croissance et developpement iv/ regulation et facteurs de...
TRANSCRIPT

SUPPORT DU COURS MODULE DE PHYSIOLOGIEVEGETALE
Mr. I. SADKI
CROISSANCE ET DEVELOPPEMENTPLAN :
I/ DEFINITION .II/ PHASES DE LA CROISSANCE .1°/ Prolifération cellulaire.2°/ Elongation cellulaire.3°/ Différentiation cellulaire.
III/ CENTRES GENERATEURS DE LA CROISSANCE .1°/ Méristèmes caulinaires.
a/Organisationb/Initiation foliaire
2°/ Méristème racinaire.
IV/ REGULATION ET FACTEURS DE LA CROISSANCE A/ REGULATION DE LA CROISSANCE
*La notion de PolaritéB/ PHYTOHORMONES1°/ Auxines.a/ Historique et découverte.b/ Polarité et transport.c/ Propriétés physiologiques.
2°/ Cytokinines.a/ Découverte.b/ Propriétés physiologiques.
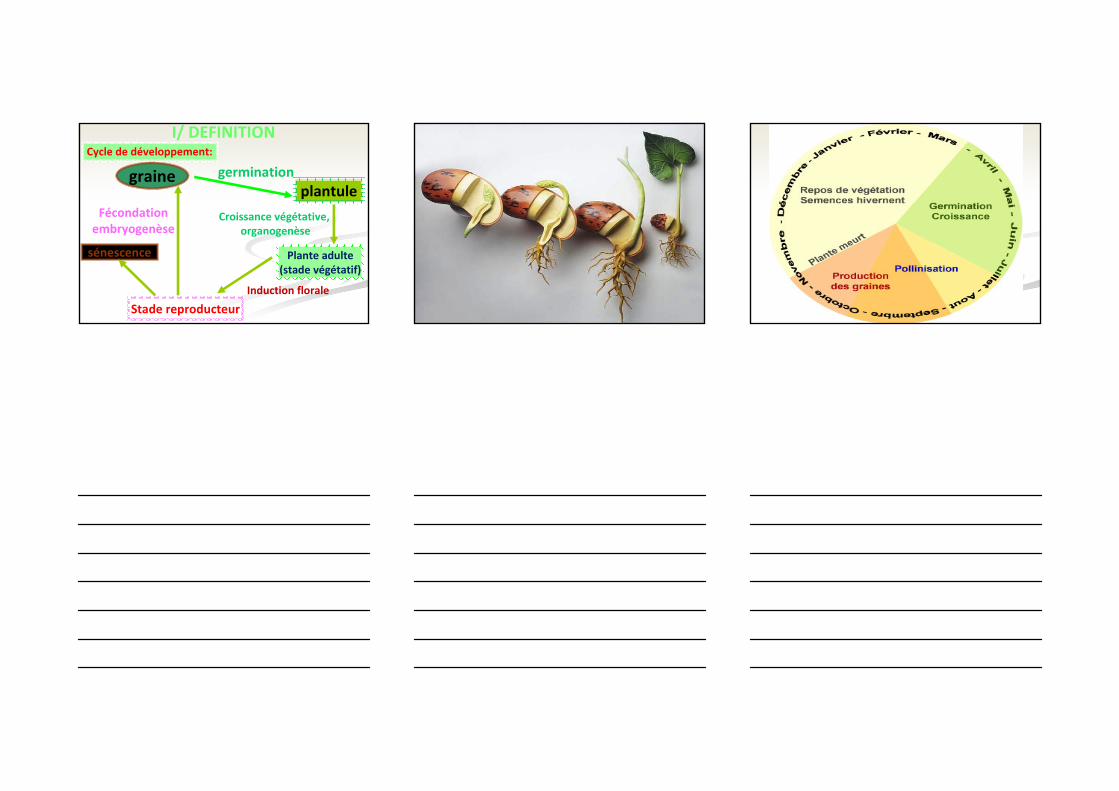
I/ DEFINITIONCycle de développement:
graineplantule
germination
Croissance végétative, organogenèse
Plante adulte(stade végétatif)
Induction florale
Stade reproducteur
Fécondationembryogenèse
sénescence

LA CROISSANCE EST UN PHENOMENE QUANTITATIF.LE DEVELLOPEMENT EST UN PHENOMENE QUALITATIF.ILS SONT TRES LIES ET SE DEROULENT EN MEME TEMPS. ON PARLE DONC DE CROISSANCE.CHEZ LES VEGETAUX LA CEROISSANCE EST INDEFINIE ET LOCALISEE.
INDEFINIE : ELLE MANIFESTE DES PHENOMENES DE CROISSANCE TANT Q’ELLE VIT. ON PARLE D’EMBRYOGENESE INDEFINIE.LOCALISEE : DANS LES CENTRES GENERATEURS DE LA CROISSANCE ( LES MERISTEMES LOCALISES AUX EXTREMITES) .
¥ LES MERISTEMES SONT LOCALISES AUX EXTREMITES DES TIGES ET DES RACINES.
¥ ILS SONT CONSTITUES DE CELLULES MERISTEMATIQUES.
¥ ILS CONTRIBUENT A L’APPARITION D’ORGANES :
tiges ; racines ; feuilles .
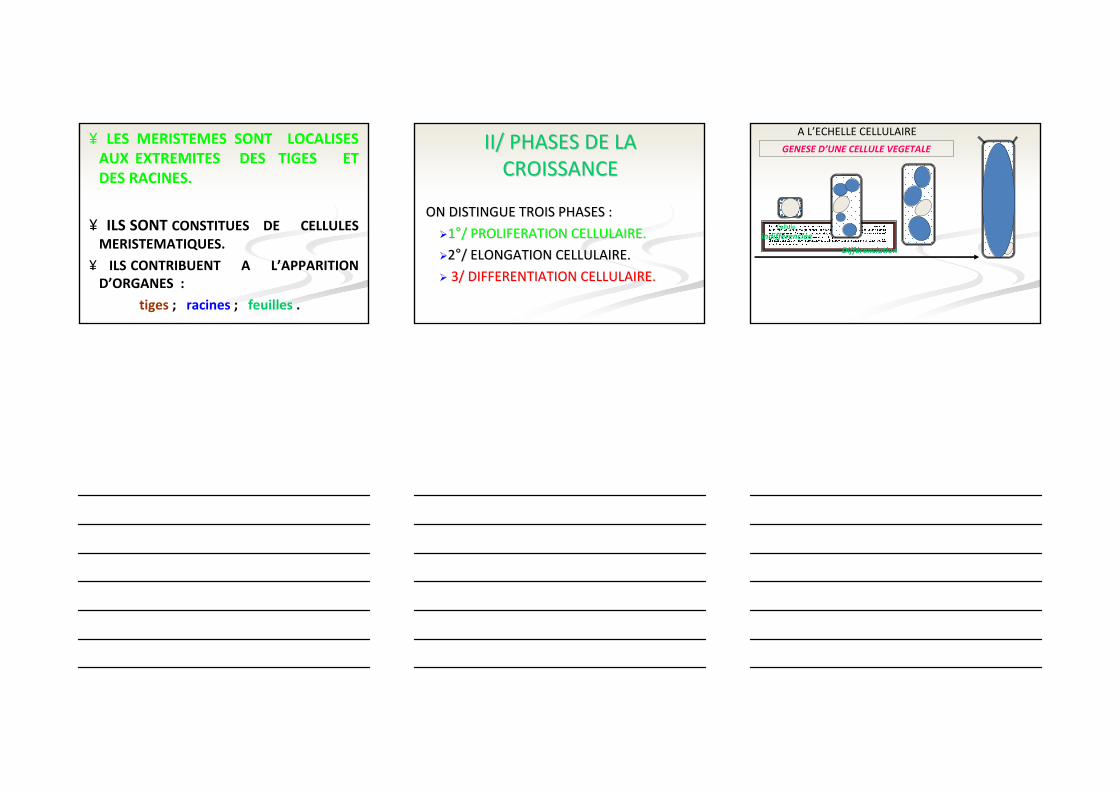
II/ PHASES DE LA II/ PHASES DE LA CROISSANCECROISSANCE
ON DISTINGUE TROIS PHASES :ON DISTINGUE TROIS PHASES :
11°°/ PROLIFERATION CELLULAIRE./ PROLIFERATION CELLULAIRE.
22°°/ ELONGATION CELLULAIRE./ ELONGATION CELLULAIRE.
3/ DIFFERENTIATION CELLULAIRE.3/ DIFFERENTIATION CELLULAIRE.
GENESE D’UNE CELLULE VEGETALE
Cellule
indifférenciée
Différenciation
A L’ECHELLE CELLULAIRE

1°/ PROLIFERATION CELLULAIRE.
a/ DEFINITION :
1er PHENOMENE AU NIVEAU DU MERISTEME.
MULTIPLICATION INTENSE DES CELLULES.
LE RESULTAT EST UNE MASSE CELLULAIRE.
QU’EST CE QUI CE PASSEAU NIVEAU CELLULAIRE ?
b/ PHASES DU CYCLE.LES CELLULES SE DIVISENT PAR
MITOSE .UNE CELLULE DONNERA DEUX CELLULES FILLES IDENTIQUES.
ENTRE DEUX MITOSES ON DISTINGUE UNE PHASE DE REPOS APPELEE :
INTERPHASE .
ON DISTINGUE DONC :
EVENEMENTS LIES A LA MITOSE :
DIVISION DU GENOME.
EVENEMENTS LIES A L’INTERPHASE :
DEDOUBLEMENTS DE TOUS LES
CONSTITUANTS CELLULAIRES.

MITOSE :Dédoublement du génome.INTERPHASE :Comprend :
2 phases de repos (Gap: G1 et G2)
1 phase de synthèse d’ADN ( S) .
CYCLE CELLULAIRE
G1
MITOSE
DébutInterphase
Fin Interphase
G2
S
c/ ETUDE DU CYCLE CELLULAIRE*LA DUREE DU CYCLE.
DUREES DIFFERENTES DUCYCLE CHEZ LE MAIS
G1G1 S S G2G2 MM TotTot
Cellules Cellules ààcroissance croissance
rapide rapide
00 88 55 22 1515
Cellules Cellules ààcroissance croissance
lentelente
151151 99 1111 33 174174
b) INFLUENCE DE FACTEURS EXTERNES SUR LA DUREE DU
CYCLE CELLULAIRE
TT°°CC G1G1 SS G2G2 MM TOTTOT
3030°° 22 1010 22 22 16h16h
2121°° 66 1111 22 22 21h21h
1313°° 1515 2323 88 55 51h51h
CELLULES DE TRADESCANTIA.

2°/ ELONGATION CELLULAIREGERMINATION DE PETIT POIS PAR SACHS
LE DIAGRAMME DE CROISSANCE MONTRE QUE L’ELONGATIONEST NULLE AU NIVEAU DES PARTIES APICALES ET BASALES IMPORTANTE AU MILIEU DE L’ORGANE.
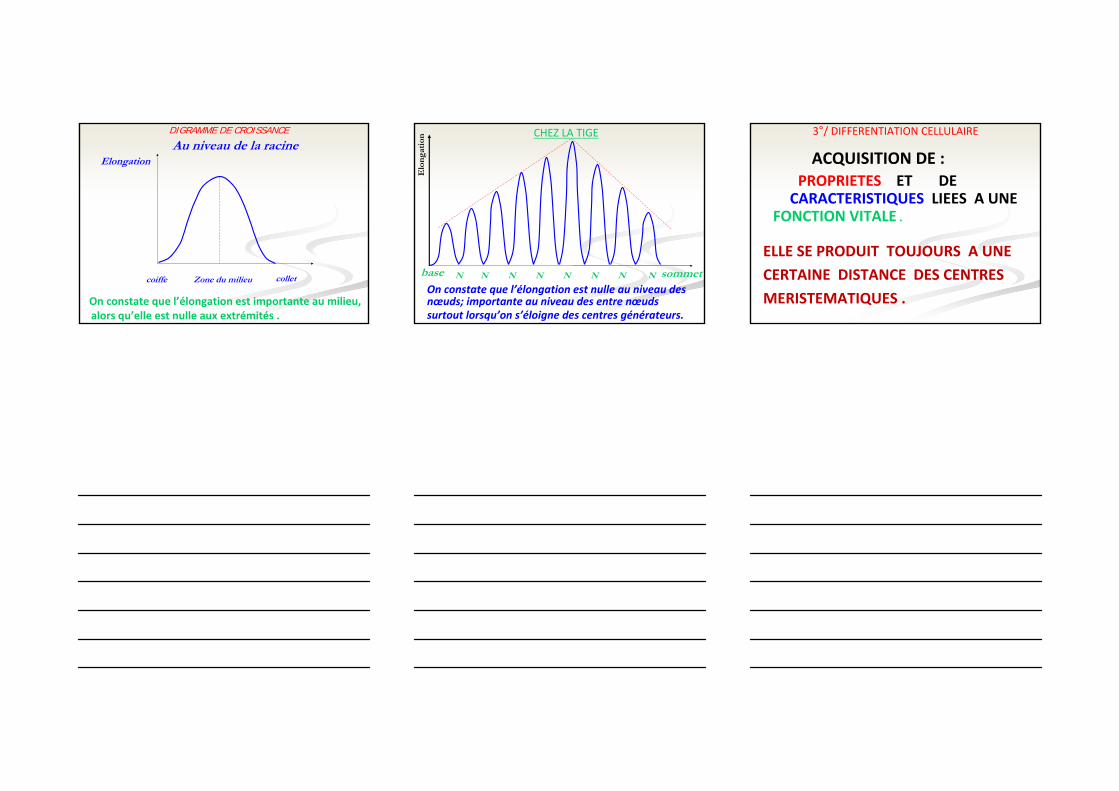
DIGRAMME DE CROISSANCE
On constate que l’élongation est importante au milieu, alors qu’elle est nulle aux extrémités .
Zone du milieu colletcoiffe
ElongationAu niveau de la racine
CHEZ LA TIGE
On constate que l’élongation est nulle au niveau desnœuds; importante au niveau des entre nœudssurtout lorsqu’on s’éloigne des centres générateurs.
base sommetN N N N N N NN
Elo
nga
tion
3°/ DIFFERENTIATION CELLULAIRE
ACQUISITION DE :PROPRIETES ET DE CARACTERISTIQUES LIEES A UNE
FONCTION VITALE .
ELLE SE PRODUIT TOUJOURS A UNE CERTAINE DISTANCE DES CENTRES
MERISTEMATIQUES .

