
BIOL 1392 : Biologie cellulaire
et Génétique 2
1ère Année Pharmacie
2019
Ousmane KOITA, PharmD, PhD
Professeur de Biologie Moléculaire

Contenu du cours
• Structure générale des cellules
eucaryotes; étude des principaux
organites et les machineries cellulaires:
structures, Propriétés et Fonctions.
• Génie génétique, Evolution et diversité
génétique des cellules animales.

Définition de la Biologie cellulaire
• La biologie cellulaire (anciennement
appelée cytologie) est une discipline
scientifique qui étudie les cellules, du point
de vue structural et fonctionnel, et les utilise
pour des applications en biotechnologie.
Elle s'intéresse à l'écosystème cellulaire,
c'est-à-dire à l'équilibre dynamique et auto-
régulé des fonctions cellulaires, dans un
contexte normal ou perturbé.

Définition de la Biologie Cellulaire
• Le champ de la biologie cellulaire concerne
une multitude de réactions chimiques
coordonnées et de mécanismes fins de
régulation entre des millions de constituants
micro et nanoscopiques.
• Ces constituants assurent durablement
l'architecture et le fonctionnement de la
cellule.

Définition de la Biologie Cellulaire
• La biologie cellulaire était née avec
l'invention du premier microscope optique
(photonique) par Antoni van Leeuwenhoek.
• L'étude des microorganismes (dont les
bactéries) ne devint réellement accessible
qu'avec le développement d'un microscope
optique composé (multilentilles) efficace
vers les années 1825.

Définition de la Biologie Cellulaire
• Rudolf Virchow (1821-1902), physiologiste
allemand est l'auteur de l'adage « omni
cellula e cellula », ou comme il le publie en
1858 dans Cellularpathologie «Là où
apparaît une cellule, il doit y avoir eu une
autre cellule auparavant » «Tout animal
apparaît comme la somme d'unités vitales
dont chacune porte en elle tous les
caractères de la vie.»

Définition de la Biologie Cellulaire
• La cellule est donc une enceinte séparée
de l'extérieur par une membrane capable
de filtrer sélectivement les échanges.
• Jusqu'au XIXe siècle, les organismes
vivants étaient classés comme animaux
ou végétaux selon des différences
évidentes de forme et de constitution, qui
dérivent de différences fondamentales
dans leur mode de nutrition.

Epigénétique
• Il s’agit des mécanismes moléculaires qui
permettent de changer de manière
héritable, l’expression des gènes sans
altérer la séquence d’ADN.
• Des anomalies épigénétiques participent
au développement de certaines maladies,
en particulier de tumeurs.

Epigénétique
• Les processus épigénétiques interviennent
dans la régulation de nombreux événements,
tels que la division cellulaire ou spécialisation
des cellules: l’altération de ces mécanismes
peut favoriser la transformation des cellules
saines en cellules cancéreuses.

Organisation de l’ADN dans la Cellule

Mécanismes épigénétiques et
développement d’une cellule cancéreuse

LA CELLULE
• La cellule représente à la fois l’unité
structurale et fonctionnelle de tous les
êtres vivants. C’est la plus petite portion
de matière vivante capable de vivre isolée
et de se reproduire.
• Il y a 2 classes de cellules:
Unicellulaires ou Protistes (Protophytes et protozoaires)
Pluricellulaires (Métaphytes ou Métazoaires)
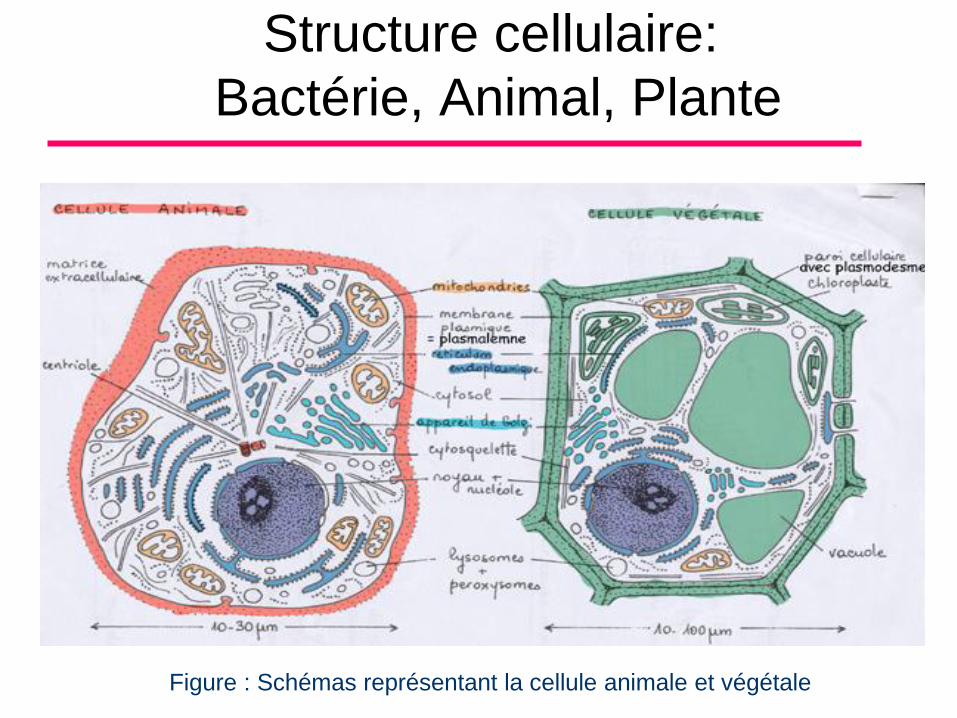
Structure cellulaire:
Bactérie, Animal, Plante
Figure : Schémas représentant la cellule animale et végétale

Structure et Composition de la
membrane plasmique
La tête hydrophile est orientée vers les versants
extérieur et intérieur de la membrane, tandis que la
queue hydrophobe est intramembranaire.

La membrane plasmique
• Elle est considérée comme filtre sélectif:
• Elle maintient des concentrations
différences d’ions entre l’intérieur et
l’extérieur de la cellule,
• Elle assure les échanges avec
l’extérieur de la cellule,
• Elle possède des molécules qui
permettent la reconnaissance des
cellules entre elles.

Membrane comme mosaique fluide
• Dans la membrane, les molécules sont
organisées en bicouche, ces molécules sont
fluides, c’est à dire que les molécules ont un
certain mouvement entre elles.
• Diffusion latérale: changement de place
dans la monocouche (figure)
• Le mouvement flip-flop: changement de
place entre les 2 monocouches (figure)
• Rotation autour de l’axe: la molécule tourne
sur elle même.
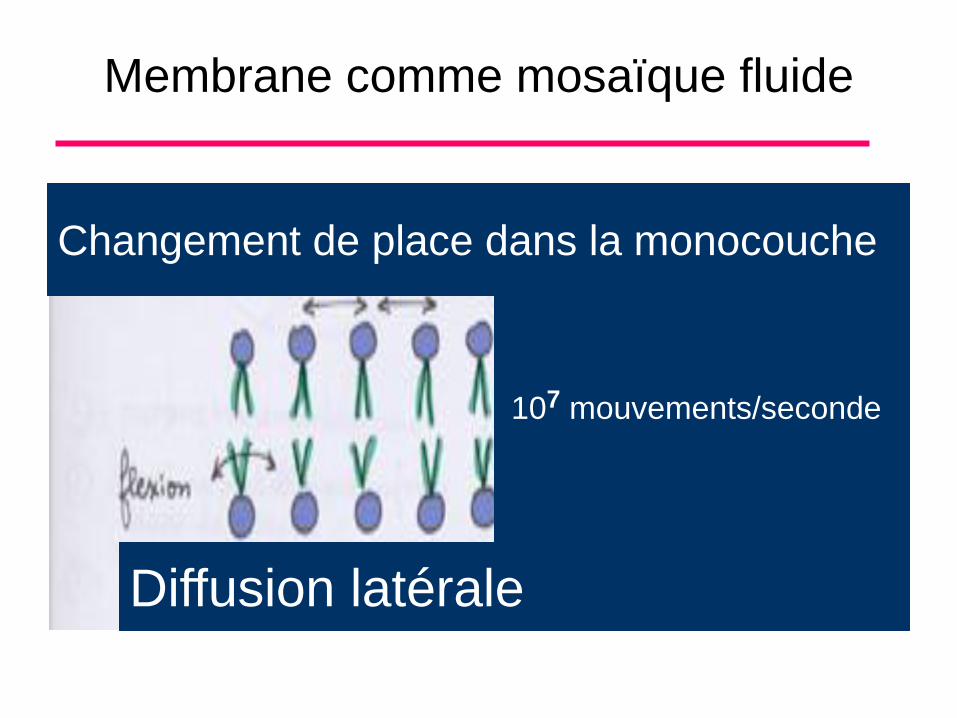
Membrane comme mosaïque fluide
107 mouvements/seconde
Diffusion latérale
Changement de place dans la monocouche

Membrane comme mosaïque fluide
la molécule
tourne sur elle même.
Rotation autour de l’Axe

Membrane comme mosaïque fluide
Changement de place entre les 2 monocouches
Mouvement Flip flop ou bascule, <1 par mois

La membrane plasmique
• Elle est considérée comme filtre sélectif:
• Elle maintient des concentrations
différences d’ions entre l’intérieur et
l’extérieur de la cellule,
• Elle assure les échanges avec
l’extérieur de la cellule,
• Elle possède des molécules qui
permettent la reconnaissance des
cellules entre elles.

Membrane comme mosaïque fluide
• Facteurs affectant la fluidité de la membrane
1. La température: une température élevée augmente la
fluidité;
2. La longueur des chaines hydrocarbonées: plus la chaine
est longue, plus la température est élevée et la fluidité
est augmentée;
3. Le nombre de doubles liaisons: plus la chaine est
insaturée, plus la température est basse, la fluidité est
faible;
4. Le contenu en cholestérol qui rigidifie la bicouche: le
cholestérol augmente la fluidité et diminue la
congélation.

Polymorphisme et spécialisation de la
membrane plasmique
• Ce sont des invaginations
en forme de doigts d’une
main qui sont des replis
qui intensifient les
échanges de l’intérieur
vers l’extérieur de la
cellule. Elles sont
appelées microvillosités
apicales.Cellule absorbante de l’intestin
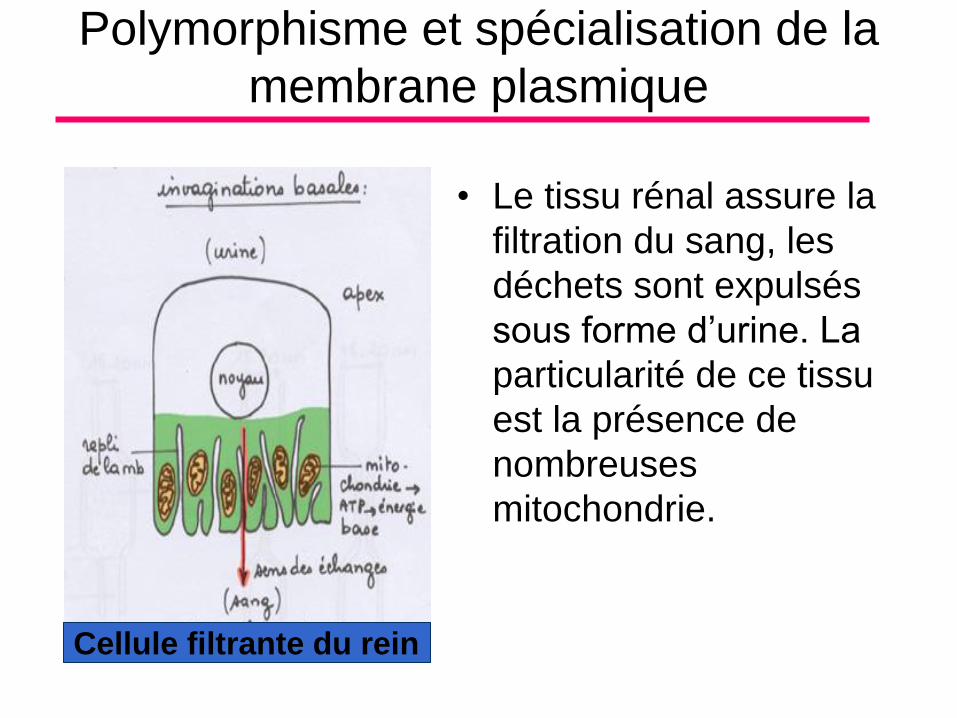
Polymorphisme et spécialisation de la
membrane plasmique
• Le tissu rénal assure la
filtration du sang, les
déchets sont expulsés
sous forme d’urine. La
particularité de ce tissu
est la présence de
nombreuses
mitochondrie.
Cellule filtrante du rein
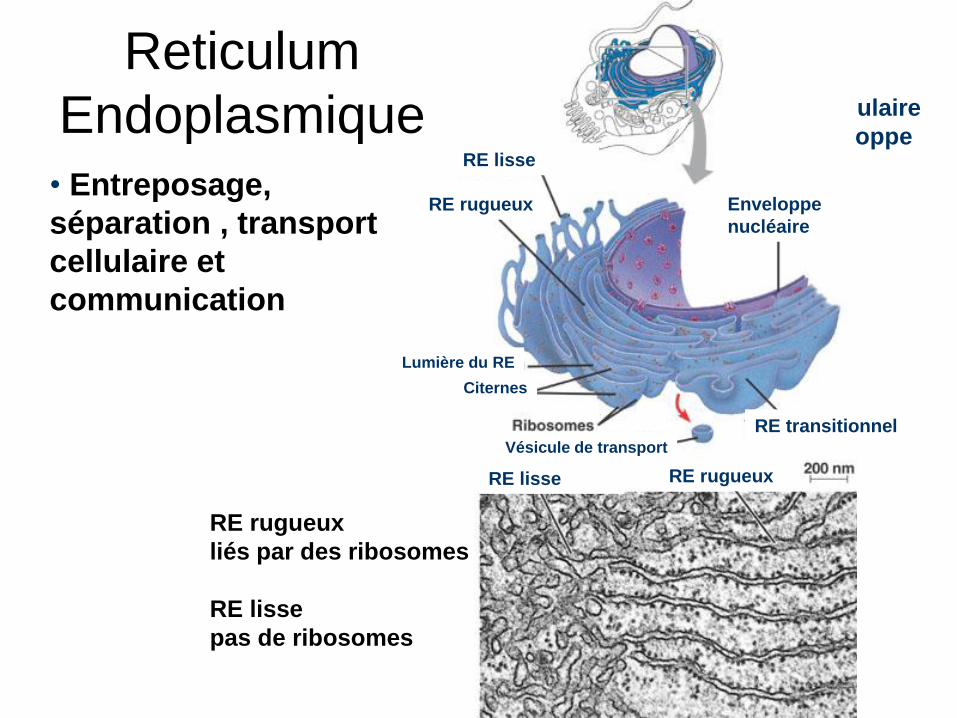
Reticulum
EndoplasmiqueRE lisse
RE rugueux Enveloppe
nucléaire
Lumière du RE
Vésicule de transport
RE lisse RE rugueux
RE transitionnel
Citernes
• Entreposage,
séparation , transport
cellulaire et
communication
RE rugueux
liés par des ribosomes
RE lisse
pas de ribosomes
Réseau tubulaire
lié à l’enveloppe
nucléaire
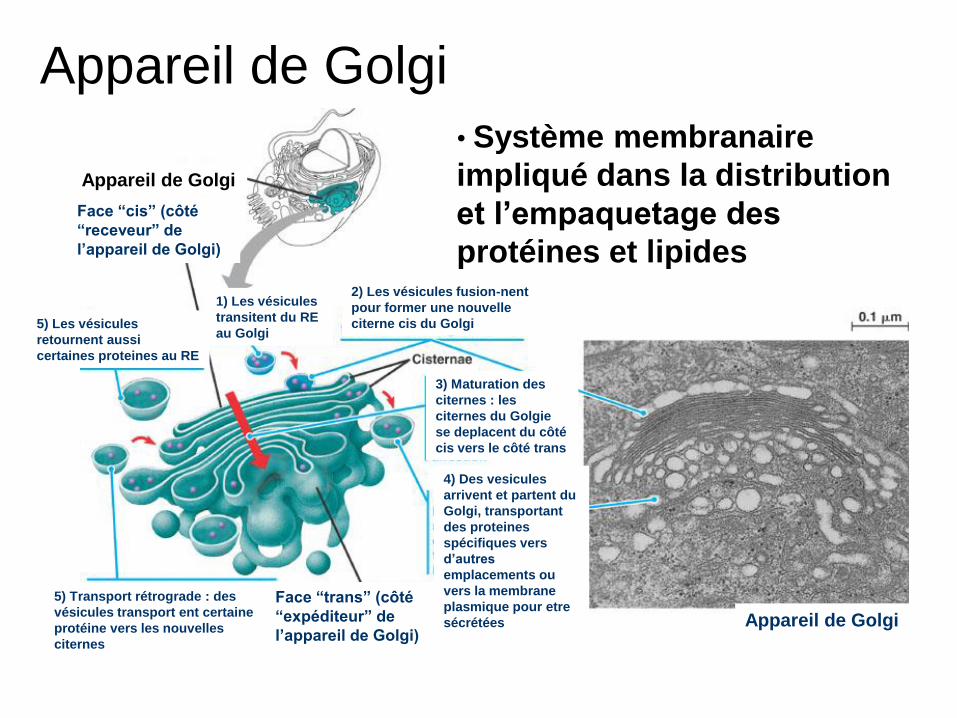
Appareil de Golgi
Appareil de Golgi
Face “cis” (côté
“receveur” de
l’appareil de Golgi)
1) Les vésicules
transitent du RE
au Golgi
2) Les vésicules fusion-nent
pour former une nouvelle
citerne cis du Golgi
3) Maturation des
citernes : les
citernes du Golgie
se deplacent du côté
cis vers le côté trans
4) Des vesicules
arrivent et partent du
Golgi, transportant
des proteines
spécifiques vers
d’autres
emplacements ou
vers la membrane
plasmique pour etre
sécrétées
Face “trans” (côté
“expéditeur” de
l’appareil de Golgi)
5) Transport rétrograde : des
vésicules transport ent certaine
protéine vers les nouvelles
citernes
5) Les vésicules
retournent aussi
certaines proteines au RE
Appareil de Golgi
• Système membranaire
impliqué dans la distribution
et l’empaquetage des
protéines et lipides

