diego minciacchi la sonification versus la composition biotique des icônes du...
TRANSCRIPT

LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DESICÔNES DU CERVEAULa musique : de la neuroscience à la performanceDiego Minciacchi ERES | Insistance 2011/2 - n° 6pages 73 à 104
ISSN 1778-7807
Article disponible en ligne à l'adresse:
--------------------------------------------------------------------------------------------------------------------http://www.cairn.info/revue-insistance-2011-2-page-73.htm
--------------------------------------------------------------------------------------------------------------------
Pour citer cet article :
--------------------------------------------------------------------------------------------------------------------Minciacchi Diego, « La sonification versus la composition biotique des icônes du cerveau » La musique : de la
neuroscience à la performance,
Insistance, 2011/2 n° 6, p. 73-104. DOI : 10.3917/insi.006.0073
--------------------------------------------------------------------------------------------------------------------
Distribution électronique Cairn.info pour ERES.
© ERES. Tous droits réservés pour tous pays.
La reproduction ou représentation de cet article, notamment par photocopie, n'est autorisée que dans les limites desconditions générales d'utilisation du site ou, le cas échéant, des conditions générales de la licence souscrite par votreétablissement. Toute autre reproduction ou représentation, en tout ou partie, sous quelque forme et de quelque manière quece soit, est interdite sauf accord préalable et écrit de l'éditeur, en dehors des cas prévus par la législation en vigueur enFrance. Il est précisé que son stockage dans une base de données est également interdit.
1 / 1
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

AVANT-PROPOS
C’est avec grand plaisir que je me souviens de l’intérêt qu’a suscité pour moi l’invitation de Jean-Michel Vivès à participer au colloque de Cerisy 2010 « L’inconscient et ses musiques ». Cela a été un vrai défi de me confronter sur ce sujet à des psychanalystes, des philosophes, des ethnologues et des sociologues.Au début des années 1990, je me suis lancé dans une approche inter-disciplinaire très personnelle et mixte de la composition musicale et de la recherche en neurosciences. Comme compositeur et spécialiste des neurosciences, j’ai commencé par établir des synergies entre ma recherche en composition musicale et celle que je mène sur le cerveau, allant jusqu’à y inclure les expériences plus significatives. J’espère que mon travail – qui est loin d’être abouti – pourra offrir un point de vue utile au développement de la musique nouvelle ainsi que de nouvelles perspectives pour la compréhension des mécanismes de fonctionne-ment du cerveau.La présente contribution décrit les bases théoriques, les résultats essentiels et les perspectives de mes recherches sur les relations
INSISTANCE N° 6 73
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAULA MUSIQUE : DE LA NEUROSCIENCE À LA PERFORMANCE
Diego Minciacchi
Diego Minciacchi, professeur de neurologie, département de sciences physiologiques, faculté de médecine, université de Florence, Italie. Compositeur, Viale Morgagni 63, I-50134 Florence, Italie. [email protected]
Insistance 6.indd 73Insistance 6.indd 73 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

74 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
entre la structure et les fonctions du cerveau et les processus de composition musicale. La première partie de cet article recense les données et les points de vue qui expliquent pourquoi la musique ne peut pas être décrite – ni définie – en termes d’universaux spécifi-ques à elle. La musique n’est pas le privilège exclusif de l’espèce humaine et sa perception est organisée selon des processus construits très tôt et largement partagés par plusieurs fonctions. Plus intéressant, peut-être, la musi-que apparaît être fondée sur des règles que nous ne pourrions décrire sans ambiguïté. En outre, pour la perception et la production de la musique, nous utilisons des réseaux céré-braux vastes et complexes qui sont largement entremêlés. Cela pose une réelle difficulté pour ce qui est de l’interprétation biologique et phénoménologique des résultats des recher-ches neurobiologiques. La deuxième partie de l’article aborde succinctement le processus de composition ainsi que le choix et la mise en œuvre des règles qui lient les paramè-tres musicaux. Je me concentrerai sur les systèmes de composition musicale qui tirent leurs informations des sciences biologiques et spécifiquement des neurosciences. Je ferai état des premières expériences et des solutions récentes de sonification biologique. Puis de quelques exemples de la stratégie que j’appelle ici biotique, et que j’ai développée en menant diverses expériences. Ils seront donnés ici pour proposer une ligne de recherche qui pourra contribuer à élaborer un point de vue unifié sur les différents paramètres du son et les aspects micro- et macro-formels de la composition
musicale 1. Une grande partie de ma produc-tion musicale est liée à l’exploration de cette perspective 2.
LA MUSIQUE N’EST PAS LE PRIVILÈGE EXCLUSIF DE L’ESPÈCE HUMAINE
Des universels spécifiques à la musique impli-queraient l’existence de propriétés musicales qui seraient communes à toutes les formes de musique. De ce point de vue, la musique peut être considérée comme une forme exclusive de conduite-production humaine et être considé-rée comme un objet d’étude particulier.Dans la perspective de cet article, en désaccord avec certains auteurs (Schwartz et coll., 2003 ; Drake et Bernard, 2001), mais en accord avec les ethnomusicologues (Parncutt, 2009), je présuppose qu’il n’y a pas d’universels spécifi-ques à la musique.Pour vérifier le caractère d’unicité de la réponse esthétique humaine à la musique, les cher-cheurs ont mis au point des méthodes pour mesurer les préférences acoustiques spontanées chez les animaux. On constate qu’ils n’ont montré aucune préférence pour les accords consonants plutôt que pour les accords disso-nants, on est amené à la conclusion que certaines des préférences essentielles sous-ten-dant la perception de la musique chez les humains ont l’air de manquer aux primates non humains (McDermott et Hauser, 2004 ; 2007). Autrement dit, il est possible que de telles préfé-rences soient uniques à nos espèces et que les
Insistance 6.indd 74Insistance 6.indd 74 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 75
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
liens essentiels à la musique soient le fait de l’espèce humaine seule. Pourtant, une étude récente a établi, en utilisant un protocole d’expéri-mentation sophistiqué, la préférence spontanée du chimpanzé pour la musique consonante plutôt que pour la musique dissonante (Sugimoto et coll., 2010). Les auteurs montrent que la préférence pour la conso-nance n’est pas propre aux humains, et soutiennent l’hypothèse que les bases de l’appréciation de la musique seraient liées à l’évolution depuis les origines.Par ailleurs, les études sur le groupement temporel produisent des résultats intéressants. Le groupement est le processus de perception des limites entre les groupes et les sous-groupes d’éléments dans une séquence musicale. Quand nous considérons la littérature sur la perception par les bébés des groupements temporels, en la rattachant aux recherches comparables faites sur les auditeurs adultes et les audi-teurs non-humains, nous pouvons voir que les processus des groupe-ments temporels ne sont pas spécifiques à des espèces, bien que les processus spécifiques à la musique puissent survenir chez les humains, de par une exposition prolongée à des morceaux complexes (Trehub et Hannon, 2006). Il peut être conclu en particulier que les capacités des bébés pour la perception de la musique sont le produit de mécanismes généraux non spécifiques à la musique, ni à l’espèce.
LA PERCEPTION DE LA MUSIQUE EST ORGANISÉE SELON DES PROCESSUS PRÉCOCES ET GÉNÉRAUX
Il est admis que le fœtus est capable de percevoir certaines informations auditives pendant les dernières semaines de la gestation, et que les principales caractéristiques de ces informations peuvent être reconnues juste après la naissance. Néanmoins, nous devons être conscients que les nouveau-nés n’ont eu fondamentalement que très peu ou pas d’exposition à la musique car l’utérus et les tissus environnants filtrent fortement le son perçu par le fœtus. Nous savons aujourd’hui que le cerveau du nouveau-né, même en absence d’une exposition réelle à la musique, possède déjà la spécialisation hémisphérique pour le trai-tement de la musique aux premières heures de vie postnatale (Perani
1. Je dois mentionner ici mes contributions de recherche spécifiques en neurosciences (Minciacchi et coll., 2008) et celles que j’ai menées sur musique et système nerveux central (Minciacchi, 2003 ; 2005).2. Les œuvres musicales pour lesquelles la présence d’informations provenant de la recherche en neurosciences est cruciale sont les suivantes : « Westbau » from Der Dom, 1992 ; Klavierstück n.4 : Vae Victis, 1996-1997 ; Quintum Desertum, 2000-2001 ; La Discipline de la Crise, 2007-2009 ; Klavierstück n.7 – Acerrima Perscrutatio et Diligentissima, 2010 ; Haß zu Wasser und Haß zu Land, 2010.
Insistance 6.indd 75Insistance 6.indd 75 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

