
Vie et mort des cellules dans les tissus
I. L'épiderme et son renouvellement par les cellules souches
II. Épithélium sensorielIII. Voies aériennes et intestinIV. Vaisseaux sanguins et cellules endothélialesV. Renouvellement par des cellules souches
multipotentes : la formation des cellules sanguinesVI. Genèse : modulation et régénération du muscle
squelettiqueVII. Les fibroblastes et leurs transformations : la famille
des cellules du tissu conjonctifVIII. Ingénierie des cellules souches

Vie et mort des cellules dans les tissus
• Être unicellulaire : individu originel• Être pluricellulaire : cellules au
service du corps tout entier• Plus de 200 types de cellules
différents dans l’organisme

3
Cells of the Adult Human Body : a Catalogue
• How many distinct cell types are there in an adult human being? In other words, how many normal adult ways are there of expressing the human genome? A large textbook of histology will mention about 200 cell types that qualify for individual names. These traditional names are not, like the names of colors, labels for parts of a continuum that has been subdivided arbitrarily: they represent, for the most part, discrete and distinctly different categories. Within a given category there is often some variation—the skeletal muscle fibers that move the eyeball are small, while those that move the leg are big; auditory hair cells in different parts of the ear may be tuned to different frequencies of sound; and so on. But there is no continuum of adult cell types intermediate in character between, say, the muscle cell and the auditory hair cell.
• The traditional histological classification is based on the shape and structure of the cell as seen in the microscope and on its chemical nature as assessed very crudely from its affinities for various stains. Subtler methods reveal new subdivisions within the traditional classification. Thus modern immunology has shown that the old category of “lymphocyte” includes more than 10 quite distinct cell types. Similarly, pharmacological and physiological tests reveal that there are many varieties of smooth muscle cell—those in the wall of the uterus, for example, are highly sensitive to estrogen, and in the later stages of pregnancy to oxytocin, while those in the wall of the gut are not. Another major type of diversity is revealed by embryological experiments of the sort discussed in Chapter 21. These show that, in many cases, apparently similar cells from different regions of the body are nonequivalent, that is, they are inherently different in their developmental capacities and in their effects on other cells. Thus, within categories such as “fibroblast” there are probably many distinct cell types, different chemically in ways that are not easy to perceive directly.
• For these reasons any classification of the cell types in the body must be somewhat arbitrary with respect to the fineness of its subdivisions. Here, we list only the adult human cell types that a histology text would recognize to be different, grouped into families roughly according to function. We have not attempted to subdivide the class of neurons of the central nervous system. Also, where a single cell type such as the keratinocyte is conventionally given a succession of different names as it matures, we give only two entries—one for the differentiating cell and one for the stem cell. With these serious provisos, the 210 varieties of cells in the catalogue represent a more or less exhaustive list of the distinctive ways in which a given mammalian genome can be expressed in the phenotype of a normal cell of the adult body.

4
http://www.garlandscience.com/textbooks/0815332181/pdfs/
appendix.pdf

5
http://www.garlandscience.com/textbooks/0815332181/pdfs/appendix.pdf

6
Tissu• Association des ces types cellulaires
qui collaborent entre elles• Forment des organes

7
Conséquences du contrôle de l’expression des gènes et des mécanismes du développement animal
• Création de la diversification cellulaire dans l’embryon par des mécanismes génétiques moléculaires
• Maintien de la diversification des cellules grâce au dialogue et à la mémoire des cellules
• Construction des tissus par la matrice extra cellulaire• Mode de vie des cellules spécialisées

8
Questions posées
• Comment les cellules collaborent entre elles pour exécuter leur tâche ?
• Comment naissent vivent et meurent les nouvelles cellules spécialisées ?
• Comment est préservée l’architecture des nouveaux tissus malgré leur perpétuel remaniement ?

9
Réponses diverses
• Exemples illustrant les principes généraux
• Intéressants par l’originalité de leurs moyens d’étude
• Nombreux problèmes non résolus

Plan
I. L'épiderme et son renouvellement par les cellules souches
II. Épithélium sensorielIII. Voies aériennes et intestinIV. Vaisseaux sanguins et cellules endothélialesV. Renouvellement par des cellules souches
multipotentes : la formation des cellules sanguinesVI. Genèse : modulation et régénération du muscle
squelettiqueVII. Les fibroblastes et leurs transformations : la famille
des cellules du tissu conjonctifVIII. Ingénierie des cellules souches

11
L'épiderme et son renouvellement par les cellules souches

12
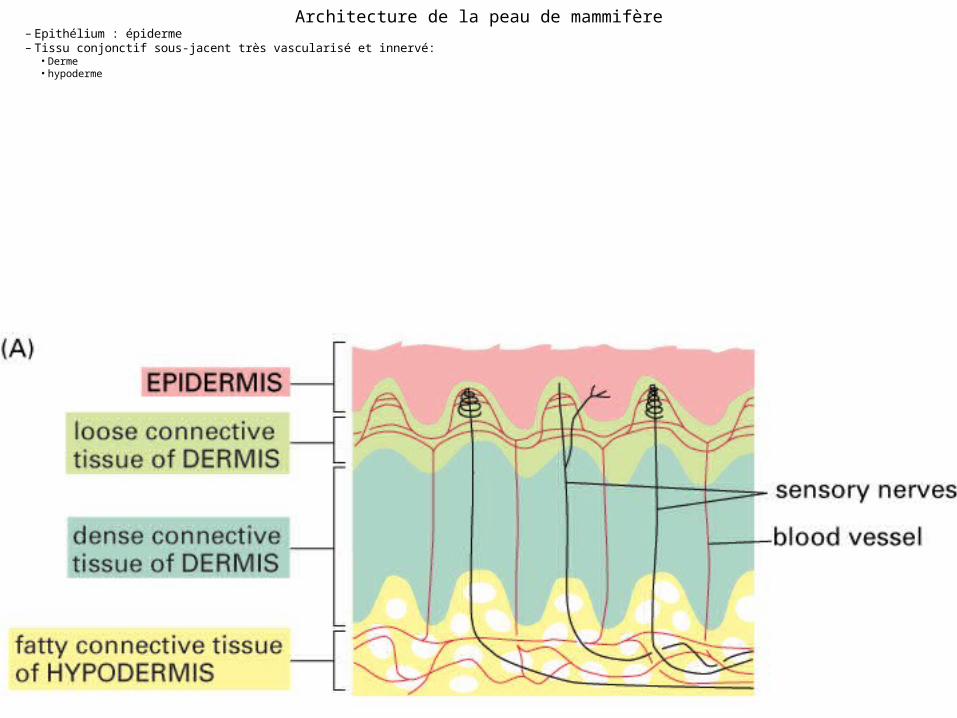
Architecture de la peau de mammifère
• Epithélium : épiderme• Tissu conjonctif sous-jacent très vascularisé et
innervé:– Dense (dont on fait le cuir) : derme – Tissu adipeux sous-cutané : hypoderme