Généralités – Définition (1)
• L’appareil de Golgi ou complexe de Golgi est
habituellement localisé près du noyau de la
cellule.
• Il est disposé près du centrosome ou au
centre de la cellule dans les cellules
animales.
• Il est composé de nombreux saccules
aplatis, limités par une membrane,
ressemblant à une pile d’assiettes.

Généralités – Définition (2)
• L'appareil de Golgi est un organite cellulaire
polymorphe constitué d'un ou plusieurs
dictyosomes (en général : un seul
dictyosome dans les cellules animales, et
plusieurs dizaines dans les cellules
végétales), empilement de saccules
membranaires de forme discoïdale et d'un
réseau de canalicules appelé réseau trans-
golgien (Trans Golgi Network ou TGN).

Généralités – Définition (3)
• Les empilements ou dictyosomes chez les
végétaux sont habituellement constitués de 4
à 6 saccules, chacun ayant un diamètre
d’environ 1µ.
• Le nombre d’empilement de Golgi par cellule
varie énormément selon le type cellulaire.
• Certaines cellules ne contiennent qu’un seul
empilement volumineux, tandis que d’autres
en contiennent de très petites, par centaine.

Généralités – Définition (4)
• Des multitudes de petites vésicules sont
associées aux empilements de Golgi,
groupées sur le côte contigu au Réticulum
endoplasmique (RE) et près des bordures
dilatées de chaque saccule.
• Ces vésicules transporteraient des
protéines et des lipides vers et depuis le
Golgi et entre les saccules de Golgi.

Généralités – Définition (5)
• Un empilement présente 2 faces distinctes:
une face cis (ou face d’entrée)
une face trans (ou face de sortie)
• La face cis est étroitement associée aux
éléments de transition du RE.
• La face trans est dilatée en un réticulum
tubulaire nommée “Réseau Trans de Golgi
(RTG).

Généralités – Définition (6)
• Les protéines et les lipides pénètrent, sur la
face cis, dans un empilement de Golgi,
enfermes dans de petites vésicules à partir
du RE et ressortent pour de multiples
destinations dans de vésicules qui se
forment sur la face trans.
• Ces molécules transportées subissent une
série ordonnée de modifications
lorsqu’elles passent d’un saccule de Golgi
à un autre.

Généralités – Définition (7)
• L’appareil de Golgi est remarquable par ses
cellules spécialisées dans la sécrétion,
telles que les cellules gobelets de
l’épithélium intestinal, qui secrètent de
grandes quantités de mucus dans l’intestin.
• Les vésicules se forment depuis la face
trans qui fait face au domaine de la
membrane plasmique ou se produit la
sécrétion.

Organisation des Saccules de Golgi (1)
• Chaque saccule est un compartiment
distinct, qui a son propre jeu d’enzymes de
transformation, de sorte que l’empilement
forme une unité de transformation
échelonnée. Les protéines sont modifiées
par étapes successives lors de leur
déplacement d’un compartiment à un
autre à travers l’empilement.

Organisation des Saccules de Golgi (2)
• Les protéines exportées du RE pénètrent
dans le 1er compartiment du Golgi (le
compartiment cis) puis se déplacent vers
le compartiment suivant (le compartiment
médian qui correspond au saccule central
de l’empilement) puis se déplacent
finalement vers le compartiment trans
(composé du dernier saccule).

Organisation des Saccules de Golgi (3)
• Les protéines se déplacent à partir de ce
compartiment trans vers le réseau trans
du Golgi (RTG). Dans ce réticulum
tubulaire, elles sont isolées dans
différentes vésicules de transport et
expédiées vers leurs destinations finales,
la membrane plasmique, les lysosomes ou
les vésicules de sécrétion.

Organisation des Saccules de Golgi (4)
• Les protéines qui pénètrent dans le Golgi
à partir du RE (exceptées celles destinées
à rester dans un compartiment du Golgi)
circulent à travers l’empilement, du
compartiment cis au compartiment
médian, puis au compartiment Trans,
permettant leur transformation
progressive.
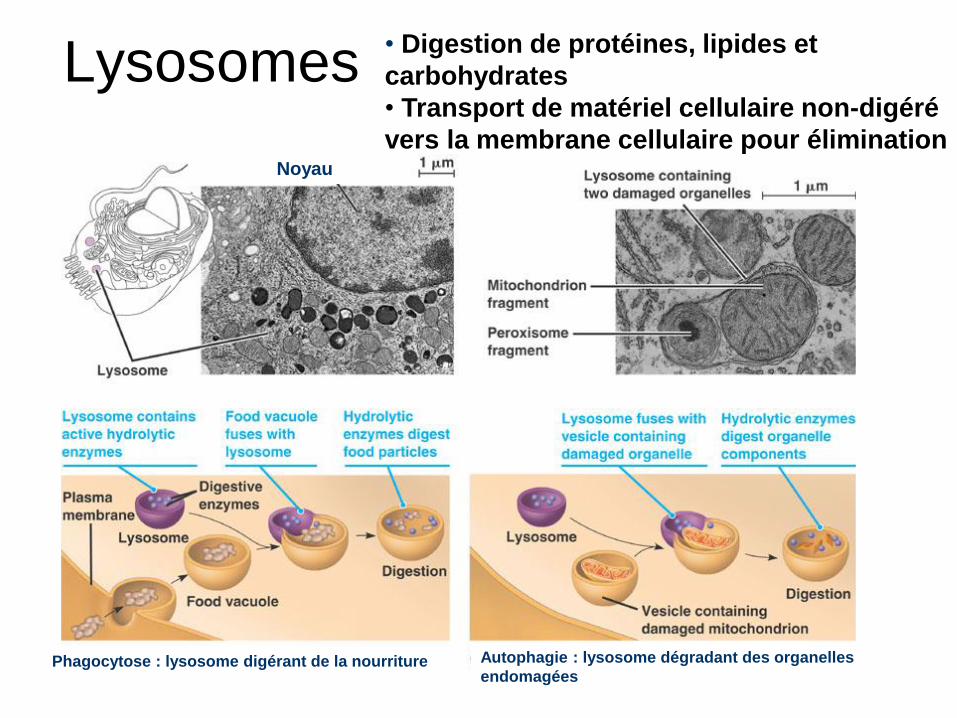
Lysosomes
Noyau
Phagocytose : lysosome digérant de la nourriture Autophagie : lysosome dégradant des organelles
endomagées
• Digestion de protéines, lipides et
carbohydrates
• Transport de matériel cellulaire non-digéré
vers la membrane cellulaire pour élimination

Transport de Protéines au niveau de
l’Appareil de Golgi (1)
1.Transport du Golgi vers les Lysosomes
1.1. Nature et localisation des lysosomes
Ce sont de volumineux sacs membraneux
contenant des enzymes hydrolytiques servant
à la digestion intracellulaire des
macromolécules.
On connait une 40taine d’enzymes
hydrolytiques contenues dans les lysosomes.

Transport de Protéines au niveau
de l’Appareil de Golgi (2)
• Ces enzymes sont les protéases, les
nucléases, glycosidases, lipases,
phospholipases, phosphatases et les
sulfatases. Ce sont toutes des hydrolases
acides dont l’activité est optimale aux environs
de pH 5, maintenu dans les lysosomes. La
membrane du lysosome retient normalement
les enzymes hors du cytosol (pH 7,2), mais la
défend de l’action des enzymes, protège le
contenu du cytosol contre les détériorations,
même si une fuite se produisait.

Transport de Protéines au niveau
de l’Appareil de Golgi (3)
• Comme les autres organites intracellulaires,
le lysosome ne contient pas seulement une
collection unique d’enzymes, mais il est
également entouré par une membrane
unique. Celle-ci contient des protéines de
transport qui permettent aux produits finaux
de la digestion de s’échapper, de sorte
qu’ils puissent être secrétés et réutilisés par
la cellule.

Transport de Protéines au niveau
de l’Appareil de Golgi (4)
• Cette membrane
contient également une
pompe à protons qui
utilise l’énergie de
l’hydrolyse de l’ATP
pour pomper des ions
H+ dans les
lysosomes, maintenant
ainsi le pH de la
lumière.
ATP ADP +Pi
pH~5
HYDROLASES
Nucleases
Proteases
Glycosidases
Lipases
Phosphatases
Sulfatases
phospholipases
pH= 7,2
H+

Transport de Protéines au niveau de
l’Appareil de Golgi (5)
1.2. Voies de pénétration des matériaux dans les
lysosomes
Il existe 3 voies possibles de formation des
lysosomes:
1. La voie la mieux étudiée est utilisée par la
digestion des matériaux inclus par
endocytose suivant le mécanisme qui conduit
à la formation des endosomes puis aux
lysosomes (1ère voie).

Transport de Protéines au niveau
de l’Appareil de Golgi (5)
• (1ère voie suite). Les matériaux endocytés
passent séquentiellement d’un compartiment
endosomal périphérique à un compartiment
endosomal périnucléaire.
• Les matériaux qui n’ont pas été
spécifiquement récupérés à partir de ces
endosomes pénètrent dans un 3ème
compartiment intermédiaire qui reçoit des
hydrolases et des protéines membranaires
lysosomales néo synthétisées dans l’AG.

Transport de Protéines au niveau
de l’Appareil de Golgi (6)
• (1ère voie suite et fin) Ce compartiment
endolysosomal étant moyennement acide, on
pense qu’il est le site de départ de la digestion
hydrolytique des matériaux endocytés.
• La conversion de l’endolysosome en un
lysosome mature nécessite la perte de ses
composants endosomaux distincts et une
baisse supplémentaire de son pH interne. On
ne sait pas comment s’effectue cette
conversion.

Transport de Protéines au niveau
de l’Appareil de Golgi (7)
• La 2ème voie est celle de l’autophagie. Le
processus de digestion implique
l’enveloppe d’un organite par des
membranes dérivées du RE, ce qui donne
naissance à une autophagosome qui
fusionnera avec un lysosome initiant la
digestion de son contenu dans une
autophagolysosome.

Transport de Protéines au niveau
de l’Appareil de Golgi (8)
• La 3ème voie qui fournit des matériaux
aux lysosomes se présente dans la
phagocytose de particules et de
microorganismes. Ainsi, des macrophages
ou des neutrophiles peuvent absorber de
gros objets pour former une phagosome.
• Les enzymes lysosomales sont triées dans
l’appareil de Golgi par un récepteur
protéique membranaire.

