génétique des populations · g en etique des populations d ecrire les g enotypes, estimer leur fr...
TRANSCRIPT

Genetique des populations
Claire Bardel
UMR CNRS 5558, Laboratoire de Biometrie et Biologie Evolutive,Equipe biostatistique - sante
Plateforme de sequencage haut debit des HCL
decembre 2015

Genetique des populations
Decrire les genotypes, estimer leur frequence et celle des alleles,determiner leur distribution au sein des individus, des populations, etentre les populations
Predire et comprendre l’evolution des frequences des genes dans lespopulations sous l’effet de differents facteurs, ou «forces evolutives»
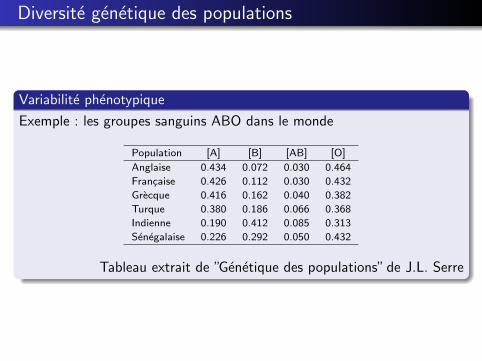
Diversite genetique des populations
Variabilite phenotypique
Exemple : les groupes sanguins ABO dans le monde
Population [A] [B] [AB] [O]
Anglaise 0.434 0.072 0.030 0.464
Francaise 0.426 0.112 0.030 0.432
Grecque 0.416 0.162 0.040 0.382
Turque 0.380 0.186 0.066 0.368
Indienne 0.190 0.412 0.085 0.313
Senegalaise 0.226 0.292 0.050 0.432
Tableau extrait de ”Genetique des populations” de J.L. Serre
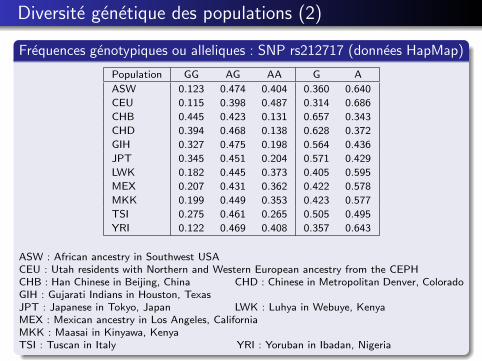
Diversite genetique des populations (2)
Frequences genotypiques ou alleliques : SNP rs212717 (donnees HapMap)
Population GG AG AA G A
ASW 0.123 0.474 0.404 0.360 0.640
CEU 0.115 0.398 0.487 0.314 0.686
CHB 0.445 0.423 0.131 0.657 0.343
CHD 0.394 0.468 0.138 0.628 0.372
GIH 0.327 0.475 0.198 0.564 0.436
JPT 0.345 0.451 0.204 0.571 0.429
LWK 0.182 0.445 0.373 0.405 0.595
MEX 0.207 0.431 0.362 0.422 0.578
MKK 0.199 0.449 0.353 0.423 0.577
TSI 0.275 0.461 0.265 0.505 0.495
YRI 0.122 0.469 0.408 0.357 0.643
ASW : African ancestry in Southwest USACEU : Utah residents with Northern and Western European ancestry from the CEPHCHB : Han Chinese in Beijing, China CHD : Chinese in Metropolitan Denver, ColoradoGIH : Gujarati Indians in Houston, TexasJPT : Japanese in Tokyo, Japan LWK : Luhya in Webuye, KenyaMEX : Mexican ancestry in Los Angeles, CaliforniaMKK : Maasai in Kinyawa, KenyaTSI : Tuscan in Italy YRI : Yoruban in Ibadan, Nigeria

Composition genetique d’une population
Au niveau d’une population, on a les frequences des phenotypes
Passage aux frequences genotypiques ou allelique dans le casd’alleles co-dominants
Frequences genotypiques egales aux frequences phenotypiques
Frequences alleliquescomptage des alleles
Phenotype [A] [Aa] [a]
Genotype AA Aa aa
Effectif 350 500 150
f (A) =350× 2 + 500
2× 1000= 0.6
f (a) =150× 2 + 500
2× 1000= 0.4 = 1−f (A)
Frequences alleliquesmethode probabiliste
Phenotype [A] [Aa] [a]
Genotype AA Aa aa
Frequence D H R
f (A) = D + H × 1
2
f (a) = R + H × 1
2
Cas d’alleles dominants ou recessifs : Loi de Hardy-Weinberg

Equilibre de Hardy-Weinberg

La loi de Hardy-Weinberg
Etablissement par le modele de l’urne gametiqueSoit un locus bi-allelique, f (A) = p, f (a) = qCondition de panmixie (choix du partenaire aleatoires) et pangamie(union aleatoire des gametes) → modelisation par deux urnes
malegameteUrne Urne
gametefemelle
f(a)=q
f(A)=p
f(a)=q
f(A)=p
p2 pq
pq q2
A a
A
a
gametemale
gametefemelle
f (AA) = p2 f (Aa) = 2pq f (aa) = q2
RemarqueLe modele de l’urne gametique correspond a la realite biologique pourcertaines especes vegetales ou de champignons