ON PEUT DISTINGUER:UNE SEPARATION DANS LE TEMPS ET DANS L’ESPACEDES DIFFERENTES ZONES
DE LA RACINE.
SI ON COUPE LABASE DE LARACINE. LA ZONE PILIFERE S’ETTENDERA JUSQU’AU NIVEAU DE LA SECTION.
LE MERISTEME INHIBE DONC LA DIFFERENTIATION.
CONTRÔLE DE LA DIFFERENTIATION DANS UNE EXPERIENCE « IN VITRO »
CALL
PHLOEME XYLEME
CAMBIUM
CHAQUE TISSU A ENGENDRE A SA PROXIMITE LE MEME TISSU D’ORIGINE.
CECI ATTESTE D’UN CONTROLE D’ORIGINE CHIMIQUE DE LA DIFFERENTIATION.

EN CONCLUSION :
• LA DIFFERENTIATION EST DONC BIEN CONTROLEE .
• ON ASSISTE A UN CONTRÔLE DANS LE TEMPS ET DANS L’ESPACE .
• LE CONTRÔLE EST DIRIGE PAR LES CENTRES GENERATEURS . IL EST D’ORDRE CHIMIQUE .
III/ CENTRES GENERATEURSDIFFERENTE TYPES :
• LES MERISTEMES TERMINAUX : D’ORIGINE PRIMAIRE . ILS SONT SOIT CAULINAIRES OU RACINAIRES .
• LES MERISTEMES SECONDAIRES (LES CAMBIUMS): ILS APPARAISSENT PLUS TARD . ON NOTE :– L’ASSISE LIBERO‐LIGNEUSE QUI DONNERA LE LIBER ET LE BOIS .
– L’ASSISE SUBERO‐PHELLODERMIQUE QUI DONNERA LE LIEGE.
Bourgeon apical
Bourgeon axillaire
Méristème racinaire

III/ CENTRES GENERATEURS DE LA CROISSANCE .
1°/ MERISTEMES CAULINAIRES
PLUSIEURES THEORIES
* AU 19° SIECLE NARGELI A MONTRE SUR DES MERISTEMES DE PTERIDPHYTES QUE TOUTES LES CELLULES ET DONC TOUS LES TISSUS SONT ISSUS A PARTIR D’UNE CELLULE UNIQUE.
a/ ORGANISATION :
* HANSTEIN EN 1870 A DEVELOPPE LA THEORIE DES HISTOGENES
DERMATOGENE (cellules superficielles)
PERIBLEME (cellules périphériques)
PLEROME (cellules du centre)
ORGANISATION MERISTEMATIQUE EN TROIS ZONES
* EN 1890 SMITH A DEVELOPPE LA THEORIE DU TUNICA ET DU CORPUS
Zone superficielle= TUNICA
Division latérale
Zone profonde= CORPUS
Division vers la profondeur
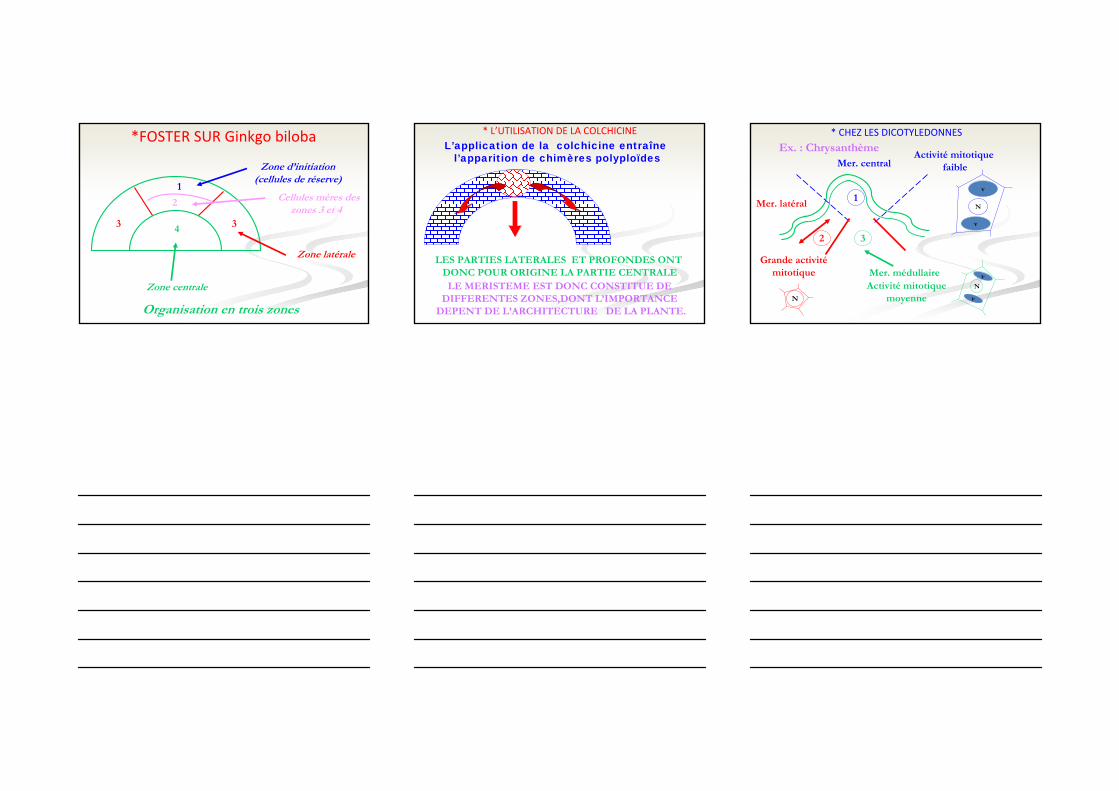
*FOSTER SUR Ginkgo biloba
1
2
33 4
Zone d’initiation(cellules de réserve)
Cellules mères deszones 3 et 4
Zone latérale
Zone centrale
Organisation en trois zones
* L’UTILISATION DE LA COLCHICINEL’application de la colchicine entraîne
l’apparition de chimères polyploïdes
LES PARTIES LATERALES ET PROFONDES ONT DONC POUR ORIGINE LA PARTIE CENTRALE
LE MERISTEME EST DONC CONSTITUE DE DIFFERENTES ZONES,DONT L’IMPORTANCE
DEPENT DE L’ARCHITECTURE DE LA PLANTE.
* CHEZ LES DICOTYLEDONNES
1
2 3
Grande activitémitotique
Mer. latéral
Mer. médullaireActivité mitotique
moyenne
Mer. centralActivité mitotique
faible
Ex. : Chrysanthème
N
N
N
v
v
v
v

* CHEZ LES MONOCOTYLEDONES
* CHEZ LES GYMNOSPERMES
Même organisation , avec : Un méristème axial plus étroit. Un méristème latéral plus développé (très large).
Le méristème axial et le méristème latéral ont la même importante.
ORGANISATIONS DIFFERENTES = ARCHITECTURES DIFFERENTES .
? Mer. foliaire
Sens des divisions
Mer.axial
Procambium
(Sys; vasculaires)
Mer. latéral
Mer. Laissé à labase de la feuille
PERICLINES
AN
TIC
LIN
ES
b/ Initiation foliaire

- LE MERISTEME FOLIAIRE PERSISTERATRES LONGTEMPS AUX EXTREMITES DES FEUILLES POUR ACTIVER LA CROISSANCE.
- DIFFERENTIATION D’UN ORGANE A LABASE DE LA FEUILLE , LE PETIOLE QUIRAPPELLE LA TIGE.
- LE RESULTAT DE CETTE CROISSANCEEST DONC UNE TIGE AVEC DESFEUILLES.
*CHEZ LES DICOTYLEDONESMOELLE
CORDONS PROVASCULAIRES
SECTION
EPIDERME
F.C.V. CROISSANCE EN EPAISSEUR
* CHEZ LES MONOCOTYLEDONES
Cordons Provasculaires
section
Moelle trèsréduite
Les feuilles naissent au niveau de l’épiderme; la zone latérale est très large.
La croissance en épaisseur se fait par étalement du méristème latéral.

* CHEZ LES PLANTES AQUATIQUES
Zone axiale
Zone latérale
Croissance de la plante
Système foliaireTissu conducteur
Lacunes qui remplace une partie duparenchyme
CONCLUSIONS
• L’ORGANISATION DE LA PARTIE APICALE A UN DETERMINISME SUR:
– L’ONTOGENESE DE LA TIGE .– LA PHYLLOTAXIE FOLIAIRE (en donnant le
nombre d’hélices foliaires et la disposition des feuilles sur cette hélice foliaire).
* NOTION DE BOURGEON
• L’APEX PLUS UN CERTAIN NOMBRE D’EBAUCHES FOLIAIRES = BOURGEON .
• ON DISTINGUE :– BOURGEONS TERMINAUX ..– BOURGEONS AXILLAIRES .– BOURGEONS ADVENTIFS . (Méristèmes qui
restent très longtemps quiescents et qui donnent des tiges sur des racines ou des racines sur des tiges .


* Méristème reproducteurMer. Ax.
Mer. Lat.
Mer. Méd.
MERISTEMEVEGETATIF
S + PPièces florales
Réceptacle
MERISTEME REPRODUCTEUR
* TRANSFORMATION FLORALE
Gynécée
Réceptacle floral
Etamines
Sépales +Pétales
CETTE ORGANISATION EST PLUS AU MOINS COMPLEXESELON LES ESPECES ET SURTOUT LORSQU’IL S’AGIT D’UNE INFLORESCENCE
* MERISTÈME REPRODUCTEURIl est généralement issu d’un
méristème végétatif.Chez les plantes annuelles tous
les méristèmes végétatifs passent en méristèmes reproducteurs.
Chez les plantes vivaces une partie seulement passe en méristèmes reproducteurs.

CQ
Ecorce
Cylindre central Tissus conducteurs (zone de
prolifération)
Assise pilifère
zone méristèmatique
zone méristèmatique
Coiffe
2°/ MERISTEME RACINAIRE On distingue trois zones méritèmatiques:
*Un centre quiescent à très faible activité.
*Un méristème qui alimente la coiffe et l’assise pilifère.
*Un méristème qui alimente les autres zones.

*ACTIVITE CELLULAIRE AU NIVEAU DU MERISTEME RACINAIRE
mamaïïss moutardemoutarde
Centre quiescentCentre quiescent 200h200h 500h500h
CoiffeCoiffe 12h12h 35h35h
Zone de Zone de prolifproliféérationration
25h25h 40h40h
*RECAPITULATION
CQ
coiffe
Mer. Vég.
Mer. Rep.
Fleur
Fruit
Graine
Méristème
Systèmevasculaire
Cambium
Croissance en épaisseur
Procambium

IV/ REGULATION ET FACTEURS DE LA CROISSANCE .
A/ REGULATION DE LA CROISSANCE
1
2
3
PA
PB
* Dans une expérience :
Culture sur milieu nutritif.
INTRODUCTION :
1
MILIEU DE CULTURE
QUELQUES JOURS
ON OBTIENT UNE PLANTE ENTIERE ENRACINEE.
2
MILIEU DE CULTURE
QUELQUES JOURS
ON OBTIENT UN CALL ( AMAS DE CELLULES DESORGANISEES).
DEDIFFERENTIATION .CELLULES MERISTEMATIQUES .PAS D’ORGANOGENESE .