Transport de Protéines au niveau
de l’Appareil de Golgi

Transport de Protéines au niveau
de l’Appareil de Golgi (9)
2. Transport de l’Appareil de Golgi vers les
vésicules sécrétrices.
• Dans les cellules, la sécrétion se produit
en réponse à un signal extracellulaire. Les
protéines secrétées sont concentrées et
stockées dans des vésicules de sécrétion
(granules de sécrétion), d’où elles sont
libérées par exocytose en réponse à un
signal.

Transport de Protéines au niveau
de l’Appareil de Golgi (10)
• Les vésicules de sécrétion se forment
par bourgeonnement à partir du RTG,
on pense que les protéines destinées
aux vésicules de sécrétion (protéines de
sécrétion) sont triées et empaquetées
dans les vésicules appropriées dans le
RTG.

Transport de Protéines au niveau de
l’Appareil de Golgi (11)
De nombreuses vésicules sécrétrices, telles
que celles des cellules acineuses
pancréatiques sont polarisées et l’exocytose
ne se produit qu’au niveau de la surface
apicale, qui fait généralement face à la
lumière d’un système de canaux qui
recueillent les secrétions. Lorsqu’une vésicule
de sécrétion fusionne avec la membrane
plasmique, son contenu est secrété loin de la
cellule par exocytose et sa membrane devient
une partie de la membrane plasmatique.

Transport de Protéines au niveau
de l’Appareil de Golgi (12)
• Cette augmentation n’est que transitoire
car les composants membranaires sont
éliminés de la surface par endocytose à
une vitesse sensiblement égale à celle de
leur incorporation par exocytose. On dit
que les composants membranaires des
vésicules sont recyclés.

Transport de Protéines au niveau
de l’Appareil de Golgi

Fonction de l’Appareil de Golgi (1)
• L'appareil de Golgi reçoit en permanence
du matériel provenant du réticulum, il le
modifie puis l'exporte vers la membrane
ou les lysosomes dans un flux vectoriel
permanent.

Fonction de l’Appareil de Golgi (2)
activation de protéines :
• certaines protéines sont sécrétées par le
réticulum sous forme inactive ; elles
s’activent en perdant une séquence
peptidique (rôle d’une endopeptidase et
d’une carboxy-peptidase).
• Les protéines glycosylées sont largement
modifiées au niveau de leur partie
glucidique dans le Golgi.

Fonction de l’Appareil de Golgi (3)
concentration des protéines
Concentration des proenzymes dans les
cellules des acini pancréatiques (trypsinogène,
proamylase, prolipase, DNAse) dans les
vésicules de sécrétion.
• ces vésicules sont stockées puis secrétées
lors d’une stimulation chimique ou nerveuse.
• La concentration des protéines peut aboutir à
leur cristallisation ou à leur précipitation dans
les granules.
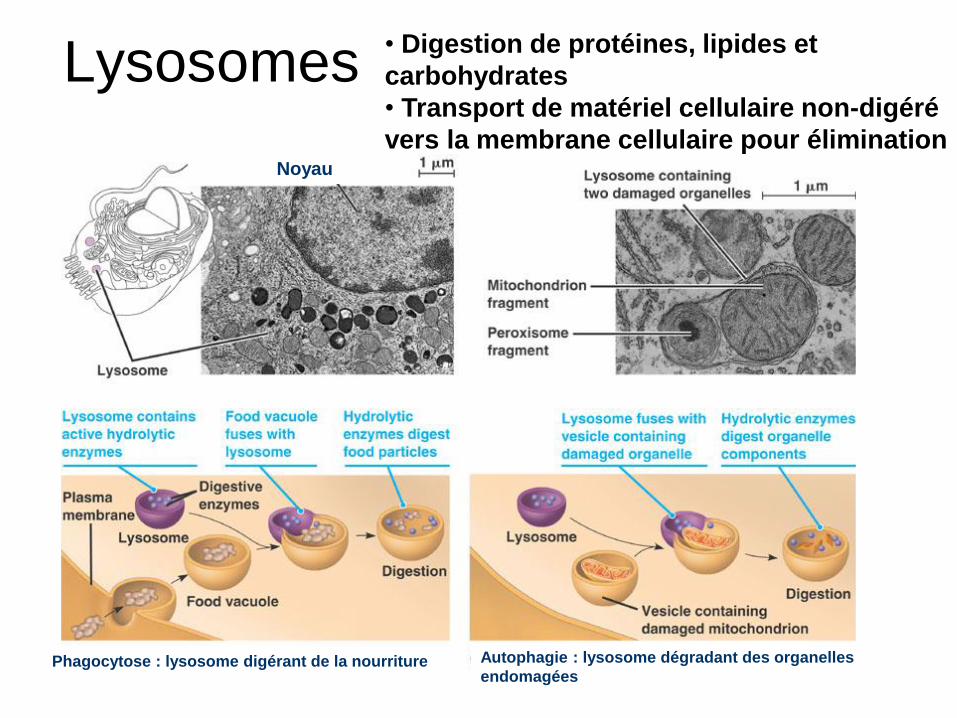
Lysosomes
Noyau
Phagocytose : lysosome digérant de la nourriture Autophagie : lysosome dégradant des organelles
endomagées
• Digestion de protéines, lipides et
carbohydrates
• Transport de matériel cellulaire non-digéré
vers la membrane cellulaire pour élimination
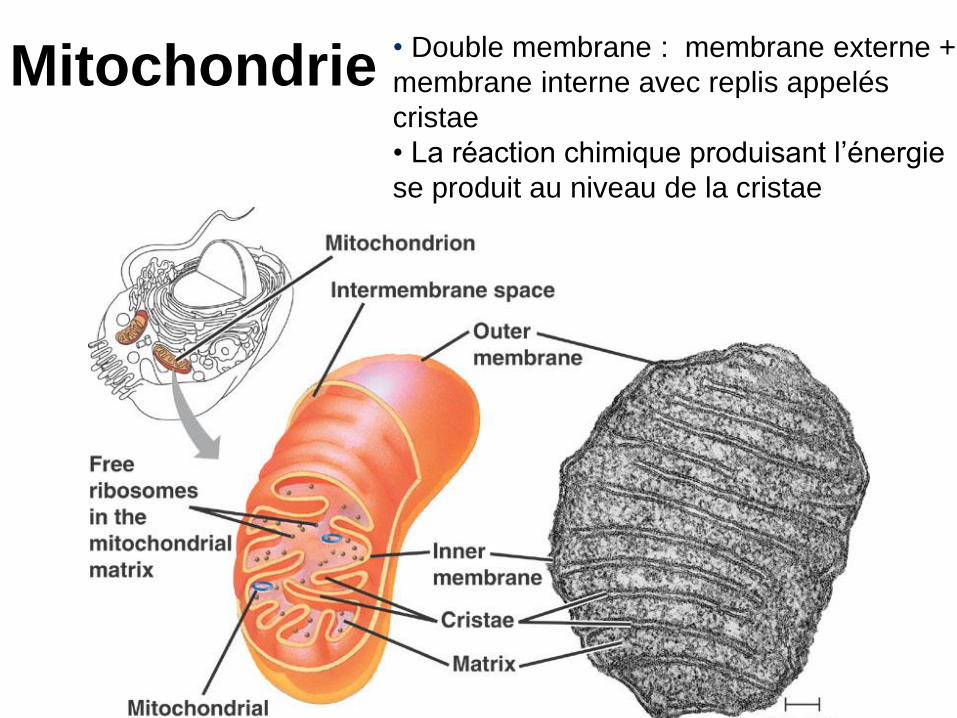
Mitochondrie• Double membrane : membrane externe +
membrane interne avec replis appelés
cristae
• La réaction chimique produisant l’énergie
se produit au niveau de la cristae

A. Généralités
• Elles occupent une part importante du
cytoplasme des eucaryotes.
• Elles se présentent comme des cylindres
allongées et rigides, ressemblent à des
bactéries. Malgré leur apparence, elles sont
malléables et changent constamment de forme
et peuvent même se fusioner pour ensuite de
dissocier.
• Elles peuvent être particulièrement nombreuses
à certaines régions telles que les myofribilles,
flagelles des spermatozoides.

A. Généralités (suite)
I. La mitochondrie possède une double membrane d’enveloppe
• L’organisation structurale de la mitochondrie est identique chez tous les eucaryotes, animaux et végétaux (figure 7/1). Les bactéries ne possèdent pas de mitochondrie (voir chapitre « la cellule »).
• La mitochondrie ne fait pas partie de système endomembranaire.
• La caractéristique de sa membrane interne est d’être spécialisée dans le transport d’ions et d’électrons.
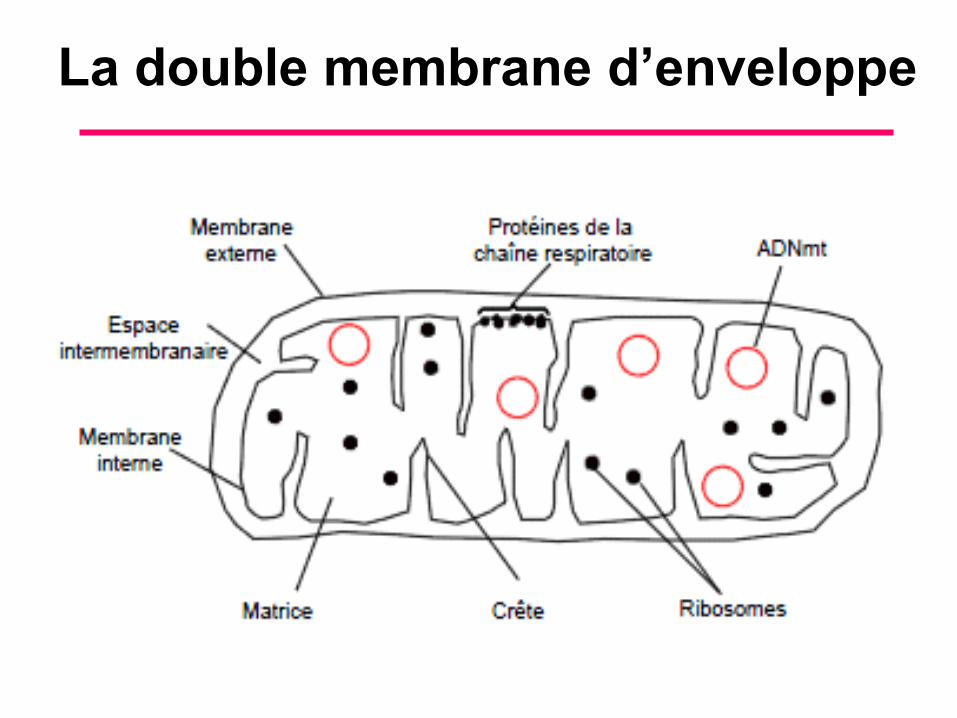
La double membrane d’enveloppe

A. Généralités (Suite)
II. Elle est responsable de la production, sous forme d’ATP, de la majeure partie de l’énergie nécessaire à la cellule par le phénomène de la phosphorylation oxydative.
• De l’ADP est phosphorylé en ATP
• Ce phénomène nécessite de l’oxygène (aérobiose). La mitochondrie est l’un des deux organites qui utilise l’oxygène. Mais c’est le seul organite qui participe à la respiration cellulaire (consommation d’O2 et production de CO2).
• Le deuxième organite consommateur d’O2 est le peroxysome, qui fabrique de l’H2O2 toutefois, d’autres réactions utilisant l’O2 dissout se déroulent en dehors de ces deux organites.

A. Généralités (Suite)
III. Elle possède son propre génome
• L’ADN mitochondrial lui permet de synthétiser, via des ARN mitochondriaux, un nombre de ses propres protéines (environ 13), les autres protéines étant synthétisées sous le contrôle du génome nucléaire.
• Il est généralement admis que les mitochondries ont pour origine une bactérie aérobie absorbée par l’ancêtre des cellules eucaryotes. Au cours de l’évolution, une parties des gènes de la bactérie ancestrale aurait migré vers le noyau de la cellule hôte alors que les autres gènes sont restés localisés dans la bactérie, d’où la notion d’un génome mitochondrie semi-autonome. C’est la théorie endosymbiotique.

A. Généralités (Suite)
• Le génome de la bactérie Rickettsia
prowasekii, responsable du typhus, est le
génome bactérien qui se rapproche le plus
de celui de la mitochondrie.
• Dans les cellules végétales, les
chloroplastes possèdent aussi leur propre
génome et exercent une fonction
comparable, la production d’ATP, en
consommant du CO2 et en produisant de
l’oxygène.

A. Généralités (Fin)
IV. La mitochondrie participe à d’autres
voies métaboliques
• Par exemple, la synthèse des stéroïdes et
de certains phospholipides membranaires,
les mouvements des ions calcium, avec
l’intervention du réticulum endoplasmique
lisse.

B. Morphologie et Composition Chimique
En microscopie optique
1. Les mitochondries peuvent être observéedirectement à l’intérieur des cellules vivantes encultures, après coloration vitale utilisant le vertJanus B (en lumière visible) ou la rhodamine 123 (enmicroscopie de fluorescence),
2. Elles se présentent sous la forme de bâtonnetsallongés et parfois ramifiés, ressemblant à desbactéries la cellule hépatique renferme environ 1700mitochondries, de volume moyen 0,3 3 (longueur ≤ 7 ,diamètre ≤ 1 ) et qui occupent environ 22% du volumecellulaire.

B. Morphologie et Composition Chimique (2)
3. Dans certains types cellulaires, les mitochondries
peuvent être immobiles dans des régions déterminées
du cytoplasme (ou elles répondent à une demande
locale importante d’énergie), par exemple le pôle
basal des cellules impliquées dans les transports
hydro-minéraux (voir chapitre « Membrane
plasmique »), le flagelle des spermatozoïdes.
4. Dans d’autres cellules, les mitochondries sont
très mobiles à l’intérieur de cytosol et présente
d’importantes modifications de forces et de taille
Leurs mouvements font intervenir le cytosquelette
(micro-filaments d’actine, microtubules et leurs
protéines associées).