Conditions de validite de la loi de Hardy-Weinberg
Contenu de l’urne
La composition de l’urne (=gametes) doit refleter les frequencesalleliques des reproducteurs
pas de selection en matiere de fertilite (meme nombre de gametespour tous les individus reproducteurs)
pas de mutation de novo
pas de selection gametique
Tirage aleatoire dans l’urne
panmixie et pangamie
Egalite des probabilites genotypes obtenus et de leurs frequences
Population de taille infinie
Stabilite de la population entre la fecondation et la reproduction
pas de selection zygotique pas de migration

L’equilibre de Hardy-Weinberg
Maintien des frequences alleliques au cours des generations
f (A) = D +H
2= p2 +
2pq
2= p(p + q) = p
f (a) = R +H
2= q2 +
2pq
2= q(p + q) = q
Bilan sur les conditions de validites
Population panmictique et pangamique→∼ vrai pour la majeure partie des genes
population de taille quasi-infinie→ peut poser probleme, phenomene de derive
mutation, migration, selection negligeables
mutation : condition acceptable sur une duree courte (qqsgenerations)selection : idem que pour les mutations (attention, generalement fauxen agronomie)migration : peut poser probleme dans les populations humaines

Generalisation de la loi de Hardy-Weinberg : genepluri-allelique
Soit un gene a n alleles A1,A2, . . . ,An de frequences p1, p2, . . . , pn
A1 A2 . . . Ai . . . An
A1 p21 p1p2 . . . p1pi . . . p1pn
A2 p2p1 p22 . . . p2pi . . . p2pn
. . . . . . . . . . . . . . . . . . . . .
Ai pip1 pip2 . . . p2i . . . pipn
. . . . . . . . . . . . . . . . . . . . .
An pnp1 pnp2 . . . pnpi . . . p2n
(p1 + p2 + . . .+ pn)2 =n∑
i=1
p2i +
n∑i=1
n∑j=i+1
2pipj
Consequence : calcul du taux d’heterozygotie theorique :
Ht = 1−n∑
i=1
p2i

Exemple de calcul des frequences alleliques
Calcul des fréquences pour des allèles multiples
Isoagglutine I : 3 allèles A (p), B (q), O (r)
6 génotypes possibles : AA, BB, 00, AB, AO, BOA et B sont codominants l’un par rapport à l’autreA et B sont dominants sur O
p + q + r = 1
HW
p²+q²+r²+ 2pq + 2pr + 2qr = 1
A partir des fréquences phénotypiqueson peut déterminer les fréquences des3 allèles dans une population
Ex : Groupe A : 0,53 Groupe B : 0,13 Groupe O : 0,26
Calculer p, q et r
r²rqrpO
qrq²qpB
prpqp²A
OBA
0,30, 0,11, 0,51

Exemple de calcul des frequences alleliques (2)
Notations
f([A]) : frequence duphenotype A
f(AA) : frequence dugenotype AA
Enonce
f([A])=0,53
f([B])=0,13
f([0])=0,26
On cherche f(A)=p, f(B)=q et f(O)=r
Hardy-Weinberg + Dominance de A et B
f([A])=f(AA)+f(AO)=p2+2pr
f([B])=f(BB)+f(BO)=q2+2qr
f([O])=f(OO)=r 2
Remarque
On pourrait calculer r directement a partir de la 3e equation, puis en injectant la valeurs de r
dans les 2 autres equations, on trouverait p et q respectivement. Mais cela impliquerait de
resoudre des equations du 2e degre. . .⇒ Utilisation d’une astuce pour simplfier les calculs !

Exemple de calcul des frequences alleliques (3)
Calcul de q
f([A])+f([O])=p2+2pr+r 2=(p + r)2
identite remarquable classique :(a + b)2 = a2 + b2 + 2ab
Or, p+q+r=1, donc p+r=1-q
Finalement, on a : f([A])+f([O])=0,53+0,26=0,79=(1− q)2
1-q=√
0, 79 = 0, 89
q=0,11
Calcul de p (meme principe)
f([B])+f([O])=q2+2qr+r 2=(q + r)2
f([B])+f([O])=0,13+0,26=0,39=(1− p)2
1-p=√
0, 39 = 0, 62
p=0,38
Calcul de r
f([O])=r 2
r=√
0, 26
r=0,51
On verifie bien que0,51+0,38+0,11=1

Comment tester l’equilibre de HW ?
Exemple d’utilisation
Test lors des etapes de controle qualite dans les etudesd’association (a verifier sur les temoins)
L’equilibre de HW est teste dans une population donnee, a un locusdonne
Exemple
Donnees de Lanfear DE, et al J Am Coll Cardiol. 2004 Jul 7 ;44(1) :165-7Genotype de 95 individus d’origine Afro-Americaine au locus C10009T dugene GJA-4 (gap junction protein alpha-4)
Genotype CC CT TT
Effectif 24 52 19
Frequence 25.3% 54.9% 19.8%
⇒ Test du χ2 de conformite a une distribution de reference

