3 - les mÉcanismes Évolutifs - manuel de … · possède un ph de 5,2, les acides aminés dont le...
TRANSCRIPT

3 - LES MÉCANISMES ÉVOLUTIFS
Ce chapitre aborde les mécanismes qui sont à l'origine de l'évolution : les
phénomènes génétiques, la sélection naturelle, et leur conséquence immédiate, la
spéciation.
3.1 - La variabilité génétique
La variabilité génétique est l’une des principales sources de l'évolution ; les
méthodes d'estimation et les mécanismes qui président à son maintien et à son
développement sont exposés dans cette section.
3.1.1 - Les méthodes d'appréciation de la variabilité
Trois méthodes principales sont utilisées :
- séquençage des acides nucléiques (voir la section 2.1.2),
- séquençage des protéines (voir la section 2.1.2),
- électrophorèse.
L'électrophorèse utilise la polarité d'un certain nombre de molécules telles que
les acides nucléiques ou les protéines. Lorsque l'on veut identifier les molécules d'un
mélange, on les sépare en les plaçant dans un champ électrique. Selon leur polarité,
leur charge électrique globale et leur conformation, elles vont migrer vers l’un des
pôles à une vitesse qui leur est propre. On utilise également leurs propriétés
amphotères. Les acides aminés, par exemple, peuvent être chargés positivement ou
négativement suivant les valeurs du pH. Si le pH est bas, ils sont chargés positivement
(+) ; pour une valeur précise du pH (pH isoélectrique ou pHi), ils sont chargés
positivement (+) et négativement (-) ; enfin, lorsque le pH est élevé, ils sont chargés
négativement (-). Par exemple, si la solution tampon utilisée pour l'électrophorèse
possède un pH de 5,2, les acides aminés dont le pHi est supérieur à 5,2 deviennent
des anions (-), ceux dont le pHi est inférieur à 5,2 deviennent des cations (+). Les
acides terminaux et les radicaux (R), comprenant parfois une ou plusieurs fonctions
carboxyles et/ou amines libres, déterminent la polarité totale du polypeptide, les
fonctions carboxyles (COOH) et amines (NH2) caractéristiques des acides aminés
d’une chaîne peptidique sont engagées dans les liaisons peptidiques et elles ne
peuvent intervenir dans la polarité de la molécule.
149

Dans la solution tampon, les protéines ont des charges résultantes différentes.
Placées dans un champ électrique, elles migrent à des vitesses spécifiques soit vers le
pôle (+), soit vers le pôle (-) . Des réactifs colorés révèlent leur emplacement.
Très sensible, l'électrophorèse peut séparer ainsi des protéines qui ne diffèrent parfois
que par un acide aminé.
Fig 3.1
Mais elle possède deux limites :
1) À la suite d'une mutation, l'acide normal et l'acide substitué n’ont pas toujours
des propriétés électriques différentes. Selon les biochimistes :
- Glu et Asp ont les mêmes propriétés chimiques.
- Leu, Ileu, Val et Met en ont de presque identiques.
- Leu, Thr, Lys et Arg en ont de différentes.
2) Les mutations synonymes ne sont pas décelées.
Quoi qu'il en soit, cette technique rapide permet d'apprécier le nombre d'allèles
présents dans une population avec une bonne précision si l’échantillonnage est
important ; aussi est-elle largement utilisée par les biologistes pour estimer plusieurs
paramètres :
- le taux d'hétérozygotie, c'est-à-dire le pourcentage des gènes hétérozygotes dans
une population ; ce taux est peu sensible à l'effectif de l'échantillon ;
Fig. 3.2
150

- le taux de polymorphisme, qui est le rapport du nombre de gènes polymorphes au
nombre total des gènes examinés ;
- le nombre moyen d'allèles, qui se calcule par le rapport du nombre total d'allèles
au nombre total de gènes étudiés ; son estimation dépend de l'effectif de
l'échantillon.
3.1.2 - Les origines de la variabilité.
L'évolution sous-entend l'apparition de nouveautés héréditaires. Par conséquent
il ne peut y avoir de changements sans mutations, c'est-à-dire sans modifications
brusques et aléatoires du matériel génétique ; c'est l’une des raisons pour laquelle le
hasard tient une place à part entière dans les phénomènes évolutifs. La variabilité
génétique concerne les mutations responsables de modifications soit géniques, soit
chromosomiques.
Différents types de mutations sont responsables d'une variabilité génétique
suffisante pour engendrer des nouveautés évolutives. Les mutations affectent aussi
bien l'ADN codant que l'ADN non-codant. Les premières peuvent avoir des
répercussions immédiates sur le phénotype, les secondes semblent souvent sans
conséquence, mais elles sont trop peu connues pour généraliser.
Les mutations ponctuelles
Elles concernent aussi bien les gènes de structure gouvernant, par exemple, la
synthèse d'enzymes, de récepteurs membranaires, d'anticorps... que les gènes de
régulation - parmi lesquels les gènes du développement - dont le fonctionnement,
beaucoup plus discret, n'en est pas moins fondamental. Elles n'affectent que des
nucléotides en nombre limité ou de courtes séquences d'ADN.
- Les mutations des gènes de structure ou gènes réalisateurs
Ce sont les mutations géniques les plus classiques. Elles consistent en
l'insertion, la délétion ou la substitution d'un ou de quelques nucléotides.
Deux exemples illustreront ce type de mutations. Le premier concerne la
synthèse d'une enzyme, la tyrosinase, responsable, par exemple chez le Chat, de la
pigmentation du pelage ; le deuxième affecte la synthèse du polypeptide β.
Le gène de la tyrosinase a subi, chez le Chat siamois, une mutation qui la rend
thermosensible. Elle ne fonctionne que pour des températures légèrement inférieures
à celles du corps. C'est pourquoi seules les régions de la tête, de la queue et de
l'extrémité des pattes sont pigmentées ; le corps, plus chaud, inhibe cette enzyme et
reste dépigmenté.
151

Le gène du polypeptide β de l’hémoglobine est sujet à des mutations
responsables de l'anémie falciforme et de maladies regroupées sous le nom de
thalassémies, maladies plus ou moins graves de l'hémoglobine, fréquentes chez les
populations du bassin méditerranéen. La figure 3.3 présente quelques exemples de
ces mutations, ainsi que la carte de l'ARN messager sur laquelle sont portées les
mutations les plus fréquentes.
Fig. 3.3
152

La formation d'allèles très proches les uns des autres augmente la variabilité
génétique. Ces mutations alléliques ne semblent pas à l'origine de grandes
nouveautés évolutives. Par sommation de plusieurs mutations, elles pourront sans
doute engendrer progressivement de nouvelles variétés et quelquefois de nouvelles
espèces. Provoquant l'apparition de nouveaux allèles, ces mutations ponctuelles sont
responsables d'une augmentation du taux d'hétérozygotie, dont l'importance sera
précisée avec l’étude de la diploïdie. Si le nouvel allèle muté s'exprime, un nouveau
phénotype apparaît ; et, s’il est toléré par la sélection, le polymorphisme de l'espèce,
c'est-à-dire le nombre d'individus reconnaissables par un caractère de leur phénotype,
augmente.
- Les mutations des gènes de régulation
Des gènes dirigent la croissance de l'individu grâce à la synthèse de protéines
régulatrices. Même si leur rôle dans les phénomènes évolutifs est encore mal connu,
on commence à comprendre qu'il est prépondérant pour certains d'entre eux.
Tel est le cas des gènes homéotiques, qui contrôlent le développement
embryonnaire des Invertébrés comme celui des Vertébrés. Leur importance est telle
que le prix Nobel de médecine 1995 a été attribué à trois chercheurs américains,
Edward LEWIS, Christiane NUESSLEIN-VOLHARD et Eric WIESCHAUS, pour leurs travaux sur
l'identification des gènes homéotiques qui déterminent l'organisation antéro-
postérieure et dorso-ventrale des Animaux. Le terme « homéotique » est dû à
William BATESON qui, à la fin du XIXe siècle, a qualifié d'homeosis le remplacement d'un
organe par un autre au cours de l'ontogenèse. Quelle que soit l'espèce, ces gènes
homéotiques assurent la synthèse de protéines de régulation qui vont se fixer sur des
gènes cibles ; certaines protéines homéotiques se terminent par une même séquence
de 60 acides aminés : l'homéodomaine, responsable de la fixation de la protéine sur
l'ADN. Le domaine d'une protéine est une région interne de la molécule qui assure une
fonction spécifique, site de fixation d'un substrat, par exemple, ou bien qui constitue
une unité structurelle facilement reconnaissable. On parle dans le premier cas d'un
domaine fonctionnel et dans le deuxième cas d'un domaine structurel.
L'homéodomaine fonctionnel de la protéine homéotique est codé par une homéobox,
portion génique terminale de 180 paires de nucléotides, dont les séquences de
plusieurs espèces sont comparées ci-après dans la section : « Le brassage génique,
la duplication génique ».
Deux observations s'imposent :
- Les homéobox des espèces étudiées (Drosophile, Xénope, Souris, Homme...)
sont très semblables les unes aux autres.
153