B. Morphologie et Composition Chimique (4)
• Les deux membranes d’enveloppe
délimitent deux compartiments, l’espace
inter membranaire et la matrice
mitochondrie. Les deux membranes
d’enveloppe présentent des zones
d’accolement transitoire au niveau desquels
se déroulent des échanges entre le cytosol
et la mitochondrie.
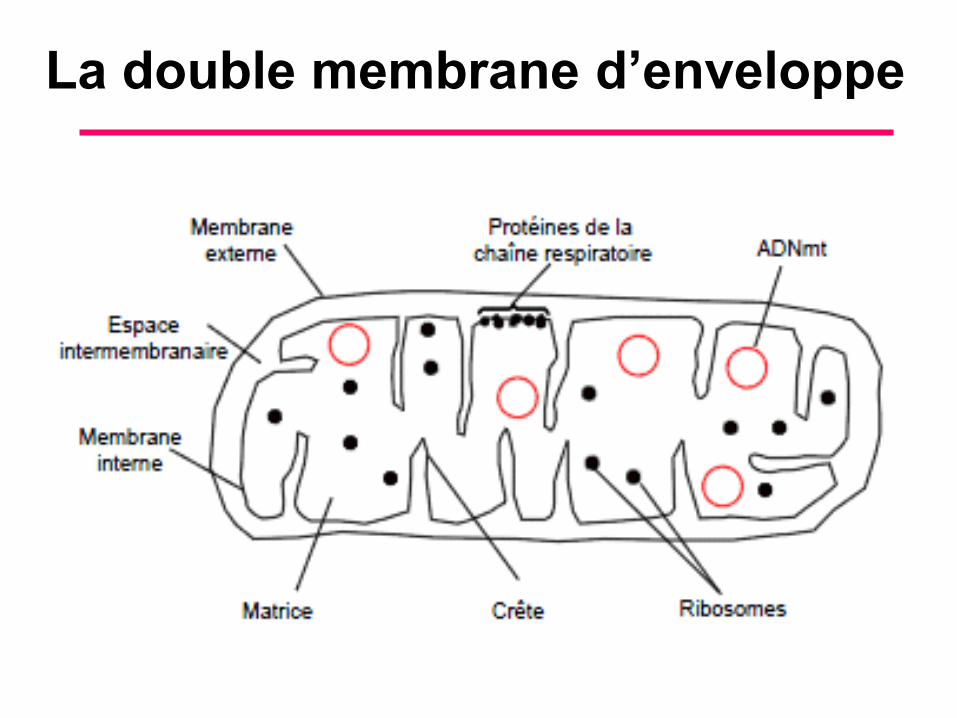
La double membrane d’enveloppe
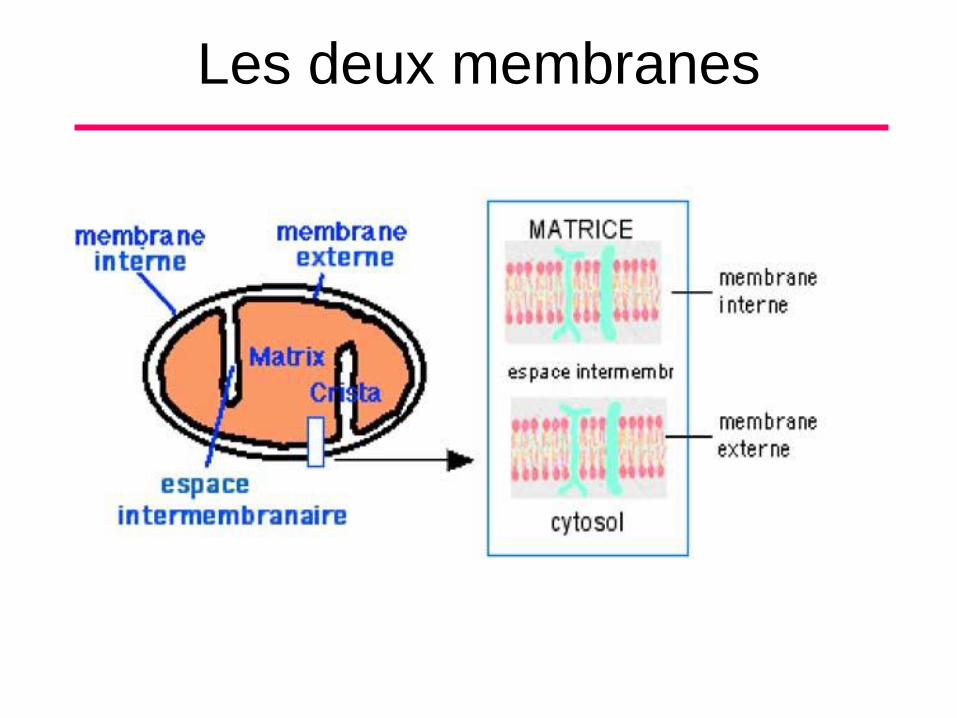
Les deux membranes

B. Morphologie et Composition Chimique (5)
• Les techniques de fractionnement
couplées à l’action de milieux hyper et
hypotonique permettent de séparer les
deux domaines membranaires et les deux
compartiments mitochondriaux et d’étudier
leur composition chimique.

B. Morphologie et Composition Chimique (6)
La membrane externe est une bicouche
lipidique renfermant des protéines en
quantité plus grande que la membrane
plasmique (3500 particules intra
membranaires /observées en microscopie
électronique en transmission après
cryofracture). Plusieurs types de protéines et
de complexes protéiques interviennent lors du
transport de molécules du cytosol vers les
deux compartiments mitochondriaux.

B. Morphologie et Composition Chimique (7)
a)Des perméases non glycosylées appelée
porines (de poids moléculaire 30000 Da) :
• Elles forment des pores aqueux au travers de
la bicouche lipidique, et permettent le
transport passif de molécules de taille
inférieure à 10 000 Da du cytosol dans
l’espace inter membranaire. De nombreuses
copies de cette protéine sont insérées dans
la membrane externe.

B. Morphologie et Composition Chimique (8)
• Les porines interagissent avec des protéines
associées aux microtubules (MAP2 pour
Microtubule Associated Protein), ce qui
pourrait rendre compte des mouvements de
ces organites.
b) Plusieurs autres protéines constituent 3
complexes au niveau de zones
d’accolement transitoire entre les
membranes mitochondriales externes

B. Morphologie et Composition Chimique (9)
L’espace inter membranaire
Très étroit, c’est un lieu de transit pour toutes
les molécules de tailles inférieures à 10 000
daltons (ions, protéines ...), qui traversent de
manière passive la membrane externe grâce
aux porines.
Il contient également les protons H+
• Transport de la matrice mitochondriale vers
l’espace inter membranaire grâce a l’action de
3 des 4 complexes transporteurs d’électrons
constituant la chaine respiratoire.

B. Morphologie et Composition Chimique (10)
• Transport de l’espace inter membranaires
vers le matrice par l’ATP-synthase et les
co-transporteurs.
• Il contient aussi une hémoprotéine, le
cytochrome C, qui circule entre les deux
membranes.
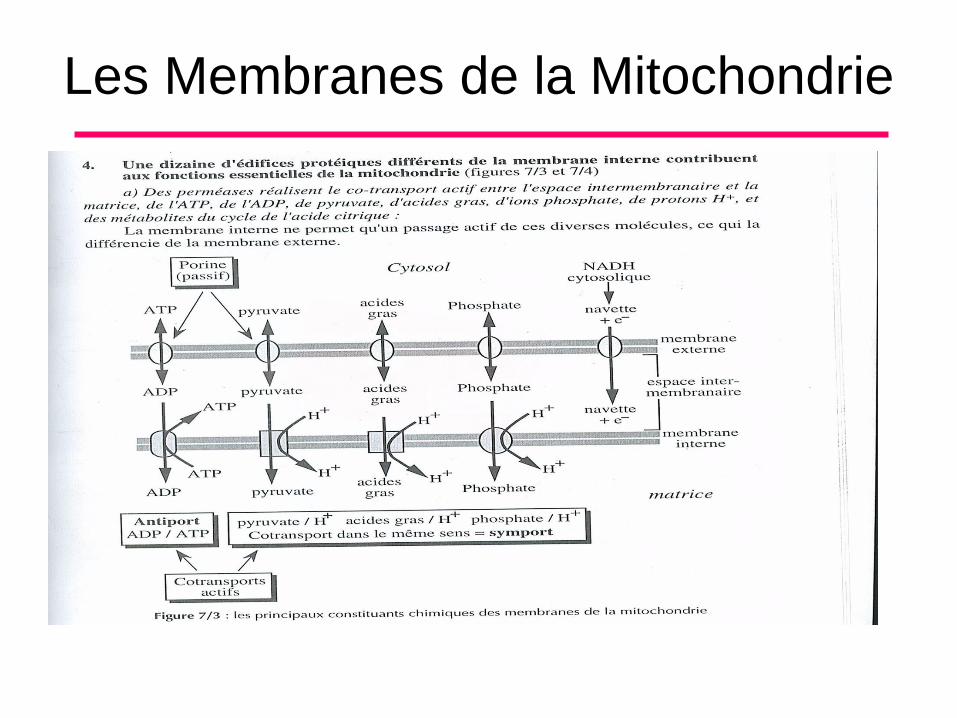
Les Membranes de la Mitochondrie

Transport d’ions au Travers de la
membrane interne
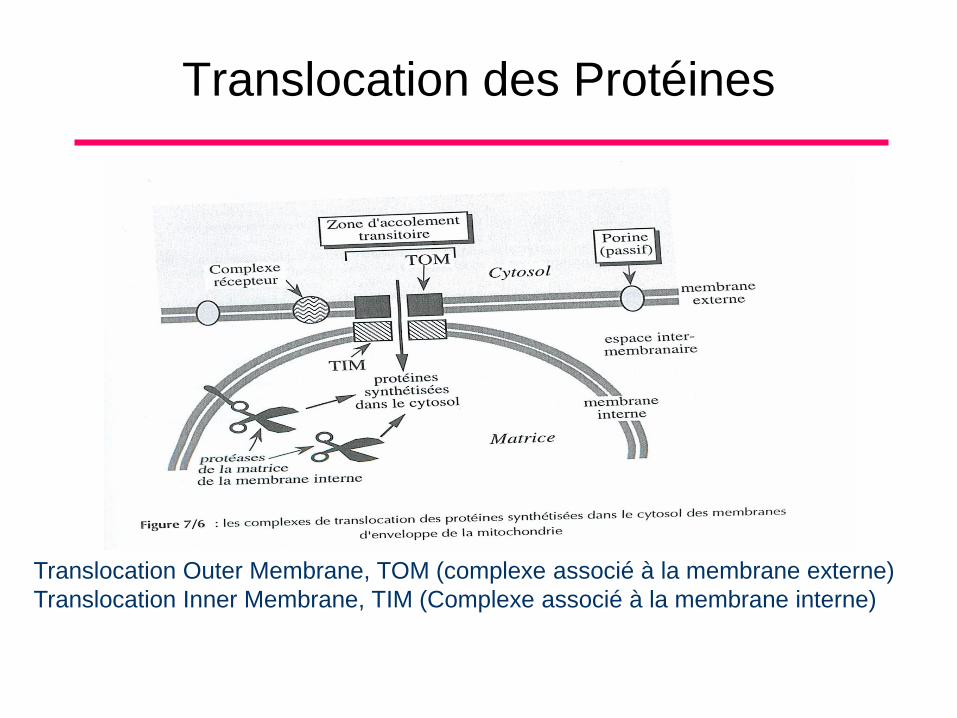
Translocation des Protéines
Translocation Outer Membrane, TOM (complexe associé à la membrane externe)
Translocation Inner Membrane, TIM (Complexe associé à la membrane interne)

Activité de l’Espace matriciel (1)
• Les acides gras et le pyruvate produit par
la glycolyse dans le cytosol sont
selectivement transportés vers la matrice
mitochondriale. A ce niveau, ils sont scindés
pour former le groupement acétyle à 2
carbones de l’ACETYL CoA. Le groupement
acétyle alimente ensuite le cycle de l’acide
citrique pour y subir une dégradation
ultérieure et l’opération se termine par le
passage des e- à haute énergie dans la
chaine respiratoire.

Activité de l’Espace matriciel (2)
• Afin d’assurer un apport continu de
combustible nécéssaire au métabolisme
oxydatif, les cellules animales stockent les
acides gras sous la forme de graisses et le
glucose sous la forme de glycogène qui peut
être dégradé pour produire du pyruvate.(Les
graisses sont quantitativement plus importantes que le glucogène parce
que leur oxydation libère 6 fois plus d’énergie que l’oxydation d’une
masse égale de glucogène).
• Un homme adulte moyen a une reserve
suffisante pour un jour d’activite normale. Il a
assez de graisses pour survivre un (1) mois.

Activité de l’Espace Matriciel (3)
Les lipides et et Acetyle CoA.
• Une molécule lipidique est constituée de 3
molécules d’acide gras réliés au glycérol par
des liaisons ester: ce sont des trialcools, Ils
n’ont aucune charge. Insolubles dans l’eau.
• Les enzymes localisés dans les membranes
interne et externe de la mitochondrie
interviennent dans le transport des acides
gras (dérivés de molécules lipidiques) vers
la matrice mitochondriale.

Activité de l’Espace matriciel (4)
• Dans la matrice, chaque molécule d’acide
gras (comme Acétyle CoA à longue chaine)
est completement dégradée suivant un
cycle de réactions qui élague 2 à 2 les
carbones de son extrémité carboxylée, et
produit une molécule d’Acetyle CoA à
chaque tour de cycle.
• L’Acetyle CoA produit est introduit dans le
cycle de l’acide citrique pour être oxydé.

Activité de l’Espace matriciel (5)
• Le cycle est catalysé par une série de 4
enzymes dans la matrice mitochondriale.
• Chaque tour de cycle raccourcit la chaine
d’acides gras de 2 carbones et produit une
molécule d’Acétyle CoA, une molécule de
NADH et de FADH2.