Comment tester l’equilibre de HW ?
Exemple d’utilisation
Test lors des etapes de controle qualite dans les etudesd’association (a verifier sur les temoins)
L’equilibre de HW est teste dans une population donnee, a un locusdonne
Exemple
Donnees de Lanfear DE, et al J Am Coll Cardiol. 2004 Jul 7 ;44(1) :165-7Genotype de 95 individus d’origine Afro-Americaine au locus C10009T dugene GJA-4 (gap junction protein alpha-4)
Genotype CC CT TT
Effectif 24 52 19
Frequence 25.3% 54.9% 19.8%
⇒ Test du χ2 de conformite a une distribution de reference

Test de l’equilibre de HW
Calcul des frequences alleliques :f (C ) = 24×2+52
2×95 ' 0.53
f (T ) = 19×2+522×95 ' 0.47
Calcul des frequences theoriques selon HWf (CC ) = 0.532 = 0.28 f (CT ) = 2× 0.53× 0.47 =0.50 f (TT ) = 0.472 = 0.22
Calcul des effectifs theoriques selon HWnCC = 95× 0.28 = 26.6 nCT = 47.5 nTT = 20.9
Table : χ2c =
∑i
(oi−ti )2
ti
Genotype CC CT TT
n observe 24 52 19
n theorique 26.6 47.5 20.9
Resultat du test :> chisq.test(x=c(24, 52, 19), p=c(0.28,0.50,0.22))
Chi-squared test for given probabilities
data: c(24, 52, 19)
X-squared = 0.8532, df = 2, p-value = 0.6527

Desequilibre de liaison

Cas de l’etude simultanee de 2 genes autosomiques
Soit 2 genes A et B avec chacun 2 alleles :f (A1) = p f (B1) = u
f (A2) = q f (B2) = v4 gametes possibles : A1B1,A2B2,A1B2 et A2,B1
Si les locus sont independants :f (A1B1) = f (A1)× f (B1) = puf (A2B2) = f (A2)× f (B2) = qvf (A1B2) = f (A1)× f (B2) = pvf (A2B1) = f (A2)× f (B1) = qu
⇒ Equilibre gametiqueSi les locus ne sont pas independants :D1,1 = f (A1B1)− f (A1)× f (B1)D2,2 = f (A2B2)− f (A2)× f (B2)D1,2 = f (A1B2)− f (A1)× f (B2)D2,1 = f (A2B1)− f (A2)× f (B1)
⇒ Desequilibre gametique : D1,1 = D2,2 = −D1,2 = −D2,1
Interpretation :
Si D1,1 > 0, A1 et B1 sont plus souvent associe qu’attendu sousl’hypothese d’equilibreSi D1,1 < 0, A1 et B1 sont moins souvent associes qu’attendu

Exemple de genese d’un desequilibre gametique : migration
Gene B :
f (B2) = v = 1f (B1) = u = 0
Gametes :f (A1B1) = pu = 0f (A2B2) = qv = 1f (A1B2) = pv = 0f (A2B1) = qu = 0
100% d’individus A1B1/A1B1
Gene A :f (A1) = p = 1f (A2) = q = 0
Gene B :f (B1) = u = 1f (B2) = v = 0
Gametes :f (A1B1) = pu = 1f (A2B2) = qv = 0f (A1B2) = pv = 0f (A2B1) = qu = 0
100% d’individus A2B2/A2B2
Gene A :f (A1) = p = 0f (A2) = q = 1
Population BPopulation A
50% d’individus A2B2/A2B2
Gene A :f (A1) = p = 0.5f (A2) = q = 0.5
Gene B :f (B1) = u = 0.5f (B2) = v = 0.5
Population C
50%50%
50% d’individus A1B1/A1B1
Gametes :f (A1B1) = 0.5 (> pu)f (A2B2) = 0.5 (> qv)f (A1B2) = 0 (< pv)f (A2B1) = 0 (< qu)
D2,2 = 0.5− 0.25 = 0.25D1,2 = 0− 0.25 = −0.25D2,1 = 0− 0.25 = −0.25
D1,1 = 0.5− 0.25 = 0.25

Exemple de genese d’un desequilibre gametique : mutation
B
B
B
B
Evolution de la populationMutation sur un gamete
portant l’allele B
A
A A
A
B
B A
A
B
A
A
A A
A
B
B A
A
A
B
A
A A
A
B
A
A
A
B A
m
f (A,m) = 0 6= f (A)× f (m)
m m
m
m
La recombinaison peut “casser” l’association entre B et m
Ici, le desequilibre gametique se maintient a cause d’une liaisongenetique entre les locus⇒ On parle de desequilibre de liaison
Autres origines possibles du desequilibre gametique
SelectionEffet fondateur (migration)

