2,5 cm dans le cas du bouleau. La figure 13 illustre bien le faible espacement
entre les verticilles d’un sapin retrouvé dans une zone non clôturée, ce qui indique la forte
pression de broutement sur ce semis. Comme suggéré par Stoeckeler et al. (1957) et

39
Whitney (1984), le recrutement des tiges de classes de hauteur intermédiaires semble donc
problématique. Par ailleurs, le cerf ne semble pas avoir d’effet sur la croissance de
l’épinette blanche, la différence de hauteur moyenne entre les zones clôturées et les zones
non clôturées n’étant pas significativement différente.
Figure 13: Semis de sapin baumier présentant un faible espacement entre les différents verticilles dans une zone non clôturée.
Les résultats obtenus dans la présente étude concordent avec ceux de la littérature. En effet,
une expérience avec le cerf muntjac de Reeve (Muntiacus reevesi Ogilby) a aussi démontré
que les espèces ayant une croissance faible ou nulle dans les parcelles non clôturées
continuaient à croître à l'intérieur des clôtures (Cooke et Farrell, 2001). De plus, Horsley et
al. (2003) ont observé qu’une augmentation de la densité de cerf induisait une diminution
linéaire de la hauteur de bouleau, d'érable rouge et de hêtre à la suite de coupes à blanc.
Pour leur part, Kullberg et Bergström (2001) ont quantifié une réduction de la hauteur des
semis de 32 % attribuable au broutement par les grands herbivores à la suite d’une coupe à

40
blanc. Finalement, Tilghman (1989) a observé une baisse substantielle de semis d'espèces
sensibles au broutement dans les classes de hauteur les plus grandes à des densités de cerf
très élevées cinq ans après une coupe à blanc.
Les résultats obtenus sous un couvert arborescent sont sensiblement les mêmes puisqu’il a
été démontré à maintes occasions que le broutement par le cerf retarde grandement la
croissance en hauteur des semis (Stoeckeler et al., 1957 ; Alverson et Waller, 1997; Cornett
et al., 2000; Opperman et Merenlender, 2000; Harmer, 2001) et parfois même la succession
forestière (Gill et Beardall, 2001). L’orignal peut aussi affecter la croissance des semis de
sapin baumier comme il a été observé sur l’île Royale (Brandner et al., 1990). En fait, la
réduction de la croissance en hauteur des semis désirés due au broutement pourrait les
amener à être dépassés par la végétation compétitive, ce qui pourrait éventuellement
produire un fort taux de mortalité (Gill et Beardall, 2001).
De leur côté, Reyes et Vasseur (2003) ont démontré que la croissance en hauteur des semis
broutés avait été significativement réduite par le cerf, mais que la différence était petite.
Cela est probablement dû au fait que la densité de cerf, d’environ 1,9 cerfs/km2, était trop
peu élevée pour provoquer une différence marquée de la hauteur.
Impact des débris ligneux L’année suivant la coupe, les analyses font ressortir que l’augmentation du pourcentage de
recouvrement de branches entraîne une diminution du coefficient de distribution et du
nombre de semis à l’hectare de sapin baumier. Ce résultat est probablement davantage le
reflet d’une difficulté à observer les semis étant donné un très grand couvert de branches au
sol, plutôt qu’une réelle tendance à la diminution des semis en présence de branches. En
effet, l’épaisseur des débris s’amenuise dans le temps (Bergquist et Örlander, 1998a), ce
qui permet une meilleure observation des semis. Cela se traduit dans le cas présent par
l’absence d’impact de la présence de branches sur le nombre et la distribution des semis.

41
Par ailleurs, 8,5 ans après la coupe, le nombre de semis à l’hectare de sapin augmentait
avec une augmentation du pourcentage d’obstacles sur le parterre de coupe. Plusieurs
auteurs ont aussi observé un effet positif des débris ligneux sur la régénération forestière
(Tilghman, 1989; Kay, 1993; Bergquist et al., 1999). Les souches, les chicots tombés au sol
et les grosses roches permettent donc de protéger les semis par obstruction visuelle jusqu’à
ce que ces derniers dépassent éventuellement la couche de débris. Probablement que les
branches n’ont pas eu d’effet 8,5 ans après la coupe puisqu’elles sont en grande partie
décomposées. Cette protection visuelle des semis fournie par les débris a été mentionnée
par Bergquist et Örlander (1998b). La plus grande présence de semis près des débris peut
aussi être le résultat de meilleures conditions d’humidité fournies par les obstacles qui
favorisent l’établissement et la croissance de ces derniers. Par ailleurs, étant donné que des
résultats statistiquement significatifs n’ont été observés que 8,5 ans après la coupe, des
mesures à plus long terme seraient nécessaires pour tester si les obstacles vont continuer à
protéger les semis du broutement dans le temps.
Des résultats contraires ont aussi été observés. Selon Putman (1994), les débris sont d’une
utilité limitée pour les espèces sensibles au broutement. Ce dernier résultat peut
possiblement être expliqué par le fait que les données étaient récoltées par des volontaires
non professionnels qui ne pouvaient faire la différence entre les formes de dommages de
broutement causés par différents mammifères (Putman, 1994). Ces résultats sont donc le
reflet de dommages causés par tous les mammifères herbivores présents sur le site d’étude
et non seulement par le cerf. L’auteur souligne aussi que l’expérience était effectuée dans
de petits blocs de coupe et que de plus grandes coupes devraient permettre d’avoir un
niveau de dommages moins élevé.
Diversité végétale
Effet des lisières boisées sur la diversité végétale La diversité végétale, en général, n’était pas influencée par la proximité de la bordure
forestière. Ainsi, ni le nombre d’espèces, ni le recouvrement total des espèces n’étaient