- L'ordre des gènes homéotiques de la Drosophile situés sur son chromosome 3
correspond à celui de leur activation ; la même concordance a été retrouvée chez les
Vertébrés (Souris et Homme).
Les gènes à homéobox des Invertébrés et des Vertébrés sont également
homologues, comme l'ont confirmé diverses expériences :
- Rebecca QUIRING (1988) découvre chez la Drosophile le gène non homéotique
eyeless, homologue du gène PAX 6 des Mammifères ; ces deux gènes gouvernent la
formation de l'œil. Le gène PAX 6, découvert chez la Souris et l'Homme (1986), est
pourvu de deux homéobox, la deuxième s'appelle la paired box, d'où son abréviation
PAX. Eyeless et PAX 6 ont des homéodomaines identiques à 94 %, et les domaines
paired identiques à 90 %. Un même gène semble capable de gouverner le
développement de l'œil chez les Insectes et les Mammifères.
- Des gènes eyeless ont été trouvés chez des animaux très éloignés
phylogénétiquement : une Planaire (Dugesia tigrina), un Ver Némerte (Lineus
sanguineus) et une Ascidie (Phallusia mammillata).
- Bill (William) McGINNIS (1990) greffe le gène homéotique HOX D3 (HOX pour
Homéobox) de Souris chez la Drosophile, dont les antennes sont remplacées par des
pattes. Les cellules de cette Mouche peuvent lire (transcrire) et comprendre (traduire)
le gène d'une espèce très éloignée.
- L'équipe de Walter GEHRING (1994) réussit à activer le gène eyeless en différents
endroits de l'Asticot de Drosophile : des yeux normaux se développent aux endroits
correspondants chez l'adulte. W. GEHRING réalise la même expérience en greffant et en
activant chez la Drosophile le gène PAX 6 des Mammifères : des yeux normaux de
Mouche se développent.
W. GERHING montre ainsi que les gènes eyeless et PAX 6 sont homologues, bien
que la séparation des Insectes et des Vertébrés datent au moins de 500 Ma ; au cours
de l'évolution, les séquences géniques qui gouvernent l'organisation de l'œil ont eu
une excellente conservation. Il prouve également que ces deux gènes sont des gènes-
maîtres, c'est-à-dire des gènes régulateurs directeurs « contrôlant » toute une série de
gènes dont les derniers affectés sont les gènes de structure. Le gène eyeless semble
diriger à peu près 2 500 gènes dont la mise en jeu régulée est nécessaire au
développement de l'œil. Il semble donc possible qu'une seule mutation ponctuelle
modifie le développement d'un organe complexe.
Cependant, il convient de signaler que, dans le développement d’un organe, un gène-
maître intervient conjointement avec un ensemble de gènes-maîtres différents ; c’est
donc leur mise en jeu ordonnée dans le temps qui est fonctionnelle. Ce
fonctionnement « collégial » spécifique explique pourquoi un même gène-maître -
154

PAX-6, par exemple - est impliqué dans l’ontogenèse d’organes divers et non pas
uniquement, ici, dans celle de l’œil.
Sur l'importance évolutive des mutations ponctuelles, responsables d'une
évolution spécifique ou micro-évolution, les avis sont partagés. Des biologistes dont
James VALENTINE doutent de l’importance évolutive des mutations ponctuelles, seules
responsables d’une évolution graduelle ou micro-évolutive ; c’est pourquoi il écrit : « La
micro-évolution prolonge et diversifie l'évolution à l'intérieur des phyla eux-mêmes ... (Elle)
est capable de créer des espèces nouvelles, mais ne crée pas de grandes percées évolutives ...
Par le seul mécanisme de la micro-évolution, on estime que la différenciation d'une seule
espèce nouvelle nécessiterait plusieurs millions d'années. Dans ces conditions, élaborer un
nouveau phylum prendrait plus de temps qu'il n'y en a de disponible au cours de toute l'histoire
de la Terre » (J. VALENTINE, La Recherche, n° 112, juin 1980 : « L'origine des grands
groupes d'animaux », p. 671). Cela est vrai sans doute pour les gènes de structure,
mais ne l'est pas obligatoirement pour les gènes de régulation.
D’autres chercheurs soutiennent que les mutations ponctuelles sont à l'origine de
l'évolution, qui est par nature graduelle. Mais on peut supposer qu'une mutation
ponctuelle affectant un gène-maître provoque immédiatement de profonds
bouleversements dans l'ontogenèse d'un animal.
Les mutations géantes semblent également capables de créer des nouveautés
évolutives dans un intervalle de temps compatible avec l'échelle des temps
géologiques.
Les mutations géantes
Contrairement aux mutations ponctuelles, elles affectent de longues portions
d'ADN. Ces mutations concernent aussi bien des transpositions, des mutations
chromosomiques que des duplications géniques. Ces recombinaisons, parfois
importantes, risquent d'entraîner de brusques modifications tant du génotype que du
phénotype. L'hémoglobine en offre un exemple. Elle est composée de quatre
tétramères : deux chaînes polypeptidiques α et deux chaînes β. Le gène d'un des
polypeptides est parfois limité chez l'Homme par deux séquences répétitives dites
« alu ». Assez peu fréquemment, ces séquences se rapprochent, éliminent le gène du
polypeptide et provoquent une grave déficience.
155
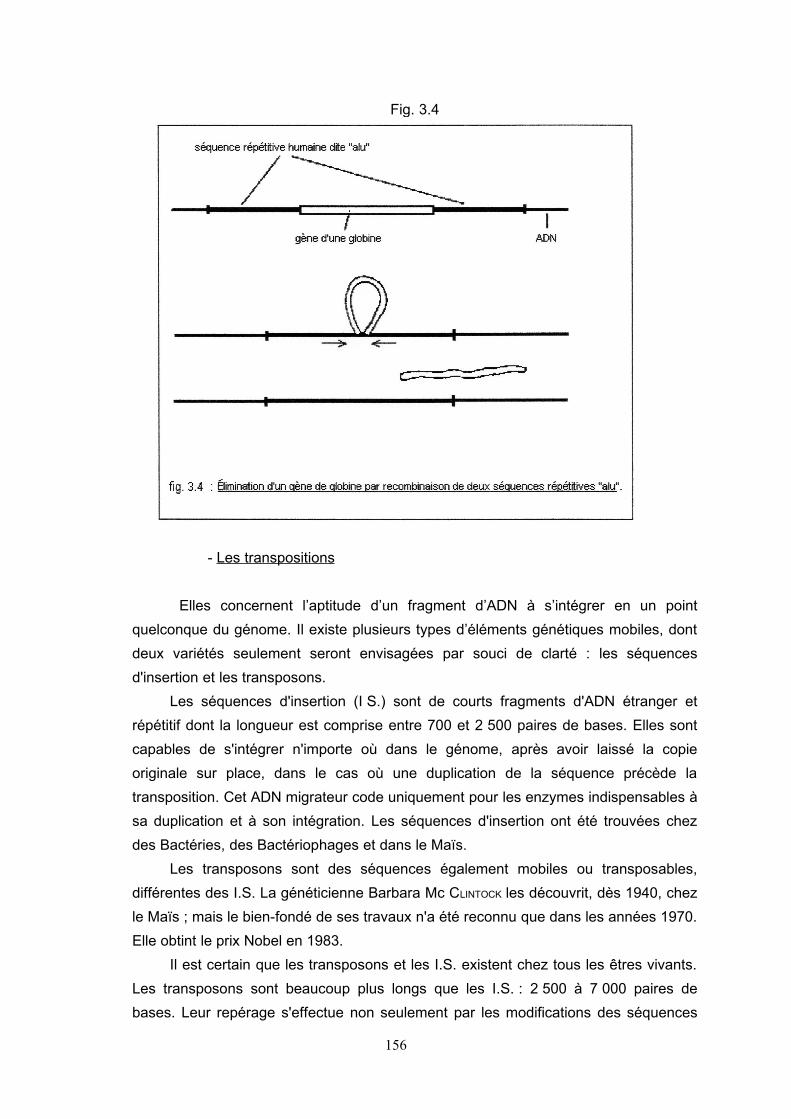
Fig. 3.4
- Les transpositions
Elles concernent l’aptitude d’un fragment d’ADN à s’intégrer en un point
quelconque du génome. Il existe plusieurs types d’éléments génétiques mobiles, dont
deux variétés seulement seront envisagées par souci de clarté : les séquences
d'insertion et les transposons.
Les séquences d'insertion (I S.) sont de courts fragments d'ADN étranger et
répétitif dont la longueur est comprise entre 700 et 2 500 paires de bases. Elles sont
capables de s'intégrer n'importe où dans le génome, après avoir laissé la copie
originale sur place, dans le cas où une duplication de la séquence précède la
transposition. Cet ADN migrateur code uniquement pour les enzymes indispensables à
sa duplication et à son intégration. Les séquences d'insertion ont été trouvées chez
des Bactéries, des Bactériophages et dans le Maïs.
Les transposons sont des séquences également mobiles ou transposables,
différentes des I.S. La généticienne Barbara Mc CLINTOCK les découvrit, dès 1940, chez
le Maïs ; mais le bien-fondé de ses travaux n'a été reconnu que dans les années 1970.
Elle obtint le prix Nobel en 1983.
Il est certain que les transposons et les I.S. existent chez tous les êtres vivants.
Les transposons sont beaucoup plus longs que les I.S. : 2 500 à 7 000 paires de
bases. Leur repérage s'effectue non seulement par les modifications des séquences
156