Evolution d’un desequilibre gametique au cours du temps
Soit θ le taux de recombinaison entre les locus A et B
Exprimons ft(A1,B1) en fonction de ft−1(A1,B1)
A la generation t, les gametes A1B1 proviennent :
des gametes A1B1 a t − 1 qui n’ont pas recombines→ ft−1(A1,B1)× (1− θ))des nouveaux gametes A1B1 formes par recombinaison→ θpu
ft(A1,B1) = ft−1(A1,B1)× (1− θ) + θpu
Exprimons D1,1(t) en fonction du tempsOn retranche pu aux deux membres et on reorganise :
ft(A1,B1)− pu = ft−1(A1,B1)× (1− θ)− pu × (1− θ)
D1,1(t) = (1− θ)D1,1(t − 1)
D1,1(t) = (1− θ)tD1,1(0)
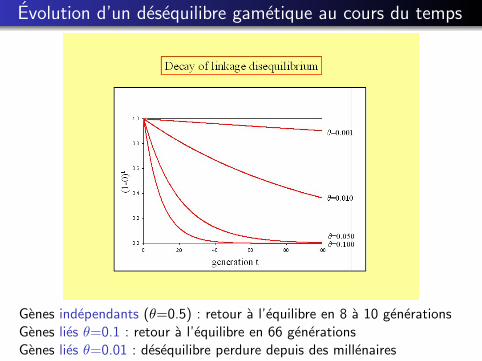
Evolution d’un desequilibre gametique au cours du temps
Genes independants (θ=0.5) : retour a l’equilibre en 8 a 10 generationsGenes lies θ=0.1 : retour a l’equilibre en 66 generationsGenes lies θ=0.01 : desequilibre perdure depuis des millenaires

D’autres mesures du desequilibre
Le D’ (Lewontin et Kojima, 1960){D ′ = D
min(pu,qv) si D < 0
D ′ = Dmin(pv ,qu) si D ≥ 0
Depend moins des frequences alleliques que DCompris entre -1 et 1Si |D’|=1, seules 3 combinaisons alleliques existent (ou 2 si lesfrequences sont egales)
Le coefficient de correlation r
r =D
√pquv
Compris entre -1 et 1si |r|=1, seules 2 combinaisons alleliques existent

Importance du desequilibre gametique en genetiqueepidemiologique
Exemple des analyses d’association en gene candidat
But : Rechercher si un gene candidat est implique dans ledeterminisme d’une maladie
gene candidat : gene dont la fonction suggere qu’il pourrait etreimplique dans la maladie
Principe : comparer les distributions alleliques ou genotypiques chezdes cas et des temoins pour differents marqueurs du gene
AA Aa aa
malades mAA mAa maa
temoins tAA tAa taa
Si allele a risque pour la maladie → plus frequent chez les malades

Importance du deqequilibre gametique en genet. epi. (2)
Test d’association indirect
Souvent, le locus de susceptibilite est inconnu → test indirect
Test d’association avec un marqueur M (alleles M1/M2) situe aproximite du polymorphisme fonctionnel S et dont l’expression y estliee
Lien entre le polymorphisme fonctionnel et le marqueur :desequilibre de liaison (DL)
Si l’allele M1 est en desequilibre gametique avec S, on observera plussouvent l’allele M1 chez les malades
⇒ Mise en evidence d’une association entre la maladie et l’allele M1
polymorphismemarqueur
associationindirecte
associationdirecte
PHENOTYPE
DL
fonctionnel

Notion d’haplotypes
Haplotype : ensemble des alleles des differents marqueurs situes surun meme chromosome
Pour trois marqueurs bi-alleliques MA (A, a), MB (B, b) etMC (C, c), en theorie 23 = 8 configurations possibles :
ABC ABc AbC Abc
aBC aBc abC abc
En pratique, on trouve beaucoup moins de configurations du fait dudesequilibre de liaison
Interet : etude simultanee de plusieurs marqueurs
Possibilite de detecter des effets dus a plusieurs locus en interaction
Un probleme : les haplotypes ne sont pas connus et doivent etrereconstruits par des methodes statistiques
Le projet HapMap (http://www.hapmap.org/)
Montpetit et Chagnon, Medecine-Sciences, 2006, vol 22, no12
”La Carte d’haplotype du genome humain : une revolution en genetique des
maladies a heredite complexe”
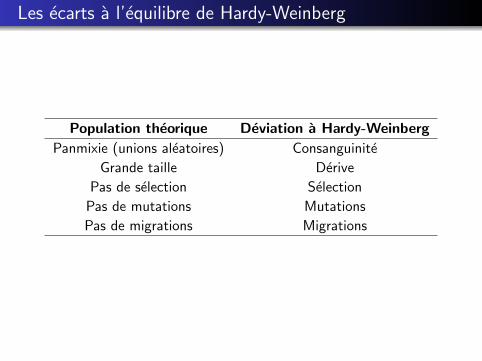
Les ecarts a l’equilibre de Hardy-Weinberg
Population theorique Deviation a Hardy-Weinberg
Panmixie (unions aleatoires) Consanguinite
Grande taille Derive
Pas de selection Selection
Pas de mutations Mutations
Pas de migrations Migrations
Ecart a la panmixie