Activité de l’Espace matriciel (6)
Le Glycogene et l’Acétyl CoA. L’hydrolyse du
glucogène libère du glucose 1- Phosphate.
• La glycolyse fait passer le glucose à 6 carbones
à 2 molécules de pyruvate à 3 carbones
• La rencontre du pyruvate et du complexe
multienzymatique dont le pyruvate
deshydrogenase assure la conversion rapide du
pyruvate en Acétyl CoA et du CO2.
• Cet Aceyl CoA rejoint celui produit à partir des
acides gras pour alimenter le cycle de l’acide
citrique

Activité de l’Espace matriciel (7)
Le Cycle de l’acide citrique
• Il est synomyme de cycle de Krebs ou cycle
des acides tricarboxyliques
• Les principaux produits finaux dans
l’oxydation totale des compossé carbonés
sont:CO2 et NADH
• Le CO2 est libéré comme déchét
• Les molécules de NADH transmettent leurs
e- à haute énergie à la chaine respiratoire. A
la fin de la chaîne, ces e- sont utilisés pour
réduire 02 en H2O
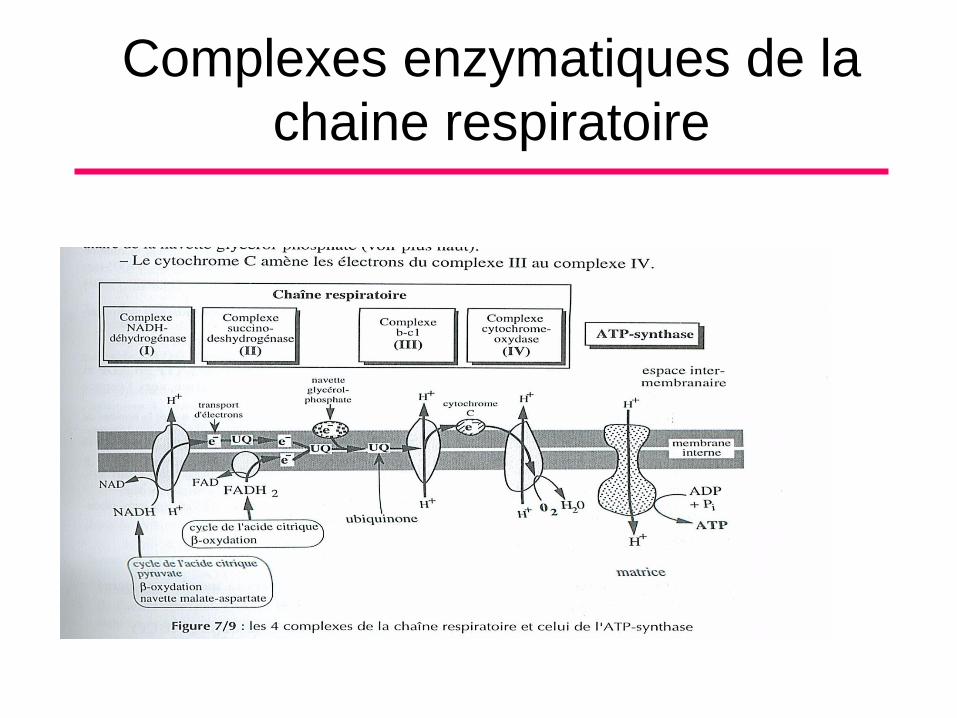
Complexes enzymatiques de la
chaine respiratoire

Activité de l’Espace matriciel (8)Le Cycle de l’acide citrique (continue)
• Chacune des étapes est catalysée par une
enzyme différente, localisée dans la matrice
mitochondriale.
• Les 2 atomes de carbones provenant de
l’Acetyl CoA qui entrent dans le cycle seront
transformés en CO2 lors des cycles ultérieurs.
• Trois molécules de NADH seront fournies.
• La molécule de GTP produite peut être
transformée en ATP par la réaction d’échange

Activité de l’Espace matriciel (9)Cycle de l’acide citrique (continue)
• La molécule de FADH2 formée reste liée à la
protéine (Acétyl CoA deshydrogenase) comme
élement du complexe succinate deshydrogenase
dans la membrame interne. Ce complexe
transmet les e- acquis par le FADH2 directement
à l’ubiquinone.
• Dans le processus de Phosphorylation
oxydative, principale conversion d’énergie, la
membrane interne sert de machine transformant
une partie de l’énergie du NADH (et du FADH2)
en énergie de liaison dans l’ATP

Activité de l’Espace matriciel (10)
• La chaîne respiratoire (aussi appelée
chaîne de transport d'électrons) est
constituée d'un ensemble complexe de
protéines membranaires de la mitochondrie
des cellules eucaryotes qui servent à
réoxyder les coenzymes NADH et
ubiquinone (CoQ) qui ont été réduits en
particulier au cours du cycle de Krebs. Cette
réoxydation s'accompagne de la création
d'un gradient transmembranaire de protons.

Activité de l’Espace matriciel (11)
• Ce gradient est une forme de stockage de
l'énergie contenue dans les coenzymes,
qui dérive elle-même de l'énergie
contenue dans les molécules dégradées
au cours du catabolisme. Le gradient de
proton va servir à fabriquer de l'ATP,
molécule énergétique universellement
utilisable, au niveau de l'ATP synthétase,
une protéine membranaire mitochondriale.

Activité de l’Espace matriciel (12)
• Ce mécanisme de phosphorylation oxydative
a été découvert par Peter Mitchell, ce qui lui
a valu le prix Nobel de chimie en 19781. Ce
mécanisme est aussi connu sous le nom de
Théorie chimiosmotique (de Mitchell).
Additionné à la synthèse d'ATP par les ATP
synthetase on la nomme phosphorylation
oxydative. (nb: le terme phosphorylation
oxydative est impropre, il faut lui préférer
celui d'oxydations phosphorylantes.)

Activité de l’Espace matriciel (13)
• La chaine respiratoire. L’ATP synthetase
ne transporte pas normalement H+ hors
de l’espace matriciel à travers la
membrane mitochondriale interne. Ce
travail sera celui de la chaine respiratoire.
• Il se crée alors un gradient
électrochimique de protons qui active la
synthèse d’ATP.
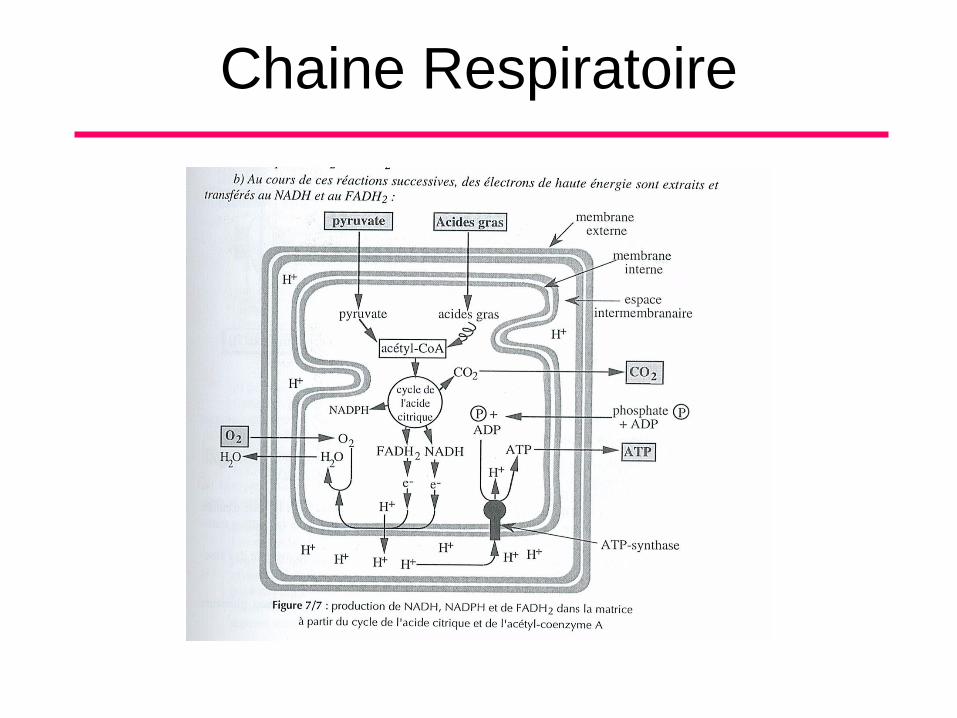
Chaine Respiratoire
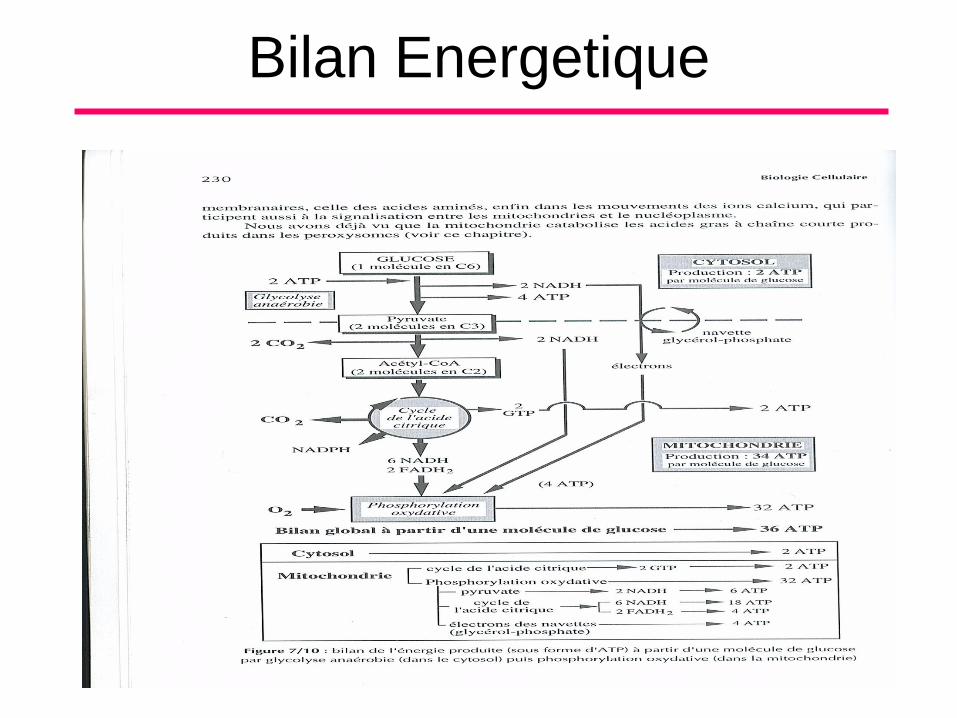
Bilan Energetique

Cycle de Krebs
• Le cycle de Krebs inverse est une voie
métabolique cyclique par laquelle certaines
bactéries fixent le dioxyde de carbone CO2 et
l'eau H2O sous forme d'acétyl-CoA en
utilisant l'hydrogène H2, l'ion sulfure S2- ou
encore l'ion thiosulfate S2O32- comme
donneurs d'électrons1,2, selon un mécanisme
reprenant le cycle de Krebs en sens inverse
pour constituer une alternative marginale au
cycle de Calvin.

Cycle de Krebs
Les principales étapes de ce cycle sont les
suivantes :
• Une première molécule de CO2 est fixée
sur la succinyl-CoA par l'alpha-
cétoglutarate synthase (EC 1.2.7.3) pour
former de l'α-cétoglutarate, sur lequel une
seconde molécule de CO2 est fixée pour
former de l'isocitrate avec réduction au
NADPH + H+.

Cycle de Krebs
• L'isocitrate est ensuite isomérisé en citrate,
lequel est clivé en oxaloacétate et acétyl-
CoA par l'ATP citrate lyase (EC 2.3.3.8), une
enzyme importante de la biosynthèse des
acides gras.
• L'oxaloacétate est ensuite converti en
succinyl-CoA par une série de réactions qui
ferment le cycle en passant par le malate, le
fumarate et le succinate

Cycle de Krebs
• La conversion du fumarate en succinate est
catalysée par une fumarate réductase à
ménaquinone, qui catalyse la réaction
réciproque de la succinate déshydrogénase
dans le cycle de Krebs.
• Ce cycle de réactions pourrait avoir joué un
rôle dans l'apparition de la vie sur Terre en
offrant une voie anabolique chimiotrophe
alternative à la phototrophie, certaines de
ses étapes pouvant être catalysées par des
minéraux3.

L’ADN Mitochondrial (1)
• Il est circulaire, comme l’ADN bactérien. Il
contient chez l’homme 16 000 paires de
bases (figure 7/12). Chaque mitochondrie
en contient plusieurs copies (de 5 a 10
selon les espèces).
• l’existence de l’ADN mitochondrial et sa
forme circulaire sont aussi des arguments
en faveur de la théorie endosymbiotique.

L’ADN Mitochondrial (2)
• Les produits du génome mitochondrial ne lui
confèrent qu’une autonomie très partielle par
rapport au génome nucléaire.
• Le code génétique mitochondrial est différent
du code génétique « universel » qui est celui
de l’ADN nucléaire des Eucaryotes :
Le code génétique est l’ensemble des
associations de 3 bases, chaque triple de
bases correspondant à un acide aminé.

L’ADN Mitochondrial (3)
• Des ARN ribosomaux propres aux
mitochondries (au nombre de 2)
• Des ARN de transfert (plus de 20)
• 13 protéines mitochondriales (dont
certaines sous-unités de la chaîne
respiratoire) codées par ARN messagers
d’origine mitochondriale.

L’ADN Mitochondrial (4)
• Par exemple, les mitochondries humaines
lisent UGA comme un codon pour le
tryptophane et non pas comme un signal
stop, comme c’est le cas pour le génome
nucléaire.

L’ADN Mitochondrial (5)
• On connait des mutants de levure chez
lesquels des anomalies de l’ADN
mitochondrial interdisent toute synthèse
protéique mitochondriale. Pourtant des
mitochondries de petite talle sont
observées chez ces mutants, ce qui
prouve l’importance des synthèses
protéiques contrôlées par le noyau dans la
genèse des mitochondries.

L’ADN Mitochondrial (6)
• Les autres protéines mitochondriales
(environ 500 dont environ 90 sont connues)
sont donc synthétisées dans le cytosol,
puis importées dans la mitochondrie :
• Parmi ces protéines, les ARN polymérases
et les ADN polymérases, ce qui rend la
réplication comme la transcription de l’ADN
mitochondrial totalement dépendantes du
génome nucléaire.

L’ADN Mitochondrial (7)
• Chez l’Homme, le génome
mitochondrial est transmis par la mère
• Il est le support de l’hérédité
dite « cytoplasmique », qualifiée de non-
mendélienne, puisque sa transmission est
monoparentale, ce qui distingue le
génome mitochondrial du génome
nucléaire.

L’ADN Mitochondrial (8)
• Cette caractéristique a été utilisée par les
généticiens qui ont tenté d’établir, à partir
des mutations observées au niveau de
l’ADN mitochondrial et de son évolution,
une carte de la filiation des espèces et
d’estimer la date de l’apparition de
l’espèce humaine.

L’ADN Mitochondrial (9)
• Toutefois, bien que l’hypothèse de base de ces
expériences soit séduisante, les données
obtenues doivent être corrigées en fonction de
deux facteurs : le fait que, au cours de
l’évolution, des fragment d’ADN mitochondrial
on été inclus dans le génome nucléaire, dont la
fréquence de mutation est environ 40 fois plus
élevée que celle du génome mitochondrial; le
fait enfin que des fragments minoritaires de
génome mitochondrial d’origine paternelle
contaminent l’ADN mitochondrial d’origine
maternelle.