3
MILIEU DE CULTURE
QUELQUES JOURS
ON OBTIENT UN CAL PLUS PETIT QUE PRECEDEMMENT .
LA CROISSANCE EST DE PLUS EN PLUS DIFFICILELOSQU’ON S’ELOIGNE DE L’APEX .
1
ZL ZL
ZA
Excision de la zone axiale
Après cicatrisation et divisions
On obtient deux apex qui naissent au niveau des zones latérales .
Le méristème latéral setransforme en méristème axial .
CC. : LA ZONE AXIALE INHIBE LES PARTIES LATERALES .
* DANS D’AUTRES EXPERIENCES *PELLIGRINI EN 1970
PROCEDE AU DECOUPAGE DELA ZONE AXIALE ET DES DEUXZONES LATERALES
PAS DE REORGANISATION DE L’APEX . LES FEUILLES OBTENUES ONT UN ASPECTANORMAL .
L’APEX ASSURE LE BON DEVELOP-PEMENT DES FEUILLES .

SI ON LAISSE LA PARTIE DUMERISTEME RESPONSABLEDE L’INITIATION FOLIAIRE .
Après cicatrisation et divisions
ON ASSISTE A LA REORGANISATION D’UN APEX
ET QUE LES FEUILLES SEDEVELOPPENT NORMALEMENT .
* LOISEAU SUR LA BALSAMINE :3 CENTRES FOLIAIRES
LORSQU’ON ENLEVE UN CENTRE
LA PLANTE FONCTIONNE AVEC LES DEUX AUTRES CENTRES
MERISTEME APICALE
LORSQU’ON ENLEVE LA PARTIE CENTRALE
LA PLANTE FONCTIONNE NORMALEMENT.REGENERATION DU MERISTEME CENTRAL .
LORSQU’ON COUPE LES TROIS CENTRES
LA REGION CENTRALE REGENERELES TROIS CENTRES FOLIAIRES
CONCLUSION :
LES CENTRES FOLIAIRES SONT INDEPENDANT LES UNS DES AUTRES . LA REGION CENTRALE PEUT REGENERER LES AUTRES CENTRES . LA PLANTE EST CAPABLE DE REGENERER LA ZONE CENTRALE .

* MERISTEME RACINAIRE
DANS TOUS LES CAS IL Y A DIVISIONS ET REGENERATIONDES CELLULES. ELLES SONT ANARCHIQUES AU DEPART PUIS SE REORGANISENT. LA ZONATION AU NIVEAU DE LA RACINE EST MOINS STRICTE QUE CHEZ LA TIGE .
DECOUPAGES AU NIVEAU DE LA RACINE DE LA FEVE PAR CLOWES EN 1953-55.
C’EST UNE CARACTERISTIQUE QUI SE TROUVE A LA BASE DE LA MORPHOGENESE ET AUSSI DE L’ORGANOGENESE. ELLE EST A L’ECHELLE DE L’ORGANE MAIS AUSSI DE LA CELLULE .
L’ŒUF EST GENERALEMENT OVALE, MAIS JUSTE AVANT LA DIVISION, ON NOTE UNE DISTRIBUTION INEGALE DU CYTOPLASME. LA POLARITE S’INSTALLE DONC DES L’ORIGINE DE LA VIE .
A L’ECHELLE CELLULAIRE :
* LA NOTION DE POLARITE EXEMPLES :
CHEZ L’ŒUF DE FUCUS :Avant la 1ere
Division
PARTIE QUI DONNERALE THALLE PARTIE QUI DONNERA
LES RHIZOIDESCHEZ LE SPORE D’Equisetum :
n nAvant la 1ere
Division
CES MODIFICATIONS CYTOPLASMIQUES SONT POSSIBLE GRACE A L’ACTION DE STRUCTURES MICROTUBULAIRES .

* AU NIVEAU ORGANOGENESE :CHEZ LA RACINE DE CHICOREE :
P.R. P.F.DEBITAGE EN TRONCONS
BOURGEONS
RACINES
CES TRONCONS SUR MILIEU DE CULTURE DONNERONT TOUJOURS DES BOURGOENS VERS LA PARTIE P.F. ETDES RACINES VERS LA PARTIE P.R.
LA POLARITE EST CONSERVEE.ON NOTE QUE LES BOURGEONS APPARAISSENT
EN PREMIER .
SI ON ELIMINE LES BOURGEONSDES LEURS APPARITION .
PAS DE RACINES
SI ON COUPE LES RACINES ; ELLESREAPPARAISSENT TOUJOURS MEME SION REPETE CELA PLUSIEURS FOIS .
ON NOTE UNE INDUCTION POLARISEE DE LA RHIZOGENESE
PAR LES BOURGEONS.
IL EXISTE DONC DES SUBSTANCES DONT LE CENTRE DE
SYNTHESE EST L’APEX, CIRCULANT DU HAUT VERS LE BAS .
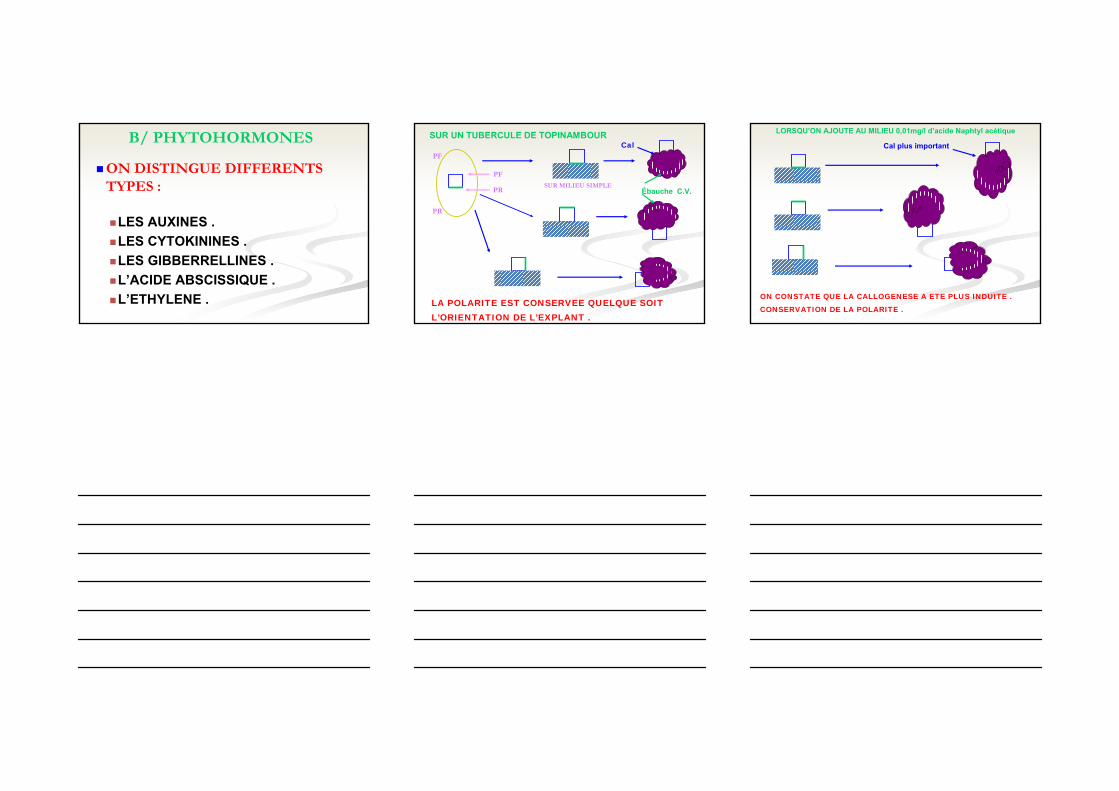
B/ PHYTOHORMONES
ON DISTINGUE DIFFERENTS TYPES :
LES AUXINES .LES CYTOKININES .LES GIBBERRELLINES .L’ACIDE ABSCISSIQUE .L’ETHYLENE .
SUR UN TUBERCULE DE TOPINAMBOUR
PF
PR
PF
PR
LA POLARITE EST CONSERVEE QUELQUE SOIT L’ORIENTATION DE L’EXPLANT .
SUR MILIEU SIMPLE
Cal
Ébauche C.V.
Cal plus important
LORSQU’ON AJOUTE AU MILIEU 0,01mg/l d’acide Naphtyl acétique
ON CONSTATE QUE LA CALLOGENESE A ETE PLUS INDUITE .CONSERVATION DE LA POLARITE .

LORSQU’ON AJOUTE AU MILIEU 1mg/l d’acide Naphtyl acétique :
LA POLARITE EXISTE , MAIS ELLE EST MASQUEE PAR L’APPORT D’UNE SUBSTANCE DE CROISSANCE .
12 racines
Quelques racines.L’ANA a changée la
polarité
1°/ LES AUXINES
C’EST GRACE A BEIJERINK ET A DARWIN QUI S’EST INTERESSE AU PHOTOTROPISME CHEZ Phalaris canariensis DEPUIS 1880 QU’ON A PU INTRODUIRE LA NOTION DE PHYTOHORMONEOU HORMONE VEGETALE .
Zone d’allongement
Croissance verticale silumière symétrique
Mise en évidence duphototropisme du
coléoptile avec unelumière asymétrique
Lumière symétrique
Lumière asymétrique
a/ Histoire et découverte.
IL REALISA L’EXPERIENCE SUIVANTE :
1ere feuille
Pointe ducoléoptile DARWIN A CONSTATE :
QUE LORSQU’ON INTERPOSE UN ECRAN DE FACON A EXPOSER LA POINTE DU COLEOPTILESEULEMENT A LA LUMIERE CELUICI SE COURBE .
IL MONTRE AUSSI QUE LORSQUE LA POINTE EST CACHEE LE COLEOPTILE NE SE COURBE PAS.
Écran
DARWIN S’EST INTERESSE ENSUITE A LA PARTIE APICALE DU COEOPTILE QUI SEMBLE L’INSTIGATEUR DU PHOTOTROPISME :

DARWIN A ENSUITE CONSTATE
LORSQU’ON DECAPITE LE SOMMET
PLUS DE COURBURE
DONC C’EST LE SOMMET QUI EST LE RESPONSABLE DE LA COURBURE

FEFER ET BOYSEN 1910-1914 :
LORSQU’ON DEPLACE LE SOMMET SOMMET
AGAR
LAME DE MICA
COURBURE
PAS DE COURBURE
COURBURE
CONCLUSION:• LA CONTINUITE CELLULAIRE N’EST PAS NECESSAIRE.• LE STIMULUS PEUT FRANCHIR L’AGAR MAIS NON LA LAME DE MICA
.
ENSUITE :
LAME DE MICA
COURBURE PAS DECOURBURE
FEFER N’EN TIRE AUCUNE CONCLUSION DE CES EXPERIENCES .

WENT EN 1928 ; EN REPRENANT CES EXPERIENCES A PU MONTRE :
COUPE LE SOMMET
BLOC DE GELOSE
&
ON OBTIENT UNE COURBURE QUI EST UN ALLONGEMENT DU COLEOPTILE DÛ A UNECROISSANCE INEGALE ENTRE LES DEUXMOITIES DU COLEOPTILE .
EN CONCLUSION
N
H
CH2 COOH
Acide indole acétique (AIA)
WENT ET SELON L’EXPERIENCE DE FEFER PENSE QUE LA SUBSTANCE RESPONSABLE DU PHENOMENE CIRCULE DANS LA MOITIE GAUCHE SEULEMENT . DONNE LE NOM D’AUXINE A CETTE SUBSTANCE, QUI VIENT DU NOM AUXESE .
SUBSTANCE IDENRIFIEE PAR KÔGL EN 1934 :
Coléoptile phototropisme auxine
Zone d’élongation
transport
productionAuxine
tissu cible tissu cible
Auxine
Lumière perception
production
Zone d’élongation
transport

CETTE SUBSTANCE A ETE DEJA IDENTIFIEE EN 1885 PAR SALKOWSKI ET MEME SYNTHETISEE PAR ELLINGER EN 1905, SANS SOUPÇONNER QUE C’EST UNE PHYTOHORMONE.
ON LA TROUVE DANS TOUS LE REGNE VEGETAL ( DANS TOUS LES ORGANES A DES DOSES VARIABLES ); SON TAUX EST INFLUENCE PAR L’AGE (IMPORTANT AU NIVEAU DES JEUNES ORGANES ) ET PAR LE MILIEU.
DISTRIBUTION DE L’AIA AU NIVEAU DE LA PLANTE
Apex collet racines
ON CONSTATE : UNE GRANDE QUANTITE D’AUXINE AU
NIVEAU DE L’APEX. ACCUMULATION AU NIVEAU DE LA RACINE.
QuantitéD’Auxine
b/ Polarité et transport.
PA
PB
PA
PB
BLOC DE GELOSEDONNEUR D’AIA
BLOC DE GELOSERCEVEUR D’AIA
COLEOPTILETRANSPORT
DU HAUTVERS LE
BAS
SI ON INVERSE LE SENS DU COLEOPTILE
PAS DE MIGRATION
L’AIA CIRCULE DONC DELA PARTIE APICALE VERS LA PARTIE BASALE

• LA POLARITE EST PLUS AU MOINS STRICTE CHEZ LES TISSUS AGES.
• LA VITESSE EST DE 10 A 12 mn / SECTION DE 2 mm A UNE T°DE 25°C ( v de 15mm/h) .
• LA TEMPERATURE INFLUE SUR LA VITESSE.25°
16°10°
6°
2°
0°
L’ABAISSEMENT DELA TEMPERATUREREDUIT LA VITESSE DE MIGRATION DEL’AIA .
LA VITESSE DEPEND AUSSI DE LA TAILLE DE L’EXPLANT .ELLE EST IMPORTANTE CHEZ LES PETITS SEGMENTS .
10
5
% AIA
100 Unité d’AIA200
* NATURE DU TRANSPORT DES AUXINES
PA
PB
BLOC DE GELOSEDONNEUR D’AIA
BLOC DE GELOSERCEVEUR D’AIA
100 U après
2h
13 U
87 UO U
PA
PB
BLOC DE GELOSEDONNEUR D’AIA
100 U
BLOC DE GELOSEDONNEUR D’AIA
200 U
après2h
14 U
280 U
LE TRANSPORT DE L’AIA EST ACTIF. IL EST INDEPENDANT DU GRADIENT DE CONCENTRATION .
C’est la seule hormone qui subit un transport actif et polarisé de cellule à cellule. Ce n’est pas un transport vasculaire, par la sève brute ou élaborée. Elle est transportée du haut vers le bas. On ne connaît pas très bien encore son mode d’action précis.
Transport actif polarisée du haut vers le bas de cellule à cellule .