Ecarts a la panmixie : consanguinite et homogamie
ConsanguiniteChoix du conjoint fonde sur les relations de parentes
Populations humaines : tabou de l’inceste, mariage entre cousinsgermains en Afrique du Nord, etc.Autofecondation chez certains plantes
⇒ La consanguinite a des consequences sur l’ensemble des locus dugenome
Homogamie ou heterogamieChoix du conjoint fonde sur la similitude (homogamie) ou ladissemblance (heterogamie) des individus pour certains caracteresindividuels
Populations humaines : la tailleSysteme d’autoincompatiblite chez les vegetaux
⇒ Homogamie et heterogamie n’ont de consequences que sur leslocus qui gouvernent les caracteres impliques
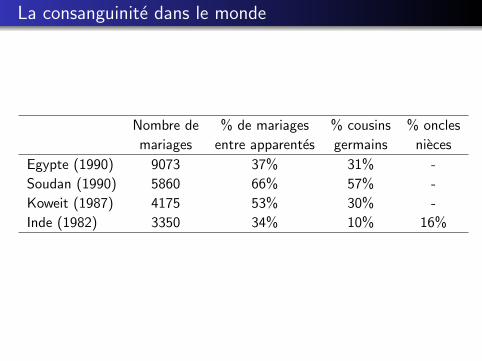
La consanguinite dans le monde
Nombre de % de mariages % cousins % oncles
mariages entre apparentes germains nieces
Egypte (1990) 9073 37% 31% -
Soudan (1990) 5860 66% 57% -
Koweit (1987) 4175 53% 30% -
Inde (1982) 3350 34% 10% 16%
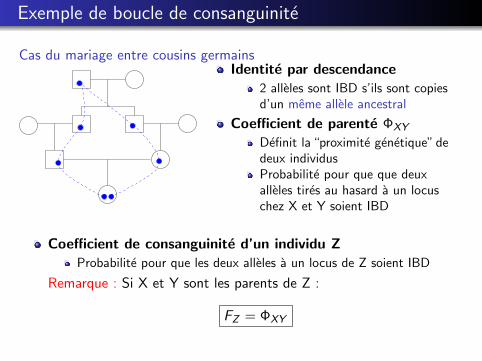
Exemple de boucle de consanguinite
Cas du mariage entre cousins germainsIdentite par descendance
2 alleles sont IBD s’ils sont copiesd’un meme allele ancestral
Coefficient de parente ΦXY
Definit la “proximite genetique” dedeux individusProbabilite pour que que deuxalleles tires au hasard a un locuschez X et Y soient IBD
Coefficient de consanguinite d’un individu ZProbabilite pour que les deux alleles a un locus de Z soient IBD
Remarque : Si X et Y sont les parents de Z :
FZ = ΦXY

Calcul du coefficient de consanguinite
Cas du mariage entre cousins germains, ancetres non consanguins
G
F
D
BA
C
E
1 G peut recevoir deux alleles IBD :
de l’arriere grand-pere Ade l’arriere grand-mere B
2 Si on considere les transmissions apartir de A
probabilite ( 12 ) pour que le meme
allele de A soit transmis a C et DSachant que le meme allele de A esttransmis a C et D, la probabiliteque G soit IBD pour cet allele estalors ( 1
2 )4
→ Probabilite pour que G recoivent deux alleles IBD de A : ( 12 )× ( 1
2 )4 = ( 12 )5
Meme raisonnement pour B et donc, au total :
FG =
(1
2
)5
+
(1
2
)5
=1
16= ΦEF

Calcul du coefficient de consanguinite
Cas du mariage entre cousins germains, ancetres consanguins
Si A est consanguin avec un coefficient de consanguinite FA, lesdeux alleles presents chez A sont IBD avec la probabilite FA
G
F
D
BA
C
E
1 Transmissions a partir de A :
probabilite 12 pour que le meme
allele de A soit transmis a C et Dprobabilite 1
2 d’avoir des allelesdifferents mais alors probabilite FA
pour qu’ils soient IBD
2 Sachant que C et D ont recu desalleles IBD de A : probabilite que G
recoive ces alleles =(
12
)4
⇒ Probabilite pour que G recoivent deux alleles IBD de A :(1
2
)(1 + FA)×
(1
2
)4
=
(1
2
)5
(1 + FA)
Meme raisonnement a partir de B

Composition genetique des populations consanguines
Soient 2 alleles A et a de frequences p et q a la generation n
F : moyenne des coefficients de parente de tous les couples de la generation n
Calculs des frequences genotypiques a la generation n+1 :
Individus avec alleles IBD (probabilite F)AA : p × Faa : q × F
Individus avec allele non IBD (probabilite (1-F)AA : p2 × (1− F )aa : q2 × (1− F )Aa : 2pq × (1− F )
f (AA) = p2(1−F ) + pF = p2 + Fp(1−p)f (aa) = q2(1− F ) + qF = q2 + Fq(1− q)f (Aa) = 2pq(1− F ) = 2pq − 2Fpq
Deux conclusions :
Augmentation des homozygotes et baisse des heterozygotes
Pas de changement des frequences alleliques, f (A) = p et f (a) = q