géniques, parce qu’ils abandonnent parfois sur place des fragments de séquence qui
leur sont spécifiques, mais aussi parce qu'ils confèrent souvent aux Bactéries une
résistance à un antibiotique. À la différence des I S., ils contiennent donc des gènes
ayant d'autres fonctions que la réplication et la transposition. Les transposons comme
les I.S. se déplacent quelquefois en laissant in situ la copie originale ; ils possèdent
alors tous deux un caractère envahissant. Au moment de la transposition, certains
transposons, les rétrotransposons, copient leur ADN en ARN, qui est lui-même copié
en ADN au moyen d'une transcriptase inverse. Cet ADN est alors capable de s'intégrer
dans quelques régions privilégiées du génome. Cette suite d'événements rappelle
celle qui caractérise l'infection d'une cellule par un Rétrovirus ; l'ADN viral intégré
constitue un type de Provirus. Ainsi, soit les rétrotransposons et les Rétrovirus ont une
origine commune, soit ils dérivent les uns des autres. Les séquences correspondant à
des Provirus sont nombreuses, transposables et codantes. On sait maintenant que le
génome des Souris utilisées en laboratoire contient des milliers de Provirus. La
transposition n'est pas un événement rare.
Dans certaines cellules comme les lymphocytes ou dans des organismes
comme les Trypanosomes, il existe des remaniements de l'ADN même des cellules,
dont les effets sont semblables à ceux des transpositions. Dans le premier cas, Ils
expliqueraient la synthèse possible d'innombrables anticorps différents et, dans le
deuxième cas, la grande variabilité de la couverture antigénique de ce parasite (voir la
section 3.2.2 : « Les adaptations immunologiques »). Les lymphocytes synthétisent les
anticorps, dont le nombre très élevé dépend de la recombinaison aléatoire de
séquences géniques différentes, recombinaison qui rappelle la transposition.
Les transposons s'intègrent en quelques points particuliers du matériel génétique,
sans que l’on sache pourquoi. Cette intégration dans les parties codantes de l'ADN
peut avoir de lourdes conséquences : inactivation du gène, fonctionnement partiel du
gène ou formation d'un gène hybride fonctionnel si le transposon, au moment de son
déplacement, entraîne un morceau du gène qui l'hébergeait.
- Les mutations chromosomiques
Elles constituent un deuxième type de recombinaison. Elles affectent non plus
les gènes mais les chromosomes, qui subissent des remaniements à la suite de
cassure, perte ou échange de segments. Elles peuvent être spontanées ou
consécutives à des radiations, à des agents chimiques mutagènes, à l'âge. Elles
modifient l'aspect des chromosomes et parfois aussi leur nombre.
157

- La perte ou le gain de fragments comprenant des séquences répétitives non
codantes ne semble pas avoir de conséquences importantes.
- La perte ou le gain d'un fragment codant de chromosome, voire d'un chromosome
entier, est souvent à l'origine de maladies héréditaires. Les médecins estiment qu'en
moyenne 50 % des œufs fécondés sont anormaux et porteurs de malformations
chromosomiques, mais qu'ils avortent spontanément dans la grande majorité des cas.
Ces remaniements sont des fissions, des inversions ou des translocations (fig. 3.5).
Dans ce dernier cas, un morceau de chromosome vient se souder sur un chromosome
d'une paire différente. Parfois, il s'agit d'un chromosome entier : si la translocation
s’effectue par les centromères, la translocation est dite alors robertsonienne. Ces
accidents rares, dont la fréquence est de 1/1 000, se transmettent difficilement, car
certains gamètes hétérozygotes, porteurs de l'anomalie, ne sont pas viables.
Fig. 3.5
158

Des cytogénéticiens soutiennent que les remaniements chromosomiques sont
suffisants pour donner naissance à deux espèces, puis pour séparer définitivement les
deux lignées issues d’un rameau commun. Les Chimpanzés et les Hommes
possèdent en commun 99 % de leurs protéines. Cela signifie une grande
ressemblance génétique : 99 % de l'ADN codant serait identique. Une telle similitude
est surprenante, car elle ne s'observe que chez des espèces jumelles à peine
différentiables, c'est-à-dire très proches génétiquement et morphologiquement, qui ne
s'hybrident pas. Or l'Homme et le Chimpanzé ne sont classés ni dans la même
espèce, ni dans le même genre. La comparaison des caryotypes montre des
similitudes, mais aussi des différences, dont 9 importantes, qui pourraient être à
l’origine de la séparation des deux lignées. Quelques-unes des différences sont
présentées sur la figure 3.6.
Fig. 3.6
159

Dans le cas de l'Homme et du Chimpanzé, les similitudes encore nombreuses
sont la preuve d'une proche parenté. Mais actuellement, selon presque tous les
biologistes, des remaniements du caryotype ne sont pas uniquement à l'origine de la
séparation Homme-Chimpanzé. Pour la comprendre, il reste à considérer le 1 %
d’ADN codant différent ; si ce 1 % concerne des gènes de régulation, le rameau
humain a pu se détacher du rameau conduisant aux Chimpanzés.
Cependant, les anomalies chromosomiques interviennent parfois dans la
création de nouvelles espèces, car elles favorisent l'isolement sexuel qui s'avère
absolument nécessaire à toute spéciation. Selon Jean GÉNERMONT, « certains
remaniements chromosomiques peuvent jouer un rôle important dans la spéciation. Ce sont
ceux qui causent une stérilité partielle lorsqu’ils sont présents à l’état hétérozygote. Il est
cependant impossible d’affirmer qu’il n’y a pas de spéciation sans remaniement
chromosomique » (J. GÉNERMONT, Les critères caryologiques de l’espèce, dans Les
problèmes de l’espèce dans le règne animal, tome III, Société Zoologique de France,
Paris, 1980).
À la suite de la méiose, des gènes surnuméraires apparaissent. Ce phénomène
général, qui correspond à la duplication génique, entraîne quelquefois des
modifications importantes dans le génotype et dans son expression phénotypique.
- La duplication génique
D'après Susumu OHNO, la duplication génique, c'est-à-dire l'apparition sur des
chromosomes de séquences géniques surnuméraires identiques ou très proches, est
une source de nouveautés évolutives ; c’est le cas, par exemple, des gènes en
tandem. « Quand un gène existe en plusieurs exemplaires dans une cellule ou un gamète, il se
trouve affranchi des contraintes imposées par la sélection naturelle. Les mutations peuvent
donc s’y accumuler plus ou moins librement et donner naissance à des structures nouvelles
dont certaines peuvent, le cas échéant, remplir des fonctions nouvelles » (article
« Bricolage » de François JACOB, Dictionnaire du darwinisme et de l’évolution, Paris,
PUF, 1996). Élément moteur de l'innovation génétique, la duplication génique se
rapporte à deux mécanismes : la polyploïdie et le crossing-over inégal.
La polyploïdie
160