76 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
et coll., 2010). Le cerveau du nouveau-né est sensible aux changements tonals et fait la diffé-rence entre consonance et dissonance.Nous pouvons affirmer que l’architecture neuro-nale au service du traitement de la musique est déjà effective à la naissance malgré le peu d’ex-position antérieure à la musique. On pourrait donc en conclure que des circuits neuronaux du nouveau-né, qui ne sont pas spécifiquement liés à l’expérience musicale, sont cependant capables d’accomplir les actes fondamentaux du processus de perception musicale. Ce que nous pouvons alors assumer en définitive, c’est que ces capacités de perception ne sont pas dédiées à la musique mais plutôt que le message musi-cal s’accorde à tirer profit de leurs potentialités.C’est donc dans cette direction que nous pouvons commencer à imaginer comment l’in-formation musicale peut circuler à travers les circuits neuronaux en utilisant des architectu-res générales du cerveau qui existent pour le traitement de perceptions complexes, mais pas nécessairement spécifiques à la musique. Pour expliquer et suivre cette voie de la pensée je vais utiliser maintenant, en tant que métaphore, les argumentations formalisées comme « darwi-nisme neuronal ».Le darwinisme neuronal est une théorie générale de l’organisation et du fonctionnement cérébral élaborée à l’origine par Gerald Edelman (Edelman et Mountcastle, 1978 ; Edelman 1987 ; voir aussi : Fernando et coll., 2010), intégrant biolo-gie, anatomie et psychologie. Edelman pose qu’il existe des dispositions innées qui sont les premières valeurs de base dont nous sommes pourvus. Ces valeurs – simples motivations,
instincts, intentionnalités – servent comme des outils dont nous avons besoin pour nous adapter et survivre. Certains ont été dévelop-pés (par sélection) au cours de l’évolution ; ils constituent le répertoire primaire. D’autres sont acquis (aussi par sélection) pendant les proces-sus d’exploration et d’expérience, constituant un répertoire secondaire (Edelman et Gally, 2001). Les circuits neuronaux de ce type sont encore à un bas niveau et Edelman présente les concepts de « cartes neuronales » et de « signa-lisation réentrante ».Les cartes sont conçues comme des groupes de neurones qui répondent sélectivement à certaines catégories élémentaires de signaux et sont large-ment et réciproquement interconnectés. Cette architecture permet une communication conti-nue entre les cartes cérébrales et rend possible lors de nouvelles expériences d’entrer, de modi-fier et de coordonner les cartes entre elles dans l’espace et le temps, ce qu’on appelle signalisation réentrante. La confirmation expérimentale de cette théorie de la fonction cérébrale est extrê-mement riche. Par exemple, pour des études sur les valeurs sélectionnées par l’évolution nous mentionnons celles sur les champs corticaux qui sont communs à différentes espèces anima-les et dont les ratios d’agrandissements des représentations spatiales sont propres à chaque espèce. Ces recherches ont permis d’établir un arbre phylogénétique des principaux groupes de mammifères, de visualiser leurs relations et de postuler l’existence d’un ancêtre commun à tous les mammifères (Krubitzer, 2007).Parmi les études sur les valeurs fondamentales déterminées par l’exploration et l’expérience il
Insistance 6.indd 76Insistance 6.indd 76 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 77
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
vaut la peine de mentionner ici les enquêtes sur les propriétés plasti-ques trans-modales pour le traitement auditif. Chez les individus qui ont perdu la vue pendant la première enfance, les régions visuelles occipitales montrent une impressionnante plasticité lors d’expériences liées au traitement spatial de sons (Collignon et coll., 2009 ; Leclerc et coll., 2005).Finalement, en partant de Hebb (1949), qui a proposé « un assem-blage de cellules » pour représenter une pensée, et en passant par la découverte de l’organisation colonnaire du cortex (Mountcastle, 1957 ; Hubel et Wiesel, 1959), différentes théories ont été développées pour expliquer le fonctionnement cortical jusqu’à la fascinante descrip-tion du cerveau comme machine de Darwin (Calvin, 1987, 1996). Sa formulation suit le développement logique suivant : il y a un dessin, le dessin est copié, pendant ce processus des variations se produisent, les populations de dessins différents rivalisent pour l’espace, la sélection naturelle fait que quelques dessins font mieux que d’autres et le cycle suivant de variations est centré sur les variations les plus réussies de la génération précédente.Pour étayer sa construction Calvin a utilisé les résultats de beaucoup de recherches structurelles et fonctionnelles sur le cortex et a finalement développé un modèle spatial du fonctionnement cortical. Le modèle est composé d’un assemblage minimal de cellules, ou « point chaud », arrangé selon un dessin hexagonal. Ce dessin peut être cloné et, sans information redondante, être confiné dans un hexagone de 0,5 milli-mètres. Les hexagones dans le cortex ne sont pas une hypothèse : ils sont dus aux propriétés des longueurs axonales et aux accouplements synaptiques que les neurones entretiennent entre eux. L’hexagone est l’assemblage Hebbian minimal de cellules, potentiellement capa-ble d’enregistrer les caractéristiques d’un objet ou les détails d’un programme de mouvement. Donc l’hexagone est potentiellement un code cérébral : ce qui représente un objet ou une idée (Figure 1).On peut évoquer différents exemples pour illustrer des processus évolu-tifs darwiniens dans la musique : l’avènement du tempérament égal prenant le pas sur les autres systèmes d’intonation comme la gamme pythagoricienne fondée sur le cycle des quintes ou les gammes natu-relles fondées sur les sons harmoniques, ou l’établissement de simples
Insistance 6.indd 77Insistance 6.indd 77 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S
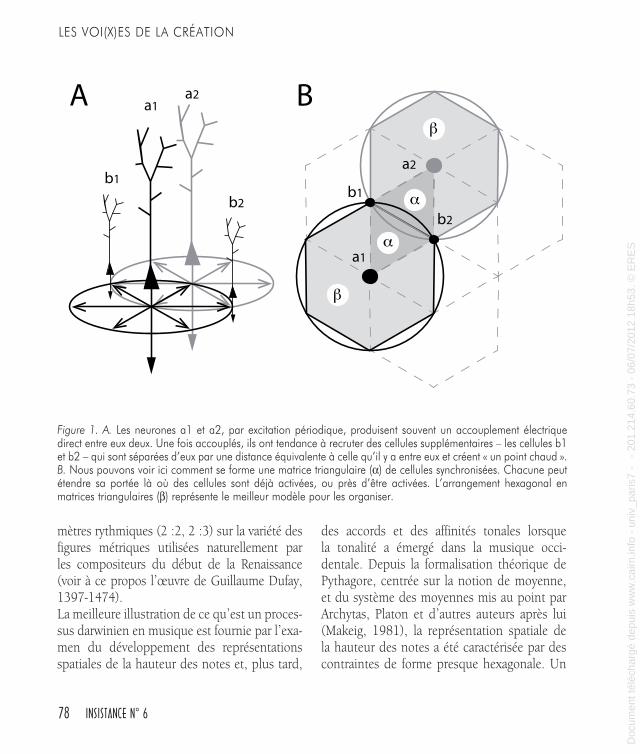
78 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
mètres rythmiques (2 :2, 2 :3) sur la variété des figures métriques utilisées naturellement par les compositeurs du début de la Renaissance (voir à ce propos l’œuvre de Guillaume Dufay, 1397-1474).La meilleure illustration de ce qu’est un proces-sus darwinien en musique est fournie par l’exa-men du développement des représentations spatiales de la hauteur des notes et, plus tard,
des accords et des affinités tonales lorsque la tonalité a émergé dans la musique occi-dentale. Depuis la formalisation théorique de Pythagore, centrée sur la notion de moyenne, et du système des moyennes mis au point par Archytas, Platon et d’autres auteurs après lui (Makeig, 1981), la représentation spatiale de la hauteur des notes a été caractérisée par des contraintes de forme presque hexagonale. Un
A Ba1
a1
b1b1
a2
a2
b2b2
Figure 1. A. Les neurones a1 et a2, par excitation périodique, produisent souvent un accouplement électrique direct entre eux deux. Une fois accouplés, ils ont tendance à recruter des cellules supplémentaires – les cellules b1 et b2 – qui sont séparées d’eux par une distance équivalente à celle qu’il y a entre eux et créent « un point chaud ». B. Nous pouvons voir ici comment se forme une matrice triangulaire (α) de cellules synchronisées. Chacune peut étendre sa portée là où des cellules sont déjà activées, ou près d’être activées. L’arrangement hexagonal en matrices triangulaires (β) représente le meilleur modèle pour les organiser.
Insistance 6.indd 78Insistance 6.indd 78 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 79
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
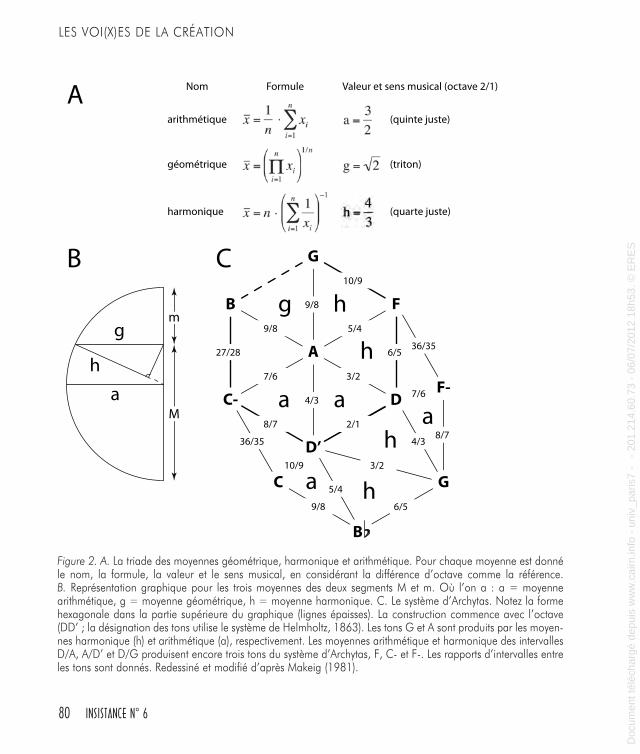
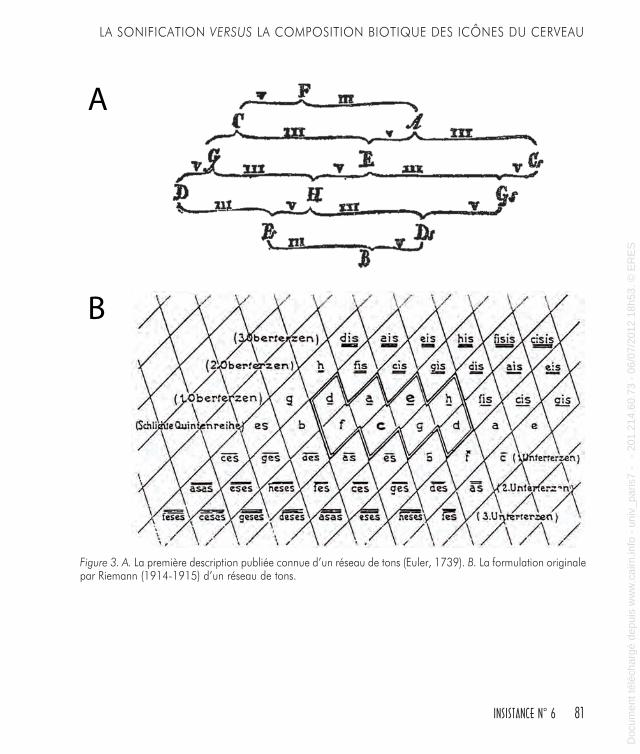
bon exemple est donné par le système d’Archytas qui a généré son ensemble de tons par l’utilisation de la triade des moyennes arith-métique, harmonique, et géométrique (Figure 2A, B, C). Plus tard, la représentation spatiale de l’espace musical a été concentrée sur les relations entre les groupes des notes, comme les triades, et les centres tonals.La gamme majeure et la gamme mineure du système tonal découlent des modes anciens médiévaux qui ont disparu progressivement. Sur les huit modes grégoriens seulement deux ont survécu : le mode ionien et le mode éolien, correspondant à nos gammes majeure et mineure, respectivement. Dans ce processus, l’intérêt musical s’est déplacé de la séquence d’intervalles, qui a caractérisé la modalité médiévale, vers le mouvement des centres tonals. Dans la musique tonale la séquence d’intervalles est plutôt invariante (pratiquement, on n’utilise que les deux gammes majeure et mineure), mais cela permet de passer d’un centre tonal à un autre avec beaucoup de souplesse.En se déplaçant entre les centres tonals il faut respecter les caractéris-tiques auditives fondamentales, la plus importante étant l’affinité de ton. Affinité signifie que des tons peuvent être perçus comme étant les mêmes, sous certains aspects, que sous certaines conditions un ton peut-être remplacé par un autre, et qu’un ton peut même être confondu avec un autre. Les plus fortes affinités reconnues sont l’équi-valence d’octave et les similarités de quinte ou de quarte (Helmholtz, 1863 ; Stumpf, 1890 ; Terhardt, 2006).Des modèles ont été développés par les théoriciens de la musique et les compositeurs pour arranger des centres tonals du point de vue des similarités, mais depuis la première description par Euler (1739) il était clair que pour représenter l’espace tonal la voie la plus efficace était un réseau triangulaire (Figure 3A). Plus récemment, les théoriciens du XIXe siècle ont exploré un réseau de tons, selon un schéma spatial, pour établir la cartographie des accords consonants (Hyer, 1995 ; Hostinsky, 1879 ; Riemann, 1914-1915 ; Figure 3B). Ici de nouveau, et jusqu’a présent (Figure 4A, B), un treillage de triades émerge dans une organi-sation spatiale composée de formes hexagonales.Après cette description on peut penser que les systèmes de relations développés pour illustrer, expliquer ou explorer les paramètres perti-
Insistance 6.indd 79Insistance 6.indd 79 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S
80 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
h
h
h
h
a
a
aa
g
G
F
G
B
A
B
C-
10/9
10/9
36/35
36/35
9/8
4/3
4/3
5/4
5/49/8
9/8
6/5
6/5
3/2
3/2
7/6
7/6
8/78/7
2/1
27/28
D’
C
DF-
Nom
(quinte juste)
(triton)
arithmétique
géométrique
harmonique (quarte juste)
Formule Valeur et sens musical (octave 2/1)A
B C
h
a
gm
M
Figure 2. A. La triade des moyennes géométrique, harmonique et arithmétique. Pour chaque moyenne est donné le nom, la formule, la valeur et le sens musical, en considérant la différence d’octave comme la référence. B. Représentation graphique pour les trois moyennes des deux segments M et m. Où l’on a : a = moyenne arithmétique, g = moyenne géométrique, h = moyenne harmonique. C. Le système d’Archytas. Notez la forme hexagonale dans la partie supérieure du graphique (lignes épaisses). La construction commence avec l’octave (DD’ ; la désignation des tons utilise le système de Helmholtz, 1863). Les tons G et A sont produits par les moyen-nes harmonique (h) et arithmétique (a), respectivement. Les moyennes arithmétique et harmonique des intervalles D/A, A/D’ et D/G produisent encore trois tons du système d’Archytas, F, C- et F-. Les rapports d’intervalles entre les tons sont donnés. Redessiné et modifié d’après Makeig (1981).
Insistance 6.indd 80Insistance 6.indd 80 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S
INSISTANCE N° 6 81
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
B
A
Figure 3. A. La première description publiée connue d’un réseau de tons (Euler, 1739). B. La formulation originale par Riemann (1914-1915) d’un réseau de tons.
Insistance 6.indd 81Insistance 6.indd 81 3/12/11 9:56:313/12/11 9:56:31
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S
82 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
g a
d
cb
e f
G
E F
A
C
D
B
g
c
f F
d
A
D
Bb
EB
A
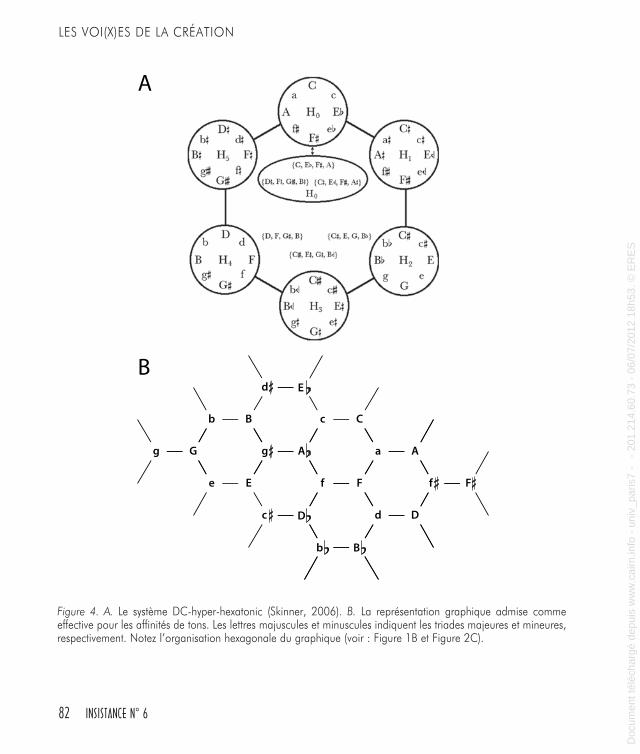
Figure 4. A. Le système DC-hyper-hexatonic (Skinner, 2006). B. La représentation graphique admise comme effective pour les affinités de tons. Les lettres majuscules et minuscules indiquent les triades majeures et mineures, respectivement. Notez l’organisation hexagonale du graphique (voir : Figure 1B et Figure 2C).
Insistance 6.indd 82Insistance 6.indd 82 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 83
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
nents d’écoulement musical d’information sont plutôt invariants. Si on utilise de nouveau la métaphore spatiale, la conformation générale des graphiques que les théoriciens et les compositeurs ont dévelop-pés au cours des siècles, en suivant des buts musicaux radicalement différents et avec des matériaux profondément divergents, emprunte constamment une forme hexagonale-triangulaire.Nous avons pu constater qu’il existe des similarités déconcertantes entre les graphiques représentant des formulations récentes de l’affinité tonale et ceux des modèles des unités fonctionnelles corticales (voir Figures 1B et 4A, B). Nous avons également précisé qu’une notion comme celle de la perception de primitives musicales n’est pas exclu-sive aux humains, qu’on peut la discerner de bonne heure, même si notre exposition préalable à la musique a été négligeable. Il nous faut préciser davantage.L’image que génèrent ces concepts une fois réunis, est tout à fait non équivoque : la perception de la musique n’est pas nécessairement accomplie par des processus spécifiques « orientés musicalement » et résidant dans notre cerveau. Ce n’est pas tout. On est en droit d’ima-giner puis de proposer une autre approche : le monde musical peut être considéré comme un des effets de la conformation architecturale élaborée de circuits du cerveau qui se trouvent là pour des ensembles de raisons complètement différentes ; la musique les emprunte, simple-ment. Nous apporterons des arguments supplémentaires dans le sens de cette idée plus loin.
DES RÉSEAUX NEURONAUX COMPLEXES ET ENTREMÊLÉS SONT IMPLIQUÉS LORS DE LA PERCEPTION ET LA PRODUCTION DE LA MUSIQUE
Bien qu’il soit généralement admis que les neurosciences doivent tendre à l’intégration, les travaux récents sur les fonctions des entités biolo-giques posent souvent des limites qui ne permettent pas d’y arriver. Il devient nécessaire d’établir une cartographie des fonctions à travers les différents systèmes neuronaux. Il faudrait utiliser pour ce faire des
Insistance 6.indd 83Insistance 6.indd 83 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