Consequences de la consanguinite sur les traits recessifs
Au niveau de la populationSoit une maladie recessive due a un allele d de frequence q
dd Dd et DD
population panmictique q2 p2 + 2pq
taux de consanguinite F q2 + Fpq p2 + 2pq − Fpq
Augmentation de la frequence des individus atteints dans lapopulation consanguine
Parmi les malades, forte frequence d’individus consanguins
Dans une population consanguine, pour estimer la frequence del’allele d, il est necessaire de connaıtre le taux moyen deconsanguinite F

Consequences de la consanguinite sur les traits recessifs
Au niveau individuelAugmentation du risque de naissance d’un enfant atteint d’une maladierecessiveExemple pour un couple de cousins germains (Φ = 1
16)
Frequence Risque Supplement Accroissement
Maladie de la mutation panmictique de risque relatif du
pathogene (q) (q2) (Φq(1− q)) risque
Mucoviscidose 2% 12500
32500 ×4
Phenylcetonurie 0.8% 116000
816000 ×9
Galactosemie 0.5% 140000
1340000 ×14
Tableau extrait de ”Genetique des populations” de J.L. Serre
Maladie tres rare : atteints quasiment tous consanguins !

La selection

La selection
Concept de selection naturelle introduit par Darwin et Wallace(1859)
Fixation des caracteres favorables et elimination des autres
En genetique des populationsIl y a selection lorsque les differents genotypes contribuentinegalement a la reproduction
Esperance de vie plus faibleFertilite plus faible
Notion de valeur selective (= valeur adaptative = fitness)Absolue : nombre moyen de descendants viables fournis par unecategorie d’individus
Relative :fitness d’un genotype
fitness maximale dans la populationsouvent notee ω
Genotypes AA Aa aa
Fitness ω1 ω2 ω3
⇒ Selon l’ordre des valeurs de fitness, consequences tres differentes surla composition genetique des populations !

Cas simple : effet deletere d’un allele recessif a
Frequences alleliques des parents : f (A) = p et f (a) = q
Genotypes AA Aa aa Somme
Frequences des zygotes p2 2pq q2
Viabilite 1 1 1-s
Frequencedes adultes
p2 2pq q2(1− s) 1− sq2
Frequence normaliseep2
1− sq2
2pq
1− sq2
q2(1− s)
1− sq21
Nouvelles frequences alleliques :
q′ =q2(1− s)
1− sq2+
1
2× 2pq
1− sq2=
q2 − sq2 + pq
1− sq2
p′ = 1− q′

Cas simple : effet deletere d’un allele recessif a (2)
Calcul de ∆q = q′ − q
∆q =q2 − sq2 + pq
1− sq2− q
∆q =q2 − sq2 + pq−q + sq3
1− sq2(reduction au meme denominateur)
Or, q2 − q = q(q − 1) = q(1− q)(−1) = −pq
∆q =−pq − sq2 + pq + sq3
1− sq2=−sq2(1− q)
1− sq2
∆q < 0
Conclusion
La frequence de a diminue au cours du temps et a terme, a disparaıt
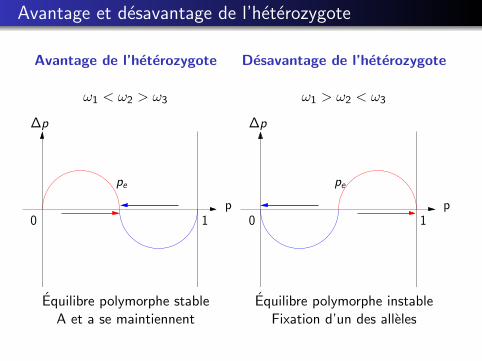
Avantage et desavantage de l’heterozygote
Avantage de l’heterozygote
ω1 < ω2 > ω3
p0
pe
∆p
1
Equilibre polymorphe stableA et a se maintiennent
Desavantage de l’heterozygote
ω1 > ω2 < ω3
p0
pe
∆p
1
Equilibre polymorphe instableFixation d’un des alleles
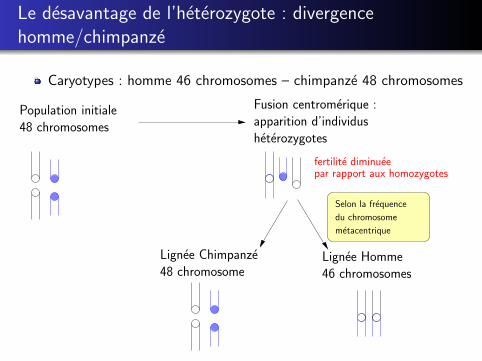
Le desavantage de l’heterozygote : divergencehomme/chimpanze
Caryotypes : homme 46 chromosomes – chimpanze 48 chromosomes
Lignee Homme46 chromosomes
Selon la frequence
du chromosome
metacentrique
Population initiale48 chromosomes
Fusion centromerique :apparition d’individusheterozygotes
Lignee Chimpanze48 chromosome
fertilite diminueepar rapport aux homozygotes
Les mutations