13
Caractères généraux de l’épiderme(communs à tous les tissus)
1. Fibroblastes2. Vascularisation3. Fibres nerveuses

14
1 - Fibroblastes
• force mécanique• Grâce à la matrice extra-cellulaire
(sécrétée par les fibroblastes)

15
2 - Vascularisation
• Cellules endothéliales
• Échanges O2, CO2, nutriments, déchets
• Défense par apport de– Macrophages– Cellules dendritiques
• Phagocytose des agents infectieux• Activation des lymphocytes

16
3 - Fibres nerveuses
• Information sensorielle au système nerveux central
• Sécrétion glandulaire• Contraction musculaire
17
Fig 22-1 A
Architecture de la peau de mammifère– Epithélium : épiderme– Tissu conjonctif sous-jacent très vascularisé et innervé:
• Derme• hypoderme

18
Fig 22-1 B
• Coupe microscopique d’une plante de pied humain (Hématoxyline - Éosine)

19
Fig 22-1 B
Coupe microscopique d’une plante de pied humain (HE)

20
Épiderme
•Spécificité de la peau•Organisation simple•Beau modèle de renouvellement des tissus comme chez l’embryon

21
1 - Structure de l’épiderme

22
L’épiderme•Très exposé aux agressions
extérieures •Nécessité de réparation et
renouvellement permanent•Constitué de kératinocytes (+
quelques autres types cellulaires)•Épithélium pavimenteux
pluristratifié (=malpighien)

23
Le kératinocyte
• LA cellule de l’épiderme

24
Autres types cellulaires
• Cellules dendritiques– = cellules de Langerhans– Proviennent de la moelle
• Mélanocytes– Cellules pigmentaires– Proviennent des crêtes neurales
• Cellules de Merkel– Associées aux terminaisons
nerveuses

25
Merkel Cells
• Modified epidermal cells located in the stratum basale.
• They are found mostly in areas where sensory perception is acute, such as the fingertips.
• Merkel cells are closely associated with an expanded terminal bulb of an afferent myelinated nerve fiber.
• Do not confuse with Merkel's corpuscle which is a combination of a neuron and an epidermal cell.

26
Human Merkel cellsMoll I, Roessler M, Brandner JM, Eispert AC, Houdek P, Moll R.
Human Merkel cells - aspects of cell biology, distribution and functions.Eur J Cell Biol. 2005;84(2-3):259-71.
• Human Merkel cells were first described by Friedrich S. Merkel in 1875 and named “Tastzellen” (touch cells) assuming a sensory touch function within the skin. Only ultrastructural research revealed their characteristics such as dense-core granules, plasma membrane spines and dendrites as well as a loosely arranged cytoskeleton. Biochemical analysis identified the expression of very specific cytokeratins (most notably CK 20) allowing the immunohistochemical detection of Merkel cells.
• In humans, they occur within the basal epidermis, being concentrated in eccrine glandular ridges of glabrous skin and in Haarscheiben of hairy skin, within belt-like clusters of hair follicles, and in certain mucosal tissues. Within the human skin, the dense-core granules contain heterogeneously distributed neuropeptides, some of which might work as neurotransmitters through which Merkel cells and their associated nerves exert their classical function as slowly adapting mechanoreceptors type I. This is the case in the Haarscheiben, small sensory organs containing keratinocytes with a special program of differentiation that includes the expression of CK 17 and Ber-EP4. Other peptides may act as growth factors and thus might participate in growth, differentiation and homeostasis of cutaneous structures. It is not yet clear whether the Merkel cell carcinomas, aggressive skin carcinomas, indeed arise from Merkel cells. We summarize and discuss data on the distribution, function and heterogeneity of human Merkel cells in normal and diseased skin.

27
Gurdip S. Sidhu , Pranil Chandra , Nicholas D. Cassai,Merkel Cells, Normal and Neoplastic: An Update.
Ultrastructural Pathology Volume 29, Number 3-4, 2005 p287 - 294• Merkel cells (MC) occur in the basal epidermal layer, hair
follicles, and oral mucosa, as complexes with sensory axons. The axons transduce slowly adapting type I mechanoreception, and MC modulate their sensitivity. MC also determine and maintain the 3-dimensional epidermal structure. They have neuroendocrine granules, rigid spinous processes, and desmosomal junctions with each other and with keratinocytes. Rare MC are dermaWl. Current evidence supports a basal cell origin.
• Merkel cell carcinomas (MCC) occur mostly in sun-exposed skin in old age. Trabecular, intermediate, or small cell in pattern, MCC have neuroendocrine granules, intercellular junctions, rigid spinous processes, and a paranuclear collection of intermediate filaments staining for cytokeratin 20.
• Most MCC behave indolently, but those with the small cell pattern, and some with the intermediate pattern, are aggressive and rapidly fatal.