84 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
méthodes novatrices afin d’identifier les compo-santes qui contribuent à une fonction, mais aussi repenser profondément la façon dont les chercheurs formulent problèmes et plans d’ex-périences. Une des premières tentatives vers la compréhension des fonctions du cerveau a été la localisation. Les neurologues ont beaucoup travaillé pour repérer les voies spinales, la zone de production du langage, établir la topographie de la motricité, celle de la mémoire et son enco-dage dans l’espace en mémoire à court terme et mémoire à long terme.Vers le milieu du XIXe siècle l’approche loca-lisationniste – ou approche phrénologique – prétendait discerner des zones correspondant à certains états mentaux ou à certaines conduites. Avec l’arrivée des techniques d’imagerie du cerveau, en particulier la résonance magnétique fonctionnelle (IRMf) et la tomographie par émis-sion de positrons (TEP), une nouvelle impulsion a été donnée aux recherches sur la topographie fine des fonctions du cerveau. D’un côté l’IRMf s’appuie sur la détection et la représentation de la consommation locale d’oxygène, de l’autre la TEP permet de visualiser en trois dimensions l’utilisation de glucose ou le flux sanguin. La consommation d’oxygène, le flux sanguin, ou l’utilisation de glucose sont des paramètres trop indirects et lents pour permettre de mesurer l’activité des neurones intervenant dans le traite-ment de l’information. Le développement de la magnétoencéphalographie (MEG), qui augmente la résolution temporelle en mesurant les champs magnétiques induits par l’activité électrique des neurones, est plus attrayant. On a pu discuter de la validité de la combinaison de la MEG avec
l’IRMf et la TEP, mais de fait, seuls quelques cher-cheurs ont publié les résultats des expériences menées en combinant les trois techniques (Horwitz et Poeppel, 2002 ; Kosik, 2003).Dans le domaine de la recherche musicale, les études du cerveau par imagerie fonctionnelle, menées suivant différents paradigmes expé-rimentaux, ont montré pléthore de régions corticales dans les lobes frontal, temporel et pariétal, dans l’insula et les circuits limbiques, impliquées dans le décodage des paramè-tres musicaux et dans leur mise en œuvre musicale ultérieure. Les études par imagerie cérébrale montrent que les aires auditives primaires et secondaires, de même que le planum temporale (en particulier la région à la frontière de la fissure sylvian avec le lobe temporel, la région SPT), mais aussi les aires motrices secondaires (comme l’aire prémo-trice dorsale et ventrale, les régions motrices supplémentaires et présupplémentaires et la région du Broca), retiennent des associations fonctionnelles importantes pour le traitement et la réception de la musique (Koelsch et coll., 2006 ; Warren, 2008 ; Fadiga et coll., 2009 ; Grand et Snyder, 2009 ; Chen et coll., 2009). On peut voir aussi que beaucoup de structu-res sous-corticales comme l’hippocampe, le complexe amygdalien, les ganglions de la base, l’épithalamus, l’aire tegmentale ventrale et le cervelet sont impliquées dans les processus liés à la musique (Mueller et coll., 2011 ; Fritz et coll., 2009 ; Blood et Zatorre, 2001).Ces processus s’entremêlent profondément avec d’autres procédures fonctionnelles, ils sont remarquablement semblables à ceux qui
Insistance 6.indd 84Insistance 6.indd 84 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 85
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
concernent le langage. Cette approche se base sur l’hypothèse que la musique et le langage sont intimement joints au début de la vie, que les éléments musicaux tiennent lieu de compétence linguistique avant que les éléments phonétiques ne se développent (Fernald, 1989 ; Jusczyk et Krumhansl, 1993). Il devient évident désormais que le cerveau humain élabore la musique et le langage en utilisant des mécanismes cognitifs qui s’interpénètrent, au sein de structures cérébrales qui elles-mêmes s’interpénètrent largement. Les structures du cerveau impliquées dans la perception de la syntaxe (le cortex frontal latéral inférieur, le cortex prémoteur ventral et la partie anté-rieure du gyrus temporel supérieur) et celle de la sémantique dans la musique (les régions temporelles postérieures) ont considérablement à voir avec celles qui sont impliquées dans la perception du langage, et nous soulignerons donc ici les liens intimes qu’il y a entre musique et langage dans le cerveau humain (Koelsch, 2009).La cartographie fonctionnelle du cerveau a récemment remporté de grands succès, mais il reste encore à évaluer les composantes temporelles selon lesquelles l’activation peut se produire, et les fonctions intrinsèques de plasticité qui permettent l’acquisition de nouvelles capacités dans des régions où il existe déjà des fonctions établies. La représentation que nous nous faisons du cerveau musical devient cohérente mais elle est complexe et évolue sans cesse. Au-delà de la vieille idée d’une élabora-tion de la musique confinée à une seule région du cerveau ou à un hémis-phère, on arrive au concept de musique comme phénomène mettant en jeu le cerveau tout entier (Altenmüller, 2001 ; Figure 5). Si c’est le cas, pour dépasser les notions induites par les premières études de locali-sation et les hypothèses tendancieuses de la phrénologie en musique, il nous faudra forger une représentation fonctionnelle inclusive dans laquelle l’écoulement logique du processus serait élégamment, à certains niveaux, d’une manière non-locationniste, et selon laquelle le système pourrait fonctionner de façon cohérente suivant des paradigmes mais aussi des échelles très différents. Dans cette approche, il faut se souvenir que les concepts classiques de champs réceptifs en électrophysiologie, de spécialisation en neuro-imagerie cérébrale et de connexionisme pour les modèles cognitifs ne rendent compte que de façon incomplète de l’architecture réelle du cerveau et de ses fonctions.
Insistance 6.indd 85Insistance 6.indd 85 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