Mutations
Modification de la sequence d’ADN qui peut avoir ou non desconsequences fonctionnelles
Exemple d’un locus avec deux alleles A1 et A2
A1 A2
u
v
Si u=v → a terme, alleles equifrequents
Si u = 0 ou v = 0 → a terme 1 seul allele dans la population
Sinon, quelles sont les frequences alleliques a l’equilibre ?Soient pt et qt les frequences alleliques a la generation t :
pt+1 = (1− u)pt + vqt
La frequence d’equilibre pe verifie cette relationd’ou pe = (1− u)pe + v(1− pe)pe = v
u+vDe meme, on montre que qe = u
u+v

Vitesse d’evolution des frequences alleliques par mutations
Ordre de grandeur des taux de mutation : 10−5 a 10−10
Les vitesses d’evolution dues aux mutations sont tres lentes !
Quantification :Soit T le temps qu’il faut pour reduire de moitie l’ecart a la limiteOn montre que T = 0.7
u+v
Exemples :u = 3× 10−6 et v = 2× 10−6
→ T = 140000 generations, soit 2.8 millions d’annees pour l’hommeu = 3× 10−6 et v = 0→ T = 233000 generations soit 4,6 millions d’annees pour l’homme
Conclusions
Legitime de negliger les mutations sur quelques generations dansl’application du modele de Hardy-Weinberg
Equilibre rarement atteint au cours de l’evolution des especes
Lenteur du phenomeneApparition de nouveaux alleles

Les equilibres selection/mutation

Les equilibres selection/mutation
Cas d’un allele defavorable dominant
Effet de la selectionSoit A l’allele defavorable, f(A)=p et f(a)=q
Genotypes AA Aa aa Somme
Frequences p2 2pq q2
valeurs selectives 1-s 1-s 1
Frequencegen n+1
p2(1− s) 2pq(1− s) q2 1− sp2 − 2spq = D
Frequencenormalisee
p2(1− s)
D
2pq(1− s)
D
q2
D1
De meme que precedemment, on peut calculer :
p′ =p2(1− s)
D+
1
2× 2pq(1− s)
D
∆pS = p′ − p = . . . =pq × (−sq)
1− sp2 − 2spq

Les equilibres selection/mutation (2)
Effet des mutations
A au
vRappel : p′ = (1− u)p + vqD’ou ∆pM = p′ − p = v − (u + v)p
Existe-t-il un point d’equilibre selection/mutation ?
Existe-t-il pe telle que ∆pM + ∆pS = 0 ?Resolution graphique de l’equation ∆pS = −∆pM
p0 1
∆p
0 1
∆pS
−∆pMpe

Les equilibres selection/mutation (2)
Effet des mutations
A au
vRappel : p′ = (1− u)p + vqD’ou ∆pM = p′ − p = v − (u + v)p
Existe-t-il un point d’equilibre selection/mutation ?
Existe-t-il pe telle que ∆pM + ∆pS = 0 ?Resolution graphique de l’equation ∆pS = −∆pM
Conclusion
Une solution au voisinage de p = 0
Existence d’un equilibre entre l’effet de la selection et l’effet desmutations
Maintien de la diversite genetique

Les equilibres selection/mutation (3)
Relation entre pe , s, u et v
Effet de la selection :
∆pS =−p(1− p)× s(1− p)
1− sp2 − 2sp(1− p)
Au voisinage de p = 0, on peut simplifier l’expression de ∆p :on neglige les termes en p de degre ≥ 2
∆pS = [−sp + o(p)]× 1
1− 2sp + o(p)
Or, quand x → 0,1
1− x= 1 + x + o(x)
∆pS = [−sp + o(p)]× [1 + 2sp + o(p)] = −sp + o(p)
Conclusion : au voisinage de p = 0, ∆pS ∼ −sp

Les equilibres selection/mutation (4)
Effet des mutations :∆pM = p′ − p = v − (u + v)p
Conclusion : au voisinage de p = 0, ∆pM ∼ v
Equilibre mutation/selection :A l’equilibre, on a ∆pS + ∆pM = 0Au voisinage de zero : v = spe → estimation de v possible
Exemples :
Maladie frequence p valeur selective taux de mutation
de la mutation (1-s) calcule (v)
Maladie de 1100 000 0.8 0.2× 10−5
Huntington
Achondroplasie 120 000 0.2 4× 10−5
Aniridie 1160 000 0.8 1.25× 10−6
⇒ Taux de mutation calcules plausibles