La duplication de l'ensemble des chromosomes est abordée dans ce paragraphe, car
certains biologistes l'incluent dans la duplication génique. Elle est fréquente chez les
Plantes, mais restreinte chez les Animaux. Elle est connue chez les Vertébrés de la
famille des Cyprinidés (Carpes à 4n) et des Salmonidés, dont la Truite qui possède
parfois des allèles multiples de 4, codant pour différentes enzymes dont la lactate
déshydrogénase ; ces nombres multiples seraient l’indice d’une succession de deux
duplications, elles-mêmes issues peut-être d’une tétraploïdie, comme le pensent des
spécialistes des Salmonidés. La polyploïdie est banale également chez les
Amphibiens des genres Rana (R. delalandii ou R. occipitalis), Bufo, Hyla
(H. versicolor) et Xenopus, et chez quelques Invertébrés dont les Annélides
oligochètes du genre Enchytraeus et des Gastéropodes du genre Bulinus.
Fig 3.7
La polyploïdie possède un intérêt certain. Des hybrides, issus du croisement de deux
espèces, sont souvent stériles, car, au moment de la méiose, les chromosomes
homologues ne s’apparient pas. La polyploïdie, en doublant le nombre des
chromosomes, lève cet écueil : les paires de chromosomes homologues sont
reconstituées, c'est l'allopolyploïdie, et les hybrides deviennent fertiles.
Chez les autres Vertébrés (Oiseaux et Mammifères), la sélection naturelle
semble avoir éliminé la polyploïdie pour ne laisser que les recombinaisons géniques,
bien que ces phénomènes ne soient pas exclusifs.
La polyploïdie apparaît comme un procédé évolutif commun, mais qui a rarement
été conservé, sans doute parce qu'elle est plus souvent délétère que bénéfique :
allongement du temps de la division cellulaire, augmentation du nombre des non-
appariements chromosomiques durant la méiose, ou bien encore modification de
l'équilibre génétique entre les chromosomes. (voir la section 3.3.2 : "La spéciation
sympatrique instantanée par polyploïdie").
161

Le crossing-over inégal
C'est un échange partiel d’une séquence d’ADN soit entre les deux
chromatides sœurs d'un chromosome au cours d'une mitose (crossing-over mitotique),
soit entre deux chromatides homologues au cours de la méiose (crossing-over
méiotique). Dans les deux cas, il y a délétion d'un segment d’ADN ou d'un fragment
génique sur l’une des chromatides sœurs et duplication correspondante sur la 2e
chromatide homologue (fig. 3.8) ; un ou plusieurs gènes entiers sont donc parfois
dupliqués.
Fig 3.8
La formation de nombreuses familles multigéniques implique des duplications. Si ces
remaniements apparaissent en dehors de toute pression sélective, ils sont successifs.
Une succession de crossing-over inégaux peut être responsable de la multiplication
d’un gène sur une des chromatides.
162

Fig 3.9
Si ces remaniements apparaissent en dehors de toute pression sélective, ils sont
ensuite soumis à la sélection naturelle, qui élimine rapidement tous les gènes
délétères. Postulant une évolution graduelle, S. OHNO résout le problème des
intermédiaires par l'existence, très fréquente chez les Vertébrés, des isozymes ou
isoenzymes ; ce sont des enzymes catalysant des réactions identiques, mais différant
par leur sensibilité à des facteurs extérieurs - pH, température...-, par leur régulation,
leurs propriétés biochimiques ou leur situation tissulaire. On suppose que les
séquences codantes des isozymes sont issues d'une série de duplications. Soumise à
une forte pression de sélection, l’une des séquences originelles continuerait d'assurer
la même fonction, tandis que, sous l'accumulation des mutations, les autres
séquences subiraient une évolution divergente et donneraient des protéines
apparentées, les isozymes, dont la fonction passerait insensiblement à une autre, sans
que cette première soit brusquement détruite ou complètement perturbée. La
tyrosinase « sauvage » et la tyrosinase thermosensible, déjà citée au sujet des Chats
siamois, sont un exemple d'isozymes ; les lactates déshydrogénases (LDH), toutes
capables de convertir le lactate en pyruvate, en sont un deuxième exemple. La LDH
est tétramérique ; les sous-unités sont codées par trois gènes (A), (B) et (C) ; si l’on
163

considère les deux premiers gènes, leur fonctionnement alternatif ou isolé permet
d’obtenir cinq isozymes : A4, A3B, A2B2, AB3 et B4.
Des chercheurs, dont Dan GRAUR, Wen-Hsiung LI ou Philippe KOURILSKY,
considèrent que la conversion génique est un mécanisme aussi important que le
crossing-over inégal. Avant d’être convertis d’une chromatide sur l’autre, les gènes
concernés sont dupliqués ; à la différence du crossing-over inégal, la conversion
génique ne s’accompagne pas de la perte d’un segment d’ADN.
Une conséquence : le brassage génique
Les recombinaisons issues des duplications qui affectent des séquences d'ADN
homologues ou très semblables entraînent soit un brassage de gènes, soit un
brassage d'exons.
1) Le brassage de gènes complets crée des familles multigéniques. Le système
HLA (Human Leucocyte Antigens), impliqué dans les réactions immunologiques, en
est un exemple, ainsi que les gènes codants pour les hémoglobines et les gènes des
globines humaines étudiées ci-dessous. Chez l'Homme, il existe, en effet, plusieurs
gènes codants pour des sous-unités de l’hémoglobine différentes, situés sur les
chromosomes 11 et 16. Après avoir étudié leur séquence, les biologistes pensent que
ces gènes dérivent d'un gène ancestral commun. Ces gènes homologues constituent
une famille multigénique. Lorsqu'ils restent voisins sur un même chromosome et avec
la même orientation, ils sont appelés gènes en tandem.
Fig 3.10
164

