Étude de la rÉgÉnÉration de la membrane basilaire au cours de la ... · enrichissantes et leur...
TRANSCRIPT

VÉRONIQUE FORTIN
ÉTUDE DE LA RÉGÉNÉRATION DE LA MEMBRANE BASILAIRE AU COURS DE LA
GUÉRISON DE PLAIES CUTANÉES HUMAINES
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en Biologie Cellulaire et Moléculaire pour l’obtention du grade de maître ès sciences (M.Sc.)
Biologie cellulaire et moléculaire Département de chirurgie
FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL
QUÉBEC
JUILLET 2005 © Véronique Fortin, 2005

Résumé
La membrane basilaire est très importante dans la peau humaine. Elle permet la diffusion
des nutriments des vaisseaux sanguins jusqu’à l’épiderme et elle assure une bonne jonction
entre le derme et l’épiderme. Durant la guérison, les kératinocytes de l’épiderme migrent
sur le derme et synthétisent les différents composants nécessaires à la formation d’une
membrane basilaire fonctionnelle.
L’objectif de cette étude est d’analyser la régénération de la membrane basilaire lors de la
guérison de plaies cutanées. L’emphase est aussi mis sur l’expression de la laminine 5
puisqu’elle joue un rôle particulier dans la migration des kératinocytes selon qu’elle est
sous sa forme longue ou courte.
Les résultats obtenus montrent la reconstruction progressive d’une membrane basilaire dans
une peau en cours de régénération. Ces analyses permettent de mieux comprendre les
mécanismes reliés à la guérison des plaies cutanées. La compréhension de ces notions
constitue la base pour une recherche plus approfondie.

iii
Avant-Propos
La réalisation des ces travaux n’aurait pas été possible sans la collaboration d’un nombre
important de personnes.
Je voudrais tout d’abord remercier le Dr Lucie Germain, ma directrice de recherche, pour
m’avoir accueillie dans son équipe et pour m’avoir guidée tout au long de ma maîtrise. Je
voudrais dire aussi un gros merci au Dr Véronique Moulin, ma co-directrice, pour m’avoir
aidée à démarrer mon projet et à analyser mes résultats.
Toute l’équipe du LOEX est d’une aide inestimable. Je tiens à dire un merci spécial à tous
les assistants de recherche qui travaillent fort pour la préparation du matériel commun. Sans
vous, notre tâche serait décuplée. Un gros merci aussi aux étudiants pour les discussions
enrichissantes et leur collaboration dans les cours et la réalisation des travaux.
Pour leur aide dans la réalisation directe de mes travaux, je voudrais remercier Céline
Langlois, qui a été un support technique et moral très important au tout début de ma
maîtrise, Alexandre Deschambeault, qui m’a beaucoup aidée pour la réalisation des
expériences au Time Lapse, et Hubert Robitaille, qui m’a montré comment « illuminer »
des kératinocytes à la GFP.
La vie au LOEX ne serait pas aussi agréable sans la présence de gens attachants. Je
voudrais donc remercier Hubert Robitaille, Patrick Carrier, Alexandre Deschambeault et
Jean Dubé pour les nombreux rires qu’ils provoquent quotidiennement au sein de l’équipe.
Je tiens aussi à remercier ma mère pour son soutien constant depuis le début de mes études.
Avec son amour et ses encouragements, j’ai pu garder confiance et persévérer dans mes
études.
Un dernier merci, mais non le moindre. Je tiens à remercier sincèrement Dany, mon mari,
qui vit mes joies et mes peines depuis déjà sept ans. Merci pour ta présence et tes
encouragements si précieux.

À mon mari, Dany, pour son amour, son soutien et ses nombreux encouragements. Je t’aime.
Tout ce qu’on découvre après avoir gravi une haute colline, c’est qu’il en reste beaucoup
d’autres à gravir.
Nelson Mandela

Table des matières
Résumé.................................................................................................................................. ii Avant-Propos....................................................................................................................... iii Table des matières ................................................................................................................v Liste des figures et des tableaux ....................................................................................... vii Chapitre 1 : Introduction...................................................................................................1
1.1 Peau normale humaine............................................................................................2
1.1.1 Épiderme.........................................................................................................3 1.1.1.1 Couche basale .............................................................................................4 1.1.1.2 Couche épineuse .........................................................................................5 1.1.1.3 Couche granuleuse......................................................................................5 1.1.1.4 Couche transitoire .......................................................................................5 1.1.1.5 Couche cornée.............................................................................................6
1.1.2 Derme..............................................................................................................6 1.1.3 Hypoderme......................................................................................................7 1.1.4 Membrane basilaire.........................................................................................8
1.1.4.1 Structure générale .......................................................................................8 1.1.4.1.1 Laminines..............................................................................................12 1.1.4.1.2 Collagènes.............................................................................................15 1.1.4.1.3 Nidogène...............................................................................................17 1.1.4.1.4 Perlécan.................................................................................................18 1.1.4.1.5 Fibronectine ..........................................................................................18 1.1.4.1.6 Intégrines ..............................................................................................19 1.1.4.2 Organisation et assemblage ......................................................................21
1.2 Guérison de plaies cutanées..................................................................................23 1.2.1 Phase inflammatoire .....................................................................................25 1.2.2 Phase de migration et de prolifération ..........................................................27 1.2.3 Phase de remodelage.....................................................................................31
1.3 Approche et objectifs des travaux de recherche ...................................................32 Chapitre 2 : Matériel et méthodes ..................................................................................35
2.1 Isolement et culture cellulaire...............................................................................36 2.2 Modèle de cicatrisation cutanée............................................................................37 2.3 Histologie..............................................................................................................38 2.4 Marquage en immunofluorescence.......................................................................38 2.5 Obtention du plasma et du sérum humain ............................................................40 2.6 Tests de migration cellulaire.................................................................................40

vi
Chapitre 3 : Résultats.......................................................................................................42 3.1 Apparence macroscopique des peaux reconstruites..............................................43 3.2 Expression des constituants de la membrane basilaire .........................................44 3.3 Régénération de la membrane basilaire ................................................................48 3.4 Migration des kératinocytes d’une plaie superficielle ..........................................55
Chapitre 4 : Discussion ....................................................................................................58
4.1 Comparaison entre la peau normale humaine et la peau reconstruite...................59 4.2 Visualisation de la reconstruction de la membrane basilaire................................61 4.3 Observation de la migration des kératinocytes en monocouche...........................66
Chapitre 5 : Conclusion et perspectives .........................................................................69 Bibliographie .......................................................................................................................72

Liste des figures et des tableaux
Figure 1 : La peau normale humaine .....................................................................................2 Figure 2 : La différenciation de l’épiderme ...........................................................................3 Figure 3 : La membrane basilaire ........................................................................................10 Figure 4 : L’hémidesmosome ..............................................................................................11 Figure 5 : Les laminines.......................................................................................................13 Figure 6 : Les clivages de la laminine 5 ..............................................................................15 Figure 7 : La structure d’une fibre de collagène ..................................................................16 Figure 8 : Le nidogène .........................................................................................................17 Figure 9 : Le perlécan ..........................................................................................................18 Figure 10 : La fibronectine ..................................................................................................19 Figure 11 : L’intégrine .........................................................................................................20 Figure 12 : Les différentes intégrines ..................................................................................21 Figure 13 : L’assemblage de la membrane basilaire............................................................23 Figure 14 : La guérison d’une plaie superficielle ................................................................24 Figure 15 : La guérison d’une plaie profonde......................................................................25 Figure 16 : Les facteurs intervenant dans l’inflammation ...................................................26 Figure 17 : Les étapes du déplacement cellulaire ................................................................30 Figure 18 : La production de peau reconstruite ...................................................................37 Figure 19 : L’induction d’une plaie dans un tissu reconstruit .............................................38 Figure 20 : Le principe de l’immunofluorescence...............................................................40 Figure 21 : L’induction de la plaie en monocouche ............................................................41 Figure 22 : La peau reconstruite ..........................................................................................44 Figure 23 : Comparaison entre la peau reconstruite et la peau normale humaine ...............45 Figure 24 : Expression de la laminine 5 et des intégrines associées....................................49 Figure 25 : Expression de l’intégrine α2β1, du collagène VII et de la fibronectine.............52 Figure 26 : Expression du nidogène, du perlécan et du collagène IV..................................53 Figure 27 : Migration cellulaire ...........................................................................................56 Tableau 1 : Liste des anticorps utilisés ................................................................................39 Tableau 2 : Résumé des marquages en immunofluorescence .............................................54

CHAPITRE 1 : INTRODUCTION

2
1.1 Peau normale humaine La peau est un organe complexe recouvrant le corps en entier. Son poids totalise environ
15 % du poids total du corps adulte, ce qui lui vaut le titre du plus grand et du plus
important organe du corps humain. Elle assure plusieurs fonctions nécessaires à la survie de
l’organisme comme la protection contre les agressions physiques, chimiques et biologiques
extérieures (Kanitakis 2002; Wysocki 1999). Elle a aussi un rôle dans la régulation
thermique, l’excrétion, l’immunité, la synthèse de la vitamine D et elle constitue un
excellent capteur d’informations extérieures grâce aux milliers de terminaisons nerveuses
qu’elle contient. De plus, les nombreux vaisseaux sanguins qui traversent le derme
transportent de 8 à 10 % du sang en circulation dans le corps, ce qui fait de la peau un
important réservoir sanguin (Tortora et al. 1994). Le développement adéquat de la peau est
nécessaire même au stade embryonnaire puisque des mutations bloquant la spécification, la
stratification ou la différenciation de l’épiderme peuvent avoir des conséquences graves sur
la survie ou la santé du fœtus. Plusieurs épidermolyses bulleuses sont d’ailleurs associées à
de telles mutations (Loomis 2001). Il est donc évident que la peau représente un facteur
important pour la santé et même la survie d’un organisme.
Figure 1 : La peau normale humaine
Source : (Geras 1990)

3
La peau normale humaine (figure 1) est constituée de trois couches principales dont les
cellules interagissent ensemble afin d’assurer les différentes fonctions de la peau. La partie
la plus superficielle, l’épiderme, est mince et composée d’un épithélium pavimenteux
stratifié et kératinisé. Elle est attachée à une couche interne plus épaisse, formée de tissu
conjonctif, le derme. La dernière couche, l’hypoderme, est la couche sous-cutanée,
composée de tissus conjonctifs auréolaires et adipeux. Les fibres du derme s’y rattachent et
fixent ainsi la peau puisque l’hypoderme est lui-même fermement attaché aux tissus et
organes sous-jacents (Gartner et al. 1992; Tortora et al. 1994).
1.1.1 Épiderme L’épiderme (figure 2) est la couche en contact direct avec l’environnement extérieur et elle
protège l’organisme contre celui-ci. Elle est en fait la couche protectrice de la peau qui
empêche les agents pathogènes d’envahir l’organisme et qui maintient l’eau et les
nutriments à l’intérieur. Elle a une épaisseur moyenne de 100 µm, mais celle-ci peut varier
considérablement selon la région du corps. Par exemple, l’épiderme des paupières n’a que
50 µm d’épaisseur tandis que l’épiderme de la plante des pieds peut avoisiner le
millimètre (Kanitakis 2002).
Figure 2 : La différenciation de l’épiderme
Source : Alain Laplante, LOEX

4
Les cellules qui composent l’épiderme forment un épithélium pavimenteux stratifié et sont
à 95 % des kératinocytes. Ils produisent de la kératine, qui imperméabilise et protège la
peau et les tissus sous-jacents (Gartner et al. 1992; Tortora et al. 1994). Le 5 % restant est
composé de mélanocytes, de cellules de Langerhans et de cellules de Merkel. Les
mélanocytes donnent la pigmentation à la peau en produisant de la mélanine synthétisée
dans leurs organites spécialisés, les mélanosomes. Ces cellules sont réparties à travers les
kératinocytes de la couche basale, mais elles se retrouvent aussi dans les follicules pileux et
dans le derme. Les cellules de Langherans jouent un rôle important dans l’immunité. Elles
sont des cellules lymphatiques dérivant de la moelle osseuse qui modifieraient les antigènes
et les transporteraient aux ganglions lymphatiques. Les cellules de Merkel se trouvent aussi
dans la couche basale. Elles sont associées à des terminaisons nerveuses afférentes et
agissent comme mécanorécepteurs (Gartner et al. 1992; Kanitakis 1998; Kanitakis 2002;
Wysocki 1999). Ces cellules sont organisées en plusieurs couches ayant chacune un rôle
dans le maintien des fonctions de la peau.
1.1.1.1 Couche basale Cette couche est composée de cellules en prolifération constante. Elle contient les cellules
souches épidermiques qui subissent des divisions mitotiques afin de produire des cellules
filles qui assureront le renouvellement de la peau en se différenciant et en migrant vers la
surface pour aller peupler les couches plus superficielles (Tortora et al. 1994; Wysocki
1999). Ce processus de différenciation s’étale sur une durée de 28 jours et est stimulé par
des facteurs de croissance comme l’EGF (epidermal growth factor) et le TGF-α
(transforming growth factor alpha) et des hormones comme l’œstrogène, la progestérone et
l’épinéphrine (Wysocki 1999). Les cellules de la couche basale ont une forme cuboïdale et
un arrangement très serré. Elles sont attachées au derme sous-jacent par des
hémidesmosomes, des structures d’attachement spéciales (Borradori et al. 1999; Kanitakis
2002). La jonction dermo-épidermique se situe sous cette couche et forme des creux et des
pics appelés crêtes épidermiques qui ont comme rôle principal d’améliorer l’attachement de
l’épiderme au derme pour éviter un décollement de l’épiderme lors de frottements

5
(Wysocki 1999). Les cellules de la couche basale vont être responsables de la production
d’une grande partie des molécules formant la membrane basilaire.
1.1.1.2 Couche épineuse Cette couche est formée de cellules présentant de nombreux desmosomes tout le long de la
bordure cellulaire qui servent à lier les cellules entre elles. Cette caractéristique donne lieu
à une apparence épineuse, d’où le nom de cette couche. Les cellules qui forment cette
couche sont plus grosses que celles de la couche basale, sont polyédriques et s’agencent en
huit à dix rangées. Ces cellules synthétisent de l’involucrine, une protéine soluble
précurseur du composé formant l’enveloppe cornifiée. La loricrine, un autre composant de
l’enveloppe cornifiée, est aussi produite par les cellules de cette couche (Wysocki 1999).
1.1.1.3 Couche granuleuse Cette couche est composée de trois à cinq rangées de cellules plus aplaties et étirées,
s’alignant parallèlement à la surface de la peau. Elles contiennent dans leur cytoplasme des
granules de kératohyaline (Tortora et al. 1994). Ces granules contiennent de la
profilaggrine qui est clivée pour devenir de la filaggrine. Cette dernière est une protéine
hautement phosphorylée qui favorise l’agrégation des filaments de kératine (Wysocki
1999). De plus, les noyaux des cellules de la couche granuleuse sont en dégénérescence,
mais toujours actifs. Cette couche constitue la dernière couche nucléée de la peau.
1.1.1.4 Couche transitoire Cette couche constitue une transition vers la couche cornée et n’est présente que dans la
peau épaisse, comme celle de la plante des pieds et de la paume des mains. Elle est
constituée de trois à cinq rangées de cellules mortes, aplaties et transparentes (Tortora et al.
1994). Les cellules de la couche transitoire ne contiennent qu’un noyau inactif en
dégradation, mais elles présentent une forte activité enzymatique où des enzymes
lysosomales, comme la Dnase et la Rnase, dégradent le noyau et les organelles. Ces cellules

6
relâchent aussi une petite quantité de matériels lipidiques qui forment une pellicule
enveloppante les protégeant contre la dégradation (Wysocki 1999).
1.1.1.5 Couche cornée Cette couche est composée de 25 à 30 rangées de cornéocytes, des cellules mortes de forme
hexagonale très aplaties et qui n’ont plus de noyau ni d’organelle. Elles sont faites d’une
matrice dense de kératines filamenteuses. Les cellules de cette couche desquament
graduellement durant le processus de différenciation et sont remplacées par des cellules de
la couche inférieure. La couche cornée est hautement résistante à la digestion enzymatique
et aux changements de température et de pH (Wysocki 1999). Elle contribue ainsi à la
fonction de barrière de la peau, la protégeant contre la lumière, la chaleur, les bactéries et
plusieurs produits chimiques.
1.1.2 Derme Le derme est la couche qui donne la souplesse et la résistance à la peau. Il est composé
principalement de tissu conjonctif qui le rend compressible et élastique. Il constitue un
support pour les différentes annexes cutanées : les vaisseaux sanguins et lymphatiques, les
poils, les terminaisons nerveuses et les glandes sébacées et sudoripares. Ces annexes sont
en fait entourées de fibres majoritairement constituées de collagènes de types I et III,
arrangées en filet pour assurer souplesse et solidité. La matrice est aussi constituée de fibres
élastiques, formées par l’élastine, une protéine insoluble. Plusieurs cellules peuplent cette
matrice. Les fibroblastes sont majoritaires et synthétisent tous les types de fibres et
quelques constituants de la membrane basilaire. Les fibrocytes, les fibroclastes et les
myofibroblastes sont des dérivés des fibroblastes ayant des fonctions plus spécifiques qui
se trouvent aussi dans le derme. D’autres types cellulaires peuvent aussi être présents en
moindre quantité : des dendrocytes, des mastocytes, des macrophages et des lymphocytes
(Kanitakis 1998; Kanitakis 2002).
L’épaisseur du derme varie considérablement selon la localisation anatomique. Par
exemple, il est beaucoup plus épais dans la plante des pieds que dans les paupières. En

7
moyenne, il a une épaisseur de deux millimètres, mais peut atteindre quatre millimètres
(Wysocki 1999). Le derme peut être divisé en deux parties distinctes : le derme papillaire
(superficiel) et le derme réticulaire (profond).
Le derme papillaire constitue le cinquième de l’épaisseur totale du derme. Il forme des
crêtes, ou projections, qui alternent avec d’autres projections formées par l’épiderme. Ces
papilles dermiques augmentent beaucoup la surface de contact entre le derme et l’épiderme,
ce qui permet une meilleure adhésion entre ces deux couches. Il contient plusieurs cellules,
des vaisseaux sanguins et des terminaisons nerveuses sensibles au toucher appelées
corpuscules tactiles de Meissner (Kanitakis 2002; Tortora et al. 1994). Il est composé de
tissu conjonctif lâche et de minces fibres élastiques.
Le derme réticulaire est la partie la plus profonde du derme. Il est fait de tissu conjonctif
dense et irrégulier composé de faisceaux de fibres de collagène I et III et de grosses fibres
élastiques. C’est cette organisation de fibres en filet qui donne force, extensibilité et
élasticité à la peau. Les espaces entre les fibres sont occupés par des follicules pileux, des
nerfs, des glandes sébacées et des canaux de glandes sudoripares (Tortora et al. 1994).
1.1.3 Hypoderme L’hypoderme est la couche la plus profonde de la peau. Il est constitué de tissus adipeux
qui ont pour fonction d’emmagasiner l’énergie sous forme de graisse. En fait, environ la
moitié de tout le gras emmagasiné dans le corps humain est retrouvée dans cette couche.
L’hypoderme joue aussi un rôle de thermorégulation et de protection contre les agressions
mécaniques. Les cellules qui peuplent l’hypoderme s’appellent les adipocytes, des cellules
rondes remplies d’acides gras et de triglycérides comprimant le noyau contre la membrane
plasmique (Kanitakis 2002). Les adipocytes sont organisés en lobules primaires et
secondaires et leur morphologie varie selon la région du corps et la race de la personne.
Cette couche contient aussi des fibroblastes et des macrophages, en plus de vaisseaux
sanguins et des terminaisons nerveuses. Ces dernières sont aussi appelées corpuscules de
Pacini ou corpuscules lamellaires et sont sensibles à la pression (Wysocki 1999).

8
1.1.4 Membrane basilaire L’épiderme et le derme sont séparés physiquement par une structure en feuillet faite de
matrice extracellulaire appelée membrane basilaire. Elle joue plusieurs rôles essentiels au
bon fonctionnement de la peau (Fleischmajer et al. 1998; Kanitakis 2002; Uitto et al.
1996). Cette structure est en effet très importante pour le maintien de la cohésion dermo-
épidermique puisqu’elle assure la jonction solide entre les deux couches principales de la
peau. Elle offre une matrice d’ancrage pour les cellules de la couche basale de l’épiderme,
ce qui lui confère aussi un rôle dans la polarité de l’épiderme et dans la régulation de la
migration cellulaire (Nishiyama et al. 2000). De plus, la membrane basilaire permet la
diffusion contrôlée des nutriments provenant des vaisseaux sanguins présents dans le derme
jusqu’aux kératinocytes de l’épiderme. La membrane basilaire joue aussi un rôle important
dans la compartimentation cellulaire, contrôlant ainsi l’invasion cellulaire. Cette dernière
fonction de barrière physique est d’ailleurs compromise dans certains types de cancers
malins (Lodish et al. 1997). Durant le processus inflammatoire, la membrane basilaire
permet aux lymphocytes et autres intervenants du système immunitaire de migrer des
vaisseaux sanguins jusqu’au site lésé (Kanitakis 2002). La membrane basilaire est aussi
impliquée dans l’apport d’informations extérieures puisqu’elle contient certains récepteurs
(Fleischmajer et al. 1998).
Les quelques maladies associées à une membrane basilaire dysfonctionelle démontrent bien
l’importance des fonctions de cette structure. L’épidermolyse bulleuse, les pemphigoïdes et
les dermatites herpétiformes sont quelques exemples de maladies dans lesquelles la
structure de la membrane basilaire est altérée, ce qui résulte en une séparation du derme et
de l’épiderme menant à la formation de bulles sous l’épiderme (Moll et al. 1998; Stevens et
al. 1997; Uitto et al. 1996).
1.1.4.1 Structure générale La membrane basilaire est composée de plusieurs composants qui ont chacun leur rôle à
jouer. L’architecture exacte de la jonction dermo-épidermique a longtemps été un mystère,
mais elle est aujourd’hui mieux connue sans être toutefois à cent pour cent élucidée. Les
progrès ont été relativement lents en raison de la rareté et de l’insolubilité du matériel

9
biologique accessible pour des analyses biochimiques détaillées (Uitto et al. 1996).
L’utilisation du microscope électronique et les études par clonage de gènes ont
radicalement changé la perception qu’avaient les scientifiques de la membrane basilaire
(Eady 1988; Uitto et al. 1996). Elle est maintenant vue comme un ensemble de matériel
complexe dont chaque composant pris individuellement est bien trop petit pour être observé
en microscopie optique (Eady 1988). Le nombre total de composants n’est pas connu
précisément, mais le nombre estimé est d’environ 50. La proportion de ceux qui sont très
bien caractérisés est encore faible, mais elle inclut la plupart des constituants majeurs
(Timpl et al. 1986).
La structure générale de la membrane basilaire est formée de trois couches principales qui
se distinguent très bien au microscope électronique (figure 3). La lamina lucida est la
couche en contact direct avec la membrane plasmique des cellules de la couche basale de
l’épiderme. D’une épaisseur d’environ 60 nm, elle est claire au microscope, comme son
nom le laisse supposer (Stevens et al. 1997). Il est toutefois possible de distinguer des
filaments d’ancrage très fins qui traversent la lamina lucida jusqu’à la membrane
plasmique. C’est, entre autres, grâce à ces filaments que les cellules de la couche basale de
l’épiderme peuvent s’attacher solidement à la matrice. L’apparence vide de cette couche est
trompeuse car elle contient des laminines et d’autres macromolécules moins connues (Eady
1988).
La lamina densa est la deuxième couche de la membrane basilaire. Elle est parallèle à la
lamina lucida et est beaucoup plus dense en électrons, ce qui lui donne l’apparence d’une
ligne d’une épaisseur qui varie entre 50 et 70 nm (Kanitakis 2002). Elle contient en
majorité du collagène de type IV qui confère à la membrane basilaire sa force naturelle,
mais aussi de la laminine 5, du perlécan et du nidogène (Eady 1988; Kanitakis 1998).
La troisième couche de la membrane basilaire est la sub-lamina densa, qui est en contact et
en continuité avec le derme. Elle constitue une zone fibreuse qui contient une grande
variété de composants dont les fibres élastiques, qui sont formées d’élastine et d’oxytalane.
Des fines fibres interstitielles de collagène peuvent aussi être retrouvées dans cette région.
Elles sont faites principalement de collagène de type III, aussi appelé réticuline (Eady
1988). Le composant majeur de la sub-lamina densa est sans contredit les fibres d’ancrage,