28
Kératinocyte•Cellule de l’épiderme•Synthétise des filaments
intermédiaires de kératine•Donne sa dureté à l’épiderme•A une forme différente en
fonction de sa hauteur dans l’épithélium

29
Fig. 22-2
• Épiderme de souris (colonnes de cellules hexagonales)cellules dendritiques (=de Langerhans), mélanocytes, Merckel ne sont pas représentées
Mitoses présentes que dans la couche basale quand l’épithélium est fin

30
Les couches de l’épiderme
1.Couche basale2.Couche des cellules à épines3.Couche granuleuse4.Couche claire5.Couche squameuse6.Couche desquamante

31
1 - Couche basale•Stratum basal•Stratum germinativum•La plus interne•En contact avec la lame basale
•La seule à se diviser

32
2 - Couche des cellules à épines
•Stratum spinosum•Nombreux desmosomes•Nombreux filaments intermédiaires de kératine
épines visibles en microscopie optique

33
Couche des cellules à épines

34
Couche des cellules à épines

35
• http://www.biocarta.com/pathfiles/h_keratinocytePathway.asp• The epidermis, which provides a protective barrier that undergoes a
constant renewal, is a multi-layered tissue with the proliferating cells located in the basal layer. As cells leave the basal layer the underog significant differentiation, biochemical and morphological remodeling. The final differentiation results in the formation of corneocytes. In vitro keratinocytes mimic this process. Several genes mark keratinocyte specific differentiation. Among the most frequently tracked markers are Transglutaminase, Cystatin and Involucrin.
• The keratinocyte differentiation studies have identified and provided significant detail regarding the involvement of three of the 4 major MAP kinase pathways (see MAPKinase Signaling Pathway ) from several diverse stimuli such as EGF, FAS, TNF and Calicium influx. The p38 cascade is represented twice since both p38alpha (p38) and p38delta (MAPK13) are involved. The keratinocyte differentiation cascased also provide for detailed study of the functions of individual PKC isoforms. It is interesting to note the contrasting functions of the PKC isoforms in this process. In recent studies it has been determined that the cPKC (conventional/classical Protein Kinase C) isoforms, which are calcium-, phospholipid-, and diacylglycerol-dependent are inhibitory where as the nPKC (novel Protein Kinase C) isoforms which are calcium independent are stimulatory for keratinocyte differentiation markers.
• On the right hand side is an earlier step showing the upregulation loop of TRAF2. This step occurs prior to the activation os ASK1 and the p38 cascade.
Différenciation du kératinocyte

36
Cellules à épines

37
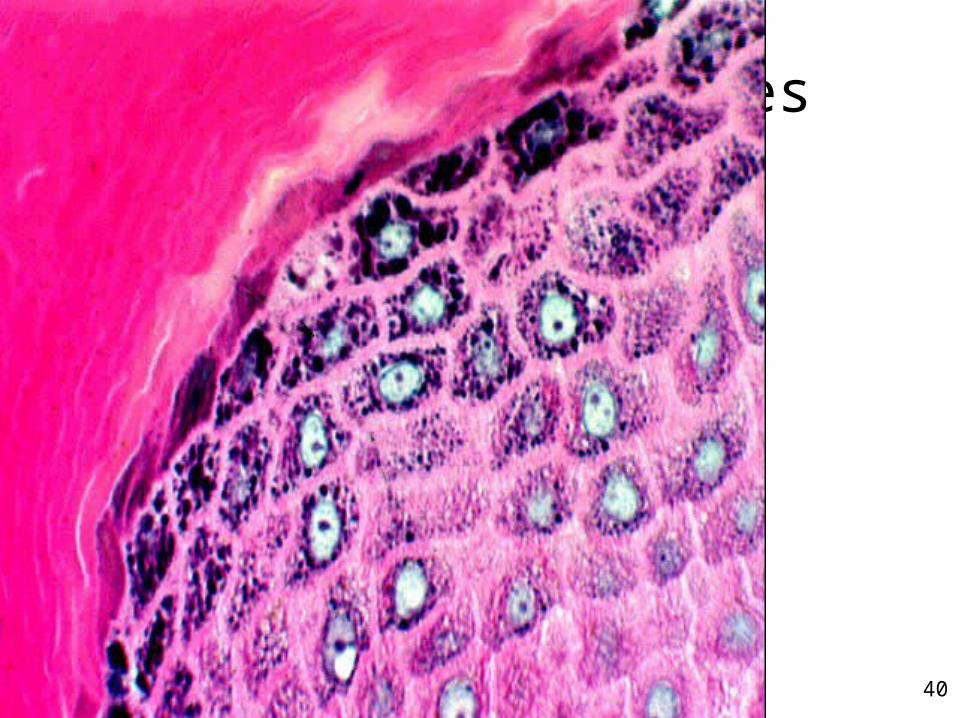
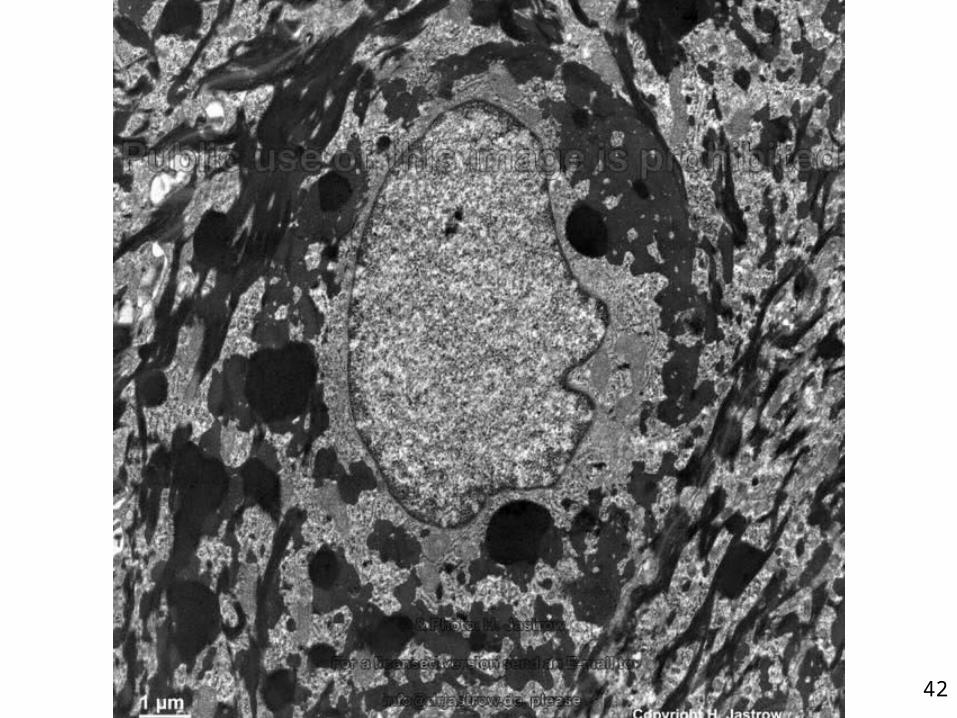
3 - Couche granuleuse• Stratum granulosum• Fine couche de cellules sombres• Présence de grains dans la cellule• Soudure des cellules entre elles • Étanchéité de la peau• Frontière entre les couches internes
métaboliquement actives et les couches plus superficielles mortes et sans organites