2) Le brassage d'exons, séquences codantes des gènes morcelés, est à l'origine de
nombreuses nouveautés évolutives. Il se rapporte soit à la duplication interne d'un seul
ou de plusieurs exons d'un même gène, soit à l'insertion d'exons étrangers. L'étude de
la composition de quelques protéines impliquées dans la coagulation du sang illustre le
brassage d'exons et elle permet de comprendre comment la nature économe fabrique,
à partir d'un nombre limité de matériaux (les exons), des nouveautés physiologiques
telles que de nouvelles enzymes. Ces remaniements ont été qualifiés par F. JACOB
(1977, 1996) de bricolage évolutif.
Les gènes homéotiques à homéobox forment une autre famille multigénique
avec des gènes en tandem. Les séquences correspondant aux homéobox sont très
peu variables, alors que celles des autres segments d'un même complexe sont très
différentes ; cette faible variabilité traduit l'importance fonctionnelle de l'homéodomaine
: les homéobox conservent d'un gène à l'autre des séquences-clés (fig.3.11). Chez la
Souris, comme chez l'Homme dont elle est, à cet égard, très proche, les homéogènes
sont regroupés en quatre complexes de 5 à 10 gènes : HOX 1, 2, 3 et 5. Chaque
complexe est porté par un chromosome différent :
HOX 1 sur le chromosome 6
HOX 2 sur le chromosome 11
HOX 3 sur le chromosome 15
HOX 5 sur le chromosome 2
La figure 3.11-A met en évidence les homologies entre des homéodomaines qui
appartiennent à deux espèces différentes, Souris et Drosophile.
165
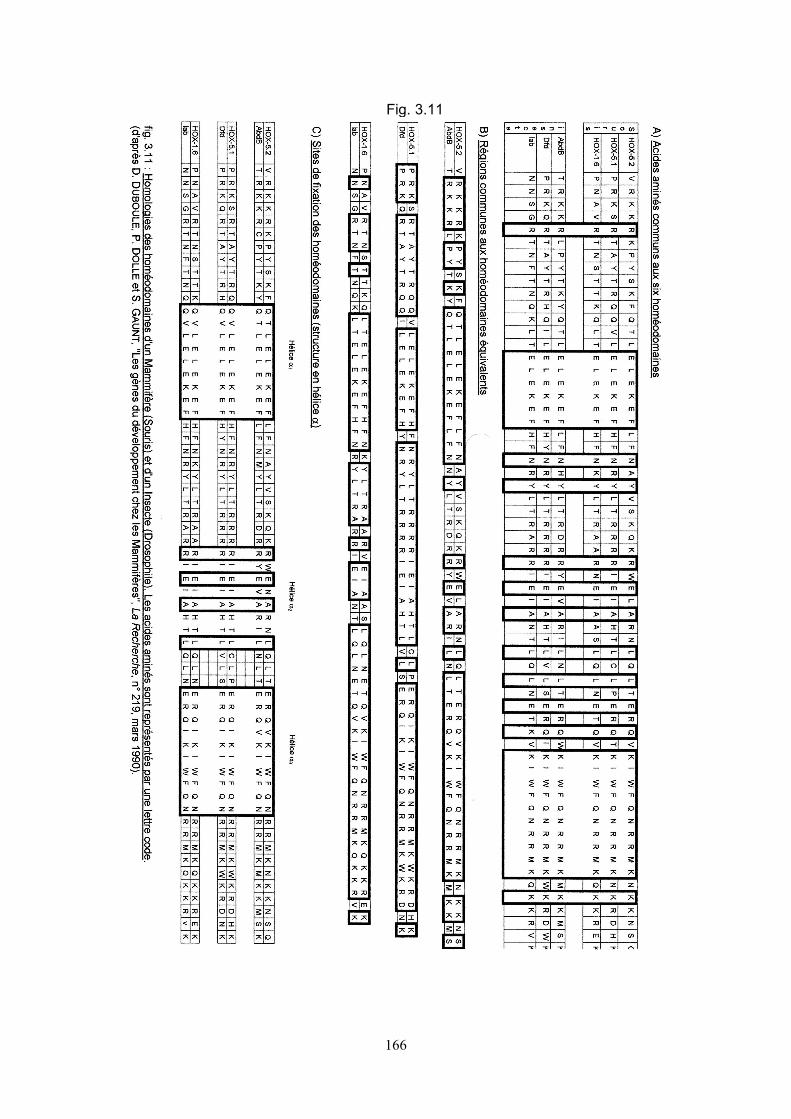
Fig. 3.11
166

Depuis 1984, les généticiens ont constaté que les homéobox des Amphibiens et des
Mammifères sont très semblables à celles de la Drosophile. Selon leurs recherches,
une telle ressemblance ne peut être rationnellement due au seul hasard. La
figure 3.11-B montre que les homéodomaines d'une même espèce, assez
dissemblables entre eux, présentent cependant d'une espèce à l'autre de fortes
similitudes. Cela laisse supposer que ces homéogènes dérivent d'un gène ancestral
qui s'est dupliqué un certain nombre de fois, donnant naissance à des gènes en
tandem qui forment les complexes ; puis ceux-ci ont évolué séparément ; enfin, deux
duplications intégrales de ces complexes, voire une polyploïdie, ont donné d'abord
deux complexes puis quatre, nombre maximum observé actuellement chez les
Mammifères. La figure 3.11-C souligne la remarquable identité et la conservation des
sites de fixation de la protéine sur l'ADN chez les deux espèces. Le bricolage de
l'évolution est toujours d'actualité ; F. JACOB souligne que l’évolution « travaille sur ce qui
existe déjà, soit qu’elle transforme un système ancien pour lui donner une fonction nouvelle,
soit qu’elle combine plusieurs systèmes pour en échafauder un autre plus complexe... Il faut
dire que l’évolution opère non à la manière d’un ingénieur, mais d’un bricoleur » (article
« Bricolage », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996).
De cette section sur les mutations géantes, quelques points sont à retenir :
- L'apparition de nouveautés évolutives ne dépend pas tant des mutations
ponctuelles des gènes de structure que de la réorganisation du matériel génétique
par des phénomènes complexes de recombinaisons.
- Les mutations des gènes de régulation et les duplications géniques semblent
constituer deux sources abondantes d'innovations évolutives.
- La diploïdie procure un avantage sélectif important qui favorise une variabilité
génétique considérable.
Les innovations et la stabilité génétiques
On peut se demander comment le patrimoine génétique demeure aussi fiable,
alors que les gènes des Eucaryotes, représentant seulement 20 % de l’ADN, sont
coupés par des introns, noyés au milieu de séquences répétitives et de pseudogènes.
L’expression « ADN poubelle » traduit ce fouillis qu’est l’ADN des Eucaryotes ; l’ADN
des Procaryotes ne comporte qu’un très petit nombre de séquences non codantes et il
semble, de ce fait, plus apte à conserver les caractéristiques génétiques de l’espèce.
C’est pourquoi certains biologistes opposent « l’ADN poubelle » des Eucaryotes et
leurs innovations génétiques à l’ADN compact des Procaryotes et leur stabilité
évolutive. J. GÉNERMONT juge cette position très discutable ; les Bactéries sont toujours
167

capables, en effet, de s’adapter rapidement à de nouvelles conditions de vie par le
fonctionnement, chez certaines d’entre elles, de gènes mutateurs qui ont pour effet
d’augmenter le taux de mutations de tous les gènes, dans une proportion de 1 à 1 000.
Une stabilité morphologique et structurale n'implique pas une adaptabilité figée.
Le vivant possède la faculté d'acquérir des nouveautés génétiques, fruits de
mutations, de duplications, fusions et amplifications géniques qui altèrent ou
perturbent l'information génétique transmise d'une génération à l'autre.
La fixation des innovations est bien discrète, ainsi que l'explique Michel GILLOIS à
propos des enzymes. La liaison enzyme-substrat est spécifique, mais cette spécificité
n'est pas absolue ; des molécules semblables au substrat peuvent également se lier
au site actif enzymatique : « Cette propriété est celle du bruit de fond phénotypique »
(M. GILLOIS, « L'information génétique : du code génétique au message héréditaire »,
Génét. Sél. Évol., 15(2), 1983, p. 315). De nouvelles molécules peuvent ainsi
s'introduire dans une chaîne métabolique, puis, par le biais des mutations, de la
sélection et des duplications, détourner des gènes de leur fonction originelle.
Cependant la stabilité de l'information génétique portée par l'ADN prouve que la
cellule est capable de résister à l'implantation de nouveautés évolutives. Dans le
matériel génétique, deux phénomènes contradictoires s’affrontent en permanence :
modifications par mutations, transpositions ou duplications du patrimoine génétique,
indispensables à l’évolution, et maintien de la permanence du message génétique,
indispensable à la stabilité de l’espèce. Deux mécanismes de résistance atténuent les
modifications de l’ADN, sans pour autant bloquer son évolution :
- Dégénérescence du code génétique (M. GILLOIS, 1983 et 1996) : plusieurs triplets
correspondent parfois à un seul acide aminé. Le nombre total des mutations modifiant
le sens du message génétique est ainsi réduit.
- Constance de la structure tertiaire des protéines malgré des modifications de la
séquence peptidique (M. GILLOIS, 1983 et 1996, et Jean-Loup RISLER, 1996). Cette
caractéristique est universelle, car le repliement d'une chaîne peptidique obéit plus à
des contraintes stéréochimiques qu'à une séquence bien déterminée d'acides aminés.
Le repliement a pour fonction de placer les acides aminés du site actif au bon endroit
de l'espace ; la situation des autres acides aminés dans la séquence polypeptidique
apparaît secondaire, comme l'ont montré diverses expériences qui modifiaient
artificiellement cette séquence : la structure tertiaire résiste à de nombreuses
substitutions, mais aussi à des délétions et à des insertions. La similarité des
propriétés chimiques de certains acides aminés, qui sont, dans une certaine mesure,
interchangeables (cf. supra la section 3.1.1 : « Les méthodes d’appréciation de la
variabilité »), renforce la stabilité de la structure tertiaire.
168