86 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
Pour ce qui est des modèles générateurs de fonctions du cerveau, les systèmes de niveau supérieur peuvent fournir une prédiction des données aux régions de niveau inférieur. La spécialisation de n’importe quelle région du cerveau serait alors également déterminée par le concours des flux de données ascendants de « bas niveau » et des prédictions descendantes
supérieures (Friston, 2002). La spécialisation ne serait donc pas le résultat d’une propriété intrin-sèque d’une quelconque région du cerveau, mais serait plutôt déterminée par la densité des flux de connexions ascendantes et descendan-tes (connexions feed-forward et feed-back) qui la relient à d’autres régions (Felleman et Van Essen, 1991).
Airesauditivesprimaires
Cortexfrontaltemporelpariétal
Cortex entier,ganglionsde la base,cervelet
Augmentation de complexitédes réseaux neuronaux
Auditive
SomatosensorielleReprésentation
MotriceSymbolique
Visuelle
Augmentationde la complexitédes informationsauditives
Systèmesous-cortical auditif
Plasticité
Apprentissage
Traitementauditifde base
Perceptionde lamusique
Comportementgénéré par lamusique
Organisationdu tondu timbredu tempo
Figure 5. Représentation illustrant l’interdépendance entre l’augmentation de complexité du traitement de l’infor-mation auditive et l’augmentation de complexité des réseaux neuronaux (redessiné et modifié de : Altenmüller, 2001).
Insistance 6.indd 86Insistance 6.indd 86 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 87
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
D’un côté nous avons les modèles localisationistes, ou néo-phrénolo-gistes, qui nous disent que les régions du cerveau dédiées à la percep-tion de musique et à ses productions impliquent la présence de réseaux neuronaux complexes abondamment entremêlés, ébauchant ainsi peut-être une sorte de modèle « cerveau tout-entier ». De l’autre côté nous trouvons les modèles prédictifs, générateurs, qui fonction-neraient sans supervision, en ce sens que les causes n’entreraient pas dans leurs règles d’apprentissage (Friston, 2002). Ces derniers modèles, pour représenter les fonctions, se conforment aux mêmes principes parallèle et distribué du traitement de l’information, qui sous-tendent la théorie du connexionisme (Rumelhart et McClelland, 1986). Ces modèles de représentation s’influencent et se soutiennent mutuellement, en suivant tous les niveaux de la hiérarchie sensorielle, en mettant en jeu les dynamiques de la signalisation réentrante, comme l’a autrefois proposé la théorie du Darwinisme neuronal (Edelman, 1993).Finalement, si nous expurgeons les résultats localisationnistes de l’en-semble de ceux pour lesquels la musique a été prise en considération pour corroborer des notions qui se rapportent à des fonctions plus génériques comme la codification auditive, l’intégration motrice-audi-tive, la formation, le stockage et le rappel de mémoire, et les interactions auditives avec les processus de récompense/motivation, d’émotion et d’éveil, on peut dire que ce que nous connaissons vraiment du traite-ment de la musique par le cerveau se réduit à peu de choses.On est un peu plus efficace avec les modèles générateurs non supervisés, selon lesquels le principe de traitement distribué de l’in-formation produit et maintient des représentations et pour lesquels l’architecture fonctionnelle du système émane des interactions continues entre le cerveau et l’environnement. Les modèles généra-teurs sont là pour « apprendre des représentations faciles à décrire mais qui permettent de reconstruire précisément ce qui les a susci-tées » (les « entrées », Hinton et coll., 1995). En ce sens la musique devrait se conformer – comme d’ailleurs elle l’a toujours fait – aux processus archétypaux du cerveau, mais il lui faudra aussi fournir des informations nouvelles afin que le cerveau puisse « apprendre quelque chose de nouveau ».
Insistance 6.indd 87Insistance 6.indd 87 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