42
affectés par l’éloignement du couvert de fuite. L’hypothèse de départ prédisant que le
centre de grandes coupes soit épargné du broutement du cerf est donc invalidée non
seulement en ce qui a trait à la régénération forestière, mais aussi du point de vue de la
diversité végétale. Cet aspect de l’étude a été discuté dans la section de la régénération
forestière.
Impact du cerf sur la richesse en espèces végétales Contrairement à notre hypothèse de départ, le cerf n’a pas eu d’effet significatif à long
terme sur la richesse en espèces végétales, le nombre d’espèces étant sensiblement le même
dans les zones clôturées et dans les zones non clôturées (5,5 et 8,5 ans après la coupe).
Cependant, immédiatement après la coupe (an 0), le nombre d’espèces était
significativement plus grand dans les exclusions du cerf que dans les zones non clôturées.
Ainsi, le cerf a pu rapidement brouter certaines espèces favorites à la suite de la coupe
(Horsley et al., 2003), alors que les espèces associées au cerf et à sa perturbation sur le
milieu n’auraient pas eu le temps de s’installer. Ce n’est que plus tard que l’impact du cerf
se serait fait sentir en favorisant l’établissement des graminées et des chardons, entre autres,
dans les zones non clôturées.
Le nombre d’espèces était en général moins élevé, autant dans les zones clôturées que dans
celles non clôturées, immédiatement après la coupe (an 0) qu’à 5,5 et 8,5 ans après la
coupe. La circulation de la machinerie forestière a certainement pu affecter les espèces
végétales au sol, ce qui pourrait expliquer le nombre d’espèces moins élevé immédiatement
après les opérations de récolte. De plus, comme il a été mentionné précédemment, la grande
quantité de débris ligneux rendait difficile l’observation de la végétation, ce qui pourrait
aussi expliquer le nombre moins élevé d’espèces recensées.
Par ailleurs, à plus long terme (5,5 et 8,5 ans après la coupe), le cerf n’a pas eu d’impact sur
le nombre d’espèces dans le milieu. Ces résultats sont en accord avec ceux démontrant
qu’en milieux ouverts ou coupés à blanc, le nombre moyen d’espèces de plantes ne varie

43
pas significativement entre les zones avec et sans exposition au broutement du cerf lorsque
ce dernier est présent en fortes densités (Bowers, 1997; Bergquist et al., 1999). Le même
type de résultat a été observé dans le sous-bois de sapinières matures de l’île d’Anticosti en
ce qui a trait à l’occurrence des espèces (Potvin et Poirier, 2004). Aussi, sur l’île Royale, le
broutement par l’orignal n’altérait pas le nombre d’espèces de plantes, d’arbustes et
d’arbres (McInnes et al., 1992). Schmitz (2005) a même observé une légère tendance pour
les parcelles exposées à l'herbivorie d'avoir une plus grande richesse d'espèces en sous
étage que les parcelles à l’abri des herbivores. Le fait que le nombre d’espèces ne varie pas
significativement entre les zones clôturées et non clôturées peut s’expliquer par
l’avènement de nouvelles espèces en présence du cerf. Ainsi, le nombre d’espèces
éliminées des placettes non clôturées est compensé par d’autres espèces que l’on ne
retrouve pas dans les zones clôturées, puisqu’elles sont associées à la perturbation
occasionnée par le cerf sur le milieu. La quasi-disparition de certaines espèces dans les
zones non clôturées, telles le framboisier et l’épilobe à feuilles étroites, est donc compensée
par le chardon des champs et les graminées qui sont des espèces fortement favorisées par la
présence du cerf. L’indice de similarité de Sørensen obtenu étant élevé (88,2 %), nous
pouvons déduire que le cerf affecte peu la composition en espèces entre les deux zones.
Cependant, 11,8 % des espèces ne se retrouvent que dans l’une ou l’autre des zones
clôturées et non clôturées. En somme, près de 12 % des espèces diffèrent, mais le nombre
total d’espèces présentes ne varie pas significativement. Ainsi, le nombre d’espèces est un
facteur important à considérer, mais il ne permet pas de démontrer l’impact d’un traitement
sur la diversité végétale lorsqu’il est utilisé seul. Une évaluation de l’impact sur les
différentes espèces permet d’approfondir l’information obtenue par le nombre d’espèces.
Par ailleurs, certains auteurs ont démontré que le broutement du cerf sous un couvert
forestier réduit la richesse en espèces herbacées, arbustives (Cockayne, 1909 dans
Bellingham and Allan, 2003) et arborescentes (Stewart et Burrows, 1989; deCalesta, 1997;
Gill et Beardall, 2001). Cette perte de richesse était plus grande lorsque la végétation était
soumise à de fortes densités de cerf (Gill et Beardall, 2001). Des résultats similaires ont
aussi été observés après l’application de coupes à blanc (Tilghman, 1989 ; Horsley et al.,