Les mutations et les recombinaisons sont une source inépuisable de nouveautés
géniques qui deviendront des nouveautés évolutives à part entière lors d’une
éventuelle sélection. Mais les mutations et les recombinaisons n’ont pas le monopole
des innovations ; à ce sujet, M. GILLOIS et Noël BONNEUIL écrivent dans leur article
« Évolution acquisitive et dynamiques darwiniennes » (Dictionnaire du darwinisme et
de l'évolution, Paris, PUF, 1996, p. 1469-1470) : « Cette interprétation génique de
l'hérédité, même complétée par les derniers apports de la biologie moléculaire, n'est pas en
mesure de rendre compte de l'hérédité d'une forme, de la morphologie, de l'embryogenèse, de
l'ontogenèse, de la convergence fonctionnelle, d'une fonction physiologique, de la permanence
des structures et de leur flexibilité dans et entre les populations au cours du temps.
Un gène seul n'a pas de sens, il n'en acquiert un que par rapport au système dynamique
phénotypique dans lequel sa fonction s'insère. » (Se reporter également à l’article de
M. GILLOIS : « Les modèles dynamiques de l’évolution », Pour Darwin (dir. P. Tort),
Paris, PUF,1997).
Les nouveautés évolutives apparaissent parfois sans aucune mutation génique
classique. Il suffit que l’ordre de lecture des gènes intervenant dans le développement
ontogénique soit inversé pour que naissent un génotype et un phénotype inconnus.
Phénomènes assez rares, les mutations entraînent à long terme une variabilité
génétique qui est amplifiée rapidement, à chaque génération, par la reproduction
sexuée. L'apparition de celle-ci a constitué une révolution génétique, car elle entretient
une extraordinaire variabilité.
La reproduction sexuée.
Un certain nombre d'organismes notamment des Procaryotes (Bactéries)
ou des Protozoaires (Amibes) se reproduisent uniquement par voie asexuée ; d'autres
comme les Rotifères Bdelloïdes, chez lesquels les mâles n'existent pas ou sont
inconnus, utilisent indéfiniment la parthénogenèse. Cela n'empêche pas ces espèces
d'évoluer. Si la reproduction sexuée n'est pas une nécessité évolutive, elle possède en
tout cas une remarquable efficacité, car elle maintient dans les populations une
extraordinaire variabilité génétique.
La reproduction sexuée se caractérise par trois particularités : l’alternance
de deux générations, une polarité (+)/(-) mâle/femelle qui implique des mécanismes de
reconnaissance et des échanges génétiques.
- L’alternance des générations
169

Chaque cellule de l’une des générations qui renferme un seul jeu de
chromosomes correspond à la phase haploïde, dite à n chromosomes. L’autre
génération comprend deux jeux de chromosomes ; cette phase diploïde est dite à 2n
chromosomes.
Vraisemblablement, les premiers organismes ont été haploïdes ; toute mutation
était soit immédiatement fixée, soit éliminée. La sélection naturelle a favorisé le
passage à la diploïdie : avec deux gènes, les mutations même létales à l'état
hétérozygote sont supportées sans pour autant être éliminées. L'allèle non muté
assure seul, en effet, un fonctionnement normal ou à peu près normal du gène. Il
arrive parfois que l'allèle mutant conservé procure même un bénéfice indirect, comme
dans le cas de l'anémie falciforme ou drépanocytose ou encore sicklémie (sickle
désigne, en anglais, une faucille), caractérisée par des hématies en forme de faucille.
Maladie héréditaire endémique des pays tropicaux, la drépanocytose frappe
surtout les populations d'Afrique noire ou d'origine africaine ; elle est responsable
d'une mortalité infantile estimée à 3 %. Les déformations de l'hématie proviennent
d'une hémoglobine anormale, dite S, incapable d'assurer le transport du dioxygène : la
drépanocytose est donc mortelle à l'état homozygote. L'hémoglobine Hb-S est due à
une mutation génique récessive du polypeptide β, qui ne diffère de la globine normale
que par un acide aminé en position 6 : la glutamine (Q ) remplace la valine (V). La
transmission du gène muté Sk s'effectue comme l'indique le tableau de la figure 3.15.
L'hétérozygotie apporte sans doute certains avantages, car le gène létal Sk ne
disparaît pas ; or, des calculs montrent qu'un gène délétère est éliminé rapidement en
un peu plus d'une dizaine de générations. Les médecins ont trouvé la réponse en
constatant que, dans ces régions intertropicales, les personnes hétérozygotes (A/Sk)
sont plus résistantes à une autre maladie endémique : la malaria ou paludisme, que
les personnes homozygotes (A/A) et, bien sûr, les personnes (Sk/Sk). Le gène Sk
procure donc un avantage sélectif qui est à l’origine de son maintien.
Cette explication classique de l'avantage du diploïde sur l'haploïde est
séduisante, mais elle n'est pas suffisante. Malgré ses « désavantages », l'haploïdie
s'est maintenue chez les Bactéries, mais aussi chez un grand nombre de Végétaux
(Algues, Végétaux non vasculaires), dont le cycle de reproduction comprend une
phase haploïde dominante. Il est difficile de considérer ces Végétaux comme des
exceptions, voire des anomalies.
L’acquisition d’une phase diploïde dominante a deux conséquences :
- Le retour à une phase haploïde dominante est impossible si l’accumulation des
mutations létales est trop importante ; les gènes létaux vont s’exprimer directement,
provoquant la disparition des individus qui les portent.
170

- La viabilité de l’autofécondation est diminuée.
- L’hérédité et la sexualité
La biologie de la reproduction sexuée a préoccupé les lamarckiens et les
évolutionnistes darwiniens du XIXe siècle.
1) Les naturalistes pensaient qu'au cours de la fécondation les caractères paternels
et maternels se mélangeaient pour en former de nouveaux (hérédité mélangeante).
Pour les évolutionnistes, cette conception renfermait un paradoxe gênant, révélé par
Ronald FISHER : le mélange des caractères paternels et maternels aurait dû diminuer
l'hétérogénéité des populations de moitié à chaque génération ; or aucun fait ne le
prouvait. Les biologistes savaient pertinemment que les populations naturelles sont
extrêmement polymorphes et que la variabilité est élevée. Grégor MENDEL, en
démontrant que les déterminants des caractères paternels et maternels restent
individualisés chez les descendants, a résolu le paradoxe. Mais ses travaux n'ont été
repris et connus de tous qu'au début du XXe siècle.
2) Dès 1883, August WEISMANN, zoologiste allemand, a énoncé l’hypothèse de
l'inaltérabilité et de la continuité du plasma germinatif : dès les premiers jours de la vie
embryonnaire, les cellules germinales s'isolent des autres cellules. Cet isolement
précoce empêche tout « caractère acquis », pendant la vie embryonnaire comme
pendant la vie adulte, de franchir la barrière qui protège le plasma germinal. Les
lamarckiens, pour qui le hasard ne peut présider à notre devenir, et les darwiniens de
la première heure se trouvaient ainsi privés d'un de leurs arguments principaux. Mais
le darwinisme, en intégrant d’abord l’hypothèse de A. WEISMANN (rejet de l’hérédité des
caractères acquis, 1883), puis la découverte des travaux de G. MENDEL au début du
XXe siècle, donnera naissance à deux néo-darwinismes successifs et
complémentaires.
- Le brassage allélique
Le brassage allélique interchromosomique concerne, au moment de la migration
polaire, lors des anaphases I et II de méiose, la répartition et la composition de deux
lots chromosomiques. Soit une espèce à 2n = 6 ; selon un calcul simple, mâle et
femelle peuvent produire respectivement 23 spermatozoïdes et 23 ovules différents. Un
couple de cette espèce peut donc avoir 23x23, soit 26 descendants différents. Le
nombre de combinaisons est ici réduit, mais il augmente très vite quand le nombre 2n
croît. Ainsi, dans l'espèce humaine où 2n est égal à 46, où les gamètes sont porteurs
de 23 chromosomes, le nombre de combinaisons est alors de 223x223, soit 246 ou 1014.
171