88 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
Figure 6. La « Tabula Mirifica » (Kircher, 1650, p. 363).
Insistance 6.indd 88Insistance 6.indd 88 3/12/11 9:56:323/12/11 9:56:32
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 89
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
Homunculus : … …Was künstlich ist, verlangt geschlossenen Raum.
« But man’s creations need to be confined. »« Mais il faut que les créations de l’homme soient contenues. »
(Johann Wolfgang von Gœthe, Faust, Partie II, Acte II. Scène dans le laboratoire. Traduit en Anglais par Philip Wayne, Penguin Books,
Baltimore, 1959)
LA COMPOSITION COMME CHOIX ET IMPLÉMENTATION DE RÈGLES RELATIONNELLES
Une fois qu’ils eurent développé un système de notation fiable, et qu’ils furent capables d’éditer des partitions complexes, les musiciens n’ont eu de cesse de se donner des contraintes pour la mise en œuvre leur musique. Depuis le début ils cherchaient à produire des systèmes logico-numériques basés sur les rapports entre les notes qui permet-traient de rendre compte et aussi de produire de façon fiable, presque mécanique, des séries de sons compatibles.Des musiciens exploraient les propriétés du son, suivant un parcours histo-rique parsemé de tétracordes, d’échelles, de tonalités et de séries, jusqu’à l’avènement des musiques microtonale et spectrale (Wyschnegradsky, 1937 ; Anderson, 2000 ; Radulescu, 2003). D’autres se sont centrés sur des systèmes numériques plus symboliques. Les systèmes logico-numéri-ques développés pour la composition de contrepoints comme le « Tabula Mirifica » (Kircher, 1650 ; Figure 6) et ses extensions, ou le « Goldberg machina », s’ouvraient à des possibilités compositionnelles multiples, même si elles suivaient la logique musicale. Beaucoup de compositeurs, incluant Johannes Ockeghem et Johann Sebastian Bach, ont utilisé ces systèmes pour réaliser les structures compositionelles les plus élaborées de la musique occidentale (Bizzi, 1982).D’autres systèmes de nombres ont été empruntés à des champs non-musicaux : la plupart d’eux sont des tables combinatoires. L’histoire de tels systèmes numériques nous ramène à l’utilisation de ces tables, carrés magiques ou matrices, qui ont été utilisées depuis longtemps pour composer de la musique et ont encore été employées au siècle
Insistance 6.indd 89Insistance 6.indd 89 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

90 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
passé par des compositeurs comme Pierre Boulez et Peter Maxwell Davies (Cross, 2003). Certains poursuivent encore cette approche de manière personnelle et intéressante. Pour n’en nommer que deux, nous citerons l’uti-lisation de la théorie des ensembles et celle de la théorie des fractales pour composer de la musique (Cross, 2003 ; Sherlaw Johnson, 2003), et nous arriverons enfin à la compo-sition algorithmique avec fonctions chaoti-ques et computation évolutive (Chapel, 2003 ; Miranda, 2004).D’autres compositeurs, dans la recherche de règles compositionnelles suffisamment riches et flexibles pour s’adapter à leur imagination créatrice, ont tourné leurs efforts vers des systè-mes de relations obtenues de domaines de la nature plus tangibles. Ils essayaient finalement de transformer en musique des informations qui venaient du monde biologique.
SONIFICATION ET BIOFEEDBACK
Avec la possibilité récente de faire des enre-gistrements physiques directs d’événements biologiques, comme la production électrique des activités du cerveau, on a pu aborder un nouveau et révolutionnaire champ de recher-ches pour ce qui est de l’établissement de règles pour la sonification. Nous posons que la soni-fication est l’utilisation d’une structure sonore dépourvue d’éléments langagiers, qui sert à transmettre un contenu informatif généré par des données biologiques. Historiquement, on a
pu observer un grand engouement pour les enre-gistrements d’évènements biologiques pendant la période des recherches menées autour du biofeedback. Ces expériences originelles furent extrêmement prometteuses et impliquèrent des musiciens comme John Cage et John Lennon (Rosenboom, 1975). Plusieurs chercheurs ont commencé alors à développer des systèmes d’interfaces ordinateur-cerveau de plus en plus sophistiqués, visant à raccorder l’expérience intérieure représentée par les événements élec-triques cérébraux, et des appareils externes, conçus pour produire des sons.On a pu utiliser des signaux physiologiques comme ceux de l’électroencéphalogramme, l’électromyogramme, et des pulsations cardia-ques pour contrôler des algorithmes de synthèse sonore ainsi que construire des instruments de musique pilotés biologiquement (Arslan et coll., 2006). Les chercheurs continuent à produire des rapports sur de nouvelles tentati-ves expérimentales et techniques afin de mettre en évidence l’électroencéphalogramme associé aux tâches musicales cognitives (Hinterberger et Baier, 2005 ; Miranda et Brouse, 2005). Au cours de toutes ces années on a développé des environnements de synthèse de musique en temps réel et des algorithmes de traitement pour agir sur les sons avec les signaux biologiques et on a pu entendre des concerts de bio-musiciens capables de commander des instruments musi-caux numériques à partir de leur corps, avec l’interface spécifique.À l’opposé, tenant compte de la similarité entre les ondes cérébrales et les rythmes musicaux, certains scientifiques proposent des règles de
Insistance 6.indd 90Insistance 6.indd 90 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 91
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
sonification qui, par l’identification des activités électriques du cerveau ou des données de mouvement, pourraient fournir des stratégies en temps réel pour contrôler des activités biologiques. Ces dernières recherches, mêmes si elles ne sont pas directement liées à la création de musique nouvelle, sont extrêmement intéressantes puisqu’elles peuvent être utiles pour la thérapie auditive par bio-feedback (Baier et coll., 2007 ; Giansanti et coll., 2009 ; Pauletto et Hunt, 2009 ; Wu et coll., 2009).Chez les auteurs qui ont travaillé sur la sonification, l’idée fondamen-tale (du moins pour ceux qui se sont spécialement concentrés sur la production de musique), émane de la prémisse selon laquelle les informations créées par le monde biologique peuvent être transférées à l’auditeur par un processus qui implique un traducteur géré par un ensemble de règles.On comprend à quel point la sonification dépend des règles de traduc-tion car, si elles sont médiocres, elles peuvent produire des résultats musicalement insignifiants comme on l’a vu avec les premières expé-riences où des séquences d’ADN ont été converties en musique (Ohno et Ohno, 1986). Le génome et la composition musicale, sont basés sur des principes de réapparition répétitive. La musique s’étant dévelop-pée dès l’origine comme un dispositif métronomique chronométrant, les séquences codantes de l’ADN pourraient être transformées elles aussi en partitions en utilisant un ensemble simple des règles. Ohno n’était pas vraiment intéressé à créer une musique nouvelle, mais il a pu en s’inspirant de cette similitude, d’une façon très convaincante, présenter des échantillons qui paraissent très proches de la musique des compositeurs du premier Baroque jusqu’au dernier Romantisme (Ohno, 1987 ; Figure 7).La limitation la plus importante à la sonification est que les solutions proposées pour la composition de la musique ne sont pas radicalement différentes de celles qui sont envisagées par les autres systèmes logico-numériques, surtout ceux qui sont basés sur des algorithmes numéri-ques. On a l’impression que finalement la sonification, si elle s’appuie avec confiance sur la solidité du milieu biologique dont elle tire sa matière de départ, se retrouve en fin de compte à dépendre beaucoup des règles choisies pour mettre en œuvre le système de traduction.
Insistance 6.indd 91Insistance 6.indd 91 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