10
qui sont formées de collagène de type VII. Elles forment des structures solides ayant pour
rôle fonctionnel de relier la lamina densa au derme papillaire où elles s’accrochent aux
fibres de collagène dermiques faites de collagène de type I, III et V (Uitto et al. 1996).
Figure 3 : La membrane basilaire
Source : sprojects.mmi.mcgill.ca/dermatology/epidermis2.jpg Une autre structure très importante dans l’organisation de la membrane basilaire est
l’hémidesmosome (figure 4). Quoiqu’il ne soit pas considéré comme un composant
structurel de la membrane basilaire, il y est fermement associé et est nécessaire à son
intégrité. Il est un complexe jonctionnel spécialisé qui relie les cellules basales à la lamina
densa par des protéines d’ancrage qui traversent la lamina lucida (Moll et al. 1998; Stevens
et al. 1997). Il assure une bonne interaction cellules-matrice en fournissant aux cellules des
signaux critiques pour leur polarisation, leur organisation spatiale et pour l’organisation
architecturale des tissus (Borradori et al. 1999). L’hémidesmosome est formé de quatre
constituants majeurs qui, une fois associés, forment à leur tour des structures visibles au
microscope électronique. Ces constituants sont la plectine, le BP230 (Bullous pemphigoid
antigen 230), le BP180 (Bullous pemphigoid antigen 180) et l’intégrine α6β4 (Moll et al.
1998; Uitto et al. 1996). L’organisation moléculaire des hémidesmosomes est basée sur
trois classes de protéines : les protéines des plaques cytoplasmiques, les protéines trans-
membranaires et les protéines de la matrice extracellulaire associées à la membrane
basilaire (Borradori et al. 1999).
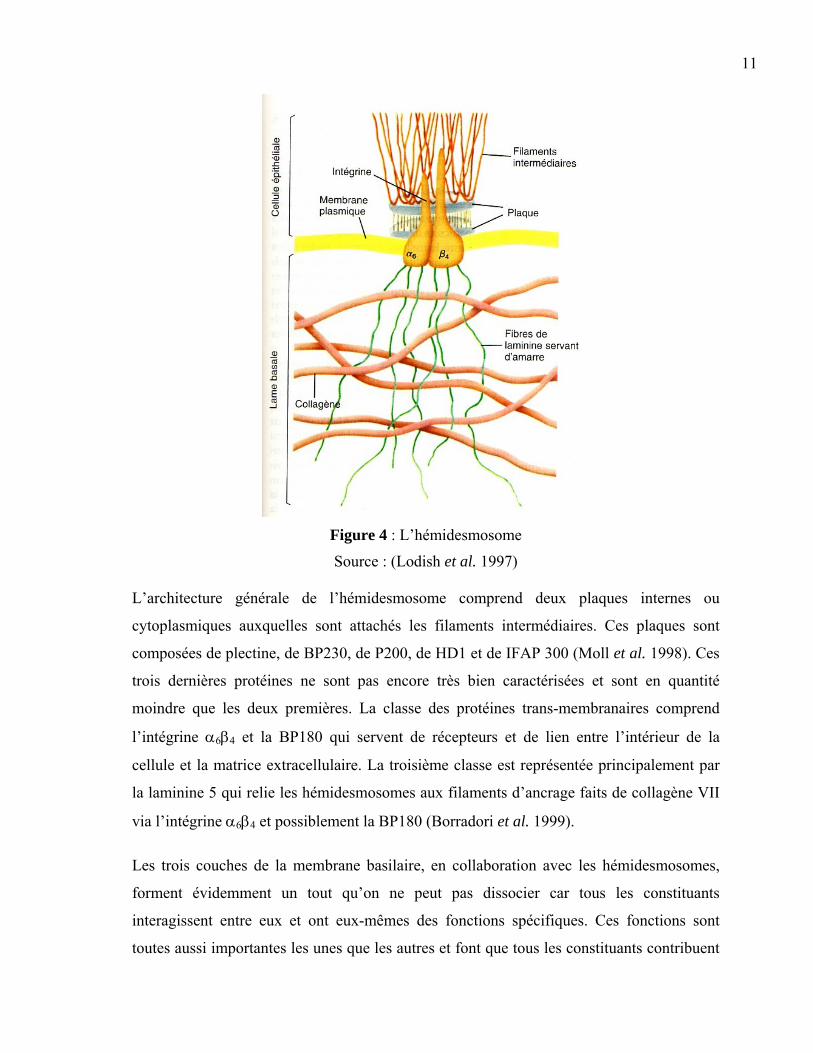
11
Figure 4 : L’hémidesmosome
Source : (Lodish et al. 1997) L’architecture générale de l’hémidesmosome comprend deux plaques internes ou
cytoplasmiques auxquelles sont attachés les filaments intermédiaires. Ces plaques sont
composées de plectine, de BP230, de P200, de HD1 et de IFAP 300 (Moll et al. 1998). Ces
trois dernières protéines ne sont pas encore très bien caractérisées et sont en quantité
moindre que les deux premières. La classe des protéines trans-membranaires comprend
l’intégrine α6β4 et la BP180 qui servent de récepteurs et de lien entre l’intérieur de la
cellule et la matrice extracellulaire. La troisième classe est représentée principalement par
la laminine 5 qui relie les hémidesmosomes aux filaments d’ancrage faits de collagène VII
via l’intégrine α6β4 et possiblement la BP180 (Borradori et al. 1999).
Les trois couches de la membrane basilaire, en collaboration avec les hémidesmosomes,
forment évidemment un tout qu’on ne peut pas dissocier car tous les constituants
interagissent entre eux et ont eux-mêmes des fonctions spécifiques. Ces fonctions sont
toutes aussi importantes les unes que les autres et font que tous les constituants contribuent

12
au maintien de l’intégrité structurale et fonctionnelle de la membrane basilaire. Les
constituants principaux sont la laminine 5, le collagène IV, le nidogène et le perlécan
(Fleischmajer et al. 1998; Mohan et al. 1986; Stevens et al. 1997). D’autres types de
collagène et d’autres isoformes de laminine font aussi partie de la structure de la membrane
basilaire. De plus, des molécules importantes pour sa formation comme des intégrines et la
fibronectine y sont aussi associées. Chacun de ces composants a ses particularités
fonctionnelles et structurales qui sont importantes à préciser.
1.1.4.1.1 Laminines Les laminines sont des molécules en forme de croix faisant partie d’une famille de
glycoprotéines constituées par l’assemblage de trois différentes chaînes polypeptidiques,
les chaînes α, β et γ. Il existe cinq différentes chaînes α, deux chaînes β et trois chaînes γ
dont l’assemblage forme jusqu’à 15 isoformes différentes simplement appelées lamimine 1
à laminine 15 (figure 5) (Ekblom et al. 2003). Toutes les chaînes de laminine ont une
région d’environ 600 résidus formée de répétitions de peptides heptad qui se torsadent pour
former les domaines I et II de la tige principale des différentes isoformes de laminine.
Chaque tige principale est donc formée d’une triple-hélice comprenant une chaîne α, une
chaîne β et une chaîne γ. À l’extrémité C-terminale de la tige principale, une séquence de 5
domaines globulaires est présente en continuité avec la chaîne α. Ces domaines globulaires
sont appelés LG1 à LG5 et sont constitués de 180 à 200 résidus. L’extrémité N-terminale
est beaucoup plus variable d’une isoforme à l’autre puisqu’elle contient trois motifs
différents, le LE, le L4 et le LN. Les chaînes de laminine varient d’ailleurs par le nombre et
l’emplacement de ces trois motifs (Aumailley et al. 1999). L’expression des différentes
chaînes de laminine dépend du type de tissu et du stade développemental. Par exemple, la
laminine 1 est détectable dans les embryons de souris à un stade très précoce de 16 cellules.
Par leur interaction avec des intégrines et d’autres molécules de surface, les laminines
contrôlent les activités cellulaires comme la migration, la différenciation, la polarité, la
prolifération, l’apoptose ou l’expression de gènes (Aumailley et al. 1999; Ryan et al. 1996).

13
Figure 5 : Les laminines
Source : (Ekblom et al. 2003)
Dans la peau normale humaine, les laminines présentes sont les laminines 5, 6 et 10, mais
l’isoforme 5 est de loin la plus abondante (Aumailley et al. 1999). La laminine 1 serait
aussi présente dans la peau humaine, mais il existe une controverse à ce sujet : certains
prétendent qu’elle n’est présente seulement qu’au stade embryonnaire où elle est nécessaire
au développement de l’épiblaste (Ekblom et al. 2003; Virtanen et al. 2000) tandis que
d’autres la détectent par immunofluorescence dans des cultures de peau humaine
(Fleischmajer et al. 1998; Fleischmajer et al. 1998; Smola et al. 1998; Sollberg et al. 1992).
Toutes les isoformes de laminine de la peau ont un rôle structural important pour la
membrane basilaire, mais la laminine 5 est particulière : l’absence de cette isoforme ou
l’altération de son expression cause une perte de la cohésion dermo-épidermique
importante (Aumailley et al. 1996; Christiano et al. 1996). La laminine 5, précédemment
appelée nicéine, épiligrine et kalinine, est formée des chaînes α3β3γ2 et est en fait
considérée comme le composant clé du complexe d’ancrage de l’épiderme et comme étant
la protéine contribuant le plus à la stabilité de la membrane basilaire. La grande importance
de cette protéine s’explique par le fait qu’elle forme les structures appelées filaments

14
d’ancrage (Rousselle et al. 1991) qui s’attachent aux hémidesmosomes via l’intégrine α6β4
et aux fibres d’ancrage constituées de collagène VII en passant au travers de la lamina
lucida et de la lamina densa (Nishiyama et al. 2000). De plus, une étude récente a démontré
que la laminine 5 est orientée à un angle de 27° par rapport à la jonction dermo-
épidermique contrairement à une orientation perpendiculaire (McMillan et al. 2003). Une
autre particularité attribuable à la laminine 5 est qu’elle subit des clivages extracellulaires
qui ont un impact sur ses fonctions (figure 6). C’est en fait la forme courte de la laminine 5
qui est présente dans la membrane basilaire de la peau normale humaine et qui permet
l’ancrage des cellules épithéliales. La forme non-clivée est sécrétée seulement par les
kératinocytes des épithéliums en formation ou lorsque la membrane basilaire doit être
réparée (Marinkovich et al. 1992; Rousselle et al. 1991).
La laminine 5 est synthétisée sous forme de précurseur d’une longueur de 460 kDa qui
permet la migration des kératinocytes qui la produisent. Dans les conditions
physiologiques, ce sont les sous-unités α3 et γ2 qui subissent des clivages post-
traductionnels et extracellulaires (Rousselle 2002). Ces clivages ont pour conséquence de
réduire la chaîne α3 une première fois de 200 kDa à 165 kDa à son extrémité carboxy-
terminale entre le domaine LG3 et LG4, une deuxième fois de 165 kDa à 145 kDa à son
extrémité amino-terminale dans le domaine IIIA et la chaîne γ2 de 155 kDa à 105 kDa à son
extrémité amino-terminale dans le domaine III (Amano et al. 2000; Marinkovich et al.
1992; Rousselle 2002). Les molécules possiblement responsables de ces clivages sont
nombreuses : la plasmine (Goldfinger et al. 1998), la BMP-1 (Bone morphogenetic protein-
1) (Amano et al. 2000), la mTLD (Métalloprotéinase Tolloïde mammaire) (Veitch et al.
2003) et une enzyme de la famille des MMP (métalloprotéases matricielles) (Tsubota et al.
2000) ont été les candidates principales pour le clivage de la chaîne α3, tandis que deux
candidates, la BMP-1 (Amano et al. 2000; Sasaki et al. 2001), et la mTLD (Veitch et al.
2003) ont été identifiées comme étant les responsables de la coupure de la chaîne γ2. Les
coupures que subit la laminine 5 surviennent presque immédiatement après sa sécrétion, ce
qui donne naissance à sa forme courte qui est alors compétente pour l’assemblage des
hémidesmosomes et donc pour l’attachement solide des cellules basales à la matrice
(Decline et al. 2000; Goldfinger et al. 1998). La nouvelle compétence de la forme courte

15
pour l’attachement s’explique par le fait que la coupure de l’extrémité carboxy-terminale de
la chaîne α3 laisse à nu le domaine globulaire LG3 sur lequel sont situés les sites
d’adhésion pour les intégrines α3β1 et α6β4 (Hirosaki et al. 2000). De la même façon, la
coupure de l’extrémité amino-terminale de la chaîne α3 pourrait améliorer l’attachement de
la forme courte à la laminine 6 et au collagène VII, mais rien n’a encore été prouvé de ce
côté (Tunggal et al. 2002). Pour ce qui est de la fonction de la coupure de la chaîne γ2, une
étude récente a démontré que la forme longue de cette chaîne était nécessaire pour
l’intégration de la laminine 5 dans la matrice extracellulaire, mais aucune fonction n’a été
attribuée à la forme courte à ce jour (Gagnoux-Palacios et al. 2001). Quoi qu’il en soit, la
laminine 5 est sans contredit un composant majeur et déterminant dans la membrane
basilaire qui, selon sa forme, assure un double rôle fascinant à étudier.
Figure 6 : Les clivages de la laminine 5
Source : Véronique Fortin, LOEX
1.1.4.1.2 Collagènes Les collagènes sont des molécules faisant partie d’une famille comptant au moins 20
différents types. Ce sont des protéines fibreuses insolubles retrouvées dans la matrice
extracellulaire et les tissus conjonctifs. Les plus abondants chez les animaux sont les types
I, II et III qui forment des longues fibrilles. L’ultrastructure des collagènes est faite d’un
module protéique en hélice triple (figure 7). Les caractéristiques structurales permettant de

16
différencier les types de collagènes sont dues principalement aux segments qui
interrompent la triple hélice pour se replier en d’autres structures tridimensionnelles. La
structure propre à chaque type de collagène lui assure une fonction particulière. Les
collagènes sont non seulement produits par les cellules des tissus conjonctifs comme les
fibroblastes, mais aussi par plusieurs types de cellules épithéliales (Lodish et al. 1997).
Dans la membrane basilaire, les deux types de collagènes principaux sont les collagène IV
et VII (Timpl et al. 1996).
Figure 7 : La structure d’une fibre de collagène
Source : www2.mcdaniel.edu/chemistry/ch332/jpgs/proteins/collagen.jpg Le collagène IV constitue la trame laminaire de la membrane basilaire. Il est le composant
le plus répandu et il est responsable de la stabilité mécanique. Trois chaînes de collagènes
IV se regroupent et forment une hélice tricaténaire longue de 400 nm dont le bout
carboxyle est replié en un grand domaine globulaire. Ces chaînes se disposent ensuite en un
réseau bi-dimensionnel formant ainsi un treillis qui assure solidité et flexibilité à la
membrane (Lodish et al. 1997; Timpl et al. 1996).
Le collagène VII, qui est synthétisé principalement par les kératinocytes, est le constituant
des fibres d’ancrage de la sub-lamina densa. À son extrémité N-terminale, chaque triple

17
hélice de collagène VII est flanquée d’un domaine globulaire NC1. Deux de ces chaînes
s’associent par leur bout C-terminal pour former un dimère. Ces dimères sont des courtes
fibrilles striées qui s’agencent latéralement pour former les fibres d’ancrage (Moll et al.
1998; Stevens et al. 1997). Le collagène VII interagit avec les autres constituants de la
matrice extracellulaires via son domaine NC1. Il s’attache ainsi à la laminine 5 par laquelle
il ancre la membrane basilaire au derme où il se lie aux autres types de collagènes (I et III)
(Moll et al. 1998; Osawa et al. 2003).
1.1.4.1.3 Nidogène Le nidogène est une molécule de forme allongée de 30 nm qui contient trois domaines
globulaires (G1 à G3) reliés par un lien flexible entre les domaines G1 et G2 et par un lien
plus rigide entre les domaines G2 et G3 (figure 8). Il est un composant de la membrane
basilaire situé dans la lamina densa (Timpl et al. 1986). Il a pour fonction de contrôler la
formation d’un complexe stable entre le collagène IV, les laminines et le perlécan (Kohfeldt
et al. 1998; Timpl et al. 1996). Ce sont surtout les épitopes localisés sur les domaines G2 et
G3 qui sont impliqués dans cette liaison (Kohfeldt et al. 1998). Le nidogène pourrait aussi
se lier à l’intégrine α3β1 par le biais du domaine G2 (Dedhar et al. 1992; Gresham et al.
1996). Le nidogène joue un rôle essentiel dans la membrane basilaire : en plus d’être
nécessaire à sa formation, il a un rôle de stabilisation et assure le maintien de son
architecture (Fleischmajer et al. 1998; Timpl et al. 1996). Des essais en laboratoire ont
d’ailleurs démontré que l’inhibition de l’attachement du nidogène aux laminines par
l’utilisation d’anticorps empêchait la formation de la membrane basilaire dans plusieurs
cultures d’organes embryonnaires (Ekblom et al. 1994).
Figure 8 : Le nidogène

18
Source : (Lodish et al. 1997)
1.1.4.1.4 Perlécan Le perlécan est le plus abondant des nombreux héparane sulfate protéoglycans présents
dans la membrane basilaire. Il est constitué d’un noyau protéique de 80 nm de long dans
lequel s’alternent des domaines globulaires et des tiges courtes (figure 9). Trois chaînes
héparane sulfate ou chondroitine sulfate se trouvent à son extrémité N-terminale (Timpl et
al. 1996). Le perlécan peut se lier au nidogène, aux laminines, au collagène IV et à la
fibronectine (Groffen et al. 1999). Ces interactions l’ancrent à la structure de la membrane
basilaire, ce qui augmente sa résistance. Il forme aussi des régions anioniques essentielles
pour la propriété filtrante de la membrane basilaire (Groffen et al. 1999) et agit en tant que
barrière (Timpl et al. 1986) grâce à ses propriétés liantes qui lui permettent de s’accrocher à
plusieurs molécules physiologiquement importantes comme des facteurs de croissance, des
agents infectieux, des composants de la matrice extracellulaire et des anticoagulants (Fuki
et al. 2000).
Figure 9 : Le perlécan
Source : http://www.glycoforum.gr.jp/science/word/proteoglycan/pga09.gif
1.1.4.1.5 Fibronectine La fibronectine est une glycoprotéine de 240 kDa ne faisant pas partie intégrante de la
structure de la membrane basilaire, mais elle aide à sa formation. Elle consiste en deux
chaînes polypeptidiques similaires connectées une à l’autre par des ponts disulfure à leur
extrémité C-terminale. Elle adopte une forme en « V » dont chaque côté atteint 63 nm de
long (figure 10). D’autres formes plus compactes peuvent aussi être observées selon la
force ionique et le pH de l’environnement. Les changements dans la forme de la

19
fibronectine sont de plus modulés par son interaction avec d’autres composants de la
matrice (Timpl et al. 1986). Elle est retrouvée à la surface des fibroblastes et de façon très
abondante dans le sérum et le plasma. Elle est donc très répandue dans le derme, mais
absente de l’épiderme (Woodley 1987). Une étude in vitro a déterminé que la fibronectine
favorisait la synthèse des composants de la membrane basilaire et par le fait même, sa
formation. Cette étude a aussi suggéré que cet effet était attribuable à des événements de
signalisation transmembranaire médiés par les intégrines (Austria et al. 1991). La
fibronectine permet aussi la migration cellulaire (Austria et al. 1991; Woodley 1987) en
s’associant à d’autres constituants structuraux de la matrice extracellulaire comme des
fibres de collagène et des protéoglycans particuliers, formant ainsi des zones fibrillaires
éparses qui offrent des espaces par où les cellules peuvent transiter (Lodish et al. 1997).
Figure 10 : La fibronectine
Source : bio.winona.msus.edu/berg/illust/fibronectin.gif
1.1.4.1.6 Intégrines Les intégrines sont des récepteurs cellulaires membranaires qui se lient à divers composants
de la matrice extracellulaire. Elles ont donc comme rôle premier de fixer solidement la
cellule à la matrice, mais certaines jouent aussi un rôle dans la communication inter-
cellulaire, dans la communication intracellulaire et dans la migration cellulaire (Ffrench-
Constant et al. 2004). Dans la membrane basilaire, elles fixent les kératinocytes de la
couche basale à cette membrane et à la matrice sous-jacente. Les intégrines sont constituées
de deux polypeptides transmembranaires, les protomères α et β, de masse variant entre 100
et 140 kDa selon la molécule formée (figure 11).

20
Figure 11 : L’intégrine
Source : ntri.tamuk.edu/homepage-ntri/lectures/protein/junction1.html Chez les mammifères, 24 hétérodimères d’intégrines sont connus. Ils sont composés de 18
types de sous-unités α et de 8 types de sous-unités β (figure 12) (Hynes 2002). Dans la
membrane basilaire, les intégrines les plus abondantes sont l’α6β4, l’α2β1 et l’α3β1. La
première, l’α6β4, fait partie intégrante de l’hémidesmosome dont il a été question dans la
section 1.1.4.1 (Borradori et al. 1999). Elle représente aussi le récepteur principal des
laminines 1 et 5 (Moll et al. 1998). L’α2β1 est un excellent récepteur du collagène IV et se
situe dans les plaques d’adhésion focale. Cette intégrine jouerait aussi un rôle important
dans la migration cellulaire (Decline et al. 2000). Finalement, l’α3β1 est un récepteur de la
laminine 5 (Groffen et al. 1999; Moll et al. 1998), du nidogène et de la fibronectine et se
situe aussi dans les plaques d’adhésion focale. De plus, elle est nécessaire à l’organisation
de la laminine 5 dans la matrice extracellulaire (deHart et al. 2003). Ces trois intégrines
contribuent donc au maintien d’une structure solide et d’une bonne organisation de la
membrane basilaire.