38
Couche des cellules granuleuses

39
Couche des cellules granuleuses
40
Couche des cellules granuleuses

41
Couche des cellules granuleuses
42
Cellule granuleuse

43
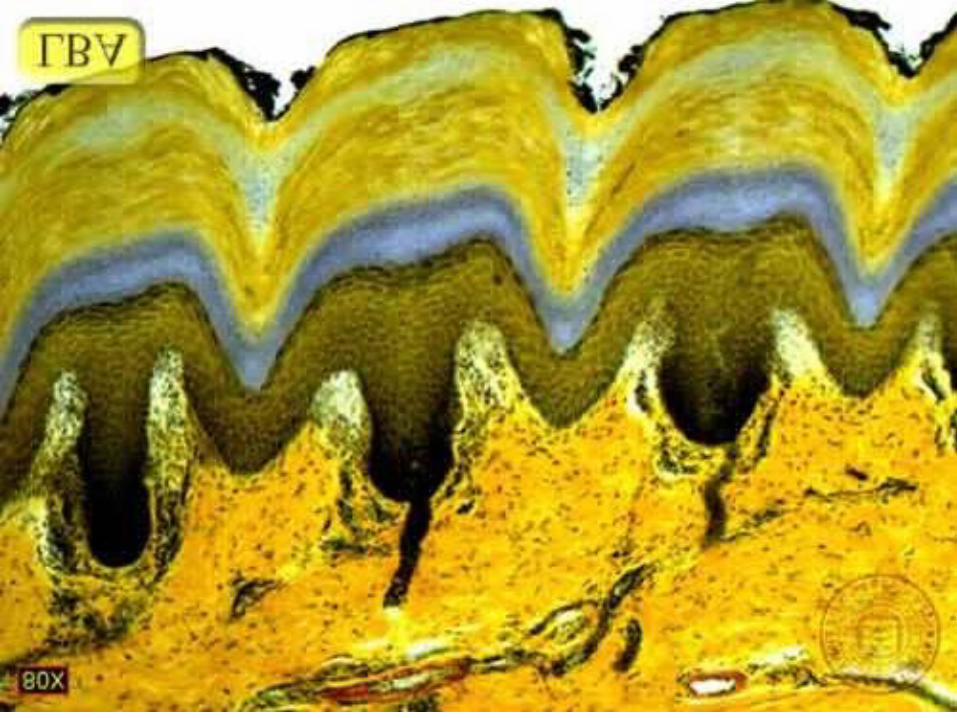
4 - Couche claire• Stratum lucidum
44
Couche claire

45
Cellule de la couche claire

46
5 - Couche cornée• Stratum corneum• Squames de cellules très aplaties• Très tassées les unes contres les autres• Remplies de kératine dense et compacte• Couche de protéines qui double la face
cytosolique de la membrane plasmique– Dure et fine (12 nm) – Contient de l’involucrine cytosolique
• Limites intercellulaires presque invisibles en microscopie optique

47
Couche cornée

48
Couche cornée

49
Couche cornée

50
6 - Couche desquamante
• Stratum disjunctum

51
Couche desquamante

52
Couche desquamante

53
Aspects dynamiques de l’épiderme
• Production de nouvelles cellules dans la couche basale– Restent dans la couche basale– Commencent le voyage vers l’extérieur
• Arrivées à la couche granulaire– Perdent leur noyau et leurs organites cytoplasmiques– Par activation partielle de l’apoptose
• Deviennent des squames de kératine• Tombent dans la poussière• 1 mois entre la naissance et la desquamation

54
Modifications moléculaires
• Kératines– Présentes en grande quantité dans toutes les
couches de l’épiderme– Grande famille de gènes homologues– Nombre de variétés encore augmenté par
épissage alternatif– Bascule d’un type de kératine à un autre au
cours du voyage de la cellule vers l’extérieur
• Involucrine : synthétisée en même temps, dans le cadre d’une différenciation cellulaire terminale