44
2003). Tel que proposé par Gill et Beardall (2001), l’effet du cerf sur la végétation n’est pas
constant et dépend des conditions écologiques locales. Il est aussi important selon eux de
comprendre les changements causés par les cerfs comme un processus qui est
continuellement affecté par les conditions changeantes du milieu. Ces résultats
contradictoires à la présente étude peuvent donc possiblement s’expliquer par des
différences de conditions écologiques des milieux. D’abord, les résultats issus d’études
effectuées sous couvert ne sont pas comparables puisque les cerfs sont moins affectés par
l’exposition aux prédateurs dans cette situation. Pour sa part, Tilghman (1989) soulignait
que le recouvrement dense de fougères en présence de fortes densités de cerf dans les
coupes à blanc pouvait affecter la croissance et la survie des semis, ce qui pourrait
expliquer la diminution de la richesse en espèces observée. L’aire d’étude de Horsley et al.
(2003) présentait aussi des conditions d’augmentation du couvert de fougères en présence
du cerf. Cette problématique reliée à l’abondance de fougères après la coupe avec de fortes
densités de cerf ne s’applique pas à l’île d’Anticosti et peut donc expliquer les divergences
de résultats obtenus.
Impact du cerf sur le pourcentage de recouvrement total des espèces végétales Pour toutes les années inventoriées, le cerf a significativement réduit le pourcentage de
recouvrement de l’ensemble des espèces végétales au sol dans les zones non clôturées
comparativement aux zones clôturées. Il apparaît donc que même si le nombre d’espèces
n’est pas différent, le recouvrement de ces espèces est grandement affecté par la présence
du cerf dans le milieu. Le même résultat avait été obtenu par Potvin et Poirier (2004) dans
une sapinière mature de l’île d’Anticosti.
L’impact du cerf sur le recouvrement peut certainement s’effectuer à plusieurs niveaux. En
effet, le broutement peut s’ajouter au piétinement qui peut avoir pour effet de compacter le
sol ou de mener plusieurs plantes à la mort (Vare et al., 1996; Persson et al., 2000; Kirby,
2001; Watkinson et al., 2001).

45
Modification de la composition végétale par le cerf Les résultats démontrent que le cerf modifie grandement l’arrangement des espèces
végétales dans les grandes coupes. À la figure 11, l’axe 1 (horizontal) est manifestement
associé à la densité de cerf, la zone clôturée et la zone non clôturée étant fortement
associées à cet axe. Par ailleurs, il semble que l’axe 2 (vertical) corresponde à un gradient
de semenciers des espèces secondaires présentes dans les peuplements avant la coupe. En
effet, les blocs situés au bas de l’axe 2 avaient davantage de semenciers de bouleau à
papier, alors que le haut de l’axe 2 serait associé à une présence plus forte de peuplier faux-
tremble sur le parterre de coupe. Ce gradient de composition en essences feuillues auquel
semble correspondre l’axe 2 s’exprime surtout dans les zones clôturées étant donné le fort
broutement dans les placettes non clôturées. Ainsi, ce gradient ne semble pas être associé à
des variations de drainage ou de qualité de station, mais plutôt à l’historique de
composition des peuplements échantillonnés. Ces différences de composition en espèces
entre des zones d’exclusion du cerf et des espaces où le cerf a libre accès ont été
mentionnées par plusieurs auteurs (Anderson et Loucks, 1979; Anderson et Katz, 1993;
Gill et Beardall, 2001; Morecroft et al., 2001). Si certaines espèces étaient présentes autant
dans les zones clôturées que dans celles non clôturées, d’autres espèces étaient plus
fortement associées à l’une ou l’autre de ces zones. Selon ces résultats, certaines espèces
tireraient profit de la présence du cerf, alors que d’autres seraient très négativement
affectées par sa présence. Il a été observé à maintes reprises que les espèces préférées du
cerf disparaissaient au profit d’espèces moins recherchées pour leur alimentation (Case et
McCullough, 1987; Stewart et Burrows, 1989; deCalesta, 1997; Cooke et Farrell, 2001;
Reyes et Vasseur, 2003; Rooney et Waller, 2003) et que la diète du cerf changeait en
fonction de cette végétation modifiée (Veblen et al., 1989; Cross, 1998). Dans la présente
étude, les espèces sensibles à la présence du cerf, et donc appelées à disparaître si les
densités de cerf ne sont pas diminuées dans le temps, sont le bouleau à papier, l’épilobe à
feuilles étroites et le framboisier (Figure 11). Certains auteurs avaient aussi noté l’impact
négatif marqué des cerfs sur l’une ou l’autre de ces espèces (Cross, 1998; Kirby, 2001;
Horsley et al., 2003 ; Potvin et Poirier, 2004). Plusieurs autres espèces, qui se retrouvent
aussi sur l’île d’Anticosti, se sont montrées très sensibles à la présence du cerf et à son
impact sur le milieu. C’est le cas du sapin baumier (Cornett et al., 2000), de la maïanthème