• CHEZ LES ORGANES JEUNES L’AIA EMPRUNTE LE CAMBIUM ET LE XYLEME EN COURS DE DIFFERENTIATION .
• CHEZ LES ORGANES AGES LA POLARITE EST MOINS STRICTE, LE PHLOEME EST LA VOIE PREFERENTIELLE, LA VITESSE EST DIX FOIS PLUS IMPORTANTE .
MAIS ON PENSE, QUE :c/ Propriétés Physiologiques
• La prolifération cellulaire est liée à la [ AIA].• Un morceau de tige badigeonné dans une patte d’AIA donne un cal important.
• On pense que l’AIA agit au niveau de la phase S.
• NITSCH a montré que l’AIA augmente le nombre de ribosomes et le volume du nucléole (gros nucléole).
* Prolifération cellulaire On pense aussi que l’AIA agit sur la synthèse
de l’ARN en agissant sur les enzymes.
On a remarqué aussi que l’AIA entraîne des modifications notables sur la membrane plasmique :
Ext. Int.
Vacuoles depinocytose
L’AIA favorisel’absorption de l’eau .
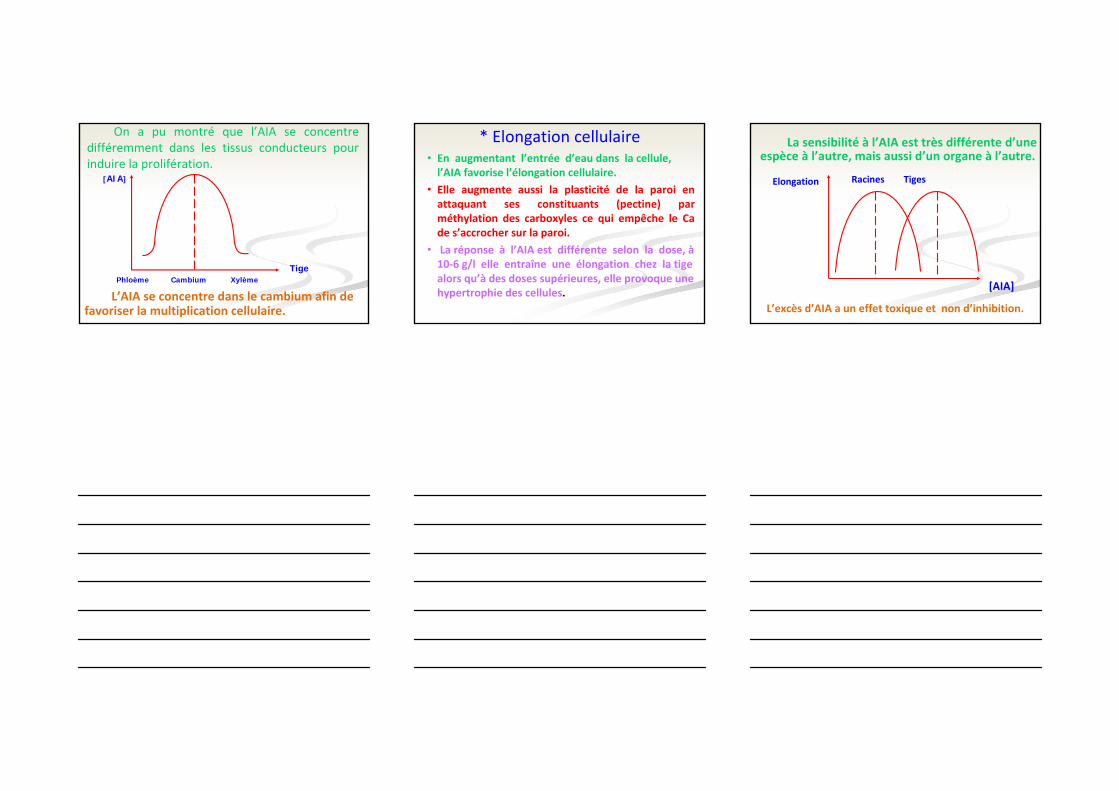
On a pu montré que l’AIA se concentre différemment dans les tissus conducteurs pour induire la prolifération.
L’AIA se concentre dans le cambium afin de favoriser la multiplication cellulaire.
Phloème Cambium Xylème
[AIA]
Tige
* Elongation cellulaire• En augmentant l’entrée d’eau dans la cellule, l’AIA favorise l’élongation cellulaire.
• Elle augmente aussi la plasticité de la paroi en attaquant ses constituants (pectine) par méthylation des carboxyles ce qui empêche le Ca de s’accrocher sur la paroi.
• La réponse à l’AIA est différente selon la dose, à10‐6 g/l elle entraîne une élongation chez la tige alors qu’à des doses supérieures, elle provoque une hypertrophie des cellules. [AIA]
Elongation Racines Tiges
L’excès d’AIA a un effet toxique et non d’inhibition.
La sensibilité à l’AIA est très différente d’uneespèce à l’autre, mais aussi d’un organe à l’autre.

*DIFFERENTIATION CELLULAIRE
Pas de racines
racinesAIA
eau
Sur des boutures de citronnier:
L’AIA favorise la Rhizogénèse (la formation de racines), elle est utilisée comme hormone de bouturage. Par ailleurs elle inhibe le bourgeonnement.
* DOMINANCE APICALE
Morceau de géloseContenant de l’AIA
THIMAN a remplacé l’apex par un morceau d’agar contenant de l’AIA :
Si on décapite le bourgeon apical:
La dominance apicaleest conservée
Bourgeons axillaires
Les bourgeons axillaires démarrent et petit à petit ladominance s’installe
ON A CONSTATE AUSSI
LES BOURGEONS NE SONT PAS LES SEULS CENTRES DE BIOSYNTHESE DES AUXINES.
ELLE SE FAIT AU NIVEAU DES CAMBIUMS, DU GRAIN DE POLLEN ET AUSSI AU NIVEAU DE L’OVAIRE APRES FECONDATION.
L’INTENSITE DE CROISSANCE D’UN TISSU DEPEND DE SA QUANTITE DE SYNTHESE DE L’AUXINE.
ON A PU DECELER UNE SYNTHESE D’AUXINE CHEZ LES BACTERIES.

22°°/CYTOKININES /CYTOKININES
Découvertes depuis 1940, en effet BLAKSLEE à constaté que l’addition de lait de coco facilite la germination d’embryons immatures .
DUHAMET a montré que le lait de coco entraîne une prolifération des tissus cultivés « in vitro ».
SKOOG a démontré que le lait de coco donne une prolifération sur la moelle de la tige de tabac « in vitro ».
a/ DécouverteMILLER à isoler d’un hydrolysat d’ADN de la
moelle de tabac une substance la Kinétine (6‐furfuryl‐aminopurine). [ substance non naturelle de fait de la dénaturation lors de l’hydrolyse ] et qui peut remplacer le lait de coco.
Par ailleurs deux substances naturelles ayantles mêmes propriétés que la Kinétine ont étédécouvertes, la zéatine (caryopse de maïs) et l’IPA (isopentyl adénine) à partir d’une culture bactérienne.
Ensuite ces composés ont pu être détecter dans la plupart des tissus végétaux à l’état libres ou liées.
En culture « in vitro », on utilise des analogues structuraux de synthèse ex.: la BA (benzyl adénine) et la BAP (9 pyranobenzyl adénine). On note aussi que l’adénine présente certaines propriétés de ces substances.

SKOOG a donné le nom de CYTOKININES àces substances qui permettent la prolifération cellulaire.
La biosynthèse semble se faire dans les extrémités racinaires et les jeunes fruits. Migrent difficilement et ont généralement un effet très localisé.
b/ Propriétés Physiologiques
*Division cellulaire : l’addition de Cytokinines entraîne la prolifération des cellules de la moelle de tige de tabac. On note cependant des exigences différentes entre les végétaux en effet :– La moelle de tige de tabac nécessite de l’AIA et des Cytokinines.
– la Ronce se multiplie sans AIA ni Cytokinines.– Le topinambour à besoin d’AIA seulement.– Le Choux navet à besoin de Cytokinines seulement.
Donc l’AIA et les Cytokinines sont complémentaires et que les doses exigées dépendent fortement des quantités exogènes dans les plantes.
*Différentiation cellulaire : les cellules de moelle de tabac peuvent verdir grâce à l’addition de cytokinines. Indispensables dans les phénomènes de proplastes, ainsi que pour le développement des cellules cambiales en xylème.
*Organogenèse : rôle important dans la formation des bourgeons. Sur la molle de tabac :
– AIA + faible dose de cytokinines racines.
– AIA + dose moyenne de cytokinines bourgeons.
– AIA + très forte dose de cytokinines pas d’organogenèse.

• Les Cytokinines inhibent les racines.
• Pour SKOOG il n’y a pas de substances spécifiques. C’est Le rapport [ AIA / CYTOKINIES ] qui détermine l’organogenèse.
AIA production de racines.
CYTOKININE
AIA Faible production de bourgeons.
CYTOKININE
Levée de dormance: ex. : des graines de laitue trompées dans des Cytokinines peuvent germer, alors qu’elles ne germaient qu’après passage à l’UV.
*Application de Cytokinines sur des germinations de pois chiche.
5,2 mm
7 mm
3,9 mm
2,8 mm
6,4 mm34,4 mm
39,1 mm
13,5 mm
12 mm
34,8 mm
50,2 mm
23,1 mm
0,2 mg 50 mg
On constate une augmentation de la longueur desbourgeons. Il y a donc levée de la dominance apicale.
*Effets sur le métabolisme de la plante : on a constaté que l’application de Cytokinines à des feuilles coupées, retarde le phénomène de sénescence ( jaunissement ). RICHMOND en 1957 a montré que l’application de cette hormone retarde le vieillissement des feuilles traitées mais accélèrent celui des feuilles adjacentes non traitées. En effet MOTHES sur des feuilles de tabac a prouvé que les Cytokinines retardent la dégradation des protéines mais stimulent aussi leurs synthèse. PENOT utilisant des produits radioactifs a montré que les Cytokinines dévient le transport des métabolites, ce qui explique le jaunissement des feuilles non traitées.

Expérience de PENOTacides
aminés 14C
Application de Cytokininessur un côté.
déviation des acidesaminéesvers la zoneTraitée.
LA GERMINATION.
I/ INTRODUCTION
1°/ La Graine.la graine est un organe résultant de la double
fécondation de l’ovule.
Il contient un embryon, un tissu de réserve et des téguments.
Caractéristique des Spermaphytes.
(plantes à graines : angiospermes, gymnospermes).

*On distingue Différents types de graines :‐ Les graines à albumen (céréales).‐ Les graines à cotylédons (légumineuses) pour lesquelles
les cotylédons ont digéré l’albumen.‐ Les graines à périsperme (peu nombreuses (caféier)) dans
lesquelles le nucelle tissu entourant l’ovule persiste.
* La taille des graines est très variable :‐ Chez une variété de Palmier (Lodoicea maldivica) qui se forme pendant environ 7 ans la graine pèse environ 10 kg.
‐D’autres graines sont de très petite taille. Celle‐ci n’a aucune relation avec la taille finale de l’individu (Carotte, tabac).
‐ D’autres sont de véritables poussières (Orchidées).

Graine de Lodoicea maldivicaGraine d’orchidées
Le grain de céréale .C'est un fruit sec (caryopse),
la paroi du fruitLe tégument
riche en protéines
Le tissu de réserve de la graine. Il contient une fraction protéique, le gluten, grains d'amidon.