55
Différenciation cellulaire terminale
• Programme de différenciation cellulaire coordonné
• Une cellule précurseur acquière ses caractéristiques spécifiques ultimes et cesse de se diviser
• Ce programme est initié dans la couche basale de l’épiderme

56
Rôle des cellules souches de la couche basale dans le renouvellement de
l’épiderme• Il faut remplacer l’épiderme un millier de fois
au cours de la vie• Des cellules de la couche basale
– Doivent rester indifférenciées– Doivent s’auto-renouveler– Doivent continuer à se diviser pour maintenir le
réservoir de cellules souches– Doivent donner une descendance qui se différencie
jusqu'à la fin– Contiennent des cellules dont la descendance
cellules indifférenciées comme les parents ET cellules différenciées
– C’est ce qu’on appelle des cellules souches

57
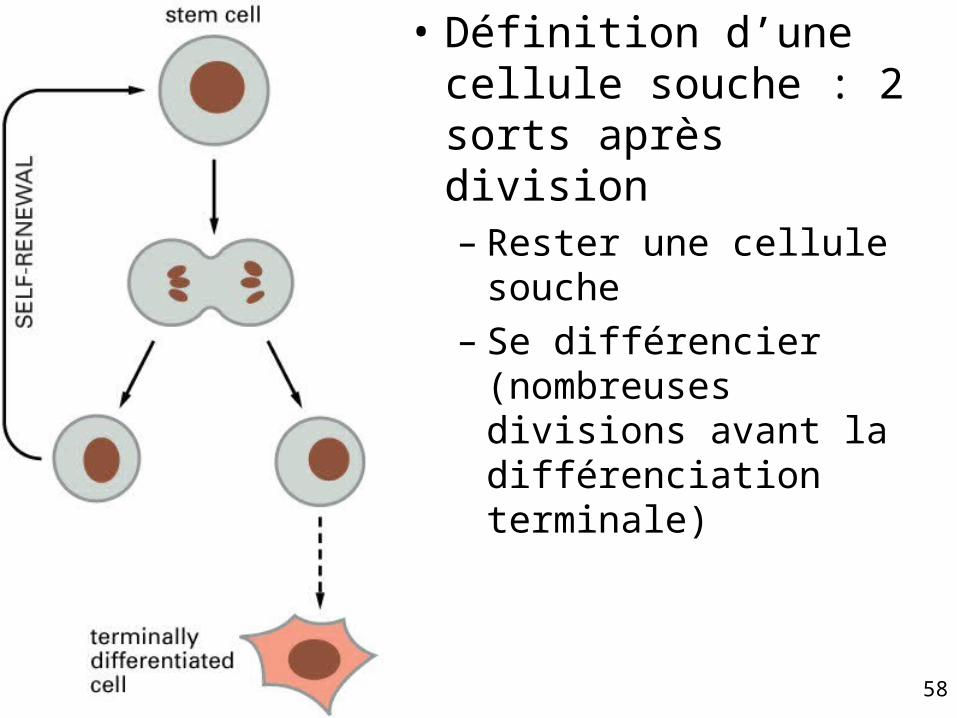
Propriétés permettant de définir une cellule souche
• Elle n’est pas dans un état de différenciation terminale
• Elle peut se diviser indéfiniment (tout au moins pendant la durée de vie de l’individu) pas nécessairement rapidement
• Quand elle se divise, chaque cellule a un choix– Rester une cellule souche– Entrer dans un cursus de différenciation
terminale
58
fig. 22-4
• Définition d’une cellule souche : 2 sorts après division– Rester une cellule
souche– Se différencier
(nombreuses divisions avant la différenciation terminale)

59
Division des cellules souches
• Doivent se diviser• Pas nécessairement rapidement• En général lentement• Se divisent quand il faut remplacer des
cellules en différenciation terminale qui ne peuvent pas elles-mêmes se diviser
• Il y en a de nombreux types– Cellules souches épidermiques– Cellules souches intestinales– Cellules souches sanguines– Cellules souches….

60
Problématique des cellules souches
• Qu’est ce qui détermine si elle doivent rester quiescentes ou se diviser ?
• Qui décide si une cellule fille doit rester cellule souche ou entrer dans une voie de différenciation terminale ?
• Quelle voie de différenciation suivre lorsqu’il y en a plusieurs (comme c’est souvent le cas)

61
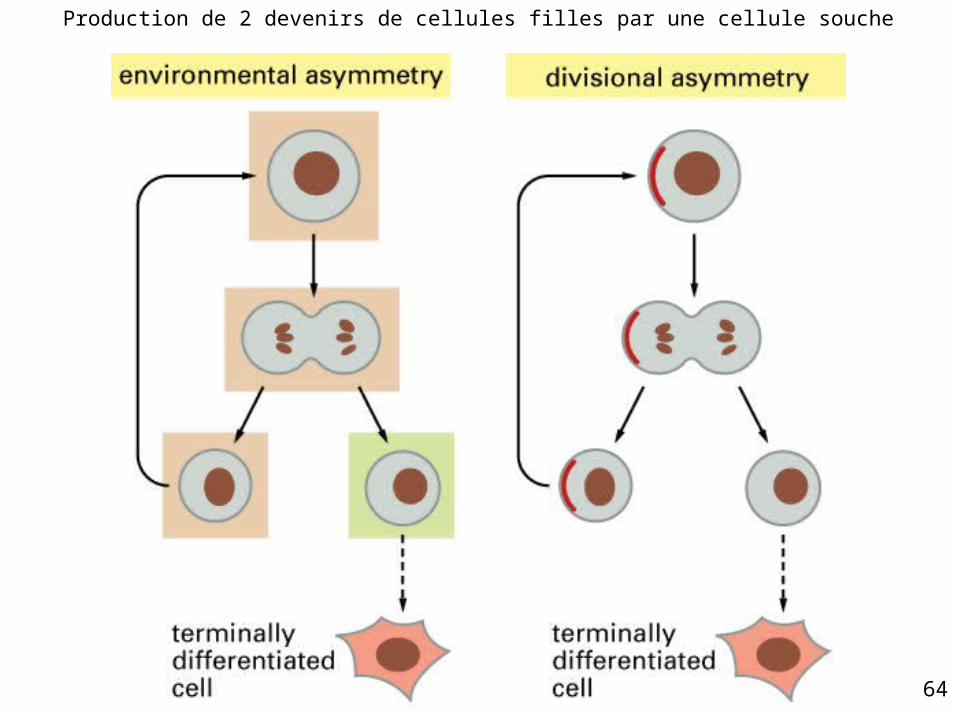
Maintien d’une population stable de cellules souches• Exactement 50 % de la
descendance de la population de cellules souches doit rester cellules souches à l’état de repos
• Deux modes1. Asymétrie environnementale2. Asymétrie de la division