46
du Canada (Waller et Alverson, 1997; Rooney, 2001), de la ronce sp. (Rubus spp.)
(Tilghman, 1989), de l’aralie à tige nue (Aralia nudicaulis L.) et de la clintonie boréale
(Clintonia borealis (Ait.) Raf.) (Waller et Alverson, 1997).
Par ailleurs, certaines espèces ont profité de la présence du cerf dans les grandes coupes
étudiées pour augmenter leur abondance. C’est le cas, entre autres, des graminées et du
chardon des champs qui étaient présents en grande quantité. Ces résultats sont en accord
avec ceux de Horsley et al. (2003) qui ont observé une augmentation linéaire du
pourcentage de recouvrement des graminées avec la densité de cerf dans les coupes à blanc.
Selon Kirby (2001), les graminées, qui ont un méristème basal et un taux de silice élevé,
sont souvent plus présentes aux endroits accessibles au cerf. En fait, étant donné que le
méristème est situé à la base de la feuille, les graminées sont les espèces les mieux
adaptées, parmi les plantes supérieures, pour supporter un broutage continu (Marie-Victorin
et al., 1995). Les graminées peuvent tirer profit de l’élimination de plusieurs individus
appartenant à des espèces fortement broutées par le cerf pour s’étendre et occuper de
grandes superficies. Par contre, la végétation dense dans les zones clôturées ne permet pas
cet étalement des graminées qui se retrouvent donc en moindre quantité. Le chardon des
champs, pour sa part, est une espèce héliophile qui se développe sur des sols à drainage de
moyen à bon. Une fois établi, son étalement s’effectue à l’aide de ses rhizomes (Marie-
Victorin et al., 1995). Ainsi, la présence d’une végétation assez dense dans les aires
clôturées entraînerait des conditions de lumière non propices à l’installation de l’espèce.
Par contre, la grande disponibilité de la lumière dans les zones fortement broutées par le
cerf favoriserait la prolifération du chardon. La violette est aussi associée aux zones non
clôturées, quoique son recouvrement soit très peu élevé. Une espèce de violette, Viola
riviniana Rchb., est réputée toxique et est donc peu broutée (Grime et al., 1988 dans Kirby,
2001), ce qui pourrait contribuer à expliquer la présence de la violette dans les zones
accessibles au cerf. Par ailleurs, la faible présence de violettes dans les zones clôturées
pourrait s’expliquer par sa faible capacité à compétitionner avec des espèces agressives
comme l’épilobe à feuilles étroites, le bouleau à papier, le framboisier ou le peuplier faux-
tremble (Tableau 8).

47
Réversibilité et extinction locale des espèces végétales Étant donné que le bouleau à papier, l’épilobe à feuilles étroites et le framboisier sont
fortement affectés par la présence du cerf, il est légitime de se demander si leur disparition
est réversible, c’est-à-dire s’il leur sera possible de recoloniser le territoire à la suite d’une
éventuelle diminution des densités de cerf. En ce qui concerne le bouleau à papier et
l’épilobe à feuilles étroites, les graines peuvent parcourir plusieurs kilomètres, ce qui
devrait possiblement leur permettre de recoloniser le territoire dans la mesure où certains
individus sont toujours présents sur l’île. Par ailleurs, la viabilité des graines du framboisier
est très longue, de l’ordre de 50 à 100 ans, et la reproduction asexuée lui permet de
coloniser rapidement une station (Whittle et al., 1997). Conséquemment, à condition qu’il y
ait présence de semenciers ayant survécus aux cerfs, nous pouvons spéculer que ces espèces
devraient pouvoir recoloniser le territoire. Horsley et al. (2003) soulignaient que là où une
source de graines est présente dans les coupes totales et les coupes partielles, des réductions
de la densité de cerf pourraient être associées à la restauration de la diversité en espèces. La
présence de ces espèces dans les zones clôturées représente de bons semenciers protégés du
broutement du cerf. De plus, il a déjà été noté que la disparition du framboisier apparaît être
facilement réversible (Horsley et Marquis, 1984 dans Gill, 1992). Selon Anderson et Katz
(1993), le temps requis pour rétablir la biodiversité initiale d’un peuplement nouvellement
protégé du cerf est proportionnel au temps durant lequel la forêt a été sujette à une pression
de broutement intensive, ce qui représente une période de plusieurs décennies sur l’île
d’Anticosti.
Dans le cas du sapin baumier, Hughes et Bechtel (1997) ont observé que la régénération
diminuait, dû à une densité de semences plus faible, à mesure que l’on s’éloignait de la
lisière boisée dans les coupes totales. Brandner et al. (1990) soulignaient que les distances
de dispersion du sapin baumier sont courtes et que les arbres matures ou surannés
fournissent la seule source de graines. Étant donnée cette faible distance effective de
dispersion des graines et que les arbres matures de sapin baumier ont été récoltées lors de la
coupe, la recolonisation du sapin à grande échelle pourrait possiblement être difficile. Si le

48
cas se présentait, le reboisement artificiel devrait être envisagé pour compenser le possible
manque de régénération naturelle sur le territoire.
Par ailleurs, si les densités de cerf ne sont pas diminuées à court terme, une extinction
localisée de certaines espèces pourrait survenir. Des exemples de ce phénomène ont été
mentionnés dans la littérature, entre autres, pour la trille (Augustine et Frelich, 1998) et la
pruche (Case et McCullough, 1987). Lors de cette expérience, le retour des espèces suite à
une diminution des densités de cerf n’a pas été quantifié et les éléments présentés dans la
présente section sont spéculatifs.