21
Figure 12 : Les différentes intégrines
Source : integrines.hypermart.net/wallpaper.htm
1.1.4.2 Organisation et assemblage La membrane basilaire est une structure macromoléculaire complexe contenant plusieurs
composants qui interagissent ensemble pour assurer solidité et stabilité. La production des
constituants de la membrane basilaire est le résultat de la coopération des kératinocytes de
l’épiderme et des fibroblastes du derme (Fleischmajer et al. 1998). La membrane basilaire
est formée par un processus d’auto-assemblage, mais chaque constituant n’interagit pas
avec tous les autres ; il existe des liens spécifiques qui permettent une organisation précise
et adéquate. De plus, un équilibre entre la production des constituants par les cellules et la
dégradation de ceux qui sont en excès par les MMP est nécessaire pour l’organisation
adéquate de la membrane (Amano et al. 2001). La structure de la membrane basilaire est
assurée par la formation de deux filets, un de collagène IV et l’autre de laminines (figure
13) (Timpl et al. 1986; Yurchenco et al. 1990). Le premier filet est formé par l’auto-
assemblage de molécules de collagène IV qui ont la capacité de se grouper en tétramères
qui s’assemblent à leur tour pour former de plus grands agrégats. Les fibres de collagènes
sont stabilisées par des ponts disulfure, ce qui donne une certaine rigidité au réseau. Le

22
deuxième filet est constitué de plusieurs molécules de laminine qui s’attachent une à
l’autre, formant ainsi un treillis parallèle à celui fait de collagène IV (Timpl et al. 1986).
Les deux filets ainsi formés n’interagissent pas directement ensemble : ils sont reliés par le
nidogène sur lequel se trouvent des sites d’attachement pour le collagène IV et les
laminines autres que la laminine 5. À l’opposé des autres laminines présentes dans la
membrane basilaire, la laminine 5 ne peut pas se fixer au nidogène puisqu’elle n’a pas la
composition en acides aminés sur la chaîne γ2 qui est nécessaire pour s’y lier normalement
(Mayer et al. 1995). Toutefois, la laminine 5 a la capacité de s’attacher à la laminine 6, ce
qui lui permet de faire partie intégrante de la structure en filet. De plus, la laminine 5 forme
les filaments d’ancrage (Rousselle et al. 1991) qui sont fixés aux hémidesmosomes, grâce
aux intégrines α3β1 et α6β4, et au domaine NC1 du collagène VII (Borradori et al. 1999).
La laminine 5 traverse donc les deux filets structuraux de part en part à un angle de 27º et
contribue par le fait même à augmenter la solidité de la membrane basilaire. Le perlécan
s’intègre à la structure de façon à être bien distribué dans la membrane afin de former des
régions anioniques nécessaires pour les propriétés filtrantes de la membrane basilaire. Ce
constituant peut se lier au nidogène, aux laminines, au collagène IV et à la fibronectine, ce
qui l’ancre dans la membrane basilaire (Groffen et al. 1999). Vu son interaction avec
plusieurs constituants, il est connu pour être requis à l’intégrité de la membrane en
participant à l’assemblage du feuillet de laminines (Henry et al. 2001). Pour ce qui est de la
fibronectine, elle favorise l’assemblage des constituants de la membrane basilaire de deux
manières : par des interactions directes avec la surface cellulaire et par son attachement à
plusieurs autres constituants de la membrane basilaire, ce qui favorise la nucléation et la
stabilisation des complexes formés par ceux-ci (Austria et al. 1991). L’organisation
macromoléculaire de la membrane basilaire semble ainsi assez simple à la base, mais elle
implique des interactions spécifiques entre les différents composants qui sont essentielles à
sa structure.

23
Figure 13 : L’assemblage de la membrane basilaire
Source : cella.cn/book/10/01.htm
1.2 Guérison de plaies cutanées Une perturbation de l’intégrité de la peau peut survenir dans plusieurs contextes. La peau
peut subir des dommages lors de chirurgies, de brûlures, de radiations, de coupures, de
déchirures, d’éraflures, d’abrasions, de frottements, de pincements et de pressions. Le degré
de gravité de la blessure varie selon certains facteurs comme l’étendue, la profondeur et la
nature. Afin de maintenir les fonctions essentielles de la peau, il est très important de la
réparer lorsqu’un tel événement survient. La guérison de plaies cutanées est un événement
dynamique coordonné qui implique la multiplication cellulaire, la migration cellulaire
active et la production de matrice extracellulaire (Li et al. 2005; Smith et al. 1998). Elle
implique donc différents processus qui doivent être respectés pour assurer la réparation.
Chez un individu en santé, toutes les étapes sont franchies, ce qui permet une guérison
adéquate. Une plaie normale est définie comme une interruption dans la continuité d’un
tissu du corps. Il existe aussi des plaies qui sont dites chroniques, qui sont définies comme
étant une interruption de la continuité d’un tissu du corps qui nécessite plus de temps pour
guérir, qui ne guérit pas ou qui est récurrente. Ce dernier type de plaie est souvent associé à

24
un âge avancé ou à des pathologies comme le diabète, l’anémie et la malnutrition (Smith et
al. 1998; Wysocki 1999).
Il existe deux sortes de plaies principales, les plaies superficielles et les plaies profondes.
Les plaies superficielles atteignent seulement les cellules de l’épiderme, ce qui rend la
guérison très facile et rapide (figure 14). En réaction au traumatisme, les kératinocytes
basaux répondent à des signaux d’alarme et brisent leurs liens avec la membrane basilaire,
ce qui les rend capables de migrer pour couvrir la surface de la plaie. Leur trajectoire sera
arrêtée lorsqu’ils rencontreront les cellules provenant de l’autre côté de la plaie. Il
surviendra alors une inhibition de contact. Pendant la migration des cellules activées, les
cellules stationnaires de la couche basale se divisent afin de combler les espaces créés par
les kératinocytes migrateurs. Une fois la plaie refermée, les cellules migratoires se divisent
à leur tour et forment de nouvelles couches selon le processus de différenciation. La
guérison d’une plaie superficielle peut prendre de 24 à 48 heures (Li et al. 2005; Tortora et
al. 1994).
Figure 14 : La guérison d’une plaie superficielle
Source : (Tortora et al. 1994)
La réparation de plaies profondes demande beaucoup plus de temps et nécessite la mise en
jeu de plus de tissus. La lésion s’étend souvent profondément dans le derme, ce qui rend la
guérison plus complexe et laisse une cicatrice (figure 15). Ce type de guérison comprend
trois phases essentielles qui devront s’effectuer pour assurer la cicatrisation complète : la
phase inflammatoire, la phase de migration et de prolifération et la phase de remodelage
(Hunt et al. 2000; Tortora et al. 1994; Wysocki 1999).
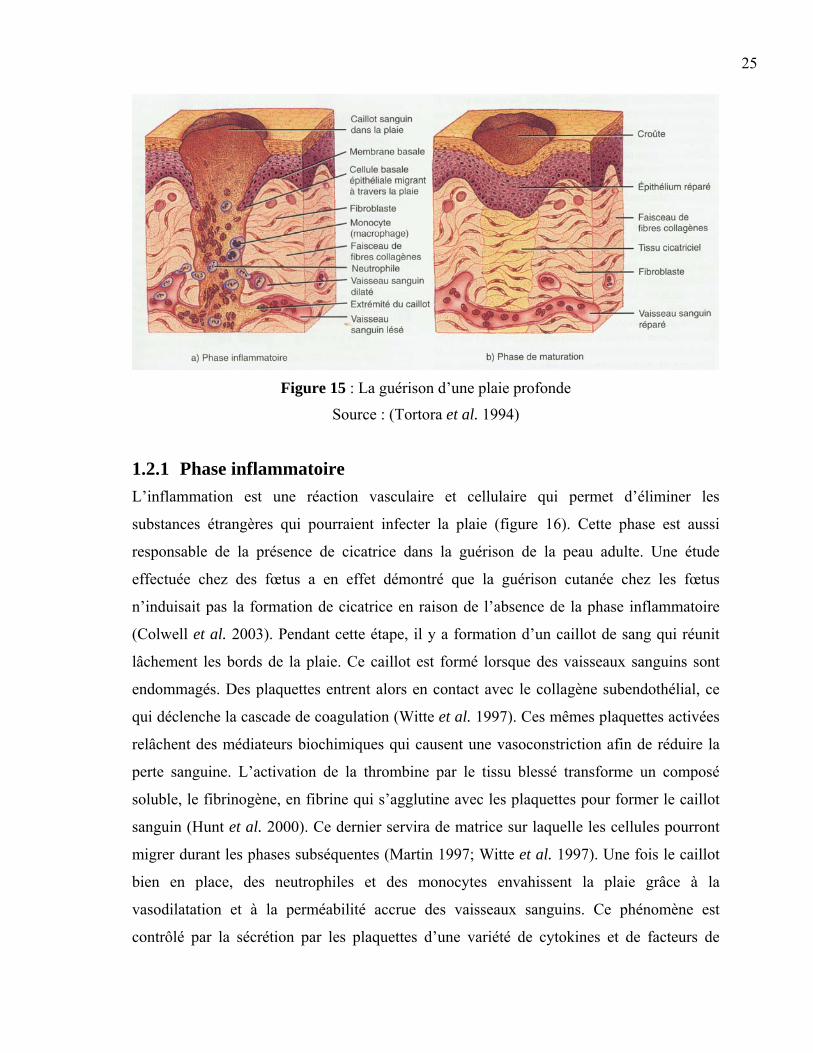
25
Figure 15 : La guérison d’une plaie profonde
Source : (Tortora et al. 1994)
1.2.1 Phase inflammatoire L’inflammation est une réaction vasculaire et cellulaire qui permet d’éliminer les
substances étrangères qui pourraient infecter la plaie (figure 16). Cette phase est aussi
responsable de la présence de cicatrice dans la guérison de la peau adulte. Une étude
effectuée chez des fœtus a en effet démontré que la guérison cutanée chez les fœtus
n’induisait pas la formation de cicatrice en raison de l’absence de la phase inflammatoire
(Colwell et al. 2003). Pendant cette étape, il y a formation d’un caillot de sang qui réunit
lâchement les bords de la plaie. Ce caillot est formé lorsque des vaisseaux sanguins sont
endommagés. Des plaquettes entrent alors en contact avec le collagène subendothélial, ce
qui déclenche la cascade de coagulation (Witte et al. 1997). Ces mêmes plaquettes activées
relâchent des médiateurs biochimiques qui causent une vasoconstriction afin de réduire la
perte sanguine. L’activation de la thrombine par le tissu blessé transforme un composé
soluble, le fibrinogène, en fibrine qui s’agglutine avec les plaquettes pour former le caillot
sanguin (Hunt et al. 2000). Ce dernier servira de matrice sur laquelle les cellules pourront
migrer durant les phases subséquentes (Martin 1997; Witte et al. 1997). Une fois le caillot
bien en place, des neutrophiles et des monocytes envahissent la plaie grâce à la
vasodilatation et à la perméabilité accrue des vaisseaux sanguins. Ce phénomène est
contrôlé par la sécrétion par les plaquettes d’une variété de cytokines et de facteurs de
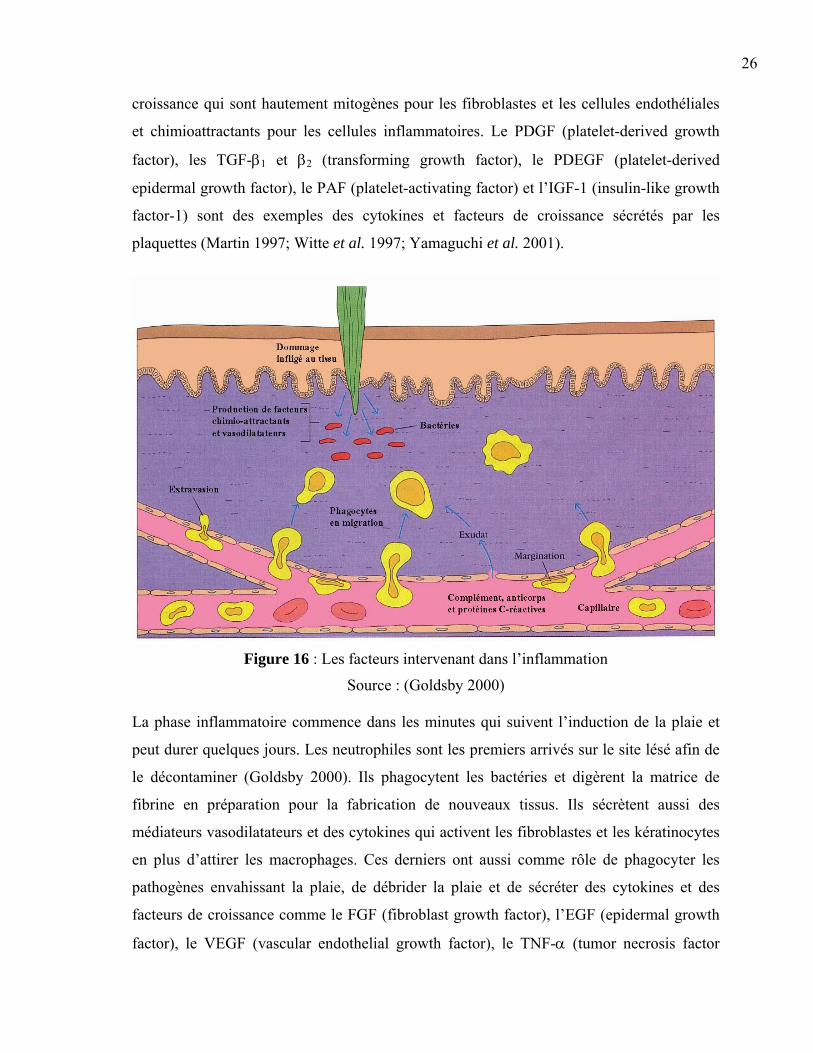
26
croissance qui sont hautement mitogènes pour les fibroblastes et les cellules endothéliales
et chimioattractants pour les cellules inflammatoires. Le PDGF (platelet-derived growth
factor), les TGF-β1 et β2 (transforming growth factor), le PDEGF (platelet-derived
epidermal growth factor), le PAF (platelet-activating factor) et l’IGF-1 (insulin-like growth
factor-1) sont des exemples des cytokines et facteurs de croissance sécrétés par les
plaquettes (Martin 1997; Witte et al. 1997; Yamaguchi et al. 2001).
Figure 16 : Les facteurs intervenant dans l’inflammation
Source : (Goldsby 2000)
La phase inflammatoire commence dans les minutes qui suivent l’induction de la plaie et
peut durer quelques jours. Les neutrophiles sont les premiers arrivés sur le site lésé afin de
le décontaminer (Goldsby 2000). Ils phagocytent les bactéries et digèrent la matrice de
fibrine en préparation pour la fabrication de nouveaux tissus. Ils sécrètent aussi des
médiateurs vasodilatateurs et des cytokines qui activent les fibroblastes et les kératinocytes
en plus d’attirer les macrophages. Ces derniers ont aussi comme rôle de phagocyter les
pathogènes envahissant la plaie, de débrider la plaie et de sécréter des cytokines et des
facteurs de croissance comme le FGF (fibroblast growth factor), l’EGF (epidermal growth
factor), le VEGF (vascular endothelial growth factor), le TNF-α (tumor necrosis factor

27
alpha), l’interleukine-1 et l’INF-γ (interféron-gamma) (Yamaguchi et al. 2001). Ces
messagers chimiques stimulent l’infiltration, la prolifération et la migration des fibroblastes
et des cellules endothéliales (Goldsby 2000; Hunt et al. 2000; Martin 1997; Witte et al.
1997). En somme, la phase inflammatoire constitue la phase de défense contre les
infections et le début de la croissance des nouvelles cellules.
1.2.2 Phase de migration et de prolifération Durant cette étape, qui commence dans les quelques heures suivant la blessure, il se produit
une organisation de l’exsudat inflammatoire où les cellules recrutées au site lésé entrent en
mitose puis s’activent afin de définir grossièrement la structure de la cicatrice. Il survient
alors un équilibre entre la production de matrice et la destruction des tissus endommagés et
irréparables. Le tissu temporaire alors formé s’appelle le tissu de granulation (Tortora et al.
1994). Des myofibroblastes participent à la réparation en rapprochant les bords de la plaie,
facilitant ainsi la fermeture (Gabbiani 1998; Grinnell 1994; Serini et al. 1999). Toutefois, la
cicatrisation est meilleure lorsque la matrice continue de s’accumuler sans se contracter
davantage (Hunt et al. 2000) puisque des contractures sévères de la peau, empêchant une
bonne mobilité, peuvent survenir (Smith et al. 1998). Même si les lymphocytes et les
macrophages sont impliqués dans le processus, ce sont les kératinocytes, les fibroblastes et
les cellules endothéliales qui vont proliférer et migrer afin d’assurer une guérison complète
des tissus. Les fibroblastes vont agir en premier en synthétisant des fibres de collagène et
des polyosides en suivant les filaments de fibrine, ce qui constituera le tissu de granulation.
Les cellules endothéliales vont être responsables de la réparation des vaisseaux sanguins
endommagés et de la croissance de ceux-ci afin de nourrir les nouveaux tissus formés. Les
kératinocytes commencent à migrer vers le centre de la plaie, et à se différencier afin de
reconstituer la barrière que constituait l’épiderme avant la blessure (Hunt et al. 2000; Smith
et al. 1998). Ils migrent alors entre le caillot sanguin et la matrice de collagène du derme.
La production de la collagénase-1 et l’activation de la plasmine par des activateurs
plasminogènes sécrétés par les kératinocytes sont nécessaires à la dégradation de la fibrine
et du collagène-1 qui forment le caillot sanguin (Yamaguchi et al. 2001). Le processus par
lequel les kératinocytes migrent pour couvrir la plaie est appelé ré-épithélialisation et

28
provoque la production de matrice extracellulaire, de facteurs de croissance et de cytokines
en plus de favoriser l’angiogenèse. Les facteurs stimulants sécrétés par les kératinocytes
sont le FGF, le PDGF, le VEGF et le TGF-α (Hunt et al. 2000). Les kératinocytes basaux
produisent aussi un haut niveau d’intégrines α5β1 et d’intégrines contenant la chaîne αv.
L’induction de celles-ci serait nécessaire à la migration des kératinocytes grâce à leur
activité liante qui favorise la formation de nouveaux points d’adhérence (Gailit et al. 1994).
Les constituants de la membrane basilaire sont aussi sécrétés par les kératinocytes et par les
fibroblastes durant cette phase pour restaurer la continuité et la solidité de la jonction
dermo-épidermique. Toutefois, très peu de littérature est disponible à ce sujet.
La migration des cellules épithéliales a lieu dans la plaie par un processus appelé épibolie
(Martin et al. 1992). Si les follicules pileux sont intacts, leurs kératinocytes vont migrer et
ainsi contribuer à la régénération du nouvel épiderme. Les cellules basales de l’épiderme
répondent généralement au traumatisme par activation. En effet, grâce aux « signaux de
détresse » envoyés par les cellules endommagées, les cellules basales s’aplatissent,
s’allongent et brisent leurs liens avec les cellules adjacentes et avec la membrane basilaire.
Les cellules épithéliales sont alors libres et en bonne position pour refermer la plaie en
migrant sur la structure de fibrine et de collagène qui se trouve en son centre. La migration
cesse lorsque les kératinocytes d’un côté de la plaie rencontrent ceux de l’autre côté. Il
survient alors une inhibition de contact. La population cellulaire est augmentée par les
nombreuses mitoses des kératinocytes et la structure et l’épaisseur originales de l’épiderme
sont restaurées par la différenciation de ces cellules (Smith et al. 1998).
La migration cellulaire peut être influencée par plusieurs molécules tels des facteurs de
croissance ou des protéines sécrétés par les cellules. Par exemple, le TGF-β1 induit la
migration des kératinocytes et diminue leur adhésion à la laminine 5. Il joue aussi un rôle
important dans le contrôle du remodelage de la matrice en régulant la transcription d’un
grand nombre de protéines de la matrice extracellulaire (Decline et al. 2000). De plus, dans
la guérison de plaie, certains constituants de la membrane basilaire et de la matrice
extracellulaire orientent les mouvements des kératinocytes et favorisent ainsi leur migration
(Lampe et al. 1998; Nguyen et al. 2000; Utani et al. 2001). C’est pourquoi l’interaction
cellule-matrice est importante pour la migration lors de la guérison de plaie. En effet, la

29
matrice extracellulaire contient des substances adhésives et des fibres qui guident les
cellules migratrices. De la même façon, des molécules présentes dans le sang peuvent
contribuer à la migration cellulaire. Par exemple, la fibrine et la fibronectine, toutes deux
présentes dans le sang, sont connues pour augmenter la migration cellulaire durant la
guérison : elles s’attachent à la matrice provisoire et forment une structure sur laquelle les
kératinocytes peuvent migrer (Campbell 1995; Woodley 1987; Yamaguchi et al. 2001). De
plus, le collagène, qui fait partie intégrante de la matrice, favorise la migration des
kératinocytes lors de la guérison (O'Toole et al. 1997).
Le mouvement cellulaire fait intervenir trois propriétés des cellules : l’extension,
l’adhérence et la contraction (figure 17). Ces déformations morphologiques impliquent un
réarrangement du cytosquelette et des molécules faisant partie des complexes d’adhésion de
la cellule (Campbell 1995; Lodish et al. 1997). Ces molécules sont souvent des intégrines
et interagissent avec d’autres molécules de la matrice qui sont présentes sous les cellules en
migration. La plupart de ces molécules sont en fait des constituants de la membrane
basilaire. Le processus de migration implique des changements dans l’adhésion cellulaire.
De la structure stable d’ancrage basée principalement sur le complexe laminine 5/intégrine
α6β4, la cellule doit transiter vers un système d’adhésion dynamique impliquant les
intégrines contenant la sous-unité β1 pour permettre la migration (Gailit et al. 1994;
Nguyen et al. 2001). Dans la guérison d’une plaie, les kératinocytes des bords migrent sur
la matrice riche en fibronectine et en laminine 5. Il survient alors une perte de la
polarisation des intégrines et les kératinocytes augmentent leur répertoire d’intégrines et
modifient leur distribution : les intégrines α3β1 et α2β1 changent leur localisation latérale à
basale pour être en contact avec la matrice extracellulaire. De plus, l’intégrine α6β4 devient
distribuée de façon plus uniforme et apparaît tout le long de la surface des kératinocytes en
migration qui ne présentent aucun hémidesmosome (Goldfinger et al. 1999; Kurpakus et al.
1991; Larjava et al. 1993).