62
1 - Asymétrie environnementale
• Cellule souche deux cellules filles identiques dont l’environnement gouverne le sort ultérieur– 50 % cellules souches– 50 % cellules qui vont se différencier
• Souvent cellule souche deux cellules filles à devenir identique

63
2 - Asymétrie de la division
• Cellule souche deux cellules filles toujours asymétriques– 1 cellule = caractère de cellule souche– 1 cellule = entrée dans la voie de
différenciation pas de possibilité d’augmentation du
nombre de cellules souches toute perte de cellule souche est irréparable
64
Fig 22-5
Production de 2 devenirs de cellules filles par une cellule souche

65
Réparation d’une perte de substance épidermique
• Migration et prolifération des cellules épidermiques avoisinantes et
• Augmentation du nombre de cellules souches =
• Divisions symétriques– 1 cellule souche 2 cellules
souches…

66
Régulation du nombre de cellules de la population de
cellules souches• Par la lame basale ?• Perte de contact déclenchant la
différenciation terminale• Maintien du contact pour préserver
le stock de cellules souches

67
Kératinocytes basaux en culture
Nouvelles cellules basales… Cellules en différenciation
terminale

68
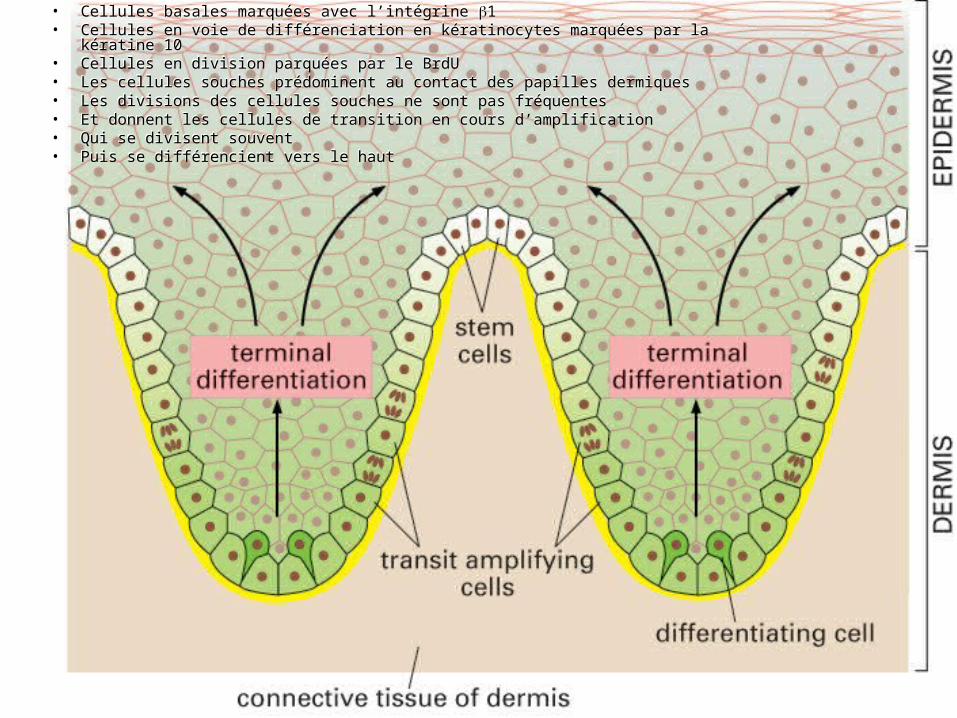
… nouvelles cellules basales
• Paraissent toutes indifférenciées• Mais grandes variations de capacité de
prolifération : une cellule basale en culture Incapable de se diviser Quelques divisions Grandes colonies
• En fonction de l’expression d’intégrine 1• Intégrine 1 élevée = cellules de la couche
basale = cellules souches
69
Fig 22-6
• Cellules basales marquées avec l’intégrine 1• Cellules en voie de différenciation en kératinocytes marquées par la kératine 10• Cellules en division parquées par le BrdU• Les cellules souches prédominent au contact des papilles dermiques• Les divisions des cellules souches ne sont pas fréquentes• Et donnent les cellules de transition en cours d’amplification• Qui se divisent souvent• Puis se différencient vers le haut
• Cellules basales marquées avec l’intégrine 1• Cellules en voie de différenciation en kératinocytes marquées par la kératine 10• Cellules en division parquées par le BrdU• Les cellules souches prédominent au contact des papilles dermiques• Les divisions des cellules souches ne sont pas fréquentes• Et donnent les cellules de transition en cours d’amplification• Qui se divisent souvent• Puis se différencient vers le haut