Conclusion
Cette étude démontre que les grandes CPRS ne représentent pas une solution dans le
contexte actuel de densité de cerf pour régénérer les sapinières et les autres espèces
associées à ces peuplements. En effet, l’impact du cerf est très marqué même à des
distances importantes de la lisière boisée. En limitant la régénération et la croissance du
sapin et du bouleau à papier au profit de l’épinette blanche, et en modifiant l’assemblage
des espèces végétales à sa portée, le cerf modifie profondément l’écosystème. La forte
population de cerf, l’absence de prédateurs et le brout relativement peu abondant
contribuent à expliquer l’échec des grandes coupes à régénérer les sapinières de l’île
d’Anticosti. Les densités de cerf devront être diminuées pour assurer le maintien des
sapinières et des espèces végétales initialement retrouvées sur l’île. Il est possible qu’une
diminution des densités de cerf puisse rendre les grandes CPRS efficaces pour
l’établissement et la croissance des semis de sapin et des espèces végétales associées. À cet
effet, une autre étude effectuée sur l’île d’Anticosti évaluait qu’une densité de cerf entre 7
et 15 cerfs/km2 en moins de 3 ans suivant la coupe serait nécessaire pour maintenir
l’assemblage naturel de plantes au sol se retrouvant dans les sapinières (Tremblay et al.,
2005). Ces auteurs sont d’avis que des densités approchant sept cerfs/km2 seraient
cependant nécessaires pour maintenir la diversité des communautés végétales
caractéristiques de la sapinière à bouleau blanc de l’Est.

Propositions d’aménagement
Les densités de cerf de l’île d’Anticosti devront être diminuées assez rapidement pour ne
pas atteindre un seuil d’irréversibilité des sapinières originales et des espèces qui y sont
associées. Le seuil d’irréversibilité correspond à l’atteinte d’un point de non retour à partir
duquel les espèces seraient incapables de réoccuper le territoire. Il est probable que seule
une chasse accrue et stratégique, telle que la récolte de cerfs sans bois ou la chasse en
enclos, pourra permettre de remplir ce mandat. À cet effet, des études sont présentement en
cours sur l’île pour tester différentes méthodes de chasse. L’importance de la chasse se
situe autant sur le plan des impacts plus faibles du cerf dans les zones chassées, puisque le
cerf fréquente moins ces endroits (Martin et Baltzinger, 2002), que sur le plan de limitation
des tailles de population de cerf (Rooney et Waller, 2003).
Pour l’instant et jusqu’à ce que les densités de cerf soient en deçà de 15 bêtes/km2, le
clôturage systématique des aires de coupe devra être maintenu. Les coupes dans lesquelles
des semenciers sont présents pourront se régénérer naturellement. Par contre, la plantation
devrait être envisagée si la régénération au sol n’est pas suffisante et qu’il n’y a pas la
présence de semenciers. Selon Sage et al. (2003), les peuplements aménagés doivent être
protégés du broutement jusqu'à ce que la régénération soit hors d’atteinte des cerfs. Sur l’île
d’Anticosti, il est prévu d’enlever les clôtures dès que la régénération aura atteint trois
mètres.
Des espèces affectées par la présence du cerf pourraient agir comme espèces indicatrices
pour caractériser le niveau de perturbation du cerf occasionné sur le milieu. Ainsi, les
espèces associées aux zones clôturées ou non clôturées pourraient indiquer si les cerfs sont
trop nombreux ou si les densités permettent de préserver les espèces sensibles au
broutement. Par exemple, une forte présence de graminées (Waller et Alverson, 1997) et de

51
chardons dans les CPRS indiquerait que le cerf est en grande densité dans le milieu. De la
même façon, la présence du bouleau à papier, du framboisier et de l’épilobe à feuilles
étroites dans les CPRS pourrait signifier que les densités de cerf sont adéquates pour
préserver les espèces sensibles dans le milieu. Rooney (2001) propose que les espèces
indicatrices doivent être sélectionnées sur la base de leur sensibilité aux changements des
densités de cerf. Par exemple, cet auteur suggère qu’une diminution des liliacées indique de
fortes densités de cerf. Par ailleurs, Rooney et Waller (2003) sont d’avis que l'approche
espèce par espèce pour déterminer les gagnants et les perdants face au broutement du cerf
est inefficace et offre peu de pouvoir de prédiction. Ces derniers proposent de regrouper les
plantes selon des caractéristiques communes pour augmenter leur capacité à anticiper la
réaction des organismes non étudiés à des augmentations des densités d'ongulés. Une
analyse plus poussée serait nécessaire afin de déterminer les groupes de plantes indicatrices
permettant de mesurer l’effet du cerf sur le milieu à l’île d’Anticosti.