30
Figure 17 : Les étapes du déplacement cellulaire
Source : (Lodish et al. 1997) La laminine 5 et les intégrines ont en effet un rôle très important dans la régulation de la
migration des kératinocytes. Il est connu depuis quelques années que la laminine 5 subit des
clivages extracellulaires et que ceux-ci ont une incidence sur ses fonctions. Ces coupures
représentent un régulateur important de l’adhésion et de la mobilité des cellules dans la
guérison de plaie (Goldfinger et al. 1999; Utani et al. 2001). Selon sa forme, la laminine 5
n’interagit pas avec les mêmes molécules, ce qui a une incidence sur la migration cellulaire.
Par exemple, la forme longue est réputée pour permettre la migration cellulaire et joue donc
un rôle dans le processus de fermeture d’une plaie. Les kératinocytes en migration sécrètent
d’ailleurs cette forme en grande quantité (Larjava et al. 1993). Il est aussi à remarquer que
dans la peau normale, il n’y a peu ou pas de marquage de la laminine 5 dans le cytoplasme
des kératinocytes basaux tandis que dans une plaie en guérison, la laminine 5 est détectée
dans les cellules basales (Kainulainen et al. 1998). La forme longue de la laminine 5
interagit avec les intégrines α3β1 et α2β1 qui sont deux récepteurs retrouvés dans les
plaques d’adhésion focale utiles pour le mouvement cellulaire (Decline et al. 2000;
Kainulainen et al. 1998; Nguyen et al. 2001). L’association de la laminine 5 et de
l’intégrine α3β1 améliore aussi la communication intercellulaire en facilitant l’assemblage
des jonctions Gap. Ces jonctions participent à une communication bidirectionnelle pour

31
réguler la réponse pré et post-migratoire dans la plaie (Lampe et al. 1998; Nguyen et al.
2000). Le clivage de la laminine 5 le plus important est celui de la chaîne α3 et il survient
entre les domaines G3 et G4, comme expliqué dans la section 1.1.4.1.1. Il a été suggéré que
le domaine G5, qui est retrouvé à l’extrémité de la chaîne α3, stimulerait la migration alors
que le domaine G3, qui est mis à nu par le clivage, serait responsable des propriétés
d’adhésion de la laminine 5, tout en permettant la migration dans certains cas (Hirosaki et
al. 2000). Une fois clivée, la laminine 5 interagit avec l’intégrine α6β4 par le biais du
domaine G3 pour former les hémidesmosomes et ainsi assurer la solidité de la jonction
dermo-épidermique (Borradori et al. 1999; Nguyen et al. 2001). Une expérimentation a
démontré que des souris ayant une délétion dans le gène de la chaîne α3 de la laminine 5
développaient des épidermolyses bulleuses sévères et des hémidesmosomes rudimentaires.
Les kératinocytes de ces souris présentaient également une habileté réduite pour la
migration (Ryan et al. 1999). La régulation de l’expression de la laminine 5 est médiée par
le TGF-β et l’INF-γ. Il a été suggéré que la production de laminine 5 pourrait être activée
par ces deux molécules présentes dans les fluides délivrés lors de l’induction de la plaie
(Kainulainen et al. 1998). Les hémidesmosomes jouent aussi un rôle dans la régulation de
l’adhésion cellulaire puisqu’ils y participent activement (Moll et al. 1998). Il importe donc
de réguler leur assemblage pour permettre la migration. Cette régulation est principalement
contrôlée par des protéines de la matrice extracellulaire et par des facteurs de croissance
(Borradori et al. 1999). Il en est de même pour l’intégrine α6β4 qui est un transmetteur
important de messages pour la cellule. Elle interagit aussi avec des éléments du
cytosquelette pour promouvoir la migration cellulaire (Mercurio et al. 2001). Une étude a
d’ailleurs démontré qu’un anticorps dirigé contre la chaîne α6 inhibait la motilité de cellules
intestinales et leur potentiel à former des lamellipodes (Rabinovitz et al. 1997).
1.2.3 Phase de remodelage Le remodelage des tissus cicatriciels peut durer jusqu’à un an et parfois plus. Les facteurs
qui influencent la durée de la phase de remodelage sont l’âge, les antécédents génétiques, la
localisation de la plaie, la longueur et l’intensité de la phase inflammatoire et la gravité de
la plaie. Au début de cette phase, l’épiderme a retrouvé son épaisseur normale et la croûte

32
formée à partir du caillot sanguin tombe. Les fibres de collagène commencent à être mieux
organisées : initialement fines et désordonnées, elles deviennent plus épaisses et alignées
avec les autres fibres des marges de la plaie (Hunt et al. 2000; Smith et al. 1998). En plus
des fibres de collagène, d’autres composants comme des glycosaminoglycans et des
protéoglycans viennent s’ajouter à la matrice extracellulaire. Avec le temps, la quantité de
fibroblastes diminue et les vaisseaux sanguins reprennent leur taille normale. L’apoptose
des fibroblastes et des cellules endothéliales marque la transition entre le tissu de
granulation et la cicatrice proprement dite. L’excès de matrice est aussi éliminé par
l’activation de collagénases comme les MMP (métalloprotéinase matricielle) (Pilcher et al.
1999; Yamaguchi et al. 2001). Malgré toutes les modifications et le remodelage que subit
le derme durant cette phase, la peau ne retrouvera jamais ni sa fonction ni sa force ou
résistance originale (Hunt et al. 2000; Tortora et al. 1994). En effet, la force de tension de
la peau cicatrisée ne dépassera pas 80 % de sa force antérieure (Clark 1985). De la même
façon, l’élasticité de la cicatrice sera moindre qu’avant la blessure (Doillon et al. 1985).
1.3 Approche et objectifs des travaux de recherche L’objectif général de ces travaux de recherche est d’analyser la reconstruction de la
membrane basilaire au cours de la guérison de plaies cutanées. Puisque la guérison cutanée
implique nécessairement une reconstruction et une expression de novo des constituants
majeurs, l’utilisation d’un nouveau modèle de guérison de plaies cutanées humaines a été
privilégiée comme approche pour cette étude.
L’utilisation d’un nouveau modèle de peau reconstruite par génie tissulaire présente
plusieurs avantages pour ce type d’expérimentation. Il a été profondément analysé,
caractérisé et optimisé par l’équipe du Laboratoire d’Organogénèse Expérimentale (LOEX)
(Auger et al. 2002), ce qui le rend des plus intéressants. Il implique l’utilisation de matériel
humain seulement, entièrement sécrété par les cellules obtenues d’une biopsie de peau
humaine. Plusieurs autres modèles existent, mais aucun n’est formé à partir de la méthode
d’auto-assemblage exclusive au modèle du LOEX (Pomahac et al. 1998). Ils utilisent tous
des cellules humaines, mais également une matrice synthétique ou provenant d’animaux
(De et al. 2002; Ruszczak et al. 2000; Yamaguchi et al. 2001). Le fait d’utiliser du matériel

33
et des cellules totalement humains est très important dans le cadre d’une étude in vitro car
le but principal est de pouvoir appliquer les résultats au vivant. Le modèle de plaie cutanée
est construit à partir du nouveau modèle de peau reconstruite auquel sont ajoutées des
étapes d’induction de plaies et de guérison. Cette innovation a aussi été analysée et
caractérisée par l’équipe du LOEX (Laplante et al. 2001) et s’est avérée tout à fait indiquée
pour l’étude de la régénération de la membrane basilaire.
Les travaux de cette maîtrise visent tout d’abord à visualiser l’expression des différents
constituants de la membrane basilaire dans le modèle de peau reconstruite et comparer les
résultats avec ceux qui sont obtenus dans la peau normale humaine. Cette étude permet de
voir chaque constituant individuellement et de bien les situer dans la peau. Il est aussi
possible de déterminer de façon encore plus fine l’exactitude du modèle par rapport à la
peau normale humaine et donc d’approfondir sa caractérisation. Des patrons d’expression
différents selon l’origine de chaque composant et un dépôt à la jonction dermo-épidermique
sont attendus.
Par la suite, il importe d’analyser l’apparition de chaque constituant à la jonction dermo-
épidermique durant la guérison de plaies. Cet examen minutieux permet d’établir une
cinétique d’apparition de chacun des constituants, ce qui fournit de précieux
renseignements sur la constitution et le processus d’assemblage de la membrane basilaire. Il
est supposé ici que les constituants principaux soient déposés en premier afin d’assurer une
bonne structure solide et résistante qui constituera la base de l’architecture générale dès les
premiers instants de la guérison.
Plus spécifiquement, le remodelage de la laminine 5 au cours de la ré-épithélialisation des
plaies doit être étudié de façon plus approfondie en raison de la particularité des fonctions
de cette protéine dans la membrane basilaire. En effet, il a été précédemment expliqué que
la laminine 5 n’avait pas la même fonction selon qu’elle est sous sa forme courte ou sous sa
forme longue. Il est donc suggéré que la forme longue de la laminine 5 soit retrouvée dans
les kératinocytes en mouvement, et donc au front de migration, tandis que la forme courte
de la laminine 5 soit déposée à la jonction dermo-épidermique.

34
Finalement, puisque la migration des kératinocytes fait partie intégrante des processus
impliqués dans la guérison cutanée, il est intéressant de visualiser et d’analyser cette
migration dans le modèle de guérison in vitro. Des tests préliminaires ont donc été effectués
en monocouche dans différentes conditions afin de mieux comprendre quels substrats
favorisent cette migration et comment les kératinocytes migrent pour refermer une plaie.
Pour cette étude, des vidéos réalisés à partir de la technologie du Time Lapse ont été
utilisés comme approche. La migration des kératinocytes en présence de différents substrats
a donc pu être observée en temps accéléré. Une vitesse plus rapide et un déplacement plus
organisé sont attendus en présence de substrat plus riche en nutriments et en facteurs de
croissance.

CHAPITRE 2 : MATÉRIEL ET MÉTHODES

36
2.1 Isolement et culture cellulaire Isolement :
Les kératinocytes humains ont été isolés à partir de prépuces de nouveau-nés et les
fibroblastes humains, de réductions mammaires, comme décrit précédemment (Michel et
al. 1999). Brièvement, les morceaux de peaux ont été incubés pour une période de 24
heures dans une solution froide de thermolysine (Sigma, Oakville, Canada) à une
concentration de 500 μg/ml dans de l’HEPES 1X à pH 7,4 (ICN biomedicals, Saint-
Laurent, Québec, Canada). L’épiderme a ensuite été séparé mécaniquement du derme à
l’aide de pinces recourbées et incubé pendant 30 minutes dans une solution de trypsine-
EDTA (0,05 % trypsine (ICN biomedicals) et 0,01 % EDTA (J.T. Baker, Phillipsburg, NJ))
afin de dissocier les cellules épidermiques. Le derme a été incubé pendant 3 heures dans
une solution de collagénase H (0,125 U/ml (Roche Diagnostics, Laval, Québec, Canada)
dans du milieu DME pour récupérer les fibroblastes.
Culture :
Les kératinocytes ont été cultivés sur une couche nourricière de fibroblastes de souris 3T3
irradiés dans du milieu DME-HAM, une combinaison de milieu Eagle’s medium (DME) et
de Ham’s F12 dans une proportion 3 :1 (Gibco BRL, Burlington, Ontario, Canada),
supplémenté de 5 % de sérum Fetal Clone II (HyClone, Logan, UT), 10 ng/ml d’epidermal
growth factor (EGF, Austral biologicals, San Ramon, CA), 24,3μg/ml d’adénine (Sigma),
5μg/ml d’insuline (Sigma), 5μg/ml de transférine (Roche), 2 X 10-9 M de 3,3’5’ triiodo-L-
thyronine (Sigma), 0,4 μg/ml d’hydrocortisone (Calbiochem, La Jolla, CA), 10-10 M de
toxine du choléra (ICN biomedicals), 100 IU/ml de pénicilline G (Sigma) et 25μg/ml de
gentamycine (Shering, Pointe-Claire, Québec, Canada). Les fibroblastes ont été cultivés
dans du milieu DME contenant 10 % de sérum de veau fœtal (SVF, HyClone), 100 IU/ml
de pénicilline G et 25μg/ml de gentamycine.

37
2.2 Modèle de cicatrisation cutanée Une peau complètement reconstruite à partir de matériel biologique a été produite par la
méthode d’auto-assemblage comme décrit précédemment (Michel et al. 1999). Brièvement,
des feuillets dermiques manipulables ont été produits en cultivant 100 000 fibroblastes en
passage 4 à 7 pendant 28 jours en présence de 50 μg/ml d’acide ascorbique (Sigma-Aldrich
Canada, Oakville, Ontario, Canada) dans du milieu DME 10 % SVF. Deux de ces feuillets
ont ensuite été superposés pour une durée de 7 jours afin de les laisser adhérer ensemble,
après quoi 2 000 000 de kératinocytes en passage 2 à 3 ont été ensemencés dans un anneau
qui a été préalablement déposé sur le derme reconstruit. La peau reconstruite a alors été
cultivée dans du milieu DME-HAM 5 % sérum Fetal Clone II pour 7 jours, puis cultivée en
interface air/liquide pendant 14 jours dans du milieu DME-HAM 5 % sérum Fetal Clone II
sans EGF afin de permettre la différenciation des kératinocytes (figure 18). Cette peau
reconstruite a ensuite été blessée avec un poinçon à biopsie de 6 mm puis placée sur un
feuillet de fibroblastes intact pour permettre la ré-épithélialisation par les kératinocytes
(figure 19). Le tissu blessé a été cultivé pendant 3 jours puis des biopsies ont été prélevées
pour analyses ultérieures.
Figure 18 : La production de peau reconstruite
Source : Roxanne Pouliot, LOEX
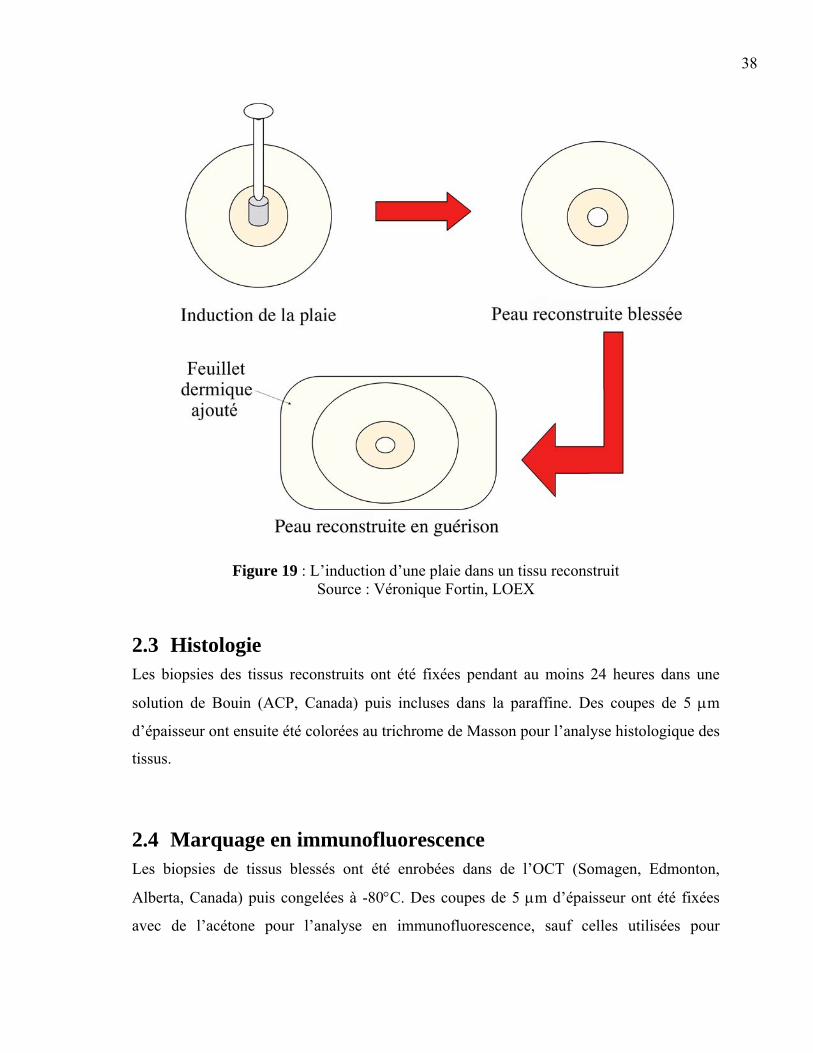
38
Figure 19 : L’induction d’une plaie dans un tissu reconstruit Source : Véronique Fortin, LOEX
2.3 Histologie Les biopsies des tissus reconstruits ont été fixées pendant au moins 24 heures dans une
solution de Bouin (ACP, Canada) puis incluses dans la paraffine. Des coupes de 5 μm
d’épaisseur ont ensuite été colorées au trichrome de Masson pour l’analyse histologique des
tissus.
2.4 Marquage en immunofluorescence Les biopsies de tissus blessés ont été enrobées dans de l’OCT (Somagen, Edmonton,
Alberta, Canada) puis congelées à -80°C. Des coupes de 5 μm d’épaisseur ont été fixées
avec de l’acétone pour l’analyse en immunofluorescence, sauf celles utilisées pour
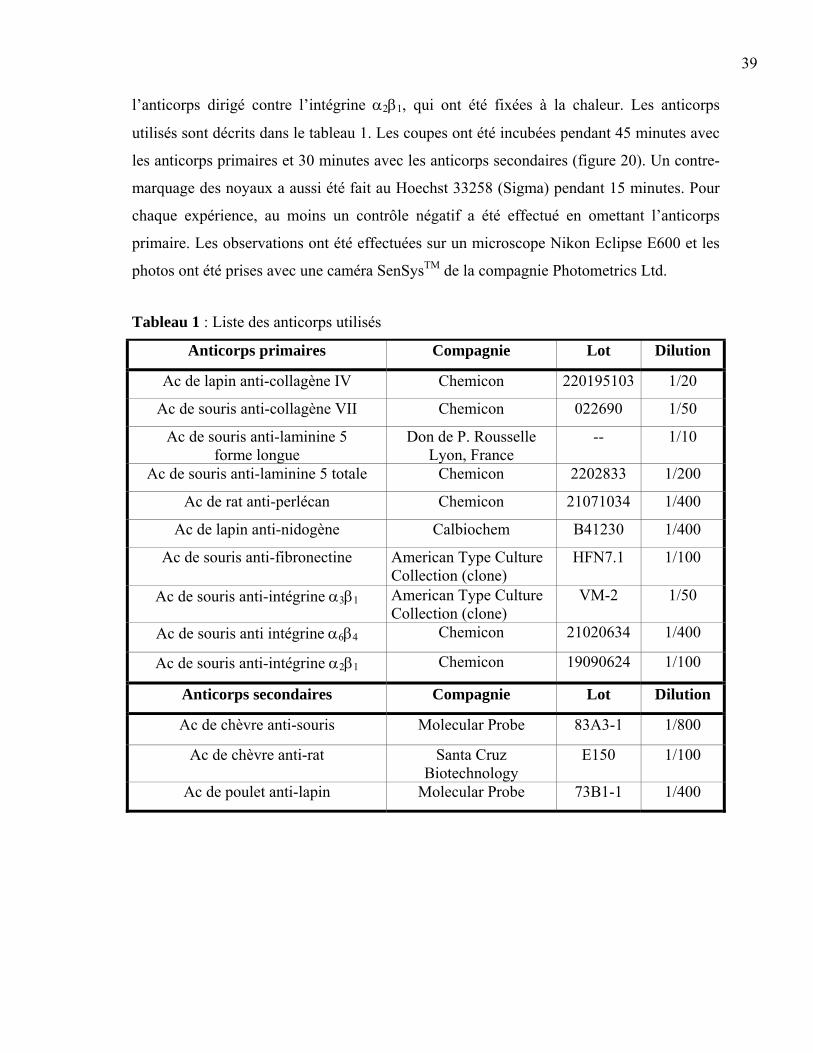
39
l’anticorps dirigé contre l’intégrine α2β1, qui ont été fixées à la chaleur. Les anticorps
utilisés sont décrits dans le tableau 1. Les coupes ont été incubées pendant 45 minutes avec
les anticorps primaires et 30 minutes avec les anticorps secondaires (figure 20). Un contre-
marquage des noyaux a aussi été fait au Hoechst 33258 (Sigma) pendant 15 minutes. Pour
chaque expérience, au moins un contrôle négatif a été effectué en omettant l’anticorps
primaire. Les observations ont été effectuées sur un microscope Nikon Eclipse E600 et les
photos ont été prises avec une caméra SenSysTM de la compagnie Photometrics Ltd.
Tableau 1 : Liste des anticorps utilisés
Anticorps primaires Compagnie Lot Dilution
Ac de lapin anti-collagène IV Chemicon 220195103 1/20
Ac de souris anti-collagène VII Chemicon 022690 1/50
Ac de souris anti-laminine 5 forme longue
Don de P. Rousselle Lyon, France
-- 1/10
Ac de souris anti-laminine 5 totale Chemicon 2202833 1/200
Ac de rat anti-perlécan Chemicon 21071034 1/400
Ac de lapin anti-nidogène Calbiochem B41230 1/400
Ac de souris anti-fibronectine American Type Culture Collection (clone)
HFN7.1 1/100
Ac de souris anti-intégrine α3β1 American Type Culture Collection (clone)
VM-2 1/50
Ac de souris anti intégrine α6β4 Chemicon 21020634 1/400
Ac de souris anti-intégrine α2β1 Chemicon 19090624 1/100
Anticorps secondaires Compagnie Lot Dilution
Ac de chèvre anti-souris Molecular Probe 83A3-1 1/800
Ac de chèvre anti-rat Santa Cruz Biotechnology
E150 1/100
Ac de poulet anti-lapin Molecular Probe 73B1-1 1/400

40
Figure 20 : Le principe de l’immunofluorescence
Source : Véronique Fortin, LOEX
2.5 Obtention du plasma et du sérum humain Le sérum et le plasma ont été obtenus à partir de sang humain prélevé sur des personnes
volontaires. Ce groupe était composé de 22 personnes, hommes et femmes dont l’intervalle
d’âge se situait entre 20 et 45 ans. Des tubes de 10 ml avec coagulants ont été utilisés pour
isoler le sérum tandis que des tubes de 10 ml contenant de l’héparine sodique, un
anticoagulant, ont été utilisés pour l’isolation du plasma. Le fournisseur de ces tubes est la
compagnie Becton Dickinson, New Jersey. Tous les tubes ont été centrifugés pendant 10
minutes à une vitesse de 3000 rpm. Le sérum et le plasma ont été prélevés avec une pipette
puis recueillis dans une bouteille afin de mélanger les échantillons de personnes différentes.
Les pools de sérum et de plasma ont été aliquotés puis congelés à -80°C.
2.6 Tests de migration cellulaire Des kératinocytes en passage 3 ont été ensemencés dans des plaques à 6 puits sur du
plastique, sur de la poly-D-lysine (20μg/ml dans de l’eau, Sigma) ou sur du collagène
(1mg/ml dans de l’acide acétique, Sigma). Les solutions de poly-D-lysine et de collagène
ont été préparées puis versées dans les puits pour une période minimale de vingt-quatre
heures. Le surplus était ensuite aspiré stérilement et les plaques fermées hermétiquement
puis entreposées à 4°C jusqu’à leur utilisation. Trois quantités de kératinocytes (50 000, 80
000 et 100 000 cellules par puits) ont été ensemencées pour chaque expérience. Le puits

41
ayant atteint la confluence à la journée voulue était alors utilisé. Les cellules ont été
cultivées sur une couche nourricière composée de 100 000 fibroblastes de souris 3T3
irradiés jusqu’à confluence dans du milieu DME-HAM supplémenté de 10 % de 3 sérums
différents : sérum humain (SH), plasma humain (PH) ou sérum de veau Fetal Clone II (SV).
Les mêmes additifs que pour la culture des kératinocytes destinés à la formation des peaux
reconstruites ont été ajoutés au milieu. Une fois la confluence atteinte, une plaie est induite
à l’aide d’un embout à pipette (figure 21). La migration est visualisée à l’aide de
photographies prises au Time Lapse puis analysée pour évaluer les vitesses de migration
pour chaque condition. La technologie du Time Lapse implique l’utilisation d’un
microscope inversé équipé d’une chambre d’incubation maintenue à une température de 37
°C. L’environnement à l’intérieur de la chambre est maintenu humide et contient aussi du
CO2. Les taux exacts d’humidité et de CO2 ne sont par contre pas connus. Les images ont
été prises à un intervalle de 10 minutes pendant 12 à 16 heures, ce qui a permis d’éviter une
exposition continue à la lumière. Les images ainsi captées ont été formatées en vidéos grâce
au logiciel Simple PCI de la compagnie Compix Inc.
Figure 21 : L’induction de la plaie en monocouche
Source : Véronique Fortin, LOEX