Ribosomes
RE
Reticulum
endoplasmique (RE)
Ribosomes libres
Ribosomes liés
Grande
sous-unité
Petite
sous-unité
Diagramme
du ribosome
Image du RE et des
ribosomes par TEM
(microscopie electronique à
balayage)
• Composés de 2 sous unités faites d’ARN et de protéines
• Traduction des ARNm en protéine
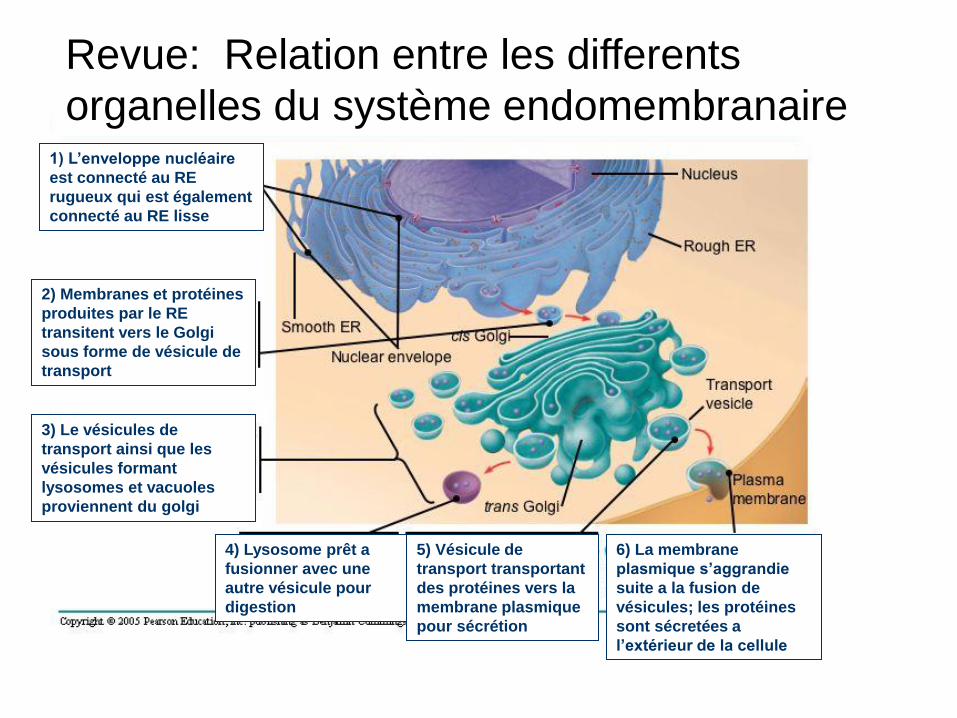
Revue: Relation entre les differents
organelles du système endomembranaire 1) L’enveloppe nucléaire
est connecté au RE
rugueux qui est également
connecté au RE lisse
2) Membranes et protéines
produites par le RE
transitent vers le Golgi
sous forme de vésicule de
transport
3) Le vésicules de
transport ainsi que les
vésicules formant
lysosomes et vacuoles
proviennent du golgi
4) Lysosome prêt a
fusionner avec une
autre vésicule pour
digestion
5) Vésicule de
transport transportant
des protéines vers la
membrane plasmique
pour sécrétion
6) La membrane
plasmique s’aggrandie
suite a la fusion de
vésicules; les protéines
sont sécretées a
l’extérieur de la cellule
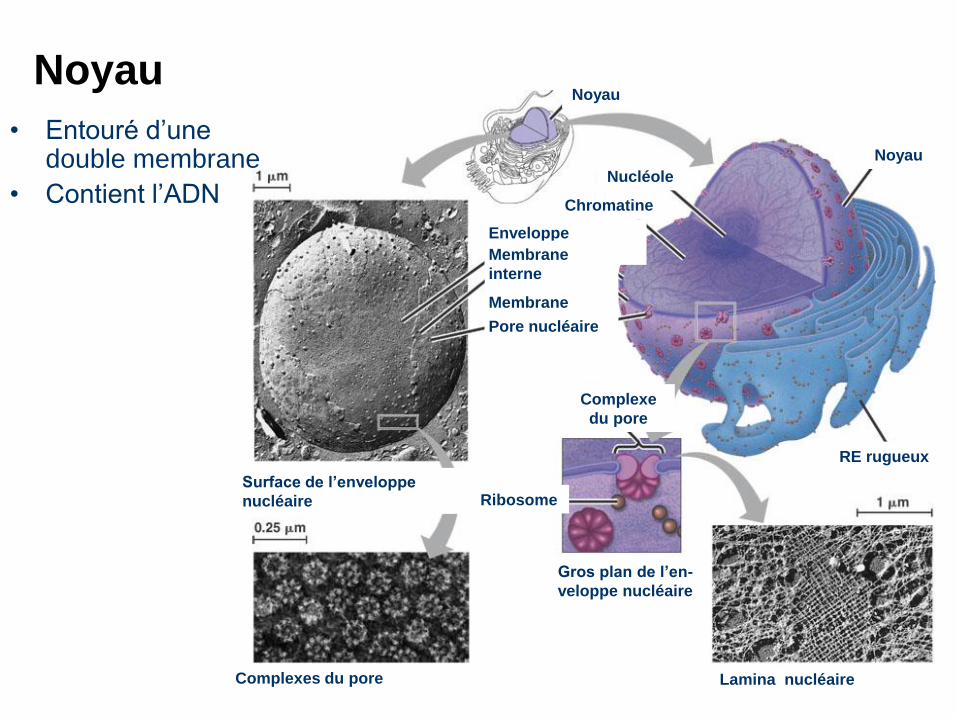
NoyauNoyau
Nucléole
Chromatine
Noyau
RE rugueux
Enveloppe
nucléaire:Membrane
interne
Membrane
externePore nucléaire
Complexe
du pore
RibosomeSurface de l’enveloppe
nucléaire
Complexes du pore
Gros plan de l’en-
veloppe nucléaire
Lamina nucléaire
• Entouré d’une double membrane
• Contient l’ADN

Dogme en Biologie
La Réplication
La Transcription
• La Traduction

Le Noyau cellulaire
• Chez les Eucaryotes,
l’ADN forme des pelotes,
les nucléosomes, ou
l’ADN est associé à des
protéines, les histones.
• Le tout est bien enroulé
et compacté sous forme
de chromosomes qui sont
localisés dans le noyau.

L’information génétique
Le matériel génétique doit posséder les propriétés suivantes:
• Il doit être capable de stocker l’information génétique et de transmettre cette information à la cellule en fonction des besoins de celle-ci.
• Il doit être capable de transférer son information aux cellules filles avec le minimum d’erreur.
• Il doit posséder une grande stabilité physique et chimique pour éviter la perte de l’information.
• Il doit être capable d’échanges génétiques mais sans perte majeure d’information.
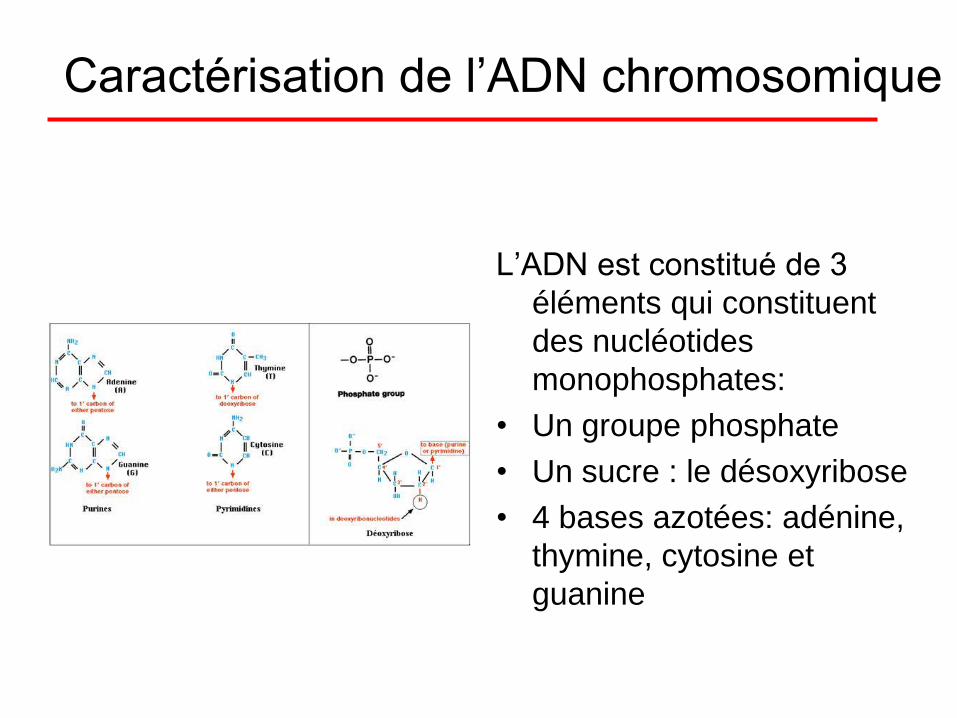
Caractérisation de l’ADN chromosomique
L’ADN est constitué de 3
éléments qui constituent
des nucléotides
monophosphates:
• Un groupe phosphate
• Un sucre : le désoxyribose
• 4 bases azotées: adénine,
thymine, cytosine et
guanine

Caractérisation de l’ADN chromosomique
•L’association de chaque base avec une molécule de sucre constitue
un nucléoside.
•L’ajout d’un groupe phosphate donne un nucléotide monophosphate.

Cycle cellulaire et Réplication
• Elle permet aux cellules
de se reproduire
identiques à elles
mêmes. Le matériel
génétique de départ de
mitose est divisé par
deux à l'arrivée.

Caractérisation de l’ADN chromosomique
• L’axe des paires de bases
ne passe pas par l’axe de la
double hélice, on aura donc
une partie plus large, le
grand sillon, et une partie
plus petite, le petit sillon.
Le grand sillon est soumis a
des interactions diverses
avec des molécules
endogènes et exogènes.

Cycle cellulaire et Réplication

La Réplication de l’ADN
• La réplication de l'ADN est semi-
conservative. À chaque mitose, la
molécule d'ADN double-brin est dupliquée
en deux molécules d'ADN double brin filles
dont chacune hérite un brin de la molécule
d'ADN initiale ou « mère » et d'un brin
synthétisé à partir de nucléotides libres.

La Réplication de l’ADN
• Lors de la réplication, les paires de bases
sont tout d'abord désappariées par la
rupture des liaisons hydrogènes de l'ADN
par une enzyme appelée ADN hélicase.
Une fourche de réplication va alors se
former donnant 2 brins d'ADN simple-brin
distincts. Chacun de ces brins va être
copié par l'action des ADN polymérases,
pour former 2 nouvelles molécules d'ADN
double brins identiques à la molécule
initiale.

La Réplication de l’ADN
• L’ ADN polymérase: cet enzyme permet la
polymérisation, c’est-à-dire l’addition des
nucléotides durant la copie, il a une fonction
d’édition ou de correction (endo ou exo
nucléaire). Il est doit être fidèle lors de la
copie.
• Les Nucléotides: ce sont l’adénine, la
cytosine, la guanine et la thymine, ils sont
ajoutés selon leur séquence sur le brin à
répliquer par l’ADN polymérase.

La Réplication de l’ADN
• L’hélicase: elle permet de dérouler le double brin
en brisant les liaisons H pour obtenir les brins
uniques.
• L’ ADN Primase ou Amorce ARN: ce sont de
courtes de séquences d’ADN ou d’ARN qui
permettent d’initier la réaction de réplication, elles
seront enlevées plus tard par l’ADN polymérase
grâce à sa fonction de correction (édition).
• Les ligases: elles fonctionnent durant la réplication
de la direction 3’ vers 5’, elles permettent de
combler les gaps crées par l’enlèvement des
amorces (ADN primase ou Amorce ARN).

La Réplication de l’ADN
• Les protéines SSB: Ce sont des protéines
qui se lient au brin d’ADN afin d’empêcher
leur appariement.
• Topoisomérase: permet d'éviter les torsions
entraînées par l'ouverture de la double-
chaîne par l'hélicase.
• Les fragments d’Okazaki: ce sont des
fragments de bases, 1000 à 2000 chez les
procaryotes, de 200 chez les eucaryotes.
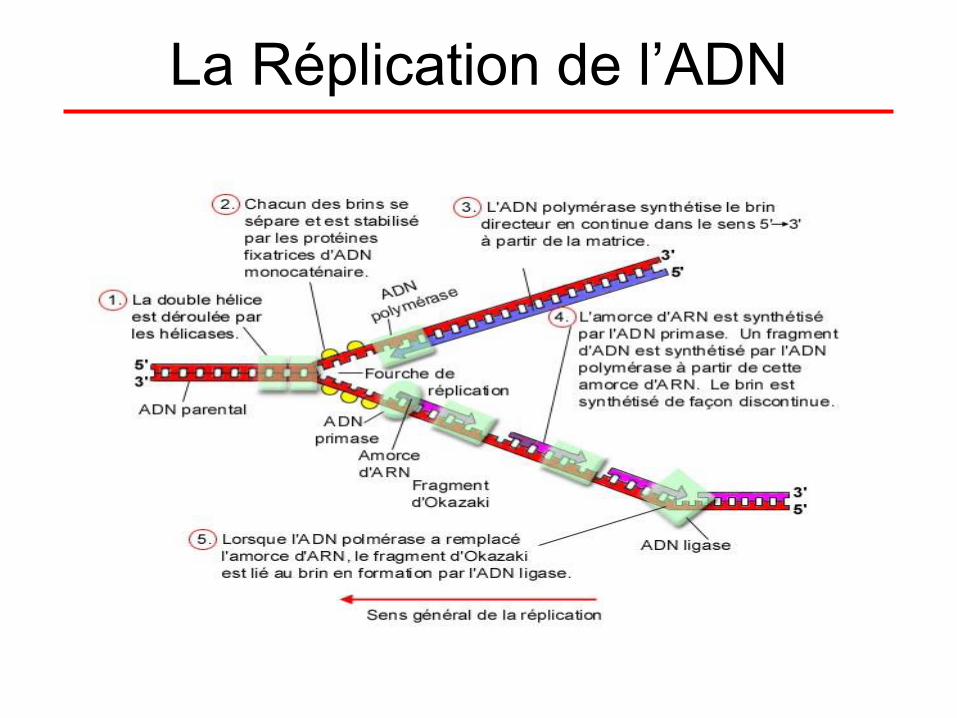
La Réplication de l’ADN

L’initiation de la Réplication (1)
• L’initiation de la réplication a lieu à l’origine de
réplication. Il existe une seule origine de
réplication dans les bactéries alors qu’il en
existe plusieurs chez les eucaryotes.
• La première étape débute par la fixation de la
protéine DnaA (protéine capable de reconnaître
ces origines) sur l'origine de réplication. Il s'en
suit un enroulement de l'ADN autour de la
protéine qui provoque une dénaturation locale
des deux brins, au niveau des 3 répétitions
consécutives de 13bp.