Différents types de graines2°/ La Notion de semence.On parle plus généralement de semences :
il s’agit de l’organe ou de la partie de l’organe que l’on sème :
‐ Qui est une graine le plus souvent.
‐Mais aussi un fruit.
ex : le caryopse de blé (grain),
fruit de betterave.
‐ Par extension ;
le terme de semences concerne :
les tubercules ( pomme de terre)
ou les bulbes.
3°/ La Formation de la graine.Trois phases sont indispensables pour l’obtention d’une graine mature et viable:
a/ l’embryogenèse ou développement de l’embryon:
On obtient deux cotylédons et un axe embryonnaire avec la radicule et l’hypocotyle.

b/ La maturation :
Phase d’accumulation des réserves soit dans l’albumen ou dans les cotylédons ou dans la nucelle (périsperme).
Ces tissus contiennent des grains d’amidon qui représentent 35% des réserves de la graine.
Les réserves des cotylédons sont de nature lipidique et protéique.
L’acide abscissique (ABA) induit la synthèse de certaines protéines de réserve et empêche la graine de germer prématurément.
c/ Une phase de dessiccation:La teneur en eau chute de 85 à 15% environ.
L’activité de l’embryon ralentit jusqu’à devenir pratiquement nulle.
L’ABA intervient dans la protection des tissus vis à vis de la déshydratation.
La sclérification intense des téguments qui se transforment en enveloppes protectrices.
C’est une protection mécanique qui s’oppose à la sortie de la radicule.
Enfin, la synthèse de substances qui inhibent la germination.
4°/ La production de semences :Les semences brutes suivent les étapes suivantes :
a/ Le Triage : Les semences sont triées et calibrées afin d’éliminer les graines anormales ou non matures.
b/ Le polissage : Le polissage permet de réduire le péricarpe, structure très dense et très lignifiée et d’éliminer des substances inhibitrices de la germination.
c/ Le lavage : Ce traitement induit la phase d’imbibition et élimine les inhibiteurs de la germination tels que les composés phénoliques présents notamment dans les téguments de la graine.

d/ L’enrobage : L’enrobage de la semence permet de la rendre sphérique, et d’augmenter ainsi sa semabilité.
De plus, il permettra de réguler l’arrivée d’eau lors de la germination de la graine pour éviter que l’embryon ne soit asphyxié. Des produits fongicides peuvent être inclus dans l’enrobage.
e/ Le pelliculage : Une fine pellicule est ensuite appliquée pour protéger l’enrobage. Ce pelliculage contient en outre des produits insecticides et une coloration pour mieux contrôler le semis.
II/ LA GERMINATION.C’est un Processus physiologique qui permet à l’embryon contenu
dans la graine de donner une jeune plantule.
1°/ Définitions‐ Physiologiquement : la germination commence avec l’imbibition de
la graine et finit avec la percée des téguments par la radicule ou par l’hypocotyle s’il sort le premier, les étapes ultérieures étant des étapes de croissance.
Il s’agit d’un stade d’hétérotrophie qui se termine à la formation des racines et des feuilles.
‐ Botaniquement : la germination s’arrête quand la radicule a percéles téguments de la graine.
‐ Agronomiquement : la germination s’arrête à la levée des semis, c’est‐à‐dire l’apparition des premières feuilles à la surface du sol.
2°/ Les Etapes de la germination:a/. Imbibition des éléments vivants déshydratés et gonflement
de la graine.
b/. Démarrage de la digestion des réserves.
c/. Grandissement des cellules de la radicule déjà formée dans
l’embryon puis prolifération des cellules du méristème radiculaire.
d/. Eclatement des téguments et sortie de la radicule.
e/. Développement de la partie aérienne et libération des
téguments

2°/ Les types de germination.
le cotylédon reste dans le sol (exemple des pois).
* La germination hypogée :
Le cotylédon sort du sol (exemple des haricots) .
* La germination épigée :

* Cas des plantes monocotylédones céréales et des graminées:
- La Coléorhize : protège la radicule.
- Le Coléoptile : protège les premières feuilles côté apical.
3°/ Les conditions de la germination
3‐1°/ facteurs internes :
La graine doit être mature, viable et non dormante.
3‐2°/ facteurs externes :Les facteurs environnementaux : (météorologiques)
l’eau, l’oxygène et la température.
les résistances mécaniques qu’elle va rencontrer (mottes de terre, croûtes de battance).
(la profondeur et l’état structural du lit de semences).
Enfin, la présence d’adventices et une activité microbienne importante peuvent créer des carences d’oxygène.
3‐1°/ facteurs internes :
La viabilité des semences varie selon les espèces.
Exemples de viabilité :
* Mimosa glomerata 221 ans.
* Lupinus arcticus 10 000 ans à l’état gelé dans le Yukon.
* Alors que Le Peuplier et Acer saccharinum meurent en quelques semaines à la T° ambiante.
Les basses T°, l’atmosphère sèche prolongent la survie.
Alors que la péroxydation et l’oxydation des acides gras insaturés des lipides accélèrent le vieillissement des graines en causant des dégâts aux membranes, aux enzymes et à la chromatine.
a/ La viabilité :

Lupinus arcticus
Mimosa glomerata
Peuplier Acer saccharinum

b/ La dormance :
Chez de nombreuses plantes, les graines, les bourgeons passent par des phases de repos.
Ainsi certaines semences mûres ne germent pas, même si les conditions externes sont favorables. (température tiède, milieu humide et aéré).
On parle alors de « dormance »: C’est l’inaptitude à croître même si les conditions externes sont favorables.
Les causes de la dormance sont très variées.
c/ Les différents types de dormance.
On distingue :
‐ la dormance tégumentaire (dite dormance exogène).
‐ la dormance embryonnaire (dite dormance endogène).
‐ la dormance combinée (où interviennent en même temps la
dormance tégumentaire et la dormance embryonnaire).
* La dormance tégumentaire :
Elle est liée aux diverses enveloppes de la graine. On distingue :
** La dormance tégumentaire chimique : se manifeste grâce aux substances inhibitrices de la germination soit dans le fruit, ou secrétées par les téguments (acide abscissique, substances phénoliques , qui inhibent aussi la croissance des microorganismes pathogènes.
** La dormance photosensible.
Elle est lié à la présence d’un pigment sensible à la lumière dans le tégument (photosensibilité positive).
‐ la lumière prolongée.
‐ Exposition courte à la lumière rouge (660 nm).
‐ Effet annulé par à la lumière rouge lointaine (730 nm).
Il existe donc dans la semence une chromoprotéine, le phytochrome, sous deux formes photoconvertibles:
Forme inactive, absorbant la lumière rouge (P660),
Forme active, absorbant la lumière rouge lointaine (P730).

** La dormance tégumentaire physique.
Elle est liée à la présence de téguments ou de péricarpes durs imperméables à l’eau ou à l’air.
la scarification et le trempage sont alors nécessaires.
Due à une alternance de saisons sèches et de saisons des pluies, et notamment chez plusieurs genres de légumineuses: Acacia, Robinia, Albizzia et Cassia.
Un cas particulier « la dormance mécanique » :
L’enveloppe est épaisse et coriace. Elle empêche l’embryon de se développer (même quand l’eau parvient à s’infiltrer).
la levée est possible grâce à un traitement à la “chaleur humide”, dont la durée varie selon les essences.
Acacia
Albizia Robinia
Cassia
* la dormance embryonnaire.
On distingue :
** la dormance embryonnaire (chimique) :
due à la présence d’inhibiteurs de germination (ex: l’ABA).
Ex: le pommier (Malus pumila), le chêne rouge (Quercus rubra). ** l’immaturité physiologique (ou dormance
morphologique): le développement de l’embryon non achevé.
Une croissance additionnelle de l’embryon est alors nécessaire,
Ex: chez les frênes européens et plusieurs sapins qui poussent en haute altitude.

* La levée de la dormance tégumentaire.Consiste à ramollir, percer, user ou fendre le tégument de
manière à le rendre perméable.
(sans endommager l’embryon et l’endosperme).
Plusieurs techniques :
+ Trempage dans l’eau : Tremper dans de l’eau àtempérature ambiante pendant 24 à 48 H.
+ Trempage dans l’eau chaude : trempage dans de l’eau très chaude, voire bouillante pendant 12H.
+ Traitement mécanique (scarification) : Une des méthodes physiques les plus simples qui consiste à couper, percer ou limer le tégument de chaque graine avant semis.
+ Traitement à l’acide :
l’acide sulfurique concentré (appelé « vitriol »).
+ Traitement spécifique à certaines espèces :
¤ le traitement biologique : la digestion par des animaux (chaleur et/ou enzymes digestives) ou par les micro‐organismes.
¤ le traitement par le feu et ou une chaleur sèche : Dans les régions tropicales, le feu est un puissant moyen naturel d’interruption de la dormance tégumentaire.
Alors qu’un feu violent tue les graines.
REMARQUE:
lorsque la coque est dure mais perméable aux liquides comme Crataegus, Carpinus ou Elaeagnus.
Le trempage dans l’eau bouillante ou dans l’acide peut entraîner la mort des embryons.
Carpinus Betulus
Crataegus monogyna