CHAPITRE 3 : RÉSULTATS

43
3.1 Apparence macroscopique des peaux reconstruites Les tissus reconstruits utilisés pour cette étude sont entièrement formés à partir de cellules
humaines obtenues de biopsies de peau. La première étape de la production d’une peau
reconstruite est la formation d’un équivalent dermique. Pour ce faire, des fibroblastes sont
cultivés dans un flacon de 25 cm2 et, grâce à la production de matrice extracellulaire,
forment un feuillet dermique après 28 jours de culture. Un derme reconstruit facilement
manipulable est obtenu en superposant deux de ces feuillets. La figure 22A montre un
feuillet dermique renforcé par un anneau de papier. Un derme reconstruit, quoique constitué
de deux feuillets de fibroblastes, demeure très mince et translucide. L’anneau de papier
permet une manipulation plus aisée et prévient la contraction du tissu durant les transferts
de pétri.
Après la période de maturation d’une semaine, des kératinocytes sont ensemencés à
l’intérieur d’un anneau métallique préalablement déposé sur le derme reconstruit. Après 14
jours à l’interface air/liquide, les kératinocytes s’organisent et forment un épiderme opaque,
différencié et d’une couleur jaunâtre (figure 22B). Cette zone épithélialisée ne couvre pas
entièrement la surface du tissu reconstruit ; son contour se restreint aux bords de l’anneau
métallique ayant servi à l’ensemencement. Cette marge coïncide presque parfaitement avec
la circonférence du trou de l’anneau de papier servant de support. La zone non
épithélialisée demeure transparente et mince, épousant la forme de l’anneau de papier.
Une fois la peau reconstruite bien formée et différenciée, une plaie est induite avec un
poinçon à biopsie de 6 mm de diamètre au centre de la zone épithélialisée. Cette
manipulation donne des marges très nettes, sans déchirure (figure 22C). Le tissu blessé est
déposé sur un feuillet de fibroblastes intact immédiatement après l’induction de la plaie.
Cette étape est essentielle puisque le feuillet servira de support pour la migration des
kératinocytes. La période de guérison dure trois jours, ce qui permet aux kératinocytes de
ré-épithélialiser la plaie en migrant vers le centre de celle-ci. Ce mouvement des cellules
épithéliales forme une langue de migration qui est visible à la figure 22D, un
agrandissement de la zone épithélialisée. Dans le trou de la plaie, cette zone de guérison est
semi-opaque et forme une bande circulaire grisâtre mesurant entre un et deux millimètres
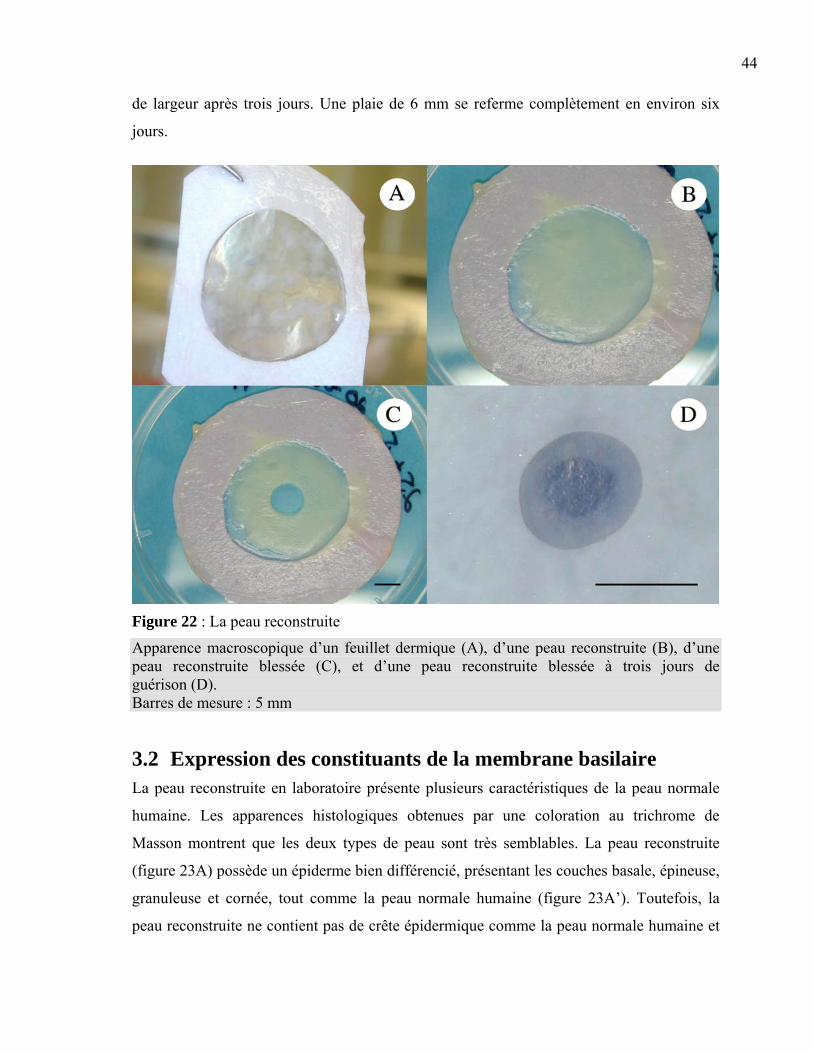
44
de largeur après trois jours. Une plaie de 6 mm se referme complètement en environ six
jours.
Figure 22 : La peau reconstruite
Apparence macroscopique d’un feuillet dermique (A), d’une peau reconstruite (B), d’une peau reconstruite blessée (C), et d’une peau reconstruite blessée à trois jours de guérison (D). Barres de mesure : 5 mm
3.2 Expression des constituants de la membrane basilaire La peau reconstruite en laboratoire présente plusieurs caractéristiques de la peau normale
humaine. Les apparences histologiques obtenues par une coloration au trichrome de
Masson montrent que les deux types de peau sont très semblables. La peau reconstruite
(figure 23A) possède un épiderme bien différencié, présentant les couches basale, épineuse,
granuleuse et cornée, tout comme la peau normale humaine (figure 23A’). Toutefois, la
peau reconstruite ne contient pas de crête épidermique comme la peau normale humaine et
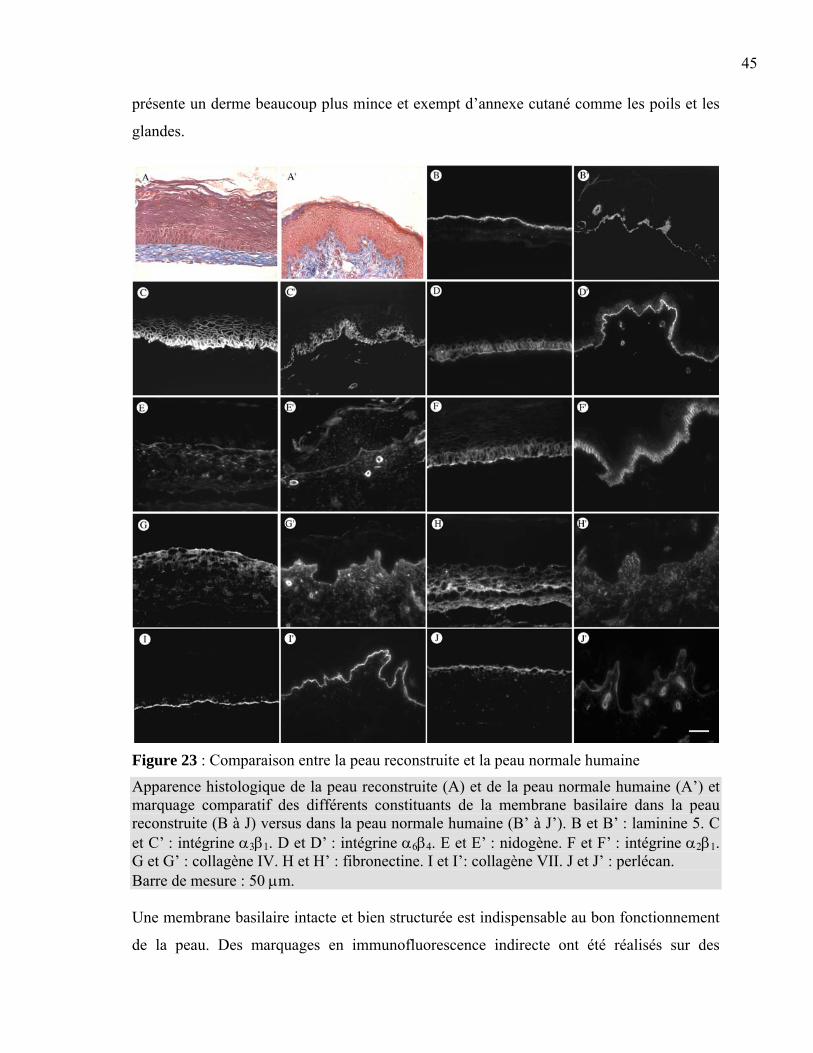
45
présente un derme beaucoup plus mince et exempt d’annexe cutané comme les poils et les
glandes.
Figure 23 : Comparaison entre la peau reconstruite et la peau normale humaine
Apparence histologique de la peau reconstruite (A) et de la peau normale humaine (A’) et marquage comparatif des différents constituants de la membrane basilaire dans la peau reconstruite (B à J) versus dans la peau normale humaine (B’ à J’). B et B’ : laminine 5. C et C’ : intégrine α3β1. D et D’ : intégrine α6β4. E et E’ : nidogène. F et F’ : intégrine α2β1. G et G’ : collagène IV. H et H’ : fibronectine. I et I’: collagène VII. J et J’ : perlécan. Barre de mesure : 50 μm. Une membrane basilaire intacte et bien structurée est indispensable au bon fonctionnement
de la peau. Des marquages en immunofluorescence indirecte ont été réalisés sur des

46
biopsies de peau normale humaine et sur des biopsies de peau reconstruite en laboratoire
afin de vérifier si la membrane basilaire de la peau reconstruite était similaire à celle de la
peau normale humaine. La distribution des composants majeurs de la membrane basilaire et
des intégrines qui y sont associées peut être visualisée à la figure 23.
La laminine 5 est une protéine structurale qui traverse la membrane basilaire de part en
part. Sa détection à l’aide d’un anticorps dirigé contre la forme totale montre un patron
d’expression très similaire dans les deux types de peaux (B et B’). La laminine 5 est
présente exclusivement à la jonction dermo-épidermique en une ligne bien nette. Dans la
peau normale humaine, le marquage suit de façon continue les crêtes épidermiques. Aucune
trace de cette protéine n’est détectée dans l’épiderme et le derme des deux types de peaux.
L’intégrine α3β1 est une molécule associée à la laminine 5 dans la membrane basilaire. Sa
détection montre qu’elle est distribuée dans toutes les cellules de la couche basale de
l’épiderme. Cette intégrine est aussi retrouvée dans les couches supérieures, mais en
moindre quantité. Le marquage est en effet moins intense à partir de la couche épineuse.
Cette présence dans les couches supérieures de l’épiderme est toutefois limitée aux
contours des cellules seulement et est légèrement plus étendue dans la peau reconstruite (C)
en comparaison à la peau normale humaine (C’). Cette intégrine, quoique principalement
retrouvée dans l’épiderme, est aussi détectée autour des vaisseaux sanguins dans le derme
de la peau normale humaine.
L’intégrine α6β4 est une molécule faisant partie intégrante de l’hémidesmosome et elle est
aussi associée à la laminine 5 dans la membrane basilaire. Son marquage en
immunofluorescence est présent de façon régulière dans toutes les cellules de la couche
basale, autant dans la peau reconstruite (D) que dans la peau normale (D’). La détection de
cette intégrine est aussi très intense au niveau de la jonction dermo-épidermique, surtout
dans la peau normale humaine. Cette molécule n’est toutefois pas détectée dans le reste de
l’épiderme ni dans le derme, sauf autour des vaisseaux sanguins, comme l’intégrine α3β1.
Le nidogène présente un patron d’expression légèrement différent dans la peau normale
(E’) par rapport à la peau reconstruite (E). Il est présent dans les cellules de l’épiderme de
la peau normale humaine de façon très claire, mais dans la peau reconstruite, le marquage

47
est très faible et incomparable à celui visible dans la peau normale humaine. Toutefois, le
marquage dans le derme est semblable dans les deux cas, et une fine ligne est visible dans
la membrane basilaire. Encore une fois, des vaisseaux sanguins sont détectés dans la peau
normale.
L’intégrine α2β1 est le récepteur principal du collagène IV et sa distribution ressemble
beaucoup à celle de l’intégrine α6β4. En effet, le marquage par immunofluorescence (F et
F’) démontre que cette intégrine se situe principalement dans les cellules de la couche
basale de l’épiderme, ainsi qu’à la membrane basilaire. Sa présence n’a été détectée nulle
part ailleurs dans les coupes des deux types de tissus.
Le collagène IV est la protéine de soutien de la membrane basilaire. Sa distribution est
semblable dans les deux types de peaux (G et G’). Il se concentre principalement dans le
derme papillaire, la portion du derme la plus rapprochée de l’épiderme. Plus en profondeur,
l’intensité du marquage diminue dramatiquement. Au niveau de la membrane basilaire,
l’intensité est à son maximum, là où le collagène IV remplit sa fonction structurale. Aucune
trace de cette molécule n’est visible dans l’épiderme, mais un vaisseau sanguin positif est
détecté dans la peau normale.
La fibronectine n’est pas directement présente dans la membrane basilaire ; elle agit plutôt
comme protéine associée. Sa détection en immunofluorescence (H et H’) montre donc une
distribution restreinte au derme sans aucune trace à la jonction dermo-épidermique. La
présence de la fibronectine de façon uniforme dans tout le derme permet de visualiser un
phénomène fréquent dans la production de peau reconstruite : il est en effet possible de voir
la séparation entre les deux feuillets de fibroblastes qui ont été superposés pour former le
derme reconstruit (H). Il arrive parfois que l’adhésion ne soit pas parfaite et que la coupe au
cryostat fasse décoller légèrement les feuillets.
Le collagène VII est une protéine essentielle à l’ancrage de la membrane basilaire au derme
sous-jacent. Son marquage en immunofluorescence (I et I’) montre une expression très
nette au niveau de la membrane basilaire. Une fine ligne positive longe en effet la jonction
dermo-épidermique des deux types de peau. Aucun marquage n’est visible dans le derme

48
dans les deux cas, mais une légère expression dans quelques cellules de la couche basale de
la peau reconstruite peut être observée.
Finalement, le perlécan, en J et J’, est détecté lui aussi à la jonction dermo-épidermique,
mais la ligne de marquage est moins bien définie et plus diffuse. Cette caractéristique du
marquage du perlécan met en évidence sa distribution aléatoire à travers la membrane
basilaire afin de former des régions anioniques essentielles aux fonctions filtrantes de la
membrane basilaire. Dans le reste du tissu, aucune trace de perlécan n’a été détectée, sauf
dans le derme de la peau normale humaine, où des vaisseaux sanguins positifs sont visibles.
3.3 Régénération de la membrane basilaire La guérison d’une plaie cutanée implique l’expression de novo de plusieurs molécules
nécessaires à la régénération des tissus et plus particulièrement à la formation de la
membrane basilaire. L’expression de ces constituants peut être visualisée par leur marquage
en immunofluorescence sur les coupes d’une langue de migration visible après trois jours
de guérison. Les figures 24, 25 et 26 illustrent l’expression des différentes molécules
contribuant à l’architecture de la membrane basilaire.
L’apparence histologique d’une langue de migration de trois jours peut être visualisée grâce
à une coloration au trichrome de Masson (figure 24). La marge de la plaie, là où le poinçon
à biopsie a sectionné la peau reconstruite, est clairement visible : il s’agit de la portion
délimitée par la barre verticale, là où le derme est plus épais. Le feuillet de fibroblastes,
ajouté après la création de la plaie, est aussi bien visible. Celui-ci ne s’est pas encore
complètement fusionné avec le derme de la peau reconstruite ; une couche de fibroblastes
est d’ailleurs présente entre les deux structures. La langue de migration est formée par les
kératinocytes des marges de la plaie qui migrent et se divisent pour refermer le trou formé
par le poinçon à biopsie. Le feuillet de migration ajouté après l’induction offre un support
pour leur migration. En fait, ce modèle mime la réparation d’une plaie superficielle, et non
la réparation d’une plaie profonde, puisque le temps alloué (trois jours) pour la guérison
n’est pas suffisant pour permettre au derme de se réparer. Les kératinocytes se déplacent
donc sur un nouveau derme avec lequel ils devront interagir afin de reformer une
membrane basilaire intacte et fonctionnelle.
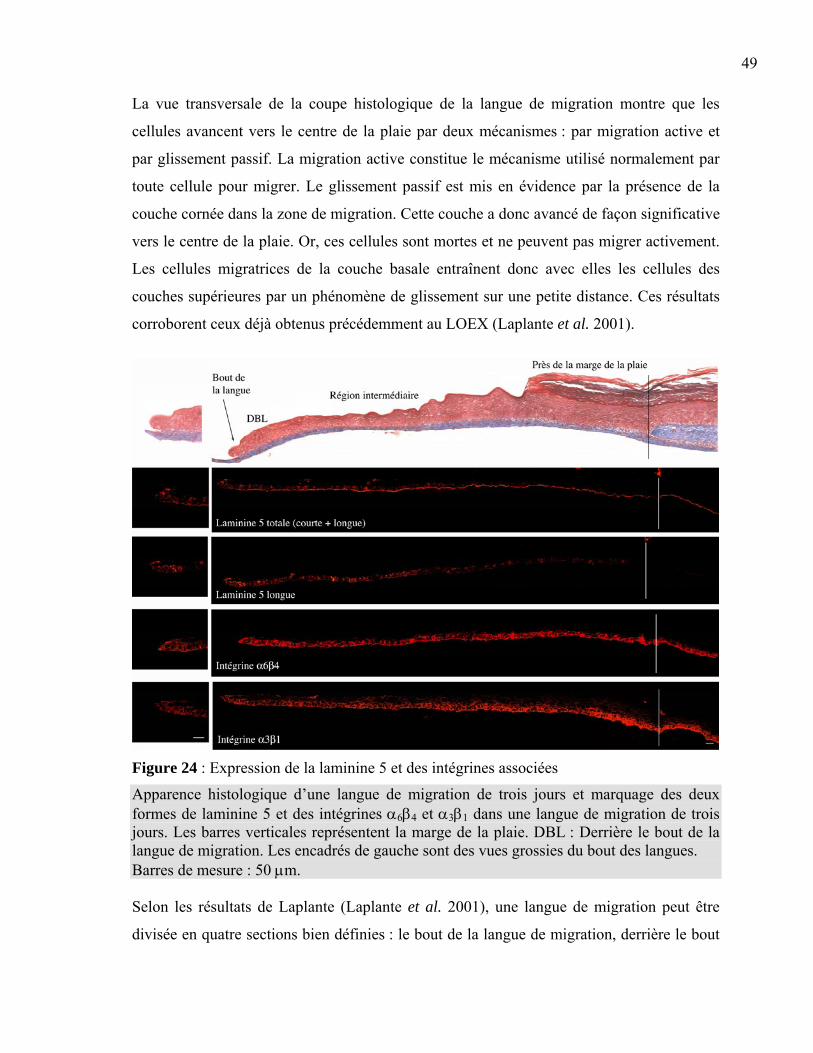
49
La vue transversale de la coupe histologique de la langue de migration montre que les
cellules avancent vers le centre de la plaie par deux mécanismes : par migration active et
par glissement passif. La migration active constitue le mécanisme utilisé normalement par
toute cellule pour migrer. Le glissement passif est mis en évidence par la présence de la
couche cornée dans la zone de migration. Cette couche a donc avancé de façon significative
vers le centre de la plaie. Or, ces cellules sont mortes et ne peuvent pas migrer activement.
Les cellules migratrices de la couche basale entraînent donc avec elles les cellules des
couches supérieures par un phénomène de glissement sur une petite distance. Ces résultats
corroborent ceux déjà obtenus précédemment au LOEX (Laplante et al. 2001).
Figure 24 : Expression de la laminine 5 et des intégrines associées
Apparence histologique d’une langue de migration de trois jours et marquage des deux formes de laminine 5 et des intégrines α6β4 et α3β1 dans une langue de migration de trois jours. Les barres verticales représentent la marge de la plaie. DBL : Derrière le bout de la langue de migration. Les encadrés de gauche sont des vues grossies du bout des langues. Barres de mesure : 50 μm. Selon les résultats de Laplante (Laplante et al. 2001), une langue de migration peut être
divisée en quatre sections bien définies : le bout de la langue de migration, derrière le bout

50
de la langue de migration, la région intermédiaire, et près de la marge de la plaie. Ces
régions sont bien identifiées à la figure 24. Cette subdivision d’une langue de migration
permet de mieux décrire les caractéristiques morphologiques et physiologiques des cellules
migratrices. Après trois jours, les kératinocytes qui sont près de la marge de la plaie tendent
vers un état stationnaire et ont les mêmes caractéristiques que les cellules épithéliales au
niveau de la marge de la plaie. Cette section de la langue de migration est d’ailleurs plus
épaisse que le reste de la langue et contient donc plus de couches cellulaires. Les cellules
présentes dans cette section ont donc commencé à se différencier pour former un épiderme
mature, contrairement aux cellules épithéliales qui se situent au bout des langues de
migration et derrière le bout des langues de migration qui sont dans la phase de migration.
Finalement, les kératinocytes qui se situent dans la région intermédiaire sont dans un état de
transition entre la migration et l’arrêt total.
Grâce à des marquages en immunofluorescence effectués sur des coupes de langues de
migration, il est possible de déterminer quels types cellulaires synthétisent les différents
composants de la membrane basilaire. En effet, au niveau de l’épiderme, les cellules au
bout de la langue de migration sont celles qui étaient en train de migrer au moment de la
biopsie. Elles expriment donc les constituants nécessaires à la régénération de la membrane
basilaire. Les immunofluorescences effectuées pour chacun des constituants permettent
aussi de visualiser leur apparition dans une membrane basilaire en formation. Un résumé
des résultats obtenus grâce à ces immunofluorescences est présenté dans le tableau 2 afin de
faciliter la compréhension et l’analyse des résultats.
La présence des deux formes de laminine 5 (longue et courte) est illustrée à la figure 24. Un
anticorps dirigé contre la forme courte de la laminine 5 n’étant pas disponible, l’étude de
l’expression de cette forme est seulement possible en comparant le résultat du marquage
réalisé avec l’anticorps dirigé contre la forme longue de celui réalisé avec l’anticorps dirigé
contre la forme totale (courte + longue) de la laminine 5. L’expression de la forme longue
de la laminine 5 est restreinte au cytoplasme des kératinocytes basaux de la langue de
migration alors que la forme courte est retrouvée à l’extérieur des cellules, formant une fine
ligne très nette dans la membrane basilaire nouvellement formée ainsi que dans la partie
non lésée de la peau. L’expression de la forme longue de la laminine 5 est particulière : elle