70
« Cellules de transition en amplification »
• Cellules basales• Exprimant peu l’intégrine 1• Capable de se diviser un peu• Puis de quitter la lame basale• Pour suivre la voie de différenciation terminale
• « de transition » : entre caractère de cellule souche et caractère de cellule différenciée
• « en amplification » : amplification de la descendance résultant de la division d’une cellule souche

71
Fig 22-7
« Cellules de transition en amplification »– La cellule souche se divise peu– mais donne des cellules destinées à se différencier– et qui se diviseront plus rapidement.– Ici chaque cellule souche donnera 8 descendants
différenciés

72
Rôle de la lame basale
• Il existe aussi quelques cellules restant en contact avec la lame basale- qui ne se divisent plus- qui commencent à se différencier
• La lame basale n’est pas suffisante pour contrôler le devenir d’une cellule basale épidermique

73
Rôle de la lame basale
• Dans une culture en suspension des kératinocytes de la couche basale arrêtent de se diviser et de se différencier
• Nécessité d’un support (lame basale ou matrice extra-cellulaire)
• La lame basale représente une limite à la croissance infinie des cellules souches (application au cancer)

74
Renouvellement de l’épiderme
• Apparemment simple• En fait nombreux points de
contrôle

75
Points de contrôle du renouvellement de l’épiderme
• Le taux de multiplication des cellules souches• La probabilité qu’une cellules fille reste une
cellule souche• Le nombre de divisions des cellules de
transition en amplification• Le temps mis pour quitter la lame basale• Le temps mis pour achever le programme de
différenciation• Le temps mis pour desquamer

76
Résultat du contrôle
• Réponse aux agressions• Augmentation de l’épaisseur de
l’épiderme• Réparation des blessures• Follicules pileux

77
Mécanismes de régulation
• Très nombreux• Mécanismes de communication
cellulaire• Signalisation intercellulaire entre
les cellules de l’épiderme• Signalisation entre épiderme et
derme

78
Mécanismes de régulation
• EGF• FGF• Wnt (mutation dans la voie cancer)• Voie Hedgehog (mutation dans la
voie cancer)• Voie Notch• BMP/TGF

79
Les voies de signalisation
• Activation de Wnt• Favorise le maintien des caractères de cellules souches• Inhibe le passage vers les « cellules de transition en
amplification »
• Activation de Notch (fait l’inverse)• Inhibe le maintien des caractères de cellules souches• Active le passage vers les « cellules de transition en
amplification »
• TGF : rôle clé dans la signalisation vers le derme pour la réparation des plaies

80
Rôle des intégrines• Intégrines maintenues artificiellement
hautes dans les couches superficielles de souris transgéniques
• Augmentation de la prolifération des cellules de la couche basale
• Épaississement de l’épiderme• Élimination des kératinocytes au bout
d’une semaine• Avant leur kératinisation complète• Sorte de psoriasis humain

81
Un cas particulier de renouvellement : la glande
mammaire• À côté des kératinocytes d’autres cellules se développent à
partir de l’épiderme de l’embryon dans certaine parties du corps
• Invaginations de l’épiderme glandes enfoncées en profondeur• Sudoripares• Lacrymales• Salivaires• Mammaires
• Même origine que l’épiderme mais• Grandes différences dans
• Fonctions• Mode de renouvellement

82
• Proposed evolution of the mammary gland from a mucus-secreting epithelial gland. Mammary glands presumably evolved as mucus-secreting skin glands that similar to many mucus surface epithelia secreted antimicrobial enzymes such as XOR and lysozyme. The evolution of additional functions of XOR and lysozyme in the ancient mammary epithelium resulted in the secretion of fat droplets, α- lactalbumin and lactose. Consequently, the mammary gland evolved from a protective immune organ into a reproductive organ unique to the class mammalia.

83
Généralités sur la glande mammaire
• Glandes sécrétrices• Définissent les mammifères• Intérêts
• Nourriture des bébés• Caractère sexuel secondaire• Industrie de lait• Cancer fréquent• Biologie du développement…

84
…Intérêt en biologie du développement
•Les processus du développement continuent chez l’adulte
•Équilibre entre développement et apoptose chez l’adulte

85
Cycle de la glande mammaire
•Production de lait à la naissance du nouveau-né
•Arrêt de la production de lait au sevrage

86
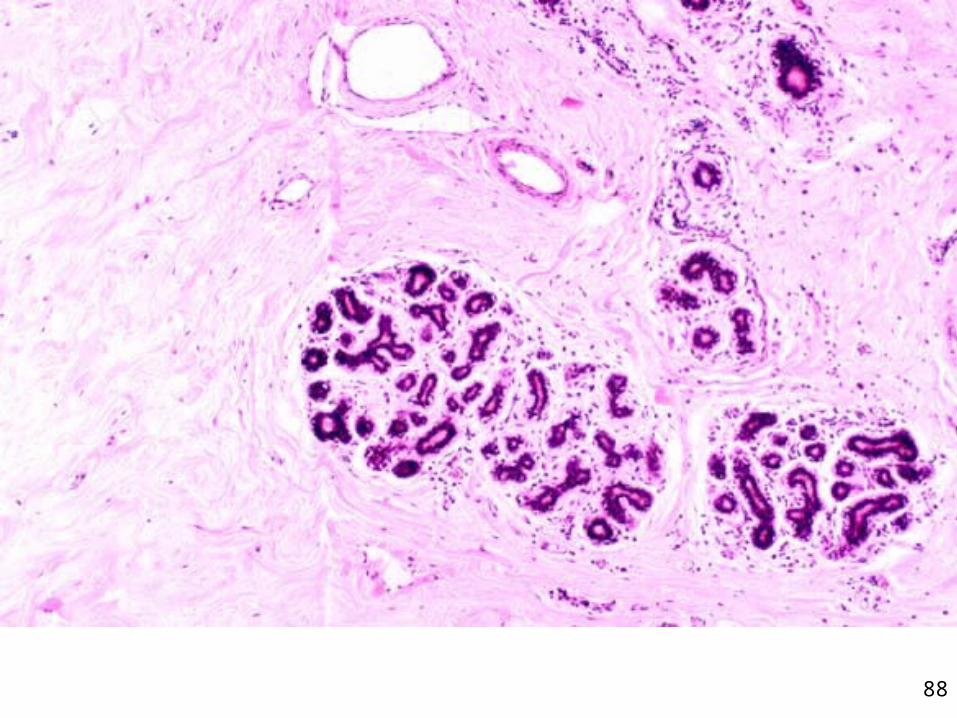
Histologie de la glande mammaire au repos
•Système de canaux ramifiés entourés de tissu adipeux