Bibliographie Alverson, W. S., et Waller, D. M. 1997. Deer populations and the widespread failure of
hemlock regeneration in northern forests. Pages 280-297 dans W. J. McShea, H. B. Underwood and J. H. Rappole, éditeurs. The Science of Overabundance: Deer Ecology and Population Management. Smithsonian Institution Press, Washington, DC, USA.
Anderson, R. C., et Katz, A. J. 1993. Recovery of browse-sensitive tree species following release from white-tailed deer Odocoileus virginianus zimmerman browsing pressure. Biological Conservation, 63: 203-208.
Anderson, R. C., et Loucks, O. L. 1979. White-tail deer (Odocoileus virginianus) influence on structure and composition of Tsuga canadensis forests. The Journal of Applied Ecology, 16: 855-861.
Andrén H. and Angelstam P. 1993. Moose browsing on Scots pine in relation to stand size and distance to forest edge. The Journal of Applied Ecology, 30: 133-142.
Augustine, D. J., et Frelich, L. E. 1998. Effects of white-tailed deer on populations of an understory forb in fragmented deciduous forests. Conservation Biology, 12: 995-1004.
Bellingham, P. J. et Allan, C. N. 2003. Forest regeneration and the influences of white-tailed deer (Odocoileus virginianus) in cool temperate New Zealand rain forests. Forest Ecology and Management, 175: 71-86.
Bergquist, J., et Örlander, G. 1998a. Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages 1. Effect of slash removal, vegetation development, and roe deer density. Forest Ecology and Management, 105: 283-293.
Bergquist, J., et Örlander, G. 1998b. Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages: 2. Effect of seedling vigour. Forest Ecology and Management, 105:295-302. Bergquist, J., Örlander, G., et Nilsson, U. 1999. Deer browsing and slash removal affect
field vegetation on south Swedish clearcuts. Forest Ecology and Management 115: 171-182.
Borlowski, J. 2000. Influence of the density of a sika deer population on activity, habitat use, and group size. Canadian Journal of Zoology, 78: 1369-1374.
Bowers, M. A. 1997. Influence of deer and other factors on an old-field plant community: An eight-year exclosure study. Pages 310-326 dans W. J. McShea, H. B. Underwood and J. H. Rappole, éditeurs. The Science of Overabundance: Deer Ecology and Population Management. Smithsonian Institution Press, Washington, DC, USA.
Brandner, T. A., Peterson, R. O., et Risenhoover, K. L. 1990. Balsam fir on Isle Royale: Effects of moose herbivory and population density. Ecology, 71: 155-164.
Byers, J. A. 1997. American pronghorn: social adaptations and the ghosts of predators past. University of Chicago Press, Chicago.
Cadenasso, M. L., et Pickett, S. T. A. 2000. Linking forest edge structure to edge function: mediation of herbivore damage. Journal of Ecology, 88: 31-44.
Case, D. J., et McCullough, D. R. 1987. The white-tailed deer of North Manitou Island. Hilgardia, 55: 1-57.

53
Conover, M. R. 1997. Monetary and intangible valuation of deer in the United States. Wildlife Society Bulletin, 25: 298-305.
Cooke, A. S., et Farrell, L. 2001. Impact of muntjac deer (Muntiacus reevesi) at Monks Wood National Nature Reserve, Cambridgeshire, eastern England. Forestry, 74: 241-250.
Cornett, M. W., Frelich, L. E., Puettmann, K. J., et Reich, P. B. 2000. Conservation implications of browsing by Odocoileus virginianus in remnant upland Thuja occidentalis forests. Biological Conservation, 93: 359-369.
Cross, P. C. 1998. The impacts of white-tailed deer and disturbance on sapling establishment and forest succession in Virginia. Undergraduate thesis, University of Virginia, Charlottesville, Virginia, USA.
deCalesta, D. S. 1997. Deer and ecosystem management. Pages 267-279 dans W. J. McShea, H. B. Underwood and J. H. Rappole, éditeurs. The Science of Overabundance: Deer Ecology and Population Management. Smithsonian Institution Press, Washington, DC, USA.
Dorais, L., Foulet, B., Chabot, M., Dupont, A., et Gagnon, R. 1996. Entomologie forestière. Ordre des ingénieurs forestiers du Québec (éd.), Manuel de foresterie. Presses de l'Université Laval, Sainte-Foy, Québec.: 1009-1043.
Drolet, C.-A. 1978. Use of forest clear-cuts by white-tailed deer in southern New Brunswick and central Nova Scotia. Canadian Field-Naturalist, 92: 275-282.
Environnement Canada. 2006. Normales et moyennes climatiques, rapports de données quotidiennes de la station de Port-Menier de 1995 à 2005 [Base de données en ligne].http://www.climate.weatheroffice.ec.gc.ca/advanceSearch/searchHistoricDataStations_f.html (Consulté le 8 juin 2006).
Frelich, L. E. et Lorimer, C. G. 1985. Current and predicted long-term effects of deer browsing in hemlock forests in Michigan, USA. Biological Conservation, 34: 99-120.
Fretwell, S. D., et Lucas, H. L. Jr. 1970. On territorial behaviour and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica, 19: 16-36.
Gill, R. M. A. 1992. A review of damage by mammals in north temperate forests: 1. Deer. Forestry, 65: 145-169.
Gill, R. M. A., et Beardall, V. 2001. The impact of deer on woodlands: the effects of browsing and seed dispersal on vegetation structure and composition. Forestry, 74: 209-218.
Grondin, P., Ansseau, C., Bélanger, L., Bergeron, J.-F., Bergeron, Y., Bouchard, A., Brisson, J., De Grandpré, L., Gagnon, G., Lavoie, C., Lessard, G., Payette, S., Richard, P. J. H., Saucier, J.-P., Sirois, L., et Vasseur, L. 1996. Écologie forestière. Pages 134-279 dans Ordre des ingénieurs forestiers du Québec, éditeur. Manuel de foresterie. Presses de l'Université Laval, Ordre des ingénieurs forestiers du Québec, Québec, Qc, Canada.
Harmer, R. 2001. The effect of plant competition and simulated summer browsing by deer on tree regeneration. The Journal of Applied Ecology, 38: 1094-1103.
Healy, W. M. 1997. Influence of deer on the structure and composition of oak forests in central Massachusetts. Pages 201-223 dans W. J. McShea, H. B. Underwood and J. H. Rappole, éditeurs. The Science of Overabundance: Deer Ecology and Population Management. Smithsonian Institution Press, Washington, DC, USA.