51
est très intense dans la région du bout de la langue, mais diminue vers la région près de la
marge de la plaie, pour enfin devenir nulle dans celle-ci.
Les marquages en immunofluorescence des deux intégrines associées à la laminine 5 sont
aussi présentés à la figure 24. L’expression de l’intégrine α6β4 est constante tout le long de
la coupe. Cette intégrine est présente dans les kératinocytes basaux, mais se concentre aussi
à la membrane basilaire, où elle participe à la formation des hémidesmosomes.
L’expression de l’intégrine α3β1 est un peu différente. Son marquage en
immunofluorescence est aussi constant tout le long de la coupe, mais diminue légèrement
d’intensité vers le bout de la langue. L’expression de cette intégrine ne se limite pas
seulement à la couche basale : quoiqu’elle soit beaucoup plus intense dans celle-ci, elle
s’étend aussi vers les couches supérieures, surtout dans la langue de migration.
Les agrandissements à la gauche de la figure 24 montrent le bout des langues de migration
grossi deux fois. Cette vision permet d’examiner l’expression des constituants de façon plus
précise et de constater que les marquages sont aussi présents à la surface du bout des
langues de migration. Cette particularité, déjà observée par Laplante (Laplante et al. 2001),
suggère que les cellules du bout de la langue migrent par roulement sur elles-mêmes. Selon
cette hypothèse, les cellules du dessus seront les prochaines à être en contact avec le derme
et à contribuer à la régénération de la membrane basilaire. L’agrandissement du bout de la
langue marquée avec l’anticorps dirigé contre l’intégrine α6β4 montre en effet une cellule
du dessus s’étirant afin de glisser par-dessus une cellule basale.
La figure 25 montre l’expression de trois autres constituants importants dans l’architecture
de la membrane basilaire. Tout d’abord, l’intégrine α2β1 est distribuée dans les cellules de
la couche basale de l’épiderme de façon régulière. Aucune différence n’est observée dans la
marge de la plaie par rapport à la langue de migration. De plus, il est évident que la
synthèse de cette intégrine est exclusive aux kératinocytes basaux puisque aucun marquage
n’est présent dans le derme et dans les autres couches de l’épiderme. Il est aussi intéressant
de remarquer que le marquage de cette intégrine est plus granuleux et moins net que celui
des deux autres intégrines décrites précédemment.
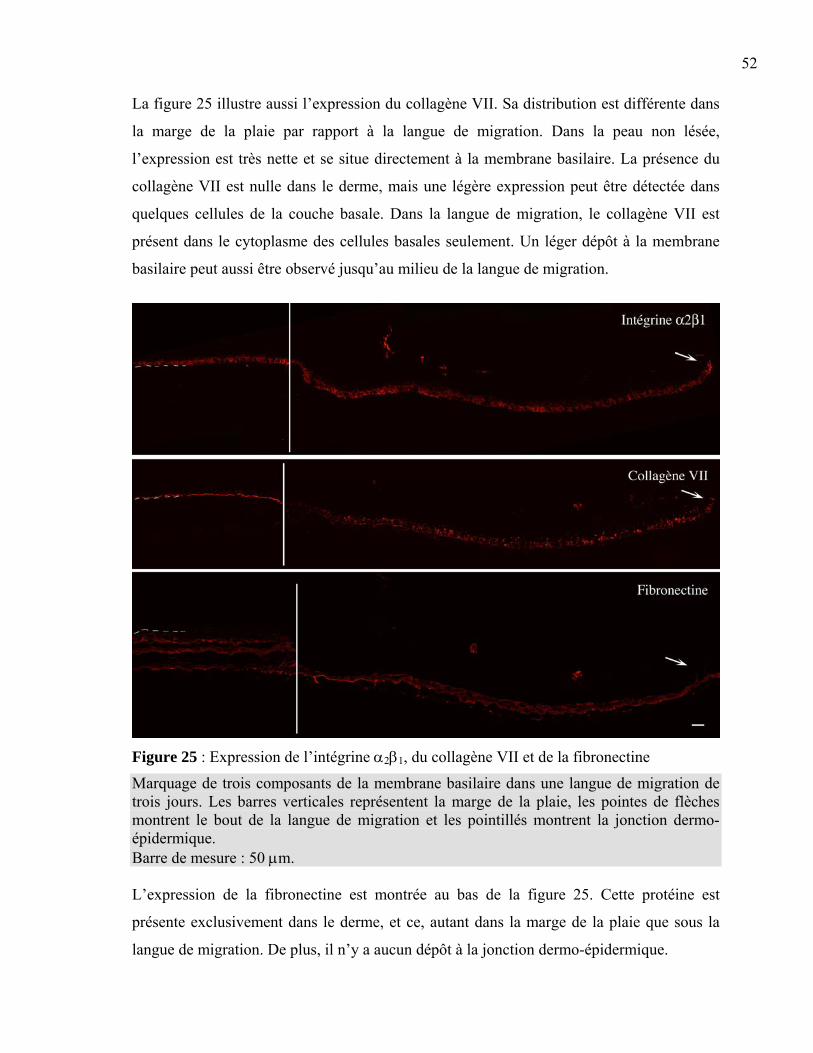
52
La figure 25 illustre aussi l’expression du collagène VII. Sa distribution est différente dans
la marge de la plaie par rapport à la langue de migration. Dans la peau non lésée,
l’expression est très nette et se situe directement à la membrane basilaire. La présence du
collagène VII est nulle dans le derme, mais une légère expression peut être détectée dans
quelques cellules de la couche basale. Dans la langue de migration, le collagène VII est
présent dans le cytoplasme des cellules basales seulement. Un léger dépôt à la membrane
basilaire peut aussi être observé jusqu’au milieu de la langue de migration.
Figure 25 : Expression de l’intégrine α2β1, du collagène VII et de la fibronectine
Marquage de trois composants de la membrane basilaire dans une langue de migration de trois jours. Les barres verticales représentent la marge de la plaie, les pointes de flèches montrent le bout de la langue de migration et les pointillés montrent la jonction dermo-épidermique. Barre de mesure : 50 μm. L’expression de la fibronectine est montrée au bas de la figure 25. Cette protéine est
présente exclusivement dans le derme, et ce, autant dans la marge de la plaie que sous la
langue de migration. De plus, il n’y a aucun dépôt à la jonction dermo-épidermique.
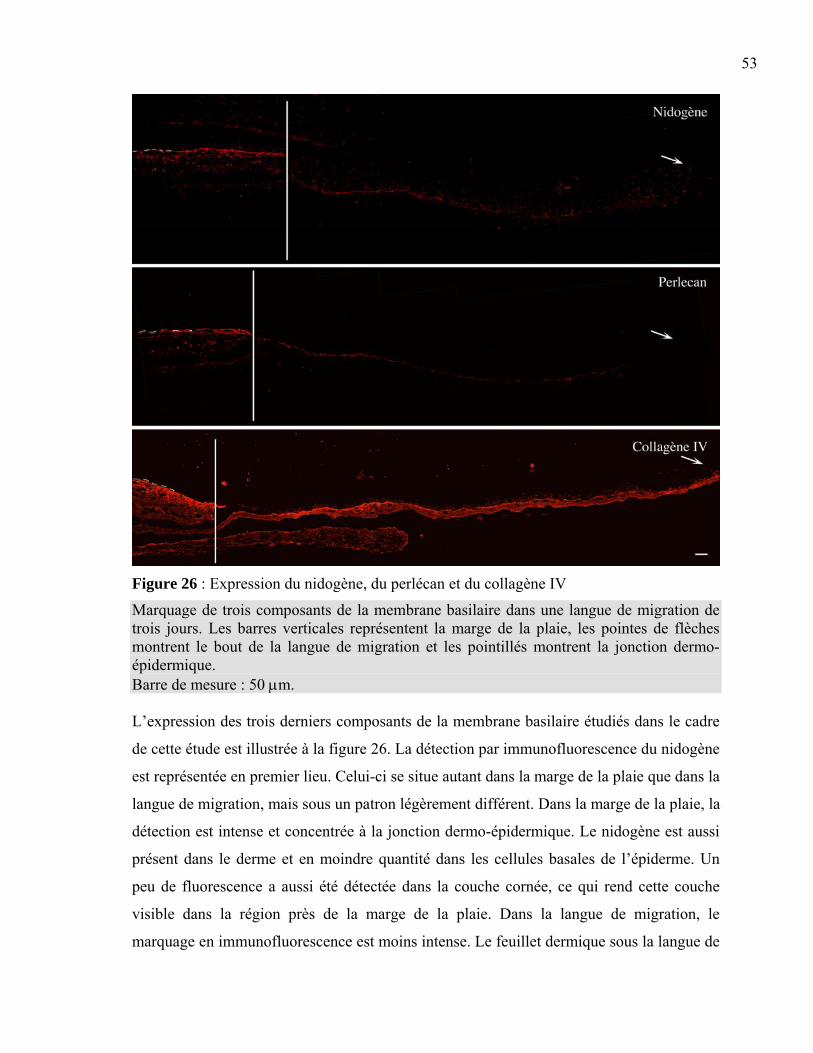
53
Figure 26 : Expression du nidogène, du perlécan et du collagène IV
Marquage de trois composants de la membrane basilaire dans une langue de migration de trois jours. Les barres verticales représentent la marge de la plaie, les pointes de flèches montrent le bout de la langue de migration et les pointillés montrent la jonction dermo-épidermique. Barre de mesure : 50 μm. L’expression des trois derniers composants de la membrane basilaire étudiés dans le cadre
de cette étude est illustrée à la figure 26. La détection par immunofluorescence du nidogène
est représentée en premier lieu. Celui-ci se situe autant dans la marge de la plaie que dans la
langue de migration, mais sous un patron légèrement différent. Dans la marge de la plaie, la
détection est intense et concentrée à la jonction dermo-épidermique. Le nidogène est aussi
présent dans le derme et en moindre quantité dans les cellules basales de l’épiderme. Un
peu de fluorescence a aussi été détectée dans la couche cornée, ce qui rend cette couche
visible dans la région près de la marge de la plaie. Dans la langue de migration, le
marquage en immunofluorescence est moins intense. Le feuillet dermique sous la langue de
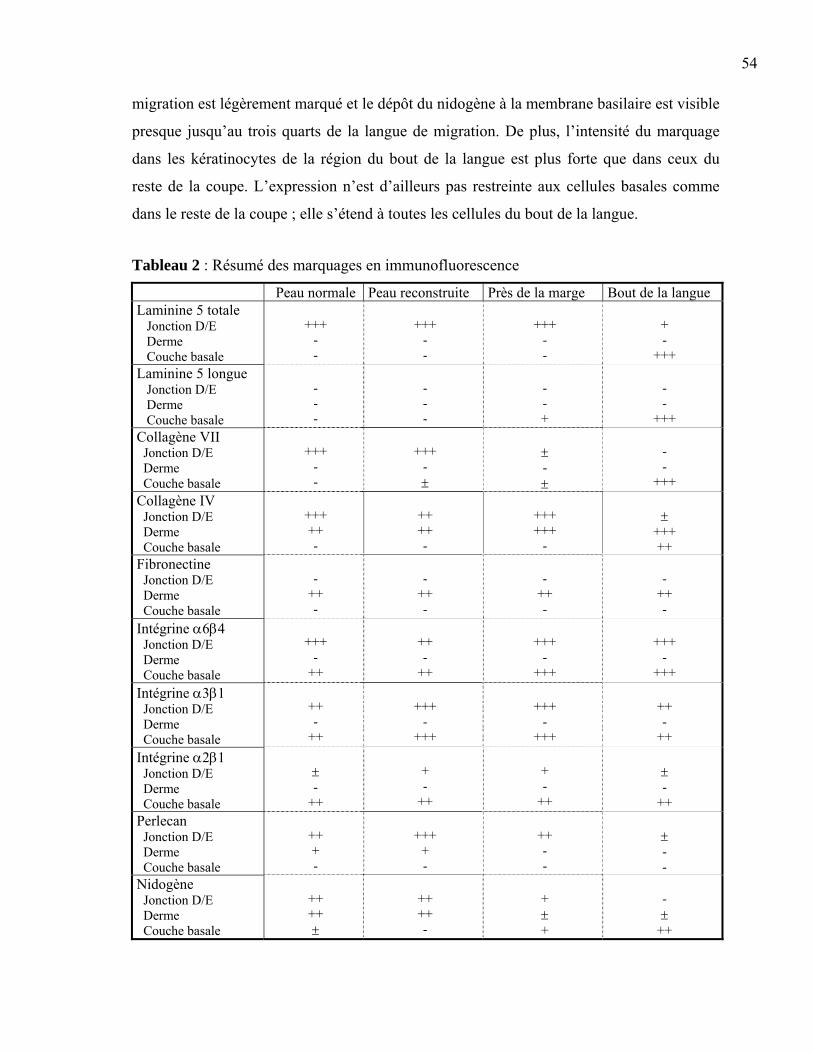
54
migration est légèrement marqué et le dépôt du nidogène à la membrane basilaire est visible
presque jusqu’au trois quarts de la langue de migration. De plus, l’intensité du marquage
dans les kératinocytes de la région du bout de la langue est plus forte que dans ceux du
reste de la coupe. L’expression n’est d’ailleurs pas restreinte aux cellules basales comme
dans le reste de la coupe ; elle s’étend à toutes les cellules du bout de la langue.
Tableau 2 : Résumé des marquages en immunofluorescence
Peau normale Peau reconstruite Près de la marge Bout de la langue Laminine 5 totale Jonction D/E Derme Couche basale
+++
- -
+++
- -
+++
- -
+ -
+++ Laminine 5 longue Jonction D/E Derme Couche basale
- - -
- - -
- - +
- -
+++ Collagène VII Jonction D/E Derme Couche basale
+++
- -
+++
- ±
± - ±
- -
+++ Collagène IV Jonction D/E Derme Couche basale
+++ ++ -
++ ++ -
+++ +++
-
±
+++ ++
Fibronectine Jonction D/E Derme Couche basale
-
++ -
-
++ -
-
++ -
-
++ -
Intégrine α6β4 Jonction D/E Derme Couche basale
+++
- ++
++ -
++
+++
- +++
+++
- +++
Intégrine α3β1 Jonction D/E Derme Couche basale
++ -
++
+++
- +++
+++
- +++
++ -
++ Intégrine α2β1 Jonction D/E Derme Couche basale
± -
++
+ -
++
+ -
++
± -
++ Perlecan Jonction D/E Derme Couche basale
++ + -
+++
+ -
++ - -
± - -
Nidogène Jonction D/E Derme Couche basale
++ ++ ±
++ ++ -
+ ± +
- ±
++

55
Le perlécan est exprimé surtout à la jonction dermo-épidermique, autant dans la marge de
la plaie que dans la langue de migration. La ligne de marquage est bien définie, mais non
continue vers le bout de la langue. Ce protéoglycan est aussi présent dans le derme, surtout
dans la marge de la plaie, mais jamais dans l’épiderme. Le dépôt dans la membrane
basilaire est présent presque jusqu’au bout de la langue de migration et diminue d’intensité
vers celle-ci.
L’expression du collagène IV est particulière. Elle est très intense dans le derme, ce qui
rend son analyse difficile. Il est toutefois possible de constater que l’intensité est plus forte
près de la membrane basilaire, et ce, presque jusqu’au bout de la langue de migration. La
présence du collagène IV dans les cellules de la couche basale est exclusive à la langue de
migration, mais elle perd de son intensité près de la marge de la plaie.
3.4 Migration des kératinocytes d’une plaie superficielle La guérison de plaies cutanées comporte plusieurs étapes essentielles. L’une d’elles est la
migration des cellules vers le centre de la plaie. Ce phénomène peut être visualisé grâce à la
technologie du Time Lapse. Cette technologie permet de prendre des photographies à des
intervalles réguliers, puis de les visualiser de façon accélérée sous forme de vidéos. Ces
vidéos peuvent ensuite être analysés afin de calculer une vitesse de migration pour chaque
condition. Dans la présente expérience, neuf conditions différentes ont été réalisées. Dans
chaque cas, du milieu DME-HAM a été utilisé, mais avec trois substrats nutritifs
différents : du sérum de veau Fetal Clone II (SV), du sérum humain (SH), ou de plasma
humain (PH). De plus, trois surfaces de migration différentes ont aussi été utilisées : le
plastique, la poly-D-lysine et le collagène.
Ces tests préliminaires ont été basés sur les expérimentations de l’équipe d’Henry (Henry et
al. 2003) afin de les appliquer éventuellement au modèle de peau reconstruite. Ces tests
constituent donc des essais en vue de la réalisation de tests plus poussés sur un modèle en
trois dimensions. Les résultats obtenus sont représentés sous forme graphique à la figure
27.
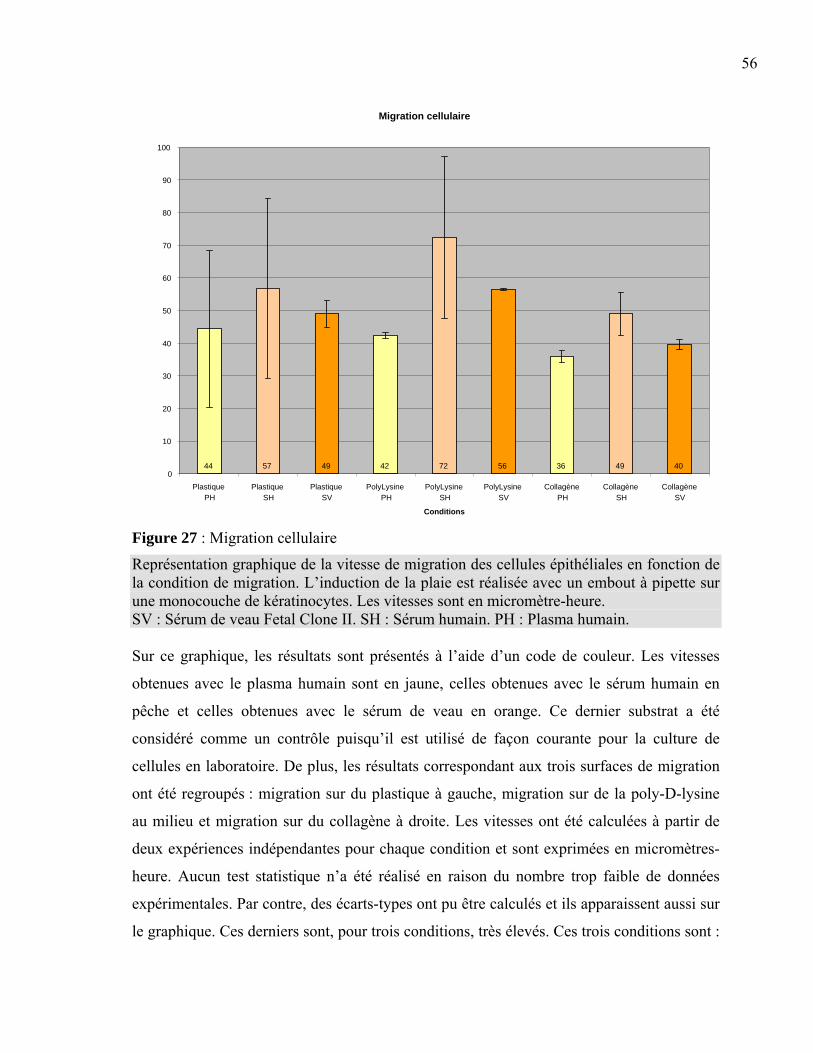
56
Migration cellulaire
44 57 49 42 72 56 36 49 400
10
20
30
40
50
60
70
80
90
100
PlastiquePH
PlastiqueSH
PlastiqueSV
PolyLysinePH
PolyLysineSH
PolyLysineSV
CollagènePH
CollagèneSH
CollagèneSV
Conditions
Figure 27 : Migration cellulaire
Représentation graphique de la vitesse de migration des cellules épithéliales en fonction de la condition de migration. L’induction de la plaie est réalisée avec un embout à pipette sur une monocouche de kératinocytes. Les vitesses sont en micromètre-heure. SV : Sérum de veau Fetal Clone II. SH : Sérum humain. PH : Plasma humain. Sur ce graphique, les résultats sont présentés à l’aide d’un code de couleur. Les vitesses
obtenues avec le plasma humain sont en jaune, celles obtenues avec le sérum humain en
pêche et celles obtenues avec le sérum de veau en orange. Ce dernier substrat a été
considéré comme un contrôle puisqu’il est utilisé de façon courante pour la culture de
cellules en laboratoire. De plus, les résultats correspondant aux trois surfaces de migration
ont été regroupés : migration sur du plastique à gauche, migration sur de la poly-D-lysine
au milieu et migration sur du collagène à droite. Les vitesses ont été calculées à partir de
deux expériences indépendantes pour chaque condition et sont exprimées en micromètres-
heure. Aucun test statistique n’a été réalisé en raison du nombre trop faible de données
expérimentales. Par contre, des écarts-types ont pu être calculés et ils apparaissent aussi sur
le graphique. Ces derniers sont, pour trois conditions, très élevés. Ces trois conditions sont :

57
en présence de plasma humain sur du plastique, en présence de sérum humain sur du
plastique, et en présence de sérum humain sur de la poly-D-lysine. Ces grands écarts-types
rendent ces résultats non significatifs et donc plus difficilement analysables. Il est toutefois
possible de constater que la migration est généralement plus rapide en présence de sérum
humain et plus lente en présence de plasma humain, et ce, pour les trois surfaces. Le sérum
de veau, qui constitue le contrôle, permet donc une vitesse de migration intermédiaire.
Pour ce qui est des surfaces de migration, un rang exact est plus difficile à définir. Aucune
différence significative n’a été observée entre les trois surfaces, quoique le collagène
semble ralentir un peu la migration.