L’initiation de la Réplication (2)
• Une hélicase s'y engouffre et sépare les
brins dans les deux sens en rompant les
liaisons hydrogène (ou "ponts hydrogènes")
entre les deux brins de la double hélice
d’ADN. Deux fourches de réplication sont
ainsi créées, délimitant l'œillet de
réplication. Des protéines SSB se lient à
l’ADN simple brin (monocaténaire) ainsi
formé et évitent la reformation de la double
hélice.

Elongation (1)
• L’élongation de l’ADN progresse toujours dans
le sens 5' vers 3' pour le brin en création. C’est
l'ADN polymérase, qui ajoute à l'extrémité 3' de
la molécule en formation,
des désoxyribonucléotides.
• Pour le brin complémentaire du brin parental
orienté 5' vers 3’ (le « brin indirect » est donc
orienté 3’ vers 5’),Il est créé de façon
discontinue, sous forme de fragments
d’Okazaki, dans le sens 5’ vers 3’.
• L'ADN polymérase a besoin d'une amorce pour
fonctionner, elle crée une extrémité 3'OH Libre

Elongation (2)
• L'amorce ARN fournira cette extrémité libre.
• La présence ici d'ARN est expliquée par le fait
que seules deux enzymes peuvent synthétiser
les chaînes nucléotidiques : L'ARN Polymérase
et L'ADN Polymérase.
• L'ADN Polymérase ne pouvant fonctionner
sans amorce, c'est L'ARN qui prend le relai
pour fournir l'amorce nécessaire. La primase va
en effet créer ces amorces d'ARN.

Terminaison
• Cette phase correspond à l’arrêt de la
réplication lorsque deux fourches de
réplication se rencontrent ou lorsqu’une
fourche rencontre un signal de terminaison
de la réplication. Il y a « ter » : terA terD
terB terC qui freinent les fourches de
réplication.

Le mécanisme de correction
• La fidélité de réplication est très grande et
elle est en très grande partie due à l'ADN
polymérase, qui incorpore les bases
nucléiques en fonction de la
complémentarité des appariements.
• Les ADN polymérases réplicatives
disposent d'une activité de relecture qui
leur permet de vérifier que le dernier
nucléotide incorporé serait le bon.
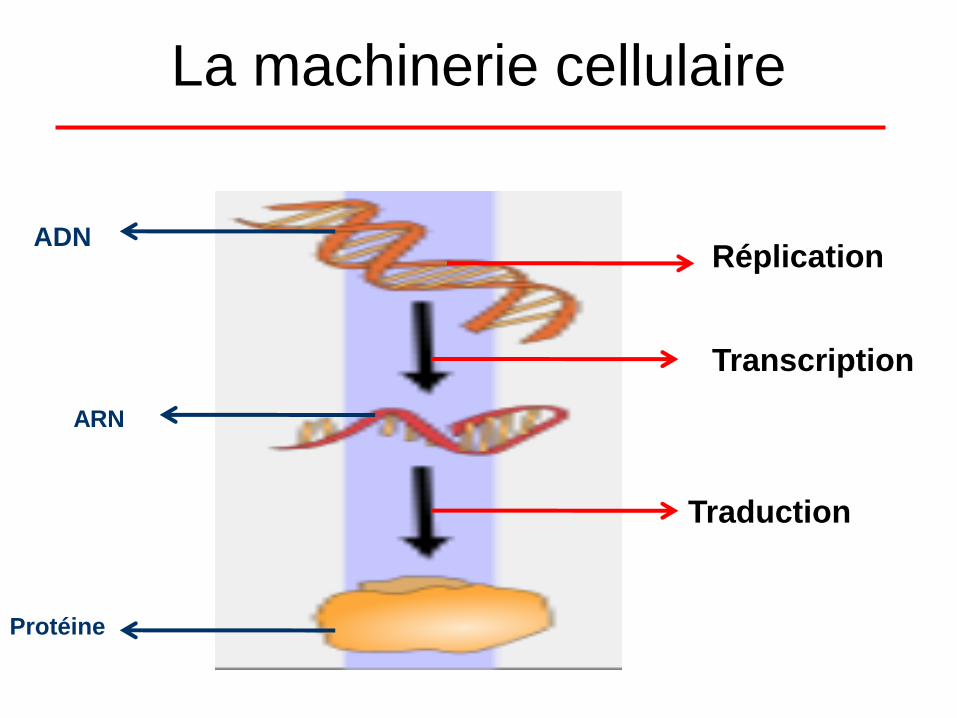
La machinerie cellulaire
ADN
ARN
Protéine
Réplication
Transcription
Traduction

Les ARN
• Les ARN sont des molécules constituées
par l'assemblage de ribonucléotides, et qui
possèdent de très nombreuses fonctions
dans la cellule.
• L'ARN est une molécule constituée d'un
enchaînement de ribonucléotides
(adénine, cytosine, guanine, uracile) reliés
entre eux par des liaisons nucléotidiques.

Les ARN
• L'ordre est dicté par la séquence des
désoxyribonucléotides portés par l'ADN. En
effet, les ARN sont issus de
la transcription de l'ADN par
une enzyme (l'ARN polymérase) qui recopie
en quelque sorte la séquence.
• Les ribonucléotides sont différents des
désoxynucléotides par la présence d'un
groupement OH en 2' du ribose, mais aussi
par le fait que la thymine (T) est remplacée
par l'uracile (U).

Les ARN
• À l'inverse de l'ADN qui est la plupart du
temps structuré en double hélice, l'ARN
peut adopter des conformations très
différentes, étroitement liées à sa fonction.
Ainsi, certaines molécules d'ARN sont
simple brin, en tige boucle (grâce à un
appariement des bases complémentaires),
en feuille de trèfle (l'ARN de transfert), etc

Les ARN
• Il existe de nombreuses familles d'ARN
(ARNr, ARNm, ARNt, ARNsi, ARNmi,
snARN...), dont chacune possède une
structure ou une fonction particulière :
• les ARN messagers (ARNm) serviront de
matrice pour la synthèse des protéines ;
• les ARN ribosomiques (ARNr) entrent
dans la composition des ribosomes, avec
les protéines ribosomiques.

Les ARN
• les ARN de transfert (ARNt) portent
des acides aminés et permettent leur
incorporation dans les protéines ;
• les ARN interférents (ARNsi, ARNmi...)
régulent l'expression des gènes en ciblant
la dégradation des ARN messagers
spécifiques ou en inhibant la traduction
des protéines.

Les ARN
• D'autres familles existent et de nouvelles
classes (ARNsno, ARNnc,...) sont
régulièrement découvertes.
• Chez certains virus (le virus de la
mosaïque du tabac, le VIH...), l'ARN
constitue le génome (alors que chez la
grande majorité des organismes, c'est
l'ADN qui remplit cette fonction).
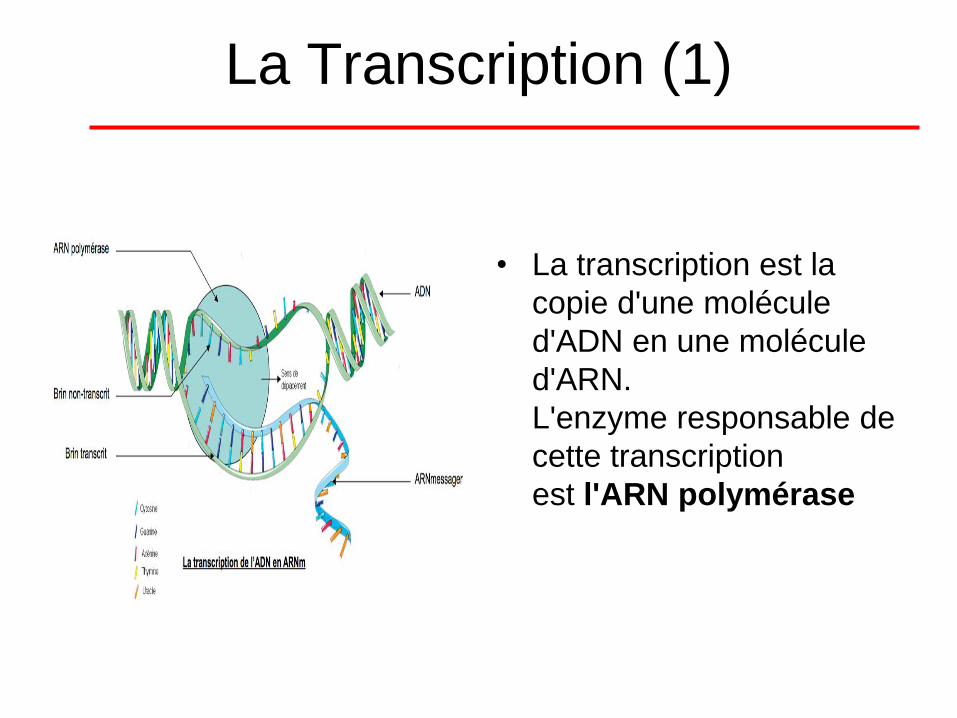
La Transcription (1)
• La transcription est la
copie d'une molécule
d'ADN en une molécule
d'ARN.
L'enzyme responsable de
cette transcription
est l'ARN polymérase

La Transcription (2)
• Les gènes des cellules eucaryotes (non
bactériennes), ont leurs séquences
codantes (appelées exons) interrompues
par des séquences non codantes
(appelées introns).

La Transcription (3)
• La molécule d'ARN directement
synthétisée à partir du modèle ADN,
ou transcrit primaire reste dans le noyau
et est traitée par un complexe
enzymatique qui enlève tous les introns.
C'est ce que l'on appelle l'épissage.
L'ARN produit est plus court, passe dans
le cytoplasme et devient un ARN
messager (ARNm).

La Transcription (4)
• L'ARNm est alors traduit en protéine à
partir des acides aminés en présence des
ribosomes et des ARN de transfert.
• La transcription se déroule dans le noyau
cellulaire. La chromatine doit au préalable
avoir été décompactée (euchromatine)
pour permettre à la machinerie protéique
d'accéder à l'ADN.

La Transcription (5)
• Contrairement aux procaryotes, l'ARN
produit par la transcription n'est pas
directement utilisable par les ribosomes pour
la traduction et devra subir plusieurs étapes
de maturation post-transcriptionnelle.

La Transcription (6)
• L'ARN polymérase ne peut se fixer seule
au promoteur du brin matrice d'ADN : chez
les eucaryotes, elle nécessite des facteurs
de transcription, protéines qui servent
d'intermédiaires à la liaison de l'ARN
polymérase sur le promoteur.

La Transcription (7)
• Cette association entre le promoteur, les
facteurs de transcription et l'ARN polymérase
forme le complexe d'initiation de la
transcription, nécessaire au commencement de
la transcription.
• Il existe 3 types d'ARN polymérase différentes
au lieu de l'unique pour les procaryotes. Mais
seules, ces ARN polymérases ne peuvent rien
faire et elles doivent être accompagnées de
facteurs de transcription généraux (protéines).

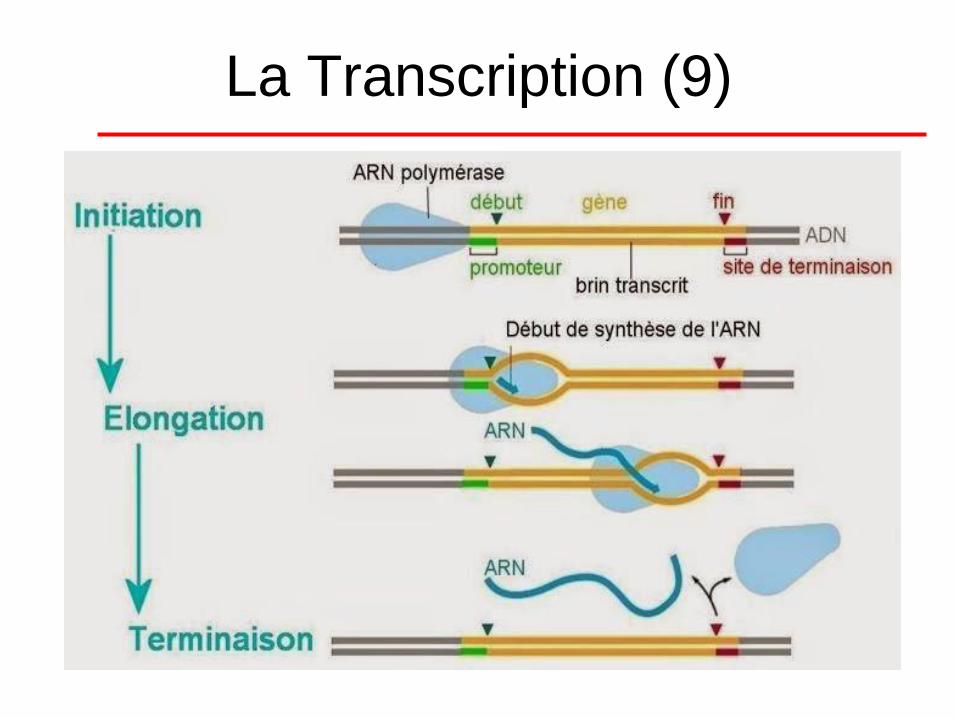
La Transcription (8)
• Ces facteurs se nomment TFI, TFII, TFIII…
ce qui constitue un complexe protéique
constitué de 8 à 14 sous-unités.
• Comme chez les procaryotes, on retrouve
les 3 phases distinctes dans le processus
de transcription : l'initiation, l'élongation et
la terminaison.
La Transcription (9)

Initiation de la Transcription (1)
• L'équivalent chez les eucaryotes de la
« boîte de Pribnow » est la « boîte TATA »
située environ 30 paires de bases avant
l'origine de transcription ; celle-ci joue un
rôle prépondérant puisque c'est à elle que
va se fixer l'ARN polymérase II.