+ La stratification à froid (vernalisation.)
Les espèces de climat tempéré ne germent pas immédiatement pour des raisons physiologiques. Le prétraitement le plus indiqué pour lever cette dormance physiologique consiste à reproduire les conditions d’hivernage auxquelles les graines sont soumises dans la nature.
la durée de stratification des graines fraîches est inférieure à celle des graines qui ont étés préalablement stockées.
+ Traitement chimique de la dormance physiologique
Utilisation d’agents chimiques tels que de l’eau oxygénée, de l’acide citrique et des gibbérellines.
+ Autres traitements .
Les rayons X, gamma, le rayonnement lumineux (spectre rouge) et les ondes sonores à haute fréquence.
L’apparition de mutations des chromosomes, et des anomalies) rendent ces techniques inutilisables.
1 : téguments ; 2 : endosperme ; 3 : hile ; 4 : cotylédons ; 5 : gemmule ; 6 : tigelle ; 7 : radicule. L’embryon est constitué de l’ensemble des pièces 4, 5, 6 et 7.
Stratification
sable
graine
e/ La dormance combinée.
Certaines espèces possèdent des graines combinant plusieurs formes de dormance.
Par exemple, les graines imperméables du Tilleul d’Amérique (Tilia americana) doivent d’abord être scarifiées avant d’être stratifiées.
f/ La double dormance.
C’est un état dans lequel il y a une deux dormances embryonnaires distinctes (du radicule et de l’épicotyle, par exemple), chacune nécessitant un traitement différent pour être levé.
Ce type de dormance est rare.
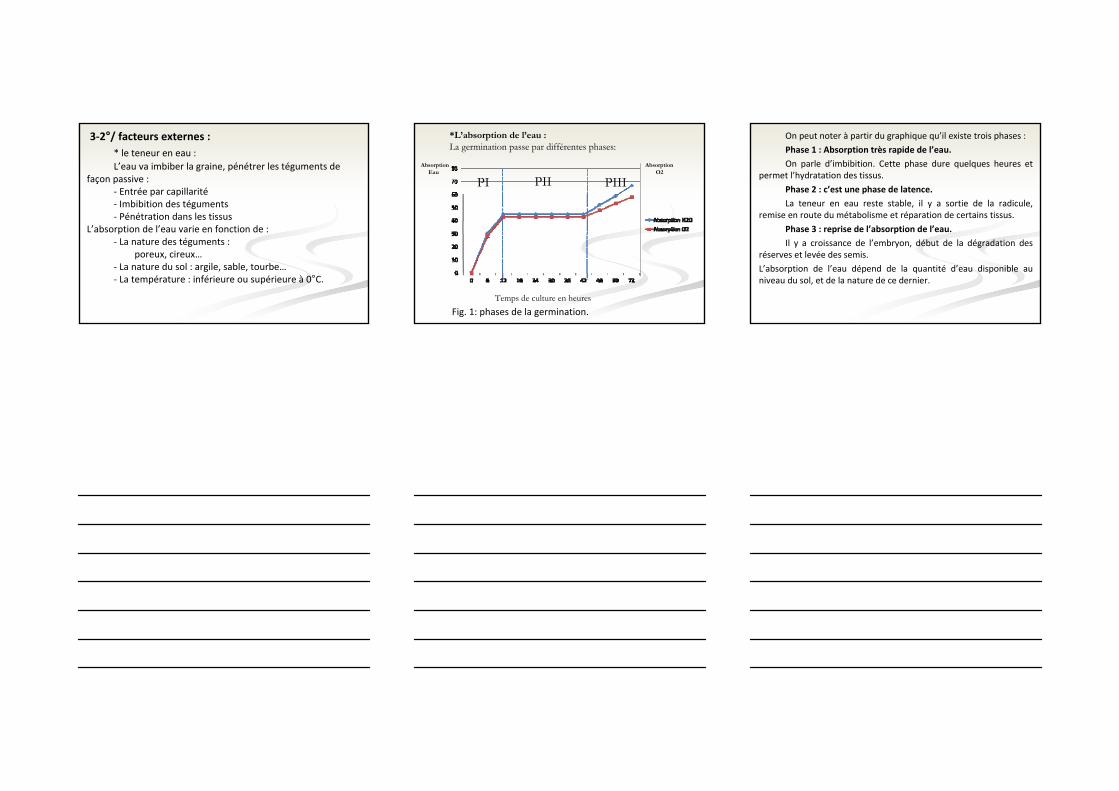
3‐2°/ facteurs externes :* le teneur en eau :L’eau va imbiber la graine, pénétrer les téguments de
façon passive :‐ Entrée par capillarité‐ Imbibition des téguments‐ Pénétration dans les tissus
L’absorption de l’eau varie en fonction de :‐ La nature des téguments :
poreux, cireux…‐ La nature du sol : argile, sable, tourbe…‐ La température : inférieure ou supérieure à 0°C.
*L’absorption de l’eau :La germination passe par différentes phases:
Temps de culture en heures
Absorption Eau
Absorption O2
PI PIIIPII
Fig. 1: phases de la germination.
On peut noter à partir du graphique qu’il existe trois phases :
Phase 1 : Absorption très rapide de l’eau.
On parle d’imbibition. Cette phase dure quelques heures et permet l’hydratation des tissus.
Phase 2 : c’est une phase de latence.
La teneur en eau reste stable, il y a sortie de la radicule, remise en route du métabolisme et réparation de certains tissus.
Phase 3 : reprise de l’absorption de l’eau.
Il y a croissance de l’embryon, début de la dégradation des réserves et levée des semis.
L’absorption de l’eau dépend de la quantité d’eau disponible au niveau du sol, et de la nature de ce dernier.
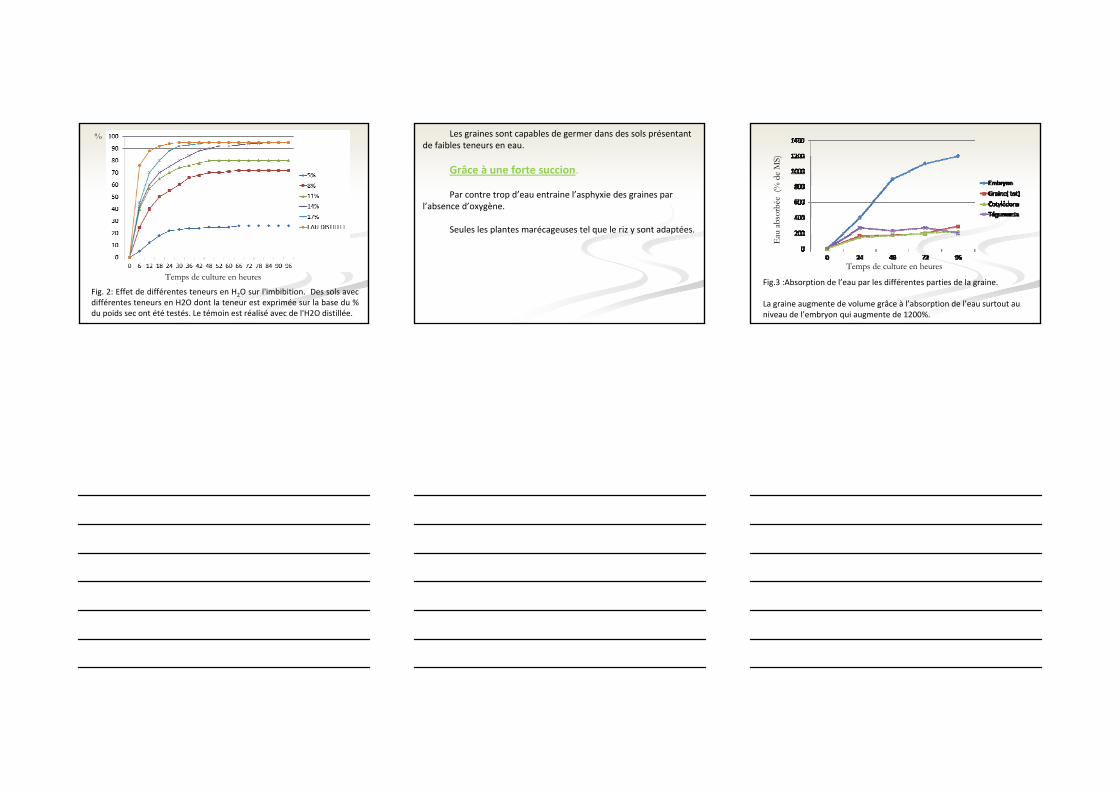
Fig. 2: Effet de différentes teneurs en H2O sur l'imbibition. Des sols avec différentes teneurs en H2O dont la teneur est exprimée sur la base du % du poids sec ont été testés. Le témoin est réalisé avec de l'H2O distillée.
%
Temps de culture en heures
Les graines sont capables de germer dans des sols présentant de faibles teneurs en eau.
Grâce à une forte succion.
Par contre trop d’eau entraine l’asphyxie des graines par l’absence d’oxygène.
Seules les plantes marécageuses tel que le riz y sont adaptées.
Fig.3 :Absorption de l’eau par les différentes parties de la graine.
La graine augmente de volume grâce à l’absorption de l’eau surtout au niveau de l’embryon qui augmente de 1200%.
Temps de culture en heures
Eau
abs
orbé
e (%
de
MS)
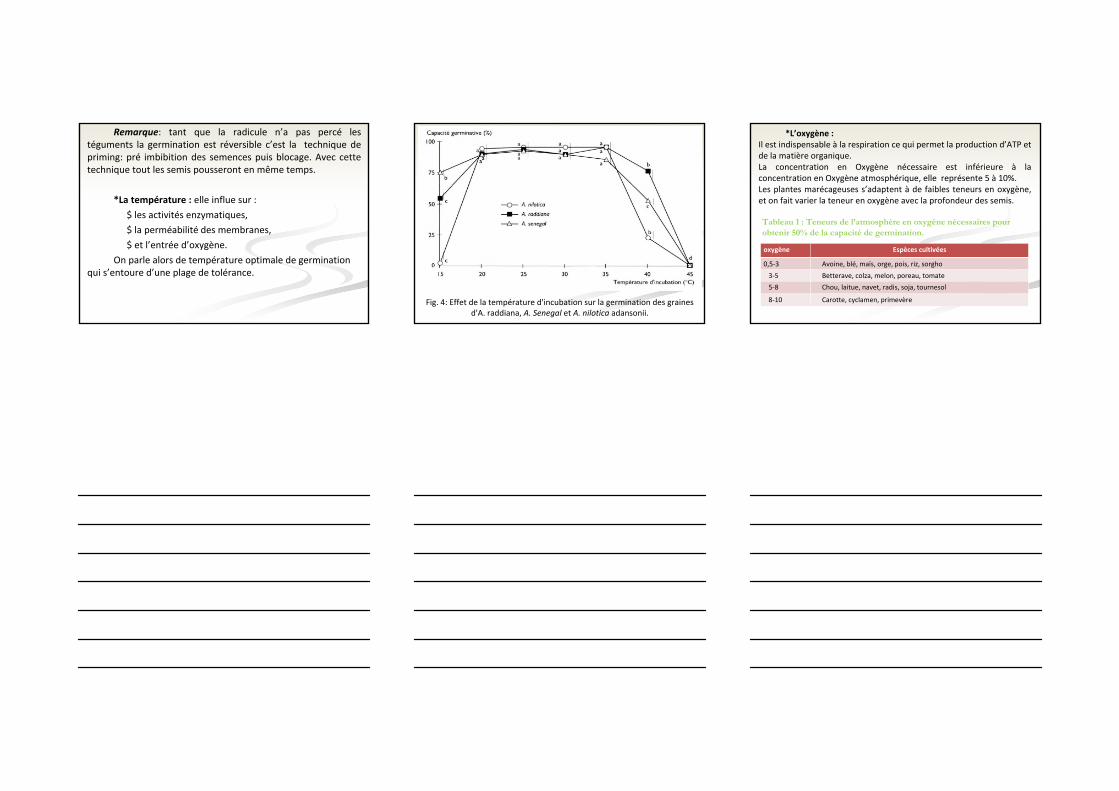
Remarque: tant que la radicule n’a pas percé les téguments la germination est réversible c’est la technique de priming: pré imbibition des semences puis blocage. Avec cette technique tout les semis pousseront en même temps.
*La température : elle influe sur :
$ les activités enzymatiques,
$ la perméabilité des membranes,
$ et l’entrée d’oxygène.
On parle alors de température optimale de germination qui s’entoure d’une plage de tolérance.
Fig. 4: Effet de la température d'incubation sur la germination des graines d'A. raddiana, A. Senegal et A. nilotica adansonii.
*L’oxygène :Il est indispensable à la respiration ce qui permet la production d’ATP et de la matière organique.La concentration en Oxygène nécessaire est inférieure à la concentration en Oxygène atmosphérique, elle représente 5 à 10%.Les plantes marécageuses s’adaptent à de faibles teneurs en oxygène, et on fait varier la teneur en oxygène avec la profondeur des semis.
oxygène Espèces cultivées
0,5‐3 Avoine, blé, maïs, orge, pois, riz, sorgho
3‐5 Betterave, colza, melon, poreau, tomate
5‐8 Chou, laitue, navet, radis, soja, tournesol
8‐10 Carotte, cyclamen, primevère
Tableau 1 : Teneurs de l’atmosphère en oxygène nécessaires pour obtenir 50% de la capacité de germination.

La consommation d’oxygène est similaire à la consommation en eau.
On a une augmentation de la respiration avec une forte consommation d’oxygène car les enzymes des mitochondries sont activées par la présence d’eau.
Parfois, on observe une diminution de la consommation d’oxygène due à une répartition non homogène de celui‐ci dans la graine àcause d’un lent développement des mitochondries ou une déficience en substrat.
Toutes ces étapes aboutissent à l’élongation et au grandissement de la radicule et à la consommation des réserves.
La chaîne respiratoire est impliquée dans la consommation d’oxygène. En effet si on utilise du cyanure, celui‐ci bloque la chaîne respiratoire et, il n’y a pas de consommation d’oxygène.
Temps en minutes de la germination d’un embryon
Con
som
mat
ion
en o
xyg
ène
Fig. 5 : Consommation d’oxygène au cours de la germination.
4°/ Aspects biochimiques de la germination.
4‐1°/ La nature des réserves.
La nature biochimique des réserves des graines est très diversifiée.
Ces réserves ont une grande importance pour l’ embryon en cours de germination ce qui lui permet d’atteindre l’autotrophie.
Ces réserves sont également importantes pour la nutrition de
l’homme et des animaux.
Ex: Céréales Tournesol, Colza, Soja, Sorgho, Féverole, Pois, Haricot. Parmi les graines celles contenant une forte proportion de protéines dites semences protéagineuses.

a/ les protéines.On distingue 4 groupes de protéines : Albumine,
Globulines, Glutélines, Prolamines.Ces protéines sont appelées grains d’aleurones répartis généralement dans tout l’organe de réserves, ou concentrés àla périphérie de la graine chez les céréales par exemple (couche de cellules à aleurone).
b/ les glucides. *L’amidon : constitue la forme principale des réserves
glucidiques, notamment chez les Graminées dont il forme presque tout l’albumen.
*Les hémicelluloses : constituent les albumens cornés ou indurés type datte (polymères de pentoses et hexoses).
*Les sucres solubles : sont en petites quantités dans la graine au repos (saccharose chez l’Amande ou le Ricin).
c/ les lipides.
les lipides constituent la forme de réserve la plus répandue, dans 9/10 des plantes.
La plus grande partie de ces réserves est constitué d’ester, de glycérol et d’acides oléique et palmitique, présents en gouttelettes de différentes tailles appelées oléosomes.
4‐2°/ Les enzymes d’hydrolyse des réserves. Chaque catégorie de graines est dotée d’enzymes en relation
avec son contenu en réserve.
a/ Réserves amylacées :
amylase, maltase, phosphorylase
amidon + P1 glucose 1 phosphate
b/ Réserves lipidiques :
Les triglycérides sont d’abord hydrolysés par des lipases qui donnent du glycérol et des acides gras. Les acides gras sont ensuite oxydés en acétyl CoA puis transformés en glucides par le cycle glycoxylique ou intégrés dans le cycle de KREBS.
c/ Réserves protéiques :
Protéases peptides, ac. aminés utilisés dans la synthèse protéique ou intégrés dans le cycle de KREBS après transamination.
4‐3°/ Les enzymes du cycle respiratoire. Elles vont fournir de l’ATP à partir de substrats libérés par les
enzymes d’hydrolyse. L’intensité respiratoire s’accroît très fortement au cours des premiers stades de la germination et s’accompagne parfois d’un dégagement de chaleur.