CHAPITRE 4 : DISCUSSION

59
La membrane basilaire est essentielle à la cohésion dermo-épidermique dans la peau
humaine. Elle a une structure complexe qui est de plus en plus connue, mais il en reste
encore beaucoup à apprendre. Elle contient environ 50 différents constituants qui forment
trois couches bien distinctes et visibles au microscope électronique. L’arrivée de cette
technologie a d’ailleurs favorisé la caractérisation de la membrane basilaire et fait avancer
la recherche à son sujet. La majorité des études parues à ce jour analyse la membrane
basilaire dans la peau normale. La membrane basilaire est donc, dans ces cas-là, construite
depuis longtemps et son architecture est stable.
Une tout autre approche est suggérée ici. Il s’agit de l’utilisation d’un modèle de guérison
de plaie cutanée humaine fait à partir de la méthode d’auto-assemblage développée au
LOEX. Le contexte de guérison de plaie implique la régénération de la membrane basilaire,
ce qui constitue un modèle unique pour l’étude de sa structure et pour la compréhension de
sa formation. Grâce à ce modèle, il est possible de visualiser une membrane basilaire en
reconstruction par l’utilisation d’anticorps dirigés contre les principaux constituants. Cette
analyse est fascinante et permet d’en apprendre plus sur la membrane basilaire en plus
d’approfondir la compréhension des mécanismes impliqués dans la guérison des plaies.
4.1 Comparaison entre la peau normale humaine et la peau reconstruite
La peau reconstruite en laboratoire constitue un très bon modèle puisqu’elle est très
semblable à la peau normale humaine. Elle a déjà été caractérisée autant du point de vue
morphologique que moléculaire, ce qui a permis de constater plusieurs points de
ressemblance (Black et al. 1998; Laplante et al. 2001; Michel et al. 1999). Tout d’abord,
l’apparence macroscopique de la peau reconstruite est semblable à une peau humaine : la
zone épithélialisée est opaque et présente une texture comparable à celle de la peau
normale. De plus, les deux couches principales d’une peau normale, l’épiderme et le derme,
sont présentes. Il est important de noter que ces deux couches ne contiennent aucune
substance synthétique ou artificielle. Toute la matrice est en effet produite par les cellules
elles-mêmes, tout comme dans la peau normale humaine. De plus, l’épiderme de la peau
reconstruite est bien différencié : comme la peau normale, il contient une couche basale,

60
une couche épineuse, une couche granuleuse et une couche cornée. Une étude moléculaire a
d’ailleurs bien caractérisé l’expression de différents marqueurs de la différenciation dans ce
modèle de peau reconstruite (Michel et al. 1999). Le derme du modèle reconstruit est
moins épais que celui de la peau normale, mais il est composé de tissu conjonctif bien
organisé mimant un derme normal. L’épaisseur du derme dans le tissu reconstruit dépend
seulement du nombre de feuillets superposés lors de son montage. Il est donc possible de
produire une peau reconstruite ayant un derme plus épais.
Une différence majeure entre la peau normale humaine et la peau reconstruite est l’absence
d’annexes cutanées. Il n’y a en effet aucun poil, ni glande, ni vaisseau sanguin dans le tissu
reconstruit. L’absence de ces structures ne nuit apparemment pas à la formation de la peau
puisque la différenciation se fait normalement. Malgré tout, des poils ont déjà été implantés
avec succès dans ce modèle de peau reconstruite (Michel et al. 1999). De plus, des cellules
endothéliales ajoutées au modèle ont aussi mené à la formation de réseaux de vaisseaux
sanguins tridimensionnels dans le derme (Black et al. 1998; Rochon et al. 2004). Ces
structures ont évidemment des rôles majeurs à jouer dans le contexte métabolique d’un être
vivant, mais elles sont moins utiles pour l’analyse in vitro. Dans le cas des vaisseaux
sanguins, par exemple, un réseau de capillaires n’est pas nécessaire dans un modèle in vitro
puisqu’il n’y a pas de sang à transporter. La peau reconstruite est nourrie par le dessous,
puisqu’elle baigne sur un milieu nutritif. L’interface air/liquide permet ainsi de mimer
l’environnement d’une vraie peau, c’est-à-dire, sec sur le dessus et humide en dessous.
Cette technique permet donc une différenciation adéquate et un apport en nutriments
suffisant.
In vivo, les nutriments nécessaires aux kératinocytes diffusent de façon sélective des
vaisseaux sanguins, qui sont dans le derme, jusqu’à l’épiderme. Cette sélection est assurée
par la membrane basilaire, qui est perméable seulement pour les bons nutriments. Ce sont
surtout les protéoglycans, dont le perlécan est le plus représenté, formant des régions
anioniques dans la membrane qui sont responsables de sa perméabilité sélective. Vu les
fonctions majeures de la membrane basilaire dans la peau, il importe de vérifier si sa
composition est la même dans la peau reconstruite que dans la peau normale humaine. Dans
les études antérieures de caractérisation du modèle de peau reconstruite (Laplante et al.

61
2001; Michel et al. 1999), la membrane basilaire n’a été caractérisée que sommairement, en
ne considérant que peu de composants. En effet, seulement trois molécules, la laminine 5,
le collagène IV et le collagène VII, ont été observées pour la caractérisation du modèle.
Toutefois, une analyse complète en microscopie électronique a été réalisée, démontrant la
qualité structurale de la membrane basilaire du modèle de peau reconstruite : ses différentes
couches sont visibles de façon continue et elle contient de nombreux hémidesmosomes
(Laplante et al. 2001). Dans la présente étude, des marquages comparatifs des principaux
composants de la membrane basilaire ont été réalisés afin de compléter la caractérisation de
la peau reconstruite.
Les immunofluorescences effectuées sur les deux types de peau (figure 23) ont permis de
constater que la membrane basilaire de la peau reconstruite contenait les mêmes
composants que celle de la peau normale humaine. De plus, leur patron d’expression est
semblable dans tous les cas. Ainsi, les intégrines α2β1, α6β4 et α3β1, la laminine 5, les
collagènes IV et VII, le nidogène, le perlécan et la fibronectine sont exprimés dans le
modèle de peau reconstruite de façon similaire à la peau normale humaine. Cette
constatation suggère que la membrane basilaire des tissus reconstruits est fonctionnelle et
bien structurée. Cette analyse complète aussi la caractérisation du modèle de peau
reconstruite en laboratoire en confirmant un autre point de ressemblance qu’est la
composition de la membrane basilaire.
L’assurance que la membrane basilaire des peaux reconstruites possède une composition
moléculaire à l’image de la peau normale humaine est encourageante en raison de la grande
importance de cette structure dans la peau. De plus, l’expression de tous les composants
majeurs était nécessaire pour l’utilisation du modèle dans la poursuite des travaux.
4.2 Visualisation de la reconstruction de la membrane basilaire Pendant la guérison d’une plaie, il se produit une série d’événements menant à la fermeture
de l’espace créé et à la réorganisation des tissus. Ces événements nécessitent la production
de nouveau matériel afin de remplacer celui qui a été endommagé. La régénération de la
membrane basilaire n’échappe pas à ce phénomène, impliquant une augmentation de la

62
production de certains composants par les kératinocytes et les fibroblastes. La collaboration
entre ces deux types cellulaires pour la formation de la membrane basilaire est d’ailleurs
très importante. Une étude a effectivement démontré que l’interaction entre les cellules
épithéliales et mésenchymateuses était complexe et dynamique, mais surtout indispensable
à la régulation de la synthèse et à l’assemblage des constituants de la membrane basilaire
(Smola et al. 1998). Chaque type cellulaire contribue donc à la production de certains
composants nécessaires à la régénération de la membrane basilaire. En effectuant des
marquages de ces composants dans une langue de migration, il est fascinant de visualiser
cette nouvelle synthèse.
Tous les constituants ne sont pas néo-synthétisés lors de la guérison. Certains sont produits
de façon continue par les cellules, durant toute la vie de la peau. C’est le cas des intégrines
α2β1, α6β4 et α3β1, de la fibronectine, du perlécan et du nidogène. Il est cependant
important de noter que ce dernier est quand même produit de façon plus intense au bout de
la langue de migration par rapport au reste de la coupe, indiquant un besoin plus grand qu’à
l’habitude (figure26). Il est aussi produit par les fibroblastes du derme, mais sans variation
d’intensité, suggérant que ce type cellulaire ne répond pas nécessairement au besoin
grandissant. Les trois intégrines étudiées sont associées à des constituants de la membrane
basilaire. Elles sont toutes les trois exprimées dans l’épiderme seulement, indiquant une
production exclusive aux kératinocytes. Quant à la fibronectine (figure 25), elle est produite
par les fibroblastes du derme seulement et n’est pas présente à la jonction dermo-
épidermique puisqu’elle n’est pas un constituant comme tel de la membrane basilaire. Pour
ce qui est du perlécan (figure 26), il est lui aussi produit seulement par les fibroblastes et
aucune modulation de l’expression n’est observée. En résumé, à part le nidogène, qui
semble avoir une expression plus soutenue par les kératinocytes du bout de la langue de
migration, tous ces constituants sont synthétisés continuellement dans la peau et ne
semblent pas répondre de façon particulière au traumatisme engendré par l’induction de la
plaie.
En fait, les constituants nouvellement synthétisés sont les collagène IV et VII et la laminine
5. Il n’est donc pas surprenant qu’ils soient les trois constituants majeurs de la membrane
basilaire, en ce sens qu’ils forment les trois structures de base. En effet, le collagène IV

63
forme une des deux structures en filet responsable de la stabilité mécanique, le collagène
VII est le constituant principal des fibres d’ancrage et la laminine 5 fait partie de la
deuxième structure en filet en plus d’être le constituant principal des filaments d’ancrage
(Paulsson 1992). Ces trois constituants sont donc synthétisés lors de la formation de la
membrane basilaire et n’ont pas besoin d’être produits de façon continue puisqu’ils forment
des structures de base stables.
En considérant tout d’abord le collagène VII (figure 25), il est en effet possible de noter que
les kératinocytes de la langue de migration expriment cette protéine contrairement à ceux
de la marge de la plaie. Les kératinocytes en migration, et donc en processus de réparation
de la plaie, contribuent donc à la reconstruction de la membrane basilaire. Il est d’ailleurs
possible de voir un léger dépôt de collagène VII au niveau de la membrane basilaire dans la
partie qui est près de la marge de la plaie. La diminution de l’intensité du marquage dans
les cellules vers la marge signifie que le besoin en collagène VII est de moins en moins
important. Les cellules épithéliales répondent donc à cette baisse de la demande en
collagène VII en cessant graduellement de produire cette protéine.
Il en est de même pour le collagène IV (figure 26), qui n’est exprimé lui aussi que par les
kératinocytes de la langue de migration. Il est toutefois fortement exprimé dans le derme et
surtout dans la région près de la jonction dermo-épidermique. Ceci signifie que les
fibroblastes contribuent aussi à la production de collagène IV, mais de façon continue
puisqu’il est aussi retrouvé dans le derme de la marge de la plaie. Tout comme pour le
collagène VII, la même diminution d’intensité du marquage dans les kératinocytes basaux
est observée vers la marge de la plaie. Ce gradient coïncide avec l’augmentation de
l’intensité du dépôt du collagène IV à la jonction dermo-épidermique. Le dépôt est donc
fort dans la marge de la plaie et près de celle-ci et va en diminuant environ jusqu’à la moitié
de la langue de migration.
Pour ce qui est de la laminine 5, le marquage en immunofluorescence des deux formes
permet de mieux comprendre ses fonctions particulières dans la migration cellulaire (figure
24). L’expression de la laminine 5 totale a tout d’abord été observée grâce à un anticorps
dirigé contre une région ne subissant aucun clivage dans la structure de la protéine.
L’expression de la forme longue de la laminine 5 a été observée grâce à l’utilisation d’un

64
nouvel anticorps dirigé contre la partie clivée de la chaîne alpha-3 de cette protéine.
L’utilisation de ces deux anticorps est en effet très utile. Elle permet de distinguer la forme
longue de la forme totale, ce qui donne, par élimination, la localisation de la forme courte,
et ce, même si aucun anticorps n’est disponible pour celle-ci.
Ces deux marquages permettent d’en apprendre beaucoup sur les fonctions des clivages de
la laminine 5. Il apparaît en effet ici que la forme longue de la laminine 5 est produite
exclusivement par les kératinocytes en migration. Elle est effectivement retrouvée dans le
cytoplasme des cellules du début de la langue de migration. L’intensité du marquage
cellulaire diminue vers la marge, pour devenir nulle dans cette dernière, là où les cellules
sont stationnaires. Les cellules migratrices sécrètent donc la forme longue de la laminine 5,
ce qui permet leur migration, et cessent de la produire lorsqu’elles n’ont plus besoin de
migrer. Quant à la forme courte, elle n’est visible que de façon extracellulaire, à la jonction
dermo-épidermique. Ce résultat concorde avec ceux de l’équipe de Rousselle (Rousselle
2002) qui démontrent bien que les clivages que subit la laminine 5 surviennent une fois la
protéine sécrétée dans la matrice. Il est donc logique de retrouver la forme longue dans le
cytoplasme des cellules et de voir le dépôt de la forme courte dans la membrane basilaire.
Des études montrent d’ailleurs que la forme courte de la laminine 5 favorise l’état
stationnaire des kératinocytes en les ancrant fermement au derme (Goldfinger et al. 1998;
Hirosaki et al. 2000; Marinkovich et al. 1992).
De plus, le dépôt de la laminine 5 se fait très tôt dans le processus de guérison. Il est
effectivement visible presque jusqu’au bout de la langue de migration. En analysant le
dépôt des autres constituants de la même manière, il est possible d’établir une séquence
d’apparition de ceux-ci. L’obtention d’une telle séquence fournit des renseignements
précieux sur le processus par lequel la membrane basilaire se régénère. Ainsi, les
constituants présents à la jonction dermo-épidermique près du bout de la langue arrivent les
premiers dans la structure de la membrane basilaire, tandis que ceux qui sont présents à la
jonction de façon plus éloignée du bout apparaissent plus tardivement dans la structure de
la membrane. Il est donc possible d’affirmer que le premier constituant à contribuer à la
formation de la membrane basilaire est la laminine 5, vu sa présence dans toute la langue de
migration et même presque jusqu’au bout de celle-ci. La laminine 5 serait suivie de près par

65
le perlécan, qui est lui-même suivi par le collagène IV, ensuite par le nidogène et
finalement, le collagène VII. Cette séquence n’est évidemment pas totalement précise
puisqu’elle est basée sur une interprétation visuelle des marquages. De plus, chaque
marquage est fait sur une coupe différente, ce qui peut faire varier la longueur de la langue.
Un fort marquage dans le derme peut aussi rendre difficile sa distinction du marquage dans
la jonction dermo-épidermique. Cette étude donne tout de même une très bonne idée de
l’ordre d’assemblage de la membrane basilaire et les résultats obtenus sont en accord avec
ceux d’une autre étude ayant un but semblable (Fine et al. 1984).
En effet, cette étude effectuée sur des fœtus avortés a permis d’établir à quel stade
embryonnaire apparaissaient certains constituants analysés dans la présente étude (Fine et
al. 1984). Ces constituants sont les collagène IV et VII, la fibronectine et la laminine 5.
Cette équipe a déterminé que ces constituants apparaissaient tôt dans la peau des fœtus. En
effet, ces quatre protéines étaient détectées dès le premier stade de gestation. De plus, la
laminine 5, le collagène IV et la fibronectine étaient détectés dès la sixième semaine de
gestation, alors que le collagène VII n’était visible qu’à la dixième semaine. Les résultats
de cette étude donnent des indices intéressants quant à la chronologie de la formation de la
membrane basilaire de la peau humaine. Ils n’offrent par contre aucune précision sur
l’apparition des autres constituants étudiés dans la présente étude, ni sur la séquence exacte
des quatre premières protéines à être détectées. Il est aussi important de préciser que la
fibronectine n’était pas présente directement à la jonction dermo-épidermique, mais bien
sous la membrane basilaire, dans le derme, tout comme les résultats présentés ici (figure
25).
Les résultats obtenus grâce au marquage des constituants de la membrane basilaire
permettent aussi d’affirmer que la membrane basilaire se régénère très rapidement.
L’observation de la section de la langue de migration près de la marge de la plaie montre,
en effet, un marquage presque identique à celui de la marge de la plaie, et ce, pour tous les
constituants. Dans certains cas, le marquage est identique en tous points. C’est le cas des
trois intégrines, de la laminine 5 et du collagène IV. Pour le perlécan et le nidogène, le
marquage est semblable, mais l’intensité est plus faible dans la langue de migration. Pour
ce qui est du collagène VII, son marquage tarde un peu à s’intensifier. Ces résultats

66
signifient que la membrane basilaire est formée complètement après seulement trois jours
de guérison. Les quelques différences entre l’expression dans la zone près de la marge et
l’expression dans la marge elle-même suggèrent toutefois que l’assemblage et la finition
dans la réorganisation de la membrane basilaire ne sont pas encore tout à fait achevés. Il est
donc possible que, même si tous les constituants sont présents après trois jours, la structure
ne soit pas encore très solide. Il est tout de même évident que la régénération de la
membrane basilaire se fait très rapidement, ce qui est très positif, considérant l’importance
de cette structure.
4.3 Observation de la migration des kératinocytes en monocouche
La migration cellulaire est une étape majeure dans la guérison cutanée. L’utilisation du
modèle de peau reconstruite constitue un outil idéal pour l’étude des mécanismes physiques
impliqués dans la migration cellulaire. Des tests préliminaires ont donc été effectués afin
d’évaluer les conditions optimales pour l’utilisation de la technologie du Time Lapse. Ces
tests ont été basés sur une étude effectuée en monocouche avec des kératinocytes humains
soumis à différentes conditions de culture (Henry et al. 2003). Le choix des conditions a été
basé sur l’environnement d’une plaie cutanée : lors de l’induction d’une plaie, les
kératinocytes sont pour la première fois exposés et entourés par du sérum plutôt que par du
plasma. L’équipe d’Henry suggère donc que le sérum contiendrait des promoteurs de la
migration cellulaire qui sont absents du plasma. Neuf conditions différentes ont été utilisées
afin de pouvoir analyser la migration des cellules épithéliales.
Les kératinocytes ont donc été exposés à trois milieux nutritifs différents et à trois surfaces
de migration différentes. Le sérum bovin a été utilisé à titre de substrat contrôle, de même
que le plastique comme surface contrôle. Les trois différentes surfaces ont été utilisées
seulement à titre comparatif pour les substrats et comme matrice pour faciliter la migration.
Les résultats obtenus sont représentatifs de deux expériences indépendantes, mais d’autres
essais seraient nécessaires afin de préciser les données. En effet, pour trois conditions, les
résultats présentent un écart-type très élevé (figure 27). Il s’agit de la migration en présence
de sérum humain sur du plastique, en présence de plasma humain sur du plastique et en

67
présence de sérum humain sur de la poly-D-lysine. Ces variations dans les vitesses
obtenues peuvent être expliquées par la difficulté d’obtenir un environnement stable à
l’intérieur de la chambre d’incubation du microscope. Il est effectivement impossible pour
l’instant de contrôler de façon précise le taux de dioxyde de carbone et le taux d’humidité.
Ces variations d’environnement peuvent certainement modifier le comportement des
cellules et ainsi changer leur réaction face à l’induction de la plaie. La vitesse de migration
s’en trouve alors changée elle aussi, menant à des écarts-type plus grands que prévu.
L’utilisation de substrats non commerciaux a aussi pu être un facteur de variabilité. Les
grands écarts-type ont en effet été observés seulement dans les cas où du sérum et du
plasma humain ont été utilisés dans le milieu. Or, ce plasma et ce sérum ont été obtenus à
partir du sang de donneurs volontaires. Quoique tous les échantillons aient été mis en
commun pour rendre les substrats homogènes, aucune analyse particulière n’a été effectuée
pour assurer la qualité de ces substrats.
Il est tout de même possible d’établir une tendance générale à partir des résultats obtenus. Il
était attendu que le sérum humain, étant plus riche que le plasma, favoriserait la migration
des kératinocytes. C’est d’ailleurs ce substrat qui permettait la meilleure migration dans
l’étude d’Henry (Henry et al. 2003). Dans la présente étude, le sérum humain semble
effectivement avoir permis une migration plus rapide et ce, pour toutes les surfaces, tandis
que le plasma humain semble avoir ralenti la migration cellulaire. Le sérum bovin, qui
représentait le contrôle, a permis aux cellules de migrer à des vitesses intermédiaires entre
le sérum humain et le plasma humain. Ce dernier résultat est très révélateur puisqu’il
indique que les cellules préféreraient être nourries par du sérum, mais préfèreraient encore
plus un substrat d’origine humaine. Cette constatation est importante puisque la majorité
des cultures en laboratoire est faite avec du sérum bovin. Il est évident que la transition des
cultures à grande échelle vers l’utilisation de sérum humain est pratiquement impossible,
considérant le coût élevé du matériel humain. Cette transition pourrait toutefois être très
utile dans le cas de cultures de cellules humaines plus capricieuses ou encore pour la
confirmation finale de résultats. Un des grands avantages du modèle de peau reconstruite
utilisé au LOEX est sa composition entièrement humaine et biologique. La seule source
animale de ce modèle constitue le sérum bovin utilisé dans le milieu de culture.

68
L’utilisation de sérum humain dans certaines études pourrait assurément augmenter la
crédibilité déjà bien établie du modèle.
Une légère variabilité a aussi été observée pour les trois différentes surfaces de migration.
Aucune différence majeure n’était attendue, mais il s’est avéré que le collagène était le
substrat permettant la moins grande vitesse. Quoique la différence soit mineure, ce résultat
est plutôt surprenant, considérant que le collagène est une surface couramment utilisée dans
les essais de migration cellulaire (Drukala et al. 2001; Li et al. 2001; Voroteliak et al.
2002).
Ces tests préliminaires sont prometteurs pour la réalisation des expérimentations en trois
dimensions. Ils ont permis d’établir les conditions optimales et les changements nécessaires
à apporter à la chambre d’incubation. Des études plus approfondies qui permettront de
visualiser la fermeture d’une plaie en trois dimensions sont donc à prévoir. Les vidéos
réalisées au Time Lapse permettent bien sûr de voir la migration des kératinocytes, mais la
monocouche ne permet pas de voir le comportement des cellules dans un épithélium
stratifié. L’analyse des coupes des langues de migration a en effet permis de constater que
les cellules se déplaçaient par deux mécanismes : par glissement et par roulement. Cette
caractéristique des kératinocytes a d’ailleurs été étudiée dans une étude antérieure (Laplante
et al. 2001). L’étude de ces mécanismes par la technologie du Time Lapse fournirait enfin
les réponses aux incertitudes à ce sujet. Une étude en trois dimensions a d’ailleurs donné
des résultats concluants dans le cas de la cornée (Zhao et al. 2003). Les résultats
préliminaires présentés ici fournissent donc tous les outils afin d’entreprendre une étude
plus poussée.

CHAPITRE 5 : CONCLUSION ET PERSPECTIVES

70
La membrane basilaire est sans contredit une structure très importante. Dans la peau
humaine, elle assure non seulement l’attachement solide de l’épiderme au derme, mais elle
permet aussi la diffusion des bons nutriments des vaisseaux sanguins jusqu’aux
kératinocytes. La peau humaine peut être endommagée de différentes façons, ce qui rend
nécessaire l’existence d’un mécanisme efficace de réparation. La peau doit en effet être
réparée rapidement pour rétablir sa structure et, par le fait même, sa fonctionnalité. La
membrane basilaire n’échappe pas à ce processus. C’est pourquoi l’utilisation d’un modèle
de peau reconstruite en guérison constitue un excellent support pour l’étude de sa structure.
Le modèle de peau reconstruite développé au LOEX a déjà fait ses preuves : il ressemble
beaucoup à la peau normale humaine, il représente le modèle de peau reconstruite en
laboratoire qui se rapproche le plus de la réalité en ne contenant aucun matériel exogène et
non humain, en plus de produire tous les constituants de la membrane basilaire. Son
utilisation pour l’étude de la régénération de la membrane basilaire était donc tout à fait
appropriée.
Dans le cadre de cette étude, il a donc été possible d’utiliser une peau reconstruite par la
méthode d’auto-assemblage qui représente un modèle pratique et efficace pour visualiser la
formation de la membrane basilaire. Les résultats obtenus ont permis de finaliser la
caractérisation du modèle, en poussant plus loin les investigations sur la structure de la
membrane basilaire.
L’analyse microscopique de la membrane basilaire en formation a aussi rendu possible
l’établissement d’une séquence d’apparition des constituants. Cette séquence enrichit les
minces connaissances acquises au cours des dernières années au sujet de la manière dont la
membrane basilaire se régénère. L’étude plus précise de la laminine 5 a aussi permis de
constater que son remodelage survenait tôt dans le processus de guérison. De plus, la
découverte que la forme longue de cette protéine n’était présente que dans les cellules en
migration et que la forme courte était déposée rapidement dans la membrane basilaire a
fourni la confirmation que l’ancrage des cellules était possible grâce à la laminine 5 clivée.
Il a ainsi été possible de conclure que le clivage de la laminine 5 contribuait à l’ancrage
solide des cellules épithéliales et permettait l’arrêt de la migration.