Le brassage allélique intrachromosomique a lieu au cours de la gamétogenèse
chez les Métazoaires et de la tétrasporogenèse chez les Plantes.
La première division cellulaire entraîne le passage à n chromosomes, réduction
nécessaire pour que, lors de la reproduction sexuée, le nombre de chromosomes
demeure constant d'une génération à l'autre. C'est au début de cette division
réductionnelle (prophase I) que les chromosomes homologues réalisent des échanges
géniques par l'intermédiaire des crossing-over qui forment des figures facilement
reconnaissables en microscopie photonique : les chiasmas.
Les recombinaisons génétiques sont nombreuses et pseudo-aléatoires, car elles
se réalisent toujours dans des zones privilégiées. Il est possible d'estimer le nombre
des recombinaisons au moment de la méiose, c'est-à-dire d'estimer le nombre de
gamètes différents qu'un organisme peut produire. Par exemple, chez l'Homme, le
nombre de gènes est estimé aujourd'hui à 35 000. L'hétérozygotie, évaluée à 30%,
implique que 30% x 35 000 soit 10 500 gènes possèdent au moins deux allèles. Si le
nombre de combinaisons entre deux gènes ayant deux allèles chacun est de 22, avec
n gènes il est de 2n. Si l’hétérozygotie, très faible, est de 10%, il y a donc 10% x 3 500
soit 3 500 gènes qui possède au moins deux allèles ; dan ce cas, le nombre total de
génotypes susceptibles d'exister pour un seul gamète produit par un même individu
est 23 500 soit 101 050, en admettant que 210 (1 024) est à peu près égal à 103. Ce chiffre
dépasse toute imagination : il serait supérieur au nombre estimé d'atomes qui
composent l'univers, évalué « seulement » aux alentours de 1080 ou 1090.
De plus, dans les brassages génétiques, le hasard préside à la rencontre des
gamètes. La loterie génétique, qui gouverne la constitution de notre génotype, se
résume dans la phrase suivante : les possibles sont infinis, le réel est unique. Dans
l’hypothèse la plus basse (3 500 gènes hétérozygotes), la probabilité d'existence de
chacun d'entre nous est à peu près égale à 1/101 050 x 1/101 050 soit 1/102 100, c'est-à-dire
statistiquement quasi nulle.
L'hérédité mendélienne sous-estime le rôle des interactions géniques. Mais elle
connaît les rapports de dominance, qui sont une interaction entre allèles homologues,
et les relations épistatiques, qui concernent des interactions entre allèles non
homologues. Un exemple historique de dominance est fourni par les Pois verts et
jaunes étudiés par G. MENDEL : le phénotype vert des hybrides masque le phénotype
jaune. Un exemple d'épistasie existe chez le Chat, qui présente deux allèles, « a »
récessif et autosomal et « O » dominant et lié au chromosome X : le génotype (a/a)
détermine un pelage de couleur uniforme non rayé, l'allèle « O » détermine un pelage
roux toujours tigré. Si l'animal possède, par exemple, le génotype (a/a, O/O), son
pelage sera roux et rayé, l'allèle « O » masquant l'allèle non homologue « a ».
172

- La sélection sexuelle
Elle se rapporte à l’avantage reproductif acquis par certains individus, souvent
des mâles, porteurs de caractères sexuels secondaires attractifs pour ceux du sexe
opposé.
La sélection sexuelle porte sur les caractères exprimés généralement chez le
mâle : « L’égale transmission des caractères aux deux sexes est la forme d’hérédité la plus
courante, du moins chez les animaux qui ne présentent pas de différences sexuelles fortement
marquées, et en fait chez un grand nombre de ces derniers. Mais, assez communément, des
caractères se transmettent exclusivement au sexe chez lequel ils sont d’abord apparus. »
(DARWIN, La filiation de l’homme et la sélection liée au sexe, Paris, Syllepse, 1999,
p. 327).
« La sélection sexuelle agit d’une manière moins rigoureuse que la sélection naturelle »
(ibid., p. 323). Elle favorise la reproduction d’individus particuliers, sans provoquer la
disparition des autres, « puisque des mâles sans armes, sans ornements et sans attraits,
réussiraient tout aussi bien dans la bataille pour la vie, et parviendraient à laisser une
nombreuse progéniture, sans la présence de mâles mieux doués » (ibid., p. 307). « Pour ce
qui est des structures acquises par l’action de la Sélection ordinaire ou Naturelle, il y a dans
la plupart des cas, tant que les conditions de vie restent les mêmes, une limite à la quantité de
modifications avantageuses liées à certaines fins spéciales… » (ibid., p. 323). La sélection
naturelle ne favorise pas une variabilité aussi étendue, car « il y a, dans la plupart des
cas, tant que les conditions d’existence restent les mêmes, une limite à l’étendue des
modifications avantageuses qui peuvent se produire dans un but déterminé » (ibid., p. 323).
La sélection sexuelle et la sélection naturelle ne sont pas globalement
antinomiques, tout en pouvant l’être dans certains cas (volume et masse de la parure
de noce empêchant le vol chez les mâles de certains Oiseaux livrés ainsi aux
prédateurs). Les rapports entre les deux types de sélection peuvent engendrer trois
cas principaux :
- Les caractères sont uniquement liés à la sexualité : ils donnent peu de prise à la
sélection naturelle.
- Les caractères sexuels « sont nécessaires pour les autres fins de la vie, comme c’est
généralement le cas, ils auront été développés par la Sélection Naturelle » (ibid., p. 305). Ils
sont adaptatifs ; l’action de la sélection sexuelle s’ajoute à celle de la sélection
naturelle. « Mais dans la plupart des cas de ce genre, il est impossible de distinguer les effets
de la Sélection Sexuelle de ceux de la Sélection Naturelle » (ibid., p. 306).
- Les caractères sexuels atteignent un tel développement, comme les bois
démesurés du Cerf fossile Cervus megaceros, qu’ils entraînent un handicap dans la
173

lutte pour l’existence. La sélection sexuelle est anti-adaptative ; la sélection naturelle
s’oppose en principe à ce que les caractères sexuels subissent un développement tel
qu’ils deviennent préjudiciables aux individus qui les portent, car « ils doivent leur être
hautement préjudiciables en les portant à une dépense excessive de leurs capacités vitales ou
en les exposant à quelque grand danger » (ibid., p. 323).
L’énorme variabilité génétique peut être restreinte par un phénomène, la dérive
génique fortuite, pressentie dès le XIXe siècle et mise en évidence par des généticiens-
mathématiciens de la première moitié du XXe siècle.
3.1.3 - La variabilité et la dérive génique fortuite
De temps en temps, des sous-populations locales présentent des caractères
originaux sans que les mutations, la sélection naturelle ou les migrations en soient
responsables ; un nouveau facteur intervient dans l'évolution des populations. Ce
facteur avait déjà été pressenti, dès 1872, par J. GULIK lorsqu'il avait découvert que
l'importante diversité d'Escargots des îles Hawaï du genre Achatinella était
indépendante des variations des facteurs de l'environnement. Maxime LAMOTTE (1951)
et C. GOODHART (1962) retrouvent le même phénomène dans des populations
d'Escargots des haies, Cepaea nemoralis, dont de nombreuses colonies sont parfois
installées le long des berges d'une rivière. Étant donné leur proximité, ces colonies
vivent dans un environnement homogène. Mais elles sont isolées génétiquement et
elles présentent les unes par rapport aux autres des phénotypes particuliers, dus au
fonctionnement d'une dizaine de couples d'allèles différents. Cette variabilité se
rapporte principalement à des caractéristiques moléculaires, et non pas seulement à
des caractères morpho-anatomiques.
En 1922, Ronald FISHER a publié un article consacré aux chances de survie d'un
gène dans une population naturelle. Dans sa conclusion, l'intervention du hasard,
future dérive génique fortuite, joue un rôle mineur dans l'évolution des populations ; un
nouveau gène a beaucoup plus de chances de disparaître que de s'implanter.
Sewall WRIGHT (1929) a reconnu l'existence de cette dérive génique. Il montre que son
action est d'autant plus importante que l'effectif de la population est réduit, le taux de
mutations faible et les migrations réduites : la dérive génique fortuite aboutit à la perte
imprévisible d'un certain nombre de gènes et à l'homogénéisation génétique de la
population. Cette homogénéisation peut être à l'origine de phénotypes originaux qui
risquent de disparaître, à moins qu'il n'y ait migration. C'est pourquoi l'étude des
petites populations et des isolats est fondamentale dans ses travaux (voir les sections
3 3 2 : « La spéciation par révolution génétique », et 4 2 3 : « La théorie synthétique,
174