54
Horsley, S. B., Stout, S. L., et deCalesta, D. S. 2003. White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecological Applications, 13: 98-118.
Hughes, J. W., et Bechtel, D. A. 1997. Effect of distance from forest edge on regeneration of red spruce and balsam fir in clearcuts. Canadian Journal of Forest Research, 27: 2088-2096.
Huot, J. 1982. Body condition and food resources of white-tailed deer on Anticosti Island, Québec. Ph.D. thesis, University of Alaska, Fairbanks, USA.
Jane, G. T. 1994. The impact of browsing animals on the stand dynamics of monotypic mountain beech (Nothofagus solandri) forests in Canterbury, New Zealand. Australian Journal of Botany, 42: 113-124.
Kay, S. 1993. Factors affecting severity of deer browsing damage within coppiced woodlands in the south of England. Biological Conservation, 63: 217-222.
Kirby, K. J. 2001. The impact of deer on the ground flora of British broadleaved woodland. Forestry, 74: 219-229.
Kullberg, Y., et Bergström, R. 2001. Winter browsing by large herbivores on planted deciduous seedlings in southern Sweden. Scandinavian Journal of Forest Research, 16: 371-378.
Legendre, L. et Legendre, P. 1984. Écologie numérique : 2. La structure des données écologiques. Deuxième édition. Presses de l’Université du Québec, Québec, Canada.
Lepš, J., et Šmilauer, P. 1999. Multivariate Analysis of Ecological Data. Faculty of Biological Sciences, University of South Bohemia. České Budějovice. 110 p.
Magurran, A. E. 2004. Measuring biological diversity. Malden, MA, Oxford, Blackwell Pub. 256 p.
Marie-Victorin, f.é.c., Brouillet, L., Goulet, I., et Rouleau, E. 1995. Flore Laurentienne, 3e édition. Les presses de l’Université de Montréal, Montréal, Qc, Canada. 1093 p.
Martin, J.-L., et Baltzinger, C. 2002. Interaction among deer browsing, hunting, and tree regeneration. Canadian Journal of Forest Research, 32: 1254-1264.
McInnes, P. F., Naiman, R. J., Pastor, J., et Cohen, Y. 1992. Effects of moose browsing on vegetation and litter of the boreal forest, Isle Royale, Michigan, USA. Ecology, 73: 2059-2075.
Milliken, G. A., et Johnson, D. E. 2002. Analysis of Messy Data, Volume III: Analysis of Covariance. Boca Raton: Chapman and Hall/CRC.
Morecroft, M. D., Taylor, M. E., Ellwood, S. A., et Quinn, S. A. 2001. Impacts of deer herbivory on ground vegetation at Wytham Woods, central England. Forestry, 74: 251-257.
Mueller-Dombois, D. et Ellenberg, H. 1974. Aims and Methods of Vegetation Ecology. Wiley, New York.
Opperman, J. J., et Merenlender, A. M. 2000. Deer herbivory as an ecological constraint to restoration of degraded riparian corridors. Restoration Ecology, 8: 41-47.
Parent, S., et Ruel, J-C. 2002. Chronologie de la croissance chez des semis de sapin baumier (Abies balsamea (L.) Mill.) après une coupe à blanc avec protection de la régénération. The Forestry Chronicle, 78: 876.
Persson, I.-L., Danell, K., et Bergström, R. 2000. Disturbance by large herbivores in boreal forests with special reference to moose. Annales Zoologici Fennici, 37: 251-263.