71
Tous ces résultats encouragent la poursuite de ces recherches. Il serait en effet intéressant
de pousser un peu plus loin l’analyse du remodelage de la laminine 5. Il subsiste encore
plusieurs interrogations quant aux responsables des clivages et à la manière dont ils sont
faits. Plusieurs hypothèses ont été élaborées, mais aucune réponse définitive n’a été trouvée
à ce jour. Plusieurs autres constituants de la jonction dermo-épidermique restent encore
dans l’ombre. Les constituants principaux ont bien sûr été analysés, mais il serait
intéressant d’approfondir l’étude des composants mineurs comme, par exemple, le
syndécan et la tenascine-C. Ils sont en effet mal caractérisés et leurs fonctions mal connues.
Une recherche plus précise pourrait toutefois leur attribuer des rôles importants dans les
fonctions de la peau humaine.
L’étude de la migration des kératinocytes en monocouche a eu comme conséquence de
mieux définir les conditions idéales pour la migration. Ainsi, quoique les résultats n’aient
pas été significativement différents, une tendance générale a pu être observée. Il a donc été
possible de constater que le sérum humain pourrait permettre une meilleure migration que
le sérum bovin, ce qui pourrait rendre possible la réalisation d’études plus difficiles. Des
corrections dans les installations pourront aussi être apportées afin d’optimiser les résultats
futurs. Par exemple, il serait essentiel de pouvoir contrôler tous les paramètres à l’intérieur
de la chambre d’incubation : le taux d’humidité et de dioxyde de carbone devraient être
calculés de la même manière que la température. Cet ajustement stabiliserait les conditions
à l’intérieur de la chambre d’incubation et permettrait d’obtenir des résultats moins
variables. La prochaine étape est bien sûr d’utiliser le modèle de peau reconstruite pour la
visualisation de la fermeture d’une plaie. Les résultats qui s’en suivront seront très
révélateurs et apporteront des réponses à des questions restées trop longtemps sans réponse.
Globalement, l’analyse microscopique de la régénération d’une membrane basilaire durant
une guérison de plaie a eu comme résultat l’approfondissement des connaissances quant à
la fonction des clivages de la laminine 5. Ces résultats, mis en commun, aboutissent
finalement à une meilleure compréhension des mécanismes impliqués dans la guérison de
plaies cutanées humaines.

Bibliographie
Amano, S., Scott, I. C., Takahara, K., Koch, M., Champliaud, M. F., Gerecke, D. R., Keene, D. R., Hudson, D. L., Nishiyama, T., Lee, S., Greenspan, D. S. et Burgeson, R. E., Bone morphogenetic protein 1 is an extracellular processing enzyme of the laminin 5 gamma 2 chain. J Biol Chem, 2000. 275(30): 22728-22735.
Amano, S., Akutsu, N., Matsunaga, Y., Kadoya, K., Nishiyama, T., Champliaud, M.-F., Burgeson, R. E. et Adachi, E., Importance of balance between extracellular matrix synthesis and degradation in basement membrane formation. Exp Cell Res, 2001. 271: 249-262.
Auger, F. A., Remy-Zolghadri, M., Grenier, G. et Germain, L., A truly new approach for tissue engineering: the LOEX self-assembly technique, in Stem cell transplantation and tissue engineering, A. Haverich H. Graf, Editors, Springer-Verlag, Berlin. 2002, Chapître 6 : 73-88.
Aumailley, M. et Krieg, T., Laminins: a family of diverse multifunctional molecules of basement membranes. J Invest Dermatol, 1996. 106: 209-214.
Aumailley, M. et Rousselle, P., Laminins of the dermo-epidermal junction. Matrix Biol, 1999. 18: 19-28.
Austria, M. R. et Couchman, J. R., Enhanced assembly of basement membrane matrix by endodermal cells in response to fibronectin substrata. J Cell Sci, 1991. 99: 443-451.
Black, A. F., Berthod, F., L'Heureux, N., Germain, L. et Auger, F. A., In vitro reconstruction of a human capillary-like network in a tissue-engineered skin equivalent. FASEB J, 1998. 12: 1331-1340.
Borradori, L. et Sonnenberg, A., Structure and function of hemidesmosomes: more than simple adhesion complexes. J Invest Dermatol, 1999. 112: 411-418.
Campbell, N. A., Biologie. Saint-Laurent: Éditions du Renouveau Pédagogique Inc. 1995, 1190 pages.
Christiano, A. M. et Uitto, J., Molecular complexity of the cutaneous basement membrane zone. Revelations from the paradigms of epidermolysis bullosa. Exp Dermatol, 1996. 5(1): 1-11.
Clark, R. A. F., Cutaneous tissue repair: Basic biologic considerations. I. J Am Acad Dermatol, 1985. 13(5): 701-725.
Colwell, A. S., Longaker, M. T. et Lorenz, H. P., Fetal wound healing. Front Biosci, 2003. 8: s1240-1248.
De, S. K., Reis, E. D. et Kerstein, M. D., Wound treatment with human skin equivalent. J Am Podiatr Med Assoc, 2002. 92(1): 19-23.

73
Decline, F. et Rousselle, P., Keratinocyte migration require a3b1 integrin-mediated interaction with the laminin 5 gamma-2 chain. J Cell Sci, 2000. 114: 811-823.
Dedhar, S., Jewell, K., Rojiani, M. et Gray, V., The receptor for the basement membrane glycoprotein entactin is the integrin alpha 3/beta 1. J Biol Chem, 1992. 267(26): 18908-18914.
deHart, G. W., Healy, K. E. et Jones, J. C. R., The role of α3β1 integrin in determining the supramolecular organization of laminin-5 in the extracellular matrix of keratinocytes. Exp Cell Res, 2003. 283: 67-79.
Doillon, C. J., Dunn, M. G., Berg, R. A. et Silver, F. H., Collagen deposition during wound repair. Scanning Elect Microsc, 1985. 11: 897-903.
Drukala, J., Bandura, L., Cieslik, K. et Korohoda, W., Locomotion of human skin keratinocytes on polystyrene, fibrin, and collagen substrata and its modification by cell-to-cell contacts. Cell Transplant, 2001. 10(8): 765-771.
Eady, R. A. J., The basement membrane. Interface between the epithelium and the dermis: structural features. Arch Dermatol, 1988. 124: 709-712.
Ekblom, P., Ekblom, M., Fecker, L., Klein, G., Zhang, H. Y., Kadoya, Y., Chu, M. L., Mayer, U. et Timpl, R., Role of mesenchymal nidogen for epithelial morphogenesis in vitro. Development, 1994. 120(7): 2003-2014.
Ekblom, P., Lonai, P. et Talts, J. F., Expression and biological role of laminin-1. Matrix Biol, 2003. 22(1): 35-47.
Ffrench-Constant, C. et Colognato, H., Integrins: versatile integrators of extracellular signals. Trends Cell Biol, 2004. 14(12): 678-686.
Fine, J.-D., Smith, L. T., Holbrook, K. A. et Katz, S. I., The appearance of four basement membrane zone antigens in developing human fetal skin. J Invest Dermatol, 1984. 83: 66-69.
Fleischmajer, R., Perlish, J. S., Macdonald II, E. D., Schechter, A., Murdoch, A. D., Iozzo, R. V. et Yamada, Y., There is binding of collagen IV to β1 integrin during early skin basement membrane assembly. Ann N Y Acad Sci, 1998. 857: 212-227.
Fleischmajer, R., Utani, A., MacDonald, E. D., Perlish, J. S., Pan, T. C., Chu, M. L., Nomizu, M., Ninomiya, Y. et Yamada, Y., Initiation of skin basement membrane formation at the epidermo-dermal interface involves assembly of laminins through binding to cell membrane receptors. J Cell Sci, 1998. 111(Pt 14): 1929-1940.
Fuki, I. V., Iozzo, R. V. et Williams, K. J., Perlecan heparan sulfate proteoglycan: a novel receptor that mediates a distinct pathway for ligand catabolism. J Biol Chem, 2000. 275(33): 25742-25750.

74
Gabbiani, G., Evolution and clinical implications of the myofibroblast concept. Cardiovasc Res, 1998. 38(3): 545-548.
Gagnoux-Palacios, L., Allegra, M., Spirito, F., Pommeret, O., Romero, C., Ortonne, J.-P. et Meneguzzi, G., The short arm of the laminin γ2 chain plays a pivotal role in the incorporation of laminin 5 into the extracellular matrix and in cell adhesion. J Cell Biol, 2001. 153(4): 835-849.
Gailit, J. et Clark, R. A. F., Wound repair in the context of extracellular matrix. Curr Opin Cell Biol, 1994. 6: 717-725.
Gartner, L. P. et Hiatt, J. L., Atlas d'histologie. 2e ed. Bruxelles: De Boeck-Wesmael. 1992, 320 pages.
Geras, A. J., Dermatology, a medical artist's interpretation. Basle: Sandoz Medical Publications. 1990, 139 pages.
Goldfinger, L. E., Stack, M. S. et Jones, J. C. R., Processing of laminini-5 and its functional consequences: role of plasmain and tissue-type plasminogen activator. J Cell Biol, 1998. 141(1): 255-265.
Goldfinger, L. E., Hopkinson, S. B., deHart, G. W., Collawn, S., Couchman, J. R. et Jones, J. C. R., The α3 laminin subunit, α6β4 and α3β1 integrin coordinately regulate wound healing in cultured epithelial cells and in the skin. J Cell Sci, 1999. 112: 2615-2629.
Goldsby, R. A., Kindt, T. J., Osborne, B. A., Kuby Immunology. 4e ed. New York: W. H. Freeman and company. 2000, 670 pages.
Gresham, H. D., Graham, I. L., Griffin, G. L., Hsieh, J. C., Dong, L. J., Chung, A. E. et Senior, R. M., Domain-specific interactions between entactin and neutrophil integrins. G2 domain ligation of integrin alpha3beta1 and E domain ligation of the leukocyte response integrin signal for different responses. J Biol Chem, 1996. 271(48): 30587-30594.
Grinnell, F., Fibroblasts, myofibroblasts, and wound contraction. J Cell Biol, 1994. 124(4): 401-404.
Groffen, A. J., Veerkamp, J. H., Monnens, L. A. et van den Heuvel, L. P., Recent insights into the structure and functions of heparan sulfate proteoglycans in the human glomerular basement membrane. Nephrol Dial Transplant, 1999. 14(9): 2119-2129.
Henry, G., Li, W., Garner, W. et Woodley, D. T., Migration of human keratinocytes in plasma and serum and wound re-epithelialisation. Lancet, 2003. 361(9357): 574-576.
Henry, M. D., Satz, J. S., Brakebusch, C., Costell, M., Gustafsson, E., Fassler, R. et Campbell, K. P., Distinct roles for dystroglycan, beta1 integrin and perlecan in cell surface laminin organization. J Cell Sci, 2001. 114(Pt 6): 1137-1144.

75
Hirosaki, T., Mizushima, H., Tsubota, Y., Moriyama, K. et Miyazaki, K., Structural requirement of carboxyl-terminal globular domains of laminin alpha 3 chain for promotion of rapid cell adhesion and migration by laminin-5. J Biol Chem, 2000. 275(29): 22495-22502.
Hunt, T. K., Hopf, H. et Hussain, Z., Physiology of wound healing. Adv Skin Wound Care, 2000. 13(2 Suppl): 6-11.
Hynes, R. O., Integrins: bidirectional, allosteric signaling machines. Cell, 2002. 110(6): 673-87.
Kainulainen, T., Hakkinen, L., Hamidi, S., Larjava, K., Kallioinen, M., Peltonen, J., Salo, T., Larjava, H. et Oikarinen, A., Laminin-5 expression is independent of the injury and the microenvironment during reepithelialization of wounds. J Histochem Cytochem, 1998. 46(3): 353-360.
Kanitakis, J., Immunohistochemistry of normal human skin. Eur J Dermatol, 1998. 8(8): 539-547.
Kanitakis, J., Anatomy, histology and immunohistochemistry of normal human skin. Eur J Dermatol, 2002. 12(4): 390-401.
Kohfeldt, E., Sasaki, T., Gohring, W. et Timpl, R., Nidogen-2: a new basement membrane protein with diverse binding properties. J Mol Biol, 1998. 282(1): 99-109.
Kurpakus, M. A., Quaranta, V. et Jones, J. C. R., Surface relocation of alpha6beta4 integrins and assembly of hemidesmosomes in an in vitro model of wound healing. J Cell Biol, 1991. 115(6): 1737-1750.
Lampe, P. D., Mguyen, B. P., Gil, S., Usui, M., Olerud, J., Takada, Y. et Carter, W. G., Cellular interaction of integrin α3β1 with laminin 5 promotes Gap junctional communication. J Cell Biol, 1998. 143(6): 1735-1747.
Laplante, A., Germain, L., Auger, F. et Moulin, V., Mechanisms of wound reepithelialization: hints from a tissue-engineered reconstructed skin to long-standing questions. FASEB J, 2001. 15: 2377-2389.
Larjava, H., Salo, T., Haapasalmi, K., Kramer, R. H. et Heino, J., Expression of integrins and basement membrane components by wound keratinocytes. J Clin Invest, 1993. 92(3): 1425-1435.
Li, W., Nadelman, C., Henry, G., Fan, J., Muellenhoff, M., Medina, E., Gratch, N. S., Chen, M., Han, J. et Woodley, D., The p38-MAPK/SAPK pathway is required for human keratinocyte migration on dermal collagen. J Invest Dermatol, 2001. 117(6): 1601-1611.
Li, W., Dasgeb, B., Phillips, T., Li, Y., Chen, M., Garner, W. et Woodley, D. T., Wound-healing perspectives. Dermatol Clin, 2005. 23(2): 181-192.

76
Lodish, H., Baltimore, D., Bark, A., Zipursky, S. L., Matsudaira, P. et Darnell, J., Biologie moléculaire de la cellule. 3e ed. Bruxelles: De Boeck Université. 1997, 1344 pages.
Loomis, C. A., Development and morphogenesis of the skin. Adv Dermatol, 2001. 17: 183-210.
Marinkovich, M. P., Lunstrum, G. P. et Burgeson, R. E., The anchoring filament protein kalinin is syntheiszed and secreted as a high molecular weight precursor. J Biol Chem, 1992. 267(25): 17900-17906.
Martin, P., Hopkinson-Woolley, J. et McCluskey, J., Growth factors and cutaneous wound repair. Prog Growth Factor Res, 1992. 4(1): 25-44.
Martin, P., Wound healing--aiming for perfect skin regeneration. Science, 1997. 276(5309): 75-81.
Mayer, U., Poschl, E., Gerecke, D. R., Wagman, D. W., Burgeson, R. E. et Timpl, R., Low nidogen affinity of laminin-5 can be attributed to two serine residues in EGF-like motif gamma 2III4. FEBS Lett, 1995. 365(2-3): 129-132.
McMillan, J. R., Akiyama, M. et Shimizu, H., Ultrastructural orientation of laminin 5 in the epidermal basement membrane: an updated model for basement membrane organization. J Histochem Cytochem, 2003. 51(10): 1299-1306.
Mercurio, A. M., Rabinovitz, I. et Shaw, L. M., The alpha 6 beta 4 integrin and epithelial cell migration. Curr Opin Cell Biol, 2001. 13(5): 541-545.
Michel, M., L'Heureux, N., Pouliot, R., Xu, W., Auger, F. A. et Germain, L., Characterization of a new tissue-engineered human skin equivalent with hair. In Vitro Cell Dev Biol. Animal, 1999. 35: 318-326.
Mohan, P., S. et Spiro, R., G., Macromolecular Organisation of Basement Membranes. J Biol Chem, 1986. 261(9): 4328-4336.
Moll, R. et Moll, I., Epidermal adhesion molecules and basement membrane components as target structure of autoimmunity. Virchows Arch, 1998. 432: 487-504.
Nguyen, B. P., Gil, S. G. et Carter, W. G., Deposition of laminin 5 by keratinocytes regulates integrin adhesion and signaling. J Biol Chem, 2000. 275(41): 31896-31907.
Nguyen, B. P., Ren, X.-D., Schwartz, M. A. et Carter, W. G., Ligation of integrin a3b1 by laminin 5 at the wound edge activates rho-dependent adhesion of leading keratinocytes on collagen. J Biol Chem, 2001. 276(47): 43860-43870.
Nishiyama, T., Amano, S., Tsunenaga, M., Kadoya, K., Takeda, A., Adachi, E. et Burgeson, R. E., The importance of laminin 5 in the dermal-epidermal basement membrane. J Dermatol Sci, 2000. 25(suppl. 1): S51-S59.

77
O'Toole, E. A., Marinkovich, M. P., Hoeffler, W. K., Furthmayr, H. et Woodley, D. T., Laminin-5 inhibits human keratinocyte migration. Exp Cell Res, 1997. 233: 330-339.
Osawa, T., Feng, X.-Y. et Nozaka, Y., Scanning electron microscopic observations of the basement membranes with dithiothreitol separation. Med Elect Micros, 2003. 2003(36): 132-138.
Paulsson, M., Basement membrane proteins: structure, assembly, and cellular interactions. Crit Rev Biochem Mol Biol, 1992. 27(1-2): 93-127.
Pilcher, B. K., Wang, M., Qin, X.-J., Parks, W. C., Senior, R. M. et Welgus, H. G., Role of matrix metalloproteinases and their inhibition in cutaneous wound healing and allergic contact hypersensitivity. Ann N Y Acad Sci, 1999. 878: 12-24.
Pomahac, B., Svenjö, T., Yao, F., Brown, H. et Erikson, E., Tissu engineering of skin. Cri Rev Oral Biol Med, 1998. 9(3): 333-344.
Rabinovitz, I. et Mercurio, A. M., The integrin alpha6beta4 functions in carcinoma cell migration on laminin-1 by mediating the formation and stabilization of actin-containing motility structures. J Cell Biol, 1997. 139(7): 1873-1884.
Rochon, M. H., Fortin, V., Roberge, C. J., Auger, F. A. et Germain, L., Normal human epithelial cells regulate the size of capillaries through the production of VEGF. J Clin Invest (Soumis), 2004.
Rousselle, P., Lunstrum, G. P., Keene, D. R. et Burgeson, R. E., Kalinin: an epithelium-specific basement membrane adhesion molecule that is a component of anchoring filaments. J Cell Biol, 1991. 114: 567-576.
Rousselle, P., Laminine 5, migration cellulaire et cancer. Med Sci, 2002. 18: 989-994.
Ruszczak, Z. et Schwartz, R. A., Modern aspects of wound healing: an update. Dermatol Surg, 2000. 26(3): 219-229.
Ryan, M. C. et Christiano, A. M., The functions of laminins: lessons from in vivo studies. Matrix Biol, 1996. 15(6): 369-381.
Ryan, M. C., Lee, K., Miyashita, Y. et Carter, W. G., Targeted disruption of the LAMA3 gene in mice reveals abnormalities in survival and late stage differentiation of epithelial cells. J Cell Biol, 1999. 145(6): 1309-1323.
Sasaki, T., Gohring, W., Mann, K., Brakebusch, C., Yamada, Y., Fassler, R. et Timpl, R., Short arm region of laminin-5 gamma2 chain: structure, mechanism of processing and binding to heparin and proteins. J Mol Biol, 2001. 314(4): 751-763.
Serini, G. et Gabbiani, G., Mechanisms of myofibroblast activity and phenotypic modulation. Exp Cell Res, 1999. 250(2): 273-283.

78
Smith, K. L. et Dean, S. J., Tissue repair of the epidermis and dermis. J Hand Ther, 1998: 95-104.
Smola, H., Stark, H.-J., Thiekötter, G., Mirancea, N., Krieg, T. et Fusenig, N. E., Dynamics of basement membrane formation by keratinocyte-fibroblast interactions in organotypic skin culture. Exp Cell Res, 1998. 239: 399-410.
Sollberg, S., Peltonen, J. et Uitto, J., Differential expression of laminin isoforms and b4 integrin epitopes in the basement membrane zone of normal human skin and basal cell carcinomas. J Invest Dermatol, 1992. 98: 864-870.
Stevens, A. et Lowe, J., Histologie humaine. 2e ed. Paris: De Boeck et Larcier. 1997, 408 pages.
Timpl, R. et Dziadek, M., Structure, development, and molecular pathology of basement membranes. Int Rev Exp Pathol, 1986. 29: 1-112.
Timpl, R. et Brown, J. C., Supramolecular assembly of basement membranes. BioEssays, 1996. 18(2): 123-132.
Tortora, G. J., Grabowski, S. R. et Parent, J., Principes d'anatomie et de physiologie. Anjou: Centre Éducatif et Culturel inc. 1994, 1203 pages.
Tsubota, Y., Mizushima, H., Hirosaki, T., Higashi, S., Yasumitsu, H. et Miyazaki, K., Isolation and activity of proteolytic fragment of laminin-5 alpha3 chain. Biochem Biophys Res Commun, 2000. 278(3): 614-620.
Tunggal, L., Ravaux, J., Pesch, M., Smola, H., Krieg, T., Gaill, F., Sasaki, T., Timpl, R., Mauch, C. et Aumailley, M., Defective laminin 5 processing in cylindroma cells. Am J Pathol, 2002. 160(2): 459-468.
Uitto, J. et Pulkkinen, L., Molecular complexity of the cutaneous basement membrane zone. Mol Biol Rep, 1996. 23: 35-46.
Utani, A., Nomizu, M., Matsuura, H., Kato, K., Kobayashi, T., Takeda, U., Aota, S., Nielsen, P. K. et Shinkai, H., A unique sequence of the laminine α3 G domain binds to hepatin and promotes cell adhesion through syndecan-2 and -4. J Biol Chem, 2001. 276(31): 28779-28788.
Veitch, D. P., Nokelainen, P., McGowan, K. A., Nguyen, T. T., Nguyen, N. E., Stephenson, R., Pappano, W. N., Keene, D. R., Spong, S. M., Greenspan, D. S., Findell, P. R. et Marinkovich, M. P., Mammalian tolloid metalloproteinase, and not matrix metalloprotease 2 or membrane type 1 metalloprotease, processes laminin-5 in keratinocytes and skin. J Biol Chem, 2003. 278(18): 15661-15668.
Virtanen, I., Gullberg, D., Rissanen, J., Kivilaakso, E., Kiviluoto, T., Laitinen, L. A., Lehto, V. P. et Ekblom, P., Laminin alpha1-chain shows a restricted distribution in epithelial

79
basement membranes of fetal and adult human tissues. Exp Cell Res, 2000. 257(2): 298-309.
Voroteliak, E. A., Shikhverdieva, A., Vasil'ev, A. V. et Terskikh, V. V., Simulation of the migration process of human epidermal keratinocytes over three-dimensional collagen gel. Izv Akad Nauk Ser Biol, 2002(1): 30-37.
Witte, M. B. et Barbul, A., General principles of wound healing. Surg Clin North Am, 1997. 77(3): 509-528.
Woodley, D. T., Importance of the dermal-epidermal junction and recent advances. Dermatologica, 1987. 174: 1-10.
Wysocki, A. B., Skin anatomy, physiology, and pathophysiology. Nurs Clin North Am, 1999. 34(4): 777-797.
Yamaguchi, Y. et Yoshikawa, K., Cutaneous wound healing: an update. J Dermatol, 2001. 28(10): 521-534.
Yurchenco, P. D. et Schittny, J. C., Molecular architecture of basement membranes. FASEB J, 1990. 4(6): 1577-1590.
Zhao, M., Song, B., Pu, J., Forrester, J. V. et McCaig, C. D., Direct visualization of a stratified epithelium reveals that wounds heal by unified sliding of cell sheets. FASEB J, 2003. 17(3): 397-406.