4‐4°/ Mode de formation des enzymes :Soit ces enzymes sont déjà présentes dans la graine et l’imbibition et
la réhydratation des tissus permet leur activité
(cas d’enzymes de la respiration).
Soit encore on peut assister à une activation d’enzymes préexistantes sous une forme inactive
(action de protéases par exemple).
Cependant dans le cas le plus fréquent les enzymes sont synthétisées « de novo » dès le début de la germination. Dans cette synthèse d’enzymes deux mécanismes ont été mis en évidence qui peuvent d’ailleurs intervenir séquentiellement:
* Stimulation de la transcription avec synthèse de nouveaux ARNm.
* Stimulation de la traduction par une transition monosomes polysomes (l’activité de traduction est beaucoup plus importante quand plusieurs ribosomes se déplacent sur une même molécule de RNA formant plusieurs copies du polypeptide dans un même temps).
Dans ce second cas il faut considérer que des ARN préexistants sont dans les graines pour permettre la reprise rapide de la synthèse protéique (on a utilisé le terme d’informosomes pour les caractériser).
5°/ Mobilisation des réserves.
a/ Le catabolisme glucidique: L’amidon est dégradé par trois enzymes selon le schéma suivant : On parle de complexe amylasique.
Amylopectine Amylose MaltoseIso Amylase α/β Amylase
Mutase
Glucose
Enzyme Orge Riz Petit pois
α‐Amylase 34,4 31,8 19
β‐ Amylase 11,4 120 Faible
Tableau 2 :Activités de deux enzymes d’hydrolyse de l’amidon chez deux céréales et une légumineuse.

b/ Le catabolisme lipidique : Pendant la germination des graines oléagineuses (ricin, tournesol), les réserves lipidiques de l'albumen ou des cotylédons des semences disparaissent. Ces lipides sont convertis en glucides, principalement du saccharose ; ainsi un flux de carbone dérivédes lipides pourra alimenter la plantule en croissance, en lui fournissant les composés carbonés indispensables à son métabolisme hétérotrophe.
Cette conversion s'effectue par l'intermédiaire d'un cycle métabolique particulier, le cycle glyoxylique, qui se déroule entièrement à l'intérieur d'un petit organite cellulaire spécialisé dans ce travail : le glyoxysome. Cycle glyoxylique et glyoxysome ont été découverts par le biologiste H. Beevers de l'université de Californie, vers 1965, dans l'albumen des graines de ricin.
La dégradation des triglycérides conduit à la formation de glycérol et d’acides gras. Le glycérol donnera ensuite du 3phosphodihydroxyacétone (DHAP) tandis que les acides gras donneront de l’acétylCoA et du succinate par le cycle de Linen. Le DHAP servira lors de la glycolyse et l’acétylCoA servira aux mitochondries.
Les courbes sont presque superposables.
g/10
0gra
ines
Temps en J.Fig 6. Dosage des quantités des glucides et des lipides au cours de la germination.
c/ Le catabolisme protéique : ce processus est alimenté par les protéines de réserves qui sont de différents types : albumine, globuline, glutéline…
La dégradation a lieu dans les tissus de réserves et favorise la libération d’acides aminés par action d’enzymes protéiques :
‐ Thiolendopeptidase pour les ponts S‐S
‐Métalloendopeptidase pour les cofacteurs
‐ Carboxyexopeptidase pour COOH
‐ Aminoexopeptidase pour NH2
Leur activation se fait lors de l’imbibition :
‐ Libération d’acides aminés simples ou deux ou trois peptides.
‐ 80% des transports d’acides aminés du site de dégradation vers l’embryon se font avec la glutamine et l’asparagine.
Les catabolismes conduisent à la production d’énergie et de pouvoir réducteur.
Les intermédiaires produits lors de la glycolyse et Krebs servent de substrats pour de nombreuses voies de biosynthèse :
‐ Synthèse d’acides aminés, de lipides, acides nucléiques, de constituants de la paroi…
Temps en heures après l’imbibition
% p
ar r
appo
rt u
n em
bry
on s
ec
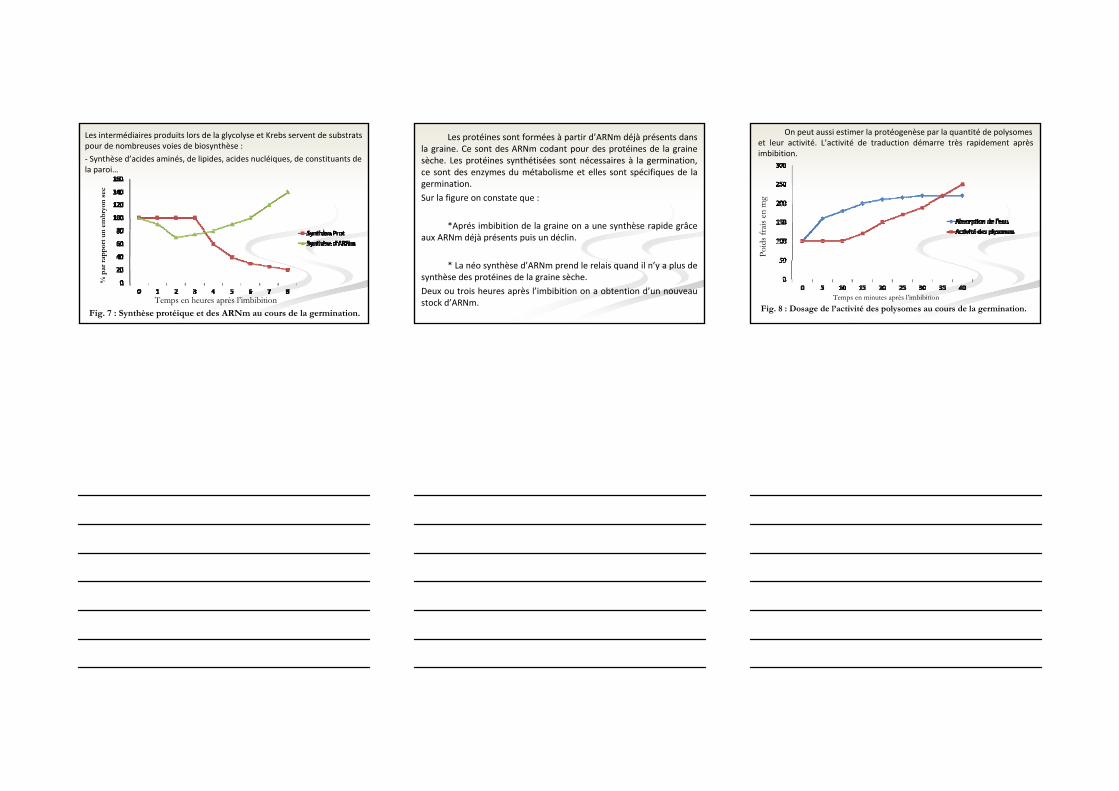
Fig. 7 : Synthèse protéique et des ARNm au cours de la germination.
Les protéines sont formées à partir d’ARNm déjà présents dans la graine. Ce sont des ARNm codant pour des protéines de la graine sèche. Les protéines synthétisées sont nécessaires à la germination, ce sont des enzymes du métabolisme et elles sont spécifiques de la germination.
Sur la figure on constate que :
*Aprés imbibition de la graine on a une synthèse rapide grâce aux ARNm déjà présents puis un déclin.
* La néo synthèse d’ARNm prend le relais quand il n’y a plus de synthèse des protéines de la graine sèche.
Deux ou trois heures après l’imbibition on a obtention d’un nouveau stock d’ARNm.
On peut aussi estimer la protéogenèse par la quantité de polysomes et leur activité. L’activité de traduction démarre très rapidement après imbibition.
Temps en minutes après l’imbibition
Poid
s fra
is en
mg
Fig. 8 : Dosage de l’activité des polysomes au cours de la germination.

Fig. 9: dosage de protéines dans la graine entière et l’embryon isolé (A), dosage de l’activité des enzymes de types protéinase(B) , teneur en acides aminés(C).
Jours après l’imbibition
Pro
téin
es(m
g/gr
aine
)A
cid
es a
min
és(µ
mol
es/
grai
ne)
Pro
téin
ases
(uni
té/
grai
ne)
A
C
B
6°/ Régulation des enzymes de dégradation des réserves.
Il existe deux types de régulation :
‐ Activation.
‐ Biosynthèse.
Il existe une interaction hormonale entre l’embryon et les tissus de réserves. On a pu noter une action des gibbérellines sur la germination du grain d’orge.
Cette corrélation est très répandue, en effet chez le Pois l’ablation de l’embryon empêche la synthèse de protéases au niveau des cotylédons.
Il y a augmentation du taux d’acide gibbérellique avant l’initiation de la synthèse d’α‐amylase.
Fig. 10: Taux d’AG3 et de la synthèse d’α‐amylase au cours de la germination.
Temps en jours
Activité
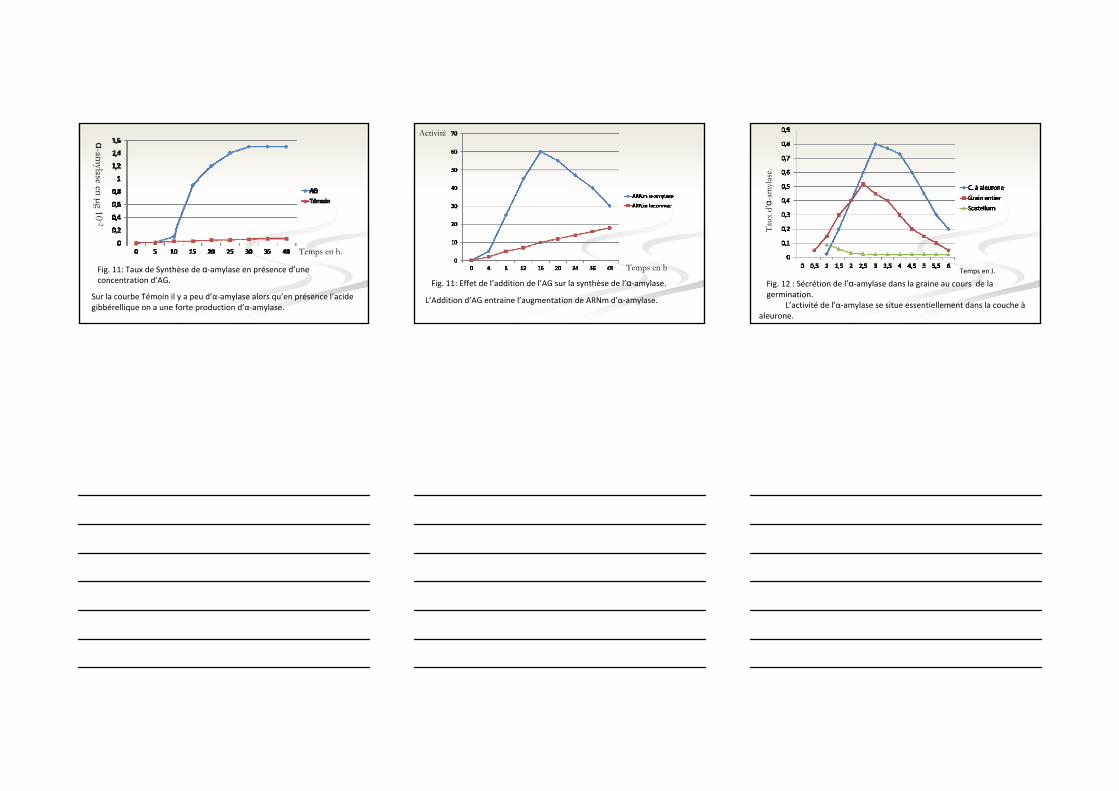
Sur la courbe Témoin il y a peu d’α‐amylase alors qu’en présence l’acide gibbérellique on a une forte production d’α‐amylase.
α-amylase en µg 10
-2
Fig. 11: Taux de Synthèse de α‐amylase en présence d’une concentration d’AG.
Temps en h.
L’Addition d’AG entraine l’augmentation de ARNm d’α‐amylase.
Fig. 11: Effet de l’addition de l’AG sur la synthèse de l’α‐amylase.
Temps en h
Activité
L’activité de l’α‐amylase se situe essentiellement dans la couche àaleurone.
Fig. 12 : Sécrétion de l’α‐amylase dans la graine au cours de la germination.
Temps en J.
Taux
d’α
-am
ylas
e.