87
Canaux
•Épithélium comprenant une sous population de cellules souches
88
Glande mammaire au repos

89
Glande mammaire au repos

90
Glande mammaire prépubère

91
Pendant la grossesse• Hormones circulantes
• Prolifération des cellules des canaux• Nombre des canaux X 10 ou X 20• Croissance et ramifications des
extrémités des canaux dilatations =
• Petits alvéoles appelés acini

92
Glande mammaire

93
Pendant la lactation
•Protéines : exocytose•Lipides: gouttelettes
entourées de membrane plasmique

94
D. B. Shennan and M. Peaker Transport of Milk Constituents by the Mammary GlandPhysiological Reviews, Vol. 80, No. 3, July 2000, pp. 925-951
Shennan, D. B. et al. Physiol. Rev. 80: 925-951 2000 Copyright ©2000 American Physiological Society
• Les 5 principales voies de sécrétion à travers ‘épithélium sécrétoire du sang vers le lait (4 trans cellulaires et 1 para cellulaire)1) Voie membranaire2) Voie golgienne3) milk fat route4) Transcytose5) Voie para cellulaire

95
Glande mammaire en lactation

96
Glande mammaire en lactation

97
Glande mammaire en lactation

98
Glande mammaire en lactation

99
La lactation•Déclenchement par une
combinaison d’hormones•Succion du mamelon par libération
d’ocytocine par l’hypothalamus•Ocytocine agit sur les cellules
myoépithéliales (même origine épithéliale que les cellules sécrétrices)

100
Glande mammaire en lactation

101
Sevrage du nouveau né
•Mort des cellules sécrétrices par apoptose
•Disparition des acini•Élimination des cellules mortes par
les macrophages•Retour à l’état de repos : fin de la
lactation…

102
Fin de la lactation•Brutale• Induite par l’accumulation de lait
(pas hormonal)•Si un canal se bouche pas
d’excrétion de lait mort des cellules sécrétrices par apoptose
•Les autres lobules continuent à fonctionner

103
Apoptose des cellules sécrétrices
•Déclenchement par des facteurs dont le TGF 3
•Qui s’accumule où la sécrétion est bloquée

104
fig. 22-9
• Mort des cellules sécrétrices à l’arrêt de la succion
A. Lactation normale (TGF 3 -)B. Arrêt de la succion depuis 9
heures (TGF 3 -)C. Canal obstrué depuis 3 jours
(mort des cellules par apoptose)

105
Rôle du TGF 3•Arrêt de la succion•Pas de drainage du lait•Production de TGF 3•Apoptose•Régression des canaux

106
Andrew V. Nguyen and Jeffrey W. PollardTransforming growth factor 3 induces cell death during the first stage of mammary gland involutionDevelopment 127, 3107-3118 (2000)
• In situ hybridization and immunohistochemistry for TGFb3 in mammary glands at D1PP.
• (A,B) Sagittal sections of Csfmop/Csfmop involuting mammary glands hybridized with (A) antisense or (B) senseTGFb3 probe.
• (C,D) Sections of +/Csfmop mammary gland isolated from a mother that had not suckled for 9 hours (C) or still feeding (D) reacted with antibody specific for TGFb3 protein.
• (E) Sections of +/Csfmop mammary gland from a mother that had not suckled for 9 hours treated with IgG as a control.
• (F) +/Csfmop mammary gland not suckled for 9 hours treated with the antibody specific for TGFb3 protein but that had been immunodepleted with excess TGFb3 peptide. Arrowheads in A and C indicate TGFb3 positive epithelial cells.

107
Andrew V. Nguyen and Jeffrey W. PollardTransforming growth factor 3 induces cell death during the first stage of mammary gland involutionDevelopment 127, 3107-3118 (2000)
• Apoptosis in mammary glands at day 1 postpartum. Sagittal sections were analyzed for apoptosis using TUNEL staining. (A) +/Csfmop; (B) Csfmop/Csfmop; (C) +/Csfmop not suckled for 9 hours; (D) sealed mammary gland from +/Csfmop feeding dam for 1 day; (E) sealed mammary gland from +/Csfm op feeding dam for 3 days; (F) contralateral gland from +/Csfmop feeding dam for 3 days. Note that the Csfmop/Csfmop mammary gland is fully in the process of involution. Arrowheads point to apoptotic cells. Inset (B,E) shows a higher magnification of apoptotic cells.

108
Signalisation dans la glande mammaire en croissance
• Hormones• Signaux
– Entre les cellules épithéliales– Entre le tissu conjonctif et l’épithélium
(= stroma)– Les mêmes que dans l’épiderme
•Intégrines : si on supprime la lame basale pas d’activation de la voie de l’intégrine la cellule épithéliale ne répond plus aux stimulations hormonales
•Tous les autres facteurs…

109
Application•Cancer