Les migrations

Les migrations
Melange de populations qui tend a homogeneiser les populationsconcernees.
Etudes genetiques complexes → cas simple du modele de l’ıle
migrationsIle
Continent
m=taux de migration
Quelles sont les frequences alleliques a l’equilibre ?Soient pt et qt les frequences alleliques dans l’ıle a la generation tSoient P et Q les frequences alleliques dans le continent
pt+1 = (1−m)pt + mPLa frequence d’equilibre pe verifie cette relation, d’ou :pe = (1−m)pe + mPpe = PDe meme, on montre que qe = Q

Vitesse d’evolution des frequences alleliques par migration
Ordre de grandeur des taux de migration : 10−1 a 10−5
Quantification :Soit T le temps qu’il faut pour reduire de moitie l’ecart a la limiteOn montre que T = 0.7
m
Exemples :
Migrations Mutations
Parametre 10−1 10−4 2.10−5 2.10−7
T (generations) 7 7000 35 000 3 500 000
En annees 140 140 000 700 000 70 000 000
Conclusions
A l’equilibre memes frequences alleliques pour l’ıle et le continent
Vitesses d’evolution liees aux migrations plus rapides que celles lieesaux mutations→ Les migrations ont deja joue un grand role dans histoiregenetique de l’homme (ex : pop noire Americaine)

La derive genetique
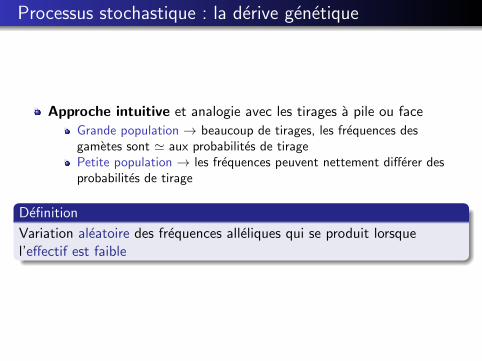
Processus stochastique : la derive genetique
Approche intuitive et analogie avec les tirages a pile ou face
Grande population → beaucoup de tirages, les frequences desgametes sont ' aux probabilites de tiragePetite population → les frequences peuvent nettement differer desprobabilites de tirage
Definition
Variation aleatoire des frequences alleliques qui se produit lorsquel’effectif est faible
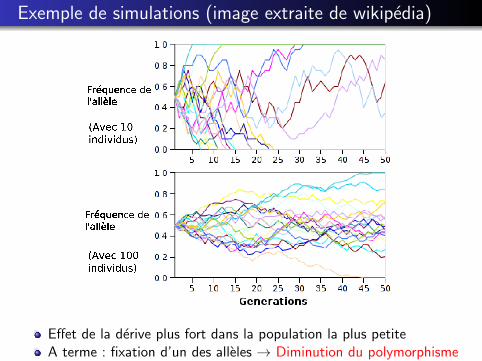
Exemple de simulations (image extraite de wikipedia)
Effet de la derive plus fort dans la population la plus petiteA terme : fixation d’un des alleles → Diminution du polymorphisme

Effet fondateur et goulot d’etranglement
Effet fondateur
Colonisation d’un nouveau milieupar un petit nombre de fondateurs
Creation de nouveaux groupesdont la composition genetique esttres differente de celle de la po-pulation d’origineExemple : Population du Sague-nay - Lac St Jean
Goulot d’etranglement =bottleneckAbaissement brutal de l’effectifd’une population
Diminution des ressourcesPredateurs, guerres, epidemies
reduction
de l’effectif
croissance
demographique
Diminution de la diversitegenetique (si baisse suffisante del’effectif !)
La destruction des individus n’estpas forcement aleatoire(resistances → favorise certainsalleles)

Conclusion

Conclusion (1) : bilan
Concept de population et mesure de sa diversite
L’equilibre de Hardy-Weinberg : theorique mais applicable
sur quelques generationsdans un espace limitepour des caracteres phenotypiques determines par 1 ou 2 genes
Les deviations a Hardy-Weinbergconsanguiniteprocessus deterministes : mutations, selection, migration
Si la nature et l’intensite de l’effet sont connues→ composition allelique finale et vitesse d’evolution sont connues
processus stochastique : derive
Phenomene aleatoire → l’etat final de la population ne peut etreprevu qu’en terme de probabilite
Dans la nature, tous ces facteurs interagissent !

Conclusion (2) : interaction de plusieurs facteurs
Interaction mutation/selectionLa selection, en general, elimine les alleles defavorablesLes mutations s’opposent au monoallelisme → maintiennent lesalleles defavorables (mais frequence faible)
Interaction selection/deriveDes alleles favorables peuvent etre perdus par deriveDes alleles defavorables peuvent etre fixes
Definition d’une ”petite population” : la derive est capable decontrecarrer l’effet de la selection
Interaction derive/mutationApparition de polymorphismes transitoires
Interaction selection/migrationmigration → nouveau contexte genetique (nouveaux alleles . . . )modification possible des valeurs selectives dans ce nouveau contexte
. . .
De plus, taux de mutation, de migration et valeurs selectives peuventvarier au cours du temps et en fonction du lieu . . .