55
Pominville, P., et Ruel, J.-C. 1995. Effets de la coupe à blanc et de la coupe par bandes sur la régénération obtenue après 5 ans dans des pessières noires du Québec. Canadian Journal of Forest Research, 25: 329-342.
Potvin, F. 1992. L'habitat du cerf à Anticosti de 1978 à 1988: suivi quinquennal. Rapport SP 1925-02-92, Direction générale de la ressource faunique, Ministère du Loisir, de la Chasse et de la Pêche, Québec, Qc, Canada. 27 p.
Potvin, F., Beaupré, P., Gingras, A., et Pothier, D. 2000. Le cerf et les sapinières de l'île d'Anticosti. Direction de la faune et des habitats, Société de la faune et des parcs du Québec, Qc, Canada. 35 p.
Potvin, F., Beaupré, P., et Laprise, G. 2003a. The eradication of balsam fir stands by white-
tailed deer on Anticosti Island, Québec: a 150-year process. Écoscience, 10: 487-495.
Potvin, F., Gingras, A., Laprise, G., Morin, D., et Rochette, B. 2003b. Réaction hivernale du cerf à de grandes coupes sur l'île d'Anticosti: une expérience de perforation et de fragmentation de l'habitat. Société de la faune et des parcs du Québec, Québec, Qc, Canada. 36 p.
Potvin, F., et Poirier, S. 2004. L'île d'Anticosti, un paradis? L'influence du cerf de Virginie sur la végétation des sapinières. Le Naturaliste Canadien, 128: 52-60.
Putman, R. J. 1994. Deer damage in coppice woodlands: an analysis of factors affecting the severity of damage and options for management. Quarterly Journal of Forestry, 88: 45-54.
Putman, R. J., et Moore, N. P. 1998. Impact of deer in lowland Britain on agriculture, forestry and conservation habitats. Mammal Review, 28: 141-164.
Reyes, G., et Vasseur, L. 2003. Factors influencing deer browsing damage to red spruce (Picea rubens) seedlings in coastal red spruce-balsam fir stands of southwestern Nova Scotia. Forest Ecology and Management, 186: 349-357.
Rooney, T. P. 2001. Deer impacts on forest ecosystems: a North American perspective. Forestry, 74: 201-208.
Rooney, T. P., et Waller, D. M. 2003. Direct and indirect effects of white-tailed deer in forest ecosystems. Forest Ecology and Management, 181: 165-176.
Russell, F. L., et Fowler, N. L. 2004. Effects of white-tailed deer on the population dynamics of acorns, seedlings and small saplings of Quercus buckleyi. Plant Ecology, 173: 59-72.
Sage, J. R.W., Porter, W. F., et Underwood, H. B. 2003. Window of opportunity: white-tailed deer and the dynamics of northern hardwood forests of the northeastern US. Journal for Nature Conservation, 10: 213-220.
Schmitz, O. J. 2005. Scaling from plot experiments to landscapes: studying grasshoppers to inform forest ecosystem management. Oecologia, 145: 225-234.
Stewart, G. H., et Burrows, L. E. 1989. The impact of white-tailed deer Odocoileus virginianus on regeneration in the coastal forests of Stewart Island, New Zealand. Biological Conservation, 49: 275-293.
Stoeckeler, J. H., Strothmann, R. O., et Krefting, L. W. 1957. Effect of deer browsing on reproduction in the northern hardwood-hemlock type in northeastern Wisconsin. Journal of Wildlife Management, 21: 75-80.
Tilghman, N. G. 1989. Impacts of white-tailed deer on forest regeneration in northwestern Pennsylvania. Journal of Wildlife Management, 53: 524-532.

56
Tremblay, J.-P., Huot, J., et Potvin, F. 2005. Relations entre les perturbations induites par les cervidés et la dynamique de régénération des écosystèmes forestiers boréaux : 4. Functional relationships linking cervids density to the ecological integrity of ground plant communities in a boreal forest. Ph.D. thesis, Université Laval, Sainte-Foy, Qc, Canada.
Vare, H., Ohtonen, R., et Mikkola, K. 1996. The effect and extent of heavy grazing by reindeer in oligotrophic pine heaths in northeastern Fennoscandia. Ecography, 19: 245-253.
Veblen, T. T., Mermoz, M., Martin, C., et Ramilo, E. 1989. Effects of exotic deer on forest regeneration and composition in northern Patagonia. The Journal of Applied Ecology, 26: 711-724.
Waller, D. M., et Alverson, W. S. 1997. The white-tailed deer: a keystone herbivore. Wildlife Society Bulletin, 25: 217-226.
Watkinson, A. R., Riding, A. E., et Cowie, N. R. 2001. A community and population perspective of the possible role of grazing in determining the ground flora of ancient woodlands. Forestry, 74: 231-239.
Welch, D., Staines, B. W., Scott, D., French, D. D., et Catt, D. C. 1991. Leader browsing by Red and Roe deer on young Sitka spruce trees in western Scotland: I. Damage rates and the influence of habitat factors. Forestry, 64: 61-82.
Whitney, G. G. 1984. Fifty years of change in the arboreal vegetation heart's content, an old-growth helmlock-white pine-northern harwood stand. Ecology, 65: 403-408.
Whittle, C. A., Duchesne, L. C., et Needham, T. 1997. The importance of buried seeds and vegetative propagation in the development of postfire plant communities. Environmental Reviews, 5: 79-87.
Williamson, S. J., et Hirth, D. H. 1985. An evaluation of edge use by white-tailed deer. Wildlife Society Bulletin, 13: 252-257.
Witmer, G. W., et deCalesta, D. S. 1991. The need and difficulty of bringing the Pennsylvania deer herd under control. Proceedings of the Eastern Wildlife Damage Control Conference: 5. Fifth Eastern Wildlife Damage Control Conference, Ithaca, NY, USA 5:130-137.1.