92 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
MUSIQUE BIOTIQUELe terme biotique signifie littéralement « se rapportant à la vie », il est donc tout à fait générique, mais il est souvent utilisé dans le domaine de l’écologie pour qualifier une composante vivante qui affecte un autre facteur et modèle un écosystème. Ce mot est particu-lièrement approprié, dans ce contexte, pour
souligner les concepts : 1) de dérivation à partir du monde biologique ; 2) de capacité à affecter d’autres parties du système ; 3) d’in-clusion dans un système interdépendant où les informations circulent à travers des éléments qui communiquent entre eux (cette réciprocité étant fondamentale pour le fonctionnement et la survie).
Figure 7. La transposition musicale du cœur du segment déterminant le code du domaine de la tyrosine kinase de la p-chaîne du récepteur de l’insuline humaine. On voit les résidus d’aminoacide des deux segments de site actifs. La transformation musicale est en mi mineur, à 4/4 (de : Ohno, 1987).
Insistance 6.indd 92Insistance 6.indd 92 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 93
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
Ainsi nous considérerons que la musique biotique est avant tout une production musicale, accompagnée des procédures de composition utilisées pour la créer, dont les données initiales sont à l’origine biolo-giques. J’ai présenté dans une précédente contribution une approche de la composition où la construction musicale était entièrement extra-polée des informations que nous avons sur la structure et les propriétés fonctionnelles du cerveau (Minciacchi, 2003).J’ai fait état des expériences dans lesquelles j’ai essayé de transformer des données du cerveau directement en musique, comme l’a récemment remarqué Massey (2009), et pour lesquelles les données du cerveau ont été utilisées à la production de tous les paramètres pertinents conve-nant à des objets sonores. Depuis, mon expérience de la composition musicale à partir de données du cerveau s’est accrue, comme je l’illus-trerai dans les paragraphes suivants, mais certains des points que j’ai appris de mes premières expériences sont toujours valides.D’abord, pour être pertinentes pour la composition, les informations que nous pouvons obtenir du cerveau n’ont pas besoin de venir de ceux des circuits du cerveau qui sont rattachés à l’audition ou corrélés à la musi-que. De tout ce que nous avons dit jusqu’à maintenant il est clair que l’architecture du cerveau, et aussi les processus fonctionnels qu’il utilise pour produire les informations, ne tiennent pas compte des catégories comme « l’audition » ou « la musique ». Deuxièmement, le processus de modelage d’un objet sonore devrait se conformer aux mêmes principes pendant le traitement de tous les paramètres de la forme musicale. Cette uniformité dans les règles de traitement devrait induire la construction tout entière du son.Troisièmement, au cœur de chaque paramètre sonore, il faut que l’ar-chitecture de traitement tienne compte de la nature essentiellement relationnelle des processus d’information sonore et de perception musicale. L’essentiel de mes dernières activités aura été d’appliquer et développer ces principes généraux aux procédures de composition.Maintenant, pour mettre en évidence les caractéristiques pertinen-tes de ce que je considère comme des procédures pour créer de la musique biotique, j’illustrerai brièvement mon propos en prenant pour exemples trois de mes travaux récents, ayant une caractéristique particulière commune : les données biologiques originales sont obte-
Insistance 6.indd 93Insistance 6.indd 93 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S
94 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
15
fffff
fpff
ff
7
77
6
6
6
55
3
A
B
a b
c d
1-b
1-b
1-b
1-b/8b
1-b
1-a
1-a1-a
1-p, 1-C,1-a
1-p, 2-C,1-a, 1-b
1-C
2-C
1-p
C
p = flûte piccolo, C = grande flûte en ut, a = flûte alto, b = flûte basse,8b = flûte octobasse, Cond = chef d'orchestre,Perc = percussion, Sp = comédien, Tu = accordeur en bois
Salle Alfred Cortot,78 rue Cardinet,Paris, France
A B
D
E FG
H I
J
1-8b
1-8b 1-8b
Sp2
Sp1
Cond
Perc
Tuner
Sp3
“La Disciplina della Crisi”
– “La Discipline de la Crise”
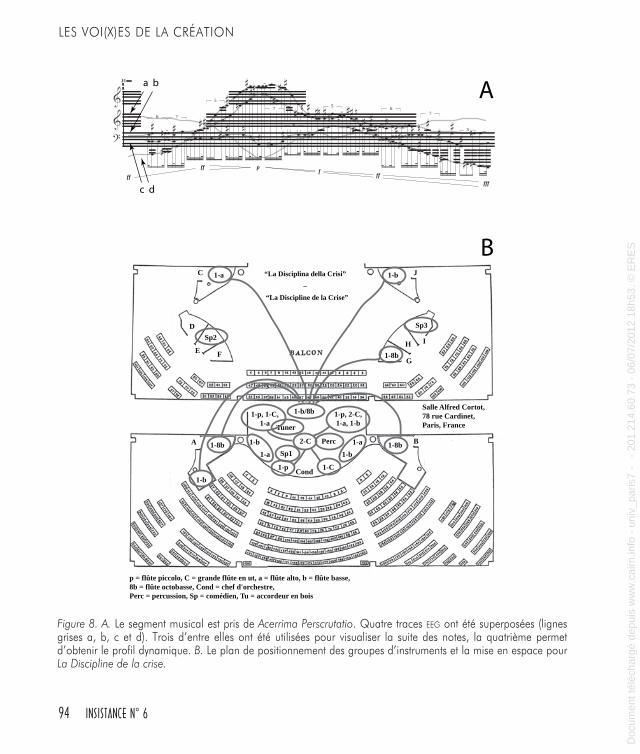
Figure 8. A. Le segment musical est pris de Acerrima Perscrutatio. Quatre traces EEG ont été superposées (lignes grises a, b, c et d). Trois d’entre elles ont été utilisées pour visualiser la suite des notes, la quatrième permet d’obtenir le profil dynamique. B. Le plan de positionnement des groupes d’instruments et la mise en espace pour La Discipline de la crise.
Insistance 6.indd 94Insistance 6.indd 94 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 95
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
A
B
5 1767
31 3773
43 2959
79101103
53107109
2347
1189
13 7 1961 714183
397
2
F8b
Fb
Fb
Fa
G1
G2
G3
Solo
S0a
S0b
5’20"5’10"5’00"
Fa
F
Fp
F2
F1
Fp
f
mp p p mf
mpmp f
mp f
f
p mf
p mf p
p mf mp
33
3
33
3
3
3
35
5
5
5 6
3
3
3
3
3
3
33
3
5
5
3 3
( )
mp / mf
Figure 9. A. Le deux gammes en quarts de tons développées comme répertoire de notes pour La Discipline de la crise. B. Un court extrait de la partition de La Discipline de la crise.
Insistance 6.indd 95Insistance 6.indd 95 3/12/11 9:56:333/12/11 9:56:33
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

96 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
= 30
(*)
(*) Diaphragmatic vibrato(speed and amplitude are indicated graphically)
incolore incolore
nulla nulla mpp pppp
= 30= 114
(§)
(§) The highest possible pitch
cantatoprecipitato!
as - ppp - as possiblemp p / mf
sfz
= 30= 76
solenne
un evento diverso,più presente, ora
mp mf / fffmf mf
= 114= 114= 76 = 76
concitato e precipitoso,continuo, senza una direzione
mp / f mp / f
Figure 10. Échantillon de quatre sections de Haß zu Wasser und Haß zu Land.
Insistance 6.indd 96Insistance 6.indd 96 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 97
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
nues de la même source 3 : un enregistrement EEG. Le choix de cette matière commune a été déterminé par le souhait de réaliser trois objets sonores radicalement différents mais partageant des affinités esthéti-ques profondes et significatives. Cette dernière considération, au-delà des connotations artistiques qui en découlent, permet cependant une grande flexibilité technique et répond à des exigences de composition très dissemblables.Le premier exemple (Figure 8A), illustre une procédure directe utilisée pour reporter des données biologiques à des paramètres musicaux. Dans cette section, extraite de la partition d’Acerrima Perscrutatio, les trois lignes de notes sont construites d’après le contour mélodique individualisé de trois traces EEG (b, c et d, dans la Figure 8A). Les notes se rapportent à des échelles développées d’après l’analyse statistique des traces EEG. Les figures rythmiques locales sont aussi tirées d’une trace EEG supplémentaire (non montrée ici). On a utilisé une dernière trace EEG (a, dans la Figure 8A) pour obtenir le profil des dynamiques. Cette section apparaît ici comme sur la partition finale ; elle est plutôt à prendre comme un matériel de référence, la partition a été modifiée en plusieurs endroits. Pour cet exemple les points-clés sont les suivants : a) les informations des traces EEG ont été utilisées pour en tirer des paramètres sonores pertinents ; b) les différentes traces créent des para-mètres différents comme les notes et leur hauteur d’octave, les figures rythmiques et le contour dynamique ; c) la section sonore obtenue est considérée comme matériel de référence puis est réorganisée plusieurs fois par la suite. Cet exemple peut être considéré comme un compte rendu assez linéaire d’une suite d’informations biologiques, et pour utiliser une métaphore linguistique, le travail est ici concentré sur le vocabulaire et la syntaxe de la musique.Le deuxième exemple est tiré du plan de scène des instruments et de mise en espace pour La Discipline de la crise (Figure 8B). Les choix concernant les groupes d’instruments et leur placement dans l’espace de la salle de concert correspondent au positionnement adéquat sur le cuir chevelu des capteurs qui génèrent les EEG. Ici les informations biologiques ont été employées pour placer le son dans l’environnement de sa performance en concert. Le développement du répertoire des hauteurs de notes, deux échelles différentes en quarts de tons, a été
3. Les trois travaux dont j’extrairai les exemples sont : La Discipline de la crise, pour orchestre de flûtes, trois speakers et deux percussions, textes d’Antonella Anedda, 2007-2009, création par l’Orchestre français de flûtes, dir : Pierre-Alain Biget ; Klavierstück n.7 – Acerrima Perscrutatio et Diligentissima, pour piano solo, 2010, création par James Clapperton ; Haß zu Wasser und Haß zu Land, pour alto flûte solo, 2010, création par Pierre-Yves Artaud.
Insistance 6.indd 97Insistance 6.indd 97 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