les interactions géniques »). Dans les isolats, le nombre total des différents génotypes
est limité à celui des individus fondateurs, toujours peu nombreux. Ces génotypes ne
sont pas le fidèle reflet de ceux de la population dont ils se sont séparés. Cette
différence due au hasard peut très bien s'accentuer par le seul fait de la dérive
génique fortuite ; au bout d'un temps variable, la colonie devient alors de plus en plus
homogène et se stabilise autour d'un type donné en perdant progressivement sa
variabilité génétique. L'ensemble de ces phénomènes contingents, non liés à la
sélection naturelle, caractérise la dérive génique fortuite. La diminution du nombre de
fécondations exogames, c’est-à-dire réalisées en dehors du groupe, baisse également
et elle participe à la perte de variantes génétiques spécifiques.
La population des Kel Kummer, Touaregs du sud saharien, à l'est de la
république du Mali, offre un exemple typique d'isolat qui s'est détaché de la société
Touareg. Les Kel Kummer présentent une rare homogénéité génétique, car leur taux
de consanguinité est très élevé. De surcroît, le nombre d'individus fondateurs,
fournissant le patrimoine génétique originel de la tribu, a été très faible : vingt-deux
individus ont apporté les 9/10es des gènes du groupe qui vit actuellement.
La dynamique des phénomènes génétiques constitue le mécanisme fondamental
qui est à l’origine des nouveautés évolutives, mais les phénomènes génétiques et les
innovations sont soumis à un deuxième mécanisme évolutif, la sélection naturelle.
Bibliographie de la section 3.1
Livres
BLANC M. , Les Héritiers de Darwin, Paris, Seuil, 1990.
BONIS (de) L. , L'Évolution dans sa réalité et ses diverses modalités, Paris, Masson, 1988.
DARWIN Ch. : La filiation de l’homme et la sélection liée au sexe, traduction de Michel Prum, publié sous la direction de Patrick Tort, Paris, Syllepse, 1999.
DEVILLERS Ch. et MAHÉ J. , Mécanismes de l'évolution animale, Paris, Masson, 1980.
DOBZHANSKY T. , L'Hérédité et la nature humaine, Paris, Flammarion, 1969.L'Homme en évolution, Paris, Flammarion, 1969.
GÉNERMONT J. , Les Mécanismes de l'évolution, Paris, Dunod, 1979.
LEWONTIN R. , Nous ne sommes pas programmés : génétique, hérédité, idéogie, Paris, La découverte, 1985.
175

LI W. et GRAUR D. , Fundamentals of Molecular evolution ; Sunderland (Massachusetts, USA), Sinauer Associates Inc., 1991.
MAYR E. , Histoire de la biologie. Diversité, évolution et hérédité, Paris, Flammarion, 1989.
OHNO S. , Evolution by gene duplication, Berlin, Springer-Verlag, 1970.
OHTA T. , Evolution and Variation of Multigene Families, Sunderland (Massachussetts, USA), Sinauer Associates Inc., 1983.
PETIT C. et PRÉVOST G. , Génétique et évolution, Paris, Hermann, 1970.
PETIT C. et ZUCKERKANDL É. , Evolution, Paris, Hermann, 1976.
SOLIGNAC M. , PÉRIQUET G. , ANXOLABÉHÈRE D. et PETIT C. , Génétique et évolution (t1 : La variation, les gènes dans les populations, t2 : L’espèce, l’évolution moléculaire), Paris, Hermann, 1995.
Articles
ANXOLABÉHÈRE D., KIDWELL M. et PÉRIQUET G. , « L'histoire d'une invasion génétique », La Recherche, n° 215, novembre 1989.
AYALA F. , « Les mécanismes de l'évolution », Pour La Science, n° spécial : L'évolution, Paris, Belin, 1980.
AYALA F. , « Variation génétique et évolution », La Recherche, n° 81, septembre 1977.
BARRIEL V. , « Mythes et réalités de l'approche génétique », La Recherche, n° 277, juin 1995.
BENNE R. et VAN der SPEK H. , « L'editing des ARN », La Recherche, n° 245, juillet-août 1992.
BLANC M. , « Grégoire Mendel : la légende du génie méconnu », La Recherche, n° 151, janvier 1984.
CHOURROUT D. , « L'amélioration génétique des poissons », La Recherche, n° 180, septembre 1986.
DANCHIN A. et SLONIMSKI P. , « Les gènes en morceaux », La Recherche, n° 155, mai 1984.
DELSOL M. , « Évolution en mosaïque », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
DELSOL M. et FLATIN J. , « Coaptation », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
DEUTSCH J. , « Les gènes homéotiques, traceurs de l’évolution chez les Métazoaires », Pour Darwin (dir. P. TORT), Paris, PUF, 1997.
176

DEVILLERS Ch. , « Atavisme », « Hérédité mélangeante », « Hérédité particulaire », « Mutation homéotique », « Mutation systémique », « Orthogenèse », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
DREUIL D. , GENERMONT J. et GILLOIS M. , « Supergène », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
DUBOULE D., DOLLE P. et GAUNT S. , « Les gènes du développement des Mammifères », La Recherche, n° 219, mars 1990
DUTRILLAUX B. , « Les chromosomes des primates », La Recherche, n° 127, novembre 1981.
GASC J.-P. , « Évolution des organismes », « Tendances évolutives et progrès », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GASSER F. : « Polymorphisme dans les populations animales », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GEHRING W. , « De la mouche à l'homme, un même supergène pour œil », La Recherche, n° 280, octobre 1995.
GÉNERMONT J. , « Les critères caryologiques de l’espèce », dans Les Problèmes de l’espèce, Paris, Société Zoologique de France, tome III, 1980 ; « Mutation réverse », « Transposons », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996, « Les yeux sont-ils des miracles de l’évolution ? », Pour Darwin (dir. P. TORT), Paris, PUF, 1997.
GÉNERMONT J. et DEVILLERS Ch. , « Allèle », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GILLOIS M. , « L'information génétique : du code génétique au message héréditaire », Génétique, Sélection, Évolution, 15(2), p. 299-322, 1983, « Consanguinité II », « Crossing-over », « Évolution du chromosome », « Héritabilité », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996 ; « Les modèles dynamiques de l’évolution », Pour Darwin (dir. P.TORT), Paris, PUF, 1997.
GILLOIS M. et BONNEUIL N. , « Évolution acquisitive et dynamiques darwiniennes », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GILLOIS M. et GÉNERMONT J. , « Épistasie », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GOUYON P.-H., MAURICE S., REBOUD X. et TILL-BOTTAND I. , « Le sexe, pour quoi faire ? », La Recherche, n° 250, janvier 1993.
GUILLE-ESCURET G. , « Orthogenèse et sélection naturelle », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
GUY Y. et DEVILLERS Ch. , « Dérive génétique », « Mutation », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
HÉLÈNE C. , « Les structures de l'ADN », La Recherche, n° 155, mai 1984.
ISRAËL A. , « Contrôle de l'activité des gènes », La Recherche, n° 164, mai 1984.
177

JACOB F. , « Bricolage de l'évolution », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
LOVTRUP S. , « Macromutation », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
LOWENSTEIN J. , « La génétique des fossiles », La Recherche, n° 148, octobre 1983.
MONDELLA F. : « Hérédité », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
MONTALENTI G. , « Génétique », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
OHNO S. , « L'évolution des gènes », La Recherche, n° 107, janvier 1980.
PERRIER J.-J. , « Les déguisements de la plasmodie », La Recherche, n° 281, novembre 1995.
PETIT M.-A. , « L'édition des ARN, un message bien caché », La Recherche, n° 217, janvier 1990
RISLER J.-L. , « Évolution des protéines », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
SCUDO F. , « Sexualité et évolution », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
SHAPIRO J. , « Les gènes sauteurs », La Recherche, n° 81, septembre 1977.
TADDEI F. et RADMAN M. , « L'évolution de l'ADN », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
TAVLITZKI J. , « Des pois de Mendel à la génétique moléculaire », La Recherche, n° 155, mai 1984.
TEMIN H. , « L'origine des provirus », La Recherche, n° 152, février 1984.
TORT P. , « Atavisme (théorie darwinienne de l') », « Femelles (choix exercé par les) », « Variation/variabilité dans l'oeuvre de Darwin », Dictionnaire du darwinisme et de l'évolution, Paris, PUF, 1996.
178