Initiation de la Transcription (2)
• Deux autres « boîtes » font aussi partie des
séquences consensus ; parmi elles se trouvent
la boîte CAAT (située à environ 70 paires de
bases en amont du site d'initiation de la
transcription), qui est un site modulateur de la
transcription, et la boîte GC.
• Des amplificateurs peuvent stimuler la
transcription à plusieurs centaines de paires de
bases du lieu de la transcription.

Initiation de la Transcription (3)
• L'initiation par l'ARN polymérase
commence par la protéine TFII D, elle-même
constituée de la protéine de liaison TBP qui
va se fixer sur la boîte TATA, ce qui va
constituer le cœur du complexe d'initiation.
• Ensuite les différents facteurs généraux de
transcription viennent s'assembler sur ce
« noyau ».

Initiation de la Transcription (4)
• La TFII A vient stabiliser le complexe TFII D-
ADN, la TFII B et TFII E se lient à l'ADN, la TFII
F qui est une hélicase ATP-dépendante et
enfin l'ARN polymérase II. Cependant ce
complexe ne peut déclencher la transcription
qu'à une faible fréquence.
• Des facteurs de transition supplémentaires
doivent intervenir. Parmi eux le CTF (NF1) se
lie à la boîte CAAT, le Sp 1 se lie aux boîtes
GC : ce sont des trans-activateurs.

Initiation de la Transcription (5)
• Les séquences « enhancers » (amplificateurs)
et « cis activateurs » seront elles-mêmes
activées par des protéines activatrices : ces
séquences vont entrer en contact avec la
boîte TATA grâce à la courbure de l'ADN qui
les rapprocheront du promoteur.
• Une fois fixée à la boîte TATA, l'ARN
polymérase déroule les deux brins d'ADN et
commence la transcription.

Elongation de la Transcription (1)
• L'élément central de l'élongation est
la phosphorylation du domaine CTD (Carboxy
Terminal Domain), domaine spécifique de la
sous unité de 220 kDa de l'ARN polymérase II.
• Celle-ci est riche en sérine et en thréonine qui
sont deux acides aminés pouvant être
phoshorylés sur leur groupement hydroxyle.

Elongation de la Transcription (2)
• La phosphorylation par TFII H (du domaine CTD)
qui est une protéine Kinase en présence
d'ATP va déplacer l'ARN polymérase jusqu'au
lieu d'origine de la transcription.
• L'addition séquentielle des ribonucléotides peut
alors démarrer. Chez les eucaryotes, la vitesse
de transcription est d'environ 40 nucléotides par
seconde.

Terminaison de la transcription (1)
• La terminaison est assurée par des signaux
spécifiques dont le signal
de polyadénylation AAUAAA.
• L'ARN polymérase continue sa transcription
un peu après ce motif puis est libérée sous
l'action de divers facteurs.
• La transcription proprement dite est terminée
mais l'ARN obtenu n'est pas fonctionnel pour
autant et doit subir 3 étapes de maturation.

Terminaison de la transcription (2)
• L'ARN est clivé au niveau du signal de
polyadénylation et une polymérase
spécifique (la polyA Polymérase ou PAP)
ajoute de nombreux résidus Adénine (50
chez les levures, 200 chez les eucaryotes
supérieurs) à l'extrémité 3' du brin d'ARN.
• Cette queue polyA est essentielle à la
stabilité de l'ARN. Il est à noter que cette
partie de l'ARN n'est pas codée dans l'ADN
sous forme de polyT.

Terminaison de la transcription (3)
• À l'autre extrémité 5', l'addition d'une coiffe
méthylguanosine est nécessaire pour la
reconnaissance par les ribosomes lors de
l'étape de traduction. Il faut malgré tout
noter que les SnRNA qui sont aussi
synthétisés par l'ARN polymérase II ont
une coiffe mais ne passent pas dans les
ribosomes pour être traduits !

Terminaison de la transcription (4)
• L'ARN des eucaryotes est d'abord produit
sous forme de pré-ARNm qui contient toute
la séquence du gène (introns + exons). Il
subit ensuite une opération d'épissage : un
complexe nucléoprotéique (le spliceosome)
reconnaît les introns et les élimine.

Epissage
• Les eucaryotes (organismes à noyau),
l’épissage est un processus par lequel
les ARN transcrits à partir de
l'ADN génomique peuvent subir des étapes
de coupure et ligature qui conduisent à
l'élimination de certaines régions dans l’ARN
final. Les segments conservés s’appellent
des exons et ceux qui sont éliminés
s’appellent des introns.

Epissage
• Les gènes sont donc constitués d’une
suite d’exons et d’introns alternés ; ceci
s’observe principalement dans les gènes
codant des protéines, mais aussi dans
certains gènes d’ARN non codants,
comme ceux des ARNt.

Epissage
• L’ARN pré-messager est synthétisé puis
est épissé dans le noyau de la cellule pour
donner lieu à l’ARN messager dit mature.
• L’ARNm mature, constitué des seuls
exons, est alors exporté vers
le cytoplasme pour être traduit en
protéine.

La Traduction
• Le code génétique est un code qui permet la
conversion d’une séquence de nucléotides
(ADN puis ARN) en séquence d’acides aminés
(protéines). Le code implique les bases A, C, T
et G ainsi que les 20 acides aminés.
• Les codons sont des triplets de nucléotides
et ils codent pour un acide aminé.
• La séquence du gène et la séquence de la
protéine codée sont colinéaires, c’est-à-dire
que la longueur du gène et la longueur de la
structure primaire de la protéine finale sont
proportionnelles.

La traduction
• Le code génétique est universel. En effet
chaque acide aminé dispose d’un ou plusieurs
codons et ceci au niveau d’une multitude
d’organismes vivants procaryote et eucaryote.
• Le code génétique est redondant (ou
dégénéré). Plusieurs codons codent pour un
même acide-aminé : on trouve 64 codons et
20 acides aminés. Souvent se sont les deux
premiers nucléotides du codon qui définissent
l’acide aminé, la redondance est donc due au
troisième nucléotide du codon.

La Traduction
• Le code génétique est non-chevauchant. Les
nucléotides d’un codon ne participe qu’au code
d’un seul acide aminé, ainsi le prochain acide-
aminé sera codé par le prochain codon présent sur
l’ARNm. On parle du cadre de lecture (ou reading
frame).
• Le code possède un système de ponctuation.
Le codon d’initiation est le codon AUG (GUG pour
la mitochondrie) et les codons de terminaison sont
les codons UAA (ocre), UAG (ambre) et UGA
(opale). Le codon UGA (opale) n’est pas présent au
niveau de la mitochondrie.

Les acteurs de la traduction
• Les ribosomes sont constitués d’ARN
ribosomiques (ARNr) et de protéines et
sont structurés sous forme de deux sous-
unités que ce soit chez les procaryotes ou
chez les eucaryotes (cf. également
chapitre généralités). Leur taille est définie
en unité Svedberg.

Les acteurs de la traduction
• Les ribosomes procaryotes (70S) sont
constitués d’une petite sous-unité 30S et
d’une grande sous-unité 50S.
– La sous-unité 30S est constituée d’un ARNr 16S
(1541 nucléotides) et de 21 protéines.
– La sous-unité 50S est constituée des ARNr 23S
(2904 nucléotides) et 5S (120 nucléotides) ainsi
que de 32 protéines.

Les acteurs de la Traduction
• Les ribosomes eucaryotes (80S) sont
constitués d’une petite sous-unité 40S et
d’une grande sous-unité 60S.
– La sous-unité 40S est constituée d’un ARNr
18S et de 33 protéines.
– La sous-unité 60S est constituée des ARNr
28S, 5,8S et 5S ainsi que de 49 protéines.

Les acteurs de la Traduction
Le ribosome bactérien comporte des sites
spécifiques:
• Site A :(site Acide-aminé/Accepteur) fixation des
acides aminés.
• Site P :(site Peptidique/Donneur) fixation de f-Met.
• Site E :(site Exit) sortie de l’ARN de transfert.
• Site EF-G: présent au niveau de la grande sous-
unité.
• Site EF-Tu: présent au niveau de la petite sous-
unité.

Les acteurs de la Traduction
• Chez les eucaryotes le premier acide aminé est la
méthionine et non pas la f-Met présent chez les
procaryotes.
• L’enchaînement des ribosomes sur l’ARNm forme le
polysome, il permet d’augmenter l’efficacité de la
traduction. La distance minimale qui sépare deux
ribosomes est de 100 nucléotides.
• Au niveau des ribosomes associés au réticulum
endoplasmique les protéines en voie de synthèse
pénètrent dans les vésicules du réticulum directement
après le site E.

Les acteurs de la Traduction
Les ARNt ont une structure secondaire en
forme de trèfle à 3 feuilles et une structure
tertiaire en forme de L à l’envers. Lors du
mécanisme de traduction il y a un
appariement antiparallèle entre l’ARNm et
l’ARNt : reconnaissance codon-anticodon
au niveau de la boucle de l’anticodon.

Les acteurs de la Traduction
Les ARNt possèdent également un bras de l’acide
aminé qui le fixe en 3’ (CCA) sur le ribose, il s’agit
d’une liaison covalente : liaison ester riche en
énergie. Les acides aminés ne vont ainsi par
arriver libre sur le ribosome mais associés à leurs
ARNt respectifs. On trouve 40 à 60 ARNt
différents par cellule, il existe donc plusieurs
ARNt différents pour un acide aminé, on les
appelle ARNt iso-accepteurs.

Les acteurs de la Traduction
• La formation du complexe amino-acyl-tARN (aa-
tARN) nécessite une Amino-acyl-tRNA-
synthétase spécifique de l’acide aminé, qui doit
ainsi reconnaître toutes les formes de codon de
cet acide aminé. Le chargement correct de l’ARNt
est un élément important dans la fidélité de la
traduction.
• L’acide aminé (aa) est tout d’abord activé et cette
activation nécessite de l’énergie sous forme
d’ATP pour permettre la formation d’aa-AMP
(liaison anhydride mixte).

Les acteurs de la Traduction
La liaison formée entre l’ARNt et l’acide
aminé est une liaison covalente de type
carboxy-ester. Les Amino-acyl-tRNA-
synthétase sont au nombre de 20 dans la
cellule, autant qu’il y a d’acides aminés qui
rentrent en compte dans la traduction.
L’acide aminé complexé peut ainsi
s’associer à la chaîne.

Les acteurs de la Traduction
Les autres acteurs sont:
• les acides aminés,
• les amino-acyl tRNA synthétases,
• le Mg2+,
• le GTP et l’ATP.

Initiation de la traduction
• Un ribosome reconnaît le début de la
séquence codante, il utilise des signaux
d’adressage en amont entre -8 et-13 du
codon initiateur (AUG) qui correspond à la
séquence de Shine-Dalgarno ou RBS
(AGGAGG). Il y a appariement
antiparallèle de bases entre l’ARNm et la
petite sous-unité (30S) du ribosome, dû à
une complémentarité de séquences entre
l’ARNm et l’ARNr 16S.
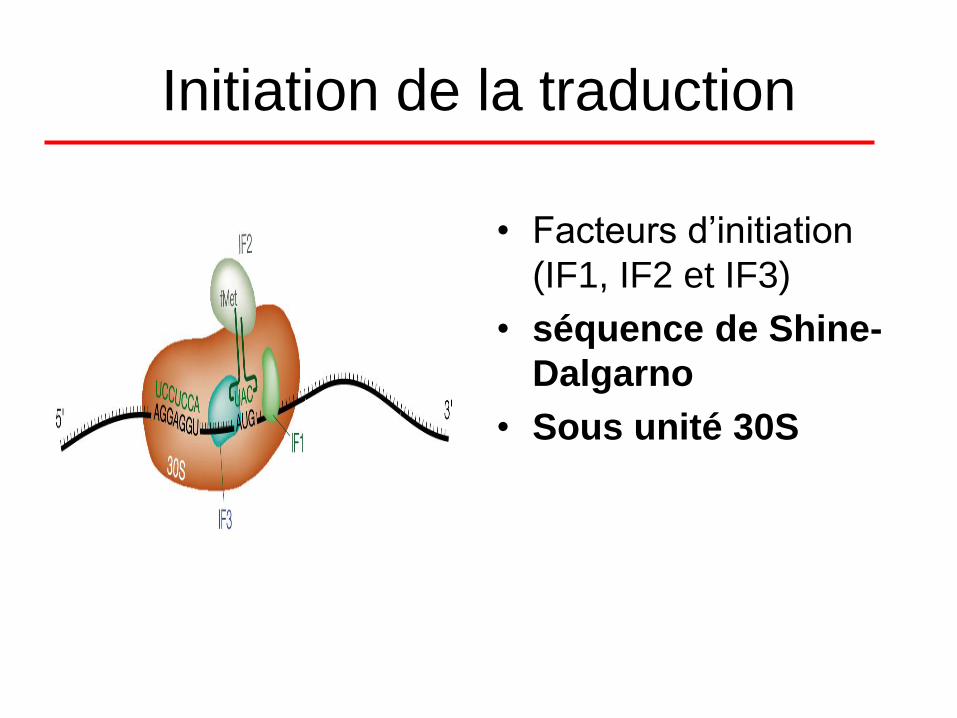
Initiation de la traduction
• Facteurs d’initiation
(IF1, IF2 et IF3)
• séquence de Shine-
Dalgarno
• Sous unité 30S

Initiation de la traduction
• L’initiation est permise grâce à la présence de
facteurs d’initiation:
• IF 1 est le facteur de dissociation du ribosome 70S.
• IF 2 est un facteur assurant la fidélité de
reconnaissance entre l’ARNt et l’acide aminé. Il
possède également une activité GTPasique (c’est-à-
dire d’hydrolyse du GTP).
• IF 3 est un facteur nécessaire à la fixation spécifique
de 30S sur l’ARNm et de contrôle de l’équilibre entre
la forme associé et dissocié du ribosome (facteur
anti-réassociation).

Initiation de la traduction
Par la suite le complexe 70S est reformé :
lorsque l’ARNt fixé à la formyl-méthionine
est fixé à la petite sous-unité de l’ARNr, il
y a hydrolyse du GTP et la grande SU se
fixe sur le complexe.

Les enzymes de Restriction
• Les enzymes de restriction sont des
protéines synthétisées par des bactéries
pour se protéger des infections de virus
(bactériophages). Ces enzymes coupent
l’ADN viral à des endroits spécifiques. Ce
mécanisme de résistance aux
bactériophages, dénommé restriction, fut
étudié par W. Arber à l’Université de
Genève dans les années 60.