98 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
obtenu à partir de traces EEG (Figure 9A). Un petit passage du score de La Discipline de la crise (Figure 9B) est aussi proposé ici pour illustrer le fait que différents plans de composition cohabitent en même temps. À nouveau, toutes les informations proviennent de traces EEG. Pourtant, pendant que dans la partie supérieure (G1, G2, G3 et Solo) la procédure suivie vise à obtenir un résultat quasi-contrapuntique pour privilégier l’interaction des interprètes, dans la partie inférieure de l’extrait (groupes S0a et S0b) les instruments sont traités individuellement sans interactions, en utilisant une procédure semblable à celle décrite dans l’exemple tiré d’Acerrima Perscrutatio (Figure 8A).Finalement, on propose (Figure 10) une collec-tion de sections tirées de Haß zu Wasser und Haß zu Land ici donnée pour montrer que l’on peut élaborer un répertoire compositionnel à partir d’une simple trace EEG. Ici les différentes sections se réfèrent plus ou moins précisément aux données originales et le flux musical a été composé en fonction d’un méta-paramètre de plus (ou paramètre latent) d’abord conçu puis utilisé. Ce méta-paramètre peut être défini comme le degré de congruité entre l’ensemble des données originales (en incluant le prétendu sens biologique) et le son final résultant du processus de composition. Dans ce dernier exemple les diverses solutions illustrées donnent une mesure de la flexibilité extrême avec laquelle nous pouvons obtenir des informations biologi-ques que nous pourrons ensuite utiliser comme nomenclature de cellules compositionnelles.Ce bref compte rendu de la méthode biotique ici décrite (sur laquelle nous avons encore beau-
coup à travailler), peut contribuer à unifier notre vue sur les différents paramètres sonores et les aspects micro- et macro-formels de la composi-tion musicale et peut-être ouvrir ainsi de nouvel-les perspectives à l’élaboration du son.Il ne me reste plus maintenant qu’à laisser à John Cage le mot de la fin (dans : Duckworth, 1995) : méthode, structure, intention, disci-pline, notation, indétermination, interprétation, imitation, dévotion et circonstances ; « ce sont les choses les plus importantes pour moi, et ils omettent, assez curieusement, les mots inven-tion et non-intention, ou des choses comme chance etc., qui relèvent surtout du mot indis-cipline ! »
BIBLIOGRAPHIE
ALTENMÜLLER, E.O. 2001. « How many music centers are in the brain ? » Ann. NY Acad. Sci., 930 : 273-280.
ANDERSON, J. 2000. « A provisional history of spectral music », Cont. Mus. Rev., 19 : 7-22.
ARSLAN, B. ; BROUSE, A. ; CASTET, J. ; LEHEMBRE, R. ; SIMON, C. ; FILATRIAU, J.J. ; NOIRHOMME, Q. 2006. « A real time music synthe-sis environment driven with biological signals. Proceedings », IEEE Int. Conf. Acoustics, Speech and Signal Proc., 2 : II.
BAIER, G. ; HERMANN, T. ; STEPHANI, U. 2007. « Event-based sonification of EEG rhythms in real time », Clin. Neurophysiol., 118 : 1377-1386.
Insistance 6.indd 98Insistance 6.indd 98 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 99
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
BIZZI, G. 1982. Specchi invisibili dei suoni. Centro Internazionale per la divulgazione della Musica Italiana, Rome, Italy.
BLOOD, A.J. ; ZATORRE, R.J. 2001. « Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion », Proc. Natl. Acad. Sci. USA, 98 : 11818-11823.
CALVIN, W.H. 1987. « The brain as a Darwin machine », Nature, 330 : 33-34.
CALVIN, W.H. 1996. The Cerebral Code, Cambridge, MIT Press, Cambridge, MA, USA.
CHAPEL, R.H. 2003. Realtime Algorithmic Music Systems from Fractals and Chaotic Functions : toward an Active Musical Instrument, Barcelona, Universitat Pompeu Fabra, Spain.
CHEN, J.L. ; PENHUNE, V.B. ; ZATORRE, R.J. 2009. « The role of auditory and premotor cortex in sensorimotor transformations », Ann. NY Acad. Sci., 1169 : 15-34.
COLLIGNON, O. ; VOSS, P. ; LASSONDE, M. ; LEPORE, F. 2009. « Cross-modal plasticity for the spatial processing of sounds in visually deprived subjects », Exp. Brain Res., 192 : 343-358.
CROSS, R. 2003. « Composing with numbers : sets, rows and magic squares », dans J. Fauvel, R. Glood, R. Wilson (eds), Music and Mathematics. From Pythagoras to Fractals, Oxford Univ. Press, Oxford, p. 131-146.
DRAKE, C. ; BERTRAND, D. 2001. « The quest for universals in temporal processing in music », Ann. NY Acad. Sci., 930 : 17-27.
DUCKWORTH, W. 1995. « Experimentalists. 1. John Cage », in Talking Music. Schirmer Books, New York, p. 3-28.
DUFAY, G. 1397-1474. « Missa super “Se la face ay pale” », dans H. Besseler (sous la direction de), Chor-Archiv., Bärenreiter Verlag, Kassel, Germany.
EDELMAN, G.M. 1987. Neural Darwinism. The Theory of Neuronal Group Selection, Basic Books, New York, USA.
EDELMAN, G.M. 1993. « Neural Darwinism : selection and reentrant signaling in higher brain function », Neuron 10 : 115-125.
EDELMAN, G.M. ; GALLY, J.A. 2001. « Degeneracy and complexity in biological systems », Proc. Natl. Acad. Sci. USA, 98 : 13763-13768.
Insistance 6.indd 99Insistance 6.indd 99 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

100 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
EDELMAN, G.M. ; MOUNTCASTLE, V.B. 1978. Mindful Brain – Cortical Organization and the Group-Selective Theory of Higher Brain Function, MIT press, Cambridge, MA, USA.
EULER, L. 1739. Tentamen novae theoriae musi-cae ex certissimis harmoniae principiis dilu-cide expositae, St. Petersburg, Typographia Academiae Scientiarum.
FADIGA, L. ; CRAIGHERO, L. ; D’AUSILIO, A. 2009. « Broca’s area in language, action, and music », Ann. NY Acad. Sci., 1169 : 448-458.
FELLEMAN, D.J. ; VAN ESSEN, D.C. 1991. « Distributed hierarchical processing in the primate cerebral cortex », Cereb. Cortex., 1 : 1-47.
FERNALD, A. 1989. « Intonation and communica-tive intent in mothers’ speech to infants : is the melody the message ? », Child Dev., 60 : 1497-1510.
FERNANDO, C. ; GOLDSTEIN, R. ; SZATHMÁRY, E. 2010. « The neuronal replicator hypoth-esis », Neural Comput., 22 : 2809-2857.
FRISTON, K. 2002. « Beyond phrenology : what can neuroimaging tell us about distrib-uted circuitry ? », Ann. Rev. Neurosci., 25 : 221-250.
FRITZ, T.H. ; MÜLLER, K. ; OTT, D. ; TURNER, R. ; KOELSCH, S. 2009. « Investigation of a distinct valence dimension from neutral to pleasant as opposed to a continuous valence dimension from unpleasant to pleasant », NeuroImage, 47 S1 : S192.
GIANSANTI, D. ; DOZZA, M. ; CHIARI, L. ; MACCIONI, G. ; CAPPELLO, A. 2009. « Energetic assess-ment of trunk postural modifications
induced by a wearable audio-biofeedback system », Med. Eng. Phys., 31 : 48-54.
HEBB, D.O. 1949. The Organisation of Behavior, John Wiley, New York, USA.
HELMHOLTZ, H. 1863. Die Lehre von den Tonempfindungen als Physiologische Grundlage für die Theorie der Musik, Braunschweig : F. Vieweg und Sohn.
HINTERBERGER, T. ; BAIER, G. 2005. Parametric Orchestral Sonification of EEG in Real Time, IEEE MultiMedia, 12 : 70-79.
HINTON, G.E. ; DAYAN, P. ; FREY, B.J. ; NEAL, R.M. 1995. « The “Wake-Sleep” algorithm for unsupervised neural networks », Science, 268 : 1158-1161.
HORWITZ, B. ; POEPPEL, D. 2002. « How can EEG/MEG and fMRI/PET data be combined ? », Hum. Brain Mapp., 17 : 1-3.
HOSTINSKY, O. 1879. Die Lehre von den musikalis-chen Klängen, H. Dominicus, Prague.
HUBEL, D.H. ; WIESEL, T.N. 1959. « Receptive fields of single neurones in the cat’s striate cortex », J. Physiol., 148 : 574-591.
HYER, B. 1995. « Re-imagining riemann », J. Music Theory, 39 : 101-138.
JUSCZYK, P.W. ; KRUMHANSL, C.L. 1993. « Pitch and rhythmic patterns affecting infants sensitivity to musical phrase structure », J. Exp. Psychol. Hum. Percept. Perform., 19 : 627-640.
KIRCHER, A. 1650. Musurgia universalis sive ars magna consoni et dissoni [tome 1]. ex typographia Haeredum Francisci Corbelletti.
KOELSCH, S. 2009. « Neural substrates of process-ing syntax and semantics in music », in
Insistance 6.indd 100Insistance 6.indd 100 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 101
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
Music that Works, Haas R. Brandes V. (eds), Springer-Verlag, Wien, p. 143-153.
KOELSCH, S. ; FRITZ, T. ; VON CRAMON, D.Y. ; MÜLLER, K. ; FRIEDERICI, A.D. 2006. « Investigating emotion with music : An fMRI study », Hum. Brain Mapp., 27 : 239-250.
KOSIK, K.S. 2003. « Beyond phrenology, at last », Nat. Rev. Neurosci., 4 : 234-239.
KRUBITZER, L. 2007. « The magnificent compromise : cortical field evolu-tion in mammals », Neuron, 56 : 201-208.
LARGE, E.W. ; SNYDER, J.S. 2009. « Pulse and meter as neural reso-nance », Ann. NY Acad. Sci., 1169 : 46-57.
LECLERC, C. ; SEGALOWITZ, S.J. ; DESJARDINS, J. ; LASSONDE, M. ; LEPORE, F. 2005. « EEG coherence in early-blind humans during sound local-ization », Neurosci. Lett., 376 : 154-159.
MAKEIG, S. 1981. Means, Meaning and Music : Pythagoras, Archytas and Plato. Ex tempore, 1-2.
MASSEY, I. 2009. The Neural Imagination : Aesthetic and Neuroscientific Approaches to the Arts, Univ. Texas Press, Austin, USA.
MCDERMOTT, J. ; HAUSER, M.D. 2004. « Are consonant intervals music to their ears ? Spontaneous acoustic preferences in a nonhuman primate », Cognition, 94 : B11-B21.
MCDERMOTT, J. ; HAUSER, M.D. 2007. « Nonhuman primates prefer slow tempos but dislike music overall », Cognition, 104 : 654-668.
MINCIACCHI, D. 2003. « Translation from neurobiological data to music parameters », Ann. NY Acad. Sci., 999 : 282-301.
MINCIACCHI, D. 2005. « Part VI : music performance. Introduction », Ann. NY Acad. Sci., 1060 : 346-349.
MINCIACCHI, D. ; DEL TONGO, C. ; TOGNETTI, A. ; DALLE MURA, G. 2008. « Has music a specific effect on temporal control of movements ? An auditory-motor task of synchronization », Soc. Neurosci. Abst. Washington : 102479-102480.
MIRANDA, E.R. 2004. « At the crossroads of evolutionary computation and music : self-programming synthesizers, swarm orchestras and the origins of melody », Evol. Comput., 12 : 137-158.
Insistance 6.indd 101Insistance 6.indd 101 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