Mode d'action de l’AG.Les hypothèses concernant le mode d'action des
gibbérellines sur la croissance découlent d'expériences réalisées sur les grains d'orge. Ces caryopses sont constitués par un embryon àun cotylédon (le scutellum) et par l'albumen ; l'ensemble est entouré de trois assises de cellules riches en grains d'aleurone et des enveloppes (téguments et péricarpe soudés). Durant la germination, l'amidon de l'albumen est hydrolysé en sucres solubles réducteurs sous l'action de l'α‐amylase, qui est une enzyme localisée dans la « couche à aleurone » (travaux de Haberland, 1890). Cependant, si un grain est coupé en deux transversalement, seule la moitié contenant l'embryon produit des sucres réducteurs à partir de l'amidon ; l'embryon est nécessaire àla production d'α‐amylase par la « couche à aleurone ».Les gibbérellines seraient le principe inducteur, car en solution elles induisent l'hydrolyse de l'amidon (donc la synthèse d'α‐amylase) dans la partie du grain dépourvue d'embryon (Yomo et Paleg, 1960)
Les gibbérellines, synthétisées, ou libérées à partir de composés inactifs après l'imbibition de l'embryon, induisent une néosynthèse d'α‐amylase et non pas une activation de molécules enzymatiques préexistantes. Cette néosynthèse s'étend àl'ensemble des enzymes hydrolytiques : protéases, nucléases.Pour expliquer l'influence des gibbérellines sur la croissance, plusieurs hypothèses ont été proposées :– stimulation de la synthèse des protéases ce qui entraîne une augmentation de la teneur en acides aminés (en particulier le tryptophane, substance mère de l'AIA) ;– stimulation de la synthèse d'enzymes hydrolytiques telles que les cellulases dont l'action pourrait provoquer une augmentation de la plasticité de la membrane ; – stimulation de la synthèse d'enzymes hydrolytiques, ce qui conduit à une hydrolyse importante des réserves, donc à une augmentation de la succion des cellules et par conséquent à un appel d'eau.
Germination d’orge
Eau
Imbibition
cotylédon
Apex
Radicelle
Tégument
Endosperme
Couche d’aleuroneReserve de Protéine
Hyd
roly
se
Acides aminées
α amylase
exocytoseHydrolys
e
Sucre
Embryon
Cro
issa
nce

L’ imbibition de la graine déclenchera la libération de l’acide gibbérellique par l’embryon ce qui va activer l’expression de l’α‐amylase via la synthèse d’une ARNm dans la couche à aleurone. L’α‐amylase est ensuite rejeté par exocytose dans l’albumen.
L’acide gibbérellique induit aussi l’expression d’une protéase qui active la β‐amylase.
Ce qui a pour effet la dégradation de l’amidon en sucre assimilable par l’embryon. Celui ci fournira l’énergie nécessaire pour la croissance . Ainsi la radicule émergera en premier suivi de l’apex caulinaire.
La graine a germée. La multiplication, l’élongation et la différentiation cellulaire aboutiront à la croissance d’une plantule.
Phase 1 : Elongation cellulaire, croissance de la radicule.
‐ On a une diminution de la résistance de la paroi grâce aux enzymes synthétisées qui l’hydrolysent et la fragilisent.
‐ Activation des pompes à protons, le milieu extérieur est plus acide, les liaisons hydrogènes sont coupées et les fibres de cellulose sont fragilisées.
‐ Augmentation de la pression osmotique du suc vacuolaire.
Dans ces étapes on trouve deux phytohormones : ABA et les Gibbérellines.
‐ L’ABA s’oppose à la germination en inhibant la synthèse des hydrolases.
‐ Les Gibbérellines stimulent la synthèse des enzymes qui dégradent la paroi.
Phase 2 : Division cellulaire :
‐ L’ABA favorise l’élongation.
‐ Les Gibbérellines favorisent la synthèse de mannoses.
‐ Les Cytokinines favorise la division cellulaire (mitose).
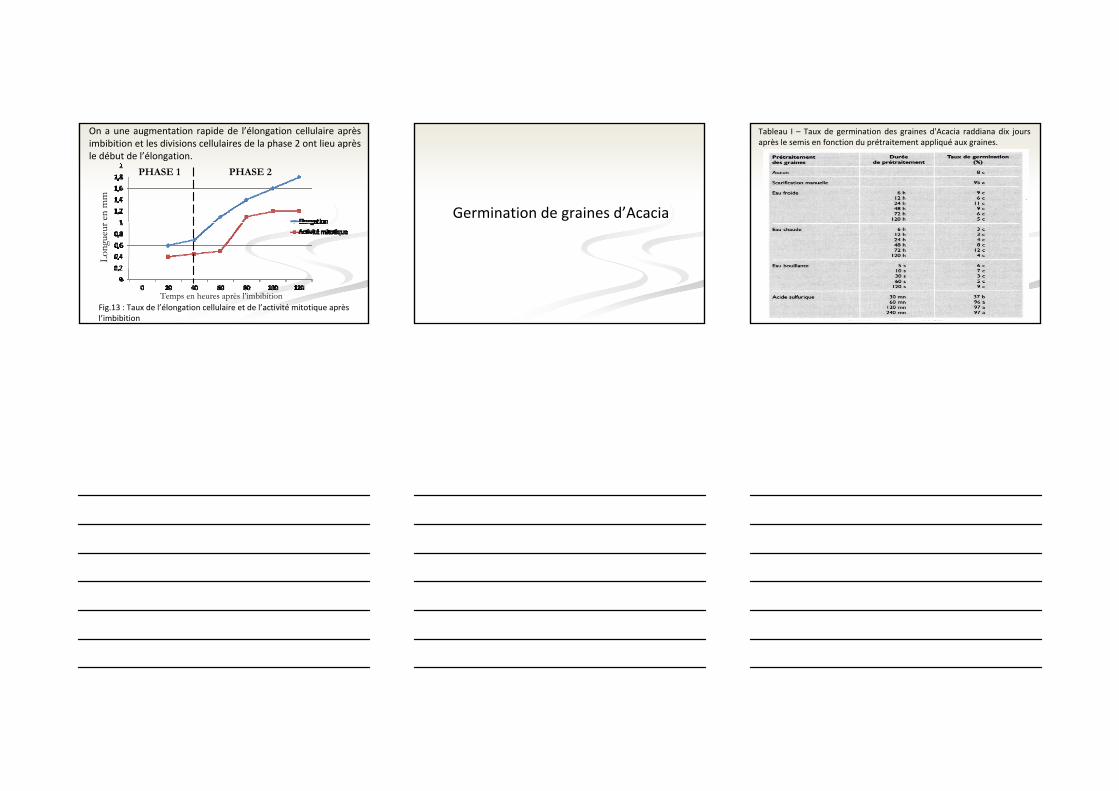
On a une augmentation rapide de l’élongation cellulaire après imbibition et les divisions cellulaires de la phase 2 ont lieu après le début de l’élongation.
Temps en heures après l’imbibition
Long
ueur
en
mm
PHASE 1 PHASE 2
Fig.13 : Taux de l’élongation cellulaire et de l’activité mitotique après l’imbibition
Germination de graines d’Acacia
Tableau I – Taux de germination des graines d'Acacia raddiana dix jours après le semis en fonction du prétraitement appliqué aux graines.
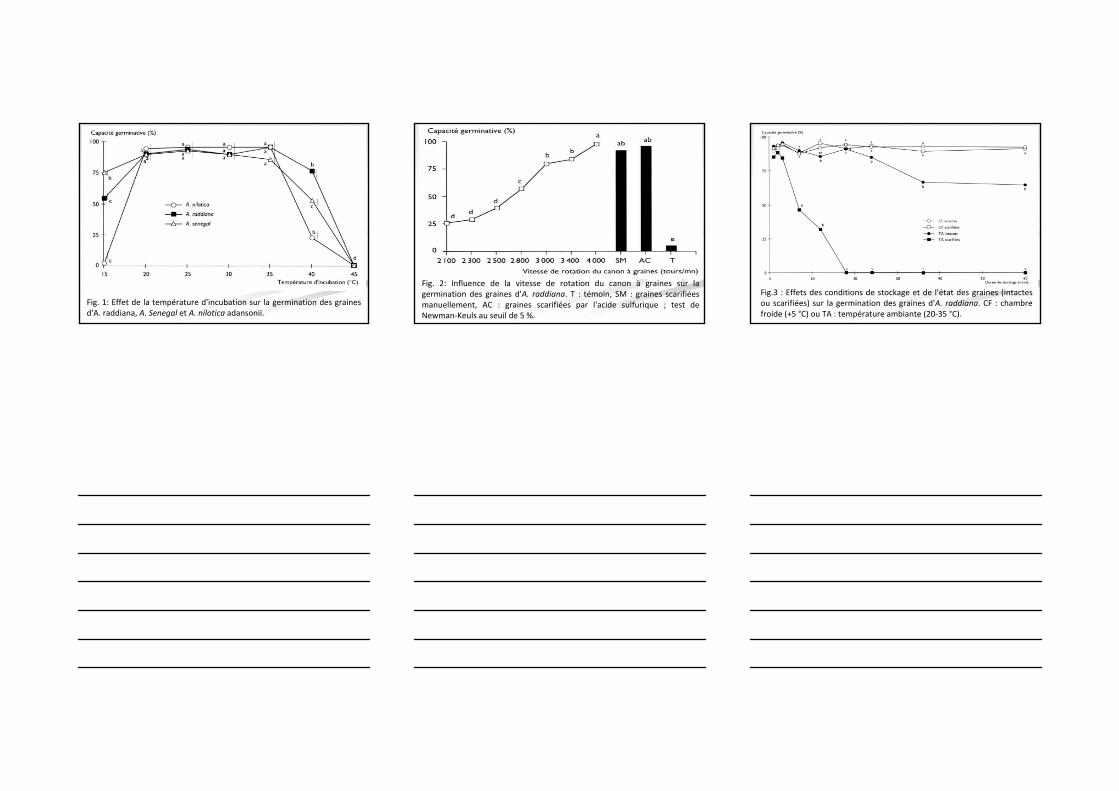
Fig. 1: Effet de la température d'incubation sur la germination des graines d'A. raddiana, A. Senegal et A. nilotica adansonii.
Fig. 2: Influence de la vitesse de rotation du canon à graines sur la germination des graines d'A. raddiana. T : témoin, SM : graines scarifiées manuellement, AC : graines scarifiées par l'acide sulfurique ; test de Newman‐Keuls au seuil de 5 %.
Fig.3 : Effets des conditions de stockage et de l'état des graines (intactes ou scarifiées) sur la germination des graines d'A. raddiana. CF : chambre froide (+5 °C) ou TA : température ambiante (20‐35 °C).
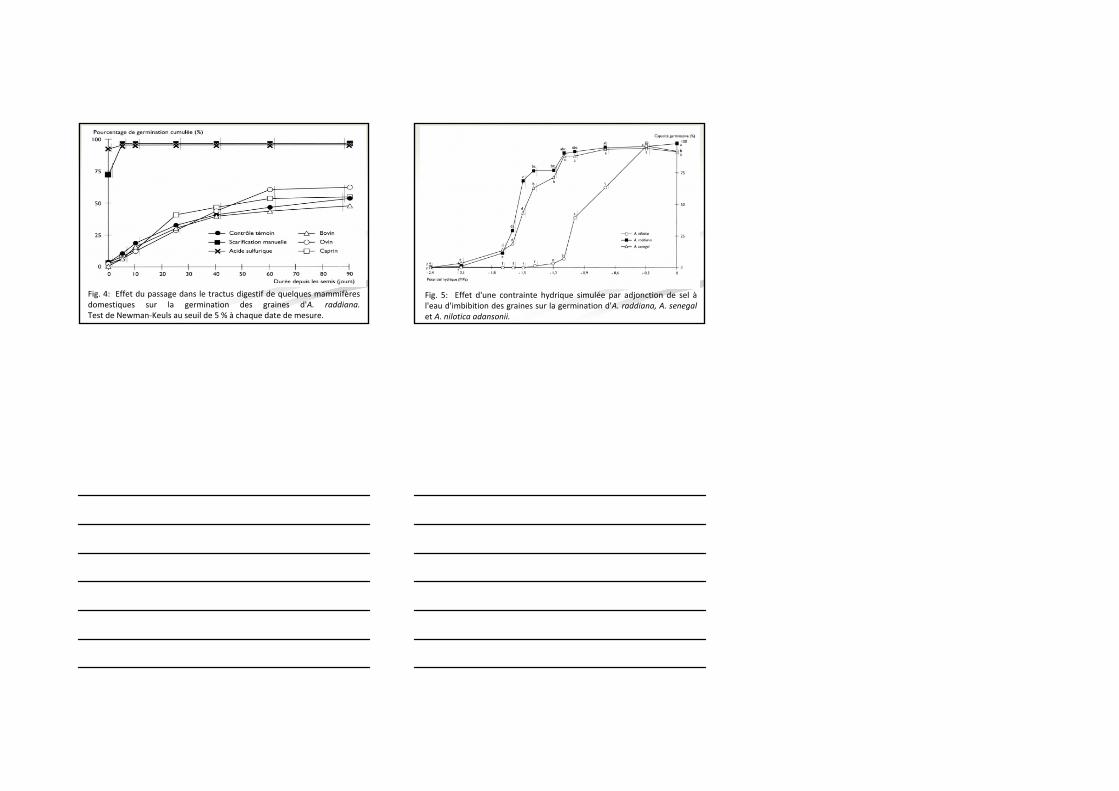
Fig. 4: Effet du passage dans le tractus digestif de quelques mammifères domestiques sur la germination des graines d'A. raddiana.Test de Newman‐Keuls au seuil de 5 % à chaque date de mesure.
Fig. 5: Effet d'une contrainte hydrique simulée par adjonction de sel àl'eau d'imbibition des graines sur la germination d'A. raddiana, A. senegalet A. nilotica adansonii.