102 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
MIRANDA, E.R. ; BROUSE, A. 2005. « Interfacing the Brain Directly with Musical Systems : On Developing Systems for Making Music with Brain Signals », Leonardo, 38 : 331-336.
MOUNTCASTLE, V.B. 1957. « Modality and topo-graphic properties of single neurons of cat’s somatic sensory cortex », J. Neurophysiol., 20 : 408-434.
MUELLER, K. ; MILDNER, T. ; FRITZ, T. ; LEPSIEN, J. ; SCHWARZBAUER, C. ; SCHROETER, M.L. ; MÖLLER, H.E. 2011. « Investigating brain response to music : a comparison of differ-ent fMRI acquisition schemes », Neuroimage, 54 : 337-343.
PARNCUTT, R. 2009. « Prenatal “experience” and the phylogenesis and ontogenesis of music », in Music that Works, Haas R., Brandes V. (eds), Springer-Verlag, Wien, 185-194.
OHNO, S. 1987. « Repetition as the essence of life on this earth : music and genes », Haematol. Blood Transfus., 31 : 511-518.
OHNO, S. ; OHNO, M. 1986. « The all pervasive principle of repetitious recurrence governs not only coding sequence construction but also human endeavor in musical composi-tion », Immunogenetics, 24 : 71-78.
PAULETTO, S. ; HUNT, A. 2009. « Interactive soni-fication of complex data », Int. J. Human-Comput. Studies, 67 : 923-933.
PERANI, D. ; SACCUMAN, M.C. ; SCIFO, P. ; SPADA, D. ; ANDREOLLI, G. ; ROVELLI, R. ; BALDOLI, C. ; KOELSCH, S. 2010. « Functional special-izations for music processing in the human
newborn brain », Proc. Natl. Acad. Sci. USA, 107 : 4758-4763.
RADULESCU, H. 2003. « Brain and sound reso-nance : The world of self-generative func-tions as a basis of the spectral language of music », Ann. NY Acad. Sci., 999 : 322-363.
RIEMANN, VON H. 1914-1915. « Ideen zu einer “Lehre von den Tonvorstellungen” », Jahrbuch der Bibliothek Peters, 21-22 : 20. Dans B. Hyer (1995). « Re-Imagining Riemann », J. Music Theory, 39 : 101-138.
ROSENBOOM, D. 1975. Biofeedback and the Arts : Results of Early Experiments (ed), Vancouver : Aesthetic Research Centre of Canada, Canada.
RUMELHART, D. ; MCCLELLAND, J. 1986. Parallel Distributed Processing : Explorations in the Microstructure of Cognition, Cambridge. MIT Press, Cambridge, MA, USA.
SCHWARTZ, D.A., HOWE, C.Q., PURVES, D. 2003. « The statistical structure of human speech sounds predicts musical universals », J. Neurosci., 23 : 7160-7168.
SHERLAW JOHNSON, J. 2003. « Composing with fractals », in J. Fauvel, R. Glood (eds), Music and Mathematics. From Pythagoras to Fractals, Oxford Univ. Press, Oxford, p. 131-146.
SKINNER, M. 2006. « Toward a quarter-tone syntax : selected analyses of works by Blackwood, Hába, Ives, and Wyschnegradsky », http://www.tierceron.com/, 194-232.
STUMPF, C. 1890. Tonpsychologie, Verlag S. Hirzel, Leipzig, Germany.
Insistance 6.indd 102Insistance 6.indd 102 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

INSISTANCE N° 6 103
LA SONIFICATION VERSUS LA COMPOSITION BIOTIQUE DES ICÔNES DU CERVEAU
SUGIMOTO, T. ; KOBAYASHI, H. ; NOBUYOSHI, N. ; KIRIYAMA, Y. ; TAKESHITA, H. ; NAKAMURA, T. ; HASHIYA, K. 2010. « Preference for consonant music over dissonant music by an infant chimpanzee », Primates, 51 : 7-12.
TERHARDT, E. 2006. Die psychoakustischen Grundlagen von Tonverwandtschaft und Harmonie. DAGA’06, Braunschweig. CD-ROM Deutsche Gesellschaft für Akustik.
TREHUB, S.E., HANNON, E.E. 2006. « Infant music perception : domain-general or domain-specific mechanisms ? », Cognition, 100 : 73-99.
WU, D. ; LI, C.Y. ; YAO, D.Z. 2009. « Scale-free music of the brain », PLoS One, 15 : e5915.
WYSCHNEGRADSKY, I. 1937. « La musique à quarts de ton et sa réalisation pratique », Rev. Mus., 171 : 26-33.
Résumé : La présente contribution retrace les bases théoriques, les résultats essen-tiels et les perspectives de mes recherches sur les relations entre la structure et les fonctions du cerveau et les processus de composition musicale. La première partie recense les données et les points de vue qui expliquent pourquoi la musique ne peut pas être décrite en termes d’universaux spécifiques : elle n’est pas le privilège exclusif de l’espèce humaine et sa perception est un processus construit très tôt et amplement partagé. En outre, pour la perception et la production de la musique, nous utilisons des réseaux cérébraux vastes, complexes, et largement entremêlés. La deuxième partie décrit le processus de composition en se concentrant sur les systèmes de sonification et de la musique biotique qui tirent des neurosciences leurs informations et les lient aux paramètres musicaux. La sonification est l’utili-sation d’une structure sonore dépourvue d’éléments langagiers pour transmettre un contenu informatif dérivé des données biologiques. Inversement, la musique biotique peut être considérée comme une production musicale dont les données d’origine sont biologiques. On présentera ici les enjeux théoriques et quelques exemples des stratégies de transformation des données du cerveau aux objets sonores. La méthode biotique décrite ici peut contribuer à une vue d’ensemble des différents paramètres du son et des aspects micro- et macro-formels de la composition musicale.
Mots clés : Universaux musicaux, Darwinisme musical.
Summary : The present contribution redraws the theoretical foundations, essential results, and perspectives, of my research on the relations between the structure
Insistance 6.indd 103Insistance 6.indd 103 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S

104 INSISTANCE N° 6
LES VOI(X)ES DE LA CRÉATION
and functions of the brain and the processes of musical composition. The first part illustrates data and views to clarify why music cannot be represented in terms of specific universals ; music is not the exclusive privilege of mankind and its perception is a process constructed very early and broadly shared. Besides, for perception and production of music, we use complex and huge cerebral networks, which are largely mixed. The second part illustrates the process of composition by concen-trating on sonification and biotic music procedures that derive information from neurosciences and link it to musical parameters. Sonification is the use of a sound structure devoid of linguistic elements to transmit an informative content originated by biological data. Conversely, biotic music can be considered as a musi-cal production where the data of origin are biologi-cal. Theoretical stakes and examples of strategies to convert brain data into sound objects are presented. The biotic method here described can contribute to a unitary view of the different sound parameters and of the micro- and macro-formal aspects of music composition.
Keywords : Music universals, musical Darwinism.
Resumen : La presente contribución rediseña los fundamentos teóricos, los resultados esenciales y las perspectivas de mi estudio sobre las relaciones entre la estructura y las funciones del cerebro y los proce-
sos de la composición musical. La primera parte ilustra datos y puntos de vista para explicar porqué la música no puede ser representada con términos de universales específicos ; la música no es privilegio exclusivo de la humanidad, y sus percepción es un proceso construido muy pronto y repartido extensa-mente. Además, para la percepción y la producción de la música, nos utilizamos redes cerebrales comple-jas y enormes, que son ampliamente mezcladas. La segunda parte ilustra el proceso de composición por medio de la atención particular sobre la sonificatión y los procedimientos de la música biótica que obtie-nen informaciones de las neurociencias y la conecta con los parámetros musicales. La sonificatión es la utilización de una estructura sonora carente de los elementos lingüísticos para transmitir un conte-nido informativo originado de datos biológicos. Sin embargo, la música biótica puede ser considerada como una producción musical en la que los datos del origen son biológicos. Son aquí presentados los cimientos teóricos y los ejemplos de las estrategias para convertir los datos cerebrales en objetos sono-ros. El método biótico aquí trazado puede contribuir a crear un punto de vista unitario de los diferentes parámetros musicales y de los aspectos micro- y macro-formales de la composición musical.
Palabras clave : Universales musicales, Darwinismo musical.
Insistance 6.indd 104Insistance 6.indd 104 3/12/11 9:56:343/12/11 9:56:34
Doc
umen
t tél
écha
rgé
depu
is w
ww
.cai
rn.in
fo -
uni
v_pa
ris7
- -
201
.214
.60.
73 -
06/
07/2
012
18h5
3. ©
ER
ES
D
ocument téléchargé depuis w
ww
.cairn.info - univ_paris7 - - 201.214.60.73 - 06/07/2012 18h53. © E
RE
S